Tonina e nocicepção
59
UNIVERSIDADE DE MOGI DAS CRUZES ALEXANDRO DE LIMA SOUZA TONINA E NOCICEPÇÃO Mogi das Cruzes, SP 2007
-
Upload
alexandro-lima -
Category
Documents
-
view
9 -
download
4
description
fisiologia da dor em camundongos transgênicos que superespressam tonina de rato.
Transcript of Tonina e nocicepção
-
UNIVERSIDADE DE MOGI DAS CRUZES ALEXANDRO DE LIMA SOUZA
TONINA E NOCICEPO
Mogi das Cruzes, SP 2007
-
UNIVERSIDADE DE MOGI DAS CRUZES ALEXANDRO DE LIMA SOUZA
TONINA E NOCICEPO Dissertao apresentada ao mestrado em Biotecnologia da Universidade de Mogi das Cruzes como parte dos requisitos para obteno do ttulo de mestre.
rea de Concentrao: Biolgica
Orientador: Prof. Dr. Jorge Luiz Pesquero
Mogi das Cruzes, SP 2007
-
FINANCIAMENTO
Os laboratrios de Biofsica e de Farmacologia da Inflamao da UFMG contam com toda a infra-estrutura para desenvolver o projeto proposto. Alm disso, o projeto conta com apoio financeiro da Fapesp e do CNPq.
Ficha Catalogrfica elaborada por Cntia de Azevedo Loureno Bibliotecria e Professora do Curso de Biblioteconomia da UFMG
612 S719t Souza, Alexandro de Lima
Tonina e nocicepo / Alexandro de Lima Souza. 2007.
46 f. : il. ; 30 cm.
Dissertao (mestrado) Universidade de Mogi das Cruzes, Programa de Ps-graduao em Biotecnologia, 2007.
Orientao: Prof. Dr. Jorge Luiz Pesquero.
1. Fisiologia da dor. 2. Tonina. 3. Nocicepo. 4. Angiotensina II. 5. Opiides. I. Ttulo.
-
DEDICATRIA
Dedico este trabalho Isabel de Lima Souza (em memria)
-
Em todo verdadeiro pesquisador da natureza mora uma espcie de reverncia religiosa, pois ele se d conta de que possvel imaginar-se como sendo o primeiro a ter conscincia da trama incomparavelmente delicada que une suas percepes. A manifestao de conhecimento que no havia ainda sido posta a nu produz no investigador sensao semelhante surgida na criana que procura alcanar o nvel de domnio com que os adultos manipulam as coisas.
(Albert Einstein)
-
AGRADECIMENTOS
Ao prof. Jorge Luiz Pesquero, pela orientao, dedicao e perseverana, por ser antes de qualquer coisa, um grande amigo, que mesmo tendo pontos de vistas diferentes, tem me apoiado nos caminhos da academia e da vida. Aos professores, estagirios e tcnicos da UFMG pelo apoio e colaborao nos momentos de dvida e necessidade de apoio. Dra. Cntia de Azevedo Loureno, por tornar menos dolorosa essa via sacra, como esposa e como amiga.
-
RESUMO
A angiotensina II (Ang II), o principal efetor do sistema renina-angiotensina (SRA) um peptdeo hormonal com potente atividade vasoconstritora, sendo tambm reconhecido como um peptdeo regulatrio multifuncional, com ao em diversos tecidos. As aes da Ang II no sistema nervoso central incluem: elevao da presso arterial, regulao do balano eletroltico, aumento da liberao de vasopressina, o aumento do apetite ao sdio e da ingesto de gua, a natriurese, possivelmente funes cognitivas. Tambm tem sido proposto o envolvimento do SRA no processamento central da informao sensorial. A tonina uma serina-proteinase capaz de formar Ang II a partir da angiotensina I (Ang I), a partir do angiotensinognio (AG) e de peptdeos sintticos correspondentes poro tetradecapeptdica N-terminal do (AG(1-14)). Recentemente, geramos camundongos transgnicos que apresentam superexpresso da tonina de rato [TGM(rTon)]. Este animal vem sendo estudado quanto ao fentipo em termos bioqumicos, fisiolgicos e morfolgicos. O animal apresenta pequeno, mas significativo aumento nos nveis pressricos, apesar de os nveis teciduais e circulantes de angiotensina II no serem diferentes dos nveis do animal selvagem. O TGM(rTon) provavelmente apresenta alteraes a nvel renal. Em relao ao volume urinrio no observamos diferenas significativas entre o TGM(rTon) e seu controle, porm para a gua ingerida observamos aumento na quantidade para o animal transgnico e este animal apresentou uma reduo da osmolaridade da urina. Neste trabalho estudou-se a resposta nociceptiva do animal TGM(rTon). Utilizamos para isso o teste das contores abdominais ao cido actico, teste de formalina e ainda o teste de rota-rod. Os resultados apresentam evidencias de que o animal TGM(rTon) apresenta reduo da capacidade motora, observada no teste do rota-rod. Esta reduo da capacidade motora dos animais TGM(rTon) pode, em parte, ser explicada por um possvel aumento nos nveis de opiides, uma vez que tonina capaz de hidrolisar beta-LPH e liberar peptdeos com ao opiide. No entanto, com relao a nocicepo, o TGM(rTon) apresentou significativo aumento na sensibilidade a dor, especificamente na segunda fase do teste da formalina, fase na qual so liberados os agentes inflamatrios. Em concluso, apesar do indcio da presena de opiides endgenos, percebidos atravs do estudo de motricidade, realizado pelo teste de rota rod, houve resposta hiperalgsica por parte dos animais TGM(rTon) na fase inflamatria do teste da formalina.
Palavras-chave: Tonina, Nocicepo, Angiotensina II, Opiides
-
ABSTRACT
Angiotesin II (Ang II), main trigger of the renin-angiotensin (SRA), is a hormonal peptide with potent vasoconstrictor activity, also being recognized as a multifunctional regulatory peptide, with action in several woven. Ang II's actions in the central nervous system include: elevation of the blood pressure, regulation of the swinging electrolytic, increase of the vasopressin liberation, the increase of sodium appetite and the increase of water ingestion, called natriurese, a process that possibly interferes in cognitive functions. There has also been proposed the involvement of SRA in the central processing of sensorial information. Tonin is a serine-proteinase capable of forming Ang II starting from angiotensin I (Ang I), or starting from angiotensinogen (AG) and of synthetic peptides corresponding to the N-terminal tetradecapeptidic portion of the (AG(1-14)). Recently we generated transgenic mice that presented super expression of the mouse tonin [TGM(rTon)]. This animal has been studied in relation to the phenotype in biochemical, physiological and morphological terms. The animal presents a small but significant increase in the pressure levels, although the tissue and circulation levels of angiotesin II are no different of the ones found in wild animals. The mouse tonin probably presents changes in the kidneys function. In relation to the urine volume we have not observed any significant changes between the TGM(rTon) and the control group, although we observed an increase in the amount of water ingested by the transgenic animal and this animals kidney presented a reduction of its filtration capacity. In this thesis we have studied the nociceptive answer of the animal TGM(rTon). For that, we used a test measuring the amount of abdominal contortion in response to acetic acid, a formalin test and a rota-rod test. The results show evidence that the TGM(rTon) animal presents a reduction of the motor skill, observed in the rota-rod test. This reduction may, in part, be explained by a possible increase of opioids, seeing that the tonine is capable of hydrolyzing beta-LPH and releasing peptides with opioid action. However, in relation to the nociceptivity, the TGM(rTon) presented a significant increase in its pain sensitivity, especially in the second fase of the formalin test, when the inflammatory agents are released. In conclusion, although there was a trace of endogenous opioids, noticed through the motricity study, realized through the rota rod test , there was an hiperalgesic response from the TGM(rTon) animals in the inflammatory fase of the formalin test.
Keywords: Tonin, Nociception, Angiotesin II, Opioids
-
LISTA DE ILUSTRAES
Figura 1. Sistema renina angiotensina .................................................... 12 Figura 2. Vias de formao de angiotensina II envolvendo o sistema
tonina angiotensina ................................................................
13
Figura 3. Organizao das fibras aferentes primrias envolvidas na conduo de um estmulo nociceptivo desde a superfcie cutnea (terminal perifrico) at o como dorsal da medula espinal. NE, fibra nociceptiva especfica; NP, fibra nociceptiva polimodal; no-N, fibra no nociceptiva. Adaptado de Millan (1999) por Castro (2000) ............................................................
18 Figura 4. Seqncia da construo gnica utilizada na gerao do
camundongo transgnico que expressa tonina de rato ...........
21 Figura 5. Aparelho de rota-rod da marca Ugo Basile .............................. 27 Figura 6. Caixa utilizada no experimento de contoro pelo cido
actico .....................................................................................
29 Figura 7. Camundongo TGM(rTon), aps experimento de contoro
abdominal, j disponibilizado para ser submetido ao processo de eutansia .............................................................
30 Figura 8. Tempo de permanncia, expresso em minutos, dos diferentes
grupos, Swiss, selvagem e transgnico no teste do rota-rod. *p < 0,05 vs TGM(rTon). As colunas representam a mdia epm (n = 6) ..............................................................................
32 Figura 9. Contores abdominais induzidas por cido actico em
camundongos selvagens e TGM(rTon) em sistema monocego. As colunas representam a mdia epm (n = 6) ...
33
Figura 10. Teste de formalina. Nmero de lambidas induzidas pela injeo de formalina em camundongos selvagens (S) e transgnicos (T). A primeira fase corresponde ao tempo de 0 - 5 minutos aps a injeao e a segunda fase 15 - 30 minutos. *p < 0,05. As colunas representam a mdia epm (n = 6) .....
34
-
LISTA DE ABREVIATURAS
AG Angiotensinognio Ang I Angiotensina I Ang II Angiotensina II Arg. Xxx Arginina + qualquer radical AT1 Receptor ligado a protena-G AT2 Receptor ligado a protena-G CD Corno distal DNA cido desoxirribonuclico ECA Enzima conversora de angiotensina FAP Promotor para insero de seqncia de gene Fen. Xxx Fenilalanina + qualquer radical IASP Associao Internacional para o Estudo da Dor Icv Intracerebroventricular RPM Rotaes por minuto SRA Sistema renina-angiotensina SV40poliA Fator de superexpresso de gene inserido TGM Transgenic mouse
-
LISTA DE ANEXOS
Tabela - Analise estatstica das contores abdominais em cido actico .....................................................................................
Comit de tica ........................................................................................
-
SUMRIO
1 INTRODUO ................................................................................... 11 1.1 Sistema renina-angiotensina ..................................................... 11 1.2 Sistema tonina-angiotensina II .................................................. 12 1.3 Dor e Nocicepo ...................................................................... 14
1.3.1 Modulao da nocicepo ............................................... 17 1.3.2 Analgesia ......................................................................... 18 1.3.3 Dor e o sistema renina-angiotensina ............................... 20
1.4 Camundongo transgnico que expressa tonina de rato ............ 21 2 OBJETIVOS ....................................................................................... 23
3 MTODOS ......................................................................................... 24 3.1 Animais ...................................................................................... 25
3.2 Mtodos para avaliar dor ........................................................... 25
3.2.1 Teste do "rota-rod ........................................................... 26
3.2.2 Teste de contoro abdominal ........................................ 27 3.2.3 Teste da formalina ........................................................... 30
3.4 Anlise dos dados ..................................................................... 30
4 RESULTADOS ................................................................................... 31
4.1 Teste de rota-rod ....................................................................... 31
4.2 Teste de contoro abdominal .................................................. 32 4.3 Teste de formalina ..................................................................... 33
5 DISCUSSO ...................................................................................... 35 6 CONCLUSES .................................................................................. 38 7 REFERNCIAS .................................................................................. 39 ANEXOS ...............................................................................................
-
1 INTRODUO 1.1 SISTEMA RENINA-ANGIOTENSINA
A angiotensina II (Ang II), o principal efetor do sistema renina-angiotensina (SRA, Figura 1) um peptdeo hormonal com potente atividade vasoconstritora (PEACH, 1977), sendo tambm reconhecido como um peptdeo regulatrio multifuncional, com ao em diversos tecidos incluindo o crebro, o
corao, a adrenal, o rim, e as gnadas (DZAU et al., 1986; DZAU et al., 1987; KUNAPULI et al., 1987; LINDPAINTER et al., 1987; KUMAR et al., 1989; LOPES et
al., 1997). As aes da Ang II no sistema nervoso central incluem: elevao da
presso arterial, regulao do balano eletroltico, aumento da liberao de
vasopressina, aumento do apetite ao sdio e da ingesto de gua, denominada
natriurese (FITZSIMONS, 1980; GANONG et al., 1982; OLSEN et al., 1985), e possivelmente funes cognitivas (PHILLIPS, 1987). Tambm tem sido proposto o envolvimento do SRA no processamento central da informao sensorial. Outros
trabalhos mostram que a angiotensina II es t ligada nocicepo (Guasti et al., 1998).
Pelo SRA, o angiotensinognio (AG) clivado pela renina, levando formao da angiotensina I (Ang I). A Ang I, pela ao da enzima conversora de
angiotensina (ECA), hidrolisada na ligao Fen8-His9, com conseqente liberao do octapeptdeo Ang II. Outras vias enzimticas podem contribuir para a formao
da Ang I e da Ang II (BOUCHER et al., 1974; GENEST, 1983). No se conhece ainda a importncia relativa de cada via formadora de Ang II que est presente nos
diferentes tecidos, porm, existem indcios de que, em alguns neurnios cerebrais, a
-
maior parte da Ang II possa ser formada por outras enzimas que no a ECA e a
renina (BUNNEMAN et al., 1993).
Figura 1: Sistema renina angiotensina e efeitos mediados pelos receptores de angiotensina
A presena do sistema renina-angiotensina j reconhecida em quase todos os tecidos (PHILLIPS et al.,1992). Isto particularmente significante no corao e na parede dos vasos sangneos, nos casos de arteriosclerose (PHILLIPS et al.,2002),( BRASIER et al.,2002), e falhas cardacas (DZAU, 1984), (NAKAMURA, M. et al.,1985),( DZAU,1986). A Ang II produzida em vrios tecidos de relevncia cardiovascular como a glndula adrenal (YOSIPIV,1996), corao e vasos sangneos (MOFFET, R. B. , BUMPUS, F. M. , HUSAIN, A.,1989), rins(CHIU, A. T. et al.,1989) e crebro (HEIN, L. et al. HEIN, L. et al.,1995). A Ang II a molcula chave do sistema, sendo um potente regulador da presso sangnea e do balano
Asp.Arg.Val.Tir.Ile.His.Pro.Fen.His.Leu.Val.Ile ... Angiotensinognio humano
Asp.Arg.Val.Tir.Ile.His.Pro.Fen.His.Leu Angiotensina I
Asp.Arg.Val.Tir.Ile.His.Pro.Fen Angiotensina II
Renina
ECA
receptor AT2
Rim reteno de sdio fibrose
Glndula adrenal liberao de aldosterona Corao
inotropia cronotropia hipertrofia fibrose
Vasos constrico hipertrofia
Crebro sede apetite ao sal ativao simptica liberao de vasopressina
-
de gua e eletrlitos atravs da dipsognese e ativao do sistema nervoso
simptico. Outras atividades relacionadas a Ang II incluem a modulao da liberao
de hormnios da pituitria, interferncia com a aprendizagem e a atividade motora e
regulao da temperatura corporal e analgesia. A ao da Ang II se d aps
interao com um dos dois receptores ligados a protena-G, e denominados AT1 e
AT2 (ICHIKI, T. et al.,1995),( WRIGHT, J. W. , HARDING, J. W.,1994),. Os principais efeitos da Ang II so mediados atravs do receptor AT1( MARRERO, M. B. et al.,1995). O receptor AT2 expresso durante a vida fetal em ncleos cerebrais distintos (GRADY, E. F. et al.,1991),(TSUTUMI, K. , SAAVEDRA, J. M.,1991) sugerindo possveis papis no crescimento, desenvolvimento e funes neuroniais.
Estudos em camundongo transgnico, no qual o gene do receptor AT2 foi
nocauteado, sugerem que este receptor pode estar envolvido no crescimento,
desenvolvimento e comportamento explorador(HEIN, L. et al.,1995),( ICHIKI, T. et al.1995) . Os efeitos da Ang II, resultantes de sua interao com estes receptores especficos esto mostrados na figura 1.
Enzimas semelhantes renina, proteinases como a catepsina D, a
pepsina e outras aspartil-proteinases, so capazes de produzir Ang I a partir do AG.
Alm disso, outras enzimas que no a ECA, so capazes de produzir Ang II, tanto a
partir de Ang I como diretamente do AG, entre elas a catepsina G e a tonina.
1.2 SISTEMA TONINA-ANGIOTENSINA II
Tonina uma enzima da famlia das serina-proteinases, capaz de formar
-
Ang II tanto a partir da Ang I, como do AG (GRIS et al., 1981) e de peptdeos sintticos correspondentes poro tetradecapeptdica N-terminal do AG (AG (1-14)) (SCHILLER et al., 1976; PESQUERO et al., 1982) (Figura 2). Alm de liberar Ang II, foi demonstrado que a tonina capaz de hidrolisar pro-opiomelanocortina
para produzir o hormnio adrenocorticotropina (ACTH) e hormnios peptdeos beta-lipotrpicos (SEIDAH et al., 1979), alm de degradar a substncia P (CHRTIEN et al., 1980, PESQUERO et al., 1982), clivando ligaes do tipo Arg. Xxx e Fen.Xxx, mostrando uma enzima com atividade trptica-quimotrptica.
Figura 2. Vias de formao de angiotensina II envolvendo o sistema tonina angiotensina.
Tonina est presente em vrios tecidos de rato, estando especialmente
concentrada na glndula submandibular, onde representa 8% do contedo total de
protena. liberada pela glndula na saliva e na corrente venosa (GARCIA et al., 1976). A atividade tonina no plasma de difcil determinao, pois a enzima encontra-se ligada a alfa2-macroglobulina, um inibidor proteico (Trembley et al., 1981). Tonina est presente em menores concentraes no pncreas, fgado, bao, partida, testculos, prstata e crebro (BOUCHER et al., 1974; JOHANSEN et al., 1987; WOODLEY-MILLER et al., 1987; ARAUJO et al., 1991).
ECA
Angiotensinognio
Angiotensina I
Angiotensina II
Renina Catepsina D
Quimase Catepsina G
TONINA
Catepsina G TONINA
-
Em trabalho realizado por Lopes et al. (1997), foram determinados os nveis de atividade tonina em diferentes regies do encfalo de rato. Os maiores
nveis de atividade, encontrados utilizando-se o AG(1-14) como substrato, foram no arquicerebelo (flculo nodular) e na neurohipfise e os menores nveis no hipocampo e adenohipfise. No foi detectada atividade no crtex cerebelar e no
plexo coride. Em trabalho desenvolvido por Araujo et al. (2002), foi mostrado que a injeo intracerebroventricular (icv) de tonina pode produzir efeito pressor em rato acordado. Observou-se aumento da ingesto de gua causado pela injeo icv de tonina, aumento que foi bloqueado completamente pela injeo prvia de losartan um antagonista de receptores AT1 de angiotensina.
3. Dor e nocicepo
Os antigos gregos, dentre eles Aristteles, acreditavam que a dor era um
"desgosto", sentimento oposto ao prazer experimentado no corao, e Willian
Harvey, descobridor da circulao, acreditava que o corao e no o crebro fosse o
"sensorium communum" onde a dor era sentida (BONICA, 1976). Em 1644, Descartes descreveu a conduo das sensaes, inclusive dor, via "sensores
delicados" contidos em nervos que conectava os tecidos ao crebro, onde a
sensao era produzida.
A dor um componente sintomtico de diversas desordens clnicas, afetando
grande parte da populao (BESSON e DICKENSON, 1997). De um modo geral, a sua funo operar como mecanismo de auto-preservao do corpo, ocorrendo
quando um tecido lesado quer seja por mtodos mecnicos ou por mtodos qumicos, fazendo com que o indivduo reaja com o intuito de remover a causa do
-
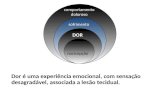
estmulo (DRAY, 1997). Segundo a Associao Internacional para o Estudo da Dor (IASP), a dor pode ser definida como uma experincia sensorial e emocional desagradvel, associada a dano tecidual ou potencial, ou descrita em termo de tal
leso (MELZACK e LOESER, 1999; ALMEIDA et al., 2004). O estmulo da dor pode ocorrer mesmo em nveis sutis, nas atividades
cotidianas onde pode ser acionado por causas muito diversas. Um exemplo que
pode ser usado para descrever esse processo o da isquemia nos squios (ossos da bacia sobre os quais o peso do corpo depositado ao se sentar, quando uma
pessoa permanece por muito tempo sentada e/ou devido a permanncia por um
tempo prolongado numa mesma posio. Este comportamento leva a diminuio do
fluxo sangneo, fazendo com que o indivduo sinta a necessidade de mudar de
posio, movendo o peso corporal inconscientemente para permanecer sentado por
um perodo mais prolongado (GUYTON, 2002; ALMEIDA et al., 2004). A dor uma experincia universal, cotidiana e quase sempre necessria.
Pessoas com doenas congnitas ou adquiridas e que devido a isso so privadas de
sensibilidade dolorosa, geralmente so acometidas por diversos tipos de ferimentos,
sem perceber, correndo srios riscos de injrias letais (MELZACK e LOESER, 1999; ALMEIDA et al., 2004).
Como exemplo de dessensibilizao dos nociceptores, podemos citar o caso
dos diabticos, os quais perdem a sensibilidade nas extremidades, sendo isto
denominado neuropatia decorrente dos nveis de glicemia alterados. Como este
um mal secundrio decorrente do diabetes, os enfermos ferem-se nos ps com
bastante freqncia sem perceberem, levando a necrose e eventualmente a uma
amputao (SCHIMID, 2007). A hansenase outro exemplo de dessensibilizao dos nociceptores e, devido a isso, ao exporem uma leso causada pela doena a
-
estmulos mecnicos ou trmicos e qumicos, no se observa qualquer reao
devido s leses causadas pela molstia (PEREIRA et al., 2006). Existe diferena entre os termos nocicepo e dor. O termo nocicepo
refere-se a manifestaes neurolgicas, geradas por estmulos nocivos. A dor
envolve a percepo de um estmulo aversivo e requer abstrao e elaborao do
impulso sensorial (MELZACK e LOESER, 1999; ALMEIDA et al., 2004). Em 1906, Sherrington props o termo nociceptivo, referindo-se a um estmulo
capaz de comprometer a integridade do organismo. Em animais esses estmulos
produzem respostas reflexas (aumento das atividades respiratria e cardaca, dilatao da pupila) e comportamentais (reao de escape, localizao) integradas em algum local. Em humanos, a sensao produzida por esses estmulos, algumas
vezes era considerada como dor aguda. Atualmente, a descrio da dor baseia-se
em dois tipos diferentes: dor rpida e dor lenta (GUYTON, 2002; ALMEIDA et al., 2004). A dor rpida caracterizada pelo estmulo e resposta dentro do tempo de aproximadamente 0,1 segundo, enquanto a dor lenta a resposta pode vir a partir de
um segundo ou mais. A dor rpida descrita de modos diferentes como dor sbita,
dor picada, dor aguda e dor eltrica. Geralmente esse tipo de dor sentido quando a
pele cortada, espetada por agulhas ou agudamente queimada. Importante
ressaltar que esse tipo de dor aparece geralmente de maneira superficial, no sendo
caracterstica em tecidos profundos (GUYTON, 2002; ALMEIDA et al., 2004). A dor lenta tambm chamada por nomes bastante diversos como, por exemplo, dor
surda, dor nauseante e dor crnica. Geralmente essa dor associada destruio
de tecidos, se prolongada pode se tornar insuportvel. Esse tipo de dor pode ocorrer
na pele ou em tecidos profundos (GUYTON, 2002; ALMEIDA et al., 2004).
-
3.1. Modulao da nocicepo
A ao pr-nociceptiva de substncias algognicas reflete a capacidade de
mobilizao das fibras aferentes primrias (FAPs) e a transmisso ortodrmica do impulso nervoso em direo ao corno dorsal (CD) da medula espinhal (figura 3). As fibras de um modo geral participam nos efeitos em cascata que transmitem os
estmulos de dor ao sistema nervoso central. De acordo com Castro (2000), as fibras aferentes primrias so:
1. Fibras C: dimetro pequeno (0,4 - 1,2 m), mielinizadas e de conjuno lenta (0,5 2,0 m/seg), localizam-se ao longo das paredes das veias desecadeando uma resposta de dor surda.
2. Fibras A: dimetro mdio (2 - 6 m), mielinizadas e de velocidade de conjuno intermediria (12 - 30 m/seg), desencadeando uma resposta de dor de natureza rpida e lancinante.
3. Fibras A: dimetro grande (>10 m), mielinizadas e de conjuno rpida (30 - 100 m/seg). Este grupo de fibras participa muito pouco de processos nociceptivos agudos, estando mais envolvidos em casos
de alodnia e em dores crnicas.
A liberao de mediadores, em tecidos inflamados pode sensibilizar fibras
nociceptivas perifricas, promovendo a facilitao central da transmisso nociceptiva
(SYRYATOWICS et al., 1999; VANEGAS, 2004), levando a uma resposta dolorosa aumentada a um estmulo nocivo, chamado de hiperalgesia (TJLSEN e HOLE, 1997; SOMER e CRESS, 2004; VANEGAS 2004; COUTAX et al., 2005). Alteraes na percepo sensorial, envolvendo sensibilizao central, pode levar alodnia,
-
que est relacionada a dor evocada por um estmulo no nocivo (MILLAN, 1999).
Figura 3. Organizao das fibras aferentes primrias envolvidas na conduo de um estmulo
nociceptivo desde a superfcie cutnea (terminal perifrico) at o como dorsal da medula espinal. NE, fibra nociceptiva especfica; NP, fibra nociceptiva polimodal; no-N, fibra no
nociceptiva. Adaptado de Millan (1999) por Castro (2000).
Levando em considerao as aes diretas dos diferentes fatores
relacionados nocicepo, algumas substncias agem modificando o limiar de
excitabilidade dos nociceptores, causando analgesia ou hiperalgesia como a
bradicinina e os prostanides (SHEWOOD e TOLIVER-KINSKY,2004). A infuso intradrmica de BK, sua aplicao em superfcies desepitelializadas ou a infuso
endovenosa desencadeiam dor intensa e imediata, em paralelo com a estimulao
de fibras C polimodais e mecanossensveis presentes na pela ou em outros tecidos
(KHAN, 1992; LANG,1990; MANNING,1991,).
3.2. Analgesia
-
A analgesia uma reduo nas percepes e no tempo de resposta do
processo nociceptivo (MILLAN, 1999). Vrios fatores podem contribuir para alterar a resposta, quer seja por ao farmacolgica, quer seja por ao de opiides endgenos.
Os hipoalgsicos so os frmacos que deprimem o sistema nervoso central
(COLLIER et al., 1961), eliminando ou reduzindo a sensao de dor, sem causar perda de conscincia. Agem complexando mltiplos receptores estereoespecficos,
em diversos locais do sistema nervoso central. So divididos entre endgenos e
exgenos.
Os principais agentes e utilizados na terapia da dor so os opiceos
(derivados do pio) e os opiides derivados sintticos com ao semelhante ao da morfina. As encefalinas (opiides endgenos), que so o grupo de opiides que envolvem os processos de analgesia produzidos fisiologicamente, so basicamente
produzidas pelo sistema nervoso central. Os representantes dessa classe,
denominados como opiceos, apresentam alta potncia analgsica, chegando em
alguns casos a ser 1000 vezes maior do que a morfina. Cerca de uma dzia dessas
substncias opiceas foram encontradas em diferentes pontos do sistema nervoso;
basicamente sendo produtos de degradao de trs grandes molculas proticas: a
pr-encefalina, a pr-opiomelacortina, a pr-dinorfina (DREY, 1997; MASOM, 1999; JULIUS e BASBAUM, 2001; VANEGAS, 2004; VANEGAS e SCHAIBLE, 2004). Mltiplas reas do crebro apresentam receptores opiceos, especialmente nas
reas do sistema de analgesia. Entre as diferentes substncias as mais importantes
so: beta-endorfina, metaencefalina, leu-encefalina e a dinorfina. As duas
encefalinas so encontradas em maior quantidade no tronco enceflico e na medula
-
espinhal; j a beta-endorfina est presente tanto no hipotlamo quanto na glndula hipfise. A dinorfina encontrada em quantidade muito menor, sobretudo nas
mesmas reas que as encefalinas. Todos os detalhes mais finos do sistema opiceo
no sistema cerebral no esto completamente compreendidos.
3.3. Dor e o sistema renina-angiotensina
De um modo geral os nociceptores no demonstram capacidade de se
adaptar, se adaptam to pouco ou no se adaptam aos estmulos, que em certas
condies agem de modo que a estimulao das fibras pode ser constante. Alguns
autores demonstraram que ratos espontaneamente hipertensos tm uma
sensibilidade reduzida dor (RODNEY et al., 1995). Entretanto o mecanismo que envolve as diferenas na sensibilidade a dor e a elevao da presso arterial ainda
no esto bem identificados. Administrao de renina e Ang II intracerebroventricular
(icv) exercem efeito analgsico em ratos, atravs da inibio da transmisso da informao nociceptiva trmica, produzindo desta forma, um aumento no tempo de
latncia no teste do tail-flick (GEORGIEVA et al., 1998). Esse efeito bloqueado pela saralasina que um antagonista da Ang II e pela naloxana que um
antagonista opiide, o que sugere a participao da Ang II como neuromodulador
central da nocicepo atravs da liberao de opiides endgenos (TOMA et al., 1997). A Tonina capaz de liberar opiides endgenos (SEIDAH et al., 1979), podendo desta forma participar do processamento da informao sensorial
juntamente com a Ang II como tem sido proposto por diversos autores (HAULICA et al., 1986; TOMA et al., 1997).
s diferentes terminaes nervosas distribudas anatomicamente pela pele
-
podem ser atribudas funes nociceptivas especficas. O conceito de uma
modalidade especfica para dor no pode ser comprovado por enquanto. A
transmisso de estmulos nociceptivos ao sistema nervoso central ocorre atravs de
fibras neuronais de pequeno dimetro no mielinizadas (C) ou finalmente que originam de pequenos neurnios bipolares dos gnglios da raiz dorsal. Em sua
maioria existem como terminaes livres na periferia. A substncia P, a angiotensina
II e a somatostatina so encontradas em seus corpos celulares (BULLINGHAM, 1985). Estas clulas projetam-se centralmente, predominantemente como dorsal da medula, sendo dividido em seis lminas que se desfazem na sinapse com
receptores especficos e /ou no especficos.
4. Camundongo transgnico que expressa tonina de rato
A tecnologia dos transgenes proporciona a oportunidade de entender in vivo o
papel de sistemas peptidrgicos. Utilizando-se do promotor gfap (protena cida fibrilar glial, 2163 pb, sequncia humana), do cDNA da tonina de glndula submandibular de rato e do SV40poliA (859 pb) (figura 4), foi gerado, aqui no Brasil, em colaborao com o Dr Bader do laboratrio de hipertenso do Max-Delbruk
Center (MDC), Berlim, Alemanha, camundongo transgnico que apresenta expresso de tonina de rato TGM(rTon).
Figura 4. Seqncia da construo gnica utilizada na gerao do camundongo transgnico
que expressa tonina de rato.
gfap ton SV40
-
O transgene est expresso em vrios tecidos deste animal sendo que o maior
nvel foi encontrado no encfalo, seguido do corao, rim, fgado, estmago, bao,
tecido adiposo e aorta. O animal apresenta pequeno aumento da presso arterial e
nenhuma diferena de freqncia cardaca quando comparado ao grupo controle
(selvagem). Para esta pesquisa, props-se caracterizar o animal TGM(rTon) quanto resposta nociceptiva.
-
II. OBJETIVOS
a) Geral Avaliar a capacidade nociceptiva dos camundongos TGM(rTon). Executar
experimentos que possam mostrar a possvel participao da tonina em processos
de nocicepo.
b) Especficos
Avaliar a resposta motricial do animal TGM(rTon) utilizando para isso os testes de rota-rod para avaliar a capacidade de
coordenao motora;
Avaliar a resposta nociceptiva dos animais TGM (rTon) atravs do nmero de contores induzidas pelo cido actico, responsvel
por resposta nociceptiva e inflamatria simultaneamente;
Avaliar as respostas nociceptiva e inflamatria em fases separadas
utilizando-se o teste de edema de pata induzido por formalina.
-
III. MATERIAL e MTODOS
1. Comit de tica
Conforme legislao em vigor, os procedimentos experimentais sero
submetidos a um comit de tica local, para garantir a legitimidade do projeto relacionado dor, garantindo assim que no ocorrero procedimentos que sejam eticamente inaceitveis ou cruis aos animais. Oportunamente, o documento de
aprovao ser anexado dissertao.
2. Material
Para a presente pesquisa proposta, sero utilizados os seguintes materiais e
equipamentos:
Caixa de vidro ou acrlico com suporte, equipada com espelhos;
cido actico glacial (ch2H4O2 ) P.A 99,7% marca comercial Vetec
Formaldedo 0,92%;
Aparelho de Rota-rod da marca Ugo-basile;
Pipetas de preciso;
Balana de preciso (para pesagem dos solutos);
Balana para pesar os animais;
-
3. Animais
Para os testes de avaliao da resposta nociceptiva foram utilizados
camundongos transgnicos que apresentam expresso da tonina de rato TGM(rTon) e camundongos controle, da linhagem C57 selvagens (S), pesando entre 25 e 30 g, mantidos em ciclo claro e escuro de 12 h, com gua comum ad libitum. Estes
animais foram provenientes do biotrio da Universidade de Mogi das Cruzes.
No dia anterior ao teste, os animais so pesados, marcados e alojados em caixas plsticas com cama de maravalha (8 animais no mximo por caixa). A rao retirada 15 horas antes dos testes, mantendo-se a gua ad libitum. No dia do
experimento os animais so levados para a sala de experimentao com
antecedncia mnima de duas horas. As condies ambientais da sala onde se
realizaram os experimentos so mantidas mais constante possvel, evitando-se
principalmente o barulho excessivo ou brusco, movimentao de pessoas na sala,
variaes de temperatura, de luminosidade e de odores. Os testes so realizados
entre 10 e 17 horas, trmino das observaes no mximo s 17h30min.
4. Mtodos para avaliar dor
Os mtodos utilizados para se estudar dor e analgesia variam, mas de um
modo geral, eles ocorrem por meios de experincias subjetivas de dor, sob condies controladas experimentais. No entanto, as diferentes manifestaes em
nocicepo, medulares ou supra-medulares, podem ser associadas dor
experimental e podem ser utilizadas na avaliao de substncias com propriedades
-
analgsicas.
Rotineiramente os testes nociceptivos utilizados em animais requerem algum
movimento para mostrar que um estmulo nocivo est tendo efeito. A ausncia ou
diminuio de uma resposta no pode ser tomada como evidncia de ao anti-
nociceptiva, a menos que seja excluda a possibilidade de que o animal no tenha a habilidade motora para executar o requerido movimento.
Colier (1970) preconiza a utilizao de um teste de coordenao motora em combinao com o teste nociceptivo rotineiro, a fim de comprovar que os animais
usados em testes so aptos a responder em situaes onde so requeridos
movimentos em resposta ao estmulo aplicado. Sua teoria foi mais tarde corroborada
por diversos pesquisadores do tema, como Millan (1999), Santos et al. (1999) e Le Bars et al. (2001).
4.1. Teste do "rota-rod
O mtodo de "rota-rod" foi desenvolvido por Kinnard e Carr (1957), com o objetivo de medir a atividade motora coordenada forada. De acordo com estes autores certos agentes deprimem a atividade motora de maneira que os animais so
capazes de mostrar movimentos normais quando estimulados, porm, com os
mtodos utilizados at ento, no podia se avaliar "atividade potencial" desses
animais deprimidos. Por isso, eles propuseram forar continuamente os movimentos,
fazendo com que os camundongos caminhassem sobre um eixo horizontal em
movimento (rota-rod), medindo a habilidade motora pelo tempo de permanncia no eixo girando a 30 RPMs. Os testes realizados para estudar a resposta nociceptiva
envolvem uma resposta motora, seja ela a retirada da cauda, o ato de lamber a pata
-
injetada, ou contores, entre outras. O teste do "rota-rod" detecta a presena de falta de coordenao motora, permitindo uma interpretao mais acurada dos
resultados obtidos nos testes de nocicepo e anti-nocicepo.
O aparelho utilizado consta de uma barra giratria horizontal, de 2,5 cm de
dimetro, revestida de plstico anti-deslizante, suspensa a uma altura de 25 cm e
com rotao constante de 32 rpm (figura 5). Os animais so pr-selecionados, sendo descartados aqueles que no permanecem na barra giratria durante pelo
menos um minuto.
Figura 5. Aparelho de rota-rod da marca Ugo Basile.
Os testes foram realizados no laboratrio da Profa Dra. Janetti Nogueira de
Francischi, do Depto de Farmacologia do Instituto de Cincias Biolgicas da
Universidade Federal de Minas Gerais.
-
4.2. Teste de contoro abdominal
O teste de contoro abdominal um dos mtodos nociceptivos de maior
sensibilidade, captando o possvel efeito analgsico dos opiides endgenos que
eventualmente possam ser produzidos pela tonina. A sndrome de contoro, na
qual ocorre contrao da parede abdominal, rotao do tronco e extenso dos
membros posteriores, foi primeiramente descrita por Vander-Wende e Margolin
(1956), que observaram ao injetar compostos orgnicos iodados, intraperitonealmente em ratos.
A injeo subcutnea de cido actico, via peritoneal, em camundongos, determina o aparecimento de uma gama de respostas motoras bem caracterizadas,
cuja quantificao permite que se avalie a intensidade da resposta nociceptiva. Esta resposta desenvolve-se bem caracterizada, sendo basicamente de origem
neurognica.
Foi utilizado o teste de contoro induzido por cido actico descrito por
Koster (1956), modificado por Castro (2000), no qual considerado o nmero de contores abdominais dos membros posteriores ocorridas entre 5 e 20 minutos
aps a administrao i.p (intraperitonealmente) de 10 mg/kg de cido actico 0,6%. Em cubas de vidro ou acrlico, medindo 12x12x12 cm, so observados dois animais
de cada vez, sendo considerados somente os camundongos que apresentam a
contoro abdominal imediatamente aps a injeo do cido. O cido actico injetado na regio abdominal do camundongo. Espelhos
colocados na parede posterior da caixa, em ngulo de 45, com piso transparente
permitem a visualizao do camundongo durante todo o tempo (figura 6).
-
Figura 6. Caixa utilizada no experimento de contoro pelo cido actico.
A soluo de cido actico 0,6% para a injeo intraperitoneal aplicada cuidadosamente atravs do abdmen com o animal imobilizado e levemente
inclinado de cabea para baixo. Esse procedimento possibilita que as vsceras do
animal, por ao gravitacional, se contraiam levemente evitando-se assim a
perfurao das mesmas. A visualizao da agulha atravs da pele permite que cido
actico seja injetado sempre na rea desejada, levando-se em considerao que a localizao da agulha deve estar sempre um pouco acima da virilha, para garantir a
insero correta da agulha na regional peritoneal.
A avaliao da resposta nociceptiva, imediatamente aps a injeo de cido actico, foi determinada computando-se o nmero de contores que o animal
apresentava, sendo as contores identificadas atravs dos movimentos de
encolhimento e de alongamento do animal na regio abdominal, seguido por
estiramento deu uma ou duas patas. Os clculos envolvendo a mdia ponderada de
cada grupo foram feitos com auxlio de programa de computador (Graph pad prism).
-
4.3. Teste da formalina
Neste teste foi realizada uma avaliao da resposta nociceptiva induzida
pelo formaldedo 0,92% nos camundongos. O formaldedo (20 L) foi injetado pela via subcutnea (sc) no dorso da pata direita posterior. O perodo no qual o animal lambe a pata injetada foi determinado entre 0 e 5 min (1 fase) e entre 15 e 30 min (2 fase) aps a injeo de formaldedo. Imediatamente aps o trmino desse experimento, os animais foram submetidos eutansia em cmara de
CO2.(BENSON,1999), (LE BARS,2001).
Figura 7. Camundongo TGM(rTon), aps experimento de contoro abdominal, j disponibilizado para ser submetido ao processo de eutansia.
-
5. Anlise dos dados
Os resultados foram apresentados como a mdia erro padro da mdia.
As comparaes estatsticas entre vrios grupos foram feitas por ANOVA one way
seguida de Newman-Keuls. Para comparaes entre dois grupos foi utilizado o teste
t de student. O nvel de significncia adotado foi de p < 0,05.
O processamento dos dados coletados e anlises estatsticas foram realizados
atravs do programa GRAPH PAD PRISM 4 for Windows.
-
IV. RESULTADOS
Um dos desafios da transgenia garantir que o animal que recebe genes de
outro animal que no de sua linhagem ir apresentar comportamento e reaes
diferenciadas dos outros de sua espcie. Entretanto, comportamentos e reaes
podem ser dos mais diversos e diferentes tipos, sendo necessrio que se teste cada
possibilidade antes de garantir a eficcia da transgenia no melhoramento gentico
das espcies. Desta forma, na presente pesquisa, o objetivo foi testar se a presena da tonina, protena existente no rato, altera ou no significativamente a resistncia a
dor.
Levando-se em considerao que os camundongos transgnicos
expressam tonina em nveis maiores que os do animal selvagem, foram realizados
testes de rota-rod, para determinar se estes animais apresentam comportamento
matricial diferenciado. Tambm foram feito testes de contoro abdominal para
identificar possveis modificaes nas respostas a dor nesses camundongos. E
finalmente foi realizado o teste de formalina para avaliar possveis modificaes nas
respostas nociceptivas em fase inflamatria.
1. Teste de rota-rod
O teste de rota-rod mede o tempo de permanncia na haste giratria a 32
RPM, ou seja, a capacidade motora dos camundongos. Inicialmente foram feitos testes de rota-rod com camundongos Swiss. Os camundongos Swiss so os
modelos mais freqentes para esse tipo de teste, portanto, so os animais que
-
serviram de parmetro controle para fins de validao do mtodo.
Ao considerar a mdia de permanncia, em minutos, os camundongos
com maior capacidade e coordenao motora foram os selvagens com uma mdia
de permanncia de 4,28 minutos. Os camundongos transgnicos apresentaram uma
mdia de permanncia de 4,12 minutos enquanto os camundongos Swiss
apresentaram uma mdia de permanncia de 3,23 minutos, como mostra a Figura 8.
Swiss Selvagem TGM (rTon)0
1
2
3
4
5
Tem
po de
pe
rman
nci
a
Figura 8. Tempo de permanncia, expresso em minutos, dos diferentes grupos, Swiss,
selvagem e transgnico no teste do rota-rod. *p < 0,05 vs TGM(rTon). As colunas representam a mdia epm (n = 6).
Observou-se uma reduo na capacidade motora dos camundongos
transgnicos quando comparados ao grupo selvagem.
2. Teste de contoro abdominal
O teste com os camundongos foi realizado em esquema monocego, ou seja,
-
o pesquisador no sabia de ante mo quais eram os animais selvagem e quais eram
TGM(rTon). Os animais foram marcados previamente por outra pessoa, que s revelou as identificaes aps o teste ter sido realizado. Nossos resultados mostram
que o TGM(rTon) apresentou tendncia de maior sensibilidade embora esta diferena no seja estatisticamente significante (figura 9).
Selvagem TGM (rTon)0
10
20
30
Nm
ero
de
co
nto
re
s
Figura 9. Contores abdominais induzidas por cido actico em camundongos selvagens e
TGM(rTon) em perodo de 20 minutos.A contagem feita em 4 tempos de 5 minutos consecutivos e em sistema monocego. As colunas representam a mdia epm Mann
Whitney test (n = 6).
3. Teste de formalina
Nesse teste foi observado o comportamento de lambida nas duas fases que
so desenvolvidas, sendo que na primeira fase temos a resposta nociceptiva, ligada
aos efeitos em cascata que so desencadeados por ao da formalina, que provoca
mobilizao e edema de pata. J na segunda fase tem-se como resposta uma
-
somatria de processo inflamatrio e nocicepo (Figura 10).
Figura 10. Teste de formalina. Nmero de lambidas induzidas pela injeo de formalina em camundongos selvagens (S) e transgnicos (T). A primeira fase corresponde ao tempo de 0 - 5 minutos aps a injeao e a segunda fase 15 - 30 minutos. *p < 0,05. As colunas representam a mdia epm (n = 6).
S T S T0
50
100
150
200
250
1 fase 2 fase
n de
la
mbi
das
-
V. DISCUSSO
Antes de analizar os resultados obtidos devemos considerar que segundo
Guasti et al. (1997), pacientes hipertensos so menos sensveis estmulos
nociceptivos, independente do mtodo de aplicao do estmulo. Neste mesmo
trabalho apresentado um dado interessante, que diz que com a administrao
de anti-hipertensivos a pacientes, quer sejam humanos ou animais, os nveis
de sensibilidade dor foram restabelecidos, o que sugere que a angiotensina II
participa no processo de nocicepo.
Estes dados no fornecem evidencia direta da participao da
angiotensina II, que seriam obtidas utilizando-se antagonistas especficos de
receptores. Existe a possibilidade de que alteraes na sensibilidade em
individuo hipertenso seja desencadeado pelos pressoreceptores mais do que
pela interao angiotensina II-receptor. Considerando que tonina uma enzima
capaz de liberar angiotensina II, ela tambm pode estar envolvida neste
processo.
O TGM(rTon) apresenta ligeiro aumento nos nveis pressricos quando
comparado com o seu controle, o WT, no entanto nenhuma diferena entre os
dois grupos foi observada nos nveis de angiotensina II tanto as nvel
plasmtico como em diversos tecidos de importncia cardiovascular. Desta
forma, conclumos que o aumento nos nveis pressricos observados para o
TGM(rTon) deve se dar por algum mecanismo que no envolve a angiotensina
II.
Analisando os dados do teste de rota-rod observamos que, os
camundongos selvagens apresentaram resposta motricial superior. Certamente
-
esta diferena no est relacionada aos nveis de angiotensina II. Outros
possveis mecanismos precisam ser pesquisados. J no teste de contoro
abdominal, nenhuma diferena significativa foi observada entre o TGM(rTon) e
seu controle, o WT. Entretanto, os resultados mostram uma tendncia de os
animais transgnicos serem mais susceptveis a dor. Acreditamos que com um
numero maior de animais poderemos demonstrar definitivamente esta
diferena entre o TGM(rTon) e o WT.
A injeo intraperitoneal de cido actico provoca reao de
nocicepo e inflamao simultaneamente. Considerando que tonina pode
hidrolisar beta-lipotropina e liberar peptdeos com ao opiide (Seidah et al.,
1979), seria de se esperar que o TGM(rTon) apresentasse menor sensibilidade
a dor. Pelo fato de o teste de contoro ao cido actico avaliar aspectos
nociceptivos e inflamatrios, torna-se um modelo menos preciso, porm mais
sensvel e com respostas mais claras. No nosso caso, alm de se fazer
necessrio aumentar o numero de animais neste modelo de estudo, faz-se
necessrio um segundo modelo a fim de confirmar os resultados obtidos.
O teste da formalina serve de salvaguarda para cobrir pelo menos
dois aspectos de nocicepo, sendo que na primeira fase a resposta est
ligada cascata nociceptiva, e na segunda fase ao processo nociceptivo e
inflamatrio. Os dados obtidos mostram que na primeira fase do teste no
houve diferena entre os dois grupos. Por outro lado, os dados referentes
segunda fase do teste mostram que os animais transgnicos mostraram ser
mais sensveis dor, concordando com a tendncia apontada pelos dados
obtidos com o cido actico.
O processo inflamatrio resulta na liberao de vrios mediadores,
-
alguns deles podendo ativar ou sensibilizar nociceptores locais das fibras
aferentes. Como j comentamos, a possvel liberao de opiides pela tonina
poderia levar a uma reduo da sensibilidade a dor. No temos no momento
uma explicao para o fato. Ao momento em que aumenta a circulao
perifrica, opiides endgenos tem sua atividade reduzida a nveis subclnicos,
no sendo suficientes para reduzir a resposta nociceptiva e consequentemente
de dor (LEVINE et al., 1993; DRAY, 1994; BESSON e DICKENSON, 1997;
BARROS et al., 2004; COUTAUX et al., 2005).
No temos ainda informaes a respeito do mecanismo pelo qual
tonina est interferindo na fisiologia do animal, por qual mecanismo estaria
envolvida, no s no aumento da presso sistmica, mas tambm na
sensibilidade a dor do animal. Certamente, estes dados abrem novas
possibilidades de estudos com esta enzima, para a qual inicialmente foi
atribudo como principal papel o de liberar angiotensina II.
-
VI. CONCLUSES
1. A introduo de transgene no camundongo levou a reduo da capacidade
motora do animal, conforme observado pelo teste de rota-rod indicando uma
possvel presena dos opiides endgenos;
2. A introduo de transgene no camundongo, diferentemente do esperado,
parece aumentar a sensibilidade do animal dor, conforme observado pelo
teste de contoro abdominal induzida pelo cido actico. Este dado dever
ser confirmado em experimento envolvendo nmero maior de animais, uma
vez que a diferena na sensibilidade no foi diferente estatisticamente;
3. Tonina pode estar envolvida no processo de nocicepo, uma vez que, no
teste da formalina, os animais mostraram diferena significativa na
sensibilidade dor, sendo esta sensibilidade aumentada na segunda fase do
processo, que envolve dor e inflamao, desta maneira sendo necessrios
maiores estudos sobre o tema, utilizando uma diversidade maior de modelos
e uma anlise com grupos maiores.
-
VII. REFERNCIAS
ABBOTT, F. V. ; FRANKLIN, K. B. J. ; WESTBROOK, R. F. The formalin test: scoring
properties of the first and second phases of the pain response in rats. Pain, v. 60, p.
91-102, 1995.
ABBOTT, F. V. et al. Improving the efficiency of the formalin test. Pain, v. 83, p. 561-
569, 1999.
ARAJO, G. W. et al. Identification of serine proteinases with tonin-like activity in the rat submandibular and prostate glands. Biochimica et Biophysica Acta, v. 1074, p.
167171, 1991.
ARAUJO, G. W. et al. Tonin-like activity in the rat submandibular and prostate
glands. Biochem Biophys Acta, v. 1074, p. 167-171, 1991.
ARAJO, R. C. et al. Tonin expression in rat brain and tonin-mediated central production of angiotensin II. Physiology and Behaviour, v. 76, p. 327333, 2002.
ARAUJO, R. C. et al. Tonin expression in the rat brain and tonin-mediated central
production of angiotensin II. Physiol Behav, v. 76, p. 327-333, 2002.
BADER, M. Transgenic animal models for the functional analysis of vasoactive
peptides. Braz J Med Biol Res, v. 31, p. 1171-1183, 1998.
BALTATU, O. et al. High levels of human chymase expression in the pineal and
pitutary gland. Brain Res, v. 759, p. 269-278, 1997.
BLEY, K. B. ; HUNTER, J. C. ; EGLEN, R. M. ; SMITH, J. A. M. The role of
prostanoid receptors in inflammatory pain. TIPS, v. 19, p. 141-147, 1998.
BOHLENDER, J. et al. High human renin hypertension in transgenic rats.
Hypertension, v. 29, p. 428-434, 1997.
BONICA, J.J. Mecanismos da dor. Revista Brasileira de Anestesiologia, v.26, n.
-
4, p. 477-502, 1976
BORGES, J. C. et al. Tonin in rat heart with experimental hypertrophy. Am J Physiol
Heart Circ Physiol, v. 284, p. H2263-2268, 2003.
BOUCHER, R. , ASSELIN, J. , GENEST, J. A new enzyme leading to the direct
formation of angiotensin II. Circ Res, n. 34/35, p. I203-I209, 1974.
BOUCHER, R. ; ASSELIN, J. ; GENEST, J. A new enzyme leading to the direct
formation of angiotensin II. Circulation Research, v.34/35, suppl. 1, p. 203-209,
1974.
BOUCHER, R. ; SAIDI, M. ; GENEST, J. A new "antiogensin I converting enzyme"
system. In: Hypertension / edited by J. Genest e E. Koiw. Heidelberg, New York :
Springer-Verlag, p. 512-523, 1972.
BOUCHER, R. et al. Tonin-angiotensin II system in hypertension. Clin Sci Mol Med
Suppl, v. 4, p. 183S-186S, 1978.
BRASIER, A. R. , RECINOS, A. , ELEDRISI, M. S. Vascular inflammation and the
renin-angiotensin system. Arterioscler Throm Vasc Biol, v. 22, p. 1257-1266, 2002.
BULLINGHAM, R. E. S. Physiological mechanisms in pain. In: Smith G. ; COVINO,
B. G. Acute pain London: Butterworths, 1985. p. 1-21.
BUNNEMAN, B. , FUXE, K. , GANTEN, D. The renin angiotensin system in the brain.
Regul Pept, v. 46, p. 487-509, 1993.
BUNNEMAN, B. ; FUXE, K. ; GANTEN, D. The renin-angiotensin system in the
brain: an up date 1993. Regulatory Peptides, v. 46, p. 487509, 1993.
CADET, R. ; AIGOUY, L. ; WODA, A. Enhanced nociceptive behaviour following
conditioning injection of formalin in the perioral area of the rat. Brain Research, v. 676, p.189-195, 1995.
CARDOSO, Cibele Campos. Gerao e caracterizao de rato e camundongo
-
transgnico apresentando superexpresso de tonina de rato. 2002. 87 f. Tese
(Doutorado em Biologia) - Curso de Ps-graduao em Cincias Biolgicas, Universidade Federal de Minas Gerais, Belo Horizonte, 2002.
CARVALHO, Wilson Andrade ; CARVALHO, Rosemary Duarte Sales ; RIOS-
SANTOS, Fabrcio. Analgsicos Inibidores especficos da Ciclooxigenase-2:
avanos teraputicos. Revista Brasileira de Anestesiologia, v. 54, n. 3, p. 448-
464, maio/jun. 2004. CASTRO, Maria Salete de Abreu. Mecanismos envolvidos no efeito
antinociceptivo do 3-O-glicosil-dihidrocanferol, flavonide estrado dos
rizomas de Cochlospermum regium (algodozinho). 2000. 155f. Tese (Doutorado em Farmacologia). Universidade Federal de So Paulo Escola Paulista de Medicina, So Paulo, 2000.
CHIU, A. T. et al. Identification of angiotensin II receptor subtypes. Biochem
Biophys Res Commun, v. 165, p. 196-203, 1989.
CHRTIEN, M. et al. Substrates and inhibitors of angiotensin converting enzyme. The Journal of Biological Chemistry, v. 255, p. 401-407, 1980.
CLAVELOU. P. et al. Application of the formalin test to the study of orofacial pain in
the rat. Neuroscience Letters, v. 103, p.349-353, 1989.
CLAVELOU. P. et al. The orofacial formalin test in rats: effects of different formalin
concentrations. Pain, v. 62, p.295-301, 1995.
CLOUSTON, W. M. , LYONS, I. G. , RICHARDS, R. I. Tissue-specific and hormonal
regulation of angiotensinogen minigenes in transgenic mice. EMBO Journal, v. 8, p.
3337-3343, 1989.
CODERRE, T. J. et al. The formalin test: a validation of the weighted-scores method
of behavioural pain rating. Pain, v. 54, p.43-50, 1995.
-
COLE, J. M. et al. New approaches to genetic manipulation of mice: tissue-specific
expression of ACE. Am J Physiol Renal Physiol, v. 284, p. F599-607, 2003.
COLLIER, H. O. Endogenous broncho-active substances and their antagonism.
Advanced Drug Research, v. 5. p. 95-107, 1970.
DALLEL, R. et al. Evidence for a peripheral origin of the tonic nociceptive response
to subcutaneous formalin. Pain, v. 61, p.11-16, 1995
DICKENSON, A. Mechanismms of central hypersensitivity: excitatory amino acids
mechanisnms and their control. In: BESSON, M. J. ; DICKENSON, A. The
farmacology of pain. Berlin : Springer-Verlag, 1997.
DRAY, A. Peripheral mediators of pain. In: BESSON, M. J. ; DICKENSON, A. The
farmacology of pain. Berlin : Springer-Verlag, 1997.
DZAU, V. J. et al. A comparative study of the distribution of renin and
angiotensinogen messenger ribonucleic acids in rat and mouse heart.
Endocrinology, v. 120, p. 23342338, 1987.
DZAU, V. J. Significance of the vascular renin-angiotensin pathway. Hipertensin, v.
8, p. 553-559, 1986.
DZAU, V. J. Vascular renin-angiotensin: a possible autocrine or paracrine system in
control of vascular function. J Cardiovasc Pharmacol, v. 6, p. S377-382, 1984.
DZAU, V.J.; INGELFINGER, P.R.E.; ELLISON, K.E. Identification of mesenger RNA
sequences in mouse and rat brains. Hypertension, v. 8, p. 544-548,1986.
FIELDS, H. L. ; BASBAUM, A. I. Endorphin-mediated pain control systems. In:
WALL, P. D. ; MELZACK, R. Textbook of Pain. London : Churchill-Livingston, 1984.
p. 142-152.
FITZSIMONS, J. T. Angiotensin stimulation of the central nervous system. Review
Physiologic Biochemical Pharmacology, v. 87, p. 117-67, 1980.
-
GANONG, W. F. et al. Role of vasopressin in the renin and ACTH responses to
intraventricular angiotensin II. Annual New York Academy Science , v. 394, p. 619-
24, 1982.
GARCIA, R. ; BOUCHER, R. ; GENEST, J. Tonin activity in rat saliva: effect of
sympathomimetic and parasympathomimetic drugs. Canadian Journal of
Physiology and Pharmacology , v. 54, p. 443-445, 1976.
GARCIA, R. et al. Effect in vivo of -adrenergic stimulation, angiotensin II, dibutyryl
cyclic AMP, and theophylline on tonin concentration in rat saliva and submaxillary
gland. Canadian Journal of Physiology and Pharmacology, v. 55, p. 983989,
1976.
GARCIA, R. et al. Effect of antitonin on blood pressure in the one-kidney
hypertension rat. Clin Sci Mol Med, v. 54, p. 457-461, 1978.
GARCIA, R. et al. Release of tonin and kallikrein by perfused rat submaxillary gland.
Am J Physiol, v. 244, p. R228-R234, 1983.
GENEST, J. Angioten-forming enzymes from extrarenal sources. In: Hypertension
and the Angiotensin System: Therapeutic Approaches. NY: Raven Press, 1983. p.
93-107
GEORGIEVA, D. ; GEORGIEV, V. The role of angiotensin II and of its receptor
subtypes in the acetic acid-induced abdominal constriction test. Pharmacology and
Biochemical Behaviour, v. 62, n. 2, p. 229-32, Feb./1999
GLOWINSKI, J. , IVERSEN, L. L. Regional studies of cathecolamines in the rat brain-
I. The disposition of (3H) norepinefrine, (3H) dopamine (3H) DOPA in various regions of the brain. J Neurochem, v. 13, p. 655-669, 1966.
GRADY, E. F. et al. Expression of AT2 receptors in the developing rat fetus. J Clin
Invest, v. 88, p. 921-933, 1991.
-
GRIS, C. et al. Formation of angiotensin II by tonin from partially purified human angiotensinogen. Can J Biochem, v. 59, p. 250-255, 1981.
GRIS, C. et al. Formation of angiotensin II by tonin from partially purified human angiotensinogen. Canadian Journal Biochemical, v. 59, p. 250-255, 1981.
GUALBERTO, M. P. , NUNES, R. L. , PESQUERO, J. L. Tonin-like activity present
in the human submandibular gland. Agents and Actions Suppl, v. 38, p. 392-400,
1992.
GUTKOWSKA, J. et al. Tonin as activator of renin. Can J Biochem, v. 60, p. 843-
846, 1982.
GUYTON, Arthur C.; HALL, John Edward. Tratado de fisiologia mdica 10. ed. Rio
de Janeiro : Guanabara Koogan, 2002. 973 p.
HAULICA, I. et al. Evidence for the involvement of cerebral renin-angiotensin system
(RAS) in stress analgesia. Pain, v. 27, n. 2, p. 237-45, Nov./1986. HOLE, K. ; TJOLSEN, A. The tail-flick and formalin tests in rodents: changes in skin
temperature as a confounding factor. Pain, v. 53, p. 247-254, 1993.
HEIN, L. et al. Behavioural and cardiovascular effects of disrupting the angiotensin
II type-2 receptor gene in mice. Nature, v. 377, p. 744-747, 1995.
HEIN, L. et al. Overexpression of angiotensin AT1 receptor transgene in the mouse
myocardium produces a lethal phenotype associated with myocyte hyperplasia and
heart block. Proc Nat Acad Sci, USA, v. 94, p. 6391-6396, 1997.
ICHIKI, T. et al. Effects on blood pressure and exploratory behaviour of mice lacking
angiotensin II type-2 receptor. Nature, v. 377, p. 748-750, 1995.
JAFFE, J. H. ; O'KEEFFE, C. From morphine clinics to buprenorphine: regulating
opioid agonist treatment of addiction in the United States. Drug and Alcohol
Dependence-1, v. 70, Suppl. 2, p. S3-S11, 21 May 2003.
-
JOHANSEN, L. et al. Excess antibody immunoassay for the measurement of tonin in
rat tissues and plasma. Journal of Immunological Methods : JIM , v. 98, p. 257-
265, 1987.
KHAN, A. A. et al. The efects of bradykinin and sequence-ralated analogs on the
response properties of cutneos nociceptors in monkeys. Somatosens Molecular
Researcj, v. 9, p. 97-106, 1992 KIM, H. S. et al. Genetic control of blood pressure and the angiotensinogen locus.
Proc Nat Acad Sci, USA, v. 92, p. 2735-2739, 1995.
KIM, S. , IWAO, H. Molecular and cellular mechanisms of angiotensin IImediated
cardiovascular and renal diseases. Pharmacol Rev, v. 52, p. 11-34, 2000.
KIMURA, S. et al. High blood pressure in transgenic mice carrying the rat
angiotensinogen gene. EMBO Journal, v. 11, p. 821-827, 1992.
KIMURA, S. et al. High blood pressure in transgenic mice carrying the rat
angiotensinogen gene. EMBO Journal, v. 11, p. 821-827, 1992.
KINNARD JR., W. J. ; CARR, C. J. A preliminary procedure for the evaluation of
central nervous system depressants. Journal of Pharmacology and Experimental
and Therapeutics, v. 120, p. 354361, 1957.
KKS, S. et al.. Cholecystokinin2 receptor-deficient mice display altered function of brain dopaminergic system. Psychopharmacology, v.158, p.198-204, 2001.
KONDO, K. et al. The effects of intracerebroventricular administration of tonin on
water drinking and blood pressure in rat. Brain Res, v. 200, p. 437-441, 1980.
KREGE, J. H. et al. Male-female differences in fertility and blood pressure in ACE-
deficient mice. Nature, v. 375, p. 146-148, 1995.
KUGAWA, F. ; MATSUMOTO, K. ; AOKI, M. Apoptosis-like cell death of human
breast cancer cell line MCF-7 induced by buprenorphine hydrochloride. Life
-
Science, v. 75, n. 3, p. 287-99, 4 Jun. 2004.
KUMAR, R. S. et al. Structure of testicular angiotensin converting enzyme: a
segmental mosaic. The Journal of Biological Chemistry, v. 264, p. 16754-8, 1989.
KUNAPULI, S. P. et al., Molecular cloning of human angiotensinogen cDNA and
evidence for the presence of its in rat hearth. Circulation Research, v. 60, p. 786-
90, 1987.
LANG, E. et al. Chemosensitivity of fine afferents from rat skin in vitro.
Neurofisiology, v. 63, p. 887-901, 1990.
LAPA, Fernanda da Rocha. Avaliao da atividade antinoceptva,
antiinflamatria e protetora gstrica do extrato hidroalcolico bruo da Polygala
paniculata L 2006. 119f. Dissertao (Mestrado em Farmacologia). Universidade Federal de Santa Catarina, Florianpolis, 2006.
LAZURE, C. et al. Aminoacid sequence of rat submaxillary tonin reveals similarities
to serine proteases. Nature, v. 307, p. 555-558, 1984.
LE BARS, D. ; GOZARIU, M. ; CADDEN, S.W. Animal models of nociception.
Pharmacological Review, v. 53, p. 597-652, 2001.
LEHMANN, Klaus A. Introductory remarks. In: PARNHAM, Michael J. Non-opioids
in pain management. Vancouver (Canad): Birkhuser, 1997. Disponvel em: Acesso em: 29/05/2007.
LINDBERG, B. F. et al. Angiotensin I is converted to angiotensin II by a serine
protease in human destrusor smooth muscle. Am J Physiol, v. 266, p. R1861-1867,
1994.
LINDPAINTER, K. et al. Tissue renin-angiotensin system in focus on the heart.
-
Journal of the Hypertension, v. 5, p. S33-S38, 1987.
LIS, M. et al. Dependence of tonin activity in rat submaxillary gland on growth
hormone and testosterone. Am J Physiol, v. 232, p. E522-525, 1977.
LOMEZ, E. S. et al. Tonin and kallikrein in the brain of transgenic rat line expressing
human tissue kallikrein. Hypertension, v. 39, p. 229-232, 2002.
LOPES, E. S. et al. Distribution of tonin- and kallikrein-like activities in the rat brain.
Brain Res, v. 769, p. 152-157, 1997.
LOPES, E. S. et al. Distribution of tonin- and kallikrein-like activities in rat brain.
Brain Research, v. 759, p. 152157, 1997.
MANNING, D. C. et al. Pain and hiperalgesia after intradermical injection of bradykinin in humans. Clinical Pharmacological Therapy, v. 50, p. 721-729, 1991.
MARRERO, M. B. et al. Direct stimulation of Jak/STAT pathway by the angiotensin II
AT1 receptor. Nature, v. 375, p. 247-250, 1995.
MELZACK, R. ; WALL, P. D. Pain mechanisms: a new theory. Science, v. 150, p.
971-979, 1965.
MILAN, Mark J. The induction of pain: an integrative review. Progress in
Neurobiology, v. 57, p. 1-164, 1999.
MOBINI, S. et al. Comparison of the effects of clozapine, haloperidol, chlorpromazine
and d-amphetamine on performance on a time-constrained progressive ratio
schedule and on locomotor behaviour in the rat. Psychopharmacology, v.152, p.47-
54, 2000.
MOFFET, R. B. , BUMPUS, F. M. , HUSAIN, A. Minireview: celular organization of
the brain renin-angiotensin system. Life Sci, v. 41, p. 1867-1979, 1987.
MULLINS, J. J. , PETERS, J. , GANTEN, D. Fulminant hypertension in transgenic
rats harbouring the mouse Ren-2 gene. Nature, v. 344, p. 541-544, 1990.
-
NAKAMURA, M. et al. A role for adrenal renin-angiotensin system in the regulation of
potassium-stimulated aldosterone production. Endocrinology, v. 117, p. 1772-1778,
1985.
NIIMURA, F. et al. Gene targeting in mice reveals a requirement for angiotensin in
the development and maintenance of kidney morphology and growth factor
regulation. J Clin Invest, v. 96, p. 2947-2954, 1995.
OHKUBO, H. et al. Generation of transgenic mice with elevated blood pressure by
introduction of the rat renin and angiotensinogen genes. Proc Nat Acad Sci, USA, v.
87, p. 5153-5157, 1990.
OLSEN, M. E. et al. Mechanisms of angiotensin II natriuriesis and antinatriuresis.
Annual Review of Physiology, v. 249, p. F299-307, 1985.
PEACH, M. J. Renin-angiotensin system: biochemistry and mechanisms of action.
Physiological Reviews, v. 57, p. 313370, 1977.
PELTO, C. T. , KERN, M. D. Inhibition of tonin activity by growth hormone and
testosterone depletion in one-kidney, one-clip hypertensive rats. Curr Surg, v. 43, p.
474-475, 1986.
PEREIRA, Helena Lcia Alves et al. Avaliao por imagem do comprometimento
osteoarticular e de nervos perifricos na hansenase. Revista Brasileira de
Reumatologia, So Paulo, v. 46, supl. 1, jun. 2006. Disponvel em: http://www.scielo.br Acesso em: 23 jun. 2007. PESQUERO, J. L. et all. Effect of substrate size on tonin activity. Biochem Biophys
Res. Commun, v. 108, n. 4, p. 1441-6, 29 Oct. 1982.
PHILLIPS, M. I. , KAGIYAMA, S. Angiotensin II as a pro-inflammatory mediator.
Curr Opin Investig Drugs, v. 3, p. 569-577, 2002.
PHILLIPS, M. I. , SPEAKMAN, E. A. , KIMURA, B. Levels of angiotensin and
-
molecular biology of the tissue renin-angiotensin systems. Regul Pept, v. 43, p. 1-
20, 1992.
PHILLIPS, M.I. Functions of brain angiotensin. Annual Review of Physiology, v.
49, p. 413-435, 1987.
RANG, H. P.; DALE, M. M. Analgesic drugs. In: Pharmacology. London: Churchil
Livingstone, 1985. p. 547-567.
RANG, H. P.; DALE, M. M. ; RITTER, J. M. Farmacologia. 4a ed. Rio de Janeiro:
Guanabara Koogan, 2001. 704 p.
ROSENTHAL, J. et al. Renin-like enzymes in human vasculature. Hypertension, v.
15, p. 848-853, 1990.
ROVERONI, R. C. et al. Development of a behavioral model of TMJ pain in rats: the
TMJ formalin test. Pain, v. 94, p.185-191, 2001.
SAYE, J. A. et al. Localization of angiotensin peptide-forming enzymes of 3T3-F442A
adipocytes. Am J Physiol, v. 264, p. C1570-C1576, 1993.
SCHIFFRIN, E. L. et al. Effects of tonin on the adrenal secretion in the rat.
Hypertension, v. 3. p. 119-13, 1981.
SCHIFFRIN, E. L. et al. Role of the pituitary in the effects of tonin on adrenal
secretion of the rat. Can J Physiol Pharmacol, v. 61, p. 201-206, 1983.
SCHILLER, P. W.; DEMASSIEUX, S.; BOUCHER, R. Substrate specificity of tonin
from rat submaxillary gland. Circulation Research, v. 39, p. 629-632, 1976.
SCHINKE, M. et al. Permanent inhibition of angiotensinogen synthesis by antisense
RNA expression. Hypertension, v. 27, p. 508-513, 1996.
SCHMID, Helena. Impacto cardiovascular da neuropata autonmica do diabetes
mellitas. Arquivos Brasileiros de Endocrinologia & Metabologia, So Paulo, v.
51, n. 2, mar. 2007. Disponvel em: Acesso em: 23 jun. 2007.
-
SEIDAH, N.G. et al. Specific cleavagr of beta-LPH and ACTH by tonin: realesed of
an opiate-like peptide beta LPH (61-78). Biochemical and Biophysical Research Community, v. 86, p. 1002-1013, 1979.
SHIBATA, M. et al. Modified formalin test: characterisitc biphasic pain response.
Pain, v. 38, p.347-352, 1989.
SHIH, H. C. , CHAO, L. , CHAO, J. Age and hormonal dependence of tonin levels in
rat submandibular gland as determined by a new direct radioimmunoassay.
Biochem J, v. 238, p.145-149, 1986.
STEINHOFF B. J. et al. The ideal characteristics of antiepileptic therapy: an overview
of old and new AEDs. Acta Neurological Scandinavian, v.107, p.87-95, 2003.
TABER, R. I. et al. Agonist and antagonist interactions of opioids on acetic acid
induced abdominal stretching in mice. Journal of Pharmacology and Experimental
and Therapeutics, v. 169, n. 1, p. 29-38, 1969.
TAKAHASHI, S. et al. Expression of the human angiotensinogen gene in transgenic
mice and transfected cells. Biochem Biophys Res Commun, v. 180, p. 1103-1109,
1991.
TANG, A. H. et al. Anxiolytic-like effects of PNU-101017, a partial agonist at the
benzodiazepine receptor. Psychopharmacology, v.131, p. 255-263, 1997.
TANIMOTO, K. et al. Angiotensinogen-deficient mice with hypotension. J Biol Chem,
v. 269, p. 31334-31337, 1994.
THOMPSON, M. W. , SMITH, S. B. , SIGMUND, C. D. Regulation of human renin
mRNA expression and protein release in transgenic mice. Hypertension, v. 28, p.
290-296, 1996.
TIAN, X. L. et al. A new transgenic rat model overexpressing human angiotensin-
converting enzyme in the heart. Hypertension, v. 28, p. 520 (Abstract), 1996.
-
Tremblay J, Thibault G, Gutkowska J, Boucher R, Genest J. Purification and partial
characterization of plasma inhibitor of tonin. Canadian Journal Biochemical, v. 59,
p. 256-261, 1981.
TREMBLAY, J. et al. Purification and partial characterization of a plasma inhibitor of
tonin. Can J Biochem, v. 59, p. 256-261, 1981.
TSUTUMI, K. , SAAVEDRA, J. M. Characterization of AT2 angiotensin II receptors in
rat anterior cerebral arteries. Am J Physiol, v. 261, p. H667-670, 1991.
VANDER-WENDE, C.; MARGOLIN, S. Analgesic tests based upon experimentally
induced abdominal pain in rats. Federal Proceedings, v. 15, p. 494, 1956.
VILELA, Hugo ; FERREIRA, Daniel. Analgesia, sedao e relaxamento
neuromuscular no doente ventilado em cuidados intensivos cardacos. Parte I:
analgesia. Revista Portuguesa de Cardiologia, Portugal, v. 25, n. 1, p. 89-98,
2006.
WRIGHT, J. W. , HARDING, J. W. Brain angiotensin receptor subtypes in the control
of physiological and behavioural responses. Neurosci Biobehav Rev, v. 18, p. 21-
53, 1994.
YOSIPIV, I. V. , EL-DAHR, S. S. Activation of angiotensin-generating system in the developing rat kidney. Hypertension, v. 27, p. 281-286, 1996. ZACHARATOS, D. T. et al. Immunohistochemistry of tonin in rat submandibular
gland during development, lactation and secretion. Histochenistry, v. 79, p. 433-
442, 1983.
-
ANEXO 1 TABELAS
Analise estatstica das contores abdominais em c. Actico
( Graph Pad Prism)
-
Anlise estatstica no teste de formalina ( Graph Pad Prism)