
ogia N 2010 -...
93
Programa de Pós-Graduação em Ecologia Universidade Federal do Rio Grande do Norte ESEC - Seridó 2010 Curso de Campo PPG Ecologia UFRN
Transcript of ogia N 2010 -...

Programa de Pós-Graduação em Ecologia Universidade Federal do Rio Grande do Norte
ES
EC
- S
eri
dó
20
10
Cu
rso
de
Ca
mp
o P
PG
Eco
log
ia
UF
RN

2 Curso de Campo PPG Ecologia, UFRN, 2010. Caatinga, ESEC Seridó
Conteúdo
Conteúdo Página
Agradecimentos 4
Sobre o Curso de Campo 5
A Caatinga da ESEC Seridó 9
Parte I Projetos com orientação
Projetos orientados 1
VIGOR DE Bauhinia cheilantha E SUA VULNERABILIDADE A HERBÍVOROS EM
UMA ÁREA DE CAATINGA
12
A ATIVIDADE DE TERMORREGULAÇÃO DE DUAS ESPÉCIES SINTÓPICAS DE
Troidurus (SQUAMATA, TROPIDURIDAE) EM UMA ÁREA DE CAATINGA NO
RIO GRANDE DO NORTE
15
INFLUÊNCIA DE RIO TEMPORÁRIO SOBRE A TAXOCENOSE DE COLLEMBOLA
EM UMA ÁREA DE CAATINGA DO NORDESTE BRASILEIRO
19
CONVERGÊNCIAS ADAPTATIVAS EM RESPOSTA À PLUVIOSIDADE ENTRE
PLANTAS BRASILEIRAS E AUSTRALIANAS
22
Projetos orientados 2
CARACTERIZAÇÃO DE COMUNIDADES DE COLÊMBOLOS EM ÁREAS COM
DIFERENTES USOS DO SOLO DA CAATINGA
27
FUGA ANTI-PREDADOR DE Tropidurus hispidus E Tropidurus semitaeniatus
(TROPIDURIDAE) EM UMA ÁREA DE CAATINGA
32
INFLUÊNCIA DO EFEITO DE BORDA NA RIQUEZA E ESTRUTURA DE PLANTAS
LENHOSAS EM UMA ÁREA DE CAATINGA NO RIO GRANDE DO NORTE
36
INFLUÊNCIA DO USO DO SOLO SOBRE A COMPOSIÇÃO DA TAXOCENOSE DE
FORMIGAS EM DUAS ÁREAS DE CAATINGA
41
Projetos orientados 3
[Digite uma
citação do
documento
ou o resumo
de uma
questão
interessante.
Você pode
posicionar a
caixa de
texto em
qualquer
lugar do
documento.
Use a guia
Ferramentas
de Caixa de
Texto para
alterar a
formatação
da caixa de
texto da
citação.]

3 Curso de Campo PPG Ecologia, UFRN, 2010. Caatinga, ESEC Seridó
INTERAÇÃO TRITRÓFICA EM Mimosa hostilis: HERBÍVOROS PODEM SER
BENÉFICOS PARA AS PLANTAS?
46
NINHOS ABANDONADOS DE TÉRMITAS POSSUEM UMA FAUNA PECULIAR DE
INVERTEBRADOS: UM ESTUDO COM Constrictotermes cyphergaster NA
CAATINGA
51
ODOR DA FUGA: EFEITO DA DETECÇÃO DE PREDADORES POR Seira sp.
(COLLEMBOLA; ENTOMOBRYIDAE) 55
SELEÇÃO DE MICROHABITAT POR Cnemidophorus ocellifer EM ÁREA DE
CAATINGA, SERRA NEGRA DO NORTE, RN 58
Parte II - Projetos livres
DEFESA DE ESTRUTURAS REPRODUTIVAS DE Macroptilium lathyroides (L.)
(FABACEAE) POR FORMIGAS Camponotus SP. EM UMA ÁREA DE CAATINGA,
NORDESTE DO BRASIL
65
PADRÃO DE DISTRIBUIÇÃO ESPACIAL DE Caesalpinia pyramidalis Tul.
(FABACEAE) e Aspidosperma pyrifolium Mart. (APOCYNACEAE) EM UMA
ÁREA DE CAATINGA, NORDESTE DO BRASIL
70
A ATIVIDADE DE ODONATOS (ARTHROPODA: HEXAPODA: INSECTA) VARIA
AO LONGO DO DIA EM UMA CAATINGA NO NORDESTE DO BRASIL? 75
EFEITO DA ALTITUDE SOBRE A DIVERSIDADE DE COLLEMBOLA
(ARTHROPODA: HEXAPODA) EM UMA ÁREA DE CAATINGA NO RIO GRANDE
DO NORTE
81
EFEITO DA QUALIDADE DO HABITAT E DENSIDADE DE PREDADORES SOBRE A
TAXOCENOSE DE COLLEMBOLA (ARTHROPODA: HEXAPODA)
87

4 Curso de Campo PPG Ecologia, UFRN, 2010. Caatinga, ESEC Seridó
AGRADECIMENTOS
Aos professores André Megali e José Luiz de Attayde, Coordenadores do Programa de
Pós-Graduação em Ecologia (PPGE) da Universidade Federal do Rio Grande do Norte,
por todo apoio dispensado para a realização desta segunda versão da disciplina
Ecologia de Campo; Ao Sr Juan Marcelo de Oliveira e ao Sr George S. Batista, Gestores
da Estação Ecológica do Seridó-RN, pelo apoio e hospitalidade na Unidade; Ao Prof
Adalberto Varela e ao Sr Carlos Varela Freire, pelo auxílio nas várias etapas da
realização deste Curso; Ao SISBIO pela rapidez na liberação da autorização (número:
23310-1) para a realização da disciplina; À Sra Ana Rúbia da Silva pelas deliciosas
refeições e compreensão.

5 Curso de Campo PPG Ecologia, UFRN, 2010. Caatinga, ESEC Seridó
O CURSO DE CAMPO
O Curso de Campo de Ecologia da
Caatinga é parte da disciplina Ecologia de
Campo do Programa de Pós-Graduação em
Ecologia da Universidade Federal do Rio
Grande do Norte.
O objetivo do curso é treinar
alunos de Pós-Graduação em atividades
acadêmicas in loco. Os alunos são
introduzidos a técnicas de coleta e design
de experimentos, assim como análise de
dados e confecção de artigos científicos.
O curso tem a duração de oito dias
e é organizado em projetos de 24 horas
com orientação, onde professores sugerem
um problema que deve ser solucionado em
24h. Nesta atividade, os alunos são
apresentados a um problema ecológico e
devem coletar, analisar e apresentar os
resultados obtidos em um prazo de 24
horas. Uma segunda atividade é o projeto
sem orientação de 48 horas. Nesta
atividade os alunos elaboram uma questão
ecológica e possuem um prazo de 48 horas
para a coleta de dados necessária para
respondê-la.
Respeitando a organização do
curso, este livro é organizado de forma que
os projetos orientados (três por grupo em
2010) são seguidos dos projetos sem
orientação.
Esperamos que o levantamento de
dados obtido ao longo dos cursos pelos
alunos ajude a conhecer melhor o bioma
da Caatinga e a ESEC Seridó.

6 Curso de Campo PPG Ecologia, UFRN, 2010. Caatinga, ESEC Seridó
DOCENTES
Adriana M. Almeida Alexandre Vaconcellos
Carlos Roberto Fonseca
PROFESSORES CONVIDADOS
Bruno Bellini Gindomar Gomes Santana
DOUTORANDOS
Cristiane Gouvêa Fajardo Vinícius de Avelar São Pedro

7 Curso de Campo PPG Ecologia, UFRN, 2010. Caatinga, ESEC Seridó
MESTRANDOS
Allan Torrecilla Batista Brunno Freire D. Oliveira Daniel de Oliveira
Diego de Medeiros Bento Fabrício R. Camacho Françoise Dantas de Lima
Gina Rodo Mantilla Heitor Bruno A. Souza Igor Galvão de Britto

8 Curso de Campo PPG Ecologia, UFRN, 2010. Caatinga, ESEC Seridó
Izabel Maria M. Souza Karol Lyncoln B. O. França Leonardo H. Teixeira Pinto
Maíra Fontes Manzan Monalisa R. O. Silva Priscila Lins P. F. Amaral
Rodrigo Xavier Soares Uirandé Oliveira Costa

9 Curso de Campo PPG Ecologia, UFRN, 2010. Caatinga, ESEC Seridó
A CAATINGA DA ESEC SERIDÓ
O bioma da Caatinga é considerado
o único bioma exclusivo do Brasil, cobrindo
734.478 km2 na região Nordeste e
ocorrendo em manchas no norte de Minas
Gerais (Scardua, 2004). O bioma Caatinga é
caracterizado por um clima quente e semi-
árido, fortemente sazonal, com menos de
1000mm de chuva anuais, concentrados
fortemente em um período de três a seis
meses. Os totais de chuva variam muito de
ano para ano e, em intervalos de dez a
vinte anos, caem a menos de metade da
média (podendo levar até cinco anos),
fenômeno conhecido como “seca” (Velloso
et al., 2002).
Os ambientes da caatinga são
extremamente diversos, proporcionados
por um mosaico de tipos de vegetação, em
geral caducifólia, xerófila e espinhosa,
dependendo do mosaico de solos e
disponibilidade hídrica (Velloso et al.,
2002). Os diversos tipos de Caatinga
estendem-se também por regiões mais
altas e de relevo variado, incluindo
caatinga arbustivo arbórea, mata seca,
mata úmida, carrasco e formações abertas
com dominância de cactáceas e
bromeliáceas.
A Estação Ecológica do Seridó
encontra-se localizada na ecorregião
caracterizada como Depressão Sertaneja
Setrentional (Velloso et al., 2002), região
essa que engloba a maior parte dos
estados da Paraíba, Rio Grande do Norte e
Ceará e prolonga-se até uma faixa no
estado do Piauí. Dentre as áreas do bioma
Caatinga, esta ecorregião é uma das mais
impactadas pela ação antrópica, mas
entretanto possui poucas áreas protegidas
em termos de número e área total.
A região do Seridó apresenta
relevo suave, com solos pedregosos e mais
raros que o restante da região. A altitude
varia de 100 a 400m, com elevações de 400
a 700m. A principal característica desta
região é a irregularidade pluviométrica,
com pluviosidade altamente concentrada e
podendo chegar a 10 meses secos ao ano.
É uma área com longa história de
perturbação, usada há longo tempo por
pecuária, assim como mineração (Velloso,
2002).
A região do Seridó é considerada
um dos centros críticos de desertificação
no país. A região do Seridó tem longo
histórico de degradação, com relatos de
queimadas feitas por fazendeiros para
defender suas terras e abrir caminho na
caatinga fechada na década de 1920
(Scardua, 2004). Atualmente existe uma
grande pressão de olarias na região que
demandam uma grande quantidade de
lenha.
A Estação Ecológica do Seridó
possui 1,163 ha e encontra-se no município
de Serra Negra do Norte, vizinho à Caicó, a
terceira cidade em tamanho e número de
habitantes do estado do Rio Grande do
norte. A ESEC-Seridó foi criada em 1982 em
uma área antes pertencente à fazenda
Solidão. Em sua área eram desenvolvidas
atividades de pecuária e cultivo de
algodão. Dentre as perturbações anteriores
à instalação da reserva encontram-se a
construção de um açude, extração de
madeira e introdução de espécies exóticas
como algaroba e leucena (Simon, 2004).

10 Curso de Campo PPG Ecologia, UFRN, 2010. Caatinga, ESEC Seridó
Referências
Scardua, F.P. (coord) 2004. Plano de manejo da
ESEC Seridó. MMA.
Simon, M.F. 2004. Avaliação ecológica rápida
para o plano de manejo da Estação
Ecológica do Seridó, RN. MMA.
Velloso, A.L., Sampaio, E.V.S.B., Pareyn, F.G.C.
2002. Ecorregiões – Proposta para o
bioma caatinga. Resultado do
Seminário de Planejamento
Ecorregional da Caatinga, Aldeia-PE.
MMA. Nature Conservancy do Brasil e
PNE.

11 Curso de Campo PPG Ecologia, UFRN, 2010. Caatinga, ESEC Seridó
Pro
jeto
s co
m o
rie
nta
ção

12 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
VIGOR DE Bauhinia cheilantha E SUA VULNERABILIDADE A HERBÍVOROS
EM UMA ÁREA DE CAATINGA
BRUNNO FREIRE DANTAS DE OLIVEIRA, CRISTIANE GOUVÊA FAJARDO, FABRÍCIO RODRIGUES
CAMACHO, UIRANDÉ OLIVEIRA COSTA
Introdução
A relação entre herbívoros e suas
hospedeiras é um assunto de bastante
interesse nas ciências ecológicas (Coley, 1983;
Strong et al., 1984; Lewinsohn et al., 2005). Os
fatores que influenciam a escolha e
preferência de plantas por herbívoros
estimulam um dos campos mais férteis nos
estudos de interação inseto-planta (Williams
& Cronin, 2004).
Algumas hipóteses se destacam e são
bastante utilizadas para explicar a distribuição
de herbívoros entre plantas, como a hipótese
do estresse (White, 1984) e a do vigor das
plantas (Price, 1991 e 1992). A primeira
hipótese propõe que plantas sob estresse são
mais preferidas por herbívoros devido a uma
maior disponibilidade de nutrientes e a um
menor investimento em metabólitos anti-
herbivoria. Já a hipótese do vigor da planta,
proposta por Price (1991), sugere que
herbívoros são mais abundantes em plantas
mais vigorosas devido a sua maior qualidade
nutricional. O vigor é definido como qualquer
individuo ou parte da planta que cresce mais
rápido ou é maior (Gonçalves-Alvin et al.
1999).
O presente trabalho objetivou analisar
se o vigor de Bauhinia cheilantha influencia as
taxas de herbivoria entre indivíduos ou no
mesmo individuo, utilizando como indicativos
de vigor o tamanho, o número de folhas e a
área foliar média.
Material e Métodos
O trabalho foi realizado na Estação
Ecológica do Seridó (6º35’S e 37º15’W),
localizada na ecorregião da Depressão
Sertaneja Setentrional do estado do Rio
Grande do Norte. Foi delimitada uma área de
100 m², próxima a trilha da caveira, em um
local onde Bauhinia cheilantha ocorre em
grande abundância. Nesse local, todas as
plantas maiores que um metro (classificadas
como maduras) foram selecionadas para
análise, bem como a plântula que estava mais
próxima a cada uma destas. Dessa maneira, 17
plantas maduras e 17 plântulas foram
amostradas e medidas suas alturas utilizando
trenas.
Foram escolhidas 15 folhas das
plantas maduras, aleatoriamente, e todas as
folhas das plântulas para análise das variáveis
indicativas de vigor (área foliar e número de
folhas). Com o auxílio de réguas foram
medidas a largura e o comprimento (cm) de
cada folha. A estimativa da área foliar foi
obtida através da multiplicação da largura
pelo comprimento da nervura central da
folha. Para estimar o número de folhas por
planta madura sorteamos aleatoriamente um
dos galhos e contamos o número de folhas
presentes e multiplicamos pelo número de
galhos total. Para as plântulas todas as folhas
foram contadas. O grau de herbivoria foi
estimado em 5 classes: classe 1 (0%), classe 2
(1-25%), classe 3 (26-50%), classe 4 (51-75%),
classe 5 (mais de 76 %). Para análises, foi
tirada a média aritmética das classes de

13 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
herbivoria, do número de folhas e da área
foliar para cada indivíduo.
Os dados foram analisados utilizando
o módulo GLM (General Linear Model) onde
foi verificado o efeito da média da altura,
número de folhas e área da folha sobre a taxa
de herbivoria. Todas as análises foram feitas
no SYSTAT v.12.
Resultados
A altura média + DP das plantas
analisadas foi de 3,37 + 1,1 m para plantas
maduras e de 0,13 + 0,03 m para plântulas. A
média de folhas por planta foi de 255 + 454,61
nas plantas maduras e de 43 + 1,28 nas
plântulas.
Comparando entre indivíduos, as taxas
de herbivoria não foram influenciadas pelo
tamanho das plantas, número de suas folhas e
tamanho das folhas (F = 0,161; gl = 3,29; P =
0,992).
Entre plantas maduras e plântulas,
também não houve diferenças significativas
nas taxas de herbivoria (F = 0,678; gl = 1,32; P
= 0,416) (Figura 1). As taxas de herbivoria
entre plantas maduras e plântulas não foram
afetadas pelo tamanho das plantas, número
de suas folhas e tamanho das folhas (F =
0,874; gl = 3,12; P = 0,484).
As taxas de herbivoria foram
diferentes entre as folhas do mesmo indivíduo
(F = 3,517 ; gl = 33,264 ; P < 0,05). Ou seja,
dentro do mesmo indivíduo existem folhas
que são menos consumidas que outras.
Entretanto, o tamanho das folhas não foi
determinante na variação das taxas de
herbivoria (F = 2,526 ; gl = 1,287 ; P = 0,113)
(Figura 2).
Figura 1. Média e desvio padrão de herbivoria para
plantas maduras e plântulas de Bauhinia
cheilantha na ESEC-Seridó.
Figura 2. Relação entre taxa de herbivoria (LOG10)
e área foliar de Bauhinia cheliantha na ESEC-
Seridó.
Discussão
Neste estudo há evidências de que os
indicativos de vigor analisados não explicam
as variações nas taxas de herbivoria sofridas
Plântula Madura

14 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
por plantas e plântulas de Bauhinia cheilantha
na área estudada. Assim, a hipótese do vigor
da planta não foi corroborada. Estes
resultados foram contrastantes com um
estudo utilizando uma espécie de herbácea na
mesma área (Viana et al. 2009). No entanto,
um outro estudo efetuado na caatinga de
Pernambuco testando vigor de uma
Euphobiacea e a performance de um inseto
galhador também não corroborou esta
hipótese (Magalhães, 2008).
Várias pesquisas suportam as
predições propostas pela hipótese do vigor da
planta, mas muitos outros encontraram fracas
evidências que há preferência por plantas
mais vigorosas, havendo alguns grupos de
insetos que corroboram mais fortemente a
estas predições que outros (Cornelissen et al
2008). Santos & Silveira (2007) observaram o
efeito do vigor plantas do gênero Bauhinia sob
a guilda de insetos galhadores.
Portanto, torna-se necessário a
execução de mais estudos que testem esta
hipótese considerando o vigor de Bauhinia
cheilantha sobre grupos específicos de insetos
herbívoros.
Agradecimentos
Agradecemos a todos aos professores e colegas do
PPG em Ecologia da UFRN, especialmente a
professora Adriana Monteiro de Almeida e Carlos
Roberto Fonseca pelos insights e contribuições ao
nosso desenvolvimento intelectual científico. À
equipe da ESEC-Seridó pelo acolhimento e apoio
durante o curso de campo.
Referências
Coley P. D. 1983. Herbivory and defensive
characteristics of tree species in a lowland
tropical forest. Ecological monographs 53,
209–33
Cornelissen, T. G., Fernandes, G. W., Vasconcellos-
Neto, J. 2008. Size does matter: variation
in herbivory between and within plants
and the plant vigour hypotesis. Oikos. 117,
1121-1130.
Gonçalves-Alvim, S. J., Faria, M. L., Fernandes,
G.W. 1999. Relationships between four
neotropical species of galling insects and
shoot vigor. � Anais da Sociedade
Entomológica do Brasil 28, 147-155.
Lewinsohn, T.M. 2005. Insects on Plants: Diversity
of herbivore assemblages revisited.
Annual Review of Ecology and Systematics
36, 597-620.
Magalhães, C.H.P. 2008. Vigor de Cnidoscolus
quercifolius Pohl. (Euphorbiaceae) e
performance de um inseto galhador em
uma área de Caatinga, Parnamirim, PE. In:
Leal, I.R., Almeida-Cortez, J. e Santos, J.C.
Ecologia de Caatinga – Curso de Campo
2008. Ed. UFPB.
Santos, J. C., Silveira, F. A. O., Fernandes, G. W.
2008. Long term oviposition preference
and larval performance of Schizomyia
crocapillata (Diptera: Cecidomyiidae) on
larger shoots of its host plant Bauhinia
brevipes (Fabaceae). Evolutionary Ecology
22, 123-137.
Viana, M., Oliveira, S. R., Torres, D.F., Maciel, L. V.
B., Vasconcelos, V. H. F. 2009. A hipótese
do vigor de planta (HPV) é válida para uma
herbácea do semi-árido brasileiro? In:
Vasconcellos, A. (ed.) Ecologia de Campo
Caatinga 2009 – PPG Ecologia, UFRN.

15 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
A ATIVIDADE DE TERMORREGULAÇÃO DE DUAS ESPÉCIES SINTÓPICAS DE
TROPIDURUS (SQUAMATA, TROPIDURIDAE) EM UMA ÁREA DE CAATINGA NO
RIO GRANDE DO NORTE
ALLAN TORRECILLA BATISTA; FRANÇOISE DANTAS DE LIMA; VINÍCIUS AVELAR SÃO PEDRO; MONALISA
RODRIGUES OLIVEIRA DA SILVA & RODRIGO XAVIER SOARES
Introdução
Grande parte da atividade diária de
várias espécies de lagartos é gasta em
interações com o ambiente térmico, pois a
temperatura é um dos fatores limitantes na
ecologia desses animais ectotérmicos (Rocha
et al. 2009). Em termos comportamentais, a
regulação da temperatura corpórea envolve
deslocamentos entre locais mais frescos e
mais quentes (Vitt et al. 2005).
Espécies de lagartos que ocupam um
mesmo local podem repartir um sítio de
termorregulação ocupando diferentes porções
desse sítio ou utilizando-o em períodos
distintos (Pianka 1973).
Tropidurus hispidus (Spix 1825) e T.
semitaeniatus (Spix 1825) são espécies de
lagartos heliófilos que ocorrem em sintopia na
Caatinga do nordeste brasileiro (Vanzolini et
al. 1980; Vitt 1995). Ambas podem ser
encontradas na Estação Ecológica do Seridó,
no Rio Grande do Norte, onde partilham
sobretudo áreas de afloramentos rochosos
(Kolodiuk 2008).
Neste trabalho avaliou-se a atividade
de termorregulação dessas espécies de
Tropidurus, buscando responder às seguintes
questões: 1) A atividade de indivíduos dessas
espécies varia ao longo do tempo? 2) A
atividade está relacionada às variações de
temperatura do ambiente? 3) A atividade de
uma espécie influencia a atividade da outra?
Material e Métodos
O projeto foi realizado na Estação
Ecológica do Seridó (ESEC-RN), localizada no
município de Serra Negra do Norte. As
observações foram realizadas em quatro
afloramentos rochosos nas imediações da
lagoa do Junco, distante, aproximadamente, 5
km da sede da estação.
Na área de estudo foram registradas a
freqüência de uso do afloramento pelas duas
espécies e a temperatura ambiente. Essas
variáveis foram medidas, respectivamente, a
cada 10 e 30 minutos (no período de 7 às 11h
da manhã) com o auxílio de um
termohidrômetro.
Além disso, quando os indivíduos das
duas espécies ocorriam simultaneamente, foi
registrada a presença ou ausência de
comportamento agonístico, no qual uma
espécie expulsava a outra do local. Durante as
observações, o observador se posicionava a
uma distância que não interferia na atividade
das espécies Tropidurus hispidus e T.
semitaeniatus.
Realizou-se o teste de MANOVA de
medidas repetidas em bloco para analisar a
variação do número de registros entre duas as
espécies nos 4 afloramentos ao longo do dia e
uma regressão linear para observar como a
quantidade de registros varia em relação à
temperatura. Uma tabela de contingência
two-way foi utilizada para análise da co-
ocorrência das espécies ao longo do tempo.

16 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Resultados
Foram realizados 100 registros da
freqüência de uso dos afloramentos para a
atividade de termorregulação no período de
quatro horas durante a manhã. O número de
registros das duas espécies de Tropidurus
variou significativamente durante o período
observado (F = 2,306; gl = 24; p < 0,05).
Entretanto, as duas espécies apresentaram
uma dinâmica temporal semelhante ao longo
da manhã (F = 1,32; gl = 24 ; p > 0,05) (Fig. 1).
Figura 1 – Médias e desvios dos números de
registros de T. hispidus (---) e T. semitaeniatus (__)
nos afloramentos ao longo de quatro horas de
observação (25 observações realizadas de 10 em
10 minutos).
A temperatura não influenciou o
número de registros de lagartos presentes nos
afloramentos (F = 1,786; p > 0,05), assim como
não houve diferença significativa nas formas
em que as duas espécies respondem a essa
variação de temperatura ao longo do dia( F=
0,216; p > 0,05) (Fig. 2).
O número de registros de co-
ocorrência das duas espécies é
significativamente superior ao valor esperado
(p < 0,05) (Tabela 1), sugerindo que as duas
espécies partilham o mesmo recurso, no caso
do estudo, a utilização do afloramento.
Figura 2 - Variação do número de registros das
espécies de lagartos em relação a temperatura. H =
Tropidurus hispidus (---); S = T. semitaeniatus (__).
Tabela 1 – Valores de registros de co-ocorrência
observados são significativamente diferentes dos
valores esperados ao acaso.
0 1 Total
0 Observado 12 16 28
Esperado -7.28 -20.72
1 Observado 14 58 72
Esperado -18.72 -53.28
Total 26 74 100
Discussão
Apesar de Tropidurus semitaeniatus
ser geralmente mais frequente que T. hispidus
(Kolodiuk 2008), a predominância de registros
de T. semitaeniatus foi marginalmente
significativa, sugerindo que a diferença entre a
freqüência de registros das espécies não é tão
evidente e pode ter ocorrido ao acaso.

17 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Alguns lagartos podem manter
temperaturas corporais muito acima da
temperatura do ar. Por isso, a presença do sol
normalmente é mais importante que a
temperatura do ar na determinação do
padrão de atividade de lagartos (Vitt et al.
2008), justificando a ausência de relação
observada entre esta variável e o número de
registros de ambas espécies de Tropidurus.
Durante as observações realizadas neste
trabalho, o céu esteve nublado na maior parte
do tempo. Variações sutis na intensidade da
insolação podem ter sido o principal fator a
influenciar a atividade dos Tropidurus.
As diferentes preferências térmicas
das espécies limitam o uso dos habitats pelos
lagartos e as horas do dia em que estão ativos
(Vitt et al. 2008). No entanto, sabe-se que
espécies de um mesmo gênero tendem a ter
temperaturas corpóreas semelhantes (Rocha
et al. 2009). Consequentemente espera-se
que também tenham estratégias semelhantes
de termorregulação, o que explicaria a
sobreposição de Tropidurus hispidus e T.
semitaeniatus no tempo e espaço. A análise
da tabela de contingência (Tab. 01) corrobora
esta idéia, indicando uma tendência das
espécies ocuparem os afloramentos ao
mesmo tempo.
Segundo Kolodiuk (2008), há grande
sobreposição quanto à composição das dietas
dessas duas espécies de Tropidurus, com
diferença apenas no tamanho médio das
presas, que é maior para T. hispidus. Apesar
disso, o número de interações agonísticas
interespecíficas observadas foi relativamente
baixo (13% dos registros), indicando que a
competição não é um fator limitante na
utilização de recursos pelas duas espécies.
Tropidurus hispidus e T. semitaeniatus
ocupam dimensões de nicho muito
semelhantes. No entanto, as observações
realizadas mostram que essas espécies, além
de não se evitarem no tempo e espaço,
demonstram pouca agressividade
interespecífica. Possivelmente, há na área de
estudo uma grande disponibilidade de
recursos, permitindo a co-ocorrência das duas
espécies. Uma análise refinada da distribuição
espacial das espécies, relacionando-as a
diferenças microclimáticas dentro de cada
afloramento, talvez revele preferências mais
sutis na escolha dos sítios de termorregulação
pelas espécies de Tropidurus.
Agradecimentos
Ao professor Gindomar Santana pela
orientação no projeto. Aos professores do
Programa de Pós Graduação em Ecologia – PPGE,
em especial a Carlos Fonseca e Alexandre
Vasconcellos pela ajuda nos testes estatísticos.
Agradecimentos especiais aos funcionários da ESEC
pela hospitalidade e por permitir o uso de toda a
infraestrutura da reserva.
Referências
Kolodiuk, M. F. 2008. Dieta e comportamento de
forrageio de duas espécies de Tropidurus
(Squamata, Tropiduridae) em uma área de
Caatinga do Nordeste do Brasil. Dissertação de
mestrado. Universidade Federal do Rio Grande
do Norte.
Pianka, E. R., 1973. The structure of lizard
communities. Annual Review of Ecology and
Systematics 4, 53-74.
Rocha, C. F. D, Sluys, M. V., Vrcibradic, D., Kiefer,
M. C., Menezes, V. A., Siqueira, C. C., 2009.
Comportamento de termorregulação em
lagartos brasileiros. Oecologia Brasiliensis. 13,
115 – 131.
Vanzolini, P. E., Ramos-Costa, A. M. M., Vitt, L. J.,
1980. Répteis das Caatingas. Academia
Brasileira de Ciências, Rio de Janeiro.

18 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Vitt, L. J., 1995. The ecology of tropical lizards in
the caatinga of northeast Brazil. Occasional
Papers of the Oklahoma Museum of Natural
History. 1, 1-29.
Vitt, L. J., Zani, P. A., Caldwell, J. P., Sartorius, S. S.,
Cooper Jr, W. E., Baird, T. A., Baird, T. D., Pérez-
Mellado, V, 2005. Pushing the edge: extended
activity as an alternative to risky body
temperatures in a herbivorous teiid lizard
(Cnemidophorus murinus: Squamata).
Functional Ecology 152-158.
Foto. Jararaca fotografada pelo grupo de trabalho na área de estudo.

19 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
INFLUÊNCIA DE RIO TEMPORÁRIO SOBRE A TAXOCENOSE DE COLLEMBOLA EM
UMA ÁREA DE CAATINGA DO NORDESTE BRASILEIRO
HEITOR B. A. SOUZA, IZABEL M. M. SOUZA, KAROL L. B. O. FRANÇA, LEONARDO H. T. PINTO,
MAÍRA F. MANZAN
Introdução
Collembola são pequenos artrópodes
que medem, em geral, de 0,2 a 3mm de
comprimento (Buzzi, 2005). São
ocasionalmente identificados como insetos
apterigotos, embora sejam hexápodas
endognatos (Zeppelini & Belllini, 2004). Os
colêmbolos são organismos bastante
abundantes, atingindo densidades de dezenas
a centenas de milhares/m² nas camadas
epiedáficas, isto é, vivem na serrapilheira, nas
primeiras camadas do solo (Zeppelimi &
Bellini, 2004).
Os colêmbolos são, mais
especificamente, encontrados no solo
humoso, no meio de musgos, sobre o solo,
folhas caídas, bainha de folhas e no meio de
detritos vegetais em decomposição. São raros
em regiões secas, mas estão presentes em
todas as partes do mundo (Buzzi, 2005).
Os Collembola são extremamente
importantes, pois estão na base da cadeia
alimentar e de ciclagem de nutrientes. Além
de servirem como presas para diversos
animais em diferentes etapas de seus
desenvolvimentos, eles atuam na
decomposição da matéria orgânica animal e
vegetal de forma determinante. Juntamente
com fungos e bactérias, os colêmbolos estão
envolvidos na formação e enriquecimento do
solo tanto em florestas como em
agroecossistemas. (Zeppelini & Bellini, 2004).
Existem mais de 7500 espécies
descritas de colêmbolos, sendo a região
Neotropical um dos lugares com maior
diversidade estimada em todo mundo. Nessa
região há enorme carência de especialistas em
taxonomia e ecologia (Zeppelini & Bellini,
2004). A Caatinga, por exemplo, não
apresenta dados publicados sobre aspectos
ecológicos de Collembola.
O domínio Caatinga abrange cerca de
900.000 Km², correspondendo
aproximadamente a 54% da região nordeste e
11% do território brasileiro, ocupando áreas
que ocorrem do Ceará ao estado de Minas
Gerais (Andrade et al, 2005). A Caatinga
apresenta pluviosidade média anual variando
entre 240 a 1500 mm, com mais da metade da
região recebendo menos de 700 mm anuais.
O presente trabalho teve o objetivo de
investigar a influência de um rio temporário
sobre a taxocenose de Collembola em uma
área de caatinga situada na ecorregião da
Depressão Sertaneja Setentrional.
Material e Métodos
O estudo foi realizado na Estação
Ecológica do Seridó – ESEC do Seridó,
localizada no município de Serra Negra do
Norte, no estado do Rio Grande do Norte,
Brasil. Com uma área de 1.166,38 ha e
localizada entre as coordenadas geográficas
06° 36’ 40’’ e 06° 33’ 50’’ Sul e 37° 17’ 10’’ e
37° 14’ 20’’ Oeste, a ESEC do Seridó possui
clima semi-árido com temperatura média
anual variando entre 33° e 22°. Há ocorrência

20 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
de precipitações irregulares, com chuvas no
período de Janeiro a Maio e variações
pluviométricas de 127 mm a 916 mm,
atingindo uma média de 497 mm por
ano.(Ibama, 2004)
Foram traçados três transectos
paralelos de 20m de comprimento distando,
respectivamente, 0m, 1m e 4m da margem de
um rio temporário. Cada transecto foi divido
em 10 parcelas, com distância mínima de 1m
entre as margens das parcelas adjacentes.
Para a coleta dos colêmbolos, foram sorteadas
5 parcelas de 1m² em cada transecto. Em cada
parcela, estabeleceu-se um esforço amostral
de 30 minutos por pessoa. Durante esse
período, foram dispostas amostras de folhiço
numa bandejas plásticas onde os colêmbolos
eram procurados ativamente e, com o auxílio
de um aspirador entomológico, coletados.
Os indivíduos amostrados foram
imediatamente fixados em álcool 70% para
posterior separação em morfoespécies com a
utilização de estereomicroscópio Olympus
SZ40.
Os dados de riqueza e abundância de
espécies, assim como as variáveis abióticas pH
e umidade foram submetidas a um
tratamento estatístico utilizando a ferramenta
General Linear Model do software Systat 12.
As medidas de umidade e pH do solo
foram realizadas com a utilização de um
medidor analógico Instrutherm pH2500 em
cada parcela amostrada.
Resultados
Foram encontradas sete
morfoespécies pertencentes a quatro gêneros
distintos (Seira, Lepidocyrtus, Branchystomella
e Sminthurides), sendo o gênero Lepidocyrtus
o com maior número de indivíduos
encontrados, com 64% das ocorrências (tabela
01).
A análise realizada através do Modelo
Linear Geral não mostrou relação significativa
da riqueza de espécies com nenhuma das
variáveis explanatórias analisadas, conforme a
tabela 02.
Tabela 2: Resultado da Análise de Variância para as
variáveis bióticas e ambientais.
Tratamentos gl F p-value
TRANSECTO 2 1,786 0,260
COLETOR 4 0,979 0,494
UMIDADE 1 0,049 0,834
PH 1 0,199 0,674
ABUNDANCIA 1 1,432 0,285
Erro 5
Discussão
Farkas et al (1998) demonstraram que
a riqueza de espécies de Collembola em um
ambiente constituído por um mosaico de
habitas e pluviosidade média anual variando
de 2000 a 2500mm, varia bastante em função
da umidade estando a similaridade entre
habitats relacionada fortemente com essa
variável ambiental. Nesse trabalho, habitats
Tabela 1: Lista de espécies e abundância de
Collembola
Abundância
Espécies Transecto 1 Transecto 2 Transecto 3 Total
Seira sp.1 2 3 0 5
Seira sp.2 1 1 0 2
Seira sp.3 0 5 0 5
Lepidocyrtus sp.1 14 7 15 36
Lepidocyrtus sp.2 0 1 0 1
Branchystomella sp. 4 1 3 8
Sminthurides sp. 0 0 1 1

21 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
mais secos apresentaram uma riqueza de
espécies muito que em áreas mais úmidas.
Eles também verificaram a ocorrência de
espécies que preferem ambientes mais secos.
A condição de maior riqueza em ambientes
mais úmidos também foi verificado para
comparações dentro de uma mesma área
entre diferentes épocas do ano em um
ambiente desértico situado no continente
norte-americano (Santos & Whitford, 1981). A
umidade apresenta elevada importância para
os colêmbolos, uma vez que, eles podem
perder elevadas quantidades de água para o
meio através de uma estrutura que realiza
trocas aquosas com o meio chamada
colenóforo situada na porção ventral de seu
corpo (Zeppelini & Bellini, 2004).
O presente estudo não encontrou uma
relação significativa do efeito da umidade e do
pH do solo sobre a riqueza de colêmbolos e
concluiu que, dentro do gradiente estudado
no entorno de um rio temporário situado num
remanescente de Caatinga na Depressão
Sertaneja Setentrional, a riqueza e abundância
de colêmbolos, diferentemente de outros
trabalhos realizados sobre a ecologia desse
táxon, não varia em função da umidade nem
em função do pH do solo.
Agradecimentos
Ao Doutor Bruno Bellini pelo
desenvolvimento e apoio dado para a realização
desse trabalho; aos Professores Doutores
Alexandre Vasconcellos e Carlos Fonseca pelo
auxílio fornecido nas análises estatísticas realizadas
nesse estudo.
Referências
Andrade L. A., Pereira I. M., Leite U. T., Barbosa M.
R. V. 2005. Análise da cobertura de duas
fitofisionomias de Caatinga, com diferentes
históricos de uso no município de São João do
Cariri, Estado da Paraíba. Cerne, Lavras, vol 11,
253-262.
Buzzi, J.Z. 2005. Entomologia Didática. Editora
UFPR, Curitiba, pp 125-127.
Farkas, J., Szathmáry, k., Strenner, A. 1998.
Ecofaunistical investigation of Collembola,
Aranae and Coleoptera in mosaic-like habitats
in the Cinege Valey, Hungary. Opusc. Zoo.
Budapest 31, 49-61.
Ibama. 2004. Plano de Manejo da ESEC do Seridó.
Santos, P.F,. Whitford, W.G. 1981. The effects of
microarthropods on litter decomposition in
Chihuahuan Desert Ecossistem. Ecology, vol 63,
654-663.
Zeppelini Filho, D., Bellini, B.C. 2004. Introdução ao
Estudo dos Collembola. Editora Universitária,
João Pessoa.

22 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
CONVERGÊNCIAS ADAPTATIVAS EM RESPOSTA À PLUVIOSIDADE ENTRE
PLANTAS BRASILEIRAS E AUSTRALIANAS
DANIEL DE OLIVEIRA; DIEGO BENTO; GINA MANTILLA; IGOR GALVÃO; PRISCILA DO AMARAL;
CARLOS ROBERTO FONSECA (ORIENTADOR)
Introdução
A separação do antigo continente
Gondwana, há 180 milhões de anos, isolou as
floras da América do Sul e da Austrália
(Teixeira et al., 2000). O isolamenteo
geográfico ocorrido levou a divergências
filogenéticas, de forma que, atualmente, as
floras do Brasil e da Austrália diferem
bastante, principalmente ao nível de gênero e
táxons inferiores.
Por outro lado, condições ambientais
semlhantes nas duas regiões podem levar a
convergências adaptativas (Burbidge, 1960;
Begon et al., 2006), trazendo semelhanças
entre as floras dos países. Assim, é possível
que plantas na Austrália e no Brasil, sujeitas a
pressões seletivas similares, apresentem
características morfo-funcionais semelhantes.
A diversidade de estratégias
ecológicas em plantas pode ser descrita a
partir de algumas características-chave.
Características morfo-funcionais como altura
da copa, tamanho da semente, área efetiva da
folha, área específica da folha, dentre outros
podem ser utilizados para identificar espécies
que têm estratégias ecológicas similares.
A largura da folha, medida como o
diâmetro do maior círculo que pode ser
inscrito no perímetro foliar, reflete a relação
entre transpiração e captação de carbono. Por
sua vez, a altura máxima que uma planta pode
atingir está relacionada com sua estratégia
para a captação de luz em relação ao custo
para a obtenção e transporte de água
(Fonseca et al., 2000).
Diante disso, este trabalho buscou
verificar se há convergência nas estratégias
adaptativas entre comunidades vegetais
brasileiras e australianas quando submetidas a
condições ambientais semelhantes, em
particular, a pluviosidade.
Material e Métodos
Dados da Austrália
Usando como base os dados contidos
na tabela 1 de Fonseca et al. (2000), buscou-se
o modelo, linear ou não-linear, que melhor
descrevesse a relação entre a largura foliar
média da comunidade, em centímetros, e a
pluviosidade anual média, em milímetros.
Também se procurou o modelo que melhor
descrevesse a relação entre a altura média da
copa da comunidade vegetal, em
centímentros, e a pluviosidade anual, em
milímetros.
As regressões lineares geralmente
serão boas representantes de correlações cuja
associação não oscile muito ao longo dos
eixos. As não-lineares, por sua vez,
representam uma mudança na amplitude da
relação das variáveis ao longo dos eixos.
Dentro desse último grupo, dois tipos
particulares foram testados: quadrático e de
potência. O primeiro representa uma
associação de forma parábola, apresentando
um ponto de inflexão (Zar, 1999). Ali, os

23 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
padrões crescentes ou decrescentes das
associações se invertem. Já o último, de
potência, representa uma associação
continuamente crescente ou decrescente, mas
em graus progressivamente mais intensos.
Dados do Brasil
O estudo foi desenvolvido na Estação
Ecolóica do Seridó (ESEC), localizada no
município de Serra Negra do Norte – RN. É
uma região de clima semi-úmido, com uma
pluviosidade média anual de 497 mm (IBAMA,
2004).
Foi escolhido um plote de 20 x 50
metros em área bastante conservada (Fig. 1),
dado que esta representaria melhor a flora
nativa da região.
Figura 1. Mapa da ESEC do Seridó (esquerda) com
destaque para o plot amostrado (direita). FONTE:
ICMBio
Dentro desse plote, todas as plantas
presentes foram morfotipadas e as alturas
máximas fora estimadas. Para cada espécie, a
largura de cada folha foi determinada com
0,001 cm de precisão, com auxílio de um
paquímetro. A largura foliar média foi
calculada a partir de 5 folhas de um ramo
representativo do morfotipo.
Para as análises, calculou-se a largura
foliar média (LFm) e a altura média da copa
(Am) entre todas as espécies, assim como seus
respectivos coeficientes de variância (CV).
Estes dados foram comparados às
previsões dos modelos australianos para
testar a hipótese da convergência morfo-
funcional.
Todas as análises foram feitas
utilizando o programa Systat 12.0.
Resultados
Dados os ajustes elevados das
regressões linear (R2 = 0,960), quadrática
(0,963) e de potência (0,961) entre P e LFm,
optou-se pelo modelo de potência, por se
acreditar que é o mais próximo ao esperado
biologicamente. A equação do modelo é a
seguinte:
(Eq. 1) LFm = 0,006 * P4,675
Para a relação entre P e Am, o modelo
quadrático foi o que apresentou o melhor
ajuste (R2 = 0,563), seguindo a equação 2:
(Eq. 2) Am = -1,643 * P2 + 9,866 * P – 14,480
Considerando-se a pluviosidade média
anual de 497 mm, o valor estimado de largura
foliar, utilizando a equação 1, foi de 0,619 cm,
enquanto o de altura, utilizando a equação 2,
foi de 0,177 m.
Foram encontrados 19 morfotipos na
área amostrada. O log da LFm foi 0,408cm
(0,210-0,606; 95% de confiança) e o da Am foi
de 0,255 (-0,003-0,514; 95% de confiança)
(Fig. 2).
O valor para largura foliar média está
ligeiramente acima do lmite superior do
intervalo observado para a área amostrada,
enquanto o segundo situa-se dentro do seu
intervalo de confiança (Fig. 3).

24 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
a.
b.
Figura 2. Distribuição das (a) larguras foliares e (b)
alturas máximas (log) para os morfotipos na área
amostrada. Os coeficientes de variação foram
1,006 e 2,095, respectivamente.
b.
Figura 3. Modelos de regressão não-lineares para
os dados de Fonseca et al. (2000). (a) Modelo de
potência entre os logs de LFm e P. (b) Modelo
quadrático entre os logs de Am e P. Os traços
vermelhos representam os limites do intervalo de
confiança para os dados observados. A seta azul
representa o valor estimado.
Discussão
Com base nos coeficientes de variação
(Fig.2), pode-se afirmar que há, na
comunidade, maior diversidade de estratégias
da característica altura máxima, quando
comparado a largura foliar.
Considerando-se que o valor estimado
para o log LFm também possui um erro
associado, mas que não foi representado, não
se pode rejeitar a hipótese de convergência
adaptativa entre as regiões analisadas, uma
vez que é possível a sobreposição entre as
margens de erro dos valores estimado e
observado. Além disso, o valor estimado para

25 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Am está contido no intervalo de confiança
observado e distante dos limites,
evidenciando a convergência adaptativa
também para essa característica.
Dessa maneira, há uma forte
convergência adaptativa dessas características
morfo-funcionais entre plantas brasileiras e
australianas submetidas a semelhante
pluviosidade.
Agradecimentos
Somos gratos ao nosso orientador, Carlos
Roberto Fonseca, pelo amplo apoio em todo o
desenvolvimento do projeto. Também
agradecemos aos funcionários da ESEC por todo o
seu apoio operacional e colaboração para nosso
bem-estar.
Referências
Begon, M., Townsend, C. R., Harper, J. L. 2006.
Ecologia: de indivíduos a ecossistemas. Artmed.
São Paulo.
Burbidge, N. T. 1960. The phytogeography of the
Australian region. Australian Journal of Botany.
8, 75-211.
Ibama. 2004. Plano de Manejo da Estação
Ecológica do Seridó.
Fonseca, C. R., Overton, J. McC., Westoby, M.
2000. Shifts in trait-combinations along rainfall
and phosphorus gradient. Journal of Ecollogy.
88, 954-977.
Teixeira, W., Toledo, M. C. N., Fairchild, T. R., Taioli,
F. 2000. Decifrando a Terra. Oficina de Textos.
São Paulo.
Zar, J. H. 1999. Biostatistical Analysis. Prentice Hall.
New Jersey.

26 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010

27 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
CARACTERIZAÇÃO DE COMUNIDADES DE COLÊMBOLOS EM ÁREAS COM
DIFERENTES USOS DO SOLO DA CAATINGA
DANIEL DE OLIVEIRA; DIEGO BENTO; GINA MANTILLA; IGOR DE BRITTO; PRISCILA DO AMARAL
Introdução
Colêmbolos (Collembola: Ellipura:
Hexapoda) são hexápodes entognatos com
um tamanho que pode variar de 0,2 a 3 mm
(Zeppelini & Bellini, 2004). É o segundo grupo
de invertebrados mais abundantes do solo,
ocorrendo em serrapilheira, árvores, litoral
marinho e água doce (Barreta et al.,2007).
Assim como outros invertebrados do
solo, os colêmbolos estão intimamente
associados aos processos ecossistêmicos que
ocorrem no compartimento serrapilheira-solo
refletindo padrões de funcionamento do
ecossistema. Vários estudos demonstraram a
contribuição direta ou indireta desse grupo
nos processos ecológicos do solo, como
mineralização de nitrogênio, respiração do
solo ou lixiviamento de carbono orgânico
dissolvido (Filser, 2002)
Mudanças no uso da terra provocam
alterações nas propriedades químicas, físicas e
biológicas dos solos, reduzindo a quantidade
de resíduos vegetais e alterando
completamente as características do solo
(Sautter et al., 1999). Como a diversidade de
colêmbolos tem sido usada como bioindicador
de distúrbios e da qualidade do solo (Barreta
et al.,2007), este estudo objetivou comparar
comunidades de colêmbolos em duas áreas
com diferentes regimes de uso do solo: uma
área preservada e outra de pastagem
extensiva.
Material e Métodos
O estudo foi desenvolvido na Estação
Ecológica do Seridó e em uma área contígua,
na Fazenda Solidão, localizadas no município
de Serra Negra do Norte-RN.
Foram estabelecidos dois transectos
de 100m paralelos entre si, sendo um em cada
área (Fig. 1). Em cada transecto sorteou-se 10
plots de 1m², com permanência de 20 minutos
em cada plot, nos quais foram coletadas
amostras de folhiço, colocadas em bandejas e
revolvidas para a captura com auxílio de
aspiradores entomológicos. Cada coletor
amostrou os mesmos pontos em ambos os
transectos (blocos) no sentido de minimizar o
erro no esforço amostral. Os plots sorteados
ficaram nas distâncias de 1, 7, 19, 22, 30, 32,
79, 87, 89, 92 m ao longo do transecto.
Figura 4. Desenho amostral. A linha tracejada
representa a cerca divisória entre as áreas
estudadas; as linhas contínuas os transectos com
os plots amostrados; as caixas claras as estradas. A
distância entre os transectos foi de 14 m, sendo
2m da cerca para a estrada e 5 m desta para os
mesmos. Os plots sorteados ficaram nas distâncias
de 1, 7, 19, 22, 30, 32, 79, 87, 89, 92 m.
As amostras foram identificadas ao
nível de gênero e posteriormente
morfotipadas de forma a possibilitar a

28 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
avaliação dos índices biológicos (riqueza,
diversidade e equitabilidade), da composição
e a análise comparativa das comunidades
(ANOVA, Agrupamento e PCA).
Resultados
Foi coletado um total de 86
indivíduos, representados principalmente
pelos grupos Poduromorpha,
Entomobryomorpha e Symphypleona,
classificados e 10 morfotipos pertecentes a 8
gêneros e 7 famílias.
Os índices de diversidade e
equitabilidade das áreas apresentaram valores
próximos (Tab. 1). O índice Bray-Curtis
quantificou apenas 33% de similaridade entre
as comunidades.
A distribuição da abundância dos
morfotipos encontrada entre as áreas com
diferentes tipos de uso de solo foi desigual.
Alguns táxons foram encontrados apenas em
uma das áreas, e os comuns a ambas
apresentaram abundâncias diferentes (Fig. 2).
A análise de variância entre as áreas
amostradas, considerando os blocos, indicou
que não há diferenças significativas entre as
riquezas (Fig. 3).
Para um melhor resultado do
agrupamento, não foram considerados os
plots em que nenhum indíviduo foi observado
(Plots E30 e S79). Essa análise gerou uma
árvore que separa as áreas amostradas em
dois grandes grupos, um contendo todos os
plots da ESEC do Seridó e alguns da Fazenda
Solidão e o outro contendo a maioria dos plots
da Fazenda Solidão (Fig. 4).
Tabela 1. Índices biológicos das comunidades da
Esec do Seridó e da Fazenda Solidão.
Índice ESEC Solidão
Shannon 1,576 1,125
Margalef 1,616 1,051
Pielou 0,81 0,7
Bray-Curtis 33%
A análise de componentes principais
demonstrou que a variabilidade dos dados foi
explicada por quatro eixos totalizando 67,25%,
com os eixos 1, 2 e 3 contribuindo com
50,76% (Tab. 2). A relação entre os três
principais eixos separou as áreas em duas
nuvens de pontos, que equivalem às duas
áreas amostradas (Fig. 5).

29 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Figura 5. Abundância de cada morfotipo (eixo X) na ESEC do Seridó e na Fazenda Solidão. Houve 5 espécies
ocorrendo apenas na fazenda Solidão e 7 apenas na ESEC. Nas duas em comum, encontrou-se abundâncias
distintas.
Figura 6. As riquezas médias das áreas
amostradas não foram diferentes (F = 1,018;
g.l. = 19; P = 0,490).
Figura 7. Análise de agrupamento das áreas
amostradas utilizando a como métrica de
distância Euclidiana e método Ward. Áreas
foram, em geral, mais similares dentro de cada
uso de terra do que entre eles, havendo alguma
sobreposição entre algumas áreas da Solidão e
da ESEC.

30 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Fig. 8. Análise de componente principal para as
áreas amostradas. Áreas da ESEC e da Solidão
representadas como pontos vermelhos e azuis,
respectivamente. Os três eixos apresentados
explicam 50,76% da variação total.
Tabela 2. Eixos da análise de componentes
principais para as áreas amostradas. Os 4 eixos
explicam 67,25% da variação a partir de
relações positivas ou negativas de cada
espécie com os eixos.
Táxon 1 2 3 4
NEANURA 0,74 0,16 0,45 0,30
SEIRA_2 -0,71 0,24 0,28 0,13
SMINTHURIDES1 -0,58 0,01 0,27 0,29
ALLACIMA 0,06 -0,85 -0,24 -0,32
DICYRTOMA 0,06 -0,80 0,09 0,22
LEPIDOCYRTUS 0,13 0,19 0,78 0,13
RASTRIOPES 0,00 -0,23 -0,66 0,02
SEIRA_1 -0,21 0,02 0,64 0,39
SMINTHURIDES2 0,01 -0,26 0,19 -0,80
BRACHYSTOMELLA 0,12 0,24 -0,16 -0,70
1 2 3 4
14,648 16,718 19,389 16,494
Discussão
Florestas nativas, de uma maneira
geral, possuem maior heterogeneidade de
hábitat e são capazes de suportar maior
diversidade de Colêmbolos (Barreta, 2007),
no entanto, a riqueza de espécies entre as
áreas estudadas foi semelhante. Não
obstante, é preciso reconhecer que hoje os
padrões de diversidade frequentemente
exibem um “fantasma do antigo uso da
terra”, refletindo anteriores atividades de
manejo da terra (Harding et al. 1998).
Como a ESEC do Seridó era parte da
Fazenda Solidão, apesar de estar livre de
maiores impactos antrópicos desde sua
implantação (1982), não pode ser
considerada uma área primária.
Apesar disso, encontraram-se
comunidades com composição bastante
diferenciada, refletindo que mudanças no
uso da terra podem não afetar a riqueza.
Todavia, ao alterar a composição da
riqueza, o funcionamento do ecossistema
também é modificado.
Devido à maior cobertura vegetal
da ESEC do Seridó, foi observada uma
maior quantidade de serrapilheira e
pressupõe-se que a umidade do folhiço é

31 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
mais elevada. A partir disso pode-se inferir
que também há uma maior diversidade da
microflora do solo, principal recurso
alimentar dos colêmbolos. (Zeppelini &
Bellini, 2004).
A análise das características morfo-
funcionais dos grupos encontrados sugere
diferentes níveis de resistência à
dessecação. Neanura sp.1 pertence ao
grupo (Poduromorpha) com menor
colóforo (superfície extra para a perda de
umidade corporal) e cutícula mais espessa,
características que reduzem a perda de
água para o meio (Bellini, comunicação
pessoal), e foi encontrado apenas na
Fazenda Solidão. Por outro lado, quatro
dos cinco morfotipos de Symphypleona
(Sminthurides sp.1, Sminthurides sp.2,
Allacma sp.1, Rastriopes sp.1 e Dicyrtoma
sp.1) foram encontrados apenas na ESEC.
Esse grupo apresenta o maior colóforo
entre os Collembola, sendo mais
dependentes de alta umidade (Bellini,
comunicação pessoal).
Diante disso, pode-se afirmar que
há uma relação entre a composição das
comunidades de Collembola e o uso dado
ao solo.
Agradecimentos
Somos gratos ao professor doutor
Bruno Bellini pela orientação, pela realização da
triagem e pelo nosso aprendizado. Ao professor
Carlos Fonseca pelo apoio nas análises
estatísticas. Aos funcionários da ESEC.
Referências
Andrade-Lima D., 1981. The caatinga
dominium. Revista Brasileira de Botânica 4,
149-63.
Barreta, D., Sousa, J.P., Ferreira, C.S., Cardoso,
E.J.B.N. 2007. Colêmbolos (Hexapoda:
Collembola) como indicadores da qualidade
do solo em áreas com Araucaria
angustifolia. XXXI Congresso Brasileiero de
Ciência do Solo. Gramado-RS.
Filser, J. 2002. The role of Collembola in carbon
and nitrogen cycling in soil. Pedobiologia,
46: 234-245.
Harding, J. S. et al. 1998. Stream biodiversity:
the ghost of land use past. Proc. Nat. Acad.
Sci. USA, 95: 14843-14847.
Sampaio, E. V. S. B., 1995. Overview of the
Brazilian Caatinga. In: Bullock, S.H.,
Mooney, H. A., Medina, E. (Eds.), Seasonally
dry forests. Cambridge University Press,
Cambridge, pp. 35-58.
Sautter, K.D., Santos, H.R., Júnior, P.J.R. 1999.
Comparação das comudades de
Entomobryidae e Isotomidae (Collembola)
entre plantio direto em três níveis de
fertilidade, plantio convencional e um
ecossistema natural (campo nativo) em
Ponta Grossa, Paraná, Brasil. Revista
Brasileira de Zoologia. 16 (1), 117-124.
Zeppelini, D., Bellini, B.C. 2004. Introdução ao
estudo dos Collembola. Editora
Universitária, João Pessoa

32 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
FUGA ANTI-PREDADOR DE Tropidurus hispidus E Tropidurus semitaeniatus
(TROPIDURIDAE) EM UMA ÁREA DE CAATINGA
FABRÍCIO RODRIGUES CAMACHO, BRUNNO FREIRE DANTAS DE OLIVEIRA, UIRANDÉ OLIVEIRA COSTA E
CRISTIANE GOUVÊA FAJARDO
Introdução
Os lagartos constituem o grupo
mais diverso dos répteis, com cerca de
5000 espécies (Uetz et al., 2008). No Brasil
encontram-se 234 espécies de lagartos, das
quais 31% dessas espécies ocorrem no
bioma Caatinga. Em algumas regiões desse
bioma duas espécies são bastante
frequentes, Tropiturus semitaeniatus e
Tropidurus hispidus, sendo a primeira
endêmica.
Apesar dessas espécies de lagartos
ocorrerem em sintopia, elas são
territorialistas. Alguns autores sugerem
que o aumento no risco de predação reduz
a freqüência de encontros agressivos entre
lagartos (Baker et al., 1999; Brick, 1999;
Díaz–Uriate, 1999), o que pode afetar
diretamente as taxas de disputa por
território e indiretamente o sucesso
reprodutivo das espécies. Dessa forma,
estudos que focam a distância de fuga anti-
predador são relevantes para o
entendimento do comportamento de
defesa de território, especialmente no se
refere ao trade-off entre forragear e
proteger o território.
Fatores como o tamanho do
animal, temperatura do ambiente e
divergências morfológicas ou
comportamentais entre espécies podem
influenciar as distâncias de fuga entre
indivíduos (Díaz–Uriate, 2001).
Assim, os objetivos do presente
trabalho foram verificar as diferenças da
distância de fuga anti-predador entre T.
semitaeniatus e T. hispidus. Além disso,
averiguar se a temperatura do ambiente e
diferenças no tamanho entre coespecíficos
influenciam na distância de fuga.
Material e Métodos
O estudo foi realizado em quatro
afloramentos rochosos, próximos à Lagoa
do Junco na Estação Ecológica do Seridó -
ESEC. Em cada afloramento foram
simuladas situações de ataque as espécies
Tropidurus semitaeniatus (Figura 1A) e T.
hispidus (Figura 1B). Os ataques foram
intervalados por um período mínimo de 15
minutos para que uma situação natural
fosse restabelecida. As distâncias de fuga
anti-predador foram medidas de acordo
com o método de Cooper (2000) pelo
mesmo observador distanciado 5 metros
do lajedo.
Figura 1. (A) Indivíduo de Tropidurus
semitaeniatus, (B) indivíduo de T. hispidus.
Para avaliar as diferenças nas
distâncias de fuga de acordo com os
tamanhos dos indivíduos de cada espécie,
foram categorizadas duas classes de
tamanho. O tamanho o indivíduo é um
fator que pode afetar seu potencial de
A
C
A B B

33 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
defesa do território (Fox et al, 1990). Na
nossa metodologia classificamos como
grandes os indivíduos maiores que 20 cm e
pequenos os menores que 20 cm, para T.
hispidus, e pequenos os indivíduos maiores
que 15 cm e pequenos os menores que 15
cm para T. semiteniatus.
As medidas de temperatura
ambiente foram registradas a cada 30
minutos, com termohigrômetro digital
INSTRUMENTHERM HT-300®.
Os dados foram analisados no
módulo GLM do programa Systat 12.0, no
qual foi analisado o efeito das variáveis
categóricas: espécie, tamanho, e a
interação entre estas; e da variável
contínua temperatura sobre a variável
dependente distância de fuga.
Resultados
Existem diferenças significativas
nas distancias de fuga entre as espécies de
T. semitaeniatus e T. hispidus (F = 7,54 ; GL
= 1,54 ; P < 0,01), com a primeira espécie
apresentando menor distância de fuga
anti-predador (Figura 2).
Não foi observado efeito das
diferenças de tamanhos entre espécies
sobre as distâncias de fuga (F = 6,28 ; GL =
1,59 ; P = 0,108). No entanto, constatamos
uma diferença aparente, onde em T.
hispidus os indivíduos grandes
apresentaram distancias de fuga menores
que os pequenos, e o inverso foi notado
para T. semiteniatus (Figura 3).
Apesar de as espécies estudadas
serem pecilotérmicas, as variações na
temperatura ambiente não surtiram efeito
sobre as distâncias de fuga das duas
espécies (F = 0,27 ; GL = 1,54 ; P = 0,605)
(Figura 3).
Figura2: Gráfico relaciona as médias e desvios
padrão nas distâncias de fuga de anti-predador
para as espécies Tropidurus hispidus (H) e T.
semitaeniatus (S).
Figura 3: Tamanho (pequeno = P ; grande = G)
dos indivíduos das espécies Tropidurus
hispidus.(H) e T. semitaeniatus (S) com relação
à distância de fuga anti-predador.

34 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Figura 4. Relacionamento da temperatura (ºC) e
distância de fuga anti-predador em metros (m)
das espécies T. hispidus (H) e T. semitaeniatus
(S).
Discussão
Kolodiuk (2008) observou que as
duas espécies estudadas apresentam
diferenças morfológicas, entretanto, não se
distinguem significativamente com relação
à dieta e comportamento.
Os resultados obtidos com o
presente trabalho indicam que há
diferenças significativas no
comportamento de fuga de entre as
espécies, sendo Tropiduros semitaeniatus a
espécie que se expõe mais tempo ao
predador.
Para espécies territorialistas o
tamanho corporal é um fator importante
durante o confronto por território (Fox et
al., 2003). No entanto, não observamos
diferenças significativas entre os tamanhos
dos indivíduos para as espécies estudadas,
mostrando que o tamanho do corpo, não é
importante para determinar diferenças na
fuga do predador.
Cooper (2000) não observou
influencia da temperatura na distância de
fuga anti-predador. Os resultados deste
trabalho corroboram os obtidos por
Cooper (2000). Apesar das espécies
estudadas serem ectotérmicas, as
distâncias de fuga parecem não serem
afetadas pela temperatura ambiente
parece não influenciar dentro da variação
climática que foi possível observar em
campo (mínima de 25,3 e máxima de 33,5
ºC).
Este trabalho revela diferenças
significativas na distância de fuga entre
Tropidurus semitaeniatus e T. hispidus, isso
pode surtir efeitos positivos sob o fitness
de T. semitaeniatus. Entretanto, tais
efeitos ainda devem ser testados.
Agradecimentos
Aos funcionários da ESEC-Seridó, ao Professor
Doutor Gindomar Gomes Santana pela
orientação, aos Professores, especialmente ao
Prof. Carlos Fonseca pelo auxílio com as
análises estatísticas, e ao Programa de Pós
Graduação em Ecologia da UFRN.
Referências
Baker, R. L., Elkin, C. M. & Brennan, H. A. 1999.
Aggressive interaction and risk of fish
predation for larval damselflies. Journal of
Insect Behaviour 12, 213-223.
Brick, O. 1999. A test of the sequential
assessment game: The effect of increased
cost of sampling. Behaviour Ecology 10,
726-732.
Coopper, W. E. 2000. Effect of temperature on
escape behaviour by na ectothermic
vertebrate, the keeled earess lizard
(Holbrookia propinqua) Behaviour. 137,
1299-1315.
Díaz-Uriart, R. 2001. Territorial intrusion risk
and antipredator behavior: a mathematical

35 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
model. Proceedings of the Royal Society of
London B 268, 1165-1173.
Fox, S. F.; McCoy, J. K.; Baird, T. A. 2003. Lizard
social behavior. The Jonhs Hopkins
University Press. 410p.
Kolodiuk, M. F. 2008. Dieta e comportamento
de forrageio de duas espécies de Tropidurus
(Squamata, Tropiduridae) área de caatinga
do nordeste do Brasil. Dissertação de
Mestrado em Psicobiologia na Universidade
Federal do Rio Grande do
Norte/UFRN.31pp.
Uetz, P. Etzold, T. & Chenna, R. 2008. The EMBL
repitale Data Base, htt://www.reptile-
database.org/ acessado em 24/06/2010

36 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
INFLUÊNCIA DO EFEITO DE BORDA NA RIQUEZA E ESTRUTURA DE PLANTAS
LENHOSAS EM UMA ÁREA DE CAATINGA NO RIO GRANDE DO NORTE
ALLAN T. BATISTA; FRANÇOISE. D. LIMA; VINÍCIUS A. SÃO PEDRO; MONALISA R. O. DA SILVA &
RODRIGO X. SOARES
Introdução
A Caatinga engloba um número
elevado de formações e tipos
vegetacionais, principalmente, pela grande
variação ambiental associada ao clima, solo
e relevo (Prado, 2003). Essa peculiaridade
ecossistêmica pode resultar em grande
variação fisionômica, tanto na densidade
quanto no porte das plantas. Contudo, a
pressão antrópica sobre esse bioma vem
sendo intensificada ao longo dos anos.
Essas atividades humanas têm
intensificado o processo de fragmentação
deste bioma, o que pode resultar na
formação de bordas nas comunidades
vegetacionais. Em paisagens fragmentadas,
as alterações biológicas são intensificadas
pelo efeito de borda (Santos & Santos,
2008).
A criação de bordas pode alterar
características da vegetação,
principalmente, devido às mudanças
microclimáticas, como maior penetração
de luz (Laurance et al., 2002). No entanto,
essas alterações ocorrem de forma distinta
dependendo do tipo de ambiente.
Para o bioma Caatinga,
representado por florestas com dossel
baixo e bem aberto, é provável que as
condições abióticas da borda e do interior
não apresentem grandes diferenças, desta
forma, o efeito de borda pode ser menos
pronunciado (Silva, 2009).
Com o intuito de avaliar o efeito de
borda, este estudo pretende comparar a
riqueza de espécies de plantas lenhosas em
dois ambientes: borda e interior da
Caatinga.
Metodologia
O estudo foi realizado na Estação
Ecológica do Seridó, localizada no
município de Serra Negra do Norte, Rio
Grande do Norte. A área encontra-se no
bioma da caatinga e está entre as mais
impactadas pela ação antrópica (IBAMA,
2004).
Na amostragem foram utilizados
dois transectos, paralelos a estrada e
distantes 50 m um do outro, divididos em
10 quadrados de 10m². Para a coleta de
dados foram selecionados
randomicamente três quadrados para cada
ambiente(fig. 1).
Em cada quadrado amostrado
foram consideradas apenas as plantas
maiores que um metro de altura, com o
intuito de selecionar as adultas. Os
indivíduos foram identificados em
morfotipos com base no guia de campo de
árvores da Caatinga (Santos et al, 2009) e,
posteriormente, contados.
Para analisar a estrutura da
vegetação nos dois ambientes foi medido o
diâmetro do tronco à altura solo (DAS) e
estimada a altura da copa de dois

37 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
indivíduos de cada espécie para cada
quadrado.
Figura 1. Delineamento amostral destacando os três quadrados (cinza claro) selecionados para estudo na borda e no interior da Caatinga.
Aplicou-se análises de variância
(ANOVA) para comparar as médias de
riqueza, abundância, dominância,
equitabilidade e diversidade (índice de
Shannon-Wiener) entre os ambientes. A
análise de covariância (ANCOVA) foi
utilizada para testar a relação do DAS com
altura para a espécie dominante Croton
sonderianus (família Euphorbiaceae) nos
dois ambientes e semelhança entre os
quadrados. Todas as análises foram feitas
através do programa Systat 12.0.
Resultados
A vegetação do local amostrado é
formada por uma floresta baixa com dossel
médio de cinco metros. As árvores e
arbustos são freqüentes e não há
vegetação rasteira. A vegetação
acompanha um córrego temporário com
leito rochoso de dois metros de largura,
porém, o dossel não é totalmente fechado.
Foram identificados 15 morfotipos
de plantas lenhosas (Tabela 1), das quais
três foram exclusivas da área de borda,
duas exclusivas do interior e 10 foram
comuns em ambos os ambientes (Figura 2).
Figura 2. Diagrama de Venn mostrando a riqueza e similaridade de espécies entre as áreas de borda e interior.
Tabela 1. Lista de morfotipos e abundância em
cada ambiente.
ESPÉCIES
(ou morfotipo) BORDA INTERIOR
Croton sonderianus 151 263
Bauhinia cheilantha 20 0
Peludinha 14 1
Mimosa hostilis 13 6
Cheirosinha 11 3
Mufumbo 9 8
Aspidosperma
pyrifolium 7 6
Caesalpinia
pyramidalis 6 13
Piptadenia
stipulaceae 6 5
Mônica 3 0
P. piauhiensis 3 1
Acerola da Caatinga 2 0
Folha dura 1 4
Commiphora
leptopholeus 0 2
Angico 0 2
A
INTERIOR BORDA

38 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
BORDA INTERIOR
ÁREA
0
1
2
3
4
5
6
AL
TU
RA
(m
)
BORDA INTERIOR
ÁREA
-50
0
50
100
150
200
DA
S (
mm
)
Não houve diferenças significativas
entre as duas áreas analisadas quanto às
variáveis, riqueza, abundância, diversidade,
dominância e equitabilidade (Tabela 2).
Em relação à estrutura da
comunidade, foi constatado que as
espécies do interior atingem tamanhos
significativamente maiores do que as da
borda (F = 19,48; gl = 1,8; p < 0,01). No
entanto, o DAS não apresentou diferenças
significativas entre os dois sítios analisados
(F = 0,005; gl = 1,82; p = 0,9) (Figuras 3 e
04).
Tabela 02. Resultados das ANOVAS para as
variáveis estudadas entre as duas áreas.
F gl p-valor
Riqueza 2,000 1 0,230
Abundância 2,035 1 0,227
Diversidade 3,415 1 0,138
Dominância 3,724 1 0,126
Equitabilidade 3,924 1 0,119
A relação positiva entre a altura da
planta e DAS foi marginalmente
significativa. A análise de covariância
(ANCOVA) realizada com a espécie mais
abundante e mais frequente nos dois
ambientes, o Croton sonderianus (Tabela
1), mostrou que a relação entre altura da
planta e DAS não foi significativamente
diferente (F = 0,258; gl = 1; p = 1,0624)
entre as duas regiões (Figura 5).
Figura 3. Comparação das médias da altura
entre as duas áreas analisadas.
Figura 4. Comparação da média do DAS
entre as duas áreas analisadas.
Figura 5. Relação entre altura e DAS do
Marmeleiro para as áreas de borda e
interior.

39 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Discussão
A criação de áreas de
bordas, segundo Laurance et al. (2002),
pode afetar a estrutura da comunidade
dentro do ecossistema. No entanto, este
comportamento não foi observado para a
área de Caatinga estudada, sugerindo que
o efeito de borda não influencia a
composição da comunidade ao longo do
gradiente estudado.
Para Prado (2003), características
peculiares deste bioma modelam a vida
animal e vegetal. Assim, a formação de
bordas pode não ter influência nas
condições microclimáticas.
Foi observado que a altura
das plantas, o diâmetro do caule ao nível
do solo, a riqueza e a abundância de
espécies não foram influenciadas pela
proximidade com a borda. Este padrão
também foi encontrado em um estudo
anterior na Caatinga (Santos & Santos,
2008) que encontrou baixa alteração na
estrutura e composição da vegetação
arbustiva neste bioma.
Apesar da similaridade
entre as espécies nos dois ambientes, em
cada um deles foram encontradas espécies
exclusivas (ver Figura 2). Este padrão pode
ser explicado pela heterogeneidade
ambiental criada pela criação da borda, o
que pode favorecer o crescimento de
espécies pioneiras e adaptadas a
ambientes perturbados. Bem como, pelo
acaso já que as amostras são pequenas.
Adicionalmente, mudanças em escala local
podem ser associadas a uma alteração na
condição ambiental, como em um
gradiente de umidade em direção a um
corpo d’água (Amorim et al., 2005).
A maior abundância para o
Croton sonderianus pode estar relacionada
ao grau de perturbação da área de estudo,
pois sabe-se que espécies de
Euphorbiaceae são apontadas como um
importante grupo colonizador desse tipo
de ambiente (Araújo et al., 2005 apud Leal,
2008).
A altura e o DAS dos
indivíduos de Croton sonderianus não
mostraram diferenças entre os dois
ambientes. O mesmo foi observado por
Leal (2008) em um estudo do efeito de
borda sobre a família Euphorbiaceae em
áreas de Caatinga. Este padrão corrobora a
hipótese da ausência do efeito de borda
neste bioma, sendo a estrutura vegetal
mais afetada por outras variáveis
ambientais, como por exemplo, a
disponibilidade de água.
Agradecimentos
À professora Adriana Monteiro pela
orientação no projeto. Aos professores do
Programa de Pós- Graduação em Ecologia –
PPGE, em especial a Carlos Fonseca pela ajuda
nos testes estatísticos. Agradecimentos
especiais aos funcionários da ESEC pela
hospitalidade e por permitir o uso de toda a
infra-estrutura da reserva.
Referências
AMORIM, I.L.; SAMPAIO, E.V.S.B.; ARAUJO, E.L.
2005. Flora e estrutura da vegetação
arbustivo-arbórea de uma área de caatinga
do Seridó, RN, Brasil. Acta Botânica
Brasílica, 19 (3): 615 – 623.
IBAMA – Instituto Brasileiro do Meio Ambiente
e dos Recursos Naturais Renováveis. 2004.
Plano de Manejo da Estação Ecológica do
Seridó. Relatório Técnico não publicado.
LAURANCE, W.F.; LOVEJOY, T.E.;
VASCONCELOS, H.L.; BRUNA, E.M.;
DIDHAM, R.K.; STOUER, P.C.; GASCON, C.;

40 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
BIERRE-GAARD, R.O.; LAURANCE, S.G.;
SAMPAIO, E. 2002. Ecosystem decay of
Amazonian forest fragments: A 22 - year
investigation. Conservation Biology, 16: 605
– 618.
LEAL, L.C. 2008. As bordas influenciam a
densidade e a riqueza de Euphorbiaceae em
áreas de Caatinga? In: LEAL, I.R.; ALMEIDA-
CORTEZ, J.; SANTOS, J.C. Ecologia da
Caatinga. Curso de Campo 2008, 332p.
PRADO, D. E. 2003. As Caatingas da América do
Sul. In: LEAL, I.R.; TABARELLI, M.; SILVA,
J.M.C. Ecologia e Conservação da Caatinga,
Recife: Ed. Universitária da UFPE, 2003. p. 3-
73.
SANTOS, A.M.M.; SANTOS, B.A. 2008. Are
the vegetation structure and composition of
the shrubby Caatinga free from edge in
fluence? Acta Botânica Brasílica, 22: 1077 –
1084.
SANTOS, A.P.B.; NASCIMENTO, M.F.S.; SANTO,
F.S.E. 2009. Guia de Campo de árvores da
Caatinga. Vol. 1. Petrolina: Editora e Gráfica
Franciscana Ltda. 64 p.
SILVA, E. E. S. 2009. Existe efeito de borda
sobre a comunidade de plântulas na
Caatinga? In: Congresso de Ecologia do
Brasil, 1 ed., 2009. Anais do IX Congresso de
Ecologia do Brasil. São Lourenço.
SYSTAT 12 © Copyright 2007, SYSTAT Software,
Inc. All Rights Reserved.

41 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
INFLUÊNCIA DO USO DO SOLO SOBRE A COMPOSIÇÃO DA TAXOCENOSE
DE FORMIGAS EM DUAS ÁREAS DE CAATINGA
HEITOR B.A. SOUZA, IZABEL M. M. SOUZA, KAROL L. B.O. FRANÇA, LEONARDO H.T. PINTO,
MAÍRA F. MANZAN
Introdução
Pertencentes à classe
Hymenoptera e à família Formicidae, as
formigas são encontradas na maioria dos
ecossistemas terrestres do globo. Apesar
de representarem apenas 1,5% da fauna de
insetos descrita, somam mais de 10% da
biomassa total de animais em diversos
habitats importantes do planeta (Agosti et
al., 2000), influenciando diversos
organismos de diferentes níveis tróficos e
participando ativamente da ciclagem de
nutrientes tanto através da ação de
herbivoria (Wirth et al., 1997 ) quanto pela
disponibilização de nutrientes para as
plantas (Hölldobler & Wilson, 1990).
Ademais, em relação a outros
animais, as formigas são organismos
especialmente apropriados para serem
usados como bioindicadores da qualidade
ambiental, pois são sensíveis às mudanças
nas condições do ambiente, apresentam
alta abundância e riqueza, além de
apresentarem muitos táxons
especializados, distribuição geográfica
ampla e serem facilmente amostrados e
separados em morfo-espécies (Silva &
Brandão, 1999). Neste sentido, o estudo de
comunidades locais de formigas tem se
mostrado uma valiosa ferramenta de
avaliação das condições ambientais em
estudos de acompanhamento de áreas
degradadas e dos diferentes padrões de
uso do solo.
Sendo assim, esse trabalho
procurou avaliar o potencial bioindicador
da taxocenose de formigas na qualidade de
solos. Para isso, testou-se a hipótese de
que a riqueza de formigas seria maior em
áreas com menor impacto sobre o solo,
buscando compreender quais fatores
estariam relacionados a esse padrão.
Material e métodos
Área de estudo
A área de coleta se divide em duas
subáreas de fragmentos de Caatinga com
diferentes estados de conservação, cada
uma com 1 ha. A primeira localiza-se
dentro da ESEC – Seridó e foi tomada como
controle por estar preservada desde a
criação da Estação Ecológica em 1982. A
segunda área localiza-se na fazenda
Solidão, onde ocorre pecuária extensiva.
Dentro de cada área foram sorteados cinco
plots, cada qual com 10m² para a
realização da coleta dos parâmetros
ambientais e bióticos a serem analisados.
Coleta de dados
Em cada plot foram medidos o pH
e a umidade do solo com a utilização de
um medidor analógico Instrutherm
pH2500, bem como a compactação, com o
auxílio de um compactômetro. Mediu-se o
DAP (diâmetro da altura do peito) para os
ramos individuais de todas as plantas com
diâmetro superior a 5 cm que,
posteriormente, foram substituídos na

42 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
equação desenvolvida por Sampaio & Silva
(2005) para o cálculo da biomassa vegetal
aérea. Também foi determinado o peso da
necromassa vegetal em uma área de
0,25m2 em cada um dos cinco plots das
áreas amostrais.
Para a amostragem de formigas
foram utilizados quatro métodos de coleta:
i. Guarda-chuva entomológico, para a
coleta de organismos da parte aérea de
três árvores de cada plot; ii. Sweep, para
coleta de organismos no extrato herbáceo
com três amostras e duas repetições; iii.
Isca, composta de água e açúcar, para a
coleta de organismos no solo, disposta
durante 20 minutos; e iv. Coleta de
necromassa vegetal para triagem das
formigas presentes na liteira.
Análise dos dados
Para verificar se há diferenças na
riqueza de espécies entre as duas áreas
estudadas, os dados foram submetidos a
uma ANOVA univariada. Para testar se
diferenças na riqueza de espécies entre as
áreas ocorria em decorrência das variáveis
abióticas avaliadas foi utilizada a
ferramenta Modelo Linear Geral (GLM) no
software Systat 12.
Resultados
Foram encontradas seis espécies
de formigas, das quais todas ocorreram na
área controle e apenas quatro foram
encontradas na área onde é praticada
pecuária extensiva (Tabela 01).
Foi constatado que há diferença
significativa na riqueza de espécies (F=
16,9; gl= 1,8; p<0,05).
Verificou-se que a área controle
apresentou uma riqueza de espécies maior
que na área da fazenda, provavelmente
pelo maior pisoteio, conforme pode ser
visualizado na figura 1.
Tabela 1. Espécies encontradas nas áreas
amostradas
Espécies
Fazenda
Solidão
ESEC
Seridó
Formicidae sp. 4 19
Crematogaster sp. 10 29
Solenopsis sp. 3 9
Camponotus sp1 13 5
Camponotus sp1 0 8
Cephalotes sp. 0 8
Total 30 58
Figura 1. Riqueza média de espécies de
formigas nas áreas em estudo. As barras
verticais representam erro padrão.
Não houve relação significativa
entre variáveis abióticas e a riqueza de
espécies, conforme visualizado na tabela
02.
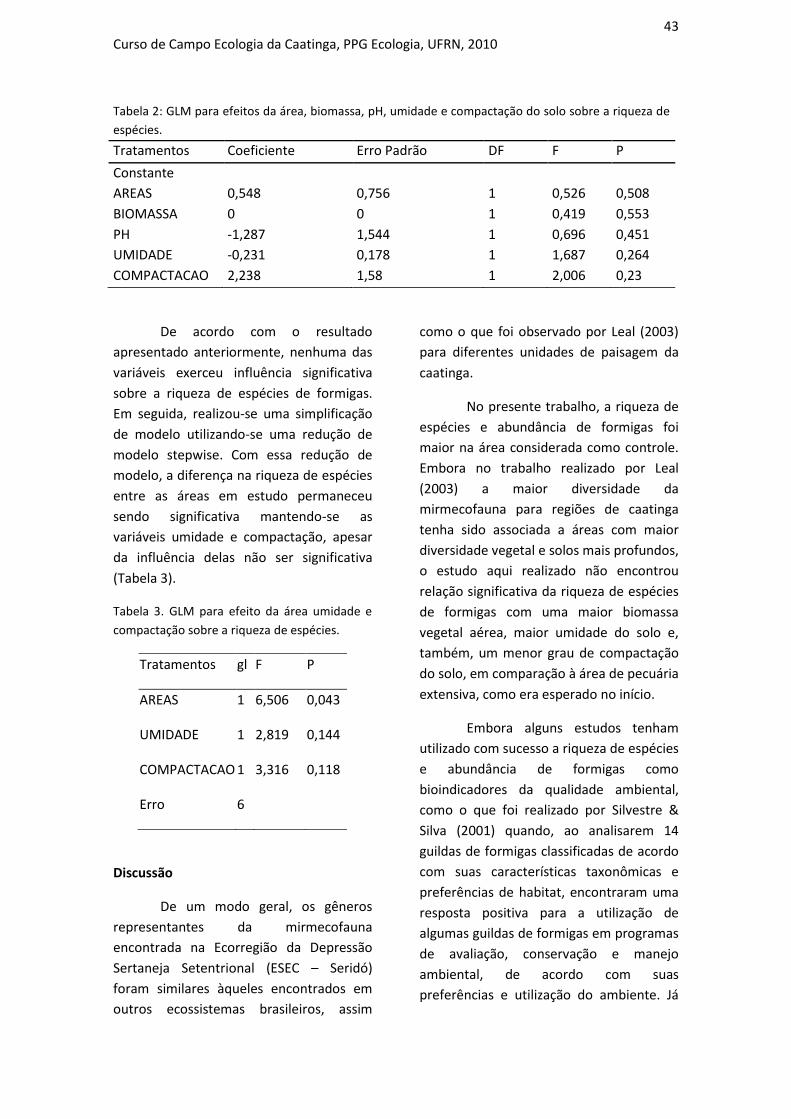
43 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
De acordo com o resultado
apresentado anteriormente, nenhuma das
variáveis exerceu influência significativa
sobre a riqueza de espécies de formigas.
Em seguida, realizou-se uma simplificação
de modelo utilizando-se uma redução de
modelo stepwise. Com essa redução de
modelo, a diferença na riqueza de espécies
entre as áreas em estudo permaneceu
sendo significativa mantendo-se as
variáveis umidade e compactação, apesar
da influência delas não ser significativa
(Tabela 3).
Tabela 3. GLM para efeito da área umidade e
compactação sobre a riqueza de espécies.
Tratamentos gl F P
AREAS 1 6,506 0,043
UMIDADE 1 2,819 0,144
COMPACTACAO 1 3,316 0,118
Erro 6
Discussão
De um modo geral, os gêneros
representantes da mirmecofauna
encontrada na Ecorregião da Depressão
Sertaneja Setentrional (ESEC – Seridó)
foram similares àqueles encontrados em
outros ecossistemas brasileiros, assim
como o que foi observado por Leal (2003)
para diferentes unidades de paisagem da
caatinga.
No presente trabalho, a riqueza de
espécies e abundância de formigas foi
maior na área considerada como controle.
Embora no trabalho realizado por Leal
(2003) a maior diversidade da
mirmecofauna para regiões de caatinga
tenha sido associada a áreas com maior
diversidade vegetal e solos mais profundos,
o estudo aqui realizado não encontrou
relação significativa da riqueza de espécies
de formigas com uma maior biomassa
vegetal aérea, maior umidade do solo e,
também, um menor grau de compactação
do solo, em comparação à área de pecuária
extensiva, como era esperado no início.
Embora alguns estudos tenham
utilizado com sucesso a riqueza de espécies
e abundância de formigas como
bioindicadores da qualidade ambiental,
como o que foi realizado por Silvestre &
Silva (2001) quando, ao analisarem 14
guildas de formigas classificadas de acordo
com suas características taxonômicas e
preferências de habitat, encontraram uma
resposta positiva para a utilização de
algumas guildas de formigas em programas
de avaliação, conservação e manejo
ambiental, de acordo com suas
preferências e utilização do ambiente. Já
Tabela 2: GLM para efeitos da área, biomassa, pH, umidade e compactação do solo sobre a riqueza de
espécies.
Tratamentos Coeficiente Erro Padrão DF F P
Constante
AREAS 0,548 0,756 1 0,526 0,508
BIOMASSA 0 0 1 0,419 0,553
PH -1,287 1,544 1 0,696 0,451
UMIDADE -0,231 0,178 1 1,687 0,264
COMPACTACAO 2,238 1,58 1 2,006 0,23

44 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Silva & Brandão (1999), revisando a
literatura sobre o uso de diferentes
gêneros de formigas como bioindicadores
e indicadores de biodiversidade concluíram
que existe correlação entre as variações
nas condições ambientais e os padrões de
resposta dos organismos estudados.
Contudo, no presente trabalho não
foi possível analisar a resposta da
taxocenose de formigas às variações nas
condições ambientais.
Sendo assim, dentro do universo
abordado, a riqueza de espécies de
formigas é significativamente diferente
entre as duas áreas com diferentes
estratégias de uso do solo, corroborando a
hipótese teórica inicial e sugerindo o
potencial de utilização da mirmecofauna
como possível bioindicadora das variações
no uso do solo. Além disso, conclui-se que
nenhum dos fatores ambientais abordados
foi responsável para a ocorrência dessa
diferença na riqueza de espécies.
Agradecimentos
Ao professor Dr. Alexandre
Vasconcellos pelo desenvolvimento e apoio
para a realização desse trabalho e ao professor
Dr. Carlos Fonseca pelo auxílio fornecido nas
análises estatísticas. E, por fim, aos
funcionários da ESEC pela cordialidade e
hospitalidade com a qual nos receberam.
Referências
AGOSTI, D. J.D. MAJER, ALONSO, L. E. &
T.SCHULTZ. 2000. Standard methods for
measuring and monitoring biodiversity.
Smithsonian institution Press, Washington.
HÖLLDOBLER, B. & WILSON, E. O. 1990. The
Ants. Havard University Press, Cambridge.
LEAL, I. R. Diversidade de formigas em
diferentes unidades de paisagem da
caatinga 2003 in LEAL, I. R. TABARELLI , M. &
SILVA, J. M. C. Ecologia e Conservação da
Caatinga. Editora Universitária da UFPE,
Recife/PE, Pp: 804.
SAMPAIO, E. V. S. B. & SILVA, G. C. 2005.
Biomass equations for Brazilian semiarid
caatinga plants. Acta botanica brasilica
19(4): 935-943.
SILVA, R. R. & BRANDÃO, C. R. F. 1999.
Formigas (Hymenoptera: Formicidae)como
indicadores da qualidade ambiental e da
biodiversidade de outros invertebrados
terrestres. Biotemas 12(2) : 55 – 73.
SILVESTRE, R. & SILVA, R. R. Guildas de formigas
da Estação Ecológica Jataí, Luiz Antônio – SP
– sugestões para aplicação do modelo de
guildas como bio-indicadores ambientais.
Biotemas, 14 (1): 37 – 69, 2001.
WIRTH R, W. BEYSCHLAG, R. J. RYEL & B.
HÖLLDOBLER. 1997. Annual foraging of the
leaf-cutting ant Atta colombic in a
semideciduos rain forest in Panama. Journal
of Tropical Ecology 13: 741-757.

45 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010

46 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
INTERAÇÃO TRITRÓFICA EM Mimosa hostilis: HERBÍVOROS PODEM SER
BENÉFICOS PARA AS PLANTAS?
HEITOR B. SOUZA, IZABEL M.M. SOUZA, KAROL L. B.O. FRANÇA, LEONARDO H.T. PINTO,
MAÍRA F. MANZAN
Introdução
De acordo com Begon (2007), os
organismos mutualistas têm sido muitas
vezes negligenciados, mesmo constituindo
a maior parte da biomassa do mundo. O
mutualismo se caracteriza pela interação
entre organismos de espécies diferentes
que se beneficiam mutuamente. É errado,
no entanto, pensar nesta interação como
uma relação sem conflitos. Estudos
recentes têm mostrado que esta interação
é, na realidade, mais relacionada com uma
exploração recíproca.
A variação espaço-temporal em
relações de mutualismo entre plantas e
formigas tem sido principalmente atribuída
as mudanças na interação entre fatores
bióticos. Por exemplo, o aumento das
relações formiga-planta em uma planta
Amazônica do gênero Tachigali determina
um padrão ontogenético de sucessão, que
involve oito diferentes formigas parceiras,
gerando um número de interações bióticas
indiretas (Fonseca & Benson 2003).
Alguns trabalhos têm mostrado
relações mutualísticas bastante evidentes
entre formigas e plantas e entre formigas e
homópteros. A presença de nectários atrai
formigas, por representarem uma fonte
rica de açúcares. Dessa forma, as formigas
defendem o seu recurso de potenciais
competidores, culminando em defesa
indireta contra herbívoros para a planta. A
presença de homópteros, os quais são
“cultivados” pelas formigas em troca de
secreções ricas em açúcar, também pode
promover uma relação positiva indireta
formiga-planta. Assim, na defesa do seu
recurso, as formigas protegem os afídeos
contra predação e a planta contra
herbivoria (Begon 2007). Nesse contexto,
Fonseca (2004) verificou que o aumento no
número de formigas faz com que ocorra a
diminuição da herbivoria e aumento da
estimativa de vida da planta.
Nesse sentido, o objetivo do
presente estudo foi verificar se a presença
dos herbívoros homópteros promove
algum benefício para a planta Mimosa
hostilis, na presença de uma formiga
predadora do gênero Camponotus.
Material e Métodos
Foram sorteadas 25 árvores ao
longo da estrada de acesso da Estação
Ecológica do Seridó – ESEC, em 15 das
quais foi realizado o experimento de
indução da resposta condicional da
formiga, enquanto a partir das demais
realizou-se o censo de estruturas
vegetativas e componentes da fauna
freqüentadora.
Coleta de dados
O censo foi realizado a partir do
sorteio de três ramos localizados na parte
inferior do extrato arbóreo das 10 árvores
citadas. Foram registrados o número de
folhas e o nível de herbivoria, classificado
em 4 categorias: 0) sem perda de folíolos ;
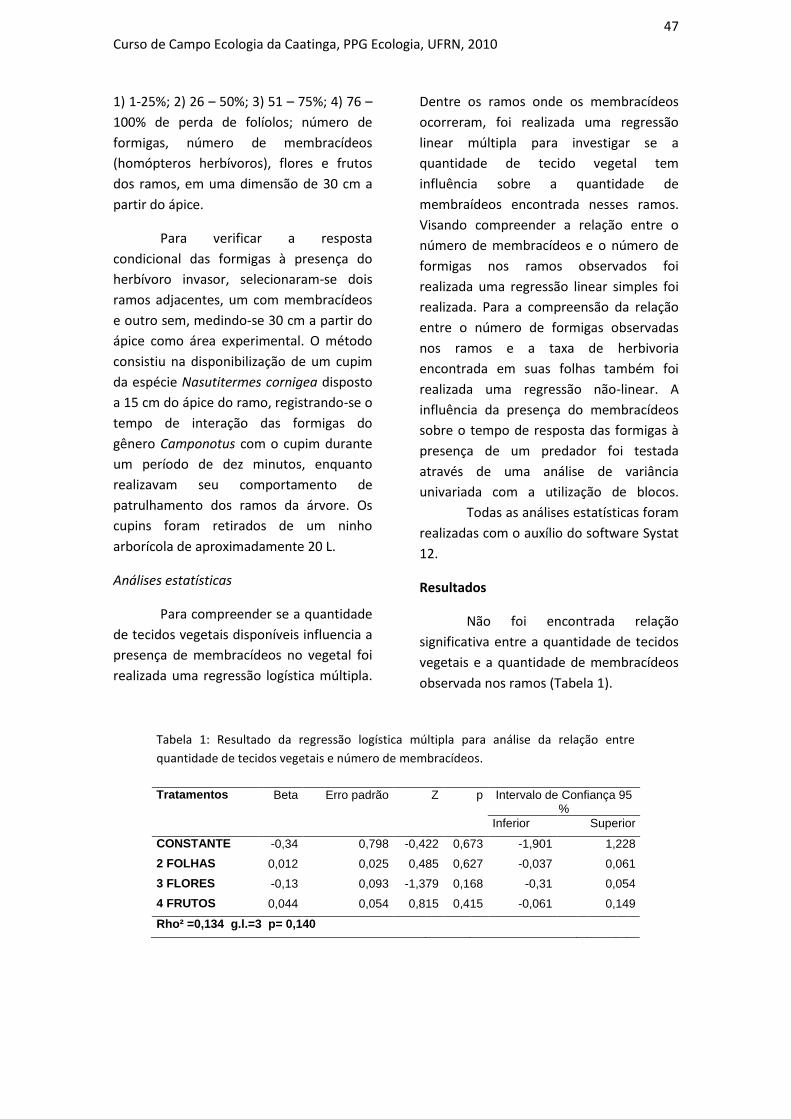
47 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
1) 1-25%; 2) 26 – 50%; 3) 51 – 75%; 4) 76 –
100% de perda de folíolos; número de
formigas, número de membracídeos
(homópteros herbívoros), flores e frutos
dos ramos, em uma dimensão de 30 cm a
partir do ápice.
Para verificar a resposta
condicional das formigas à presença do
herbívoro invasor, selecionaram-se dois
ramos adjacentes, um com membracídeos
e outro sem, medindo-se 30 cm a partir do
ápice como área experimental. O método
consistiu na disponibilização de um cupim
da espécie Nasutitermes cornigea disposto
a 15 cm do ápice do ramo, registrando-se o
tempo de interação das formigas do
gênero Camponotus com o cupim durante
um período de dez minutos, enquanto
realizavam seu comportamento de
patrulhamento dos ramos da árvore. Os
cupins foram retirados de um ninho
arborícola de aproximadamente 20 L.
Análises estatísticas
Para compreender se a quantidade
de tecidos vegetais disponíveis influencia a
presença de membracídeos no vegetal foi
realizada uma regressão logística múltipla.
Dentre os ramos onde os membracídeos
ocorreram, foi realizada uma regressão
linear múltipla para investigar se a
quantidade de tecido vegetal tem
influência sobre a quantidade de
membraídeos encontrada nesses ramos.
Visando compreender a relação entre o
número de membracídeos e o número de
formigas nos ramos observados foi
realizada uma regressão linear simples foi
realizada. Para a compreensão da relação
entre o número de formigas observadas
nos ramos e a taxa de herbivoria
encontrada em suas folhas também foi
realizada uma regressão não-linear. A
influência da presença do membracídeos
sobre o tempo de resposta das formigas à
presença de um predador foi testada
através de uma análise de variância
univariada com a utilização de blocos.
Todas as análises estatísticas foram
realizadas com o auxílio do software Systat
12.
Resultados
Não foi encontrada relação
significativa entre a quantidade de tecidos
vegetais e a quantidade de membracídeos
observada nos ramos (Tabela 1).
Tabela 1: Resultado da regressão logística múltipla para análise da relação entre
quantidade de tecidos vegetais e número de membracídeos.
Tratamentos Beta Erro padrão Z p Intervalo de Confiança 95 %
Inferior Superior
CONSTANTE -0,34 0,798 -0,422 0,673 -1,901 1,228
2 FOLHAS 0,012 0,025 0,485 0,627 -0,037 0,061
3 FLORES -0,13 0,093 -1,379 0,168 -0,31 0,054
4 FRUTOS 0,044 0,054 0,815 0,415 -0,061 0,149
Rho² =0,134 g.l.=3 p= 0,140

48 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Tabela 2: Resultado da regressão linear múltipla para análise da relação entre quantidade de
tecidos vegetais e número de membracídeos.
Tratamentos Coeficientes Erro Padrão t p
CONSTANTE 4,534 1,862 2,435 0,022
FOLHAS -0,017 0,056 -0,302 0,765
FRUTOS 0,02 0,094 0,21 0,835
FLORES -0,286 0,169 -1,69 0,1
R²=0,105
Dentre os ramos que apresentaram
membracídeos, não houve relação
significativa entre a quantidade de tecidos
vegetais e o número de membracídeos
(tabela 2).
Foi encontrada relação significativa
entre o número de formigas e o número de
membracídeos nos ramos observados
(Tabela 3).
A relação entre o número de
formigas e a taxa de herbivoria apresentou
um comportamento onde, quando não
havia formigas ou quando o número de
formigas era muito pequeno, a taxa de
herbivoria continha bastante variação.
Quando o número de formigas foi maior, a
taxa de herbivoria foi reduzida
progressivamente (Figura 1). A regressão
não-linear, utilizando um modelo
polinomial apresentou um valor de F=58,53
p<0,001
Tabela 3: Análise de regressão linear simples da
relação entre o número de formigas e o número de
membracídeos.
Tratamentos Coeficientes Erro
padrão t p
CONSTANTE 0,143 0,298 0,481 0,635
MEMBRACIDAE 0,444 0,05 8,921 0
R²= 0,74
Figura 1: Variação da taxa de herbivoria em
função do número de formigas.
A análise de variância univariada
em blocos revelou que as formigas
encontravam o herbívoro num tempo
significativamente menor quando havia a
presença do Membracidae, conforme pode
ser verificado na figura 2 F=7,601; ( g.l. = 1;
p=0,010).

49 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Figura 2: Tempo de resposta da formiga (Log10)
à presença do cupim.
Discussão
A falta de significância na relação
entre o número de tecidos vegetais e a
presença ou abundância de membracídeos
nos ramos de Mimosa hostilis parece
indicar que os membracídeos não são
afetados pela maior disponibilidade de
tecidos vegetais passíveis de serem
parasitados.
O número de formigas em cada
ramo de Mimosa hostilis, por sua vez, foi
positivamente afetado pela abundância de
membracídeos, evidenciando a
importância desses organismos na dieta
das formigas do gênero Camponotus.
Contudo, um estudo realizado por Bentley
(1976) mostrou que os nectários
extraflorais são importantes componentes
na dieta de uma comunidade de formigas
visitantes de plantas em uma área de
floresta tropical na Costa Rica, onde a
presença das formigas exerceu um efeito
de redução da herbivoria, que foi mais
acentuado em áreas perturbadas. Por
outro lado, Lima Jr. (2004) comparando as
diferenças de atração de formigas entre
nectários extraflorais e herbívoros
homópteros, não encontrou diferenças
significativas entre ambos os recursos
ofertados às formigas em um experimento
realizado na área do Projeto Dinâmica
Biológica de Fragmentos Florestais em
Manaus, AM, durante um curso de campo.
No presente estudo, por sua vez,
não foram contabilizados os números de
nectários extraflorais presentes nas folhas
de Mimosa hostilis, portanto não há
possibilidade de se fazer uma comparação
mais aprofundada com os resultados de
Bentley (1976), buscando compreender
quais vegetais influenciam a presença ou
abundância de membracídeos em plantas
de Mimosa hostilis, entretanto, foi
observada uma forte correlação entre o
número de formigas e o número de
membracídeos, assim como, uma redução
progressiva nas taxas de herbivoria a partir
de uma maior abundância de formigas.
Trabalhos experimentais
manipulando a presença de formigas têm
demonstrado um profundo impacto sobre
a sobrevivência de plantas a sobre o
destino dos herbívoros que as consomem
(Fonseca 1994, Kersch & Fonseca 2005). O
presente estudo encontrou uma relação
significativamente positiva entre a
presença de membracídeos nos ramos de
Mimosa hostilis e o tempo da resposta
condicional das formigas, sendo esse
tempo menor nos ramos com
membracídeos, assim como foi esperado
inicialmente.
Essa interação indica que as
formigas do gênero Camponotus atuam de
forma a proteger os membracídeos contra
possíveis predadores, devido à sua
importância como fonte de açúcares e,
indiretamente, são benéficas para as
plantas analisadas, participando de duas
interações mutualísticas, sendo a primeira

50 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
formiga-membracídeo e, após interagir
agonisticamente com os organismos
herbívoros invasores, formiga-planta.
Sendo assim, pode-se concluir que
dependendo das interações existentes, a
presença do herbívoro membracídeo
(Homoptera) será benéfica para Mimosa
hostilis.
Agradecimentos
Ao professores, Dr. Carlos Fonseca e Dra.
Adriana Almeida por toda orientação e apoio
para a realização desse trabalho. E, por fim, aos
funcionários da ESEC pela cordialidade e
hospitalidade com a qual nos receberam.
Referências
Begon, M., Townsend, C.R. & Harper, J.L., 2007.
Ecologia: de indivíduos ecossistemas. Cap
13, pp. 381.
Bentley, B. L., 1976. Plants bearing extrafloral
nectaries and the associated ant
community: interhabitat diferences in the
reduction of herbivore damage. Ecology, 57:
pp. 815-820.
Fonseca, C. R., 1994. Herbivory and long-lived
leaves of in Amazonian ant-tree. Journal of
Ecology 82, 833-842.
Fonseca, C. R. & Benson, W. W., 2003.
Ontogenetic sucession in Amazonian ant
trees. Oikos, 102:2.
Kersch, M. F. & Fonseca, C. R., 2005. Abiotic
factors and conditional outcome of an ant-
plant mutualism. Ecology, 86(8), pp. 2117-
2126.
Lima Jr., D. P., 2004. Curso de Campo em
Biologia da Conservação,
PDBFF/INPA/Smithsonian Institution.
Foto. Camponotus sp. e Membracidae em ramo. Foto do grupo de trabalho.

51 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
NINHOS ABANDONADOS DE TÉRMITAS POSSUEM UMA FAUNA PECULIAR
DE INVERTEBRADOS: UM ESTUDO COM Constrictotermes cyphergaster NA
CAATINGA
ALLAN T. BATISTA; FRANÇOISE. D. LIMA; VINÍCIUS A. SÃO PEDRO; MONALISA R. O. DA SILVA &
RODRIGO X. SOARES
Introdução
Os térmitas estão entre os grupos de
invertebrados mais abundantes, sendo
amplamente distribuídos nos trópicos e
subtrópicos (Araújo, 1970). Eles são
considerados espécies “engenheiras de
ecossistemas”, pois constroem ninhos
formados por partículas do solo,
excrementos e restos vegetais, os quais
influenciam a composição química e física
do solo, alterando a disponibilidade de
recursos para outros organismos, (Jones,
1994). Além disso, os ninhos abandonados
de térmitas podem ser ocupados por
outros animais conhecidos como
inquilinos. Nesse sentido, eles são
importantes sítios de abrigo, reprodução e
alimentação para outros invertebrados,
como formigas, abelhas e aranhas, e até
mesmo vertebrados, como lagartos e aves
(Moura et al., 2006 ).
A espécie Constrictotermes cyphergaster
ocorre no Paraguai, Bolívia, nordeste da
Argentina (Constantino, 1998) e no Brasil é
encontrado no cerrado e na caatinga, onde
constroem ninhos conspícuos (Lima-
Ribeiro, 2006; Mélo & Bandeira 2004).
Nesses ambientes, os ninhos podem atuar
como um elemento importante de
heterogeneidade do habitat, funcionando,
dessa forma, como fator de aumento de
diversidade de invertebrados de solos.
Assim, sua importância fica evidenciada
principalmente em regiões que vêm
sofrendo impactos constantes, como
muitas áreas de Caatinga (IBAMA, 2004).
Tendo em vista a importância dos ninhos
abandonados como um fator de
heterogeneidade do ambiente, este estudo
propõe uma comparação na composição
de invertebrados de solo em quatro tipos
de micro-habitats em uma região de
caatinga: ninhos abandonados de cupins,
serrapilheira, troncos caídos e sob pedras.
Metodologia
O estudo foi realizado na Estação Ecológica
do Seridó, localizada no município de Serra
Negra do Norte, no Rio Grande do Norte. A
área encontra-se na Caatinga e está entre
as mais impactadas pela ação antrópica na
região (IBAMA, 2004). A amostragem foi
realizada na Serra Verde, localizada na
porção norte da ESEC Seridó, a uma
altitude aproximada de 320 metros.
Foram selecionados quatro ninhos inativos
da espécie Constrictotermes cyphergaster
com volume aproximado de 30L cada.
Buscou-se por invertebrados habitando os
ninhos (N) e em três outros micro-habitats
que se encontravam localizados a pelo
menos 10 m do cupinzeiro abandonado:
serrapilheira (S), troncos caídos (T) e sob
pedras (P) (Figura 1).

52 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
10m
Figura 1. Desenho amostral em cada ninho de
cupim selecionado para coleta de invertebrados
nos micro-habitats.
As buscas por invertebrados foram
realizadas simultaneamente nos quatro
micro-habitats por um período de 20
minutos.
Os espécimes encontrados foram
analisados em laboratório com o auxílio de
um microscópio binocular e identificados
em morfoespécies.
Foi realizada uma análise de
Cluster (Bray-Curtis) para agrupar os micro-
habitats com composição de espécies
semelhantes; uma análise de
escalonamento multidimensional para
dados não paramétricos (nMDS), em um
corte 20% de similaridade, para melhor
visualizar o agrupamento dos micro-
habitats e um ANOSIM para diferir os
agrupamentos propostos pelo cluster. Para
tal foi usado o programa de estatística
denominado Primer.
Resultados
Foram obtidas 34 morfoespécies,
pertencentes a 12 grupos de
invertebrados, ocupando os quatro micro-
habitats (Tabela 1).
Tabela 1. Número de morfoespécies de
invertebrados registrados por grupo.
Taxon N. de espécies
Araneae 7 Blattaria 3
Chilopoda 2
Coleoptera 1
Embioptera 1
Formicidae 7
Gastropoda 5
Isopoda 2
Isoptera 3
Orthoptera 1
Scorpiones 1
Thysanura 1
Total 12 34
Figura 2. Riqueza de espécies de cada grupo de
invertebrados em cada micro-habitat.
Grupos distintos foram dominantes em
cada micro-habitat. Nos ninhos houve
predomínio de espécies de Araneae.
O grupo Formicidae foi mais rico na
serrapilheira e, juntamente com Isoptera,
dominou também nos troncos caídos. Sob
pedras o grupo com o maior número de
espécies foi Gastropoda (Figura 2).
Os quatro ninhos analisados mostraram
uma composição de morfoespécies similar,
Araneae

53 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
sendo agrupados dentro de um grupo
único, enquanto os demais micro-habitats,
serrapilheira, troncos caídos e sob pedras,
não mostraram a mesma tendência de se
agruparem, sendo assim mais
heterogêneos. O micro-habitat T1 (tronco
caído) mostrou uma composição de
espécies similar àquela dos ninhos, mas no
corte de 20% de similaridade, formou um
grupo separado dos demais (Figuras 3 e 4).
Figura 3. Análise de agrupamento da similaridade entre os micro-habitats. N = ninho; S = serrapilheira; T = tronco e P = pedra.
Figura 4. Análise nMDS mostrando agrupamento dos pontos de coleta em um corte de 20% de similaridade.
A composição de espécies dos
ninhos é significativamente diferente da
composição da serrapilheira (R = 0,849; p <
0,05) e da de pedras (R = 0,541; p < 0,05).
Esta diferença foi marginalmente
significativa (R = 0,474; p = 0,057) entre
ninhos e troncos.
Discussão
Sabe-se que os cupins atuam como
engenheiros de ecossistemas, pois
transformam a paisagem, aumentando a
disponibilidade de recursos para outras
espécies (Jones et al., 1994). Corroborando
essa perspectiva, os ninhos abandonados
de Constrictotermes cyphergaster
analisados, mostraram-se especiais em
relação aos demais micro-habitats, pois
possuíam uma composição peculiar de
invertebrados de solo. Isso mostra a
importância desses ninhos como um fator
adicional da heterogeneidade espacial.
Essa variedade de micro-habitat propicia
uma amplitude do espectro de recursos
disponibilizado para a comunidade desses
ambientes, como fontes de alimentação,
locais de refúgio e reprodução, microclimas
(Begon et al., 2007). Tal fato indica que os
ninhos abandonados são essenciais no
aumento e manutenção da diversidade
local.
Cada micro-habitat estudado
mostrou abrigar grupos relativamente
distintos de invertebrados. Isso se deve ao
fato de que as características de cada
micro-habitat favorecem determinados
grupos, conforme suas necessidades e
adaptações (Townsend, et al., 2008). Os
Gastropoda, por exemplo, estão associados
a ambientes mais úmidos e por isso são
predominantes sob as pedras. De alguma
forma, pode-se inferir que os ninhos
inativos de Constrictotermes cyphergaster
proporcionam condições favoráveis a
várias espécies de aracnídeos no ambiente
de caatinga. No entanto, estudos adicionais
relacionados às peculiaridades dos
organismos presentes nesses ninhos
Riq
uez
a d
e es
péc
ies

54 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
devem ser realizados a fim de comprovar
tal inferência.
Agradecimentos
Ao professor Dr. Alexandre
Vasconcellos pelo desenvolvimento e apoio
para a realização desse trabalho e ao professor
Dr. Carlos Fonseca pelo auxílio fornecido nas
análises estatísticas. E, por fim, aos
funcionários da ESEC pela cordialidade e
hospitalidade com a qual nos receberam.
Referências
Araújo, R.L. 1970. Termites of the Neotropical
Region, 527-576p. In K. Krishna & F. R.
Weesner (eds.), Biology of termites, v.
II. New York, Academic Press, 643p.
Begon M., Townsend C.R., Harper J.L. 2007.
Padrão na riqueza de species. In:
Ecologia de indivíduos a Ecossistemas.
4 ed. 740p.
Constantino, R. 1998. 2010. Catalog of the
living termites of the New World
(Insecta: Isoptera). Arq. Zool. (São
Paulo) 35: 135-231.
IBAMA – Instituto Brasileiro do Meio Ambiente
e dos Recursos Naturais Renováveis.
2004. Plano de Manejo da Estação
Ecológica do Seridó. Relatório Técnico
não publicado.
Jones C.G., Lawton, J.H., Sachak, M., 1994.
Organisms as a ecossystem engineers.
Oikos 69: 373-386.
Lima-Ribeiro, M.S.., Pinto, M.P., Costa, S. S.,
Nabout, J.C., Rangel, T., Melo, T.I.,
Moura, I., 2006. Associação de
Constrictotermes cyphergaster Silvestri
(Isoptera: Termitidae) com Espécies
Arbóreas do Cerrado Brasileiro.
Neotropical Entomology 35(1): 049-
055.
Melo, A.C.S., Bandeira, A.G., 2004. A qualitative
and quantitative survey of termites
(Isoptera) in an open shrubby Caatinga
in Northeast Brazil. Sociobiologia, 44:
707-716.
Moura F.M.S., Vasconcellos A. , Araffljo V.F.P.,
Bandeira A.G. 2006. Seasonality in
foraging behaviour of
Constrictotermes cyphergaster
(Termitidae, Nasutitermitinae) in the
Caatinga of Northeastern Brazil. Insect.
Soc. 53: 472 – 479.
Townsend C.R., Begon M., Harper J.L. 2008.
Padrão na riqueza de espécies. In:
Fundamentos em Ecologia. 3 ed. 576p
.

55 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
ODOR DA FUGA: EFEITO DA DETECÇÃO DE PREDADORES POR Seira sp.
(COLLEMBOLA; ENTOMOBRYIDAE)
CRISTIANE GOUVÊA FAJARDO, BRUNNO FREIRE DANTAS DE OLIVEIRA, FABRÍCIO RODRIGUES
CAMACHO, UIRANDÉ OLIVEIRA COSTA
Introdução
A injuria ou a morte de outro
indivíduo pode liberam sinais químicos
(Rollo et al., 1995), ou necromônios (Rollo
et al., 1994). Animais de solo podem
apresentar sensibilidade química para
reconhecer esses odores.
Presas podem responder a sinais
de predadores paralisando, reduzindo a
sua mobilidade ou escapar para se
defender (Dicke & Grostal, 2001). Nilson e
Bengtsson (2004a) observaram que
indivíduos de uma espécie de Collembola,
Protaphorura armata, podem reconhecer
sinais da presença de um predador se este
se alimentou de um coespecífico,
sugerindo sensibilidade a necromônios.
Os objetivos do presente estudo
foram testar se indivíduos de uma espécie
de Seira: (a) reconhecem e evitam locais
onde encontra-se um predador ou
coespecífico morto, distanciando-se deste
local e, (b) se a intensidade desta resposta
varia com o tempo de exposição ao
estímulo.
Materiais e Métodos
Neste trabalho foram utilizadas
espécimes de Seira sp. e um espécime de
aracnídeo, que foram coletados em uma
região de caatinga localizada dentro da
Estação Ecológica do Seridó – ESEC.
Dentro de placas de Petri foram
testados os efeitos de dois tratamentos
sobre a movimentação de Seira sp. no
tempo: um com um espécime coespecífico
morto e outro com um espécime de
predador morto. Cada espécime morto foi
posicionado no centro de uma placa de
Petri de 7 cm de raio localizada sobre um
painel com 14 círculos concêntricos,
espaçados 0,5cm entre si, de forma que
era possível visualizar as linhas do painel
através do vidro da placa (Figura 1). Cada
linha do circulo foi numerada para
observação da posição do espécime de
Seira sp. no decorrer do tempo. Também
foi realizado um terceiro tratamento
(controle), sem coespecífico morto ou
predador, para se comparar diferenças no
no distanciamento ao centro da placa por
Seria sp na ausência de estímulo. Cada
indivíduo testado passou somente por uma
vez pelo tratamento sendo, logo em
seguida, foi devolvido no campo.
Figura 1. Arena experimental utilizada para os
tratamentos.

56 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Para cada tratamento 10 réplicas
sequeciadas temporalmente foram
realizadas. A distância do indivíduo vivo em
relação ao centro foi registrada a cada 10
segundos, observando-se a posição do
animal em relação às linhas circulares
enumeradas do painel. Antes do início do
experimento o colêmbolo passou por um
tempo de aclimatação na placa de Petri de
20 segundos.
Os dados obtidos foram analisados
por meio de uma ANOVA de Medidas
repetidas, rodada no software Systat 12.
Resultados
A distância média de Seira sp. até o
centro da arena experimental foi similar
para os três tratamentos (F = 0,625 ; GL =
2,27 ; P = 0,543). O padrão de
distanciamento do centro foi similar no
decorrer do tempo do experimento (F=
1,268 ; GL = 29,783 ; P = 0,158), mesmo se
comparado entre tratamentos (F =0,877;
GL = 58,783; P = 0,729). A dinâmica
temporal de distanciamento do centro de
Seira sp. não foi significativo (Figura 2), de
modo que as observações em todos os
experimentos foram concentrados na
borda da placa de Petri (Figura 3), ou seja,
posição 14.
Figura 2: Dinâmica temporal das posições de
Seria sp. no tempo de acordo com os
tratamentos. Linha vermelha para tratamento
com predador, azul para coespecífico morto e
preto para controle.
Figura 3: Distribuição do número de
observações constatadas para cada posição do
colêmbolo na placa de Petri.
Discussão
Algumas espécies de colêmbolos
são capazes de identificar a presença de
predadores e coespecíficos mortos através
da detecção de feromônios, fugindo ou
evitando regiões onde o odor está
presente (Nilsson & Bengtsson, 2004a,b).
Entretanto, os resultados do presente
trabalho sugerem que Seira sp. não reage
da mesma forma, pois seu comportamento
não diferiu do comportamento do
colêmbolo no tratamento controle.

57 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Nilsson & Bengtsson (2004)
observaram que uma espécie de
colêmbolo, Protaphotuts armata, não
reconhecem o odor do predador. O
presente trabalho corrobora esse estudo,
demonstrando que o comportamento do
colêmbolo no tratamento com predador
morto foi semelhante ao encontrado no
controle.
Seira sp. não conseguiu identificar
a presença do predador diante das
condições testadas neste experimento. Isso
pode significar que Seira sp. detecta
predadores de outra forma, sem a
utilização do olfato, ou que este
experimento não foi sensível para
detectar.este efeito. O aumento do tempo
de aclimatação na placa de Petri antes do
início do experimento pode reduzir o
estresse do colêmbolo e tornar os
resultados ainda mais confiáveis.
Agradecimentos
Agradecemos aos funcionários da ESEC, aos
caros professores, colegas do PPG Ecologia da
UFRN e em especial a Bruno Belline na ajuda
com os Collembolas e a Carlos Fonseca pela
ajuda no delineamento experimental análise
dos dados.
Referências
Andrade-Lima D., 1981. The caatinga
dominium. Revista Brasileira de
Botânica 4, 149-63.
Dicke, M. & Grostal, P. 2001. Chemical
detection of natural enemies by
arthropods: an ecological perspective.
Annu.Rev. Syst. 32: 1-23.
Lavelle, P., Lattaud, D. T.& Barois, I., 1995.
Mutualism and biodiversity in soils.
Plant Soil. 170, 23–33.
Nilsson, E. & Bentsson, G., 2004a. Death odour
changes movement pattern of
Collembola. Oikos. 104. 509-517.
Nilsson, E. & Bentsson, G., 2004b. Endogenous
free fatty acids repel and attract
Collembola. Journal of Chemical
Ecology. 30, 1431-1443.
Sampaio, E. V. S. B., 1995. Overview of the
Brazilian Caatinga. In: Bullock, S.H.,
Mooney, H. A., Medina, E. (Eds.),
Seasonally dry forests. Cambridge
University Press, Cambridge, pp. 35-
58.
Swift, M. J., Heal, 0. W., Anderson. J. M., 1979.
Decomposition in terrestrial
ecosystems. Blackwell Scientific
Publications, London.
Rollo, C. D., Borden, J. H. e Casey, I. B. 1995.
Endogenously produced repellent from
American cockroach (Blattaria:
Blattidae): function in death
recognition. Environ. Entomol. 24:
116-124.
Rollo, C. D., Czyewska, E. e Borden, J. H. 1994.
Fatty acid necromones for
cockroaches. Naturwissenschaften 81:
409-410.

58 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
SELEÇÃO DE MICROHABITAT POR Cnemidophorus ocellifer EM ÁREA DE
CAATINGA, SERRA NEGRA DO NORTE, RN
GINA MANTILLA; DANIEL DE OLIVEIRA; DIEGO BENTO; PRISCILA DO AMARAL; IGOR GALVÃO
Introdução
Dentro do seu habitat, os indivíduos
optam pela utilização de microhabitats de
acordo com características relacionadas à sua
história de vida, tais como alimentação,
segurança, reprodução e termorregulação,
fator fundamental para a manutenção da
homeostase dos répteis (Dawson, 1975). Na
maioria dos lagartos, a seleção de
microhabitats depende primariamente de
características estruturais do ambiente (Vitt et
al., 2007) e a sua forma de uso varia sobre
diferentes condições de habitat (Veja et al.,
2000).
Cnemidophorus ocellifer é um lagarto
forrageador ativo comum às regiões tropicais
e subtropicais da América do Sul, distribuindo-
se no Nordeste, Centro e Sudeste do Brasil. É
abundante nos campos, cerrados e caatingas
(Vanzonlini et al., 1980). Na Caatinga, seu
período de atividade máxima ocorre nas horas
mais quentes do dia, ou seja, entre 10:00 e
12:00, com início às 08:00 e permanecendo
ativo até 13:00-14:00 (Teixeira-Filho et al,
1995).
A dieta é generalista, envolvendo
insetos de várias ordens (principalmente
Isoptera, Orthoptera, Coleoptera, Homoptera
e Hymenoptera, larvas de Lepidoptera), além
de aranhas e partes vegetais (frutos e flores)
(Dias & Rocha, 2005).
De acordo com as características da
biologia de C. ocellifer anteriormente
descritas, espera-se a preferência por
determinados ambientes, se estes estiverem
disponíveis, dentro da heterogeneidade
ambiental disponível em seu habitat. Dessa
forma, entender a associação entre a
abundância da espécie e as características de
microhabitats tem um valor considerável em
conservação, pois isso permite fazer previsões
sobre as respostas a mudanças naturais e
antropogênicas no habitat (Law & Dickman,
1998).
Deste modo, este estudo objetivou
analisar a seleção de microhabitats por C.
ocellifer em uma área de Caatinga do Seridó
do Rio Grande do Norte, bem como identificar
as características ambientais que determinam
a preferência do lagarto por determinados
microambientes.
Material e Métodos
O estudo foi desenvolvido na Estação
Ecológica do Seridó localizada no município de
Serra Negra do Norte-RN. Determinou-se um
transecto de 1440 metros (iniciado em
6o35’49,5”N, 37o14’56,95”W e finalizando em
06o36’22,26”N e 37o15’20,12”W) por 13
metros, com abrangência de 5 metros em
cada lateral da estrada (Fig. 1).
Em um sentido do transecto
registraram-se as observações de C. ocellifer
através de procura visual, enquanto no
sentido inverso sorteou-se o mesmo número
de observações para caracterizar diferentes
microhabitats da área. Em todos os pontos
mediu-se 11 parâmetros ambientais que
refletem características de habitat (Tabela 1).
Os dados foram analisados através de
regressão logística simples e tabela de

59 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
contingência, utilizando o programa Systat
12.0.
Figura 9. Delineamento amostral da área de coleta
As linhas cinza representam as faixas sem
vegetação da estrada, as verdes margens à direita
(9-13), esquerda (1-5) e centro da estrada (7). Cada
número representa 1 m.
Tabela 3. Parâmetros medidos para caracterização de microhabitat no transecto selecionado.
Parâmetro Fator de história de vida
relacionado Medição
Abundância de itens alimentares
Recurso alimentar Número de artrópodes de solo contados em plot de 50x50 cm por 2 minutos
Porcentagem de cobertura do solo
Estratégia anti-predador, recurso alimentar e termorregulação
Estimativa visual da proporção de solo coberto por folhiço e colheita em pé, categorizada em três classes
Profundidade do folhiço Recurso alimentar Média de quatro medidas do material orgânico do solo num raio 1 m, a partir do ponto
Distância ao afloramento mais próximo
Termorregulação Medida em metros da menor distância do ponto ao afloramento
Distância ao tronco mais próximo
Estratégia anti-predador e recurso alimentar
Medida em metros da menor distância do ponto ao tronco no solo
Ocorrência de toca Estratégia anti-predador Presença e ausência de toca
Ocorrência de touceira Estratégia anti-predador e recurso alimentar
Presença e ausência de touceira
Altura do estrato herbáceo Estratégia anti-predador e termorregulação
Média de quatro medidas de altura máxima de espécimes herbácea num raio 1 m, a partir do ponto
Número de plantas com Diâmetro Altura do Solo (DAS) > 5 cm
Termorregulação e recurso alimentar
Contagem de plantas com DAS > 5 cm num raio de 5 m, a partir do ponto
Riqueza da flora Termorregulação e recurso alimentar
Contagem de morfotipos de flora num raio de 5 m, a partir do ponto
Cobertura vegetal predominante
Termorregulação e recurso alimentar
Categorização da vegetação predominante num raio de 5 m, a partir do ponto, em herbácea e não herbácea

60 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Resultados
Foi amostrado um total de 40
pontos ao longo do transecto, dos quais 20
são registros de avistamento e os outros
20, não associados aos avistamentos,
sorteados (Fig. 2).
Figura 10. Área estudada com destaque para o
transecto amostrado com os registros de
visualização da espécie (pontos verdes) e os
sorteados (pontos vermelhos).
Dentre os parâmetros medidos,
somente quatro, profundidade de folhiço,
número de árvores com DAS > 5 cm,
riqueza da flora e ocorrência de touceira,
apresentaram relação significativa com a
probabilidade de ocorrência de C. ocellifer.
A regressão logística entre
probabilidade de ocorrência e o log do
número de árvores com DAS maior que 5
cm mostrou uma relação negativa e
significativa (β = -2,618; p = 0,04) (Fig. 3).
Houve relação significativa (P <
0,01) também na regressão logística entre
a probabilidade de ocorrência e o log da
profundidade do folhiço em centímetros.
Desta vez, a associação foi positiva (β =
7,541), sendo possível ver um grande
agrupamento de ocorrências nas situações
de folhiço mais profundo e um grande
agrupamento de não-ocorrências de C.
ocellifer nas situações de folhiço mais raso
(Fig. 4).
Figura 11. Regressão logística entre a
probabilidade de ocorrência de C. ocellifer e o
log da quantidade de árvores com DAS > 5 cm
(Z = -2,052; g.l. = 39; P < 0,05).
Figura 12. Regressão logística entre a
probabilidade de ocorrência de C. ocellifer e log
da profundidade de folhiço (Z = 2,148; g.l. = 39;
P = 0,032).
A regressão entre a probabilidade de
ocorrência e a riqueza vegetal também se
mostrou positiva e significativa (β = 0,530;

61 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
P = 0,002). Os encontros foram mais
freqüentes em situações de maior riqueza.
Figura 13. Regressão logística entre
probabilidade de ocorrência e riqueza vegetal.
As ocorrências se concentraram onde houve
maior riqueza de espécies vegetais. Z = 3,052;
g.l. = 39; p = 0,002.
Figura 14. Quantidades de ocorrência e não-
ocorrência de C. ocellifer relacionadas com
ausência e presença de touceira(Likelihood =
11,872; g.l. = 1; P < 0,01).
A tabela de contingência
relacionando presença ou ausência de
touceiras com avistamento e não
avistamento da C. ocellifer também foi
significativa (Likelihood = 11,872; P < 0,01)
(Fig. 6). O número de registros na presença
de touceira foi bastante maior que na
ausência, ocorrendo o comportamento
contrário para o não registro C. ocellifer.
Discussão
É consistente a significância da
relação positiva de profundidade de folhiço
e da riqueza da flora com a probabilidade
de ocorrência de C. ocellifer, uma vez que
esses parâmetros estão associados,
respectivamente, à abundância e a
diversidade de recursos alimentares. O
número de árvores com DAS maior que 5
centímetros, apesar de poder contribuir
com folhiço, pode ter uma maior influência
com o aumento do sombreamento, dado o
saldo de sua relação ser negativo com a
probabilidade de ocorrência do animal.
O hábito do animal de buscar ficar
próximo a refúgios vegetais (Teixeira-Filho
et al., 1995) pode explicar as relações
positivas entre a probabilidade de
ocorrência e a presença de touceiras.
Os resultados obtidos concordam
com a bibliografia em relação à maior
freqüência de uso de borda, arbustos e
áreas iluminadas (Teixeira-Filho et al.,
1995). Por outro lado, há divergências
quanto à preferência pelo tipo de solo, pois
o presente estudo revelou preferência por
folhiço em área de Caatinga, enquanto
Mesquita & Coli (2003) afirmam haver
preferência por terra nua e rochas para
esse tipo de ecorregião.
Sabe-se haver três grandes fontes
de calor que este ((lagarto utiliza para
termorregulação: radiação direta do sol,

62 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
calor do ar e o calor do substrato.
Entretanto, há indícios de que a radiação
direta seja a mais importante (Teixeira-
Filho et al., 1995). Essa previsão é
suportada pelos dados obtidos, uma vez
que não houve relações significativas com
os afloramentos rochosos, mas uma
relação negativa com o número de árvores,
que podem sombrear a área e impedir o
acesso à radiação direta.
Assim, conclui-se que há seleção de
microhabitat pelo C. ocellifer e que a
mesma está associada a peculiaridades
importantes de sua história de vida.
Agradecimentos
Somos gratos a Gindomar Santana por
sua orientação e grande auxílio na preparação e
execução do projeto. Agradecemos a Carlos
Roberto Fonseca pelos insights no
delineamento do projeto e auxílio no
tratamento estatístico dos dados. Agradecemos
também aos funcionários da ESEC Seridó pelo
acolhimento e auxílio logístico.
Referências
Dawson, W.R., 1975. On the physiological
significance of the preferred body
temperatures of reptiles, In. Gates,
D.M. & Schmerl, R.B. (eds.).
Perspectives of biophysical ecology.
Ecology studies. New York, Springer-
Verlag. Vol. 12. P. 433-473.
Dias, E.J.R., Rocha, C.F.D., 2005. Niche
differences between two sympatric
whiptail lizards (Cnemidophorus
abaetensis and C. ocellifer, Teiidae) in
the resting habitat of northeastern
Brazil. Braz. J. Biol., 67(1), 41-46.
Law, B.S., Dickman, C.R., 1998. The use of
habitat mosaic by terrestrial
vertebrate fauna: implications for
conservation and management.
Biodivers. Conserv. 7, 323- 333.
Mesquita, D., Colli, G.R., 2003. The ecology of
Cnemidophorus occellifer (Squamata:
Teiidae) in a neotropical savanna. J.
Herpetol. 37, 498-509.
Teixeira-Filho, P.F.; Rocha, C.F.D.; Ribas, S.C.
1995. Aspectos da ecologia termal e
uso do habitat por Cnemidophorus
ocellifer (Sauria: Teiidae) na restinga
da Barra de Maricá, RJ. Oecologia
Brasiliensis. V1. P.155-165. UFRJ.
Vanzolini, P.E., Ramos-Costa, A.M.M., Vitt, L.J.,
1980. Répteis das caatingas. Rio de
Janeiro, Acad. Bras. Ciênc. P. 111-114.
Vega, L.E., Bellagamba, P.J., Fitzgerald, L.A.,
2000. Longterm effects of
anthropogenic habitat disturbance on
a lizard assemblage inhabiting coastal
dunes in Argentina. Can. J. Zool. 78,
1653-1660.

63 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010

64 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Pro
jeto
s se
m o
rie
nta
ção

65 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
DEFESA DE ESTRUTURAS REPRODUTIVAS DE Macroptilium lathyroides (L.)
(FABACEAE) POR FORMIGAS Camponotus SP. EM UMA ÁREA DE
CAATINGA, NORDESTE DO BRASIL
IGOR GALVÃO DE BRITTO; VINÍCIUS DE AVELAR SÃO PEDRO; IZABEL MARIA MATOS DE SOUZA
Introdução
Em uma gama diversa de interações, as
plantas podem ser alimentadas, protegidas
ou transportadas por formigas (Beattie,
1985). Sabe-se que o mutualismo formiga-
planta é um importante componente das
comunidades tropicais, envolvendo mais
de 100 gêneros de angiospermas e 40
gêneros de formigas (Heil & Mckey, 2003).
Algumas espécies vegetais
possuem nectários extraflorais (NEFs), que
são glândulas secretoras de néctar não
envolvidas diretamente com a polinização
(Fiala & Maschwitz, 1991). Formigas que
visitam os NEFs podem beneficiar a planta
detendo herbívoros interessados em suas
folhas ou estruturas reprodutivas (Oliveira,
1997).
A espécie anual herbácea
Macroptilium. lathyroides (L.) (Fabaceae),
conhecida como feijão-de-rola, possui NEFs
associados às suas estruturas reprodutivas.
Esses NEFs atraem uma série de
invertebrados, predominantemente
formigas do gênero Camponotus sp.
(Hymenoptera: Formicidae).
Neste trabalho, investigou-se a
interação entre a herbácea M. lathyroides
e as formigas Camponotus sp. em uma
área de caatinga no Rio Grande do Norte,
visando responder às seguintes questões:
1) Quais características da planta mais
favorecem a presença das formigas? 2)
Como a planta se beneficia com a presença
das formigas? Suspeita-se que a formiga
proteja a planta. Caso isso se confirme, 3)
Quais partes da planta são efetivamente
protegidas pelas formigas?.
Material e Métodos
O estudo foi desenvolvido na
Estação Ecológica do Seridó, localizada no
município de Serra Negra do Norte-RN.
Traçou-se um transecto de 20 metros a
partir do começo da trilha que leva ao
açude e 3 metros para cada lado da trilha,
dentro do qual foi realizado o censo dos
indivíduos de M. lathyroides. As plantas
foram marcadas com uma fita
identificadora e, para cada uma, contou-se:
formigas nos ramos reprodutivos, formigas
nos ramos vegetativos, total de formigas,
hastes, ramos, folhas, frutos, flores, brotos,
folhas com sinais de herbivoria. O censo foi
realizado entre as 08:30 e 13:30.
No dia seguinte, checou-se, para as
plantas que apresentavam frutos, folhas e
flores, o tempo de resposta de
Camponotus sp. a um térmita
(Nasutitermis corniger (Motschulsky,
1855)) (Isoptera: Termitidae:
Nasutitermitinae) vivo aderido, com cola
branca, respectivamente, à folha mais alta
do ramo vegetativo mais próximo ao
reprodutivo utilizado, ao meio do maior
ramo vegetativo, ao meio de um fruto
nesse ramo e ao meio de uma flor no
mesmo ramo. As observações foram feitas

66 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
sucessivamente, uma só iniciando após
término da outra, na sequência folha >
haste > fruto > flor e por um período
máximo de 10 minutos. Por fim, observou-
se, quando as formigas tocaram os
invasores com suas antenas por mais dois
minutos para avaliar qual seria a reação
final da formiga: ignorá-lo ou tentar
removê-lo.
Testou-se a diferença no número
de formigas entre plantas com e sem
estruturas reprodutivas através de uma
ANOVA. Também se testou a diferença na
distribuição das formigas entre ramos
reprodutivos e vegetativos das plantas com
estruturas reprodutivas através de uma
ANOVA. Fez-se ainda uma regressão
múltipla para relacionar o número de
formigas ao número de folhas, ramos
vegetativos e estruturas com nectários;
outra relacionando o número de frutos
com o de folhas, ramos e formigas. Por fim,
testou-se a diferença nos tempos de
resposta das formigas nas diferentes
estruturas vegetais através de uma
ANOVA. Análises conduzidas com
programa Systat 12.
Resultados
Foram observados 32 indivíduos na
área de estudo, dos quais 16 apresentaram
folhas, frutos e flores e, nos demais, uma
ou mais das estruturas não estava
presente. Observou-se que as plantas com
estruturas reprodutivas apresentaram mais
formigas que as sem (F = 6,581; g.l. = 1,30;
P < 0,02; Fig 1.).
Realizou-se também o teste não
paramétrico de Kruskal-Wallis e verificou-
se que, apesar da grande variação no
número de formigas em plantas sem
estruturas reprodutivas, essa diferença
ainda é significativa (P < 0,05).
Figura 15. Média e erro padrão do número de
formigas em plantas com e sem estruturas
reprodutivas. (F = 6,581; g.l. = 1,30; P < 0,02).
Nas plantas com estruturas
reprodutivas, as formigas também
tenderam a se concentrar em ramos
vegetativos (F= 16,69; g.l. = 1,62; P < 0,01;
Fig. 2).
O número de formigas também foi
significativamente relacionado com o
número de folhas (β = 1,042; P < 0,01),
ramos (β = -0,774; P < 0,02) e estruturas
reprodutivas (β = 0,423; P < 0,01; R2 =
0,662; Fig. 3).
O número de frutos apresentou
relações significativas apenas com o
número de formigas (β = 0,621; P < 0,01).
Não foram significativas as relações com
ramos vegetativos (β = 0,783; p = 0,099) e
folhas (β = -0,119; p = 0,799) (Fig. 4).
Indivíduos de Camponotus sp.
também reagiram mais rapidamente aos
cupins colocados em flores, frutos e ramos
vegetativos que em folhas (F = 7,39; g.l. =

67 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
3,60; P < 0,01). Contudo, não houve
diferenças significativas entre os três
primeiros (Fig. 5).
Figura 16. Média e erro padrão do número de
formigas em ramos vegetativos e reprodutivos
nas plantas com estruturas reprodutivas.
Figura 17. Número de indivíduos de
Camponotus sp (Log) em resposta ao número
de estruturas reprodutivas (Log) em cada
indivíduo observado de M. lathyroides.
Figura 18. Relação entre o número de frutos e o
número de formigas. A relação é positiva e
significativa β = 0,621; P < 0,01) .
Figura 19. Média e erro padrão do tempo de
resposta, em segundos, das formigas aos
térmitas colocados em diferentes estruturas da
mesma. Resposta mais lenta para folhas e não
significativamente diferente entre flores, frutos
e hastes.
Reprodutivo Vegetativo
Tipo de ramo
0,0
0,5
1,0
1,5
2,0
Nú
me
ro d
e fo
rmig
as [L
og
]

68 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Discussão
Sabe-se que, para algumas
espécies vegetais, altas taxas de predação
floral podem resultar em uma menor taxa
de produção de sementes e
conseqüentemente de descendentes
(Nascimento & Del-Claro, 2007). Nesse
sentido, as relações encontradas apontam
que as formigas podem ter um papel
fundamental na reprodução de M.
lathyroides.
Sabendo-se haver espécies
endoparasitas dos ovários e que se
alimentam das sementes que surgem
depois (Elias, 2008), o investimento em
proteção floral pode apresentar relações
diretas e indiretas com o número de
sementes viáveis.
O fato das formigas se
concentrarem nas áreas com nectários
indica que sejam essas as estruturas que
mais as beneficiam. Isso pode estar
associado tanto ao suprimento de glicídeos
pela planta como a atração de possíveis
presas, de maior conteúdo protéico
(Bentley, 1977).
O padrão de concentrar os esforços
de proteção em suas estruturas
reprodutivas pode estar ocorrendo em
função dos elevados gastos com a
produção de descendentes (Schulze et al.,
2002), que podem estar potencializados
nessa planta pelas condições em que vive.
A aridez local, a escassez de corpos d’água
próximos e a concentração das chuvas
podem limitar a atividade fotossintética
(Schulze et al., 2002) e o encontro de
parceiros. Assim, para indivíduos pouco
abundantes e semelparos ou com ciclos de
vida curtos o suficiente para que a perda
de folhas durante ou após o período
reprodutivo não seja grave, surgem
vantagens aos que protegem seus
descendentes mais do que suas folhas.
Além disso, evidenciou-se que o
número de elementos reprodutivos
pareceu mais relacionado com o número
de formigas na planta do que com o
tamanho da mesma. Esse padrão parece
reforçar a idéia de que M. lathyroides é
bastante sensível a predação em suas
estruturas reprodutivas.
Assim, pode-se concluir que M.
lathyroides se beneficia pela proteção de
suas estruturas vegetativas e,
principalmente, reprodutivas pela ação de
Camponotus sp.. Estudos futuros podem
testar mais a fundo a redução do fitness de
plantas com menos formigas, verificando
se há redução no número de sementes nas
plantas e se as formigas teriam diferenças
nos tempos de respostas caso o agente
estranho na planta lhe seja palatável (uma
presa conhecida) ou impalatável (algum
fator abiótico ou indivíduo que se saiba
não ser presa de Camponotus sp.).
Agradecimentos
Agradecemos a Carlos Roberto Fonseca, pelo
incansável apoio, madrugada a dentro, nas
análises estatísticas e valiosas sugestões sobre
a elaboração do projeto. Agradecemos também
aos funcionários da ESEC Seridó pelo
acolhimento e auxílio logístico.
Referências
Beattie, A. J. 1985. The Evolutionary Ecology of
Ant-Plant Mutualisms. Cambridge, UK:
Cambridge University Press.
Bentley, B. 1977. Extrafloral nectarines and
protection by pugnacious bodyguards.

69 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Annual Review of Ecology and
Systematics. Vol. 8. P 407-427.
Elias, L.G. 1975. Estratégias de utilização de
recursos em vespas parasitas do
mutualismo da figueira – vespas de
figo. Dissertação apresentada à
Faculdade de Filosofia, Ciências e
Letras de Ribeião Preto da USP.
Fiala, B., Maschwitz, U., 1991. Extrafloral
nectaries in the genus Macaranga
(Euphorbiaceae) in Malaysia:
comparative studies of their possible
significance as predispositions for
myrmecophytism. Biological Journal of
Linnean Society. 44: 287 - 305.
Heil, M., Mckey, D., 2003. Protective ant-plant
interactions as model systems in
ecological and evolutionary research.
Annual Review of Ecology, Evolution
and Systematics. 34: 425-553.
Nascimento, E. A., Del-Claro, K., 2007. Floral
visitors of Chamaecrista debilis (Vogel)
H.S.Irwin & Barneby (Fabaceae-
Caesalpinoideae) at Cerrado of Estação
Ecológica do Jataí, São Paulo State,
Brazil. Neotropical Entomology. 36 (4):
619-624.
Oliveira, P. S. 1997. The ecological function of
extrafloral nectaries: herbivore
deterrence by visiting ants and
reproductive output in Caryocar
brasiliense (Caryocaraceae). Functional
Ecology. 11: 323 – 330.
Schulze, E., Beck, E., Müller-hohenstein, K.
2002. Plant Ecology. Berlin. Springer
Berlin.
Foto. Detalhe do ramo florido com as formigas. Foto do grupo de trabalho.

70 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
PADRÃO DE DISTRIBUIÇÃO ESPACIAL DE Caesalpinia pyramidalis Tul.
(FABACEAE) e Aspidosperma pyrifolium Mart. (APOCYNACEAE) EM
UMA ÁREA DE CAATINGA, NORDESTE DO BRASIL
CRISTIANE GOUVÊA FAJARDO, DIEGO BENTO, MAIRA MANZAN, KAROL LYNCOLN
Introdução
A Caatinga ocupa 11% do território
nacional com uma área de
aproximadamente 800.000 km2.
Corresponde a 70% da região Nordeste,
ocorrendo nos Estados do Piauí, Ceará, Rio
Grande do Norte, Paraíba, Pernambuco,
Alagoas, Sergipe e Bahia. A caatinga ocorre
em áreas com marcada sazonalidade e
baixos índices pluviométricos. É um bioma
exclusivamente brasileiro e apresenta uma
fitofisionomia arbustivo-arbórea composta
principalmente por espécies xerofíticas
(Rodal & Melo, 1999).
Somente recentemente a biota da
Caatinga vem sendo mais bem estudada
(Leal et al., 2003). Quanto à flora, por
exemplo, há registro de 932 espécies
vegetais, sendo 380 endêmicas (Giulietti et
al., 2004).
Dentre as espécies vegetais com
ocorrência na Caatinga, Caesalpinia
pyramidalis Tul., popularmente conhecida
como “catingueira”, é bastante
característica e endêmica deste bioma
(Leite & Machado, 2009). É uma espécie
pioneira (Santana & Souto, 2006) com
dispersão autocórica.
Aspidosperma pyrifolium Mart.
também ocorre na caatinga e é conhecida
como pereiro, pau-de-coaru e pequiá-da-
mata. Possui sementes aladas,
características de espécies de dispersão
anemocórica. A árvore apresenta porte
regular, podendo atingir até 5m de altura.
Além disso, é uma espécie heliófila e
secundária (Andrade et al., 2005).
Sendo C. pyramidalis e A.
pyrifolium espécies que coexistem em um
mesmo habitat na Caatinga, buscou-se
investigar interações intra e
interespecíficas entre as plantas por meio
de análises do padrão espacial, pois os
padrões observados são frequentemente
reflexos dessas interações.
O objetivo do presente trabalho foi
analisar a distribuição espacial e estrutural
de Caesalpinia pyramidalis e Aspidosperma
pyrifolium, considerando-se os indivíduos
adultos, jovens e plântulas. Além disso,
averiguar se há associação espacial
interespecífica.
Material e Métodos
Local do estudo e amostragem
Delimitou-se uma área de estudo
de 2.500 m2 dentro da Estação Ecológica
do Seridó (Figura 1), região de caatinga
pertencente ao município de Serra Negra
do Norte, estado do Rio Grande do Norte,
Brasil. A região da ESEC Seridó possui uma
área de 1.166,38 ha, clima semi-árido e
localiza-se entre as coordenadas
geográficas 06° 36’ 40’’ e 06° 33’ 50’’ Sul e
37° 17’ 10’’ e 37° 14’ 20’’ Oeste.

71 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Figura 1: Área de estudo indicada pela seta
dentro do mapa da ESEC Seridó.
Dentro da parcela, mapearam-se
todos os indivíduos das espécies
Aspidosperma pyrifolium e Caesalpinia
pyramidalis e foram registradas as suas
posições x e y. Adicionamente, foram
medidos o PAS (Perímetro à Altura do Solo,
posteriormente convertido em DAS –
Diâmetro à Altura do Solo) e a altura dos
indivíduos, com o objetivo de avaliar a
maturidade.
Análise do padrão de espacial
Para determinar se o padrão de
distribuição espacial para uma única
espécie é agregado (análise univariada) e
se há associação espacial entre as espécies
(análise bivariada), utilizou-se a função de
segunda ordem da densidade de vizinhos
ou NDF (Neighbourhood Density Function)
(Condit et al., 2000). A estatística NDF é
similar à função K de Ripley, entretanto
não é cumulativa (Perry et al., 2006).
Foram utilizadas classes de distância (t)
entre 2 e 24 m, pois, como sugerido por
Haase (1995) elas não devem exceder
aproximadamente ½ do menor
comprimento da parcela. Adicionalmente,
foi calculada a correção do efeito de borda,
baseado em Goreaud & Pelissier (1999). Os
valores da estatística NDF (t) foram
plotados nos correlogramas em função da
distância t e comparados a “envelopes” de
completa aleatoriedade, obtidos a partir de
499 simulações randômicas de Monte
Carlo (α = 0,01). Essas análises foram
realizadas por meio do programa SpPack
1.38 (Perry, 2004).
Correlação entre as distribuições espaciais
das duas espécies
Para analisar se as distribuições
espaciais das duas espécies estão
correlacionadas, a área amostrada foi
subdividida em 100 parcelas de 5x5 metros
e contaram-se quantos indivíduos de cada
espécie ocupavam cada parcela. Foi
efetuado um teste de correlação não-
paramétrica (utilizando o coeficiente de
correlação de Spearman) no programa
Statística 7.0.
Resultados
Demografia
Foram amostrados no total 196
indivíduos de Caesalpinia pyramidalis
(Figura 2), sendo esses 126 adultos, 61
juvenis e 9 plântulas. Para Aspidosperma
pyrifolium (Figura 3) foram encontrados 89
indivíduos, sendo 83 adultos e 6 jovens.

72 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Figura 2: Distribuição espacial de plântulas (PAS = 0
a 50), jovens (PAS= 51 a 100) e adultos (PAS = 101 a
150) de C. pyramidalis na área amostrada de
50x50m.
Figura 3: Distribuição espacial de jovens (PAS= 0 a
60) e adultos (PAS = 61 a 120) de A. pyrifolium na
área amostrada de 50x50m.
Padrão espacial
A análise do padrão espacial dos
indivíduos de C. pyramidalis (N = 196)
mostrou significantes níveis de agregação
até um raio de 10 m do indivíduo focal e
padrão aleatório em distâncias superiores
(Figura 4 A). Para a espécie A. pyrifolium (N
= 89) a densidade de vizinhos até um raio
de 4 m e entre 8 e 12 m foi maior do que o
esperado para a hipótese de completa
aleatoriedade, já nas demais classes
observou-se padrão espacial aleatório
(Figura 4 B).
Figura 4: Padrão espacial obtido por meio da análise
univariada da densidade de vizinhos (NDF) de
indivíduos adultos de Caesalpinia pyramidalis (A) e
Aspidosperma pyrifolium (B). Linhas pontilhadas
correspondem ao intervalo de confiança de 99%
para a hipótese nula de padrão espacial
completamente aleatório.
Os indivíduos jovens e plântulas
(N=70) e os adultos (N=126) de C.
pyramidalis apresentaram padrão espacial
agregado semelhante num raio de 9m do
indivíduo focal (Figuras 5 A e B).
O padrão espacial de indivíduos
adultos de Caesalpinia pyramidalis e
Aspidosperma pyrifolium foi agregado até
o raio de 6m do indivíduo focal e padrão
aleatório em distâncias superiores (Figura
6).
Foi encontrada uma correlação
significativa (r=0.4645; p <0.05) entre as
distribuições espaciais da Caesalpinia
pyramidalis e Aspidosperma pyrifolium
(Figura 7).
0
50
100
150
PAS
0 10 20 30 40 50
X
0
10
20
30
40
50
Y 0.01.02.03.04.05.0
2 8 14 20
ND
F
Distância t (m)
NDF
IC
IC
0.00.51.01.52.0
2 8 14 20
ND
F
Distância t (m)
NDF
IC
IC
A A
B

73 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Figura 5: Padrão espacial obtido por meio da análise
univariada da densidade de vizinhos (NDF) para
jovens (A) e indivíduos e adultos (B) de Caesalpinia
pyramidalis. Linhas pontilhadas correspondem ao
intervalo de confiança de 99% para a hipótese nula
de padrão espacial completamente aleatório.
Figura 6. Padrão espacial obtido por meio da análise
bivariada da densidade de vizinhos (NDF) adultos de
Caesalpinia pyramidalis e Aspidosperma pyrifolium.
Linhas pontilhadas correspondem ao intervalo de
confiança de 99% para a hipótese nula de padrão
espacial completamente aleatório.
Figura 7: Correlação não-paramétrica entre as
distribuições espaciais da e catingueira (C.
pyramidalis) e do pereiro (A. pyrifolium) na área
amostrada.
Discussão
O padrão de distribuição espacial
encontrado provavelmente reflete as
estratégias reprodutivas das espécies e o
histórico do local. Devido à síndrome de
dispersão autocórica, esperava-se uma
distribuição mais agregada em C.
pyramidalis do que em A. pyrifolium, que
possui dispersão anemocórica. Entretanto,
os resultados mostraram que tanto os
indivíduos adultos de C. pyramidalis quanto
os de A. pyrifolium apresentam elevada
densidade de vizinhos em curtas distâncias,
por volta de 6m.
Quanto ao padrão espacial das
faixas etárias, não foi observada diferença
entre os indivíduos jovens e adultos da
espécie C. pyramidalis. Esse resultado
diverge da literatura, que relata que a
maioria das espécies arbóreas apresenta
padrão agregado nos primeiros estágios de
vida e padrão aleatório com o aumento da
idade (Hamrick et al., 1993; Epperson &
Alvarez-Buylla, 1997). Provavelmente, os
valores similares da densidade de
indivíduos jovens e adultos em C.
pyramidalis estão relacionados a fatores
antrópicos. Segundo Meyer et al. (1961), a
0.0
1.0
2.0
3.0
4.0
3 9 15 21
ND
F
Distância t ( m)
NDF
IC
IC
0.00.51.01.52.02.53.03.54.0
3 9 15 21 27
ND
F
Distância t ( m)
NDF
IC
IC
0.00.51.01.52.02.53.0
3 9 15 21
NDF
Distância t ( m)
NDF
IC
IC
A
B

74 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
estrutura demográfica reflete o histórico
da floresta, bem como a ocorrência, no
passado, de distúrbios como fogo, corte
seletivo e herbivoria. Mais
especificamente, a herbivoria por animais
domesticados está associada à redução do
recrutamento e crescimento das plantas
(Oba, 1998), assim como na redução da
capacidade de regeneração da vegetação
(Perevolottsky & Haimov, 1992).
Além de C. pyramidalis e A.
pyrifolium apresentarem padrão espacial
agregado em curtas distâncias, há
considerável coexistência, conforme
observado na análise bivariada da
densidade de vizinhos e na correlação
positiva de Spearman. De acordo com
Paine et al. (2008), a magnitude da
competição interespecífica ainda é pouco
conhecida, mas geralmente é fraca a
competição entre algumas espécies
arbóreas Neotropicais. Segundo Byrne et
al. (2007), a intensidade com a qual os
indivíduos estão agregados afeta o sistema
de reprodução e também como as plantas
usam os recursos (Getzin et al., 2008).
Dessa forma, a agregação das
espécies poderia ser justificada pelo
processo de interação interespecífica
positiva, como a facilitação. Processo esse
comum em regiões semi-áridas sob
condições estressantes (Smit et al., 2009),
ou simplesmente, porque na área
amostrada existe uma heterogeneidade na
distribuição de nutrientes no solo,
proporcionando regiões mais férteis do
que outras, e portanto, contribuindo na
associação de C. pyramidalis e A.
pyrifolium. Entretanto, tais interações
carecem de estudos para a comprovação, e
poderão ser elucidados em futuros
trabalhos.
Agradecimentos
Aos professores Dr. Alexandre Vasconcellos e
Carlos Fonseca pelo auxílio fornecido no
presente estudo. Aos funcionários da ESEC
Seridó pela hospitalidade dedicada durante os
dias de pesquisa.
Referências
Andrade, L. A.; Pereira, I. M.; Leite, U. T.;
Barbosa, M. R. 2005. Análise de duas
coberturas fitofisionomias de Caatinga,
com diferentes históricos de uso, no
município de São João do Cariri, Estado
da Paraíba. Cerne v. 11, n. 3, pp. 253-
262.
Byrne, M.; Elliott, C. P.; Yates, C.; Coates, D. J.
2007.Extensive pollen dispersal in a
bird-pollinated shrub, Calothamnus
quadrifidus, in a fragmented
landscape. Molecular Ecology, v. 16, p.
1303-1314.
Condit, R.; Ashton, P. S.; Aker, P.;
Bunyavejchewin, S.; Gunatilleke,
S.;Gunatilleke, n.; Hubbell, S. P.; Losos,
E.; Manokaran, N.; Sukumar,
R.;Yamakura, T. 2000. Spatial patterns
in the distribution of tropical tree
species. Science, v. 288, p. 1414-1418.
Epperson, B.K.; Alvarez Buylla, E.R. 1997.
Limited seed dispersal and genetic
structure in life stages of Cecropia
obtusifolia. Evolution, v. 51, p. 275–
282.
Getzin, S.; Wiegand, T.; Wiegand, K.; He, F.
2008.Heterogeneity influences spatial
patterns and demographics in forest
stands. Journal of Ecology, v. 96, p.
807-820.
Giulietti, A. M., Du Bocage Neta, A.L., Castro,
A.A.J.F., Gamarra-ROJAS, C.F.L.,
Sampaio, E.V.S.B., Virgínio, J.F.,
Queiroz, L.P., Figueiredo, M.A., Rodal,
M.J.N., Barbosa, M.R.V. & Harley, R.M.
2004. Diagnóstico da vegetação nativa
do bioma Caatinga. In Biodiversidade

75 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
da Caatinga: áreas e ações prioritárias
para a conservação (J.M.C. Silva, M.
Tabarelli, M.T. Fonseca & L.V. Lins,
orgs.). MMA,UFPE, Conservation
International do Brasil, Fundação
Biodiversitas, Embrapa Semi-Árido,
Brasília, p.48-90.
Goreaud, F.; Pelissier, R. 1999. On explicit
formulas of edge effect correction for
Ripley’s K-function. Journal of
Vegetation Science, v. 10, p. 433-438.
Haase, P. 1995.Spatial pattern analysis in
ecology based on Ripley’s K-function:
introduction and methods of edge
correction. Journal of Vegetation
Science, v. 6, p. 575-582.
Hamrick, J.L.; Murawski, D.A.; Nason, J.D. 1993.
The influence of seed dispersal
mechanisms on the genetic-tructure of
tropical tree populations. Vegetatio, v.
108, p. 281-297.
Leal, I.R., TABARELLI, M. & SILVA, J.M.C. 2003.
Ecologiae conservação da Caatinga.
Editora Universitária, UFPE, Recife.
Leite, A. V.; Machado, I. C. 2009. Biologia
reprodutiva da catingueira (Caesalpinia
pyramidalis Tul., Leguminosae-
Caesalpinioideae), uma espécie
endêmica da caatinga. Revista
Brasileira de Botânica, v. 32, p. 79-88.
Meyer, H. A.; Recknagel, A. B.; Stevenson, D. D.;
Bartoo, R. A. , 1961. Forest
management. 2. ed. New York: Ronald,
282p
Oba, G. 1998.Effects of excluding goat
herbivory on Acacia tortilis woodland
around pastoralist settlements in
northwest Kenya. Acta Oecologica, v.
19, p. 395-404.
Paine, C. E. T.; Harms, K. E.; Schnitzer, S. A.;
Carson, W. P. 2008. Weak competition
among tropical tree seedlings:
implications for species coexistence.
Biotropica, v. 40, n. 4, p. 432-440.
Perevolotsky, A.; Haimov, Y. 1992. The effect of
thinning and goat browsing on the
structure and development of
mediterranean woodland in Israel.
Forest Ecology and Management, v.
49, p. 61-74.
Perry, G. L. W. 2004. SpPack: spatial point
pattern analysis in Excel using Visual
Basic for Applications (VBA).
Environmental Modelling & Software,
v. 19, p. 559-569.
Perry, G. L. W.; Miller, B. P.; Enright, N.J. 2006.A
comparison of methods for the
statistical analysis of spatial point
patterns in plant ecology. Plant
Ecology, v. 187, p. 59-82.
Rodal, M. J. N.; Melo, A. L. 1999. Levantamento
preliminar das espécies lenhosas da
Caatinga de Pernambuco. In Plantas do
Nordeste. Anais do I Workshop Geral
(F.D. Araujo, H.D.V. Prendergast & S.J.
Mayo, eds.). Royal Botanical Garden,
Kew, p.53-62.
Santana, J. A. S., Souto. J. S. 2006. Diversidade e
Estrutura Fitossociológica da Caatinga
na EstaçãoEcológica do Seridó-RN
Revista de Biologia E Ciências da Terra,
v. 6,n. 2, pp 232- 242.
Smit. C.; Rietkerk, M.; Martin J. Wassen, M. J.
2009. Inclusion of biotic stress
(consumer pressure) alters predictions
from the stress gradient hypothesis.
Journal of Ecology, v.97 n.6, pp.1215-
1219.

76 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
A ATIVIDADE DE ODONATOS (ARTHROPODA: HEXAPODA: INSECTA) VARIA
AO LONGO DO DIA EM UMA CAATINGA NO NORDESTE DO BRASIL?
ALLAN BATISTA; FRANÇOISE LIMA; HEITOR SOUZA; PRISCILA DO AMARAL
Introdução
Odonatos, também conhecidos
como lavadeiras, libélulas ou zigue-zagues,
constituem uma ordem amplamente
distribuída de insetos composta por
aproximadamente 5000 espécies
conhecidas e agrupadas em cerca de 500
gêneros, distribuídos principalmente nas
regiões neotropicais (BUZZI, 2002).
A ordem Odonata se divide em três
subordens: Anisozygoptera, contendo
apenas duas espécies conhecidas de
Epiophlebia encontradas no Japão e na
Índia, Zygoptera e Anisoptera (CORBET,
1980). Os indivíduos da subordem
Zygoptera possuem olhos compostos
afastados um do outro, vôo lento e,
quando em repouso, asas elevadas sobre o
abdômen. Já os indivíduos da subordem
Anisoptera possuem olhos aproximados ou
contíguos, vôo rápido, chegando a atingir
80km/hora, e, quando em repouso,
mantém asas abertas. A subordem
Anisozygoptera é considerada uma
assembleia parafilética morfologicamente
primitiva de Anisoptera. (BUZZI, 2002).
Os indivíduos adultos de Odonata
são tipicamente ativos durante o dia,
sendo encontrados próximos a lagoas e
rios (CORBET, 1980). A maioria dos adultos
se alimenta de insetos capturados em
pleno vôo. Alguns pousam para comer a
presa, enquanto outros o fazem durante o
vôo (BUZZI 2002).
Os odonatos vêm sendo
amplamente utilizados como
biondicadores (IMOOBE & OHIOZEBAU,
2010; SILVA et al., 2010; YOON et al., 2010)
devido a algumas características como (i)
estarem presentes em todos os ambientes
aquáticos de água doce; (ii) suas ninfas
possuírem baixa tolerância a distúrbios
ambientais; (iii) as ninfas serem
relativamente sedentárias; e (iv) pelos
adultos poderem ser facilmente
observados devido ao seu comportamento
territorial (SILVA et al., 2010). Além disso,
os odonatos possuem grande importância
nas relações tróficas dos ecossistemas,
uma vez que são importantes predadores
de topo (McCAULEY, 2010).
A atividade destes organismos
pode variar de acordo com a sazonalidade
anual, e diariamente em função das
variações de temperatura. Tais variações
podem também ocorrer em função das
características intrísecas de cada espécies
(LUTZ & PITTMAN, 1970).
Apesar de sua importância nas
redes tróficas, seu potencial uso como
bioindicador e de estar distribuído
principalmente nas regiões neotropicais,
não existe nenhum estudo publicado sobre
a biologia ou ecologia de Odonata no
bioma Caatinga. Sendo assim, esse estudo
visa compreender a variação na freqüência
de atividade de Odonata ao longo de 12
horas em uma área de Caatinga.

77 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Material e Métodos
O estudo foi desenvolvido nas
proximidades do açude da Estação
Ecológica do Seridó (6°15’S e 37°16’O),
localizada no município de Serra Negra do
Norte, RN. Durante um dia (28/06/2010),
no período de 6 h da manhã às 18 h da
tarde, registrou-se o número de
ocorrências dos morfotipos de Odonata em
três parcelas de 10 m², sorteados ao longo
de um transecto de 150 m (Fig. 1). As
observações ocorriam simultaneamente
nas parcelas por 10 min, a cada 30 minutos
decorridos, totalizando 4,16 h de esforço
amostral.
Figura 1. Esquema ilustrativo da área amostrada limítrofe ao açude da ESEC do Seridó no município de Serra Negra do Norte, RN. Na forma malhada as parcelas sorteadas, no transecto de 150 m, estão em destaque (4, 19 e 29 m). A forma em azul ilustra o açude.
Um indivíduo de cada morfotipo foi
coletado com o auxílio de uma rede
entomológica para posterior identificação
ao nível do menor táxon possível e para
medição de índice do tamanho corporal
(Largura tórax/Comprimento total).
Os dados foram analisados de
forma descritiva, para verificar a ocorrência
ao longo do dia, assim como por meio de
Análise de Agrupamentos, para verificar
horários similares, e nMDS, para verificar
quais morfotipos possuem
comportamentos semelhantes. Em todas
as análises o software Systat 12.0 foi
utilizado.
RESULTADOS
Foram identificadas sete
morfoespécies da ordem Odonata,
pertencentes a duas subordens: Anisoptera
e Zygoptera. Dentro dessas subordens,
foram identificadas três famílias
(Libellulidae, Coenagrionidae e Aeshnidae)
(Tab. 1).
As morfoespécies analisadas
mostraram variação em relação à
frequência e abundância de acordo com os
diferentes horários de atividade
observados durante o dia (Fig. 2). Pôde-se
verificar que as morfoespécies Bombom e
Bambu apresentaram uma frequência de
atividade similar ao longo do dia, assim
como maiores números de registros,
enquanto a Dourada e Olho Verde se
mostraram pouco frequentes, e surgiam
em menores quantidades. Além disso, a
morfoespécie Azul surgiu em quase todas
as observações, porém em baixas
quantidades.
Tabela 1. Morfoespécies de Odonata identificadas dentro de duas subordens e três famílias.
MORFOTIPO SUBORDEM FAMÍLIA
Sp 1 (Bombom) ANISOPTERA LIBELLULIDAE Sp 2 (Vermelha) ANISOPTERA LIBELLULIDAE Sp 3 (Bambu) ANISOPTERA LIBELLULIDAE Sp 4 (Olho verde) ANISOPTERA LIBELLULIDAE Sp 5 (Roxa) ANISOPTERA AESHNIDAE Sp 6 (Dourada) ZYGOPTERA COENAGRIONIDAE Sp 7 (Azul) ZYGOPTERA COENAGRIONIDAE

78 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Figura 2. Número de registros das sete
morfoespécies de Odonata, observados ao longo
dos horários do dia, mostrando frequências de atividades diferentes.
Foi constatado que as
morfoespécies possuem similaridade em
relação à frequência de atividades
durantes horários do dia (Fig. 3). Algumas
preferem horários nos quais as
temperaturas são mais amenas, como é o
caso da Dourada e da Olho Verde que são
mais frequentes durante as horas com
temperaturas amenas, no amanhecer e no
crepúsculo. Enquanto isso, outras
morfoespécies como a Bombom e a Bambu
são mais frequentes nos horários mais
quentes do dia.
As morfoespécies aparentemente
mostraram se agrupar também em relação
ao tamanho, pois Bombom e Bambu são as
maiores em tamanho e mais frequentes,
enquanto Dourada e Olho Verde são as
menores e menos frequentes. Vermelha,
Azul e Roxa são intermediárias em relação
à frequência, tamanho e número de
registros. Porém, não houve evidências de
que o tamanho estaria influenciando o
período de atividade das Odonata (R2 = -
0,49; P = 0,3) (Fig. 4).
Figura 3. A: Gráfico de Similaridade de Bray-Curtis
agrupando as morfoespécies que apresentaram
horários de atividades semelhantes. B: Análise do
nMDS com corte no nível de similaridade 16 e 35%
mostrando quais morfoespécies compartilham
padrões semelhantes relacionados a freqüência de
atividade.
A
B

79 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Figura 4. Tamanho médio da comunidade de
Odonata plotado em relação a um dos eixos gerados pelo nMDS.
DISCUSSÃO
É conhecido que fatores como
tamanho do corpo, comportamento de
movimento e fatores ambientais, como
temperatura e exposição solar local,
influenciam na temperatura destes
organismos ectotérmicos (SAMEJIMA &
TSUBAKI, 2010). Como algumas destas
características variam de espécie para
espécie, provavelmente o padrão de
ocorrência de odonatos constatado está
ocorrendo em função dessas
características intrínsecas e das variações
diárias do ambiente.
Relacionando este conhecimento
com o presente estudo, temos que o fato
de os morfotipos Bambu e Bombom terem
sido as mais abundantes e terem ocorrido
durante a faixa de horários com
temperaturas mais elevadas pode estar
relacionado com o maior tamanho corporal
desses indivíduos, uma vez que, a
capacidade de manter a temperatura
corporal relativamente constante pode ser
associada a essa característica (CORBET,
1980), apesar de a análise de correlação
não ter se apresentado significativa. Para
uma maior certeza dessa hipótese se faz
necessária uma nova análise, onde seriam
utilizados valores referentes à temperatura
mensurados com a utilização de um
termômetro e medições de índice de
tamanho corporal em vários indivíduos por
morfotipo.
Notou-se que a parcela localizada
aproximadamente no centro do transecto
apresentou uma frequência de ocorrência
de odonatos bem maior que as outras duas
parcelas. De acordo com Corbet (1980),
microhabitats sombreados por arvores já
apresentam grandes diferenças na
composição das espécies de Odonata em
relação aos habitats não sombreados.
Estando a primeira e a terceira parcelas
perto de estratos arbóreos, pode estar aí a
justificativa para essa diferença encontrada
entre as parcelas.
Conclui-se que existe variação na
freqüência de ocorrência de Odonata ao
longo de um dia na área estudada e que
essa variação pode estar relacionada à
capacidade das diversas espécies lidarem
com as diferentes temperaturas ao longo
do dia, sugerindo que a comunidade está
estruturada através de uma divisão de
nicho circadiano. São necessários estudos
futuros mais detalhados, com um esforço
amostral maior, verificando não somente
variações diárias como também anuais da
ocorrência das espécies de Odonata,
podendo assim inferir sobre partição de
nicho espacial e temporal.
Agradecimentos
Nossa gratidão aos professores
Alexandre Vasconcelos e Carlos Fonseca
pela orientação nas análises estatísticas, e
ao Adalberto Varella pela identificação dos
Odonata. Também não podemos deixar de
agradecer à equipe da ESEC do Seridó por

80 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
todo suporte oferecido durante nossa
estadia.
Referências
Andrade-Lima D., 1981. The caatinga
dominium. Revista Brasileira de Botânica 4,
149-63.
Buzzi, J.Z. 2005. Entomologia Didática. Editora
UFPR, Curitiba, pp 125-127.
Corbet, P. S. 1980. Biology of Odonata. Annual
Review of Entomology 25, 189-217.
Imoobe, T. O. T. and E. Ohiozebau. 2010.
Pollution status of a tropical forest river
using aquatic insects as indicators. African
Journal of Ecology 48(1), 232-238.
Lavelle, P., Lattaud, D. T., Barois, I., 1995.
Mutualism and biodiversity in soils. Plant
Soil 170, 23–33.
Lutz, P. E., Pittman, A. R. 1970. Some ecological
factors influencing a community of adult
Odonata. Ecology 51 (2), 279-284.
McCauley, S. J. 2010. Body size and social
dominance influence breeding dispersal in
male Pachydiplax longipennis (Odonata).
Ecological Entomology 35(3), 377-385.
Sampaio, E. V. S. B., 1995. Overview of the
Brazilian Caatinga. In: Bullock, S.H.,
Mooney, H. A., Medina, E. (Eds.), Seasonally
dry forests. Cambridge University Press,
Cambridge, pp. 35-58.
Silva, D. D., P. De Marco, et al. (2010). Adult
odonate abundance and community
assemblage measures as indicators of
stream ecological integrity: A case study.
Ecological Indicators 10(3), 744-752.
Swift, M. J., Heal, 0. W., Anderson. J. M., 1979.
Decomposition in terrestrial ecosystems.
Blackwell Scientific Publications, London.
Samejima, Y. & Tsubaki, Y. 2010. Body
temperature and body size affect flight
performance in a damselfly. Behav. Ecol.
Sociobiol. 64, 685–692.
Yoon, J., J. M. Nam, et al. 2010. Nannophya
pygmaea (Odonata: Libellulidae), an
Endangered Dragonfly in Korea, Prefers
Abandoned Paddy Fields in the Early Seral
Stage. Environmental Entomology 39(2),
278-285.

81 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
EFEITO DA ALTITUDE SOBRE A DIVERSIDADE DE COLLEMBOLA
(ARTHROPODA: HEXAPODA) EM UMA ÁREA DE CAATINGA NO RIO
GRANDE DO NORTE
DANIEL OLIVEIRA; GINA MANTILLA; MONALISA RODRIGUES O. DA SILVA & UIRANDÉ O.
COSTA
Introdução
O padrão de riqueza de espécies
nas comunidades biológicas pode ser
explicado por uma série de fatores
classificados em diferentes categorias. Um
dos padrões mais difundidos e aceitos na
ecologia aponta para uma diminuição no
número de espécies diante do aumento
altitudinal (Begon et al., 2007). Stevens
(1992) observou para plantas e
vertebrados que a riqueza de espécies nos
topos das montanhas é geralmente menor
do que em áreas mais baixas. No entanto,
há um crescente número de estudos
contrários a essa constatação, sugerindo
que a configuração dos fatores climáticos,
biológicos, geográficos e históricos pode
ser determinante para explicar a variação
na riqueza de espécies ao longo de
gradientes altitudinais (Lomolino, 2001).
Carsten (1999) ressaltou que
muitos grupos de animais, incluindo alguns
táxons de insetos, apresentam uma maior
riqueza em altitudes mais elevadas, por
exemplo, formigas em um ecossistema
árido. Em ecossistemas áridos, a riqueza de
espécies para algumas ordens de insetos
pode ser maior em maiores altitudes, onde
temperaturas mais baixas e maior
precipitação podem suportar maiores
níveis de produção primária e causar
menores níveis de estresses fisiológicos a
determinados grupos (Sanders et al.,
2003).
Colêmbolos são organismos
pequenos e semelhantes aos insetos,
podem estar associados a ambientes
terrestres, aquáticos e arborícolas
(Zeppelini & Bellini, 2004). Estes
organismos estão intimamente associados
aos processos ecossistêmicos que ocorrem
no compartimento serrapilheira-solo.
Vários estudos demonstraram sua
contribuição direta ou indireta nos
processos ecológicos do solo, como
mineralização de nitrogênio, respiração do
solo ou lixiviamento de carbono orgânico
dissolvido (Filser, 2002). Assim, conhecer a
diversidade e distribuição destes
organismos torna-se essencial em estudos
para a conservação, os quais englobam as
características e funcionalidade das
espécies.
O bioma Caatinga possui
características peculiares com grande
variação ambiental e climática. A
vegetação é um mosaico de florestas
arbóreas e arbustivas que compreende
diversas adaptações necessárias à
sobrevivência num ambiente de alta
deficiência hídrica e de elevadas
temperaturas, característico do clima do
semi-árido quente (Prado, 2003). Diante de
um clima sazonal bastante pronunciado
esperam-se efeitos significativos na
comunidade de invertebrados.

82 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Assim, é válido considerar a
hipótese de que na Caatinga, o padrão de
diversidade, através de um gradiente
altitudinal, pode ser o inverso para alguns
grupos, visto que ambientes mais elevados
podem oferecer condições diferenciadas as
encontradas normalmente nas regiões
mais baixas.
Com o intuito de testar essa
hipótese, o estudo teve como objetivo
investigar o padrão de diversidade de
espécies de Collembola ao longo de um
gradiente altitudinal. Assim como,
examinar algumas variáveis ambientais que
podem influenciar a diversidade desse
grupo no gradiente em questão, em uma
área de Caatinga no Rio Grande do Norte.
Metodologia
O estudo foi realizado na Serra
Verde, área de Caatinga localizada na
Estação Ecológica do Seridó - ESEC, no
município de Serra Negra do Norte, Rio
Grande do Norte. A região do Seridó está
entre as mais impactadas pela ação
antrópica, mas a ESEC encontra-se
conservada desde 1982. Nosso ambiente
de estudo, a Serra Verde, é considerada
uma das áreas mais conservadas da
estação (IBAMA, 2004).
A amostragem ocorreu em um
gradiente altitudinal correspondendo a 120
metros, sendo o primeiro ponto localizado
a 227 do nível do mar e o último a 347
metros.
Na coleta de dados foram
utilizados cinco transectos horizontais de
10 metros de comprimento,
perpendiculares à trilha e distantes 30m
(altitude) entre si. Os transectos foram
divididos em 10 retângulos de 1m² e para a
coleta de dados foram selecionados
randomicamente seis retângulos para cada
transecto (Fig. 1).
Figura 20: Delineamento amostral destacando
os transectos e seus seis retângulos
selecionados para estudo de gradiente
altitudinal.
A coleta de colêmbolos foi
realizada com aspirador entomológico e
armadilha de queda (pit-fall). No primeiro
método o folhiço presente no retângulo é
analisado em bandejas durante 20 minutos
e os indivíduos de collembola são
aspirados. Três coletores foram
responsáveis pela aspiração entomológica
em todo gradiente, totalizando dois
retângulos por transecto para cada coletor.
Ao longo do gradiente foram mantidos os
coletores e seus respectivos pontos.
Colocou-se armadilhas de queda (pit-fall),
com álcool a 70%, ao longo do gradiente
altitudinal. Para cada retângulo foi
colocado uma armadilha, totalizando 30
armadilhas, com tempo de exposição de
24h.
Devido problemas operacionais a
coleta ativa com aspirador entomológico
não foi amostrada no último ponto do
gradiente.

83 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Figura 21: Métodos de coleta para collembola:
A – Armadilha de pit-fall; B – Aspirador
entomológico.
Os indivíduos foram mantidos em
álcool a 70% e, após a triagem em campo,
os espécimes foram contados e
identificados em morfoespécies. A
identificação ocorreu em laboratório sob
microscópio estereoscópico, de acordo
com seus gêneros.
Utilizou-se um termohigrômetro na
medição da temperatura e pHmetro para
medir o pH e a umidade do solo ao longo
do gradiente altitudinal. A serrapilheira
utilizada para coleta de colêmbolos, por
meio do aspirador, foi pesada para
estimativa do esforço amostral entre os
transectos. A compactação do solo foi
estimada após soltar, na altura do peito,
um facão em direção ao solo e com o
paquímetro mediu-se quanto o facão
penetrou no solo.
O índice de diversidade de
Shannon-Wiener foi calculado a partir da
riqueza e abundância de cada transecto, e
para tal utilizou-se o programa PRIMER 6
Beta. Foi realizada uma análise de
regressão linear para verificar a relação da
diversidade e altitude nos transectos. A
análise de variância (ANOVA) foi utilizada
para testar diferenças entre os transectos
em relação aos métodos de coleta.
Aplicou-se o modelo linear geral (GLM)
para verificar diferenças das variáveis entre
os transectos analisados.
Todas as análises estatísticas foram
feitas através do programa Systat 12.0.
Resultados
Foram obtidos 67 indivíduos,
pertencentes a seis gêneros e classificados
em sete morfoespécies (Tabela 1), das
quais duas ocorreram em todos os
transectos e duas em apenas em um
transecto.
Tabela 4: Abundância e riqueza de
morfoespécies de Collembola em cada ponto
altitudinal analisado.
MORFOESPÉCIE P1 227m
P2 257m
P3 287m
P4 317m
P5 347m
Seira sp. 14 8 1 19 2 Allacma sp.1 4 3 2 0 4 Allacma sp.2 2 0 0 1 0 Allacma sp.3 1 0 0 0 0 Brachystomella sp. 1 2 1 2 0 Sphaeridae sp.1 1 1 3 1 2
Cryptopygus sp. 2 0 0 0 0
Abundância 25 14 7 23 8 Riqueza 7 5 5 5 4
Não houve diferenças significativas
entre os dois métodos, aspirador
entomológico e pit-fall, na riqueza (F =
1,227; gl 3= ; p = 0,320) e na abundância (F
= 1,387; gl = 5; p = 0,264) (Fig. 3).
Ao analisar o índice de diversidade
Shannon-Wiener de colêmbolos diante do
gradiente trabalhado, observou-se uma
relação negativa e significativa (R2 = -0,92;
p = 0,023) entre diversidade e altitude (Fig.
4).
Nenhuma das variáveis abióticas de
solo avaliadas mostraram relação
significativa com riqueza (F = 2,505; gl = 4;
p = 0,109) e abundância (F = 1,427; gl = 4; p
= 0,295) (Tabela 2) de Collembola ao longo
do gradiente estudado.
A
A
A
B
A
A

84 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
Figura 22: Gráficos comparando a riqueza (A) e
abundância (B) por método de coleta em
diferentes altitudes, nos transectos
amostrados. ■ Aspirador entomológico; □ Pit-
fall.
Figura 23: Variação da diversidade de
Collembola ao longo do gradiente altitudinal
analisado. Valor de diversidade é o índice de
Shannon-Wiener (R2 = -0,92; p = 0,023).
Tabela 5: P-valores para as variáveis
relacionadas à riqueza e abundância
Variáveis
p – valor
(riqueza)
p – valor
(abundância)
Compactação (solo) 0,980 0,699
Peso (serrapilheira) 0,061 0,092
Umidade (solo) 0,359 0,422
Temperatura
(transecto)
0,334 0,954
Discussão
Foi verificado que a distribuição
das espécies de colêmbolos variou diante
do gradiente de altitude estudado, como
evidenciado na tabela 1.
Observou-se que as morfoespécies
Seira sp., Sphaeridae sp.1, Allacma sp.1 e
Brachystomella sp. foram encontrados na
maioria dos níveis altitudinais. No entanto,
outras morfoespécies foram peculiares a
um único transecto, como por exemplo
Allacma sp.3 e Cryptopygus sp. presentes
apenas na parte mais baixa do transecto
altitudinal. Nesse sentido, Begon et. al.
(2007), observou que os limites de
distribuição para algumas espécies não
terminam bruscamente, mas de forma
lenta em análises de gradiente, o que é
corroborado pelos resultados obtidos
nesse trabalho.
A distribuição das morfoespécies
de colêmbolos ao longo do gradiente
altitudinal estudado pode sugerir que
algumas dessas entidades são mais
ocorrentes, possuindo maior amplitude em
algumas dimensões do seu nicho ecológico,
197 227 257 287 317 347 377Altitude (m)
0
1
2
3
4
5
6
7
Nú
me
ro d
e in
div
ídu
os
B
A

85 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
permitindo assim uma maior distribuição
altitudinal. Por outro lado, outras
morfoespécies como a Allacma sp.3 e
Cryptopygus sp., de menor largura de
nicho, foram mais específicas em apenas
um dos níveis altitudinais, ou podem ser
consideradas espécies raras pois não foram
observadas nos outros pontos do
gradiente.
Percebeu-se que existiu uma
variação na diversidade de colêmbolos
inversamente proporcional ao aumento da
altitude na área estudada. Esse resultado
corrobora a hipótese mais ampla (Stevens,
1992), contrastando com a hipótese que
motivou o estudo (Carsten, 1999), e
confirmando um padrão de declínio da
diversidade diante do aumento altitude.
McCain (2004), ao trabalhar com
pequenos mamíferos em gradientes
altitudinais, sugeriu que as diferenças na
diversidade desses grupos em diversas
altitudes são refletidas pela disponibilidade
de micro-habitats e/ou por diferenças
causadas na geomorfologia do relevo.
Dessa forma, o padrão de diversidade
encontrado pode estar relacionado com as
diferenças entre a maior heterogeneidade
espacial e maior disponibilidade de micro-
habitats nas altitudes mais baixas. Bem
como, a maior homogeneização espacial e
menor disponibilidade de micro-habitat
próximo ao topo da Serra.
É importante ressaltar que a coleta
ativa, através do aspirador entomológico,
não foi realizada no último ponto
altitudinal. No entanto, como não houve
diferenças significativas entre as duas
técnicas de coleta, as análises não foram
afetadas e o padrão encontrado na análise
de regressão também não foi prejudicado.
As variáveis ambientais
temperatura, compactação e umidade do
solo não afetaram significativamente a
riqueza e abundância dos colêmbolos.
Consequentemente, o padrão de
diversidade encontrado não pode ser
explicado pela alteração dessas variáveis. O
peso da serrapilheira foi marginalmente
significativo, indicando que essa variável
pode estar afetando uma parte da variação
na diversidade de colêmbolos. Desse
modo, a camada de serrapilheira pode
estar permitindo uma maior variação de
condições e recursos ambientais que
podem ser essenciais para estes
organismos. No entanto, outros fatores
podem estar influenciando a diversidade
desse grupo na Serra Verde. Conforme
Sanders et al. (2003), os fatores ambientais
que influenciam a diversidade em um
gradiente altitudinal são complexos e
diferem entre espécies ao longo do
gradiente.
Por fim, diante da alta relevância
biológica da Caatinga e das intensas
ameaças à sua integridade e permanência,
ainda há uma grande carência de
interesses e políticas para a conservação da
biodiversidade, das paisagens e demais
recursos naturais da Caatinga.
Dessa forma, estudos como este,
que consideram padrões de diversidade
das espécies e que ressaltam a importância
de alguns locais peculiares como refúgios
de diversidade presentes na paisagem,
devem ser repetidos e fazer parte das
iniciativas de conservação e proteção do
bioma.
Agradecimentos
Aos professores do Programa de Pós-
Graduação em Ecologia – PPGE pelas
contribuições no projeto. Agradecimentos

86 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
especiais ao Bruno Bellini e Alexandre
Vasconcellos pelo fornecimento de material e
atenção dedicada ao grupo, e aos funcionários
da ESEC pela hospitalidade e por permitir o uso
de toda a infra-estrutura da reserva.
Referências
BEGON, M.; TOWNSEND, C. R.; HARPER, J. L.
2007. Padrão na riqueza de species. In:
Ecologia de indivíduos a Ecossistemas.
4 ed. 740 p.
CARSTEN, A. B.; MOHAMED, M.; LINSENMAIR,
K. E. 1999. Altitudinal distribution of
leaf litter ants along a transect in
primary forests on Mount Kinabalu,
Sabah, Malaysia. Journal of Tropical
Ecology. Cambridge University Press.
1999. 15, 265-277.
FILSER, J. 2002. The role of Collembola in
carbon and nitrogen cycling in soil.
Pedobiologia. 2002. 46, 234-245.
IBAMA – Instituto Brasileiro do Meio Ambiente
e dos Recursos Naturais Renováveis.
2004. Plano de Manejo da Estação
Ecológica do Seridó. Relatório Técnico
não publicado.
LOMOLINO, M. V. 2001. Elevation gradients of
species-density: historical and
prospective views. Global Ecology &
Biogeography. 2001. 10, 3-13.
McCAIN, C. M. 2004. The mid-domain effect
applied to elevational gradients:
species richness of small mammals in
Costa. Journal of Biogeography. 2004.
31, 19–31.
PRADO, D. E. 2003. As Caatingas da América do
Sul. In: LEAL, I.R.; TABARELLI, M.;
SILVA, J.M.C. Ecologia e Conservação
da Caatinga, Recife: Ed. Universitária
da UFPE, 2003. p. 3-73.
SANDERS, N. J.; WAGNER, J. M. D. 2003.
Patterns of ant species richness along
elevational gradients in an arid
ecosystem. Global Ecology &
Biogeography. 2003. 12, 93–102.
STEVENS, G. C. 1992. The elevational gradient
in altitudinal range: an extension of
rapoport’s latitudinal rule to altitude.
The American naturalist. 1992. 140,
893-911.
ZEPPELINI, D.; BELLINI, B. C. 2004. Introdução
ao estudo dos Collembola. João
Pessoa: Editora Universitária.

87 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
EFEITO DA QUALIDADE DO HABITAT E DENSIDADE DE PREDADORES SOBRE
A TAXOCENOSE DE COLLEMBOLA (ARTHROPODA: HEXAPODA)
BRUNNO FREIRE DANTAS DE OLIVEIRA, FABRICIO RODRIGUES CAMACHO, LEONARDO H. TEIXEIRA
PINTO & RODRIGO XAVIER SOARES
Introdução
A densidade de microorganismos
de solo é conhecida por ser controlada por
efeitos ascendentes resultantes da
quantidade e qualidade da serrapilheira
(Scheu & Schaefer, 1998; Rantalainen et
al., 2004). Existem evidências que sugerem
que a densidade de microartrópodes de
solo é regulada não apenas pela
quantidade e qualidade da serrapilheira,
mas também pela disponibilidade de
fungos (Hairston et al.,1960; Scheu &
Schaefer, 1998; Rantalainen et al., 2004).
Colêmbolos são bons
representantes da biodiversidade
faunística do solo (Zepelline et al., 2008),
compõem a guilda de fungívoros e
detritívoros da microfauna, e executam um
importante papel no consumo de detritos e
ciclagem de nutrientes (Zinckler & Stecken,
1986). São organismos sensíveis a fatores
exógenos como temperatura e umidade
(Klironomos & Kendrick, 1995). Além dos
fatores exógenos, a densidade de
colêmbolos pode ser controlada por
predação (Hagvar, 1995; Schaefer, 1995;
Ferguson, 2001). Estes organismos
compõem um grupo diverso de artrópodes
de solo e compartilham uma diversa fauna
de predadores na serrapilheira (Schneider
& Maraun, 2009).
O objetivo do presente estudo foi
avaliar o efeito da qualidade do habitat,
em particular a umidade e disponibilidade
de fungo (alimento) no solo, e a densidade
de predadores sobre a densidade de
colêmbolos. As hipóteses testadas são: (a)
a densidade de colêmbolos responde
positivamente à umidade e disponibilidade
de fungos (efeito ascendente); e (b) a
densidade de colêmbolos responde
negativamente a densidade de predadores
(efeito descendente).
Material e Métodos
O presente trabalho foi realizado
na Estação Ecológica do Seridó (6º35’S e
37º15’O), uma área de caatinga localizada
na ecorregião da Depressão Sertaneja
Setentrional, município de Serra Negra do
Norte.
O delineamento experimental
adotado foi uma análise de variância
fatorial em bloco. Estabeleceu-se sete
blocos de 9m² (3x3m), dispostos de
maneira aleatória, em uma área próxima à
jusante do açude da Sede da ESEC-Seridó.
Cada bloco era composto por quatro
parcelas internas de 1m², dispostas em
cada vértice do bloco. A área restante no

88 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
interior do bloco foi utilizada para
deslocamento, facilitando a coleta em cada
parcela, como apresentado na figura 1.
Cada bloco continha todos os
tratamentos. Dentro de cada bloco, os
tratamentos foram aleatorizados e
dispostos de forma que houvesse parcela
somente com água; somente com fungo;
com água e fungo; e um controle. Assim,
para cada tratamento existiam sete
réplicas (parcelas), uma em cada bloco.
Figura 1: Representação esquemática da
distribuição dos blocos na área de estudo. “A”
representa as parcelas onde foram dispostos os
tratamentos; “B” é a área utilizada para
deslocamento.
Uma mistura de 150g de fermento
biológico (Saccharomyces cerevisiae)
desidratado em pó, juntamente com água
e farinha de trigo foi produzida para ser
utilizada nos tratamentos com adição de
fungo. Alíquotas de 20ml dessa mistura
foram depositadas no centro das parcelas.
Um litro de água foi distribuido da mesma
forma para o tratamento com adição de
água.
Após 18 horas do início do
experimento, coletas de 10 minutos foram
conduzidas em cada parcela. O folhiço da
serrapilheira e solo foram depositados em
bandejas de plástico e os animais
observados foram coletados com aspirador
entomológico. Todos os colêmbolos e
pequenos artrópodes predadores, que
passassem pelo tubo do aparelho, foram
coletados e fixados com álcool a 70%. Toda
a serrapilheira triada durante o tempo de
coleta foi pesada para o cálculo da
densidade de indivíduos (densidade =
indivíduos/g. serrapilheira). Os indivíduos
coletados foram levados ao laboratório
para identificação em nível de gênero para
colêmbolos e de menor táxon possível para
predadores, sob microscópio
estereoscópico.
O efeito dos tratamentos e da
densidade de predadores sobre a
densidade de colêmbolos foi analisada
através de General Linear Model (GLM),
utilizando o software estatístico SYSTAT 12.
Resultados
Foram coletados 46 indivíduos de
Collembola, classificados em 4 espécies; e
229 indivíduos de predadores,
pertencentes a 22 espécies. Em média 2,14
indivíduos de Collembola e 5 indivíduos de
predadores foram coletados nas parcelas
controle, e 1,52 indivíduos de Collembola e
9,48 indivíduos de predadores nas parcelas
manipuladas pelos tratamentos.
A densidade de colêmbolos foi
similar entre blocos (F=0,54; gl = 6,17 ; p =
0,769); não respondeu aos tratamentos de
adição de água (F=0,653 ; gl = 1,17 ; p =
0,43); adição de fermento (F = 0,815 ; 1,17
; p = 0,38); e também não foi influenciada
pela interação entre estes (F = 0,003 ; gl =
1,17 ; p = 0,958 – Figura 2). Por fim, a
densidade de colêmbolos não respondeu à
densidade de predadores (F = 0,182 ; gl =
1,17 ; p = 0,675 – Figura 3).

89 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
A riqueza de colêmbolos, assim
como a densidade, também foi similar
entre blocos (F=0,667; gl = 6,17 ; p = 0,677)
e não respondeu aos tratamentos de
adição de água (F=0,595 ; gl = 1,17 ; p =
0,45) ou de fermento (F = 0,791 ; 1,17 ; p =
0,38), bem como à interação entre
tratamentos (F = 1,303 ; gl = 1,17 ; p = 0,27
– Figura 4). Ainda, a riqueza de colêmbolos
não respondeu a riqueza de predadores (F
= 0,182 ; gl = 1,17 ; p = 0,675)
Figura 2: Densidade de colêmbolos (ind./g. serrapilheira) e erro padrão nos tratamentos de adição de água e fungo.
Figura 3: Relação entre densidade de colêmbolos e densidade de predadores [Log(ind./g. serrapilheira)] na área de estudo.
.
Figura 4: Riqueza de colêmbolos e erro padrão nos tratamentos de adição de água e fungo.
Tabela 1: Riqueza e abundância de colêmbolos e predadores. Táxon N
Collembola Brachystomella sp1 6
Dicyrtoma sp 2 Lepidocyrtus sp1 17 Seira sp1 21
Total 46
Predadores Acari sp1 14
Acari sp2 1 Acari sp3 1 Acari sp4 4 Aranae sp1 4 Aranae sp2 1 Aranae sp3 2 Aranae sp4 1 Aranae sp5 2 Formicidae sp1 91 Formicidae sp2 9 Formicidae sp3 45 Formicidae sp4 8 Formicidae sp5 15 Formicidae sp6 1 Formicidae sp7 1 Opiliones sp1 2 Opiliones sp2 1 Pseudoescorpiones sp1 13 Pseudoescorpiones sp2 1 Pseudoescorpiones sp3 10 Pseudoescorpiones sp4 1 Reduviidae sp1 1
Total 229
Discussão
O presente trabalho mostra que a
densidade e riqueza de colêmbolos foi
indiferente aos tratamentos de adição de
fungo (alimento) e água e, também, à
R² = 0,009 ; p = 0,63

90 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
densidade e riqueza de predadores, após
um período de 18 horas do início do
experimento.
Trabalhos com adição de água
sugerem que colêmbolos respondem
positivamente a tratamentos similares ao
aqui executado após um período de uma
(Ferguson & Joly, 2002) e duas semanas
(Chikoski et al., 2006). Colêmbolos são
sensíveis a fatores exógenos como
temperatura e umidade (Klironomos &
Kendrick, 1995) podendo responder
imediatamente a variações nas condições
locais (Bellini, comunicação pessoal). O
presente estudo sugere que os organismos
analisados estão adaptados ao ambiente
de caatinga, não respondendo tão
rapidamente a um tratamento que simulou
uma precipitação momentânea de
1000mm³.m-2.
Colêmbolos são organismos
conhecidos por se alimentar de fungos
(Zinckler & Stecken, 1986; Klironomos &
Kendrick, 1995; Scheu & Schaefer, 1998).
Entretanto, no presente trabalho, a
densidade de colêmbolos não respondeu
ao tratamento de adição de fungo.
Contudo, já foi observado que este grupo
pode ser seletivo quanto à preferência em
forragear em determinadas espécies de
fungos (Ngosong et al, 2009). Neste
trabalho, a assembléia de colêmbolos não
foi capaz de responder rapidamente a
adição de alimento (Saccharomyces
cerevisiae), mantendo densidades
populacionais e riquezas de espécies
similares entre tratamentos e grupos
controle.
Resultados contrastantes podem
ser obtidos para os tratamentos realizados
neste experimento caso a comunidade de
colêmbolos tivesse mais tempo para
responder as variações ambientais aqui
manipuladas. Talvez exista um lag para se
perceber uma resposta numérica na
densidade e/ou riqueza de colêmbolos,
como resposta aos tratamentos aqui
realizados, como sugerem os trabalhos de
Ferguson & Joly (2002) e Chikoski e
colaboradores (2006).
Alguns autores observaram que
colêmbolos sofrem grande pressão de
predação por serem superabundantes em
detritos e massas de fungo (Hairston et al.
1960; Schneider & Maraun, 2009). Essa
tendência, no entanto, não foi evidenciada
pelo experimento realizado na ESEC –
Seridó. Isto, por sua vez, sugere que a
fauna de colêmbolos de solo parece ser
regulada por fatores exógenos à interação
predador-presa ou que o período
experimental adotado não foi sensível para
evidenciar tal relação.
Agradecimentos
Somos imensamente gratos aos funcionários da
ESEC-Seridó pelo carinho e recepção durante o
curso de campo de 2010. Aos professores do
Programa de Pós graduação em Ecologia da
UFRN pela paciência e compartilhamento
intelectual, em especial aos sempre solícitos,
Carlos Fonseca e Bruno Bellini. Também, aos
alunos deste PPG pelo companheirismo e
afabilidade.
Referências
Chikoski, J.M., Fergusonb, S.H. e Meyer, L.
2006. Effects of water addition on soil
arthropods and soil characteristics in a
precipitation-limited environment.
Acta Ecologica. 30. 203-211.
Ferguson, S.H. & Joly, D.O. 2002. Dynamics of
springtail and mite populations: the

91 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
role of density dependence, predation,
and weather. 27. 565-573.
Hairston, N.G., Smith, F.E., Slobodkin, L.B. 1960.
Community structure, population
control and competition. American
Naturalist. 94. 421–425.
Ngosong, C., Raupp, J., Scheu, S. e Ruess, L.
2009. Low importance for a fungal
based food web in arable soils under
mineral and organic fertilization
indicated by Collembola grazers. Soil
biology and biochemistry. 41. 2308-
2317.
Scheu, S. & Schaefer, M., 1998. Bottom-up
control of the soil macrofauna
community in a beechwood on
limestone: manipulation of food
resources. Ecology. 79. 1573–1585.
Schneider, K. & Maraun,M. 2009. Top-down
control of soil microarthropods.
Evidence from a laboratory
experiment. Soil Biology and
Biochemistry. 41. 170-175.
Rantalainen, M.L., Kontiola, L., Haimi, J., Fritze,
H. e Setala, H. 2004. Influence of
resource quality on the composition of
soil decomposer community in
fragmented and continuous habitat.
Soil Biology & Biochemistry 36, 1983–
1996.
Zinkler, R.D. & Stecken, R. 1986. Digestion of
leaf litter by the spring tail Tomocerus
flavescens (Insecta, Collembola). Soil
Fauna and Soil Fertility. Ed. B. R.
Striganova. Pp. 730-734.
Klironomos, J.N & Kendrick, B. 1995.
Relationships among microarthropods,
fungi, and their environment. Plant
soil. 170. 183-197.
Zeppelini, D., Belline, B.C., Creão-Duarte, A.J. e
Hernández, M.I.M. 2008. Collembola
as bioindicators of restoration in
mined sand dunes of Northeast Brazil.
Biodivers Conserv. 18. 1161-1170.

92 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010

93 Curso de Campo Ecologia da Caatinga, PPG Ecologia, UFRN, 2010
TURMA ECOLOGIA DE CAMPO 2010
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA-UFRN








