
Eletrofisiologia 5A. C. Cassola1 Excitabilidade Elétrica das Células.

UNIVERSIDADE FEDERAL DE GOIÁS
ESCOLA DE VETERINÁRIA E ZOOTECNIA PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA ANIMAL
Disciplina: SEMINÁRIOS APLICADOS
TÉCNICA DE ELETROFISIOLOGIA EM CARRAPATOS
Lorena Lopes Ferreira
Orientadora: Profª Drª Lígia Miranda Ferreira Borges
GOIÂNIA 2011

ii
LORENA LOPES FERREIRA
TÉCNICA DE ELETROFISIOLOGIA EM CARRAPATOS
Seminário apresentado junto à
Disciplina Seminários Aplicados do Programa de Pós-Graduação em
Ciência Animal da Escola de Veterinária e Zootecnia da Universidade Federal de Goiás.
Nível: Mestrado
Área de concentração: Sanidade Animal, Higiene e Tecnologia de Alimentos
Linha de Pesquisa: Parasitos e doenças parasitárias dos animais
Orientadora: Profª Drª Lígia Miranda Ferreira Borges – IPTSP/UFG Comitê de orientação: Profª Drª Andréa Caetano da Silva – IPTSP/UFG Drª Carla Cristina Braz Louly – PNPD/EVZ/UFG
GOIÂNIA 2011

iii
SUMÁRIO
1 INTRODUÇÃO .................................................................................................... 1
2 REVISÃO DA LITERATURA ............................................................................... 2
2.1 Sistema nervoso, sensilas e órgãos sensoriais dos carrapatos ....................... 2
2.1.1 Sistema nervoso ............................................................................................ 2
2.1.2 Sensilas e órgãos sensoriais ......................................................................... 3
2.2 Eletrofisiologia .................................................................................................. 8
2.2.1 Eletroantenograma (EAG) x Registro de sensila única................................11
2.2.2 Percepção do estímulo e disparo do potencial de ação .............................. 11
2.2.3 Eletrofisiologia gustativa e olfativa em carrapatos ....................................... 13
2.2.4 Resultados da técnica de RSU .................................................................... 18
3 CONSIDERAÇÕES FINAIS .............................................................................. 20
REFERÊNCIAS .................................................................................................... 21

1 INTRODUÇÃO
Assim como os demais animais, os carrapatos utilizam seus órgãos
sensoriais para a busca de hospedeiros, exploração do ambiente e encontro de
parceiros. Os primeiros estímulos emitidos pelo hospedeiro são sentidos por
receptores (sensilas) mecânicos, térmicos e olfativos localizados principalmente
no primeiro par de patas do carrapato. Para selecionar o hospedeiro e o sítio de
fixação, o odor, a temperatura e os estímulos gustativos são fatores decisivos, e
os receptores que captam estes sentidos estão localizados no tarso e peças
bucais, especialmente nos palpos e quelíceras.
A eletrofisiologia consiste no estudo das propriedades elétricas em
células e tecidos que envolvem o disparo de potenciais de ação quando
estimulados. Nos artrópodes a eletrofisiologia tem uma grande contribuição na
detecção de feromônios e demais substâncias capazes de atuar no
comportamento dos mesmos. Para o registro de sensilas dos carrapatos adota-se
a eletrofisiologia em sensila única (ESU), no qual as sensilas presentes nos
órgãos sensoriais são expostas a estímulos voláteis ou a compostos gustativos.
Os carrapatos causam inúmeros prejuízos aos seus hospedeiros,
desde a espoliação sanguínea e irritabilidade até transmissão de doenças. A
busca por métodos alternativos de controle é constante e desejável, já que o uso
indiscriminado de acaricidas ocasionou o surgimento de cepas cada vez mais
resistentes as bases químicas disponíveis no mercado. A técnica de
eletrofisiologia pode permitir o encontro de substâncias que participam da
ecologia química dos carrapatos. Tais conhecimentos poderiam embasar novas
alternativas viáveis para o controle destes.
O objetivo deste seminário é comentar sobre as quimiossensilas
presentes nos órgãos sensoriais dos carrapatos e descrever os aspectos da
técnica de eletrofisiologia olfativa e gustativa em carrapatos.

2
2 REVISÃO DA LITERATURA
2.1 Sistema nervoso, sensilas e órgãos sensoriais dos carrapatos
Os carrapatos, assim como os insetos, se comunicam através de
estímulos químicos e físicos. Compostos que carregam informações químicas
são conhecidos como semioquímicos. Em carrapatos os principais semioquímicos
que participam da sua ecologia são os feromônios (reunião; sexual; agregação;
atração/agregação e fixação) e os cairomônios. Os feromônios são substâncias
que servem para guiar o comportamento de organismos facilitando atividades
vitais e o desenvolvimento fisiológico, secretadas por um indivíduo para o exterior
e recebido por um segundo indivíduo da mesma espécie. Os cairomônios são
substâncias produzidas pelos hospedeiros e percebidas pelos carrapatos para
sua localização (VILELA & DELLA LUCIA, 2001; SONENSHINE, 2006). Sendo
assim, os ectoparasitas desenvolveram quimiossensores no corpo e adaptações
comportamentais essenciais para a busca de seu hospedeiro, alimentação e
encontro de parceiros para a cópula (GUERIN et al., 2000).
2.1.1 Sistema nervoso
O sistema nervoso é o aparelho que regula as relações entre os
organismos e o ambiente e que coordena as ações dos demais órgãos (IVANOV,
1983). Assim como nos demais aracnídeos, os gânglios que compõem o sistema
nervoso central (SNC) do carrapato estão condensados em uma massa
denominada singânglio (BINNINGTON & OBENCHAIN, 1982). Do singânglio
partem nervos que inervam órgãos alvos como, por exemplo, as quelíceras,
palpos e glândulas salivares (SONENSHINE, 1991).

3
2.1.2 Sensilas
Carrapatos possuem uma variedade de órgãos sensoriais (Figura 1)
que monitoram o ambiente externo e interno. Os órgãos possuem receptores
celulares, mais conhecidos como sensilas, que estão distribuídos por todo corpo e
apêndices e são classificados de acordo com suas funções (quimiossensila,
mecanossensila, fotossensila e termossensila), sendo que na maioria das vezes
são multifuncionais. Das várias sensilas existentes que detectam uma diversidade
de sinais químicos vindos do ambiente, as mais importantes são as sensilas do
órgão de Haller, as sensilas palpais e as sensilas do dígito da quelícera.
(SONENSHINE, 1991).
FIGURA 1: Localização do órgão de Haller, palpos e quelíceras em um ixodídeo.
Fonte: http://www.nhm.ac.uk/nature-online/species-of-the-day/scientific-
advances/disease/ixodes-ricinus/index.html
As células receptoras correspondem a neurônios bipolares, com dois
processos que se estendem a partir do corpo celular: axônio (processo central)
que se estende para as regiões correspondentes do SNC e o dendrito (processo
periférico) que se estende para a superfície do corpo. Sendo que dendrito possui
um cílio que constitui o aparato perceptivo propriamente dito da célula (SLIFER,
1970; MCIVER, 1975).

4
As quimiossensilas estão envolvidas com a percepção do cheiro
(olfação) e gosto (gustação), são determinantes nos comportamentos de busca
pelo hospedeiro e de encontro para a cópula, uma vez que permitem que o
carrapato identifique substâncias que sinalizem a presença do hospedeiro ou do
parceiro sexual. Os estímulos olfativos são mais específicos, enquanto a gustação
é de fundamental importância no comportamento de exploração do hospedeiro,
alimentação e fixação, ou seja, nas etapas que precedem o ingurgitamento e
também na identificação de substâncias tóxicas que poderão interferir no
desenvolvimento do carrapato (WALADDE, 1982).
O sistema gustativo atua com a finalidade de dar o aval para a aceitação
ou rejeição do alimento (SCOTT, 2005). O suor e sebo dos hospedeiros
vertebrados afetam o comportamento de fixação do ectoparasita, e os receptores
que identificam estes produtos situam nas patas e peça bucal (GUERIN et al.,
2000). Cada espécie animal possui uma composição individual dos fluidos
corpóreos. A partir do momento que os carrapatos introduzem as peças bucais no
tegumento de seus hospedeiros, estes quimiorreceptores conseguem identificar
se a composição dos fluidos corresponde à de seu hospedeiro adequado. Deste
modo, as sensilas das quelíceras estão envolvidas especialmente nas fases de
fixação e ingurgitamento do carrapato (WALADDE & RICE, 1982; SONENSHINE,
1991).
As sensilas dos carrapatos seguem o modelo geral dos artrópodes e
podem ser divididas em quatro categorias de acordo com suas características
estruturais: sensila multiporosa com parede simples (SM-PS) e sensila
multiporosa com parede dupla (SM-PD), sensila com poro terminal (SPT) e
sensila sem poros (SSP) (HESS & VLIMANT, 1982, 1986).
As SM-PS apresentam cutícula delgada perfurada por inúmeros poros,
são olfativas mono ou multifuncionais com funções adicionais de termo e
higrossensibilidade. As SM-PD possuem cutícula dupla, função olfativa e podem
ser mono ou multifuncionais. As SPT possuem um único poro terminal ou
subterminal, são receptores mecanogustativos e podem ser sensíveis a
temperatura e água. As SSP não possuem poros ou fendas, são sensilas mono
ou multifuncionais, com possíveis funções mecano, termo e higrossensibilidade
(HESS & VLIMANT, 1982, 1986).

5
A principal diferença entre sensilas gustativas e olfativas é quanto a
estrutura e o modo de percepção. As sensilas gustativas usualmente possuem um
único poro em sua ponta, respondem a soluções e requerem grandes
concentrações para detectar a substância. Já as sensilas olfativas possuem
vários poros que facilitam a entrada dos odores e respondem a concentrações
pequenas, diferentemente das sensilas gustativas (WALADDE & RICE, 1982;
SONENSHINE 1991).
2.1.2 Órgãos sensoriais dos carrapatos
O órgão de Haller (Figura 2) está localizado no dorso do tarso no
primeiro par de patas e é englobado por uma cápsula. As principais regiões
sensoriais deste órgão são a cápsula posterior que é exposta através de uma
fenda e uma depressão distal. Possui um pequeno grupo de sensilas olfativas
(Figura 3), cujo número varia entre espécies. A função principal desse órgão
complexo é a olfação, apresenta também mecanossensilas, termossensilas e
sensilas gustativas (SONENSHINE, 1991).
FIGURA 2: órgão de Haller de Ixodes ricinus. Fonte:
http://www.flickr.com/photos/28088928@N07/2668482530/
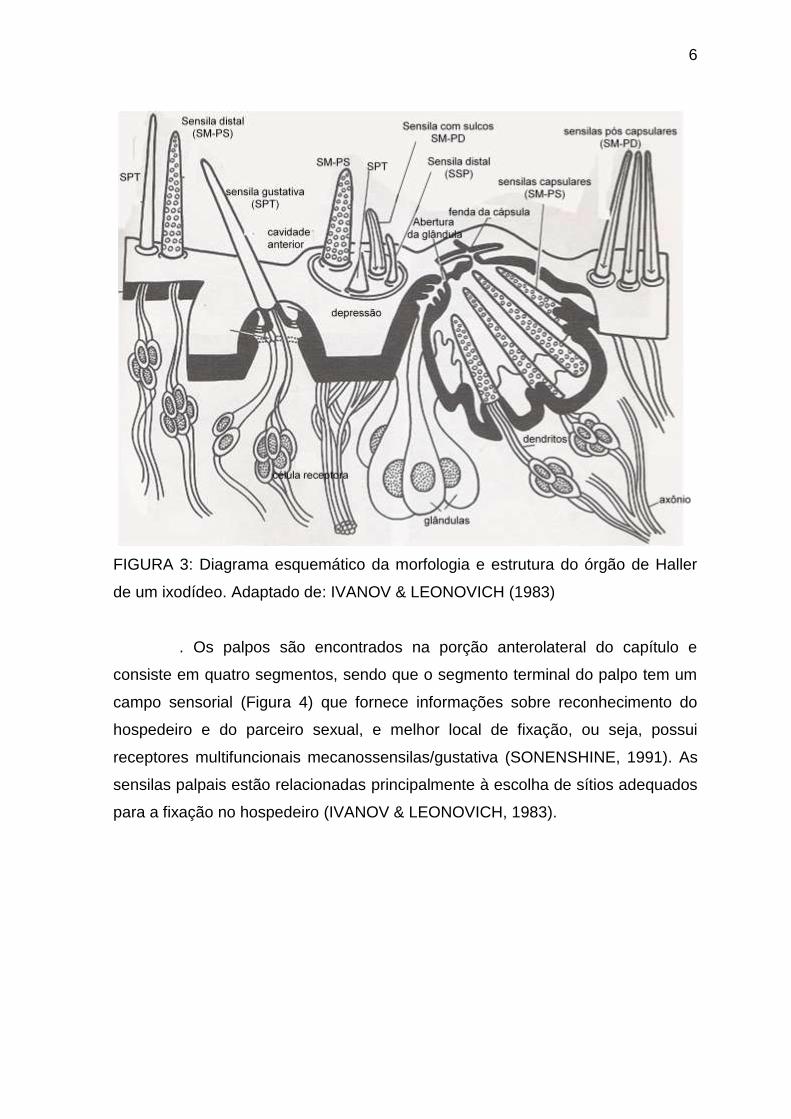
6
FIGURA 3: Diagrama esquemático da morfologia e estrutura do órgão de Haller
de um ixodídeo. Adaptado de: IVANOV & LEONOVICH (1983)
. Os palpos são encontrados na porção anterolateral do capítulo e
consiste em quatro segmentos, sendo que o segmento terminal do palpo tem um
campo sensorial (Figura 4) que fornece informações sobre reconhecimento do
hospedeiro e do parceiro sexual, e melhor local de fixação, ou seja, possui
receptores multifuncionais mecanossensilas/gustativa (SONENSHINE, 1991). As
sensilas palpais estão relacionadas principalmente à escolha de sítios adequados
para a fixação no hospedeiro (IVANOV & LEONOVICH, 1983).
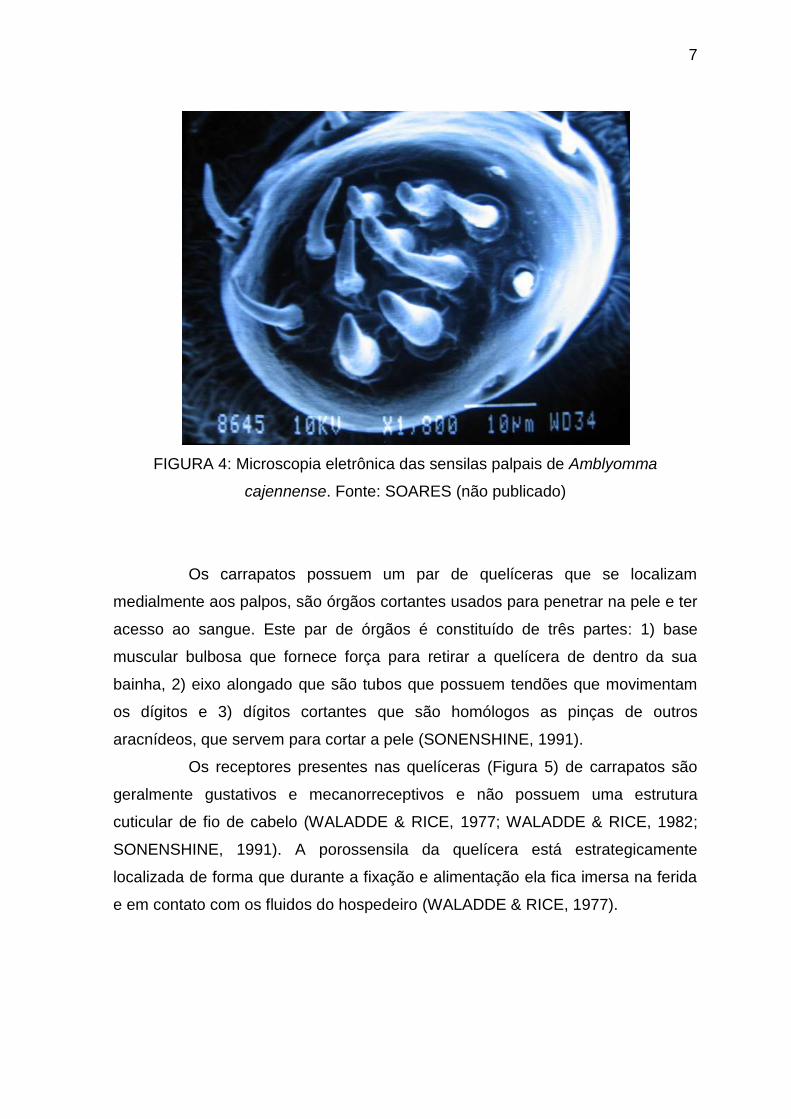
7
FIGURA 4: Microscopia eletrônica das sensilas palpais de Amblyomma
cajennense. Fonte: SOARES (não publicado)
Os carrapatos possuem um par de quelíceras que se localizam
medialmente aos palpos, são órgãos cortantes usados para penetrar na pele e ter
acesso ao sangue. Este par de órgãos é constituído de três partes: 1) base
muscular bulbosa que fornece força para retirar a quelícera de dentro da sua
bainha, 2) eixo alongado que são tubos que possuem tendões que movimentam
os dígitos e 3) dígitos cortantes que são homólogos as pinças de outros
aracnídeos, que servem para cortar a pele (SONENSHINE, 1991).
Os receptores presentes nas quelíceras (Figura 5) de carrapatos são
geralmente gustativos e mecanorreceptivos e não possuem uma estrutura
cuticular de fio de cabelo (WALADDE & RICE, 1977; WALADDE & RICE, 1982;
SONENSHINE, 1991). A porossensila da quelícera está estrategicamente
localizada de forma que durante a fixação e alimentação ela fica imersa na ferida
e em contato com os fluidos do hospedeiro (WALADDE & RICE, 1977).
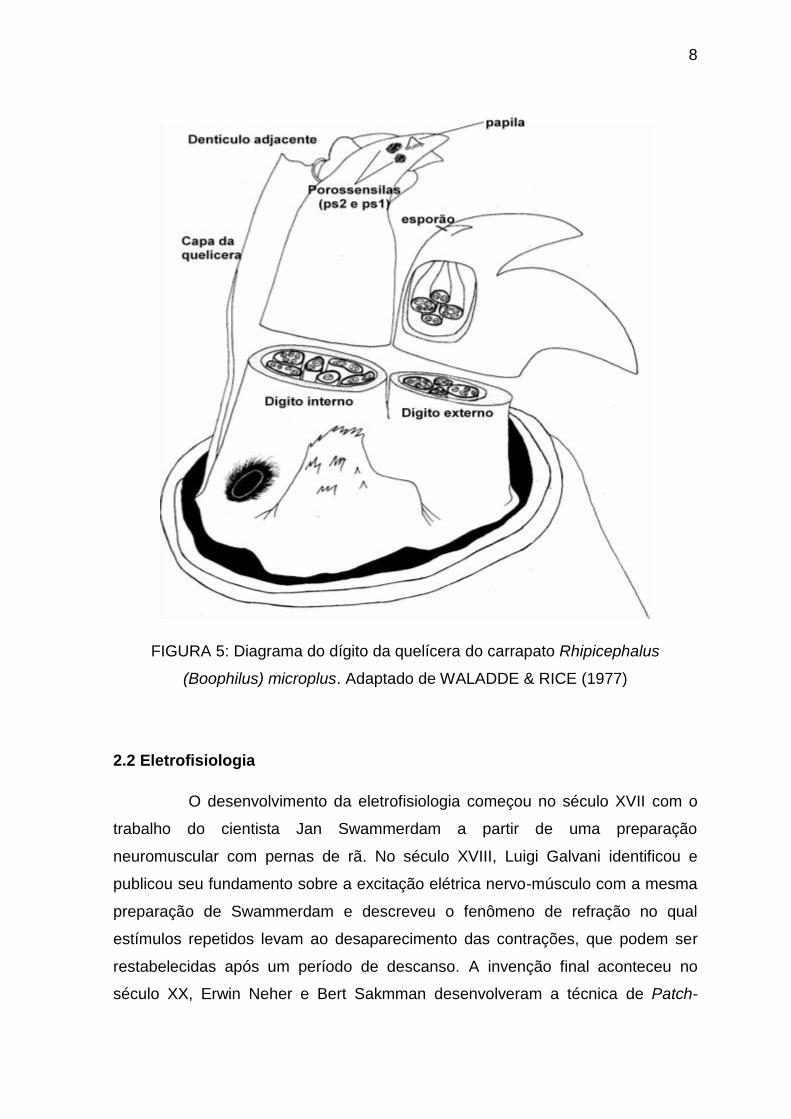
8
FIGURA 5: Diagrama do dígito da quelícera do carrapato Rhipicephalus
(Boophilus) microplus. Adaptado de WALADDE & RICE (1977)
2.2 Eletrofisiologia
O desenvolvimento da eletrofisiologia começou no século XVII com o
trabalho do cientista Jan Swammerdam a partir de uma preparação
neuromuscular com pernas de rã. No século XVIII, Luigi Galvani identificou e
publicou seu fundamento sobre a excitação elétrica nervo-músculo com a mesma
preparação de Swammerdam e descreveu o fenômeno de refração no qual
estímulos repetidos levam ao desaparecimento das contrações, que podem ser
restabelecidas após um período de descanso. A invenção final aconteceu no
século XX, Erwin Neher e Bert Sakmman desenvolveram a técnica de Patch-

9
Clamp que mensura o potencial de células excitáveis, ou seja, analisa a
desempenho de um simples canal de íon utilizando um eletrodo de prata e uma
micropipeta de vidro (VERKHRATSKY et al., 2006)
Eletrofisiologia é o estudo das propriedades elétricas de células e
tecidos (bioeletricidade) que envolve variação da voltagem ou da corrente elétrica
em uma ampla variedade de escalas, que pode ser de um único canal iônico até
de tecidos como o cardíaco e inclui a avaliação da atividade elétrica dos
neurônios (atividade dos potenciais de ação) (KANDEL et al., 2000). Ou seja, é a
ciência e a técnica que estuda o fenômeno elétrico de plantas e animais, envolve
o potencial de ação das células que podem sofrer variações lentas ou rápidas
devido a uma mudança de concentração de uma substância química e estas
mudanças geram picos que podem ser chamados de “potenciais de ação” ou
“spikes” (BRETSCHNEIDER & DE WEILLE, 2006).
A eletrofisiologia em artrópodes é eficaz em identificar a fisiologia
sensitiva dos mesmos, tem importância na descoberta de feromônios (sexual e
agregação), sítios atrativos de oviposição, compostos naturalmente repelentes ou
estimulantes (HOLSCHER et al., 1982). É uma técnica de triagem que tem como
objetivo determinar com mais precisão as respostas das sensilas olfativas e
gustativas dos artrópodes, detectando substâncias possivelmente repelentes ou
atrativas. Essas substâncias posteriormente são submetidas a testes
comportamentais para a determinação de sua identidade e dependendo dos
resultados podem ser usadas no controle dos artrópodes (BJOSTAD, 1998;
BORGES, 2001).
A eletrofisiologia em artrópodes iniciou-se com o eletroantenograma
(EAG) (Figura 6) em insetos para a detecção de feromônios, o princípio do EAG
baseia-se no registro da voltagem entre a ponta e a base da antena de um inseto
durante um estímulo volátil (WIBE, 2004). Desde então, outras técnicas têm sido
desenvolvidas, como a técnica de eletrofisiologia em sensila única que envolve o
registro do potencial elétrico característico de cada célula através da implantação
de eletrodos na base da sensila, seguida de sua estimulação com substâncias
específicas (SONENSHINE, 1991).

10
FIGURA 6: Esquema de um EAG. Fonte: BJOSTAD (1998)
A técnica de registro de sensila única ou de ponta (RSU) para contato
em sensilas quimiorreceptoras foi introduzida por HODGSTON et al. em 1955
(Figura 7). Compostos solúveis em água (açúcar, sal, ácidos, álcoois) foram
dissolvidos em uma solução salina no eletrodo de registro, e quando este eletrodo
entrou em contato com a sensila do labelo do inseto causou uma resposta de
duas séries de spikes (BJOSTAD, 1998).
FIGURA 7: Diagrama da preparação para a técnica de RSU. Adaptado de
HODGSON et al. (1955)

11
2.2.1 Eletroantenograma (EAG) vs Registro de sensila única (RSU)
Os receptores olfativos de insetos e carrapatos localizados na antena e
no primeiro par de patas, respectivamente, permitem que estes artrópodes
encontrem hospedeiros à distância (GUERIN et al., 2000). O primeiro par de
patas dos carrapatos seria análogo as antenas dos insetos na parte de busca de
hospedeiros (IVANOV & LEONOVICH, 1983).
As sensilas dos insetos estão distribuídas por todo o corpo, estão
concentradas nas patas, abdome, palpos e especialmente nas antenas onde há
inúmeras sensilas que respondem para um mesmo composto. Estas sensilas
possuem receptores para CO2, umidade, temperatura, estímulos gustativos e
olfativos (KAISSLING, 1995). Os carrapatos possuem um número bem menor de
sensilas e não possuem antenas, porém possuem um órgão similar, que é o
órgão de Haller. Há cerca de 20 quimiossensilas no órgão de Haller, enquanto na
antena de insetos há inúmeras (HESS & VLIMANT, 1983; IVANOV &
LEONOVICH, 1983; STEULLET, 1993). Sendo assim, o EAG por ser uma técnica
menos sensível não pode ser usado para carrapatos (WIBE, 2004).
No EAG uma única grande despolarização é observada após a
estimulação, devido á soma de baixos potenciais de ação de milhares de
neurônios. Já no ESU a principal característica é a „explosão‟ de potenciais de
ação após a estimulação, ou seja, os compostos químicos geram spikes mais
rápidos e com alta freqüência sem alteração da amplitude por englobar apenas
uma sensila (BJOSTAD, 1998).
2.2.2 Percepção do estímulo e disparo do potencial de ação
O sistema nervoso periférico dos carrapatos detecta uma variedade de
sinais químicos do ambiente que transmitem informações para sua sobrevivência
e o SNC recebe e integra estas informações. As sensilas mais importantes para
tal ação são as sensilas do órgão de Haller, do dígito das quelíceras e palpais,
lembrando que sensilas com estruturas diferentes são sensibilizadas por
estímulos diferentes (SONENSHINE, 1991).

12
O objetivo de experimentos eletrofisiológicos no geral é estudar os
potenciais de membrana e suas eventuais alterações (BRETSCHNEIDER & DE
WEILLE, 2006). As células são estruturas separadas por uma membrana que
apresenta seletividade a diversos íons (principalmente sódio (Na+) e potássio (K+))
e a diferença de potencial elétrico entre o meio intra e extracelular, ou seja, a
diferença de potencial entre o exterior e interior de uma célula, é denominado
potencial de membrana. A concentração de íons K+ é maior dentro da célula,
enquanto a concentração de Na+ é maior no meio extracelular (DELATTRE,
2007).
Os sinais nervosos são transmitidos por potenciais de ação, que por
sua vez são variações rápidas do potencial de membrana que viajam da
extremidade das fibras nervosas até o SNC devido a vibrações, luminosidade,
temperatura e/ou presença de alguma substância química (RASHBASS, 1967).
O mecanismo é a bomba de sódio e potássio (Na+/ K+ / ATPase), no
qual os íons fluem pelos canais iônicos que são responsáveis pela transmissão
elétrica em todo o sistema nervoso (DELATTRE, 2007). No repouso o neurônio
encontra-se polarizado, o estímulo altera a permeabilidade dos íons. A
despolarização da membrana promove a abertura dos canais de Na+, com isso
íons de Na+ começam a entrar na célula despolarizando mais ainda a membrana,
há então mais abertura de canais de Na+. Os canais de Na+ se fecham
espontaneamente e os de K+ começam a se abrir, com isso os íons de K+ fluem
para fora da célula. O K+ tem uma maior capacidade de difusão que o Na+, então
ele leva sua carga positiva deixando o meio intracelular negativo em relação ao
meio extracelular. Causando a repolarização do neurônio de volta ao potencial de
repouso (DELATTRE, 2007).

13
2.2.3 Eletrofisiologia olfativa e gustativa em carrapatos
Para a realização da técnica de eletrofisiologia olfativa e gustativa são
necessários uma variedade de equipamentos e materiais (Figura 8), como
eletrodos de vidro, micropipetas, fio de prata, micromanipuladores, amplificadores,
microscópio esterioscópio, fabricador de eletrodos, estimulador de sinais (inseto
artificial), controlador de estímulos (Stimulus Controller CS 55 – Syntech®), fitas
crepe e dupla-face, pinças, tesoura. (BJOSTAD, 1998; JORGENSEN et al., 2007;
SOARES, não publicado).
FIGURA 8: Equipamentos de uma estação de eletrofisiologia. A: TastePROBE; B:
IDAC-4; C: inseto artificial (Antennal Signal Stimulator); D: probe; E: holders; F:
micromanipuladores; G: fabricador de eletrodos (PC-10 PULLER); H: controlador
de estímulos (Stimulus Controller). Fonte: http://www.syntech.nl/; http://narishige-
group.com/. Acesso em: 25 out. 2011
A IDAC-4 é um amplificador com multicanais desenvolvido para a
gravação, armazenamento e análise de sinais fisiológicos. Durante o

14
desenvolvimento da técnica de eletrofisiologia as oscilações dos sinais são
mostradas na tela de um computador em uma base de tempo ajustável, com isso
facilita um ajuste rápido e fácil dos níveis de sinais permitindo o controle de
amplificação, configurações de filtro e amostragem. O software Autospike que é
apresentado junto com a IDAC-4 grava em tempo real os potenciais de ação e
permite uma série de adaptações para a avaliação dos mesmos (SYNTECH,
2004a). O controlador de estímulos produz uma corrente de ar constante e bem
definida para aplicação em pesquisa de quimiorrecepção em uma estação de
eletrofisiologia olfativa (SYNTECH, 2004b). A TastePROBE é um pré amplificador
que grava os potenciais de ação das sensilas gustativas, permite a gravação de
baixos potenciais de ação (spikes), melhora o sinal de ruído e preserva as formas
dos spikes (MARION-POLL & VAN DER PERS, 1996).
Para que a técnica seja realizada a estação de eletrofisiologia (Figura
6) deve estar aterrada. O aterramento protege de uma eletrocussão fatal, além de
auxiliar no controle de interferências que outros aparelhos presentes no local
possam causar que prejudicam a gravação do registro e posterior interpretação.
(PURVES, 1998). Para a verificação dos sinais e o adequado funcionamento dos
equipamentos utiliza-se um inseto artificial (Antennal Signal Stimulator –
Syntech®) que simula estímulos elétricos (SYNTECH, 2002).
Os eletrodos de vidro para eletrofisiologia possuem um diâmetro
microscópico e sua estrutura é dividida em três partes: ponta, cone e eixo (Figura
9). São fabricados a partir de capilares de vidro com parede fina de 1 a 2 mm por
meio de um puxador de pipetas (PC PULLER – Narishige®) com diferentes
propriedades para suprir a necessidade de diferentes tipos de técnica/registro
(BRETSCHNEIDER & DE WEILLE, 2006).
FIGURA 9: Estrutura de um eletrodo de vidro. Fonte: BRETSCHNEIDER & DE
WEILLE (2006)

15
A confecção de um eletrodo de vidro demanda tempo para se acertar a
temperatura que deve ser atingida e a forma de preenchimento. O diâmetro da
ponta deve ser capaz de englobar a sensila do carrapato. O preenchimento dos
eletrodos com soluções é dificultado devido à tensão de superfície em um
diâmetro muito pequeno (PURVES, 1998).
Os eletrodos devem ser preenchidos inicialmente por uma solução de
ensaio um pouco antes do registro para que se evite a cristalização e as
mudanças de concentração na ponta do eletrodo de vidro (JORGENSEN, 2007).
As substâncias iniciais, de registro e referência são eletrólitos, ou seja, são
substâncias que quando misturadas em um soluto (comumente água) se
dissociam em partículas ou íons transportando cargas (BRETSCHNEIDER & DE
WEILLE, 2006). Quando preenchidos por uma solução salina, a mesma será
responsável pela condução elétrica da sensila até o fio de prata (BJOSTAD,
1998).
As substâncias utilizadas para o estímulo olfativo devem ser
preparadas, impregnadas em papel filtro e colocadas na pipeta de vidro para
posteriormente serem acopladas no controlador de estímulos. Já as substâncias
gustativas são dissolvidas e colocadas no eletrodo de vidro (BJOSTAD, 1998;
SOARES, não publicado).
Para eletrofisiologia olfativa, o carrapato vivo é fixado ventralmente com fita
dupla-face, o primeiro par de patas deve ser estendido para frente com a intenção
de facilitar o acesso ao órgão de Haller e os demais pares de patas imobilizados
(Figura 10). Insere-se um eletrodo de vidro indiferente com solução salina em
alguma parte do carrapato (escudo, coxa) para fechar o circuito. A ponta de uma
sensila multiporosa é cortada e um eletrodo acoplado à abertura. O controlador de
estímulos emite um fluxo de ar constante e quando acionado por um pedal libera
o odor na concentração que se deseja testar. A umidade deve estar sempre
acima de 90%. O eletrodo de registro deve ser posicionado bem na ponta da
sensila, para que os poros fiquem livres para a percepção do estímulo volátil. Os
sinais são captados pelo amplificador e analisados pelo software (STEULLET &
GUERIN, 1994; BJOSTAD, 1998).

16
FIGURA 10: Preparação do carrapato para realização da técnica de
eletrofisiologia olfativa. Fonte: SOARES, não publicado
Para a eletrofisiologia gustativa, um espécime vivo de carrapato na
fase adulta é fixado em um suporte imantado com fita dupla-face pela região
dorsal, os quatro pares de patas devem ser presos com fita crepe (Figura 11)
(WALADDE & RICE, 1977). Realiza-se a introdução do eletrodo de
referência/indiferente já preenchido com solução salina no orifício anal e o
direciona a região anterior do carrapato. Após, o eletrodo de registro deve ser
preenchido com solução que se deseja testar, coloca-se no holder da
TASTEprobe, já com o fio de prata no seu interior, que por sua vez é apoiada por
um micromanipulador. O eletrodo de registro deve entrar em contato com a
sensila, para realizar um novo contato deve aguardar dois minutos para que se
restabeleça o potencial de ação. Os sinais são captados pelo pré amplificador e
vai para o amplificador e é analisado pelo software (JORGENSEN, 2007;
SOARES, não publicado). Em ambos os tipos, olfativa e gustativa, o eletrodo de
registro pode ser inserido na base da sensila, mas neste caso usa-se eletrodo de
tungstênio com a ponta afinada.
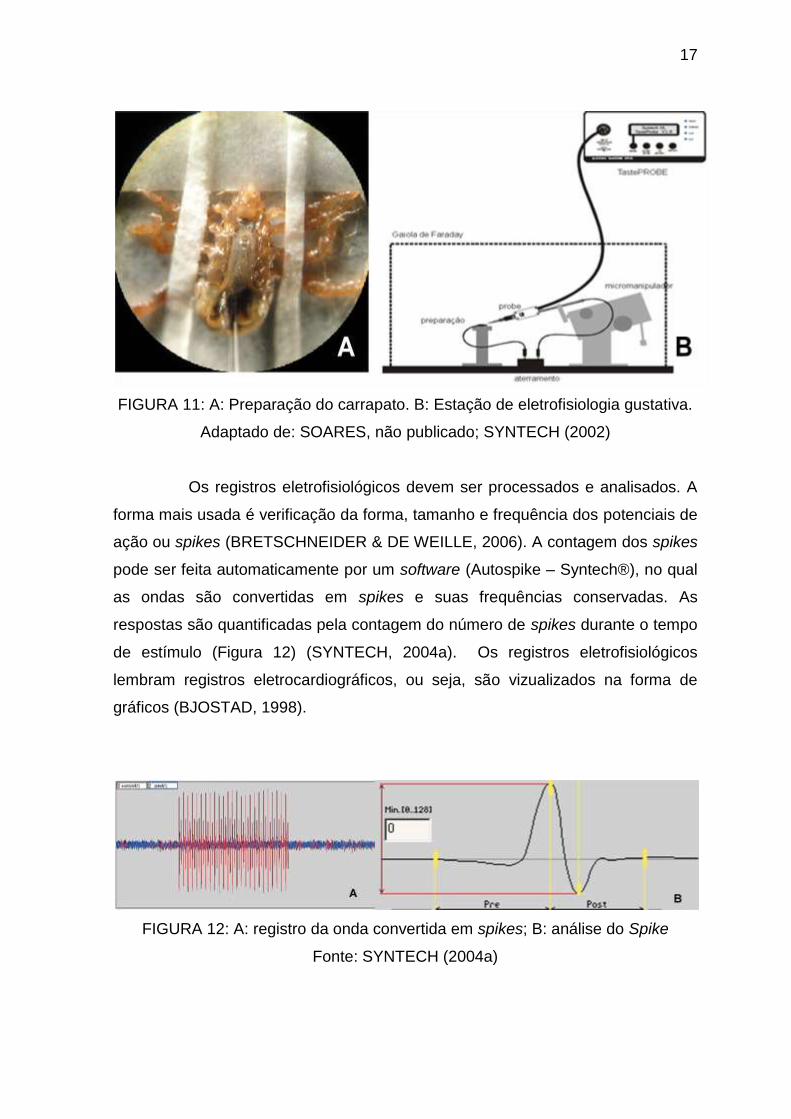
17
FIGURA 11: A: Preparação do carrapato. B: Estação de eletrofisiologia gustativa.
Adaptado de: SOARES, não publicado; SYNTECH (2002)
Os registros eletrofisiológicos devem ser processados e analisados. A
forma mais usada é verificação da forma, tamanho e frequência dos potenciais de
ação ou spikes (BRETSCHNEIDER & DE WEILLE, 2006). A contagem dos spikes
pode ser feita automaticamente por um software (Autospike – Syntech®), no qual
as ondas são convertidas em spikes e suas frequências conservadas. As
respostas são quantificadas pela contagem do número de spikes durante o tempo
de estímulo (Figura 12) (SYNTECH, 2004a). Os registros eletrofisiológicos
lembram registros eletrocardiográficos, ou seja, são vizualizados na forma de
gráficos (BJOSTAD, 1998).
FIGURA 12: A: registro da onda convertida em spikes; B: análise do Spike
Fonte: SYNTECH (2004a)

18
Circuitos eletrônicos são afetados por diversos fatores que causam a
diminuição do desempenho dos equipamentos, estes fatores podem ser
controlados, mas não eliminados por inteiro (WHITAKER, 2005). A presença de
ruídos (flutuações espontâneas imprevisíveis resultantes da física das
substâncias que fazem parte do circuito elétrico) no registro prejudica a técnica de
eletrofisiologia, porém são facilmente detectáveis (BRETSCHNEIDER & DE
WEILLE, 2006).
Falhas no equipamento de eletrofisiologia são bastante comuns, o
sucesso de um diagnóstico da falha requer conhecimento sobre o funcionamento
normal de cada item da estação de eletrofisiologia, reposição/modificação das
fontes de sinais e de alguns dispositivos (eletrodo de prata, eletrodo de vidro,
aterramento) e o uso da lógica (PURVES, 1998).
2.2.4 Resultados da técnica de ESU em carrapatos
Receptores fora da cápsula do órgão de Haller de Amblyomma
variegatum respondem a quatro compostos ativos de odores de bovinos, sendo
dois altamente estimulantes e identificados como 2-nitrofenol e 4-metil-2-nitrofenol
(STEULLET & GUERIN, 1994). As sensilas de Ixodes ricinus também possuem
receptores para fenol e lactona (LEONOVICH, 2004). Compostos normalmente
presentes no ambiente, como amônia e CO2, também são detectados pelos
carrapatos dependendo da concentração indica a proximidade do hospedeiro
(HAGGART et al., 1980; HOLSCHER et al., 1980). Outros sinalizadores de
hospedeiros detectados por carrapatos (A. variegatum, A. hebraeum, I. ricinus, I.
persulcatus, I. scapulares) são substâncias presentes no hálito e pele humana e
substâncias da eructação dos bovinos (STEULLET & GUERIN, 1992; DONZÉ,
2004),
Algumas espécies de carrapatos, como Rhipicephalus (Boophilus)
microplus, R. sanguineus e Ornithodorus tholozani respondem a
hematofagoestimulantes (compostos que estimulam a alimentação à medida que
são ingeridos) tais como cloreto de sódio (NaCl), adenosina trifosfato (ATP) e
glutationa (GT), que são componentes normais do plasma sanguíneo. Também
respondem ao plasma bovino quando estimulados (GALUN & KINDLER, 1965;

19
GALUN & KINDLER, 1968; WALADDE & RICE, 1977). R. sanguineus responde a
glicose, ATP, GT e alta concentração de sais (SOARES et al., 2011). Sensilas
palpais de machos de R. microplus são ativas para feromônios sexuais (BRUYNE
& GUERIN, 1998).

20
3 CONSIDERAÇÕES FINAIS
O estudo das estruturas dos órgãos sensoriais dos carrapatos,
principalmente do órgão de Haller, palpos e quelíceras, permite a identificação
dos receptores presentes em cada um deles, assim como suas funções. Funções
estas importantes na sobrevivência e perpetuação do ectoparasita (identificação e
localização do hospedeiro, fixação, alimentação, busca de parceiros para a
cópula).
A técnica de eletrofisiologia em artrópodes é uma técnica de triagem
minuciosa, laboriosa e demanda treino, porém bastante aplicável, uma vez que a
partir disso é possível identificar compostos potencialmente repelentes ou não aos
artrópodes, que serão confirmados a partir da realização de um teste
comportamental. O manuseio dos equipamentos, a preparação do carrapato,
realização dos testes e a análise dos registros devem ser executados da maneira
mais correta de modo que falhas sejam evitadas.
Atualmente, o uso de acaricidas para o combate de carrapatos é
bastante preocupante devido ao impacto causado na saúde humana, no meio
ambiente e até mesmo nos animais. Apesar das técnicas olfativa e gustativa em
carrapatos serem iniciantes, com poucos estudos e a estação de eletrofisiologia
do CPV/EVZ/UFG ser a única no Brasil os registros eletrofisiológicos podem ser o
início para o desenvolvimento de uma forma alternativa de controle de carrapatos,
pois a partir da identificação de uma substância, seja ela capaz de inibir ou
estimular o parasitismo, pode-se criar um produto que promova a repelência ou o
destacamento do carrapato do hospedeiro.

21
REFERÊNCIAS
1. BINNINGTON, K. C.; OBENCHAIN, F. D. Structure and function of the circulatory, nervous, and neuroendocrine systems of ticks. In: OBENCHAIN, F. G.; GALUN, R. Physiology of ticks. 1. ed. Oxford: Pergamon Press, 1982. cap. 10, p. 351-398. 2. BJOSTAD, L. B. Electrophysiological methods. In: MILLAR, J. G.; HAYNES, K. F. Methods in chemical ecology. vol. 2. Norwell: Kluwer Academic Publishers, 1998, cap. 9, p. 339-375. 3. BORGES, M. Os semioquímicos no controle de pragas e vetores de doenças: estudos biológicos comportamentais e químicos para o emprego de semioquímicos em agroecossistemas. Biotecnologia Ciência & Desenvolvimento, Brasília, n. 22, p. 4-7, 2001. 4. BRETSCHNEIDER, F.; DE WEILLE, J. R. Introduction to electrophysiological methods and instrumentation. [online]. 1.ed. Netherlands:Elsevier, 2006. Disponível em: http://pt.scribd.com/doc/8239928/Introduction-to-Electrophysiological-Methods-and-Instrumentation. Acesso em: 25 out. 2011. 5. BRUYNE, M.; GUERIN, P.M. Contact chemostimuli in the mating behaviour of the cattle tick, Boophilus microplus. Archives of insect biochemistry and physiology, v. 39, n. 2, p. 65-80. 1998. 6. DELATTRE, E. Fundamentos e eletrofisiologia: potenciais de membrana. Medicina, Ribeirão Preto, v.40, n.3, p. 378-393, 2007. 7. DONZÉ, G.; MAMAHON, C.; GERIN, P. M. Rumen metabolites serve ticks exploit large mammals. The Journal of Experimental Biology, London, v. 207, n. 5, p. 4283-4289, 2004. 8. GALUN, R.; KINDLER, S. H. Chemical basis of feeding in the tick Ornithodoros tholozani. Journal of Insect Physiology, Oxford, v. 14, n.10, p. 1409-1421, 1968. 9. GALUN, R.; KINDLER, S. H. Glutathione as an inducer of feeding in ticks.Science, Washington, v. 147, n. 3654, p. 166-167, 1965. 10. GUERIN, P. M.; KROBER, T.; MCMAHON, C.; GUERENSTEIN, P.; GRENACHER, S.; VLIMANT, M.; DIEHL, P.A.; STEULLET, P.; SYED, Z. Chemosensory and behavioural adaptations of ectoparasitic arthropods. Nova Acta Leopoldina, Stuttgart, v. 83, n. 316, p. 213-229, 2000. 11. HAGGART, D. A.; DAVIS, E. E. Ammonia-sensitive neurons on the first tarsi of the tick, Rhipicephalus sanguineus. Journal of Insect Physiology, Oxford, vol. 26, p. 517-523, 1980

22
12. HESS, E.; VLIMANT, M. The tarsal sensory system of Amblyomma variegatum Fabricius (Ixodidae, Metastriata). I. Wall pore and terminal pore sensila. Review Suisse of Zoology, Geneve, v. 89, n. 3, p. 713-729, 1982. 13. HESS, E.; VLIMANT, M. The tarsal sensory system of Amblyomma variegatum Fabricius (Ixodidae, Metastriata). III. Mapping of sensory hairs and evolution of the relative importance of sensory modalities during post-embryonic development. Review Suisse of Zoology, Geneve, v. 90, n. 4, p. 887-897, 1983. 14. HODGSON, E. S.; LETTVIN, J. Y.; ROEDER, K. D. Physiology of a primary chemoreceptor unit. Science, Washington, v. 122, p. 417-418, 1955.
15. HOLSCHER, K. H.; BARKER, R. W; STAATS, J. J. Selected bibliography of sensory systems in hematophagous arthropods, using electrophysiological methods. Entomological Society of America, Lanham, v. 28, n. 1, p. 27-30, 1982. 16. IVANOV, V. P. Central nervous system. In: BALASHOV, Y. S. An Atlas of Ixodid Tick Ultrastructure, Leningard: Naka Publishers, 1983, cap. 7, p. 175-190. 17. IVANOV, V. P.; LEONOVICH, S. A. Sensory organs. In: BALASHOV, Y. S. An Atlas of Ixodid Tick Ultrastructure, Leningard: Naka Publishers, 1983, cap. 8, p. 191-220. 18. JORGENSEN, K.; ALMAAS, T. J.; MARION POLL, F.; MUSTAPARTA, H. Electrophysiological Characterization of responses from gustatory receptor neurons of sensilla chaetica in the Moth Heliothis virescens. Chemical senses, Oxford, v. 32, p. 863-879, 2007. 19. KAISSLING, K. E. Single unit and electroantennogram recordings in insect olfactory organs. In: SPIELMAN, A. I.; BRAND, J. G. Experimental cell biology of taste and olfaction:current techniques and protocols. CRC Press: Boca Raton, 1995, cap. 4, p. 361-377. 20. KANDEL, E.; SCHWARTZ, J. H.; JESSEL, T. M. Principles of neural science. New York: McGraw-Hill Professional, 2000. 1568 p. 21. LEONOVICH, S. A. Phenol and lactone receptors in the distal sensila of the Haller‟s organ in Ixodes ricinus ticks and their possible role in host perception. Experimental and Applied Acarology, Netherlands, v. 32, p. 89-102, 2004. 22. MARION-POLL, F.; VAN DER PERS, J. Un-filtered recordings from insect taste sensilla. Entomologia Experimentalis et Applicata, Bélgica, v. 80, p. 113-115, 1996. 23. MCIVER, S. B. Structure of cuticular mechanoreceptors of arthropods. Annual Review of Entomology, Palo Alto, v. 20, p. 381-397, 1975.

23
24. PURVES, R. D. Microelectrodes methods for intracellular recordings and ionophoresis (Biological Techniques Series).Academic Press, 1998. 146 p. 25. RASHBASS, C. Sense organs – transducers of the environment. Reports on Progress in Physics, London, v. 30, n. 2, p. 375-407, 1967. 26. SCOTT, K. Taste recognition: food for thought. Neuron, Cambridge, v. 48, p. 455-464, 2005. 27. SLIEFER, E. H. The structure of arthropod chemoreceptors. Annual Review of Entomology, Palo Alto, v. 15, p. 121-142, 1970. 28. SOARES, S. F.; LOULY, C. C. B.; MARION-POLL, F.; BORGES, L. M. F. Electrophysiology on contact chemoreceptors in chelicerae. In: TICK AND TICK-BORNE DISEASE CONFERENCE, 7., 2011. Zaragoza. Anais eletrônicos… [CD-ROM], Zaragoza: Universidad Zaragoza, 2011. [nutm 46]. 29. SONENSHINE, D. E. Biology of ticks. vol. 1. New York: Oxford University Press, 1991. 447 p. 30. SONENSHINE, D. Tick pheromones and their use in tick control. Annual Review of Entomology, Palo Alto, v. 51, p. 557-580, 2006. 31. STEULLET, P.; GUERIN, P. M. Identification of vertebrate volatiles stimulating olfactory receptors on tarsus I of the tick Amblyomma variegatum Fabricius (Ixodidae) II. Receptors outside the Haller‟s organ capsule. Journal of Comparative Physiology A Sensory, Neural and Behavioral Physiology,Berlin, v. 174, p. 39-47, 1994. 32. STEULLET, P.; GUERIN, P. M. Perception of breath components by the tropical bont tick, Amblyomma variegatum Fabricius (ixodidae). II. Sulfide receptors. Journal of Comparative Physiology A Sensory, Neural and Behavioral Physiology,Berlin, v. 170, n. 6, p. 677-685, 1992.
33. STEULLET, P. Perception of vertebrate volatiles in the tropical bont tick, Amblyomma variegatum Fabricius. 1993. 122 f. Tese (Doutorado em Ciências) - Faculté des Sciences, Université de Neuchâtel, Neuchâtel. 34. SYNTECH. IDAC-4 Intelligent Data Acquisition Controller with programmable output control, AUTOSPIKE Software for signal recording and programmable output control: user‟s guide. Netherlands, 2004a. 55 p. 35. SYNTECH. Stimulus Controller, Type CS – 55: user‟s guide. Netherlands 2004b. 12p. 36. SYNTECH. TastePROBE type DTP-1 pre-amplifier for recording from contact chemosensilla: user‟s guide. Netherlands, 2002. 18 p.

24
37. VERKHRATSKY, A.; KRISHTAL, O. A.; PETERSEN, O. H. From Galvani to patch clamp: the development of electrophysiology. European Journal of Physiology, v. 453, p. 233-247, 2006. 38. VILELA, E. F.; DELLA LUCIA, T. M. C. Feromônios de insetos: biologia, química e emprego no manejo de pragas. 2. ed. São Paulo: Holos, 2001, 206 p. 39. WALADDE, S. M. Tip-recording from ixodid tick olfactory sensila: responses to tick relates odours. Journal of comparative physiology A Sensory, Neural and Behavioral Physiology, Berlin, v. 148, n. 4, p. 441-418, 1982. 40. WALADDE, S. M.; RICE, M. J. The sensory basis of tick feeding behaviour. In: OBENCHAIN, F. D.; GALUN, R. Physiology of Ticks. Oxford: Pergamon, 1982. p. 71-118. 41. WALADDE, S. M.; RICE, M. J. The sensory nervous system of the adult cattle tick Boophilus microplus (Canestrini) Ixodidae. Part III. Ultra-structure and electrophysiology of the cheliceral receptors. Australian Journal of Entomology, Camberra, v. 16, n. 4, p. 441-453, 1977. 42. WIBE, A. How the choice of method influence on the results in electrophysiological studies of insect olfaction. Journal of Insect Physiology, Oxford, v. 50, n. 6, p. 497-503, 2004. 43. WHITAKER, J. C. The electronics handbook. [on line]. 2. ed. Boca Raton: Taylor & Francis Group, 2005. Disponível em: http://pt.scribd.com/doc/59438138/ALL-TEXT. Acesso em: 01 nov. 2011.


















