
Toca que o cérebro também dança: Efeito da ritmicidade de ...
110
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS PROGRAMA DE PÓS-GRADUAÇÃO EM PSICOBIOLOGIA LABORATÓRIO DE NEUROBIOLOGIA E RITMICIDADE BIOLÓGICA JOÃO CARLOS NASCIMENTO DE ALENCAR Toca que o cérebro também dança: Efeito da ritmicidade de tons auditivos sobre parâmetros comportamentais e eletrofisiológicos da execução de uma tarefa de soma aritmética NATAL/RN 2017
Transcript of Toca que o cérebro também dança: Efeito da ritmicidade de ...

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM PSICOBIOLOGIA
LABORATÓRIO DE NEUROBIOLOGIA E RITMICIDADE BIOLÓGICA
JOÃO CARLOS NASCIMENTO DE ALENCAR
Toca que o cérebro também dança: Efeito da ritmicidade de tons
auditivos sobre parâmetros comportamentais e eletrofisiológicos da
execução de uma tarefa de soma aritmética
NATAL/RN
2017

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM PSICOBIOLOGIA
LABORATÓRIO DE NEUROBIOLOGIA E RITMICIDADE BIOLÓGICA
JOÃO CARLOS NASCIMENTO DE ALENCAR
Toca que o cérebro também dança: Efeito da ritmicidade de tons
auditivos sobre parâmetros comportamentais e eletrofisiológicos da
execução de uma tarefa de soma aritmética
Dissertação apresentada ao Programa de Pós-
Graduação em Psicobiologia, como requisito a
obtenção do título de Mestre em Psicobiologia
(Área de concentração: Psicologia Fisiológica)
Orientador: Prof. Dr. John Fontenele-Araújo
NATAL/RN
2017

Universidade Federal do Rio Grande do Norte - UFRN
Sistema de Bibliotecas - SISBI
Catalogação de Publicação na Fonte. UFRN - Biblioteca Setorial Prof. Leopoldo Nelson - Centro de Biociências - CB
Alencar, João Carlos Nascimento de.
Toca que o cérebro também dança: efeito da ritmicidade de tons
auditivos sobre parâmetros comportamentais e eletrofisiológicos
da execução de uma tarefa de soma aritmética / Joao Carlos
Nascimento de Alencar. - Natal, 2017.
110 f.: il.
Dissertação (Mestrado) - Universidade Federal do Rio Grande do
Norte. Centro de Biociências. Programa de Pós-Graduação em
Psicobiologia.
Orientador: Prof. Dr. John Fontenele-Araújo.
1. Funções executivas - Dissertação. 2. Córtex pré-frontal -
Dissertação. 3. Dynamic Attending Theory - Dissertação. 4.
Acoplamento fase-amplitude - Dissertação. 5. Integração sensório-
motora - Dissertação. I. Fontenele-Araújo, John. II. Universidade
Federal do Rio Grande do Norte. III. Título.
RN/UF/BSE-CB CDU 573

JOÃO CARLOS NASCIMENTO DE ALENCAR
Toca que o cérebro também dança: Efeito da ritmicidade de tons
auditivos sobre parâmetros comportamentais e eletrofisiológicos da
execução de uma tarefa de soma aritmética
Dissertação apresentada ao Programa de Pós-
Graduação em Psicobiologia, como requisito a
obtenção do título de Mestre em Psicobiologia
(Área de concentração: Psicologia Fisiológica)
Aprovado em: ____/____/_________.
BANCA EXAMINADORA
____________________________________
Prof. Dr. John Fontenele-Araújo (Orientador)
Departamento de Fisiologia - Universidade Federal do Rio Grande do Norte
____________________________________
Prof. Dr. Diego Andrés Laplagne
Instituto do Cérebro - Universidade Federal do Rio Grande do Norte
____________________________________
Prof. Dr. Bernardino Fernández Calvo
Departamento de Psicologia - Universidade Federal da Paraíba

AGRADECIMENTOS
Uma das coisas que mais me encantam nas crianças é o encanto com a novidade. Este
encanto reflete uma genuína curiosidade com o funcionamento do mundo e da natureza. Se
olharmos bem, a criança mais pequena já desenvolve explicações e suposições sobre o
mundo, e, muitas vezes, a despeito do controle e da vigilância, testa suas hipóteses. Não sei
bem onde, mas em algum ponto, aprendemos que o estabelecido deve ser introjetado, e o
pensamento atípico, o diferente, suprimido. Neste ponto, este trabalho materializa a
oportunidade, há muito aguardada, de me encantar com a natureza, de cogitar, de pensar
modelos explicativos e, como a criança, testá-los. Durante os dois anos de mestrado, aprendi
muita coisa, mas, acima de tudo, descobri que se há algo de nobre no ofício do cientista, este
“algo” é o encanto pela natureza.
Em 2001, cheguei a Natal, como um “retirante sudestino”. E como foi receptiva esta
cidade! Assim, tenho muito a agradecer, especialmente ao governo federal e à minha segunda
(primeira) casa, a UFRN. Desde meu ingresso no curso de Psicologia, em 2010, tive contato
com pessoas e ideias fascinantes, com uma estrutura física privilegiada, com equipamentos,
graças ao investimento público em educação de qualidade. Este trabalho tem o proposito de
retornar este investimento, e ser mais uma, dentre tantas outras, demonstrações de que
precisamos reafirmar o compromisso com políticas que ampliem ainda mais os caminhos que
trazem à universidade e à equidade em nossa sociedade.
Agradeço à meu pai, Carlos, minha mãe, Cleide, sobretudo por terem sempre e
incansavelmente feito seu melhor para que eu chegasse até aqui. À minha companheira,
Janaína, agradeço pela força, pelo companheirismo, pelo carinho, pelo incentivo para seguir
sempre em frente. Vocês poderão sempre contar comigo, assim como eu conto sempre com
vocês.
Pela recepção e orientação, agradeço ao Professor John Fontenele Araújo. Com John
aprendi que esta curiosidade que citei é o motor da ciência. Agradeço ainda ao Professor
Mário Miguel, pela confiança depositada nos diversos projetos. Descobri que a experiência
que têm não impede que possamos trabalharmos como pares, em pé de igualdade. E esta
motivação foi muito importante para a conclusão desta etapa.
Agradeço aos meus chefes do Departamento de História, Roberto, Margarida, Nonato e
Aurinete pela compreensão e por, dentro daquilo que a legislação permite, terem possibilitado

a realização deste mestrado em conciliação com minhas atividades como técnico-
administrativo.
À Pedro Henrique e à Prof. Mahayana Godoy, por terem auxiliado no recrutamento dos
voluntários. Devo confessar que esta foi uma etapa crítica de meu trabalho. Assim, deixo meu
muito obrigado a vocês e outros que contribuíram indicando pessoas para participar de meu
experimento.
Aos meus colegas de laboratório, especialmente Cleanto, Laura, Júlio, Breno, Patrícia,
Iara, Ringo. Agradeço pelas conversas, pela recepção no grupo. Desejo-lhes toda a sorte,
onde quer que nossa vida acadêmica nos leve.
Aos meus voluntários, cujo nome não posso citar por questões éticas, agradeço
especialmente pelo tempo e confiança. Sei como o tempo é um fator caro e raro hoje em dia.
Espero tê-los retribuído com um tratamento adequado dos resultados e com a construção de
conhecimento.
Que essas palavras sempre os encontrem bem. Avante!
João

RESUMO
Sequências de estímulos rítmicos apresentados na faixa 125-2000ms (0.3-8Hz) podem
ter efeitos importantes sobre a percepção, motricidade e a cognição. A "Dynamic Attending" é
uma teoria que pressupõe a existência de oscilações atencionais flexíveis à sincronização com
eventos ambientais rítmicos, fenômeno denominado arrastamento. A percepção de ritmo tem
sido implicada a áreas classicamente de controle motor, notadamente os núcleos da base.
Estes núcleos têm sido atrelados também ao funcionamento executivo, através de laços
córtico-estriatais-talâmico-corticais. Considerando as funções executivas sediadas no córtex
pré-frontal como integrantes desse circuito, o objetivo deste estudo é caracterizar o efeito de
estímulos auditivos rítmicos sobre parâmetros de comportamento motor, desempenho
cognitivo executivo e sinais eletrofisiológicos. A amostra (n=24) foi calculada a partir do
software G*Power 3.1. Os participantes foram submetidos a tarefas de tempo motor
espontâneo e tarefas de processamento executivo (soma sequencial de dígitos), com ou sem a
presença de pulsos sonoros isocrônicos de três diferentes intervalos. Os resultados mostram
que há sincronização do período motor aos pulsos com ISI (Intervalo entre estímulos, do
inglês, “Inter Stimulus Interval”) igual a 500 ms, e que ISI com 350 ms ou 650 ms aumentam
a variabilidade das batidas. Não foi encontrada diferença entre as manipulações
experimentais no que concerne à ativação autonômica, exceto por diminuição da frequência
cardíaca durante a realização da tarefa com som Não-Sincronizado 350 ms. Com relação aos
dados eletroencefalográficos, diferentes padrões de potência espectral estão vinculados ao
intervalo de estimulação. O índice de modulação delta-gama está negativamente
correlacionado à taxa de acertos na condição 350 ms, e positivamente correlacionado na
condição Sincronizada. O número de acertos está atrelado ao ISI, mas o Tempo de reação
parece ter maior modulação do ISI e do som. Nossos resultados de tempo motor espontâneo
sugerem a existência de uma “região ótima de arrastamento”, com centro em 500 ms, no qual
há maior suscetibilidade de sincronização motora involuntária. Além disso, os dados indicam
independência entre as funções executivas “quentes” e “frias”, e que o sistema nervoso
parece acionar diferentes estratégias de processamento para diferentes contextos temporais do
ambiente. Isto é consistente com um modo rítmico de processamento, que, no entanto, pode
implicar em redução do desempenho cognitivo mediante de alta exigência por velocidade de
processamento.
Palavras-chave: funções executivas; córtex pré-frontal; Dynamic Attending Theory;
acoplamento fase amplitude; integração sensório-motora.

ABSTRACT
Sequences of stimuli presented between 125-2000 ms (0.3-8Hz) may impact on perception,
motor responses and cognition. Dynamic Attending Theory postulates oscillations of
attention, that may synchronize by entrainment to rhythmic events from environment.
Rhythm perception has been shown to involve classical motor areas, notably basal ganglia.
These sub-cortical nuclei has been also linked to executive functioning, due to existence of
cortical-striatum-thalamic-cortical loops. Since executive function is based on prefrontal
cortex activity, this work aims to characterize effect of auditory rhythmic stimuli on motor
behavior, executive function and electrophysiological responses. Sample (n=24) was
previously calculated by G*Power 3.1 software. Participants were submitted to finger tapping
and executive tasks (sequential arithmetic sum), while auditory tones were played or not. Our
results show motor synchronization for ISI 500 ms, and increased variability for ISI 350 ms
and 650 ms. Autonomic electrophysiological responses did not show any changes, except for
a decreased heart rate at ISI 350 ms, Non-Synchronized Condition. Delta-Gama Modulation
Index is negatively correlated with hitrate for ISI 350 ms, but positively correlated with
hitrate at Synchronized Condition. ISI impacts on hitrate, but a larger modulation of both ISI
and auditory stimuli over Reaction Time is shown. Our results suggest an “optimal
entrainment range”, centered at 500 ms, and an increased likelihood for involuntary
synchronization at this point. Besides, results suggest so-called cold and hot executive
functions are independent processes, and also that nervous system may activate different
processing ways for different temporal contexts. These findings also corroborate the
existence of a rhythmic processing mode, however, with decreased performance through high
speed processing request.
Keywords: executive function; prefrontal cortex; Dynamic Attending Theory; phase-
amplitude coupling; sensorimotor integration.

Sumário
APRESENTAÇÃO ................................................................................................................ 10
CAPÍTULO 1 – COMO A RITMICIDADE INFLUENCIA O SISTEMA NERVOSO E
COMPORTAMENTO ........................................................................................................... 11
1.1 RITMO, EXCITABILIDADE CORTICAL E COMPORTAMENTO
SENSÓRIOMOTOR ...................................................................................................... 12
1.2 MODELOS TEÓRICOS DA PERCEPÇÃO DE RITMO E SINCRONIZAÇÃO .. 15
1.2.1 “Dynamic Attending Theory” e sua relação com oscilações dos sistemas
biológicos .................................................................................................................. 17
1.2.1.1 O período endógeno das oscilações atencionais é de 500 ms ................... 20
1.3 ATIVIDADE CEREBRAL E RITMICIDADE AMBIENTAL ............................... 22
CAPÍTULO 2 – COMO O CÓRTEX PRÉ-FRONTAL ATUA NO CONTROLE
EXECUTIVO DO COMPORTAMENTO .......................................................................... 27
2.1 O CÓRTEX PRÉ-FRONTAL .................................................................................. 28
2.1.1 Função Executiva ............................................................................................ 33
2.2 VIAS CORTICOESTRIATAIS E O CONTROLE DO COMPORTAMENTO E DA
COGNIÇÃO ................................................................................................................... 35
CAPÍTULO 3 - EXPERIMENTO ........................................................................................... 39
3.1 JUSTIFICATIVA ..................................................................................................... 39
3.2 OBJETIVOS ............................................................................................................. 41
3.2.1 Objetivo Geral ................................................................................................. 41
3.2.2 Objetivos específicos ....................................................................................... 41
3.4 MATERIAIS E MÉTODOS..................................................................................... 43
3.4.1 Participantes .................................................................................................... 43
3.4.2 Aspectos Legais e Éticos ................................................................................. 43
3.4.3 Procedimentos ................................................................................................. 44
3.4.3.1 Instrumentos .............................................................................................. 44
3.4.3.2 Registro dos Sinais Biológicos ................................................................. 45
3.4.4 Estímulos ......................................................................................................... 45
3.4.4.1 Estímulos Auditivos .................................................................................. 45
3.4.4.2 Estímulos Visuais ...................................................................................... 46
3.4.5 Tarefas ............................................................................................................. 46

3.4.5.1 Tarefa de Tempo Motor Espontâneo ........................................................ 46
3.4.5.2 Tarefa de soma aritmética ......................................................................... 46
3.5 PROTOCOLO EXPERIMENTAL .......................................................................... 47
3.6 ANÁLISE DE DADOS ............................................................................................ 49
3.6.1 Dados Comportamentais ................................................................................ 50
3.6.2 Condutância de pele ....................................................................................... 50
3.6.3 Eletrocardiografia (ECG) .............................................................................. 51
3.6.4 Dados Eletroencefalográficos (EEG) ............................................................ 51
3.6.5 Análise estatística ............................................................................................ 54
3.7 RESULTADOS ........................................................................................................ 56
Tarefa de Tempo Motor Espontâneo ..................................................................... 56
Tarefa de soma de dígitos – Condutância de pele ................................................. 62
Tarefa de soma de dígitos - Dados Eletroencefalográficos (EEG) ...................... 64
Acoplamento de delta aos estímulos ....................................................................... 74
Modulação de delta a gama .................................................................................... 78
Tarefa de processamento executivo (Soma de dígitos) – Acertos e Tempo de
reação ........................................................................................................................ 79
3.8 DISCUSSÃO ............................................................................................................ 83
CONCLUSÃO ........................................................................................................................ 95
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................. 97
APENDICE 1 – Termo de consentimento Livre e esclarecido (TCLE) .......................... 104
Anexo 1 – Parecer do Comitê de Ética ao projeto ............................................................ 106

10
APRESENTAÇÃO
Ubíquo é o termo que caracteriza aquilo que está presente todo o tempo, o onipresente.
Nesse aspecto, podemos pensar que nada seja mais ubíquo que o tempo. No campo da
psicologia e das neurociências, talvez a área que mais precocemente tenha tomado o tempo e
o ritmo como objeto de pesquisa seja a psicofísica, sob a qual se registram já no fim do
século XIX e início do século XX as tentativas de estudar empiricamente a produção e
percepção de intervalos de tempo (Wearden, 2005) e de ritmos motores (Fraisse, 1982). É
claro que nós percebemos o tempo, mas que este não pode ser visto como um sistema
sensorial no sentido usual: ainda hoje não foram descritos receptores para o tempo (Miguel,
2012).
Além disso, o tempo possui uma sinestesia intrínseca: vemos um filme, ouvimos uma
música, mas, ao mesmo tempo, sentimos sua duração. Estas talvez sejam expressões
paradoxais da desafiadora missão que é estudar a percepção temporal, já que, apesar de dois
séculos de investigação, como bem coloca Grondin (2010), sequer estão claramente
estabelecidos os parâmetros e aspectos relevantes à uma ciência da experiência sensorial ou
perceptual do tempo.
Neste trabalho, será discutida a existência de oscilações rápidas da atenção, como tais
oscilações podem se ajustar ao ambiente e, dessa forma, otimizar o processamento cognitivo
geral em uma tarefa com alta demanda das funções pré-frontais do cérebro (funcionamento
executivo). Tentaremos propor uma abordagem experimental para entender como a
organização temporal de sequências de estímulos influencia o desempenho do sistema
nervoso, especificamente sobre a função pré-frontal. Além da resposta comportamental,
dados de eletroencefalograma (EEG), eletrocardiograma (ECG) e condutância de pele serão
utilizados como ferramentas na busca dessas respostas.
No capítulo 1, apresentamos os principais conceitos e achados relativos à percepção e
sincronização à ritmos. No capítulo 2, fazemos uma explanação sobre a função do córtex pré-
frontal no controle executivo do comportamento e da cognição. O capítulo 3 apresenta o
experimento e os resultados obtidos.

11
CAPÍTULO 1 – COMO A RITMICIDADE INFLUENCIA O SISTEMA NERVOSO
E COMPORTAMENTO
A etimologia da palavra “ritmo” sugere que os gregos antigos a utilizavam para
descrever “aquilo que flui no tempo”. Na contemporaneidade, duas das suas várias acepções
conhecidas são particularmente interessantes a este trabalho: ‘ritmo musicológico’ e ‘ritmo
biológico’. Enquanto a primeira permite referir a organização temporal de estímulos, a
segunda, mais complexa, carrega em si um conjunto de pressupostos que descrevem como
oscilações dos sistemas biológicos podem se ajustar à estímulos ambientais.
Em musicologia, o ritmo é formalmente concebido como configuração das sequências
temporais regulares entre estímulos. Um ritmo, portanto, pode ser descrito a partir de duas
características fundamentais: um ‘tempo’ (ou pelo sinônimo: ‘batida’) e uma ‘métrica’.
Enquanto a batida emerge durante a apresentação dos estímulos, como um pulso percebido
que marca pontos igualmente espaçados no tempo, a métrica são os possíveis múltiplos níveis
de periodicidade que podem simultaneamente emergir na estrutura rítmica. É percebida, via
de regra, em frações ou múltiplos inteiros dos intervalos de tempo da batida fundamental
(Patel, Iversen, Chen, & Repp, 2005). O compasso musical ocidental, por exemplo, constitui-
se como um elemento métrico do ritmo em que a cada duas, três ou quatro batidas ocorre um
pulso forte (compasso binário, ternário ou quaternário, respectivamente).
Tabela 1.1 - Figuras musicais
Nota: Notar que todas as figuras são múltiplos ou frações de inteiros da semínima, com tempo =1,
denotando diversos níveis de ritmicidade.
Fonte: http://magiadamusica.webnode.pt/figuras-musicais/.
Entre as diversas áreas que pesquisam a percepção de ritmo (musicologia, neurociência,
psicologia experimental, etc.), não existe consenso geral quanto à unidade de medição da

12
batida. Entre musicólogos, é comum a notação em batidas por minuto (em bpm), ao passo
que nos estudos em psicologia e neurociências se utiliza mais comumente o Intervalo Entre
Estímulos (ISI, do inglês, “Inter-Stimulus Interval”, em ms) ou ainda a frequência (em Hz).
Para estímulos auditivos, por exemplo, o psicólogo francês Paul Fraisse (1911-1996) propôs
que, sequências de estímulos auditivos com ISI inferior a 125ms (480 bpm, ou 8 Hz) passam
a ser percebidos como um som constante, e que sequências com ISI superior a 2000ms (30
bpm, ou 0.5 Hz) são percebidas como estímulos isolados, constituindo esses os limites
sensoriais para a percepção de ritmos sonoros em humanos (Fraisse, 1982; Fraisse, 1984).
A música e a dança representam atividades humanas que conferem grande importância
à capacidade de perceber ritmos. No entanto, a filogenia do sistema nervoso demonstra que
uma de suas mais primeiras funções foi o processamento de estímulos ambientais e a resposta
comportamental através de efetores. Obter previsibilidade sobre os eventos ambientais dos
mais diversos permite a alocação mais efetiva dos recursos fisiológicos, e, por conseguinte,
uma resposta antecipatória e mais adaptada (Campos, Santos, & Xavier, 1997). Nesse
aspecto, é possível cogitar que nossa capacidade percepção de ritmos esteja ligada a um
incremento da previsibilidade ambiental, assim, à otimização de processos desempenhados
pelo sistema nervoso.
Nenhuma outra interação com a percepção de ritmo é mais óbvia e bem estudada que a
existente com o comportamento motor. Basta dizer que qualquer um de nós sente que dança
ou canta melhor enquanto ouve a música. Isto dá uma importante pista de como nossa
percepção de ritmicidade em estímulos e nossa capacidade de sincronizar nosso
comportamento a eles também está ligada intimamente à coordenação temporal de sequências
de estímulos aferentes ao sistema nervoso e respostas motoras mais adequadas.
1.1 RITMO, EXCITABILIDADE CORTICAL E COMPORTAMENTO
SENSÓRIOMOTOR
Através de que mecanismos estes estímulos percebidos tem influência sobre nosso
comportamento motor? Para responder essa pergunta, vamos pensar em uma situação
comum: quem nunca sentiu ao menos por um instante o impulso involuntário de bater o pé ao
ouvir uma música? Este comportamento é conhecido como “foot-tapping”. Alguns
experimentos interessantes relacionados ao “foot-tapping” dão sugestões de como isso
ocorre. Wilson & Davey (2002) investigaram a possível modulação de estímulos sonoros

13
isocrônicos sobre o comportamento motor, avaliando as respostas de potenciais musculares
evocados no Gastrocnêmio e Tíbia (respectivamente, músculos extensor e flexor para o
comportamento de “foot-tapping”). Através de Estimulação Magnética Transcraniana no
córtex motor contralateral, foi possível excitar de forma controlada e simultânea ambos os
músculos, e verificar a resposta muscular evocada. Considerando a intensidade controlada da
estimulação, seria esperado que a resposta de ambos os músculos fosse idêntica. No entanto,
durante a apresentação de música, a correlação entre o potencial evocado dos dois músculos
no momento da batida desapareceu em 66% dos indivíduos, indicando que a estimulação
interagiu com diferentes padrões de excitabilidade cortical que favoreceram a resposta em um
músculo, e a reduziram no músculo antagonista.
Cameron, Stewart, Pearce, Grube, & Muggleton (2012) também submeteram
voluntários a este protocolo, com objetivo de verificar se a saliência rítmica1 das batidas
possui influência sobre o resultado encontrado por Wilson e Davey. Para tanto, aplicaram o
mesmo protocolo, diante da apresentação de tons com baixa, alta saliência ou música. Além
de corroborar a influência da sincronia com a batida sonora, os resultados indicam que pulsos
isocrônicos de alta saliência rítmica são ainda mais efetivos que música para elicitar respostas
motoras. Isto demonstra que o componente rítmico é o principal fator para a facilitação
motora, e explica o efeito facilitador da música com alta saliência rítmica sobre o exercício,
utilizado intuitivamente pelas academias, por exemplo (Wilson & Davey, 2002). Ao que
parece, a facilitação motora em fase aos estímulos isocrônicos ou à batida percebida está
ligada à maior excitabilidade cortical nesses instantes (Michaelis et al., 2014).
1.1.1 Filogenia e percepção da batida e sincronização em animais não-humanos
Nesse aspecto, os achados de maior excitabilidade cortical sincronizada à batida
explicam a facilitação motora e, portanto, a extraordinária capacidade e predisposição dos
seres humanos para sincronizar seus movimentos a estímulos rítmicos ambientais variados.
Esta habilidade é ainda mais surpreendente se considerarmos que muitas espécies de animais
não-humanos expressam comportamento motor rítmico (danças, cantos, e comportamentos
estereotipados), mas, interessantemente, eles não parecem ser capazes de sincronizar a fase
1 Saliência rítmica, aqui, representa a capacidade da sequência de estímulos para induzir ritmicidade.

14
ou frequência desses comportamentos à eventos externos, como nós fazemos tão facilmente
com música, por exemplo (Patel, Iversen, Bregman, & Schulz, 2009).
Uma demonstração disso são as evidências de que animais não-humanos podem ser
condicionados a estimar o intervalo entre dois estímulos, mas não a sincronizar
eficientemente a sequências de estímulos distintos níveis de ritmicidade (Bizo, Chu, Sanabria,
& Killeen, 2006; Hattori, Tomonaga, & Matsuzawa, 2015). O estudo mais recentemente
publicado relata que uma fração dos chimpanzés amostrados consegue, mas com enorme
dificuldade, sincronizar a tons isocrônicos altamente salientes (Hattori et al., 2015). Em um
interessante estudo de caso, que vem se tornando um clássico da área da percepção de ritmo e
sincronização, Patel et al. (2009) validaram a intrigante cracatoa “Snowball” como um dos
únicos animais reconhecidamente capazes de sincronizar espontaneamente movimentos do
corpo a batidas musicais variadas. Ainda assim, não são conhecidos outros indivíduos da
mesma espécie com tal capacidade, sendo a percepção de ritmos variados fora da espécie
humana um fenômeno raro.
Do ponto de vista evolutivo, o surgimento de uma característica normalmente possui
homólogos dentro do clado monofilético, o que não ocorre para a capacidade de perceber
ritmos. Merker, Madison, & Eckerdal (2009), em uma interessante revisão, exploraram
atentamente este paradoxo. Segundo esses autores, uma possível explicação seria o pequeno
número de circunstâncias ecológicas nas quais um comportamento sincronizado possa
representar maior sucesso evolutivo aos indivíduos de espécies. Os mais bem reconhecidos
exemplos seriam o canto sincronizado emitido por machos de certas espécies de insetos
(Buck & Buck, 1968) e anfíbios anuros, com fins reprodutivos. Ainda assim, a sincronização
comportamental dessas espécies é limitada ao canto, pouquíssimo flexível, e ocorre somente
em resposta à um estímulo bastante particular (ex.: canto de outro co-específico), o que
sugere a evolução independente de tais características.
Em conjunto, há ainda poucas evidências empíricas que apoie qualquer especulação
sobre o aparecimento filogenético da capacidade (e predisposição) para inferir e sincronizar a
batidas regulares de natureza ampla e variada, que no homem é tão facilmente observada.
Alguns pesquisadores cogitam que seu aparecimento possa ter ocorrido em espécies
ancestrais do gênero Homo, já extintas (Merker et al., 2009).
Assim sendo, considerando a pequena pressão evolutiva para o surgimento de áreas
encefálicas dedicadas à percepção de ritmos, alguns teóricos cogitam que a captação de
padrões temporais rítmicos do ambiente – e seus desdobramentos - na verdade recrute de

15
forma secundária áreas originalmente dedicadas a funções de outros domínios, como a
linguagem. Patel (2014), representante da chamada “hipótese da sincronização e aprendizado
vocal”, sugere que a aprendizagem da linguagem vocal complexa passa pela capacidade de
emitir sinais vocais baseados na experiência auditiva e na retroalimentação sensorial, um
processo “online” em que ocorre a integração dos sistemas auditivos e motores para produzir
sequências de vocalizações conforme um modelo desejado (Patel, 2006).
Portanto, a capacidade para sincronização motora pode ser filogeneticamente derivada
de áreas encefálicas cunhadas no processo evolutivo pela pressão evolutiva para a aquisição
da linguagem vocal, o que poderia explicar porque a capacidade de perceber e sincronizar à
ritmos externos, assim como a linguagem vocal complexa, é tão rara na natureza (Merker et
al., 2009; Patel, 2006, 2014).
1.2 MODELOS TEÓRICOS DA PERCEPÇÃO DE RITMO E SINCRONIZAÇÃO
Alguns modelos teóricos se consolidaram como possíveis mecanismos da percepção
de tempo, ritmo e sincronização em humanos. Grondin (2010), em estado da arte sobre o
tema, categorizou esses modelos em dois grandes grupos fundamentais: os que se apoiam e
os que não se apoiam na noção de um relógio temporizador endógeno como mecanismo para
perceber o tempo.
A ideia de um relógio temporizador é considerada como desnecessária para muitos
estudiosos que defendem os chamados modelos intrínsecos da percepção de tempo. O
conceito de relógio temporizador se refere a um marca-passo ou oscilador endógeno, que, do
ponto de vista funcional, gera pulsos ou períodos que serviriam de base para o sistema
nervoso computar a passagem do tempo (maiores detalhes adiante). A principal afirmação
dos questionadores desses modelos é que o tempo pode percebido por um subproduto da
atividade neural que processa o estímulo. Por exemplo, a duração de um pulso luminoso ou
auditivo, por exemplo, seriam subprodutos do processamento perceptual do estímulo pelo
córtex visual ou auditivo, respectivamente. Enquanto para alguns modelos intrínsecos, como
o chamado “state-dependent network model”, isto implica que a duração percebida depende
da diferença entre o estado da rede neural entre o início e o fim do estímulo (Goel &
Buonomano, 2014), para outros, a duração percebida do estímulo depende da magnitude da
atividade neural recrutada para processá-lo (Ivry & Schlerf, 2008).

16
Apesar dessas críticas, os modelos baseados em relógio endógeno, possuem maior
tradição experimental, e um corpo sólido de evidências comportamentais que os amparam.
Por exemplo, a capacidade de estimar o tempo transcorrido entre dois estímulos – sem
qualquer estímulo entre eles - pode ser parcimoniosamente explicado através de algum
mecanismo temporizador endógeno, mas não a partir de mecanismos intrínsecos. Além disso,
existe alta correlação entre medidas de tempo percebido e tempo produzido (Michaelis et al.,
2014), o que dá suporte à ideia de centralidade do mecanismo perceptivo de tempo.
O agrupamento de uma diversidade de modelos teóricos que se apoiam na ideia de
relógio temporizador necessita detalhe. Uma divisão importante concerne aos modelos
intervalares e modelos de arrastamento. Com relação aos modelos intervalares, entre os quais
se destaca a “Scalar Expectancy Theory” (SET), são dominantes e de maior tradição
psicofísica na pesquisa de percepção de tempo, e conceituam o relógio como um marca-passo
contador endógeno, cujo número de pulsos acumulados ao longo de um intervalo de tempo
determinaria a extensão percebida (Treisman, 1963). Os modelos de arrastamento, tal qual a
“Dynamic Attending Theory” (DAT), por sua vez, pressupõem oscilações endógenas que
capazes de sincronizar a eventos externos através de arrastamento, e daí obter estimativas de
tempo implícitas que guiam o comportamento, (Jones, 1976; Jones & Boltz, 1989).
McAuley & Jones (2003) examinaram a adequação da SET e da DAT a diferentes
contextos rítmicos, através de quatro experimentos em que diferentes estímulos sonoros com
eram apresentados aos participantes. A tarefa era avaliar se o intervalo de tempo dois últimos
estímulos de uma sequência era igual ao intervalo das sequências rítmicas precedentes, cujos
arranjos temporais foram controlados para induzir maior ou menor percepção da batida. Os
autores propuseram modelos matemáticos baseados na SET e na DAT, e compararam as
simulações matemáticas aos dados empiricamente obtidos. Os resultados demonstram que as
previsões teóricas propostas pela DAT são robustas, ao passo que nenhum dos modelos
matemáticos propostos baseados na SET mantém seu poder preditivo para os diferentes
arranjos temporais rítmicos.
Naturalmente, é necessário que exista uma sequência de estímulos prévios aos quais
oscilador proposto pela DAT se sintonize para gerar qualquer estimativa de tempo, o que
parece apontar para a melhor adequação teórica da SET para estimativas de tempo discreto e
explícitas, e da DAT para contextos rítmicos, onde a tarefa de temporização é implícita e é
somente um elemento entre tarefas não-temporais (Coull, Cheng, & Meck, 2011; Grondin,

17
2010). Isto faz com a que a DAT seja um modelo teórico adequado ao estudo da percepção de
ritmo e sincronização.
Na seção a seguir, são esclarecidos os principais conceitos e pressupostos da DAT
como um modelo teórico da percepção de ritmo e sincronização.
1.2.1 “Dynamic Attending Theory” e sua relação com oscilações dos sistemas
biológicos
A “Dynamic Attending Theory” foi proposta em 1976 por Mari Jones, como “uma
teoria da percepção e da atenção que enfatiza a natureza relacional entre perceptuais não-
variantes” (Jones, 1976). Basicamente, o modelo cogita a existência de oscilações, ou ritmos
atencionais, com a propriedade de sincronizar a estímulos externos relativamente isocrônicos.
Essas oscilações são auto-sustentadas, com período relativamente estável, mas, diante de
estimulações regulares no tempo (rítmicas), tem a propriedade de ajustar-se por meio de
arrastamento. O acoplamento das oscilações endógenas à ritmicidade ambiental resulta em
um gradativo aumento da expectância: uma antecipação ao próximo estímulo da sequência,
caracterizada pela maior alocação dos recursos atencionais e perceptuais disponíveis para este
momento em que o estímulo é esperado (Large & Jones, 1999).
Figura 1.1 – “Dynamic Attenting Theory”

18
Nota: A partir de tons auditivos regulares no tempo, um arrastamento dos chamados “pulsos atencionais”
implicaria na alocação temporal da atenção, com consequente aumento da expectância e da acurácia perceptual
para o momento em que o estímulo é esperado.
Fonte: Adaptado pelo autor, de Jones, Moynihan, MacKenzie, & Puente (2002).
O reconhecimento visual de casas ou rostos humanos é melhorado quando as imagens
são apresentadas em sincronia ao tempo forte de um ritmo musical ao qual não havia
qualquer instrução explícita para prestar atenção na batida (Escoffier, Sheng, & Schirmer,
2010). Brochard, Tassin, & Zagar (2013), adaptando o mesmo protocolo, demonstraram que
o reconhecimento visual de palavras também é facilitado pelo ritmo musical. Além de
apresentar evidências empíricas para o conceito de expectância, esses estudos demonstram a
capacidade de estímulos auditivos para afetarem a expectância a estímulos aferentes por
outras vias sensoriais.
Para fins de simplificação, o modelo é normalmente apresentado a partir de um único
oscilador, embora seja mais aceitável a existência de múltiplos osciladores, atuando
simultaneamente em vários níveis temporais, hierarquicamente acoplados (ou aninhados) e

19
que permitem, por exemplo, explicar a capacidade de processar e reconhecer as múltiplas
escalas de tempo da métrica musical, ou ainda, as diversas periodicidades embutidas na fala
(Henry & Herrmann, 2014; Jones, Moynihan, MacKenzie, & Puente, 2002).
Um dos sistemas biológicos oscilatórios funcional e anatomicamente mais bem
conhecidos é o oscilador gerador dos ritmos circadianos, o Núcleo Supraquiasmático do
hipotálamo. Embora não haja qualquer evidência de relação neurofisiológica entre os ritmos
circadianos e os ritmos atencionais propostos pela DAT, como demonstra a Fig. 1, o
arrastamento das oscilações atencionais por estímulos externos possui similaridades
funcionais com a sincronização circadiana à luz solar (McAuley & Jones, 2003).
Figura 1.2 –Similaridade funcional entre o arrastamento de osciladores circadianos e do oscilador
atencional proposto pela DAT
Nota: Note-se o gradativo arrastamento de fase/período a partir dos estímulos sincronizadores.
Fonte: Autor
Roenneberg e colaboradores (Roenneberg, Dragovic, & Merrow, 2005) discorreram
sobre as algumas características do arrastamento de oscilações biológicas a padrões cíclicos
ambientais: o período de um oscilador biológico, ou a média do período de vários ciclos,
tende à ajustar-se ao período dos eventos externos aos quais se arrasta; pode levar vários
ciclos até que o oscilador atinja estabilidade; redução no período do ciclo ambiental acarreta
atraso de fase, e vice-versa; além da função de arrastamento do oscilador, o ciclo externo tem
efeito indireto sobre os ritmos controlados pelo oscilador (efeito de mascaramento). É sabido
também que, na ausência de qualquer estímulo, os osciladores funcionam “em livre curso”,
isto é, em seu período endógeno basal.

20
Mas, como se pode medir o período dos osciladores biológicos? Na escala circadiana,
se infere este período a partir da mensuração de um ritmo subordinado ao oscilador de
interesse (normalmente atividade-repouso). Na ausência de um ritmo arrastador exógeno
(zeitgeber, normalmente o claro-escuro), podemos obter o período espontâneo do oscilador
em livre curso. Da mesma forma, a grande maioria dos pesquisadores acredita que o período
das oscilações atencionais propostas pela DAT podem ser acessadas através de medidas de
ritmos comportamentais espontâneos a ele subordinados.
A mais popular dessas medidas é o ‘tempo motor espontâneo’ (SMT, do inglês,
“Spontaneous Motor Tempo”), que corresponde ao intervalo médio entre toques do dedo
indicador (“finger-tapping”), quando sujeitos são instruídos a manter a uma cadência motora
mais confortável para si, “nem rápido, nem lento”, e na ausência de estimulação externa.
Outra medida, conhecida como ‘tempo perceptual preferido’ é obtido apresentando ao sujeito
estímulos isocrônicos, e instruindo-o a relatar a velocidade de apresentação dos estímulos
mais confortável para si, “nem rápido, nem lento”. Sendo as oscilações atencionais um
mecanismo regulatório central, pressupõe-se que haja alta correlação entre as duas medidas, o
que é corroborado por estudos empíricos (McAuley, Jones, Holub, Johnston, & Miller, 2006;
Michaelis et al., 2014).
1.2.1.1 O período endógeno das oscilações atencionais é de 500 ms
Como já citado, nossa percepção de ritmos ocorre em dentro de certos limites
perceptuais, a saber, frequências de apresentação dos estímulos em que somos capazes de
associar as batidas isoladas à ritmicidade (0.5-8 Hz, que equivale a 30-480 bpm ou ISIs entre
2000-125 ms). Os osciladores biológicos podem sincronizar a estímulos externos, e a
dificuldade de sincronizar eficientemente pode ser medida pelo coeficiente de variação dos
períodos produzidos numa tarefa de “finger tapping”, na qual se instruiu o sujeito a bater o
dedo em sincronia a estímulos regulares de eixo do tempo. Cabe salientar, portanto, que
nossa percepção da isocronia, ou nossa capacidade de realizar “finger tapping” mantendo a
ritmicidade do movimento nesses intervalos é bastante precária, marcada por coeficientes de
variação elevados.
A sincronização diferenciada a ritmos de diferentes ISI contraria a lei geral dos
processos psicofísicos de Weber-Fechner, que estabelece que o coeficiente de variação da

21
percepção subjetiva do estímulo permanece estável conforme cresce sua magnitude objetiva
(Bizo et al., 2006). Isto levou McAuley & Jones (2006) a cunhar o termo “região de
arrastamento”, para referir a uma faixa dentro da qual estímulos externos podem efetivamente
se sincronizar oscilações atencionais. Fora dessa região, os coeficientes de variação em uma
tarefa de sincronização são tão grandes que considerada nula.
Se nos extremos desse intervalo apresentamos grande dificuldade de sincronizar nosso
comportamento, deve haver um ponto central, onde há uma percepção ótima da ritmicidade
dos estímulos, e somos capazes de realizar “finger-tapping” sincronizado ao som associado
com acurácia máxima (Moelants, 2002). Para referir-se a este constructo, o psicólogo francês
Paul Fraisse cunhou simultaneamente os termos Tempo preferido, Tempo natural e tempo
espontâneo (Fraisse, 1982). Fraisse propôs que este período estaria na faixa do ISI igual a
600 ms, mas, tem havido consenso quanto à período mais rápidos, de aproximadamente 500
ms, ou 2Hz (120bpm) (Moelants, 2002). Curiosamente, não apenas na produção de intervalos
através do comportamento motor espontâneo, mas também no tempo das batidas da maior
parte da música ocidental expressamos nosso viés pelo tempo preferido na faixa de 500 ms
(Figura 1.3)
Figura 1.3 – Tempo motor espontâneo e tempo preferido
Nota: . À esquerda, histograma dos tempos motores espontâneos de 68 voluntários (em ms). À direita, a
distribuição do tempo encontrado em músicas de listas de maiores sucessos, músicas que mais tocaram em
rádios, músicas populares, e músicas de diferentes estilos. Note-se que em ambos os gráficos ocorre uma
concentração dos valores em torno de 500 ms, ou 120 bpm.
Fonte: Adaptado pelo autor, de Moelants (2002)

22
Há ainda suporte para a afirmação de que comportamentos cotidianos imbuídos de
ritmicidade também expressam tempo espontâneo com período na faixa de 500 ms (2 Hz).
MacDougall & Moore (2005), por exemplo, registraram a aceleração da cabeça de 20
voluntários, durante 10h, em suas rotinas habituais. O espectro das frequências mostra a
presença de uma frequência de 2Hz, que prepondera não apenas durante o caminhar, mas
também durante diversas atividades locomotoras, como descer escadas, limpar a casa ou
pedalar durante a prática de exercício em bicicletas ergométricas (MacDougall & Moore,
2005). Em outro estudo, sem qualquer relação explícita com o conceito de tempo motor
espontâneo, Néda, Ravasz, Brechet, Vicsek, & Barabási (2000) relataram que os aplausos do
público ao final de um espetáculo tendem a sincronizar entre si, de forma espontânea, em
2Hz. Assim, a frequência de 2Hz (que equivale a ISI igual a 500 ms ou 120 bpm) parece uma
promissora proposta para a frequência de repouso do oscilador endógeno proposto pela DAT.
Nesse aspecto, seria a frequência ótima na qual ocorre a melhor sincronização, e, por
conseguinte, melhor previsibilidade e melhor processamento dos estímulos.
Embora diversas evidências se coadunem para estender o efeito da percepção de
ritmicidade à processos perceptuais e cognitivos, certamente é a psicomotricidade a função na
qual esta relação é mais extensamente documentada. Na seção a seguir, veremos o que isto
tem a ver com o que acontece dentro do encéfalo.
1.3 ATIVIDADE CEREBRAL E RITMICIDADE AMBIENTAL
Estudos de imagem observaram a ativação de áreas encefálicas de humanos diante de
tarefas de temporização. À parte das diferenças metodológicas entre cada estudo, sobretudo
considerando que nos protocolos experimentais muitas vezes a diferença entre temporização
explícita e implícita não é levada em consideração, está clara a grande influência de áreas
classicamente envolvidas com funções motoras sobre o processamento temporal na escala
dos milisegundos, especialmente estriado, o cerebelo, a área motora suplementar, a área pré-
motora, além de regiões do córtex frontal (Dreher & Grafman, 2002; J. a. Grahn & McAuley,
2009; Ivry & Schlerf, 2008; Kornysheva, Von Cramon, Jacobsen, & Schubotz, 2010; Mello,
Soares, & Paton, 2015).

23
Figura 1.4 – Áreas encefálicas ativadas durante percepção de ritmos.
Nota: PMd – Córtex pré-motor dorsal; SMA – Área motora suplementar; STG – Giro Temporal
Superior (cortex auditivo primário); basal ganglia – núcleos da base.
Fonte: Grahn & Brett (2007).
No entanto, num interessante protocolo experimental utilizando Imageamento por
Ressonância Magnética Funcional, Grahn & McAuley (2009) conseguiram criar um
protocolo de fraca ritmicidade, e assim separar participantes cuja estratégia utilizada pelos
para julgar o intervalo entre dois estímulos apresentados 1200ms após uma sequência rítmica.
Neste protocolo foi possível separar os participantes que tomavam o intervalo de tempo
discreto entre os últimos estímulos (consistente com a SET) ou a batida percebida global
(consistente com a DAT) para estimar o tempo, e comparar a ativação de áreas entre os
grupos. Em suma, os resultados indicam a ativação de áreas encefálicas da área pré-motora,
área motora suplementar (AMS), além do estriado e do cerebelo são comuns às duas
estratégias de temporização, sendo a ativação superior no grupo que inferiu fortemente a
batida (Grahn & McAuley, 2009). Teki, Grube, Kumar, & Griffiths (2011) relataram que o
sistema olivo-cerebelar esteja implicado à temporização discreta e o estriado e áreas motoras
corticais estejam mais diretamente implicado à inferência da batida em estímulos auditivos.

24
Ainda que os resultados relativos à diferenças nas áreas de ativação específicas para
cada mecanismo de temporização ainda careçam de maior replicabilidade, a participação de
áreas sabidamente relacionadas ao controle do movimento é notável, e justifica o grande
efeito da ritmicidade sobre os processos psicomotores.
Além de técnicas de imageamento cerebral por ressonância magnética funcional, se
tem tentado elucidar os fenômenos relacionados à percepção de ritmicidade e sincronização
através de métodos eletrofisiológicos, como a eletroencefalografia (EEG).
Nesse aspecto, diversos estudos têm buscado elucidar o papel das oscilações
eletroencefalográficas nos fenômenos atencionais, em contextos com ou sem ritmicidade
explícita. As evidências sugerem fortemente que ondas rápidas do tipo gama (30-100Hz)
podem constituir um modelo fidedigno para a seleção de estímulos externos a serem
processados (i.e., a atenção). A partir de resultados empíricos, está demonstrado que a
sincronização entre disparos de unidades neuronais codificadoras de um estímulo em
particular favorece o processamento desse estímulo em detrimento dos demais, construindo
assim a possibilidade do cérebro aumentar a relação sinal-ruído de um entre diversos
estímulos aferentes simultaneamente, e assim dirigir a ele (Jensen, Kaiser, & Lachaux,
2007).
O ritmo alfa (8-13Hz), classicamente atrelado à um repouso do córtex durante a vigília,
tem sido cada vez mais implicado nos fenômenos atencionais, especialmente na primeira
supressão do estímulo interveniente, e sua redução parece necessária para aparecimento do
ritmo gama que caracterizaria efetivamente o processamento do estímulo alvo (Ward, 2003).
Cabe destacar, no entanto, que nenhuma dessas bandas de frequência está na ordem dos
estímulos passíveis de arrastar as oscilações atencionais (~0.5-8Hz). Estes estudos, no
entanto, não se preocupavam em apontar o papel da ritmicidade dos estímulos sobre o
funcionamento cognitivo, assim sendo, não ficam neles elucidados possíveis mecanismos
para o acoplamento ‘ambiente-cérebro’ proposto pela DAT.
Além do espectro das frequências, outros parâmetros podem ser derivados do sinal de
EEG, como a fase, envelope de amplitude, medidas de coerência entre frequências distintas,
entre outras. Já foi mostrado que a fase de oscilações neurais podem promover ou reduzir a
detecção de um estímulo externo (Calderone, Lakatos, Butler, & Castellanos, 2014), o que
sugere que a fase de tais oscilações correspondem a correlatos neurais de processos
atencionais.

25
De fato, estudos utilizando estimulação rítmica sonora ou visual tem encontrado
consistentemente que o arrastamento e sincronização de fase de oscilações
eletroencefalográficas mais lentas - tipo delta e teta - aos estímulos, e também,
processamento perceptual e cognitivo mais rápido nessas condições. Stefanics et al. (2010)
administraram tons auditivos com ISI entre 1200 e 1500ms (0.83-0.66Hz) e encontraram
arrastamento das ondas delta, concomitantemente a melhores tempos de reação para
estímulos alvo apresentados em fase com os tons precedentes. Ondas delta no lobo occipital
também apresentaram sincronia de fase a estímulos visuais com ISI igual a 400ms, com
melhora da sensibilidade perceptual (Cravo, Rohenkohl, Wyart, & Nobre, 2013).
Aparentemente, conforme diminui o ISI (i.e., os estímulos ficam mais rápidos), as ondas delta
(0.5-4Hz) são menos capazes de sincronizar, e ondas teta (4-7Hz), mais rápidas, passam a
arrastar para sincronizar aos estímulos, sendo as oscilações atencionais provavelmente
emergentes de interações entre diversas bandas de frequências lentas e rápidas (Calderone et
al., 2014).
Com vistas à proposição de correlatos eletrofisiológicos da modulação atencional por
estímulos externos rítmicos proposta pela DAT, Henry & Herrmann (2014) hipotetizaram
distintos modos de funcionamento do sistema nervoso, a depender do contexto temporal dos
estímulos ambientais. Nesta proposta, o sistema nervoso escolheria o mais vantajoso (i.e.,
com menor custo energético) entre dois modos de funcionamento: contínuo ou rítmico. O
primeiro estaria atrelado a contextos em que a chegada dos estímulos é temporalmente
aleatória, portanto, com nenhuma ou pouca previsibilidade temporal. Ao contrário, o segundo
emergiria a partir da possibilidade de antecipar a ocorrência dos estímulos a serem
processados, e assim, permitiria ao sistema uma otimização dos recursos de processamento
para esses instantes. Eletrofisiologicamente, a modulação dos estímulos ambientais se daria
pela modulação dos níveis de excitabilidade cortical, expressos por oscilações
eletroencefalográficas de baixa frequência, em especial delta (1-4 Hz) e teta (4-7 Hz) . Estas
bandas teriam papel modulatório sobre áreas corticais geradoras das bandas rápidas do EEG
através de acoplamentos fase-amplitude, permitindo assim o controle temporal do
processamento perceptual (Figura 4) (Calderone et al., 2014; Henry, Herrmann, & Obleser,
2014).
Figura 1.5 – Processamento cortical contínuo e processamento rítmico.

26
Nota: Os retângulos acima do eixo horizontal da imagem (tempo) representam estímulos auditivos.
Quando os estímulos não tem regularidade temporal (imagem à direita), bandas lentas do sinal
eletroencefalográficos não tem efeito sobre bandas mais altas, como gama. À esquerda, vê-se que a regularidade
temporal dos estímulos induz o funcionamento em modo rítmico, ou seja, maior potência das bandas de baixa
frequência, e, mais importante, o aninhamento da amplitude das oscilações mais rápidas pela fase dessas
oscilações mais lentas (acoplamento fase-amplitude), devido à modulação da excitabilidade cortical pelas
bandas eletroencefalográficas mais lentas (ex.: delta e teta).
Fonte: Adaptado pelo autor, de Henry & Herrmann (2014).

27
CAPÍTULO 2 – COMO O CÓRTEX PRÉ-FRONTAL ATUA NO CONTROLE
EXECUTIVO DO COMPORTAMENTO
O estudo da nossa capacidade de sincronização temporal a ritmos externos é um
exemplo de como nosso sistema nervoso permite que nosso comportamento ocorra integrado
aos eventos e demandas ambientais, através da integração sensório-motora.
Integração sensório-motora pode ser definida como a forma pela qual o sistema nervoso
integra uma série de informações perceptuais registradas por diferentes modalidades
sensoriais a uma resposta motora voluntária adequada às demandas ambientais. Além de ser
um processo extremamente dinâmico, implica na comunicação funcional entre uma grande
variedade de estruturas encefálicas. Assim, fica visto que nosso extenso repertório
comportamental está subjacente à um alto nível de processamento medular-encefálico,
online2
,que envolve desde setores sensoriais primários até as vias finais comuns que
conduzem aos efetores, passando por uma gama de regiões associativas que permitem o
monitoramento, refinamento e modulação contínua da saída motora.
Machado et al. (2010) propõem três níveis, hierarquicamente organizados, e que
permitem a miríade comportamental humana. À nível medular, no extremo mais primitivo,
ocorre a integração dos estímulos sensoriais aferentes, provenientes de estruturas que
transduzem sinais do exterior (ex. tato) ou do interior do organismo (propriocepção da
musculatura), para produzir saídas motoras estereotipadas. Neste nível, estão os arcos
reflexos, ou esquemas motores de locomoção. Em segundo nível, a nível sub-cortical, áreas
do tronco encefálico, como o núcleo vestibular, os colículos superiores e a formação reticular
atuam para estabelecer o controle e correção postural axial, com a participação de regiões
cerebelares e de núcleos da base. Por último, no mais alto nível hierárquico, estão áreas
corticais associativas, recebem aferências não apenas motoras e sensoriais, mas também
cognitivas, que, através de conexões funcionais com regiões sub-corticais, permitem integrar
aspectos voluntários, mnemônicos e afetivos ao comportamento. Durante muitas de nossas
tarefas diárias, a integração realizada nos níveis primário e secundário recebe modulação
2 Define-se aqui como processo online aquele que ocorre em tempo real, continuamente, durante a
produção/execução da percepção ou ação.

28
“top-down” dos níveis hierarquicamente superiores, permitindo maior flexibilidade e
coordenação geral das respostas do indivíduo.
Via de regra, a integração sensório-motora de alto nível inclui a recepção dos sinais
aferentes pelas áreas corticais sensoriais primárias, que processam aspectos específicos de
modalidade (por exemplo, no córtex somatossenssorial: pressão, temperatura, nocicepção; ou
ainda: cor, movimento no córtex visual), sendo a seguir conduzidos à áreas associativas
unimodais, que integram essas diversas características (Machado et al., 2010). O sinal
unimodal é integrado a outras modalidades perceptivas, por uma das três áreas corticais
associativas multimodais conhecidas: área associativa posterior (córtex parietal), área
associativa límbica (córtex temporal), ou ainda, a área de associação anterior (córtex pré-
frontal). O resultado dessas operações é enviado a áreas corticais motoras com nível
associativo decrescente (área motora suplementar, área pré-motora), até chegar ao córtex
motor primário, via final comum à musculatura esquelética. Cabe salientar que este
processamento ocorre simultaneamente em paralelo e em série, de forma que a saída
comportamental final conta com participação, ainda que eventualmente desigual, de todas as
três áreas associativas multimodais (Machado et al., 2010).
Dentre as três áreas multimodais citadas, o córtex pré-frontal se destaca como aquela
responsável pela incorporação dos aspectos voluntários ao comportamento. Além de
responder de forma planejada a estímulos, o processamento pré-frontal permite que possamos
proceder à execução de ações apropriadamente dirigidas a objetivos, ou mesmo, gerar o
comportamento espontaneamente em função de motivações internas. Nesse aspecto, a seção a
seguir apresenta de forma mais detalhada a contribuição de regiões pré-frontais - e das
funções por elas subsidiadas - para a consecução de nossas atividades diárias.
2.1 O CÓRTEX PRÉ-FRONTAL
O córtex pré-frontal está localizado no lobo frontal encefálico. A definição exata das
fronteiras entre o córtex pré-frontal e áreas motoras, também sediadas no lobo frontal, é ainda
atualmente alvo de debate acadêmico, embora haja maior consenso que o córtex pré-frontal
equivale às áreas frontais com citoarquitetura histológica de alta densidade neuronal na

29
camada granular interna (IV)3 (Passimgham & Wise, 2012). Outros estudiosos, buscando
uma definição mais ampla e válida, definem o córtex pré-frontal como as áreas alvo das
projeções do núcleo médio-dorsal talâmico (Fuster, 2008).
Do ponto de vista anatômico, estão estabelecidas três principais subdivisões do córtex
pré-frontal: lateral, medial e orbitofrontal, Cada região participa de uma série de diferentes
processos cognitivos, e pode ser subdividida ainda com base na citoarquitetura, dando origem
às diversas áreas do mapa de Brodmann, por exemplo (Figura 2.1) (Fuster, 2002). Neste
aspecto, por sua implicação em uma série de processos cognitivos que serão tratados adiante
neste trabalho, damos detalhe para as áreas 9 e 46, que integram o córtex pré-frontal
dorsolateral (DLPFC, do inglês, “dorsolateral prefrontal cortex”) e áreas 45 e 12/47 de
Brodmann, que localizam o córtex pré-frontal ventromedial (VMPFC, do inglês,
“ventromedial prefrontal cortex") (Passingham & Wise, 2012). As regiões pré-frontais
possuem intensa conectividade estrutural e funcional entre si (Fuster, 2004; Fuster, 2008).
FIGURA 2.1 – Áreas do córtex frontal. Note-se a proximidade de regiões pré-frontais a regiões
motoras.
Fonte: Badre (2008)
3 Uma exceção à esta regra geral são porções orbitofrontais e mediais do córtex pré-frontal que tem
citoarquitetura disgranular.

30
Como já citado, todas as áreas pré-frontais recebem aferências do núcleo médio-dorsal
do tálamo, o que não esgota as todas as aferências que recebe de áreas sub-corticais e
corticais (Fuster, 2008). Em última análise, córtex pré-frontal recebe aferências de diversos
outros núcleos talâmicas, dos núcleos da base, cerebelo, regiões límbicas, córtices
associativos temporal e parietal posterior, e, via de regra, há reciprocidade nestas conexões
(Fuster, 2001).
Portanto, as conexões com áreas sensoriais e motoras primárias ocorrem somente de
modo indireto, através das regiões citadas. Além disso, os três núcleos pré-frontais também
conectam-se entre si, e recebem projeções das regiões produtoras dos principais sistemas de
neurotransmissores (acetilcolina, noradrelina, serotonina e dopamina), sediadas no tronco
encefálico. Em especial, o córtex pré-frontal lateral (onde está no DLPFC) é alvo direto ou
indireto de aferências sensoriais visuais, auditivas, somatossensoriais provenientes dos
córtices temporal, parietal e occipital, e em sua porção dorsolateral, possui projeções da área
46 até áreas associativas unimodais motoras (córtex pré-motor e área motora suplementar),
cerebelo, colículo superior do mesencéfalo, além dos núcleos da base, que recebem sinais de
praticamente todo o encéfalo (Miller & Cohen, 2001). Este alto nível de conectividade com
outras áreas corticais e sub-corticais denotam sua posição privilegiada como centro de
processamento e modulação da codificação perceptual, motora, cognitiva e emocional que
ocorre em outras porções do encéfalo.

31
Figura 2.2 – Principais aferências (direita) e eferências (esquerda) ao córtex pré-frontal.
Fonte: Adaptado pelo autor, de Fuster (2008)
As primeiras pistas da função do córtex pré-frontal para o comportamento são
provenientes de estudos a partir de lesões em pacientes clínicos. São exemplos famosos o do
paciente Phineas Gage (1823-1860, relatado em 1848 por seu médico, John Harlow), bem
como dos diversos pacientes que, a partir da década de 1930, foram submetidos ao
procedimento de lobotomia - secção das vias talâmicas ao córtex pré-frontal. Um número
maior de evidências, a partir de estudos com humanos e animais, e com maior refinamento
das lesões, valida três síndromes provenientes de lesões pré-frontais, correspondentes à lesão
das três grandes divisões anatômicas do córtex pré-frontal (Fuster, 2004; Negrón-Oyarzo,
Aboitiz, & Fuentealba, 2016).
Phineas Gage é um exemplo característico das lesões orbitofrontais: o paciente se torna
impulsivo, e desinibido perante as regras morais e sociais de conduta. Por isso, envolve-se
constantemente em problemas legais, e, ao realizar atividades, distrai-se facilmente perante
estímulos irrelevantes. Lesões do córtex pré-frontal medial também estão associadas a falhas
na atenção e nas reações emocionais, no entanto, são marcadas por apatia e aparente perda da
espontaneidade. As falhas na atenção decorrem da falta de motivação para empreender-se
continuamente nas tarefas. Lesões do córtex pré-frontal lateral, por sua vez, são as mais
características: o paciente passa a ter grande dificuldade para planejar ações ou lidar com a

32
sequência de atos necessários para concluir uma tarefa com sucesso. Estes déficits podem,
inclusive, acarretar dificuldade para articulação da comunicação verbal, especialmente para a
exposição de ideias novas ou de maior complexidade (Fuster, 2004). Obviamente, pacientes
clínicos podem apresentar uma mescla complexa desses sintomas, em virtude dos limites da
lesão.
A partir de uma série de evidências anatômicas e funcionais, o médico espanhol Joaquín
Fuster (1930-) tem tentado validar a tese de que a função última do córtex pré-frontal é a
organização temporal da ação (Fuster, 2008). Assim, o córtex pré-frontal como um todo não
está associado primordialmente à nenhum processo mental específico, mas sim, à
coordenação temporal de conjuntos de processos, visando executar o ciclo percepção-ação
através de respostas comportamentais adequadas e direcionadas a objetivos, especialmente
nos momentos em que esquemas previamente consolidados nos sistemas de memória não
podem totalmente responder à demanda externa (Fuster, 2001; Negrón-Oyarzo et al., 2016).
O ciclo percepção-ação concebido por Fuster ocorre através da construção de um mapa
cognitivo, ou seja, o engajamento de representações perceptuais do ambiente, bem como de
representações mnemônicas, e de repertórios de comportamentais esparsamente localizados
em diversas áreas do encéfalo que serão utilizados para compor uma resposta adaptada. Essas
representações são instáveis, devido à dinamicidade que lhes é intrínseca, inclusive para que
novos fluxos de percepção-ação sejam continuamente elaborados, em tempo real. Como já
implicitamente colocado, isto deriva da interação entre uma grande parte de estruturas
encefálicas, corticais e subcorticais, e, notadamente, da interação entre o córtex pré-frontal e
o córtex parietal posterior (Fuster, 2008).
A figura 2.3 apresenta um esquema ilustrativo simplificado desse processo, assim como
uma representação esquemática das grandes regiões corticais envolvidas.

33
Figura 2.3 – Ciclo ação percepção.
Fonte: Adaptado pelo autor, a partir de Negrón-Oyarzo et al. (2016); Fuster (2001).
Neste aspecto, do ponto de vista funcional, diversos pesquisadores têm delimitar uma
série de constructos, funções cognitivas que servem ao controle do comportamento
objetivado. Em conjunto, formam a chamada Função ou Controle executivo (FE).
2.1.1 Função Executiva
Diversos modelos neuropsicológicos tem tentado descrever o funcionamento executivo.
Cada modelo conta com volumosa documentação. Antes de uma exposição exaustiva,
Diamond (2014) sugere a existência de relativo consenso em torno de três grupos de funções
executivas fundamentais.

34
A inibição corresponde à nossa capacidade de controlar a atenção, o comportamento,
pensamentos e emoções para superar uma série de predisposições internas ou distratores
externos e realizar o que é mais apropriado ao contexto. A ausência de controle inibitório faz
com que estejamos totalmente passíveis a desejos internos ou simplesmente nos
comportemos de forma reflexa aos estímulos perceptivos mais salientes. Assim, o controle
inibitório está associado ao auto-controle, à inibição comportamental e cognitiva.
Uma segunda função fundamental, a memória de trabalho, diz respeito a nossa
capacidade de manter e manipular informações, mesmo na ausência do estímulo físico. Isto
permite representar o mundo, categorizá-lo, construir suposições, prever ou antever
acontecimentos e relacionar entre si objetos sem que haja real proximidade física ou
conceitual entre eles. Por seu conteúdo, pode ser classificada como verbal ou não-verbal.
Por fim, a flexibilidade cognitiva diz respeito a nossa capacidade de mudar
perspectivas cognitivas ou modos de resposta diante de alterações das contingencias do
ambiente. É uma função que se embasa no controle inibitório para inibir representações e
carregar outras representações na memória de trabalho, permitindo que sejamos capazes de
mudar as regras pelas conduzimos comportamento diante de mudanças das contingências do
ambiente.
A partir dessas três funções fundamentais, funções executivas superiores são
constituídas: planejamento, resolução de problemas, tomada de decisão, raciocínio lógico-
abstrato, iniciação e monitoramento da ação. No geral, o funcionamento executivo está
amplamente associado à capacidade de manejar a própria vida e obter sucesso acadêmico e
profissional (Diamond, 2014) .
O nível de fracionamento e interdependência desses constructos e a busca de regiões
discretas do córtex pré-frontal que subsidiam cada função é ainda alvo de debate. Enquanto
alguns pesquisadores utilizam as fracas correlações entre resultados de testes de diferentes
funções executivas e as associações de áreas lesionadas a déficits executivos específicos para
argumentar que é possível tratar independentemente as funções executivas (Gilbert &
Burgess, 2008), para outros, não se deve jamais perder de vista que, em indivíduos saudáveis,
a função executiva resulta das interações das diversas conexões entre áreas pré-frontais
laterais, mediais e orbitofrontais (para não falar de áreas fora do córtex pré-frontal), e,
portanto, sempre ocorre a interação e intermodulação entre as funções pré-frontais, ainda que
os níveis de contribuição de cada uma para consecução de uma tarefa específica possa ser

35
desigual (Fuster, 2008). Apesar desse debate, há ampla aceitação quanto ao refinamento
conceitual das funções executivas tratado acima.
No tocante à função do córtex pré-frontal para o comportamento, apesar da ampla
variedade de modelos cognitivistas do funcionamento executivo, a proposta de Fuster implica
que a função primordial dessa região é servir ao laço sensorial-motor, ao contrário de servir à
cognição per se. Além disso, a visão do córtex pré-frontal como integrador temporal das
sequências de processos cognitivos com vistas à conclusão de um objetivo permite supor
como somos capazes de responder online ao ambiente durante a execução de tarefas
cotidianas.
2.2 VIAS CORTICOESTRIATAIS E O CONTROLE DO COMPORTAMENTO E DA
COGNIÇÃO
Alguns pesquisadores acusam a volumosa literatura que estuda a função pré-frontal de
supervalorizar a importância das estruturas corticais no processamento sensorial e motor, em
detrimento de estruturas subcorticais, como o cerebelo e os núcleos da base, cuja filogenia
mostra inegável e conservada importância entre espécies (Koziol & Lutz, 2013). Para eles, a
visão córtico-cêntrica é especialmente perigosa, tendo em vista que, especialmente, as
questões centrais da função pré-frontal e executiva permanecem não respondidas: como o
córtex sabe quais ações ou planos selecionar para conduzir o comportamento? Que
características específicas do córtex pré-frontal tornam possível que faça isso, e em que ele
difere de regiões não executivas? Esses questionamentos em aberto dão a entender que a
função pré-frontal córtico-cêntrica se assemelha a um homúnculo (pequeno homem), vivendo
em nosso córtex pré-frontal e controlando nossas ações, o que, do ponto de vista científico
formal, não explica parcimoniosamente o controle executivo (Hazy, Frank, & O’reilly, 2007).
Em lugar do córtico-centrismo, o neuropsicólogo norte-americano Leonard Koziol vem
sugerindo uma abordagem fronto-estriatal da função executiva. A premissão fundamental do
modelo é que aquilo que se classifica como função executiva pré-frontal tem surgimento
filogenético do desenvolvimento do sistema motor e de estruturas para o controle da ação, de
forma que a dualidade entre processos motores e cognitivos neste ponto de vista é inexistente
(Koziol & Lutz, 2013). Assim, o modelo dá grande ênfase sobre a comunicação de regiões
corticais e sub-corticais, notadamente os núcleos da base.

36
A figura 2.4 demonstra as ligações (“loops”) entre o córtex e os núcleos da base. O que
se conhece por núcleos da base é uma série de quatro principais núcleos neuronais
subcorticais, bilaterais: estriado (composto pelo caudado e putâmen), pálido (composto pelo
pálido, globo pálido interno e globo pálido externo), substância negra (pars compacta e pars
reticulata) e núcleo sub-talâmico (Purves et al., 2004).
Figura 2.4 – Vias córtico-estriatais-talâmico-corticais implicadas no controle do movimento e da
cognição.
Nota: Put: putâmen; Gpe: Globo pálido externo;Gpi: Globo pálido interno; Cd: caudado; STN: Núcleo
Subtalâmico; VA/VL: Núcleos Ventroanteriores e Ventrolaterais do tálamo; MD: Núcleo Mediodorsal do
tálamo.
Fonte: Adaptado pelo autor, a partir de Koziol & Budding (2010)

37
As primeiras funções da via córtico-estriatal descritas foram as classicamente
associadas ao controle motor, especificamente na iniciação do movimento e na inibição de
movimentos indesejados. Basicamente, a ação do circuito ocorre por três vias4. Antes de
tudo, é importante saber que, apesar de receber aferentes diretos do córtex, a via de saída às
regiões motoras corticais dos núcleos da base ocorre através de núcleos talâmicos. A
comunicação desses núcleos aos núcleos da base ocorre através do Globo Pálido interno
(Gpi). Os neurônios do Gpi tem baixo limiar de disparo, o que se junta à sua transmissão
gabaérgica e resulta na inibição tônica do tálamo pelo Gpi. Por conseguinte, nesta condição
de repouso há inibição do movimento pelo circuito.
A primeira via pela qual o estriado influencia o movimento é a chamada via direta.
Aferências corticais ao putâmen causam o disparo de neurônios inibitórios ligados ao Gpi.
Isso inibe a atividade inibitória tônica sobre o tálamo e o movimento é desimpedido.
A segunda e a terceira vias resultam na inibição de movimentos indesejados. Quando
ocorre com a excitação de áreas do putâmen não ligadas ao Gpi, mas sim ao Globo pálido
externo (Gpe), há excitação da estrutura denominada Núcleo Sub-talâmico (NST). O NST
possui conexões excitatórias com o Gpi, assim, ocorre o fortalecimento da inibição tônica
deste sobre o tálamo e logo, do movimento. Tendo em vista que a ligação putâmen-Gpi é
realizada através do circuito Putamen-Gpe-NST-Gpi, esta é a chamada via indireta. A terceira
via também resulta na inibição de movimentos indesejados. Sua característica fundamental é
a ligação de regiões frontais diretamente ao STN (via hiper-direta), permitindo uma
interrupção de movimentos indesejados mais rapidamente que aquela realizada pela via
indireta (Koziol & Budding, 2010).
Koziol se apóia em evidências anatomo-histólógicas que sugerem a alta conectividade
do neocórtex ao estriado para justificar a afirmação de que a função dos núcleos da base é a
seleção ou o chaveamento de um conjunto de movimentos (representações motoras) a ser
executado. A taxa de entrada de informação cortical ao estriado é estimada entre 10 a 20 mil
células corticais por célula estriatal. O sinal converge ainda mais na ligação do putâmen ao
4 As três vias indicadas aqui são aquelas que dizem respeito à participação das vias corticoestriatais no controle
executivo/motor. Pode-se apontar ainda: a via de ligação de áreas corticais orbitofrontais e límbicas à substância
negra pars compacta, com projeções ao núcleo accumbens, e permite a avaliação de recompensas e motivação;
a via de ligação de controle óculo-motor, que conecta córtex, núcleos da base, substância negra pars reticulata e
os colículos inferior e superior do tronco encefálico. A descrição pormenorizada foge ao escopo atual, mas pode
ser facilmente encontrada em livros-texto de neurociência.

38
Gpi (aproximadamente 300:1). Isto proporciona que os núcleos da base recebam uma ampla
variedade de informações do contexto ambiental, e, caso haja similaridade das características
fundamentais do contexto com situações já vivenciadas anteriormente, pode ocorrer a
ativação dos mesmos neurônios estriatais ligados à representações corticais (neste caso,
esquemas motores) previamente estabelecidas (Koziol & Budding, 2010).
De forma idêntica ao que se aplica para o controle motor, o circuito córtico-estriatal (i.e,
pré-frontal-estriatal) teria a propriedade de selecionar ou chavear representações cognitivas
pertinentes à conclusão de objetivos, gerando a função pré-frontal. Em última análise, o
cérebro não distingue entre representações cognitivas ou motoras, aplicando a ambos os
mesmos princípios: a filogenia mostra uma conservada alocação das áreas dorsais do sistema
nervoso à sensação e áreas ventrais à motricidade, assim, pela sua localização anatômica, “o
sistema de controle cognitivo é provavelmente uma extensão evolucionária do sistema de
controle motor desempenhado pelo circuito córtex frontal-núcleos da base” (Koziol &
Budding, 2010, p.50). Dessa forma, a via córtico-estriatal direta atuaria no carregamento da
informação na memória de trabalho, e a via indireta impediria que representações distratoras
ascendam até o córtex.
Mas, o que acontece quando uma situação é entendida como nova, ou inesperada? Cabe
salientar que a utilização adequada de representações previamente consolidadas é
extremamente vantajosa: em primeiro, porque economiza recursos gastos no processamento
de informações pelo córtex; em segundo, porque, com base na primeira vantagem apontada,
ativar esquemas consolidados previamente permite maior rapidez na resposta. Assim, a
ausência de similaridade contextual com representações motoras ou cognitivas anteriormente
consolidadas faz com que seja necessário ao estriado efetivamente recrutar o córtex pré-
frontal para estabelecer novos modelos de ação.
Na verdade, esta é uma abstração: mesmo em situações novas, recorremos às nossas
experiências anteriores. Assim, é mais razoável pensar na dinâmica de um processo contínuo
e online, em que áreas pré-frontais e o estriado comunicam-se mutuamente, para recrutar um
ou mais esquemas anteriores (difusamente espalhados pelo encefálo), carregá-los na memória
de trabalho, modificá-los com base nas funções executivas fundamentais e superiores
descritas, monitorar continuamente os resultados e consolidar novos esquemas de ação.

39
CAPÍTULO 3 - EXPERIMENTO
Toca que o cérebro também dança: Efeito da ritmicidade de tons auditivos
sobre parâmetros comportamentais e eletrofisiológicos da execução de uma tarefa
de soma aritmética
3.1 JUSTIFICATIVA
A partir do exposto no capítulo 1, se pode afirmar que a ritmicidade dos estímulos
ambientais efetivamente desempenhe um papel sincronizador sobre diversos processos
encefálicos. A organização temporal do comportamento é extremamente importante
para a execução de tarefas visando atingir metas e objetivos necessários à
sobrevivência, como vimos no capítulo 2.
A coordenação temporal da atividade paralela de diversos sítios encefálicos é
executada pela interação contínua de áreas do córtex pré-frontal, especialmente o córtex
pré-frontal dorsolateral, e núcleos da base, responsáveis não apenas pelos aspectos
motores mas também cognitivos associados à tarefa. Estas duas áreas, pode-se dizer,
regulam continuamente a atividade de áreas encefálicas em função da meta a ser obtida,
com similaridade ao maestro que conduz os diversos instrumentos de uma orquestra.
Esta modulação tem ainda um importante papel de núcleos talâmicos, até então
conhecidos como um importante centro integrador das informações aferentes e eferentes
do sistema.
Os achados acumulados evidenciam ainda que os núcleos da base têm importante
para a captação ativa dos padrões temporais presentes no ambiente pelo sistema
nervoso. No tocante ao ritmo, já está demonstrada influência positiva da ritmicidade
sobre processos motores, perceptivos e cognitivos, como reconhecimento de faces
(Escoffier et al., 2010). Esta melhora do processamento se daria pelo aumento da
previsibilidade ambiental em contexto temporal rítmico.
As ligações entre os núcleos da base e o córtex pré-frontal sugerem que a
capacidade de acoplarmos nosso comportamento motor a estímulos externos rítmicos
possa surgir também em tarefas complexas de processamento executivo superior. À data
desse trabalho, no entanto, não foram encontrados relatos de melhor desempenho em

40
tarefas cognitivas superiores atrelado ao arrastamento de oscilações atencionais. Nesse
sentido, com base nas interações cortiço-talâmico-estriatais que envolvem tanto a
sincronização à ritmos como o controle cognitivo da ação pelo córtex pré-frontal
(Figura 3.1), este estudo se justifica pela necessidade de aprofundamento do
conhecimento da interação do funcionamento do sistema nervoso com padrões
temporais ambientais.
Figura 3.1 – Conexões das áreas do córtex frontal.
Nota: Notar as conexões aferentes do córtex pré-frontal dorso-lateral com áreas talâmicas (sinais
sensoriais) e eferentes a áreas estriatais/motoras (envolvidas com a percepção de ritmos).
Fonte: Adaptado pelo autor, a partir de <http://brmlab.cz/project/brain_hacking/tdcs/pfc>.

41
3.2 OBJETIVOS
3.2.1 Objetivo Geral
Caracterizar o efeito de estímulos auditivos rítmicos sobre parâmetros de
comportamento motor, desempenho cognitivo e sinais eletrofisiológicos.
3.2.2 Objetivos específicos
Descrever a existência de arrastamento do tempo motor por estímulos auditivos
rítmicos;
Descrever alterações de medidas eletrofisiológicas do sistema nervoso central e
autonômo associadas a diferentes padrões de estimulação rítmica;
Verificar se acoplamento fase-amplitude de delta a gama está associado à melhoria
de desempenho cognitivo executivo;
Verificar se a apresentação de uma demanda cognitiva executiva (soma sequencial
de dígitos) em diferentes intervalos ou em sincronia de fase à estimulação sonora
rítmica implica em processamento mais rápido ou mais acurado.

42
3.3 HIPÓTESES E PREDIÇÕES
H1 – O tempo motor espontâneo é suscetível à ritmicidade de estímulos auditivos
externos.
P1 – O tempo motor será arrastado pelo ISI dos estímulos auditivos rítmicos,
sobretudo na estimulação com ISI igual a 500 ms.
H2 – O ISI dos estímulos em uma tarefa cognitiva influencia a ativação do sistema
nervoso autônomo.
P2 – O aumento da demanda cognitiva, representado em nosso estudo pelo ISI de
apresentação dos estímulos, estará associado a maior ativação do sistema nervoso
autônomo simpático.
H3 – O contexto temporal de influxo dos estímulos está associado a diferentes perfis de
processamento cortical.
P3 – Haverá diferentes perfis na potência espectral das bandas entre diferentes
contextos temporais de nosso experimento.
H4 – Estímulos auditivos rítmicos provocam o arrastamento de fase das ondas
eletroencefalográficas da banda delta (1-4 Hz).
P4 – Uma concentração das fases de delta no instante do áudio será encontrada,
tanto na condição Sincronizada como na condição Não-Sincronizada.
H5 – O arrastamento de fase de delta representa um correlato do aumento da
excitabilidade cortical nos momentos esperados para a ocorrência do próximo estímulo.
P5 – Considerando que os instantes de maior excitabilidade cortical estão
associados aos vales da onda delta, encontraremos a concentração das fases de
delta no instante do áudio em 180º.
H6 – Melhoria do desempenho cognitivo em função do contexto rítmico dos estímulos é
mediado pelo acoplamento delta-gama.
P6 – O acoplamento fase-amplitude delta-gama estará positivamente
correlacionado ao desempenho na condição Sincronizada. Na condição Não-
Sincronizada haverá uma correlação negativa com o desempenho cognitivo, visto
que o estímulo a ser processado ocorre fora do pico atencional.
H7 – Ocorre melhor processamento cognitivo executivo quando um padrão rítmico
induzido pelos tons auditivos é apresentado em sincronia ao estímulo a ser processado.
P7 – A condição Sincronizada terá melhor número de acertos e menores tempos
de reação em relação às demais condições experimentais.

43
3.4 MATERIAIS E MÉTODOS
Os experimentos foram realizados no Laboratório de Neurobiologia e Ritmicidade
Biológica do Departamento de Fisiologia no Centro de Biociências da Universidade
Federal do Rio Grande do Norte, em sala especialmente equipada de nosso laboratório.
3.4.1 Participantes
Foi realizado cálculo amostral através do software G*Power 3.1 (Universität
Düsseldorf, 2014). Considerando o delineamento metodológico proposto (descrito
adiante), o cálculo foi realizado com base na Análise de Variância (ANOVA) de
medidas repetidas, e se obteve 24 sujeitos como tamanho amostral previsto. Os
parâmetros para a obtenção desse tamanho amostral foram: Número de grupos
independentes=1; Número de medidas repetidas = 4; Correlação entre medidas = 0.5;
Epsilon (corretor de não-esferidade) = 1 (espera-se esfericidade); alfa = 0.05; poder
estatístico = 0.8; Efeito (f) = 0.25, equivalente a η2 = 0.06 (médio efeito). Foram
critérios de inclusão: idade entre 18 e 35 anos e aceitar participar voluntariamente da
pesquisa através da assinatura do Termo de Consentimento Livre e Esclarecido (TCLE,
Apêndice 1). Foram critérios de exclusão: acuidade visual ou auditiva insuficiente para
perceber os estímulos, apresentar déficits motores centrais ou periféricos que
impossibilitem a resposta comportamental, relatar proficiência musical (ex.: saber tocar
um instrumento), estar sob uso de medicamentos com ação sobre o sistema nervoso
central (psicotrópicos), relato de uso de drogas de abuso psico-estimulantes no dia do
experimento, e não ser capaz de compreender adequadamente as instruções da tarefa.
3.4.2 Aspectos Legais e Éticos
O estudo passou por apreciação ética pelo Comitê de Ética em Pesquisa (CEP) da
UFRN, e aprovado sob o CAAE 53301516.7.0000.5537 (Anexo I). Após
esclarecimento dos objetivos e procedimentos, os voluntários expressaram
consentimento em participar da pesquisa através da assinatura do Termo de
Consentimento Livre e Esclarecido (TCLE – Apêndice 1).

44
3.4.3 Procedimentos
Os voluntários foram abordados diretamente das dependências do Centro de
Biociências, esclarecidos sobre a temática e os procedimentos da pesquisa, e conduzidos
ao laboratório para leitura e assinatura do Termo de consentimento Livre e Esclarecido
(TCLE), dando-se início à coleta de dados logo a seguir. Caso houvesse interesse em
participar da pesquisa mas não fosse possível na ocasião, era realizado o agendamento
da coleta para momento posterior. A coleta dos dados requereu a implantação de
eletrodos para coleta de dados encefalográficos, eletrocardiográficos e de condutância
de pele, a primeira mediante limpeza da pele do couro cabeludo com pasta abrasiva
específica. Os eletrodos foram implantados sobre o couro cabeludo, com impedância
inferior a sete Kilohms, com o uso de gel condutor específico para esta finalidade.
Foi realizado um balanceamento da sequência de apresentação dos blocos (350
ms, 500 ms e 650 ms) e da ordem de apresentação da tarefa de soma de dígitos
(Silêncio, Sincronizada e Não-Sincronizada). Esta randomização ocorreu por sorteio da
ordem ao qual cada participante deveria se submeter, sem reposição. A seguir, as
instruções da tarefa eram fornecidas e três rodadas de teste com ISI igual a 1500 ms
eram realizadas. Todos os participantes compreenderam adequadamente a tarefa ao final
das três rodadas de teste.
3.4.3.1 Instrumentos
A apresentação dos estímulos foi realizada através do software Superlab 5.0,
conectado à caixa de resposta RB-740 (Cedrus Corporation). O registro de EEG foi
realizado pelo sistema de aquisição de dados biológicos Powerlab 8/30 e Bio-
amplificador Octal ML138 (ADInstruments), com 8 canais. Visando a confiabilidade
temporal do momento de apresentação dos estímulos, o marcador de eventos Cedrus
Stimtracker foi utilizado para marcar o começo dos estímulos visuais e auditivos
diretamente no traçado eletrofisiológico. O registro da atividade eletrocardiográfica
(ECG) e eletroculográfica (EOG) foi realizado em um segundo módulo Powerlab 26T,
com dois canais e bioamplificador integrado. O módulo GSR Amp (ADInstruments) foi
utilizado para registro da condutância de pele. Todo o registro eletrofisiológico descrito
foi efetuado através do software Labchart 7.

45
3.4.3.2 Registro dos Sinais Biológicos
O registro eletroencefalográfico contínuo foi realizado com eletrodos Ag-AgCl, à
taxa de amostragem de 1000Hz, com filtragem analógica passa-banda entre 0.1 e
100Hz, e filtro notch para minimizar interferência da rede elétrica (60Hz). Os eletrodos
foram posicionados no escalpo conforme o Sistema Internacional 10-20. Neste sistema,
é medida a distância entre a depressão anterior formada pela intersecção do osso frontal
e dos ossos nasais (nasion) e a protuberância posterior do osso occipital do crânio
(inion), e intervalos de 10% ou 20% dessa distância localizam cada ponto. Sabidamente,
o EEG possui pequena resolução espacial, no entanto, a localização dos eletrodos sobre
áreas corticais de interesse possibilita melhor razão sinal/ruído. Assim, foram eleitas as
localizações Fz,F3,F4,Cz,C3,C4, com derivação unipolar, referência no processo
mastoide e terra na fronte (Fpz), para o monitoramento da atividade do córtex pré-
frontal, áreas motoras e associativas parietais. O registro de ECG também foi realizado
de forma contínua, com uso de eletrodos descartáveis e derivação bipolar DII. A
condutância de pele foi registrada através do posicionamento de eletrodos Ag-AgCl
para registro de GSR nas falanges dos dedos da mão não-dominante.
Os sujeitos foram instruídos evitar movimentos não relacionados ao desempenho
da tarefa, o que poderia introduzir no registro artefatos decorrentes de potenciais
musculares ou movimentação dos eletrodos.
3.4.4 Estímulos
3.4.4.1 Estímulos Auditivos
Os estímulos auditivos (pulsos sonoros) foram ondas senoidais com frequência de
660Hz, duração de 50ms, apresentados através de fones de ouvido, com intensidade de
aproximadamente 60db. Para a criação desses tons, foi utilizado o editor de áudio
Audacity 2.1.2. O intervalo entre estímulos em cada etapa é isocrônico (igualmente
espaçado no tempo), com intervalo de apresentação dependente do bloco, conforme
indicado na seção protocolo experimental.

46
3.4.4.2 Estímulos Visuais
Os estímulos visuais doravante denominados “dígitos” consistiram em imagens
dos algarismos “1”, “2” e “3”, fonte “Arial Rounded”, tamanho 120, em branco sobre
tela negra. Estes estímulos foram randomicamente exibidos a cada dois pulsos auditivos
isocrônicos, em sincronia ou não com a ocorrência do som, o que depende da condição
experimental. Os estímulos visuais denominado “limiar da soma”, correspondem à
imagens dos números “12”, “13”,”14”, fonte “Arial Rounded”, tamanho 120, em
branco sobre tela negra logo abaixo do dígito. Este estímulo mudava randomicamente a
cada rodada de soma.
3.4.5 Tarefas
Cada bloco experimental será composto pelas tarefas de: (1) Tempo motor
espontâneo; (2) soma aritmética sequencial – doravante denominada “Tarefa de soma”,
descritas abaixo em detalhe.
3.4.5.1 Tarefa de Tempo Motor Espontâneo
Ao início de um dos três cada blocos experimentais, uma tarefa de “finger-
tapping” será utilizada para acessar o tempo motor espontâneo e a capacidade do
participante para sincronizar e inferir a batida dos pulsos sonoros. O participante será
instruído a pressionar regularmente o botão de resposta, com a mão dominante, na
velocidade que lhes for mais confortável, ou seja, “nem rápido, nem lento”.
Ao final de 31 sequências motoras (gerando 30 intervalos de tempo espontâneo),
pulsos sonoros isocrônicos com intervalos de tempo controlado (ver a seção protocolo
experimental) serão apresentados, enquanto o participante continua a realizar “finger-
tapping”. Não haverá instrução explícita ao participante para sincronizar a taxa ou a
fase de “finger-tapping” aos estímulos sonoros. Com isso, desejamos verificar se
ocorrerá sincronização espontânea, e qual a estratégia de sincronização utilizada pelos
participantes.
3.4.5.2 Tarefa de soma aritmética

47
Após a tarefa de tempo motor espontâneo, os participantes receberão, na tela, uma
dica visual (asterisco - *), indicando a preparação para o inicio da tarefa de soma. Esta
tarefa foi adaptada a partir do protocolo clínico de avaliação neuropsicológica “Paced
Auditory Serial Addition Test” (PASAT). A versão original do teste apresenta dois
dígitos aleatórios (0 a 9), com intervalo de 3 ou 2 segundos entre eles, e compete ao
participante fornecer ao avaliador o resultado da soma entre os dígitos. Sabidamente,
menor intervalo de tempo entre os dígitos dificulta a tarefa
(http://www.pasat.us/PDF/PASAT_Manual.pdf).
O participante foi instruído a somar mentalmente os dígitos, que seriam
sequencialmente apresentados. Quando a soma acumulativa fosse igual ou superior ao
estímulo “limiar da soma” , deveria pressionar o botão de respostas o mais rápido
possível. A mão dominante foi utilizada para esta resposta.
Um esquema ilustrativo dos estímulos e da tarefa estão representados na figura
3.2.
Figura 3.2 – Estímulos visuais da tarefa de processamento executivo (exemplo).
Nota: Um asteristico (*) sinaliza início da rodada de adição. O número superior refere-se ao dígito
(entre 1 e 3, aleatoriamente apresentado a cada dois pulsos sonoros), o número inferior refere-se ao limiar
da soma (entre 12 e 14, aleatório a cada rodada). No momento em que o somatório acumulado dos dígitos
for igual ou superior ao limiar da soma, o participante deve pressionar o botão e preparar-se para nova
rodada.
Fonte: Autor
3.5 PROTOCOLO EXPERIMENTAL

48
O experimento em três grandes blocos experimentais (Figura 3.3), com ISI igual a
350 ms, 500 ms ou 650 ms. Um balanceamento dessas condições foi realizada mediante
sorteio da ordem de apresentação dos estímulos a cada participante. Tendo em vista o
esforço mental requerido pela tarefa, um intervalo de tempo entre os blocos, com até 3
minutos, poderá ser concedido ao participante que assim desejar.
Figura 3.3 – Protocolo experimental do estudo.
Nota: A porção superior da figura apresenta dos três blocos experimentais, definidos pelo
Intervalo entre estímulos (ISI). Todos os participantes foram submetidos aos três blocos mostrados, de
forma balanceada (randomizada). A porção inferior apresenta em detalhe a sequência das tarefas dentro
do bloco. Note-se que os blocos se diferenciam tão somente pelo intervalo entre estímulos.
Fonte: Autor
Conforme a figura 3.3, cada bloco se constitui pela realização da tarefa de tempo
motor espontâneo sem som, seguida da tarefa de tempo motor na presença dos
estímulos com ISI referente ao bloco (350 ms, 500 ms ou 650 ms). O ponto de
exclamação indica a advertência dada aos participantes, para encerrar o tapping e
preparar para a tarefa de soma.
Visando estudar o efeito da fase dos estímulos auditivos sobre o processamento
executivo, os tons foram apresentados em três condições distintas. Condição

49
Sincronizada: apresentação de estímulos auditivos e visuais coincidem (Figura 3.4,
esquerda); Condição Não-Sincronizada: apresentação de estímulos auditivos e visuais
tem um deslocamento de fase entre si (Figura 3.4, direita); Condição Silêncio:
apresentação dos estímulos visuais ocorre sem estimulação auditiva.
Figura 3.4 – Esquema ilustrativo da Tarefa Sincronizada e Tarefa Não-Sincronizada
Fonte: Autor
3.6 ANÁLISE DE DADOS
Os dados comportamentais e eletrofisiológicos de cada sujeito foram registrados
pelos aplicativos Superlab e Labchart, respectivamente. Esses arquivos foram
processados para correção dos acertos e análise de sinais, através de rotinas
computacionais desenvolvidas no software Matlab 2013a (Mathworks Inc.). Estas
rotinas podem ser obtidas mediante solicitação ao autor, pelo e-mail
Os detalhes para cada um dos parâmetros obtidos está exposto nas seções a seguir.

50
3.6.1 Dados Comportamentais
Os dados brutos registrados para cada participante são: os 30 intervalos entre
preensões do botão de resposta da tarefa de tempo motor espontâneo, os 30 intervalos
entre preensões do botão de resposta da tarefa de sincronização à batida, os estímulos
apresentados durante a tarefa de soma, e o tempo de reação (TR) desde o começo do
último “dígito” apresentado ao sujeito.
A partir dos dados brutos, o tempo motor espontâneo de cada sujeito (Período) foi
definido como a média aritmética dos intervalos entre preensões do botão de resposta,
excluindo-se o primeiro intervalo, ou seja:
∑ (1)
Onde: SMT é o tempo motor espontâneo médio, n é o número total de intervalos
de tempo motor espontâneo a serem promediados (30), ΔSMTi é o intervalo de tempo
entre a i-ésima preensão do botão de resposta e a preensão anterior.
Além do período motor, alterações da fase do comportamento motor provocadas
pela ritmicidade dos eventos externos foi medida pela Assincronia Média, a diferença
entre o instante do estímulo externo (tons auditivos) e o instante do “tapping” (Repp &
Su, 2013). O desvio-padrão dos “taps” indica a variabilidade atrelada ao
comportamento motor. O coeficiente de variação (CV) é também uma medida usual em
estudos com “finger-tapping”, e representa a dificuldade do sujeito para sincronizar a
um estímulo externo. É definido neste trabalho como a razão entre a média e o desvio
padrão dos 30 ΔSMT da formula anteriormente citada.
Com relação à tarefa de soma, foi definido como acerto a preensão do botão entre
100 e 800 ms após o aparecimento do estímulo “dígitos” cuja soma acumulada fosse
maior ou igual ao “limiar da soma”. A partir disso, a taxa de acertos e a média dos
tempos de reação foram obtidos para cada condição experimental.
3.6.2 Condutância de pele

51
O sinal bruto de condutância de pele foi processado através de rotinas
customizadas em MATLAB, através do toolbox Ledalab para análise de dados de
condutância de pele, v.4.3.9 (Benedek & Kaernbach, 2010). O sinal teve a frequência de
amostragem reduzida para 10 Hz e foi corrigido para artefatos através do método de
Gauss, ambas as funções embutidas no pacote Ledalab. O parâmetro de saída utilizado
foi a condutância tônica da pele (CDA.Tonic), que evidencia a ativação do sistema
nervoso autônomo simpático em dentro de cada condição experimental (Boucsein,
2012).
3.6.3 Eletrocardiografia (ECG)
Com relação ao sinal de eletrocardiograma, utilizamos o módulo HRV do
Labchart 7 para calcular os intervalos R-R durante todo o experimento. Estes valores
foram exportados para formato .csv, e importados para tratamento em MATLAB. Com
base em rotinas customizadas, os intervalos foram separados de acordo com a condição
experimental, e foram removidos os intervalos ectópicos, definidos como aqueles
maiores que 1000 ms e menores que 600 ms. Estes intervalos foram substituídos através
de interpolação por “spline” cúbico (função interp1).
Com base nos dados pré-processados, os seguintes parâmetros de saída foram
calculados para cada condição experimental: média dos intervalos R-R; desvio-padrão
dos intervalos R-R (SDNN); raiz quadrada dos quadrados das diferenças entre R-R
sucessivos (RMSSD).
3.6.4 Dados Eletroencefalográficos (EEG)
O sinal de EEG foi inicialmente importado para MATLAB através do
ADInstruments (LabChart) SDK
(https://www.mathworks.com/matlabcentral/fileexchange/50520-adinstruments--
labchart--sdk).
O sinal de EEG, particularmente nas regiões frontais, está sujeito à captação de
artefatos biológicos não provenientes do córtex, tais como o movimento ocular, ou sinal
cardíaco. Assim, os dados foram submetidos à remoção de artefatos oculares e cardíacos

52
por Análise de componentes independentes (ICA, do inglês Independent Component
Analysis) através do toolbox EEGLAB (Delorme & Makeig, 2004). Para a realização da
ICA, foram seguidas as orientações do roteiro de pré-processamento de Makoto
(https://sccn.ucsd.edu/wiki/Makoto%27s_preprocessing_pipeline). O sinal foi filtrado
entre 1 e 100 Hz, e teve a frequência de amostragem reduzida para 250 Hz. A seguir, foi
feita a inspeção visual e a rejeição de janelas com artefatos não estereotipados, que
podem prejudicar a correta identificação dos artefatos. O plugin CleanLine
(https://www.nitrc.org/projects/cleanline/) foi utilizado para remover eventual
contaminação de sinal de 60 Hz proveniente da rede elétrica. Finalmente, a ICA foi
realizada, com o algoritmo “binica.m”.
Os componentes foram rejeitados a partir da inspeção visual dos componentes
com espectro de frequências suavemente decrescente, da distribuição topográfica
frontal e da identificação de alta amplitude do sinal compatível com piscadas ou
movimento ocular. Outros componentes suspeitos não foram excluídos, tendo em vista a
manutenção das características fundamentais do sinal.
A partir do sinal pré-processado, para cada condição e bloco, as potências
espectrais foram calculadas através do método de Welch. Este método consiste em
dividir a série temporal em estudo em m segmentos, e realizar a promediação das
potencias espectrais obtidas pela Transformada Rápida de Fourier de cada segmento
(Figura 3.5). Se comparado à simples execução da transformada em toda a série
temporal, este método resulta em menor ruído no espectro de potências resultantes.
Figura 3.5 – Ilustração do método de Welch para cálculo das potências espectrais.

53
Fonte: Krejcar & Frischer (2011)
A obtenção do sinal filtrado por banda se deu através da utilização da função
eegfilt. Delta foi definido como a banda entre 1 a 4 Hz, e Gama como 30 a 50 Hz. Com
vistas unicamente à obtenção dos potenciais evocados pelo som, foi realizada a
filtragem do sinal bruto entre 1 e 20 Hz, e a posterior promediação intra-sujeito, seguida
da promediação entre sujeitos.
O arrastamento da fase de delta pelos estímulos auditivos foi estimado pela
obtenção da fase instantânea dessa banda no momento dos pulsos auditivos, através da
transformada de Hilbert. Estes valores foram apresentados através de diagramas de
roseta, e comparados estatisticamente através do teste de Rayleigh para uniformidade,
disponível no toolbox CircStat (Berens, 2009).
O acoplamento fase-amplitude entre baixas e altas frequências do sinal de EEG é
um importante aspecto dos objetivos desse trabalho. Existem diversos métodos para
quantificar este acoplamento, mas ainda não há coesão quanto ao melhor deles (Tort,
Komorowski, Eichenbaum, & Kopell, 2010). No entanto, todos os métodos iniciam com
a decomposição do sinal original de EEG nas suas bandas constituintes, e extrair a fase
da série temporal de baixa frequência (representada pela letra grega Phi: Φ) e o
envelope de amplitude do série temporal de alta frequência (A), e plotar um histograma
de A em relação a Φ . Assim, é possível observar visualmente o acoplamento entre Φ e
A (Figura 3.6E).

54
Figura 3.6 – Mensuração do Acoplamento fase-amplitude
Nota: Primeiramente, extrai-se a fase instantânea de um sinal de baixa frequência, teta (Fig. C,
Φfp) e do envelope de amplitude de um sinal de alta frequência, baixo-gama (Fig. D, Afa), a partir de sinal
original (Fig.A, xraw). A plotagem do histograma de Afa contra Φfp (Fig. E) demonstra a existência de
maior amplitude de gama em torno da fase 180º de teta.
Fonte: Adaptado pelo autor, de Tort, Komorowski, Eichenbaum, & Kopell (2010)
A fase e a amplitude dos sinais filtrados podem ser definidas como a fase angular
e a magnitude da representação analítica do sinal pela transformada Hilbert. Além da
visualização do acoplamento pelo histograma (Fig. 3.6E), Tort, Komorowski,
Eichenbaum, & Kopell (2010), após revisão dos métodos de avaliação do acoplamento
fase-amplitude entre dois sinais, propuseram o cálculo de um Índice de Modulação,
entre 0 e 1, para quantificar o acoplamento fase-amplitude. Considerando que este
índice depende do tamanho da série temporal, estabelecemos seu valor através de z-
scores, considerando distribuições randômicas do Índice de Modulação com
acoplamento nulo em séries de mesma extensão (“Surrogate tests”). Através desse
procedimento, esperamos estimar o quanto a fase de delta modula a amplitude das ondas
gama na presença dos estímulos isocrônicos.
3.6.5 Análise estatística
Os parâmetros computados em Matlab foram exportados para comparação
estatística no SPSS for Windows 20. As variáveis foram submetidas ao teste de
normalidade de Shapiro-Wilk, constatando-se a quebra do parâmetro de normalidade

55
(p<0,05) para todas elas, exceto para os índices comportamentais de acertos e tempo de
reação.
Assim, optamos pelo uso de estatísticas não paramétricas na maioria de nossas
análises. A mediana será utilizada como medida de tendência central, e a Distância
Inter-quartil (IQR, do inglês, “Inter Quartile Range”) como medida de dispersão. O
intervalo de confiança de 95 % da mediana foi definido como a 1,57 x IQR/ raiz
quadrada (n). Especificamente para os dados de acertos e tempo de reação durante a
tarefa de soma, a média, o desvio-padrão e o Intervalo de confiança de 95 % da média
serão priorizados.
Os dados obtidos foram comparadas através do teste não-paramétrico de
Friedman, e submetidos a comparações planejadas pelo teste de Wilcoxon, sob correção
de Šidák. A correlação de Spearman foi utilizada para buscar associações entre o Índice
de Modulação e o desempenho cognitivo. Especificamente para os dados paramétricos
relativos ao desempenho, foi utilizado um modelo misto de Análise de variâncias, com
os fatores Sexo e Condição experimental, seguido por testes posthocs (correção de
Šidák). O teste de Kruskal-Wallis foi pontualmente utilizado para realizar comparações
entre participantes submetidos a diferentes ordens experimentais.

56
3.7 RESULTADOS
Participaram do estudo 24 indivíduos saudáveis, homens e mulheres com idade
entre 18 e 28 anos. A tabela 3.1 apresenta uma caracterização dos participantes com
relação à idade, sexo, lateralidade relatada e consumo de cafeína nas 4 horas prévias ao
experimento.
Tabela 3.1 – Caracterização da amostra do estudo
Masculino Feminino Total
n (%) 14 (58,3%) 10(41,7%) 24
Idade média (DP) 20,42 (1,82) 22,4(3,09) 21,21(2,57)
Lateralidade relatada (canhotos) 3 (21,4%) 1 (10%) 4(16,7%)
Consumo de cafeína até 4 horas
antes do experimento (%)
1 (7,1%) - 1 (4,1%)
Tarefa de Tempo Motor Espontâneo
Conforme descrito na seção Método, ao início de cada bloco experimental os
sujeitos foram instruídos a executar uma tarefa de “finger tapping”, sendo os primeiros
31 toques realizados sem qualquer estímulo e os 31 toques restantes realizados na
presença de tons auditivos isocrônicos, com ISI igual a 350ms, 500ms ou 650ms, a
depender do bloco experimental. A tabela 3.2 apresenta os principais parâmetros que
caracterizam a resposta dos voluntários com relação ao período e variabilidade das
respostas motoras, assim como indicadores estatísticos de eventuais diferenças entre o
“tapping” concomitante ou não concomitante à a apresentação de tons isocrônicos com
ISI igual a 350ms.
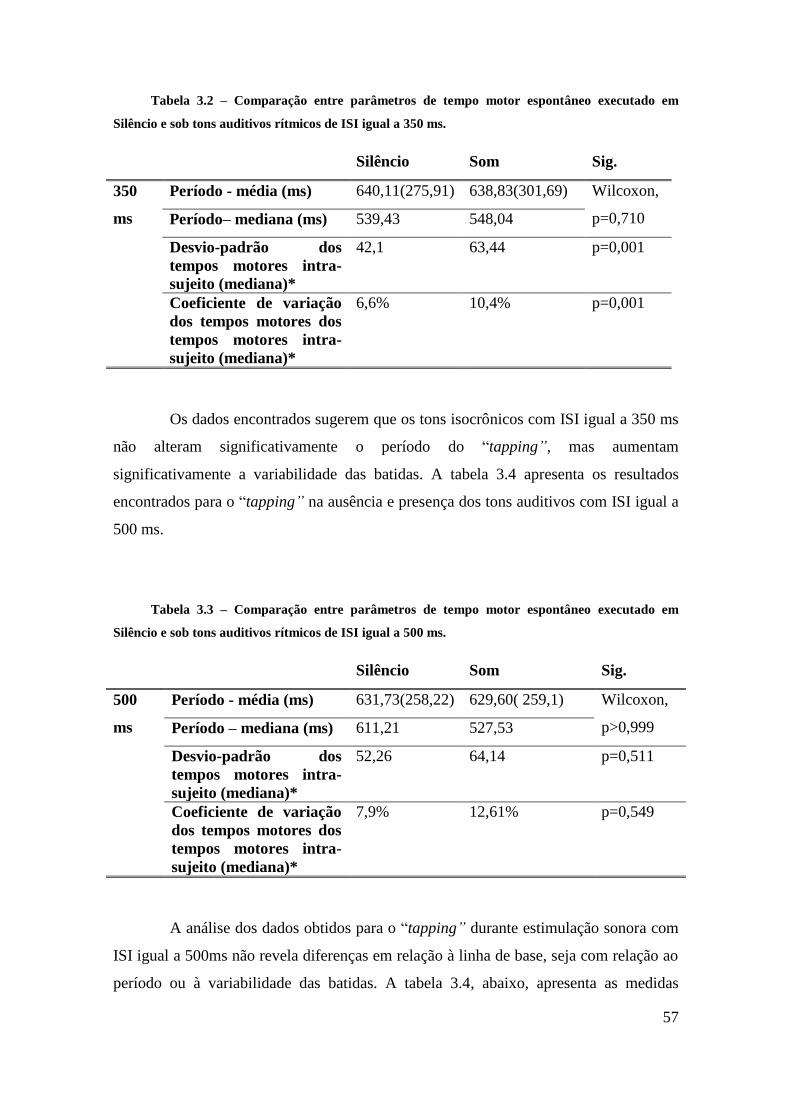
57
Tabela 3.2 – Comparação entre parâmetros de tempo motor espontâneo executado em
Silêncio e sob tons auditivos rítmicos de ISI igual a 350 ms.
Silêncio Som Sig.
350
ms
Período - média (ms) 640,11(275,91) 638,83(301,69) Wilcoxon,
p=0,710 Período– mediana (ms) 539,43 548,04
Desvio-padrão dos
tempos motores intra-
sujeito (mediana)*
42,1 63,44 p=0,001
Coeficiente de variação
dos tempos motores dos
tempos motores intra-
sujeito (mediana)*
6,6% 10,4% p=0,001
Os dados encontrados sugerem que os tons isocrônicos com ISI igual a 350 ms
não alteram significativamente o período do “tapping”, mas aumentam
significativamente a variabilidade das batidas. A tabela 3.4 apresenta os resultados
encontrados para o “tapping” na ausência e presença dos tons auditivos com ISI igual a
500 ms.
Tabela 3.3 – Comparação entre parâmetros de tempo motor espontâneo executado em
Silêncio e sob tons auditivos rítmicos de ISI igual a 500 ms.
Silêncio Som Sig.
500
ms
Período - média (ms) 631,73(258,22) 629,60( 259,1) Wilcoxon,
p>0,999 Período – mediana (ms) 611,21 527,53
Desvio-padrão dos
tempos motores intra-
sujeito (mediana)*
52,26 64,14 p=0,511
Coeficiente de variação
dos tempos motores dos
tempos motores intra-
sujeito (mediana)*
7,9% 12,61% p=0,549
A análise dos dados obtidos para o “tapping” durante estimulação sonora com
ISI igual a 500ms não revela diferenças em relação à linha de base, seja com relação ao
período ou à variabilidade das batidas. A tabela 3.4, abaixo, apresenta as medidas

58
estatísticas para o “tapping” na condição 650 ms. Nota-se que, assim como para os
blocos anteriores, as pistas temporais auditivas com ISI igual a 650 ms não tiveram
impacto significativo sobre o período médio. Quanto às medidas de variabilidade,
embora a significância estatística para o desvio-padrão e o coeficiente de variação
tenham sido limítrofes em relação valor de significância estabelecido, diferença foi
encontrada apenas para o desvio-padrão médio.
Tabela 3.4 – Comparação entre parâmetros de tempo motor espontâneo executado em
Silêncio e sob tons auditivos rítmicos de ISI igual a 650 ms.
Silêncio Som Sig.
650
ms
Período - média (ms) 658,58 666,72 Wilcoxon,
p=0,493 Período– mediana (ms) 550,23 588,58
Desvio-padrão dos
tempos motores intra-
sujeito (mediana)*
48,44 86,95 p=0,049
Coeficiente de variação
dos tempos motores dos
tempos motores intra-
sujeito (mediana)*
9% 12,77% p=0,043
A média encontrada para os períodos motores espontâneos nas três sessões de
“finger tapping” sem som 566 ms, e coeficiente de variação igual a 7,83%. Não foram
encontradas diferenças nos parâmetros de “finger tapping” descritos acima entre
diferentes tempos de estimulação (ISI 350,500 ou 650) (Friedman, p>0,05). Realizar
“finger tapping” no primeiro, segundo ou terceiro bloco experimental não tem efeito
sobre o período motor (Friedman, p>0,05). Para a tarefa de “finger tapping”, não havia
qualquer estímulo que diferenciasse os blocos. Coerentemente, não foram encontradas
diferenças entre sujeitos com ordem de apresentação diferenciada (Kruskal-Wallis,
p>0,05) (Tabela 3.5).
Tabela 3.5 - Comparação entre a primeira, segunda e terceira sessão de “finger tapping”
realizada pelos sujeitos (mediana).

59
Ordem de apresentação
1 2 3 p (Kruskal-Wallis)
ISI 350 492,55 644,38 725,03 0,88
ISI 500 519,55 677,03 644,38 0,88
ISI 650 813,56 521,03 491,01 0,41
p (Friedman) 0,43 0,92 0,65
Nota: Note-se que não há estímulo atrelado aos tempos motores apresentados, sendo os ISI
indicados apenas para referir ao bloco experimental em que o “tapping“ se insere.
A diferença entre o “tapping” realizado com a introdução das pistas auditivas
sonoras também pode ser apresentada através da estimativa da razão média entre esta
condição e o “tapping” espontâneo, que antecedeu o início do som. A figura 3.6
apresenta a mediana e o IC 95% da estimativa, assim como o resultado dos testes de
significância realizados.
Figura 3.6 – Razão entre parâmetros de tempo motor na condição com som rítmico e em silêncio.
Nota: Razão entre o desvio-padrão médio encontrado para a condição de “tapping” espontâneo e
o desvio-padrão encontrado para a condição Experimental (A); Razão entre o Coeficiente de variação
médio encontrado para a condição de “tapping” espontâneo e o Coeficiente de variação médio
encontrado para a condição Experimental (B); Razão entre o período das batidas para a condição de
“tapping” espontâneo e para a condição Experimental (C). Asteriscos indicam diferença significativa em
relação ao valor neutro, 1.
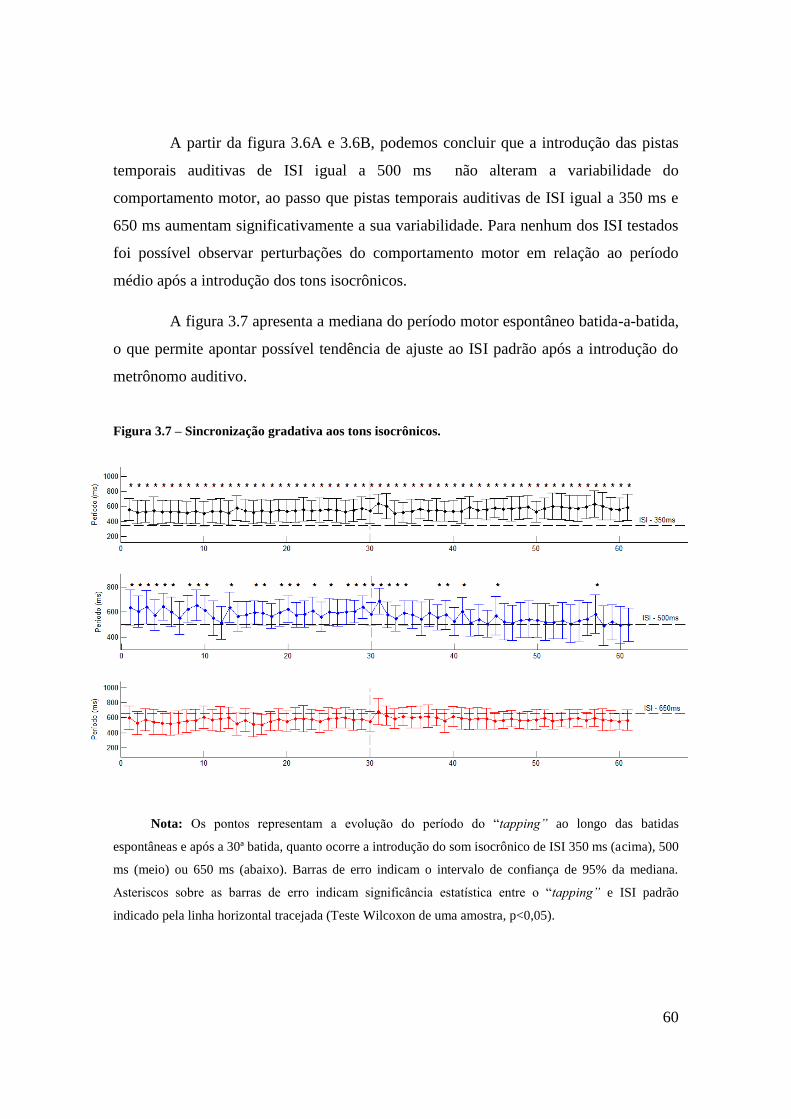
60
A partir da figura 3.6A e 3.6B, podemos concluir que a introdução das pistas
temporais auditivas de ISI igual a 500 ms não alteram a variabilidade do
comportamento motor, ao passo que pistas temporais auditivas de ISI igual a 350 ms e
650 ms aumentam significativamente a sua variabilidade. Para nenhum dos ISI testados
foi possível observar perturbações do comportamento motor em relação ao período
médio após a introdução dos tons isocrônicos.
A figura 3.7 apresenta a mediana do período motor espontâneo batida-a-batida,
o que permite apontar possível tendência de ajuste ao ISI padrão após a introdução do
metrônomo auditivo.
Figura 3.7 – Sincronização gradativa aos tons isocrônicos.
Nota: Os pontos representam a evolução do período do “tapping” ao longo das batidas
espontâneas e após a 30ª batida, quanto ocorre a introdução do som isocrônico de ISI 350 ms (acima), 500
ms (meio) ou 650 ms (abaixo). Barras de erro indicam o intervalo de confiança de 95% da mediana.
Asteriscos sobre as barras de erro indicam significância estatística entre o “tapping” e ISI padrão
indicado pela linha horizontal tracejada (Teste Wilcoxon de uma amostra, p<0,05).

61
A partir da figura 3.7A, podemos observar que não existe qualquer indício de
ajustamento do período 350ms, o que é indicado pela diferença estatística entre as
medianas e este valor. O mesmo pode ser visto até a 30ª batida da figura 16B, quando
são introduzidos tons isocrônicos com ISI igual a 500 ms. A partir de então, observamos
que a estimativa tende e se torna consistentemente igual a 500 ms. Os dados obtidos
com relação ao ISI igual a 650ms (Figura 3.7) não permitem um resultado conclusivo
quanto à sincronização ao som, já que tanto o “tapping” espontâneo (sem qualquer
som) quanto sob estimulação são estatisticamente iguais ao período do som de ISI 650
ms. O mesmo ocorreu quanto ao possível ajustamento do comportamento motor à 2ª
harmônica da condição 350 ms (700ms) também foi testado, também sem um resultado
conclusivo do ponto de vista estatístico, embora a sequência da série indique que esta
sincronização ocorra para um número maior de batidas que as estudadas nesse trabalho.
Este resultado parece indicar que o ISI igual a 500 ms tem maior força como
sincronização do “tapping”.
Além do período, podemos verificar alterações da fase do comportamento motor
provocadas pela ritmicidade dos eventos externos, através da assincronia média. A
figura 3.8 apresenta os resultados encontrados.
Figura 3.8 – Assincronia Média entre “tapping” e estímulos auditivos rítmicos
Nota: À esquerda, a assincronia média (instante do “tapping”- instante do evento externo (tom
auditivo rítmico), cujo valor 0 indica “tapping” coincidente ao instante do estímulos auditivos. À direita, a

62
média do módulo dessas diferenças, que expressa a diferença temporal entre o “tapping” e o instante dos
estímulos auditivos. Os altos valores apontados na batida 30 resultam da alteração do som em virtude do
início do próximo bloco experimental .
Tarefa de soma de dígitos – Condutância de pele
Com relação à segunda tarefa que compunha cada bloco experimental, (tarefa de
soma de dígitos), a figura 3.9 demonstra as medianas e o intervalo de confiança de 95
%, através dos boxplots abaixo. Tendo em vista a quebra do parâmetro de normalidade
dos dados (Shapiro-Wilk, p>0,05), foi utilizado o teste de Friedman para abordar
eventuais efeitos do ISI e das condições de estimulação auditiva rítmica (Silêncio,
Sincronizada, Não-Sincronizada). A análise não permite rejeitar a hipótese nula de
medianas iguais para todas as condições testadas (p>0,05).
Figura 3.9 – Níveis tônicos de condutância da pele durante as nove condições experimentais
de realização da tarefa de soma.
Nota: Os entalhes representam o Intervalo de confiança de 95% da mediana.
Com relação ao sinal de EcG, três parâmetros foram estudados: Intervalo R-R,
SDNN, e RMSSD. A comparação estatística mostra um aumento do Intervalo RR na
condição Não-Sincronizada em relação à condição Sincronizada, sem outras alterações
na frequência e variabilidade cardíaca (Friedman, p>0,05) (Figura 3.10, 3.11 e 3.12).

63
Figura 3.10 – Intervalo R-R durante as nove condições experimentais de realização da tarefa
de processamento executivo (soma de dígitos).
Nota: Os entalhes representam o Intervalo de confiança de 95% da mediana. Chave e asterisco
ligando dois box-plots indicam significância estatística.
Figura 3.11 – Desvio-padrão dos intervalos RR normais (SDNN) durante as nove condições
experimentais de realização da tarefa de processamento executivo (soma de dígitos). Os entalhes
representam o Intervalo de confiança de 95% da mediana.

64
Figura 3.12 – Raiz da média dos quadrados da diferença sucessiva entre os intervalos RR
(RMSSD) durante as nove condições experimentais de realização da tarefa de processamento
executivo (soma de dígitos). Os entalhes representam o Intervalo de confiança de 95% da mediana.
Tarefa de soma de dígitos - Dados Eletroencefalográficos (EEG)
Durante a tarefa, foi realizado o registro da atividade cortical através de sete
eletrodos eletroencefalográficos. As figuras 3.13, 3.14 e 3.15 apresentam os potenciais
evocados pelo som (Média geral) durante as condições Sincronizada e Não-
Sincronizada. A amplitude máxima encontrada para as respostas evocadas não superou
1.1 uV.

65
Figura 3.13 – Respostas evocadas pelo áudio e componentes evocados propostos – Canais Frontais
Nota: As áreas sombreadas em amarelo representam significância estatística (teste t dependente, correção de Šidák).

66
Figura 3.14 – Respostas evocadas pelo áudio e componentes evocados propostos – Canais Centrais
Nota: As áreas sombreadas em amarelo representam significância estatística (teste t dependente, correção de Šidák).

67
Figura 3.15 – Respostas evocadas pelo áudio e componentes evocados propostos – Canal Parietal
Nota: As áreas sombreadas em amarelo representam significância estatística (teste t dependente,
correção de Šidák).
As figuras 3.16, 3.17 e 3.18 apresentam a mediana entre-sujeitos dos
periodogramas, representações dos sinais registrados durante a tarefa executiva no
domínio da frequência em todas as condições estudadas. Além da tendência 1/f, que
caracteriza o sinal eletroencefalográfico representado no domínio da frequência, a
análise visual não sugere picos de frequência que suportem diferenças no perfil
eletroencefalográfico entre nenhum dos ISI e manipulações experimentais testadas. Um
pico saliente na frequência 60 Hz é visto no canal Fz em todas as condições testadas,
provavelmente proveniente da rede elétrica que não pode ser dilapidado através das
técnicas de filtragem e redução de ruído utilizadas. É notável ainda o surgimento de um
pico nos canais centrais e parietal, com frequência 10Hz, que sugere a existência de
oscilações na banda de alfa (8-13 Hz) durante todas as sessões experimentais (incluindo
a linha de base).
A análise visual do periodograma, ainda que importante, não permite um bom
dimensionamento do efeito de diferentes fatores sobre a atividade eletroencefalográfica.
Para tanto, foi realizada a análise do incremento da potência espectral de cada banda em
cada condição, ou seja, o quociente entre a potência espectral de cada banda na
condição experimental e a potência espectral na condição linha de base. Os resultados
estão apresentados por canal, nas figuras 3.19, 3.20 e 3.21.
Os resultados encontrados nas figuras 3.19, 3.20 e 3.21 apontam diferenças no
padrão eletroencefalográfico durante a realização da tarefa executiva, em comparação à
linha de base. Realizar a tarefa sob ISI igual a 350 ms implica aumento de atividade da
banda gama (30-50 Hz) no porção central esquerda (C3) do encefálo, ao passo que

68
realizar a tarefa sob ISI igual a 500 ms tem efeito positivo sobre a banda delta (1-4 Hz)
em Cz ,C4 e Pz. Sob ISI igual a 650 ms, há um efeito negativo sobre delta em ambos
hemisférios do lobo frontal (F3 e F4), e no eletrodo central do lado direito (C4).

69
Figura 3.16 – Mediana das potências espectrais dos sete eletrodos eletroencefalográficos (F3,Fz,F4,C3,Cz,C3,Pz), durante as três condições experimentais da
tarefa executiva com ISI igual a 350 ms. Para efeito de comparação, um bloco de base também é mostrado.

70
Figura 3.17 – Mediana das potências espectrais dos sete eletrodos eletroencefalográficos (F3,Fz,F4,C3,Cz,C3,Pz), durante as três condições experimentais da
tarefa executiva com ISI igual a 500 ms. Para efeito de comparação, um bloco de base também é mostrado.

71
Figura 3.18 – Mediana das potência espectrais dos sete eletrodos eletroencefalográficos (F3,Fz,F4,C3,Cz,C3,Pz), durante as três condições experimentais da
tarefa executiva com ISI igual a 650 ms. Para efeito de comparação, um bloco de base também é mostrado.

72
Figura 3.19 – Incremento de potência das bandas eletroencefalográficas em relação à condição basal – canais frontais.
Nota: Valores superiores a 1 indicam aumento, valores inferiores a 1 indicam diminuição. Os entalhes indicam a aproximação do intervalo de confiança de 95% da
mediana (+- 1,57*IQR/sqrt(n)). A presença do símbolo # sobre a barra indica diferença estatística em relação ao valor de nulo incremento, 1, indicado pela linha horizontal
tracejada.

73
Figura 3.20 – Incremento de potência das bandas eletroencefalográficas em relação à condição basal – canais centrais.
Nota: Valores superiores a 1 indicam aumento, valores inferiores a 1 indicam diminuição. Os entalhes indicam a aproximação do intervalo de confiança de 95% da
mediana (+- 1,57*IQR/sqrt(n)). A presença do símbolo # sobre a barra indica diferença estatística em relação ao valor de nulo incremento, 1, indicado pela linha horizontal
tracejada.

74
Figura 3.21 – Incremento de potência das bandas eletroencefalográficas em relação à condição
basal – canal parietal.
Nota: Valores superiores a 1 indicam aumento, valores inferiores a 1 indicam diminuição. Os
entalhes indicam a aproximação do intervalo de confiança de 95% da mediana (+- 1,57*IQR/sqrt(n)). A
presença do símbolo # sobre a barra indica diferença estatística em relação ao valor de nulo incremento,
1, indicado pela linha horizontal tracejada.
Acoplamento de delta aos estímulos
O arrastamento da fase de delta aos estímulos foi testado através de medidas de
estatística circular. Um trecho demonstrativo do sinal registrado está apresentado na
figura 3.22.

75
Figura 3.22 – Época demonstrativa do sinal delta, gama, e da fase de delta (1-4 Hz) e envelope
de amplitude de gama (30-50 Hz) extraídos do sinal bruto (~6s, Condição Sincronizada 500 ms)
As figuras 3.23 e 3.24 apresentam a distribuição uniforme encontrada nas fases de
delta no momento do áudio, e os p-valores associados (teste de Rayleigh para
uniformidade). Não foi encontrada orientação das fases de delta para o instante dos
pulsos sonoros rítmicos em nenhum dos sítios ou condições testadas (p>0.05).

76
Figura 3.24 – Fase de delta no momento do áudio.
Nota: Os diagramas de roseta apresentam o histograma da fase de delta no instante dos tons isocrônicos na condição Sincronizada, para cada sítio e ISI testados.

77
Figura 3.24 – Fase de delta no momento do áudio.
Nota: Os diagramas de roseta apresentam o histograma da fase de delta no instante dos tons isocrônicos na condição não-Sincronizada, para cada sítio e ISI testados.
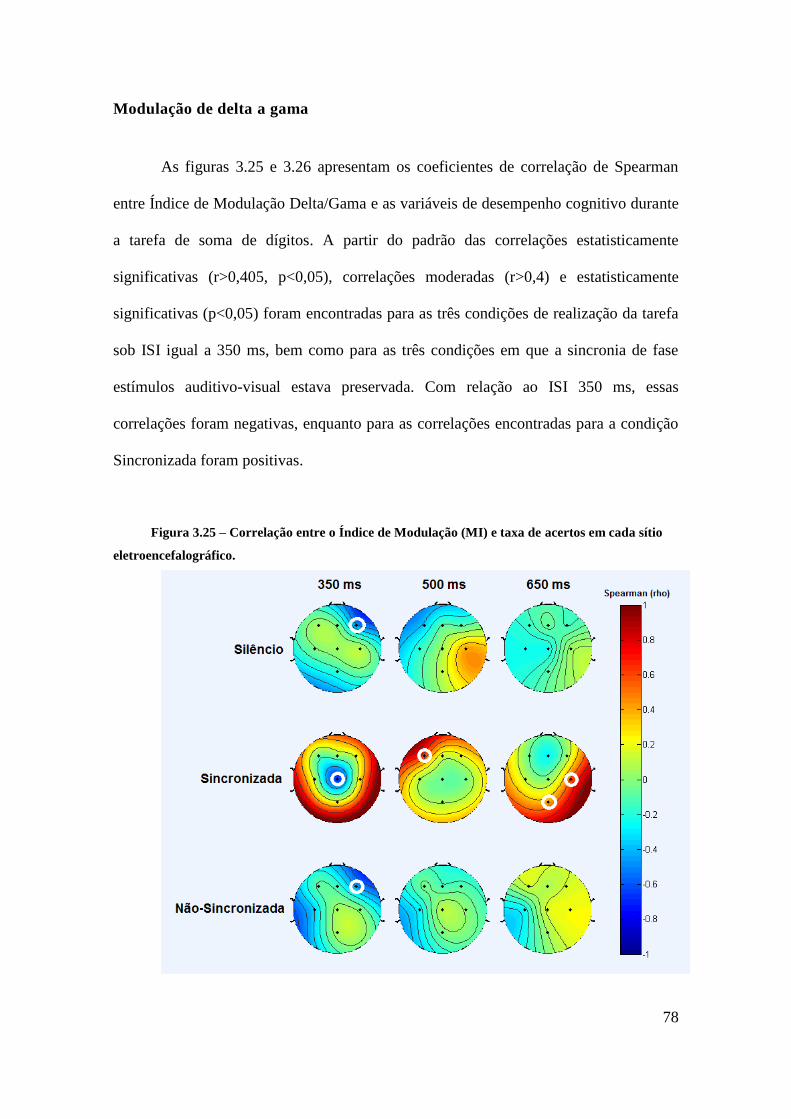
78
Modulação de delta a gama
As figuras 3.25 e 3.26 apresentam os coeficientes de correlação de Spearman
entre Índice de Modulação Delta/Gama e as variáveis de desempenho cognitivo durante
a tarefa de soma de dígitos. A partir do padrão das correlações estatisticamente
significativas (r>0,405, p<0,05), correlações moderadas (r>0,4) e estatisticamente
significativas (p<0,05) foram encontradas para as três condições de realização da tarefa
sob ISI igual a 350 ms, bem como para as três condições em que a sincronia de fase
estímulos auditivo-visual estava preservada. Com relação ao ISI 350 ms, essas
correlações foram negativas, enquanto para as correlações encontradas para a condição
Sincronizada foram positivas.
Figura 3.25 – Correlação entre o Índice de Modulação (MI) e taxa de acertos em cada sítio
eletroencefalográfico.

79
Figura 3.26 – Correlação entre o Índice de Modulação (MI) e o tempo de reação (RT) em
cada sítio eletroencefalográfico
Tarefa de processamento executivo (Soma de dígitos) – Acertos e Tempo de
reação
Conforme descrito em detalhes na seção Método, cada um dos três blocos
experimentais do estudo (ISI=350 ms, ISI=500 ms e ISI =650 ms) era iniciado pela
tarefa de “finger tapping” e seguido por uma tarefa de processamento executivo, a soma
de dígitos sequencialmente apresentados. Os voluntários foram expostos à realização da
tarefa sob três diferentes condições: “Silêncio”, na qual era desempenhada sem qualquer
estímulo sonoro; “Sincronizados”, na qual tons isocrônicos com o ISI padrão do bloco
era apresentado em fase com o dígito para o qual deve ocorrer a adição mental; “Não-
sincronizados”, quando o estímulo auditivo (tons isocrônicos) e visual (dígito) eram
apresentados em completa assincronia de fase (para maiores detalhes, ver seção
Método). O efeito do fator Condição (Silêncio/ Sincronizados/Não-sincronizados) e

80
Sexo (M/F) foi testado através de um modelo de ANOVA fatorial mista. O teste de
Mauchly indica que a hipótese de esferidade não foi violada (χ2 (35)=41,89, p=0,21), e
no que couber os resultados serão relativos à esfericidade considerada. Os resultados
mostram não haver efeito do Sexo (F(1,20)=0,84, p=0,775,η² parcial=0,004) ou da
Interação Sexo*Condição sobre a taxa de acertos (F(1,20)=1,2,p=0,3, η² parcial=0,052).
A Condição experimental possui efeito sobre a taxa de acertos (F(8,176)=20,64,
p<0,001,η² parcial=0,484). A figura 3.27 apresenta os resultados no que tange à taxa de
acertos à tarefa em cada condição testada e os resultados obtidos para as comparações
par-a-par.
Figura 3.27 – Taxa de acertos na tarefa de processamento executivo (soma sequência de
dígitos), nas condições Silêncio, Sincronizada e Não-Sincronizada e ISI 350 ms, 500 ms e 650 ms.
Nota: Barras de erro apresentam o Intervalo de confiança (95%). Letras diferentes sobre as
barras indicam diferença estatística entre elas (ANOVA mista, testes posthoc com correção de Šidák e
p<0,05).
A partir da figura, nota-se que a taxa de acertos na tarefa com ISI igual a 500
ms ou 650 ms não diferem entre si, em nenhuma das condições, mas que a taxa de
acertos desses blocos foram significamente superiores às do bloco de ISI igual a 350
ms. Para este bloco, a condição Silêncio foi aquela em que os sujeitos obtiveram o
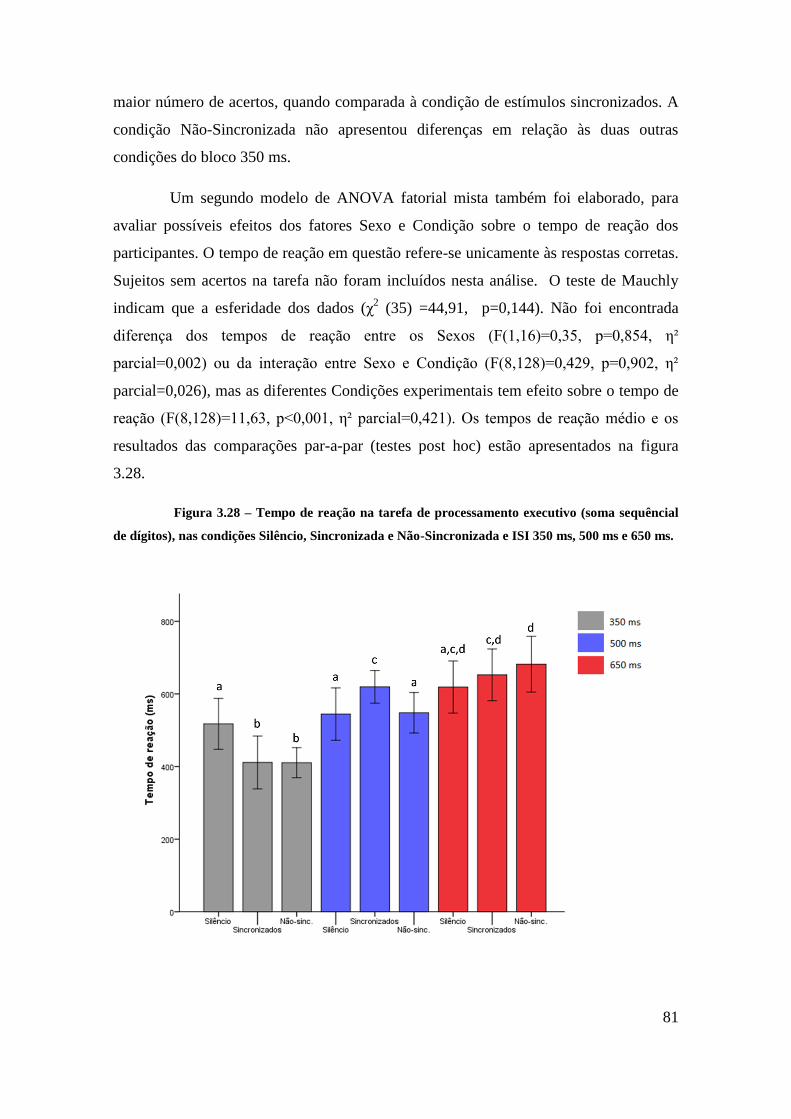
81
maior número de acertos, quando comparada à condição de estímulos sincronizados. A
condição Não-Sincronizada não apresentou diferenças em relação às duas outras
condições do bloco 350 ms.
Um segundo modelo de ANOVA fatorial mista também foi elaborado, para
avaliar possíveis efeitos dos fatores Sexo e Condição sobre o tempo de reação dos
participantes. O tempo de reação em questão refere-se unicamente às respostas corretas.
Sujeitos sem acertos na tarefa não foram incluídos nesta análise. O teste de Mauchly
indicam que a esferidade dos dados (χ2 (35) =44,91, p=0,144). Não foi encontrada
diferença dos tempos de reação entre os Sexos (F(1,16)=0,35, p=0,854, η²
parcial=0,002) ou da interação entre Sexo e Condição (F(8,128)=0,429, p=0,902, η²
parcial=0,026), mas as diferentes Condições experimentais tem efeito sobre o tempo de
reação (F(8,128)=11,63, p<0,001, η² parcial=0,421). Os tempos de reação médio e os
resultados das comparações par-a-par (testes post hoc) estão apresentados na figura
3.28.
Figura 3.28 – Tempo de reação na tarefa de processamento executivo (soma sequêncial
de dígitos), nas condições Silêncio, Sincronizada e Não-Sincronizada e ISI 350 ms, 500 ms e 650 ms.

82
Nota: Barras de erro apresentam o Intervalo de confiança (95%). Letras diferentes sobre as barras
indicam diferença estatística entre elas (ANOVA mista, testes posthoc com correção de Šidák e p<0,05).

83
3.8 DISCUSSÃO
Neste trabalho, buscamos entender como padrões de estimulação rítmica influenciam o
sistema nervoso. Para tanto, o experimento foi composto por duas tarefas, executadas sob três
períodos de apresentação entre estímulos: 350 ms, 500 ms, 650 ms.
A primeira tarefa, “finger tapping”, foi inicialmente realizada espontaneamente pelos
voluntários (sem nenhum estímulo). Após 31 toques, tons auditivos rítmicos com ISI
controlado eram apresentados ao voluntário, que não recebia qualquer instrução para
pressionar o botão de resposta sincronizado ao som. O principal objetivo desta tarefa foi
avaliar alterações no padrão motor associadas ao som. A segunda tarefa, destinada a avaliar
possíveis alterações no processamento cognitivo associadas à ritmicidade dos estímulos
auditivos, consistia numa versão alterada de uma tarefa clínica de avaliação da memória
operacional (PASAT), uma soma sequencial de dígitos numéricos. Dígitos de 1 a 3 eram
apresentados na tela, em intervalos controlados (700 ms, 1000 ms ou 1300 ms), e os sujeitos
eram instruídos a pressionar o botão de resposta quando a soma atingisse um valor pré-
determinado. Esta tarefa foi executada pelos participantes em três condições experimentais
distintas, e eram registrados os acertos e tempo de reação. Em duas, tons rítmicos isocrônicos
de intervalo controlado (350 ms, 500 ms e 750 ms) eram apresentados juntamente aos dígitos.
Contudo, a condição “Sincronizada” implicava que o estímulo auditivo (som rítmico) e visual
(dígitos) tinham sincronização temporal, ou seja, eram entregues ao sujeito no mesmo
instante; a Condição “Não-Sincronizada”, por sua vez, implicava que o estímulo auditivo e
visual possuíam deslocamento de fase entre si. Uma última condição, denominada “Silêncio”,
consistia na apresentação dos dígitos sem qualquer som.
A média dos três períodos motores espontâneos (i.e., sem som) encontrada é de 566
ms, com ampla variação entre sujeitos, mas conservada estabilidade intra-sujeito nas 3
sessões de “finger tapping”. Este resultado é consistente com em diversos achados que
situam o tempo preferido na casa dos 500 ms, ou 2 Hz, (Delevoye-Turrell, Dione, &
Agneray, 2014; MacDougall & Moore, 2005; McAuley et al., 2006; Moelants, 2002).
Não foram encontradas diferenças significativas entre o desvio-padrão ou coeficiente
de variação das batidas nas três sessões experimentais, o que a reforça a afirmação de

84
estabilidade intra-sujeito da medida. A variabilidade das batidas é constantemente atrelada
aos aspectos motores e atencionais subsidiadas pelo circuito córtico-estriatal (Schwartze,
Keller, Patel, & Kotz, 2011), e diversos estudos descrevem coeficientes de variação na casa
de 4 a 7 % para sujeitos normais (Janzen, 2009; McAuley et al., 2006; Schwartze et al.,
2011), 7% para pacientes com lesões nos núcleos da base (Schwartze et al., 2011) e 6% para
pacientes bipolares (Bolbecker et al., 2011). Nossos resultados sugerem coeficientes de
variação mais elevados, entre 6,6 a 9%, com média nas três sessões igual a 7,83%.
Assim, o alto coeficiente de variação encontrado em nossos resultados parece mostrar
lentificação psicomotora ou baixos níveis de atenção associados à participação no
experimento. Considerando que o período motor espontâneo encontrado é consistente com
achados anteriores, conferimos maior plausibilidade à última proposição. Nesta mesma linha,
cabe salientar que a coleta dos dados tomou lugar precisamente entre 15h e 19h, fase do dia
sabidamente marcada por baixos níveis de atenção e alerta (Valdez et al., 2005).
A introdução do som não teve efeito sobre a mediana dos períodos motores antes e
após o som, mas aumentou significativamente a variabilidade das batidas nas condições 350
ms e 650 ms. Aparentemente, os estímulos auditivos rítmicos com periodicidade diferente do
tempo preferido (500 ms) tendem a exercer papel como estímulos distratores da atenção.
O estudo do período motor ao longo da tarefa demonstra efeito sincronizador dos tons
com ISI igual a 500 ms sobre o tempo motor. Obviamente, o presente resultado não exclui
que o efeito sincronizador dos períodos 350 e 650 ms venha a tona após um maior número de
batidas na presença do som. No entanto, fica evidenciado que o período 500 ms possui maior
força de sincronização sobre o “finger tapping”. Conforme exposto na figura 3.8, no entanto,
observamos que a diferença temporal entre o instante do áudio e do “tapping” se mantém
constante ao longo do experimento, assim, este efeito não foi observado com relação à fase
do ”tapping”.
Cabe salientar que, havendo arrastamento do tempo motor pelos estímulos, duas
estratégias poderiam ser utilizadas para sincronizar a ação à ritmicidade ambiental: reset de
fase ou arrastamento. Enquanto o primeiro pode ser observado por uma abrupta mudança no
padrão de ritmicidade para acoplar ao evento externo, o conceito de arrastamento pressupõe a
adaptação gradual do sistema oscilatório ao evento externo, o que pode levar vários ciclos
(Roenneberg et al., 2005). O efeito de arrastamento encontrado para a estimulação com ISI

85
igual a 500 ms é consistente com a proposta de oscilador endógeno proposto pela “Dynamic
Attending Theory” (Jones & Boltz, 1989; E. Large & Jones, 1999).
Com relação à essa capacidade de sincronização motora, McAuley et al. (2006)
sugerem a existência de uma “região de arrastamento”, uma faixa de intervalos entre
estímulos na qual somos capazes de sincronizar de forma ótima, i.e., com pequenos
coeficientes de variação. Os três intervalos abordados no presente estudo estão dentro dessa
faixa. Assim, a partir do aumento da variabilidade das batidas após o início dos tons com ISI
igual a 350 e 650 ms (não observado para 500 ms), e da ausência de efeito de arrastamento
desses intervalos sobre o comportamento motor (presente para ISI igual a 500 ms), propomos
que a região de arrastamento é ainda mais estreita se excluído o componente voluntário da
sincronização, e tem centro em aproximadamente 500 ms (2 Hz).
Antes de passar a uma análise pormenorizada dos resultados eletrofisiológicos e
comportamentais obtidos para a tarefa de soma de dígitos, faz-se necessário abordar os
aspectos neuropsicológicos associados à sua realização. A tarefa requer que os sujeitos sejam
capazes de manter, de 5 a 10 segundos por rodada, simultaneamente ativas a representação da
soma no instante atual, do limiar da soma da rodada, e, além disso, operar continuamente
sobre a representação da soma no eixo do tempo, quando ocorre a chegada de um novo
dígito. Considerando todo um bloco experimental, ou seja, as condições Sincronizada, não-
Sincronizada e Silêncio, temos a realização sustentada destas operações por
aproximadamente 30 rodadas, equivalentes a 1 a 3 minutos, a depender do ISI de cada bloco.
Obviamente, os sujeitos poderiam empregar diferentes estratégias cognitivas para
cumprir essa tarefa. Operações aditivas simples, envolvendo dois valores de dígito único,
como “1+3”, não estão calcadas em processamento executivo, mas sim em sistemas
mnemônicos, o que evidencia que esse tipo de operação aritmética simples incorre na
consulta aos chamados “fatos matemáticos” (Grabner et al., 2009). Embora a tarefa de soma
utilizada em nosso experimento seja baseada na adição de dígitos simples, esta era
acumulativa, com limite entre 12 e 14, e, portanto, implica na capacidade de operar adições
entre valores de um e dois dígitos. Assim, acredita-se que a tarefa não poderia ser resolvida
através da recuperação mnemônica de fatos matemáticos, mas sim, esteja associada ao
funcionamento de circuitos fronto-parietais (Grabner et al., 2009; Mihulowicz, Willmes,
Karnath, & Klein, 2014), consistente com a utilização de processos cognitivos de controle
executivo, em especial, memória de trabalho (Cragg & Gilmore, 2014).

86
Erros nesta tarefa também poderiam resultar comportamentos impulsivos, como
pressionar o botão antes de realizar a soma. Estes são resultado de falhas no controle
inibitório, e, do ponto de vista qualitativo, normalmente acompanhavam expressões de
frustração do participante, tão logo o erro era percebido. Embora não tenhamos abordado
medidas objetivas deste fenômeno, a maior parte desses comportamentos ocorreu durante a
realização da tarefa com ISI igual a 350 ms.
Em nosso estudo, utilizamos parâmetros da atividade eletrodermal (também conhecida
como condutância da pele), frequência e variabilidade cardíaca para estimar o impacto da
demanda cognitiva sobre a ativação de sistemas autonômicos (sistema nervoso simpático e
parassimpático) em diferentes contextos temporais. Nossos resultados indicam e que os
parâmetros de condutância tônica da pele e variabilidade cardíaca (SDNN,RMSSD) se
mantém estáveis ao longo das condições testadas, no entanto, há um aumento do intervalo R-
R entre as condições Sincronizada e Não-Sincronizada durante a realização da tarefa com ISI
igual a 350 ms.
Com exceção da diminuição da frequência observada entre a tarefa Sincronizada e Não
sincronizada, os dados sugerem que não há impacto do intervalo de apresentação dos
estímulos ou da presença de som (sincronizado às estímulos visuais ou não) sobre o sistema
nervoso autônomo simpático e parassimpático. Embora já tenha sido aventada a possibilidade
de interação do tempo musical com índices de variabilidade cardíaca como correlatos da
capacidade da música induzir emoção (Wang & Huang, 2014), nossos resultados não
suportam essa afirmação.
A participação de sistemas autonômicos em tarefas de cunho cognitivo é também uma
possível perspectiva para estes resultados. Especificamente com relação ao controle
executivo, alguns teóricos fazem a distinção entre funções executivas “frias” e “quentes”,
sendo as primeiras associadas às funções cognitivas associadas ao córtex pré-frontal
dorsolateral, e as últimas, aos córtices pré-frontal ventromedial e orbitofrontal (Chan, Shum,
Toulopoulou, & Chen, 2008). Nossos resultados parecem corroborar a independência dos
componentes “frios” e “quentes” do controle executivo, já apontada por outros estudos
(Zimmerman, Ownsworth, O’Donovan, Roberts, & Gullo, 2016).
A baixa amplitude das respostas evocadas pelo som (<1.1 uV) indica que a ritmicidade
dos estímulos auditivos teve pequeno efeito sobre a amplitude das bandas, incluindo delta.

87
Além disso, foram encontradas diferenças estatisticamente significativas entre a condição
Sincronizada e Não-Sincronizada, sugerindo a existência de polaridades opostas na resposta
evocada pelo som nas duas condições. Em F3, sob ISI igual a 350 ms, por exemplo,
sugerimos o aparecimento dos componentes P1 e N2 atrelados à condição Sincronizada, e
N1-P2 atrelados à condição Não-Sincronizada. Acreditamos que o protocolo do estudo, no
qual dois estímulos – em vez de apenas um estímulo, como é comum em protocolos
“oddball” típicos de estudos de potenciais evocados - são entregues para processamento
cortical explique esse achado. No entanto, a replicação e elucidação desse fenômeno é digna
de melhor aprofundamento experimental.
Os resultados encontrados apontam diferenças no padrão eletroencefalográfico
durante a realização da tarefa executiva, em comparação à linha de base. Os dados sugerem
que realizar a tarefa sob ISI igual a 350 ms implica aumento de atividade da banda gama (30-
50 Hz) no porção central esquerda (C3) do encefálo, ao passo que realizar a tarefa sob ISI
igual a 500 ms tem efeito positivo sobre a banda delta (1-4 Hz) em Cz ,C4 e Pz, e sob ISI
igual a 650 ms tem efeito negativo sobre delta em ambos hemisférios do lobo frontal (F3 e
F4), assim como em C4.
Embora o EEG possua baixa resolução espacial, a localização dos eletrodos
eletroencefalográficos faz prevalecer o sinal proveniente de certas áreas encefálicas, em
detrimento de áreas mais distantes (Burle et al., 2015). Dessa forma, a localização topográfica
e o sentido das diferenças (aumento ou diminuição) encontrada para as bandas delta e gama
pode ser utilizada para inferir suposições quanto ao processamento cortical durante a
realização da tarefa. Nesse sentido, os sítios eletroencefalográficos frontais laterais utilizados
neste estudo (F3 e F4) estão anatomicamente posicionados à porção média do giro frontal, ao
passo que os eletrodos centrais (C3 e C4) sob o giro pós-central, enquanto Fz, Cz, Pz
respectivamente sobre a linha média entre os giros superiores frontais, os giros centrais e os
giros superiores parietais (Kabdebon et al., 2014).
Assim, os resultados encontrados parecem sugerir que haja diferenças no
processamento cortical diante dos três intervalos de estimulação rítmica. Nesse aspecto, cabe
salientar que os parâmetros para estabelecer a linha de base foram os mesmos utilizados para
a tarefa em si, exceto pelo fato que, nesta última, os estímulos visuais apresentados ao sujeito
foram dígitos para processamento executivo em vez de estímulos inócuos de sentido (ex.:
“&”,”%”, ou ”@”). Portanto, creditamos as diferenças no padrão cortical à realização da

88
tarefa sob diferentes contextos rítmicos (ISI e sincronia dos estímulos auditivos e visuais), e
não à ritmicidade dos estímulos auditivos ou visuais em si mesmos.
A ativação da banda gama tem sido associada a diferentes processos perceptuais,
motores e cognitivos. Embora não haja consenso quanto aos limites precisos, há relativo
consenso que frequências acima de 30 Hz constituem oscilações gama. Propõe-se que o
surgimento dessas oscilações rápidas é transitório, e está ligado ao funcionamento de grupos
de assembleias neuronais locais envolvidas na interligação de diferentes características dos
estímulos como um todo coerente. Esta propriedade fundamental coloca a atividade gama
induzida pelo estímulo como possível correlato da preparação de áreas sensório motoras ao
movimento, a percepção de estímulos e a atenção (Tallon-Baudry & Bertrand, 1999). Já foi
demonstrado aumento transitório da atividade gama (20-60 Hz) em eletrodos fronto-centrais
a partir de estímulos auditivos rítmicos com ISI igual a 390 ms (Snyder & Large, 2005).
Embora este fator não tenha sido objeto de análise estatística, a atividade induzida parece
estar restrita ao hemisfério esquerdo, consistente com nossos resultados. Neste aspecto, o
aumento de gama nos eletrodos na porção central esquerda, isolado à apresentação dos
estímulos em ISI 350 ms, parece sugerir que há uma alocação de recursos para antecipar o
tempo de processamento sensório-motor em condições nas quais é necessária uma rápida
resposta do sistema.
Nossos resultados demonstram ainda incremento da atividade da banda delta (1-4 Hz)
atrelada à tarefa de soma com ISI igual a 500 ms e decréscimo da atividade desta banda
durante a realização da tarefa sob ISI 650 ms. A distribuição topográfica diverge nos dois
casos, sendo significativa nos eletrodos C4, Cz e Pz para a tarefa em 500 ms e F3,F4,C4 para
650 ms.
Como descrito em nossos resultados relativos ao tempo motor nos três ISI
investigados, o intervalo de 500 ms é aquele com maior força de arrastamento sobre do
comportamento motor. O aumento de delta na porção central-parietal direita do encefálo
durante a realização da tarefa executiva sob intervalo indica aumento dessa banda em áreas
sensoriomotoras uni e multimodais (Machado et al., 2010; Purves et al., 2004).
Aumento da potência de delta nos lobos frontais é associado a diversos processos
cognitivos, em especial, a inibição de estímulos intervenientes à tarefa (Harmony, 2013).
Assim, a diminuição bilateral de delta em áreas pré-frontais parece indicar que realizar a
soma aritmética sob ISI igual a 650 ms – portanto, sob menor demanda de velocidade de

89
processamento - requer menor controle inibitório cognitivo. Este resultado é coerente com a
maior variabilidade do tapping após a introdução das pistas auditivas, relatada acima.
Sob ISI 500 ms, especificamente na região parietal o aumento foi observado somente
na condição Silêncio. O aumento de potência espectral de delta é comumente associado a
tarefas cognitivas, aspectos motivacionais e de recompensa, mas há um maior volume de
evidências de sua função em áreas frontais (Harmony, 2013; Knyazev, 2007, 2012) e poucas
evidências quanto ao papel ou aos correlatos comportamentais, cognitivos ou perceptuais de
delta no córtex parietal. Conforme Knyazev (2007, 2012), este papel pode ser inferido pela
contribuição de oscilações na frequência delta sobre o potencial evocado P300. O potencial
evocado P300, ou P3b, é resposta elétrica cortical positiva, com latência entre 300 e 400 ms e
máxima amplitude no córtex parietal, evocado por estímulos inesperados ou salientes.
Portanto, a maior atividade delta em Pz, encontrada neste estudo, pode estar vinculada à ação
de circuitos fronto-parietais ligados ao deslocamento da atenção para eventos considerados
relevantes e o consequente carregamento desses na memória de trabalho (Knyazev, 2012; S.
Patel & Azzam, 2005).
Assim como para o córtex parietal, há poucos relatos do papel funcional de delta em
áreas motoras ou somatossensoriais que permitam uma discussão com o aumento dessa banda
nos eletrodos C4 e Cz durante o ISI igual a 500 ms. É necessário notar que no hemisfério
encefálico direito, este aumento está associado unicamente às condições em que o estímulo
auditivo estava presente. Considerando que a maior parte de nossos voluntários é destro
(83%), e que a tarefa exigiu a resposta com a mão dominante, o incremento no hemisfério
direito parece não ter associação direta com a resposta motora da tarefa. Nesse aspecto,
propomos que os estímulos auditivos rítmicos com ISI igual a 500 ms (2 Hz) tenham atuado
sobre o sistema nervoso aumentando a potência espectral da banda delta no hemisfério motor
não-dominante. Esta proposição, em linhas gerais, corrobora a hipótese de “ressonância
neural” como codificadora da percepção de ritmos (Large & Snyder, 2009). Ademais, nossos
resultados parecem mostrar que a demanda cognitiva imposta pela tarefa de soma exerce
papel modulatório sobre este fenômeno.
Os resultados referentes à análise espectral do sinal eletroencefalográfico durante a
tarefa de soma de dígitos sob 3 diferentes ISI e 3 diferentes relações de fase entre os
estímulos visuais e auditivos rítmicos parece evidenciar que diferentes estratégias são postas
em ação, a depender da velocidade de processamento que é demandada ao sistema nervoso. O

90
aumento de delta em áreas motoras não-dominantes, observado durante as condições
Sincronizada e Não-Sincronizada de ISI 500 ms é um possível correlato da codificação neural
da ritmicidade dos estímulos. Os resultados referentes à banda delta nestas áreas parecem
indicar um padrão em ‘U invertido”, com incremento máximo de delta no ponto ótimo
próximo 500 ms.
Lakatos, Chen, O’Connell, Mills, & Schroeder (2007) propõem que o sinal
eletroencefalográfico possui uma organização hierárquica de suas bandas constituintes, na
qual a amplitude de bandas mais rápidas é modulada pela fase de bandas
eletroencefalográficas mais lentas. A fase de delta está associada aos níveis de excitabilidade
cortical, que exercem papel modulatório sobre a atividade de assembleias neuronais locais,
responsáveis por aspectos específicos do processamento do estímulo. Assim, bandas de baixa
frequência, notadamente delta e teta, têm sido associadas à coordenação temporal do
processamento neural paralelamente realizado por diferentes áreas encefálicas (Canolty &
Knight, 2010).
Conforme a “Dynamic Attending Theory” (DAT), estímulos periódicos do ambiente
atuam no arrastamento de oscilações atencionais (Jones & Boltz, 1989). Mais recentemente,
tem sido proposto que oscilações na escala temporal desses estímulos (delta e teta) seriam
prováveis correlatos eletrofisiológicos da DAT (Henry & Herrmann, 2014). Neste aspecto,
uma de nossas hipóteses experimentais é que a fase de delta seria um parâmetro através do
qual o sistema nervoso poderia codificar a ritmicidade ambiental, e assim, acoplar a seu
funcionamento a diferentes taxas temporais de aferências sensoriais do ambiente. Isto poderia
explicar não só nossa capacidade de previsibilidade e expectância temporal, mas também
nosso acoplamento psicomotor e cognitivo, que permite responder adaptativamente às
demandas temporais ambientais.
Para tanto, verificamos se há acoplamento desta banda aos estímulos auditivos
rítmicos. Nossos resultados indicam que a fase de delta não está relacionada aos estímulos
auditivos rítmicos, para nenhum dos ISI ou das condições testadas (Sincronizada, Não-
Sincronizada). Assim, se o ISI parece exercer impacto sobre o aumento de delta, o que,
conforme nossa proposição, ocorre em virtude de ressonância neuronal da banda delta ao
ritmo auditivo, a fase não parece ser um parâmetro eletrofisiológico implicado neste
fenômeno.

91
Este resultado não corrobora dados anteriores. Estímulos auditivos com frequências
mais agudas (com ISI igual a 1350 ms (0,74 Hz) provocaram significativo alinhamento de
fase de delta em Cz (Stefanics et al., 2010). Estímulos visuais com ISI dentro da faixa de
nossos resultados também provocaram arrastamento de delta occipital encefalográfico em
humanos (Breska & Deouell, 2017; Cravo et al., 2013) e intracortical em macacos (Lakatos,
Karmos, Mehta, Ulbert, & Schroeder, 2008). Uma característica desses estudos que diverge
do presente trabalho é que a fase das oscilações foi estudada na ausência do estímulo. Ou
seja, enquanto realizamos a análise da fase nos momentos dos estímulos isocrônicos, outros
estudos tomaram a fase de delta no momento em que o estímulo era esperado, mas não
ocorria. Infelizmente, não foram encontrados protocolos experimentais idênticos a este estudo
e que tenham analisado este parâmetro. Assim, uma possibilidade é que os resultados
mostrem que não há acoplamento da fase de delta aos estímulos rítmicos, o que, em conjunto
com resultados anteriores, reforça que a fase de delta esteja envolvida na expectativa
temporal da ocorrência do estímulo, mas não na própria codificação temporal da ritmicidade
(Breska & Deouell, 2017).
Ainda que a fase não tenha sido um parâmetro relevante, a comparação das potências
espectrais na tarefa e na condição linha de base sugere que as bandas delta e gama
desempenham papel significativo na execução do protocolo ao qual submetemos os
participantes. Assim, o estudo de uma possível correlação da modulação de delta a gama com
o desempenho poderia ser uma via para um melhor entendimento da relação dessas bandas
eletroencefalográficas com o funcionamento cortical.
As correlações entre variáveis de desempenho na tarefa executiva e o Índice de
Modulação sugerem que o acoplamento fase-amplitude de delta a gama em alguns sítios
como fator explicativo do desempenho na tarefa, bem corroboram a proposição de que o
sistema nervoso faz uso de diferentes estratégias de processamento encefálico, a depender das
exigências temporais ambientais. Nesse aspecto, chama atenção que as correlações para a
condição 350 ms são todas negativas, e indicam que o acoplamento de delta a gama no córtex
pré-frontal do hemisfério direito foi deletério para um maior número de acertos. Com relação
às correlações da condição Sincronizada, parece haver modulação do intervalo entre
estímulos na topografia das correlações, já que, como citado, em 350 ms se obteve correlação
negativa em Cz, mas em 500 ms se obteve correlação positiva em F3, e em 650 ms

92
correlações positivas em C4 e Pz. Estas últimas parecem associar acoplamento delta-gama a
melhor desempenho cognitivo executivo.
Obviamente, a medida de acoplamento entre duas bandas de frequência que
utilizamos neste trabalho tem caráter exploratório. Considerando apenas os resultados
coletados no presente trabalho, seria possível realizar o estudo de acoplamento delta-gama no
mesmo sítio, como fizemos, mas também entre diferentes sítios. Elegemos este protocolo de
análise para verificar especificamente se diferentes níveis de excitabilidade cortical locais,
medidos por delta, poderiam influenciar a potência de oscilações implicadas com fenômenos
atencionais (gama). A existência de tal associação poderia demonstrar empiricamente que o
acoplamento fase-amplitude delta-gama é um plausível correlato eletrofisiológicos da DAT
(Henry & Herrmann (2014).
Portanto, os dois parâmetros de ritmicidade estudados neste trabalho, ISI e
Sincronização de fase, parecem desempenhar um importante papel para o desvelamento desse
fenômeno. Nossa proposta é o acoplamento fase-amplitude delta-gama tenha uma importante
participação no aumento da eficácia cognitiva, mas que, diante de contextos com maior
demanda por velocidade de processamento, o acoplamento de fase desempenhado um papel
negativo dada a complexidade da tarefa e a necessidade de manter os níveis de atenção e
processamento elevados durante toda a tarefa (modo contínuo). Isto equivale dizer que a
economia de recursos proporcionada pelo modo rítmico de processamento cortical neste
contexto ocorre às custas de menor eficácia do sistema. Em ISI menos exigentes do ponto de
vista temporal, aqui representados pelas condições 500 ms e 650 ms, o modo rítmico de
processamento seria não apenas econômico, mas também favorável ao desempenho.
Com relação às correlações encontradas entre Índice de modulação e Tempo de
reação, foram encontradas correlações positivas apenas nas condições Sincronizada 350 ms,
Silêncio 500 ms e Silêncio 650 ms. A distribuição topográfica não guarda relações com
outros achados deste estudo. No entanto, uma observação pertinente é que, diferentemente da
taxa de acertos, temos que maiores Índices de Modulação estiveram atrelados a maior tempo
de reação (i.e., menor eficiência) em todas as condições apontadas.
Nossos resultados relativos ao desempenho na tarefa executiva de soma de dígitos
demonstram que o ISI tem forte influência sobre a taxa de acertos. As condições Silêncio,
Sincronizada e Não-Sincronizada com ISI 350 ms apresentaram taxas de acerto

93
significativamente inferiores que as demais condições experimentais. As taxas de acerto para
este intervalo foram de aproximadamente 40%, contra aproximadamente 70% das condições
500 ms e 650 ms. Isto sugere maior dificuldade dessa condição perante as demais. Por outro
lado, a condição 350 ms apresenta tempos de reação médio aproximadamente 100 ms mais
rápidos que as demais, se tomadas as condições Sincronizada e Não-Sincronizada (em que o
som esteve presente).
As diferenças entre as manipulações experimentais relacionadas ao som
demonstraram efeito significativo apenas sob ISI 350 ms, com menor taxa de acertos para a
condição Sincronizada. Este achado evidencia que pulsos auditivos isocrônicos apresentam
efeito negativo para o número de acertos em nosso protocolo de desempenho executivo.
Com relação ao tempo de reação, os resultados indicam efeito tanto do ISI como
também da condição de apresentação dos estímulos. Na condição 350 ms, executar a tarefa
em Silêncio implica maiores tempos de reação em relação às condições Sincronizada e Não-
Sincronizada. Para 500 ms, a condição Sincronizada apresenta maiores tempos de reação em
relação à Silêncio e Não-Sincronizada. Para 650 ms, não houve efeito. Comparando o
desempenho entre a mesma condição e diferentes ISI (ex.: Silencio 350 ms vs Silêncio 500
ms), observamos que para as condições Sincronizada e não-Sincronizada há modulação do
ISI no tempo de reação, sempre com menor tempo de reação atrelados a menor ISI.
O resultado referente à taxa de acertos parece ir de encontro à melhoria da atenção
cogitada pela DAT. Primeiramente, os dados referentes à 350 ms indicam que, na única
condição em que foi observado efeito do som, o estímulo auditivo Sincronizado ao som
exerceu efeito distrator, e não sincronizador. No entanto, com base nos dados
eletrofisiológicos apresentados nesse trabalho para a condição 350 ms, fornecemos a
explicação que alternativa que, em situações de grande demanda por velocidade cognitiva,
diferentes estratégias podem ser postas em ação pelo sistema nervoso. Assim sendo, também
do ponto de vista comportamental, a menor taxa de acertos para a condição Sincronizada
pode representar o impacto negativo do funcionamento cognitivo em modo rítmico, quando
há alta demanda por alta velocidade de processamento. Em segundo, não houve qualquer
efeito do som sobre a taxa de acertos nas condições 500 ms e 650 ms. Uma possibilidade é
que, tendo em vista a alta taxa de acertos para esses dois intervalos, a tarefa não tenha
permitido distinguir adequadamente o efeito do som entre as condições.

94
Diferentemente da taxa de acertos, foi observada grande modulação do ISI e da
sincronia de fase entre estímulos auditivos e visuais sobre o tempo de reação. Enquanto o
som, Sincronizado ou não, parece ter efeito positivo na tarefa em 350 ms, em 500 ms a
Sincronia parece ter sido prejudicial. Cabe salientar que diversos estudos já apontaram efeito
positivo da sincronização temporal de tons auditivos rítmicos a estímulos alvo de
processamento (Brochard et al., 2013; Cravo et al., 2013; Escoffier et al., 2010). Em todos
eles, somente o tempo de reação foi melhorado.
Uma limitação deste estudo diz respeito ao protocolo utilizado não ser conclusivo
quanto à sincronização motora na condição 650 ms, já que o tempo motor anterior ao som era
já estatisticamente igual a este intervalo. Com relação à tarefa de soma, não foi possível
diferenciar respostas à tarefa de soma que fossem decorrentes de erros de controle inibitório.
Ademais, em nosso protocolo, nenhum tipo de reforçamento – nem mesmo o feedback
quanto às respostas certas ou erradas - era concedido aos participantes. Uma possibilidade é
que este contexto pouco ecológico tenha evocado pequena ativação de áreas límbicas, e,
consequente, menor ativação dos sistemas autonômicos.

95
CONCLUSÃO
Neste estudo, buscamos estudar como a ritmicidade ambiental afeta respostas motoras e
cognitivas, e, especificamente, averiguar indícios de acoplamento temporal entre o
funcionamento do sistema nervoso e o ambiente externo. Em suma, encontramos indícios de
arrastamento involuntário do período motor espontâneo por estímulos auditivos rítmicos com
intervalo de 500 ms, e que intervalos distantes desse ponto aumentam a variabilidade da
atividade motora, sem favorecer a sincronização ao evento externo. Isso sugere a existência
de uma “região de arrastamento”, na qual somos mais suscetíveis a sincronização
involuntária. Os limites dessa região de arrastamento podem ser objeto de investigações
futuras.
Demonstramos que a demanda cognitiva executiva teve pequeno impacto sobre
medidas fisiológicas do sistema nervoso simpático e parassimpático. Acreditamos que os
resultados espelham a independência funcional entre processos subsidiados por circuitos
dorsolaterais, diretamente envolvidos com as chamadas funções executivas “frias”, e circuitos
ventromediais/orbitofrontais, envolvidos com as chamadas funções executivas “quentes”. No
entanto, a extrapolação dessa conclusão faz jus à estudos em que estejam em jogo
contingências de reforçamento diante da tarefa, que poderiam resultar em maior controle
cognitivo da emoção pelo córtex pré-frontal.
Nossos dados eletroencefalográficos demonstram que o sistema nervoso utiliza
diferentes meios para cumprir demandas cognitivas de diferentes exigências temporais. Em
conjunto, os dados demonstram que um modo de funcionamento “rítmico” emerge no sistema
nervoso central, a partir da inferência de regularidade ambiental. Isto nem sempre representa
melhora do desempenho cognitivo, já que diante de tarefas complexas e que exigem
processamento cortical intenso a economia de recursos proporcionada pelo modo de
processamento “rítmico” leva a menor eficácia do sistema. Uma importante perspectiva é o
estudo da associação entre o desempenho cognitivo e o acoplamento fase-amplitude, uma vez
que, do ponto de vista formal, os resultados apresentados são apenas correlacionais.
Por fim, os dados relativos ao desempenho na tarefa sugerem que o ISI e o
arrastamento atencional pelos tons auditivos rítmicos tem efeito apenas sobre o tempo de
reação das respostas corretas. Nossos dados mostram que o intervalo entre estímulos tem

96
relação direta com o tempo de reação. Uma questão que merece aprofundamento, portanto, é
como este tempo de reação se relaciona precisamente com o intervalo dos estímulos. No
entanto, entendemos que estes achados dão credibilidade à afirmação de acoplamento do
funcionamento cognitivo à estrutura temporal ambiental.

97
REFERÊNCIAS BIBLIOGRÁFICAS
Badre, D. (2008). Cognitive control, hierarchy, and the rostro-caudal organization of the
frontal lobes. Trends in Cognitive Sciences, 12(5), 193–200.
https://doi.org/10.1016/j.tics.2008.02.004
Benedek, M., & Kaernbach, C. (2010). Decomposition of skin conductance data by means of
nonnegative deconvolution. Psychophysiology, 47(4), 647–658.
https://doi.org/10.1111/j.1469-8986.2009.00972.x
Berens, P. (2009). CircStat: A MATLAB toolbox for circular statistics. Journal of Statistical
Software, 31(10), 1–21. https://doi.org/10.1002/wics.10
Bizo, L. a, Chu, J. Y. M., Sanabria, F., & Killeen, P. R. (2006). The failure of Weber’s law in
time perception and production. Behavioural Processes, 71(2–3), 201–10.
https://doi.org/10.1016/j.beproc.2005.11.006
Bolbecker, A. R., Hong, S. L., Kent, J. S., Forsyth, J. K., Klaunig, M. J., Lazar, E. K., …
Hetrick, W. P. (2011). Paced finger-tapping abnormalities in bipolar disorder indicate
timing dysfunction. Bipolar Disorders, 13(1), 99–110. https://doi.org/10.1111/j.1399-
5618.2011.00895.x
Boucsein, W. (2012). Electrodermal activity. Springer (Second ed.). New York.
https://doi.org/10.1007/978-1-4614-1126-0
Breska, A., & Deouell, L. Y. (2017). Neural mechanisms of rhythm-based temporal
prediction: Delta phase-locking reflects temporal predictability but not rhythmic
entrainment. PLOS Biology, 15(2), e2001665.
https://doi.org/10.1371/journal.pbio.2001665
Brochard, R., Tassin, M., & Zagar, D. (2013). Got rhythm... for better and for worse. Cross-
modal effects of auditory rhythm on visual word recognition. Cognition, 127(2), 214–
219. https://doi.org/10.1016/j.cognition.2013.01.007
Buck, J., & Buck, E. (1968). Mechanism of Rhythmic Synchronous Flashing of Fireflies.
Fireflies of Southeast Asia may use anticipatory time-measuring in synchronizing their
flashing. Science, 159(3821), 1319–27.
Burle, B., Spieser, L., Roger, C., Casini, L., Hasbroucq, T., & Vidal, F. (2015). Spatial and
temporal resolutions of EEG: Is it really black and white? A scalp current density view.
International Journal of Psychophysiology, 97(3), 210–220.
https://doi.org/10.1016/j.ijpsycho.2015.05.004
Calderone, D. J., Lakatos, P., Butler, P. D., & Castellanos, F. X. (2014). Entrainment of
neural oscillations as a modifiable substrate of attention. Trends in Cognitive Sciences,
18(6), 300–309. https://doi.org/10.1016/j.tics.2014.02.005
Cameron, D. J., Stewart, L., Pearce, M. T., Grube, M., & Muggleton, N. G. (2012).
Modulation of Motor Excitability by Metricality of Tone Sequences. Psychomusicology:
Music, Mind and Brain, 22(2), 122–128. https://doi.org/10.1037/a0031229
Campos, A. de, Santos, A. M. G., & Xavier, G. F. (1997). A Consciência como fruto da
evolução e do funcionamento do sistema nervoso. Psicologia USP, 8(2), 181–226.

98
https://doi.org/http://dx.doi.org/10.1590/S0103-65641997000200010
Canolty, R. T., & Knight, R. T. (2010). The functional role of cross-frequency coupling.
Trends in Cognitive Sciences, 14(11), 506–515.
https://doi.org/10.1016/j.tics.2010.09.001.The
Chan, R. C. K., Shum, D., Toulopoulou, T., & Chen, E. (2008). Assessment of executive
functions: Review of instruments and identification of critical issues. Archives of
Clinical Neuropsychology, 23(2), 201–216. https://doi.org/10.1016/j.acn.2007.08.010
Coull, J. T., Cheng, R.-K., & Meck, W. H. (2011). Neuroanatomical and Neurochemical
Substrates of Timing. Neuropsychopharmacology, 36(1), 3–25.
https://doi.org/10.1038/npp.2010.113
Cragg, L., & Gilmore, C. (2014). Skills underlying mathematics: The role of executive
function in the development of mathematics proficiency. Trends in Neuroscience and
Education, 3(2), 63–68. https://doi.org/10.1016/j.tine.2013.12.001
Cravo, A. M., Rohenkohl, G., Wyart, V., & Nobre, A. C. (2013). Temporal expectation
enhances contrast sensitivity by phase entrainment of low-frequency oscillations in
visual cortex. The Journal of Neuroscience : The Official Journal of the Society for
Neuroscience, 33(9), 4002–10. https://doi.org/10.1523/JNEUROSCI.4675-12.2013
Delevoye-Turrell, Y., Dione, M., & Agneray, G. (2014). Spontaneous Motor Tempo is the
Easiest Pace to Act Upon for Both the Emergent and the Predictive Timing Modes.
Procedia - Social and Behavioral Sciences, 126, 121–122.
https://doi.org/10.1016/j.sbspro.2014.02.338
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source toolbox for analysis of single-
trial EEG dynamics including independent component analysis. Journal of Neuroscience
Methods, 134(1), 9–21. https://doi.org/10.1016/j.jneumeth.2003.10.009
Diamond, A. (2014). Executive Functions. Annual Review of Clinical psychologyPsychol.,
64, 135–168. https://doi.org/10.1146/annurev-psych-113011-143750.Executive
Dreher, J. C., & Grafman, J. (2002). The roles of the cerebellum and basal ganglia in timing
and error prediction. European Journal of Neuroscience, 16(8), 1609–1619.
https://doi.org/10.1046/j.1460-9568.2002.02212.x
Escoffier, N., Sheng, D. Y. J., & Schirmer, A. (2010). Unattended musical beats enhance
visual processing. Acta Psychologica, 135(1), 12–16.
https://doi.org/10.1016/j.actpsy.2010.04.005
Fraisse, P. (1982). Rhythm and Tempo. In The Psychology of Music (pp. 149–180).
https://doi.org/10.1007/978-1-4419-6114-3
Fraisse, P. (1984). Perception and estimation of time. October, 35, 1–36. Retrieved from
http://www.annualreviews.org/doi/abs/10.1146/annurev.ms.10.080180.000245
Fuster, J. (2004). Upper processing stages of the perception-action cycle. Trends in Cognitive
Sciences, 8(4), 143–145. https://doi.org/10.1016/j.tics.2004.02.004
Fuster, J. (2008). The Prefrontal Cortex (Fourth ed.). London, UK: Elsevier.
Fuster, J. M. (2001). The prefrontal cortex - An update: Time is of the essence. Neuron,
30(2), 319–333. https://doi.org/10.1016/S0896-6273(01)00285-9

99
Fuster, J. M. (2002). Frontal lobe and cognitive development. J Neurocytol, 31, 373–385.
https://doi.org/10.1023/A:1024190429920
Gilbert, S. J., & Burgess, P. W. (2008). Executive function. Current Biology, 18(3), R110–
R114. https://doi.org/10.1037/a0013399
Goel, A., & Buonomano, D. V. (2014). Timing as an intrinsic property of neural networks:
evidence from in vivo and in vitro experiments. Philosophical Transactions of the Royal
Society of London. Series B, Biological Sciences, 369(1637), 20120460.
https://doi.org/10.1098/rstb.2012.0460
Grabner, R. H., Ansari, D., Koschutnig, K., Reishofer, G., Ebner, F., & Neuper, C. (2009). To
retrieve or to calculate? Left angular gyrus mediates the retrieval of arithmetic facts
during problem solving. Neuropsychologia, 47(2), 604–608.
https://doi.org/10.1016/j.neuropsychologia.2008.10.013
Grahn, J. A., & Brett, M. (2007). Rhythm and beat perception in motor areas of the brain.
Journal of Cognitive Neuroscience, 19(5), 893–906.
https://doi.org/10.1162/jocn.2007.19.5.893
Grahn, J. a., & McAuley, J. D. (2009). Neural bases of individual differences in beat
perception. NeuroImage, 47(4), 1894–1903.
https://doi.org/10.1016/j.neuroimage.2009.04.039
Grondin, S. (2010). Timing and time perception: a Review of recent behavioral and
neuroscience findings and theoretical directions. Attention, Perception & Psychophysics,
72(3), 561–582. https://doi.org/10.3758
Harmony, T. (2013). The functional significance of delta oscillations in cognitive processing.
Frontiers in Integrative Neuroscience, 7(December), 83.
https://doi.org/10.3389/fnint.2013.00083
Hattori, Y., Tomonaga, M., & Matsuzawa, T. (2015). Distractor Effect of Auditory Rhythms
on Self-Paced Tapping in Chimpanzees and Humans. Plos, 1–17.
https://doi.org/10.5061/dryad.1618r.Funding
Hazy, T. E., Frank, M. J., & O’reilly, R. C. (2007). Towards an executive without a
homunculus: computational models of the prefrontal cortex/basal ganglia system.
Philosophical Transactions of the Royal Society of London. Series B, Biological
Sciences, 362(1485), 1601–13. https://doi.org/10.1098/rstb.2007.2055
Henry, M. J., & Herrmann, B. (2014). Low-Frequency Neural Oscillations Support Dynamic
Attending in Temporal Context. Timing & Time Perception, 2(1), 62–86.
https://doi.org/10.1163/22134468-00002011
Henry, M. J., Herrmann, B., & Obleser, J. (2014). Entrained neural oscillations in multiple
frequency bands comodulate behavior. Proceedings of the National Academy of
Sciences, 111(41), 14935–14940. https://doi.org/10.1073/pnas.1408741111
Ivry, R. B., & Schlerf, J. E. (2008). Dedicated and intrinsic models of time perception. Trends
in Cognitive Sciences, 12(7), 273–280.
https://doi.org/10.1016/j.tics.2008.04.002.Dedicated
Janzen, T. B. (2009). Análise psicofísica de medidas subjetivas de tempo em contexto rítmico.
Universidade de São Paulo.

100
Jensen, O., Kaiser, J., & Lachaux, J. (2007). Human gamma-frequency oscillations associated
with attention and memory. Trends in Neurosciences, 30(7), 317–324.
https://doi.org/10.1016/j.tins.2007.05.001
Jones, M. R. (1976). Time, our lost dimension: toward a new theory of perception, attention,
and memory. Psychological Review, 83(5), 323–355. https://doi.org/10.1037/0033-
295X.83.5.323
Jones, M. R., & Boltz, M. (1989). Dynamic attending and responses to time. Psychological
Review, 96(3), 459–491. https://doi.org/10.1037/0033-295X.96.3.459
Jones, M. R., Moynihan, H., MacKenzie, N., & Puente, J. (2002). Temporal aspects of
stimulus-driven attending in dynamic arrays. Psychological Science : A Journal of the
American Psychological Society / APS, 13(4), 313–319. https://doi.org/10.1111/j.0956-
7976.2002.00458.x
Kabdebon, C., Leroy, F., Simmonet, H., Perrot, M., Dubois, J., & Dehaene-Lambertz, G.
(2014). Anatomical correlations of the international 10-20 sensor placement system in
infants. NeuroImage, 99, 342–356. https://doi.org/10.1016/j.neuroimage.2014.05.046
Knyazev, G. G. (2007). Motivation, emotion, and their inhibitory control mirrored in brain
oscillations. Neuroscience and Biobehavioral Reviews, 31(3), 377–395.
https://doi.org/10.1016/j.neubiorev.2006.10.004
Knyazev, G. G. (2012). EEG delta oscillations as a correlate of basic homeostatic and
motivational processes. Neuroscience and Biobehavioral Reviews, 36(1), 677–695.
https://doi.org/10.1016/j.neubiorev.2011.10.002
Kornysheva, K., Von Cramon, D. Y., Jacobsen, T., & Schubotz, R. I. (2010). Tuning-in to the
beat: Aesthetic appreciation of musical rhythms correlates with a premotor activity
boost. Human Brain Mapping, 31(1), 48–64. https://doi.org/10.1002/hbm.20844
Koziol, L. F., & Budding, D. E. (2010). Subcortical structures and Cognition: Implications
for neuropsychological assessment. Park Ridge, USA: Springer.
Koziol, L. F., & Lutz, J. T. (2013). From Movement to Thought: The Development of
Executive Function. Applied Neuropsychology: Child, 2, 104–115.
https://doi.org/10.1080/21622965.2013.748386
Krejcar, O., & Frischer, R. (2011). Non destructive defect detection by spectral density
analysis. Sensors, 11(3), 2334–2346. https://doi.org/10.3390/s110302334
Lakatos, P., Chen, C. M., O’Connell, M. N., Mills, A., & Schroeder, C. E. (2007). Neuronal
Oscillations and Multisensory Interaction in Primary Auditory Cortex. Neuron, 53(2),
279–292. https://doi.org/10.1016/j.neuron.2006.12.011
Lakatos, P., Karmos, G., Mehta, A. D., Ulbert, I., & Schroeder, C. E. (2008). Entrainment of
neuronal oscillations as a mechanism of attentional selection. Science (New York, N.Y.).
https://doi.org/10.1126/science.1154735
Large, E., & Jones, M. R. (1999). The Dinamics of attending: how people Track time
Varying Events. Psychological Review, 106(1), 119–159.
Large, E. W., & Snyder, J. S. (2009). Pulse and meter as neural resonance. Annals of the New
York Academy of Sciences, 1169, 46–57. https://doi.org/10.1111/j.1749-
6632.2009.04550.x

101
MacDougall, H. G., & Moore, S. T. (2005). Marching to the beat of the same drummer: the
spontaneous tempo of human locomotion. Journal of Applied Physiology (Bethesda,
Md. : 1985). https://doi.org/10.1152/japplphysiol.00138.2005
Machado, S. E. D. C., Cunha, M., Velasques, B., Minc, D., Teixeira, S., Domingues, C. A.,
… Ribeiro, P. (2010). Sensorimotor integration: Basic concepts, abnormalities related to
movement disorders and sensorimotor training-induced cortical reorganization |
Integración sensitivomotora: Conceptos básicos, anomalías relacionadas con trastornos
del movimiento y reorga. Revista de Neurologia, 51(7).
McAuley, J. D., & Jones, M. R. (2003). Modeling Effects of Rhythmic Context on Perceived
Duration: A Comparison of Interval and Entrainment Approaches to Short-Interval
Timing. Journal of Experimental Psychology: Human Perception and Performance,
29(6), 1102–1125. https://doi.org/10.1037/0096-1523.29.6.1102
McAuley, J. D., Jones, M. R., Holub, S., Johnston, H. M., & Miller, N. S. (2006). The time of
our lives: life span development of timing and event tracking. Journal of Experimental
Psychology. General, 135(3), 348–367. https://doi.org/10.1037/0096-3445.135.3.348
Mello, G. B. M., Soares, S., & Paton, J. J. (2015). A Scalable Population Code for Time in
the Striatum. Current Biology, 25(9), 1113–1122.
https://doi.org/10.1016/j.cub.2015.02.036
Merker, B. H., Madison, G. S., & Eckerdal, P. (2009). On the role and origin of isochrony in
human rhythmic entrainment. Cortex, 45(1), 4–17.
https://doi.org/10.1016/j.cortex.2008.06.011
Michaelis, K., Wiener, M., & Thompson, J. C. (2014). Passive listening to preferred motor
tempo modulates corticospinal excitability. Frontiers in Human Neuroscience, 8(April),
252. https://doi.org/10.3389/fnhum.2014.00252
Mihulowicz, U., Willmes, K., Karnath, H.-O., & Klein, E. (2014). Single-digit arithmetic
processing-anatomical evidence from statistical voxel-based lesion analysis. Frontiers in
Human Neuroscience, 8(May), 286. https://doi.org/10.3389/fnhum.2014.00286
Miller, E. K., & Cohen, J. D. (2001). AN INTEGRATIVE THEORY OF PREFRONTAL
CORTEX FUNCTION. Annual Review of Neuroscience, 24, 167–202.
Moelants, D. (2002). Preferred tempo reconsidered. Proceedings of the 7th International
Conference on Music Perception and Cognition, 580–583.
https://doi.org/10.1080/00140138208924946
Néda, Z., Ravasz, E., Brechet, Y., Vicsek, T., & Barabási, a L. (2000). The sound of many
hands clapping. Nature, 403(6772), 849–850. https://doi.org/10.1038/35002660
Negrón-Oyarzo, I., Aboitiz, F., & Fuentealba, P. (2016). Impaired functional connectivity in
the prefrontal cortex: A mechanism for chronic stress-induced neuropsychiatric
disorders. Neural Plasticity, 2016, 21–24. https://doi.org/10.1155/2016/7539065
Passimgham, R., & Wise, S. (2012). Neurobiology of prefrontal cortex. Oxford: Oxford
University Press.
Passingham, R., & Wise, S. (2012). Neurobiology of prefrontal cortex. Oxford: Oxford
University Press.
Patel, A. D. (2006). Musical Rhythm, linguistic Rhythm, and human evolution. Music

102
Perception, 24(1), 99–104.
Patel, A. D. (2014). The Evolutionary Biology of Musical Rhythm: Was Darwin Wrong?
PLoS Biology, 12(3), 1–6. https://doi.org/10.1371/journal.pbio.1001821
Patel, A. D., Iversen, J. R., Bregman, M. R., & Schulz, I. (2009). Studying synchronization to
a musical beat in nonhuman animals. Annals of the New York Academy of Sciences,
1169(10), 459–469. https://doi.org/10.1111/j.1749-6632.2009.04581.x
Patel, A. D., Iversen, J. R., Chen, Y., & Repp, B. H. (2005). The influence of metricality and
modality on synchronization with a beat. Experimental Brain Research, 163(2), 226–
238. https://doi.org/10.1007/s00221-004-2159-8
Patel, S. H., & Azzam, P. N. (2005). Characterization of N200 and P300: Selected studies of
the Event-Related Potential. International Journal of Medical Sciences.
https://doi.org/10.7150/ijms.2.147
Purves, D., Augustine, G. J., Fitzpatrick, D., Hall, W. C., Lamantia, A.-S., Mcnamara, J. O.,
& Willians, S. M. (2004). Neuroscience. Sunderland (Vol. 3). https://doi.org/978-
0878937257
Repp, B. H., & Su, Y.-H. (2013). Sensorimotor synchronization: A review of recent research
(2006–2012). Psychonomic Bulletin & Review, 20(3), 403–452.
https://doi.org/10.3758/s13423-012-0371-2
Roenneberg, T., Dragovic, Z., & Merrow, M. (2005). Demasking biological oscillators:
properties and principles of entrainment exemplified by the Neurospora circadian clock.
Proceedings of the National Academy of Sciences of the United States of America,
102(21), 7742–7747. https://doi.org/10.1073/pnas.0501884102
Schwartze, M., Keller, P. E., Patel, A. D., & Kotz, S. A. (2011). The impact of basal ganglia
lesions on sensorimotor synchronization, spontaneous motor tempo, and the detection of
tempo changes. Behavioural Brain Research, 216(2), 685–691.
https://doi.org/10.1016/j.bbr.2010.09.015
Snyder, J. S., & Large, E. W. (2005). Gamma-band activity reflects the metric structure of
rhythmic tone sequences. Cognitive Brain Research, 24(1), 117–126.
https://doi.org/10.1016/j.cogbrainres.2004.12.014
Stefanics, G., Hangya, B., Hernádi, I., Winkler, I., Lakatos, P., & Ulbert, I. (2010). Phase
entrainment of human delta oscillations can mediate the effects of expectation on
reaction speed. The Journal of Neuroscience : The Official Journal of the Society for
Neuroscience, 30(41), 13578–13585. https://doi.org/10.1523/JNEUROSCI.0703-
10.2010
Tallon-Baudry, C., & Bertrand, O. (1999). Oscillatory gamma activity in humans and its role
in object representation. Trends in Cognitive Sciences, 3(4), 151–162.
https://doi.org/10.1016/S1364-6613(99)01299-1
Teki, S., Grube, M., Kumar, S., & Griffiths, T. D. (2011). Distinct neural substrates of
duration-based and beat-based auditory timing. The Journal of Neuroscience : The
Official Journal of the Society for Neuroscience, 31(10), 3805–3812.
https://doi.org/10.1523/JNEUROSCI.5561-10.2011
Tort, A. B. L., Komorowski, R., Eichenbaum, H., & Kopell, N. (2010). Measuring Phase-
Amplitude Coupling Between Neuronal Oscillations of Different Frequencies. Journal

103
of Neurophysiology, 104(2), 1195–1210. https://doi.org/10.1152/jn.00106.2010
Treisman, M. (1963). THE TEMPORAL DISCRIMINATION AND INDIFFERENCE
INTERVAL : IMPLICATIONS FOR A MODEL OF THE “ INTERNAL CLOCK ” :
Psychological Monographs : General and Applied, 77(13).
Valdez, P., Ramírez, C., García, A., Talamantes, J., Armijo, P., & Borrani, J. (2005).
Circadian rhythms in components of attention. Biological Rhythm Research, 36(1–2),
57–65. https://doi.org/10.1080/09291010400028633
Wang, H., & Huang, S. (2014). Musical Rhythms Affect Heart Rate Variability. Advances in
Electrical Engineering, 2014, 14. https://doi.org/10.1155/2014/851796
Ward, L. M. (2003). Synchronous neural oscillations and cognitive processes. Trends in
Cognitive Sciences, 7(12), 553–559. https://doi.org/10.1016/j.tics.2003.10.012
Wearden, J. H. (2005). Origins and development of internal clock theories of time.
Psychologie Française, 50(1), 7–25.
https://doi.org/http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.116.1006&rep
=rep1&type=pdf
Wilson, E. M., & Davey, N. J. (2002). Musical beat influences corticospinal drive to ankle
flexor and extensor muscles in man. International Journal of Psychophysiology :
Official Journal of the International Organization of Psychophysiology, 44(2), 177–184.
https://doi.org/10.1016/S0167-8760(01)00203-3
Zimmerman, D. L., Ownsworth, T., O’Donovan, A., Roberts, J., & Gullo, M. J. (2016).
Independence of Hot and Cold Executive Function Deficits in High-Functioning Adults
with Autism Spectrum Disorder. Frontiers in Human Neuroscience, 10(February), 24.
https://doi.org/10.3389/fnhum.2016.00024

104
APENDICE 1 – Termo de consentimento Livre e esclarecido (TCLE)
TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO – TCLE
Esclarecimentos
Este é um convite para você participar da pesquisa: “Toca que o cérebro também
dança: a sincronização de oscilações atencionais e neurais à estímulos externos melhora o
processamento cognitivo?”, que tem como pesquisador responsável o Prof. Dr. John
Fontenelle Araújo. Esta pesquisa pretende caracterizar o arrastamento (sincronização) de
oscilações das funções executivas à estímulos auditivos. O motivo que nos leva a fazer este
estudo são evidências dessa sincronização em diversos fenômenos cognitivos, no entanto,
nenhum estudo até o momento abordou as funções executivas, extremamente importantes em
nosso dia-a-dia, ao executarmos tarefas cotidianas.
Caso você decida participar, você deverá responder à uma tarefa baseada em adições
matemáticas simples ouvindo tons auditivos de intensidade mediana (60db), ao mesmo tempo
em que ocorre o registro de sinais eletroencefalográficos e eletrocardiográficos. Estima-se
que as tarefas durem cerca de 20 minutos, e que a duração total do experimento seja de 50
minutos, incluído aí a assinatura deste termo e o tempo para implantação e retirada dos
eletrodos para coleta dos sinais eletrofisiológicos.
Durante a realização, você deverá permitir a implantação dos eletrodos em seu couro
cabeludo e abdômen, e executar as tarefas sob a execução dos estímulos auditivos. A previsão
de riscos é mínima, ou seja, o risco que você corre é semelhante àquele sentido num exame
físico ou psicológico de rotina. Pode acontecer um desconforto com relação ao esforço
mental requerido na tarefa, que pode ser minimizado com intervalos entre as etapas do
estudo, e com os eletrodos, que poderá ser minimizado pela utilização de pasta condutora
convencional, largamente utilizada tanto na pesquisa básica quanto na prática clínica de
eletrofisiologia. De qualquer forma, o estudo será imediatamente interrompido caso qualquer
desconforto seja relatado por você.
Durante todo o período da pesquisa você poderá tirar suas dúvidas ligando para John
Fontenele Araújo, a través do fone: 3215-3409.
Você tem o direito de se recursar a participar ou retirar seu consentimento.
Os dados que você irá nos fornecer serão confidenciais e serão divulgados apenas em
congressos ou publicações científicas, não havendo divulgação de nenhum dado que possa
lhe identificar.
Esses dados serão guardados pelo pesquisador responsável por essa pesquisa em local
seguro e por um período de 5 anos. Se você tiver algum gasto pela sua participação nessa
pesquisa, ele será assumido pelo pesquisador e reembolsado para você. Se você sofrer algum
dano comprovadamente decorrente desta pesquisa, você será indenizado.

105
Qualquer dúvida sobre a ética desta pesquisa você deverá ligar para o Comitê de Ética
em Pesquisa da Universidade Federal do Rio Grande do Norte, telefone: 3215-3135.
Este documento foi impresso em duas vias, uma ficará com você e a outra com o
pesquisador responsável John Fontenele Araújo.
Consentimento Livre e Esclarecido
Após ter sido esclarecido sobre os objetivos, importância e o modo como os
dados serão coletados nessa pesquisa, além de conhecer os riscos, desconfortos e benefícios
que ela trará para mim e ter ficado ciente de todos os meus direitos, concordo em participar
da pesquisa “Toca que o cérebro também dança: a sincronização de oscilações atencionais e
neurais à estímulos externos melhora o processamento cognitivo?”, e autorizo a divulgação
das informações por mim fornecidas em congressos e/ou publicações científicas desde que
nenhum dado possa me identificar.
Natal, 02 de fevereiro de 2016.
__________________________________
Assinatura do participante da pesquisa
Declaração do pesquisador responsável
Como pesquisador responsável pelo estudo “Toca que o cérebro também dança: a
sincronização de oscilações atencionais e neurais à estímulos externos melhora o
processamento cognitivo”, declaro que assumo a inteira responsabilidade de cumprir
fielmente os procedimentos metodologicamente e direitos que foram esclarecidos e
assegurados ao participante desse estudo, assim como manter sigilo e confidencialidade sobre
a identidade do mesmo.
Declaro ainda estar ciente que na inobservância do compromisso ora assumido estarei
infringindo as normas e diretrizes propostas pela Resolução 466/12 do Conselho Nacional de
Saúde – CNS, que regulamenta as pesquisas envolvendo o ser humano.
Natal, 02 de fevereiro de 2016.
__________________________________
Assinatura do pesquisador responsável
Impressão
datiloscópica do participante

106
Anexo 1 – Parecer do Comitê de Ética ao projeto

107

108
109


















