
Otimização do Meio de Cultura para a Produção de Quitosanase … · 2017. 11. 2. ·...
1000
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE TECNOLOGIA DEPARTAMENTO DE ENGENHARIA QUÍMICA PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA TESE DE DOUTORADO Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo Descontínuo Submerso Raimundo Cosme da Silva Filho Natal/RN Abril/2013
Transcript of Otimização do Meio de Cultura para a Produção de Quitosanase … · 2017. 11. 2. ·...

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE TECNOLOGIA
DEPARTAMENTO DE ENGENHARIA QUÍMICA PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA
TESE DE DOUTORADO
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em
Cultivo Descontínuo Submerso
Raimundo Cosme da Silva Filho
Natal/RN
Abril/2013

Raimundo Cosme da Silva Filho
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo
Descontínuo Submerso
Tese apresentada ao Programa de Pós-Graduação em Engenharia Química – PPGEQ, da Universidade Federal do Rio Grande do Norte, como parte dos requisitos para obtenção do título de Doutor em Engenharia Química, sob a orientação do Prof. Dr. Everaldo Silvino dos Santos e a coorientação da Profª. Drª. Gorete Ribeiro de Macedo.
Natal/RN
Abril/2013

Catalogação da Publicação na Fonte. UFRN / CT / PPGEQ
Biblioteca Setorial “Professor Horácio Nícolas Sólimo”.
Silva Filho, Raimundo Cosme da. Otimização do meio de cultura para a produção de quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso / Raimundo Cosme da Silva Filho. - Natal, 2013. 98 f.: il.
Orientador: Everaldo Silvino dos Santos. Co-orientador: Gorete Ribeiro de Macedo. Tese (Doutorado) - Universidade Federal do Rio Grande do Norte.
Centro de Tecnologia. Departamento de Engenharia Química. Programa de Pós-Graduação em Engenharia Química.
1. Quitosana - Tese. 2. Quitina - Tese. 3. Quitosanase – Tese. 4. Metarhizium
anisopliae - Tese. I. Santos, Everaldo Silvino dos. II. Macedo, Gorete Ribeiro de. III. Universidade Federal do Rio Grande do Norte. IV. Título. RN/UF/BSEQ CDU 547.458 (043.2)


Ao meu pai, minha mãe e minha
família, em especial aos meus
irmãos e sobrinhos

AGRADECIMENTOS
A Deus, por estar presente em todos os momentos da minha vida e por me ceder à
oportunidade de iniciar e concluir este trabalho.
A minha mãe Cândida Inocência da Silva que esteve comigo em todos os momentos da
minha vida me educando, me amando e sempre me dando muito carinho.
Ao meu pai Raimundo Cosme da Silva, que não está mais aqui para ver este trabalho,
mas que foi a pessoa que mais me apoiou e me incentivou, em todos os momentos da minha vida,
para que eu pudesse chegar o mais longe possível na minha carreira acadêmica. Pai muito
obrigado por toda atenção, educação, amor e carinho que recebi durante toda a minha vida.
Aos meus irmãos e familiares que sempre estiveram comigo dividindo momentos de
grande alegria e às vezes de tristeza, mas sempre nos mantendo unidos para superarmos todos os
obstáculos.
A minha namorada Rochelly Caroliny por todo amor e carinho que me deu e que me
entendeu nos momentos que estive ausente para a conclusão deste trabalho.
Ao meu orientador Professor Doutor Everaldo Silvino dos Santos por confiar em mim e
me aceitar como orientando além de toda a amizade, orientação e dedicação em todas as etapas de
desenvolvimento e conclusão deste trabalho.
À Professora Doutora Gorete Ribeiro de Macedo, “pessoa que admiro muito como
profissional e principalmente, como pessoa, um exemplo a ser seguido”, pelos conselhos e críticas
quando necessários ao desenvolvimento deste trabalho. Professora a senhora é considerada uma
mãe para de todos os alunos que tiveram a honra de conhecê-la e trabalhar com a senhora.
Aos meus amigos Engenheiros Químicos, Ana Carmen, Ana Katerine, Andréa Farias,
Anita Lima, Sirtys Lessa, Alexandre, Sérgio, Márcio Bezerra e ao Biólogo Daniel Souza, que
sempre me ouviram e que me ajudaram de alguma forma nesse trabalho.
A Eusamar Coelho de Lima, Mazinha, secretária do PPGEQ, pelo apoio, pela paciência e
orientação nas questões burocráticas do processo.
A Carlos e Jardelson, pela grande colaboração e dedicação nos experimentos do
laboratório.
A Sérgio Santana pela grande colaboração e dedicação, o qual não poupou esforços nem
tempo em me ajudar na realização dos experimentos em laboratório.

Ao meu amigo Ubiratan Nazaré por me ajudar na organização do texto e na tradução para
o Abstract.
Finalizando, agradeço ao Laboratório de Engenharia Bioquímica da UFRN por ter cedido
toda a sua estrutura para realização dos experimentos e ao Programa de Pós-Graduação em
Engenharia Química, assim como a todos os professores do Departamento de Engenharia Química
da UFRN que trabalham em pró de uma educação digna e ensino de qualidade para todos os
alunos.

SILVA FILHO, Raimundo Cosme da – Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo Descontínuo Submerso. Tese de Doutorado. UFRN - Programa de Pós-Graduação em Engenharia Química. Área de Concentração: Pesquisa e Desenvolvimento de Tecnologias Regionais – Subárea: Engenharia de Processos – Alimentos e Biotecnologia, Natal/RN, Brasil.
Orientação: Prof. Dr. Everaldo Silvino dos Santos Coorientação: Profª. Drª. Gorete Ribeiro de Macedo ___________________________________________________________________________
RESUMO: No presente trabalho utilizou-se um Planejamento Plackett & Burman, com 8 fatores e 12 ensaios em 2 níveis e mais 3 repetições na condição do ponto central, para se investigar a influência das concentrações de quitosana, peptona, extrato de levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O e FeSO4 na produção da enzima quitosanase por Metarhizium
anisopliae. Os ensaios para produção da enzima foram realizados em cultivo descontínuo submerso. Os resultados para o Planejamento Plackett & Burman mostraram que a atividade quitosanolítica foi favorecida pelo aumento da concentração de substrato (quitosana) e de sulfato ferroso (FeSO4), enquanto que o aumento da concentração dos outros fatores não contribuiu de forma significativa para a atividade quitosanolítica. A otimização do meio de cultura para a produção da enzima foi realizado por meio de um Planejamento Composto Central, com os dois fatores que mais influenciaram a atividade quitosanolítica (quitosana e FeSO4), conforme Planejamento Plackett & Burman, mantendo-se os outros nutrientes em seus valores mínimos. Nesse planejamento, para o fator FeSO4, tomou-se o limite inferior (-1) como sendo o limite superior do Planejamento Plackett & Burman. Os resultados mostraram que a produção da enzima foi favorecida pelo aumento da concentração quitosana e pela diminuição da concentração de FeSO4. A produção máxima de atividade quitosanolítica foi da ordem de 70,0 U/L e foi atingida em apenas 18 h de fermentação, resultado esse vinte e oito vezes maior aos obtidos anteriormente para o mesmo microrganismo que foi 2,5 U/L em 48h.
Palavras-chave: quitosana; quitosanase; Metarhizium anisopliae; Plackett & Burman.

ABSTRACT
In this work a Plackett-Burman Design with 8 factors and 12 trials in 2 levels with 3 repetitions at the center point was used in order to investigate the influence of the concentration of chitosan, peptone, yeast extract, NaNO3, K2HPO4, KCl, MgSO4.7H2O and FeSO4 on chitosanase production by Metarhizium anisopliae. Runs were carried out using submerged discontinuous cultivation for enzyme production. The results of the Plackett & Burman Design showed that only two factors, chitosan concentration as well as FeSO4 had influence on chitosanolytic activity, while the increase in concentration of other factors not contributed significantly to the quitosanolítica activity. Cultivation medium optimization for enzyme production was carried out using a Composite Central Design, with the most important factors for chitosanolytic activity (chitosan and FeSO4), in accordance with Plackett & Burman Design, and keeping the other nutrients in their minimum values. On this other design, it was taken the highest limit in Plackett & Burman Design as the lowest limit (-1) to FeSO4 factor. The results showed that the enzyme production was favoured by increasing the chitosan concentration and by decreasing FeSO4. Maximum production for chitosanolytic activity was about 70.0 U/L and was reached in only 18 h of fermentation, a result about twenty-eight times greater than a former study using the same microorganism (about 2.5 U/L at 48 h).
Key-words: Chitosan; Chitosanase; Metarhizium anisopliae; Plackett & Burman

Sumário LISTA DE FIGURAS
LISTA DE TABELAS
NOMENCLATURA
INTRODUÇÃO...............................................................................................................................15
REVISÃO BIBLIOGRÁFICA........................................................................................................18
2.1. Enzimas................................................................................................................................18
2.1.1. Quitosana e seus derivados........................................................................................19
2.1.2. Quitosanases...............................................................................................................24
2.2. Fungos..................................................................................................................................29
2.2.1. Características gerais..................................................................................................29
2.2.2. Reprodução dos Fungos.............................................................................................32
2.2.3. Metabolismo de fungos..............................................................................................33
2.2.4. Nutrição de fungos.....................................................................................................34
2.2.5. O Fungo Metarhizium anisopliae ..............................................................................36
2.3. Fermentação.........................................................................................................................40
2.3.1. Preparo do inóculo......................................................................................................41
2.3.2. Produção de enzimas em cultivo submerso descontínuo...........................................42
2.4. Planejamento experimental..................................................................................................42
2.4.1. Planejamento fatorial..................................................................................................43
2.4.2. Planejamento Plackett & Burman..............................................................................43
2.4.3. Aplicações do Planejamento Plackett & Burman.......................................................44
METODOLOGIA EXPERIMENTAL.............................................................................................50
3.1. Quitosana..............................................................................................................................50
3.2. Microrganismo.....................................................................................................................50
3.2.1. Escolha da cepa produtora de quitosanase.................................................................50

3.2.2. Manutenção da cepa...................................................................................................50
3.3. Produção de enzima.............................................................................................................51
3.4. Determinação da Proteína Total...........................................................................................51
3.5. Determinação da Atividade Quitosanolítica e Atividade Específica...................................51
3.6. Planejamento Experimental.................................................................................................52
3.6.1. Planejamento de seleção de fatores Plackett Burman................................................52
3.6.2. Planejamento Composto Central para Otimização do meio de cultura para a
Produção da Enzima em Cultivo Submerso.....................................................................................53
3.7. Perfil Cinético durante o Cultivo.........................................................................................56
RESULTADOS E DISCUSSÃO.....................................................................................................58
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA ..........................................58
4.2. Planejamento Plackett & Burman........................................................................................59
4.2.1. Análise do Plackett & Burman com 8 fatores e 12 ensaios .......................................59
4.2.2. Análise do Plackett & Burman com 12 ensaios e mais 3 repetições na condição do
ponto central ....................................................................................................................................61
4.3. Planejamento Composto Central para Produção de Quitosanase por Metarhizium
anisopliae em Cultivo Descontínuo Submerso................................................................................64
4.4. Perfil Cinético – Melhor Condição......................................................................................69
CONCLUSÃO.................................................................................................................................75
REFERÊNCIAS BIBLIOGRÁFICAS.............................................................................................78
APÊNDICE I...................................................................................................................................93
Metodologia para Repique da Cepa ................................................................................................93
APÊNDICE II.................................................................................................................................94
Montagem da Curva de Calibração Usando o Método DNS..........................................................94
APÊNDICE III................................................................................................................................96
Método de Determinação de Proteína Utilizando o Corante Comassie Brilhante Blue G 250
(Método modificado de Sedmak e Grossberg)................................................................................96

Lista de Figuras
Figura 1 – Estrutura da quitosana..................................................................................................20
Figura 2 – Estrutura da glicosamina..............................................................................................23
Figura 3 – Estrutura do sulfato de glicosamina.............................................................................23
Figura 4 – Representação da estrutura geral de uma célula de fungo..........................................30
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas...........................................................31
Figura 6 – Conídios de Aspergillus e Penicillium.........................................................................33
Figura 7 – Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA ...................58
Figura 8 – Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados............................................................................................................................60
Figura 9 – Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto central
mostrando a influência dos fatores estudados................................................................................63
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos
fatores estudados............................................................................................................................67
Figura 11 – Valores preditos em função dos valores observados..................................................68
Figura 12 – Atividade quitosanolítica em função da concentração de quitosana e da concentração
de FeSO4 para os fatores não significativos no ponto central .......................................................68
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana ........70
Figura 14 – Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana....................................70
Figura 15 – Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de
quitosana........................................................................................................................................71
Figura 16 – Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana..........................................................................................................72
Figura 17 – Curva padrão de glicosamina ....................................................................................95
Figura 18 – Curva Padrão de BSA ................................................................................................98

Lista de Tabelas
Tabela 1 – Comparação das enzimas com os catalisadores químicos..............................................19
Tabela 2 – Fontes naturais de quitina e quitosana ...........................................................................20
Tabela 3 – Níveis para os fatores e seus valores codificados...........................................................53
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central .........................................................................................53
Tabela 5 – Matriz do planejamento Composto Central...................................................................54
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central......................................................................................................55
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.......................59
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman...............................................60
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.......................61
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central...............................................................................................................................................62
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da atividade
enzimática........................................................................................................................................65
Tabela 12 – Efeitos Estimados do planejamento Composto Central...............................................65
Tabela 13 – Resumo da ANOVA....................................................................................................66
Tabela 14 – Dados para construção da curva de calibração usando o Método DNS ......................94
Tabela 15 – Medidas de absorbância a 600 nm para construção da curva de calibração................94
Tabela 16 – Dados para construção da curva padrão de BSA ........................................................97

NOMENCLATURA
A – Atividade quitosanolítica (U/L)
AE – Atividade específica (U/mg)
ANN-GA – algoritmo genético de rede neural artificial
ANOVA – Análise de variância (Analysis of Variance)
BSA – Albumina de soro bovino (Bovine Sereum Albumin)
CCD – Planejamento Composto Central
CDA – Chitosan Dector Agar
Cs – Concentração de substrato (%)
CP – Concentração de KCl (%)
Da – Dalton
DEQ – Departamento de Engenharia Química
DNS – Ácido Dinitrosalissílico
E – Enzima
Ea – Energia de ativação
EC – Comissão para nomenclatura e classificação de enzimas
EL – Concentração de Extrato de Levedura (%)
FP – Concentração de K2HPO4 (%)
NS – Concentração de NaNO3 (%)
GDA – Grânulos dispersíveis em água
k – Número de fatores, variáveis
LEB – Laboratório de Engenharia Bioquímica
mRNA – RNA mensageiro
n – Número de níveis de um determinado fator
N – Número de ensaios de um planejamento
P – Pressão
PP – Concentração de Peptona (%)
PDA – Batata Dextrose Agar
pH – Potencial hidrogeniônico
Q – Concentração de Quitosana (%)
Qm – Quadrado médio
r – Agitação (rpm)
R2 – Coeficiente de determinação

RA – Razão de aeração
rpm – Rotações por minuto
RSM – Metodologia de superfície de resposta
S – Molécula de substrato
SF – Concentração de FeSO4 (%)
SM – Concentração de MgSO4.7H2O (%)
SQ – Soma de quadrados
SSF – Fermentação de estado sólido
T – Temperatura
TMC – Temperatura de cultivo
V11,V12,V13 – Variáveis inertes para cálculo do erro
VHG – Fermentação de alta densidade
WP – Pó molhável
X – Concentração celular
Xi – variável independente
Xj – variável independente
xi – Fator independente real
x0 – Valor de xi no ponto central
∆xi – Variação de valor
Y – Resposta do modelo de primeira ordem
Yp – Resposta do modelo polinomial de segunda ordem
β0 – Interseção dos modelos de primeira ou segunda ordem
βi – Coeficiente linear dos modelos de primeira ou segunda ordem
βii – Coeficiente quadrático
βij – Coeficiente de interação

Capítulo 1 Introdução

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
15
1. Introdução As quitosanases foram descobertas simultaneamente há quase quarenta anos por
Monaghan et al. (1973), como pelo grupo de Ramirez-León & Ruiz-Herrera (1972), e
são enzimas obtidas a partir do metabolismo de bactérias, fungos e extraídas de plantas.
Desde então, vários estudos vêm apresentando a obtenção de quitosanase deste
microrganismos, na forma intracelular ou extracelular (Osswald et al., 1994;
Somashekar & Joseph, 1996), como a do presente estudo. A quitosanase (EC 3.2.1.132)
representa uma classe de enzimas hidrolíticas que catalisam a hidrólise de ligação β-
(1→ 4) glicosídica da quitosana, gerando oligômeros chamados quito-oligossacarídeos
(Reddy et al., 2008).
A quitosana é um biopolímero hidrofílico obtido a partir da quitina, um
polissacarídeo abundante na natureza. Na literatura, é considerado quitosana o produto
da desacetilação da quitina para um grau de desacetilação superior a 75% e solúveis em
ácidos como o acético e o fórmico. A quitosana é conhecida por exibir uma ampla
variedade de atividade fisiológica como atividade antitumoral, mas o peso molecular e a
insolubilidade em água da quitosana são desvantagens para várias aplicações.
Atualmente, há interesse nas quitosanas quando as mesmas são convertidas em quito-
oligossacarídeos porque eles não apenas são solúveis em água e possuem baixo peso
molecular, mas também têm atividades biológicas bem notáveis, como atividade
anticancerígena atóxica e biocompatível, incluindo efeito antimetastático, atividade anti-
HIV, hepatoprotetora, antioxidante, hipocolesterolêmica, antimicrobiana,
imunoestimulate, antitumoral, anti-inflamatória e atividade aceleradora da absorção de
cálcio e ferro. No entanto, estes quito-oligossacarídeos são obtidos tradicionalmente na
indústria por reação de hidrólise usando ácidos, que apresenta muitos problemas como
dificuldade em controlar a hidrólise, produzindo uma grande quantidade de
oligossacarídeos de cadeia pequena e oligossacarídeos de baixo rendimento, alto custo
de separação e, também, poluição ambiental, uma vez que utiliza um ácido como
catalisador. Como alternativa, com suas vantagens em termos de compatibilidade
ambiental, baixo custo e reprodutibilidade, a obtenção destes quito-oligossacarídeos por
hidrólise com quitosanase se tornou cada vez mais popular nos últimos anos (Azevedo
et al., 2007).

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
16
Devido a todos estes aspectos positivos é que, em geral, as enzimas vêm sendo
estudadas em diferentes aplicações, mostrando que esta é uma tendência na indústria.
No presente trabalho, foi desenvolvido um processo fermentativo visando a
otimização do meio de cultura para a produção da enzima quitosanase em cultivo
descontínuo submerso, por Metarhizium anisopliae, capaz de sintetizar oligômeros com
aplicações na indústria farmacêutica e de química fina, partindo de quitosana como
única fonte de carbono.
Nesse caso, avaliou-se, através de um planejamento Plackett & Burman (método
de delineamento de seleção de fatores) e de um planejamento Composto Central as
melhores condições para a produção da enzima. A técnica do planejamento
experimental Plackett & Burman para realizar triagem de fatores na otimização de um
determinado processo foi utilizada neste trabalho para identificar quais os fatores que
influenciam de forma significativa na produção da enzima quitosanase. A influência
destes fatores (concentração de quitosana e concentração de FeSO4) foi então
investigada pelo planejamento Composto Central com 22 experimentos na parte fatorial,
repetição em triplicata no ponto central e quatro experimentos na parte axial. A
repetição do ponto central permite também a determinação do erro puro e da
repetibilidade do processo, sendo uma forma de identificar melhor se o processo está
sob controle.
Do ponto de vista industrial é de relevante importância a realização de estudos
para a otimização da produção da quitosanase de forma contínua ou em grandes
bateladas.

Capítulo 2 Revisão Bibliográfica

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
18
2. Revisão Bibliográfica 2.1. Enzimas
As enzimas são catalisadores biológicos constituídos por longas cadeias de
aminoácidos que atuam em substratos reduzindo a energia de ativação, acelerando assim
a velocidade da reação, e transformando-os em produtos. Sua atuação catalítica visa
facilitar a atividade das células através da quebra de moléculas ou junção das mesmas
para formação de novos produtos. Podem apresentar alto grau de especificidade relativa
a apenas um substrato ou a vários substratos ao mesmo tempo, como a certos tipos de
ligações glicosídicas α-1,4 das moléculas de amido, ou para determinado tipo particular
de isômero óptico como ocorre na oxidação da β-D-glicose pela glicose oxidase (Furigo
Junior & Pereira, 2001).
Enzimas são proteínas que apresentam em sua estrutura um centro ativo
(apoenzima) e, em alguns casos, um grupo prostético denominado cofator (coenzima ou
íon metálico ativo) e sua atividade depende de alguns fatores como estrutura da
proteína, número de cadeias peptídicas, arranjo dessas cadeias na molécula, natureza do
substrato e natureza do grupo prostético (Politzer & Bon, 2006).
Todos os organismos vivos produzem, em pequena ou grande quantidade, certa
variedade de enzimas, sejam elas intra ou extracelulares. As enzimas extracelulares são
excretadas para o meio externo e podem decompor compostos insolúveis como
celulose, quitina e quitosana.
Algumas destas enzimas podem ser usadas tanto nas indústrias de detergentes,
têxteis e farmacêuticas, como também de alimentos, de bebidas, etc., sendo utilizadas
em processos biotecnológicos industriais em substituição aos processos químicos que
muitas vezes causam grandes prejuízos ao meio ambiente. A produção comercial das
enzimas pode ser realizada através do cultivo de microrganismos como fungos e
bactérias.
Na Tabela 1 mostra-se uma comparação das enzimas com os catalisadores
químicos, levando-se em conta algumas das características dos dois catalisadores.
Destaca-se que alguns parâmetros da Tabela 1 demonstram a maior importância das
enzimas, quando comparadas com os catalisadores químicos, principalmente a
especificidade ao substrato, as condições de reação, o consumo de energia e a

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
19
velocidade de reação, como desvantagem cita-se o custo de obtenção (isolamento e
purificação).
Tabela 1 – Comparação das enzimas com os catalisadores químicos (Adaptado de
Furigo Junior & Pereira, 2001).
Característica Enzimas Catalisadores Químicos
Especificidade ao substrato Alta Baixa
Natureza da estrutura Complexa Simples
Sensibilidade à T e pH Alta Baixa
Condições de reação (T, P e pH) Suaves Drástica (geralmente)
Custo de obtenção (isolamento e purificação) Alto Moderado
Natureza do processo Batelada Contínuo
Consumo de energia Baixo Alto
Formação de subprodutos Baixa Alto
Separação catalisador/produtos Difícil/cara Simples
Atividade catalítica (temperatura ambiente) Alta Baixa
Presença de cofatores Sim Não
Estabilidade do preparado Baixa Alta
Energia de Ativação Baixa Alta
Velocidade de reação Alta Baixa
2.1.1. Quitosana e seus derivados
A quitosana e seus derivados apresentam grande importância devido ao
conhecimento de suas funcionalidades biológicas, médicas, tecnológicas, etc
(Shimosaka et al, 1993; Shimosaka et al, 1995; Mitsutomi et al., 1995). A quitosana é
obtida da quitina, polissacarídeo abundante na natureza, extraído principalmente das
carapaças de caranguejos, lagosta e camarões. A quitosana é um polímero natural,
biodegradável, não tóxico e de alta massa molecular obtido a partir de animais
marinhos, insetos e microrganismos (Tabela 2).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
20
Tabela 2 – Fontes naturais de quitina e quitosana. (Adaptado de Polymar, 2003).
Animais
Marinhos
Insetos Aracnídeos Microrganismos
Anelídios Formigas Escorpiões Leveduras
Moluscos Besouros Aranhas Fungos
Lagosta Esporos
Camarão
Caranguejo
Krill
A Figura 1 abaixo mostra a estrutura química da quitosana
Figura 1 – Estrutura da quitosana (Fonte: Polymar, 2003)
Quitosana totalmente desacetilada não existe na natureza. Graus de acetilação
(GA) nferior a 50% a torna mais solúvel em soluções ácidas (Muzzarelli, 1993;
Muzzarelli et al., 1994; Dung et al., 1994; Stoyachenko et al.,1994). Pelletier &
Sygusch (1990) enfatizaram que grande parte dos resíduos sólidos da indústria
pesqueira, principalmente da carcinicultura, é jogada ao mar ocasionando degradação
ambiental. Logo, a utilização desses resíduos como matéria-prima para formação de
novos produtos, não só reduz de forma drástica o impacto ambiental, mas também daria
maior valor agregado à indústria pesqueira.
A demanda cada vez maior de consumidores por alimentos mais saudáveis, sem
conservantes químicos, tem motivado pesquisas à descoberta de novos agentes
antimicrobianos naturais. Sendo assim, a atividade antimicrobiana incomum da

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
21
quitosana e seus derivados contra microrganismos, tais como bactérias, bolores e
leveduras tem tido maior atenção ultimamente.
Devido à interação entre a quitosana, que é positivamente carregada, e as
membranas celulares dos microrganismos, negativamente carregadas, ocorre a formação
de verdadeiros poros na membrana, que são canais de vazamento de conteúdo protéico e
outros componentes intracelulares. A quitosana tem também a propriedade de agir como
agente quelante que se liga de forma seletiva a traços de metais, inibindo, assim, a
produção de toxinas e o crescimento microbiano. Ainda é capaz de ativar mecanismos
de defesa em tecidos hospedeiros, atuando como agente sequestrante de água e inibindo
várias enzimas. A ligação da quitosana com DNA e a inibição da síntese do mRNA
ocorre via penetração da quitosana no núcleo dos microrganismos e interferência na
síntese do mRNA e proteínas, (Craveiro et al., 2004).
Em um estudo clínico conduzido por Giustina & Ventura (1995) com 100
pessoas com sobrepeso de 10 a 25%, ao final de 4 semanas as pessoas que receberam
quitosana demonstraram perda de peso significativa (7,3 Kg contra 3,0 Kg nos grupos
controle). As pessoas que tomaram quitosana também demonstraram redução na pressão
sanguínea sistólica (de 145 para 135 mm Hg) e diastólica (92.6 para 84.2 mm Hg),
assim como uma redução na taxa respiratória (27.6/min a 21.2/min).
Kondo & Osada (1996) pesquisaram a influência da quitina e da quitosana na
biodisponibilidade do zinco em ratos alimentados com uma dieta com suplementos de
5% ou sem fibra dietética (quitina, quitosana, celulose, pectina e agar-agar) durante 31
dias. Observaram que os ratos alimentados com dietas contendo 5% de fibras, exceto
quitosana, o consumo de alimento foi maior do que o controle. O ganho de peso em
todos os grupos alimentados com fibra foi maior do que o controle, exceto para o grupo
suplementado com quitosana (5%) onde a ingestão de alimento e o ganho de peso foram
menores do que os grupos controle e das demais fibras.
Segundo Craveiro et al. (2004), derivados da quitosana obtidos pela reação com
ácidos carboxílicos, podem promover ao polímero uma solubilidade especial em água.
Esses compostos possuem estrutura química semelhante ao ácido hialurônico e
fornecem uma excelente compatibilidade com o tecido epitelial. As soluções obtidas
com esses derivados podem ser facilmente incorporadas a formulações cosméticas,
mostrando melhor compatibilidade com surfactantes iônicos e emulsificantes,
apresentando estabilidade em uma larga faixa de pH.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
22
Han et al. (1999) examinaram os efeitos da quitosana sobre a atividade da lipase
pancreática in vitro e sobre o grau de armazenamento de gorduras induzido em ratos
pela administração oral de uma dieta rica em gorduras por nove semanas. Como
resultado, observou-se que os animais alimentados com quitosana, reduziram o peso
corporal, a hiperlipidemia e a gordura hepática. Como conclusão, os autores propõem
que os efeitos anti-obesidade da quitosana em ratos alimentados com alto teor de
gordura se devem parcialmente a inibição da absorção intestinal das gorduras dietéticas.
Em outro ensaio, Zahorska-Markiewicz et al. (2002) realizaram um estudo para
avaliar o efeito da quitosana no tratamento da obesidade. Segundo os autores do estudo,
a quitosana pode ser usada como um valioso e seguro coadjuvante em tratamentos para
a obesidade em longo prazo. A quitosana parece acentuar a redução da pressão arterial,
juntamente com a perda de peso.
Assis (2010) estudou a produção e caracterização de quitooligossacarídeos
produzidos pelo fungo Metarhizium anisopliae e avaliação da citotoxicidade em células
tumorais. A hidrólise da quitosana foi realizada em tempos de 10 a 60 min para a
produção de quitooligossacarídeos. A avaliação da citotoxicidade dos oligômeros de
quitosana foi realizada em células tumorais (HepG2 e HeLa) e não tumoral (3T3). A
produção de oligômeros de quitosana teve maiores rendimentos durante 10 minutos de
hidrólise, os pentâmeros apresentaram concentração de 0,15 mg/mL, no entanto os
hexâmeros, que apresentam maior interesse por causa de suas propriedades biológicas,
só foram detectados com 30 minutos de hidrólise com uma concentração de 0,004
mg/mL.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o planejamento Plackett-Burmam e a metodologia de superfície de resposta.
Os resultados mostraram que a eficiência máxima de extração da quitina foi conseguida
nas seguintes condições: concentração de casca de camarão 50g/L, glicose 50g/L, tempo
de incubação de 5 dias e inoculo como 0,05 OD (densidade óptica).
A glicosamina, unidade monomérica básica da quitosana, possui atividade
antiinflamatória e tem grande importância na construção e manutenção da cartilagem,
no tratamento de enfermidades como: febre reumática, artrose e principalmente a artrite
e osteoartrite na forma aguda ou crônica (Polymar, 2003). A Figura 2 apresenta a
estrutura química da glicosamina.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
23
Figura 2 – Estrutura da glicosamina (Fonte: Polymar, 2003)
O sulfato de glicosamina é um derivado da quitosana sendo utilizado no
tratamento da osteoartrite sintomática dos joelhos, quadris, coluna, mãos e outras
regiões (Craveiro et al., 2004). O sulfato de glicosamina estimula a síntese de
proteoglicanos necessários para o bom funcionamento das articulações, estimula a
regeneração das cartilagens e ainda incorpora enxofre ao tecido cartilaginoso. Estudos
de longa duração, acima de 3 anos, mostram que o sulfatos de glicosamina evita a
progressão de danos estruturais nas articulações e induz melhora significativa nas dores
e funções físicas (Craveiro et al., 2004). A Figura 3 apresenta a estrutura química do
sulfato de glicosamina.
Figura 3 – Estrutura do sulfato de glicosamina (Fonte: Polymar, 2003)

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
24
2.1.2. Quitosanases
Dependendo do tipo de quitosanase obtida, ocorrem diferentes mecanismos de
hidrólise da quitosana, isso devido a diferentes microrganismos produzirem enzimas
com diferenças nas suas estruturas (Kurita et al., 1977; Seino et al., 1991).
Quitosanases (EC 3.2.1.132) são enzimas que catalisam a reação de hidrólise de
quitosana em oligossacarídeos. As enzimas quitosanases são produzidas por vários
microrganismos, bactérias, actinomicetos e fungos, e até por algumas plantas. Estas
enzimas agem em polímeros com grau de acetilação de 30% a 60%. Os monômeros
resultantes da hidrólise da quitosana por estas enzimas são melhores digeridos, ao serem
ingeridos, como é o caso da glicosamina. Estudos efetuados até o presente momento
classificam as quitosanases em cinco famílias: 5, 8, 46, 75, 80, sendo que a família 46 é
a mais estudada. As quitosanases tem grande aplicação na geração de oligômeros de
quitosana (Somashekar & Joseph, 1996; Chen et al., 2005).
Nogawa et al. (1998) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida pelo fungo Trichoderma reseei PC-3-7. Estes pesquisadores
inocularam 106 conídios em 100 mL de um meio basal contendo 0,3% de N-
acetilglicosamina em incubator rotativo com agitação de 220 rpm a 28oC. Destaca-se
que o fungo não produziu quitosanase em meio contendo quitosana ou quitina.
Entretanto, o microrganismo produziu quitosanase em meio contendo glicosamina e N-
acetilglicosamina, sendo que para este último obteve-se uma atividade específica
máxima de 1,5 U/mg após 72 horas de incubação. É importante ressaltar que uma
agitação vigorosa favoreceu a produção da enzima. Esse artigo ilustra a presença de
exo-β-D-glicosaminidase e exo-β-D-N-acetilglicosaminidase.
Piza et al. (1999) estudaram a produção, purificação e caracterização parcial de
uma quitosanase produzida por Bacillus cereus. Nesse caso, investigou-se através de um
planejamento fatorial fracionário a influência dos fatores: concentração de sulfato de
amônio (0,4; 2,2 e 4,0%), razão de aeração (2; 6 e 7), tempo de fermentação (16; 24 e
32 horas), pH (5; 6 e 6,5) e concentração de quitosana (1,0; 1,5 e 2,0%). Os resultados
mostraram que os fatores mais significativos para esse planejamento foram a
concentração de sulfato de amônio, a aeração, o pH e a interação entre sulfato de
amônio e a aeração.
Cheng & Li (2000) estudaram a preparação de oligossacarídeos utilizando
quitosanase produzida por Aspergillus sp e constataram que a enzima foi largamente

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
25
induzida em meio contendo quitosana como única fonte de carbono e também na
presença de quitina coloidal. A indução da enzima foi realizada em temperaturas
variando na faixa de 28 a 30ºC, pois essa espécie não cresceu bem em temperaturas
superiores a 37ºC.
Zhang et al. (2000) estudaram a produção, purificação e caracterização de
quitosanase e Exo-β-D-Glucosaminidase por Aspergillus oryzae IAM2660. Os
microrganismos utilizados neste estudo foram mantidos em placas de Petri contendo
agar para contagem em placas (PDA). Destaca-se que para produzir a enzima, conídios
foram retirados das placas e inoculados em meio Czapeq-dox com os seguintes
componentes: 1,0g K2HPO4, 0,5g KCl, 2,0g NaNO3, 0,5g MgSO4.7H2O, 0,02g
FeSO4.7H2O, 20g de sacarose e 1,0L de água deionizada, com pH do meio igual a 5,5.
Foi adicionado ao meio 0,5 g de peptona e 0,5g de extrato de levedura com
concentração de esporos da ordem de 1,0x106 esporos.mL-1. Após a incubação, feita em
incubador rotativo a 26ºC por um período de 60h, micélios foram coletados e após
filtração e secagem a 80ºC foram utilizados para ensaio de quitosanase.
Kuroiwa et al. (2002) estudaram os fatores que afetam a composição de
oligossacarídeos produzidos através da hidrólise de quitosana por quitosanase originária
de Bacillus pumilus BN-262. Neste experimento foi utilizado quitosana 100%
desacetilada dissolvida em ácido acético 1M. O pH do meio foi ajustado em 5,6. Neste
estudo a quitosanase foi imobilizada em agar e Sepharose. Os resultados mostraram que
a concentração de enzima, a temperatura, a velocidade de agitação, e concentração de
substrato influenciaram de forma significativa na composição dos oligossacarídeos
produzidos.
Ichikawa et al. (2002) estudaram a imobilização e estabilização da ligação
multiponto da quitosanase tendo agar como suporte e concluíram que a imobilização
aumentou a termoestabilidade da quitosanase. Os oligossacarídeos de quitosana foram
continuamente produzidos usando um reator coluna contendo quitosanase imobilizada.
A percentagem de quitosana imobilizada após 28 dias de reação foi de 44%.
Hung et al. (2002) estudaram a purificação, através de troca iônica, e
caracterização da hidrólise sobre ação da quitinase e quitosanase purificadas do meio
contendo Bromelina comercial. Destaca-se que a quitinase purificada apresentou pH,
temperatura e Km ótimo para ação catalítica em meio contendo quitina de 4,0, 60ºC e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
26
0,2mg.mL-1, respectivamente. Enquanto que quitosanase apresentou o pH, temperatura e
Km ótimo de 3,0, 50ºC e 0,88mg.mL-1, respectivamente.
Zhu et al. (2003) estudaram as melhores condições fermentativas e propriedades
de uma quitosanase produzida por Acinetobacter sp. C-17. A espécie de bactéria
utilizada neste ensaio foi isolada do solo e identificada como sendo Acinetobacter sp C-
17 e capaz de produzir quitosanase. O microrganismo foi mantido em placas contendo o
meio agar com os seguintes componentes: 0,5 % de quitosana, 0,2 % K2HPO4, 0,1 %
KH2PO4, 0,07 % MgSO4, 0,05 % NaCl, 0,05 % KCl, 0,01 % CaCl2, 0,05 % de extrato
de levedura e 2 % agar, o pH do meio de manutenção foi mantido em 7,2 e a
temperatura foi mantida em 30ºC durante 3 a 5 dias, para identificar a ação da enzima
quitosanase. A ação da quitosanase foi identificada através da observação de uma
colônia de microrganismo existente na placa. A bactéria foi então isolada da placa e
colocada em fermentador rotativo a 200 rpm durante 72h em meio contendo: 0,07 %
K2HPO4, 0,03 % KH2PO4, 0,05 % MgSO4, 0,03 % peptona, 0,03 % de extrato de
levedura e 1,0 % de quitosana. O pH foi de 7,0 e a temperatura 30ºC. Como fontes de
carbono foram utilizadas além de 1,0 % de quitosana, 1,0 % de quitina e 1,0 % de
glicose. Os resultados mostraram que as melhores condições para produção da
quitosanase foram alcançadas quando se utilizou 1,0 % de solução de quitosana a 30ºC,
pH 7,0 durante 24 horas de incubação.
Liu et al. (2003) estudaram a produção de quitinase por Verticillium lecanii
F091 usando fermentação submersa, a atividade de quitinase foi de 9,95 mU/mL em um
meio de cultura otimizado, com um volume de cultura de 200 mL, agitação de 150 rpm
e temperatura de 24ºC cultivado em incubador rotativo e concluíram que a taxa de
agitação e o pH foram os fatores mais significativos para a produção de quitinase, no
entanto, as taxas de agitação e aeração poderiam modificar a concentração de oxigênio
dissolvido afetando diretamente no crescimento do Verticillium lecanii e na produção
de quitinase.
Donzelli et al. (2003) estudaram a hidrólise enzimática avançada de quitina de
casca de caranguejo langostino com misturas de enzimas de bactérias e fungos e
concluiram que a combinação de enzimas de Trichoderma atroviride e Serratia
marcescens foram capazes de degradar completamente altas concentrações de quitina
(100g/mL) a partir de carapaças de caranguejo langostino convertendo a N-
Acetilglicosamina (78%), glucosamina (2%) e quitobiose (10%). Enzimas de fonte

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
27
procariótica apresentaram atividades menores que as enzimas de T. atroviride, para
catalisar a reação com quitosana.
Jo et al. (2003) estudaram a caracterização e a avaliação cinética de quitosanase
produzida por Bacillus sp. Destaca-se que na produção da enzima utilizada neste ensaio
o meio continha 0,5% de concentração de quitosana, 1% de triptona e 1% de NaCl, o
pH do meio foi ajustado em 5,5 e a temperatura foi mantida em 37ºC com agitação de
180 rpm em incubador rotativo.
Fu et al (2003) estudaram a caracterização de três iso-enzimas de quitosanase
isolada de pepsina comercial e denominado como PSC-I, PSC-II, PSC-III,
respectivamente, em relação a hidrólise enzimática da quitosana, o meio ótimo para a
produção destas iso-enzimas continha as seguintes características: pH 5,0; 5,0 e 4,0 e
temperaturas de 40ºC, 40ºC e 30ºC, o valor de Km foi de 5,2, 4,0 e 5,6 mg.mL-1,
respectivamente. As massas moleculares das três iso-enzimas foram de 40 kDa. Os
resultados mostraram que as três iso-enzimas apresentaram atividade em meio contendo
quitosana, produzindo oligômeros da ordem de 68 – 88% desacetilada.
Choi et al. (2004) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida por Bacillus sp KCTC 0377BP. Nesse caso, com relação à
produção da enzima foi possível obter um aumento de 1,2 U/ml em incubator rotativo
para 100,0 U/ml em fermentador piloto de 500 L, utilizando uma concentração de
substrato de 0,5% de quitosana e como fonte de nitrogênio polipeptona (2,5%) em pH
igual a 6,8, com 100 rpm de 100,0 rpm a 30oC.
Shimosaka et al. (2003) estudaram a produção, purificação e caracterização total
de duas quitosanases produzida por Acinetobacter sp CHB101. Nesse caso, com relação
à produção da enzima os pesquisadores usaram dois meios, um contendo glicose e outro
contendo quitosana. Com relação à cinética, o meio contendo quitosana fornece um
tempo de geração de 280 minutos, enquanto que no meio contendo glicose a taxa de
crescimento foi mais elevada e o tempo de geração foi de apenas 110 minutos. Os
autores observaram que após os dois primeiros dias de cultivo a atividade
quitosanolítica permaneceu baixa (na ordem 0,10 a 0,20 U/mL). Durante a fase de
crescimento exponencial o valor da atividade permaneceu com valor inferior a 0,01
U/mL, começando a aumentar ao atingir a fase estacionária, indicando assim que a
produção da enzima é não associada ao crescimento.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
28
Kim et al. (2004) estudaram a produção, purificação e caracterização de uma
quitosanase produzida pelo Bacillus sp e identificado pelo número 1299. Destaca-se que
na produção da enzima quitosanase foi utilizado um meio contendo os seguintes
componentes: 0,5 % de quitosana, 0,42 % (NH4)2SO4, 0,2 % K2HPO4, 0,02 % uréia,
0,03 % CaCl2.2H2O, 0,03% MgSO4.7H2O, 0,1% peptona, 0,2% Tween 80, 0,2 % da
solução (0,5% FeSO4.7H2O, 0,16% MnSO4.H2O, 0,14% ZnSO4.7H2O, 0,2% CoCl2) e
1,7% agar. O pH do meio para este ensaio foi mantido em 7,0. Este microrganismo foi
escolhido por ser grande produtor de quitosanase.
Chen et al. (2005) estudaram a purificação e a caracterização de dois tipos de
quitosanase produzidas por Aspergillus sp. CJ22-326 em cultivo submerso. Na
produção da enzima foi utilizado um meio de cultivo contendo 1,0% de quitosana, 2,0%
de gérmen de trigo, 0,2% de sulfato de amônio, 0,2 % de fosfato de potássio e 0,05% de
sulfato de magnésio por litro de solução em pH 5,6. O cultivo foi realizado em frasco de
500 mL contendo 150 mL de meio de cultura a 30ºC por 96 horas com uma agitação de
150 rpm. Os autores apresentam um gel de eletroforese com as duas enzimas purificadas
com massa molar de 29 kDa e 109 kDa.
Silva Filho et al. (2005) estudaram, através de planejamento fatorial 24 com
repetição em triplicata no ponto central, a produção de quitosanase por Aspergillus
ochraceus em cultivo descontínuo submerso. Os resultados mostraram que foi possível
produzir quitosanase com atividade na ordem de aproximadamente 5,9 U/mL utilizando
Aspergillus ochraceus e que a atividade foi favorecida pelo aumento da agitação (r), da
razão de aeração (RA) e da concentração de substrato (Cs), enquanto que o aumento da
temperatura de cultivo (TMC) não favoreceu a resposta (atividade quitosanolítica).
Pode-se classificar as quitosanases em endoquitosanases e exoquitosanases. As
primeiras diferenciam-se por catalisar a hidrólise aleatoriamente, no interior do
biopolímero, gerando diversos tamanhos de oligossacarídeos. As exoquitosanases são
capazes de hidrolisar as terminações não redutoras, produzindo assim unidades
redutoras (Peter, 2005).
Palma-Guerrero et al. (2008) estudaram o biocontrole de fungos patogênicos e
demonstraram a lise da membrana plasmática e a inibição da germinação de esporos de
fungos. Eles observaram que os fungos entomopatogênicos e nematófagos foram menos
inibidos pela quitosana que os fitopatogênicos e micoparasíticos provavelmente porque
apresentam a enzima quitosanase que degrada a quitosana. Mesmo assim, a quitosana

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
29
pode ajudar a desenvolver novas estratégias para o biocontrole em geral de fungos
patogênicos, evitando o uso de fungicidas que prejudicam o meio ambiente e a saúde
humana.
Pagnoncelli (2008) estudou a produção de quitooligossacarídeos com
propriedades nutracêuticas a partir da quitosana por hidrólise enzimática, utilizando
processo fermentativo simultâneo. Duas cepas produtoras de quitosanaes foram
selecionadas, Paenibacillus chitinolyticus e Paenibacillus ehimensis, e avaliadas quanto
ao comportamento em meio de cultivo contendo açúcares simples e em relação às
variações de pH do meio. Os quitooligossacarídeos produzidos a partir do complexo
enzimático da primeira cepa apresentaram maior pico após 9h de hidrólise, enquanto
para a segunda cepa avaliada, após 20min, observou-se quitooligossacarídeos com grau
de polimerização entre 3 e 6 unidades.
2.2. Fungos
2.2.1. Características gerais
Desde o final dos anos 60 que os fungos deixaram de pertencer ao reino das
plantas e passaram a ter um reino próprio chamado de Reino Fungi. Foi a partir de
estudos morfológicos, bioquímicos e citológicos onde se concluiu que eles são tão
diferentes dos vegetais como dos animais. Distinguem-se de outros organismos
eucariotas por serem seres quimiorganoheterotróficos e, ainda, por apresentarem parede
celular rígida que é composta por quitina e glucano e uma membrana celular em que o
ergosterol substitui o colesterol (Trabulsi & Toledo, 1996; Murray et al., 2005).
Fazem parte do Reino Fungi cerca de 100 mil espécies, desde grandes como
os cogumelos, até microscópicas como os bolores e leveduras. Ainda hoje é utilizada
essa classificação, mas os avanços da biologia molecular e estudos bioquímicos levaram
a outras propostas de classificação em que os fungos e organismos afins foram incluídos
em três diferentes reinos: Protozoa, Chromista e Fungi (Kirk et al., 2008).
Abaixo, são apresentadas características que diferenciam os fungos das plantas:
• são incapazes de sintetizar clorofila, como também não apresentar celulose em
sua parede celular, com exceções como fungos aquáticos;
• apresentam algumas características de células animais (capacidade de depositar
glicogênio e a presença de substâncias quitinosas, em sua parede celular);

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
30
• podem ser eucarióticos (as leveduras), ou multinucleados (os bolores), e
apresentar retículo endoplasmático rugoso e mitocôndrias em seu citoplasma;
• são heterotróficos, alimentam-se de matéria orgânica viva (fungos parasitários),
ou morta (fungos saprofíticos);
Os fungos são amplamente encontrados na natureza, crescem rapidamente no
solo, na água, na atmosfera, bem como nos animais, nos vegetais, resíduos orgânicos e
desempenham um papel importante na vida do homem, quer de uma maneira benéfica,
quer de um modo prejudicial. Os fungos são um dos principais microrganismos
responsáveis pela decomposição da matéria orgânica, interferindo no ciclo do carbono,
do nitrogênio e de outros nutrientes da biosfera. São capazes de deteriorarem produtos e
bens de consumo, tais como alimentos, tecidos, papel, madeira. Os alimentos
armazenados representam um vasto meio para a proliferação dos fungos, principalmente
quando não são consideradas as condições básicas de armazenamento correto (Trabulsi
& Toledo, 1996).
Estão presentes funcionalmente em vários processos industriais, tais como, de
fabricação de pão, cervejas, vinhos e determinados tipos de queijos, sendo também
utilizados na produção comercial de muitos ácidos orgânicos, de alguns fármacos, como
a ergometrina e a cortisona, na obtenção de diferentes antibióticos, como a penicilina, e
de substâncias imunossupressoras, como a ciclosporina (Esteves et al., 1990). A Figura
4 ilustra a estrutura geral de uma célula de fungo.
Figura 4 – Representação da estrutura geral de uma célula de fungo.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
31
Os fungos podem se desenvolver em meios de cultivo especiais formando
colônias de dois tipos, leveduriformes e filamentosas, que se diferenciam pela
macromorfologia e micromorfologia. O primeiro tipo apresenta, em geral, característica
de consistência cremosa de cor branca a creme, brilhantes ou opacas, podendo
apresentar às vezes coloração escura ou alaranjada, e são formados por microrganismos
unicelulares que apresentam as funções vegetativas e reprodutivas. As filamentosas
apresentam características granulares, cotonosas, aveludadas ou pulverulentas, são
multinucleadas e apresentam-se em forma de tubos denominadas de hifas. Essas hifas
podem ser não septadas (cenocíticas) ou septadas (tabicadas). O conjunto de hifas é
denominado micélio.
As hifas asseptadas têm aparência anastomosada (concisa), formada por um
citoplasma estendido e polinucleado, enquanto que as hifas septadas contêm células
individualizadas, cada uma contendo o seu núcleo. Na Figura 5 são mostradas as
estruturas das hifas septadas e não septadas.
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas. (Adaptado de Guia.Bio,
2012)
O micélio que se desenvolve no interior do substrato e que é responsável pela
sustentação e absorção de nutrientes é denominado micélio vegetativo. O micélio que
cresce na superfície do meio de cultura é denominado micélio aéreo. Este pode ser
denominado também de micélio reprodutivo, quando o mesmo se diferencia para
sustentar os corpos de frutificação ou propágulos (Putzke & Putzke, 1998). Os
propágulos classificam-se de acordo com a origem em externos e internos, sexuados e
assexuados.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
32
2.2.2. Reprodução dos Fungos
A reprodução dos fungos ocorre em ciclos assexuais, sexuais e parassexuais. Em
fungos, a reprodução assexuada é observada com mais freqüência do que a reprodução
sexuada. A reprodução assexuada é normalmente a reprodução mais importante para a
propagação da espécie, feita através de fragmentação de artroconídios, fissão de células
somáticas, brotamento ou gemulação do blastoconídios-mãe e produção de conídios. A
reprodução sexuada envolve a união de duas células ou de dois órgãos sexuais
sexualmente compatíveis. A reprodução sexuada culmina na produção de basidiósporos,
no caso de basidiomicetos. A reprodução parassexuada ocorre com a fusão de hifas e
formação de um heterocário, que consiste na existência de núcleos haplóides
geneticamente distintos num mesmo citoplasma, os quais, após a fusão, originam
núcleos diplóides heterozigotos sujeitos a sucessivas divisões mitóticas. Apesar da
raridade dessas, a reprodução parassexual é de grande importância na evolução de
alguns fungos (Pelczar et al., 1996).
A reprodução assexuada é feita por conídios, esporos assexuados formados no
ápice de hifas modificadas, os conidióforos. Os conidióforos não possuem esporângios
de forma que os conídios ficam expostos. Na reprodução sexuada, com o encontro de
duas hifas haplóides distintas, ocorre a plasmogamia e um novo conjunto de hifas
dicarióticas é formado.
Os conídios podem ter formas esféricas, fusiformes, cilíndricas, piriformes, entre
outras. Apresentam-se como hialinos ou pigmentados, podem estar isolados ou
agrupados, a parede celular pode ser lisa ou rugosa e podem ser formados por apenas
uma célula ou apresentarem septos em mais de um plano.
Os conídios também podem originar-se em qualquer parte do micélio vegetativo
e são denominados conídios sésseis como é o caso do Trichophyton rubrum.
Alguns fungos patogênicos são identificados a partir de estruturas formadas pela
célula conidiogênica e o conidióforo. Essas estruturas são denominadas: aparelho de
frutificação ou conidiação. Os conídios podem formar cadeias sobre fiálides em volta de
uma vesícula, dilatando a extremidade do conidióforo. Essas estruturas ocorrem em
Aspergillus, apresentando-se em forma de cabeça ao redor de uma vesícula, e em
Penicillium agrupados em forma de pincel, como é ilustrada na Figura 6.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
33
Figura 6 – Conídios de Aspergillus e Penicillium
2.2.3. Metabolismo de fungos
Os fungos são seres aeróbicos e heterotróficos, podendo se desenvolver em
ambientes com baixa quantidade de oxigênio, como ocorre no crescimento vegetativo e
na reprodução assexuada, como também em ambientes com grande quantidade de
oxigênio como é o caso da reprodução sexuada.
Os fungos são capazes de produzir certas enzimas como lipases, invertases,
lactases, quitosanases, proteases, amilases, etc., que fazem a hidrólise do substrato
facilitando o mecanismo de transporte ativo e passivo. Esses substratos induzem a
formação de enzimas degradativas. Algumas espécies de fungos desenvolvem-se em
meios que apresentam amônia ou nitrilos como única fonte de nitrogênio. As melhores
fontes orgânicas para o seu desenvolvimento são carboidratos, como a D-glicose, e
alguns sais minerais como sulfatos e fosfatos.
A água é uma necessidade de todos os seres vivos, não sendo diferente para os
fungos que precisam de água para o seu desenvolvimento. Certos fungos apresentam
características halofílicas podendo crescer nos ambientes com grandes concentrações de
sal.
A razão de não existirem ambientes livres de espécies fúngicas deve-se ao facto
destes microrganismos conseguirem sobreviver em situações extremas como, por
exemplo, temperaturas que variam entre os - 5 e os 60 ºC, com pH de 1 a 9 e com baixas

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
34
concentrações de oxigênio, apesar de cada espécie possuir condições ideais para o seu
desenvolvimento (Chao et al., 2002).
Devido a grande faixa de temperatura ótima para o seu crescimento, os fungos
apresentam espécies psicrófilas, mesófilas e termófilas. Para a maior parte das espécies
fúngicas, a temperatura de desenvolvimento situa-se entre os 20 e 24 ºC e não crescem
acima dos 29 ºC. Em relação à umidade relativa, necessitam de pelo menos 50 %, mas
geralmente necessitam de mais de 65 % (Colakoglu, 2001).
O pH ideal para o desenvolvimento dos fungos varia na faixa entre 5 e 7.
Destaca-se, entretanto, que a maioria dos fungos suportam grandes variações de pH
como ocorre com os fungos filamentosos que podem crescer em pH variando entre 1,5 e
11. Por outro lado, as leveduras não suportam pH alcalino.
Os fungos crescem lentamente, se comparados com as bactérias, em meio de
incubação para o seu desenvolvimento. Para evitar contaminação por bactérias, que
podem se sobrepor ou inibir o seu crescimento, pode-se introduzir no meio de cultura
um antibacteriano de largo espectro como o cloranfenicol.
Algumas espécies de fungos se desenvolvem na presença de luz, outras são
inibidas e algumas podem crescer tanto na presença quanto na ausência deste agente. No
entanto a irradiação solar, em geral, é prejudicial ao seu desenvolvimento, por causa da
radiação ultravioleta.
2.2.4. Nutrição de fungos
Quanto ao aspecto da nutrição, os fungos são heterótrofos, ou seja, incapazes de
produzir seu próprio alimento. Entretanto, a forma de assimilar matéria orgânica é
diferente daquela utilizada pela maioria dos animais, que são heterótrofos por ingestão.
Nos fungos, a digestão é extracorpórea, ou seja, é realizada fora do corpo. O fungo
lança no ambiente, enzimas que degradam as moléculas orgânicas complexas e, depois,
absorve moléculas menores, mais simples. Por essa razão, são designados heterótrofos
por absorção. É devido a este processo que os fungos podem crescer dentro ou sobre os
alimentos. Desenvolve-se geralmente em meios contendo um pH baixo, uma fonte de
carbono, uma fonte de nitrogênio orgânico ou inorgânico e alguns minerais. Alguns
necessitam de vitaminas. Quanto aos modos de vida, os fungos podem ser
decompositores, parasitas, mutualísticos e predadores (Raven, 2001).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
35
Os fungos decompositores, saprófagos (do grego, saprós, “podre”, e phagos,
“comedor”), juntamente com as bactérias, são os principais decompositores da biosfera,
participando intensamente do processo de degradação da matéria orgânica morta, o que
promove a reciclagem dos elementos químicos constituintes dos seres vivos. A maioria
deles vive no solo, obtendo nutrientes de seres mortos. Em florestas, onde a matéria
orgânica vegetal é abundante, os micélios dos fungos decompositores absorvem os
nutrientes de folhas e galhos caídos, decompondo-os. Os fungos decompositores
também são responsáveis pelo apodrecimento de alimentos e de outros materiais, como
a madeira.
Os fungos parasitas vivem à custa de outros organismos vivos, prejudicando-os.
Ao parasitar o corpo de um ser vivo, animal ou vegetal, o fungo pode até provocar sua
morte. Várias doenças são causadas por fungos parasitas. Nas plantas, cita-se a
ferrugem do café, que, no Brasil, é motivo de grande preocupação por parte dos
cafeicultores, devido aos prejuízos que acarreta. As micoses que atacam a pele de seres
humanos e animais são provocadas por fungos.
Existem fungos que se associam a outros organismos, estabelecendo uma relação
em que há benefício mútuo para os indivíduos envolvidos. Esses fungos são
denominados mutualísticos. A maioria vive associada a seres fotossintetizantes, como
as plantas, cedendo-lhes parte da água e dos nutrientes que as hifas absorvem do solo.
As plantas, por sua vez, cedem ao fungo certos açúcares e aminoácidos. Duas
associações mutualísticas formam estruturas bem características: as micorrizas,
associação que envolve fungos e raízes de plantas, e os líquens, formados por fungos e
algumas variedades de algas e cianobactérias.
Certos fungos atuam como predadores. Na maioria dos casos, as hifas secretam
substâncias aderentes que aprisionam os organismos que tocam os fungos. Dessa
maneira, as hifas penetram o corpo da presa, crescem e se ramificam, espalhando-se no
interior do organismo e absorvendo seus nutrientes, causando-lhe a morte. O gênero
Arthrobotrys apresenta uma forma mais elaborada de predação. O fungo vive no solo e
captura nematódeos microscópicos que habitam o mesmo ambiente. As hifas
apresentam pequenos anéis que se estreitam quando a presa passa por eles. As hifas do
fungo logo invadem o corpo do nematódeo, digerindo-o.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
36
2.2.5. O fungo Metarhizium anisopliae
Existem três espécies de fungos do gênero Metarhizium, divididas em dez
variedades: M. anisopliae, variedades anisopliae, majus, lepidiotum e acridum; M.
flavoviride, variedades tipo E, flavoviride, minus, novazealandicum e pemphigum; e M.
album (Driver et al. 2000). E, mais recentemente, foi descrita na China uma nova
variedade de M. anisopliae, dcjhyium (Dong et al. 2009).
Data de 1879 quando o russo Ilya Metchnikoff aplicou o fungo para combater
larvas de um curculionídeo, praga de beterraba, e foi classificado como Metarhizium
anisopliae por Sorokin em 1883. É um fungicida natural para mais de 300 espécies de
insetos, especialmente pragas da agricultura e pecuária. No Brasil, o fungo Metarhizium
anisopliae teve a sua aplicação com sucesso pela primeira vez em 1965, no controle da
cigarrinha-da-cana Mahanarva posticata (Alves, 1998).
O fungo Metarhizium anisopliae é um importante agente entomopatogênico
utilizado no controle de pragas, como cupins, gafanhotos, cigarrinhas e besouros, sendo
amplamente estudado em todo o mundo. Existe amplamente na natureza e é facilmente
encontrado no solo, onde sobrevive por longos períodos e seu desenvolvimento se dá
geralmente entre 15 e 32 °C, sendo ideal entre 24 e 30 °C e com pH ótimo igual a 6,9.
Apresenta micélio hialino e septado, com conidióforos característicos, sobre os quais
surgem conídios cilíndricos organizados em colunas. Este fungo ao atacar os insetos
provoca rigidez e os recobre por uma camada pulverulenta de conídios, resultando em
colorações que variam do verde claro ao escuro, cinza ou branco com pontos verdes
(Tinline, 1971; Alves, 1998, Driver et al. 2000; Arruda, 2005).
Quanto a sua morfologia, apresenta-se como um fungo filamentoso, com corpo
de frutificação equivalente a um esporodóquio agregado a hifas intimamente
entrelaçadas, com massa compactada de conidióforos característicos, simples ou
ramificados, formando células esporogênicas denominadas fiálides, resultando nos
fialosporos (Wang et al., 2002).
O Metarhizium anisopliae é um fungo de cultivo simples, sendo necessário
quase que exclusivamente o nutriente a base de amido. Um dos fungos mais estudados
em programas de manejo de pragas já que apresenta grande potencialidade
entomopatogênica (Onofre et al., 2002). O Metarhizium anisopliae é um microrganismo
pouco exigente, pois pode se desenvolver em diversos meios de cultura como: amido,
glicose, glicerina, levulose, maltose, sacarose e quitina (Jabor et al., 2003).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
37
Em geral, os fungos produzem enzimas que convertem os tecidos dos insetos em
nutrientes. Porém, a cutícula esclerotizada dos insetos não é bem utilizada pelos fungos,
excetos pelos entomopatogênicos que desenvolveram enzimas eficientes para degradar
essa camada protetora. Destaca-se a enzima Pr1, produzida por Metarhizium anisopliae,
que tem um poder de degradação bem superior a qualquer outra enzima sintetizada por
esse grupo de fungos (Dias, 2005). Desta forma, o Metarhizium anisopliae (fungo
entomopatogênico) ao atacar a célula dos insetos, produz grande quantidade de enzimas
que degradam a membrana citoplasmática e inativa a célula e, consequentemente, mata
o inseto, logo é evidente a relação entre o controle de pragas e produção de enzimas.
No momento, sobre a comercialização de fungos entomopatogênicos no Brasil,
em sua maioria, são formulados em grânulos, compostas do fungo mais o substrato
(arroz + fungo), como também na forma de pó molhável (WP), resultado da moagem do
fungo com o substrato. Esses produtos representam 90% e 8,5%, respectivamente, da
produção de fungos no Brasil, e sendo suas concentrações finais ficam em torno de
5x108 conídios/g para Metarhizium anisopliae e 1x109 conídios/g para Beauveria
bassiana (Alves et al., 2008).
Segundo Batista Filho et al. (2002), Garcia et al. (2005) e Mello et al. (2006), o
Metarhizium anisopliae tem sido bastante utilizado para o controle de diferentes pragas
no Brasil, como a broca da bananeira, pragas de grãos armazenados, carrapatos, cupim
de montículo em pastagens, larvas de escarabeídeos que atacam a cana de açúcar e
cigarrinha da cana-de-açúcar.
É comum a ocorrência natural do fungo Metarhizium anisopliae atacando
cigarrinha-das-raízes de canaviais pelo país, o que incentiva a utilização dele como
agente de controle biológico em áreas infestadas por esse tipo de praga. A aplicação de
recursos no uso de programa de manejo integrado da cigarrinha tem sido reforçado por
razões ambientais e econômicas (Dinardo-Miranda, 2001).
Faria & Magalhães (2001) fizeram uma avaliação do uso de fungos
entomopatogênicos no Brasil, na qual observaram que a qualidade dos micoinseticidas
disponíveis no Brasil poderia ser incrementada de forma considerável quanto a sua
formulação, ou seja, antes da venda, poderiam realizar melhorias no tratamento
posterior, adição de substâncias que lhes assegurem melhorias na eficiência de controle,
capacidade de armazenamento ou praticidade de manuseio, ou de qualquer outro critério
que resulte em vantagem em relação ao produto bruto. Ressaltaram a pouca praticidade

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
38
em alguns casos, como exigência de lavagem, e em outros podendo causar o
entupimento de bicos dos pulverizadores devido à elevada proporção de inertes,
principalmente quando são empregados baixos volumes de aplicação. Outro problema
evidenciado com os inseticidas nacionais diz respeito à pequena sobrevida, devendo a
ser usados em, no máximo, 30 dias depois de produzidos, quando armazenados à
temperatura ambiente e em local sombreado, fazendo as vendas ocorrerem quase que,
exclusivamente, sob encomenda. Comparativamente, os bons inseticidas sob
comercialização em outros países apresentam maior concentração de ingrediente ativo,
maior sobrevida (alguns produtos podem ser armazenados por mais de 8 meses à
temperatura ambiente) e praticidade (são formulados na forma de GDA (grânulos
dispersíveis em água) ou de óleos emulsionáveis, por exemplo, podendo ser adicionados
diretamente ao tanque do pulverizador.
Rangel et al. (2007), instalaram uma Unidade de Observação para o controle
microbiano das cigarrinhas das pastagens Distrito de Itahum, em Dourados/MS, devido
as necessidades de agricultores familiares de produção de leite. As aplicações do fungo
Metarhizium anisopliae foram realizadas por pulverização com equipamento de tração
animal. As condições do ambiente de estudo foram favoráveis ao desenvolvimento da
população das cigarrinhas e à incidência do fungo. Como resultados, observaram que os
níveis de espumas, ninfas e adultos reduziram significativamente após as aplicações,
levando os Agricultores Familiares a total satisfação com os ganhos na produção de
todos os envolvidos.
Segundo Vieira et al. (2007), o fungo entomopatogênico Metarhizium anisopliae
var. acridum tem sido utilizado no controle de pragas, e assim, visando este fim,
analisaram em laboratório a esporulação, a germinação dos conídios e as características
morfológicas peculiares desse fungo em diferentes temperaturas. Esperou-se o
desenvolvimento das colônias em períodos de 1 a 15 dias. Foi observado que
temperaturas de 25, 28 e 30 ºC proporcionaram maiores taxas de esporulação e elevadas
porcentagens de germinação dos conídios. A 25, 28 e 30 ºC observou-se um
desenvolvimento mais rápido das microestruturas somáticas e reprodutivas (hifas,
anastomoses, apressórios, conidióforos e conídios elipsóides). Nas temperaturas
extremas de 40 e 45 ºC não foi detectado esporulação, germinação, nem
desenvolvimento de nenhuma microestrutura.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
39
Lopes et al. (2007) avaliaram o efeito de formulações de Beauveria bassiana e
Metarhizium anisopliae sobre ninfas de Amblyomma cajennense (transmissor da febre
maculosa ao homem) em laboratório. Ninfas não alimentadas foram pulverizadas com
suspensões contendo 2x107 e 4x107 conídios/mL de B. bassiana, e 1x107, 1,5x107,
2x107, 3x107 e 5x107 conídios/mL de M. anisopliae. A mortalidade de ninfas,
decorridos três dias da aplicação, foi significativa nas concentrações de 3x107 e 5x107
conídios/mL de M. anisopliae, com 57,1 e 56,2 %, respectivamente. Após seis dias a
mortalidade foi de 100 % nessas duas concentrações. A mortalidade foi
significativamente maior que a observada para B. bassiana em concentrações
semelhantes.
Guirado et al. (2009) realizaram estudos para controlar cupins de montículo em
área de pastagem, utilizando fungos entomopatogênicos Metarhizium anisopliae e
Beauveria bassiana em diferentes doses e formulações. A avaliação foi realizada após
30 dias da aplicação dos produtos. Os melhores resultados em ordem decrescente de
eficiência foram: M. anisopliae 30 mL + 1000 mL de água com 100 % de eficiência; B.
bassiana 10g + M. anisopliae 20 mL + 1000 mL de água com 85,71 % de eficiência; M.
anisopliae 20 mL + 1000 mL de água com 71,42 % de eficiência. Os demais
tratamentos não diferiram da testemunha.
Loureiro et al. (2012) compararam a eficiência de isolados de Metarhizium
anisopliae em controlar populações naturais de Mahanarva fimbriolata (cigarrinha-da-
raiz) em cultivos comerciais de cana-de-açúcar colhida mecanicamente. Foram
aplicados os isolados IBCB 348, IBCB 408, IBCB 410 e IBCB 425 do fungo em
concentração de 1,5×1012 conídios/ha. Aplicou-se o planejamento experimental em
blocos casualizados com 5 tratamentos e 4 repetições, sendo cada repetição composta
por 7 linhas de 100 m de comprimento e espaçamento de 1,5 m. Após 30 dias de
pulverização foi observado para os isolados IBCB 408 e IBCB 425 uma eficiência de
controle de ninfas de 63 e 62%, respectivamente, já no caso dos adultos chegou a 100%.
Após 60 dias o isolado IBCB 425 foi o mais eficiente para controlar as ninfas com
48,4%.
Assis et al. (2012) estudaram a citotoxidade de quitoolissacarídeos (QOS) e
oligômeros A, B e C (soluções compostas por diferentes quantidades de QOS)
produzidos por hidrólise enzimática pelo fungo Metarhrizium anisopliae. Foi analisado
o efeito de antiproliferativo destas moléculas em células tumorais (HepG2 e HeLa) e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
40
também em célula normal (3T3). A atividade antioxidante foi analisada em vários
experimentos in vitro. A glicosamina apresentou maior toxicidade (cerca de 92%) para
todas as células estudadas. No entanto, os oligômeros obtidos após hidrólise não
demonstraram efeito tóxico sobre a célula normal (3T3). Além disso, mostraram que
uma pequena quantidade de outros COS pode diminuir o efeito citotóxico da
glicosamina contra células 3T3, indicando que glicosamina poderia ser usada como uma
droga antitumoral na presença de outros COS.
2.3. Fermentação
Os processos fermentativos são caracterizados por processos nos quais
microrganismos realizam a conversão catalítica de uma dada substância num
determinado produto. Estes processos podem ser classificados como processos
descontínuos, contínuos e descontínuos alimentados. A classificação destes processos
está diretamente ligada à forma de como o substrato é adicionado e como o produto é
retirado. No processo de fermentação descontínuo o substrato é colocado no
fermentador, em seguida é inoculado e após o término do processo o produto é retirado.
No processo contínuo, o substrato é colocado num reator com vazão constante e o meio
fermentado é retirado com a mesma vazão de entrada do reator (Faccioti, 2001). Nos
processo descontínuos alimentados à adição de substrato e a retirada do mesmo deve
ocorrer de forma intermitente (Carvalho & Sato, 2001).
De certa forma, os processos fermentativos representam um elo entre as antigas
artes de elaboração de alimentos, como queijos e vinho, a partir de uma flora
microbiana natural, com a moderna indústria de fermentação de alimentos, onde são
utilisados cultivos puros e equipamentos sofisticados para o controle do processo
(Assis, 2010).
Atualmente, a maioria dos processos industriais se desenvolve em cubas de
vidro ou de aço de até 20 litros. Os agentes biológicos se encontram submersos no meio de
cultivo que ocupa, aproximadamente, 75% da cuba. As vezes é necessário injetar ar, e em muitas
fermentações se forma espuma. O desenho do biorreator deve se adequar ao objetivo do
processo, respondendo eventualmente a diversos imperativos, tais como a esterilização do
sistema, a aeração e homogeneização do meio, o acréscimo de nutrientes e de aditivos
antiespuma, a manutenção do pH, etc. Os modelos de fermentadores mais utilizados com
microrganismos contam com aeração e agitação mecânica. Esta facilita a distribuição dos

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
41
nutrientes na cuba, mas gera calor que deve ser eliminado mediante a circulação de água
fria. Em outros tipos de biorreatores, em coluna ou torre, a homogeneização depende da injeção de
ar. Os sistemas submersos são apropriados para o cultivo de microrganismos livres, mas resultam
pouco econômicos quando se trabalha com células ou enzimas caras. A imobilização em
fermentadores menores, seja por adesão a um suporte inerte, seja por inclusão dentro de um
polímero que permita o contato com o meio de cultura, além de simplificar a purificação do
produto permite a reutilização das células ou das enzimas, que permanecem dentro do
biorreator (Malajovich, 2009).
As enzimas são produtos de alto valor agregado, amplamente empregadas em
uma gama de indústrias, e entre as fontes para sua obtenção, destacam-se as de origem
microbiana, principalmente, devido a elevada seletividades dos substratos utilizados e
produtos formados. A principal forma de obtenção de enzimas, em escala industrial, é a
fermentação submersa, por esta apresentar vantagens em relação a fermentação
semisólida, como: maior controle do processo fermentativo e consequente reprodução
da cinética microbiana, além de maiores rendimentos (Wanderley, 2011).
2.3.1. Preparo do inóculo
Denomina-se inóculo um volume de suspensão de microrganismo de
concentração adequada, capaz de garantir, em condições econômicas, a fermentação de
uma dada quantidade de meio. O volume do inóculo, assim como sua concentração de
células, vai depender tanto do processo de fermentação c omo da quantidade de meio a
ser fermentado (Carvalho & Sato, 2001).
A partir de uma cultura pura, inocula-se um volume relativamente pequeno de
meio nutriente, que vai depender do tipo de microrganismo cultivado (pois cada espécie
microbiana possui velocidades diferentes de crescimento), sendo, em seguida, incubado
em condições favoráveis ao desenvolvimento do microrganismo. A suspensão obtida é
transferida para um frasco maior em agitador rotativo ou recíproco (shakers), que por
sua vez é adequadamente incubado, e a seguir, transferido para um frasco contendo um
volume ainda maior deste meio, até se conseguir alguns litros de suspensão microbiana.
Os cuidados com assepsia, volumes de meio utilizado, numero de transferência bem
como as condições de incubação, podem variar de acordo com o tipo de processo de
fermentação que se deseja realizar.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
42
2.3.2. Produção de enzimas em cultivo submerso descontínuo
A produção de enzimas, usando processos fermentativos, é constituída
basicamente por duas etapas: a primeira caracteriza-se pela preparação do inóculo e a
segunda, pela fermentação do substrato (Carvalho & Sato, 2001). A finalidade da
primeira etapa é o preparo do microrganismo em condições adequadas para que possa
ocorrer o desenvolvimento da segunda etapa. Diferentemente do cultivo em estado
sólido, no cultivo submerso os microrganismos encontram-se em um meio com elevada
atividade de água.
2.4. Planejamento experimental
Um bom planejamento tem como fundamental projetar um experimento de
forma que ele seja capaz de fornecer exatamente o tipo de informação que procuramos.
Para isso é necessário saber, em primeiro lugar, o que é mesmo que se deseja procurar.
Planejamento experimental, ou delineamento experimental, é um conjunto de
ensaios estabelecido com critérios científicos e estatísticos, objetivando determinar a
influência de diversas variáveis nos resultados de um dado sistema ou processo (Barros
Neto et al., 2001; Rodrigues & Iemma, 2005). De acordo com o propósito dos ensaios,
esse objetivo maior pode ser mais detalhado em:
– determinar quais variáveis são mais influentes nos resultados;
– atribuir valores às variáveis influentes de modo a otimizar os resultados;
– atribuir valores às variáveis influentes de modo a minimizar a variabilidade dos
resultados;
– atribuir valores às variáveis influentes de modo a minimizar a influência de variáveis
incontroláveis.
O planejamento experimental, baseado nos fundamentos estatísticos é, sem
dúvida alguma, uma ferramenta poderosa para se chegar às condições otimizadas de um
processo, desenvolvimento da formulação de produtos dentro das especificações
desejadas ou simplesmente para avaliar os efeitos ou impactos que os fatores (variáveis
independentes) têm nas respostas (variáveis dependentes) desejadas (Barros Neto et al.,
2001; Rodrigues & Iemma, 2005). Da utilização das técnicas estatísticas de
planejamento experimental, é possível chegar benefícios importantes, como:

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
43
– reduzir o número de ensaios sem prejuízo da qualidade da informação;
– estudar simultaneamente diversas variáveis, separando seus efeitos;
– determinar a confiabilidade dos resultados;
– realizar a pesquisa em etapas, num processo iterativo de acréscimo de novos ensaios;
– selecionar as variáveis que influem num processo com número reduzido de ensaios;
– representar o processo estudado através de expressões matemáticas;
– elaborar conclusões a partir de resultados qualitativos.
2.4.1. Planejamento fatorial
Em um planejamento fatorial é investigado as combinações de níveis de estudo
de cada fator. Para tanto, inicialmente, é necessário especificar esses níveis em cada
fator, quer dizer, os valores dos fatores que serão utilizados nos experimentos. Um
ensaio experimental é definido como um experimento constituído de um conjunto de
níveis definido. Geralmente, quando se tem n1 níveis de um determinado fator 1, n2 do
fator 2, ..., e nk do fator k, o planejamento será descrito por um fatorial n1 x n2 x ... x nk,
que corresponde ao número mínimo de experimentos para se ter um planejamento
fatorial completo.
É necessário fazer cada fator variar e observar o resultado dessa variação para se
estudar o efeito sobre a resposta. Assim, os ensaios precisam ter um mínimo de dois
níveis por fator para a realização desses ensaios. Para k fatores, ou seja, k variáveis de
controle, o planejamento de dois níveis precisará de 2 x 2 x ... x 2 = 2k, para o número
de ensaios diferentes, sendo conhecido como planejamento fatorial 2k.
Para o planejamento fatorial, são especificados os valores dos fatores em cada
experimento, e estes são classificados em níveis, variando-se, por exemplo, de -1
(mínimo) a +1 (máximo). A partir de uma matriz serão apresentadas todas as
combinações entre o número de fatores e de níveis nos experimentos a serem realizados
(Barros Neto et al., 2001; Rodrigues & Iemma, 2005).
2.4.2. Planejamento Plackett & Burman
A maioria dos planejamentos do tipo “Screening” (Delineamentos de Seleção de
fatores) foi descrito originalmente por Plackett & Burman em 1946. Os planejamentos
experimentais de Plackett & Burman são eficientes em situações que envolvem uma

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
44
grande quantidade de variáveis a serem exploradas em um processo de otimização de
fatores (Reddy et al., 2008).
As matrizes Plackett & Burman consistem de planejamentos fatoriais de dois
níveis para estudar k = N – 1 variáveis em N ensaios. São construídas para ensaios
múltiplos de 4, sendo essas matrizes para ensaios de 8, 12, 16,..., 100. Cada variável
independente em uma matriz assume uma configuração de dois níveis, o inferior (-1) e o
superior (+1), para o nível intermediário é estabelecido (0). Para cada vez que a variável
aparece no nível inferior, terá o par equivalente no nível superior (Rodrigues & Iemma,
2005).
Para evitar dúvidas nos resultados dos efeitos de algum fator no planejamento de
Plackett & Burman, o número de ensaios a ser realizado deve ser no mínimo o número
de variáveis mais 4. No caso de um fatorial completo o número de ensaios a ser
realizado é bem maior (Rodrigues & Iemma, 2005).
Mesmo já determinado o planejamento Plackett & Burman, torna-se importante
à realização de experimentos relativos aos pontos centrais de modo a se avaliar melhor
as possíveis interferências analíticas que podem ocorrer durante os diferentes
experimentos. A repetição do ponto central permite também a determinação do erro
puro e da repetibilidade do processo. Na prática, esta é uma ferramenta que permite
identificar melhor se o processo em estudo está sob controle (Rodrigues & Iemma,
2005).
É possível que no planejamento Plackett & Burman não tenha sido considerado
a codificação do ponto central (0) para repetição, pois, matematicamente, a estimativa
do valor desse efeito codificado (zero) vezes, teria como resposta zero. Porém, no
resultado prático do experimento, consegue-se informações valiosas com essas
repetições no ponto central (Rodrigues & Iemma, 2005).
2.4.3. Aplicações do Planejamento Plackett & Burman
Ghanem et al. (2000) produziram xilanase por Aspergillus terreus cultivadas
sobre palha de trigo finamente moída em fermentação de estado sólido. A composição
ideal do meio foi estabelecida aplicando-se o planejamento experimental de Plackett &
Burman. A melhor atividade enzimática foi obtida em um meio contendo 10 g/L de

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
45
palha de trigo por frasco umedecido com uma solução concentrada de sal com teor de
água inicial de 75 % e incubadas durante 4 dias a 30 °C. Observou-se que íons
metálicos como Hg2+, Cu2+, Co2+, Fe3+ e Pb2+ inibiram fortemente a xilanase, enquanto
que Ca2+ ativou a enzima.
Mu et al. (2005) estudaram as otimização de meio de fermentação para a reação
de estereoinversão por Candida parapsilosis CCTCC M203011 utilizando o
planejamento Plackett & Burman. Os resultados mostraram que dentre os fatores
estudados, os fatores mais significativos foram: a Concentração de MgSO4, ZnSO4 e
KH2PO4 e que após a utilização da metodologia de superfície de resposta foi constatado
que o ponto ótimo para a otimização foi conseguido com as concentrações dos fatores
MgSO4, ZnSO4 e KH2PO4 nos valores em: 1,17 g/L, 0,10 g/L e 9,41 g/L,
respectivamente, elevando a taxa de conversão enantiosseletiva de 75,2 % para 96 %.
Naveena et al. (2005) empregaram o planejamento Plackett & Burman para a
seleção de 15 parâmetros para produção de ácido láctico L(+) de farelo de trigo,
substrato de baixo custo e suporte sólido, por Lactobacillus amylophilus GV6 em
fermentação de estado sólido (SSF). Onze nutrientes pertencentes a duas categorias,
fontes de nitrogênio e fontes de sal, junto com três parâmetros físicos e um tampão
foram os parâmetros considerados. Os autores verificaram influência na produtividade
devido às fontes de nitrogênio, peptona, extrato de levedura e tri-citrato de amônio,
juntamente com o NaH2PO4.2H2O e Tween 80, que podem ainda serem otimizados para
o aumento na produção de ácido lático. Esses pesquisadores ainda chamam atenção para
o uso escasso deste planejamento em fermentação em estado sólido e que o mesmo
ainda não havia sido considerado anteriormente para a conversão direta de amido em
ácido láctico L (+) usando um sistema bacteriano.
Khosravi-Darani et al. (2008) estudaram a aplicação de Plackett & Burman na
análise dos fatores significativos para a produção de ácido cítrico por Aspergillus niger
ATCC 9142 utilizando palha de trigo como substrato. Os resultados mostraram que a
produção, com a palha de trigo pré-tratada com HCl, NaOH e Uréia, foi mais eficiente e
que os fatores mais significativos na produção foram: Solvente, Teor de Umidade,
Temperatura, Tempo de Incubação, Concentração Inicial de Açúcar e Concentração de
Metanol.
Ji et al. (2009) estudaram o desenvolvimento de um meio de cultura econômico
para a produção do 2,3-butanodiol através da fermentação da glicose e xilose por

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
46
Klebsiella oxytoca utilizando o planejamento Plackett & Burman. Os resultados
mostraram que a uréia e a maceração de milho foram os fatores mais significativos para
a produção do 2,3-butanodiol e que através de planejamento experimental com repetição
em triplicata no ponto central para determinar os níveis ótimos de produção. Os autores
constaram que o rendimento atingido foi 85,6% do valor teórico.
Paula & Arruda. (2009) observaram a necessidade da busca por alternativas mais
eficientes ao método termomecânico utilizado para a degradação de milhões de
toneladas de penas de aves provenientes de frigoríficos e aviários, cuja rígida estrutura é
composta por mais de 90 % de queratina. Para esse fim, utilizaram de comunidades
microbianas para a finalidade proposta, testando o planejamento experimental Plackett
& Burman para verificar a influência de diferentes microrganismos simultaneamente,
com um número reduzido de experimentos. Testaram 32 bactérias isoladas, inoculadas
em meio contendo somente uma pena como única fonte de carbono, durante sete dias,
de onde foi selecionado as cepas para os consórcios microbianos. Experimentos unindo
as cepas NP5, FCA7, NP4 e S14 demonstraram diferentes atividades queratinolíticas,
aumentando o nível de degradação, até 87,8%, comparada com as cepas
individualmente, variando entre 26,4% e 78,0%.
Rajendran & Thangavelu (2009) obtiveram valores máximos da atividade de
produção de lipase em 3,98 U/mL e concentração de massa celular em 5,62 g.L-1 com R.
arrhizus em cultivo submerso descontínuo otimizado pelo planejamento Plackett &
Burman. As variáveis mais importantes que afetaram a produção da lipase foram azeite
de oliva, peptona, KH2PO4, CaCl2.2H2O e MgSO4.7H2O.
Li et al. (2010) estudaram a otimização de meio de cultura para a produção de
Streptolydigin (um antibiótico) por Streptomyces lydicus AS 4,2501 utilizando Plackett
& Burman e a metodologia de superfície de resposta. Os resultados mostraram que
dentre os fatores estudados, os fatores mais significativos para a produção de
Streptolydigin e otimização do meio de cultura foram: Concentração de Amido,
Concentração de Peptona e a Concentração de K2HPO4.
Chen et al. (2010) estudaram a otimização de meio de cultura para a produção da
Luciferase – que são enzimas que catalisam reações biológicas transformando energia
química em energia luminosa - por Bacillus Subtilis em fermentação descontínua
utilizando planejamento Plackett & Burmam. Os resultados mostraram que dentre os
componentes estudados, os fatores mais significativos para a produção e otimização do

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
47
meio de cultura para a produção da Luciferase foram: Concentração de Glicose 1,17 %,
Extrato de Levedura 2,27 % e Glutamato 0,55 %.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o projeto Plackett & Burman e a metodologia de superfície de resposta. Os
resultados mostraram que a eficiência máxima de extração da quitina foi conseguida nas
seguintes condições: Concentração de casca de camarão (50 g/L), glicose (50 g/L),
tempo de incubação de 5 dias e inoculo com 0,05 de OD (densidade óptica).
Gao et al. (2010) desenvolveram um meio para a fermentação do extrato bruto
de inulina de tubérculos de alcachofra de Jerusalém por Paenibacillus polymyxa ZJ-9
para produzir R,R-2,3-butanodiol (R-R,2,3-BD). Inulina, K2HPO4 e NH4Cl foram os
parâmetros principais na fermentação de acordo com os resultados obtidos com o
planejamento experimental Plackett & Burman. A concentração ideal dos três fatores,
77,14 g/L, 3,09 g/L e 0,93 g/L, respectivamente, foi obtida pelo planejamento Box-
Behnken e pela superfície de resposta. Sob as condições ideais, a concentração do R-
R,2,3-BD obtida foi 36,92 g/L, em mais de 98 % de pureza óptica. Comparado com
outras fontes de carbono investigadas, a fermentação do extrato bruto de inulina obteve
o maior rendimento de R-R,2,3-BD.
Uma metodologia de superfície de resposta foi empregada com sucesso para
otimizar um meio de fermentação chamada VHG (Very High Gravity), referência à alta
concentração de açúcares e densidade dos mostos, baseado na água maceração de milho
CSL (Corn Steep Liquor) e outros nutrientes de baixo custo para a produção eficiente de
etanol a partir de glicose por S. cerevisiae. Usando a composição média otimizada (g/L:
CSL 44,3; uréia 2,3; MgSO4.7H2O 3,8; CuSO4.5H2O 0,03), a levedura industrial PE-2
foi capaz de fermentar até 330 g/L de glicose e produzir 18,6 % (v/v) de etanol, com
uma produção descontínua de 2,4 g/L/h e um rendimento em etanol de 93 % (Pereira et
al., 2010).
O planejamento Plackett & Burman juntamente com o Planejamento Composto
Central (CCD) foram aplicados para otimizar o meio de cultura para produção de etanol
por Clostridium autoethanogenum com monóxido de carbono (CO) como fonte única de
carbono e um meio contendo (g/L): 1,0 de NaCl, 0,1 de KH2PO4, 0,02 de CaCl2, 0,15 de
extrato de levedura, 0,116 de MgSO4, 1,694 de NH4Cl e pH 4,74. O rendimento ótimo
de etanol previsto pela metodologia de superfície de resposta (RSM) e um algoritmo

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
48
genético de rede neural artificial (ANN-GA) foram 247,48 e 261,48 mg/L,
respectivamente. Valores semelhantes aos obtidos experimentalmente sob as condições
ideais sugeridas pelos métodos estatísticos (254,26 e 259,64 mg/L). A adequação do
modelo ANN-GA foi maior do que o modelo RSM. Os rendimentos obtidos foram
substancialmente superiores aos relatados anteriormente (60-70 mg/L) com essa cultura
(Guo et al., 2010).
Rigueira (2010) avaliou a influência das variáveis do procedimento de extração
na porcentagem de recuperação das aminas encontradas no leite cru utilizando um
planejamento para “screening” do tipo Plackett & Burman, para os ácidos
tricloroacético (TCA) e sulfosalicílico (ASS).
Quines (2010) estudou a necessidade ou não de se fazer um pré-tratamento da
biomassa antes da extração. Um planejamento experimental Plackett & Burman de doze
ensaios mais três pontos centrais foi utilizado para selecionar as variáveis mais
significativas na extração de Poli(3-hidroxibutirato) (P(3HB)) por 1,2-carbonato de
propileno. Foram realizados dois testes de recuperação do solvente 1,2-carbonato de
propileno, após a extração, sendo um com aplicação de aquecimento e o outro com
aquecimento e vácuo. Os resultados mostraram que o pré-tratamento térmico levou a
uma maior porcentagem de recuperação e de pureza do polímero.

Capítulo 3 Metodologia Experimental

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
50
3. Metodologia Experimental
O presente capítulo apresenta os matérias e métodos utilizados na produção de
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso.
3.1. Quitosana
Neste trabalho utilizou-se uma quitosana (Lote 067) obtida junto a POLYMAR/CE,
com no mínimo 80% de desacetilação. Esse substrato foi utilizado como fonte de carbono e
energia tanto no estudo cinético como nos ensaios do planejamento de seleção de fatores
Plackett & Burman e Planejamento Composto Central, para a produção de quitosanase pelo
Metarhizium anisopliae, por fermentação submersa.
3.2. Microrganismo
O fungo Metarhizium anisopliae estirpe (CG374), utilizado neste estudo, foi cedido
pela Embrapa Recursos Genéticos e Biotecnologia (Brasília / DF-Brasil) e foi mantido em
placas de Petri contendo o meio Batata Dextrose Agar (PDA). As placas foram incubadas em
estufa a 25 ºC. O pH do meio de manutenção foi mantido entre 6,0 e 6,5.
3.2.1. Escolha da cepa produtora de quitosanase
A escolha do microrganismo Metarhizium anisopliae foi feita com base na capacidade
de crescer em meio contendo quitosana como única fonte de carbono, além de apresentar
grande tolerância a pH variando a uma taxa de 2,0 a 8,5, sendo 6,9 a melhor condição para o
crescimento vegetativo e esporução, de acordo com Assis (2010).
3.2.2. Manutenção da cepa
A manutenção da cultura foi realizada em placas petri contendo o meio Batata
Dextrose Agar (PDA). A renovação das células foi realizada a cada trinta dias através de
subcultivos das células no meio PDA.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
51
3.3. Produção de enzima
No preparo do inóculo, utilizou-se 7 placas cultivadas e mantidas, com o fungo
Metarhizium anisopliae, em meio PDA a 25 ºC durante 5 dias em estufa. O meio de cultura
para a produção da enzima quitosanase apresentou os seguintes componentes: quitosana;
peptona; extrato de levedura; NaNO3; K2HPO4; KCl; MgSO4.7H2O e FeSO4, com
concentrações de acordo as matrizes dos planejamentos: Plackett & Burman e composto
central. Para a produção da enzima no cultivo, a biomassa foi transferida através de água
estéril, para um Béquer de 500 mL que, em seguida, foi colocado em Erlenmeyers de 250 mL
na proporção de 20 mL de biomassa para 180 mL do meio de cultura. Os Erlenmeyers
cultivados foram então incubados em incubador rotativo a 25 ºC e 150 rpm durante 24 h.
Alíquotas foram retiradas e centrifugadas a 8000xg por 5 min, a cada 6 horas, para se
determinar a atividade quitosanolítica, teores de açúcares redutores e de proteína total.
Ressalta-se que todos os experimentos foram feitos em triplicata.
3.4. Determinação da Proteína Total
A determinação da proteína total foi efetuada de acordo com o método modificado de
Sedmak & Grossberg (1977), que se baseia na diminuição da absorbância do corante azul de
Coomassie, devido à ligação proteína-corante, a 595 nm, comparado a absorção máxima a
465 nm, conforme Santos (2001). Nesse caso, após centrifugação para retirada da biomassa,
transferia-se para um eppendorf 1,5 mL de amostra (caldo fermentativo) e adicionava-se 1,5
mL de solução azul de Coomassie. Deixava-se a solução em repouso por 5 minutos,
transferindo-se em seguida para um cubeta de quartzo e efetuando-se as leituras de
absorbância nos comprimentos de ondas 465 nm e 595 nm, respectivamente. O
espectrofotômetro (Thermo Spectronic modelo Genesys 10UV) foi zerado com água
destilada. O conteúdo protéico foi obtido a partir de uma curva padrão na qual graficava-se a
relação de absorbância A595/A465 para diferentes concentrações de Albumina de Soro
Bovino (Fração V – Lote 110881/Sigma Aldrich-Ohio-USA).
3.5. Determinação da Atividade Quitosanolítica e da Atividade Específica
A atividade quitosanolítica foi obtida monitorando-se o aumento dos açúcares
redutores totais, utilizando-se o método DNS a partir de uma curva padrão de glicosamina.
Para a determinação da atividade quitosanolítica adicionaram-se, aos tubos de ensaios 500 µL

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
52
de substrato (solução 0,2 % de quitosana solubilizada em ácido clorídrico 1N) e 500 µL de
caldo bruto (contendo enzima), controlando-se o pH entre 6,0 e 6,5 e incubando-se o sistema
em um banho termostatizado com temperaturas da ordem de 55 ºC. Após o tempo de
incubação de 30 minutos no banho foi adicionado aos tubos de ensaio 2,5 mL de DNS,
aquecendo-se à ebulição em banho-maria por 10 minutos, resfriando-se, em seguida, em
banho de gelo. Após essa etapa, o sistema foi centrifugado e com o sobrenadante efetuou-se a
leitura no espectrofotômetro a 600 nm. Destaca-se, que antes da leitura da amostra zerava-se o
equipamento (branco do espectro) com 1,0 mL de água destilada acrescidos de 2,5 mL de
DNS e realizava-se o branco da enzima que consistia da adição de 500 µL de solução de
quitosana, completando-se até um 1,0 mL com água destilada. Uma unidade de atividade
quitosanolítica (A) foi definida como a quantidade de enzima necessária para produzir 1
µmol.min-1 de glicosamina por mL de extrato enzimático, nas condições de temperatura e pH
citadas, sendo o resultado expresso em (U.mL-1). A Atividade específica foi determinada pela
divisão entre a atividade da enzima (U.mL-1) e a concentração de proteína total (µg.mL-1),
sendo expressa em (U. µg-1).
3.6. Planejamento experimental
3.6.1. Planejamento de seleção de fatores Plackett & Burman
O planejamento experimental Plackett & Burman é uma técnica muito usada, para se
realizar triagem de fatores na otimização num determinado processo (Reddy et al., 2008), e
foi utilizado neste trabalho para identificar os fatores que influenciam de forma significativa
na produção da enzima quitosanase.
A Tabela 3 ilustra os níveis de cada variável (fator) utilizada no planejamento Plackett
& Burman, sendo: Concentração de Quitosana (Q), Peptona (PP), Extrato de Levedura (EL),
NaNO3 (NS), K2HPO4 (FP), KCl (CP), MgSO4.7H2O (SM), FeSO4 (SF). Já a Tabela 4 mostra
a matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3 repetições
na condição do ponto central.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
53
Tabela 3 – Níveis para os fatores e seus valores codificados.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1 0,1 0,2 0,2 0,1 0,05 0,025 0,025 0,0005
0 0,2 0,5 0,3 0,2 0,10 0,050 0,050 0,0010
1 0,3 0,8 0,4 0,3 0,15 0,075 0,075 0,0015
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1
2 1 1 -1 1 -1 -1 -1 1 -1 1 1
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1
5 1 1 -1 1 1 -1 1 -1 1 -1 1
6 1 1 1 -1 1 1 -1 1 1 -1 -1
7 -1 1 1 1 -1 1 1 -1 -1 1 1
8 -1 -1 1 1 1 -1 1 1 1 1 1
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1
11 -1 1 -1 -1 -1 1 1 1 1 1 -1
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1
13 0 0 0 0 0 0 0 0 0 0 0
14 0 0 0 0 0 0 0 0 0 0 0
15 0 0 0 0 0 0 0 0 0 0 0
Sendo: V11,V12,V13 = Variáveis inertes para cálculo do erro.
3.6.2. Planejamento Composto Central para a Otimização do Meio de Cultura para a
Produção da Enzima em Cultivo Submerso
Um dos problemas mais comuns para quem realiza experimentos é determinar a
influência de uma ou mais variáveis sobre outra variável de interesse (Lobato, 2003). Este

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
54
problema pode ser minimizado utilizando-se um planejamento experimental de modo a se
obter uma função, ou uma aproximação satisfatória para a mesma, de forma que se operando
sobre as variáveis de entrada produza-se como saída a resposta ou as respostas observadas
(Barros Neto, 2001; Silva, 2004).
Em um planejamento, inicialmente, deve-se definir os fatores e as respostas de
interesse. Tendo-se identificado os fatores e respostas define-se o objetivo que se pretende
alcançar com os experimentos para que se possa escolher o planejamento mais apropriado.
(Silva, 2004).
No presente trabalho foi utilizado um Planejamento Composto Central (CCD) com 22
experimentos na parte fatorial, repetição em triplicata no ponto central e quatro experimentos
na parte axial (planejamento do tipo estrela), para se investigar a influência dos fatores:
concentração de quitosana e concentração de FeSO4 que foram os dois fatores que
influenciaram a produção de quitosanase, conforme o planejamento Plackett & Burman de 12
ensaios e mais 3 repetições na condição do ponto central. Estes dois fatores foram estudados
em cinco diferentes níveis (-1,41; -1; 0; 1; 1,41) e um conjunto de 11 experimentos foi
realizado, conforme Tabela 5.
Tabela 5 – Matriz do planejamento Composto Central.
Ensaio Q SF
1 -1 -1
2 -1 1
3 1 -1
4 1 1
5 -1,41 0
6 1,41 0
7 0 -1,41
8 0 1,41
9 0 0
10 0 0
11 0 0
Os outros fatores foram mantidos nos níveis mais baixos (para evitar custos). Uma que
não influenciaram de forma significativa na produção da enzima quitosanase.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
55
A codificação dos fatores foi realizada de acordo com a seguinte equação (Ji et al.,
2009):
Xi = (xi – x0)/∆xi , i = 1, 2, 3, ..., k (2)
Onde Xi é o fator independente codificado, xi é o fator independente real, x0 é o valor de xi no
ponto central e ∆xi é a variação de valor. O comportamento do sistema foi explicado pela
seguinte equação polinomial de segunda ordem:
Yp = β0 + ∑ βiXi + ∑ βiiXi2 + ∑ βijXiXj, i, j = 1, 2, 3,..., k (3)
Onde Yp foi a resposta esperada, β0 foi a interseção, Xi e Xj
foram os fatores independentes codificados, βi foi o coeficiente linear, βii
foi o coeficiente quadrático e βij foi o coeficiente de interação. Destaca-se que os outros
fatores não significativos foram mantidos no nível mais baixo (-1).
A Tabela 6 apresenta os níveis para os fatores e seus valores codificados, para o
Planejamento Composto Central, com triplicata no ponto central.
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1,41 0,218 0,2 0,2 0,1 0,05 0,025 0,025 0,0009
-1 0,3 0,2 0,2 0,1 0,05 0,025 0,025 0,0015
0 0,5 0,2 0,2 0,1 0,05 0,025 0,025 0,0030
1 0,7 0,2 0,2 0,1 0,05 0,025 0,025 0,0045
1,41 0,782 0,2 0,2 0,1 0,05 0,025 0,025 0,0051
Na análise do planejamento de seleção de fatores Plackett & Burman e do
planejamento experimental fatorial foi utilizado um pacote computacional STATISTICA for
Windows versão 7.0.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
56
3.7. Perfil cinético durante o cultivo
Um perfil cinético na melhor condição de cultivo, conforme planejamento
apresentado, foi realizado em frascos Erlenmyer de 250 mL contendo 200 mL de meio de
cultivo contendo os seguintes componentes, em 1 Litro: 2,0g de quitosana; 5,0g de peptona;
3,0g de extrato de levedura; 2,0g de NaNO3; 1,0g de K2HPO4; 0,5g de KCl; 0,5g de
MgSO4.7H2O; 0,01g de FeSO4, que foram inoculadas com 10% (vol/vol) de inóculo contendo
o fungo Metarhizium anisopliae e cultivadas em incubador rotativo durante 5 dias a 150 rpm
e 25 ºC. Alíquotas foram retiradas a cada seis horas e analisadas quanto à proteína total no
sobredanante, atividade enzimática, atividade específica e açúcares redutores. Os ensaios
foram realizados em triplicata (Silva Filho, 2005).

Capítulo 4 Resultados e
Discussão

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
58
4. Resultados e discussão
Neste capítulo são apresentados e discutidos os resultados obtidos no presente
trabalho. Inicialmente, é apresentada imagem do MEV do Metarhizium anisopliae
mantidos em meio PDA. Em seguida, apresenta-se e discute-se os resultados da avaliação
dos fatores significativos, que podem influenciar na produção da quitosanase, no
planejamento de seleção de fatores, conforme planejamento Plackett & Burman bem como
são apresentados e discutidos os resultados obtidos para o planejamento composto central.
Por último, apresenta-se o perfil cinético, na melhor condição, para a produção de
quitosanase em cultivo submerso descontínuo usando o fungo Metarhizium anisopliae.
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA.
A Figura 7 ilustra a imagem da Microscopia Eletrônica de Varredura (MEV) do micélio de Metarhizium anisopliae mantido em meio PDA.
Figura 7. Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA.
Observa-se na Figura 7, quanto à morfologia que o Metarhizium anisopliae, apresenta-
se como um fungo filamentoso com as hifas intimamente entrelaçadas conforme citado por
Wang et al. 2002.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
59
4.2. Planejamento Plackett & Burman
4.2.1 Análise do Plackett & Burman com 8 fatores e 12 ensaios
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis, foi utilizado para avaliar a influência dos fatores: Concentração de
Quitosana (Q), Peptona (PP), Extrato de Levedura (EL), NaNO3 (NS), K2HPO4 (FP), KCl
(CP), MgSO4.7H2O (SM), FeSO4 (SF) na produção da enzima quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso. Ressalta-se que os experimentos foram
realizados aleatoriamente.
A Tabela 7 apresenta a matriz do modelo Plackett & Burman e os respectivos valores
obtidos para a variável resposta (Atividade Quitosanalítica).
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.
Ensaio Q PP EL NS FP CP SM SF A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 0,0195
2 1 1 -1 1 -1 -1 -1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 0,0109
4 1 -1 1 1 -1 1 -1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 0,0136
6 1 1 1 -1 1 1 -1 1 0,0156
7 -1 1 1 1 -1 1 1 -1 0,0109
8 -1 -1 1 1 1 -1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 0,0085
A Tabela 8 apresenta os efeitos estimados para o planejamento Plackett & Burman.
Observa-se na Tabela 8 que o valor do coeficiente de determinação R2 foi de
aproximadamente 90%, indicando que um modelo linear poderia representar bem a relação
entre o fator significativo, concentração de quitosana (1), e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
60
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman.
Efeitos Estimados; R2=0,8996; Ajuste:0,63178 MS Erro Residual =0,0000054
Desvio Padrão t(3) p
Lim. Conf. Lim. Conf.
Efeito Erro Residual
-95,% +95
Média/Interação 0,0127 0,0007 18,903 0,0003 0,0105 0,0149 (1) Quitosana 0,0055 0,0013 4,0628 0,0268 0,0011 0,0098
(2) Peptona 0,0015 0,0013 1,1080 0,3486 -0,0028 0,0058 (3) Ext. de leved. 0,0009 0,0013 0,7387 0,5136 -0,0033 0,0053
(4) NaNO3 0,0002 0,0013 0,1758 0,8715 -0,0040 0,0045 (5) K2HPO4 -0,0014 0,0013 -1,0552 0,3687 -0,0057 0,0028
(6) KCl -0,0022 0,0013 -1,6708 0,1933 -0,0065 0,0020 (7) MgSO4.7H2O -0,0003 0,0013 -0,2286 0,8338 -0,0046 0,0039
(8) FeSO4 0,0029 0,0013 2,1457 0,1211 -0,0014 0,0071
A Figura 8 mostra o gráfico de Pareto, uma das formas de se avaliar visualmente a
influência dos fatores estudados na resposta. A magnitude dos efeitos é representada pelas
colunas enquanto que a linha transversal às colunas representa a magnitude dos efeitos com
significado estatístico para p=0,05, ou seja, os fatores que são estatisticamente significativos
ao nível de 95% de confiança.
Figura 8. Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
61
Analisando-se o gráfico de Pareto (Figura 8) observa-se que o fator concentração de
quitosana (1) foi o único fator que influenciou de forma mais significativa na atividade
enzimática atingindo efeitos estimados de aproximadamente 4,06. O sinal positivo indica que
o aumento da concentração de quitosana aumentará à atividade enzimática. Por outro lado, os
outros fatores parecem desprezíveis estatisticamente conforme o gráfico de Pareto.
4.2.2 Análise do Plackett & Burman com 12 Ensaios e mais 3 Repetições na condição do
ponto central.
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis com repetição em triplicata no ponto central, foi utilizado para se avaliar a
influência dos mesmos fatores descritos na Seção 4.2.1 para a produção da enzima
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso. Ressalta-se que os
experimentos foram realizados aleatoriamente. A Tabela 9 apresenta a matriz do modelo
Plackett & Burman e os respectivos valores obtidos para a variável resposta (Atividade
Quitosanalítica).
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13 A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1 0,0195
2 1 1 -1 1 -1 -1 -1 1 -1 1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1 0,0109
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 1 -1 1 0,0136
6 1 1 1 -1 1 1 -1 1 1 -1 -1 0,0156
7 -1 1 1 1 -1 1 1 -1 -1 1 1 0,0109
8 -1 -1 1 1 1 -1 1 1 1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 1 1 -1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1 0,0085
13 0 0 0 0 0 0 0 0 0 0 0 0,0704
14 0 0 0 0 0 0 0 0 0 0 0 0,0688
15 0 0 0 0 0 0 0 0 0 0 0 0,0710

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
62
A Tabela 10 a seguir apresenta os efeitos estimados para planejamento Plackett &
Burman com 3 repetições na condição do poto central. Observa-se na Tabela 10 que o valor
do coeficiente de determinação R2 foi de aproximadamente 2,0%, indicando que um modelo
linear não poderia representar a relação entre os fatores significativos concentração de
quitosana (1), concentração de FeSO4 (8) e a resposta.
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central.
Efeitos Estimados; R2=0,01811; Ajuste:0,0 MS Erro Puro =0,0000012
Desvio Padrão t(2) P
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0242 0,0003 85,509 0,0001 0,0229 0,0254 (1) Quitosana 0,0055 0,0006 8,6432 0,0131 0,0027 0,0082
(2) Peptona 0,0015 0,0006 2,3572 0,1425 -0,0012 0,0042 (3) Ext. de leved. 0,0009 0,0006 1,5715 0,2567 -0,0017 0,0037
(4) NaNO3 0,0002 0,0006 0,3742 0,7442 -0,0024 0,0029 (5) K2HPO4 -0,0014 0,0006 -2,2450 0,1538 -0,0041 0,0013
(6) KCl -0,0022 0,0006 -3,5546 0,0708 -0,0049 0,0005 (7) MgSO4.7H2O -0,0003 0,0006 -0,4864 0,6747 -0,0030 0,0024
(8) FeSO4 0,0029 0,0006 4,5648 0,0448 0,0002 0,0056
A Figura 9 ilustra o gráfico de Pareto para o planejamento Plackett & Burman, com
repetição em triplicata no ponto central, uma das formas de se avaliar visualmente a influência
dos fatores estudados na resposta, onde é mostrado a magnitude dos efeitos para 95 % de
confiança.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
63
Figura 9. Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto
central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto, observa-se que os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma mais
significativa na atividade enzimática atingindo efeitos estimados de aproximadamente 8,84 e
4,67, respectivamente. O sinal positivo indica que o aumento das concentrações de quitosana
e de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores parecem
desprezíveis estatisticamente conforme o gráfico de Pareto.
Este comportamento do FeSO4 está de acordo com o observado por Hagag et al.
(2012) que utilizando o Aspergillus fumigatus na produção de protease mostraram que um
fator de transcrição PrtT regula positivamente o controle da produção de protease, da
assimilação de Ferro e da síntese de ergosterol e que a produção de protease foi favorecida
pelo aumento da concentração de ferro.
Rajendran & Thangavelu (2009) utilizaram um planejamento Plackett & Burman para
identificar quais os fatores significativos na produção de lipase por Rhizopus arrhizus MTCC
2233 e observaram o efeito significativo médio do FeSO4, apresentando um nível de
confiança para esse fator de 48,92% (para um máximo de 98,7 % de MgSO4), com uma
produção de lipase da ordem de 2,98 U/mL em 84 horas de fermentação e que ocorreu uma
produção considerável de protease no meio de cultivo da ordem de 1,92 U/mL em 96 horas de
cultivo.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
64
Mu et al. (2005) também observaram um efeito positivo do FeSO4, da ordem de 1,48
para um fator máximo encontrado de 3,11 de MgSO4, no aumento da atividade da
estereoinversão de S-fenil-1,2-etanodiol por Candida parapsilosis CCTCC M203011 na
otimização da fermentação utilizando um planejamento Plackett & Burman.
Dessa forma, observa-se que o FeSO4 é importante uma que vez o mesmo está
diretamente associado a síntese de proteases (principalmente metalloproteases), conforme
citado por Hagag et al. (2012).
4.3. Planejamento Composto Central para Produção de Quitosanase por
Metarhizium anisopliae em Cultivo Descontínuo Submerso
Um planejamento experimentos 22 com repetição em triplicata no ponto central e mais
quatro ensaios axiais, foram utilizados para se chegar a uma combinação ideal de quitosana e
FeSO4 na produção da enzima quitosanase (Tabela 11). A partir do modelo Plackett &
Burman (Tabela 12), que mostrou que o aumento das concentrações de quitosana e FeSO4,
favoreceram a produção de quitosanase, os valores utilizados neste modelo para estes
componentes nos níveis mais altos, serão considerados como os níveis mais baixos no
planejamento experimental do composto central. Os outros fatores foram fixados em todos os
ensaios em seus níveis mais baixos para a redução de custos (Tabela 11).

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
65
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da
atividade enzimática.
Ensaio Q SF A (U/mL)
1 -1 -1 0,0195
2 -1 1 0,0202
3 1 -1 0,0109
4 1 1 0,0121
5 -1,41 0 0,0136
6 1,41 0 0,0156
7 0 -1,41 0,0108
8 0 1,41 0,0103
9 0 0 0,0099
10 0 0 0,0116
11 0 0 0,0094
A Tabela 12 apresenta os efeitos estimados para o planejamento composto central.
Tabela 12 – Efeitos Estimados do planejamento Composto Central.
Efeitos Estimados; R2=0,629; Ajuste:0,25836 MS Erro Puro =0,0000014
Desvio Padrão t(2) p
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0103 0,0006 15,305 0,0042 0,0074 0,0132 (1) Quitosana (L) 0,0011 0,0008 1,4316 0,2885 -0,0023 0,0047
Quitosana (Q) 0,0058 0,0009 5,9637 0,0269 0,0016 0,0101 (2) FeSO4 (L) -0,0043 0,0008 -5,3037 0,0337 -0,0079 -0,0008 (2) FeSO4 (Q) 0,0018 0,0009 1,8366 0,2076 -0,0024 0,0060
1L x 2L 0,0001 0,0011 0,1520 0,8931 -0,0048 0,0052
Observa-se na Tabela 12 que o valor do coeficiente de determinação R2 foi de
aproximadamente 63%, indicando que um modelo linear representaria bem a relação entre os
fatores significativos concentração de quitosana (1), concentração de FeSO4 (8) e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
66
A Tabela 13 mostra um resumo da ANOVA para o planejamento composto central
com repetição em triplicata no ponto central.
Tabela 13 – Resumo da ANOVA.
Fonte de
Variação
Soma dos
Quadrados
Grau de
Liberdade
Quadrado
Médio
Fcalculado Ftabelado
Regressão
Resíduos
0,000086 2 0,000043 6,14 4,46
0,000056 8 0,000007
Falta de
Ajuste
Erro puro
0,000053 6 0,000009 6 19,33
0,000003
2
0,0000015
Total 0,000142 10
De acordo com a Tabela 13, partindo-se dos efeitos significativos propõe-se o
modelo:
� = 0,010316 + 0,002942� � − 0,002193�� (8)
% de variação explicável: 60,5
% máxima de variação explicável: 97,9
A análise da ANOVA mostra que o modelo proposto possui significância estatística, ao
nível de 95% de confiança, uma vez que o valor de F (calculado) é maior que F (tabelado),
entretanto a ANOVA mostra também que o modelo proposto pode ser útil para se fazer
previsões, uma vez que a relação entre F (calculado) e F (tabelado) para a falta de ajuste e o
erro puro é menor que 1. O coeficiente de determinação R2 foi de aproximadamente 63%,
indicando que o modelo consegue explicar 63% da variação total em torno da média.
Os cálculos para variação explicável e para máxima variação explicável são dados pelas
Equações (9) e (10):
% de variação explicável = (SQ da regressão) / (SQ Total) (9)
% máxima de variação explicável = (SQ Total – SQ Erro puro) / (SQ Total) (10)
Sendo:
SQ – Soma dos quadrados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
67
A Figura 10 mostra o gráfico de Pareto para o planejamento Composto Central
utilizando erro puro. A magnitude dos efeitos é representada pelas colunas. Já a linha
transversal às colunas representa a magnitude dos efeitos com significado estatístico para 95%
de confiança.
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto (Figura 10) observa-se que os fatores concentração
de quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma
mais significativa na atividade enzimática atingindo efeitos estimados de aproximadamente
5,96 e - 5,30, respectivamente. O sinal positivo indica que o aumento das concentrações de
quitosana aumentará à atividade enzimática. O sinal negativo indica que o decréscimo da
concentração de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores
parecem desprezíveis estatisticamente no gráfico de Pareto.
A avaliação do modelo também pode ser feita através da observação do gráfico dos
valores preditos em função os valores observados que são mostrados na Figura 11. Os valores
preditos pelo modelo são representados pela reta, enquanto que os valores observados
representam-se pelos pontos.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
68
Figura 11 – Valores preditos em função dos valores observados.
Observa-se que alguns valores preditos apresentam um afastamento dos valores
observados. A Figura 12 ilustra a superfície de resposta da atividade quitosanolítica em
função dos dois fatores significativos, concentração de quitosana e concentração de FeSO4.
Figura 12. Atividade quitosanolítica em função da concentração de quitosana e da
concentração de FeSO4 para os fatores não significativos no ponto central.
Observa-se a existência de um ponto de mínimo, ocorrendo para concentração de
quitosana no ponto central (0) e para maiores concentrações de FeSO4 igual a (1,5). Essa

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
69
região deve ser evitada uma vez que a atividade quitosanolítica atinge níveis tão reduzidos
quanto 0,0094 U.mL-1. Destaca-se que nesse planejamento o maior nível da concentração de
FeSO4 (α=+1,41) equivale a 0,0051% (Tabela 6) que é aproximadamente cinco vezes maior
que o nível desse fator no ponto central do planejamento Plackett & Burman (0,001%),
conforme Tabela 3. Dessa forma, ao se utilizar o nível mínimo da concentração de FeSO4 no
Planejamento Composto Central (α=-1,41), que equivale a aproximadamente 0,001%,
aproxima-se do ponto central do do planejamento Plackett & Burman (0,001%), condição
essa que apresenta os melhores resultados de atividade quitosanolítica usando quitosana como
fonte de carbono e energia e fungo Metarhizium anisopliae.
4.4. Perfil Cinético – Melhor Condição
A seguir, apresenta-se um perfil cinético, na melhor condição, para produção da
quitosanase usando o fungo Metarhizium anisopliae, em que foram monitorados durante 24
horas os seguintes parâmetros: Concentração de proteínas totais (µg/mL), atividade
quitosanolítica (U/mL), atividade específica (U/µg) e açúcares redutores (mg/mL), ilustradas
nas Figuras 13, 14, 15 e 16, respectivamente.
Com relação à proteína extracelular, observa-se na Figura 13 que, inicialmente, ocorre
um aumento da mesma para o seu nível máximo (6,2 µg/mL), durante as 6 primeiras horas de
cultivo, seguido de uma redução para 5,7 µg/mL em 18 horas de cultivo, possivelmente
devido à presença de proteases no meio. Observa-se que a partir de então ocorre uma redução
e aumento da proteína total, em uma quantidade inferior, sugerindo um mecanismo de
controle.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
70
0 4 8 12 16 20 245,0
5,2
5,4
5,6
5,8
6,0
6,2
6,4
Pro
tein
a (
µg
/mL
)
Tempo (h)
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana.
Com relação à atividade quitosanolítica, conforme Figura 14, a atividade máxima
ocorreu em 18 horas de produção em que se obteve aproximadamente 70,0 U/L, o que foi 28
vezes maior que o valor (2,5 U/L em 48 horas) relatado por Assis (2010) para o mesmo fungo,
e em apenas um terço do tempo de produção.
0 4 8 12 16 20 240,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,10
Ativid
ade
(U
/mL
)
Tempo (h)
Figura 14. Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
71
Além disso, destaca-se o considerável aumento da produção de quitosanase a partir das
12 horas até 18 horas de cultivo onde atingiu o pico máximo de produção (70,0 U/L). Após o
pico em 18 horas, a produção começa a cair rapidamente, comportamento esse observado
durante todos os ensaios cinéticos. Destaca-se que é importante do ponto de vista industrial
manter a produção em alto nível por muitas horas, onde se espera uma produção contínua ou
em grandes bateladas.
Ak et al. (1998) utilizaram o fungo Penicillium spinulosum para produzir a quitosanase
usando um meio de cultura contendo 0,1% de K2HPO4; 0,05 % de MgSO4.7H2O; 0,05% de
KCl; 0,001 % de FeSO4, 0,5% NH4NO3 e como fonte de carbono e energia, os autores
usaram quitosana e parede de Rhizopus. A produção foi realizada em cultivo submerso a 28
oC. Os autores apresentam atividade quitosanolítica máxima de aproximadamente 47,0 U/mL,
ao se utilizar a parede do fungo como fonte de carbono e energia, e 30,0 U/mL ao se utilizar
quitosana como fonte de carbono e energia. Destaca-se que essas atividades ocorreram após 4
dias de cultivo. Os perfís com o tempo da atividade quitosanolítica e da atividade específica
apresentaram tendência de crescimento, com o pico máximo em 4 dias, seguindo com uma
tendência de queda da atividade. Neste tempo, a atividade específica máxima da quitosanase
foi de aproximadamente 60,0 U/mg de proteína. Essas tendências também foram observadas
nos resultados apresentados nos perfís de atividade das Figuras 14 e 15.
0 4 8 12 16 20 24
-0,010
-0,005
0,000
0,005
0,010
0,015
0,020
0,025
0,030
Ativid
ad
e E
spe
cific
a (
U/µ
g)
Tempo (h)
Figura 15. Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
72
Observa-se na Figura 15 que a atividade específica máxima da quitosanase segue a
mesma tendência da atividade quitosanolítica apresentada na Figura 14, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
O perfil para os açúcares redutores durante o cultivo de Metarhizium anisopliae
cultivado em meio contendo 0,2% de quitosana é ilustrado na Figura 16.
0 4 8 12 16 20 240,004
0,008
0,012
0,016
0,020
0,024
0,028
Acuca
res R
ed
uto
res (
mg/m
L)
Tempo (h)
Figura 16. Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana.
Observa-se, na Figura 16, que taxa máxima de redução dos açúcares redutores
(aproximadamente 0,021 mg/mL) ocorre logo após a produção máxima da enzima, em 18
horas de cultivo, conforme Figura 14. A quantidade de açúcares redutores foi a
aproximadamente 0,012 mg/mL após 24 h de cultivo. O fungo excreta para o meio enzimas
com atividade quitosanolítica, essas enzimas hidrolisam a quitosana aumentando a quantidade
de açúcares redutores para o meio, sendo que esses açúcares também são consumidos pelo
fungo durante seu crescimento.
Foi demonstrado por Assis (2010) que os açúcares redutores produzidos através da
quitossana, ocasionada pelas enzimas quitossanases metabolizadas por Metarhizium
anisopliae, com quitosana como sua única fonte de carbono e energia, atingiu um pico de 0,53
mg/mL em 48 horas. Apresentaram como quitooligossacarídeos, constituídos principalmente
por dímeros e pentâmeros, formados pela hidrólise enzimática de quitosanase em quantidades
maiores do que o consumido pelos microrganismos. Mostraram, também, que essa produção

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
73
segue a mesma tendência da sua atividade enzimática, corroborando com os resultados
apresentados no presente trabalho.
Os resultados do presente trabalho mostram a relevância de se adotar novas condições
para a produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter quitooligossacarídeo
com atividades biológicas. Esta pesquisa, portanto, sugere base para novos estudos que
permitam alcançar rapidamente a produção de quitosanase e quitooligossacarídeo com todos
os seus benefícios.

Capítulo 5 Conclusão

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
75
5. Conclusão
Após o desenvolvimento desse trabalho conclui-se:
- Com relação ao Planejamento de Seleção de Fatores Plackett & Burman:
Os resultados das análises do Planejamento de Seleção de Fatores Plackett &
Burman com 8 fatores e 12 ensaios em 2 níveis, mais 3 repetições no ponto central, para
se investigar a influência dos fatores: Concentração de Quitosana, Peptona, Extrato de
Levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O, FeSO4 na produção da enzima
quitosanase por Metarhizium anisopliae, mostraram que a atividade foi favorecida pelo
aumento da concentração de substrato (quitosana) e sulfato ferroso (FeSO4), enquanto
que o aumento da concentração dos outros fatores não contribuíram de forma
significativa para a resposta (atividade quitosanolítica). Os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) atingiram efeitos estimados de
aproximadamente 8,84 e 4,67, respectivamente. O sinal positivo indica que o aumento
das concentrações de quitosana e de FeSO4 aumentará a atividade enzimática. Conforme
visto na literatura, observa-se importância do FeSO4, já que o mesmo está diretamente
associado a síntese de proteases (principalmente metalloproteases),
- Com relação ao Planejamento Composto Central:
A análise do planejamento composto central para avaliar a influência dos
fatores, concentração de substrato (quitosana) e concentração de FeSO4, na produção da
enzima quitosanase por Metarhizium anisopliae (a partir do planejamento Plackett &
Burman) mostrou que os fatores concentração de quitosana e concentração de FeSO4
foram os fatores que influenciaram de forma mais significativa na atividade enzimática
atingindo efeitos estimados de aproximadamente 5,96 e - 5,30, respectivamente. O sinal
positivo indica que o aumento das concentrações de quitosana aumentará a atividade
enzimática. O sinal negativo indica que o decréscimo da concentração de FeSO4
aumentará a atividade enzimática. Observa-se a existência de um ponto de mínimo,
ocorrendo para concentração de quitosana (X1) no ponto central (0) e para maiores

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
76
concentrações de FeSO4 (X2) igual a (1,5) com a atividade quitosanolítica de 0,0094
U.mL-1.
- Com relação ao cultivo na melhor condição:
O cultivo na melhor condição mostrou uma produção máxima de quitosanase em
18 horas de cultivo, com a atividade enzimática de 70,0 U/L, o que foi 28 vezes maior
que o valor (2,5 U/L em 48 horas) relatado na literatura (Assis, 2010) para o mesmo
fungo e em apenas um terço do tempo de produção.
Além disso, observou-se que um elevado incremento na produção de
quitosanase a partir das 12 horas de cultivo, antes de se atingir a atividade
quitosanolítica máxima, 18 horas de cultivo. Essa produção em alto nível por muitas
horas é importante do ponto de vista industrial, onde se espera uma produção contínua
ou em grandes bateladas. Observou-se também que a atividade específica máxima da
quitosanase segue a mesma tendência da atividade quitosanolítica, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
- Com relação aos açúcares redutores:
Foi observado que os açúcares redutores seguem a mesma tendência da atividade
enzimática, com uma produção máxima de cerca de 0,21 mg/mL em 18 horas de
cultivo, produção essa que coincinde com o máximo pico de atividade enzimática.
Estes resultados mostram a relevância de se adotar novas condições para a
produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter
quitooligossacarídeo com atividades biológicas. Esta pesquisa, portanto, sugere base
para novos estudos que permitam alcançar rapidamente a produção de quitosanase e
quitooligossacarídeo com todos os seus benefícios.

Referências Bibliográficas

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
78
Referências bibliográficas
AK, Ö.; BAKIR, U.; GÜRAY, T. Isolation of a chitosan degrading fungus, Penicillium
spinulosum, and chitosanase production by the isolate. In: SYMPOSIUM - NEW
FRONTIERS IN SCREENING FOR MICROBIAL BIOCATALYSTS, 1996, Ede, The
Netherlands. Anais, Editado por K. Kieslich, C. P. van der Beek, J. A. M. de Bont e W.
J. J. van den Tweel, Elsevier Science B. V., 1998, v.53, p.265-268.
ALVES, S. B. Patologia e controle microbiano: Vantagens e desvantagens. In: ALVES,
S. B. Controle Microbiano de Insetos. 2ed., Piracicaba, FEALQ, 1163p, 1998, v.4, p.21-
37.
ALVES, S. B.; LEITE, L. G.; FILHO, A. B.; ALMEIDA, F. E. M.; MARQUES, E. J.
Produção massal de fungos entomopatogênicos na América Latina, In: ALVES, S. B.;
LOPES, R. B. Controle microbiano de pragas na América Latina: avanços e desafios.
Piracicaba, FEALQ, 414p, 2008, v.14, p.215-237.
ARRUDA, W.; LUBECK, I.; SCHRANK, A.; VAINSTEIN, M. H. Morphological
alterations of Metarhizium anisopliae during penetration of Boophilus microplus ticks.
Experimental and Applied Acarology, v. 37, p.231-244, 2005.
ASSIS, C. F. Produção e caracterização de quitooligossacarídeos produzidos pelo fungo
Metarhizium anisopliae e avaliação da citotoxicidade em células tumorais. 2010. 110p.
Tese (Doutorado em Biotecnologia) – Renorbio - Programa de Pós-Graduação em
Biotecnologia. Universidade Federal do Rio Grande do Norte, Natal.
ASSIS, C. F.; COSTA, L. S.; MELO-SILVEIRA, R. F.; OLIVEIRA, R. M.;
PAGNONCELLI, M. G. B.; ROCHA, H. A. O.; MACEDO, G. R.; SANTOS, E. S.,
Chitooligosaccharides antagonizes the cytotoxic effect of glucosamine. World J.
Microbiol. Biotechnol., v.28, n.3, p.1097-1105, 2012.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
79
AZEVEDO, V. V. C.; CHAVES, S. A.; BEZERRA, D. C.; FOOK, M. V. L.; COSTA,
A. C. F. M. Quitina e Quitosana: aplicações como biomateriais. Revista Eletrônica de
Materiais e Processos, v.2.3, p.27-34, 2007.
BARROS NETO, B.; SCARMINIO, I. S.; BRUNS, R. Como fazer experimentos:
pesquisa e desenvolvimento na ciência e na indústria. 2. ed. Campinas: Unicamp, 2001.
401p.
BATISTA FILHO, A.; ALMEIDA, J. E. M.; SANTOS, A. S.; MACHADO, L. A.;
ALVES, S. B. Eficiência de isolados de Metarhizium anisopliae no controle de
cigarrinha-da-raiz de cana-de-açúcar Mahanarva fimbriolata (Hom.: Cercopidae). STAB
– Açúcar, Álcool e Subprodutos, v.21, p.84-89, 2002.
CARVALHO, J. C. M.; SATO, S. Fermentação Descontínua, In: SCHMIDELL, W.;
LIMA, U. A.; AQUARONE, E.; BORZANI, W. Biotecnologia Industrial. São Paulo,
Ed. Edgard Blücher, v.2, p.193-204, 2001.
CHAO, H. J.; SCHWARTZ, J.; MILTON, D. K. Populations and determinats of
airborne fungi in large office buildings. Environmental Health Perspectives, v.110, n.8,
p.777-782, 2002.
CHEN, X.; XIA, W.; YU, X. Purification and characterization of two types of
chitosanase from Aspergillus sp. CJ22-326. Food Research International, v.38, n.3,
p.315-322, 2005.
CHEN, P. T.; CHIANG, C.-J.; CHAO, Y.-P. Medium optimization and production of
secreted Renilla luciferase in Bacillus subtilis by fed-batch fermentation. Biochemical
Engineering Journal, v.49, p.395-400, 2010.
CHENG, C. Y.; LI, Y. An Aspergillus chitosanase with potential for large-ecale
preparation of chitosan oligosaccharides. Biotechnol. Appl. Biochem., v.32, p.197-203,
2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
80
CHOI, Y. J.; Kim, E. J.; Piao, Z.; Yun, Y. C.; Shin, Y. C. Purification and
characterization of chitosanase from Bacillus sp. Strain KCTC 0377BP and its
application for the production of chitosan oligosaccharides. Applied and Environmental
Microbiology, v.70, n.8, p.4522-4531, 2004.
COLAKOGLU, G. The chemical and physical environment for fungal growth. Fak
Matbaa Birimi, Istanbul, 149-166, 2001.
CRAVEIRO, A. C.; CRAVEIRO, A. A.; QUEIROZ, D. C. Quitosana a fibra do futuro.
2. ed. Fortaleza: PADETEC, 2004. V.1, 218p.
DIAS, B. A. Estudo de proteases degradadoras de cutícula produzidas pelo fungo
entomopatogênico Beauveria bassiana. 2005. 70p. Dissertação (Mestrado em
Microbiologia) – Centro de Ciências Biológicas, Programa de Pós-Graduação em
Microbiologia, Universidade Estadual de Londrina, Londrina.
DINARDO-MIRANDA, L. L.; FERREIRA, J. M. G.; CARVALHO, P. A. M.
Influência da época de colheita e do genótipo de cana-de-açúcar sobre a infestação de
Mahanarva fimbriolata (Stal) (Hemiptera: Cercopidae). Neotrop. Entomol, v.30, n.1,
p.145-149, 200.
DONG, C.; ZHANG, J.; HUANG, H.; CHEN, H.; HU, Y. Pathogenicity of a new China
variety of Metarhizium anisopliae (M. anisopliae var. Dcjhyium) to subterranean termite
Odontotermes formosanus. Microbiological Research, v.164, p.27-35, 2009.
DONZELLI, B. G. G.; OSTROFF, G.; HARMAN, G. E. Enhanced enzymatic
hydrolysis of langostino shell chitin with mixtures of enzymes from bacterial and fungal
sources. Carbohydrate Research, v.338, p.1823-1833, 2003.
DRIVER, F.; MILNER, R. J.; TRUEMAN, J. W. H. A taxonomic revision of
Metarhizium based on phylogenetic analysis of rDNA sequence data. Mycological
Research, v.104, n.2, p.134-150, 2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
81
DUNG, P.; MILAS, M.; RINAUDO, M.; DESBRIÉRES, J. Water soluble derivatives
obtained by controlled chemical modifications of chitosan, Carbohydrate Polymers,
v.24, p.209-214, 1994.
ESTEVES, J. A.; CABRITA, J. D.; NOBRE, G. N. Micologia médica, 2. ed. Lisboa:
Fundação Calouste Gulbenkian, 1990. 1058p.
FACCIOTI, M. C. R. Fermentação Contínua, In: Biotecnologia Industrial. Editado por:
SCHMIDELL, W.; LIMA, U. A.; AQUARONE, E.; BORZANI, W., São Paulo, Ed.
Edgard Blücher, v.2, p.541, 2001.
FARIA, M. R.; MAGALHÃES, B. P. O uso de fungos entomopatogênicos no Brasil -
Situação atual e perspectivas. Biotecnologia Ciência & Desenvolvimento, n.22., p.18-
21, setembro/outubro 2001.
FU, J.-Y.; WU, S.-M.; CHANG, C.-T.; SUNG, H.-Y. Characterization of Three
Chitosanase Isozymes Isolated from a Commercial Crude Porcine Pepsin Preparation. J.
Agric. Food Chem., v.51, p.1042-1048, 2003.
FURIGO JUNIOR, A.; PEREIRA, E. P. Enzimas e suas aplicações cinética enzimática.
2001. 39f. Apostila (Graduação de Engenharia Química e Engenharia de Alimentos) –
Centro de Tecnologia, Departamento de Engenharia Química e Engenharia de
Alimentos, Universidade Federal de Santa Catarina, Florianópolis.
GAO, J.; XU, H.; LI, Q.-J.; FENG, X.-H.; LI, S. Optimization of medium for one-step
fermentation of inulin extract from Jerusalem artichoke tubers using Paenibacillus
polymyxa ZJ-9 to produce R,R-2,3-butanediol, Bioresource Technology, v.101, p.7076-
7082, 2010.
GARCIA, M. V.; MONTEIROI, A. C.; SZABO, M. J. P.; PRETTE, N.; BECHARA, G.
H. Mechanism of infection and colonization of Rhipicephalus sanguineus eggs by
Metarhizium anisopliae as revealed by scanning electron microscopy and
histopathology. Brazilian Journal of Microbiology, São Paulo, v.36, n.4, p.368-372,
2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
82
GHANEM, N. B.; YUSEF, H. H.; MAHROUSE, H. K. Production of Aspergillus
terreus xylanase in solid-state cultures: application of the Plackett & Burman
experimental design to evaluate nutritional requirements, Bioresource Technology,
v.73, p.113-121, 2000.
GHORBEL-BELLAJ, O.; HMIDET, N.; JELLOULI, K.; YOUNES, I.; MAALEJ, H.;
HACHICHA, R.; NASRI, M. Shrimp waste fermentation with Pseudomonas aeruginosa
A2: Optimization of chitin extraction conditions through Plackett–Burman and response
surface methodology approaches, International Journal of Biological Macromolecules,
v.48, p.596–602, 2010.
GIUSTINA, A.; VENTURA, P. Weight-reducing regimens in obese subjects: Effects of
a new dietary fiber integrator. Acta Toxicol Ther, 16, 4, p.199-214, 1995.
GUIA.BIO, Reino dos fungos, 2012. Disponível em:
<http://www.http://guia.bio.br/reino-dos-fungos>. Acesso em: 12 novembro 2012.
GUIRADO, N.; AMBROSANO, E. J.; ROSSI, F.; DIAS, F. L. F. Controle de Cupins
de Montículo com Metarhizium anisopliae e Beauveria bassiana, Rev. Bras. de
Agroecologia, v.4, n.2, 2009.
GUO, Y.; XU, J.; ZHANG, Y.; XU, H.; YUAN, Z.; LI, D. Medium optimization for
ethanol production with Clostridium autoethanogenum with carbon monoxide as sole
carbon source, Bioresource Technology, v.101, p.8784–8789, 2010.
HAGAG, S.; KUBITSCHEK-BARREIRA, P.; NEVES, G. W. P.; AMAR, D.;
NIERMAN, W.; SHALIT, I.; SHAMIR, R.; LOPES-BEZERRA, L.; OSHEROV, N.
Transcriptional and proteomic analysis of the Aspergillus fumigatus ∆prtT protease-
deficient mutant. PLoS One, v.7, n.4, e33604, 2012.
HAN, L.-K.; KIMURA, Y.; OKUDA, H. Reduction in fat storage during chitin-chitosan
treatment in mice fed a high-fat diet, International Journal of Obesity, 23:174-179,1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
83
HUNG, T.; CHANG, Y.; SUNG, H.; CHANG, C. Purification and Characterization of
Hydrolase with Chitinase and Chitosanase Activity from Commercial Stem Bromelain.
J. Agric. Food Chem., 50, 4666-4673, 2002.
ICHIKAWA, S.; TAKANO, K.; TAKASHIKUROIWA, HIRUTA, O.; SATO, S.;
MUKATAKA, A. S. Immobilization and stabilization of chitosanase by multipoint
attachment to agar gel support. Journal of Bioscience and Bioengineering, v.93, n.2,
p.201-206, 2002.
JABOR, I. A. S.; PAMPHILE, J. A.; RODRIGUES, S. B.; MARQUES-SILVA, G. G.;
ROCHA, C. L. M. S. Análise do desenvolvimento do fungo entomopatogênico
Metarhizium anisopliae em resposta a fatores nutricionais. Acta Scientiarum.
Agronomy, Maringá, v.25, n.2, p.497-501, 2003.
JI, X.-J.; HUANG, H.; DU, J.; ZHU, J.-G.; REN, L.-J.; LI, S.; NIE, Z.-K. Development
of an industrial medium for economical 2,3-butanediol production through co-
fermentation of glucose and xylose by Klebsiella oxytoca, Bioresource Technology,
v.100, p.5214–5218, 2009.
JO, Y.; JO, K.; JIN, Y.; KIM, K.; SHIN, J.; KIM, Y.; PARK, R. Characterization and
kinetics of 45 kDa chitosanase from bacillus sp. P16. Biosci. Biotechnol. Biochem.,
v.67, n.9, 1875 – 1882, 2003.
KHOSRAVI-DARANI, K.; ZOGHI, A.; ALAVI, S. A.; FATEMI, S. S. A. Application
of Plackett Burman Design for Citric Acid Production from Pretreated and Untreated
Wheat Straw, Iran. J. Chem. Chem. Eng.. v.27, n.1, p.91-104, 2008.
KIM, P.; KANG, T. H.; CHUNG, K. J.; KIM, I. S.; CHUNG, K. Purification of a
constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and
expression of the gene. FEMS Microbiolog.y Letters 240, 31–39, 2004.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
84
KIM, S.-K.; RAJAPAKSE, N. Enzymatic production and biological activities of
chitosan oligosaccharides (COS): A review. Carbohydrate Polymers, v.62, n.4, p.357-
368, 2005.
KIRK, P. M.; CANON, P.F.; MINTER, D.W.; STALPERS, J.A. Dictionary of the
fungi. CAB International, Wallingford, 2008.
KONDO, H.; OSADA, A. Influence of dietary fiber on the bioavailability of zinc in
rats, Biomed Environ Sci, 9, 204-8, 1996.
KUMAR, S.; KOH, J. Physiochemical, Optical and Biological Activity of Chitosan-
Chromone Derivative for Biomedical Applications. Int. J. Mol. Sci., v.13, p.6102-6116,
2012.
KURITA, K.; SANNAN, T.; IWAKURA, Y. Studies on chitin, 4. Evidence for
formation of block and random copolymers of N-acetyl-o-glucosamine and
oglucosamine by hetero and homogeneous hydrolyses. Die Makromolecular Chemie,
v.178, p.3197-3202, 1977.
KUROIWA, T.; ICHIKAWA, S.; HIRUTA, O.; SATO, S.; MUKATAKA, S. Factors
Affecting the Composition of Oligosaccharides Produced in Chitosan Hydrolysis Using
Immobilized Chitosanases. Biotechnol. Prog., 18, 969-974, 2002.
LI, L.-Z.; HUI, Z.; MO, X. Optimization of the medium for streptolydigin production
by Streptomyces lydicus AS 4.2501 with precursor feeding using response surface
methodology, Journal of the Taiwan Institute of Chemical Engineers, 41, 252–258,
2010.
LIU, B.-L.; KAO, P.-M.; TZENG, Y.-M.; FENG, K.-C. Production of chitinase from
Verticillium lecanii F091 using submerged fermentation. Enzyme and Microbial
Technology, v.33, p.410-415, 2003.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
85
LOBATO, A. K. C. L. Estudo da produção de biosurfactantes por micro-organismos
isolados de poços de petróleo. 2003. 108p. Dissertação (Mestrado) – Programa de Pós-
Graduação em Engenharia Química. Universidade Federal do Rio Grande do Norte.
Natal.
LOPES, R. B.; ALVES, S. B.; PADULLA, L. F. L.; PÉREZ, C. A. Eficiência de
formulações de Beauveria bassiana e Metarhizium anisopliae para o controle de ninfas
de Amblyomma cajennense (Fabricius, 1787). Revista Brasileira de Parasitologia
Veterinária, v.16, n.1, p.27-31, 2007.
LOUREIRO, E. S.; BATISTA FILHO, A.; ALMEIDA, J. E. M.; MENDES, J. M.;
PESSOA, L. G. A. Eficiência de isolados de Metarhizium anisopliae (metsch.) sorok.
no controle da cigarrinha-da-raiz da cana-de-açúcar, Mahanarva fimbriolata (stal, 1854)
(hemiptera: cercopidae), em condições de campo. Arq. Inst. Biol., São Paulo, v.79, n.1,
p. 47-53, 2012.
MELLO, D. R.; REIS, R. C. S.; BITTENCOURT, V. R. E. P. Patogenicidade in vitro
do fungo Metarhizium anisopliae (Metschnikoff, 1879) Sorokin, 1883, sobre o carrapato
Boophilus microplus (Canestrini, 1887). Revista Brasileira de Parasitologia Veterinária,
São Paulo, v.15, p.157-162, 2006.
MITSUTOMI, M.; KIDOH, H.; TOMITA, H.; WATANABE, T. The action of Bacillus
circulans WL-12 chitinases on partially N-acetylated chitosan, Bioscience
Biotechnology and Biochemistry, v.59, p.529-531, 1995.
MALAJOVICH, M. A. Biotecnologia: fundamentos. Ed. biblioteca Max Feffer do
Instituto de Tecnologia ORT, Rio de Janeiro, 2009. 104p.
MONAGHAN, R. L.; EVELEIGH, D. E.; TEWARI, R. P.; REESE, E. T. Chitosanase,
a novel enzyme. Nature - New Biology, 245, p.78-80, 1973.
MU, X. Q.; XU, Y.; NIE, Y.; OUYANG, J.; SUN, Z. H. Candida parapsilosis CCTCC
M203011 and the optimization of fermentation medium for stereoinversion of (S)-1-
phenyl-1,2-ethanediol, Process Biochemistry, v.40, p.2345–2350, 2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
86
MURRAY, P. R.; ROSENTHAL, K. S.; PFALLER, M. A. Medical microbiology. 5th
ed. London, Elsevier Mosby, 2005.
MUZZARELLI, R. A. A. Biochemical significance of exogenous chitins and chitosans
in animals and patients, Carbohydrate Polymers, v.20, p.7-16, 1993.
MUZZARELLI, R. A. A.; TOMASETTI, M.; ILARI, P. Depolymerization of chitosan
with the aid of papain, Enzyme Microbiology and Technology, v.16, p.110-114, 1994.
NAVEENA, B. J.; ALTAF, Md.; BHADRIAH, K.; REDDY, G. Selection of medium
components by Plackett–Burman design for production of L(+) lactic acid by
Lactobacillus amylophilus GV6 in SSF using wheat bran, Bioresource Technology,
v.96, p.485-490, 2005.
NOGAWA, M.; TAKAHASHI, H.; KASHIWAGI, A.; OHSHIMA, K.; OKADA, H.;
MORIKAWA, Y. Purification and Characterization of Exo--D-Glucosaminidase from
a Cellulolytic Fungus, Trichoderma reesei PC-3-7, Appl. Environ. Microbiol., v.64, n.3,
p.890-895, 1998.
ONOFRE, S. B.; VARGAS; L. R. B.; ROSSATO, M.; BARROS, N. M.; BOLDO, J.
T.; NUNES, A. R. F.; AZEVEDO, J. L. Controle biológico de pragas na agropecuária
por meio de fungos entomopatogênicos. In: SERAFINI, L. A.; BARROS, N. M.;
AZEVEDO, J. L. (Org.). Biotecnologia – avanços na agricultura e na agroindústria.
Caxias do Sul: EDUCS, p.295-328, cap.8, 2002.
OSSWALD, W. F.; SHAPIRO, J. P.; DOOSTDAR, H.; MCDONALD, R. E.; NIEDZ,
R. P.; NAIRN, C. J.; HEARN, C. J.; MAYER, R. T. Identification and Characterization
of Acidic Hydrolases with Chitinase and Chitosanase Activities from Sweet Orange
Callus Tissue, Plant Cell Physiol. 35, p.811-820, 1994.
PAGNONCELLI, M. G. B. Estudo do mecanismo de produção de oligossacarídeos com
atividades nutracêuticas a partir da quitosana por hidrólise enzimática com processo

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
87
fermentativo simultâneo. 2008. 118p. Tese (Doutorado) – Centro de Tecnologia,
Programa de Pós-Graduação em Engenharia Química. Universidade Federal do Rio
Grande do Norte. Natal.
PALMA-GUERRERO, J., JANSSON, H.-B., SALINAS, J.; LOPEZ-LLORCA, L. V.
Effect of chitosan on hyphal growth and spore germination of plant pathogenic and
biocontrol fungi, J. Appl. Microbiol., v.104, p.541-553, 2008.
PAULA, F. A.; ARRUDA, D. C. Análise do método de planejamento experimental
Plackett Burman na otimização de degradação de penas de aves por comunidades
microbianas, XXI Salão de Iniciação Científica, 19-23, outubro, 2009, UFRGS, Porto
Alegre.
PELCZAR, M. J.; CHANG, E. C. S.; KRIEG, N. R. Microbiologia: conceitos e
aplicações, v.2, 2.ed. São Paulo: Makron BooKs, 1996.
PELLETIER, A.; SYGUSCH, J. Purification and characterization of three chitosanase
activities from Bacillus megaterium P1, Applied and Environmental Microbiology, v.
56, n.4, p.844-848, 1990.
PEREIRA, F. B.; GUIMARÃES, P. M. R.; TEIXEIRA, J. A.; DOMINGUES, L.
Optimization of low-cost medium for very high gravity ethanol fermentations by
Saccharomyces cerevisiae using statistical experimental designs, Bioresource
Technology, v.101, p.7856-7863, 2010.
PETER, M. G. Chitin and chitosan from aninal sources. In: STEINBUCHEL, A.;
RHEE, S. K. Polysaccharides and Polyamides in the Food Industry. Weinheim:
Wileyvch, v.1, p.115-208, 2005.
PIZA, F. A. T.; SILOTO, A. P.; CARVALHO, C. V.; FRANCO, T. T. Purification,
characterization and purification of chitosanase from Bacillus cereus, Brazilian Journal
of Chemical Engineering., v.16, n 2, p.185-192, 1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
88
POLITZER, K.; BON, E. P. S. Enzimas Industriais e Especiais. C.G.E.E. Ciência,
Tecnologia e Inovação. Rio de Janeiro, 554p, 2006.
POLYMAR, Ciência é a nossa Natureza, 2003. Disponível em:
<http://www.polymar.com.br>. Acesso em: 6 setembro 2012.
PUTZKE, J.; PUTZKE, M. T. L. Os reinos dos fungos. Santa Cruz do Sul, RS:
EDUNISC, 1998.
QUINES, L. K. M. Extração de poli(3-hidroxibutirato) produzido por Cupriavidus
necator dsm 545 com 1,2-carbonato de propileno. 2010. 99p. Dissertação (Mestrado em
Engenharia de Alimentos) – Centro Tecnológico, Departamento de Engenharia Química
e Engenharia de Alimentos, Programa de Pós-Graduação em Engenharia de Alimentos,
Universidade Federal de Santa Catarina, Florianópolis.
RAJENDRAN, A.; THANGAVELU, V. Statistical experimental design for evaluation
of medium components for lipase production by Rhizopus arrhizus MTCC 2233, LWT -
Food Science and Technology, v.42, p.985–992, 2009.
RAMIREZ-LEÓN, L. F.; RUIZ-HERRERA, J. Hydrolysis of walls and formation of
spheroplasts in Mucor rouxii. J. Gen. Microbiol., v.72, p.281-290, 1972.
RANGEL, M. A. S.; LOUREIRO, E. S.; ALMEIDA, A. S.; ALVES, H. C.; ONO, F.
B.; LOBO, P. M.; MARIANI, M. A.; PESSOA, L. G. A.; SILVA, A. J. Avaliação
participativa do controle de cigarrinhas das pastagens com a aplicação do fungo
Metarhizium anisopliae (Metsch.) Sorok. Rev. Bras. de Agroecologia, v.2, n.2, 731-734,
2007.
RAVEN, P. H.; EVERT, R.F.; EICHHORN, S.E. Biologia Vegetal, 6ª Ed, Rio Janeiro,
Editora Guanabara Koogan, 2001.
REDDY, L. V. A.; WEE, Y.-J.; YUN, J.-S.; RYU, H.-W. Optimization of alkaline
protease production by batch culture of Bacillus sp. RKY3 through Plackett–Burman

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
89
and response surface methodological approaches, Bioresource Technology, v.99, p.242–
2249, 2008.
RIGUEIRA, J. C. S. Influência da contagem de células somáticas no perfil e teores de
aminas bioativas e na qualidade de leite cru e de queijo mussarela. 2010. 147p. Tese
(Doutorado em Ciência de Alimentos) - Faculdade de Farmácia, Departamento de
Alimentos, Programa de Pós-Graduação em Ciência de Alimentos, Faculdade de
Farmácia da Universidade Federal de Minas Gerais, Belo Horizonte.
RODRIGUES, M. I.; IEMMA, A. F. Planejamento de Experimentos e Otimização de
Processos: uma estratégia sequencial de planejamentos, Campinas, SP, Casa do Pão
Editora, 2005.
SANTOS, E. S. Recuperação e Purificação de Enzimas usando Adsorção em Leito
Expandido, Tese de Doutorado, Universidade Estadual de Campinas, Campinas/SP,
p.152, 2001.
SEDMAK, J. J.; GROSSBERG, S. E. A rapid, sensitive and versatile assay for protein
using coomassie brilliant blue G250, Analytical Biochemistry, v.79, p. 544-552, 1977.
SEINO, H.; TSUKUDA, K.; SHIMASUE, Y. Properties and action pattern of a
chitosanase from Bacillus sp. PI- 7S. Agricultural Biological Chemistry, 55, p.2421-
2423, 1991.
SHIMOSAKA, M.; NOGAWA, M.; OHNO, Y.; OKAZAKI, M. Chitosanase from the
plant pathogenic fungus, Fusarium solani f. sp. Phaseoli – purification and some
properties, Bioscience Biotechnolgy and Biochemistry, v.57, n.2, p.231-235, 1993.
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Production of two chitosanases from a chitosan-assimilating bacterium, Acinetobacter
sp. Strain CHB101, Applied and Environmental Microbiology, v.61, n.2, p.438-442,
1995.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
90
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Characterization of Three Chitosanase Isozymes Isolated from a Commercial Crude
Porcine Pepsin Preparation, J. Agric. Food Chem. 51, p.1042-1048, 2003.
SILVA FILHO, R. C.; COSTA, A. S. S.; CHAGAS, B. M. E.; SANTOS, F. N.;
MACEDO, G. R.; SANTOS, E. S. Avaliação do teste de atividade quitosanolítica
produzida por fungo. In: XV SIMPÓSIO NACIONAL DE BIOPROCESSOS, 15,
Recife, 2005, 02 a 05 de agosto.
SILVA, E. P. Avaliação Preliminar do Potencial de Aplicação da Tecnologia de
Biopilhas para a Biorremediação do Solo de Guamaré-RN. 2004. 119p. Dissertação
(Mestrado) – Centro de Tecnologia, Programa de Pós-Graduação em Engenharia
Química. Universidade Federal do Rio Grande do Norte. Natal.
SOMASHEKAR, D.; JOSEPH, R. Chitosanases – Properties and apllications: A review.
Bioresource Technology, v.55, p.35-45, 1996.
STOYACHENKO, I. A.; VARLAMOV, V. P.; DAVANKOV, V. A. Chitinases of
Streptomyces kurssanovii: purification and some properties, Carbohydrate Polymers,
v.24, p.47-54, 1994.
TINLINE, R. D. Nuclear distribuition in Metarhizium anisopliae. Mycologia, v. 63, p.
713-721, 1971.
TRABULSI, L. R.; TOLEDO, M. R. F. Microbiologia. 2. Ed., São Paulo: Atheneu,
1996.
VIEIRA, P. D. S.; SILVA, W. M. T.; PAIVA, L. M.; LIMA, E. A. L. A.;
CAVALCANTI, V. L. B. Estudo da caracterização morfológica, esporulação e
germinação dos conídiosde Metarhizium anisopliae var. acridum em diferentes
temperaturas. Biológico, São Paulo, v.69, n.1, p.17-21, 2007.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
91
WANDERLEY, M. D.; NEVES, E.; ANDRADE, C. J. Aspectos da produção industrial
de enzimas. Revista Citino, v.1, n.1, p.30-36, 2011.
WANG, C.; TYPAS, M. A.; BUTT, T. M. Detection and characterization of pr1
virulent gene deficiencies in the insect pathogenic fungus Metarhizium anisopliae.
FEMS Microbiology Letters, v. 213, p. 251-255, 2002.
ZAHORSKA-MARKIEWICZ, B.; KROTKIEWSKI, M.; OLSZANECKA-
GLINIANOWICZ, M.; ZURAKOWSKI, A. Effect of chitosan in complex management
of obesity, Pol Merkur Lekarski, v.13, n.74, p.129-132, 2002.
ZHANG, X. Y.; DAI, A. L.; ZHANG, X. K.; KUROIWA, K.; KODAIRA, R.;
SHIMOSAKA, M.; OKAZAKI, M. Purification and Characterization of and Exo--D-
glucosaminidase from a koji mold, Aspergillus oryzae IAM2660. Biosci. Biotechnol.
Biochem., v.64, n.9, 1896-1902, 2000.
ZHU, X.; WU, X.; DAI, Y. Fermentation conditions of a chitosanase from Acinetobcter
sp C-17. Biosci. Biotechnol. Biochem., v.67, n.2, p.284-290, 2003.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
92
APÊNDICE

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
93
APÊNDICE I
Metodologia para repique da cepa
Em Erlenmeyer de 1000 mL colocava-se 500 mL do meio de manutenção
PDA. Este Erlenmeyer, com o meio de manutenção, era autoclavado junto com placas
de Petri a 1,5 kgf/cm2 por 15 minutos, em seguida, era levado para uma câmara de fluxo
laminar e transferido para as placas de Petri já autoclavadas, onde eram mantidas numa
estufa a 27 ºC durante 24 h. Após este período, as placas de Petri eram levadas a câmara
de fluxo laminar, utilizando bico de Bunsen para esterilizar a alça de platina. Procedia-
se o repique utilizando uma placa de Petri contendo a cepa pura. Cada placa de Petri
contendo a cepa pura era transferida para duas placas de Petri contendo o meio de
manutenção. As placas eram mantidas na estufa a 27 ºC durante 30 dias onde após esse
período, repetia-se este procedimento.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
94
APÊNDICE II
Montagem da Curva de Calibração Usando o Método DNS
Soluções:
Glicosamina = 0,01 mol/mL (10000,0 µmol/mL)
Tabela 14. Dados para construção da curva de calibração usando o Método DNS.
Tubo de
Ensaio
Solução de
glicosamina (mL)
Água Destilada
(mL)
DNS (mL)
Volume Total
Massa (µµµµmol)
1 0 1,0 1,5 2,5 0
2 0,2 0,8 1,5 2,5 0,2
3 0,4 0,6 1,5 2,5 0,4
4 0,6 0,4 1,5 2,5 0,6
5 0,8 0,2 1,5 2,5 0,8
6 1,0 0 1,5 2,5 1,0
Tabela 15 - Medidas de absorbância a 600nm
Dados adicionais:
Peso Molecular D-Glicosamina = 215,64 (recipiente SIGMA)
Conc. Sol. D-Glicosamina = 0,05 g/50 mL
Massa calc.(µmol) = conc. Sol. D-Glic. x Vol. Utiliz. D-Glics. =
= 0,05 x 1000 x Vol.util./50*215,64
Tempo atividade enzimática = 30 min
1 U/mL = 1 µmol/(min.mL)
Vol. Total (Glicosamina+caldo) = 1 mL
Medições Adsorbância a 600 nm Vol. Utiliz. Massa Massa Atividade
II III Média(Abs)(mL) calc. (mg) calc. (µg) Enzim. (U/mL)
0 0 0 0 0 0 0
0,133 0,128 0,1307 0,2 0,2 200 0,0309
0,279 0,277 0,2787 0,4 0,4 400 0,0618
0,433 0,428 0,4297 0,6 0,6 600 0,0927
0,587 0,583 0,5797 0,8 0,8 800 0,1237
0,727 0,716 0,7157 1 1 1000 0,1546

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
95
Figura 17 – Curva padrão de glicosamina.
y = 1900ralx + 1900ral
R² = 1900ral
00,000
00,020
00,040
00,060
00,080
00,100
00,120
00,140
00,160
00,180
00,000 00,100 00,200 00,300 00,400 00,500 00,600 00,700 00,800
Ati
v.
En
zim
áti
ca (
U/m
L)
Absorbância (Abs) a 600 nm
Curva-Padrão D-Glicosamina

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
96
APÊNDICE III
Método de Determinação de Proteína Utilizando o Corante Comassie
Brilhante Blue G 250 (Método modificado de Sedmak e Grossberg)
Micrométodo
Preparar uma solução 0,06% de Comassie Blue G250 em 1,5 % HCl (w/V). Filtrar e
papel de filtro Whatman nº1.
Curva Padrão (1 a 20 µµµµg)
a) Preparar soluções estoque de BSA de 10 a 200 µg/mL.
b) Colocar em tubos de ensaio 100 µL da solução estoque, 1400 µL de água e
1500µL do Comassie e agitar.
c) Preparar um tubo contendo 1500 µL de água e 1500 µL do corante que
corresponde ao zero de proteína.
d) Após 5 minutos (padronizar o tempo) fazer as leituras das absorbâncias.
e) As leituras das amostras devem ser feitas em dois comprimentos de onda 595 e
465 contra a água.
f) Plotar a razão das absorbâncias a 595/465 como uma função da massa de BSA e
calcular o melhor ajuste linear dos dados.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
97
Micrométodo (1 a 20 µµµµg de BSA)
Solução Estoque de BSA = 0,2 mg/mL
Tabela 16. Dados para construção da curva padrão de BSA.
Tubo de
Ensaio
H2O (µµµµL)
BSA (µµµµg)
Coomassie (µµµµL)
BSA (µµµµg)
1 1500 0 1500 0
2 1490 10 1500 2
3 1480 20 1500 4
4 1470 30 1500 6
5 1460 40 1500 8
6 1450 50 1500 10
7 1440 60 1500 12
8 1430 70 1500 14
9 1420 80 1500 16
10 1410 90 1500 18
11 1400 100 1500 20
Dados adicionais:
Concentração Sol. Estoque
BSA = 0,2 mg/mL = 0,2 µg/µL
Vol. total sol. = vol. H2O + vol. BSA + vol.
Coomassie = 3000 µL = 3 mL
Concentração Sol. BSA (mg/mL) = (massa BSA mg) / 3 mL
.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
98
Figura 18 – Curva Padrão de BSA
y = 1900ralx - 1900ral
R² = 1900ral
000,000
001,000
002,000
003,000
004,000
005,000
006,000
007,000
000,400 000,500 000,600 000,700 000,800 000,900 001,000 001,100
µg
BS
A
Razão Absorbâncias (595nm/465nm)
Curva-Padrão BSA

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE TECNOLOGIA
DEPARTAMENTO DE ENGENHARIA QUÍMICA PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA
TESE DE DOUTORADO
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em
Cultivo Descontínuo Submerso
Raimundo Cosme da Silva Filho
Natal/RN
Abril/2013

Raimundo Cosme da Silva Filho
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo
Descontínuo Submerso
Tese apresentada ao Programa de Pós-Graduação em Engenharia Química – PPGEQ, da Universidade Federal do Rio Grande do Norte, como parte dos requisitos para obtenção do título de Doutor em Engenharia Química, sob a orientação do Prof. Dr. Everaldo Silvino dos Santos e a coorientação da Profª. Drª. Gorete Ribeiro de Macedo.
Natal/RN
Abril/2013

Catalogação da Publicação na Fonte. UFRN / CT / PPGEQ
Biblioteca Setorial “Professor Horácio Nícolas Sólimo”.
Silva Filho, Raimundo Cosme da. Otimização do meio de cultura para a produção de quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso / Raimundo Cosme da Silva Filho. - Natal, 2013. 98 f.: il.
Orientador: Everaldo Silvino dos Santos. Co-orientador: Gorete Ribeiro de Macedo. Tese (Doutorado) - Universidade Federal do Rio Grande do Norte.
Centro de Tecnologia. Departamento de Engenharia Química. Programa de Pós-Graduação em Engenharia Química.
1. Quitosana - Tese. 2. Quitina - Tese. 3. Quitosanase – Tese. 4. Metarhizium
anisopliae - Tese. I. Santos, Everaldo Silvino dos. II. Macedo, Gorete Ribeiro de. III. Universidade Federal do Rio Grande do Norte. IV. Título. RN/UF/BSEQ CDU 547.458 (043.2)

SILVA FILHO, Raimundo Cosme da – Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo Descontínuo Submerso. Tese de Doutorado. UFRN - Programa de Pós-Graduação em Engenharia Química. Área de Concentração: Pesquisa e Desenvolvimento de Tecnologias Regionais – Subárea: Engenharia de Processos – Alimentos e Biotecnologia, Natal/RN, Brasil.
Orientação: Prof. Dr. Everaldo Silvino dos Santos Coorientação: Profª. Drª. Gorete Ribeiro de Macedo ___________________________________________________________________________
RESUMO: No presente trabalho utilizou-se um Planejamento Plackett & Burman, com 8 fatores e 12 ensaios em 2 níveis e mais 3 repetições na condição do ponto central, para se investigar a influência das concentrações de quitosana, peptona, extrato de levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O e FeSO4 na produção da enzima quitosanase por Metarhizium
anisopliae. Os ensaios para produção da enzima foram realizados em cultivo descontínuo submerso. Os resultados para o Planejamento Plackett & Burman mostraram que a atividade quitosanolítica foi favorecida pelo aumento da concentração de substrato (quitosana) e de sulfato ferroso (FeSO4), enquanto que o aumento da concentração dos outros fatores não contribuiu de forma significativa para a atividade quitosanolítica. A otimização do meio de cultura para a produção da enzima foi realizado por meio de um Planejamento Composto Central, com os dois fatores que mais influenciaram a atividade quitosanolítica (quitosana e FeSO4), conforme Planejamento Plackett & Burman, mantendo-se os outros nutrientes em seus valores mínimos. Nesse planejamento, para o fator FeSO4, tomou-se o limite inferior (-1) como sendo o limite superior do Planejamento Plackett & Burman. Os resultados mostraram que a produção da enzima foi favorecida pelo aumento da concentração quitosana e pela diminuição da concentração de FeSO4. A produção máxima de atividade quitosanolítica foi da ordem de 70,0 U/L e foi atingida em apenas 18 h de fermentação, resultado esse vinte e oito vezes maior aos obtidos anteriormente para o mesmo microrganismo que foi 2,5 U/L em 48h.
Palavras-chave: quitosana; quitosanase; Metarhizium anisopliae; Plackett & Burman.


ABSTRACT
In this work a Plackett-Burman Design with 8 factors and 12 trials in 2 levels with 3 repetitions at the center point was used in order to investigate the influence of the concentration of chitosan, peptone, yeast extract, NaNO3, K2HPO4, KCl, MgSO4.7H2O and FeSO4 on chitosanase production by Metarhizium anisopliae. Runs were carried out using submerged discontinuous cultivation for enzyme production. The results of the Plackett & Burman Design showed that only two factors, chitosan concentration as well as FeSO4 had influence on chitosanolytic activity, while the increase in concentration of other factors not contributed significantly to the quitosanolítica activity. Cultivation medium optimization for enzyme production was carried out using a Composite Central Design, with the most important factors for chitosanolytic activity (chitosan and FeSO4), in accordance with Plackett & Burman Design, and keeping the other nutrients in their minimum values. On this other design, it was taken the highest limit in Plackett & Burman Design as the lowest limit (-1) to FeSO4 factor. The results showed that the enzyme production was favoured by increasing the chitosan concentration and by decreasing FeSO4. Maximum production for chitosanolytic activity was about 70.0 U/L and was reached in only 18 h of fermentation, a result about twenty-eight times greater than a former study using the same microorganism (about 2.5 U/L at 48 h).
Key-words: Chitosan; Chitosanase; Metarhizium anisopliae; Plackett & Burman

Ao meu pai, minha mãe e minha
família, em especial aos meus
irmãos e sobrinhos

AGRADECIMENTOS
A Deus, por estar presente em todos os momentos da minha vida e por me ceder à
oportunidade de iniciar e concluir este trabalho.
A minha mãe Cândida Inocência da Silva que esteve comigo em todos os momentos da
minha vida me educando, me amando e sempre me dando muito carinho.
Ao meu pai Raimundo Cosme da Silva, que não está mais aqui para ver este trabalho,
mas que foi a pessoa que mais me apoiou e me incentivou, em todos os momentos da minha vida,
para que eu pudesse chegar o mais longe possível na minha carreira acadêmica. Pai muito
obrigado por toda atenção, educação, amor e carinho que recebi durante toda a minha vida.
Aos meus irmãos e familiares que sempre estiveram comigo dividindo momentos de
grande alegria e às vezes de tristeza, mas sempre nos mantendo unidos para superarmos todos os
obstáculos.
A minha namorada Rochelly Caroliny por todo amor e carinho que me deu e que me
entendeu nos momentos que estive ausente para a conclusão deste trabalho.
Ao meu orientador Professor Doutor Everaldo Silvino dos Santos por confiar em mim e
me aceitar como orientando além de toda a amizade, orientação e dedicação em todas as etapas de
desenvolvimento e conclusão deste trabalho.
À Professora Doutora Gorete Ribeiro de Macedo, “pessoa que admiro muito como
profissional e principalmente, como pessoa, um exemplo a ser seguido”, pelos conselhos e críticas
quando necessários ao desenvolvimento deste trabalho. Professora a senhora é considerada uma
mãe para de todos os alunos que tiveram a honra de conhecê-la e trabalhar com a senhora.
Aos meus amigos Engenheiros Químicos, Ana Carmen, Ana Katerine, Andréa Farias,
Anita Lima, Sirtys Lessa, Alexandre, Sérgio, Márcio Bezerra e ao Biólogo Daniel Souza, que
sempre me ouviram e que me ajudaram de alguma forma nesse trabalho.
A Eusamar Coelho de Lima, Mazinha, secretária do PPGEQ, pelo apoio, pela paciência e
orientação nas questões burocráticas do processo.
A Carlos e Jardelson, pela grande colaboração e dedicação nos experimentos do
laboratório.
A Sérgio Santana pela grande colaboração e dedicação, o qual não poupou esforços nem
tempo em me ajudar na realização dos experimentos em laboratório.

Ao meu amigo Ubiratan Nazaré por me ajudar na organização do texto e na tradução para
o Abstract.
Finalizando, agradeço ao Laboratório de Engenharia Bioquímica da UFRN por ter cedido
toda a sua estrutura para realização dos experimentos e ao Programa de Pós-Graduação em
Engenharia Química, assim como a todos os professores do Departamento de Engenharia Química
da UFRN que trabalham em pró de uma educação digna e ensino de qualidade para todos os
alunos.

Sumário LISTA DE FIGURAS
LISTA DE TABELAS
NOMENCLATURA
INTRODUÇÃO...............................................................................................................................15
REVISÃO BIBLIOGRÁFICA........................................................................................................18
2.1. Enzimas................................................................................................................................18
2.1.1. Quitosana e seus derivados........................................................................................19
2.1.2. Quitosanases...............................................................................................................24
2.2. Fungos..................................................................................................................................29
2.2.1. Características gerais..................................................................................................29
2.2.2. Reprodução dos Fungos.............................................................................................32
2.2.3. Metabolismo de fungos..............................................................................................33
2.2.4. Nutrição de fungos.....................................................................................................34
2.2.5. O Fungo Metarhizium anisopliae ..............................................................................36
2.3. Fermentação.........................................................................................................................40
2.3.1. Preparo do inóculo......................................................................................................41
2.3.2. Produção de enzimas em cultivo submerso descontínuo...........................................42
2.4. Planejamento experimental..................................................................................................42
2.4.1. Planejamento fatorial..................................................................................................43
2.4.2. Planejamento Plackett & Burman..............................................................................43
2.4.3. Aplicações do Planejamento Plackett & Burman.......................................................44
METODOLOGIA EXPERIMENTAL.............................................................................................50
3.1. Quitosana..............................................................................................................................50
3.2. Microrganismo.....................................................................................................................50
3.2.1. Escolha da cepa produtora de quitosanase.................................................................50

3.2.2. Manutenção da cepa...................................................................................................50
3.3. Produção de enzima.............................................................................................................51
3.4. Determinação da Proteína Total...........................................................................................51
3.5. Determinação da Atividade Quitosanolítica e Atividade Específica...................................51
3.6. Planejamento Experimental.................................................................................................52
3.6.1. Planejamento de seleção de fatores Plackett Burman................................................52
3.6.2. Planejamento Composto Central para Otimização do meio de cultura para a
Produção da Enzima em Cultivo Submerso.....................................................................................53
3.7. Perfil Cinético durante o Cultivo.........................................................................................56
RESULTADOS E DISCUSSÃO.....................................................................................................58
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA ..........................................58
4.2. Planejamento Plackett & Burman........................................................................................59
4.2.1. Análise do Plackett & Burman com 8 fatores e 12 ensaios .......................................59
4.2.2. Análise do Plackett & Burman com 12 ensaios e mais 3 repetições na condição do
ponto central ....................................................................................................................................61
4.3. Planejamento Composto Central para Produção de Quitosanase por Metarhizium
anisopliae em Cultivo Descontínuo Submerso................................................................................64
4.4. Perfil Cinético – Melhor Condição......................................................................................69
CONCLUSÃO.................................................................................................................................75
REFERÊNCIAS BIBLIOGRÁFICAS.............................................................................................78
APÊNDICE I...................................................................................................................................93
Metodologia para Repique da Cepa ................................................................................................93
APÊNDICE II.................................................................................................................................94
Montagem da Curva de Calibração Usando o Método DNS..........................................................94
APÊNDICE III................................................................................................................................96
Método de Determinação de Proteína Utilizando o Corante Comassie Brilhante Blue G 250
(Método modificado de Sedmak e Grossberg)................................................................................96

Lista de Figuras
Figura 1 – Estrutura da quitosana..................................................................................................20
Figura 2 – Estrutura da glicosamina..............................................................................................23
Figura 3 – Estrutura do sulfato de glicosamina.............................................................................23
Figura 4 – Representação da estrutura geral de uma célula de fungo..........................................30
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas...........................................................31
Figura 6 – Conídios de Aspergillus e Penicillium.........................................................................33
Figura 7 – Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA ...................58
Figura 8 – Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados............................................................................................................................60
Figura 9 – Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto central
mostrando a influência dos fatores estudados................................................................................63
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos
fatores estudados............................................................................................................................67
Figura 11 – Valores preditos em função dos valores observados..................................................68
Figura 12 – Atividade quitosanolítica em função da concentração de quitosana e da concentração
de FeSO4 para os fatores não significativos no ponto central .......................................................68
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana ........70
Figura 14 – Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana....................................70
Figura 15 – Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de
quitosana........................................................................................................................................71
Figura 16 – Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana..........................................................................................................72
Figura 17 – Curva padrão de glicosamina ....................................................................................95
Figura 18 – Curva Padrão de BSA ................................................................................................98

Lista de Tabelas
Tabela 1 – Comparação das enzimas com os catalisadores químicos..............................................19
Tabela 2 – Fontes naturais de quitina e quitosana ...........................................................................20
Tabela 3 – Níveis para os fatores e seus valores codificados...........................................................53
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central .........................................................................................53
Tabela 5 – Matriz do planejamento Composto Central...................................................................54
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central......................................................................................................55
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.......................59
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman...............................................60
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.......................61
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central...............................................................................................................................................62
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da atividade
enzimática........................................................................................................................................65
Tabela 12 – Efeitos Estimados do planejamento Composto Central...............................................65
Tabela 13 – Resumo da ANOVA....................................................................................................66
Tabela 14 – Dados para construção da curva de calibração usando o Método DNS ......................94
Tabela 15 – Medidas de absorbância a 600 nm para construção da curva de calibração................94
Tabela 16 – Dados para construção da curva padrão de BSA ........................................................97

NOMENCLATURA
A – Atividade quitosanolítica (U/L)
AE – Atividade específica (U/mg)
ANN-GA – algoritmo genético de rede neural artificial
ANOVA – Análise de variância (Analysis of Variance)
BSA – Albumina de soro bovino (Bovine Sereum Albumin)
CCD – Planejamento Composto Central
CDA – Chitosan Dector Agar
Cs – Concentração de substrato (%)
CP – Concentração de KCl (%)
Da – Dalton
DEQ – Departamento de Engenharia Química
DNS – Ácido Dinitrosalissílico
E – Enzima
Ea – Energia de ativação
EC – Comissão para nomenclatura e classificação de enzimas
EL – Concentração de Extrato de Levedura (%)
FP – Concentração de K2HPO4 (%)
NS – Concentração de NaNO3 (%)
GDA – Grânulos dispersíveis em água
k – Número de fatores, variáveis
LEB – Laboratório de Engenharia Bioquímica
mRNA – RNA mensageiro
n – Número de níveis de um determinado fator
N – Número de ensaios de um planejamento
P – Pressão
PP – Concentração de Peptona (%)
PDA – Batata Dextrose Agar
pH – Potencial hidrogeniônico
Q – Concentração de Quitosana (%)
Qm – Quadrado médio
r – Agitação (rpm)
R2 – Coeficiente de determinação

RA – Razão de aeração
rpm – Rotações por minuto
RSM – Metodologia de superfície de resposta
S – Molécula de substrato
SF – Concentração de FeSO4 (%)
SM – Concentração de MgSO4.7H2O (%)
SQ – Soma de quadrados
SSF – Fermentação de estado sólido
T – Temperatura
TMC – Temperatura de cultivo
V11,V12,V13 – Variáveis inertes para cálculo do erro
VHG – Fermentação de alta densidade
WP – Pó molhável
X – Concentração celular
Xi – variável independente
Xj – variável independente
xi – Fator independente real
x0 – Valor de xi no ponto central
∆xi – Variação de valor
Y – Resposta do modelo de primeira ordem
Yp – Resposta do modelo polinomial de segunda ordem
β0 – Interseção dos modelos de primeira ou segunda ordem
βi – Coeficiente linear dos modelos de primeira ou segunda ordem
βii – Coeficiente quadrático
βij – Coeficiente de interação

Capítulo 1 Introdução

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
15
1. Introdução As quitosanases foram descobertas simultaneamente há quase quarenta anos por
Monaghan et al. (1973), como pelo grupo de Ramirez-León & Ruiz-Herrera (1972), e
são enzimas obtidas a partir do metabolismo de bactérias, fungos e extraídas de plantas.
Desde então, vários estudos vêm apresentando a obtenção de quitosanase deste
microrganismos, na forma intracelular ou extracelular (Osswald et al., 1994;
Somashekar & Joseph, 1996), como a do presente estudo. A quitosanase (EC 3.2.1.132)
representa uma classe de enzimas hidrolíticas que catalisam a hidrólise de ligação β-
(1→ 4) glicosídica da quitosana, gerando oligômeros chamados quito-oligossacarídeos
(Reddy et al., 2008).
A quitosana é um biopolímero hidrofílico obtido a partir da quitina, um
polissacarídeo abundante na natureza. Na literatura, é considerado quitosana o produto
da desacetilação da quitina para um grau de desacetilação superior a 75% e solúveis em
ácidos como o acético e o fórmico. A quitosana é conhecida por exibir uma ampla
variedade de atividade fisiológica como atividade antitumoral, mas o peso molecular e a
insolubilidade em água da quitosana são desvantagens para várias aplicações.
Atualmente, há interesse nas quitosanas quando as mesmas são convertidas em quito-
oligossacarídeos porque eles não apenas são solúveis em água e possuem baixo peso
molecular, mas também têm atividades biológicas bem notáveis, como atividade
anticancerígena atóxica e biocompatível, incluindo efeito antimetastático, atividade anti-
HIV, hepatoprotetora, antioxidante, hipocolesterolêmica, antimicrobiana,
imunoestimulate, antitumoral, anti-inflamatória e atividade aceleradora da absorção de
cálcio e ferro. No entanto, estes quito-oligossacarídeos são obtidos tradicionalmente na
indústria por reação de hidrólise usando ácidos, que apresenta muitos problemas como
dificuldade em controlar a hidrólise, produzindo uma grande quantidade de
oligossacarídeos de cadeia pequena e oligossacarídeos de baixo rendimento, alto custo
de separação e, também, poluição ambiental, uma vez que utiliza um ácido como
catalisador. Como alternativa, com suas vantagens em termos de compatibilidade
ambiental, baixo custo e reprodutibilidade, a obtenção destes quito-oligossacarídeos por
hidrólise com quitosanase se tornou cada vez mais popular nos últimos anos (Azevedo
et al., 2007).

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
16
Devido a todos estes aspectos positivos é que, em geral, as enzimas vêm sendo
estudadas em diferentes aplicações, mostrando que esta é uma tendência na indústria.
No presente trabalho, foi desenvolvido um processo fermentativo visando a
otimização do meio de cultura para a produção da enzima quitosanase em cultivo
descontínuo submerso, por Metarhizium anisopliae, capaz de sintetizar oligômeros com
aplicações na indústria farmacêutica e de química fina, partindo de quitosana como
única fonte de carbono.
Nesse caso, avaliou-se, através de um planejamento Plackett & Burman (método
de delineamento de seleção de fatores) e de um planejamento Composto Central as
melhores condições para a produção da enzima. A técnica do planejamento
experimental Plackett & Burman para realizar triagem de fatores na otimização de um
determinado processo foi utilizada neste trabalho para identificar quais os fatores que
influenciam de forma significativa na produção da enzima quitosanase. A influência
destes fatores (concentração de quitosana e concentração de FeSO4) foi então
investigada pelo planejamento Composto Central com 22 experimentos na parte fatorial,
repetição em triplicata no ponto central e quatro experimentos na parte axial. A
repetição do ponto central permite também a determinação do erro puro e da
repetibilidade do processo, sendo uma forma de identificar melhor se o processo está
sob controle.
Do ponto de vista industrial é de relevante importância a realização de estudos
para a otimização da produção da quitosanase de forma contínua ou em grandes
bateladas.

Capítulo 2 Revisão Bibliográfica

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
18
2. Revisão Bibliográfica 2.1. Enzimas
As enzimas são catalisadores biológicos constituídos por longas cadeias de
aminoácidos que atuam em substratos reduzindo a energia de ativação, acelerando assim
a velocidade da reação, e transformando-os em produtos. Sua atuação catalítica visa
facilitar a atividade das células através da quebra de moléculas ou junção das mesmas
para formação de novos produtos. Podem apresentar alto grau de especificidade relativa
a apenas um substrato ou a vários substratos ao mesmo tempo, como a certos tipos de
ligações glicosídicas α-1,4 das moléculas de amido, ou para determinado tipo particular
de isômero óptico como ocorre na oxidação da β-D-glicose pela glicose oxidase (Furigo
Junior & Pereira, 2001).
Enzimas são proteínas que apresentam em sua estrutura um centro ativo
(apoenzima) e, em alguns casos, um grupo prostético denominado cofator (coenzima ou
íon metálico ativo) e sua atividade depende de alguns fatores como estrutura da
proteína, número de cadeias peptídicas, arranjo dessas cadeias na molécula, natureza do
substrato e natureza do grupo prostético (Politzer & Bon, 2006).
Todos os organismos vivos produzem, em pequena ou grande quantidade, certa
variedade de enzimas, sejam elas intra ou extracelulares. As enzimas extracelulares são
excretadas para o meio externo e podem decompor compostos insolúveis como
celulose, quitina e quitosana.
Algumas destas enzimas podem ser usadas tanto nas indústrias de detergentes,
têxteis e farmacêuticas, como também de alimentos, de bebidas, etc., sendo utilizadas
em processos biotecnológicos industriais em substituição aos processos químicos que
muitas vezes causam grandes prejuízos ao meio ambiente. A produção comercial das
enzimas pode ser realizada através do cultivo de microrganismos como fungos e
bactérias.
Na Tabela 1 mostra-se uma comparação das enzimas com os catalisadores
químicos, levando-se em conta algumas das características dos dois catalisadores.
Destaca-se que alguns parâmetros da Tabela 1 demonstram a maior importância das
enzimas, quando comparadas com os catalisadores químicos, principalmente a
especificidade ao substrato, as condições de reação, o consumo de energia e a

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
19
velocidade de reação, como desvantagem cita-se o custo de obtenção (isolamento e
purificação).
Tabela 1 – Comparação das enzimas com os catalisadores químicos (Adaptado de
Furigo Junior & Pereira, 2001).
Característica Enzimas Catalisadores Químicos
Especificidade ao substrato Alta Baixa
Natureza da estrutura Complexa Simples
Sensibilidade à T e pH Alta Baixa
Condições de reação (T, P e pH) Suaves Drástica (geralmente)
Custo de obtenção (isolamento e purificação) Alto Moderado
Natureza do processo Batelada Contínuo
Consumo de energia Baixo Alto
Formação de subprodutos Baixa Alto
Separação catalisador/produtos Difícil/cara Simples
Atividade catalítica (temperatura ambiente) Alta Baixa
Presença de cofatores Sim Não
Estabilidade do preparado Baixa Alta
Energia de Ativação Baixa Alta
Velocidade de reação Alta Baixa
2.1.1. Quitosana e seus derivados
A quitosana e seus derivados apresentam grande importância devido ao
conhecimento de suas funcionalidades biológicas, médicas, tecnológicas, etc
(Shimosaka et al, 1993; Shimosaka et al, 1995; Mitsutomi et al., 1995). A quitosana é
obtida da quitina, polissacarídeo abundante na natureza, extraído principalmente das
carapaças de caranguejos, lagosta e camarões. A quitosana é um polímero natural,
biodegradável, não tóxico e de alta massa molecular obtido a partir de animais
marinhos, insetos e microrganismos (Tabela 2).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
20
Tabela 2 – Fontes naturais de quitina e quitosana. (Adaptado de Polymar, 2003).
Animais
Marinhos
Insetos Aracnídeos Microrganismos
Anelídios Formigas Escorpiões Leveduras
Moluscos Besouros Aranhas Fungos
Lagosta Esporos
Camarão
Caranguejo
Krill
A Figura 1 abaixo mostra a estrutura química da quitosana
Figura 1 – Estrutura da quitosana (Fonte: Polymar, 2003)
Quitosana totalmente desacetilada não existe na natureza. Graus de acetilação
(GA) nferior a 50% a torna mais solúvel em soluções ácidas (Muzzarelli, 1993;
Muzzarelli et al., 1994; Dung et al., 1994; Stoyachenko et al.,1994). Pelletier &
Sygusch (1990) enfatizaram que grande parte dos resíduos sólidos da indústria
pesqueira, principalmente da carcinicultura, é jogada ao mar ocasionando degradação
ambiental. Logo, a utilização desses resíduos como matéria-prima para formação de
novos produtos, não só reduz de forma drástica o impacto ambiental, mas também daria
maior valor agregado à indústria pesqueira.
A demanda cada vez maior de consumidores por alimentos mais saudáveis, sem
conservantes químicos, tem motivado pesquisas à descoberta de novos agentes
antimicrobianos naturais. Sendo assim, a atividade antimicrobiana incomum da

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
21
quitosana e seus derivados contra microrganismos, tais como bactérias, bolores e
leveduras tem tido maior atenção ultimamente.
Devido à interação entre a quitosana, que é positivamente carregada, e as
membranas celulares dos microrganismos, negativamente carregadas, ocorre a formação
de verdadeiros poros na membrana, que são canais de vazamento de conteúdo protéico e
outros componentes intracelulares. A quitosana tem também a propriedade de agir como
agente quelante que se liga de forma seletiva a traços de metais, inibindo, assim, a
produção de toxinas e o crescimento microbiano. Ainda é capaz de ativar mecanismos
de defesa em tecidos hospedeiros, atuando como agente sequestrante de água e inibindo
várias enzimas. A ligação da quitosana com DNA e a inibição da síntese do mRNA
ocorre via penetração da quitosana no núcleo dos microrganismos e interferência na
síntese do mRNA e proteínas, (Craveiro et al., 2004).
Em um estudo clínico conduzido por Giustina & Ventura (1995) com 100
pessoas com sobrepeso de 10 a 25%, ao final de 4 semanas as pessoas que receberam
quitosana demonstraram perda de peso significativa (7,3 Kg contra 3,0 Kg nos grupos
controle). As pessoas que tomaram quitosana também demonstraram redução na pressão
sanguínea sistólica (de 145 para 135 mm Hg) e diastólica (92.6 para 84.2 mm Hg),
assim como uma redução na taxa respiratória (27.6/min a 21.2/min).
Kondo & Osada (1996) pesquisaram a influência da quitina e da quitosana na
biodisponibilidade do zinco em ratos alimentados com uma dieta com suplementos de
5% ou sem fibra dietética (quitina, quitosana, celulose, pectina e agar-agar) durante 31
dias. Observaram que os ratos alimentados com dietas contendo 5% de fibras, exceto
quitosana, o consumo de alimento foi maior do que o controle. O ganho de peso em
todos os grupos alimentados com fibra foi maior do que o controle, exceto para o grupo
suplementado com quitosana (5%) onde a ingestão de alimento e o ganho de peso foram
menores do que os grupos controle e das demais fibras.
Segundo Craveiro et al. (2004), derivados da quitosana obtidos pela reação com
ácidos carboxílicos, podem promover ao polímero uma solubilidade especial em água.
Esses compostos possuem estrutura química semelhante ao ácido hialurônico e
fornecem uma excelente compatibilidade com o tecido epitelial. As soluções obtidas
com esses derivados podem ser facilmente incorporadas a formulações cosméticas,
mostrando melhor compatibilidade com surfactantes iônicos e emulsificantes,
apresentando estabilidade em uma larga faixa de pH.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
22
Han et al. (1999) examinaram os efeitos da quitosana sobre a atividade da lipase
pancreática in vitro e sobre o grau de armazenamento de gorduras induzido em ratos
pela administração oral de uma dieta rica em gorduras por nove semanas. Como
resultado, observou-se que os animais alimentados com quitosana, reduziram o peso
corporal, a hiperlipidemia e a gordura hepática. Como conclusão, os autores propõem
que os efeitos anti-obesidade da quitosana em ratos alimentados com alto teor de
gordura se devem parcialmente a inibição da absorção intestinal das gorduras dietéticas.
Em outro ensaio, Zahorska-Markiewicz et al. (2002) realizaram um estudo para
avaliar o efeito da quitosana no tratamento da obesidade. Segundo os autores do estudo,
a quitosana pode ser usada como um valioso e seguro coadjuvante em tratamentos para
a obesidade em longo prazo. A quitosana parece acentuar a redução da pressão arterial,
juntamente com a perda de peso.
Assis (2010) estudou a produção e caracterização de quitooligossacarídeos
produzidos pelo fungo Metarhizium anisopliae e avaliação da citotoxicidade em células
tumorais. A hidrólise da quitosana foi realizada em tempos de 10 a 60 min para a
produção de quitooligossacarídeos. A avaliação da citotoxicidade dos oligômeros de
quitosana foi realizada em células tumorais (HepG2 e HeLa) e não tumoral (3T3). A
produção de oligômeros de quitosana teve maiores rendimentos durante 10 minutos de
hidrólise, os pentâmeros apresentaram concentração de 0,15 mg/mL, no entanto os
hexâmeros, que apresentam maior interesse por causa de suas propriedades biológicas,
só foram detectados com 30 minutos de hidrólise com uma concentração de 0,004
mg/mL.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o planejamento Plackett-Burmam e a metodologia de superfície de resposta.
Os resultados mostraram que a eficiência máxima de extração da quitina foi conseguida
nas seguintes condições: concentração de casca de camarão 50g/L, glicose 50g/L, tempo
de incubação de 5 dias e inoculo como 0,05 OD (densidade óptica).
A glicosamina, unidade monomérica básica da quitosana, possui atividade
antiinflamatória e tem grande importância na construção e manutenção da cartilagem,
no tratamento de enfermidades como: febre reumática, artrose e principalmente a artrite
e osteoartrite na forma aguda ou crônica (Polymar, 2003). A Figura 2 apresenta a
estrutura química da glicosamina.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
23
Figura 2 – Estrutura da glicosamina (Fonte: Polymar, 2003)
O sulfato de glicosamina é um derivado da quitosana sendo utilizado no
tratamento da osteoartrite sintomática dos joelhos, quadris, coluna, mãos e outras
regiões (Craveiro et al., 2004). O sulfato de glicosamina estimula a síntese de
proteoglicanos necessários para o bom funcionamento das articulações, estimula a
regeneração das cartilagens e ainda incorpora enxofre ao tecido cartilaginoso. Estudos
de longa duração, acima de 3 anos, mostram que o sulfatos de glicosamina evita a
progressão de danos estruturais nas articulações e induz melhora significativa nas dores
e funções físicas (Craveiro et al., 2004). A Figura 3 apresenta a estrutura química do
sulfato de glicosamina.
Figura 3 – Estrutura do sulfato de glicosamina (Fonte: Polymar, 2003)

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
24
2.1.2. Quitosanases
Dependendo do tipo de quitosanase obtida, ocorrem diferentes mecanismos de
hidrólise da quitosana, isso devido a diferentes microrganismos produzirem enzimas
com diferenças nas suas estruturas (Kurita et al., 1977; Seino et al., 1991).
Quitosanases (EC 3.2.1.132) são enzimas que catalisam a reação de hidrólise de
quitosana em oligossacarídeos. As enzimas quitosanases são produzidas por vários
microrganismos, bactérias, actinomicetos e fungos, e até por algumas plantas. Estas
enzimas agem em polímeros com grau de acetilação de 30% a 60%. Os monômeros
resultantes da hidrólise da quitosana por estas enzimas são melhores digeridos, ao serem
ingeridos, como é o caso da glicosamina. Estudos efetuados até o presente momento
classificam as quitosanases em cinco famílias: 5, 8, 46, 75, 80, sendo que a família 46 é
a mais estudada. As quitosanases tem grande aplicação na geração de oligômeros de
quitosana (Somashekar & Joseph, 1996; Chen et al., 2005).
Nogawa et al. (1998) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida pelo fungo Trichoderma reseei PC-3-7. Estes pesquisadores
inocularam 106 conídios em 100 mL de um meio basal contendo 0,3% de N-
acetilglicosamina em incubator rotativo com agitação de 220 rpm a 28oC. Destaca-se
que o fungo não produziu quitosanase em meio contendo quitosana ou quitina.
Entretanto, o microrganismo produziu quitosanase em meio contendo glicosamina e N-
acetilglicosamina, sendo que para este último obteve-se uma atividade específica
máxima de 1,5 U/mg após 72 horas de incubação. É importante ressaltar que uma
agitação vigorosa favoreceu a produção da enzima. Esse artigo ilustra a presença de
exo-β-D-glicosaminidase e exo-β-D-N-acetilglicosaminidase.
Piza et al. (1999) estudaram a produção, purificação e caracterização parcial de
uma quitosanase produzida por Bacillus cereus. Nesse caso, investigou-se através de um
planejamento fatorial fracionário a influência dos fatores: concentração de sulfato de
amônio (0,4; 2,2 e 4,0%), razão de aeração (2; 6 e 7), tempo de fermentação (16; 24 e
32 horas), pH (5; 6 e 6,5) e concentração de quitosana (1,0; 1,5 e 2,0%). Os resultados
mostraram que os fatores mais significativos para esse planejamento foram a
concentração de sulfato de amônio, a aeração, o pH e a interação entre sulfato de
amônio e a aeração.
Cheng & Li (2000) estudaram a preparação de oligossacarídeos utilizando
quitosanase produzida por Aspergillus sp e constataram que a enzima foi largamente

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
25
induzida em meio contendo quitosana como única fonte de carbono e também na
presença de quitina coloidal. A indução da enzima foi realizada em temperaturas
variando na faixa de 28 a 30ºC, pois essa espécie não cresceu bem em temperaturas
superiores a 37ºC.
Zhang et al. (2000) estudaram a produção, purificação e caracterização de
quitosanase e Exo-β-D-Glucosaminidase por Aspergillus oryzae IAM2660. Os
microrganismos utilizados neste estudo foram mantidos em placas de Petri contendo
agar para contagem em placas (PDA). Destaca-se que para produzir a enzima, conídios
foram retirados das placas e inoculados em meio Czapeq-dox com os seguintes
componentes: 1,0g K2HPO4, 0,5g KCl, 2,0g NaNO3, 0,5g MgSO4.7H2O, 0,02g
FeSO4.7H2O, 20g de sacarose e 1,0L de água deionizada, com pH do meio igual a 5,5.
Foi adicionado ao meio 0,5 g de peptona e 0,5g de extrato de levedura com
concentração de esporos da ordem de 1,0x106 esporos.mL-1. Após a incubação, feita em
incubador rotativo a 26ºC por um período de 60h, micélios foram coletados e após
filtração e secagem a 80ºC foram utilizados para ensaio de quitosanase.
Kuroiwa et al. (2002) estudaram os fatores que afetam a composição de
oligossacarídeos produzidos através da hidrólise de quitosana por quitosanase originária
de Bacillus pumilus BN-262. Neste experimento foi utilizado quitosana 100%
desacetilada dissolvida em ácido acético 1M. O pH do meio foi ajustado em 5,6. Neste
estudo a quitosanase foi imobilizada em agar e Sepharose. Os resultados mostraram que
a concentração de enzima, a temperatura, a velocidade de agitação, e concentração de
substrato influenciaram de forma significativa na composição dos oligossacarídeos
produzidos.
Ichikawa et al. (2002) estudaram a imobilização e estabilização da ligação
multiponto da quitosanase tendo agar como suporte e concluíram que a imobilização
aumentou a termoestabilidade da quitosanase. Os oligossacarídeos de quitosana foram
continuamente produzidos usando um reator coluna contendo quitosanase imobilizada.
A percentagem de quitosana imobilizada após 28 dias de reação foi de 44%.
Hung et al. (2002) estudaram a purificação, através de troca iônica, e
caracterização da hidrólise sobre ação da quitinase e quitosanase purificadas do meio
contendo Bromelina comercial. Destaca-se que a quitinase purificada apresentou pH,
temperatura e Km ótimo para ação catalítica em meio contendo quitina de 4,0, 60ºC e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
26
0,2mg.mL-1, respectivamente. Enquanto que quitosanase apresentou o pH, temperatura e
Km ótimo de 3,0, 50ºC e 0,88mg.mL-1, respectivamente.
Zhu et al. (2003) estudaram as melhores condições fermentativas e propriedades
de uma quitosanase produzida por Acinetobacter sp. C-17. A espécie de bactéria
utilizada neste ensaio foi isolada do solo e identificada como sendo Acinetobacter sp C-
17 e capaz de produzir quitosanase. O microrganismo foi mantido em placas contendo o
meio agar com os seguintes componentes: 0,5 % de quitosana, 0,2 % K2HPO4, 0,1 %
KH2PO4, 0,07 % MgSO4, 0,05 % NaCl, 0,05 % KCl, 0,01 % CaCl2, 0,05 % de extrato
de levedura e 2 % agar, o pH do meio de manutenção foi mantido em 7,2 e a
temperatura foi mantida em 30ºC durante 3 a 5 dias, para identificar a ação da enzima
quitosanase. A ação da quitosanase foi identificada através da observação de uma
colônia de microrganismo existente na placa. A bactéria foi então isolada da placa e
colocada em fermentador rotativo a 200 rpm durante 72h em meio contendo: 0,07 %
K2HPO4, 0,03 % KH2PO4, 0,05 % MgSO4, 0,03 % peptona, 0,03 % de extrato de
levedura e 1,0 % de quitosana. O pH foi de 7,0 e a temperatura 30ºC. Como fontes de
carbono foram utilizadas além de 1,0 % de quitosana, 1,0 % de quitina e 1,0 % de
glicose. Os resultados mostraram que as melhores condições para produção da
quitosanase foram alcançadas quando se utilizou 1,0 % de solução de quitosana a 30ºC,
pH 7,0 durante 24 horas de incubação.
Liu et al. (2003) estudaram a produção de quitinase por Verticillium lecanii
F091 usando fermentação submersa, a atividade de quitinase foi de 9,95 mU/mL em um
meio de cultura otimizado, com um volume de cultura de 200 mL, agitação de 150 rpm
e temperatura de 24ºC cultivado em incubador rotativo e concluíram que a taxa de
agitação e o pH foram os fatores mais significativos para a produção de quitinase, no
entanto, as taxas de agitação e aeração poderiam modificar a concentração de oxigênio
dissolvido afetando diretamente no crescimento do Verticillium lecanii e na produção
de quitinase.
Donzelli et al. (2003) estudaram a hidrólise enzimática avançada de quitina de
casca de caranguejo langostino com misturas de enzimas de bactérias e fungos e
concluiram que a combinação de enzimas de Trichoderma atroviride e Serratia
marcescens foram capazes de degradar completamente altas concentrações de quitina
(100g/mL) a partir de carapaças de caranguejo langostino convertendo a N-
Acetilglicosamina (78%), glucosamina (2%) e quitobiose (10%). Enzimas de fonte

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
27
procariótica apresentaram atividades menores que as enzimas de T. atroviride, para
catalisar a reação com quitosana.
Jo et al. (2003) estudaram a caracterização e a avaliação cinética de quitosanase
produzida por Bacillus sp. Destaca-se que na produção da enzima utilizada neste ensaio
o meio continha 0,5% de concentração de quitosana, 1% de triptona e 1% de NaCl, o
pH do meio foi ajustado em 5,5 e a temperatura foi mantida em 37ºC com agitação de
180 rpm em incubador rotativo.
Fu et al (2003) estudaram a caracterização de três iso-enzimas de quitosanase
isolada de pepsina comercial e denominado como PSC-I, PSC-II, PSC-III,
respectivamente, em relação a hidrólise enzimática da quitosana, o meio ótimo para a
produção destas iso-enzimas continha as seguintes características: pH 5,0; 5,0 e 4,0 e
temperaturas de 40ºC, 40ºC e 30ºC, o valor de Km foi de 5,2, 4,0 e 5,6 mg.mL-1,
respectivamente. As massas moleculares das três iso-enzimas foram de 40 kDa. Os
resultados mostraram que as três iso-enzimas apresentaram atividade em meio contendo
quitosana, produzindo oligômeros da ordem de 68 – 88% desacetilada.
Choi et al. (2004) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida por Bacillus sp KCTC 0377BP. Nesse caso, com relação à
produção da enzima foi possível obter um aumento de 1,2 U/ml em incubator rotativo
para 100,0 U/ml em fermentador piloto de 500 L, utilizando uma concentração de
substrato de 0,5% de quitosana e como fonte de nitrogênio polipeptona (2,5%) em pH
igual a 6,8, com 100 rpm de 100,0 rpm a 30oC.
Shimosaka et al. (2003) estudaram a produção, purificação e caracterização total
de duas quitosanases produzida por Acinetobacter sp CHB101. Nesse caso, com relação
à produção da enzima os pesquisadores usaram dois meios, um contendo glicose e outro
contendo quitosana. Com relação à cinética, o meio contendo quitosana fornece um
tempo de geração de 280 minutos, enquanto que no meio contendo glicose a taxa de
crescimento foi mais elevada e o tempo de geração foi de apenas 110 minutos. Os
autores observaram que após os dois primeiros dias de cultivo a atividade
quitosanolítica permaneceu baixa (na ordem 0,10 a 0,20 U/mL). Durante a fase de
crescimento exponencial o valor da atividade permaneceu com valor inferior a 0,01
U/mL, começando a aumentar ao atingir a fase estacionária, indicando assim que a
produção da enzima é não associada ao crescimento.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
28
Kim et al. (2004) estudaram a produção, purificação e caracterização de uma
quitosanase produzida pelo Bacillus sp e identificado pelo número 1299. Destaca-se que
na produção da enzima quitosanase foi utilizado um meio contendo os seguintes
componentes: 0,5 % de quitosana, 0,42 % (NH4)2SO4, 0,2 % K2HPO4, 0,02 % uréia,
0,03 % CaCl2.2H2O, 0,03% MgSO4.7H2O, 0,1% peptona, 0,2% Tween 80, 0,2 % da
solução (0,5% FeSO4.7H2O, 0,16% MnSO4.H2O, 0,14% ZnSO4.7H2O, 0,2% CoCl2) e
1,7% agar. O pH do meio para este ensaio foi mantido em 7,0. Este microrganismo foi
escolhido por ser grande produtor de quitosanase.
Chen et al. (2005) estudaram a purificação e a caracterização de dois tipos de
quitosanase produzidas por Aspergillus sp. CJ22-326 em cultivo submerso. Na
produção da enzima foi utilizado um meio de cultivo contendo 1,0% de quitosana, 2,0%
de gérmen de trigo, 0,2% de sulfato de amônio, 0,2 % de fosfato de potássio e 0,05% de
sulfato de magnésio por litro de solução em pH 5,6. O cultivo foi realizado em frasco de
500 mL contendo 150 mL de meio de cultura a 30ºC por 96 horas com uma agitação de
150 rpm. Os autores apresentam um gel de eletroforese com as duas enzimas purificadas
com massa molar de 29 kDa e 109 kDa.
Silva Filho et al. (2005) estudaram, através de planejamento fatorial 24 com
repetição em triplicata no ponto central, a produção de quitosanase por Aspergillus
ochraceus em cultivo descontínuo submerso. Os resultados mostraram que foi possível
produzir quitosanase com atividade na ordem de aproximadamente 5,9 U/mL utilizando
Aspergillus ochraceus e que a atividade foi favorecida pelo aumento da agitação (r), da
razão de aeração (RA) e da concentração de substrato (Cs), enquanto que o aumento da
temperatura de cultivo (TMC) não favoreceu a resposta (atividade quitosanolítica).
Pode-se classificar as quitosanases em endoquitosanases e exoquitosanases. As
primeiras diferenciam-se por catalisar a hidrólise aleatoriamente, no interior do
biopolímero, gerando diversos tamanhos de oligossacarídeos. As exoquitosanases são
capazes de hidrolisar as terminações não redutoras, produzindo assim unidades
redutoras (Peter, 2005).
Palma-Guerrero et al. (2008) estudaram o biocontrole de fungos patogênicos e
demonstraram a lise da membrana plasmática e a inibição da germinação de esporos de
fungos. Eles observaram que os fungos entomopatogênicos e nematófagos foram menos
inibidos pela quitosana que os fitopatogênicos e micoparasíticos provavelmente porque
apresentam a enzima quitosanase que degrada a quitosana. Mesmo assim, a quitosana

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
29
pode ajudar a desenvolver novas estratégias para o biocontrole em geral de fungos
patogênicos, evitando o uso de fungicidas que prejudicam o meio ambiente e a saúde
humana.
Pagnoncelli (2008) estudou a produção de quitooligossacarídeos com
propriedades nutracêuticas a partir da quitosana por hidrólise enzimática, utilizando
processo fermentativo simultâneo. Duas cepas produtoras de quitosanaes foram
selecionadas, Paenibacillus chitinolyticus e Paenibacillus ehimensis, e avaliadas quanto
ao comportamento em meio de cultivo contendo açúcares simples e em relação às
variações de pH do meio. Os quitooligossacarídeos produzidos a partir do complexo
enzimático da primeira cepa apresentaram maior pico após 9h de hidrólise, enquanto
para a segunda cepa avaliada, após 20min, observou-se quitooligossacarídeos com grau
de polimerização entre 3 e 6 unidades.
2.2. Fungos
2.2.1. Características gerais
Desde o final dos anos 60 que os fungos deixaram de pertencer ao reino das
plantas e passaram a ter um reino próprio chamado de Reino Fungi. Foi a partir de
estudos morfológicos, bioquímicos e citológicos onde se concluiu que eles são tão
diferentes dos vegetais como dos animais. Distinguem-se de outros organismos
eucariotas por serem seres quimiorganoheterotróficos e, ainda, por apresentarem parede
celular rígida que é composta por quitina e glucano e uma membrana celular em que o
ergosterol substitui o colesterol (Trabulsi & Toledo, 1996; Murray et al., 2005).
Fazem parte do Reino Fungi cerca de 100 mil espécies, desde grandes como
os cogumelos, até microscópicas como os bolores e leveduras. Ainda hoje é utilizada
essa classificação, mas os avanços da biologia molecular e estudos bioquímicos levaram
a outras propostas de classificação em que os fungos e organismos afins foram incluídos
em três diferentes reinos: Protozoa, Chromista e Fungi (Kirk et al., 2008).
Abaixo, são apresentadas características que diferenciam os fungos das plantas:
• são incapazes de sintetizar clorofila, como também não apresentar celulose em
sua parede celular, com exceções como fungos aquáticos;
• apresentam algumas características de células animais (capacidade de depositar
glicogênio e a presença de substâncias quitinosas, em sua parede celular);

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
30
• podem ser eucarióticos (as leveduras), ou multinucleados (os bolores), e
apresentar retículo endoplasmático rugoso e mitocôndrias em seu citoplasma;
• são heterotróficos, alimentam-se de matéria orgânica viva (fungos parasitários),
ou morta (fungos saprofíticos);
Os fungos são amplamente encontrados na natureza, crescem rapidamente no
solo, na água, na atmosfera, bem como nos animais, nos vegetais, resíduos orgânicos e
desempenham um papel importante na vida do homem, quer de uma maneira benéfica,
quer de um modo prejudicial. Os fungos são um dos principais microrganismos
responsáveis pela decomposição da matéria orgânica, interferindo no ciclo do carbono,
do nitrogênio e de outros nutrientes da biosfera. São capazes de deteriorarem produtos e
bens de consumo, tais como alimentos, tecidos, papel, madeira. Os alimentos
armazenados representam um vasto meio para a proliferação dos fungos, principalmente
quando não são consideradas as condições básicas de armazenamento correto (Trabulsi
& Toledo, 1996).
Estão presentes funcionalmente em vários processos industriais, tais como, de
fabricação de pão, cervejas, vinhos e determinados tipos de queijos, sendo também
utilizados na produção comercial de muitos ácidos orgânicos, de alguns fármacos, como
a ergometrina e a cortisona, na obtenção de diferentes antibióticos, como a penicilina, e
de substâncias imunossupressoras, como a ciclosporina (Esteves et al., 1990). A Figura
4 ilustra a estrutura geral de uma célula de fungo.
Figura 4 – Representação da estrutura geral de uma célula de fungo.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
31
Os fungos podem se desenvolver em meios de cultivo especiais formando
colônias de dois tipos, leveduriformes e filamentosas, que se diferenciam pela
macromorfologia e micromorfologia. O primeiro tipo apresenta, em geral, característica
de consistência cremosa de cor branca a creme, brilhantes ou opacas, podendo
apresentar às vezes coloração escura ou alaranjada, e são formados por microrganismos
unicelulares que apresentam as funções vegetativas e reprodutivas. As filamentosas
apresentam características granulares, cotonosas, aveludadas ou pulverulentas, são
multinucleadas e apresentam-se em forma de tubos denominadas de hifas. Essas hifas
podem ser não septadas (cenocíticas) ou septadas (tabicadas). O conjunto de hifas é
denominado micélio.
As hifas asseptadas têm aparência anastomosada (concisa), formada por um
citoplasma estendido e polinucleado, enquanto que as hifas septadas contêm células
individualizadas, cada uma contendo o seu núcleo. Na Figura 5 são mostradas as
estruturas das hifas septadas e não septadas.
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas. (Adaptado de Guia.Bio,
2012)
O micélio que se desenvolve no interior do substrato e que é responsável pela
sustentação e absorção de nutrientes é denominado micélio vegetativo. O micélio que
cresce na superfície do meio de cultura é denominado micélio aéreo. Este pode ser
denominado também de micélio reprodutivo, quando o mesmo se diferencia para
sustentar os corpos de frutificação ou propágulos (Putzke & Putzke, 1998). Os
propágulos classificam-se de acordo com a origem em externos e internos, sexuados e
assexuados.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
32
2.2.2. Reprodução dos Fungos
A reprodução dos fungos ocorre em ciclos assexuais, sexuais e parassexuais. Em
fungos, a reprodução assexuada é observada com mais freqüência do que a reprodução
sexuada. A reprodução assexuada é normalmente a reprodução mais importante para a
propagação da espécie, feita através de fragmentação de artroconídios, fissão de células
somáticas, brotamento ou gemulação do blastoconídios-mãe e produção de conídios. A
reprodução sexuada envolve a união de duas células ou de dois órgãos sexuais
sexualmente compatíveis. A reprodução sexuada culmina na produção de basidiósporos,
no caso de basidiomicetos. A reprodução parassexuada ocorre com a fusão de hifas e
formação de um heterocário, que consiste na existência de núcleos haplóides
geneticamente distintos num mesmo citoplasma, os quais, após a fusão, originam
núcleos diplóides heterozigotos sujeitos a sucessivas divisões mitóticas. Apesar da
raridade dessas, a reprodução parassexual é de grande importância na evolução de
alguns fungos (Pelczar et al., 1996).
A reprodução assexuada é feita por conídios, esporos assexuados formados no
ápice de hifas modificadas, os conidióforos. Os conidióforos não possuem esporângios
de forma que os conídios ficam expostos. Na reprodução sexuada, com o encontro de
duas hifas haplóides distintas, ocorre a plasmogamia e um novo conjunto de hifas
dicarióticas é formado.
Os conídios podem ter formas esféricas, fusiformes, cilíndricas, piriformes, entre
outras. Apresentam-se como hialinos ou pigmentados, podem estar isolados ou
agrupados, a parede celular pode ser lisa ou rugosa e podem ser formados por apenas
uma célula ou apresentarem septos em mais de um plano.
Os conídios também podem originar-se em qualquer parte do micélio vegetativo
e são denominados conídios sésseis como é o caso do Trichophyton rubrum.
Alguns fungos patogênicos são identificados a partir de estruturas formadas pela
célula conidiogênica e o conidióforo. Essas estruturas são denominadas: aparelho de
frutificação ou conidiação. Os conídios podem formar cadeias sobre fiálides em volta de
uma vesícula, dilatando a extremidade do conidióforo. Essas estruturas ocorrem em
Aspergillus, apresentando-se em forma de cabeça ao redor de uma vesícula, e em
Penicillium agrupados em forma de pincel, como é ilustrada na Figura 6.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
33
Figura 6 – Conídios de Aspergillus e Penicillium
2.2.3. Metabolismo de fungos
Os fungos são seres aeróbicos e heterotróficos, podendo se desenvolver em
ambientes com baixa quantidade de oxigênio, como ocorre no crescimento vegetativo e
na reprodução assexuada, como também em ambientes com grande quantidade de
oxigênio como é o caso da reprodução sexuada.
Os fungos são capazes de produzir certas enzimas como lipases, invertases,
lactases, quitosanases, proteases, amilases, etc., que fazem a hidrólise do substrato
facilitando o mecanismo de transporte ativo e passivo. Esses substratos induzem a
formação de enzimas degradativas. Algumas espécies de fungos desenvolvem-se em
meios que apresentam amônia ou nitrilos como única fonte de nitrogênio. As melhores
fontes orgânicas para o seu desenvolvimento são carboidratos, como a D-glicose, e
alguns sais minerais como sulfatos e fosfatos.
A água é uma necessidade de todos os seres vivos, não sendo diferente para os
fungos que precisam de água para o seu desenvolvimento. Certos fungos apresentam
características halofílicas podendo crescer nos ambientes com grandes concentrações de
sal.
A razão de não existirem ambientes livres de espécies fúngicas deve-se ao facto
destes microrganismos conseguirem sobreviver em situações extremas como, por
exemplo, temperaturas que variam entre os - 5 e os 60 ºC, com pH de 1 a 9 e com baixas

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
34
concentrações de oxigênio, apesar de cada espécie possuir condições ideais para o seu
desenvolvimento (Chao et al., 2002).
Devido a grande faixa de temperatura ótima para o seu crescimento, os fungos
apresentam espécies psicrófilas, mesófilas e termófilas. Para a maior parte das espécies
fúngicas, a temperatura de desenvolvimento situa-se entre os 20 e 24 ºC e não crescem
acima dos 29 ºC. Em relação à umidade relativa, necessitam de pelo menos 50 %, mas
geralmente necessitam de mais de 65 % (Colakoglu, 2001).
O pH ideal para o desenvolvimento dos fungos varia na faixa entre 5 e 7.
Destaca-se, entretanto, que a maioria dos fungos suportam grandes variações de pH
como ocorre com os fungos filamentosos que podem crescer em pH variando entre 1,5 e
11. Por outro lado, as leveduras não suportam pH alcalino.
Os fungos crescem lentamente, se comparados com as bactérias, em meio de
incubação para o seu desenvolvimento. Para evitar contaminação por bactérias, que
podem se sobrepor ou inibir o seu crescimento, pode-se introduzir no meio de cultura
um antibacteriano de largo espectro como o cloranfenicol.
Algumas espécies de fungos se desenvolvem na presença de luz, outras são
inibidas e algumas podem crescer tanto na presença quanto na ausência deste agente. No
entanto a irradiação solar, em geral, é prejudicial ao seu desenvolvimento, por causa da
radiação ultravioleta.
2.2.4. Nutrição de fungos
Quanto ao aspecto da nutrição, os fungos são heterótrofos, ou seja, incapazes de
produzir seu próprio alimento. Entretanto, a forma de assimilar matéria orgânica é
diferente daquela utilizada pela maioria dos animais, que são heterótrofos por ingestão.
Nos fungos, a digestão é extracorpórea, ou seja, é realizada fora do corpo. O fungo
lança no ambiente, enzimas que degradam as moléculas orgânicas complexas e, depois,
absorve moléculas menores, mais simples. Por essa razão, são designados heterótrofos
por absorção. É devido a este processo que os fungos podem crescer dentro ou sobre os
alimentos. Desenvolve-se geralmente em meios contendo um pH baixo, uma fonte de
carbono, uma fonte de nitrogênio orgânico ou inorgânico e alguns minerais. Alguns
necessitam de vitaminas. Quanto aos modos de vida, os fungos podem ser
decompositores, parasitas, mutualísticos e predadores (Raven, 2001).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
35
Os fungos decompositores, saprófagos (do grego, saprós, “podre”, e phagos,
“comedor”), juntamente com as bactérias, são os principais decompositores da biosfera,
participando intensamente do processo de degradação da matéria orgânica morta, o que
promove a reciclagem dos elementos químicos constituintes dos seres vivos. A maioria
deles vive no solo, obtendo nutrientes de seres mortos. Em florestas, onde a matéria
orgânica vegetal é abundante, os micélios dos fungos decompositores absorvem os
nutrientes de folhas e galhos caídos, decompondo-os. Os fungos decompositores
também são responsáveis pelo apodrecimento de alimentos e de outros materiais, como
a madeira.
Os fungos parasitas vivem à custa de outros organismos vivos, prejudicando-os.
Ao parasitar o corpo de um ser vivo, animal ou vegetal, o fungo pode até provocar sua
morte. Várias doenças são causadas por fungos parasitas. Nas plantas, cita-se a
ferrugem do café, que, no Brasil, é motivo de grande preocupação por parte dos
cafeicultores, devido aos prejuízos que acarreta. As micoses que atacam a pele de seres
humanos e animais são provocadas por fungos.
Existem fungos que se associam a outros organismos, estabelecendo uma relação
em que há benefício mútuo para os indivíduos envolvidos. Esses fungos são
denominados mutualísticos. A maioria vive associada a seres fotossintetizantes, como
as plantas, cedendo-lhes parte da água e dos nutrientes que as hifas absorvem do solo.
As plantas, por sua vez, cedem ao fungo certos açúcares e aminoácidos. Duas
associações mutualísticas formam estruturas bem características: as micorrizas,
associação que envolve fungos e raízes de plantas, e os líquens, formados por fungos e
algumas variedades de algas e cianobactérias.
Certos fungos atuam como predadores. Na maioria dos casos, as hifas secretam
substâncias aderentes que aprisionam os organismos que tocam os fungos. Dessa
maneira, as hifas penetram o corpo da presa, crescem e se ramificam, espalhando-se no
interior do organismo e absorvendo seus nutrientes, causando-lhe a morte. O gênero
Arthrobotrys apresenta uma forma mais elaborada de predação. O fungo vive no solo e
captura nematódeos microscópicos que habitam o mesmo ambiente. As hifas
apresentam pequenos anéis que se estreitam quando a presa passa por eles. As hifas do
fungo logo invadem o corpo do nematódeo, digerindo-o.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
36
2.2.5. O fungo Metarhizium anisopliae
Existem três espécies de fungos do gênero Metarhizium, divididas em dez
variedades: M. anisopliae, variedades anisopliae, majus, lepidiotum e acridum; M.
flavoviride, variedades tipo E, flavoviride, minus, novazealandicum e pemphigum; e M.
album (Driver et al. 2000). E, mais recentemente, foi descrita na China uma nova
variedade de M. anisopliae, dcjhyium (Dong et al. 2009).
Data de 1879 quando o russo Ilya Metchnikoff aplicou o fungo para combater
larvas de um curculionídeo, praga de beterraba, e foi classificado como Metarhizium
anisopliae por Sorokin em 1883. É um fungicida natural para mais de 300 espécies de
insetos, especialmente pragas da agricultura e pecuária. No Brasil, o fungo Metarhizium
anisopliae teve a sua aplicação com sucesso pela primeira vez em 1965, no controle da
cigarrinha-da-cana Mahanarva posticata (Alves, 1998).
O fungo Metarhizium anisopliae é um importante agente entomopatogênico
utilizado no controle de pragas, como cupins, gafanhotos, cigarrinhas e besouros, sendo
amplamente estudado em todo o mundo. Existe amplamente na natureza e é facilmente
encontrado no solo, onde sobrevive por longos períodos e seu desenvolvimento se dá
geralmente entre 15 e 32 °C, sendo ideal entre 24 e 30 °C e com pH ótimo igual a 6,9.
Apresenta micélio hialino e septado, com conidióforos característicos, sobre os quais
surgem conídios cilíndricos organizados em colunas. Este fungo ao atacar os insetos
provoca rigidez e os recobre por uma camada pulverulenta de conídios, resultando em
colorações que variam do verde claro ao escuro, cinza ou branco com pontos verdes
(Tinline, 1971; Alves, 1998, Driver et al. 2000; Arruda, 2005).
Quanto a sua morfologia, apresenta-se como um fungo filamentoso, com corpo
de frutificação equivalente a um esporodóquio agregado a hifas intimamente
entrelaçadas, com massa compactada de conidióforos característicos, simples ou
ramificados, formando células esporogênicas denominadas fiálides, resultando nos
fialosporos (Wang et al., 2002).
O Metarhizium anisopliae é um fungo de cultivo simples, sendo necessário
quase que exclusivamente o nutriente a base de amido. Um dos fungos mais estudados
em programas de manejo de pragas já que apresenta grande potencialidade
entomopatogênica (Onofre et al., 2002). O Metarhizium anisopliae é um microrganismo
pouco exigente, pois pode se desenvolver em diversos meios de cultura como: amido,
glicose, glicerina, levulose, maltose, sacarose e quitina (Jabor et al., 2003).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
37
Em geral, os fungos produzem enzimas que convertem os tecidos dos insetos em
nutrientes. Porém, a cutícula esclerotizada dos insetos não é bem utilizada pelos fungos,
excetos pelos entomopatogênicos que desenvolveram enzimas eficientes para degradar
essa camada protetora. Destaca-se a enzima Pr1, produzida por Metarhizium anisopliae,
que tem um poder de degradação bem superior a qualquer outra enzima sintetizada por
esse grupo de fungos (Dias, 2005). Desta forma, o Metarhizium anisopliae (fungo
entomopatogênico) ao atacar a célula dos insetos, produz grande quantidade de enzimas
que degradam a membrana citoplasmática e inativa a célula e, consequentemente, mata
o inseto, logo é evidente a relação entre o controle de pragas e produção de enzimas.
No momento, sobre a comercialização de fungos entomopatogênicos no Brasil,
em sua maioria, são formulados em grânulos, compostas do fungo mais o substrato
(arroz + fungo), como também na forma de pó molhável (WP), resultado da moagem do
fungo com o substrato. Esses produtos representam 90% e 8,5%, respectivamente, da
produção de fungos no Brasil, e sendo suas concentrações finais ficam em torno de
5x108 conídios/g para Metarhizium anisopliae e 1x109 conídios/g para Beauveria
bassiana (Alves et al., 2008).
Segundo Batista Filho et al. (2002), Garcia et al. (2005) e Mello et al. (2006), o
Metarhizium anisopliae tem sido bastante utilizado para o controle de diferentes pragas
no Brasil, como a broca da bananeira, pragas de grãos armazenados, carrapatos, cupim
de montículo em pastagens, larvas de escarabeídeos que atacam a cana de açúcar e
cigarrinha da cana-de-açúcar.
É comum a ocorrência natural do fungo Metarhizium anisopliae atacando
cigarrinha-das-raízes de canaviais pelo país, o que incentiva a utilização dele como
agente de controle biológico em áreas infestadas por esse tipo de praga. A aplicação de
recursos no uso de programa de manejo integrado da cigarrinha tem sido reforçado por
razões ambientais e econômicas (Dinardo-Miranda, 2001).
Faria & Magalhães (2001) fizeram uma avaliação do uso de fungos
entomopatogênicos no Brasil, na qual observaram que a qualidade dos micoinseticidas
disponíveis no Brasil poderia ser incrementada de forma considerável quanto a sua
formulação, ou seja, antes da venda, poderiam realizar melhorias no tratamento
posterior, adição de substâncias que lhes assegurem melhorias na eficiência de controle,
capacidade de armazenamento ou praticidade de manuseio, ou de qualquer outro critério
que resulte em vantagem em relação ao produto bruto. Ressaltaram a pouca praticidade

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
38
em alguns casos, como exigência de lavagem, e em outros podendo causar o
entupimento de bicos dos pulverizadores devido à elevada proporção de inertes,
principalmente quando são empregados baixos volumes de aplicação. Outro problema
evidenciado com os inseticidas nacionais diz respeito à pequena sobrevida, devendo a
ser usados em, no máximo, 30 dias depois de produzidos, quando armazenados à
temperatura ambiente e em local sombreado, fazendo as vendas ocorrerem quase que,
exclusivamente, sob encomenda. Comparativamente, os bons inseticidas sob
comercialização em outros países apresentam maior concentração de ingrediente ativo,
maior sobrevida (alguns produtos podem ser armazenados por mais de 8 meses à
temperatura ambiente) e praticidade (são formulados na forma de GDA (grânulos
dispersíveis em água) ou de óleos emulsionáveis, por exemplo, podendo ser adicionados
diretamente ao tanque do pulverizador.
Rangel et al. (2007), instalaram uma Unidade de Observação para o controle
microbiano das cigarrinhas das pastagens Distrito de Itahum, em Dourados/MS, devido
as necessidades de agricultores familiares de produção de leite. As aplicações do fungo
Metarhizium anisopliae foram realizadas por pulverização com equipamento de tração
animal. As condições do ambiente de estudo foram favoráveis ao desenvolvimento da
população das cigarrinhas e à incidência do fungo. Como resultados, observaram que os
níveis de espumas, ninfas e adultos reduziram significativamente após as aplicações,
levando os Agricultores Familiares a total satisfação com os ganhos na produção de
todos os envolvidos.
Segundo Vieira et al. (2007), o fungo entomopatogênico Metarhizium anisopliae
var. acridum tem sido utilizado no controle de pragas, e assim, visando este fim,
analisaram em laboratório a esporulação, a germinação dos conídios e as características
morfológicas peculiares desse fungo em diferentes temperaturas. Esperou-se o
desenvolvimento das colônias em períodos de 1 a 15 dias. Foi observado que
temperaturas de 25, 28 e 30 ºC proporcionaram maiores taxas de esporulação e elevadas
porcentagens de germinação dos conídios. A 25, 28 e 30 ºC observou-se um
desenvolvimento mais rápido das microestruturas somáticas e reprodutivas (hifas,
anastomoses, apressórios, conidióforos e conídios elipsóides). Nas temperaturas
extremas de 40 e 45 ºC não foi detectado esporulação, germinação, nem
desenvolvimento de nenhuma microestrutura.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
39
Lopes et al. (2007) avaliaram o efeito de formulações de Beauveria bassiana e
Metarhizium anisopliae sobre ninfas de Amblyomma cajennense (transmissor da febre
maculosa ao homem) em laboratório. Ninfas não alimentadas foram pulverizadas com
suspensões contendo 2x107 e 4x107 conídios/mL de B. bassiana, e 1x107, 1,5x107,
2x107, 3x107 e 5x107 conídios/mL de M. anisopliae. A mortalidade de ninfas,
decorridos três dias da aplicação, foi significativa nas concentrações de 3x107 e 5x107
conídios/mL de M. anisopliae, com 57,1 e 56,2 %, respectivamente. Após seis dias a
mortalidade foi de 100 % nessas duas concentrações. A mortalidade foi
significativamente maior que a observada para B. bassiana em concentrações
semelhantes.
Guirado et al. (2009) realizaram estudos para controlar cupins de montículo em
área de pastagem, utilizando fungos entomopatogênicos Metarhizium anisopliae e
Beauveria bassiana em diferentes doses e formulações. A avaliação foi realizada após
30 dias da aplicação dos produtos. Os melhores resultados em ordem decrescente de
eficiência foram: M. anisopliae 30 mL + 1000 mL de água com 100 % de eficiência; B.
bassiana 10g + M. anisopliae 20 mL + 1000 mL de água com 85,71 % de eficiência; M.
anisopliae 20 mL + 1000 mL de água com 71,42 % de eficiência. Os demais
tratamentos não diferiram da testemunha.
Loureiro et al. (2012) compararam a eficiência de isolados de Metarhizium
anisopliae em controlar populações naturais de Mahanarva fimbriolata (cigarrinha-da-
raiz) em cultivos comerciais de cana-de-açúcar colhida mecanicamente. Foram
aplicados os isolados IBCB 348, IBCB 408, IBCB 410 e IBCB 425 do fungo em
concentração de 1,5×1012 conídios/ha. Aplicou-se o planejamento experimental em
blocos casualizados com 5 tratamentos e 4 repetições, sendo cada repetição composta
por 7 linhas de 100 m de comprimento e espaçamento de 1,5 m. Após 30 dias de
pulverização foi observado para os isolados IBCB 408 e IBCB 425 uma eficiência de
controle de ninfas de 63 e 62%, respectivamente, já no caso dos adultos chegou a 100%.
Após 60 dias o isolado IBCB 425 foi o mais eficiente para controlar as ninfas com
48,4%.
Assis et al. (2012) estudaram a citotoxidade de quitoolissacarídeos (QOS) e
oligômeros A, B e C (soluções compostas por diferentes quantidades de QOS)
produzidos por hidrólise enzimática pelo fungo Metarhrizium anisopliae. Foi analisado
o efeito de antiproliferativo destas moléculas em células tumorais (HepG2 e HeLa) e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
40
também em célula normal (3T3). A atividade antioxidante foi analisada em vários
experimentos in vitro. A glicosamina apresentou maior toxicidade (cerca de 92%) para
todas as células estudadas. No entanto, os oligômeros obtidos após hidrólise não
demonstraram efeito tóxico sobre a célula normal (3T3). Além disso, mostraram que
uma pequena quantidade de outros COS pode diminuir o efeito citotóxico da
glicosamina contra células 3T3, indicando que glicosamina poderia ser usada como uma
droga antitumoral na presença de outros COS.
2.3. Fermentação
Os processos fermentativos são caracterizados por processos nos quais
microrganismos realizam a conversão catalítica de uma dada substância num
determinado produto. Estes processos podem ser classificados como processos
descontínuos, contínuos e descontínuos alimentados. A classificação destes processos
está diretamente ligada à forma de como o substrato é adicionado e como o produto é
retirado. No processo de fermentação descontínuo o substrato é colocado no
fermentador, em seguida é inoculado e após o término do processo o produto é retirado.
No processo contínuo, o substrato é colocado num reator com vazão constante e o meio
fermentado é retirado com a mesma vazão de entrada do reator (Faccioti, 2001). Nos
processo descontínuos alimentados à adição de substrato e a retirada do mesmo deve
ocorrer de forma intermitente (Carvalho & Sato, 2001).
De certa forma, os processos fermentativos representam um elo entre as antigas
artes de elaboração de alimentos, como queijos e vinho, a partir de uma flora
microbiana natural, com a moderna indústria de fermentação de alimentos, onde são
utilisados cultivos puros e equipamentos sofisticados para o controle do processo
(Assis, 2010).
Atualmente, a maioria dos processos industriais se desenvolve em cubas de
vidro ou de aço de até 20 litros. Os agentes biológicos se encontram submersos no meio de
cultivo que ocupa, aproximadamente, 75% da cuba. As vezes é necessário injetar ar, e em muitas
fermentações se forma espuma. O desenho do biorreator deve se adequar ao objetivo do
processo, respondendo eventualmente a diversos imperativos, tais como a esterilização do
sistema, a aeração e homogeneização do meio, o acréscimo de nutrientes e de aditivos
antiespuma, a manutenção do pH, etc. Os modelos de fermentadores mais utilizados com
microrganismos contam com aeração e agitação mecânica. Esta facilita a distribuição dos

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
41
nutrientes na cuba, mas gera calor que deve ser eliminado mediante a circulação de água
fria. Em outros tipos de biorreatores, em coluna ou torre, a homogeneização depende da injeção de
ar. Os sistemas submersos são apropriados para o cultivo de microrganismos livres, mas resultam
pouco econômicos quando se trabalha com células ou enzimas caras. A imobilização em
fermentadores menores, seja por adesão a um suporte inerte, seja por inclusão dentro de um
polímero que permita o contato com o meio de cultura, além de simplificar a purificação do
produto permite a reutilização das células ou das enzimas, que permanecem dentro do
biorreator (Malajovich, 2009).
As enzimas são produtos de alto valor agregado, amplamente empregadas em
uma gama de indústrias, e entre as fontes para sua obtenção, destacam-se as de origem
microbiana, principalmente, devido a elevada seletividades dos substratos utilizados e
produtos formados. A principal forma de obtenção de enzimas, em escala industrial, é a
fermentação submersa, por esta apresentar vantagens em relação a fermentação
semisólida, como: maior controle do processo fermentativo e consequente reprodução
da cinética microbiana, além de maiores rendimentos (Wanderley, 2011).
2.3.1. Preparo do inóculo
Denomina-se inóculo um volume de suspensão de microrganismo de
concentração adequada, capaz de garantir, em condições econômicas, a fermentação de
uma dada quantidade de meio. O volume do inóculo, assim como sua concentração de
células, vai depender tanto do processo de fermentação c omo da quantidade de meio a
ser fermentado (Carvalho & Sato, 2001).
A partir de uma cultura pura, inocula-se um volume relativamente pequeno de
meio nutriente, que vai depender do tipo de microrganismo cultivado (pois cada espécie
microbiana possui velocidades diferentes de crescimento), sendo, em seguida, incubado
em condições favoráveis ao desenvolvimento do microrganismo. A suspensão obtida é
transferida para um frasco maior em agitador rotativo ou recíproco (shakers), que por
sua vez é adequadamente incubado, e a seguir, transferido para um frasco contendo um
volume ainda maior deste meio, até se conseguir alguns litros de suspensão microbiana.
Os cuidados com assepsia, volumes de meio utilizado, numero de transferência bem
como as condições de incubação, podem variar de acordo com o tipo de processo de
fermentação que se deseja realizar.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
42
2.3.2. Produção de enzimas em cultivo submerso descontínuo
A produção de enzimas, usando processos fermentativos, é constituída
basicamente por duas etapas: a primeira caracteriza-se pela preparação do inóculo e a
segunda, pela fermentação do substrato (Carvalho & Sato, 2001). A finalidade da
primeira etapa é o preparo do microrganismo em condições adequadas para que possa
ocorrer o desenvolvimento da segunda etapa. Diferentemente do cultivo em estado
sólido, no cultivo submerso os microrganismos encontram-se em um meio com elevada
atividade de água.
2.4. Planejamento experimental
Um bom planejamento tem como fundamental projetar um experimento de
forma que ele seja capaz de fornecer exatamente o tipo de informação que procuramos.
Para isso é necessário saber, em primeiro lugar, o que é mesmo que se deseja procurar.
Planejamento experimental, ou delineamento experimental, é um conjunto de
ensaios estabelecido com critérios científicos e estatísticos, objetivando determinar a
influência de diversas variáveis nos resultados de um dado sistema ou processo (Barros
Neto et al., 2001; Rodrigues & Iemma, 2005). De acordo com o propósito dos ensaios,
esse objetivo maior pode ser mais detalhado em:
– determinar quais variáveis são mais influentes nos resultados;
– atribuir valores às variáveis influentes de modo a otimizar os resultados;
– atribuir valores às variáveis influentes de modo a minimizar a variabilidade dos
resultados;
– atribuir valores às variáveis influentes de modo a minimizar a influência de variáveis
incontroláveis.
O planejamento experimental, baseado nos fundamentos estatísticos é, sem
dúvida alguma, uma ferramenta poderosa para se chegar às condições otimizadas de um
processo, desenvolvimento da formulação de produtos dentro das especificações
desejadas ou simplesmente para avaliar os efeitos ou impactos que os fatores (variáveis
independentes) têm nas respostas (variáveis dependentes) desejadas (Barros Neto et al.,
2001; Rodrigues & Iemma, 2005). Da utilização das técnicas estatísticas de
planejamento experimental, é possível chegar benefícios importantes, como:

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
43
– reduzir o número de ensaios sem prejuízo da qualidade da informação;
– estudar simultaneamente diversas variáveis, separando seus efeitos;
– determinar a confiabilidade dos resultados;
– realizar a pesquisa em etapas, num processo iterativo de acréscimo de novos ensaios;
– selecionar as variáveis que influem num processo com número reduzido de ensaios;
– representar o processo estudado através de expressões matemáticas;
– elaborar conclusões a partir de resultados qualitativos.
2.4.1. Planejamento fatorial
Em um planejamento fatorial é investigado as combinações de níveis de estudo
de cada fator. Para tanto, inicialmente, é necessário especificar esses níveis em cada
fator, quer dizer, os valores dos fatores que serão utilizados nos experimentos. Um
ensaio experimental é definido como um experimento constituído de um conjunto de
níveis definido. Geralmente, quando se tem n1 níveis de um determinado fator 1, n2 do
fator 2, ..., e nk do fator k, o planejamento será descrito por um fatorial n1 x n2 x ... x nk,
que corresponde ao número mínimo de experimentos para se ter um planejamento
fatorial completo.
É necessário fazer cada fator variar e observar o resultado dessa variação para se
estudar o efeito sobre a resposta. Assim, os ensaios precisam ter um mínimo de dois
níveis por fator para a realização desses ensaios. Para k fatores, ou seja, k variáveis de
controle, o planejamento de dois níveis precisará de 2 x 2 x ... x 2 = 2k, para o número
de ensaios diferentes, sendo conhecido como planejamento fatorial 2k.
Para o planejamento fatorial, são especificados os valores dos fatores em cada
experimento, e estes são classificados em níveis, variando-se, por exemplo, de -1
(mínimo) a +1 (máximo). A partir de uma matriz serão apresentadas todas as
combinações entre o número de fatores e de níveis nos experimentos a serem realizados
(Barros Neto et al., 2001; Rodrigues & Iemma, 2005).
2.4.2. Planejamento Plackett & Burman
A maioria dos planejamentos do tipo “Screening” (Delineamentos de Seleção de
fatores) foi descrito originalmente por Plackett & Burman em 1946. Os planejamentos
experimentais de Plackett & Burman são eficientes em situações que envolvem uma

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
44
grande quantidade de variáveis a serem exploradas em um processo de otimização de
fatores (Reddy et al., 2008).
As matrizes Plackett & Burman consistem de planejamentos fatoriais de dois
níveis para estudar k = N – 1 variáveis em N ensaios. São construídas para ensaios
múltiplos de 4, sendo essas matrizes para ensaios de 8, 12, 16,..., 100. Cada variável
independente em uma matriz assume uma configuração de dois níveis, o inferior (-1) e o
superior (+1), para o nível intermediário é estabelecido (0). Para cada vez que a variável
aparece no nível inferior, terá o par equivalente no nível superior (Rodrigues & Iemma,
2005).
Para evitar dúvidas nos resultados dos efeitos de algum fator no planejamento de
Plackett & Burman, o número de ensaios a ser realizado deve ser no mínimo o número
de variáveis mais 4. No caso de um fatorial completo o número de ensaios a ser
realizado é bem maior (Rodrigues & Iemma, 2005).
Mesmo já determinado o planejamento Plackett & Burman, torna-se importante
à realização de experimentos relativos aos pontos centrais de modo a se avaliar melhor
as possíveis interferências analíticas que podem ocorrer durante os diferentes
experimentos. A repetição do ponto central permite também a determinação do erro
puro e da repetibilidade do processo. Na prática, esta é uma ferramenta que permite
identificar melhor se o processo em estudo está sob controle (Rodrigues & Iemma,
2005).
É possível que no planejamento Plackett & Burman não tenha sido considerado
a codificação do ponto central (0) para repetição, pois, matematicamente, a estimativa
do valor desse efeito codificado (zero) vezes, teria como resposta zero. Porém, no
resultado prático do experimento, consegue-se informações valiosas com essas
repetições no ponto central (Rodrigues & Iemma, 2005).
2.4.3. Aplicações do Planejamento Plackett & Burman
Ghanem et al. (2000) produziram xilanase por Aspergillus terreus cultivadas
sobre palha de trigo finamente moída em fermentação de estado sólido. A composição
ideal do meio foi estabelecida aplicando-se o planejamento experimental de Plackett &
Burman. A melhor atividade enzimática foi obtida em um meio contendo 10 g/L de

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
45
palha de trigo por frasco umedecido com uma solução concentrada de sal com teor de
água inicial de 75 % e incubadas durante 4 dias a 30 °C. Observou-se que íons
metálicos como Hg2+, Cu2+, Co2+, Fe3+ e Pb2+ inibiram fortemente a xilanase, enquanto
que Ca2+ ativou a enzima.
Mu et al. (2005) estudaram as otimização de meio de fermentação para a reação
de estereoinversão por Candida parapsilosis CCTCC M203011 utilizando o
planejamento Plackett & Burman. Os resultados mostraram que dentre os fatores
estudados, os fatores mais significativos foram: a Concentração de MgSO4, ZnSO4 e
KH2PO4 e que após a utilização da metodologia de superfície de resposta foi constatado
que o ponto ótimo para a otimização foi conseguido com as concentrações dos fatores
MgSO4, ZnSO4 e KH2PO4 nos valores em: 1,17 g/L, 0,10 g/L e 9,41 g/L,
respectivamente, elevando a taxa de conversão enantiosseletiva de 75,2 % para 96 %.
Naveena et al. (2005) empregaram o planejamento Plackett & Burman para a
seleção de 15 parâmetros para produção de ácido láctico L(+) de farelo de trigo,
substrato de baixo custo e suporte sólido, por Lactobacillus amylophilus GV6 em
fermentação de estado sólido (SSF). Onze nutrientes pertencentes a duas categorias,
fontes de nitrogênio e fontes de sal, junto com três parâmetros físicos e um tampão
foram os parâmetros considerados. Os autores verificaram influência na produtividade
devido às fontes de nitrogênio, peptona, extrato de levedura e tri-citrato de amônio,
juntamente com o NaH2PO4.2H2O e Tween 80, que podem ainda serem otimizados para
o aumento na produção de ácido lático. Esses pesquisadores ainda chamam atenção para
o uso escasso deste planejamento em fermentação em estado sólido e que o mesmo
ainda não havia sido considerado anteriormente para a conversão direta de amido em
ácido láctico L (+) usando um sistema bacteriano.
Khosravi-Darani et al. (2008) estudaram a aplicação de Plackett & Burman na
análise dos fatores significativos para a produção de ácido cítrico por Aspergillus niger
ATCC 9142 utilizando palha de trigo como substrato. Os resultados mostraram que a
produção, com a palha de trigo pré-tratada com HCl, NaOH e Uréia, foi mais eficiente e
que os fatores mais significativos na produção foram: Solvente, Teor de Umidade,
Temperatura, Tempo de Incubação, Concentração Inicial de Açúcar e Concentração de
Metanol.
Ji et al. (2009) estudaram o desenvolvimento de um meio de cultura econômico
para a produção do 2,3-butanodiol através da fermentação da glicose e xilose por

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
46
Klebsiella oxytoca utilizando o planejamento Plackett & Burman. Os resultados
mostraram que a uréia e a maceração de milho foram os fatores mais significativos para
a produção do 2,3-butanodiol e que através de planejamento experimental com repetição
em triplicata no ponto central para determinar os níveis ótimos de produção. Os autores
constaram que o rendimento atingido foi 85,6% do valor teórico.
Paula & Arruda. (2009) observaram a necessidade da busca por alternativas mais
eficientes ao método termomecânico utilizado para a degradação de milhões de
toneladas de penas de aves provenientes de frigoríficos e aviários, cuja rígida estrutura é
composta por mais de 90 % de queratina. Para esse fim, utilizaram de comunidades
microbianas para a finalidade proposta, testando o planejamento experimental Plackett
& Burman para verificar a influência de diferentes microrganismos simultaneamente,
com um número reduzido de experimentos. Testaram 32 bactérias isoladas, inoculadas
em meio contendo somente uma pena como única fonte de carbono, durante sete dias,
de onde foi selecionado as cepas para os consórcios microbianos. Experimentos unindo
as cepas NP5, FCA7, NP4 e S14 demonstraram diferentes atividades queratinolíticas,
aumentando o nível de degradação, até 87,8%, comparada com as cepas
individualmente, variando entre 26,4% e 78,0%.
Rajendran & Thangavelu (2009) obtiveram valores máximos da atividade de
produção de lipase em 3,98 U/mL e concentração de massa celular em 5,62 g.L-1 com R.
arrhizus em cultivo submerso descontínuo otimizado pelo planejamento Plackett &
Burman. As variáveis mais importantes que afetaram a produção da lipase foram azeite
de oliva, peptona, KH2PO4, CaCl2.2H2O e MgSO4.7H2O.
Li et al. (2010) estudaram a otimização de meio de cultura para a produção de
Streptolydigin (um antibiótico) por Streptomyces lydicus AS 4,2501 utilizando Plackett
& Burman e a metodologia de superfície de resposta. Os resultados mostraram que
dentre os fatores estudados, os fatores mais significativos para a produção de
Streptolydigin e otimização do meio de cultura foram: Concentração de Amido,
Concentração de Peptona e a Concentração de K2HPO4.
Chen et al. (2010) estudaram a otimização de meio de cultura para a produção da
Luciferase – que são enzimas que catalisam reações biológicas transformando energia
química em energia luminosa - por Bacillus Subtilis em fermentação descontínua
utilizando planejamento Plackett & Burmam. Os resultados mostraram que dentre os
componentes estudados, os fatores mais significativos para a produção e otimização do

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
47
meio de cultura para a produção da Luciferase foram: Concentração de Glicose 1,17 %,
Extrato de Levedura 2,27 % e Glutamato 0,55 %.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o projeto Plackett & Burman e a metodologia de superfície de resposta. Os
resultados mostraram que a eficiência máxima de extração da quitina foi conseguida nas
seguintes condições: Concentração de casca de camarão (50 g/L), glicose (50 g/L),
tempo de incubação de 5 dias e inoculo com 0,05 de OD (densidade óptica).
Gao et al. (2010) desenvolveram um meio para a fermentação do extrato bruto
de inulina de tubérculos de alcachofra de Jerusalém por Paenibacillus polymyxa ZJ-9
para produzir R,R-2,3-butanodiol (R-R,2,3-BD). Inulina, K2HPO4 e NH4Cl foram os
parâmetros principais na fermentação de acordo com os resultados obtidos com o
planejamento experimental Plackett & Burman. A concentração ideal dos três fatores,
77,14 g/L, 3,09 g/L e 0,93 g/L, respectivamente, foi obtida pelo planejamento Box-
Behnken e pela superfície de resposta. Sob as condições ideais, a concentração do R-
R,2,3-BD obtida foi 36,92 g/L, em mais de 98 % de pureza óptica. Comparado com
outras fontes de carbono investigadas, a fermentação do extrato bruto de inulina obteve
o maior rendimento de R-R,2,3-BD.
Uma metodologia de superfície de resposta foi empregada com sucesso para
otimizar um meio de fermentação chamada VHG (Very High Gravity), referência à alta
concentração de açúcares e densidade dos mostos, baseado na água maceração de milho
CSL (Corn Steep Liquor) e outros nutrientes de baixo custo para a produção eficiente de
etanol a partir de glicose por S. cerevisiae. Usando a composição média otimizada (g/L:
CSL 44,3; uréia 2,3; MgSO4.7H2O 3,8; CuSO4.5H2O 0,03), a levedura industrial PE-2
foi capaz de fermentar até 330 g/L de glicose e produzir 18,6 % (v/v) de etanol, com
uma produção descontínua de 2,4 g/L/h e um rendimento em etanol de 93 % (Pereira et
al., 2010).
O planejamento Plackett & Burman juntamente com o Planejamento Composto
Central (CCD) foram aplicados para otimizar o meio de cultura para produção de etanol
por Clostridium autoethanogenum com monóxido de carbono (CO) como fonte única de
carbono e um meio contendo (g/L): 1,0 de NaCl, 0,1 de KH2PO4, 0,02 de CaCl2, 0,15 de
extrato de levedura, 0,116 de MgSO4, 1,694 de NH4Cl e pH 4,74. O rendimento ótimo
de etanol previsto pela metodologia de superfície de resposta (RSM) e um algoritmo

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
48
genético de rede neural artificial (ANN-GA) foram 247,48 e 261,48 mg/L,
respectivamente. Valores semelhantes aos obtidos experimentalmente sob as condições
ideais sugeridas pelos métodos estatísticos (254,26 e 259,64 mg/L). A adequação do
modelo ANN-GA foi maior do que o modelo RSM. Os rendimentos obtidos foram
substancialmente superiores aos relatados anteriormente (60-70 mg/L) com essa cultura
(Guo et al., 2010).
Rigueira (2010) avaliou a influência das variáveis do procedimento de extração
na porcentagem de recuperação das aminas encontradas no leite cru utilizando um
planejamento para “screening” do tipo Plackett & Burman, para os ácidos
tricloroacético (TCA) e sulfosalicílico (ASS).
Quines (2010) estudou a necessidade ou não de se fazer um pré-tratamento da
biomassa antes da extração. Um planejamento experimental Plackett & Burman de doze
ensaios mais três pontos centrais foi utilizado para selecionar as variáveis mais
significativas na extração de Poli(3-hidroxibutirato) (P(3HB)) por 1,2-carbonato de
propileno. Foram realizados dois testes de recuperação do solvente 1,2-carbonato de
propileno, após a extração, sendo um com aplicação de aquecimento e o outro com
aquecimento e vácuo. Os resultados mostraram que o pré-tratamento térmico levou a
uma maior porcentagem de recuperação e de pureza do polímero.

Capítulo 3 Metodologia Experimental

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
50
3. Metodologia Experimental
O presente capítulo apresenta os matérias e métodos utilizados na produção de
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso.
3.1. Quitosana
Neste trabalho utilizou-se uma quitosana (Lote 067) obtida junto a POLYMAR/CE,
com no mínimo 80% de desacetilação. Esse substrato foi utilizado como fonte de carbono e
energia tanto no estudo cinético como nos ensaios do planejamento de seleção de fatores
Plackett & Burman e Planejamento Composto Central, para a produção de quitosanase pelo
Metarhizium anisopliae, por fermentação submersa.
3.2. Microrganismo
O fungo Metarhizium anisopliae estirpe (CG374), utilizado neste estudo, foi cedido
pela Embrapa Recursos Genéticos e Biotecnologia (Brasília / DF-Brasil) e foi mantido em
placas de Petri contendo o meio Batata Dextrose Agar (PDA). As placas foram incubadas em
estufa a 25 ºC. O pH do meio de manutenção foi mantido entre 6,0 e 6,5.
3.2.1. Escolha da cepa produtora de quitosanase
A escolha do microrganismo Metarhizium anisopliae foi feita com base na capacidade
de crescer em meio contendo quitosana como única fonte de carbono, além de apresentar
grande tolerância a pH variando a uma taxa de 2,0 a 8,5, sendo 6,9 a melhor condição para o
crescimento vegetativo e esporução, de acordo com Assis (2010).
3.2.2. Manutenção da cepa
A manutenção da cultura foi realizada em placas petri contendo o meio Batata
Dextrose Agar (PDA). A renovação das células foi realizada a cada trinta dias através de
subcultivos das células no meio PDA.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
51
3.3. Produção de enzima
No preparo do inóculo, utilizou-se 7 placas cultivadas e mantidas, com o fungo
Metarhizium anisopliae, em meio PDA a 25 ºC durante 5 dias em estufa. O meio de cultura
para a produção da enzima quitosanase apresentou os seguintes componentes: quitosana;
peptona; extrato de levedura; NaNO3; K2HPO4; KCl; MgSO4.7H2O e FeSO4, com
concentrações de acordo as matrizes dos planejamentos: Plackett & Burman e composto
central. Para a produção da enzima no cultivo, a biomassa foi transferida através de água
estéril, para um Béquer de 500 mL que, em seguida, foi colocado em Erlenmeyers de 250 mL
na proporção de 20 mL de biomassa para 180 mL do meio de cultura. Os Erlenmeyers
cultivados foram então incubados em incubador rotativo a 25 ºC e 150 rpm durante 24 h.
Alíquotas foram retiradas e centrifugadas a 8000xg por 5 min, a cada 6 horas, para se
determinar a atividade quitosanolítica, teores de açúcares redutores e de proteína total.
Ressalta-se que todos os experimentos foram feitos em triplicata.
3.4. Determinação da Proteína Total
A determinação da proteína total foi efetuada de acordo com o método modificado de
Sedmak & Grossberg (1977), que se baseia na diminuição da absorbância do corante azul de
Coomassie, devido à ligação proteína-corante, a 595 nm, comparado a absorção máxima a
465 nm, conforme Santos (2001). Nesse caso, após centrifugação para retirada da biomassa,
transferia-se para um eppendorf 1,5 mL de amostra (caldo fermentativo) e adicionava-se 1,5
mL de solução azul de Coomassie. Deixava-se a solução em repouso por 5 minutos,
transferindo-se em seguida para um cubeta de quartzo e efetuando-se as leituras de
absorbância nos comprimentos de ondas 465 nm e 595 nm, respectivamente. O
espectrofotômetro (Thermo Spectronic modelo Genesys 10UV) foi zerado com água
destilada. O conteúdo protéico foi obtido a partir de uma curva padrão na qual graficava-se a
relação de absorbância A595/A465 para diferentes concentrações de Albumina de Soro
Bovino (Fração V – Lote 110881/Sigma Aldrich-Ohio-USA).
3.5. Determinação da Atividade Quitosanolítica e da Atividade Específica
A atividade quitosanolítica foi obtida monitorando-se o aumento dos açúcares
redutores totais, utilizando-se o método DNS a partir de uma curva padrão de glicosamina.
Para a determinação da atividade quitosanolítica adicionaram-se, aos tubos de ensaios 500 µL

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
52
de substrato (solução 0,2 % de quitosana solubilizada em ácido clorídrico 1N) e 500 µL de
caldo bruto (contendo enzima), controlando-se o pH entre 6,0 e 6,5 e incubando-se o sistema
em um banho termostatizado com temperaturas da ordem de 55 ºC. Após o tempo de
incubação de 30 minutos no banho foi adicionado aos tubos de ensaio 2,5 mL de DNS,
aquecendo-se à ebulição em banho-maria por 10 minutos, resfriando-se, em seguida, em
banho de gelo. Após essa etapa, o sistema foi centrifugado e com o sobrenadante efetuou-se a
leitura no espectrofotômetro a 600 nm. Destaca-se, que antes da leitura da amostra zerava-se o
equipamento (branco do espectro) com 1,0 mL de água destilada acrescidos de 2,5 mL de
DNS e realizava-se o branco da enzima que consistia da adição de 500 µL de solução de
quitosana, completando-se até um 1,0 mL com água destilada. Uma unidade de atividade
quitosanolítica (A) foi definida como a quantidade de enzima necessária para produzir 1
µmol.min-1 de glicosamina por mL de extrato enzimático, nas condições de temperatura e pH
citadas, sendo o resultado expresso em (U.mL-1). A Atividade específica foi determinada pela
divisão entre a atividade da enzima (U.mL-1) e a concentração de proteína total (µg.mL-1),
sendo expressa em (U. µg-1).
3.6. Planejamento experimental
3.6.1. Planejamento de seleção de fatores Plackett & Burman
O planejamento experimental Plackett & Burman é uma técnica muito usada, para se
realizar triagem de fatores na otimização num determinado processo (Reddy et al., 2008), e
foi utilizado neste trabalho para identificar os fatores que influenciam de forma significativa
na produção da enzima quitosanase.
A Tabela 3 ilustra os níveis de cada variável (fator) utilizada no planejamento Plackett
& Burman, sendo: Concentração de Quitosana (Q), Peptona (PP), Extrato de Levedura (EL),
NaNO3 (NS), K2HPO4 (FP), KCl (CP), MgSO4.7H2O (SM), FeSO4 (SF). Já a Tabela 4 mostra
a matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3 repetições
na condição do ponto central.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
53
Tabela 3 – Níveis para os fatores e seus valores codificados.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1 0,1 0,2 0,2 0,1 0,05 0,025 0,025 0,0005
0 0,2 0,5 0,3 0,2 0,10 0,050 0,050 0,0010
1 0,3 0,8 0,4 0,3 0,15 0,075 0,075 0,0015
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1
2 1 1 -1 1 -1 -1 -1 1 -1 1 1
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1
5 1 1 -1 1 1 -1 1 -1 1 -1 1
6 1 1 1 -1 1 1 -1 1 1 -1 -1
7 -1 1 1 1 -1 1 1 -1 -1 1 1
8 -1 -1 1 1 1 -1 1 1 1 1 1
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1
11 -1 1 -1 -1 -1 1 1 1 1 1 -1
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1
13 0 0 0 0 0 0 0 0 0 0 0
14 0 0 0 0 0 0 0 0 0 0 0
15 0 0 0 0 0 0 0 0 0 0 0
Sendo: V11,V12,V13 = Variáveis inertes para cálculo do erro.
3.6.2. Planejamento Composto Central para a Otimização do Meio de Cultura para a
Produção da Enzima em Cultivo Submerso
Um dos problemas mais comuns para quem realiza experimentos é determinar a
influência de uma ou mais variáveis sobre outra variável de interesse (Lobato, 2003). Este

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
54
problema pode ser minimizado utilizando-se um planejamento experimental de modo a se
obter uma função, ou uma aproximação satisfatória para a mesma, de forma que se operando
sobre as variáveis de entrada produza-se como saída a resposta ou as respostas observadas
(Barros Neto, 2001; Silva, 2004).
Em um planejamento, inicialmente, deve-se definir os fatores e as respostas de
interesse. Tendo-se identificado os fatores e respostas define-se o objetivo que se pretende
alcançar com os experimentos para que se possa escolher o planejamento mais apropriado.
(Silva, 2004).
No presente trabalho foi utilizado um Planejamento Composto Central (CCD) com 22
experimentos na parte fatorial, repetição em triplicata no ponto central e quatro experimentos
na parte axial (planejamento do tipo estrela), para se investigar a influência dos fatores:
concentração de quitosana e concentração de FeSO4 que foram os dois fatores que
influenciaram a produção de quitosanase, conforme o planejamento Plackett & Burman de 12
ensaios e mais 3 repetições na condição do ponto central. Estes dois fatores foram estudados
em cinco diferentes níveis (-1,41; -1; 0; 1; 1,41) e um conjunto de 11 experimentos foi
realizado, conforme Tabela 5.
Tabela 5 – Matriz do planejamento Composto Central.
Ensaio Q SF
1 -1 -1
2 -1 1
3 1 -1
4 1 1
5 -1,41 0
6 1,41 0
7 0 -1,41
8 0 1,41
9 0 0
10 0 0
11 0 0
Os outros fatores foram mantidos nos níveis mais baixos (para evitar custos). Uma que
não influenciaram de forma significativa na produção da enzima quitosanase.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
55
A codificação dos fatores foi realizada de acordo com a seguinte equação (Ji et al.,
2009):
Xi = (xi – x0)/∆xi , i = 1, 2, 3, ..., k (2)
Onde Xi é o fator independente codificado, xi é o fator independente real, x0 é o valor de xi no
ponto central e ∆xi é a variação de valor. O comportamento do sistema foi explicado pela
seguinte equação polinomial de segunda ordem:
Yp = β0 + ∑ βiXi + ∑ βiiXi2 + ∑ βijXiXj, i, j = 1, 2, 3,..., k (3)
Onde Yp foi a resposta esperada, β0 foi a interseção, Xi e Xj
foram os fatores independentes codificados, βi foi o coeficiente linear, βii
foi o coeficiente quadrático e βij foi o coeficiente de interação. Destaca-se que os outros
fatores não significativos foram mantidos no nível mais baixo (-1).
A Tabela 6 apresenta os níveis para os fatores e seus valores codificados, para o
Planejamento Composto Central, com triplicata no ponto central.
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1,41 0,218 0,2 0,2 0,1 0,05 0,025 0,025 0,0009
-1 0,3 0,2 0,2 0,1 0,05 0,025 0,025 0,0015
0 0,5 0,2 0,2 0,1 0,05 0,025 0,025 0,0030
1 0,7 0,2 0,2 0,1 0,05 0,025 0,025 0,0045
1,41 0,782 0,2 0,2 0,1 0,05 0,025 0,025 0,0051
Na análise do planejamento de seleção de fatores Plackett & Burman e do
planejamento experimental fatorial foi utilizado um pacote computacional STATISTICA for
Windows versão 7.0.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
56
3.7. Perfil cinético durante o cultivo
Um perfil cinético na melhor condição de cultivo, conforme planejamento
apresentado, foi realizado em frascos Erlenmyer de 250 mL contendo 200 mL de meio de
cultivo contendo os seguintes componentes, em 1 Litro: 2,0g de quitosana; 5,0g de peptona;
3,0g de extrato de levedura; 2,0g de NaNO3; 1,0g de K2HPO4; 0,5g de KCl; 0,5g de
MgSO4.7H2O; 0,01g de FeSO4, que foram inoculadas com 10% (vol/vol) de inóculo contendo
o fungo Metarhizium anisopliae e cultivadas em incubador rotativo durante 5 dias a 150 rpm
e 25 ºC. Alíquotas foram retiradas a cada seis horas e analisadas quanto à proteína total no
sobredanante, atividade enzimática, atividade específica e açúcares redutores. Os ensaios
foram realizados em triplicata (Silva Filho, 2005).

Capítulo 4 Resultados e
Discussão

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
58
4. Resultados e discussão
Neste capítulo são apresentados e discutidos os resultados obtidos no presente
trabalho. Inicialmente, é apresentada imagem do MEV do Metarhizium anisopliae
mantidos em meio PDA. Em seguida, apresenta-se e discute-se os resultados da avaliação
dos fatores significativos, que podem influenciar na produção da quitosanase, no
planejamento de seleção de fatores, conforme planejamento Plackett & Burman bem como
são apresentados e discutidos os resultados obtidos para o planejamento composto central.
Por último, apresenta-se o perfil cinético, na melhor condição, para a produção de
quitosanase em cultivo submerso descontínuo usando o fungo Metarhizium anisopliae.
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA.
A Figura 7 ilustra a imagem da Microscopia Eletrônica de Varredura (MEV) do micélio de Metarhizium anisopliae mantido em meio PDA.
Figura 7. Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA.
Observa-se na Figura 7, quanto à morfologia que o Metarhizium anisopliae, apresenta-
se como um fungo filamentoso com as hifas intimamente entrelaçadas conforme citado por
Wang et al. 2002.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
59
4.2. Planejamento Plackett & Burman
4.2.1 Análise do Plackett & Burman com 8 fatores e 12 ensaios
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis, foi utilizado para avaliar a influência dos fatores: Concentração de
Quitosana (Q), Peptona (PP), Extrato de Levedura (EL), NaNO3 (NS), K2HPO4 (FP), KCl
(CP), MgSO4.7H2O (SM), FeSO4 (SF) na produção da enzima quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso. Ressalta-se que os experimentos foram
realizados aleatoriamente.
A Tabela 7 apresenta a matriz do modelo Plackett & Burman e os respectivos valores
obtidos para a variável resposta (Atividade Quitosanalítica).
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.
Ensaio Q PP EL NS FP CP SM SF A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 0,0195
2 1 1 -1 1 -1 -1 -1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 0,0109
4 1 -1 1 1 -1 1 -1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 0,0136
6 1 1 1 -1 1 1 -1 1 0,0156
7 -1 1 1 1 -1 1 1 -1 0,0109
8 -1 -1 1 1 1 -1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 0,0085
A Tabela 8 apresenta os efeitos estimados para o planejamento Plackett & Burman.
Observa-se na Tabela 8 que o valor do coeficiente de determinação R2 foi de
aproximadamente 90%, indicando que um modelo linear poderia representar bem a relação
entre o fator significativo, concentração de quitosana (1), e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
60
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman.
Efeitos Estimados; R2=0,8996; Ajuste:0,63178 MS Erro Residual =0,0000054
Desvio Padrão t(3) p
Lim. Conf. Lim. Conf.
Efeito Erro Residual
-95,% +95
Média/Interação 0,0127 0,0007 18,903 0,0003 0,0105 0,0149 (1) Quitosana 0,0055 0,0013 4,0628 0,0268 0,0011 0,0098
(2) Peptona 0,0015 0,0013 1,1080 0,3486 -0,0028 0,0058 (3) Ext. de leved. 0,0009 0,0013 0,7387 0,5136 -0,0033 0,0053
(4) NaNO3 0,0002 0,0013 0,1758 0,8715 -0,0040 0,0045 (5) K2HPO4 -0,0014 0,0013 -1,0552 0,3687 -0,0057 0,0028
(6) KCl -0,0022 0,0013 -1,6708 0,1933 -0,0065 0,0020 (7) MgSO4.7H2O -0,0003 0,0013 -0,2286 0,8338 -0,0046 0,0039
(8) FeSO4 0,0029 0,0013 2,1457 0,1211 -0,0014 0,0071
A Figura 8 mostra o gráfico de Pareto, uma das formas de se avaliar visualmente a
influência dos fatores estudados na resposta. A magnitude dos efeitos é representada pelas
colunas enquanto que a linha transversal às colunas representa a magnitude dos efeitos com
significado estatístico para p=0,05, ou seja, os fatores que são estatisticamente significativos
ao nível de 95% de confiança.
Figura 8. Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
61
Analisando-se o gráfico de Pareto (Figura 8) observa-se que o fator concentração de
quitosana (1) foi o único fator que influenciou de forma mais significativa na atividade
enzimática atingindo efeitos estimados de aproximadamente 4,06. O sinal positivo indica que
o aumento da concentração de quitosana aumentará à atividade enzimática. Por outro lado, os
outros fatores parecem desprezíveis estatisticamente conforme o gráfico de Pareto.
4.2.2 Análise do Plackett & Burman com 12 Ensaios e mais 3 Repetições na condição do
ponto central.
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis com repetição em triplicata no ponto central, foi utilizado para se avaliar a
influência dos mesmos fatores descritos na Seção 4.2.1 para a produção da enzima
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso. Ressalta-se que os
experimentos foram realizados aleatoriamente. A Tabela 9 apresenta a matriz do modelo
Plackett & Burman e os respectivos valores obtidos para a variável resposta (Atividade
Quitosanalítica).
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13 A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1 0,0195
2 1 1 -1 1 -1 -1 -1 1 -1 1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1 0,0109
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 1 -1 1 0,0136
6 1 1 1 -1 1 1 -1 1 1 -1 -1 0,0156
7 -1 1 1 1 -1 1 1 -1 -1 1 1 0,0109
8 -1 -1 1 1 1 -1 1 1 1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 1 1 -1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1 0,0085
13 0 0 0 0 0 0 0 0 0 0 0 0,0704
14 0 0 0 0 0 0 0 0 0 0 0 0,0688
15 0 0 0 0 0 0 0 0 0 0 0 0,0710

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
62
A Tabela 10 a seguir apresenta os efeitos estimados para planejamento Plackett &
Burman com 3 repetições na condição do poto central. Observa-se na Tabela 10 que o valor
do coeficiente de determinação R2 foi de aproximadamente 2,0%, indicando que um modelo
linear não poderia representar a relação entre os fatores significativos concentração de
quitosana (1), concentração de FeSO4 (8) e a resposta.
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central.
Efeitos Estimados; R2=0,01811; Ajuste:0,0 MS Erro Puro =0,0000012
Desvio Padrão t(2) P
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0242 0,0003 85,509 0,0001 0,0229 0,0254 (1) Quitosana 0,0055 0,0006 8,6432 0,0131 0,0027 0,0082
(2) Peptona 0,0015 0,0006 2,3572 0,1425 -0,0012 0,0042 (3) Ext. de leved. 0,0009 0,0006 1,5715 0,2567 -0,0017 0,0037
(4) NaNO3 0,0002 0,0006 0,3742 0,7442 -0,0024 0,0029 (5) K2HPO4 -0,0014 0,0006 -2,2450 0,1538 -0,0041 0,0013
(6) KCl -0,0022 0,0006 -3,5546 0,0708 -0,0049 0,0005 (7) MgSO4.7H2O -0,0003 0,0006 -0,4864 0,6747 -0,0030 0,0024
(8) FeSO4 0,0029 0,0006 4,5648 0,0448 0,0002 0,0056
A Figura 9 ilustra o gráfico de Pareto para o planejamento Plackett & Burman, com
repetição em triplicata no ponto central, uma das formas de se avaliar visualmente a influência
dos fatores estudados na resposta, onde é mostrado a magnitude dos efeitos para 95 % de
confiança.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
63
Figura 9. Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto
central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto, observa-se que os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma mais
significativa na atividade enzimática atingindo efeitos estimados de aproximadamente 8,84 e
4,67, respectivamente. O sinal positivo indica que o aumento das concentrações de quitosana
e de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores parecem
desprezíveis estatisticamente conforme o gráfico de Pareto.
Este comportamento do FeSO4 está de acordo com o observado por Hagag et al.
(2012) que utilizando o Aspergillus fumigatus na produção de protease mostraram que um
fator de transcrição PrtT regula positivamente o controle da produção de protease, da
assimilação de Ferro e da síntese de ergosterol e que a produção de protease foi favorecida
pelo aumento da concentração de ferro.
Rajendran & Thangavelu (2009) utilizaram um planejamento Plackett & Burman para
identificar quais os fatores significativos na produção de lipase por Rhizopus arrhizus MTCC
2233 e observaram o efeito significativo médio do FeSO4, apresentando um nível de
confiança para esse fator de 48,92% (para um máximo de 98,7 % de MgSO4), com uma
produção de lipase da ordem de 2,98 U/mL em 84 horas de fermentação e que ocorreu uma
produção considerável de protease no meio de cultivo da ordem de 1,92 U/mL em 96 horas de
cultivo.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
64
Mu et al. (2005) também observaram um efeito positivo do FeSO4, da ordem de 1,48
para um fator máximo encontrado de 3,11 de MgSO4, no aumento da atividade da
estereoinversão de S-fenil-1,2-etanodiol por Candida parapsilosis CCTCC M203011 na
otimização da fermentação utilizando um planejamento Plackett & Burman.
Dessa forma, observa-se que o FeSO4 é importante uma que vez o mesmo está
diretamente associado a síntese de proteases (principalmente metalloproteases), conforme
citado por Hagag et al. (2012).
4.3. Planejamento Composto Central para Produção de Quitosanase por
Metarhizium anisopliae em Cultivo Descontínuo Submerso
Um planejamento experimentos 22 com repetição em triplicata no ponto central e mais
quatro ensaios axiais, foram utilizados para se chegar a uma combinação ideal de quitosana e
FeSO4 na produção da enzima quitosanase (Tabela 11). A partir do modelo Plackett &
Burman (Tabela 12), que mostrou que o aumento das concentrações de quitosana e FeSO4,
favoreceram a produção de quitosanase, os valores utilizados neste modelo para estes
componentes nos níveis mais altos, serão considerados como os níveis mais baixos no
planejamento experimental do composto central. Os outros fatores foram fixados em todos os
ensaios em seus níveis mais baixos para a redução de custos (Tabela 11).

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
65
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da
atividade enzimática.
Ensaio Q SF A (U/mL)
1 -1 -1 0,0195
2 -1 1 0,0202
3 1 -1 0,0109
4 1 1 0,0121
5 -1,41 0 0,0136
6 1,41 0 0,0156
7 0 -1,41 0,0108
8 0 1,41 0,0103
9 0 0 0,0099
10 0 0 0,0116
11 0 0 0,0094
A Tabela 12 apresenta os efeitos estimados para o planejamento composto central.
Tabela 12 – Efeitos Estimados do planejamento Composto Central.
Efeitos Estimados; R2=0,629; Ajuste:0,25836 MS Erro Puro =0,0000014
Desvio Padrão t(2) p
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0103 0,0006 15,305 0,0042 0,0074 0,0132 (1) Quitosana (L) 0,0011 0,0008 1,4316 0,2885 -0,0023 0,0047
Quitosana (Q) 0,0058 0,0009 5,9637 0,0269 0,0016 0,0101 (2) FeSO4 (L) -0,0043 0,0008 -5,3037 0,0337 -0,0079 -0,0008 (2) FeSO4 (Q) 0,0018 0,0009 1,8366 0,2076 -0,0024 0,0060
1L x 2L 0,0001 0,0011 0,1520 0,8931 -0,0048 0,0052
Observa-se na Tabela 12 que o valor do coeficiente de determinação R2 foi de
aproximadamente 63%, indicando que um modelo linear representaria bem a relação entre os
fatores significativos concentração de quitosana (1), concentração de FeSO4 (8) e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
66
A Tabela 13 mostra um resumo da ANOVA para o planejamento composto central
com repetição em triplicata no ponto central.
Tabela 13 – Resumo da ANOVA.
Fonte de
Variação
Soma dos
Quadrados
Grau de
Liberdade
Quadrado
Médio
Fcalculado Ftabelado
Regressão
Resíduos
0,000086 2 0,000043 6,14 4,46
0,000056 8 0,000007
Falta de
Ajuste
Erro puro
0,000053 6 0,000009 6 19,33
0,000003
2
0,0000015
Total 0,000142 10
De acordo com a Tabela 13, partindo-se dos efeitos significativos propõe-se o
modelo:
� = 0,010316 + 0,002942� � − 0,002193�� (8)
% de variação explicável: 60,5
% máxima de variação explicável: 97,9
A análise da ANOVA mostra que o modelo proposto possui significância estatística, ao
nível de 95% de confiança, uma vez que o valor de F (calculado) é maior que F (tabelado),
entretanto a ANOVA mostra também que o modelo proposto pode ser útil para se fazer
previsões, uma vez que a relação entre F (calculado) e F (tabelado) para a falta de ajuste e o
erro puro é menor que 1. O coeficiente de determinação R2 foi de aproximadamente 63%,
indicando que o modelo consegue explicar 63% da variação total em torno da média.
Os cálculos para variação explicável e para máxima variação explicável são dados pelas
Equações (9) e (10):
% de variação explicável = (SQ da regressão) / (SQ Total) (9)
% máxima de variação explicável = (SQ Total – SQ Erro puro) / (SQ Total) (10)
Sendo:
SQ – Soma dos quadrados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
67
A Figura 10 mostra o gráfico de Pareto para o planejamento Composto Central
utilizando erro puro. A magnitude dos efeitos é representada pelas colunas. Já a linha
transversal às colunas representa a magnitude dos efeitos com significado estatístico para 95%
de confiança.
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto (Figura 10) observa-se que os fatores concentração
de quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma
mais significativa na atividade enzimática atingindo efeitos estimados de aproximadamente
5,96 e - 5,30, respectivamente. O sinal positivo indica que o aumento das concentrações de
quitosana aumentará à atividade enzimática. O sinal negativo indica que o decréscimo da
concentração de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores
parecem desprezíveis estatisticamente no gráfico de Pareto.
A avaliação do modelo também pode ser feita através da observação do gráfico dos
valores preditos em função os valores observados que são mostrados na Figura 11. Os valores
preditos pelo modelo são representados pela reta, enquanto que os valores observados
representam-se pelos pontos.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
68
Figura 11 – Valores preditos em função dos valores observados.
Observa-se que alguns valores preditos apresentam um afastamento dos valores
observados. A Figura 12 ilustra a superfície de resposta da atividade quitosanolítica em
função dos dois fatores significativos, concentração de quitosana e concentração de FeSO4.
Figura 12. Atividade quitosanolítica em função da concentração de quitosana e da
concentração de FeSO4 para os fatores não significativos no ponto central.
Observa-se a existência de um ponto de mínimo, ocorrendo para concentração de
quitosana no ponto central (0) e para maiores concentrações de FeSO4 igual a (1,5). Essa

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
69
região deve ser evitada uma vez que a atividade quitosanolítica atinge níveis tão reduzidos
quanto 0,0094 U.mL-1. Destaca-se que nesse planejamento o maior nível da concentração de
FeSO4 (α=+1,41) equivale a 0,0051% (Tabela 6) que é aproximadamente cinco vezes maior
que o nível desse fator no ponto central do planejamento Plackett & Burman (0,001%),
conforme Tabela 3. Dessa forma, ao se utilizar o nível mínimo da concentração de FeSO4 no
Planejamento Composto Central (α=-1,41), que equivale a aproximadamente 0,001%,
aproxima-se do ponto central do do planejamento Plackett & Burman (0,001%), condição
essa que apresenta os melhores resultados de atividade quitosanolítica usando quitosana como
fonte de carbono e energia e fungo Metarhizium anisopliae.
4.4. Perfil Cinético – Melhor Condição
A seguir, apresenta-se um perfil cinético, na melhor condição, para produção da
quitosanase usando o fungo Metarhizium anisopliae, em que foram monitorados durante 24
horas os seguintes parâmetros: Concentração de proteínas totais (µg/mL), atividade
quitosanolítica (U/mL), atividade específica (U/µg) e açúcares redutores (mg/mL), ilustradas
nas Figuras 13, 14, 15 e 16, respectivamente.
Com relação à proteína extracelular, observa-se na Figura 13 que, inicialmente, ocorre
um aumento da mesma para o seu nível máximo (6,2 µg/mL), durante as 6 primeiras horas de
cultivo, seguido de uma redução para 5,7 µg/mL em 18 horas de cultivo, possivelmente
devido à presença de proteases no meio. Observa-se que a partir de então ocorre uma redução
e aumento da proteína total, em uma quantidade inferior, sugerindo um mecanismo de
controle.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
70
0 4 8 12 16 20 245,0
5,2
5,4
5,6
5,8
6,0
6,2
6,4
Pro
tein
a (
µg
/mL
)
Tempo (h)
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana.
Com relação à atividade quitosanolítica, conforme Figura 14, a atividade máxima
ocorreu em 18 horas de produção em que se obteve aproximadamente 70,0 U/L, o que foi 28
vezes maior que o valor (2,5 U/L em 48 horas) relatado por Assis (2010) para o mesmo fungo,
e em apenas um terço do tempo de produção.
0 4 8 12 16 20 240,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,10
Ativid
ade
(U
/mL
)
Tempo (h)
Figura 14. Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
71
Além disso, destaca-se o considerável aumento da produção de quitosanase a partir das
12 horas até 18 horas de cultivo onde atingiu o pico máximo de produção (70,0 U/L). Após o
pico em 18 horas, a produção começa a cair rapidamente, comportamento esse observado
durante todos os ensaios cinéticos. Destaca-se que é importante do ponto de vista industrial
manter a produção em alto nível por muitas horas, onde se espera uma produção contínua ou
em grandes bateladas.
Ak et al. (1998) utilizaram o fungo Penicillium spinulosum para produzir a quitosanase
usando um meio de cultura contendo 0,1% de K2HPO4; 0,05 % de MgSO4.7H2O; 0,05% de
KCl; 0,001 % de FeSO4, 0,5% NH4NO3 e como fonte de carbono e energia, os autores
usaram quitosana e parede de Rhizopus. A produção foi realizada em cultivo submerso a 28
oC. Os autores apresentam atividade quitosanolítica máxima de aproximadamente 47,0 U/mL,
ao se utilizar a parede do fungo como fonte de carbono e energia, e 30,0 U/mL ao se utilizar
quitosana como fonte de carbono e energia. Destaca-se que essas atividades ocorreram após 4
dias de cultivo. Os perfís com o tempo da atividade quitosanolítica e da atividade específica
apresentaram tendência de crescimento, com o pico máximo em 4 dias, seguindo com uma
tendência de queda da atividade. Neste tempo, a atividade específica máxima da quitosanase
foi de aproximadamente 60,0 U/mg de proteína. Essas tendências também foram observadas
nos resultados apresentados nos perfís de atividade das Figuras 14 e 15.
0 4 8 12 16 20 24
-0,010
-0,005
0,000
0,005
0,010
0,015
0,020
0,025
0,030
Ativid
ad
e E
spe
cific
a (
U/µ
g)
Tempo (h)
Figura 15. Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.
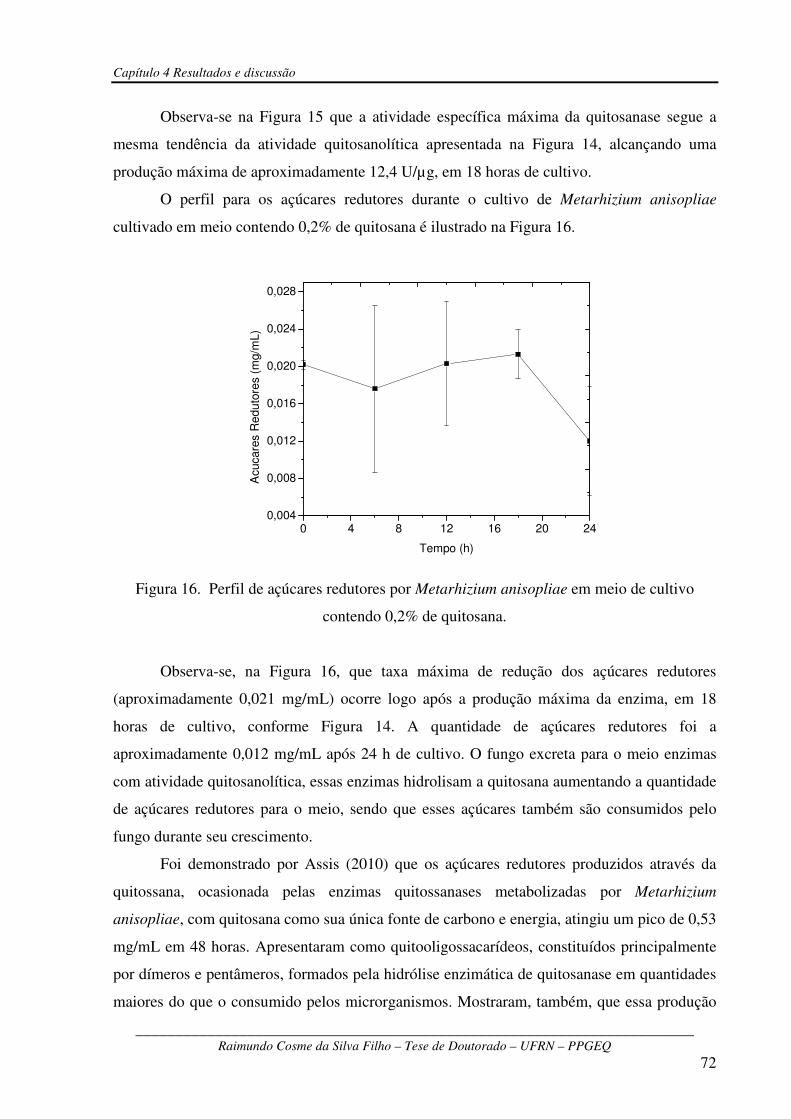
Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
72
Observa-se na Figura 15 que a atividade específica máxima da quitosanase segue a
mesma tendência da atividade quitosanolítica apresentada na Figura 14, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
O perfil para os açúcares redutores durante o cultivo de Metarhizium anisopliae
cultivado em meio contendo 0,2% de quitosana é ilustrado na Figura 16.
0 4 8 12 16 20 240,004
0,008
0,012
0,016
0,020
0,024
0,028
Acuca
res R
ed
uto
res (
mg/m
L)
Tempo (h)
Figura 16. Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana.
Observa-se, na Figura 16, que taxa máxima de redução dos açúcares redutores
(aproximadamente 0,021 mg/mL) ocorre logo após a produção máxima da enzima, em 18
horas de cultivo, conforme Figura 14. A quantidade de açúcares redutores foi a
aproximadamente 0,012 mg/mL após 24 h de cultivo. O fungo excreta para o meio enzimas
com atividade quitosanolítica, essas enzimas hidrolisam a quitosana aumentando a quantidade
de açúcares redutores para o meio, sendo que esses açúcares também são consumidos pelo
fungo durante seu crescimento.
Foi demonstrado por Assis (2010) que os açúcares redutores produzidos através da
quitossana, ocasionada pelas enzimas quitossanases metabolizadas por Metarhizium
anisopliae, com quitosana como sua única fonte de carbono e energia, atingiu um pico de 0,53
mg/mL em 48 horas. Apresentaram como quitooligossacarídeos, constituídos principalmente
por dímeros e pentâmeros, formados pela hidrólise enzimática de quitosanase em quantidades
maiores do que o consumido pelos microrganismos. Mostraram, também, que essa produção

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
73
segue a mesma tendência da sua atividade enzimática, corroborando com os resultados
apresentados no presente trabalho.
Os resultados do presente trabalho mostram a relevância de se adotar novas condições
para a produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter quitooligossacarídeo
com atividades biológicas. Esta pesquisa, portanto, sugere base para novos estudos que
permitam alcançar rapidamente a produção de quitosanase e quitooligossacarídeo com todos
os seus benefícios.

Capítulo 5 Conclusão

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
75
5. Conclusão
Após o desenvolvimento desse trabalho conclui-se:
- Com relação ao Planejamento de Seleção de Fatores Plackett & Burman:
Os resultados das análises do Planejamento de Seleção de Fatores Plackett &
Burman com 8 fatores e 12 ensaios em 2 níveis, mais 3 repetições no ponto central, para
se investigar a influência dos fatores: Concentração de Quitosana, Peptona, Extrato de
Levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O, FeSO4 na produção da enzima
quitosanase por Metarhizium anisopliae, mostraram que a atividade foi favorecida pelo
aumento da concentração de substrato (quitosana) e sulfato ferroso (FeSO4), enquanto
que o aumento da concentração dos outros fatores não contribuíram de forma
significativa para a resposta (atividade quitosanolítica). Os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) atingiram efeitos estimados de
aproximadamente 8,84 e 4,67, respectivamente. O sinal positivo indica que o aumento
das concentrações de quitosana e de FeSO4 aumentará a atividade enzimática. Conforme
visto na literatura, observa-se importância do FeSO4, já que o mesmo está diretamente
associado a síntese de proteases (principalmente metalloproteases),
- Com relação ao Planejamento Composto Central:
A análise do planejamento composto central para avaliar a influência dos
fatores, concentração de substrato (quitosana) e concentração de FeSO4, na produção da
enzima quitosanase por Metarhizium anisopliae (a partir do planejamento Plackett &
Burman) mostrou que os fatores concentração de quitosana e concentração de FeSO4
foram os fatores que influenciaram de forma mais significativa na atividade enzimática
atingindo efeitos estimados de aproximadamente 5,96 e - 5,30, respectivamente. O sinal
positivo indica que o aumento das concentrações de quitosana aumentará a atividade
enzimática. O sinal negativo indica que o decréscimo da concentração de FeSO4
aumentará a atividade enzimática. Observa-se a existência de um ponto de mínimo,
ocorrendo para concentração de quitosana (X1) no ponto central (0) e para maiores

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
76
concentrações de FeSO4 (X2) igual a (1,5) com a atividade quitosanolítica de 0,0094
U.mL-1.
- Com relação ao cultivo na melhor condição:
O cultivo na melhor condição mostrou uma produção máxima de quitosanase em
18 horas de cultivo, com a atividade enzimática de 70,0 U/L, o que foi 28 vezes maior
que o valor (2,5 U/L em 48 horas) relatado na literatura (Assis, 2010) para o mesmo
fungo e em apenas um terço do tempo de produção.
Além disso, observou-se que um elevado incremento na produção de
quitosanase a partir das 12 horas de cultivo, antes de se atingir a atividade
quitosanolítica máxima, 18 horas de cultivo. Essa produção em alto nível por muitas
horas é importante do ponto de vista industrial, onde se espera uma produção contínua
ou em grandes bateladas. Observou-se também que a atividade específica máxima da
quitosanase segue a mesma tendência da atividade quitosanolítica, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
- Com relação aos açúcares redutores:
Foi observado que os açúcares redutores seguem a mesma tendência da atividade
enzimática, com uma produção máxima de cerca de 0,21 mg/mL em 18 horas de
cultivo, produção essa que coincinde com o máximo pico de atividade enzimática.
Estes resultados mostram a relevância de se adotar novas condições para a
produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter
quitooligossacarídeo com atividades biológicas. Esta pesquisa, portanto, sugere base
para novos estudos que permitam alcançar rapidamente a produção de quitosanase e
quitooligossacarídeo com todos os seus benefícios.

Referências Bibliográficas

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
78
Referências bibliográficas
AK, Ö.; BAKIR, U.; GÜRAY, T. Isolation of a chitosan degrading fungus, Penicillium
spinulosum, and chitosanase production by the isolate. In: SYMPOSIUM - NEW
FRONTIERS IN SCREENING FOR MICROBIAL BIOCATALYSTS, 1996, Ede, The
Netherlands. Anais, Editado por K. Kieslich, C. P. van der Beek, J. A. M. de Bont e W.
J. J. van den Tweel, Elsevier Science B. V., 1998, v.53, p.265-268.
ALVES, S. B. Patologia e controle microbiano: Vantagens e desvantagens. In: ALVES,
S. B. Controle Microbiano de Insetos. 2ed., Piracicaba, FEALQ, 1163p, 1998, v.4, p.21-
37.
ALVES, S. B.; LEITE, L. G.; FILHO, A. B.; ALMEIDA, F. E. M.; MARQUES, E. J.
Produção massal de fungos entomopatogênicos na América Latina, In: ALVES, S. B.;
LOPES, R. B. Controle microbiano de pragas na América Latina: avanços e desafios.
Piracicaba, FEALQ, 414p, 2008, v.14, p.215-237.
ARRUDA, W.; LUBECK, I.; SCHRANK, A.; VAINSTEIN, M. H. Morphological
alterations of Metarhizium anisopliae during penetration of Boophilus microplus ticks.
Experimental and Applied Acarology, v. 37, p.231-244, 2005.
ASSIS, C. F. Produção e caracterização de quitooligossacarídeos produzidos pelo fungo
Metarhizium anisopliae e avaliação da citotoxicidade em células tumorais. 2010. 110p.
Tese (Doutorado em Biotecnologia) – Renorbio - Programa de Pós-Graduação em
Biotecnologia. Universidade Federal do Rio Grande do Norte, Natal.
ASSIS, C. F.; COSTA, L. S.; MELO-SILVEIRA, R. F.; OLIVEIRA, R. M.;
PAGNONCELLI, M. G. B.; ROCHA, H. A. O.; MACEDO, G. R.; SANTOS, E. S.,
Chitooligosaccharides antagonizes the cytotoxic effect of glucosamine. World J.
Microbiol. Biotechnol., v.28, n.3, p.1097-1105, 2012.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
79
AZEVEDO, V. V. C.; CHAVES, S. A.; BEZERRA, D. C.; FOOK, M. V. L.; COSTA,
A. C. F. M. Quitina e Quitosana: aplicações como biomateriais. Revista Eletrônica de
Materiais e Processos, v.2.3, p.27-34, 2007.
BARROS NETO, B.; SCARMINIO, I. S.; BRUNS, R. Como fazer experimentos:
pesquisa e desenvolvimento na ciência e na indústria. 2. ed. Campinas: Unicamp, 2001.
401p.
BATISTA FILHO, A.; ALMEIDA, J. E. M.; SANTOS, A. S.; MACHADO, L. A.;
ALVES, S. B. Eficiência de isolados de Metarhizium anisopliae no controle de
cigarrinha-da-raiz de cana-de-açúcar Mahanarva fimbriolata (Hom.: Cercopidae). STAB
– Açúcar, Álcool e Subprodutos, v.21, p.84-89, 2002.
CARVALHO, J. C. M.; SATO, S. Fermentação Descontínua, In: SCHMIDELL, W.;
LIMA, U. A.; AQUARONE, E.; BORZANI, W. Biotecnologia Industrial. São Paulo,
Ed. Edgard Blücher, v.2, p.193-204, 2001.
CHAO, H. J.; SCHWARTZ, J.; MILTON, D. K. Populations and determinats of
airborne fungi in large office buildings. Environmental Health Perspectives, v.110, n.8,
p.777-782, 2002.
CHEN, X.; XIA, W.; YU, X. Purification and characterization of two types of
chitosanase from Aspergillus sp. CJ22-326. Food Research International, v.38, n.3,
p.315-322, 2005.
CHEN, P. T.; CHIANG, C.-J.; CHAO, Y.-P. Medium optimization and production of
secreted Renilla luciferase in Bacillus subtilis by fed-batch fermentation. Biochemical
Engineering Journal, v.49, p.395-400, 2010.
CHENG, C. Y.; LI, Y. An Aspergillus chitosanase with potential for large-ecale
preparation of chitosan oligosaccharides. Biotechnol. Appl. Biochem., v.32, p.197-203,
2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
80
CHOI, Y. J.; Kim, E. J.; Piao, Z.; Yun, Y. C.; Shin, Y. C. Purification and
characterization of chitosanase from Bacillus sp. Strain KCTC 0377BP and its
application for the production of chitosan oligosaccharides. Applied and Environmental
Microbiology, v.70, n.8, p.4522-4531, 2004.
COLAKOGLU, G. The chemical and physical environment for fungal growth. Fak
Matbaa Birimi, Istanbul, 149-166, 2001.
CRAVEIRO, A. C.; CRAVEIRO, A. A.; QUEIROZ, D. C. Quitosana a fibra do futuro.
2. ed. Fortaleza: PADETEC, 2004. V.1, 218p.
DIAS, B. A. Estudo de proteases degradadoras de cutícula produzidas pelo fungo
entomopatogênico Beauveria bassiana. 2005. 70p. Dissertação (Mestrado em
Microbiologia) – Centro de Ciências Biológicas, Programa de Pós-Graduação em
Microbiologia, Universidade Estadual de Londrina, Londrina.
DINARDO-MIRANDA, L. L.; FERREIRA, J. M. G.; CARVALHO, P. A. M.
Influência da época de colheita e do genótipo de cana-de-açúcar sobre a infestação de
Mahanarva fimbriolata (Stal) (Hemiptera: Cercopidae). Neotrop. Entomol, v.30, n.1,
p.145-149, 200.
DONG, C.; ZHANG, J.; HUANG, H.; CHEN, H.; HU, Y. Pathogenicity of a new China
variety of Metarhizium anisopliae (M. anisopliae var. Dcjhyium) to subterranean termite
Odontotermes formosanus. Microbiological Research, v.164, p.27-35, 2009.
DONZELLI, B. G. G.; OSTROFF, G.; HARMAN, G. E. Enhanced enzymatic
hydrolysis of langostino shell chitin with mixtures of enzymes from bacterial and fungal
sources. Carbohydrate Research, v.338, p.1823-1833, 2003.
DRIVER, F.; MILNER, R. J.; TRUEMAN, J. W. H. A taxonomic revision of
Metarhizium based on phylogenetic analysis of rDNA sequence data. Mycological
Research, v.104, n.2, p.134-150, 2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
81
DUNG, P.; MILAS, M.; RINAUDO, M.; DESBRIÉRES, J. Water soluble derivatives
obtained by controlled chemical modifications of chitosan, Carbohydrate Polymers,
v.24, p.209-214, 1994.
ESTEVES, J. A.; CABRITA, J. D.; NOBRE, G. N. Micologia médica, 2. ed. Lisboa:
Fundação Calouste Gulbenkian, 1990. 1058p.
FACCIOTI, M. C. R. Fermentação Contínua, In: Biotecnologia Industrial. Editado por:
SCHMIDELL, W.; LIMA, U. A.; AQUARONE, E.; BORZANI, W., São Paulo, Ed.
Edgard Blücher, v.2, p.541, 2001.
FARIA, M. R.; MAGALHÃES, B. P. O uso de fungos entomopatogênicos no Brasil -
Situação atual e perspectivas. Biotecnologia Ciência & Desenvolvimento, n.22., p.18-
21, setembro/outubro 2001.
FU, J.-Y.; WU, S.-M.; CHANG, C.-T.; SUNG, H.-Y. Characterization of Three
Chitosanase Isozymes Isolated from a Commercial Crude Porcine Pepsin Preparation. J.
Agric. Food Chem., v.51, p.1042-1048, 2003.
FURIGO JUNIOR, A.; PEREIRA, E. P. Enzimas e suas aplicações cinética enzimática.
2001. 39f. Apostila (Graduação de Engenharia Química e Engenharia de Alimentos) –
Centro de Tecnologia, Departamento de Engenharia Química e Engenharia de
Alimentos, Universidade Federal de Santa Catarina, Florianópolis.
GAO, J.; XU, H.; LI, Q.-J.; FENG, X.-H.; LI, S. Optimization of medium for one-step
fermentation of inulin extract from Jerusalem artichoke tubers using Paenibacillus
polymyxa ZJ-9 to produce R,R-2,3-butanediol, Bioresource Technology, v.101, p.7076-
7082, 2010.
GARCIA, M. V.; MONTEIROI, A. C.; SZABO, M. J. P.; PRETTE, N.; BECHARA, G.
H. Mechanism of infection and colonization of Rhipicephalus sanguineus eggs by
Metarhizium anisopliae as revealed by scanning electron microscopy and
histopathology. Brazilian Journal of Microbiology, São Paulo, v.36, n.4, p.368-372,
2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
82
GHANEM, N. B.; YUSEF, H. H.; MAHROUSE, H. K. Production of Aspergillus
terreus xylanase in solid-state cultures: application of the Plackett & Burman
experimental design to evaluate nutritional requirements, Bioresource Technology,
v.73, p.113-121, 2000.
GHORBEL-BELLAJ, O.; HMIDET, N.; JELLOULI, K.; YOUNES, I.; MAALEJ, H.;
HACHICHA, R.; NASRI, M. Shrimp waste fermentation with Pseudomonas aeruginosa
A2: Optimization of chitin extraction conditions through Plackett–Burman and response
surface methodology approaches, International Journal of Biological Macromolecules,
v.48, p.596–602, 2010.
GIUSTINA, A.; VENTURA, P. Weight-reducing regimens in obese subjects: Effects of
a new dietary fiber integrator. Acta Toxicol Ther, 16, 4, p.199-214, 1995.
GUIA.BIO, Reino dos fungos, 2012. Disponível em:
<http://www.http://guia.bio.br/reino-dos-fungos>. Acesso em: 12 novembro 2012.
GUIRADO, N.; AMBROSANO, E. J.; ROSSI, F.; DIAS, F. L. F. Controle de Cupins
de Montículo com Metarhizium anisopliae e Beauveria bassiana, Rev. Bras. de
Agroecologia, v.4, n.2, 2009.
GUO, Y.; XU, J.; ZHANG, Y.; XU, H.; YUAN, Z.; LI, D. Medium optimization for
ethanol production with Clostridium autoethanogenum with carbon monoxide as sole
carbon source, Bioresource Technology, v.101, p.8784–8789, 2010.
HAGAG, S.; KUBITSCHEK-BARREIRA, P.; NEVES, G. W. P.; AMAR, D.;
NIERMAN, W.; SHALIT, I.; SHAMIR, R.; LOPES-BEZERRA, L.; OSHEROV, N.
Transcriptional and proteomic analysis of the Aspergillus fumigatus ∆prtT protease-
deficient mutant. PLoS One, v.7, n.4, e33604, 2012.
HAN, L.-K.; KIMURA, Y.; OKUDA, H. Reduction in fat storage during chitin-chitosan
treatment in mice fed a high-fat diet, International Journal of Obesity, 23:174-179,1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
83
HUNG, T.; CHANG, Y.; SUNG, H.; CHANG, C. Purification and Characterization of
Hydrolase with Chitinase and Chitosanase Activity from Commercial Stem Bromelain.
J. Agric. Food Chem., 50, 4666-4673, 2002.
ICHIKAWA, S.; TAKANO, K.; TAKASHIKUROIWA, HIRUTA, O.; SATO, S.;
MUKATAKA, A. S. Immobilization and stabilization of chitosanase by multipoint
attachment to agar gel support. Journal of Bioscience and Bioengineering, v.93, n.2,
p.201-206, 2002.
JABOR, I. A. S.; PAMPHILE, J. A.; RODRIGUES, S. B.; MARQUES-SILVA, G. G.;
ROCHA, C. L. M. S. Análise do desenvolvimento do fungo entomopatogênico
Metarhizium anisopliae em resposta a fatores nutricionais. Acta Scientiarum.
Agronomy, Maringá, v.25, n.2, p.497-501, 2003.
JI, X.-J.; HUANG, H.; DU, J.; ZHU, J.-G.; REN, L.-J.; LI, S.; NIE, Z.-K. Development
of an industrial medium for economical 2,3-butanediol production through co-
fermentation of glucose and xylose by Klebsiella oxytoca, Bioresource Technology,
v.100, p.5214–5218, 2009.
JO, Y.; JO, K.; JIN, Y.; KIM, K.; SHIN, J.; KIM, Y.; PARK, R. Characterization and
kinetics of 45 kDa chitosanase from bacillus sp. P16. Biosci. Biotechnol. Biochem.,
v.67, n.9, 1875 – 1882, 2003.
KHOSRAVI-DARANI, K.; ZOGHI, A.; ALAVI, S. A.; FATEMI, S. S. A. Application
of Plackett Burman Design for Citric Acid Production from Pretreated and Untreated
Wheat Straw, Iran. J. Chem. Chem. Eng.. v.27, n.1, p.91-104, 2008.
KIM, P.; KANG, T. H.; CHUNG, K. J.; KIM, I. S.; CHUNG, K. Purification of a
constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and
expression of the gene. FEMS Microbiolog.y Letters 240, 31–39, 2004.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
84
KIM, S.-K.; RAJAPAKSE, N. Enzymatic production and biological activities of
chitosan oligosaccharides (COS): A review. Carbohydrate Polymers, v.62, n.4, p.357-
368, 2005.
KIRK, P. M.; CANON, P.F.; MINTER, D.W.; STALPERS, J.A. Dictionary of the
fungi. CAB International, Wallingford, 2008.
KONDO, H.; OSADA, A. Influence of dietary fiber on the bioavailability of zinc in
rats, Biomed Environ Sci, 9, 204-8, 1996.
KUMAR, S.; KOH, J. Physiochemical, Optical and Biological Activity of Chitosan-
Chromone Derivative for Biomedical Applications. Int. J. Mol. Sci., v.13, p.6102-6116,
2012.
KURITA, K.; SANNAN, T.; IWAKURA, Y. Studies on chitin, 4. Evidence for
formation of block and random copolymers of N-acetyl-o-glucosamine and
oglucosamine by hetero and homogeneous hydrolyses. Die Makromolecular Chemie,
v.178, p.3197-3202, 1977.
KUROIWA, T.; ICHIKAWA, S.; HIRUTA, O.; SATO, S.; MUKATAKA, S. Factors
Affecting the Composition of Oligosaccharides Produced in Chitosan Hydrolysis Using
Immobilized Chitosanases. Biotechnol. Prog., 18, 969-974, 2002.
LI, L.-Z.; HUI, Z.; MO, X. Optimization of the medium for streptolydigin production
by Streptomyces lydicus AS 4.2501 with precursor feeding using response surface
methodology, Journal of the Taiwan Institute of Chemical Engineers, 41, 252–258,
2010.
LIU, B.-L.; KAO, P.-M.; TZENG, Y.-M.; FENG, K.-C. Production of chitinase from
Verticillium lecanii F091 using submerged fermentation. Enzyme and Microbial
Technology, v.33, p.410-415, 2003.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
85
LOBATO, A. K. C. L. Estudo da produção de biosurfactantes por micro-organismos
isolados de poços de petróleo. 2003. 108p. Dissertação (Mestrado) – Programa de Pós-
Graduação em Engenharia Química. Universidade Federal do Rio Grande do Norte.
Natal.
LOPES, R. B.; ALVES, S. B.; PADULLA, L. F. L.; PÉREZ, C. A. Eficiência de
formulações de Beauveria bassiana e Metarhizium anisopliae para o controle de ninfas
de Amblyomma cajennense (Fabricius, 1787). Revista Brasileira de Parasitologia
Veterinária, v.16, n.1, p.27-31, 2007.
LOUREIRO, E. S.; BATISTA FILHO, A.; ALMEIDA, J. E. M.; MENDES, J. M.;
PESSOA, L. G. A. Eficiência de isolados de Metarhizium anisopliae (metsch.) sorok.
no controle da cigarrinha-da-raiz da cana-de-açúcar, Mahanarva fimbriolata (stal, 1854)
(hemiptera: cercopidae), em condições de campo. Arq. Inst. Biol., São Paulo, v.79, n.1,
p. 47-53, 2012.
MELLO, D. R.; REIS, R. C. S.; BITTENCOURT, V. R. E. P. Patogenicidade in vitro
do fungo Metarhizium anisopliae (Metschnikoff, 1879) Sorokin, 1883, sobre o carrapato
Boophilus microplus (Canestrini, 1887). Revista Brasileira de Parasitologia Veterinária,
São Paulo, v.15, p.157-162, 2006.
MITSUTOMI, M.; KIDOH, H.; TOMITA, H.; WATANABE, T. The action of Bacillus
circulans WL-12 chitinases on partially N-acetylated chitosan, Bioscience
Biotechnology and Biochemistry, v.59, p.529-531, 1995.
MALAJOVICH, M. A. Biotecnologia: fundamentos. Ed. biblioteca Max Feffer do
Instituto de Tecnologia ORT, Rio de Janeiro, 2009. 104p.
MONAGHAN, R. L.; EVELEIGH, D. E.; TEWARI, R. P.; REESE, E. T. Chitosanase,
a novel enzyme. Nature - New Biology, 245, p.78-80, 1973.
MU, X. Q.; XU, Y.; NIE, Y.; OUYANG, J.; SUN, Z. H. Candida parapsilosis CCTCC
M203011 and the optimization of fermentation medium for stereoinversion of (S)-1-
phenyl-1,2-ethanediol, Process Biochemistry, v.40, p.2345–2350, 2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
86
MURRAY, P. R.; ROSENTHAL, K. S.; PFALLER, M. A. Medical microbiology. 5th
ed. London, Elsevier Mosby, 2005.
MUZZARELLI, R. A. A. Biochemical significance of exogenous chitins and chitosans
in animals and patients, Carbohydrate Polymers, v.20, p.7-16, 1993.
MUZZARELLI, R. A. A.; TOMASETTI, M.; ILARI, P. Depolymerization of chitosan
with the aid of papain, Enzyme Microbiology and Technology, v.16, p.110-114, 1994.
NAVEENA, B. J.; ALTAF, Md.; BHADRIAH, K.; REDDY, G. Selection of medium
components by Plackett–Burman design for production of L(+) lactic acid by
Lactobacillus amylophilus GV6 in SSF using wheat bran, Bioresource Technology,
v.96, p.485-490, 2005.
NOGAWA, M.; TAKAHASHI, H.; KASHIWAGI, A.; OHSHIMA, K.; OKADA, H.;
MORIKAWA, Y. Purification and Characterization of Exo--D-Glucosaminidase from
a Cellulolytic Fungus, Trichoderma reesei PC-3-7, Appl. Environ. Microbiol., v.64, n.3,
p.890-895, 1998.
ONOFRE, S. B.; VARGAS; L. R. B.; ROSSATO, M.; BARROS, N. M.; BOLDO, J.
T.; NUNES, A. R. F.; AZEVEDO, J. L. Controle biológico de pragas na agropecuária
por meio de fungos entomopatogênicos. In: SERAFINI, L. A.; BARROS, N. M.;
AZEVEDO, J. L. (Org.). Biotecnologia – avanços na agricultura e na agroindústria.
Caxias do Sul: EDUCS, p.295-328, cap.8, 2002.
OSSWALD, W. F.; SHAPIRO, J. P.; DOOSTDAR, H.; MCDONALD, R. E.; NIEDZ,
R. P.; NAIRN, C. J.; HEARN, C. J.; MAYER, R. T. Identification and Characterization
of Acidic Hydrolases with Chitinase and Chitosanase Activities from Sweet Orange
Callus Tissue, Plant Cell Physiol. 35, p.811-820, 1994.
PAGNONCELLI, M. G. B. Estudo do mecanismo de produção de oligossacarídeos com
atividades nutracêuticas a partir da quitosana por hidrólise enzimática com processo

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
87
fermentativo simultâneo. 2008. 118p. Tese (Doutorado) – Centro de Tecnologia,
Programa de Pós-Graduação em Engenharia Química. Universidade Federal do Rio
Grande do Norte. Natal.
PALMA-GUERRERO, J., JANSSON, H.-B., SALINAS, J.; LOPEZ-LLORCA, L. V.
Effect of chitosan on hyphal growth and spore germination of plant pathogenic and
biocontrol fungi, J. Appl. Microbiol., v.104, p.541-553, 2008.
PAULA, F. A.; ARRUDA, D. C. Análise do método de planejamento experimental
Plackett Burman na otimização de degradação de penas de aves por comunidades
microbianas, XXI Salão de Iniciação Científica, 19-23, outubro, 2009, UFRGS, Porto
Alegre.
PELCZAR, M. J.; CHANG, E. C. S.; KRIEG, N. R. Microbiologia: conceitos e
aplicações, v.2, 2.ed. São Paulo: Makron BooKs, 1996.
PELLETIER, A.; SYGUSCH, J. Purification and characterization of three chitosanase
activities from Bacillus megaterium P1, Applied and Environmental Microbiology, v.
56, n.4, p.844-848, 1990.
PEREIRA, F. B.; GUIMARÃES, P. M. R.; TEIXEIRA, J. A.; DOMINGUES, L.
Optimization of low-cost medium for very high gravity ethanol fermentations by
Saccharomyces cerevisiae using statistical experimental designs, Bioresource
Technology, v.101, p.7856-7863, 2010.
PETER, M. G. Chitin and chitosan from aninal sources. In: STEINBUCHEL, A.;
RHEE, S. K. Polysaccharides and Polyamides in the Food Industry. Weinheim:
Wileyvch, v.1, p.115-208, 2005.
PIZA, F. A. T.; SILOTO, A. P.; CARVALHO, C. V.; FRANCO, T. T. Purification,
characterization and purification of chitosanase from Bacillus cereus, Brazilian Journal
of Chemical Engineering., v.16, n 2, p.185-192, 1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
88
POLITZER, K.; BON, E. P. S. Enzimas Industriais e Especiais. C.G.E.E. Ciência,
Tecnologia e Inovação. Rio de Janeiro, 554p, 2006.
POLYMAR, Ciência é a nossa Natureza, 2003. Disponível em:
<http://www.polymar.com.br>. Acesso em: 6 setembro 2012.
PUTZKE, J.; PUTZKE, M. T. L. Os reinos dos fungos. Santa Cruz do Sul, RS:
EDUNISC, 1998.
QUINES, L. K. M. Extração de poli(3-hidroxibutirato) produzido por Cupriavidus
necator dsm 545 com 1,2-carbonato de propileno. 2010. 99p. Dissertação (Mestrado em
Engenharia de Alimentos) – Centro Tecnológico, Departamento de Engenharia Química
e Engenharia de Alimentos, Programa de Pós-Graduação em Engenharia de Alimentos,
Universidade Federal de Santa Catarina, Florianópolis.
RAJENDRAN, A.; THANGAVELU, V. Statistical experimental design for evaluation
of medium components for lipase production by Rhizopus arrhizus MTCC 2233, LWT -
Food Science and Technology, v.42, p.985–992, 2009.
RAMIREZ-LEÓN, L. F.; RUIZ-HERRERA, J. Hydrolysis of walls and formation of
spheroplasts in Mucor rouxii. J. Gen. Microbiol., v.72, p.281-290, 1972.
RANGEL, M. A. S.; LOUREIRO, E. S.; ALMEIDA, A. S.; ALVES, H. C.; ONO, F.
B.; LOBO, P. M.; MARIANI, M. A.; PESSOA, L. G. A.; SILVA, A. J. Avaliação
participativa do controle de cigarrinhas das pastagens com a aplicação do fungo
Metarhizium anisopliae (Metsch.) Sorok. Rev. Bras. de Agroecologia, v.2, n.2, 731-734,
2007.
RAVEN, P. H.; EVERT, R.F.; EICHHORN, S.E. Biologia Vegetal, 6ª Ed, Rio Janeiro,
Editora Guanabara Koogan, 2001.
REDDY, L. V. A.; WEE, Y.-J.; YUN, J.-S.; RYU, H.-W. Optimization of alkaline
protease production by batch culture of Bacillus sp. RKY3 through Plackett–Burman

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
89
and response surface methodological approaches, Bioresource Technology, v.99, p.242–
2249, 2008.
RIGUEIRA, J. C. S. Influência da contagem de células somáticas no perfil e teores de
aminas bioativas e na qualidade de leite cru e de queijo mussarela. 2010. 147p. Tese
(Doutorado em Ciência de Alimentos) - Faculdade de Farmácia, Departamento de
Alimentos, Programa de Pós-Graduação em Ciência de Alimentos, Faculdade de
Farmácia da Universidade Federal de Minas Gerais, Belo Horizonte.
RODRIGUES, M. I.; IEMMA, A. F. Planejamento de Experimentos e Otimização de
Processos: uma estratégia sequencial de planejamentos, Campinas, SP, Casa do Pão
Editora, 2005.
SANTOS, E. S. Recuperação e Purificação de Enzimas usando Adsorção em Leito
Expandido, Tese de Doutorado, Universidade Estadual de Campinas, Campinas/SP,
p.152, 2001.
SEDMAK, J. J.; GROSSBERG, S. E. A rapid, sensitive and versatile assay for protein
using coomassie brilliant blue G250, Analytical Biochemistry, v.79, p. 544-552, 1977.
SEINO, H.; TSUKUDA, K.; SHIMASUE, Y. Properties and action pattern of a
chitosanase from Bacillus sp. PI- 7S. Agricultural Biological Chemistry, 55, p.2421-
2423, 1991.
SHIMOSAKA, M.; NOGAWA, M.; OHNO, Y.; OKAZAKI, M. Chitosanase from the
plant pathogenic fungus, Fusarium solani f. sp. Phaseoli – purification and some
properties, Bioscience Biotechnolgy and Biochemistry, v.57, n.2, p.231-235, 1993.
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Production of two chitosanases from a chitosan-assimilating bacterium, Acinetobacter
sp. Strain CHB101, Applied and Environmental Microbiology, v.61, n.2, p.438-442,
1995.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
90
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Characterization of Three Chitosanase Isozymes Isolated from a Commercial Crude
Porcine Pepsin Preparation, J. Agric. Food Chem. 51, p.1042-1048, 2003.
SILVA FILHO, R. C.; COSTA, A. S. S.; CHAGAS, B. M. E.; SANTOS, F. N.;
MACEDO, G. R.; SANTOS, E. S. Avaliação do teste de atividade quitosanolítica
produzida por fungo. In: XV SIMPÓSIO NACIONAL DE BIOPROCESSOS, 15,
Recife, 2005, 02 a 05 de agosto.
SILVA, E. P. Avaliação Preliminar do Potencial de Aplicação da Tecnologia de
Biopilhas para a Biorremediação do Solo de Guamaré-RN. 2004. 119p. Dissertação
(Mestrado) – Centro de Tecnologia, Programa de Pós-Graduação em Engenharia
Química. Universidade Federal do Rio Grande do Norte. Natal.
SOMASHEKAR, D.; JOSEPH, R. Chitosanases – Properties and apllications: A review.
Bioresource Technology, v.55, p.35-45, 1996.
STOYACHENKO, I. A.; VARLAMOV, V. P.; DAVANKOV, V. A. Chitinases of
Streptomyces kurssanovii: purification and some properties, Carbohydrate Polymers,
v.24, p.47-54, 1994.
TINLINE, R. D. Nuclear distribuition in Metarhizium anisopliae. Mycologia, v. 63, p.
713-721, 1971.
TRABULSI, L. R.; TOLEDO, M. R. F. Microbiologia. 2. Ed., São Paulo: Atheneu,
1996.
VIEIRA, P. D. S.; SILVA, W. M. T.; PAIVA, L. M.; LIMA, E. A. L. A.;
CAVALCANTI, V. L. B. Estudo da caracterização morfológica, esporulação e
germinação dos conídiosde Metarhizium anisopliae var. acridum em diferentes
temperaturas. Biológico, São Paulo, v.69, n.1, p.17-21, 2007.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
91
WANDERLEY, M. D.; NEVES, E.; ANDRADE, C. J. Aspectos da produção industrial
de enzimas. Revista Citino, v.1, n.1, p.30-36, 2011.
WANG, C.; TYPAS, M. A.; BUTT, T. M. Detection and characterization of pr1
virulent gene deficiencies in the insect pathogenic fungus Metarhizium anisopliae.
FEMS Microbiology Letters, v. 213, p. 251-255, 2002.
ZAHORSKA-MARKIEWICZ, B.; KROTKIEWSKI, M.; OLSZANECKA-
GLINIANOWICZ, M.; ZURAKOWSKI, A. Effect of chitosan in complex management
of obesity, Pol Merkur Lekarski, v.13, n.74, p.129-132, 2002.
ZHANG, X. Y.; DAI, A. L.; ZHANG, X. K.; KUROIWA, K.; KODAIRA, R.;
SHIMOSAKA, M.; OKAZAKI, M. Purification and Characterization of and Exo--D-
glucosaminidase from a koji mold, Aspergillus oryzae IAM2660. Biosci. Biotechnol.
Biochem., v.64, n.9, 1896-1902, 2000.
ZHU, X.; WU, X.; DAI, Y. Fermentation conditions of a chitosanase from Acinetobcter
sp C-17. Biosci. Biotechnol. Biochem., v.67, n.2, p.284-290, 2003.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
92
APÊNDICE

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
93
APÊNDICE I
Metodologia para repique da cepa
Em Erlenmeyer de 1000 mL colocava-se 500 mL do meio de manutenção
PDA. Este Erlenmeyer, com o meio de manutenção, era autoclavado junto com placas
de Petri a 1,5 kgf/cm2 por 15 minutos, em seguida, era levado para uma câmara de fluxo
laminar e transferido para as placas de Petri já autoclavadas, onde eram mantidas numa
estufa a 27 ºC durante 24 h. Após este período, as placas de Petri eram levadas a câmara
de fluxo laminar, utilizando bico de Bunsen para esterilizar a alça de platina. Procedia-
se o repique utilizando uma placa de Petri contendo a cepa pura. Cada placa de Petri
contendo a cepa pura era transferida para duas placas de Petri contendo o meio de
manutenção. As placas eram mantidas na estufa a 27 ºC durante 30 dias onde após esse
período, repetia-se este procedimento.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
94
APÊNDICE II
Montagem da Curva de Calibração Usando o Método DNS
Soluções:
Glicosamina = 0,01 mol/mL (10000,0 µmol/mL)
Tabela 14. Dados para construção da curva de calibração usando o Método DNS.
Tubo de
Ensaio
Solução de
glicosamina (mL)
Água Destilada
(mL)
DNS (mL)
Volume Total
Massa (µµµµmol)
1 0 1,0 1,5 2,5 0
2 0,2 0,8 1,5 2,5 0,2
3 0,4 0,6 1,5 2,5 0,4
4 0,6 0,4 1,5 2,5 0,6
5 0,8 0,2 1,5 2,5 0,8
6 1,0 0 1,5 2,5 1,0
Tabela 15 - Medidas de absorbância a 600nm
Dados adicionais:
Peso Molecular D-Glicosamina = 215,64 (recipiente SIGMA)
Conc. Sol. D-Glicosamina = 0,05 g/50 mL
Massa calc.(µmol) = conc. Sol. D-Glic. x Vol. Utiliz. D-Glics. =
= 0,05 x 1000 x Vol.util./50*215,64
Tempo atividade enzimática = 30 min
1 U/mL = 1 µmol/(min.mL)
Vol. Total (Glicosamina+caldo) = 1 mL
Medições Adsorbância a 600 nm Vol. Utiliz. Massa Massa Atividade
II III Média(Abs)(mL) calc. (mg) calc. (µg) Enzim. (U/mL)
0 0 0 0 0 0 0
0,133 0,128 0,1307 0,2 0,2 200 0,0309
0,279 0,277 0,2787 0,4 0,4 400 0,0618
0,433 0,428 0,4297 0,6 0,6 600 0,0927
0,587 0,583 0,5797 0,8 0,8 800 0,1237
0,727 0,716 0,7157 1 1 1000 0,1546

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
95
Figura 17 – Curva padrão de glicosamina.
y = 1900ralx + 1900ral
R² = 1900ral
00,000
00,020
00,040
00,060
00,080
00,100
00,120
00,140
00,160
00,180
00,000 00,100 00,200 00,300 00,400 00,500 00,600 00,700 00,800
Ati
v.
En
zim
áti
ca (
U/m
L)
Absorbância (Abs) a 600 nm
Curva-Padrão D-Glicosamina

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
96
APÊNDICE III
Método de Determinação de Proteína Utilizando o Corante Comassie
Brilhante Blue G 250 (Método modificado de Sedmak e Grossberg)
Micrométodo
Preparar uma solução 0,06% de Comassie Blue G250 em 1,5 % HCl (w/V). Filtrar e
papel de filtro Whatman nº1.
Curva Padrão (1 a 20 µµµµg)
a) Preparar soluções estoque de BSA de 10 a 200 µg/mL.
b) Colocar em tubos de ensaio 100 µL da solução estoque, 1400 µL de água e
1500µL do Comassie e agitar.
c) Preparar um tubo contendo 1500 µL de água e 1500 µL do corante que
corresponde ao zero de proteína.
d) Após 5 minutos (padronizar o tempo) fazer as leituras das absorbâncias.
e) As leituras das amostras devem ser feitas em dois comprimentos de onda 595 e
465 contra a água.
f) Plotar a razão das absorbâncias a 595/465 como uma função da massa de BSA e
calcular o melhor ajuste linear dos dados.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
97
Micrométodo (1 a 20 µµµµg de BSA)
Solução Estoque de BSA = 0,2 mg/mL
Tabela 16. Dados para construção da curva padrão de BSA.
Tubo de
Ensaio
H2O (µµµµL)
BSA (µµµµg)
Coomassie (µµµµL)
BSA (µµµµg)
1 1500 0 1500 0
2 1490 10 1500 2
3 1480 20 1500 4
4 1470 30 1500 6
5 1460 40 1500 8
6 1450 50 1500 10
7 1440 60 1500 12
8 1430 70 1500 14
9 1420 80 1500 16
10 1410 90 1500 18
11 1400 100 1500 20
Dados adicionais:
Concentração Sol. Estoque
BSA = 0,2 mg/mL = 0,2 µg/µL
Vol. total sol. = vol. H2O + vol. BSA + vol.
Coomassie = 3000 µL = 3 mL
Concentração Sol. BSA (mg/mL) = (massa BSA mg) / 3 mL
.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
98
Figura 18 – Curva Padrão de BSA
y = 1900ralx - 1900ral
R² = 1900ral
000,000
001,000
002,000
003,000
004,000
005,000
006,000
007,000
000,400 000,500 000,600 000,700 000,800 000,900 001,000 001,100
µg
BS
A
Razão Absorbâncias (595nm/465nm)
Curva-Padrão BSA

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE TECNOLOGIA
DEPARTAMENTO DE ENGENHARIA QUÍMICA PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA
TESE DE DOUTORADO
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em
Cultivo Descontínuo Submerso
Raimundo Cosme da Silva Filho
Natal/RN
Abril/2013

Raimundo Cosme da Silva Filho
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo
Descontínuo Submerso
Tese apresentada ao Programa de Pós-Graduação em Engenharia Química – PPGEQ, da Universidade Federal do Rio Grande do Norte, como parte dos requisitos para obtenção do título de Doutor em Engenharia Química, sob a orientação do Prof. Dr. Everaldo Silvino dos Santos e a coorientação da Profª. Drª. Gorete Ribeiro de Macedo.
Natal/RN
Abril/2013

Catalogação da Publicação na Fonte. UFRN / CT / PPGEQ
Biblioteca Setorial “Professor Horácio Nícolas Sólimo”.
Silva Filho, Raimundo Cosme da. Otimização do meio de cultura para a produção de quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso / Raimundo Cosme da Silva Filho. - Natal, 2013. 98 f.: il.
Orientador: Everaldo Silvino dos Santos. Co-orientador: Gorete Ribeiro de Macedo. Tese (Doutorado) - Universidade Federal do Rio Grande do Norte.
Centro de Tecnologia. Departamento de Engenharia Química. Programa de Pós-Graduação em Engenharia Química.
1. Quitosana - Tese. 2. Quitina - Tese. 3. Quitosanase – Tese. 4. Metarhizium
anisopliae - Tese. I. Santos, Everaldo Silvino dos. II. Macedo, Gorete Ribeiro de. III. Universidade Federal do Rio Grande do Norte. IV. Título. RN/UF/BSEQ CDU 547.458 (043.2)

SILVA FILHO, Raimundo Cosme da – Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo Descontínuo Submerso. Tese de Doutorado. UFRN - Programa de Pós-Graduação em Engenharia Química. Área de Concentração: Pesquisa e Desenvolvimento de Tecnologias Regionais – Subárea: Engenharia de Processos – Alimentos e Biotecnologia, Natal/RN, Brasil.
Orientação: Prof. Dr. Everaldo Silvino dos Santos Coorientação: Profª. Drª. Gorete Ribeiro de Macedo ___________________________________________________________________________
RESUMO: No presente trabalho utilizou-se um Planejamento Plackett & Burman, com 8 fatores e 12 ensaios em 2 níveis e mais 3 repetições na condição do ponto central, para se investigar a influência das concentrações de quitosana, peptona, extrato de levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O e FeSO4 na produção da enzima quitosanase por Metarhizium
anisopliae. Os ensaios para produção da enzima foram realizados em cultivo descontínuo submerso. Os resultados para o Planejamento Plackett & Burman mostraram que a atividade quitosanolítica foi favorecida pelo aumento da concentração de substrato (quitosana) e de sulfato ferroso (FeSO4), enquanto que o aumento da concentração dos outros fatores não contribuiu de forma significativa para a atividade quitosanolítica. A otimização do meio de cultura para a produção da enzima foi realizado por meio de um Planejamento Composto Central, com os dois fatores que mais influenciaram a atividade quitosanolítica (quitosana e FeSO4), conforme Planejamento Plackett & Burman, mantendo-se os outros nutrientes em seus valores mínimos. Nesse planejamento, para o fator FeSO4, tomou-se o limite inferior (-1) como sendo o limite superior do Planejamento Plackett & Burman. Os resultados mostraram que a produção da enzima foi favorecida pelo aumento da concentração quitosana e pela diminuição da concentração de FeSO4. A produção máxima de atividade quitosanolítica foi da ordem de 70,0 U/L e foi atingida em apenas 18 h de fermentação, resultado esse vinte e oito vezes maior aos obtidos anteriormente para o mesmo microrganismo que foi 2,5 U/L em 48h.
Palavras-chave: quitosana; quitosanase; Metarhizium anisopliae; Plackett & Burman.


ABSTRACT
In this work a Plackett-Burman Design with 8 factors and 12 trials in 2 levels with 3 repetitions at the center point was used in order to investigate the influence of the concentration of chitosan, peptone, yeast extract, NaNO3, K2HPO4, KCl, MgSO4.7H2O and FeSO4 on chitosanase production by Metarhizium anisopliae. Runs were carried out using submerged discontinuous cultivation for enzyme production. The results of the Plackett & Burman Design showed that only two factors, chitosan concentration as well as FeSO4 had influence on chitosanolytic activity, while the increase in concentration of other factors not contributed significantly to the quitosanolítica activity. Cultivation medium optimization for enzyme production was carried out using a Composite Central Design, with the most important factors for chitosanolytic activity (chitosan and FeSO4), in accordance with Plackett & Burman Design, and keeping the other nutrients in their minimum values. On this other design, it was taken the highest limit in Plackett & Burman Design as the lowest limit (-1) to FeSO4 factor. The results showed that the enzyme production was favoured by increasing the chitosan concentration and by decreasing FeSO4. Maximum production for chitosanolytic activity was about 70.0 U/L and was reached in only 18 h of fermentation, a result about twenty-eight times greater than a former study using the same microorganism (about 2.5 U/L at 48 h).
Key-words: Chitosan; Chitosanase; Metarhizium anisopliae; Plackett & Burman

Ao meu pai, minha mãe e minha
família, em especial aos meus
irmãos e sobrinhos

AGRADECIMENTOS
A Deus, por estar presente em todos os momentos da minha vida e por me ceder à
oportunidade de iniciar e concluir este trabalho.
A minha mãe Cândida Inocência da Silva que esteve comigo em todos os momentos da
minha vida me educando, me amando e sempre me dando muito carinho.
Ao meu pai Raimundo Cosme da Silva, que não está mais aqui para ver este trabalho,
mas que foi a pessoa que mais me apoiou e me incentivou, em todos os momentos da minha vida,
para que eu pudesse chegar o mais longe possível na minha carreira acadêmica. Pai muito
obrigado por toda atenção, educação, amor e carinho que recebi durante toda a minha vida.
Aos meus irmãos e familiares que sempre estiveram comigo dividindo momentos de
grande alegria e às vezes de tristeza, mas sempre nos mantendo unidos para superarmos todos os
obstáculos.
A minha namorada Rochelly Caroliny por todo amor e carinho que me deu e que me
entendeu nos momentos que estive ausente para a conclusão deste trabalho.
Ao meu orientador Professor Doutor Everaldo Silvino dos Santos por confiar em mim e
me aceitar como orientando além de toda a amizade, orientação e dedicação em todas as etapas de
desenvolvimento e conclusão deste trabalho.
À Professora Doutora Gorete Ribeiro de Macedo, “pessoa que admiro muito como
profissional e principalmente, como pessoa, um exemplo a ser seguido”, pelos conselhos e críticas
quando necessários ao desenvolvimento deste trabalho. Professora a senhora é considerada uma
mãe para de todos os alunos que tiveram a honra de conhecê-la e trabalhar com a senhora.
Aos meus amigos Engenheiros Químicos, Ana Carmen, Ana Katerine, Andréa Farias,
Anita Lima, Sirtys Lessa, Alexandre, Sérgio, Márcio Bezerra e ao Biólogo Daniel Souza, que
sempre me ouviram e que me ajudaram de alguma forma nesse trabalho.
A Eusamar Coelho de Lima, Mazinha, secretária do PPGEQ, pelo apoio, pela paciência e
orientação nas questões burocráticas do processo.
A Carlos e Jardelson, pela grande colaboração e dedicação nos experimentos do
laboratório.
A Sérgio Santana pela grande colaboração e dedicação, o qual não poupou esforços nem
tempo em me ajudar na realização dos experimentos em laboratório.

Ao meu amigo Ubiratan Nazaré por me ajudar na organização do texto e na tradução para
o Abstract.
Finalizando, agradeço ao Laboratório de Engenharia Bioquímica da UFRN por ter cedido
toda a sua estrutura para realização dos experimentos e ao Programa de Pós-Graduação em
Engenharia Química, assim como a todos os professores do Departamento de Engenharia Química
da UFRN que trabalham em pró de uma educação digna e ensino de qualidade para todos os
alunos.

Sumário LISTA DE FIGURAS
LISTA DE TABELAS
NOMENCLATURA
INTRODUÇÃO...............................................................................................................................15
REVISÃO BIBLIOGRÁFICA........................................................................................................18
2.1. Enzimas................................................................................................................................18
2.1.1. Quitosana e seus derivados........................................................................................19
2.1.2. Quitosanases...............................................................................................................24
2.2. Fungos..................................................................................................................................29
2.2.1. Características gerais..................................................................................................29
2.2.2. Reprodução dos Fungos.............................................................................................32
2.2.3. Metabolismo de fungos..............................................................................................33
2.2.4. Nutrição de fungos.....................................................................................................34
2.2.5. O Fungo Metarhizium anisopliae ..............................................................................36
2.3. Fermentação.........................................................................................................................40
2.3.1. Preparo do inóculo......................................................................................................41
2.3.2. Produção de enzimas em cultivo submerso descontínuo...........................................42
2.4. Planejamento experimental..................................................................................................42
2.4.1. Planejamento fatorial..................................................................................................43
2.4.2. Planejamento Plackett & Burman..............................................................................43
2.4.3. Aplicações do Planejamento Plackett & Burman.......................................................44
METODOLOGIA EXPERIMENTAL.............................................................................................50
3.1. Quitosana..............................................................................................................................50
3.2. Microrganismo.....................................................................................................................50
3.2.1. Escolha da cepa produtora de quitosanase.................................................................50

3.2.2. Manutenção da cepa...................................................................................................50
3.3. Produção de enzima.............................................................................................................51
3.4. Determinação da Proteína Total...........................................................................................51
3.5. Determinação da Atividade Quitosanolítica e Atividade Específica...................................51
3.6. Planejamento Experimental.................................................................................................52
3.6.1. Planejamento de seleção de fatores Plackett Burman................................................52
3.6.2. Planejamento Composto Central para Otimização do meio de cultura para a
Produção da Enzima em Cultivo Submerso.....................................................................................53
3.7. Perfil Cinético durante o Cultivo.........................................................................................56
RESULTADOS E DISCUSSÃO.....................................................................................................58
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA ..........................................58
4.2. Planejamento Plackett & Burman........................................................................................59
4.2.1. Análise do Plackett & Burman com 8 fatores e 12 ensaios .......................................59
4.2.2. Análise do Plackett & Burman com 12 ensaios e mais 3 repetições na condição do
ponto central ....................................................................................................................................61
4.3. Planejamento Composto Central para Produção de Quitosanase por Metarhizium
anisopliae em Cultivo Descontínuo Submerso................................................................................64
4.4. Perfil Cinético – Melhor Condição......................................................................................69
CONCLUSÃO.................................................................................................................................75
REFERÊNCIAS BIBLIOGRÁFICAS.............................................................................................78
APÊNDICE I...................................................................................................................................93
Metodologia para Repique da Cepa ................................................................................................93
APÊNDICE II.................................................................................................................................94
Montagem da Curva de Calibração Usando o Método DNS..........................................................94
APÊNDICE III................................................................................................................................96
Método de Determinação de Proteína Utilizando o Corante Comassie Brilhante Blue G 250
(Método modificado de Sedmak e Grossberg)................................................................................96

Lista de Figuras
Figura 1 – Estrutura da quitosana..................................................................................................20
Figura 2 – Estrutura da glicosamina..............................................................................................23
Figura 3 – Estrutura do sulfato de glicosamina.............................................................................23
Figura 4 – Representação da estrutura geral de uma célula de fungo..........................................30
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas...........................................................31
Figura 6 – Conídios de Aspergillus e Penicillium.........................................................................33
Figura 7 – Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA ...................58
Figura 8 – Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados............................................................................................................................60
Figura 9 – Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto central
mostrando a influência dos fatores estudados................................................................................63
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos
fatores estudados............................................................................................................................67
Figura 11 – Valores preditos em função dos valores observados..................................................68
Figura 12 – Atividade quitosanolítica em função da concentração de quitosana e da concentração
de FeSO4 para os fatores não significativos no ponto central .......................................................68
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana ........70
Figura 14 – Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana....................................70
Figura 15 – Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de
quitosana........................................................................................................................................71
Figura 16 – Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana..........................................................................................................72
Figura 17 – Curva padrão de glicosamina ....................................................................................95
Figura 18 – Curva Padrão de BSA ................................................................................................98

Lista de Tabelas
Tabela 1 – Comparação das enzimas com os catalisadores químicos..............................................19
Tabela 2 – Fontes naturais de quitina e quitosana ...........................................................................20
Tabela 3 – Níveis para os fatores e seus valores codificados...........................................................53
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central .........................................................................................53
Tabela 5 – Matriz do planejamento Composto Central...................................................................54
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central......................................................................................................55
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.......................59
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman...............................................60
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.......................61
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central...............................................................................................................................................62
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da atividade
enzimática........................................................................................................................................65
Tabela 12 – Efeitos Estimados do planejamento Composto Central...............................................65
Tabela 13 – Resumo da ANOVA....................................................................................................66
Tabela 14 – Dados para construção da curva de calibração usando o Método DNS ......................94
Tabela 15 – Medidas de absorbância a 600 nm para construção da curva de calibração................94
Tabela 16 – Dados para construção da curva padrão de BSA ........................................................97

NOMENCLATURA
A – Atividade quitosanolítica (U/L)
AE – Atividade específica (U/mg)
ANN-GA – algoritmo genético de rede neural artificial
ANOVA – Análise de variância (Analysis of Variance)
BSA – Albumina de soro bovino (Bovine Sereum Albumin)
CCD – Planejamento Composto Central
CDA – Chitosan Dector Agar
Cs – Concentração de substrato (%)
CP – Concentração de KCl (%)
Da – Dalton
DEQ – Departamento de Engenharia Química
DNS – Ácido Dinitrosalissílico
E – Enzima
Ea – Energia de ativação
EC – Comissão para nomenclatura e classificação de enzimas
EL – Concentração de Extrato de Levedura (%)
FP – Concentração de K2HPO4 (%)
NS – Concentração de NaNO3 (%)
GDA – Grânulos dispersíveis em água
k – Número de fatores, variáveis
LEB – Laboratório de Engenharia Bioquímica
mRNA – RNA mensageiro
n – Número de níveis de um determinado fator
N – Número de ensaios de um planejamento
P – Pressão
PP – Concentração de Peptona (%)
PDA – Batata Dextrose Agar
pH – Potencial hidrogeniônico
Q – Concentração de Quitosana (%)
Qm – Quadrado médio
r – Agitação (rpm)
R2 – Coeficiente de determinação

RA – Razão de aeração
rpm – Rotações por minuto
RSM – Metodologia de superfície de resposta
S – Molécula de substrato
SF – Concentração de FeSO4 (%)
SM – Concentração de MgSO4.7H2O (%)
SQ – Soma de quadrados
SSF – Fermentação de estado sólido
T – Temperatura
TMC – Temperatura de cultivo
V11,V12,V13 – Variáveis inertes para cálculo do erro
VHG – Fermentação de alta densidade
WP – Pó molhável
X – Concentração celular
Xi – variável independente
Xj – variável independente
xi – Fator independente real
x0 – Valor de xi no ponto central
∆xi – Variação de valor
Y – Resposta do modelo de primeira ordem
Yp – Resposta do modelo polinomial de segunda ordem
β0 – Interseção dos modelos de primeira ou segunda ordem
βi – Coeficiente linear dos modelos de primeira ou segunda ordem
βii – Coeficiente quadrático
βij – Coeficiente de interação

Capítulo 1 Introdução

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
15
1. Introdução As quitosanases foram descobertas simultaneamente há quase quarenta anos por
Monaghan et al. (1973), como pelo grupo de Ramirez-León & Ruiz-Herrera (1972), e
são enzimas obtidas a partir do metabolismo de bactérias, fungos e extraídas de plantas.
Desde então, vários estudos vêm apresentando a obtenção de quitosanase deste
microrganismos, na forma intracelular ou extracelular (Osswald et al., 1994;
Somashekar & Joseph, 1996), como a do presente estudo. A quitosanase (EC 3.2.1.132)
representa uma classe de enzimas hidrolíticas que catalisam a hidrólise de ligação β-
(1→ 4) glicosídica da quitosana, gerando oligômeros chamados quito-oligossacarídeos
(Reddy et al., 2008).
A quitosana é um biopolímero hidrofílico obtido a partir da quitina, um
polissacarídeo abundante na natureza. Na literatura, é considerado quitosana o produto
da desacetilação da quitina para um grau de desacetilação superior a 75% e solúveis em
ácidos como o acético e o fórmico. A quitosana é conhecida por exibir uma ampla
variedade de atividade fisiológica como atividade antitumoral, mas o peso molecular e a
insolubilidade em água da quitosana são desvantagens para várias aplicações.
Atualmente, há interesse nas quitosanas quando as mesmas são convertidas em quito-
oligossacarídeos porque eles não apenas são solúveis em água e possuem baixo peso
molecular, mas também têm atividades biológicas bem notáveis, como atividade
anticancerígena atóxica e biocompatível, incluindo efeito antimetastático, atividade anti-
HIV, hepatoprotetora, antioxidante, hipocolesterolêmica, antimicrobiana,
imunoestimulate, antitumoral, anti-inflamatória e atividade aceleradora da absorção de
cálcio e ferro. No entanto, estes quito-oligossacarídeos são obtidos tradicionalmente na
indústria por reação de hidrólise usando ácidos, que apresenta muitos problemas como
dificuldade em controlar a hidrólise, produzindo uma grande quantidade de
oligossacarídeos de cadeia pequena e oligossacarídeos de baixo rendimento, alto custo
de separação e, também, poluição ambiental, uma vez que utiliza um ácido como
catalisador. Como alternativa, com suas vantagens em termos de compatibilidade
ambiental, baixo custo e reprodutibilidade, a obtenção destes quito-oligossacarídeos por
hidrólise com quitosanase se tornou cada vez mais popular nos últimos anos (Azevedo
et al., 2007).

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
16
Devido a todos estes aspectos positivos é que, em geral, as enzimas vêm sendo
estudadas em diferentes aplicações, mostrando que esta é uma tendência na indústria.
No presente trabalho, foi desenvolvido um processo fermentativo visando a
otimização do meio de cultura para a produção da enzima quitosanase em cultivo
descontínuo submerso, por Metarhizium anisopliae, capaz de sintetizar oligômeros com
aplicações na indústria farmacêutica e de química fina, partindo de quitosana como
única fonte de carbono.
Nesse caso, avaliou-se, através de um planejamento Plackett & Burman (método
de delineamento de seleção de fatores) e de um planejamento Composto Central as
melhores condições para a produção da enzima. A técnica do planejamento
experimental Plackett & Burman para realizar triagem de fatores na otimização de um
determinado processo foi utilizada neste trabalho para identificar quais os fatores que
influenciam de forma significativa na produção da enzima quitosanase. A influência
destes fatores (concentração de quitosana e concentração de FeSO4) foi então
investigada pelo planejamento Composto Central com 22 experimentos na parte fatorial,
repetição em triplicata no ponto central e quatro experimentos na parte axial. A
repetição do ponto central permite também a determinação do erro puro e da
repetibilidade do processo, sendo uma forma de identificar melhor se o processo está
sob controle.
Do ponto de vista industrial é de relevante importância a realização de estudos
para a otimização da produção da quitosanase de forma contínua ou em grandes
bateladas.

Capítulo 2 Revisão Bibliográfica

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
18
2. Revisão Bibliográfica 2.1. Enzimas
As enzimas são catalisadores biológicos constituídos por longas cadeias de
aminoácidos que atuam em substratos reduzindo a energia de ativação, acelerando assim
a velocidade da reação, e transformando-os em produtos. Sua atuação catalítica visa
facilitar a atividade das células através da quebra de moléculas ou junção das mesmas
para formação de novos produtos. Podem apresentar alto grau de especificidade relativa
a apenas um substrato ou a vários substratos ao mesmo tempo, como a certos tipos de
ligações glicosídicas α-1,4 das moléculas de amido, ou para determinado tipo particular
de isômero óptico como ocorre na oxidação da β-D-glicose pela glicose oxidase (Furigo
Junior & Pereira, 2001).
Enzimas são proteínas que apresentam em sua estrutura um centro ativo
(apoenzima) e, em alguns casos, um grupo prostético denominado cofator (coenzima ou
íon metálico ativo) e sua atividade depende de alguns fatores como estrutura da
proteína, número de cadeias peptídicas, arranjo dessas cadeias na molécula, natureza do
substrato e natureza do grupo prostético (Politzer & Bon, 2006).
Todos os organismos vivos produzem, em pequena ou grande quantidade, certa
variedade de enzimas, sejam elas intra ou extracelulares. As enzimas extracelulares são
excretadas para o meio externo e podem decompor compostos insolúveis como
celulose, quitina e quitosana.
Algumas destas enzimas podem ser usadas tanto nas indústrias de detergentes,
têxteis e farmacêuticas, como também de alimentos, de bebidas, etc., sendo utilizadas
em processos biotecnológicos industriais em substituição aos processos químicos que
muitas vezes causam grandes prejuízos ao meio ambiente. A produção comercial das
enzimas pode ser realizada através do cultivo de microrganismos como fungos e
bactérias.
Na Tabela 1 mostra-se uma comparação das enzimas com os catalisadores
químicos, levando-se em conta algumas das características dos dois catalisadores.
Destaca-se que alguns parâmetros da Tabela 1 demonstram a maior importância das
enzimas, quando comparadas com os catalisadores químicos, principalmente a
especificidade ao substrato, as condições de reação, o consumo de energia e a

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
19
velocidade de reação, como desvantagem cita-se o custo de obtenção (isolamento e
purificação).
Tabela 1 – Comparação das enzimas com os catalisadores químicos (Adaptado de
Furigo Junior & Pereira, 2001).
Característica Enzimas Catalisadores Químicos
Especificidade ao substrato Alta Baixa
Natureza da estrutura Complexa Simples
Sensibilidade à T e pH Alta Baixa
Condições de reação (T, P e pH) Suaves Drástica (geralmente)
Custo de obtenção (isolamento e purificação) Alto Moderado
Natureza do processo Batelada Contínuo
Consumo de energia Baixo Alto
Formação de subprodutos Baixa Alto
Separação catalisador/produtos Difícil/cara Simples
Atividade catalítica (temperatura ambiente) Alta Baixa
Presença de cofatores Sim Não
Estabilidade do preparado Baixa Alta
Energia de Ativação Baixa Alta
Velocidade de reação Alta Baixa
2.1.1. Quitosana e seus derivados
A quitosana e seus derivados apresentam grande importância devido ao
conhecimento de suas funcionalidades biológicas, médicas, tecnológicas, etc
(Shimosaka et al, 1993; Shimosaka et al, 1995; Mitsutomi et al., 1995). A quitosana é
obtida da quitina, polissacarídeo abundante na natureza, extraído principalmente das
carapaças de caranguejos, lagosta e camarões. A quitosana é um polímero natural,
biodegradável, não tóxico e de alta massa molecular obtido a partir de animais
marinhos, insetos e microrganismos (Tabela 2).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
20
Tabela 2 – Fontes naturais de quitina e quitosana. (Adaptado de Polymar, 2003).
Animais
Marinhos
Insetos Aracnídeos Microrganismos
Anelídios Formigas Escorpiões Leveduras
Moluscos Besouros Aranhas Fungos
Lagosta Esporos
Camarão
Caranguejo
Krill
A Figura 1 abaixo mostra a estrutura química da quitosana
Figura 1 – Estrutura da quitosana (Fonte: Polymar, 2003)
Quitosana totalmente desacetilada não existe na natureza. Graus de acetilação
(GA) nferior a 50% a torna mais solúvel em soluções ácidas (Muzzarelli, 1993;
Muzzarelli et al., 1994; Dung et al., 1994; Stoyachenko et al.,1994). Pelletier &
Sygusch (1990) enfatizaram que grande parte dos resíduos sólidos da indústria
pesqueira, principalmente da carcinicultura, é jogada ao mar ocasionando degradação
ambiental. Logo, a utilização desses resíduos como matéria-prima para formação de
novos produtos, não só reduz de forma drástica o impacto ambiental, mas também daria
maior valor agregado à indústria pesqueira.
A demanda cada vez maior de consumidores por alimentos mais saudáveis, sem
conservantes químicos, tem motivado pesquisas à descoberta de novos agentes
antimicrobianos naturais. Sendo assim, a atividade antimicrobiana incomum da

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
21
quitosana e seus derivados contra microrganismos, tais como bactérias, bolores e
leveduras tem tido maior atenção ultimamente.
Devido à interação entre a quitosana, que é positivamente carregada, e as
membranas celulares dos microrganismos, negativamente carregadas, ocorre a formação
de verdadeiros poros na membrana, que são canais de vazamento de conteúdo protéico e
outros componentes intracelulares. A quitosana tem também a propriedade de agir como
agente quelante que se liga de forma seletiva a traços de metais, inibindo, assim, a
produção de toxinas e o crescimento microbiano. Ainda é capaz de ativar mecanismos
de defesa em tecidos hospedeiros, atuando como agente sequestrante de água e inibindo
várias enzimas. A ligação da quitosana com DNA e a inibição da síntese do mRNA
ocorre via penetração da quitosana no núcleo dos microrganismos e interferência na
síntese do mRNA e proteínas, (Craveiro et al., 2004).
Em um estudo clínico conduzido por Giustina & Ventura (1995) com 100
pessoas com sobrepeso de 10 a 25%, ao final de 4 semanas as pessoas que receberam
quitosana demonstraram perda de peso significativa (7,3 Kg contra 3,0 Kg nos grupos
controle). As pessoas que tomaram quitosana também demonstraram redução na pressão
sanguínea sistólica (de 145 para 135 mm Hg) e diastólica (92.6 para 84.2 mm Hg),
assim como uma redução na taxa respiratória (27.6/min a 21.2/min).
Kondo & Osada (1996) pesquisaram a influência da quitina e da quitosana na
biodisponibilidade do zinco em ratos alimentados com uma dieta com suplementos de
5% ou sem fibra dietética (quitina, quitosana, celulose, pectina e agar-agar) durante 31
dias. Observaram que os ratos alimentados com dietas contendo 5% de fibras, exceto
quitosana, o consumo de alimento foi maior do que o controle. O ganho de peso em
todos os grupos alimentados com fibra foi maior do que o controle, exceto para o grupo
suplementado com quitosana (5%) onde a ingestão de alimento e o ganho de peso foram
menores do que os grupos controle e das demais fibras.
Segundo Craveiro et al. (2004), derivados da quitosana obtidos pela reação com
ácidos carboxílicos, podem promover ao polímero uma solubilidade especial em água.
Esses compostos possuem estrutura química semelhante ao ácido hialurônico e
fornecem uma excelente compatibilidade com o tecido epitelial. As soluções obtidas
com esses derivados podem ser facilmente incorporadas a formulações cosméticas,
mostrando melhor compatibilidade com surfactantes iônicos e emulsificantes,
apresentando estabilidade em uma larga faixa de pH.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
22
Han et al. (1999) examinaram os efeitos da quitosana sobre a atividade da lipase
pancreática in vitro e sobre o grau de armazenamento de gorduras induzido em ratos
pela administração oral de uma dieta rica em gorduras por nove semanas. Como
resultado, observou-se que os animais alimentados com quitosana, reduziram o peso
corporal, a hiperlipidemia e a gordura hepática. Como conclusão, os autores propõem
que os efeitos anti-obesidade da quitosana em ratos alimentados com alto teor de
gordura se devem parcialmente a inibição da absorção intestinal das gorduras dietéticas.
Em outro ensaio, Zahorska-Markiewicz et al. (2002) realizaram um estudo para
avaliar o efeito da quitosana no tratamento da obesidade. Segundo os autores do estudo,
a quitosana pode ser usada como um valioso e seguro coadjuvante em tratamentos para
a obesidade em longo prazo. A quitosana parece acentuar a redução da pressão arterial,
juntamente com a perda de peso.
Assis (2010) estudou a produção e caracterização de quitooligossacarídeos
produzidos pelo fungo Metarhizium anisopliae e avaliação da citotoxicidade em células
tumorais. A hidrólise da quitosana foi realizada em tempos de 10 a 60 min para a
produção de quitooligossacarídeos. A avaliação da citotoxicidade dos oligômeros de
quitosana foi realizada em células tumorais (HepG2 e HeLa) e não tumoral (3T3). A
produção de oligômeros de quitosana teve maiores rendimentos durante 10 minutos de
hidrólise, os pentâmeros apresentaram concentração de 0,15 mg/mL, no entanto os
hexâmeros, que apresentam maior interesse por causa de suas propriedades biológicas,
só foram detectados com 30 minutos de hidrólise com uma concentração de 0,004
mg/mL.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o planejamento Plackett-Burmam e a metodologia de superfície de resposta.
Os resultados mostraram que a eficiência máxima de extração da quitina foi conseguida
nas seguintes condições: concentração de casca de camarão 50g/L, glicose 50g/L, tempo
de incubação de 5 dias e inoculo como 0,05 OD (densidade óptica).
A glicosamina, unidade monomérica básica da quitosana, possui atividade
antiinflamatória e tem grande importância na construção e manutenção da cartilagem,
no tratamento de enfermidades como: febre reumática, artrose e principalmente a artrite
e osteoartrite na forma aguda ou crônica (Polymar, 2003). A Figura 2 apresenta a
estrutura química da glicosamina.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
23
Figura 2 – Estrutura da glicosamina (Fonte: Polymar, 2003)
O sulfato de glicosamina é um derivado da quitosana sendo utilizado no
tratamento da osteoartrite sintomática dos joelhos, quadris, coluna, mãos e outras
regiões (Craveiro et al., 2004). O sulfato de glicosamina estimula a síntese de
proteoglicanos necessários para o bom funcionamento das articulações, estimula a
regeneração das cartilagens e ainda incorpora enxofre ao tecido cartilaginoso. Estudos
de longa duração, acima de 3 anos, mostram que o sulfatos de glicosamina evita a
progressão de danos estruturais nas articulações e induz melhora significativa nas dores
e funções físicas (Craveiro et al., 2004). A Figura 3 apresenta a estrutura química do
sulfato de glicosamina.
Figura 3 – Estrutura do sulfato de glicosamina (Fonte: Polymar, 2003)

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
24
2.1.2. Quitosanases
Dependendo do tipo de quitosanase obtida, ocorrem diferentes mecanismos de
hidrólise da quitosana, isso devido a diferentes microrganismos produzirem enzimas
com diferenças nas suas estruturas (Kurita et al., 1977; Seino et al., 1991).
Quitosanases (EC 3.2.1.132) são enzimas que catalisam a reação de hidrólise de
quitosana em oligossacarídeos. As enzimas quitosanases são produzidas por vários
microrganismos, bactérias, actinomicetos e fungos, e até por algumas plantas. Estas
enzimas agem em polímeros com grau de acetilação de 30% a 60%. Os monômeros
resultantes da hidrólise da quitosana por estas enzimas são melhores digeridos, ao serem
ingeridos, como é o caso da glicosamina. Estudos efetuados até o presente momento
classificam as quitosanases em cinco famílias: 5, 8, 46, 75, 80, sendo que a família 46 é
a mais estudada. As quitosanases tem grande aplicação na geração de oligômeros de
quitosana (Somashekar & Joseph, 1996; Chen et al., 2005).
Nogawa et al. (1998) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida pelo fungo Trichoderma reseei PC-3-7. Estes pesquisadores
inocularam 106 conídios em 100 mL de um meio basal contendo 0,3% de N-
acetilglicosamina em incubator rotativo com agitação de 220 rpm a 28oC. Destaca-se
que o fungo não produziu quitosanase em meio contendo quitosana ou quitina.
Entretanto, o microrganismo produziu quitosanase em meio contendo glicosamina e N-
acetilglicosamina, sendo que para este último obteve-se uma atividade específica
máxima de 1,5 U/mg após 72 horas de incubação. É importante ressaltar que uma
agitação vigorosa favoreceu a produção da enzima. Esse artigo ilustra a presença de
exo-β-D-glicosaminidase e exo-β-D-N-acetilglicosaminidase.
Piza et al. (1999) estudaram a produção, purificação e caracterização parcial de
uma quitosanase produzida por Bacillus cereus. Nesse caso, investigou-se através de um
planejamento fatorial fracionário a influência dos fatores: concentração de sulfato de
amônio (0,4; 2,2 e 4,0%), razão de aeração (2; 6 e 7), tempo de fermentação (16; 24 e
32 horas), pH (5; 6 e 6,5) e concentração de quitosana (1,0; 1,5 e 2,0%). Os resultados
mostraram que os fatores mais significativos para esse planejamento foram a
concentração de sulfato de amônio, a aeração, o pH e a interação entre sulfato de
amônio e a aeração.
Cheng & Li (2000) estudaram a preparação de oligossacarídeos utilizando
quitosanase produzida por Aspergillus sp e constataram que a enzima foi largamente

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
25
induzida em meio contendo quitosana como única fonte de carbono e também na
presença de quitina coloidal. A indução da enzima foi realizada em temperaturas
variando na faixa de 28 a 30ºC, pois essa espécie não cresceu bem em temperaturas
superiores a 37ºC.
Zhang et al. (2000) estudaram a produção, purificação e caracterização de
quitosanase e Exo-β-D-Glucosaminidase por Aspergillus oryzae IAM2660. Os
microrganismos utilizados neste estudo foram mantidos em placas de Petri contendo
agar para contagem em placas (PDA). Destaca-se que para produzir a enzima, conídios
foram retirados das placas e inoculados em meio Czapeq-dox com os seguintes
componentes: 1,0g K2HPO4, 0,5g KCl, 2,0g NaNO3, 0,5g MgSO4.7H2O, 0,02g
FeSO4.7H2O, 20g de sacarose e 1,0L de água deionizada, com pH do meio igual a 5,5.
Foi adicionado ao meio 0,5 g de peptona e 0,5g de extrato de levedura com
concentração de esporos da ordem de 1,0x106 esporos.mL-1. Após a incubação, feita em
incubador rotativo a 26ºC por um período de 60h, micélios foram coletados e após
filtração e secagem a 80ºC foram utilizados para ensaio de quitosanase.
Kuroiwa et al. (2002) estudaram os fatores que afetam a composição de
oligossacarídeos produzidos através da hidrólise de quitosana por quitosanase originária
de Bacillus pumilus BN-262. Neste experimento foi utilizado quitosana 100%
desacetilada dissolvida em ácido acético 1M. O pH do meio foi ajustado em 5,6. Neste
estudo a quitosanase foi imobilizada em agar e Sepharose. Os resultados mostraram que
a concentração de enzima, a temperatura, a velocidade de agitação, e concentração de
substrato influenciaram de forma significativa na composição dos oligossacarídeos
produzidos.
Ichikawa et al. (2002) estudaram a imobilização e estabilização da ligação
multiponto da quitosanase tendo agar como suporte e concluíram que a imobilização
aumentou a termoestabilidade da quitosanase. Os oligossacarídeos de quitosana foram
continuamente produzidos usando um reator coluna contendo quitosanase imobilizada.
A percentagem de quitosana imobilizada após 28 dias de reação foi de 44%.
Hung et al. (2002) estudaram a purificação, através de troca iônica, e
caracterização da hidrólise sobre ação da quitinase e quitosanase purificadas do meio
contendo Bromelina comercial. Destaca-se que a quitinase purificada apresentou pH,
temperatura e Km ótimo para ação catalítica em meio contendo quitina de 4,0, 60ºC e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
26
0,2mg.mL-1, respectivamente. Enquanto que quitosanase apresentou o pH, temperatura e
Km ótimo de 3,0, 50ºC e 0,88mg.mL-1, respectivamente.
Zhu et al. (2003) estudaram as melhores condições fermentativas e propriedades
de uma quitosanase produzida por Acinetobacter sp. C-17. A espécie de bactéria
utilizada neste ensaio foi isolada do solo e identificada como sendo Acinetobacter sp C-
17 e capaz de produzir quitosanase. O microrganismo foi mantido em placas contendo o
meio agar com os seguintes componentes: 0,5 % de quitosana, 0,2 % K2HPO4, 0,1 %
KH2PO4, 0,07 % MgSO4, 0,05 % NaCl, 0,05 % KCl, 0,01 % CaCl2, 0,05 % de extrato
de levedura e 2 % agar, o pH do meio de manutenção foi mantido em 7,2 e a
temperatura foi mantida em 30ºC durante 3 a 5 dias, para identificar a ação da enzima
quitosanase. A ação da quitosanase foi identificada através da observação de uma
colônia de microrganismo existente na placa. A bactéria foi então isolada da placa e
colocada em fermentador rotativo a 200 rpm durante 72h em meio contendo: 0,07 %
K2HPO4, 0,03 % KH2PO4, 0,05 % MgSO4, 0,03 % peptona, 0,03 % de extrato de
levedura e 1,0 % de quitosana. O pH foi de 7,0 e a temperatura 30ºC. Como fontes de
carbono foram utilizadas além de 1,0 % de quitosana, 1,0 % de quitina e 1,0 % de
glicose. Os resultados mostraram que as melhores condições para produção da
quitosanase foram alcançadas quando se utilizou 1,0 % de solução de quitosana a 30ºC,
pH 7,0 durante 24 horas de incubação.
Liu et al. (2003) estudaram a produção de quitinase por Verticillium lecanii
F091 usando fermentação submersa, a atividade de quitinase foi de 9,95 mU/mL em um
meio de cultura otimizado, com um volume de cultura de 200 mL, agitação de 150 rpm
e temperatura de 24ºC cultivado em incubador rotativo e concluíram que a taxa de
agitação e o pH foram os fatores mais significativos para a produção de quitinase, no
entanto, as taxas de agitação e aeração poderiam modificar a concentração de oxigênio
dissolvido afetando diretamente no crescimento do Verticillium lecanii e na produção
de quitinase.
Donzelli et al. (2003) estudaram a hidrólise enzimática avançada de quitina de
casca de caranguejo langostino com misturas de enzimas de bactérias e fungos e
concluiram que a combinação de enzimas de Trichoderma atroviride e Serratia
marcescens foram capazes de degradar completamente altas concentrações de quitina
(100g/mL) a partir de carapaças de caranguejo langostino convertendo a N-
Acetilglicosamina (78%), glucosamina (2%) e quitobiose (10%). Enzimas de fonte

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
27
procariótica apresentaram atividades menores que as enzimas de T. atroviride, para
catalisar a reação com quitosana.
Jo et al. (2003) estudaram a caracterização e a avaliação cinética de quitosanase
produzida por Bacillus sp. Destaca-se que na produção da enzima utilizada neste ensaio
o meio continha 0,5% de concentração de quitosana, 1% de triptona e 1% de NaCl, o
pH do meio foi ajustado em 5,5 e a temperatura foi mantida em 37ºC com agitação de
180 rpm em incubador rotativo.
Fu et al (2003) estudaram a caracterização de três iso-enzimas de quitosanase
isolada de pepsina comercial e denominado como PSC-I, PSC-II, PSC-III,
respectivamente, em relação a hidrólise enzimática da quitosana, o meio ótimo para a
produção destas iso-enzimas continha as seguintes características: pH 5,0; 5,0 e 4,0 e
temperaturas de 40ºC, 40ºC e 30ºC, o valor de Km foi de 5,2, 4,0 e 5,6 mg.mL-1,
respectivamente. As massas moleculares das três iso-enzimas foram de 40 kDa. Os
resultados mostraram que as três iso-enzimas apresentaram atividade em meio contendo
quitosana, produzindo oligômeros da ordem de 68 – 88% desacetilada.
Choi et al. (2004) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida por Bacillus sp KCTC 0377BP. Nesse caso, com relação à
produção da enzima foi possível obter um aumento de 1,2 U/ml em incubator rotativo
para 100,0 U/ml em fermentador piloto de 500 L, utilizando uma concentração de
substrato de 0,5% de quitosana e como fonte de nitrogênio polipeptona (2,5%) em pH
igual a 6,8, com 100 rpm de 100,0 rpm a 30oC.
Shimosaka et al. (2003) estudaram a produção, purificação e caracterização total
de duas quitosanases produzida por Acinetobacter sp CHB101. Nesse caso, com relação
à produção da enzima os pesquisadores usaram dois meios, um contendo glicose e outro
contendo quitosana. Com relação à cinética, o meio contendo quitosana fornece um
tempo de geração de 280 minutos, enquanto que no meio contendo glicose a taxa de
crescimento foi mais elevada e o tempo de geração foi de apenas 110 minutos. Os
autores observaram que após os dois primeiros dias de cultivo a atividade
quitosanolítica permaneceu baixa (na ordem 0,10 a 0,20 U/mL). Durante a fase de
crescimento exponencial o valor da atividade permaneceu com valor inferior a 0,01
U/mL, começando a aumentar ao atingir a fase estacionária, indicando assim que a
produção da enzima é não associada ao crescimento.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
28
Kim et al. (2004) estudaram a produção, purificação e caracterização de uma
quitosanase produzida pelo Bacillus sp e identificado pelo número 1299. Destaca-se que
na produção da enzima quitosanase foi utilizado um meio contendo os seguintes
componentes: 0,5 % de quitosana, 0,42 % (NH4)2SO4, 0,2 % K2HPO4, 0,02 % uréia,
0,03 % CaCl2.2H2O, 0,03% MgSO4.7H2O, 0,1% peptona, 0,2% Tween 80, 0,2 % da
solução (0,5% FeSO4.7H2O, 0,16% MnSO4.H2O, 0,14% ZnSO4.7H2O, 0,2% CoCl2) e
1,7% agar. O pH do meio para este ensaio foi mantido em 7,0. Este microrganismo foi
escolhido por ser grande produtor de quitosanase.
Chen et al. (2005) estudaram a purificação e a caracterização de dois tipos de
quitosanase produzidas por Aspergillus sp. CJ22-326 em cultivo submerso. Na
produção da enzima foi utilizado um meio de cultivo contendo 1,0% de quitosana, 2,0%
de gérmen de trigo, 0,2% de sulfato de amônio, 0,2 % de fosfato de potássio e 0,05% de
sulfato de magnésio por litro de solução em pH 5,6. O cultivo foi realizado em frasco de
500 mL contendo 150 mL de meio de cultura a 30ºC por 96 horas com uma agitação de
150 rpm. Os autores apresentam um gel de eletroforese com as duas enzimas purificadas
com massa molar de 29 kDa e 109 kDa.
Silva Filho et al. (2005) estudaram, através de planejamento fatorial 24 com
repetição em triplicata no ponto central, a produção de quitosanase por Aspergillus
ochraceus em cultivo descontínuo submerso. Os resultados mostraram que foi possível
produzir quitosanase com atividade na ordem de aproximadamente 5,9 U/mL utilizando
Aspergillus ochraceus e que a atividade foi favorecida pelo aumento da agitação (r), da
razão de aeração (RA) e da concentração de substrato (Cs), enquanto que o aumento da
temperatura de cultivo (TMC) não favoreceu a resposta (atividade quitosanolítica).
Pode-se classificar as quitosanases em endoquitosanases e exoquitosanases. As
primeiras diferenciam-se por catalisar a hidrólise aleatoriamente, no interior do
biopolímero, gerando diversos tamanhos de oligossacarídeos. As exoquitosanases são
capazes de hidrolisar as terminações não redutoras, produzindo assim unidades
redutoras (Peter, 2005).
Palma-Guerrero et al. (2008) estudaram o biocontrole de fungos patogênicos e
demonstraram a lise da membrana plasmática e a inibição da germinação de esporos de
fungos. Eles observaram que os fungos entomopatogênicos e nematófagos foram menos
inibidos pela quitosana que os fitopatogênicos e micoparasíticos provavelmente porque
apresentam a enzima quitosanase que degrada a quitosana. Mesmo assim, a quitosana

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
29
pode ajudar a desenvolver novas estratégias para o biocontrole em geral de fungos
patogênicos, evitando o uso de fungicidas que prejudicam o meio ambiente e a saúde
humana.
Pagnoncelli (2008) estudou a produção de quitooligossacarídeos com
propriedades nutracêuticas a partir da quitosana por hidrólise enzimática, utilizando
processo fermentativo simultâneo. Duas cepas produtoras de quitosanaes foram
selecionadas, Paenibacillus chitinolyticus e Paenibacillus ehimensis, e avaliadas quanto
ao comportamento em meio de cultivo contendo açúcares simples e em relação às
variações de pH do meio. Os quitooligossacarídeos produzidos a partir do complexo
enzimático da primeira cepa apresentaram maior pico após 9h de hidrólise, enquanto
para a segunda cepa avaliada, após 20min, observou-se quitooligossacarídeos com grau
de polimerização entre 3 e 6 unidades.
2.2. Fungos
2.2.1. Características gerais
Desde o final dos anos 60 que os fungos deixaram de pertencer ao reino das
plantas e passaram a ter um reino próprio chamado de Reino Fungi. Foi a partir de
estudos morfológicos, bioquímicos e citológicos onde se concluiu que eles são tão
diferentes dos vegetais como dos animais. Distinguem-se de outros organismos
eucariotas por serem seres quimiorganoheterotróficos e, ainda, por apresentarem parede
celular rígida que é composta por quitina e glucano e uma membrana celular em que o
ergosterol substitui o colesterol (Trabulsi & Toledo, 1996; Murray et al., 2005).
Fazem parte do Reino Fungi cerca de 100 mil espécies, desde grandes como
os cogumelos, até microscópicas como os bolores e leveduras. Ainda hoje é utilizada
essa classificação, mas os avanços da biologia molecular e estudos bioquímicos levaram
a outras propostas de classificação em que os fungos e organismos afins foram incluídos
em três diferentes reinos: Protozoa, Chromista e Fungi (Kirk et al., 2008).
Abaixo, são apresentadas características que diferenciam os fungos das plantas:
• são incapazes de sintetizar clorofila, como também não apresentar celulose em
sua parede celular, com exceções como fungos aquáticos;
• apresentam algumas características de células animais (capacidade de depositar
glicogênio e a presença de substâncias quitinosas, em sua parede celular);

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
30
• podem ser eucarióticos (as leveduras), ou multinucleados (os bolores), e
apresentar retículo endoplasmático rugoso e mitocôndrias em seu citoplasma;
• são heterotróficos, alimentam-se de matéria orgânica viva (fungos parasitários),
ou morta (fungos saprofíticos);
Os fungos são amplamente encontrados na natureza, crescem rapidamente no
solo, na água, na atmosfera, bem como nos animais, nos vegetais, resíduos orgânicos e
desempenham um papel importante na vida do homem, quer de uma maneira benéfica,
quer de um modo prejudicial. Os fungos são um dos principais microrganismos
responsáveis pela decomposição da matéria orgânica, interferindo no ciclo do carbono,
do nitrogênio e de outros nutrientes da biosfera. São capazes de deteriorarem produtos e
bens de consumo, tais como alimentos, tecidos, papel, madeira. Os alimentos
armazenados representam um vasto meio para a proliferação dos fungos, principalmente
quando não são consideradas as condições básicas de armazenamento correto (Trabulsi
& Toledo, 1996).
Estão presentes funcionalmente em vários processos industriais, tais como, de
fabricação de pão, cervejas, vinhos e determinados tipos de queijos, sendo também
utilizados na produção comercial de muitos ácidos orgânicos, de alguns fármacos, como
a ergometrina e a cortisona, na obtenção de diferentes antibióticos, como a penicilina, e
de substâncias imunossupressoras, como a ciclosporina (Esteves et al., 1990). A Figura
4 ilustra a estrutura geral de uma célula de fungo.
Figura 4 – Representação da estrutura geral de uma célula de fungo.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
31
Os fungos podem se desenvolver em meios de cultivo especiais formando
colônias de dois tipos, leveduriformes e filamentosas, que se diferenciam pela
macromorfologia e micromorfologia. O primeiro tipo apresenta, em geral, característica
de consistência cremosa de cor branca a creme, brilhantes ou opacas, podendo
apresentar às vezes coloração escura ou alaranjada, e são formados por microrganismos
unicelulares que apresentam as funções vegetativas e reprodutivas. As filamentosas
apresentam características granulares, cotonosas, aveludadas ou pulverulentas, são
multinucleadas e apresentam-se em forma de tubos denominadas de hifas. Essas hifas
podem ser não septadas (cenocíticas) ou septadas (tabicadas). O conjunto de hifas é
denominado micélio.
As hifas asseptadas têm aparência anastomosada (concisa), formada por um
citoplasma estendido e polinucleado, enquanto que as hifas septadas contêm células
individualizadas, cada uma contendo o seu núcleo. Na Figura 5 são mostradas as
estruturas das hifas septadas e não septadas.
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas. (Adaptado de Guia.Bio,
2012)
O micélio que se desenvolve no interior do substrato e que é responsável pela
sustentação e absorção de nutrientes é denominado micélio vegetativo. O micélio que
cresce na superfície do meio de cultura é denominado micélio aéreo. Este pode ser
denominado também de micélio reprodutivo, quando o mesmo se diferencia para
sustentar os corpos de frutificação ou propágulos (Putzke & Putzke, 1998). Os
propágulos classificam-se de acordo com a origem em externos e internos, sexuados e
assexuados.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
32
2.2.2. Reprodução dos Fungos
A reprodução dos fungos ocorre em ciclos assexuais, sexuais e parassexuais. Em
fungos, a reprodução assexuada é observada com mais freqüência do que a reprodução
sexuada. A reprodução assexuada é normalmente a reprodução mais importante para a
propagação da espécie, feita através de fragmentação de artroconídios, fissão de células
somáticas, brotamento ou gemulação do blastoconídios-mãe e produção de conídios. A
reprodução sexuada envolve a união de duas células ou de dois órgãos sexuais
sexualmente compatíveis. A reprodução sexuada culmina na produção de basidiósporos,
no caso de basidiomicetos. A reprodução parassexuada ocorre com a fusão de hifas e
formação de um heterocário, que consiste na existência de núcleos haplóides
geneticamente distintos num mesmo citoplasma, os quais, após a fusão, originam
núcleos diplóides heterozigotos sujeitos a sucessivas divisões mitóticas. Apesar da
raridade dessas, a reprodução parassexual é de grande importância na evolução de
alguns fungos (Pelczar et al., 1996).
A reprodução assexuada é feita por conídios, esporos assexuados formados no
ápice de hifas modificadas, os conidióforos. Os conidióforos não possuem esporângios
de forma que os conídios ficam expostos. Na reprodução sexuada, com o encontro de
duas hifas haplóides distintas, ocorre a plasmogamia e um novo conjunto de hifas
dicarióticas é formado.
Os conídios podem ter formas esféricas, fusiformes, cilíndricas, piriformes, entre
outras. Apresentam-se como hialinos ou pigmentados, podem estar isolados ou
agrupados, a parede celular pode ser lisa ou rugosa e podem ser formados por apenas
uma célula ou apresentarem septos em mais de um plano.
Os conídios também podem originar-se em qualquer parte do micélio vegetativo
e são denominados conídios sésseis como é o caso do Trichophyton rubrum.
Alguns fungos patogênicos são identificados a partir de estruturas formadas pela
célula conidiogênica e o conidióforo. Essas estruturas são denominadas: aparelho de
frutificação ou conidiação. Os conídios podem formar cadeias sobre fiálides em volta de
uma vesícula, dilatando a extremidade do conidióforo. Essas estruturas ocorrem em
Aspergillus, apresentando-se em forma de cabeça ao redor de uma vesícula, e em
Penicillium agrupados em forma de pincel, como é ilustrada na Figura 6.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
33
Figura 6 – Conídios de Aspergillus e Penicillium
2.2.3. Metabolismo de fungos
Os fungos são seres aeróbicos e heterotróficos, podendo se desenvolver em
ambientes com baixa quantidade de oxigênio, como ocorre no crescimento vegetativo e
na reprodução assexuada, como também em ambientes com grande quantidade de
oxigênio como é o caso da reprodução sexuada.
Os fungos são capazes de produzir certas enzimas como lipases, invertases,
lactases, quitosanases, proteases, amilases, etc., que fazem a hidrólise do substrato
facilitando o mecanismo de transporte ativo e passivo. Esses substratos induzem a
formação de enzimas degradativas. Algumas espécies de fungos desenvolvem-se em
meios que apresentam amônia ou nitrilos como única fonte de nitrogênio. As melhores
fontes orgânicas para o seu desenvolvimento são carboidratos, como a D-glicose, e
alguns sais minerais como sulfatos e fosfatos.
A água é uma necessidade de todos os seres vivos, não sendo diferente para os
fungos que precisam de água para o seu desenvolvimento. Certos fungos apresentam
características halofílicas podendo crescer nos ambientes com grandes concentrações de
sal.
A razão de não existirem ambientes livres de espécies fúngicas deve-se ao facto
destes microrganismos conseguirem sobreviver em situações extremas como, por
exemplo, temperaturas que variam entre os - 5 e os 60 ºC, com pH de 1 a 9 e com baixas

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
34
concentrações de oxigênio, apesar de cada espécie possuir condições ideais para o seu
desenvolvimento (Chao et al., 2002).
Devido a grande faixa de temperatura ótima para o seu crescimento, os fungos
apresentam espécies psicrófilas, mesófilas e termófilas. Para a maior parte das espécies
fúngicas, a temperatura de desenvolvimento situa-se entre os 20 e 24 ºC e não crescem
acima dos 29 ºC. Em relação à umidade relativa, necessitam de pelo menos 50 %, mas
geralmente necessitam de mais de 65 % (Colakoglu, 2001).
O pH ideal para o desenvolvimento dos fungos varia na faixa entre 5 e 7.
Destaca-se, entretanto, que a maioria dos fungos suportam grandes variações de pH
como ocorre com os fungos filamentosos que podem crescer em pH variando entre 1,5 e
11. Por outro lado, as leveduras não suportam pH alcalino.
Os fungos crescem lentamente, se comparados com as bactérias, em meio de
incubação para o seu desenvolvimento. Para evitar contaminação por bactérias, que
podem se sobrepor ou inibir o seu crescimento, pode-se introduzir no meio de cultura
um antibacteriano de largo espectro como o cloranfenicol.
Algumas espécies de fungos se desenvolvem na presença de luz, outras são
inibidas e algumas podem crescer tanto na presença quanto na ausência deste agente. No
entanto a irradiação solar, em geral, é prejudicial ao seu desenvolvimento, por causa da
radiação ultravioleta.
2.2.4. Nutrição de fungos
Quanto ao aspecto da nutrição, os fungos são heterótrofos, ou seja, incapazes de
produzir seu próprio alimento. Entretanto, a forma de assimilar matéria orgânica é
diferente daquela utilizada pela maioria dos animais, que são heterótrofos por ingestão.
Nos fungos, a digestão é extracorpórea, ou seja, é realizada fora do corpo. O fungo
lança no ambiente, enzimas que degradam as moléculas orgânicas complexas e, depois,
absorve moléculas menores, mais simples. Por essa razão, são designados heterótrofos
por absorção. É devido a este processo que os fungos podem crescer dentro ou sobre os
alimentos. Desenvolve-se geralmente em meios contendo um pH baixo, uma fonte de
carbono, uma fonte de nitrogênio orgânico ou inorgânico e alguns minerais. Alguns
necessitam de vitaminas. Quanto aos modos de vida, os fungos podem ser
decompositores, parasitas, mutualísticos e predadores (Raven, 2001).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
35
Os fungos decompositores, saprófagos (do grego, saprós, “podre”, e phagos,
“comedor”), juntamente com as bactérias, são os principais decompositores da biosfera,
participando intensamente do processo de degradação da matéria orgânica morta, o que
promove a reciclagem dos elementos químicos constituintes dos seres vivos. A maioria
deles vive no solo, obtendo nutrientes de seres mortos. Em florestas, onde a matéria
orgânica vegetal é abundante, os micélios dos fungos decompositores absorvem os
nutrientes de folhas e galhos caídos, decompondo-os. Os fungos decompositores
também são responsáveis pelo apodrecimento de alimentos e de outros materiais, como
a madeira.
Os fungos parasitas vivem à custa de outros organismos vivos, prejudicando-os.
Ao parasitar o corpo de um ser vivo, animal ou vegetal, o fungo pode até provocar sua
morte. Várias doenças são causadas por fungos parasitas. Nas plantas, cita-se a
ferrugem do café, que, no Brasil, é motivo de grande preocupação por parte dos
cafeicultores, devido aos prejuízos que acarreta. As micoses que atacam a pele de seres
humanos e animais são provocadas por fungos.
Existem fungos que se associam a outros organismos, estabelecendo uma relação
em que há benefício mútuo para os indivíduos envolvidos. Esses fungos são
denominados mutualísticos. A maioria vive associada a seres fotossintetizantes, como
as plantas, cedendo-lhes parte da água e dos nutrientes que as hifas absorvem do solo.
As plantas, por sua vez, cedem ao fungo certos açúcares e aminoácidos. Duas
associações mutualísticas formam estruturas bem características: as micorrizas,
associação que envolve fungos e raízes de plantas, e os líquens, formados por fungos e
algumas variedades de algas e cianobactérias.
Certos fungos atuam como predadores. Na maioria dos casos, as hifas secretam
substâncias aderentes que aprisionam os organismos que tocam os fungos. Dessa
maneira, as hifas penetram o corpo da presa, crescem e se ramificam, espalhando-se no
interior do organismo e absorvendo seus nutrientes, causando-lhe a morte. O gênero
Arthrobotrys apresenta uma forma mais elaborada de predação. O fungo vive no solo e
captura nematódeos microscópicos que habitam o mesmo ambiente. As hifas
apresentam pequenos anéis que se estreitam quando a presa passa por eles. As hifas do
fungo logo invadem o corpo do nematódeo, digerindo-o.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
36
2.2.5. O fungo Metarhizium anisopliae
Existem três espécies de fungos do gênero Metarhizium, divididas em dez
variedades: M. anisopliae, variedades anisopliae, majus, lepidiotum e acridum; M.
flavoviride, variedades tipo E, flavoviride, minus, novazealandicum e pemphigum; e M.
album (Driver et al. 2000). E, mais recentemente, foi descrita na China uma nova
variedade de M. anisopliae, dcjhyium (Dong et al. 2009).
Data de 1879 quando o russo Ilya Metchnikoff aplicou o fungo para combater
larvas de um curculionídeo, praga de beterraba, e foi classificado como Metarhizium
anisopliae por Sorokin em 1883. É um fungicida natural para mais de 300 espécies de
insetos, especialmente pragas da agricultura e pecuária. No Brasil, o fungo Metarhizium
anisopliae teve a sua aplicação com sucesso pela primeira vez em 1965, no controle da
cigarrinha-da-cana Mahanarva posticata (Alves, 1998).
O fungo Metarhizium anisopliae é um importante agente entomopatogênico
utilizado no controle de pragas, como cupins, gafanhotos, cigarrinhas e besouros, sendo
amplamente estudado em todo o mundo. Existe amplamente na natureza e é facilmente
encontrado no solo, onde sobrevive por longos períodos e seu desenvolvimento se dá
geralmente entre 15 e 32 °C, sendo ideal entre 24 e 30 °C e com pH ótimo igual a 6,9.
Apresenta micélio hialino e septado, com conidióforos característicos, sobre os quais
surgem conídios cilíndricos organizados em colunas. Este fungo ao atacar os insetos
provoca rigidez e os recobre por uma camada pulverulenta de conídios, resultando em
colorações que variam do verde claro ao escuro, cinza ou branco com pontos verdes
(Tinline, 1971; Alves, 1998, Driver et al. 2000; Arruda, 2005).
Quanto a sua morfologia, apresenta-se como um fungo filamentoso, com corpo
de frutificação equivalente a um esporodóquio agregado a hifas intimamente
entrelaçadas, com massa compactada de conidióforos característicos, simples ou
ramificados, formando células esporogênicas denominadas fiálides, resultando nos
fialosporos (Wang et al., 2002).
O Metarhizium anisopliae é um fungo de cultivo simples, sendo necessário
quase que exclusivamente o nutriente a base de amido. Um dos fungos mais estudados
em programas de manejo de pragas já que apresenta grande potencialidade
entomopatogênica (Onofre et al., 2002). O Metarhizium anisopliae é um microrganismo
pouco exigente, pois pode se desenvolver em diversos meios de cultura como: amido,
glicose, glicerina, levulose, maltose, sacarose e quitina (Jabor et al., 2003).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
37
Em geral, os fungos produzem enzimas que convertem os tecidos dos insetos em
nutrientes. Porém, a cutícula esclerotizada dos insetos não é bem utilizada pelos fungos,
excetos pelos entomopatogênicos que desenvolveram enzimas eficientes para degradar
essa camada protetora. Destaca-se a enzima Pr1, produzida por Metarhizium anisopliae,
que tem um poder de degradação bem superior a qualquer outra enzima sintetizada por
esse grupo de fungos (Dias, 2005). Desta forma, o Metarhizium anisopliae (fungo
entomopatogênico) ao atacar a célula dos insetos, produz grande quantidade de enzimas
que degradam a membrana citoplasmática e inativa a célula e, consequentemente, mata
o inseto, logo é evidente a relação entre o controle de pragas e produção de enzimas.
No momento, sobre a comercialização de fungos entomopatogênicos no Brasil,
em sua maioria, são formulados em grânulos, compostas do fungo mais o substrato
(arroz + fungo), como também na forma de pó molhável (WP), resultado da moagem do
fungo com o substrato. Esses produtos representam 90% e 8,5%, respectivamente, da
produção de fungos no Brasil, e sendo suas concentrações finais ficam em torno de
5x108 conídios/g para Metarhizium anisopliae e 1x109 conídios/g para Beauveria
bassiana (Alves et al., 2008).
Segundo Batista Filho et al. (2002), Garcia et al. (2005) e Mello et al. (2006), o
Metarhizium anisopliae tem sido bastante utilizado para o controle de diferentes pragas
no Brasil, como a broca da bananeira, pragas de grãos armazenados, carrapatos, cupim
de montículo em pastagens, larvas de escarabeídeos que atacam a cana de açúcar e
cigarrinha da cana-de-açúcar.
É comum a ocorrência natural do fungo Metarhizium anisopliae atacando
cigarrinha-das-raízes de canaviais pelo país, o que incentiva a utilização dele como
agente de controle biológico em áreas infestadas por esse tipo de praga. A aplicação de
recursos no uso de programa de manejo integrado da cigarrinha tem sido reforçado por
razões ambientais e econômicas (Dinardo-Miranda, 2001).
Faria & Magalhães (2001) fizeram uma avaliação do uso de fungos
entomopatogênicos no Brasil, na qual observaram que a qualidade dos micoinseticidas
disponíveis no Brasil poderia ser incrementada de forma considerável quanto a sua
formulação, ou seja, antes da venda, poderiam realizar melhorias no tratamento
posterior, adição de substâncias que lhes assegurem melhorias na eficiência de controle,
capacidade de armazenamento ou praticidade de manuseio, ou de qualquer outro critério
que resulte em vantagem em relação ao produto bruto. Ressaltaram a pouca praticidade

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
38
em alguns casos, como exigência de lavagem, e em outros podendo causar o
entupimento de bicos dos pulverizadores devido à elevada proporção de inertes,
principalmente quando são empregados baixos volumes de aplicação. Outro problema
evidenciado com os inseticidas nacionais diz respeito à pequena sobrevida, devendo a
ser usados em, no máximo, 30 dias depois de produzidos, quando armazenados à
temperatura ambiente e em local sombreado, fazendo as vendas ocorrerem quase que,
exclusivamente, sob encomenda. Comparativamente, os bons inseticidas sob
comercialização em outros países apresentam maior concentração de ingrediente ativo,
maior sobrevida (alguns produtos podem ser armazenados por mais de 8 meses à
temperatura ambiente) e praticidade (são formulados na forma de GDA (grânulos
dispersíveis em água) ou de óleos emulsionáveis, por exemplo, podendo ser adicionados
diretamente ao tanque do pulverizador.
Rangel et al. (2007), instalaram uma Unidade de Observação para o controle
microbiano das cigarrinhas das pastagens Distrito de Itahum, em Dourados/MS, devido
as necessidades de agricultores familiares de produção de leite. As aplicações do fungo
Metarhizium anisopliae foram realizadas por pulverização com equipamento de tração
animal. As condições do ambiente de estudo foram favoráveis ao desenvolvimento da
população das cigarrinhas e à incidência do fungo. Como resultados, observaram que os
níveis de espumas, ninfas e adultos reduziram significativamente após as aplicações,
levando os Agricultores Familiares a total satisfação com os ganhos na produção de
todos os envolvidos.
Segundo Vieira et al. (2007), o fungo entomopatogênico Metarhizium anisopliae
var. acridum tem sido utilizado no controle de pragas, e assim, visando este fim,
analisaram em laboratório a esporulação, a germinação dos conídios e as características
morfológicas peculiares desse fungo em diferentes temperaturas. Esperou-se o
desenvolvimento das colônias em períodos de 1 a 15 dias. Foi observado que
temperaturas de 25, 28 e 30 ºC proporcionaram maiores taxas de esporulação e elevadas
porcentagens de germinação dos conídios. A 25, 28 e 30 ºC observou-se um
desenvolvimento mais rápido das microestruturas somáticas e reprodutivas (hifas,
anastomoses, apressórios, conidióforos e conídios elipsóides). Nas temperaturas
extremas de 40 e 45 ºC não foi detectado esporulação, germinação, nem
desenvolvimento de nenhuma microestrutura.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
39
Lopes et al. (2007) avaliaram o efeito de formulações de Beauveria bassiana e
Metarhizium anisopliae sobre ninfas de Amblyomma cajennense (transmissor da febre
maculosa ao homem) em laboratório. Ninfas não alimentadas foram pulverizadas com
suspensões contendo 2x107 e 4x107 conídios/mL de B. bassiana, e 1x107, 1,5x107,
2x107, 3x107 e 5x107 conídios/mL de M. anisopliae. A mortalidade de ninfas,
decorridos três dias da aplicação, foi significativa nas concentrações de 3x107 e 5x107
conídios/mL de M. anisopliae, com 57,1 e 56,2 %, respectivamente. Após seis dias a
mortalidade foi de 100 % nessas duas concentrações. A mortalidade foi
significativamente maior que a observada para B. bassiana em concentrações
semelhantes.
Guirado et al. (2009) realizaram estudos para controlar cupins de montículo em
área de pastagem, utilizando fungos entomopatogênicos Metarhizium anisopliae e
Beauveria bassiana em diferentes doses e formulações. A avaliação foi realizada após
30 dias da aplicação dos produtos. Os melhores resultados em ordem decrescente de
eficiência foram: M. anisopliae 30 mL + 1000 mL de água com 100 % de eficiência; B.
bassiana 10g + M. anisopliae 20 mL + 1000 mL de água com 85,71 % de eficiência; M.
anisopliae 20 mL + 1000 mL de água com 71,42 % de eficiência. Os demais
tratamentos não diferiram da testemunha.
Loureiro et al. (2012) compararam a eficiência de isolados de Metarhizium
anisopliae em controlar populações naturais de Mahanarva fimbriolata (cigarrinha-da-
raiz) em cultivos comerciais de cana-de-açúcar colhida mecanicamente. Foram
aplicados os isolados IBCB 348, IBCB 408, IBCB 410 e IBCB 425 do fungo em
concentração de 1,5×1012 conídios/ha. Aplicou-se o planejamento experimental em
blocos casualizados com 5 tratamentos e 4 repetições, sendo cada repetição composta
por 7 linhas de 100 m de comprimento e espaçamento de 1,5 m. Após 30 dias de
pulverização foi observado para os isolados IBCB 408 e IBCB 425 uma eficiência de
controle de ninfas de 63 e 62%, respectivamente, já no caso dos adultos chegou a 100%.
Após 60 dias o isolado IBCB 425 foi o mais eficiente para controlar as ninfas com
48,4%.
Assis et al. (2012) estudaram a citotoxidade de quitoolissacarídeos (QOS) e
oligômeros A, B e C (soluções compostas por diferentes quantidades de QOS)
produzidos por hidrólise enzimática pelo fungo Metarhrizium anisopliae. Foi analisado
o efeito de antiproliferativo destas moléculas em células tumorais (HepG2 e HeLa) e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
40
também em célula normal (3T3). A atividade antioxidante foi analisada em vários
experimentos in vitro. A glicosamina apresentou maior toxicidade (cerca de 92%) para
todas as células estudadas. No entanto, os oligômeros obtidos após hidrólise não
demonstraram efeito tóxico sobre a célula normal (3T3). Além disso, mostraram que
uma pequena quantidade de outros COS pode diminuir o efeito citotóxico da
glicosamina contra células 3T3, indicando que glicosamina poderia ser usada como uma
droga antitumoral na presença de outros COS.
2.3. Fermentação
Os processos fermentativos são caracterizados por processos nos quais
microrganismos realizam a conversão catalítica de uma dada substância num
determinado produto. Estes processos podem ser classificados como processos
descontínuos, contínuos e descontínuos alimentados. A classificação destes processos
está diretamente ligada à forma de como o substrato é adicionado e como o produto é
retirado. No processo de fermentação descontínuo o substrato é colocado no
fermentador, em seguida é inoculado e após o término do processo o produto é retirado.
No processo contínuo, o substrato é colocado num reator com vazão constante e o meio
fermentado é retirado com a mesma vazão de entrada do reator (Faccioti, 2001). Nos
processo descontínuos alimentados à adição de substrato e a retirada do mesmo deve
ocorrer de forma intermitente (Carvalho & Sato, 2001).
De certa forma, os processos fermentativos representam um elo entre as antigas
artes de elaboração de alimentos, como queijos e vinho, a partir de uma flora
microbiana natural, com a moderna indústria de fermentação de alimentos, onde são
utilisados cultivos puros e equipamentos sofisticados para o controle do processo
(Assis, 2010).
Atualmente, a maioria dos processos industriais se desenvolve em cubas de
vidro ou de aço de até 20 litros. Os agentes biológicos se encontram submersos no meio de
cultivo que ocupa, aproximadamente, 75% da cuba. As vezes é necessário injetar ar, e em muitas
fermentações se forma espuma. O desenho do biorreator deve se adequar ao objetivo do
processo, respondendo eventualmente a diversos imperativos, tais como a esterilização do
sistema, a aeração e homogeneização do meio, o acréscimo de nutrientes e de aditivos
antiespuma, a manutenção do pH, etc. Os modelos de fermentadores mais utilizados com
microrganismos contam com aeração e agitação mecânica. Esta facilita a distribuição dos

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
41
nutrientes na cuba, mas gera calor que deve ser eliminado mediante a circulação de água
fria. Em outros tipos de biorreatores, em coluna ou torre, a homogeneização depende da injeção de
ar. Os sistemas submersos são apropriados para o cultivo de microrganismos livres, mas resultam
pouco econômicos quando se trabalha com células ou enzimas caras. A imobilização em
fermentadores menores, seja por adesão a um suporte inerte, seja por inclusão dentro de um
polímero que permita o contato com o meio de cultura, além de simplificar a purificação do
produto permite a reutilização das células ou das enzimas, que permanecem dentro do
biorreator (Malajovich, 2009).
As enzimas são produtos de alto valor agregado, amplamente empregadas em
uma gama de indústrias, e entre as fontes para sua obtenção, destacam-se as de origem
microbiana, principalmente, devido a elevada seletividades dos substratos utilizados e
produtos formados. A principal forma de obtenção de enzimas, em escala industrial, é a
fermentação submersa, por esta apresentar vantagens em relação a fermentação
semisólida, como: maior controle do processo fermentativo e consequente reprodução
da cinética microbiana, além de maiores rendimentos (Wanderley, 2011).
2.3.1. Preparo do inóculo
Denomina-se inóculo um volume de suspensão de microrganismo de
concentração adequada, capaz de garantir, em condições econômicas, a fermentação de
uma dada quantidade de meio. O volume do inóculo, assim como sua concentração de
células, vai depender tanto do processo de fermentação c omo da quantidade de meio a
ser fermentado (Carvalho & Sato, 2001).
A partir de uma cultura pura, inocula-se um volume relativamente pequeno de
meio nutriente, que vai depender do tipo de microrganismo cultivado (pois cada espécie
microbiana possui velocidades diferentes de crescimento), sendo, em seguida, incubado
em condições favoráveis ao desenvolvimento do microrganismo. A suspensão obtida é
transferida para um frasco maior em agitador rotativo ou recíproco (shakers), que por
sua vez é adequadamente incubado, e a seguir, transferido para um frasco contendo um
volume ainda maior deste meio, até se conseguir alguns litros de suspensão microbiana.
Os cuidados com assepsia, volumes de meio utilizado, numero de transferência bem
como as condições de incubação, podem variar de acordo com o tipo de processo de
fermentação que se deseja realizar.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
42
2.3.2. Produção de enzimas em cultivo submerso descontínuo
A produção de enzimas, usando processos fermentativos, é constituída
basicamente por duas etapas: a primeira caracteriza-se pela preparação do inóculo e a
segunda, pela fermentação do substrato (Carvalho & Sato, 2001). A finalidade da
primeira etapa é o preparo do microrganismo em condições adequadas para que possa
ocorrer o desenvolvimento da segunda etapa. Diferentemente do cultivo em estado
sólido, no cultivo submerso os microrganismos encontram-se em um meio com elevada
atividade de água.
2.4. Planejamento experimental
Um bom planejamento tem como fundamental projetar um experimento de
forma que ele seja capaz de fornecer exatamente o tipo de informação que procuramos.
Para isso é necessário saber, em primeiro lugar, o que é mesmo que se deseja procurar.
Planejamento experimental, ou delineamento experimental, é um conjunto de
ensaios estabelecido com critérios científicos e estatísticos, objetivando determinar a
influência de diversas variáveis nos resultados de um dado sistema ou processo (Barros
Neto et al., 2001; Rodrigues & Iemma, 2005). De acordo com o propósito dos ensaios,
esse objetivo maior pode ser mais detalhado em:
– determinar quais variáveis são mais influentes nos resultados;
– atribuir valores às variáveis influentes de modo a otimizar os resultados;
– atribuir valores às variáveis influentes de modo a minimizar a variabilidade dos
resultados;
– atribuir valores às variáveis influentes de modo a minimizar a influência de variáveis
incontroláveis.
O planejamento experimental, baseado nos fundamentos estatísticos é, sem
dúvida alguma, uma ferramenta poderosa para se chegar às condições otimizadas de um
processo, desenvolvimento da formulação de produtos dentro das especificações
desejadas ou simplesmente para avaliar os efeitos ou impactos que os fatores (variáveis
independentes) têm nas respostas (variáveis dependentes) desejadas (Barros Neto et al.,
2001; Rodrigues & Iemma, 2005). Da utilização das técnicas estatísticas de
planejamento experimental, é possível chegar benefícios importantes, como:

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
43
– reduzir o número de ensaios sem prejuízo da qualidade da informação;
– estudar simultaneamente diversas variáveis, separando seus efeitos;
– determinar a confiabilidade dos resultados;
– realizar a pesquisa em etapas, num processo iterativo de acréscimo de novos ensaios;
– selecionar as variáveis que influem num processo com número reduzido de ensaios;
– representar o processo estudado através de expressões matemáticas;
– elaborar conclusões a partir de resultados qualitativos.
2.4.1. Planejamento fatorial
Em um planejamento fatorial é investigado as combinações de níveis de estudo
de cada fator. Para tanto, inicialmente, é necessário especificar esses níveis em cada
fator, quer dizer, os valores dos fatores que serão utilizados nos experimentos. Um
ensaio experimental é definido como um experimento constituído de um conjunto de
níveis definido. Geralmente, quando se tem n1 níveis de um determinado fator 1, n2 do
fator 2, ..., e nk do fator k, o planejamento será descrito por um fatorial n1 x n2 x ... x nk,
que corresponde ao número mínimo de experimentos para se ter um planejamento
fatorial completo.
É necessário fazer cada fator variar e observar o resultado dessa variação para se
estudar o efeito sobre a resposta. Assim, os ensaios precisam ter um mínimo de dois
níveis por fator para a realização desses ensaios. Para k fatores, ou seja, k variáveis de
controle, o planejamento de dois níveis precisará de 2 x 2 x ... x 2 = 2k, para o número
de ensaios diferentes, sendo conhecido como planejamento fatorial 2k.
Para o planejamento fatorial, são especificados os valores dos fatores em cada
experimento, e estes são classificados em níveis, variando-se, por exemplo, de -1
(mínimo) a +1 (máximo). A partir de uma matriz serão apresentadas todas as
combinações entre o número de fatores e de níveis nos experimentos a serem realizados
(Barros Neto et al., 2001; Rodrigues & Iemma, 2005).
2.4.2. Planejamento Plackett & Burman
A maioria dos planejamentos do tipo “Screening” (Delineamentos de Seleção de
fatores) foi descrito originalmente por Plackett & Burman em 1946. Os planejamentos
experimentais de Plackett & Burman são eficientes em situações que envolvem uma

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
44
grande quantidade de variáveis a serem exploradas em um processo de otimização de
fatores (Reddy et al., 2008).
As matrizes Plackett & Burman consistem de planejamentos fatoriais de dois
níveis para estudar k = N – 1 variáveis em N ensaios. São construídas para ensaios
múltiplos de 4, sendo essas matrizes para ensaios de 8, 12, 16,..., 100. Cada variável
independente em uma matriz assume uma configuração de dois níveis, o inferior (-1) e o
superior (+1), para o nível intermediário é estabelecido (0). Para cada vez que a variável
aparece no nível inferior, terá o par equivalente no nível superior (Rodrigues & Iemma,
2005).
Para evitar dúvidas nos resultados dos efeitos de algum fator no planejamento de
Plackett & Burman, o número de ensaios a ser realizado deve ser no mínimo o número
de variáveis mais 4. No caso de um fatorial completo o número de ensaios a ser
realizado é bem maior (Rodrigues & Iemma, 2005).
Mesmo já determinado o planejamento Plackett & Burman, torna-se importante
à realização de experimentos relativos aos pontos centrais de modo a se avaliar melhor
as possíveis interferências analíticas que podem ocorrer durante os diferentes
experimentos. A repetição do ponto central permite também a determinação do erro
puro e da repetibilidade do processo. Na prática, esta é uma ferramenta que permite
identificar melhor se o processo em estudo está sob controle (Rodrigues & Iemma,
2005).
É possível que no planejamento Plackett & Burman não tenha sido considerado
a codificação do ponto central (0) para repetição, pois, matematicamente, a estimativa
do valor desse efeito codificado (zero) vezes, teria como resposta zero. Porém, no
resultado prático do experimento, consegue-se informações valiosas com essas
repetições no ponto central (Rodrigues & Iemma, 2005).
2.4.3. Aplicações do Planejamento Plackett & Burman
Ghanem et al. (2000) produziram xilanase por Aspergillus terreus cultivadas
sobre palha de trigo finamente moída em fermentação de estado sólido. A composição
ideal do meio foi estabelecida aplicando-se o planejamento experimental de Plackett &
Burman. A melhor atividade enzimática foi obtida em um meio contendo 10 g/L de

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
45
palha de trigo por frasco umedecido com uma solução concentrada de sal com teor de
água inicial de 75 % e incubadas durante 4 dias a 30 °C. Observou-se que íons
metálicos como Hg2+, Cu2+, Co2+, Fe3+ e Pb2+ inibiram fortemente a xilanase, enquanto
que Ca2+ ativou a enzima.
Mu et al. (2005) estudaram as otimização de meio de fermentação para a reação
de estereoinversão por Candida parapsilosis CCTCC M203011 utilizando o
planejamento Plackett & Burman. Os resultados mostraram que dentre os fatores
estudados, os fatores mais significativos foram: a Concentração de MgSO4, ZnSO4 e
KH2PO4 e que após a utilização da metodologia de superfície de resposta foi constatado
que o ponto ótimo para a otimização foi conseguido com as concentrações dos fatores
MgSO4, ZnSO4 e KH2PO4 nos valores em: 1,17 g/L, 0,10 g/L e 9,41 g/L,
respectivamente, elevando a taxa de conversão enantiosseletiva de 75,2 % para 96 %.
Naveena et al. (2005) empregaram o planejamento Plackett & Burman para a
seleção de 15 parâmetros para produção de ácido láctico L(+) de farelo de trigo,
substrato de baixo custo e suporte sólido, por Lactobacillus amylophilus GV6 em
fermentação de estado sólido (SSF). Onze nutrientes pertencentes a duas categorias,
fontes de nitrogênio e fontes de sal, junto com três parâmetros físicos e um tampão
foram os parâmetros considerados. Os autores verificaram influência na produtividade
devido às fontes de nitrogênio, peptona, extrato de levedura e tri-citrato de amônio,
juntamente com o NaH2PO4.2H2O e Tween 80, que podem ainda serem otimizados para
o aumento na produção de ácido lático. Esses pesquisadores ainda chamam atenção para
o uso escasso deste planejamento em fermentação em estado sólido e que o mesmo
ainda não havia sido considerado anteriormente para a conversão direta de amido em
ácido láctico L (+) usando um sistema bacteriano.
Khosravi-Darani et al. (2008) estudaram a aplicação de Plackett & Burman na
análise dos fatores significativos para a produção de ácido cítrico por Aspergillus niger
ATCC 9142 utilizando palha de trigo como substrato. Os resultados mostraram que a
produção, com a palha de trigo pré-tratada com HCl, NaOH e Uréia, foi mais eficiente e
que os fatores mais significativos na produção foram: Solvente, Teor de Umidade,
Temperatura, Tempo de Incubação, Concentração Inicial de Açúcar e Concentração de
Metanol.
Ji et al. (2009) estudaram o desenvolvimento de um meio de cultura econômico
para a produção do 2,3-butanodiol através da fermentação da glicose e xilose por

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
46
Klebsiella oxytoca utilizando o planejamento Plackett & Burman. Os resultados
mostraram que a uréia e a maceração de milho foram os fatores mais significativos para
a produção do 2,3-butanodiol e que através de planejamento experimental com repetição
em triplicata no ponto central para determinar os níveis ótimos de produção. Os autores
constaram que o rendimento atingido foi 85,6% do valor teórico.
Paula & Arruda. (2009) observaram a necessidade da busca por alternativas mais
eficientes ao método termomecânico utilizado para a degradação de milhões de
toneladas de penas de aves provenientes de frigoríficos e aviários, cuja rígida estrutura é
composta por mais de 90 % de queratina. Para esse fim, utilizaram de comunidades
microbianas para a finalidade proposta, testando o planejamento experimental Plackett
& Burman para verificar a influência de diferentes microrganismos simultaneamente,
com um número reduzido de experimentos. Testaram 32 bactérias isoladas, inoculadas
em meio contendo somente uma pena como única fonte de carbono, durante sete dias,
de onde foi selecionado as cepas para os consórcios microbianos. Experimentos unindo
as cepas NP5, FCA7, NP4 e S14 demonstraram diferentes atividades queratinolíticas,
aumentando o nível de degradação, até 87,8%, comparada com as cepas
individualmente, variando entre 26,4% e 78,0%.
Rajendran & Thangavelu (2009) obtiveram valores máximos da atividade de
produção de lipase em 3,98 U/mL e concentração de massa celular em 5,62 g.L-1 com R.
arrhizus em cultivo submerso descontínuo otimizado pelo planejamento Plackett &
Burman. As variáveis mais importantes que afetaram a produção da lipase foram azeite
de oliva, peptona, KH2PO4, CaCl2.2H2O e MgSO4.7H2O.
Li et al. (2010) estudaram a otimização de meio de cultura para a produção de
Streptolydigin (um antibiótico) por Streptomyces lydicus AS 4,2501 utilizando Plackett
& Burman e a metodologia de superfície de resposta. Os resultados mostraram que
dentre os fatores estudados, os fatores mais significativos para a produção de
Streptolydigin e otimização do meio de cultura foram: Concentração de Amido,
Concentração de Peptona e a Concentração de K2HPO4.
Chen et al. (2010) estudaram a otimização de meio de cultura para a produção da
Luciferase – que são enzimas que catalisam reações biológicas transformando energia
química em energia luminosa - por Bacillus Subtilis em fermentação descontínua
utilizando planejamento Plackett & Burmam. Os resultados mostraram que dentre os
componentes estudados, os fatores mais significativos para a produção e otimização do

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
47
meio de cultura para a produção da Luciferase foram: Concentração de Glicose 1,17 %,
Extrato de Levedura 2,27 % e Glutamato 0,55 %.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o projeto Plackett & Burman e a metodologia de superfície de resposta. Os
resultados mostraram que a eficiência máxima de extração da quitina foi conseguida nas
seguintes condições: Concentração de casca de camarão (50 g/L), glicose (50 g/L),
tempo de incubação de 5 dias e inoculo com 0,05 de OD (densidade óptica).
Gao et al. (2010) desenvolveram um meio para a fermentação do extrato bruto
de inulina de tubérculos de alcachofra de Jerusalém por Paenibacillus polymyxa ZJ-9
para produzir R,R-2,3-butanodiol (R-R,2,3-BD). Inulina, K2HPO4 e NH4Cl foram os
parâmetros principais na fermentação de acordo com os resultados obtidos com o
planejamento experimental Plackett & Burman. A concentração ideal dos três fatores,
77,14 g/L, 3,09 g/L e 0,93 g/L, respectivamente, foi obtida pelo planejamento Box-
Behnken e pela superfície de resposta. Sob as condições ideais, a concentração do R-
R,2,3-BD obtida foi 36,92 g/L, em mais de 98 % de pureza óptica. Comparado com
outras fontes de carbono investigadas, a fermentação do extrato bruto de inulina obteve
o maior rendimento de R-R,2,3-BD.
Uma metodologia de superfície de resposta foi empregada com sucesso para
otimizar um meio de fermentação chamada VHG (Very High Gravity), referência à alta
concentração de açúcares e densidade dos mostos, baseado na água maceração de milho
CSL (Corn Steep Liquor) e outros nutrientes de baixo custo para a produção eficiente de
etanol a partir de glicose por S. cerevisiae. Usando a composição média otimizada (g/L:
CSL 44,3; uréia 2,3; MgSO4.7H2O 3,8; CuSO4.5H2O 0,03), a levedura industrial PE-2
foi capaz de fermentar até 330 g/L de glicose e produzir 18,6 % (v/v) de etanol, com
uma produção descontínua de 2,4 g/L/h e um rendimento em etanol de 93 % (Pereira et
al., 2010).
O planejamento Plackett & Burman juntamente com o Planejamento Composto
Central (CCD) foram aplicados para otimizar o meio de cultura para produção de etanol
por Clostridium autoethanogenum com monóxido de carbono (CO) como fonte única de
carbono e um meio contendo (g/L): 1,0 de NaCl, 0,1 de KH2PO4, 0,02 de CaCl2, 0,15 de
extrato de levedura, 0,116 de MgSO4, 1,694 de NH4Cl e pH 4,74. O rendimento ótimo
de etanol previsto pela metodologia de superfície de resposta (RSM) e um algoritmo

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
48
genético de rede neural artificial (ANN-GA) foram 247,48 e 261,48 mg/L,
respectivamente. Valores semelhantes aos obtidos experimentalmente sob as condições
ideais sugeridas pelos métodos estatísticos (254,26 e 259,64 mg/L). A adequação do
modelo ANN-GA foi maior do que o modelo RSM. Os rendimentos obtidos foram
substancialmente superiores aos relatados anteriormente (60-70 mg/L) com essa cultura
(Guo et al., 2010).
Rigueira (2010) avaliou a influência das variáveis do procedimento de extração
na porcentagem de recuperação das aminas encontradas no leite cru utilizando um
planejamento para “screening” do tipo Plackett & Burman, para os ácidos
tricloroacético (TCA) e sulfosalicílico (ASS).
Quines (2010) estudou a necessidade ou não de se fazer um pré-tratamento da
biomassa antes da extração. Um planejamento experimental Plackett & Burman de doze
ensaios mais três pontos centrais foi utilizado para selecionar as variáveis mais
significativas na extração de Poli(3-hidroxibutirato) (P(3HB)) por 1,2-carbonato de
propileno. Foram realizados dois testes de recuperação do solvente 1,2-carbonato de
propileno, após a extração, sendo um com aplicação de aquecimento e o outro com
aquecimento e vácuo. Os resultados mostraram que o pré-tratamento térmico levou a
uma maior porcentagem de recuperação e de pureza do polímero.

Capítulo 3 Metodologia Experimental

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
50
3. Metodologia Experimental
O presente capítulo apresenta os matérias e métodos utilizados na produção de
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso.
3.1. Quitosana
Neste trabalho utilizou-se uma quitosana (Lote 067) obtida junto a POLYMAR/CE,
com no mínimo 80% de desacetilação. Esse substrato foi utilizado como fonte de carbono e
energia tanto no estudo cinético como nos ensaios do planejamento de seleção de fatores
Plackett & Burman e Planejamento Composto Central, para a produção de quitosanase pelo
Metarhizium anisopliae, por fermentação submersa.
3.2. Microrganismo
O fungo Metarhizium anisopliae estirpe (CG374), utilizado neste estudo, foi cedido
pela Embrapa Recursos Genéticos e Biotecnologia (Brasília / DF-Brasil) e foi mantido em
placas de Petri contendo o meio Batata Dextrose Agar (PDA). As placas foram incubadas em
estufa a 25 ºC. O pH do meio de manutenção foi mantido entre 6,0 e 6,5.
3.2.1. Escolha da cepa produtora de quitosanase
A escolha do microrganismo Metarhizium anisopliae foi feita com base na capacidade
de crescer em meio contendo quitosana como única fonte de carbono, além de apresentar
grande tolerância a pH variando a uma taxa de 2,0 a 8,5, sendo 6,9 a melhor condição para o
crescimento vegetativo e esporução, de acordo com Assis (2010).
3.2.2. Manutenção da cepa
A manutenção da cultura foi realizada em placas petri contendo o meio Batata
Dextrose Agar (PDA). A renovação das células foi realizada a cada trinta dias através de
subcultivos das células no meio PDA.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
51
3.3. Produção de enzima
No preparo do inóculo, utilizou-se 7 placas cultivadas e mantidas, com o fungo
Metarhizium anisopliae, em meio PDA a 25 ºC durante 5 dias em estufa. O meio de cultura
para a produção da enzima quitosanase apresentou os seguintes componentes: quitosana;
peptona; extrato de levedura; NaNO3; K2HPO4; KCl; MgSO4.7H2O e FeSO4, com
concentrações de acordo as matrizes dos planejamentos: Plackett & Burman e composto
central. Para a produção da enzima no cultivo, a biomassa foi transferida através de água
estéril, para um Béquer de 500 mL que, em seguida, foi colocado em Erlenmeyers de 250 mL
na proporção de 20 mL de biomassa para 180 mL do meio de cultura. Os Erlenmeyers
cultivados foram então incubados em incubador rotativo a 25 ºC e 150 rpm durante 24 h.
Alíquotas foram retiradas e centrifugadas a 8000xg por 5 min, a cada 6 horas, para se
determinar a atividade quitosanolítica, teores de açúcares redutores e de proteína total.
Ressalta-se que todos os experimentos foram feitos em triplicata.
3.4. Determinação da Proteína Total
A determinação da proteína total foi efetuada de acordo com o método modificado de
Sedmak & Grossberg (1977), que se baseia na diminuição da absorbância do corante azul de
Coomassie, devido à ligação proteína-corante, a 595 nm, comparado a absorção máxima a
465 nm, conforme Santos (2001). Nesse caso, após centrifugação para retirada da biomassa,
transferia-se para um eppendorf 1,5 mL de amostra (caldo fermentativo) e adicionava-se 1,5
mL de solução azul de Coomassie. Deixava-se a solução em repouso por 5 minutos,
transferindo-se em seguida para um cubeta de quartzo e efetuando-se as leituras de
absorbância nos comprimentos de ondas 465 nm e 595 nm, respectivamente. O
espectrofotômetro (Thermo Spectronic modelo Genesys 10UV) foi zerado com água
destilada. O conteúdo protéico foi obtido a partir de uma curva padrão na qual graficava-se a
relação de absorbância A595/A465 para diferentes concentrações de Albumina de Soro
Bovino (Fração V – Lote 110881/Sigma Aldrich-Ohio-USA).
3.5. Determinação da Atividade Quitosanolítica e da Atividade Específica
A atividade quitosanolítica foi obtida monitorando-se o aumento dos açúcares
redutores totais, utilizando-se o método DNS a partir de uma curva padrão de glicosamina.
Para a determinação da atividade quitosanolítica adicionaram-se, aos tubos de ensaios 500 µL

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
52
de substrato (solução 0,2 % de quitosana solubilizada em ácido clorídrico 1N) e 500 µL de
caldo bruto (contendo enzima), controlando-se o pH entre 6,0 e 6,5 e incubando-se o sistema
em um banho termostatizado com temperaturas da ordem de 55 ºC. Após o tempo de
incubação de 30 minutos no banho foi adicionado aos tubos de ensaio 2,5 mL de DNS,
aquecendo-se à ebulição em banho-maria por 10 minutos, resfriando-se, em seguida, em
banho de gelo. Após essa etapa, o sistema foi centrifugado e com o sobrenadante efetuou-se a
leitura no espectrofotômetro a 600 nm. Destaca-se, que antes da leitura da amostra zerava-se o
equipamento (branco do espectro) com 1,0 mL de água destilada acrescidos de 2,5 mL de
DNS e realizava-se o branco da enzima que consistia da adição de 500 µL de solução de
quitosana, completando-se até um 1,0 mL com água destilada. Uma unidade de atividade
quitosanolítica (A) foi definida como a quantidade de enzima necessária para produzir 1
µmol.min-1 de glicosamina por mL de extrato enzimático, nas condições de temperatura e pH
citadas, sendo o resultado expresso em (U.mL-1). A Atividade específica foi determinada pela
divisão entre a atividade da enzima (U.mL-1) e a concentração de proteína total (µg.mL-1),
sendo expressa em (U. µg-1).
3.6. Planejamento experimental
3.6.1. Planejamento de seleção de fatores Plackett & Burman
O planejamento experimental Plackett & Burman é uma técnica muito usada, para se
realizar triagem de fatores na otimização num determinado processo (Reddy et al., 2008), e
foi utilizado neste trabalho para identificar os fatores que influenciam de forma significativa
na produção da enzima quitosanase.
A Tabela 3 ilustra os níveis de cada variável (fator) utilizada no planejamento Plackett
& Burman, sendo: Concentração de Quitosana (Q), Peptona (PP), Extrato de Levedura (EL),
NaNO3 (NS), K2HPO4 (FP), KCl (CP), MgSO4.7H2O (SM), FeSO4 (SF). Já a Tabela 4 mostra
a matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3 repetições
na condição do ponto central.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
53
Tabela 3 – Níveis para os fatores e seus valores codificados.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1 0,1 0,2 0,2 0,1 0,05 0,025 0,025 0,0005
0 0,2 0,5 0,3 0,2 0,10 0,050 0,050 0,0010
1 0,3 0,8 0,4 0,3 0,15 0,075 0,075 0,0015
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1
2 1 1 -1 1 -1 -1 -1 1 -1 1 1
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1
5 1 1 -1 1 1 -1 1 -1 1 -1 1
6 1 1 1 -1 1 1 -1 1 1 -1 -1
7 -1 1 1 1 -1 1 1 -1 -1 1 1
8 -1 -1 1 1 1 -1 1 1 1 1 1
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1
11 -1 1 -1 -1 -1 1 1 1 1 1 -1
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1
13 0 0 0 0 0 0 0 0 0 0 0
14 0 0 0 0 0 0 0 0 0 0 0
15 0 0 0 0 0 0 0 0 0 0 0
Sendo: V11,V12,V13 = Variáveis inertes para cálculo do erro.
3.6.2. Planejamento Composto Central para a Otimização do Meio de Cultura para a
Produção da Enzima em Cultivo Submerso
Um dos problemas mais comuns para quem realiza experimentos é determinar a
influência de uma ou mais variáveis sobre outra variável de interesse (Lobato, 2003). Este

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
54
problema pode ser minimizado utilizando-se um planejamento experimental de modo a se
obter uma função, ou uma aproximação satisfatória para a mesma, de forma que se operando
sobre as variáveis de entrada produza-se como saída a resposta ou as respostas observadas
(Barros Neto, 2001; Silva, 2004).
Em um planejamento, inicialmente, deve-se definir os fatores e as respostas de
interesse. Tendo-se identificado os fatores e respostas define-se o objetivo que se pretende
alcançar com os experimentos para que se possa escolher o planejamento mais apropriado.
(Silva, 2004).
No presente trabalho foi utilizado um Planejamento Composto Central (CCD) com 22
experimentos na parte fatorial, repetição em triplicata no ponto central e quatro experimentos
na parte axial (planejamento do tipo estrela), para se investigar a influência dos fatores:
concentração de quitosana e concentração de FeSO4 que foram os dois fatores que
influenciaram a produção de quitosanase, conforme o planejamento Plackett & Burman de 12
ensaios e mais 3 repetições na condição do ponto central. Estes dois fatores foram estudados
em cinco diferentes níveis (-1,41; -1; 0; 1; 1,41) e um conjunto de 11 experimentos foi
realizado, conforme Tabela 5.
Tabela 5 – Matriz do planejamento Composto Central.
Ensaio Q SF
1 -1 -1
2 -1 1
3 1 -1
4 1 1
5 -1,41 0
6 1,41 0
7 0 -1,41
8 0 1,41
9 0 0
10 0 0
11 0 0
Os outros fatores foram mantidos nos níveis mais baixos (para evitar custos). Uma que
não influenciaram de forma significativa na produção da enzima quitosanase.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
55
A codificação dos fatores foi realizada de acordo com a seguinte equação (Ji et al.,
2009):
Xi = (xi – x0)/∆xi , i = 1, 2, 3, ..., k (2)
Onde Xi é o fator independente codificado, xi é o fator independente real, x0 é o valor de xi no
ponto central e ∆xi é a variação de valor. O comportamento do sistema foi explicado pela
seguinte equação polinomial de segunda ordem:
Yp = β0 + ∑ βiXi + ∑ βiiXi2 + ∑ βijXiXj, i, j = 1, 2, 3,..., k (3)
Onde Yp foi a resposta esperada, β0 foi a interseção, Xi e Xj
foram os fatores independentes codificados, βi foi o coeficiente linear, βii
foi o coeficiente quadrático e βij foi o coeficiente de interação. Destaca-se que os outros
fatores não significativos foram mantidos no nível mais baixo (-1).
A Tabela 6 apresenta os níveis para os fatores e seus valores codificados, para o
Planejamento Composto Central, com triplicata no ponto central.
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1,41 0,218 0,2 0,2 0,1 0,05 0,025 0,025 0,0009
-1 0,3 0,2 0,2 0,1 0,05 0,025 0,025 0,0015
0 0,5 0,2 0,2 0,1 0,05 0,025 0,025 0,0030
1 0,7 0,2 0,2 0,1 0,05 0,025 0,025 0,0045
1,41 0,782 0,2 0,2 0,1 0,05 0,025 0,025 0,0051
Na análise do planejamento de seleção de fatores Plackett & Burman e do
planejamento experimental fatorial foi utilizado um pacote computacional STATISTICA for
Windows versão 7.0.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
56
3.7. Perfil cinético durante o cultivo
Um perfil cinético na melhor condição de cultivo, conforme planejamento
apresentado, foi realizado em frascos Erlenmyer de 250 mL contendo 200 mL de meio de
cultivo contendo os seguintes componentes, em 1 Litro: 2,0g de quitosana; 5,0g de peptona;
3,0g de extrato de levedura; 2,0g de NaNO3; 1,0g de K2HPO4; 0,5g de KCl; 0,5g de
MgSO4.7H2O; 0,01g de FeSO4, que foram inoculadas com 10% (vol/vol) de inóculo contendo
o fungo Metarhizium anisopliae e cultivadas em incubador rotativo durante 5 dias a 150 rpm
e 25 ºC. Alíquotas foram retiradas a cada seis horas e analisadas quanto à proteína total no
sobredanante, atividade enzimática, atividade específica e açúcares redutores. Os ensaios
foram realizados em triplicata (Silva Filho, 2005).

Capítulo 4 Resultados e
Discussão

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
58
4. Resultados e discussão
Neste capítulo são apresentados e discutidos os resultados obtidos no presente
trabalho. Inicialmente, é apresentada imagem do MEV do Metarhizium anisopliae
mantidos em meio PDA. Em seguida, apresenta-se e discute-se os resultados da avaliação
dos fatores significativos, que podem influenciar na produção da quitosanase, no
planejamento de seleção de fatores, conforme planejamento Plackett & Burman bem como
são apresentados e discutidos os resultados obtidos para o planejamento composto central.
Por último, apresenta-se o perfil cinético, na melhor condição, para a produção de
quitosanase em cultivo submerso descontínuo usando o fungo Metarhizium anisopliae.
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA.
A Figura 7 ilustra a imagem da Microscopia Eletrônica de Varredura (MEV) do micélio de Metarhizium anisopliae mantido em meio PDA.
Figura 7. Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA.
Observa-se na Figura 7, quanto à morfologia que o Metarhizium anisopliae, apresenta-
se como um fungo filamentoso com as hifas intimamente entrelaçadas conforme citado por
Wang et al. 2002.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
59
4.2. Planejamento Plackett & Burman
4.2.1 Análise do Plackett & Burman com 8 fatores e 12 ensaios
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis, foi utilizado para avaliar a influência dos fatores: Concentração de
Quitosana (Q), Peptona (PP), Extrato de Levedura (EL), NaNO3 (NS), K2HPO4 (FP), KCl
(CP), MgSO4.7H2O (SM), FeSO4 (SF) na produção da enzima quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso. Ressalta-se que os experimentos foram
realizados aleatoriamente.
A Tabela 7 apresenta a matriz do modelo Plackett & Burman e os respectivos valores
obtidos para a variável resposta (Atividade Quitosanalítica).
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.
Ensaio Q PP EL NS FP CP SM SF A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 0,0195
2 1 1 -1 1 -1 -1 -1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 0,0109
4 1 -1 1 1 -1 1 -1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 0,0136
6 1 1 1 -1 1 1 -1 1 0,0156
7 -1 1 1 1 -1 1 1 -1 0,0109
8 -1 -1 1 1 1 -1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 0,0085
A Tabela 8 apresenta os efeitos estimados para o planejamento Plackett & Burman.
Observa-se na Tabela 8 que o valor do coeficiente de determinação R2 foi de
aproximadamente 90%, indicando que um modelo linear poderia representar bem a relação
entre o fator significativo, concentração de quitosana (1), e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
60
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman.
Efeitos Estimados; R2=0,8996; Ajuste:0,63178 MS Erro Residual =0,0000054
Desvio Padrão t(3) p
Lim. Conf. Lim. Conf.
Efeito Erro Residual
-95,% +95
Média/Interação 0,0127 0,0007 18,903 0,0003 0,0105 0,0149 (1) Quitosana 0,0055 0,0013 4,0628 0,0268 0,0011 0,0098
(2) Peptona 0,0015 0,0013 1,1080 0,3486 -0,0028 0,0058 (3) Ext. de leved. 0,0009 0,0013 0,7387 0,5136 -0,0033 0,0053
(4) NaNO3 0,0002 0,0013 0,1758 0,8715 -0,0040 0,0045 (5) K2HPO4 -0,0014 0,0013 -1,0552 0,3687 -0,0057 0,0028
(6) KCl -0,0022 0,0013 -1,6708 0,1933 -0,0065 0,0020 (7) MgSO4.7H2O -0,0003 0,0013 -0,2286 0,8338 -0,0046 0,0039
(8) FeSO4 0,0029 0,0013 2,1457 0,1211 -0,0014 0,0071
A Figura 8 mostra o gráfico de Pareto, uma das formas de se avaliar visualmente a
influência dos fatores estudados na resposta. A magnitude dos efeitos é representada pelas
colunas enquanto que a linha transversal às colunas representa a magnitude dos efeitos com
significado estatístico para p=0,05, ou seja, os fatores que são estatisticamente significativos
ao nível de 95% de confiança.
Figura 8. Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
61
Analisando-se o gráfico de Pareto (Figura 8) observa-se que o fator concentração de
quitosana (1) foi o único fator que influenciou de forma mais significativa na atividade
enzimática atingindo efeitos estimados de aproximadamente 4,06. O sinal positivo indica que
o aumento da concentração de quitosana aumentará à atividade enzimática. Por outro lado, os
outros fatores parecem desprezíveis estatisticamente conforme o gráfico de Pareto.
4.2.2 Análise do Plackett & Burman com 12 Ensaios e mais 3 Repetições na condição do
ponto central.
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis com repetição em triplicata no ponto central, foi utilizado para se avaliar a
influência dos mesmos fatores descritos na Seção 4.2.1 para a produção da enzima
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso. Ressalta-se que os
experimentos foram realizados aleatoriamente. A Tabela 9 apresenta a matriz do modelo
Plackett & Burman e os respectivos valores obtidos para a variável resposta (Atividade
Quitosanalítica).
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13 A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1 0,0195
2 1 1 -1 1 -1 -1 -1 1 -1 1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1 0,0109
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 1 -1 1 0,0136
6 1 1 1 -1 1 1 -1 1 1 -1 -1 0,0156
7 -1 1 1 1 -1 1 1 -1 -1 1 1 0,0109
8 -1 -1 1 1 1 -1 1 1 1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 1 1 -1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1 0,0085
13 0 0 0 0 0 0 0 0 0 0 0 0,0704
14 0 0 0 0 0 0 0 0 0 0 0 0,0688
15 0 0 0 0 0 0 0 0 0 0 0 0,0710

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
62
A Tabela 10 a seguir apresenta os efeitos estimados para planejamento Plackett &
Burman com 3 repetições na condição do poto central. Observa-se na Tabela 10 que o valor
do coeficiente de determinação R2 foi de aproximadamente 2,0%, indicando que um modelo
linear não poderia representar a relação entre os fatores significativos concentração de
quitosana (1), concentração de FeSO4 (8) e a resposta.
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central.
Efeitos Estimados; R2=0,01811; Ajuste:0,0 MS Erro Puro =0,0000012
Desvio Padrão t(2) P
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0242 0,0003 85,509 0,0001 0,0229 0,0254 (1) Quitosana 0,0055 0,0006 8,6432 0,0131 0,0027 0,0082
(2) Peptona 0,0015 0,0006 2,3572 0,1425 -0,0012 0,0042 (3) Ext. de leved. 0,0009 0,0006 1,5715 0,2567 -0,0017 0,0037
(4) NaNO3 0,0002 0,0006 0,3742 0,7442 -0,0024 0,0029 (5) K2HPO4 -0,0014 0,0006 -2,2450 0,1538 -0,0041 0,0013
(6) KCl -0,0022 0,0006 -3,5546 0,0708 -0,0049 0,0005 (7) MgSO4.7H2O -0,0003 0,0006 -0,4864 0,6747 -0,0030 0,0024
(8) FeSO4 0,0029 0,0006 4,5648 0,0448 0,0002 0,0056
A Figura 9 ilustra o gráfico de Pareto para o planejamento Plackett & Burman, com
repetição em triplicata no ponto central, uma das formas de se avaliar visualmente a influência
dos fatores estudados na resposta, onde é mostrado a magnitude dos efeitos para 95 % de
confiança.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
63
Figura 9. Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto
central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto, observa-se que os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma mais
significativa na atividade enzimática atingindo efeitos estimados de aproximadamente 8,84 e
4,67, respectivamente. O sinal positivo indica que o aumento das concentrações de quitosana
e de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores parecem
desprezíveis estatisticamente conforme o gráfico de Pareto.
Este comportamento do FeSO4 está de acordo com o observado por Hagag et al.
(2012) que utilizando o Aspergillus fumigatus na produção de protease mostraram que um
fator de transcrição PrtT regula positivamente o controle da produção de protease, da
assimilação de Ferro e da síntese de ergosterol e que a produção de protease foi favorecida
pelo aumento da concentração de ferro.
Rajendran & Thangavelu (2009) utilizaram um planejamento Plackett & Burman para
identificar quais os fatores significativos na produção de lipase por Rhizopus arrhizus MTCC
2233 e observaram o efeito significativo médio do FeSO4, apresentando um nível de
confiança para esse fator de 48,92% (para um máximo de 98,7 % de MgSO4), com uma
produção de lipase da ordem de 2,98 U/mL em 84 horas de fermentação e que ocorreu uma
produção considerável de protease no meio de cultivo da ordem de 1,92 U/mL em 96 horas de
cultivo.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
64
Mu et al. (2005) também observaram um efeito positivo do FeSO4, da ordem de 1,48
para um fator máximo encontrado de 3,11 de MgSO4, no aumento da atividade da
estereoinversão de S-fenil-1,2-etanodiol por Candida parapsilosis CCTCC M203011 na
otimização da fermentação utilizando um planejamento Plackett & Burman.
Dessa forma, observa-se que o FeSO4 é importante uma que vez o mesmo está
diretamente associado a síntese de proteases (principalmente metalloproteases), conforme
citado por Hagag et al. (2012).
4.3. Planejamento Composto Central para Produção de Quitosanase por
Metarhizium anisopliae em Cultivo Descontínuo Submerso
Um planejamento experimentos 22 com repetição em triplicata no ponto central e mais
quatro ensaios axiais, foram utilizados para se chegar a uma combinação ideal de quitosana e
FeSO4 na produção da enzima quitosanase (Tabela 11). A partir do modelo Plackett &
Burman (Tabela 12), que mostrou que o aumento das concentrações de quitosana e FeSO4,
favoreceram a produção de quitosanase, os valores utilizados neste modelo para estes
componentes nos níveis mais altos, serão considerados como os níveis mais baixos no
planejamento experimental do composto central. Os outros fatores foram fixados em todos os
ensaios em seus níveis mais baixos para a redução de custos (Tabela 11).

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
65
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da
atividade enzimática.
Ensaio Q SF A (U/mL)
1 -1 -1 0,0195
2 -1 1 0,0202
3 1 -1 0,0109
4 1 1 0,0121
5 -1,41 0 0,0136
6 1,41 0 0,0156
7 0 -1,41 0,0108
8 0 1,41 0,0103
9 0 0 0,0099
10 0 0 0,0116
11 0 0 0,0094
A Tabela 12 apresenta os efeitos estimados para o planejamento composto central.
Tabela 12 – Efeitos Estimados do planejamento Composto Central.
Efeitos Estimados; R2=0,629; Ajuste:0,25836 MS Erro Puro =0,0000014
Desvio Padrão t(2) p
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0103 0,0006 15,305 0,0042 0,0074 0,0132 (1) Quitosana (L) 0,0011 0,0008 1,4316 0,2885 -0,0023 0,0047
Quitosana (Q) 0,0058 0,0009 5,9637 0,0269 0,0016 0,0101 (2) FeSO4 (L) -0,0043 0,0008 -5,3037 0,0337 -0,0079 -0,0008 (2) FeSO4 (Q) 0,0018 0,0009 1,8366 0,2076 -0,0024 0,0060
1L x 2L 0,0001 0,0011 0,1520 0,8931 -0,0048 0,0052
Observa-se na Tabela 12 que o valor do coeficiente de determinação R2 foi de
aproximadamente 63%, indicando que um modelo linear representaria bem a relação entre os
fatores significativos concentração de quitosana (1), concentração de FeSO4 (8) e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
66
A Tabela 13 mostra um resumo da ANOVA para o planejamento composto central
com repetição em triplicata no ponto central.
Tabela 13 – Resumo da ANOVA.
Fonte de
Variação
Soma dos
Quadrados
Grau de
Liberdade
Quadrado
Médio
Fcalculado Ftabelado
Regressão
Resíduos
0,000086 2 0,000043 6,14 4,46
0,000056 8 0,000007
Falta de
Ajuste
Erro puro
0,000053 6 0,000009 6 19,33
0,000003
2
0,0000015
Total 0,000142 10
De acordo com a Tabela 13, partindo-se dos efeitos significativos propõe-se o
modelo:
� = 0,010316 + 0,002942� � − 0,002193�� (8)
% de variação explicável: 60,5
% máxima de variação explicável: 97,9
A análise da ANOVA mostra que o modelo proposto possui significância estatística, ao
nível de 95% de confiança, uma vez que o valor de F (calculado) é maior que F (tabelado),
entretanto a ANOVA mostra também que o modelo proposto pode ser útil para se fazer
previsões, uma vez que a relação entre F (calculado) e F (tabelado) para a falta de ajuste e o
erro puro é menor que 1. O coeficiente de determinação R2 foi de aproximadamente 63%,
indicando que o modelo consegue explicar 63% da variação total em torno da média.
Os cálculos para variação explicável e para máxima variação explicável são dados pelas
Equações (9) e (10):
% de variação explicável = (SQ da regressão) / (SQ Total) (9)
% máxima de variação explicável = (SQ Total – SQ Erro puro) / (SQ Total) (10)
Sendo:
SQ – Soma dos quadrados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
67
A Figura 10 mostra o gráfico de Pareto para o planejamento Composto Central
utilizando erro puro. A magnitude dos efeitos é representada pelas colunas. Já a linha
transversal às colunas representa a magnitude dos efeitos com significado estatístico para 95%
de confiança.
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto (Figura 10) observa-se que os fatores concentração
de quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma
mais significativa na atividade enzimática atingindo efeitos estimados de aproximadamente
5,96 e - 5,30, respectivamente. O sinal positivo indica que o aumento das concentrações de
quitosana aumentará à atividade enzimática. O sinal negativo indica que o decréscimo da
concentração de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores
parecem desprezíveis estatisticamente no gráfico de Pareto.
A avaliação do modelo também pode ser feita através da observação do gráfico dos
valores preditos em função os valores observados que são mostrados na Figura 11. Os valores
preditos pelo modelo são representados pela reta, enquanto que os valores observados
representam-se pelos pontos.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
68
Figura 11 – Valores preditos em função dos valores observados.
Observa-se que alguns valores preditos apresentam um afastamento dos valores
observados. A Figura 12 ilustra a superfície de resposta da atividade quitosanolítica em
função dos dois fatores significativos, concentração de quitosana e concentração de FeSO4.
Figura 12. Atividade quitosanolítica em função da concentração de quitosana e da
concentração de FeSO4 para os fatores não significativos no ponto central.
Observa-se a existência de um ponto de mínimo, ocorrendo para concentração de
quitosana no ponto central (0) e para maiores concentrações de FeSO4 igual a (1,5). Essa

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
69
região deve ser evitada uma vez que a atividade quitosanolítica atinge níveis tão reduzidos
quanto 0,0094 U.mL-1. Destaca-se que nesse planejamento o maior nível da concentração de
FeSO4 (α=+1,41) equivale a 0,0051% (Tabela 6) que é aproximadamente cinco vezes maior
que o nível desse fator no ponto central do planejamento Plackett & Burman (0,001%),
conforme Tabela 3. Dessa forma, ao se utilizar o nível mínimo da concentração de FeSO4 no
Planejamento Composto Central (α=-1,41), que equivale a aproximadamente 0,001%,
aproxima-se do ponto central do do planejamento Plackett & Burman (0,001%), condição
essa que apresenta os melhores resultados de atividade quitosanolítica usando quitosana como
fonte de carbono e energia e fungo Metarhizium anisopliae.
4.4. Perfil Cinético – Melhor Condição
A seguir, apresenta-se um perfil cinético, na melhor condição, para produção da
quitosanase usando o fungo Metarhizium anisopliae, em que foram monitorados durante 24
horas os seguintes parâmetros: Concentração de proteínas totais (µg/mL), atividade
quitosanolítica (U/mL), atividade específica (U/µg) e açúcares redutores (mg/mL), ilustradas
nas Figuras 13, 14, 15 e 16, respectivamente.
Com relação à proteína extracelular, observa-se na Figura 13 que, inicialmente, ocorre
um aumento da mesma para o seu nível máximo (6,2 µg/mL), durante as 6 primeiras horas de
cultivo, seguido de uma redução para 5,7 µg/mL em 18 horas de cultivo, possivelmente
devido à presença de proteases no meio. Observa-se que a partir de então ocorre uma redução
e aumento da proteína total, em uma quantidade inferior, sugerindo um mecanismo de
controle.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
70
0 4 8 12 16 20 245,0
5,2
5,4
5,6
5,8
6,0
6,2
6,4
Pro
tein
a (
µg
/mL
)
Tempo (h)
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana.
Com relação à atividade quitosanolítica, conforme Figura 14, a atividade máxima
ocorreu em 18 horas de produção em que se obteve aproximadamente 70,0 U/L, o que foi 28
vezes maior que o valor (2,5 U/L em 48 horas) relatado por Assis (2010) para o mesmo fungo,
e em apenas um terço do tempo de produção.
0 4 8 12 16 20 240,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,10
Ativid
ade
(U
/mL
)
Tempo (h)
Figura 14. Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
71
Além disso, destaca-se o considerável aumento da produção de quitosanase a partir das
12 horas até 18 horas de cultivo onde atingiu o pico máximo de produção (70,0 U/L). Após o
pico em 18 horas, a produção começa a cair rapidamente, comportamento esse observado
durante todos os ensaios cinéticos. Destaca-se que é importante do ponto de vista industrial
manter a produção em alto nível por muitas horas, onde se espera uma produção contínua ou
em grandes bateladas.
Ak et al. (1998) utilizaram o fungo Penicillium spinulosum para produzir a quitosanase
usando um meio de cultura contendo 0,1% de K2HPO4; 0,05 % de MgSO4.7H2O; 0,05% de
KCl; 0,001 % de FeSO4, 0,5% NH4NO3 e como fonte de carbono e energia, os autores
usaram quitosana e parede de Rhizopus. A produção foi realizada em cultivo submerso a 28
oC. Os autores apresentam atividade quitosanolítica máxima de aproximadamente 47,0 U/mL,
ao se utilizar a parede do fungo como fonte de carbono e energia, e 30,0 U/mL ao se utilizar
quitosana como fonte de carbono e energia. Destaca-se que essas atividades ocorreram após 4
dias de cultivo. Os perfís com o tempo da atividade quitosanolítica e da atividade específica
apresentaram tendência de crescimento, com o pico máximo em 4 dias, seguindo com uma
tendência de queda da atividade. Neste tempo, a atividade específica máxima da quitosanase
foi de aproximadamente 60,0 U/mg de proteína. Essas tendências também foram observadas
nos resultados apresentados nos perfís de atividade das Figuras 14 e 15.
0 4 8 12 16 20 24
-0,010
-0,005
0,000
0,005
0,010
0,015
0,020
0,025
0,030
Ativid
ad
e E
spe
cific
a (
U/µ
g)
Tempo (h)
Figura 15. Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
72
Observa-se na Figura 15 que a atividade específica máxima da quitosanase segue a
mesma tendência da atividade quitosanolítica apresentada na Figura 14, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
O perfil para os açúcares redutores durante o cultivo de Metarhizium anisopliae
cultivado em meio contendo 0,2% de quitosana é ilustrado na Figura 16.
0 4 8 12 16 20 240,004
0,008
0,012
0,016
0,020
0,024
0,028
Acuca
res R
ed
uto
res (
mg/m
L)
Tempo (h)
Figura 16. Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana.
Observa-se, na Figura 16, que taxa máxima de redução dos açúcares redutores
(aproximadamente 0,021 mg/mL) ocorre logo após a produção máxima da enzima, em 18
horas de cultivo, conforme Figura 14. A quantidade de açúcares redutores foi a
aproximadamente 0,012 mg/mL após 24 h de cultivo. O fungo excreta para o meio enzimas
com atividade quitosanolítica, essas enzimas hidrolisam a quitosana aumentando a quantidade
de açúcares redutores para o meio, sendo que esses açúcares também são consumidos pelo
fungo durante seu crescimento.
Foi demonstrado por Assis (2010) que os açúcares redutores produzidos através da
quitossana, ocasionada pelas enzimas quitossanases metabolizadas por Metarhizium
anisopliae, com quitosana como sua única fonte de carbono e energia, atingiu um pico de 0,53
mg/mL em 48 horas. Apresentaram como quitooligossacarídeos, constituídos principalmente
por dímeros e pentâmeros, formados pela hidrólise enzimática de quitosanase em quantidades
maiores do que o consumido pelos microrganismos. Mostraram, também, que essa produção

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
73
segue a mesma tendência da sua atividade enzimática, corroborando com os resultados
apresentados no presente trabalho.
Os resultados do presente trabalho mostram a relevância de se adotar novas condições
para a produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter quitooligossacarídeo
com atividades biológicas. Esta pesquisa, portanto, sugere base para novos estudos que
permitam alcançar rapidamente a produção de quitosanase e quitooligossacarídeo com todos
os seus benefícios.

Capítulo 5 Conclusão

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
75
5. Conclusão
Após o desenvolvimento desse trabalho conclui-se:
- Com relação ao Planejamento de Seleção de Fatores Plackett & Burman:
Os resultados das análises do Planejamento de Seleção de Fatores Plackett &
Burman com 8 fatores e 12 ensaios em 2 níveis, mais 3 repetições no ponto central, para
se investigar a influência dos fatores: Concentração de Quitosana, Peptona, Extrato de
Levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O, FeSO4 na produção da enzima
quitosanase por Metarhizium anisopliae, mostraram que a atividade foi favorecida pelo
aumento da concentração de substrato (quitosana) e sulfato ferroso (FeSO4), enquanto
que o aumento da concentração dos outros fatores não contribuíram de forma
significativa para a resposta (atividade quitosanolítica). Os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) atingiram efeitos estimados de
aproximadamente 8,84 e 4,67, respectivamente. O sinal positivo indica que o aumento
das concentrações de quitosana e de FeSO4 aumentará a atividade enzimática. Conforme
visto na literatura, observa-se importância do FeSO4, já que o mesmo está diretamente
associado a síntese de proteases (principalmente metalloproteases),
- Com relação ao Planejamento Composto Central:
A análise do planejamento composto central para avaliar a influência dos
fatores, concentração de substrato (quitosana) e concentração de FeSO4, na produção da
enzima quitosanase por Metarhizium anisopliae (a partir do planejamento Plackett &
Burman) mostrou que os fatores concentração de quitosana e concentração de FeSO4
foram os fatores que influenciaram de forma mais significativa na atividade enzimática
atingindo efeitos estimados de aproximadamente 5,96 e - 5,30, respectivamente. O sinal
positivo indica que o aumento das concentrações de quitosana aumentará a atividade
enzimática. O sinal negativo indica que o decréscimo da concentração de FeSO4
aumentará a atividade enzimática. Observa-se a existência de um ponto de mínimo,
ocorrendo para concentração de quitosana (X1) no ponto central (0) e para maiores

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
76
concentrações de FeSO4 (X2) igual a (1,5) com a atividade quitosanolítica de 0,0094
U.mL-1.
- Com relação ao cultivo na melhor condição:
O cultivo na melhor condição mostrou uma produção máxima de quitosanase em
18 horas de cultivo, com a atividade enzimática de 70,0 U/L, o que foi 28 vezes maior
que o valor (2,5 U/L em 48 horas) relatado na literatura (Assis, 2010) para o mesmo
fungo e em apenas um terço do tempo de produção.
Além disso, observou-se que um elevado incremento na produção de
quitosanase a partir das 12 horas de cultivo, antes de se atingir a atividade
quitosanolítica máxima, 18 horas de cultivo. Essa produção em alto nível por muitas
horas é importante do ponto de vista industrial, onde se espera uma produção contínua
ou em grandes bateladas. Observou-se também que a atividade específica máxima da
quitosanase segue a mesma tendência da atividade quitosanolítica, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
- Com relação aos açúcares redutores:
Foi observado que os açúcares redutores seguem a mesma tendência da atividade
enzimática, com uma produção máxima de cerca de 0,21 mg/mL em 18 horas de
cultivo, produção essa que coincinde com o máximo pico de atividade enzimática.
Estes resultados mostram a relevância de se adotar novas condições para a
produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter
quitooligossacarídeo com atividades biológicas. Esta pesquisa, portanto, sugere base
para novos estudos que permitam alcançar rapidamente a produção de quitosanase e
quitooligossacarídeo com todos os seus benefícios.

Referências Bibliográficas

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
78
Referências bibliográficas
AK, Ö.; BAKIR, U.; GÜRAY, T. Isolation of a chitosan degrading fungus, Penicillium
spinulosum, and chitosanase production by the isolate. In: SYMPOSIUM - NEW
FRONTIERS IN SCREENING FOR MICROBIAL BIOCATALYSTS, 1996, Ede, The
Netherlands. Anais, Editado por K. Kieslich, C. P. van der Beek, J. A. M. de Bont e W.
J. J. van den Tweel, Elsevier Science B. V., 1998, v.53, p.265-268.
ALVES, S. B. Patologia e controle microbiano: Vantagens e desvantagens. In: ALVES,
S. B. Controle Microbiano de Insetos. 2ed., Piracicaba, FEALQ, 1163p, 1998, v.4, p.21-
37.
ALVES, S. B.; LEITE, L. G.; FILHO, A. B.; ALMEIDA, F. E. M.; MARQUES, E. J.
Produção massal de fungos entomopatogênicos na América Latina, In: ALVES, S. B.;
LOPES, R. B. Controle microbiano de pragas na América Latina: avanços e desafios.
Piracicaba, FEALQ, 414p, 2008, v.14, p.215-237.
ARRUDA, W.; LUBECK, I.; SCHRANK, A.; VAINSTEIN, M. H. Morphological
alterations of Metarhizium anisopliae during penetration of Boophilus microplus ticks.
Experimental and Applied Acarology, v. 37, p.231-244, 2005.
ASSIS, C. F. Produção e caracterização de quitooligossacarídeos produzidos pelo fungo
Metarhizium anisopliae e avaliação da citotoxicidade em células tumorais. 2010. 110p.
Tese (Doutorado em Biotecnologia) – Renorbio - Programa de Pós-Graduação em
Biotecnologia. Universidade Federal do Rio Grande do Norte, Natal.
ASSIS, C. F.; COSTA, L. S.; MELO-SILVEIRA, R. F.; OLIVEIRA, R. M.;
PAGNONCELLI, M. G. B.; ROCHA, H. A. O.; MACEDO, G. R.; SANTOS, E. S.,
Chitooligosaccharides antagonizes the cytotoxic effect of glucosamine. World J.
Microbiol. Biotechnol., v.28, n.3, p.1097-1105, 2012.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
79
AZEVEDO, V. V. C.; CHAVES, S. A.; BEZERRA, D. C.; FOOK, M. V. L.; COSTA,
A. C. F. M. Quitina e Quitosana: aplicações como biomateriais. Revista Eletrônica de
Materiais e Processos, v.2.3, p.27-34, 2007.
BARROS NETO, B.; SCARMINIO, I. S.; BRUNS, R. Como fazer experimentos:
pesquisa e desenvolvimento na ciência e na indústria. 2. ed. Campinas: Unicamp, 2001.
401p.
BATISTA FILHO, A.; ALMEIDA, J. E. M.; SANTOS, A. S.; MACHADO, L. A.;
ALVES, S. B. Eficiência de isolados de Metarhizium anisopliae no controle de
cigarrinha-da-raiz de cana-de-açúcar Mahanarva fimbriolata (Hom.: Cercopidae). STAB
– Açúcar, Álcool e Subprodutos, v.21, p.84-89, 2002.
CARVALHO, J. C. M.; SATO, S. Fermentação Descontínua, In: SCHMIDELL, W.;
LIMA, U. A.; AQUARONE, E.; BORZANI, W. Biotecnologia Industrial. São Paulo,
Ed. Edgard Blücher, v.2, p.193-204, 2001.
CHAO, H. J.; SCHWARTZ, J.; MILTON, D. K. Populations and determinats of
airborne fungi in large office buildings. Environmental Health Perspectives, v.110, n.8,
p.777-782, 2002.
CHEN, X.; XIA, W.; YU, X. Purification and characterization of two types of
chitosanase from Aspergillus sp. CJ22-326. Food Research International, v.38, n.3,
p.315-322, 2005.
CHEN, P. T.; CHIANG, C.-J.; CHAO, Y.-P. Medium optimization and production of
secreted Renilla luciferase in Bacillus subtilis by fed-batch fermentation. Biochemical
Engineering Journal, v.49, p.395-400, 2010.
CHENG, C. Y.; LI, Y. An Aspergillus chitosanase with potential for large-ecale
preparation of chitosan oligosaccharides. Biotechnol. Appl. Biochem., v.32, p.197-203,
2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
80
CHOI, Y. J.; Kim, E. J.; Piao, Z.; Yun, Y. C.; Shin, Y. C. Purification and
characterization of chitosanase from Bacillus sp. Strain KCTC 0377BP and its
application for the production of chitosan oligosaccharides. Applied and Environmental
Microbiology, v.70, n.8, p.4522-4531, 2004.
COLAKOGLU, G. The chemical and physical environment for fungal growth. Fak
Matbaa Birimi, Istanbul, 149-166, 2001.
CRAVEIRO, A. C.; CRAVEIRO, A. A.; QUEIROZ, D. C. Quitosana a fibra do futuro.
2. ed. Fortaleza: PADETEC, 2004. V.1, 218p.
DIAS, B. A. Estudo de proteases degradadoras de cutícula produzidas pelo fungo
entomopatogênico Beauveria bassiana. 2005. 70p. Dissertação (Mestrado em
Microbiologia) – Centro de Ciências Biológicas, Programa de Pós-Graduação em
Microbiologia, Universidade Estadual de Londrina, Londrina.
DINARDO-MIRANDA, L. L.; FERREIRA, J. M. G.; CARVALHO, P. A. M.
Influência da época de colheita e do genótipo de cana-de-açúcar sobre a infestação de
Mahanarva fimbriolata (Stal) (Hemiptera: Cercopidae). Neotrop. Entomol, v.30, n.1,
p.145-149, 200.
DONG, C.; ZHANG, J.; HUANG, H.; CHEN, H.; HU, Y. Pathogenicity of a new China
variety of Metarhizium anisopliae (M. anisopliae var. Dcjhyium) to subterranean termite
Odontotermes formosanus. Microbiological Research, v.164, p.27-35, 2009.
DONZELLI, B. G. G.; OSTROFF, G.; HARMAN, G. E. Enhanced enzymatic
hydrolysis of langostino shell chitin with mixtures of enzymes from bacterial and fungal
sources. Carbohydrate Research, v.338, p.1823-1833, 2003.
DRIVER, F.; MILNER, R. J.; TRUEMAN, J. W. H. A taxonomic revision of
Metarhizium based on phylogenetic analysis of rDNA sequence data. Mycological
Research, v.104, n.2, p.134-150, 2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
81
DUNG, P.; MILAS, M.; RINAUDO, M.; DESBRIÉRES, J. Water soluble derivatives
obtained by controlled chemical modifications of chitosan, Carbohydrate Polymers,
v.24, p.209-214, 1994.
ESTEVES, J. A.; CABRITA, J. D.; NOBRE, G. N. Micologia médica, 2. ed. Lisboa:
Fundação Calouste Gulbenkian, 1990. 1058p.
FACCIOTI, M. C. R. Fermentação Contínua, In: Biotecnologia Industrial. Editado por:
SCHMIDELL, W.; LIMA, U. A.; AQUARONE, E.; BORZANI, W., São Paulo, Ed.
Edgard Blücher, v.2, p.541, 2001.
FARIA, M. R.; MAGALHÃES, B. P. O uso de fungos entomopatogênicos no Brasil -
Situação atual e perspectivas. Biotecnologia Ciência & Desenvolvimento, n.22., p.18-
21, setembro/outubro 2001.
FU, J.-Y.; WU, S.-M.; CHANG, C.-T.; SUNG, H.-Y. Characterization of Three
Chitosanase Isozymes Isolated from a Commercial Crude Porcine Pepsin Preparation. J.
Agric. Food Chem., v.51, p.1042-1048, 2003.
FURIGO JUNIOR, A.; PEREIRA, E. P. Enzimas e suas aplicações cinética enzimática.
2001. 39f. Apostila (Graduação de Engenharia Química e Engenharia de Alimentos) –
Centro de Tecnologia, Departamento de Engenharia Química e Engenharia de
Alimentos, Universidade Federal de Santa Catarina, Florianópolis.
GAO, J.; XU, H.; LI, Q.-J.; FENG, X.-H.; LI, S. Optimization of medium for one-step
fermentation of inulin extract from Jerusalem artichoke tubers using Paenibacillus
polymyxa ZJ-9 to produce R,R-2,3-butanediol, Bioresource Technology, v.101, p.7076-
7082, 2010.
GARCIA, M. V.; MONTEIROI, A. C.; SZABO, M. J. P.; PRETTE, N.; BECHARA, G.
H. Mechanism of infection and colonization of Rhipicephalus sanguineus eggs by
Metarhizium anisopliae as revealed by scanning electron microscopy and
histopathology. Brazilian Journal of Microbiology, São Paulo, v.36, n.4, p.368-372,
2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
82
GHANEM, N. B.; YUSEF, H. H.; MAHROUSE, H. K. Production of Aspergillus
terreus xylanase in solid-state cultures: application of the Plackett & Burman
experimental design to evaluate nutritional requirements, Bioresource Technology,
v.73, p.113-121, 2000.
GHORBEL-BELLAJ, O.; HMIDET, N.; JELLOULI, K.; YOUNES, I.; MAALEJ, H.;
HACHICHA, R.; NASRI, M. Shrimp waste fermentation with Pseudomonas aeruginosa
A2: Optimization of chitin extraction conditions through Plackett–Burman and response
surface methodology approaches, International Journal of Biological Macromolecules,
v.48, p.596–602, 2010.
GIUSTINA, A.; VENTURA, P. Weight-reducing regimens in obese subjects: Effects of
a new dietary fiber integrator. Acta Toxicol Ther, 16, 4, p.199-214, 1995.
GUIA.BIO, Reino dos fungos, 2012. Disponível em:
<http://www.http://guia.bio.br/reino-dos-fungos>. Acesso em: 12 novembro 2012.
GUIRADO, N.; AMBROSANO, E. J.; ROSSI, F.; DIAS, F. L. F. Controle de Cupins
de Montículo com Metarhizium anisopliae e Beauveria bassiana, Rev. Bras. de
Agroecologia, v.4, n.2, 2009.
GUO, Y.; XU, J.; ZHANG, Y.; XU, H.; YUAN, Z.; LI, D. Medium optimization for
ethanol production with Clostridium autoethanogenum with carbon monoxide as sole
carbon source, Bioresource Technology, v.101, p.8784–8789, 2010.
HAGAG, S.; KUBITSCHEK-BARREIRA, P.; NEVES, G. W. P.; AMAR, D.;
NIERMAN, W.; SHALIT, I.; SHAMIR, R.; LOPES-BEZERRA, L.; OSHEROV, N.
Transcriptional and proteomic analysis of the Aspergillus fumigatus ∆prtT protease-
deficient mutant. PLoS One, v.7, n.4, e33604, 2012.
HAN, L.-K.; KIMURA, Y.; OKUDA, H. Reduction in fat storage during chitin-chitosan
treatment in mice fed a high-fat diet, International Journal of Obesity, 23:174-179,1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
83
HUNG, T.; CHANG, Y.; SUNG, H.; CHANG, C. Purification and Characterization of
Hydrolase with Chitinase and Chitosanase Activity from Commercial Stem Bromelain.
J. Agric. Food Chem., 50, 4666-4673, 2002.
ICHIKAWA, S.; TAKANO, K.; TAKASHIKUROIWA, HIRUTA, O.; SATO, S.;
MUKATAKA, A. S. Immobilization and stabilization of chitosanase by multipoint
attachment to agar gel support. Journal of Bioscience and Bioengineering, v.93, n.2,
p.201-206, 2002.
JABOR, I. A. S.; PAMPHILE, J. A.; RODRIGUES, S. B.; MARQUES-SILVA, G. G.;
ROCHA, C. L. M. S. Análise do desenvolvimento do fungo entomopatogênico
Metarhizium anisopliae em resposta a fatores nutricionais. Acta Scientiarum.
Agronomy, Maringá, v.25, n.2, p.497-501, 2003.
JI, X.-J.; HUANG, H.; DU, J.; ZHU, J.-G.; REN, L.-J.; LI, S.; NIE, Z.-K. Development
of an industrial medium for economical 2,3-butanediol production through co-
fermentation of glucose and xylose by Klebsiella oxytoca, Bioresource Technology,
v.100, p.5214–5218, 2009.
JO, Y.; JO, K.; JIN, Y.; KIM, K.; SHIN, J.; KIM, Y.; PARK, R. Characterization and
kinetics of 45 kDa chitosanase from bacillus sp. P16. Biosci. Biotechnol. Biochem.,
v.67, n.9, 1875 – 1882, 2003.
KHOSRAVI-DARANI, K.; ZOGHI, A.; ALAVI, S. A.; FATEMI, S. S. A. Application
of Plackett Burman Design for Citric Acid Production from Pretreated and Untreated
Wheat Straw, Iran. J. Chem. Chem. Eng.. v.27, n.1, p.91-104, 2008.
KIM, P.; KANG, T. H.; CHUNG, K. J.; KIM, I. S.; CHUNG, K. Purification of a
constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and
expression of the gene. FEMS Microbiolog.y Letters 240, 31–39, 2004.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
84
KIM, S.-K.; RAJAPAKSE, N. Enzymatic production and biological activities of
chitosan oligosaccharides (COS): A review. Carbohydrate Polymers, v.62, n.4, p.357-
368, 2005.
KIRK, P. M.; CANON, P.F.; MINTER, D.W.; STALPERS, J.A. Dictionary of the
fungi. CAB International, Wallingford, 2008.
KONDO, H.; OSADA, A. Influence of dietary fiber on the bioavailability of zinc in
rats, Biomed Environ Sci, 9, 204-8, 1996.
KUMAR, S.; KOH, J. Physiochemical, Optical and Biological Activity of Chitosan-
Chromone Derivative for Biomedical Applications. Int. J. Mol. Sci., v.13, p.6102-6116,
2012.
KURITA, K.; SANNAN, T.; IWAKURA, Y. Studies on chitin, 4. Evidence for
formation of block and random copolymers of N-acetyl-o-glucosamine and
oglucosamine by hetero and homogeneous hydrolyses. Die Makromolecular Chemie,
v.178, p.3197-3202, 1977.
KUROIWA, T.; ICHIKAWA, S.; HIRUTA, O.; SATO, S.; MUKATAKA, S. Factors
Affecting the Composition of Oligosaccharides Produced in Chitosan Hydrolysis Using
Immobilized Chitosanases. Biotechnol. Prog., 18, 969-974, 2002.
LI, L.-Z.; HUI, Z.; MO, X. Optimization of the medium for streptolydigin production
by Streptomyces lydicus AS 4.2501 with precursor feeding using response surface
methodology, Journal of the Taiwan Institute of Chemical Engineers, 41, 252–258,
2010.
LIU, B.-L.; KAO, P.-M.; TZENG, Y.-M.; FENG, K.-C. Production of chitinase from
Verticillium lecanii F091 using submerged fermentation. Enzyme and Microbial
Technology, v.33, p.410-415, 2003.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
85
LOBATO, A. K. C. L. Estudo da produção de biosurfactantes por micro-organismos
isolados de poços de petróleo. 2003. 108p. Dissertação (Mestrado) – Programa de Pós-
Graduação em Engenharia Química. Universidade Federal do Rio Grande do Norte.
Natal.
LOPES, R. B.; ALVES, S. B.; PADULLA, L. F. L.; PÉREZ, C. A. Eficiência de
formulações de Beauveria bassiana e Metarhizium anisopliae para o controle de ninfas
de Amblyomma cajennense (Fabricius, 1787). Revista Brasileira de Parasitologia
Veterinária, v.16, n.1, p.27-31, 2007.
LOUREIRO, E. S.; BATISTA FILHO, A.; ALMEIDA, J. E. M.; MENDES, J. M.;
PESSOA, L. G. A. Eficiência de isolados de Metarhizium anisopliae (metsch.) sorok.
no controle da cigarrinha-da-raiz da cana-de-açúcar, Mahanarva fimbriolata (stal, 1854)
(hemiptera: cercopidae), em condições de campo. Arq. Inst. Biol., São Paulo, v.79, n.1,
p. 47-53, 2012.
MELLO, D. R.; REIS, R. C. S.; BITTENCOURT, V. R. E. P. Patogenicidade in vitro
do fungo Metarhizium anisopliae (Metschnikoff, 1879) Sorokin, 1883, sobre o carrapato
Boophilus microplus (Canestrini, 1887). Revista Brasileira de Parasitologia Veterinária,
São Paulo, v.15, p.157-162, 2006.
MITSUTOMI, M.; KIDOH, H.; TOMITA, H.; WATANABE, T. The action of Bacillus
circulans WL-12 chitinases on partially N-acetylated chitosan, Bioscience
Biotechnology and Biochemistry, v.59, p.529-531, 1995.
MALAJOVICH, M. A. Biotecnologia: fundamentos. Ed. biblioteca Max Feffer do
Instituto de Tecnologia ORT, Rio de Janeiro, 2009. 104p.
MONAGHAN, R. L.; EVELEIGH, D. E.; TEWARI, R. P.; REESE, E. T. Chitosanase,
a novel enzyme. Nature - New Biology, 245, p.78-80, 1973.
MU, X. Q.; XU, Y.; NIE, Y.; OUYANG, J.; SUN, Z. H. Candida parapsilosis CCTCC
M203011 and the optimization of fermentation medium for stereoinversion of (S)-1-
phenyl-1,2-ethanediol, Process Biochemistry, v.40, p.2345–2350, 2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
86
MURRAY, P. R.; ROSENTHAL, K. S.; PFALLER, M. A. Medical microbiology. 5th
ed. London, Elsevier Mosby, 2005.
MUZZARELLI, R. A. A. Biochemical significance of exogenous chitins and chitosans
in animals and patients, Carbohydrate Polymers, v.20, p.7-16, 1993.
MUZZARELLI, R. A. A.; TOMASETTI, M.; ILARI, P. Depolymerization of chitosan
with the aid of papain, Enzyme Microbiology and Technology, v.16, p.110-114, 1994.
NAVEENA, B. J.; ALTAF, Md.; BHADRIAH, K.; REDDY, G. Selection of medium
components by Plackett–Burman design for production of L(+) lactic acid by
Lactobacillus amylophilus GV6 in SSF using wheat bran, Bioresource Technology,
v.96, p.485-490, 2005.
NOGAWA, M.; TAKAHASHI, H.; KASHIWAGI, A.; OHSHIMA, K.; OKADA, H.;
MORIKAWA, Y. Purification and Characterization of Exo--D-Glucosaminidase from
a Cellulolytic Fungus, Trichoderma reesei PC-3-7, Appl. Environ. Microbiol., v.64, n.3,
p.890-895, 1998.
ONOFRE, S. B.; VARGAS; L. R. B.; ROSSATO, M.; BARROS, N. M.; BOLDO, J.
T.; NUNES, A. R. F.; AZEVEDO, J. L. Controle biológico de pragas na agropecuária
por meio de fungos entomopatogênicos. In: SERAFINI, L. A.; BARROS, N. M.;
AZEVEDO, J. L. (Org.). Biotecnologia – avanços na agricultura e na agroindústria.
Caxias do Sul: EDUCS, p.295-328, cap.8, 2002.
OSSWALD, W. F.; SHAPIRO, J. P.; DOOSTDAR, H.; MCDONALD, R. E.; NIEDZ,
R. P.; NAIRN, C. J.; HEARN, C. J.; MAYER, R. T. Identification and Characterization
of Acidic Hydrolases with Chitinase and Chitosanase Activities from Sweet Orange
Callus Tissue, Plant Cell Physiol. 35, p.811-820, 1994.
PAGNONCELLI, M. G. B. Estudo do mecanismo de produção de oligossacarídeos com
atividades nutracêuticas a partir da quitosana por hidrólise enzimática com processo

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
87
fermentativo simultâneo. 2008. 118p. Tese (Doutorado) – Centro de Tecnologia,
Programa de Pós-Graduação em Engenharia Química. Universidade Federal do Rio
Grande do Norte. Natal.
PALMA-GUERRERO, J., JANSSON, H.-B., SALINAS, J.; LOPEZ-LLORCA, L. V.
Effect of chitosan on hyphal growth and spore germination of plant pathogenic and
biocontrol fungi, J. Appl. Microbiol., v.104, p.541-553, 2008.
PAULA, F. A.; ARRUDA, D. C. Análise do método de planejamento experimental
Plackett Burman na otimização de degradação de penas de aves por comunidades
microbianas, XXI Salão de Iniciação Científica, 19-23, outubro, 2009, UFRGS, Porto
Alegre.
PELCZAR, M. J.; CHANG, E. C. S.; KRIEG, N. R. Microbiologia: conceitos e
aplicações, v.2, 2.ed. São Paulo: Makron BooKs, 1996.
PELLETIER, A.; SYGUSCH, J. Purification and characterization of three chitosanase
activities from Bacillus megaterium P1, Applied and Environmental Microbiology, v.
56, n.4, p.844-848, 1990.
PEREIRA, F. B.; GUIMARÃES, P. M. R.; TEIXEIRA, J. A.; DOMINGUES, L.
Optimization of low-cost medium for very high gravity ethanol fermentations by
Saccharomyces cerevisiae using statistical experimental designs, Bioresource
Technology, v.101, p.7856-7863, 2010.
PETER, M. G. Chitin and chitosan from aninal sources. In: STEINBUCHEL, A.;
RHEE, S. K. Polysaccharides and Polyamides in the Food Industry. Weinheim:
Wileyvch, v.1, p.115-208, 2005.
PIZA, F. A. T.; SILOTO, A. P.; CARVALHO, C. V.; FRANCO, T. T. Purification,
characterization and purification of chitosanase from Bacillus cereus, Brazilian Journal
of Chemical Engineering., v.16, n 2, p.185-192, 1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
88
POLITZER, K.; BON, E. P. S. Enzimas Industriais e Especiais. C.G.E.E. Ciência,
Tecnologia e Inovação. Rio de Janeiro, 554p, 2006.
POLYMAR, Ciência é a nossa Natureza, 2003. Disponível em:
<http://www.polymar.com.br>. Acesso em: 6 setembro 2012.
PUTZKE, J.; PUTZKE, M. T. L. Os reinos dos fungos. Santa Cruz do Sul, RS:
EDUNISC, 1998.
QUINES, L. K. M. Extração de poli(3-hidroxibutirato) produzido por Cupriavidus
necator dsm 545 com 1,2-carbonato de propileno. 2010. 99p. Dissertação (Mestrado em
Engenharia de Alimentos) – Centro Tecnológico, Departamento de Engenharia Química
e Engenharia de Alimentos, Programa de Pós-Graduação em Engenharia de Alimentos,
Universidade Federal de Santa Catarina, Florianópolis.
RAJENDRAN, A.; THANGAVELU, V. Statistical experimental design for evaluation
of medium components for lipase production by Rhizopus arrhizus MTCC 2233, LWT -
Food Science and Technology, v.42, p.985–992, 2009.
RAMIREZ-LEÓN, L. F.; RUIZ-HERRERA, J. Hydrolysis of walls and formation of
spheroplasts in Mucor rouxii. J. Gen. Microbiol., v.72, p.281-290, 1972.
RANGEL, M. A. S.; LOUREIRO, E. S.; ALMEIDA, A. S.; ALVES, H. C.; ONO, F.
B.; LOBO, P. M.; MARIANI, M. A.; PESSOA, L. G. A.; SILVA, A. J. Avaliação
participativa do controle de cigarrinhas das pastagens com a aplicação do fungo
Metarhizium anisopliae (Metsch.) Sorok. Rev. Bras. de Agroecologia, v.2, n.2, 731-734,
2007.
RAVEN, P. H.; EVERT, R.F.; EICHHORN, S.E. Biologia Vegetal, 6ª Ed, Rio Janeiro,
Editora Guanabara Koogan, 2001.
REDDY, L. V. A.; WEE, Y.-J.; YUN, J.-S.; RYU, H.-W. Optimization of alkaline
protease production by batch culture of Bacillus sp. RKY3 through Plackett–Burman

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
89
and response surface methodological approaches, Bioresource Technology, v.99, p.242–
2249, 2008.
RIGUEIRA, J. C. S. Influência da contagem de células somáticas no perfil e teores de
aminas bioativas e na qualidade de leite cru e de queijo mussarela. 2010. 147p. Tese
(Doutorado em Ciência de Alimentos) - Faculdade de Farmácia, Departamento de
Alimentos, Programa de Pós-Graduação em Ciência de Alimentos, Faculdade de
Farmácia da Universidade Federal de Minas Gerais, Belo Horizonte.
RODRIGUES, M. I.; IEMMA, A. F. Planejamento de Experimentos e Otimização de
Processos: uma estratégia sequencial de planejamentos, Campinas, SP, Casa do Pão
Editora, 2005.
SANTOS, E. S. Recuperação e Purificação de Enzimas usando Adsorção em Leito
Expandido, Tese de Doutorado, Universidade Estadual de Campinas, Campinas/SP,
p.152, 2001.
SEDMAK, J. J.; GROSSBERG, S. E. A rapid, sensitive and versatile assay for protein
using coomassie brilliant blue G250, Analytical Biochemistry, v.79, p. 544-552, 1977.
SEINO, H.; TSUKUDA, K.; SHIMASUE, Y. Properties and action pattern of a
chitosanase from Bacillus sp. PI- 7S. Agricultural Biological Chemistry, 55, p.2421-
2423, 1991.
SHIMOSAKA, M.; NOGAWA, M.; OHNO, Y.; OKAZAKI, M. Chitosanase from the
plant pathogenic fungus, Fusarium solani f. sp. Phaseoli – purification and some
properties, Bioscience Biotechnolgy and Biochemistry, v.57, n.2, p.231-235, 1993.
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Production of two chitosanases from a chitosan-assimilating bacterium, Acinetobacter
sp. Strain CHB101, Applied and Environmental Microbiology, v.61, n.2, p.438-442,
1995.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
90
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Characterization of Three Chitosanase Isozymes Isolated from a Commercial Crude
Porcine Pepsin Preparation, J. Agric. Food Chem. 51, p.1042-1048, 2003.
SILVA FILHO, R. C.; COSTA, A. S. S.; CHAGAS, B. M. E.; SANTOS, F. N.;
MACEDO, G. R.; SANTOS, E. S. Avaliação do teste de atividade quitosanolítica
produzida por fungo. In: XV SIMPÓSIO NACIONAL DE BIOPROCESSOS, 15,
Recife, 2005, 02 a 05 de agosto.
SILVA, E. P. Avaliação Preliminar do Potencial de Aplicação da Tecnologia de
Biopilhas para a Biorremediação do Solo de Guamaré-RN. 2004. 119p. Dissertação
(Mestrado) – Centro de Tecnologia, Programa de Pós-Graduação em Engenharia
Química. Universidade Federal do Rio Grande do Norte. Natal.
SOMASHEKAR, D.; JOSEPH, R. Chitosanases – Properties and apllications: A review.
Bioresource Technology, v.55, p.35-45, 1996.
STOYACHENKO, I. A.; VARLAMOV, V. P.; DAVANKOV, V. A. Chitinases of
Streptomyces kurssanovii: purification and some properties, Carbohydrate Polymers,
v.24, p.47-54, 1994.
TINLINE, R. D. Nuclear distribuition in Metarhizium anisopliae. Mycologia, v. 63, p.
713-721, 1971.
TRABULSI, L. R.; TOLEDO, M. R. F. Microbiologia. 2. Ed., São Paulo: Atheneu,
1996.
VIEIRA, P. D. S.; SILVA, W. M. T.; PAIVA, L. M.; LIMA, E. A. L. A.;
CAVALCANTI, V. L. B. Estudo da caracterização morfológica, esporulação e
germinação dos conídiosde Metarhizium anisopliae var. acridum em diferentes
temperaturas. Biológico, São Paulo, v.69, n.1, p.17-21, 2007.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
91
WANDERLEY, M. D.; NEVES, E.; ANDRADE, C. J. Aspectos da produção industrial
de enzimas. Revista Citino, v.1, n.1, p.30-36, 2011.
WANG, C.; TYPAS, M. A.; BUTT, T. M. Detection and characterization of pr1
virulent gene deficiencies in the insect pathogenic fungus Metarhizium anisopliae.
FEMS Microbiology Letters, v. 213, p. 251-255, 2002.
ZAHORSKA-MARKIEWICZ, B.; KROTKIEWSKI, M.; OLSZANECKA-
GLINIANOWICZ, M.; ZURAKOWSKI, A. Effect of chitosan in complex management
of obesity, Pol Merkur Lekarski, v.13, n.74, p.129-132, 2002.
ZHANG, X. Y.; DAI, A. L.; ZHANG, X. K.; KUROIWA, K.; KODAIRA, R.;
SHIMOSAKA, M.; OKAZAKI, M. Purification and Characterization of and Exo--D-
glucosaminidase from a koji mold, Aspergillus oryzae IAM2660. Biosci. Biotechnol.
Biochem., v.64, n.9, 1896-1902, 2000.
ZHU, X.; WU, X.; DAI, Y. Fermentation conditions of a chitosanase from Acinetobcter
sp C-17. Biosci. Biotechnol. Biochem., v.67, n.2, p.284-290, 2003.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
92
APÊNDICE

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
93
APÊNDICE I
Metodologia para repique da cepa
Em Erlenmeyer de 1000 mL colocava-se 500 mL do meio de manutenção
PDA. Este Erlenmeyer, com o meio de manutenção, era autoclavado junto com placas
de Petri a 1,5 kgf/cm2 por 15 minutos, em seguida, era levado para uma câmara de fluxo
laminar e transferido para as placas de Petri já autoclavadas, onde eram mantidas numa
estufa a 27 ºC durante 24 h. Após este período, as placas de Petri eram levadas a câmara
de fluxo laminar, utilizando bico de Bunsen para esterilizar a alça de platina. Procedia-
se o repique utilizando uma placa de Petri contendo a cepa pura. Cada placa de Petri
contendo a cepa pura era transferida para duas placas de Petri contendo o meio de
manutenção. As placas eram mantidas na estufa a 27 ºC durante 30 dias onde após esse
período, repetia-se este procedimento.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
94
APÊNDICE II
Montagem da Curva de Calibração Usando o Método DNS
Soluções:
Glicosamina = 0,01 mol/mL (10000,0 µmol/mL)
Tabela 14. Dados para construção da curva de calibração usando o Método DNS.
Tubo de
Ensaio
Solução de
glicosamina (mL)
Água Destilada
(mL)
DNS (mL)
Volume Total
Massa (µµµµmol)
1 0 1,0 1,5 2,5 0
2 0,2 0,8 1,5 2,5 0,2
3 0,4 0,6 1,5 2,5 0,4
4 0,6 0,4 1,5 2,5 0,6
5 0,8 0,2 1,5 2,5 0,8
6 1,0 0 1,5 2,5 1,0
Tabela 15 - Medidas de absorbância a 600nm
Dados adicionais:
Peso Molecular D-Glicosamina = 215,64 (recipiente SIGMA)
Conc. Sol. D-Glicosamina = 0,05 g/50 mL
Massa calc.(µmol) = conc. Sol. D-Glic. x Vol. Utiliz. D-Glics. =
= 0,05 x 1000 x Vol.util./50*215,64
Tempo atividade enzimática = 30 min
1 U/mL = 1 µmol/(min.mL)
Vol. Total (Glicosamina+caldo) = 1 mL
Medições Adsorbância a 600 nm Vol. Utiliz. Massa Massa Atividade
II III Média(Abs)(mL) calc. (mg) calc. (µg) Enzim. (U/mL)
0 0 0 0 0 0 0
0,133 0,128 0,1307 0,2 0,2 200 0,0309
0,279 0,277 0,2787 0,4 0,4 400 0,0618
0,433 0,428 0,4297 0,6 0,6 600 0,0927
0,587 0,583 0,5797 0,8 0,8 800 0,1237
0,727 0,716 0,7157 1 1 1000 0,1546

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
95
Figura 17 – Curva padrão de glicosamina.
y = 1900ralx + 1900ral
R² = 1900ral
00,000
00,020
00,040
00,060
00,080
00,100
00,120
00,140
00,160
00,180
00,000 00,100 00,200 00,300 00,400 00,500 00,600 00,700 00,800
Ati
v.
En
zim
áti
ca (
U/m
L)
Absorbância (Abs) a 600 nm
Curva-Padrão D-Glicosamina

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
96
APÊNDICE III
Método de Determinação de Proteína Utilizando o Corante Comassie
Brilhante Blue G 250 (Método modificado de Sedmak e Grossberg)
Micrométodo
Preparar uma solução 0,06% de Comassie Blue G250 em 1,5 % HCl (w/V). Filtrar e
papel de filtro Whatman nº1.
Curva Padrão (1 a 20 µµµµg)
a) Preparar soluções estoque de BSA de 10 a 200 µg/mL.
b) Colocar em tubos de ensaio 100 µL da solução estoque, 1400 µL de água e
1500µL do Comassie e agitar.
c) Preparar um tubo contendo 1500 µL de água e 1500 µL do corante que
corresponde ao zero de proteína.
d) Após 5 minutos (padronizar o tempo) fazer as leituras das absorbâncias.
e) As leituras das amostras devem ser feitas em dois comprimentos de onda 595 e
465 contra a água.
f) Plotar a razão das absorbâncias a 595/465 como uma função da massa de BSA e
calcular o melhor ajuste linear dos dados.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
97
Micrométodo (1 a 20 µµµµg de BSA)
Solução Estoque de BSA = 0,2 mg/mL
Tabela 16. Dados para construção da curva padrão de BSA.
Tubo de
Ensaio
H2O (µµµµL)
BSA (µµµµg)
Coomassie (µµµµL)
BSA (µµµµg)
1 1500 0 1500 0
2 1490 10 1500 2
3 1480 20 1500 4
4 1470 30 1500 6
5 1460 40 1500 8
6 1450 50 1500 10
7 1440 60 1500 12
8 1430 70 1500 14
9 1420 80 1500 16
10 1410 90 1500 18
11 1400 100 1500 20
Dados adicionais:
Concentração Sol. Estoque
BSA = 0,2 mg/mL = 0,2 µg/µL
Vol. total sol. = vol. H2O + vol. BSA + vol.
Coomassie = 3000 µL = 3 mL
Concentração Sol. BSA (mg/mL) = (massa BSA mg) / 3 mL
.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
98
Figura 18 – Curva Padrão de BSA
y = 1900ralx - 1900ral
R² = 1900ral
000,000
001,000
002,000
003,000
004,000
005,000
006,000
007,000
000,400 000,500 000,600 000,700 000,800 000,900 001,000 001,100
µg
BS
A
Razão Absorbâncias (595nm/465nm)
Curva-Padrão BSA

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE TECNOLOGIA
DEPARTAMENTO DE ENGENHARIA QUÍMICA PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA
TESE DE DOUTORADO
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em
Cultivo Descontínuo Submerso
Raimundo Cosme da Silva Filho
Natal/RN
Abril/2013

Raimundo Cosme da Silva Filho
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo
Descontínuo Submerso
Tese apresentada ao Programa de Pós-Graduação em Engenharia Química – PPGEQ, da Universidade Federal do Rio Grande do Norte, como parte dos requisitos para obtenção do título de Doutor em Engenharia Química, sob a orientação do Prof. Dr. Everaldo Silvino dos Santos e a coorientação da Profª. Drª. Gorete Ribeiro de Macedo.
Natal/RN
Abril/2013

Catalogação da Publicação na Fonte. UFRN / CT / PPGEQ
Biblioteca Setorial “Professor Horácio Nícolas Sólimo”.
Silva Filho, Raimundo Cosme da. Otimização do meio de cultura para a produção de quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso / Raimundo Cosme da Silva Filho. - Natal, 2013. 98 f.: il.
Orientador: Everaldo Silvino dos Santos. Co-orientador: Gorete Ribeiro de Macedo. Tese (Doutorado) - Universidade Federal do Rio Grande do Norte.
Centro de Tecnologia. Departamento de Engenharia Química. Programa de Pós-Graduação em Engenharia Química.
1. Quitosana - Tese. 2. Quitina - Tese. 3. Quitosanase – Tese. 4. Metarhizium
anisopliae - Tese. I. Santos, Everaldo Silvino dos. II. Macedo, Gorete Ribeiro de. III. Universidade Federal do Rio Grande do Norte. IV. Título. RN/UF/BSEQ CDU 547.458 (043.2)

SILVA FILHO, Raimundo Cosme da – Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo Descontínuo Submerso. Tese de Doutorado. UFRN - Programa de Pós-Graduação em Engenharia Química. Área de Concentração: Pesquisa e Desenvolvimento de Tecnologias Regionais – Subárea: Engenharia de Processos – Alimentos e Biotecnologia, Natal/RN, Brasil.
Orientação: Prof. Dr. Everaldo Silvino dos Santos Coorientação: Profª. Drª. Gorete Ribeiro de Macedo ___________________________________________________________________________
RESUMO: No presente trabalho utilizou-se um Planejamento Plackett & Burman, com 8 fatores e 12 ensaios em 2 níveis e mais 3 repetições na condição do ponto central, para se investigar a influência das concentrações de quitosana, peptona, extrato de levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O e FeSO4 na produção da enzima quitosanase por Metarhizium
anisopliae. Os ensaios para produção da enzima foram realizados em cultivo descontínuo submerso. Os resultados para o Planejamento Plackett & Burman mostraram que a atividade quitosanolítica foi favorecida pelo aumento da concentração de substrato (quitosana) e de sulfato ferroso (FeSO4), enquanto que o aumento da concentração dos outros fatores não contribuiu de forma significativa para a atividade quitosanolítica. A otimização do meio de cultura para a produção da enzima foi realizado por meio de um Planejamento Composto Central, com os dois fatores que mais influenciaram a atividade quitosanolítica (quitosana e FeSO4), conforme Planejamento Plackett & Burman, mantendo-se os outros nutrientes em seus valores mínimos. Nesse planejamento, para o fator FeSO4, tomou-se o limite inferior (-1) como sendo o limite superior do Planejamento Plackett & Burman. Os resultados mostraram que a produção da enzima foi favorecida pelo aumento da concentração quitosana e pela diminuição da concentração de FeSO4. A produção máxima de atividade quitosanolítica foi da ordem de 70,0 U/L e foi atingida em apenas 18 h de fermentação, resultado esse vinte e oito vezes maior aos obtidos anteriormente para o mesmo microrganismo que foi 2,5 U/L em 48h.
Palavras-chave: quitosana; quitosanase; Metarhizium anisopliae; Plackett & Burman.


ABSTRACT
In this work a Plackett-Burman Design with 8 factors and 12 trials in 2 levels with 3 repetitions at the center point was used in order to investigate the influence of the concentration of chitosan, peptone, yeast extract, NaNO3, K2HPO4, KCl, MgSO4.7H2O and FeSO4 on chitosanase production by Metarhizium anisopliae. Runs were carried out using submerged discontinuous cultivation for enzyme production. The results of the Plackett & Burman Design showed that only two factors, chitosan concentration as well as FeSO4 had influence on chitosanolytic activity, while the increase in concentration of other factors not contributed significantly to the quitosanolítica activity. Cultivation medium optimization for enzyme production was carried out using a Composite Central Design, with the most important factors for chitosanolytic activity (chitosan and FeSO4), in accordance with Plackett & Burman Design, and keeping the other nutrients in their minimum values. On this other design, it was taken the highest limit in Plackett & Burman Design as the lowest limit (-1) to FeSO4 factor. The results showed that the enzyme production was favoured by increasing the chitosan concentration and by decreasing FeSO4. Maximum production for chitosanolytic activity was about 70.0 U/L and was reached in only 18 h of fermentation, a result about twenty-eight times greater than a former study using the same microorganism (about 2.5 U/L at 48 h).
Key-words: Chitosan; Chitosanase; Metarhizium anisopliae; Plackett & Burman

Ao meu pai, minha mãe e minha
família, em especial aos meus
irmãos e sobrinhos

AGRADECIMENTOS
A Deus, por estar presente em todos os momentos da minha vida e por me ceder à
oportunidade de iniciar e concluir este trabalho.
A minha mãe Cândida Inocência da Silva que esteve comigo em todos os momentos da
minha vida me educando, me amando e sempre me dando muito carinho.
Ao meu pai Raimundo Cosme da Silva, que não está mais aqui para ver este trabalho,
mas que foi a pessoa que mais me apoiou e me incentivou, em todos os momentos da minha vida,
para que eu pudesse chegar o mais longe possível na minha carreira acadêmica. Pai muito
obrigado por toda atenção, educação, amor e carinho que recebi durante toda a minha vida.
Aos meus irmãos e familiares que sempre estiveram comigo dividindo momentos de
grande alegria e às vezes de tristeza, mas sempre nos mantendo unidos para superarmos todos os
obstáculos.
A minha namorada Rochelly Caroliny por todo amor e carinho que me deu e que me
entendeu nos momentos que estive ausente para a conclusão deste trabalho.
Ao meu orientador Professor Doutor Everaldo Silvino dos Santos por confiar em mim e
me aceitar como orientando além de toda a amizade, orientação e dedicação em todas as etapas de
desenvolvimento e conclusão deste trabalho.
À Professora Doutora Gorete Ribeiro de Macedo, “pessoa que admiro muito como
profissional e principalmente, como pessoa, um exemplo a ser seguido”, pelos conselhos e críticas
quando necessários ao desenvolvimento deste trabalho. Professora a senhora é considerada uma
mãe para de todos os alunos que tiveram a honra de conhecê-la e trabalhar com a senhora.
Aos meus amigos Engenheiros Químicos, Ana Carmen, Ana Katerine, Andréa Farias,
Anita Lima, Sirtys Lessa, Alexandre, Sérgio, Márcio Bezerra e ao Biólogo Daniel Souza, que
sempre me ouviram e que me ajudaram de alguma forma nesse trabalho.
A Eusamar Coelho de Lima, Mazinha, secretária do PPGEQ, pelo apoio, pela paciência e
orientação nas questões burocráticas do processo.
A Carlos e Jardelson, pela grande colaboração e dedicação nos experimentos do
laboratório.
A Sérgio Santana pela grande colaboração e dedicação, o qual não poupou esforços nem
tempo em me ajudar na realização dos experimentos em laboratório.

Ao meu amigo Ubiratan Nazaré por me ajudar na organização do texto e na tradução para
o Abstract.
Finalizando, agradeço ao Laboratório de Engenharia Bioquímica da UFRN por ter cedido
toda a sua estrutura para realização dos experimentos e ao Programa de Pós-Graduação em
Engenharia Química, assim como a todos os professores do Departamento de Engenharia Química
da UFRN que trabalham em pró de uma educação digna e ensino de qualidade para todos os
alunos.

Sumário LISTA DE FIGURAS
LISTA DE TABELAS
NOMENCLATURA
INTRODUÇÃO...............................................................................................................................15
REVISÃO BIBLIOGRÁFICA........................................................................................................18
2.1. Enzimas................................................................................................................................18
2.1.1. Quitosana e seus derivados........................................................................................19
2.1.2. Quitosanases...............................................................................................................24
2.2. Fungos..................................................................................................................................29
2.2.1. Características gerais..................................................................................................29
2.2.2. Reprodução dos Fungos.............................................................................................32
2.2.3. Metabolismo de fungos..............................................................................................33
2.2.4. Nutrição de fungos.....................................................................................................34
2.2.5. O Fungo Metarhizium anisopliae ..............................................................................36
2.3. Fermentação.........................................................................................................................40
2.3.1. Preparo do inóculo......................................................................................................41
2.3.2. Produção de enzimas em cultivo submerso descontínuo...........................................42
2.4. Planejamento experimental..................................................................................................42
2.4.1. Planejamento fatorial..................................................................................................43
2.4.2. Planejamento Plackett & Burman..............................................................................43
2.4.3. Aplicações do Planejamento Plackett & Burman.......................................................44
METODOLOGIA EXPERIMENTAL.............................................................................................50
3.1. Quitosana..............................................................................................................................50
3.2. Microrganismo.....................................................................................................................50
3.2.1. Escolha da cepa produtora de quitosanase.................................................................50

3.2.2. Manutenção da cepa...................................................................................................50
3.3. Produção de enzima.............................................................................................................51
3.4. Determinação da Proteína Total...........................................................................................51
3.5. Determinação da Atividade Quitosanolítica e Atividade Específica...................................51
3.6. Planejamento Experimental.................................................................................................52
3.6.1. Planejamento de seleção de fatores Plackett Burman................................................52
3.6.2. Planejamento Composto Central para Otimização do meio de cultura para a
Produção da Enzima em Cultivo Submerso.....................................................................................53
3.7. Perfil Cinético durante o Cultivo.........................................................................................56
RESULTADOS E DISCUSSÃO.....................................................................................................58
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA ..........................................58
4.2. Planejamento Plackett & Burman........................................................................................59
4.2.1. Análise do Plackett & Burman com 8 fatores e 12 ensaios .......................................59
4.2.2. Análise do Plackett & Burman com 12 ensaios e mais 3 repetições na condição do
ponto central ....................................................................................................................................61
4.3. Planejamento Composto Central para Produção de Quitosanase por Metarhizium
anisopliae em Cultivo Descontínuo Submerso................................................................................64
4.4. Perfil Cinético – Melhor Condição......................................................................................69
CONCLUSÃO.................................................................................................................................75
REFERÊNCIAS BIBLIOGRÁFICAS.............................................................................................78
APÊNDICE I...................................................................................................................................93
Metodologia para Repique da Cepa ................................................................................................93
APÊNDICE II.................................................................................................................................94
Montagem da Curva de Calibração Usando o Método DNS..........................................................94
APÊNDICE III................................................................................................................................96
Método de Determinação de Proteína Utilizando o Corante Comassie Brilhante Blue G 250
(Método modificado de Sedmak e Grossberg)................................................................................96

Lista de Figuras
Figura 1 – Estrutura da quitosana..................................................................................................20
Figura 2 – Estrutura da glicosamina..............................................................................................23
Figura 3 – Estrutura do sulfato de glicosamina.............................................................................23
Figura 4 – Representação da estrutura geral de uma célula de fungo..........................................30
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas...........................................................31
Figura 6 – Conídios de Aspergillus e Penicillium.........................................................................33
Figura 7 – Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA ...................58
Figura 8 – Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados............................................................................................................................60
Figura 9 – Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto central
mostrando a influência dos fatores estudados................................................................................63
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos
fatores estudados............................................................................................................................67
Figura 11 – Valores preditos em função dos valores observados..................................................68
Figura 12 – Atividade quitosanolítica em função da concentração de quitosana e da concentração
de FeSO4 para os fatores não significativos no ponto central .......................................................68
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana ........70
Figura 14 – Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana....................................70
Figura 15 – Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de
quitosana........................................................................................................................................71
Figura 16 – Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana..........................................................................................................72
Figura 17 – Curva padrão de glicosamina ....................................................................................95
Figura 18 – Curva Padrão de BSA ................................................................................................98

Lista de Tabelas
Tabela 1 – Comparação das enzimas com os catalisadores químicos..............................................19
Tabela 2 – Fontes naturais de quitina e quitosana ...........................................................................20
Tabela 3 – Níveis para os fatores e seus valores codificados...........................................................53
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central .........................................................................................53
Tabela 5 – Matriz do planejamento Composto Central...................................................................54
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central......................................................................................................55
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.......................59
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman...............................................60
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.......................61
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central...............................................................................................................................................62
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da atividade
enzimática........................................................................................................................................65
Tabela 12 – Efeitos Estimados do planejamento Composto Central...............................................65
Tabela 13 – Resumo da ANOVA....................................................................................................66
Tabela 14 – Dados para construção da curva de calibração usando o Método DNS ......................94
Tabela 15 – Medidas de absorbância a 600 nm para construção da curva de calibração................94
Tabela 16 – Dados para construção da curva padrão de BSA ........................................................97

NOMENCLATURA
A – Atividade quitosanolítica (U/L)
AE – Atividade específica (U/mg)
ANN-GA – algoritmo genético de rede neural artificial
ANOVA – Análise de variância (Analysis of Variance)
BSA – Albumina de soro bovino (Bovine Sereum Albumin)
CCD – Planejamento Composto Central
CDA – Chitosan Dector Agar
Cs – Concentração de substrato (%)
CP – Concentração de KCl (%)
Da – Dalton
DEQ – Departamento de Engenharia Química
DNS – Ácido Dinitrosalissílico
E – Enzima
Ea – Energia de ativação
EC – Comissão para nomenclatura e classificação de enzimas
EL – Concentração de Extrato de Levedura (%)
FP – Concentração de K2HPO4 (%)
NS – Concentração de NaNO3 (%)
GDA – Grânulos dispersíveis em água
k – Número de fatores, variáveis
LEB – Laboratório de Engenharia Bioquímica
mRNA – RNA mensageiro
n – Número de níveis de um determinado fator
N – Número de ensaios de um planejamento
P – Pressão
PP – Concentração de Peptona (%)
PDA – Batata Dextrose Agar
pH – Potencial hidrogeniônico
Q – Concentração de Quitosana (%)
Qm – Quadrado médio
r – Agitação (rpm)
R2 – Coeficiente de determinação

RA – Razão de aeração
rpm – Rotações por minuto
RSM – Metodologia de superfície de resposta
S – Molécula de substrato
SF – Concentração de FeSO4 (%)
SM – Concentração de MgSO4.7H2O (%)
SQ – Soma de quadrados
SSF – Fermentação de estado sólido
T – Temperatura
TMC – Temperatura de cultivo
V11,V12,V13 – Variáveis inertes para cálculo do erro
VHG – Fermentação de alta densidade
WP – Pó molhável
X – Concentração celular
Xi – variável independente
Xj – variável independente
xi – Fator independente real
x0 – Valor de xi no ponto central
∆xi – Variação de valor
Y – Resposta do modelo de primeira ordem
Yp – Resposta do modelo polinomial de segunda ordem
β0 – Interseção dos modelos de primeira ou segunda ordem
βi – Coeficiente linear dos modelos de primeira ou segunda ordem
βii – Coeficiente quadrático
βij – Coeficiente de interação

Capítulo 1 Introdução

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
15
1. Introdução As quitosanases foram descobertas simultaneamente há quase quarenta anos por
Monaghan et al. (1973), como pelo grupo de Ramirez-León & Ruiz-Herrera (1972), e
são enzimas obtidas a partir do metabolismo de bactérias, fungos e extraídas de plantas.
Desde então, vários estudos vêm apresentando a obtenção de quitosanase deste
microrganismos, na forma intracelular ou extracelular (Osswald et al., 1994;
Somashekar & Joseph, 1996), como a do presente estudo. A quitosanase (EC 3.2.1.132)
representa uma classe de enzimas hidrolíticas que catalisam a hidrólise de ligação β-
(1→ 4) glicosídica da quitosana, gerando oligômeros chamados quito-oligossacarídeos
(Reddy et al., 2008).
A quitosana é um biopolímero hidrofílico obtido a partir da quitina, um
polissacarídeo abundante na natureza. Na literatura, é considerado quitosana o produto
da desacetilação da quitina para um grau de desacetilação superior a 75% e solúveis em
ácidos como o acético e o fórmico. A quitosana é conhecida por exibir uma ampla
variedade de atividade fisiológica como atividade antitumoral, mas o peso molecular e a
insolubilidade em água da quitosana são desvantagens para várias aplicações.
Atualmente, há interesse nas quitosanas quando as mesmas são convertidas em quito-
oligossacarídeos porque eles não apenas são solúveis em água e possuem baixo peso
molecular, mas também têm atividades biológicas bem notáveis, como atividade
anticancerígena atóxica e biocompatível, incluindo efeito antimetastático, atividade anti-
HIV, hepatoprotetora, antioxidante, hipocolesterolêmica, antimicrobiana,
imunoestimulate, antitumoral, anti-inflamatória e atividade aceleradora da absorção de
cálcio e ferro. No entanto, estes quito-oligossacarídeos são obtidos tradicionalmente na
indústria por reação de hidrólise usando ácidos, que apresenta muitos problemas como
dificuldade em controlar a hidrólise, produzindo uma grande quantidade de
oligossacarídeos de cadeia pequena e oligossacarídeos de baixo rendimento, alto custo
de separação e, também, poluição ambiental, uma vez que utiliza um ácido como
catalisador. Como alternativa, com suas vantagens em termos de compatibilidade
ambiental, baixo custo e reprodutibilidade, a obtenção destes quito-oligossacarídeos por
hidrólise com quitosanase se tornou cada vez mais popular nos últimos anos (Azevedo
et al., 2007).

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
16
Devido a todos estes aspectos positivos é que, em geral, as enzimas vêm sendo
estudadas em diferentes aplicações, mostrando que esta é uma tendência na indústria.
No presente trabalho, foi desenvolvido um processo fermentativo visando a
otimização do meio de cultura para a produção da enzima quitosanase em cultivo
descontínuo submerso, por Metarhizium anisopliae, capaz de sintetizar oligômeros com
aplicações na indústria farmacêutica e de química fina, partindo de quitosana como
única fonte de carbono.
Nesse caso, avaliou-se, através de um planejamento Plackett & Burman (método
de delineamento de seleção de fatores) e de um planejamento Composto Central as
melhores condições para a produção da enzima. A técnica do planejamento
experimental Plackett & Burman para realizar triagem de fatores na otimização de um
determinado processo foi utilizada neste trabalho para identificar quais os fatores que
influenciam de forma significativa na produção da enzima quitosanase. A influência
destes fatores (concentração de quitosana e concentração de FeSO4) foi então
investigada pelo planejamento Composto Central com 22 experimentos na parte fatorial,
repetição em triplicata no ponto central e quatro experimentos na parte axial. A
repetição do ponto central permite também a determinação do erro puro e da
repetibilidade do processo, sendo uma forma de identificar melhor se o processo está
sob controle.
Do ponto de vista industrial é de relevante importância a realização de estudos
para a otimização da produção da quitosanase de forma contínua ou em grandes
bateladas.

Capítulo 2 Revisão Bibliográfica

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
18
2. Revisão Bibliográfica 2.1. Enzimas
As enzimas são catalisadores biológicos constituídos por longas cadeias de
aminoácidos que atuam em substratos reduzindo a energia de ativação, acelerando assim
a velocidade da reação, e transformando-os em produtos. Sua atuação catalítica visa
facilitar a atividade das células através da quebra de moléculas ou junção das mesmas
para formação de novos produtos. Podem apresentar alto grau de especificidade relativa
a apenas um substrato ou a vários substratos ao mesmo tempo, como a certos tipos de
ligações glicosídicas α-1,4 das moléculas de amido, ou para determinado tipo particular
de isômero óptico como ocorre na oxidação da β-D-glicose pela glicose oxidase (Furigo
Junior & Pereira, 2001).
Enzimas são proteínas que apresentam em sua estrutura um centro ativo
(apoenzima) e, em alguns casos, um grupo prostético denominado cofator (coenzima ou
íon metálico ativo) e sua atividade depende de alguns fatores como estrutura da
proteína, número de cadeias peptídicas, arranjo dessas cadeias na molécula, natureza do
substrato e natureza do grupo prostético (Politzer & Bon, 2006).
Todos os organismos vivos produzem, em pequena ou grande quantidade, certa
variedade de enzimas, sejam elas intra ou extracelulares. As enzimas extracelulares são
excretadas para o meio externo e podem decompor compostos insolúveis como
celulose, quitina e quitosana.
Algumas destas enzimas podem ser usadas tanto nas indústrias de detergentes,
têxteis e farmacêuticas, como também de alimentos, de bebidas, etc., sendo utilizadas
em processos biotecnológicos industriais em substituição aos processos químicos que
muitas vezes causam grandes prejuízos ao meio ambiente. A produção comercial das
enzimas pode ser realizada através do cultivo de microrganismos como fungos e
bactérias.
Na Tabela 1 mostra-se uma comparação das enzimas com os catalisadores
químicos, levando-se em conta algumas das características dos dois catalisadores.
Destaca-se que alguns parâmetros da Tabela 1 demonstram a maior importância das
enzimas, quando comparadas com os catalisadores químicos, principalmente a
especificidade ao substrato, as condições de reação, o consumo de energia e a

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
19
velocidade de reação, como desvantagem cita-se o custo de obtenção (isolamento e
purificação).
Tabela 1 – Comparação das enzimas com os catalisadores químicos (Adaptado de
Furigo Junior & Pereira, 2001).
Característica Enzimas Catalisadores Químicos
Especificidade ao substrato Alta Baixa
Natureza da estrutura Complexa Simples
Sensibilidade à T e pH Alta Baixa
Condições de reação (T, P e pH) Suaves Drástica (geralmente)
Custo de obtenção (isolamento e purificação) Alto Moderado
Natureza do processo Batelada Contínuo
Consumo de energia Baixo Alto
Formação de subprodutos Baixa Alto
Separação catalisador/produtos Difícil/cara Simples
Atividade catalítica (temperatura ambiente) Alta Baixa
Presença de cofatores Sim Não
Estabilidade do preparado Baixa Alta
Energia de Ativação Baixa Alta
Velocidade de reação Alta Baixa
2.1.1. Quitosana e seus derivados
A quitosana e seus derivados apresentam grande importância devido ao
conhecimento de suas funcionalidades biológicas, médicas, tecnológicas, etc
(Shimosaka et al, 1993; Shimosaka et al, 1995; Mitsutomi et al., 1995). A quitosana é
obtida da quitina, polissacarídeo abundante na natureza, extraído principalmente das
carapaças de caranguejos, lagosta e camarões. A quitosana é um polímero natural,
biodegradável, não tóxico e de alta massa molecular obtido a partir de animais
marinhos, insetos e microrganismos (Tabela 2).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
20
Tabela 2 – Fontes naturais de quitina e quitosana. (Adaptado de Polymar, 2003).
Animais
Marinhos
Insetos Aracnídeos Microrganismos
Anelídios Formigas Escorpiões Leveduras
Moluscos Besouros Aranhas Fungos
Lagosta Esporos
Camarão
Caranguejo
Krill
A Figura 1 abaixo mostra a estrutura química da quitosana
Figura 1 – Estrutura da quitosana (Fonte: Polymar, 2003)
Quitosana totalmente desacetilada não existe na natureza. Graus de acetilação
(GA) nferior a 50% a torna mais solúvel em soluções ácidas (Muzzarelli, 1993;
Muzzarelli et al., 1994; Dung et al., 1994; Stoyachenko et al.,1994). Pelletier &
Sygusch (1990) enfatizaram que grande parte dos resíduos sólidos da indústria
pesqueira, principalmente da carcinicultura, é jogada ao mar ocasionando degradação
ambiental. Logo, a utilização desses resíduos como matéria-prima para formação de
novos produtos, não só reduz de forma drástica o impacto ambiental, mas também daria
maior valor agregado à indústria pesqueira.
A demanda cada vez maior de consumidores por alimentos mais saudáveis, sem
conservantes químicos, tem motivado pesquisas à descoberta de novos agentes
antimicrobianos naturais. Sendo assim, a atividade antimicrobiana incomum da

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
21
quitosana e seus derivados contra microrganismos, tais como bactérias, bolores e
leveduras tem tido maior atenção ultimamente.
Devido à interação entre a quitosana, que é positivamente carregada, e as
membranas celulares dos microrganismos, negativamente carregadas, ocorre a formação
de verdadeiros poros na membrana, que são canais de vazamento de conteúdo protéico e
outros componentes intracelulares. A quitosana tem também a propriedade de agir como
agente quelante que se liga de forma seletiva a traços de metais, inibindo, assim, a
produção de toxinas e o crescimento microbiano. Ainda é capaz de ativar mecanismos
de defesa em tecidos hospedeiros, atuando como agente sequestrante de água e inibindo
várias enzimas. A ligação da quitosana com DNA e a inibição da síntese do mRNA
ocorre via penetração da quitosana no núcleo dos microrganismos e interferência na
síntese do mRNA e proteínas, (Craveiro et al., 2004).
Em um estudo clínico conduzido por Giustina & Ventura (1995) com 100
pessoas com sobrepeso de 10 a 25%, ao final de 4 semanas as pessoas que receberam
quitosana demonstraram perda de peso significativa (7,3 Kg contra 3,0 Kg nos grupos
controle). As pessoas que tomaram quitosana também demonstraram redução na pressão
sanguínea sistólica (de 145 para 135 mm Hg) e diastólica (92.6 para 84.2 mm Hg),
assim como uma redução na taxa respiratória (27.6/min a 21.2/min).
Kondo & Osada (1996) pesquisaram a influência da quitina e da quitosana na
biodisponibilidade do zinco em ratos alimentados com uma dieta com suplementos de
5% ou sem fibra dietética (quitina, quitosana, celulose, pectina e agar-agar) durante 31
dias. Observaram que os ratos alimentados com dietas contendo 5% de fibras, exceto
quitosana, o consumo de alimento foi maior do que o controle. O ganho de peso em
todos os grupos alimentados com fibra foi maior do que o controle, exceto para o grupo
suplementado com quitosana (5%) onde a ingestão de alimento e o ganho de peso foram
menores do que os grupos controle e das demais fibras.
Segundo Craveiro et al. (2004), derivados da quitosana obtidos pela reação com
ácidos carboxílicos, podem promover ao polímero uma solubilidade especial em água.
Esses compostos possuem estrutura química semelhante ao ácido hialurônico e
fornecem uma excelente compatibilidade com o tecido epitelial. As soluções obtidas
com esses derivados podem ser facilmente incorporadas a formulações cosméticas,
mostrando melhor compatibilidade com surfactantes iônicos e emulsificantes,
apresentando estabilidade em uma larga faixa de pH.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
22
Han et al. (1999) examinaram os efeitos da quitosana sobre a atividade da lipase
pancreática in vitro e sobre o grau de armazenamento de gorduras induzido em ratos
pela administração oral de uma dieta rica em gorduras por nove semanas. Como
resultado, observou-se que os animais alimentados com quitosana, reduziram o peso
corporal, a hiperlipidemia e a gordura hepática. Como conclusão, os autores propõem
que os efeitos anti-obesidade da quitosana em ratos alimentados com alto teor de
gordura se devem parcialmente a inibição da absorção intestinal das gorduras dietéticas.
Em outro ensaio, Zahorska-Markiewicz et al. (2002) realizaram um estudo para
avaliar o efeito da quitosana no tratamento da obesidade. Segundo os autores do estudo,
a quitosana pode ser usada como um valioso e seguro coadjuvante em tratamentos para
a obesidade em longo prazo. A quitosana parece acentuar a redução da pressão arterial,
juntamente com a perda de peso.
Assis (2010) estudou a produção e caracterização de quitooligossacarídeos
produzidos pelo fungo Metarhizium anisopliae e avaliação da citotoxicidade em células
tumorais. A hidrólise da quitosana foi realizada em tempos de 10 a 60 min para a
produção de quitooligossacarídeos. A avaliação da citotoxicidade dos oligômeros de
quitosana foi realizada em células tumorais (HepG2 e HeLa) e não tumoral (3T3). A
produção de oligômeros de quitosana teve maiores rendimentos durante 10 minutos de
hidrólise, os pentâmeros apresentaram concentração de 0,15 mg/mL, no entanto os
hexâmeros, que apresentam maior interesse por causa de suas propriedades biológicas,
só foram detectados com 30 minutos de hidrólise com uma concentração de 0,004
mg/mL.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o planejamento Plackett-Burmam e a metodologia de superfície de resposta.
Os resultados mostraram que a eficiência máxima de extração da quitina foi conseguida
nas seguintes condições: concentração de casca de camarão 50g/L, glicose 50g/L, tempo
de incubação de 5 dias e inoculo como 0,05 OD (densidade óptica).
A glicosamina, unidade monomérica básica da quitosana, possui atividade
antiinflamatória e tem grande importância na construção e manutenção da cartilagem,
no tratamento de enfermidades como: febre reumática, artrose e principalmente a artrite
e osteoartrite na forma aguda ou crônica (Polymar, 2003). A Figura 2 apresenta a
estrutura química da glicosamina.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
23
Figura 2 – Estrutura da glicosamina (Fonte: Polymar, 2003)
O sulfato de glicosamina é um derivado da quitosana sendo utilizado no
tratamento da osteoartrite sintomática dos joelhos, quadris, coluna, mãos e outras
regiões (Craveiro et al., 2004). O sulfato de glicosamina estimula a síntese de
proteoglicanos necessários para o bom funcionamento das articulações, estimula a
regeneração das cartilagens e ainda incorpora enxofre ao tecido cartilaginoso. Estudos
de longa duração, acima de 3 anos, mostram que o sulfatos de glicosamina evita a
progressão de danos estruturais nas articulações e induz melhora significativa nas dores
e funções físicas (Craveiro et al., 2004). A Figura 3 apresenta a estrutura química do
sulfato de glicosamina.
Figura 3 – Estrutura do sulfato de glicosamina (Fonte: Polymar, 2003)

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
24
2.1.2. Quitosanases
Dependendo do tipo de quitosanase obtida, ocorrem diferentes mecanismos de
hidrólise da quitosana, isso devido a diferentes microrganismos produzirem enzimas
com diferenças nas suas estruturas (Kurita et al., 1977; Seino et al., 1991).
Quitosanases (EC 3.2.1.132) são enzimas que catalisam a reação de hidrólise de
quitosana em oligossacarídeos. As enzimas quitosanases são produzidas por vários
microrganismos, bactérias, actinomicetos e fungos, e até por algumas plantas. Estas
enzimas agem em polímeros com grau de acetilação de 30% a 60%. Os monômeros
resultantes da hidrólise da quitosana por estas enzimas são melhores digeridos, ao serem
ingeridos, como é o caso da glicosamina. Estudos efetuados até o presente momento
classificam as quitosanases em cinco famílias: 5, 8, 46, 75, 80, sendo que a família 46 é
a mais estudada. As quitosanases tem grande aplicação na geração de oligômeros de
quitosana (Somashekar & Joseph, 1996; Chen et al., 2005).
Nogawa et al. (1998) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida pelo fungo Trichoderma reseei PC-3-7. Estes pesquisadores
inocularam 106 conídios em 100 mL de um meio basal contendo 0,3% de N-
acetilglicosamina em incubator rotativo com agitação de 220 rpm a 28oC. Destaca-se
que o fungo não produziu quitosanase em meio contendo quitosana ou quitina.
Entretanto, o microrganismo produziu quitosanase em meio contendo glicosamina e N-
acetilglicosamina, sendo que para este último obteve-se uma atividade específica
máxima de 1,5 U/mg após 72 horas de incubação. É importante ressaltar que uma
agitação vigorosa favoreceu a produção da enzima. Esse artigo ilustra a presença de
exo-β-D-glicosaminidase e exo-β-D-N-acetilglicosaminidase.
Piza et al. (1999) estudaram a produção, purificação e caracterização parcial de
uma quitosanase produzida por Bacillus cereus. Nesse caso, investigou-se através de um
planejamento fatorial fracionário a influência dos fatores: concentração de sulfato de
amônio (0,4; 2,2 e 4,0%), razão de aeração (2; 6 e 7), tempo de fermentação (16; 24 e
32 horas), pH (5; 6 e 6,5) e concentração de quitosana (1,0; 1,5 e 2,0%). Os resultados
mostraram que os fatores mais significativos para esse planejamento foram a
concentração de sulfato de amônio, a aeração, o pH e a interação entre sulfato de
amônio e a aeração.
Cheng & Li (2000) estudaram a preparação de oligossacarídeos utilizando
quitosanase produzida por Aspergillus sp e constataram que a enzima foi largamente

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
25
induzida em meio contendo quitosana como única fonte de carbono e também na
presença de quitina coloidal. A indução da enzima foi realizada em temperaturas
variando na faixa de 28 a 30ºC, pois essa espécie não cresceu bem em temperaturas
superiores a 37ºC.
Zhang et al. (2000) estudaram a produção, purificação e caracterização de
quitosanase e Exo-β-D-Glucosaminidase por Aspergillus oryzae IAM2660. Os
microrganismos utilizados neste estudo foram mantidos em placas de Petri contendo
agar para contagem em placas (PDA). Destaca-se que para produzir a enzima, conídios
foram retirados das placas e inoculados em meio Czapeq-dox com os seguintes
componentes: 1,0g K2HPO4, 0,5g KCl, 2,0g NaNO3, 0,5g MgSO4.7H2O, 0,02g
FeSO4.7H2O, 20g de sacarose e 1,0L de água deionizada, com pH do meio igual a 5,5.
Foi adicionado ao meio 0,5 g de peptona e 0,5g de extrato de levedura com
concentração de esporos da ordem de 1,0x106 esporos.mL-1. Após a incubação, feita em
incubador rotativo a 26ºC por um período de 60h, micélios foram coletados e após
filtração e secagem a 80ºC foram utilizados para ensaio de quitosanase.
Kuroiwa et al. (2002) estudaram os fatores que afetam a composição de
oligossacarídeos produzidos através da hidrólise de quitosana por quitosanase originária
de Bacillus pumilus BN-262. Neste experimento foi utilizado quitosana 100%
desacetilada dissolvida em ácido acético 1M. O pH do meio foi ajustado em 5,6. Neste
estudo a quitosanase foi imobilizada em agar e Sepharose. Os resultados mostraram que
a concentração de enzima, a temperatura, a velocidade de agitação, e concentração de
substrato influenciaram de forma significativa na composição dos oligossacarídeos
produzidos.
Ichikawa et al. (2002) estudaram a imobilização e estabilização da ligação
multiponto da quitosanase tendo agar como suporte e concluíram que a imobilização
aumentou a termoestabilidade da quitosanase. Os oligossacarídeos de quitosana foram
continuamente produzidos usando um reator coluna contendo quitosanase imobilizada.
A percentagem de quitosana imobilizada após 28 dias de reação foi de 44%.
Hung et al. (2002) estudaram a purificação, através de troca iônica, e
caracterização da hidrólise sobre ação da quitinase e quitosanase purificadas do meio
contendo Bromelina comercial. Destaca-se que a quitinase purificada apresentou pH,
temperatura e Km ótimo para ação catalítica em meio contendo quitina de 4,0, 60ºC e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
26
0,2mg.mL-1, respectivamente. Enquanto que quitosanase apresentou o pH, temperatura e
Km ótimo de 3,0, 50ºC e 0,88mg.mL-1, respectivamente.
Zhu et al. (2003) estudaram as melhores condições fermentativas e propriedades
de uma quitosanase produzida por Acinetobacter sp. C-17. A espécie de bactéria
utilizada neste ensaio foi isolada do solo e identificada como sendo Acinetobacter sp C-
17 e capaz de produzir quitosanase. O microrganismo foi mantido em placas contendo o
meio agar com os seguintes componentes: 0,5 % de quitosana, 0,2 % K2HPO4, 0,1 %
KH2PO4, 0,07 % MgSO4, 0,05 % NaCl, 0,05 % KCl, 0,01 % CaCl2, 0,05 % de extrato
de levedura e 2 % agar, o pH do meio de manutenção foi mantido em 7,2 e a
temperatura foi mantida em 30ºC durante 3 a 5 dias, para identificar a ação da enzima
quitosanase. A ação da quitosanase foi identificada através da observação de uma
colônia de microrganismo existente na placa. A bactéria foi então isolada da placa e
colocada em fermentador rotativo a 200 rpm durante 72h em meio contendo: 0,07 %
K2HPO4, 0,03 % KH2PO4, 0,05 % MgSO4, 0,03 % peptona, 0,03 % de extrato de
levedura e 1,0 % de quitosana. O pH foi de 7,0 e a temperatura 30ºC. Como fontes de
carbono foram utilizadas além de 1,0 % de quitosana, 1,0 % de quitina e 1,0 % de
glicose. Os resultados mostraram que as melhores condições para produção da
quitosanase foram alcançadas quando se utilizou 1,0 % de solução de quitosana a 30ºC,
pH 7,0 durante 24 horas de incubação.
Liu et al. (2003) estudaram a produção de quitinase por Verticillium lecanii
F091 usando fermentação submersa, a atividade de quitinase foi de 9,95 mU/mL em um
meio de cultura otimizado, com um volume de cultura de 200 mL, agitação de 150 rpm
e temperatura de 24ºC cultivado em incubador rotativo e concluíram que a taxa de
agitação e o pH foram os fatores mais significativos para a produção de quitinase, no
entanto, as taxas de agitação e aeração poderiam modificar a concentração de oxigênio
dissolvido afetando diretamente no crescimento do Verticillium lecanii e na produção
de quitinase.
Donzelli et al. (2003) estudaram a hidrólise enzimática avançada de quitina de
casca de caranguejo langostino com misturas de enzimas de bactérias e fungos e
concluiram que a combinação de enzimas de Trichoderma atroviride e Serratia
marcescens foram capazes de degradar completamente altas concentrações de quitina
(100g/mL) a partir de carapaças de caranguejo langostino convertendo a N-
Acetilglicosamina (78%), glucosamina (2%) e quitobiose (10%). Enzimas de fonte

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
27
procariótica apresentaram atividades menores que as enzimas de T. atroviride, para
catalisar a reação com quitosana.
Jo et al. (2003) estudaram a caracterização e a avaliação cinética de quitosanase
produzida por Bacillus sp. Destaca-se que na produção da enzima utilizada neste ensaio
o meio continha 0,5% de concentração de quitosana, 1% de triptona e 1% de NaCl, o
pH do meio foi ajustado em 5,5 e a temperatura foi mantida em 37ºC com agitação de
180 rpm em incubador rotativo.
Fu et al (2003) estudaram a caracterização de três iso-enzimas de quitosanase
isolada de pepsina comercial e denominado como PSC-I, PSC-II, PSC-III,
respectivamente, em relação a hidrólise enzimática da quitosana, o meio ótimo para a
produção destas iso-enzimas continha as seguintes características: pH 5,0; 5,0 e 4,0 e
temperaturas de 40ºC, 40ºC e 30ºC, o valor de Km foi de 5,2, 4,0 e 5,6 mg.mL-1,
respectivamente. As massas moleculares das três iso-enzimas foram de 40 kDa. Os
resultados mostraram que as três iso-enzimas apresentaram atividade em meio contendo
quitosana, produzindo oligômeros da ordem de 68 – 88% desacetilada.
Choi et al. (2004) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida por Bacillus sp KCTC 0377BP. Nesse caso, com relação à
produção da enzima foi possível obter um aumento de 1,2 U/ml em incubator rotativo
para 100,0 U/ml em fermentador piloto de 500 L, utilizando uma concentração de
substrato de 0,5% de quitosana e como fonte de nitrogênio polipeptona (2,5%) em pH
igual a 6,8, com 100 rpm de 100,0 rpm a 30oC.
Shimosaka et al. (2003) estudaram a produção, purificação e caracterização total
de duas quitosanases produzida por Acinetobacter sp CHB101. Nesse caso, com relação
à produção da enzima os pesquisadores usaram dois meios, um contendo glicose e outro
contendo quitosana. Com relação à cinética, o meio contendo quitosana fornece um
tempo de geração de 280 minutos, enquanto que no meio contendo glicose a taxa de
crescimento foi mais elevada e o tempo de geração foi de apenas 110 minutos. Os
autores observaram que após os dois primeiros dias de cultivo a atividade
quitosanolítica permaneceu baixa (na ordem 0,10 a 0,20 U/mL). Durante a fase de
crescimento exponencial o valor da atividade permaneceu com valor inferior a 0,01
U/mL, começando a aumentar ao atingir a fase estacionária, indicando assim que a
produção da enzima é não associada ao crescimento.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
28
Kim et al. (2004) estudaram a produção, purificação e caracterização de uma
quitosanase produzida pelo Bacillus sp e identificado pelo número 1299. Destaca-se que
na produção da enzima quitosanase foi utilizado um meio contendo os seguintes
componentes: 0,5 % de quitosana, 0,42 % (NH4)2SO4, 0,2 % K2HPO4, 0,02 % uréia,
0,03 % CaCl2.2H2O, 0,03% MgSO4.7H2O, 0,1% peptona, 0,2% Tween 80, 0,2 % da
solução (0,5% FeSO4.7H2O, 0,16% MnSO4.H2O, 0,14% ZnSO4.7H2O, 0,2% CoCl2) e
1,7% agar. O pH do meio para este ensaio foi mantido em 7,0. Este microrganismo foi
escolhido por ser grande produtor de quitosanase.
Chen et al. (2005) estudaram a purificação e a caracterização de dois tipos de
quitosanase produzidas por Aspergillus sp. CJ22-326 em cultivo submerso. Na
produção da enzima foi utilizado um meio de cultivo contendo 1,0% de quitosana, 2,0%
de gérmen de trigo, 0,2% de sulfato de amônio, 0,2 % de fosfato de potássio e 0,05% de
sulfato de magnésio por litro de solução em pH 5,6. O cultivo foi realizado em frasco de
500 mL contendo 150 mL de meio de cultura a 30ºC por 96 horas com uma agitação de
150 rpm. Os autores apresentam um gel de eletroforese com as duas enzimas purificadas
com massa molar de 29 kDa e 109 kDa.
Silva Filho et al. (2005) estudaram, através de planejamento fatorial 24 com
repetição em triplicata no ponto central, a produção de quitosanase por Aspergillus
ochraceus em cultivo descontínuo submerso. Os resultados mostraram que foi possível
produzir quitosanase com atividade na ordem de aproximadamente 5,9 U/mL utilizando
Aspergillus ochraceus e que a atividade foi favorecida pelo aumento da agitação (r), da
razão de aeração (RA) e da concentração de substrato (Cs), enquanto que o aumento da
temperatura de cultivo (TMC) não favoreceu a resposta (atividade quitosanolítica).
Pode-se classificar as quitosanases em endoquitosanases e exoquitosanases. As
primeiras diferenciam-se por catalisar a hidrólise aleatoriamente, no interior do
biopolímero, gerando diversos tamanhos de oligossacarídeos. As exoquitosanases são
capazes de hidrolisar as terminações não redutoras, produzindo assim unidades
redutoras (Peter, 2005).
Palma-Guerrero et al. (2008) estudaram o biocontrole de fungos patogênicos e
demonstraram a lise da membrana plasmática e a inibição da germinação de esporos de
fungos. Eles observaram que os fungos entomopatogênicos e nematófagos foram menos
inibidos pela quitosana que os fitopatogênicos e micoparasíticos provavelmente porque
apresentam a enzima quitosanase que degrada a quitosana. Mesmo assim, a quitosana

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
29
pode ajudar a desenvolver novas estratégias para o biocontrole em geral de fungos
patogênicos, evitando o uso de fungicidas que prejudicam o meio ambiente e a saúde
humana.
Pagnoncelli (2008) estudou a produção de quitooligossacarídeos com
propriedades nutracêuticas a partir da quitosana por hidrólise enzimática, utilizando
processo fermentativo simultâneo. Duas cepas produtoras de quitosanaes foram
selecionadas, Paenibacillus chitinolyticus e Paenibacillus ehimensis, e avaliadas quanto
ao comportamento em meio de cultivo contendo açúcares simples e em relação às
variações de pH do meio. Os quitooligossacarídeos produzidos a partir do complexo
enzimático da primeira cepa apresentaram maior pico após 9h de hidrólise, enquanto
para a segunda cepa avaliada, após 20min, observou-se quitooligossacarídeos com grau
de polimerização entre 3 e 6 unidades.
2.2. Fungos
2.2.1. Características gerais
Desde o final dos anos 60 que os fungos deixaram de pertencer ao reino das
plantas e passaram a ter um reino próprio chamado de Reino Fungi. Foi a partir de
estudos morfológicos, bioquímicos e citológicos onde se concluiu que eles são tão
diferentes dos vegetais como dos animais. Distinguem-se de outros organismos
eucariotas por serem seres quimiorganoheterotróficos e, ainda, por apresentarem parede
celular rígida que é composta por quitina e glucano e uma membrana celular em que o
ergosterol substitui o colesterol (Trabulsi & Toledo, 1996; Murray et al., 2005).
Fazem parte do Reino Fungi cerca de 100 mil espécies, desde grandes como
os cogumelos, até microscópicas como os bolores e leveduras. Ainda hoje é utilizada
essa classificação, mas os avanços da biologia molecular e estudos bioquímicos levaram
a outras propostas de classificação em que os fungos e organismos afins foram incluídos
em três diferentes reinos: Protozoa, Chromista e Fungi (Kirk et al., 2008).
Abaixo, são apresentadas características que diferenciam os fungos das plantas:
• são incapazes de sintetizar clorofila, como também não apresentar celulose em
sua parede celular, com exceções como fungos aquáticos;
• apresentam algumas características de células animais (capacidade de depositar
glicogênio e a presença de substâncias quitinosas, em sua parede celular);

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
30
• podem ser eucarióticos (as leveduras), ou multinucleados (os bolores), e
apresentar retículo endoplasmático rugoso e mitocôndrias em seu citoplasma;
• são heterotróficos, alimentam-se de matéria orgânica viva (fungos parasitários),
ou morta (fungos saprofíticos);
Os fungos são amplamente encontrados na natureza, crescem rapidamente no
solo, na água, na atmosfera, bem como nos animais, nos vegetais, resíduos orgânicos e
desempenham um papel importante na vida do homem, quer de uma maneira benéfica,
quer de um modo prejudicial. Os fungos são um dos principais microrganismos
responsáveis pela decomposição da matéria orgânica, interferindo no ciclo do carbono,
do nitrogênio e de outros nutrientes da biosfera. São capazes de deteriorarem produtos e
bens de consumo, tais como alimentos, tecidos, papel, madeira. Os alimentos
armazenados representam um vasto meio para a proliferação dos fungos, principalmente
quando não são consideradas as condições básicas de armazenamento correto (Trabulsi
& Toledo, 1996).
Estão presentes funcionalmente em vários processos industriais, tais como, de
fabricação de pão, cervejas, vinhos e determinados tipos de queijos, sendo também
utilizados na produção comercial de muitos ácidos orgânicos, de alguns fármacos, como
a ergometrina e a cortisona, na obtenção de diferentes antibióticos, como a penicilina, e
de substâncias imunossupressoras, como a ciclosporina (Esteves et al., 1990). A Figura
4 ilustra a estrutura geral de uma célula de fungo.
Figura 4 – Representação da estrutura geral de uma célula de fungo.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
31
Os fungos podem se desenvolver em meios de cultivo especiais formando
colônias de dois tipos, leveduriformes e filamentosas, que se diferenciam pela
macromorfologia e micromorfologia. O primeiro tipo apresenta, em geral, característica
de consistência cremosa de cor branca a creme, brilhantes ou opacas, podendo
apresentar às vezes coloração escura ou alaranjada, e são formados por microrganismos
unicelulares que apresentam as funções vegetativas e reprodutivas. As filamentosas
apresentam características granulares, cotonosas, aveludadas ou pulverulentas, são
multinucleadas e apresentam-se em forma de tubos denominadas de hifas. Essas hifas
podem ser não septadas (cenocíticas) ou septadas (tabicadas). O conjunto de hifas é
denominado micélio.
As hifas asseptadas têm aparência anastomosada (concisa), formada por um
citoplasma estendido e polinucleado, enquanto que as hifas septadas contêm células
individualizadas, cada uma contendo o seu núcleo. Na Figura 5 são mostradas as
estruturas das hifas septadas e não septadas.
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas. (Adaptado de Guia.Bio,
2012)
O micélio que se desenvolve no interior do substrato e que é responsável pela
sustentação e absorção de nutrientes é denominado micélio vegetativo. O micélio que
cresce na superfície do meio de cultura é denominado micélio aéreo. Este pode ser
denominado também de micélio reprodutivo, quando o mesmo se diferencia para
sustentar os corpos de frutificação ou propágulos (Putzke & Putzke, 1998). Os
propágulos classificam-se de acordo com a origem em externos e internos, sexuados e
assexuados.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
32
2.2.2. Reprodução dos Fungos
A reprodução dos fungos ocorre em ciclos assexuais, sexuais e parassexuais. Em
fungos, a reprodução assexuada é observada com mais freqüência do que a reprodução
sexuada. A reprodução assexuada é normalmente a reprodução mais importante para a
propagação da espécie, feita através de fragmentação de artroconídios, fissão de células
somáticas, brotamento ou gemulação do blastoconídios-mãe e produção de conídios. A
reprodução sexuada envolve a união de duas células ou de dois órgãos sexuais
sexualmente compatíveis. A reprodução sexuada culmina na produção de basidiósporos,
no caso de basidiomicetos. A reprodução parassexuada ocorre com a fusão de hifas e
formação de um heterocário, que consiste na existência de núcleos haplóides
geneticamente distintos num mesmo citoplasma, os quais, após a fusão, originam
núcleos diplóides heterozigotos sujeitos a sucessivas divisões mitóticas. Apesar da
raridade dessas, a reprodução parassexual é de grande importância na evolução de
alguns fungos (Pelczar et al., 1996).
A reprodução assexuada é feita por conídios, esporos assexuados formados no
ápice de hifas modificadas, os conidióforos. Os conidióforos não possuem esporângios
de forma que os conídios ficam expostos. Na reprodução sexuada, com o encontro de
duas hifas haplóides distintas, ocorre a plasmogamia e um novo conjunto de hifas
dicarióticas é formado.
Os conídios podem ter formas esféricas, fusiformes, cilíndricas, piriformes, entre
outras. Apresentam-se como hialinos ou pigmentados, podem estar isolados ou
agrupados, a parede celular pode ser lisa ou rugosa e podem ser formados por apenas
uma célula ou apresentarem septos em mais de um plano.
Os conídios também podem originar-se em qualquer parte do micélio vegetativo
e são denominados conídios sésseis como é o caso do Trichophyton rubrum.
Alguns fungos patogênicos são identificados a partir de estruturas formadas pela
célula conidiogênica e o conidióforo. Essas estruturas são denominadas: aparelho de
frutificação ou conidiação. Os conídios podem formar cadeias sobre fiálides em volta de
uma vesícula, dilatando a extremidade do conidióforo. Essas estruturas ocorrem em
Aspergillus, apresentando-se em forma de cabeça ao redor de uma vesícula, e em
Penicillium agrupados em forma de pincel, como é ilustrada na Figura 6.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
33
Figura 6 – Conídios de Aspergillus e Penicillium
2.2.3. Metabolismo de fungos
Os fungos são seres aeróbicos e heterotróficos, podendo se desenvolver em
ambientes com baixa quantidade de oxigênio, como ocorre no crescimento vegetativo e
na reprodução assexuada, como também em ambientes com grande quantidade de
oxigênio como é o caso da reprodução sexuada.
Os fungos são capazes de produzir certas enzimas como lipases, invertases,
lactases, quitosanases, proteases, amilases, etc., que fazem a hidrólise do substrato
facilitando o mecanismo de transporte ativo e passivo. Esses substratos induzem a
formação de enzimas degradativas. Algumas espécies de fungos desenvolvem-se em
meios que apresentam amônia ou nitrilos como única fonte de nitrogênio. As melhores
fontes orgânicas para o seu desenvolvimento são carboidratos, como a D-glicose, e
alguns sais minerais como sulfatos e fosfatos.
A água é uma necessidade de todos os seres vivos, não sendo diferente para os
fungos que precisam de água para o seu desenvolvimento. Certos fungos apresentam
características halofílicas podendo crescer nos ambientes com grandes concentrações de
sal.
A razão de não existirem ambientes livres de espécies fúngicas deve-se ao facto
destes microrganismos conseguirem sobreviver em situações extremas como, por
exemplo, temperaturas que variam entre os - 5 e os 60 ºC, com pH de 1 a 9 e com baixas

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
34
concentrações de oxigênio, apesar de cada espécie possuir condições ideais para o seu
desenvolvimento (Chao et al., 2002).
Devido a grande faixa de temperatura ótima para o seu crescimento, os fungos
apresentam espécies psicrófilas, mesófilas e termófilas. Para a maior parte das espécies
fúngicas, a temperatura de desenvolvimento situa-se entre os 20 e 24 ºC e não crescem
acima dos 29 ºC. Em relação à umidade relativa, necessitam de pelo menos 50 %, mas
geralmente necessitam de mais de 65 % (Colakoglu, 2001).
O pH ideal para o desenvolvimento dos fungos varia na faixa entre 5 e 7.
Destaca-se, entretanto, que a maioria dos fungos suportam grandes variações de pH
como ocorre com os fungos filamentosos que podem crescer em pH variando entre 1,5 e
11. Por outro lado, as leveduras não suportam pH alcalino.
Os fungos crescem lentamente, se comparados com as bactérias, em meio de
incubação para o seu desenvolvimento. Para evitar contaminação por bactérias, que
podem se sobrepor ou inibir o seu crescimento, pode-se introduzir no meio de cultura
um antibacteriano de largo espectro como o cloranfenicol.
Algumas espécies de fungos se desenvolvem na presença de luz, outras são
inibidas e algumas podem crescer tanto na presença quanto na ausência deste agente. No
entanto a irradiação solar, em geral, é prejudicial ao seu desenvolvimento, por causa da
radiação ultravioleta.
2.2.4. Nutrição de fungos
Quanto ao aspecto da nutrição, os fungos são heterótrofos, ou seja, incapazes de
produzir seu próprio alimento. Entretanto, a forma de assimilar matéria orgânica é
diferente daquela utilizada pela maioria dos animais, que são heterótrofos por ingestão.
Nos fungos, a digestão é extracorpórea, ou seja, é realizada fora do corpo. O fungo
lança no ambiente, enzimas que degradam as moléculas orgânicas complexas e, depois,
absorve moléculas menores, mais simples. Por essa razão, são designados heterótrofos
por absorção. É devido a este processo que os fungos podem crescer dentro ou sobre os
alimentos. Desenvolve-se geralmente em meios contendo um pH baixo, uma fonte de
carbono, uma fonte de nitrogênio orgânico ou inorgânico e alguns minerais. Alguns
necessitam de vitaminas. Quanto aos modos de vida, os fungos podem ser
decompositores, parasitas, mutualísticos e predadores (Raven, 2001).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
35
Os fungos decompositores, saprófagos (do grego, saprós, “podre”, e phagos,
“comedor”), juntamente com as bactérias, são os principais decompositores da biosfera,
participando intensamente do processo de degradação da matéria orgânica morta, o que
promove a reciclagem dos elementos químicos constituintes dos seres vivos. A maioria
deles vive no solo, obtendo nutrientes de seres mortos. Em florestas, onde a matéria
orgânica vegetal é abundante, os micélios dos fungos decompositores absorvem os
nutrientes de folhas e galhos caídos, decompondo-os. Os fungos decompositores
também são responsáveis pelo apodrecimento de alimentos e de outros materiais, como
a madeira.
Os fungos parasitas vivem à custa de outros organismos vivos, prejudicando-os.
Ao parasitar o corpo de um ser vivo, animal ou vegetal, o fungo pode até provocar sua
morte. Várias doenças são causadas por fungos parasitas. Nas plantas, cita-se a
ferrugem do café, que, no Brasil, é motivo de grande preocupação por parte dos
cafeicultores, devido aos prejuízos que acarreta. As micoses que atacam a pele de seres
humanos e animais são provocadas por fungos.
Existem fungos que se associam a outros organismos, estabelecendo uma relação
em que há benefício mútuo para os indivíduos envolvidos. Esses fungos são
denominados mutualísticos. A maioria vive associada a seres fotossintetizantes, como
as plantas, cedendo-lhes parte da água e dos nutrientes que as hifas absorvem do solo.
As plantas, por sua vez, cedem ao fungo certos açúcares e aminoácidos. Duas
associações mutualísticas formam estruturas bem características: as micorrizas,
associação que envolve fungos e raízes de plantas, e os líquens, formados por fungos e
algumas variedades de algas e cianobactérias.
Certos fungos atuam como predadores. Na maioria dos casos, as hifas secretam
substâncias aderentes que aprisionam os organismos que tocam os fungos. Dessa
maneira, as hifas penetram o corpo da presa, crescem e se ramificam, espalhando-se no
interior do organismo e absorvendo seus nutrientes, causando-lhe a morte. O gênero
Arthrobotrys apresenta uma forma mais elaborada de predação. O fungo vive no solo e
captura nematódeos microscópicos que habitam o mesmo ambiente. As hifas
apresentam pequenos anéis que se estreitam quando a presa passa por eles. As hifas do
fungo logo invadem o corpo do nematódeo, digerindo-o.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
36
2.2.5. O fungo Metarhizium anisopliae
Existem três espécies de fungos do gênero Metarhizium, divididas em dez
variedades: M. anisopliae, variedades anisopliae, majus, lepidiotum e acridum; M.
flavoviride, variedades tipo E, flavoviride, minus, novazealandicum e pemphigum; e M.
album (Driver et al. 2000). E, mais recentemente, foi descrita na China uma nova
variedade de M. anisopliae, dcjhyium (Dong et al. 2009).
Data de 1879 quando o russo Ilya Metchnikoff aplicou o fungo para combater
larvas de um curculionídeo, praga de beterraba, e foi classificado como Metarhizium
anisopliae por Sorokin em 1883. É um fungicida natural para mais de 300 espécies de
insetos, especialmente pragas da agricultura e pecuária. No Brasil, o fungo Metarhizium
anisopliae teve a sua aplicação com sucesso pela primeira vez em 1965, no controle da
cigarrinha-da-cana Mahanarva posticata (Alves, 1998).
O fungo Metarhizium anisopliae é um importante agente entomopatogênico
utilizado no controle de pragas, como cupins, gafanhotos, cigarrinhas e besouros, sendo
amplamente estudado em todo o mundo. Existe amplamente na natureza e é facilmente
encontrado no solo, onde sobrevive por longos períodos e seu desenvolvimento se dá
geralmente entre 15 e 32 °C, sendo ideal entre 24 e 30 °C e com pH ótimo igual a 6,9.
Apresenta micélio hialino e septado, com conidióforos característicos, sobre os quais
surgem conídios cilíndricos organizados em colunas. Este fungo ao atacar os insetos
provoca rigidez e os recobre por uma camada pulverulenta de conídios, resultando em
colorações que variam do verde claro ao escuro, cinza ou branco com pontos verdes
(Tinline, 1971; Alves, 1998, Driver et al. 2000; Arruda, 2005).
Quanto a sua morfologia, apresenta-se como um fungo filamentoso, com corpo
de frutificação equivalente a um esporodóquio agregado a hifas intimamente
entrelaçadas, com massa compactada de conidióforos característicos, simples ou
ramificados, formando células esporogênicas denominadas fiálides, resultando nos
fialosporos (Wang et al., 2002).
O Metarhizium anisopliae é um fungo de cultivo simples, sendo necessário
quase que exclusivamente o nutriente a base de amido. Um dos fungos mais estudados
em programas de manejo de pragas já que apresenta grande potencialidade
entomopatogênica (Onofre et al., 2002). O Metarhizium anisopliae é um microrganismo
pouco exigente, pois pode se desenvolver em diversos meios de cultura como: amido,
glicose, glicerina, levulose, maltose, sacarose e quitina (Jabor et al., 2003).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
37
Em geral, os fungos produzem enzimas que convertem os tecidos dos insetos em
nutrientes. Porém, a cutícula esclerotizada dos insetos não é bem utilizada pelos fungos,
excetos pelos entomopatogênicos que desenvolveram enzimas eficientes para degradar
essa camada protetora. Destaca-se a enzima Pr1, produzida por Metarhizium anisopliae,
que tem um poder de degradação bem superior a qualquer outra enzima sintetizada por
esse grupo de fungos (Dias, 2005). Desta forma, o Metarhizium anisopliae (fungo
entomopatogênico) ao atacar a célula dos insetos, produz grande quantidade de enzimas
que degradam a membrana citoplasmática e inativa a célula e, consequentemente, mata
o inseto, logo é evidente a relação entre o controle de pragas e produção de enzimas.
No momento, sobre a comercialização de fungos entomopatogênicos no Brasil,
em sua maioria, são formulados em grânulos, compostas do fungo mais o substrato
(arroz + fungo), como também na forma de pó molhável (WP), resultado da moagem do
fungo com o substrato. Esses produtos representam 90% e 8,5%, respectivamente, da
produção de fungos no Brasil, e sendo suas concentrações finais ficam em torno de
5x108 conídios/g para Metarhizium anisopliae e 1x109 conídios/g para Beauveria
bassiana (Alves et al., 2008).
Segundo Batista Filho et al. (2002), Garcia et al. (2005) e Mello et al. (2006), o
Metarhizium anisopliae tem sido bastante utilizado para o controle de diferentes pragas
no Brasil, como a broca da bananeira, pragas de grãos armazenados, carrapatos, cupim
de montículo em pastagens, larvas de escarabeídeos que atacam a cana de açúcar e
cigarrinha da cana-de-açúcar.
É comum a ocorrência natural do fungo Metarhizium anisopliae atacando
cigarrinha-das-raízes de canaviais pelo país, o que incentiva a utilização dele como
agente de controle biológico em áreas infestadas por esse tipo de praga. A aplicação de
recursos no uso de programa de manejo integrado da cigarrinha tem sido reforçado por
razões ambientais e econômicas (Dinardo-Miranda, 2001).
Faria & Magalhães (2001) fizeram uma avaliação do uso de fungos
entomopatogênicos no Brasil, na qual observaram que a qualidade dos micoinseticidas
disponíveis no Brasil poderia ser incrementada de forma considerável quanto a sua
formulação, ou seja, antes da venda, poderiam realizar melhorias no tratamento
posterior, adição de substâncias que lhes assegurem melhorias na eficiência de controle,
capacidade de armazenamento ou praticidade de manuseio, ou de qualquer outro critério
que resulte em vantagem em relação ao produto bruto. Ressaltaram a pouca praticidade

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
38
em alguns casos, como exigência de lavagem, e em outros podendo causar o
entupimento de bicos dos pulverizadores devido à elevada proporção de inertes,
principalmente quando são empregados baixos volumes de aplicação. Outro problema
evidenciado com os inseticidas nacionais diz respeito à pequena sobrevida, devendo a
ser usados em, no máximo, 30 dias depois de produzidos, quando armazenados à
temperatura ambiente e em local sombreado, fazendo as vendas ocorrerem quase que,
exclusivamente, sob encomenda. Comparativamente, os bons inseticidas sob
comercialização em outros países apresentam maior concentração de ingrediente ativo,
maior sobrevida (alguns produtos podem ser armazenados por mais de 8 meses à
temperatura ambiente) e praticidade (são formulados na forma de GDA (grânulos
dispersíveis em água) ou de óleos emulsionáveis, por exemplo, podendo ser adicionados
diretamente ao tanque do pulverizador.
Rangel et al. (2007), instalaram uma Unidade de Observação para o controle
microbiano das cigarrinhas das pastagens Distrito de Itahum, em Dourados/MS, devido
as necessidades de agricultores familiares de produção de leite. As aplicações do fungo
Metarhizium anisopliae foram realizadas por pulverização com equipamento de tração
animal. As condições do ambiente de estudo foram favoráveis ao desenvolvimento da
população das cigarrinhas e à incidência do fungo. Como resultados, observaram que os
níveis de espumas, ninfas e adultos reduziram significativamente após as aplicações,
levando os Agricultores Familiares a total satisfação com os ganhos na produção de
todos os envolvidos.
Segundo Vieira et al. (2007), o fungo entomopatogênico Metarhizium anisopliae
var. acridum tem sido utilizado no controle de pragas, e assim, visando este fim,
analisaram em laboratório a esporulação, a germinação dos conídios e as características
morfológicas peculiares desse fungo em diferentes temperaturas. Esperou-se o
desenvolvimento das colônias em períodos de 1 a 15 dias. Foi observado que
temperaturas de 25, 28 e 30 ºC proporcionaram maiores taxas de esporulação e elevadas
porcentagens de germinação dos conídios. A 25, 28 e 30 ºC observou-se um
desenvolvimento mais rápido das microestruturas somáticas e reprodutivas (hifas,
anastomoses, apressórios, conidióforos e conídios elipsóides). Nas temperaturas
extremas de 40 e 45 ºC não foi detectado esporulação, germinação, nem
desenvolvimento de nenhuma microestrutura.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
39
Lopes et al. (2007) avaliaram o efeito de formulações de Beauveria bassiana e
Metarhizium anisopliae sobre ninfas de Amblyomma cajennense (transmissor da febre
maculosa ao homem) em laboratório. Ninfas não alimentadas foram pulverizadas com
suspensões contendo 2x107 e 4x107 conídios/mL de B. bassiana, e 1x107, 1,5x107,
2x107, 3x107 e 5x107 conídios/mL de M. anisopliae. A mortalidade de ninfas,
decorridos três dias da aplicação, foi significativa nas concentrações de 3x107 e 5x107
conídios/mL de M. anisopliae, com 57,1 e 56,2 %, respectivamente. Após seis dias a
mortalidade foi de 100 % nessas duas concentrações. A mortalidade foi
significativamente maior que a observada para B. bassiana em concentrações
semelhantes.
Guirado et al. (2009) realizaram estudos para controlar cupins de montículo em
área de pastagem, utilizando fungos entomopatogênicos Metarhizium anisopliae e
Beauveria bassiana em diferentes doses e formulações. A avaliação foi realizada após
30 dias da aplicação dos produtos. Os melhores resultados em ordem decrescente de
eficiência foram: M. anisopliae 30 mL + 1000 mL de água com 100 % de eficiência; B.
bassiana 10g + M. anisopliae 20 mL + 1000 mL de água com 85,71 % de eficiência; M.
anisopliae 20 mL + 1000 mL de água com 71,42 % de eficiência. Os demais
tratamentos não diferiram da testemunha.
Loureiro et al. (2012) compararam a eficiência de isolados de Metarhizium
anisopliae em controlar populações naturais de Mahanarva fimbriolata (cigarrinha-da-
raiz) em cultivos comerciais de cana-de-açúcar colhida mecanicamente. Foram
aplicados os isolados IBCB 348, IBCB 408, IBCB 410 e IBCB 425 do fungo em
concentração de 1,5×1012 conídios/ha. Aplicou-se o planejamento experimental em
blocos casualizados com 5 tratamentos e 4 repetições, sendo cada repetição composta
por 7 linhas de 100 m de comprimento e espaçamento de 1,5 m. Após 30 dias de
pulverização foi observado para os isolados IBCB 408 e IBCB 425 uma eficiência de
controle de ninfas de 63 e 62%, respectivamente, já no caso dos adultos chegou a 100%.
Após 60 dias o isolado IBCB 425 foi o mais eficiente para controlar as ninfas com
48,4%.
Assis et al. (2012) estudaram a citotoxidade de quitoolissacarídeos (QOS) e
oligômeros A, B e C (soluções compostas por diferentes quantidades de QOS)
produzidos por hidrólise enzimática pelo fungo Metarhrizium anisopliae. Foi analisado
o efeito de antiproliferativo destas moléculas em células tumorais (HepG2 e HeLa) e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
40
também em célula normal (3T3). A atividade antioxidante foi analisada em vários
experimentos in vitro. A glicosamina apresentou maior toxicidade (cerca de 92%) para
todas as células estudadas. No entanto, os oligômeros obtidos após hidrólise não
demonstraram efeito tóxico sobre a célula normal (3T3). Além disso, mostraram que
uma pequena quantidade de outros COS pode diminuir o efeito citotóxico da
glicosamina contra células 3T3, indicando que glicosamina poderia ser usada como uma
droga antitumoral na presença de outros COS.
2.3. Fermentação
Os processos fermentativos são caracterizados por processos nos quais
microrganismos realizam a conversão catalítica de uma dada substância num
determinado produto. Estes processos podem ser classificados como processos
descontínuos, contínuos e descontínuos alimentados. A classificação destes processos
está diretamente ligada à forma de como o substrato é adicionado e como o produto é
retirado. No processo de fermentação descontínuo o substrato é colocado no
fermentador, em seguida é inoculado e após o término do processo o produto é retirado.
No processo contínuo, o substrato é colocado num reator com vazão constante e o meio
fermentado é retirado com a mesma vazão de entrada do reator (Faccioti, 2001). Nos
processo descontínuos alimentados à adição de substrato e a retirada do mesmo deve
ocorrer de forma intermitente (Carvalho & Sato, 2001).
De certa forma, os processos fermentativos representam um elo entre as antigas
artes de elaboração de alimentos, como queijos e vinho, a partir de uma flora
microbiana natural, com a moderna indústria de fermentação de alimentos, onde são
utilisados cultivos puros e equipamentos sofisticados para o controle do processo
(Assis, 2010).
Atualmente, a maioria dos processos industriais se desenvolve em cubas de
vidro ou de aço de até 20 litros. Os agentes biológicos se encontram submersos no meio de
cultivo que ocupa, aproximadamente, 75% da cuba. As vezes é necessário injetar ar, e em muitas
fermentações se forma espuma. O desenho do biorreator deve se adequar ao objetivo do
processo, respondendo eventualmente a diversos imperativos, tais como a esterilização do
sistema, a aeração e homogeneização do meio, o acréscimo de nutrientes e de aditivos
antiespuma, a manutenção do pH, etc. Os modelos de fermentadores mais utilizados com
microrganismos contam com aeração e agitação mecânica. Esta facilita a distribuição dos

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
41
nutrientes na cuba, mas gera calor que deve ser eliminado mediante a circulação de água
fria. Em outros tipos de biorreatores, em coluna ou torre, a homogeneização depende da injeção de
ar. Os sistemas submersos são apropriados para o cultivo de microrganismos livres, mas resultam
pouco econômicos quando se trabalha com células ou enzimas caras. A imobilização em
fermentadores menores, seja por adesão a um suporte inerte, seja por inclusão dentro de um
polímero que permita o contato com o meio de cultura, além de simplificar a purificação do
produto permite a reutilização das células ou das enzimas, que permanecem dentro do
biorreator (Malajovich, 2009).
As enzimas são produtos de alto valor agregado, amplamente empregadas em
uma gama de indústrias, e entre as fontes para sua obtenção, destacam-se as de origem
microbiana, principalmente, devido a elevada seletividades dos substratos utilizados e
produtos formados. A principal forma de obtenção de enzimas, em escala industrial, é a
fermentação submersa, por esta apresentar vantagens em relação a fermentação
semisólida, como: maior controle do processo fermentativo e consequente reprodução
da cinética microbiana, além de maiores rendimentos (Wanderley, 2011).
2.3.1. Preparo do inóculo
Denomina-se inóculo um volume de suspensão de microrganismo de
concentração adequada, capaz de garantir, em condições econômicas, a fermentação de
uma dada quantidade de meio. O volume do inóculo, assim como sua concentração de
células, vai depender tanto do processo de fermentação c omo da quantidade de meio a
ser fermentado (Carvalho & Sato, 2001).
A partir de uma cultura pura, inocula-se um volume relativamente pequeno de
meio nutriente, que vai depender do tipo de microrganismo cultivado (pois cada espécie
microbiana possui velocidades diferentes de crescimento), sendo, em seguida, incubado
em condições favoráveis ao desenvolvimento do microrganismo. A suspensão obtida é
transferida para um frasco maior em agitador rotativo ou recíproco (shakers), que por
sua vez é adequadamente incubado, e a seguir, transferido para um frasco contendo um
volume ainda maior deste meio, até se conseguir alguns litros de suspensão microbiana.
Os cuidados com assepsia, volumes de meio utilizado, numero de transferência bem
como as condições de incubação, podem variar de acordo com o tipo de processo de
fermentação que se deseja realizar.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
42
2.3.2. Produção de enzimas em cultivo submerso descontínuo
A produção de enzimas, usando processos fermentativos, é constituída
basicamente por duas etapas: a primeira caracteriza-se pela preparação do inóculo e a
segunda, pela fermentação do substrato (Carvalho & Sato, 2001). A finalidade da
primeira etapa é o preparo do microrganismo em condições adequadas para que possa
ocorrer o desenvolvimento da segunda etapa. Diferentemente do cultivo em estado
sólido, no cultivo submerso os microrganismos encontram-se em um meio com elevada
atividade de água.
2.4. Planejamento experimental
Um bom planejamento tem como fundamental projetar um experimento de
forma que ele seja capaz de fornecer exatamente o tipo de informação que procuramos.
Para isso é necessário saber, em primeiro lugar, o que é mesmo que se deseja procurar.
Planejamento experimental, ou delineamento experimental, é um conjunto de
ensaios estabelecido com critérios científicos e estatísticos, objetivando determinar a
influência de diversas variáveis nos resultados de um dado sistema ou processo (Barros
Neto et al., 2001; Rodrigues & Iemma, 2005). De acordo com o propósito dos ensaios,
esse objetivo maior pode ser mais detalhado em:
– determinar quais variáveis são mais influentes nos resultados;
– atribuir valores às variáveis influentes de modo a otimizar os resultados;
– atribuir valores às variáveis influentes de modo a minimizar a variabilidade dos
resultados;
– atribuir valores às variáveis influentes de modo a minimizar a influência de variáveis
incontroláveis.
O planejamento experimental, baseado nos fundamentos estatísticos é, sem
dúvida alguma, uma ferramenta poderosa para se chegar às condições otimizadas de um
processo, desenvolvimento da formulação de produtos dentro das especificações
desejadas ou simplesmente para avaliar os efeitos ou impactos que os fatores (variáveis
independentes) têm nas respostas (variáveis dependentes) desejadas (Barros Neto et al.,
2001; Rodrigues & Iemma, 2005). Da utilização das técnicas estatísticas de
planejamento experimental, é possível chegar benefícios importantes, como:

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
43
– reduzir o número de ensaios sem prejuízo da qualidade da informação;
– estudar simultaneamente diversas variáveis, separando seus efeitos;
– determinar a confiabilidade dos resultados;
– realizar a pesquisa em etapas, num processo iterativo de acréscimo de novos ensaios;
– selecionar as variáveis que influem num processo com número reduzido de ensaios;
– representar o processo estudado através de expressões matemáticas;
– elaborar conclusões a partir de resultados qualitativos.
2.4.1. Planejamento fatorial
Em um planejamento fatorial é investigado as combinações de níveis de estudo
de cada fator. Para tanto, inicialmente, é necessário especificar esses níveis em cada
fator, quer dizer, os valores dos fatores que serão utilizados nos experimentos. Um
ensaio experimental é definido como um experimento constituído de um conjunto de
níveis definido. Geralmente, quando se tem n1 níveis de um determinado fator 1, n2 do
fator 2, ..., e nk do fator k, o planejamento será descrito por um fatorial n1 x n2 x ... x nk,
que corresponde ao número mínimo de experimentos para se ter um planejamento
fatorial completo.
É necessário fazer cada fator variar e observar o resultado dessa variação para se
estudar o efeito sobre a resposta. Assim, os ensaios precisam ter um mínimo de dois
níveis por fator para a realização desses ensaios. Para k fatores, ou seja, k variáveis de
controle, o planejamento de dois níveis precisará de 2 x 2 x ... x 2 = 2k, para o número
de ensaios diferentes, sendo conhecido como planejamento fatorial 2k.
Para o planejamento fatorial, são especificados os valores dos fatores em cada
experimento, e estes são classificados em níveis, variando-se, por exemplo, de -1
(mínimo) a +1 (máximo). A partir de uma matriz serão apresentadas todas as
combinações entre o número de fatores e de níveis nos experimentos a serem realizados
(Barros Neto et al., 2001; Rodrigues & Iemma, 2005).
2.4.2. Planejamento Plackett & Burman
A maioria dos planejamentos do tipo “Screening” (Delineamentos de Seleção de
fatores) foi descrito originalmente por Plackett & Burman em 1946. Os planejamentos
experimentais de Plackett & Burman são eficientes em situações que envolvem uma

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
44
grande quantidade de variáveis a serem exploradas em um processo de otimização de
fatores (Reddy et al., 2008).
As matrizes Plackett & Burman consistem de planejamentos fatoriais de dois
níveis para estudar k = N – 1 variáveis em N ensaios. São construídas para ensaios
múltiplos de 4, sendo essas matrizes para ensaios de 8, 12, 16,..., 100. Cada variável
independente em uma matriz assume uma configuração de dois níveis, o inferior (-1) e o
superior (+1), para o nível intermediário é estabelecido (0). Para cada vez que a variável
aparece no nível inferior, terá o par equivalente no nível superior (Rodrigues & Iemma,
2005).
Para evitar dúvidas nos resultados dos efeitos de algum fator no planejamento de
Plackett & Burman, o número de ensaios a ser realizado deve ser no mínimo o número
de variáveis mais 4. No caso de um fatorial completo o número de ensaios a ser
realizado é bem maior (Rodrigues & Iemma, 2005).
Mesmo já determinado o planejamento Plackett & Burman, torna-se importante
à realização de experimentos relativos aos pontos centrais de modo a se avaliar melhor
as possíveis interferências analíticas que podem ocorrer durante os diferentes
experimentos. A repetição do ponto central permite também a determinação do erro
puro e da repetibilidade do processo. Na prática, esta é uma ferramenta que permite
identificar melhor se o processo em estudo está sob controle (Rodrigues & Iemma,
2005).
É possível que no planejamento Plackett & Burman não tenha sido considerado
a codificação do ponto central (0) para repetição, pois, matematicamente, a estimativa
do valor desse efeito codificado (zero) vezes, teria como resposta zero. Porém, no
resultado prático do experimento, consegue-se informações valiosas com essas
repetições no ponto central (Rodrigues & Iemma, 2005).
2.4.3. Aplicações do Planejamento Plackett & Burman
Ghanem et al. (2000) produziram xilanase por Aspergillus terreus cultivadas
sobre palha de trigo finamente moída em fermentação de estado sólido. A composição
ideal do meio foi estabelecida aplicando-se o planejamento experimental de Plackett &
Burman. A melhor atividade enzimática foi obtida em um meio contendo 10 g/L de

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
45
palha de trigo por frasco umedecido com uma solução concentrada de sal com teor de
água inicial de 75 % e incubadas durante 4 dias a 30 °C. Observou-se que íons
metálicos como Hg2+, Cu2+, Co2+, Fe3+ e Pb2+ inibiram fortemente a xilanase, enquanto
que Ca2+ ativou a enzima.
Mu et al. (2005) estudaram as otimização de meio de fermentação para a reação
de estereoinversão por Candida parapsilosis CCTCC M203011 utilizando o
planejamento Plackett & Burman. Os resultados mostraram que dentre os fatores
estudados, os fatores mais significativos foram: a Concentração de MgSO4, ZnSO4 e
KH2PO4 e que após a utilização da metodologia de superfície de resposta foi constatado
que o ponto ótimo para a otimização foi conseguido com as concentrações dos fatores
MgSO4, ZnSO4 e KH2PO4 nos valores em: 1,17 g/L, 0,10 g/L e 9,41 g/L,
respectivamente, elevando a taxa de conversão enantiosseletiva de 75,2 % para 96 %.
Naveena et al. (2005) empregaram o planejamento Plackett & Burman para a
seleção de 15 parâmetros para produção de ácido láctico L(+) de farelo de trigo,
substrato de baixo custo e suporte sólido, por Lactobacillus amylophilus GV6 em
fermentação de estado sólido (SSF). Onze nutrientes pertencentes a duas categorias,
fontes de nitrogênio e fontes de sal, junto com três parâmetros físicos e um tampão
foram os parâmetros considerados. Os autores verificaram influência na produtividade
devido às fontes de nitrogênio, peptona, extrato de levedura e tri-citrato de amônio,
juntamente com o NaH2PO4.2H2O e Tween 80, que podem ainda serem otimizados para
o aumento na produção de ácido lático. Esses pesquisadores ainda chamam atenção para
o uso escasso deste planejamento em fermentação em estado sólido e que o mesmo
ainda não havia sido considerado anteriormente para a conversão direta de amido em
ácido láctico L (+) usando um sistema bacteriano.
Khosravi-Darani et al. (2008) estudaram a aplicação de Plackett & Burman na
análise dos fatores significativos para a produção de ácido cítrico por Aspergillus niger
ATCC 9142 utilizando palha de trigo como substrato. Os resultados mostraram que a
produção, com a palha de trigo pré-tratada com HCl, NaOH e Uréia, foi mais eficiente e
que os fatores mais significativos na produção foram: Solvente, Teor de Umidade,
Temperatura, Tempo de Incubação, Concentração Inicial de Açúcar e Concentração de
Metanol.
Ji et al. (2009) estudaram o desenvolvimento de um meio de cultura econômico
para a produção do 2,3-butanodiol através da fermentação da glicose e xilose por

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
46
Klebsiella oxytoca utilizando o planejamento Plackett & Burman. Os resultados
mostraram que a uréia e a maceração de milho foram os fatores mais significativos para
a produção do 2,3-butanodiol e que através de planejamento experimental com repetição
em triplicata no ponto central para determinar os níveis ótimos de produção. Os autores
constaram que o rendimento atingido foi 85,6% do valor teórico.
Paula & Arruda. (2009) observaram a necessidade da busca por alternativas mais
eficientes ao método termomecânico utilizado para a degradação de milhões de
toneladas de penas de aves provenientes de frigoríficos e aviários, cuja rígida estrutura é
composta por mais de 90 % de queratina. Para esse fim, utilizaram de comunidades
microbianas para a finalidade proposta, testando o planejamento experimental Plackett
& Burman para verificar a influência de diferentes microrganismos simultaneamente,
com um número reduzido de experimentos. Testaram 32 bactérias isoladas, inoculadas
em meio contendo somente uma pena como única fonte de carbono, durante sete dias,
de onde foi selecionado as cepas para os consórcios microbianos. Experimentos unindo
as cepas NP5, FCA7, NP4 e S14 demonstraram diferentes atividades queratinolíticas,
aumentando o nível de degradação, até 87,8%, comparada com as cepas
individualmente, variando entre 26,4% e 78,0%.
Rajendran & Thangavelu (2009) obtiveram valores máximos da atividade de
produção de lipase em 3,98 U/mL e concentração de massa celular em 5,62 g.L-1 com R.
arrhizus em cultivo submerso descontínuo otimizado pelo planejamento Plackett &
Burman. As variáveis mais importantes que afetaram a produção da lipase foram azeite
de oliva, peptona, KH2PO4, CaCl2.2H2O e MgSO4.7H2O.
Li et al. (2010) estudaram a otimização de meio de cultura para a produção de
Streptolydigin (um antibiótico) por Streptomyces lydicus AS 4,2501 utilizando Plackett
& Burman e a metodologia de superfície de resposta. Os resultados mostraram que
dentre os fatores estudados, os fatores mais significativos para a produção de
Streptolydigin e otimização do meio de cultura foram: Concentração de Amido,
Concentração de Peptona e a Concentração de K2HPO4.
Chen et al. (2010) estudaram a otimização de meio de cultura para a produção da
Luciferase – que são enzimas que catalisam reações biológicas transformando energia
química em energia luminosa - por Bacillus Subtilis em fermentação descontínua
utilizando planejamento Plackett & Burmam. Os resultados mostraram que dentre os
componentes estudados, os fatores mais significativos para a produção e otimização do

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
47
meio de cultura para a produção da Luciferase foram: Concentração de Glicose 1,17 %,
Extrato de Levedura 2,27 % e Glutamato 0,55 %.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o projeto Plackett & Burman e a metodologia de superfície de resposta. Os
resultados mostraram que a eficiência máxima de extração da quitina foi conseguida nas
seguintes condições: Concentração de casca de camarão (50 g/L), glicose (50 g/L),
tempo de incubação de 5 dias e inoculo com 0,05 de OD (densidade óptica).
Gao et al. (2010) desenvolveram um meio para a fermentação do extrato bruto
de inulina de tubérculos de alcachofra de Jerusalém por Paenibacillus polymyxa ZJ-9
para produzir R,R-2,3-butanodiol (R-R,2,3-BD). Inulina, K2HPO4 e NH4Cl foram os
parâmetros principais na fermentação de acordo com os resultados obtidos com o
planejamento experimental Plackett & Burman. A concentração ideal dos três fatores,
77,14 g/L, 3,09 g/L e 0,93 g/L, respectivamente, foi obtida pelo planejamento Box-
Behnken e pela superfície de resposta. Sob as condições ideais, a concentração do R-
R,2,3-BD obtida foi 36,92 g/L, em mais de 98 % de pureza óptica. Comparado com
outras fontes de carbono investigadas, a fermentação do extrato bruto de inulina obteve
o maior rendimento de R-R,2,3-BD.
Uma metodologia de superfície de resposta foi empregada com sucesso para
otimizar um meio de fermentação chamada VHG (Very High Gravity), referência à alta
concentração de açúcares e densidade dos mostos, baseado na água maceração de milho
CSL (Corn Steep Liquor) e outros nutrientes de baixo custo para a produção eficiente de
etanol a partir de glicose por S. cerevisiae. Usando a composição média otimizada (g/L:
CSL 44,3; uréia 2,3; MgSO4.7H2O 3,8; CuSO4.5H2O 0,03), a levedura industrial PE-2
foi capaz de fermentar até 330 g/L de glicose e produzir 18,6 % (v/v) de etanol, com
uma produção descontínua de 2,4 g/L/h e um rendimento em etanol de 93 % (Pereira et
al., 2010).
O planejamento Plackett & Burman juntamente com o Planejamento Composto
Central (CCD) foram aplicados para otimizar o meio de cultura para produção de etanol
por Clostridium autoethanogenum com monóxido de carbono (CO) como fonte única de
carbono e um meio contendo (g/L): 1,0 de NaCl, 0,1 de KH2PO4, 0,02 de CaCl2, 0,15 de
extrato de levedura, 0,116 de MgSO4, 1,694 de NH4Cl e pH 4,74. O rendimento ótimo
de etanol previsto pela metodologia de superfície de resposta (RSM) e um algoritmo

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
48
genético de rede neural artificial (ANN-GA) foram 247,48 e 261,48 mg/L,
respectivamente. Valores semelhantes aos obtidos experimentalmente sob as condições
ideais sugeridas pelos métodos estatísticos (254,26 e 259,64 mg/L). A adequação do
modelo ANN-GA foi maior do que o modelo RSM. Os rendimentos obtidos foram
substancialmente superiores aos relatados anteriormente (60-70 mg/L) com essa cultura
(Guo et al., 2010).
Rigueira (2010) avaliou a influência das variáveis do procedimento de extração
na porcentagem de recuperação das aminas encontradas no leite cru utilizando um
planejamento para “screening” do tipo Plackett & Burman, para os ácidos
tricloroacético (TCA) e sulfosalicílico (ASS).
Quines (2010) estudou a necessidade ou não de se fazer um pré-tratamento da
biomassa antes da extração. Um planejamento experimental Plackett & Burman de doze
ensaios mais três pontos centrais foi utilizado para selecionar as variáveis mais
significativas na extração de Poli(3-hidroxibutirato) (P(3HB)) por 1,2-carbonato de
propileno. Foram realizados dois testes de recuperação do solvente 1,2-carbonato de
propileno, após a extração, sendo um com aplicação de aquecimento e o outro com
aquecimento e vácuo. Os resultados mostraram que o pré-tratamento térmico levou a
uma maior porcentagem de recuperação e de pureza do polímero.

Capítulo 3 Metodologia Experimental

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
50
3. Metodologia Experimental
O presente capítulo apresenta os matérias e métodos utilizados na produção de
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso.
3.1. Quitosana
Neste trabalho utilizou-se uma quitosana (Lote 067) obtida junto a POLYMAR/CE,
com no mínimo 80% de desacetilação. Esse substrato foi utilizado como fonte de carbono e
energia tanto no estudo cinético como nos ensaios do planejamento de seleção de fatores
Plackett & Burman e Planejamento Composto Central, para a produção de quitosanase pelo
Metarhizium anisopliae, por fermentação submersa.
3.2. Microrganismo
O fungo Metarhizium anisopliae estirpe (CG374), utilizado neste estudo, foi cedido
pela Embrapa Recursos Genéticos e Biotecnologia (Brasília / DF-Brasil) e foi mantido em
placas de Petri contendo o meio Batata Dextrose Agar (PDA). As placas foram incubadas em
estufa a 25 ºC. O pH do meio de manutenção foi mantido entre 6,0 e 6,5.
3.2.1. Escolha da cepa produtora de quitosanase
A escolha do microrganismo Metarhizium anisopliae foi feita com base na capacidade
de crescer em meio contendo quitosana como única fonte de carbono, além de apresentar
grande tolerância a pH variando a uma taxa de 2,0 a 8,5, sendo 6,9 a melhor condição para o
crescimento vegetativo e esporução, de acordo com Assis (2010).
3.2.2. Manutenção da cepa
A manutenção da cultura foi realizada em placas petri contendo o meio Batata
Dextrose Agar (PDA). A renovação das células foi realizada a cada trinta dias através de
subcultivos das células no meio PDA.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
51
3.3. Produção de enzima
No preparo do inóculo, utilizou-se 7 placas cultivadas e mantidas, com o fungo
Metarhizium anisopliae, em meio PDA a 25 ºC durante 5 dias em estufa. O meio de cultura
para a produção da enzima quitosanase apresentou os seguintes componentes: quitosana;
peptona; extrato de levedura; NaNO3; K2HPO4; KCl; MgSO4.7H2O e FeSO4, com
concentrações de acordo as matrizes dos planejamentos: Plackett & Burman e composto
central. Para a produção da enzima no cultivo, a biomassa foi transferida através de água
estéril, para um Béquer de 500 mL que, em seguida, foi colocado em Erlenmeyers de 250 mL
na proporção de 20 mL de biomassa para 180 mL do meio de cultura. Os Erlenmeyers
cultivados foram então incubados em incubador rotativo a 25 ºC e 150 rpm durante 24 h.
Alíquotas foram retiradas e centrifugadas a 8000xg por 5 min, a cada 6 horas, para se
determinar a atividade quitosanolítica, teores de açúcares redutores e de proteína total.
Ressalta-se que todos os experimentos foram feitos em triplicata.
3.4. Determinação da Proteína Total
A determinação da proteína total foi efetuada de acordo com o método modificado de
Sedmak & Grossberg (1977), que se baseia na diminuição da absorbância do corante azul de
Coomassie, devido à ligação proteína-corante, a 595 nm, comparado a absorção máxima a
465 nm, conforme Santos (2001). Nesse caso, após centrifugação para retirada da biomassa,
transferia-se para um eppendorf 1,5 mL de amostra (caldo fermentativo) e adicionava-se 1,5
mL de solução azul de Coomassie. Deixava-se a solução em repouso por 5 minutos,
transferindo-se em seguida para um cubeta de quartzo e efetuando-se as leituras de
absorbância nos comprimentos de ondas 465 nm e 595 nm, respectivamente. O
espectrofotômetro (Thermo Spectronic modelo Genesys 10UV) foi zerado com água
destilada. O conteúdo protéico foi obtido a partir de uma curva padrão na qual graficava-se a
relação de absorbância A595/A465 para diferentes concentrações de Albumina de Soro
Bovino (Fração V – Lote 110881/Sigma Aldrich-Ohio-USA).
3.5. Determinação da Atividade Quitosanolítica e da Atividade Específica
A atividade quitosanolítica foi obtida monitorando-se o aumento dos açúcares
redutores totais, utilizando-se o método DNS a partir de uma curva padrão de glicosamina.
Para a determinação da atividade quitosanolítica adicionaram-se, aos tubos de ensaios 500 µL

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
52
de substrato (solução 0,2 % de quitosana solubilizada em ácido clorídrico 1N) e 500 µL de
caldo bruto (contendo enzima), controlando-se o pH entre 6,0 e 6,5 e incubando-se o sistema
em um banho termostatizado com temperaturas da ordem de 55 ºC. Após o tempo de
incubação de 30 minutos no banho foi adicionado aos tubos de ensaio 2,5 mL de DNS,
aquecendo-se à ebulição em banho-maria por 10 minutos, resfriando-se, em seguida, em
banho de gelo. Após essa etapa, o sistema foi centrifugado e com o sobrenadante efetuou-se a
leitura no espectrofotômetro a 600 nm. Destaca-se, que antes da leitura da amostra zerava-se o
equipamento (branco do espectro) com 1,0 mL de água destilada acrescidos de 2,5 mL de
DNS e realizava-se o branco da enzima que consistia da adição de 500 µL de solução de
quitosana, completando-se até um 1,0 mL com água destilada. Uma unidade de atividade
quitosanolítica (A) foi definida como a quantidade de enzima necessária para produzir 1
µmol.min-1 de glicosamina por mL de extrato enzimático, nas condições de temperatura e pH
citadas, sendo o resultado expresso em (U.mL-1). A Atividade específica foi determinada pela
divisão entre a atividade da enzima (U.mL-1) e a concentração de proteína total (µg.mL-1),
sendo expressa em (U. µg-1).
3.6. Planejamento experimental
3.6.1. Planejamento de seleção de fatores Plackett & Burman
O planejamento experimental Plackett & Burman é uma técnica muito usada, para se
realizar triagem de fatores na otimização num determinado processo (Reddy et al., 2008), e
foi utilizado neste trabalho para identificar os fatores que influenciam de forma significativa
na produção da enzima quitosanase.
A Tabela 3 ilustra os níveis de cada variável (fator) utilizada no planejamento Plackett
& Burman, sendo: Concentração de Quitosana (Q), Peptona (PP), Extrato de Levedura (EL),
NaNO3 (NS), K2HPO4 (FP), KCl (CP), MgSO4.7H2O (SM), FeSO4 (SF). Já a Tabela 4 mostra
a matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3 repetições
na condição do ponto central.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
53
Tabela 3 – Níveis para os fatores e seus valores codificados.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1 0,1 0,2 0,2 0,1 0,05 0,025 0,025 0,0005
0 0,2 0,5 0,3 0,2 0,10 0,050 0,050 0,0010
1 0,3 0,8 0,4 0,3 0,15 0,075 0,075 0,0015
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1
2 1 1 -1 1 -1 -1 -1 1 -1 1 1
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1
5 1 1 -1 1 1 -1 1 -1 1 -1 1
6 1 1 1 -1 1 1 -1 1 1 -1 -1
7 -1 1 1 1 -1 1 1 -1 -1 1 1
8 -1 -1 1 1 1 -1 1 1 1 1 1
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1
11 -1 1 -1 -1 -1 1 1 1 1 1 -1
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1
13 0 0 0 0 0 0 0 0 0 0 0
14 0 0 0 0 0 0 0 0 0 0 0
15 0 0 0 0 0 0 0 0 0 0 0
Sendo: V11,V12,V13 = Variáveis inertes para cálculo do erro.
3.6.2. Planejamento Composto Central para a Otimização do Meio de Cultura para a
Produção da Enzima em Cultivo Submerso
Um dos problemas mais comuns para quem realiza experimentos é determinar a
influência de uma ou mais variáveis sobre outra variável de interesse (Lobato, 2003). Este

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
54
problema pode ser minimizado utilizando-se um planejamento experimental de modo a se
obter uma função, ou uma aproximação satisfatória para a mesma, de forma que se operando
sobre as variáveis de entrada produza-se como saída a resposta ou as respostas observadas
(Barros Neto, 2001; Silva, 2004).
Em um planejamento, inicialmente, deve-se definir os fatores e as respostas de
interesse. Tendo-se identificado os fatores e respostas define-se o objetivo que se pretende
alcançar com os experimentos para que se possa escolher o planejamento mais apropriado.
(Silva, 2004).
No presente trabalho foi utilizado um Planejamento Composto Central (CCD) com 22
experimentos na parte fatorial, repetição em triplicata no ponto central e quatro experimentos
na parte axial (planejamento do tipo estrela), para se investigar a influência dos fatores:
concentração de quitosana e concentração de FeSO4 que foram os dois fatores que
influenciaram a produção de quitosanase, conforme o planejamento Plackett & Burman de 12
ensaios e mais 3 repetições na condição do ponto central. Estes dois fatores foram estudados
em cinco diferentes níveis (-1,41; -1; 0; 1; 1,41) e um conjunto de 11 experimentos foi
realizado, conforme Tabela 5.
Tabela 5 – Matriz do planejamento Composto Central.
Ensaio Q SF
1 -1 -1
2 -1 1
3 1 -1
4 1 1
5 -1,41 0
6 1,41 0
7 0 -1,41
8 0 1,41
9 0 0
10 0 0
11 0 0
Os outros fatores foram mantidos nos níveis mais baixos (para evitar custos). Uma que
não influenciaram de forma significativa na produção da enzima quitosanase.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
55
A codificação dos fatores foi realizada de acordo com a seguinte equação (Ji et al.,
2009):
Xi = (xi – x0)/∆xi , i = 1, 2, 3, ..., k (2)
Onde Xi é o fator independente codificado, xi é o fator independente real, x0 é o valor de xi no
ponto central e ∆xi é a variação de valor. O comportamento do sistema foi explicado pela
seguinte equação polinomial de segunda ordem:
Yp = β0 + ∑ βiXi + ∑ βiiXi2 + ∑ βijXiXj, i, j = 1, 2, 3,..., k (3)
Onde Yp foi a resposta esperada, β0 foi a interseção, Xi e Xj
foram os fatores independentes codificados, βi foi o coeficiente linear, βii
foi o coeficiente quadrático e βij foi o coeficiente de interação. Destaca-se que os outros
fatores não significativos foram mantidos no nível mais baixo (-1).
A Tabela 6 apresenta os níveis para os fatores e seus valores codificados, para o
Planejamento Composto Central, com triplicata no ponto central.
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1,41 0,218 0,2 0,2 0,1 0,05 0,025 0,025 0,0009
-1 0,3 0,2 0,2 0,1 0,05 0,025 0,025 0,0015
0 0,5 0,2 0,2 0,1 0,05 0,025 0,025 0,0030
1 0,7 0,2 0,2 0,1 0,05 0,025 0,025 0,0045
1,41 0,782 0,2 0,2 0,1 0,05 0,025 0,025 0,0051
Na análise do planejamento de seleção de fatores Plackett & Burman e do
planejamento experimental fatorial foi utilizado um pacote computacional STATISTICA for
Windows versão 7.0.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
56
3.7. Perfil cinético durante o cultivo
Um perfil cinético na melhor condição de cultivo, conforme planejamento
apresentado, foi realizado em frascos Erlenmyer de 250 mL contendo 200 mL de meio de
cultivo contendo os seguintes componentes, em 1 Litro: 2,0g de quitosana; 5,0g de peptona;
3,0g de extrato de levedura; 2,0g de NaNO3; 1,0g de K2HPO4; 0,5g de KCl; 0,5g de
MgSO4.7H2O; 0,01g de FeSO4, que foram inoculadas com 10% (vol/vol) de inóculo contendo
o fungo Metarhizium anisopliae e cultivadas em incubador rotativo durante 5 dias a 150 rpm
e 25 ºC. Alíquotas foram retiradas a cada seis horas e analisadas quanto à proteína total no
sobredanante, atividade enzimática, atividade específica e açúcares redutores. Os ensaios
foram realizados em triplicata (Silva Filho, 2005).

Capítulo 4 Resultados e
Discussão

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
58
4. Resultados e discussão
Neste capítulo são apresentados e discutidos os resultados obtidos no presente
trabalho. Inicialmente, é apresentada imagem do MEV do Metarhizium anisopliae
mantidos em meio PDA. Em seguida, apresenta-se e discute-se os resultados da avaliação
dos fatores significativos, que podem influenciar na produção da quitosanase, no
planejamento de seleção de fatores, conforme planejamento Plackett & Burman bem como
são apresentados e discutidos os resultados obtidos para o planejamento composto central.
Por último, apresenta-se o perfil cinético, na melhor condição, para a produção de
quitosanase em cultivo submerso descontínuo usando o fungo Metarhizium anisopliae.
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA.
A Figura 7 ilustra a imagem da Microscopia Eletrônica de Varredura (MEV) do micélio de Metarhizium anisopliae mantido em meio PDA.
Figura 7. Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA.
Observa-se na Figura 7, quanto à morfologia que o Metarhizium anisopliae, apresenta-
se como um fungo filamentoso com as hifas intimamente entrelaçadas conforme citado por
Wang et al. 2002.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
59
4.2. Planejamento Plackett & Burman
4.2.1 Análise do Plackett & Burman com 8 fatores e 12 ensaios
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis, foi utilizado para avaliar a influência dos fatores: Concentração de
Quitosana (Q), Peptona (PP), Extrato de Levedura (EL), NaNO3 (NS), K2HPO4 (FP), KCl
(CP), MgSO4.7H2O (SM), FeSO4 (SF) na produção da enzima quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso. Ressalta-se que os experimentos foram
realizados aleatoriamente.
A Tabela 7 apresenta a matriz do modelo Plackett & Burman e os respectivos valores
obtidos para a variável resposta (Atividade Quitosanalítica).
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.
Ensaio Q PP EL NS FP CP SM SF A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 0,0195
2 1 1 -1 1 -1 -1 -1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 0,0109
4 1 -1 1 1 -1 1 -1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 0,0136
6 1 1 1 -1 1 1 -1 1 0,0156
7 -1 1 1 1 -1 1 1 -1 0,0109
8 -1 -1 1 1 1 -1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 0,0085
A Tabela 8 apresenta os efeitos estimados para o planejamento Plackett & Burman.
Observa-se na Tabela 8 que o valor do coeficiente de determinação R2 foi de
aproximadamente 90%, indicando que um modelo linear poderia representar bem a relação
entre o fator significativo, concentração de quitosana (1), e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
60
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman.
Efeitos Estimados; R2=0,8996; Ajuste:0,63178 MS Erro Residual =0,0000054
Desvio Padrão t(3) p
Lim. Conf. Lim. Conf.
Efeito Erro Residual
-95,% +95
Média/Interação 0,0127 0,0007 18,903 0,0003 0,0105 0,0149 (1) Quitosana 0,0055 0,0013 4,0628 0,0268 0,0011 0,0098
(2) Peptona 0,0015 0,0013 1,1080 0,3486 -0,0028 0,0058 (3) Ext. de leved. 0,0009 0,0013 0,7387 0,5136 -0,0033 0,0053
(4) NaNO3 0,0002 0,0013 0,1758 0,8715 -0,0040 0,0045 (5) K2HPO4 -0,0014 0,0013 -1,0552 0,3687 -0,0057 0,0028
(6) KCl -0,0022 0,0013 -1,6708 0,1933 -0,0065 0,0020 (7) MgSO4.7H2O -0,0003 0,0013 -0,2286 0,8338 -0,0046 0,0039
(8) FeSO4 0,0029 0,0013 2,1457 0,1211 -0,0014 0,0071
A Figura 8 mostra o gráfico de Pareto, uma das formas de se avaliar visualmente a
influência dos fatores estudados na resposta. A magnitude dos efeitos é representada pelas
colunas enquanto que a linha transversal às colunas representa a magnitude dos efeitos com
significado estatístico para p=0,05, ou seja, os fatores que são estatisticamente significativos
ao nível de 95% de confiança.
Figura 8. Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
61
Analisando-se o gráfico de Pareto (Figura 8) observa-se que o fator concentração de
quitosana (1) foi o único fator que influenciou de forma mais significativa na atividade
enzimática atingindo efeitos estimados de aproximadamente 4,06. O sinal positivo indica que
o aumento da concentração de quitosana aumentará à atividade enzimática. Por outro lado, os
outros fatores parecem desprezíveis estatisticamente conforme o gráfico de Pareto.
4.2.2 Análise do Plackett & Burman com 12 Ensaios e mais 3 Repetições na condição do
ponto central.
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis com repetição em triplicata no ponto central, foi utilizado para se avaliar a
influência dos mesmos fatores descritos na Seção 4.2.1 para a produção da enzima
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso. Ressalta-se que os
experimentos foram realizados aleatoriamente. A Tabela 9 apresenta a matriz do modelo
Plackett & Burman e os respectivos valores obtidos para a variável resposta (Atividade
Quitosanalítica).
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13 A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1 0,0195
2 1 1 -1 1 -1 -1 -1 1 -1 1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1 0,0109
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 1 -1 1 0,0136
6 1 1 1 -1 1 1 -1 1 1 -1 -1 0,0156
7 -1 1 1 1 -1 1 1 -1 -1 1 1 0,0109
8 -1 -1 1 1 1 -1 1 1 1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 1 1 -1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1 0,0085
13 0 0 0 0 0 0 0 0 0 0 0 0,0704
14 0 0 0 0 0 0 0 0 0 0 0 0,0688
15 0 0 0 0 0 0 0 0 0 0 0 0,0710

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
62
A Tabela 10 a seguir apresenta os efeitos estimados para planejamento Plackett &
Burman com 3 repetições na condição do poto central. Observa-se na Tabela 10 que o valor
do coeficiente de determinação R2 foi de aproximadamente 2,0%, indicando que um modelo
linear não poderia representar a relação entre os fatores significativos concentração de
quitosana (1), concentração de FeSO4 (8) e a resposta.
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central.
Efeitos Estimados; R2=0,01811; Ajuste:0,0 MS Erro Puro =0,0000012
Desvio Padrão t(2) P
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0242 0,0003 85,509 0,0001 0,0229 0,0254 (1) Quitosana 0,0055 0,0006 8,6432 0,0131 0,0027 0,0082
(2) Peptona 0,0015 0,0006 2,3572 0,1425 -0,0012 0,0042 (3) Ext. de leved. 0,0009 0,0006 1,5715 0,2567 -0,0017 0,0037
(4) NaNO3 0,0002 0,0006 0,3742 0,7442 -0,0024 0,0029 (5) K2HPO4 -0,0014 0,0006 -2,2450 0,1538 -0,0041 0,0013
(6) KCl -0,0022 0,0006 -3,5546 0,0708 -0,0049 0,0005 (7) MgSO4.7H2O -0,0003 0,0006 -0,4864 0,6747 -0,0030 0,0024
(8) FeSO4 0,0029 0,0006 4,5648 0,0448 0,0002 0,0056
A Figura 9 ilustra o gráfico de Pareto para o planejamento Plackett & Burman, com
repetição em triplicata no ponto central, uma das formas de se avaliar visualmente a influência
dos fatores estudados na resposta, onde é mostrado a magnitude dos efeitos para 95 % de
confiança.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
63
Figura 9. Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto
central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto, observa-se que os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma mais
significativa na atividade enzimática atingindo efeitos estimados de aproximadamente 8,84 e
4,67, respectivamente. O sinal positivo indica que o aumento das concentrações de quitosana
e de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores parecem
desprezíveis estatisticamente conforme o gráfico de Pareto.
Este comportamento do FeSO4 está de acordo com o observado por Hagag et al.
(2012) que utilizando o Aspergillus fumigatus na produção de protease mostraram que um
fator de transcrição PrtT regula positivamente o controle da produção de protease, da
assimilação de Ferro e da síntese de ergosterol e que a produção de protease foi favorecida
pelo aumento da concentração de ferro.
Rajendran & Thangavelu (2009) utilizaram um planejamento Plackett & Burman para
identificar quais os fatores significativos na produção de lipase por Rhizopus arrhizus MTCC
2233 e observaram o efeito significativo médio do FeSO4, apresentando um nível de
confiança para esse fator de 48,92% (para um máximo de 98,7 % de MgSO4), com uma
produção de lipase da ordem de 2,98 U/mL em 84 horas de fermentação e que ocorreu uma
produção considerável de protease no meio de cultivo da ordem de 1,92 U/mL em 96 horas de
cultivo.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
64
Mu et al. (2005) também observaram um efeito positivo do FeSO4, da ordem de 1,48
para um fator máximo encontrado de 3,11 de MgSO4, no aumento da atividade da
estereoinversão de S-fenil-1,2-etanodiol por Candida parapsilosis CCTCC M203011 na
otimização da fermentação utilizando um planejamento Plackett & Burman.
Dessa forma, observa-se que o FeSO4 é importante uma que vez o mesmo está
diretamente associado a síntese de proteases (principalmente metalloproteases), conforme
citado por Hagag et al. (2012).
4.3. Planejamento Composto Central para Produção de Quitosanase por
Metarhizium anisopliae em Cultivo Descontínuo Submerso
Um planejamento experimentos 22 com repetição em triplicata no ponto central e mais
quatro ensaios axiais, foram utilizados para se chegar a uma combinação ideal de quitosana e
FeSO4 na produção da enzima quitosanase (Tabela 11). A partir do modelo Plackett &
Burman (Tabela 12), que mostrou que o aumento das concentrações de quitosana e FeSO4,
favoreceram a produção de quitosanase, os valores utilizados neste modelo para estes
componentes nos níveis mais altos, serão considerados como os níveis mais baixos no
planejamento experimental do composto central. Os outros fatores foram fixados em todos os
ensaios em seus níveis mais baixos para a redução de custos (Tabela 11).
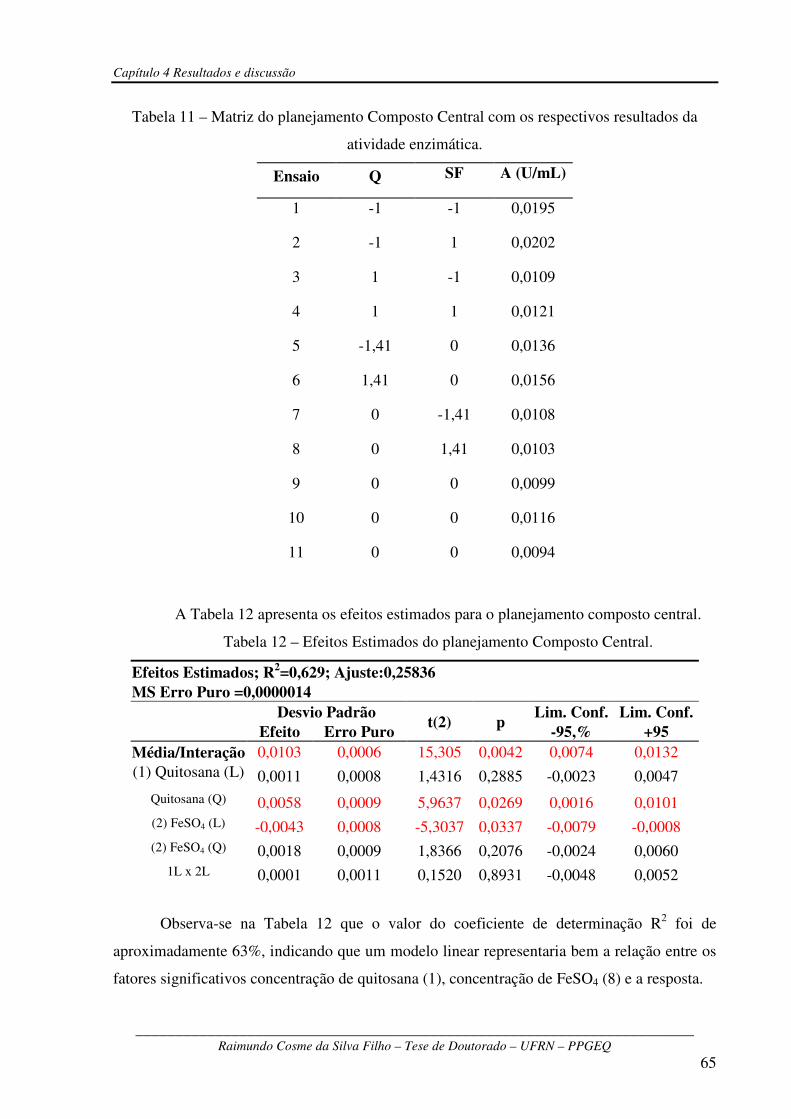
Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
65
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da
atividade enzimática.
Ensaio Q SF A (U/mL)
1 -1 -1 0,0195
2 -1 1 0,0202
3 1 -1 0,0109
4 1 1 0,0121
5 -1,41 0 0,0136
6 1,41 0 0,0156
7 0 -1,41 0,0108
8 0 1,41 0,0103
9 0 0 0,0099
10 0 0 0,0116
11 0 0 0,0094
A Tabela 12 apresenta os efeitos estimados para o planejamento composto central.
Tabela 12 – Efeitos Estimados do planejamento Composto Central.
Efeitos Estimados; R2=0,629; Ajuste:0,25836 MS Erro Puro =0,0000014
Desvio Padrão t(2) p
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0103 0,0006 15,305 0,0042 0,0074 0,0132 (1) Quitosana (L) 0,0011 0,0008 1,4316 0,2885 -0,0023 0,0047
Quitosana (Q) 0,0058 0,0009 5,9637 0,0269 0,0016 0,0101 (2) FeSO4 (L) -0,0043 0,0008 -5,3037 0,0337 -0,0079 -0,0008 (2) FeSO4 (Q) 0,0018 0,0009 1,8366 0,2076 -0,0024 0,0060
1L x 2L 0,0001 0,0011 0,1520 0,8931 -0,0048 0,0052
Observa-se na Tabela 12 que o valor do coeficiente de determinação R2 foi de
aproximadamente 63%, indicando que um modelo linear representaria bem a relação entre os
fatores significativos concentração de quitosana (1), concentração de FeSO4 (8) e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
66
A Tabela 13 mostra um resumo da ANOVA para o planejamento composto central
com repetição em triplicata no ponto central.
Tabela 13 – Resumo da ANOVA.
Fonte de
Variação
Soma dos
Quadrados
Grau de
Liberdade
Quadrado
Médio
Fcalculado Ftabelado
Regressão
Resíduos
0,000086 2 0,000043 6,14 4,46
0,000056 8 0,000007
Falta de
Ajuste
Erro puro
0,000053 6 0,000009 6 19,33
0,000003
2
0,0000015
Total 0,000142 10
De acordo com a Tabela 13, partindo-se dos efeitos significativos propõe-se o
modelo:
� = 0,010316 + 0,002942� � − 0,002193�� (8)
% de variação explicável: 60,5
% máxima de variação explicável: 97,9
A análise da ANOVA mostra que o modelo proposto possui significância estatística, ao
nível de 95% de confiança, uma vez que o valor de F (calculado) é maior que F (tabelado),
entretanto a ANOVA mostra também que o modelo proposto pode ser útil para se fazer
previsões, uma vez que a relação entre F (calculado) e F (tabelado) para a falta de ajuste e o
erro puro é menor que 1. O coeficiente de determinação R2 foi de aproximadamente 63%,
indicando que o modelo consegue explicar 63% da variação total em torno da média.
Os cálculos para variação explicável e para máxima variação explicável são dados pelas
Equações (9) e (10):
% de variação explicável = (SQ da regressão) / (SQ Total) (9)
% máxima de variação explicável = (SQ Total – SQ Erro puro) / (SQ Total) (10)
Sendo:
SQ – Soma dos quadrados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
67
A Figura 10 mostra o gráfico de Pareto para o planejamento Composto Central
utilizando erro puro. A magnitude dos efeitos é representada pelas colunas. Já a linha
transversal às colunas representa a magnitude dos efeitos com significado estatístico para 95%
de confiança.
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto (Figura 10) observa-se que os fatores concentração
de quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma
mais significativa na atividade enzimática atingindo efeitos estimados de aproximadamente
5,96 e - 5,30, respectivamente. O sinal positivo indica que o aumento das concentrações de
quitosana aumentará à atividade enzimática. O sinal negativo indica que o decréscimo da
concentração de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores
parecem desprezíveis estatisticamente no gráfico de Pareto.
A avaliação do modelo também pode ser feita através da observação do gráfico dos
valores preditos em função os valores observados que são mostrados na Figura 11. Os valores
preditos pelo modelo são representados pela reta, enquanto que os valores observados
representam-se pelos pontos.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
68
Figura 11 – Valores preditos em função dos valores observados.
Observa-se que alguns valores preditos apresentam um afastamento dos valores
observados. A Figura 12 ilustra a superfície de resposta da atividade quitosanolítica em
função dos dois fatores significativos, concentração de quitosana e concentração de FeSO4.
Figura 12. Atividade quitosanolítica em função da concentração de quitosana e da
concentração de FeSO4 para os fatores não significativos no ponto central.
Observa-se a existência de um ponto de mínimo, ocorrendo para concentração de
quitosana no ponto central (0) e para maiores concentrações de FeSO4 igual a (1,5). Essa

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
69
região deve ser evitada uma vez que a atividade quitosanolítica atinge níveis tão reduzidos
quanto 0,0094 U.mL-1. Destaca-se que nesse planejamento o maior nível da concentração de
FeSO4 (α=+1,41) equivale a 0,0051% (Tabela 6) que é aproximadamente cinco vezes maior
que o nível desse fator no ponto central do planejamento Plackett & Burman (0,001%),
conforme Tabela 3. Dessa forma, ao se utilizar o nível mínimo da concentração de FeSO4 no
Planejamento Composto Central (α=-1,41), que equivale a aproximadamente 0,001%,
aproxima-se do ponto central do do planejamento Plackett & Burman (0,001%), condição
essa que apresenta os melhores resultados de atividade quitosanolítica usando quitosana como
fonte de carbono e energia e fungo Metarhizium anisopliae.
4.4. Perfil Cinético – Melhor Condição
A seguir, apresenta-se um perfil cinético, na melhor condição, para produção da
quitosanase usando o fungo Metarhizium anisopliae, em que foram monitorados durante 24
horas os seguintes parâmetros: Concentração de proteínas totais (µg/mL), atividade
quitosanolítica (U/mL), atividade específica (U/µg) e açúcares redutores (mg/mL), ilustradas
nas Figuras 13, 14, 15 e 16, respectivamente.
Com relação à proteína extracelular, observa-se na Figura 13 que, inicialmente, ocorre
um aumento da mesma para o seu nível máximo (6,2 µg/mL), durante as 6 primeiras horas de
cultivo, seguido de uma redução para 5,7 µg/mL em 18 horas de cultivo, possivelmente
devido à presença de proteases no meio. Observa-se que a partir de então ocorre uma redução
e aumento da proteína total, em uma quantidade inferior, sugerindo um mecanismo de
controle.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
70
0 4 8 12 16 20 245,0
5,2
5,4
5,6
5,8
6,0
6,2
6,4
Pro
tein
a (
µg
/mL
)
Tempo (h)
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana.
Com relação à atividade quitosanolítica, conforme Figura 14, a atividade máxima
ocorreu em 18 horas de produção em que se obteve aproximadamente 70,0 U/L, o que foi 28
vezes maior que o valor (2,5 U/L em 48 horas) relatado por Assis (2010) para o mesmo fungo,
e em apenas um terço do tempo de produção.
0 4 8 12 16 20 240,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,10
Ativid
ade
(U
/mL
)
Tempo (h)
Figura 14. Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
71
Além disso, destaca-se o considerável aumento da produção de quitosanase a partir das
12 horas até 18 horas de cultivo onde atingiu o pico máximo de produção (70,0 U/L). Após o
pico em 18 horas, a produção começa a cair rapidamente, comportamento esse observado
durante todos os ensaios cinéticos. Destaca-se que é importante do ponto de vista industrial
manter a produção em alto nível por muitas horas, onde se espera uma produção contínua ou
em grandes bateladas.
Ak et al. (1998) utilizaram o fungo Penicillium spinulosum para produzir a quitosanase
usando um meio de cultura contendo 0,1% de K2HPO4; 0,05 % de MgSO4.7H2O; 0,05% de
KCl; 0,001 % de FeSO4, 0,5% NH4NO3 e como fonte de carbono e energia, os autores
usaram quitosana e parede de Rhizopus. A produção foi realizada em cultivo submerso a 28
oC. Os autores apresentam atividade quitosanolítica máxima de aproximadamente 47,0 U/mL,
ao se utilizar a parede do fungo como fonte de carbono e energia, e 30,0 U/mL ao se utilizar
quitosana como fonte de carbono e energia. Destaca-se que essas atividades ocorreram após 4
dias de cultivo. Os perfís com o tempo da atividade quitosanolítica e da atividade específica
apresentaram tendência de crescimento, com o pico máximo em 4 dias, seguindo com uma
tendência de queda da atividade. Neste tempo, a atividade específica máxima da quitosanase
foi de aproximadamente 60,0 U/mg de proteína. Essas tendências também foram observadas
nos resultados apresentados nos perfís de atividade das Figuras 14 e 15.
0 4 8 12 16 20 24
-0,010
-0,005
0,000
0,005
0,010
0,015
0,020
0,025
0,030
Ativid
ad
e E
spe
cific
a (
U/µ
g)
Tempo (h)
Figura 15. Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
72
Observa-se na Figura 15 que a atividade específica máxima da quitosanase segue a
mesma tendência da atividade quitosanolítica apresentada na Figura 14, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
O perfil para os açúcares redutores durante o cultivo de Metarhizium anisopliae
cultivado em meio contendo 0,2% de quitosana é ilustrado na Figura 16.
0 4 8 12 16 20 240,004
0,008
0,012
0,016
0,020
0,024
0,028
Acuca
res R
ed
uto
res (
mg/m
L)
Tempo (h)
Figura 16. Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana.
Observa-se, na Figura 16, que taxa máxima de redução dos açúcares redutores
(aproximadamente 0,021 mg/mL) ocorre logo após a produção máxima da enzima, em 18
horas de cultivo, conforme Figura 14. A quantidade de açúcares redutores foi a
aproximadamente 0,012 mg/mL após 24 h de cultivo. O fungo excreta para o meio enzimas
com atividade quitosanolítica, essas enzimas hidrolisam a quitosana aumentando a quantidade
de açúcares redutores para o meio, sendo que esses açúcares também são consumidos pelo
fungo durante seu crescimento.
Foi demonstrado por Assis (2010) que os açúcares redutores produzidos através da
quitossana, ocasionada pelas enzimas quitossanases metabolizadas por Metarhizium
anisopliae, com quitosana como sua única fonte de carbono e energia, atingiu um pico de 0,53
mg/mL em 48 horas. Apresentaram como quitooligossacarídeos, constituídos principalmente
por dímeros e pentâmeros, formados pela hidrólise enzimática de quitosanase em quantidades
maiores do que o consumido pelos microrganismos. Mostraram, também, que essa produção

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
73
segue a mesma tendência da sua atividade enzimática, corroborando com os resultados
apresentados no presente trabalho.
Os resultados do presente trabalho mostram a relevância de se adotar novas condições
para a produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter quitooligossacarídeo
com atividades biológicas. Esta pesquisa, portanto, sugere base para novos estudos que
permitam alcançar rapidamente a produção de quitosanase e quitooligossacarídeo com todos
os seus benefícios.

Capítulo 5 Conclusão

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
75
5. Conclusão
Após o desenvolvimento desse trabalho conclui-se:
- Com relação ao Planejamento de Seleção de Fatores Plackett & Burman:
Os resultados das análises do Planejamento de Seleção de Fatores Plackett &
Burman com 8 fatores e 12 ensaios em 2 níveis, mais 3 repetições no ponto central, para
se investigar a influência dos fatores: Concentração de Quitosana, Peptona, Extrato de
Levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O, FeSO4 na produção da enzima
quitosanase por Metarhizium anisopliae, mostraram que a atividade foi favorecida pelo
aumento da concentração de substrato (quitosana) e sulfato ferroso (FeSO4), enquanto
que o aumento da concentração dos outros fatores não contribuíram de forma
significativa para a resposta (atividade quitosanolítica). Os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) atingiram efeitos estimados de
aproximadamente 8,84 e 4,67, respectivamente. O sinal positivo indica que o aumento
das concentrações de quitosana e de FeSO4 aumentará a atividade enzimática. Conforme
visto na literatura, observa-se importância do FeSO4, já que o mesmo está diretamente
associado a síntese de proteases (principalmente metalloproteases),
- Com relação ao Planejamento Composto Central:
A análise do planejamento composto central para avaliar a influência dos
fatores, concentração de substrato (quitosana) e concentração de FeSO4, na produção da
enzima quitosanase por Metarhizium anisopliae (a partir do planejamento Plackett &
Burman) mostrou que os fatores concentração de quitosana e concentração de FeSO4
foram os fatores que influenciaram de forma mais significativa na atividade enzimática
atingindo efeitos estimados de aproximadamente 5,96 e - 5,30, respectivamente. O sinal
positivo indica que o aumento das concentrações de quitosana aumentará a atividade
enzimática. O sinal negativo indica que o decréscimo da concentração de FeSO4
aumentará a atividade enzimática. Observa-se a existência de um ponto de mínimo,
ocorrendo para concentração de quitosana (X1) no ponto central (0) e para maiores

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
76
concentrações de FeSO4 (X2) igual a (1,5) com a atividade quitosanolítica de 0,0094
U.mL-1.
- Com relação ao cultivo na melhor condição:
O cultivo na melhor condição mostrou uma produção máxima de quitosanase em
18 horas de cultivo, com a atividade enzimática de 70,0 U/L, o que foi 28 vezes maior
que o valor (2,5 U/L em 48 horas) relatado na literatura (Assis, 2010) para o mesmo
fungo e em apenas um terço do tempo de produção.
Além disso, observou-se que um elevado incremento na produção de
quitosanase a partir das 12 horas de cultivo, antes de se atingir a atividade
quitosanolítica máxima, 18 horas de cultivo. Essa produção em alto nível por muitas
horas é importante do ponto de vista industrial, onde se espera uma produção contínua
ou em grandes bateladas. Observou-se também que a atividade específica máxima da
quitosanase segue a mesma tendência da atividade quitosanolítica, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
- Com relação aos açúcares redutores:
Foi observado que os açúcares redutores seguem a mesma tendência da atividade
enzimática, com uma produção máxima de cerca de 0,21 mg/mL em 18 horas de
cultivo, produção essa que coincinde com o máximo pico de atividade enzimática.
Estes resultados mostram a relevância de se adotar novas condições para a
produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter
quitooligossacarídeo com atividades biológicas. Esta pesquisa, portanto, sugere base
para novos estudos que permitam alcançar rapidamente a produção de quitosanase e
quitooligossacarídeo com todos os seus benefícios.

Referências Bibliográficas

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
78
Referências bibliográficas
AK, Ö.; BAKIR, U.; GÜRAY, T. Isolation of a chitosan degrading fungus, Penicillium
spinulosum, and chitosanase production by the isolate. In: SYMPOSIUM - NEW
FRONTIERS IN SCREENING FOR MICROBIAL BIOCATALYSTS, 1996, Ede, The
Netherlands. Anais, Editado por K. Kieslich, C. P. van der Beek, J. A. M. de Bont e W.
J. J. van den Tweel, Elsevier Science B. V., 1998, v.53, p.265-268.
ALVES, S. B. Patologia e controle microbiano: Vantagens e desvantagens. In: ALVES,
S. B. Controle Microbiano de Insetos. 2ed., Piracicaba, FEALQ, 1163p, 1998, v.4, p.21-
37.
ALVES, S. B.; LEITE, L. G.; FILHO, A. B.; ALMEIDA, F. E. M.; MARQUES, E. J.
Produção massal de fungos entomopatogênicos na América Latina, In: ALVES, S. B.;
LOPES, R. B. Controle microbiano de pragas na América Latina: avanços e desafios.
Piracicaba, FEALQ, 414p, 2008, v.14, p.215-237.
ARRUDA, W.; LUBECK, I.; SCHRANK, A.; VAINSTEIN, M. H. Morphological
alterations of Metarhizium anisopliae during penetration of Boophilus microplus ticks.
Experimental and Applied Acarology, v. 37, p.231-244, 2005.
ASSIS, C. F. Produção e caracterização de quitooligossacarídeos produzidos pelo fungo
Metarhizium anisopliae e avaliação da citotoxicidade em células tumorais. 2010. 110p.
Tese (Doutorado em Biotecnologia) – Renorbio - Programa de Pós-Graduação em
Biotecnologia. Universidade Federal do Rio Grande do Norte, Natal.
ASSIS, C. F.; COSTA, L. S.; MELO-SILVEIRA, R. F.; OLIVEIRA, R. M.;
PAGNONCELLI, M. G. B.; ROCHA, H. A. O.; MACEDO, G. R.; SANTOS, E. S.,
Chitooligosaccharides antagonizes the cytotoxic effect of glucosamine. World J.
Microbiol. Biotechnol., v.28, n.3, p.1097-1105, 2012.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
79
AZEVEDO, V. V. C.; CHAVES, S. A.; BEZERRA, D. C.; FOOK, M. V. L.; COSTA,
A. C. F. M. Quitina e Quitosana: aplicações como biomateriais. Revista Eletrônica de
Materiais e Processos, v.2.3, p.27-34, 2007.
BARROS NETO, B.; SCARMINIO, I. S.; BRUNS, R. Como fazer experimentos:
pesquisa e desenvolvimento na ciência e na indústria. 2. ed. Campinas: Unicamp, 2001.
401p.
BATISTA FILHO, A.; ALMEIDA, J. E. M.; SANTOS, A. S.; MACHADO, L. A.;
ALVES, S. B. Eficiência de isolados de Metarhizium anisopliae no controle de
cigarrinha-da-raiz de cana-de-açúcar Mahanarva fimbriolata (Hom.: Cercopidae). STAB
– Açúcar, Álcool e Subprodutos, v.21, p.84-89, 2002.
CARVALHO, J. C. M.; SATO, S. Fermentação Descontínua, In: SCHMIDELL, W.;
LIMA, U. A.; AQUARONE, E.; BORZANI, W. Biotecnologia Industrial. São Paulo,
Ed. Edgard Blücher, v.2, p.193-204, 2001.
CHAO, H. J.; SCHWARTZ, J.; MILTON, D. K. Populations and determinats of
airborne fungi in large office buildings. Environmental Health Perspectives, v.110, n.8,
p.777-782, 2002.
CHEN, X.; XIA, W.; YU, X. Purification and characterization of two types of
chitosanase from Aspergillus sp. CJ22-326. Food Research International, v.38, n.3,
p.315-322, 2005.
CHEN, P. T.; CHIANG, C.-J.; CHAO, Y.-P. Medium optimization and production of
secreted Renilla luciferase in Bacillus subtilis by fed-batch fermentation. Biochemical
Engineering Journal, v.49, p.395-400, 2010.
CHENG, C. Y.; LI, Y. An Aspergillus chitosanase with potential for large-ecale
preparation of chitosan oligosaccharides. Biotechnol. Appl. Biochem., v.32, p.197-203,
2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
80
CHOI, Y. J.; Kim, E. J.; Piao, Z.; Yun, Y. C.; Shin, Y. C. Purification and
characterization of chitosanase from Bacillus sp. Strain KCTC 0377BP and its
application for the production of chitosan oligosaccharides. Applied and Environmental
Microbiology, v.70, n.8, p.4522-4531, 2004.
COLAKOGLU, G. The chemical and physical environment for fungal growth. Fak
Matbaa Birimi, Istanbul, 149-166, 2001.
CRAVEIRO, A. C.; CRAVEIRO, A. A.; QUEIROZ, D. C. Quitosana a fibra do futuro.
2. ed. Fortaleza: PADETEC, 2004. V.1, 218p.
DIAS, B. A. Estudo de proteases degradadoras de cutícula produzidas pelo fungo
entomopatogênico Beauveria bassiana. 2005. 70p. Dissertação (Mestrado em
Microbiologia) – Centro de Ciências Biológicas, Programa de Pós-Graduação em
Microbiologia, Universidade Estadual de Londrina, Londrina.
DINARDO-MIRANDA, L. L.; FERREIRA, J. M. G.; CARVALHO, P. A. M.
Influência da época de colheita e do genótipo de cana-de-açúcar sobre a infestação de
Mahanarva fimbriolata (Stal) (Hemiptera: Cercopidae). Neotrop. Entomol, v.30, n.1,
p.145-149, 200.
DONG, C.; ZHANG, J.; HUANG, H.; CHEN, H.; HU, Y. Pathogenicity of a new China
variety of Metarhizium anisopliae (M. anisopliae var. Dcjhyium) to subterranean termite
Odontotermes formosanus. Microbiological Research, v.164, p.27-35, 2009.
DONZELLI, B. G. G.; OSTROFF, G.; HARMAN, G. E. Enhanced enzymatic
hydrolysis of langostino shell chitin with mixtures of enzymes from bacterial and fungal
sources. Carbohydrate Research, v.338, p.1823-1833, 2003.
DRIVER, F.; MILNER, R. J.; TRUEMAN, J. W. H. A taxonomic revision of
Metarhizium based on phylogenetic analysis of rDNA sequence data. Mycological
Research, v.104, n.2, p.134-150, 2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
81
DUNG, P.; MILAS, M.; RINAUDO, M.; DESBRIÉRES, J. Water soluble derivatives
obtained by controlled chemical modifications of chitosan, Carbohydrate Polymers,
v.24, p.209-214, 1994.
ESTEVES, J. A.; CABRITA, J. D.; NOBRE, G. N. Micologia médica, 2. ed. Lisboa:
Fundação Calouste Gulbenkian, 1990. 1058p.
FACCIOTI, M. C. R. Fermentação Contínua, In: Biotecnologia Industrial. Editado por:
SCHMIDELL, W.; LIMA, U. A.; AQUARONE, E.; BORZANI, W., São Paulo, Ed.
Edgard Blücher, v.2, p.541, 2001.
FARIA, M. R.; MAGALHÃES, B. P. O uso de fungos entomopatogênicos no Brasil -
Situação atual e perspectivas. Biotecnologia Ciência & Desenvolvimento, n.22., p.18-
21, setembro/outubro 2001.
FU, J.-Y.; WU, S.-M.; CHANG, C.-T.; SUNG, H.-Y. Characterization of Three
Chitosanase Isozymes Isolated from a Commercial Crude Porcine Pepsin Preparation. J.
Agric. Food Chem., v.51, p.1042-1048, 2003.
FURIGO JUNIOR, A.; PEREIRA, E. P. Enzimas e suas aplicações cinética enzimática.
2001. 39f. Apostila (Graduação de Engenharia Química e Engenharia de Alimentos) –
Centro de Tecnologia, Departamento de Engenharia Química e Engenharia de
Alimentos, Universidade Federal de Santa Catarina, Florianópolis.
GAO, J.; XU, H.; LI, Q.-J.; FENG, X.-H.; LI, S. Optimization of medium for one-step
fermentation of inulin extract from Jerusalem artichoke tubers using Paenibacillus
polymyxa ZJ-9 to produce R,R-2,3-butanediol, Bioresource Technology, v.101, p.7076-
7082, 2010.
GARCIA, M. V.; MONTEIROI, A. C.; SZABO, M. J. P.; PRETTE, N.; BECHARA, G.
H. Mechanism of infection and colonization of Rhipicephalus sanguineus eggs by
Metarhizium anisopliae as revealed by scanning electron microscopy and
histopathology. Brazilian Journal of Microbiology, São Paulo, v.36, n.4, p.368-372,
2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
82
GHANEM, N. B.; YUSEF, H. H.; MAHROUSE, H. K. Production of Aspergillus
terreus xylanase in solid-state cultures: application of the Plackett & Burman
experimental design to evaluate nutritional requirements, Bioresource Technology,
v.73, p.113-121, 2000.
GHORBEL-BELLAJ, O.; HMIDET, N.; JELLOULI, K.; YOUNES, I.; MAALEJ, H.;
HACHICHA, R.; NASRI, M. Shrimp waste fermentation with Pseudomonas aeruginosa
A2: Optimization of chitin extraction conditions through Plackett–Burman and response
surface methodology approaches, International Journal of Biological Macromolecules,
v.48, p.596–602, 2010.
GIUSTINA, A.; VENTURA, P. Weight-reducing regimens in obese subjects: Effects of
a new dietary fiber integrator. Acta Toxicol Ther, 16, 4, p.199-214, 1995.
GUIA.BIO, Reino dos fungos, 2012. Disponível em:
<http://www.http://guia.bio.br/reino-dos-fungos>. Acesso em: 12 novembro 2012.
GUIRADO, N.; AMBROSANO, E. J.; ROSSI, F.; DIAS, F. L. F. Controle de Cupins
de Montículo com Metarhizium anisopliae e Beauveria bassiana, Rev. Bras. de
Agroecologia, v.4, n.2, 2009.
GUO, Y.; XU, J.; ZHANG, Y.; XU, H.; YUAN, Z.; LI, D. Medium optimization for
ethanol production with Clostridium autoethanogenum with carbon monoxide as sole
carbon source, Bioresource Technology, v.101, p.8784–8789, 2010.
HAGAG, S.; KUBITSCHEK-BARREIRA, P.; NEVES, G. W. P.; AMAR, D.;
NIERMAN, W.; SHALIT, I.; SHAMIR, R.; LOPES-BEZERRA, L.; OSHEROV, N.
Transcriptional and proteomic analysis of the Aspergillus fumigatus ∆prtT protease-
deficient mutant. PLoS One, v.7, n.4, e33604, 2012.
HAN, L.-K.; KIMURA, Y.; OKUDA, H. Reduction in fat storage during chitin-chitosan
treatment in mice fed a high-fat diet, International Journal of Obesity, 23:174-179,1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
83
HUNG, T.; CHANG, Y.; SUNG, H.; CHANG, C. Purification and Characterization of
Hydrolase with Chitinase and Chitosanase Activity from Commercial Stem Bromelain.
J. Agric. Food Chem., 50, 4666-4673, 2002.
ICHIKAWA, S.; TAKANO, K.; TAKASHIKUROIWA, HIRUTA, O.; SATO, S.;
MUKATAKA, A. S. Immobilization and stabilization of chitosanase by multipoint
attachment to agar gel support. Journal of Bioscience and Bioengineering, v.93, n.2,
p.201-206, 2002.
JABOR, I. A. S.; PAMPHILE, J. A.; RODRIGUES, S. B.; MARQUES-SILVA, G. G.;
ROCHA, C. L. M. S. Análise do desenvolvimento do fungo entomopatogênico
Metarhizium anisopliae em resposta a fatores nutricionais. Acta Scientiarum.
Agronomy, Maringá, v.25, n.2, p.497-501, 2003.
JI, X.-J.; HUANG, H.; DU, J.; ZHU, J.-G.; REN, L.-J.; LI, S.; NIE, Z.-K. Development
of an industrial medium for economical 2,3-butanediol production through co-
fermentation of glucose and xylose by Klebsiella oxytoca, Bioresource Technology,
v.100, p.5214–5218, 2009.
JO, Y.; JO, K.; JIN, Y.; KIM, K.; SHIN, J.; KIM, Y.; PARK, R. Characterization and
kinetics of 45 kDa chitosanase from bacillus sp. P16. Biosci. Biotechnol. Biochem.,
v.67, n.9, 1875 – 1882, 2003.
KHOSRAVI-DARANI, K.; ZOGHI, A.; ALAVI, S. A.; FATEMI, S. S. A. Application
of Plackett Burman Design for Citric Acid Production from Pretreated and Untreated
Wheat Straw, Iran. J. Chem. Chem. Eng.. v.27, n.1, p.91-104, 2008.
KIM, P.; KANG, T. H.; CHUNG, K. J.; KIM, I. S.; CHUNG, K. Purification of a
constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and
expression of the gene. FEMS Microbiolog.y Letters 240, 31–39, 2004.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
84
KIM, S.-K.; RAJAPAKSE, N. Enzymatic production and biological activities of
chitosan oligosaccharides (COS): A review. Carbohydrate Polymers, v.62, n.4, p.357-
368, 2005.
KIRK, P. M.; CANON, P.F.; MINTER, D.W.; STALPERS, J.A. Dictionary of the
fungi. CAB International, Wallingford, 2008.
KONDO, H.; OSADA, A. Influence of dietary fiber on the bioavailability of zinc in
rats, Biomed Environ Sci, 9, 204-8, 1996.
KUMAR, S.; KOH, J. Physiochemical, Optical and Biological Activity of Chitosan-
Chromone Derivative for Biomedical Applications. Int. J. Mol. Sci., v.13, p.6102-6116,
2012.
KURITA, K.; SANNAN, T.; IWAKURA, Y. Studies on chitin, 4. Evidence for
formation of block and random copolymers of N-acetyl-o-glucosamine and
oglucosamine by hetero and homogeneous hydrolyses. Die Makromolecular Chemie,
v.178, p.3197-3202, 1977.
KUROIWA, T.; ICHIKAWA, S.; HIRUTA, O.; SATO, S.; MUKATAKA, S. Factors
Affecting the Composition of Oligosaccharides Produced in Chitosan Hydrolysis Using
Immobilized Chitosanases. Biotechnol. Prog., 18, 969-974, 2002.
LI, L.-Z.; HUI, Z.; MO, X. Optimization of the medium for streptolydigin production
by Streptomyces lydicus AS 4.2501 with precursor feeding using response surface
methodology, Journal of the Taiwan Institute of Chemical Engineers, 41, 252–258,
2010.
LIU, B.-L.; KAO, P.-M.; TZENG, Y.-M.; FENG, K.-C. Production of chitinase from
Verticillium lecanii F091 using submerged fermentation. Enzyme and Microbial
Technology, v.33, p.410-415, 2003.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
85
LOBATO, A. K. C. L. Estudo da produção de biosurfactantes por micro-organismos
isolados de poços de petróleo. 2003. 108p. Dissertação (Mestrado) – Programa de Pós-
Graduação em Engenharia Química. Universidade Federal do Rio Grande do Norte.
Natal.
LOPES, R. B.; ALVES, S. B.; PADULLA, L. F. L.; PÉREZ, C. A. Eficiência de
formulações de Beauveria bassiana e Metarhizium anisopliae para o controle de ninfas
de Amblyomma cajennense (Fabricius, 1787). Revista Brasileira de Parasitologia
Veterinária, v.16, n.1, p.27-31, 2007.
LOUREIRO, E. S.; BATISTA FILHO, A.; ALMEIDA, J. E. M.; MENDES, J. M.;
PESSOA, L. G. A. Eficiência de isolados de Metarhizium anisopliae (metsch.) sorok.
no controle da cigarrinha-da-raiz da cana-de-açúcar, Mahanarva fimbriolata (stal, 1854)
(hemiptera: cercopidae), em condições de campo. Arq. Inst. Biol., São Paulo, v.79, n.1,
p. 47-53, 2012.
MELLO, D. R.; REIS, R. C. S.; BITTENCOURT, V. R. E. P. Patogenicidade in vitro
do fungo Metarhizium anisopliae (Metschnikoff, 1879) Sorokin, 1883, sobre o carrapato
Boophilus microplus (Canestrini, 1887). Revista Brasileira de Parasitologia Veterinária,
São Paulo, v.15, p.157-162, 2006.
MITSUTOMI, M.; KIDOH, H.; TOMITA, H.; WATANABE, T. The action of Bacillus
circulans WL-12 chitinases on partially N-acetylated chitosan, Bioscience
Biotechnology and Biochemistry, v.59, p.529-531, 1995.
MALAJOVICH, M. A. Biotecnologia: fundamentos. Ed. biblioteca Max Feffer do
Instituto de Tecnologia ORT, Rio de Janeiro, 2009. 104p.
MONAGHAN, R. L.; EVELEIGH, D. E.; TEWARI, R. P.; REESE, E. T. Chitosanase,
a novel enzyme. Nature - New Biology, 245, p.78-80, 1973.
MU, X. Q.; XU, Y.; NIE, Y.; OUYANG, J.; SUN, Z. H. Candida parapsilosis CCTCC
M203011 and the optimization of fermentation medium for stereoinversion of (S)-1-
phenyl-1,2-ethanediol, Process Biochemistry, v.40, p.2345–2350, 2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
86
MURRAY, P. R.; ROSENTHAL, K. S.; PFALLER, M. A. Medical microbiology. 5th
ed. London, Elsevier Mosby, 2005.
MUZZARELLI, R. A. A. Biochemical significance of exogenous chitins and chitosans
in animals and patients, Carbohydrate Polymers, v.20, p.7-16, 1993.
MUZZARELLI, R. A. A.; TOMASETTI, M.; ILARI, P. Depolymerization of chitosan
with the aid of papain, Enzyme Microbiology and Technology, v.16, p.110-114, 1994.
NAVEENA, B. J.; ALTAF, Md.; BHADRIAH, K.; REDDY, G. Selection of medium
components by Plackett–Burman design for production of L(+) lactic acid by
Lactobacillus amylophilus GV6 in SSF using wheat bran, Bioresource Technology,
v.96, p.485-490, 2005.
NOGAWA, M.; TAKAHASHI, H.; KASHIWAGI, A.; OHSHIMA, K.; OKADA, H.;
MORIKAWA, Y. Purification and Characterization of Exo--D-Glucosaminidase from
a Cellulolytic Fungus, Trichoderma reesei PC-3-7, Appl. Environ. Microbiol., v.64, n.3,
p.890-895, 1998.
ONOFRE, S. B.; VARGAS; L. R. B.; ROSSATO, M.; BARROS, N. M.; BOLDO, J.
T.; NUNES, A. R. F.; AZEVEDO, J. L. Controle biológico de pragas na agropecuária
por meio de fungos entomopatogênicos. In: SERAFINI, L. A.; BARROS, N. M.;
AZEVEDO, J. L. (Org.). Biotecnologia – avanços na agricultura e na agroindústria.
Caxias do Sul: EDUCS, p.295-328, cap.8, 2002.
OSSWALD, W. F.; SHAPIRO, J. P.; DOOSTDAR, H.; MCDONALD, R. E.; NIEDZ,
R. P.; NAIRN, C. J.; HEARN, C. J.; MAYER, R. T. Identification and Characterization
of Acidic Hydrolases with Chitinase and Chitosanase Activities from Sweet Orange
Callus Tissue, Plant Cell Physiol. 35, p.811-820, 1994.
PAGNONCELLI, M. G. B. Estudo do mecanismo de produção de oligossacarídeos com
atividades nutracêuticas a partir da quitosana por hidrólise enzimática com processo

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
87
fermentativo simultâneo. 2008. 118p. Tese (Doutorado) – Centro de Tecnologia,
Programa de Pós-Graduação em Engenharia Química. Universidade Federal do Rio
Grande do Norte. Natal.
PALMA-GUERRERO, J., JANSSON, H.-B., SALINAS, J.; LOPEZ-LLORCA, L. V.
Effect of chitosan on hyphal growth and spore germination of plant pathogenic and
biocontrol fungi, J. Appl. Microbiol., v.104, p.541-553, 2008.
PAULA, F. A.; ARRUDA, D. C. Análise do método de planejamento experimental
Plackett Burman na otimização de degradação de penas de aves por comunidades
microbianas, XXI Salão de Iniciação Científica, 19-23, outubro, 2009, UFRGS, Porto
Alegre.
PELCZAR, M. J.; CHANG, E. C. S.; KRIEG, N. R. Microbiologia: conceitos e
aplicações, v.2, 2.ed. São Paulo: Makron BooKs, 1996.
PELLETIER, A.; SYGUSCH, J. Purification and characterization of three chitosanase
activities from Bacillus megaterium P1, Applied and Environmental Microbiology, v.
56, n.4, p.844-848, 1990.
PEREIRA, F. B.; GUIMARÃES, P. M. R.; TEIXEIRA, J. A.; DOMINGUES, L.
Optimization of low-cost medium for very high gravity ethanol fermentations by
Saccharomyces cerevisiae using statistical experimental designs, Bioresource
Technology, v.101, p.7856-7863, 2010.
PETER, M. G. Chitin and chitosan from aninal sources. In: STEINBUCHEL, A.;
RHEE, S. K. Polysaccharides and Polyamides in the Food Industry. Weinheim:
Wileyvch, v.1, p.115-208, 2005.
PIZA, F. A. T.; SILOTO, A. P.; CARVALHO, C. V.; FRANCO, T. T. Purification,
characterization and purification of chitosanase from Bacillus cereus, Brazilian Journal
of Chemical Engineering., v.16, n 2, p.185-192, 1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
88
POLITZER, K.; BON, E. P. S. Enzimas Industriais e Especiais. C.G.E.E. Ciência,
Tecnologia e Inovação. Rio de Janeiro, 554p, 2006.
POLYMAR, Ciência é a nossa Natureza, 2003. Disponível em:
<http://www.polymar.com.br>. Acesso em: 6 setembro 2012.
PUTZKE, J.; PUTZKE, M. T. L. Os reinos dos fungos. Santa Cruz do Sul, RS:
EDUNISC, 1998.
QUINES, L. K. M. Extração de poli(3-hidroxibutirato) produzido por Cupriavidus
necator dsm 545 com 1,2-carbonato de propileno. 2010. 99p. Dissertação (Mestrado em
Engenharia de Alimentos) – Centro Tecnológico, Departamento de Engenharia Química
e Engenharia de Alimentos, Programa de Pós-Graduação em Engenharia de Alimentos,
Universidade Federal de Santa Catarina, Florianópolis.
RAJENDRAN, A.; THANGAVELU, V. Statistical experimental design for evaluation
of medium components for lipase production by Rhizopus arrhizus MTCC 2233, LWT -
Food Science and Technology, v.42, p.985–992, 2009.
RAMIREZ-LEÓN, L. F.; RUIZ-HERRERA, J. Hydrolysis of walls and formation of
spheroplasts in Mucor rouxii. J. Gen. Microbiol., v.72, p.281-290, 1972.
RANGEL, M. A. S.; LOUREIRO, E. S.; ALMEIDA, A. S.; ALVES, H. C.; ONO, F.
B.; LOBO, P. M.; MARIANI, M. A.; PESSOA, L. G. A.; SILVA, A. J. Avaliação
participativa do controle de cigarrinhas das pastagens com a aplicação do fungo
Metarhizium anisopliae (Metsch.) Sorok. Rev. Bras. de Agroecologia, v.2, n.2, 731-734,
2007.
RAVEN, P. H.; EVERT, R.F.; EICHHORN, S.E. Biologia Vegetal, 6ª Ed, Rio Janeiro,
Editora Guanabara Koogan, 2001.
REDDY, L. V. A.; WEE, Y.-J.; YUN, J.-S.; RYU, H.-W. Optimization of alkaline
protease production by batch culture of Bacillus sp. RKY3 through Plackett–Burman

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
89
and response surface methodological approaches, Bioresource Technology, v.99, p.242–
2249, 2008.
RIGUEIRA, J. C. S. Influência da contagem de células somáticas no perfil e teores de
aminas bioativas e na qualidade de leite cru e de queijo mussarela. 2010. 147p. Tese
(Doutorado em Ciência de Alimentos) - Faculdade de Farmácia, Departamento de
Alimentos, Programa de Pós-Graduação em Ciência de Alimentos, Faculdade de
Farmácia da Universidade Federal de Minas Gerais, Belo Horizonte.
RODRIGUES, M. I.; IEMMA, A. F. Planejamento de Experimentos e Otimização de
Processos: uma estratégia sequencial de planejamentos, Campinas, SP, Casa do Pão
Editora, 2005.
SANTOS, E. S. Recuperação e Purificação de Enzimas usando Adsorção em Leito
Expandido, Tese de Doutorado, Universidade Estadual de Campinas, Campinas/SP,
p.152, 2001.
SEDMAK, J. J.; GROSSBERG, S. E. A rapid, sensitive and versatile assay for protein
using coomassie brilliant blue G250, Analytical Biochemistry, v.79, p. 544-552, 1977.
SEINO, H.; TSUKUDA, K.; SHIMASUE, Y. Properties and action pattern of a
chitosanase from Bacillus sp. PI- 7S. Agricultural Biological Chemistry, 55, p.2421-
2423, 1991.
SHIMOSAKA, M.; NOGAWA, M.; OHNO, Y.; OKAZAKI, M. Chitosanase from the
plant pathogenic fungus, Fusarium solani f. sp. Phaseoli – purification and some
properties, Bioscience Biotechnolgy and Biochemistry, v.57, n.2, p.231-235, 1993.
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Production of two chitosanases from a chitosan-assimilating bacterium, Acinetobacter
sp. Strain CHB101, Applied and Environmental Microbiology, v.61, n.2, p.438-442,
1995.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
90
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Characterization of Three Chitosanase Isozymes Isolated from a Commercial Crude
Porcine Pepsin Preparation, J. Agric. Food Chem. 51, p.1042-1048, 2003.
SILVA FILHO, R. C.; COSTA, A. S. S.; CHAGAS, B. M. E.; SANTOS, F. N.;
MACEDO, G. R.; SANTOS, E. S. Avaliação do teste de atividade quitosanolítica
produzida por fungo. In: XV SIMPÓSIO NACIONAL DE BIOPROCESSOS, 15,
Recife, 2005, 02 a 05 de agosto.
SILVA, E. P. Avaliação Preliminar do Potencial de Aplicação da Tecnologia de
Biopilhas para a Biorremediação do Solo de Guamaré-RN. 2004. 119p. Dissertação
(Mestrado) – Centro de Tecnologia, Programa de Pós-Graduação em Engenharia
Química. Universidade Federal do Rio Grande do Norte. Natal.
SOMASHEKAR, D.; JOSEPH, R. Chitosanases – Properties and apllications: A review.
Bioresource Technology, v.55, p.35-45, 1996.
STOYACHENKO, I. A.; VARLAMOV, V. P.; DAVANKOV, V. A. Chitinases of
Streptomyces kurssanovii: purification and some properties, Carbohydrate Polymers,
v.24, p.47-54, 1994.
TINLINE, R. D. Nuclear distribuition in Metarhizium anisopliae. Mycologia, v. 63, p.
713-721, 1971.
TRABULSI, L. R.; TOLEDO, M. R. F. Microbiologia. 2. Ed., São Paulo: Atheneu,
1996.
VIEIRA, P. D. S.; SILVA, W. M. T.; PAIVA, L. M.; LIMA, E. A. L. A.;
CAVALCANTI, V. L. B. Estudo da caracterização morfológica, esporulação e
germinação dos conídiosde Metarhizium anisopliae var. acridum em diferentes
temperaturas. Biológico, São Paulo, v.69, n.1, p.17-21, 2007.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
91
WANDERLEY, M. D.; NEVES, E.; ANDRADE, C. J. Aspectos da produção industrial
de enzimas. Revista Citino, v.1, n.1, p.30-36, 2011.
WANG, C.; TYPAS, M. A.; BUTT, T. M. Detection and characterization of pr1
virulent gene deficiencies in the insect pathogenic fungus Metarhizium anisopliae.
FEMS Microbiology Letters, v. 213, p. 251-255, 2002.
ZAHORSKA-MARKIEWICZ, B.; KROTKIEWSKI, M.; OLSZANECKA-
GLINIANOWICZ, M.; ZURAKOWSKI, A. Effect of chitosan in complex management
of obesity, Pol Merkur Lekarski, v.13, n.74, p.129-132, 2002.
ZHANG, X. Y.; DAI, A. L.; ZHANG, X. K.; KUROIWA, K.; KODAIRA, R.;
SHIMOSAKA, M.; OKAZAKI, M. Purification and Characterization of and Exo--D-
glucosaminidase from a koji mold, Aspergillus oryzae IAM2660. Biosci. Biotechnol.
Biochem., v.64, n.9, 1896-1902, 2000.
ZHU, X.; WU, X.; DAI, Y. Fermentation conditions of a chitosanase from Acinetobcter
sp C-17. Biosci. Biotechnol. Biochem., v.67, n.2, p.284-290, 2003.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
92
APÊNDICE

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
93
APÊNDICE I
Metodologia para repique da cepa
Em Erlenmeyer de 1000 mL colocava-se 500 mL do meio de manutenção
PDA. Este Erlenmeyer, com o meio de manutenção, era autoclavado junto com placas
de Petri a 1,5 kgf/cm2 por 15 minutos, em seguida, era levado para uma câmara de fluxo
laminar e transferido para as placas de Petri já autoclavadas, onde eram mantidas numa
estufa a 27 ºC durante 24 h. Após este período, as placas de Petri eram levadas a câmara
de fluxo laminar, utilizando bico de Bunsen para esterilizar a alça de platina. Procedia-
se o repique utilizando uma placa de Petri contendo a cepa pura. Cada placa de Petri
contendo a cepa pura era transferida para duas placas de Petri contendo o meio de
manutenção. As placas eram mantidas na estufa a 27 ºC durante 30 dias onde após esse
período, repetia-se este procedimento.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
94
APÊNDICE II
Montagem da Curva de Calibração Usando o Método DNS
Soluções:
Glicosamina = 0,01 mol/mL (10000,0 µmol/mL)
Tabela 14. Dados para construção da curva de calibração usando o Método DNS.
Tubo de
Ensaio
Solução de
glicosamina (mL)
Água Destilada
(mL)
DNS (mL)
Volume Total
Massa (µµµµmol)
1 0 1,0 1,5 2,5 0
2 0,2 0,8 1,5 2,5 0,2
3 0,4 0,6 1,5 2,5 0,4
4 0,6 0,4 1,5 2,5 0,6
5 0,8 0,2 1,5 2,5 0,8
6 1,0 0 1,5 2,5 1,0
Tabela 15 - Medidas de absorbância a 600nm
Dados adicionais:
Peso Molecular D-Glicosamina = 215,64 (recipiente SIGMA)
Conc. Sol. D-Glicosamina = 0,05 g/50 mL
Massa calc.(µmol) = conc. Sol. D-Glic. x Vol. Utiliz. D-Glics. =
= 0,05 x 1000 x Vol.util./50*215,64
Tempo atividade enzimática = 30 min
1 U/mL = 1 µmol/(min.mL)
Vol. Total (Glicosamina+caldo) = 1 mL
Medições Adsorbância a 600 nm Vol. Utiliz. Massa Massa Atividade
II III Média(Abs)(mL) calc. (mg) calc. (µg) Enzim. (U/mL)
0 0 0 0 0 0 0
0,133 0,128 0,1307 0,2 0,2 200 0,0309
0,279 0,277 0,2787 0,4 0,4 400 0,0618
0,433 0,428 0,4297 0,6 0,6 600 0,0927
0,587 0,583 0,5797 0,8 0,8 800 0,1237
0,727 0,716 0,7157 1 1 1000 0,1546

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
95
Figura 17 – Curva padrão de glicosamina.
y = 1900ralx + 1900ral
R² = 1900ral
00,000
00,020
00,040
00,060
00,080
00,100
00,120
00,140
00,160
00,180
00,000 00,100 00,200 00,300 00,400 00,500 00,600 00,700 00,800
Ati
v.
En
zim
áti
ca (
U/m
L)
Absorbância (Abs) a 600 nm
Curva-Padrão D-Glicosamina

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
96
APÊNDICE III
Método de Determinação de Proteína Utilizando o Corante Comassie
Brilhante Blue G 250 (Método modificado de Sedmak e Grossberg)
Micrométodo
Preparar uma solução 0,06% de Comassie Blue G250 em 1,5 % HCl (w/V). Filtrar e
papel de filtro Whatman nº1.
Curva Padrão (1 a 20 µµµµg)
a) Preparar soluções estoque de BSA de 10 a 200 µg/mL.
b) Colocar em tubos de ensaio 100 µL da solução estoque, 1400 µL de água e
1500µL do Comassie e agitar.
c) Preparar um tubo contendo 1500 µL de água e 1500 µL do corante que
corresponde ao zero de proteína.
d) Após 5 minutos (padronizar o tempo) fazer as leituras das absorbâncias.
e) As leituras das amostras devem ser feitas em dois comprimentos de onda 595 e
465 contra a água.
f) Plotar a razão das absorbâncias a 595/465 como uma função da massa de BSA e
calcular o melhor ajuste linear dos dados.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
97
Micrométodo (1 a 20 µµµµg de BSA)
Solução Estoque de BSA = 0,2 mg/mL
Tabela 16. Dados para construção da curva padrão de BSA.
Tubo de
Ensaio
H2O (µµµµL)
BSA (µµµµg)
Coomassie (µµµµL)
BSA (µµµµg)
1 1500 0 1500 0
2 1490 10 1500 2
3 1480 20 1500 4
4 1470 30 1500 6
5 1460 40 1500 8
6 1450 50 1500 10
7 1440 60 1500 12
8 1430 70 1500 14
9 1420 80 1500 16
10 1410 90 1500 18
11 1400 100 1500 20
Dados adicionais:
Concentração Sol. Estoque
BSA = 0,2 mg/mL = 0,2 µg/µL
Vol. total sol. = vol. H2O + vol. BSA + vol.
Coomassie = 3000 µL = 3 mL
Concentração Sol. BSA (mg/mL) = (massa BSA mg) / 3 mL
.
Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
98
Figura 18 – Curva Padrão de BSA
y = 1900ralx - 1900ral
R² = 1900ral
000,000
001,000
002,000
003,000
004,000
005,000
006,000
007,000
000,400 000,500 000,600 000,700 000,800 000,900 001,000 001,100
µg
BS
A
Razão Absorbâncias (595nm/465nm)
Curva-Padrão BSA

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE TECNOLOGIA
DEPARTAMENTO DE ENGENHARIA QUÍMICA PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA
TESE DE DOUTORADO
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em
Cultivo Descontínuo Submerso
Raimundo Cosme da Silva Filho
Natal/RN
Abril/2013

Raimundo Cosme da Silva Filho
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo
Descontínuo Submerso
Tese apresentada ao Programa de Pós-Graduação em Engenharia Química – PPGEQ, da Universidade Federal do Rio Grande do Norte, como parte dos requisitos para obtenção do título de Doutor em Engenharia Química, sob a orientação do Prof. Dr. Everaldo Silvino dos Santos e a coorientação da Profª. Drª. Gorete Ribeiro de Macedo.
Natal/RN
Abril/2013

Catalogação da Publicação na Fonte. UFRN / CT / PPGEQ
Biblioteca Setorial “Professor Horácio Nícolas Sólimo”.
Silva Filho, Raimundo Cosme da. Otimização do meio de cultura para a produção de quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso / Raimundo Cosme da Silva Filho. - Natal, 2013. 98 f.: il.
Orientador: Everaldo Silvino dos Santos. Co-orientador: Gorete Ribeiro de Macedo. Tese (Doutorado) - Universidade Federal do Rio Grande do Norte.
Centro de Tecnologia. Departamento de Engenharia Química. Programa de Pós-Graduação em Engenharia Química.
1. Quitosana - Tese. 2. Quitina - Tese. 3. Quitosanase – Tese. 4. Metarhizium
anisopliae - Tese. I. Santos, Everaldo Silvino dos. II. Macedo, Gorete Ribeiro de. III. Universidade Federal do Rio Grande do Norte. IV. Título. RN/UF/BSEQ CDU 547.458 (043.2)

SILVA FILHO, Raimundo Cosme da – Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo Descontínuo Submerso. Tese de Doutorado. UFRN - Programa de Pós-Graduação em Engenharia Química. Área de Concentração: Pesquisa e Desenvolvimento de Tecnologias Regionais – Subárea: Engenharia de Processos – Alimentos e Biotecnologia, Natal/RN, Brasil.
Orientação: Prof. Dr. Everaldo Silvino dos Santos Coorientação: Profª. Drª. Gorete Ribeiro de Macedo ___________________________________________________________________________
RESUMO: No presente trabalho utilizou-se um Planejamento Plackett & Burman, com 8 fatores e 12 ensaios em 2 níveis e mais 3 repetições na condição do ponto central, para se investigar a influência das concentrações de quitosana, peptona, extrato de levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O e FeSO4 na produção da enzima quitosanase por Metarhizium
anisopliae. Os ensaios para produção da enzima foram realizados em cultivo descontínuo submerso. Os resultados para o Planejamento Plackett & Burman mostraram que a atividade quitosanolítica foi favorecida pelo aumento da concentração de substrato (quitosana) e de sulfato ferroso (FeSO4), enquanto que o aumento da concentração dos outros fatores não contribuiu de forma significativa para a atividade quitosanolítica. A otimização do meio de cultura para a produção da enzima foi realizado por meio de um Planejamento Composto Central, com os dois fatores que mais influenciaram a atividade quitosanolítica (quitosana e FeSO4), conforme Planejamento Plackett & Burman, mantendo-se os outros nutrientes em seus valores mínimos. Nesse planejamento, para o fator FeSO4, tomou-se o limite inferior (-1) como sendo o limite superior do Planejamento Plackett & Burman. Os resultados mostraram que a produção da enzima foi favorecida pelo aumento da concentração quitosana e pela diminuição da concentração de FeSO4. A produção máxima de atividade quitosanolítica foi da ordem de 70,0 U/L e foi atingida em apenas 18 h de fermentação, resultado esse vinte e oito vezes maior aos obtidos anteriormente para o mesmo microrganismo que foi 2,5 U/L em 48h.
Palavras-chave: quitosana; quitosanase; Metarhizium anisopliae; Plackett & Burman.


ABSTRACT
In this work a Plackett-Burman Design with 8 factors and 12 trials in 2 levels with 3 repetitions at the center point was used in order to investigate the influence of the concentration of chitosan, peptone, yeast extract, NaNO3, K2HPO4, KCl, MgSO4.7H2O and FeSO4 on chitosanase production by Metarhizium anisopliae. Runs were carried out using submerged discontinuous cultivation for enzyme production. The results of the Plackett & Burman Design showed that only two factors, chitosan concentration as well as FeSO4 had influence on chitosanolytic activity, while the increase in concentration of other factors not contributed significantly to the quitosanolítica activity. Cultivation medium optimization for enzyme production was carried out using a Composite Central Design, with the most important factors for chitosanolytic activity (chitosan and FeSO4), in accordance with Plackett & Burman Design, and keeping the other nutrients in their minimum values. On this other design, it was taken the highest limit in Plackett & Burman Design as the lowest limit (-1) to FeSO4 factor. The results showed that the enzyme production was favoured by increasing the chitosan concentration and by decreasing FeSO4. Maximum production for chitosanolytic activity was about 70.0 U/L and was reached in only 18 h of fermentation, a result about twenty-eight times greater than a former study using the same microorganism (about 2.5 U/L at 48 h).
Key-words: Chitosan; Chitosanase; Metarhizium anisopliae; Plackett & Burman

Ao meu pai, minha mãe e minha
família, em especial aos meus
irmãos e sobrinhos

AGRADECIMENTOS
A Deus, por estar presente em todos os momentos da minha vida e por me ceder à
oportunidade de iniciar e concluir este trabalho.
A minha mãe Cândida Inocência da Silva que esteve comigo em todos os momentos da
minha vida me educando, me amando e sempre me dando muito carinho.
Ao meu pai Raimundo Cosme da Silva, que não está mais aqui para ver este trabalho,
mas que foi a pessoa que mais me apoiou e me incentivou, em todos os momentos da minha vida,
para que eu pudesse chegar o mais longe possível na minha carreira acadêmica. Pai muito
obrigado por toda atenção, educação, amor e carinho que recebi durante toda a minha vida.
Aos meus irmãos e familiares que sempre estiveram comigo dividindo momentos de
grande alegria e às vezes de tristeza, mas sempre nos mantendo unidos para superarmos todos os
obstáculos.
A minha namorada Rochelly Caroliny por todo amor e carinho que me deu e que me
entendeu nos momentos que estive ausente para a conclusão deste trabalho.
Ao meu orientador Professor Doutor Everaldo Silvino dos Santos por confiar em mim e
me aceitar como orientando além de toda a amizade, orientação e dedicação em todas as etapas de
desenvolvimento e conclusão deste trabalho.
À Professora Doutora Gorete Ribeiro de Macedo, “pessoa que admiro muito como
profissional e principalmente, como pessoa, um exemplo a ser seguido”, pelos conselhos e críticas
quando necessários ao desenvolvimento deste trabalho. Professora a senhora é considerada uma
mãe para de todos os alunos que tiveram a honra de conhecê-la e trabalhar com a senhora.
Aos meus amigos Engenheiros Químicos, Ana Carmen, Ana Katerine, Andréa Farias,
Anita Lima, Sirtys Lessa, Alexandre, Sérgio, Márcio Bezerra e ao Biólogo Daniel Souza, que
sempre me ouviram e que me ajudaram de alguma forma nesse trabalho.
A Eusamar Coelho de Lima, Mazinha, secretária do PPGEQ, pelo apoio, pela paciência e
orientação nas questões burocráticas do processo.
A Carlos e Jardelson, pela grande colaboração e dedicação nos experimentos do
laboratório.
A Sérgio Santana pela grande colaboração e dedicação, o qual não poupou esforços nem
tempo em me ajudar na realização dos experimentos em laboratório.

Ao meu amigo Ubiratan Nazaré por me ajudar na organização do texto e na tradução para
o Abstract.
Finalizando, agradeço ao Laboratório de Engenharia Bioquímica da UFRN por ter cedido
toda a sua estrutura para realização dos experimentos e ao Programa de Pós-Graduação em
Engenharia Química, assim como a todos os professores do Departamento de Engenharia Química
da UFRN que trabalham em pró de uma educação digna e ensino de qualidade para todos os
alunos.

Sumário LISTA DE FIGURAS
LISTA DE TABELAS
NOMENCLATURA
INTRODUÇÃO...............................................................................................................................15
REVISÃO BIBLIOGRÁFICA........................................................................................................18
2.1. Enzimas................................................................................................................................18
2.1.1. Quitosana e seus derivados........................................................................................19
2.1.2. Quitosanases...............................................................................................................24
2.2. Fungos..................................................................................................................................29
2.2.1. Características gerais..................................................................................................29
2.2.2. Reprodução dos Fungos.............................................................................................32
2.2.3. Metabolismo de fungos..............................................................................................33
2.2.4. Nutrição de fungos.....................................................................................................34
2.2.5. O Fungo Metarhizium anisopliae ..............................................................................36
2.3. Fermentação.........................................................................................................................40
2.3.1. Preparo do inóculo......................................................................................................41
2.3.2. Produção de enzimas em cultivo submerso descontínuo...........................................42
2.4. Planejamento experimental..................................................................................................42
2.4.1. Planejamento fatorial..................................................................................................43
2.4.2. Planejamento Plackett & Burman..............................................................................43
2.4.3. Aplicações do Planejamento Plackett & Burman.......................................................44
METODOLOGIA EXPERIMENTAL.............................................................................................50
3.1. Quitosana..............................................................................................................................50
3.2. Microrganismo.....................................................................................................................50
3.2.1. Escolha da cepa produtora de quitosanase.................................................................50

3.2.2. Manutenção da cepa...................................................................................................50
3.3. Produção de enzima.............................................................................................................51
3.4. Determinação da Proteína Total...........................................................................................51
3.5. Determinação da Atividade Quitosanolítica e Atividade Específica...................................51
3.6. Planejamento Experimental.................................................................................................52
3.6.1. Planejamento de seleção de fatores Plackett Burman................................................52
3.6.2. Planejamento Composto Central para Otimização do meio de cultura para a
Produção da Enzima em Cultivo Submerso.....................................................................................53
3.7. Perfil Cinético durante o Cultivo.........................................................................................56
RESULTADOS E DISCUSSÃO.....................................................................................................58
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA ..........................................58
4.2. Planejamento Plackett & Burman........................................................................................59
4.2.1. Análise do Plackett & Burman com 8 fatores e 12 ensaios .......................................59
4.2.2. Análise do Plackett & Burman com 12 ensaios e mais 3 repetições na condição do
ponto central ....................................................................................................................................61
4.3. Planejamento Composto Central para Produção de Quitosanase por Metarhizium
anisopliae em Cultivo Descontínuo Submerso................................................................................64
4.4. Perfil Cinético – Melhor Condição......................................................................................69
CONCLUSÃO.................................................................................................................................75
REFERÊNCIAS BIBLIOGRÁFICAS.............................................................................................78
APÊNDICE I...................................................................................................................................93
Metodologia para Repique da Cepa ................................................................................................93
APÊNDICE II.................................................................................................................................94
Montagem da Curva de Calibração Usando o Método DNS..........................................................94
APÊNDICE III................................................................................................................................96
Método de Determinação de Proteína Utilizando o Corante Comassie Brilhante Blue G 250
(Método modificado de Sedmak e Grossberg)................................................................................96

Lista de Figuras
Figura 1 – Estrutura da quitosana..................................................................................................20
Figura 2 – Estrutura da glicosamina..............................................................................................23
Figura 3 – Estrutura do sulfato de glicosamina.............................................................................23
Figura 4 – Representação da estrutura geral de uma célula de fungo..........................................30
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas...........................................................31
Figura 6 – Conídios de Aspergillus e Penicillium.........................................................................33
Figura 7 – Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA ...................58
Figura 8 – Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados............................................................................................................................60
Figura 9 – Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto central
mostrando a influência dos fatores estudados................................................................................63
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos
fatores estudados............................................................................................................................67
Figura 11 – Valores preditos em função dos valores observados..................................................68
Figura 12 – Atividade quitosanolítica em função da concentração de quitosana e da concentração
de FeSO4 para os fatores não significativos no ponto central .......................................................68
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana ........70
Figura 14 – Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana....................................70
Figura 15 – Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de
quitosana........................................................................................................................................71
Figura 16 – Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana..........................................................................................................72
Figura 17 – Curva padrão de glicosamina ....................................................................................95
Figura 18 – Curva Padrão de BSA ................................................................................................98

Lista de Tabelas
Tabela 1 – Comparação das enzimas com os catalisadores químicos..............................................19
Tabela 2 – Fontes naturais de quitina e quitosana ...........................................................................20
Tabela 3 – Níveis para os fatores e seus valores codificados...........................................................53
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central .........................................................................................53
Tabela 5 – Matriz do planejamento Composto Central...................................................................54
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central......................................................................................................55
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.......................59
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman...............................................60
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.......................61
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central...............................................................................................................................................62
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da atividade
enzimática........................................................................................................................................65
Tabela 12 – Efeitos Estimados do planejamento Composto Central...............................................65
Tabela 13 – Resumo da ANOVA....................................................................................................66
Tabela 14 – Dados para construção da curva de calibração usando o Método DNS ......................94
Tabela 15 – Medidas de absorbância a 600 nm para construção da curva de calibração................94
Tabela 16 – Dados para construção da curva padrão de BSA ........................................................97

NOMENCLATURA
A – Atividade quitosanolítica (U/L)
AE – Atividade específica (U/mg)
ANN-GA – algoritmo genético de rede neural artificial
ANOVA – Análise de variância (Analysis of Variance)
BSA – Albumina de soro bovino (Bovine Sereum Albumin)
CCD – Planejamento Composto Central
CDA – Chitosan Dector Agar
Cs – Concentração de substrato (%)
CP – Concentração de KCl (%)
Da – Dalton
DEQ – Departamento de Engenharia Química
DNS – Ácido Dinitrosalissílico
E – Enzima
Ea – Energia de ativação
EC – Comissão para nomenclatura e classificação de enzimas
EL – Concentração de Extrato de Levedura (%)
FP – Concentração de K2HPO4 (%)
NS – Concentração de NaNO3 (%)
GDA – Grânulos dispersíveis em água
k – Número de fatores, variáveis
LEB – Laboratório de Engenharia Bioquímica
mRNA – RNA mensageiro
n – Número de níveis de um determinado fator
N – Número de ensaios de um planejamento
P – Pressão
PP – Concentração de Peptona (%)
PDA – Batata Dextrose Agar
pH – Potencial hidrogeniônico
Q – Concentração de Quitosana (%)
Qm – Quadrado médio
r – Agitação (rpm)
R2 – Coeficiente de determinação

RA – Razão de aeração
rpm – Rotações por minuto
RSM – Metodologia de superfície de resposta
S – Molécula de substrato
SF – Concentração de FeSO4 (%)
SM – Concentração de MgSO4.7H2O (%)
SQ – Soma de quadrados
SSF – Fermentação de estado sólido
T – Temperatura
TMC – Temperatura de cultivo
V11,V12,V13 – Variáveis inertes para cálculo do erro
VHG – Fermentação de alta densidade
WP – Pó molhável
X – Concentração celular
Xi – variável independente
Xj – variável independente
xi – Fator independente real
x0 – Valor de xi no ponto central
∆xi – Variação de valor
Y – Resposta do modelo de primeira ordem
Yp – Resposta do modelo polinomial de segunda ordem
β0 – Interseção dos modelos de primeira ou segunda ordem
βi – Coeficiente linear dos modelos de primeira ou segunda ordem
βii – Coeficiente quadrático
βij – Coeficiente de interação

Capítulo 1 Introdução

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
15
1. Introdução As quitosanases foram descobertas simultaneamente há quase quarenta anos por
Monaghan et al. (1973), como pelo grupo de Ramirez-León & Ruiz-Herrera (1972), e
são enzimas obtidas a partir do metabolismo de bactérias, fungos e extraídas de plantas.
Desde então, vários estudos vêm apresentando a obtenção de quitosanase deste
microrganismos, na forma intracelular ou extracelular (Osswald et al., 1994;
Somashekar & Joseph, 1996), como a do presente estudo. A quitosanase (EC 3.2.1.132)
representa uma classe de enzimas hidrolíticas que catalisam a hidrólise de ligação β-
(1→ 4) glicosídica da quitosana, gerando oligômeros chamados quito-oligossacarídeos
(Reddy et al., 2008).
A quitosana é um biopolímero hidrofílico obtido a partir da quitina, um
polissacarídeo abundante na natureza. Na literatura, é considerado quitosana o produto
da desacetilação da quitina para um grau de desacetilação superior a 75% e solúveis em
ácidos como o acético e o fórmico. A quitosana é conhecida por exibir uma ampla
variedade de atividade fisiológica como atividade antitumoral, mas o peso molecular e a
insolubilidade em água da quitosana são desvantagens para várias aplicações.
Atualmente, há interesse nas quitosanas quando as mesmas são convertidas em quito-
oligossacarídeos porque eles não apenas são solúveis em água e possuem baixo peso
molecular, mas também têm atividades biológicas bem notáveis, como atividade
anticancerígena atóxica e biocompatível, incluindo efeito antimetastático, atividade anti-
HIV, hepatoprotetora, antioxidante, hipocolesterolêmica, antimicrobiana,
imunoestimulate, antitumoral, anti-inflamatória e atividade aceleradora da absorção de
cálcio e ferro. No entanto, estes quito-oligossacarídeos são obtidos tradicionalmente na
indústria por reação de hidrólise usando ácidos, que apresenta muitos problemas como
dificuldade em controlar a hidrólise, produzindo uma grande quantidade de
oligossacarídeos de cadeia pequena e oligossacarídeos de baixo rendimento, alto custo
de separação e, também, poluição ambiental, uma vez que utiliza um ácido como
catalisador. Como alternativa, com suas vantagens em termos de compatibilidade
ambiental, baixo custo e reprodutibilidade, a obtenção destes quito-oligossacarídeos por
hidrólise com quitosanase se tornou cada vez mais popular nos últimos anos (Azevedo
et al., 2007).

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
16
Devido a todos estes aspectos positivos é que, em geral, as enzimas vêm sendo
estudadas em diferentes aplicações, mostrando que esta é uma tendência na indústria.
No presente trabalho, foi desenvolvido um processo fermentativo visando a
otimização do meio de cultura para a produção da enzima quitosanase em cultivo
descontínuo submerso, por Metarhizium anisopliae, capaz de sintetizar oligômeros com
aplicações na indústria farmacêutica e de química fina, partindo de quitosana como
única fonte de carbono.
Nesse caso, avaliou-se, através de um planejamento Plackett & Burman (método
de delineamento de seleção de fatores) e de um planejamento Composto Central as
melhores condições para a produção da enzima. A técnica do planejamento
experimental Plackett & Burman para realizar triagem de fatores na otimização de um
determinado processo foi utilizada neste trabalho para identificar quais os fatores que
influenciam de forma significativa na produção da enzima quitosanase. A influência
destes fatores (concentração de quitosana e concentração de FeSO4) foi então
investigada pelo planejamento Composto Central com 22 experimentos na parte fatorial,
repetição em triplicata no ponto central e quatro experimentos na parte axial. A
repetição do ponto central permite também a determinação do erro puro e da
repetibilidade do processo, sendo uma forma de identificar melhor se o processo está
sob controle.
Do ponto de vista industrial é de relevante importância a realização de estudos
para a otimização da produção da quitosanase de forma contínua ou em grandes
bateladas.

Capítulo 2 Revisão Bibliográfica

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
18
2. Revisão Bibliográfica 2.1. Enzimas
As enzimas são catalisadores biológicos constituídos por longas cadeias de
aminoácidos que atuam em substratos reduzindo a energia de ativação, acelerando assim
a velocidade da reação, e transformando-os em produtos. Sua atuação catalítica visa
facilitar a atividade das células através da quebra de moléculas ou junção das mesmas
para formação de novos produtos. Podem apresentar alto grau de especificidade relativa
a apenas um substrato ou a vários substratos ao mesmo tempo, como a certos tipos de
ligações glicosídicas α-1,4 das moléculas de amido, ou para determinado tipo particular
de isômero óptico como ocorre na oxidação da β-D-glicose pela glicose oxidase (Furigo
Junior & Pereira, 2001).
Enzimas são proteínas que apresentam em sua estrutura um centro ativo
(apoenzima) e, em alguns casos, um grupo prostético denominado cofator (coenzima ou
íon metálico ativo) e sua atividade depende de alguns fatores como estrutura da
proteína, número de cadeias peptídicas, arranjo dessas cadeias na molécula, natureza do
substrato e natureza do grupo prostético (Politzer & Bon, 2006).
Todos os organismos vivos produzem, em pequena ou grande quantidade, certa
variedade de enzimas, sejam elas intra ou extracelulares. As enzimas extracelulares são
excretadas para o meio externo e podem decompor compostos insolúveis como
celulose, quitina e quitosana.
Algumas destas enzimas podem ser usadas tanto nas indústrias de detergentes,
têxteis e farmacêuticas, como também de alimentos, de bebidas, etc., sendo utilizadas
em processos biotecnológicos industriais em substituição aos processos químicos que
muitas vezes causam grandes prejuízos ao meio ambiente. A produção comercial das
enzimas pode ser realizada através do cultivo de microrganismos como fungos e
bactérias.
Na Tabela 1 mostra-se uma comparação das enzimas com os catalisadores
químicos, levando-se em conta algumas das características dos dois catalisadores.
Destaca-se que alguns parâmetros da Tabela 1 demonstram a maior importância das
enzimas, quando comparadas com os catalisadores químicos, principalmente a
especificidade ao substrato, as condições de reação, o consumo de energia e a

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
19
velocidade de reação, como desvantagem cita-se o custo de obtenção (isolamento e
purificação).
Tabela 1 – Comparação das enzimas com os catalisadores químicos (Adaptado de
Furigo Junior & Pereira, 2001).
Característica Enzimas Catalisadores Químicos
Especificidade ao substrato Alta Baixa
Natureza da estrutura Complexa Simples
Sensibilidade à T e pH Alta Baixa
Condições de reação (T, P e pH) Suaves Drástica (geralmente)
Custo de obtenção (isolamento e purificação) Alto Moderado
Natureza do processo Batelada Contínuo
Consumo de energia Baixo Alto
Formação de subprodutos Baixa Alto
Separação catalisador/produtos Difícil/cara Simples
Atividade catalítica (temperatura ambiente) Alta Baixa
Presença de cofatores Sim Não
Estabilidade do preparado Baixa Alta
Energia de Ativação Baixa Alta
Velocidade de reação Alta Baixa
2.1.1. Quitosana e seus derivados
A quitosana e seus derivados apresentam grande importância devido ao
conhecimento de suas funcionalidades biológicas, médicas, tecnológicas, etc
(Shimosaka et al, 1993; Shimosaka et al, 1995; Mitsutomi et al., 1995). A quitosana é
obtida da quitina, polissacarídeo abundante na natureza, extraído principalmente das
carapaças de caranguejos, lagosta e camarões. A quitosana é um polímero natural,
biodegradável, não tóxico e de alta massa molecular obtido a partir de animais
marinhos, insetos e microrganismos (Tabela 2).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
20
Tabela 2 – Fontes naturais de quitina e quitosana. (Adaptado de Polymar, 2003).
Animais
Marinhos
Insetos Aracnídeos Microrganismos
Anelídios Formigas Escorpiões Leveduras
Moluscos Besouros Aranhas Fungos
Lagosta Esporos
Camarão
Caranguejo
Krill
A Figura 1 abaixo mostra a estrutura química da quitosana
Figura 1 – Estrutura da quitosana (Fonte: Polymar, 2003)
Quitosana totalmente desacetilada não existe na natureza. Graus de acetilação
(GA) nferior a 50% a torna mais solúvel em soluções ácidas (Muzzarelli, 1993;
Muzzarelli et al., 1994; Dung et al., 1994; Stoyachenko et al.,1994). Pelletier &
Sygusch (1990) enfatizaram que grande parte dos resíduos sólidos da indústria
pesqueira, principalmente da carcinicultura, é jogada ao mar ocasionando degradação
ambiental. Logo, a utilização desses resíduos como matéria-prima para formação de
novos produtos, não só reduz de forma drástica o impacto ambiental, mas também daria
maior valor agregado à indústria pesqueira.
A demanda cada vez maior de consumidores por alimentos mais saudáveis, sem
conservantes químicos, tem motivado pesquisas à descoberta de novos agentes
antimicrobianos naturais. Sendo assim, a atividade antimicrobiana incomum da

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
21
quitosana e seus derivados contra microrganismos, tais como bactérias, bolores e
leveduras tem tido maior atenção ultimamente.
Devido à interação entre a quitosana, que é positivamente carregada, e as
membranas celulares dos microrganismos, negativamente carregadas, ocorre a formação
de verdadeiros poros na membrana, que são canais de vazamento de conteúdo protéico e
outros componentes intracelulares. A quitosana tem também a propriedade de agir como
agente quelante que se liga de forma seletiva a traços de metais, inibindo, assim, a
produção de toxinas e o crescimento microbiano. Ainda é capaz de ativar mecanismos
de defesa em tecidos hospedeiros, atuando como agente sequestrante de água e inibindo
várias enzimas. A ligação da quitosana com DNA e a inibição da síntese do mRNA
ocorre via penetração da quitosana no núcleo dos microrganismos e interferência na
síntese do mRNA e proteínas, (Craveiro et al., 2004).
Em um estudo clínico conduzido por Giustina & Ventura (1995) com 100
pessoas com sobrepeso de 10 a 25%, ao final de 4 semanas as pessoas que receberam
quitosana demonstraram perda de peso significativa (7,3 Kg contra 3,0 Kg nos grupos
controle). As pessoas que tomaram quitosana também demonstraram redução na pressão
sanguínea sistólica (de 145 para 135 mm Hg) e diastólica (92.6 para 84.2 mm Hg),
assim como uma redução na taxa respiratória (27.6/min a 21.2/min).
Kondo & Osada (1996) pesquisaram a influência da quitina e da quitosana na
biodisponibilidade do zinco em ratos alimentados com uma dieta com suplementos de
5% ou sem fibra dietética (quitina, quitosana, celulose, pectina e agar-agar) durante 31
dias. Observaram que os ratos alimentados com dietas contendo 5% de fibras, exceto
quitosana, o consumo de alimento foi maior do que o controle. O ganho de peso em
todos os grupos alimentados com fibra foi maior do que o controle, exceto para o grupo
suplementado com quitosana (5%) onde a ingestão de alimento e o ganho de peso foram
menores do que os grupos controle e das demais fibras.
Segundo Craveiro et al. (2004), derivados da quitosana obtidos pela reação com
ácidos carboxílicos, podem promover ao polímero uma solubilidade especial em água.
Esses compostos possuem estrutura química semelhante ao ácido hialurônico e
fornecem uma excelente compatibilidade com o tecido epitelial. As soluções obtidas
com esses derivados podem ser facilmente incorporadas a formulações cosméticas,
mostrando melhor compatibilidade com surfactantes iônicos e emulsificantes,
apresentando estabilidade em uma larga faixa de pH.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
22
Han et al. (1999) examinaram os efeitos da quitosana sobre a atividade da lipase
pancreática in vitro e sobre o grau de armazenamento de gorduras induzido em ratos
pela administração oral de uma dieta rica em gorduras por nove semanas. Como
resultado, observou-se que os animais alimentados com quitosana, reduziram o peso
corporal, a hiperlipidemia e a gordura hepática. Como conclusão, os autores propõem
que os efeitos anti-obesidade da quitosana em ratos alimentados com alto teor de
gordura se devem parcialmente a inibição da absorção intestinal das gorduras dietéticas.
Em outro ensaio, Zahorska-Markiewicz et al. (2002) realizaram um estudo para
avaliar o efeito da quitosana no tratamento da obesidade. Segundo os autores do estudo,
a quitosana pode ser usada como um valioso e seguro coadjuvante em tratamentos para
a obesidade em longo prazo. A quitosana parece acentuar a redução da pressão arterial,
juntamente com a perda de peso.
Assis (2010) estudou a produção e caracterização de quitooligossacarídeos
produzidos pelo fungo Metarhizium anisopliae e avaliação da citotoxicidade em células
tumorais. A hidrólise da quitosana foi realizada em tempos de 10 a 60 min para a
produção de quitooligossacarídeos. A avaliação da citotoxicidade dos oligômeros de
quitosana foi realizada em células tumorais (HepG2 e HeLa) e não tumoral (3T3). A
produção de oligômeros de quitosana teve maiores rendimentos durante 10 minutos de
hidrólise, os pentâmeros apresentaram concentração de 0,15 mg/mL, no entanto os
hexâmeros, que apresentam maior interesse por causa de suas propriedades biológicas,
só foram detectados com 30 minutos de hidrólise com uma concentração de 0,004
mg/mL.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o planejamento Plackett-Burmam e a metodologia de superfície de resposta.
Os resultados mostraram que a eficiência máxima de extração da quitina foi conseguida
nas seguintes condições: concentração de casca de camarão 50g/L, glicose 50g/L, tempo
de incubação de 5 dias e inoculo como 0,05 OD (densidade óptica).
A glicosamina, unidade monomérica básica da quitosana, possui atividade
antiinflamatória e tem grande importância na construção e manutenção da cartilagem,
no tratamento de enfermidades como: febre reumática, artrose e principalmente a artrite
e osteoartrite na forma aguda ou crônica (Polymar, 2003). A Figura 2 apresenta a
estrutura química da glicosamina.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
23
Figura 2 – Estrutura da glicosamina (Fonte: Polymar, 2003)
O sulfato de glicosamina é um derivado da quitosana sendo utilizado no
tratamento da osteoartrite sintomática dos joelhos, quadris, coluna, mãos e outras
regiões (Craveiro et al., 2004). O sulfato de glicosamina estimula a síntese de
proteoglicanos necessários para o bom funcionamento das articulações, estimula a
regeneração das cartilagens e ainda incorpora enxofre ao tecido cartilaginoso. Estudos
de longa duração, acima de 3 anos, mostram que o sulfatos de glicosamina evita a
progressão de danos estruturais nas articulações e induz melhora significativa nas dores
e funções físicas (Craveiro et al., 2004). A Figura 3 apresenta a estrutura química do
sulfato de glicosamina.
Figura 3 – Estrutura do sulfato de glicosamina (Fonte: Polymar, 2003)

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
24
2.1.2. Quitosanases
Dependendo do tipo de quitosanase obtida, ocorrem diferentes mecanismos de
hidrólise da quitosana, isso devido a diferentes microrganismos produzirem enzimas
com diferenças nas suas estruturas (Kurita et al., 1977; Seino et al., 1991).
Quitosanases (EC 3.2.1.132) são enzimas que catalisam a reação de hidrólise de
quitosana em oligossacarídeos. As enzimas quitosanases são produzidas por vários
microrganismos, bactérias, actinomicetos e fungos, e até por algumas plantas. Estas
enzimas agem em polímeros com grau de acetilação de 30% a 60%. Os monômeros
resultantes da hidrólise da quitosana por estas enzimas são melhores digeridos, ao serem
ingeridos, como é o caso da glicosamina. Estudos efetuados até o presente momento
classificam as quitosanases em cinco famílias: 5, 8, 46, 75, 80, sendo que a família 46 é
a mais estudada. As quitosanases tem grande aplicação na geração de oligômeros de
quitosana (Somashekar & Joseph, 1996; Chen et al., 2005).
Nogawa et al. (1998) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida pelo fungo Trichoderma reseei PC-3-7. Estes pesquisadores
inocularam 106 conídios em 100 mL de um meio basal contendo 0,3% de N-
acetilglicosamina em incubator rotativo com agitação de 220 rpm a 28oC. Destaca-se
que o fungo não produziu quitosanase em meio contendo quitosana ou quitina.
Entretanto, o microrganismo produziu quitosanase em meio contendo glicosamina e N-
acetilglicosamina, sendo que para este último obteve-se uma atividade específica
máxima de 1,5 U/mg após 72 horas de incubação. É importante ressaltar que uma
agitação vigorosa favoreceu a produção da enzima. Esse artigo ilustra a presença de
exo-β-D-glicosaminidase e exo-β-D-N-acetilglicosaminidase.
Piza et al. (1999) estudaram a produção, purificação e caracterização parcial de
uma quitosanase produzida por Bacillus cereus. Nesse caso, investigou-se através de um
planejamento fatorial fracionário a influência dos fatores: concentração de sulfato de
amônio (0,4; 2,2 e 4,0%), razão de aeração (2; 6 e 7), tempo de fermentação (16; 24 e
32 horas), pH (5; 6 e 6,5) e concentração de quitosana (1,0; 1,5 e 2,0%). Os resultados
mostraram que os fatores mais significativos para esse planejamento foram a
concentração de sulfato de amônio, a aeração, o pH e a interação entre sulfato de
amônio e a aeração.
Cheng & Li (2000) estudaram a preparação de oligossacarídeos utilizando
quitosanase produzida por Aspergillus sp e constataram que a enzima foi largamente

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
25
induzida em meio contendo quitosana como única fonte de carbono e também na
presença de quitina coloidal. A indução da enzima foi realizada em temperaturas
variando na faixa de 28 a 30ºC, pois essa espécie não cresceu bem em temperaturas
superiores a 37ºC.
Zhang et al. (2000) estudaram a produção, purificação e caracterização de
quitosanase e Exo-β-D-Glucosaminidase por Aspergillus oryzae IAM2660. Os
microrganismos utilizados neste estudo foram mantidos em placas de Petri contendo
agar para contagem em placas (PDA). Destaca-se que para produzir a enzima, conídios
foram retirados das placas e inoculados em meio Czapeq-dox com os seguintes
componentes: 1,0g K2HPO4, 0,5g KCl, 2,0g NaNO3, 0,5g MgSO4.7H2O, 0,02g
FeSO4.7H2O, 20g de sacarose e 1,0L de água deionizada, com pH do meio igual a 5,5.
Foi adicionado ao meio 0,5 g de peptona e 0,5g de extrato de levedura com
concentração de esporos da ordem de 1,0x106 esporos.mL-1. Após a incubação, feita em
incubador rotativo a 26ºC por um período de 60h, micélios foram coletados e após
filtração e secagem a 80ºC foram utilizados para ensaio de quitosanase.
Kuroiwa et al. (2002) estudaram os fatores que afetam a composição de
oligossacarídeos produzidos através da hidrólise de quitosana por quitosanase originária
de Bacillus pumilus BN-262. Neste experimento foi utilizado quitosana 100%
desacetilada dissolvida em ácido acético 1M. O pH do meio foi ajustado em 5,6. Neste
estudo a quitosanase foi imobilizada em agar e Sepharose. Os resultados mostraram que
a concentração de enzima, a temperatura, a velocidade de agitação, e concentração de
substrato influenciaram de forma significativa na composição dos oligossacarídeos
produzidos.
Ichikawa et al. (2002) estudaram a imobilização e estabilização da ligação
multiponto da quitosanase tendo agar como suporte e concluíram que a imobilização
aumentou a termoestabilidade da quitosanase. Os oligossacarídeos de quitosana foram
continuamente produzidos usando um reator coluna contendo quitosanase imobilizada.
A percentagem de quitosana imobilizada após 28 dias de reação foi de 44%.
Hung et al. (2002) estudaram a purificação, através de troca iônica, e
caracterização da hidrólise sobre ação da quitinase e quitosanase purificadas do meio
contendo Bromelina comercial. Destaca-se que a quitinase purificada apresentou pH,
temperatura e Km ótimo para ação catalítica em meio contendo quitina de 4,0, 60ºC e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
26
0,2mg.mL-1, respectivamente. Enquanto que quitosanase apresentou o pH, temperatura e
Km ótimo de 3,0, 50ºC e 0,88mg.mL-1, respectivamente.
Zhu et al. (2003) estudaram as melhores condições fermentativas e propriedades
de uma quitosanase produzida por Acinetobacter sp. C-17. A espécie de bactéria
utilizada neste ensaio foi isolada do solo e identificada como sendo Acinetobacter sp C-
17 e capaz de produzir quitosanase. O microrganismo foi mantido em placas contendo o
meio agar com os seguintes componentes: 0,5 % de quitosana, 0,2 % K2HPO4, 0,1 %
KH2PO4, 0,07 % MgSO4, 0,05 % NaCl, 0,05 % KCl, 0,01 % CaCl2, 0,05 % de extrato
de levedura e 2 % agar, o pH do meio de manutenção foi mantido em 7,2 e a
temperatura foi mantida em 30ºC durante 3 a 5 dias, para identificar a ação da enzima
quitosanase. A ação da quitosanase foi identificada através da observação de uma
colônia de microrganismo existente na placa. A bactéria foi então isolada da placa e
colocada em fermentador rotativo a 200 rpm durante 72h em meio contendo: 0,07 %
K2HPO4, 0,03 % KH2PO4, 0,05 % MgSO4, 0,03 % peptona, 0,03 % de extrato de
levedura e 1,0 % de quitosana. O pH foi de 7,0 e a temperatura 30ºC. Como fontes de
carbono foram utilizadas além de 1,0 % de quitosana, 1,0 % de quitina e 1,0 % de
glicose. Os resultados mostraram que as melhores condições para produção da
quitosanase foram alcançadas quando se utilizou 1,0 % de solução de quitosana a 30ºC,
pH 7,0 durante 24 horas de incubação.
Liu et al. (2003) estudaram a produção de quitinase por Verticillium lecanii
F091 usando fermentação submersa, a atividade de quitinase foi de 9,95 mU/mL em um
meio de cultura otimizado, com um volume de cultura de 200 mL, agitação de 150 rpm
e temperatura de 24ºC cultivado em incubador rotativo e concluíram que a taxa de
agitação e o pH foram os fatores mais significativos para a produção de quitinase, no
entanto, as taxas de agitação e aeração poderiam modificar a concentração de oxigênio
dissolvido afetando diretamente no crescimento do Verticillium lecanii e na produção
de quitinase.
Donzelli et al. (2003) estudaram a hidrólise enzimática avançada de quitina de
casca de caranguejo langostino com misturas de enzimas de bactérias e fungos e
concluiram que a combinação de enzimas de Trichoderma atroviride e Serratia
marcescens foram capazes de degradar completamente altas concentrações de quitina
(100g/mL) a partir de carapaças de caranguejo langostino convertendo a N-
Acetilglicosamina (78%), glucosamina (2%) e quitobiose (10%). Enzimas de fonte

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
27
procariótica apresentaram atividades menores que as enzimas de T. atroviride, para
catalisar a reação com quitosana.
Jo et al. (2003) estudaram a caracterização e a avaliação cinética de quitosanase
produzida por Bacillus sp. Destaca-se que na produção da enzima utilizada neste ensaio
o meio continha 0,5% de concentração de quitosana, 1% de triptona e 1% de NaCl, o
pH do meio foi ajustado em 5,5 e a temperatura foi mantida em 37ºC com agitação de
180 rpm em incubador rotativo.
Fu et al (2003) estudaram a caracterização de três iso-enzimas de quitosanase
isolada de pepsina comercial e denominado como PSC-I, PSC-II, PSC-III,
respectivamente, em relação a hidrólise enzimática da quitosana, o meio ótimo para a
produção destas iso-enzimas continha as seguintes características: pH 5,0; 5,0 e 4,0 e
temperaturas de 40ºC, 40ºC e 30ºC, o valor de Km foi de 5,2, 4,0 e 5,6 mg.mL-1,
respectivamente. As massas moleculares das três iso-enzimas foram de 40 kDa. Os
resultados mostraram que as três iso-enzimas apresentaram atividade em meio contendo
quitosana, produzindo oligômeros da ordem de 68 – 88% desacetilada.
Choi et al. (2004) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida por Bacillus sp KCTC 0377BP. Nesse caso, com relação à
produção da enzima foi possível obter um aumento de 1,2 U/ml em incubator rotativo
para 100,0 U/ml em fermentador piloto de 500 L, utilizando uma concentração de
substrato de 0,5% de quitosana e como fonte de nitrogênio polipeptona (2,5%) em pH
igual a 6,8, com 100 rpm de 100,0 rpm a 30oC.
Shimosaka et al. (2003) estudaram a produção, purificação e caracterização total
de duas quitosanases produzida por Acinetobacter sp CHB101. Nesse caso, com relação
à produção da enzima os pesquisadores usaram dois meios, um contendo glicose e outro
contendo quitosana. Com relação à cinética, o meio contendo quitosana fornece um
tempo de geração de 280 minutos, enquanto que no meio contendo glicose a taxa de
crescimento foi mais elevada e o tempo de geração foi de apenas 110 minutos. Os
autores observaram que após os dois primeiros dias de cultivo a atividade
quitosanolítica permaneceu baixa (na ordem 0,10 a 0,20 U/mL). Durante a fase de
crescimento exponencial o valor da atividade permaneceu com valor inferior a 0,01
U/mL, começando a aumentar ao atingir a fase estacionária, indicando assim que a
produção da enzima é não associada ao crescimento.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
28
Kim et al. (2004) estudaram a produção, purificação e caracterização de uma
quitosanase produzida pelo Bacillus sp e identificado pelo número 1299. Destaca-se que
na produção da enzima quitosanase foi utilizado um meio contendo os seguintes
componentes: 0,5 % de quitosana, 0,42 % (NH4)2SO4, 0,2 % K2HPO4, 0,02 % uréia,
0,03 % CaCl2.2H2O, 0,03% MgSO4.7H2O, 0,1% peptona, 0,2% Tween 80, 0,2 % da
solução (0,5% FeSO4.7H2O, 0,16% MnSO4.H2O, 0,14% ZnSO4.7H2O, 0,2% CoCl2) e
1,7% agar. O pH do meio para este ensaio foi mantido em 7,0. Este microrganismo foi
escolhido por ser grande produtor de quitosanase.
Chen et al. (2005) estudaram a purificação e a caracterização de dois tipos de
quitosanase produzidas por Aspergillus sp. CJ22-326 em cultivo submerso. Na
produção da enzima foi utilizado um meio de cultivo contendo 1,0% de quitosana, 2,0%
de gérmen de trigo, 0,2% de sulfato de amônio, 0,2 % de fosfato de potássio e 0,05% de
sulfato de magnésio por litro de solução em pH 5,6. O cultivo foi realizado em frasco de
500 mL contendo 150 mL de meio de cultura a 30ºC por 96 horas com uma agitação de
150 rpm. Os autores apresentam um gel de eletroforese com as duas enzimas purificadas
com massa molar de 29 kDa e 109 kDa.
Silva Filho et al. (2005) estudaram, através de planejamento fatorial 24 com
repetição em triplicata no ponto central, a produção de quitosanase por Aspergillus
ochraceus em cultivo descontínuo submerso. Os resultados mostraram que foi possível
produzir quitosanase com atividade na ordem de aproximadamente 5,9 U/mL utilizando
Aspergillus ochraceus e que a atividade foi favorecida pelo aumento da agitação (r), da
razão de aeração (RA) e da concentração de substrato (Cs), enquanto que o aumento da
temperatura de cultivo (TMC) não favoreceu a resposta (atividade quitosanolítica).
Pode-se classificar as quitosanases em endoquitosanases e exoquitosanases. As
primeiras diferenciam-se por catalisar a hidrólise aleatoriamente, no interior do
biopolímero, gerando diversos tamanhos de oligossacarídeos. As exoquitosanases são
capazes de hidrolisar as terminações não redutoras, produzindo assim unidades
redutoras (Peter, 2005).
Palma-Guerrero et al. (2008) estudaram o biocontrole de fungos patogênicos e
demonstraram a lise da membrana plasmática e a inibição da germinação de esporos de
fungos. Eles observaram que os fungos entomopatogênicos e nematófagos foram menos
inibidos pela quitosana que os fitopatogênicos e micoparasíticos provavelmente porque
apresentam a enzima quitosanase que degrada a quitosana. Mesmo assim, a quitosana

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
29
pode ajudar a desenvolver novas estratégias para o biocontrole em geral de fungos
patogênicos, evitando o uso de fungicidas que prejudicam o meio ambiente e a saúde
humana.
Pagnoncelli (2008) estudou a produção de quitooligossacarídeos com
propriedades nutracêuticas a partir da quitosana por hidrólise enzimática, utilizando
processo fermentativo simultâneo. Duas cepas produtoras de quitosanaes foram
selecionadas, Paenibacillus chitinolyticus e Paenibacillus ehimensis, e avaliadas quanto
ao comportamento em meio de cultivo contendo açúcares simples e em relação às
variações de pH do meio. Os quitooligossacarídeos produzidos a partir do complexo
enzimático da primeira cepa apresentaram maior pico após 9h de hidrólise, enquanto
para a segunda cepa avaliada, após 20min, observou-se quitooligossacarídeos com grau
de polimerização entre 3 e 6 unidades.
2.2. Fungos
2.2.1. Características gerais
Desde o final dos anos 60 que os fungos deixaram de pertencer ao reino das
plantas e passaram a ter um reino próprio chamado de Reino Fungi. Foi a partir de
estudos morfológicos, bioquímicos e citológicos onde se concluiu que eles são tão
diferentes dos vegetais como dos animais. Distinguem-se de outros organismos
eucariotas por serem seres quimiorganoheterotróficos e, ainda, por apresentarem parede
celular rígida que é composta por quitina e glucano e uma membrana celular em que o
ergosterol substitui o colesterol (Trabulsi & Toledo, 1996; Murray et al., 2005).
Fazem parte do Reino Fungi cerca de 100 mil espécies, desde grandes como
os cogumelos, até microscópicas como os bolores e leveduras. Ainda hoje é utilizada
essa classificação, mas os avanços da biologia molecular e estudos bioquímicos levaram
a outras propostas de classificação em que os fungos e organismos afins foram incluídos
em três diferentes reinos: Protozoa, Chromista e Fungi (Kirk et al., 2008).
Abaixo, são apresentadas características que diferenciam os fungos das plantas:
• são incapazes de sintetizar clorofila, como também não apresentar celulose em
sua parede celular, com exceções como fungos aquáticos;
• apresentam algumas características de células animais (capacidade de depositar
glicogênio e a presença de substâncias quitinosas, em sua parede celular);

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
30
• podem ser eucarióticos (as leveduras), ou multinucleados (os bolores), e
apresentar retículo endoplasmático rugoso e mitocôndrias em seu citoplasma;
• são heterotróficos, alimentam-se de matéria orgânica viva (fungos parasitários),
ou morta (fungos saprofíticos);
Os fungos são amplamente encontrados na natureza, crescem rapidamente no
solo, na água, na atmosfera, bem como nos animais, nos vegetais, resíduos orgânicos e
desempenham um papel importante na vida do homem, quer de uma maneira benéfica,
quer de um modo prejudicial. Os fungos são um dos principais microrganismos
responsáveis pela decomposição da matéria orgânica, interferindo no ciclo do carbono,
do nitrogênio e de outros nutrientes da biosfera. São capazes de deteriorarem produtos e
bens de consumo, tais como alimentos, tecidos, papel, madeira. Os alimentos
armazenados representam um vasto meio para a proliferação dos fungos, principalmente
quando não são consideradas as condições básicas de armazenamento correto (Trabulsi
& Toledo, 1996).
Estão presentes funcionalmente em vários processos industriais, tais como, de
fabricação de pão, cervejas, vinhos e determinados tipos de queijos, sendo também
utilizados na produção comercial de muitos ácidos orgânicos, de alguns fármacos, como
a ergometrina e a cortisona, na obtenção de diferentes antibióticos, como a penicilina, e
de substâncias imunossupressoras, como a ciclosporina (Esteves et al., 1990). A Figura
4 ilustra a estrutura geral de uma célula de fungo.
Figura 4 – Representação da estrutura geral de uma célula de fungo.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
31
Os fungos podem se desenvolver em meios de cultivo especiais formando
colônias de dois tipos, leveduriformes e filamentosas, que se diferenciam pela
macromorfologia e micromorfologia. O primeiro tipo apresenta, em geral, característica
de consistência cremosa de cor branca a creme, brilhantes ou opacas, podendo
apresentar às vezes coloração escura ou alaranjada, e são formados por microrganismos
unicelulares que apresentam as funções vegetativas e reprodutivas. As filamentosas
apresentam características granulares, cotonosas, aveludadas ou pulverulentas, são
multinucleadas e apresentam-se em forma de tubos denominadas de hifas. Essas hifas
podem ser não septadas (cenocíticas) ou septadas (tabicadas). O conjunto de hifas é
denominado micélio.
As hifas asseptadas têm aparência anastomosada (concisa), formada por um
citoplasma estendido e polinucleado, enquanto que as hifas septadas contêm células
individualizadas, cada uma contendo o seu núcleo. Na Figura 5 são mostradas as
estruturas das hifas septadas e não septadas.
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas. (Adaptado de Guia.Bio,
2012)
O micélio que se desenvolve no interior do substrato e que é responsável pela
sustentação e absorção de nutrientes é denominado micélio vegetativo. O micélio que
cresce na superfície do meio de cultura é denominado micélio aéreo. Este pode ser
denominado também de micélio reprodutivo, quando o mesmo se diferencia para
sustentar os corpos de frutificação ou propágulos (Putzke & Putzke, 1998). Os
propágulos classificam-se de acordo com a origem em externos e internos, sexuados e
assexuados.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
32
2.2.2. Reprodução dos Fungos
A reprodução dos fungos ocorre em ciclos assexuais, sexuais e parassexuais. Em
fungos, a reprodução assexuada é observada com mais freqüência do que a reprodução
sexuada. A reprodução assexuada é normalmente a reprodução mais importante para a
propagação da espécie, feita através de fragmentação de artroconídios, fissão de células
somáticas, brotamento ou gemulação do blastoconídios-mãe e produção de conídios. A
reprodução sexuada envolve a união de duas células ou de dois órgãos sexuais
sexualmente compatíveis. A reprodução sexuada culmina na produção de basidiósporos,
no caso de basidiomicetos. A reprodução parassexuada ocorre com a fusão de hifas e
formação de um heterocário, que consiste na existência de núcleos haplóides
geneticamente distintos num mesmo citoplasma, os quais, após a fusão, originam
núcleos diplóides heterozigotos sujeitos a sucessivas divisões mitóticas. Apesar da
raridade dessas, a reprodução parassexual é de grande importância na evolução de
alguns fungos (Pelczar et al., 1996).
A reprodução assexuada é feita por conídios, esporos assexuados formados no
ápice de hifas modificadas, os conidióforos. Os conidióforos não possuem esporângios
de forma que os conídios ficam expostos. Na reprodução sexuada, com o encontro de
duas hifas haplóides distintas, ocorre a plasmogamia e um novo conjunto de hifas
dicarióticas é formado.
Os conídios podem ter formas esféricas, fusiformes, cilíndricas, piriformes, entre
outras. Apresentam-se como hialinos ou pigmentados, podem estar isolados ou
agrupados, a parede celular pode ser lisa ou rugosa e podem ser formados por apenas
uma célula ou apresentarem septos em mais de um plano.
Os conídios também podem originar-se em qualquer parte do micélio vegetativo
e são denominados conídios sésseis como é o caso do Trichophyton rubrum.
Alguns fungos patogênicos são identificados a partir de estruturas formadas pela
célula conidiogênica e o conidióforo. Essas estruturas são denominadas: aparelho de
frutificação ou conidiação. Os conídios podem formar cadeias sobre fiálides em volta de
uma vesícula, dilatando a extremidade do conidióforo. Essas estruturas ocorrem em
Aspergillus, apresentando-se em forma de cabeça ao redor de uma vesícula, e em
Penicillium agrupados em forma de pincel, como é ilustrada na Figura 6.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
33
Figura 6 – Conídios de Aspergillus e Penicillium
2.2.3. Metabolismo de fungos
Os fungos são seres aeróbicos e heterotróficos, podendo se desenvolver em
ambientes com baixa quantidade de oxigênio, como ocorre no crescimento vegetativo e
na reprodução assexuada, como também em ambientes com grande quantidade de
oxigênio como é o caso da reprodução sexuada.
Os fungos são capazes de produzir certas enzimas como lipases, invertases,
lactases, quitosanases, proteases, amilases, etc., que fazem a hidrólise do substrato
facilitando o mecanismo de transporte ativo e passivo. Esses substratos induzem a
formação de enzimas degradativas. Algumas espécies de fungos desenvolvem-se em
meios que apresentam amônia ou nitrilos como única fonte de nitrogênio. As melhores
fontes orgânicas para o seu desenvolvimento são carboidratos, como a D-glicose, e
alguns sais minerais como sulfatos e fosfatos.
A água é uma necessidade de todos os seres vivos, não sendo diferente para os
fungos que precisam de água para o seu desenvolvimento. Certos fungos apresentam
características halofílicas podendo crescer nos ambientes com grandes concentrações de
sal.
A razão de não existirem ambientes livres de espécies fúngicas deve-se ao facto
destes microrganismos conseguirem sobreviver em situações extremas como, por
exemplo, temperaturas que variam entre os - 5 e os 60 ºC, com pH de 1 a 9 e com baixas

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
34
concentrações de oxigênio, apesar de cada espécie possuir condições ideais para o seu
desenvolvimento (Chao et al., 2002).
Devido a grande faixa de temperatura ótima para o seu crescimento, os fungos
apresentam espécies psicrófilas, mesófilas e termófilas. Para a maior parte das espécies
fúngicas, a temperatura de desenvolvimento situa-se entre os 20 e 24 ºC e não crescem
acima dos 29 ºC. Em relação à umidade relativa, necessitam de pelo menos 50 %, mas
geralmente necessitam de mais de 65 % (Colakoglu, 2001).
O pH ideal para o desenvolvimento dos fungos varia na faixa entre 5 e 7.
Destaca-se, entretanto, que a maioria dos fungos suportam grandes variações de pH
como ocorre com os fungos filamentosos que podem crescer em pH variando entre 1,5 e
11. Por outro lado, as leveduras não suportam pH alcalino.
Os fungos crescem lentamente, se comparados com as bactérias, em meio de
incubação para o seu desenvolvimento. Para evitar contaminação por bactérias, que
podem se sobrepor ou inibir o seu crescimento, pode-se introduzir no meio de cultura
um antibacteriano de largo espectro como o cloranfenicol.
Algumas espécies de fungos se desenvolvem na presença de luz, outras são
inibidas e algumas podem crescer tanto na presença quanto na ausência deste agente. No
entanto a irradiação solar, em geral, é prejudicial ao seu desenvolvimento, por causa da
radiação ultravioleta.
2.2.4. Nutrição de fungos
Quanto ao aspecto da nutrição, os fungos são heterótrofos, ou seja, incapazes de
produzir seu próprio alimento. Entretanto, a forma de assimilar matéria orgânica é
diferente daquela utilizada pela maioria dos animais, que são heterótrofos por ingestão.
Nos fungos, a digestão é extracorpórea, ou seja, é realizada fora do corpo. O fungo
lança no ambiente, enzimas que degradam as moléculas orgânicas complexas e, depois,
absorve moléculas menores, mais simples. Por essa razão, são designados heterótrofos
por absorção. É devido a este processo que os fungos podem crescer dentro ou sobre os
alimentos. Desenvolve-se geralmente em meios contendo um pH baixo, uma fonte de
carbono, uma fonte de nitrogênio orgânico ou inorgânico e alguns minerais. Alguns
necessitam de vitaminas. Quanto aos modos de vida, os fungos podem ser
decompositores, parasitas, mutualísticos e predadores (Raven, 2001).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
35
Os fungos decompositores, saprófagos (do grego, saprós, “podre”, e phagos,
“comedor”), juntamente com as bactérias, são os principais decompositores da biosfera,
participando intensamente do processo de degradação da matéria orgânica morta, o que
promove a reciclagem dos elementos químicos constituintes dos seres vivos. A maioria
deles vive no solo, obtendo nutrientes de seres mortos. Em florestas, onde a matéria
orgânica vegetal é abundante, os micélios dos fungos decompositores absorvem os
nutrientes de folhas e galhos caídos, decompondo-os. Os fungos decompositores
também são responsáveis pelo apodrecimento de alimentos e de outros materiais, como
a madeira.
Os fungos parasitas vivem à custa de outros organismos vivos, prejudicando-os.
Ao parasitar o corpo de um ser vivo, animal ou vegetal, o fungo pode até provocar sua
morte. Várias doenças são causadas por fungos parasitas. Nas plantas, cita-se a
ferrugem do café, que, no Brasil, é motivo de grande preocupação por parte dos
cafeicultores, devido aos prejuízos que acarreta. As micoses que atacam a pele de seres
humanos e animais são provocadas por fungos.
Existem fungos que se associam a outros organismos, estabelecendo uma relação
em que há benefício mútuo para os indivíduos envolvidos. Esses fungos são
denominados mutualísticos. A maioria vive associada a seres fotossintetizantes, como
as plantas, cedendo-lhes parte da água e dos nutrientes que as hifas absorvem do solo.
As plantas, por sua vez, cedem ao fungo certos açúcares e aminoácidos. Duas
associações mutualísticas formam estruturas bem características: as micorrizas,
associação que envolve fungos e raízes de plantas, e os líquens, formados por fungos e
algumas variedades de algas e cianobactérias.
Certos fungos atuam como predadores. Na maioria dos casos, as hifas secretam
substâncias aderentes que aprisionam os organismos que tocam os fungos. Dessa
maneira, as hifas penetram o corpo da presa, crescem e se ramificam, espalhando-se no
interior do organismo e absorvendo seus nutrientes, causando-lhe a morte. O gênero
Arthrobotrys apresenta uma forma mais elaborada de predação. O fungo vive no solo e
captura nematódeos microscópicos que habitam o mesmo ambiente. As hifas
apresentam pequenos anéis que se estreitam quando a presa passa por eles. As hifas do
fungo logo invadem o corpo do nematódeo, digerindo-o.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
36
2.2.5. O fungo Metarhizium anisopliae
Existem três espécies de fungos do gênero Metarhizium, divididas em dez
variedades: M. anisopliae, variedades anisopliae, majus, lepidiotum e acridum; M.
flavoviride, variedades tipo E, flavoviride, minus, novazealandicum e pemphigum; e M.
album (Driver et al. 2000). E, mais recentemente, foi descrita na China uma nova
variedade de M. anisopliae, dcjhyium (Dong et al. 2009).
Data de 1879 quando o russo Ilya Metchnikoff aplicou o fungo para combater
larvas de um curculionídeo, praga de beterraba, e foi classificado como Metarhizium
anisopliae por Sorokin em 1883. É um fungicida natural para mais de 300 espécies de
insetos, especialmente pragas da agricultura e pecuária. No Brasil, o fungo Metarhizium
anisopliae teve a sua aplicação com sucesso pela primeira vez em 1965, no controle da
cigarrinha-da-cana Mahanarva posticata (Alves, 1998).
O fungo Metarhizium anisopliae é um importante agente entomopatogênico
utilizado no controle de pragas, como cupins, gafanhotos, cigarrinhas e besouros, sendo
amplamente estudado em todo o mundo. Existe amplamente na natureza e é facilmente
encontrado no solo, onde sobrevive por longos períodos e seu desenvolvimento se dá
geralmente entre 15 e 32 °C, sendo ideal entre 24 e 30 °C e com pH ótimo igual a 6,9.
Apresenta micélio hialino e septado, com conidióforos característicos, sobre os quais
surgem conídios cilíndricos organizados em colunas. Este fungo ao atacar os insetos
provoca rigidez e os recobre por uma camada pulverulenta de conídios, resultando em
colorações que variam do verde claro ao escuro, cinza ou branco com pontos verdes
(Tinline, 1971; Alves, 1998, Driver et al. 2000; Arruda, 2005).
Quanto a sua morfologia, apresenta-se como um fungo filamentoso, com corpo
de frutificação equivalente a um esporodóquio agregado a hifas intimamente
entrelaçadas, com massa compactada de conidióforos característicos, simples ou
ramificados, formando células esporogênicas denominadas fiálides, resultando nos
fialosporos (Wang et al., 2002).
O Metarhizium anisopliae é um fungo de cultivo simples, sendo necessário
quase que exclusivamente o nutriente a base de amido. Um dos fungos mais estudados
em programas de manejo de pragas já que apresenta grande potencialidade
entomopatogênica (Onofre et al., 2002). O Metarhizium anisopliae é um microrganismo
pouco exigente, pois pode se desenvolver em diversos meios de cultura como: amido,
glicose, glicerina, levulose, maltose, sacarose e quitina (Jabor et al., 2003).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
37
Em geral, os fungos produzem enzimas que convertem os tecidos dos insetos em
nutrientes. Porém, a cutícula esclerotizada dos insetos não é bem utilizada pelos fungos,
excetos pelos entomopatogênicos que desenvolveram enzimas eficientes para degradar
essa camada protetora. Destaca-se a enzima Pr1, produzida por Metarhizium anisopliae,
que tem um poder de degradação bem superior a qualquer outra enzima sintetizada por
esse grupo de fungos (Dias, 2005). Desta forma, o Metarhizium anisopliae (fungo
entomopatogênico) ao atacar a célula dos insetos, produz grande quantidade de enzimas
que degradam a membrana citoplasmática e inativa a célula e, consequentemente, mata
o inseto, logo é evidente a relação entre o controle de pragas e produção de enzimas.
No momento, sobre a comercialização de fungos entomopatogênicos no Brasil,
em sua maioria, são formulados em grânulos, compostas do fungo mais o substrato
(arroz + fungo), como também na forma de pó molhável (WP), resultado da moagem do
fungo com o substrato. Esses produtos representam 90% e 8,5%, respectivamente, da
produção de fungos no Brasil, e sendo suas concentrações finais ficam em torno de
5x108 conídios/g para Metarhizium anisopliae e 1x109 conídios/g para Beauveria
bassiana (Alves et al., 2008).
Segundo Batista Filho et al. (2002), Garcia et al. (2005) e Mello et al. (2006), o
Metarhizium anisopliae tem sido bastante utilizado para o controle de diferentes pragas
no Brasil, como a broca da bananeira, pragas de grãos armazenados, carrapatos, cupim
de montículo em pastagens, larvas de escarabeídeos que atacam a cana de açúcar e
cigarrinha da cana-de-açúcar.
É comum a ocorrência natural do fungo Metarhizium anisopliae atacando
cigarrinha-das-raízes de canaviais pelo país, o que incentiva a utilização dele como
agente de controle biológico em áreas infestadas por esse tipo de praga. A aplicação de
recursos no uso de programa de manejo integrado da cigarrinha tem sido reforçado por
razões ambientais e econômicas (Dinardo-Miranda, 2001).
Faria & Magalhães (2001) fizeram uma avaliação do uso de fungos
entomopatogênicos no Brasil, na qual observaram que a qualidade dos micoinseticidas
disponíveis no Brasil poderia ser incrementada de forma considerável quanto a sua
formulação, ou seja, antes da venda, poderiam realizar melhorias no tratamento
posterior, adição de substâncias que lhes assegurem melhorias na eficiência de controle,
capacidade de armazenamento ou praticidade de manuseio, ou de qualquer outro critério
que resulte em vantagem em relação ao produto bruto. Ressaltaram a pouca praticidade

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
38
em alguns casos, como exigência de lavagem, e em outros podendo causar o
entupimento de bicos dos pulverizadores devido à elevada proporção de inertes,
principalmente quando são empregados baixos volumes de aplicação. Outro problema
evidenciado com os inseticidas nacionais diz respeito à pequena sobrevida, devendo a
ser usados em, no máximo, 30 dias depois de produzidos, quando armazenados à
temperatura ambiente e em local sombreado, fazendo as vendas ocorrerem quase que,
exclusivamente, sob encomenda. Comparativamente, os bons inseticidas sob
comercialização em outros países apresentam maior concentração de ingrediente ativo,
maior sobrevida (alguns produtos podem ser armazenados por mais de 8 meses à
temperatura ambiente) e praticidade (são formulados na forma de GDA (grânulos
dispersíveis em água) ou de óleos emulsionáveis, por exemplo, podendo ser adicionados
diretamente ao tanque do pulverizador.
Rangel et al. (2007), instalaram uma Unidade de Observação para o controle
microbiano das cigarrinhas das pastagens Distrito de Itahum, em Dourados/MS, devido
as necessidades de agricultores familiares de produção de leite. As aplicações do fungo
Metarhizium anisopliae foram realizadas por pulverização com equipamento de tração
animal. As condições do ambiente de estudo foram favoráveis ao desenvolvimento da
população das cigarrinhas e à incidência do fungo. Como resultados, observaram que os
níveis de espumas, ninfas e adultos reduziram significativamente após as aplicações,
levando os Agricultores Familiares a total satisfação com os ganhos na produção de
todos os envolvidos.
Segundo Vieira et al. (2007), o fungo entomopatogênico Metarhizium anisopliae
var. acridum tem sido utilizado no controle de pragas, e assim, visando este fim,
analisaram em laboratório a esporulação, a germinação dos conídios e as características
morfológicas peculiares desse fungo em diferentes temperaturas. Esperou-se o
desenvolvimento das colônias em períodos de 1 a 15 dias. Foi observado que
temperaturas de 25, 28 e 30 ºC proporcionaram maiores taxas de esporulação e elevadas
porcentagens de germinação dos conídios. A 25, 28 e 30 ºC observou-se um
desenvolvimento mais rápido das microestruturas somáticas e reprodutivas (hifas,
anastomoses, apressórios, conidióforos e conídios elipsóides). Nas temperaturas
extremas de 40 e 45 ºC não foi detectado esporulação, germinação, nem
desenvolvimento de nenhuma microestrutura.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
39
Lopes et al. (2007) avaliaram o efeito de formulações de Beauveria bassiana e
Metarhizium anisopliae sobre ninfas de Amblyomma cajennense (transmissor da febre
maculosa ao homem) em laboratório. Ninfas não alimentadas foram pulverizadas com
suspensões contendo 2x107 e 4x107 conídios/mL de B. bassiana, e 1x107, 1,5x107,
2x107, 3x107 e 5x107 conídios/mL de M. anisopliae. A mortalidade de ninfas,
decorridos três dias da aplicação, foi significativa nas concentrações de 3x107 e 5x107
conídios/mL de M. anisopliae, com 57,1 e 56,2 %, respectivamente. Após seis dias a
mortalidade foi de 100 % nessas duas concentrações. A mortalidade foi
significativamente maior que a observada para B. bassiana em concentrações
semelhantes.
Guirado et al. (2009) realizaram estudos para controlar cupins de montículo em
área de pastagem, utilizando fungos entomopatogênicos Metarhizium anisopliae e
Beauveria bassiana em diferentes doses e formulações. A avaliação foi realizada após
30 dias da aplicação dos produtos. Os melhores resultados em ordem decrescente de
eficiência foram: M. anisopliae 30 mL + 1000 mL de água com 100 % de eficiência; B.
bassiana 10g + M. anisopliae 20 mL + 1000 mL de água com 85,71 % de eficiência; M.
anisopliae 20 mL + 1000 mL de água com 71,42 % de eficiência. Os demais
tratamentos não diferiram da testemunha.
Loureiro et al. (2012) compararam a eficiência de isolados de Metarhizium
anisopliae em controlar populações naturais de Mahanarva fimbriolata (cigarrinha-da-
raiz) em cultivos comerciais de cana-de-açúcar colhida mecanicamente. Foram
aplicados os isolados IBCB 348, IBCB 408, IBCB 410 e IBCB 425 do fungo em
concentração de 1,5×1012 conídios/ha. Aplicou-se o planejamento experimental em
blocos casualizados com 5 tratamentos e 4 repetições, sendo cada repetição composta
por 7 linhas de 100 m de comprimento e espaçamento de 1,5 m. Após 30 dias de
pulverização foi observado para os isolados IBCB 408 e IBCB 425 uma eficiência de
controle de ninfas de 63 e 62%, respectivamente, já no caso dos adultos chegou a 100%.
Após 60 dias o isolado IBCB 425 foi o mais eficiente para controlar as ninfas com
48,4%.
Assis et al. (2012) estudaram a citotoxidade de quitoolissacarídeos (QOS) e
oligômeros A, B e C (soluções compostas por diferentes quantidades de QOS)
produzidos por hidrólise enzimática pelo fungo Metarhrizium anisopliae. Foi analisado
o efeito de antiproliferativo destas moléculas em células tumorais (HepG2 e HeLa) e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
40
também em célula normal (3T3). A atividade antioxidante foi analisada em vários
experimentos in vitro. A glicosamina apresentou maior toxicidade (cerca de 92%) para
todas as células estudadas. No entanto, os oligômeros obtidos após hidrólise não
demonstraram efeito tóxico sobre a célula normal (3T3). Além disso, mostraram que
uma pequena quantidade de outros COS pode diminuir o efeito citotóxico da
glicosamina contra células 3T3, indicando que glicosamina poderia ser usada como uma
droga antitumoral na presença de outros COS.
2.3. Fermentação
Os processos fermentativos são caracterizados por processos nos quais
microrganismos realizam a conversão catalítica de uma dada substância num
determinado produto. Estes processos podem ser classificados como processos
descontínuos, contínuos e descontínuos alimentados. A classificação destes processos
está diretamente ligada à forma de como o substrato é adicionado e como o produto é
retirado. No processo de fermentação descontínuo o substrato é colocado no
fermentador, em seguida é inoculado e após o término do processo o produto é retirado.
No processo contínuo, o substrato é colocado num reator com vazão constante e o meio
fermentado é retirado com a mesma vazão de entrada do reator (Faccioti, 2001). Nos
processo descontínuos alimentados à adição de substrato e a retirada do mesmo deve
ocorrer de forma intermitente (Carvalho & Sato, 2001).
De certa forma, os processos fermentativos representam um elo entre as antigas
artes de elaboração de alimentos, como queijos e vinho, a partir de uma flora
microbiana natural, com a moderna indústria de fermentação de alimentos, onde são
utilisados cultivos puros e equipamentos sofisticados para o controle do processo
(Assis, 2010).
Atualmente, a maioria dos processos industriais se desenvolve em cubas de
vidro ou de aço de até 20 litros. Os agentes biológicos se encontram submersos no meio de
cultivo que ocupa, aproximadamente, 75% da cuba. As vezes é necessário injetar ar, e em muitas
fermentações se forma espuma. O desenho do biorreator deve se adequar ao objetivo do
processo, respondendo eventualmente a diversos imperativos, tais como a esterilização do
sistema, a aeração e homogeneização do meio, o acréscimo de nutrientes e de aditivos
antiespuma, a manutenção do pH, etc. Os modelos de fermentadores mais utilizados com
microrganismos contam com aeração e agitação mecânica. Esta facilita a distribuição dos

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
41
nutrientes na cuba, mas gera calor que deve ser eliminado mediante a circulação de água
fria. Em outros tipos de biorreatores, em coluna ou torre, a homogeneização depende da injeção de
ar. Os sistemas submersos são apropriados para o cultivo de microrganismos livres, mas resultam
pouco econômicos quando se trabalha com células ou enzimas caras. A imobilização em
fermentadores menores, seja por adesão a um suporte inerte, seja por inclusão dentro de um
polímero que permita o contato com o meio de cultura, além de simplificar a purificação do
produto permite a reutilização das células ou das enzimas, que permanecem dentro do
biorreator (Malajovich, 2009).
As enzimas são produtos de alto valor agregado, amplamente empregadas em
uma gama de indústrias, e entre as fontes para sua obtenção, destacam-se as de origem
microbiana, principalmente, devido a elevada seletividades dos substratos utilizados e
produtos formados. A principal forma de obtenção de enzimas, em escala industrial, é a
fermentação submersa, por esta apresentar vantagens em relação a fermentação
semisólida, como: maior controle do processo fermentativo e consequente reprodução
da cinética microbiana, além de maiores rendimentos (Wanderley, 2011).
2.3.1. Preparo do inóculo
Denomina-se inóculo um volume de suspensão de microrganismo de
concentração adequada, capaz de garantir, em condições econômicas, a fermentação de
uma dada quantidade de meio. O volume do inóculo, assim como sua concentração de
células, vai depender tanto do processo de fermentação c omo da quantidade de meio a
ser fermentado (Carvalho & Sato, 2001).
A partir de uma cultura pura, inocula-se um volume relativamente pequeno de
meio nutriente, que vai depender do tipo de microrganismo cultivado (pois cada espécie
microbiana possui velocidades diferentes de crescimento), sendo, em seguida, incubado
em condições favoráveis ao desenvolvimento do microrganismo. A suspensão obtida é
transferida para um frasco maior em agitador rotativo ou recíproco (shakers), que por
sua vez é adequadamente incubado, e a seguir, transferido para um frasco contendo um
volume ainda maior deste meio, até se conseguir alguns litros de suspensão microbiana.
Os cuidados com assepsia, volumes de meio utilizado, numero de transferência bem
como as condições de incubação, podem variar de acordo com o tipo de processo de
fermentação que se deseja realizar.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
42
2.3.2. Produção de enzimas em cultivo submerso descontínuo
A produção de enzimas, usando processos fermentativos, é constituída
basicamente por duas etapas: a primeira caracteriza-se pela preparação do inóculo e a
segunda, pela fermentação do substrato (Carvalho & Sato, 2001). A finalidade da
primeira etapa é o preparo do microrganismo em condições adequadas para que possa
ocorrer o desenvolvimento da segunda etapa. Diferentemente do cultivo em estado
sólido, no cultivo submerso os microrganismos encontram-se em um meio com elevada
atividade de água.
2.4. Planejamento experimental
Um bom planejamento tem como fundamental projetar um experimento de
forma que ele seja capaz de fornecer exatamente o tipo de informação que procuramos.
Para isso é necessário saber, em primeiro lugar, o que é mesmo que se deseja procurar.
Planejamento experimental, ou delineamento experimental, é um conjunto de
ensaios estabelecido com critérios científicos e estatísticos, objetivando determinar a
influência de diversas variáveis nos resultados de um dado sistema ou processo (Barros
Neto et al., 2001; Rodrigues & Iemma, 2005). De acordo com o propósito dos ensaios,
esse objetivo maior pode ser mais detalhado em:
– determinar quais variáveis são mais influentes nos resultados;
– atribuir valores às variáveis influentes de modo a otimizar os resultados;
– atribuir valores às variáveis influentes de modo a minimizar a variabilidade dos
resultados;
– atribuir valores às variáveis influentes de modo a minimizar a influência de variáveis
incontroláveis.
O planejamento experimental, baseado nos fundamentos estatísticos é, sem
dúvida alguma, uma ferramenta poderosa para se chegar às condições otimizadas de um
processo, desenvolvimento da formulação de produtos dentro das especificações
desejadas ou simplesmente para avaliar os efeitos ou impactos que os fatores (variáveis
independentes) têm nas respostas (variáveis dependentes) desejadas (Barros Neto et al.,
2001; Rodrigues & Iemma, 2005). Da utilização das técnicas estatísticas de
planejamento experimental, é possível chegar benefícios importantes, como:

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
43
– reduzir o número de ensaios sem prejuízo da qualidade da informação;
– estudar simultaneamente diversas variáveis, separando seus efeitos;
– determinar a confiabilidade dos resultados;
– realizar a pesquisa em etapas, num processo iterativo de acréscimo de novos ensaios;
– selecionar as variáveis que influem num processo com número reduzido de ensaios;
– representar o processo estudado através de expressões matemáticas;
– elaborar conclusões a partir de resultados qualitativos.
2.4.1. Planejamento fatorial
Em um planejamento fatorial é investigado as combinações de níveis de estudo
de cada fator. Para tanto, inicialmente, é necessário especificar esses níveis em cada
fator, quer dizer, os valores dos fatores que serão utilizados nos experimentos. Um
ensaio experimental é definido como um experimento constituído de um conjunto de
níveis definido. Geralmente, quando se tem n1 níveis de um determinado fator 1, n2 do
fator 2, ..., e nk do fator k, o planejamento será descrito por um fatorial n1 x n2 x ... x nk,
que corresponde ao número mínimo de experimentos para se ter um planejamento
fatorial completo.
É necessário fazer cada fator variar e observar o resultado dessa variação para se
estudar o efeito sobre a resposta. Assim, os ensaios precisam ter um mínimo de dois
níveis por fator para a realização desses ensaios. Para k fatores, ou seja, k variáveis de
controle, o planejamento de dois níveis precisará de 2 x 2 x ... x 2 = 2k, para o número
de ensaios diferentes, sendo conhecido como planejamento fatorial 2k.
Para o planejamento fatorial, são especificados os valores dos fatores em cada
experimento, e estes são classificados em níveis, variando-se, por exemplo, de -1
(mínimo) a +1 (máximo). A partir de uma matriz serão apresentadas todas as
combinações entre o número de fatores e de níveis nos experimentos a serem realizados
(Barros Neto et al., 2001; Rodrigues & Iemma, 2005).
2.4.2. Planejamento Plackett & Burman
A maioria dos planejamentos do tipo “Screening” (Delineamentos de Seleção de
fatores) foi descrito originalmente por Plackett & Burman em 1946. Os planejamentos
experimentais de Plackett & Burman são eficientes em situações que envolvem uma

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
44
grande quantidade de variáveis a serem exploradas em um processo de otimização de
fatores (Reddy et al., 2008).
As matrizes Plackett & Burman consistem de planejamentos fatoriais de dois
níveis para estudar k = N – 1 variáveis em N ensaios. São construídas para ensaios
múltiplos de 4, sendo essas matrizes para ensaios de 8, 12, 16,..., 100. Cada variável
independente em uma matriz assume uma configuração de dois níveis, o inferior (-1) e o
superior (+1), para o nível intermediário é estabelecido (0). Para cada vez que a variável
aparece no nível inferior, terá o par equivalente no nível superior (Rodrigues & Iemma,
2005).
Para evitar dúvidas nos resultados dos efeitos de algum fator no planejamento de
Plackett & Burman, o número de ensaios a ser realizado deve ser no mínimo o número
de variáveis mais 4. No caso de um fatorial completo o número de ensaios a ser
realizado é bem maior (Rodrigues & Iemma, 2005).
Mesmo já determinado o planejamento Plackett & Burman, torna-se importante
à realização de experimentos relativos aos pontos centrais de modo a se avaliar melhor
as possíveis interferências analíticas que podem ocorrer durante os diferentes
experimentos. A repetição do ponto central permite também a determinação do erro
puro e da repetibilidade do processo. Na prática, esta é uma ferramenta que permite
identificar melhor se o processo em estudo está sob controle (Rodrigues & Iemma,
2005).
É possível que no planejamento Plackett & Burman não tenha sido considerado
a codificação do ponto central (0) para repetição, pois, matematicamente, a estimativa
do valor desse efeito codificado (zero) vezes, teria como resposta zero. Porém, no
resultado prático do experimento, consegue-se informações valiosas com essas
repetições no ponto central (Rodrigues & Iemma, 2005).
2.4.3. Aplicações do Planejamento Plackett & Burman
Ghanem et al. (2000) produziram xilanase por Aspergillus terreus cultivadas
sobre palha de trigo finamente moída em fermentação de estado sólido. A composição
ideal do meio foi estabelecida aplicando-se o planejamento experimental de Plackett &
Burman. A melhor atividade enzimática foi obtida em um meio contendo 10 g/L de

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
45
palha de trigo por frasco umedecido com uma solução concentrada de sal com teor de
água inicial de 75 % e incubadas durante 4 dias a 30 °C. Observou-se que íons
metálicos como Hg2+, Cu2+, Co2+, Fe3+ e Pb2+ inibiram fortemente a xilanase, enquanto
que Ca2+ ativou a enzima.
Mu et al. (2005) estudaram as otimização de meio de fermentação para a reação
de estereoinversão por Candida parapsilosis CCTCC M203011 utilizando o
planejamento Plackett & Burman. Os resultados mostraram que dentre os fatores
estudados, os fatores mais significativos foram: a Concentração de MgSO4, ZnSO4 e
KH2PO4 e que após a utilização da metodologia de superfície de resposta foi constatado
que o ponto ótimo para a otimização foi conseguido com as concentrações dos fatores
MgSO4, ZnSO4 e KH2PO4 nos valores em: 1,17 g/L, 0,10 g/L e 9,41 g/L,
respectivamente, elevando a taxa de conversão enantiosseletiva de 75,2 % para 96 %.
Naveena et al. (2005) empregaram o planejamento Plackett & Burman para a
seleção de 15 parâmetros para produção de ácido láctico L(+) de farelo de trigo,
substrato de baixo custo e suporte sólido, por Lactobacillus amylophilus GV6 em
fermentação de estado sólido (SSF). Onze nutrientes pertencentes a duas categorias,
fontes de nitrogênio e fontes de sal, junto com três parâmetros físicos e um tampão
foram os parâmetros considerados. Os autores verificaram influência na produtividade
devido às fontes de nitrogênio, peptona, extrato de levedura e tri-citrato de amônio,
juntamente com o NaH2PO4.2H2O e Tween 80, que podem ainda serem otimizados para
o aumento na produção de ácido lático. Esses pesquisadores ainda chamam atenção para
o uso escasso deste planejamento em fermentação em estado sólido e que o mesmo
ainda não havia sido considerado anteriormente para a conversão direta de amido em
ácido láctico L (+) usando um sistema bacteriano.
Khosravi-Darani et al. (2008) estudaram a aplicação de Plackett & Burman na
análise dos fatores significativos para a produção de ácido cítrico por Aspergillus niger
ATCC 9142 utilizando palha de trigo como substrato. Os resultados mostraram que a
produção, com a palha de trigo pré-tratada com HCl, NaOH e Uréia, foi mais eficiente e
que os fatores mais significativos na produção foram: Solvente, Teor de Umidade,
Temperatura, Tempo de Incubação, Concentração Inicial de Açúcar e Concentração de
Metanol.
Ji et al. (2009) estudaram o desenvolvimento de um meio de cultura econômico
para a produção do 2,3-butanodiol através da fermentação da glicose e xilose por

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
46
Klebsiella oxytoca utilizando o planejamento Plackett & Burman. Os resultados
mostraram que a uréia e a maceração de milho foram os fatores mais significativos para
a produção do 2,3-butanodiol e que através de planejamento experimental com repetição
em triplicata no ponto central para determinar os níveis ótimos de produção. Os autores
constaram que o rendimento atingido foi 85,6% do valor teórico.
Paula & Arruda. (2009) observaram a necessidade da busca por alternativas mais
eficientes ao método termomecânico utilizado para a degradação de milhões de
toneladas de penas de aves provenientes de frigoríficos e aviários, cuja rígida estrutura é
composta por mais de 90 % de queratina. Para esse fim, utilizaram de comunidades
microbianas para a finalidade proposta, testando o planejamento experimental Plackett
& Burman para verificar a influência de diferentes microrganismos simultaneamente,
com um número reduzido de experimentos. Testaram 32 bactérias isoladas, inoculadas
em meio contendo somente uma pena como única fonte de carbono, durante sete dias,
de onde foi selecionado as cepas para os consórcios microbianos. Experimentos unindo
as cepas NP5, FCA7, NP4 e S14 demonstraram diferentes atividades queratinolíticas,
aumentando o nível de degradação, até 87,8%, comparada com as cepas
individualmente, variando entre 26,4% e 78,0%.
Rajendran & Thangavelu (2009) obtiveram valores máximos da atividade de
produção de lipase em 3,98 U/mL e concentração de massa celular em 5,62 g.L-1 com R.
arrhizus em cultivo submerso descontínuo otimizado pelo planejamento Plackett &
Burman. As variáveis mais importantes que afetaram a produção da lipase foram azeite
de oliva, peptona, KH2PO4, CaCl2.2H2O e MgSO4.7H2O.
Li et al. (2010) estudaram a otimização de meio de cultura para a produção de
Streptolydigin (um antibiótico) por Streptomyces lydicus AS 4,2501 utilizando Plackett
& Burman e a metodologia de superfície de resposta. Os resultados mostraram que
dentre os fatores estudados, os fatores mais significativos para a produção de
Streptolydigin e otimização do meio de cultura foram: Concentração de Amido,
Concentração de Peptona e a Concentração de K2HPO4.
Chen et al. (2010) estudaram a otimização de meio de cultura para a produção da
Luciferase – que são enzimas que catalisam reações biológicas transformando energia
química em energia luminosa - por Bacillus Subtilis em fermentação descontínua
utilizando planejamento Plackett & Burmam. Os resultados mostraram que dentre os
componentes estudados, os fatores mais significativos para a produção e otimização do

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
47
meio de cultura para a produção da Luciferase foram: Concentração de Glicose 1,17 %,
Extrato de Levedura 2,27 % e Glutamato 0,55 %.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o projeto Plackett & Burman e a metodologia de superfície de resposta. Os
resultados mostraram que a eficiência máxima de extração da quitina foi conseguida nas
seguintes condições: Concentração de casca de camarão (50 g/L), glicose (50 g/L),
tempo de incubação de 5 dias e inoculo com 0,05 de OD (densidade óptica).
Gao et al. (2010) desenvolveram um meio para a fermentação do extrato bruto
de inulina de tubérculos de alcachofra de Jerusalém por Paenibacillus polymyxa ZJ-9
para produzir R,R-2,3-butanodiol (R-R,2,3-BD). Inulina, K2HPO4 e NH4Cl foram os
parâmetros principais na fermentação de acordo com os resultados obtidos com o
planejamento experimental Plackett & Burman. A concentração ideal dos três fatores,
77,14 g/L, 3,09 g/L e 0,93 g/L, respectivamente, foi obtida pelo planejamento Box-
Behnken e pela superfície de resposta. Sob as condições ideais, a concentração do R-
R,2,3-BD obtida foi 36,92 g/L, em mais de 98 % de pureza óptica. Comparado com
outras fontes de carbono investigadas, a fermentação do extrato bruto de inulina obteve
o maior rendimento de R-R,2,3-BD.
Uma metodologia de superfície de resposta foi empregada com sucesso para
otimizar um meio de fermentação chamada VHG (Very High Gravity), referência à alta
concentração de açúcares e densidade dos mostos, baseado na água maceração de milho
CSL (Corn Steep Liquor) e outros nutrientes de baixo custo para a produção eficiente de
etanol a partir de glicose por S. cerevisiae. Usando a composição média otimizada (g/L:
CSL 44,3; uréia 2,3; MgSO4.7H2O 3,8; CuSO4.5H2O 0,03), a levedura industrial PE-2
foi capaz de fermentar até 330 g/L de glicose e produzir 18,6 % (v/v) de etanol, com
uma produção descontínua de 2,4 g/L/h e um rendimento em etanol de 93 % (Pereira et
al., 2010).
O planejamento Plackett & Burman juntamente com o Planejamento Composto
Central (CCD) foram aplicados para otimizar o meio de cultura para produção de etanol
por Clostridium autoethanogenum com monóxido de carbono (CO) como fonte única de
carbono e um meio contendo (g/L): 1,0 de NaCl, 0,1 de KH2PO4, 0,02 de CaCl2, 0,15 de
extrato de levedura, 0,116 de MgSO4, 1,694 de NH4Cl e pH 4,74. O rendimento ótimo
de etanol previsto pela metodologia de superfície de resposta (RSM) e um algoritmo

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
48
genético de rede neural artificial (ANN-GA) foram 247,48 e 261,48 mg/L,
respectivamente. Valores semelhantes aos obtidos experimentalmente sob as condições
ideais sugeridas pelos métodos estatísticos (254,26 e 259,64 mg/L). A adequação do
modelo ANN-GA foi maior do que o modelo RSM. Os rendimentos obtidos foram
substancialmente superiores aos relatados anteriormente (60-70 mg/L) com essa cultura
(Guo et al., 2010).
Rigueira (2010) avaliou a influência das variáveis do procedimento de extração
na porcentagem de recuperação das aminas encontradas no leite cru utilizando um
planejamento para “screening” do tipo Plackett & Burman, para os ácidos
tricloroacético (TCA) e sulfosalicílico (ASS).
Quines (2010) estudou a necessidade ou não de se fazer um pré-tratamento da
biomassa antes da extração. Um planejamento experimental Plackett & Burman de doze
ensaios mais três pontos centrais foi utilizado para selecionar as variáveis mais
significativas na extração de Poli(3-hidroxibutirato) (P(3HB)) por 1,2-carbonato de
propileno. Foram realizados dois testes de recuperação do solvente 1,2-carbonato de
propileno, após a extração, sendo um com aplicação de aquecimento e o outro com
aquecimento e vácuo. Os resultados mostraram que o pré-tratamento térmico levou a
uma maior porcentagem de recuperação e de pureza do polímero.

Capítulo 3 Metodologia Experimental

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
50
3. Metodologia Experimental
O presente capítulo apresenta os matérias e métodos utilizados na produção de
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso.
3.1. Quitosana
Neste trabalho utilizou-se uma quitosana (Lote 067) obtida junto a POLYMAR/CE,
com no mínimo 80% de desacetilação. Esse substrato foi utilizado como fonte de carbono e
energia tanto no estudo cinético como nos ensaios do planejamento de seleção de fatores
Plackett & Burman e Planejamento Composto Central, para a produção de quitosanase pelo
Metarhizium anisopliae, por fermentação submersa.
3.2. Microrganismo
O fungo Metarhizium anisopliae estirpe (CG374), utilizado neste estudo, foi cedido
pela Embrapa Recursos Genéticos e Biotecnologia (Brasília / DF-Brasil) e foi mantido em
placas de Petri contendo o meio Batata Dextrose Agar (PDA). As placas foram incubadas em
estufa a 25 ºC. O pH do meio de manutenção foi mantido entre 6,0 e 6,5.
3.2.1. Escolha da cepa produtora de quitosanase
A escolha do microrganismo Metarhizium anisopliae foi feita com base na capacidade
de crescer em meio contendo quitosana como única fonte de carbono, além de apresentar
grande tolerância a pH variando a uma taxa de 2,0 a 8,5, sendo 6,9 a melhor condição para o
crescimento vegetativo e esporução, de acordo com Assis (2010).
3.2.2. Manutenção da cepa
A manutenção da cultura foi realizada em placas petri contendo o meio Batata
Dextrose Agar (PDA). A renovação das células foi realizada a cada trinta dias através de
subcultivos das células no meio PDA.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
51
3.3. Produção de enzima
No preparo do inóculo, utilizou-se 7 placas cultivadas e mantidas, com o fungo
Metarhizium anisopliae, em meio PDA a 25 ºC durante 5 dias em estufa. O meio de cultura
para a produção da enzima quitosanase apresentou os seguintes componentes: quitosana;
peptona; extrato de levedura; NaNO3; K2HPO4; KCl; MgSO4.7H2O e FeSO4, com
concentrações de acordo as matrizes dos planejamentos: Plackett & Burman e composto
central. Para a produção da enzima no cultivo, a biomassa foi transferida através de água
estéril, para um Béquer de 500 mL que, em seguida, foi colocado em Erlenmeyers de 250 mL
na proporção de 20 mL de biomassa para 180 mL do meio de cultura. Os Erlenmeyers
cultivados foram então incubados em incubador rotativo a 25 ºC e 150 rpm durante 24 h.
Alíquotas foram retiradas e centrifugadas a 8000xg por 5 min, a cada 6 horas, para se
determinar a atividade quitosanolítica, teores de açúcares redutores e de proteína total.
Ressalta-se que todos os experimentos foram feitos em triplicata.
3.4. Determinação da Proteína Total
A determinação da proteína total foi efetuada de acordo com o método modificado de
Sedmak & Grossberg (1977), que se baseia na diminuição da absorbância do corante azul de
Coomassie, devido à ligação proteína-corante, a 595 nm, comparado a absorção máxima a
465 nm, conforme Santos (2001). Nesse caso, após centrifugação para retirada da biomassa,
transferia-se para um eppendorf 1,5 mL de amostra (caldo fermentativo) e adicionava-se 1,5
mL de solução azul de Coomassie. Deixava-se a solução em repouso por 5 minutos,
transferindo-se em seguida para um cubeta de quartzo e efetuando-se as leituras de
absorbância nos comprimentos de ondas 465 nm e 595 nm, respectivamente. O
espectrofotômetro (Thermo Spectronic modelo Genesys 10UV) foi zerado com água
destilada. O conteúdo protéico foi obtido a partir de uma curva padrão na qual graficava-se a
relação de absorbância A595/A465 para diferentes concentrações de Albumina de Soro
Bovino (Fração V – Lote 110881/Sigma Aldrich-Ohio-USA).
3.5. Determinação da Atividade Quitosanolítica e da Atividade Específica
A atividade quitosanolítica foi obtida monitorando-se o aumento dos açúcares
redutores totais, utilizando-se o método DNS a partir de uma curva padrão de glicosamina.
Para a determinação da atividade quitosanolítica adicionaram-se, aos tubos de ensaios 500 µL

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
52
de substrato (solução 0,2 % de quitosana solubilizada em ácido clorídrico 1N) e 500 µL de
caldo bruto (contendo enzima), controlando-se o pH entre 6,0 e 6,5 e incubando-se o sistema
em um banho termostatizado com temperaturas da ordem de 55 ºC. Após o tempo de
incubação de 30 minutos no banho foi adicionado aos tubos de ensaio 2,5 mL de DNS,
aquecendo-se à ebulição em banho-maria por 10 minutos, resfriando-se, em seguida, em
banho de gelo. Após essa etapa, o sistema foi centrifugado e com o sobrenadante efetuou-se a
leitura no espectrofotômetro a 600 nm. Destaca-se, que antes da leitura da amostra zerava-se o
equipamento (branco do espectro) com 1,0 mL de água destilada acrescidos de 2,5 mL de
DNS e realizava-se o branco da enzima que consistia da adição de 500 µL de solução de
quitosana, completando-se até um 1,0 mL com água destilada. Uma unidade de atividade
quitosanolítica (A) foi definida como a quantidade de enzima necessária para produzir 1
µmol.min-1 de glicosamina por mL de extrato enzimático, nas condições de temperatura e pH
citadas, sendo o resultado expresso em (U.mL-1). A Atividade específica foi determinada pela
divisão entre a atividade da enzima (U.mL-1) e a concentração de proteína total (µg.mL-1),
sendo expressa em (U. µg-1).
3.6. Planejamento experimental
3.6.1. Planejamento de seleção de fatores Plackett & Burman
O planejamento experimental Plackett & Burman é uma técnica muito usada, para se
realizar triagem de fatores na otimização num determinado processo (Reddy et al., 2008), e
foi utilizado neste trabalho para identificar os fatores que influenciam de forma significativa
na produção da enzima quitosanase.
A Tabela 3 ilustra os níveis de cada variável (fator) utilizada no planejamento Plackett
& Burman, sendo: Concentração de Quitosana (Q), Peptona (PP), Extrato de Levedura (EL),
NaNO3 (NS), K2HPO4 (FP), KCl (CP), MgSO4.7H2O (SM), FeSO4 (SF). Já a Tabela 4 mostra
a matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3 repetições
na condição do ponto central.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
53
Tabela 3 – Níveis para os fatores e seus valores codificados.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1 0,1 0,2 0,2 0,1 0,05 0,025 0,025 0,0005
0 0,2 0,5 0,3 0,2 0,10 0,050 0,050 0,0010
1 0,3 0,8 0,4 0,3 0,15 0,075 0,075 0,0015
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1
2 1 1 -1 1 -1 -1 -1 1 -1 1 1
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1
5 1 1 -1 1 1 -1 1 -1 1 -1 1
6 1 1 1 -1 1 1 -1 1 1 -1 -1
7 -1 1 1 1 -1 1 1 -1 -1 1 1
8 -1 -1 1 1 1 -1 1 1 1 1 1
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1
11 -1 1 -1 -1 -1 1 1 1 1 1 -1
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1
13 0 0 0 0 0 0 0 0 0 0 0
14 0 0 0 0 0 0 0 0 0 0 0
15 0 0 0 0 0 0 0 0 0 0 0
Sendo: V11,V12,V13 = Variáveis inertes para cálculo do erro.
3.6.2. Planejamento Composto Central para a Otimização do Meio de Cultura para a
Produção da Enzima em Cultivo Submerso
Um dos problemas mais comuns para quem realiza experimentos é determinar a
influência de uma ou mais variáveis sobre outra variável de interesse (Lobato, 2003). Este

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
54
problema pode ser minimizado utilizando-se um planejamento experimental de modo a se
obter uma função, ou uma aproximação satisfatória para a mesma, de forma que se operando
sobre as variáveis de entrada produza-se como saída a resposta ou as respostas observadas
(Barros Neto, 2001; Silva, 2004).
Em um planejamento, inicialmente, deve-se definir os fatores e as respostas de
interesse. Tendo-se identificado os fatores e respostas define-se o objetivo que se pretende
alcançar com os experimentos para que se possa escolher o planejamento mais apropriado.
(Silva, 2004).
No presente trabalho foi utilizado um Planejamento Composto Central (CCD) com 22
experimentos na parte fatorial, repetição em triplicata no ponto central e quatro experimentos
na parte axial (planejamento do tipo estrela), para se investigar a influência dos fatores:
concentração de quitosana e concentração de FeSO4 que foram os dois fatores que
influenciaram a produção de quitosanase, conforme o planejamento Plackett & Burman de 12
ensaios e mais 3 repetições na condição do ponto central. Estes dois fatores foram estudados
em cinco diferentes níveis (-1,41; -1; 0; 1; 1,41) e um conjunto de 11 experimentos foi
realizado, conforme Tabela 5.
Tabela 5 – Matriz do planejamento Composto Central.
Ensaio Q SF
1 -1 -1
2 -1 1
3 1 -1
4 1 1
5 -1,41 0
6 1,41 0
7 0 -1,41
8 0 1,41
9 0 0
10 0 0
11 0 0
Os outros fatores foram mantidos nos níveis mais baixos (para evitar custos). Uma que
não influenciaram de forma significativa na produção da enzima quitosanase.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
55
A codificação dos fatores foi realizada de acordo com a seguinte equação (Ji et al.,
2009):
Xi = (xi – x0)/∆xi , i = 1, 2, 3, ..., k (2)
Onde Xi é o fator independente codificado, xi é o fator independente real, x0 é o valor de xi no
ponto central e ∆xi é a variação de valor. O comportamento do sistema foi explicado pela
seguinte equação polinomial de segunda ordem:
Yp = β0 + ∑ βiXi + ∑ βiiXi2 + ∑ βijXiXj, i, j = 1, 2, 3,..., k (3)
Onde Yp foi a resposta esperada, β0 foi a interseção, Xi e Xj
foram os fatores independentes codificados, βi foi o coeficiente linear, βii
foi o coeficiente quadrático e βij foi o coeficiente de interação. Destaca-se que os outros
fatores não significativos foram mantidos no nível mais baixo (-1).
A Tabela 6 apresenta os níveis para os fatores e seus valores codificados, para o
Planejamento Composto Central, com triplicata no ponto central.
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1,41 0,218 0,2 0,2 0,1 0,05 0,025 0,025 0,0009
-1 0,3 0,2 0,2 0,1 0,05 0,025 0,025 0,0015
0 0,5 0,2 0,2 0,1 0,05 0,025 0,025 0,0030
1 0,7 0,2 0,2 0,1 0,05 0,025 0,025 0,0045
1,41 0,782 0,2 0,2 0,1 0,05 0,025 0,025 0,0051
Na análise do planejamento de seleção de fatores Plackett & Burman e do
planejamento experimental fatorial foi utilizado um pacote computacional STATISTICA for
Windows versão 7.0.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
56
3.7. Perfil cinético durante o cultivo
Um perfil cinético na melhor condição de cultivo, conforme planejamento
apresentado, foi realizado em frascos Erlenmyer de 250 mL contendo 200 mL de meio de
cultivo contendo os seguintes componentes, em 1 Litro: 2,0g de quitosana; 5,0g de peptona;
3,0g de extrato de levedura; 2,0g de NaNO3; 1,0g de K2HPO4; 0,5g de KCl; 0,5g de
MgSO4.7H2O; 0,01g de FeSO4, que foram inoculadas com 10% (vol/vol) de inóculo contendo
o fungo Metarhizium anisopliae e cultivadas em incubador rotativo durante 5 dias a 150 rpm
e 25 ºC. Alíquotas foram retiradas a cada seis horas e analisadas quanto à proteína total no
sobredanante, atividade enzimática, atividade específica e açúcares redutores. Os ensaios
foram realizados em triplicata (Silva Filho, 2005).

Capítulo 4 Resultados e
Discussão

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
58
4. Resultados e discussão
Neste capítulo são apresentados e discutidos os resultados obtidos no presente
trabalho. Inicialmente, é apresentada imagem do MEV do Metarhizium anisopliae
mantidos em meio PDA. Em seguida, apresenta-se e discute-se os resultados da avaliação
dos fatores significativos, que podem influenciar na produção da quitosanase, no
planejamento de seleção de fatores, conforme planejamento Plackett & Burman bem como
são apresentados e discutidos os resultados obtidos para o planejamento composto central.
Por último, apresenta-se o perfil cinético, na melhor condição, para a produção de
quitosanase em cultivo submerso descontínuo usando o fungo Metarhizium anisopliae.
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA.
A Figura 7 ilustra a imagem da Microscopia Eletrônica de Varredura (MEV) do micélio de Metarhizium anisopliae mantido em meio PDA.
Figura 7. Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA.
Observa-se na Figura 7, quanto à morfologia que o Metarhizium anisopliae, apresenta-
se como um fungo filamentoso com as hifas intimamente entrelaçadas conforme citado por
Wang et al. 2002.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
59
4.2. Planejamento Plackett & Burman
4.2.1 Análise do Plackett & Burman com 8 fatores e 12 ensaios
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis, foi utilizado para avaliar a influência dos fatores: Concentração de
Quitosana (Q), Peptona (PP), Extrato de Levedura (EL), NaNO3 (NS), K2HPO4 (FP), KCl
(CP), MgSO4.7H2O (SM), FeSO4 (SF) na produção da enzima quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso. Ressalta-se que os experimentos foram
realizados aleatoriamente.
A Tabela 7 apresenta a matriz do modelo Plackett & Burman e os respectivos valores
obtidos para a variável resposta (Atividade Quitosanalítica).
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.
Ensaio Q PP EL NS FP CP SM SF A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 0,0195
2 1 1 -1 1 -1 -1 -1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 0,0109
4 1 -1 1 1 -1 1 -1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 0,0136
6 1 1 1 -1 1 1 -1 1 0,0156
7 -1 1 1 1 -1 1 1 -1 0,0109
8 -1 -1 1 1 1 -1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 0,0085
A Tabela 8 apresenta os efeitos estimados para o planejamento Plackett & Burman.
Observa-se na Tabela 8 que o valor do coeficiente de determinação R2 foi de
aproximadamente 90%, indicando que um modelo linear poderia representar bem a relação
entre o fator significativo, concentração de quitosana (1), e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
60
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman.
Efeitos Estimados; R2=0,8996; Ajuste:0,63178 MS Erro Residual =0,0000054
Desvio Padrão t(3) p
Lim. Conf. Lim. Conf.
Efeito Erro Residual
-95,% +95
Média/Interação 0,0127 0,0007 18,903 0,0003 0,0105 0,0149 (1) Quitosana 0,0055 0,0013 4,0628 0,0268 0,0011 0,0098
(2) Peptona 0,0015 0,0013 1,1080 0,3486 -0,0028 0,0058 (3) Ext. de leved. 0,0009 0,0013 0,7387 0,5136 -0,0033 0,0053
(4) NaNO3 0,0002 0,0013 0,1758 0,8715 -0,0040 0,0045 (5) K2HPO4 -0,0014 0,0013 -1,0552 0,3687 -0,0057 0,0028
(6) KCl -0,0022 0,0013 -1,6708 0,1933 -0,0065 0,0020 (7) MgSO4.7H2O -0,0003 0,0013 -0,2286 0,8338 -0,0046 0,0039
(8) FeSO4 0,0029 0,0013 2,1457 0,1211 -0,0014 0,0071
A Figura 8 mostra o gráfico de Pareto, uma das formas de se avaliar visualmente a
influência dos fatores estudados na resposta. A magnitude dos efeitos é representada pelas
colunas enquanto que a linha transversal às colunas representa a magnitude dos efeitos com
significado estatístico para p=0,05, ou seja, os fatores que são estatisticamente significativos
ao nível de 95% de confiança.
Figura 8. Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados.
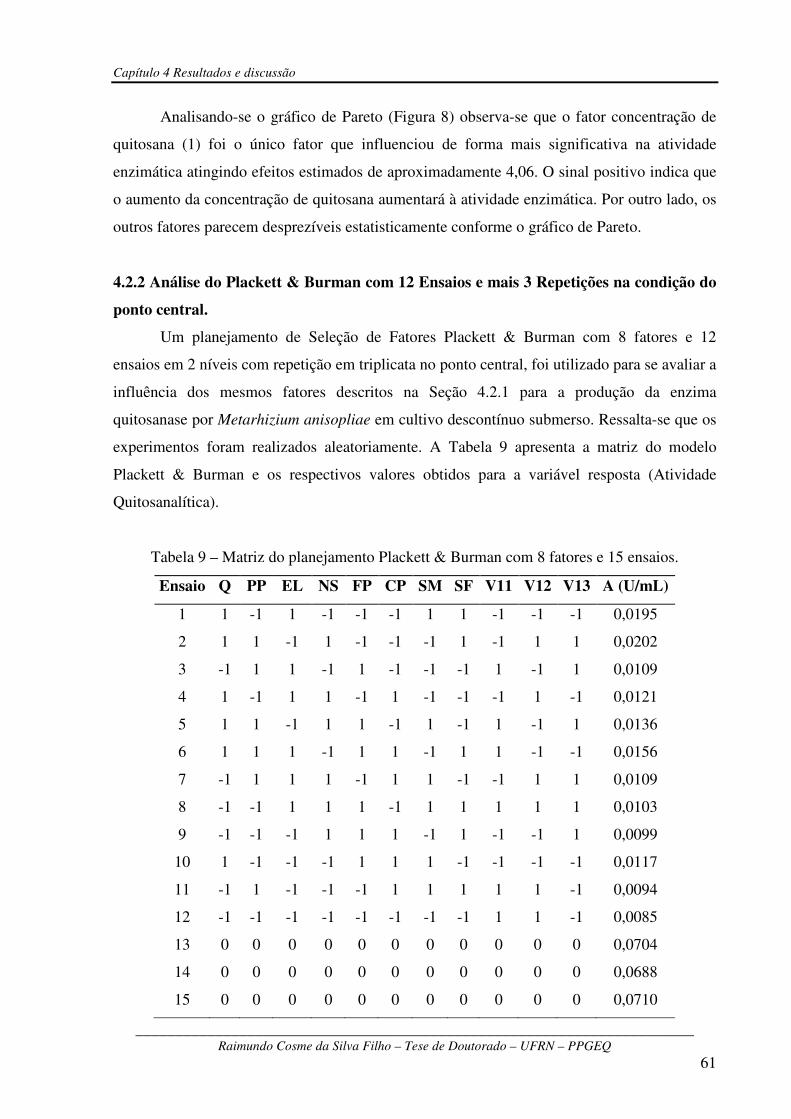
Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
61
Analisando-se o gráfico de Pareto (Figura 8) observa-se que o fator concentração de
quitosana (1) foi o único fator que influenciou de forma mais significativa na atividade
enzimática atingindo efeitos estimados de aproximadamente 4,06. O sinal positivo indica que
o aumento da concentração de quitosana aumentará à atividade enzimática. Por outro lado, os
outros fatores parecem desprezíveis estatisticamente conforme o gráfico de Pareto.
4.2.2 Análise do Plackett & Burman com 12 Ensaios e mais 3 Repetições na condição do
ponto central.
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis com repetição em triplicata no ponto central, foi utilizado para se avaliar a
influência dos mesmos fatores descritos na Seção 4.2.1 para a produção da enzima
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso. Ressalta-se que os
experimentos foram realizados aleatoriamente. A Tabela 9 apresenta a matriz do modelo
Plackett & Burman e os respectivos valores obtidos para a variável resposta (Atividade
Quitosanalítica).
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13 A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1 0,0195
2 1 1 -1 1 -1 -1 -1 1 -1 1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1 0,0109
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 1 -1 1 0,0136
6 1 1 1 -1 1 1 -1 1 1 -1 -1 0,0156
7 -1 1 1 1 -1 1 1 -1 -1 1 1 0,0109
8 -1 -1 1 1 1 -1 1 1 1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 1 1 -1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1 0,0085
13 0 0 0 0 0 0 0 0 0 0 0 0,0704
14 0 0 0 0 0 0 0 0 0 0 0 0,0688
15 0 0 0 0 0 0 0 0 0 0 0 0,0710
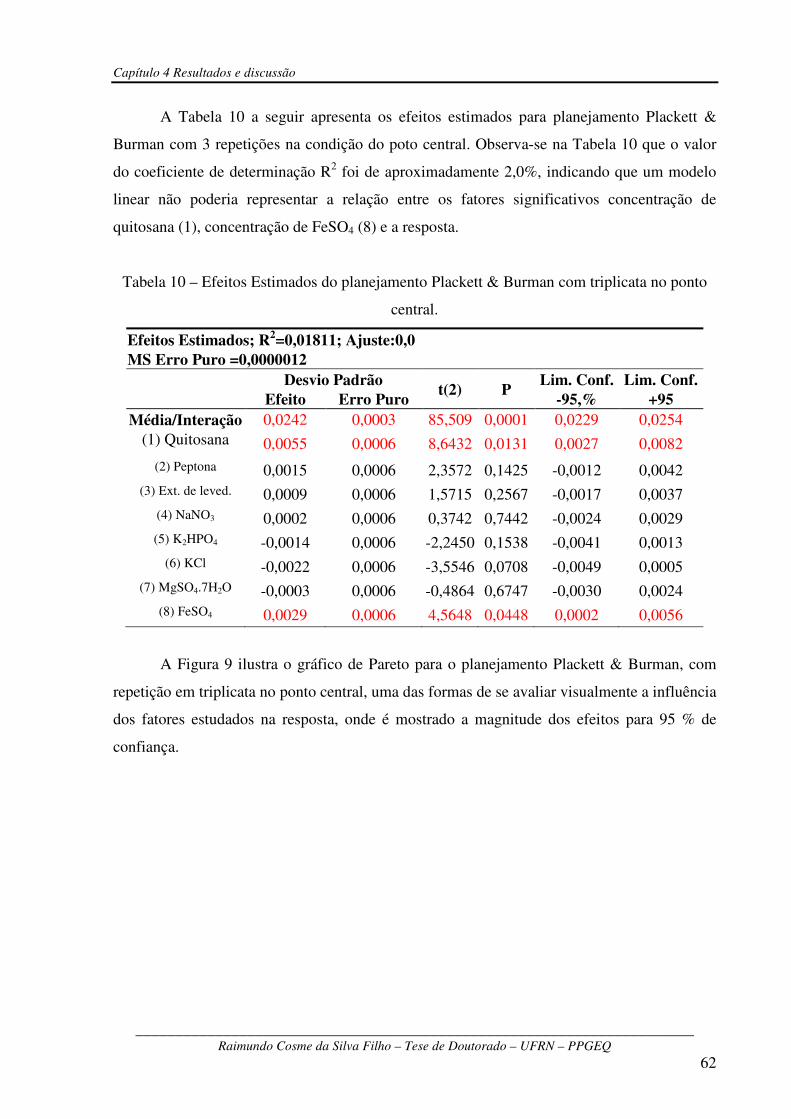
Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
62
A Tabela 10 a seguir apresenta os efeitos estimados para planejamento Plackett &
Burman com 3 repetições na condição do poto central. Observa-se na Tabela 10 que o valor
do coeficiente de determinação R2 foi de aproximadamente 2,0%, indicando que um modelo
linear não poderia representar a relação entre os fatores significativos concentração de
quitosana (1), concentração de FeSO4 (8) e a resposta.
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central.
Efeitos Estimados; R2=0,01811; Ajuste:0,0 MS Erro Puro =0,0000012
Desvio Padrão t(2) P
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0242 0,0003 85,509 0,0001 0,0229 0,0254 (1) Quitosana 0,0055 0,0006 8,6432 0,0131 0,0027 0,0082
(2) Peptona 0,0015 0,0006 2,3572 0,1425 -0,0012 0,0042 (3) Ext. de leved. 0,0009 0,0006 1,5715 0,2567 -0,0017 0,0037
(4) NaNO3 0,0002 0,0006 0,3742 0,7442 -0,0024 0,0029 (5) K2HPO4 -0,0014 0,0006 -2,2450 0,1538 -0,0041 0,0013
(6) KCl -0,0022 0,0006 -3,5546 0,0708 -0,0049 0,0005 (7) MgSO4.7H2O -0,0003 0,0006 -0,4864 0,6747 -0,0030 0,0024
(8) FeSO4 0,0029 0,0006 4,5648 0,0448 0,0002 0,0056
A Figura 9 ilustra o gráfico de Pareto para o planejamento Plackett & Burman, com
repetição em triplicata no ponto central, uma das formas de se avaliar visualmente a influência
dos fatores estudados na resposta, onde é mostrado a magnitude dos efeitos para 95 % de
confiança.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
63
Figura 9. Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto
central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto, observa-se que os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma mais
significativa na atividade enzimática atingindo efeitos estimados de aproximadamente 8,84 e
4,67, respectivamente. O sinal positivo indica que o aumento das concentrações de quitosana
e de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores parecem
desprezíveis estatisticamente conforme o gráfico de Pareto.
Este comportamento do FeSO4 está de acordo com o observado por Hagag et al.
(2012) que utilizando o Aspergillus fumigatus na produção de protease mostraram que um
fator de transcrição PrtT regula positivamente o controle da produção de protease, da
assimilação de Ferro e da síntese de ergosterol e que a produção de protease foi favorecida
pelo aumento da concentração de ferro.
Rajendran & Thangavelu (2009) utilizaram um planejamento Plackett & Burman para
identificar quais os fatores significativos na produção de lipase por Rhizopus arrhizus MTCC
2233 e observaram o efeito significativo médio do FeSO4, apresentando um nível de
confiança para esse fator de 48,92% (para um máximo de 98,7 % de MgSO4), com uma
produção de lipase da ordem de 2,98 U/mL em 84 horas de fermentação e que ocorreu uma
produção considerável de protease no meio de cultivo da ordem de 1,92 U/mL em 96 horas de
cultivo.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
64
Mu et al. (2005) também observaram um efeito positivo do FeSO4, da ordem de 1,48
para um fator máximo encontrado de 3,11 de MgSO4, no aumento da atividade da
estereoinversão de S-fenil-1,2-etanodiol por Candida parapsilosis CCTCC M203011 na
otimização da fermentação utilizando um planejamento Plackett & Burman.
Dessa forma, observa-se que o FeSO4 é importante uma que vez o mesmo está
diretamente associado a síntese de proteases (principalmente metalloproteases), conforme
citado por Hagag et al. (2012).
4.3. Planejamento Composto Central para Produção de Quitosanase por
Metarhizium anisopliae em Cultivo Descontínuo Submerso
Um planejamento experimentos 22 com repetição em triplicata no ponto central e mais
quatro ensaios axiais, foram utilizados para se chegar a uma combinação ideal de quitosana e
FeSO4 na produção da enzima quitosanase (Tabela 11). A partir do modelo Plackett &
Burman (Tabela 12), que mostrou que o aumento das concentrações de quitosana e FeSO4,
favoreceram a produção de quitosanase, os valores utilizados neste modelo para estes
componentes nos níveis mais altos, serão considerados como os níveis mais baixos no
planejamento experimental do composto central. Os outros fatores foram fixados em todos os
ensaios em seus níveis mais baixos para a redução de custos (Tabela 11).

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
65
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da
atividade enzimática.
Ensaio Q SF A (U/mL)
1 -1 -1 0,0195
2 -1 1 0,0202
3 1 -1 0,0109
4 1 1 0,0121
5 -1,41 0 0,0136
6 1,41 0 0,0156
7 0 -1,41 0,0108
8 0 1,41 0,0103
9 0 0 0,0099
10 0 0 0,0116
11 0 0 0,0094
A Tabela 12 apresenta os efeitos estimados para o planejamento composto central.
Tabela 12 – Efeitos Estimados do planejamento Composto Central.
Efeitos Estimados; R2=0,629; Ajuste:0,25836 MS Erro Puro =0,0000014
Desvio Padrão t(2) p
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0103 0,0006 15,305 0,0042 0,0074 0,0132 (1) Quitosana (L) 0,0011 0,0008 1,4316 0,2885 -0,0023 0,0047
Quitosana (Q) 0,0058 0,0009 5,9637 0,0269 0,0016 0,0101 (2) FeSO4 (L) -0,0043 0,0008 -5,3037 0,0337 -0,0079 -0,0008 (2) FeSO4 (Q) 0,0018 0,0009 1,8366 0,2076 -0,0024 0,0060
1L x 2L 0,0001 0,0011 0,1520 0,8931 -0,0048 0,0052
Observa-se na Tabela 12 que o valor do coeficiente de determinação R2 foi de
aproximadamente 63%, indicando que um modelo linear representaria bem a relação entre os
fatores significativos concentração de quitosana (1), concentração de FeSO4 (8) e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
66
A Tabela 13 mostra um resumo da ANOVA para o planejamento composto central
com repetição em triplicata no ponto central.
Tabela 13 – Resumo da ANOVA.
Fonte de
Variação
Soma dos
Quadrados
Grau de
Liberdade
Quadrado
Médio
Fcalculado Ftabelado
Regressão
Resíduos
0,000086 2 0,000043 6,14 4,46
0,000056 8 0,000007
Falta de
Ajuste
Erro puro
0,000053 6 0,000009 6 19,33
0,000003
2
0,0000015
Total 0,000142 10
De acordo com a Tabela 13, partindo-se dos efeitos significativos propõe-se o
modelo:
� = 0,010316 + 0,002942� � − 0,002193�� (8)
% de variação explicável: 60,5
% máxima de variação explicável: 97,9
A análise da ANOVA mostra que o modelo proposto possui significância estatística, ao
nível de 95% de confiança, uma vez que o valor de F (calculado) é maior que F (tabelado),
entretanto a ANOVA mostra também que o modelo proposto pode ser útil para se fazer
previsões, uma vez que a relação entre F (calculado) e F (tabelado) para a falta de ajuste e o
erro puro é menor que 1. O coeficiente de determinação R2 foi de aproximadamente 63%,
indicando que o modelo consegue explicar 63% da variação total em torno da média.
Os cálculos para variação explicável e para máxima variação explicável são dados pelas
Equações (9) e (10):
% de variação explicável = (SQ da regressão) / (SQ Total) (9)
% máxima de variação explicável = (SQ Total – SQ Erro puro) / (SQ Total) (10)
Sendo:
SQ – Soma dos quadrados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
67
A Figura 10 mostra o gráfico de Pareto para o planejamento Composto Central
utilizando erro puro. A magnitude dos efeitos é representada pelas colunas. Já a linha
transversal às colunas representa a magnitude dos efeitos com significado estatístico para 95%
de confiança.
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto (Figura 10) observa-se que os fatores concentração
de quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma
mais significativa na atividade enzimática atingindo efeitos estimados de aproximadamente
5,96 e - 5,30, respectivamente. O sinal positivo indica que o aumento das concentrações de
quitosana aumentará à atividade enzimática. O sinal negativo indica que o decréscimo da
concentração de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores
parecem desprezíveis estatisticamente no gráfico de Pareto.
A avaliação do modelo também pode ser feita através da observação do gráfico dos
valores preditos em função os valores observados que são mostrados na Figura 11. Os valores
preditos pelo modelo são representados pela reta, enquanto que os valores observados
representam-se pelos pontos.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
68
Figura 11 – Valores preditos em função dos valores observados.
Observa-se que alguns valores preditos apresentam um afastamento dos valores
observados. A Figura 12 ilustra a superfície de resposta da atividade quitosanolítica em
função dos dois fatores significativos, concentração de quitosana e concentração de FeSO4.
Figura 12. Atividade quitosanolítica em função da concentração de quitosana e da
concentração de FeSO4 para os fatores não significativos no ponto central.
Observa-se a existência de um ponto de mínimo, ocorrendo para concentração de
quitosana no ponto central (0) e para maiores concentrações de FeSO4 igual a (1,5). Essa

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
69
região deve ser evitada uma vez que a atividade quitosanolítica atinge níveis tão reduzidos
quanto 0,0094 U.mL-1. Destaca-se que nesse planejamento o maior nível da concentração de
FeSO4 (α=+1,41) equivale a 0,0051% (Tabela 6) que é aproximadamente cinco vezes maior
que o nível desse fator no ponto central do planejamento Plackett & Burman (0,001%),
conforme Tabela 3. Dessa forma, ao se utilizar o nível mínimo da concentração de FeSO4 no
Planejamento Composto Central (α=-1,41), que equivale a aproximadamente 0,001%,
aproxima-se do ponto central do do planejamento Plackett & Burman (0,001%), condição
essa que apresenta os melhores resultados de atividade quitosanolítica usando quitosana como
fonte de carbono e energia e fungo Metarhizium anisopliae.
4.4. Perfil Cinético – Melhor Condição
A seguir, apresenta-se um perfil cinético, na melhor condição, para produção da
quitosanase usando o fungo Metarhizium anisopliae, em que foram monitorados durante 24
horas os seguintes parâmetros: Concentração de proteínas totais (µg/mL), atividade
quitosanolítica (U/mL), atividade específica (U/µg) e açúcares redutores (mg/mL), ilustradas
nas Figuras 13, 14, 15 e 16, respectivamente.
Com relação à proteína extracelular, observa-se na Figura 13 que, inicialmente, ocorre
um aumento da mesma para o seu nível máximo (6,2 µg/mL), durante as 6 primeiras horas de
cultivo, seguido de uma redução para 5,7 µg/mL em 18 horas de cultivo, possivelmente
devido à presença de proteases no meio. Observa-se que a partir de então ocorre uma redução
e aumento da proteína total, em uma quantidade inferior, sugerindo um mecanismo de
controle.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
70
0 4 8 12 16 20 245,0
5,2
5,4
5,6
5,8
6,0
6,2
6,4
Pro
tein
a (
µg
/mL
)
Tempo (h)
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana.
Com relação à atividade quitosanolítica, conforme Figura 14, a atividade máxima
ocorreu em 18 horas de produção em que se obteve aproximadamente 70,0 U/L, o que foi 28
vezes maior que o valor (2,5 U/L em 48 horas) relatado por Assis (2010) para o mesmo fungo,
e em apenas um terço do tempo de produção.
0 4 8 12 16 20 240,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,10
Ativid
ade
(U
/mL
)
Tempo (h)
Figura 14. Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
71
Além disso, destaca-se o considerável aumento da produção de quitosanase a partir das
12 horas até 18 horas de cultivo onde atingiu o pico máximo de produção (70,0 U/L). Após o
pico em 18 horas, a produção começa a cair rapidamente, comportamento esse observado
durante todos os ensaios cinéticos. Destaca-se que é importante do ponto de vista industrial
manter a produção em alto nível por muitas horas, onde se espera uma produção contínua ou
em grandes bateladas.
Ak et al. (1998) utilizaram o fungo Penicillium spinulosum para produzir a quitosanase
usando um meio de cultura contendo 0,1% de K2HPO4; 0,05 % de MgSO4.7H2O; 0,05% de
KCl; 0,001 % de FeSO4, 0,5% NH4NO3 e como fonte de carbono e energia, os autores
usaram quitosana e parede de Rhizopus. A produção foi realizada em cultivo submerso a 28
oC. Os autores apresentam atividade quitosanolítica máxima de aproximadamente 47,0 U/mL,
ao se utilizar a parede do fungo como fonte de carbono e energia, e 30,0 U/mL ao se utilizar
quitosana como fonte de carbono e energia. Destaca-se que essas atividades ocorreram após 4
dias de cultivo. Os perfís com o tempo da atividade quitosanolítica e da atividade específica
apresentaram tendência de crescimento, com o pico máximo em 4 dias, seguindo com uma
tendência de queda da atividade. Neste tempo, a atividade específica máxima da quitosanase
foi de aproximadamente 60,0 U/mg de proteína. Essas tendências também foram observadas
nos resultados apresentados nos perfís de atividade das Figuras 14 e 15.
0 4 8 12 16 20 24
-0,010
-0,005
0,000
0,005
0,010
0,015
0,020
0,025
0,030
Ativid
ad
e E
spe
cific
a (
U/µ
g)
Tempo (h)
Figura 15. Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
72
Observa-se na Figura 15 que a atividade específica máxima da quitosanase segue a
mesma tendência da atividade quitosanolítica apresentada na Figura 14, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
O perfil para os açúcares redutores durante o cultivo de Metarhizium anisopliae
cultivado em meio contendo 0,2% de quitosana é ilustrado na Figura 16.
0 4 8 12 16 20 240,004
0,008
0,012
0,016
0,020
0,024
0,028
Acuca
res R
ed
uto
res (
mg/m
L)
Tempo (h)
Figura 16. Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana.
Observa-se, na Figura 16, que taxa máxima de redução dos açúcares redutores
(aproximadamente 0,021 mg/mL) ocorre logo após a produção máxima da enzima, em 18
horas de cultivo, conforme Figura 14. A quantidade de açúcares redutores foi a
aproximadamente 0,012 mg/mL após 24 h de cultivo. O fungo excreta para o meio enzimas
com atividade quitosanolítica, essas enzimas hidrolisam a quitosana aumentando a quantidade
de açúcares redutores para o meio, sendo que esses açúcares também são consumidos pelo
fungo durante seu crescimento.
Foi demonstrado por Assis (2010) que os açúcares redutores produzidos através da
quitossana, ocasionada pelas enzimas quitossanases metabolizadas por Metarhizium
anisopliae, com quitosana como sua única fonte de carbono e energia, atingiu um pico de 0,53
mg/mL em 48 horas. Apresentaram como quitooligossacarídeos, constituídos principalmente
por dímeros e pentâmeros, formados pela hidrólise enzimática de quitosanase em quantidades
maiores do que o consumido pelos microrganismos. Mostraram, também, que essa produção

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
73
segue a mesma tendência da sua atividade enzimática, corroborando com os resultados
apresentados no presente trabalho.
Os resultados do presente trabalho mostram a relevância de se adotar novas condições
para a produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter quitooligossacarídeo
com atividades biológicas. Esta pesquisa, portanto, sugere base para novos estudos que
permitam alcançar rapidamente a produção de quitosanase e quitooligossacarídeo com todos
os seus benefícios.

Capítulo 5 Conclusão

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
75
5. Conclusão
Após o desenvolvimento desse trabalho conclui-se:
- Com relação ao Planejamento de Seleção de Fatores Plackett & Burman:
Os resultados das análises do Planejamento de Seleção de Fatores Plackett &
Burman com 8 fatores e 12 ensaios em 2 níveis, mais 3 repetições no ponto central, para
se investigar a influência dos fatores: Concentração de Quitosana, Peptona, Extrato de
Levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O, FeSO4 na produção da enzima
quitosanase por Metarhizium anisopliae, mostraram que a atividade foi favorecida pelo
aumento da concentração de substrato (quitosana) e sulfato ferroso (FeSO4), enquanto
que o aumento da concentração dos outros fatores não contribuíram de forma
significativa para a resposta (atividade quitosanolítica). Os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) atingiram efeitos estimados de
aproximadamente 8,84 e 4,67, respectivamente. O sinal positivo indica que o aumento
das concentrações de quitosana e de FeSO4 aumentará a atividade enzimática. Conforme
visto na literatura, observa-se importância do FeSO4, já que o mesmo está diretamente
associado a síntese de proteases (principalmente metalloproteases),
- Com relação ao Planejamento Composto Central:
A análise do planejamento composto central para avaliar a influência dos
fatores, concentração de substrato (quitosana) e concentração de FeSO4, na produção da
enzima quitosanase por Metarhizium anisopliae (a partir do planejamento Plackett &
Burman) mostrou que os fatores concentração de quitosana e concentração de FeSO4
foram os fatores que influenciaram de forma mais significativa na atividade enzimática
atingindo efeitos estimados de aproximadamente 5,96 e - 5,30, respectivamente. O sinal
positivo indica que o aumento das concentrações de quitosana aumentará a atividade
enzimática. O sinal negativo indica que o decréscimo da concentração de FeSO4
aumentará a atividade enzimática. Observa-se a existência de um ponto de mínimo,
ocorrendo para concentração de quitosana (X1) no ponto central (0) e para maiores

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
76
concentrações de FeSO4 (X2) igual a (1,5) com a atividade quitosanolítica de 0,0094
U.mL-1.
- Com relação ao cultivo na melhor condição:
O cultivo na melhor condição mostrou uma produção máxima de quitosanase em
18 horas de cultivo, com a atividade enzimática de 70,0 U/L, o que foi 28 vezes maior
que o valor (2,5 U/L em 48 horas) relatado na literatura (Assis, 2010) para o mesmo
fungo e em apenas um terço do tempo de produção.
Além disso, observou-se que um elevado incremento na produção de
quitosanase a partir das 12 horas de cultivo, antes de se atingir a atividade
quitosanolítica máxima, 18 horas de cultivo. Essa produção em alto nível por muitas
horas é importante do ponto de vista industrial, onde se espera uma produção contínua
ou em grandes bateladas. Observou-se também que a atividade específica máxima da
quitosanase segue a mesma tendência da atividade quitosanolítica, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
- Com relação aos açúcares redutores:
Foi observado que os açúcares redutores seguem a mesma tendência da atividade
enzimática, com uma produção máxima de cerca de 0,21 mg/mL em 18 horas de
cultivo, produção essa que coincinde com o máximo pico de atividade enzimática.
Estes resultados mostram a relevância de se adotar novas condições para a
produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter
quitooligossacarídeo com atividades biológicas. Esta pesquisa, portanto, sugere base
para novos estudos que permitam alcançar rapidamente a produção de quitosanase e
quitooligossacarídeo com todos os seus benefícios.

Referências Bibliográficas

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
78
Referências bibliográficas
AK, Ö.; BAKIR, U.; GÜRAY, T. Isolation of a chitosan degrading fungus, Penicillium
spinulosum, and chitosanase production by the isolate. In: SYMPOSIUM - NEW
FRONTIERS IN SCREENING FOR MICROBIAL BIOCATALYSTS, 1996, Ede, The
Netherlands. Anais, Editado por K. Kieslich, C. P. van der Beek, J. A. M. de Bont e W.
J. J. van den Tweel, Elsevier Science B. V., 1998, v.53, p.265-268.
ALVES, S. B. Patologia e controle microbiano: Vantagens e desvantagens. In: ALVES,
S. B. Controle Microbiano de Insetos. 2ed., Piracicaba, FEALQ, 1163p, 1998, v.4, p.21-
37.
ALVES, S. B.; LEITE, L. G.; FILHO, A. B.; ALMEIDA, F. E. M.; MARQUES, E. J.
Produção massal de fungos entomopatogênicos na América Latina, In: ALVES, S. B.;
LOPES, R. B. Controle microbiano de pragas na América Latina: avanços e desafios.
Piracicaba, FEALQ, 414p, 2008, v.14, p.215-237.
ARRUDA, W.; LUBECK, I.; SCHRANK, A.; VAINSTEIN, M. H. Morphological
alterations of Metarhizium anisopliae during penetration of Boophilus microplus ticks.
Experimental and Applied Acarology, v. 37, p.231-244, 2005.
ASSIS, C. F. Produção e caracterização de quitooligossacarídeos produzidos pelo fungo
Metarhizium anisopliae e avaliação da citotoxicidade em células tumorais. 2010. 110p.
Tese (Doutorado em Biotecnologia) – Renorbio - Programa de Pós-Graduação em
Biotecnologia. Universidade Federal do Rio Grande do Norte, Natal.
ASSIS, C. F.; COSTA, L. S.; MELO-SILVEIRA, R. F.; OLIVEIRA, R. M.;
PAGNONCELLI, M. G. B.; ROCHA, H. A. O.; MACEDO, G. R.; SANTOS, E. S.,
Chitooligosaccharides antagonizes the cytotoxic effect of glucosamine. World J.
Microbiol. Biotechnol., v.28, n.3, p.1097-1105, 2012.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
79
AZEVEDO, V. V. C.; CHAVES, S. A.; BEZERRA, D. C.; FOOK, M. V. L.; COSTA,
A. C. F. M. Quitina e Quitosana: aplicações como biomateriais. Revista Eletrônica de
Materiais e Processos, v.2.3, p.27-34, 2007.
BARROS NETO, B.; SCARMINIO, I. S.; BRUNS, R. Como fazer experimentos:
pesquisa e desenvolvimento na ciência e na indústria. 2. ed. Campinas: Unicamp, 2001.
401p.
BATISTA FILHO, A.; ALMEIDA, J. E. M.; SANTOS, A. S.; MACHADO, L. A.;
ALVES, S. B. Eficiência de isolados de Metarhizium anisopliae no controle de
cigarrinha-da-raiz de cana-de-açúcar Mahanarva fimbriolata (Hom.: Cercopidae). STAB
– Açúcar, Álcool e Subprodutos, v.21, p.84-89, 2002.
CARVALHO, J. C. M.; SATO, S. Fermentação Descontínua, In: SCHMIDELL, W.;
LIMA, U. A.; AQUARONE, E.; BORZANI, W. Biotecnologia Industrial. São Paulo,
Ed. Edgard Blücher, v.2, p.193-204, 2001.
CHAO, H. J.; SCHWARTZ, J.; MILTON, D. K. Populations and determinats of
airborne fungi in large office buildings. Environmental Health Perspectives, v.110, n.8,
p.777-782, 2002.
CHEN, X.; XIA, W.; YU, X. Purification and characterization of two types of
chitosanase from Aspergillus sp. CJ22-326. Food Research International, v.38, n.3,
p.315-322, 2005.
CHEN, P. T.; CHIANG, C.-J.; CHAO, Y.-P. Medium optimization and production of
secreted Renilla luciferase in Bacillus subtilis by fed-batch fermentation. Biochemical
Engineering Journal, v.49, p.395-400, 2010.
CHENG, C. Y.; LI, Y. An Aspergillus chitosanase with potential for large-ecale
preparation of chitosan oligosaccharides. Biotechnol. Appl. Biochem., v.32, p.197-203,
2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
80
CHOI, Y. J.; Kim, E. J.; Piao, Z.; Yun, Y. C.; Shin, Y. C. Purification and
characterization of chitosanase from Bacillus sp. Strain KCTC 0377BP and its
application for the production of chitosan oligosaccharides. Applied and Environmental
Microbiology, v.70, n.8, p.4522-4531, 2004.
COLAKOGLU, G. The chemical and physical environment for fungal growth. Fak
Matbaa Birimi, Istanbul, 149-166, 2001.
CRAVEIRO, A. C.; CRAVEIRO, A. A.; QUEIROZ, D. C. Quitosana a fibra do futuro.
2. ed. Fortaleza: PADETEC, 2004. V.1, 218p.
DIAS, B. A. Estudo de proteases degradadoras de cutícula produzidas pelo fungo
entomopatogênico Beauveria bassiana. 2005. 70p. Dissertação (Mestrado em
Microbiologia) – Centro de Ciências Biológicas, Programa de Pós-Graduação em
Microbiologia, Universidade Estadual de Londrina, Londrina.
DINARDO-MIRANDA, L. L.; FERREIRA, J. M. G.; CARVALHO, P. A. M.
Influência da época de colheita e do genótipo de cana-de-açúcar sobre a infestação de
Mahanarva fimbriolata (Stal) (Hemiptera: Cercopidae). Neotrop. Entomol, v.30, n.1,
p.145-149, 200.
DONG, C.; ZHANG, J.; HUANG, H.; CHEN, H.; HU, Y. Pathogenicity of a new China
variety of Metarhizium anisopliae (M. anisopliae var. Dcjhyium) to subterranean termite
Odontotermes formosanus. Microbiological Research, v.164, p.27-35, 2009.
DONZELLI, B. G. G.; OSTROFF, G.; HARMAN, G. E. Enhanced enzymatic
hydrolysis of langostino shell chitin with mixtures of enzymes from bacterial and fungal
sources. Carbohydrate Research, v.338, p.1823-1833, 2003.
DRIVER, F.; MILNER, R. J.; TRUEMAN, J. W. H. A taxonomic revision of
Metarhizium based on phylogenetic analysis of rDNA sequence data. Mycological
Research, v.104, n.2, p.134-150, 2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
81
DUNG, P.; MILAS, M.; RINAUDO, M.; DESBRIÉRES, J. Water soluble derivatives
obtained by controlled chemical modifications of chitosan, Carbohydrate Polymers,
v.24, p.209-214, 1994.
ESTEVES, J. A.; CABRITA, J. D.; NOBRE, G. N. Micologia médica, 2. ed. Lisboa:
Fundação Calouste Gulbenkian, 1990. 1058p.
FACCIOTI, M. C. R. Fermentação Contínua, In: Biotecnologia Industrial. Editado por:
SCHMIDELL, W.; LIMA, U. A.; AQUARONE, E.; BORZANI, W., São Paulo, Ed.
Edgard Blücher, v.2, p.541, 2001.
FARIA, M. R.; MAGALHÃES, B. P. O uso de fungos entomopatogênicos no Brasil -
Situação atual e perspectivas. Biotecnologia Ciência & Desenvolvimento, n.22., p.18-
21, setembro/outubro 2001.
FU, J.-Y.; WU, S.-M.; CHANG, C.-T.; SUNG, H.-Y. Characterization of Three
Chitosanase Isozymes Isolated from a Commercial Crude Porcine Pepsin Preparation. J.
Agric. Food Chem., v.51, p.1042-1048, 2003.
FURIGO JUNIOR, A.; PEREIRA, E. P. Enzimas e suas aplicações cinética enzimática.
2001. 39f. Apostila (Graduação de Engenharia Química e Engenharia de Alimentos) –
Centro de Tecnologia, Departamento de Engenharia Química e Engenharia de
Alimentos, Universidade Federal de Santa Catarina, Florianópolis.
GAO, J.; XU, H.; LI, Q.-J.; FENG, X.-H.; LI, S. Optimization of medium for one-step
fermentation of inulin extract from Jerusalem artichoke tubers using Paenibacillus
polymyxa ZJ-9 to produce R,R-2,3-butanediol, Bioresource Technology, v.101, p.7076-
7082, 2010.
GARCIA, M. V.; MONTEIROI, A. C.; SZABO, M. J. P.; PRETTE, N.; BECHARA, G.
H. Mechanism of infection and colonization of Rhipicephalus sanguineus eggs by
Metarhizium anisopliae as revealed by scanning electron microscopy and
histopathology. Brazilian Journal of Microbiology, São Paulo, v.36, n.4, p.368-372,
2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
82
GHANEM, N. B.; YUSEF, H. H.; MAHROUSE, H. K. Production of Aspergillus
terreus xylanase in solid-state cultures: application of the Plackett & Burman
experimental design to evaluate nutritional requirements, Bioresource Technology,
v.73, p.113-121, 2000.
GHORBEL-BELLAJ, O.; HMIDET, N.; JELLOULI, K.; YOUNES, I.; MAALEJ, H.;
HACHICHA, R.; NASRI, M. Shrimp waste fermentation with Pseudomonas aeruginosa
A2: Optimization of chitin extraction conditions through Plackett–Burman and response
surface methodology approaches, International Journal of Biological Macromolecules,
v.48, p.596–602, 2010.
GIUSTINA, A.; VENTURA, P. Weight-reducing regimens in obese subjects: Effects of
a new dietary fiber integrator. Acta Toxicol Ther, 16, 4, p.199-214, 1995.
GUIA.BIO, Reino dos fungos, 2012. Disponível em:
<http://www.http://guia.bio.br/reino-dos-fungos>. Acesso em: 12 novembro 2012.
GUIRADO, N.; AMBROSANO, E. J.; ROSSI, F.; DIAS, F. L. F. Controle de Cupins
de Montículo com Metarhizium anisopliae e Beauveria bassiana, Rev. Bras. de
Agroecologia, v.4, n.2, 2009.
GUO, Y.; XU, J.; ZHANG, Y.; XU, H.; YUAN, Z.; LI, D. Medium optimization for
ethanol production with Clostridium autoethanogenum with carbon monoxide as sole
carbon source, Bioresource Technology, v.101, p.8784–8789, 2010.
HAGAG, S.; KUBITSCHEK-BARREIRA, P.; NEVES, G. W. P.; AMAR, D.;
NIERMAN, W.; SHALIT, I.; SHAMIR, R.; LOPES-BEZERRA, L.; OSHEROV, N.
Transcriptional and proteomic analysis of the Aspergillus fumigatus ∆prtT protease-
deficient mutant. PLoS One, v.7, n.4, e33604, 2012.
HAN, L.-K.; KIMURA, Y.; OKUDA, H. Reduction in fat storage during chitin-chitosan
treatment in mice fed a high-fat diet, International Journal of Obesity, 23:174-179,1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
83
HUNG, T.; CHANG, Y.; SUNG, H.; CHANG, C. Purification and Characterization of
Hydrolase with Chitinase and Chitosanase Activity from Commercial Stem Bromelain.
J. Agric. Food Chem., 50, 4666-4673, 2002.
ICHIKAWA, S.; TAKANO, K.; TAKASHIKUROIWA, HIRUTA, O.; SATO, S.;
MUKATAKA, A. S. Immobilization and stabilization of chitosanase by multipoint
attachment to agar gel support. Journal of Bioscience and Bioengineering, v.93, n.2,
p.201-206, 2002.
JABOR, I. A. S.; PAMPHILE, J. A.; RODRIGUES, S. B.; MARQUES-SILVA, G. G.;
ROCHA, C. L. M. S. Análise do desenvolvimento do fungo entomopatogênico
Metarhizium anisopliae em resposta a fatores nutricionais. Acta Scientiarum.
Agronomy, Maringá, v.25, n.2, p.497-501, 2003.
JI, X.-J.; HUANG, H.; DU, J.; ZHU, J.-G.; REN, L.-J.; LI, S.; NIE, Z.-K. Development
of an industrial medium for economical 2,3-butanediol production through co-
fermentation of glucose and xylose by Klebsiella oxytoca, Bioresource Technology,
v.100, p.5214–5218, 2009.
JO, Y.; JO, K.; JIN, Y.; KIM, K.; SHIN, J.; KIM, Y.; PARK, R. Characterization and
kinetics of 45 kDa chitosanase from bacillus sp. P16. Biosci. Biotechnol. Biochem.,
v.67, n.9, 1875 – 1882, 2003.
KHOSRAVI-DARANI, K.; ZOGHI, A.; ALAVI, S. A.; FATEMI, S. S. A. Application
of Plackett Burman Design for Citric Acid Production from Pretreated and Untreated
Wheat Straw, Iran. J. Chem. Chem. Eng.. v.27, n.1, p.91-104, 2008.
KIM, P.; KANG, T. H.; CHUNG, K. J.; KIM, I. S.; CHUNG, K. Purification of a
constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and
expression of the gene. FEMS Microbiolog.y Letters 240, 31–39, 2004.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
84
KIM, S.-K.; RAJAPAKSE, N. Enzymatic production and biological activities of
chitosan oligosaccharides (COS): A review. Carbohydrate Polymers, v.62, n.4, p.357-
368, 2005.
KIRK, P. M.; CANON, P.F.; MINTER, D.W.; STALPERS, J.A. Dictionary of the
fungi. CAB International, Wallingford, 2008.
KONDO, H.; OSADA, A. Influence of dietary fiber on the bioavailability of zinc in
rats, Biomed Environ Sci, 9, 204-8, 1996.
KUMAR, S.; KOH, J. Physiochemical, Optical and Biological Activity of Chitosan-
Chromone Derivative for Biomedical Applications. Int. J. Mol. Sci., v.13, p.6102-6116,
2012.
KURITA, K.; SANNAN, T.; IWAKURA, Y. Studies on chitin, 4. Evidence for
formation of block and random copolymers of N-acetyl-o-glucosamine and
oglucosamine by hetero and homogeneous hydrolyses. Die Makromolecular Chemie,
v.178, p.3197-3202, 1977.
KUROIWA, T.; ICHIKAWA, S.; HIRUTA, O.; SATO, S.; MUKATAKA, S. Factors
Affecting the Composition of Oligosaccharides Produced in Chitosan Hydrolysis Using
Immobilized Chitosanases. Biotechnol. Prog., 18, 969-974, 2002.
LI, L.-Z.; HUI, Z.; MO, X. Optimization of the medium for streptolydigin production
by Streptomyces lydicus AS 4.2501 with precursor feeding using response surface
methodology, Journal of the Taiwan Institute of Chemical Engineers, 41, 252–258,
2010.
LIU, B.-L.; KAO, P.-M.; TZENG, Y.-M.; FENG, K.-C. Production of chitinase from
Verticillium lecanii F091 using submerged fermentation. Enzyme and Microbial
Technology, v.33, p.410-415, 2003.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
85
LOBATO, A. K. C. L. Estudo da produção de biosurfactantes por micro-organismos
isolados de poços de petróleo. 2003. 108p. Dissertação (Mestrado) – Programa de Pós-
Graduação em Engenharia Química. Universidade Federal do Rio Grande do Norte.
Natal.
LOPES, R. B.; ALVES, S. B.; PADULLA, L. F. L.; PÉREZ, C. A. Eficiência de
formulações de Beauveria bassiana e Metarhizium anisopliae para o controle de ninfas
de Amblyomma cajennense (Fabricius, 1787). Revista Brasileira de Parasitologia
Veterinária, v.16, n.1, p.27-31, 2007.
LOUREIRO, E. S.; BATISTA FILHO, A.; ALMEIDA, J. E. M.; MENDES, J. M.;
PESSOA, L. G. A. Eficiência de isolados de Metarhizium anisopliae (metsch.) sorok.
no controle da cigarrinha-da-raiz da cana-de-açúcar, Mahanarva fimbriolata (stal, 1854)
(hemiptera: cercopidae), em condições de campo. Arq. Inst. Biol., São Paulo, v.79, n.1,
p. 47-53, 2012.
MELLO, D. R.; REIS, R. C. S.; BITTENCOURT, V. R. E. P. Patogenicidade in vitro
do fungo Metarhizium anisopliae (Metschnikoff, 1879) Sorokin, 1883, sobre o carrapato
Boophilus microplus (Canestrini, 1887). Revista Brasileira de Parasitologia Veterinária,
São Paulo, v.15, p.157-162, 2006.
MITSUTOMI, M.; KIDOH, H.; TOMITA, H.; WATANABE, T. The action of Bacillus
circulans WL-12 chitinases on partially N-acetylated chitosan, Bioscience
Biotechnology and Biochemistry, v.59, p.529-531, 1995.
MALAJOVICH, M. A. Biotecnologia: fundamentos. Ed. biblioteca Max Feffer do
Instituto de Tecnologia ORT, Rio de Janeiro, 2009. 104p.
MONAGHAN, R. L.; EVELEIGH, D. E.; TEWARI, R. P.; REESE, E. T. Chitosanase,
a novel enzyme. Nature - New Biology, 245, p.78-80, 1973.
MU, X. Q.; XU, Y.; NIE, Y.; OUYANG, J.; SUN, Z. H. Candida parapsilosis CCTCC
M203011 and the optimization of fermentation medium for stereoinversion of (S)-1-
phenyl-1,2-ethanediol, Process Biochemistry, v.40, p.2345–2350, 2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
86
MURRAY, P. R.; ROSENTHAL, K. S.; PFALLER, M. A. Medical microbiology. 5th
ed. London, Elsevier Mosby, 2005.
MUZZARELLI, R. A. A. Biochemical significance of exogenous chitins and chitosans
in animals and patients, Carbohydrate Polymers, v.20, p.7-16, 1993.
MUZZARELLI, R. A. A.; TOMASETTI, M.; ILARI, P. Depolymerization of chitosan
with the aid of papain, Enzyme Microbiology and Technology, v.16, p.110-114, 1994.
NAVEENA, B. J.; ALTAF, Md.; BHADRIAH, K.; REDDY, G. Selection of medium
components by Plackett–Burman design for production of L(+) lactic acid by
Lactobacillus amylophilus GV6 in SSF using wheat bran, Bioresource Technology,
v.96, p.485-490, 2005.
NOGAWA, M.; TAKAHASHI, H.; KASHIWAGI, A.; OHSHIMA, K.; OKADA, H.;
MORIKAWA, Y. Purification and Characterization of Exo--D-Glucosaminidase from
a Cellulolytic Fungus, Trichoderma reesei PC-3-7, Appl. Environ. Microbiol., v.64, n.3,
p.890-895, 1998.
ONOFRE, S. B.; VARGAS; L. R. B.; ROSSATO, M.; BARROS, N. M.; BOLDO, J.
T.; NUNES, A. R. F.; AZEVEDO, J. L. Controle biológico de pragas na agropecuária
por meio de fungos entomopatogênicos. In: SERAFINI, L. A.; BARROS, N. M.;
AZEVEDO, J. L. (Org.). Biotecnologia – avanços na agricultura e na agroindústria.
Caxias do Sul: EDUCS, p.295-328, cap.8, 2002.
OSSWALD, W. F.; SHAPIRO, J. P.; DOOSTDAR, H.; MCDONALD, R. E.; NIEDZ,
R. P.; NAIRN, C. J.; HEARN, C. J.; MAYER, R. T. Identification and Characterization
of Acidic Hydrolases with Chitinase and Chitosanase Activities from Sweet Orange
Callus Tissue, Plant Cell Physiol. 35, p.811-820, 1994.
PAGNONCELLI, M. G. B. Estudo do mecanismo de produção de oligossacarídeos com
atividades nutracêuticas a partir da quitosana por hidrólise enzimática com processo

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
87
fermentativo simultâneo. 2008. 118p. Tese (Doutorado) – Centro de Tecnologia,
Programa de Pós-Graduação em Engenharia Química. Universidade Federal do Rio
Grande do Norte. Natal.
PALMA-GUERRERO, J., JANSSON, H.-B., SALINAS, J.; LOPEZ-LLORCA, L. V.
Effect of chitosan on hyphal growth and spore germination of plant pathogenic and
biocontrol fungi, J. Appl. Microbiol., v.104, p.541-553, 2008.
PAULA, F. A.; ARRUDA, D. C. Análise do método de planejamento experimental
Plackett Burman na otimização de degradação de penas de aves por comunidades
microbianas, XXI Salão de Iniciação Científica, 19-23, outubro, 2009, UFRGS, Porto
Alegre.
PELCZAR, M. J.; CHANG, E. C. S.; KRIEG, N. R. Microbiologia: conceitos e
aplicações, v.2, 2.ed. São Paulo: Makron BooKs, 1996.
PELLETIER, A.; SYGUSCH, J. Purification and characterization of three chitosanase
activities from Bacillus megaterium P1, Applied and Environmental Microbiology, v.
56, n.4, p.844-848, 1990.
PEREIRA, F. B.; GUIMARÃES, P. M. R.; TEIXEIRA, J. A.; DOMINGUES, L.
Optimization of low-cost medium for very high gravity ethanol fermentations by
Saccharomyces cerevisiae using statistical experimental designs, Bioresource
Technology, v.101, p.7856-7863, 2010.
PETER, M. G. Chitin and chitosan from aninal sources. In: STEINBUCHEL, A.;
RHEE, S. K. Polysaccharides and Polyamides in the Food Industry. Weinheim:
Wileyvch, v.1, p.115-208, 2005.
PIZA, F. A. T.; SILOTO, A. P.; CARVALHO, C. V.; FRANCO, T. T. Purification,
characterization and purification of chitosanase from Bacillus cereus, Brazilian Journal
of Chemical Engineering., v.16, n 2, p.185-192, 1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
88
POLITZER, K.; BON, E. P. S. Enzimas Industriais e Especiais. C.G.E.E. Ciência,
Tecnologia e Inovação. Rio de Janeiro, 554p, 2006.
POLYMAR, Ciência é a nossa Natureza, 2003. Disponível em:
<http://www.polymar.com.br>. Acesso em: 6 setembro 2012.
PUTZKE, J.; PUTZKE, M. T. L. Os reinos dos fungos. Santa Cruz do Sul, RS:
EDUNISC, 1998.
QUINES, L. K. M. Extração de poli(3-hidroxibutirato) produzido por Cupriavidus
necator dsm 545 com 1,2-carbonato de propileno. 2010. 99p. Dissertação (Mestrado em
Engenharia de Alimentos) – Centro Tecnológico, Departamento de Engenharia Química
e Engenharia de Alimentos, Programa de Pós-Graduação em Engenharia de Alimentos,
Universidade Federal de Santa Catarina, Florianópolis.
RAJENDRAN, A.; THANGAVELU, V. Statistical experimental design for evaluation
of medium components for lipase production by Rhizopus arrhizus MTCC 2233, LWT -
Food Science and Technology, v.42, p.985–992, 2009.
RAMIREZ-LEÓN, L. F.; RUIZ-HERRERA, J. Hydrolysis of walls and formation of
spheroplasts in Mucor rouxii. J. Gen. Microbiol., v.72, p.281-290, 1972.
RANGEL, M. A. S.; LOUREIRO, E. S.; ALMEIDA, A. S.; ALVES, H. C.; ONO, F.
B.; LOBO, P. M.; MARIANI, M. A.; PESSOA, L. G. A.; SILVA, A. J. Avaliação
participativa do controle de cigarrinhas das pastagens com a aplicação do fungo
Metarhizium anisopliae (Metsch.) Sorok. Rev. Bras. de Agroecologia, v.2, n.2, 731-734,
2007.
RAVEN, P. H.; EVERT, R.F.; EICHHORN, S.E. Biologia Vegetal, 6ª Ed, Rio Janeiro,
Editora Guanabara Koogan, 2001.
REDDY, L. V. A.; WEE, Y.-J.; YUN, J.-S.; RYU, H.-W. Optimization of alkaline
protease production by batch culture of Bacillus sp. RKY3 through Plackett–Burman

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
89
and response surface methodological approaches, Bioresource Technology, v.99, p.242–
2249, 2008.
RIGUEIRA, J. C. S. Influência da contagem de células somáticas no perfil e teores de
aminas bioativas e na qualidade de leite cru e de queijo mussarela. 2010. 147p. Tese
(Doutorado em Ciência de Alimentos) - Faculdade de Farmácia, Departamento de
Alimentos, Programa de Pós-Graduação em Ciência de Alimentos, Faculdade de
Farmácia da Universidade Federal de Minas Gerais, Belo Horizonte.
RODRIGUES, M. I.; IEMMA, A. F. Planejamento de Experimentos e Otimização de
Processos: uma estratégia sequencial de planejamentos, Campinas, SP, Casa do Pão
Editora, 2005.
SANTOS, E. S. Recuperação e Purificação de Enzimas usando Adsorção em Leito
Expandido, Tese de Doutorado, Universidade Estadual de Campinas, Campinas/SP,
p.152, 2001.
SEDMAK, J. J.; GROSSBERG, S. E. A rapid, sensitive and versatile assay for protein
using coomassie brilliant blue G250, Analytical Biochemistry, v.79, p. 544-552, 1977.
SEINO, H.; TSUKUDA, K.; SHIMASUE, Y. Properties and action pattern of a
chitosanase from Bacillus sp. PI- 7S. Agricultural Biological Chemistry, 55, p.2421-
2423, 1991.
SHIMOSAKA, M.; NOGAWA, M.; OHNO, Y.; OKAZAKI, M. Chitosanase from the
plant pathogenic fungus, Fusarium solani f. sp. Phaseoli – purification and some
properties, Bioscience Biotechnolgy and Biochemistry, v.57, n.2, p.231-235, 1993.
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Production of two chitosanases from a chitosan-assimilating bacterium, Acinetobacter
sp. Strain CHB101, Applied and Environmental Microbiology, v.61, n.2, p.438-442,
1995.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
90
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Characterization of Three Chitosanase Isozymes Isolated from a Commercial Crude
Porcine Pepsin Preparation, J. Agric. Food Chem. 51, p.1042-1048, 2003.
SILVA FILHO, R. C.; COSTA, A. S. S.; CHAGAS, B. M. E.; SANTOS, F. N.;
MACEDO, G. R.; SANTOS, E. S. Avaliação do teste de atividade quitosanolítica
produzida por fungo. In: XV SIMPÓSIO NACIONAL DE BIOPROCESSOS, 15,
Recife, 2005, 02 a 05 de agosto.
SILVA, E. P. Avaliação Preliminar do Potencial de Aplicação da Tecnologia de
Biopilhas para a Biorremediação do Solo de Guamaré-RN. 2004. 119p. Dissertação
(Mestrado) – Centro de Tecnologia, Programa de Pós-Graduação em Engenharia
Química. Universidade Federal do Rio Grande do Norte. Natal.
SOMASHEKAR, D.; JOSEPH, R. Chitosanases – Properties and apllications: A review.
Bioresource Technology, v.55, p.35-45, 1996.
STOYACHENKO, I. A.; VARLAMOV, V. P.; DAVANKOV, V. A. Chitinases of
Streptomyces kurssanovii: purification and some properties, Carbohydrate Polymers,
v.24, p.47-54, 1994.
TINLINE, R. D. Nuclear distribuition in Metarhizium anisopliae. Mycologia, v. 63, p.
713-721, 1971.
TRABULSI, L. R.; TOLEDO, M. R. F. Microbiologia. 2. Ed., São Paulo: Atheneu,
1996.
VIEIRA, P. D. S.; SILVA, W. M. T.; PAIVA, L. M.; LIMA, E. A. L. A.;
CAVALCANTI, V. L. B. Estudo da caracterização morfológica, esporulação e
germinação dos conídiosde Metarhizium anisopliae var. acridum em diferentes
temperaturas. Biológico, São Paulo, v.69, n.1, p.17-21, 2007.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
91
WANDERLEY, M. D.; NEVES, E.; ANDRADE, C. J. Aspectos da produção industrial
de enzimas. Revista Citino, v.1, n.1, p.30-36, 2011.
WANG, C.; TYPAS, M. A.; BUTT, T. M. Detection and characterization of pr1
virulent gene deficiencies in the insect pathogenic fungus Metarhizium anisopliae.
FEMS Microbiology Letters, v. 213, p. 251-255, 2002.
ZAHORSKA-MARKIEWICZ, B.; KROTKIEWSKI, M.; OLSZANECKA-
GLINIANOWICZ, M.; ZURAKOWSKI, A. Effect of chitosan in complex management
of obesity, Pol Merkur Lekarski, v.13, n.74, p.129-132, 2002.
ZHANG, X. Y.; DAI, A. L.; ZHANG, X. K.; KUROIWA, K.; KODAIRA, R.;
SHIMOSAKA, M.; OKAZAKI, M. Purification and Characterization of and Exo--D-
glucosaminidase from a koji mold, Aspergillus oryzae IAM2660. Biosci. Biotechnol.
Biochem., v.64, n.9, 1896-1902, 2000.
ZHU, X.; WU, X.; DAI, Y. Fermentation conditions of a chitosanase from Acinetobcter
sp C-17. Biosci. Biotechnol. Biochem., v.67, n.2, p.284-290, 2003.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
92
APÊNDICE

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
93
APÊNDICE I
Metodologia para repique da cepa
Em Erlenmeyer de 1000 mL colocava-se 500 mL do meio de manutenção
PDA. Este Erlenmeyer, com o meio de manutenção, era autoclavado junto com placas
de Petri a 1,5 kgf/cm2 por 15 minutos, em seguida, era levado para uma câmara de fluxo
laminar e transferido para as placas de Petri já autoclavadas, onde eram mantidas numa
estufa a 27 ºC durante 24 h. Após este período, as placas de Petri eram levadas a câmara
de fluxo laminar, utilizando bico de Bunsen para esterilizar a alça de platina. Procedia-
se o repique utilizando uma placa de Petri contendo a cepa pura. Cada placa de Petri
contendo a cepa pura era transferida para duas placas de Petri contendo o meio de
manutenção. As placas eram mantidas na estufa a 27 ºC durante 30 dias onde após esse
período, repetia-se este procedimento.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
94
APÊNDICE II
Montagem da Curva de Calibração Usando o Método DNS
Soluções:
Glicosamina = 0,01 mol/mL (10000,0 µmol/mL)
Tabela 14. Dados para construção da curva de calibração usando o Método DNS.
Tubo de
Ensaio
Solução de
glicosamina (mL)
Água Destilada
(mL)
DNS (mL)
Volume Total
Massa (µµµµmol)
1 0 1,0 1,5 2,5 0
2 0,2 0,8 1,5 2,5 0,2
3 0,4 0,6 1,5 2,5 0,4
4 0,6 0,4 1,5 2,5 0,6
5 0,8 0,2 1,5 2,5 0,8
6 1,0 0 1,5 2,5 1,0
Tabela 15 - Medidas de absorbância a 600nm
Dados adicionais:
Peso Molecular D-Glicosamina = 215,64 (recipiente SIGMA)
Conc. Sol. D-Glicosamina = 0,05 g/50 mL
Massa calc.(µmol) = conc. Sol. D-Glic. x Vol. Utiliz. D-Glics. =
= 0,05 x 1000 x Vol.util./50*215,64
Tempo atividade enzimática = 30 min
1 U/mL = 1 µmol/(min.mL)
Vol. Total (Glicosamina+caldo) = 1 mL
Medições Adsorbância a 600 nm Vol. Utiliz. Massa Massa Atividade
II III Média(Abs)(mL) calc. (mg) calc. (µg) Enzim. (U/mL)
0 0 0 0 0 0 0
0,133 0,128 0,1307 0,2 0,2 200 0,0309
0,279 0,277 0,2787 0,4 0,4 400 0,0618
0,433 0,428 0,4297 0,6 0,6 600 0,0927
0,587 0,583 0,5797 0,8 0,8 800 0,1237
0,727 0,716 0,7157 1 1 1000 0,1546
Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
95
Figura 17 – Curva padrão de glicosamina.
y = 1900ralx + 1900ral
R² = 1900ral
00,000
00,020
00,040
00,060
00,080
00,100
00,120
00,140
00,160
00,180
00,000 00,100 00,200 00,300 00,400 00,500 00,600 00,700 00,800
Ati
v.
En
zim
áti
ca (
U/m
L)
Absorbância (Abs) a 600 nm
Curva-Padrão D-Glicosamina

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
96
APÊNDICE III
Método de Determinação de Proteína Utilizando o Corante Comassie
Brilhante Blue G 250 (Método modificado de Sedmak e Grossberg)
Micrométodo
Preparar uma solução 0,06% de Comassie Blue G250 em 1,5 % HCl (w/V). Filtrar e
papel de filtro Whatman nº1.
Curva Padrão (1 a 20 µµµµg)
a) Preparar soluções estoque de BSA de 10 a 200 µg/mL.
b) Colocar em tubos de ensaio 100 µL da solução estoque, 1400 µL de água e
1500µL do Comassie e agitar.
c) Preparar um tubo contendo 1500 µL de água e 1500 µL do corante que
corresponde ao zero de proteína.
d) Após 5 minutos (padronizar o tempo) fazer as leituras das absorbâncias.
e) As leituras das amostras devem ser feitas em dois comprimentos de onda 595 e
465 contra a água.
f) Plotar a razão das absorbâncias a 595/465 como uma função da massa de BSA e
calcular o melhor ajuste linear dos dados.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
97
Micrométodo (1 a 20 µµµµg de BSA)
Solução Estoque de BSA = 0,2 mg/mL
Tabela 16. Dados para construção da curva padrão de BSA.
Tubo de
Ensaio
H2O (µµµµL)
BSA (µµµµg)
Coomassie (µµµµL)
BSA (µµµµg)
1 1500 0 1500 0
2 1490 10 1500 2
3 1480 20 1500 4
4 1470 30 1500 6
5 1460 40 1500 8
6 1450 50 1500 10
7 1440 60 1500 12
8 1430 70 1500 14
9 1420 80 1500 16
10 1410 90 1500 18
11 1400 100 1500 20
Dados adicionais:
Concentração Sol. Estoque
BSA = 0,2 mg/mL = 0,2 µg/µL
Vol. total sol. = vol. H2O + vol. BSA + vol.
Coomassie = 3000 µL = 3 mL
Concentração Sol. BSA (mg/mL) = (massa BSA mg) / 3 mL
.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
98
Figura 18 – Curva Padrão de BSA
y = 1900ralx - 1900ral
R² = 1900ral
000,000
001,000
002,000
003,000
004,000
005,000
006,000
007,000
000,400 000,500 000,600 000,700 000,800 000,900 001,000 001,100
µg
BS
A
Razão Absorbâncias (595nm/465nm)
Curva-Padrão BSA

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE TECNOLOGIA
DEPARTAMENTO DE ENGENHARIA QUÍMICA PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA
TESE DE DOUTORADO
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em
Cultivo Descontínuo Submerso
Raimundo Cosme da Silva Filho
Natal/RN
Abril/2013

Raimundo Cosme da Silva Filho
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo
Descontínuo Submerso
Tese apresentada ao Programa de Pós-Graduação em Engenharia Química – PPGEQ, da Universidade Federal do Rio Grande do Norte, como parte dos requisitos para obtenção do título de Doutor em Engenharia Química, sob a orientação do Prof. Dr. Everaldo Silvino dos Santos e a coorientação da Profª. Drª. Gorete Ribeiro de Macedo.
Natal/RN
Abril/2013

Catalogação da Publicação na Fonte. UFRN / CT / PPGEQ
Biblioteca Setorial “Professor Horácio Nícolas Sólimo”.
Silva Filho, Raimundo Cosme da. Otimização do meio de cultura para a produção de quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso / Raimundo Cosme da Silva Filho. - Natal, 2013. 98 f.: il.
Orientador: Everaldo Silvino dos Santos. Co-orientador: Gorete Ribeiro de Macedo. Tese (Doutorado) - Universidade Federal do Rio Grande do Norte.
Centro de Tecnologia. Departamento de Engenharia Química. Programa de Pós-Graduação em Engenharia Química.
1. Quitosana - Tese. 2. Quitina - Tese. 3. Quitosanase – Tese. 4. Metarhizium
anisopliae - Tese. I. Santos, Everaldo Silvino dos. II. Macedo, Gorete Ribeiro de. III. Universidade Federal do Rio Grande do Norte. IV. Título. RN/UF/BSEQ CDU 547.458 (043.2)

SILVA FILHO, Raimundo Cosme da – Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo Descontínuo Submerso. Tese de Doutorado. UFRN - Programa de Pós-Graduação em Engenharia Química. Área de Concentração: Pesquisa e Desenvolvimento de Tecnologias Regionais – Subárea: Engenharia de Processos – Alimentos e Biotecnologia, Natal/RN, Brasil.
Orientação: Prof. Dr. Everaldo Silvino dos Santos Coorientação: Profª. Drª. Gorete Ribeiro de Macedo ___________________________________________________________________________
RESUMO: No presente trabalho utilizou-se um Planejamento Plackett & Burman, com 8 fatores e 12 ensaios em 2 níveis e mais 3 repetições na condição do ponto central, para se investigar a influência das concentrações de quitosana, peptona, extrato de levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O e FeSO4 na produção da enzima quitosanase por Metarhizium
anisopliae. Os ensaios para produção da enzima foram realizados em cultivo descontínuo submerso. Os resultados para o Planejamento Plackett & Burman mostraram que a atividade quitosanolítica foi favorecida pelo aumento da concentração de substrato (quitosana) e de sulfato ferroso (FeSO4), enquanto que o aumento da concentração dos outros fatores não contribuiu de forma significativa para a atividade quitosanolítica. A otimização do meio de cultura para a produção da enzima foi realizado por meio de um Planejamento Composto Central, com os dois fatores que mais influenciaram a atividade quitosanolítica (quitosana e FeSO4), conforme Planejamento Plackett & Burman, mantendo-se os outros nutrientes em seus valores mínimos. Nesse planejamento, para o fator FeSO4, tomou-se o limite inferior (-1) como sendo o limite superior do Planejamento Plackett & Burman. Os resultados mostraram que a produção da enzima foi favorecida pelo aumento da concentração quitosana e pela diminuição da concentração de FeSO4. A produção máxima de atividade quitosanolítica foi da ordem de 70,0 U/L e foi atingida em apenas 18 h de fermentação, resultado esse vinte e oito vezes maior aos obtidos anteriormente para o mesmo microrganismo que foi 2,5 U/L em 48h.
Palavras-chave: quitosana; quitosanase; Metarhizium anisopliae; Plackett & Burman.


ABSTRACT
In this work a Plackett-Burman Design with 8 factors and 12 trials in 2 levels with 3 repetitions at the center point was used in order to investigate the influence of the concentration of chitosan, peptone, yeast extract, NaNO3, K2HPO4, KCl, MgSO4.7H2O and FeSO4 on chitosanase production by Metarhizium anisopliae. Runs were carried out using submerged discontinuous cultivation for enzyme production. The results of the Plackett & Burman Design showed that only two factors, chitosan concentration as well as FeSO4 had influence on chitosanolytic activity, while the increase in concentration of other factors not contributed significantly to the quitosanolítica activity. Cultivation medium optimization for enzyme production was carried out using a Composite Central Design, with the most important factors for chitosanolytic activity (chitosan and FeSO4), in accordance with Plackett & Burman Design, and keeping the other nutrients in their minimum values. On this other design, it was taken the highest limit in Plackett & Burman Design as the lowest limit (-1) to FeSO4 factor. The results showed that the enzyme production was favoured by increasing the chitosan concentration and by decreasing FeSO4. Maximum production for chitosanolytic activity was about 70.0 U/L and was reached in only 18 h of fermentation, a result about twenty-eight times greater than a former study using the same microorganism (about 2.5 U/L at 48 h).
Key-words: Chitosan; Chitosanase; Metarhizium anisopliae; Plackett & Burman

Ao meu pai, minha mãe e minha
família, em especial aos meus
irmãos e sobrinhos

AGRADECIMENTOS
A Deus, por estar presente em todos os momentos da minha vida e por me ceder à
oportunidade de iniciar e concluir este trabalho.
A minha mãe Cândida Inocência da Silva que esteve comigo em todos os momentos da
minha vida me educando, me amando e sempre me dando muito carinho.
Ao meu pai Raimundo Cosme da Silva, que não está mais aqui para ver este trabalho,
mas que foi a pessoa que mais me apoiou e me incentivou, em todos os momentos da minha vida,
para que eu pudesse chegar o mais longe possível na minha carreira acadêmica. Pai muito
obrigado por toda atenção, educação, amor e carinho que recebi durante toda a minha vida.
Aos meus irmãos e familiares que sempre estiveram comigo dividindo momentos de
grande alegria e às vezes de tristeza, mas sempre nos mantendo unidos para superarmos todos os
obstáculos.
A minha namorada Rochelly Caroliny por todo amor e carinho que me deu e que me
entendeu nos momentos que estive ausente para a conclusão deste trabalho.
Ao meu orientador Professor Doutor Everaldo Silvino dos Santos por confiar em mim e
me aceitar como orientando além de toda a amizade, orientação e dedicação em todas as etapas de
desenvolvimento e conclusão deste trabalho.
À Professora Doutora Gorete Ribeiro de Macedo, “pessoa que admiro muito como
profissional e principalmente, como pessoa, um exemplo a ser seguido”, pelos conselhos e críticas
quando necessários ao desenvolvimento deste trabalho. Professora a senhora é considerada uma
mãe para de todos os alunos que tiveram a honra de conhecê-la e trabalhar com a senhora.
Aos meus amigos Engenheiros Químicos, Ana Carmen, Ana Katerine, Andréa Farias,
Anita Lima, Sirtys Lessa, Alexandre, Sérgio, Márcio Bezerra e ao Biólogo Daniel Souza, que
sempre me ouviram e que me ajudaram de alguma forma nesse trabalho.
A Eusamar Coelho de Lima, Mazinha, secretária do PPGEQ, pelo apoio, pela paciência e
orientação nas questões burocráticas do processo.
A Carlos e Jardelson, pela grande colaboração e dedicação nos experimentos do
laboratório.
A Sérgio Santana pela grande colaboração e dedicação, o qual não poupou esforços nem
tempo em me ajudar na realização dos experimentos em laboratório.

Ao meu amigo Ubiratan Nazaré por me ajudar na organização do texto e na tradução para
o Abstract.
Finalizando, agradeço ao Laboratório de Engenharia Bioquímica da UFRN por ter cedido
toda a sua estrutura para realização dos experimentos e ao Programa de Pós-Graduação em
Engenharia Química, assim como a todos os professores do Departamento de Engenharia Química
da UFRN que trabalham em pró de uma educação digna e ensino de qualidade para todos os
alunos.

Sumário LISTA DE FIGURAS
LISTA DE TABELAS
NOMENCLATURA
INTRODUÇÃO...............................................................................................................................15
REVISÃO BIBLIOGRÁFICA........................................................................................................18
2.1. Enzimas................................................................................................................................18
2.1.1. Quitosana e seus derivados........................................................................................19
2.1.2. Quitosanases...............................................................................................................24
2.2. Fungos..................................................................................................................................29
2.2.1. Características gerais..................................................................................................29
2.2.2. Reprodução dos Fungos.............................................................................................32
2.2.3. Metabolismo de fungos..............................................................................................33
2.2.4. Nutrição de fungos.....................................................................................................34
2.2.5. O Fungo Metarhizium anisopliae ..............................................................................36
2.3. Fermentação.........................................................................................................................40
2.3.1. Preparo do inóculo......................................................................................................41
2.3.2. Produção de enzimas em cultivo submerso descontínuo...........................................42
2.4. Planejamento experimental..................................................................................................42
2.4.1. Planejamento fatorial..................................................................................................43
2.4.2. Planejamento Plackett & Burman..............................................................................43
2.4.3. Aplicações do Planejamento Plackett & Burman.......................................................44
METODOLOGIA EXPERIMENTAL.............................................................................................50
3.1. Quitosana..............................................................................................................................50
3.2. Microrganismo.....................................................................................................................50
3.2.1. Escolha da cepa produtora de quitosanase.................................................................50

3.2.2. Manutenção da cepa...................................................................................................50
3.3. Produção de enzima.............................................................................................................51
3.4. Determinação da Proteína Total...........................................................................................51
3.5. Determinação da Atividade Quitosanolítica e Atividade Específica...................................51
3.6. Planejamento Experimental.................................................................................................52
3.6.1. Planejamento de seleção de fatores Plackett Burman................................................52
3.6.2. Planejamento Composto Central para Otimização do meio de cultura para a
Produção da Enzima em Cultivo Submerso.....................................................................................53
3.7. Perfil Cinético durante o Cultivo.........................................................................................56
RESULTADOS E DISCUSSÃO.....................................................................................................58
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA ..........................................58
4.2. Planejamento Plackett & Burman........................................................................................59
4.2.1. Análise do Plackett & Burman com 8 fatores e 12 ensaios .......................................59
4.2.2. Análise do Plackett & Burman com 12 ensaios e mais 3 repetições na condição do
ponto central ....................................................................................................................................61
4.3. Planejamento Composto Central para Produção de Quitosanase por Metarhizium
anisopliae em Cultivo Descontínuo Submerso................................................................................64
4.4. Perfil Cinético – Melhor Condição......................................................................................69
CONCLUSÃO.................................................................................................................................75
REFERÊNCIAS BIBLIOGRÁFICAS.............................................................................................78
APÊNDICE I...................................................................................................................................93
Metodologia para Repique da Cepa ................................................................................................93
APÊNDICE II.................................................................................................................................94
Montagem da Curva de Calibração Usando o Método DNS..........................................................94
APÊNDICE III................................................................................................................................96
Método de Determinação de Proteína Utilizando o Corante Comassie Brilhante Blue G 250
(Método modificado de Sedmak e Grossberg)................................................................................96

Lista de Figuras
Figura 1 – Estrutura da quitosana..................................................................................................20
Figura 2 – Estrutura da glicosamina..............................................................................................23
Figura 3 – Estrutura do sulfato de glicosamina.............................................................................23
Figura 4 – Representação da estrutura geral de uma célula de fungo..........................................30
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas...........................................................31
Figura 6 – Conídios de Aspergillus e Penicillium.........................................................................33
Figura 7 – Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA ...................58
Figura 8 – Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados............................................................................................................................60
Figura 9 – Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto central
mostrando a influência dos fatores estudados................................................................................63
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos
fatores estudados............................................................................................................................67
Figura 11 – Valores preditos em função dos valores observados..................................................68
Figura 12 – Atividade quitosanolítica em função da concentração de quitosana e da concentração
de FeSO4 para os fatores não significativos no ponto central .......................................................68
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana ........70
Figura 14 – Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana....................................70
Figura 15 – Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de
quitosana........................................................................................................................................71
Figura 16 – Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana..........................................................................................................72
Figura 17 – Curva padrão de glicosamina ....................................................................................95
Figura 18 – Curva Padrão de BSA ................................................................................................98

Lista de Tabelas
Tabela 1 – Comparação das enzimas com os catalisadores químicos..............................................19
Tabela 2 – Fontes naturais de quitina e quitosana ...........................................................................20
Tabela 3 – Níveis para os fatores e seus valores codificados...........................................................53
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central .........................................................................................53
Tabela 5 – Matriz do planejamento Composto Central...................................................................54
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central......................................................................................................55
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.......................59
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman...............................................60
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.......................61
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central...............................................................................................................................................62
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da atividade
enzimática........................................................................................................................................65
Tabela 12 – Efeitos Estimados do planejamento Composto Central...............................................65
Tabela 13 – Resumo da ANOVA....................................................................................................66
Tabela 14 – Dados para construção da curva de calibração usando o Método DNS ......................94
Tabela 15 – Medidas de absorbância a 600 nm para construção da curva de calibração................94
Tabela 16 – Dados para construção da curva padrão de BSA ........................................................97

NOMENCLATURA
A – Atividade quitosanolítica (U/L)
AE – Atividade específica (U/mg)
ANN-GA – algoritmo genético de rede neural artificial
ANOVA – Análise de variância (Analysis of Variance)
BSA – Albumina de soro bovino (Bovine Sereum Albumin)
CCD – Planejamento Composto Central
CDA – Chitosan Dector Agar
Cs – Concentração de substrato (%)
CP – Concentração de KCl (%)
Da – Dalton
DEQ – Departamento de Engenharia Química
DNS – Ácido Dinitrosalissílico
E – Enzima
Ea – Energia de ativação
EC – Comissão para nomenclatura e classificação de enzimas
EL – Concentração de Extrato de Levedura (%)
FP – Concentração de K2HPO4 (%)
NS – Concentração de NaNO3 (%)
GDA – Grânulos dispersíveis em água
k – Número de fatores, variáveis
LEB – Laboratório de Engenharia Bioquímica
mRNA – RNA mensageiro
n – Número de níveis de um determinado fator
N – Número de ensaios de um planejamento
P – Pressão
PP – Concentração de Peptona (%)
PDA – Batata Dextrose Agar
pH – Potencial hidrogeniônico
Q – Concentração de Quitosana (%)
Qm – Quadrado médio
r – Agitação (rpm)
R2 – Coeficiente de determinação

RA – Razão de aeração
rpm – Rotações por minuto
RSM – Metodologia de superfície de resposta
S – Molécula de substrato
SF – Concentração de FeSO4 (%)
SM – Concentração de MgSO4.7H2O (%)
SQ – Soma de quadrados
SSF – Fermentação de estado sólido
T – Temperatura
TMC – Temperatura de cultivo
V11,V12,V13 – Variáveis inertes para cálculo do erro
VHG – Fermentação de alta densidade
WP – Pó molhável
X – Concentração celular
Xi – variável independente
Xj – variável independente
xi – Fator independente real
x0 – Valor de xi no ponto central
∆xi – Variação de valor
Y – Resposta do modelo de primeira ordem
Yp – Resposta do modelo polinomial de segunda ordem
β0 – Interseção dos modelos de primeira ou segunda ordem
βi – Coeficiente linear dos modelos de primeira ou segunda ordem
βii – Coeficiente quadrático
βij – Coeficiente de interação

Capítulo 1 Introdução

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
15
1. Introdução As quitosanases foram descobertas simultaneamente há quase quarenta anos por
Monaghan et al. (1973), como pelo grupo de Ramirez-León & Ruiz-Herrera (1972), e
são enzimas obtidas a partir do metabolismo de bactérias, fungos e extraídas de plantas.
Desde então, vários estudos vêm apresentando a obtenção de quitosanase deste
microrganismos, na forma intracelular ou extracelular (Osswald et al., 1994;
Somashekar & Joseph, 1996), como a do presente estudo. A quitosanase (EC 3.2.1.132)
representa uma classe de enzimas hidrolíticas que catalisam a hidrólise de ligação β-
(1→ 4) glicosídica da quitosana, gerando oligômeros chamados quito-oligossacarídeos
(Reddy et al., 2008).
A quitosana é um biopolímero hidrofílico obtido a partir da quitina, um
polissacarídeo abundante na natureza. Na literatura, é considerado quitosana o produto
da desacetilação da quitina para um grau de desacetilação superior a 75% e solúveis em
ácidos como o acético e o fórmico. A quitosana é conhecida por exibir uma ampla
variedade de atividade fisiológica como atividade antitumoral, mas o peso molecular e a
insolubilidade em água da quitosana são desvantagens para várias aplicações.
Atualmente, há interesse nas quitosanas quando as mesmas são convertidas em quito-
oligossacarídeos porque eles não apenas são solúveis em água e possuem baixo peso
molecular, mas também têm atividades biológicas bem notáveis, como atividade
anticancerígena atóxica e biocompatível, incluindo efeito antimetastático, atividade anti-
HIV, hepatoprotetora, antioxidante, hipocolesterolêmica, antimicrobiana,
imunoestimulate, antitumoral, anti-inflamatória e atividade aceleradora da absorção de
cálcio e ferro. No entanto, estes quito-oligossacarídeos são obtidos tradicionalmente na
indústria por reação de hidrólise usando ácidos, que apresenta muitos problemas como
dificuldade em controlar a hidrólise, produzindo uma grande quantidade de
oligossacarídeos de cadeia pequena e oligossacarídeos de baixo rendimento, alto custo
de separação e, também, poluição ambiental, uma vez que utiliza um ácido como
catalisador. Como alternativa, com suas vantagens em termos de compatibilidade
ambiental, baixo custo e reprodutibilidade, a obtenção destes quito-oligossacarídeos por
hidrólise com quitosanase se tornou cada vez mais popular nos últimos anos (Azevedo
et al., 2007).

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
16
Devido a todos estes aspectos positivos é que, em geral, as enzimas vêm sendo
estudadas em diferentes aplicações, mostrando que esta é uma tendência na indústria.
No presente trabalho, foi desenvolvido um processo fermentativo visando a
otimização do meio de cultura para a produção da enzima quitosanase em cultivo
descontínuo submerso, por Metarhizium anisopliae, capaz de sintetizar oligômeros com
aplicações na indústria farmacêutica e de química fina, partindo de quitosana como
única fonte de carbono.
Nesse caso, avaliou-se, através de um planejamento Plackett & Burman (método
de delineamento de seleção de fatores) e de um planejamento Composto Central as
melhores condições para a produção da enzima. A técnica do planejamento
experimental Plackett & Burman para realizar triagem de fatores na otimização de um
determinado processo foi utilizada neste trabalho para identificar quais os fatores que
influenciam de forma significativa na produção da enzima quitosanase. A influência
destes fatores (concentração de quitosana e concentração de FeSO4) foi então
investigada pelo planejamento Composto Central com 22 experimentos na parte fatorial,
repetição em triplicata no ponto central e quatro experimentos na parte axial. A
repetição do ponto central permite também a determinação do erro puro e da
repetibilidade do processo, sendo uma forma de identificar melhor se o processo está
sob controle.
Do ponto de vista industrial é de relevante importância a realização de estudos
para a otimização da produção da quitosanase de forma contínua ou em grandes
bateladas.

Capítulo 2 Revisão Bibliográfica

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
18
2. Revisão Bibliográfica 2.1. Enzimas
As enzimas são catalisadores biológicos constituídos por longas cadeias de
aminoácidos que atuam em substratos reduzindo a energia de ativação, acelerando assim
a velocidade da reação, e transformando-os em produtos. Sua atuação catalítica visa
facilitar a atividade das células através da quebra de moléculas ou junção das mesmas
para formação de novos produtos. Podem apresentar alto grau de especificidade relativa
a apenas um substrato ou a vários substratos ao mesmo tempo, como a certos tipos de
ligações glicosídicas α-1,4 das moléculas de amido, ou para determinado tipo particular
de isômero óptico como ocorre na oxidação da β-D-glicose pela glicose oxidase (Furigo
Junior & Pereira, 2001).
Enzimas são proteínas que apresentam em sua estrutura um centro ativo
(apoenzima) e, em alguns casos, um grupo prostético denominado cofator (coenzima ou
íon metálico ativo) e sua atividade depende de alguns fatores como estrutura da
proteína, número de cadeias peptídicas, arranjo dessas cadeias na molécula, natureza do
substrato e natureza do grupo prostético (Politzer & Bon, 2006).
Todos os organismos vivos produzem, em pequena ou grande quantidade, certa
variedade de enzimas, sejam elas intra ou extracelulares. As enzimas extracelulares são
excretadas para o meio externo e podem decompor compostos insolúveis como
celulose, quitina e quitosana.
Algumas destas enzimas podem ser usadas tanto nas indústrias de detergentes,
têxteis e farmacêuticas, como também de alimentos, de bebidas, etc., sendo utilizadas
em processos biotecnológicos industriais em substituição aos processos químicos que
muitas vezes causam grandes prejuízos ao meio ambiente. A produção comercial das
enzimas pode ser realizada através do cultivo de microrganismos como fungos e
bactérias.
Na Tabela 1 mostra-se uma comparação das enzimas com os catalisadores
químicos, levando-se em conta algumas das características dos dois catalisadores.
Destaca-se que alguns parâmetros da Tabela 1 demonstram a maior importância das
enzimas, quando comparadas com os catalisadores químicos, principalmente a
especificidade ao substrato, as condições de reação, o consumo de energia e a

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
19
velocidade de reação, como desvantagem cita-se o custo de obtenção (isolamento e
purificação).
Tabela 1 – Comparação das enzimas com os catalisadores químicos (Adaptado de
Furigo Junior & Pereira, 2001).
Característica Enzimas Catalisadores Químicos
Especificidade ao substrato Alta Baixa
Natureza da estrutura Complexa Simples
Sensibilidade à T e pH Alta Baixa
Condições de reação (T, P e pH) Suaves Drástica (geralmente)
Custo de obtenção (isolamento e purificação) Alto Moderado
Natureza do processo Batelada Contínuo
Consumo de energia Baixo Alto
Formação de subprodutos Baixa Alto
Separação catalisador/produtos Difícil/cara Simples
Atividade catalítica (temperatura ambiente) Alta Baixa
Presença de cofatores Sim Não
Estabilidade do preparado Baixa Alta
Energia de Ativação Baixa Alta
Velocidade de reação Alta Baixa
2.1.1. Quitosana e seus derivados
A quitosana e seus derivados apresentam grande importância devido ao
conhecimento de suas funcionalidades biológicas, médicas, tecnológicas, etc
(Shimosaka et al, 1993; Shimosaka et al, 1995; Mitsutomi et al., 1995). A quitosana é
obtida da quitina, polissacarídeo abundante na natureza, extraído principalmente das
carapaças de caranguejos, lagosta e camarões. A quitosana é um polímero natural,
biodegradável, não tóxico e de alta massa molecular obtido a partir de animais
marinhos, insetos e microrganismos (Tabela 2).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
20
Tabela 2 – Fontes naturais de quitina e quitosana. (Adaptado de Polymar, 2003).
Animais
Marinhos
Insetos Aracnídeos Microrganismos
Anelídios Formigas Escorpiões Leveduras
Moluscos Besouros Aranhas Fungos
Lagosta Esporos
Camarão
Caranguejo
Krill
A Figura 1 abaixo mostra a estrutura química da quitosana
Figura 1 – Estrutura da quitosana (Fonte: Polymar, 2003)
Quitosana totalmente desacetilada não existe na natureza. Graus de acetilação
(GA) nferior a 50% a torna mais solúvel em soluções ácidas (Muzzarelli, 1993;
Muzzarelli et al., 1994; Dung et al., 1994; Stoyachenko et al.,1994). Pelletier &
Sygusch (1990) enfatizaram que grande parte dos resíduos sólidos da indústria
pesqueira, principalmente da carcinicultura, é jogada ao mar ocasionando degradação
ambiental. Logo, a utilização desses resíduos como matéria-prima para formação de
novos produtos, não só reduz de forma drástica o impacto ambiental, mas também daria
maior valor agregado à indústria pesqueira.
A demanda cada vez maior de consumidores por alimentos mais saudáveis, sem
conservantes químicos, tem motivado pesquisas à descoberta de novos agentes
antimicrobianos naturais. Sendo assim, a atividade antimicrobiana incomum da

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
21
quitosana e seus derivados contra microrganismos, tais como bactérias, bolores e
leveduras tem tido maior atenção ultimamente.
Devido à interação entre a quitosana, que é positivamente carregada, e as
membranas celulares dos microrganismos, negativamente carregadas, ocorre a formação
de verdadeiros poros na membrana, que são canais de vazamento de conteúdo protéico e
outros componentes intracelulares. A quitosana tem também a propriedade de agir como
agente quelante que se liga de forma seletiva a traços de metais, inibindo, assim, a
produção de toxinas e o crescimento microbiano. Ainda é capaz de ativar mecanismos
de defesa em tecidos hospedeiros, atuando como agente sequestrante de água e inibindo
várias enzimas. A ligação da quitosana com DNA e a inibição da síntese do mRNA
ocorre via penetração da quitosana no núcleo dos microrganismos e interferência na
síntese do mRNA e proteínas, (Craveiro et al., 2004).
Em um estudo clínico conduzido por Giustina & Ventura (1995) com 100
pessoas com sobrepeso de 10 a 25%, ao final de 4 semanas as pessoas que receberam
quitosana demonstraram perda de peso significativa (7,3 Kg contra 3,0 Kg nos grupos
controle). As pessoas que tomaram quitosana também demonstraram redução na pressão
sanguínea sistólica (de 145 para 135 mm Hg) e diastólica (92.6 para 84.2 mm Hg),
assim como uma redução na taxa respiratória (27.6/min a 21.2/min).
Kondo & Osada (1996) pesquisaram a influência da quitina e da quitosana na
biodisponibilidade do zinco em ratos alimentados com uma dieta com suplementos de
5% ou sem fibra dietética (quitina, quitosana, celulose, pectina e agar-agar) durante 31
dias. Observaram que os ratos alimentados com dietas contendo 5% de fibras, exceto
quitosana, o consumo de alimento foi maior do que o controle. O ganho de peso em
todos os grupos alimentados com fibra foi maior do que o controle, exceto para o grupo
suplementado com quitosana (5%) onde a ingestão de alimento e o ganho de peso foram
menores do que os grupos controle e das demais fibras.
Segundo Craveiro et al. (2004), derivados da quitosana obtidos pela reação com
ácidos carboxílicos, podem promover ao polímero uma solubilidade especial em água.
Esses compostos possuem estrutura química semelhante ao ácido hialurônico e
fornecem uma excelente compatibilidade com o tecido epitelial. As soluções obtidas
com esses derivados podem ser facilmente incorporadas a formulações cosméticas,
mostrando melhor compatibilidade com surfactantes iônicos e emulsificantes,
apresentando estabilidade em uma larga faixa de pH.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
22
Han et al. (1999) examinaram os efeitos da quitosana sobre a atividade da lipase
pancreática in vitro e sobre o grau de armazenamento de gorduras induzido em ratos
pela administração oral de uma dieta rica em gorduras por nove semanas. Como
resultado, observou-se que os animais alimentados com quitosana, reduziram o peso
corporal, a hiperlipidemia e a gordura hepática. Como conclusão, os autores propõem
que os efeitos anti-obesidade da quitosana em ratos alimentados com alto teor de
gordura se devem parcialmente a inibição da absorção intestinal das gorduras dietéticas.
Em outro ensaio, Zahorska-Markiewicz et al. (2002) realizaram um estudo para
avaliar o efeito da quitosana no tratamento da obesidade. Segundo os autores do estudo,
a quitosana pode ser usada como um valioso e seguro coadjuvante em tratamentos para
a obesidade em longo prazo. A quitosana parece acentuar a redução da pressão arterial,
juntamente com a perda de peso.
Assis (2010) estudou a produção e caracterização de quitooligossacarídeos
produzidos pelo fungo Metarhizium anisopliae e avaliação da citotoxicidade em células
tumorais. A hidrólise da quitosana foi realizada em tempos de 10 a 60 min para a
produção de quitooligossacarídeos. A avaliação da citotoxicidade dos oligômeros de
quitosana foi realizada em células tumorais (HepG2 e HeLa) e não tumoral (3T3). A
produção de oligômeros de quitosana teve maiores rendimentos durante 10 minutos de
hidrólise, os pentâmeros apresentaram concentração de 0,15 mg/mL, no entanto os
hexâmeros, que apresentam maior interesse por causa de suas propriedades biológicas,
só foram detectados com 30 minutos de hidrólise com uma concentração de 0,004
mg/mL.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o planejamento Plackett-Burmam e a metodologia de superfície de resposta.
Os resultados mostraram que a eficiência máxima de extração da quitina foi conseguida
nas seguintes condições: concentração de casca de camarão 50g/L, glicose 50g/L, tempo
de incubação de 5 dias e inoculo como 0,05 OD (densidade óptica).
A glicosamina, unidade monomérica básica da quitosana, possui atividade
antiinflamatória e tem grande importância na construção e manutenção da cartilagem,
no tratamento de enfermidades como: febre reumática, artrose e principalmente a artrite
e osteoartrite na forma aguda ou crônica (Polymar, 2003). A Figura 2 apresenta a
estrutura química da glicosamina.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
23
Figura 2 – Estrutura da glicosamina (Fonte: Polymar, 2003)
O sulfato de glicosamina é um derivado da quitosana sendo utilizado no
tratamento da osteoartrite sintomática dos joelhos, quadris, coluna, mãos e outras
regiões (Craveiro et al., 2004). O sulfato de glicosamina estimula a síntese de
proteoglicanos necessários para o bom funcionamento das articulações, estimula a
regeneração das cartilagens e ainda incorpora enxofre ao tecido cartilaginoso. Estudos
de longa duração, acima de 3 anos, mostram que o sulfatos de glicosamina evita a
progressão de danos estruturais nas articulações e induz melhora significativa nas dores
e funções físicas (Craveiro et al., 2004). A Figura 3 apresenta a estrutura química do
sulfato de glicosamina.
Figura 3 – Estrutura do sulfato de glicosamina (Fonte: Polymar, 2003)

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
24
2.1.2. Quitosanases
Dependendo do tipo de quitosanase obtida, ocorrem diferentes mecanismos de
hidrólise da quitosana, isso devido a diferentes microrganismos produzirem enzimas
com diferenças nas suas estruturas (Kurita et al., 1977; Seino et al., 1991).
Quitosanases (EC 3.2.1.132) são enzimas que catalisam a reação de hidrólise de
quitosana em oligossacarídeos. As enzimas quitosanases são produzidas por vários
microrganismos, bactérias, actinomicetos e fungos, e até por algumas plantas. Estas
enzimas agem em polímeros com grau de acetilação de 30% a 60%. Os monômeros
resultantes da hidrólise da quitosana por estas enzimas são melhores digeridos, ao serem
ingeridos, como é o caso da glicosamina. Estudos efetuados até o presente momento
classificam as quitosanases em cinco famílias: 5, 8, 46, 75, 80, sendo que a família 46 é
a mais estudada. As quitosanases tem grande aplicação na geração de oligômeros de
quitosana (Somashekar & Joseph, 1996; Chen et al., 2005).
Nogawa et al. (1998) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida pelo fungo Trichoderma reseei PC-3-7. Estes pesquisadores
inocularam 106 conídios em 100 mL de um meio basal contendo 0,3% de N-
acetilglicosamina em incubator rotativo com agitação de 220 rpm a 28oC. Destaca-se
que o fungo não produziu quitosanase em meio contendo quitosana ou quitina.
Entretanto, o microrganismo produziu quitosanase em meio contendo glicosamina e N-
acetilglicosamina, sendo que para este último obteve-se uma atividade específica
máxima de 1,5 U/mg após 72 horas de incubação. É importante ressaltar que uma
agitação vigorosa favoreceu a produção da enzima. Esse artigo ilustra a presença de
exo-β-D-glicosaminidase e exo-β-D-N-acetilglicosaminidase.
Piza et al. (1999) estudaram a produção, purificação e caracterização parcial de
uma quitosanase produzida por Bacillus cereus. Nesse caso, investigou-se através de um
planejamento fatorial fracionário a influência dos fatores: concentração de sulfato de
amônio (0,4; 2,2 e 4,0%), razão de aeração (2; 6 e 7), tempo de fermentação (16; 24 e
32 horas), pH (5; 6 e 6,5) e concentração de quitosana (1,0; 1,5 e 2,0%). Os resultados
mostraram que os fatores mais significativos para esse planejamento foram a
concentração de sulfato de amônio, a aeração, o pH e a interação entre sulfato de
amônio e a aeração.
Cheng & Li (2000) estudaram a preparação de oligossacarídeos utilizando
quitosanase produzida por Aspergillus sp e constataram que a enzima foi largamente

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
25
induzida em meio contendo quitosana como única fonte de carbono e também na
presença de quitina coloidal. A indução da enzima foi realizada em temperaturas
variando na faixa de 28 a 30ºC, pois essa espécie não cresceu bem em temperaturas
superiores a 37ºC.
Zhang et al. (2000) estudaram a produção, purificação e caracterização de
quitosanase e Exo-β-D-Glucosaminidase por Aspergillus oryzae IAM2660. Os
microrganismos utilizados neste estudo foram mantidos em placas de Petri contendo
agar para contagem em placas (PDA). Destaca-se que para produzir a enzima, conídios
foram retirados das placas e inoculados em meio Czapeq-dox com os seguintes
componentes: 1,0g K2HPO4, 0,5g KCl, 2,0g NaNO3, 0,5g MgSO4.7H2O, 0,02g
FeSO4.7H2O, 20g de sacarose e 1,0L de água deionizada, com pH do meio igual a 5,5.
Foi adicionado ao meio 0,5 g de peptona e 0,5g de extrato de levedura com
concentração de esporos da ordem de 1,0x106 esporos.mL-1. Após a incubação, feita em
incubador rotativo a 26ºC por um período de 60h, micélios foram coletados e após
filtração e secagem a 80ºC foram utilizados para ensaio de quitosanase.
Kuroiwa et al. (2002) estudaram os fatores que afetam a composição de
oligossacarídeos produzidos através da hidrólise de quitosana por quitosanase originária
de Bacillus pumilus BN-262. Neste experimento foi utilizado quitosana 100%
desacetilada dissolvida em ácido acético 1M. O pH do meio foi ajustado em 5,6. Neste
estudo a quitosanase foi imobilizada em agar e Sepharose. Os resultados mostraram que
a concentração de enzima, a temperatura, a velocidade de agitação, e concentração de
substrato influenciaram de forma significativa na composição dos oligossacarídeos
produzidos.
Ichikawa et al. (2002) estudaram a imobilização e estabilização da ligação
multiponto da quitosanase tendo agar como suporte e concluíram que a imobilização
aumentou a termoestabilidade da quitosanase. Os oligossacarídeos de quitosana foram
continuamente produzidos usando um reator coluna contendo quitosanase imobilizada.
A percentagem de quitosana imobilizada após 28 dias de reação foi de 44%.
Hung et al. (2002) estudaram a purificação, através de troca iônica, e
caracterização da hidrólise sobre ação da quitinase e quitosanase purificadas do meio
contendo Bromelina comercial. Destaca-se que a quitinase purificada apresentou pH,
temperatura e Km ótimo para ação catalítica em meio contendo quitina de 4,0, 60ºC e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
26
0,2mg.mL-1, respectivamente. Enquanto que quitosanase apresentou o pH, temperatura e
Km ótimo de 3,0, 50ºC e 0,88mg.mL-1, respectivamente.
Zhu et al. (2003) estudaram as melhores condições fermentativas e propriedades
de uma quitosanase produzida por Acinetobacter sp. C-17. A espécie de bactéria
utilizada neste ensaio foi isolada do solo e identificada como sendo Acinetobacter sp C-
17 e capaz de produzir quitosanase. O microrganismo foi mantido em placas contendo o
meio agar com os seguintes componentes: 0,5 % de quitosana, 0,2 % K2HPO4, 0,1 %
KH2PO4, 0,07 % MgSO4, 0,05 % NaCl, 0,05 % KCl, 0,01 % CaCl2, 0,05 % de extrato
de levedura e 2 % agar, o pH do meio de manutenção foi mantido em 7,2 e a
temperatura foi mantida em 30ºC durante 3 a 5 dias, para identificar a ação da enzima
quitosanase. A ação da quitosanase foi identificada através da observação de uma
colônia de microrganismo existente na placa. A bactéria foi então isolada da placa e
colocada em fermentador rotativo a 200 rpm durante 72h em meio contendo: 0,07 %
K2HPO4, 0,03 % KH2PO4, 0,05 % MgSO4, 0,03 % peptona, 0,03 % de extrato de
levedura e 1,0 % de quitosana. O pH foi de 7,0 e a temperatura 30ºC. Como fontes de
carbono foram utilizadas além de 1,0 % de quitosana, 1,0 % de quitina e 1,0 % de
glicose. Os resultados mostraram que as melhores condições para produção da
quitosanase foram alcançadas quando se utilizou 1,0 % de solução de quitosana a 30ºC,
pH 7,0 durante 24 horas de incubação.
Liu et al. (2003) estudaram a produção de quitinase por Verticillium lecanii
F091 usando fermentação submersa, a atividade de quitinase foi de 9,95 mU/mL em um
meio de cultura otimizado, com um volume de cultura de 200 mL, agitação de 150 rpm
e temperatura de 24ºC cultivado em incubador rotativo e concluíram que a taxa de
agitação e o pH foram os fatores mais significativos para a produção de quitinase, no
entanto, as taxas de agitação e aeração poderiam modificar a concentração de oxigênio
dissolvido afetando diretamente no crescimento do Verticillium lecanii e na produção
de quitinase.
Donzelli et al. (2003) estudaram a hidrólise enzimática avançada de quitina de
casca de caranguejo langostino com misturas de enzimas de bactérias e fungos e
concluiram que a combinação de enzimas de Trichoderma atroviride e Serratia
marcescens foram capazes de degradar completamente altas concentrações de quitina
(100g/mL) a partir de carapaças de caranguejo langostino convertendo a N-
Acetilglicosamina (78%), glucosamina (2%) e quitobiose (10%). Enzimas de fonte

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
27
procariótica apresentaram atividades menores que as enzimas de T. atroviride, para
catalisar a reação com quitosana.
Jo et al. (2003) estudaram a caracterização e a avaliação cinética de quitosanase
produzida por Bacillus sp. Destaca-se que na produção da enzima utilizada neste ensaio
o meio continha 0,5% de concentração de quitosana, 1% de triptona e 1% de NaCl, o
pH do meio foi ajustado em 5,5 e a temperatura foi mantida em 37ºC com agitação de
180 rpm em incubador rotativo.
Fu et al (2003) estudaram a caracterização de três iso-enzimas de quitosanase
isolada de pepsina comercial e denominado como PSC-I, PSC-II, PSC-III,
respectivamente, em relação a hidrólise enzimática da quitosana, o meio ótimo para a
produção destas iso-enzimas continha as seguintes características: pH 5,0; 5,0 e 4,0 e
temperaturas de 40ºC, 40ºC e 30ºC, o valor de Km foi de 5,2, 4,0 e 5,6 mg.mL-1,
respectivamente. As massas moleculares das três iso-enzimas foram de 40 kDa. Os
resultados mostraram que as três iso-enzimas apresentaram atividade em meio contendo
quitosana, produzindo oligômeros da ordem de 68 – 88% desacetilada.
Choi et al. (2004) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida por Bacillus sp KCTC 0377BP. Nesse caso, com relação à
produção da enzima foi possível obter um aumento de 1,2 U/ml em incubator rotativo
para 100,0 U/ml em fermentador piloto de 500 L, utilizando uma concentração de
substrato de 0,5% de quitosana e como fonte de nitrogênio polipeptona (2,5%) em pH
igual a 6,8, com 100 rpm de 100,0 rpm a 30oC.
Shimosaka et al. (2003) estudaram a produção, purificação e caracterização total
de duas quitosanases produzida por Acinetobacter sp CHB101. Nesse caso, com relação
à produção da enzima os pesquisadores usaram dois meios, um contendo glicose e outro
contendo quitosana. Com relação à cinética, o meio contendo quitosana fornece um
tempo de geração de 280 minutos, enquanto que no meio contendo glicose a taxa de
crescimento foi mais elevada e o tempo de geração foi de apenas 110 minutos. Os
autores observaram que após os dois primeiros dias de cultivo a atividade
quitosanolítica permaneceu baixa (na ordem 0,10 a 0,20 U/mL). Durante a fase de
crescimento exponencial o valor da atividade permaneceu com valor inferior a 0,01
U/mL, começando a aumentar ao atingir a fase estacionária, indicando assim que a
produção da enzima é não associada ao crescimento.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
28
Kim et al. (2004) estudaram a produção, purificação e caracterização de uma
quitosanase produzida pelo Bacillus sp e identificado pelo número 1299. Destaca-se que
na produção da enzima quitosanase foi utilizado um meio contendo os seguintes
componentes: 0,5 % de quitosana, 0,42 % (NH4)2SO4, 0,2 % K2HPO4, 0,02 % uréia,
0,03 % CaCl2.2H2O, 0,03% MgSO4.7H2O, 0,1% peptona, 0,2% Tween 80, 0,2 % da
solução (0,5% FeSO4.7H2O, 0,16% MnSO4.H2O, 0,14% ZnSO4.7H2O, 0,2% CoCl2) e
1,7% agar. O pH do meio para este ensaio foi mantido em 7,0. Este microrganismo foi
escolhido por ser grande produtor de quitosanase.
Chen et al. (2005) estudaram a purificação e a caracterização de dois tipos de
quitosanase produzidas por Aspergillus sp. CJ22-326 em cultivo submerso. Na
produção da enzima foi utilizado um meio de cultivo contendo 1,0% de quitosana, 2,0%
de gérmen de trigo, 0,2% de sulfato de amônio, 0,2 % de fosfato de potássio e 0,05% de
sulfato de magnésio por litro de solução em pH 5,6. O cultivo foi realizado em frasco de
500 mL contendo 150 mL de meio de cultura a 30ºC por 96 horas com uma agitação de
150 rpm. Os autores apresentam um gel de eletroforese com as duas enzimas purificadas
com massa molar de 29 kDa e 109 kDa.
Silva Filho et al. (2005) estudaram, através de planejamento fatorial 24 com
repetição em triplicata no ponto central, a produção de quitosanase por Aspergillus
ochraceus em cultivo descontínuo submerso. Os resultados mostraram que foi possível
produzir quitosanase com atividade na ordem de aproximadamente 5,9 U/mL utilizando
Aspergillus ochraceus e que a atividade foi favorecida pelo aumento da agitação (r), da
razão de aeração (RA) e da concentração de substrato (Cs), enquanto que o aumento da
temperatura de cultivo (TMC) não favoreceu a resposta (atividade quitosanolítica).
Pode-se classificar as quitosanases em endoquitosanases e exoquitosanases. As
primeiras diferenciam-se por catalisar a hidrólise aleatoriamente, no interior do
biopolímero, gerando diversos tamanhos de oligossacarídeos. As exoquitosanases são
capazes de hidrolisar as terminações não redutoras, produzindo assim unidades
redutoras (Peter, 2005).
Palma-Guerrero et al. (2008) estudaram o biocontrole de fungos patogênicos e
demonstraram a lise da membrana plasmática e a inibição da germinação de esporos de
fungos. Eles observaram que os fungos entomopatogênicos e nematófagos foram menos
inibidos pela quitosana que os fitopatogênicos e micoparasíticos provavelmente porque
apresentam a enzima quitosanase que degrada a quitosana. Mesmo assim, a quitosana

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
29
pode ajudar a desenvolver novas estratégias para o biocontrole em geral de fungos
patogênicos, evitando o uso de fungicidas que prejudicam o meio ambiente e a saúde
humana.
Pagnoncelli (2008) estudou a produção de quitooligossacarídeos com
propriedades nutracêuticas a partir da quitosana por hidrólise enzimática, utilizando
processo fermentativo simultâneo. Duas cepas produtoras de quitosanaes foram
selecionadas, Paenibacillus chitinolyticus e Paenibacillus ehimensis, e avaliadas quanto
ao comportamento em meio de cultivo contendo açúcares simples e em relação às
variações de pH do meio. Os quitooligossacarídeos produzidos a partir do complexo
enzimático da primeira cepa apresentaram maior pico após 9h de hidrólise, enquanto
para a segunda cepa avaliada, após 20min, observou-se quitooligossacarídeos com grau
de polimerização entre 3 e 6 unidades.
2.2. Fungos
2.2.1. Características gerais
Desde o final dos anos 60 que os fungos deixaram de pertencer ao reino das
plantas e passaram a ter um reino próprio chamado de Reino Fungi. Foi a partir de
estudos morfológicos, bioquímicos e citológicos onde se concluiu que eles são tão
diferentes dos vegetais como dos animais. Distinguem-se de outros organismos
eucariotas por serem seres quimiorganoheterotróficos e, ainda, por apresentarem parede
celular rígida que é composta por quitina e glucano e uma membrana celular em que o
ergosterol substitui o colesterol (Trabulsi & Toledo, 1996; Murray et al., 2005).
Fazem parte do Reino Fungi cerca de 100 mil espécies, desde grandes como
os cogumelos, até microscópicas como os bolores e leveduras. Ainda hoje é utilizada
essa classificação, mas os avanços da biologia molecular e estudos bioquímicos levaram
a outras propostas de classificação em que os fungos e organismos afins foram incluídos
em três diferentes reinos: Protozoa, Chromista e Fungi (Kirk et al., 2008).
Abaixo, são apresentadas características que diferenciam os fungos das plantas:
• são incapazes de sintetizar clorofila, como também não apresentar celulose em
sua parede celular, com exceções como fungos aquáticos;
• apresentam algumas características de células animais (capacidade de depositar
glicogênio e a presença de substâncias quitinosas, em sua parede celular);

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
30
• podem ser eucarióticos (as leveduras), ou multinucleados (os bolores), e
apresentar retículo endoplasmático rugoso e mitocôndrias em seu citoplasma;
• são heterotróficos, alimentam-se de matéria orgânica viva (fungos parasitários),
ou morta (fungos saprofíticos);
Os fungos são amplamente encontrados na natureza, crescem rapidamente no
solo, na água, na atmosfera, bem como nos animais, nos vegetais, resíduos orgânicos e
desempenham um papel importante na vida do homem, quer de uma maneira benéfica,
quer de um modo prejudicial. Os fungos são um dos principais microrganismos
responsáveis pela decomposição da matéria orgânica, interferindo no ciclo do carbono,
do nitrogênio e de outros nutrientes da biosfera. São capazes de deteriorarem produtos e
bens de consumo, tais como alimentos, tecidos, papel, madeira. Os alimentos
armazenados representam um vasto meio para a proliferação dos fungos, principalmente
quando não são consideradas as condições básicas de armazenamento correto (Trabulsi
& Toledo, 1996).
Estão presentes funcionalmente em vários processos industriais, tais como, de
fabricação de pão, cervejas, vinhos e determinados tipos de queijos, sendo também
utilizados na produção comercial de muitos ácidos orgânicos, de alguns fármacos, como
a ergometrina e a cortisona, na obtenção de diferentes antibióticos, como a penicilina, e
de substâncias imunossupressoras, como a ciclosporina (Esteves et al., 1990). A Figura
4 ilustra a estrutura geral de uma célula de fungo.
Figura 4 – Representação da estrutura geral de uma célula de fungo.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
31
Os fungos podem se desenvolver em meios de cultivo especiais formando
colônias de dois tipos, leveduriformes e filamentosas, que se diferenciam pela
macromorfologia e micromorfologia. O primeiro tipo apresenta, em geral, característica
de consistência cremosa de cor branca a creme, brilhantes ou opacas, podendo
apresentar às vezes coloração escura ou alaranjada, e são formados por microrganismos
unicelulares que apresentam as funções vegetativas e reprodutivas. As filamentosas
apresentam características granulares, cotonosas, aveludadas ou pulverulentas, são
multinucleadas e apresentam-se em forma de tubos denominadas de hifas. Essas hifas
podem ser não septadas (cenocíticas) ou septadas (tabicadas). O conjunto de hifas é
denominado micélio.
As hifas asseptadas têm aparência anastomosada (concisa), formada por um
citoplasma estendido e polinucleado, enquanto que as hifas septadas contêm células
individualizadas, cada uma contendo o seu núcleo. Na Figura 5 são mostradas as
estruturas das hifas septadas e não septadas.
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas. (Adaptado de Guia.Bio,
2012)
O micélio que se desenvolve no interior do substrato e que é responsável pela
sustentação e absorção de nutrientes é denominado micélio vegetativo. O micélio que
cresce na superfície do meio de cultura é denominado micélio aéreo. Este pode ser
denominado também de micélio reprodutivo, quando o mesmo se diferencia para
sustentar os corpos de frutificação ou propágulos (Putzke & Putzke, 1998). Os
propágulos classificam-se de acordo com a origem em externos e internos, sexuados e
assexuados.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
32
2.2.2. Reprodução dos Fungos
A reprodução dos fungos ocorre em ciclos assexuais, sexuais e parassexuais. Em
fungos, a reprodução assexuada é observada com mais freqüência do que a reprodução
sexuada. A reprodução assexuada é normalmente a reprodução mais importante para a
propagação da espécie, feita através de fragmentação de artroconídios, fissão de células
somáticas, brotamento ou gemulação do blastoconídios-mãe e produção de conídios. A
reprodução sexuada envolve a união de duas células ou de dois órgãos sexuais
sexualmente compatíveis. A reprodução sexuada culmina na produção de basidiósporos,
no caso de basidiomicetos. A reprodução parassexuada ocorre com a fusão de hifas e
formação de um heterocário, que consiste na existência de núcleos haplóides
geneticamente distintos num mesmo citoplasma, os quais, após a fusão, originam
núcleos diplóides heterozigotos sujeitos a sucessivas divisões mitóticas. Apesar da
raridade dessas, a reprodução parassexual é de grande importância na evolução de
alguns fungos (Pelczar et al., 1996).
A reprodução assexuada é feita por conídios, esporos assexuados formados no
ápice de hifas modificadas, os conidióforos. Os conidióforos não possuem esporângios
de forma que os conídios ficam expostos. Na reprodução sexuada, com o encontro de
duas hifas haplóides distintas, ocorre a plasmogamia e um novo conjunto de hifas
dicarióticas é formado.
Os conídios podem ter formas esféricas, fusiformes, cilíndricas, piriformes, entre
outras. Apresentam-se como hialinos ou pigmentados, podem estar isolados ou
agrupados, a parede celular pode ser lisa ou rugosa e podem ser formados por apenas
uma célula ou apresentarem septos em mais de um plano.
Os conídios também podem originar-se em qualquer parte do micélio vegetativo
e são denominados conídios sésseis como é o caso do Trichophyton rubrum.
Alguns fungos patogênicos são identificados a partir de estruturas formadas pela
célula conidiogênica e o conidióforo. Essas estruturas são denominadas: aparelho de
frutificação ou conidiação. Os conídios podem formar cadeias sobre fiálides em volta de
uma vesícula, dilatando a extremidade do conidióforo. Essas estruturas ocorrem em
Aspergillus, apresentando-se em forma de cabeça ao redor de uma vesícula, e em
Penicillium agrupados em forma de pincel, como é ilustrada na Figura 6.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
33
Figura 6 – Conídios de Aspergillus e Penicillium
2.2.3. Metabolismo de fungos
Os fungos são seres aeróbicos e heterotróficos, podendo se desenvolver em
ambientes com baixa quantidade de oxigênio, como ocorre no crescimento vegetativo e
na reprodução assexuada, como também em ambientes com grande quantidade de
oxigênio como é o caso da reprodução sexuada.
Os fungos são capazes de produzir certas enzimas como lipases, invertases,
lactases, quitosanases, proteases, amilases, etc., que fazem a hidrólise do substrato
facilitando o mecanismo de transporte ativo e passivo. Esses substratos induzem a
formação de enzimas degradativas. Algumas espécies de fungos desenvolvem-se em
meios que apresentam amônia ou nitrilos como única fonte de nitrogênio. As melhores
fontes orgânicas para o seu desenvolvimento são carboidratos, como a D-glicose, e
alguns sais minerais como sulfatos e fosfatos.
A água é uma necessidade de todos os seres vivos, não sendo diferente para os
fungos que precisam de água para o seu desenvolvimento. Certos fungos apresentam
características halofílicas podendo crescer nos ambientes com grandes concentrações de
sal.
A razão de não existirem ambientes livres de espécies fúngicas deve-se ao facto
destes microrganismos conseguirem sobreviver em situações extremas como, por
exemplo, temperaturas que variam entre os - 5 e os 60 ºC, com pH de 1 a 9 e com baixas

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
34
concentrações de oxigênio, apesar de cada espécie possuir condições ideais para o seu
desenvolvimento (Chao et al., 2002).
Devido a grande faixa de temperatura ótima para o seu crescimento, os fungos
apresentam espécies psicrófilas, mesófilas e termófilas. Para a maior parte das espécies
fúngicas, a temperatura de desenvolvimento situa-se entre os 20 e 24 ºC e não crescem
acima dos 29 ºC. Em relação à umidade relativa, necessitam de pelo menos 50 %, mas
geralmente necessitam de mais de 65 % (Colakoglu, 2001).
O pH ideal para o desenvolvimento dos fungos varia na faixa entre 5 e 7.
Destaca-se, entretanto, que a maioria dos fungos suportam grandes variações de pH
como ocorre com os fungos filamentosos que podem crescer em pH variando entre 1,5 e
11. Por outro lado, as leveduras não suportam pH alcalino.
Os fungos crescem lentamente, se comparados com as bactérias, em meio de
incubação para o seu desenvolvimento. Para evitar contaminação por bactérias, que
podem se sobrepor ou inibir o seu crescimento, pode-se introduzir no meio de cultura
um antibacteriano de largo espectro como o cloranfenicol.
Algumas espécies de fungos se desenvolvem na presença de luz, outras são
inibidas e algumas podem crescer tanto na presença quanto na ausência deste agente. No
entanto a irradiação solar, em geral, é prejudicial ao seu desenvolvimento, por causa da
radiação ultravioleta.
2.2.4. Nutrição de fungos
Quanto ao aspecto da nutrição, os fungos são heterótrofos, ou seja, incapazes de
produzir seu próprio alimento. Entretanto, a forma de assimilar matéria orgânica é
diferente daquela utilizada pela maioria dos animais, que são heterótrofos por ingestão.
Nos fungos, a digestão é extracorpórea, ou seja, é realizada fora do corpo. O fungo
lança no ambiente, enzimas que degradam as moléculas orgânicas complexas e, depois,
absorve moléculas menores, mais simples. Por essa razão, são designados heterótrofos
por absorção. É devido a este processo que os fungos podem crescer dentro ou sobre os
alimentos. Desenvolve-se geralmente em meios contendo um pH baixo, uma fonte de
carbono, uma fonte de nitrogênio orgânico ou inorgânico e alguns minerais. Alguns
necessitam de vitaminas. Quanto aos modos de vida, os fungos podem ser
decompositores, parasitas, mutualísticos e predadores (Raven, 2001).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
35
Os fungos decompositores, saprófagos (do grego, saprós, “podre”, e phagos,
“comedor”), juntamente com as bactérias, são os principais decompositores da biosfera,
participando intensamente do processo de degradação da matéria orgânica morta, o que
promove a reciclagem dos elementos químicos constituintes dos seres vivos. A maioria
deles vive no solo, obtendo nutrientes de seres mortos. Em florestas, onde a matéria
orgânica vegetal é abundante, os micélios dos fungos decompositores absorvem os
nutrientes de folhas e galhos caídos, decompondo-os. Os fungos decompositores
também são responsáveis pelo apodrecimento de alimentos e de outros materiais, como
a madeira.
Os fungos parasitas vivem à custa de outros organismos vivos, prejudicando-os.
Ao parasitar o corpo de um ser vivo, animal ou vegetal, o fungo pode até provocar sua
morte. Várias doenças são causadas por fungos parasitas. Nas plantas, cita-se a
ferrugem do café, que, no Brasil, é motivo de grande preocupação por parte dos
cafeicultores, devido aos prejuízos que acarreta. As micoses que atacam a pele de seres
humanos e animais são provocadas por fungos.
Existem fungos que se associam a outros organismos, estabelecendo uma relação
em que há benefício mútuo para os indivíduos envolvidos. Esses fungos são
denominados mutualísticos. A maioria vive associada a seres fotossintetizantes, como
as plantas, cedendo-lhes parte da água e dos nutrientes que as hifas absorvem do solo.
As plantas, por sua vez, cedem ao fungo certos açúcares e aminoácidos. Duas
associações mutualísticas formam estruturas bem características: as micorrizas,
associação que envolve fungos e raízes de plantas, e os líquens, formados por fungos e
algumas variedades de algas e cianobactérias.
Certos fungos atuam como predadores. Na maioria dos casos, as hifas secretam
substâncias aderentes que aprisionam os organismos que tocam os fungos. Dessa
maneira, as hifas penetram o corpo da presa, crescem e se ramificam, espalhando-se no
interior do organismo e absorvendo seus nutrientes, causando-lhe a morte. O gênero
Arthrobotrys apresenta uma forma mais elaborada de predação. O fungo vive no solo e
captura nematódeos microscópicos que habitam o mesmo ambiente. As hifas
apresentam pequenos anéis que se estreitam quando a presa passa por eles. As hifas do
fungo logo invadem o corpo do nematódeo, digerindo-o.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
36
2.2.5. O fungo Metarhizium anisopliae
Existem três espécies de fungos do gênero Metarhizium, divididas em dez
variedades: M. anisopliae, variedades anisopliae, majus, lepidiotum e acridum; M.
flavoviride, variedades tipo E, flavoviride, minus, novazealandicum e pemphigum; e M.
album (Driver et al. 2000). E, mais recentemente, foi descrita na China uma nova
variedade de M. anisopliae, dcjhyium (Dong et al. 2009).
Data de 1879 quando o russo Ilya Metchnikoff aplicou o fungo para combater
larvas de um curculionídeo, praga de beterraba, e foi classificado como Metarhizium
anisopliae por Sorokin em 1883. É um fungicida natural para mais de 300 espécies de
insetos, especialmente pragas da agricultura e pecuária. No Brasil, o fungo Metarhizium
anisopliae teve a sua aplicação com sucesso pela primeira vez em 1965, no controle da
cigarrinha-da-cana Mahanarva posticata (Alves, 1998).
O fungo Metarhizium anisopliae é um importante agente entomopatogênico
utilizado no controle de pragas, como cupins, gafanhotos, cigarrinhas e besouros, sendo
amplamente estudado em todo o mundo. Existe amplamente na natureza e é facilmente
encontrado no solo, onde sobrevive por longos períodos e seu desenvolvimento se dá
geralmente entre 15 e 32 °C, sendo ideal entre 24 e 30 °C e com pH ótimo igual a 6,9.
Apresenta micélio hialino e septado, com conidióforos característicos, sobre os quais
surgem conídios cilíndricos organizados em colunas. Este fungo ao atacar os insetos
provoca rigidez e os recobre por uma camada pulverulenta de conídios, resultando em
colorações que variam do verde claro ao escuro, cinza ou branco com pontos verdes
(Tinline, 1971; Alves, 1998, Driver et al. 2000; Arruda, 2005).
Quanto a sua morfologia, apresenta-se como um fungo filamentoso, com corpo
de frutificação equivalente a um esporodóquio agregado a hifas intimamente
entrelaçadas, com massa compactada de conidióforos característicos, simples ou
ramificados, formando células esporogênicas denominadas fiálides, resultando nos
fialosporos (Wang et al., 2002).
O Metarhizium anisopliae é um fungo de cultivo simples, sendo necessário
quase que exclusivamente o nutriente a base de amido. Um dos fungos mais estudados
em programas de manejo de pragas já que apresenta grande potencialidade
entomopatogênica (Onofre et al., 2002). O Metarhizium anisopliae é um microrganismo
pouco exigente, pois pode se desenvolver em diversos meios de cultura como: amido,
glicose, glicerina, levulose, maltose, sacarose e quitina (Jabor et al., 2003).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
37
Em geral, os fungos produzem enzimas que convertem os tecidos dos insetos em
nutrientes. Porém, a cutícula esclerotizada dos insetos não é bem utilizada pelos fungos,
excetos pelos entomopatogênicos que desenvolveram enzimas eficientes para degradar
essa camada protetora. Destaca-se a enzima Pr1, produzida por Metarhizium anisopliae,
que tem um poder de degradação bem superior a qualquer outra enzima sintetizada por
esse grupo de fungos (Dias, 2005). Desta forma, o Metarhizium anisopliae (fungo
entomopatogênico) ao atacar a célula dos insetos, produz grande quantidade de enzimas
que degradam a membrana citoplasmática e inativa a célula e, consequentemente, mata
o inseto, logo é evidente a relação entre o controle de pragas e produção de enzimas.
No momento, sobre a comercialização de fungos entomopatogênicos no Brasil,
em sua maioria, são formulados em grânulos, compostas do fungo mais o substrato
(arroz + fungo), como também na forma de pó molhável (WP), resultado da moagem do
fungo com o substrato. Esses produtos representam 90% e 8,5%, respectivamente, da
produção de fungos no Brasil, e sendo suas concentrações finais ficam em torno de
5x108 conídios/g para Metarhizium anisopliae e 1x109 conídios/g para Beauveria
bassiana (Alves et al., 2008).
Segundo Batista Filho et al. (2002), Garcia et al. (2005) e Mello et al. (2006), o
Metarhizium anisopliae tem sido bastante utilizado para o controle de diferentes pragas
no Brasil, como a broca da bananeira, pragas de grãos armazenados, carrapatos, cupim
de montículo em pastagens, larvas de escarabeídeos que atacam a cana de açúcar e
cigarrinha da cana-de-açúcar.
É comum a ocorrência natural do fungo Metarhizium anisopliae atacando
cigarrinha-das-raízes de canaviais pelo país, o que incentiva a utilização dele como
agente de controle biológico em áreas infestadas por esse tipo de praga. A aplicação de
recursos no uso de programa de manejo integrado da cigarrinha tem sido reforçado por
razões ambientais e econômicas (Dinardo-Miranda, 2001).
Faria & Magalhães (2001) fizeram uma avaliação do uso de fungos
entomopatogênicos no Brasil, na qual observaram que a qualidade dos micoinseticidas
disponíveis no Brasil poderia ser incrementada de forma considerável quanto a sua
formulação, ou seja, antes da venda, poderiam realizar melhorias no tratamento
posterior, adição de substâncias que lhes assegurem melhorias na eficiência de controle,
capacidade de armazenamento ou praticidade de manuseio, ou de qualquer outro critério
que resulte em vantagem em relação ao produto bruto. Ressaltaram a pouca praticidade

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
38
em alguns casos, como exigência de lavagem, e em outros podendo causar o
entupimento de bicos dos pulverizadores devido à elevada proporção de inertes,
principalmente quando são empregados baixos volumes de aplicação. Outro problema
evidenciado com os inseticidas nacionais diz respeito à pequena sobrevida, devendo a
ser usados em, no máximo, 30 dias depois de produzidos, quando armazenados à
temperatura ambiente e em local sombreado, fazendo as vendas ocorrerem quase que,
exclusivamente, sob encomenda. Comparativamente, os bons inseticidas sob
comercialização em outros países apresentam maior concentração de ingrediente ativo,
maior sobrevida (alguns produtos podem ser armazenados por mais de 8 meses à
temperatura ambiente) e praticidade (são formulados na forma de GDA (grânulos
dispersíveis em água) ou de óleos emulsionáveis, por exemplo, podendo ser adicionados
diretamente ao tanque do pulverizador.
Rangel et al. (2007), instalaram uma Unidade de Observação para o controle
microbiano das cigarrinhas das pastagens Distrito de Itahum, em Dourados/MS, devido
as necessidades de agricultores familiares de produção de leite. As aplicações do fungo
Metarhizium anisopliae foram realizadas por pulverização com equipamento de tração
animal. As condições do ambiente de estudo foram favoráveis ao desenvolvimento da
população das cigarrinhas e à incidência do fungo. Como resultados, observaram que os
níveis de espumas, ninfas e adultos reduziram significativamente após as aplicações,
levando os Agricultores Familiares a total satisfação com os ganhos na produção de
todos os envolvidos.
Segundo Vieira et al. (2007), o fungo entomopatogênico Metarhizium anisopliae
var. acridum tem sido utilizado no controle de pragas, e assim, visando este fim,
analisaram em laboratório a esporulação, a germinação dos conídios e as características
morfológicas peculiares desse fungo em diferentes temperaturas. Esperou-se o
desenvolvimento das colônias em períodos de 1 a 15 dias. Foi observado que
temperaturas de 25, 28 e 30 ºC proporcionaram maiores taxas de esporulação e elevadas
porcentagens de germinação dos conídios. A 25, 28 e 30 ºC observou-se um
desenvolvimento mais rápido das microestruturas somáticas e reprodutivas (hifas,
anastomoses, apressórios, conidióforos e conídios elipsóides). Nas temperaturas
extremas de 40 e 45 ºC não foi detectado esporulação, germinação, nem
desenvolvimento de nenhuma microestrutura.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
39
Lopes et al. (2007) avaliaram o efeito de formulações de Beauveria bassiana e
Metarhizium anisopliae sobre ninfas de Amblyomma cajennense (transmissor da febre
maculosa ao homem) em laboratório. Ninfas não alimentadas foram pulverizadas com
suspensões contendo 2x107 e 4x107 conídios/mL de B. bassiana, e 1x107, 1,5x107,
2x107, 3x107 e 5x107 conídios/mL de M. anisopliae. A mortalidade de ninfas,
decorridos três dias da aplicação, foi significativa nas concentrações de 3x107 e 5x107
conídios/mL de M. anisopliae, com 57,1 e 56,2 %, respectivamente. Após seis dias a
mortalidade foi de 100 % nessas duas concentrações. A mortalidade foi
significativamente maior que a observada para B. bassiana em concentrações
semelhantes.
Guirado et al. (2009) realizaram estudos para controlar cupins de montículo em
área de pastagem, utilizando fungos entomopatogênicos Metarhizium anisopliae e
Beauveria bassiana em diferentes doses e formulações. A avaliação foi realizada após
30 dias da aplicação dos produtos. Os melhores resultados em ordem decrescente de
eficiência foram: M. anisopliae 30 mL + 1000 mL de água com 100 % de eficiência; B.
bassiana 10g + M. anisopliae 20 mL + 1000 mL de água com 85,71 % de eficiência; M.
anisopliae 20 mL + 1000 mL de água com 71,42 % de eficiência. Os demais
tratamentos não diferiram da testemunha.
Loureiro et al. (2012) compararam a eficiência de isolados de Metarhizium
anisopliae em controlar populações naturais de Mahanarva fimbriolata (cigarrinha-da-
raiz) em cultivos comerciais de cana-de-açúcar colhida mecanicamente. Foram
aplicados os isolados IBCB 348, IBCB 408, IBCB 410 e IBCB 425 do fungo em
concentração de 1,5×1012 conídios/ha. Aplicou-se o planejamento experimental em
blocos casualizados com 5 tratamentos e 4 repetições, sendo cada repetição composta
por 7 linhas de 100 m de comprimento e espaçamento de 1,5 m. Após 30 dias de
pulverização foi observado para os isolados IBCB 408 e IBCB 425 uma eficiência de
controle de ninfas de 63 e 62%, respectivamente, já no caso dos adultos chegou a 100%.
Após 60 dias o isolado IBCB 425 foi o mais eficiente para controlar as ninfas com
48,4%.
Assis et al. (2012) estudaram a citotoxidade de quitoolissacarídeos (QOS) e
oligômeros A, B e C (soluções compostas por diferentes quantidades de QOS)
produzidos por hidrólise enzimática pelo fungo Metarhrizium anisopliae. Foi analisado
o efeito de antiproliferativo destas moléculas em células tumorais (HepG2 e HeLa) e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
40
também em célula normal (3T3). A atividade antioxidante foi analisada em vários
experimentos in vitro. A glicosamina apresentou maior toxicidade (cerca de 92%) para
todas as células estudadas. No entanto, os oligômeros obtidos após hidrólise não
demonstraram efeito tóxico sobre a célula normal (3T3). Além disso, mostraram que
uma pequena quantidade de outros COS pode diminuir o efeito citotóxico da
glicosamina contra células 3T3, indicando que glicosamina poderia ser usada como uma
droga antitumoral na presença de outros COS.
2.3. Fermentação
Os processos fermentativos são caracterizados por processos nos quais
microrganismos realizam a conversão catalítica de uma dada substância num
determinado produto. Estes processos podem ser classificados como processos
descontínuos, contínuos e descontínuos alimentados. A classificação destes processos
está diretamente ligada à forma de como o substrato é adicionado e como o produto é
retirado. No processo de fermentação descontínuo o substrato é colocado no
fermentador, em seguida é inoculado e após o término do processo o produto é retirado.
No processo contínuo, o substrato é colocado num reator com vazão constante e o meio
fermentado é retirado com a mesma vazão de entrada do reator (Faccioti, 2001). Nos
processo descontínuos alimentados à adição de substrato e a retirada do mesmo deve
ocorrer de forma intermitente (Carvalho & Sato, 2001).
De certa forma, os processos fermentativos representam um elo entre as antigas
artes de elaboração de alimentos, como queijos e vinho, a partir de uma flora
microbiana natural, com a moderna indústria de fermentação de alimentos, onde são
utilisados cultivos puros e equipamentos sofisticados para o controle do processo
(Assis, 2010).
Atualmente, a maioria dos processos industriais se desenvolve em cubas de
vidro ou de aço de até 20 litros. Os agentes biológicos se encontram submersos no meio de
cultivo que ocupa, aproximadamente, 75% da cuba. As vezes é necessário injetar ar, e em muitas
fermentações se forma espuma. O desenho do biorreator deve se adequar ao objetivo do
processo, respondendo eventualmente a diversos imperativos, tais como a esterilização do
sistema, a aeração e homogeneização do meio, o acréscimo de nutrientes e de aditivos
antiespuma, a manutenção do pH, etc. Os modelos de fermentadores mais utilizados com
microrganismos contam com aeração e agitação mecânica. Esta facilita a distribuição dos

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
41
nutrientes na cuba, mas gera calor que deve ser eliminado mediante a circulação de água
fria. Em outros tipos de biorreatores, em coluna ou torre, a homogeneização depende da injeção de
ar. Os sistemas submersos são apropriados para o cultivo de microrganismos livres, mas resultam
pouco econômicos quando se trabalha com células ou enzimas caras. A imobilização em
fermentadores menores, seja por adesão a um suporte inerte, seja por inclusão dentro de um
polímero que permita o contato com o meio de cultura, além de simplificar a purificação do
produto permite a reutilização das células ou das enzimas, que permanecem dentro do
biorreator (Malajovich, 2009).
As enzimas são produtos de alto valor agregado, amplamente empregadas em
uma gama de indústrias, e entre as fontes para sua obtenção, destacam-se as de origem
microbiana, principalmente, devido a elevada seletividades dos substratos utilizados e
produtos formados. A principal forma de obtenção de enzimas, em escala industrial, é a
fermentação submersa, por esta apresentar vantagens em relação a fermentação
semisólida, como: maior controle do processo fermentativo e consequente reprodução
da cinética microbiana, além de maiores rendimentos (Wanderley, 2011).
2.3.1. Preparo do inóculo
Denomina-se inóculo um volume de suspensão de microrganismo de
concentração adequada, capaz de garantir, em condições econômicas, a fermentação de
uma dada quantidade de meio. O volume do inóculo, assim como sua concentração de
células, vai depender tanto do processo de fermentação c omo da quantidade de meio a
ser fermentado (Carvalho & Sato, 2001).
A partir de uma cultura pura, inocula-se um volume relativamente pequeno de
meio nutriente, que vai depender do tipo de microrganismo cultivado (pois cada espécie
microbiana possui velocidades diferentes de crescimento), sendo, em seguida, incubado
em condições favoráveis ao desenvolvimento do microrganismo. A suspensão obtida é
transferida para um frasco maior em agitador rotativo ou recíproco (shakers), que por
sua vez é adequadamente incubado, e a seguir, transferido para um frasco contendo um
volume ainda maior deste meio, até se conseguir alguns litros de suspensão microbiana.
Os cuidados com assepsia, volumes de meio utilizado, numero de transferência bem
como as condições de incubação, podem variar de acordo com o tipo de processo de
fermentação que se deseja realizar.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
42
2.3.2. Produção de enzimas em cultivo submerso descontínuo
A produção de enzimas, usando processos fermentativos, é constituída
basicamente por duas etapas: a primeira caracteriza-se pela preparação do inóculo e a
segunda, pela fermentação do substrato (Carvalho & Sato, 2001). A finalidade da
primeira etapa é o preparo do microrganismo em condições adequadas para que possa
ocorrer o desenvolvimento da segunda etapa. Diferentemente do cultivo em estado
sólido, no cultivo submerso os microrganismos encontram-se em um meio com elevada
atividade de água.
2.4. Planejamento experimental
Um bom planejamento tem como fundamental projetar um experimento de
forma que ele seja capaz de fornecer exatamente o tipo de informação que procuramos.
Para isso é necessário saber, em primeiro lugar, o que é mesmo que se deseja procurar.
Planejamento experimental, ou delineamento experimental, é um conjunto de
ensaios estabelecido com critérios científicos e estatísticos, objetivando determinar a
influência de diversas variáveis nos resultados de um dado sistema ou processo (Barros
Neto et al., 2001; Rodrigues & Iemma, 2005). De acordo com o propósito dos ensaios,
esse objetivo maior pode ser mais detalhado em:
– determinar quais variáveis são mais influentes nos resultados;
– atribuir valores às variáveis influentes de modo a otimizar os resultados;
– atribuir valores às variáveis influentes de modo a minimizar a variabilidade dos
resultados;
– atribuir valores às variáveis influentes de modo a minimizar a influência de variáveis
incontroláveis.
O planejamento experimental, baseado nos fundamentos estatísticos é, sem
dúvida alguma, uma ferramenta poderosa para se chegar às condições otimizadas de um
processo, desenvolvimento da formulação de produtos dentro das especificações
desejadas ou simplesmente para avaliar os efeitos ou impactos que os fatores (variáveis
independentes) têm nas respostas (variáveis dependentes) desejadas (Barros Neto et al.,
2001; Rodrigues & Iemma, 2005). Da utilização das técnicas estatísticas de
planejamento experimental, é possível chegar benefícios importantes, como:

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
43
– reduzir o número de ensaios sem prejuízo da qualidade da informação;
– estudar simultaneamente diversas variáveis, separando seus efeitos;
– determinar a confiabilidade dos resultados;
– realizar a pesquisa em etapas, num processo iterativo de acréscimo de novos ensaios;
– selecionar as variáveis que influem num processo com número reduzido de ensaios;
– representar o processo estudado através de expressões matemáticas;
– elaborar conclusões a partir de resultados qualitativos.
2.4.1. Planejamento fatorial
Em um planejamento fatorial é investigado as combinações de níveis de estudo
de cada fator. Para tanto, inicialmente, é necessário especificar esses níveis em cada
fator, quer dizer, os valores dos fatores que serão utilizados nos experimentos. Um
ensaio experimental é definido como um experimento constituído de um conjunto de
níveis definido. Geralmente, quando se tem n1 níveis de um determinado fator 1, n2 do
fator 2, ..., e nk do fator k, o planejamento será descrito por um fatorial n1 x n2 x ... x nk,
que corresponde ao número mínimo de experimentos para se ter um planejamento
fatorial completo.
É necessário fazer cada fator variar e observar o resultado dessa variação para se
estudar o efeito sobre a resposta. Assim, os ensaios precisam ter um mínimo de dois
níveis por fator para a realização desses ensaios. Para k fatores, ou seja, k variáveis de
controle, o planejamento de dois níveis precisará de 2 x 2 x ... x 2 = 2k, para o número
de ensaios diferentes, sendo conhecido como planejamento fatorial 2k.
Para o planejamento fatorial, são especificados os valores dos fatores em cada
experimento, e estes são classificados em níveis, variando-se, por exemplo, de -1
(mínimo) a +1 (máximo). A partir de uma matriz serão apresentadas todas as
combinações entre o número de fatores e de níveis nos experimentos a serem realizados
(Barros Neto et al., 2001; Rodrigues & Iemma, 2005).
2.4.2. Planejamento Plackett & Burman
A maioria dos planejamentos do tipo “Screening” (Delineamentos de Seleção de
fatores) foi descrito originalmente por Plackett & Burman em 1946. Os planejamentos
experimentais de Plackett & Burman são eficientes em situações que envolvem uma

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
44
grande quantidade de variáveis a serem exploradas em um processo de otimização de
fatores (Reddy et al., 2008).
As matrizes Plackett & Burman consistem de planejamentos fatoriais de dois
níveis para estudar k = N – 1 variáveis em N ensaios. São construídas para ensaios
múltiplos de 4, sendo essas matrizes para ensaios de 8, 12, 16,..., 100. Cada variável
independente em uma matriz assume uma configuração de dois níveis, o inferior (-1) e o
superior (+1), para o nível intermediário é estabelecido (0). Para cada vez que a variável
aparece no nível inferior, terá o par equivalente no nível superior (Rodrigues & Iemma,
2005).
Para evitar dúvidas nos resultados dos efeitos de algum fator no planejamento de
Plackett & Burman, o número de ensaios a ser realizado deve ser no mínimo o número
de variáveis mais 4. No caso de um fatorial completo o número de ensaios a ser
realizado é bem maior (Rodrigues & Iemma, 2005).
Mesmo já determinado o planejamento Plackett & Burman, torna-se importante
à realização de experimentos relativos aos pontos centrais de modo a se avaliar melhor
as possíveis interferências analíticas que podem ocorrer durante os diferentes
experimentos. A repetição do ponto central permite também a determinação do erro
puro e da repetibilidade do processo. Na prática, esta é uma ferramenta que permite
identificar melhor se o processo em estudo está sob controle (Rodrigues & Iemma,
2005).
É possível que no planejamento Plackett & Burman não tenha sido considerado
a codificação do ponto central (0) para repetição, pois, matematicamente, a estimativa
do valor desse efeito codificado (zero) vezes, teria como resposta zero. Porém, no
resultado prático do experimento, consegue-se informações valiosas com essas
repetições no ponto central (Rodrigues & Iemma, 2005).
2.4.3. Aplicações do Planejamento Plackett & Burman
Ghanem et al. (2000) produziram xilanase por Aspergillus terreus cultivadas
sobre palha de trigo finamente moída em fermentação de estado sólido. A composição
ideal do meio foi estabelecida aplicando-se o planejamento experimental de Plackett &
Burman. A melhor atividade enzimática foi obtida em um meio contendo 10 g/L de

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
45
palha de trigo por frasco umedecido com uma solução concentrada de sal com teor de
água inicial de 75 % e incubadas durante 4 dias a 30 °C. Observou-se que íons
metálicos como Hg2+, Cu2+, Co2+, Fe3+ e Pb2+ inibiram fortemente a xilanase, enquanto
que Ca2+ ativou a enzima.
Mu et al. (2005) estudaram as otimização de meio de fermentação para a reação
de estereoinversão por Candida parapsilosis CCTCC M203011 utilizando o
planejamento Plackett & Burman. Os resultados mostraram que dentre os fatores
estudados, os fatores mais significativos foram: a Concentração de MgSO4, ZnSO4 e
KH2PO4 e que após a utilização da metodologia de superfície de resposta foi constatado
que o ponto ótimo para a otimização foi conseguido com as concentrações dos fatores
MgSO4, ZnSO4 e KH2PO4 nos valores em: 1,17 g/L, 0,10 g/L e 9,41 g/L,
respectivamente, elevando a taxa de conversão enantiosseletiva de 75,2 % para 96 %.
Naveena et al. (2005) empregaram o planejamento Plackett & Burman para a
seleção de 15 parâmetros para produção de ácido láctico L(+) de farelo de trigo,
substrato de baixo custo e suporte sólido, por Lactobacillus amylophilus GV6 em
fermentação de estado sólido (SSF). Onze nutrientes pertencentes a duas categorias,
fontes de nitrogênio e fontes de sal, junto com três parâmetros físicos e um tampão
foram os parâmetros considerados. Os autores verificaram influência na produtividade
devido às fontes de nitrogênio, peptona, extrato de levedura e tri-citrato de amônio,
juntamente com o NaH2PO4.2H2O e Tween 80, que podem ainda serem otimizados para
o aumento na produção de ácido lático. Esses pesquisadores ainda chamam atenção para
o uso escasso deste planejamento em fermentação em estado sólido e que o mesmo
ainda não havia sido considerado anteriormente para a conversão direta de amido em
ácido láctico L (+) usando um sistema bacteriano.
Khosravi-Darani et al. (2008) estudaram a aplicação de Plackett & Burman na
análise dos fatores significativos para a produção de ácido cítrico por Aspergillus niger
ATCC 9142 utilizando palha de trigo como substrato. Os resultados mostraram que a
produção, com a palha de trigo pré-tratada com HCl, NaOH e Uréia, foi mais eficiente e
que os fatores mais significativos na produção foram: Solvente, Teor de Umidade,
Temperatura, Tempo de Incubação, Concentração Inicial de Açúcar e Concentração de
Metanol.
Ji et al. (2009) estudaram o desenvolvimento de um meio de cultura econômico
para a produção do 2,3-butanodiol através da fermentação da glicose e xilose por

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
46
Klebsiella oxytoca utilizando o planejamento Plackett & Burman. Os resultados
mostraram que a uréia e a maceração de milho foram os fatores mais significativos para
a produção do 2,3-butanodiol e que através de planejamento experimental com repetição
em triplicata no ponto central para determinar os níveis ótimos de produção. Os autores
constaram que o rendimento atingido foi 85,6% do valor teórico.
Paula & Arruda. (2009) observaram a necessidade da busca por alternativas mais
eficientes ao método termomecânico utilizado para a degradação de milhões de
toneladas de penas de aves provenientes de frigoríficos e aviários, cuja rígida estrutura é
composta por mais de 90 % de queratina. Para esse fim, utilizaram de comunidades
microbianas para a finalidade proposta, testando o planejamento experimental Plackett
& Burman para verificar a influência de diferentes microrganismos simultaneamente,
com um número reduzido de experimentos. Testaram 32 bactérias isoladas, inoculadas
em meio contendo somente uma pena como única fonte de carbono, durante sete dias,
de onde foi selecionado as cepas para os consórcios microbianos. Experimentos unindo
as cepas NP5, FCA7, NP4 e S14 demonstraram diferentes atividades queratinolíticas,
aumentando o nível de degradação, até 87,8%, comparada com as cepas
individualmente, variando entre 26,4% e 78,0%.
Rajendran & Thangavelu (2009) obtiveram valores máximos da atividade de
produção de lipase em 3,98 U/mL e concentração de massa celular em 5,62 g.L-1 com R.
arrhizus em cultivo submerso descontínuo otimizado pelo planejamento Plackett &
Burman. As variáveis mais importantes que afetaram a produção da lipase foram azeite
de oliva, peptona, KH2PO4, CaCl2.2H2O e MgSO4.7H2O.
Li et al. (2010) estudaram a otimização de meio de cultura para a produção de
Streptolydigin (um antibiótico) por Streptomyces lydicus AS 4,2501 utilizando Plackett
& Burman e a metodologia de superfície de resposta. Os resultados mostraram que
dentre os fatores estudados, os fatores mais significativos para a produção de
Streptolydigin e otimização do meio de cultura foram: Concentração de Amido,
Concentração de Peptona e a Concentração de K2HPO4.
Chen et al. (2010) estudaram a otimização de meio de cultura para a produção da
Luciferase – que são enzimas que catalisam reações biológicas transformando energia
química em energia luminosa - por Bacillus Subtilis em fermentação descontínua
utilizando planejamento Plackett & Burmam. Os resultados mostraram que dentre os
componentes estudados, os fatores mais significativos para a produção e otimização do

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
47
meio de cultura para a produção da Luciferase foram: Concentração de Glicose 1,17 %,
Extrato de Levedura 2,27 % e Glutamato 0,55 %.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o projeto Plackett & Burman e a metodologia de superfície de resposta. Os
resultados mostraram que a eficiência máxima de extração da quitina foi conseguida nas
seguintes condições: Concentração de casca de camarão (50 g/L), glicose (50 g/L),
tempo de incubação de 5 dias e inoculo com 0,05 de OD (densidade óptica).
Gao et al. (2010) desenvolveram um meio para a fermentação do extrato bruto
de inulina de tubérculos de alcachofra de Jerusalém por Paenibacillus polymyxa ZJ-9
para produzir R,R-2,3-butanodiol (R-R,2,3-BD). Inulina, K2HPO4 e NH4Cl foram os
parâmetros principais na fermentação de acordo com os resultados obtidos com o
planejamento experimental Plackett & Burman. A concentração ideal dos três fatores,
77,14 g/L, 3,09 g/L e 0,93 g/L, respectivamente, foi obtida pelo planejamento Box-
Behnken e pela superfície de resposta. Sob as condições ideais, a concentração do R-
R,2,3-BD obtida foi 36,92 g/L, em mais de 98 % de pureza óptica. Comparado com
outras fontes de carbono investigadas, a fermentação do extrato bruto de inulina obteve
o maior rendimento de R-R,2,3-BD.
Uma metodologia de superfície de resposta foi empregada com sucesso para
otimizar um meio de fermentação chamada VHG (Very High Gravity), referência à alta
concentração de açúcares e densidade dos mostos, baseado na água maceração de milho
CSL (Corn Steep Liquor) e outros nutrientes de baixo custo para a produção eficiente de
etanol a partir de glicose por S. cerevisiae. Usando a composição média otimizada (g/L:
CSL 44,3; uréia 2,3; MgSO4.7H2O 3,8; CuSO4.5H2O 0,03), a levedura industrial PE-2
foi capaz de fermentar até 330 g/L de glicose e produzir 18,6 % (v/v) de etanol, com
uma produção descontínua de 2,4 g/L/h e um rendimento em etanol de 93 % (Pereira et
al., 2010).
O planejamento Plackett & Burman juntamente com o Planejamento Composto
Central (CCD) foram aplicados para otimizar o meio de cultura para produção de etanol
por Clostridium autoethanogenum com monóxido de carbono (CO) como fonte única de
carbono e um meio contendo (g/L): 1,0 de NaCl, 0,1 de KH2PO4, 0,02 de CaCl2, 0,15 de
extrato de levedura, 0,116 de MgSO4, 1,694 de NH4Cl e pH 4,74. O rendimento ótimo
de etanol previsto pela metodologia de superfície de resposta (RSM) e um algoritmo

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
48
genético de rede neural artificial (ANN-GA) foram 247,48 e 261,48 mg/L,
respectivamente. Valores semelhantes aos obtidos experimentalmente sob as condições
ideais sugeridas pelos métodos estatísticos (254,26 e 259,64 mg/L). A adequação do
modelo ANN-GA foi maior do que o modelo RSM. Os rendimentos obtidos foram
substancialmente superiores aos relatados anteriormente (60-70 mg/L) com essa cultura
(Guo et al., 2010).
Rigueira (2010) avaliou a influência das variáveis do procedimento de extração
na porcentagem de recuperação das aminas encontradas no leite cru utilizando um
planejamento para “screening” do tipo Plackett & Burman, para os ácidos
tricloroacético (TCA) e sulfosalicílico (ASS).
Quines (2010) estudou a necessidade ou não de se fazer um pré-tratamento da
biomassa antes da extração. Um planejamento experimental Plackett & Burman de doze
ensaios mais três pontos centrais foi utilizado para selecionar as variáveis mais
significativas na extração de Poli(3-hidroxibutirato) (P(3HB)) por 1,2-carbonato de
propileno. Foram realizados dois testes de recuperação do solvente 1,2-carbonato de
propileno, após a extração, sendo um com aplicação de aquecimento e o outro com
aquecimento e vácuo. Os resultados mostraram que o pré-tratamento térmico levou a
uma maior porcentagem de recuperação e de pureza do polímero.

Capítulo 3 Metodologia Experimental

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
50
3. Metodologia Experimental
O presente capítulo apresenta os matérias e métodos utilizados na produção de
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso.
3.1. Quitosana
Neste trabalho utilizou-se uma quitosana (Lote 067) obtida junto a POLYMAR/CE,
com no mínimo 80% de desacetilação. Esse substrato foi utilizado como fonte de carbono e
energia tanto no estudo cinético como nos ensaios do planejamento de seleção de fatores
Plackett & Burman e Planejamento Composto Central, para a produção de quitosanase pelo
Metarhizium anisopliae, por fermentação submersa.
3.2. Microrganismo
O fungo Metarhizium anisopliae estirpe (CG374), utilizado neste estudo, foi cedido
pela Embrapa Recursos Genéticos e Biotecnologia (Brasília / DF-Brasil) e foi mantido em
placas de Petri contendo o meio Batata Dextrose Agar (PDA). As placas foram incubadas em
estufa a 25 ºC. O pH do meio de manutenção foi mantido entre 6,0 e 6,5.
3.2.1. Escolha da cepa produtora de quitosanase
A escolha do microrganismo Metarhizium anisopliae foi feita com base na capacidade
de crescer em meio contendo quitosana como única fonte de carbono, além de apresentar
grande tolerância a pH variando a uma taxa de 2,0 a 8,5, sendo 6,9 a melhor condição para o
crescimento vegetativo e esporução, de acordo com Assis (2010).
3.2.2. Manutenção da cepa
A manutenção da cultura foi realizada em placas petri contendo o meio Batata
Dextrose Agar (PDA). A renovação das células foi realizada a cada trinta dias através de
subcultivos das células no meio PDA.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
51
3.3. Produção de enzima
No preparo do inóculo, utilizou-se 7 placas cultivadas e mantidas, com o fungo
Metarhizium anisopliae, em meio PDA a 25 ºC durante 5 dias em estufa. O meio de cultura
para a produção da enzima quitosanase apresentou os seguintes componentes: quitosana;
peptona; extrato de levedura; NaNO3; K2HPO4; KCl; MgSO4.7H2O e FeSO4, com
concentrações de acordo as matrizes dos planejamentos: Plackett & Burman e composto
central. Para a produção da enzima no cultivo, a biomassa foi transferida através de água
estéril, para um Béquer de 500 mL que, em seguida, foi colocado em Erlenmeyers de 250 mL
na proporção de 20 mL de biomassa para 180 mL do meio de cultura. Os Erlenmeyers
cultivados foram então incubados em incubador rotativo a 25 ºC e 150 rpm durante 24 h.
Alíquotas foram retiradas e centrifugadas a 8000xg por 5 min, a cada 6 horas, para se
determinar a atividade quitosanolítica, teores de açúcares redutores e de proteína total.
Ressalta-se que todos os experimentos foram feitos em triplicata.
3.4. Determinação da Proteína Total
A determinação da proteína total foi efetuada de acordo com o método modificado de
Sedmak & Grossberg (1977), que se baseia na diminuição da absorbância do corante azul de
Coomassie, devido à ligação proteína-corante, a 595 nm, comparado a absorção máxima a
465 nm, conforme Santos (2001). Nesse caso, após centrifugação para retirada da biomassa,
transferia-se para um eppendorf 1,5 mL de amostra (caldo fermentativo) e adicionava-se 1,5
mL de solução azul de Coomassie. Deixava-se a solução em repouso por 5 minutos,
transferindo-se em seguida para um cubeta de quartzo e efetuando-se as leituras de
absorbância nos comprimentos de ondas 465 nm e 595 nm, respectivamente. O
espectrofotômetro (Thermo Spectronic modelo Genesys 10UV) foi zerado com água
destilada. O conteúdo protéico foi obtido a partir de uma curva padrão na qual graficava-se a
relação de absorbância A595/A465 para diferentes concentrações de Albumina de Soro
Bovino (Fração V – Lote 110881/Sigma Aldrich-Ohio-USA).
3.5. Determinação da Atividade Quitosanolítica e da Atividade Específica
A atividade quitosanolítica foi obtida monitorando-se o aumento dos açúcares
redutores totais, utilizando-se o método DNS a partir de uma curva padrão de glicosamina.
Para a determinação da atividade quitosanolítica adicionaram-se, aos tubos de ensaios 500 µL

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
52
de substrato (solução 0,2 % de quitosana solubilizada em ácido clorídrico 1N) e 500 µL de
caldo bruto (contendo enzima), controlando-se o pH entre 6,0 e 6,5 e incubando-se o sistema
em um banho termostatizado com temperaturas da ordem de 55 ºC. Após o tempo de
incubação de 30 minutos no banho foi adicionado aos tubos de ensaio 2,5 mL de DNS,
aquecendo-se à ebulição em banho-maria por 10 minutos, resfriando-se, em seguida, em
banho de gelo. Após essa etapa, o sistema foi centrifugado e com o sobrenadante efetuou-se a
leitura no espectrofotômetro a 600 nm. Destaca-se, que antes da leitura da amostra zerava-se o
equipamento (branco do espectro) com 1,0 mL de água destilada acrescidos de 2,5 mL de
DNS e realizava-se o branco da enzima que consistia da adição de 500 µL de solução de
quitosana, completando-se até um 1,0 mL com água destilada. Uma unidade de atividade
quitosanolítica (A) foi definida como a quantidade de enzima necessária para produzir 1
µmol.min-1 de glicosamina por mL de extrato enzimático, nas condições de temperatura e pH
citadas, sendo o resultado expresso em (U.mL-1). A Atividade específica foi determinada pela
divisão entre a atividade da enzima (U.mL-1) e a concentração de proteína total (µg.mL-1),
sendo expressa em (U. µg-1).
3.6. Planejamento experimental
3.6.1. Planejamento de seleção de fatores Plackett & Burman
O planejamento experimental Plackett & Burman é uma técnica muito usada, para se
realizar triagem de fatores na otimização num determinado processo (Reddy et al., 2008), e
foi utilizado neste trabalho para identificar os fatores que influenciam de forma significativa
na produção da enzima quitosanase.
A Tabela 3 ilustra os níveis de cada variável (fator) utilizada no planejamento Plackett
& Burman, sendo: Concentração de Quitosana (Q), Peptona (PP), Extrato de Levedura (EL),
NaNO3 (NS), K2HPO4 (FP), KCl (CP), MgSO4.7H2O (SM), FeSO4 (SF). Já a Tabela 4 mostra
a matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3 repetições
na condição do ponto central.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
53
Tabela 3 – Níveis para os fatores e seus valores codificados.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1 0,1 0,2 0,2 0,1 0,05 0,025 0,025 0,0005
0 0,2 0,5 0,3 0,2 0,10 0,050 0,050 0,0010
1 0,3 0,8 0,4 0,3 0,15 0,075 0,075 0,0015
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1
2 1 1 -1 1 -1 -1 -1 1 -1 1 1
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1
5 1 1 -1 1 1 -1 1 -1 1 -1 1
6 1 1 1 -1 1 1 -1 1 1 -1 -1
7 -1 1 1 1 -1 1 1 -1 -1 1 1
8 -1 -1 1 1 1 -1 1 1 1 1 1
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1
11 -1 1 -1 -1 -1 1 1 1 1 1 -1
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1
13 0 0 0 0 0 0 0 0 0 0 0
14 0 0 0 0 0 0 0 0 0 0 0
15 0 0 0 0 0 0 0 0 0 0 0
Sendo: V11,V12,V13 = Variáveis inertes para cálculo do erro.
3.6.2. Planejamento Composto Central para a Otimização do Meio de Cultura para a
Produção da Enzima em Cultivo Submerso
Um dos problemas mais comuns para quem realiza experimentos é determinar a
influência de uma ou mais variáveis sobre outra variável de interesse (Lobato, 2003). Este

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
54
problema pode ser minimizado utilizando-se um planejamento experimental de modo a se
obter uma função, ou uma aproximação satisfatória para a mesma, de forma que se operando
sobre as variáveis de entrada produza-se como saída a resposta ou as respostas observadas
(Barros Neto, 2001; Silva, 2004).
Em um planejamento, inicialmente, deve-se definir os fatores e as respostas de
interesse. Tendo-se identificado os fatores e respostas define-se o objetivo que se pretende
alcançar com os experimentos para que se possa escolher o planejamento mais apropriado.
(Silva, 2004).
No presente trabalho foi utilizado um Planejamento Composto Central (CCD) com 22
experimentos na parte fatorial, repetição em triplicata no ponto central e quatro experimentos
na parte axial (planejamento do tipo estrela), para se investigar a influência dos fatores:
concentração de quitosana e concentração de FeSO4 que foram os dois fatores que
influenciaram a produção de quitosanase, conforme o planejamento Plackett & Burman de 12
ensaios e mais 3 repetições na condição do ponto central. Estes dois fatores foram estudados
em cinco diferentes níveis (-1,41; -1; 0; 1; 1,41) e um conjunto de 11 experimentos foi
realizado, conforme Tabela 5.
Tabela 5 – Matriz do planejamento Composto Central.
Ensaio Q SF
1 -1 -1
2 -1 1
3 1 -1
4 1 1
5 -1,41 0
6 1,41 0
7 0 -1,41
8 0 1,41
9 0 0
10 0 0
11 0 0
Os outros fatores foram mantidos nos níveis mais baixos (para evitar custos). Uma que
não influenciaram de forma significativa na produção da enzima quitosanase.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
55
A codificação dos fatores foi realizada de acordo com a seguinte equação (Ji et al.,
2009):
Xi = (xi – x0)/∆xi , i = 1, 2, 3, ..., k (2)
Onde Xi é o fator independente codificado, xi é o fator independente real, x0 é o valor de xi no
ponto central e ∆xi é a variação de valor. O comportamento do sistema foi explicado pela
seguinte equação polinomial de segunda ordem:
Yp = β0 + ∑ βiXi + ∑ βiiXi2 + ∑ βijXiXj, i, j = 1, 2, 3,..., k (3)
Onde Yp foi a resposta esperada, β0 foi a interseção, Xi e Xj
foram os fatores independentes codificados, βi foi o coeficiente linear, βii
foi o coeficiente quadrático e βij foi o coeficiente de interação. Destaca-se que os outros
fatores não significativos foram mantidos no nível mais baixo (-1).
A Tabela 6 apresenta os níveis para os fatores e seus valores codificados, para o
Planejamento Composto Central, com triplicata no ponto central.
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1,41 0,218 0,2 0,2 0,1 0,05 0,025 0,025 0,0009
-1 0,3 0,2 0,2 0,1 0,05 0,025 0,025 0,0015
0 0,5 0,2 0,2 0,1 0,05 0,025 0,025 0,0030
1 0,7 0,2 0,2 0,1 0,05 0,025 0,025 0,0045
1,41 0,782 0,2 0,2 0,1 0,05 0,025 0,025 0,0051
Na análise do planejamento de seleção de fatores Plackett & Burman e do
planejamento experimental fatorial foi utilizado um pacote computacional STATISTICA for
Windows versão 7.0.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
56
3.7. Perfil cinético durante o cultivo
Um perfil cinético na melhor condição de cultivo, conforme planejamento
apresentado, foi realizado em frascos Erlenmyer de 250 mL contendo 200 mL de meio de
cultivo contendo os seguintes componentes, em 1 Litro: 2,0g de quitosana; 5,0g de peptona;
3,0g de extrato de levedura; 2,0g de NaNO3; 1,0g de K2HPO4; 0,5g de KCl; 0,5g de
MgSO4.7H2O; 0,01g de FeSO4, que foram inoculadas com 10% (vol/vol) de inóculo contendo
o fungo Metarhizium anisopliae e cultivadas em incubador rotativo durante 5 dias a 150 rpm
e 25 ºC. Alíquotas foram retiradas a cada seis horas e analisadas quanto à proteína total no
sobredanante, atividade enzimática, atividade específica e açúcares redutores. Os ensaios
foram realizados em triplicata (Silva Filho, 2005).

Capítulo 4 Resultados e
Discussão

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
58
4. Resultados e discussão
Neste capítulo são apresentados e discutidos os resultados obtidos no presente
trabalho. Inicialmente, é apresentada imagem do MEV do Metarhizium anisopliae
mantidos em meio PDA. Em seguida, apresenta-se e discute-se os resultados da avaliação
dos fatores significativos, que podem influenciar na produção da quitosanase, no
planejamento de seleção de fatores, conforme planejamento Plackett & Burman bem como
são apresentados e discutidos os resultados obtidos para o planejamento composto central.
Por último, apresenta-se o perfil cinético, na melhor condição, para a produção de
quitosanase em cultivo submerso descontínuo usando o fungo Metarhizium anisopliae.
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA.
A Figura 7 ilustra a imagem da Microscopia Eletrônica de Varredura (MEV) do micélio de Metarhizium anisopliae mantido em meio PDA.
Figura 7. Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA.
Observa-se na Figura 7, quanto à morfologia que o Metarhizium anisopliae, apresenta-
se como um fungo filamentoso com as hifas intimamente entrelaçadas conforme citado por
Wang et al. 2002.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
59
4.2. Planejamento Plackett & Burman
4.2.1 Análise do Plackett & Burman com 8 fatores e 12 ensaios
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis, foi utilizado para avaliar a influência dos fatores: Concentração de
Quitosana (Q), Peptona (PP), Extrato de Levedura (EL), NaNO3 (NS), K2HPO4 (FP), KCl
(CP), MgSO4.7H2O (SM), FeSO4 (SF) na produção da enzima quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso. Ressalta-se que os experimentos foram
realizados aleatoriamente.
A Tabela 7 apresenta a matriz do modelo Plackett & Burman e os respectivos valores
obtidos para a variável resposta (Atividade Quitosanalítica).
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.
Ensaio Q PP EL NS FP CP SM SF A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 0,0195
2 1 1 -1 1 -1 -1 -1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 0,0109
4 1 -1 1 1 -1 1 -1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 0,0136
6 1 1 1 -1 1 1 -1 1 0,0156
7 -1 1 1 1 -1 1 1 -1 0,0109
8 -1 -1 1 1 1 -1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 0,0085
A Tabela 8 apresenta os efeitos estimados para o planejamento Plackett & Burman.
Observa-se na Tabela 8 que o valor do coeficiente de determinação R2 foi de
aproximadamente 90%, indicando que um modelo linear poderia representar bem a relação
entre o fator significativo, concentração de quitosana (1), e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
60
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman.
Efeitos Estimados; R2=0,8996; Ajuste:0,63178 MS Erro Residual =0,0000054
Desvio Padrão t(3) p
Lim. Conf. Lim. Conf.
Efeito Erro Residual
-95,% +95
Média/Interação 0,0127 0,0007 18,903 0,0003 0,0105 0,0149 (1) Quitosana 0,0055 0,0013 4,0628 0,0268 0,0011 0,0098
(2) Peptona 0,0015 0,0013 1,1080 0,3486 -0,0028 0,0058 (3) Ext. de leved. 0,0009 0,0013 0,7387 0,5136 -0,0033 0,0053
(4) NaNO3 0,0002 0,0013 0,1758 0,8715 -0,0040 0,0045 (5) K2HPO4 -0,0014 0,0013 -1,0552 0,3687 -0,0057 0,0028
(6) KCl -0,0022 0,0013 -1,6708 0,1933 -0,0065 0,0020 (7) MgSO4.7H2O -0,0003 0,0013 -0,2286 0,8338 -0,0046 0,0039
(8) FeSO4 0,0029 0,0013 2,1457 0,1211 -0,0014 0,0071
A Figura 8 mostra o gráfico de Pareto, uma das formas de se avaliar visualmente a
influência dos fatores estudados na resposta. A magnitude dos efeitos é representada pelas
colunas enquanto que a linha transversal às colunas representa a magnitude dos efeitos com
significado estatístico para p=0,05, ou seja, os fatores que são estatisticamente significativos
ao nível de 95% de confiança.
Figura 8. Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
61
Analisando-se o gráfico de Pareto (Figura 8) observa-se que o fator concentração de
quitosana (1) foi o único fator que influenciou de forma mais significativa na atividade
enzimática atingindo efeitos estimados de aproximadamente 4,06. O sinal positivo indica que
o aumento da concentração de quitosana aumentará à atividade enzimática. Por outro lado, os
outros fatores parecem desprezíveis estatisticamente conforme o gráfico de Pareto.
4.2.2 Análise do Plackett & Burman com 12 Ensaios e mais 3 Repetições na condição do
ponto central.
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis com repetição em triplicata no ponto central, foi utilizado para se avaliar a
influência dos mesmos fatores descritos na Seção 4.2.1 para a produção da enzima
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso. Ressalta-se que os
experimentos foram realizados aleatoriamente. A Tabela 9 apresenta a matriz do modelo
Plackett & Burman e os respectivos valores obtidos para a variável resposta (Atividade
Quitosanalítica).
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13 A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1 0,0195
2 1 1 -1 1 -1 -1 -1 1 -1 1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1 0,0109
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 1 -1 1 0,0136
6 1 1 1 -1 1 1 -1 1 1 -1 -1 0,0156
7 -1 1 1 1 -1 1 1 -1 -1 1 1 0,0109
8 -1 -1 1 1 1 -1 1 1 1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 1 1 -1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1 0,0085
13 0 0 0 0 0 0 0 0 0 0 0 0,0704
14 0 0 0 0 0 0 0 0 0 0 0 0,0688
15 0 0 0 0 0 0 0 0 0 0 0 0,0710

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
62
A Tabela 10 a seguir apresenta os efeitos estimados para planejamento Plackett &
Burman com 3 repetições na condição do poto central. Observa-se na Tabela 10 que o valor
do coeficiente de determinação R2 foi de aproximadamente 2,0%, indicando que um modelo
linear não poderia representar a relação entre os fatores significativos concentração de
quitosana (1), concentração de FeSO4 (8) e a resposta.
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central.
Efeitos Estimados; R2=0,01811; Ajuste:0,0 MS Erro Puro =0,0000012
Desvio Padrão t(2) P
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0242 0,0003 85,509 0,0001 0,0229 0,0254 (1) Quitosana 0,0055 0,0006 8,6432 0,0131 0,0027 0,0082
(2) Peptona 0,0015 0,0006 2,3572 0,1425 -0,0012 0,0042 (3) Ext. de leved. 0,0009 0,0006 1,5715 0,2567 -0,0017 0,0037
(4) NaNO3 0,0002 0,0006 0,3742 0,7442 -0,0024 0,0029 (5) K2HPO4 -0,0014 0,0006 -2,2450 0,1538 -0,0041 0,0013
(6) KCl -0,0022 0,0006 -3,5546 0,0708 -0,0049 0,0005 (7) MgSO4.7H2O -0,0003 0,0006 -0,4864 0,6747 -0,0030 0,0024
(8) FeSO4 0,0029 0,0006 4,5648 0,0448 0,0002 0,0056
A Figura 9 ilustra o gráfico de Pareto para o planejamento Plackett & Burman, com
repetição em triplicata no ponto central, uma das formas de se avaliar visualmente a influência
dos fatores estudados na resposta, onde é mostrado a magnitude dos efeitos para 95 % de
confiança.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
63
Figura 9. Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto
central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto, observa-se que os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma mais
significativa na atividade enzimática atingindo efeitos estimados de aproximadamente 8,84 e
4,67, respectivamente. O sinal positivo indica que o aumento das concentrações de quitosana
e de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores parecem
desprezíveis estatisticamente conforme o gráfico de Pareto.
Este comportamento do FeSO4 está de acordo com o observado por Hagag et al.
(2012) que utilizando o Aspergillus fumigatus na produção de protease mostraram que um
fator de transcrição PrtT regula positivamente o controle da produção de protease, da
assimilação de Ferro e da síntese de ergosterol e que a produção de protease foi favorecida
pelo aumento da concentração de ferro.
Rajendran & Thangavelu (2009) utilizaram um planejamento Plackett & Burman para
identificar quais os fatores significativos na produção de lipase por Rhizopus arrhizus MTCC
2233 e observaram o efeito significativo médio do FeSO4, apresentando um nível de
confiança para esse fator de 48,92% (para um máximo de 98,7 % de MgSO4), com uma
produção de lipase da ordem de 2,98 U/mL em 84 horas de fermentação e que ocorreu uma
produção considerável de protease no meio de cultivo da ordem de 1,92 U/mL em 96 horas de
cultivo.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
64
Mu et al. (2005) também observaram um efeito positivo do FeSO4, da ordem de 1,48
para um fator máximo encontrado de 3,11 de MgSO4, no aumento da atividade da
estereoinversão de S-fenil-1,2-etanodiol por Candida parapsilosis CCTCC M203011 na
otimização da fermentação utilizando um planejamento Plackett & Burman.
Dessa forma, observa-se que o FeSO4 é importante uma que vez o mesmo está
diretamente associado a síntese de proteases (principalmente metalloproteases), conforme
citado por Hagag et al. (2012).
4.3. Planejamento Composto Central para Produção de Quitosanase por
Metarhizium anisopliae em Cultivo Descontínuo Submerso
Um planejamento experimentos 22 com repetição em triplicata no ponto central e mais
quatro ensaios axiais, foram utilizados para se chegar a uma combinação ideal de quitosana e
FeSO4 na produção da enzima quitosanase (Tabela 11). A partir do modelo Plackett &
Burman (Tabela 12), que mostrou que o aumento das concentrações de quitosana e FeSO4,
favoreceram a produção de quitosanase, os valores utilizados neste modelo para estes
componentes nos níveis mais altos, serão considerados como os níveis mais baixos no
planejamento experimental do composto central. Os outros fatores foram fixados em todos os
ensaios em seus níveis mais baixos para a redução de custos (Tabela 11).

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
65
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da
atividade enzimática.
Ensaio Q SF A (U/mL)
1 -1 -1 0,0195
2 -1 1 0,0202
3 1 -1 0,0109
4 1 1 0,0121
5 -1,41 0 0,0136
6 1,41 0 0,0156
7 0 -1,41 0,0108
8 0 1,41 0,0103
9 0 0 0,0099
10 0 0 0,0116
11 0 0 0,0094
A Tabela 12 apresenta os efeitos estimados para o planejamento composto central.
Tabela 12 – Efeitos Estimados do planejamento Composto Central.
Efeitos Estimados; R2=0,629; Ajuste:0,25836 MS Erro Puro =0,0000014
Desvio Padrão t(2) p
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0103 0,0006 15,305 0,0042 0,0074 0,0132 (1) Quitosana (L) 0,0011 0,0008 1,4316 0,2885 -0,0023 0,0047
Quitosana (Q) 0,0058 0,0009 5,9637 0,0269 0,0016 0,0101 (2) FeSO4 (L) -0,0043 0,0008 -5,3037 0,0337 -0,0079 -0,0008 (2) FeSO4 (Q) 0,0018 0,0009 1,8366 0,2076 -0,0024 0,0060
1L x 2L 0,0001 0,0011 0,1520 0,8931 -0,0048 0,0052
Observa-se na Tabela 12 que o valor do coeficiente de determinação R2 foi de
aproximadamente 63%, indicando que um modelo linear representaria bem a relação entre os
fatores significativos concentração de quitosana (1), concentração de FeSO4 (8) e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
66
A Tabela 13 mostra um resumo da ANOVA para o planejamento composto central
com repetição em triplicata no ponto central.
Tabela 13 – Resumo da ANOVA.
Fonte de
Variação
Soma dos
Quadrados
Grau de
Liberdade
Quadrado
Médio
Fcalculado Ftabelado
Regressão
Resíduos
0,000086 2 0,000043 6,14 4,46
0,000056 8 0,000007
Falta de
Ajuste
Erro puro
0,000053 6 0,000009 6 19,33
0,000003
2
0,0000015
Total 0,000142 10
De acordo com a Tabela 13, partindo-se dos efeitos significativos propõe-se o
modelo:
� = 0,010316 + 0,002942� � − 0,002193�� (8)
% de variação explicável: 60,5
% máxima de variação explicável: 97,9
A análise da ANOVA mostra que o modelo proposto possui significância estatística, ao
nível de 95% de confiança, uma vez que o valor de F (calculado) é maior que F (tabelado),
entretanto a ANOVA mostra também que o modelo proposto pode ser útil para se fazer
previsões, uma vez que a relação entre F (calculado) e F (tabelado) para a falta de ajuste e o
erro puro é menor que 1. O coeficiente de determinação R2 foi de aproximadamente 63%,
indicando que o modelo consegue explicar 63% da variação total em torno da média.
Os cálculos para variação explicável e para máxima variação explicável são dados pelas
Equações (9) e (10):
% de variação explicável = (SQ da regressão) / (SQ Total) (9)
% máxima de variação explicável = (SQ Total – SQ Erro puro) / (SQ Total) (10)
Sendo:
SQ – Soma dos quadrados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
67
A Figura 10 mostra o gráfico de Pareto para o planejamento Composto Central
utilizando erro puro. A magnitude dos efeitos é representada pelas colunas. Já a linha
transversal às colunas representa a magnitude dos efeitos com significado estatístico para 95%
de confiança.
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto (Figura 10) observa-se que os fatores concentração
de quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma
mais significativa na atividade enzimática atingindo efeitos estimados de aproximadamente
5,96 e - 5,30, respectivamente. O sinal positivo indica que o aumento das concentrações de
quitosana aumentará à atividade enzimática. O sinal negativo indica que o decréscimo da
concentração de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores
parecem desprezíveis estatisticamente no gráfico de Pareto.
A avaliação do modelo também pode ser feita através da observação do gráfico dos
valores preditos em função os valores observados que são mostrados na Figura 11. Os valores
preditos pelo modelo são representados pela reta, enquanto que os valores observados
representam-se pelos pontos.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
68
Figura 11 – Valores preditos em função dos valores observados.
Observa-se que alguns valores preditos apresentam um afastamento dos valores
observados. A Figura 12 ilustra a superfície de resposta da atividade quitosanolítica em
função dos dois fatores significativos, concentração de quitosana e concentração de FeSO4.
Figura 12. Atividade quitosanolítica em função da concentração de quitosana e da
concentração de FeSO4 para os fatores não significativos no ponto central.
Observa-se a existência de um ponto de mínimo, ocorrendo para concentração de
quitosana no ponto central (0) e para maiores concentrações de FeSO4 igual a (1,5). Essa

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
69
região deve ser evitada uma vez que a atividade quitosanolítica atinge níveis tão reduzidos
quanto 0,0094 U.mL-1. Destaca-se que nesse planejamento o maior nível da concentração de
FeSO4 (α=+1,41) equivale a 0,0051% (Tabela 6) que é aproximadamente cinco vezes maior
que o nível desse fator no ponto central do planejamento Plackett & Burman (0,001%),
conforme Tabela 3. Dessa forma, ao se utilizar o nível mínimo da concentração de FeSO4 no
Planejamento Composto Central (α=-1,41), que equivale a aproximadamente 0,001%,
aproxima-se do ponto central do do planejamento Plackett & Burman (0,001%), condição
essa que apresenta os melhores resultados de atividade quitosanolítica usando quitosana como
fonte de carbono e energia e fungo Metarhizium anisopliae.
4.4. Perfil Cinético – Melhor Condição
A seguir, apresenta-se um perfil cinético, na melhor condição, para produção da
quitosanase usando o fungo Metarhizium anisopliae, em que foram monitorados durante 24
horas os seguintes parâmetros: Concentração de proteínas totais (µg/mL), atividade
quitosanolítica (U/mL), atividade específica (U/µg) e açúcares redutores (mg/mL), ilustradas
nas Figuras 13, 14, 15 e 16, respectivamente.
Com relação à proteína extracelular, observa-se na Figura 13 que, inicialmente, ocorre
um aumento da mesma para o seu nível máximo (6,2 µg/mL), durante as 6 primeiras horas de
cultivo, seguido de uma redução para 5,7 µg/mL em 18 horas de cultivo, possivelmente
devido à presença de proteases no meio. Observa-se que a partir de então ocorre uma redução
e aumento da proteína total, em uma quantidade inferior, sugerindo um mecanismo de
controle.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
70
0 4 8 12 16 20 245,0
5,2
5,4
5,6
5,8
6,0
6,2
6,4
Pro
tein
a (
µg
/mL
)
Tempo (h)
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana.
Com relação à atividade quitosanolítica, conforme Figura 14, a atividade máxima
ocorreu em 18 horas de produção em que se obteve aproximadamente 70,0 U/L, o que foi 28
vezes maior que o valor (2,5 U/L em 48 horas) relatado por Assis (2010) para o mesmo fungo,
e em apenas um terço do tempo de produção.
0 4 8 12 16 20 240,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,10
Ativid
ade
(U
/mL
)
Tempo (h)
Figura 14. Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
71
Além disso, destaca-se o considerável aumento da produção de quitosanase a partir das
12 horas até 18 horas de cultivo onde atingiu o pico máximo de produção (70,0 U/L). Após o
pico em 18 horas, a produção começa a cair rapidamente, comportamento esse observado
durante todos os ensaios cinéticos. Destaca-se que é importante do ponto de vista industrial
manter a produção em alto nível por muitas horas, onde se espera uma produção contínua ou
em grandes bateladas.
Ak et al. (1998) utilizaram o fungo Penicillium spinulosum para produzir a quitosanase
usando um meio de cultura contendo 0,1% de K2HPO4; 0,05 % de MgSO4.7H2O; 0,05% de
KCl; 0,001 % de FeSO4, 0,5% NH4NO3 e como fonte de carbono e energia, os autores
usaram quitosana e parede de Rhizopus. A produção foi realizada em cultivo submerso a 28
oC. Os autores apresentam atividade quitosanolítica máxima de aproximadamente 47,0 U/mL,
ao se utilizar a parede do fungo como fonte de carbono e energia, e 30,0 U/mL ao se utilizar
quitosana como fonte de carbono e energia. Destaca-se que essas atividades ocorreram após 4
dias de cultivo. Os perfís com o tempo da atividade quitosanolítica e da atividade específica
apresentaram tendência de crescimento, com o pico máximo em 4 dias, seguindo com uma
tendência de queda da atividade. Neste tempo, a atividade específica máxima da quitosanase
foi de aproximadamente 60,0 U/mg de proteína. Essas tendências também foram observadas
nos resultados apresentados nos perfís de atividade das Figuras 14 e 15.
0 4 8 12 16 20 24
-0,010
-0,005
0,000
0,005
0,010
0,015
0,020
0,025
0,030
Ativid
ad
e E
spe
cific
a (
U/µ
g)
Tempo (h)
Figura 15. Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
72
Observa-se na Figura 15 que a atividade específica máxima da quitosanase segue a
mesma tendência da atividade quitosanolítica apresentada na Figura 14, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
O perfil para os açúcares redutores durante o cultivo de Metarhizium anisopliae
cultivado em meio contendo 0,2% de quitosana é ilustrado na Figura 16.
0 4 8 12 16 20 240,004
0,008
0,012
0,016
0,020
0,024
0,028
Acuca
res R
ed
uto
res (
mg/m
L)
Tempo (h)
Figura 16. Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana.
Observa-se, na Figura 16, que taxa máxima de redução dos açúcares redutores
(aproximadamente 0,021 mg/mL) ocorre logo após a produção máxima da enzima, em 18
horas de cultivo, conforme Figura 14. A quantidade de açúcares redutores foi a
aproximadamente 0,012 mg/mL após 24 h de cultivo. O fungo excreta para o meio enzimas
com atividade quitosanolítica, essas enzimas hidrolisam a quitosana aumentando a quantidade
de açúcares redutores para o meio, sendo que esses açúcares também são consumidos pelo
fungo durante seu crescimento.
Foi demonstrado por Assis (2010) que os açúcares redutores produzidos através da
quitossana, ocasionada pelas enzimas quitossanases metabolizadas por Metarhizium
anisopliae, com quitosana como sua única fonte de carbono e energia, atingiu um pico de 0,53
mg/mL em 48 horas. Apresentaram como quitooligossacarídeos, constituídos principalmente
por dímeros e pentâmeros, formados pela hidrólise enzimática de quitosanase em quantidades
maiores do que o consumido pelos microrganismos. Mostraram, também, que essa produção

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
73
segue a mesma tendência da sua atividade enzimática, corroborando com os resultados
apresentados no presente trabalho.
Os resultados do presente trabalho mostram a relevância de se adotar novas condições
para a produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter quitooligossacarídeo
com atividades biológicas. Esta pesquisa, portanto, sugere base para novos estudos que
permitam alcançar rapidamente a produção de quitosanase e quitooligossacarídeo com todos
os seus benefícios.

Capítulo 5 Conclusão

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
75
5. Conclusão
Após o desenvolvimento desse trabalho conclui-se:
- Com relação ao Planejamento de Seleção de Fatores Plackett & Burman:
Os resultados das análises do Planejamento de Seleção de Fatores Plackett &
Burman com 8 fatores e 12 ensaios em 2 níveis, mais 3 repetições no ponto central, para
se investigar a influência dos fatores: Concentração de Quitosana, Peptona, Extrato de
Levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O, FeSO4 na produção da enzima
quitosanase por Metarhizium anisopliae, mostraram que a atividade foi favorecida pelo
aumento da concentração de substrato (quitosana) e sulfato ferroso (FeSO4), enquanto
que o aumento da concentração dos outros fatores não contribuíram de forma
significativa para a resposta (atividade quitosanolítica). Os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) atingiram efeitos estimados de
aproximadamente 8,84 e 4,67, respectivamente. O sinal positivo indica que o aumento
das concentrações de quitosana e de FeSO4 aumentará a atividade enzimática. Conforme
visto na literatura, observa-se importância do FeSO4, já que o mesmo está diretamente
associado a síntese de proteases (principalmente metalloproteases),
- Com relação ao Planejamento Composto Central:
A análise do planejamento composto central para avaliar a influência dos
fatores, concentração de substrato (quitosana) e concentração de FeSO4, na produção da
enzima quitosanase por Metarhizium anisopliae (a partir do planejamento Plackett &
Burman) mostrou que os fatores concentração de quitosana e concentração de FeSO4
foram os fatores que influenciaram de forma mais significativa na atividade enzimática
atingindo efeitos estimados de aproximadamente 5,96 e - 5,30, respectivamente. O sinal
positivo indica que o aumento das concentrações de quitosana aumentará a atividade
enzimática. O sinal negativo indica que o decréscimo da concentração de FeSO4
aumentará a atividade enzimática. Observa-se a existência de um ponto de mínimo,
ocorrendo para concentração de quitosana (X1) no ponto central (0) e para maiores

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
76
concentrações de FeSO4 (X2) igual a (1,5) com a atividade quitosanolítica de 0,0094
U.mL-1.
- Com relação ao cultivo na melhor condição:
O cultivo na melhor condição mostrou uma produção máxima de quitosanase em
18 horas de cultivo, com a atividade enzimática de 70,0 U/L, o que foi 28 vezes maior
que o valor (2,5 U/L em 48 horas) relatado na literatura (Assis, 2010) para o mesmo
fungo e em apenas um terço do tempo de produção.
Além disso, observou-se que um elevado incremento na produção de
quitosanase a partir das 12 horas de cultivo, antes de se atingir a atividade
quitosanolítica máxima, 18 horas de cultivo. Essa produção em alto nível por muitas
horas é importante do ponto de vista industrial, onde se espera uma produção contínua
ou em grandes bateladas. Observou-se também que a atividade específica máxima da
quitosanase segue a mesma tendência da atividade quitosanolítica, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
- Com relação aos açúcares redutores:
Foi observado que os açúcares redutores seguem a mesma tendência da atividade
enzimática, com uma produção máxima de cerca de 0,21 mg/mL em 18 horas de
cultivo, produção essa que coincinde com o máximo pico de atividade enzimática.
Estes resultados mostram a relevância de se adotar novas condições para a
produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter
quitooligossacarídeo com atividades biológicas. Esta pesquisa, portanto, sugere base
para novos estudos que permitam alcançar rapidamente a produção de quitosanase e
quitooligossacarídeo com todos os seus benefícios.

Referências Bibliográficas

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
78
Referências bibliográficas
AK, Ö.; BAKIR, U.; GÜRAY, T. Isolation of a chitosan degrading fungus, Penicillium
spinulosum, and chitosanase production by the isolate. In: SYMPOSIUM - NEW
FRONTIERS IN SCREENING FOR MICROBIAL BIOCATALYSTS, 1996, Ede, The
Netherlands. Anais, Editado por K. Kieslich, C. P. van der Beek, J. A. M. de Bont e W.
J. J. van den Tweel, Elsevier Science B. V., 1998, v.53, p.265-268.
ALVES, S. B. Patologia e controle microbiano: Vantagens e desvantagens. In: ALVES,
S. B. Controle Microbiano de Insetos. 2ed., Piracicaba, FEALQ, 1163p, 1998, v.4, p.21-
37.
ALVES, S. B.; LEITE, L. G.; FILHO, A. B.; ALMEIDA, F. E. M.; MARQUES, E. J.
Produção massal de fungos entomopatogênicos na América Latina, In: ALVES, S. B.;
LOPES, R. B. Controle microbiano de pragas na América Latina: avanços e desafios.
Piracicaba, FEALQ, 414p, 2008, v.14, p.215-237.
ARRUDA, W.; LUBECK, I.; SCHRANK, A.; VAINSTEIN, M. H. Morphological
alterations of Metarhizium anisopliae during penetration of Boophilus microplus ticks.
Experimental and Applied Acarology, v. 37, p.231-244, 2005.
ASSIS, C. F. Produção e caracterização de quitooligossacarídeos produzidos pelo fungo
Metarhizium anisopliae e avaliação da citotoxicidade em células tumorais. 2010. 110p.
Tese (Doutorado em Biotecnologia) – Renorbio - Programa de Pós-Graduação em
Biotecnologia. Universidade Federal do Rio Grande do Norte, Natal.
ASSIS, C. F.; COSTA, L. S.; MELO-SILVEIRA, R. F.; OLIVEIRA, R. M.;
PAGNONCELLI, M. G. B.; ROCHA, H. A. O.; MACEDO, G. R.; SANTOS, E. S.,
Chitooligosaccharides antagonizes the cytotoxic effect of glucosamine. World J.
Microbiol. Biotechnol., v.28, n.3, p.1097-1105, 2012.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
79
AZEVEDO, V. V. C.; CHAVES, S. A.; BEZERRA, D. C.; FOOK, M. V. L.; COSTA,
A. C. F. M. Quitina e Quitosana: aplicações como biomateriais. Revista Eletrônica de
Materiais e Processos, v.2.3, p.27-34, 2007.
BARROS NETO, B.; SCARMINIO, I. S.; BRUNS, R. Como fazer experimentos:
pesquisa e desenvolvimento na ciência e na indústria. 2. ed. Campinas: Unicamp, 2001.
401p.
BATISTA FILHO, A.; ALMEIDA, J. E. M.; SANTOS, A. S.; MACHADO, L. A.;
ALVES, S. B. Eficiência de isolados de Metarhizium anisopliae no controle de
cigarrinha-da-raiz de cana-de-açúcar Mahanarva fimbriolata (Hom.: Cercopidae). STAB
– Açúcar, Álcool e Subprodutos, v.21, p.84-89, 2002.
CARVALHO, J. C. M.; SATO, S. Fermentação Descontínua, In: SCHMIDELL, W.;
LIMA, U. A.; AQUARONE, E.; BORZANI, W. Biotecnologia Industrial. São Paulo,
Ed. Edgard Blücher, v.2, p.193-204, 2001.
CHAO, H. J.; SCHWARTZ, J.; MILTON, D. K. Populations and determinats of
airborne fungi in large office buildings. Environmental Health Perspectives, v.110, n.8,
p.777-782, 2002.
CHEN, X.; XIA, W.; YU, X. Purification and characterization of two types of
chitosanase from Aspergillus sp. CJ22-326. Food Research International, v.38, n.3,
p.315-322, 2005.
CHEN, P. T.; CHIANG, C.-J.; CHAO, Y.-P. Medium optimization and production of
secreted Renilla luciferase in Bacillus subtilis by fed-batch fermentation. Biochemical
Engineering Journal, v.49, p.395-400, 2010.
CHENG, C. Y.; LI, Y. An Aspergillus chitosanase with potential for large-ecale
preparation of chitosan oligosaccharides. Biotechnol. Appl. Biochem., v.32, p.197-203,
2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
80
CHOI, Y. J.; Kim, E. J.; Piao, Z.; Yun, Y. C.; Shin, Y. C. Purification and
characterization of chitosanase from Bacillus sp. Strain KCTC 0377BP and its
application for the production of chitosan oligosaccharides. Applied and Environmental
Microbiology, v.70, n.8, p.4522-4531, 2004.
COLAKOGLU, G. The chemical and physical environment for fungal growth. Fak
Matbaa Birimi, Istanbul, 149-166, 2001.
CRAVEIRO, A. C.; CRAVEIRO, A. A.; QUEIROZ, D. C. Quitosana a fibra do futuro.
2. ed. Fortaleza: PADETEC, 2004. V.1, 218p.
DIAS, B. A. Estudo de proteases degradadoras de cutícula produzidas pelo fungo
entomopatogênico Beauveria bassiana. 2005. 70p. Dissertação (Mestrado em
Microbiologia) – Centro de Ciências Biológicas, Programa de Pós-Graduação em
Microbiologia, Universidade Estadual de Londrina, Londrina.
DINARDO-MIRANDA, L. L.; FERREIRA, J. M. G.; CARVALHO, P. A. M.
Influência da época de colheita e do genótipo de cana-de-açúcar sobre a infestação de
Mahanarva fimbriolata (Stal) (Hemiptera: Cercopidae). Neotrop. Entomol, v.30, n.1,
p.145-149, 200.
DONG, C.; ZHANG, J.; HUANG, H.; CHEN, H.; HU, Y. Pathogenicity of a new China
variety of Metarhizium anisopliae (M. anisopliae var. Dcjhyium) to subterranean termite
Odontotermes formosanus. Microbiological Research, v.164, p.27-35, 2009.
DONZELLI, B. G. G.; OSTROFF, G.; HARMAN, G. E. Enhanced enzymatic
hydrolysis of langostino shell chitin with mixtures of enzymes from bacterial and fungal
sources. Carbohydrate Research, v.338, p.1823-1833, 2003.
DRIVER, F.; MILNER, R. J.; TRUEMAN, J. W. H. A taxonomic revision of
Metarhizium based on phylogenetic analysis of rDNA sequence data. Mycological
Research, v.104, n.2, p.134-150, 2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
81
DUNG, P.; MILAS, M.; RINAUDO, M.; DESBRIÉRES, J. Water soluble derivatives
obtained by controlled chemical modifications of chitosan, Carbohydrate Polymers,
v.24, p.209-214, 1994.
ESTEVES, J. A.; CABRITA, J. D.; NOBRE, G. N. Micologia médica, 2. ed. Lisboa:
Fundação Calouste Gulbenkian, 1990. 1058p.
FACCIOTI, M. C. R. Fermentação Contínua, In: Biotecnologia Industrial. Editado por:
SCHMIDELL, W.; LIMA, U. A.; AQUARONE, E.; BORZANI, W., São Paulo, Ed.
Edgard Blücher, v.2, p.541, 2001.
FARIA, M. R.; MAGALHÃES, B. P. O uso de fungos entomopatogênicos no Brasil -
Situação atual e perspectivas. Biotecnologia Ciência & Desenvolvimento, n.22., p.18-
21, setembro/outubro 2001.
FU, J.-Y.; WU, S.-M.; CHANG, C.-T.; SUNG, H.-Y. Characterization of Three
Chitosanase Isozymes Isolated from a Commercial Crude Porcine Pepsin Preparation. J.
Agric. Food Chem., v.51, p.1042-1048, 2003.
FURIGO JUNIOR, A.; PEREIRA, E. P. Enzimas e suas aplicações cinética enzimática.
2001. 39f. Apostila (Graduação de Engenharia Química e Engenharia de Alimentos) –
Centro de Tecnologia, Departamento de Engenharia Química e Engenharia de
Alimentos, Universidade Federal de Santa Catarina, Florianópolis.
GAO, J.; XU, H.; LI, Q.-J.; FENG, X.-H.; LI, S. Optimization of medium for one-step
fermentation of inulin extract from Jerusalem artichoke tubers using Paenibacillus
polymyxa ZJ-9 to produce R,R-2,3-butanediol, Bioresource Technology, v.101, p.7076-
7082, 2010.
GARCIA, M. V.; MONTEIROI, A. C.; SZABO, M. J. P.; PRETTE, N.; BECHARA, G.
H. Mechanism of infection and colonization of Rhipicephalus sanguineus eggs by
Metarhizium anisopliae as revealed by scanning electron microscopy and
histopathology. Brazilian Journal of Microbiology, São Paulo, v.36, n.4, p.368-372,
2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
82
GHANEM, N. B.; YUSEF, H. H.; MAHROUSE, H. K. Production of Aspergillus
terreus xylanase in solid-state cultures: application of the Plackett & Burman
experimental design to evaluate nutritional requirements, Bioresource Technology,
v.73, p.113-121, 2000.
GHORBEL-BELLAJ, O.; HMIDET, N.; JELLOULI, K.; YOUNES, I.; MAALEJ, H.;
HACHICHA, R.; NASRI, M. Shrimp waste fermentation with Pseudomonas aeruginosa
A2: Optimization of chitin extraction conditions through Plackett–Burman and response
surface methodology approaches, International Journal of Biological Macromolecules,
v.48, p.596–602, 2010.
GIUSTINA, A.; VENTURA, P. Weight-reducing regimens in obese subjects: Effects of
a new dietary fiber integrator. Acta Toxicol Ther, 16, 4, p.199-214, 1995.
GUIA.BIO, Reino dos fungos, 2012. Disponível em:
<http://www.http://guia.bio.br/reino-dos-fungos>. Acesso em: 12 novembro 2012.
GUIRADO, N.; AMBROSANO, E. J.; ROSSI, F.; DIAS, F. L. F. Controle de Cupins
de Montículo com Metarhizium anisopliae e Beauveria bassiana, Rev. Bras. de
Agroecologia, v.4, n.2, 2009.
GUO, Y.; XU, J.; ZHANG, Y.; XU, H.; YUAN, Z.; LI, D. Medium optimization for
ethanol production with Clostridium autoethanogenum with carbon monoxide as sole
carbon source, Bioresource Technology, v.101, p.8784–8789, 2010.
HAGAG, S.; KUBITSCHEK-BARREIRA, P.; NEVES, G. W. P.; AMAR, D.;
NIERMAN, W.; SHALIT, I.; SHAMIR, R.; LOPES-BEZERRA, L.; OSHEROV, N.
Transcriptional and proteomic analysis of the Aspergillus fumigatus ∆prtT protease-
deficient mutant. PLoS One, v.7, n.4, e33604, 2012.
HAN, L.-K.; KIMURA, Y.; OKUDA, H. Reduction in fat storage during chitin-chitosan
treatment in mice fed a high-fat diet, International Journal of Obesity, 23:174-179,1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
83
HUNG, T.; CHANG, Y.; SUNG, H.; CHANG, C. Purification and Characterization of
Hydrolase with Chitinase and Chitosanase Activity from Commercial Stem Bromelain.
J. Agric. Food Chem., 50, 4666-4673, 2002.
ICHIKAWA, S.; TAKANO, K.; TAKASHIKUROIWA, HIRUTA, O.; SATO, S.;
MUKATAKA, A. S. Immobilization and stabilization of chitosanase by multipoint
attachment to agar gel support. Journal of Bioscience and Bioengineering, v.93, n.2,
p.201-206, 2002.
JABOR, I. A. S.; PAMPHILE, J. A.; RODRIGUES, S. B.; MARQUES-SILVA, G. G.;
ROCHA, C. L. M. S. Análise do desenvolvimento do fungo entomopatogênico
Metarhizium anisopliae em resposta a fatores nutricionais. Acta Scientiarum.
Agronomy, Maringá, v.25, n.2, p.497-501, 2003.
JI, X.-J.; HUANG, H.; DU, J.; ZHU, J.-G.; REN, L.-J.; LI, S.; NIE, Z.-K. Development
of an industrial medium for economical 2,3-butanediol production through co-
fermentation of glucose and xylose by Klebsiella oxytoca, Bioresource Technology,
v.100, p.5214–5218, 2009.
JO, Y.; JO, K.; JIN, Y.; KIM, K.; SHIN, J.; KIM, Y.; PARK, R. Characterization and
kinetics of 45 kDa chitosanase from bacillus sp. P16. Biosci. Biotechnol. Biochem.,
v.67, n.9, 1875 – 1882, 2003.
KHOSRAVI-DARANI, K.; ZOGHI, A.; ALAVI, S. A.; FATEMI, S. S. A. Application
of Plackett Burman Design for Citric Acid Production from Pretreated and Untreated
Wheat Straw, Iran. J. Chem. Chem. Eng.. v.27, n.1, p.91-104, 2008.
KIM, P.; KANG, T. H.; CHUNG, K. J.; KIM, I. S.; CHUNG, K. Purification of a
constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and
expression of the gene. FEMS Microbiolog.y Letters 240, 31–39, 2004.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
84
KIM, S.-K.; RAJAPAKSE, N. Enzymatic production and biological activities of
chitosan oligosaccharides (COS): A review. Carbohydrate Polymers, v.62, n.4, p.357-
368, 2005.
KIRK, P. M.; CANON, P.F.; MINTER, D.W.; STALPERS, J.A. Dictionary of the
fungi. CAB International, Wallingford, 2008.
KONDO, H.; OSADA, A. Influence of dietary fiber on the bioavailability of zinc in
rats, Biomed Environ Sci, 9, 204-8, 1996.
KUMAR, S.; KOH, J. Physiochemical, Optical and Biological Activity of Chitosan-
Chromone Derivative for Biomedical Applications. Int. J. Mol. Sci., v.13, p.6102-6116,
2012.
KURITA, K.; SANNAN, T.; IWAKURA, Y. Studies on chitin, 4. Evidence for
formation of block and random copolymers of N-acetyl-o-glucosamine and
oglucosamine by hetero and homogeneous hydrolyses. Die Makromolecular Chemie,
v.178, p.3197-3202, 1977.
KUROIWA, T.; ICHIKAWA, S.; HIRUTA, O.; SATO, S.; MUKATAKA, S. Factors
Affecting the Composition of Oligosaccharides Produced in Chitosan Hydrolysis Using
Immobilized Chitosanases. Biotechnol. Prog., 18, 969-974, 2002.
LI, L.-Z.; HUI, Z.; MO, X. Optimization of the medium for streptolydigin production
by Streptomyces lydicus AS 4.2501 with precursor feeding using response surface
methodology, Journal of the Taiwan Institute of Chemical Engineers, 41, 252–258,
2010.
LIU, B.-L.; KAO, P.-M.; TZENG, Y.-M.; FENG, K.-C. Production of chitinase from
Verticillium lecanii F091 using submerged fermentation. Enzyme and Microbial
Technology, v.33, p.410-415, 2003.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
85
LOBATO, A. K. C. L. Estudo da produção de biosurfactantes por micro-organismos
isolados de poços de petróleo. 2003. 108p. Dissertação (Mestrado) – Programa de Pós-
Graduação em Engenharia Química. Universidade Federal do Rio Grande do Norte.
Natal.
LOPES, R. B.; ALVES, S. B.; PADULLA, L. F. L.; PÉREZ, C. A. Eficiência de
formulações de Beauveria bassiana e Metarhizium anisopliae para o controle de ninfas
de Amblyomma cajennense (Fabricius, 1787). Revista Brasileira de Parasitologia
Veterinária, v.16, n.1, p.27-31, 2007.
LOUREIRO, E. S.; BATISTA FILHO, A.; ALMEIDA, J. E. M.; MENDES, J. M.;
PESSOA, L. G. A. Eficiência de isolados de Metarhizium anisopliae (metsch.) sorok.
no controle da cigarrinha-da-raiz da cana-de-açúcar, Mahanarva fimbriolata (stal, 1854)
(hemiptera: cercopidae), em condições de campo. Arq. Inst. Biol., São Paulo, v.79, n.1,
p. 47-53, 2012.
MELLO, D. R.; REIS, R. C. S.; BITTENCOURT, V. R. E. P. Patogenicidade in vitro
do fungo Metarhizium anisopliae (Metschnikoff, 1879) Sorokin, 1883, sobre o carrapato
Boophilus microplus (Canestrini, 1887). Revista Brasileira de Parasitologia Veterinária,
São Paulo, v.15, p.157-162, 2006.
MITSUTOMI, M.; KIDOH, H.; TOMITA, H.; WATANABE, T. The action of Bacillus
circulans WL-12 chitinases on partially N-acetylated chitosan, Bioscience
Biotechnology and Biochemistry, v.59, p.529-531, 1995.
MALAJOVICH, M. A. Biotecnologia: fundamentos. Ed. biblioteca Max Feffer do
Instituto de Tecnologia ORT, Rio de Janeiro, 2009. 104p.
MONAGHAN, R. L.; EVELEIGH, D. E.; TEWARI, R. P.; REESE, E. T. Chitosanase,
a novel enzyme. Nature - New Biology, 245, p.78-80, 1973.
MU, X. Q.; XU, Y.; NIE, Y.; OUYANG, J.; SUN, Z. H. Candida parapsilosis CCTCC
M203011 and the optimization of fermentation medium for stereoinversion of (S)-1-
phenyl-1,2-ethanediol, Process Biochemistry, v.40, p.2345–2350, 2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
86
MURRAY, P. R.; ROSENTHAL, K. S.; PFALLER, M. A. Medical microbiology. 5th
ed. London, Elsevier Mosby, 2005.
MUZZARELLI, R. A. A. Biochemical significance of exogenous chitins and chitosans
in animals and patients, Carbohydrate Polymers, v.20, p.7-16, 1993.
MUZZARELLI, R. A. A.; TOMASETTI, M.; ILARI, P. Depolymerization of chitosan
with the aid of papain, Enzyme Microbiology and Technology, v.16, p.110-114, 1994.
NAVEENA, B. J.; ALTAF, Md.; BHADRIAH, K.; REDDY, G. Selection of medium
components by Plackett–Burman design for production of L(+) lactic acid by
Lactobacillus amylophilus GV6 in SSF using wheat bran, Bioresource Technology,
v.96, p.485-490, 2005.
NOGAWA, M.; TAKAHASHI, H.; KASHIWAGI, A.; OHSHIMA, K.; OKADA, H.;
MORIKAWA, Y. Purification and Characterization of Exo--D-Glucosaminidase from
a Cellulolytic Fungus, Trichoderma reesei PC-3-7, Appl. Environ. Microbiol., v.64, n.3,
p.890-895, 1998.
ONOFRE, S. B.; VARGAS; L. R. B.; ROSSATO, M.; BARROS, N. M.; BOLDO, J.
T.; NUNES, A. R. F.; AZEVEDO, J. L. Controle biológico de pragas na agropecuária
por meio de fungos entomopatogênicos. In: SERAFINI, L. A.; BARROS, N. M.;
AZEVEDO, J. L. (Org.). Biotecnologia – avanços na agricultura e na agroindústria.
Caxias do Sul: EDUCS, p.295-328, cap.8, 2002.
OSSWALD, W. F.; SHAPIRO, J. P.; DOOSTDAR, H.; MCDONALD, R. E.; NIEDZ,
R. P.; NAIRN, C. J.; HEARN, C. J.; MAYER, R. T. Identification and Characterization
of Acidic Hydrolases with Chitinase and Chitosanase Activities from Sweet Orange
Callus Tissue, Plant Cell Physiol. 35, p.811-820, 1994.
PAGNONCELLI, M. G. B. Estudo do mecanismo de produção de oligossacarídeos com
atividades nutracêuticas a partir da quitosana por hidrólise enzimática com processo

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
87
fermentativo simultâneo. 2008. 118p. Tese (Doutorado) – Centro de Tecnologia,
Programa de Pós-Graduação em Engenharia Química. Universidade Federal do Rio
Grande do Norte. Natal.
PALMA-GUERRERO, J., JANSSON, H.-B., SALINAS, J.; LOPEZ-LLORCA, L. V.
Effect of chitosan on hyphal growth and spore germination of plant pathogenic and
biocontrol fungi, J. Appl. Microbiol., v.104, p.541-553, 2008.
PAULA, F. A.; ARRUDA, D. C. Análise do método de planejamento experimental
Plackett Burman na otimização de degradação de penas de aves por comunidades
microbianas, XXI Salão de Iniciação Científica, 19-23, outubro, 2009, UFRGS, Porto
Alegre.
PELCZAR, M. J.; CHANG, E. C. S.; KRIEG, N. R. Microbiologia: conceitos e
aplicações, v.2, 2.ed. São Paulo: Makron BooKs, 1996.
PELLETIER, A.; SYGUSCH, J. Purification and characterization of three chitosanase
activities from Bacillus megaterium P1, Applied and Environmental Microbiology, v.
56, n.4, p.844-848, 1990.
PEREIRA, F. B.; GUIMARÃES, P. M. R.; TEIXEIRA, J. A.; DOMINGUES, L.
Optimization of low-cost medium for very high gravity ethanol fermentations by
Saccharomyces cerevisiae using statistical experimental designs, Bioresource
Technology, v.101, p.7856-7863, 2010.
PETER, M. G. Chitin and chitosan from aninal sources. In: STEINBUCHEL, A.;
RHEE, S. K. Polysaccharides and Polyamides in the Food Industry. Weinheim:
Wileyvch, v.1, p.115-208, 2005.
PIZA, F. A. T.; SILOTO, A. P.; CARVALHO, C. V.; FRANCO, T. T. Purification,
characterization and purification of chitosanase from Bacillus cereus, Brazilian Journal
of Chemical Engineering., v.16, n 2, p.185-192, 1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
88
POLITZER, K.; BON, E. P. S. Enzimas Industriais e Especiais. C.G.E.E. Ciência,
Tecnologia e Inovação. Rio de Janeiro, 554p, 2006.
POLYMAR, Ciência é a nossa Natureza, 2003. Disponível em:
<http://www.polymar.com.br>. Acesso em: 6 setembro 2012.
PUTZKE, J.; PUTZKE, M. T. L. Os reinos dos fungos. Santa Cruz do Sul, RS:
EDUNISC, 1998.
QUINES, L. K. M. Extração de poli(3-hidroxibutirato) produzido por Cupriavidus
necator dsm 545 com 1,2-carbonato de propileno. 2010. 99p. Dissertação (Mestrado em
Engenharia de Alimentos) – Centro Tecnológico, Departamento de Engenharia Química
e Engenharia de Alimentos, Programa de Pós-Graduação em Engenharia de Alimentos,
Universidade Federal de Santa Catarina, Florianópolis.
RAJENDRAN, A.; THANGAVELU, V. Statistical experimental design for evaluation
of medium components for lipase production by Rhizopus arrhizus MTCC 2233, LWT -
Food Science and Technology, v.42, p.985–992, 2009.
RAMIREZ-LEÓN, L. F.; RUIZ-HERRERA, J. Hydrolysis of walls and formation of
spheroplasts in Mucor rouxii. J. Gen. Microbiol., v.72, p.281-290, 1972.
RANGEL, M. A. S.; LOUREIRO, E. S.; ALMEIDA, A. S.; ALVES, H. C.; ONO, F.
B.; LOBO, P. M.; MARIANI, M. A.; PESSOA, L. G. A.; SILVA, A. J. Avaliação
participativa do controle de cigarrinhas das pastagens com a aplicação do fungo
Metarhizium anisopliae (Metsch.) Sorok. Rev. Bras. de Agroecologia, v.2, n.2, 731-734,
2007.
RAVEN, P. H.; EVERT, R.F.; EICHHORN, S.E. Biologia Vegetal, 6ª Ed, Rio Janeiro,
Editora Guanabara Koogan, 2001.
REDDY, L. V. A.; WEE, Y.-J.; YUN, J.-S.; RYU, H.-W. Optimization of alkaline
protease production by batch culture of Bacillus sp. RKY3 through Plackett–Burman

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
89
and response surface methodological approaches, Bioresource Technology, v.99, p.242–
2249, 2008.
RIGUEIRA, J. C. S. Influência da contagem de células somáticas no perfil e teores de
aminas bioativas e na qualidade de leite cru e de queijo mussarela. 2010. 147p. Tese
(Doutorado em Ciência de Alimentos) - Faculdade de Farmácia, Departamento de
Alimentos, Programa de Pós-Graduação em Ciência de Alimentos, Faculdade de
Farmácia da Universidade Federal de Minas Gerais, Belo Horizonte.
RODRIGUES, M. I.; IEMMA, A. F. Planejamento de Experimentos e Otimização de
Processos: uma estratégia sequencial de planejamentos, Campinas, SP, Casa do Pão
Editora, 2005.
SANTOS, E. S. Recuperação e Purificação de Enzimas usando Adsorção em Leito
Expandido, Tese de Doutorado, Universidade Estadual de Campinas, Campinas/SP,
p.152, 2001.
SEDMAK, J. J.; GROSSBERG, S. E. A rapid, sensitive and versatile assay for protein
using coomassie brilliant blue G250, Analytical Biochemistry, v.79, p. 544-552, 1977.
SEINO, H.; TSUKUDA, K.; SHIMASUE, Y. Properties and action pattern of a
chitosanase from Bacillus sp. PI- 7S. Agricultural Biological Chemistry, 55, p.2421-
2423, 1991.
SHIMOSAKA, M.; NOGAWA, M.; OHNO, Y.; OKAZAKI, M. Chitosanase from the
plant pathogenic fungus, Fusarium solani f. sp. Phaseoli – purification and some
properties, Bioscience Biotechnolgy and Biochemistry, v.57, n.2, p.231-235, 1993.
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Production of two chitosanases from a chitosan-assimilating bacterium, Acinetobacter
sp. Strain CHB101, Applied and Environmental Microbiology, v.61, n.2, p.438-442,
1995.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
90
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Characterization of Three Chitosanase Isozymes Isolated from a Commercial Crude
Porcine Pepsin Preparation, J. Agric. Food Chem. 51, p.1042-1048, 2003.
SILVA FILHO, R. C.; COSTA, A. S. S.; CHAGAS, B. M. E.; SANTOS, F. N.;
MACEDO, G. R.; SANTOS, E. S. Avaliação do teste de atividade quitosanolítica
produzida por fungo. In: XV SIMPÓSIO NACIONAL DE BIOPROCESSOS, 15,
Recife, 2005, 02 a 05 de agosto.
SILVA, E. P. Avaliação Preliminar do Potencial de Aplicação da Tecnologia de
Biopilhas para a Biorremediação do Solo de Guamaré-RN. 2004. 119p. Dissertação
(Mestrado) – Centro de Tecnologia, Programa de Pós-Graduação em Engenharia
Química. Universidade Federal do Rio Grande do Norte. Natal.
SOMASHEKAR, D.; JOSEPH, R. Chitosanases – Properties and apllications: A review.
Bioresource Technology, v.55, p.35-45, 1996.
STOYACHENKO, I. A.; VARLAMOV, V. P.; DAVANKOV, V. A. Chitinases of
Streptomyces kurssanovii: purification and some properties, Carbohydrate Polymers,
v.24, p.47-54, 1994.
TINLINE, R. D. Nuclear distribuition in Metarhizium anisopliae. Mycologia, v. 63, p.
713-721, 1971.
TRABULSI, L. R.; TOLEDO, M. R. F. Microbiologia. 2. Ed., São Paulo: Atheneu,
1996.
VIEIRA, P. D. S.; SILVA, W. M. T.; PAIVA, L. M.; LIMA, E. A. L. A.;
CAVALCANTI, V. L. B. Estudo da caracterização morfológica, esporulação e
germinação dos conídiosde Metarhizium anisopliae var. acridum em diferentes
temperaturas. Biológico, São Paulo, v.69, n.1, p.17-21, 2007.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
91
WANDERLEY, M. D.; NEVES, E.; ANDRADE, C. J. Aspectos da produção industrial
de enzimas. Revista Citino, v.1, n.1, p.30-36, 2011.
WANG, C.; TYPAS, M. A.; BUTT, T. M. Detection and characterization of pr1
virulent gene deficiencies in the insect pathogenic fungus Metarhizium anisopliae.
FEMS Microbiology Letters, v. 213, p. 251-255, 2002.
ZAHORSKA-MARKIEWICZ, B.; KROTKIEWSKI, M.; OLSZANECKA-
GLINIANOWICZ, M.; ZURAKOWSKI, A. Effect of chitosan in complex management
of obesity, Pol Merkur Lekarski, v.13, n.74, p.129-132, 2002.
ZHANG, X. Y.; DAI, A. L.; ZHANG, X. K.; KUROIWA, K.; KODAIRA, R.;
SHIMOSAKA, M.; OKAZAKI, M. Purification and Characterization of and Exo--D-
glucosaminidase from a koji mold, Aspergillus oryzae IAM2660. Biosci. Biotechnol.
Biochem., v.64, n.9, 1896-1902, 2000.
ZHU, X.; WU, X.; DAI, Y. Fermentation conditions of a chitosanase from Acinetobcter
sp C-17. Biosci. Biotechnol. Biochem., v.67, n.2, p.284-290, 2003.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
92
APÊNDICE

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
93
APÊNDICE I
Metodologia para repique da cepa
Em Erlenmeyer de 1000 mL colocava-se 500 mL do meio de manutenção
PDA. Este Erlenmeyer, com o meio de manutenção, era autoclavado junto com placas
de Petri a 1,5 kgf/cm2 por 15 minutos, em seguida, era levado para uma câmara de fluxo
laminar e transferido para as placas de Petri já autoclavadas, onde eram mantidas numa
estufa a 27 ºC durante 24 h. Após este período, as placas de Petri eram levadas a câmara
de fluxo laminar, utilizando bico de Bunsen para esterilizar a alça de platina. Procedia-
se o repique utilizando uma placa de Petri contendo a cepa pura. Cada placa de Petri
contendo a cepa pura era transferida para duas placas de Petri contendo o meio de
manutenção. As placas eram mantidas na estufa a 27 ºC durante 30 dias onde após esse
período, repetia-se este procedimento.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
94
APÊNDICE II
Montagem da Curva de Calibração Usando o Método DNS
Soluções:
Glicosamina = 0,01 mol/mL (10000,0 µmol/mL)
Tabela 14. Dados para construção da curva de calibração usando o Método DNS.
Tubo de
Ensaio
Solução de
glicosamina (mL)
Água Destilada
(mL)
DNS (mL)
Volume Total
Massa (µµµµmol)
1 0 1,0 1,5 2,5 0
2 0,2 0,8 1,5 2,5 0,2
3 0,4 0,6 1,5 2,5 0,4
4 0,6 0,4 1,5 2,5 0,6
5 0,8 0,2 1,5 2,5 0,8
6 1,0 0 1,5 2,5 1,0
Tabela 15 - Medidas de absorbância a 600nm
Dados adicionais:
Peso Molecular D-Glicosamina = 215,64 (recipiente SIGMA)
Conc. Sol. D-Glicosamina = 0,05 g/50 mL
Massa calc.(µmol) = conc. Sol. D-Glic. x Vol. Utiliz. D-Glics. =
= 0,05 x 1000 x Vol.util./50*215,64
Tempo atividade enzimática = 30 min
1 U/mL = 1 µmol/(min.mL)
Vol. Total (Glicosamina+caldo) = 1 mL
Medições Adsorbância a 600 nm Vol. Utiliz. Massa Massa Atividade
II III Média(Abs)(mL) calc. (mg) calc. (µg) Enzim. (U/mL)
0 0 0 0 0 0 0
0,133 0,128 0,1307 0,2 0,2 200 0,0309
0,279 0,277 0,2787 0,4 0,4 400 0,0618
0,433 0,428 0,4297 0,6 0,6 600 0,0927
0,587 0,583 0,5797 0,8 0,8 800 0,1237
0,727 0,716 0,7157 1 1 1000 0,1546

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
95
Figura 17 – Curva padrão de glicosamina.
y = 1900ralx + 1900ral
R² = 1900ral
00,000
00,020
00,040
00,060
00,080
00,100
00,120
00,140
00,160
00,180
00,000 00,100 00,200 00,300 00,400 00,500 00,600 00,700 00,800
Ati
v.
En
zim
áti
ca (
U/m
L)
Absorbância (Abs) a 600 nm
Curva-Padrão D-Glicosamina

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
96
APÊNDICE III
Método de Determinação de Proteína Utilizando o Corante Comassie
Brilhante Blue G 250 (Método modificado de Sedmak e Grossberg)
Micrométodo
Preparar uma solução 0,06% de Comassie Blue G250 em 1,5 % HCl (w/V). Filtrar e
papel de filtro Whatman nº1.
Curva Padrão (1 a 20 µµµµg)
a) Preparar soluções estoque de BSA de 10 a 200 µg/mL.
b) Colocar em tubos de ensaio 100 µL da solução estoque, 1400 µL de água e
1500µL do Comassie e agitar.
c) Preparar um tubo contendo 1500 µL de água e 1500 µL do corante que
corresponde ao zero de proteína.
d) Após 5 minutos (padronizar o tempo) fazer as leituras das absorbâncias.
e) As leituras das amostras devem ser feitas em dois comprimentos de onda 595 e
465 contra a água.
f) Plotar a razão das absorbâncias a 595/465 como uma função da massa de BSA e
calcular o melhor ajuste linear dos dados.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
97
Micrométodo (1 a 20 µµµµg de BSA)
Solução Estoque de BSA = 0,2 mg/mL
Tabela 16. Dados para construção da curva padrão de BSA.
Tubo de
Ensaio
H2O (µµµµL)
BSA (µµµµg)
Coomassie (µµµµL)
BSA (µµµµg)
1 1500 0 1500 0
2 1490 10 1500 2
3 1480 20 1500 4
4 1470 30 1500 6
5 1460 40 1500 8
6 1450 50 1500 10
7 1440 60 1500 12
8 1430 70 1500 14
9 1420 80 1500 16
10 1410 90 1500 18
11 1400 100 1500 20
Dados adicionais:
Concentração Sol. Estoque
BSA = 0,2 mg/mL = 0,2 µg/µL
Vol. total sol. = vol. H2O + vol. BSA + vol.
Coomassie = 3000 µL = 3 mL
Concentração Sol. BSA (mg/mL) = (massa BSA mg) / 3 mL
.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
98
Figura 18 – Curva Padrão de BSA
y = 1900ralx - 1900ral
R² = 1900ral
000,000
001,000
002,000
003,000
004,000
005,000
006,000
007,000
000,400 000,500 000,600 000,700 000,800 000,900 001,000 001,100
µg
BS
A
Razão Absorbâncias (595nm/465nm)
Curva-Padrão BSA

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE TECNOLOGIA
DEPARTAMENTO DE ENGENHARIA QUÍMICA PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA
TESE DE DOUTORADO
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em
Cultivo Descontínuo Submerso
Raimundo Cosme da Silva Filho
Natal/RN
Abril/2013

Raimundo Cosme da Silva Filho
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo
Descontínuo Submerso
Tese apresentada ao Programa de Pós-Graduação em Engenharia Química – PPGEQ, da Universidade Federal do Rio Grande do Norte, como parte dos requisitos para obtenção do título de Doutor em Engenharia Química, sob a orientação do Prof. Dr. Everaldo Silvino dos Santos e a coorientação da Profª. Drª. Gorete Ribeiro de Macedo.
Natal/RN
Abril/2013

Catalogação da Publicação na Fonte. UFRN / CT / PPGEQ
Biblioteca Setorial “Professor Horácio Nícolas Sólimo”.
Silva Filho, Raimundo Cosme da. Otimização do meio de cultura para a produção de quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso / Raimundo Cosme da Silva Filho. - Natal, 2013. 98 f.: il.
Orientador: Everaldo Silvino dos Santos. Co-orientador: Gorete Ribeiro de Macedo. Tese (Doutorado) - Universidade Federal do Rio Grande do Norte.
Centro de Tecnologia. Departamento de Engenharia Química. Programa de Pós-Graduação em Engenharia Química.
1. Quitosana - Tese. 2. Quitina - Tese. 3. Quitosanase – Tese. 4. Metarhizium
anisopliae - Tese. I. Santos, Everaldo Silvino dos. II. Macedo, Gorete Ribeiro de. III. Universidade Federal do Rio Grande do Norte. IV. Título. RN/UF/BSEQ CDU 547.458 (043.2)

SILVA FILHO, Raimundo Cosme da – Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo Descontínuo Submerso. Tese de Doutorado. UFRN - Programa de Pós-Graduação em Engenharia Química. Área de Concentração: Pesquisa e Desenvolvimento de Tecnologias Regionais – Subárea: Engenharia de Processos – Alimentos e Biotecnologia, Natal/RN, Brasil.
Orientação: Prof. Dr. Everaldo Silvino dos Santos Coorientação: Profª. Drª. Gorete Ribeiro de Macedo ___________________________________________________________________________
RESUMO: No presente trabalho utilizou-se um Planejamento Plackett & Burman, com 8 fatores e 12 ensaios em 2 níveis e mais 3 repetições na condição do ponto central, para se investigar a influência das concentrações de quitosana, peptona, extrato de levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O e FeSO4 na produção da enzima quitosanase por Metarhizium
anisopliae. Os ensaios para produção da enzima foram realizados em cultivo descontínuo submerso. Os resultados para o Planejamento Plackett & Burman mostraram que a atividade quitosanolítica foi favorecida pelo aumento da concentração de substrato (quitosana) e de sulfato ferroso (FeSO4), enquanto que o aumento da concentração dos outros fatores não contribuiu de forma significativa para a atividade quitosanolítica. A otimização do meio de cultura para a produção da enzima foi realizado por meio de um Planejamento Composto Central, com os dois fatores que mais influenciaram a atividade quitosanolítica (quitosana e FeSO4), conforme Planejamento Plackett & Burman, mantendo-se os outros nutrientes em seus valores mínimos. Nesse planejamento, para o fator FeSO4, tomou-se o limite inferior (-1) como sendo o limite superior do Planejamento Plackett & Burman. Os resultados mostraram que a produção da enzima foi favorecida pelo aumento da concentração quitosana e pela diminuição da concentração de FeSO4. A produção máxima de atividade quitosanolítica foi da ordem de 70,0 U/L e foi atingida em apenas 18 h de fermentação, resultado esse vinte e oito vezes maior aos obtidos anteriormente para o mesmo microrganismo que foi 2,5 U/L em 48h.
Palavras-chave: quitosana; quitosanase; Metarhizium anisopliae; Plackett & Burman.


ABSTRACT
In this work a Plackett-Burman Design with 8 factors and 12 trials in 2 levels with 3 repetitions at the center point was used in order to investigate the influence of the concentration of chitosan, peptone, yeast extract, NaNO3, K2HPO4, KCl, MgSO4.7H2O and FeSO4 on chitosanase production by Metarhizium anisopliae. Runs were carried out using submerged discontinuous cultivation for enzyme production. The results of the Plackett & Burman Design showed that only two factors, chitosan concentration as well as FeSO4 had influence on chitosanolytic activity, while the increase in concentration of other factors not contributed significantly to the quitosanolítica activity. Cultivation medium optimization for enzyme production was carried out using a Composite Central Design, with the most important factors for chitosanolytic activity (chitosan and FeSO4), in accordance with Plackett & Burman Design, and keeping the other nutrients in their minimum values. On this other design, it was taken the highest limit in Plackett & Burman Design as the lowest limit (-1) to FeSO4 factor. The results showed that the enzyme production was favoured by increasing the chitosan concentration and by decreasing FeSO4. Maximum production for chitosanolytic activity was about 70.0 U/L and was reached in only 18 h of fermentation, a result about twenty-eight times greater than a former study using the same microorganism (about 2.5 U/L at 48 h).
Key-words: Chitosan; Chitosanase; Metarhizium anisopliae; Plackett & Burman

Ao meu pai, minha mãe e minha
família, em especial aos meus
irmãos e sobrinhos

AGRADECIMENTOS
A Deus, por estar presente em todos os momentos da minha vida e por me ceder à
oportunidade de iniciar e concluir este trabalho.
A minha mãe Cândida Inocência da Silva que esteve comigo em todos os momentos da
minha vida me educando, me amando e sempre me dando muito carinho.
Ao meu pai Raimundo Cosme da Silva, que não está mais aqui para ver este trabalho,
mas que foi a pessoa que mais me apoiou e me incentivou, em todos os momentos da minha vida,
para que eu pudesse chegar o mais longe possível na minha carreira acadêmica. Pai muito
obrigado por toda atenção, educação, amor e carinho que recebi durante toda a minha vida.
Aos meus irmãos e familiares que sempre estiveram comigo dividindo momentos de
grande alegria e às vezes de tristeza, mas sempre nos mantendo unidos para superarmos todos os
obstáculos.
A minha namorada Rochelly Caroliny por todo amor e carinho que me deu e que me
entendeu nos momentos que estive ausente para a conclusão deste trabalho.
Ao meu orientador Professor Doutor Everaldo Silvino dos Santos por confiar em mim e
me aceitar como orientando além de toda a amizade, orientação e dedicação em todas as etapas de
desenvolvimento e conclusão deste trabalho.
À Professora Doutora Gorete Ribeiro de Macedo, “pessoa que admiro muito como
profissional e principalmente, como pessoa, um exemplo a ser seguido”, pelos conselhos e críticas
quando necessários ao desenvolvimento deste trabalho. Professora a senhora é considerada uma
mãe para de todos os alunos que tiveram a honra de conhecê-la e trabalhar com a senhora.
Aos meus amigos Engenheiros Químicos, Ana Carmen, Ana Katerine, Andréa Farias,
Anita Lima, Sirtys Lessa, Alexandre, Sérgio, Márcio Bezerra e ao Biólogo Daniel Souza, que
sempre me ouviram e que me ajudaram de alguma forma nesse trabalho.
A Eusamar Coelho de Lima, Mazinha, secretária do PPGEQ, pelo apoio, pela paciência e
orientação nas questões burocráticas do processo.
A Carlos e Jardelson, pela grande colaboração e dedicação nos experimentos do
laboratório.
A Sérgio Santana pela grande colaboração e dedicação, o qual não poupou esforços nem
tempo em me ajudar na realização dos experimentos em laboratório.

Ao meu amigo Ubiratan Nazaré por me ajudar na organização do texto e na tradução para
o Abstract.
Finalizando, agradeço ao Laboratório de Engenharia Bioquímica da UFRN por ter cedido
toda a sua estrutura para realização dos experimentos e ao Programa de Pós-Graduação em
Engenharia Química, assim como a todos os professores do Departamento de Engenharia Química
da UFRN que trabalham em pró de uma educação digna e ensino de qualidade para todos os
alunos.

Sumário LISTA DE FIGURAS
LISTA DE TABELAS
NOMENCLATURA
INTRODUÇÃO...............................................................................................................................15
REVISÃO BIBLIOGRÁFICA........................................................................................................18
2.1. Enzimas................................................................................................................................18
2.1.1. Quitosana e seus derivados........................................................................................19
2.1.2. Quitosanases...............................................................................................................24
2.2. Fungos..................................................................................................................................29
2.2.1. Características gerais..................................................................................................29
2.2.2. Reprodução dos Fungos.............................................................................................32
2.2.3. Metabolismo de fungos..............................................................................................33
2.2.4. Nutrição de fungos.....................................................................................................34
2.2.5. O Fungo Metarhizium anisopliae ..............................................................................36
2.3. Fermentação.........................................................................................................................40
2.3.1. Preparo do inóculo......................................................................................................41
2.3.2. Produção de enzimas em cultivo submerso descontínuo...........................................42
2.4. Planejamento experimental..................................................................................................42
2.4.1. Planejamento fatorial..................................................................................................43
2.4.2. Planejamento Plackett & Burman..............................................................................43
2.4.3. Aplicações do Planejamento Plackett & Burman.......................................................44
METODOLOGIA EXPERIMENTAL.............................................................................................50
3.1. Quitosana..............................................................................................................................50
3.2. Microrganismo.....................................................................................................................50
3.2.1. Escolha da cepa produtora de quitosanase.................................................................50

3.2.2. Manutenção da cepa...................................................................................................50
3.3. Produção de enzima.............................................................................................................51
3.4. Determinação da Proteína Total...........................................................................................51
3.5. Determinação da Atividade Quitosanolítica e Atividade Específica...................................51
3.6. Planejamento Experimental.................................................................................................52
3.6.1. Planejamento de seleção de fatores Plackett Burman................................................52
3.6.2. Planejamento Composto Central para Otimização do meio de cultura para a
Produção da Enzima em Cultivo Submerso.....................................................................................53
3.7. Perfil Cinético durante o Cultivo.........................................................................................56
RESULTADOS E DISCUSSÃO.....................................................................................................58
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA ..........................................58
4.2. Planejamento Plackett & Burman........................................................................................59
4.2.1. Análise do Plackett & Burman com 8 fatores e 12 ensaios .......................................59
4.2.2. Análise do Plackett & Burman com 12 ensaios e mais 3 repetições na condição do
ponto central ....................................................................................................................................61
4.3. Planejamento Composto Central para Produção de Quitosanase por Metarhizium
anisopliae em Cultivo Descontínuo Submerso................................................................................64
4.4. Perfil Cinético – Melhor Condição......................................................................................69
CONCLUSÃO.................................................................................................................................75
REFERÊNCIAS BIBLIOGRÁFICAS.............................................................................................78
APÊNDICE I...................................................................................................................................93
Metodologia para Repique da Cepa ................................................................................................93
APÊNDICE II.................................................................................................................................94
Montagem da Curva de Calibração Usando o Método DNS..........................................................94
APÊNDICE III................................................................................................................................96
Método de Determinação de Proteína Utilizando o Corante Comassie Brilhante Blue G 250
(Método modificado de Sedmak e Grossberg)................................................................................96

Lista de Figuras
Figura 1 – Estrutura da quitosana..................................................................................................20
Figura 2 – Estrutura da glicosamina..............................................................................................23
Figura 3 – Estrutura do sulfato de glicosamina.............................................................................23
Figura 4 – Representação da estrutura geral de uma célula de fungo..........................................30
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas...........................................................31
Figura 6 – Conídios de Aspergillus e Penicillium.........................................................................33
Figura 7 – Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA ...................58
Figura 8 – Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados............................................................................................................................60
Figura 9 – Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto central
mostrando a influência dos fatores estudados................................................................................63
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos
fatores estudados............................................................................................................................67
Figura 11 – Valores preditos em função dos valores observados..................................................68
Figura 12 – Atividade quitosanolítica em função da concentração de quitosana e da concentração
de FeSO4 para os fatores não significativos no ponto central .......................................................68
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana ........70
Figura 14 – Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana....................................70
Figura 15 – Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de
quitosana........................................................................................................................................71
Figura 16 – Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana..........................................................................................................72
Figura 17 – Curva padrão de glicosamina ....................................................................................95
Figura 18 – Curva Padrão de BSA ................................................................................................98

Lista de Tabelas
Tabela 1 – Comparação das enzimas com os catalisadores químicos..............................................19
Tabela 2 – Fontes naturais de quitina e quitosana ...........................................................................20
Tabela 3 – Níveis para os fatores e seus valores codificados...........................................................53
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central .........................................................................................53
Tabela 5 – Matriz do planejamento Composto Central...................................................................54
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central......................................................................................................55
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.......................59
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman...............................................60
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.......................61
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central...............................................................................................................................................62
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da atividade
enzimática........................................................................................................................................65
Tabela 12 – Efeitos Estimados do planejamento Composto Central...............................................65
Tabela 13 – Resumo da ANOVA....................................................................................................66
Tabela 14 – Dados para construção da curva de calibração usando o Método DNS ......................94
Tabela 15 – Medidas de absorbância a 600 nm para construção da curva de calibração................94
Tabela 16 – Dados para construção da curva padrão de BSA ........................................................97

NOMENCLATURA
A – Atividade quitosanolítica (U/L)
AE – Atividade específica (U/mg)
ANN-GA – algoritmo genético de rede neural artificial
ANOVA – Análise de variância (Analysis of Variance)
BSA – Albumina de soro bovino (Bovine Sereum Albumin)
CCD – Planejamento Composto Central
CDA – Chitosan Dector Agar
Cs – Concentração de substrato (%)
CP – Concentração de KCl (%)
Da – Dalton
DEQ – Departamento de Engenharia Química
DNS – Ácido Dinitrosalissílico
E – Enzima
Ea – Energia de ativação
EC – Comissão para nomenclatura e classificação de enzimas
EL – Concentração de Extrato de Levedura (%)
FP – Concentração de K2HPO4 (%)
NS – Concentração de NaNO3 (%)
GDA – Grânulos dispersíveis em água
k – Número de fatores, variáveis
LEB – Laboratório de Engenharia Bioquímica
mRNA – RNA mensageiro
n – Número de níveis de um determinado fator
N – Número de ensaios de um planejamento
P – Pressão
PP – Concentração de Peptona (%)
PDA – Batata Dextrose Agar
pH – Potencial hidrogeniônico
Q – Concentração de Quitosana (%)
Qm – Quadrado médio
r – Agitação (rpm)
R2 – Coeficiente de determinação

RA – Razão de aeração
rpm – Rotações por minuto
RSM – Metodologia de superfície de resposta
S – Molécula de substrato
SF – Concentração de FeSO4 (%)
SM – Concentração de MgSO4.7H2O (%)
SQ – Soma de quadrados
SSF – Fermentação de estado sólido
T – Temperatura
TMC – Temperatura de cultivo
V11,V12,V13 – Variáveis inertes para cálculo do erro
VHG – Fermentação de alta densidade
WP – Pó molhável
X – Concentração celular
Xi – variável independente
Xj – variável independente
xi – Fator independente real
x0 – Valor de xi no ponto central
∆xi – Variação de valor
Y – Resposta do modelo de primeira ordem
Yp – Resposta do modelo polinomial de segunda ordem
β0 – Interseção dos modelos de primeira ou segunda ordem
βi – Coeficiente linear dos modelos de primeira ou segunda ordem
βii – Coeficiente quadrático
βij – Coeficiente de interação

Capítulo 1 Introdução

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
15
1. Introdução As quitosanases foram descobertas simultaneamente há quase quarenta anos por
Monaghan et al. (1973), como pelo grupo de Ramirez-León & Ruiz-Herrera (1972), e
são enzimas obtidas a partir do metabolismo de bactérias, fungos e extraídas de plantas.
Desde então, vários estudos vêm apresentando a obtenção de quitosanase deste
microrganismos, na forma intracelular ou extracelular (Osswald et al., 1994;
Somashekar & Joseph, 1996), como a do presente estudo. A quitosanase (EC 3.2.1.132)
representa uma classe de enzimas hidrolíticas que catalisam a hidrólise de ligação β-
(1→ 4) glicosídica da quitosana, gerando oligômeros chamados quito-oligossacarídeos
(Reddy et al., 2008).
A quitosana é um biopolímero hidrofílico obtido a partir da quitina, um
polissacarídeo abundante na natureza. Na literatura, é considerado quitosana o produto
da desacetilação da quitina para um grau de desacetilação superior a 75% e solúveis em
ácidos como o acético e o fórmico. A quitosana é conhecida por exibir uma ampla
variedade de atividade fisiológica como atividade antitumoral, mas o peso molecular e a
insolubilidade em água da quitosana são desvantagens para várias aplicações.
Atualmente, há interesse nas quitosanas quando as mesmas são convertidas em quito-
oligossacarídeos porque eles não apenas são solúveis em água e possuem baixo peso
molecular, mas também têm atividades biológicas bem notáveis, como atividade
anticancerígena atóxica e biocompatível, incluindo efeito antimetastático, atividade anti-
HIV, hepatoprotetora, antioxidante, hipocolesterolêmica, antimicrobiana,
imunoestimulate, antitumoral, anti-inflamatória e atividade aceleradora da absorção de
cálcio e ferro. No entanto, estes quito-oligossacarídeos são obtidos tradicionalmente na
indústria por reação de hidrólise usando ácidos, que apresenta muitos problemas como
dificuldade em controlar a hidrólise, produzindo uma grande quantidade de
oligossacarídeos de cadeia pequena e oligossacarídeos de baixo rendimento, alto custo
de separação e, também, poluição ambiental, uma vez que utiliza um ácido como
catalisador. Como alternativa, com suas vantagens em termos de compatibilidade
ambiental, baixo custo e reprodutibilidade, a obtenção destes quito-oligossacarídeos por
hidrólise com quitosanase se tornou cada vez mais popular nos últimos anos (Azevedo
et al., 2007).

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
16
Devido a todos estes aspectos positivos é que, em geral, as enzimas vêm sendo
estudadas em diferentes aplicações, mostrando que esta é uma tendência na indústria.
No presente trabalho, foi desenvolvido um processo fermentativo visando a
otimização do meio de cultura para a produção da enzima quitosanase em cultivo
descontínuo submerso, por Metarhizium anisopliae, capaz de sintetizar oligômeros com
aplicações na indústria farmacêutica e de química fina, partindo de quitosana como
única fonte de carbono.
Nesse caso, avaliou-se, através de um planejamento Plackett & Burman (método
de delineamento de seleção de fatores) e de um planejamento Composto Central as
melhores condições para a produção da enzima. A técnica do planejamento
experimental Plackett & Burman para realizar triagem de fatores na otimização de um
determinado processo foi utilizada neste trabalho para identificar quais os fatores que
influenciam de forma significativa na produção da enzima quitosanase. A influência
destes fatores (concentração de quitosana e concentração de FeSO4) foi então
investigada pelo planejamento Composto Central com 22 experimentos na parte fatorial,
repetição em triplicata no ponto central e quatro experimentos na parte axial. A
repetição do ponto central permite também a determinação do erro puro e da
repetibilidade do processo, sendo uma forma de identificar melhor se o processo está
sob controle.
Do ponto de vista industrial é de relevante importância a realização de estudos
para a otimização da produção da quitosanase de forma contínua ou em grandes
bateladas.

Capítulo 2 Revisão Bibliográfica

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
18
2. Revisão Bibliográfica 2.1. Enzimas
As enzimas são catalisadores biológicos constituídos por longas cadeias de
aminoácidos que atuam em substratos reduzindo a energia de ativação, acelerando assim
a velocidade da reação, e transformando-os em produtos. Sua atuação catalítica visa
facilitar a atividade das células através da quebra de moléculas ou junção das mesmas
para formação de novos produtos. Podem apresentar alto grau de especificidade relativa
a apenas um substrato ou a vários substratos ao mesmo tempo, como a certos tipos de
ligações glicosídicas α-1,4 das moléculas de amido, ou para determinado tipo particular
de isômero óptico como ocorre na oxidação da β-D-glicose pela glicose oxidase (Furigo
Junior & Pereira, 2001).
Enzimas são proteínas que apresentam em sua estrutura um centro ativo
(apoenzima) e, em alguns casos, um grupo prostético denominado cofator (coenzima ou
íon metálico ativo) e sua atividade depende de alguns fatores como estrutura da
proteína, número de cadeias peptídicas, arranjo dessas cadeias na molécula, natureza do
substrato e natureza do grupo prostético (Politzer & Bon, 2006).
Todos os organismos vivos produzem, em pequena ou grande quantidade, certa
variedade de enzimas, sejam elas intra ou extracelulares. As enzimas extracelulares são
excretadas para o meio externo e podem decompor compostos insolúveis como
celulose, quitina e quitosana.
Algumas destas enzimas podem ser usadas tanto nas indústrias de detergentes,
têxteis e farmacêuticas, como também de alimentos, de bebidas, etc., sendo utilizadas
em processos biotecnológicos industriais em substituição aos processos químicos que
muitas vezes causam grandes prejuízos ao meio ambiente. A produção comercial das
enzimas pode ser realizada através do cultivo de microrganismos como fungos e
bactérias.
Na Tabela 1 mostra-se uma comparação das enzimas com os catalisadores
químicos, levando-se em conta algumas das características dos dois catalisadores.
Destaca-se que alguns parâmetros da Tabela 1 demonstram a maior importância das
enzimas, quando comparadas com os catalisadores químicos, principalmente a
especificidade ao substrato, as condições de reação, o consumo de energia e a

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
19
velocidade de reação, como desvantagem cita-se o custo de obtenção (isolamento e
purificação).
Tabela 1 – Comparação das enzimas com os catalisadores químicos (Adaptado de
Furigo Junior & Pereira, 2001).
Característica Enzimas Catalisadores Químicos
Especificidade ao substrato Alta Baixa
Natureza da estrutura Complexa Simples
Sensibilidade à T e pH Alta Baixa
Condições de reação (T, P e pH) Suaves Drástica (geralmente)
Custo de obtenção (isolamento e purificação) Alto Moderado
Natureza do processo Batelada Contínuo
Consumo de energia Baixo Alto
Formação de subprodutos Baixa Alto
Separação catalisador/produtos Difícil/cara Simples
Atividade catalítica (temperatura ambiente) Alta Baixa
Presença de cofatores Sim Não
Estabilidade do preparado Baixa Alta
Energia de Ativação Baixa Alta
Velocidade de reação Alta Baixa
2.1.1. Quitosana e seus derivados
A quitosana e seus derivados apresentam grande importância devido ao
conhecimento de suas funcionalidades biológicas, médicas, tecnológicas, etc
(Shimosaka et al, 1993; Shimosaka et al, 1995; Mitsutomi et al., 1995). A quitosana é
obtida da quitina, polissacarídeo abundante na natureza, extraído principalmente das
carapaças de caranguejos, lagosta e camarões. A quitosana é um polímero natural,
biodegradável, não tóxico e de alta massa molecular obtido a partir de animais
marinhos, insetos e microrganismos (Tabela 2).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
20
Tabela 2 – Fontes naturais de quitina e quitosana. (Adaptado de Polymar, 2003).
Animais
Marinhos
Insetos Aracnídeos Microrganismos
Anelídios Formigas Escorpiões Leveduras
Moluscos Besouros Aranhas Fungos
Lagosta Esporos
Camarão
Caranguejo
Krill
A Figura 1 abaixo mostra a estrutura química da quitosana
Figura 1 – Estrutura da quitosana (Fonte: Polymar, 2003)
Quitosana totalmente desacetilada não existe na natureza. Graus de acetilação
(GA) nferior a 50% a torna mais solúvel em soluções ácidas (Muzzarelli, 1993;
Muzzarelli et al., 1994; Dung et al., 1994; Stoyachenko et al.,1994). Pelletier &
Sygusch (1990) enfatizaram que grande parte dos resíduos sólidos da indústria
pesqueira, principalmente da carcinicultura, é jogada ao mar ocasionando degradação
ambiental. Logo, a utilização desses resíduos como matéria-prima para formação de
novos produtos, não só reduz de forma drástica o impacto ambiental, mas também daria
maior valor agregado à indústria pesqueira.
A demanda cada vez maior de consumidores por alimentos mais saudáveis, sem
conservantes químicos, tem motivado pesquisas à descoberta de novos agentes
antimicrobianos naturais. Sendo assim, a atividade antimicrobiana incomum da

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
21
quitosana e seus derivados contra microrganismos, tais como bactérias, bolores e
leveduras tem tido maior atenção ultimamente.
Devido à interação entre a quitosana, que é positivamente carregada, e as
membranas celulares dos microrganismos, negativamente carregadas, ocorre a formação
de verdadeiros poros na membrana, que são canais de vazamento de conteúdo protéico e
outros componentes intracelulares. A quitosana tem também a propriedade de agir como
agente quelante que se liga de forma seletiva a traços de metais, inibindo, assim, a
produção de toxinas e o crescimento microbiano. Ainda é capaz de ativar mecanismos
de defesa em tecidos hospedeiros, atuando como agente sequestrante de água e inibindo
várias enzimas. A ligação da quitosana com DNA e a inibição da síntese do mRNA
ocorre via penetração da quitosana no núcleo dos microrganismos e interferência na
síntese do mRNA e proteínas, (Craveiro et al., 2004).
Em um estudo clínico conduzido por Giustina & Ventura (1995) com 100
pessoas com sobrepeso de 10 a 25%, ao final de 4 semanas as pessoas que receberam
quitosana demonstraram perda de peso significativa (7,3 Kg contra 3,0 Kg nos grupos
controle). As pessoas que tomaram quitosana também demonstraram redução na pressão
sanguínea sistólica (de 145 para 135 mm Hg) e diastólica (92.6 para 84.2 mm Hg),
assim como uma redução na taxa respiratória (27.6/min a 21.2/min).
Kondo & Osada (1996) pesquisaram a influência da quitina e da quitosana na
biodisponibilidade do zinco em ratos alimentados com uma dieta com suplementos de
5% ou sem fibra dietética (quitina, quitosana, celulose, pectina e agar-agar) durante 31
dias. Observaram que os ratos alimentados com dietas contendo 5% de fibras, exceto
quitosana, o consumo de alimento foi maior do que o controle. O ganho de peso em
todos os grupos alimentados com fibra foi maior do que o controle, exceto para o grupo
suplementado com quitosana (5%) onde a ingestão de alimento e o ganho de peso foram
menores do que os grupos controle e das demais fibras.
Segundo Craveiro et al. (2004), derivados da quitosana obtidos pela reação com
ácidos carboxílicos, podem promover ao polímero uma solubilidade especial em água.
Esses compostos possuem estrutura química semelhante ao ácido hialurônico e
fornecem uma excelente compatibilidade com o tecido epitelial. As soluções obtidas
com esses derivados podem ser facilmente incorporadas a formulações cosméticas,
mostrando melhor compatibilidade com surfactantes iônicos e emulsificantes,
apresentando estabilidade em uma larga faixa de pH.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
22
Han et al. (1999) examinaram os efeitos da quitosana sobre a atividade da lipase
pancreática in vitro e sobre o grau de armazenamento de gorduras induzido em ratos
pela administração oral de uma dieta rica em gorduras por nove semanas. Como
resultado, observou-se que os animais alimentados com quitosana, reduziram o peso
corporal, a hiperlipidemia e a gordura hepática. Como conclusão, os autores propõem
que os efeitos anti-obesidade da quitosana em ratos alimentados com alto teor de
gordura se devem parcialmente a inibição da absorção intestinal das gorduras dietéticas.
Em outro ensaio, Zahorska-Markiewicz et al. (2002) realizaram um estudo para
avaliar o efeito da quitosana no tratamento da obesidade. Segundo os autores do estudo,
a quitosana pode ser usada como um valioso e seguro coadjuvante em tratamentos para
a obesidade em longo prazo. A quitosana parece acentuar a redução da pressão arterial,
juntamente com a perda de peso.
Assis (2010) estudou a produção e caracterização de quitooligossacarídeos
produzidos pelo fungo Metarhizium anisopliae e avaliação da citotoxicidade em células
tumorais. A hidrólise da quitosana foi realizada em tempos de 10 a 60 min para a
produção de quitooligossacarídeos. A avaliação da citotoxicidade dos oligômeros de
quitosana foi realizada em células tumorais (HepG2 e HeLa) e não tumoral (3T3). A
produção de oligômeros de quitosana teve maiores rendimentos durante 10 minutos de
hidrólise, os pentâmeros apresentaram concentração de 0,15 mg/mL, no entanto os
hexâmeros, que apresentam maior interesse por causa de suas propriedades biológicas,
só foram detectados com 30 minutos de hidrólise com uma concentração de 0,004
mg/mL.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o planejamento Plackett-Burmam e a metodologia de superfície de resposta.
Os resultados mostraram que a eficiência máxima de extração da quitina foi conseguida
nas seguintes condições: concentração de casca de camarão 50g/L, glicose 50g/L, tempo
de incubação de 5 dias e inoculo como 0,05 OD (densidade óptica).
A glicosamina, unidade monomérica básica da quitosana, possui atividade
antiinflamatória e tem grande importância na construção e manutenção da cartilagem,
no tratamento de enfermidades como: febre reumática, artrose e principalmente a artrite
e osteoartrite na forma aguda ou crônica (Polymar, 2003). A Figura 2 apresenta a
estrutura química da glicosamina.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
23
Figura 2 – Estrutura da glicosamina (Fonte: Polymar, 2003)
O sulfato de glicosamina é um derivado da quitosana sendo utilizado no
tratamento da osteoartrite sintomática dos joelhos, quadris, coluna, mãos e outras
regiões (Craveiro et al., 2004). O sulfato de glicosamina estimula a síntese de
proteoglicanos necessários para o bom funcionamento das articulações, estimula a
regeneração das cartilagens e ainda incorpora enxofre ao tecido cartilaginoso. Estudos
de longa duração, acima de 3 anos, mostram que o sulfatos de glicosamina evita a
progressão de danos estruturais nas articulações e induz melhora significativa nas dores
e funções físicas (Craveiro et al., 2004). A Figura 3 apresenta a estrutura química do
sulfato de glicosamina.
Figura 3 – Estrutura do sulfato de glicosamina (Fonte: Polymar, 2003)

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
24
2.1.2. Quitosanases
Dependendo do tipo de quitosanase obtida, ocorrem diferentes mecanismos de
hidrólise da quitosana, isso devido a diferentes microrganismos produzirem enzimas
com diferenças nas suas estruturas (Kurita et al., 1977; Seino et al., 1991).
Quitosanases (EC 3.2.1.132) são enzimas que catalisam a reação de hidrólise de
quitosana em oligossacarídeos. As enzimas quitosanases são produzidas por vários
microrganismos, bactérias, actinomicetos e fungos, e até por algumas plantas. Estas
enzimas agem em polímeros com grau de acetilação de 30% a 60%. Os monômeros
resultantes da hidrólise da quitosana por estas enzimas são melhores digeridos, ao serem
ingeridos, como é o caso da glicosamina. Estudos efetuados até o presente momento
classificam as quitosanases em cinco famílias: 5, 8, 46, 75, 80, sendo que a família 46 é
a mais estudada. As quitosanases tem grande aplicação na geração de oligômeros de
quitosana (Somashekar & Joseph, 1996; Chen et al., 2005).
Nogawa et al. (1998) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida pelo fungo Trichoderma reseei PC-3-7. Estes pesquisadores
inocularam 106 conídios em 100 mL de um meio basal contendo 0,3% de N-
acetilglicosamina em incubator rotativo com agitação de 220 rpm a 28oC. Destaca-se
que o fungo não produziu quitosanase em meio contendo quitosana ou quitina.
Entretanto, o microrganismo produziu quitosanase em meio contendo glicosamina e N-
acetilglicosamina, sendo que para este último obteve-se uma atividade específica
máxima de 1,5 U/mg após 72 horas de incubação. É importante ressaltar que uma
agitação vigorosa favoreceu a produção da enzima. Esse artigo ilustra a presença de
exo-β-D-glicosaminidase e exo-β-D-N-acetilglicosaminidase.
Piza et al. (1999) estudaram a produção, purificação e caracterização parcial de
uma quitosanase produzida por Bacillus cereus. Nesse caso, investigou-se através de um
planejamento fatorial fracionário a influência dos fatores: concentração de sulfato de
amônio (0,4; 2,2 e 4,0%), razão de aeração (2; 6 e 7), tempo de fermentação (16; 24 e
32 horas), pH (5; 6 e 6,5) e concentração de quitosana (1,0; 1,5 e 2,0%). Os resultados
mostraram que os fatores mais significativos para esse planejamento foram a
concentração de sulfato de amônio, a aeração, o pH e a interação entre sulfato de
amônio e a aeração.
Cheng & Li (2000) estudaram a preparação de oligossacarídeos utilizando
quitosanase produzida por Aspergillus sp e constataram que a enzima foi largamente

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
25
induzida em meio contendo quitosana como única fonte de carbono e também na
presença de quitina coloidal. A indução da enzima foi realizada em temperaturas
variando na faixa de 28 a 30ºC, pois essa espécie não cresceu bem em temperaturas
superiores a 37ºC.
Zhang et al. (2000) estudaram a produção, purificação e caracterização de
quitosanase e Exo-β-D-Glucosaminidase por Aspergillus oryzae IAM2660. Os
microrganismos utilizados neste estudo foram mantidos em placas de Petri contendo
agar para contagem em placas (PDA). Destaca-se que para produzir a enzima, conídios
foram retirados das placas e inoculados em meio Czapeq-dox com os seguintes
componentes: 1,0g K2HPO4, 0,5g KCl, 2,0g NaNO3, 0,5g MgSO4.7H2O, 0,02g
FeSO4.7H2O, 20g de sacarose e 1,0L de água deionizada, com pH do meio igual a 5,5.
Foi adicionado ao meio 0,5 g de peptona e 0,5g de extrato de levedura com
concentração de esporos da ordem de 1,0x106 esporos.mL-1. Após a incubação, feita em
incubador rotativo a 26ºC por um período de 60h, micélios foram coletados e após
filtração e secagem a 80ºC foram utilizados para ensaio de quitosanase.
Kuroiwa et al. (2002) estudaram os fatores que afetam a composição de
oligossacarídeos produzidos através da hidrólise de quitosana por quitosanase originária
de Bacillus pumilus BN-262. Neste experimento foi utilizado quitosana 100%
desacetilada dissolvida em ácido acético 1M. O pH do meio foi ajustado em 5,6. Neste
estudo a quitosanase foi imobilizada em agar e Sepharose. Os resultados mostraram que
a concentração de enzima, a temperatura, a velocidade de agitação, e concentração de
substrato influenciaram de forma significativa na composição dos oligossacarídeos
produzidos.
Ichikawa et al. (2002) estudaram a imobilização e estabilização da ligação
multiponto da quitosanase tendo agar como suporte e concluíram que a imobilização
aumentou a termoestabilidade da quitosanase. Os oligossacarídeos de quitosana foram
continuamente produzidos usando um reator coluna contendo quitosanase imobilizada.
A percentagem de quitosana imobilizada após 28 dias de reação foi de 44%.
Hung et al. (2002) estudaram a purificação, através de troca iônica, e
caracterização da hidrólise sobre ação da quitinase e quitosanase purificadas do meio
contendo Bromelina comercial. Destaca-se que a quitinase purificada apresentou pH,
temperatura e Km ótimo para ação catalítica em meio contendo quitina de 4,0, 60ºC e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
26
0,2mg.mL-1, respectivamente. Enquanto que quitosanase apresentou o pH, temperatura e
Km ótimo de 3,0, 50ºC e 0,88mg.mL-1, respectivamente.
Zhu et al. (2003) estudaram as melhores condições fermentativas e propriedades
de uma quitosanase produzida por Acinetobacter sp. C-17. A espécie de bactéria
utilizada neste ensaio foi isolada do solo e identificada como sendo Acinetobacter sp C-
17 e capaz de produzir quitosanase. O microrganismo foi mantido em placas contendo o
meio agar com os seguintes componentes: 0,5 % de quitosana, 0,2 % K2HPO4, 0,1 %
KH2PO4, 0,07 % MgSO4, 0,05 % NaCl, 0,05 % KCl, 0,01 % CaCl2, 0,05 % de extrato
de levedura e 2 % agar, o pH do meio de manutenção foi mantido em 7,2 e a
temperatura foi mantida em 30ºC durante 3 a 5 dias, para identificar a ação da enzima
quitosanase. A ação da quitosanase foi identificada através da observação de uma
colônia de microrganismo existente na placa. A bactéria foi então isolada da placa e
colocada em fermentador rotativo a 200 rpm durante 72h em meio contendo: 0,07 %
K2HPO4, 0,03 % KH2PO4, 0,05 % MgSO4, 0,03 % peptona, 0,03 % de extrato de
levedura e 1,0 % de quitosana. O pH foi de 7,0 e a temperatura 30ºC. Como fontes de
carbono foram utilizadas além de 1,0 % de quitosana, 1,0 % de quitina e 1,0 % de
glicose. Os resultados mostraram que as melhores condições para produção da
quitosanase foram alcançadas quando se utilizou 1,0 % de solução de quitosana a 30ºC,
pH 7,0 durante 24 horas de incubação.
Liu et al. (2003) estudaram a produção de quitinase por Verticillium lecanii
F091 usando fermentação submersa, a atividade de quitinase foi de 9,95 mU/mL em um
meio de cultura otimizado, com um volume de cultura de 200 mL, agitação de 150 rpm
e temperatura de 24ºC cultivado em incubador rotativo e concluíram que a taxa de
agitação e o pH foram os fatores mais significativos para a produção de quitinase, no
entanto, as taxas de agitação e aeração poderiam modificar a concentração de oxigênio
dissolvido afetando diretamente no crescimento do Verticillium lecanii e na produção
de quitinase.
Donzelli et al. (2003) estudaram a hidrólise enzimática avançada de quitina de
casca de caranguejo langostino com misturas de enzimas de bactérias e fungos e
concluiram que a combinação de enzimas de Trichoderma atroviride e Serratia
marcescens foram capazes de degradar completamente altas concentrações de quitina
(100g/mL) a partir de carapaças de caranguejo langostino convertendo a N-
Acetilglicosamina (78%), glucosamina (2%) e quitobiose (10%). Enzimas de fonte

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
27
procariótica apresentaram atividades menores que as enzimas de T. atroviride, para
catalisar a reação com quitosana.
Jo et al. (2003) estudaram a caracterização e a avaliação cinética de quitosanase
produzida por Bacillus sp. Destaca-se que na produção da enzima utilizada neste ensaio
o meio continha 0,5% de concentração de quitosana, 1% de triptona e 1% de NaCl, o
pH do meio foi ajustado em 5,5 e a temperatura foi mantida em 37ºC com agitação de
180 rpm em incubador rotativo.
Fu et al (2003) estudaram a caracterização de três iso-enzimas de quitosanase
isolada de pepsina comercial e denominado como PSC-I, PSC-II, PSC-III,
respectivamente, em relação a hidrólise enzimática da quitosana, o meio ótimo para a
produção destas iso-enzimas continha as seguintes características: pH 5,0; 5,0 e 4,0 e
temperaturas de 40ºC, 40ºC e 30ºC, o valor de Km foi de 5,2, 4,0 e 5,6 mg.mL-1,
respectivamente. As massas moleculares das três iso-enzimas foram de 40 kDa. Os
resultados mostraram que as três iso-enzimas apresentaram atividade em meio contendo
quitosana, produzindo oligômeros da ordem de 68 – 88% desacetilada.
Choi et al. (2004) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida por Bacillus sp KCTC 0377BP. Nesse caso, com relação à
produção da enzima foi possível obter um aumento de 1,2 U/ml em incubator rotativo
para 100,0 U/ml em fermentador piloto de 500 L, utilizando uma concentração de
substrato de 0,5% de quitosana e como fonte de nitrogênio polipeptona (2,5%) em pH
igual a 6,8, com 100 rpm de 100,0 rpm a 30oC.
Shimosaka et al. (2003) estudaram a produção, purificação e caracterização total
de duas quitosanases produzida por Acinetobacter sp CHB101. Nesse caso, com relação
à produção da enzima os pesquisadores usaram dois meios, um contendo glicose e outro
contendo quitosana. Com relação à cinética, o meio contendo quitosana fornece um
tempo de geração de 280 minutos, enquanto que no meio contendo glicose a taxa de
crescimento foi mais elevada e o tempo de geração foi de apenas 110 minutos. Os
autores observaram que após os dois primeiros dias de cultivo a atividade
quitosanolítica permaneceu baixa (na ordem 0,10 a 0,20 U/mL). Durante a fase de
crescimento exponencial o valor da atividade permaneceu com valor inferior a 0,01
U/mL, começando a aumentar ao atingir a fase estacionária, indicando assim que a
produção da enzima é não associada ao crescimento.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
28
Kim et al. (2004) estudaram a produção, purificação e caracterização de uma
quitosanase produzida pelo Bacillus sp e identificado pelo número 1299. Destaca-se que
na produção da enzima quitosanase foi utilizado um meio contendo os seguintes
componentes: 0,5 % de quitosana, 0,42 % (NH4)2SO4, 0,2 % K2HPO4, 0,02 % uréia,
0,03 % CaCl2.2H2O, 0,03% MgSO4.7H2O, 0,1% peptona, 0,2% Tween 80, 0,2 % da
solução (0,5% FeSO4.7H2O, 0,16% MnSO4.H2O, 0,14% ZnSO4.7H2O, 0,2% CoCl2) e
1,7% agar. O pH do meio para este ensaio foi mantido em 7,0. Este microrganismo foi
escolhido por ser grande produtor de quitosanase.
Chen et al. (2005) estudaram a purificação e a caracterização de dois tipos de
quitosanase produzidas por Aspergillus sp. CJ22-326 em cultivo submerso. Na
produção da enzima foi utilizado um meio de cultivo contendo 1,0% de quitosana, 2,0%
de gérmen de trigo, 0,2% de sulfato de amônio, 0,2 % de fosfato de potássio e 0,05% de
sulfato de magnésio por litro de solução em pH 5,6. O cultivo foi realizado em frasco de
500 mL contendo 150 mL de meio de cultura a 30ºC por 96 horas com uma agitação de
150 rpm. Os autores apresentam um gel de eletroforese com as duas enzimas purificadas
com massa molar de 29 kDa e 109 kDa.
Silva Filho et al. (2005) estudaram, através de planejamento fatorial 24 com
repetição em triplicata no ponto central, a produção de quitosanase por Aspergillus
ochraceus em cultivo descontínuo submerso. Os resultados mostraram que foi possível
produzir quitosanase com atividade na ordem de aproximadamente 5,9 U/mL utilizando
Aspergillus ochraceus e que a atividade foi favorecida pelo aumento da agitação (r), da
razão de aeração (RA) e da concentração de substrato (Cs), enquanto que o aumento da
temperatura de cultivo (TMC) não favoreceu a resposta (atividade quitosanolítica).
Pode-se classificar as quitosanases em endoquitosanases e exoquitosanases. As
primeiras diferenciam-se por catalisar a hidrólise aleatoriamente, no interior do
biopolímero, gerando diversos tamanhos de oligossacarídeos. As exoquitosanases são
capazes de hidrolisar as terminações não redutoras, produzindo assim unidades
redutoras (Peter, 2005).
Palma-Guerrero et al. (2008) estudaram o biocontrole de fungos patogênicos e
demonstraram a lise da membrana plasmática e a inibição da germinação de esporos de
fungos. Eles observaram que os fungos entomopatogênicos e nematófagos foram menos
inibidos pela quitosana que os fitopatogênicos e micoparasíticos provavelmente porque
apresentam a enzima quitosanase que degrada a quitosana. Mesmo assim, a quitosana

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
29
pode ajudar a desenvolver novas estratégias para o biocontrole em geral de fungos
patogênicos, evitando o uso de fungicidas que prejudicam o meio ambiente e a saúde
humana.
Pagnoncelli (2008) estudou a produção de quitooligossacarídeos com
propriedades nutracêuticas a partir da quitosana por hidrólise enzimática, utilizando
processo fermentativo simultâneo. Duas cepas produtoras de quitosanaes foram
selecionadas, Paenibacillus chitinolyticus e Paenibacillus ehimensis, e avaliadas quanto
ao comportamento em meio de cultivo contendo açúcares simples e em relação às
variações de pH do meio. Os quitooligossacarídeos produzidos a partir do complexo
enzimático da primeira cepa apresentaram maior pico após 9h de hidrólise, enquanto
para a segunda cepa avaliada, após 20min, observou-se quitooligossacarídeos com grau
de polimerização entre 3 e 6 unidades.
2.2. Fungos
2.2.1. Características gerais
Desde o final dos anos 60 que os fungos deixaram de pertencer ao reino das
plantas e passaram a ter um reino próprio chamado de Reino Fungi. Foi a partir de
estudos morfológicos, bioquímicos e citológicos onde se concluiu que eles são tão
diferentes dos vegetais como dos animais. Distinguem-se de outros organismos
eucariotas por serem seres quimiorganoheterotróficos e, ainda, por apresentarem parede
celular rígida que é composta por quitina e glucano e uma membrana celular em que o
ergosterol substitui o colesterol (Trabulsi & Toledo, 1996; Murray et al., 2005).
Fazem parte do Reino Fungi cerca de 100 mil espécies, desde grandes como
os cogumelos, até microscópicas como os bolores e leveduras. Ainda hoje é utilizada
essa classificação, mas os avanços da biologia molecular e estudos bioquímicos levaram
a outras propostas de classificação em que os fungos e organismos afins foram incluídos
em três diferentes reinos: Protozoa, Chromista e Fungi (Kirk et al., 2008).
Abaixo, são apresentadas características que diferenciam os fungos das plantas:
• são incapazes de sintetizar clorofila, como também não apresentar celulose em
sua parede celular, com exceções como fungos aquáticos;
• apresentam algumas características de células animais (capacidade de depositar
glicogênio e a presença de substâncias quitinosas, em sua parede celular);

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
30
• podem ser eucarióticos (as leveduras), ou multinucleados (os bolores), e
apresentar retículo endoplasmático rugoso e mitocôndrias em seu citoplasma;
• são heterotróficos, alimentam-se de matéria orgânica viva (fungos parasitários),
ou morta (fungos saprofíticos);
Os fungos são amplamente encontrados na natureza, crescem rapidamente no
solo, na água, na atmosfera, bem como nos animais, nos vegetais, resíduos orgânicos e
desempenham um papel importante na vida do homem, quer de uma maneira benéfica,
quer de um modo prejudicial. Os fungos são um dos principais microrganismos
responsáveis pela decomposição da matéria orgânica, interferindo no ciclo do carbono,
do nitrogênio e de outros nutrientes da biosfera. São capazes de deteriorarem produtos e
bens de consumo, tais como alimentos, tecidos, papel, madeira. Os alimentos
armazenados representam um vasto meio para a proliferação dos fungos, principalmente
quando não são consideradas as condições básicas de armazenamento correto (Trabulsi
& Toledo, 1996).
Estão presentes funcionalmente em vários processos industriais, tais como, de
fabricação de pão, cervejas, vinhos e determinados tipos de queijos, sendo também
utilizados na produção comercial de muitos ácidos orgânicos, de alguns fármacos, como
a ergometrina e a cortisona, na obtenção de diferentes antibióticos, como a penicilina, e
de substâncias imunossupressoras, como a ciclosporina (Esteves et al., 1990). A Figura
4 ilustra a estrutura geral de uma célula de fungo.
Figura 4 – Representação da estrutura geral de uma célula de fungo.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
31
Os fungos podem se desenvolver em meios de cultivo especiais formando
colônias de dois tipos, leveduriformes e filamentosas, que se diferenciam pela
macromorfologia e micromorfologia. O primeiro tipo apresenta, em geral, característica
de consistência cremosa de cor branca a creme, brilhantes ou opacas, podendo
apresentar às vezes coloração escura ou alaranjada, e são formados por microrganismos
unicelulares que apresentam as funções vegetativas e reprodutivas. As filamentosas
apresentam características granulares, cotonosas, aveludadas ou pulverulentas, são
multinucleadas e apresentam-se em forma de tubos denominadas de hifas. Essas hifas
podem ser não septadas (cenocíticas) ou septadas (tabicadas). O conjunto de hifas é
denominado micélio.
As hifas asseptadas têm aparência anastomosada (concisa), formada por um
citoplasma estendido e polinucleado, enquanto que as hifas septadas contêm células
individualizadas, cada uma contendo o seu núcleo. Na Figura 5 são mostradas as
estruturas das hifas septadas e não septadas.
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas. (Adaptado de Guia.Bio,
2012)
O micélio que se desenvolve no interior do substrato e que é responsável pela
sustentação e absorção de nutrientes é denominado micélio vegetativo. O micélio que
cresce na superfície do meio de cultura é denominado micélio aéreo. Este pode ser
denominado também de micélio reprodutivo, quando o mesmo se diferencia para
sustentar os corpos de frutificação ou propágulos (Putzke & Putzke, 1998). Os
propágulos classificam-se de acordo com a origem em externos e internos, sexuados e
assexuados.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
32
2.2.2. Reprodução dos Fungos
A reprodução dos fungos ocorre em ciclos assexuais, sexuais e parassexuais. Em
fungos, a reprodução assexuada é observada com mais freqüência do que a reprodução
sexuada. A reprodução assexuada é normalmente a reprodução mais importante para a
propagação da espécie, feita através de fragmentação de artroconídios, fissão de células
somáticas, brotamento ou gemulação do blastoconídios-mãe e produção de conídios. A
reprodução sexuada envolve a união de duas células ou de dois órgãos sexuais
sexualmente compatíveis. A reprodução sexuada culmina na produção de basidiósporos,
no caso de basidiomicetos. A reprodução parassexuada ocorre com a fusão de hifas e
formação de um heterocário, que consiste na existência de núcleos haplóides
geneticamente distintos num mesmo citoplasma, os quais, após a fusão, originam
núcleos diplóides heterozigotos sujeitos a sucessivas divisões mitóticas. Apesar da
raridade dessas, a reprodução parassexual é de grande importância na evolução de
alguns fungos (Pelczar et al., 1996).
A reprodução assexuada é feita por conídios, esporos assexuados formados no
ápice de hifas modificadas, os conidióforos. Os conidióforos não possuem esporângios
de forma que os conídios ficam expostos. Na reprodução sexuada, com o encontro de
duas hifas haplóides distintas, ocorre a plasmogamia e um novo conjunto de hifas
dicarióticas é formado.
Os conídios podem ter formas esféricas, fusiformes, cilíndricas, piriformes, entre
outras. Apresentam-se como hialinos ou pigmentados, podem estar isolados ou
agrupados, a parede celular pode ser lisa ou rugosa e podem ser formados por apenas
uma célula ou apresentarem septos em mais de um plano.
Os conídios também podem originar-se em qualquer parte do micélio vegetativo
e são denominados conídios sésseis como é o caso do Trichophyton rubrum.
Alguns fungos patogênicos são identificados a partir de estruturas formadas pela
célula conidiogênica e o conidióforo. Essas estruturas são denominadas: aparelho de
frutificação ou conidiação. Os conídios podem formar cadeias sobre fiálides em volta de
uma vesícula, dilatando a extremidade do conidióforo. Essas estruturas ocorrem em
Aspergillus, apresentando-se em forma de cabeça ao redor de uma vesícula, e em
Penicillium agrupados em forma de pincel, como é ilustrada na Figura 6.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
33
Figura 6 – Conídios de Aspergillus e Penicillium
2.2.3. Metabolismo de fungos
Os fungos são seres aeróbicos e heterotróficos, podendo se desenvolver em
ambientes com baixa quantidade de oxigênio, como ocorre no crescimento vegetativo e
na reprodução assexuada, como também em ambientes com grande quantidade de
oxigênio como é o caso da reprodução sexuada.
Os fungos são capazes de produzir certas enzimas como lipases, invertases,
lactases, quitosanases, proteases, amilases, etc., que fazem a hidrólise do substrato
facilitando o mecanismo de transporte ativo e passivo. Esses substratos induzem a
formação de enzimas degradativas. Algumas espécies de fungos desenvolvem-se em
meios que apresentam amônia ou nitrilos como única fonte de nitrogênio. As melhores
fontes orgânicas para o seu desenvolvimento são carboidratos, como a D-glicose, e
alguns sais minerais como sulfatos e fosfatos.
A água é uma necessidade de todos os seres vivos, não sendo diferente para os
fungos que precisam de água para o seu desenvolvimento. Certos fungos apresentam
características halofílicas podendo crescer nos ambientes com grandes concentrações de
sal.
A razão de não existirem ambientes livres de espécies fúngicas deve-se ao facto
destes microrganismos conseguirem sobreviver em situações extremas como, por
exemplo, temperaturas que variam entre os - 5 e os 60 ºC, com pH de 1 a 9 e com baixas

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
34
concentrações de oxigênio, apesar de cada espécie possuir condições ideais para o seu
desenvolvimento (Chao et al., 2002).
Devido a grande faixa de temperatura ótima para o seu crescimento, os fungos
apresentam espécies psicrófilas, mesófilas e termófilas. Para a maior parte das espécies
fúngicas, a temperatura de desenvolvimento situa-se entre os 20 e 24 ºC e não crescem
acima dos 29 ºC. Em relação à umidade relativa, necessitam de pelo menos 50 %, mas
geralmente necessitam de mais de 65 % (Colakoglu, 2001).
O pH ideal para o desenvolvimento dos fungos varia na faixa entre 5 e 7.
Destaca-se, entretanto, que a maioria dos fungos suportam grandes variações de pH
como ocorre com os fungos filamentosos que podem crescer em pH variando entre 1,5 e
11. Por outro lado, as leveduras não suportam pH alcalino.
Os fungos crescem lentamente, se comparados com as bactérias, em meio de
incubação para o seu desenvolvimento. Para evitar contaminação por bactérias, que
podem se sobrepor ou inibir o seu crescimento, pode-se introduzir no meio de cultura
um antibacteriano de largo espectro como o cloranfenicol.
Algumas espécies de fungos se desenvolvem na presença de luz, outras são
inibidas e algumas podem crescer tanto na presença quanto na ausência deste agente. No
entanto a irradiação solar, em geral, é prejudicial ao seu desenvolvimento, por causa da
radiação ultravioleta.
2.2.4. Nutrição de fungos
Quanto ao aspecto da nutrição, os fungos são heterótrofos, ou seja, incapazes de
produzir seu próprio alimento. Entretanto, a forma de assimilar matéria orgânica é
diferente daquela utilizada pela maioria dos animais, que são heterótrofos por ingestão.
Nos fungos, a digestão é extracorpórea, ou seja, é realizada fora do corpo. O fungo
lança no ambiente, enzimas que degradam as moléculas orgânicas complexas e, depois,
absorve moléculas menores, mais simples. Por essa razão, são designados heterótrofos
por absorção. É devido a este processo que os fungos podem crescer dentro ou sobre os
alimentos. Desenvolve-se geralmente em meios contendo um pH baixo, uma fonte de
carbono, uma fonte de nitrogênio orgânico ou inorgânico e alguns minerais. Alguns
necessitam de vitaminas. Quanto aos modos de vida, os fungos podem ser
decompositores, parasitas, mutualísticos e predadores (Raven, 2001).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
35
Os fungos decompositores, saprófagos (do grego, saprós, “podre”, e phagos,
“comedor”), juntamente com as bactérias, são os principais decompositores da biosfera,
participando intensamente do processo de degradação da matéria orgânica morta, o que
promove a reciclagem dos elementos químicos constituintes dos seres vivos. A maioria
deles vive no solo, obtendo nutrientes de seres mortos. Em florestas, onde a matéria
orgânica vegetal é abundante, os micélios dos fungos decompositores absorvem os
nutrientes de folhas e galhos caídos, decompondo-os. Os fungos decompositores
também são responsáveis pelo apodrecimento de alimentos e de outros materiais, como
a madeira.
Os fungos parasitas vivem à custa de outros organismos vivos, prejudicando-os.
Ao parasitar o corpo de um ser vivo, animal ou vegetal, o fungo pode até provocar sua
morte. Várias doenças são causadas por fungos parasitas. Nas plantas, cita-se a
ferrugem do café, que, no Brasil, é motivo de grande preocupação por parte dos
cafeicultores, devido aos prejuízos que acarreta. As micoses que atacam a pele de seres
humanos e animais são provocadas por fungos.
Existem fungos que se associam a outros organismos, estabelecendo uma relação
em que há benefício mútuo para os indivíduos envolvidos. Esses fungos são
denominados mutualísticos. A maioria vive associada a seres fotossintetizantes, como
as plantas, cedendo-lhes parte da água e dos nutrientes que as hifas absorvem do solo.
As plantas, por sua vez, cedem ao fungo certos açúcares e aminoácidos. Duas
associações mutualísticas formam estruturas bem características: as micorrizas,
associação que envolve fungos e raízes de plantas, e os líquens, formados por fungos e
algumas variedades de algas e cianobactérias.
Certos fungos atuam como predadores. Na maioria dos casos, as hifas secretam
substâncias aderentes que aprisionam os organismos que tocam os fungos. Dessa
maneira, as hifas penetram o corpo da presa, crescem e se ramificam, espalhando-se no
interior do organismo e absorvendo seus nutrientes, causando-lhe a morte. O gênero
Arthrobotrys apresenta uma forma mais elaborada de predação. O fungo vive no solo e
captura nematódeos microscópicos que habitam o mesmo ambiente. As hifas
apresentam pequenos anéis que se estreitam quando a presa passa por eles. As hifas do
fungo logo invadem o corpo do nematódeo, digerindo-o.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
36
2.2.5. O fungo Metarhizium anisopliae
Existem três espécies de fungos do gênero Metarhizium, divididas em dez
variedades: M. anisopliae, variedades anisopliae, majus, lepidiotum e acridum; M.
flavoviride, variedades tipo E, flavoviride, minus, novazealandicum e pemphigum; e M.
album (Driver et al. 2000). E, mais recentemente, foi descrita na China uma nova
variedade de M. anisopliae, dcjhyium (Dong et al. 2009).
Data de 1879 quando o russo Ilya Metchnikoff aplicou o fungo para combater
larvas de um curculionídeo, praga de beterraba, e foi classificado como Metarhizium
anisopliae por Sorokin em 1883. É um fungicida natural para mais de 300 espécies de
insetos, especialmente pragas da agricultura e pecuária. No Brasil, o fungo Metarhizium
anisopliae teve a sua aplicação com sucesso pela primeira vez em 1965, no controle da
cigarrinha-da-cana Mahanarva posticata (Alves, 1998).
O fungo Metarhizium anisopliae é um importante agente entomopatogênico
utilizado no controle de pragas, como cupins, gafanhotos, cigarrinhas e besouros, sendo
amplamente estudado em todo o mundo. Existe amplamente na natureza e é facilmente
encontrado no solo, onde sobrevive por longos períodos e seu desenvolvimento se dá
geralmente entre 15 e 32 °C, sendo ideal entre 24 e 30 °C e com pH ótimo igual a 6,9.
Apresenta micélio hialino e septado, com conidióforos característicos, sobre os quais
surgem conídios cilíndricos organizados em colunas. Este fungo ao atacar os insetos
provoca rigidez e os recobre por uma camada pulverulenta de conídios, resultando em
colorações que variam do verde claro ao escuro, cinza ou branco com pontos verdes
(Tinline, 1971; Alves, 1998, Driver et al. 2000; Arruda, 2005).
Quanto a sua morfologia, apresenta-se como um fungo filamentoso, com corpo
de frutificação equivalente a um esporodóquio agregado a hifas intimamente
entrelaçadas, com massa compactada de conidióforos característicos, simples ou
ramificados, formando células esporogênicas denominadas fiálides, resultando nos
fialosporos (Wang et al., 2002).
O Metarhizium anisopliae é um fungo de cultivo simples, sendo necessário
quase que exclusivamente o nutriente a base de amido. Um dos fungos mais estudados
em programas de manejo de pragas já que apresenta grande potencialidade
entomopatogênica (Onofre et al., 2002). O Metarhizium anisopliae é um microrganismo
pouco exigente, pois pode se desenvolver em diversos meios de cultura como: amido,
glicose, glicerina, levulose, maltose, sacarose e quitina (Jabor et al., 2003).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
37
Em geral, os fungos produzem enzimas que convertem os tecidos dos insetos em
nutrientes. Porém, a cutícula esclerotizada dos insetos não é bem utilizada pelos fungos,
excetos pelos entomopatogênicos que desenvolveram enzimas eficientes para degradar
essa camada protetora. Destaca-se a enzima Pr1, produzida por Metarhizium anisopliae,
que tem um poder de degradação bem superior a qualquer outra enzima sintetizada por
esse grupo de fungos (Dias, 2005). Desta forma, o Metarhizium anisopliae (fungo
entomopatogênico) ao atacar a célula dos insetos, produz grande quantidade de enzimas
que degradam a membrana citoplasmática e inativa a célula e, consequentemente, mata
o inseto, logo é evidente a relação entre o controle de pragas e produção de enzimas.
No momento, sobre a comercialização de fungos entomopatogênicos no Brasil,
em sua maioria, são formulados em grânulos, compostas do fungo mais o substrato
(arroz + fungo), como também na forma de pó molhável (WP), resultado da moagem do
fungo com o substrato. Esses produtos representam 90% e 8,5%, respectivamente, da
produção de fungos no Brasil, e sendo suas concentrações finais ficam em torno de
5x108 conídios/g para Metarhizium anisopliae e 1x109 conídios/g para Beauveria
bassiana (Alves et al., 2008).
Segundo Batista Filho et al. (2002), Garcia et al. (2005) e Mello et al. (2006), o
Metarhizium anisopliae tem sido bastante utilizado para o controle de diferentes pragas
no Brasil, como a broca da bananeira, pragas de grãos armazenados, carrapatos, cupim
de montículo em pastagens, larvas de escarabeídeos que atacam a cana de açúcar e
cigarrinha da cana-de-açúcar.
É comum a ocorrência natural do fungo Metarhizium anisopliae atacando
cigarrinha-das-raízes de canaviais pelo país, o que incentiva a utilização dele como
agente de controle biológico em áreas infestadas por esse tipo de praga. A aplicação de
recursos no uso de programa de manejo integrado da cigarrinha tem sido reforçado por
razões ambientais e econômicas (Dinardo-Miranda, 2001).
Faria & Magalhães (2001) fizeram uma avaliação do uso de fungos
entomopatogênicos no Brasil, na qual observaram que a qualidade dos micoinseticidas
disponíveis no Brasil poderia ser incrementada de forma considerável quanto a sua
formulação, ou seja, antes da venda, poderiam realizar melhorias no tratamento
posterior, adição de substâncias que lhes assegurem melhorias na eficiência de controle,
capacidade de armazenamento ou praticidade de manuseio, ou de qualquer outro critério
que resulte em vantagem em relação ao produto bruto. Ressaltaram a pouca praticidade

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
38
em alguns casos, como exigência de lavagem, e em outros podendo causar o
entupimento de bicos dos pulverizadores devido à elevada proporção de inertes,
principalmente quando são empregados baixos volumes de aplicação. Outro problema
evidenciado com os inseticidas nacionais diz respeito à pequena sobrevida, devendo a
ser usados em, no máximo, 30 dias depois de produzidos, quando armazenados à
temperatura ambiente e em local sombreado, fazendo as vendas ocorrerem quase que,
exclusivamente, sob encomenda. Comparativamente, os bons inseticidas sob
comercialização em outros países apresentam maior concentração de ingrediente ativo,
maior sobrevida (alguns produtos podem ser armazenados por mais de 8 meses à
temperatura ambiente) e praticidade (são formulados na forma de GDA (grânulos
dispersíveis em água) ou de óleos emulsionáveis, por exemplo, podendo ser adicionados
diretamente ao tanque do pulverizador.
Rangel et al. (2007), instalaram uma Unidade de Observação para o controle
microbiano das cigarrinhas das pastagens Distrito de Itahum, em Dourados/MS, devido
as necessidades de agricultores familiares de produção de leite. As aplicações do fungo
Metarhizium anisopliae foram realizadas por pulverização com equipamento de tração
animal. As condições do ambiente de estudo foram favoráveis ao desenvolvimento da
população das cigarrinhas e à incidência do fungo. Como resultados, observaram que os
níveis de espumas, ninfas e adultos reduziram significativamente após as aplicações,
levando os Agricultores Familiares a total satisfação com os ganhos na produção de
todos os envolvidos.
Segundo Vieira et al. (2007), o fungo entomopatogênico Metarhizium anisopliae
var. acridum tem sido utilizado no controle de pragas, e assim, visando este fim,
analisaram em laboratório a esporulação, a germinação dos conídios e as características
morfológicas peculiares desse fungo em diferentes temperaturas. Esperou-se o
desenvolvimento das colônias em períodos de 1 a 15 dias. Foi observado que
temperaturas de 25, 28 e 30 ºC proporcionaram maiores taxas de esporulação e elevadas
porcentagens de germinação dos conídios. A 25, 28 e 30 ºC observou-se um
desenvolvimento mais rápido das microestruturas somáticas e reprodutivas (hifas,
anastomoses, apressórios, conidióforos e conídios elipsóides). Nas temperaturas
extremas de 40 e 45 ºC não foi detectado esporulação, germinação, nem
desenvolvimento de nenhuma microestrutura.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
39
Lopes et al. (2007) avaliaram o efeito de formulações de Beauveria bassiana e
Metarhizium anisopliae sobre ninfas de Amblyomma cajennense (transmissor da febre
maculosa ao homem) em laboratório. Ninfas não alimentadas foram pulverizadas com
suspensões contendo 2x107 e 4x107 conídios/mL de B. bassiana, e 1x107, 1,5x107,
2x107, 3x107 e 5x107 conídios/mL de M. anisopliae. A mortalidade de ninfas,
decorridos três dias da aplicação, foi significativa nas concentrações de 3x107 e 5x107
conídios/mL de M. anisopliae, com 57,1 e 56,2 %, respectivamente. Após seis dias a
mortalidade foi de 100 % nessas duas concentrações. A mortalidade foi
significativamente maior que a observada para B. bassiana em concentrações
semelhantes.
Guirado et al. (2009) realizaram estudos para controlar cupins de montículo em
área de pastagem, utilizando fungos entomopatogênicos Metarhizium anisopliae e
Beauveria bassiana em diferentes doses e formulações. A avaliação foi realizada após
30 dias da aplicação dos produtos. Os melhores resultados em ordem decrescente de
eficiência foram: M. anisopliae 30 mL + 1000 mL de água com 100 % de eficiência; B.
bassiana 10g + M. anisopliae 20 mL + 1000 mL de água com 85,71 % de eficiência; M.
anisopliae 20 mL + 1000 mL de água com 71,42 % de eficiência. Os demais
tratamentos não diferiram da testemunha.
Loureiro et al. (2012) compararam a eficiência de isolados de Metarhizium
anisopliae em controlar populações naturais de Mahanarva fimbriolata (cigarrinha-da-
raiz) em cultivos comerciais de cana-de-açúcar colhida mecanicamente. Foram
aplicados os isolados IBCB 348, IBCB 408, IBCB 410 e IBCB 425 do fungo em
concentração de 1,5×1012 conídios/ha. Aplicou-se o planejamento experimental em
blocos casualizados com 5 tratamentos e 4 repetições, sendo cada repetição composta
por 7 linhas de 100 m de comprimento e espaçamento de 1,5 m. Após 30 dias de
pulverização foi observado para os isolados IBCB 408 e IBCB 425 uma eficiência de
controle de ninfas de 63 e 62%, respectivamente, já no caso dos adultos chegou a 100%.
Após 60 dias o isolado IBCB 425 foi o mais eficiente para controlar as ninfas com
48,4%.
Assis et al. (2012) estudaram a citotoxidade de quitoolissacarídeos (QOS) e
oligômeros A, B e C (soluções compostas por diferentes quantidades de QOS)
produzidos por hidrólise enzimática pelo fungo Metarhrizium anisopliae. Foi analisado
o efeito de antiproliferativo destas moléculas em células tumorais (HepG2 e HeLa) e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
40
também em célula normal (3T3). A atividade antioxidante foi analisada em vários
experimentos in vitro. A glicosamina apresentou maior toxicidade (cerca de 92%) para
todas as células estudadas. No entanto, os oligômeros obtidos após hidrólise não
demonstraram efeito tóxico sobre a célula normal (3T3). Além disso, mostraram que
uma pequena quantidade de outros COS pode diminuir o efeito citotóxico da
glicosamina contra células 3T3, indicando que glicosamina poderia ser usada como uma
droga antitumoral na presença de outros COS.
2.3. Fermentação
Os processos fermentativos são caracterizados por processos nos quais
microrganismos realizam a conversão catalítica de uma dada substância num
determinado produto. Estes processos podem ser classificados como processos
descontínuos, contínuos e descontínuos alimentados. A classificação destes processos
está diretamente ligada à forma de como o substrato é adicionado e como o produto é
retirado. No processo de fermentação descontínuo o substrato é colocado no
fermentador, em seguida é inoculado e após o término do processo o produto é retirado.
No processo contínuo, o substrato é colocado num reator com vazão constante e o meio
fermentado é retirado com a mesma vazão de entrada do reator (Faccioti, 2001). Nos
processo descontínuos alimentados à adição de substrato e a retirada do mesmo deve
ocorrer de forma intermitente (Carvalho & Sato, 2001).
De certa forma, os processos fermentativos representam um elo entre as antigas
artes de elaboração de alimentos, como queijos e vinho, a partir de uma flora
microbiana natural, com a moderna indústria de fermentação de alimentos, onde são
utilisados cultivos puros e equipamentos sofisticados para o controle do processo
(Assis, 2010).
Atualmente, a maioria dos processos industriais se desenvolve em cubas de
vidro ou de aço de até 20 litros. Os agentes biológicos se encontram submersos no meio de
cultivo que ocupa, aproximadamente, 75% da cuba. As vezes é necessário injetar ar, e em muitas
fermentações se forma espuma. O desenho do biorreator deve se adequar ao objetivo do
processo, respondendo eventualmente a diversos imperativos, tais como a esterilização do
sistema, a aeração e homogeneização do meio, o acréscimo de nutrientes e de aditivos
antiespuma, a manutenção do pH, etc. Os modelos de fermentadores mais utilizados com
microrganismos contam com aeração e agitação mecânica. Esta facilita a distribuição dos

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
41
nutrientes na cuba, mas gera calor que deve ser eliminado mediante a circulação de água
fria. Em outros tipos de biorreatores, em coluna ou torre, a homogeneização depende da injeção de
ar. Os sistemas submersos são apropriados para o cultivo de microrganismos livres, mas resultam
pouco econômicos quando se trabalha com células ou enzimas caras. A imobilização em
fermentadores menores, seja por adesão a um suporte inerte, seja por inclusão dentro de um
polímero que permita o contato com o meio de cultura, além de simplificar a purificação do
produto permite a reutilização das células ou das enzimas, que permanecem dentro do
biorreator (Malajovich, 2009).
As enzimas são produtos de alto valor agregado, amplamente empregadas em
uma gama de indústrias, e entre as fontes para sua obtenção, destacam-se as de origem
microbiana, principalmente, devido a elevada seletividades dos substratos utilizados e
produtos formados. A principal forma de obtenção de enzimas, em escala industrial, é a
fermentação submersa, por esta apresentar vantagens em relação a fermentação
semisólida, como: maior controle do processo fermentativo e consequente reprodução
da cinética microbiana, além de maiores rendimentos (Wanderley, 2011).
2.3.1. Preparo do inóculo
Denomina-se inóculo um volume de suspensão de microrganismo de
concentração adequada, capaz de garantir, em condições econômicas, a fermentação de
uma dada quantidade de meio. O volume do inóculo, assim como sua concentração de
células, vai depender tanto do processo de fermentação c omo da quantidade de meio a
ser fermentado (Carvalho & Sato, 2001).
A partir de uma cultura pura, inocula-se um volume relativamente pequeno de
meio nutriente, que vai depender do tipo de microrganismo cultivado (pois cada espécie
microbiana possui velocidades diferentes de crescimento), sendo, em seguida, incubado
em condições favoráveis ao desenvolvimento do microrganismo. A suspensão obtida é
transferida para um frasco maior em agitador rotativo ou recíproco (shakers), que por
sua vez é adequadamente incubado, e a seguir, transferido para um frasco contendo um
volume ainda maior deste meio, até se conseguir alguns litros de suspensão microbiana.
Os cuidados com assepsia, volumes de meio utilizado, numero de transferência bem
como as condições de incubação, podem variar de acordo com o tipo de processo de
fermentação que se deseja realizar.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
42
2.3.2. Produção de enzimas em cultivo submerso descontínuo
A produção de enzimas, usando processos fermentativos, é constituída
basicamente por duas etapas: a primeira caracteriza-se pela preparação do inóculo e a
segunda, pela fermentação do substrato (Carvalho & Sato, 2001). A finalidade da
primeira etapa é o preparo do microrganismo em condições adequadas para que possa
ocorrer o desenvolvimento da segunda etapa. Diferentemente do cultivo em estado
sólido, no cultivo submerso os microrganismos encontram-se em um meio com elevada
atividade de água.
2.4. Planejamento experimental
Um bom planejamento tem como fundamental projetar um experimento de
forma que ele seja capaz de fornecer exatamente o tipo de informação que procuramos.
Para isso é necessário saber, em primeiro lugar, o que é mesmo que se deseja procurar.
Planejamento experimental, ou delineamento experimental, é um conjunto de
ensaios estabelecido com critérios científicos e estatísticos, objetivando determinar a
influência de diversas variáveis nos resultados de um dado sistema ou processo (Barros
Neto et al., 2001; Rodrigues & Iemma, 2005). De acordo com o propósito dos ensaios,
esse objetivo maior pode ser mais detalhado em:
– determinar quais variáveis são mais influentes nos resultados;
– atribuir valores às variáveis influentes de modo a otimizar os resultados;
– atribuir valores às variáveis influentes de modo a minimizar a variabilidade dos
resultados;
– atribuir valores às variáveis influentes de modo a minimizar a influência de variáveis
incontroláveis.
O planejamento experimental, baseado nos fundamentos estatísticos é, sem
dúvida alguma, uma ferramenta poderosa para se chegar às condições otimizadas de um
processo, desenvolvimento da formulação de produtos dentro das especificações
desejadas ou simplesmente para avaliar os efeitos ou impactos que os fatores (variáveis
independentes) têm nas respostas (variáveis dependentes) desejadas (Barros Neto et al.,
2001; Rodrigues & Iemma, 2005). Da utilização das técnicas estatísticas de
planejamento experimental, é possível chegar benefícios importantes, como:

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
43
– reduzir o número de ensaios sem prejuízo da qualidade da informação;
– estudar simultaneamente diversas variáveis, separando seus efeitos;
– determinar a confiabilidade dos resultados;
– realizar a pesquisa em etapas, num processo iterativo de acréscimo de novos ensaios;
– selecionar as variáveis que influem num processo com número reduzido de ensaios;
– representar o processo estudado através de expressões matemáticas;
– elaborar conclusões a partir de resultados qualitativos.
2.4.1. Planejamento fatorial
Em um planejamento fatorial é investigado as combinações de níveis de estudo
de cada fator. Para tanto, inicialmente, é necessário especificar esses níveis em cada
fator, quer dizer, os valores dos fatores que serão utilizados nos experimentos. Um
ensaio experimental é definido como um experimento constituído de um conjunto de
níveis definido. Geralmente, quando se tem n1 níveis de um determinado fator 1, n2 do
fator 2, ..., e nk do fator k, o planejamento será descrito por um fatorial n1 x n2 x ... x nk,
que corresponde ao número mínimo de experimentos para se ter um planejamento
fatorial completo.
É necessário fazer cada fator variar e observar o resultado dessa variação para se
estudar o efeito sobre a resposta. Assim, os ensaios precisam ter um mínimo de dois
níveis por fator para a realização desses ensaios. Para k fatores, ou seja, k variáveis de
controle, o planejamento de dois níveis precisará de 2 x 2 x ... x 2 = 2k, para o número
de ensaios diferentes, sendo conhecido como planejamento fatorial 2k.
Para o planejamento fatorial, são especificados os valores dos fatores em cada
experimento, e estes são classificados em níveis, variando-se, por exemplo, de -1
(mínimo) a +1 (máximo). A partir de uma matriz serão apresentadas todas as
combinações entre o número de fatores e de níveis nos experimentos a serem realizados
(Barros Neto et al., 2001; Rodrigues & Iemma, 2005).
2.4.2. Planejamento Plackett & Burman
A maioria dos planejamentos do tipo “Screening” (Delineamentos de Seleção de
fatores) foi descrito originalmente por Plackett & Burman em 1946. Os planejamentos
experimentais de Plackett & Burman são eficientes em situações que envolvem uma

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
44
grande quantidade de variáveis a serem exploradas em um processo de otimização de
fatores (Reddy et al., 2008).
As matrizes Plackett & Burman consistem de planejamentos fatoriais de dois
níveis para estudar k = N – 1 variáveis em N ensaios. São construídas para ensaios
múltiplos de 4, sendo essas matrizes para ensaios de 8, 12, 16,..., 100. Cada variável
independente em uma matriz assume uma configuração de dois níveis, o inferior (-1) e o
superior (+1), para o nível intermediário é estabelecido (0). Para cada vez que a variável
aparece no nível inferior, terá o par equivalente no nível superior (Rodrigues & Iemma,
2005).
Para evitar dúvidas nos resultados dos efeitos de algum fator no planejamento de
Plackett & Burman, o número de ensaios a ser realizado deve ser no mínimo o número
de variáveis mais 4. No caso de um fatorial completo o número de ensaios a ser
realizado é bem maior (Rodrigues & Iemma, 2005).
Mesmo já determinado o planejamento Plackett & Burman, torna-se importante
à realização de experimentos relativos aos pontos centrais de modo a se avaliar melhor
as possíveis interferências analíticas que podem ocorrer durante os diferentes
experimentos. A repetição do ponto central permite também a determinação do erro
puro e da repetibilidade do processo. Na prática, esta é uma ferramenta que permite
identificar melhor se o processo em estudo está sob controle (Rodrigues & Iemma,
2005).
É possível que no planejamento Plackett & Burman não tenha sido considerado
a codificação do ponto central (0) para repetição, pois, matematicamente, a estimativa
do valor desse efeito codificado (zero) vezes, teria como resposta zero. Porém, no
resultado prático do experimento, consegue-se informações valiosas com essas
repetições no ponto central (Rodrigues & Iemma, 2005).
2.4.3. Aplicações do Planejamento Plackett & Burman
Ghanem et al. (2000) produziram xilanase por Aspergillus terreus cultivadas
sobre palha de trigo finamente moída em fermentação de estado sólido. A composição
ideal do meio foi estabelecida aplicando-se o planejamento experimental de Plackett &
Burman. A melhor atividade enzimática foi obtida em um meio contendo 10 g/L de

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
45
palha de trigo por frasco umedecido com uma solução concentrada de sal com teor de
água inicial de 75 % e incubadas durante 4 dias a 30 °C. Observou-se que íons
metálicos como Hg2+, Cu2+, Co2+, Fe3+ e Pb2+ inibiram fortemente a xilanase, enquanto
que Ca2+ ativou a enzima.
Mu et al. (2005) estudaram as otimização de meio de fermentação para a reação
de estereoinversão por Candida parapsilosis CCTCC M203011 utilizando o
planejamento Plackett & Burman. Os resultados mostraram que dentre os fatores
estudados, os fatores mais significativos foram: a Concentração de MgSO4, ZnSO4 e
KH2PO4 e que após a utilização da metodologia de superfície de resposta foi constatado
que o ponto ótimo para a otimização foi conseguido com as concentrações dos fatores
MgSO4, ZnSO4 e KH2PO4 nos valores em: 1,17 g/L, 0,10 g/L e 9,41 g/L,
respectivamente, elevando a taxa de conversão enantiosseletiva de 75,2 % para 96 %.
Naveena et al. (2005) empregaram o planejamento Plackett & Burman para a
seleção de 15 parâmetros para produção de ácido láctico L(+) de farelo de trigo,
substrato de baixo custo e suporte sólido, por Lactobacillus amylophilus GV6 em
fermentação de estado sólido (SSF). Onze nutrientes pertencentes a duas categorias,
fontes de nitrogênio e fontes de sal, junto com três parâmetros físicos e um tampão
foram os parâmetros considerados. Os autores verificaram influência na produtividade
devido às fontes de nitrogênio, peptona, extrato de levedura e tri-citrato de amônio,
juntamente com o NaH2PO4.2H2O e Tween 80, que podem ainda serem otimizados para
o aumento na produção de ácido lático. Esses pesquisadores ainda chamam atenção para
o uso escasso deste planejamento em fermentação em estado sólido e que o mesmo
ainda não havia sido considerado anteriormente para a conversão direta de amido em
ácido láctico L (+) usando um sistema bacteriano.
Khosravi-Darani et al. (2008) estudaram a aplicação de Plackett & Burman na
análise dos fatores significativos para a produção de ácido cítrico por Aspergillus niger
ATCC 9142 utilizando palha de trigo como substrato. Os resultados mostraram que a
produção, com a palha de trigo pré-tratada com HCl, NaOH e Uréia, foi mais eficiente e
que os fatores mais significativos na produção foram: Solvente, Teor de Umidade,
Temperatura, Tempo de Incubação, Concentração Inicial de Açúcar e Concentração de
Metanol.
Ji et al. (2009) estudaram o desenvolvimento de um meio de cultura econômico
para a produção do 2,3-butanodiol através da fermentação da glicose e xilose por

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
46
Klebsiella oxytoca utilizando o planejamento Plackett & Burman. Os resultados
mostraram que a uréia e a maceração de milho foram os fatores mais significativos para
a produção do 2,3-butanodiol e que através de planejamento experimental com repetição
em triplicata no ponto central para determinar os níveis ótimos de produção. Os autores
constaram que o rendimento atingido foi 85,6% do valor teórico.
Paula & Arruda. (2009) observaram a necessidade da busca por alternativas mais
eficientes ao método termomecânico utilizado para a degradação de milhões de
toneladas de penas de aves provenientes de frigoríficos e aviários, cuja rígida estrutura é
composta por mais de 90 % de queratina. Para esse fim, utilizaram de comunidades
microbianas para a finalidade proposta, testando o planejamento experimental Plackett
& Burman para verificar a influência de diferentes microrganismos simultaneamente,
com um número reduzido de experimentos. Testaram 32 bactérias isoladas, inoculadas
em meio contendo somente uma pena como única fonte de carbono, durante sete dias,
de onde foi selecionado as cepas para os consórcios microbianos. Experimentos unindo
as cepas NP5, FCA7, NP4 e S14 demonstraram diferentes atividades queratinolíticas,
aumentando o nível de degradação, até 87,8%, comparada com as cepas
individualmente, variando entre 26,4% e 78,0%.
Rajendran & Thangavelu (2009) obtiveram valores máximos da atividade de
produção de lipase em 3,98 U/mL e concentração de massa celular em 5,62 g.L-1 com R.
arrhizus em cultivo submerso descontínuo otimizado pelo planejamento Plackett &
Burman. As variáveis mais importantes que afetaram a produção da lipase foram azeite
de oliva, peptona, KH2PO4, CaCl2.2H2O e MgSO4.7H2O.
Li et al. (2010) estudaram a otimização de meio de cultura para a produção de
Streptolydigin (um antibiótico) por Streptomyces lydicus AS 4,2501 utilizando Plackett
& Burman e a metodologia de superfície de resposta. Os resultados mostraram que
dentre os fatores estudados, os fatores mais significativos para a produção de
Streptolydigin e otimização do meio de cultura foram: Concentração de Amido,
Concentração de Peptona e a Concentração de K2HPO4.
Chen et al. (2010) estudaram a otimização de meio de cultura para a produção da
Luciferase – que são enzimas que catalisam reações biológicas transformando energia
química em energia luminosa - por Bacillus Subtilis em fermentação descontínua
utilizando planejamento Plackett & Burmam. Os resultados mostraram que dentre os
componentes estudados, os fatores mais significativos para a produção e otimização do

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
47
meio de cultura para a produção da Luciferase foram: Concentração de Glicose 1,17 %,
Extrato de Levedura 2,27 % e Glutamato 0,55 %.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o projeto Plackett & Burman e a metodologia de superfície de resposta. Os
resultados mostraram que a eficiência máxima de extração da quitina foi conseguida nas
seguintes condições: Concentração de casca de camarão (50 g/L), glicose (50 g/L),
tempo de incubação de 5 dias e inoculo com 0,05 de OD (densidade óptica).
Gao et al. (2010) desenvolveram um meio para a fermentação do extrato bruto
de inulina de tubérculos de alcachofra de Jerusalém por Paenibacillus polymyxa ZJ-9
para produzir R,R-2,3-butanodiol (R-R,2,3-BD). Inulina, K2HPO4 e NH4Cl foram os
parâmetros principais na fermentação de acordo com os resultados obtidos com o
planejamento experimental Plackett & Burman. A concentração ideal dos três fatores,
77,14 g/L, 3,09 g/L e 0,93 g/L, respectivamente, foi obtida pelo planejamento Box-
Behnken e pela superfície de resposta. Sob as condições ideais, a concentração do R-
R,2,3-BD obtida foi 36,92 g/L, em mais de 98 % de pureza óptica. Comparado com
outras fontes de carbono investigadas, a fermentação do extrato bruto de inulina obteve
o maior rendimento de R-R,2,3-BD.
Uma metodologia de superfície de resposta foi empregada com sucesso para
otimizar um meio de fermentação chamada VHG (Very High Gravity), referência à alta
concentração de açúcares e densidade dos mostos, baseado na água maceração de milho
CSL (Corn Steep Liquor) e outros nutrientes de baixo custo para a produção eficiente de
etanol a partir de glicose por S. cerevisiae. Usando a composição média otimizada (g/L:
CSL 44,3; uréia 2,3; MgSO4.7H2O 3,8; CuSO4.5H2O 0,03), a levedura industrial PE-2
foi capaz de fermentar até 330 g/L de glicose e produzir 18,6 % (v/v) de etanol, com
uma produção descontínua de 2,4 g/L/h e um rendimento em etanol de 93 % (Pereira et
al., 2010).
O planejamento Plackett & Burman juntamente com o Planejamento Composto
Central (CCD) foram aplicados para otimizar o meio de cultura para produção de etanol
por Clostridium autoethanogenum com monóxido de carbono (CO) como fonte única de
carbono e um meio contendo (g/L): 1,0 de NaCl, 0,1 de KH2PO4, 0,02 de CaCl2, 0,15 de
extrato de levedura, 0,116 de MgSO4, 1,694 de NH4Cl e pH 4,74. O rendimento ótimo
de etanol previsto pela metodologia de superfície de resposta (RSM) e um algoritmo

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
48
genético de rede neural artificial (ANN-GA) foram 247,48 e 261,48 mg/L,
respectivamente. Valores semelhantes aos obtidos experimentalmente sob as condições
ideais sugeridas pelos métodos estatísticos (254,26 e 259,64 mg/L). A adequação do
modelo ANN-GA foi maior do que o modelo RSM. Os rendimentos obtidos foram
substancialmente superiores aos relatados anteriormente (60-70 mg/L) com essa cultura
(Guo et al., 2010).
Rigueira (2010) avaliou a influência das variáveis do procedimento de extração
na porcentagem de recuperação das aminas encontradas no leite cru utilizando um
planejamento para “screening” do tipo Plackett & Burman, para os ácidos
tricloroacético (TCA) e sulfosalicílico (ASS).
Quines (2010) estudou a necessidade ou não de se fazer um pré-tratamento da
biomassa antes da extração. Um planejamento experimental Plackett & Burman de doze
ensaios mais três pontos centrais foi utilizado para selecionar as variáveis mais
significativas na extração de Poli(3-hidroxibutirato) (P(3HB)) por 1,2-carbonato de
propileno. Foram realizados dois testes de recuperação do solvente 1,2-carbonato de
propileno, após a extração, sendo um com aplicação de aquecimento e o outro com
aquecimento e vácuo. Os resultados mostraram que o pré-tratamento térmico levou a
uma maior porcentagem de recuperação e de pureza do polímero.

Capítulo 3 Metodologia Experimental

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
50
3. Metodologia Experimental
O presente capítulo apresenta os matérias e métodos utilizados na produção de
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso.
3.1. Quitosana
Neste trabalho utilizou-se uma quitosana (Lote 067) obtida junto a POLYMAR/CE,
com no mínimo 80% de desacetilação. Esse substrato foi utilizado como fonte de carbono e
energia tanto no estudo cinético como nos ensaios do planejamento de seleção de fatores
Plackett & Burman e Planejamento Composto Central, para a produção de quitosanase pelo
Metarhizium anisopliae, por fermentação submersa.
3.2. Microrganismo
O fungo Metarhizium anisopliae estirpe (CG374), utilizado neste estudo, foi cedido
pela Embrapa Recursos Genéticos e Biotecnologia (Brasília / DF-Brasil) e foi mantido em
placas de Petri contendo o meio Batata Dextrose Agar (PDA). As placas foram incubadas em
estufa a 25 ºC. O pH do meio de manutenção foi mantido entre 6,0 e 6,5.
3.2.1. Escolha da cepa produtora de quitosanase
A escolha do microrganismo Metarhizium anisopliae foi feita com base na capacidade
de crescer em meio contendo quitosana como única fonte de carbono, além de apresentar
grande tolerância a pH variando a uma taxa de 2,0 a 8,5, sendo 6,9 a melhor condição para o
crescimento vegetativo e esporução, de acordo com Assis (2010).
3.2.2. Manutenção da cepa
A manutenção da cultura foi realizada em placas petri contendo o meio Batata
Dextrose Agar (PDA). A renovação das células foi realizada a cada trinta dias através de
subcultivos das células no meio PDA.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
51
3.3. Produção de enzima
No preparo do inóculo, utilizou-se 7 placas cultivadas e mantidas, com o fungo
Metarhizium anisopliae, em meio PDA a 25 ºC durante 5 dias em estufa. O meio de cultura
para a produção da enzima quitosanase apresentou os seguintes componentes: quitosana;
peptona; extrato de levedura; NaNO3; K2HPO4; KCl; MgSO4.7H2O e FeSO4, com
concentrações de acordo as matrizes dos planejamentos: Plackett & Burman e composto
central. Para a produção da enzima no cultivo, a biomassa foi transferida através de água
estéril, para um Béquer de 500 mL que, em seguida, foi colocado em Erlenmeyers de 250 mL
na proporção de 20 mL de biomassa para 180 mL do meio de cultura. Os Erlenmeyers
cultivados foram então incubados em incubador rotativo a 25 ºC e 150 rpm durante 24 h.
Alíquotas foram retiradas e centrifugadas a 8000xg por 5 min, a cada 6 horas, para se
determinar a atividade quitosanolítica, teores de açúcares redutores e de proteína total.
Ressalta-se que todos os experimentos foram feitos em triplicata.
3.4. Determinação da Proteína Total
A determinação da proteína total foi efetuada de acordo com o método modificado de
Sedmak & Grossberg (1977), que se baseia na diminuição da absorbância do corante azul de
Coomassie, devido à ligação proteína-corante, a 595 nm, comparado a absorção máxima a
465 nm, conforme Santos (2001). Nesse caso, após centrifugação para retirada da biomassa,
transferia-se para um eppendorf 1,5 mL de amostra (caldo fermentativo) e adicionava-se 1,5
mL de solução azul de Coomassie. Deixava-se a solução em repouso por 5 minutos,
transferindo-se em seguida para um cubeta de quartzo e efetuando-se as leituras de
absorbância nos comprimentos de ondas 465 nm e 595 nm, respectivamente. O
espectrofotômetro (Thermo Spectronic modelo Genesys 10UV) foi zerado com água
destilada. O conteúdo protéico foi obtido a partir de uma curva padrão na qual graficava-se a
relação de absorbância A595/A465 para diferentes concentrações de Albumina de Soro
Bovino (Fração V – Lote 110881/Sigma Aldrich-Ohio-USA).
3.5. Determinação da Atividade Quitosanolítica e da Atividade Específica
A atividade quitosanolítica foi obtida monitorando-se o aumento dos açúcares
redutores totais, utilizando-se o método DNS a partir de uma curva padrão de glicosamina.
Para a determinação da atividade quitosanolítica adicionaram-se, aos tubos de ensaios 500 µL

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
52
de substrato (solução 0,2 % de quitosana solubilizada em ácido clorídrico 1N) e 500 µL de
caldo bruto (contendo enzima), controlando-se o pH entre 6,0 e 6,5 e incubando-se o sistema
em um banho termostatizado com temperaturas da ordem de 55 ºC. Após o tempo de
incubação de 30 minutos no banho foi adicionado aos tubos de ensaio 2,5 mL de DNS,
aquecendo-se à ebulição em banho-maria por 10 minutos, resfriando-se, em seguida, em
banho de gelo. Após essa etapa, o sistema foi centrifugado e com o sobrenadante efetuou-se a
leitura no espectrofotômetro a 600 nm. Destaca-se, que antes da leitura da amostra zerava-se o
equipamento (branco do espectro) com 1,0 mL de água destilada acrescidos de 2,5 mL de
DNS e realizava-se o branco da enzima que consistia da adição de 500 µL de solução de
quitosana, completando-se até um 1,0 mL com água destilada. Uma unidade de atividade
quitosanolítica (A) foi definida como a quantidade de enzima necessária para produzir 1
µmol.min-1 de glicosamina por mL de extrato enzimático, nas condições de temperatura e pH
citadas, sendo o resultado expresso em (U.mL-1). A Atividade específica foi determinada pela
divisão entre a atividade da enzima (U.mL-1) e a concentração de proteína total (µg.mL-1),
sendo expressa em (U. µg-1).
3.6. Planejamento experimental
3.6.1. Planejamento de seleção de fatores Plackett & Burman
O planejamento experimental Plackett & Burman é uma técnica muito usada, para se
realizar triagem de fatores na otimização num determinado processo (Reddy et al., 2008), e
foi utilizado neste trabalho para identificar os fatores que influenciam de forma significativa
na produção da enzima quitosanase.
A Tabela 3 ilustra os níveis de cada variável (fator) utilizada no planejamento Plackett
& Burman, sendo: Concentração de Quitosana (Q), Peptona (PP), Extrato de Levedura (EL),
NaNO3 (NS), K2HPO4 (FP), KCl (CP), MgSO4.7H2O (SM), FeSO4 (SF). Já a Tabela 4 mostra
a matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3 repetições
na condição do ponto central.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
53
Tabela 3 – Níveis para os fatores e seus valores codificados.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1 0,1 0,2 0,2 0,1 0,05 0,025 0,025 0,0005
0 0,2 0,5 0,3 0,2 0,10 0,050 0,050 0,0010
1 0,3 0,8 0,4 0,3 0,15 0,075 0,075 0,0015
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1
2 1 1 -1 1 -1 -1 -1 1 -1 1 1
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1
5 1 1 -1 1 1 -1 1 -1 1 -1 1
6 1 1 1 -1 1 1 -1 1 1 -1 -1
7 -1 1 1 1 -1 1 1 -1 -1 1 1
8 -1 -1 1 1 1 -1 1 1 1 1 1
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1
11 -1 1 -1 -1 -1 1 1 1 1 1 -1
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1
13 0 0 0 0 0 0 0 0 0 0 0
14 0 0 0 0 0 0 0 0 0 0 0
15 0 0 0 0 0 0 0 0 0 0 0
Sendo: V11,V12,V13 = Variáveis inertes para cálculo do erro.
3.6.2. Planejamento Composto Central para a Otimização do Meio de Cultura para a
Produção da Enzima em Cultivo Submerso
Um dos problemas mais comuns para quem realiza experimentos é determinar a
influência de uma ou mais variáveis sobre outra variável de interesse (Lobato, 2003). Este

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
54
problema pode ser minimizado utilizando-se um planejamento experimental de modo a se
obter uma função, ou uma aproximação satisfatória para a mesma, de forma que se operando
sobre as variáveis de entrada produza-se como saída a resposta ou as respostas observadas
(Barros Neto, 2001; Silva, 2004).
Em um planejamento, inicialmente, deve-se definir os fatores e as respostas de
interesse. Tendo-se identificado os fatores e respostas define-se o objetivo que se pretende
alcançar com os experimentos para que se possa escolher o planejamento mais apropriado.
(Silva, 2004).
No presente trabalho foi utilizado um Planejamento Composto Central (CCD) com 22
experimentos na parte fatorial, repetição em triplicata no ponto central e quatro experimentos
na parte axial (planejamento do tipo estrela), para se investigar a influência dos fatores:
concentração de quitosana e concentração de FeSO4 que foram os dois fatores que
influenciaram a produção de quitosanase, conforme o planejamento Plackett & Burman de 12
ensaios e mais 3 repetições na condição do ponto central. Estes dois fatores foram estudados
em cinco diferentes níveis (-1,41; -1; 0; 1; 1,41) e um conjunto de 11 experimentos foi
realizado, conforme Tabela 5.
Tabela 5 – Matriz do planejamento Composto Central.
Ensaio Q SF
1 -1 -1
2 -1 1
3 1 -1
4 1 1
5 -1,41 0
6 1,41 0
7 0 -1,41
8 0 1,41
9 0 0
10 0 0
11 0 0
Os outros fatores foram mantidos nos níveis mais baixos (para evitar custos). Uma que
não influenciaram de forma significativa na produção da enzima quitosanase.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
55
A codificação dos fatores foi realizada de acordo com a seguinte equação (Ji et al.,
2009):
Xi = (xi – x0)/∆xi , i = 1, 2, 3, ..., k (2)
Onde Xi é o fator independente codificado, xi é o fator independente real, x0 é o valor de xi no
ponto central e ∆xi é a variação de valor. O comportamento do sistema foi explicado pela
seguinte equação polinomial de segunda ordem:
Yp = β0 + ∑ βiXi + ∑ βiiXi2 + ∑ βijXiXj, i, j = 1, 2, 3,..., k (3)
Onde Yp foi a resposta esperada, β0 foi a interseção, Xi e Xj
foram os fatores independentes codificados, βi foi o coeficiente linear, βii
foi o coeficiente quadrático e βij foi o coeficiente de interação. Destaca-se que os outros
fatores não significativos foram mantidos no nível mais baixo (-1).
A Tabela 6 apresenta os níveis para os fatores e seus valores codificados, para o
Planejamento Composto Central, com triplicata no ponto central.
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1,41 0,218 0,2 0,2 0,1 0,05 0,025 0,025 0,0009
-1 0,3 0,2 0,2 0,1 0,05 0,025 0,025 0,0015
0 0,5 0,2 0,2 0,1 0,05 0,025 0,025 0,0030
1 0,7 0,2 0,2 0,1 0,05 0,025 0,025 0,0045
1,41 0,782 0,2 0,2 0,1 0,05 0,025 0,025 0,0051
Na análise do planejamento de seleção de fatores Plackett & Burman e do
planejamento experimental fatorial foi utilizado um pacote computacional STATISTICA for
Windows versão 7.0.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
56
3.7. Perfil cinético durante o cultivo
Um perfil cinético na melhor condição de cultivo, conforme planejamento
apresentado, foi realizado em frascos Erlenmyer de 250 mL contendo 200 mL de meio de
cultivo contendo os seguintes componentes, em 1 Litro: 2,0g de quitosana; 5,0g de peptona;
3,0g de extrato de levedura; 2,0g de NaNO3; 1,0g de K2HPO4; 0,5g de KCl; 0,5g de
MgSO4.7H2O; 0,01g de FeSO4, que foram inoculadas com 10% (vol/vol) de inóculo contendo
o fungo Metarhizium anisopliae e cultivadas em incubador rotativo durante 5 dias a 150 rpm
e 25 ºC. Alíquotas foram retiradas a cada seis horas e analisadas quanto à proteína total no
sobredanante, atividade enzimática, atividade específica e açúcares redutores. Os ensaios
foram realizados em triplicata (Silva Filho, 2005).

Capítulo 4 Resultados e
Discussão

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
58
4. Resultados e discussão
Neste capítulo são apresentados e discutidos os resultados obtidos no presente
trabalho. Inicialmente, é apresentada imagem do MEV do Metarhizium anisopliae
mantidos em meio PDA. Em seguida, apresenta-se e discute-se os resultados da avaliação
dos fatores significativos, que podem influenciar na produção da quitosanase, no
planejamento de seleção de fatores, conforme planejamento Plackett & Burman bem como
são apresentados e discutidos os resultados obtidos para o planejamento composto central.
Por último, apresenta-se o perfil cinético, na melhor condição, para a produção de
quitosanase em cultivo submerso descontínuo usando o fungo Metarhizium anisopliae.
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA.
A Figura 7 ilustra a imagem da Microscopia Eletrônica de Varredura (MEV) do micélio de Metarhizium anisopliae mantido em meio PDA.
Figura 7. Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA.
Observa-se na Figura 7, quanto à morfologia que o Metarhizium anisopliae, apresenta-
se como um fungo filamentoso com as hifas intimamente entrelaçadas conforme citado por
Wang et al. 2002.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
59
4.2. Planejamento Plackett & Burman
4.2.1 Análise do Plackett & Burman com 8 fatores e 12 ensaios
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis, foi utilizado para avaliar a influência dos fatores: Concentração de
Quitosana (Q), Peptona (PP), Extrato de Levedura (EL), NaNO3 (NS), K2HPO4 (FP), KCl
(CP), MgSO4.7H2O (SM), FeSO4 (SF) na produção da enzima quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso. Ressalta-se que os experimentos foram
realizados aleatoriamente.
A Tabela 7 apresenta a matriz do modelo Plackett & Burman e os respectivos valores
obtidos para a variável resposta (Atividade Quitosanalítica).
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.
Ensaio Q PP EL NS FP CP SM SF A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 0,0195
2 1 1 -1 1 -1 -1 -1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 0,0109
4 1 -1 1 1 -1 1 -1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 0,0136
6 1 1 1 -1 1 1 -1 1 0,0156
7 -1 1 1 1 -1 1 1 -1 0,0109
8 -1 -1 1 1 1 -1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 0,0085
A Tabela 8 apresenta os efeitos estimados para o planejamento Plackett & Burman.
Observa-se na Tabela 8 que o valor do coeficiente de determinação R2 foi de
aproximadamente 90%, indicando que um modelo linear poderia representar bem a relação
entre o fator significativo, concentração de quitosana (1), e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
60
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman.
Efeitos Estimados; R2=0,8996; Ajuste:0,63178 MS Erro Residual =0,0000054
Desvio Padrão t(3) p
Lim. Conf. Lim. Conf.
Efeito Erro Residual
-95,% +95
Média/Interação 0,0127 0,0007 18,903 0,0003 0,0105 0,0149 (1) Quitosana 0,0055 0,0013 4,0628 0,0268 0,0011 0,0098
(2) Peptona 0,0015 0,0013 1,1080 0,3486 -0,0028 0,0058 (3) Ext. de leved. 0,0009 0,0013 0,7387 0,5136 -0,0033 0,0053
(4) NaNO3 0,0002 0,0013 0,1758 0,8715 -0,0040 0,0045 (5) K2HPO4 -0,0014 0,0013 -1,0552 0,3687 -0,0057 0,0028
(6) KCl -0,0022 0,0013 -1,6708 0,1933 -0,0065 0,0020 (7) MgSO4.7H2O -0,0003 0,0013 -0,2286 0,8338 -0,0046 0,0039
(8) FeSO4 0,0029 0,0013 2,1457 0,1211 -0,0014 0,0071
A Figura 8 mostra o gráfico de Pareto, uma das formas de se avaliar visualmente a
influência dos fatores estudados na resposta. A magnitude dos efeitos é representada pelas
colunas enquanto que a linha transversal às colunas representa a magnitude dos efeitos com
significado estatístico para p=0,05, ou seja, os fatores que são estatisticamente significativos
ao nível de 95% de confiança.
Figura 8. Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
61
Analisando-se o gráfico de Pareto (Figura 8) observa-se que o fator concentração de
quitosana (1) foi o único fator que influenciou de forma mais significativa na atividade
enzimática atingindo efeitos estimados de aproximadamente 4,06. O sinal positivo indica que
o aumento da concentração de quitosana aumentará à atividade enzimática. Por outro lado, os
outros fatores parecem desprezíveis estatisticamente conforme o gráfico de Pareto.
4.2.2 Análise do Plackett & Burman com 12 Ensaios e mais 3 Repetições na condição do
ponto central.
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis com repetição em triplicata no ponto central, foi utilizado para se avaliar a
influência dos mesmos fatores descritos na Seção 4.2.1 para a produção da enzima
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso. Ressalta-se que os
experimentos foram realizados aleatoriamente. A Tabela 9 apresenta a matriz do modelo
Plackett & Burman e os respectivos valores obtidos para a variável resposta (Atividade
Quitosanalítica).
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13 A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1 0,0195
2 1 1 -1 1 -1 -1 -1 1 -1 1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1 0,0109
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 1 -1 1 0,0136
6 1 1 1 -1 1 1 -1 1 1 -1 -1 0,0156
7 -1 1 1 1 -1 1 1 -1 -1 1 1 0,0109
8 -1 -1 1 1 1 -1 1 1 1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 1 1 -1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1 0,0085
13 0 0 0 0 0 0 0 0 0 0 0 0,0704
14 0 0 0 0 0 0 0 0 0 0 0 0,0688
15 0 0 0 0 0 0 0 0 0 0 0 0,0710

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
62
A Tabela 10 a seguir apresenta os efeitos estimados para planejamento Plackett &
Burman com 3 repetições na condição do poto central. Observa-se na Tabela 10 que o valor
do coeficiente de determinação R2 foi de aproximadamente 2,0%, indicando que um modelo
linear não poderia representar a relação entre os fatores significativos concentração de
quitosana (1), concentração de FeSO4 (8) e a resposta.
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central.
Efeitos Estimados; R2=0,01811; Ajuste:0,0 MS Erro Puro =0,0000012
Desvio Padrão t(2) P
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0242 0,0003 85,509 0,0001 0,0229 0,0254 (1) Quitosana 0,0055 0,0006 8,6432 0,0131 0,0027 0,0082
(2) Peptona 0,0015 0,0006 2,3572 0,1425 -0,0012 0,0042 (3) Ext. de leved. 0,0009 0,0006 1,5715 0,2567 -0,0017 0,0037
(4) NaNO3 0,0002 0,0006 0,3742 0,7442 -0,0024 0,0029 (5) K2HPO4 -0,0014 0,0006 -2,2450 0,1538 -0,0041 0,0013
(6) KCl -0,0022 0,0006 -3,5546 0,0708 -0,0049 0,0005 (7) MgSO4.7H2O -0,0003 0,0006 -0,4864 0,6747 -0,0030 0,0024
(8) FeSO4 0,0029 0,0006 4,5648 0,0448 0,0002 0,0056
A Figura 9 ilustra o gráfico de Pareto para o planejamento Plackett & Burman, com
repetição em triplicata no ponto central, uma das formas de se avaliar visualmente a influência
dos fatores estudados na resposta, onde é mostrado a magnitude dos efeitos para 95 % de
confiança.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
63
Figura 9. Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto
central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto, observa-se que os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma mais
significativa na atividade enzimática atingindo efeitos estimados de aproximadamente 8,84 e
4,67, respectivamente. O sinal positivo indica que o aumento das concentrações de quitosana
e de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores parecem
desprezíveis estatisticamente conforme o gráfico de Pareto.
Este comportamento do FeSO4 está de acordo com o observado por Hagag et al.
(2012) que utilizando o Aspergillus fumigatus na produção de protease mostraram que um
fator de transcrição PrtT regula positivamente o controle da produção de protease, da
assimilação de Ferro e da síntese de ergosterol e que a produção de protease foi favorecida
pelo aumento da concentração de ferro.
Rajendran & Thangavelu (2009) utilizaram um planejamento Plackett & Burman para
identificar quais os fatores significativos na produção de lipase por Rhizopus arrhizus MTCC
2233 e observaram o efeito significativo médio do FeSO4, apresentando um nível de
confiança para esse fator de 48,92% (para um máximo de 98,7 % de MgSO4), com uma
produção de lipase da ordem de 2,98 U/mL em 84 horas de fermentação e que ocorreu uma
produção considerável de protease no meio de cultivo da ordem de 1,92 U/mL em 96 horas de
cultivo.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
64
Mu et al. (2005) também observaram um efeito positivo do FeSO4, da ordem de 1,48
para um fator máximo encontrado de 3,11 de MgSO4, no aumento da atividade da
estereoinversão de S-fenil-1,2-etanodiol por Candida parapsilosis CCTCC M203011 na
otimização da fermentação utilizando um planejamento Plackett & Burman.
Dessa forma, observa-se que o FeSO4 é importante uma que vez o mesmo está
diretamente associado a síntese de proteases (principalmente metalloproteases), conforme
citado por Hagag et al. (2012).
4.3. Planejamento Composto Central para Produção de Quitosanase por
Metarhizium anisopliae em Cultivo Descontínuo Submerso
Um planejamento experimentos 22 com repetição em triplicata no ponto central e mais
quatro ensaios axiais, foram utilizados para se chegar a uma combinação ideal de quitosana e
FeSO4 na produção da enzima quitosanase (Tabela 11). A partir do modelo Plackett &
Burman (Tabela 12), que mostrou que o aumento das concentrações de quitosana e FeSO4,
favoreceram a produção de quitosanase, os valores utilizados neste modelo para estes
componentes nos níveis mais altos, serão considerados como os níveis mais baixos no
planejamento experimental do composto central. Os outros fatores foram fixados em todos os
ensaios em seus níveis mais baixos para a redução de custos (Tabela 11).

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
65
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da
atividade enzimática.
Ensaio Q SF A (U/mL)
1 -1 -1 0,0195
2 -1 1 0,0202
3 1 -1 0,0109
4 1 1 0,0121
5 -1,41 0 0,0136
6 1,41 0 0,0156
7 0 -1,41 0,0108
8 0 1,41 0,0103
9 0 0 0,0099
10 0 0 0,0116
11 0 0 0,0094
A Tabela 12 apresenta os efeitos estimados para o planejamento composto central.
Tabela 12 – Efeitos Estimados do planejamento Composto Central.
Efeitos Estimados; R2=0,629; Ajuste:0,25836 MS Erro Puro =0,0000014
Desvio Padrão t(2) p
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0103 0,0006 15,305 0,0042 0,0074 0,0132 (1) Quitosana (L) 0,0011 0,0008 1,4316 0,2885 -0,0023 0,0047
Quitosana (Q) 0,0058 0,0009 5,9637 0,0269 0,0016 0,0101 (2) FeSO4 (L) -0,0043 0,0008 -5,3037 0,0337 -0,0079 -0,0008 (2) FeSO4 (Q) 0,0018 0,0009 1,8366 0,2076 -0,0024 0,0060
1L x 2L 0,0001 0,0011 0,1520 0,8931 -0,0048 0,0052
Observa-se na Tabela 12 que o valor do coeficiente de determinação R2 foi de
aproximadamente 63%, indicando que um modelo linear representaria bem a relação entre os
fatores significativos concentração de quitosana (1), concentração de FeSO4 (8) e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
66
A Tabela 13 mostra um resumo da ANOVA para o planejamento composto central
com repetição em triplicata no ponto central.
Tabela 13 – Resumo da ANOVA.
Fonte de
Variação
Soma dos
Quadrados
Grau de
Liberdade
Quadrado
Médio
Fcalculado Ftabelado
Regressão
Resíduos
0,000086 2 0,000043 6,14 4,46
0,000056 8 0,000007
Falta de
Ajuste
Erro puro
0,000053 6 0,000009 6 19,33
0,000003
2
0,0000015
Total 0,000142 10
De acordo com a Tabela 13, partindo-se dos efeitos significativos propõe-se o
modelo:
� = 0,010316 + 0,002942� � − 0,002193�� (8)
% de variação explicável: 60,5
% máxima de variação explicável: 97,9
A análise da ANOVA mostra que o modelo proposto possui significância estatística, ao
nível de 95% de confiança, uma vez que o valor de F (calculado) é maior que F (tabelado),
entretanto a ANOVA mostra também que o modelo proposto pode ser útil para se fazer
previsões, uma vez que a relação entre F (calculado) e F (tabelado) para a falta de ajuste e o
erro puro é menor que 1. O coeficiente de determinação R2 foi de aproximadamente 63%,
indicando que o modelo consegue explicar 63% da variação total em torno da média.
Os cálculos para variação explicável e para máxima variação explicável são dados pelas
Equações (9) e (10):
% de variação explicável = (SQ da regressão) / (SQ Total) (9)
% máxima de variação explicável = (SQ Total – SQ Erro puro) / (SQ Total) (10)
Sendo:
SQ – Soma dos quadrados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
67
A Figura 10 mostra o gráfico de Pareto para o planejamento Composto Central
utilizando erro puro. A magnitude dos efeitos é representada pelas colunas. Já a linha
transversal às colunas representa a magnitude dos efeitos com significado estatístico para 95%
de confiança.
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto (Figura 10) observa-se que os fatores concentração
de quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma
mais significativa na atividade enzimática atingindo efeitos estimados de aproximadamente
5,96 e - 5,30, respectivamente. O sinal positivo indica que o aumento das concentrações de
quitosana aumentará à atividade enzimática. O sinal negativo indica que o decréscimo da
concentração de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores
parecem desprezíveis estatisticamente no gráfico de Pareto.
A avaliação do modelo também pode ser feita através da observação do gráfico dos
valores preditos em função os valores observados que são mostrados na Figura 11. Os valores
preditos pelo modelo são representados pela reta, enquanto que os valores observados
representam-se pelos pontos.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
68
Figura 11 – Valores preditos em função dos valores observados.
Observa-se que alguns valores preditos apresentam um afastamento dos valores
observados. A Figura 12 ilustra a superfície de resposta da atividade quitosanolítica em
função dos dois fatores significativos, concentração de quitosana e concentração de FeSO4.
Figura 12. Atividade quitosanolítica em função da concentração de quitosana e da
concentração de FeSO4 para os fatores não significativos no ponto central.
Observa-se a existência de um ponto de mínimo, ocorrendo para concentração de
quitosana no ponto central (0) e para maiores concentrações de FeSO4 igual a (1,5). Essa

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
69
região deve ser evitada uma vez que a atividade quitosanolítica atinge níveis tão reduzidos
quanto 0,0094 U.mL-1. Destaca-se que nesse planejamento o maior nível da concentração de
FeSO4 (α=+1,41) equivale a 0,0051% (Tabela 6) que é aproximadamente cinco vezes maior
que o nível desse fator no ponto central do planejamento Plackett & Burman (0,001%),
conforme Tabela 3. Dessa forma, ao se utilizar o nível mínimo da concentração de FeSO4 no
Planejamento Composto Central (α=-1,41), que equivale a aproximadamente 0,001%,
aproxima-se do ponto central do do planejamento Plackett & Burman (0,001%), condição
essa que apresenta os melhores resultados de atividade quitosanolítica usando quitosana como
fonte de carbono e energia e fungo Metarhizium anisopliae.
4.4. Perfil Cinético – Melhor Condição
A seguir, apresenta-se um perfil cinético, na melhor condição, para produção da
quitosanase usando o fungo Metarhizium anisopliae, em que foram monitorados durante 24
horas os seguintes parâmetros: Concentração de proteínas totais (µg/mL), atividade
quitosanolítica (U/mL), atividade específica (U/µg) e açúcares redutores (mg/mL), ilustradas
nas Figuras 13, 14, 15 e 16, respectivamente.
Com relação à proteína extracelular, observa-se na Figura 13 que, inicialmente, ocorre
um aumento da mesma para o seu nível máximo (6,2 µg/mL), durante as 6 primeiras horas de
cultivo, seguido de uma redução para 5,7 µg/mL em 18 horas de cultivo, possivelmente
devido à presença de proteases no meio. Observa-se que a partir de então ocorre uma redução
e aumento da proteína total, em uma quantidade inferior, sugerindo um mecanismo de
controle.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
70
0 4 8 12 16 20 245,0
5,2
5,4
5,6
5,8
6,0
6,2
6,4
Pro
tein
a (
µg
/mL
)
Tempo (h)
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana.
Com relação à atividade quitosanolítica, conforme Figura 14, a atividade máxima
ocorreu em 18 horas de produção em que se obteve aproximadamente 70,0 U/L, o que foi 28
vezes maior que o valor (2,5 U/L em 48 horas) relatado por Assis (2010) para o mesmo fungo,
e em apenas um terço do tempo de produção.
0 4 8 12 16 20 240,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,10
Ativid
ade
(U
/mL
)
Tempo (h)
Figura 14. Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
71
Além disso, destaca-se o considerável aumento da produção de quitosanase a partir das
12 horas até 18 horas de cultivo onde atingiu o pico máximo de produção (70,0 U/L). Após o
pico em 18 horas, a produção começa a cair rapidamente, comportamento esse observado
durante todos os ensaios cinéticos. Destaca-se que é importante do ponto de vista industrial
manter a produção em alto nível por muitas horas, onde se espera uma produção contínua ou
em grandes bateladas.
Ak et al. (1998) utilizaram o fungo Penicillium spinulosum para produzir a quitosanase
usando um meio de cultura contendo 0,1% de K2HPO4; 0,05 % de MgSO4.7H2O; 0,05% de
KCl; 0,001 % de FeSO4, 0,5% NH4NO3 e como fonte de carbono e energia, os autores
usaram quitosana e parede de Rhizopus. A produção foi realizada em cultivo submerso a 28
oC. Os autores apresentam atividade quitosanolítica máxima de aproximadamente 47,0 U/mL,
ao se utilizar a parede do fungo como fonte de carbono e energia, e 30,0 U/mL ao se utilizar
quitosana como fonte de carbono e energia. Destaca-se que essas atividades ocorreram após 4
dias de cultivo. Os perfís com o tempo da atividade quitosanolítica e da atividade específica
apresentaram tendência de crescimento, com o pico máximo em 4 dias, seguindo com uma
tendência de queda da atividade. Neste tempo, a atividade específica máxima da quitosanase
foi de aproximadamente 60,0 U/mg de proteína. Essas tendências também foram observadas
nos resultados apresentados nos perfís de atividade das Figuras 14 e 15.
0 4 8 12 16 20 24
-0,010
-0,005
0,000
0,005
0,010
0,015
0,020
0,025
0,030
Ativid
ad
e E
spe
cific
a (
U/µ
g)
Tempo (h)
Figura 15. Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
72
Observa-se na Figura 15 que a atividade específica máxima da quitosanase segue a
mesma tendência da atividade quitosanolítica apresentada na Figura 14, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
O perfil para os açúcares redutores durante o cultivo de Metarhizium anisopliae
cultivado em meio contendo 0,2% de quitosana é ilustrado na Figura 16.
0 4 8 12 16 20 240,004
0,008
0,012
0,016
0,020
0,024
0,028
Acuca
res R
ed
uto
res (
mg/m
L)
Tempo (h)
Figura 16. Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana.
Observa-se, na Figura 16, que taxa máxima de redução dos açúcares redutores
(aproximadamente 0,021 mg/mL) ocorre logo após a produção máxima da enzima, em 18
horas de cultivo, conforme Figura 14. A quantidade de açúcares redutores foi a
aproximadamente 0,012 mg/mL após 24 h de cultivo. O fungo excreta para o meio enzimas
com atividade quitosanolítica, essas enzimas hidrolisam a quitosana aumentando a quantidade
de açúcares redutores para o meio, sendo que esses açúcares também são consumidos pelo
fungo durante seu crescimento.
Foi demonstrado por Assis (2010) que os açúcares redutores produzidos através da
quitossana, ocasionada pelas enzimas quitossanases metabolizadas por Metarhizium
anisopliae, com quitosana como sua única fonte de carbono e energia, atingiu um pico de 0,53
mg/mL em 48 horas. Apresentaram como quitooligossacarídeos, constituídos principalmente
por dímeros e pentâmeros, formados pela hidrólise enzimática de quitosanase em quantidades
maiores do que o consumido pelos microrganismos. Mostraram, também, que essa produção

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
73
segue a mesma tendência da sua atividade enzimática, corroborando com os resultados
apresentados no presente trabalho.
Os resultados do presente trabalho mostram a relevância de se adotar novas condições
para a produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter quitooligossacarídeo
com atividades biológicas. Esta pesquisa, portanto, sugere base para novos estudos que
permitam alcançar rapidamente a produção de quitosanase e quitooligossacarídeo com todos
os seus benefícios.

Capítulo 5 Conclusão

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
75
5. Conclusão
Após o desenvolvimento desse trabalho conclui-se:
- Com relação ao Planejamento de Seleção de Fatores Plackett & Burman:
Os resultados das análises do Planejamento de Seleção de Fatores Plackett &
Burman com 8 fatores e 12 ensaios em 2 níveis, mais 3 repetições no ponto central, para
se investigar a influência dos fatores: Concentração de Quitosana, Peptona, Extrato de
Levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O, FeSO4 na produção da enzima
quitosanase por Metarhizium anisopliae, mostraram que a atividade foi favorecida pelo
aumento da concentração de substrato (quitosana) e sulfato ferroso (FeSO4), enquanto
que o aumento da concentração dos outros fatores não contribuíram de forma
significativa para a resposta (atividade quitosanolítica). Os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) atingiram efeitos estimados de
aproximadamente 8,84 e 4,67, respectivamente. O sinal positivo indica que o aumento
das concentrações de quitosana e de FeSO4 aumentará a atividade enzimática. Conforme
visto na literatura, observa-se importância do FeSO4, já que o mesmo está diretamente
associado a síntese de proteases (principalmente metalloproteases),
- Com relação ao Planejamento Composto Central:
A análise do planejamento composto central para avaliar a influência dos
fatores, concentração de substrato (quitosana) e concentração de FeSO4, na produção da
enzima quitosanase por Metarhizium anisopliae (a partir do planejamento Plackett &
Burman) mostrou que os fatores concentração de quitosana e concentração de FeSO4
foram os fatores que influenciaram de forma mais significativa na atividade enzimática
atingindo efeitos estimados de aproximadamente 5,96 e - 5,30, respectivamente. O sinal
positivo indica que o aumento das concentrações de quitosana aumentará a atividade
enzimática. O sinal negativo indica que o decréscimo da concentração de FeSO4
aumentará a atividade enzimática. Observa-se a existência de um ponto de mínimo,
ocorrendo para concentração de quitosana (X1) no ponto central (0) e para maiores

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
76
concentrações de FeSO4 (X2) igual a (1,5) com a atividade quitosanolítica de 0,0094
U.mL-1.
- Com relação ao cultivo na melhor condição:
O cultivo na melhor condição mostrou uma produção máxima de quitosanase em
18 horas de cultivo, com a atividade enzimática de 70,0 U/L, o que foi 28 vezes maior
que o valor (2,5 U/L em 48 horas) relatado na literatura (Assis, 2010) para o mesmo
fungo e em apenas um terço do tempo de produção.
Além disso, observou-se que um elevado incremento na produção de
quitosanase a partir das 12 horas de cultivo, antes de se atingir a atividade
quitosanolítica máxima, 18 horas de cultivo. Essa produção em alto nível por muitas
horas é importante do ponto de vista industrial, onde se espera uma produção contínua
ou em grandes bateladas. Observou-se também que a atividade específica máxima da
quitosanase segue a mesma tendência da atividade quitosanolítica, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
- Com relação aos açúcares redutores:
Foi observado que os açúcares redutores seguem a mesma tendência da atividade
enzimática, com uma produção máxima de cerca de 0,21 mg/mL em 18 horas de
cultivo, produção essa que coincinde com o máximo pico de atividade enzimática.
Estes resultados mostram a relevância de se adotar novas condições para a
produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter
quitooligossacarídeo com atividades biológicas. Esta pesquisa, portanto, sugere base
para novos estudos que permitam alcançar rapidamente a produção de quitosanase e
quitooligossacarídeo com todos os seus benefícios.

Referências Bibliográficas

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
78
Referências bibliográficas
AK, Ö.; BAKIR, U.; GÜRAY, T. Isolation of a chitosan degrading fungus, Penicillium
spinulosum, and chitosanase production by the isolate. In: SYMPOSIUM - NEW
FRONTIERS IN SCREENING FOR MICROBIAL BIOCATALYSTS, 1996, Ede, The
Netherlands. Anais, Editado por K. Kieslich, C. P. van der Beek, J. A. M. de Bont e W.
J. J. van den Tweel, Elsevier Science B. V., 1998, v.53, p.265-268.
ALVES, S. B. Patologia e controle microbiano: Vantagens e desvantagens. In: ALVES,
S. B. Controle Microbiano de Insetos. 2ed., Piracicaba, FEALQ, 1163p, 1998, v.4, p.21-
37.
ALVES, S. B.; LEITE, L. G.; FILHO, A. B.; ALMEIDA, F. E. M.; MARQUES, E. J.
Produção massal de fungos entomopatogênicos na América Latina, In: ALVES, S. B.;
LOPES, R. B. Controle microbiano de pragas na América Latina: avanços e desafios.
Piracicaba, FEALQ, 414p, 2008, v.14, p.215-237.
ARRUDA, W.; LUBECK, I.; SCHRANK, A.; VAINSTEIN, M. H. Morphological
alterations of Metarhizium anisopliae during penetration of Boophilus microplus ticks.
Experimental and Applied Acarology, v. 37, p.231-244, 2005.
ASSIS, C. F. Produção e caracterização de quitooligossacarídeos produzidos pelo fungo
Metarhizium anisopliae e avaliação da citotoxicidade em células tumorais. 2010. 110p.
Tese (Doutorado em Biotecnologia) – Renorbio - Programa de Pós-Graduação em
Biotecnologia. Universidade Federal do Rio Grande do Norte, Natal.
ASSIS, C. F.; COSTA, L. S.; MELO-SILVEIRA, R. F.; OLIVEIRA, R. M.;
PAGNONCELLI, M. G. B.; ROCHA, H. A. O.; MACEDO, G. R.; SANTOS, E. S.,
Chitooligosaccharides antagonizes the cytotoxic effect of glucosamine. World J.
Microbiol. Biotechnol., v.28, n.3, p.1097-1105, 2012.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
79
AZEVEDO, V. V. C.; CHAVES, S. A.; BEZERRA, D. C.; FOOK, M. V. L.; COSTA,
A. C. F. M. Quitina e Quitosana: aplicações como biomateriais. Revista Eletrônica de
Materiais e Processos, v.2.3, p.27-34, 2007.
BARROS NETO, B.; SCARMINIO, I. S.; BRUNS, R. Como fazer experimentos:
pesquisa e desenvolvimento na ciência e na indústria. 2. ed. Campinas: Unicamp, 2001.
401p.
BATISTA FILHO, A.; ALMEIDA, J. E. M.; SANTOS, A. S.; MACHADO, L. A.;
ALVES, S. B. Eficiência de isolados de Metarhizium anisopliae no controle de
cigarrinha-da-raiz de cana-de-açúcar Mahanarva fimbriolata (Hom.: Cercopidae). STAB
– Açúcar, Álcool e Subprodutos, v.21, p.84-89, 2002.
CARVALHO, J. C. M.; SATO, S. Fermentação Descontínua, In: SCHMIDELL, W.;
LIMA, U. A.; AQUARONE, E.; BORZANI, W. Biotecnologia Industrial. São Paulo,
Ed. Edgard Blücher, v.2, p.193-204, 2001.
CHAO, H. J.; SCHWARTZ, J.; MILTON, D. K. Populations and determinats of
airborne fungi in large office buildings. Environmental Health Perspectives, v.110, n.8,
p.777-782, 2002.
CHEN, X.; XIA, W.; YU, X. Purification and characterization of two types of
chitosanase from Aspergillus sp. CJ22-326. Food Research International, v.38, n.3,
p.315-322, 2005.
CHEN, P. T.; CHIANG, C.-J.; CHAO, Y.-P. Medium optimization and production of
secreted Renilla luciferase in Bacillus subtilis by fed-batch fermentation. Biochemical
Engineering Journal, v.49, p.395-400, 2010.
CHENG, C. Y.; LI, Y. An Aspergillus chitosanase with potential for large-ecale
preparation of chitosan oligosaccharides. Biotechnol. Appl. Biochem., v.32, p.197-203,
2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
80
CHOI, Y. J.; Kim, E. J.; Piao, Z.; Yun, Y. C.; Shin, Y. C. Purification and
characterization of chitosanase from Bacillus sp. Strain KCTC 0377BP and its
application for the production of chitosan oligosaccharides. Applied and Environmental
Microbiology, v.70, n.8, p.4522-4531, 2004.
COLAKOGLU, G. The chemical and physical environment for fungal growth. Fak
Matbaa Birimi, Istanbul, 149-166, 2001.
CRAVEIRO, A. C.; CRAVEIRO, A. A.; QUEIROZ, D. C. Quitosana a fibra do futuro.
2. ed. Fortaleza: PADETEC, 2004. V.1, 218p.
DIAS, B. A. Estudo de proteases degradadoras de cutícula produzidas pelo fungo
entomopatogênico Beauveria bassiana. 2005. 70p. Dissertação (Mestrado em
Microbiologia) – Centro de Ciências Biológicas, Programa de Pós-Graduação em
Microbiologia, Universidade Estadual de Londrina, Londrina.
DINARDO-MIRANDA, L. L.; FERREIRA, J. M. G.; CARVALHO, P. A. M.
Influência da época de colheita e do genótipo de cana-de-açúcar sobre a infestação de
Mahanarva fimbriolata (Stal) (Hemiptera: Cercopidae). Neotrop. Entomol, v.30, n.1,
p.145-149, 200.
DONG, C.; ZHANG, J.; HUANG, H.; CHEN, H.; HU, Y. Pathogenicity of a new China
variety of Metarhizium anisopliae (M. anisopliae var. Dcjhyium) to subterranean termite
Odontotermes formosanus. Microbiological Research, v.164, p.27-35, 2009.
DONZELLI, B. G. G.; OSTROFF, G.; HARMAN, G. E. Enhanced enzymatic
hydrolysis of langostino shell chitin with mixtures of enzymes from bacterial and fungal
sources. Carbohydrate Research, v.338, p.1823-1833, 2003.
DRIVER, F.; MILNER, R. J.; TRUEMAN, J. W. H. A taxonomic revision of
Metarhizium based on phylogenetic analysis of rDNA sequence data. Mycological
Research, v.104, n.2, p.134-150, 2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
81
DUNG, P.; MILAS, M.; RINAUDO, M.; DESBRIÉRES, J. Water soluble derivatives
obtained by controlled chemical modifications of chitosan, Carbohydrate Polymers,
v.24, p.209-214, 1994.
ESTEVES, J. A.; CABRITA, J. D.; NOBRE, G. N. Micologia médica, 2. ed. Lisboa:
Fundação Calouste Gulbenkian, 1990. 1058p.
FACCIOTI, M. C. R. Fermentação Contínua, In: Biotecnologia Industrial. Editado por:
SCHMIDELL, W.; LIMA, U. A.; AQUARONE, E.; BORZANI, W., São Paulo, Ed.
Edgard Blücher, v.2, p.541, 2001.
FARIA, M. R.; MAGALHÃES, B. P. O uso de fungos entomopatogênicos no Brasil -
Situação atual e perspectivas. Biotecnologia Ciência & Desenvolvimento, n.22., p.18-
21, setembro/outubro 2001.
FU, J.-Y.; WU, S.-M.; CHANG, C.-T.; SUNG, H.-Y. Characterization of Three
Chitosanase Isozymes Isolated from a Commercial Crude Porcine Pepsin Preparation. J.
Agric. Food Chem., v.51, p.1042-1048, 2003.
FURIGO JUNIOR, A.; PEREIRA, E. P. Enzimas e suas aplicações cinética enzimática.
2001. 39f. Apostila (Graduação de Engenharia Química e Engenharia de Alimentos) –
Centro de Tecnologia, Departamento de Engenharia Química e Engenharia de
Alimentos, Universidade Federal de Santa Catarina, Florianópolis.
GAO, J.; XU, H.; LI, Q.-J.; FENG, X.-H.; LI, S. Optimization of medium for one-step
fermentation of inulin extract from Jerusalem artichoke tubers using Paenibacillus
polymyxa ZJ-9 to produce R,R-2,3-butanediol, Bioresource Technology, v.101, p.7076-
7082, 2010.
GARCIA, M. V.; MONTEIROI, A. C.; SZABO, M. J. P.; PRETTE, N.; BECHARA, G.
H. Mechanism of infection and colonization of Rhipicephalus sanguineus eggs by
Metarhizium anisopliae as revealed by scanning electron microscopy and
histopathology. Brazilian Journal of Microbiology, São Paulo, v.36, n.4, p.368-372,
2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
82
GHANEM, N. B.; YUSEF, H. H.; MAHROUSE, H. K. Production of Aspergillus
terreus xylanase in solid-state cultures: application of the Plackett & Burman
experimental design to evaluate nutritional requirements, Bioresource Technology,
v.73, p.113-121, 2000.
GHORBEL-BELLAJ, O.; HMIDET, N.; JELLOULI, K.; YOUNES, I.; MAALEJ, H.;
HACHICHA, R.; NASRI, M. Shrimp waste fermentation with Pseudomonas aeruginosa
A2: Optimization of chitin extraction conditions through Plackett–Burman and response
surface methodology approaches, International Journal of Biological Macromolecules,
v.48, p.596–602, 2010.
GIUSTINA, A.; VENTURA, P. Weight-reducing regimens in obese subjects: Effects of
a new dietary fiber integrator. Acta Toxicol Ther, 16, 4, p.199-214, 1995.
GUIA.BIO, Reino dos fungos, 2012. Disponível em:
<http://www.http://guia.bio.br/reino-dos-fungos>. Acesso em: 12 novembro 2012.
GUIRADO, N.; AMBROSANO, E. J.; ROSSI, F.; DIAS, F. L. F. Controle de Cupins
de Montículo com Metarhizium anisopliae e Beauveria bassiana, Rev. Bras. de
Agroecologia, v.4, n.2, 2009.
GUO, Y.; XU, J.; ZHANG, Y.; XU, H.; YUAN, Z.; LI, D. Medium optimization for
ethanol production with Clostridium autoethanogenum with carbon monoxide as sole
carbon source, Bioresource Technology, v.101, p.8784–8789, 2010.
HAGAG, S.; KUBITSCHEK-BARREIRA, P.; NEVES, G. W. P.; AMAR, D.;
NIERMAN, W.; SHALIT, I.; SHAMIR, R.; LOPES-BEZERRA, L.; OSHEROV, N.
Transcriptional and proteomic analysis of the Aspergillus fumigatus ∆prtT protease-
deficient mutant. PLoS One, v.7, n.4, e33604, 2012.
HAN, L.-K.; KIMURA, Y.; OKUDA, H. Reduction in fat storage during chitin-chitosan
treatment in mice fed a high-fat diet, International Journal of Obesity, 23:174-179,1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
83
HUNG, T.; CHANG, Y.; SUNG, H.; CHANG, C. Purification and Characterization of
Hydrolase with Chitinase and Chitosanase Activity from Commercial Stem Bromelain.
J. Agric. Food Chem., 50, 4666-4673, 2002.
ICHIKAWA, S.; TAKANO, K.; TAKASHIKUROIWA, HIRUTA, O.; SATO, S.;
MUKATAKA, A. S. Immobilization and stabilization of chitosanase by multipoint
attachment to agar gel support. Journal of Bioscience and Bioengineering, v.93, n.2,
p.201-206, 2002.
JABOR, I. A. S.; PAMPHILE, J. A.; RODRIGUES, S. B.; MARQUES-SILVA, G. G.;
ROCHA, C. L. M. S. Análise do desenvolvimento do fungo entomopatogênico
Metarhizium anisopliae em resposta a fatores nutricionais. Acta Scientiarum.
Agronomy, Maringá, v.25, n.2, p.497-501, 2003.
JI, X.-J.; HUANG, H.; DU, J.; ZHU, J.-G.; REN, L.-J.; LI, S.; NIE, Z.-K. Development
of an industrial medium for economical 2,3-butanediol production through co-
fermentation of glucose and xylose by Klebsiella oxytoca, Bioresource Technology,
v.100, p.5214–5218, 2009.
JO, Y.; JO, K.; JIN, Y.; KIM, K.; SHIN, J.; KIM, Y.; PARK, R. Characterization and
kinetics of 45 kDa chitosanase from bacillus sp. P16. Biosci. Biotechnol. Biochem.,
v.67, n.9, 1875 – 1882, 2003.
KHOSRAVI-DARANI, K.; ZOGHI, A.; ALAVI, S. A.; FATEMI, S. S. A. Application
of Plackett Burman Design for Citric Acid Production from Pretreated and Untreated
Wheat Straw, Iran. J. Chem. Chem. Eng.. v.27, n.1, p.91-104, 2008.
KIM, P.; KANG, T. H.; CHUNG, K. J.; KIM, I. S.; CHUNG, K. Purification of a
constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and
expression of the gene. FEMS Microbiolog.y Letters 240, 31–39, 2004.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
84
KIM, S.-K.; RAJAPAKSE, N. Enzymatic production and biological activities of
chitosan oligosaccharides (COS): A review. Carbohydrate Polymers, v.62, n.4, p.357-
368, 2005.
KIRK, P. M.; CANON, P.F.; MINTER, D.W.; STALPERS, J.A. Dictionary of the
fungi. CAB International, Wallingford, 2008.
KONDO, H.; OSADA, A. Influence of dietary fiber on the bioavailability of zinc in
rats, Biomed Environ Sci, 9, 204-8, 1996.
KUMAR, S.; KOH, J. Physiochemical, Optical and Biological Activity of Chitosan-
Chromone Derivative for Biomedical Applications. Int. J. Mol. Sci., v.13, p.6102-6116,
2012.
KURITA, K.; SANNAN, T.; IWAKURA, Y. Studies on chitin, 4. Evidence for
formation of block and random copolymers of N-acetyl-o-glucosamine and
oglucosamine by hetero and homogeneous hydrolyses. Die Makromolecular Chemie,
v.178, p.3197-3202, 1977.
KUROIWA, T.; ICHIKAWA, S.; HIRUTA, O.; SATO, S.; MUKATAKA, S. Factors
Affecting the Composition of Oligosaccharides Produced in Chitosan Hydrolysis Using
Immobilized Chitosanases. Biotechnol. Prog., 18, 969-974, 2002.
LI, L.-Z.; HUI, Z.; MO, X. Optimization of the medium for streptolydigin production
by Streptomyces lydicus AS 4.2501 with precursor feeding using response surface
methodology, Journal of the Taiwan Institute of Chemical Engineers, 41, 252–258,
2010.
LIU, B.-L.; KAO, P.-M.; TZENG, Y.-M.; FENG, K.-C. Production of chitinase from
Verticillium lecanii F091 using submerged fermentation. Enzyme and Microbial
Technology, v.33, p.410-415, 2003.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
85
LOBATO, A. K. C. L. Estudo da produção de biosurfactantes por micro-organismos
isolados de poços de petróleo. 2003. 108p. Dissertação (Mestrado) – Programa de Pós-
Graduação em Engenharia Química. Universidade Federal do Rio Grande do Norte.
Natal.
LOPES, R. B.; ALVES, S. B.; PADULLA, L. F. L.; PÉREZ, C. A. Eficiência de
formulações de Beauveria bassiana e Metarhizium anisopliae para o controle de ninfas
de Amblyomma cajennense (Fabricius, 1787). Revista Brasileira de Parasitologia
Veterinária, v.16, n.1, p.27-31, 2007.
LOUREIRO, E. S.; BATISTA FILHO, A.; ALMEIDA, J. E. M.; MENDES, J. M.;
PESSOA, L. G. A. Eficiência de isolados de Metarhizium anisopliae (metsch.) sorok.
no controle da cigarrinha-da-raiz da cana-de-açúcar, Mahanarva fimbriolata (stal, 1854)
(hemiptera: cercopidae), em condições de campo. Arq. Inst. Biol., São Paulo, v.79, n.1,
p. 47-53, 2012.
MELLO, D. R.; REIS, R. C. S.; BITTENCOURT, V. R. E. P. Patogenicidade in vitro
do fungo Metarhizium anisopliae (Metschnikoff, 1879) Sorokin, 1883, sobre o carrapato
Boophilus microplus (Canestrini, 1887). Revista Brasileira de Parasitologia Veterinária,
São Paulo, v.15, p.157-162, 2006.
MITSUTOMI, M.; KIDOH, H.; TOMITA, H.; WATANABE, T. The action of Bacillus
circulans WL-12 chitinases on partially N-acetylated chitosan, Bioscience
Biotechnology and Biochemistry, v.59, p.529-531, 1995.
MALAJOVICH, M. A. Biotecnologia: fundamentos. Ed. biblioteca Max Feffer do
Instituto de Tecnologia ORT, Rio de Janeiro, 2009. 104p.
MONAGHAN, R. L.; EVELEIGH, D. E.; TEWARI, R. P.; REESE, E. T. Chitosanase,
a novel enzyme. Nature - New Biology, 245, p.78-80, 1973.
MU, X. Q.; XU, Y.; NIE, Y.; OUYANG, J.; SUN, Z. H. Candida parapsilosis CCTCC
M203011 and the optimization of fermentation medium for stereoinversion of (S)-1-
phenyl-1,2-ethanediol, Process Biochemistry, v.40, p.2345–2350, 2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
86
MURRAY, P. R.; ROSENTHAL, K. S.; PFALLER, M. A. Medical microbiology. 5th
ed. London, Elsevier Mosby, 2005.
MUZZARELLI, R. A. A. Biochemical significance of exogenous chitins and chitosans
in animals and patients, Carbohydrate Polymers, v.20, p.7-16, 1993.
MUZZARELLI, R. A. A.; TOMASETTI, M.; ILARI, P. Depolymerization of chitosan
with the aid of papain, Enzyme Microbiology and Technology, v.16, p.110-114, 1994.
NAVEENA, B. J.; ALTAF, Md.; BHADRIAH, K.; REDDY, G. Selection of medium
components by Plackett–Burman design for production of L(+) lactic acid by
Lactobacillus amylophilus GV6 in SSF using wheat bran, Bioresource Technology,
v.96, p.485-490, 2005.
NOGAWA, M.; TAKAHASHI, H.; KASHIWAGI, A.; OHSHIMA, K.; OKADA, H.;
MORIKAWA, Y. Purification and Characterization of Exo--D-Glucosaminidase from
a Cellulolytic Fungus, Trichoderma reesei PC-3-7, Appl. Environ. Microbiol., v.64, n.3,
p.890-895, 1998.
ONOFRE, S. B.; VARGAS; L. R. B.; ROSSATO, M.; BARROS, N. M.; BOLDO, J.
T.; NUNES, A. R. F.; AZEVEDO, J. L. Controle biológico de pragas na agropecuária
por meio de fungos entomopatogênicos. In: SERAFINI, L. A.; BARROS, N. M.;
AZEVEDO, J. L. (Org.). Biotecnologia – avanços na agricultura e na agroindústria.
Caxias do Sul: EDUCS, p.295-328, cap.8, 2002.
OSSWALD, W. F.; SHAPIRO, J. P.; DOOSTDAR, H.; MCDONALD, R. E.; NIEDZ,
R. P.; NAIRN, C. J.; HEARN, C. J.; MAYER, R. T. Identification and Characterization
of Acidic Hydrolases with Chitinase and Chitosanase Activities from Sweet Orange
Callus Tissue, Plant Cell Physiol. 35, p.811-820, 1994.
PAGNONCELLI, M. G. B. Estudo do mecanismo de produção de oligossacarídeos com
atividades nutracêuticas a partir da quitosana por hidrólise enzimática com processo

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
87
fermentativo simultâneo. 2008. 118p. Tese (Doutorado) – Centro de Tecnologia,
Programa de Pós-Graduação em Engenharia Química. Universidade Federal do Rio
Grande do Norte. Natal.
PALMA-GUERRERO, J., JANSSON, H.-B., SALINAS, J.; LOPEZ-LLORCA, L. V.
Effect of chitosan on hyphal growth and spore germination of plant pathogenic and
biocontrol fungi, J. Appl. Microbiol., v.104, p.541-553, 2008.
PAULA, F. A.; ARRUDA, D. C. Análise do método de planejamento experimental
Plackett Burman na otimização de degradação de penas de aves por comunidades
microbianas, XXI Salão de Iniciação Científica, 19-23, outubro, 2009, UFRGS, Porto
Alegre.
PELCZAR, M. J.; CHANG, E. C. S.; KRIEG, N. R. Microbiologia: conceitos e
aplicações, v.2, 2.ed. São Paulo: Makron BooKs, 1996.
PELLETIER, A.; SYGUSCH, J. Purification and characterization of three chitosanase
activities from Bacillus megaterium P1, Applied and Environmental Microbiology, v.
56, n.4, p.844-848, 1990.
PEREIRA, F. B.; GUIMARÃES, P. M. R.; TEIXEIRA, J. A.; DOMINGUES, L.
Optimization of low-cost medium for very high gravity ethanol fermentations by
Saccharomyces cerevisiae using statistical experimental designs, Bioresource
Technology, v.101, p.7856-7863, 2010.
PETER, M. G. Chitin and chitosan from aninal sources. In: STEINBUCHEL, A.;
RHEE, S. K. Polysaccharides and Polyamides in the Food Industry. Weinheim:
Wileyvch, v.1, p.115-208, 2005.
PIZA, F. A. T.; SILOTO, A. P.; CARVALHO, C. V.; FRANCO, T. T. Purification,
characterization and purification of chitosanase from Bacillus cereus, Brazilian Journal
of Chemical Engineering., v.16, n 2, p.185-192, 1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
88
POLITZER, K.; BON, E. P. S. Enzimas Industriais e Especiais. C.G.E.E. Ciência,
Tecnologia e Inovação. Rio de Janeiro, 554p, 2006.
POLYMAR, Ciência é a nossa Natureza, 2003. Disponível em:
<http://www.polymar.com.br>. Acesso em: 6 setembro 2012.
PUTZKE, J.; PUTZKE, M. T. L. Os reinos dos fungos. Santa Cruz do Sul, RS:
EDUNISC, 1998.
QUINES, L. K. M. Extração de poli(3-hidroxibutirato) produzido por Cupriavidus
necator dsm 545 com 1,2-carbonato de propileno. 2010. 99p. Dissertação (Mestrado em
Engenharia de Alimentos) – Centro Tecnológico, Departamento de Engenharia Química
e Engenharia de Alimentos, Programa de Pós-Graduação em Engenharia de Alimentos,
Universidade Federal de Santa Catarina, Florianópolis.
RAJENDRAN, A.; THANGAVELU, V. Statistical experimental design for evaluation
of medium components for lipase production by Rhizopus arrhizus MTCC 2233, LWT -
Food Science and Technology, v.42, p.985–992, 2009.
RAMIREZ-LEÓN, L. F.; RUIZ-HERRERA, J. Hydrolysis of walls and formation of
spheroplasts in Mucor rouxii. J. Gen. Microbiol., v.72, p.281-290, 1972.
RANGEL, M. A. S.; LOUREIRO, E. S.; ALMEIDA, A. S.; ALVES, H. C.; ONO, F.
B.; LOBO, P. M.; MARIANI, M. A.; PESSOA, L. G. A.; SILVA, A. J. Avaliação
participativa do controle de cigarrinhas das pastagens com a aplicação do fungo
Metarhizium anisopliae (Metsch.) Sorok. Rev. Bras. de Agroecologia, v.2, n.2, 731-734,
2007.
RAVEN, P. H.; EVERT, R.F.; EICHHORN, S.E. Biologia Vegetal, 6ª Ed, Rio Janeiro,
Editora Guanabara Koogan, 2001.
REDDY, L. V. A.; WEE, Y.-J.; YUN, J.-S.; RYU, H.-W. Optimization of alkaline
protease production by batch culture of Bacillus sp. RKY3 through Plackett–Burman

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
89
and response surface methodological approaches, Bioresource Technology, v.99, p.242–
2249, 2008.
RIGUEIRA, J. C. S. Influência da contagem de células somáticas no perfil e teores de
aminas bioativas e na qualidade de leite cru e de queijo mussarela. 2010. 147p. Tese
(Doutorado em Ciência de Alimentos) - Faculdade de Farmácia, Departamento de
Alimentos, Programa de Pós-Graduação em Ciência de Alimentos, Faculdade de
Farmácia da Universidade Federal de Minas Gerais, Belo Horizonte.
RODRIGUES, M. I.; IEMMA, A. F. Planejamento de Experimentos e Otimização de
Processos: uma estratégia sequencial de planejamentos, Campinas, SP, Casa do Pão
Editora, 2005.
SANTOS, E. S. Recuperação e Purificação de Enzimas usando Adsorção em Leito
Expandido, Tese de Doutorado, Universidade Estadual de Campinas, Campinas/SP,
p.152, 2001.
SEDMAK, J. J.; GROSSBERG, S. E. A rapid, sensitive and versatile assay for protein
using coomassie brilliant blue G250, Analytical Biochemistry, v.79, p. 544-552, 1977.
SEINO, H.; TSUKUDA, K.; SHIMASUE, Y. Properties and action pattern of a
chitosanase from Bacillus sp. PI- 7S. Agricultural Biological Chemistry, 55, p.2421-
2423, 1991.
SHIMOSAKA, M.; NOGAWA, M.; OHNO, Y.; OKAZAKI, M. Chitosanase from the
plant pathogenic fungus, Fusarium solani f. sp. Phaseoli – purification and some
properties, Bioscience Biotechnolgy and Biochemistry, v.57, n.2, p.231-235, 1993.
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Production of two chitosanases from a chitosan-assimilating bacterium, Acinetobacter
sp. Strain CHB101, Applied and Environmental Microbiology, v.61, n.2, p.438-442,
1995.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
90
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Characterization of Three Chitosanase Isozymes Isolated from a Commercial Crude
Porcine Pepsin Preparation, J. Agric. Food Chem. 51, p.1042-1048, 2003.
SILVA FILHO, R. C.; COSTA, A. S. S.; CHAGAS, B. M. E.; SANTOS, F. N.;
MACEDO, G. R.; SANTOS, E. S. Avaliação do teste de atividade quitosanolítica
produzida por fungo. In: XV SIMPÓSIO NACIONAL DE BIOPROCESSOS, 15,
Recife, 2005, 02 a 05 de agosto.
SILVA, E. P. Avaliação Preliminar do Potencial de Aplicação da Tecnologia de
Biopilhas para a Biorremediação do Solo de Guamaré-RN. 2004. 119p. Dissertação
(Mestrado) – Centro de Tecnologia, Programa de Pós-Graduação em Engenharia
Química. Universidade Federal do Rio Grande do Norte. Natal.
SOMASHEKAR, D.; JOSEPH, R. Chitosanases – Properties and apllications: A review.
Bioresource Technology, v.55, p.35-45, 1996.
STOYACHENKO, I. A.; VARLAMOV, V. P.; DAVANKOV, V. A. Chitinases of
Streptomyces kurssanovii: purification and some properties, Carbohydrate Polymers,
v.24, p.47-54, 1994.
TINLINE, R. D. Nuclear distribuition in Metarhizium anisopliae. Mycologia, v. 63, p.
713-721, 1971.
TRABULSI, L. R.; TOLEDO, M. R. F. Microbiologia. 2. Ed., São Paulo: Atheneu,
1996.
VIEIRA, P. D. S.; SILVA, W. M. T.; PAIVA, L. M.; LIMA, E. A. L. A.;
CAVALCANTI, V. L. B. Estudo da caracterização morfológica, esporulação e
germinação dos conídiosde Metarhizium anisopliae var. acridum em diferentes
temperaturas. Biológico, São Paulo, v.69, n.1, p.17-21, 2007.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
91
WANDERLEY, M. D.; NEVES, E.; ANDRADE, C. J. Aspectos da produção industrial
de enzimas. Revista Citino, v.1, n.1, p.30-36, 2011.
WANG, C.; TYPAS, M. A.; BUTT, T. M. Detection and characterization of pr1
virulent gene deficiencies in the insect pathogenic fungus Metarhizium anisopliae.
FEMS Microbiology Letters, v. 213, p. 251-255, 2002.
ZAHORSKA-MARKIEWICZ, B.; KROTKIEWSKI, M.; OLSZANECKA-
GLINIANOWICZ, M.; ZURAKOWSKI, A. Effect of chitosan in complex management
of obesity, Pol Merkur Lekarski, v.13, n.74, p.129-132, 2002.
ZHANG, X. Y.; DAI, A. L.; ZHANG, X. K.; KUROIWA, K.; KODAIRA, R.;
SHIMOSAKA, M.; OKAZAKI, M. Purification and Characterization of and Exo--D-
glucosaminidase from a koji mold, Aspergillus oryzae IAM2660. Biosci. Biotechnol.
Biochem., v.64, n.9, 1896-1902, 2000.
ZHU, X.; WU, X.; DAI, Y. Fermentation conditions of a chitosanase from Acinetobcter
sp C-17. Biosci. Biotechnol. Biochem., v.67, n.2, p.284-290, 2003.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
92
APÊNDICE

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
93
APÊNDICE I
Metodologia para repique da cepa
Em Erlenmeyer de 1000 mL colocava-se 500 mL do meio de manutenção
PDA. Este Erlenmeyer, com o meio de manutenção, era autoclavado junto com placas
de Petri a 1,5 kgf/cm2 por 15 minutos, em seguida, era levado para uma câmara de fluxo
laminar e transferido para as placas de Petri já autoclavadas, onde eram mantidas numa
estufa a 27 ºC durante 24 h. Após este período, as placas de Petri eram levadas a câmara
de fluxo laminar, utilizando bico de Bunsen para esterilizar a alça de platina. Procedia-
se o repique utilizando uma placa de Petri contendo a cepa pura. Cada placa de Petri
contendo a cepa pura era transferida para duas placas de Petri contendo o meio de
manutenção. As placas eram mantidas na estufa a 27 ºC durante 30 dias onde após esse
período, repetia-se este procedimento.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
94
APÊNDICE II
Montagem da Curva de Calibração Usando o Método DNS
Soluções:
Glicosamina = 0,01 mol/mL (10000,0 µmol/mL)
Tabela 14. Dados para construção da curva de calibração usando o Método DNS.
Tubo de
Ensaio
Solução de
glicosamina (mL)
Água Destilada
(mL)
DNS (mL)
Volume Total
Massa (µµµµmol)
1 0 1,0 1,5 2,5 0
2 0,2 0,8 1,5 2,5 0,2
3 0,4 0,6 1,5 2,5 0,4
4 0,6 0,4 1,5 2,5 0,6
5 0,8 0,2 1,5 2,5 0,8
6 1,0 0 1,5 2,5 1,0
Tabela 15 - Medidas de absorbância a 600nm
Dados adicionais:
Peso Molecular D-Glicosamina = 215,64 (recipiente SIGMA)
Conc. Sol. D-Glicosamina = 0,05 g/50 mL
Massa calc.(µmol) = conc. Sol. D-Glic. x Vol. Utiliz. D-Glics. =
= 0,05 x 1000 x Vol.util./50*215,64
Tempo atividade enzimática = 30 min
1 U/mL = 1 µmol/(min.mL)
Vol. Total (Glicosamina+caldo) = 1 mL
Medições Adsorbância a 600 nm Vol. Utiliz. Massa Massa Atividade
II III Média(Abs)(mL) calc. (mg) calc. (µg) Enzim. (U/mL)
0 0 0 0 0 0 0
0,133 0,128 0,1307 0,2 0,2 200 0,0309
0,279 0,277 0,2787 0,4 0,4 400 0,0618
0,433 0,428 0,4297 0,6 0,6 600 0,0927
0,587 0,583 0,5797 0,8 0,8 800 0,1237
0,727 0,716 0,7157 1 1 1000 0,1546

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
95
Figura 17 – Curva padrão de glicosamina.
y = 1900ralx + 1900ral
R² = 1900ral
00,000
00,020
00,040
00,060
00,080
00,100
00,120
00,140
00,160
00,180
00,000 00,100 00,200 00,300 00,400 00,500 00,600 00,700 00,800
Ati
v.
En
zim
áti
ca (
U/m
L)
Absorbância (Abs) a 600 nm
Curva-Padrão D-Glicosamina

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
96
APÊNDICE III
Método de Determinação de Proteína Utilizando o Corante Comassie
Brilhante Blue G 250 (Método modificado de Sedmak e Grossberg)
Micrométodo
Preparar uma solução 0,06% de Comassie Blue G250 em 1,5 % HCl (w/V). Filtrar e
papel de filtro Whatman nº1.
Curva Padrão (1 a 20 µµµµg)
a) Preparar soluções estoque de BSA de 10 a 200 µg/mL.
b) Colocar em tubos de ensaio 100 µL da solução estoque, 1400 µL de água e
1500µL do Comassie e agitar.
c) Preparar um tubo contendo 1500 µL de água e 1500 µL do corante que
corresponde ao zero de proteína.
d) Após 5 minutos (padronizar o tempo) fazer as leituras das absorbâncias.
e) As leituras das amostras devem ser feitas em dois comprimentos de onda 595 e
465 contra a água.
f) Plotar a razão das absorbâncias a 595/465 como uma função da massa de BSA e
calcular o melhor ajuste linear dos dados.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
97
Micrométodo (1 a 20 µµµµg de BSA)
Solução Estoque de BSA = 0,2 mg/mL
Tabela 16. Dados para construção da curva padrão de BSA.
Tubo de
Ensaio
H2O (µµµµL)
BSA (µµµµg)
Coomassie (µµµµL)
BSA (µµµµg)
1 1500 0 1500 0
2 1490 10 1500 2
3 1480 20 1500 4
4 1470 30 1500 6
5 1460 40 1500 8
6 1450 50 1500 10
7 1440 60 1500 12
8 1430 70 1500 14
9 1420 80 1500 16
10 1410 90 1500 18
11 1400 100 1500 20
Dados adicionais:
Concentração Sol. Estoque
BSA = 0,2 mg/mL = 0,2 µg/µL
Vol. total sol. = vol. H2O + vol. BSA + vol.
Coomassie = 3000 µL = 3 mL
Concentração Sol. BSA (mg/mL) = (massa BSA mg) / 3 mL
.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
98
Figura 18 – Curva Padrão de BSA
y = 1900ralx - 1900ral
R² = 1900ral
000,000
001,000
002,000
003,000
004,000
005,000
006,000
007,000
000,400 000,500 000,600 000,700 000,800 000,900 001,000 001,100
µg
BS
A
Razão Absorbâncias (595nm/465nm)
Curva-Padrão BSA

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE TECNOLOGIA
DEPARTAMENTO DE ENGENHARIA QUÍMICA PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA
TESE DE DOUTORADO
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em
Cultivo Descontínuo Submerso
Raimundo Cosme da Silva Filho
Natal/RN
Abril/2013

Raimundo Cosme da Silva Filho
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo
Descontínuo Submerso
Tese apresentada ao Programa de Pós-Graduação em Engenharia Química – PPGEQ, da Universidade Federal do Rio Grande do Norte, como parte dos requisitos para obtenção do título de Doutor em Engenharia Química, sob a orientação do Prof. Dr. Everaldo Silvino dos Santos e a coorientação da Profª. Drª. Gorete Ribeiro de Macedo.
Natal/RN
Abril/2013

Catalogação da Publicação na Fonte. UFRN / CT / PPGEQ
Biblioteca Setorial “Professor Horácio Nícolas Sólimo”.
Silva Filho, Raimundo Cosme da. Otimização do meio de cultura para a produção de quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso / Raimundo Cosme da Silva Filho. - Natal, 2013. 98 f.: il.
Orientador: Everaldo Silvino dos Santos. Co-orientador: Gorete Ribeiro de Macedo. Tese (Doutorado) - Universidade Federal do Rio Grande do Norte.
Centro de Tecnologia. Departamento de Engenharia Química. Programa de Pós-Graduação em Engenharia Química.
1. Quitosana - Tese. 2. Quitina - Tese. 3. Quitosanase – Tese. 4. Metarhizium
anisopliae - Tese. I. Santos, Everaldo Silvino dos. II. Macedo, Gorete Ribeiro de. III. Universidade Federal do Rio Grande do Norte. IV. Título. RN/UF/BSEQ CDU 547.458 (043.2)

SILVA FILHO, Raimundo Cosme da – Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo Descontínuo Submerso. Tese de Doutorado. UFRN - Programa de Pós-Graduação em Engenharia Química. Área de Concentração: Pesquisa e Desenvolvimento de Tecnologias Regionais – Subárea: Engenharia de Processos – Alimentos e Biotecnologia, Natal/RN, Brasil.
Orientação: Prof. Dr. Everaldo Silvino dos Santos Coorientação: Profª. Drª. Gorete Ribeiro de Macedo ___________________________________________________________________________
RESUMO: No presente trabalho utilizou-se um Planejamento Plackett & Burman, com 8 fatores e 12 ensaios em 2 níveis e mais 3 repetições na condição do ponto central, para se investigar a influência das concentrações de quitosana, peptona, extrato de levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O e FeSO4 na produção da enzima quitosanase por Metarhizium
anisopliae. Os ensaios para produção da enzima foram realizados em cultivo descontínuo submerso. Os resultados para o Planejamento Plackett & Burman mostraram que a atividade quitosanolítica foi favorecida pelo aumento da concentração de substrato (quitosana) e de sulfato ferroso (FeSO4), enquanto que o aumento da concentração dos outros fatores não contribuiu de forma significativa para a atividade quitosanolítica. A otimização do meio de cultura para a produção da enzima foi realizado por meio de um Planejamento Composto Central, com os dois fatores que mais influenciaram a atividade quitosanolítica (quitosana e FeSO4), conforme Planejamento Plackett & Burman, mantendo-se os outros nutrientes em seus valores mínimos. Nesse planejamento, para o fator FeSO4, tomou-se o limite inferior (-1) como sendo o limite superior do Planejamento Plackett & Burman. Os resultados mostraram que a produção da enzima foi favorecida pelo aumento da concentração quitosana e pela diminuição da concentração de FeSO4. A produção máxima de atividade quitosanolítica foi da ordem de 70,0 U/L e foi atingida em apenas 18 h de fermentação, resultado esse vinte e oito vezes maior aos obtidos anteriormente para o mesmo microrganismo que foi 2,5 U/L em 48h.
Palavras-chave: quitosana; quitosanase; Metarhizium anisopliae; Plackett & Burman.


ABSTRACT
In this work a Plackett-Burman Design with 8 factors and 12 trials in 2 levels with 3 repetitions at the center point was used in order to investigate the influence of the concentration of chitosan, peptone, yeast extract, NaNO3, K2HPO4, KCl, MgSO4.7H2O and FeSO4 on chitosanase production by Metarhizium anisopliae. Runs were carried out using submerged discontinuous cultivation for enzyme production. The results of the Plackett & Burman Design showed that only two factors, chitosan concentration as well as FeSO4 had influence on chitosanolytic activity, while the increase in concentration of other factors not contributed significantly to the quitosanolítica activity. Cultivation medium optimization for enzyme production was carried out using a Composite Central Design, with the most important factors for chitosanolytic activity (chitosan and FeSO4), in accordance with Plackett & Burman Design, and keeping the other nutrients in their minimum values. On this other design, it was taken the highest limit in Plackett & Burman Design as the lowest limit (-1) to FeSO4 factor. The results showed that the enzyme production was favoured by increasing the chitosan concentration and by decreasing FeSO4. Maximum production for chitosanolytic activity was about 70.0 U/L and was reached in only 18 h of fermentation, a result about twenty-eight times greater than a former study using the same microorganism (about 2.5 U/L at 48 h).
Key-words: Chitosan; Chitosanase; Metarhizium anisopliae; Plackett & Burman

Ao meu pai, minha mãe e minha
família, em especial aos meus
irmãos e sobrinhos

AGRADECIMENTOS
A Deus, por estar presente em todos os momentos da minha vida e por me ceder à
oportunidade de iniciar e concluir este trabalho.
A minha mãe Cândida Inocência da Silva que esteve comigo em todos os momentos da
minha vida me educando, me amando e sempre me dando muito carinho.
Ao meu pai Raimundo Cosme da Silva, que não está mais aqui para ver este trabalho,
mas que foi a pessoa que mais me apoiou e me incentivou, em todos os momentos da minha vida,
para que eu pudesse chegar o mais longe possível na minha carreira acadêmica. Pai muito
obrigado por toda atenção, educação, amor e carinho que recebi durante toda a minha vida.
Aos meus irmãos e familiares que sempre estiveram comigo dividindo momentos de
grande alegria e às vezes de tristeza, mas sempre nos mantendo unidos para superarmos todos os
obstáculos.
A minha namorada Rochelly Caroliny por todo amor e carinho que me deu e que me
entendeu nos momentos que estive ausente para a conclusão deste trabalho.
Ao meu orientador Professor Doutor Everaldo Silvino dos Santos por confiar em mim e
me aceitar como orientando além de toda a amizade, orientação e dedicação em todas as etapas de
desenvolvimento e conclusão deste trabalho.
À Professora Doutora Gorete Ribeiro de Macedo, “pessoa que admiro muito como
profissional e principalmente, como pessoa, um exemplo a ser seguido”, pelos conselhos e críticas
quando necessários ao desenvolvimento deste trabalho. Professora a senhora é considerada uma
mãe para de todos os alunos que tiveram a honra de conhecê-la e trabalhar com a senhora.
Aos meus amigos Engenheiros Químicos, Ana Carmen, Ana Katerine, Andréa Farias,
Anita Lima, Sirtys Lessa, Alexandre, Sérgio, Márcio Bezerra e ao Biólogo Daniel Souza, que
sempre me ouviram e que me ajudaram de alguma forma nesse trabalho.
A Eusamar Coelho de Lima, Mazinha, secretária do PPGEQ, pelo apoio, pela paciência e
orientação nas questões burocráticas do processo.
A Carlos e Jardelson, pela grande colaboração e dedicação nos experimentos do
laboratório.
A Sérgio Santana pela grande colaboração e dedicação, o qual não poupou esforços nem
tempo em me ajudar na realização dos experimentos em laboratório.

Ao meu amigo Ubiratan Nazaré por me ajudar na organização do texto e na tradução para
o Abstract.
Finalizando, agradeço ao Laboratório de Engenharia Bioquímica da UFRN por ter cedido
toda a sua estrutura para realização dos experimentos e ao Programa de Pós-Graduação em
Engenharia Química, assim como a todos os professores do Departamento de Engenharia Química
da UFRN que trabalham em pró de uma educação digna e ensino de qualidade para todos os
alunos.

Sumário LISTA DE FIGURAS
LISTA DE TABELAS
NOMENCLATURA
INTRODUÇÃO...............................................................................................................................15
REVISÃO BIBLIOGRÁFICA........................................................................................................18
2.1. Enzimas................................................................................................................................18
2.1.1. Quitosana e seus derivados........................................................................................19
2.1.2. Quitosanases...............................................................................................................24
2.2. Fungos..................................................................................................................................29
2.2.1. Características gerais..................................................................................................29
2.2.2. Reprodução dos Fungos.............................................................................................32
2.2.3. Metabolismo de fungos..............................................................................................33
2.2.4. Nutrição de fungos.....................................................................................................34
2.2.5. O Fungo Metarhizium anisopliae ..............................................................................36
2.3. Fermentação.........................................................................................................................40
2.3.1. Preparo do inóculo......................................................................................................41
2.3.2. Produção de enzimas em cultivo submerso descontínuo...........................................42
2.4. Planejamento experimental..................................................................................................42
2.4.1. Planejamento fatorial..................................................................................................43
2.4.2. Planejamento Plackett & Burman..............................................................................43
2.4.3. Aplicações do Planejamento Plackett & Burman.......................................................44
METODOLOGIA EXPERIMENTAL.............................................................................................50
3.1. Quitosana..............................................................................................................................50
3.2. Microrganismo.....................................................................................................................50
3.2.1. Escolha da cepa produtora de quitosanase.................................................................50

3.2.2. Manutenção da cepa...................................................................................................50
3.3. Produção de enzima.............................................................................................................51
3.4. Determinação da Proteína Total...........................................................................................51
3.5. Determinação da Atividade Quitosanolítica e Atividade Específica...................................51
3.6. Planejamento Experimental.................................................................................................52
3.6.1. Planejamento de seleção de fatores Plackett Burman................................................52
3.6.2. Planejamento Composto Central para Otimização do meio de cultura para a
Produção da Enzima em Cultivo Submerso.....................................................................................53
3.7. Perfil Cinético durante o Cultivo.........................................................................................56
RESULTADOS E DISCUSSÃO.....................................................................................................58
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA ..........................................58
4.2. Planejamento Plackett & Burman........................................................................................59
4.2.1. Análise do Plackett & Burman com 8 fatores e 12 ensaios .......................................59
4.2.2. Análise do Plackett & Burman com 12 ensaios e mais 3 repetições na condição do
ponto central ....................................................................................................................................61
4.3. Planejamento Composto Central para Produção de Quitosanase por Metarhizium
anisopliae em Cultivo Descontínuo Submerso................................................................................64
4.4. Perfil Cinético – Melhor Condição......................................................................................69
CONCLUSÃO.................................................................................................................................75
REFERÊNCIAS BIBLIOGRÁFICAS.............................................................................................78
APÊNDICE I...................................................................................................................................93
Metodologia para Repique da Cepa ................................................................................................93
APÊNDICE II.................................................................................................................................94
Montagem da Curva de Calibração Usando o Método DNS..........................................................94
APÊNDICE III................................................................................................................................96
Método de Determinação de Proteína Utilizando o Corante Comassie Brilhante Blue G 250
(Método modificado de Sedmak e Grossberg)................................................................................96

Lista de Figuras
Figura 1 – Estrutura da quitosana..................................................................................................20
Figura 2 – Estrutura da glicosamina..............................................................................................23
Figura 3 – Estrutura do sulfato de glicosamina.............................................................................23
Figura 4 – Representação da estrutura geral de uma célula de fungo..........................................30
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas...........................................................31
Figura 6 – Conídios de Aspergillus e Penicillium.........................................................................33
Figura 7 – Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA ...................58
Figura 8 – Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados............................................................................................................................60
Figura 9 – Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto central
mostrando a influência dos fatores estudados................................................................................63
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos
fatores estudados............................................................................................................................67
Figura 11 – Valores preditos em função dos valores observados..................................................68
Figura 12 – Atividade quitosanolítica em função da concentração de quitosana e da concentração
de FeSO4 para os fatores não significativos no ponto central .......................................................68
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana ........70
Figura 14 – Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana....................................70
Figura 15 – Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de
quitosana........................................................................................................................................71
Figura 16 – Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana..........................................................................................................72
Figura 17 – Curva padrão de glicosamina ....................................................................................95
Figura 18 – Curva Padrão de BSA ................................................................................................98

Lista de Tabelas
Tabela 1 – Comparação das enzimas com os catalisadores químicos..............................................19
Tabela 2 – Fontes naturais de quitina e quitosana ...........................................................................20
Tabela 3 – Níveis para os fatores e seus valores codificados...........................................................53
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central .........................................................................................53
Tabela 5 – Matriz do planejamento Composto Central...................................................................54
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central......................................................................................................55
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.......................59
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman...............................................60
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.......................61
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central...............................................................................................................................................62
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da atividade
enzimática........................................................................................................................................65
Tabela 12 – Efeitos Estimados do planejamento Composto Central...............................................65
Tabela 13 – Resumo da ANOVA....................................................................................................66
Tabela 14 – Dados para construção da curva de calibração usando o Método DNS ......................94
Tabela 15 – Medidas de absorbância a 600 nm para construção da curva de calibração................94
Tabela 16 – Dados para construção da curva padrão de BSA ........................................................97

NOMENCLATURA
A – Atividade quitosanolítica (U/L)
AE – Atividade específica (U/mg)
ANN-GA – algoritmo genético de rede neural artificial
ANOVA – Análise de variância (Analysis of Variance)
BSA – Albumina de soro bovino (Bovine Sereum Albumin)
CCD – Planejamento Composto Central
CDA – Chitosan Dector Agar
Cs – Concentração de substrato (%)
CP – Concentração de KCl (%)
Da – Dalton
DEQ – Departamento de Engenharia Química
DNS – Ácido Dinitrosalissílico
E – Enzima
Ea – Energia de ativação
EC – Comissão para nomenclatura e classificação de enzimas
EL – Concentração de Extrato de Levedura (%)
FP – Concentração de K2HPO4 (%)
NS – Concentração de NaNO3 (%)
GDA – Grânulos dispersíveis em água
k – Número de fatores, variáveis
LEB – Laboratório de Engenharia Bioquímica
mRNA – RNA mensageiro
n – Número de níveis de um determinado fator
N – Número de ensaios de um planejamento
P – Pressão
PP – Concentração de Peptona (%)
PDA – Batata Dextrose Agar
pH – Potencial hidrogeniônico
Q – Concentração de Quitosana (%)
Qm – Quadrado médio
r – Agitação (rpm)
R2 – Coeficiente de determinação

RA – Razão de aeração
rpm – Rotações por minuto
RSM – Metodologia de superfície de resposta
S – Molécula de substrato
SF – Concentração de FeSO4 (%)
SM – Concentração de MgSO4.7H2O (%)
SQ – Soma de quadrados
SSF – Fermentação de estado sólido
T – Temperatura
TMC – Temperatura de cultivo
V11,V12,V13 – Variáveis inertes para cálculo do erro
VHG – Fermentação de alta densidade
WP – Pó molhável
X – Concentração celular
Xi – variável independente
Xj – variável independente
xi – Fator independente real
x0 – Valor de xi no ponto central
∆xi – Variação de valor
Y – Resposta do modelo de primeira ordem
Yp – Resposta do modelo polinomial de segunda ordem
β0 – Interseção dos modelos de primeira ou segunda ordem
βi – Coeficiente linear dos modelos de primeira ou segunda ordem
βii – Coeficiente quadrático
βij – Coeficiente de interação

Capítulo 1 Introdução

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
15
1. Introdução As quitosanases foram descobertas simultaneamente há quase quarenta anos por
Monaghan et al. (1973), como pelo grupo de Ramirez-León & Ruiz-Herrera (1972), e
são enzimas obtidas a partir do metabolismo de bactérias, fungos e extraídas de plantas.
Desde então, vários estudos vêm apresentando a obtenção de quitosanase deste
microrganismos, na forma intracelular ou extracelular (Osswald et al., 1994;
Somashekar & Joseph, 1996), como a do presente estudo. A quitosanase (EC 3.2.1.132)
representa uma classe de enzimas hidrolíticas que catalisam a hidrólise de ligação β-
(1→ 4) glicosídica da quitosana, gerando oligômeros chamados quito-oligossacarídeos
(Reddy et al., 2008).
A quitosana é um biopolímero hidrofílico obtido a partir da quitina, um
polissacarídeo abundante na natureza. Na literatura, é considerado quitosana o produto
da desacetilação da quitina para um grau de desacetilação superior a 75% e solúveis em
ácidos como o acético e o fórmico. A quitosana é conhecida por exibir uma ampla
variedade de atividade fisiológica como atividade antitumoral, mas o peso molecular e a
insolubilidade em água da quitosana são desvantagens para várias aplicações.
Atualmente, há interesse nas quitosanas quando as mesmas são convertidas em quito-
oligossacarídeos porque eles não apenas são solúveis em água e possuem baixo peso
molecular, mas também têm atividades biológicas bem notáveis, como atividade
anticancerígena atóxica e biocompatível, incluindo efeito antimetastático, atividade anti-
HIV, hepatoprotetora, antioxidante, hipocolesterolêmica, antimicrobiana,
imunoestimulate, antitumoral, anti-inflamatória e atividade aceleradora da absorção de
cálcio e ferro. No entanto, estes quito-oligossacarídeos são obtidos tradicionalmente na
indústria por reação de hidrólise usando ácidos, que apresenta muitos problemas como
dificuldade em controlar a hidrólise, produzindo uma grande quantidade de
oligossacarídeos de cadeia pequena e oligossacarídeos de baixo rendimento, alto custo
de separação e, também, poluição ambiental, uma vez que utiliza um ácido como
catalisador. Como alternativa, com suas vantagens em termos de compatibilidade
ambiental, baixo custo e reprodutibilidade, a obtenção destes quito-oligossacarídeos por
hidrólise com quitosanase se tornou cada vez mais popular nos últimos anos (Azevedo
et al., 2007).

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
16
Devido a todos estes aspectos positivos é que, em geral, as enzimas vêm sendo
estudadas em diferentes aplicações, mostrando que esta é uma tendência na indústria.
No presente trabalho, foi desenvolvido um processo fermentativo visando a
otimização do meio de cultura para a produção da enzima quitosanase em cultivo
descontínuo submerso, por Metarhizium anisopliae, capaz de sintetizar oligômeros com
aplicações na indústria farmacêutica e de química fina, partindo de quitosana como
única fonte de carbono.
Nesse caso, avaliou-se, através de um planejamento Plackett & Burman (método
de delineamento de seleção de fatores) e de um planejamento Composto Central as
melhores condições para a produção da enzima. A técnica do planejamento
experimental Plackett & Burman para realizar triagem de fatores na otimização de um
determinado processo foi utilizada neste trabalho para identificar quais os fatores que
influenciam de forma significativa na produção da enzima quitosanase. A influência
destes fatores (concentração de quitosana e concentração de FeSO4) foi então
investigada pelo planejamento Composto Central com 22 experimentos na parte fatorial,
repetição em triplicata no ponto central e quatro experimentos na parte axial. A
repetição do ponto central permite também a determinação do erro puro e da
repetibilidade do processo, sendo uma forma de identificar melhor se o processo está
sob controle.
Do ponto de vista industrial é de relevante importância a realização de estudos
para a otimização da produção da quitosanase de forma contínua ou em grandes
bateladas.

Capítulo 2 Revisão Bibliográfica

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
18
2. Revisão Bibliográfica 2.1. Enzimas
As enzimas são catalisadores biológicos constituídos por longas cadeias de
aminoácidos que atuam em substratos reduzindo a energia de ativação, acelerando assim
a velocidade da reação, e transformando-os em produtos. Sua atuação catalítica visa
facilitar a atividade das células através da quebra de moléculas ou junção das mesmas
para formação de novos produtos. Podem apresentar alto grau de especificidade relativa
a apenas um substrato ou a vários substratos ao mesmo tempo, como a certos tipos de
ligações glicosídicas α-1,4 das moléculas de amido, ou para determinado tipo particular
de isômero óptico como ocorre na oxidação da β-D-glicose pela glicose oxidase (Furigo
Junior & Pereira, 2001).
Enzimas são proteínas que apresentam em sua estrutura um centro ativo
(apoenzima) e, em alguns casos, um grupo prostético denominado cofator (coenzima ou
íon metálico ativo) e sua atividade depende de alguns fatores como estrutura da
proteína, número de cadeias peptídicas, arranjo dessas cadeias na molécula, natureza do
substrato e natureza do grupo prostético (Politzer & Bon, 2006).
Todos os organismos vivos produzem, em pequena ou grande quantidade, certa
variedade de enzimas, sejam elas intra ou extracelulares. As enzimas extracelulares são
excretadas para o meio externo e podem decompor compostos insolúveis como
celulose, quitina e quitosana.
Algumas destas enzimas podem ser usadas tanto nas indústrias de detergentes,
têxteis e farmacêuticas, como também de alimentos, de bebidas, etc., sendo utilizadas
em processos biotecnológicos industriais em substituição aos processos químicos que
muitas vezes causam grandes prejuízos ao meio ambiente. A produção comercial das
enzimas pode ser realizada através do cultivo de microrganismos como fungos e
bactérias.
Na Tabela 1 mostra-se uma comparação das enzimas com os catalisadores
químicos, levando-se em conta algumas das características dos dois catalisadores.
Destaca-se que alguns parâmetros da Tabela 1 demonstram a maior importância das
enzimas, quando comparadas com os catalisadores químicos, principalmente a
especificidade ao substrato, as condições de reação, o consumo de energia e a

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
19
velocidade de reação, como desvantagem cita-se o custo de obtenção (isolamento e
purificação).
Tabela 1 – Comparação das enzimas com os catalisadores químicos (Adaptado de
Furigo Junior & Pereira, 2001).
Característica Enzimas Catalisadores Químicos
Especificidade ao substrato Alta Baixa
Natureza da estrutura Complexa Simples
Sensibilidade à T e pH Alta Baixa
Condições de reação (T, P e pH) Suaves Drástica (geralmente)
Custo de obtenção (isolamento e purificação) Alto Moderado
Natureza do processo Batelada Contínuo
Consumo de energia Baixo Alto
Formação de subprodutos Baixa Alto
Separação catalisador/produtos Difícil/cara Simples
Atividade catalítica (temperatura ambiente) Alta Baixa
Presença de cofatores Sim Não
Estabilidade do preparado Baixa Alta
Energia de Ativação Baixa Alta
Velocidade de reação Alta Baixa
2.1.1. Quitosana e seus derivados
A quitosana e seus derivados apresentam grande importância devido ao
conhecimento de suas funcionalidades biológicas, médicas, tecnológicas, etc
(Shimosaka et al, 1993; Shimosaka et al, 1995; Mitsutomi et al., 1995). A quitosana é
obtida da quitina, polissacarídeo abundante na natureza, extraído principalmente das
carapaças de caranguejos, lagosta e camarões. A quitosana é um polímero natural,
biodegradável, não tóxico e de alta massa molecular obtido a partir de animais
marinhos, insetos e microrganismos (Tabela 2).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
20
Tabela 2 – Fontes naturais de quitina e quitosana. (Adaptado de Polymar, 2003).
Animais
Marinhos
Insetos Aracnídeos Microrganismos
Anelídios Formigas Escorpiões Leveduras
Moluscos Besouros Aranhas Fungos
Lagosta Esporos
Camarão
Caranguejo
Krill
A Figura 1 abaixo mostra a estrutura química da quitosana
Figura 1 – Estrutura da quitosana (Fonte: Polymar, 2003)
Quitosana totalmente desacetilada não existe na natureza. Graus de acetilação
(GA) nferior a 50% a torna mais solúvel em soluções ácidas (Muzzarelli, 1993;
Muzzarelli et al., 1994; Dung et al., 1994; Stoyachenko et al.,1994). Pelletier &
Sygusch (1990) enfatizaram que grande parte dos resíduos sólidos da indústria
pesqueira, principalmente da carcinicultura, é jogada ao mar ocasionando degradação
ambiental. Logo, a utilização desses resíduos como matéria-prima para formação de
novos produtos, não só reduz de forma drástica o impacto ambiental, mas também daria
maior valor agregado à indústria pesqueira.
A demanda cada vez maior de consumidores por alimentos mais saudáveis, sem
conservantes químicos, tem motivado pesquisas à descoberta de novos agentes
antimicrobianos naturais. Sendo assim, a atividade antimicrobiana incomum da

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
21
quitosana e seus derivados contra microrganismos, tais como bactérias, bolores e
leveduras tem tido maior atenção ultimamente.
Devido à interação entre a quitosana, que é positivamente carregada, e as
membranas celulares dos microrganismos, negativamente carregadas, ocorre a formação
de verdadeiros poros na membrana, que são canais de vazamento de conteúdo protéico e
outros componentes intracelulares. A quitosana tem também a propriedade de agir como
agente quelante que se liga de forma seletiva a traços de metais, inibindo, assim, a
produção de toxinas e o crescimento microbiano. Ainda é capaz de ativar mecanismos
de defesa em tecidos hospedeiros, atuando como agente sequestrante de água e inibindo
várias enzimas. A ligação da quitosana com DNA e a inibição da síntese do mRNA
ocorre via penetração da quitosana no núcleo dos microrganismos e interferência na
síntese do mRNA e proteínas, (Craveiro et al., 2004).
Em um estudo clínico conduzido por Giustina & Ventura (1995) com 100
pessoas com sobrepeso de 10 a 25%, ao final de 4 semanas as pessoas que receberam
quitosana demonstraram perda de peso significativa (7,3 Kg contra 3,0 Kg nos grupos
controle). As pessoas que tomaram quitosana também demonstraram redução na pressão
sanguínea sistólica (de 145 para 135 mm Hg) e diastólica (92.6 para 84.2 mm Hg),
assim como uma redução na taxa respiratória (27.6/min a 21.2/min).
Kondo & Osada (1996) pesquisaram a influência da quitina e da quitosana na
biodisponibilidade do zinco em ratos alimentados com uma dieta com suplementos de
5% ou sem fibra dietética (quitina, quitosana, celulose, pectina e agar-agar) durante 31
dias. Observaram que os ratos alimentados com dietas contendo 5% de fibras, exceto
quitosana, o consumo de alimento foi maior do que o controle. O ganho de peso em
todos os grupos alimentados com fibra foi maior do que o controle, exceto para o grupo
suplementado com quitosana (5%) onde a ingestão de alimento e o ganho de peso foram
menores do que os grupos controle e das demais fibras.
Segundo Craveiro et al. (2004), derivados da quitosana obtidos pela reação com
ácidos carboxílicos, podem promover ao polímero uma solubilidade especial em água.
Esses compostos possuem estrutura química semelhante ao ácido hialurônico e
fornecem uma excelente compatibilidade com o tecido epitelial. As soluções obtidas
com esses derivados podem ser facilmente incorporadas a formulações cosméticas,
mostrando melhor compatibilidade com surfactantes iônicos e emulsificantes,
apresentando estabilidade em uma larga faixa de pH.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
22
Han et al. (1999) examinaram os efeitos da quitosana sobre a atividade da lipase
pancreática in vitro e sobre o grau de armazenamento de gorduras induzido em ratos
pela administração oral de uma dieta rica em gorduras por nove semanas. Como
resultado, observou-se que os animais alimentados com quitosana, reduziram o peso
corporal, a hiperlipidemia e a gordura hepática. Como conclusão, os autores propõem
que os efeitos anti-obesidade da quitosana em ratos alimentados com alto teor de
gordura se devem parcialmente a inibição da absorção intestinal das gorduras dietéticas.
Em outro ensaio, Zahorska-Markiewicz et al. (2002) realizaram um estudo para
avaliar o efeito da quitosana no tratamento da obesidade. Segundo os autores do estudo,
a quitosana pode ser usada como um valioso e seguro coadjuvante em tratamentos para
a obesidade em longo prazo. A quitosana parece acentuar a redução da pressão arterial,
juntamente com a perda de peso.
Assis (2010) estudou a produção e caracterização de quitooligossacarídeos
produzidos pelo fungo Metarhizium anisopliae e avaliação da citotoxicidade em células
tumorais. A hidrólise da quitosana foi realizada em tempos de 10 a 60 min para a
produção de quitooligossacarídeos. A avaliação da citotoxicidade dos oligômeros de
quitosana foi realizada em células tumorais (HepG2 e HeLa) e não tumoral (3T3). A
produção de oligômeros de quitosana teve maiores rendimentos durante 10 minutos de
hidrólise, os pentâmeros apresentaram concentração de 0,15 mg/mL, no entanto os
hexâmeros, que apresentam maior interesse por causa de suas propriedades biológicas,
só foram detectados com 30 minutos de hidrólise com uma concentração de 0,004
mg/mL.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o planejamento Plackett-Burmam e a metodologia de superfície de resposta.
Os resultados mostraram que a eficiência máxima de extração da quitina foi conseguida
nas seguintes condições: concentração de casca de camarão 50g/L, glicose 50g/L, tempo
de incubação de 5 dias e inoculo como 0,05 OD (densidade óptica).
A glicosamina, unidade monomérica básica da quitosana, possui atividade
antiinflamatória e tem grande importância na construção e manutenção da cartilagem,
no tratamento de enfermidades como: febre reumática, artrose e principalmente a artrite
e osteoartrite na forma aguda ou crônica (Polymar, 2003). A Figura 2 apresenta a
estrutura química da glicosamina.
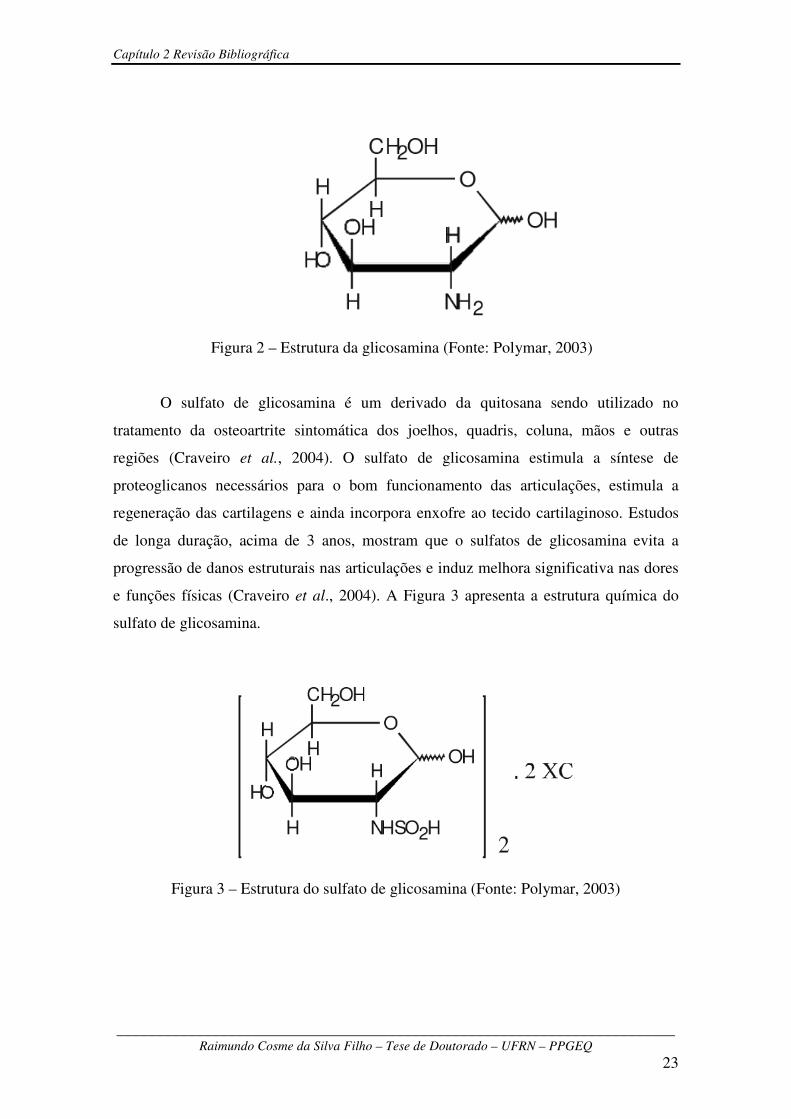
Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
23
Figura 2 – Estrutura da glicosamina (Fonte: Polymar, 2003)
O sulfato de glicosamina é um derivado da quitosana sendo utilizado no
tratamento da osteoartrite sintomática dos joelhos, quadris, coluna, mãos e outras
regiões (Craveiro et al., 2004). O sulfato de glicosamina estimula a síntese de
proteoglicanos necessários para o bom funcionamento das articulações, estimula a
regeneração das cartilagens e ainda incorpora enxofre ao tecido cartilaginoso. Estudos
de longa duração, acima de 3 anos, mostram que o sulfatos de glicosamina evita a
progressão de danos estruturais nas articulações e induz melhora significativa nas dores
e funções físicas (Craveiro et al., 2004). A Figura 3 apresenta a estrutura química do
sulfato de glicosamina.
Figura 3 – Estrutura do sulfato de glicosamina (Fonte: Polymar, 2003)

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
24
2.1.2. Quitosanases
Dependendo do tipo de quitosanase obtida, ocorrem diferentes mecanismos de
hidrólise da quitosana, isso devido a diferentes microrganismos produzirem enzimas
com diferenças nas suas estruturas (Kurita et al., 1977; Seino et al., 1991).
Quitosanases (EC 3.2.1.132) são enzimas que catalisam a reação de hidrólise de
quitosana em oligossacarídeos. As enzimas quitosanases são produzidas por vários
microrganismos, bactérias, actinomicetos e fungos, e até por algumas plantas. Estas
enzimas agem em polímeros com grau de acetilação de 30% a 60%. Os monômeros
resultantes da hidrólise da quitosana por estas enzimas são melhores digeridos, ao serem
ingeridos, como é o caso da glicosamina. Estudos efetuados até o presente momento
classificam as quitosanases em cinco famílias: 5, 8, 46, 75, 80, sendo que a família 46 é
a mais estudada. As quitosanases tem grande aplicação na geração de oligômeros de
quitosana (Somashekar & Joseph, 1996; Chen et al., 2005).
Nogawa et al. (1998) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida pelo fungo Trichoderma reseei PC-3-7. Estes pesquisadores
inocularam 106 conídios em 100 mL de um meio basal contendo 0,3% de N-
acetilglicosamina em incubator rotativo com agitação de 220 rpm a 28oC. Destaca-se
que o fungo não produziu quitosanase em meio contendo quitosana ou quitina.
Entretanto, o microrganismo produziu quitosanase em meio contendo glicosamina e N-
acetilglicosamina, sendo que para este último obteve-se uma atividade específica
máxima de 1,5 U/mg após 72 horas de incubação. É importante ressaltar que uma
agitação vigorosa favoreceu a produção da enzima. Esse artigo ilustra a presença de
exo-β-D-glicosaminidase e exo-β-D-N-acetilglicosaminidase.
Piza et al. (1999) estudaram a produção, purificação e caracterização parcial de
uma quitosanase produzida por Bacillus cereus. Nesse caso, investigou-se através de um
planejamento fatorial fracionário a influência dos fatores: concentração de sulfato de
amônio (0,4; 2,2 e 4,0%), razão de aeração (2; 6 e 7), tempo de fermentação (16; 24 e
32 horas), pH (5; 6 e 6,5) e concentração de quitosana (1,0; 1,5 e 2,0%). Os resultados
mostraram que os fatores mais significativos para esse planejamento foram a
concentração de sulfato de amônio, a aeração, o pH e a interação entre sulfato de
amônio e a aeração.
Cheng & Li (2000) estudaram a preparação de oligossacarídeos utilizando
quitosanase produzida por Aspergillus sp e constataram que a enzima foi largamente

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
25
induzida em meio contendo quitosana como única fonte de carbono e também na
presença de quitina coloidal. A indução da enzima foi realizada em temperaturas
variando na faixa de 28 a 30ºC, pois essa espécie não cresceu bem em temperaturas
superiores a 37ºC.
Zhang et al. (2000) estudaram a produção, purificação e caracterização de
quitosanase e Exo-β-D-Glucosaminidase por Aspergillus oryzae IAM2660. Os
microrganismos utilizados neste estudo foram mantidos em placas de Petri contendo
agar para contagem em placas (PDA). Destaca-se que para produzir a enzima, conídios
foram retirados das placas e inoculados em meio Czapeq-dox com os seguintes
componentes: 1,0g K2HPO4, 0,5g KCl, 2,0g NaNO3, 0,5g MgSO4.7H2O, 0,02g
FeSO4.7H2O, 20g de sacarose e 1,0L de água deionizada, com pH do meio igual a 5,5.
Foi adicionado ao meio 0,5 g de peptona e 0,5g de extrato de levedura com
concentração de esporos da ordem de 1,0x106 esporos.mL-1. Após a incubação, feita em
incubador rotativo a 26ºC por um período de 60h, micélios foram coletados e após
filtração e secagem a 80ºC foram utilizados para ensaio de quitosanase.
Kuroiwa et al. (2002) estudaram os fatores que afetam a composição de
oligossacarídeos produzidos através da hidrólise de quitosana por quitosanase originária
de Bacillus pumilus BN-262. Neste experimento foi utilizado quitosana 100%
desacetilada dissolvida em ácido acético 1M. O pH do meio foi ajustado em 5,6. Neste
estudo a quitosanase foi imobilizada em agar e Sepharose. Os resultados mostraram que
a concentração de enzima, a temperatura, a velocidade de agitação, e concentração de
substrato influenciaram de forma significativa na composição dos oligossacarídeos
produzidos.
Ichikawa et al. (2002) estudaram a imobilização e estabilização da ligação
multiponto da quitosanase tendo agar como suporte e concluíram que a imobilização
aumentou a termoestabilidade da quitosanase. Os oligossacarídeos de quitosana foram
continuamente produzidos usando um reator coluna contendo quitosanase imobilizada.
A percentagem de quitosana imobilizada após 28 dias de reação foi de 44%.
Hung et al. (2002) estudaram a purificação, através de troca iônica, e
caracterização da hidrólise sobre ação da quitinase e quitosanase purificadas do meio
contendo Bromelina comercial. Destaca-se que a quitinase purificada apresentou pH,
temperatura e Km ótimo para ação catalítica em meio contendo quitina de 4,0, 60ºC e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
26
0,2mg.mL-1, respectivamente. Enquanto que quitosanase apresentou o pH, temperatura e
Km ótimo de 3,0, 50ºC e 0,88mg.mL-1, respectivamente.
Zhu et al. (2003) estudaram as melhores condições fermentativas e propriedades
de uma quitosanase produzida por Acinetobacter sp. C-17. A espécie de bactéria
utilizada neste ensaio foi isolada do solo e identificada como sendo Acinetobacter sp C-
17 e capaz de produzir quitosanase. O microrganismo foi mantido em placas contendo o
meio agar com os seguintes componentes: 0,5 % de quitosana, 0,2 % K2HPO4, 0,1 %
KH2PO4, 0,07 % MgSO4, 0,05 % NaCl, 0,05 % KCl, 0,01 % CaCl2, 0,05 % de extrato
de levedura e 2 % agar, o pH do meio de manutenção foi mantido em 7,2 e a
temperatura foi mantida em 30ºC durante 3 a 5 dias, para identificar a ação da enzima
quitosanase. A ação da quitosanase foi identificada através da observação de uma
colônia de microrganismo existente na placa. A bactéria foi então isolada da placa e
colocada em fermentador rotativo a 200 rpm durante 72h em meio contendo: 0,07 %
K2HPO4, 0,03 % KH2PO4, 0,05 % MgSO4, 0,03 % peptona, 0,03 % de extrato de
levedura e 1,0 % de quitosana. O pH foi de 7,0 e a temperatura 30ºC. Como fontes de
carbono foram utilizadas além de 1,0 % de quitosana, 1,0 % de quitina e 1,0 % de
glicose. Os resultados mostraram que as melhores condições para produção da
quitosanase foram alcançadas quando se utilizou 1,0 % de solução de quitosana a 30ºC,
pH 7,0 durante 24 horas de incubação.
Liu et al. (2003) estudaram a produção de quitinase por Verticillium lecanii
F091 usando fermentação submersa, a atividade de quitinase foi de 9,95 mU/mL em um
meio de cultura otimizado, com um volume de cultura de 200 mL, agitação de 150 rpm
e temperatura de 24ºC cultivado em incubador rotativo e concluíram que a taxa de
agitação e o pH foram os fatores mais significativos para a produção de quitinase, no
entanto, as taxas de agitação e aeração poderiam modificar a concentração de oxigênio
dissolvido afetando diretamente no crescimento do Verticillium lecanii e na produção
de quitinase.
Donzelli et al. (2003) estudaram a hidrólise enzimática avançada de quitina de
casca de caranguejo langostino com misturas de enzimas de bactérias e fungos e
concluiram que a combinação de enzimas de Trichoderma atroviride e Serratia
marcescens foram capazes de degradar completamente altas concentrações de quitina
(100g/mL) a partir de carapaças de caranguejo langostino convertendo a N-
Acetilglicosamina (78%), glucosamina (2%) e quitobiose (10%). Enzimas de fonte

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
27
procariótica apresentaram atividades menores que as enzimas de T. atroviride, para
catalisar a reação com quitosana.
Jo et al. (2003) estudaram a caracterização e a avaliação cinética de quitosanase
produzida por Bacillus sp. Destaca-se que na produção da enzima utilizada neste ensaio
o meio continha 0,5% de concentração de quitosana, 1% de triptona e 1% de NaCl, o
pH do meio foi ajustado em 5,5 e a temperatura foi mantida em 37ºC com agitação de
180 rpm em incubador rotativo.
Fu et al (2003) estudaram a caracterização de três iso-enzimas de quitosanase
isolada de pepsina comercial e denominado como PSC-I, PSC-II, PSC-III,
respectivamente, em relação a hidrólise enzimática da quitosana, o meio ótimo para a
produção destas iso-enzimas continha as seguintes características: pH 5,0; 5,0 e 4,0 e
temperaturas de 40ºC, 40ºC e 30ºC, o valor de Km foi de 5,2, 4,0 e 5,6 mg.mL-1,
respectivamente. As massas moleculares das três iso-enzimas foram de 40 kDa. Os
resultados mostraram que as três iso-enzimas apresentaram atividade em meio contendo
quitosana, produzindo oligômeros da ordem de 68 – 88% desacetilada.
Choi et al. (2004) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida por Bacillus sp KCTC 0377BP. Nesse caso, com relação à
produção da enzima foi possível obter um aumento de 1,2 U/ml em incubator rotativo
para 100,0 U/ml em fermentador piloto de 500 L, utilizando uma concentração de
substrato de 0,5% de quitosana e como fonte de nitrogênio polipeptona (2,5%) em pH
igual a 6,8, com 100 rpm de 100,0 rpm a 30oC.
Shimosaka et al. (2003) estudaram a produção, purificação e caracterização total
de duas quitosanases produzida por Acinetobacter sp CHB101. Nesse caso, com relação
à produção da enzima os pesquisadores usaram dois meios, um contendo glicose e outro
contendo quitosana. Com relação à cinética, o meio contendo quitosana fornece um
tempo de geração de 280 minutos, enquanto que no meio contendo glicose a taxa de
crescimento foi mais elevada e o tempo de geração foi de apenas 110 minutos. Os
autores observaram que após os dois primeiros dias de cultivo a atividade
quitosanolítica permaneceu baixa (na ordem 0,10 a 0,20 U/mL). Durante a fase de
crescimento exponencial o valor da atividade permaneceu com valor inferior a 0,01
U/mL, começando a aumentar ao atingir a fase estacionária, indicando assim que a
produção da enzima é não associada ao crescimento.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
28
Kim et al. (2004) estudaram a produção, purificação e caracterização de uma
quitosanase produzida pelo Bacillus sp e identificado pelo número 1299. Destaca-se que
na produção da enzima quitosanase foi utilizado um meio contendo os seguintes
componentes: 0,5 % de quitosana, 0,42 % (NH4)2SO4, 0,2 % K2HPO4, 0,02 % uréia,
0,03 % CaCl2.2H2O, 0,03% MgSO4.7H2O, 0,1% peptona, 0,2% Tween 80, 0,2 % da
solução (0,5% FeSO4.7H2O, 0,16% MnSO4.H2O, 0,14% ZnSO4.7H2O, 0,2% CoCl2) e
1,7% agar. O pH do meio para este ensaio foi mantido em 7,0. Este microrganismo foi
escolhido por ser grande produtor de quitosanase.
Chen et al. (2005) estudaram a purificação e a caracterização de dois tipos de
quitosanase produzidas por Aspergillus sp. CJ22-326 em cultivo submerso. Na
produção da enzima foi utilizado um meio de cultivo contendo 1,0% de quitosana, 2,0%
de gérmen de trigo, 0,2% de sulfato de amônio, 0,2 % de fosfato de potássio e 0,05% de
sulfato de magnésio por litro de solução em pH 5,6. O cultivo foi realizado em frasco de
500 mL contendo 150 mL de meio de cultura a 30ºC por 96 horas com uma agitação de
150 rpm. Os autores apresentam um gel de eletroforese com as duas enzimas purificadas
com massa molar de 29 kDa e 109 kDa.
Silva Filho et al. (2005) estudaram, através de planejamento fatorial 24 com
repetição em triplicata no ponto central, a produção de quitosanase por Aspergillus
ochraceus em cultivo descontínuo submerso. Os resultados mostraram que foi possível
produzir quitosanase com atividade na ordem de aproximadamente 5,9 U/mL utilizando
Aspergillus ochraceus e que a atividade foi favorecida pelo aumento da agitação (r), da
razão de aeração (RA) e da concentração de substrato (Cs), enquanto que o aumento da
temperatura de cultivo (TMC) não favoreceu a resposta (atividade quitosanolítica).
Pode-se classificar as quitosanases em endoquitosanases e exoquitosanases. As
primeiras diferenciam-se por catalisar a hidrólise aleatoriamente, no interior do
biopolímero, gerando diversos tamanhos de oligossacarídeos. As exoquitosanases são
capazes de hidrolisar as terminações não redutoras, produzindo assim unidades
redutoras (Peter, 2005).
Palma-Guerrero et al. (2008) estudaram o biocontrole de fungos patogênicos e
demonstraram a lise da membrana plasmática e a inibição da germinação de esporos de
fungos. Eles observaram que os fungos entomopatogênicos e nematófagos foram menos
inibidos pela quitosana que os fitopatogênicos e micoparasíticos provavelmente porque
apresentam a enzima quitosanase que degrada a quitosana. Mesmo assim, a quitosana

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
29
pode ajudar a desenvolver novas estratégias para o biocontrole em geral de fungos
patogênicos, evitando o uso de fungicidas que prejudicam o meio ambiente e a saúde
humana.
Pagnoncelli (2008) estudou a produção de quitooligossacarídeos com
propriedades nutracêuticas a partir da quitosana por hidrólise enzimática, utilizando
processo fermentativo simultâneo. Duas cepas produtoras de quitosanaes foram
selecionadas, Paenibacillus chitinolyticus e Paenibacillus ehimensis, e avaliadas quanto
ao comportamento em meio de cultivo contendo açúcares simples e em relação às
variações de pH do meio. Os quitooligossacarídeos produzidos a partir do complexo
enzimático da primeira cepa apresentaram maior pico após 9h de hidrólise, enquanto
para a segunda cepa avaliada, após 20min, observou-se quitooligossacarídeos com grau
de polimerização entre 3 e 6 unidades.
2.2. Fungos
2.2.1. Características gerais
Desde o final dos anos 60 que os fungos deixaram de pertencer ao reino das
plantas e passaram a ter um reino próprio chamado de Reino Fungi. Foi a partir de
estudos morfológicos, bioquímicos e citológicos onde se concluiu que eles são tão
diferentes dos vegetais como dos animais. Distinguem-se de outros organismos
eucariotas por serem seres quimiorganoheterotróficos e, ainda, por apresentarem parede
celular rígida que é composta por quitina e glucano e uma membrana celular em que o
ergosterol substitui o colesterol (Trabulsi & Toledo, 1996; Murray et al., 2005).
Fazem parte do Reino Fungi cerca de 100 mil espécies, desde grandes como
os cogumelos, até microscópicas como os bolores e leveduras. Ainda hoje é utilizada
essa classificação, mas os avanços da biologia molecular e estudos bioquímicos levaram
a outras propostas de classificação em que os fungos e organismos afins foram incluídos
em três diferentes reinos: Protozoa, Chromista e Fungi (Kirk et al., 2008).
Abaixo, são apresentadas características que diferenciam os fungos das plantas:
• são incapazes de sintetizar clorofila, como também não apresentar celulose em
sua parede celular, com exceções como fungos aquáticos;
• apresentam algumas características de células animais (capacidade de depositar
glicogênio e a presença de substâncias quitinosas, em sua parede celular);

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
30
• podem ser eucarióticos (as leveduras), ou multinucleados (os bolores), e
apresentar retículo endoplasmático rugoso e mitocôndrias em seu citoplasma;
• são heterotróficos, alimentam-se de matéria orgânica viva (fungos parasitários),
ou morta (fungos saprofíticos);
Os fungos são amplamente encontrados na natureza, crescem rapidamente no
solo, na água, na atmosfera, bem como nos animais, nos vegetais, resíduos orgânicos e
desempenham um papel importante na vida do homem, quer de uma maneira benéfica,
quer de um modo prejudicial. Os fungos são um dos principais microrganismos
responsáveis pela decomposição da matéria orgânica, interferindo no ciclo do carbono,
do nitrogênio e de outros nutrientes da biosfera. São capazes de deteriorarem produtos e
bens de consumo, tais como alimentos, tecidos, papel, madeira. Os alimentos
armazenados representam um vasto meio para a proliferação dos fungos, principalmente
quando não são consideradas as condições básicas de armazenamento correto (Trabulsi
& Toledo, 1996).
Estão presentes funcionalmente em vários processos industriais, tais como, de
fabricação de pão, cervejas, vinhos e determinados tipos de queijos, sendo também
utilizados na produção comercial de muitos ácidos orgânicos, de alguns fármacos, como
a ergometrina e a cortisona, na obtenção de diferentes antibióticos, como a penicilina, e
de substâncias imunossupressoras, como a ciclosporina (Esteves et al., 1990). A Figura
4 ilustra a estrutura geral de uma célula de fungo.
Figura 4 – Representação da estrutura geral de uma célula de fungo.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
31
Os fungos podem se desenvolver em meios de cultivo especiais formando
colônias de dois tipos, leveduriformes e filamentosas, que se diferenciam pela
macromorfologia e micromorfologia. O primeiro tipo apresenta, em geral, característica
de consistência cremosa de cor branca a creme, brilhantes ou opacas, podendo
apresentar às vezes coloração escura ou alaranjada, e são formados por microrganismos
unicelulares que apresentam as funções vegetativas e reprodutivas. As filamentosas
apresentam características granulares, cotonosas, aveludadas ou pulverulentas, são
multinucleadas e apresentam-se em forma de tubos denominadas de hifas. Essas hifas
podem ser não septadas (cenocíticas) ou septadas (tabicadas). O conjunto de hifas é
denominado micélio.
As hifas asseptadas têm aparência anastomosada (concisa), formada por um
citoplasma estendido e polinucleado, enquanto que as hifas septadas contêm células
individualizadas, cada uma contendo o seu núcleo. Na Figura 5 são mostradas as
estruturas das hifas septadas e não septadas.
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas. (Adaptado de Guia.Bio,
2012)
O micélio que se desenvolve no interior do substrato e que é responsável pela
sustentação e absorção de nutrientes é denominado micélio vegetativo. O micélio que
cresce na superfície do meio de cultura é denominado micélio aéreo. Este pode ser
denominado também de micélio reprodutivo, quando o mesmo se diferencia para
sustentar os corpos de frutificação ou propágulos (Putzke & Putzke, 1998). Os
propágulos classificam-se de acordo com a origem em externos e internos, sexuados e
assexuados.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
32
2.2.2. Reprodução dos Fungos
A reprodução dos fungos ocorre em ciclos assexuais, sexuais e parassexuais. Em
fungos, a reprodução assexuada é observada com mais freqüência do que a reprodução
sexuada. A reprodução assexuada é normalmente a reprodução mais importante para a
propagação da espécie, feita através de fragmentação de artroconídios, fissão de células
somáticas, brotamento ou gemulação do blastoconídios-mãe e produção de conídios. A
reprodução sexuada envolve a união de duas células ou de dois órgãos sexuais
sexualmente compatíveis. A reprodução sexuada culmina na produção de basidiósporos,
no caso de basidiomicetos. A reprodução parassexuada ocorre com a fusão de hifas e
formação de um heterocário, que consiste na existência de núcleos haplóides
geneticamente distintos num mesmo citoplasma, os quais, após a fusão, originam
núcleos diplóides heterozigotos sujeitos a sucessivas divisões mitóticas. Apesar da
raridade dessas, a reprodução parassexual é de grande importância na evolução de
alguns fungos (Pelczar et al., 1996).
A reprodução assexuada é feita por conídios, esporos assexuados formados no
ápice de hifas modificadas, os conidióforos. Os conidióforos não possuem esporângios
de forma que os conídios ficam expostos. Na reprodução sexuada, com o encontro de
duas hifas haplóides distintas, ocorre a plasmogamia e um novo conjunto de hifas
dicarióticas é formado.
Os conídios podem ter formas esféricas, fusiformes, cilíndricas, piriformes, entre
outras. Apresentam-se como hialinos ou pigmentados, podem estar isolados ou
agrupados, a parede celular pode ser lisa ou rugosa e podem ser formados por apenas
uma célula ou apresentarem septos em mais de um plano.
Os conídios também podem originar-se em qualquer parte do micélio vegetativo
e são denominados conídios sésseis como é o caso do Trichophyton rubrum.
Alguns fungos patogênicos são identificados a partir de estruturas formadas pela
célula conidiogênica e o conidióforo. Essas estruturas são denominadas: aparelho de
frutificação ou conidiação. Os conídios podem formar cadeias sobre fiálides em volta de
uma vesícula, dilatando a extremidade do conidióforo. Essas estruturas ocorrem em
Aspergillus, apresentando-se em forma de cabeça ao redor de uma vesícula, e em
Penicillium agrupados em forma de pincel, como é ilustrada na Figura 6.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
33
Figura 6 – Conídios de Aspergillus e Penicillium
2.2.3. Metabolismo de fungos
Os fungos são seres aeróbicos e heterotróficos, podendo se desenvolver em
ambientes com baixa quantidade de oxigênio, como ocorre no crescimento vegetativo e
na reprodução assexuada, como também em ambientes com grande quantidade de
oxigênio como é o caso da reprodução sexuada.
Os fungos são capazes de produzir certas enzimas como lipases, invertases,
lactases, quitosanases, proteases, amilases, etc., que fazem a hidrólise do substrato
facilitando o mecanismo de transporte ativo e passivo. Esses substratos induzem a
formação de enzimas degradativas. Algumas espécies de fungos desenvolvem-se em
meios que apresentam amônia ou nitrilos como única fonte de nitrogênio. As melhores
fontes orgânicas para o seu desenvolvimento são carboidratos, como a D-glicose, e
alguns sais minerais como sulfatos e fosfatos.
A água é uma necessidade de todos os seres vivos, não sendo diferente para os
fungos que precisam de água para o seu desenvolvimento. Certos fungos apresentam
características halofílicas podendo crescer nos ambientes com grandes concentrações de
sal.
A razão de não existirem ambientes livres de espécies fúngicas deve-se ao facto
destes microrganismos conseguirem sobreviver em situações extremas como, por
exemplo, temperaturas que variam entre os - 5 e os 60 ºC, com pH de 1 a 9 e com baixas

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
34
concentrações de oxigênio, apesar de cada espécie possuir condições ideais para o seu
desenvolvimento (Chao et al., 2002).
Devido a grande faixa de temperatura ótima para o seu crescimento, os fungos
apresentam espécies psicrófilas, mesófilas e termófilas. Para a maior parte das espécies
fúngicas, a temperatura de desenvolvimento situa-se entre os 20 e 24 ºC e não crescem
acima dos 29 ºC. Em relação à umidade relativa, necessitam de pelo menos 50 %, mas
geralmente necessitam de mais de 65 % (Colakoglu, 2001).
O pH ideal para o desenvolvimento dos fungos varia na faixa entre 5 e 7.
Destaca-se, entretanto, que a maioria dos fungos suportam grandes variações de pH
como ocorre com os fungos filamentosos que podem crescer em pH variando entre 1,5 e
11. Por outro lado, as leveduras não suportam pH alcalino.
Os fungos crescem lentamente, se comparados com as bactérias, em meio de
incubação para o seu desenvolvimento. Para evitar contaminação por bactérias, que
podem se sobrepor ou inibir o seu crescimento, pode-se introduzir no meio de cultura
um antibacteriano de largo espectro como o cloranfenicol.
Algumas espécies de fungos se desenvolvem na presença de luz, outras são
inibidas e algumas podem crescer tanto na presença quanto na ausência deste agente. No
entanto a irradiação solar, em geral, é prejudicial ao seu desenvolvimento, por causa da
radiação ultravioleta.
2.2.4. Nutrição de fungos
Quanto ao aspecto da nutrição, os fungos são heterótrofos, ou seja, incapazes de
produzir seu próprio alimento. Entretanto, a forma de assimilar matéria orgânica é
diferente daquela utilizada pela maioria dos animais, que são heterótrofos por ingestão.
Nos fungos, a digestão é extracorpórea, ou seja, é realizada fora do corpo. O fungo
lança no ambiente, enzimas que degradam as moléculas orgânicas complexas e, depois,
absorve moléculas menores, mais simples. Por essa razão, são designados heterótrofos
por absorção. É devido a este processo que os fungos podem crescer dentro ou sobre os
alimentos. Desenvolve-se geralmente em meios contendo um pH baixo, uma fonte de
carbono, uma fonte de nitrogênio orgânico ou inorgânico e alguns minerais. Alguns
necessitam de vitaminas. Quanto aos modos de vida, os fungos podem ser
decompositores, parasitas, mutualísticos e predadores (Raven, 2001).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
35
Os fungos decompositores, saprófagos (do grego, saprós, “podre”, e phagos,
“comedor”), juntamente com as bactérias, são os principais decompositores da biosfera,
participando intensamente do processo de degradação da matéria orgânica morta, o que
promove a reciclagem dos elementos químicos constituintes dos seres vivos. A maioria
deles vive no solo, obtendo nutrientes de seres mortos. Em florestas, onde a matéria
orgânica vegetal é abundante, os micélios dos fungos decompositores absorvem os
nutrientes de folhas e galhos caídos, decompondo-os. Os fungos decompositores
também são responsáveis pelo apodrecimento de alimentos e de outros materiais, como
a madeira.
Os fungos parasitas vivem à custa de outros organismos vivos, prejudicando-os.
Ao parasitar o corpo de um ser vivo, animal ou vegetal, o fungo pode até provocar sua
morte. Várias doenças são causadas por fungos parasitas. Nas plantas, cita-se a
ferrugem do café, que, no Brasil, é motivo de grande preocupação por parte dos
cafeicultores, devido aos prejuízos que acarreta. As micoses que atacam a pele de seres
humanos e animais são provocadas por fungos.
Existem fungos que se associam a outros organismos, estabelecendo uma relação
em que há benefício mútuo para os indivíduos envolvidos. Esses fungos são
denominados mutualísticos. A maioria vive associada a seres fotossintetizantes, como
as plantas, cedendo-lhes parte da água e dos nutrientes que as hifas absorvem do solo.
As plantas, por sua vez, cedem ao fungo certos açúcares e aminoácidos. Duas
associações mutualísticas formam estruturas bem características: as micorrizas,
associação que envolve fungos e raízes de plantas, e os líquens, formados por fungos e
algumas variedades de algas e cianobactérias.
Certos fungos atuam como predadores. Na maioria dos casos, as hifas secretam
substâncias aderentes que aprisionam os organismos que tocam os fungos. Dessa
maneira, as hifas penetram o corpo da presa, crescem e se ramificam, espalhando-se no
interior do organismo e absorvendo seus nutrientes, causando-lhe a morte. O gênero
Arthrobotrys apresenta uma forma mais elaborada de predação. O fungo vive no solo e
captura nematódeos microscópicos que habitam o mesmo ambiente. As hifas
apresentam pequenos anéis que se estreitam quando a presa passa por eles. As hifas do
fungo logo invadem o corpo do nematódeo, digerindo-o.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
36
2.2.5. O fungo Metarhizium anisopliae
Existem três espécies de fungos do gênero Metarhizium, divididas em dez
variedades: M. anisopliae, variedades anisopliae, majus, lepidiotum e acridum; M.
flavoviride, variedades tipo E, flavoviride, minus, novazealandicum e pemphigum; e M.
album (Driver et al. 2000). E, mais recentemente, foi descrita na China uma nova
variedade de M. anisopliae, dcjhyium (Dong et al. 2009).
Data de 1879 quando o russo Ilya Metchnikoff aplicou o fungo para combater
larvas de um curculionídeo, praga de beterraba, e foi classificado como Metarhizium
anisopliae por Sorokin em 1883. É um fungicida natural para mais de 300 espécies de
insetos, especialmente pragas da agricultura e pecuária. No Brasil, o fungo Metarhizium
anisopliae teve a sua aplicação com sucesso pela primeira vez em 1965, no controle da
cigarrinha-da-cana Mahanarva posticata (Alves, 1998).
O fungo Metarhizium anisopliae é um importante agente entomopatogênico
utilizado no controle de pragas, como cupins, gafanhotos, cigarrinhas e besouros, sendo
amplamente estudado em todo o mundo. Existe amplamente na natureza e é facilmente
encontrado no solo, onde sobrevive por longos períodos e seu desenvolvimento se dá
geralmente entre 15 e 32 °C, sendo ideal entre 24 e 30 °C e com pH ótimo igual a 6,9.
Apresenta micélio hialino e septado, com conidióforos característicos, sobre os quais
surgem conídios cilíndricos organizados em colunas. Este fungo ao atacar os insetos
provoca rigidez e os recobre por uma camada pulverulenta de conídios, resultando em
colorações que variam do verde claro ao escuro, cinza ou branco com pontos verdes
(Tinline, 1971; Alves, 1998, Driver et al. 2000; Arruda, 2005).
Quanto a sua morfologia, apresenta-se como um fungo filamentoso, com corpo
de frutificação equivalente a um esporodóquio agregado a hifas intimamente
entrelaçadas, com massa compactada de conidióforos característicos, simples ou
ramificados, formando células esporogênicas denominadas fiálides, resultando nos
fialosporos (Wang et al., 2002).
O Metarhizium anisopliae é um fungo de cultivo simples, sendo necessário
quase que exclusivamente o nutriente a base de amido. Um dos fungos mais estudados
em programas de manejo de pragas já que apresenta grande potencialidade
entomopatogênica (Onofre et al., 2002). O Metarhizium anisopliae é um microrganismo
pouco exigente, pois pode se desenvolver em diversos meios de cultura como: amido,
glicose, glicerina, levulose, maltose, sacarose e quitina (Jabor et al., 2003).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
37
Em geral, os fungos produzem enzimas que convertem os tecidos dos insetos em
nutrientes. Porém, a cutícula esclerotizada dos insetos não é bem utilizada pelos fungos,
excetos pelos entomopatogênicos que desenvolveram enzimas eficientes para degradar
essa camada protetora. Destaca-se a enzima Pr1, produzida por Metarhizium anisopliae,
que tem um poder de degradação bem superior a qualquer outra enzima sintetizada por
esse grupo de fungos (Dias, 2005). Desta forma, o Metarhizium anisopliae (fungo
entomopatogênico) ao atacar a célula dos insetos, produz grande quantidade de enzimas
que degradam a membrana citoplasmática e inativa a célula e, consequentemente, mata
o inseto, logo é evidente a relação entre o controle de pragas e produção de enzimas.
No momento, sobre a comercialização de fungos entomopatogênicos no Brasil,
em sua maioria, são formulados em grânulos, compostas do fungo mais o substrato
(arroz + fungo), como também na forma de pó molhável (WP), resultado da moagem do
fungo com o substrato. Esses produtos representam 90% e 8,5%, respectivamente, da
produção de fungos no Brasil, e sendo suas concentrações finais ficam em torno de
5x108 conídios/g para Metarhizium anisopliae e 1x109 conídios/g para Beauveria
bassiana (Alves et al., 2008).
Segundo Batista Filho et al. (2002), Garcia et al. (2005) e Mello et al. (2006), o
Metarhizium anisopliae tem sido bastante utilizado para o controle de diferentes pragas
no Brasil, como a broca da bananeira, pragas de grãos armazenados, carrapatos, cupim
de montículo em pastagens, larvas de escarabeídeos que atacam a cana de açúcar e
cigarrinha da cana-de-açúcar.
É comum a ocorrência natural do fungo Metarhizium anisopliae atacando
cigarrinha-das-raízes de canaviais pelo país, o que incentiva a utilização dele como
agente de controle biológico em áreas infestadas por esse tipo de praga. A aplicação de
recursos no uso de programa de manejo integrado da cigarrinha tem sido reforçado por
razões ambientais e econômicas (Dinardo-Miranda, 2001).
Faria & Magalhães (2001) fizeram uma avaliação do uso de fungos
entomopatogênicos no Brasil, na qual observaram que a qualidade dos micoinseticidas
disponíveis no Brasil poderia ser incrementada de forma considerável quanto a sua
formulação, ou seja, antes da venda, poderiam realizar melhorias no tratamento
posterior, adição de substâncias que lhes assegurem melhorias na eficiência de controle,
capacidade de armazenamento ou praticidade de manuseio, ou de qualquer outro critério
que resulte em vantagem em relação ao produto bruto. Ressaltaram a pouca praticidade

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
38
em alguns casos, como exigência de lavagem, e em outros podendo causar o
entupimento de bicos dos pulverizadores devido à elevada proporção de inertes,
principalmente quando são empregados baixos volumes de aplicação. Outro problema
evidenciado com os inseticidas nacionais diz respeito à pequena sobrevida, devendo a
ser usados em, no máximo, 30 dias depois de produzidos, quando armazenados à
temperatura ambiente e em local sombreado, fazendo as vendas ocorrerem quase que,
exclusivamente, sob encomenda. Comparativamente, os bons inseticidas sob
comercialização em outros países apresentam maior concentração de ingrediente ativo,
maior sobrevida (alguns produtos podem ser armazenados por mais de 8 meses à
temperatura ambiente) e praticidade (são formulados na forma de GDA (grânulos
dispersíveis em água) ou de óleos emulsionáveis, por exemplo, podendo ser adicionados
diretamente ao tanque do pulverizador.
Rangel et al. (2007), instalaram uma Unidade de Observação para o controle
microbiano das cigarrinhas das pastagens Distrito de Itahum, em Dourados/MS, devido
as necessidades de agricultores familiares de produção de leite. As aplicações do fungo
Metarhizium anisopliae foram realizadas por pulverização com equipamento de tração
animal. As condições do ambiente de estudo foram favoráveis ao desenvolvimento da
população das cigarrinhas e à incidência do fungo. Como resultados, observaram que os
níveis de espumas, ninfas e adultos reduziram significativamente após as aplicações,
levando os Agricultores Familiares a total satisfação com os ganhos na produção de
todos os envolvidos.
Segundo Vieira et al. (2007), o fungo entomopatogênico Metarhizium anisopliae
var. acridum tem sido utilizado no controle de pragas, e assim, visando este fim,
analisaram em laboratório a esporulação, a germinação dos conídios e as características
morfológicas peculiares desse fungo em diferentes temperaturas. Esperou-se o
desenvolvimento das colônias em períodos de 1 a 15 dias. Foi observado que
temperaturas de 25, 28 e 30 ºC proporcionaram maiores taxas de esporulação e elevadas
porcentagens de germinação dos conídios. A 25, 28 e 30 ºC observou-se um
desenvolvimento mais rápido das microestruturas somáticas e reprodutivas (hifas,
anastomoses, apressórios, conidióforos e conídios elipsóides). Nas temperaturas
extremas de 40 e 45 ºC não foi detectado esporulação, germinação, nem
desenvolvimento de nenhuma microestrutura.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
39
Lopes et al. (2007) avaliaram o efeito de formulações de Beauveria bassiana e
Metarhizium anisopliae sobre ninfas de Amblyomma cajennense (transmissor da febre
maculosa ao homem) em laboratório. Ninfas não alimentadas foram pulverizadas com
suspensões contendo 2x107 e 4x107 conídios/mL de B. bassiana, e 1x107, 1,5x107,
2x107, 3x107 e 5x107 conídios/mL de M. anisopliae. A mortalidade de ninfas,
decorridos três dias da aplicação, foi significativa nas concentrações de 3x107 e 5x107
conídios/mL de M. anisopliae, com 57,1 e 56,2 %, respectivamente. Após seis dias a
mortalidade foi de 100 % nessas duas concentrações. A mortalidade foi
significativamente maior que a observada para B. bassiana em concentrações
semelhantes.
Guirado et al. (2009) realizaram estudos para controlar cupins de montículo em
área de pastagem, utilizando fungos entomopatogênicos Metarhizium anisopliae e
Beauveria bassiana em diferentes doses e formulações. A avaliação foi realizada após
30 dias da aplicação dos produtos. Os melhores resultados em ordem decrescente de
eficiência foram: M. anisopliae 30 mL + 1000 mL de água com 100 % de eficiência; B.
bassiana 10g + M. anisopliae 20 mL + 1000 mL de água com 85,71 % de eficiência; M.
anisopliae 20 mL + 1000 mL de água com 71,42 % de eficiência. Os demais
tratamentos não diferiram da testemunha.
Loureiro et al. (2012) compararam a eficiência de isolados de Metarhizium
anisopliae em controlar populações naturais de Mahanarva fimbriolata (cigarrinha-da-
raiz) em cultivos comerciais de cana-de-açúcar colhida mecanicamente. Foram
aplicados os isolados IBCB 348, IBCB 408, IBCB 410 e IBCB 425 do fungo em
concentração de 1,5×1012 conídios/ha. Aplicou-se o planejamento experimental em
blocos casualizados com 5 tratamentos e 4 repetições, sendo cada repetição composta
por 7 linhas de 100 m de comprimento e espaçamento de 1,5 m. Após 30 dias de
pulverização foi observado para os isolados IBCB 408 e IBCB 425 uma eficiência de
controle de ninfas de 63 e 62%, respectivamente, já no caso dos adultos chegou a 100%.
Após 60 dias o isolado IBCB 425 foi o mais eficiente para controlar as ninfas com
48,4%.
Assis et al. (2012) estudaram a citotoxidade de quitoolissacarídeos (QOS) e
oligômeros A, B e C (soluções compostas por diferentes quantidades de QOS)
produzidos por hidrólise enzimática pelo fungo Metarhrizium anisopliae. Foi analisado
o efeito de antiproliferativo destas moléculas em células tumorais (HepG2 e HeLa) e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
40
também em célula normal (3T3). A atividade antioxidante foi analisada em vários
experimentos in vitro. A glicosamina apresentou maior toxicidade (cerca de 92%) para
todas as células estudadas. No entanto, os oligômeros obtidos após hidrólise não
demonstraram efeito tóxico sobre a célula normal (3T3). Além disso, mostraram que
uma pequena quantidade de outros COS pode diminuir o efeito citotóxico da
glicosamina contra células 3T3, indicando que glicosamina poderia ser usada como uma
droga antitumoral na presença de outros COS.
2.3. Fermentação
Os processos fermentativos são caracterizados por processos nos quais
microrganismos realizam a conversão catalítica de uma dada substância num
determinado produto. Estes processos podem ser classificados como processos
descontínuos, contínuos e descontínuos alimentados. A classificação destes processos
está diretamente ligada à forma de como o substrato é adicionado e como o produto é
retirado. No processo de fermentação descontínuo o substrato é colocado no
fermentador, em seguida é inoculado e após o término do processo o produto é retirado.
No processo contínuo, o substrato é colocado num reator com vazão constante e o meio
fermentado é retirado com a mesma vazão de entrada do reator (Faccioti, 2001). Nos
processo descontínuos alimentados à adição de substrato e a retirada do mesmo deve
ocorrer de forma intermitente (Carvalho & Sato, 2001).
De certa forma, os processos fermentativos representam um elo entre as antigas
artes de elaboração de alimentos, como queijos e vinho, a partir de uma flora
microbiana natural, com a moderna indústria de fermentação de alimentos, onde são
utilisados cultivos puros e equipamentos sofisticados para o controle do processo
(Assis, 2010).
Atualmente, a maioria dos processos industriais se desenvolve em cubas de
vidro ou de aço de até 20 litros. Os agentes biológicos se encontram submersos no meio de
cultivo que ocupa, aproximadamente, 75% da cuba. As vezes é necessário injetar ar, e em muitas
fermentações se forma espuma. O desenho do biorreator deve se adequar ao objetivo do
processo, respondendo eventualmente a diversos imperativos, tais como a esterilização do
sistema, a aeração e homogeneização do meio, o acréscimo de nutrientes e de aditivos
antiespuma, a manutenção do pH, etc. Os modelos de fermentadores mais utilizados com
microrganismos contam com aeração e agitação mecânica. Esta facilita a distribuição dos

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
41
nutrientes na cuba, mas gera calor que deve ser eliminado mediante a circulação de água
fria. Em outros tipos de biorreatores, em coluna ou torre, a homogeneização depende da injeção de
ar. Os sistemas submersos são apropriados para o cultivo de microrganismos livres, mas resultam
pouco econômicos quando se trabalha com células ou enzimas caras. A imobilização em
fermentadores menores, seja por adesão a um suporte inerte, seja por inclusão dentro de um
polímero que permita o contato com o meio de cultura, além de simplificar a purificação do
produto permite a reutilização das células ou das enzimas, que permanecem dentro do
biorreator (Malajovich, 2009).
As enzimas são produtos de alto valor agregado, amplamente empregadas em
uma gama de indústrias, e entre as fontes para sua obtenção, destacam-se as de origem
microbiana, principalmente, devido a elevada seletividades dos substratos utilizados e
produtos formados. A principal forma de obtenção de enzimas, em escala industrial, é a
fermentação submersa, por esta apresentar vantagens em relação a fermentação
semisólida, como: maior controle do processo fermentativo e consequente reprodução
da cinética microbiana, além de maiores rendimentos (Wanderley, 2011).
2.3.1. Preparo do inóculo
Denomina-se inóculo um volume de suspensão de microrganismo de
concentração adequada, capaz de garantir, em condições econômicas, a fermentação de
uma dada quantidade de meio. O volume do inóculo, assim como sua concentração de
células, vai depender tanto do processo de fermentação c omo da quantidade de meio a
ser fermentado (Carvalho & Sato, 2001).
A partir de uma cultura pura, inocula-se um volume relativamente pequeno de
meio nutriente, que vai depender do tipo de microrganismo cultivado (pois cada espécie
microbiana possui velocidades diferentes de crescimento), sendo, em seguida, incubado
em condições favoráveis ao desenvolvimento do microrganismo. A suspensão obtida é
transferida para um frasco maior em agitador rotativo ou recíproco (shakers), que por
sua vez é adequadamente incubado, e a seguir, transferido para um frasco contendo um
volume ainda maior deste meio, até se conseguir alguns litros de suspensão microbiana.
Os cuidados com assepsia, volumes de meio utilizado, numero de transferência bem
como as condições de incubação, podem variar de acordo com o tipo de processo de
fermentação que se deseja realizar.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
42
2.3.2. Produção de enzimas em cultivo submerso descontínuo
A produção de enzimas, usando processos fermentativos, é constituída
basicamente por duas etapas: a primeira caracteriza-se pela preparação do inóculo e a
segunda, pela fermentação do substrato (Carvalho & Sato, 2001). A finalidade da
primeira etapa é o preparo do microrganismo em condições adequadas para que possa
ocorrer o desenvolvimento da segunda etapa. Diferentemente do cultivo em estado
sólido, no cultivo submerso os microrganismos encontram-se em um meio com elevada
atividade de água.
2.4. Planejamento experimental
Um bom planejamento tem como fundamental projetar um experimento de
forma que ele seja capaz de fornecer exatamente o tipo de informação que procuramos.
Para isso é necessário saber, em primeiro lugar, o que é mesmo que se deseja procurar.
Planejamento experimental, ou delineamento experimental, é um conjunto de
ensaios estabelecido com critérios científicos e estatísticos, objetivando determinar a
influência de diversas variáveis nos resultados de um dado sistema ou processo (Barros
Neto et al., 2001; Rodrigues & Iemma, 2005). De acordo com o propósito dos ensaios,
esse objetivo maior pode ser mais detalhado em:
– determinar quais variáveis são mais influentes nos resultados;
– atribuir valores às variáveis influentes de modo a otimizar os resultados;
– atribuir valores às variáveis influentes de modo a minimizar a variabilidade dos
resultados;
– atribuir valores às variáveis influentes de modo a minimizar a influência de variáveis
incontroláveis.
O planejamento experimental, baseado nos fundamentos estatísticos é, sem
dúvida alguma, uma ferramenta poderosa para se chegar às condições otimizadas de um
processo, desenvolvimento da formulação de produtos dentro das especificações
desejadas ou simplesmente para avaliar os efeitos ou impactos que os fatores (variáveis
independentes) têm nas respostas (variáveis dependentes) desejadas (Barros Neto et al.,
2001; Rodrigues & Iemma, 2005). Da utilização das técnicas estatísticas de
planejamento experimental, é possível chegar benefícios importantes, como:

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
43
– reduzir o número de ensaios sem prejuízo da qualidade da informação;
– estudar simultaneamente diversas variáveis, separando seus efeitos;
– determinar a confiabilidade dos resultados;
– realizar a pesquisa em etapas, num processo iterativo de acréscimo de novos ensaios;
– selecionar as variáveis que influem num processo com número reduzido de ensaios;
– representar o processo estudado através de expressões matemáticas;
– elaborar conclusões a partir de resultados qualitativos.
2.4.1. Planejamento fatorial
Em um planejamento fatorial é investigado as combinações de níveis de estudo
de cada fator. Para tanto, inicialmente, é necessário especificar esses níveis em cada
fator, quer dizer, os valores dos fatores que serão utilizados nos experimentos. Um
ensaio experimental é definido como um experimento constituído de um conjunto de
níveis definido. Geralmente, quando se tem n1 níveis de um determinado fator 1, n2 do
fator 2, ..., e nk do fator k, o planejamento será descrito por um fatorial n1 x n2 x ... x nk,
que corresponde ao número mínimo de experimentos para se ter um planejamento
fatorial completo.
É necessário fazer cada fator variar e observar o resultado dessa variação para se
estudar o efeito sobre a resposta. Assim, os ensaios precisam ter um mínimo de dois
níveis por fator para a realização desses ensaios. Para k fatores, ou seja, k variáveis de
controle, o planejamento de dois níveis precisará de 2 x 2 x ... x 2 = 2k, para o número
de ensaios diferentes, sendo conhecido como planejamento fatorial 2k.
Para o planejamento fatorial, são especificados os valores dos fatores em cada
experimento, e estes são classificados em níveis, variando-se, por exemplo, de -1
(mínimo) a +1 (máximo). A partir de uma matriz serão apresentadas todas as
combinações entre o número de fatores e de níveis nos experimentos a serem realizados
(Barros Neto et al., 2001; Rodrigues & Iemma, 2005).
2.4.2. Planejamento Plackett & Burman
A maioria dos planejamentos do tipo “Screening” (Delineamentos de Seleção de
fatores) foi descrito originalmente por Plackett & Burman em 1946. Os planejamentos
experimentais de Plackett & Burman são eficientes em situações que envolvem uma

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
44
grande quantidade de variáveis a serem exploradas em um processo de otimização de
fatores (Reddy et al., 2008).
As matrizes Plackett & Burman consistem de planejamentos fatoriais de dois
níveis para estudar k = N – 1 variáveis em N ensaios. São construídas para ensaios
múltiplos de 4, sendo essas matrizes para ensaios de 8, 12, 16,..., 100. Cada variável
independente em uma matriz assume uma configuração de dois níveis, o inferior (-1) e o
superior (+1), para o nível intermediário é estabelecido (0). Para cada vez que a variável
aparece no nível inferior, terá o par equivalente no nível superior (Rodrigues & Iemma,
2005).
Para evitar dúvidas nos resultados dos efeitos de algum fator no planejamento de
Plackett & Burman, o número de ensaios a ser realizado deve ser no mínimo o número
de variáveis mais 4. No caso de um fatorial completo o número de ensaios a ser
realizado é bem maior (Rodrigues & Iemma, 2005).
Mesmo já determinado o planejamento Plackett & Burman, torna-se importante
à realização de experimentos relativos aos pontos centrais de modo a se avaliar melhor
as possíveis interferências analíticas que podem ocorrer durante os diferentes
experimentos. A repetição do ponto central permite também a determinação do erro
puro e da repetibilidade do processo. Na prática, esta é uma ferramenta que permite
identificar melhor se o processo em estudo está sob controle (Rodrigues & Iemma,
2005).
É possível que no planejamento Plackett & Burman não tenha sido considerado
a codificação do ponto central (0) para repetição, pois, matematicamente, a estimativa
do valor desse efeito codificado (zero) vezes, teria como resposta zero. Porém, no
resultado prático do experimento, consegue-se informações valiosas com essas
repetições no ponto central (Rodrigues & Iemma, 2005).
2.4.3. Aplicações do Planejamento Plackett & Burman
Ghanem et al. (2000) produziram xilanase por Aspergillus terreus cultivadas
sobre palha de trigo finamente moída em fermentação de estado sólido. A composição
ideal do meio foi estabelecida aplicando-se o planejamento experimental de Plackett &
Burman. A melhor atividade enzimática foi obtida em um meio contendo 10 g/L de

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
45
palha de trigo por frasco umedecido com uma solução concentrada de sal com teor de
água inicial de 75 % e incubadas durante 4 dias a 30 °C. Observou-se que íons
metálicos como Hg2+, Cu2+, Co2+, Fe3+ e Pb2+ inibiram fortemente a xilanase, enquanto
que Ca2+ ativou a enzima.
Mu et al. (2005) estudaram as otimização de meio de fermentação para a reação
de estereoinversão por Candida parapsilosis CCTCC M203011 utilizando o
planejamento Plackett & Burman. Os resultados mostraram que dentre os fatores
estudados, os fatores mais significativos foram: a Concentração de MgSO4, ZnSO4 e
KH2PO4 e que após a utilização da metodologia de superfície de resposta foi constatado
que o ponto ótimo para a otimização foi conseguido com as concentrações dos fatores
MgSO4, ZnSO4 e KH2PO4 nos valores em: 1,17 g/L, 0,10 g/L e 9,41 g/L,
respectivamente, elevando a taxa de conversão enantiosseletiva de 75,2 % para 96 %.
Naveena et al. (2005) empregaram o planejamento Plackett & Burman para a
seleção de 15 parâmetros para produção de ácido láctico L(+) de farelo de trigo,
substrato de baixo custo e suporte sólido, por Lactobacillus amylophilus GV6 em
fermentação de estado sólido (SSF). Onze nutrientes pertencentes a duas categorias,
fontes de nitrogênio e fontes de sal, junto com três parâmetros físicos e um tampão
foram os parâmetros considerados. Os autores verificaram influência na produtividade
devido às fontes de nitrogênio, peptona, extrato de levedura e tri-citrato de amônio,
juntamente com o NaH2PO4.2H2O e Tween 80, que podem ainda serem otimizados para
o aumento na produção de ácido lático. Esses pesquisadores ainda chamam atenção para
o uso escasso deste planejamento em fermentação em estado sólido e que o mesmo
ainda não havia sido considerado anteriormente para a conversão direta de amido em
ácido láctico L (+) usando um sistema bacteriano.
Khosravi-Darani et al. (2008) estudaram a aplicação de Plackett & Burman na
análise dos fatores significativos para a produção de ácido cítrico por Aspergillus niger
ATCC 9142 utilizando palha de trigo como substrato. Os resultados mostraram que a
produção, com a palha de trigo pré-tratada com HCl, NaOH e Uréia, foi mais eficiente e
que os fatores mais significativos na produção foram: Solvente, Teor de Umidade,
Temperatura, Tempo de Incubação, Concentração Inicial de Açúcar e Concentração de
Metanol.
Ji et al. (2009) estudaram o desenvolvimento de um meio de cultura econômico
para a produção do 2,3-butanodiol através da fermentação da glicose e xilose por

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
46
Klebsiella oxytoca utilizando o planejamento Plackett & Burman. Os resultados
mostraram que a uréia e a maceração de milho foram os fatores mais significativos para
a produção do 2,3-butanodiol e que através de planejamento experimental com repetição
em triplicata no ponto central para determinar os níveis ótimos de produção. Os autores
constaram que o rendimento atingido foi 85,6% do valor teórico.
Paula & Arruda. (2009) observaram a necessidade da busca por alternativas mais
eficientes ao método termomecânico utilizado para a degradação de milhões de
toneladas de penas de aves provenientes de frigoríficos e aviários, cuja rígida estrutura é
composta por mais de 90 % de queratina. Para esse fim, utilizaram de comunidades
microbianas para a finalidade proposta, testando o planejamento experimental Plackett
& Burman para verificar a influência de diferentes microrganismos simultaneamente,
com um número reduzido de experimentos. Testaram 32 bactérias isoladas, inoculadas
em meio contendo somente uma pena como única fonte de carbono, durante sete dias,
de onde foi selecionado as cepas para os consórcios microbianos. Experimentos unindo
as cepas NP5, FCA7, NP4 e S14 demonstraram diferentes atividades queratinolíticas,
aumentando o nível de degradação, até 87,8%, comparada com as cepas
individualmente, variando entre 26,4% e 78,0%.
Rajendran & Thangavelu (2009) obtiveram valores máximos da atividade de
produção de lipase em 3,98 U/mL e concentração de massa celular em 5,62 g.L-1 com R.
arrhizus em cultivo submerso descontínuo otimizado pelo planejamento Plackett &
Burman. As variáveis mais importantes que afetaram a produção da lipase foram azeite
de oliva, peptona, KH2PO4, CaCl2.2H2O e MgSO4.7H2O.
Li et al. (2010) estudaram a otimização de meio de cultura para a produção de
Streptolydigin (um antibiótico) por Streptomyces lydicus AS 4,2501 utilizando Plackett
& Burman e a metodologia de superfície de resposta. Os resultados mostraram que
dentre os fatores estudados, os fatores mais significativos para a produção de
Streptolydigin e otimização do meio de cultura foram: Concentração de Amido,
Concentração de Peptona e a Concentração de K2HPO4.
Chen et al. (2010) estudaram a otimização de meio de cultura para a produção da
Luciferase – que são enzimas que catalisam reações biológicas transformando energia
química em energia luminosa - por Bacillus Subtilis em fermentação descontínua
utilizando planejamento Plackett & Burmam. Os resultados mostraram que dentre os
componentes estudados, os fatores mais significativos para a produção e otimização do

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
47
meio de cultura para a produção da Luciferase foram: Concentração de Glicose 1,17 %,
Extrato de Levedura 2,27 % e Glutamato 0,55 %.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o projeto Plackett & Burman e a metodologia de superfície de resposta. Os
resultados mostraram que a eficiência máxima de extração da quitina foi conseguida nas
seguintes condições: Concentração de casca de camarão (50 g/L), glicose (50 g/L),
tempo de incubação de 5 dias e inoculo com 0,05 de OD (densidade óptica).
Gao et al. (2010) desenvolveram um meio para a fermentação do extrato bruto
de inulina de tubérculos de alcachofra de Jerusalém por Paenibacillus polymyxa ZJ-9
para produzir R,R-2,3-butanodiol (R-R,2,3-BD). Inulina, K2HPO4 e NH4Cl foram os
parâmetros principais na fermentação de acordo com os resultados obtidos com o
planejamento experimental Plackett & Burman. A concentração ideal dos três fatores,
77,14 g/L, 3,09 g/L e 0,93 g/L, respectivamente, foi obtida pelo planejamento Box-
Behnken e pela superfície de resposta. Sob as condições ideais, a concentração do R-
R,2,3-BD obtida foi 36,92 g/L, em mais de 98 % de pureza óptica. Comparado com
outras fontes de carbono investigadas, a fermentação do extrato bruto de inulina obteve
o maior rendimento de R-R,2,3-BD.
Uma metodologia de superfície de resposta foi empregada com sucesso para
otimizar um meio de fermentação chamada VHG (Very High Gravity), referência à alta
concentração de açúcares e densidade dos mostos, baseado na água maceração de milho
CSL (Corn Steep Liquor) e outros nutrientes de baixo custo para a produção eficiente de
etanol a partir de glicose por S. cerevisiae. Usando a composição média otimizada (g/L:
CSL 44,3; uréia 2,3; MgSO4.7H2O 3,8; CuSO4.5H2O 0,03), a levedura industrial PE-2
foi capaz de fermentar até 330 g/L de glicose e produzir 18,6 % (v/v) de etanol, com
uma produção descontínua de 2,4 g/L/h e um rendimento em etanol de 93 % (Pereira et
al., 2010).
O planejamento Plackett & Burman juntamente com o Planejamento Composto
Central (CCD) foram aplicados para otimizar o meio de cultura para produção de etanol
por Clostridium autoethanogenum com monóxido de carbono (CO) como fonte única de
carbono e um meio contendo (g/L): 1,0 de NaCl, 0,1 de KH2PO4, 0,02 de CaCl2, 0,15 de
extrato de levedura, 0,116 de MgSO4, 1,694 de NH4Cl e pH 4,74. O rendimento ótimo
de etanol previsto pela metodologia de superfície de resposta (RSM) e um algoritmo

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
48
genético de rede neural artificial (ANN-GA) foram 247,48 e 261,48 mg/L,
respectivamente. Valores semelhantes aos obtidos experimentalmente sob as condições
ideais sugeridas pelos métodos estatísticos (254,26 e 259,64 mg/L). A adequação do
modelo ANN-GA foi maior do que o modelo RSM. Os rendimentos obtidos foram
substancialmente superiores aos relatados anteriormente (60-70 mg/L) com essa cultura
(Guo et al., 2010).
Rigueira (2010) avaliou a influência das variáveis do procedimento de extração
na porcentagem de recuperação das aminas encontradas no leite cru utilizando um
planejamento para “screening” do tipo Plackett & Burman, para os ácidos
tricloroacético (TCA) e sulfosalicílico (ASS).
Quines (2010) estudou a necessidade ou não de se fazer um pré-tratamento da
biomassa antes da extração. Um planejamento experimental Plackett & Burman de doze
ensaios mais três pontos centrais foi utilizado para selecionar as variáveis mais
significativas na extração de Poli(3-hidroxibutirato) (P(3HB)) por 1,2-carbonato de
propileno. Foram realizados dois testes de recuperação do solvente 1,2-carbonato de
propileno, após a extração, sendo um com aplicação de aquecimento e o outro com
aquecimento e vácuo. Os resultados mostraram que o pré-tratamento térmico levou a
uma maior porcentagem de recuperação e de pureza do polímero.

Capítulo 3 Metodologia Experimental

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
50
3. Metodologia Experimental
O presente capítulo apresenta os matérias e métodos utilizados na produção de
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso.
3.1. Quitosana
Neste trabalho utilizou-se uma quitosana (Lote 067) obtida junto a POLYMAR/CE,
com no mínimo 80% de desacetilação. Esse substrato foi utilizado como fonte de carbono e
energia tanto no estudo cinético como nos ensaios do planejamento de seleção de fatores
Plackett & Burman e Planejamento Composto Central, para a produção de quitosanase pelo
Metarhizium anisopliae, por fermentação submersa.
3.2. Microrganismo
O fungo Metarhizium anisopliae estirpe (CG374), utilizado neste estudo, foi cedido
pela Embrapa Recursos Genéticos e Biotecnologia (Brasília / DF-Brasil) e foi mantido em
placas de Petri contendo o meio Batata Dextrose Agar (PDA). As placas foram incubadas em
estufa a 25 ºC. O pH do meio de manutenção foi mantido entre 6,0 e 6,5.
3.2.1. Escolha da cepa produtora de quitosanase
A escolha do microrganismo Metarhizium anisopliae foi feita com base na capacidade
de crescer em meio contendo quitosana como única fonte de carbono, além de apresentar
grande tolerância a pH variando a uma taxa de 2,0 a 8,5, sendo 6,9 a melhor condição para o
crescimento vegetativo e esporução, de acordo com Assis (2010).
3.2.2. Manutenção da cepa
A manutenção da cultura foi realizada em placas petri contendo o meio Batata
Dextrose Agar (PDA). A renovação das células foi realizada a cada trinta dias através de
subcultivos das células no meio PDA.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
51
3.3. Produção de enzima
No preparo do inóculo, utilizou-se 7 placas cultivadas e mantidas, com o fungo
Metarhizium anisopliae, em meio PDA a 25 ºC durante 5 dias em estufa. O meio de cultura
para a produção da enzima quitosanase apresentou os seguintes componentes: quitosana;
peptona; extrato de levedura; NaNO3; K2HPO4; KCl; MgSO4.7H2O e FeSO4, com
concentrações de acordo as matrizes dos planejamentos: Plackett & Burman e composto
central. Para a produção da enzima no cultivo, a biomassa foi transferida através de água
estéril, para um Béquer de 500 mL que, em seguida, foi colocado em Erlenmeyers de 250 mL
na proporção de 20 mL de biomassa para 180 mL do meio de cultura. Os Erlenmeyers
cultivados foram então incubados em incubador rotativo a 25 ºC e 150 rpm durante 24 h.
Alíquotas foram retiradas e centrifugadas a 8000xg por 5 min, a cada 6 horas, para se
determinar a atividade quitosanolítica, teores de açúcares redutores e de proteína total.
Ressalta-se que todos os experimentos foram feitos em triplicata.
3.4. Determinação da Proteína Total
A determinação da proteína total foi efetuada de acordo com o método modificado de
Sedmak & Grossberg (1977), que se baseia na diminuição da absorbância do corante azul de
Coomassie, devido à ligação proteína-corante, a 595 nm, comparado a absorção máxima a
465 nm, conforme Santos (2001). Nesse caso, após centrifugação para retirada da biomassa,
transferia-se para um eppendorf 1,5 mL de amostra (caldo fermentativo) e adicionava-se 1,5
mL de solução azul de Coomassie. Deixava-se a solução em repouso por 5 minutos,
transferindo-se em seguida para um cubeta de quartzo e efetuando-se as leituras de
absorbância nos comprimentos de ondas 465 nm e 595 nm, respectivamente. O
espectrofotômetro (Thermo Spectronic modelo Genesys 10UV) foi zerado com água
destilada. O conteúdo protéico foi obtido a partir de uma curva padrão na qual graficava-se a
relação de absorbância A595/A465 para diferentes concentrações de Albumina de Soro
Bovino (Fração V – Lote 110881/Sigma Aldrich-Ohio-USA).
3.5. Determinação da Atividade Quitosanolítica e da Atividade Específica
A atividade quitosanolítica foi obtida monitorando-se o aumento dos açúcares
redutores totais, utilizando-se o método DNS a partir de uma curva padrão de glicosamina.
Para a determinação da atividade quitosanolítica adicionaram-se, aos tubos de ensaios 500 µL

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
52
de substrato (solução 0,2 % de quitosana solubilizada em ácido clorídrico 1N) e 500 µL de
caldo bruto (contendo enzima), controlando-se o pH entre 6,0 e 6,5 e incubando-se o sistema
em um banho termostatizado com temperaturas da ordem de 55 ºC. Após o tempo de
incubação de 30 minutos no banho foi adicionado aos tubos de ensaio 2,5 mL de DNS,
aquecendo-se à ebulição em banho-maria por 10 minutos, resfriando-se, em seguida, em
banho de gelo. Após essa etapa, o sistema foi centrifugado e com o sobrenadante efetuou-se a
leitura no espectrofotômetro a 600 nm. Destaca-se, que antes da leitura da amostra zerava-se o
equipamento (branco do espectro) com 1,0 mL de água destilada acrescidos de 2,5 mL de
DNS e realizava-se o branco da enzima que consistia da adição de 500 µL de solução de
quitosana, completando-se até um 1,0 mL com água destilada. Uma unidade de atividade
quitosanolítica (A) foi definida como a quantidade de enzima necessária para produzir 1
µmol.min-1 de glicosamina por mL de extrato enzimático, nas condições de temperatura e pH
citadas, sendo o resultado expresso em (U.mL-1). A Atividade específica foi determinada pela
divisão entre a atividade da enzima (U.mL-1) e a concentração de proteína total (µg.mL-1),
sendo expressa em (U. µg-1).
3.6. Planejamento experimental
3.6.1. Planejamento de seleção de fatores Plackett & Burman
O planejamento experimental Plackett & Burman é uma técnica muito usada, para se
realizar triagem de fatores na otimização num determinado processo (Reddy et al., 2008), e
foi utilizado neste trabalho para identificar os fatores que influenciam de forma significativa
na produção da enzima quitosanase.
A Tabela 3 ilustra os níveis de cada variável (fator) utilizada no planejamento Plackett
& Burman, sendo: Concentração de Quitosana (Q), Peptona (PP), Extrato de Levedura (EL),
NaNO3 (NS), K2HPO4 (FP), KCl (CP), MgSO4.7H2O (SM), FeSO4 (SF). Já a Tabela 4 mostra
a matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3 repetições
na condição do ponto central.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
53
Tabela 3 – Níveis para os fatores e seus valores codificados.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1 0,1 0,2 0,2 0,1 0,05 0,025 0,025 0,0005
0 0,2 0,5 0,3 0,2 0,10 0,050 0,050 0,0010
1 0,3 0,8 0,4 0,3 0,15 0,075 0,075 0,0015
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1
2 1 1 -1 1 -1 -1 -1 1 -1 1 1
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1
5 1 1 -1 1 1 -1 1 -1 1 -1 1
6 1 1 1 -1 1 1 -1 1 1 -1 -1
7 -1 1 1 1 -1 1 1 -1 -1 1 1
8 -1 -1 1 1 1 -1 1 1 1 1 1
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1
11 -1 1 -1 -1 -1 1 1 1 1 1 -1
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1
13 0 0 0 0 0 0 0 0 0 0 0
14 0 0 0 0 0 0 0 0 0 0 0
15 0 0 0 0 0 0 0 0 0 0 0
Sendo: V11,V12,V13 = Variáveis inertes para cálculo do erro.
3.6.2. Planejamento Composto Central para a Otimização do Meio de Cultura para a
Produção da Enzima em Cultivo Submerso
Um dos problemas mais comuns para quem realiza experimentos é determinar a
influência de uma ou mais variáveis sobre outra variável de interesse (Lobato, 2003). Este

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
54
problema pode ser minimizado utilizando-se um planejamento experimental de modo a se
obter uma função, ou uma aproximação satisfatória para a mesma, de forma que se operando
sobre as variáveis de entrada produza-se como saída a resposta ou as respostas observadas
(Barros Neto, 2001; Silva, 2004).
Em um planejamento, inicialmente, deve-se definir os fatores e as respostas de
interesse. Tendo-se identificado os fatores e respostas define-se o objetivo que se pretende
alcançar com os experimentos para que se possa escolher o planejamento mais apropriado.
(Silva, 2004).
No presente trabalho foi utilizado um Planejamento Composto Central (CCD) com 22
experimentos na parte fatorial, repetição em triplicata no ponto central e quatro experimentos
na parte axial (planejamento do tipo estrela), para se investigar a influência dos fatores:
concentração de quitosana e concentração de FeSO4 que foram os dois fatores que
influenciaram a produção de quitosanase, conforme o planejamento Plackett & Burman de 12
ensaios e mais 3 repetições na condição do ponto central. Estes dois fatores foram estudados
em cinco diferentes níveis (-1,41; -1; 0; 1; 1,41) e um conjunto de 11 experimentos foi
realizado, conforme Tabela 5.
Tabela 5 – Matriz do planejamento Composto Central.
Ensaio Q SF
1 -1 -1
2 -1 1
3 1 -1
4 1 1
5 -1,41 0
6 1,41 0
7 0 -1,41
8 0 1,41
9 0 0
10 0 0
11 0 0
Os outros fatores foram mantidos nos níveis mais baixos (para evitar custos). Uma que
não influenciaram de forma significativa na produção da enzima quitosanase.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
55
A codificação dos fatores foi realizada de acordo com a seguinte equação (Ji et al.,
2009):
Xi = (xi – x0)/∆xi , i = 1, 2, 3, ..., k (2)
Onde Xi é o fator independente codificado, xi é o fator independente real, x0 é o valor de xi no
ponto central e ∆xi é a variação de valor. O comportamento do sistema foi explicado pela
seguinte equação polinomial de segunda ordem:
Yp = β0 + ∑ βiXi + ∑ βiiXi2 + ∑ βijXiXj, i, j = 1, 2, 3,..., k (3)
Onde Yp foi a resposta esperada, β0 foi a interseção, Xi e Xj
foram os fatores independentes codificados, βi foi o coeficiente linear, βii
foi o coeficiente quadrático e βij foi o coeficiente de interação. Destaca-se que os outros
fatores não significativos foram mantidos no nível mais baixo (-1).
A Tabela 6 apresenta os níveis para os fatores e seus valores codificados, para o
Planejamento Composto Central, com triplicata no ponto central.
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1,41 0,218 0,2 0,2 0,1 0,05 0,025 0,025 0,0009
-1 0,3 0,2 0,2 0,1 0,05 0,025 0,025 0,0015
0 0,5 0,2 0,2 0,1 0,05 0,025 0,025 0,0030
1 0,7 0,2 0,2 0,1 0,05 0,025 0,025 0,0045
1,41 0,782 0,2 0,2 0,1 0,05 0,025 0,025 0,0051
Na análise do planejamento de seleção de fatores Plackett & Burman e do
planejamento experimental fatorial foi utilizado um pacote computacional STATISTICA for
Windows versão 7.0.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
56
3.7. Perfil cinético durante o cultivo
Um perfil cinético na melhor condição de cultivo, conforme planejamento
apresentado, foi realizado em frascos Erlenmyer de 250 mL contendo 200 mL de meio de
cultivo contendo os seguintes componentes, em 1 Litro: 2,0g de quitosana; 5,0g de peptona;
3,0g de extrato de levedura; 2,0g de NaNO3; 1,0g de K2HPO4; 0,5g de KCl; 0,5g de
MgSO4.7H2O; 0,01g de FeSO4, que foram inoculadas com 10% (vol/vol) de inóculo contendo
o fungo Metarhizium anisopliae e cultivadas em incubador rotativo durante 5 dias a 150 rpm
e 25 ºC. Alíquotas foram retiradas a cada seis horas e analisadas quanto à proteína total no
sobredanante, atividade enzimática, atividade específica e açúcares redutores. Os ensaios
foram realizados em triplicata (Silva Filho, 2005).

Capítulo 4 Resultados e
Discussão

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
58
4. Resultados e discussão
Neste capítulo são apresentados e discutidos os resultados obtidos no presente
trabalho. Inicialmente, é apresentada imagem do MEV do Metarhizium anisopliae
mantidos em meio PDA. Em seguida, apresenta-se e discute-se os resultados da avaliação
dos fatores significativos, que podem influenciar na produção da quitosanase, no
planejamento de seleção de fatores, conforme planejamento Plackett & Burman bem como
são apresentados e discutidos os resultados obtidos para o planejamento composto central.
Por último, apresenta-se o perfil cinético, na melhor condição, para a produção de
quitosanase em cultivo submerso descontínuo usando o fungo Metarhizium anisopliae.
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA.
A Figura 7 ilustra a imagem da Microscopia Eletrônica de Varredura (MEV) do micélio de Metarhizium anisopliae mantido em meio PDA.
Figura 7. Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA.
Observa-se na Figura 7, quanto à morfologia que o Metarhizium anisopliae, apresenta-
se como um fungo filamentoso com as hifas intimamente entrelaçadas conforme citado por
Wang et al. 2002.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
59
4.2. Planejamento Plackett & Burman
4.2.1 Análise do Plackett & Burman com 8 fatores e 12 ensaios
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis, foi utilizado para avaliar a influência dos fatores: Concentração de
Quitosana (Q), Peptona (PP), Extrato de Levedura (EL), NaNO3 (NS), K2HPO4 (FP), KCl
(CP), MgSO4.7H2O (SM), FeSO4 (SF) na produção da enzima quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso. Ressalta-se que os experimentos foram
realizados aleatoriamente.
A Tabela 7 apresenta a matriz do modelo Plackett & Burman e os respectivos valores
obtidos para a variável resposta (Atividade Quitosanalítica).
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.
Ensaio Q PP EL NS FP CP SM SF A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 0,0195
2 1 1 -1 1 -1 -1 -1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 0,0109
4 1 -1 1 1 -1 1 -1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 0,0136
6 1 1 1 -1 1 1 -1 1 0,0156
7 -1 1 1 1 -1 1 1 -1 0,0109
8 -1 -1 1 1 1 -1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 0,0085
A Tabela 8 apresenta os efeitos estimados para o planejamento Plackett & Burman.
Observa-se na Tabela 8 que o valor do coeficiente de determinação R2 foi de
aproximadamente 90%, indicando que um modelo linear poderia representar bem a relação
entre o fator significativo, concentração de quitosana (1), e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
60
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman.
Efeitos Estimados; R2=0,8996; Ajuste:0,63178 MS Erro Residual =0,0000054
Desvio Padrão t(3) p
Lim. Conf. Lim. Conf.
Efeito Erro Residual
-95,% +95
Média/Interação 0,0127 0,0007 18,903 0,0003 0,0105 0,0149 (1) Quitosana 0,0055 0,0013 4,0628 0,0268 0,0011 0,0098
(2) Peptona 0,0015 0,0013 1,1080 0,3486 -0,0028 0,0058 (3) Ext. de leved. 0,0009 0,0013 0,7387 0,5136 -0,0033 0,0053
(4) NaNO3 0,0002 0,0013 0,1758 0,8715 -0,0040 0,0045 (5) K2HPO4 -0,0014 0,0013 -1,0552 0,3687 -0,0057 0,0028
(6) KCl -0,0022 0,0013 -1,6708 0,1933 -0,0065 0,0020 (7) MgSO4.7H2O -0,0003 0,0013 -0,2286 0,8338 -0,0046 0,0039
(8) FeSO4 0,0029 0,0013 2,1457 0,1211 -0,0014 0,0071
A Figura 8 mostra o gráfico de Pareto, uma das formas de se avaliar visualmente a
influência dos fatores estudados na resposta. A magnitude dos efeitos é representada pelas
colunas enquanto que a linha transversal às colunas representa a magnitude dos efeitos com
significado estatístico para p=0,05, ou seja, os fatores que são estatisticamente significativos
ao nível de 95% de confiança.
Figura 8. Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
61
Analisando-se o gráfico de Pareto (Figura 8) observa-se que o fator concentração de
quitosana (1) foi o único fator que influenciou de forma mais significativa na atividade
enzimática atingindo efeitos estimados de aproximadamente 4,06. O sinal positivo indica que
o aumento da concentração de quitosana aumentará à atividade enzimática. Por outro lado, os
outros fatores parecem desprezíveis estatisticamente conforme o gráfico de Pareto.
4.2.2 Análise do Plackett & Burman com 12 Ensaios e mais 3 Repetições na condição do
ponto central.
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis com repetição em triplicata no ponto central, foi utilizado para se avaliar a
influência dos mesmos fatores descritos na Seção 4.2.1 para a produção da enzima
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso. Ressalta-se que os
experimentos foram realizados aleatoriamente. A Tabela 9 apresenta a matriz do modelo
Plackett & Burman e os respectivos valores obtidos para a variável resposta (Atividade
Quitosanalítica).
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13 A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1 0,0195
2 1 1 -1 1 -1 -1 -1 1 -1 1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1 0,0109
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 1 -1 1 0,0136
6 1 1 1 -1 1 1 -1 1 1 -1 -1 0,0156
7 -1 1 1 1 -1 1 1 -1 -1 1 1 0,0109
8 -1 -1 1 1 1 -1 1 1 1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 1 1 -1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1 0,0085
13 0 0 0 0 0 0 0 0 0 0 0 0,0704
14 0 0 0 0 0 0 0 0 0 0 0 0,0688
15 0 0 0 0 0 0 0 0 0 0 0 0,0710

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
62
A Tabela 10 a seguir apresenta os efeitos estimados para planejamento Plackett &
Burman com 3 repetições na condição do poto central. Observa-se na Tabela 10 que o valor
do coeficiente de determinação R2 foi de aproximadamente 2,0%, indicando que um modelo
linear não poderia representar a relação entre os fatores significativos concentração de
quitosana (1), concentração de FeSO4 (8) e a resposta.
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central.
Efeitos Estimados; R2=0,01811; Ajuste:0,0 MS Erro Puro =0,0000012
Desvio Padrão t(2) P
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0242 0,0003 85,509 0,0001 0,0229 0,0254 (1) Quitosana 0,0055 0,0006 8,6432 0,0131 0,0027 0,0082
(2) Peptona 0,0015 0,0006 2,3572 0,1425 -0,0012 0,0042 (3) Ext. de leved. 0,0009 0,0006 1,5715 0,2567 -0,0017 0,0037
(4) NaNO3 0,0002 0,0006 0,3742 0,7442 -0,0024 0,0029 (5) K2HPO4 -0,0014 0,0006 -2,2450 0,1538 -0,0041 0,0013
(6) KCl -0,0022 0,0006 -3,5546 0,0708 -0,0049 0,0005 (7) MgSO4.7H2O -0,0003 0,0006 -0,4864 0,6747 -0,0030 0,0024
(8) FeSO4 0,0029 0,0006 4,5648 0,0448 0,0002 0,0056
A Figura 9 ilustra o gráfico de Pareto para o planejamento Plackett & Burman, com
repetição em triplicata no ponto central, uma das formas de se avaliar visualmente a influência
dos fatores estudados na resposta, onde é mostrado a magnitude dos efeitos para 95 % de
confiança.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
63
Figura 9. Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto
central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto, observa-se que os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma mais
significativa na atividade enzimática atingindo efeitos estimados de aproximadamente 8,84 e
4,67, respectivamente. O sinal positivo indica que o aumento das concentrações de quitosana
e de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores parecem
desprezíveis estatisticamente conforme o gráfico de Pareto.
Este comportamento do FeSO4 está de acordo com o observado por Hagag et al.
(2012) que utilizando o Aspergillus fumigatus na produção de protease mostraram que um
fator de transcrição PrtT regula positivamente o controle da produção de protease, da
assimilação de Ferro e da síntese de ergosterol e que a produção de protease foi favorecida
pelo aumento da concentração de ferro.
Rajendran & Thangavelu (2009) utilizaram um planejamento Plackett & Burman para
identificar quais os fatores significativos na produção de lipase por Rhizopus arrhizus MTCC
2233 e observaram o efeito significativo médio do FeSO4, apresentando um nível de
confiança para esse fator de 48,92% (para um máximo de 98,7 % de MgSO4), com uma
produção de lipase da ordem de 2,98 U/mL em 84 horas de fermentação e que ocorreu uma
produção considerável de protease no meio de cultivo da ordem de 1,92 U/mL em 96 horas de
cultivo.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
64
Mu et al. (2005) também observaram um efeito positivo do FeSO4, da ordem de 1,48
para um fator máximo encontrado de 3,11 de MgSO4, no aumento da atividade da
estereoinversão de S-fenil-1,2-etanodiol por Candida parapsilosis CCTCC M203011 na
otimização da fermentação utilizando um planejamento Plackett & Burman.
Dessa forma, observa-se que o FeSO4 é importante uma que vez o mesmo está
diretamente associado a síntese de proteases (principalmente metalloproteases), conforme
citado por Hagag et al. (2012).
4.3. Planejamento Composto Central para Produção de Quitosanase por
Metarhizium anisopliae em Cultivo Descontínuo Submerso
Um planejamento experimentos 22 com repetição em triplicata no ponto central e mais
quatro ensaios axiais, foram utilizados para se chegar a uma combinação ideal de quitosana e
FeSO4 na produção da enzima quitosanase (Tabela 11). A partir do modelo Plackett &
Burman (Tabela 12), que mostrou que o aumento das concentrações de quitosana e FeSO4,
favoreceram a produção de quitosanase, os valores utilizados neste modelo para estes
componentes nos níveis mais altos, serão considerados como os níveis mais baixos no
planejamento experimental do composto central. Os outros fatores foram fixados em todos os
ensaios em seus níveis mais baixos para a redução de custos (Tabela 11).

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
65
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da
atividade enzimática.
Ensaio Q SF A (U/mL)
1 -1 -1 0,0195
2 -1 1 0,0202
3 1 -1 0,0109
4 1 1 0,0121
5 -1,41 0 0,0136
6 1,41 0 0,0156
7 0 -1,41 0,0108
8 0 1,41 0,0103
9 0 0 0,0099
10 0 0 0,0116
11 0 0 0,0094
A Tabela 12 apresenta os efeitos estimados para o planejamento composto central.
Tabela 12 – Efeitos Estimados do planejamento Composto Central.
Efeitos Estimados; R2=0,629; Ajuste:0,25836 MS Erro Puro =0,0000014
Desvio Padrão t(2) p
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0103 0,0006 15,305 0,0042 0,0074 0,0132 (1) Quitosana (L) 0,0011 0,0008 1,4316 0,2885 -0,0023 0,0047
Quitosana (Q) 0,0058 0,0009 5,9637 0,0269 0,0016 0,0101 (2) FeSO4 (L) -0,0043 0,0008 -5,3037 0,0337 -0,0079 -0,0008 (2) FeSO4 (Q) 0,0018 0,0009 1,8366 0,2076 -0,0024 0,0060
1L x 2L 0,0001 0,0011 0,1520 0,8931 -0,0048 0,0052
Observa-se na Tabela 12 que o valor do coeficiente de determinação R2 foi de
aproximadamente 63%, indicando que um modelo linear representaria bem a relação entre os
fatores significativos concentração de quitosana (1), concentração de FeSO4 (8) e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
66
A Tabela 13 mostra um resumo da ANOVA para o planejamento composto central
com repetição em triplicata no ponto central.
Tabela 13 – Resumo da ANOVA.
Fonte de
Variação
Soma dos
Quadrados
Grau de
Liberdade
Quadrado
Médio
Fcalculado Ftabelado
Regressão
Resíduos
0,000086 2 0,000043 6,14 4,46
0,000056 8 0,000007
Falta de
Ajuste
Erro puro
0,000053 6 0,000009 6 19,33
0,000003
2
0,0000015
Total 0,000142 10
De acordo com a Tabela 13, partindo-se dos efeitos significativos propõe-se o
modelo:
� = 0,010316 + 0,002942� � − 0,002193�� (8)
% de variação explicável: 60,5
% máxima de variação explicável: 97,9
A análise da ANOVA mostra que o modelo proposto possui significância estatística, ao
nível de 95% de confiança, uma vez que o valor de F (calculado) é maior que F (tabelado),
entretanto a ANOVA mostra também que o modelo proposto pode ser útil para se fazer
previsões, uma vez que a relação entre F (calculado) e F (tabelado) para a falta de ajuste e o
erro puro é menor que 1. O coeficiente de determinação R2 foi de aproximadamente 63%,
indicando que o modelo consegue explicar 63% da variação total em torno da média.
Os cálculos para variação explicável e para máxima variação explicável são dados pelas
Equações (9) e (10):
% de variação explicável = (SQ da regressão) / (SQ Total) (9)
% máxima de variação explicável = (SQ Total – SQ Erro puro) / (SQ Total) (10)
Sendo:
SQ – Soma dos quadrados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
67
A Figura 10 mostra o gráfico de Pareto para o planejamento Composto Central
utilizando erro puro. A magnitude dos efeitos é representada pelas colunas. Já a linha
transversal às colunas representa a magnitude dos efeitos com significado estatístico para 95%
de confiança.
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto (Figura 10) observa-se que os fatores concentração
de quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma
mais significativa na atividade enzimática atingindo efeitos estimados de aproximadamente
5,96 e - 5,30, respectivamente. O sinal positivo indica que o aumento das concentrações de
quitosana aumentará à atividade enzimática. O sinal negativo indica que o decréscimo da
concentração de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores
parecem desprezíveis estatisticamente no gráfico de Pareto.
A avaliação do modelo também pode ser feita através da observação do gráfico dos
valores preditos em função os valores observados que são mostrados na Figura 11. Os valores
preditos pelo modelo são representados pela reta, enquanto que os valores observados
representam-se pelos pontos.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
68
Figura 11 – Valores preditos em função dos valores observados.
Observa-se que alguns valores preditos apresentam um afastamento dos valores
observados. A Figura 12 ilustra a superfície de resposta da atividade quitosanolítica em
função dos dois fatores significativos, concentração de quitosana e concentração de FeSO4.
Figura 12. Atividade quitosanolítica em função da concentração de quitosana e da
concentração de FeSO4 para os fatores não significativos no ponto central.
Observa-se a existência de um ponto de mínimo, ocorrendo para concentração de
quitosana no ponto central (0) e para maiores concentrações de FeSO4 igual a (1,5). Essa

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
69
região deve ser evitada uma vez que a atividade quitosanolítica atinge níveis tão reduzidos
quanto 0,0094 U.mL-1. Destaca-se que nesse planejamento o maior nível da concentração de
FeSO4 (α=+1,41) equivale a 0,0051% (Tabela 6) que é aproximadamente cinco vezes maior
que o nível desse fator no ponto central do planejamento Plackett & Burman (0,001%),
conforme Tabela 3. Dessa forma, ao se utilizar o nível mínimo da concentração de FeSO4 no
Planejamento Composto Central (α=-1,41), que equivale a aproximadamente 0,001%,
aproxima-se do ponto central do do planejamento Plackett & Burman (0,001%), condição
essa que apresenta os melhores resultados de atividade quitosanolítica usando quitosana como
fonte de carbono e energia e fungo Metarhizium anisopliae.
4.4. Perfil Cinético – Melhor Condição
A seguir, apresenta-se um perfil cinético, na melhor condição, para produção da
quitosanase usando o fungo Metarhizium anisopliae, em que foram monitorados durante 24
horas os seguintes parâmetros: Concentração de proteínas totais (µg/mL), atividade
quitosanolítica (U/mL), atividade específica (U/µg) e açúcares redutores (mg/mL), ilustradas
nas Figuras 13, 14, 15 e 16, respectivamente.
Com relação à proteína extracelular, observa-se na Figura 13 que, inicialmente, ocorre
um aumento da mesma para o seu nível máximo (6,2 µg/mL), durante as 6 primeiras horas de
cultivo, seguido de uma redução para 5,7 µg/mL em 18 horas de cultivo, possivelmente
devido à presença de proteases no meio. Observa-se que a partir de então ocorre uma redução
e aumento da proteína total, em uma quantidade inferior, sugerindo um mecanismo de
controle.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
70
0 4 8 12 16 20 245,0
5,2
5,4
5,6
5,8
6,0
6,2
6,4
Pro
tein
a (
µg
/mL
)
Tempo (h)
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana.
Com relação à atividade quitosanolítica, conforme Figura 14, a atividade máxima
ocorreu em 18 horas de produção em que se obteve aproximadamente 70,0 U/L, o que foi 28
vezes maior que o valor (2,5 U/L em 48 horas) relatado por Assis (2010) para o mesmo fungo,
e em apenas um terço do tempo de produção.
0 4 8 12 16 20 240,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,10
Ativid
ade
(U
/mL
)
Tempo (h)
Figura 14. Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
71
Além disso, destaca-se o considerável aumento da produção de quitosanase a partir das
12 horas até 18 horas de cultivo onde atingiu o pico máximo de produção (70,0 U/L). Após o
pico em 18 horas, a produção começa a cair rapidamente, comportamento esse observado
durante todos os ensaios cinéticos. Destaca-se que é importante do ponto de vista industrial
manter a produção em alto nível por muitas horas, onde se espera uma produção contínua ou
em grandes bateladas.
Ak et al. (1998) utilizaram o fungo Penicillium spinulosum para produzir a quitosanase
usando um meio de cultura contendo 0,1% de K2HPO4; 0,05 % de MgSO4.7H2O; 0,05% de
KCl; 0,001 % de FeSO4, 0,5% NH4NO3 e como fonte de carbono e energia, os autores
usaram quitosana e parede de Rhizopus. A produção foi realizada em cultivo submerso a 28
oC. Os autores apresentam atividade quitosanolítica máxima de aproximadamente 47,0 U/mL,
ao se utilizar a parede do fungo como fonte de carbono e energia, e 30,0 U/mL ao se utilizar
quitosana como fonte de carbono e energia. Destaca-se que essas atividades ocorreram após 4
dias de cultivo. Os perfís com o tempo da atividade quitosanolítica e da atividade específica
apresentaram tendência de crescimento, com o pico máximo em 4 dias, seguindo com uma
tendência de queda da atividade. Neste tempo, a atividade específica máxima da quitosanase
foi de aproximadamente 60,0 U/mg de proteína. Essas tendências também foram observadas
nos resultados apresentados nos perfís de atividade das Figuras 14 e 15.
0 4 8 12 16 20 24
-0,010
-0,005
0,000
0,005
0,010
0,015
0,020
0,025
0,030
Ativid
ad
e E
spe
cific
a (
U/µ
g)
Tempo (h)
Figura 15. Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
72
Observa-se na Figura 15 que a atividade específica máxima da quitosanase segue a
mesma tendência da atividade quitosanolítica apresentada na Figura 14, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
O perfil para os açúcares redutores durante o cultivo de Metarhizium anisopliae
cultivado em meio contendo 0,2% de quitosana é ilustrado na Figura 16.
0 4 8 12 16 20 240,004
0,008
0,012
0,016
0,020
0,024
0,028
Acuca
res R
ed
uto
res (
mg/m
L)
Tempo (h)
Figura 16. Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana.
Observa-se, na Figura 16, que taxa máxima de redução dos açúcares redutores
(aproximadamente 0,021 mg/mL) ocorre logo após a produção máxima da enzima, em 18
horas de cultivo, conforme Figura 14. A quantidade de açúcares redutores foi a
aproximadamente 0,012 mg/mL após 24 h de cultivo. O fungo excreta para o meio enzimas
com atividade quitosanolítica, essas enzimas hidrolisam a quitosana aumentando a quantidade
de açúcares redutores para o meio, sendo que esses açúcares também são consumidos pelo
fungo durante seu crescimento.
Foi demonstrado por Assis (2010) que os açúcares redutores produzidos através da
quitossana, ocasionada pelas enzimas quitossanases metabolizadas por Metarhizium
anisopliae, com quitosana como sua única fonte de carbono e energia, atingiu um pico de 0,53
mg/mL em 48 horas. Apresentaram como quitooligossacarídeos, constituídos principalmente
por dímeros e pentâmeros, formados pela hidrólise enzimática de quitosanase em quantidades
maiores do que o consumido pelos microrganismos. Mostraram, também, que essa produção

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
73
segue a mesma tendência da sua atividade enzimática, corroborando com os resultados
apresentados no presente trabalho.
Os resultados do presente trabalho mostram a relevância de se adotar novas condições
para a produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter quitooligossacarídeo
com atividades biológicas. Esta pesquisa, portanto, sugere base para novos estudos que
permitam alcançar rapidamente a produção de quitosanase e quitooligossacarídeo com todos
os seus benefícios.

Capítulo 5 Conclusão

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
75
5. Conclusão
Após o desenvolvimento desse trabalho conclui-se:
- Com relação ao Planejamento de Seleção de Fatores Plackett & Burman:
Os resultados das análises do Planejamento de Seleção de Fatores Plackett &
Burman com 8 fatores e 12 ensaios em 2 níveis, mais 3 repetições no ponto central, para
se investigar a influência dos fatores: Concentração de Quitosana, Peptona, Extrato de
Levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O, FeSO4 na produção da enzima
quitosanase por Metarhizium anisopliae, mostraram que a atividade foi favorecida pelo
aumento da concentração de substrato (quitosana) e sulfato ferroso (FeSO4), enquanto
que o aumento da concentração dos outros fatores não contribuíram de forma
significativa para a resposta (atividade quitosanolítica). Os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) atingiram efeitos estimados de
aproximadamente 8,84 e 4,67, respectivamente. O sinal positivo indica que o aumento
das concentrações de quitosana e de FeSO4 aumentará a atividade enzimática. Conforme
visto na literatura, observa-se importância do FeSO4, já que o mesmo está diretamente
associado a síntese de proteases (principalmente metalloproteases),
- Com relação ao Planejamento Composto Central:
A análise do planejamento composto central para avaliar a influência dos
fatores, concentração de substrato (quitosana) e concentração de FeSO4, na produção da
enzima quitosanase por Metarhizium anisopliae (a partir do planejamento Plackett &
Burman) mostrou que os fatores concentração de quitosana e concentração de FeSO4
foram os fatores que influenciaram de forma mais significativa na atividade enzimática
atingindo efeitos estimados de aproximadamente 5,96 e - 5,30, respectivamente. O sinal
positivo indica que o aumento das concentrações de quitosana aumentará a atividade
enzimática. O sinal negativo indica que o decréscimo da concentração de FeSO4
aumentará a atividade enzimática. Observa-se a existência de um ponto de mínimo,
ocorrendo para concentração de quitosana (X1) no ponto central (0) e para maiores

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
76
concentrações de FeSO4 (X2) igual a (1,5) com a atividade quitosanolítica de 0,0094
U.mL-1.
- Com relação ao cultivo na melhor condição:
O cultivo na melhor condição mostrou uma produção máxima de quitosanase em
18 horas de cultivo, com a atividade enzimática de 70,0 U/L, o que foi 28 vezes maior
que o valor (2,5 U/L em 48 horas) relatado na literatura (Assis, 2010) para o mesmo
fungo e em apenas um terço do tempo de produção.
Além disso, observou-se que um elevado incremento na produção de
quitosanase a partir das 12 horas de cultivo, antes de se atingir a atividade
quitosanolítica máxima, 18 horas de cultivo. Essa produção em alto nível por muitas
horas é importante do ponto de vista industrial, onde se espera uma produção contínua
ou em grandes bateladas. Observou-se também que a atividade específica máxima da
quitosanase segue a mesma tendência da atividade quitosanolítica, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
- Com relação aos açúcares redutores:
Foi observado que os açúcares redutores seguem a mesma tendência da atividade
enzimática, com uma produção máxima de cerca de 0,21 mg/mL em 18 horas de
cultivo, produção essa que coincinde com o máximo pico de atividade enzimática.
Estes resultados mostram a relevância de se adotar novas condições para a
produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter
quitooligossacarídeo com atividades biológicas. Esta pesquisa, portanto, sugere base
para novos estudos que permitam alcançar rapidamente a produção de quitosanase e
quitooligossacarídeo com todos os seus benefícios.

Referências Bibliográficas

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
78
Referências bibliográficas
AK, Ö.; BAKIR, U.; GÜRAY, T. Isolation of a chitosan degrading fungus, Penicillium
spinulosum, and chitosanase production by the isolate. In: SYMPOSIUM - NEW
FRONTIERS IN SCREENING FOR MICROBIAL BIOCATALYSTS, 1996, Ede, The
Netherlands. Anais, Editado por K. Kieslich, C. P. van der Beek, J. A. M. de Bont e W.
J. J. van den Tweel, Elsevier Science B. V., 1998, v.53, p.265-268.
ALVES, S. B. Patologia e controle microbiano: Vantagens e desvantagens. In: ALVES,
S. B. Controle Microbiano de Insetos. 2ed., Piracicaba, FEALQ, 1163p, 1998, v.4, p.21-
37.
ALVES, S. B.; LEITE, L. G.; FILHO, A. B.; ALMEIDA, F. E. M.; MARQUES, E. J.
Produção massal de fungos entomopatogênicos na América Latina, In: ALVES, S. B.;
LOPES, R. B. Controle microbiano de pragas na América Latina: avanços e desafios.
Piracicaba, FEALQ, 414p, 2008, v.14, p.215-237.
ARRUDA, W.; LUBECK, I.; SCHRANK, A.; VAINSTEIN, M. H. Morphological
alterations of Metarhizium anisopliae during penetration of Boophilus microplus ticks.
Experimental and Applied Acarology, v. 37, p.231-244, 2005.
ASSIS, C. F. Produção e caracterização de quitooligossacarídeos produzidos pelo fungo
Metarhizium anisopliae e avaliação da citotoxicidade em células tumorais. 2010. 110p.
Tese (Doutorado em Biotecnologia) – Renorbio - Programa de Pós-Graduação em
Biotecnologia. Universidade Federal do Rio Grande do Norte, Natal.
ASSIS, C. F.; COSTA, L. S.; MELO-SILVEIRA, R. F.; OLIVEIRA, R. M.;
PAGNONCELLI, M. G. B.; ROCHA, H. A. O.; MACEDO, G. R.; SANTOS, E. S.,
Chitooligosaccharides antagonizes the cytotoxic effect of glucosamine. World J.
Microbiol. Biotechnol., v.28, n.3, p.1097-1105, 2012.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
79
AZEVEDO, V. V. C.; CHAVES, S. A.; BEZERRA, D. C.; FOOK, M. V. L.; COSTA,
A. C. F. M. Quitina e Quitosana: aplicações como biomateriais. Revista Eletrônica de
Materiais e Processos, v.2.3, p.27-34, 2007.
BARROS NETO, B.; SCARMINIO, I. S.; BRUNS, R. Como fazer experimentos:
pesquisa e desenvolvimento na ciência e na indústria. 2. ed. Campinas: Unicamp, 2001.
401p.
BATISTA FILHO, A.; ALMEIDA, J. E. M.; SANTOS, A. S.; MACHADO, L. A.;
ALVES, S. B. Eficiência de isolados de Metarhizium anisopliae no controle de
cigarrinha-da-raiz de cana-de-açúcar Mahanarva fimbriolata (Hom.: Cercopidae). STAB
– Açúcar, Álcool e Subprodutos, v.21, p.84-89, 2002.
CARVALHO, J. C. M.; SATO, S. Fermentação Descontínua, In: SCHMIDELL, W.;
LIMA, U. A.; AQUARONE, E.; BORZANI, W. Biotecnologia Industrial. São Paulo,
Ed. Edgard Blücher, v.2, p.193-204, 2001.
CHAO, H. J.; SCHWARTZ, J.; MILTON, D. K. Populations and determinats of
airborne fungi in large office buildings. Environmental Health Perspectives, v.110, n.8,
p.777-782, 2002.
CHEN, X.; XIA, W.; YU, X. Purification and characterization of two types of
chitosanase from Aspergillus sp. CJ22-326. Food Research International, v.38, n.3,
p.315-322, 2005.
CHEN, P. T.; CHIANG, C.-J.; CHAO, Y.-P. Medium optimization and production of
secreted Renilla luciferase in Bacillus subtilis by fed-batch fermentation. Biochemical
Engineering Journal, v.49, p.395-400, 2010.
CHENG, C. Y.; LI, Y. An Aspergillus chitosanase with potential for large-ecale
preparation of chitosan oligosaccharides. Biotechnol. Appl. Biochem., v.32, p.197-203,
2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
80
CHOI, Y. J.; Kim, E. J.; Piao, Z.; Yun, Y. C.; Shin, Y. C. Purification and
characterization of chitosanase from Bacillus sp. Strain KCTC 0377BP and its
application for the production of chitosan oligosaccharides. Applied and Environmental
Microbiology, v.70, n.8, p.4522-4531, 2004.
COLAKOGLU, G. The chemical and physical environment for fungal growth. Fak
Matbaa Birimi, Istanbul, 149-166, 2001.
CRAVEIRO, A. C.; CRAVEIRO, A. A.; QUEIROZ, D. C. Quitosana a fibra do futuro.
2. ed. Fortaleza: PADETEC, 2004. V.1, 218p.
DIAS, B. A. Estudo de proteases degradadoras de cutícula produzidas pelo fungo
entomopatogênico Beauveria bassiana. 2005. 70p. Dissertação (Mestrado em
Microbiologia) – Centro de Ciências Biológicas, Programa de Pós-Graduação em
Microbiologia, Universidade Estadual de Londrina, Londrina.
DINARDO-MIRANDA, L. L.; FERREIRA, J. M. G.; CARVALHO, P. A. M.
Influência da época de colheita e do genótipo de cana-de-açúcar sobre a infestação de
Mahanarva fimbriolata (Stal) (Hemiptera: Cercopidae). Neotrop. Entomol, v.30, n.1,
p.145-149, 200.
DONG, C.; ZHANG, J.; HUANG, H.; CHEN, H.; HU, Y. Pathogenicity of a new China
variety of Metarhizium anisopliae (M. anisopliae var. Dcjhyium) to subterranean termite
Odontotermes formosanus. Microbiological Research, v.164, p.27-35, 2009.
DONZELLI, B. G. G.; OSTROFF, G.; HARMAN, G. E. Enhanced enzymatic
hydrolysis of langostino shell chitin with mixtures of enzymes from bacterial and fungal
sources. Carbohydrate Research, v.338, p.1823-1833, 2003.
DRIVER, F.; MILNER, R. J.; TRUEMAN, J. W. H. A taxonomic revision of
Metarhizium based on phylogenetic analysis of rDNA sequence data. Mycological
Research, v.104, n.2, p.134-150, 2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
81
DUNG, P.; MILAS, M.; RINAUDO, M.; DESBRIÉRES, J. Water soluble derivatives
obtained by controlled chemical modifications of chitosan, Carbohydrate Polymers,
v.24, p.209-214, 1994.
ESTEVES, J. A.; CABRITA, J. D.; NOBRE, G. N. Micologia médica, 2. ed. Lisboa:
Fundação Calouste Gulbenkian, 1990. 1058p.
FACCIOTI, M. C. R. Fermentação Contínua, In: Biotecnologia Industrial. Editado por:
SCHMIDELL, W.; LIMA, U. A.; AQUARONE, E.; BORZANI, W., São Paulo, Ed.
Edgard Blücher, v.2, p.541, 2001.
FARIA, M. R.; MAGALHÃES, B. P. O uso de fungos entomopatogênicos no Brasil -
Situação atual e perspectivas. Biotecnologia Ciência & Desenvolvimento, n.22., p.18-
21, setembro/outubro 2001.
FU, J.-Y.; WU, S.-M.; CHANG, C.-T.; SUNG, H.-Y. Characterization of Three
Chitosanase Isozymes Isolated from a Commercial Crude Porcine Pepsin Preparation. J.
Agric. Food Chem., v.51, p.1042-1048, 2003.
FURIGO JUNIOR, A.; PEREIRA, E. P. Enzimas e suas aplicações cinética enzimática.
2001. 39f. Apostila (Graduação de Engenharia Química e Engenharia de Alimentos) –
Centro de Tecnologia, Departamento de Engenharia Química e Engenharia de
Alimentos, Universidade Federal de Santa Catarina, Florianópolis.
GAO, J.; XU, H.; LI, Q.-J.; FENG, X.-H.; LI, S. Optimization of medium for one-step
fermentation of inulin extract from Jerusalem artichoke tubers using Paenibacillus
polymyxa ZJ-9 to produce R,R-2,3-butanediol, Bioresource Technology, v.101, p.7076-
7082, 2010.
GARCIA, M. V.; MONTEIROI, A. C.; SZABO, M. J. P.; PRETTE, N.; BECHARA, G.
H. Mechanism of infection and colonization of Rhipicephalus sanguineus eggs by
Metarhizium anisopliae as revealed by scanning electron microscopy and
histopathology. Brazilian Journal of Microbiology, São Paulo, v.36, n.4, p.368-372,
2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
82
GHANEM, N. B.; YUSEF, H. H.; MAHROUSE, H. K. Production of Aspergillus
terreus xylanase in solid-state cultures: application of the Plackett & Burman
experimental design to evaluate nutritional requirements, Bioresource Technology,
v.73, p.113-121, 2000.
GHORBEL-BELLAJ, O.; HMIDET, N.; JELLOULI, K.; YOUNES, I.; MAALEJ, H.;
HACHICHA, R.; NASRI, M. Shrimp waste fermentation with Pseudomonas aeruginosa
A2: Optimization of chitin extraction conditions through Plackett–Burman and response
surface methodology approaches, International Journal of Biological Macromolecules,
v.48, p.596–602, 2010.
GIUSTINA, A.; VENTURA, P. Weight-reducing regimens in obese subjects: Effects of
a new dietary fiber integrator. Acta Toxicol Ther, 16, 4, p.199-214, 1995.
GUIA.BIO, Reino dos fungos, 2012. Disponível em:
<http://www.http://guia.bio.br/reino-dos-fungos>. Acesso em: 12 novembro 2012.
GUIRADO, N.; AMBROSANO, E. J.; ROSSI, F.; DIAS, F. L. F. Controle de Cupins
de Montículo com Metarhizium anisopliae e Beauveria bassiana, Rev. Bras. de
Agroecologia, v.4, n.2, 2009.
GUO, Y.; XU, J.; ZHANG, Y.; XU, H.; YUAN, Z.; LI, D. Medium optimization for
ethanol production with Clostridium autoethanogenum with carbon monoxide as sole
carbon source, Bioresource Technology, v.101, p.8784–8789, 2010.
HAGAG, S.; KUBITSCHEK-BARREIRA, P.; NEVES, G. W. P.; AMAR, D.;
NIERMAN, W.; SHALIT, I.; SHAMIR, R.; LOPES-BEZERRA, L.; OSHEROV, N.
Transcriptional and proteomic analysis of the Aspergillus fumigatus ∆prtT protease-
deficient mutant. PLoS One, v.7, n.4, e33604, 2012.
HAN, L.-K.; KIMURA, Y.; OKUDA, H. Reduction in fat storage during chitin-chitosan
treatment in mice fed a high-fat diet, International Journal of Obesity, 23:174-179,1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
83
HUNG, T.; CHANG, Y.; SUNG, H.; CHANG, C. Purification and Characterization of
Hydrolase with Chitinase and Chitosanase Activity from Commercial Stem Bromelain.
J. Agric. Food Chem., 50, 4666-4673, 2002.
ICHIKAWA, S.; TAKANO, K.; TAKASHIKUROIWA, HIRUTA, O.; SATO, S.;
MUKATAKA, A. S. Immobilization and stabilization of chitosanase by multipoint
attachment to agar gel support. Journal of Bioscience and Bioengineering, v.93, n.2,
p.201-206, 2002.
JABOR, I. A. S.; PAMPHILE, J. A.; RODRIGUES, S. B.; MARQUES-SILVA, G. G.;
ROCHA, C. L. M. S. Análise do desenvolvimento do fungo entomopatogênico
Metarhizium anisopliae em resposta a fatores nutricionais. Acta Scientiarum.
Agronomy, Maringá, v.25, n.2, p.497-501, 2003.
JI, X.-J.; HUANG, H.; DU, J.; ZHU, J.-G.; REN, L.-J.; LI, S.; NIE, Z.-K. Development
of an industrial medium for economical 2,3-butanediol production through co-
fermentation of glucose and xylose by Klebsiella oxytoca, Bioresource Technology,
v.100, p.5214–5218, 2009.
JO, Y.; JO, K.; JIN, Y.; KIM, K.; SHIN, J.; KIM, Y.; PARK, R. Characterization and
kinetics of 45 kDa chitosanase from bacillus sp. P16. Biosci. Biotechnol. Biochem.,
v.67, n.9, 1875 – 1882, 2003.
KHOSRAVI-DARANI, K.; ZOGHI, A.; ALAVI, S. A.; FATEMI, S. S. A. Application
of Plackett Burman Design for Citric Acid Production from Pretreated and Untreated
Wheat Straw, Iran. J. Chem. Chem. Eng.. v.27, n.1, p.91-104, 2008.
KIM, P.; KANG, T. H.; CHUNG, K. J.; KIM, I. S.; CHUNG, K. Purification of a
constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and
expression of the gene. FEMS Microbiolog.y Letters 240, 31–39, 2004.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
84
KIM, S.-K.; RAJAPAKSE, N. Enzymatic production and biological activities of
chitosan oligosaccharides (COS): A review. Carbohydrate Polymers, v.62, n.4, p.357-
368, 2005.
KIRK, P. M.; CANON, P.F.; MINTER, D.W.; STALPERS, J.A. Dictionary of the
fungi. CAB International, Wallingford, 2008.
KONDO, H.; OSADA, A. Influence of dietary fiber on the bioavailability of zinc in
rats, Biomed Environ Sci, 9, 204-8, 1996.
KUMAR, S.; KOH, J. Physiochemical, Optical and Biological Activity of Chitosan-
Chromone Derivative for Biomedical Applications. Int. J. Mol. Sci., v.13, p.6102-6116,
2012.
KURITA, K.; SANNAN, T.; IWAKURA, Y. Studies on chitin, 4. Evidence for
formation of block and random copolymers of N-acetyl-o-glucosamine and
oglucosamine by hetero and homogeneous hydrolyses. Die Makromolecular Chemie,
v.178, p.3197-3202, 1977.
KUROIWA, T.; ICHIKAWA, S.; HIRUTA, O.; SATO, S.; MUKATAKA, S. Factors
Affecting the Composition of Oligosaccharides Produced in Chitosan Hydrolysis Using
Immobilized Chitosanases. Biotechnol. Prog., 18, 969-974, 2002.
LI, L.-Z.; HUI, Z.; MO, X. Optimization of the medium for streptolydigin production
by Streptomyces lydicus AS 4.2501 with precursor feeding using response surface
methodology, Journal of the Taiwan Institute of Chemical Engineers, 41, 252–258,
2010.
LIU, B.-L.; KAO, P.-M.; TZENG, Y.-M.; FENG, K.-C. Production of chitinase from
Verticillium lecanii F091 using submerged fermentation. Enzyme and Microbial
Technology, v.33, p.410-415, 2003.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
85
LOBATO, A. K. C. L. Estudo da produção de biosurfactantes por micro-organismos
isolados de poços de petróleo. 2003. 108p. Dissertação (Mestrado) – Programa de Pós-
Graduação em Engenharia Química. Universidade Federal do Rio Grande do Norte.
Natal.
LOPES, R. B.; ALVES, S. B.; PADULLA, L. F. L.; PÉREZ, C. A. Eficiência de
formulações de Beauveria bassiana e Metarhizium anisopliae para o controle de ninfas
de Amblyomma cajennense (Fabricius, 1787). Revista Brasileira de Parasitologia
Veterinária, v.16, n.1, p.27-31, 2007.
LOUREIRO, E. S.; BATISTA FILHO, A.; ALMEIDA, J. E. M.; MENDES, J. M.;
PESSOA, L. G. A. Eficiência de isolados de Metarhizium anisopliae (metsch.) sorok.
no controle da cigarrinha-da-raiz da cana-de-açúcar, Mahanarva fimbriolata (stal, 1854)
(hemiptera: cercopidae), em condições de campo. Arq. Inst. Biol., São Paulo, v.79, n.1,
p. 47-53, 2012.
MELLO, D. R.; REIS, R. C. S.; BITTENCOURT, V. R. E. P. Patogenicidade in vitro
do fungo Metarhizium anisopliae (Metschnikoff, 1879) Sorokin, 1883, sobre o carrapato
Boophilus microplus (Canestrini, 1887). Revista Brasileira de Parasitologia Veterinária,
São Paulo, v.15, p.157-162, 2006.
MITSUTOMI, M.; KIDOH, H.; TOMITA, H.; WATANABE, T. The action of Bacillus
circulans WL-12 chitinases on partially N-acetylated chitosan, Bioscience
Biotechnology and Biochemistry, v.59, p.529-531, 1995.
MALAJOVICH, M. A. Biotecnologia: fundamentos. Ed. biblioteca Max Feffer do
Instituto de Tecnologia ORT, Rio de Janeiro, 2009. 104p.
MONAGHAN, R. L.; EVELEIGH, D. E.; TEWARI, R. P.; REESE, E. T. Chitosanase,
a novel enzyme. Nature - New Biology, 245, p.78-80, 1973.
MU, X. Q.; XU, Y.; NIE, Y.; OUYANG, J.; SUN, Z. H. Candida parapsilosis CCTCC
M203011 and the optimization of fermentation medium for stereoinversion of (S)-1-
phenyl-1,2-ethanediol, Process Biochemistry, v.40, p.2345–2350, 2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
86
MURRAY, P. R.; ROSENTHAL, K. S.; PFALLER, M. A. Medical microbiology. 5th
ed. London, Elsevier Mosby, 2005.
MUZZARELLI, R. A. A. Biochemical significance of exogenous chitins and chitosans
in animals and patients, Carbohydrate Polymers, v.20, p.7-16, 1993.
MUZZARELLI, R. A. A.; TOMASETTI, M.; ILARI, P. Depolymerization of chitosan
with the aid of papain, Enzyme Microbiology and Technology, v.16, p.110-114, 1994.
NAVEENA, B. J.; ALTAF, Md.; BHADRIAH, K.; REDDY, G. Selection of medium
components by Plackett–Burman design for production of L(+) lactic acid by
Lactobacillus amylophilus GV6 in SSF using wheat bran, Bioresource Technology,
v.96, p.485-490, 2005.
NOGAWA, M.; TAKAHASHI, H.; KASHIWAGI, A.; OHSHIMA, K.; OKADA, H.;
MORIKAWA, Y. Purification and Characterization of Exo--D-Glucosaminidase from
a Cellulolytic Fungus, Trichoderma reesei PC-3-7, Appl. Environ. Microbiol., v.64, n.3,
p.890-895, 1998.
ONOFRE, S. B.; VARGAS; L. R. B.; ROSSATO, M.; BARROS, N. M.; BOLDO, J.
T.; NUNES, A. R. F.; AZEVEDO, J. L. Controle biológico de pragas na agropecuária
por meio de fungos entomopatogênicos. In: SERAFINI, L. A.; BARROS, N. M.;
AZEVEDO, J. L. (Org.). Biotecnologia – avanços na agricultura e na agroindústria.
Caxias do Sul: EDUCS, p.295-328, cap.8, 2002.
OSSWALD, W. F.; SHAPIRO, J. P.; DOOSTDAR, H.; MCDONALD, R. E.; NIEDZ,
R. P.; NAIRN, C. J.; HEARN, C. J.; MAYER, R. T. Identification and Characterization
of Acidic Hydrolases with Chitinase and Chitosanase Activities from Sweet Orange
Callus Tissue, Plant Cell Physiol. 35, p.811-820, 1994.
PAGNONCELLI, M. G. B. Estudo do mecanismo de produção de oligossacarídeos com
atividades nutracêuticas a partir da quitosana por hidrólise enzimática com processo

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
87
fermentativo simultâneo. 2008. 118p. Tese (Doutorado) – Centro de Tecnologia,
Programa de Pós-Graduação em Engenharia Química. Universidade Federal do Rio
Grande do Norte. Natal.
PALMA-GUERRERO, J., JANSSON, H.-B., SALINAS, J.; LOPEZ-LLORCA, L. V.
Effect of chitosan on hyphal growth and spore germination of plant pathogenic and
biocontrol fungi, J. Appl. Microbiol., v.104, p.541-553, 2008.
PAULA, F. A.; ARRUDA, D. C. Análise do método de planejamento experimental
Plackett Burman na otimização de degradação de penas de aves por comunidades
microbianas, XXI Salão de Iniciação Científica, 19-23, outubro, 2009, UFRGS, Porto
Alegre.
PELCZAR, M. J.; CHANG, E. C. S.; KRIEG, N. R. Microbiologia: conceitos e
aplicações, v.2, 2.ed. São Paulo: Makron BooKs, 1996.
PELLETIER, A.; SYGUSCH, J. Purification and characterization of three chitosanase
activities from Bacillus megaterium P1, Applied and Environmental Microbiology, v.
56, n.4, p.844-848, 1990.
PEREIRA, F. B.; GUIMARÃES, P. M. R.; TEIXEIRA, J. A.; DOMINGUES, L.
Optimization of low-cost medium for very high gravity ethanol fermentations by
Saccharomyces cerevisiae using statistical experimental designs, Bioresource
Technology, v.101, p.7856-7863, 2010.
PETER, M. G. Chitin and chitosan from aninal sources. In: STEINBUCHEL, A.;
RHEE, S. K. Polysaccharides and Polyamides in the Food Industry. Weinheim:
Wileyvch, v.1, p.115-208, 2005.
PIZA, F. A. T.; SILOTO, A. P.; CARVALHO, C. V.; FRANCO, T. T. Purification,
characterization and purification of chitosanase from Bacillus cereus, Brazilian Journal
of Chemical Engineering., v.16, n 2, p.185-192, 1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
88
POLITZER, K.; BON, E. P. S. Enzimas Industriais e Especiais. C.G.E.E. Ciência,
Tecnologia e Inovação. Rio de Janeiro, 554p, 2006.
POLYMAR, Ciência é a nossa Natureza, 2003. Disponível em:
<http://www.polymar.com.br>. Acesso em: 6 setembro 2012.
PUTZKE, J.; PUTZKE, M. T. L. Os reinos dos fungos. Santa Cruz do Sul, RS:
EDUNISC, 1998.
QUINES, L. K. M. Extração de poli(3-hidroxibutirato) produzido por Cupriavidus
necator dsm 545 com 1,2-carbonato de propileno. 2010. 99p. Dissertação (Mestrado em
Engenharia de Alimentos) – Centro Tecnológico, Departamento de Engenharia Química
e Engenharia de Alimentos, Programa de Pós-Graduação em Engenharia de Alimentos,
Universidade Federal de Santa Catarina, Florianópolis.
RAJENDRAN, A.; THANGAVELU, V. Statistical experimental design for evaluation
of medium components for lipase production by Rhizopus arrhizus MTCC 2233, LWT -
Food Science and Technology, v.42, p.985–992, 2009.
RAMIREZ-LEÓN, L. F.; RUIZ-HERRERA, J. Hydrolysis of walls and formation of
spheroplasts in Mucor rouxii. J. Gen. Microbiol., v.72, p.281-290, 1972.
RANGEL, M. A. S.; LOUREIRO, E. S.; ALMEIDA, A. S.; ALVES, H. C.; ONO, F.
B.; LOBO, P. M.; MARIANI, M. A.; PESSOA, L. G. A.; SILVA, A. J. Avaliação
participativa do controle de cigarrinhas das pastagens com a aplicação do fungo
Metarhizium anisopliae (Metsch.) Sorok. Rev. Bras. de Agroecologia, v.2, n.2, 731-734,
2007.
RAVEN, P. H.; EVERT, R.F.; EICHHORN, S.E. Biologia Vegetal, 6ª Ed, Rio Janeiro,
Editora Guanabara Koogan, 2001.
REDDY, L. V. A.; WEE, Y.-J.; YUN, J.-S.; RYU, H.-W. Optimization of alkaline
protease production by batch culture of Bacillus sp. RKY3 through Plackett–Burman

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
89
and response surface methodological approaches, Bioresource Technology, v.99, p.242–
2249, 2008.
RIGUEIRA, J. C. S. Influência da contagem de células somáticas no perfil e teores de
aminas bioativas e na qualidade de leite cru e de queijo mussarela. 2010. 147p. Tese
(Doutorado em Ciência de Alimentos) - Faculdade de Farmácia, Departamento de
Alimentos, Programa de Pós-Graduação em Ciência de Alimentos, Faculdade de
Farmácia da Universidade Federal de Minas Gerais, Belo Horizonte.
RODRIGUES, M. I.; IEMMA, A. F. Planejamento de Experimentos e Otimização de
Processos: uma estratégia sequencial de planejamentos, Campinas, SP, Casa do Pão
Editora, 2005.
SANTOS, E. S. Recuperação e Purificação de Enzimas usando Adsorção em Leito
Expandido, Tese de Doutorado, Universidade Estadual de Campinas, Campinas/SP,
p.152, 2001.
SEDMAK, J. J.; GROSSBERG, S. E. A rapid, sensitive and versatile assay for protein
using coomassie brilliant blue G250, Analytical Biochemistry, v.79, p. 544-552, 1977.
SEINO, H.; TSUKUDA, K.; SHIMASUE, Y. Properties and action pattern of a
chitosanase from Bacillus sp. PI- 7S. Agricultural Biological Chemistry, 55, p.2421-
2423, 1991.
SHIMOSAKA, M.; NOGAWA, M.; OHNO, Y.; OKAZAKI, M. Chitosanase from the
plant pathogenic fungus, Fusarium solani f. sp. Phaseoli – purification and some
properties, Bioscience Biotechnolgy and Biochemistry, v.57, n.2, p.231-235, 1993.
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Production of two chitosanases from a chitosan-assimilating bacterium, Acinetobacter
sp. Strain CHB101, Applied and Environmental Microbiology, v.61, n.2, p.438-442,
1995.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
90
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Characterization of Three Chitosanase Isozymes Isolated from a Commercial Crude
Porcine Pepsin Preparation, J. Agric. Food Chem. 51, p.1042-1048, 2003.
SILVA FILHO, R. C.; COSTA, A. S. S.; CHAGAS, B. M. E.; SANTOS, F. N.;
MACEDO, G. R.; SANTOS, E. S. Avaliação do teste de atividade quitosanolítica
produzida por fungo. In: XV SIMPÓSIO NACIONAL DE BIOPROCESSOS, 15,
Recife, 2005, 02 a 05 de agosto.
SILVA, E. P. Avaliação Preliminar do Potencial de Aplicação da Tecnologia de
Biopilhas para a Biorremediação do Solo de Guamaré-RN. 2004. 119p. Dissertação
(Mestrado) – Centro de Tecnologia, Programa de Pós-Graduação em Engenharia
Química. Universidade Federal do Rio Grande do Norte. Natal.
SOMASHEKAR, D.; JOSEPH, R. Chitosanases – Properties and apllications: A review.
Bioresource Technology, v.55, p.35-45, 1996.
STOYACHENKO, I. A.; VARLAMOV, V. P.; DAVANKOV, V. A. Chitinases of
Streptomyces kurssanovii: purification and some properties, Carbohydrate Polymers,
v.24, p.47-54, 1994.
TINLINE, R. D. Nuclear distribuition in Metarhizium anisopliae. Mycologia, v. 63, p.
713-721, 1971.
TRABULSI, L. R.; TOLEDO, M. R. F. Microbiologia. 2. Ed., São Paulo: Atheneu,
1996.
VIEIRA, P. D. S.; SILVA, W. M. T.; PAIVA, L. M.; LIMA, E. A. L. A.;
CAVALCANTI, V. L. B. Estudo da caracterização morfológica, esporulação e
germinação dos conídiosde Metarhizium anisopliae var. acridum em diferentes
temperaturas. Biológico, São Paulo, v.69, n.1, p.17-21, 2007.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
91
WANDERLEY, M. D.; NEVES, E.; ANDRADE, C. J. Aspectos da produção industrial
de enzimas. Revista Citino, v.1, n.1, p.30-36, 2011.
WANG, C.; TYPAS, M. A.; BUTT, T. M. Detection and characterization of pr1
virulent gene deficiencies in the insect pathogenic fungus Metarhizium anisopliae.
FEMS Microbiology Letters, v. 213, p. 251-255, 2002.
ZAHORSKA-MARKIEWICZ, B.; KROTKIEWSKI, M.; OLSZANECKA-
GLINIANOWICZ, M.; ZURAKOWSKI, A. Effect of chitosan in complex management
of obesity, Pol Merkur Lekarski, v.13, n.74, p.129-132, 2002.
ZHANG, X. Y.; DAI, A. L.; ZHANG, X. K.; KUROIWA, K.; KODAIRA, R.;
SHIMOSAKA, M.; OKAZAKI, M. Purification and Characterization of and Exo--D-
glucosaminidase from a koji mold, Aspergillus oryzae IAM2660. Biosci. Biotechnol.
Biochem., v.64, n.9, 1896-1902, 2000.
ZHU, X.; WU, X.; DAI, Y. Fermentation conditions of a chitosanase from Acinetobcter
sp C-17. Biosci. Biotechnol. Biochem., v.67, n.2, p.284-290, 2003.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
92
APÊNDICE

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
93
APÊNDICE I
Metodologia para repique da cepa
Em Erlenmeyer de 1000 mL colocava-se 500 mL do meio de manutenção
PDA. Este Erlenmeyer, com o meio de manutenção, era autoclavado junto com placas
de Petri a 1,5 kgf/cm2 por 15 minutos, em seguida, era levado para uma câmara de fluxo
laminar e transferido para as placas de Petri já autoclavadas, onde eram mantidas numa
estufa a 27 ºC durante 24 h. Após este período, as placas de Petri eram levadas a câmara
de fluxo laminar, utilizando bico de Bunsen para esterilizar a alça de platina. Procedia-
se o repique utilizando uma placa de Petri contendo a cepa pura. Cada placa de Petri
contendo a cepa pura era transferida para duas placas de Petri contendo o meio de
manutenção. As placas eram mantidas na estufa a 27 ºC durante 30 dias onde após esse
período, repetia-se este procedimento.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
94
APÊNDICE II
Montagem da Curva de Calibração Usando o Método DNS
Soluções:
Glicosamina = 0,01 mol/mL (10000,0 µmol/mL)
Tabela 14. Dados para construção da curva de calibração usando o Método DNS.
Tubo de
Ensaio
Solução de
glicosamina (mL)
Água Destilada
(mL)
DNS (mL)
Volume Total
Massa (µµµµmol)
1 0 1,0 1,5 2,5 0
2 0,2 0,8 1,5 2,5 0,2
3 0,4 0,6 1,5 2,5 0,4
4 0,6 0,4 1,5 2,5 0,6
5 0,8 0,2 1,5 2,5 0,8
6 1,0 0 1,5 2,5 1,0
Tabela 15 - Medidas de absorbância a 600nm
Dados adicionais:
Peso Molecular D-Glicosamina = 215,64 (recipiente SIGMA)
Conc. Sol. D-Glicosamina = 0,05 g/50 mL
Massa calc.(µmol) = conc. Sol. D-Glic. x Vol. Utiliz. D-Glics. =
= 0,05 x 1000 x Vol.util./50*215,64
Tempo atividade enzimática = 30 min
1 U/mL = 1 µmol/(min.mL)
Vol. Total (Glicosamina+caldo) = 1 mL
Medições Adsorbância a 600 nm Vol. Utiliz. Massa Massa Atividade
II III Média(Abs)(mL) calc. (mg) calc. (µg) Enzim. (U/mL)
0 0 0 0 0 0 0
0,133 0,128 0,1307 0,2 0,2 200 0,0309
0,279 0,277 0,2787 0,4 0,4 400 0,0618
0,433 0,428 0,4297 0,6 0,6 600 0,0927
0,587 0,583 0,5797 0,8 0,8 800 0,1237
0,727 0,716 0,7157 1 1 1000 0,1546

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
95
Figura 17 – Curva padrão de glicosamina.
y = 1900ralx + 1900ral
R² = 1900ral
00,000
00,020
00,040
00,060
00,080
00,100
00,120
00,140
00,160
00,180
00,000 00,100 00,200 00,300 00,400 00,500 00,600 00,700 00,800
Ati
v.
En
zim
áti
ca (
U/m
L)
Absorbância (Abs) a 600 nm
Curva-Padrão D-Glicosamina

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
96
APÊNDICE III
Método de Determinação de Proteína Utilizando o Corante Comassie
Brilhante Blue G 250 (Método modificado de Sedmak e Grossberg)
Micrométodo
Preparar uma solução 0,06% de Comassie Blue G250 em 1,5 % HCl (w/V). Filtrar e
papel de filtro Whatman nº1.
Curva Padrão (1 a 20 µµµµg)
a) Preparar soluções estoque de BSA de 10 a 200 µg/mL.
b) Colocar em tubos de ensaio 100 µL da solução estoque, 1400 µL de água e
1500µL do Comassie e agitar.
c) Preparar um tubo contendo 1500 µL de água e 1500 µL do corante que
corresponde ao zero de proteína.
d) Após 5 minutos (padronizar o tempo) fazer as leituras das absorbâncias.
e) As leituras das amostras devem ser feitas em dois comprimentos de onda 595 e
465 contra a água.
f) Plotar a razão das absorbâncias a 595/465 como uma função da massa de BSA e
calcular o melhor ajuste linear dos dados.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
97
Micrométodo (1 a 20 µµµµg de BSA)
Solução Estoque de BSA = 0,2 mg/mL
Tabela 16. Dados para construção da curva padrão de BSA.
Tubo de
Ensaio
H2O (µµµµL)
BSA (µµµµg)
Coomassie (µµµµL)
BSA (µµµµg)
1 1500 0 1500 0
2 1490 10 1500 2
3 1480 20 1500 4
4 1470 30 1500 6
5 1460 40 1500 8
6 1450 50 1500 10
7 1440 60 1500 12
8 1430 70 1500 14
9 1420 80 1500 16
10 1410 90 1500 18
11 1400 100 1500 20
Dados adicionais:
Concentração Sol. Estoque
BSA = 0,2 mg/mL = 0,2 µg/µL
Vol. total sol. = vol. H2O + vol. BSA + vol.
Coomassie = 3000 µL = 3 mL
Concentração Sol. BSA (mg/mL) = (massa BSA mg) / 3 mL
.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
98
Figura 18 – Curva Padrão de BSA
y = 1900ralx - 1900ral
R² = 1900ral
000,000
001,000
002,000
003,000
004,000
005,000
006,000
007,000
000,400 000,500 000,600 000,700 000,800 000,900 001,000 001,100
µg
BS
A
Razão Absorbâncias (595nm/465nm)
Curva-Padrão BSA

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE TECNOLOGIA
DEPARTAMENTO DE ENGENHARIA QUÍMICA PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA
TESE DE DOUTORADO
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em
Cultivo Descontínuo Submerso
Raimundo Cosme da Silva Filho
Natal/RN
Abril/2013

Raimundo Cosme da Silva Filho
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo
Descontínuo Submerso
Tese apresentada ao Programa de Pós-Graduação em Engenharia Química – PPGEQ, da Universidade Federal do Rio Grande do Norte, como parte dos requisitos para obtenção do título de Doutor em Engenharia Química, sob a orientação do Prof. Dr. Everaldo Silvino dos Santos e a coorientação da Profª. Drª. Gorete Ribeiro de Macedo.
Natal/RN
Abril/2013

Catalogação da Publicação na Fonte. UFRN / CT / PPGEQ
Biblioteca Setorial “Professor Horácio Nícolas Sólimo”.
Silva Filho, Raimundo Cosme da. Otimização do meio de cultura para a produção de quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso / Raimundo Cosme da Silva Filho. - Natal, 2013. 98 f.: il.
Orientador: Everaldo Silvino dos Santos. Co-orientador: Gorete Ribeiro de Macedo. Tese (Doutorado) - Universidade Federal do Rio Grande do Norte.
Centro de Tecnologia. Departamento de Engenharia Química. Programa de Pós-Graduação em Engenharia Química.
1. Quitosana - Tese. 2. Quitina - Tese. 3. Quitosanase – Tese. 4. Metarhizium
anisopliae - Tese. I. Santos, Everaldo Silvino dos. II. Macedo, Gorete Ribeiro de. III. Universidade Federal do Rio Grande do Norte. IV. Título. RN/UF/BSEQ CDU 547.458 (043.2)

SILVA FILHO, Raimundo Cosme da – Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo Descontínuo Submerso. Tese de Doutorado. UFRN - Programa de Pós-Graduação em Engenharia Química. Área de Concentração: Pesquisa e Desenvolvimento de Tecnologias Regionais – Subárea: Engenharia de Processos – Alimentos e Biotecnologia, Natal/RN, Brasil.
Orientação: Prof. Dr. Everaldo Silvino dos Santos Coorientação: Profª. Drª. Gorete Ribeiro de Macedo ___________________________________________________________________________
RESUMO: No presente trabalho utilizou-se um Planejamento Plackett & Burman, com 8 fatores e 12 ensaios em 2 níveis e mais 3 repetições na condição do ponto central, para se investigar a influência das concentrações de quitosana, peptona, extrato de levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O e FeSO4 na produção da enzima quitosanase por Metarhizium
anisopliae. Os ensaios para produção da enzima foram realizados em cultivo descontínuo submerso. Os resultados para o Planejamento Plackett & Burman mostraram que a atividade quitosanolítica foi favorecida pelo aumento da concentração de substrato (quitosana) e de sulfato ferroso (FeSO4), enquanto que o aumento da concentração dos outros fatores não contribuiu de forma significativa para a atividade quitosanolítica. A otimização do meio de cultura para a produção da enzima foi realizado por meio de um Planejamento Composto Central, com os dois fatores que mais influenciaram a atividade quitosanolítica (quitosana e FeSO4), conforme Planejamento Plackett & Burman, mantendo-se os outros nutrientes em seus valores mínimos. Nesse planejamento, para o fator FeSO4, tomou-se o limite inferior (-1) como sendo o limite superior do Planejamento Plackett & Burman. Os resultados mostraram que a produção da enzima foi favorecida pelo aumento da concentração quitosana e pela diminuição da concentração de FeSO4. A produção máxima de atividade quitosanolítica foi da ordem de 70,0 U/L e foi atingida em apenas 18 h de fermentação, resultado esse vinte e oito vezes maior aos obtidos anteriormente para o mesmo microrganismo que foi 2,5 U/L em 48h.
Palavras-chave: quitosana; quitosanase; Metarhizium anisopliae; Plackett & Burman.


ABSTRACT
In this work a Plackett-Burman Design with 8 factors and 12 trials in 2 levels with 3 repetitions at the center point was used in order to investigate the influence of the concentration of chitosan, peptone, yeast extract, NaNO3, K2HPO4, KCl, MgSO4.7H2O and FeSO4 on chitosanase production by Metarhizium anisopliae. Runs were carried out using submerged discontinuous cultivation for enzyme production. The results of the Plackett & Burman Design showed that only two factors, chitosan concentration as well as FeSO4 had influence on chitosanolytic activity, while the increase in concentration of other factors not contributed significantly to the quitosanolítica activity. Cultivation medium optimization for enzyme production was carried out using a Composite Central Design, with the most important factors for chitosanolytic activity (chitosan and FeSO4), in accordance with Plackett & Burman Design, and keeping the other nutrients in their minimum values. On this other design, it was taken the highest limit in Plackett & Burman Design as the lowest limit (-1) to FeSO4 factor. The results showed that the enzyme production was favoured by increasing the chitosan concentration and by decreasing FeSO4. Maximum production for chitosanolytic activity was about 70.0 U/L and was reached in only 18 h of fermentation, a result about twenty-eight times greater than a former study using the same microorganism (about 2.5 U/L at 48 h).
Key-words: Chitosan; Chitosanase; Metarhizium anisopliae; Plackett & Burman

Ao meu pai, minha mãe e minha
família, em especial aos meus
irmãos e sobrinhos

AGRADECIMENTOS
A Deus, por estar presente em todos os momentos da minha vida e por me ceder à
oportunidade de iniciar e concluir este trabalho.
A minha mãe Cândida Inocência da Silva que esteve comigo em todos os momentos da
minha vida me educando, me amando e sempre me dando muito carinho.
Ao meu pai Raimundo Cosme da Silva, que não está mais aqui para ver este trabalho,
mas que foi a pessoa que mais me apoiou e me incentivou, em todos os momentos da minha vida,
para que eu pudesse chegar o mais longe possível na minha carreira acadêmica. Pai muito
obrigado por toda atenção, educação, amor e carinho que recebi durante toda a minha vida.
Aos meus irmãos e familiares que sempre estiveram comigo dividindo momentos de
grande alegria e às vezes de tristeza, mas sempre nos mantendo unidos para superarmos todos os
obstáculos.
A minha namorada Rochelly Caroliny por todo amor e carinho que me deu e que me
entendeu nos momentos que estive ausente para a conclusão deste trabalho.
Ao meu orientador Professor Doutor Everaldo Silvino dos Santos por confiar em mim e
me aceitar como orientando além de toda a amizade, orientação e dedicação em todas as etapas de
desenvolvimento e conclusão deste trabalho.
À Professora Doutora Gorete Ribeiro de Macedo, “pessoa que admiro muito como
profissional e principalmente, como pessoa, um exemplo a ser seguido”, pelos conselhos e críticas
quando necessários ao desenvolvimento deste trabalho. Professora a senhora é considerada uma
mãe para de todos os alunos que tiveram a honra de conhecê-la e trabalhar com a senhora.
Aos meus amigos Engenheiros Químicos, Ana Carmen, Ana Katerine, Andréa Farias,
Anita Lima, Sirtys Lessa, Alexandre, Sérgio, Márcio Bezerra e ao Biólogo Daniel Souza, que
sempre me ouviram e que me ajudaram de alguma forma nesse trabalho.
A Eusamar Coelho de Lima, Mazinha, secretária do PPGEQ, pelo apoio, pela paciência e
orientação nas questões burocráticas do processo.
A Carlos e Jardelson, pela grande colaboração e dedicação nos experimentos do
laboratório.
A Sérgio Santana pela grande colaboração e dedicação, o qual não poupou esforços nem
tempo em me ajudar na realização dos experimentos em laboratório.

Ao meu amigo Ubiratan Nazaré por me ajudar na organização do texto e na tradução para
o Abstract.
Finalizando, agradeço ao Laboratório de Engenharia Bioquímica da UFRN por ter cedido
toda a sua estrutura para realização dos experimentos e ao Programa de Pós-Graduação em
Engenharia Química, assim como a todos os professores do Departamento de Engenharia Química
da UFRN que trabalham em pró de uma educação digna e ensino de qualidade para todos os
alunos.

Sumário LISTA DE FIGURAS
LISTA DE TABELAS
NOMENCLATURA
INTRODUÇÃO...............................................................................................................................15
REVISÃO BIBLIOGRÁFICA........................................................................................................18
2.1. Enzimas................................................................................................................................18
2.1.1. Quitosana e seus derivados........................................................................................19
2.1.2. Quitosanases...............................................................................................................24
2.2. Fungos..................................................................................................................................29
2.2.1. Características gerais..................................................................................................29
2.2.2. Reprodução dos Fungos.............................................................................................32
2.2.3. Metabolismo de fungos..............................................................................................33
2.2.4. Nutrição de fungos.....................................................................................................34
2.2.5. O Fungo Metarhizium anisopliae ..............................................................................36
2.3. Fermentação.........................................................................................................................40
2.3.1. Preparo do inóculo......................................................................................................41
2.3.2. Produção de enzimas em cultivo submerso descontínuo...........................................42
2.4. Planejamento experimental..................................................................................................42
2.4.1. Planejamento fatorial..................................................................................................43
2.4.2. Planejamento Plackett & Burman..............................................................................43
2.4.3. Aplicações do Planejamento Plackett & Burman.......................................................44
METODOLOGIA EXPERIMENTAL.............................................................................................50
3.1. Quitosana..............................................................................................................................50
3.2. Microrganismo.....................................................................................................................50
3.2.1. Escolha da cepa produtora de quitosanase.................................................................50

3.2.2. Manutenção da cepa...................................................................................................50
3.3. Produção de enzima.............................................................................................................51
3.4. Determinação da Proteína Total...........................................................................................51
3.5. Determinação da Atividade Quitosanolítica e Atividade Específica...................................51
3.6. Planejamento Experimental.................................................................................................52
3.6.1. Planejamento de seleção de fatores Plackett Burman................................................52
3.6.2. Planejamento Composto Central para Otimização do meio de cultura para a
Produção da Enzima em Cultivo Submerso.....................................................................................53
3.7. Perfil Cinético durante o Cultivo.........................................................................................56
RESULTADOS E DISCUSSÃO.....................................................................................................58
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA ..........................................58
4.2. Planejamento Plackett & Burman........................................................................................59
4.2.1. Análise do Plackett & Burman com 8 fatores e 12 ensaios .......................................59
4.2.2. Análise do Plackett & Burman com 12 ensaios e mais 3 repetições na condição do
ponto central ....................................................................................................................................61
4.3. Planejamento Composto Central para Produção de Quitosanase por Metarhizium
anisopliae em Cultivo Descontínuo Submerso................................................................................64
4.4. Perfil Cinético – Melhor Condição......................................................................................69
CONCLUSÃO.................................................................................................................................75
REFERÊNCIAS BIBLIOGRÁFICAS.............................................................................................78
APÊNDICE I...................................................................................................................................93
Metodologia para Repique da Cepa ................................................................................................93
APÊNDICE II.................................................................................................................................94
Montagem da Curva de Calibração Usando o Método DNS..........................................................94
APÊNDICE III................................................................................................................................96
Método de Determinação de Proteína Utilizando o Corante Comassie Brilhante Blue G 250
(Método modificado de Sedmak e Grossberg)................................................................................96

Lista de Figuras
Figura 1 – Estrutura da quitosana..................................................................................................20
Figura 2 – Estrutura da glicosamina..............................................................................................23
Figura 3 – Estrutura do sulfato de glicosamina.............................................................................23
Figura 4 – Representação da estrutura geral de uma célula de fungo..........................................30
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas...........................................................31
Figura 6 – Conídios de Aspergillus e Penicillium.........................................................................33
Figura 7 – Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA ...................58
Figura 8 – Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados............................................................................................................................60
Figura 9 – Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto central
mostrando a influência dos fatores estudados................................................................................63
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos
fatores estudados............................................................................................................................67
Figura 11 – Valores preditos em função dos valores observados..................................................68
Figura 12 – Atividade quitosanolítica em função da concentração de quitosana e da concentração
de FeSO4 para os fatores não significativos no ponto central .......................................................68
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana ........70
Figura 14 – Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana....................................70
Figura 15 – Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de
quitosana........................................................................................................................................71
Figura 16 – Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana..........................................................................................................72
Figura 17 – Curva padrão de glicosamina ....................................................................................95
Figura 18 – Curva Padrão de BSA ................................................................................................98

Lista de Tabelas
Tabela 1 – Comparação das enzimas com os catalisadores químicos..............................................19
Tabela 2 – Fontes naturais de quitina e quitosana ...........................................................................20
Tabela 3 – Níveis para os fatores e seus valores codificados...........................................................53
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central .........................................................................................53
Tabela 5 – Matriz do planejamento Composto Central...................................................................54
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central......................................................................................................55
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.......................59
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman...............................................60
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.......................61
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central...............................................................................................................................................62
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da atividade
enzimática........................................................................................................................................65
Tabela 12 – Efeitos Estimados do planejamento Composto Central...............................................65
Tabela 13 – Resumo da ANOVA....................................................................................................66
Tabela 14 – Dados para construção da curva de calibração usando o Método DNS ......................94
Tabela 15 – Medidas de absorbância a 600 nm para construção da curva de calibração................94
Tabela 16 – Dados para construção da curva padrão de BSA ........................................................97

NOMENCLATURA
A – Atividade quitosanolítica (U/L)
AE – Atividade específica (U/mg)
ANN-GA – algoritmo genético de rede neural artificial
ANOVA – Análise de variância (Analysis of Variance)
BSA – Albumina de soro bovino (Bovine Sereum Albumin)
CCD – Planejamento Composto Central
CDA – Chitosan Dector Agar
Cs – Concentração de substrato (%)
CP – Concentração de KCl (%)
Da – Dalton
DEQ – Departamento de Engenharia Química
DNS – Ácido Dinitrosalissílico
E – Enzima
Ea – Energia de ativação
EC – Comissão para nomenclatura e classificação de enzimas
EL – Concentração de Extrato de Levedura (%)
FP – Concentração de K2HPO4 (%)
NS – Concentração de NaNO3 (%)
GDA – Grânulos dispersíveis em água
k – Número de fatores, variáveis
LEB – Laboratório de Engenharia Bioquímica
mRNA – RNA mensageiro
n – Número de níveis de um determinado fator
N – Número de ensaios de um planejamento
P – Pressão
PP – Concentração de Peptona (%)
PDA – Batata Dextrose Agar
pH – Potencial hidrogeniônico
Q – Concentração de Quitosana (%)
Qm – Quadrado médio
r – Agitação (rpm)
R2 – Coeficiente de determinação

RA – Razão de aeração
rpm – Rotações por minuto
RSM – Metodologia de superfície de resposta
S – Molécula de substrato
SF – Concentração de FeSO4 (%)
SM – Concentração de MgSO4.7H2O (%)
SQ – Soma de quadrados
SSF – Fermentação de estado sólido
T – Temperatura
TMC – Temperatura de cultivo
V11,V12,V13 – Variáveis inertes para cálculo do erro
VHG – Fermentação de alta densidade
WP – Pó molhável
X – Concentração celular
Xi – variável independente
Xj – variável independente
xi – Fator independente real
x0 – Valor de xi no ponto central
∆xi – Variação de valor
Y – Resposta do modelo de primeira ordem
Yp – Resposta do modelo polinomial de segunda ordem
β0 – Interseção dos modelos de primeira ou segunda ordem
βi – Coeficiente linear dos modelos de primeira ou segunda ordem
βii – Coeficiente quadrático
βij – Coeficiente de interação

Capítulo 1 Introdução

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
15
1. Introdução As quitosanases foram descobertas simultaneamente há quase quarenta anos por
Monaghan et al. (1973), como pelo grupo de Ramirez-León & Ruiz-Herrera (1972), e
são enzimas obtidas a partir do metabolismo de bactérias, fungos e extraídas de plantas.
Desde então, vários estudos vêm apresentando a obtenção de quitosanase deste
microrganismos, na forma intracelular ou extracelular (Osswald et al., 1994;
Somashekar & Joseph, 1996), como a do presente estudo. A quitosanase (EC 3.2.1.132)
representa uma classe de enzimas hidrolíticas que catalisam a hidrólise de ligação β-
(1→ 4) glicosídica da quitosana, gerando oligômeros chamados quito-oligossacarídeos
(Reddy et al., 2008).
A quitosana é um biopolímero hidrofílico obtido a partir da quitina, um
polissacarídeo abundante na natureza. Na literatura, é considerado quitosana o produto
da desacetilação da quitina para um grau de desacetilação superior a 75% e solúveis em
ácidos como o acético e o fórmico. A quitosana é conhecida por exibir uma ampla
variedade de atividade fisiológica como atividade antitumoral, mas o peso molecular e a
insolubilidade em água da quitosana são desvantagens para várias aplicações.
Atualmente, há interesse nas quitosanas quando as mesmas são convertidas em quito-
oligossacarídeos porque eles não apenas são solúveis em água e possuem baixo peso
molecular, mas também têm atividades biológicas bem notáveis, como atividade
anticancerígena atóxica e biocompatível, incluindo efeito antimetastático, atividade anti-
HIV, hepatoprotetora, antioxidante, hipocolesterolêmica, antimicrobiana,
imunoestimulate, antitumoral, anti-inflamatória e atividade aceleradora da absorção de
cálcio e ferro. No entanto, estes quito-oligossacarídeos são obtidos tradicionalmente na
indústria por reação de hidrólise usando ácidos, que apresenta muitos problemas como
dificuldade em controlar a hidrólise, produzindo uma grande quantidade de
oligossacarídeos de cadeia pequena e oligossacarídeos de baixo rendimento, alto custo
de separação e, também, poluição ambiental, uma vez que utiliza um ácido como
catalisador. Como alternativa, com suas vantagens em termos de compatibilidade
ambiental, baixo custo e reprodutibilidade, a obtenção destes quito-oligossacarídeos por
hidrólise com quitosanase se tornou cada vez mais popular nos últimos anos (Azevedo
et al., 2007).

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
16
Devido a todos estes aspectos positivos é que, em geral, as enzimas vêm sendo
estudadas em diferentes aplicações, mostrando que esta é uma tendência na indústria.
No presente trabalho, foi desenvolvido um processo fermentativo visando a
otimização do meio de cultura para a produção da enzima quitosanase em cultivo
descontínuo submerso, por Metarhizium anisopliae, capaz de sintetizar oligômeros com
aplicações na indústria farmacêutica e de química fina, partindo de quitosana como
única fonte de carbono.
Nesse caso, avaliou-se, através de um planejamento Plackett & Burman (método
de delineamento de seleção de fatores) e de um planejamento Composto Central as
melhores condições para a produção da enzima. A técnica do planejamento
experimental Plackett & Burman para realizar triagem de fatores na otimização de um
determinado processo foi utilizada neste trabalho para identificar quais os fatores que
influenciam de forma significativa na produção da enzima quitosanase. A influência
destes fatores (concentração de quitosana e concentração de FeSO4) foi então
investigada pelo planejamento Composto Central com 22 experimentos na parte fatorial,
repetição em triplicata no ponto central e quatro experimentos na parte axial. A
repetição do ponto central permite também a determinação do erro puro e da
repetibilidade do processo, sendo uma forma de identificar melhor se o processo está
sob controle.
Do ponto de vista industrial é de relevante importância a realização de estudos
para a otimização da produção da quitosanase de forma contínua ou em grandes
bateladas.

Capítulo 2 Revisão Bibliográfica

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
18
2. Revisão Bibliográfica 2.1. Enzimas
As enzimas são catalisadores biológicos constituídos por longas cadeias de
aminoácidos que atuam em substratos reduzindo a energia de ativação, acelerando assim
a velocidade da reação, e transformando-os em produtos. Sua atuação catalítica visa
facilitar a atividade das células através da quebra de moléculas ou junção das mesmas
para formação de novos produtos. Podem apresentar alto grau de especificidade relativa
a apenas um substrato ou a vários substratos ao mesmo tempo, como a certos tipos de
ligações glicosídicas α-1,4 das moléculas de amido, ou para determinado tipo particular
de isômero óptico como ocorre na oxidação da β-D-glicose pela glicose oxidase (Furigo
Junior & Pereira, 2001).
Enzimas são proteínas que apresentam em sua estrutura um centro ativo
(apoenzima) e, em alguns casos, um grupo prostético denominado cofator (coenzima ou
íon metálico ativo) e sua atividade depende de alguns fatores como estrutura da
proteína, número de cadeias peptídicas, arranjo dessas cadeias na molécula, natureza do
substrato e natureza do grupo prostético (Politzer & Bon, 2006).
Todos os organismos vivos produzem, em pequena ou grande quantidade, certa
variedade de enzimas, sejam elas intra ou extracelulares. As enzimas extracelulares são
excretadas para o meio externo e podem decompor compostos insolúveis como
celulose, quitina e quitosana.
Algumas destas enzimas podem ser usadas tanto nas indústrias de detergentes,
têxteis e farmacêuticas, como também de alimentos, de bebidas, etc., sendo utilizadas
em processos biotecnológicos industriais em substituição aos processos químicos que
muitas vezes causam grandes prejuízos ao meio ambiente. A produção comercial das
enzimas pode ser realizada através do cultivo de microrganismos como fungos e
bactérias.
Na Tabela 1 mostra-se uma comparação das enzimas com os catalisadores
químicos, levando-se em conta algumas das características dos dois catalisadores.
Destaca-se que alguns parâmetros da Tabela 1 demonstram a maior importância das
enzimas, quando comparadas com os catalisadores químicos, principalmente a
especificidade ao substrato, as condições de reação, o consumo de energia e a

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
19
velocidade de reação, como desvantagem cita-se o custo de obtenção (isolamento e
purificação).
Tabela 1 – Comparação das enzimas com os catalisadores químicos (Adaptado de
Furigo Junior & Pereira, 2001).
Característica Enzimas Catalisadores Químicos
Especificidade ao substrato Alta Baixa
Natureza da estrutura Complexa Simples
Sensibilidade à T e pH Alta Baixa
Condições de reação (T, P e pH) Suaves Drástica (geralmente)
Custo de obtenção (isolamento e purificação) Alto Moderado
Natureza do processo Batelada Contínuo
Consumo de energia Baixo Alto
Formação de subprodutos Baixa Alto
Separação catalisador/produtos Difícil/cara Simples
Atividade catalítica (temperatura ambiente) Alta Baixa
Presença de cofatores Sim Não
Estabilidade do preparado Baixa Alta
Energia de Ativação Baixa Alta
Velocidade de reação Alta Baixa
2.1.1. Quitosana e seus derivados
A quitosana e seus derivados apresentam grande importância devido ao
conhecimento de suas funcionalidades biológicas, médicas, tecnológicas, etc
(Shimosaka et al, 1993; Shimosaka et al, 1995; Mitsutomi et al., 1995). A quitosana é
obtida da quitina, polissacarídeo abundante na natureza, extraído principalmente das
carapaças de caranguejos, lagosta e camarões. A quitosana é um polímero natural,
biodegradável, não tóxico e de alta massa molecular obtido a partir de animais
marinhos, insetos e microrganismos (Tabela 2).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
20
Tabela 2 – Fontes naturais de quitina e quitosana. (Adaptado de Polymar, 2003).
Animais
Marinhos
Insetos Aracnídeos Microrganismos
Anelídios Formigas Escorpiões Leveduras
Moluscos Besouros Aranhas Fungos
Lagosta Esporos
Camarão
Caranguejo
Krill
A Figura 1 abaixo mostra a estrutura química da quitosana
Figura 1 – Estrutura da quitosana (Fonte: Polymar, 2003)
Quitosana totalmente desacetilada não existe na natureza. Graus de acetilação
(GA) nferior a 50% a torna mais solúvel em soluções ácidas (Muzzarelli, 1993;
Muzzarelli et al., 1994; Dung et al., 1994; Stoyachenko et al.,1994). Pelletier &
Sygusch (1990) enfatizaram que grande parte dos resíduos sólidos da indústria
pesqueira, principalmente da carcinicultura, é jogada ao mar ocasionando degradação
ambiental. Logo, a utilização desses resíduos como matéria-prima para formação de
novos produtos, não só reduz de forma drástica o impacto ambiental, mas também daria
maior valor agregado à indústria pesqueira.
A demanda cada vez maior de consumidores por alimentos mais saudáveis, sem
conservantes químicos, tem motivado pesquisas à descoberta de novos agentes
antimicrobianos naturais. Sendo assim, a atividade antimicrobiana incomum da

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
21
quitosana e seus derivados contra microrganismos, tais como bactérias, bolores e
leveduras tem tido maior atenção ultimamente.
Devido à interação entre a quitosana, que é positivamente carregada, e as
membranas celulares dos microrganismos, negativamente carregadas, ocorre a formação
de verdadeiros poros na membrana, que são canais de vazamento de conteúdo protéico e
outros componentes intracelulares. A quitosana tem também a propriedade de agir como
agente quelante que se liga de forma seletiva a traços de metais, inibindo, assim, a
produção de toxinas e o crescimento microbiano. Ainda é capaz de ativar mecanismos
de defesa em tecidos hospedeiros, atuando como agente sequestrante de água e inibindo
várias enzimas. A ligação da quitosana com DNA e a inibição da síntese do mRNA
ocorre via penetração da quitosana no núcleo dos microrganismos e interferência na
síntese do mRNA e proteínas, (Craveiro et al., 2004).
Em um estudo clínico conduzido por Giustina & Ventura (1995) com 100
pessoas com sobrepeso de 10 a 25%, ao final de 4 semanas as pessoas que receberam
quitosana demonstraram perda de peso significativa (7,3 Kg contra 3,0 Kg nos grupos
controle). As pessoas que tomaram quitosana também demonstraram redução na pressão
sanguínea sistólica (de 145 para 135 mm Hg) e diastólica (92.6 para 84.2 mm Hg),
assim como uma redução na taxa respiratória (27.6/min a 21.2/min).
Kondo & Osada (1996) pesquisaram a influência da quitina e da quitosana na
biodisponibilidade do zinco em ratos alimentados com uma dieta com suplementos de
5% ou sem fibra dietética (quitina, quitosana, celulose, pectina e agar-agar) durante 31
dias. Observaram que os ratos alimentados com dietas contendo 5% de fibras, exceto
quitosana, o consumo de alimento foi maior do que o controle. O ganho de peso em
todos os grupos alimentados com fibra foi maior do que o controle, exceto para o grupo
suplementado com quitosana (5%) onde a ingestão de alimento e o ganho de peso foram
menores do que os grupos controle e das demais fibras.
Segundo Craveiro et al. (2004), derivados da quitosana obtidos pela reação com
ácidos carboxílicos, podem promover ao polímero uma solubilidade especial em água.
Esses compostos possuem estrutura química semelhante ao ácido hialurônico e
fornecem uma excelente compatibilidade com o tecido epitelial. As soluções obtidas
com esses derivados podem ser facilmente incorporadas a formulações cosméticas,
mostrando melhor compatibilidade com surfactantes iônicos e emulsificantes,
apresentando estabilidade em uma larga faixa de pH.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
22
Han et al. (1999) examinaram os efeitos da quitosana sobre a atividade da lipase
pancreática in vitro e sobre o grau de armazenamento de gorduras induzido em ratos
pela administração oral de uma dieta rica em gorduras por nove semanas. Como
resultado, observou-se que os animais alimentados com quitosana, reduziram o peso
corporal, a hiperlipidemia e a gordura hepática. Como conclusão, os autores propõem
que os efeitos anti-obesidade da quitosana em ratos alimentados com alto teor de
gordura se devem parcialmente a inibição da absorção intestinal das gorduras dietéticas.
Em outro ensaio, Zahorska-Markiewicz et al. (2002) realizaram um estudo para
avaliar o efeito da quitosana no tratamento da obesidade. Segundo os autores do estudo,
a quitosana pode ser usada como um valioso e seguro coadjuvante em tratamentos para
a obesidade em longo prazo. A quitosana parece acentuar a redução da pressão arterial,
juntamente com a perda de peso.
Assis (2010) estudou a produção e caracterização de quitooligossacarídeos
produzidos pelo fungo Metarhizium anisopliae e avaliação da citotoxicidade em células
tumorais. A hidrólise da quitosana foi realizada em tempos de 10 a 60 min para a
produção de quitooligossacarídeos. A avaliação da citotoxicidade dos oligômeros de
quitosana foi realizada em células tumorais (HepG2 e HeLa) e não tumoral (3T3). A
produção de oligômeros de quitosana teve maiores rendimentos durante 10 minutos de
hidrólise, os pentâmeros apresentaram concentração de 0,15 mg/mL, no entanto os
hexâmeros, que apresentam maior interesse por causa de suas propriedades biológicas,
só foram detectados com 30 minutos de hidrólise com uma concentração de 0,004
mg/mL.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o planejamento Plackett-Burmam e a metodologia de superfície de resposta.
Os resultados mostraram que a eficiência máxima de extração da quitina foi conseguida
nas seguintes condições: concentração de casca de camarão 50g/L, glicose 50g/L, tempo
de incubação de 5 dias e inoculo como 0,05 OD (densidade óptica).
A glicosamina, unidade monomérica básica da quitosana, possui atividade
antiinflamatória e tem grande importância na construção e manutenção da cartilagem,
no tratamento de enfermidades como: febre reumática, artrose e principalmente a artrite
e osteoartrite na forma aguda ou crônica (Polymar, 2003). A Figura 2 apresenta a
estrutura química da glicosamina.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
23
Figura 2 – Estrutura da glicosamina (Fonte: Polymar, 2003)
O sulfato de glicosamina é um derivado da quitosana sendo utilizado no
tratamento da osteoartrite sintomática dos joelhos, quadris, coluna, mãos e outras
regiões (Craveiro et al., 2004). O sulfato de glicosamina estimula a síntese de
proteoglicanos necessários para o bom funcionamento das articulações, estimula a
regeneração das cartilagens e ainda incorpora enxofre ao tecido cartilaginoso. Estudos
de longa duração, acima de 3 anos, mostram que o sulfatos de glicosamina evita a
progressão de danos estruturais nas articulações e induz melhora significativa nas dores
e funções físicas (Craveiro et al., 2004). A Figura 3 apresenta a estrutura química do
sulfato de glicosamina.
Figura 3 – Estrutura do sulfato de glicosamina (Fonte: Polymar, 2003)

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
24
2.1.2. Quitosanases
Dependendo do tipo de quitosanase obtida, ocorrem diferentes mecanismos de
hidrólise da quitosana, isso devido a diferentes microrganismos produzirem enzimas
com diferenças nas suas estruturas (Kurita et al., 1977; Seino et al., 1991).
Quitosanases (EC 3.2.1.132) são enzimas que catalisam a reação de hidrólise de
quitosana em oligossacarídeos. As enzimas quitosanases são produzidas por vários
microrganismos, bactérias, actinomicetos e fungos, e até por algumas plantas. Estas
enzimas agem em polímeros com grau de acetilação de 30% a 60%. Os monômeros
resultantes da hidrólise da quitosana por estas enzimas são melhores digeridos, ao serem
ingeridos, como é o caso da glicosamina. Estudos efetuados até o presente momento
classificam as quitosanases em cinco famílias: 5, 8, 46, 75, 80, sendo que a família 46 é
a mais estudada. As quitosanases tem grande aplicação na geração de oligômeros de
quitosana (Somashekar & Joseph, 1996; Chen et al., 2005).
Nogawa et al. (1998) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida pelo fungo Trichoderma reseei PC-3-7. Estes pesquisadores
inocularam 106 conídios em 100 mL de um meio basal contendo 0,3% de N-
acetilglicosamina em incubator rotativo com agitação de 220 rpm a 28oC. Destaca-se
que o fungo não produziu quitosanase em meio contendo quitosana ou quitina.
Entretanto, o microrganismo produziu quitosanase em meio contendo glicosamina e N-
acetilglicosamina, sendo que para este último obteve-se uma atividade específica
máxima de 1,5 U/mg após 72 horas de incubação. É importante ressaltar que uma
agitação vigorosa favoreceu a produção da enzima. Esse artigo ilustra a presença de
exo-β-D-glicosaminidase e exo-β-D-N-acetilglicosaminidase.
Piza et al. (1999) estudaram a produção, purificação e caracterização parcial de
uma quitosanase produzida por Bacillus cereus. Nesse caso, investigou-se através de um
planejamento fatorial fracionário a influência dos fatores: concentração de sulfato de
amônio (0,4; 2,2 e 4,0%), razão de aeração (2; 6 e 7), tempo de fermentação (16; 24 e
32 horas), pH (5; 6 e 6,5) e concentração de quitosana (1,0; 1,5 e 2,0%). Os resultados
mostraram que os fatores mais significativos para esse planejamento foram a
concentração de sulfato de amônio, a aeração, o pH e a interação entre sulfato de
amônio e a aeração.
Cheng & Li (2000) estudaram a preparação de oligossacarídeos utilizando
quitosanase produzida por Aspergillus sp e constataram que a enzima foi largamente

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
25
induzida em meio contendo quitosana como única fonte de carbono e também na
presença de quitina coloidal. A indução da enzima foi realizada em temperaturas
variando na faixa de 28 a 30ºC, pois essa espécie não cresceu bem em temperaturas
superiores a 37ºC.
Zhang et al. (2000) estudaram a produção, purificação e caracterização de
quitosanase e Exo-β-D-Glucosaminidase por Aspergillus oryzae IAM2660. Os
microrganismos utilizados neste estudo foram mantidos em placas de Petri contendo
agar para contagem em placas (PDA). Destaca-se que para produzir a enzima, conídios
foram retirados das placas e inoculados em meio Czapeq-dox com os seguintes
componentes: 1,0g K2HPO4, 0,5g KCl, 2,0g NaNO3, 0,5g MgSO4.7H2O, 0,02g
FeSO4.7H2O, 20g de sacarose e 1,0L de água deionizada, com pH do meio igual a 5,5.
Foi adicionado ao meio 0,5 g de peptona e 0,5g de extrato de levedura com
concentração de esporos da ordem de 1,0x106 esporos.mL-1. Após a incubação, feita em
incubador rotativo a 26ºC por um período de 60h, micélios foram coletados e após
filtração e secagem a 80ºC foram utilizados para ensaio de quitosanase.
Kuroiwa et al. (2002) estudaram os fatores que afetam a composição de
oligossacarídeos produzidos através da hidrólise de quitosana por quitosanase originária
de Bacillus pumilus BN-262. Neste experimento foi utilizado quitosana 100%
desacetilada dissolvida em ácido acético 1M. O pH do meio foi ajustado em 5,6. Neste
estudo a quitosanase foi imobilizada em agar e Sepharose. Os resultados mostraram que
a concentração de enzima, a temperatura, a velocidade de agitação, e concentração de
substrato influenciaram de forma significativa na composição dos oligossacarídeos
produzidos.
Ichikawa et al. (2002) estudaram a imobilização e estabilização da ligação
multiponto da quitosanase tendo agar como suporte e concluíram que a imobilização
aumentou a termoestabilidade da quitosanase. Os oligossacarídeos de quitosana foram
continuamente produzidos usando um reator coluna contendo quitosanase imobilizada.
A percentagem de quitosana imobilizada após 28 dias de reação foi de 44%.
Hung et al. (2002) estudaram a purificação, através de troca iônica, e
caracterização da hidrólise sobre ação da quitinase e quitosanase purificadas do meio
contendo Bromelina comercial. Destaca-se que a quitinase purificada apresentou pH,
temperatura e Km ótimo para ação catalítica em meio contendo quitina de 4,0, 60ºC e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
26
0,2mg.mL-1, respectivamente. Enquanto que quitosanase apresentou o pH, temperatura e
Km ótimo de 3,0, 50ºC e 0,88mg.mL-1, respectivamente.
Zhu et al. (2003) estudaram as melhores condições fermentativas e propriedades
de uma quitosanase produzida por Acinetobacter sp. C-17. A espécie de bactéria
utilizada neste ensaio foi isolada do solo e identificada como sendo Acinetobacter sp C-
17 e capaz de produzir quitosanase. O microrganismo foi mantido em placas contendo o
meio agar com os seguintes componentes: 0,5 % de quitosana, 0,2 % K2HPO4, 0,1 %
KH2PO4, 0,07 % MgSO4, 0,05 % NaCl, 0,05 % KCl, 0,01 % CaCl2, 0,05 % de extrato
de levedura e 2 % agar, o pH do meio de manutenção foi mantido em 7,2 e a
temperatura foi mantida em 30ºC durante 3 a 5 dias, para identificar a ação da enzima
quitosanase. A ação da quitosanase foi identificada através da observação de uma
colônia de microrganismo existente na placa. A bactéria foi então isolada da placa e
colocada em fermentador rotativo a 200 rpm durante 72h em meio contendo: 0,07 %
K2HPO4, 0,03 % KH2PO4, 0,05 % MgSO4, 0,03 % peptona, 0,03 % de extrato de
levedura e 1,0 % de quitosana. O pH foi de 7,0 e a temperatura 30ºC. Como fontes de
carbono foram utilizadas além de 1,0 % de quitosana, 1,0 % de quitina e 1,0 % de
glicose. Os resultados mostraram que as melhores condições para produção da
quitosanase foram alcançadas quando se utilizou 1,0 % de solução de quitosana a 30ºC,
pH 7,0 durante 24 horas de incubação.
Liu et al. (2003) estudaram a produção de quitinase por Verticillium lecanii
F091 usando fermentação submersa, a atividade de quitinase foi de 9,95 mU/mL em um
meio de cultura otimizado, com um volume de cultura de 200 mL, agitação de 150 rpm
e temperatura de 24ºC cultivado em incubador rotativo e concluíram que a taxa de
agitação e o pH foram os fatores mais significativos para a produção de quitinase, no
entanto, as taxas de agitação e aeração poderiam modificar a concentração de oxigênio
dissolvido afetando diretamente no crescimento do Verticillium lecanii e na produção
de quitinase.
Donzelli et al. (2003) estudaram a hidrólise enzimática avançada de quitina de
casca de caranguejo langostino com misturas de enzimas de bactérias e fungos e
concluiram que a combinação de enzimas de Trichoderma atroviride e Serratia
marcescens foram capazes de degradar completamente altas concentrações de quitina
(100g/mL) a partir de carapaças de caranguejo langostino convertendo a N-
Acetilglicosamina (78%), glucosamina (2%) e quitobiose (10%). Enzimas de fonte

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
27
procariótica apresentaram atividades menores que as enzimas de T. atroviride, para
catalisar a reação com quitosana.
Jo et al. (2003) estudaram a caracterização e a avaliação cinética de quitosanase
produzida por Bacillus sp. Destaca-se que na produção da enzima utilizada neste ensaio
o meio continha 0,5% de concentração de quitosana, 1% de triptona e 1% de NaCl, o
pH do meio foi ajustado em 5,5 e a temperatura foi mantida em 37ºC com agitação de
180 rpm em incubador rotativo.
Fu et al (2003) estudaram a caracterização de três iso-enzimas de quitosanase
isolada de pepsina comercial e denominado como PSC-I, PSC-II, PSC-III,
respectivamente, em relação a hidrólise enzimática da quitosana, o meio ótimo para a
produção destas iso-enzimas continha as seguintes características: pH 5,0; 5,0 e 4,0 e
temperaturas de 40ºC, 40ºC e 30ºC, o valor de Km foi de 5,2, 4,0 e 5,6 mg.mL-1,
respectivamente. As massas moleculares das três iso-enzimas foram de 40 kDa. Os
resultados mostraram que as três iso-enzimas apresentaram atividade em meio contendo
quitosana, produzindo oligômeros da ordem de 68 – 88% desacetilada.
Choi et al. (2004) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida por Bacillus sp KCTC 0377BP. Nesse caso, com relação à
produção da enzima foi possível obter um aumento de 1,2 U/ml em incubator rotativo
para 100,0 U/ml em fermentador piloto de 500 L, utilizando uma concentração de
substrato de 0,5% de quitosana e como fonte de nitrogênio polipeptona (2,5%) em pH
igual a 6,8, com 100 rpm de 100,0 rpm a 30oC.
Shimosaka et al. (2003) estudaram a produção, purificação e caracterização total
de duas quitosanases produzida por Acinetobacter sp CHB101. Nesse caso, com relação
à produção da enzima os pesquisadores usaram dois meios, um contendo glicose e outro
contendo quitosana. Com relação à cinética, o meio contendo quitosana fornece um
tempo de geração de 280 minutos, enquanto que no meio contendo glicose a taxa de
crescimento foi mais elevada e o tempo de geração foi de apenas 110 minutos. Os
autores observaram que após os dois primeiros dias de cultivo a atividade
quitosanolítica permaneceu baixa (na ordem 0,10 a 0,20 U/mL). Durante a fase de
crescimento exponencial o valor da atividade permaneceu com valor inferior a 0,01
U/mL, começando a aumentar ao atingir a fase estacionária, indicando assim que a
produção da enzima é não associada ao crescimento.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
28
Kim et al. (2004) estudaram a produção, purificação e caracterização de uma
quitosanase produzida pelo Bacillus sp e identificado pelo número 1299. Destaca-se que
na produção da enzima quitosanase foi utilizado um meio contendo os seguintes
componentes: 0,5 % de quitosana, 0,42 % (NH4)2SO4, 0,2 % K2HPO4, 0,02 % uréia,
0,03 % CaCl2.2H2O, 0,03% MgSO4.7H2O, 0,1% peptona, 0,2% Tween 80, 0,2 % da
solução (0,5% FeSO4.7H2O, 0,16% MnSO4.H2O, 0,14% ZnSO4.7H2O, 0,2% CoCl2) e
1,7% agar. O pH do meio para este ensaio foi mantido em 7,0. Este microrganismo foi
escolhido por ser grande produtor de quitosanase.
Chen et al. (2005) estudaram a purificação e a caracterização de dois tipos de
quitosanase produzidas por Aspergillus sp. CJ22-326 em cultivo submerso. Na
produção da enzima foi utilizado um meio de cultivo contendo 1,0% de quitosana, 2,0%
de gérmen de trigo, 0,2% de sulfato de amônio, 0,2 % de fosfato de potássio e 0,05% de
sulfato de magnésio por litro de solução em pH 5,6. O cultivo foi realizado em frasco de
500 mL contendo 150 mL de meio de cultura a 30ºC por 96 horas com uma agitação de
150 rpm. Os autores apresentam um gel de eletroforese com as duas enzimas purificadas
com massa molar de 29 kDa e 109 kDa.
Silva Filho et al. (2005) estudaram, através de planejamento fatorial 24 com
repetição em triplicata no ponto central, a produção de quitosanase por Aspergillus
ochraceus em cultivo descontínuo submerso. Os resultados mostraram que foi possível
produzir quitosanase com atividade na ordem de aproximadamente 5,9 U/mL utilizando
Aspergillus ochraceus e que a atividade foi favorecida pelo aumento da agitação (r), da
razão de aeração (RA) e da concentração de substrato (Cs), enquanto que o aumento da
temperatura de cultivo (TMC) não favoreceu a resposta (atividade quitosanolítica).
Pode-se classificar as quitosanases em endoquitosanases e exoquitosanases. As
primeiras diferenciam-se por catalisar a hidrólise aleatoriamente, no interior do
biopolímero, gerando diversos tamanhos de oligossacarídeos. As exoquitosanases são
capazes de hidrolisar as terminações não redutoras, produzindo assim unidades
redutoras (Peter, 2005).
Palma-Guerrero et al. (2008) estudaram o biocontrole de fungos patogênicos e
demonstraram a lise da membrana plasmática e a inibição da germinação de esporos de
fungos. Eles observaram que os fungos entomopatogênicos e nematófagos foram menos
inibidos pela quitosana que os fitopatogênicos e micoparasíticos provavelmente porque
apresentam a enzima quitosanase que degrada a quitosana. Mesmo assim, a quitosana

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
29
pode ajudar a desenvolver novas estratégias para o biocontrole em geral de fungos
patogênicos, evitando o uso de fungicidas que prejudicam o meio ambiente e a saúde
humana.
Pagnoncelli (2008) estudou a produção de quitooligossacarídeos com
propriedades nutracêuticas a partir da quitosana por hidrólise enzimática, utilizando
processo fermentativo simultâneo. Duas cepas produtoras de quitosanaes foram
selecionadas, Paenibacillus chitinolyticus e Paenibacillus ehimensis, e avaliadas quanto
ao comportamento em meio de cultivo contendo açúcares simples e em relação às
variações de pH do meio. Os quitooligossacarídeos produzidos a partir do complexo
enzimático da primeira cepa apresentaram maior pico após 9h de hidrólise, enquanto
para a segunda cepa avaliada, após 20min, observou-se quitooligossacarídeos com grau
de polimerização entre 3 e 6 unidades.
2.2. Fungos
2.2.1. Características gerais
Desde o final dos anos 60 que os fungos deixaram de pertencer ao reino das
plantas e passaram a ter um reino próprio chamado de Reino Fungi. Foi a partir de
estudos morfológicos, bioquímicos e citológicos onde se concluiu que eles são tão
diferentes dos vegetais como dos animais. Distinguem-se de outros organismos
eucariotas por serem seres quimiorganoheterotróficos e, ainda, por apresentarem parede
celular rígida que é composta por quitina e glucano e uma membrana celular em que o
ergosterol substitui o colesterol (Trabulsi & Toledo, 1996; Murray et al., 2005).
Fazem parte do Reino Fungi cerca de 100 mil espécies, desde grandes como
os cogumelos, até microscópicas como os bolores e leveduras. Ainda hoje é utilizada
essa classificação, mas os avanços da biologia molecular e estudos bioquímicos levaram
a outras propostas de classificação em que os fungos e organismos afins foram incluídos
em três diferentes reinos: Protozoa, Chromista e Fungi (Kirk et al., 2008).
Abaixo, são apresentadas características que diferenciam os fungos das plantas:
• são incapazes de sintetizar clorofila, como também não apresentar celulose em
sua parede celular, com exceções como fungos aquáticos;
• apresentam algumas características de células animais (capacidade de depositar
glicogênio e a presença de substâncias quitinosas, em sua parede celular);

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
30
• podem ser eucarióticos (as leveduras), ou multinucleados (os bolores), e
apresentar retículo endoplasmático rugoso e mitocôndrias em seu citoplasma;
• são heterotróficos, alimentam-se de matéria orgânica viva (fungos parasitários),
ou morta (fungos saprofíticos);
Os fungos são amplamente encontrados na natureza, crescem rapidamente no
solo, na água, na atmosfera, bem como nos animais, nos vegetais, resíduos orgânicos e
desempenham um papel importante na vida do homem, quer de uma maneira benéfica,
quer de um modo prejudicial. Os fungos são um dos principais microrganismos
responsáveis pela decomposição da matéria orgânica, interferindo no ciclo do carbono,
do nitrogênio e de outros nutrientes da biosfera. São capazes de deteriorarem produtos e
bens de consumo, tais como alimentos, tecidos, papel, madeira. Os alimentos
armazenados representam um vasto meio para a proliferação dos fungos, principalmente
quando não são consideradas as condições básicas de armazenamento correto (Trabulsi
& Toledo, 1996).
Estão presentes funcionalmente em vários processos industriais, tais como, de
fabricação de pão, cervejas, vinhos e determinados tipos de queijos, sendo também
utilizados na produção comercial de muitos ácidos orgânicos, de alguns fármacos, como
a ergometrina e a cortisona, na obtenção de diferentes antibióticos, como a penicilina, e
de substâncias imunossupressoras, como a ciclosporina (Esteves et al., 1990). A Figura
4 ilustra a estrutura geral de uma célula de fungo.
Figura 4 – Representação da estrutura geral de uma célula de fungo.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
31
Os fungos podem se desenvolver em meios de cultivo especiais formando
colônias de dois tipos, leveduriformes e filamentosas, que se diferenciam pela
macromorfologia e micromorfologia. O primeiro tipo apresenta, em geral, característica
de consistência cremosa de cor branca a creme, brilhantes ou opacas, podendo
apresentar às vezes coloração escura ou alaranjada, e são formados por microrganismos
unicelulares que apresentam as funções vegetativas e reprodutivas. As filamentosas
apresentam características granulares, cotonosas, aveludadas ou pulverulentas, são
multinucleadas e apresentam-se em forma de tubos denominadas de hifas. Essas hifas
podem ser não septadas (cenocíticas) ou septadas (tabicadas). O conjunto de hifas é
denominado micélio.
As hifas asseptadas têm aparência anastomosada (concisa), formada por um
citoplasma estendido e polinucleado, enquanto que as hifas septadas contêm células
individualizadas, cada uma contendo o seu núcleo. Na Figura 5 são mostradas as
estruturas das hifas septadas e não septadas.
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas. (Adaptado de Guia.Bio,
2012)
O micélio que se desenvolve no interior do substrato e que é responsável pela
sustentação e absorção de nutrientes é denominado micélio vegetativo. O micélio que
cresce na superfície do meio de cultura é denominado micélio aéreo. Este pode ser
denominado também de micélio reprodutivo, quando o mesmo se diferencia para
sustentar os corpos de frutificação ou propágulos (Putzke & Putzke, 1998). Os
propágulos classificam-se de acordo com a origem em externos e internos, sexuados e
assexuados.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
32
2.2.2. Reprodução dos Fungos
A reprodução dos fungos ocorre em ciclos assexuais, sexuais e parassexuais. Em
fungos, a reprodução assexuada é observada com mais freqüência do que a reprodução
sexuada. A reprodução assexuada é normalmente a reprodução mais importante para a
propagação da espécie, feita através de fragmentação de artroconídios, fissão de células
somáticas, brotamento ou gemulação do blastoconídios-mãe e produção de conídios. A
reprodução sexuada envolve a união de duas células ou de dois órgãos sexuais
sexualmente compatíveis. A reprodução sexuada culmina na produção de basidiósporos,
no caso de basidiomicetos. A reprodução parassexuada ocorre com a fusão de hifas e
formação de um heterocário, que consiste na existência de núcleos haplóides
geneticamente distintos num mesmo citoplasma, os quais, após a fusão, originam
núcleos diplóides heterozigotos sujeitos a sucessivas divisões mitóticas. Apesar da
raridade dessas, a reprodução parassexual é de grande importância na evolução de
alguns fungos (Pelczar et al., 1996).
A reprodução assexuada é feita por conídios, esporos assexuados formados no
ápice de hifas modificadas, os conidióforos. Os conidióforos não possuem esporângios
de forma que os conídios ficam expostos. Na reprodução sexuada, com o encontro de
duas hifas haplóides distintas, ocorre a plasmogamia e um novo conjunto de hifas
dicarióticas é formado.
Os conídios podem ter formas esféricas, fusiformes, cilíndricas, piriformes, entre
outras. Apresentam-se como hialinos ou pigmentados, podem estar isolados ou
agrupados, a parede celular pode ser lisa ou rugosa e podem ser formados por apenas
uma célula ou apresentarem septos em mais de um plano.
Os conídios também podem originar-se em qualquer parte do micélio vegetativo
e são denominados conídios sésseis como é o caso do Trichophyton rubrum.
Alguns fungos patogênicos são identificados a partir de estruturas formadas pela
célula conidiogênica e o conidióforo. Essas estruturas são denominadas: aparelho de
frutificação ou conidiação. Os conídios podem formar cadeias sobre fiálides em volta de
uma vesícula, dilatando a extremidade do conidióforo. Essas estruturas ocorrem em
Aspergillus, apresentando-se em forma de cabeça ao redor de uma vesícula, e em
Penicillium agrupados em forma de pincel, como é ilustrada na Figura 6.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
33
Figura 6 – Conídios de Aspergillus e Penicillium
2.2.3. Metabolismo de fungos
Os fungos são seres aeróbicos e heterotróficos, podendo se desenvolver em
ambientes com baixa quantidade de oxigênio, como ocorre no crescimento vegetativo e
na reprodução assexuada, como também em ambientes com grande quantidade de
oxigênio como é o caso da reprodução sexuada.
Os fungos são capazes de produzir certas enzimas como lipases, invertases,
lactases, quitosanases, proteases, amilases, etc., que fazem a hidrólise do substrato
facilitando o mecanismo de transporte ativo e passivo. Esses substratos induzem a
formação de enzimas degradativas. Algumas espécies de fungos desenvolvem-se em
meios que apresentam amônia ou nitrilos como única fonte de nitrogênio. As melhores
fontes orgânicas para o seu desenvolvimento são carboidratos, como a D-glicose, e
alguns sais minerais como sulfatos e fosfatos.
A água é uma necessidade de todos os seres vivos, não sendo diferente para os
fungos que precisam de água para o seu desenvolvimento. Certos fungos apresentam
características halofílicas podendo crescer nos ambientes com grandes concentrações de
sal.
A razão de não existirem ambientes livres de espécies fúngicas deve-se ao facto
destes microrganismos conseguirem sobreviver em situações extremas como, por
exemplo, temperaturas que variam entre os - 5 e os 60 ºC, com pH de 1 a 9 e com baixas

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
34
concentrações de oxigênio, apesar de cada espécie possuir condições ideais para o seu
desenvolvimento (Chao et al., 2002).
Devido a grande faixa de temperatura ótima para o seu crescimento, os fungos
apresentam espécies psicrófilas, mesófilas e termófilas. Para a maior parte das espécies
fúngicas, a temperatura de desenvolvimento situa-se entre os 20 e 24 ºC e não crescem
acima dos 29 ºC. Em relação à umidade relativa, necessitam de pelo menos 50 %, mas
geralmente necessitam de mais de 65 % (Colakoglu, 2001).
O pH ideal para o desenvolvimento dos fungos varia na faixa entre 5 e 7.
Destaca-se, entretanto, que a maioria dos fungos suportam grandes variações de pH
como ocorre com os fungos filamentosos que podem crescer em pH variando entre 1,5 e
11. Por outro lado, as leveduras não suportam pH alcalino.
Os fungos crescem lentamente, se comparados com as bactérias, em meio de
incubação para o seu desenvolvimento. Para evitar contaminação por bactérias, que
podem se sobrepor ou inibir o seu crescimento, pode-se introduzir no meio de cultura
um antibacteriano de largo espectro como o cloranfenicol.
Algumas espécies de fungos se desenvolvem na presença de luz, outras são
inibidas e algumas podem crescer tanto na presença quanto na ausência deste agente. No
entanto a irradiação solar, em geral, é prejudicial ao seu desenvolvimento, por causa da
radiação ultravioleta.
2.2.4. Nutrição de fungos
Quanto ao aspecto da nutrição, os fungos são heterótrofos, ou seja, incapazes de
produzir seu próprio alimento. Entretanto, a forma de assimilar matéria orgânica é
diferente daquela utilizada pela maioria dos animais, que são heterótrofos por ingestão.
Nos fungos, a digestão é extracorpórea, ou seja, é realizada fora do corpo. O fungo
lança no ambiente, enzimas que degradam as moléculas orgânicas complexas e, depois,
absorve moléculas menores, mais simples. Por essa razão, são designados heterótrofos
por absorção. É devido a este processo que os fungos podem crescer dentro ou sobre os
alimentos. Desenvolve-se geralmente em meios contendo um pH baixo, uma fonte de
carbono, uma fonte de nitrogênio orgânico ou inorgânico e alguns minerais. Alguns
necessitam de vitaminas. Quanto aos modos de vida, os fungos podem ser
decompositores, parasitas, mutualísticos e predadores (Raven, 2001).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
35
Os fungos decompositores, saprófagos (do grego, saprós, “podre”, e phagos,
“comedor”), juntamente com as bactérias, são os principais decompositores da biosfera,
participando intensamente do processo de degradação da matéria orgânica morta, o que
promove a reciclagem dos elementos químicos constituintes dos seres vivos. A maioria
deles vive no solo, obtendo nutrientes de seres mortos. Em florestas, onde a matéria
orgânica vegetal é abundante, os micélios dos fungos decompositores absorvem os
nutrientes de folhas e galhos caídos, decompondo-os. Os fungos decompositores
também são responsáveis pelo apodrecimento de alimentos e de outros materiais, como
a madeira.
Os fungos parasitas vivem à custa de outros organismos vivos, prejudicando-os.
Ao parasitar o corpo de um ser vivo, animal ou vegetal, o fungo pode até provocar sua
morte. Várias doenças são causadas por fungos parasitas. Nas plantas, cita-se a
ferrugem do café, que, no Brasil, é motivo de grande preocupação por parte dos
cafeicultores, devido aos prejuízos que acarreta. As micoses que atacam a pele de seres
humanos e animais são provocadas por fungos.
Existem fungos que se associam a outros organismos, estabelecendo uma relação
em que há benefício mútuo para os indivíduos envolvidos. Esses fungos são
denominados mutualísticos. A maioria vive associada a seres fotossintetizantes, como
as plantas, cedendo-lhes parte da água e dos nutrientes que as hifas absorvem do solo.
As plantas, por sua vez, cedem ao fungo certos açúcares e aminoácidos. Duas
associações mutualísticas formam estruturas bem características: as micorrizas,
associação que envolve fungos e raízes de plantas, e os líquens, formados por fungos e
algumas variedades de algas e cianobactérias.
Certos fungos atuam como predadores. Na maioria dos casos, as hifas secretam
substâncias aderentes que aprisionam os organismos que tocam os fungos. Dessa
maneira, as hifas penetram o corpo da presa, crescem e se ramificam, espalhando-se no
interior do organismo e absorvendo seus nutrientes, causando-lhe a morte. O gênero
Arthrobotrys apresenta uma forma mais elaborada de predação. O fungo vive no solo e
captura nematódeos microscópicos que habitam o mesmo ambiente. As hifas
apresentam pequenos anéis que se estreitam quando a presa passa por eles. As hifas do
fungo logo invadem o corpo do nematódeo, digerindo-o.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
36
2.2.5. O fungo Metarhizium anisopliae
Existem três espécies de fungos do gênero Metarhizium, divididas em dez
variedades: M. anisopliae, variedades anisopliae, majus, lepidiotum e acridum; M.
flavoviride, variedades tipo E, flavoviride, minus, novazealandicum e pemphigum; e M.
album (Driver et al. 2000). E, mais recentemente, foi descrita na China uma nova
variedade de M. anisopliae, dcjhyium (Dong et al. 2009).
Data de 1879 quando o russo Ilya Metchnikoff aplicou o fungo para combater
larvas de um curculionídeo, praga de beterraba, e foi classificado como Metarhizium
anisopliae por Sorokin em 1883. É um fungicida natural para mais de 300 espécies de
insetos, especialmente pragas da agricultura e pecuária. No Brasil, o fungo Metarhizium
anisopliae teve a sua aplicação com sucesso pela primeira vez em 1965, no controle da
cigarrinha-da-cana Mahanarva posticata (Alves, 1998).
O fungo Metarhizium anisopliae é um importante agente entomopatogênico
utilizado no controle de pragas, como cupins, gafanhotos, cigarrinhas e besouros, sendo
amplamente estudado em todo o mundo. Existe amplamente na natureza e é facilmente
encontrado no solo, onde sobrevive por longos períodos e seu desenvolvimento se dá
geralmente entre 15 e 32 °C, sendo ideal entre 24 e 30 °C e com pH ótimo igual a 6,9.
Apresenta micélio hialino e septado, com conidióforos característicos, sobre os quais
surgem conídios cilíndricos organizados em colunas. Este fungo ao atacar os insetos
provoca rigidez e os recobre por uma camada pulverulenta de conídios, resultando em
colorações que variam do verde claro ao escuro, cinza ou branco com pontos verdes
(Tinline, 1971; Alves, 1998, Driver et al. 2000; Arruda, 2005).
Quanto a sua morfologia, apresenta-se como um fungo filamentoso, com corpo
de frutificação equivalente a um esporodóquio agregado a hifas intimamente
entrelaçadas, com massa compactada de conidióforos característicos, simples ou
ramificados, formando células esporogênicas denominadas fiálides, resultando nos
fialosporos (Wang et al., 2002).
O Metarhizium anisopliae é um fungo de cultivo simples, sendo necessário
quase que exclusivamente o nutriente a base de amido. Um dos fungos mais estudados
em programas de manejo de pragas já que apresenta grande potencialidade
entomopatogênica (Onofre et al., 2002). O Metarhizium anisopliae é um microrganismo
pouco exigente, pois pode se desenvolver em diversos meios de cultura como: amido,
glicose, glicerina, levulose, maltose, sacarose e quitina (Jabor et al., 2003).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
37
Em geral, os fungos produzem enzimas que convertem os tecidos dos insetos em
nutrientes. Porém, a cutícula esclerotizada dos insetos não é bem utilizada pelos fungos,
excetos pelos entomopatogênicos que desenvolveram enzimas eficientes para degradar
essa camada protetora. Destaca-se a enzima Pr1, produzida por Metarhizium anisopliae,
que tem um poder de degradação bem superior a qualquer outra enzima sintetizada por
esse grupo de fungos (Dias, 2005). Desta forma, o Metarhizium anisopliae (fungo
entomopatogênico) ao atacar a célula dos insetos, produz grande quantidade de enzimas
que degradam a membrana citoplasmática e inativa a célula e, consequentemente, mata
o inseto, logo é evidente a relação entre o controle de pragas e produção de enzimas.
No momento, sobre a comercialização de fungos entomopatogênicos no Brasil,
em sua maioria, são formulados em grânulos, compostas do fungo mais o substrato
(arroz + fungo), como também na forma de pó molhável (WP), resultado da moagem do
fungo com o substrato. Esses produtos representam 90% e 8,5%, respectivamente, da
produção de fungos no Brasil, e sendo suas concentrações finais ficam em torno de
5x108 conídios/g para Metarhizium anisopliae e 1x109 conídios/g para Beauveria
bassiana (Alves et al., 2008).
Segundo Batista Filho et al. (2002), Garcia et al. (2005) e Mello et al. (2006), o
Metarhizium anisopliae tem sido bastante utilizado para o controle de diferentes pragas
no Brasil, como a broca da bananeira, pragas de grãos armazenados, carrapatos, cupim
de montículo em pastagens, larvas de escarabeídeos que atacam a cana de açúcar e
cigarrinha da cana-de-açúcar.
É comum a ocorrência natural do fungo Metarhizium anisopliae atacando
cigarrinha-das-raízes de canaviais pelo país, o que incentiva a utilização dele como
agente de controle biológico em áreas infestadas por esse tipo de praga. A aplicação de
recursos no uso de programa de manejo integrado da cigarrinha tem sido reforçado por
razões ambientais e econômicas (Dinardo-Miranda, 2001).
Faria & Magalhães (2001) fizeram uma avaliação do uso de fungos
entomopatogênicos no Brasil, na qual observaram que a qualidade dos micoinseticidas
disponíveis no Brasil poderia ser incrementada de forma considerável quanto a sua
formulação, ou seja, antes da venda, poderiam realizar melhorias no tratamento
posterior, adição de substâncias que lhes assegurem melhorias na eficiência de controle,
capacidade de armazenamento ou praticidade de manuseio, ou de qualquer outro critério
que resulte em vantagem em relação ao produto bruto. Ressaltaram a pouca praticidade

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
38
em alguns casos, como exigência de lavagem, e em outros podendo causar o
entupimento de bicos dos pulverizadores devido à elevada proporção de inertes,
principalmente quando são empregados baixos volumes de aplicação. Outro problema
evidenciado com os inseticidas nacionais diz respeito à pequena sobrevida, devendo a
ser usados em, no máximo, 30 dias depois de produzidos, quando armazenados à
temperatura ambiente e em local sombreado, fazendo as vendas ocorrerem quase que,
exclusivamente, sob encomenda. Comparativamente, os bons inseticidas sob
comercialização em outros países apresentam maior concentração de ingrediente ativo,
maior sobrevida (alguns produtos podem ser armazenados por mais de 8 meses à
temperatura ambiente) e praticidade (são formulados na forma de GDA (grânulos
dispersíveis em água) ou de óleos emulsionáveis, por exemplo, podendo ser adicionados
diretamente ao tanque do pulverizador.
Rangel et al. (2007), instalaram uma Unidade de Observação para o controle
microbiano das cigarrinhas das pastagens Distrito de Itahum, em Dourados/MS, devido
as necessidades de agricultores familiares de produção de leite. As aplicações do fungo
Metarhizium anisopliae foram realizadas por pulverização com equipamento de tração
animal. As condições do ambiente de estudo foram favoráveis ao desenvolvimento da
população das cigarrinhas e à incidência do fungo. Como resultados, observaram que os
níveis de espumas, ninfas e adultos reduziram significativamente após as aplicações,
levando os Agricultores Familiares a total satisfação com os ganhos na produção de
todos os envolvidos.
Segundo Vieira et al. (2007), o fungo entomopatogênico Metarhizium anisopliae
var. acridum tem sido utilizado no controle de pragas, e assim, visando este fim,
analisaram em laboratório a esporulação, a germinação dos conídios e as características
morfológicas peculiares desse fungo em diferentes temperaturas. Esperou-se o
desenvolvimento das colônias em períodos de 1 a 15 dias. Foi observado que
temperaturas de 25, 28 e 30 ºC proporcionaram maiores taxas de esporulação e elevadas
porcentagens de germinação dos conídios. A 25, 28 e 30 ºC observou-se um
desenvolvimento mais rápido das microestruturas somáticas e reprodutivas (hifas,
anastomoses, apressórios, conidióforos e conídios elipsóides). Nas temperaturas
extremas de 40 e 45 ºC não foi detectado esporulação, germinação, nem
desenvolvimento de nenhuma microestrutura.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
39
Lopes et al. (2007) avaliaram o efeito de formulações de Beauveria bassiana e
Metarhizium anisopliae sobre ninfas de Amblyomma cajennense (transmissor da febre
maculosa ao homem) em laboratório. Ninfas não alimentadas foram pulverizadas com
suspensões contendo 2x107 e 4x107 conídios/mL de B. bassiana, e 1x107, 1,5x107,
2x107, 3x107 e 5x107 conídios/mL de M. anisopliae. A mortalidade de ninfas,
decorridos três dias da aplicação, foi significativa nas concentrações de 3x107 e 5x107
conídios/mL de M. anisopliae, com 57,1 e 56,2 %, respectivamente. Após seis dias a
mortalidade foi de 100 % nessas duas concentrações. A mortalidade foi
significativamente maior que a observada para B. bassiana em concentrações
semelhantes.
Guirado et al. (2009) realizaram estudos para controlar cupins de montículo em
área de pastagem, utilizando fungos entomopatogênicos Metarhizium anisopliae e
Beauveria bassiana em diferentes doses e formulações. A avaliação foi realizada após
30 dias da aplicação dos produtos. Os melhores resultados em ordem decrescente de
eficiência foram: M. anisopliae 30 mL + 1000 mL de água com 100 % de eficiência; B.
bassiana 10g + M. anisopliae 20 mL + 1000 mL de água com 85,71 % de eficiência; M.
anisopliae 20 mL + 1000 mL de água com 71,42 % de eficiência. Os demais
tratamentos não diferiram da testemunha.
Loureiro et al. (2012) compararam a eficiência de isolados de Metarhizium
anisopliae em controlar populações naturais de Mahanarva fimbriolata (cigarrinha-da-
raiz) em cultivos comerciais de cana-de-açúcar colhida mecanicamente. Foram
aplicados os isolados IBCB 348, IBCB 408, IBCB 410 e IBCB 425 do fungo em
concentração de 1,5×1012 conídios/ha. Aplicou-se o planejamento experimental em
blocos casualizados com 5 tratamentos e 4 repetições, sendo cada repetição composta
por 7 linhas de 100 m de comprimento e espaçamento de 1,5 m. Após 30 dias de
pulverização foi observado para os isolados IBCB 408 e IBCB 425 uma eficiência de
controle de ninfas de 63 e 62%, respectivamente, já no caso dos adultos chegou a 100%.
Após 60 dias o isolado IBCB 425 foi o mais eficiente para controlar as ninfas com
48,4%.
Assis et al. (2012) estudaram a citotoxidade de quitoolissacarídeos (QOS) e
oligômeros A, B e C (soluções compostas por diferentes quantidades de QOS)
produzidos por hidrólise enzimática pelo fungo Metarhrizium anisopliae. Foi analisado
o efeito de antiproliferativo destas moléculas em células tumorais (HepG2 e HeLa) e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
40
também em célula normal (3T3). A atividade antioxidante foi analisada em vários
experimentos in vitro. A glicosamina apresentou maior toxicidade (cerca de 92%) para
todas as células estudadas. No entanto, os oligômeros obtidos após hidrólise não
demonstraram efeito tóxico sobre a célula normal (3T3). Além disso, mostraram que
uma pequena quantidade de outros COS pode diminuir o efeito citotóxico da
glicosamina contra células 3T3, indicando que glicosamina poderia ser usada como uma
droga antitumoral na presença de outros COS.
2.3. Fermentação
Os processos fermentativos são caracterizados por processos nos quais
microrganismos realizam a conversão catalítica de uma dada substância num
determinado produto. Estes processos podem ser classificados como processos
descontínuos, contínuos e descontínuos alimentados. A classificação destes processos
está diretamente ligada à forma de como o substrato é adicionado e como o produto é
retirado. No processo de fermentação descontínuo o substrato é colocado no
fermentador, em seguida é inoculado e após o término do processo o produto é retirado.
No processo contínuo, o substrato é colocado num reator com vazão constante e o meio
fermentado é retirado com a mesma vazão de entrada do reator (Faccioti, 2001). Nos
processo descontínuos alimentados à adição de substrato e a retirada do mesmo deve
ocorrer de forma intermitente (Carvalho & Sato, 2001).
De certa forma, os processos fermentativos representam um elo entre as antigas
artes de elaboração de alimentos, como queijos e vinho, a partir de uma flora
microbiana natural, com a moderna indústria de fermentação de alimentos, onde são
utilisados cultivos puros e equipamentos sofisticados para o controle do processo
(Assis, 2010).
Atualmente, a maioria dos processos industriais se desenvolve em cubas de
vidro ou de aço de até 20 litros. Os agentes biológicos se encontram submersos no meio de
cultivo que ocupa, aproximadamente, 75% da cuba. As vezes é necessário injetar ar, e em muitas
fermentações se forma espuma. O desenho do biorreator deve se adequar ao objetivo do
processo, respondendo eventualmente a diversos imperativos, tais como a esterilização do
sistema, a aeração e homogeneização do meio, o acréscimo de nutrientes e de aditivos
antiespuma, a manutenção do pH, etc. Os modelos de fermentadores mais utilizados com
microrganismos contam com aeração e agitação mecânica. Esta facilita a distribuição dos

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
41
nutrientes na cuba, mas gera calor que deve ser eliminado mediante a circulação de água
fria. Em outros tipos de biorreatores, em coluna ou torre, a homogeneização depende da injeção de
ar. Os sistemas submersos são apropriados para o cultivo de microrganismos livres, mas resultam
pouco econômicos quando se trabalha com células ou enzimas caras. A imobilização em
fermentadores menores, seja por adesão a um suporte inerte, seja por inclusão dentro de um
polímero que permita o contato com o meio de cultura, além de simplificar a purificação do
produto permite a reutilização das células ou das enzimas, que permanecem dentro do
biorreator (Malajovich, 2009).
As enzimas são produtos de alto valor agregado, amplamente empregadas em
uma gama de indústrias, e entre as fontes para sua obtenção, destacam-se as de origem
microbiana, principalmente, devido a elevada seletividades dos substratos utilizados e
produtos formados. A principal forma de obtenção de enzimas, em escala industrial, é a
fermentação submersa, por esta apresentar vantagens em relação a fermentação
semisólida, como: maior controle do processo fermentativo e consequente reprodução
da cinética microbiana, além de maiores rendimentos (Wanderley, 2011).
2.3.1. Preparo do inóculo
Denomina-se inóculo um volume de suspensão de microrganismo de
concentração adequada, capaz de garantir, em condições econômicas, a fermentação de
uma dada quantidade de meio. O volume do inóculo, assim como sua concentração de
células, vai depender tanto do processo de fermentação c omo da quantidade de meio a
ser fermentado (Carvalho & Sato, 2001).
A partir de uma cultura pura, inocula-se um volume relativamente pequeno de
meio nutriente, que vai depender do tipo de microrganismo cultivado (pois cada espécie
microbiana possui velocidades diferentes de crescimento), sendo, em seguida, incubado
em condições favoráveis ao desenvolvimento do microrganismo. A suspensão obtida é
transferida para um frasco maior em agitador rotativo ou recíproco (shakers), que por
sua vez é adequadamente incubado, e a seguir, transferido para um frasco contendo um
volume ainda maior deste meio, até se conseguir alguns litros de suspensão microbiana.
Os cuidados com assepsia, volumes de meio utilizado, numero de transferência bem
como as condições de incubação, podem variar de acordo com o tipo de processo de
fermentação que se deseja realizar.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
42
2.3.2. Produção de enzimas em cultivo submerso descontínuo
A produção de enzimas, usando processos fermentativos, é constituída
basicamente por duas etapas: a primeira caracteriza-se pela preparação do inóculo e a
segunda, pela fermentação do substrato (Carvalho & Sato, 2001). A finalidade da
primeira etapa é o preparo do microrganismo em condições adequadas para que possa
ocorrer o desenvolvimento da segunda etapa. Diferentemente do cultivo em estado
sólido, no cultivo submerso os microrganismos encontram-se em um meio com elevada
atividade de água.
2.4. Planejamento experimental
Um bom planejamento tem como fundamental projetar um experimento de
forma que ele seja capaz de fornecer exatamente o tipo de informação que procuramos.
Para isso é necessário saber, em primeiro lugar, o que é mesmo que se deseja procurar.
Planejamento experimental, ou delineamento experimental, é um conjunto de
ensaios estabelecido com critérios científicos e estatísticos, objetivando determinar a
influência de diversas variáveis nos resultados de um dado sistema ou processo (Barros
Neto et al., 2001; Rodrigues & Iemma, 2005). De acordo com o propósito dos ensaios,
esse objetivo maior pode ser mais detalhado em:
– determinar quais variáveis são mais influentes nos resultados;
– atribuir valores às variáveis influentes de modo a otimizar os resultados;
– atribuir valores às variáveis influentes de modo a minimizar a variabilidade dos
resultados;
– atribuir valores às variáveis influentes de modo a minimizar a influência de variáveis
incontroláveis.
O planejamento experimental, baseado nos fundamentos estatísticos é, sem
dúvida alguma, uma ferramenta poderosa para se chegar às condições otimizadas de um
processo, desenvolvimento da formulação de produtos dentro das especificações
desejadas ou simplesmente para avaliar os efeitos ou impactos que os fatores (variáveis
independentes) têm nas respostas (variáveis dependentes) desejadas (Barros Neto et al.,
2001; Rodrigues & Iemma, 2005). Da utilização das técnicas estatísticas de
planejamento experimental, é possível chegar benefícios importantes, como:

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
43
– reduzir o número de ensaios sem prejuízo da qualidade da informação;
– estudar simultaneamente diversas variáveis, separando seus efeitos;
– determinar a confiabilidade dos resultados;
– realizar a pesquisa em etapas, num processo iterativo de acréscimo de novos ensaios;
– selecionar as variáveis que influem num processo com número reduzido de ensaios;
– representar o processo estudado através de expressões matemáticas;
– elaborar conclusões a partir de resultados qualitativos.
2.4.1. Planejamento fatorial
Em um planejamento fatorial é investigado as combinações de níveis de estudo
de cada fator. Para tanto, inicialmente, é necessário especificar esses níveis em cada
fator, quer dizer, os valores dos fatores que serão utilizados nos experimentos. Um
ensaio experimental é definido como um experimento constituído de um conjunto de
níveis definido. Geralmente, quando se tem n1 níveis de um determinado fator 1, n2 do
fator 2, ..., e nk do fator k, o planejamento será descrito por um fatorial n1 x n2 x ... x nk,
que corresponde ao número mínimo de experimentos para se ter um planejamento
fatorial completo.
É necessário fazer cada fator variar e observar o resultado dessa variação para se
estudar o efeito sobre a resposta. Assim, os ensaios precisam ter um mínimo de dois
níveis por fator para a realização desses ensaios. Para k fatores, ou seja, k variáveis de
controle, o planejamento de dois níveis precisará de 2 x 2 x ... x 2 = 2k, para o número
de ensaios diferentes, sendo conhecido como planejamento fatorial 2k.
Para o planejamento fatorial, são especificados os valores dos fatores em cada
experimento, e estes são classificados em níveis, variando-se, por exemplo, de -1
(mínimo) a +1 (máximo). A partir de uma matriz serão apresentadas todas as
combinações entre o número de fatores e de níveis nos experimentos a serem realizados
(Barros Neto et al., 2001; Rodrigues & Iemma, 2005).
2.4.2. Planejamento Plackett & Burman
A maioria dos planejamentos do tipo “Screening” (Delineamentos de Seleção de
fatores) foi descrito originalmente por Plackett & Burman em 1946. Os planejamentos
experimentais de Plackett & Burman são eficientes em situações que envolvem uma

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
44
grande quantidade de variáveis a serem exploradas em um processo de otimização de
fatores (Reddy et al., 2008).
As matrizes Plackett & Burman consistem de planejamentos fatoriais de dois
níveis para estudar k = N – 1 variáveis em N ensaios. São construídas para ensaios
múltiplos de 4, sendo essas matrizes para ensaios de 8, 12, 16,..., 100. Cada variável
independente em uma matriz assume uma configuração de dois níveis, o inferior (-1) e o
superior (+1), para o nível intermediário é estabelecido (0). Para cada vez que a variável
aparece no nível inferior, terá o par equivalente no nível superior (Rodrigues & Iemma,
2005).
Para evitar dúvidas nos resultados dos efeitos de algum fator no planejamento de
Plackett & Burman, o número de ensaios a ser realizado deve ser no mínimo o número
de variáveis mais 4. No caso de um fatorial completo o número de ensaios a ser
realizado é bem maior (Rodrigues & Iemma, 2005).
Mesmo já determinado o planejamento Plackett & Burman, torna-se importante
à realização de experimentos relativos aos pontos centrais de modo a se avaliar melhor
as possíveis interferências analíticas que podem ocorrer durante os diferentes
experimentos. A repetição do ponto central permite também a determinação do erro
puro e da repetibilidade do processo. Na prática, esta é uma ferramenta que permite
identificar melhor se o processo em estudo está sob controle (Rodrigues & Iemma,
2005).
É possível que no planejamento Plackett & Burman não tenha sido considerado
a codificação do ponto central (0) para repetição, pois, matematicamente, a estimativa
do valor desse efeito codificado (zero) vezes, teria como resposta zero. Porém, no
resultado prático do experimento, consegue-se informações valiosas com essas
repetições no ponto central (Rodrigues & Iemma, 2005).
2.4.3. Aplicações do Planejamento Plackett & Burman
Ghanem et al. (2000) produziram xilanase por Aspergillus terreus cultivadas
sobre palha de trigo finamente moída em fermentação de estado sólido. A composição
ideal do meio foi estabelecida aplicando-se o planejamento experimental de Plackett &
Burman. A melhor atividade enzimática foi obtida em um meio contendo 10 g/L de

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
45
palha de trigo por frasco umedecido com uma solução concentrada de sal com teor de
água inicial de 75 % e incubadas durante 4 dias a 30 °C. Observou-se que íons
metálicos como Hg2+, Cu2+, Co2+, Fe3+ e Pb2+ inibiram fortemente a xilanase, enquanto
que Ca2+ ativou a enzima.
Mu et al. (2005) estudaram as otimização de meio de fermentação para a reação
de estereoinversão por Candida parapsilosis CCTCC M203011 utilizando o
planejamento Plackett & Burman. Os resultados mostraram que dentre os fatores
estudados, os fatores mais significativos foram: a Concentração de MgSO4, ZnSO4 e
KH2PO4 e que após a utilização da metodologia de superfície de resposta foi constatado
que o ponto ótimo para a otimização foi conseguido com as concentrações dos fatores
MgSO4, ZnSO4 e KH2PO4 nos valores em: 1,17 g/L, 0,10 g/L e 9,41 g/L,
respectivamente, elevando a taxa de conversão enantiosseletiva de 75,2 % para 96 %.
Naveena et al. (2005) empregaram o planejamento Plackett & Burman para a
seleção de 15 parâmetros para produção de ácido láctico L(+) de farelo de trigo,
substrato de baixo custo e suporte sólido, por Lactobacillus amylophilus GV6 em
fermentação de estado sólido (SSF). Onze nutrientes pertencentes a duas categorias,
fontes de nitrogênio e fontes de sal, junto com três parâmetros físicos e um tampão
foram os parâmetros considerados. Os autores verificaram influência na produtividade
devido às fontes de nitrogênio, peptona, extrato de levedura e tri-citrato de amônio,
juntamente com o NaH2PO4.2H2O e Tween 80, que podem ainda serem otimizados para
o aumento na produção de ácido lático. Esses pesquisadores ainda chamam atenção para
o uso escasso deste planejamento em fermentação em estado sólido e que o mesmo
ainda não havia sido considerado anteriormente para a conversão direta de amido em
ácido láctico L (+) usando um sistema bacteriano.
Khosravi-Darani et al. (2008) estudaram a aplicação de Plackett & Burman na
análise dos fatores significativos para a produção de ácido cítrico por Aspergillus niger
ATCC 9142 utilizando palha de trigo como substrato. Os resultados mostraram que a
produção, com a palha de trigo pré-tratada com HCl, NaOH e Uréia, foi mais eficiente e
que os fatores mais significativos na produção foram: Solvente, Teor de Umidade,
Temperatura, Tempo de Incubação, Concentração Inicial de Açúcar e Concentração de
Metanol.
Ji et al. (2009) estudaram o desenvolvimento de um meio de cultura econômico
para a produção do 2,3-butanodiol através da fermentação da glicose e xilose por

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
46
Klebsiella oxytoca utilizando o planejamento Plackett & Burman. Os resultados
mostraram que a uréia e a maceração de milho foram os fatores mais significativos para
a produção do 2,3-butanodiol e que através de planejamento experimental com repetição
em triplicata no ponto central para determinar os níveis ótimos de produção. Os autores
constaram que o rendimento atingido foi 85,6% do valor teórico.
Paula & Arruda. (2009) observaram a necessidade da busca por alternativas mais
eficientes ao método termomecânico utilizado para a degradação de milhões de
toneladas de penas de aves provenientes de frigoríficos e aviários, cuja rígida estrutura é
composta por mais de 90 % de queratina. Para esse fim, utilizaram de comunidades
microbianas para a finalidade proposta, testando o planejamento experimental Plackett
& Burman para verificar a influência de diferentes microrganismos simultaneamente,
com um número reduzido de experimentos. Testaram 32 bactérias isoladas, inoculadas
em meio contendo somente uma pena como única fonte de carbono, durante sete dias,
de onde foi selecionado as cepas para os consórcios microbianos. Experimentos unindo
as cepas NP5, FCA7, NP4 e S14 demonstraram diferentes atividades queratinolíticas,
aumentando o nível de degradação, até 87,8%, comparada com as cepas
individualmente, variando entre 26,4% e 78,0%.
Rajendran & Thangavelu (2009) obtiveram valores máximos da atividade de
produção de lipase em 3,98 U/mL e concentração de massa celular em 5,62 g.L-1 com R.
arrhizus em cultivo submerso descontínuo otimizado pelo planejamento Plackett &
Burman. As variáveis mais importantes que afetaram a produção da lipase foram azeite
de oliva, peptona, KH2PO4, CaCl2.2H2O e MgSO4.7H2O.
Li et al. (2010) estudaram a otimização de meio de cultura para a produção de
Streptolydigin (um antibiótico) por Streptomyces lydicus AS 4,2501 utilizando Plackett
& Burman e a metodologia de superfície de resposta. Os resultados mostraram que
dentre os fatores estudados, os fatores mais significativos para a produção de
Streptolydigin e otimização do meio de cultura foram: Concentração de Amido,
Concentração de Peptona e a Concentração de K2HPO4.
Chen et al. (2010) estudaram a otimização de meio de cultura para a produção da
Luciferase – que são enzimas que catalisam reações biológicas transformando energia
química em energia luminosa - por Bacillus Subtilis em fermentação descontínua
utilizando planejamento Plackett & Burmam. Os resultados mostraram que dentre os
componentes estudados, os fatores mais significativos para a produção e otimização do

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
47
meio de cultura para a produção da Luciferase foram: Concentração de Glicose 1,17 %,
Extrato de Levedura 2,27 % e Glutamato 0,55 %.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o projeto Plackett & Burman e a metodologia de superfície de resposta. Os
resultados mostraram que a eficiência máxima de extração da quitina foi conseguida nas
seguintes condições: Concentração de casca de camarão (50 g/L), glicose (50 g/L),
tempo de incubação de 5 dias e inoculo com 0,05 de OD (densidade óptica).
Gao et al. (2010) desenvolveram um meio para a fermentação do extrato bruto
de inulina de tubérculos de alcachofra de Jerusalém por Paenibacillus polymyxa ZJ-9
para produzir R,R-2,3-butanodiol (R-R,2,3-BD). Inulina, K2HPO4 e NH4Cl foram os
parâmetros principais na fermentação de acordo com os resultados obtidos com o
planejamento experimental Plackett & Burman. A concentração ideal dos três fatores,
77,14 g/L, 3,09 g/L e 0,93 g/L, respectivamente, foi obtida pelo planejamento Box-
Behnken e pela superfície de resposta. Sob as condições ideais, a concentração do R-
R,2,3-BD obtida foi 36,92 g/L, em mais de 98 % de pureza óptica. Comparado com
outras fontes de carbono investigadas, a fermentação do extrato bruto de inulina obteve
o maior rendimento de R-R,2,3-BD.
Uma metodologia de superfície de resposta foi empregada com sucesso para
otimizar um meio de fermentação chamada VHG (Very High Gravity), referência à alta
concentração de açúcares e densidade dos mostos, baseado na água maceração de milho
CSL (Corn Steep Liquor) e outros nutrientes de baixo custo para a produção eficiente de
etanol a partir de glicose por S. cerevisiae. Usando a composição média otimizada (g/L:
CSL 44,3; uréia 2,3; MgSO4.7H2O 3,8; CuSO4.5H2O 0,03), a levedura industrial PE-2
foi capaz de fermentar até 330 g/L de glicose e produzir 18,6 % (v/v) de etanol, com
uma produção descontínua de 2,4 g/L/h e um rendimento em etanol de 93 % (Pereira et
al., 2010).
O planejamento Plackett & Burman juntamente com o Planejamento Composto
Central (CCD) foram aplicados para otimizar o meio de cultura para produção de etanol
por Clostridium autoethanogenum com monóxido de carbono (CO) como fonte única de
carbono e um meio contendo (g/L): 1,0 de NaCl, 0,1 de KH2PO4, 0,02 de CaCl2, 0,15 de
extrato de levedura, 0,116 de MgSO4, 1,694 de NH4Cl e pH 4,74. O rendimento ótimo
de etanol previsto pela metodologia de superfície de resposta (RSM) e um algoritmo

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
48
genético de rede neural artificial (ANN-GA) foram 247,48 e 261,48 mg/L,
respectivamente. Valores semelhantes aos obtidos experimentalmente sob as condições
ideais sugeridas pelos métodos estatísticos (254,26 e 259,64 mg/L). A adequação do
modelo ANN-GA foi maior do que o modelo RSM. Os rendimentos obtidos foram
substancialmente superiores aos relatados anteriormente (60-70 mg/L) com essa cultura
(Guo et al., 2010).
Rigueira (2010) avaliou a influência das variáveis do procedimento de extração
na porcentagem de recuperação das aminas encontradas no leite cru utilizando um
planejamento para “screening” do tipo Plackett & Burman, para os ácidos
tricloroacético (TCA) e sulfosalicílico (ASS).
Quines (2010) estudou a necessidade ou não de se fazer um pré-tratamento da
biomassa antes da extração. Um planejamento experimental Plackett & Burman de doze
ensaios mais três pontos centrais foi utilizado para selecionar as variáveis mais
significativas na extração de Poli(3-hidroxibutirato) (P(3HB)) por 1,2-carbonato de
propileno. Foram realizados dois testes de recuperação do solvente 1,2-carbonato de
propileno, após a extração, sendo um com aplicação de aquecimento e o outro com
aquecimento e vácuo. Os resultados mostraram que o pré-tratamento térmico levou a
uma maior porcentagem de recuperação e de pureza do polímero.

Capítulo 3 Metodologia Experimental

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
50
3. Metodologia Experimental
O presente capítulo apresenta os matérias e métodos utilizados na produção de
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso.
3.1. Quitosana
Neste trabalho utilizou-se uma quitosana (Lote 067) obtida junto a POLYMAR/CE,
com no mínimo 80% de desacetilação. Esse substrato foi utilizado como fonte de carbono e
energia tanto no estudo cinético como nos ensaios do planejamento de seleção de fatores
Plackett & Burman e Planejamento Composto Central, para a produção de quitosanase pelo
Metarhizium anisopliae, por fermentação submersa.
3.2. Microrganismo
O fungo Metarhizium anisopliae estirpe (CG374), utilizado neste estudo, foi cedido
pela Embrapa Recursos Genéticos e Biotecnologia (Brasília / DF-Brasil) e foi mantido em
placas de Petri contendo o meio Batata Dextrose Agar (PDA). As placas foram incubadas em
estufa a 25 ºC. O pH do meio de manutenção foi mantido entre 6,0 e 6,5.
3.2.1. Escolha da cepa produtora de quitosanase
A escolha do microrganismo Metarhizium anisopliae foi feita com base na capacidade
de crescer em meio contendo quitosana como única fonte de carbono, além de apresentar
grande tolerância a pH variando a uma taxa de 2,0 a 8,5, sendo 6,9 a melhor condição para o
crescimento vegetativo e esporução, de acordo com Assis (2010).
3.2.2. Manutenção da cepa
A manutenção da cultura foi realizada em placas petri contendo o meio Batata
Dextrose Agar (PDA). A renovação das células foi realizada a cada trinta dias através de
subcultivos das células no meio PDA.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
51
3.3. Produção de enzima
No preparo do inóculo, utilizou-se 7 placas cultivadas e mantidas, com o fungo
Metarhizium anisopliae, em meio PDA a 25 ºC durante 5 dias em estufa. O meio de cultura
para a produção da enzima quitosanase apresentou os seguintes componentes: quitosana;
peptona; extrato de levedura; NaNO3; K2HPO4; KCl; MgSO4.7H2O e FeSO4, com
concentrações de acordo as matrizes dos planejamentos: Plackett & Burman e composto
central. Para a produção da enzima no cultivo, a biomassa foi transferida através de água
estéril, para um Béquer de 500 mL que, em seguida, foi colocado em Erlenmeyers de 250 mL
na proporção de 20 mL de biomassa para 180 mL do meio de cultura. Os Erlenmeyers
cultivados foram então incubados em incubador rotativo a 25 ºC e 150 rpm durante 24 h.
Alíquotas foram retiradas e centrifugadas a 8000xg por 5 min, a cada 6 horas, para se
determinar a atividade quitosanolítica, teores de açúcares redutores e de proteína total.
Ressalta-se que todos os experimentos foram feitos em triplicata.
3.4. Determinação da Proteína Total
A determinação da proteína total foi efetuada de acordo com o método modificado de
Sedmak & Grossberg (1977), que se baseia na diminuição da absorbância do corante azul de
Coomassie, devido à ligação proteína-corante, a 595 nm, comparado a absorção máxima a
465 nm, conforme Santos (2001). Nesse caso, após centrifugação para retirada da biomassa,
transferia-se para um eppendorf 1,5 mL de amostra (caldo fermentativo) e adicionava-se 1,5
mL de solução azul de Coomassie. Deixava-se a solução em repouso por 5 minutos,
transferindo-se em seguida para um cubeta de quartzo e efetuando-se as leituras de
absorbância nos comprimentos de ondas 465 nm e 595 nm, respectivamente. O
espectrofotômetro (Thermo Spectronic modelo Genesys 10UV) foi zerado com água
destilada. O conteúdo protéico foi obtido a partir de uma curva padrão na qual graficava-se a
relação de absorbância A595/A465 para diferentes concentrações de Albumina de Soro
Bovino (Fração V – Lote 110881/Sigma Aldrich-Ohio-USA).
3.5. Determinação da Atividade Quitosanolítica e da Atividade Específica
A atividade quitosanolítica foi obtida monitorando-se o aumento dos açúcares
redutores totais, utilizando-se o método DNS a partir de uma curva padrão de glicosamina.
Para a determinação da atividade quitosanolítica adicionaram-se, aos tubos de ensaios 500 µL

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
52
de substrato (solução 0,2 % de quitosana solubilizada em ácido clorídrico 1N) e 500 µL de
caldo bruto (contendo enzima), controlando-se o pH entre 6,0 e 6,5 e incubando-se o sistema
em um banho termostatizado com temperaturas da ordem de 55 ºC. Após o tempo de
incubação de 30 minutos no banho foi adicionado aos tubos de ensaio 2,5 mL de DNS,
aquecendo-se à ebulição em banho-maria por 10 minutos, resfriando-se, em seguida, em
banho de gelo. Após essa etapa, o sistema foi centrifugado e com o sobrenadante efetuou-se a
leitura no espectrofotômetro a 600 nm. Destaca-se, que antes da leitura da amostra zerava-se o
equipamento (branco do espectro) com 1,0 mL de água destilada acrescidos de 2,5 mL de
DNS e realizava-se o branco da enzima que consistia da adição de 500 µL de solução de
quitosana, completando-se até um 1,0 mL com água destilada. Uma unidade de atividade
quitosanolítica (A) foi definida como a quantidade de enzima necessária para produzir 1
µmol.min-1 de glicosamina por mL de extrato enzimático, nas condições de temperatura e pH
citadas, sendo o resultado expresso em (U.mL-1). A Atividade específica foi determinada pela
divisão entre a atividade da enzima (U.mL-1) e a concentração de proteína total (µg.mL-1),
sendo expressa em (U. µg-1).
3.6. Planejamento experimental
3.6.1. Planejamento de seleção de fatores Plackett & Burman
O planejamento experimental Plackett & Burman é uma técnica muito usada, para se
realizar triagem de fatores na otimização num determinado processo (Reddy et al., 2008), e
foi utilizado neste trabalho para identificar os fatores que influenciam de forma significativa
na produção da enzima quitosanase.
A Tabela 3 ilustra os níveis de cada variável (fator) utilizada no planejamento Plackett
& Burman, sendo: Concentração de Quitosana (Q), Peptona (PP), Extrato de Levedura (EL),
NaNO3 (NS), K2HPO4 (FP), KCl (CP), MgSO4.7H2O (SM), FeSO4 (SF). Já a Tabela 4 mostra
a matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3 repetições
na condição do ponto central.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
53
Tabela 3 – Níveis para os fatores e seus valores codificados.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1 0,1 0,2 0,2 0,1 0,05 0,025 0,025 0,0005
0 0,2 0,5 0,3 0,2 0,10 0,050 0,050 0,0010
1 0,3 0,8 0,4 0,3 0,15 0,075 0,075 0,0015
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1
2 1 1 -1 1 -1 -1 -1 1 -1 1 1
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1
5 1 1 -1 1 1 -1 1 -1 1 -1 1
6 1 1 1 -1 1 1 -1 1 1 -1 -1
7 -1 1 1 1 -1 1 1 -1 -1 1 1
8 -1 -1 1 1 1 -1 1 1 1 1 1
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1
11 -1 1 -1 -1 -1 1 1 1 1 1 -1
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1
13 0 0 0 0 0 0 0 0 0 0 0
14 0 0 0 0 0 0 0 0 0 0 0
15 0 0 0 0 0 0 0 0 0 0 0
Sendo: V11,V12,V13 = Variáveis inertes para cálculo do erro.
3.6.2. Planejamento Composto Central para a Otimização do Meio de Cultura para a
Produção da Enzima em Cultivo Submerso
Um dos problemas mais comuns para quem realiza experimentos é determinar a
influência de uma ou mais variáveis sobre outra variável de interesse (Lobato, 2003). Este

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
54
problema pode ser minimizado utilizando-se um planejamento experimental de modo a se
obter uma função, ou uma aproximação satisfatória para a mesma, de forma que se operando
sobre as variáveis de entrada produza-se como saída a resposta ou as respostas observadas
(Barros Neto, 2001; Silva, 2004).
Em um planejamento, inicialmente, deve-se definir os fatores e as respostas de
interesse. Tendo-se identificado os fatores e respostas define-se o objetivo que se pretende
alcançar com os experimentos para que se possa escolher o planejamento mais apropriado.
(Silva, 2004).
No presente trabalho foi utilizado um Planejamento Composto Central (CCD) com 22
experimentos na parte fatorial, repetição em triplicata no ponto central e quatro experimentos
na parte axial (planejamento do tipo estrela), para se investigar a influência dos fatores:
concentração de quitosana e concentração de FeSO4 que foram os dois fatores que
influenciaram a produção de quitosanase, conforme o planejamento Plackett & Burman de 12
ensaios e mais 3 repetições na condição do ponto central. Estes dois fatores foram estudados
em cinco diferentes níveis (-1,41; -1; 0; 1; 1,41) e um conjunto de 11 experimentos foi
realizado, conforme Tabela 5.
Tabela 5 – Matriz do planejamento Composto Central.
Ensaio Q SF
1 -1 -1
2 -1 1
3 1 -1
4 1 1
5 -1,41 0
6 1,41 0
7 0 -1,41
8 0 1,41
9 0 0
10 0 0
11 0 0
Os outros fatores foram mantidos nos níveis mais baixos (para evitar custos). Uma que
não influenciaram de forma significativa na produção da enzima quitosanase.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
55
A codificação dos fatores foi realizada de acordo com a seguinte equação (Ji et al.,
2009):
Xi = (xi – x0)/∆xi , i = 1, 2, 3, ..., k (2)
Onde Xi é o fator independente codificado, xi é o fator independente real, x0 é o valor de xi no
ponto central e ∆xi é a variação de valor. O comportamento do sistema foi explicado pela
seguinte equação polinomial de segunda ordem:
Yp = β0 + ∑ βiXi + ∑ βiiXi2 + ∑ βijXiXj, i, j = 1, 2, 3,..., k (3)
Onde Yp foi a resposta esperada, β0 foi a interseção, Xi e Xj
foram os fatores independentes codificados, βi foi o coeficiente linear, βii
foi o coeficiente quadrático e βij foi o coeficiente de interação. Destaca-se que os outros
fatores não significativos foram mantidos no nível mais baixo (-1).
A Tabela 6 apresenta os níveis para os fatores e seus valores codificados, para o
Planejamento Composto Central, com triplicata no ponto central.
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1,41 0,218 0,2 0,2 0,1 0,05 0,025 0,025 0,0009
-1 0,3 0,2 0,2 0,1 0,05 0,025 0,025 0,0015
0 0,5 0,2 0,2 0,1 0,05 0,025 0,025 0,0030
1 0,7 0,2 0,2 0,1 0,05 0,025 0,025 0,0045
1,41 0,782 0,2 0,2 0,1 0,05 0,025 0,025 0,0051
Na análise do planejamento de seleção de fatores Plackett & Burman e do
planejamento experimental fatorial foi utilizado um pacote computacional STATISTICA for
Windows versão 7.0.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
56
3.7. Perfil cinético durante o cultivo
Um perfil cinético na melhor condição de cultivo, conforme planejamento
apresentado, foi realizado em frascos Erlenmyer de 250 mL contendo 200 mL de meio de
cultivo contendo os seguintes componentes, em 1 Litro: 2,0g de quitosana; 5,0g de peptona;
3,0g de extrato de levedura; 2,0g de NaNO3; 1,0g de K2HPO4; 0,5g de KCl; 0,5g de
MgSO4.7H2O; 0,01g de FeSO4, que foram inoculadas com 10% (vol/vol) de inóculo contendo
o fungo Metarhizium anisopliae e cultivadas em incubador rotativo durante 5 dias a 150 rpm
e 25 ºC. Alíquotas foram retiradas a cada seis horas e analisadas quanto à proteína total no
sobredanante, atividade enzimática, atividade específica e açúcares redutores. Os ensaios
foram realizados em triplicata (Silva Filho, 2005).

Capítulo 4 Resultados e
Discussão

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
58
4. Resultados e discussão
Neste capítulo são apresentados e discutidos os resultados obtidos no presente
trabalho. Inicialmente, é apresentada imagem do MEV do Metarhizium anisopliae
mantidos em meio PDA. Em seguida, apresenta-se e discute-se os resultados da avaliação
dos fatores significativos, que podem influenciar na produção da quitosanase, no
planejamento de seleção de fatores, conforme planejamento Plackett & Burman bem como
são apresentados e discutidos os resultados obtidos para o planejamento composto central.
Por último, apresenta-se o perfil cinético, na melhor condição, para a produção de
quitosanase em cultivo submerso descontínuo usando o fungo Metarhizium anisopliae.
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA.
A Figura 7 ilustra a imagem da Microscopia Eletrônica de Varredura (MEV) do micélio de Metarhizium anisopliae mantido em meio PDA.
Figura 7. Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA.
Observa-se na Figura 7, quanto à morfologia que o Metarhizium anisopliae, apresenta-
se como um fungo filamentoso com as hifas intimamente entrelaçadas conforme citado por
Wang et al. 2002.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
59
4.2. Planejamento Plackett & Burman
4.2.1 Análise do Plackett & Burman com 8 fatores e 12 ensaios
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis, foi utilizado para avaliar a influência dos fatores: Concentração de
Quitosana (Q), Peptona (PP), Extrato de Levedura (EL), NaNO3 (NS), K2HPO4 (FP), KCl
(CP), MgSO4.7H2O (SM), FeSO4 (SF) na produção da enzima quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso. Ressalta-se que os experimentos foram
realizados aleatoriamente.
A Tabela 7 apresenta a matriz do modelo Plackett & Burman e os respectivos valores
obtidos para a variável resposta (Atividade Quitosanalítica).
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.
Ensaio Q PP EL NS FP CP SM SF A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 0,0195
2 1 1 -1 1 -1 -1 -1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 0,0109
4 1 -1 1 1 -1 1 -1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 0,0136
6 1 1 1 -1 1 1 -1 1 0,0156
7 -1 1 1 1 -1 1 1 -1 0,0109
8 -1 -1 1 1 1 -1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 0,0085
A Tabela 8 apresenta os efeitos estimados para o planejamento Plackett & Burman.
Observa-se na Tabela 8 que o valor do coeficiente de determinação R2 foi de
aproximadamente 90%, indicando que um modelo linear poderia representar bem a relação
entre o fator significativo, concentração de quitosana (1), e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
60
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman.
Efeitos Estimados; R2=0,8996; Ajuste:0,63178 MS Erro Residual =0,0000054
Desvio Padrão t(3) p
Lim. Conf. Lim. Conf.
Efeito Erro Residual
-95,% +95
Média/Interação 0,0127 0,0007 18,903 0,0003 0,0105 0,0149 (1) Quitosana 0,0055 0,0013 4,0628 0,0268 0,0011 0,0098
(2) Peptona 0,0015 0,0013 1,1080 0,3486 -0,0028 0,0058 (3) Ext. de leved. 0,0009 0,0013 0,7387 0,5136 -0,0033 0,0053
(4) NaNO3 0,0002 0,0013 0,1758 0,8715 -0,0040 0,0045 (5) K2HPO4 -0,0014 0,0013 -1,0552 0,3687 -0,0057 0,0028
(6) KCl -0,0022 0,0013 -1,6708 0,1933 -0,0065 0,0020 (7) MgSO4.7H2O -0,0003 0,0013 -0,2286 0,8338 -0,0046 0,0039
(8) FeSO4 0,0029 0,0013 2,1457 0,1211 -0,0014 0,0071
A Figura 8 mostra o gráfico de Pareto, uma das formas de se avaliar visualmente a
influência dos fatores estudados na resposta. A magnitude dos efeitos é representada pelas
colunas enquanto que a linha transversal às colunas representa a magnitude dos efeitos com
significado estatístico para p=0,05, ou seja, os fatores que são estatisticamente significativos
ao nível de 95% de confiança.
Figura 8. Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
61
Analisando-se o gráfico de Pareto (Figura 8) observa-se que o fator concentração de
quitosana (1) foi o único fator que influenciou de forma mais significativa na atividade
enzimática atingindo efeitos estimados de aproximadamente 4,06. O sinal positivo indica que
o aumento da concentração de quitosana aumentará à atividade enzimática. Por outro lado, os
outros fatores parecem desprezíveis estatisticamente conforme o gráfico de Pareto.
4.2.2 Análise do Plackett & Burman com 12 Ensaios e mais 3 Repetições na condição do
ponto central.
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis com repetição em triplicata no ponto central, foi utilizado para se avaliar a
influência dos mesmos fatores descritos na Seção 4.2.1 para a produção da enzima
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso. Ressalta-se que os
experimentos foram realizados aleatoriamente. A Tabela 9 apresenta a matriz do modelo
Plackett & Burman e os respectivos valores obtidos para a variável resposta (Atividade
Quitosanalítica).
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13 A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1 0,0195
2 1 1 -1 1 -1 -1 -1 1 -1 1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1 0,0109
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 1 -1 1 0,0136
6 1 1 1 -1 1 1 -1 1 1 -1 -1 0,0156
7 -1 1 1 1 -1 1 1 -1 -1 1 1 0,0109
8 -1 -1 1 1 1 -1 1 1 1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 1 1 -1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1 0,0085
13 0 0 0 0 0 0 0 0 0 0 0 0,0704
14 0 0 0 0 0 0 0 0 0 0 0 0,0688
15 0 0 0 0 0 0 0 0 0 0 0 0,0710

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
62
A Tabela 10 a seguir apresenta os efeitos estimados para planejamento Plackett &
Burman com 3 repetições na condição do poto central. Observa-se na Tabela 10 que o valor
do coeficiente de determinação R2 foi de aproximadamente 2,0%, indicando que um modelo
linear não poderia representar a relação entre os fatores significativos concentração de
quitosana (1), concentração de FeSO4 (8) e a resposta.
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central.
Efeitos Estimados; R2=0,01811; Ajuste:0,0 MS Erro Puro =0,0000012
Desvio Padrão t(2) P
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0242 0,0003 85,509 0,0001 0,0229 0,0254 (1) Quitosana 0,0055 0,0006 8,6432 0,0131 0,0027 0,0082
(2) Peptona 0,0015 0,0006 2,3572 0,1425 -0,0012 0,0042 (3) Ext. de leved. 0,0009 0,0006 1,5715 0,2567 -0,0017 0,0037
(4) NaNO3 0,0002 0,0006 0,3742 0,7442 -0,0024 0,0029 (5) K2HPO4 -0,0014 0,0006 -2,2450 0,1538 -0,0041 0,0013
(6) KCl -0,0022 0,0006 -3,5546 0,0708 -0,0049 0,0005 (7) MgSO4.7H2O -0,0003 0,0006 -0,4864 0,6747 -0,0030 0,0024
(8) FeSO4 0,0029 0,0006 4,5648 0,0448 0,0002 0,0056
A Figura 9 ilustra o gráfico de Pareto para o planejamento Plackett & Burman, com
repetição em triplicata no ponto central, uma das formas de se avaliar visualmente a influência
dos fatores estudados na resposta, onde é mostrado a magnitude dos efeitos para 95 % de
confiança.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
63
Figura 9. Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto
central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto, observa-se que os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma mais
significativa na atividade enzimática atingindo efeitos estimados de aproximadamente 8,84 e
4,67, respectivamente. O sinal positivo indica que o aumento das concentrações de quitosana
e de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores parecem
desprezíveis estatisticamente conforme o gráfico de Pareto.
Este comportamento do FeSO4 está de acordo com o observado por Hagag et al.
(2012) que utilizando o Aspergillus fumigatus na produção de protease mostraram que um
fator de transcrição PrtT regula positivamente o controle da produção de protease, da
assimilação de Ferro e da síntese de ergosterol e que a produção de protease foi favorecida
pelo aumento da concentração de ferro.
Rajendran & Thangavelu (2009) utilizaram um planejamento Plackett & Burman para
identificar quais os fatores significativos na produção de lipase por Rhizopus arrhizus MTCC
2233 e observaram o efeito significativo médio do FeSO4, apresentando um nível de
confiança para esse fator de 48,92% (para um máximo de 98,7 % de MgSO4), com uma
produção de lipase da ordem de 2,98 U/mL em 84 horas de fermentação e que ocorreu uma
produção considerável de protease no meio de cultivo da ordem de 1,92 U/mL em 96 horas de
cultivo.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
64
Mu et al. (2005) também observaram um efeito positivo do FeSO4, da ordem de 1,48
para um fator máximo encontrado de 3,11 de MgSO4, no aumento da atividade da
estereoinversão de S-fenil-1,2-etanodiol por Candida parapsilosis CCTCC M203011 na
otimização da fermentação utilizando um planejamento Plackett & Burman.
Dessa forma, observa-se que o FeSO4 é importante uma que vez o mesmo está
diretamente associado a síntese de proteases (principalmente metalloproteases), conforme
citado por Hagag et al. (2012).
4.3. Planejamento Composto Central para Produção de Quitosanase por
Metarhizium anisopliae em Cultivo Descontínuo Submerso
Um planejamento experimentos 22 com repetição em triplicata no ponto central e mais
quatro ensaios axiais, foram utilizados para se chegar a uma combinação ideal de quitosana e
FeSO4 na produção da enzima quitosanase (Tabela 11). A partir do modelo Plackett &
Burman (Tabela 12), que mostrou que o aumento das concentrações de quitosana e FeSO4,
favoreceram a produção de quitosanase, os valores utilizados neste modelo para estes
componentes nos níveis mais altos, serão considerados como os níveis mais baixos no
planejamento experimental do composto central. Os outros fatores foram fixados em todos os
ensaios em seus níveis mais baixos para a redução de custos (Tabela 11).

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
65
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da
atividade enzimática.
Ensaio Q SF A (U/mL)
1 -1 -1 0,0195
2 -1 1 0,0202
3 1 -1 0,0109
4 1 1 0,0121
5 -1,41 0 0,0136
6 1,41 0 0,0156
7 0 -1,41 0,0108
8 0 1,41 0,0103
9 0 0 0,0099
10 0 0 0,0116
11 0 0 0,0094
A Tabela 12 apresenta os efeitos estimados para o planejamento composto central.
Tabela 12 – Efeitos Estimados do planejamento Composto Central.
Efeitos Estimados; R2=0,629; Ajuste:0,25836 MS Erro Puro =0,0000014
Desvio Padrão t(2) p
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0103 0,0006 15,305 0,0042 0,0074 0,0132 (1) Quitosana (L) 0,0011 0,0008 1,4316 0,2885 -0,0023 0,0047
Quitosana (Q) 0,0058 0,0009 5,9637 0,0269 0,0016 0,0101 (2) FeSO4 (L) -0,0043 0,0008 -5,3037 0,0337 -0,0079 -0,0008 (2) FeSO4 (Q) 0,0018 0,0009 1,8366 0,2076 -0,0024 0,0060
1L x 2L 0,0001 0,0011 0,1520 0,8931 -0,0048 0,0052
Observa-se na Tabela 12 que o valor do coeficiente de determinação R2 foi de
aproximadamente 63%, indicando que um modelo linear representaria bem a relação entre os
fatores significativos concentração de quitosana (1), concentração de FeSO4 (8) e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
66
A Tabela 13 mostra um resumo da ANOVA para o planejamento composto central
com repetição em triplicata no ponto central.
Tabela 13 – Resumo da ANOVA.
Fonte de
Variação
Soma dos
Quadrados
Grau de
Liberdade
Quadrado
Médio
Fcalculado Ftabelado
Regressão
Resíduos
0,000086 2 0,000043 6,14 4,46
0,000056 8 0,000007
Falta de
Ajuste
Erro puro
0,000053 6 0,000009 6 19,33
0,000003
2
0,0000015
Total 0,000142 10
De acordo com a Tabela 13, partindo-se dos efeitos significativos propõe-se o
modelo:
� = 0,010316 + 0,002942� � − 0,002193�� (8)
% de variação explicável: 60,5
% máxima de variação explicável: 97,9
A análise da ANOVA mostra que o modelo proposto possui significância estatística, ao
nível de 95% de confiança, uma vez que o valor de F (calculado) é maior que F (tabelado),
entretanto a ANOVA mostra também que o modelo proposto pode ser útil para se fazer
previsões, uma vez que a relação entre F (calculado) e F (tabelado) para a falta de ajuste e o
erro puro é menor que 1. O coeficiente de determinação R2 foi de aproximadamente 63%,
indicando que o modelo consegue explicar 63% da variação total em torno da média.
Os cálculos para variação explicável e para máxima variação explicável são dados pelas
Equações (9) e (10):
% de variação explicável = (SQ da regressão) / (SQ Total) (9)
% máxima de variação explicável = (SQ Total – SQ Erro puro) / (SQ Total) (10)
Sendo:
SQ – Soma dos quadrados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
67
A Figura 10 mostra o gráfico de Pareto para o planejamento Composto Central
utilizando erro puro. A magnitude dos efeitos é representada pelas colunas. Já a linha
transversal às colunas representa a magnitude dos efeitos com significado estatístico para 95%
de confiança.
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto (Figura 10) observa-se que os fatores concentração
de quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma
mais significativa na atividade enzimática atingindo efeitos estimados de aproximadamente
5,96 e - 5,30, respectivamente. O sinal positivo indica que o aumento das concentrações de
quitosana aumentará à atividade enzimática. O sinal negativo indica que o decréscimo da
concentração de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores
parecem desprezíveis estatisticamente no gráfico de Pareto.
A avaliação do modelo também pode ser feita através da observação do gráfico dos
valores preditos em função os valores observados que são mostrados na Figura 11. Os valores
preditos pelo modelo são representados pela reta, enquanto que os valores observados
representam-se pelos pontos.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
68
Figura 11 – Valores preditos em função dos valores observados.
Observa-se que alguns valores preditos apresentam um afastamento dos valores
observados. A Figura 12 ilustra a superfície de resposta da atividade quitosanolítica em
função dos dois fatores significativos, concentração de quitosana e concentração de FeSO4.
Figura 12. Atividade quitosanolítica em função da concentração de quitosana e da
concentração de FeSO4 para os fatores não significativos no ponto central.
Observa-se a existência de um ponto de mínimo, ocorrendo para concentração de
quitosana no ponto central (0) e para maiores concentrações de FeSO4 igual a (1,5). Essa

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
69
região deve ser evitada uma vez que a atividade quitosanolítica atinge níveis tão reduzidos
quanto 0,0094 U.mL-1. Destaca-se que nesse planejamento o maior nível da concentração de
FeSO4 (α=+1,41) equivale a 0,0051% (Tabela 6) que é aproximadamente cinco vezes maior
que o nível desse fator no ponto central do planejamento Plackett & Burman (0,001%),
conforme Tabela 3. Dessa forma, ao se utilizar o nível mínimo da concentração de FeSO4 no
Planejamento Composto Central (α=-1,41), que equivale a aproximadamente 0,001%,
aproxima-se do ponto central do do planejamento Plackett & Burman (0,001%), condição
essa que apresenta os melhores resultados de atividade quitosanolítica usando quitosana como
fonte de carbono e energia e fungo Metarhizium anisopliae.
4.4. Perfil Cinético – Melhor Condição
A seguir, apresenta-se um perfil cinético, na melhor condição, para produção da
quitosanase usando o fungo Metarhizium anisopliae, em que foram monitorados durante 24
horas os seguintes parâmetros: Concentração de proteínas totais (µg/mL), atividade
quitosanolítica (U/mL), atividade específica (U/µg) e açúcares redutores (mg/mL), ilustradas
nas Figuras 13, 14, 15 e 16, respectivamente.
Com relação à proteína extracelular, observa-se na Figura 13 que, inicialmente, ocorre
um aumento da mesma para o seu nível máximo (6,2 µg/mL), durante as 6 primeiras horas de
cultivo, seguido de uma redução para 5,7 µg/mL em 18 horas de cultivo, possivelmente
devido à presença de proteases no meio. Observa-se que a partir de então ocorre uma redução
e aumento da proteína total, em uma quantidade inferior, sugerindo um mecanismo de
controle.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
70
0 4 8 12 16 20 245,0
5,2
5,4
5,6
5,8
6,0
6,2
6,4
Pro
tein
a (
µg
/mL
)
Tempo (h)
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana.
Com relação à atividade quitosanolítica, conforme Figura 14, a atividade máxima
ocorreu em 18 horas de produção em que se obteve aproximadamente 70,0 U/L, o que foi 28
vezes maior que o valor (2,5 U/L em 48 horas) relatado por Assis (2010) para o mesmo fungo,
e em apenas um terço do tempo de produção.
0 4 8 12 16 20 240,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,10
Ativid
ade
(U
/mL
)
Tempo (h)
Figura 14. Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.
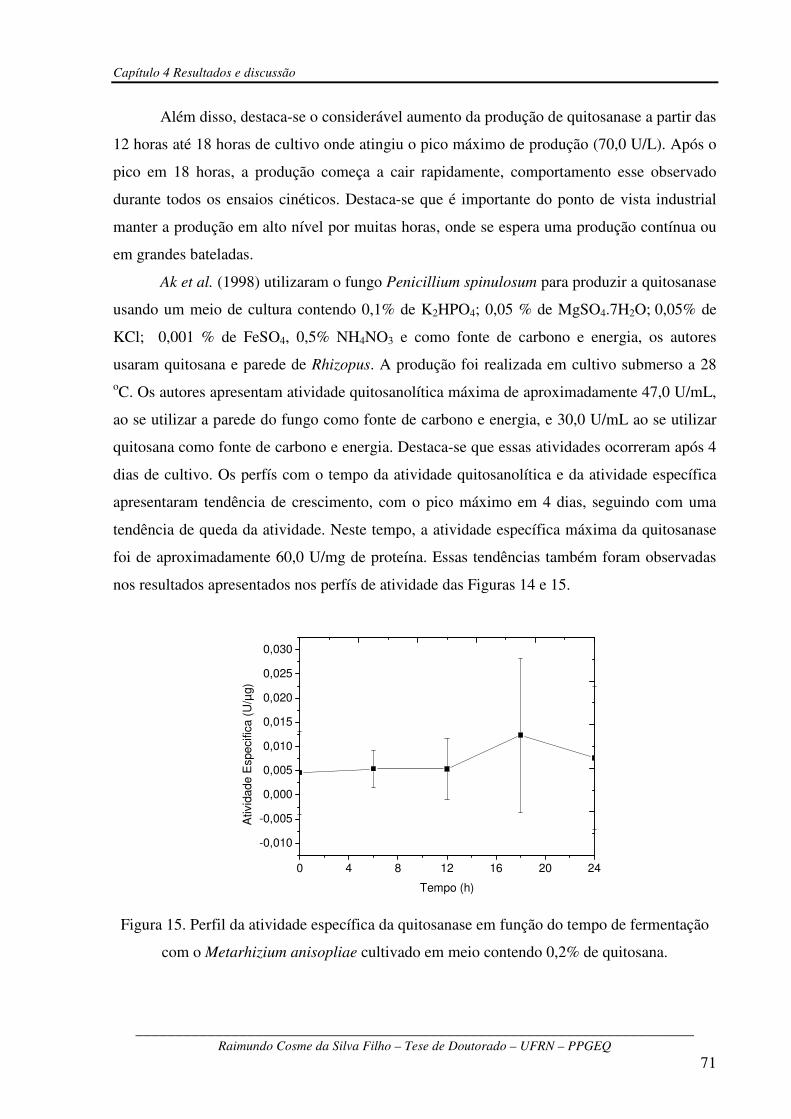
Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
71
Além disso, destaca-se o considerável aumento da produção de quitosanase a partir das
12 horas até 18 horas de cultivo onde atingiu o pico máximo de produção (70,0 U/L). Após o
pico em 18 horas, a produção começa a cair rapidamente, comportamento esse observado
durante todos os ensaios cinéticos. Destaca-se que é importante do ponto de vista industrial
manter a produção em alto nível por muitas horas, onde se espera uma produção contínua ou
em grandes bateladas.
Ak et al. (1998) utilizaram o fungo Penicillium spinulosum para produzir a quitosanase
usando um meio de cultura contendo 0,1% de K2HPO4; 0,05 % de MgSO4.7H2O; 0,05% de
KCl; 0,001 % de FeSO4, 0,5% NH4NO3 e como fonte de carbono e energia, os autores
usaram quitosana e parede de Rhizopus. A produção foi realizada em cultivo submerso a 28
oC. Os autores apresentam atividade quitosanolítica máxima de aproximadamente 47,0 U/mL,
ao se utilizar a parede do fungo como fonte de carbono e energia, e 30,0 U/mL ao se utilizar
quitosana como fonte de carbono e energia. Destaca-se que essas atividades ocorreram após 4
dias de cultivo. Os perfís com o tempo da atividade quitosanolítica e da atividade específica
apresentaram tendência de crescimento, com o pico máximo em 4 dias, seguindo com uma
tendência de queda da atividade. Neste tempo, a atividade específica máxima da quitosanase
foi de aproximadamente 60,0 U/mg de proteína. Essas tendências também foram observadas
nos resultados apresentados nos perfís de atividade das Figuras 14 e 15.
0 4 8 12 16 20 24
-0,010
-0,005
0,000
0,005
0,010
0,015
0,020
0,025
0,030
Ativid
ad
e E
spe
cific
a (
U/µ
g)
Tempo (h)
Figura 15. Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
72
Observa-se na Figura 15 que a atividade específica máxima da quitosanase segue a
mesma tendência da atividade quitosanolítica apresentada na Figura 14, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
O perfil para os açúcares redutores durante o cultivo de Metarhizium anisopliae
cultivado em meio contendo 0,2% de quitosana é ilustrado na Figura 16.
0 4 8 12 16 20 240,004
0,008
0,012
0,016
0,020
0,024
0,028
Acuca
res R
ed
uto
res (
mg/m
L)
Tempo (h)
Figura 16. Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana.
Observa-se, na Figura 16, que taxa máxima de redução dos açúcares redutores
(aproximadamente 0,021 mg/mL) ocorre logo após a produção máxima da enzima, em 18
horas de cultivo, conforme Figura 14. A quantidade de açúcares redutores foi a
aproximadamente 0,012 mg/mL após 24 h de cultivo. O fungo excreta para o meio enzimas
com atividade quitosanolítica, essas enzimas hidrolisam a quitosana aumentando a quantidade
de açúcares redutores para o meio, sendo que esses açúcares também são consumidos pelo
fungo durante seu crescimento.
Foi demonstrado por Assis (2010) que os açúcares redutores produzidos através da
quitossana, ocasionada pelas enzimas quitossanases metabolizadas por Metarhizium
anisopliae, com quitosana como sua única fonte de carbono e energia, atingiu um pico de 0,53
mg/mL em 48 horas. Apresentaram como quitooligossacarídeos, constituídos principalmente
por dímeros e pentâmeros, formados pela hidrólise enzimática de quitosanase em quantidades
maiores do que o consumido pelos microrganismos. Mostraram, também, que essa produção

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
73
segue a mesma tendência da sua atividade enzimática, corroborando com os resultados
apresentados no presente trabalho.
Os resultados do presente trabalho mostram a relevância de se adotar novas condições
para a produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter quitooligossacarídeo
com atividades biológicas. Esta pesquisa, portanto, sugere base para novos estudos que
permitam alcançar rapidamente a produção de quitosanase e quitooligossacarídeo com todos
os seus benefícios.

Capítulo 5 Conclusão

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
75
5. Conclusão
Após o desenvolvimento desse trabalho conclui-se:
- Com relação ao Planejamento de Seleção de Fatores Plackett & Burman:
Os resultados das análises do Planejamento de Seleção de Fatores Plackett &
Burman com 8 fatores e 12 ensaios em 2 níveis, mais 3 repetições no ponto central, para
se investigar a influência dos fatores: Concentração de Quitosana, Peptona, Extrato de
Levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O, FeSO4 na produção da enzima
quitosanase por Metarhizium anisopliae, mostraram que a atividade foi favorecida pelo
aumento da concentração de substrato (quitosana) e sulfato ferroso (FeSO4), enquanto
que o aumento da concentração dos outros fatores não contribuíram de forma
significativa para a resposta (atividade quitosanolítica). Os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) atingiram efeitos estimados de
aproximadamente 8,84 e 4,67, respectivamente. O sinal positivo indica que o aumento
das concentrações de quitosana e de FeSO4 aumentará a atividade enzimática. Conforme
visto na literatura, observa-se importância do FeSO4, já que o mesmo está diretamente
associado a síntese de proteases (principalmente metalloproteases),
- Com relação ao Planejamento Composto Central:
A análise do planejamento composto central para avaliar a influência dos
fatores, concentração de substrato (quitosana) e concentração de FeSO4, na produção da
enzima quitosanase por Metarhizium anisopliae (a partir do planejamento Plackett &
Burman) mostrou que os fatores concentração de quitosana e concentração de FeSO4
foram os fatores que influenciaram de forma mais significativa na atividade enzimática
atingindo efeitos estimados de aproximadamente 5,96 e - 5,30, respectivamente. O sinal
positivo indica que o aumento das concentrações de quitosana aumentará a atividade
enzimática. O sinal negativo indica que o decréscimo da concentração de FeSO4
aumentará a atividade enzimática. Observa-se a existência de um ponto de mínimo,
ocorrendo para concentração de quitosana (X1) no ponto central (0) e para maiores

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
76
concentrações de FeSO4 (X2) igual a (1,5) com a atividade quitosanolítica de 0,0094
U.mL-1.
- Com relação ao cultivo na melhor condição:
O cultivo na melhor condição mostrou uma produção máxima de quitosanase em
18 horas de cultivo, com a atividade enzimática de 70,0 U/L, o que foi 28 vezes maior
que o valor (2,5 U/L em 48 horas) relatado na literatura (Assis, 2010) para o mesmo
fungo e em apenas um terço do tempo de produção.
Além disso, observou-se que um elevado incremento na produção de
quitosanase a partir das 12 horas de cultivo, antes de se atingir a atividade
quitosanolítica máxima, 18 horas de cultivo. Essa produção em alto nível por muitas
horas é importante do ponto de vista industrial, onde se espera uma produção contínua
ou em grandes bateladas. Observou-se também que a atividade específica máxima da
quitosanase segue a mesma tendência da atividade quitosanolítica, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
- Com relação aos açúcares redutores:
Foi observado que os açúcares redutores seguem a mesma tendência da atividade
enzimática, com uma produção máxima de cerca de 0,21 mg/mL em 18 horas de
cultivo, produção essa que coincinde com o máximo pico de atividade enzimática.
Estes resultados mostram a relevância de se adotar novas condições para a
produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter
quitooligossacarídeo com atividades biológicas. Esta pesquisa, portanto, sugere base
para novos estudos que permitam alcançar rapidamente a produção de quitosanase e
quitooligossacarídeo com todos os seus benefícios.

Referências Bibliográficas

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
78
Referências bibliográficas
AK, Ö.; BAKIR, U.; GÜRAY, T. Isolation of a chitosan degrading fungus, Penicillium
spinulosum, and chitosanase production by the isolate. In: SYMPOSIUM - NEW
FRONTIERS IN SCREENING FOR MICROBIAL BIOCATALYSTS, 1996, Ede, The
Netherlands. Anais, Editado por K. Kieslich, C. P. van der Beek, J. A. M. de Bont e W.
J. J. van den Tweel, Elsevier Science B. V., 1998, v.53, p.265-268.
ALVES, S. B. Patologia e controle microbiano: Vantagens e desvantagens. In: ALVES,
S. B. Controle Microbiano de Insetos. 2ed., Piracicaba, FEALQ, 1163p, 1998, v.4, p.21-
37.
ALVES, S. B.; LEITE, L. G.; FILHO, A. B.; ALMEIDA, F. E. M.; MARQUES, E. J.
Produção massal de fungos entomopatogênicos na América Latina, In: ALVES, S. B.;
LOPES, R. B. Controle microbiano de pragas na América Latina: avanços e desafios.
Piracicaba, FEALQ, 414p, 2008, v.14, p.215-237.
ARRUDA, W.; LUBECK, I.; SCHRANK, A.; VAINSTEIN, M. H. Morphological
alterations of Metarhizium anisopliae during penetration of Boophilus microplus ticks.
Experimental and Applied Acarology, v. 37, p.231-244, 2005.
ASSIS, C. F. Produção e caracterização de quitooligossacarídeos produzidos pelo fungo
Metarhizium anisopliae e avaliação da citotoxicidade em células tumorais. 2010. 110p.
Tese (Doutorado em Biotecnologia) – Renorbio - Programa de Pós-Graduação em
Biotecnologia. Universidade Federal do Rio Grande do Norte, Natal.
ASSIS, C. F.; COSTA, L. S.; MELO-SILVEIRA, R. F.; OLIVEIRA, R. M.;
PAGNONCELLI, M. G. B.; ROCHA, H. A. O.; MACEDO, G. R.; SANTOS, E. S.,
Chitooligosaccharides antagonizes the cytotoxic effect of glucosamine. World J.
Microbiol. Biotechnol., v.28, n.3, p.1097-1105, 2012.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
79
AZEVEDO, V. V. C.; CHAVES, S. A.; BEZERRA, D. C.; FOOK, M. V. L.; COSTA,
A. C. F. M. Quitina e Quitosana: aplicações como biomateriais. Revista Eletrônica de
Materiais e Processos, v.2.3, p.27-34, 2007.
BARROS NETO, B.; SCARMINIO, I. S.; BRUNS, R. Como fazer experimentos:
pesquisa e desenvolvimento na ciência e na indústria. 2. ed. Campinas: Unicamp, 2001.
401p.
BATISTA FILHO, A.; ALMEIDA, J. E. M.; SANTOS, A. S.; MACHADO, L. A.;
ALVES, S. B. Eficiência de isolados de Metarhizium anisopliae no controle de
cigarrinha-da-raiz de cana-de-açúcar Mahanarva fimbriolata (Hom.: Cercopidae). STAB
– Açúcar, Álcool e Subprodutos, v.21, p.84-89, 2002.
CARVALHO, J. C. M.; SATO, S. Fermentação Descontínua, In: SCHMIDELL, W.;
LIMA, U. A.; AQUARONE, E.; BORZANI, W. Biotecnologia Industrial. São Paulo,
Ed. Edgard Blücher, v.2, p.193-204, 2001.
CHAO, H. J.; SCHWARTZ, J.; MILTON, D. K. Populations and determinats of
airborne fungi in large office buildings. Environmental Health Perspectives, v.110, n.8,
p.777-782, 2002.
CHEN, X.; XIA, W.; YU, X. Purification and characterization of two types of
chitosanase from Aspergillus sp. CJ22-326. Food Research International, v.38, n.3,
p.315-322, 2005.
CHEN, P. T.; CHIANG, C.-J.; CHAO, Y.-P. Medium optimization and production of
secreted Renilla luciferase in Bacillus subtilis by fed-batch fermentation. Biochemical
Engineering Journal, v.49, p.395-400, 2010.
CHENG, C. Y.; LI, Y. An Aspergillus chitosanase with potential for large-ecale
preparation of chitosan oligosaccharides. Biotechnol. Appl. Biochem., v.32, p.197-203,
2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
80
CHOI, Y. J.; Kim, E. J.; Piao, Z.; Yun, Y. C.; Shin, Y. C. Purification and
characterization of chitosanase from Bacillus sp. Strain KCTC 0377BP and its
application for the production of chitosan oligosaccharides. Applied and Environmental
Microbiology, v.70, n.8, p.4522-4531, 2004.
COLAKOGLU, G. The chemical and physical environment for fungal growth. Fak
Matbaa Birimi, Istanbul, 149-166, 2001.
CRAVEIRO, A. C.; CRAVEIRO, A. A.; QUEIROZ, D. C. Quitosana a fibra do futuro.
2. ed. Fortaleza: PADETEC, 2004. V.1, 218p.
DIAS, B. A. Estudo de proteases degradadoras de cutícula produzidas pelo fungo
entomopatogênico Beauveria bassiana. 2005. 70p. Dissertação (Mestrado em
Microbiologia) – Centro de Ciências Biológicas, Programa de Pós-Graduação em
Microbiologia, Universidade Estadual de Londrina, Londrina.
DINARDO-MIRANDA, L. L.; FERREIRA, J. M. G.; CARVALHO, P. A. M.
Influência da época de colheita e do genótipo de cana-de-açúcar sobre a infestação de
Mahanarva fimbriolata (Stal) (Hemiptera: Cercopidae). Neotrop. Entomol, v.30, n.1,
p.145-149, 200.
DONG, C.; ZHANG, J.; HUANG, H.; CHEN, H.; HU, Y. Pathogenicity of a new China
variety of Metarhizium anisopliae (M. anisopliae var. Dcjhyium) to subterranean termite
Odontotermes formosanus. Microbiological Research, v.164, p.27-35, 2009.
DONZELLI, B. G. G.; OSTROFF, G.; HARMAN, G. E. Enhanced enzymatic
hydrolysis of langostino shell chitin with mixtures of enzymes from bacterial and fungal
sources. Carbohydrate Research, v.338, p.1823-1833, 2003.
DRIVER, F.; MILNER, R. J.; TRUEMAN, J. W. H. A taxonomic revision of
Metarhizium based on phylogenetic analysis of rDNA sequence data. Mycological
Research, v.104, n.2, p.134-150, 2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
81
DUNG, P.; MILAS, M.; RINAUDO, M.; DESBRIÉRES, J. Water soluble derivatives
obtained by controlled chemical modifications of chitosan, Carbohydrate Polymers,
v.24, p.209-214, 1994.
ESTEVES, J. A.; CABRITA, J. D.; NOBRE, G. N. Micologia médica, 2. ed. Lisboa:
Fundação Calouste Gulbenkian, 1990. 1058p.
FACCIOTI, M. C. R. Fermentação Contínua, In: Biotecnologia Industrial. Editado por:
SCHMIDELL, W.; LIMA, U. A.; AQUARONE, E.; BORZANI, W., São Paulo, Ed.
Edgard Blücher, v.2, p.541, 2001.
FARIA, M. R.; MAGALHÃES, B. P. O uso de fungos entomopatogênicos no Brasil -
Situação atual e perspectivas. Biotecnologia Ciência & Desenvolvimento, n.22., p.18-
21, setembro/outubro 2001.
FU, J.-Y.; WU, S.-M.; CHANG, C.-T.; SUNG, H.-Y. Characterization of Three
Chitosanase Isozymes Isolated from a Commercial Crude Porcine Pepsin Preparation. J.
Agric. Food Chem., v.51, p.1042-1048, 2003.
FURIGO JUNIOR, A.; PEREIRA, E. P. Enzimas e suas aplicações cinética enzimática.
2001. 39f. Apostila (Graduação de Engenharia Química e Engenharia de Alimentos) –
Centro de Tecnologia, Departamento de Engenharia Química e Engenharia de
Alimentos, Universidade Federal de Santa Catarina, Florianópolis.
GAO, J.; XU, H.; LI, Q.-J.; FENG, X.-H.; LI, S. Optimization of medium for one-step
fermentation of inulin extract from Jerusalem artichoke tubers using Paenibacillus
polymyxa ZJ-9 to produce R,R-2,3-butanediol, Bioresource Technology, v.101, p.7076-
7082, 2010.
GARCIA, M. V.; MONTEIROI, A. C.; SZABO, M. J. P.; PRETTE, N.; BECHARA, G.
H. Mechanism of infection and colonization of Rhipicephalus sanguineus eggs by
Metarhizium anisopliae as revealed by scanning electron microscopy and
histopathology. Brazilian Journal of Microbiology, São Paulo, v.36, n.4, p.368-372,
2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
82
GHANEM, N. B.; YUSEF, H. H.; MAHROUSE, H. K. Production of Aspergillus
terreus xylanase in solid-state cultures: application of the Plackett & Burman
experimental design to evaluate nutritional requirements, Bioresource Technology,
v.73, p.113-121, 2000.
GHORBEL-BELLAJ, O.; HMIDET, N.; JELLOULI, K.; YOUNES, I.; MAALEJ, H.;
HACHICHA, R.; NASRI, M. Shrimp waste fermentation with Pseudomonas aeruginosa
A2: Optimization of chitin extraction conditions through Plackett–Burman and response
surface methodology approaches, International Journal of Biological Macromolecules,
v.48, p.596–602, 2010.
GIUSTINA, A.; VENTURA, P. Weight-reducing regimens in obese subjects: Effects of
a new dietary fiber integrator. Acta Toxicol Ther, 16, 4, p.199-214, 1995.
GUIA.BIO, Reino dos fungos, 2012. Disponível em:
<http://www.http://guia.bio.br/reino-dos-fungos>. Acesso em: 12 novembro 2012.
GUIRADO, N.; AMBROSANO, E. J.; ROSSI, F.; DIAS, F. L. F. Controle de Cupins
de Montículo com Metarhizium anisopliae e Beauveria bassiana, Rev. Bras. de
Agroecologia, v.4, n.2, 2009.
GUO, Y.; XU, J.; ZHANG, Y.; XU, H.; YUAN, Z.; LI, D. Medium optimization for
ethanol production with Clostridium autoethanogenum with carbon monoxide as sole
carbon source, Bioresource Technology, v.101, p.8784–8789, 2010.
HAGAG, S.; KUBITSCHEK-BARREIRA, P.; NEVES, G. W. P.; AMAR, D.;
NIERMAN, W.; SHALIT, I.; SHAMIR, R.; LOPES-BEZERRA, L.; OSHEROV, N.
Transcriptional and proteomic analysis of the Aspergillus fumigatus ∆prtT protease-
deficient mutant. PLoS One, v.7, n.4, e33604, 2012.
HAN, L.-K.; KIMURA, Y.; OKUDA, H. Reduction in fat storage during chitin-chitosan
treatment in mice fed a high-fat diet, International Journal of Obesity, 23:174-179,1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
83
HUNG, T.; CHANG, Y.; SUNG, H.; CHANG, C. Purification and Characterization of
Hydrolase with Chitinase and Chitosanase Activity from Commercial Stem Bromelain.
J. Agric. Food Chem., 50, 4666-4673, 2002.
ICHIKAWA, S.; TAKANO, K.; TAKASHIKUROIWA, HIRUTA, O.; SATO, S.;
MUKATAKA, A. S. Immobilization and stabilization of chitosanase by multipoint
attachment to agar gel support. Journal of Bioscience and Bioengineering, v.93, n.2,
p.201-206, 2002.
JABOR, I. A. S.; PAMPHILE, J. A.; RODRIGUES, S. B.; MARQUES-SILVA, G. G.;
ROCHA, C. L. M. S. Análise do desenvolvimento do fungo entomopatogênico
Metarhizium anisopliae em resposta a fatores nutricionais. Acta Scientiarum.
Agronomy, Maringá, v.25, n.2, p.497-501, 2003.
JI, X.-J.; HUANG, H.; DU, J.; ZHU, J.-G.; REN, L.-J.; LI, S.; NIE, Z.-K. Development
of an industrial medium for economical 2,3-butanediol production through co-
fermentation of glucose and xylose by Klebsiella oxytoca, Bioresource Technology,
v.100, p.5214–5218, 2009.
JO, Y.; JO, K.; JIN, Y.; KIM, K.; SHIN, J.; KIM, Y.; PARK, R. Characterization and
kinetics of 45 kDa chitosanase from bacillus sp. P16. Biosci. Biotechnol. Biochem.,
v.67, n.9, 1875 – 1882, 2003.
KHOSRAVI-DARANI, K.; ZOGHI, A.; ALAVI, S. A.; FATEMI, S. S. A. Application
of Plackett Burman Design for Citric Acid Production from Pretreated and Untreated
Wheat Straw, Iran. J. Chem. Chem. Eng.. v.27, n.1, p.91-104, 2008.
KIM, P.; KANG, T. H.; CHUNG, K. J.; KIM, I. S.; CHUNG, K. Purification of a
constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and
expression of the gene. FEMS Microbiolog.y Letters 240, 31–39, 2004.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
84
KIM, S.-K.; RAJAPAKSE, N. Enzymatic production and biological activities of
chitosan oligosaccharides (COS): A review. Carbohydrate Polymers, v.62, n.4, p.357-
368, 2005.
KIRK, P. M.; CANON, P.F.; MINTER, D.W.; STALPERS, J.A. Dictionary of the
fungi. CAB International, Wallingford, 2008.
KONDO, H.; OSADA, A. Influence of dietary fiber on the bioavailability of zinc in
rats, Biomed Environ Sci, 9, 204-8, 1996.
KUMAR, S.; KOH, J. Physiochemical, Optical and Biological Activity of Chitosan-
Chromone Derivative for Biomedical Applications. Int. J. Mol. Sci., v.13, p.6102-6116,
2012.
KURITA, K.; SANNAN, T.; IWAKURA, Y. Studies on chitin, 4. Evidence for
formation of block and random copolymers of N-acetyl-o-glucosamine and
oglucosamine by hetero and homogeneous hydrolyses. Die Makromolecular Chemie,
v.178, p.3197-3202, 1977.
KUROIWA, T.; ICHIKAWA, S.; HIRUTA, O.; SATO, S.; MUKATAKA, S. Factors
Affecting the Composition of Oligosaccharides Produced in Chitosan Hydrolysis Using
Immobilized Chitosanases. Biotechnol. Prog., 18, 969-974, 2002.
LI, L.-Z.; HUI, Z.; MO, X. Optimization of the medium for streptolydigin production
by Streptomyces lydicus AS 4.2501 with precursor feeding using response surface
methodology, Journal of the Taiwan Institute of Chemical Engineers, 41, 252–258,
2010.
LIU, B.-L.; KAO, P.-M.; TZENG, Y.-M.; FENG, K.-C. Production of chitinase from
Verticillium lecanii F091 using submerged fermentation. Enzyme and Microbial
Technology, v.33, p.410-415, 2003.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
85
LOBATO, A. K. C. L. Estudo da produção de biosurfactantes por micro-organismos
isolados de poços de petróleo. 2003. 108p. Dissertação (Mestrado) – Programa de Pós-
Graduação em Engenharia Química. Universidade Federal do Rio Grande do Norte.
Natal.
LOPES, R. B.; ALVES, S. B.; PADULLA, L. F. L.; PÉREZ, C. A. Eficiência de
formulações de Beauveria bassiana e Metarhizium anisopliae para o controle de ninfas
de Amblyomma cajennense (Fabricius, 1787). Revista Brasileira de Parasitologia
Veterinária, v.16, n.1, p.27-31, 2007.
LOUREIRO, E. S.; BATISTA FILHO, A.; ALMEIDA, J. E. M.; MENDES, J. M.;
PESSOA, L. G. A. Eficiência de isolados de Metarhizium anisopliae (metsch.) sorok.
no controle da cigarrinha-da-raiz da cana-de-açúcar, Mahanarva fimbriolata (stal, 1854)
(hemiptera: cercopidae), em condições de campo. Arq. Inst. Biol., São Paulo, v.79, n.1,
p. 47-53, 2012.
MELLO, D. R.; REIS, R. C. S.; BITTENCOURT, V. R. E. P. Patogenicidade in vitro
do fungo Metarhizium anisopliae (Metschnikoff, 1879) Sorokin, 1883, sobre o carrapato
Boophilus microplus (Canestrini, 1887). Revista Brasileira de Parasitologia Veterinária,
São Paulo, v.15, p.157-162, 2006.
MITSUTOMI, M.; KIDOH, H.; TOMITA, H.; WATANABE, T. The action of Bacillus
circulans WL-12 chitinases on partially N-acetylated chitosan, Bioscience
Biotechnology and Biochemistry, v.59, p.529-531, 1995.
MALAJOVICH, M. A. Biotecnologia: fundamentos. Ed. biblioteca Max Feffer do
Instituto de Tecnologia ORT, Rio de Janeiro, 2009. 104p.
MONAGHAN, R. L.; EVELEIGH, D. E.; TEWARI, R. P.; REESE, E. T. Chitosanase,
a novel enzyme. Nature - New Biology, 245, p.78-80, 1973.
MU, X. Q.; XU, Y.; NIE, Y.; OUYANG, J.; SUN, Z. H. Candida parapsilosis CCTCC
M203011 and the optimization of fermentation medium for stereoinversion of (S)-1-
phenyl-1,2-ethanediol, Process Biochemistry, v.40, p.2345–2350, 2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
86
MURRAY, P. R.; ROSENTHAL, K. S.; PFALLER, M. A. Medical microbiology. 5th
ed. London, Elsevier Mosby, 2005.
MUZZARELLI, R. A. A. Biochemical significance of exogenous chitins and chitosans
in animals and patients, Carbohydrate Polymers, v.20, p.7-16, 1993.
MUZZARELLI, R. A. A.; TOMASETTI, M.; ILARI, P. Depolymerization of chitosan
with the aid of papain, Enzyme Microbiology and Technology, v.16, p.110-114, 1994.
NAVEENA, B. J.; ALTAF, Md.; BHADRIAH, K.; REDDY, G. Selection of medium
components by Plackett–Burman design for production of L(+) lactic acid by
Lactobacillus amylophilus GV6 in SSF using wheat bran, Bioresource Technology,
v.96, p.485-490, 2005.
NOGAWA, M.; TAKAHASHI, H.; KASHIWAGI, A.; OHSHIMA, K.; OKADA, H.;
MORIKAWA, Y. Purification and Characterization of Exo--D-Glucosaminidase from
a Cellulolytic Fungus, Trichoderma reesei PC-3-7, Appl. Environ. Microbiol., v.64, n.3,
p.890-895, 1998.
ONOFRE, S. B.; VARGAS; L. R. B.; ROSSATO, M.; BARROS, N. M.; BOLDO, J.
T.; NUNES, A. R. F.; AZEVEDO, J. L. Controle biológico de pragas na agropecuária
por meio de fungos entomopatogênicos. In: SERAFINI, L. A.; BARROS, N. M.;
AZEVEDO, J. L. (Org.). Biotecnologia – avanços na agricultura e na agroindústria.
Caxias do Sul: EDUCS, p.295-328, cap.8, 2002.
OSSWALD, W. F.; SHAPIRO, J. P.; DOOSTDAR, H.; MCDONALD, R. E.; NIEDZ,
R. P.; NAIRN, C. J.; HEARN, C. J.; MAYER, R. T. Identification and Characterization
of Acidic Hydrolases with Chitinase and Chitosanase Activities from Sweet Orange
Callus Tissue, Plant Cell Physiol. 35, p.811-820, 1994.
PAGNONCELLI, M. G. B. Estudo do mecanismo de produção de oligossacarídeos com
atividades nutracêuticas a partir da quitosana por hidrólise enzimática com processo

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
87
fermentativo simultâneo. 2008. 118p. Tese (Doutorado) – Centro de Tecnologia,
Programa de Pós-Graduação em Engenharia Química. Universidade Federal do Rio
Grande do Norte. Natal.
PALMA-GUERRERO, J., JANSSON, H.-B., SALINAS, J.; LOPEZ-LLORCA, L. V.
Effect of chitosan on hyphal growth and spore germination of plant pathogenic and
biocontrol fungi, J. Appl. Microbiol., v.104, p.541-553, 2008.
PAULA, F. A.; ARRUDA, D. C. Análise do método de planejamento experimental
Plackett Burman na otimização de degradação de penas de aves por comunidades
microbianas, XXI Salão de Iniciação Científica, 19-23, outubro, 2009, UFRGS, Porto
Alegre.
PELCZAR, M. J.; CHANG, E. C. S.; KRIEG, N. R. Microbiologia: conceitos e
aplicações, v.2, 2.ed. São Paulo: Makron BooKs, 1996.
PELLETIER, A.; SYGUSCH, J. Purification and characterization of three chitosanase
activities from Bacillus megaterium P1, Applied and Environmental Microbiology, v.
56, n.4, p.844-848, 1990.
PEREIRA, F. B.; GUIMARÃES, P. M. R.; TEIXEIRA, J. A.; DOMINGUES, L.
Optimization of low-cost medium for very high gravity ethanol fermentations by
Saccharomyces cerevisiae using statistical experimental designs, Bioresource
Technology, v.101, p.7856-7863, 2010.
PETER, M. G. Chitin and chitosan from aninal sources. In: STEINBUCHEL, A.;
RHEE, S. K. Polysaccharides and Polyamides in the Food Industry. Weinheim:
Wileyvch, v.1, p.115-208, 2005.
PIZA, F. A. T.; SILOTO, A. P.; CARVALHO, C. V.; FRANCO, T. T. Purification,
characterization and purification of chitosanase from Bacillus cereus, Brazilian Journal
of Chemical Engineering., v.16, n 2, p.185-192, 1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
88
POLITZER, K.; BON, E. P. S. Enzimas Industriais e Especiais. C.G.E.E. Ciência,
Tecnologia e Inovação. Rio de Janeiro, 554p, 2006.
POLYMAR, Ciência é a nossa Natureza, 2003. Disponível em:
<http://www.polymar.com.br>. Acesso em: 6 setembro 2012.
PUTZKE, J.; PUTZKE, M. T. L. Os reinos dos fungos. Santa Cruz do Sul, RS:
EDUNISC, 1998.
QUINES, L. K. M. Extração de poli(3-hidroxibutirato) produzido por Cupriavidus
necator dsm 545 com 1,2-carbonato de propileno. 2010. 99p. Dissertação (Mestrado em
Engenharia de Alimentos) – Centro Tecnológico, Departamento de Engenharia Química
e Engenharia de Alimentos, Programa de Pós-Graduação em Engenharia de Alimentos,
Universidade Federal de Santa Catarina, Florianópolis.
RAJENDRAN, A.; THANGAVELU, V. Statistical experimental design for evaluation
of medium components for lipase production by Rhizopus arrhizus MTCC 2233, LWT -
Food Science and Technology, v.42, p.985–992, 2009.
RAMIREZ-LEÓN, L. F.; RUIZ-HERRERA, J. Hydrolysis of walls and formation of
spheroplasts in Mucor rouxii. J. Gen. Microbiol., v.72, p.281-290, 1972.
RANGEL, M. A. S.; LOUREIRO, E. S.; ALMEIDA, A. S.; ALVES, H. C.; ONO, F.
B.; LOBO, P. M.; MARIANI, M. A.; PESSOA, L. G. A.; SILVA, A. J. Avaliação
participativa do controle de cigarrinhas das pastagens com a aplicação do fungo
Metarhizium anisopliae (Metsch.) Sorok. Rev. Bras. de Agroecologia, v.2, n.2, 731-734,
2007.
RAVEN, P. H.; EVERT, R.F.; EICHHORN, S.E. Biologia Vegetal, 6ª Ed, Rio Janeiro,
Editora Guanabara Koogan, 2001.
REDDY, L. V. A.; WEE, Y.-J.; YUN, J.-S.; RYU, H.-W. Optimization of alkaline
protease production by batch culture of Bacillus sp. RKY3 through Plackett–Burman

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
89
and response surface methodological approaches, Bioresource Technology, v.99, p.242–
2249, 2008.
RIGUEIRA, J. C. S. Influência da contagem de células somáticas no perfil e teores de
aminas bioativas e na qualidade de leite cru e de queijo mussarela. 2010. 147p. Tese
(Doutorado em Ciência de Alimentos) - Faculdade de Farmácia, Departamento de
Alimentos, Programa de Pós-Graduação em Ciência de Alimentos, Faculdade de
Farmácia da Universidade Federal de Minas Gerais, Belo Horizonte.
RODRIGUES, M. I.; IEMMA, A. F. Planejamento de Experimentos e Otimização de
Processos: uma estratégia sequencial de planejamentos, Campinas, SP, Casa do Pão
Editora, 2005.
SANTOS, E. S. Recuperação e Purificação de Enzimas usando Adsorção em Leito
Expandido, Tese de Doutorado, Universidade Estadual de Campinas, Campinas/SP,
p.152, 2001.
SEDMAK, J. J.; GROSSBERG, S. E. A rapid, sensitive and versatile assay for protein
using coomassie brilliant blue G250, Analytical Biochemistry, v.79, p. 544-552, 1977.
SEINO, H.; TSUKUDA, K.; SHIMASUE, Y. Properties and action pattern of a
chitosanase from Bacillus sp. PI- 7S. Agricultural Biological Chemistry, 55, p.2421-
2423, 1991.
SHIMOSAKA, M.; NOGAWA, M.; OHNO, Y.; OKAZAKI, M. Chitosanase from the
plant pathogenic fungus, Fusarium solani f. sp. Phaseoli – purification and some
properties, Bioscience Biotechnolgy and Biochemistry, v.57, n.2, p.231-235, 1993.
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Production of two chitosanases from a chitosan-assimilating bacterium, Acinetobacter
sp. Strain CHB101, Applied and Environmental Microbiology, v.61, n.2, p.438-442,
1995.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
90
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Characterization of Three Chitosanase Isozymes Isolated from a Commercial Crude
Porcine Pepsin Preparation, J. Agric. Food Chem. 51, p.1042-1048, 2003.
SILVA FILHO, R. C.; COSTA, A. S. S.; CHAGAS, B. M. E.; SANTOS, F. N.;
MACEDO, G. R.; SANTOS, E. S. Avaliação do teste de atividade quitosanolítica
produzida por fungo. In: XV SIMPÓSIO NACIONAL DE BIOPROCESSOS, 15,
Recife, 2005, 02 a 05 de agosto.
SILVA, E. P. Avaliação Preliminar do Potencial de Aplicação da Tecnologia de
Biopilhas para a Biorremediação do Solo de Guamaré-RN. 2004. 119p. Dissertação
(Mestrado) – Centro de Tecnologia, Programa de Pós-Graduação em Engenharia
Química. Universidade Federal do Rio Grande do Norte. Natal.
SOMASHEKAR, D.; JOSEPH, R. Chitosanases – Properties and apllications: A review.
Bioresource Technology, v.55, p.35-45, 1996.
STOYACHENKO, I. A.; VARLAMOV, V. P.; DAVANKOV, V. A. Chitinases of
Streptomyces kurssanovii: purification and some properties, Carbohydrate Polymers,
v.24, p.47-54, 1994.
TINLINE, R. D. Nuclear distribuition in Metarhizium anisopliae. Mycologia, v. 63, p.
713-721, 1971.
TRABULSI, L. R.; TOLEDO, M. R. F. Microbiologia. 2. Ed., São Paulo: Atheneu,
1996.
VIEIRA, P. D. S.; SILVA, W. M. T.; PAIVA, L. M.; LIMA, E. A. L. A.;
CAVALCANTI, V. L. B. Estudo da caracterização morfológica, esporulação e
germinação dos conídiosde Metarhizium anisopliae var. acridum em diferentes
temperaturas. Biológico, São Paulo, v.69, n.1, p.17-21, 2007.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
91
WANDERLEY, M. D.; NEVES, E.; ANDRADE, C. J. Aspectos da produção industrial
de enzimas. Revista Citino, v.1, n.1, p.30-36, 2011.
WANG, C.; TYPAS, M. A.; BUTT, T. M. Detection and characterization of pr1
virulent gene deficiencies in the insect pathogenic fungus Metarhizium anisopliae.
FEMS Microbiology Letters, v. 213, p. 251-255, 2002.
ZAHORSKA-MARKIEWICZ, B.; KROTKIEWSKI, M.; OLSZANECKA-
GLINIANOWICZ, M.; ZURAKOWSKI, A. Effect of chitosan in complex management
of obesity, Pol Merkur Lekarski, v.13, n.74, p.129-132, 2002.
ZHANG, X. Y.; DAI, A. L.; ZHANG, X. K.; KUROIWA, K.; KODAIRA, R.;
SHIMOSAKA, M.; OKAZAKI, M. Purification and Characterization of and Exo--D-
glucosaminidase from a koji mold, Aspergillus oryzae IAM2660. Biosci. Biotechnol.
Biochem., v.64, n.9, 1896-1902, 2000.
ZHU, X.; WU, X.; DAI, Y. Fermentation conditions of a chitosanase from Acinetobcter
sp C-17. Biosci. Biotechnol. Biochem., v.67, n.2, p.284-290, 2003.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
92
APÊNDICE

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
93
APÊNDICE I
Metodologia para repique da cepa
Em Erlenmeyer de 1000 mL colocava-se 500 mL do meio de manutenção
PDA. Este Erlenmeyer, com o meio de manutenção, era autoclavado junto com placas
de Petri a 1,5 kgf/cm2 por 15 minutos, em seguida, era levado para uma câmara de fluxo
laminar e transferido para as placas de Petri já autoclavadas, onde eram mantidas numa
estufa a 27 ºC durante 24 h. Após este período, as placas de Petri eram levadas a câmara
de fluxo laminar, utilizando bico de Bunsen para esterilizar a alça de platina. Procedia-
se o repique utilizando uma placa de Petri contendo a cepa pura. Cada placa de Petri
contendo a cepa pura era transferida para duas placas de Petri contendo o meio de
manutenção. As placas eram mantidas na estufa a 27 ºC durante 30 dias onde após esse
período, repetia-se este procedimento.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
94
APÊNDICE II
Montagem da Curva de Calibração Usando o Método DNS
Soluções:
Glicosamina = 0,01 mol/mL (10000,0 µmol/mL)
Tabela 14. Dados para construção da curva de calibração usando o Método DNS.
Tubo de
Ensaio
Solução de
glicosamina (mL)
Água Destilada
(mL)
DNS (mL)
Volume Total
Massa (µµµµmol)
1 0 1,0 1,5 2,5 0
2 0,2 0,8 1,5 2,5 0,2
3 0,4 0,6 1,5 2,5 0,4
4 0,6 0,4 1,5 2,5 0,6
5 0,8 0,2 1,5 2,5 0,8
6 1,0 0 1,5 2,5 1,0
Tabela 15 - Medidas de absorbância a 600nm
Dados adicionais:
Peso Molecular D-Glicosamina = 215,64 (recipiente SIGMA)
Conc. Sol. D-Glicosamina = 0,05 g/50 mL
Massa calc.(µmol) = conc. Sol. D-Glic. x Vol. Utiliz. D-Glics. =
= 0,05 x 1000 x Vol.util./50*215,64
Tempo atividade enzimática = 30 min
1 U/mL = 1 µmol/(min.mL)
Vol. Total (Glicosamina+caldo) = 1 mL
Medições Adsorbância a 600 nm Vol. Utiliz. Massa Massa Atividade
II III Média(Abs)(mL) calc. (mg) calc. (µg) Enzim. (U/mL)
0 0 0 0 0 0 0
0,133 0,128 0,1307 0,2 0,2 200 0,0309
0,279 0,277 0,2787 0,4 0,4 400 0,0618
0,433 0,428 0,4297 0,6 0,6 600 0,0927
0,587 0,583 0,5797 0,8 0,8 800 0,1237
0,727 0,716 0,7157 1 1 1000 0,1546

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
95
Figura 17 – Curva padrão de glicosamina.
y = 1900ralx + 1900ral
R² = 1900ral
00,000
00,020
00,040
00,060
00,080
00,100
00,120
00,140
00,160
00,180
00,000 00,100 00,200 00,300 00,400 00,500 00,600 00,700 00,800
Ati
v.
En
zim
áti
ca (
U/m
L)
Absorbância (Abs) a 600 nm
Curva-Padrão D-Glicosamina

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
96
APÊNDICE III
Método de Determinação de Proteína Utilizando o Corante Comassie
Brilhante Blue G 250 (Método modificado de Sedmak e Grossberg)
Micrométodo
Preparar uma solução 0,06% de Comassie Blue G250 em 1,5 % HCl (w/V). Filtrar e
papel de filtro Whatman nº1.
Curva Padrão (1 a 20 µµµµg)
a) Preparar soluções estoque de BSA de 10 a 200 µg/mL.
b) Colocar em tubos de ensaio 100 µL da solução estoque, 1400 µL de água e
1500µL do Comassie e agitar.
c) Preparar um tubo contendo 1500 µL de água e 1500 µL do corante que
corresponde ao zero de proteína.
d) Após 5 minutos (padronizar o tempo) fazer as leituras das absorbâncias.
e) As leituras das amostras devem ser feitas em dois comprimentos de onda 595 e
465 contra a água.
f) Plotar a razão das absorbâncias a 595/465 como uma função da massa de BSA e
calcular o melhor ajuste linear dos dados.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
97
Micrométodo (1 a 20 µµµµg de BSA)
Solução Estoque de BSA = 0,2 mg/mL
Tabela 16. Dados para construção da curva padrão de BSA.
Tubo de
Ensaio
H2O (µµµµL)
BSA (µµµµg)
Coomassie (µµµµL)
BSA (µµµµg)
1 1500 0 1500 0
2 1490 10 1500 2
3 1480 20 1500 4
4 1470 30 1500 6
5 1460 40 1500 8
6 1450 50 1500 10
7 1440 60 1500 12
8 1430 70 1500 14
9 1420 80 1500 16
10 1410 90 1500 18
11 1400 100 1500 20
Dados adicionais:
Concentração Sol. Estoque
BSA = 0,2 mg/mL = 0,2 µg/µL
Vol. total sol. = vol. H2O + vol. BSA + vol.
Coomassie = 3000 µL = 3 mL
Concentração Sol. BSA (mg/mL) = (massa BSA mg) / 3 mL
.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
98
Figura 18 – Curva Padrão de BSA
y = 1900ralx - 1900ral
R² = 1900ral
000,000
001,000
002,000
003,000
004,000
005,000
006,000
007,000
000,400 000,500 000,600 000,700 000,800 000,900 001,000 001,100
µg
BS
A
Razão Absorbâncias (595nm/465nm)
Curva-Padrão BSA

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE TECNOLOGIA
DEPARTAMENTO DE ENGENHARIA QUÍMICA PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA
TESE DE DOUTORADO
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em
Cultivo Descontínuo Submerso
Raimundo Cosme da Silva Filho
Natal/RN
Abril/2013

Raimundo Cosme da Silva Filho
Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo
Descontínuo Submerso
Tese apresentada ao Programa de Pós-Graduação em Engenharia Química – PPGEQ, da Universidade Federal do Rio Grande do Norte, como parte dos requisitos para obtenção do título de Doutor em Engenharia Química, sob a orientação do Prof. Dr. Everaldo Silvino dos Santos e a coorientação da Profª. Drª. Gorete Ribeiro de Macedo.
Natal/RN
Abril/2013

Catalogação da Publicação na Fonte. UFRN / CT / PPGEQ
Biblioteca Setorial “Professor Horácio Nícolas Sólimo”.
Silva Filho, Raimundo Cosme da. Otimização do meio de cultura para a produção de quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso / Raimundo Cosme da Silva Filho. - Natal, 2013. 98 f.: il.
Orientador: Everaldo Silvino dos Santos. Co-orientador: Gorete Ribeiro de Macedo. Tese (Doutorado) - Universidade Federal do Rio Grande do Norte.
Centro de Tecnologia. Departamento de Engenharia Química. Programa de Pós-Graduação em Engenharia Química.
1. Quitosana - Tese. 2. Quitina - Tese. 3. Quitosanase – Tese. 4. Metarhizium
anisopliae - Tese. I. Santos, Everaldo Silvino dos. II. Macedo, Gorete Ribeiro de. III. Universidade Federal do Rio Grande do Norte. IV. Título. RN/UF/BSEQ CDU 547.458 (043.2)

SILVA FILHO, Raimundo Cosme da – Otimização do Meio de Cultura para a Produção de Quitosanase por Metarhizium anisopliae em Cultivo Descontínuo Submerso. Tese de Doutorado. UFRN - Programa de Pós-Graduação em Engenharia Química. Área de Concentração: Pesquisa e Desenvolvimento de Tecnologias Regionais – Subárea: Engenharia de Processos – Alimentos e Biotecnologia, Natal/RN, Brasil.
Orientação: Prof. Dr. Everaldo Silvino dos Santos Coorientação: Profª. Drª. Gorete Ribeiro de Macedo ___________________________________________________________________________
RESUMO: No presente trabalho utilizou-se um Planejamento Plackett & Burman, com 8 fatores e 12 ensaios em 2 níveis e mais 3 repetições na condição do ponto central, para se investigar a influência das concentrações de quitosana, peptona, extrato de levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O e FeSO4 na produção da enzima quitosanase por Metarhizium
anisopliae. Os ensaios para produção da enzima foram realizados em cultivo descontínuo submerso. Os resultados para o Planejamento Plackett & Burman mostraram que a atividade quitosanolítica foi favorecida pelo aumento da concentração de substrato (quitosana) e de sulfato ferroso (FeSO4), enquanto que o aumento da concentração dos outros fatores não contribuiu de forma significativa para a atividade quitosanolítica. A otimização do meio de cultura para a produção da enzima foi realizado por meio de um Planejamento Composto Central, com os dois fatores que mais influenciaram a atividade quitosanolítica (quitosana e FeSO4), conforme Planejamento Plackett & Burman, mantendo-se os outros nutrientes em seus valores mínimos. Nesse planejamento, para o fator FeSO4, tomou-se o limite inferior (-1) como sendo o limite superior do Planejamento Plackett & Burman. Os resultados mostraram que a produção da enzima foi favorecida pelo aumento da concentração quitosana e pela diminuição da concentração de FeSO4. A produção máxima de atividade quitosanolítica foi da ordem de 70,0 U/L e foi atingida em apenas 18 h de fermentação, resultado esse vinte e oito vezes maior aos obtidos anteriormente para o mesmo microrganismo que foi 2,5 U/L em 48h.
Palavras-chave: quitosana; quitosanase; Metarhizium anisopliae; Plackett & Burman.


ABSTRACT
In this work a Plackett-Burman Design with 8 factors and 12 trials in 2 levels with 3 repetitions at the center point was used in order to investigate the influence of the concentration of chitosan, peptone, yeast extract, NaNO3, K2HPO4, KCl, MgSO4.7H2O and FeSO4 on chitosanase production by Metarhizium anisopliae. Runs were carried out using submerged discontinuous cultivation for enzyme production. The results of the Plackett & Burman Design showed that only two factors, chitosan concentration as well as FeSO4 had influence on chitosanolytic activity, while the increase in concentration of other factors not contributed significantly to the quitosanolítica activity. Cultivation medium optimization for enzyme production was carried out using a Composite Central Design, with the most important factors for chitosanolytic activity (chitosan and FeSO4), in accordance with Plackett & Burman Design, and keeping the other nutrients in their minimum values. On this other design, it was taken the highest limit in Plackett & Burman Design as the lowest limit (-1) to FeSO4 factor. The results showed that the enzyme production was favoured by increasing the chitosan concentration and by decreasing FeSO4. Maximum production for chitosanolytic activity was about 70.0 U/L and was reached in only 18 h of fermentation, a result about twenty-eight times greater than a former study using the same microorganism (about 2.5 U/L at 48 h).
Key-words: Chitosan; Chitosanase; Metarhizium anisopliae; Plackett & Burman

Ao meu pai, minha mãe e minha
família, em especial aos meus
irmãos e sobrinhos

AGRADECIMENTOS
A Deus, por estar presente em todos os momentos da minha vida e por me ceder à
oportunidade de iniciar e concluir este trabalho.
A minha mãe Cândida Inocência da Silva que esteve comigo em todos os momentos da
minha vida me educando, me amando e sempre me dando muito carinho.
Ao meu pai Raimundo Cosme da Silva, que não está mais aqui para ver este trabalho,
mas que foi a pessoa que mais me apoiou e me incentivou, em todos os momentos da minha vida,
para que eu pudesse chegar o mais longe possível na minha carreira acadêmica. Pai muito
obrigado por toda atenção, educação, amor e carinho que recebi durante toda a minha vida.
Aos meus irmãos e familiares que sempre estiveram comigo dividindo momentos de
grande alegria e às vezes de tristeza, mas sempre nos mantendo unidos para superarmos todos os
obstáculos.
A minha namorada Rochelly Caroliny por todo amor e carinho que me deu e que me
entendeu nos momentos que estive ausente para a conclusão deste trabalho.
Ao meu orientador Professor Doutor Everaldo Silvino dos Santos por confiar em mim e
me aceitar como orientando além de toda a amizade, orientação e dedicação em todas as etapas de
desenvolvimento e conclusão deste trabalho.
À Professora Doutora Gorete Ribeiro de Macedo, “pessoa que admiro muito como
profissional e principalmente, como pessoa, um exemplo a ser seguido”, pelos conselhos e críticas
quando necessários ao desenvolvimento deste trabalho. Professora a senhora é considerada uma
mãe para de todos os alunos que tiveram a honra de conhecê-la e trabalhar com a senhora.
Aos meus amigos Engenheiros Químicos, Ana Carmen, Ana Katerine, Andréa Farias,
Anita Lima, Sirtys Lessa, Alexandre, Sérgio, Márcio Bezerra e ao Biólogo Daniel Souza, que
sempre me ouviram e que me ajudaram de alguma forma nesse trabalho.
A Eusamar Coelho de Lima, Mazinha, secretária do PPGEQ, pelo apoio, pela paciência e
orientação nas questões burocráticas do processo.
A Carlos e Jardelson, pela grande colaboração e dedicação nos experimentos do
laboratório.
A Sérgio Santana pela grande colaboração e dedicação, o qual não poupou esforços nem
tempo em me ajudar na realização dos experimentos em laboratório.

Ao meu amigo Ubiratan Nazaré por me ajudar na organização do texto e na tradução para
o Abstract.
Finalizando, agradeço ao Laboratório de Engenharia Bioquímica da UFRN por ter cedido
toda a sua estrutura para realização dos experimentos e ao Programa de Pós-Graduação em
Engenharia Química, assim como a todos os professores do Departamento de Engenharia Química
da UFRN que trabalham em pró de uma educação digna e ensino de qualidade para todos os
alunos.

Sumário LISTA DE FIGURAS
LISTA DE TABELAS
NOMENCLATURA
INTRODUÇÃO...............................................................................................................................15
REVISÃO BIBLIOGRÁFICA........................................................................................................18
2.1. Enzimas................................................................................................................................18
2.1.1. Quitosana e seus derivados........................................................................................19
2.1.2. Quitosanases...............................................................................................................24
2.2. Fungos..................................................................................................................................29
2.2.1. Características gerais..................................................................................................29
2.2.2. Reprodução dos Fungos.............................................................................................32
2.2.3. Metabolismo de fungos..............................................................................................33
2.2.4. Nutrição de fungos.....................................................................................................34
2.2.5. O Fungo Metarhizium anisopliae ..............................................................................36
2.3. Fermentação.........................................................................................................................40
2.3.1. Preparo do inóculo......................................................................................................41
2.3.2. Produção de enzimas em cultivo submerso descontínuo...........................................42
2.4. Planejamento experimental..................................................................................................42
2.4.1. Planejamento fatorial..................................................................................................43
2.4.2. Planejamento Plackett & Burman..............................................................................43
2.4.3. Aplicações do Planejamento Plackett & Burman.......................................................44
METODOLOGIA EXPERIMENTAL.............................................................................................50
3.1. Quitosana..............................................................................................................................50
3.2. Microrganismo.....................................................................................................................50
3.2.1. Escolha da cepa produtora de quitosanase.................................................................50

3.2.2. Manutenção da cepa...................................................................................................50
3.3. Produção de enzima.............................................................................................................51
3.4. Determinação da Proteína Total...........................................................................................51
3.5. Determinação da Atividade Quitosanolítica e Atividade Específica...................................51
3.6. Planejamento Experimental.................................................................................................52
3.6.1. Planejamento de seleção de fatores Plackett Burman................................................52
3.6.2. Planejamento Composto Central para Otimização do meio de cultura para a
Produção da Enzima em Cultivo Submerso.....................................................................................53
3.7. Perfil Cinético durante o Cultivo.........................................................................................56
RESULTADOS E DISCUSSÃO.....................................................................................................58
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA ..........................................58
4.2. Planejamento Plackett & Burman........................................................................................59
4.2.1. Análise do Plackett & Burman com 8 fatores e 12 ensaios .......................................59
4.2.2. Análise do Plackett & Burman com 12 ensaios e mais 3 repetições na condição do
ponto central ....................................................................................................................................61
4.3. Planejamento Composto Central para Produção de Quitosanase por Metarhizium
anisopliae em Cultivo Descontínuo Submerso................................................................................64
4.4. Perfil Cinético – Melhor Condição......................................................................................69
CONCLUSÃO.................................................................................................................................75
REFERÊNCIAS BIBLIOGRÁFICAS.............................................................................................78
APÊNDICE I...................................................................................................................................93
Metodologia para Repique da Cepa ................................................................................................93
APÊNDICE II.................................................................................................................................94
Montagem da Curva de Calibração Usando o Método DNS..........................................................94
APÊNDICE III................................................................................................................................96
Método de Determinação de Proteína Utilizando o Corante Comassie Brilhante Blue G 250
(Método modificado de Sedmak e Grossberg)................................................................................96

Lista de Figuras
Figura 1 – Estrutura da quitosana..................................................................................................20
Figura 2 – Estrutura da glicosamina..............................................................................................23
Figura 3 – Estrutura do sulfato de glicosamina.............................................................................23
Figura 4 – Representação da estrutura geral de uma célula de fungo..........................................30
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas...........................................................31
Figura 6 – Conídios de Aspergillus e Penicillium.........................................................................33
Figura 7 – Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA ...................58
Figura 8 – Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados............................................................................................................................60
Figura 9 – Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto central
mostrando a influência dos fatores estudados................................................................................63
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos
fatores estudados............................................................................................................................67
Figura 11 – Valores preditos em função dos valores observados..................................................68
Figura 12 – Atividade quitosanolítica em função da concentração de quitosana e da concentração
de FeSO4 para os fatores não significativos no ponto central .......................................................68
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana ........70
Figura 14 – Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana....................................70
Figura 15 – Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de
quitosana........................................................................................................................................71
Figura 16 – Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana..........................................................................................................72
Figura 17 – Curva padrão de glicosamina ....................................................................................95
Figura 18 – Curva Padrão de BSA ................................................................................................98

Lista de Tabelas
Tabela 1 – Comparação das enzimas com os catalisadores químicos..............................................19
Tabela 2 – Fontes naturais de quitina e quitosana ...........................................................................20
Tabela 3 – Níveis para os fatores e seus valores codificados...........................................................53
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central .........................................................................................53
Tabela 5 – Matriz do planejamento Composto Central...................................................................54
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central......................................................................................................55
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.......................59
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman...............................................60
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.......................61
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central...............................................................................................................................................62
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da atividade
enzimática........................................................................................................................................65
Tabela 12 – Efeitos Estimados do planejamento Composto Central...............................................65
Tabela 13 – Resumo da ANOVA....................................................................................................66
Tabela 14 – Dados para construção da curva de calibração usando o Método DNS ......................94
Tabela 15 – Medidas de absorbância a 600 nm para construção da curva de calibração................94
Tabela 16 – Dados para construção da curva padrão de BSA ........................................................97

NOMENCLATURA
A – Atividade quitosanolítica (U/L)
AE – Atividade específica (U/mg)
ANN-GA – algoritmo genético de rede neural artificial
ANOVA – Análise de variância (Analysis of Variance)
BSA – Albumina de soro bovino (Bovine Sereum Albumin)
CCD – Planejamento Composto Central
CDA – Chitosan Dector Agar
Cs – Concentração de substrato (%)
CP – Concentração de KCl (%)
Da – Dalton
DEQ – Departamento de Engenharia Química
DNS – Ácido Dinitrosalissílico
E – Enzima
Ea – Energia de ativação
EC – Comissão para nomenclatura e classificação de enzimas
EL – Concentração de Extrato de Levedura (%)
FP – Concentração de K2HPO4 (%)
NS – Concentração de NaNO3 (%)
GDA – Grânulos dispersíveis em água
k – Número de fatores, variáveis
LEB – Laboratório de Engenharia Bioquímica
mRNA – RNA mensageiro
n – Número de níveis de um determinado fator
N – Número de ensaios de um planejamento
P – Pressão
PP – Concentração de Peptona (%)
PDA – Batata Dextrose Agar
pH – Potencial hidrogeniônico
Q – Concentração de Quitosana (%)
Qm – Quadrado médio
r – Agitação (rpm)
R2 – Coeficiente de determinação

RA – Razão de aeração
rpm – Rotações por minuto
RSM – Metodologia de superfície de resposta
S – Molécula de substrato
SF – Concentração de FeSO4 (%)
SM – Concentração de MgSO4.7H2O (%)
SQ – Soma de quadrados
SSF – Fermentação de estado sólido
T – Temperatura
TMC – Temperatura de cultivo
V11,V12,V13 – Variáveis inertes para cálculo do erro
VHG – Fermentação de alta densidade
WP – Pó molhável
X – Concentração celular
Xi – variável independente
Xj – variável independente
xi – Fator independente real
x0 – Valor de xi no ponto central
∆xi – Variação de valor
Y – Resposta do modelo de primeira ordem
Yp – Resposta do modelo polinomial de segunda ordem
β0 – Interseção dos modelos de primeira ou segunda ordem
βi – Coeficiente linear dos modelos de primeira ou segunda ordem
βii – Coeficiente quadrático
βij – Coeficiente de interação

Capítulo 1 Introdução

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
15
1. Introdução As quitosanases foram descobertas simultaneamente há quase quarenta anos por
Monaghan et al. (1973), como pelo grupo de Ramirez-León & Ruiz-Herrera (1972), e
são enzimas obtidas a partir do metabolismo de bactérias, fungos e extraídas de plantas.
Desde então, vários estudos vêm apresentando a obtenção de quitosanase deste
microrganismos, na forma intracelular ou extracelular (Osswald et al., 1994;
Somashekar & Joseph, 1996), como a do presente estudo. A quitosanase (EC 3.2.1.132)
representa uma classe de enzimas hidrolíticas que catalisam a hidrólise de ligação β-
(1→ 4) glicosídica da quitosana, gerando oligômeros chamados quito-oligossacarídeos
(Reddy et al., 2008).
A quitosana é um biopolímero hidrofílico obtido a partir da quitina, um
polissacarídeo abundante na natureza. Na literatura, é considerado quitosana o produto
da desacetilação da quitina para um grau de desacetilação superior a 75% e solúveis em
ácidos como o acético e o fórmico. A quitosana é conhecida por exibir uma ampla
variedade de atividade fisiológica como atividade antitumoral, mas o peso molecular e a
insolubilidade em água da quitosana são desvantagens para várias aplicações.
Atualmente, há interesse nas quitosanas quando as mesmas são convertidas em quito-
oligossacarídeos porque eles não apenas são solúveis em água e possuem baixo peso
molecular, mas também têm atividades biológicas bem notáveis, como atividade
anticancerígena atóxica e biocompatível, incluindo efeito antimetastático, atividade anti-
HIV, hepatoprotetora, antioxidante, hipocolesterolêmica, antimicrobiana,
imunoestimulate, antitumoral, anti-inflamatória e atividade aceleradora da absorção de
cálcio e ferro. No entanto, estes quito-oligossacarídeos são obtidos tradicionalmente na
indústria por reação de hidrólise usando ácidos, que apresenta muitos problemas como
dificuldade em controlar a hidrólise, produzindo uma grande quantidade de
oligossacarídeos de cadeia pequena e oligossacarídeos de baixo rendimento, alto custo
de separação e, também, poluição ambiental, uma vez que utiliza um ácido como
catalisador. Como alternativa, com suas vantagens em termos de compatibilidade
ambiental, baixo custo e reprodutibilidade, a obtenção destes quito-oligossacarídeos por
hidrólise com quitosanase se tornou cada vez mais popular nos últimos anos (Azevedo
et al., 2007).

Capítulo 1 Introdução
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
16
Devido a todos estes aspectos positivos é que, em geral, as enzimas vêm sendo
estudadas em diferentes aplicações, mostrando que esta é uma tendência na indústria.
No presente trabalho, foi desenvolvido um processo fermentativo visando a
otimização do meio de cultura para a produção da enzima quitosanase em cultivo
descontínuo submerso, por Metarhizium anisopliae, capaz de sintetizar oligômeros com
aplicações na indústria farmacêutica e de química fina, partindo de quitosana como
única fonte de carbono.
Nesse caso, avaliou-se, através de um planejamento Plackett & Burman (método
de delineamento de seleção de fatores) e de um planejamento Composto Central as
melhores condições para a produção da enzima. A técnica do planejamento
experimental Plackett & Burman para realizar triagem de fatores na otimização de um
determinado processo foi utilizada neste trabalho para identificar quais os fatores que
influenciam de forma significativa na produção da enzima quitosanase. A influência
destes fatores (concentração de quitosana e concentração de FeSO4) foi então
investigada pelo planejamento Composto Central com 22 experimentos na parte fatorial,
repetição em triplicata no ponto central e quatro experimentos na parte axial. A
repetição do ponto central permite também a determinação do erro puro e da
repetibilidade do processo, sendo uma forma de identificar melhor se o processo está
sob controle.
Do ponto de vista industrial é de relevante importância a realização de estudos
para a otimização da produção da quitosanase de forma contínua ou em grandes
bateladas.

Capítulo 2 Revisão Bibliográfica

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
18
2. Revisão Bibliográfica 2.1. Enzimas
As enzimas são catalisadores biológicos constituídos por longas cadeias de
aminoácidos que atuam em substratos reduzindo a energia de ativação, acelerando assim
a velocidade da reação, e transformando-os em produtos. Sua atuação catalítica visa
facilitar a atividade das células através da quebra de moléculas ou junção das mesmas
para formação de novos produtos. Podem apresentar alto grau de especificidade relativa
a apenas um substrato ou a vários substratos ao mesmo tempo, como a certos tipos de
ligações glicosídicas α-1,4 das moléculas de amido, ou para determinado tipo particular
de isômero óptico como ocorre na oxidação da β-D-glicose pela glicose oxidase (Furigo
Junior & Pereira, 2001).
Enzimas são proteínas que apresentam em sua estrutura um centro ativo
(apoenzima) e, em alguns casos, um grupo prostético denominado cofator (coenzima ou
íon metálico ativo) e sua atividade depende de alguns fatores como estrutura da
proteína, número de cadeias peptídicas, arranjo dessas cadeias na molécula, natureza do
substrato e natureza do grupo prostético (Politzer & Bon, 2006).
Todos os organismos vivos produzem, em pequena ou grande quantidade, certa
variedade de enzimas, sejam elas intra ou extracelulares. As enzimas extracelulares são
excretadas para o meio externo e podem decompor compostos insolúveis como
celulose, quitina e quitosana.
Algumas destas enzimas podem ser usadas tanto nas indústrias de detergentes,
têxteis e farmacêuticas, como também de alimentos, de bebidas, etc., sendo utilizadas
em processos biotecnológicos industriais em substituição aos processos químicos que
muitas vezes causam grandes prejuízos ao meio ambiente. A produção comercial das
enzimas pode ser realizada através do cultivo de microrganismos como fungos e
bactérias.
Na Tabela 1 mostra-se uma comparação das enzimas com os catalisadores
químicos, levando-se em conta algumas das características dos dois catalisadores.
Destaca-se que alguns parâmetros da Tabela 1 demonstram a maior importância das
enzimas, quando comparadas com os catalisadores químicos, principalmente a
especificidade ao substrato, as condições de reação, o consumo de energia e a

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
19
velocidade de reação, como desvantagem cita-se o custo de obtenção (isolamento e
purificação).
Tabela 1 – Comparação das enzimas com os catalisadores químicos (Adaptado de
Furigo Junior & Pereira, 2001).
Característica Enzimas Catalisadores Químicos
Especificidade ao substrato Alta Baixa
Natureza da estrutura Complexa Simples
Sensibilidade à T e pH Alta Baixa
Condições de reação (T, P e pH) Suaves Drástica (geralmente)
Custo de obtenção (isolamento e purificação) Alto Moderado
Natureza do processo Batelada Contínuo
Consumo de energia Baixo Alto
Formação de subprodutos Baixa Alto
Separação catalisador/produtos Difícil/cara Simples
Atividade catalítica (temperatura ambiente) Alta Baixa
Presença de cofatores Sim Não
Estabilidade do preparado Baixa Alta
Energia de Ativação Baixa Alta
Velocidade de reação Alta Baixa
2.1.1. Quitosana e seus derivados
A quitosana e seus derivados apresentam grande importância devido ao
conhecimento de suas funcionalidades biológicas, médicas, tecnológicas, etc
(Shimosaka et al, 1993; Shimosaka et al, 1995; Mitsutomi et al., 1995). A quitosana é
obtida da quitina, polissacarídeo abundante na natureza, extraído principalmente das
carapaças de caranguejos, lagosta e camarões. A quitosana é um polímero natural,
biodegradável, não tóxico e de alta massa molecular obtido a partir de animais
marinhos, insetos e microrganismos (Tabela 2).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
20
Tabela 2 – Fontes naturais de quitina e quitosana. (Adaptado de Polymar, 2003).
Animais
Marinhos
Insetos Aracnídeos Microrganismos
Anelídios Formigas Escorpiões Leveduras
Moluscos Besouros Aranhas Fungos
Lagosta Esporos
Camarão
Caranguejo
Krill
A Figura 1 abaixo mostra a estrutura química da quitosana
Figura 1 – Estrutura da quitosana (Fonte: Polymar, 2003)
Quitosana totalmente desacetilada não existe na natureza. Graus de acetilação
(GA) nferior a 50% a torna mais solúvel em soluções ácidas (Muzzarelli, 1993;
Muzzarelli et al., 1994; Dung et al., 1994; Stoyachenko et al.,1994). Pelletier &
Sygusch (1990) enfatizaram que grande parte dos resíduos sólidos da indústria
pesqueira, principalmente da carcinicultura, é jogada ao mar ocasionando degradação
ambiental. Logo, a utilização desses resíduos como matéria-prima para formação de
novos produtos, não só reduz de forma drástica o impacto ambiental, mas também daria
maior valor agregado à indústria pesqueira.
A demanda cada vez maior de consumidores por alimentos mais saudáveis, sem
conservantes químicos, tem motivado pesquisas à descoberta de novos agentes
antimicrobianos naturais. Sendo assim, a atividade antimicrobiana incomum da

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
21
quitosana e seus derivados contra microrganismos, tais como bactérias, bolores e
leveduras tem tido maior atenção ultimamente.
Devido à interação entre a quitosana, que é positivamente carregada, e as
membranas celulares dos microrganismos, negativamente carregadas, ocorre a formação
de verdadeiros poros na membrana, que são canais de vazamento de conteúdo protéico e
outros componentes intracelulares. A quitosana tem também a propriedade de agir como
agente quelante que se liga de forma seletiva a traços de metais, inibindo, assim, a
produção de toxinas e o crescimento microbiano. Ainda é capaz de ativar mecanismos
de defesa em tecidos hospedeiros, atuando como agente sequestrante de água e inibindo
várias enzimas. A ligação da quitosana com DNA e a inibição da síntese do mRNA
ocorre via penetração da quitosana no núcleo dos microrganismos e interferência na
síntese do mRNA e proteínas, (Craveiro et al., 2004).
Em um estudo clínico conduzido por Giustina & Ventura (1995) com 100
pessoas com sobrepeso de 10 a 25%, ao final de 4 semanas as pessoas que receberam
quitosana demonstraram perda de peso significativa (7,3 Kg contra 3,0 Kg nos grupos
controle). As pessoas que tomaram quitosana também demonstraram redução na pressão
sanguínea sistólica (de 145 para 135 mm Hg) e diastólica (92.6 para 84.2 mm Hg),
assim como uma redução na taxa respiratória (27.6/min a 21.2/min).
Kondo & Osada (1996) pesquisaram a influência da quitina e da quitosana na
biodisponibilidade do zinco em ratos alimentados com uma dieta com suplementos de
5% ou sem fibra dietética (quitina, quitosana, celulose, pectina e agar-agar) durante 31
dias. Observaram que os ratos alimentados com dietas contendo 5% de fibras, exceto
quitosana, o consumo de alimento foi maior do que o controle. O ganho de peso em
todos os grupos alimentados com fibra foi maior do que o controle, exceto para o grupo
suplementado com quitosana (5%) onde a ingestão de alimento e o ganho de peso foram
menores do que os grupos controle e das demais fibras.
Segundo Craveiro et al. (2004), derivados da quitosana obtidos pela reação com
ácidos carboxílicos, podem promover ao polímero uma solubilidade especial em água.
Esses compostos possuem estrutura química semelhante ao ácido hialurônico e
fornecem uma excelente compatibilidade com o tecido epitelial. As soluções obtidas
com esses derivados podem ser facilmente incorporadas a formulações cosméticas,
mostrando melhor compatibilidade com surfactantes iônicos e emulsificantes,
apresentando estabilidade em uma larga faixa de pH.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
22
Han et al. (1999) examinaram os efeitos da quitosana sobre a atividade da lipase
pancreática in vitro e sobre o grau de armazenamento de gorduras induzido em ratos
pela administração oral de uma dieta rica em gorduras por nove semanas. Como
resultado, observou-se que os animais alimentados com quitosana, reduziram o peso
corporal, a hiperlipidemia e a gordura hepática. Como conclusão, os autores propõem
que os efeitos anti-obesidade da quitosana em ratos alimentados com alto teor de
gordura se devem parcialmente a inibição da absorção intestinal das gorduras dietéticas.
Em outro ensaio, Zahorska-Markiewicz et al. (2002) realizaram um estudo para
avaliar o efeito da quitosana no tratamento da obesidade. Segundo os autores do estudo,
a quitosana pode ser usada como um valioso e seguro coadjuvante em tratamentos para
a obesidade em longo prazo. A quitosana parece acentuar a redução da pressão arterial,
juntamente com a perda de peso.
Assis (2010) estudou a produção e caracterização de quitooligossacarídeos
produzidos pelo fungo Metarhizium anisopliae e avaliação da citotoxicidade em células
tumorais. A hidrólise da quitosana foi realizada em tempos de 10 a 60 min para a
produção de quitooligossacarídeos. A avaliação da citotoxicidade dos oligômeros de
quitosana foi realizada em células tumorais (HepG2 e HeLa) e não tumoral (3T3). A
produção de oligômeros de quitosana teve maiores rendimentos durante 10 minutos de
hidrólise, os pentâmeros apresentaram concentração de 0,15 mg/mL, no entanto os
hexâmeros, que apresentam maior interesse por causa de suas propriedades biológicas,
só foram detectados com 30 minutos de hidrólise com uma concentração de 0,004
mg/mL.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o planejamento Plackett-Burmam e a metodologia de superfície de resposta.
Os resultados mostraram que a eficiência máxima de extração da quitina foi conseguida
nas seguintes condições: concentração de casca de camarão 50g/L, glicose 50g/L, tempo
de incubação de 5 dias e inoculo como 0,05 OD (densidade óptica).
A glicosamina, unidade monomérica básica da quitosana, possui atividade
antiinflamatória e tem grande importância na construção e manutenção da cartilagem,
no tratamento de enfermidades como: febre reumática, artrose e principalmente a artrite
e osteoartrite na forma aguda ou crônica (Polymar, 2003). A Figura 2 apresenta a
estrutura química da glicosamina.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
23
Figura 2 – Estrutura da glicosamina (Fonte: Polymar, 2003)
O sulfato de glicosamina é um derivado da quitosana sendo utilizado no
tratamento da osteoartrite sintomática dos joelhos, quadris, coluna, mãos e outras
regiões (Craveiro et al., 2004). O sulfato de glicosamina estimula a síntese de
proteoglicanos necessários para o bom funcionamento das articulações, estimula a
regeneração das cartilagens e ainda incorpora enxofre ao tecido cartilaginoso. Estudos
de longa duração, acima de 3 anos, mostram que o sulfatos de glicosamina evita a
progressão de danos estruturais nas articulações e induz melhora significativa nas dores
e funções físicas (Craveiro et al., 2004). A Figura 3 apresenta a estrutura química do
sulfato de glicosamina.
Figura 3 – Estrutura do sulfato de glicosamina (Fonte: Polymar, 2003)

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
24
2.1.2. Quitosanases
Dependendo do tipo de quitosanase obtida, ocorrem diferentes mecanismos de
hidrólise da quitosana, isso devido a diferentes microrganismos produzirem enzimas
com diferenças nas suas estruturas (Kurita et al., 1977; Seino et al., 1991).
Quitosanases (EC 3.2.1.132) são enzimas que catalisam a reação de hidrólise de
quitosana em oligossacarídeos. As enzimas quitosanases são produzidas por vários
microrganismos, bactérias, actinomicetos e fungos, e até por algumas plantas. Estas
enzimas agem em polímeros com grau de acetilação de 30% a 60%. Os monômeros
resultantes da hidrólise da quitosana por estas enzimas são melhores digeridos, ao serem
ingeridos, como é o caso da glicosamina. Estudos efetuados até o presente momento
classificam as quitosanases em cinco famílias: 5, 8, 46, 75, 80, sendo que a família 46 é
a mais estudada. As quitosanases tem grande aplicação na geração de oligômeros de
quitosana (Somashekar & Joseph, 1996; Chen et al., 2005).
Nogawa et al. (1998) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida pelo fungo Trichoderma reseei PC-3-7. Estes pesquisadores
inocularam 106 conídios em 100 mL de um meio basal contendo 0,3% de N-
acetilglicosamina em incubator rotativo com agitação de 220 rpm a 28oC. Destaca-se
que o fungo não produziu quitosanase em meio contendo quitosana ou quitina.
Entretanto, o microrganismo produziu quitosanase em meio contendo glicosamina e N-
acetilglicosamina, sendo que para este último obteve-se uma atividade específica
máxima de 1,5 U/mg após 72 horas de incubação. É importante ressaltar que uma
agitação vigorosa favoreceu a produção da enzima. Esse artigo ilustra a presença de
exo-β-D-glicosaminidase e exo-β-D-N-acetilglicosaminidase.
Piza et al. (1999) estudaram a produção, purificação e caracterização parcial de
uma quitosanase produzida por Bacillus cereus. Nesse caso, investigou-se através de um
planejamento fatorial fracionário a influência dos fatores: concentração de sulfato de
amônio (0,4; 2,2 e 4,0%), razão de aeração (2; 6 e 7), tempo de fermentação (16; 24 e
32 horas), pH (5; 6 e 6,5) e concentração de quitosana (1,0; 1,5 e 2,0%). Os resultados
mostraram que os fatores mais significativos para esse planejamento foram a
concentração de sulfato de amônio, a aeração, o pH e a interação entre sulfato de
amônio e a aeração.
Cheng & Li (2000) estudaram a preparação de oligossacarídeos utilizando
quitosanase produzida por Aspergillus sp e constataram que a enzima foi largamente

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
25
induzida em meio contendo quitosana como única fonte de carbono e também na
presença de quitina coloidal. A indução da enzima foi realizada em temperaturas
variando na faixa de 28 a 30ºC, pois essa espécie não cresceu bem em temperaturas
superiores a 37ºC.
Zhang et al. (2000) estudaram a produção, purificação e caracterização de
quitosanase e Exo-β-D-Glucosaminidase por Aspergillus oryzae IAM2660. Os
microrganismos utilizados neste estudo foram mantidos em placas de Petri contendo
agar para contagem em placas (PDA). Destaca-se que para produzir a enzima, conídios
foram retirados das placas e inoculados em meio Czapeq-dox com os seguintes
componentes: 1,0g K2HPO4, 0,5g KCl, 2,0g NaNO3, 0,5g MgSO4.7H2O, 0,02g
FeSO4.7H2O, 20g de sacarose e 1,0L de água deionizada, com pH do meio igual a 5,5.
Foi adicionado ao meio 0,5 g de peptona e 0,5g de extrato de levedura com
concentração de esporos da ordem de 1,0x106 esporos.mL-1. Após a incubação, feita em
incubador rotativo a 26ºC por um período de 60h, micélios foram coletados e após
filtração e secagem a 80ºC foram utilizados para ensaio de quitosanase.
Kuroiwa et al. (2002) estudaram os fatores que afetam a composição de
oligossacarídeos produzidos através da hidrólise de quitosana por quitosanase originária
de Bacillus pumilus BN-262. Neste experimento foi utilizado quitosana 100%
desacetilada dissolvida em ácido acético 1M. O pH do meio foi ajustado em 5,6. Neste
estudo a quitosanase foi imobilizada em agar e Sepharose. Os resultados mostraram que
a concentração de enzima, a temperatura, a velocidade de agitação, e concentração de
substrato influenciaram de forma significativa na composição dos oligossacarídeos
produzidos.
Ichikawa et al. (2002) estudaram a imobilização e estabilização da ligação
multiponto da quitosanase tendo agar como suporte e concluíram que a imobilização
aumentou a termoestabilidade da quitosanase. Os oligossacarídeos de quitosana foram
continuamente produzidos usando um reator coluna contendo quitosanase imobilizada.
A percentagem de quitosana imobilizada após 28 dias de reação foi de 44%.
Hung et al. (2002) estudaram a purificação, através de troca iônica, e
caracterização da hidrólise sobre ação da quitinase e quitosanase purificadas do meio
contendo Bromelina comercial. Destaca-se que a quitinase purificada apresentou pH,
temperatura e Km ótimo para ação catalítica em meio contendo quitina de 4,0, 60ºC e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
26
0,2mg.mL-1, respectivamente. Enquanto que quitosanase apresentou o pH, temperatura e
Km ótimo de 3,0, 50ºC e 0,88mg.mL-1, respectivamente.
Zhu et al. (2003) estudaram as melhores condições fermentativas e propriedades
de uma quitosanase produzida por Acinetobacter sp. C-17. A espécie de bactéria
utilizada neste ensaio foi isolada do solo e identificada como sendo Acinetobacter sp C-
17 e capaz de produzir quitosanase. O microrganismo foi mantido em placas contendo o
meio agar com os seguintes componentes: 0,5 % de quitosana, 0,2 % K2HPO4, 0,1 %
KH2PO4, 0,07 % MgSO4, 0,05 % NaCl, 0,05 % KCl, 0,01 % CaCl2, 0,05 % de extrato
de levedura e 2 % agar, o pH do meio de manutenção foi mantido em 7,2 e a
temperatura foi mantida em 30ºC durante 3 a 5 dias, para identificar a ação da enzima
quitosanase. A ação da quitosanase foi identificada através da observação de uma
colônia de microrganismo existente na placa. A bactéria foi então isolada da placa e
colocada em fermentador rotativo a 200 rpm durante 72h em meio contendo: 0,07 %
K2HPO4, 0,03 % KH2PO4, 0,05 % MgSO4, 0,03 % peptona, 0,03 % de extrato de
levedura e 1,0 % de quitosana. O pH foi de 7,0 e a temperatura 30ºC. Como fontes de
carbono foram utilizadas além de 1,0 % de quitosana, 1,0 % de quitina e 1,0 % de
glicose. Os resultados mostraram que as melhores condições para produção da
quitosanase foram alcançadas quando se utilizou 1,0 % de solução de quitosana a 30ºC,
pH 7,0 durante 24 horas de incubação.
Liu et al. (2003) estudaram a produção de quitinase por Verticillium lecanii
F091 usando fermentação submersa, a atividade de quitinase foi de 9,95 mU/mL em um
meio de cultura otimizado, com um volume de cultura de 200 mL, agitação de 150 rpm
e temperatura de 24ºC cultivado em incubador rotativo e concluíram que a taxa de
agitação e o pH foram os fatores mais significativos para a produção de quitinase, no
entanto, as taxas de agitação e aeração poderiam modificar a concentração de oxigênio
dissolvido afetando diretamente no crescimento do Verticillium lecanii e na produção
de quitinase.
Donzelli et al. (2003) estudaram a hidrólise enzimática avançada de quitina de
casca de caranguejo langostino com misturas de enzimas de bactérias e fungos e
concluiram que a combinação de enzimas de Trichoderma atroviride e Serratia
marcescens foram capazes de degradar completamente altas concentrações de quitina
(100g/mL) a partir de carapaças de caranguejo langostino convertendo a N-
Acetilglicosamina (78%), glucosamina (2%) e quitobiose (10%). Enzimas de fonte

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
27
procariótica apresentaram atividades menores que as enzimas de T. atroviride, para
catalisar a reação com quitosana.
Jo et al. (2003) estudaram a caracterização e a avaliação cinética de quitosanase
produzida por Bacillus sp. Destaca-se que na produção da enzima utilizada neste ensaio
o meio continha 0,5% de concentração de quitosana, 1% de triptona e 1% de NaCl, o
pH do meio foi ajustado em 5,5 e a temperatura foi mantida em 37ºC com agitação de
180 rpm em incubador rotativo.
Fu et al (2003) estudaram a caracterização de três iso-enzimas de quitosanase
isolada de pepsina comercial e denominado como PSC-I, PSC-II, PSC-III,
respectivamente, em relação a hidrólise enzimática da quitosana, o meio ótimo para a
produção destas iso-enzimas continha as seguintes características: pH 5,0; 5,0 e 4,0 e
temperaturas de 40ºC, 40ºC e 30ºC, o valor de Km foi de 5,2, 4,0 e 5,6 mg.mL-1,
respectivamente. As massas moleculares das três iso-enzimas foram de 40 kDa. Os
resultados mostraram que as três iso-enzimas apresentaram atividade em meio contendo
quitosana, produzindo oligômeros da ordem de 68 – 88% desacetilada.
Choi et al. (2004) estudaram a produção, purificação e caracterização total de
uma quitosanase produzida por Bacillus sp KCTC 0377BP. Nesse caso, com relação à
produção da enzima foi possível obter um aumento de 1,2 U/ml em incubator rotativo
para 100,0 U/ml em fermentador piloto de 500 L, utilizando uma concentração de
substrato de 0,5% de quitosana e como fonte de nitrogênio polipeptona (2,5%) em pH
igual a 6,8, com 100 rpm de 100,0 rpm a 30oC.
Shimosaka et al. (2003) estudaram a produção, purificação e caracterização total
de duas quitosanases produzida por Acinetobacter sp CHB101. Nesse caso, com relação
à produção da enzima os pesquisadores usaram dois meios, um contendo glicose e outro
contendo quitosana. Com relação à cinética, o meio contendo quitosana fornece um
tempo de geração de 280 minutos, enquanto que no meio contendo glicose a taxa de
crescimento foi mais elevada e o tempo de geração foi de apenas 110 minutos. Os
autores observaram que após os dois primeiros dias de cultivo a atividade
quitosanolítica permaneceu baixa (na ordem 0,10 a 0,20 U/mL). Durante a fase de
crescimento exponencial o valor da atividade permaneceu com valor inferior a 0,01
U/mL, começando a aumentar ao atingir a fase estacionária, indicando assim que a
produção da enzima é não associada ao crescimento.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
28
Kim et al. (2004) estudaram a produção, purificação e caracterização de uma
quitosanase produzida pelo Bacillus sp e identificado pelo número 1299. Destaca-se que
na produção da enzima quitosanase foi utilizado um meio contendo os seguintes
componentes: 0,5 % de quitosana, 0,42 % (NH4)2SO4, 0,2 % K2HPO4, 0,02 % uréia,
0,03 % CaCl2.2H2O, 0,03% MgSO4.7H2O, 0,1% peptona, 0,2% Tween 80, 0,2 % da
solução (0,5% FeSO4.7H2O, 0,16% MnSO4.H2O, 0,14% ZnSO4.7H2O, 0,2% CoCl2) e
1,7% agar. O pH do meio para este ensaio foi mantido em 7,0. Este microrganismo foi
escolhido por ser grande produtor de quitosanase.
Chen et al. (2005) estudaram a purificação e a caracterização de dois tipos de
quitosanase produzidas por Aspergillus sp. CJ22-326 em cultivo submerso. Na
produção da enzima foi utilizado um meio de cultivo contendo 1,0% de quitosana, 2,0%
de gérmen de trigo, 0,2% de sulfato de amônio, 0,2 % de fosfato de potássio e 0,05% de
sulfato de magnésio por litro de solução em pH 5,6. O cultivo foi realizado em frasco de
500 mL contendo 150 mL de meio de cultura a 30ºC por 96 horas com uma agitação de
150 rpm. Os autores apresentam um gel de eletroforese com as duas enzimas purificadas
com massa molar de 29 kDa e 109 kDa.
Silva Filho et al. (2005) estudaram, através de planejamento fatorial 24 com
repetição em triplicata no ponto central, a produção de quitosanase por Aspergillus
ochraceus em cultivo descontínuo submerso. Os resultados mostraram que foi possível
produzir quitosanase com atividade na ordem de aproximadamente 5,9 U/mL utilizando
Aspergillus ochraceus e que a atividade foi favorecida pelo aumento da agitação (r), da
razão de aeração (RA) e da concentração de substrato (Cs), enquanto que o aumento da
temperatura de cultivo (TMC) não favoreceu a resposta (atividade quitosanolítica).
Pode-se classificar as quitosanases em endoquitosanases e exoquitosanases. As
primeiras diferenciam-se por catalisar a hidrólise aleatoriamente, no interior do
biopolímero, gerando diversos tamanhos de oligossacarídeos. As exoquitosanases são
capazes de hidrolisar as terminações não redutoras, produzindo assim unidades
redutoras (Peter, 2005).
Palma-Guerrero et al. (2008) estudaram o biocontrole de fungos patogênicos e
demonstraram a lise da membrana plasmática e a inibição da germinação de esporos de
fungos. Eles observaram que os fungos entomopatogênicos e nematófagos foram menos
inibidos pela quitosana que os fitopatogênicos e micoparasíticos provavelmente porque
apresentam a enzima quitosanase que degrada a quitosana. Mesmo assim, a quitosana

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
29
pode ajudar a desenvolver novas estratégias para o biocontrole em geral de fungos
patogênicos, evitando o uso de fungicidas que prejudicam o meio ambiente e a saúde
humana.
Pagnoncelli (2008) estudou a produção de quitooligossacarídeos com
propriedades nutracêuticas a partir da quitosana por hidrólise enzimática, utilizando
processo fermentativo simultâneo. Duas cepas produtoras de quitosanaes foram
selecionadas, Paenibacillus chitinolyticus e Paenibacillus ehimensis, e avaliadas quanto
ao comportamento em meio de cultivo contendo açúcares simples e em relação às
variações de pH do meio. Os quitooligossacarídeos produzidos a partir do complexo
enzimático da primeira cepa apresentaram maior pico após 9h de hidrólise, enquanto
para a segunda cepa avaliada, após 20min, observou-se quitooligossacarídeos com grau
de polimerização entre 3 e 6 unidades.
2.2. Fungos
2.2.1. Características gerais
Desde o final dos anos 60 que os fungos deixaram de pertencer ao reino das
plantas e passaram a ter um reino próprio chamado de Reino Fungi. Foi a partir de
estudos morfológicos, bioquímicos e citológicos onde se concluiu que eles são tão
diferentes dos vegetais como dos animais. Distinguem-se de outros organismos
eucariotas por serem seres quimiorganoheterotróficos e, ainda, por apresentarem parede
celular rígida que é composta por quitina e glucano e uma membrana celular em que o
ergosterol substitui o colesterol (Trabulsi & Toledo, 1996; Murray et al., 2005).
Fazem parte do Reino Fungi cerca de 100 mil espécies, desde grandes como
os cogumelos, até microscópicas como os bolores e leveduras. Ainda hoje é utilizada
essa classificação, mas os avanços da biologia molecular e estudos bioquímicos levaram
a outras propostas de classificação em que os fungos e organismos afins foram incluídos
em três diferentes reinos: Protozoa, Chromista e Fungi (Kirk et al., 2008).
Abaixo, são apresentadas características que diferenciam os fungos das plantas:
• são incapazes de sintetizar clorofila, como também não apresentar celulose em
sua parede celular, com exceções como fungos aquáticos;
• apresentam algumas características de células animais (capacidade de depositar
glicogênio e a presença de substâncias quitinosas, em sua parede celular);

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
30
• podem ser eucarióticos (as leveduras), ou multinucleados (os bolores), e
apresentar retículo endoplasmático rugoso e mitocôndrias em seu citoplasma;
• são heterotróficos, alimentam-se de matéria orgânica viva (fungos parasitários),
ou morta (fungos saprofíticos);
Os fungos são amplamente encontrados na natureza, crescem rapidamente no
solo, na água, na atmosfera, bem como nos animais, nos vegetais, resíduos orgânicos e
desempenham um papel importante na vida do homem, quer de uma maneira benéfica,
quer de um modo prejudicial. Os fungos são um dos principais microrganismos
responsáveis pela decomposição da matéria orgânica, interferindo no ciclo do carbono,
do nitrogênio e de outros nutrientes da biosfera. São capazes de deteriorarem produtos e
bens de consumo, tais como alimentos, tecidos, papel, madeira. Os alimentos
armazenados representam um vasto meio para a proliferação dos fungos, principalmente
quando não são consideradas as condições básicas de armazenamento correto (Trabulsi
& Toledo, 1996).
Estão presentes funcionalmente em vários processos industriais, tais como, de
fabricação de pão, cervejas, vinhos e determinados tipos de queijos, sendo também
utilizados na produção comercial de muitos ácidos orgânicos, de alguns fármacos, como
a ergometrina e a cortisona, na obtenção de diferentes antibióticos, como a penicilina, e
de substâncias imunossupressoras, como a ciclosporina (Esteves et al., 1990). A Figura
4 ilustra a estrutura geral de uma célula de fungo.
Figura 4 – Representação da estrutura geral de uma célula de fungo.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
31
Os fungos podem se desenvolver em meios de cultivo especiais formando
colônias de dois tipos, leveduriformes e filamentosas, que se diferenciam pela
macromorfologia e micromorfologia. O primeiro tipo apresenta, em geral, característica
de consistência cremosa de cor branca a creme, brilhantes ou opacas, podendo
apresentar às vezes coloração escura ou alaranjada, e são formados por microrganismos
unicelulares que apresentam as funções vegetativas e reprodutivas. As filamentosas
apresentam características granulares, cotonosas, aveludadas ou pulverulentas, são
multinucleadas e apresentam-se em forma de tubos denominadas de hifas. Essas hifas
podem ser não septadas (cenocíticas) ou septadas (tabicadas). O conjunto de hifas é
denominado micélio.
As hifas asseptadas têm aparência anastomosada (concisa), formada por um
citoplasma estendido e polinucleado, enquanto que as hifas septadas contêm células
individualizadas, cada uma contendo o seu núcleo. Na Figura 5 são mostradas as
estruturas das hifas septadas e não septadas.
Figura 5 – Estrutura das Hifas: a) septadas e b) asseptadas. (Adaptado de Guia.Bio,
2012)
O micélio que se desenvolve no interior do substrato e que é responsável pela
sustentação e absorção de nutrientes é denominado micélio vegetativo. O micélio que
cresce na superfície do meio de cultura é denominado micélio aéreo. Este pode ser
denominado também de micélio reprodutivo, quando o mesmo se diferencia para
sustentar os corpos de frutificação ou propágulos (Putzke & Putzke, 1998). Os
propágulos classificam-se de acordo com a origem em externos e internos, sexuados e
assexuados.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
32
2.2.2. Reprodução dos Fungos
A reprodução dos fungos ocorre em ciclos assexuais, sexuais e parassexuais. Em
fungos, a reprodução assexuada é observada com mais freqüência do que a reprodução
sexuada. A reprodução assexuada é normalmente a reprodução mais importante para a
propagação da espécie, feita através de fragmentação de artroconídios, fissão de células
somáticas, brotamento ou gemulação do blastoconídios-mãe e produção de conídios. A
reprodução sexuada envolve a união de duas células ou de dois órgãos sexuais
sexualmente compatíveis. A reprodução sexuada culmina na produção de basidiósporos,
no caso de basidiomicetos. A reprodução parassexuada ocorre com a fusão de hifas e
formação de um heterocário, que consiste na existência de núcleos haplóides
geneticamente distintos num mesmo citoplasma, os quais, após a fusão, originam
núcleos diplóides heterozigotos sujeitos a sucessivas divisões mitóticas. Apesar da
raridade dessas, a reprodução parassexual é de grande importância na evolução de
alguns fungos (Pelczar et al., 1996).
A reprodução assexuada é feita por conídios, esporos assexuados formados no
ápice de hifas modificadas, os conidióforos. Os conidióforos não possuem esporângios
de forma que os conídios ficam expostos. Na reprodução sexuada, com o encontro de
duas hifas haplóides distintas, ocorre a plasmogamia e um novo conjunto de hifas
dicarióticas é formado.
Os conídios podem ter formas esféricas, fusiformes, cilíndricas, piriformes, entre
outras. Apresentam-se como hialinos ou pigmentados, podem estar isolados ou
agrupados, a parede celular pode ser lisa ou rugosa e podem ser formados por apenas
uma célula ou apresentarem septos em mais de um plano.
Os conídios também podem originar-se em qualquer parte do micélio vegetativo
e são denominados conídios sésseis como é o caso do Trichophyton rubrum.
Alguns fungos patogênicos são identificados a partir de estruturas formadas pela
célula conidiogênica e o conidióforo. Essas estruturas são denominadas: aparelho de
frutificação ou conidiação. Os conídios podem formar cadeias sobre fiálides em volta de
uma vesícula, dilatando a extremidade do conidióforo. Essas estruturas ocorrem em
Aspergillus, apresentando-se em forma de cabeça ao redor de uma vesícula, e em
Penicillium agrupados em forma de pincel, como é ilustrada na Figura 6.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
33
Figura 6 – Conídios de Aspergillus e Penicillium
2.2.3. Metabolismo de fungos
Os fungos são seres aeróbicos e heterotróficos, podendo se desenvolver em
ambientes com baixa quantidade de oxigênio, como ocorre no crescimento vegetativo e
na reprodução assexuada, como também em ambientes com grande quantidade de
oxigênio como é o caso da reprodução sexuada.
Os fungos são capazes de produzir certas enzimas como lipases, invertases,
lactases, quitosanases, proteases, amilases, etc., que fazem a hidrólise do substrato
facilitando o mecanismo de transporte ativo e passivo. Esses substratos induzem a
formação de enzimas degradativas. Algumas espécies de fungos desenvolvem-se em
meios que apresentam amônia ou nitrilos como única fonte de nitrogênio. As melhores
fontes orgânicas para o seu desenvolvimento são carboidratos, como a D-glicose, e
alguns sais minerais como sulfatos e fosfatos.
A água é uma necessidade de todos os seres vivos, não sendo diferente para os
fungos que precisam de água para o seu desenvolvimento. Certos fungos apresentam
características halofílicas podendo crescer nos ambientes com grandes concentrações de
sal.
A razão de não existirem ambientes livres de espécies fúngicas deve-se ao facto
destes microrganismos conseguirem sobreviver em situações extremas como, por
exemplo, temperaturas que variam entre os - 5 e os 60 ºC, com pH de 1 a 9 e com baixas

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
34
concentrações de oxigênio, apesar de cada espécie possuir condições ideais para o seu
desenvolvimento (Chao et al., 2002).
Devido a grande faixa de temperatura ótima para o seu crescimento, os fungos
apresentam espécies psicrófilas, mesófilas e termófilas. Para a maior parte das espécies
fúngicas, a temperatura de desenvolvimento situa-se entre os 20 e 24 ºC e não crescem
acima dos 29 ºC. Em relação à umidade relativa, necessitam de pelo menos 50 %, mas
geralmente necessitam de mais de 65 % (Colakoglu, 2001).
O pH ideal para o desenvolvimento dos fungos varia na faixa entre 5 e 7.
Destaca-se, entretanto, que a maioria dos fungos suportam grandes variações de pH
como ocorre com os fungos filamentosos que podem crescer em pH variando entre 1,5 e
11. Por outro lado, as leveduras não suportam pH alcalino.
Os fungos crescem lentamente, se comparados com as bactérias, em meio de
incubação para o seu desenvolvimento. Para evitar contaminação por bactérias, que
podem se sobrepor ou inibir o seu crescimento, pode-se introduzir no meio de cultura
um antibacteriano de largo espectro como o cloranfenicol.
Algumas espécies de fungos se desenvolvem na presença de luz, outras são
inibidas e algumas podem crescer tanto na presença quanto na ausência deste agente. No
entanto a irradiação solar, em geral, é prejudicial ao seu desenvolvimento, por causa da
radiação ultravioleta.
2.2.4. Nutrição de fungos
Quanto ao aspecto da nutrição, os fungos são heterótrofos, ou seja, incapazes de
produzir seu próprio alimento. Entretanto, a forma de assimilar matéria orgânica é
diferente daquela utilizada pela maioria dos animais, que são heterótrofos por ingestão.
Nos fungos, a digestão é extracorpórea, ou seja, é realizada fora do corpo. O fungo
lança no ambiente, enzimas que degradam as moléculas orgânicas complexas e, depois,
absorve moléculas menores, mais simples. Por essa razão, são designados heterótrofos
por absorção. É devido a este processo que os fungos podem crescer dentro ou sobre os
alimentos. Desenvolve-se geralmente em meios contendo um pH baixo, uma fonte de
carbono, uma fonte de nitrogênio orgânico ou inorgânico e alguns minerais. Alguns
necessitam de vitaminas. Quanto aos modos de vida, os fungos podem ser
decompositores, parasitas, mutualísticos e predadores (Raven, 2001).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
35
Os fungos decompositores, saprófagos (do grego, saprós, “podre”, e phagos,
“comedor”), juntamente com as bactérias, são os principais decompositores da biosfera,
participando intensamente do processo de degradação da matéria orgânica morta, o que
promove a reciclagem dos elementos químicos constituintes dos seres vivos. A maioria
deles vive no solo, obtendo nutrientes de seres mortos. Em florestas, onde a matéria
orgânica vegetal é abundante, os micélios dos fungos decompositores absorvem os
nutrientes de folhas e galhos caídos, decompondo-os. Os fungos decompositores
também são responsáveis pelo apodrecimento de alimentos e de outros materiais, como
a madeira.
Os fungos parasitas vivem à custa de outros organismos vivos, prejudicando-os.
Ao parasitar o corpo de um ser vivo, animal ou vegetal, o fungo pode até provocar sua
morte. Várias doenças são causadas por fungos parasitas. Nas plantas, cita-se a
ferrugem do café, que, no Brasil, é motivo de grande preocupação por parte dos
cafeicultores, devido aos prejuízos que acarreta. As micoses que atacam a pele de seres
humanos e animais são provocadas por fungos.
Existem fungos que se associam a outros organismos, estabelecendo uma relação
em que há benefício mútuo para os indivíduos envolvidos. Esses fungos são
denominados mutualísticos. A maioria vive associada a seres fotossintetizantes, como
as plantas, cedendo-lhes parte da água e dos nutrientes que as hifas absorvem do solo.
As plantas, por sua vez, cedem ao fungo certos açúcares e aminoácidos. Duas
associações mutualísticas formam estruturas bem características: as micorrizas,
associação que envolve fungos e raízes de plantas, e os líquens, formados por fungos e
algumas variedades de algas e cianobactérias.
Certos fungos atuam como predadores. Na maioria dos casos, as hifas secretam
substâncias aderentes que aprisionam os organismos que tocam os fungos. Dessa
maneira, as hifas penetram o corpo da presa, crescem e se ramificam, espalhando-se no
interior do organismo e absorvendo seus nutrientes, causando-lhe a morte. O gênero
Arthrobotrys apresenta uma forma mais elaborada de predação. O fungo vive no solo e
captura nematódeos microscópicos que habitam o mesmo ambiente. As hifas
apresentam pequenos anéis que se estreitam quando a presa passa por eles. As hifas do
fungo logo invadem o corpo do nematódeo, digerindo-o.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
36
2.2.5. O fungo Metarhizium anisopliae
Existem três espécies de fungos do gênero Metarhizium, divididas em dez
variedades: M. anisopliae, variedades anisopliae, majus, lepidiotum e acridum; M.
flavoviride, variedades tipo E, flavoviride, minus, novazealandicum e pemphigum; e M.
album (Driver et al. 2000). E, mais recentemente, foi descrita na China uma nova
variedade de M. anisopliae, dcjhyium (Dong et al. 2009).
Data de 1879 quando o russo Ilya Metchnikoff aplicou o fungo para combater
larvas de um curculionídeo, praga de beterraba, e foi classificado como Metarhizium
anisopliae por Sorokin em 1883. É um fungicida natural para mais de 300 espécies de
insetos, especialmente pragas da agricultura e pecuária. No Brasil, o fungo Metarhizium
anisopliae teve a sua aplicação com sucesso pela primeira vez em 1965, no controle da
cigarrinha-da-cana Mahanarva posticata (Alves, 1998).
O fungo Metarhizium anisopliae é um importante agente entomopatogênico
utilizado no controle de pragas, como cupins, gafanhotos, cigarrinhas e besouros, sendo
amplamente estudado em todo o mundo. Existe amplamente na natureza e é facilmente
encontrado no solo, onde sobrevive por longos períodos e seu desenvolvimento se dá
geralmente entre 15 e 32 °C, sendo ideal entre 24 e 30 °C e com pH ótimo igual a 6,9.
Apresenta micélio hialino e septado, com conidióforos característicos, sobre os quais
surgem conídios cilíndricos organizados em colunas. Este fungo ao atacar os insetos
provoca rigidez e os recobre por uma camada pulverulenta de conídios, resultando em
colorações que variam do verde claro ao escuro, cinza ou branco com pontos verdes
(Tinline, 1971; Alves, 1998, Driver et al. 2000; Arruda, 2005).
Quanto a sua morfologia, apresenta-se como um fungo filamentoso, com corpo
de frutificação equivalente a um esporodóquio agregado a hifas intimamente
entrelaçadas, com massa compactada de conidióforos característicos, simples ou
ramificados, formando células esporogênicas denominadas fiálides, resultando nos
fialosporos (Wang et al., 2002).
O Metarhizium anisopliae é um fungo de cultivo simples, sendo necessário
quase que exclusivamente o nutriente a base de amido. Um dos fungos mais estudados
em programas de manejo de pragas já que apresenta grande potencialidade
entomopatogênica (Onofre et al., 2002). O Metarhizium anisopliae é um microrganismo
pouco exigente, pois pode se desenvolver em diversos meios de cultura como: amido,
glicose, glicerina, levulose, maltose, sacarose e quitina (Jabor et al., 2003).

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
37
Em geral, os fungos produzem enzimas que convertem os tecidos dos insetos em
nutrientes. Porém, a cutícula esclerotizada dos insetos não é bem utilizada pelos fungos,
excetos pelos entomopatogênicos que desenvolveram enzimas eficientes para degradar
essa camada protetora. Destaca-se a enzima Pr1, produzida por Metarhizium anisopliae,
que tem um poder de degradação bem superior a qualquer outra enzima sintetizada por
esse grupo de fungos (Dias, 2005). Desta forma, o Metarhizium anisopliae (fungo
entomopatogênico) ao atacar a célula dos insetos, produz grande quantidade de enzimas
que degradam a membrana citoplasmática e inativa a célula e, consequentemente, mata
o inseto, logo é evidente a relação entre o controle de pragas e produção de enzimas.
No momento, sobre a comercialização de fungos entomopatogênicos no Brasil,
em sua maioria, são formulados em grânulos, compostas do fungo mais o substrato
(arroz + fungo), como também na forma de pó molhável (WP), resultado da moagem do
fungo com o substrato. Esses produtos representam 90% e 8,5%, respectivamente, da
produção de fungos no Brasil, e sendo suas concentrações finais ficam em torno de
5x108 conídios/g para Metarhizium anisopliae e 1x109 conídios/g para Beauveria
bassiana (Alves et al., 2008).
Segundo Batista Filho et al. (2002), Garcia et al. (2005) e Mello et al. (2006), o
Metarhizium anisopliae tem sido bastante utilizado para o controle de diferentes pragas
no Brasil, como a broca da bananeira, pragas de grãos armazenados, carrapatos, cupim
de montículo em pastagens, larvas de escarabeídeos que atacam a cana de açúcar e
cigarrinha da cana-de-açúcar.
É comum a ocorrência natural do fungo Metarhizium anisopliae atacando
cigarrinha-das-raízes de canaviais pelo país, o que incentiva a utilização dele como
agente de controle biológico em áreas infestadas por esse tipo de praga. A aplicação de
recursos no uso de programa de manejo integrado da cigarrinha tem sido reforçado por
razões ambientais e econômicas (Dinardo-Miranda, 2001).
Faria & Magalhães (2001) fizeram uma avaliação do uso de fungos
entomopatogênicos no Brasil, na qual observaram que a qualidade dos micoinseticidas
disponíveis no Brasil poderia ser incrementada de forma considerável quanto a sua
formulação, ou seja, antes da venda, poderiam realizar melhorias no tratamento
posterior, adição de substâncias que lhes assegurem melhorias na eficiência de controle,
capacidade de armazenamento ou praticidade de manuseio, ou de qualquer outro critério
que resulte em vantagem em relação ao produto bruto. Ressaltaram a pouca praticidade

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
38
em alguns casos, como exigência de lavagem, e em outros podendo causar o
entupimento de bicos dos pulverizadores devido à elevada proporção de inertes,
principalmente quando são empregados baixos volumes de aplicação. Outro problema
evidenciado com os inseticidas nacionais diz respeito à pequena sobrevida, devendo a
ser usados em, no máximo, 30 dias depois de produzidos, quando armazenados à
temperatura ambiente e em local sombreado, fazendo as vendas ocorrerem quase que,
exclusivamente, sob encomenda. Comparativamente, os bons inseticidas sob
comercialização em outros países apresentam maior concentração de ingrediente ativo,
maior sobrevida (alguns produtos podem ser armazenados por mais de 8 meses à
temperatura ambiente) e praticidade (são formulados na forma de GDA (grânulos
dispersíveis em água) ou de óleos emulsionáveis, por exemplo, podendo ser adicionados
diretamente ao tanque do pulverizador.
Rangel et al. (2007), instalaram uma Unidade de Observação para o controle
microbiano das cigarrinhas das pastagens Distrito de Itahum, em Dourados/MS, devido
as necessidades de agricultores familiares de produção de leite. As aplicações do fungo
Metarhizium anisopliae foram realizadas por pulverização com equipamento de tração
animal. As condições do ambiente de estudo foram favoráveis ao desenvolvimento da
população das cigarrinhas e à incidência do fungo. Como resultados, observaram que os
níveis de espumas, ninfas e adultos reduziram significativamente após as aplicações,
levando os Agricultores Familiares a total satisfação com os ganhos na produção de
todos os envolvidos.
Segundo Vieira et al. (2007), o fungo entomopatogênico Metarhizium anisopliae
var. acridum tem sido utilizado no controle de pragas, e assim, visando este fim,
analisaram em laboratório a esporulação, a germinação dos conídios e as características
morfológicas peculiares desse fungo em diferentes temperaturas. Esperou-se o
desenvolvimento das colônias em períodos de 1 a 15 dias. Foi observado que
temperaturas de 25, 28 e 30 ºC proporcionaram maiores taxas de esporulação e elevadas
porcentagens de germinação dos conídios. A 25, 28 e 30 ºC observou-se um
desenvolvimento mais rápido das microestruturas somáticas e reprodutivas (hifas,
anastomoses, apressórios, conidióforos e conídios elipsóides). Nas temperaturas
extremas de 40 e 45 ºC não foi detectado esporulação, germinação, nem
desenvolvimento de nenhuma microestrutura.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
39
Lopes et al. (2007) avaliaram o efeito de formulações de Beauveria bassiana e
Metarhizium anisopliae sobre ninfas de Amblyomma cajennense (transmissor da febre
maculosa ao homem) em laboratório. Ninfas não alimentadas foram pulverizadas com
suspensões contendo 2x107 e 4x107 conídios/mL de B. bassiana, e 1x107, 1,5x107,
2x107, 3x107 e 5x107 conídios/mL de M. anisopliae. A mortalidade de ninfas,
decorridos três dias da aplicação, foi significativa nas concentrações de 3x107 e 5x107
conídios/mL de M. anisopliae, com 57,1 e 56,2 %, respectivamente. Após seis dias a
mortalidade foi de 100 % nessas duas concentrações. A mortalidade foi
significativamente maior que a observada para B. bassiana em concentrações
semelhantes.
Guirado et al. (2009) realizaram estudos para controlar cupins de montículo em
área de pastagem, utilizando fungos entomopatogênicos Metarhizium anisopliae e
Beauveria bassiana em diferentes doses e formulações. A avaliação foi realizada após
30 dias da aplicação dos produtos. Os melhores resultados em ordem decrescente de
eficiência foram: M. anisopliae 30 mL + 1000 mL de água com 100 % de eficiência; B.
bassiana 10g + M. anisopliae 20 mL + 1000 mL de água com 85,71 % de eficiência; M.
anisopliae 20 mL + 1000 mL de água com 71,42 % de eficiência. Os demais
tratamentos não diferiram da testemunha.
Loureiro et al. (2012) compararam a eficiência de isolados de Metarhizium
anisopliae em controlar populações naturais de Mahanarva fimbriolata (cigarrinha-da-
raiz) em cultivos comerciais de cana-de-açúcar colhida mecanicamente. Foram
aplicados os isolados IBCB 348, IBCB 408, IBCB 410 e IBCB 425 do fungo em
concentração de 1,5×1012 conídios/ha. Aplicou-se o planejamento experimental em
blocos casualizados com 5 tratamentos e 4 repetições, sendo cada repetição composta
por 7 linhas de 100 m de comprimento e espaçamento de 1,5 m. Após 30 dias de
pulverização foi observado para os isolados IBCB 408 e IBCB 425 uma eficiência de
controle de ninfas de 63 e 62%, respectivamente, já no caso dos adultos chegou a 100%.
Após 60 dias o isolado IBCB 425 foi o mais eficiente para controlar as ninfas com
48,4%.
Assis et al. (2012) estudaram a citotoxidade de quitoolissacarídeos (QOS) e
oligômeros A, B e C (soluções compostas por diferentes quantidades de QOS)
produzidos por hidrólise enzimática pelo fungo Metarhrizium anisopliae. Foi analisado
o efeito de antiproliferativo destas moléculas em células tumorais (HepG2 e HeLa) e

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
40
também em célula normal (3T3). A atividade antioxidante foi analisada em vários
experimentos in vitro. A glicosamina apresentou maior toxicidade (cerca de 92%) para
todas as células estudadas. No entanto, os oligômeros obtidos após hidrólise não
demonstraram efeito tóxico sobre a célula normal (3T3). Além disso, mostraram que
uma pequena quantidade de outros COS pode diminuir o efeito citotóxico da
glicosamina contra células 3T3, indicando que glicosamina poderia ser usada como uma
droga antitumoral na presença de outros COS.
2.3. Fermentação
Os processos fermentativos são caracterizados por processos nos quais
microrganismos realizam a conversão catalítica de uma dada substância num
determinado produto. Estes processos podem ser classificados como processos
descontínuos, contínuos e descontínuos alimentados. A classificação destes processos
está diretamente ligada à forma de como o substrato é adicionado e como o produto é
retirado. No processo de fermentação descontínuo o substrato é colocado no
fermentador, em seguida é inoculado e após o término do processo o produto é retirado.
No processo contínuo, o substrato é colocado num reator com vazão constante e o meio
fermentado é retirado com a mesma vazão de entrada do reator (Faccioti, 2001). Nos
processo descontínuos alimentados à adição de substrato e a retirada do mesmo deve
ocorrer de forma intermitente (Carvalho & Sato, 2001).
De certa forma, os processos fermentativos representam um elo entre as antigas
artes de elaboração de alimentos, como queijos e vinho, a partir de uma flora
microbiana natural, com a moderna indústria de fermentação de alimentos, onde são
utilisados cultivos puros e equipamentos sofisticados para o controle do processo
(Assis, 2010).
Atualmente, a maioria dos processos industriais se desenvolve em cubas de
vidro ou de aço de até 20 litros. Os agentes biológicos se encontram submersos no meio de
cultivo que ocupa, aproximadamente, 75% da cuba. As vezes é necessário injetar ar, e em muitas
fermentações se forma espuma. O desenho do biorreator deve se adequar ao objetivo do
processo, respondendo eventualmente a diversos imperativos, tais como a esterilização do
sistema, a aeração e homogeneização do meio, o acréscimo de nutrientes e de aditivos
antiespuma, a manutenção do pH, etc. Os modelos de fermentadores mais utilizados com
microrganismos contam com aeração e agitação mecânica. Esta facilita a distribuição dos

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
41
nutrientes na cuba, mas gera calor que deve ser eliminado mediante a circulação de água
fria. Em outros tipos de biorreatores, em coluna ou torre, a homogeneização depende da injeção de
ar. Os sistemas submersos são apropriados para o cultivo de microrganismos livres, mas resultam
pouco econômicos quando se trabalha com células ou enzimas caras. A imobilização em
fermentadores menores, seja por adesão a um suporte inerte, seja por inclusão dentro de um
polímero que permita o contato com o meio de cultura, além de simplificar a purificação do
produto permite a reutilização das células ou das enzimas, que permanecem dentro do
biorreator (Malajovich, 2009).
As enzimas são produtos de alto valor agregado, amplamente empregadas em
uma gama de indústrias, e entre as fontes para sua obtenção, destacam-se as de origem
microbiana, principalmente, devido a elevada seletividades dos substratos utilizados e
produtos formados. A principal forma de obtenção de enzimas, em escala industrial, é a
fermentação submersa, por esta apresentar vantagens em relação a fermentação
semisólida, como: maior controle do processo fermentativo e consequente reprodução
da cinética microbiana, além de maiores rendimentos (Wanderley, 2011).
2.3.1. Preparo do inóculo
Denomina-se inóculo um volume de suspensão de microrganismo de
concentração adequada, capaz de garantir, em condições econômicas, a fermentação de
uma dada quantidade de meio. O volume do inóculo, assim como sua concentração de
células, vai depender tanto do processo de fermentação c omo da quantidade de meio a
ser fermentado (Carvalho & Sato, 2001).
A partir de uma cultura pura, inocula-se um volume relativamente pequeno de
meio nutriente, que vai depender do tipo de microrganismo cultivado (pois cada espécie
microbiana possui velocidades diferentes de crescimento), sendo, em seguida, incubado
em condições favoráveis ao desenvolvimento do microrganismo. A suspensão obtida é
transferida para um frasco maior em agitador rotativo ou recíproco (shakers), que por
sua vez é adequadamente incubado, e a seguir, transferido para um frasco contendo um
volume ainda maior deste meio, até se conseguir alguns litros de suspensão microbiana.
Os cuidados com assepsia, volumes de meio utilizado, numero de transferência bem
como as condições de incubação, podem variar de acordo com o tipo de processo de
fermentação que se deseja realizar.

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
42
2.3.2. Produção de enzimas em cultivo submerso descontínuo
A produção de enzimas, usando processos fermentativos, é constituída
basicamente por duas etapas: a primeira caracteriza-se pela preparação do inóculo e a
segunda, pela fermentação do substrato (Carvalho & Sato, 2001). A finalidade da
primeira etapa é o preparo do microrganismo em condições adequadas para que possa
ocorrer o desenvolvimento da segunda etapa. Diferentemente do cultivo em estado
sólido, no cultivo submerso os microrganismos encontram-se em um meio com elevada
atividade de água.
2.4. Planejamento experimental
Um bom planejamento tem como fundamental projetar um experimento de
forma que ele seja capaz de fornecer exatamente o tipo de informação que procuramos.
Para isso é necessário saber, em primeiro lugar, o que é mesmo que se deseja procurar.
Planejamento experimental, ou delineamento experimental, é um conjunto de
ensaios estabelecido com critérios científicos e estatísticos, objetivando determinar a
influência de diversas variáveis nos resultados de um dado sistema ou processo (Barros
Neto et al., 2001; Rodrigues & Iemma, 2005). De acordo com o propósito dos ensaios,
esse objetivo maior pode ser mais detalhado em:
– determinar quais variáveis são mais influentes nos resultados;
– atribuir valores às variáveis influentes de modo a otimizar os resultados;
– atribuir valores às variáveis influentes de modo a minimizar a variabilidade dos
resultados;
– atribuir valores às variáveis influentes de modo a minimizar a influência de variáveis
incontroláveis.
O planejamento experimental, baseado nos fundamentos estatísticos é, sem
dúvida alguma, uma ferramenta poderosa para se chegar às condições otimizadas de um
processo, desenvolvimento da formulação de produtos dentro das especificações
desejadas ou simplesmente para avaliar os efeitos ou impactos que os fatores (variáveis
independentes) têm nas respostas (variáveis dependentes) desejadas (Barros Neto et al.,
2001; Rodrigues & Iemma, 2005). Da utilização das técnicas estatísticas de
planejamento experimental, é possível chegar benefícios importantes, como:

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
43
– reduzir o número de ensaios sem prejuízo da qualidade da informação;
– estudar simultaneamente diversas variáveis, separando seus efeitos;
– determinar a confiabilidade dos resultados;
– realizar a pesquisa em etapas, num processo iterativo de acréscimo de novos ensaios;
– selecionar as variáveis que influem num processo com número reduzido de ensaios;
– representar o processo estudado através de expressões matemáticas;
– elaborar conclusões a partir de resultados qualitativos.
2.4.1. Planejamento fatorial
Em um planejamento fatorial é investigado as combinações de níveis de estudo
de cada fator. Para tanto, inicialmente, é necessário especificar esses níveis em cada
fator, quer dizer, os valores dos fatores que serão utilizados nos experimentos. Um
ensaio experimental é definido como um experimento constituído de um conjunto de
níveis definido. Geralmente, quando se tem n1 níveis de um determinado fator 1, n2 do
fator 2, ..., e nk do fator k, o planejamento será descrito por um fatorial n1 x n2 x ... x nk,
que corresponde ao número mínimo de experimentos para se ter um planejamento
fatorial completo.
É necessário fazer cada fator variar e observar o resultado dessa variação para se
estudar o efeito sobre a resposta. Assim, os ensaios precisam ter um mínimo de dois
níveis por fator para a realização desses ensaios. Para k fatores, ou seja, k variáveis de
controle, o planejamento de dois níveis precisará de 2 x 2 x ... x 2 = 2k, para o número
de ensaios diferentes, sendo conhecido como planejamento fatorial 2k.
Para o planejamento fatorial, são especificados os valores dos fatores em cada
experimento, e estes são classificados em níveis, variando-se, por exemplo, de -1
(mínimo) a +1 (máximo). A partir de uma matriz serão apresentadas todas as
combinações entre o número de fatores e de níveis nos experimentos a serem realizados
(Barros Neto et al., 2001; Rodrigues & Iemma, 2005).
2.4.2. Planejamento Plackett & Burman
A maioria dos planejamentos do tipo “Screening” (Delineamentos de Seleção de
fatores) foi descrito originalmente por Plackett & Burman em 1946. Os planejamentos
experimentais de Plackett & Burman são eficientes em situações que envolvem uma

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
44
grande quantidade de variáveis a serem exploradas em um processo de otimização de
fatores (Reddy et al., 2008).
As matrizes Plackett & Burman consistem de planejamentos fatoriais de dois
níveis para estudar k = N – 1 variáveis em N ensaios. São construídas para ensaios
múltiplos de 4, sendo essas matrizes para ensaios de 8, 12, 16,..., 100. Cada variável
independente em uma matriz assume uma configuração de dois níveis, o inferior (-1) e o
superior (+1), para o nível intermediário é estabelecido (0). Para cada vez que a variável
aparece no nível inferior, terá o par equivalente no nível superior (Rodrigues & Iemma,
2005).
Para evitar dúvidas nos resultados dos efeitos de algum fator no planejamento de
Plackett & Burman, o número de ensaios a ser realizado deve ser no mínimo o número
de variáveis mais 4. No caso de um fatorial completo o número de ensaios a ser
realizado é bem maior (Rodrigues & Iemma, 2005).
Mesmo já determinado o planejamento Plackett & Burman, torna-se importante
à realização de experimentos relativos aos pontos centrais de modo a se avaliar melhor
as possíveis interferências analíticas que podem ocorrer durante os diferentes
experimentos. A repetição do ponto central permite também a determinação do erro
puro e da repetibilidade do processo. Na prática, esta é uma ferramenta que permite
identificar melhor se o processo em estudo está sob controle (Rodrigues & Iemma,
2005).
É possível que no planejamento Plackett & Burman não tenha sido considerado
a codificação do ponto central (0) para repetição, pois, matematicamente, a estimativa
do valor desse efeito codificado (zero) vezes, teria como resposta zero. Porém, no
resultado prático do experimento, consegue-se informações valiosas com essas
repetições no ponto central (Rodrigues & Iemma, 2005).
2.4.3. Aplicações do Planejamento Plackett & Burman
Ghanem et al. (2000) produziram xilanase por Aspergillus terreus cultivadas
sobre palha de trigo finamente moída em fermentação de estado sólido. A composição
ideal do meio foi estabelecida aplicando-se o planejamento experimental de Plackett &
Burman. A melhor atividade enzimática foi obtida em um meio contendo 10 g/L de

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
45
palha de trigo por frasco umedecido com uma solução concentrada de sal com teor de
água inicial de 75 % e incubadas durante 4 dias a 30 °C. Observou-se que íons
metálicos como Hg2+, Cu2+, Co2+, Fe3+ e Pb2+ inibiram fortemente a xilanase, enquanto
que Ca2+ ativou a enzima.
Mu et al. (2005) estudaram as otimização de meio de fermentação para a reação
de estereoinversão por Candida parapsilosis CCTCC M203011 utilizando o
planejamento Plackett & Burman. Os resultados mostraram que dentre os fatores
estudados, os fatores mais significativos foram: a Concentração de MgSO4, ZnSO4 e
KH2PO4 e que após a utilização da metodologia de superfície de resposta foi constatado
que o ponto ótimo para a otimização foi conseguido com as concentrações dos fatores
MgSO4, ZnSO4 e KH2PO4 nos valores em: 1,17 g/L, 0,10 g/L e 9,41 g/L,
respectivamente, elevando a taxa de conversão enantiosseletiva de 75,2 % para 96 %.
Naveena et al. (2005) empregaram o planejamento Plackett & Burman para a
seleção de 15 parâmetros para produção de ácido láctico L(+) de farelo de trigo,
substrato de baixo custo e suporte sólido, por Lactobacillus amylophilus GV6 em
fermentação de estado sólido (SSF). Onze nutrientes pertencentes a duas categorias,
fontes de nitrogênio e fontes de sal, junto com três parâmetros físicos e um tampão
foram os parâmetros considerados. Os autores verificaram influência na produtividade
devido às fontes de nitrogênio, peptona, extrato de levedura e tri-citrato de amônio,
juntamente com o NaH2PO4.2H2O e Tween 80, que podem ainda serem otimizados para
o aumento na produção de ácido lático. Esses pesquisadores ainda chamam atenção para
o uso escasso deste planejamento em fermentação em estado sólido e que o mesmo
ainda não havia sido considerado anteriormente para a conversão direta de amido em
ácido láctico L (+) usando um sistema bacteriano.
Khosravi-Darani et al. (2008) estudaram a aplicação de Plackett & Burman na
análise dos fatores significativos para a produção de ácido cítrico por Aspergillus niger
ATCC 9142 utilizando palha de trigo como substrato. Os resultados mostraram que a
produção, com a palha de trigo pré-tratada com HCl, NaOH e Uréia, foi mais eficiente e
que os fatores mais significativos na produção foram: Solvente, Teor de Umidade,
Temperatura, Tempo de Incubação, Concentração Inicial de Açúcar e Concentração de
Metanol.
Ji et al. (2009) estudaram o desenvolvimento de um meio de cultura econômico
para a produção do 2,3-butanodiol através da fermentação da glicose e xilose por

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
46
Klebsiella oxytoca utilizando o planejamento Plackett & Burman. Os resultados
mostraram que a uréia e a maceração de milho foram os fatores mais significativos para
a produção do 2,3-butanodiol e que através de planejamento experimental com repetição
em triplicata no ponto central para determinar os níveis ótimos de produção. Os autores
constaram que o rendimento atingido foi 85,6% do valor teórico.
Paula & Arruda. (2009) observaram a necessidade da busca por alternativas mais
eficientes ao método termomecânico utilizado para a degradação de milhões de
toneladas de penas de aves provenientes de frigoríficos e aviários, cuja rígida estrutura é
composta por mais de 90 % de queratina. Para esse fim, utilizaram de comunidades
microbianas para a finalidade proposta, testando o planejamento experimental Plackett
& Burman para verificar a influência de diferentes microrganismos simultaneamente,
com um número reduzido de experimentos. Testaram 32 bactérias isoladas, inoculadas
em meio contendo somente uma pena como única fonte de carbono, durante sete dias,
de onde foi selecionado as cepas para os consórcios microbianos. Experimentos unindo
as cepas NP5, FCA7, NP4 e S14 demonstraram diferentes atividades queratinolíticas,
aumentando o nível de degradação, até 87,8%, comparada com as cepas
individualmente, variando entre 26,4% e 78,0%.
Rajendran & Thangavelu (2009) obtiveram valores máximos da atividade de
produção de lipase em 3,98 U/mL e concentração de massa celular em 5,62 g.L-1 com R.
arrhizus em cultivo submerso descontínuo otimizado pelo planejamento Plackett &
Burman. As variáveis mais importantes que afetaram a produção da lipase foram azeite
de oliva, peptona, KH2PO4, CaCl2.2H2O e MgSO4.7H2O.
Li et al. (2010) estudaram a otimização de meio de cultura para a produção de
Streptolydigin (um antibiótico) por Streptomyces lydicus AS 4,2501 utilizando Plackett
& Burman e a metodologia de superfície de resposta. Os resultados mostraram que
dentre os fatores estudados, os fatores mais significativos para a produção de
Streptolydigin e otimização do meio de cultura foram: Concentração de Amido,
Concentração de Peptona e a Concentração de K2HPO4.
Chen et al. (2010) estudaram a otimização de meio de cultura para a produção da
Luciferase – que são enzimas que catalisam reações biológicas transformando energia
química em energia luminosa - por Bacillus Subtilis em fermentação descontínua
utilizando planejamento Plackett & Burmam. Os resultados mostraram que dentre os
componentes estudados, os fatores mais significativos para a produção e otimização do

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
47
meio de cultura para a produção da Luciferase foram: Concentração de Glicose 1,17 %,
Extrato de Levedura 2,27 % e Glutamato 0,55 %.
Ghorbel-Bellaaj et al. (2010) estudaram a otimização da extração de quitina a
partir de fermentação de resíduos de camarão por Pseudomonas aeruginosa A2
utilizando o projeto Plackett & Burman e a metodologia de superfície de resposta. Os
resultados mostraram que a eficiência máxima de extração da quitina foi conseguida nas
seguintes condições: Concentração de casca de camarão (50 g/L), glicose (50 g/L),
tempo de incubação de 5 dias e inoculo com 0,05 de OD (densidade óptica).
Gao et al. (2010) desenvolveram um meio para a fermentação do extrato bruto
de inulina de tubérculos de alcachofra de Jerusalém por Paenibacillus polymyxa ZJ-9
para produzir R,R-2,3-butanodiol (R-R,2,3-BD). Inulina, K2HPO4 e NH4Cl foram os
parâmetros principais na fermentação de acordo com os resultados obtidos com o
planejamento experimental Plackett & Burman. A concentração ideal dos três fatores,
77,14 g/L, 3,09 g/L e 0,93 g/L, respectivamente, foi obtida pelo planejamento Box-
Behnken e pela superfície de resposta. Sob as condições ideais, a concentração do R-
R,2,3-BD obtida foi 36,92 g/L, em mais de 98 % de pureza óptica. Comparado com
outras fontes de carbono investigadas, a fermentação do extrato bruto de inulina obteve
o maior rendimento de R-R,2,3-BD.
Uma metodologia de superfície de resposta foi empregada com sucesso para
otimizar um meio de fermentação chamada VHG (Very High Gravity), referência à alta
concentração de açúcares e densidade dos mostos, baseado na água maceração de milho
CSL (Corn Steep Liquor) e outros nutrientes de baixo custo para a produção eficiente de
etanol a partir de glicose por S. cerevisiae. Usando a composição média otimizada (g/L:
CSL 44,3; uréia 2,3; MgSO4.7H2O 3,8; CuSO4.5H2O 0,03), a levedura industrial PE-2
foi capaz de fermentar até 330 g/L de glicose e produzir 18,6 % (v/v) de etanol, com
uma produção descontínua de 2,4 g/L/h e um rendimento em etanol de 93 % (Pereira et
al., 2010).
O planejamento Plackett & Burman juntamente com o Planejamento Composto
Central (CCD) foram aplicados para otimizar o meio de cultura para produção de etanol
por Clostridium autoethanogenum com monóxido de carbono (CO) como fonte única de
carbono e um meio contendo (g/L): 1,0 de NaCl, 0,1 de KH2PO4, 0,02 de CaCl2, 0,15 de
extrato de levedura, 0,116 de MgSO4, 1,694 de NH4Cl e pH 4,74. O rendimento ótimo
de etanol previsto pela metodologia de superfície de resposta (RSM) e um algoritmo

Capítulo 2 Revisão Bibliográfica
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
48
genético de rede neural artificial (ANN-GA) foram 247,48 e 261,48 mg/L,
respectivamente. Valores semelhantes aos obtidos experimentalmente sob as condições
ideais sugeridas pelos métodos estatísticos (254,26 e 259,64 mg/L). A adequação do
modelo ANN-GA foi maior do que o modelo RSM. Os rendimentos obtidos foram
substancialmente superiores aos relatados anteriormente (60-70 mg/L) com essa cultura
(Guo et al., 2010).
Rigueira (2010) avaliou a influência das variáveis do procedimento de extração
na porcentagem de recuperação das aminas encontradas no leite cru utilizando um
planejamento para “screening” do tipo Plackett & Burman, para os ácidos
tricloroacético (TCA) e sulfosalicílico (ASS).
Quines (2010) estudou a necessidade ou não de se fazer um pré-tratamento da
biomassa antes da extração. Um planejamento experimental Plackett & Burman de doze
ensaios mais três pontos centrais foi utilizado para selecionar as variáveis mais
significativas na extração de Poli(3-hidroxibutirato) (P(3HB)) por 1,2-carbonato de
propileno. Foram realizados dois testes de recuperação do solvente 1,2-carbonato de
propileno, após a extração, sendo um com aplicação de aquecimento e o outro com
aquecimento e vácuo. Os resultados mostraram que o pré-tratamento térmico levou a
uma maior porcentagem de recuperação e de pureza do polímero.

Capítulo 3 Metodologia Experimental

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
50
3. Metodologia Experimental
O presente capítulo apresenta os matérias e métodos utilizados na produção de
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso.
3.1. Quitosana
Neste trabalho utilizou-se uma quitosana (Lote 067) obtida junto a POLYMAR/CE,
com no mínimo 80% de desacetilação. Esse substrato foi utilizado como fonte de carbono e
energia tanto no estudo cinético como nos ensaios do planejamento de seleção de fatores
Plackett & Burman e Planejamento Composto Central, para a produção de quitosanase pelo
Metarhizium anisopliae, por fermentação submersa.
3.2. Microrganismo
O fungo Metarhizium anisopliae estirpe (CG374), utilizado neste estudo, foi cedido
pela Embrapa Recursos Genéticos e Biotecnologia (Brasília / DF-Brasil) e foi mantido em
placas de Petri contendo o meio Batata Dextrose Agar (PDA). As placas foram incubadas em
estufa a 25 ºC. O pH do meio de manutenção foi mantido entre 6,0 e 6,5.
3.2.1. Escolha da cepa produtora de quitosanase
A escolha do microrganismo Metarhizium anisopliae foi feita com base na capacidade
de crescer em meio contendo quitosana como única fonte de carbono, além de apresentar
grande tolerância a pH variando a uma taxa de 2,0 a 8,5, sendo 6,9 a melhor condição para o
crescimento vegetativo e esporução, de acordo com Assis (2010).
3.2.2. Manutenção da cepa
A manutenção da cultura foi realizada em placas petri contendo o meio Batata
Dextrose Agar (PDA). A renovação das células foi realizada a cada trinta dias através de
subcultivos das células no meio PDA.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
51
3.3. Produção de enzima
No preparo do inóculo, utilizou-se 7 placas cultivadas e mantidas, com o fungo
Metarhizium anisopliae, em meio PDA a 25 ºC durante 5 dias em estufa. O meio de cultura
para a produção da enzima quitosanase apresentou os seguintes componentes: quitosana;
peptona; extrato de levedura; NaNO3; K2HPO4; KCl; MgSO4.7H2O e FeSO4, com
concentrações de acordo as matrizes dos planejamentos: Plackett & Burman e composto
central. Para a produção da enzima no cultivo, a biomassa foi transferida através de água
estéril, para um Béquer de 500 mL que, em seguida, foi colocado em Erlenmeyers de 250 mL
na proporção de 20 mL de biomassa para 180 mL do meio de cultura. Os Erlenmeyers
cultivados foram então incubados em incubador rotativo a 25 ºC e 150 rpm durante 24 h.
Alíquotas foram retiradas e centrifugadas a 8000xg por 5 min, a cada 6 horas, para se
determinar a atividade quitosanolítica, teores de açúcares redutores e de proteína total.
Ressalta-se que todos os experimentos foram feitos em triplicata.
3.4. Determinação da Proteína Total
A determinação da proteína total foi efetuada de acordo com o método modificado de
Sedmak & Grossberg (1977), que se baseia na diminuição da absorbância do corante azul de
Coomassie, devido à ligação proteína-corante, a 595 nm, comparado a absorção máxima a
465 nm, conforme Santos (2001). Nesse caso, após centrifugação para retirada da biomassa,
transferia-se para um eppendorf 1,5 mL de amostra (caldo fermentativo) e adicionava-se 1,5
mL de solução azul de Coomassie. Deixava-se a solução em repouso por 5 minutos,
transferindo-se em seguida para um cubeta de quartzo e efetuando-se as leituras de
absorbância nos comprimentos de ondas 465 nm e 595 nm, respectivamente. O
espectrofotômetro (Thermo Spectronic modelo Genesys 10UV) foi zerado com água
destilada. O conteúdo protéico foi obtido a partir de uma curva padrão na qual graficava-se a
relação de absorbância A595/A465 para diferentes concentrações de Albumina de Soro
Bovino (Fração V – Lote 110881/Sigma Aldrich-Ohio-USA).
3.5. Determinação da Atividade Quitosanolítica e da Atividade Específica
A atividade quitosanolítica foi obtida monitorando-se o aumento dos açúcares
redutores totais, utilizando-se o método DNS a partir de uma curva padrão de glicosamina.
Para a determinação da atividade quitosanolítica adicionaram-se, aos tubos de ensaios 500 µL

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
52
de substrato (solução 0,2 % de quitosana solubilizada em ácido clorídrico 1N) e 500 µL de
caldo bruto (contendo enzima), controlando-se o pH entre 6,0 e 6,5 e incubando-se o sistema
em um banho termostatizado com temperaturas da ordem de 55 ºC. Após o tempo de
incubação de 30 minutos no banho foi adicionado aos tubos de ensaio 2,5 mL de DNS,
aquecendo-se à ebulição em banho-maria por 10 minutos, resfriando-se, em seguida, em
banho de gelo. Após essa etapa, o sistema foi centrifugado e com o sobrenadante efetuou-se a
leitura no espectrofotômetro a 600 nm. Destaca-se, que antes da leitura da amostra zerava-se o
equipamento (branco do espectro) com 1,0 mL de água destilada acrescidos de 2,5 mL de
DNS e realizava-se o branco da enzima que consistia da adição de 500 µL de solução de
quitosana, completando-se até um 1,0 mL com água destilada. Uma unidade de atividade
quitosanolítica (A) foi definida como a quantidade de enzima necessária para produzir 1
µmol.min-1 de glicosamina por mL de extrato enzimático, nas condições de temperatura e pH
citadas, sendo o resultado expresso em (U.mL-1). A Atividade específica foi determinada pela
divisão entre a atividade da enzima (U.mL-1) e a concentração de proteína total (µg.mL-1),
sendo expressa em (U. µg-1).
3.6. Planejamento experimental
3.6.1. Planejamento de seleção de fatores Plackett & Burman
O planejamento experimental Plackett & Burman é uma técnica muito usada, para se
realizar triagem de fatores na otimização num determinado processo (Reddy et al., 2008), e
foi utilizado neste trabalho para identificar os fatores que influenciam de forma significativa
na produção da enzima quitosanase.
A Tabela 3 ilustra os níveis de cada variável (fator) utilizada no planejamento Plackett
& Burman, sendo: Concentração de Quitosana (Q), Peptona (PP), Extrato de Levedura (EL),
NaNO3 (NS), K2HPO4 (FP), KCl (CP), MgSO4.7H2O (SM), FeSO4 (SF). Já a Tabela 4 mostra
a matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3 repetições
na condição do ponto central.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
53
Tabela 3 – Níveis para os fatores e seus valores codificados.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1 0,1 0,2 0,2 0,1 0,05 0,025 0,025 0,0005
0 0,2 0,5 0,3 0,2 0,10 0,050 0,050 0,0010
1 0,3 0,8 0,4 0,3 0,15 0,075 0,075 0,0015
Tabela 4 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios e mais 3
repetições na condição do ponto central.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1
2 1 1 -1 1 -1 -1 -1 1 -1 1 1
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1
5 1 1 -1 1 1 -1 1 -1 1 -1 1
6 1 1 1 -1 1 1 -1 1 1 -1 -1
7 -1 1 1 1 -1 1 1 -1 -1 1 1
8 -1 -1 1 1 1 -1 1 1 1 1 1
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1
11 -1 1 -1 -1 -1 1 1 1 1 1 -1
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1
13 0 0 0 0 0 0 0 0 0 0 0
14 0 0 0 0 0 0 0 0 0 0 0
15 0 0 0 0 0 0 0 0 0 0 0
Sendo: V11,V12,V13 = Variáveis inertes para cálculo do erro.
3.6.2. Planejamento Composto Central para a Otimização do Meio de Cultura para a
Produção da Enzima em Cultivo Submerso
Um dos problemas mais comuns para quem realiza experimentos é determinar a
influência de uma ou mais variáveis sobre outra variável de interesse (Lobato, 2003). Este

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
54
problema pode ser minimizado utilizando-se um planejamento experimental de modo a se
obter uma função, ou uma aproximação satisfatória para a mesma, de forma que se operando
sobre as variáveis de entrada produza-se como saída a resposta ou as respostas observadas
(Barros Neto, 2001; Silva, 2004).
Em um planejamento, inicialmente, deve-se definir os fatores e as respostas de
interesse. Tendo-se identificado os fatores e respostas define-se o objetivo que se pretende
alcançar com os experimentos para que se possa escolher o planejamento mais apropriado.
(Silva, 2004).
No presente trabalho foi utilizado um Planejamento Composto Central (CCD) com 22
experimentos na parte fatorial, repetição em triplicata no ponto central e quatro experimentos
na parte axial (planejamento do tipo estrela), para se investigar a influência dos fatores:
concentração de quitosana e concentração de FeSO4 que foram os dois fatores que
influenciaram a produção de quitosanase, conforme o planejamento Plackett & Burman de 12
ensaios e mais 3 repetições na condição do ponto central. Estes dois fatores foram estudados
em cinco diferentes níveis (-1,41; -1; 0; 1; 1,41) e um conjunto de 11 experimentos foi
realizado, conforme Tabela 5.
Tabela 5 – Matriz do planejamento Composto Central.
Ensaio Q SF
1 -1 -1
2 -1 1
3 1 -1
4 1 1
5 -1,41 0
6 1,41 0
7 0 -1,41
8 0 1,41
9 0 0
10 0 0
11 0 0
Os outros fatores foram mantidos nos níveis mais baixos (para evitar custos). Uma que
não influenciaram de forma significativa na produção da enzima quitosanase.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
55
A codificação dos fatores foi realizada de acordo com a seguinte equação (Ji et al.,
2009):
Xi = (xi – x0)/∆xi , i = 1, 2, 3, ..., k (2)
Onde Xi é o fator independente codificado, xi é o fator independente real, x0 é o valor de xi no
ponto central e ∆xi é a variação de valor. O comportamento do sistema foi explicado pela
seguinte equação polinomial de segunda ordem:
Yp = β0 + ∑ βiXi + ∑ βiiXi2 + ∑ βijXiXj, i, j = 1, 2, 3,..., k (3)
Onde Yp foi a resposta esperada, β0 foi a interseção, Xi e Xj
foram os fatores independentes codificados, βi foi o coeficiente linear, βii
foi o coeficiente quadrático e βij foi o coeficiente de interação. Destaca-se que os outros
fatores não significativos foram mantidos no nível mais baixo (-1).
A Tabela 6 apresenta os níveis para os fatores e seus valores codificados, para o
Planejamento Composto Central, com triplicata no ponto central.
Tabela 6 – Níveis para os fatores e seus valores codificados para o cultivo submerso –
Planejamento Composto Central.
Nível Q
(%)
PP
(%)
EL
(%)
NS
(%)
FP
(%)
CP
(%)
SM
(%)
SF
(%)
-1,41 0,218 0,2 0,2 0,1 0,05 0,025 0,025 0,0009
-1 0,3 0,2 0,2 0,1 0,05 0,025 0,025 0,0015
0 0,5 0,2 0,2 0,1 0,05 0,025 0,025 0,0030
1 0,7 0,2 0,2 0,1 0,05 0,025 0,025 0,0045
1,41 0,782 0,2 0,2 0,1 0,05 0,025 0,025 0,0051
Na análise do planejamento de seleção de fatores Plackett & Burman e do
planejamento experimental fatorial foi utilizado um pacote computacional STATISTICA for
Windows versão 7.0.

Capítulo 3 Metodologia Experimental
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
56
3.7. Perfil cinético durante o cultivo
Um perfil cinético na melhor condição de cultivo, conforme planejamento
apresentado, foi realizado em frascos Erlenmyer de 250 mL contendo 200 mL de meio de
cultivo contendo os seguintes componentes, em 1 Litro: 2,0g de quitosana; 5,0g de peptona;
3,0g de extrato de levedura; 2,0g de NaNO3; 1,0g de K2HPO4; 0,5g de KCl; 0,5g de
MgSO4.7H2O; 0,01g de FeSO4, que foram inoculadas com 10% (vol/vol) de inóculo contendo
o fungo Metarhizium anisopliae e cultivadas em incubador rotativo durante 5 dias a 150 rpm
e 25 ºC. Alíquotas foram retiradas a cada seis horas e analisadas quanto à proteína total no
sobredanante, atividade enzimática, atividade específica e açúcares redutores. Os ensaios
foram realizados em triplicata (Silva Filho, 2005).

Capítulo 4 Resultados e
Discussão

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
58
4. Resultados e discussão
Neste capítulo são apresentados e discutidos os resultados obtidos no presente
trabalho. Inicialmente, é apresentada imagem do MEV do Metarhizium anisopliae
mantidos em meio PDA. Em seguida, apresenta-se e discute-se os resultados da avaliação
dos fatores significativos, que podem influenciar na produção da quitosanase, no
planejamento de seleção de fatores, conforme planejamento Plackett & Burman bem como
são apresentados e discutidos os resultados obtidos para o planejamento composto central.
Por último, apresenta-se o perfil cinético, na melhor condição, para a produção de
quitosanase em cultivo submerso descontínuo usando o fungo Metarhizium anisopliae.
4.1. Imagem do micélio de Metarhizium anisopliae em meio PDA.
A Figura 7 ilustra a imagem da Microscopia Eletrônica de Varredura (MEV) do micélio de Metarhizium anisopliae mantido em meio PDA.
Figura 7. Imagem do MEV do micélio de Metarhizium anisopliae em meio PDA.
Observa-se na Figura 7, quanto à morfologia que o Metarhizium anisopliae, apresenta-
se como um fungo filamentoso com as hifas intimamente entrelaçadas conforme citado por
Wang et al. 2002.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
59
4.2. Planejamento Plackett & Burman
4.2.1 Análise do Plackett & Burman com 8 fatores e 12 ensaios
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis, foi utilizado para avaliar a influência dos fatores: Concentração de
Quitosana (Q), Peptona (PP), Extrato de Levedura (EL), NaNO3 (NS), K2HPO4 (FP), KCl
(CP), MgSO4.7H2O (SM), FeSO4 (SF) na produção da enzima quitosanase por Metarhizium
anisopliae em cultivo descontínuo submerso. Ressalta-se que os experimentos foram
realizados aleatoriamente.
A Tabela 7 apresenta a matriz do modelo Plackett & Burman e os respectivos valores
obtidos para a variável resposta (Atividade Quitosanalítica).
Tabela 7 – Matriz do planejamento Plackett & Burman com 8 fatores e 12 ensaios.
Ensaio Q PP EL NS FP CP SM SF A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 0,0195
2 1 1 -1 1 -1 -1 -1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 0,0109
4 1 -1 1 1 -1 1 -1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 0,0136
6 1 1 1 -1 1 1 -1 1 0,0156
7 -1 1 1 1 -1 1 1 -1 0,0109
8 -1 -1 1 1 1 -1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 0,0085
A Tabela 8 apresenta os efeitos estimados para o planejamento Plackett & Burman.
Observa-se na Tabela 8 que o valor do coeficiente de determinação R2 foi de
aproximadamente 90%, indicando que um modelo linear poderia representar bem a relação
entre o fator significativo, concentração de quitosana (1), e a resposta.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
60
Tabela 8 – Efeitos Estimados do planejamento Plackett & Burman.
Efeitos Estimados; R2=0,8996; Ajuste:0,63178 MS Erro Residual =0,0000054
Desvio Padrão t(3) p
Lim. Conf. Lim. Conf.
Efeito Erro Residual
-95,% +95
Média/Interação 0,0127 0,0007 18,903 0,0003 0,0105 0,0149 (1) Quitosana 0,0055 0,0013 4,0628 0,0268 0,0011 0,0098
(2) Peptona 0,0015 0,0013 1,1080 0,3486 -0,0028 0,0058 (3) Ext. de leved. 0,0009 0,0013 0,7387 0,5136 -0,0033 0,0053
(4) NaNO3 0,0002 0,0013 0,1758 0,8715 -0,0040 0,0045 (5) K2HPO4 -0,0014 0,0013 -1,0552 0,3687 -0,0057 0,0028
(6) KCl -0,0022 0,0013 -1,6708 0,1933 -0,0065 0,0020 (7) MgSO4.7H2O -0,0003 0,0013 -0,2286 0,8338 -0,0046 0,0039
(8) FeSO4 0,0029 0,0013 2,1457 0,1211 -0,0014 0,0071
A Figura 8 mostra o gráfico de Pareto, uma das formas de se avaliar visualmente a
influência dos fatores estudados na resposta. A magnitude dos efeitos é representada pelas
colunas enquanto que a linha transversal às colunas representa a magnitude dos efeitos com
significado estatístico para p=0,05, ou seja, os fatores que são estatisticamente significativos
ao nível de 95% de confiança.
Figura 8. Gráfico de Pareto do planejamento Plackett & Burman mostrando a influência dos
fatores estudados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
61
Analisando-se o gráfico de Pareto (Figura 8) observa-se que o fator concentração de
quitosana (1) foi o único fator que influenciou de forma mais significativa na atividade
enzimática atingindo efeitos estimados de aproximadamente 4,06. O sinal positivo indica que
o aumento da concentração de quitosana aumentará à atividade enzimática. Por outro lado, os
outros fatores parecem desprezíveis estatisticamente conforme o gráfico de Pareto.
4.2.2 Análise do Plackett & Burman com 12 Ensaios e mais 3 Repetições na condição do
ponto central.
Um planejamento de Seleção de Fatores Plackett & Burman com 8 fatores e 12
ensaios em 2 níveis com repetição em triplicata no ponto central, foi utilizado para se avaliar a
influência dos mesmos fatores descritos na Seção 4.2.1 para a produção da enzima
quitosanase por Metarhizium anisopliae em cultivo descontínuo submerso. Ressalta-se que os
experimentos foram realizados aleatoriamente. A Tabela 9 apresenta a matriz do modelo
Plackett & Burman e os respectivos valores obtidos para a variável resposta (Atividade
Quitosanalítica).
Tabela 9 – Matriz do planejamento Plackett & Burman com 8 fatores e 15 ensaios.
Ensaio Q PP EL NS FP CP SM SF V11 V12 V13 A (U/mL)
1 1 -1 1 -1 -1 -1 1 1 -1 -1 -1 0,0195
2 1 1 -1 1 -1 -1 -1 1 -1 1 1 0,0202
3 -1 1 1 -1 1 -1 -1 -1 1 -1 1 0,0109
4 1 -1 1 1 -1 1 -1 -1 -1 1 -1 0,0121
5 1 1 -1 1 1 -1 1 -1 1 -1 1 0,0136
6 1 1 1 -1 1 1 -1 1 1 -1 -1 0,0156
7 -1 1 1 1 -1 1 1 -1 -1 1 1 0,0109
8 -1 -1 1 1 1 -1 1 1 1 1 1 0,0103
9 -1 -1 -1 1 1 1 -1 1 -1 -1 1 0,0099
10 1 -1 -1 -1 1 1 1 -1 -1 -1 -1 0,0117
11 -1 1 -1 -1 -1 1 1 1 1 1 -1 0,0094
12 -1 -1 -1 -1 -1 -1 -1 -1 1 1 -1 0,0085
13 0 0 0 0 0 0 0 0 0 0 0 0,0704
14 0 0 0 0 0 0 0 0 0 0 0 0,0688
15 0 0 0 0 0 0 0 0 0 0 0 0,0710

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
62
A Tabela 10 a seguir apresenta os efeitos estimados para planejamento Plackett &
Burman com 3 repetições na condição do poto central. Observa-se na Tabela 10 que o valor
do coeficiente de determinação R2 foi de aproximadamente 2,0%, indicando que um modelo
linear não poderia representar a relação entre os fatores significativos concentração de
quitosana (1), concentração de FeSO4 (8) e a resposta.
Tabela 10 – Efeitos Estimados do planejamento Plackett & Burman com triplicata no ponto
central.
Efeitos Estimados; R2=0,01811; Ajuste:0,0 MS Erro Puro =0,0000012
Desvio Padrão t(2) P
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0242 0,0003 85,509 0,0001 0,0229 0,0254 (1) Quitosana 0,0055 0,0006 8,6432 0,0131 0,0027 0,0082
(2) Peptona 0,0015 0,0006 2,3572 0,1425 -0,0012 0,0042 (3) Ext. de leved. 0,0009 0,0006 1,5715 0,2567 -0,0017 0,0037
(4) NaNO3 0,0002 0,0006 0,3742 0,7442 -0,0024 0,0029 (5) K2HPO4 -0,0014 0,0006 -2,2450 0,1538 -0,0041 0,0013
(6) KCl -0,0022 0,0006 -3,5546 0,0708 -0,0049 0,0005 (7) MgSO4.7H2O -0,0003 0,0006 -0,4864 0,6747 -0,0030 0,0024
(8) FeSO4 0,0029 0,0006 4,5648 0,0448 0,0002 0,0056
A Figura 9 ilustra o gráfico de Pareto para o planejamento Plackett & Burman, com
repetição em triplicata no ponto central, uma das formas de se avaliar visualmente a influência
dos fatores estudados na resposta, onde é mostrado a magnitude dos efeitos para 95 % de
confiança.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
63
Figura 9. Gráfico de Pareto do planejamento Plackett & Burman com triplicata no ponto
central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto, observa-se que os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma mais
significativa na atividade enzimática atingindo efeitos estimados de aproximadamente 8,84 e
4,67, respectivamente. O sinal positivo indica que o aumento das concentrações de quitosana
e de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores parecem
desprezíveis estatisticamente conforme o gráfico de Pareto.
Este comportamento do FeSO4 está de acordo com o observado por Hagag et al.
(2012) que utilizando o Aspergillus fumigatus na produção de protease mostraram que um
fator de transcrição PrtT regula positivamente o controle da produção de protease, da
assimilação de Ferro e da síntese de ergosterol e que a produção de protease foi favorecida
pelo aumento da concentração de ferro.
Rajendran & Thangavelu (2009) utilizaram um planejamento Plackett & Burman para
identificar quais os fatores significativos na produção de lipase por Rhizopus arrhizus MTCC
2233 e observaram o efeito significativo médio do FeSO4, apresentando um nível de
confiança para esse fator de 48,92% (para um máximo de 98,7 % de MgSO4), com uma
produção de lipase da ordem de 2,98 U/mL em 84 horas de fermentação e que ocorreu uma
produção considerável de protease no meio de cultivo da ordem de 1,92 U/mL em 96 horas de
cultivo.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
64
Mu et al. (2005) também observaram um efeito positivo do FeSO4, da ordem de 1,48
para um fator máximo encontrado de 3,11 de MgSO4, no aumento da atividade da
estereoinversão de S-fenil-1,2-etanodiol por Candida parapsilosis CCTCC M203011 na
otimização da fermentação utilizando um planejamento Plackett & Burman.
Dessa forma, observa-se que o FeSO4 é importante uma que vez o mesmo está
diretamente associado a síntese de proteases (principalmente metalloproteases), conforme
citado por Hagag et al. (2012).
4.3. Planejamento Composto Central para Produção de Quitosanase por
Metarhizium anisopliae em Cultivo Descontínuo Submerso
Um planejamento experimentos 22 com repetição em triplicata no ponto central e mais
quatro ensaios axiais, foram utilizados para se chegar a uma combinação ideal de quitosana e
FeSO4 na produção da enzima quitosanase (Tabela 11). A partir do modelo Plackett &
Burman (Tabela 12), que mostrou que o aumento das concentrações de quitosana e FeSO4,
favoreceram a produção de quitosanase, os valores utilizados neste modelo para estes
componentes nos níveis mais altos, serão considerados como os níveis mais baixos no
planejamento experimental do composto central. Os outros fatores foram fixados em todos os
ensaios em seus níveis mais baixos para a redução de custos (Tabela 11).

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
65
Tabela 11 – Matriz do planejamento Composto Central com os respectivos resultados da
atividade enzimática.
Ensaio Q SF A (U/mL)
1 -1 -1 0,0195
2 -1 1 0,0202
3 1 -1 0,0109
4 1 1 0,0121
5 -1,41 0 0,0136
6 1,41 0 0,0156
7 0 -1,41 0,0108
8 0 1,41 0,0103
9 0 0 0,0099
10 0 0 0,0116
11 0 0 0,0094
A Tabela 12 apresenta os efeitos estimados para o planejamento composto central.
Tabela 12 – Efeitos Estimados do planejamento Composto Central.
Efeitos Estimados; R2=0,629; Ajuste:0,25836 MS Erro Puro =0,0000014
Desvio Padrão t(2) p
Lim. Conf. Lim. Conf. Efeito Erro Puro -95,% +95
Média/Interação 0,0103 0,0006 15,305 0,0042 0,0074 0,0132 (1) Quitosana (L) 0,0011 0,0008 1,4316 0,2885 -0,0023 0,0047
Quitosana (Q) 0,0058 0,0009 5,9637 0,0269 0,0016 0,0101 (2) FeSO4 (L) -0,0043 0,0008 -5,3037 0,0337 -0,0079 -0,0008 (2) FeSO4 (Q) 0,0018 0,0009 1,8366 0,2076 -0,0024 0,0060
1L x 2L 0,0001 0,0011 0,1520 0,8931 -0,0048 0,0052
Observa-se na Tabela 12 que o valor do coeficiente de determinação R2 foi de
aproximadamente 63%, indicando que um modelo linear representaria bem a relação entre os
fatores significativos concentração de quitosana (1), concentração de FeSO4 (8) e a resposta.
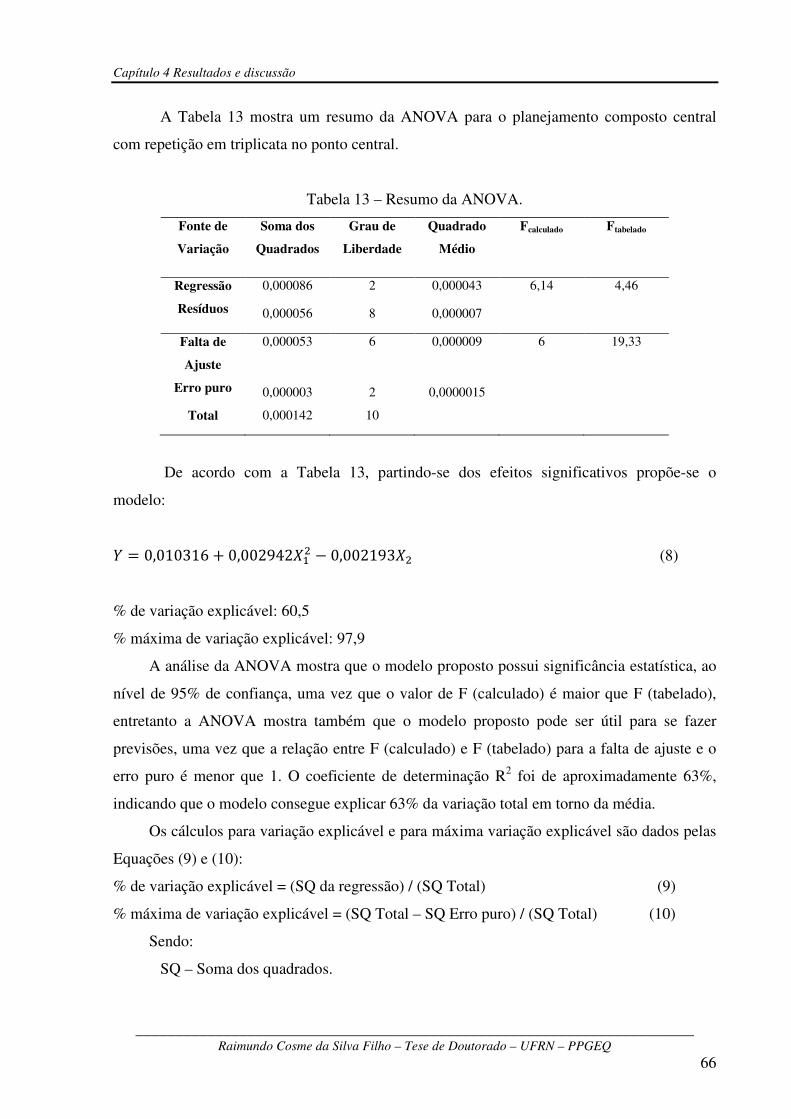
Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
66
A Tabela 13 mostra um resumo da ANOVA para o planejamento composto central
com repetição em triplicata no ponto central.
Tabela 13 – Resumo da ANOVA.
Fonte de
Variação
Soma dos
Quadrados
Grau de
Liberdade
Quadrado
Médio
Fcalculado Ftabelado
Regressão
Resíduos
0,000086 2 0,000043 6,14 4,46
0,000056 8 0,000007
Falta de
Ajuste
Erro puro
0,000053 6 0,000009 6 19,33
0,000003
2
0,0000015
Total 0,000142 10
De acordo com a Tabela 13, partindo-se dos efeitos significativos propõe-se o
modelo:
� = 0,010316 + 0,002942� � − 0,002193�� (8)
% de variação explicável: 60,5
% máxima de variação explicável: 97,9
A análise da ANOVA mostra que o modelo proposto possui significância estatística, ao
nível de 95% de confiança, uma vez que o valor de F (calculado) é maior que F (tabelado),
entretanto a ANOVA mostra também que o modelo proposto pode ser útil para se fazer
previsões, uma vez que a relação entre F (calculado) e F (tabelado) para a falta de ajuste e o
erro puro é menor que 1. O coeficiente de determinação R2 foi de aproximadamente 63%,
indicando que o modelo consegue explicar 63% da variação total em torno da média.
Os cálculos para variação explicável e para máxima variação explicável são dados pelas
Equações (9) e (10):
% de variação explicável = (SQ da regressão) / (SQ Total) (9)
% máxima de variação explicável = (SQ Total – SQ Erro puro) / (SQ Total) (10)
Sendo:
SQ – Soma dos quadrados.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
67
A Figura 10 mostra o gráfico de Pareto para o planejamento Composto Central
utilizando erro puro. A magnitude dos efeitos é representada pelas colunas. Já a linha
transversal às colunas representa a magnitude dos efeitos com significado estatístico para 95%
de confiança.
Figura 10 – Gráfico de Pareto do planejamento Composto Central mostrando a influência dos fatores estudados.
Analisando-se o gráfico de Pareto (Figura 10) observa-se que os fatores concentração
de quitosana (1) e concentração de FeSO4 (8) foram os fatores que influenciaram de forma
mais significativa na atividade enzimática atingindo efeitos estimados de aproximadamente
5,96 e - 5,30, respectivamente. O sinal positivo indica que o aumento das concentrações de
quitosana aumentará à atividade enzimática. O sinal negativo indica que o decréscimo da
concentração de FeSO4 aumentará à atividade enzimática. Por outro lado, os outros fatores
parecem desprezíveis estatisticamente no gráfico de Pareto.
A avaliação do modelo também pode ser feita através da observação do gráfico dos
valores preditos em função os valores observados que são mostrados na Figura 11. Os valores
preditos pelo modelo são representados pela reta, enquanto que os valores observados
representam-se pelos pontos.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
68
Figura 11 – Valores preditos em função dos valores observados.
Observa-se que alguns valores preditos apresentam um afastamento dos valores
observados. A Figura 12 ilustra a superfície de resposta da atividade quitosanolítica em
função dos dois fatores significativos, concentração de quitosana e concentração de FeSO4.
Figura 12. Atividade quitosanolítica em função da concentração de quitosana e da
concentração de FeSO4 para os fatores não significativos no ponto central.
Observa-se a existência de um ponto de mínimo, ocorrendo para concentração de
quitosana no ponto central (0) e para maiores concentrações de FeSO4 igual a (1,5). Essa

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
69
região deve ser evitada uma vez que a atividade quitosanolítica atinge níveis tão reduzidos
quanto 0,0094 U.mL-1. Destaca-se que nesse planejamento o maior nível da concentração de
FeSO4 (α=+1,41) equivale a 0,0051% (Tabela 6) que é aproximadamente cinco vezes maior
que o nível desse fator no ponto central do planejamento Plackett & Burman (0,001%),
conforme Tabela 3. Dessa forma, ao se utilizar o nível mínimo da concentração de FeSO4 no
Planejamento Composto Central (α=-1,41), que equivale a aproximadamente 0,001%,
aproxima-se do ponto central do do planejamento Plackett & Burman (0,001%), condição
essa que apresenta os melhores resultados de atividade quitosanolítica usando quitosana como
fonte de carbono e energia e fungo Metarhizium anisopliae.
4.4. Perfil Cinético – Melhor Condição
A seguir, apresenta-se um perfil cinético, na melhor condição, para produção da
quitosanase usando o fungo Metarhizium anisopliae, em que foram monitorados durante 24
horas os seguintes parâmetros: Concentração de proteínas totais (µg/mL), atividade
quitosanolítica (U/mL), atividade específica (U/µg) e açúcares redutores (mg/mL), ilustradas
nas Figuras 13, 14, 15 e 16, respectivamente.
Com relação à proteína extracelular, observa-se na Figura 13 que, inicialmente, ocorre
um aumento da mesma para o seu nível máximo (6,2 µg/mL), durante as 6 primeiras horas de
cultivo, seguido de uma redução para 5,7 µg/mL em 18 horas de cultivo, possivelmente
devido à presença de proteases no meio. Observa-se que a partir de então ocorre uma redução
e aumento da proteína total, em uma quantidade inferior, sugerindo um mecanismo de
controle.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
70
0 4 8 12 16 20 245,0
5,2
5,4
5,6
5,8
6,0
6,2
6,4
Pro
tein
a (
µg
/mL
)
Tempo (h)
Figura 13 – Concentração de proteínas totais em função do tempo de cultivo na produção de
quitosanase por Metarhizium anisopliae em meio de cultivo contendo 0,2% de quitosana.
Com relação à atividade quitosanolítica, conforme Figura 14, a atividade máxima
ocorreu em 18 horas de produção em que se obteve aproximadamente 70,0 U/L, o que foi 28
vezes maior que o valor (2,5 U/L em 48 horas) relatado por Assis (2010) para o mesmo fungo,
e em apenas um terço do tempo de produção.
0 4 8 12 16 20 240,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,10
Ativid
ade
(U
/mL
)
Tempo (h)
Figura 14. Perfil da atividade quitosanolítica em função do tempo de fermentação com o
Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.
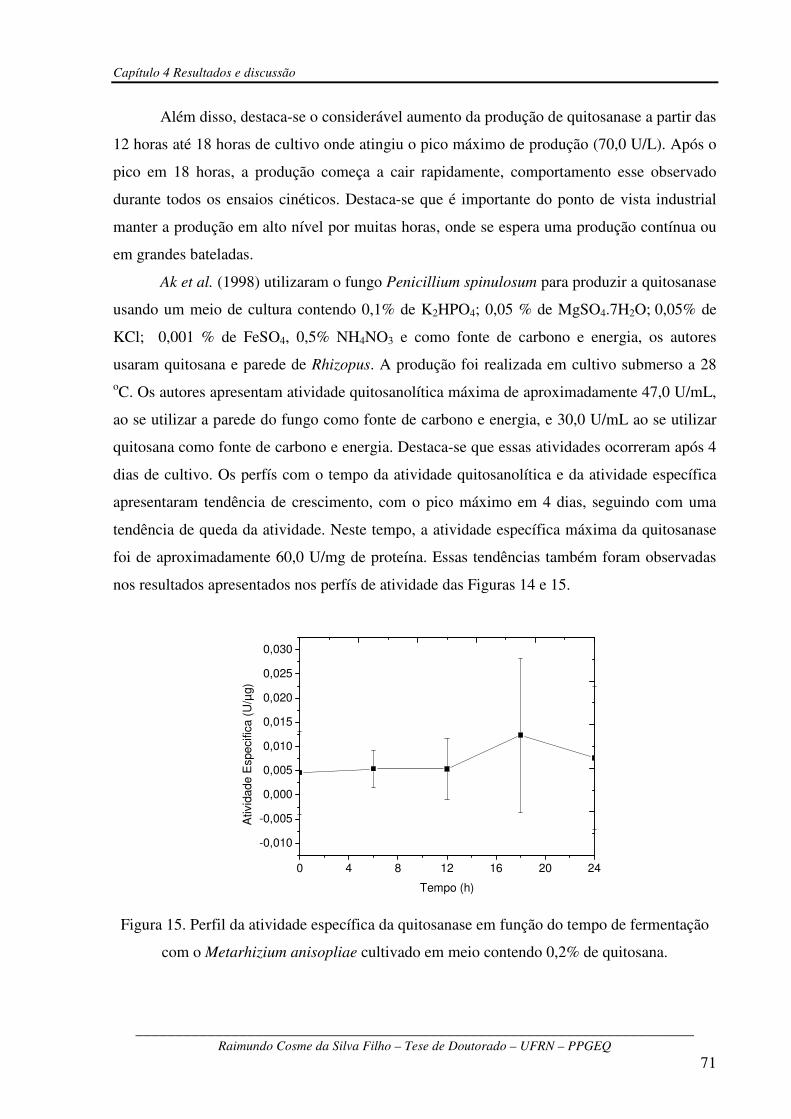
Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
71
Além disso, destaca-se o considerável aumento da produção de quitosanase a partir das
12 horas até 18 horas de cultivo onde atingiu o pico máximo de produção (70,0 U/L). Após o
pico em 18 horas, a produção começa a cair rapidamente, comportamento esse observado
durante todos os ensaios cinéticos. Destaca-se que é importante do ponto de vista industrial
manter a produção em alto nível por muitas horas, onde se espera uma produção contínua ou
em grandes bateladas.
Ak et al. (1998) utilizaram o fungo Penicillium spinulosum para produzir a quitosanase
usando um meio de cultura contendo 0,1% de K2HPO4; 0,05 % de MgSO4.7H2O; 0,05% de
KCl; 0,001 % de FeSO4, 0,5% NH4NO3 e como fonte de carbono e energia, os autores
usaram quitosana e parede de Rhizopus. A produção foi realizada em cultivo submerso a 28
oC. Os autores apresentam atividade quitosanolítica máxima de aproximadamente 47,0 U/mL,
ao se utilizar a parede do fungo como fonte de carbono e energia, e 30,0 U/mL ao se utilizar
quitosana como fonte de carbono e energia. Destaca-se que essas atividades ocorreram após 4
dias de cultivo. Os perfís com o tempo da atividade quitosanolítica e da atividade específica
apresentaram tendência de crescimento, com o pico máximo em 4 dias, seguindo com uma
tendência de queda da atividade. Neste tempo, a atividade específica máxima da quitosanase
foi de aproximadamente 60,0 U/mg de proteína. Essas tendências também foram observadas
nos resultados apresentados nos perfís de atividade das Figuras 14 e 15.
0 4 8 12 16 20 24
-0,010
-0,005
0,000
0,005
0,010
0,015
0,020
0,025
0,030
Ativid
ad
e E
spe
cific
a (
U/µ
g)
Tempo (h)
Figura 15. Perfil da atividade específica da quitosanase em função do tempo de fermentação
com o Metarhizium anisopliae cultivado em meio contendo 0,2% de quitosana.

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
72
Observa-se na Figura 15 que a atividade específica máxima da quitosanase segue a
mesma tendência da atividade quitosanolítica apresentada na Figura 14, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
O perfil para os açúcares redutores durante o cultivo de Metarhizium anisopliae
cultivado em meio contendo 0,2% de quitosana é ilustrado na Figura 16.
0 4 8 12 16 20 240,004
0,008
0,012
0,016
0,020
0,024
0,028
Acuca
res R
ed
uto
res (
mg/m
L)
Tempo (h)
Figura 16. Perfil de açúcares redutores por Metarhizium anisopliae em meio de cultivo
contendo 0,2% de quitosana.
Observa-se, na Figura 16, que taxa máxima de redução dos açúcares redutores
(aproximadamente 0,021 mg/mL) ocorre logo após a produção máxima da enzima, em 18
horas de cultivo, conforme Figura 14. A quantidade de açúcares redutores foi a
aproximadamente 0,012 mg/mL após 24 h de cultivo. O fungo excreta para o meio enzimas
com atividade quitosanolítica, essas enzimas hidrolisam a quitosana aumentando a quantidade
de açúcares redutores para o meio, sendo que esses açúcares também são consumidos pelo
fungo durante seu crescimento.
Foi demonstrado por Assis (2010) que os açúcares redutores produzidos através da
quitossana, ocasionada pelas enzimas quitossanases metabolizadas por Metarhizium
anisopliae, com quitosana como sua única fonte de carbono e energia, atingiu um pico de 0,53
mg/mL em 48 horas. Apresentaram como quitooligossacarídeos, constituídos principalmente
por dímeros e pentâmeros, formados pela hidrólise enzimática de quitosanase em quantidades
maiores do que o consumido pelos microrganismos. Mostraram, também, que essa produção

Capítulo 4 Resultados e discussão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
73
segue a mesma tendência da sua atividade enzimática, corroborando com os resultados
apresentados no presente trabalho.
Os resultados do presente trabalho mostram a relevância de se adotar novas condições
para a produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter quitooligossacarídeo
com atividades biológicas. Esta pesquisa, portanto, sugere base para novos estudos que
permitam alcançar rapidamente a produção de quitosanase e quitooligossacarídeo com todos
os seus benefícios.

Capítulo 5 Conclusão

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
75
5. Conclusão
Após o desenvolvimento desse trabalho conclui-se:
- Com relação ao Planejamento de Seleção de Fatores Plackett & Burman:
Os resultados das análises do Planejamento de Seleção de Fatores Plackett &
Burman com 8 fatores e 12 ensaios em 2 níveis, mais 3 repetições no ponto central, para
se investigar a influência dos fatores: Concentração de Quitosana, Peptona, Extrato de
Levedura, NaNO3, K2HPO4, KCl, MgSO4.7H2O, FeSO4 na produção da enzima
quitosanase por Metarhizium anisopliae, mostraram que a atividade foi favorecida pelo
aumento da concentração de substrato (quitosana) e sulfato ferroso (FeSO4), enquanto
que o aumento da concentração dos outros fatores não contribuíram de forma
significativa para a resposta (atividade quitosanolítica). Os fatores concentração de
quitosana (1) e concentração de FeSO4 (8) atingiram efeitos estimados de
aproximadamente 8,84 e 4,67, respectivamente. O sinal positivo indica que o aumento
das concentrações de quitosana e de FeSO4 aumentará a atividade enzimática. Conforme
visto na literatura, observa-se importância do FeSO4, já que o mesmo está diretamente
associado a síntese de proteases (principalmente metalloproteases),
- Com relação ao Planejamento Composto Central:
A análise do planejamento composto central para avaliar a influência dos
fatores, concentração de substrato (quitosana) e concentração de FeSO4, na produção da
enzima quitosanase por Metarhizium anisopliae (a partir do planejamento Plackett &
Burman) mostrou que os fatores concentração de quitosana e concentração de FeSO4
foram os fatores que influenciaram de forma mais significativa na atividade enzimática
atingindo efeitos estimados de aproximadamente 5,96 e - 5,30, respectivamente. O sinal
positivo indica que o aumento das concentrações de quitosana aumentará a atividade
enzimática. O sinal negativo indica que o decréscimo da concentração de FeSO4
aumentará a atividade enzimática. Observa-se a existência de um ponto de mínimo,
ocorrendo para concentração de quitosana (X1) no ponto central (0) e para maiores

Capítulo 5 Conclusão
______________________________________________________________________ Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
76
concentrações de FeSO4 (X2) igual a (1,5) com a atividade quitosanolítica de 0,0094
U.mL-1.
- Com relação ao cultivo na melhor condição:
O cultivo na melhor condição mostrou uma produção máxima de quitosanase em
18 horas de cultivo, com a atividade enzimática de 70,0 U/L, o que foi 28 vezes maior
que o valor (2,5 U/L em 48 horas) relatado na literatura (Assis, 2010) para o mesmo
fungo e em apenas um terço do tempo de produção.
Além disso, observou-se que um elevado incremento na produção de
quitosanase a partir das 12 horas de cultivo, antes de se atingir a atividade
quitosanolítica máxima, 18 horas de cultivo. Essa produção em alto nível por muitas
horas é importante do ponto de vista industrial, onde se espera uma produção contínua
ou em grandes bateladas. Observou-se também que a atividade específica máxima da
quitosanase segue a mesma tendência da atividade quitosanolítica, alcançando uma
produção máxima de aproximadamente 12,4 U/µg, em 18 horas de cultivo.
- Com relação aos açúcares redutores:
Foi observado que os açúcares redutores seguem a mesma tendência da atividade
enzimática, com uma produção máxima de cerca de 0,21 mg/mL em 18 horas de
cultivo, produção essa que coincinde com o máximo pico de atividade enzimática.
Estes resultados mostram a relevância de se adotar novas condições para a
produção de quitosanase, com suas vantagens em termos de compatibilidade ambiental,
baixo custo e reprodutibilidade com a possibilidade também de se obter
quitooligossacarídeo com atividades biológicas. Esta pesquisa, portanto, sugere base
para novos estudos que permitam alcançar rapidamente a produção de quitosanase e
quitooligossacarídeo com todos os seus benefícios.

Referências Bibliográficas

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
78
Referências bibliográficas
AK, Ö.; BAKIR, U.; GÜRAY, T. Isolation of a chitosan degrading fungus, Penicillium
spinulosum, and chitosanase production by the isolate. In: SYMPOSIUM - NEW
FRONTIERS IN SCREENING FOR MICROBIAL BIOCATALYSTS, 1996, Ede, The
Netherlands. Anais, Editado por K. Kieslich, C. P. van der Beek, J. A. M. de Bont e W.
J. J. van den Tweel, Elsevier Science B. V., 1998, v.53, p.265-268.
ALVES, S. B. Patologia e controle microbiano: Vantagens e desvantagens. In: ALVES,
S. B. Controle Microbiano de Insetos. 2ed., Piracicaba, FEALQ, 1163p, 1998, v.4, p.21-
37.
ALVES, S. B.; LEITE, L. G.; FILHO, A. B.; ALMEIDA, F. E. M.; MARQUES, E. J.
Produção massal de fungos entomopatogênicos na América Latina, In: ALVES, S. B.;
LOPES, R. B. Controle microbiano de pragas na América Latina: avanços e desafios.
Piracicaba, FEALQ, 414p, 2008, v.14, p.215-237.
ARRUDA, W.; LUBECK, I.; SCHRANK, A.; VAINSTEIN, M. H. Morphological
alterations of Metarhizium anisopliae during penetration of Boophilus microplus ticks.
Experimental and Applied Acarology, v. 37, p.231-244, 2005.
ASSIS, C. F. Produção e caracterização de quitooligossacarídeos produzidos pelo fungo
Metarhizium anisopliae e avaliação da citotoxicidade em células tumorais. 2010. 110p.
Tese (Doutorado em Biotecnologia) – Renorbio - Programa de Pós-Graduação em
Biotecnologia. Universidade Federal do Rio Grande do Norte, Natal.
ASSIS, C. F.; COSTA, L. S.; MELO-SILVEIRA, R. F.; OLIVEIRA, R. M.;
PAGNONCELLI, M. G. B.; ROCHA, H. A. O.; MACEDO, G. R.; SANTOS, E. S.,
Chitooligosaccharides antagonizes the cytotoxic effect of glucosamine. World J.
Microbiol. Biotechnol., v.28, n.3, p.1097-1105, 2012.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
79
AZEVEDO, V. V. C.; CHAVES, S. A.; BEZERRA, D. C.; FOOK, M. V. L.; COSTA,
A. C. F. M. Quitina e Quitosana: aplicações como biomateriais. Revista Eletrônica de
Materiais e Processos, v.2.3, p.27-34, 2007.
BARROS NETO, B.; SCARMINIO, I. S.; BRUNS, R. Como fazer experimentos:
pesquisa e desenvolvimento na ciência e na indústria. 2. ed. Campinas: Unicamp, 2001.
401p.
BATISTA FILHO, A.; ALMEIDA, J. E. M.; SANTOS, A. S.; MACHADO, L. A.;
ALVES, S. B. Eficiência de isolados de Metarhizium anisopliae no controle de
cigarrinha-da-raiz de cana-de-açúcar Mahanarva fimbriolata (Hom.: Cercopidae). STAB
– Açúcar, Álcool e Subprodutos, v.21, p.84-89, 2002.
CARVALHO, J. C. M.; SATO, S. Fermentação Descontínua, In: SCHMIDELL, W.;
LIMA, U. A.; AQUARONE, E.; BORZANI, W. Biotecnologia Industrial. São Paulo,
Ed. Edgard Blücher, v.2, p.193-204, 2001.
CHAO, H. J.; SCHWARTZ, J.; MILTON, D. K. Populations and determinats of
airborne fungi in large office buildings. Environmental Health Perspectives, v.110, n.8,
p.777-782, 2002.
CHEN, X.; XIA, W.; YU, X. Purification and characterization of two types of
chitosanase from Aspergillus sp. CJ22-326. Food Research International, v.38, n.3,
p.315-322, 2005.
CHEN, P. T.; CHIANG, C.-J.; CHAO, Y.-P. Medium optimization and production of
secreted Renilla luciferase in Bacillus subtilis by fed-batch fermentation. Biochemical
Engineering Journal, v.49, p.395-400, 2010.
CHENG, C. Y.; LI, Y. An Aspergillus chitosanase with potential for large-ecale
preparation of chitosan oligosaccharides. Biotechnol. Appl. Biochem., v.32, p.197-203,
2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
80
CHOI, Y. J.; Kim, E. J.; Piao, Z.; Yun, Y. C.; Shin, Y. C. Purification and
characterization of chitosanase from Bacillus sp. Strain KCTC 0377BP and its
application for the production of chitosan oligosaccharides. Applied and Environmental
Microbiology, v.70, n.8, p.4522-4531, 2004.
COLAKOGLU, G. The chemical and physical environment for fungal growth. Fak
Matbaa Birimi, Istanbul, 149-166, 2001.
CRAVEIRO, A. C.; CRAVEIRO, A. A.; QUEIROZ, D. C. Quitosana a fibra do futuro.
2. ed. Fortaleza: PADETEC, 2004. V.1, 218p.
DIAS, B. A. Estudo de proteases degradadoras de cutícula produzidas pelo fungo
entomopatogênico Beauveria bassiana. 2005. 70p. Dissertação (Mestrado em
Microbiologia) – Centro de Ciências Biológicas, Programa de Pós-Graduação em
Microbiologia, Universidade Estadual de Londrina, Londrina.
DINARDO-MIRANDA, L. L.; FERREIRA, J. M. G.; CARVALHO, P. A. M.
Influência da época de colheita e do genótipo de cana-de-açúcar sobre a infestação de
Mahanarva fimbriolata (Stal) (Hemiptera: Cercopidae). Neotrop. Entomol, v.30, n.1,
p.145-149, 200.
DONG, C.; ZHANG, J.; HUANG, H.; CHEN, H.; HU, Y. Pathogenicity of a new China
variety of Metarhizium anisopliae (M. anisopliae var. Dcjhyium) to subterranean termite
Odontotermes formosanus. Microbiological Research, v.164, p.27-35, 2009.
DONZELLI, B. G. G.; OSTROFF, G.; HARMAN, G. E. Enhanced enzymatic
hydrolysis of langostino shell chitin with mixtures of enzymes from bacterial and fungal
sources. Carbohydrate Research, v.338, p.1823-1833, 2003.
DRIVER, F.; MILNER, R. J.; TRUEMAN, J. W. H. A taxonomic revision of
Metarhizium based on phylogenetic analysis of rDNA sequence data. Mycological
Research, v.104, n.2, p.134-150, 2000.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
81
DUNG, P.; MILAS, M.; RINAUDO, M.; DESBRIÉRES, J. Water soluble derivatives
obtained by controlled chemical modifications of chitosan, Carbohydrate Polymers,
v.24, p.209-214, 1994.
ESTEVES, J. A.; CABRITA, J. D.; NOBRE, G. N. Micologia médica, 2. ed. Lisboa:
Fundação Calouste Gulbenkian, 1990. 1058p.
FACCIOTI, M. C. R. Fermentação Contínua, In: Biotecnologia Industrial. Editado por:
SCHMIDELL, W.; LIMA, U. A.; AQUARONE, E.; BORZANI, W., São Paulo, Ed.
Edgard Blücher, v.2, p.541, 2001.
FARIA, M. R.; MAGALHÃES, B. P. O uso de fungos entomopatogênicos no Brasil -
Situação atual e perspectivas. Biotecnologia Ciência & Desenvolvimento, n.22., p.18-
21, setembro/outubro 2001.
FU, J.-Y.; WU, S.-M.; CHANG, C.-T.; SUNG, H.-Y. Characterization of Three
Chitosanase Isozymes Isolated from a Commercial Crude Porcine Pepsin Preparation. J.
Agric. Food Chem., v.51, p.1042-1048, 2003.
FURIGO JUNIOR, A.; PEREIRA, E. P. Enzimas e suas aplicações cinética enzimática.
2001. 39f. Apostila (Graduação de Engenharia Química e Engenharia de Alimentos) –
Centro de Tecnologia, Departamento de Engenharia Química e Engenharia de
Alimentos, Universidade Federal de Santa Catarina, Florianópolis.
GAO, J.; XU, H.; LI, Q.-J.; FENG, X.-H.; LI, S. Optimization of medium for one-step
fermentation of inulin extract from Jerusalem artichoke tubers using Paenibacillus
polymyxa ZJ-9 to produce R,R-2,3-butanediol, Bioresource Technology, v.101, p.7076-
7082, 2010.
GARCIA, M. V.; MONTEIROI, A. C.; SZABO, M. J. P.; PRETTE, N.; BECHARA, G.
H. Mechanism of infection and colonization of Rhipicephalus sanguineus eggs by
Metarhizium anisopliae as revealed by scanning electron microscopy and
histopathology. Brazilian Journal of Microbiology, São Paulo, v.36, n.4, p.368-372,
2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
82
GHANEM, N. B.; YUSEF, H. H.; MAHROUSE, H. K. Production of Aspergillus
terreus xylanase in solid-state cultures: application of the Plackett & Burman
experimental design to evaluate nutritional requirements, Bioresource Technology,
v.73, p.113-121, 2000.
GHORBEL-BELLAJ, O.; HMIDET, N.; JELLOULI, K.; YOUNES, I.; MAALEJ, H.;
HACHICHA, R.; NASRI, M. Shrimp waste fermentation with Pseudomonas aeruginosa
A2: Optimization of chitin extraction conditions through Plackett–Burman and response
surface methodology approaches, International Journal of Biological Macromolecules,
v.48, p.596–602, 2010.
GIUSTINA, A.; VENTURA, P. Weight-reducing regimens in obese subjects: Effects of
a new dietary fiber integrator. Acta Toxicol Ther, 16, 4, p.199-214, 1995.
GUIA.BIO, Reino dos fungos, 2012. Disponível em:
<http://www.http://guia.bio.br/reino-dos-fungos>. Acesso em: 12 novembro 2012.
GUIRADO, N.; AMBROSANO, E. J.; ROSSI, F.; DIAS, F. L. F. Controle de Cupins
de Montículo com Metarhizium anisopliae e Beauveria bassiana, Rev. Bras. de
Agroecologia, v.4, n.2, 2009.
GUO, Y.; XU, J.; ZHANG, Y.; XU, H.; YUAN, Z.; LI, D. Medium optimization for
ethanol production with Clostridium autoethanogenum with carbon monoxide as sole
carbon source, Bioresource Technology, v.101, p.8784–8789, 2010.
HAGAG, S.; KUBITSCHEK-BARREIRA, P.; NEVES, G. W. P.; AMAR, D.;
NIERMAN, W.; SHALIT, I.; SHAMIR, R.; LOPES-BEZERRA, L.; OSHEROV, N.
Transcriptional and proteomic analysis of the Aspergillus fumigatus ∆prtT protease-
deficient mutant. PLoS One, v.7, n.4, e33604, 2012.
HAN, L.-K.; KIMURA, Y.; OKUDA, H. Reduction in fat storage during chitin-chitosan
treatment in mice fed a high-fat diet, International Journal of Obesity, 23:174-179,1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
83
HUNG, T.; CHANG, Y.; SUNG, H.; CHANG, C. Purification and Characterization of
Hydrolase with Chitinase and Chitosanase Activity from Commercial Stem Bromelain.
J. Agric. Food Chem., 50, 4666-4673, 2002.
ICHIKAWA, S.; TAKANO, K.; TAKASHIKUROIWA, HIRUTA, O.; SATO, S.;
MUKATAKA, A. S. Immobilization and stabilization of chitosanase by multipoint
attachment to agar gel support. Journal of Bioscience and Bioengineering, v.93, n.2,
p.201-206, 2002.
JABOR, I. A. S.; PAMPHILE, J. A.; RODRIGUES, S. B.; MARQUES-SILVA, G. G.;
ROCHA, C. L. M. S. Análise do desenvolvimento do fungo entomopatogênico
Metarhizium anisopliae em resposta a fatores nutricionais. Acta Scientiarum.
Agronomy, Maringá, v.25, n.2, p.497-501, 2003.
JI, X.-J.; HUANG, H.; DU, J.; ZHU, J.-G.; REN, L.-J.; LI, S.; NIE, Z.-K. Development
of an industrial medium for economical 2,3-butanediol production through co-
fermentation of glucose and xylose by Klebsiella oxytoca, Bioresource Technology,
v.100, p.5214–5218, 2009.
JO, Y.; JO, K.; JIN, Y.; KIM, K.; SHIN, J.; KIM, Y.; PARK, R. Characterization and
kinetics of 45 kDa chitosanase from bacillus sp. P16. Biosci. Biotechnol. Biochem.,
v.67, n.9, 1875 – 1882, 2003.
KHOSRAVI-DARANI, K.; ZOGHI, A.; ALAVI, S. A.; FATEMI, S. S. A. Application
of Plackett Burman Design for Citric Acid Production from Pretreated and Untreated
Wheat Straw, Iran. J. Chem. Chem. Eng.. v.27, n.1, p.91-104, 2008.
KIM, P.; KANG, T. H.; CHUNG, K. J.; KIM, I. S.; CHUNG, K. Purification of a
constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and
expression of the gene. FEMS Microbiolog.y Letters 240, 31–39, 2004.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
84
KIM, S.-K.; RAJAPAKSE, N. Enzymatic production and biological activities of
chitosan oligosaccharides (COS): A review. Carbohydrate Polymers, v.62, n.4, p.357-
368, 2005.
KIRK, P. M.; CANON, P.F.; MINTER, D.W.; STALPERS, J.A. Dictionary of the
fungi. CAB International, Wallingford, 2008.
KONDO, H.; OSADA, A. Influence of dietary fiber on the bioavailability of zinc in
rats, Biomed Environ Sci, 9, 204-8, 1996.
KUMAR, S.; KOH, J. Physiochemical, Optical and Biological Activity of Chitosan-
Chromone Derivative for Biomedical Applications. Int. J. Mol. Sci., v.13, p.6102-6116,
2012.
KURITA, K.; SANNAN, T.; IWAKURA, Y. Studies on chitin, 4. Evidence for
formation of block and random copolymers of N-acetyl-o-glucosamine and
oglucosamine by hetero and homogeneous hydrolyses. Die Makromolecular Chemie,
v.178, p.3197-3202, 1977.
KUROIWA, T.; ICHIKAWA, S.; HIRUTA, O.; SATO, S.; MUKATAKA, S. Factors
Affecting the Composition of Oligosaccharides Produced in Chitosan Hydrolysis Using
Immobilized Chitosanases. Biotechnol. Prog., 18, 969-974, 2002.
LI, L.-Z.; HUI, Z.; MO, X. Optimization of the medium for streptolydigin production
by Streptomyces lydicus AS 4.2501 with precursor feeding using response surface
methodology, Journal of the Taiwan Institute of Chemical Engineers, 41, 252–258,
2010.
LIU, B.-L.; KAO, P.-M.; TZENG, Y.-M.; FENG, K.-C. Production of chitinase from
Verticillium lecanii F091 using submerged fermentation. Enzyme and Microbial
Technology, v.33, p.410-415, 2003.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
85
LOBATO, A. K. C. L. Estudo da produção de biosurfactantes por micro-organismos
isolados de poços de petróleo. 2003. 108p. Dissertação (Mestrado) – Programa de Pós-
Graduação em Engenharia Química. Universidade Federal do Rio Grande do Norte.
Natal.
LOPES, R. B.; ALVES, S. B.; PADULLA, L. F. L.; PÉREZ, C. A. Eficiência de
formulações de Beauveria bassiana e Metarhizium anisopliae para o controle de ninfas
de Amblyomma cajennense (Fabricius, 1787). Revista Brasileira de Parasitologia
Veterinária, v.16, n.1, p.27-31, 2007.
LOUREIRO, E. S.; BATISTA FILHO, A.; ALMEIDA, J. E. M.; MENDES, J. M.;
PESSOA, L. G. A. Eficiência de isolados de Metarhizium anisopliae (metsch.) sorok.
no controle da cigarrinha-da-raiz da cana-de-açúcar, Mahanarva fimbriolata (stal, 1854)
(hemiptera: cercopidae), em condições de campo. Arq. Inst. Biol., São Paulo, v.79, n.1,
p. 47-53, 2012.
MELLO, D. R.; REIS, R. C. S.; BITTENCOURT, V. R. E. P. Patogenicidade in vitro
do fungo Metarhizium anisopliae (Metschnikoff, 1879) Sorokin, 1883, sobre o carrapato
Boophilus microplus (Canestrini, 1887). Revista Brasileira de Parasitologia Veterinária,
São Paulo, v.15, p.157-162, 2006.
MITSUTOMI, M.; KIDOH, H.; TOMITA, H.; WATANABE, T. The action of Bacillus
circulans WL-12 chitinases on partially N-acetylated chitosan, Bioscience
Biotechnology and Biochemistry, v.59, p.529-531, 1995.
MALAJOVICH, M. A. Biotecnologia: fundamentos. Ed. biblioteca Max Feffer do
Instituto de Tecnologia ORT, Rio de Janeiro, 2009. 104p.
MONAGHAN, R. L.; EVELEIGH, D. E.; TEWARI, R. P.; REESE, E. T. Chitosanase,
a novel enzyme. Nature - New Biology, 245, p.78-80, 1973.
MU, X. Q.; XU, Y.; NIE, Y.; OUYANG, J.; SUN, Z. H. Candida parapsilosis CCTCC
M203011 and the optimization of fermentation medium for stereoinversion of (S)-1-
phenyl-1,2-ethanediol, Process Biochemistry, v.40, p.2345–2350, 2005.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
86
MURRAY, P. R.; ROSENTHAL, K. S.; PFALLER, M. A. Medical microbiology. 5th
ed. London, Elsevier Mosby, 2005.
MUZZARELLI, R. A. A. Biochemical significance of exogenous chitins and chitosans
in animals and patients, Carbohydrate Polymers, v.20, p.7-16, 1993.
MUZZARELLI, R. A. A.; TOMASETTI, M.; ILARI, P. Depolymerization of chitosan
with the aid of papain, Enzyme Microbiology and Technology, v.16, p.110-114, 1994.
NAVEENA, B. J.; ALTAF, Md.; BHADRIAH, K.; REDDY, G. Selection of medium
components by Plackett–Burman design for production of L(+) lactic acid by
Lactobacillus amylophilus GV6 in SSF using wheat bran, Bioresource Technology,
v.96, p.485-490, 2005.
NOGAWA, M.; TAKAHASHI, H.; KASHIWAGI, A.; OHSHIMA, K.; OKADA, H.;
MORIKAWA, Y. Purification and Characterization of Exo--D-Glucosaminidase from
a Cellulolytic Fungus, Trichoderma reesei PC-3-7, Appl. Environ. Microbiol., v.64, n.3,
p.890-895, 1998.
ONOFRE, S. B.; VARGAS; L. R. B.; ROSSATO, M.; BARROS, N. M.; BOLDO, J.
T.; NUNES, A. R. F.; AZEVEDO, J. L. Controle biológico de pragas na agropecuária
por meio de fungos entomopatogênicos. In: SERAFINI, L. A.; BARROS, N. M.;
AZEVEDO, J. L. (Org.). Biotecnologia – avanços na agricultura e na agroindústria.
Caxias do Sul: EDUCS, p.295-328, cap.8, 2002.
OSSWALD, W. F.; SHAPIRO, J. P.; DOOSTDAR, H.; MCDONALD, R. E.; NIEDZ,
R. P.; NAIRN, C. J.; HEARN, C. J.; MAYER, R. T. Identification and Characterization
of Acidic Hydrolases with Chitinase and Chitosanase Activities from Sweet Orange
Callus Tissue, Plant Cell Physiol. 35, p.811-820, 1994.
PAGNONCELLI, M. G. B. Estudo do mecanismo de produção de oligossacarídeos com
atividades nutracêuticas a partir da quitosana por hidrólise enzimática com processo

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
87
fermentativo simultâneo. 2008. 118p. Tese (Doutorado) – Centro de Tecnologia,
Programa de Pós-Graduação em Engenharia Química. Universidade Federal do Rio
Grande do Norte. Natal.
PALMA-GUERRERO, J., JANSSON, H.-B., SALINAS, J.; LOPEZ-LLORCA, L. V.
Effect of chitosan on hyphal growth and spore germination of plant pathogenic and
biocontrol fungi, J. Appl. Microbiol., v.104, p.541-553, 2008.
PAULA, F. A.; ARRUDA, D. C. Análise do método de planejamento experimental
Plackett Burman na otimização de degradação de penas de aves por comunidades
microbianas, XXI Salão de Iniciação Científica, 19-23, outubro, 2009, UFRGS, Porto
Alegre.
PELCZAR, M. J.; CHANG, E. C. S.; KRIEG, N. R. Microbiologia: conceitos e
aplicações, v.2, 2.ed. São Paulo: Makron BooKs, 1996.
PELLETIER, A.; SYGUSCH, J. Purification and characterization of three chitosanase
activities from Bacillus megaterium P1, Applied and Environmental Microbiology, v.
56, n.4, p.844-848, 1990.
PEREIRA, F. B.; GUIMARÃES, P. M. R.; TEIXEIRA, J. A.; DOMINGUES, L.
Optimization of low-cost medium for very high gravity ethanol fermentations by
Saccharomyces cerevisiae using statistical experimental designs, Bioresource
Technology, v.101, p.7856-7863, 2010.
PETER, M. G. Chitin and chitosan from aninal sources. In: STEINBUCHEL, A.;
RHEE, S. K. Polysaccharides and Polyamides in the Food Industry. Weinheim:
Wileyvch, v.1, p.115-208, 2005.
PIZA, F. A. T.; SILOTO, A. P.; CARVALHO, C. V.; FRANCO, T. T. Purification,
characterization and purification of chitosanase from Bacillus cereus, Brazilian Journal
of Chemical Engineering., v.16, n 2, p.185-192, 1999.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
88
POLITZER, K.; BON, E. P. S. Enzimas Industriais e Especiais. C.G.E.E. Ciência,
Tecnologia e Inovação. Rio de Janeiro, 554p, 2006.
POLYMAR, Ciência é a nossa Natureza, 2003. Disponível em:
<http://www.polymar.com.br>. Acesso em: 6 setembro 2012.
PUTZKE, J.; PUTZKE, M. T. L. Os reinos dos fungos. Santa Cruz do Sul, RS:
EDUNISC, 1998.
QUINES, L. K. M. Extração de poli(3-hidroxibutirato) produzido por Cupriavidus
necator dsm 545 com 1,2-carbonato de propileno. 2010. 99p. Dissertação (Mestrado em
Engenharia de Alimentos) – Centro Tecnológico, Departamento de Engenharia Química
e Engenharia de Alimentos, Programa de Pós-Graduação em Engenharia de Alimentos,
Universidade Federal de Santa Catarina, Florianópolis.
RAJENDRAN, A.; THANGAVELU, V. Statistical experimental design for evaluation
of medium components for lipase production by Rhizopus arrhizus MTCC 2233, LWT -
Food Science and Technology, v.42, p.985–992, 2009.
RAMIREZ-LEÓN, L. F.; RUIZ-HERRERA, J. Hydrolysis of walls and formation of
spheroplasts in Mucor rouxii. J. Gen. Microbiol., v.72, p.281-290, 1972.
RANGEL, M. A. S.; LOUREIRO, E. S.; ALMEIDA, A. S.; ALVES, H. C.; ONO, F.
B.; LOBO, P. M.; MARIANI, M. A.; PESSOA, L. G. A.; SILVA, A. J. Avaliação
participativa do controle de cigarrinhas das pastagens com a aplicação do fungo
Metarhizium anisopliae (Metsch.) Sorok. Rev. Bras. de Agroecologia, v.2, n.2, 731-734,
2007.
RAVEN, P. H.; EVERT, R.F.; EICHHORN, S.E. Biologia Vegetal, 6ª Ed, Rio Janeiro,
Editora Guanabara Koogan, 2001.
REDDY, L. V. A.; WEE, Y.-J.; YUN, J.-S.; RYU, H.-W. Optimization of alkaline
protease production by batch culture of Bacillus sp. RKY3 through Plackett–Burman

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
89
and response surface methodological approaches, Bioresource Technology, v.99, p.242–
2249, 2008.
RIGUEIRA, J. C. S. Influência da contagem de células somáticas no perfil e teores de
aminas bioativas e na qualidade de leite cru e de queijo mussarela. 2010. 147p. Tese
(Doutorado em Ciência de Alimentos) - Faculdade de Farmácia, Departamento de
Alimentos, Programa de Pós-Graduação em Ciência de Alimentos, Faculdade de
Farmácia da Universidade Federal de Minas Gerais, Belo Horizonte.
RODRIGUES, M. I.; IEMMA, A. F. Planejamento de Experimentos e Otimização de
Processos: uma estratégia sequencial de planejamentos, Campinas, SP, Casa do Pão
Editora, 2005.
SANTOS, E. S. Recuperação e Purificação de Enzimas usando Adsorção em Leito
Expandido, Tese de Doutorado, Universidade Estadual de Campinas, Campinas/SP,
p.152, 2001.
SEDMAK, J. J.; GROSSBERG, S. E. A rapid, sensitive and versatile assay for protein
using coomassie brilliant blue G250, Analytical Biochemistry, v.79, p. 544-552, 1977.
SEINO, H.; TSUKUDA, K.; SHIMASUE, Y. Properties and action pattern of a
chitosanase from Bacillus sp. PI- 7S. Agricultural Biological Chemistry, 55, p.2421-
2423, 1991.
SHIMOSAKA, M.; NOGAWA, M.; OHNO, Y.; OKAZAKI, M. Chitosanase from the
plant pathogenic fungus, Fusarium solani f. sp. Phaseoli – purification and some
properties, Bioscience Biotechnolgy and Biochemistry, v.57, n.2, p.231-235, 1993.
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Production of two chitosanases from a chitosan-assimilating bacterium, Acinetobacter
sp. Strain CHB101, Applied and Environmental Microbiology, v.61, n.2, p.438-442,
1995.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
90
SHIMOSAKA, M.; NOGAWA, M.; WANG, X.-Y.; KUMEHARA, M.; OKAZAKI, M.
Characterization of Three Chitosanase Isozymes Isolated from a Commercial Crude
Porcine Pepsin Preparation, J. Agric. Food Chem. 51, p.1042-1048, 2003.
SILVA FILHO, R. C.; COSTA, A. S. S.; CHAGAS, B. M. E.; SANTOS, F. N.;
MACEDO, G. R.; SANTOS, E. S. Avaliação do teste de atividade quitosanolítica
produzida por fungo. In: XV SIMPÓSIO NACIONAL DE BIOPROCESSOS, 15,
Recife, 2005, 02 a 05 de agosto.
SILVA, E. P. Avaliação Preliminar do Potencial de Aplicação da Tecnologia de
Biopilhas para a Biorremediação do Solo de Guamaré-RN. 2004. 119p. Dissertação
(Mestrado) – Centro de Tecnologia, Programa de Pós-Graduação em Engenharia
Química. Universidade Federal do Rio Grande do Norte. Natal.
SOMASHEKAR, D.; JOSEPH, R. Chitosanases – Properties and apllications: A review.
Bioresource Technology, v.55, p.35-45, 1996.
STOYACHENKO, I. A.; VARLAMOV, V. P.; DAVANKOV, V. A. Chitinases of
Streptomyces kurssanovii: purification and some properties, Carbohydrate Polymers,
v.24, p.47-54, 1994.
TINLINE, R. D. Nuclear distribuition in Metarhizium anisopliae. Mycologia, v. 63, p.
713-721, 1971.
TRABULSI, L. R.; TOLEDO, M. R. F. Microbiologia. 2. Ed., São Paulo: Atheneu,
1996.
VIEIRA, P. D. S.; SILVA, W. M. T.; PAIVA, L. M.; LIMA, E. A. L. A.;
CAVALCANTI, V. L. B. Estudo da caracterização morfológica, esporulação e
germinação dos conídiosde Metarhizium anisopliae var. acridum em diferentes
temperaturas. Biológico, São Paulo, v.69, n.1, p.17-21, 2007.

Referências Bibliográficas
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
91
WANDERLEY, M. D.; NEVES, E.; ANDRADE, C. J. Aspectos da produção industrial
de enzimas. Revista Citino, v.1, n.1, p.30-36, 2011.
WANG, C.; TYPAS, M. A.; BUTT, T. M. Detection and characterization of pr1
virulent gene deficiencies in the insect pathogenic fungus Metarhizium anisopliae.
FEMS Microbiology Letters, v. 213, p. 251-255, 2002.
ZAHORSKA-MARKIEWICZ, B.; KROTKIEWSKI, M.; OLSZANECKA-
GLINIANOWICZ, M.; ZURAKOWSKI, A. Effect of chitosan in complex management
of obesity, Pol Merkur Lekarski, v.13, n.74, p.129-132, 2002.
ZHANG, X. Y.; DAI, A. L.; ZHANG, X. K.; KUROIWA, K.; KODAIRA, R.;
SHIMOSAKA, M.; OKAZAKI, M. Purification and Characterization of and Exo--D-
glucosaminidase from a koji mold, Aspergillus oryzae IAM2660. Biosci. Biotechnol.
Biochem., v.64, n.9, 1896-1902, 2000.
ZHU, X.; WU, X.; DAI, Y. Fermentation conditions of a chitosanase from Acinetobcter
sp C-17. Biosci. Biotechnol. Biochem., v.67, n.2, p.284-290, 2003.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
92
APÊNDICE

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
93
APÊNDICE I
Metodologia para repique da cepa
Em Erlenmeyer de 1000 mL colocava-se 500 mL do meio de manutenção
PDA. Este Erlenmeyer, com o meio de manutenção, era autoclavado junto com placas
de Petri a 1,5 kgf/cm2 por 15 minutos, em seguida, era levado para uma câmara de fluxo
laminar e transferido para as placas de Petri já autoclavadas, onde eram mantidas numa
estufa a 27 ºC durante 24 h. Após este período, as placas de Petri eram levadas a câmara
de fluxo laminar, utilizando bico de Bunsen para esterilizar a alça de platina. Procedia-
se o repique utilizando uma placa de Petri contendo a cepa pura. Cada placa de Petri
contendo a cepa pura era transferida para duas placas de Petri contendo o meio de
manutenção. As placas eram mantidas na estufa a 27 ºC durante 30 dias onde após esse
período, repetia-se este procedimento.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
94
APÊNDICE II
Montagem da Curva de Calibração Usando o Método DNS
Soluções:
Glicosamina = 0,01 mol/mL (10000,0 µmol/mL)
Tabela 14. Dados para construção da curva de calibração usando o Método DNS.
Tubo de
Ensaio
Solução de
glicosamina (mL)
Água Destilada
(mL)
DNS (mL)
Volume Total
Massa (µµµµmol)
1 0 1,0 1,5 2,5 0
2 0,2 0,8 1,5 2,5 0,2
3 0,4 0,6 1,5 2,5 0,4
4 0,6 0,4 1,5 2,5 0,6
5 0,8 0,2 1,5 2,5 0,8
6 1,0 0 1,5 2,5 1,0
Tabela 15 - Medidas de absorbância a 600nm
Dados adicionais:
Peso Molecular D-Glicosamina = 215,64 (recipiente SIGMA)
Conc. Sol. D-Glicosamina = 0,05 g/50 mL
Massa calc.(µmol) = conc. Sol. D-Glic. x Vol. Utiliz. D-Glics. =
= 0,05 x 1000 x Vol.util./50*215,64
Tempo atividade enzimática = 30 min
1 U/mL = 1 µmol/(min.mL)
Vol. Total (Glicosamina+caldo) = 1 mL
Medições Adsorbância a 600 nm Vol. Utiliz. Massa Massa Atividade
II III Média(Abs)(mL) calc. (mg) calc. (µg) Enzim. (U/mL)
0 0 0 0 0 0 0
0,133 0,128 0,1307 0,2 0,2 200 0,0309
0,279 0,277 0,2787 0,4 0,4 400 0,0618
0,433 0,428 0,4297 0,6 0,6 600 0,0927
0,587 0,583 0,5797 0,8 0,8 800 0,1237
0,727 0,716 0,7157 1 1 1000 0,1546
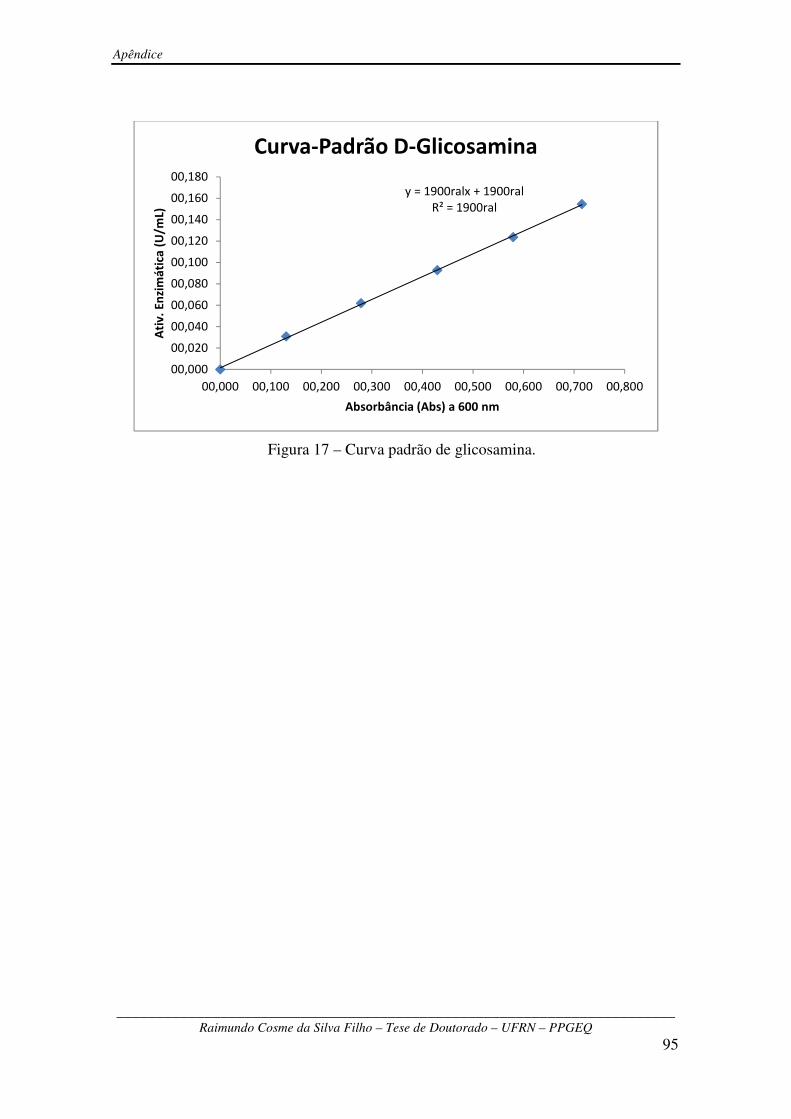
Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
95
Figura 17 – Curva padrão de glicosamina.
y = 1900ralx + 1900ral
R² = 1900ral
00,000
00,020
00,040
00,060
00,080
00,100
00,120
00,140
00,160
00,180
00,000 00,100 00,200 00,300 00,400 00,500 00,600 00,700 00,800
Ati
v.
En
zim
áti
ca (
U/m
L)
Absorbância (Abs) a 600 nm
Curva-Padrão D-Glicosamina

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
96
APÊNDICE III
Método de Determinação de Proteína Utilizando o Corante Comassie
Brilhante Blue G 250 (Método modificado de Sedmak e Grossberg)
Micrométodo
Preparar uma solução 0,06% de Comassie Blue G250 em 1,5 % HCl (w/V). Filtrar e
papel de filtro Whatman nº1.
Curva Padrão (1 a 20 µµµµg)
a) Preparar soluções estoque de BSA de 10 a 200 µg/mL.
b) Colocar em tubos de ensaio 100 µL da solução estoque, 1400 µL de água e
1500µL do Comassie e agitar.
c) Preparar um tubo contendo 1500 µL de água e 1500 µL do corante que
corresponde ao zero de proteína.
d) Após 5 minutos (padronizar o tempo) fazer as leituras das absorbâncias.
e) As leituras das amostras devem ser feitas em dois comprimentos de onda 595 e
465 contra a água.
f) Plotar a razão das absorbâncias a 595/465 como uma função da massa de BSA e
calcular o melhor ajuste linear dos dados.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
97
Micrométodo (1 a 20 µµµµg de BSA)
Solução Estoque de BSA = 0,2 mg/mL
Tabela 16. Dados para construção da curva padrão de BSA.
Tubo de
Ensaio
H2O (µµµµL)
BSA (µµµµg)
Coomassie (µµµµL)
BSA (µµµµg)
1 1500 0 1500 0
2 1490 10 1500 2
3 1480 20 1500 4
4 1470 30 1500 6
5 1460 40 1500 8
6 1450 50 1500 10
7 1440 60 1500 12
8 1430 70 1500 14
9 1420 80 1500 16
10 1410 90 1500 18
11 1400 100 1500 20
Dados adicionais:
Concentração Sol. Estoque
BSA = 0,2 mg/mL = 0,2 µg/µL
Vol. total sol. = vol. H2O + vol. BSA + vol.
Coomassie = 3000 µL = 3 mL
Concentração Sol. BSA (mg/mL) = (massa BSA mg) / 3 mL
.

Apêndice
______________________________________________________________________Raimundo Cosme da Silva Filho – Tese de Doutorado – UFRN – PPGEQ
98
Figura 18 – Curva Padrão de BSA
y = 1900ralx - 1900ral
R² = 1900ral
000,000
001,000
002,000
003,000
004,000
005,000
006,000
007,000
000,400 000,500 000,600 000,700 000,800 000,900 001,000 001,100
µg
BS
A
Razão Absorbâncias (595nm/465nm)
Curva-Padrão BSA


















