
PDF (868.42 KB)
31
Transcript of PDF (868.42 KB)


Revista da Biologia – www.ib.usp.br/revista ISSN 1984-5154
Sumário _____________________________ Volume 3
Dezembro de 2009 1
SERIA A TEORIA DA EVOLUÇÃO DARWINIANA DOMÍNIO EXCLUSIVO DOS BIÓLOGOS? IMPLICAÇÕES DA EVOLUÇÃO BIOLÓGICA PARA AS CIÊNCIAS HUMANAS Pedro José Tótora da Glória
6
INSETOS EUSOCIAIS E O DESAFIO PARA A IDÉIA DE SELEÇÃO NATURAL Pedro Leite Ribeiro
9
NEUROGÊNESE NO SISTEMA NERVOSO ADULTO DE MAMÍFEROS Ilton Santos da Silva
15
UMA INTRODUÇÃO À NEUROFILOSOFIA: O PROBLEMA MENTE-CORPO Camila Gomes Victorino
21
PROJETO GENOGRÁFICO E AS IMPLICAÇÕES DA POPULARIZAÇÃO DOS ESTUDOS DE GENEALOGIA GÊNICA Bárbara Domingues Bitarello
24
TÉCNICAS PARA O PROCESSAMENTO DE FÓSSEIS DE BIVALVES: UM ESTUDO DE CASO SOBRE A FORMAÇÃO CORUMBATAÍ, BACIA DO PARANÁ, BRASIL Rodrigo B. Salvador e Luiz R. L. Simone

Revista da Biologia – www.ib.usp.br/revista
Expediente ____________________________________ Editor Executivo:
Welington Braz Carvalho Delitti Coordenação:
Agustín Camacho Guerrero, Pedro Ribeiro e Rodrigo Pavão Secretaria Executiva: CABio USP
Gustavo Busanelli e Juliano Zequini Polidoro Comissão Científica: Docentes IB USP
Editor Botânica: Déborah Yara Alves Cursino dos Santos e Lucia Soares-Silva Editor Ecologia: Marcelo Luiz Martins Pompêo e Paulo Enrique Cardoso Peixoto Editor Fisiologia: Gilberto Fernando Xavier Editor Genética e Biologia Evolutiva: Carlos Ribeiro Vilela Editor Zoologia: Rui Cerqueira Silva e Sônia Godoy Bueno Carvalho Lopes
Consultores desse volume:
Anary Priscila Monteiro Egydio André Menezes Strauss Barbara Kazue Amaral Onishi Cintia Etsuko Yamashita Daniel José Galafasse Lahr Denise de Araujo Alves Erika Cecon Felipe Ibañez de Santi Ferrara José Eduardo Amoroso Rodriguez Marian José Eduardo Soubhia Natali Marcelo Veronesi Fukuda Marco Antonio Monteiro Granzinolli Monique Nouailhetas Simon Pedro Leite Ribeiro Renata Pereira Lima Ricardo Mendonça Neves dos Santos Rodrigo Machado Feitosa Sérgio Marinho da Silva Sérgio Nascimento Stampar

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
1
SERIA A TEORIA DA EVOLUÇÃO DARWINIANA DOMÍNIO EXCLUSIVO DOS BIÓLOGOS? IMPLICAÇÕES DA EVOLUÇÃO
BIOLÓGICA PARA AS CIÊNCIAS HUMANAS
Pedro José Tótora da Glória Department of Anthropology,The Ohio State University
Recebido 04mai09 / Aceito 28ago09 / Publicação inicial 31ago09 [email protected]
Resumo. A teoria evolutiva darwiniana tornou-se o atual paradigma nas ciências biológicas desde a
formulação da teoria sintética nas décadas de 1930 e 1940. No entanto, o alcance da teoria darwiniana tem impactado áreas do conhecimento que vão muito além da biologia. O objetivo deste artigo é fazer uma incursão histórica sobre a relação entre a teoria evolutiva biológica e as ciências humanas. A partir desta perspectiva histórica é possível fazer uma reflexão sobre o papel do biólogo nesse diálogo, tanto no âmbito acadêmico como no âmbito social.
Palavras-chave. História da biologia, Humanidades, Sociedade, Interdisciplinaridade. DOES THE DARWINIAN THEORY OF EVOLUTION BELONG ONLY TO BIOLOGISTS?
IMPLICATIONS OF THE BIOLOGICAL EVOLUTION FOR SOCIAL SCIENCES Abstract. The Darwinian theory of evolution has become the current paradigm in the biological
sciences since the formulation of the synthetic theory in the 1930s and 1940s. However, the scope of the Darwinian theory has impacted areas of knowledge that go far beyond biology. The aim of this article is to make a raid on the historical relationship between evolutionary theory and social sciences. From that historical perspective, a discussion is promoted on the role of biologists in this dialogue, both in academic and social spheres.
Keywords. History of Biology, Humanities, Society, Interdisciplinarity. A teoria da evolução darwiniana é o atual
paradigma nas ciências biológicas, tanto por sua capacidade de integração teórica de diferentes campos da biologia como por sua extensa corroboração empírica (Ridley, 2003). Por outro lado, o alcance dessa teoria biológica não é claramente definido. Diferentes correntes na área de humanidades têm usado seus princípios para gerar novas áreas do saber. Mais ainda, setores da sociedade e segmentos políticos têm se envolvido em polêmicas acerca da validade da teoria evolutiva, tais como o crescente movimento criacionista nos Estados Unidos. Dentro desse contexto, este ensaio se propõe a fornecer uma breve visão histórica da relação entre a teoria da evolução biológica e as ciências humanas, e então precisar qual exatamente deve ser o papel do biólogo e da biologia nessa comunicação entre diferentes áreas do saber.
A hipótese de evolução biológica não foi proposta pela primeira vez por Charles Darwin (1809-1882) em 1859 com a publicação de seu clássico livro “A origem das espécies”. Pensadores do século XVIII como conde de Buffon (1707-1788) e Erasmus Darwin (1731-1802) já haviam sugerido a existência de mudança biológica ao longo do tempo (Mayr, 1982). Porém, não havia ainda uma sistematização dessas ideias na forma de uma teoria da evolução. Essa sistematização ocorreu somente com Jean-Baptiste de Lamarck (1744-1829) na sua clássica publicação “Philosophie zoologique” em 1809. Baseado em parte no legado de seus predecessores, Lamarck defendia
categoricamente que organismos se transformavam ao longo do tempo. Porém, ao contrário dos anteriores, ele foi capaz de precisar dois mecanismos de mudança. O primeiro refere-se à capacidade inata dos organismos de aumentar em complexidade, iniciando-se a partir da geração espontânea de organismos simples. A ideia Lamarckiana de progresso é oriunda do conceito grego da “grande cadeia dos seres”, o qual estabelece uma ordem crescente de complexidade entre os seres. Lamarck, no entanto, transformou essa ideia de um conceito estático para uma linha temporal de descendência biológica (Mayr, 1982). O segundo refere-se à capacidade dos organismos de se transformarem por meio do uso e desuso de estruturas; ideia essa que já era difundida em sua época (Meyer e El-Hani, 2005).
Em oposição a Lamarck, George Cuvier (1769-1832) defendia a mudança biológica mediada por eventos de catástrofe, causando discretas substituições da fauna do passado. Ainda preso a uma ideia essencialista da biologia, Cuvier trabalhou com importantes conceitos anatômicos tais como homologia e analogia. A ideia do catastrofismo e da extinção de espécies divergia marcadamente da noção Lamarckiana de progresso (Mayr, 1982).
Foi somente com o estabelecimento da teoria de seleção natural de Darwin que os mecanismos de transformação biológica de Lamarck e Cuvier foram seriamente questionados dentro de uma perspectiva científica. É digno de nota, porém, que naquela época Darwin ainda
Ensaio:

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
2
considerava, por exemplo, o uso e desuso como um possível mecanismo evolutivo, ainda que relegado a uma menor importância (Darwin, 1859). Apesar de Darwin ter se tornado um símbolo da moderna teoria evolutiva biológica, Alfred Wallace (1823-1913) propôs as ideias de seleção natural simultaneamente a Darwin (Darwin e Wallace, 1958). Porém, é importante frisar que a robustez da obra de Darwin e sua centralidade na discussão evolutiva na Inglaterra do século XIX o fizeram personagem muito mais influente do que Wallace.
Todavia, a ideia de progresso, como formulada na teoria de Lamarck, não foi prontamente abandonada com o surgimento das ideias de Darwin. A ideia de progresso no século XIX estava enraizada em importantes mudanças sociais e econômicas na Europa, como exemplificado pelo estabelecimento do capitalismo industrial na Inglaterra. No âmbito ideológico, o filósofo Auguste Comte (1798-1857), por exemplo, foi um notável proponente das ideias positivistas de progresso, dando subsídios teóricos para diversas correntes do pensamento europeu do século XIX. A sociedade europeia, de fato, era vista como o ápice da evolução social humana, criando uma demanda por explicações racionais para essa crença. Herbert Spencer (1820-1903), seguindo essa tendência, foi um expoente da noção de progresso nas ciências humanas. Conhecido como o mais influente proponente da corrente atualmente chamada de “darwinismo social”, Spencer chegou mesmo a usar o termo “sobrevivência do mais adaptado” (survival of the fittest) antes mesmo que Darwin. Porém, suas ideias contrastavam com as de Darwin, uma vez que Spencer defendia a noção de progresso a partir de uma perspectiva social. Darwin, por sua vez, defendia a ocorrência de seleção natural por meio de uma perspectiva biológica, postulando ausência de necessário progresso (Freeman, 1974). Darwin, de fato, sempre foi muito reticente em abordar os seres humanos em seus escritos. Assim o fez somente no final de sua vida com os livros “A origem do Homem, e a seleção sexual” de 1871 e “A expressão das emoções no Homem e nos animais” de 1872. Ainda assim, ele evitou polêmicas sobre as implicações de sua teoria para a evolução social.
No começo do século XX, a teoria da evolução de Darwin estava longe de ser uma unanimidade. Muito pelo contrário, havia um acirrado debate entre gradualistas, defendendo a concepção darwiniana de mudança lenta e gradual, e os saltacionistas, defendendo mutações de grande efeito como principal mecanismo evolutivo (Gould, 2002). Nas ciências humanas, a teoria evolutiva darwiniana era ainda menos aceita. Na antropologia física, por exemplo, o começo do século XX foi marcado por ideias tipológicas de classificação das chamadas
raças humanas, continuando a tradição da noção progressista de evolução do século XIX. Ideias etnocêntricas, como a superioridade intelectual dos Europeus, eram baseadas em inferências sobre o formato e o tamanho do crânio humano (Gould, 1981). Como reação a essas ideias, Franz Boas (1858-1942), o fundador do atual modelo acadêmico da antropologia norte-americana, propôs o conceito de relativismo cultural. Segundo ele, era preciso entender os outros povos em seus próprios termos, evitando impor nossos moldes sobre outras culturas. Nos Estados Unidos do começo do século XX, uma postura eugênica dominava o cenário cultural e intelectual da época. Essa doutrina defendia a purificação das raças, eliminando variações humanas que não atendessem à norma por eles defendida, tais como negros e indígenas na América. Em oposição às ideias eugênicas de seu tempo, Boas fundou uma tradição de abordagem histórico-cultural do comportamento humano (Kroeber, 1956). Boas reagiu contra os evolucionistas sociais do século XIX, tais como Lewis Morgan (1818-1883), Edward Tylor (1832-1917) e James Frazer (1854-1951). Ao contrário das ideias de progresso e evolução linear, Boas defendia o particularismo histórico, considerando as características da geografia, cultura e língua local. Essa polarização entre cultura/história e biologia, acentuada ainda mais pelos alunos de Boas, pode ser mencionada como um fator para a pouca adesão da teoria biológica de evolução nas ciências humanas na primeira metade do século XX na América (Kroeber, 1946).
A teoria de Darwin foi estabelecida como paradigma nas ciências biológicas somente com o advento da teoria sintética da evolução nas décadas de 1930 e 1940. Encabeçada por Ronald Fisher (1890-1962), John Haldane (1892-1964) e Sewall Wright (1889-1988), a teoria sintética de evolução foi capaz de integrar a seleção natural de Darwin com os conceitos da genética mendeliana, usando para isso uma sólida base matemática. Notáveis contribuições também foram feitas por Ernst Mayr (1904-2005) e Theodosius Dobzhansky (1900-1975). A aceitação da teoria sintética da evolução, por um lado, teve imediatas consequências nas ciências humanas. O 15º simpósio de biologia quantitativa em Long Island, EUA (Warren, 1951), foi um marco para o estabelecimento da biologia evolutiva em diferentes áreas. O artigo de Washburn (1951), por exemplo, estabeleceu os princípios da nova antropologia física, agora sob uma perspectiva evolutiva. Por outro lado, porém, os então recentes acontecimentos da segunda guerra mundial frearam qualquer tipo de avanço no sentido de aplicar a teoria da evolução biológica para entender o comportamento humano, especialmente no cenário europeu.
Já no campo da antropologia norte-americana, a década de 1950 foi marcada pelo

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
3
surgimento dos neo-evolucionistas. Influenciados mais pelo materialismo Marxista do que pela nova síntese biológica, Leslie White (1900-1975) e Julian Steward (1902-1972) retomaram a ideia evolutiva abandonada por Boas. Steward fundou a escola da ecologia cultural, baseando-se em uma evolução multilinear das sociedades humanas. O conceito de adaptação era usado por ele para indicar a relação de ajuste entre o núcleo cultural – tecnologia e bens de produção – e o ecossistema, implicando em diferentes trajetórias de evolução social (Steward, 1955). Em oposição, White investigava o conceito de Cultura como um todo. Ele elaborou uma fórmula de mudança cultural baseada na quantidade de energia utilizada por cada habitante. Suas ideias retomaram a evolução linear do século XIX, porém usando uma perspectiva menos valorativa e mais científica (White, 1959). Os neo-evolucionistas tiveram grande influência no surgimento da antropologia ecológica e no materialismo cultural na década de 1960 (Eriksen e Nielsen, 2001).
Foi somente com os trabalhos de Hamilton (1964) e Williams (1966) que a importância da teoria evolutiva biológica para o entendimento do comportamento social humano ganhou força na biologia. De fato, esses autores foram severos críticos da ideia de seleção de grupo na evolução, proposta naquele momento por Wynne-Edwards (1962). Por outro lado, essa discussão abriu portas para o entendimento do comportamento social humano por meio dos conceitos de seleção no nível do indivíduo e “inclusive fitness”. Este último conceito, por exemplo, surgiu como forma de explicar a cooperação entre indivíduos que apresentam compartilhamento gênico. Ao agir em benefício de um parente, promove-se a sobrevivência e reprodução de parte de seu material genético. Essas publicações serviram de base para o desenvolvimento da ideia de altruísmo recíproco elaborada por Trivers (1971). Segundo esse conceito, as relações sociais seriam vantajosas para ambas as partes desde que o investimento realizado em uma atividade social pudesse ser retribuído no futuro, resultando em mais benefícios do que custos. Esses benefícios deveriam ser ponderados considerando-se também o grau de compartilhamento gênico entre as partes.
Seguindo essa abordagem evolutiva, Edward Wilson propôs o termo “Sociobiologia” em 1975, causando um enorme impacto na biologia e nas ciências humanas. Conhecido por seus estudos com insetos sociais (Wilson, 1971), ele transpôs os princípios da teoria evolutiva darwiniana para o estudo da sociedade e comportamento humanos. Segundo Wilson, assuntos longamente debatidos pelas ciências humanas, tais como violência, comportamento social, moral e religião, poderiam passar a serem entendidos usando uma abordagem genético–
evolutiva (Wilson, 1975). A reação contra suas ideias foi imediata, causando ira por parte de cientistas sociais e até mesmo por parte de alguns biólogos como Stephen Jay Gould e Richard Lewontin. A mobilização e o impacto gerado por suas ideias foi enorme, tanto no meio acadêmico como fora dele. Embora a sociobiologia tenha impulsionado novos campos de estudo do comportamento humano sob uma perspectiva biológica, o termo sociobiologia sucumbiu às ferrenhas críticas feitas a ele. Como pode ser contemplado no livro de Kitcher (1985), aspectos culturais fazem com que o comportamento humano seja dificilmente entendido somente em termos biológicos, caracterizando a sociobiologia como uma abordagem extremamente simplificada do fenômeno social humano.
De fato, as ciências humanas na década de 1980 caminharam em uma direção diametralmente oposta às ciências biológicas. O pós-modernismo e o desconstrutivismo criticavam os preceitos da ciência tradicional, questionando ideias como verdade, objetividade e neutralidade científica. Departamentos de antropologia nos Estados Unidos passaram a se polarizar entre os defensores da ciência e os seus críticos (Lett, 1997). A teoria da evolução passou a ser vista como fruto do positivismo e imperialismo do século XIX, longe de representar a realidade cultural dos diversos tipos de sociedades humanas. Porém, como uma fricção constante entre ideias opostas, as sementes da sociobiologia não tinham morrido. Campos das ciências humanas ganharam novas e provocantes abordagens a partir de uma visão evolutiva. Dois exemplos serão brevemente descritos aqui: arqueologia e psicologia evolutivas.
A arqueologia evolutiva tem Robert Dunnell como seu maior expoente, iniciando suas publicações na área em 1978. Os preceitos teóricos da arqueologia evolutiva são incorporações diretas da seleção natural darwiniana a fim de explicar mudanças no registro arqueológico. Porém, o que é defendido por seus propositores não é a origem biológica do comportamento humano, mas sim o uso da teoria evolutiva para entender a permanência de determinados traços materiais, tecnológicos ou sociais através do tempo. Para isso, Dunnell adapta conceitos da biologia evolutiva como unidade de seleção, variabilidade e transmissão, e os aplica nos vestígios arqueológicos – instrumentos, restos vegetais, cerâmica (Dunnell, 1980). Longe de ser uma abordagem dominante nas ciências humanas, a arqueologia evolutiva no entanto ainda é praticada por um pequeno grupo, como pode ser demonstrado pelos trabalhos recentes de O’Brien (1996, 2005).
A psicologia evolutiva, por outro lado, apresenta um impacto muito maior que a abordagem anterior. De forte apelo popular e

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
4
midiático (e.g. Pinker, 2002), ela surgiu em 1992 com o livro “Adapted Mind” editado por Barkow et al. O preceito básico dessa abordagem é que podemos entender o comportamento humano investigando o ambiente de adaptação evolutiva onde os nossos antepassados foram selecionados, isto é, caçando e coletando nas savanas africanas. O comportamento humano, portanto, é grandemente fruto da seleção de características especializadas para solucionar problemas ambientais e sociais do passado. Evidências da neurociência apontando para a modularidade funcional do cérebro humano (Pinker, 1997) são vistas pelos psicólogos evolutivos como o resultado de distintos e específicos processos de seleção natural. Embora sustentado sobre os alicerces da moderna teoria evolutiva, Buller (2005) tem demonstrado erros e simplificações no uso dessa teoria, além de falhas na sustentação empírica da psicologia evolutiva.
Apesar da abordagem evolutiva nas ciências humanas ainda ser muito discutível, a teoria sintética de evolução é o atual paradigma na biologia (Ridley, 2003). Por outro lado, fora das universidades, tem-se presenciado recentemente um ferrenho debate entre criacionistas e biólogos evolutivos. Por parte dos criacionistas, a corrente denominada planejamento inteligente (intelligent design) procura provar por meio de dados empíricos a inexistência da evolução. Do outro lado, biólogos evolutivos como Richard Dawkins defendem contundentemente os preceitos do darwinismo (Dawkins, 1986, 1996). Este último autor, inclusive, publicou recentemente um livro chamado “A desilusão de Deus” (Dawkins, 2006), onde argumenta sobre a impossibilidade da existência divina dentro de sua visão científico-materialista. Estes debates recentes ilustram o impacto que a evolução biológica ainda tem sobre o grande público. De fato, a biologia evolutiva é uma área do conhecimento com alcance muito mais amplo que as cercanias do meio acadêmico.
Feita esta breve incursão histórica sobre as implicações da teoria darwiniana nas ciências humanas e na sociedade, pode-se chegar a algumas conclusões. Primeiro, a palavra evolução se mistura com a noção popular de progresso, sendo amplamente utilizada no meio acadêmico do século XIX e chegando até mesmo à década de 1950 com o trabalho de White (1959) sobre evolução cultural. Por outro lado, evolução é usada por Darwin como um indicador de transformação ao longo do tempo, sem nenhuma implicação finalista ou progressista. Dessa forma, o termo evolução não é autoexplicativo, mas, muito pelo contrário, precisa ser definido com rigor pelo pesquisador, uma vez que historicamente esse termo foi utilizado em diferentes sensos. Segundo, diversas tentativas de utilização da teoria evolutiva biológica nas ciências humanas foram atacadas devido às
associações da biologia com o eugenismo e o “darwinismo social”. Nesse sentido, o contexto social e os fatores históricos não podem ser negligenciados quando avaliamos o impacto da teoria evolutiva na academia e na sociedade. O pesquisador, portanto, tem o dever de estar informado e de ser capaz de discutir esses aspectos sociais quando pertinente. Terceiro, tentativas de transposição literal da evolução biológica para o comportamento humano acabaram por ignorar fatores culturais e sociais, sendo constantemente acusadas de simplificação e determinismo biológico pelos cientistas sociais. Esse ponto nos remete à importância e ao cuidado que o biólogo deve ter ao transpor suas teorias diretamente para áreas das ciências humanas. Quarto, os apelos midiático e popular de certas correntes acadêmicas podem gerar uma super simplificação de questões que na verdade são muito mais complexas. Essas veiculações populares podem mesmo alimentar visões religiosas críticas à teoria de evolução biológica, como é o caso do planejamento inteligente. O pesquisador nesse sentido tem o dever de fornecer sua visão científica sobre o assunto, fazendo-se inteligível para um público mais amplo que seu território acadêmico.
Esses quatro pontos nos remetem ao papel do biólogo evolutivo dentro desse diálogo entre áreas do conhecimento. Melhor do que qualquer pesquisador, o biólogo é o mais capacitado para descrever com propriedade os princípios teóricos e a sustentação empírica da teoria evolutiva. Mais ainda, ele é o mais capacitado para delinear os debates internos dentro de seu próprio campo de atuação. Embora a aplicação da evolução biológica nas ciências humanas exija mais do que conhecimento biológico, o biólogo precisa ser ouvido nesse diálogo. Para isso, ele precisa adequar sua linguagem e se fazer claro para áreas do conhecimento que não detenham seu jargão, incluindo aí o grande público. Cientistas como Richard Dawkins, Stephen Jay Gould e Richard Lewontin têm realizado um importante papel de divulgação científica, porém o diálogo na academia brasileira ainda é muito precário. Visto que as ciências humanas brasileiras têm uma profunda aversão pela teoria evolutiva biológica (Strauss e Waizbort, 2008), faz-se desta comunicação um dever do biólogo evolutivo brasileiro. Mais do que uma obrigação, esse tipo de comunicação apresenta uma enorme demanda na realidade científica brasileira. Espero que a partir deste texto, os biólogos evolutivos se sensibilizem para as importantes implicações do darwinismo tanto nas ciências humanas como na opinião pública, e encarem esse diálogo como parte constituinte de seu papel profissional.
Agradecimentos. Eu sou imensamente
grato pelas intermináveis conversas com Andre Strauss ao longo dos últimos anos, e pelas

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
5
incontáveis ideias que nossas conversam me proporcionam. Qualquer erro nesse ensaio, no entanto, é de minha total responsabilidade. Agradeço também aos editores e revisores da Revista da Biologia pelas suas importantes sugestões.
Bibliografia
Barkow, J., Cosmides, L., e Tooby, J. (editores) (1992) The adapted mind: evolutionary psychology and the generation of culture. New York: Oxford University Press.
Buller, D. J. (2005) Adapting minds: evolutionary psychology and the persistent quest for human nature. Cambridge: MIT Press.
Darwin, C. (1859 [2004]). A origem das espécies. São Paulo: Editora Martin Claret.
_______. (1871 [2004]) A origem do Homem e a seleção sexual. Belo Horizonte: Editora Itatiaia.
_______. (1872 [2000]). A expressão das emoções no Homem e nos animais. São Paulo: Companhia das letras.
_______ e Wallace, A. R. (1958) Evolution by natural selection, ed. por de Beer, G. Cambridge: Cambridge University Press.
Dawkins, R. (1986 [2001]). O relojoeiro cego. São Paulo: Companhia das Letras.
_______. (1996 [1998]) A escalada do monte improvável. São Paulo: Companhia das Letras.
_______. (2006 [2007]) A desilusão de Deus. Lisboa: Casa da Letras.
Dunnell, R. C. (1980). Evolutionary theory and archaeology. In: (M.B. Schiffer, Ed.) Advances in Archaeological Method and Theory vol. 3.New York, NY: Academic Press, pp. 35-99.
Eriksen, T. H., e Nielsen, F. S. (2001 [2007]) História da antropologia. Petrópolis: Editora Vozes.
Freeman, D. (1974) The evolutionary theories of Charles Darwin and Hebert Spencer. Current Anthropology 15, 211-237.
Gould, S.J. (1981 [1991]). A falsa medida do Homem. São Paulo: Martins Fontes.
_______. (2002). The structure of evolutionary theory. Cambridge: The Belknap Press of Harvard University Press.
Hamilton, W. D. (1964) The genetical evolution of social behaviour I and II. Journal of Theoretical Biology 7, 1-52.
Kitcher, P. (1985) Vaulting ambition: sociobiology and the quest for human nature. Cambridge, Mass.: MIT Press.
Kroeber, A. L. (1946) History and evolution. Southwestern Journal of Anthropology 2, 1-15.
_______. (1956) The place of Boas in anthropology. American Anthropologist 58, 151-159.
Mayr, E. (1982 [1998]). O desenvolvimento do pensamento biológico. Brasília: Editora da Universidade de Brasília.
Meyer, D. e El-Hani, C. N. (2005) Evolução: o sentido da biologia. São Paulo: Editora Unesp.
O’Brien, M. J., editor (1996) Evolutionary archaeology: theory and application. Salt Lake City: University of Utah Press.
_______. (2005) Evolutionism and North America’s archaeological record. World Archaeology 37, 26-45.
Lamarck, J.-B. (1809) Philosophie zoologique, ou exposition des considérations relative à l’histoire naturelle des animaux, 2 vol. Paris: Dentu.
Lett, J. (1997) Science, reason and anthropology: the principles of rational inquiry. Oxford: Rowman & Littlefield Publishers, Inc.
Pinker, S. (1997 [1998]) Como a mente funciona. São Paulo: Companhia das Letras.
_______. (2002 [2004]) Tábula rasa: a negação contemporânea da natureza humana. São Paulo: Companhia das Letras.
Ridley, M. (2003 [2006]) Evolução. 3.ed. Porto Alegre: Artmed. Steward, J. (1955) The theory of culture change: the
methodology of multilinear evolution. Urbana: University of Illinois Press.
Strauss, A. e Waizbort, R. (2008) Sob o signo de Darwin? Sobre o mau uso de uma quimera. Revista Brasileira de Ciências Sociais 23, 125-134.
Trivers, R. L. (1971) The evolution of reciprocal altruism. Quarterly Review of Biology 46, 35-57.
Warren, K. B. (1951) Origin and evolution of Man. vol. 15 of Cold Spring Harbor Symposia on Quantitative Biology.
Washburn, S. L. (1951) The new physical anthropology. Transactions of the New York Academy of Science 13 (2d ser.), 298-304.
White, L. (1959) The evolution of culture. New York: McGraw Hill.
Williams, G. C. (1966) Adaptation and Natural Selection. Princeton: Princeton University Press.
Wilson, E. O. (1971) The insect societies. Cambridge, Mass.: Harvard University Press.
_______. (1975) Sociobiology: the new synthesis. Cambridge, Mass.: Harvard University Press.
Wynne-Edwards, V. C. (1962) Animal dispersion in relation to social behaviour. New York: Hafner Publishing Company.

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
6
INSETOS EUSOCIAIS E O DESAFIO PARA A IDÉIA DE SELEÇÃO NATURAL
Pedro Leite Ribeiro Departamento de Fisiologia, Instituto de Biociências, USP
Recebido 26mar09 / Aceito 24nov09 / Publicação inicial 24nov09 Pedro Leite Ribeiro
Resumo. Este texto versa sobre a clássica discussão a respeito da dificuldade que a teoria da
Seleção Natural tem em explicar o surgimento de indivíduos não férteis. Para tanto é apresentado o conceito de eusocialidade, as idéias de seleção de parentesco, altruísmo, altruísmo forçado e seleção de grupo.
Palavras-chave. Seleção natural, Seleção de parentesco, Hamilton EUSOCIAL INSECTS AND THE CHALLENGE TO THE IDEA OF NATURAL SELECTION Abstract. This article is about the classic debate about the difficulty that the theory of natural selection
has in explaining the emergence of non-fertile individuals. Therefore, it is presented the concept of eusociality, the ideas of kin selection, altruism, enforced altruism and group selection.
Keywords. Natural Selection, Kin selection, Hamilton
Darwin, ao propor a teoria da Seleção Natural em seu livro Origem das Espécies, questiona a abrangência de sua teoria quando, por exemplo, menciona os insetos sociais: “há que se admitir a existência de casos que apresentam especial dificuldade com relação à teoria da seleção natural. Um dos mais curiosos é o da existência de duas ou três castas definidas de formigas-operárias ou fêmeas estéreis na mesma comunidade de insetos” (Darwin, 1859).
Wilson, em 1971, estipula três critérios, que têm a pretensão de classificar todas as espécies animais com relação aos níveis de socialidade, a saber: 1) que indivíduos da mesma espécie ajudam de forma cooperativa na criação dos jovens; 2) que há uma divisão de tarefas reprodutivas em uma sociedade em que um grupo de indivíduos é infértil e colabora com a criação dos filhos dos indivíduos férteis da colônia, além de desempenharem outras funções de manutenção da colônia; e 3) a existência de uma sobreposição de pelo menos duas gerações, de forma que os filhos possam ajudar seus pais. Sob a perspectiva de Wilson, os animais ditos eusociais são apenas aqueles que exibem os três critérios por ele estipulados (Wilson, 1971). Então, é importante notar que existem também vários estágios considerados intermediários, como, por exemplo, os seres humanos, que são capazes de comportamentos solidários com a prole alheia, sem, no entanto, abdicarem de sua própria capacidade reprodutiva.
Portanto, a idéia de que a seleção natural agiria exclusivamente sobre o indivíduo traz uma dificuldade para a explicação de como teriam sido selecionadas as relações altruísticas das castas inférteis de uma colônia de formigas. Afinal, nesse caso, como a esterilidade de um indivíduo pode ter sido selecionada? Se um indivíduo é estéril, jamais poderá deixar descendentes e não terá qualquer aptidão, (capacidade de deixar descendentes diretos) algo que, pensando sob o olhar clássico da evolução, torna-se paradoxal. A
dificuldade torna-se ainda maior se considerarmos que estas castas inférteis apresentam marcadas diferenças morfológicas: como essas diferenças poderiam ser selecionadas num sistema no qual não há reprodução (Darwin, 1859)?
Hamilton em 1964 publicou dois trabalhos que procuravam explicar, sob a luz da seleção natural, o comportamento eusocial nas diferentes espécies animais (Hamilton, 1964a; Hamilton, 1964b) Sob essa perspectiva, ambas as pesquisas introduziram uma nova e importante idéia no contexto da evolução, a idéia da seleção de parentesco. Para entendermos a concepção desta idéia, temos, antes de tudo, que explanar outro conceito fundamental, a saber, o conceito de aptidão abrangente (fitness inclusivo, ou fitness total), que é a base da teoria de Hamilton ao definir aptidão (fitness) como a capacidade de um indivíduo transmitir seus genes para as gerações futuras.
De acordo com as idéias de Hamilton aptidão abrangente é o resultado da soma da aptidão direta com a aptidão indireta. A aptidão direta é oriunda da capacidade reprodutiva do indivíduo em questão, enquanto que a aptidão indireta é dada pela capacidade reprodutiva dos outros membros da comunidade em que o animal vive, e que guardam algum tipo de parentesco com o indivíduo de referência. Como o cálculo da aptidão indireta leva em conta, necessariamente, a consideração do grau de parentesco dos outros membros da comunidade com o indivíduo em questão, quanto maior o grau de parentesco com os parentes reprodutivamente ativos, maior seria, então, a aptidão indireta desse indivíduo. Assim, a capacidade reprodutiva de membros geneticamente relacionados (parentes) de um determinado indivíduo tem uma importante participação na aptidão abrangente (total) do indivíduo. Dessa forma, temos a aptidão abrangente de um indivíduo parcialmente desvinculada da capacidade do indivíduo de gerar
Revisão:

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
7
filhos, pois os seus genes podem ser transmitidos para as gerações futuras, por exemplo, pelos seus irmãos, através dos sobrinhos, consistindo, nisso, o principal argumento sobre o qual Hamilton teceu a sua teoria. Portanto, do ponto de vista de Hamilton, comportamentos altruísticos poderiam ser justificados pelo aumento da aptidão indireta, desde que este aumento seja maior do que o prejuízo na aptidão direta que o comportamento altruístico possa provocar no indivíduo que faz a generosidade. Ou seja, trata-se, nesse caso, de um aumento na aptidão total. Assim, em situações particulares em que exista um alto grau de parentesco entre os membros de uma comunidade, pode ser mais vantajoso, do ponto de vista da transmissão de genes para gerações futuras, abdicar da aptidão direta em prol da aptidão indireta. É em sociedades partenogênicas que este raciocínio ganha força, afinal, as irmãs compartilham, em média, 75% dos genes umas com as outras, enquanto que mães e filhas compartilham apenas 50% dos genes, o que torna a aptidão indireta potencializada. Quanto à transmissão gênica, pode tornar-se mais interessante que se tenha maior cuidado com as irmãs do que com os próprios filhos. Estas idéias parecem, portanto, acalmar as críticas com relação à espetacular contradição de que a seleção natural teria levado ao surgimento de indivíduos que não têm qualquer capacidade de transmitir os seus genes diretamente, algo que, à primeira vista, pode parecer improvável de ser selecionado. Afinal, a teoria da seleção natural versa: os indivíduos que são selecionados são aqueles que têm maior capacidade de deixar descendentes.
Desse modo, uma vez que temos uma explicação lógica e coerente sobre o status quo dos sistemas biológicos que nos propusemos a estudar, por que não tentarmos explicar o surgimento dos sistemas biológicos eusociais? Podemos, com isso, nos perguntar: quais eram – ou deveriam ser – as condições ecológicas dos ancestrais desses animais eusociais que hoje conhecemos? De fato há artigos de autores consagrados que lidam com essas questões, por exemplo, Arillo (2007) e Wilson e Holldobler (2005).
Dessa forma, a descoberta de características comuns aos ancestrais destes grupos pode ser de grande valor para o entendimento de quais são as condições necessárias ou pelo menos favoráveis ao surgimento de animais eusociais. Infelizmente, o estudo de fósseis é pouco revelador com relação ao comportamento e às condições ecológicas de um determinado momento da história evolutiva. Como há poucas informações extraídas das estruturas morfológicas que podem evidenciar algum tipo de comportamento, torna-se necessário recorrer aos estudos sobre a fossilização de grupos de insetos, que, ocorrida
em único evento, pôde eternizar um momento da vida social de uma determinada espécie. As fossilizações de formigas aladas e de outras ápteras podem revelar a existência de indivíduos tanto férteis como estéreis, o que seria um indicativo de eusocialidade. Infelizmente, as condições ecológicas em que estes animais vivem são de difícil fossilização, e os estudiosos do assunto devem construir inferências teóricas para demonstrar como deveriam ser as condições ecológicas dos ancestrais dos animais eusociais que conhecemos hoje. Tais inferências, por sua vez, podem estar apoiadas nas poucas evidências fósseis, como também nas características comportamentais e ecológicas dos animais que hoje se encontram em estágios intermediários de socialidade, ou, ainda, no próprio estudo dos animais eusociais da atualidade.
De maneira geral, acredita-se que algumas condições devam ser satisfeitas para que exista a possibilidade do surgimento da eusocialidade. Em primeiro lugar, é preciso que exista uma situação ecológica tal que grupos de indivíduos da mesma espécie sejam obrigados a viver juntos. Tanto por viverem em ninhos, ou pela necessidade de manutenção de um território que, dificilmente, possa ser mantido por um único indivíduo. Em segundo lugar, devido às necessidades alimentares restritas, a procura por alimento torne-se demasiado intensa, que acabe por inviabilizar o cumprimento das outras tarefas que este indivíduo precisa executar, como, por exemplo, o seu cuidado com a prole (Wilson, 2008). Portanto, dados todos esses fatores, parece existir uma situação em que as condições ecológicas adversas obriguem o trabalho coletivo por meio da imposição de dificuldades à vida solitária.
A idéia de seleção de parentesco pode parecer, às vezes, insuficiente, por exemplo, quando se leva em consideração o surgimento da eusocialidade em cupins (diplobiontes), como também em formigas, cujas fêmeas foram fecundadas por vários machos. Afinal, nesses casos a relação de parentesco entre os irmãos deixa de ser de 75% e pode passar para menos de 30%. Nesse contexto a aptidão indireta nunca será maior que a aptidão direta – pois cuidar da própria prole seria sempre mais vantajoso do que abdicar dos próprios filhos em favor dos irmãos. Surge, então, uma idéia alternativa ou, pelo menos, complementar às idéias de Hamilton (seleção de parentesco). Esta idéia – inicialmente apresentada por Richard Alexander (1974) e que ganha apoio mais de 20 anos depois (Foster e col., 2002; Foster e Ratnieks, 2000; Foster e Ratnieks, 2001; Ratnieks e Reeve, 1992; Ratnieks e Visscher, 1989; Ratnieks e Wenseleers, 2005; Ratnieks e Wenseleers, 2008) – introduz um novo conceito dentro do contexto

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
8
da evolução da eusocialidade: o altruísmo forçado.
Idéias e concepções como essas ganham suporte em recentes estudos que descrevem uma série de comportamentos agressivos dentro da sociedade de insetos (Boomsma e Ratnieks, 1996; Chaline e col., 2004; Foster e Ratnieks, 2000; Foster e col., 2001; Foster e col., 2006; Ratnieks, 1988; Ratnieks, 1991; Wenseleers e col., 2005; Wenseleers e Ratnieks, 2006; Wenseleers e col., 2003) que podem ser resumidos em três tipos: 1) Coerção – comportamento social agressivo, que pune e policia o comportamento individual egoísta. 2) Manipulação parental – comportamento exibido pelos pais, que visa à persuasão dos filhos para que cuidem dos irmãos. 3) Policiamento – comportamento de inibição da atividade de reprodução de determinada operários (formas não reprodutivas presentes em colônias de insetos socias) que pode acontecer pela destruição dos ovos postos ou agressão física a ela (Ratnieks e Wenseleers, 2008). Foi a descrição de tais comportamentos que levou os estudiosos do assunto a acreditarem que os comportamentos tidos como altruístas poderiam não ser voluntários como Hamilton supunha. Há, a partir desse posicionamento, uma sutil e importante diferença na compreensão de como se deu o surgimento do comportamento eusocial, já que a questão permissiva e causal do surgimento de castas inférteis não mais estaria pautada somente nas relações genéticas entre os indivíduos que vivem conjuntamente; mas, preponderantemente, esse surgimento consistiria no comportamento agressivo de membros dominantes dentro do grupo, o que levaria à esterilidade de alguns indivíduos que a ele pertencem. É claro que essas descobertas não necessariamente são contrárias às idéias de seleção de parentesco, e podem inclusive servir de substrato para uma nova classe de argumentação que suporte as idéias de Hamilton.
É evidente que as idéias de Hamilton não podem ser desconsideradas ou totalmente substituídas, afinal, a razão pela qual se deu a origem do comportamento eusocial é um assunto que ainda não está totalmente elucidado. Poderíamos argumentar do ponto de vista evolutivo que, por exemplo, a manutenção das relações altruísticas pacíficas em animais eusociais pode ter como explicação parcial – ou até mesmo total – as idéias de seleção de parentesco. De qualquer modo, uma idéia que permanece válida, com relação ao comportamento social das diferentes espécies animais, é a de que, dentro do grupo, o comportamento altruísta parece ser prejudicial para o indivíduo que pratica a generosidade, enquanto que, quando há uma comparação entre grupos, parece-nos que o grupo que exibe comportamento altruísta entre os seus membros
angaria alguma vantagem (Wilson e Wilson, 2007).
Bibliografia: Alexander, R. (1974). The evolution of social
behaviour. Annual Review of Ecology and Systematics. Arillo, A. (2007). Paleoethology: fossilized
behaviours in amber. Geologica Acta 5, 159-166. Boomsma, J. J. e Ratnieks, F. L. W. (1996).
Paternity in eusocial Hymenoptera. Philosophical Transactions Of The Royal Society Of London Series B-Biological Sciences 351, 947-975.
Chaline, N., Martin, S. J. e Ratnieks, F. L. W. (2004). Worker policing persists in a hopelessly queenless honey bee colony (Apis mellifera). Insectes Sociaux 51, 113-116.
Darwin, C. (1859). The origin of species (By Means of Natural Selection). London: John Murray.
Foster, K. R., Gulliver, J. e Ratnieks, F. L. W. (2002). Worker policing in the European hornet Vespa crabro. Insectes Sociaux 49, 41-44.
Foster, K. R. e Ratnieks, F. L. W. (2000). Social insects - Facultative worker policing in a wasp. Nature 407, 692-693.
Foster, K. R. e Ratnieks, F. L. W. (2001). Convergent evolution of worker policing by egg eating in the honeybee and common wasp. Proceedings Of The Royal Society Of London Series B-Biological Sciences 268, 169-174.
Foster, K. R., Wenseleers, T. e Ratnieks, F. L. W. (2001). Spite: Hamilton's unproven theory. Annales Zoologici Fennici 38, 229-238.
Foster, K. R., Wenseleers, T. e Ratnieks, F. L. W. (2006). Kin selection is the key to altruism. Trends In Ecology & Evolution 21, 57-60.
Hamilton, W. D. (1964a). Genetical Evolution Of Social Behaviour 2. Journal of Theoretical Biology 7, 17-&.
Hamilton, W. D. (1964b). Genetical Evolution Of Social Behaviour I. Journal of Theoretical Biology 7, 1-&.
Ratnieks, F. L. W. (1988). Reproductive Harmony Via Mutual Policing By Workers In Eusocial Hymenoptera. American Naturalist 132, 217-236.
Ratnieks, F. L. W. (1991). Evolution Of Discriminatory Aggression In Marine-Invertebrates. Journal of Theoretical Biology 152, 557-565.
Ratnieks, F. L. W. e Reeve, H. K. (1992). Conflict In Single-Queen Hymenopteran Societies - The Structure Of Conflict And Processes That Reduce Conflict In Advanced Eusocial Species. Journal of Theoretical Biology 158, 33-65.
Ratnieks, F. L. W. e Visscher, P. K. (1989). Worker Policing In The Honeybee. Nature 342, 796-797.
Ratnieks, F. L. W. e Wenseleers, T. (2005). Policing insect societies. Science 307, 54-56.
Ratnieks, F. L. W. e Wenseleers, T. (2008). Altruism in insect societies and beyond: voluntary or enforced? Trends In Ecology & Evolution 23, 45-52.
Wenseleers, T., Badcock, N. S., Erven, K., e col. (2005). A test of worker policing theory in an advanced eusocial wasp, Vespula rufa. Evolution 59, 1306-1314.
Wenseleers, T. e Ratnieks, F. L. W. (2006). Enforced altruism in insect societies. Nature 444, 50-50.
Wenseleers, T., Ratnieks, F. L. W. e Billen, J. (2003). Caste fate conflict in swarm-founding social Hymenoptera: an inclusive fitness analysis. Journal Of Evolutionary Biology 16, 647-658.
Wilson, D. S. e Wilson, E. O. (2007). Rethinking the theoretical foundation of sociobiology. Quarterly Review of Biology 82, 327-348.
Wilson, E. O. (1971). The insect societies. Cambridge, MA: Belknap Press of Harvard University Press.
Wilson, E. O. (2008). One giant leap: How insects achieved altruism and colonial life. Bioscience 58, 17-25.
Wilson, E. O. e Holldobler, B. (2005). Eusociality: Origin and consequences (vol 102, pg 13367, 2005). Proceedings of the National Academy of Sciences of the United States of America 102, 16119-16119.

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
9
NEUROGÊNESE NO SISTEMA NERVOSO ADULTO DE MAMÍFEROS
Ilton Santos da Silva
Departamento de Fisiologia, Instituto de Biociências, USP Recebido 27jul09 / Aceito 15dez09 / Publicação inicial 17dez09
[email protected] Resumo. O cientista espanhol Ramón y Cajal e seus contemporâneos defendiam a idéia de que o
sistema nervoso de indivíduos adultos fosse imutável, sem a possibilidade de que novos neurônios surgissem. No entanto, com o desenvolvimento recente de novas técnicas, foi possível confirmar a existência da neurogênese na fase adulta. Como a maior parte dos novos neurônios surge em uma estrutura crítica nas funções de memória, acredita-se que eles contribuam para o aprimoramento destas funções e não sejam apenas vestígios ontogenéticos. Essa breve revisão busca discutir algumas teorias e evidências experimentais a respeito da neurogênese e seu papel no sistema nervoso de indivíduos adultos.
Palavras-chave. Aprendizagem e memória, neurogênese adulta, giro denteado, hipocampo. NEUROGENESIS IN THE ADULT MAMMALIAN NERVOUS SYSTEM Abstract. The Spanish scientist Ramón y Cajal and his contemporaries advocated the idea that the
adult mammalian nervous systems were immutable and new neurons could not arise. However, with the recent development of new techniques, it was possible to confirm the existence of neurogenesis in adulthood. Like most of the new neurons arise in a critical structure for memory functions, it is believed that they contribute to improve these functions and not only constitute ontogenetic traces. This brief review discusses some theories and experimental evidences regarding to the neurogenesis and its role in the adult mammalian nervous system.
Keywords. Learning and memory, adult neurogenesis, dentate gyrus, hippocampus.
Breve histórico As primeiras descrições detalhadas de
células nervosas foram realizadas no final do século XIX por Camilo Golgi e Ramón y Cajal, contribuindo imensamente para a compreensão dos aspectos estruturais do sistema nervoso. Golgi desenvolveu técnicas de coloração utilizando sais de prata para corar neurônios, revelando detalhes da estrutura neuronal sob o microscópio, tais como o corpo celular, dendritos e axônio. Utilizando a técnica de coloração empregada por Golgi, o histologista espanhol Santiago Ramón y Cajal observou que o tecido nervoso é formado por uma rede de células distintas e que estas seriam os elementos fundamentais do sistema nervoso (Fig.1). Ramón y Cajal estudou as diferentes fases de desenvolvimento dos neurônios em mamíferos, observando que não havia a presença de qualquer sinal do surgimento de novas células no encéfalo adulto, além daquelas já estabelecidas ao nascimento. Outros pesquisadores da época também concluíram que a elaborada arquitetura do encéfalo de mamíferos permanece fixa e defenderam a idéia de que a adição de novas células era completamente inconcebível. Dessa forma, postulou-se que o sistema nervoso central possui conexões fixas e imutáveis, sem qualquer possibilidade de que novos neurônios surgissem.
Já na primeira metade do século XX, alguns estudos mostraram que havia o nascimento de novas células em encéfalos adultos (Hamilton, 1901; Allen, 1912; Sugita, 1918). Entretanto, a grande dificuldade era afirmar se essas novas células eram realmente
neurônios ou glia, tendo em vista a limitação dos métodos empregados na época.
Na década de 1960, Joseph Altman publicou uma série de estudos relatando a ocorrência de neurogênese em ratos jovens e adultos (Altman, 1962; Altman, 1963; Altman e Das, 1965; Altman e Das, 1966; Altman, 1966; Altman, 1969). Utilizando a técnica de auto-radiografia com [3H]-Timidina, uma substância que é incorporada ao DNA das células em divisão, Altman observou o surgimento de novas células em diversas áreas, incluindo neocórtex, giro denteado e bulbo olfatório. Esse autor sugeriu ainda que estas células, que ele chamava de “microneurônios”, possuíam axônios curtos e apresentavam forma granular ou estelar.
Figura 1 - Neurônios no cerebelo de ave.
Desenho realizado por Ramón y Cajal, mostrando os cinco tipos celulares existentes no cerebelo: células de Purkinje; células esteladas; células em forma de cesto;
Revisão:

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
10
células granulares e células de Golgi (extraído de Sotelo, 2003).
Novamente as críticas surgiram em relação
ao método empregado, que de fato não era o mais adequado para diferenciar células gliais de neurônios propriamente ditos. Outro motivo da não aceitação das observações de Altman foi a pouca credibilidade direcionada ao pesquisador, que na ocasião era pós-doutorando e trabalhava por conta própria. Dessa forma foi julgado incapaz de alterar um fato amplamente aceito pela comunidade científica contemporânea.
Com a ajuda da microscopia eletrônica, Michael Kaplan e seus colaboradores realizaram vários estudos, publicados a partir de 1977, mostrando que as células incorporadas com [3H]-Timidina no giro denteado e bulbo olfatório de ratos possuíam características ultra-estruturais de neurônios, tais como dendritos e sinapses, o que não é observado em astrócitos e oligodendrócitos (Kaplan e Hinds, 1977; Kaplan, 1984). O mesmo autor observou ainda novos neurônios no córtex cerebral de ratos adultos (Kaplan, 1981; Kaplan, 1985), confirmando assim as afirmações feitas por Altman. Os trabalhos de Kaplan receberam pouca atenção e também não foram suficientes para quebrar o “dogma” que parecia fortemente estabelecido.
O grande avanço no estudo da neurogênese ocorreu no final da década de 1980 com o emprego da 5-bromo-3’-deoxiuridina (BrdU) que é captada pelas células durante a fase S da mitose, sendo desta forma, um marcador de células em proliferação. As células marcadas com BrdU podem ser visualizadas por técnicas de imunocitoquímica, sem a necessidade de empregar auto-radiografia (Nowakowski e col., 1989). Desde então, vários estudos mostraram que a neurogênese é um processo que ocorre continuamente em certas regiões encefálicas de diversas espécies, incluindo aves (Goldman e Nottebohm, 1983), roedores (van Praag e col., 1999), macacos (Kornack e Rakic, 1999), e humanos (Eriksson e col., 1998). Portanto, depois de mais de um século de estudos e muita resistência quanto à existência do fenômeno, hoje a neurogênese em encéfalos adultos é um fato amplamente aceito pelos neurocientistas.
Como ocorre maior parte da neurogênese no hipocampo, uma estrutura nervosa reconhecidamente envolvida em processos de aprendizagem e memória (O’Keefe e Nadel, 1978), as pesquisas recentes na área tentam apontar qual seria o papel desses novos neurônios nestas funções.
O surgimento de novos neurônios pode ainda ser regulado por fatores psico-fisiológicos como estresse e complexidade ambiental a que o animal é exposto. Estas questões serão discutidas com mais detalhes nos tópicos a seguir.
Regulação da Neurogênese Diversos fatores podem interferir nos
processos de neurogênese (e.g., neurais, endócrinos e ambientais), aumentando ou diminuindo a produção de novos neurônios no indivíduo adulto. Por exemplo, a elevação nos níveis de glicocorticóides, hormônios que têm seus níveis alterados em virtude de experiências estressantes como a exposição ao odor de um predador natural, pode diminuir a taxa de proliferação de células granulares do giro denteado do hipocampo de ratos (Heale e col., 1994), possivelmente por meio de um mecanismo que envolve a liberação e o acúmulo de glutamato no hipocampo (Moghddam e col., 1994; Gould e col., 2000). Eisch e col. (2000) mostraram também que o tratamento crônico com morfina ou heroína reduz significativamente a taxa de neurogênese na camada de células granulares do giro denteado de ratos. E esse resultado parece não estar relacionado com alterações dos níveis circulantes de glicocorticóides, pois os mesmos autores observaram efeitos similares em animais que foram submetidos à adrenalectomia e posterior reposição de corticosterona.
Por outro lado, certas condições propiciam a produção de novas células no hipocampo. Tanapat e col. (1999) mostraram que ratas submetidas ao procedimento de ovariectomia, visando eliminar a produção de estrógenos, apresentam menor número de células em proliferação marcadas com BrdU. No mesmo estudo, os autores observaram que durante o proestro (fase do ciclo estral de ratas em que os níveis de estrógenos estão altos) ocorre maior produção de novas células, sendo que a maioria delas adquire características neuronais.
Ambientes que fornecem uma combinação variada de estímulos também podem aumentar a neurogênese. Roedores adultos que são mantidos em gaiolas que contém objetos diferentes, tais como pequenos brinquedos, túneis, rodas de atividade física e obstáculos, exibem significativo aumento no número de células no giro denteado do hipocampo (Fig.2), (Kempermann e col., 1997). Ou seja, parece que o aumento da atividade exploratória e novas experiências sensoriais proporcionadas por estímulos diversos do ambiente enriquecido estimulam a aprendizagem, fazendo com que esses animais aumentem a sua capacidade de desempenhar tarefas cognitivas, diferentemente daqueles que vivem em gaiolas comuns de laboratório.

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
11
Figura 2 - Parâmetros em que camundongos
mantidos em ambiente enriquecido diferem de animais controle. (a) Número de células marcadas com BrdU na camada granular do giro denteado (CCG) um dia ou 4 semanas após injeção intraperitoneal de BrdU, (b) o volume total do giro denteado 4 semanas após a injeção, (c) número absoluto de células granulares do giro denteado 4 semanas após a injeção, e (d) extensão do trajeto até encontrar a plataforma no labirinto aquático de Morris (modificado de Kempermann e col., 1997).
Esses animais mostram ainda melhor
desempenho quando submetidos a uma tarefa de aprendizagem e memória espacial no labirinto aquático de Morris (Fig. 2-d) (Kempermann e col., 1997), que consiste na busca por uma plataforma submersa ao longo de vários dias de treino (Morris, 1981). Porém, os autores deixam claro que não se pode concluir que esse desempenho melhor seja devido ao aumento do número de células no hipocampo, embora seja plausível pensar que a combinação do maior número de neurônios, sinapses e dendritos, contribua para um melhor desempenho induzido pelo ambiente enriquecido. Entretanto, tal ambiente é formado por diversos componentes, incluindo a oportunidade de interação social, atividade física e aprendizagem. Cabe questionar, então, se o aumento de células no hipocampo e o melhor desempenho na tarefa espacial (Kempermann e col., 1997) seriam decorrentes das experiências sensoriais no ambiente enriquecido ou da constante atividade física proporcionada pelas rodas de atividade, túneis e obstáculos presentes nas gaiolas.
van Praag e col. (1999) tentaram identificar quais destes fatores (atividade física voluntária ou forçada e uma tarefa que envolve aprendizagem) contribuem para o aumento da neurogênese hipocampal em camundongos adultos.
Figura 3 - Condições experimentais utilizadas
por van Praag e col. (1999). (A) animais mantidos em ambiente enriquecido; (B) em gaiola contendo a roda de exercício físico voluntário; ou (C) em gaiolas-padrão de laboratório (extraído de van Praag e col., 1999).
Grupos independentes de camundongos
foram submetidos às seguintes condições: acondicionamento em ambiente enriquecido; treino de busca pela plataforma no labirinto aquático; natação forçada; exercício voluntário em roda de atividade física, e acondicionamento em gaiolas-padrão (grupo controle) (Fig.3). A proliferação de células no giro denteado foi investigada por meio da marcação com BrdU e posterior análise imunohistoquímica. Os resultados (Fig.4-a) mostraram que o grupo submetido à roda de atividade física exibiu maior proliferação do que qualquer outro grupo avaliado. Já a avaliação da sobrevivência da progênie das células em divisão foi feita quatro semanas após a última injeção de BrdU. A análise estatística mostrou que os animais submetidos ao ambiente enriquecido e roda de atividade física apresentaram taxas maiores de sobrevivência das novas células (Fig.4-b) (85% e 56%, respectivamente), quando comparados aos grupos que passaram pelo treinamento no labirinto aquático (42%) e natação forçada (46%). Além disso, em ambos os grupos (ambiente enriquecido e roda de atividade física) a maioria das células marcadas com BrdU apresentou características neuronais, reveladas por meio do uso de marcadores específicos.
A
B C

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
12
Figura 4 – Número estimado de células
marcadas com BrdU no giro denteado de camundongos adultos. (a) número total de células em proliferação marcadas um dia após a última injeção de BrdU, (b) número estimado de células sobreviventes quatro semanas após a última injeção de BrdU (modificado de van Praag e col., 1999).
Portanto, este estudo mostra que somente
o exercício físico voluntário é suficiente para estimular a proliferação de células hipocampais em camundongos adultos. Porém, quanto à taxa de sobrevivência dessas células, as duas condições (ambiente enriquecido e exercício físico) mostraram efeito significativo, sugerindo que diferentes protocolos de manipulações comportamentais podem aumentar consideravelmente a neurogênese e manutenção dos novos neurônios formados. Embora ambos os grupos apresentem números comparáveis de células marcadas com BrdU após quatro semanas, a sobrevivência dessas células foi relativamente menor no grupo submetido ao exercício físico voluntário (56%) quando comparado ao grupo mantido em ambiente enriquecido (85%), sugerindo que estas condições apresentam efeitos diferentes a longo prazo. Os autores discutem ainda se o tempo de treino empregado no labirinto aquático teria sido suficiente para revelar eventuais efeitos sobre a neurogênese. E também considerando possíveis efeitos prejudiciais induzidos pelo estresse, a natação forçada pode ter sido um fator que prejudicou a proliferação e sobrevivência das novas células devido à elevação nos níveis de glicocorticóides.
Dessa forma, a neurogênese no hipocampo, regulada por diferentes variáveis ambientais, reforça a idéia de que a produção de novos neurônios não faz parte de vestígios dos estágios iniciais de desenvolvimento do sistema nervoso, mas constituem um recurso neural notadamente flexível e adaptativo também na idade adulta.
Possíveis funções dos novos
neurônios: aprendizagem e memória? Estudos relatando neurogênese no sistema
nervoso permitem afirmar que milhares de neurônios novos são formados todos os dias no encéfalo adulto, principalmente no giro denteado
da formação hipocampal. No entanto, diversas questões relativas à possível função (ou funções) da neurogênese em adultos têm gerado debates e muitas especulações entre os pesquisadores.
Kempermann (2002) propôs que os novos neurônios do giro denteado atuam como “comportas” na “entrada de informações” para a memória (Fig.5). Segundo o autor, os novos neurônios são adicionados em um local do circuito hipocampal determinante no processamento de informações. Como não há evidências de que o hipocampo possa armazenar memórias por um longo período de tempo, a neurogênese hipocampal adulta não estaria envolvida nesse tipo de memória “per se”, mas no processamento de informações mantidas transitoriamente. Nesse caso, se os novos neurônios forem estrategicamente introduzidos na circuitaria existente, podem aumentar significativamente a capacidade de processamento de informações e também sua complexidade.
Figura 5 – Teoria proposta por Kempermann
(2002). (A) Segundo o autor, os novos neurônios atuam como “comportas”, reforçando circuitarias pré-existentes conforme a demanda do ambiente; (B) uma analogia com um computador, mostrando que os novos neurônios podem aumentar a capacidade de processamento de informações na memória “RAM”, função atribuída ao hipocampo; e (C) localização estratégica dos novos neurônios (em vermelho) na circuitaria pré-existente (Cx=áreas corticais; DG=giro denteado; e sub-campos CA1 e CA3 da formação hipocampal) (modificado de Kempermann, 2002).
Uma vez que as novas células granulares
do giro denteado apresentam propriedades funcionais similares aos neurônios maduros, incluindo potenciais de ação, sinapses e recebem aferências de outras regiões do circuito (van Praag e col., 2002), aparentemente, elas são integradas funcionalmente à circuitaria hipocampal. Portanto, considerando que poucos dos novos neurônios sobrevivem e um número menor ainda é funcionalmente integrado ao sistema, parece que a neurogênese hipocampal adulta representa um “ajuste a longo prazo” da circuitaria para processar conteúdos em um alto nível de complexidade (Kempermann, 2002).

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
13
No entanto, a real contribuição dos novos neurônios sobre o desempenho de animais experimentais em tarefas dependentes do hipocampo permanece desconhecida, pois os resultados obtidos são divergentes. Por exemplo, alguns estudos sugerem que o aumento da neurogênese pode melhorar o desempenho em alguns tipos de tarefa espacial, como no labirinto aquático de Morris (Kempermann e col., 1997; van Praag e col., 1999); paralelamente, prejuízos são observados na ausência de novos neurônios (Kempermann & Gage, 2002). Entretanto, existe ainda relação inversa entre os fatores regulatórios da neurogênese e a aprendizagem; fatores que diminuem a produção de novos neurônios, como a elevação nos níveis de glicocorticóides, decorrentes de estresse moderado não necessariamente alteram o desempenho, mas podem até mesmo facilitar a aquisição de uma tarefa espacial (Akirav e col., 2004). Outro estudo mostra que a diminuição da neurogênese decorrente da idade também não interfere no desempenho de uma tarefa espacial no labirinto aquático (Bizon e Gallagher, 2003). Essa discrepância de resultados pode ser devido a diversos fatores. O agente antimitótico acetato de metilazoximetanol (MAM) e a radiação ionizante geralmente são utilizados para cessar a produção de precursores de células granulares (Cameron & Christie, 2007); assim, a neurogênese e seus efeitos no giro denteado podem ser estudados. Porém, estes métodos podem causar efeitos secundários e levar a interpretações viesadas sobre o comportamento dos animais. Além disso, ambiente enriquecido e a atividade física podem causar outros efeitos, como angiogênese (Isaacs e col., 1992) e alterações estruturais nos neurônios pré-existentes (Leggio e col., 2005), induzindo dessa forma a obtenção de resultados falsos positivos em relação a contribuição da neurogênese sobre o comportamento.
Embora existam fortes evidências de correlação entre neurogênese e função hipocampal, seria de grande valia o desenvolvimento de abordagens experimentais que contemplassem o papel da neurogênese hipocampal adulta por si só.
Considerações finais As pressões cotidianas exigem que
mecanismos neurais diferenciados sejam desenvolvidos, permitindo a sobrevivência dos animais, pois os mesmos não nascem com um repertório comportamental completo. Neste sentido, a plasticidade do sistema nervoso tem papel fundamental na adaptação às contingências ambientais. A neurogênese, hoje um fenômeno indiscutivelmente bem estabelecido e amplamente aceito, oferece a oportunidade de entender como o sistema nervoso desenvolveu mecanismos para suprir as demandas do ambiente. Embora pouco ainda se saiba sobre a
função da neurogênese adulta, parece que o surgimento de novas células nervosas faz parte de uma gama de recursos neurais que podem contribuir para a flexibilidade comportamental dos animais. Considerando que um número substancial de neurônios é gerado em uma região crítica para as funções de memória, é pouco provável que essas novas células não tenham qualquer função e apenas façam parte de vestígios do desenvolvimento neural.
Agradecimentos. Agradeço ao Professor
Gilberto F. Xavier pelas sugestões e comentários sobre o texto e pelas discussões acerca do tema.
Bibliografia
Akirav I., et al. (2004) A facilitative role for corticosterone in the acquisition of a spatial task under moderate stress. Learning and Memory 11:188–195.
Allen E. (1912) The cessation of mitosis in the central nervous system of the albino rat. Journal of Comparative Neurology. 19:547-68.
Altman, J. (1962) Are new neurons formed in the brains of adult mammals? Science 135, 1127–1128.
Altman, J. (1963) Autoradiographic investigation of cell proliferation in the brains of rats and cats. Postnatal growth and differentiation of the mammalian brain, with implications for a morphological theory of memory. Anatomical Record. 145, 573–591.
Altman, J. & Das, G. D. (1965) Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. Journal of Comparative Neurology. 124, 319–335.
Altman, J. & Das, G. D. (1966) Autoradiographic and histological studies of postnatal neurogenesis. I. A longitudinal investigation of the kinetics, migration and transformation of cells incorporating tritiated thymidine in neonate rats, with special reference to postnatal neurogenesis in some brain regions. Journal of Comparative Neurology. 126, 337–390.
Altman, J. (1966) Autoradiographic and histological studies of postnatal neurogenesis II. A longitudinal investigation of the kinetics, migration and transformation of cells incorporating tritiated thymidine in infant rats, with special reference to postnatal neurogenesis in some brain regions. Journal of Comparative Neurology. 128, 431–474.
Altman, J. (1969) Autoradiographic and histological studies of postnatal neurogenesis IV. Cell proliferation and migration in the anterior forebrain, with special reference to persisting neurogenesis in the olfactory bulb. Journal of Comparative Neurology. 137, 433–458.
Cameron, H.A. & Christie, B.R. (2007) Do new neurons have a functional role in the adult hippocampus? Debates in Neuroscience 1:26–32.
Eisch, A. J., Barrot, M., Schad, C. A., Self, D. W. & Nestler, E. J. (2000). Opiates inhibit neurogenesis in the adult rat hippocampus. Proceedings of the National Academy of sciences of the USA. 97, 7579–7584.
Eriksson, P. S. et al. (1998) Neurogenesis in the adult human hippocampus. Nature Medicine. 4, 1313–1317.
Goldman, S. A. & Nottebohm, F. (1983) Neuronal production, migration, and differentiation in a vocal control nucleus of the adult female canary brain. Proceedings of the National Academy of Sciences of the USA. 80, 2390–2394.
Gould, E. et al. (2000) Regulation of hippocampal neurogenesis in adulthood. Biological psychiatry 48(8):715-20.
Hamilton A. (1901) The division of differentiated cells in the central nervous system of the white rat. Journal of Comparative Neurology. 11:297-320.
Heale VR, Vanderwolf CH, Kavaliers M (1994): Components of weasel and fox odors elicit fast wave bursts in the

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
14
dentate gyrus of rats. Behavior Brain Research. 63:159 –165.
Isaacs, K. R. et al. (1992) Exercise and the brain: angiogenesis in the adult rat cerebellum after vigorous physical activity and motor skill learning. Journal of Cerebral Blood Flow and Metabolism 12, 110–119.
Kandel, E.R.; Schwartz, J.H.; Jessell, T.M. (2000) Principles of Neural Science. 4th ed. New York: McGraw-Hill.
Kaplan, M. S. & Hinds, J. W. (1977) Neurogenesis in the adult rat: electron microscopic analysis of light radioautographs. Science 197, 1092–1094.
Kaplan, M. S. (1981) Neurogenesis in the 3-month-old rat visual cortex. Journal of Comparative Neurology. 195, 323–338.
Kaplan, M. S. (1984) Mitotitic neuroblasts in the 9-day-old and 11-month-old rodent hippocampus. Journal of Neuroscience. 4,1429–1441.
Kaplan, M. S. (1985) Formation and turnover of neurons in young and senescent animals: an electron microscopic and morphometric analysis. Annals of the New York Academy of Sciences. 457, 173–192.
Kempermann G, Kuhn HG, Gage FH. (1997) More hippocampal neurons in adult mice living in an enriched environment. Nature 3;386(6624):493-5.
Kempermann G, Gage FH. (2002) Genetic determinants of adult hippocampal neurogenesis correlate with acquisition, but not probe trial performance, in the water maze task. European Journal of Neuroscience 16:129–136.
Kempermann, G. (2002) Why new neurons? Possible functions for adult hippocampal neurogenesis. The Journal of Neuroscience. 22(3):635-8.
Kornack, D. R. & Rakic, P. (1999) Continuation of neurogenesis in the hippocampus of the adult macaque monkey. Proceedings of the National Academy of Sciences of the USA. 96 5768–5773.
Leggio, M.G. (2005) Environmental enrichment promotes improved spatial abilities and enhanced dendritic growth in the rat. Behavioural Brain Research 163(1):78-90.
Moghaddam B, Bolinao ML, Stein-Behrens B, Sapolsky R (1994): Glucocorticoids mediate the stress-induced extracellular accumulation of glutamate. Brain Research. 655:251–254.
Morris, R.G.M. (1981). Spatial localization does not require the presence of local cues, Learning and Motivation 12: 239-260.
Nowakowski RS, Lewin SB, Miller MW. (1989) Bromodeoxyuridine immunohistochemical determination of the lengths of the cell cycle and the DNA-synthetic phase for an anatomically defined population. Journal of Neurocytology. 18(3):311-8.
O’Keefe J, Nadel L. (1978). The hippocampus as a cognitive map. Oxford: Clarendon Press. 570 p.
Sotelo, C. (2003) Viewing the brain through the master hand of Ramón y Cajal. Nature Reviews Neuroscience. 4(1):71-7.
Sugita N. (1918) Comparative studies on the growth of the cerebral cortex. Journal of Comparative Neurology. 29:61-117.
Tanapat P, Hastings N, Reeves AJ, Gould E (1999): Estrogen stimulates a transient increase in the number of new neurons in the dentate gyrus of the adult female rat. Journal of Neuroscience. 19:5792–5801.
van Praag H, Kempermann G, Gage FH. (1999) Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nature Neuroscience 2(3):266-70.
van Praag et al. (2002) Functional neurogenesis in the adult hippocampus. Nature 415(6875):1030-4.

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
15
UMA INTRODUÇÃO À NEUROFILOSOFIA: O PROBLEMA MENTE-CORPO
Camila Gomes Victorino
Departamento de Fisiologia, Instituto de Biociências, USP Recebido 25mai2009 / Aceito 15dez2009 / Publicação inicial 17dez2009
Resumo. A neurofilosofia tenta desvendar os mistérios da inteligência consciente, aliada a outras ciências como a neurociência e a inteligência artificial. Apesar do crescente avanço da neurofisiologia, uma filosofia da mente que se adeque a todos os resultados e eventos observados das ciências cognitivas ainda não pôde ser formulada, existindo um enorme desacordo quanto à origem dos estados mentais. Designado como problema mente-corpo, ele divide a comunidade filosófica e científica em dualistas e monistas. Se os primeiros afirmam que os estados mentais não têm como origem os estados cerebrais, os segundos afirmam que há uma correlação entre estados físicos do encéfalo e estados mentais, sem, no entanto, chegar a um consenso quanto aos detalhes da correlação entre encéfalo e mente.
Palavras-chave. neurofilosofia, filosofia da ciência, dualismo, monismo. AN INTRODUCTION TO NEUROPHILOSOPHY: THE MIND-BODY PROBLEM Abstract. Neurophilosophy, allied to other sciences, like Neuroscience and Artificial Intelligence, tries
to unravel the consciousness intelligence mysteries. In spite of the Neurophilosophy development, a Mind Philosophy which essays to conform to the cognitive science observed-events has never had the chance to be formulated, because there is an enormous disagree concerning the mental states origins. Designed as the mind-body problem, it breaks up the philosophical and scientific community in dualists and monists. If the first ones claim that mental states don´t have the brain as source, the last ones claim that there is a relation between brain physical states and mental states, without, however, being in a consensus about the details concerning the mind-brain relationship
Keywords. Neurophilosophy, Science Philosophy, dualism, monism. Quando as dúvidas relativas ao
funcionamento do encéfalo humano surgem na mente de um indivíduo, sua primeira reação é pensar nas neurociências. Ela é a linha de estudo mais bem conhecida do não-especialista, porém não é a única, fazendo parte de um grupo que contém muitas outras ciências que tentam elucidar o funcionamento do encéfalo, bem como as repercussões que elas podem trazer para a sociedade, tanto em termos filosóficos e sociais, como tecnológicos. Essas ciências, quando agrupadas, passam a fazer parte de um grande agrupamento de ciências chamado de ciências cognitivas, o qual pode ser dividido nos seguintes domínios: inteligência artificial, evolução e cognição, linguística, neurociências, neurofilosofia e psicologia (Wilson & Keil, 1999).
Todas essas ciências uniram forças para tratar o problema da natureza da inteligência consciente, tendo a neurofilosofia ou filosofia da mente um papel crucial na elucidação do estatuto do autoconhecimento da mente e na elaboração de uma concepção mais clara da natureza (Churchland, 2004). Entretanto, a neurofilosofia é pouco conhecida do público, pois ainda não existem muitos laboratórios que trabalham com o tema no Brasil. Tendo em vista sua baixa difusão, o presente ensaio visa introduzir uma das questões primordiais que ela tenta responder: o problema mente-corpo.
Para quê neurofilosofia? Quando trabalhamos com uma ciência
experimental aplicamos - na maioria das vezes de maneira inconsciente - conceitos filosóficos que influenciam nossa decisão quanto à escolha de um método ou outro, bem como a interpretação dos resultados. Um exemplo de como a filosofia da ciência influencia essa interpretação, se dá quando - ao aplicarmos erroneamente o método científico - pressupomos que os nossos resultados (a partir do momento em que eles são aceitos pela comunidade científica) podem ser generalizados para toda a natureza, apagando as possíveis exceções que possam existir. Essa generalização se baseia num pressuposto filosófico chamado de indução. Segundo Chalmers (1993), a resposta indutivista é que, desde que certas condições sejam satisfeitas, é legítimo generalizar a partir de uma lista finita de proposições de observação singulares para uma lei universal.
Para se entender melhor o principio da indução, atentemos para o exemplo do mesmo autor que escreve: suponhamos que até hoje eu tenha observado uma grande variedade de corvos sob uma ampla variedade de circunstâncias e tenha observado que todos eles são pretos e que, com base nisto, concluo: "Todos os corvos são pretos". Esta é uma inferência indutiva perfeitamente legítima. (...) Mas não há garantia lógica de que o próximo corvo que observarei não seja cor-de-rosa.
De acordo com Chalmers (1993), na posição indutivista ingênua, a ciência é baseada no princípio de indução (...). De acordo com o
Ensaio:
REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
16
indutivista ingênuo, o corpo do conhecimento científico é construído pela indução a partir da base segura fornecida pela observação. A análise até aqui constitui apenas uma explicação parcial da ciência. Pois certamente uma característica importante da ciência é sua capacidade de explicar e prever.
Tendo em vista equívocos metodológicos como esses, a filosofia da ciência visa à elucidação dos limites do fazer científico e assim ao seu aprimoramento. O filósofo não tenta ditar como a investigação e os argumentos científicos devem ser conduzidos; ao invés disso, ele procura enumerar os princípios e práticas que têm contribuído para a boa ciência (Fodor, 1981).
Da mesma maneira, a neurofilosofia trabalha os pressupostos empregados nas ciências cognitivas experimentais. Desde seu surgimento, a filosofia da mente se caracterizou como um novo esforço para retornar aos principais temas clássicos que atravessaram o pensamento na modernidade. Era preciso fazer uma nova tentativa no sentido de determinar a natureza última dos fenômenos mentais (...) sem ignorar os resultados das pesquisas sobre o cérebro humano (Teixeira, 1994). Para se ter uma idéia dos principais temas estudados pela neurofilosofia, o problema mente-corpo será apresentado, o qual não só concerne a própria filosofia da mente, mas todo o desenvolvimento das outras ciências cognitivas.
O problema mente-corpo O problema mente-corpo se levanta a partir
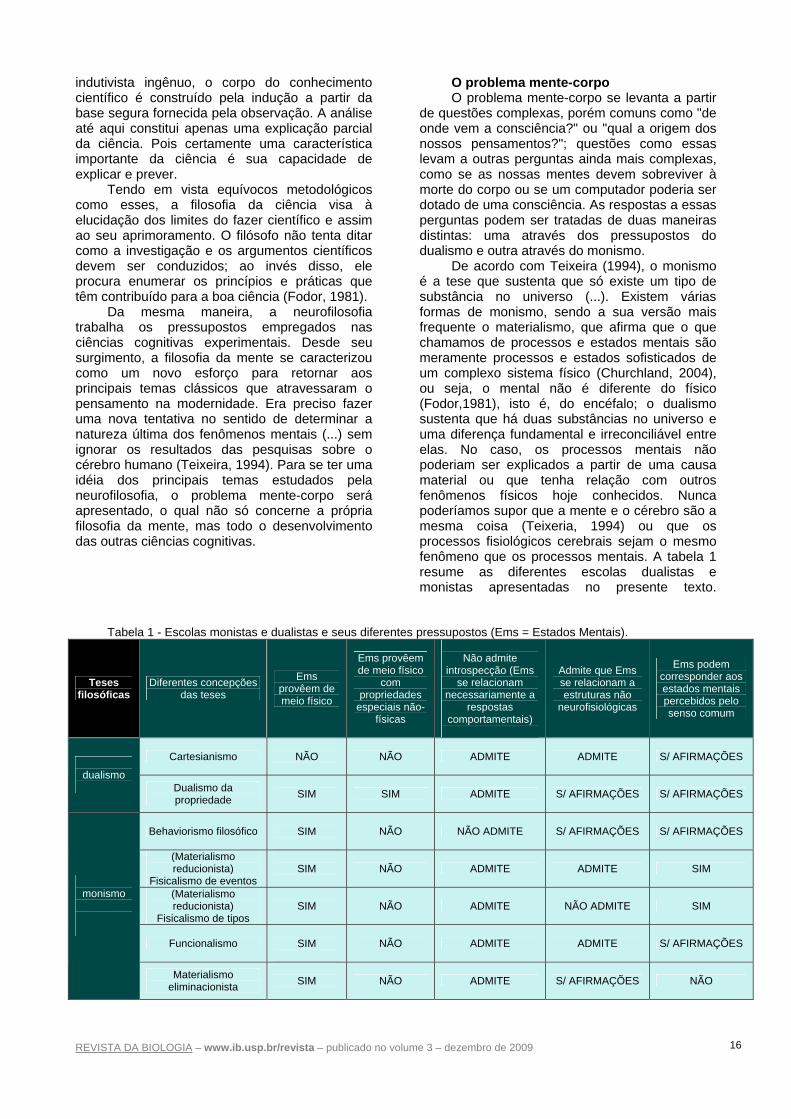
de questões complexas, porém comuns como "de onde vem a consciência?" ou "qual a origem dos nossos pensamentos?"; questões como essas levam a outras perguntas ainda mais complexas, como se as nossas mentes devem sobreviver à morte do corpo ou se um computador poderia ser dotado de uma consciência. As respostas a essas perguntas podem ser tratadas de duas maneiras distintas: uma através dos pressupostos do dualismo e outra através do monismo.
De acordo com Teixeira (1994), o monismo é a tese que sustenta que só existe um tipo de substância no universo (...). Existem várias formas de monismo, sendo a sua versão mais frequente o materialismo, que afirma que o que chamamos de processos e estados mentais são meramente processos e estados sofisticados de um complexo sistema físico (Churchland, 2004), ou seja, o mental não é diferente do físico (Fodor,1981), isto é, do encéfalo; o dualismo sustenta que há duas substâncias no universo e uma diferença fundamental e irreconciliável entre elas. No caso, os processos mentais não poderiam ser explicados a partir de uma causa material ou que tenha relação com outros fenômenos físicos hoje conhecidos. Nunca poderíamos supor que a mente e o cérebro são a mesma coisa (Teixeria, 1994) ou que os processos fisiológicos cerebrais sejam o mesmo fenômeno que os processos mentais. A tabela 1 resume as diferentes escolas dualistas e monistas apresentadas no presente texto.
Tabela 1 - Escolas monistas e dualistas e seus diferentes pressupostos (Ems = Estados Mentais).
Teses filosóficas
Diferentes concepções das teses
Ems provêem de meio físico
Ems provêem de meio físico
com propriedades
especiais não-físicas
Não admite
introspecção (Ems se relacionam
necessariamente a respostas
comportamentais)
Admite que Ems se relacionam a estruturas não
neurofisiológicas
Ems podem corresponder aos estados mentais percebidos pelo senso comum
Cartesianismo
NÃO
NÃO ADMITE ADMITE S/ AFIRMAÇÕES dualismo
Dualismo da propriedade
SIM
SIM ADMITE S/ AFIRMAÇÕES S/ AFIRMAÇÕES
Behaviorismo filosófico
SIM
NÃO NÃO ADMITE S/ AFIRMAÇÕES S/ AFIRMAÇÕES
(Materialismo reducionista)
Fisicalismo de eventos
SIM
NÃO ADMITE ADMITE SIM
(Materialismo reducionista)
Fisicalismo de tipos
SIM
NÃO ADMITE NÃO ADMITE SIM
Funcionalismo
SIM
NÃO ADMITE ADMITE S/ AFIRMAÇÕES
monismo
Materialismo eliminacionista
SIM
NÃO ADMITE S/ AFIRMAÇÕES NÃO

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
17
O dualismo não é a concepção mais amplamente defendida em meio à comunidade científica e filosófica atual, mas é a ”teoria da mente” mais difundida em meio às pessoas em geral (Churchland, 2004), tendo, aliás, importância fundamental no desenrolar histórico do desenvolvimento das “teorias da mente”, pois as primeiras teorias que tentaram de alguma maneira explicar a mente humana foram dualistas (Churchland, 2004).
Existem várias formas de dualismo, entre as quais o dualismo da substância ou cartesianismo é a mais conhecida. Ele admite que a mente possui uma natureza inteiramente independente do corpo e consequentemente que ela não é sujeita à morte (Descartes, 1973). Essa idéia é bastante popular, mas cheia de problemas quando passamos a nos questionar que se a mente provém de algo totalmente diferente da matéria, como seria possível a sua ligação ao corpo? Alguns argumentam que essa substância proveria de alguma forma de energia ainda não-descrita pela ciência, porém as provas de sua existência são extremamente parcas, o que levou à modificação desta teoria para uma forma menos radical: o dualismo da propriedade (Churchland, 2004).
Nesse tipo de dualismo, embora não haja outra substância envolvida além da matéria do encéfalo, afirma-se que o último seria dotado de um conjunto especial de propriedades (no caso, os estados mentais; nome que indica estados como sensação de cor, desejos, apreciações e outras sensações e pensamentos que proviriam de nossa mente) que nenhum outro tipo de objeto físico dispõem; tais propriedades são consideradas não-físicas, no sentido de que jamais podem ser reduzidas ou explicadas em termos dos conceitos das ciências físicas habituais (Churchland, 2004).
Pra se entender melhor, basta pensar que essas propriedades poderiam ser consideradas como emergentes (provenientes do tipo de organização complexa que a matéria física de origem dispõe), assim como a cor é uma propriedade emergente da organização específica dos átomos de uma substância. Essas propriedades são facilmente descritas na natureza, mas para o dualista, os estados mentais não se resumiriam somente a isso, pois elas iriam para além da estrutura física do encéfalo, apresentando-se como uma lei presente em todo o universo e independente da organização da matéria que as abriga. Nesse caso, a experiência consciente deve ser considerada como sendo uma característica fundamental do mundo, do mesmo jeito que massa, carga eletromagnética e espaço-tempo (Leal-Toledo & Teixeira, 2005).
De acordo com Chalmers (1996) - um dos mais conhecidos dualistas da atualidade -, não há conexão lógica entre base física ou arquitetura
funcional e consciência. (...). Segundo Teixeira (1997), o livro de D. J. Chalmers, "The Conscious Mind", é talvez a tentativa mais recente de se formular uma teoria abrangente da natureza da consciência. De acordo com o autor, sua teoria é ousada e corre na direção oposta a tudo o que os cientistas cognitivos e neurocientistas desejam: reduzir estados conscientes a uma base neurofisiológica ou física.
Esta posição é uma variedade de dualismo, na medida em que ela postula propriedades básicas além daquelas estipuladas pela física. Mas trata-se de uma variedade inocente de dualismo, inteiramente compatível com uma visão científica do mundo (Teixeira, 1997).
Enquanto que as teorias dualistas se baseiam em hipóteses que ainda não encontraram fundamento experimental ou matemático, reações baseadas nos avanços da pesquisa em neurofisiologia se desenvolveram, originando diversas teorias monistas.
O behaviorismo filosófico foi a primeira delas e afirma que um estado mental nada mais é do que o conjunto de comportamentos observáveis, desencadeados pela entrada de dados ambientais, também observáveis (Churchland, 2004). Assim, segundo Churchland (2004), o estado mental não mais se caracterizaria como algo interior e extraordinário, mas como um conjunto de operações que o individuo realiza em determinada situação.
Por exemplo, o estado mental "dor", não é visto como algo único e pessoal, mas apenas como o conjunto de reações comportamentais (o afastamento do membro da fonte de danos, o grito "ai" etc) que a ação "danos celulares" desencadearia. Uma das falhas dessa visão é que ela nega o aspecto introspectivo, isto é, único e pessoal, do estado mental, ou seja, o fato de que, ao vermos a cor vermelha, não só realizamos um conjunto de operações padrão (como o recebimento de dados ambientais e sua saída), como temos a sensação de que algo é vermelho e não laranja, um aspecto puramente introspectivo do ato de enxergar (Churchland, 2004). Essa falha levou ao desenvolvimento de outras concepções de filosofia da mente como o materialismo reducionista, o funcionalismo e o materialismo eliminacionista.
O materialismo reducionista ou teoria da identidade afirma de maneira simples que cada estado mental é idêntico a um estado físico (evento, processo) (Boyd, 1980). Seu nome vem do fato de que cada estado mental seria idêntico a um processo fisiológico interior do encéfalo, fato que seria devidamente provado com o avanço das descobertas no campo das neurociências (Churchland, 2004). Ao assumir que a neurociência pode revelar os mecanismos físicos subjacentes às funções psicológicas, assume-se que é de fato o cérebro que realiza essas funções, isto é, que as capacidades da mente

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
18
humana são, na verdade, capacidades do cérebro humano, o que é hipótese altamente provável, baseada em evidências atualmente disponíveis da física, química, neurociência e biologia evolucionária (Churchland, 1994).
A teoria da identidade reconhece que fenômenos mentais possam interagir entre si, sem que para isso, tenham que desenvolver respostas comportamentais, também designadas como "saída de dados" pelos behavioristas (Churchland, 2004). Por exemplo, quando alguém tem uma idéia e abandona a idéia no mesmo instante, sem desencadear deste modo uma ação, estaria se produzindo uma interação entre vários estados mentais, mas, sem o desencadeamento de uma ação ou de uma saída de dados. Assim, essa teoria completa mais uma lacuna deixada pela teoria anterior, a qual negava a existência desses estados mentais.
De acordo com Fodor (1981), a teoria da identidade pode ser dividida em duas doutrinas: o fisicalismo de eventos e o fisicalismo de tipos. Ambos afirmam que existe uma identidade entre os eventos fisiológicos e os processos mentais correspondentes a esses eventos, todavia, o primeiro sustenta que todos os estados mentais até então descobertos são de ordem neurofisiológica, deixando assim, a possibilidade de que existam estados mentais produzidos por qualquer outro meio físico suficientemente complexo e não-neurofisiológico; já o segundo faz a asserção mais arrebatadora de que todos os estados mentais possivelmente existentes são neurofisiológicos, excluindo a possibilidade da existência de outras formas de inteligência não-neurofisiológicas, como, por exemplo, um andróide. Resumindo, o fisicalismo de eventos não elimina a possibilidade lógica de máquinas terem propriedades mentais. O fisicalismo de tipos rejeita essa possibilidade porque as máquinas não possuem neurônios (Fodor, 1981). Para Fodor (1981) o problema com o fisicalismo de tipos é que a constituição psicológica de um sistema parece não depender de seu "hardware", ou seja, de sua composição física, mas sim de seu "software", isto é, de seu programa.
A idéia de que mentes seriam análogas a máquinas computacionais implementadas em diferentes tipos de substrato mostrava-se bastante atraente, não somente porque consistente com, e sugerida pela pesquisa empírica sobre a cognição, mas também por fortalecer a autonomia da pesquisa do mental face à pesquisa do físico (Kim 1998, citado de Abrantes & Amaral, 2002). Entretanto, para o teórico da identidade cada tipo mental é individuado por um tipo físico, engessando-se assim as relações mente-corpo e impossibilitando a múltipla realização (capacidade de estados mentais criarem outros estados mentais), situação que desvela a inépcia desta postura para lidar com o problema mente-corpo (Abrantes &
Amaral, 2002). Para responder a essa intuição da múltipla
realização do mental, vários funcionalistas caracterizaram os estados mentais de um sistema cognitivo em termos da função causal desses estados numa rede causal envolvendo outros estados mentais, “inputs” (entrada de dados) e “outputs” (saída de dados) do sistema (Abrantes & Amaral, 2002). Isto é, os estados mentais poderiam ser a causa de outros estados mentais, afirmando assim, que este estado se resumiria ao seu papel funcional, já que é o fato dele causar outros estados mentais (servindo como “inputs” ao gerar e como “outputs” ao produzir efeitos) que realmente interessa.
Apesar de parecer similar ao behaviorismo filosófico, essa escola não resume os estados mentais a entradas e saídas de dados somente, mas admite que exista uma conexão desse estado a outros estados mentais (o que admitiria a existência de estados mentais que não gerariam, obrigatoriamente, respostas externas comportamentais, como o simples fato de refletir um problema, por exemplo). Assim, a mente passou a se caracterizar como uma rede de diversas relações causais (“inputs” e “outputs”), sem que elas estivessem sempre engessadas a um meio físico.
Aparentemente, o funcionalismo seria também parecido ao materialismo reducionista, afirmando que o estado mental poderia corresponder a qualquer outra estrutura física - seja ela feita de neurônios ou de circuitos -, desde que sua organização seja tão complexa, quanto à organização do encéfalo. Entretanto, diferentemente do primeiro, o funcionalismo admite a possibilidade da criação de estados mentais por outros estados mentais.
O que querem dizer os funcionalistas quando asseveram que uma propriedade não-mental qualquer realiza uma propriedade mental? (...) O fato de ocorrências mentais causarem ocorrências físicas é razão suficiente para concluirmos que tipos mentais façam diferença causal em um mundo físico? (Abrantes & Amaral, 2002).
Mesmo que fenômenos mentais sejam diferentes de fenômenos físicos, os primeiros são necessariamente embasados pelos últimos, no sentido em que, se há alguma mudança no nível do mental, há mudança no nível que o embasa, o físico (Abrantes & Amaral, 2002). Todavia, o funcionalismo pareceu conformar-se a algumas das intuições fundamentais de doutrinas fisicalistas sem, com isso, abrir mão do não-reducionismo do mental ao físico (Abrantes & Amaral, 2002), como o faz o materialismo reducionista.
Outra reação ao materialismo reducionista foi o materialismo eliminacionista. O materialismo eliminacionista afirma que o ultimo é falso, pois uma correspondência exata dos estados mentais

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
19
percebidos pelo senso comum, aos estados físicos do encéfalo é impossível, já que a interpretação que o senso comum faria desses estados é totalmente equivocada e distorcida. Assim, de acordo com a primeira escola, as neurociências, ao invés de reiterar as correspondências de um plano a outro, destruiria, com o tempo, as concepções de senso comum, de nossos estados mentais. A importância do materialismo eliminacionista vem do fato de que seria mais adequado considerar nossas mais ternas intuições sobre a função mente/cérebro como hipóteses revisáveis, ao invés de encará-las como certezas transcendentais absolutas ou certezas introspectivamente dadas. O reconhecimento da possibilidade de uma tal revisão faz uma enorme diferença na maneira pela qual nós conduzimos experimentos psicológicos e neurobiológicos, e em como nós interpretamos seus resultados (Churchland, 1994). Desse modo, o materialismo eliminacionista parece ser mais uma forma de revisão do conceito de teoria da identidade, do que como uma teoria contraria a esta.
Como se pôde perceber, variações sobre um mesmo conjunto de problemas vêm desafiando e vencendo todos aqueles que se debruçaram sobre a análise do fenômeno mental no presente século (Vicentini, 2001), Além das controvérsias e adições postas por novas teorias, a neurofilosofia ainda se vê desafiada por tópicos que perspassam todas as escolas. Pretende-se assim, apresentar adiante os princípais dilemas com os quais ela está lidando no momento.
Desafios futuros É sensível para todo aquele que possui ao
menos um pequeno conhecimento dos preceitos da ciência atual, um aparente descompasso entre a visão resultante das descrições científicas e aquela imagem do mundo que se obtém pela experiência, por meio dos sentidos e que está baseada nas mais íntimas intuições de como o mundo é. Esse mundo de cores, odores, sabores, etc. é reduzido a um amontoado de desbotados elementos básicos e enunciados de leis que não deixa ver como tão vívidas propriedades possam ser deduzidas a partir de uma base tão elementar (Vicentini, 2001). Dentro dessa linha de pensamento, os qualia aparecem como candidatos resistentes às tentativas de lhe outorgarem uma cidadania científica (Vicentini, 2001).
Qualia é o termo empregado para denotar as características intrínsecas de nossas sensações, as quais podemos obter apenas por meio da introspecção (Churchland, 1989, citado por Vicentini, 2001). Os qualia resistem a qualquer tentativa de descrição, pois são acessíveis somente à consciência do experienciador (...) e pode-se dizer que o problema central que direciona essa investigação
é o de apurar se um fenômeno com as características que, aparentemente pertencem aos qualia, pode ser tratado a partir de uma abordagem fisicalista, como parece ser a diretriz dominante nas ciências da mente contemporâneas (...), pois diante de uma paisagem marinha, onde o céu e o mar aparecem como sendo igualmente azuis, como posso afirmar que estou percebendo uma única cor (Vicentini, 2001)? Afinal, uma vez que todos nós aprendemos as palavras que designam as cores, vendo objetos coloridos e públicos, nosso comportamento verbal concordaria mesmo se nós experienciássemos as cores subjetivamente de modos inteiramente diferentes (Dennett, 1991, citado de Vicentini, 2001).
Deste modo, o problema dos qualia se caracteriza como o principal desafio para os neurofilósofos que visam - dentro de cada concepção de filosofia da mente - construir uma abordagem teórica que se adeque a todos os conhecimentos até então adquiridos pelas ciências cognitivas. Deve-se lembrar, entretanto, que, apesar dos desafios e falhas que apresenta cada escola, a abordagem fisicalista está cada vez mais em voga, desviando a atenção outrora dada ao dualismo. São os avanços da pesquisa em neurociências, psicologia e inteligência artificial que são os responsáveis pelo fortalecimento das diversas formas de monismo; aliás, serão possivelmente os avanços futuros da pesquisa nessas áreas que poderão gerar um consenso e mesmo uma concepção de filosofia da mente unificada que possa explicar toda a complexidade da mente humana, isso, claro, se a relação entre os fenômenos físicos do encéfalo aos fenômenos mentais, afirmados pela teoria da identidade e suas complementações, estiver correta.
Dualismo ou monismo? O dualismo, apesar de ter sido deixado de
lado pelas ciências cognitivas experimentais, ainda gera fortes debates em meio à comunidade filosófica e à sociedade em geral. A existência da possibilidade de manutenção da mente após a morte do corpo físico, afirmada pelas escolas dualistas, faz com que essa teoria seja uma das preferidas da comunidade não científica, pois ela responderia às dúvidas existenciais humanas que a ciência e a filosofia não puderam até agora responder. Todavia, em termos científicos, o dualismo não parece apresentar comprovação experimental. Até o momento, o materialismo reducionista parece conseguir responder às atuais dúvidas científicas a respeito da mente, bem como contribuir na construção de um modelo físico desta. Suas complementações, dadas pelo funcionalismo e pelo materialismo eliminacionista, a fortalecem ainda mais, pois abrem respectivamente, o horizonte para a existência de mentes não neurofisiológicas e para a idéia de

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
20
que há uma ilusão ao se correlacionar os estados mentais aos estados percebidos pelo senso comum.
Assim, parece não restar dúvidas quanto à veracidade das afirmações do materialismo reducionista, porém estados mentais mais complexos, como a consciência, ainda carecem de um modelo científico maduro, o que gera uma série de reticências à afirmação de que não existem mais questões quanto à correlação estado físico / estado mental. Por enquanto, as evidências se inclinam para o lado do monismo, porém mesmo que esta escola seja a correta, o cientista cognitivo deve manter a mente aberta para as novas descobertas que poderão mudar a idéia do que seria um estado mental; somente a adoção desse espírito aberto, impedirá que o materialismo reducionista se transforme em um dogma, impedindo no futuro, o avanço rumo a um modelo da mente mais próximo da realidade.
Conclusão As questões neurofilosóficas são tão antigas
quanto o dia em que o primeiro ser humano se fez a pergunta de "quem somos nós". A natureza do pensamento e sua origem perspassou as mentes de grandes filósofos, todavia sem obter consenso. Atualmente, entretanto, os avanços da pesquisa em neurociências são um forte argumento a favor das teorias materialistas, sem que, no entanto, as teorias dualistas sejam abandonadas, pois a pouca idade das neurociências ainda não permite a construção de um modelo científico capaz de explicar completamente estados mentais mais complexos como a consciência.
Se o materialismo se mostrar correto, é fato que antigas questões de ordem filosófica serão respondidas pelas ciências cognitivas, porém isso não quer dizer que a neurofilosofia se tornará obsoleta, já que sempre existirão perguntas para serem respondidas com relação à natureza da mente e, portanto, neurofilósofos para definirem pressupostos por trás das diversas linhas experimentais que irão surgir.
Agradecimentos Agradeço a Arnaldo Cheixas-Dias e Rodrigo
Pavão pelos valiosos comentários Bibliografia
Abrantes, P.; Amaral, F. Funcionalismo e causação mental. In. International Colloquium in Philosophy of Mind, 3, 2002, Campinas, Brasil. Proceedings of the Third International Colloquium in Philosophy of Mind, Campinas, Manuscrito, 2002, 14-45;
Boyd, R. (1980 ). Materialism without reductionism. In. Readings in philosophy of psychology, I, USA, Library of Congress Cataloging in Publication Data;
Chalmers, A. F. (1993) O que é ciência, afinal? São Paulo, SP, Editora Brasiliense;
Chalmers, D. J. (1996) The conscious mind: In search of a fundamental theory. Oxford, Oxford University Press;
Churchland, P. M. (2004) Matéria e Consciência, Uma introdução contemporânea à filosofia da mente. São Paulo, SP, Ed. Unesp;
Churchland, P. S. (1994). Can neurobiology teach us anything about consciousness? Presidential Address to the American Philosophical Association, Pacific Division. Proceedings and Addresses of the American Philosophical, Association. Lancaster, PA: Lancaster Press. 67-4: 23-40 ;
Descartes (1973). Discours de la méthode. Paris, Librairie Générale Française ;
Fodor, J. (1981). The Mind/body Problem. Scientific American, 244, 124-132;
Leal-Toledo, G.; Teixeira, J. F. (2005). O dualismo e o argumento dos zumbis na filosofia da mente do século XX. Cadernos do Centro Universitário São Camilo, 11, 1;
Teixeira, J. F. (1994). O que é filosofia da mente. Coleção Primeiros Passos. São Paulo, SP, Ed. Brasiliense
Teixeira, J. F. (1997). A teoria da consciência de David Chalmers. Psicologia USP, 8, 2;
Vicentini, M. R. (2001). O critério do desempenho: do behaviorismo ao funcionalismo. Acta Scientiarum, Maringá, 23, 1, 223-230;
Wilson R. A., Keil F. (Ed.s) (1999) The MIT Encyclopedia of the Cognitive Sciences. Cambridge, MA, MIT Press

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
21
PROJETO GENOGRÁFICO E AS IMPLICAÇÕES DA POPULARIZAÇÃO DOS ESTUDOS DE GENEALOGIA GÊNICA
Bárbara Domingues Bitarello
Departamento de Genética e Biologia Evolutiva, Instituto de Biociências, USP Recebido 05ago09 / Aceito 24nov09 / Publicação inicial 30nov09
[email protected] Resumo. O Projeto Genográfico busca caracterizar a variabilidade genética da população mundial,
baseando-se majoritariamente em dados genéticos de grupos indígenas isolados e, em menor escala, na participação voluntária de indivíduos interessados em entender suas origens. No entanto, as implicações deste conhecimento merecem ressalva, assim como é importante avaliar o grau de ilusão que está sendo gerado entre os leigos que buscam conhecer sua ancestralidade genética.
Palavras-chave. Genográfico, genealogia gênica, DNA. THE GENOGRAPHIC PROJECT AND THE IMPLICATIONS OF THE POPULARIZATION OF GENE
GENALOGY STUDIES Abstract. The Genographic Project seeks to characterize the genetic variation of the global human
population, based mainly on genetic data from isolated indigenous groups and, on a smaller scale, on the voluntary participation of individuals interested in understanding their origins. However, the implications of this knowledge deserve careful consideration, and it is also important to evaluate the degree of illusion that is being created among laymen who seek knowledge about their genetic ancestry.
Keywords. Genographic, gene genealogy, DNA. Introdução Em 2005, teve início o Projeto Genográfico1
caracterizado por seu pioneirismo em duas frentes: (1) é o primeiro projeto de pesquisa genética-antropológica em escala mundial; e (2) por criar um enorme banco de dados de seqüências mitocondriais e da região não-recombinante do cromossomo Y, sendo que existem protocolos muito bem definidos para extração, análise e armazenamento de tais seqüências. O objetivo central do projeto consiste em tentar mapear, através de marcadores moleculares, as principais ondas migratórias de nossa espécie pelo planeta.
Além de coletar e analisar DNA de grupos étnicos relativamente isolados e bem definidos (por exemplo, índios da Amazônia, esquimós, aborígenes etc), um dos grandes diferenciais do projeto é justamente que qualquer pessoa do mundo pode comprar um kit para extração e enviar o material para análise pela equipe que compõe o projeto.
A descrição do resultado, no site do projeto, dispõe de mapas mostrando as rotas de migração e textos informativos sobre a importância histórica deste ou daquele haplótipo. Por fim, a pessoa pode optar por doar, ou não, sua seqüência (anônima) para o banco de dados do projeto.
É importante ressaltar que o objetivo central do projeto é a análise dos marcadores de DNA provenientes destas populações relativamente isoladas. A participação do público geral, embora encorajada, tende a ser menos informativa do ponto de vista das migrações, devido ao alto grau de miscigenação das populações em geral. 1Genographic Project – www.nationalgeographic.com/ genographic
Técnicas empregadas e implicações Basicamente, as amostras femininas
passam por uma genotipagem padrão do DNA mitocondrial, que inclui o sequenciamento direto da HVR-I2 e a tipagem de 22 sítios bi-alélicos situados em regiões codificantes. Já as amostras masculinas são analisadas em função de alguns STRs3 e SNPs4 específicos da região não-recombinante do cromossomo Y. A sequência como um todo não é avaliada e não se pode, portanto, chamá-lo de ambicioso no que tange a história demográfica humana.
Para ilustrar essa observação, basta comparar o projeto genográfico com estudos em larga escala do genoma (os populares “genome wide association studies”), que dispõem de painéis contendo milhões de SNPs “varrendo” o genoma em busca de blocos haplotípicos relacionados. Ao fazermos isso, não fica claro o que o projeto pode trazer em termos de contribuição inovadora ao atual conhecimento sobre nossa história evolutiva recente.
Por outro lado, se focarmos nas amostras que o projeto promete coletar, pode-se dizer que o ganho que poderá ser proporcionado pelo Projeto Genográfico é o armazenamento de grande quantidade de informações genômicas de alta qualidade e fáceis de serem recuperadas, reflexo do padrão bem definido com que todas as análises são feitas. Ao menos em teoria, e as 2Hipervariable region I, ou região hipervariável do DNA mitocondrial. 3Short tandem repeats, também conhecidos como microssatélites. 4Single nucleotide polymorphisms, ou polimorfismos de um nucleotídeo.
Opinião:

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009 22
recentes publicações têm mostrado muito empenho em reforçar essa preocupação, todas as etapas do projeto, da coleta de DNA às análises, são feitas de forma “penless”5. Além disso, as regiões escolhidas para análise, conforme descrito anteriormente, estão bem definidas e amplamente estudadas, de modo que protocolos foram estabelecidos de maneira a minimizar a perda de amostras. Todo esse cuidado tem gerado um vasto banco de dados, onde todas as análises seguem um mesmo padrão, facilitando estudos em escala global. Resta averiguar se essa abordagem “penless” não estaria, por outro lado, dificultando a detecção de erros sistemáticos relacionados à análise e coleta do material, que poderia ser feita por curação manual6.
Questões éticas As delicadas questões éticas que
concernem os agrupamentos étnicos isolados privaram o Projeto Genográfico de fazer as tão populares análises de miscigenação. Em troca, o projeto logrou coletar amostras de certos grupos étnicos jamais analisados antes.
Em geral, essas questões éticas estão relacionadas ao medo, justificável, de que análises de miscigenação pudessem gerar ou aumentar o grau de discriminação sofrido por certos grupos étnicos, ou mesmo tirar alguns direitos que tais grupos lutaram para conseguir. A título de exemplo, suponha que um grupo de índios em algum estado brasileiro tenha conseguido o direito a uma área que teria pertencido a seus antepassados. Se uma inocente análise de miscigenação revelasse alto grau de mistura com os europeus7, por exemplo, que implicações políticas poderiam decorrer dessa descoberta?
Para Fernando Mathias, advogado do Instituto Socioambiental (ISA), organização de defesa indígena, o fato de o projeto ter objetivos históricos, e sua equipe se comprometer a não fazer uso indevido dos dados, não significa que outros não possam utilizar seus resultados, uma vez publicados. Nesse campo, a linha que define o bom e o mau uso é difícil de ser estabelecida,
5 Literalmente, “sem caneta”. Significa que todo o processo de coleta de amostras e análise é feito de forma bastante automatizada e padronizada, de forma a evitar erros humanos, como, por exemplo, anotações ambíguas, letra ilegível, contaminação, etc. 6 Um pequeno erro em um algoritmo utilizado em uma etapa básica da identificação da qualidade da amostra, por exemplo, poderia gerar o descarte de amostras de boa qualidade, resultando em perda de informação. Por isso é importante aliar a curação manual aos métodos automatizados, em qualquer área da ciência. 7 Considerando as freqüentes ocorrências de uniões entre colonizadores e índias, não seria tão improvável detectar tal fenômeno no cromossomo Y.
segundo o advogado8. Já para muitos cientistas, a simplicidade das análises promovidas pelo Projeto Genográfico contrasta com a qualidade e quantidade de amostras de que dispõe. Muitos lamentam o pouco “uso” que será feito de tais amostras e o ganho de conteúdo que a genética de populações receberia se análises mais abrangentes fossem feitas9.
Motivação do público à adesão ao
projeto Se por um lado muitos grupos temem
repercussões éticas do projeto, existe um número cada vez maior de pessoas que têm buscado descobrir mais sobre sua ascendência genética. O Projeto Genográfico tem dado essa oportunidade. Por 9910 dólares (para mais informações, visite o site do projeto), qualquer pessoa pode adquirir um kit de extração de DNA e enviar o material coletado para ser analisado por uma equipe do projeto, tendo direito a um relatório didático sobre seu haplótipo mitocondrial ou do cromossomo Y.
Numa publicação relativamente recente, um dos grupos de pesquisa vinculados ao projeto (Behar e col., 2007) divulgou alguns resultados preliminares. Eles relataram que mais de 188 mil pessoas já haviam adquirido o kit, das quais 95% eram provenientes da Europa ou dos EUA. Se um dos grandes objetivos do projeto é atingir escala global de participação, uma questão que surge naturalmente é: por que não tentar baratear o custo da participação pública? A resposta é que a participação pública não é um objetivo central do projeto e o lucro com a venda dos kits é destinado a iniciativas de preservação cultural de grupos indígenas e suas respectivas culturas ao redor do mundo. O que o projeto realmente está buscando, declaradamente, é o DNA de grupos indígenas relativamente isolados, que contêm alto grau de informação para os propósitos do projeto.
Implicações da popularização das
genealogias gênicas Mas o que esse grande interesse pelas
genealogias gênicas reflete? Será que as pessoas encontram o que buscam ao ter seu material genético analisado pelo Projeto Genográfico ou por qualquer empresa especializada em genealogias genéticas? Podemos antecipar potenciais erros de
8 Matéria “ISA vê riscos na coleta de DNA indígena”, de 12/07/2006: http://www.comciencia.br/comciencia/ handler.php?section=3¬icia=161 9 Por exemplo, as tais análises de miscigenação, comparações de grandes trechos do genoma ou mesmo, num futuro próximo, de genomas completos, a fim de melhor compreender a diversidade humana. 10 Para um brasileiro, algo em torno de 170 reais, mais os custos de importação. Para fins de comparação, o salário mínimo brasileiro é de 465 reais.

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009 23
interpretação dos resultados do Projeto Genográfico, decorrentes de falsas expectativas acerca do mesmo?
Um bom exemplo disso foi dado por Wolinsky (2006), que relatou o memorável caso do professor Thomas Robinson. Como muitos norte-americanos, Thomas tinha uma profunda curiosidade sobre suas origens e, ao contrário de outros curiosos ao redor do mundo (fora do eixo América do Norte/Europa Ocidental), ele sabia que tinha como recorrer a uma empresa especializada em genealogias gênicas para começar a decifrar suas origens remotas, desde que pagasse bem por isso. Curiosamente, Thomas foi informado pela empresa que lhe prestou tais serviços que possuía sete de nove marcadores atribuídos ao famoso imperador mongol, Genghis Khan (1162-1227). Wolinsky comenta que o alvoroço foi tamanho em torno da notícia, que o embaixador da Mongólia nos EUA prometeu homenagear Thomas Robinson em um evento público.
A euforia se desfez pouco tempo depois, quando outra empresa especializada em genealogias gênicas esclareceu que Thomas e o grande imperador não compartilharam um ancestral comum nos últimos 30 ou 40 mil anos, muito menos nos últimos 800 anos. Mas somente mais tarde ficou clara a fonte de discordância: a segunda empresa havia analisado alguns marcadores do cromossomo Y além daqueles analisados pela primeira empresa, mostrando que a relação entre as duas amostras11 não era tão próxima quanto havia sido alardeado.
Numa busca pela internet, a descoberta de um blog12 que alardeia com entusiasmo a excitante e promissora possibilidade de descoberta de ancestrais via Projeto Genográfico é preocupante. A matéria, embora bastante adequada e baseada em informações obtidas no site oficial do projeto, gerou comentários inusitados. Muitas pessoas forneceram informações relativas a seu potencial parentesco com figuras históricas ilustres (relembrando o caso anteriormente relatado, do parentesco com Genghis Khan), além de relatos de parentes desaparecidos. Esse exemplo mostra que não só o público leigo deve ser preparado para interpretar os resultados, como faz-se necessário esclarecer abertamente que as genealogias gênicas não têm o poder de responder perguntas como essas que foram colocadas no blog.
Enquanto a maior parte das pessoas ainda procura esse tipo de empresa (especializada em
11 Nestes casos, geralmente calcula-se o MRCA (ancestral comum mais recente) entre as duas seqüências que estão sendo comparadas. 12 Matéria “À procura de antepassados”, de 15/09/2009: http://www.meusparentes.com.br/blog/2009/09/15/a-procura-de-antepassados/
análise de DNA e genealogias gênicas) com objetivos mais modestos (testes de paternidade, análises forenses etc.), a busca por suas origens tem se tornado cada vez mais popular, na medida em que as promessas das genealogias gênicas são divulgadas e tornam-se acessíveis a todas as pessoas que possam pagar por uma análise de sua ancestralidade genética. Deve haver, entretanto, por parte daqueles relacionados à divulgação científica, uma cautela redobrada acerca das expectativas dos leigos quanto às promessas da genética. Os estudos genético-populacionais buscam traçar rotas de migração de linhagens grandes de nossa espécie, e não de antepassados recentes de certo indivíduo.
Conclusão Pode-se concluir, portanto, que o grande
diferencial do Projeto Genográfico é a escala global que ele pretende abarcar, coletando e analisando criteriosamente material genético de grupos étnicos relativamente isolados ao redor do mundo. Seu objetivo secundário de encorajar a participação do público por meio da compra de kits poderia ser alavancado se o preço fosse mais acessível. Por fim, a divulgação científica depara-se com um importante desafio relativo às genealogias gênicas em geral: melhorar o entendimento da população acerca de que tipo de informação elas podem adquirir com sua participação no Projeto Genográfico e, assim, evitar interpretações fantasiosas e exageradas dos resultados fornecidos pelas análises genéticas.
Agradecimentos Agradeço a Revista da Biologia, Daniel
Lahr e Ricardo Mendonça pelas sugestões. Agradeço também o professor Dr. Diogo Meyer.
Bibliografia
Behar, D.M., Rosset, S., Blue-Smith, J., Balanovsky, O., Tzur, S., Comas, D., Mitchell, R.J., Quintana-Murci, L.,Tyler-Smith, C., Spencer Wells, R.S., The Genographic Consortium (2007). The Genographic Project Public Participation Mitochondrial DNA Database. PLoS Genet 3(6), e104.
Blog Meus Parentes: http://www.meusparentes.com.br/blog/2 Com Ciência – Revista Eletrônica de Jornalismo Científico:
www.comciencia.com.br National Geographic Project: www.nationalgeographic.com/
genographic Wolinsky, H. (2006). Genetic Genealogy Goes Global. EMBO
Reports 7(11), 1072-1074.

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009.
24
TÉCNICAS PARA O PROCESSAMENTO DE FÓSSEIS DE BIVALVES: UM ESTUDO DE CASO SOBRE A FORMAÇÃO CORUMBATAÍ, BACIA DO
PARANÁ, BRASIL
Rodrigo B. Salvador1, Luiz R. L. Simone2 Instituto de Biociências, USP
Museu de Zoologia Recebido 12set09 / Aceito 25nov09 / Publicação inicial 14dez09
[email protected]; [email protected] Resumo. A Formação Corumbataí faz parte do Grupo Passa Dois na Bacia do Paraná e data do final
da Era Paleozóica, mais precisamente do Período Permiano (Superior). Seu registro fóssil conta principalmente com moluscos bivalves silicificados e imersos em matrizes de arenito, os quais são de difícil processamento e preparo. Este trabalho descreve algumas das técnicas utilizadas no processamento desses fósseis, testando a eficácia de cada uma no tratamento dos bivalves fósseis da Formação Corumbataí. Tratamentos mecânicos e químicos foram utilizados e os resultados indicam que os primeiros são mais eficientes do que os últimos.
Palavras-chave. Arenito, fósseis silicificados, Grupo Passa Dois, moluscos, Permiano Superior. TECHNIQUES FOR PROCESSING BIVALVE MOLLUSK FOSSILS: A CASE-STUDY ON THE
CORUMBATAÍ FORMATION, PARANÁ BASIN, BRAZIL Abstract. The Corumbataí Formation belongs to the Passa Dois Group in Paraná Basin, Brazil, and
dates from the end of the Paleozoic Era, more precisely from the (Upper) Permian Period. The Formation’s fossil record is composed mainly of silicified bivalve mollusks embedded in sandstone, which are usually difficult to process and prepare. The present article describes some of the techniques commonly used in fossil processing, and tests their efficacy on fossil bivalves from the Corumbataí Formation. Mechanical and chemical treatments were performed, and the results show that the former are considerably more efficient than the latter.
Keywords. Mollusks, Passa Dois Group, sandstone, silicified fossils, Upper Permian. Introdução A Formação Corumbataí é a camada
superior do Grupo Passa Dois na Bacia do Paraná e data do final da Era Paleozóica, mais precisamente do Período Permiano (Superior). A formação se estende desde a fronteira entre os Estados do Paraná e de São Paulo até o Estado de Minas Gerais e apresenta argilitos, siltitos e folhelhos em sua seção inferior, e argilitos e arenitos finos em sua seção superior (Mezzalira, 1981). Seu registro fóssil, que conta principalmente com moluscos bivalves, vem sendo estudado desde o início do século XX. Entre os bivalves da formação, que totalizam 33 espécies distribuídas em 20 gêneros e 7 famílias, há representantes das seguintes subclasses: Anomalodesmata, que apresenta a maior diversidade de espécies; Heterodonta, que contém as duas espécies mais abundantes da Formação (Pinzonella illusa e P. neotropica); Pteriomorphia; e Palaeoheterodonta (Runnegar e Newell, 1971; Simone e Mezzalira, 1994). Acredita-se que o paleoambiente da Bacia durante o final do Permiano tenha sido um mar epicontinental salobro, de salinidade variável, clima quente, intensa evaporação e, portanto, de alto estresse ambiental (Rohn, 1994; Ghilardi e Simões, 2002).
Os fósseis da Formação Corumbataí encontram-se silicificados, isto é, o carbonato de cálcio original das conchas foi substituído por sílica durante o processo de fossilização, e
imersos em rochas arenáceas. Podem ocorrer tanto esparsos na matriz como na forma de aglomerados muito compactados (coquinas). Para que os fósseis possam ser integrados em coleções (tanto particulares quanto de museus) ou exibidos em exposições, devem ser previamente processados de modo adequado (Lichter, 1993). O tipo de material presente na Formação Corumbataí (fósseis silicificados, em arenito) é um exemplo de ocorrência comum, sendo, entretanto, de difícil processamento (Campbell, 2006).
Objetivos Tendo em vista as dificuldades do
processamento e preparo de fósseis silicificados imersos em matrizes de arenito, este trabalho teve como objetivo testar diferentes técnicas a fim de definir qual a metodologia mais eficaz para o tratamento desse tipo de fóssil. Para tal, foram utilizados os bivalves fósseis da Formação Corumbataí como modelo.
Material Utilizado Neste trabalho foram utilizadas rochas
fossilíferas provenientes da Formação Corumbataí, das regiões de Rio Claro e Tambaú, Estado de São Paulo (Figura 1). Os fósseis de bivalves podem ser encontrados silicificados, em bom estado de preservação, ou ainda na forma de moldes, em estado de preservação variável de ruim a bom.
Técnica:

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009.
25
Figura 1 – Mapa do Estado de São Paulo, Brasil, mostrando a localização dos municípios de procedência das rochas: Rio Claro e Tambaú. (modificado de http://commons.wikimedia.org).
Há dois tipos principais de rochas
sedimentares na Formação em Rio Claro: a mais comum é uma rocha arenácea com fósseis esparsos (Figura 2), sendo que a matriz (a parte da rocha sem fósseis) não é tão compacta na maioria dos casos, o que facilita o trabalho de remoção dos fósseis, mas pode apresentar-se bastante dura em alguns casos; o outro tipo é uma coquina muito compactada (Figura 3), com pouca matriz, sendo que as conchas encontram-se muito agregadas, o que torna a remoção dos fósseis difícil, por vezes impossibilitando o trabalho e freqüentemente resultando na quebra dos espécimes. O material de Tambaú é uma coquina semelhante à de Rio Claro, porém de matriz ainda mais dura.
Figura 2 – Rocha arenácea, apresentando fósseis esparsos e grande quantidade de matriz.
Preparação do Material Espécimes fósseis podem ser processados
e preparados, para a incorporação a coleções e futuros estudos, de diversas maneiras, as quais podem ser divididas em duas grandes categorias: tratamentos mecânicos e tratamentos químicos. Ambos são eficientes em seu respectivo contexto e são aplicados de modos distintos dependendo do material a ser processado. Em geral, prefere-se que a técnica seja o menos destrutiva possível, para evitar danos desnecessários aos fósseis (Nobre e Carvalho, 2004). Neste trabalho,
utilizamos técnicas de ambos os tipos de tratamentos a fim de testar sua eficácia no processamento do material da Formação Corumbataí.
Figura 3 – Coquina: rocha muito compactada, apresentando muitos fósseis aglomerados.
Antes de processar os espécimes, foi
necessário removê-los das rochas em que se encontravam. Devido ao seu grande tamanho, fez-se necessário o uso de uma morsa ou de martelo e talhadeira para quebrar as rochas em fragmentos menores, passíveis de serem processados sob estereomicroscópio. Esse procedimento é grosseiro e pode ocasionar a obliteração de alguns espécimes no momento em que a rocha se parte. No caso da Formação Corumbataí, que é relativamente abundante em fósseis, a perda de alguns exemplares não chega a ser um problema.
Tratamentos Mecânicos O tratamento mecânico básico consistiu em
limpeza utilizando-se água corrente e ferramentas para a abrasão da matriz e conseguinte remoção do sedimento próximo ao fóssil. As ferramentas utilizadas foram instrumentais de dentística (como calçador, esculpidor, brocas etc.), pincéis de diferentes tamanhos, estiletes, agulhas reforçadas, pequenas lâminas, entre outros, que variaram em seu uso dependendo do tamanho e fragilidade da amostra. O processo, sempre que possível ou necessário, foi feito sob lupa estereoscópica, de modo a evidenciar com maior nitidez a fronteira entre o fóssil e a matriz.
Os espécimes foram completamente removidos da matriz (Figura 4) ou, quando isso não foi possível, foram deixados parcialmente imersos, revelando o máximo possível do fóssil. Alguns fósseis foram propositalmente deixados com uma quantidade generosa de matriz, devido à sua importância em estudos paleontológicos mais completos (Foote e Miller, 2007). Pela sua natureza frágil, alguns fósseis fragmentaram-se durante o processamento e, sempre que tal problema ocorreu, utilizou-se cola adesiva à base

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009.
26
de éster de cianoacrilato para repará-los. Uma técnica comum, e empregada no presente trabalho, é misturar-se um pouco de pó proveniente da matriz rochosa à cola, para preencher melhor algumas lacunas deixadas pela quebra (Lichter, 1993; Nobre e Carvalho, 2004).
Figura 4 – Alguns espécimes que passaram pelo tratamento mecânico básico, prontos para serem expostos ou adicionados à uma coleção. a) Plesiocyprinella carinata, proveniente da rocha arenácea de Rio Claro. b) Pinzonella neotropica, proveniente da coquina de Rio Claro. c) Pyramus anceps, proveniente da coquina de Tambaú.
Além desse tratamento básico, utilizamos também um banho de ultrassom (Quinn, 1994; Magalhães Ribeiro, 1997), técnica útil para amolecer, desagregar parcialmente ou até mesmo remover o sedimento ao redor do fóssil. Tal tratamento é comumente utilizado na paleontologia de vertebrados, especialmente para fragmentos de cascas de ovos de dinossauros, e foram utilizados no presente trabalho esperando-se resultados similares, pois as conchas de bivalves e as cascas dos ovos são semelhantes em seus formato e processo de fossilização.
Foram utilizados dois tipos de banho de ultrassom: em um deles a amostra foi imersa em água destilada e, no outro, em peróxido de hidrogênio a 100 volumes (H2O2 100 vol.). Em ambos os casos, as amostras foram deixadas no banho por diferentes intervalos de tempo, a fim de se descobrir qual a duração mais adequada para o tratamento. Os tempos utilizados foram de 1, 5, 10, 15 e 20 minutos. Os espécimes utilizados nesse tratamento apresentavam pouca quantidade de matriz envolvente: a maior parte da matriz foi removida previamente pelo tratamento básico descrito anteriormente. Os dois tipos de sedimentos da Formação Corumbataí, o sedimento arenáceo e a coquina (tanto a de Rio Claro quanto a de Tambaú), foram utilizados, sendo 10 amostras de cada, para se definir a eficácia do tratamento para cada tipo de sedimento. O aparelho utilizado foi o Ultrassonic Cleaner (Odontobrás Ltda.), modelo T-1025, freqüência do ultrassom 25 KHz, potência ultrassônica 50 W. Após o banho, os espécimes passaram novamente pelo tratamento básico, a fim de determinar o efeito do ultrassom, ou seja, para verificar se a matriz podia ser removida com maior facilidade ou não.
Tratamentos Químicos Também se testou a eficácia do tratamento
químico descrito por Simões (1988) especificamente para os bivalves fósseis da Formação Corumbataí. Tal procedimento consistiu em três banhos de diferentes reagentes químicos. Primeiramente, os espécimes, ainda preenchidos por uma pequena quantidade de sedimento, foram imersos em banho de H2O2 100 vol. e em seguida transferidos para banho de ácido acético (CH3COOH) a 100%. Cada um desses banhos teve duração de 72 horas e, após passar por ambos, os sedimentos mais grosseiros de cada espécime foram removidos mecanicamente sob estereomicroscópio (pelo tratamento mecânico básico, descrito acima). Subseqüentemente, os espécimes foram imersos em banho de solução de ácido clorídrico (HCl) a 10% por 24 horas. Finalmente, as amostras foram lavadas em água corrente e o restante do sedimento removido mecanicamente.
Em um primeiro momento seguiu-se à risca esse protocolo, mas outros testes foram

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009.
27
efetuados, aumentando-se a duração dos dois primeiros banhos para 96 horas. Novamente, ambos os tipos de sedimentos da Formação (o sedimento arenáceo e a coquina, de Rio Claro e de Tambaú) foram utilizados em cada caso, sendo 5 amostras de cada.
Resultados Ao todo, foram obtidos 488 espécimes de
bivalves, que foram adicionados à coleção de Malacologia do MZUSP: lotes MZSP-90860 a MZSP-90911.
O tratamento mecânico básico mostrou-se eficiente para a rocha arenácea na maioria dos casos, sendo que a matriz era removida lentamente, mas com segurança e sem acarretar muitos danos aos fósseis (Figura 4). Apenas alguns sedimentos mais compactados não puderam ser satisfatoriamente removidos com as ferramentas utilizadas. As conchas das coquinas, entretanto, geralmente se fragmentavam quando se tentava removê-las. Foi preciso, portanto, escolher quais conchas pretendia-se remover e focar os esforços nas mesmas, em detrimento das conchas ao redor, que eram quebradas.
Os banhos de ultrassom, com água destilada e com H2O2, mostraram-se totalmente ineficazes para as coquinas, não importando o tempo do tratamento. O sedimento não amoleceu e a remoção da matriz não foi facilitada. Já para o sedimento arenáceo, os banhos com água e H2O2 parecem ser igualmente eficientes, ajudando a desagregar a matriz, mas mesmo assim somente nos sedimentos menos compactados (os mais compactados não foram afetados, de modo semelhante ao que aconteceu com as coquinas). Com 1 minuto de banho não houve resultados satisfatórios, mas com 5 minutos a matriz podia ser removida mais facilmente, ao menos na camada superficial da amostra. A partir de 10 minutos a remoção é bastante facilitada: toda a matriz pôde ser removida em todos os espécimes. Entretanto, esse tratamento fragiliza os fósseis, fazendo com que se quebrem mais facilmente.
O tratamento químico não forneceu resultados satisfatórios em nenhum dos casos, não desagregando a matriz rochosa nem facilitando sua remoção. Além disso, tal tratamento (especialmente o último banho, de HCl) sempre afetou as esculturas da concha, apagando-as parcial ou totalmente.
Discussão e Conclusão De um modo geral, a remoção do
sedimento com ferramentas mostrou-se a maneira mais eficiente e segura para o processamento dos bivalves fósseis da Formação Corumbataí, apesar de ser um processo lento e delicado.
O banho do ultrassom é eficiente para ajudar na remoção dos sedimentos pouco
compactados das rochas arenáceas. Tal resultado reflete a eficácia desse tratamento, uma vez que é comumente utilizado no processamento de cascas de ovos fósseis de dinossauros (Quinn, 1994; Magalhães Ribeiro, 1997). Entretanto, esse procedimento fragiliza o fóssil, tornando-o mais quebradiço, pois as conchas de um modo geral tendem a ser mais frágeis que as cascas de ovos, em especial aquelas de tamanho diminuto como muitos espécimes da Formação Corumbataí. Portanto, o banho de ultrassom é indicado apenas para as conchas maiores e mais resistentes, mas pode ainda ser utilizado para conchas menores quando o estudo de suas estruturas internas ou charneira for necessário, uma vez que há maior dificuldade para a remoção da matriz dessas regiões pelo tratamento mecânico básico.
O tratamento químico utilizado no presente estudo é completamente desaconselhado, uma vez que, além de não surtir o efeito desejado, tende a apagar as esculturas da concha, características muitas vezes importantes para a identificação dos fósseis de moluscos e estudos em sistemática. Tal resultado foi inesperado, tendo em vista que esse tratamento foi aconselhado por Simões (1988) especificamente para os bivalves da Formação Corumbataí.
Desse modo, para o processamento de fósseis de bivalves silicificados em arenitos, inclusive em coquinas, e demais moluscos nas mesmas condições, aconselha-se sempre o uso de tratamentos mecânicos, que, mesmo sendo mais demorados e trabalhosos, apresentam os melhores resultados, além de danificarem menos os espécimes.
Agradecimentos. Ao Museu de Zoologia
da USP pela oportunidade de realizarmos este trabalho e pelos recursos e instalações para tal. A Renato P. Ghilardi (Universidade Estadual Paulista Júlio de Mesquita Filho – UNESP de Bauru) por possibilitar a coleta em Rio Claro e por sua grande disposição em ajudar. A Lucas E. Fiorelli (Centro Regional de Investigaciones Científicas y Transferencia Tecnológica de La Rioja, Argentina – CRILAR) e a Luiz E. Anelli (Instituto de Geociências da USP) por todos os conselhos; a Rodrigo C. Marques e Ana Paula S. Dornellas (Museu de Zoologia da USP) pela ajuda na coleta; à Barbara M. Tomotani (Instituto de Biociências da USP) pela leitura crítica do artigo e ajuda com as imagens; aos consultores da Revista da Biologia pela revisão do artigo e valiosos comentários.
Contribuição dos autores. Concepção e
delineamento do projeto: R. B. Salvador e L. R. L. Simone. Execução: R. B. Salvador. Análise dos resultados: R. B. Salvador. Redação do artigo: R. B. Salvador e L. R. L. Simone.

REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009.
28
Referências Bibliográficas Campbell, D. (2006). Fossil mollusks. p. 197-205. In: Sturm,
C.F.; Parce, T.A.; Valdés, A. (Eds.). (2006). The Mollusks: A guide to their study, collection and preservation. American Malacological Society, Universal Publishers, Boca Raton, USA.
Foote, M. e Miller, A. I. (2007). Principles of Paleontology. 3rd ed. W. H. Freeman and Company, New York. USA.
Ghilardi, R.P. e Simões, M.G. (2002). Foram os bivalves do Grupo Passa Dois (exclusive Formação Rio do Rasto), Neopermiano, invertebrados tipicamente dulcícolas? Pesquisas em Geociências 29(1), 3-13.
Lichter, G. (1993). Fossil Collector’s Handbook: finding, identifying, preparing, displaying. Sterling Pub. Comp., Inc. New York, USA.
Magalhães Ribeiro, C.M. (1997). Descrição de caracteres morfológicos e estudo composicional de cascas de ovos de dinossauros da Formação Allen (Cretáceo Superior), do Bajo de Santa Rosa, província de Rio Negro (Argentina). Tese de Mestrado, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brasil.
Mezzalira, S. (1981). Léxico Estratigráfico do Estado de São Paulo. Governo do Estado de São Paulo, Secretaria de Agricultura e Abastecimento, Coordenadoria da Pesquisa de Recursos Naturais, Instituto Geológico.
Nobre, P.H. e Carvalho, I.S. (2004). Fósseis: Coleta e Métodos de Estudo. p. 27-42. In: Carvalho, I.S. (Ed.). (2004). Paleontologia vol. 2 (2ª Ed.). Editora Interciência, Rio de Janeiro, Brasil.
Quinn, B. 1994. Fossilized eggshell preparation. p. 146-153. In: Leiggi, P. & May, P. (Eds.) Vertebrate Paleontological Techniques. Cambridge University Press, New York, USA.
Rohn, R. (1994). Evolução ambiental da Bacia do Paraná durante o Neopermiano no leste de Santa Catarina e do Paraná. Tese de Doutoramento, Instituto de Geociências, Universidade de São Paulo, São Paulo, Brasil.
Runnegar B. e Newell, ND. (1971). Caspian-like relict molluscan fauna in South American Permian. Bulletin of the American Museum of Natural History 146, 1-66.
Simões, M.G. (1988). Técnica de preparação de bivalves silicificados, com vistas ao estudo da morfologia interna. Resumos do XV Congresso Brasileiro de Zoologia, Curitiba. São Paulo: Sociedade Brasileira de Zoologia. p. 49.
Simone, L.R.L. e Mezzalira, S. (1994). Fossil Molluscs of Brazil. Governo do Estado de São Paulo, Secretaria do Meio Ambiente, Coordenadoria de Informações Técnicas, Documentação e Pesquisa Ambiental, Instituto Geológico.















![PDF [5149 KB]](https://static.fdocumentos.com/doc/165x107/5874b9e31a28abc52a8b8543/pdf-5149-kb.jpg)


