
Universidade do Algarve - fctec.ualg.pt · 1.1 – Metabolismo de xenobióticos A maior parte dos...
83
Universidade do Algarve Faculdade de Ciências e Tecnologia Isa da Conceição Lopes Cavaco Faro Novembro, 2001
Transcript of Universidade do Algarve - fctec.ualg.pt · 1.1 – Metabolismo de xenobióticos A maior parte dos...

Universidade do Algarve Faculdade de Ciências e Tecnologia
Isa da Conceição Lopes Cavaco
Faro
Novembro, 2001

Relatório de estágio da Licenciatura em
Bioquímica, da Faculdade de Ciências e
Tecnologia, sob a orientação da Professora
Doutora Vera Ribeiro Marques

I
Agradecimentos
Quero expressar a minha gratidão para com todos aqueles que de alguma forma contribuiram para a
realização deste trabalho, assim como, para a minha formação.
Em especial gostaria de agradecer,
Ao Professor Doutor Manuel Aureliano Alves por todo empenho, dedicação e amizade que tem para
com todos os alunos de Bioquímica.
À Professora Doutora Vera Ribeiro Marques pela possibilidade de realizar o estágio sobre a sua
orientação e também pela sua amizade, permamente disponibilidade, apoio e ensinamentos
transmitidos ao longo deste trabalho.
Ao Professor Doutor José Pedro Gil pela colaboração prestada ao longo de todo o trabalho, assim
como pela amizade e boa disposição sempre presentes.
A todos os docentes da licenciatura em Bioquímica da Universidade do Algarve pelos conhecimentos
transmitidos ao longo destes anos.
À Professora Doutora Leonor Cancela e à Merck Eurolab pelas facilidades técnicas concedidas que
permitiram a realização deste trabalho.
Ao Engenheiro Eduardo Naia, meu professor de técnicas laboratorias de química do secundário, por
ter feito nascer em mim o gosto pelo trabalho em laboratório e pela investigação, e, acima de tudo,
pela amizade e por todos os conselhos que sempre me deu.
Às minhas amigas, colegas de curso e de laboratório, Orquídea e N’zola por tudo aquilo que passámos
ao longo destes cinco anos e pela amizade que nos une.
À Rosinda, minha amiga do coração, que embora por vezes longe, esteve sempre presente com a sua
amizade, e também a todos aqueles que com a sua amizade marcaram a minha vida.
Ao Manuel por ter estado sempre a meu lado, pelo carinho e por tudo o resto.
Por fim, mas em primeiro lugar, aos meus pais por tudo o que sempre fizeram por mim e que me fez
chegar até aqui. Também à minha irmã pela amizade de sempre que nos une.

II
Abreviaturas e simbologia
A maioria das siglas e abreviaturas aqui compiladas estão também definidas ao longo do texto
da tese.
A – Nucleótido contendo adenina
ABC – ATP Binding Cassete*
Bis-acrilamida – N, N’- metileno-bis-acrilamida
bp – pares de bases (base pairs)
C – Nucleótido contendo citosina
CAR – Receptor Constitutivo dos Androstanos (Constitutive Androstane Receptor)
cDNA – DNA complementar ao RNA
CI – intervalo de confiança (Confidence Intervals)
C-terminal – Extremidade da cadeia polipeptídica que apresenta um grupo carboxilo livre
CYP – citocromo P450
df – graus de liberdade (degrees of freedom)
DME – enzima metabolizadora de drogas (Drug Metabolyzing Enzyme)
DNA – Ácido desoxiribonucleico
dNTP – desoxiribonucleótido
EDTA – Ácido (etilenodiamino)tetracético
ER – Receptor de estrogénios
f – frequência
G – Nucleótido contendo guanina
g – Aceleração da gravidade
GR – Receptor dos Glucocorticóides
* Alguns termos técnicos com origem na língua inglesa são utilizados no decorrer do texto desta tese. Não são traduzidos visto que fazem parte de um glossário científico estabelecido na literatura internacional, sendo o seu sentido provavelmente desvirtuado numa tradução literal.

III
MDR – Multidrug Resistance
MRP – Multidrug Resistance Protein ou Multidrug Resistance-Related Protein
NAD(P)H – (Difosfato de) dinucleótido de adenina e nicotinamida, reduzido
N-terminal – Extremidade da cadeia polipeptídica que apresenta um grupo amino livre
p – probabilidade
PAGE – Electroforese em gel de poliacrilamida
PCR – Reacção em cadeia da polimerase de DNA (Polymerase Chain Reaction)
P-gp – P-glicoproteína (P-glycoprotein)
PSA – Persulfato de amónio
PXR – Receptor de Pregnanos (Pregnane X-Receptor)
RFLP – Restriction Fragment Length Polymorphism
RXR – Receptor de Retinóides (Retinoid X Receptor)
SD – Desvio Padrão (Standard deviation)
s – segundo
SNP – Single Nucleotide Polymorphism
SXR – Receptor de Esteróides e Xenobióticos (Steroid and Xenobiotic Receptor)
T – Nucleótido contendo timina
Taq – Thermus aquaticus
TEMED – N, N, N’, N’ - tetrametilenodiamina
Tris – tris (hidroximetil) aminometano
Tween 20 – poli-oxietileno sorbitan monolaurato
U – Unidade de actividade enzimática
χ2 – Qui quadrado
Os aminoácidos são indicados pelo código de uma letra ou de três letras.

IV
Índice
Agradecimentos I
Abreviaturas e simbologia II
Índice IV
Índice de Figuras VI
Índice de Tabelas VII
Resumo VIII
Abstract IX
1 – Introdução 1
1.1 – Metabolismo de xenobióticos 1
1.1.1 – Reacções de Fase I 2
1.1.2 – Reacções de Fase II 3
1.1.3 – Fase III – O transporte de xenobióticos 3
1.2 – O sistema citocromo P450 5
1.2.1 – História e bioquímica dos citocromos P450 5
1.2.2 – Nomenclatura 8
1.2.3 – A evolução dos citocromos P450 9
1.2.4 – A estrutura das proteínas CYP 10
1.3 – As enzimas CYP no Homem 12
1.3.1 – A subfamília CYP3A 13
1.3.1.1 – O gene CYP3A4 16
1.3.1.1 – O polimorfismo genético no CYP3A4 19
2 – Objectivos 23
3 – Material e Métodos – análise do gene CYP3A4 24
3.1 – Introdução 24

V
3.2 – Materiais 29
3.2.1 – Material Biológico - Indivíduos 29
3.2.2 – Reagentes 29
3.3 – Métodos 30
3.3.1 – Extracção de DNA genómico – Extracção rápida de DNA 30
3.3.2 – Pesquisa dos alelos do gene CYP3A4 por PCR-RFLP 31
3.3.3 – Detecção dos produtos de digestão 35
3.3.4 – Análise estatística 37
3.3.4.1 – Frequências alélicas 37
3.3.4.2 – Frequências genotípicas 37
3.3.4.3 – Desvios em relação à distribuição de Hardy-Weinberg- Teste χ2 38
4 – Resultados 39
4.1 – Identificação das variantes CYP3A4*1B, CYP3A4*2 e CYP3A4*3 39
4.2 – Análise estatística dos resultados 40
5 – Discussão dos resultados 43
6 – Conclusão 47
7 – Referências Bibliográficas 48
Anexos 58

VI
Índice de Figuras
Figura 1.1 – Fase III do metabolismo: sistema antiporter 4
Figura 1.2 – O ciclo catalítico dos citocromos P450 7
Figura 1.3 – Árvore filogenética da superfamília dos citocromos P450 9
Figura 1.4 – Estrutura tridimensional das enzimas CYP101, CYP102 e CYP2C5 11
Figura 1.5 – As familías e subfamílias de genes CYP descritas no Homem 12
Figura 1.6 – Contribuição relativa das mais importantes enzimas metabolizadoras de drogas no
fígado humano adulto
13
Figura 3.1 – Exemplo de uma reacção de PCR em que se esquematizam os diferentes passos de
cada ciclo
25
Figura 3.2 – Esquema do gene CYP3A4 e da zona de amplificação de cada par de primers.
Representação esquemática dos padrões de RFLP obtidos para a identificação do CYP3A4*1B,
CYP3A4*2 e CYP3A4*3
36
Figura 4.1 – Caracterização das variantes CYP3A4*1B, CYP3A4*2 e CYP3A4*3 por PCR-RFLP 39
Índice de Tabelas

VII
Tabela 1.1 – A evolução dos CYPs 10
Tabela 1.2 – Semelhança estrutural na sequência de aminoácidos entre os membros da
subfamília CYP3A
14
Tabela 1.3 – Substratos, indutores e inibidores da isoenzima CYP3A4 17
Tabela 1.4 – Nomenclatura das variantes descritas para o CYP3A4 22
Tabela 3.1 – Oligonucleótidos usados na reacção de PCR para a genotipagem do CYP3A4 33
Tabela 3.2 – Frequências de Hardy-Weinberg geradas pela união ao acaso de dois alelos, A e a,
cujas frequências são p e q, respectivamente
38
Tabela 4.1 – Frequências genotípicas para as variantes do CYP3A4 40
Tabela 4.2 – Frequências alélicas das mutações CYP3A4*1B, CYP3A4*2 e CYP3A4*3 na
população Caucasiana Portuguesa
41
Tabela 4.3 – Resultados do teste do ajustamento χ2 utilizados na determinação dos desvios às
frequências esperadas pelo equilíbrio de Hardy-Weinberg numa população de 131 indivíduos
portugueses de origem caucasiana
42
Tabela 5.1 – Frequências alélicas das variantes CYP3A4*1B, CYP3A4*2 e CYP3A4*3 em
populações Caucasianas
44
Resumo

VIII
Os citocromos P450 (CYP) são vitais para o mecanismo oxidativo e redutor de uma grande
variedade de xenobióticos, carcinógenios e compostos endógenos. As enzimas pertencentes à
subfamília CYP3A são as mais importantes para o metabolismo de drogas, contribuindo para a
biotransformação de pelo menos 50% de todos os fármacos usados no Homem. Vários são os
compostos de uso terapêutico que são substrato para estas enzimas, como é o caso da nifedipina,
midazolam, inibidores de proteases e drogas antimaláricas.
A actividade das enzimas CYP3A humanas pode ser o reflexo da contribuição de quatro
enzimas distintas: CYP3A4, CYP3A5, CYP3A7 e CYP3A43. Estudos de fenótipo demonstraram a
existência de uma variabilidade interindividual significativa no metabolismo mediado pelos CYP3A.
No caso da enzima CYP3A4, a forma predominante no fígado adulto, várias mutações foram
recentemente descritas que podem contribuir para a presença de enzimas com actividade variável em
cada indivíduo.
Devido à sua importância na farmacogenética, neste trabalho foram desenvolvidos métodos de
PCR-RFLP para identificar três das variantes do CYP3A4, CYP3A4*1B, CYP3A4*2 e CYP3A4*3.
Estes métodos foram depois usados para analisar a população Portuguesa. A frequência alélica
observada foi de 3,8% no caso do CYP3A4*1B, de 4,2% para o CYP3A4*2 e de 12,6% para a variante
CYP3A4*3.
Uma vez que a actividade do CYP3A4 pode determinar a eficácia dos fármacos e/ou a sua
toxicidade, a sua genotipagem pode tornar-se muito útil para a optimização das doses a usar, assim
como para a determinação da susceptibilidade de cada indivíduo à doença. Estes métodos poderão
também contribuir para que haja um melhor conhecimento das variações étnicas e geográficas dos
SNPs do CYP3A4, e para que seja aplicada uma melhor terapêutica individual.
Abstract

IX
Cytochrome P450 (CYPs) are vital for the oxidative and reductive metabolism of a large
variety of xenobiotics, carcinogens and endogenous compounds. CYP3As are the major drug
metabolizing enzymes, accounting for the biotransformation of about 50% of all drugs used in
humans. A wide range of compounds of therapeutical use are known substrates for these enzymes,
including nifedipine, midazolam, protease inhibitors and antimalarial drugs.
Human CYP3A activity may reflect the contribution of four distinct enzymes: CYP3A4,
CYP3A5, CYP3A7 and CYP3A43. Phenotyping studies have demonstrated the existence of
significant inter-individual variability in CYP3A-mediated metabolism. For CYP3A4, the major form
expressed in adult liver, several mutations were recently described, which may account for the
presence of enzymes with variable activity in each individual.
Due to its pharmacogenetic importance, we have developed a PCR-RFLP method to identify
three of the CYP3A4 variants, CYP3A4*1B, CYP3A4*2 and CYP3A4*3, and used it to analyse the
Portuguese population. The allelic frequency observed was 3,8% for CYP3A4*1B, 4,2% for
CYP3A4*2 and 12,6% for the CYP3A4*3 variant.
Since CYP3A4 activity may determine drug efficacy and/or toxicity, CYP3A4 genotyping may
become a useful tool for optimizing drug dosage, as well for assessing individual susceptibility to
disease. These methods may also contribute to a better knowledge of ethnic and geographical
variation in CYP3A4 SNPs, providing a useful tool for the improvement of therapeutic outcome.

INTRODUÇÃO
1
1 – Introdução
No dia-a-dia o corpo humano encontra-se sujeito a um grande número de xenobióticos que
necessitam de ser eliminados do organismo. Para levar a cabo esta tarefa o corpo humano possui um
sistema de destoxificação que tem como função minimizar os potenciais danos dos xenobióticos. Este
sistema de destoxificação é bastante complexo, apresentando grande variabilidade interindividual, e
dependendo de factores como o meio ambiente, estilo de vida e genética individual.
1.1 – Metabolismo de xenobióticos
A maior parte dos compostos farmacologicamente activos, assim como outros xenobióticos,
são compostos lipofílicos, e de modo a serem mais facilmente excretados do organismo estes
compostos têm que sofrer uma transformação enzimática em metabolitos com características mais
hidrofílicas, num processo denominado biotransformação (Di Giulio et al., 1995). Os metabolitos
resultantes deste processo são geralmente menos activos que o composto inicial ou mesmo inactivos,
embora possa também acontecer o caso dos produtos finais apresentarem uma actividade
farmacológica realçada. Porém, o metabolismo de xenobióticos pode produzir produtos reactivos e
tóxicos que podem ser responsáveis por várias formas de toxicidade, incluindo a iniciação e
progressão de tumores (Nelson et al., 1996). Deste modo, a biotransformação resulta normalmente
num aumento da velocidade de excreção dos xenobióticos, o que reduz o risco de acumulação destas
substâncias a níveis tóxicos no organismo, sendo por isso a biotransformação de xenobióticos o
mecanismo principal para a manutenção da homeostase durante a exposição a moléculas estranhas ao
organismo (Klaassen e Watkins, 1999).
O processo de biotransformação é levado a cabo pelas enzimas metabolizadoras de drogas
(DME’s - Drug Metabolizing Enzymes), um grupo diversificado de enzimas responsáveis pela
metabolização de um vasto leque de xenobióticos, incluindo drogas e poluentes ambientais, e
compostos endógenos como os esteróides e as prostaglandinas. As DMEs são usualmente divididas
em dois grupos de enzimas, ou seja, enzimas de Fase I e as enzimas de Fase II. Os produtos gerados

INTRODUÇÃO
2
pelas reacções de fase I são geralmente metabolitos intermediários reactivos e/ou espécies reactivas de
oxigénio, o que pode causar danos nos tecidos. As reacções de fase II envolvem geralmente a
conversão dos metabolitos intermediários da fase I em produtos finais que são depois eliminados. Os
processos da fase I e fase II são, no entanto, independentes e como tal, o composto pode sofrer apenas
reacções de fase I ou de fase II, ou as duas sequencialmente. Embora ambas as fases tenham diferentes
características, é essencial que as suas funções estejam balanceadas entre si para minimizar a presença
de metabolitos intermediários reactivos.
A concentração de compostos exógenos nas células é também condicionada pela presença de
transportadores na membrana da célula, responsáveis pela chamada Fase III do metabolismo de
drogas.
1.1.1 - Reacções de Fase I
O sistema de destoxificação de fase I é geralmente a primeira defesa enzimática contra
compostos estranhos. Os compostos tóxicos são tipicamente compostos não reactivos, e como tal não
contêm sítios reactivos que possam ligar os grupos solúveis em água que são conjugados nas reacções
de fase II. Assim, na fase I os xenobióticos são sujeitos a reacções de funcionalização ou de activação,
onde se introduz ou revela, por reacções de oxidação, redução ou hidrólise, um grupo funcional (-OH,
-NH2, -SH ou -COOH) transformando-os assim em substâncias activas prontas para o processo de fase
II.
As reacções de fase I podem ser catalisadas por diversas enzimas, de que são exemplo as
monooxigenases citocromo P450, as monooxigenases contendo flavina, as hidrolases, as
lipooxigenases, as peroxidases, as oxidases e as reductases (Klaassen e Watkins, 1999; Nebert, 1991).
De todas as famílias de enzimas de fase I a dos citocromos P450 é a mais importante, tanto a nível do
elevado número de xenobióticos que destoxificam, como da versatilidade catalítica que apresentam
(Nebert, 1991). O papel destas enzimas é relevante para o metabolismo oxidativo, peroxidativo e
redutor de numerosos e diversos compostos endógenos e exógenos (Nelson et al., 1996).

INTRODUÇÃO
3
Os metabolitos activados nesta fase podem, em certos casos, ser mais tóxicos que o substrato
original, podendo ser depois conjugados com uma molécula endógena, como o ácido glucurónico, a
glutationa ou o sulfato, nas chamadas reacções de fase II ou de conjugação.
1.1.2. - Reacções de Fase II
As enzimas desta fase participam na biotransformação de certas substâncias lipofílicas que
geralmente resultam das reacções de fase I. Nesta fase, essas moléculas são conjugadas através da
adição de um grupo solúvel em água ao sítio reactivo. As reacções de fase II são mediadas por várias
enzimas que podem pertencer a superfamílias de genes distintas, onde estão incluídas as
sulfotransferases, as transaminases, as acetiltransferases, as metiltransferases, as aciltransferases, as
aldocetoreductases, as carboxilesterases, as glicosilases, as glucuroniltransferases e várias hidrolases e
esterases (Nebert, 1991).
Nos metabolitos conjugados existe normalmente um aumento na hidrofilicidade e como tal
estes compostos são rapidamente excretados na urina e na bilis, que são as principais vias de
eliminação para a maior parte das drogas (Meyer, 1996).
1.1.3. – Fase III – O transporte de xenobióticos
Para além do metabolismo que é levado a cabo pelas enzimas de fase I e de fase II, a
disponibilidade dos medicamentos no organismo pode variar como resultado de diferenças genéticas
no mecanismo de transporte através da membrana plasmática. A esta fase de transporte para o exterior
da célula dos compostos resultantes da acção das enzimas de fase I e fase II é dado o nome de fase
III, sendo esta actividade de transporte um factor importante no metabolismo de xenobióticos (Liska,
1998; Müller, 2000). Associadas à fase III estão várias superfamílias de proteínas, sendo uma delas a
dos transportadores ABC (ATP Binding Cassette), associada aos eucariotas (Van Bambeke et al.,
2000). Esta superfamília de proteínas é responsável pelo transporte de um grande número de
substratos através da membrana celular, nos quais estão incluídos açúcares, aminoácidos, proteínas,
metais, iões orgânicos e inorgânicos e antibióticos (Raggers et al., 2000). Os transportadores ABC

INTRODUÇÃO
4
têm sido associados a diferentes manifestações clínicas no Homem, sendo uma delas a resistência a
várias drogas, onde estão envolvidas a família MDR (Multi Drug Resistance) e a família MRP
(Multidrug Resistance Protein ou Multidrug Resistance-Related Protein) (Raggers et al., 2000; Van
Bambeke et al., 2000). Ambas as famílias estão associadas à resistência das células tumorais a drogas
citotóxicas em cerca de 50% dos cancros humanos (Stavrovskaya, 2000). No entanto, estes genes
podem ser expressos em condições normais, e no caso do MDR, a proteína que é expressa, a P-
glicoproteína (P-gp), está associada à defesa do organismo de lesões tóxicas, tendo funções de
“barreira”, e como tal, protege as células de substâncias perigosas que existem, por exemplo, no cólon
(Stavrovskaya, 2000).
Foi recentemente demonstrada a existência de uma regulação coordenada entre a actividade de
transporte no intestino e a enzima de fase I CYP3A4, pelo que os transportadores podem suportar e
promover a destoxificação. Possivelmente, a sua função de bombear xenobióticos não-metabolizados
para fora da célula e de novo para o lúmen intestinal permite aumentar a probabilidade de que os
metabolitos sejam metabolizados pelas enzimas de fase I antes de voltarem à circulação, tendo sido
atribuído a esta função o nome de Fase 0 (ver figura 1.1) (Liska, 1998).
1.2 – O sistema citocromo P450
As enzimas citocromo P450 (CYP) constituem uma superfamília de proteínas que podem ser
encontradas desde as bactérias até ao Homem, e estão envolvidas no metabolismo de uma grande
Estômago
Enterocito
Fezes
Veiaportapara ofígado
Lúmen dointestino
Figura 1.1 – Fase III do metabolismo: sistema antiporter (adaptado de Liska, 1998).

INTRODUÇÃO
5
variedade de compostos (Nelson et al., 1996). Desde a sua origem há mais de 3,5 mil milhões de anos,
a família dos genes CYP diversificou-se para acomodar o metabolismo de um número crescente de
toxinas ambientais, compostos da dieta e drogas (Nelson et al., 1996; Streetman et al., 2000).
1.2.1. – História e bioquímica dos citocromos P450
A capacidade dos tecidos de mamífero para oxidar xenobióticos hidrófobos já era conhecida
antes do ano de 1950, embora as enzimas responsáveis por esta actividade fossem então
desconhecidas. Em 1955, G.R. Williams e M. Klingenberg num estudo sobre hemoproteínas
microssomais de rato e usando a espectroscopia óptica, identificaram um pigmento celular
especializado que apresentava no seu espectro diferencial um pico de Soret a 450 nm quando
complexado com monóxido de carbono (Nebert e Gonzalez, 1987), posteriormente designado por
citocromo P450 (Omura e Sato, 1964). Estudos posteriores atribuíram funções enzimáticas aos CYP e
mostraram a sua capacidade para metabolizar numerosos substratos.
Como referido, estas enzimas foram inicialmente descritas em tecidos animais, mas
posteriormente foi descrita a sua expressão em vários eucariotas, que incluem os vertebrados,
invertebrados, fungos e plantas, e ainda nos procariotas (Nelson et al., 1996). Estando presente numa
tão grande variedade de espécies, actualmente o número de membros desta superfamília excede já as
1200 isoformas (Lewis e Sheridan, 2001). Uma tão elevada variedade de isoformas traduz-se num
elevado número de substratos metabolizados por estas superfamília que incluem vários compostos
endógenos (esteróides, ácidos biliares, ácidos gordos, prostaglandinas, leucotrienos, retinóides) e
exógenos, como fármacos (debrisoquina, codeína, nifedipina, mefenitoina, ciclosporina), químicos
ambientais (hidrocarbonetos policíclicos aromáticos, nitrosaminas, hidrazinas, arilaminas) e produtos
naturais de plantas (fitoalexinas) (Nelson et al., 1996; Stegeman e Hahn, 1994).
As enzimas CYP são hemoproteínas ligadas a membranas com cerca de 48 a 60 kDa, cada uma
contendo aproximadamente 500 resíduos de aminoácidos (Lewis e Sheridan, 2001). Nos animais, a
maior parte das proteínas CYP encontram-se no retículo endoplasmático liso, embora uma minoria

INTRODUÇÃO
6
possa estar localizada na membrana interior da mitocôndria (Guengerich, 1992; Nebert e Gonzalez,
1987). A expressão das enzimas CYP foi identificada no tecido hepático, assim como nos tecidos
extrahepáticos de que são exemplos o rim, o intestino, o pulmão, as gónadas, a pituitária, a próstata, o
cérebro, a pele, os leucócitos, o baço e o sistema cardiovascular (Celander, 1999).
A maior parte do metabolismo mediado pelos CYPs baseia-se na catálise da transformação
oxidativa de um largo número de substratos, sendo a reacção básica catalisada por estas enzimas a
monooxigenação, na qual um átomo de oxigénio (derivado do O2) é incorporado no substrato, e o
outro átomo é reduzido a água com os equivalentes redutores do NAD(P)H (Josephy, 1997; Klaassen
e Watkins, 1999). Esta reacção pode ser representada pela seguinte equação:
Substrato (RH) + O2 + NAD(P)H + H+ → Produto (ROH) + H2O + NAD(P)+
Dependendo do substrato, diferentes reacções podem ocorrer, incluindo a desalogenação
oxidativa ou redutora, a sulfoxidação, a N-hidroxilação e N- e S-oxidação, a desaminação oxidativa, a
S-, N- e O-desalquilação, a redução de peróxidos e epóxidos e a hidroxilação alifática e aromática. O
ciclo catalítico das enzimas CYP é mostrado na figura 1.2 (Anzenbacher e Anzenbacherová, 2001;
Guengerich, 1999; Lewis e Sheridan, 2001; Roberts, 1999). Este é iniciado quando o substrato se liga
a um sítio perto do grupo heme para interagir com o oxigénio (passo 1). O átomo de ferro é então
reduzido para a forma ferrosa (Fe2+) por intermédio de um electrão transferido do NAD(P)H através
da oxidoredutase, tornando-se capaz de captar uma molécula de oxigénio (passo 2). O oxigénio
molecular liga-se então ao complexo CYP-substrato (passo 3), seguindo-se depois a transferência de
um segundo electrão do NAD(P)H através da oxidoredutase para o complexo (passo 4). Finalmente, o
complexo rearranja com a inserção de um átomo de oxigénio no substrato para dar um produto e água
ou peróxido de hidrogénio (passo 5 e 6).
Fe3+
Ls RH
Fe3+ RH Hs NADPH-P450 reductasered
NADPH P450 d t oxe-
Fe3+ ROH
Ls
- ROH
FeIII
H2O
1
2
6

INTRODUÇÃO
7
Figura 1.2 – O ciclo catalítico dos citocromos P450. RH, substrato; ROH, produto hidroxilado.
(adaptado de Guengerich, 1999).
1.2.2. – Nomenclatura
As evidências iniciais da multiplicidade nos CYPs surgiram cedo através da purificação de
várias enzimas e como CYPs individuais foram identificados em vários laboratórios, vários sistemas
de nomenclatura surgiram baseados nos seus pesos moleculares, especificidade de substratos ou

INTRODUÇÃO
8
mobilidade electroforética. Como tal houve a necessidade de desenvolver um sistema de nomenclatura
universal para a superfamília CYP, tendo este como fundamento a comparação das sequências de
aminoácidos e as relações evolutivas dos genes correspondentes, baseada numa evolução divergente
da superfamília (Nebert et al., 1987).
O novo sistema de nomenclatura foi proposto pela primeira vez em 1987, mas desde essa
altura diversas actualizações foram publicadas à medida que surgiram novas formas desta enzima
(Nebert et al., 1989, 1991; Nelson et al., 1993, 1996). Desde a última publicação a informação é
constantemente actualizada em sites na Internet como -
http://drnelson.utmem.edu/CytochromeP450.html e http://www.icgeb.trieste.it/p450.
As recomendações para designar um gene citocromo P450 (Nelson et al., 1996) incluem em
primeiro lugar a sigla “CYP” (“Cyp” para murganho e Drosophila), que advém de CYtochrome P450.
Em seguida, as enzimas CYP dentro da mesma família são designadas por um número e partilham
entre si mais de 40% de identidade na sequência de aminoácidos. As famílias são depois divididas em
subfamílias, sendo as enzimas dentro da mesma subfamília designadas pela mesma letra. Genes dentro
da mesma subfamília partilham mais de 55% de identidade na sua sequência de aminoácidos.
Finalmente, um número após a letra denota cada isoenzima individual, e assim um gene individual
designa-se, por exemplo, como CYP3A4. Para ser considerado um novo membro individual, uma
sequência deve diferir de uma outra mais do que 3%. No caso dos genes a notação CYP apresenta-se
em itálico (CYP), enquanto para as proteínas, mRNA e cDNAs a sigla não é apresentada em itálico e
todas as letras são maiúsculas, qualquer que seja a espécie (Nelson et al., 1996).
1.2.3. – A evolução dos citocromos P450
O sistema de classificação baseado na percentagem de homologia entre as sequências dos
CYPs facilita a construção de uma árvore filogenética para esta superfamília de enzimas, que se
encontra representada na figura 1.3.
Superfamília dos genes CYP

INTRODUÇÃO
9
Figura 1.3 – Árvore filogenética da superfamília dos citocromos P450. Em abcissas encontra-se
representada a distância evolutiva, estimada com base na divergência entre as sequências de resíduos
de aminoácidos das proteínas (Gonzalez e Nebert, 1990).
O gene ancestral CYP deverá ter aparecido há cerca de 3500 milhões de anos nas espécies
procariotas, na altura em que a atmosfera sofreu alterações e terá começado a haver oxigénio
disponível para as transformações catalíticas mediadas pelos CYPs (Lewis e Sheridan, 2001). O gene
ancestral terá depois sofrido duplicação há cerca de 1360 milhões de anos, o que deu origem a
citocromos P450 que se encontram em dois organelos celulares diferentes, a mitocôndria e o retículo
endoplasmático. Várias etapas importantes da evolução dos citocromos P450 encontram-se resumidos
na tabela 1.1.
Tabela 1.1 – A evolução dos CYPs (adaptado de Lewis e Sheridan, 2001).
Tempo (milhões de anos)
Divergências na superfamília CYP
2000 CYPs eucariotas e procariotas
1000 CYPs microssomais e mitocôndriais
800 As famílias CYP3 e CYP4 separaram-se dos outros CYPs microssomais
650 Divergência entre CYP3 e CYP4
550 As famílias CYP1 e CYP2 divergiram da CYP17 e CYP21
470 Divergência entre CYP1 e CYP2
450 Divergência entre CYP17 e CYP21
370 Divergência da subfamília CYP2D da família CYP2
300 A subfamília CYP2D expande-se
250 Divergência da família CYP2
230 Divergência das subfamílias CYP2A e CYP2B, e das CYP2C e CYP2E
120 Divergência entre CYP1A1 e CYP1A2
80 Especialização das famílias e subfamílias CYP
1.2.4. – A estrutura das proteínas CYP
Muitas são as sequências de CYPs conhecidas, mas poucas são as estruturas tridimensionais já
determinadas por cristalografia de raio-X. Até agora, as estruturas tridimensionais mais bem
conhecidas são as estruturas de CYPs de procariotas, de que são exemplo a proteína CYP101
Distância Evolutiva

INTRODUÇÃO
10
(P450cam – ver figura 1.4A) da Pseudomonas putida (Poulos et al., 1987) e a CYP102 (BM-3 – ver
figura 1.4B) do Bacillus megaterium (Ravichandran et al., 1993). Só recentemente foi resolvida uma
estrutura de um cristal de CYP de mamíferos, o CYP2C5 de coelho (ver figura 1.4C) (Williams et al.,
2000).
Nos CYPs existem sequências absolutamente conservadas como as que rodeiam dois resíduos
chave, o resíduo de cisteína C-terminal e o resíduo de treonina na hélice I. O resíduo Cys do C-
terminal faz parte de uma folha β antiparalela que começa com Phe (F) e acaba com Cys (C), parte de
uma sequência consenso altamente conservada F(G/S)XGX(H/R)XCXGX(I/L/F)A, conhecida como o
domínio de ligação ao heme no sítio activo, que pode servir como identificador de uma proteína CYP
(Gonzalez, 1989; Kalb e Loper, 1988). A Thr-252 na hélice I aponta para o sítio activo da enzima e
está envolvida na ligação ao oxigénio. Outras regiões conservadas existem, embora com funções
menos claras, como é o caso da região rica em prolina no N-terminal Pro-Pro-Gly-Pro, que pode
servir para juntar o N-terminal de ligação à membrana com a região globular da proteína.
A B
C

INTRODUÇÃO
11
Figura 1.4 – Estrutura tridimensional das enzimas CYP101 (A), CYP102 (B) e CYP2C5 (C).
1.3. – As enzimas CYP no Homem
No Homem foram descritos até à data 56 genes (Kuehl et al., 2001), inseridos em 17 famílias
(ver figura 1.5), e 21 pseudogenes (Leeder, 1996; Mackinnon, 2000). Os CYPs humanos podem ser
funcionalmente divididos em duas classes, uma em que os genes estão, por exemplo, associados à
biossíntese de esteróides (CYP4, CYP5 e famílias acima destas) e uma outra em que os genes estão
primariamente envolvidos no metabolismo de xenobióticos (CYP1, CYP2 e CYP3) (Leeder, 1996).
Figura 1.5 – As famílias e subfamílias de genes CYP descritas no Homem (adaptado de Leeder, 1996).
Cerca de metade das formas de CYP humanas conhecidas pertencem às famílias 1, 2 e 3, e
como tal estas são as famílias mais importantes do metabolismo hepático de drogas. A contribuição
Citocromos P450 Humanos
1 2 3 19 21 5 7 11 27 4 17 24 26 8 46 51 39
1A1 1A2
1B1 3A4 3A5 3A7
3A43
2A6 2A7 2A13
2B6 2C8 2C9 2C18 2C19
2D6 2E1 2F1 2R1 2J2 2S1 2W1 2V1
5A1
4F2 4F3 4F8
4F11 4F12 4F22
4A11 4A20 4A22
4B1 4V2 4X1
7A1 7B1
8B1 8A1
11A1 11B1 11B2
21A2
26A1 26B1 26C1
27A1 27B1 27C1
39A1

INTRODUÇÃO
12
relativa das enzimas CYP envolvidas na metabolização de drogas no fígado humano adulto está
apresentada na figura 1.6 (Leeder, 1996), sendo aí visível a importância da subfamília CYP3A.
����������������������������������������������������������������������������������������������������������������������������
����������������������������������������������������������������������������������������������������������������������������
������������������������������������������������������������������������������������������������������������������������������������������������������������
������������������������������������������������������������������������������������������������������������������������������������������������������������������������
����������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������
�����������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������
�������������������������������������������������������������������������������������������������������������������������������������������������
���������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������
�������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������
������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������
CYP3A (30% - 60%)
CYP2C (20% - 25%)
CYP1A2 ~ 13%
CYP2E1~ 7%
CYP2A6 ~4%
CYP2D6 ~2% CYP2B6
< 1% Outros ~2%
Figura 1.6 – Contribuição relativa das mais importantes enzimas metabolizadoras de drogas no fígado
humano adulto (adaptado de Leeder, 1996).
1.3.1 – A subfamília CYP3A
Na família CYP3 apenas está descrita uma subfamília, a CYP3A (Nelson et al., 1996). A
subfamília CYP3A está envolvida no metabolismo de um grande número de compostos endógenos e
exógenos, como o cortisol, a ciclosporina e a nifedipina. Como cerca de 50% de todas as drogas
usadas no Homem são metabolizadas, ao menos parcialmente, pela oxidação mediada pelos CYP3A,
esta parece ser a subfamília mais importante no metabolismo de drogas (Thummel e Wilkinson, 1998;
Wrighton et al., 2000).
Os genes CYP3A estão localizados num cluster no cromossoma 7q21-q22.1, apresentando uma
estrutura exão-intrão altamente conservada, com 13 exões e 12 intrões (Hashimoto et al., 1993;
Nelson et al., 1996; Ourlin et al., 1997). Esta subfamília inclui quatro genes, CYP3A4, CYP3A5,
CYP3A7 (Nelson et al., 1996) e o recentemente identificado, CYP3A43 (Domanski et al., 2001;
Gellner et al., 2001). Os genes que fazem parte desta subfamília apresentam grande semelhança
estrutural tanto em termos do cDNA, como da proteína, como se pode ver tabela 1.2 (Gelner et al.,
2001).

INTRODUÇÃO
13
Tabela 1.2 – Semelhança estrutural na sequência de aminoácidos entre os membros da subfamília
CYP3A (adaptado de Gelner et al., 2001).
CYP3A4 CYP3A5 CYP3A7 CYP3A43
CYP3A4 **** 84,1 88,1 75,8
CYP3A5 **** **** 81,9 75,8
CYP3A7 **** **** **** 71,5
CYP3A43 **** **** **** ****
O citocromo 3A4 é a enzima CYP mais importante para o metabolismo de drogas no Homem,
não só pela quantidade em que existe no fígado (que pode aumentar por indução até mais de 60%),
mas também porque participa no metabolismo da maioria das drogas.
O CYP3A5 é expresso no rim (Schuetz et al., 1992), esófago (Lechvrel et al., 1999), cólon
(Gervot et al., 1996), pulmão (Kivistö et al., 1996), intestino (Kivistö et al., 1996a), glândula
pituitária (Murray et al., 1995) e, polimorficamente, no fígado (Wrighton et al., 1989). A expressão
deste gene é detectada em apenas 10 a 30% dos fígados humanos, sendo o seu nível, quando expresso,
cerca de 25% do nível da enzima CYP3A4 (Guengerich, 1999; Paulussen et al., 2000). Este gene
apresenta variantes alélicas tanto na região do promotor, assim como na região codificante do gene
(Jounaidi et al., 1996; Kuehl et al., 2001). A região do promotor do gene CYP3A5 contém um
elemento funcional de resposta aos glucocorticóides, sendo este o único gene CYP humano que é
directamente induzido pelo receptor dos glucocorticóides (GR) (Schuetz et al., 1996). A sua
regulação, ao contrário de outras enzimas CYP3A, não tem evidências de ser efectuada pelo
PXR/SXR (para pregnane X-receptor/steroid and xenobiotic receptor) (Whighton et al., 2000).

INTRODUÇÃO
14
O CYP3A7 é a isoforma CYP3A dominante no fígado fetal, sendo encontrada a baixos níveis
de expressão no fígado adulto (Yang et al., 1994). Esta enzima é também detectada em pequenas
quantidades na placenta, endométrio, tumores e linhas celulares tumorais (Lacroix et al., 1997;
Pascussi et al., 1999). Recentemente foi descrita a sua expressão em vários orgãos no adulto, como o
rim, pâncreas e pulmão (Finta e Zaphiropoulos, 2000). A enzima CYP3A7, embora
predominantemente fetal, tem as propriedades catalíticas semelhantes às outras enzimas CYP3A,
incluindo a hidroxilação da testosterona na posição 6β. Esta isoforma é induzida por várias
substâncias, sendo esta indução mediada por um receptor nuclear, o receptor nuclear de pregnanos
(PXR), que se liga ao elemento de resposta do PXR no promotor do gene (Kuehl et al., 2001; Pascussi
et al., 1999). Também o CAR (Constitutive Androstane Receptor) parece ter um papel na resposta a
xenobióticos no caso do CYP3A7 (Bertilsson et al., 2001).
A expressão do CYP3A43 foi descrita em vários tecidos, mas na maior parte dos casos a baixos
níveis. A expressão a níveis mais elevados foi detectada na próstata, fígado, rim e pâncreas no adulto,
e ainda no fígado e músculo esquelético fetais (Domanski et al., 2001). Níveis menores de expressão
foram descritos no adulto no coração, cérebro, placenta, cólon, leucócitos, intestino delgado, baço e
testículo (Gellner et al., 2001). A contribuição deste gene para o metabolismo de drogas ainda não foi
esclarecida até à data.

INTRODUÇÃO
15
1.3.1.1 – O gene CYP3A4
A isoenzima CYP3A4 representa cerca de 30% do conteúdo total de enzimas CYP no fígado
adulto (Shimada et al., 1994), mas para além deste órgão é também expressa no intestino delgado
(Kolars et al.,1992; Zhang et al., 1999), cólon e estômago (de Waziers et al., 1990). Cerca de 60% das
drogas metabolizadas pelos CYPs são metabolizadas pelo CYP3A4. Esta enzima cataliza o
metabolismo oxidativo de uma grande variedade de químicos com diferentes características
estruturais,e assim os substratos para esta enzima incluem uma grande variedade de xenobióticos (ver
tabela 1.3), que se distribuem por várias classes como, anticoagulantes, antidepressivos,
antihistamínicos, antimaláricos, bloqueadores dos canais de cálcio, agentes quimioterapêuticos,
imunosupressores, antibióticos macrólidos, e inibidores de proteases (Michalets, 1998; Ducharme e
Farinotti, 1996). O CYP3A4 tem também um papel importante no metabolismo de esteróides
endógenos, como a testosterona, a progesterona, o cortisol e o estradiol.
A sequenciação do gene CYP3A4 mostrou que a região do promotor deste contém uma TATA
box e um elemento básico de transcrição (BTE) (Hashimoto et al., 1993). Para além destes existem
sítios de ligação para diversos factores de transcrição, tais como, a proteína octamérica (Oct-1),
proteína activadora 3 (AP-3), proteína p53, factor hepático nuclear -4 e -5 (HNF-4 e HNF-5), receptor
dos glucocorticóides (GR) e receptor de estrogénios (ER) (Guengerich, 1999; Hashimoto et al., 1993;
Jover et al., 2001). Existe ainda uma sequência na região 5’, o elemento de resposta à nifedipina (-389
a –398), denominado NFSE (P450NF specific element), que poderá estar envolvida na expressão do
CYP3A4, embora não estejam identificados os factores que são capazes de se ligar a este elemento.

INTRODUÇÃO
16
Tabela 1.3 – Substratos, indutores e inibidores da isoenzima CYP3A4 (Mackinnon e Evans, 2000;
Michalets, 1998; Piscitelli e Gallaciano, 2001).
Substratos Alfentanil Alprazolam Amitriptilina Amiodarona Amlodipina Astemizol Atorvastatine Budesonide Buprenorfina Busulfan Canabinóides Carbamazepina Cetoconazol Cisapride Claritromicina Clindamicina Clonazepam Clozapina Cocaína Cortisol Ciclobenzaprina Ciclofosfamida Ciclosporina
Dapsona Dexametasona Dextrometorfano Diazepam Digitoxina Diltiazem Disopiramida Donepezil Doxorubicina Dronabiol Efavirenz Eritromicina Estradiol Estrogénios, Contraceptivos orais Etiniloestradiol Etosuximida Etopósido Felodipina Fentanil Fexofenadina Flutamida Ifosfamida
Imipramina Indinavir Isradipina Lansoprazol Lidocaína Loratadina Losartan Lovastatina Mibefradil Miconazol Midazolam Navelbine Nefazodona Nelfinavir Nicardipina Nifedipina Nimodipina Nisoldipina Omeprazol Ondansetron Paclitaxel Pravastatina Prednisona
Propafenona Quinidina Quinino Ritonavir Saquinavir Sertraline Simvastatina Tacrolimus Tamoxifen Temazepam Tenipósido Terfenadina Testosterona Tetrahidrocanabinol Teofilina Trazadona Triazolam Troleandomicina Verapamil Vinblastina R-warfarina Zileuton
Inibidores Indutores Amiodarone Amprenavir Canabinóides Cetoconazol Cimetidine Claritromicina Clotrimazol Delavindina Diltiazem Efavirenz Eritromicina Fluconazole Fluoxetina Fluvoxamina Indinavir Inibidores de proteases (ritonavir é o mais potente)
Itraconazol Lopinavir Omeprazol Metronidazol Mibefradil Miconazol Nefazodona Nelfinavir Norfloxacina Paroxetina Quinino Saquinavir Sertralina Sumo de Toranja Troleandomicina Zafirlukast
Carbamazepina Dexametasona Etosuximida Fenobarbital Fenitoína Glucocorticóides Nevirapina Primidona Rifabutina Rifampina Rifampicina Sulfadimidina Sulfinpirazona Troglitazona

INTRODUÇÃO
17
O metabolismo de drogas é afectado pela indução e inibição das enzimas. A indução das
enzimas resulta usualmente num aumento da quantidade da enzima, normalmente devido a uma
indução da transcrição, e o período de tempo necessário para a indução da enzima torna-se
clinicamente significativo. Os níveis de CYP3A4 são induzidos pela exposição a vários agentes
estruturalmente diversos (ver tabela 1.3), que incluem a dexametasona, carbamazepina, fenobarbital e
rifampicina (Wrighton et al., 2000). A inibição das enzimas CYP, por sua vez, ocorre rapidamente,
mesmo após uma dose única de inibidor. Para o CYP3A4 existem vários inibidores (ver tabela 1.3), de
que são exemplo o itraconazol ou o indinavir.
O metabolismo de drogas efectuado pelos CYPs humanos é levada a cabo por enzimas
polimórficas que podem apresentar uma actividade reduzida ou aumentada em relação a um dado
substrato. As principais causas para a variação no metabolismo são o polimorfismo genético, a
indução ou inibição devido a terapias com drogas ou factores ambientais, o estado fisiológico e alguns
estados patológicos (Ingelman-Sundberg et al., 1999). Recentemente foi sugerido que 60% a 90% da
variabilidade interindividual nos CYP3A pode ser causada por factores genéticos (Özdemir et al.,
2000). O princípio genético do polimorfismo de enzimas que participam no metabolismo de drogas
reside em mutações no gene que codifica uma enzima em particular. O polimorfismo genético das
enzimas do metabolismo de drogas pode ser caracterizado como consequência da presença de duas ou
mais variantes de enzimas numa população etnicamente homogénea e ocorre quando a variante de
uma enzima em particular está presente em pelo menos 1% da população (≤ 3% de divergência)
(Kohút e Kalina, 1998; Nelson et al., 1996).
A variabilidade na expressão do CYP3A4 pode também ser o resultado da regulação que existe
a nível da transcrição, tanto através das variações que existem nos elementos reguladores upstream,
como nos genes que codificam para os factores de transcrição (Eichelbaum e Burk, 2001). O factor de
transcrição que parece desempenhar um papel central na expressão do CYP3A4 é o receptor orfão
PXR (também chamado de SXR) (Bertilsson et al., 1998; Blumberg et al., 1998; Lehmann et al.,
1998), na forma de heterodímero com o RXR (Retinoid X Receptor) (Waxman, 1999; Moore e

INTRODUÇÃO
18
Kliewer, 2000; Xie et al., 2000). Outros receptores parecem também estar ligados à expressão do
CYP3A4, como é o caso do CAR ou do GR, embora neste último caso a regulação pareça ser
indirecta (Wrighton et al., 2000; P. Maurel, comunicação pessoal).
1.3.1.1.1 – O polimorfismo genético no CYP3A4
O CYP3A4 exibe uma grande variação interindividual nos níveis de expressão que in vitro
pode ir até 50 vezes, enquanto que in vivo a actividade da proteína pode variar até 20 vezes (Shimada
et al., 1994; Özdemir et al., 2000). Embora vários estudos baseados na fenotipagem tenham há muito
demonstrado a existência desta variabilidade interindividual no metabolismo do CYP3A4 só
recentemente foram identificadas variações na sequência do gene que a podem explicar (Rebbeck et
al., 1998; Sata et al., 2000; Hshie et al., 2001; Eiselt et al., 2001).
A primeira evidência da existência de polimorfismo genético no gene CYP3A4 surgiu em 1998
com a descrição de uma variação na região 5’ do gene que consiste numa mutação pontual de uma
adenina para guanina (A-392G) no elemento de resposta específico da nifedipina (Rebbeck et al.,
1998). A esta mutação na região não codificante do gene foi dada a designação de CYP3A4*1B, tendo
sido descrita com maior incidência nos indivíduos negros, menor incidência nos Caucasianos e como
inexistente em populações Asiáticas (Rebbeck et al., 1998). Desde logo foi feita a associação desta
variante com uma actividade proteíca menor, que poderá estar associada ao cancro da próstata
(Rebbeck et al., 1998) e a um menor risco de leucemias relacionadas com o tratamento (Felix et al.,
1998). Outros estudos, no entanto, apresentam resultados que põem em causa os anteriormente
referenciados, uma vez que nestes não são observadas alterações significativas da expressão da
enzima ou da sua actividade catalítica (Ando et al., 1999; Ball et al., 1999; Wandel et al., 2000;
Westlind et al., 1999).
No início do ano 2000 foram descritas as primeiras variantes na região codificante do
CYP3A4, a variante CYP3A4*2 e CYP3A4*3 (Sata et al., 2000).

INTRODUÇÃO
19
A mutação no CYP3A4*2 consiste numa transição T → C que resulta numa alteração
Ser222Pro (Sata et al., 2000). Este SNP (Single Nucleotide Polymorphism) foi descrito na população
Finlandesa e dado como ausente nos sujeitos Chineses e Negros analisados (Sata et al., 2000).
Recentemente, num estudo realizado na população Alemã e Suíça, esta variante não foi detectada pelo
que foi sugerido que esta seria uma variante específica dos Finlandeses (Eiselt et al., 2001). A
expressão do CYP3A4*2 em células de mamífero mostrou que esta mutação leva a uma actividade
catalítica alterada perante o substrato nifedipina (Sata et al., 2000).
A variação T → C que encontramos no caso do CYP3A4*3 leva a uma alteração Met445Thr na
região conservada de ligação ao heme (Sata et al., 2000). Esta mutação quando descrita, foi
encontrada num único indivíduo Chinês, num universo de 72 indivíduos analisados, o que levou à sua
caracterização como sendo um alelo raro (Sata et al., 2000). Um estudo recente na população
Holandesa chegou à conclusão de que este não seria um alelo raro, uma vez que nesta população a
mutação foi descrita com uma incidência superior a 1 %, o que leva a que possa ser designada por
variante polimórfica (van Schaik et al., 2001). Um outro estudo realizado nas populações Alemã e
Suíça também encontrou esta mutação, tendo aqui sido estudadas as consequências funcionais da
mutação (Eiselt et al., 2001). O estudo do efeito da mutação Met445Thr na actividade da proteína
revelou que esta não tinha qualquer efeito a nível da testosterona e progesterona (Eiselt et al., 2001).
Para além destas variantes sobre as quais incidiu este trabalho, outras variantes foram
descritas. Hsieh e seus colaboradores (2001) descreveram, numa população Chinesa, três novas
mutações na região codificante do gene. A primeira variante (CYP3A4*4) consiste numa mutação de
A13989G que resulta numa alteração Ile118Val, o que parece levar a uma alteração da actividade da
enzima in vivo (Hsieh et al., 2001). A outra variante (CYP3A4*5) apresenta uma mutação pontual
C15820G que causa uma alteração Pro218Arg, e que leva à diminuição da actividade da proteína
(Hsieh et al., 2001). A variante CYP3A4*6 consiste na inserção de uma adenina na posição 17776, e
este frameshift devido à inserção cria um codão stop na posição 285, antes do que acontece na

INTRODUÇÃO
20
sequência de referência (Hsieh et al., 2001). Isto origina uma diminuição da actividade a valores
muito inferiores do que o que acontece com as outras variantes descritas no estudo, assim como em
relação à sequência de referência (Hsieh et al., 2001).
Muito recentemente foram descritos mais sete SNPs na região codificante, o CYP3A4*7,
CYP3A4*8, CYP3A4*9, CYP3A4*10, CYP3A4*11, CYP3A4*12 e CYP3A4*13, dos quais apenas o
CYP3A4*11 é expresso a níveis significativamente diferentes em relação à sequência de referência e o
CYP3A4*12 apresenta uma actividade alterada na metabolização da testosterona (Eiselt et al., 2001).
A busca de variantes para este gene já permitiu a localização de mais de 20 alterações na
sequência que se encontram apresentadas na tabela 1.4.
Tabela 1.4 – Nomenclatura das variantes descritas para o CYP3A4.
(adaptado de http://www.imm.ki.se/CYPalleles/cyp3a4)
Alelo Alterações na Sequência Efeito Referências

INTRODUÇÃO
21
cDNA Gene
CYP3A4*1A Nenhuma Nenhuma Gonzalez et al., 1988
CYP3A4*1B -392 A>G Rebbeck et al., 1998 Westlind et al., 1999
CYP3A4*1C -444 T>G Kuehl et al., 2001
CYP3A4*1D -62 C>A Kuehl et al., 2001
CYP3A4*1E -369 T>A Dr. H. Hamzeiy
CYP3A4*1F -747 C>G Dr. H. Hamzeiy
CYP3A4*2 S222P Sata et al., 2000
CYP3A4*3 1334 T>C M445T Sata et al., 2000
CYP3A4*4 352 A>G I118V Hsieh et al., 2001
CYP3A4*5 653 C>G P218R Hsieh et al., 2001
CYP3A4*6 831 ins A frameshift Hsieh et al., 2001
CYP3A4*7 167 G>A 6004 G>A G56D Eiselt et al., 2001
CYP3A4*8 389 G>A 13908 G>A R130Q Eiselt et al., 2001
CYP3A4*9 508 G>A 14292 G>A V170I Eiselt et al., 2001
CYP3A4*10 520 G>C 14304 G>C D174H Eiselt et al., 2001
CYP3A4*11 1088 C>T 21867 C>T T363M Eiselt et al., 2001
CYP3A4*12 1117 C>T 21896 C>T L373F Eiselt et al., 2001
CYP3A4*13 1247 C>T 22026 C>T P416L Eiselt et al., 2001
CYP3A4*14 44 T>C 44 T>C L15P Dr. J. Lamba
CYP3A4*15A 485 G>A 14269 G>A R162Q Dr. J. Lamba
CYP3A4*15B - 845 ins ATGGAGTGA;
-392 A>G; 485 G>A
- 845 ins ATGGAGTGA;
-392 A>G; 14269 G>A
R162Q
Dr. H. Hamzeiy
CYP3A4*16 554 C>G 15603 C>G T185S Dr. J. Lamba
CYP3A4*17 566 T>C 15615 T>C F189S Dr. Dai
CYP3A4*18 878 T>C 20072 T>C L293P Dr.Dai
CYP3A4*19 1399 C>T VS10+12 G>A
23237 C>T 20230 G>A
P467S Dr. Dai

OBJECTIVOS
23
2 – Objectivos
Perante a inexistência de métodos de genotipagem para as variantes do CYP3A4, assim como
de informação relativa à população Portuguesa no que respeita à incidência dessas variantes, este
trabalho teve por objectivos principais o desenvolvimento de métodos de genotipagem, baseados em
PCR-RFLP, para a detecção das mutações CYP3A4*1B, CYP3A4*2 e CYP3A4*3, assim como a
validação do método através da caracterização das respectivas frequências na população Portuguesa.

MATERIAL E MÉTODOS
24
3 - Material e Métodos – análise do gene CYP3A4
3.1 - Introdução
A abordagem experimental usada para a caracterização das variantes do CYP3A4 teve
como base a amplificação e análise de segmentos específicos de DNA por PCR (Polymerase
Chain Reaction - reacção de amplificação em cadeia da polimerase de DNA termostável) (Mullis e
Faloona, 1987). O PCR é o método experimental indicado para a amplificação exponencial in vitro
de um fragmento específico de DNA, de tamanho e sequência definidos, a partir de quantidades
mínimas de um “molde” de DNA complexo (Mullis e Faloona, 1987; White et al., 1989). De
modo a iniciar a amplificação são necessárias sequências oligonucleotídicas - primers -
complementares às regiões adjacentes (5’ e 3’) de cada uma das cadeias da dupla hélice do DNA
de origem (Sambrook et al., 1989). Os primers flanqueiam o segmento de DNA que se deseja
amplificar, uma vez que as DNA polimerases catalisam reacções de polimerização apenas a partir
de um grupo 3’-OH terminal.
Como o produto da amplificação pode servir de molde para outros primers cada ciclo irá
duplicar a quantidade de fragmentos de DNA produzidos no ciclo anterior, o que resulta numa
acumulação exponencial do fragmento de DNA alvo inicial (cerca de 2n, onde n é o número de
ciclos de amplificação realizados) (Saiki, 1990; White et al., 1989).
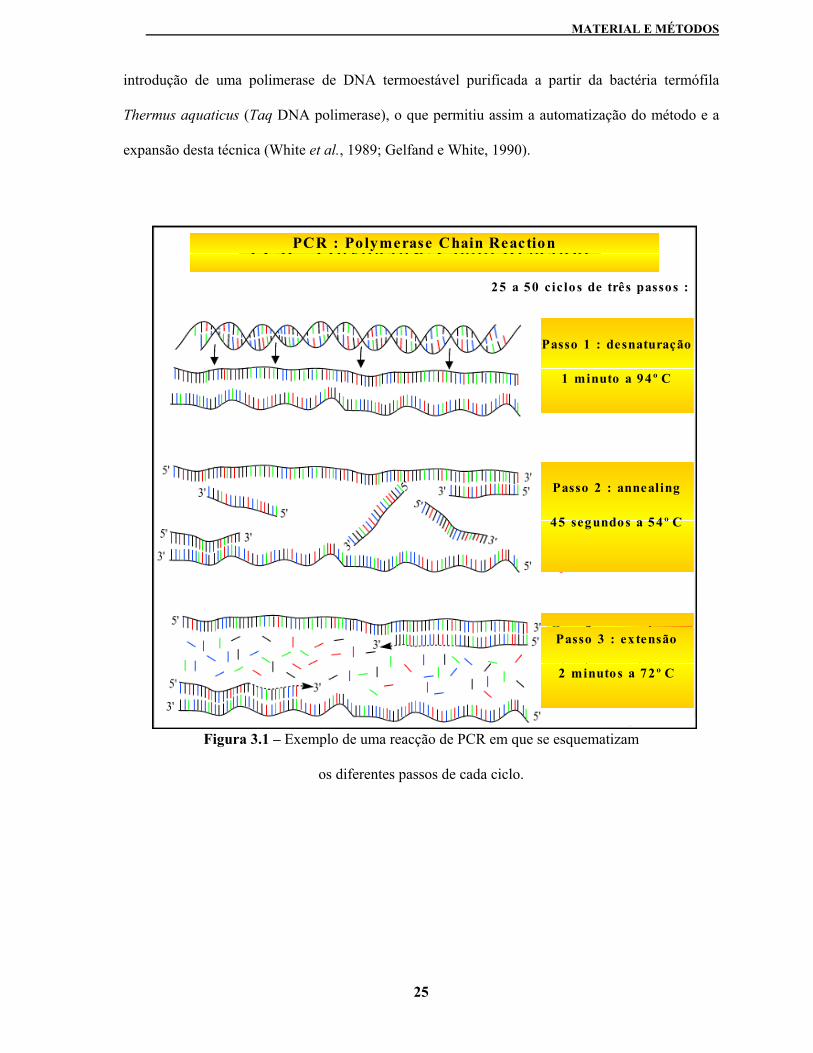
A metodologia de amplificação por PCR baseia-se na repetição de uma série de três passos
(ver figura 3.1): (1) desnaturação do DNA molde por aquecimento, (2) hibridação dos primers
com as sequências de DNA complementares por abaixamento da temperatura – passo de
annealing - e (3) elongação da cadeia de DNA a partir dos primers usando a TaqDNA polimerase
(Sambrook et al., 1989).
Os protocolos originais para as reacções de PCR (Mullis e Faloona, 1987) usavam o
fragmento Klenow da polimerase I de DNA de E. coli para catalisar a reacção de extensão dos
primers, e como esta enzima era inactivada durante o passo de desnaturação havia a necessidade
de a adicionar durante cada ciclo (Sambrook et al., 1989). Este problema foi resolvido com a
MATERIAL E MÉTODOS
25
introdução de uma polimerase de DNA termoestável purificada a partir da bactéria termófila
Thermus aquaticus (Taq DNA polimerase), o que permitiu assim a automatização do método e a
expansão desta técnica (White et al., 1989; Gelfand e White, 1990).
Figura 3.1 – Exemplo de uma reacção de PCR em que se esquematizam
os diferentes passos de cada ciclo.
25 a 50 ciclos de três passos :
Passo 1 : desnaturação
1 minuto a 94º C
Passo 2 : annealing
45 segundos a 54º C
Passo 3 : extensão
2 minutos a 72º C
PCR : Polymerase Chain Reaction

MATERIAL E MÉTODOS
26
No método de PCR geralmente existe a necessidade de optimização do processo, o que
envolve diversos parâmetros (Innis e Gelfand, 1990):
• Primers – Para uma elevada especificidade da reacção é necessária uma baixa concentração de
primers. As altas concentrações de primers podem favorecer o annealing e a produção de
produtos inespecíficos, assim como a formação de dímeros de primers, onde estes em vez de
hibridar com o DNA alvo, hibridam entre si.
• Taq polimerase – Uma alta concentração da enzima Taq polimerase leva a uma redução da
especificidade da reacção, levando à síntese de produtos indesejados. O contrário origina uma
formação ineficiente de produto, o que leva à diminuição do rendimento da reacção.
• dNTPs – Os dNTPs (dATP, dCTP, dTTP, dGTP) devem ser usados em concentrações iguais,
para que assim um deles em maior concentração não seja preferencialmente incorporado. Para
aumentar a especificidade da reacção a concentração de dNTPs deve ser reduzida, enquanto
para o aumento da eficiência é necessário um aumento destes.
• Iões magnésio – O Mg++ é um catião divalente e liga-se ao DNA molde, dNTPs, primers e
polimerase. Logo a sua concentração afecta a especificidade do produto, a hibridação dos
primers, a formação de dímeros de primers, a temperatura de dissociação e a actividade e
fidelidade da enzima. Um excesso de Mg++ pode causar reacções inespecíficas, ou seja, a
hibridação inespecífica dos primers com a cadeia molde, o que diminui a fidelidade da
reacção.
• Temperatura de desnaturação – O passo de desnaturação normalmente ocorre entre 92ºC e
96ºC, o que assegura que haja uma desnaturação completa do DNA molde.

MATERIAL E MÉTODOS
27
• Temperatura de hibridação – A temperatura requerida para a hibridação dos primers com o
DNA alvo depende da sua composição em bases, tamanho e concentração. A temperatura de
hibridação é calculada por diversas formas, dependendo do tamanho da cadeia de
oligonucleótidos. A forma mais simples consiste em fazer o somatório dos nucleótidos
contendo adenina e timina e multiplicar por 2ºC e adicionar o somatório dos nucleótidos
contendo guanina e citosina multiplicado por 4ºC:
Tm = 4 ºC x (número de G’s e C’s no primer) + 2 ºC x (número de A’s e T’s no primer)
Para aumentar a especificidade da reacção aumenta-se a temperatura de hibridação, uma vez
que assim é reduzida a possibilidade de hibridações não específicas dos primers e a
consequente amplificação de produtos inespecíficos.
• Número de ciclos de amplificação – A optimização do número de ciclos de amplificação
numa reacção, partindo do princípio que todos os outros parâmetros estão optimizados,
dependerá sobretudo da concentração inicial de DNA molde. Para aumentar a especificidade
da reacção deve reduzir-se o número de ciclos. Se esta diminuição for demasiada irá levar a
uma diminuição do rendimento da reacção.
A detecção de alterações na estrutura do gene CYP3A4 envolvem a digestão dos
fragmentos amplificados com endonucleases de restrição e a análise por separação electroforética
dos fragmentos de restrição do DNA em gel de poliacrilamida.
As endonucleases de restrição são usadas para hidrolisar o DNA de uma maneira precisa
em posições internas. Estas são membros de uma classe de enzimas que têm a propriedade de
quebrar as ligações fosfodiéster que ligam os nucleótidos adjacentes nas moléculas de DNA. As
enzimas de restrição são produzidas por bactérias como mecanismo de defesa contra os
bacteriofagos, e actuam clivando sequências específicas de nucleótidos do DNA, tornando-o então
inactivo (Griffiths et al., 1996).

MATERIAL E MÉTODOS
28
Em relação à electroforese, esta é uma técnica que tem como base a migração de uma
partícula carregada sobre a influência de um campo eléctrico. Muitas moléculas de importância
biológica, incluindo os ácidos nucleicos, possuem grupos ionizáveis e, assim, a um dado pH,
existem em solução como espécies carregadas. Sobre a influência de um campo eléctrico estas
partículas carregadas irão migrar para o cátodo ou ânodo, dependendo da natureza da sua carga. A
electroforese em gel de poliacrilamida (PAGE) utiliza um suporte em que os monómeros de
acrilamida ao polimerizarem formam longas cadeias ligadas entre si por ligações cruzadas
(crosslinking), através dos resíduos de N, N’-metilenobisacrilamida (bisacrilamida) presentes, o
que forma uma malha de tamanho controlável.
A polimerização da acrilamida é um exemplo de catálise por radicais livres iniciada pelo
persulfato de amónio (PSA) e catalisada pelo N, N, N’, N’- tetrametilenodiamina (TEMED). O
TEMED catalisa a decomposição do ião persulfato para dar um radical anião sulfato:
S2O82- + e- → SO4
− • + SO42−
Este radical (SO4− •) reage com o monómero de acrilamida transformando-a num radical
que vai reagir com outra molécula de acrilamida, ou uma de bisacrilamida, dando origem a outro
radical, e assim sucessivamente, o que dá origem ao polímero com ligações cruzadas. O número de
ligações determina o tamanho dos poros do gel, e assim a gama de massas moleculares das
moléculas que podem ser separadas no gel (Walker, 2000).
3.2 - Materiais

MATERIAL E MÉTODOS
29
3.2.1 - Material Biológico - Indivíduos
A população controlo genotipada no presente estudo foi constituída por 131 indivíduos (59
homens, idades 8 – 77, média de idades = 49,88, SD = 18,35; 72 mulheres, idades 13-89, média
de idades = 49,58, SD = 18,50). As amostras de sangue utilizadas foram obtidas em voluntários
Caucasianos normais, não relacionados e saudáveis, que foram assistidos nos Hospitais de Faro e
do Barlavento, no Algarve. Todos os protocolos experimentais foram aprovados pelos Quadros de
Ética dos Hospitais, tendo sido os indivíduos analisados neste estudo recrutados de check-ups
médicos regulares.
As amostras de sangue foram colhidas sob controlo médico e em obediência à legislação e
normas éticas vigentes. As amostras foram colhidas para tubos estéreis com o anticoagulante
K3EDTA a pH8,0, tendo sido depois congeladas a -20ºC, até a extracção de DNA.
3.2.2 - Reagentes
Tris (Riedel-de Haën), ácido bórico (Merck), EDTA (Merck), acrilamida (Pharmacia Biotech,
Riedel-de Haën), N, N’- metileno-bis-acrilamida (Sigma, Riedel-de Haën), Glicerol (Sigma), Azul
de bromofenol (Merck), Xileno cianol (Sigma), Persulfato de amónio (Sigma), TEMED (BDH
Chemicals Ltd), brometo de etídeo (Sigma), Igepal CA-630 (Sigma), Tween 20 (Merck),
Proteinase K (Sigma), cloreto de magnésio (Riedel-de Haën).
A origem de outros reagentes e dos equipamentos usados é mencionada no decorrer do
texto.
3.3 – Métodos

MATERIAL E MÉTODOS
30
3.3.1 - Extracção de DNA genómico – Extracção rápida de DNA
O método de extracção rápida de DNA é usado para extrair o DNA genómico (Innis et al.,
1990), e é um método que não implica passos de purificação, o que faz com que seja
extremamente rápido e económico, evitando o uso de solventes orgânicos.
Após a descongelação da amostra, misturou-se 100 µl de sangue com 750 µl de tampão TE
(pH 8,0)1. A mistura foi agitada no vortex (Heidolph 94323) e seguidamente sujeita a uma
centrifugação a 12000 g (Eppendorf 5415D) durante 1 minuto. Como este passo leva à lise dos
eritrócitos, o sobrenadante foi cuidadosamente removido por aspiração. Este procedimento foi
depois repetido até o sedimento se apresentar livre de hemoglobina. O sedimento de células
nucleadas foi ressuspendido em 100 µl de tampão de lise2 com o objectivo de lisar os leucócitos. A
desproteinização foi feita através da adição de 1 µl de proteinase K (20 mg/ml). Esta mistura foi
depois incubada a 55 ºC durante cerca de 16 horas, e após este tempo foi-lhe adicionado 100 µl de
água MilliQ (Millipore) estéril. A amostra foi então aquecida a 95 ºC por cerca de 15 minutos, de
modo a dar-se a desnaturação proteica, e consequente inactivação da proteínase K. As amostras de
DNA foram mantidas a –20 ºC até utilização para as reacções de PCR.
A avaliação da concentração e pureza do DNA obtido foi efectuada por espectrofotometria
(Shimadzu UV-260), determinando-se a absorvância de uma amostra diluída de DNA (diluição de
1:10) aos comprimentos de onda de 260 e 280 nm (Sambrook et al., 1989).
3.3.2 – Pesquisa dos alelos do gene CYP3A4 por PCR-RFLP
1 Tampão TE (pH 8,0): 10 mM Tris-HCl (pH 8,0), 1 mM EDTA 2 Tampão de lise: 20 mM Tris-HCl (pH 8,0), 50 mM MgCl2, Igepal CA-630 (0,45%), Tween 20 (0,45%)

MATERIAL E MÉTODOS
31
O desenvolvimento de métodos de PCR depende do desenho de primers específicos para as
amplificações. Neste processo o primeiro passo consiste na consulta de bases de dados para a
obtenção das sequências dos genes a estudar. Após a localização na sequência do gene das
mutações a estudar, começa o desenho de possíveis primers a usar na reacção de PCR. Aqui é
necessário ter em conta várias regras (Innis e Gelfand, 1990):
♦ os primers devem ter um tamanho de pelo menos entre 17 e 28 nucleótidos, de
modo a minimizar o problema de hibridação secundária;
♦ os primers devem ter uma composição em G /C entre 50 e 60%;
♦ os primers devem ter no terminal 3’ um G ou C, ou CG ou GC, o que aumenta a
eficiência dos primers;
♦ as temperaturas de dissociação entre 55 – 80ºC são as ideais;
♦ os primers não devem apresentar complementaridade interior (palindromas), ou
seja, não devem formar estruturas como hairpins;
♦ os primers não devem conter sequências de nucleótidos complementares entre
elas, uma vez que dímeros serão sintetizados preferencialmente em relação a
outros produtos;
♦ os primers escolhidos devem ter homologia específica com o gene que se quer
amplificar.
No caso do gene CYP3A4 foi necessário assegurar que os primers escolhidos não
hibridassem com formas relacionadas , como é o caso dos genes CYP3A5, CYP3A7 e CYP3A43.
Para o SNP CYP3A4*1B a amplificação foi levada a cabo numa mistura de reacção de 50
µl contendo 3 µl de DNA alvo, 0,6 µM de cada primer (1Bfw e 1Brv, ver tabela 3.1) (Eurogentec),

MATERIAL E MÉTODOS
32
200 µM de cada dNTP (Life Technologies), 1,5 mM de MgCl2 (Promega), tampão para Taq
polimerase3 (1x) (Promega), 2,5 U4 de Taq polimerase (Promega) e H2O estéril.
As condições de amplificação consistiram em um ciclo de 2 min a 94ºC, 30 ciclos de
amplificação que incluiram 45 s de desnaturação a 94 ºC, 45 s de annealing a 59 ºC e 45 s de
extensão a 72 ºC, e ciclo final de 1 min a 72ºC. O termociclador usado foi o Mastercycler personal
5332 (Eppendorf). Os produtos de amplificação obtidos foram mantidos a 4 ºC até serem
submetidos à digestão pelos enzimas de restrição.
O CYP3A4*1B corresponde a uma transição A → G na região 5’ do gene. A verificação da
presença desta mutação baseia-se na introdução de uma base sem emparelhamento (mismatch),
que neste caso é uma citosina assinalada a negrito na sequência do primer 1Bfw (ver tabela 3.1). A
presença deste mismatch, juntamente com a mutação permite a existência de um sítio de restrição
para a enzima Bst NI:
CYP3A4*1A CCAAG Sem sítio de restrição
CYP3A4*1B CCAGG Sequência de restrição Bst NI A
5’- CC↓TGG -3’ 3’- GGA↑CC -5’
T
3 10 mM Tris-HCl (pH 9,0 a 25ºC), 50 mM KCl e 0,1 % Triton®X-100 4 Uma unidade é definida como a quantidade de enzima necessária para catalizar a incorporação de 10 nmoles de dNTPs numa forma ácida, não solúvel, em 30 minutos a 74 ºC, nas seguintes condições: 50 mM Tris-HCl (pH 9,0), 50 mM NaCl, 5 mM MgCl2, 200 µM de dNTPs, 10 µg de DNA de timo de vitela e 0,1 mg/ml de BSA, num volume final de 50 µl (Promega, Madison).

MATERIAL E MÉTODOS
34
Para a análise desta variante foi introduzido um controlo interno através da amplificação, nas
mesmas condições descritas anteriormente, de um fragmento de 211 pares de bases (bp) (primers
1BCfw e 1BCrv, ver tabela 3.1) (Life Technologies), que através de sítios de restrição adicionais para
a Bst NI, que dão origem a fragmentos de 158, 31, 12 e 10 bp, permitem o controlo da eficiência da
digestão e a consequente identificação inequívoca dos homozigotas CYP3A4*1A.
O produto de cada uma das reacções de PCR com 122 e 211 bp, (10 µl de cada produto
amplificado) foi digerido com 5 U5 de Bst NI (Stratagene), num volume total de 30 µl durante 3 horas
a 60 ºC. A restrição do alelo contendo a mutação origina dois fragmentos (95 bp + 27 bp), enquanto
que o CYP3A4*1A manterá o fragmento amplificado de 122 bp (ver figura 3.2B).
No caso do CYP3A4*2 a amplificação foi em tudo semelhante ao descrito anteriormente para o
CYP3A4*1B, excepto nos primers usados (2fw e 2 rv, ver tabela 3.1) (Eurogentec) e na temperatura
de annealing que foi de 45 ºC.
O SNP CYP3A4*2 consiste numa transição T → C na região codificante. A verificação da
presença desta variante baseia-se na introdução de um mismatch, uma guanina assinalada a negrito na
sequência do primer 2fw (ver tabela 3.1). A presença do mismatch e a ausência da mutação permite a
existência de um sítio de restrição Alw26 I:
CYP3A4*1A GTCTC Sequência de restrição Alw26 I
5’- GTCTC↓ -3’ 3’- CAGAG↑ -5’
CYP3A4*2 GTCCC Perda do sítio de restrição
Deste modo, a restrição do produto amplificado (20 µl), com 5 U5 Alw26 I (Promega), num
volume total de 30 µl durante 3 horas a 37 ºC, origina para o caso do CYP3A4*1A dois fragmentos
(123 bp + 28 bp), enquanto que o CYP3A4*2 mantém o fragmento amplificado de 151 bp intacto (ver
figura 3.2C).
5 Definição de unidade: quantidade de enzima necessária para a restrição completa de 1 µg de DNA genómico de bacteriofago λ em condições óptimas de pH e temperatura (Stratagene).

MATERIAL E MÉTODOS
35
A amplificação do CYP3A4*3, usando os primers 3fw e 3rv (ver tabela 3.1) (Life
Technologies), foi realizada como descrito anteriormente, excepto para a temperatura de annealing
que foi de 52ºC. Esta variante consiste na substituição T → C que leva ao desaparecimento de um
sítio de reconhecimento para a enzima de restrição Nla III:
Alelo CYP3A4*1A CATGA Sequência de restrição Nla III
5’- CATG↓ -3’ 3’- ↑GTAC -5’
Alelo CYP3A4*3 CACGA Perda do sítio de restrição
A restrição do produto amplificado (20 µl) com 2,5 U5 de Nla III (New England BioLabs),
num volume total de 30 µl, durante 3h a 37ºC, origina para o CYP3A4*1A dois fragmentos (89 bp +
85 bp), enquanto que no caso da mutação mantêm-se o fragmento amplificado de 174 bp (ver figura
3.2D).
3.3.3 – Detecção dos produtos de digestão
A detecção dos fragmentos de restrição foi realizada em gel de poliacrilamida a 8% utilizando
um sistema de electroforese vertical Hoefer SE600 (Amersham Pharmacia Biotech). A migração
efectuou-se em tampão TBE6 (1x), à temperatura ambiente e a uma voltagem de 25 V/cm, durante
cerca de uma hora, ou em alternativa, overnight a 2 V/cm. Para a preparação da amostra a aplicar
misturou-se 25 µl de produto digerido com 2,5 µl de tampão de aplicação7. Como marcador de pesos
moleculares foram usados o marcador V ou o marcador IX (Roche Biochemicals). Os géis foram
corados numa solução de brometo de etídeo (0,5 µg/ml), visualizados por exposição a luz ultravioleta
e fotografados num sistema Gel Doc 2000 (Bio-Rad).
6 Tampão TBE: 89 mM Tris, 89 mM ácido bórico e 2 mM EDTA (pH 8,0). 7 Tampão de aplicação: 0,25 % xileno cianol e 30 % glicerol ou 0,25% azul bromofenol e 30% glicerol.

MATERIAL E MÉTODOS
36
Os padrões de restrição previstos para as variantes CYP3A4*1B, CYP3A4*2 e CYP3A4*3 estão
apresentadas na figura 3.2.
(A)
(B) (C)
(D)
Figura 3.2 – Esquema do gene CYP3A4 e da zona de amplificação de cada par de primers (A).
Representação esquemática dos padrões de RFLP obtidos para a identificação do CYP3A4*1B (B),
CYP3A4*2 (C) e CYP3A4*3 (D).
_ Nla III
+ *1A/*3 *3/*3 *1A/*1A
174 bp
89 bp 85 bp
5’
3’ 5’
3’ 1 2 3 4 5 7 8 9 10 11 12 13 6
1 *
7 *
12
_ Alw26 I
+ *1A/*2 *2/*2 *1A/*1A
151 bp
123 bp
28 bp
_ Bst NI
+ *1A/*1B *1B/*1B *1A/*1A
211 bp
122 bp 158 bp
95 bp
31 bp 27 bp 12 bp 10 bp

MATERIAL E MÉTODOS
37
3.3.4 – Análise estatística
3.3.4.1 – Frequências alélicas
A frequência alélica representa uma medida do quanto frequente é um alelo numa população,
ou seja, exprime o valor que caracteriza cada população particular de indivíduos do ponto de vista da
natureza genética, permitindo a sua posterior comparação com outras populações.
O cálculo da frequência de um alelo é feita através da contagem dos homozigotas e com a
adição de metade dos heterozigotas. A forma geral para este cálculo é (Griffiths et al., 2000):
p = fA/A + ½ fA/a = frequência de A
q = fa/a + ½ fA/a = frequência de a
em que fA/A , fA/a e fa/a são as proporções dos três genótipos num locus com dois alelos e onde p
corresponde à frequência do alelo A e q à frequência do alelo a.
3.3.4.2 – Frequências genotípicas
A frequência genotípica numa população é a proporção de organismos que têm um genótipo
em particular. O cálculo das frequências genotípicas teóricas efectua-se tendo como base o princípio
de Hardy-Weinberg. Este modelo baseia-se em cinco princípios, ou seja, de que uma população é
grande, de que não existem flutuações de genes entre populações, resultantes de migrações ou de
transferência de gâmetas, de que as mutações são negligenciáveis, de que os indivíduos casam-se
aleatóriamente (população em panmixia) e de que a selecção natural não opera na população. Se todos
estes princípios se verificarem, o genótipo da população e as frequências alélicas permanecerão
inalteradas ao longo de gerações sucessivas, e a população diz-se que se encontra em equilíbrio de
Hardy-Weinberg.

MATERIAL E MÉTODOS
38
O modelo de Hardy-Weinberg baseia-se em duas equações, uma que calcula as frequências
alélicas e outra que calcula as frequências genotípicas. A equação p + q = 1 descreve as frequências
alélicas para um gene com dois alelos (o caso mais simples). Num organismo diplóide com os alelos A
e a num dado locus existem três genótipos possíveis: AA, Aa e aa. Usando o p para representar a
frequência de A e q para representar a frequência de a temos que a equação para a frequência
genotípica é p2 + 2 pq + q2 = 1, uma equação que descreve uma distribuição binomial.
Tabela 3.2 – Frequências de Hardy-Weinberg geradas pela união ao acaso de dois alelos, A e a, cujas
frequências são p e q, respectivamente.
A p
a q
A p
AA P2
Aa pq
a q
Aa pq
aa q2
3.3.4.3 – Desvios em relação à distribuição de Hardy-Weinberg – Teste de χ2
O qui-quadrado é um teste estatístico usado geralmente para comparar resultados observados
com o seu valor esperado, segundo critérios estatísticos. A fórmula usada para calcular o χ2 é (Fowler
e Cohen, 1992):
χ2 ∑ −=
esperadovaloresperadovalorobservadovalor
_)__( 2
Isto é, o χ2 é a soma do quadrado da diferença entre o valor observado e o esperado, dividida pelo
valor esperado.
O teste de χ2 é uma forma simples de quantificar o desvio esperado se uma hipótese se mostrar
verdadeira.
Os resultados foram tratados estatisticamente utilizando o programa informático CIA (Gardner
e Altman, 1989) para o cálculo de frequências e intervalos de confiança.

MATERIAL E MÉTODOS
33
Tabela 3.1 - Oligonucleótidos usados na reacção de PCR para a genotipagem do CYP3A4.
Variantes Sequência dos primers Posição na sequência Tamanho dos fragmentos
de PCR (bp)
CYP3A4*1B
1Bfw: 5’ -AAT GAG GAC AGC CAT AGA GAC AAG GcC A-3’
1Brv: 5’ -CAA TCA ATG TTA CTG GGG AGT CCA AGG G- 3’
788 – 815 a
909 – 882 a
122
1BCfw: 5’ – ACA ATC CAA CAG CCT CAC TGA ATC AC-3’
1BCrv: 5’ – GAA CAG TTA CTC ACA GAT AGA GGA GC-3’
1081 – 1106 a
1292 – 1267 a
211
CYP3A4*2 2fw: 5’ –AGA TTT GAT TTT TTG GAT CCA TTC TTT gTC-3’
2rv: 5’ -CAA ATC ACT GAA CTG TAT ATT TTA AGT GG- 3’
15801 – 15830 b
15951 – 15923 b
151
CYP3A4*3 3fw : 5’ -CCA GAT TCA GCA AGA AGA ACA AGG- 3’
3rv: 5’ -GAC TAA CCT GTG TTT CTT TAC AAG G- 3’
23194 – 23217 b
23367 – 23343 b
174
a Genbank, número de acesso D11131 b Genbank, número de acesso AF209389
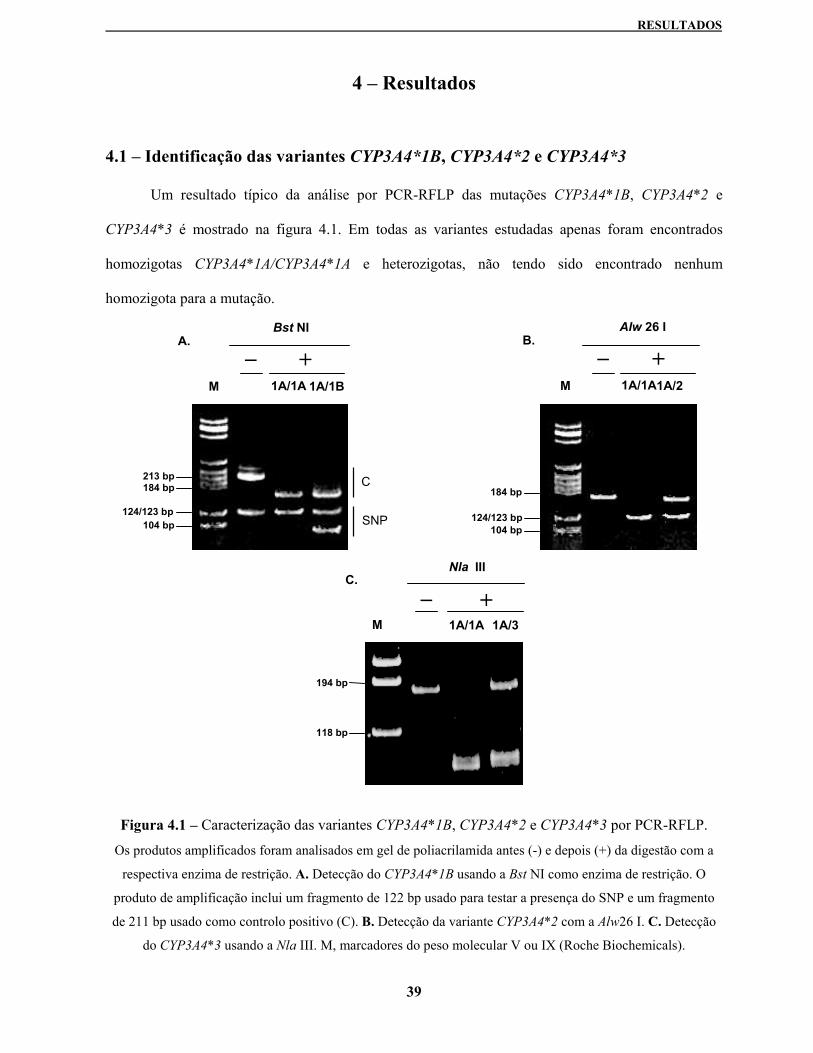
RESULTADOS
39
4 – Resultados
4.1 – Identificação das variantes CYP3A4*1B, CYP3A4*2 e CYP3A4*3
Um resultado típico da análise por PCR-RFLP das mutações CYP3A4*1B, CYP3A4*2 e
CYP3A4*3 é mostrado na figura 4.1. Em todas as variantes estudadas apenas foram encontrados
homozigotas CYP3A4*1A/CYP3A4*1A e heterozigotas, não tendo sido encontrado nenhum
homozigota para a mutação.
Figura 4.1 – Caracterização das variantes CYP3A4*1B, CYP3A4*2 e CYP3A4*3 por PCR-RFLP.
Os produtos amplificados foram analisados em gel de poliacrilamida antes (-) e depois (+) da digestão com a
respectiva enzima de restrição. A. Detecção do CYP3A4*1B usando a Bst NI como enzima de restrição. O
produto de amplificação inclui um fragmento de 122 bp usado para testar a presença do SNP e um fragmento
de 211 bp usado como controlo positivo (C). B. Detecção da variante CYP3A4*2 com a Alw26 I. C. Detecção
do CYP3A4*3 usando a Nla III. M, marcadores do peso molecular V ou IX (Roche Biochemicals).
1A/1A 1A/1B M
+ _ Bst NI
A.
213 bp
124/123 bp
184 bp
104 bp
C
SNP
1A/1A 1A/2 M
+ _ Alw 26 I
B.
124/123 bp
184 bp
104 bp
1A/1A 1A/3 M
+ _
Nla III C.
194 bp
118 bp
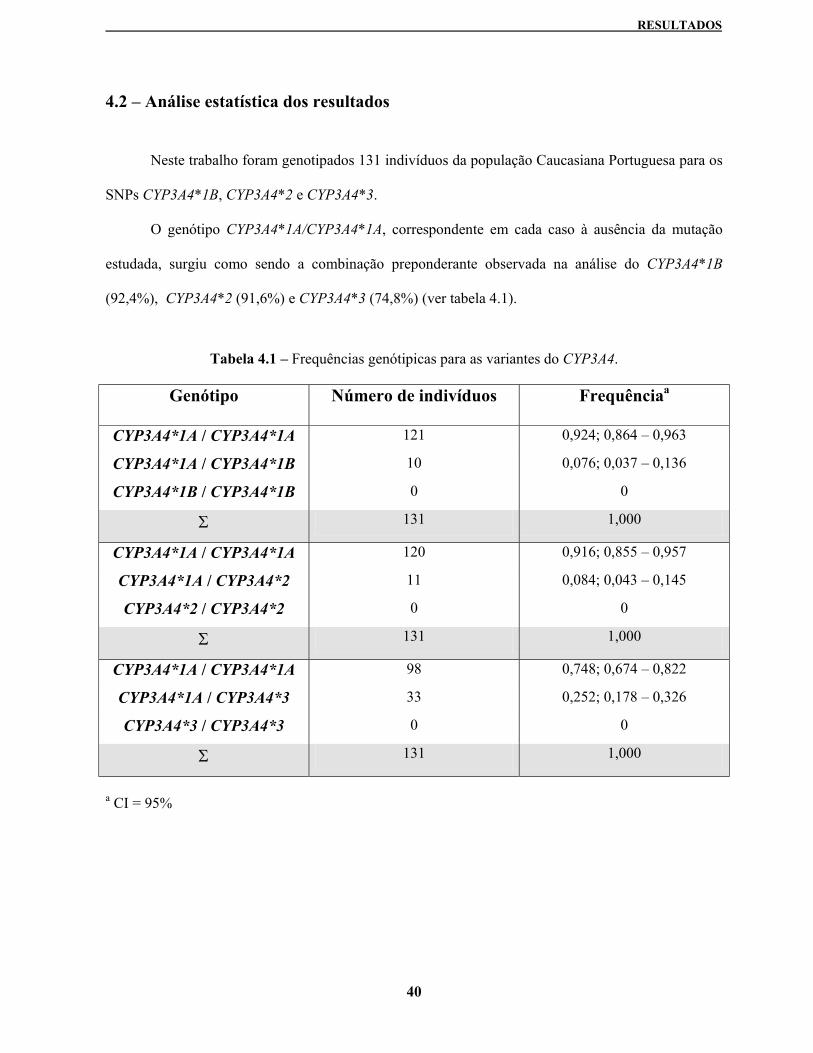
RESULTADOS
40
4.2 – Análise estatística dos resultados
Neste trabalho foram genotipados 131 indivíduos da população Caucasiana Portuguesa para os
SNPs CYP3A4*1B, CYP3A4*2 e CYP3A4*3.
O genótipo CYP3A4*1A/CYP3A4*1A, correspondente em cada caso à ausência da mutação
estudada, surgiu como sendo a combinação preponderante observada na análise do CYP3A4*1B
(92,4%), CYP3A4*2 (91,6%) e CYP3A4*3 (74,8%) (ver tabela 4.1).
Tabela 4.1 – Frequências genótipicas para as variantes do CYP3A4.
Genótipo Número de indivíduos Frequênciaa
CYP3A4*1A / CYP3A4*1A 121 0,924; 0,864 – 0,963
CYP3A4*1A / CYP3A4*1B 10 0,076; 0,037 – 0,136
CYP3A4*1B / CYP3A4*1B 0 0
Σ 131 1,000
CYP3A4*1A / CYP3A4*1A 120 0,916; 0,855 – 0,957
CYP3A4*1A / CYP3A4*2 11 0,084; 0,043 – 0,145
CYP3A4*2 / CYP3A4*2 0 0
Σ 131 1,000
CYP3A4*1A / CYP3A4*1A 98 0,748; 0,674 – 0,822
CYP3A4*1A / CYP3A4*3 33 0,252; 0,178 – 0,326
CYP3A4*3 / CYP3A4*3 0 0
Σ 131 1,000
a CI = 95%

RESULTADOS
41
Em relação às frequências alélicas encontradas os valores calculados estão apresentados na
tabela 4.2. Entre as variantes estudadas, a variante CYP3A4*3 apresentou-se como sendo a mais
frequente na população Portuguesa (12,6%), isto após a frequência correspondente à sequência de
referência CYP3A4*1A.
Tabela 4.2 – Frequências alélicas das mutações CYP3A4*1B, CYP3A4*2 e CYP3A4*3 na população
Caucasiana Portuguesa.
CYP3A4*1B CYP3A4*2 CYP3A4*3
A G T C T C
Número de
alelos
252 10 251 11 229 33
Frequência
alélica
(95% CI)
0,962 (0,931 – 0,982)
0,0382 (0,0185 – 0,096)
0,958 (0,926 – 0,979)
0,042 (0,0212 – 0,0739)
0,874 (0,834 – 0,914)
0,126 (0,0858 – 0,166)
O cálculo da frequência genotípica esperada para cada uma das variantes efectuou-se segundo
o princípio de Hardy-Weinberg e os resultados obtidos encontram-se representados na tabela 4.3. Aos
valores obtidos aplicou-se o teste de cqui-quadrado, cujos valores também se encontram apresentados
na tabela 4.3.
O conjunto dos resultados obtidos para as variantes CYP3A4*1B, CYP3A4*2 e CYP3A4*3
revela que a população estudada se encontra em equilíbrio de Hardy-Weinberg. O valor de p para o χ2
calculado é p > 0,05 (ver tabela 4.3), pelo que os desvios que possam existir em relação às frequências
esperadas serão devido ao acaso e não a outros factores (mutações, selecção natural, entre outros).

RESULTADOS
42
Tabela 4.3 – Resultados do teste do ajustamento χ2 utilizado na determinação dos desvios às
frequências esperadas pelo equilíbrio de Hardy-Weinberg numa população de 131 indivíduos
portugueses de origem caucasiana.
Frequência Genotípica
Genótipo Observada Esperada χ2 Graus de
liberdade
p
CYP3A4*1B *1A/*1A 0,924 0,9254
Transição *1A/*1B 0,076 0,0731 1,517 x 10-3 2 0,9993
A → G *1B/*1B 0 0,0014
CYP3A4*2 *1A/*1A 0,916 0,9178
Transição *1A/*2 0,084 0,0805 1,918 x 10-3 2 0,9991
T → C *2/*2 0 0,0018
CYP3A4*3 *1A/*1A 0,748 0,7639
Transição *1A/*3 0,252 0,2202 2,08 x 10-2 2 0,9909
T → C *3/*3 0 0,0159

DISCUSSÃO
43
5 – Discussão dos resultados
O CYP3A4 é o citocromo P450 mais abundante no fígado, apresentando uma grande
variabilidade interindividual atribuída quase na totalidade a factores genéticos. No entanto, só
recentemente, foram descritas variantes polimórficas para o CYP3A4 que podem estar na base das
variações existentes.
Os resultados apresentados constituem os primeiros dados sobre a caracterização da população
Portuguesa no que diz respeito ao gene polimórfico CYP3A4, assim como, representam a validação do
método de PCR-RFLP desenvolvido para as variantes CYP3A4*1B, CYP3A4*2 e CYP3A4*3. O
protocolo descrito para o CYP3A4*2 é também o primeiro apresentado até à data que se baseia nesta
aproximação experimental.
No que diz respeito ao CYP3A4*1B os dados obtidos mostram que esta é uma variante que está
presente com uma baixa incidência na população Portuguesa (3,8%). Quando comparamos a
frequência alélica obtida neste estudo com as obtidas noutras populações (ver tabela 5.1) esta
apresenta-se como sendo significativamente menor (p < 0,01) do que a observada numa população
Caucasiana Americana (Rebbeck et al., 1998), mas em relação a outras populações recentemente
caracterizadas a diferença não é estatisticamente significativa (Ball et al., 1999; Westlind et al., 1999;
Sata et al., 2000; Tayeb et al., 2000; van Schaik et al., 2000).
A variante CYP3A4*2 foi observada na população Portuguesa com uma frequência de 4,2%, o
que não é significativamente diferente do observado inicialmente na população Finlandesa (ver tabela
5.1) (Sata et al., 2000). Tendo sido apenas descrita na população Finlandesa, foi sugerido que esta
seria uma mutação específica desta população, uma vez que esta apresenta diferenças em termos de
hereditariedade genética em relação às outras populações (Eiselt et al., 2001). Os dados obtidos para a
população Portuguesa demonstram que esta hipótese não se verifica e estudos futuros noutras
populações poderão também demonstrá-lo.
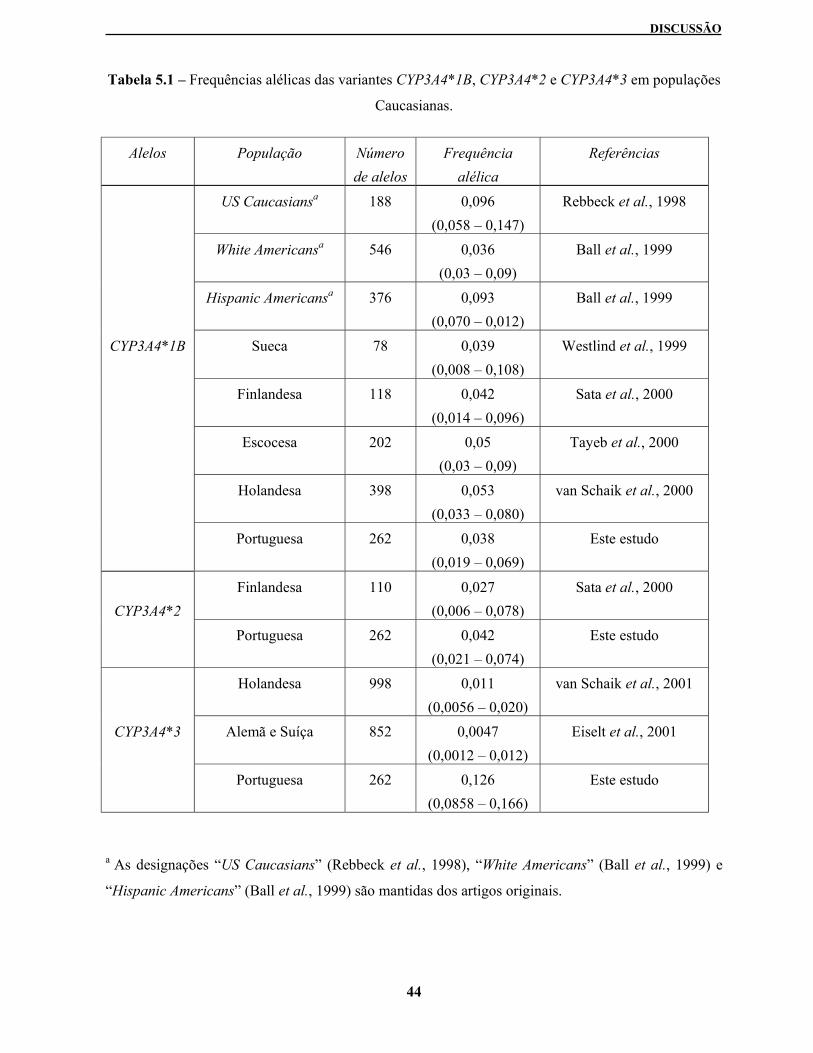
DISCUSSÃO
44
Tabela 5.1 – Frequências alélicas das variantes CYP3A4*1B, CYP3A4*2 e CYP3A4*3 em populações
Caucasianas.
Alelos População Número de alelos
Frequência alélica
Referências
US Caucasiansa 188 0,096 (0,058 – 0,147)
Rebbeck et al., 1998
White Americansa 546 0,036 (0,03 – 0,09)
Ball et al., 1999
Hispanic Americansa 376 0,093 (0,070 – 0,012)
Ball et al., 1999
CYP3A4*1B Sueca 78 0,039 (0,008 – 0,108)
Westlind et al., 1999
Finlandesa 118 0,042 (0,014 – 0,096)
Sata et al., 2000
Escocesa 202 0,05 (0,03 – 0,09)
Tayeb et al., 2000
Holandesa 398 0,053 (0,033 – 0,080)
van Schaik et al., 2000
Portuguesa 262 0,038 (0,019 – 0,069)
Este estudo
CYP3A4*2
Finlandesa 110 0,027 (0,006 – 0,078)
Sata et al., 2000
Portuguesa 262 0,042 (0,021 – 0,074)
Este estudo
Holandesa 998 0,011 (0,0056 – 0,020)
van Schaik et al., 2001
CYP3A4*3 Alemã e Suíça 852 0,0047 (0,0012 – 0,012)
Eiselt et al., 2001
Portuguesa 262 0,126 (0,0858 – 0,166)
Este estudo
a As designações “US Caucasians” (Rebbeck et al., 1998), “White Americans” (Ball et al., 1999) e
“Hispanic Americans” (Ball et al., 1999) são mantidas dos artigos originais.

DISCUSSÃO
45
No caso do CYP3A4*3 a frequência alélica obtida foi de 12,6%, um valor significativamente
maior do que aquele que foi obtido para as outras duas mutações nesta população. Quando comparada
com os valores obtidos em estudos na população Holandesa (van Schaik et al., 2001) ou na população
Alemã e Suíça (Eiselt et al., 2001), a frequência obtida na população Portuguesa não apresenta
diferenças significativas (ver tabela 5.1). Este estudo confirma também uma recente sugestão de que
esta mutação representa uma variante polimórfica e não um alelo raro, como havia sido inicialmente
considerado (Sata et al., 2000; van Schaik et al., 2001).
Assim, no conjunto as variantes CYP3A4*1B, CYP3A4*2 e CYP3A4*3 representam uma
frequência não negligenciável de cerca de 20 – 21% dos SNPs do CYP3A4 encontrados na população
Portuguesa.
No estudo realizado foi observada a existência de indivíduos que se apresentam como sendo
heterozigotas tanto para a mutação CYP3A4*1B, assim como para a CYP3A4*3. Na população
estudada, para o CYP3A4*1B em 10 indivíduos 6 apresentam também a mutação CYP3A4*3. Assim,
como a frequência alélica obtida para o CYP3A4*1B é baixa, a existência, ou não, de uma associação
entre as duas mutações deverá ser verificada através do estudo de uma população em que a incidência
da variante CYP3A4*1B seja maior, como é o caso da população negra. Se nesta população se
verificar a existência de indivíduos com ambas as mutações será necessário confirmar se ambas as
mutações se encontram localizadas no mesmo cromatídio. Isto poderá ser feito se forem encontrados
homozigotas para a mutação em qualquer uma das mutações, existindo assim a certeza de que as duas
mutações estão no mesmo cromatídio.
Deste modo, os métodos desenvolvidos neste estudo, tendo como base a metodologia de PCR-
-RFLP, poderão ser importantes na caracterização de outras populações, assim como poderão
representar ferramentas importantes para um futuro ajustamento da terapêutica, uma vez que as
variantes aqui caracterizadas poderão prejudicar o metabolismo de drogas mediado pelo CYP3A4.
Embora existam resultados contraditórios no que diz respeito ao efeito da mutação CYP3A4*1B na
expressão da proteína (Rebbeck et al., 1998; Westlind et al., 1999), a proteína expressa para o

DISCUSSÃO
46
CYP3A4*2 demonstrou ter um metabolismo alterado em relação à nifedipina (Sata et al., 2000). No
caso da enzima CYP3A4*3, esta apresenta uma actividade enzimática alterada perante a testosterona e
progesterona (Eiselt et al., 2001).
No entanto, não pode ser excluído o efeito destas mutações (S222P, M445T) na metabolização
de outros xenobióticos ou drogas de uso terapêutico. As variantes do CYP3A4 constituem, por outro
lado, potenciais factores para a susceptibilidade à doença. Esta associação a doenças foi já sugerida
para a variante CYP3A4*1B no caso da leucemia (Felix et al., 1998) e do cancro da próstata (Rebbeck
et al., 1998; Walker et al., 1998; Paris et al., 1999).
Deste modo, a genotipagem destes SNPs do CYP3A4 poderão ser uma ferramenta importante
para a optimização da dose de drogas administrada, assim como na determinação da susceptibilidade
de cada indivíduo para a doença.

CONCLUSÃO
47
6 – Conclusão
O polimorfismo genético existente no CYP3A4 poderá contribuir para a variabilidade
interindividual existente em termos de actividade enzimática, pelo que a detecção das variantes
polimórficas é importante para um melhor conhecimento da resposta individual às várias drogas. O
trabalho aqui descrito contribui para um melhor conhecimento desta variação, nomeadamente que:
♦ as variantes CYP3A4*1B, CYP3A4*2 e CYP3A4*3 estão presentes na população Portuguesa,
embora no caso das duas primeiras com baixa incidência (3,8 e 4,2%, respectivamente). A terceira
variante estudada, CYP3A4*3, apresentou-se como sendo a predominante na população
estudada(12,6%), o que confirma o facto desta mutação ser uma variante polimórfica, e não um
alelo raro.
♦ no total, as variantes estudadas representam uma frequência total de cerca de 21% na população
estudada.
♦ os métodos desenvolvidos são válidos e, deste modo, poderão ser usados para estudos noutras
populações, assim como no estudo de patologias.
♦ existe uma possível associação entre a variante CYP3A4*1B e a CYP3A4*3.
Deste modo, uma vez que é provável que as variantes do CYP3A4 possam influenciar a
metabolização de várias drogas, afectando a eficácia, assim como a toxicidade, estes são dados
importantes para o ajustamento da terapêutica.
O trabalho futuro passará pelo estudo da incidência de variantes do CYP3A4 que sejam descritas
como afectando a actividade enzimática ou o metabolismo de compostos exógenos ou endógenos de
modo significativo, e desenvolvimento de métodos para o estudos das mesmas. Outra vertente passará
também pelo estudo de possíveis associações dessas mutações com diversas patologias, sendo aqui
também importante verificar a relação destas patologias com as variações recentemente descritas nos
receptores nucleares que potencialmente regulam a expressão do CYP3A4 (Zhang et al., 2001). Por
fim, existe a necessidade de confirmar a possível associação entre a variante CYP3A4*1B e a variante
CYP3A4*3 numa população em que seja observada uma maior incidência da primeira mutação
(CYP3A4*1B).

REFERÊNCIAS BIBLIOGRÁFICAS
48
7 – Referências Bibliográficas
♦ Ando Y, Tateishi T, Sekido Y, Yamamoto T, Satoh T, Hasegawa Y, Kobayashi S, Katsumata Y,
Shimokata K, Saito H. Re: Modification od clinical presentation of prostate tumors by a novel
genetic variant in CYP3A4. J Natl Cancer Inst 1999; 91: 1587 – 1588.
♦ Anzenbacher P, Anzenbacherová E. Cytochromes P450 and metabolism of xenobiotics. Cell. Mol.
Life Sci. 2001; 58: 737 – 747.
♦ Ball SE, Scatina J, Kao J, Ferron GM, Fruncillo R, Mayer P, Weinryb I, Guida M, Hopkins PJ,
Warner N, Hall J. Population distribution and effects on drug metabolism of a genetic variant in
the 5’ promotor region of CYP3A4. Clin Pharmacol Ther 1999; 66: 288 – 294.
♦ Bertilsson G, Heidrich J, Svensson K, Åsman M, Jendeberg L, Sydow-Bäckman M, Ohlsson R,
Postlind H, Blomquist P, Berkenstam A. Identification of a human nuclear receptor defines a new
signaling pathway for CYP3A induction. Proc Natl Acad Sci USA 1998; 95: 12208 – 12213.
♦ Bertilsson G, Berkenstam A, Blomquist P. Functionally conserved xenobiotic responsive enhancer
in cytochrome P450 3A7. Biochem Biophys Res Commun 2001; 280: 139 – 144.
♦ Blumberg B, Sabbagh W, Jr., Juguilon H, Bolado J, Jr., van Meter
CM, Ong ES & Evans RM. SXR, a novel steroid and xenobiotic-sensing
nuclear receptor. Genes 1998; 12: 3195 – 3205.
♦ Celander M. Impact of stress on animal toxicology in Stress Physiology in Animals (ed. Balm
PH). USA: CRC Press, 1999.
♦ Di Giulio RT, Benson WH, Sanders BM, Van Veld PA. Biochemical Mechanisms: Metabolism,
Adaptation, and Toxicity, in Fundamentals of Aquatic Toxicology – Effects, Environmental Fate,
and Risk Assessment (ed. G. Pand). London: Taylor & Francis, 1995.
♦ Domanski TL, Finta C, Halpert JR, Zaphiropoulos PG. cDNA Cloning and Initial Characterization
of CYP3A43, a Novel Human Cytochrome P450. Mol Pharmacol 2001; 59: 386 – 392.

REFERÊNCIAS BIBLIOGRÁFICAS
49
♦ Ducharme J, Farinotti R. Clinical pharmacokinetics and metabolism of cloroquine - Focus on
recent advancements. Clin Pharmacokinet. 1996; 31:257-274.
♦ Eichelbaum M, Burk O. CYP3A genetics in drug metabolism. Nature Medicine 2001; 3: 285 –
287.
♦ Eiselt R, Domanski T, Zibat A, Mueller R, Presecan-Siedel E, Huestert E, Zanger U, Brockmoller
J, Klenk H, Meyer U, Khan K, He Y, Halpert J, Wojnowski L. Identification and functional
characterization of eight CYP3A4 protein variants. Pharmacogenetics 2001; 11: 447 – 458.
♦ Felix CA, Walker AH, Lange BJ, Williams TM, Winick NJ, Cheung NV, Lovett BD, Nowell PC,
Blair IA, Rebbeck TR. Association of CYP3A4 genotype with treatment-related leukemia. Proc
Natl Acad Sci USA 1998; 95: 13176 – 13181.
♦ Finta C, Zaphiropoulos PG. The human cytochrome P450 3A locus. Gene evolution by capture of
downstream exons. Gene 2000; 260: 13 – 23.
♦ Fowler J, Cohen L. Practical Statistics for Field Bilogy. England: John Wiley & Sons, 1992.
♦ Gardner MJ, Altman DG (eds). Statistics with confidence , London: British Med Journal, 1989.
♦ Gelfand DH, White TJ. Thermostable DNA Polymerases in PCR protocols: a guide to methods
and applications. London: Academic Press, 1990.
♦ Gellner K, Eiselt R, Hustert E, Arnold H, Koch I, Haberl M, Deglmann CJ, Burk O, Buntefuss D,
Escher S, Bishop C, Koebe H, Brinkmann U, Klenk H, Kleine K, Meyer UA, Wojnowski L.
Genomic organization of the human CYP3A locus: identification of a new, inducible CYP3A gene.
Pharmacogenetics 2001; 11: 111 – 121.
♦ Gervot L, Carriere V, Costet P, Cugnenc PH, Berger A, Beaune P, de Waziers I. CYP3A5 is the
major cytochrome P450 3A expressed in human colon and colonic cell lines. Environ Tox Pharm
1996; 2: 381 – 388.
♦ Gonzalez FJ, Schmid BJ, Umeno M, Mcbride OW, Hardwick JP, Meyer UA, Gelboin HV, Idle
JR. Human P450PCN1: sequence, chromosome localization, and direct evidence through cDNA
expression that P450PCN1 is nifedipine oxidase. DNA 1988; 7: 79 – 86.

REFERÊNCIAS BIBLIOGRÁFICAS
50
♦ Gonzalez FJ. The Molecular Biology of Cytochrome P450s. Pharmacological Reviews 1989; 40:
243 – 288.
♦ Gonzalez FJ, Nebert DW. Evolution of the P450 gene superfamily: animal-plant ‘warfare’,
molecular drive and human genetic differences in drug oxidation. TIG 1990; 6: 182 – 186.
♦ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, William M. Gelbart. An Introduction to
Genetic Analysis. 6th Edition. USA: W.H. Freeman, 1996.
♦ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, William M. Gelbart. An Introduction to
Genetic Analysis. 7th Edition. USA: W.H. Freeman, 2000.
♦ Guengerich FP. Cytochrome P450: advances and prospects. FASEB J 1992; 6: 667 – 668.
♦ Guengerich FP. CYTOCHROME P-450 3A4: Regulation and Role in Drug Metabolism. Annu.
Rev. Pharmacol. Toxicol. 1999; 39: 1 – 17.
♦ Hashimoto H, Toide K, Kitamura R, Fujita M, Tagawa S, Itoh S, Kamataki T. Gene struture of
CYP3A4, an adult-specific form of cytochrome P450 in human livers, and its transcriptional
control. Eur. J. Biochem. 1993; 218: 585 – 595.
♦ Hsieh K, Lin Y, Cheng C, Lai M, Lin M, Siest J, Huang J. Novel mutations of CYP3A4 in
Chinese. Drug Metab Dispos 2001; 29: 268 – 273.
♦ Ingelman-Sundberg M, Oscarson M, McLellan RA. Polymorphic human cytochrome P450
enzymes: an oppurtunity for individualized drug treatment. TiPS 1999; 20: 342 – 349.
♦ Innis MA, Gelfand DH, Sninsky JJ, White TJ. PCR protocols: A guide to methods and
applications. London: Academic Press, 1990.
♦ Innis MA, Gelfand DH. Optimization of PCRs in PCR protocols: A guide to methods and
applications (Innis, Gelfand, Sninsky e White, eds). London: Academic Press, 1990.
♦ Josephy PD. Molecular Toxicology. New York: Oxford University Press, 1997.
♦ Jounaïdi Y, Hyrailles V, Gervot L, Maurel P. Detection of a CYP3A5 Allelic Variant: a Candidate
for the Polymorphic Expression of the Protein? Bioch. Biophy. Res. Comm. 1996; 221: 466 – 470.

REFERÊNCIAS BIBLIOGRÁFICAS
51
♦ Jover R, Bort R, Gómez-Lechón MJ, Castell JV. Cytochrome P450 Regulation by Hepatocyte
Nuclear Factor 4 in Human Hepatocytes: A Study Using Adenovirus-Mediated Antisense
Targeting. Hepatology 2001; 33: 668 – 675.
♦ Kivistö KT, Griese E-U, Fritz P, Linder A, Hakkola J, Raunio H,
Beaune P, Kroemer HK. Expression of cytochrome P4503A enzymes in
human lung: a combined RT-PCR and immuno-histochemical analysis of
normal tissue and lung tumors. Naunyn-Schmiedeberg Arch Pharmacol
1996; 353: 207 – 212.
♦ Kivistö KT, Bookjans G, Fromm MF, Griese EU, Münzel P, Kroemer HK.
Expression of CYP3A4, CYP3A5 and CYP3A7 in human duodenal tissue.
Br J Clin Pharm 1996a; 42: 387 – 389.
♦ Klaassen CD, Watkins JB. Biotransformation of xenobiotics in Casarett & Doull’s Toxicology –
The basic science of poisons. USA: McGraw-Hill Companion Handbook, 1999.
♦ Kohút A, Kalina I. Genetic polymorphism of enzymes and changes in the effect of drugs.
Slovakofarma Revue VIII 1998; 4: 141 – 146.
♦ Kolars JC, Schmiedlin-Ren P, Schuetz JD, Fang C, Watkins PB. Identification of rifampin-
inducible P450IIIA4 (CYP3A4) in human small bowel enterocytes. J Clin Invest 1992; 90: 1871 –
1878.
♦ Kuehl P, Zhang J, Lin Y, Lamba J, Assem M, Schuetz J, Watkins PB, Dally A, Wrighton SA, Hall
SD, Maurel P, Brimer C, Yasuda K, Venkataramanan R, Strom S, Thummel K, Boguski M,
Schuetz E. Sequence diversity in CYP3A promoters and characterization of the genetic basis of
polymorphic CYP3A5 expression. Nature Genetics 2001; 27: 383 – 391.
♦ Lacroix D, Sonnier M, Moncion A, Cheron G, Cresteil T. Expression of CYP3A in the human
liver – Evidence that the shift between CYP3A7 and CYP3A4 occurs immediately after birth. Eur.
J. Biochem 1997; 247: 625 – 634.
♦ Lechevrel M, Casson AG, Wolf CR, Hardie LJ, Flinterman MB,
Montesano R, Wild CP. Characterization of cytochrome P450

REFERÊNCIAS BIBLIOGRÁFICAS
52
expression in human oesophageal mucosa. Carcinogenesis 1999; 20:
243 – 248.
♦ Leeder JS. Developmental Aspects of Drug Metabolism in Children. Drug Information Journal
1996; 30: 1135 – 1143.
♦ Lehmann JM, McKee DD, Watson MA, Willson TM, Moore JT, Kliewer SA. The human orphan
nuclear receptor PXR is activated by compounds that regulate CYP3A4 gene expression and cause
drug interactions. J Clin Invest 1998; 102: 1016 – 1023.
♦ Lewis DF, Sheridan G. Cytochrome P450, Oxygen and Evolution. The Scientific World 2001; 1:
151 – 167.
♦ Liska DJ. The Detoxification Enzyme Systems. Altern Med Rev 1998; 3: 187 – 198.
♦ McKinnon RA. Cytochrome P450 – 1. Multiplicity and Function. Aust J Hosp Pharm 2000; 30: 54
– 56.
♦ McKinnon RA, Evans AM. Cytochrome P450 – 3. Clinically Significant Drug Interactions. Aust J
Hosp Pharm 2000; 30: 146 – 149.
♦ Meyer UA. Overview of enzymes of drug metabolism. J Pharmacokinet Biopharm 1996; 24: 449-
459
♦ Michalets EL. Update: Clinically significant cytochrome P450 drug interactions. Pharmacotherapy
1998; 18:84 – 112.
♦ Moore JT, Kliewer SA. Use of the nuclear receptor PXR to predict drug interations. Toxicology
2000; 153: 1 – 10.
♦ Müller M. Transcriptional Control of Hepatocanalicular Transporter Gene Expression. Seminars in
Liver Disease 2000; 20: 323 – 337.
♦ Mullis KB, Faloona FA. Specific Synthesis of DNA in Vitro via a polymerase-catalyzed chain
reaction. Methods Enzymol. 1987; 155: 335 – 350.
♦ Murray GI, Pritchard S, Melvin WT, Burke MD. Cytochrome P450
CYP3A5 in the human anterior pituitary gland. FEBS Lett 1995; 364:
79 – 82.

REFERÊNCIAS BIBLIOGRÁFICAS
53
♦ Nebert DW, Gonzalez FJ. P450 Genes: Structure, Evolution, and Regulation. Ann. Rev. Biochem.
1987: 56: 945 – 993.
♦ Nebert DW, Adesnik M, Coon MJ, Estabrook RW, Gonzalez FJ, Guengerich FP, Gunsalus IC,
Johnson EF, Kemper B, Levin W, Phillips IR, Sato R, Waterman MR. The P450 Gene
Superfamily: Recommended Nomenclature. DNA 1987; 6: 1 – 11.
♦ Nebert DW, Nelson DR, Adesnik M, Coon MJ, Estabrook RW, Gonzalez FJ, Guengerich FP,
Gunsalus IC, Johnson EF, Kemper B, Levin W, Phillips IR, Sato R, Waterman MR. The P450
Superfamily: Updated Listing of All Genes and Recommended Nomenclature for the
Chromosomal Loci. DNA 1989; 8: 1 – 13.
♦ Nebert DW, Nelson DR, Coon MJ, Estabrook RW, Feyereisen R, Fuji-Kuriyama Y, Gonzalez FJ,
Guengerich FP, Gunsalus IC, Johnson EF, Loper JC, Sato R, Waterman MR, Waxman DJ. The
P450 Superfamily: Updated on new sequences, gene mapping, and recommended nomenclature.
DNA Cell Biol.1991; 10: 1 – 14. Corrigendum DNA Cell Biol. 1991; 10: 397 – 398.
♦ Nebert DW. Proposed Role of Drug-Metabolizing Enzymes: Regulation of Steady State Levels of
the Ligands that Effect Growth, Homeostasis, Differentiation, and Neuroendocrine Functions.
Molecular Endocrinology 1991; 5: 1203 – 1214.
♦ Nelson DR, Kamataki T, Waxman DJ, Guengerich FP, Estabrook RW, Feyereisen R, Gonzalez FJ,
Coon MJ, Gunsalus IC, Gotoh O, Okuda K, Nebert DW. P450 superfamily: update on new
sequences, gene mapping, accession numbers early trivial names of enzymes, and nomenclature.
DNA Cell Biol 1993; 12: 1 – 51.
♦ Nelson DR, Koymans L, Kamataki T, Stegeman JJ, Feyerelsen R, Waxman DJ, Waterman MR,
Gotoh O, Coon MJ, Estabrook RW, Gunsalus IC, Nebert DW. P450 superfamily: update on new
sequences, gene mapping, accession numbers and nomenclature. Pharmacogenetics 1996; 6: 1 –
42.
♦ Omura T, Sato R. The carbon monoxide-binding pigment of liver microsomes. J. Biol. Chem.
1964; 239: 2370 – 2378 e 2379 – 2385.

REFERÊNCIAS BIBLIOGRÁFICAS
54
♦ Ourlin JC, Jounaïdi Y, Maurel P, Vilarem M. Role of the liver-enriched transcription factors
C/EBPα and DBP in the expression of human CYP3A4 and CYP3A7. J Hepatology 1997; 26: 54
– 62.
♦ Özdemir V, Kalow W, Tang B, Paterson AD, Walker SE, Endrenyi L, Kashuba AD. Evaluation of
the genetic component of variability in CYP3A4 activity: a repeated drug administration method.
Pharmacogenetics 2000; 10: 373 – 388.
♦ Paris PL, Kupelian PA, Hall JM, Williams TL, Levin H, Klein EA, Casey G, Witte JS.
Association between a CYP3A4 genetic variant and clinical presentation in African-American
prostate cancer patients. Cancer Epidemiol Biomarkers Prev 1999; 8: 901 – 905.
♦ Pascussi J, Jounaidi Y, Drocourt L, Domergue J, Balabaud C, Maurel P, Vilarem M. Evidence for
the Presence of a Functional Pregnane X Receptor Response Element in the CYP3A7 Promotor
Gene. Biochem. Biophys. Res. Commun. 1999; 260: 377 – 381.
♦ Paulussen A, Lavrijsen K, Bohets H, Hendrickx J, Verhasselt P, Luyten W, Konings F, Armstrong
M. Two linked mutations in transcriptional regulatory elements of the CYP3A5 gene constitute the
major genetic determinant of polymorphic activity in humans. Pharmacogenetics 2000; 10: 415 –
424.
♦ Piscitelli SC, Gallicano KD. Interactions Among Drugs For HIV And Opportunistic Infections. N
Engl J Med 2001; 344: 984 – 996.
♦ Poulos TL, Finzel BC, Howard AJ. High-resolution crystal structure of cytochrome 450cam. J. Mol.
Biol 1987; 195: 687 - 700.
♦ Raggers RJ, Pomorski T, Holthuis JC, Kälin N, van Meer G. Lipid Traffic: The ABC of
Transbilayer Movement. Traffic 2000; 1: 226 – 234.
♦ Ravichandran KG, Boddupalli SS, Hasemann CA, Peterson JA, Deisenhofer J. Crystal structure of
hemoprotein domain of P450BM-3 , a prototype for microsomal P450s. Science 1993; 261: 731 -
736.

REFERÊNCIAS BIBLIOGRÁFICAS
55
♦ Rebbeck TR, Jaffe JM, Walker AH, Wein AJ, Malkowicz SB. Modification of clinical
presentation of prostate tumors by a novel genetic variant in CYP3A4. J Natl Cancer Inst 1998;
90: 1225 – 1229.
♦ Roberts GC. The power of evolution: accessing the synthetic potential of P450s. Chemistry &
Biology 1999; 6: R269 – R272.
♦ Saiki RK. Amplification of genomic DNA in PCR protocols: a guide to methods and applications.
London: Academic Press, 1990.
♦ Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning – A laboratory manual. 2nd Edition. Cold
Spring Harbor Laboratory Press, 1989, New York, USA.
♦ Sata F, Sapone A, Elizondo G, Stocker P, Miller VP, Zheng W, Raunio H, Crespi CL, Gonzalez
FJ. CYP3A4 allelic variants with amino acid substitutions in exons 7 and 12: evidence for an
allelic variant with altered catalytic activity. Clin Pharmacol Ther 2000; 67: 48 – 56.
♦ Schuetz EG, Schuetz JD, Grogan WM, Naray-Fejes-Toth A, Fejes-Toth
G, Raucy J, Guzelian P, Gionela K, Watlington CO. Expression of
cytochrome P450 3A in amphibian, rat, and human kidney. Arch
Biochem Biophys 1992; 294: 206 – 214.
♦ Schuetz JD, Schuetz EG, Thottassery JV, Guzelian PS, Strom S, Sun D. Identification of a novel
dexamethasone responsive enhancer in the human CYP3A5 gene and its activation in human and
rat liver cells. Mol Pharmacol 1996; 49: 63 – 72.
♦ Shimada T, Yamazaki H, Mimura M, Inui Y, Guengerich FP. Interindividual variations in human
liver cytochrome P-450 enzymes involved in the oxidation of drugs, carcinogens and toxic
chemicals: studies with liver microsomes of 30 Japanese and 30 Caucasians. J Pharmacol Exp
Ther 1994; 270: 414 – 423.
♦ Stavrovskaya AA. Cellular Mechanisms of Multidrug Resistance of Tumor Cells. Biochemistry
(Moscow) 2000; 65: 95 – 106.
♦ Stegeman JS, Hahn ME. Biochemistry and Molecular Biology of Monooxygenases: Current
Perspectives on Forms, Functions, and Regulation of Cytochrome P450 in Aquatic Species in

REFERÊNCIAS BIBLIOGRÁFICAS
56
Aquatic Toxicology – Molecular, Biochemical and Cellular Perspectives (ed. Malins DC e
Ostrander GK). Lewis Publishers; 1994.
♦ Tayeb MT, Clark C, Ameyaw M, Haites NE, Evans DA, Tariq M, Mobarek A, Ofori-Adjei D,
McLeod HL. CYP3A4 promotor variants in Saudi, Ghanaian and Scottish Caucasian populations.
Pharmacogenetics 2000; 10: 753 – 756.
♦ Thummel KE, Wilkinson GR. In Vitro and In Vivo Drug Interations Involving Human CYP3A.
Annu. Rev. Pharmacol. Toxicol. 1998; 38: 389 – 430.
♦ Van Bambeke F, Balzi E, Tulkens PM. Antibiotic Efflux Pumps. Biochemical Pharmacology
2000; 60: 457 – 470.
♦ van Schaik RH, Wildt SN, van Iperen NM, Uitterlinden AG, van den Anker JN, Lindemans J.
CYP3A4-V Polymorphism detection by PCR-Restriction Fragment Length Polymorphism
analysis and its allelic frequency among 199 dutch Caucasians. Clin Chem 2000; 46: 1834 – 1836.
♦ van Schaik RH, Wildt SN, Brosens R, van Fessem M, van der Anker JN, Lindemans J. The
CYP3A4*3 allele: Is it really rare? Clin Chem 2001; 47: 1104 - 1106.
♦ Walker AH, Jaffe JM, Gunasegaram S, Cummings SA, Huang CS, Chern HD, Olopade OI, Weber
BL, Rebbeck TR. Characterization of an allelic variant in the nifedipine-specific element of
CYP3A4: ethnic distribution and implications for prostate cancer risk. Hum Mutat 1998; 12: 289.
♦ Walker JM. Electrophoretic techiques in Principles and Techniques of Practical Biochemistry
(ed.Wilson K, Walker J). Cambridge: Cambridge University Press, 2000.
♦ Wandel C, Witte JS, Hall JM, Stein CM, Wood AJ, Wilkinson GR. CYP3A activity in African
American and European American men: population differences and functional effect of the
CYP3A4*1B 5’-promotor region polymorphism. Clin Pharmacol Ther 2000; 68: 82 – 91.
♦ Waxman DJ. P450 gene induction by structurally diverse
xenochemicals: central role of nuclear receptors CAR, PXR, and
PPAR. Arch Biochem Biophys 1999; 369: 11 – 23.

REFERÊNCIAS BIBLIOGRÁFICAS
57
♦ de Waziers I, Cugnenc PH, Yang CS, Leroux JP, Beaune PH. Cytochrome P 450 isoenzymes,
epoxide hydrolase and glutathione transferases in rat and human hepatic and extrahepatic tissues. J
Pharmacol Exp Ther 1990; 253: 387 – 394.
♦ Westlind A, Lofberg L, Tindberg N, Andersson TB, Ingelman-Sundberg M. Interindividual
differences in hepatic expression of CYP3A4: relationship to genetic polymorphism in the 5’-
upstream regulatory region. Biochem Biophys Res Commun 1999; 259: 201 – 205.
♦ Williams PA, Cosme J, Sridhar V, Johnson EF, McRee DE. Mammalian microsomal cytochrome
P450 monooxygenase: structural adaptations for membrana binding and functional diversity. Mol.
Cell 2000; 5: 121 – 131.
♦ White TJ, Arnheim N, Erlich HA. The polymerase chain reaction. TIG 1989; 5: 185 – 189.
♦ Wrighton SA, Ring BJ, Watkins PB, VandenBranden M. Identification
of a polymorphically expressed member of the human cytochrome P-
450III family. Mol Pharmacol 1989; 86: 97 – 105.
♦ Wrighton SA, Schuetz EG, Thummel KE, Shen DD, Korzerwa KR, Watkins PB. The Human
CYP3A4 Subfamily: Practical Considerations. Drug Metab. Rev. 2000; 32: 339 – 361.
♦ Xie W, Barwick JL, Downes M, Blumberg B, Simon CM, Nelson MC, Neuschwander-Tetri BA,
Brunt EM, Guzelian PS, Evans RM. Humanized xenobiotic response in mice expressing nuclear
receptor SXR. Nature 2000; 406: 435 – 439.
♦ Yang HL, Lee QP, Rettie AE, Juchau MR. Functional Cytochrome P4503A Isoforms in human
Embryonic Tissues: Expression During Organogenesis. Mol. Pharmacol. 1994; 46: 922 – 928.
♦ Zhang J, Kuehl P, Green ED, Touchman JW, Watkins PB, Daly A, Hall SD, Maurel P, Relling M,
Brimer C, Yasuda K, Wrighton SA, Hancock M, Kim RB, Strom S, Thummel K, Russell CG,
Hudson JR, Schuetz EG, Boguski MS. The human pregnane X receptor: genomic structure and
identification and functional characterization of natural allelic variants. Pharmacogenetics 2001;
11: 555 – 572.

REFERÊNCIAS BIBLIOGRÁFICAS
58
♦ Zhang Q, Dunbar D, Ostrowska A, Zeisloft S, Yang J, Kaminsky LS. Characterization of human
small intestinal cytochromes P450. Drug Metab Dispos 1999; 27: 804 – 809.


Anexo I Sequência do promotor do gene CYP3A4 gfedc 1: D11131. Human cytochrome ...[gi:219569]
LOCUS HUMCYP3A4 1345 bp DNA PRI 14-APR-2000 DEFINITION Human cytochrome P-450IIIA4 gene, partial sequence. ACCESSION D11131 VERSION D11131.1 GI:219569 KEYWORDS P-450 IIIA4; cytochrome P-450. SOURCE Homo sapiens (library: Charon 4A library of T. Maniatis.) adult liver DNA, clone p32B. ORGANISM Homo sapiens Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi; Mammalia; Eutheria; Primates; Catarrhini; Hominidae; Homo. REFERENCE 1 (bases 1 to 1345) AUTHORS Hashimoto,H., Toide,K., Kitamura,R., Fujita,M., Tagawa,S., Itoh,S. and Kamataki,T. TITLE Gene structure of CYP3A4, an adult-specific form of cytochrome P450 in human livers, and its transcriptional control JOURNAL Eur. J. Biochem. 218 (2), 585-595 (1993) MEDLINE 94094853 COMMENT Submitted (13-May-1992) to DDBJ by: Hisashi Hashimoto Department of Division of Drug Metabolism Faculty of Pharmaceutical Sciences of Hokkaido University N12 W6, Kitaku Sapporo, Hokkaido 060 Japan Phone: 011-709-6390 Fax: 011-709-6390. FEATURES Location/Qualifiers source 1..1345 /organism="Homo sapiens" /db_xref="taxon:9606" /tissue_type="Liver" /clone_lib="Charon 4A library of T. Maniatis." /dev_stage="Adult" misc_feature 810..819 /note="'P-450NF specific element (NFSE)'" CAAT_signal 843..848 enhancer 1054..1069 /note="'basic transcription element (BTE)'" TATA_signal 1076..1081 exon 1104..1278 /note="'minor'" /number=1 /evidence=experimental exon 1105..1278 /note="'major'" /number=1 /evidence=experimental exon 1106..1278 /note="'minor'" /number=1 /evidence=experimental intron 1279..>1345 /number=1 BASE COUNT 353 a 314 c 356 g 322 t

ORIGIN Chromosome 7. 1 ctgcagtgac cactgcccca tcattgctgg ctgaggtggt tggggtccat ctggctatct 61 gggcagctgt tctcttctct cctttctctc ctgtttccag acatgcagta tttccagaga 121 gaaggggcca ctctttggca aagaacctgt ctaacttgct atctatggca ggacctttga 181 agggttcaca ggaagcagca caaattgata ctattccacc aagccatcag ctccatctca 241 tccatgccct gtctctcctt taggggtccc cttgccaaca gaatcacaga ggaccagcct 301 gaaagtgcag agacagcagc tgaggcacag ccaagagctc tggctgtatt aatgacctaa 361 gaagtcacca gaaagtcaga aggatgcata gcagaggccc agcaatctca gctaagtcaa 421 ctccaccagc ctttctagtt gcccactgtg tgtacagcac cctggtaggg accagagcca 481 tgacagggaa taagactaga ctatgccctt gaggagctca cctctgttca gggaaacagg 541 cgtggaaaca caatggtggt aaagaggaaa gaggacaata ggattgcatg aaggggatgg 601 aaagtgccca ggggaggaaa tggttacatc tgtgtgagga gtttggtgag gaaagactct 661 aagagaaggc tctgtctgtc tgggtttgga aggatgtgta ggagtcttct agggggcaca 721 ggcacactcc aggcataggt aaagatctgt aggtgtggct tgttgggatg aatttcaagt 781 attttggaat gaggacagcc atagagacaa gggcaagaga gaggcgattt aatagatttt 841 atgccaatgg ctccacttga gtttctgata agaacccaga acccttggac tccccagtaa 901 cattgattga gttgtttatg atacctcata gaatatgaac tcaaaggagg tcagtgagtg 961 gtgtgtgtgt gattctttgc caacttccaa ggtggagaag cctcttccaa ctgcaggcag 1021 agcacaggtg gccctgctac tggctgcagc tccagccctg cctccttctc tagcatataa 1081 acaatccaac agcctcactg aatcactgct gtgcagggca ggaaagctcc atgcacatag 1141 cccagcaaag agcaacacag agctgaaagg aagactcaga ggagagagat aagtaaggaa 1201 agtagtgatg gctctcatcc cagacttggc catggaaacc tggcttctcc tggctgtcag 1261 cctggtgctc ctctatctgt gagtaactgt tcaggctcct cttctctgtt tcttggactt 1321 ggggtcgtaa tcaggcctct ctttt // Primers 1Bfw e 1Brv Primers 1BCfw e 1BCrv Sequência do gene CYP3A4

gfedc 1: AF209389. Homo sapiens cyto...[gi:6644371] LOCUS AF209389 26502 bp DNA PRI 26-JUL-2001 DEFINITION Homo sapiens cytochrome P450 IIIA4 (CYP3A4) gene, exons 1 through 13 and complete cds. ACCESSION AF209389 VERSION AF209389.1 GI:6644371 KEYWORDS . SOURCE human. ORGANISM Homo sapiens Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi; Mammalia; Eutheria; Primates; Catarrhini; Hominidae; Homo. REFERENCE 1 (bases 1 to 26502) AUTHORS Hsieh,K.P., Lin,Y.Y., Cheng,C.L., Lai,M.L., Lin,M.S., Siest,J.P. and Huang,J.D. TITLE Novel mutations of CYP3A4 in Chinese JOURNAL Drug Metab. Dispos. 29 (3), 268-273 (2001) MEDLINE 21098888 PUBMED 11181494 REFERENCE 2 (bases 1 to 26502) AUTHORS Hsieh,K.P., Cheng,C.L., Lai,M.D. and Huang,J.D. TITLE Direct Submission JOURNAL Submitted (30-NOV-1999) Pharmacology, College of Medicine, National Cheng Kung University, 1 University Road, Tainan, Taiwan 70101, R.O.C. FEATURES Location/Qualifiers source 1..26502 /organism="Homo sapiens" /db_xref="taxon:9606" /chromosome="7" /map="7q21-q22.1" exon <1..71 /gene="CYP3A4" /number=1 mRNA join(<1..71,4004..4097,6009..6061,11502..11601, 13956..14069,14335..14423,15689..15837,16932..17059, 17744..17810,20166..20326,21912..22138,23198..23360, 25950..>26502) /gene="CYP3A4" /product="cytochrome P450 IIIA4" CDS join(1..71,4004..4097,6009..6061,11502..11601, 13956..14069,14335..14423,15689..15837,16932..17059, 17744..17810,20166..20326,21912..22138,23198..23360, 25950..26045) /gene="CYP3A4" /codon_start=1 /product="cytochrome P450 IIIA4" /protein_id="AAF21034.1" /db_xref="GI:6644372" /translation="MALIPDLAMETWLLLAVSLVLLYLYGTHSHGLFKKLGIPGPTPL PFLGNILSYHKGFCMFDMECHKKYGKVWGFYDGQQPVLAITDPDMIKTVLVKECYSVF TNRRPFGPVGFMKSAISIAEDEEWKRLRSLLSPTFTSGKLKEMVPIIAQYGDVLVRNL RREAETGKPVTLKDVFGAYSMDVITSTSFGVNIDSLNNPQDPFVENTKKLLRFDFLDP FFLSITVFPFLIPILEVLNICVFPREVTNFLRKSVKRMKESRLEDTQKHRVDFLQLMI DSQNSKETESHKALSDLELVAQSIIFIFAGYETTSSVLSFIMYELATHPDVQQKLQEE IDAVLPNKAPPTYDTVLQMEYLDMVVNETLRLFPIAMRLERVCKKDVEINGMFIPKGV VVMIPSYALHRDPKYWTEPEKFLPERFSKKNKDNIDPYIYTPFGSGPRNCIGMRFALM NMKLALIRVLQNFSFKPCKETQIPLKLSLGGLLQPEKPVVLKVESRDGTVSGA" gene <1..>26502 /gene="CYP3A4" exon 4004..4097 /gene="CYP3A4" /number=2

exon 6009..6061 /gene="CYP3A4" /number=3 exon 11502..11601 /gene="CYP3A4" /number=4 exon 13956..14069 /gene="CYP3A4" /number=5 exon 14335..14423 /gene="CYP3A4" /number=6 exon 15689..15837 /gene="CYP3A4" /number=7 exon 16932..17059 /gene="CYP3A4" /number=8 exon 17744..17810 /gene="CYP3A4" /number=9 exon 20166..20326 /gene="CYP3A4" /number=10 exon 21912..22138 /gene="CYP3A4" /number=11 exon 23198..23360 /gene="CYP3A4" /number=12 exon 25950..>26502 /gene="CYP3A4" /number=13 BASE COUNT 7293 a 5226 c 5275 g 8708 t ORIGIN 1 atggctctca tcccagactt ggccatggaa acctggcttc tcctggctgt cagcctggtg 61 ctcctctatc tgtgagtaac tgtccaggct cctcttctct gtttccttgg acttggggcg 121 ctaatcacgc ctctctttcc cttatctgga ttgaagatca aaaaagatgt tcaggccggg 181 cgtggtggct tacacctgta atcccagcac tttgggaggc taaggcaagt ggactgcctg 241 aggtcaagag tccaagacca gcctggctaa catggcgata ctctgtctct actaaaaata 301 caaaaattag ctgggcatgg tggtgcacgc ctgtattccc agttacttgg gaggctgagg 361 caggagaatt gcttgaaccc ggcaggcgga ggttgcagtg agctgagatc atgccagtgc 421 acttcagcct gggtgacaga gtgaggctgt cttaaaaaaa aaaaaaaaaa aaaaaagatg 481 ttcaaggagc agtagcttaa gtgttggatg ctacaaacat atagaggtta ttgtagatct 541 tatgcagctc tataaaggaa taaataagca tcttccccat ccatctttag tggcaagaag 601 ggttttggga tagcattgat tgaggatgat ctacttgaca atagtttgga cccaaggagg 661 ataaggaagg aaagtagtga cggatctcat tccaaacttg gctgtggaaa cctggcttct 721 ccttactaaa ctagaatttg gattttacat tttccccttt atgttgcagt agaagaggat 781 gaatcctctc actggtggga tcctgccatc ctagagcagg tagagagaag agtcactccc 841 cactgtgggt agtggaggct tctcacatgt cacatttcac ttctacctca atttcactct 901 tactaagatt tgggaatcat aatgacagga aaatagaaaa tataaacctc attttaattc 961 tttcacagaa aggttagaaa ttcagtgagt tgtggcaaca tattttccat cttctgacct 1021 tttaacacta attgatatgg cttaaattca ttctatttta aaccagattt ttttggagat 1081 agtctatttc caacatgttc cttctaggtg acaaatgagg gctgttagtt cagtatttgt 1141 tacaataaat gtgtgtaaaa taacctcacc tttccagaat catgtcagga atatgaatct 1201 aatgcacaaa tgtataactc tatgacaaga ttgcatatat cttttaaaat ataccttccc 1261 aacgttcatt ttaatacccc tatttcaaac aaacctgctt agcaggttat gttaaacgct 1321 cagggcagag gagtaagcaa gactgtgagc cagtgatgac agcaaaagca tccaggtagg 1381 atcaaaatgg agtaagaaaa tattcctcat ccctcagggt agaactccaa agagatattc 1441 atgggtcctg gccccgtagt ggaggtcact caaaggacaa acatgtttgc atctcatctg 1501 cttgaagcct ggacacagag gcaccatctg tgtcactctg tgtgtggtct gccatgttgt 1561 ggggtggtca ctacagactc acagactagg cagctgggca gacaatacct tagccttaga 1621 tgatgctgat gcagcccagg agtcagaaac tgtagtgcag acaatgccct ccttaggcca 1681 acacaattaa gtgcaataga tgactggctt ttctgttagc ctcttcattg gaaccaaaag 1741 cagcattact ctaccaaaca gaggggagct ggaaagaaac tacacagttt gcccagccta

1801 gcctctgcct tgacacggaa ccatgtgagt ctagacattc acctagatca ttccttgggg 1861 accaatgcta ctgacacatt aactcaatag tttgtcctgg cctgagaggt catgtaactt 1921 gtagaaagtt tagaagcaga gattagtgtc atttatttgc catggctgtg acaacaaagg 1981 aaggaacagg agtgggaaaa cccaaggcca ccctggtttt ggtagatggt gcacacgctt 2041 ccactagctg ttctggggca aagatccaaa tgcactattg ggcctggcta tgctgcttct 2101 gctgggtccc ccaaacatga gcctccacgc catttctcag ttgtatttta ccacatatta 2161 tcacagtcac cggatttgta cagaatattt ggaacctata ctgtcttaag ggctaccctt 2221 taaagaagag aaaacaaggt tttaattcaa ctgtctggaa cattttatgt ttacttatgt 2281 ggaatactac atcttttgtt ataaacagga gggaatgtgg acattcgaag gcccctacct 2341 tttagctaaa agcccatatg aagcatatgg atccatttat acacaccatg cttttcagct 2401 acattttcct aatttgcctc tctggggcca accttgtggg actagcagat tcatggttga 2461 gttgaaggat ggtgacctct tccatcagct tttcttcttc ctccagtctt ccaaccctca 2521 gtaacatcag actgggaagg tcttcagaca tccagaaacc ccagttcggg gagttcatac 2581 atgacccatc aaagatgagt tgcaagcagg cctgccttag ggagcaccag ccttaatggg 2641 ttttcctaca gagatagttg atgggcagat gcaataaact gactgctttg tgattgacca 2701 ccttgagaaa aatagcatgt ctggctatgt tagtcttttc ttgcattgtt ataaagaaat 2761 accagagtct gggtatttat aaagaaaagg gatttaattg tctcatggtt ctgcaggctg 2821 tacagaaagc atagtgactt cttcctctag ggaggtctcg ggaaatttac aatcatggca 2881 gaaggtgaag gagaagcaga cagatcttac atggctggag cagaagcaag agaggctggg 2941 gagaaggcgc cacacattca aaccaccaga tctcataaaa actcattgtc acgaggacaa 3001 cacctaaggc gggatggtgt gaaacgatga gaaactgccc ctaggatcca atcatctcct 3061 accaggcccc acctccagta ttggggattg catttcaaca tgagattttg gtaggggcac 3121 agattcagac catatcactg gcactgtgct aatcagatga atatcaccag ttggaaggct 3181 agattccaca agaggaggaa tgacctggaa attggttctt tagttgtgat tcttctgcac 3241 actgtcattc agggaaatat gagtcaatca tccttcccaa taggtcaaat caaccagatc 3301 atctgatcat agagactgag gtgtagctga aagctgctca catttctatg aggtcaatgg 3361 aagccttgag cacagttgtc aatctgtaga aataaggact ctgtgactcc tccaagacct 3421 ctctgtgaat gacggtttaa gaagaaccag atcctaaaac agggtcagag cttagaggga 3481 agggaaagca taaaagcctc tgagcaaatt ctaaagacag ggtcaccata ggctctcagt 3541 gaccctctgt gactgagtgg ctgcagtgat gcaaaatctc atcatcactg cggaagacaa 3601 aaaaatgtca ccctttctac ctaggatgag aatccccaaa tttggggaga ggccacttac 3661 taaatagacg taaaggaaca aagtgacctg gaagaattcc tgcctgaacc tctcaggatc 3721 attcacattt gagaacattt atcaaatatt cattccagga ctgggaccat gaagacttca 3781 gctgctttga gctaatcatt gtaacttttt ggtgtctcat ggtggaggca ggaaaggacc 3841 tgatgaacaa atataatcat tgccgtcaga gttactgtta ttatttcttg ccttaatgtt 3901 acctcgttct cttgagcatt ccagttcctc agtcaatgac tcatcaaccc ctatatctat 3961 aaagtcacaa tcgctgtgac ttgatttctg tttcactttg tagatatgga acccattcac 4021 atggactttt taagaagctt ggaattccag ggcccacacc tctgcctttt ttgggaaata 4081 ttttgtccta ccataaggtg agtgtttttg agctcactct tttgcttctt atgattgcca 4141 agagcagctt agttccatca gtaaaaatgc ttctcctcag ggggaaggtc tgaagtttta 4201 cactttcaga aacagtgtgt aggcatcacc cagaacatgg caatgtttac ccaagggctc 4261 tcttgctaac tctcaggaac ctcaggtttg cctcagttga acagcccaaa tctcaggtag 4321 atcagcaacc tgatgctcag aacttgatgt gcaaactttg tgagcgcgcc ataaagaagg 4381 ttttcttttt gctctatgca ggttcccagg aaggtacagt catacttagt tagtattaaa 4441 agtaggaaaa ggaccccctg attgtgactt gttatctgtg tacatgagag tcaaactttc 4501 ctctctgtag caagagctct tgagtagcct tttttccctt ccttctggac agctttgaaa 4561 ctcaatttat tagggagccc aggtatttac ctttgggtca tttgcacctg cttcaatcct 4621 ctccgaaaaa tgctaaattc cctgagggta tctccatggc tctaccccag ctcttactcc 4681 ccatgacatc ttttgcagct ttgagagaca gaacaaaggg gttctgcttt atttcttttc 4741 cttcaaaggc tgcaccttat gggtgagact gatcacagac tggatataac tgagatgaga 4801 cagtggtggt caatcaaact cagatattta agcacaaaat gagtttgtgg ggtttttttt 4861 tacacaaagg cggaatcaca tgcaaaatca ctagaaaggg gatgatttga tgaaattatt 4921 aaatatttaa ttttgctgaa tagaatataa tatgtgccac gtggaccgtg aactttggcc 4981 gagcctttgt gcctaaccta gtctgatgct ctgcccaggg tctgggccct ggatggaaaa 5041 ttaggagccc atgtccacat ggccagccgc agcagtcagc tctttggctt acatcatctt 5101 tcccacatac ctgaggcttg tttctcagat tctaattctc tcaggtgagg gctttgttgt 5161 ctaattacta tccaggaatt tcatattttt tccctgtgca aaagcaataa tttccccgcc 5221 accttttcca ggtcaactct ttagtagatg tacccccaag atgcacattc ctgggacctt 5281 tgtttgcaca gttaaaatgt cacccctgaa atgtcgatac aggaaggttt gtttttaagt 5341 ttcagtgaaa actctgagca agtgttgtaa tttgctgtgt ccgatgtgta gaggggacat 5401 tttctcagaa cttccatgtt aagctggaaa actggaaagt gagttcactt tgtcattctg 5461 tcactcgttc attttctcac tcaacaacat gcctcatact tacctaaatc tgctagacta 5521 aaggagttcc ctggtgtctg tactttccaa ttctgctaga actctagagc gagctcatga 5581 aataaatgaa aaggatgaca aagagataaa acactgtgca ttctcttctg atgctaattc 5641 actttccctt ggcctcagtt tccccatgtg cccctggagg tgatcattca gggattcatg

5701 agattttcaa gacaacacat gaaaaagcaa aaagacatca gaaagacaaa gaggtactta 5761 gtatttatac acaaggataa gtcattcagt atccacaaca cttggagaga attcaagagt 5821 gattttaaat ttcccttttc aaatacctcc tctgttttct cttatttcct ttatgacgtc 5881 tccaaataag cttcctctaa ctgccagcaa gtctgatttc attggcttcg actgttttca 5941 tcccaattag aggcagggtt aagtacatta aaaataataa tcaaatatta ttttgtttct 6001 cctcccaggg cttttgtatg tttgacatgg aatgtcataa aaagtatgga aaagtgtggg 6061 ggtgagtatt ctggaaactt ccattggata gacttgtttc tatgatgagt ttaccccact 6121 gcacagagga cagtctcagc ccaaagcctc ttgggatgaa gctcttgtca acctaactac 6181 aaacagagag aagttctctg aaagaagaag atatttattt gggtgtagag tattgcaatg 6241 ggaatctgca tgcctttata aactatgtgc aaattcaggg aagtaaagca agacaaagag 6301 gctccaagga aaatatgagg aggatttctt atcagttttg aaataattat ccttcgctac 6361 aaagatcagt aacaagggtg acgcctcacc aaggttggac aggcagttgc tgggcaggtg 6421 tccttgcaga aatatttttt ttaatgttgg gatggccttt gtgcaagctt gtagttttgc 6481 ggagtctttt gtgatagttt tgttatcagg cacacaagca tgagaatcct ctcttcatag 6541 ccttctttga tttatttgtc agggttttta cacacacaca cacacacaca cacacacaca 6601 caactagtga catcattttg gttctaacaa cattcacact ggttattgta aaacttttcg 6661 aaggttgtcc taccaaggat cccatgtgtc accaggtgtc aagttctaca gtctgaacta 6721 ggctgggagc attgtgatta cttttctcca gactttggtg gcccagggac tcacagcatc 6781 atgctctgtc cagtgtctgc ctattcccct cttctttttt tttttcctta ggtgcccttt 6841 tattacatgt gttgtctcag acccttctaa tatgtgctca taaatacatc atatcatctc 6901 cttcccacat caattcactt tcaattaaaa gccaaaactc tttcatttag actttggatt 6961 taaagtgctt ttgaatgaag ggttgagaga taatagagaa atagattggc aaaccattta 7021 tactctgctg ttgttgtttt ttaattttat ctgcaagtgt ggaacttttc attctgtttt 7081 gttattaaat ttaagccaag actttttaat agaagggtat ataagcattt ctttgtctat 7141 accttcctgc tgaatttgaa gaaatgctga atattcttaa ccactggcgg gctgatggac 7201 tgtgatttta ttttattttt tatttttagt tttttaaatt ataccttaag ttctgggtta 7261 catgcataga atgtgtagtt ttgttacata ggtatacacg tgccatggtg gtttgctgca 7321 cccatcaacc tgtcacctac attaggtatt tctcctaatg ctatctctcc cctagccccc 7381 cacccaacaa caggccccag tgtgtgatgt ttccctcccc gtgtccatgt gttctcattg 7441 ttcaactcct acttaggagt gagaacatgt ggtgttgagt tttctgatct tgtgatagtt 7501 tgctgagaat gatggtttcc agcttcatcc ttgtccctgc aaaggacatg aactcattgt 7561 ttttttatgg ctgcatagta ttccatgggg tatacgtggc acattttctt tatccagtct 7621 gtcactgatg gacatttggg ttggttccaa gtctttggta ttgtgaatag ttctgcaata 7681 aacatatgcg tgcatgtgtc tttatcatag aatgatttat gctttgggta tatgcccagt 7741 aatgggattg ctgggtcaaa tggtatttct agttctagat ccttgaggaa tcaccacact 7801 gtcttccaca atggttgaac taatttacac tcccaccaac agtgtaaaag tgttcctatt 7861 tctccacatc ctctccagca tctgttgttt cctgactttt taatgatcac cataccacct 7921 ggcatgagat ggtatctcat tgtggttttg atttgcattt ctctaatgac cagtgatgat 7981 gagcattttt tcacatgtct gttggctgca tagatgtctt cttttgagaa gtgtctgttc 8041 atatcctttg cctatttttt gatggggttg tttgcttttt ttcttgtaaa tttgtttaag 8101 ttctttgtag attctggatg ttagcccttc gtcagatgga tagattgcaa aaattttctc 8161 ccattctgta ggttgcctgt ttgctctgat gatagtttct tttgctgtgt agaagctctt 8221 tagtttaatc atatcccatt tgtcaatttt ggcttttgtt gccattgctt ttggtgttat 8281 atttatgaag cctttgccca tgcctgtgtc ctgaatggta ttgcccaggt tttcttctag 8341 gatttttatg gtcctaggtc ttacatttaa gtctttaatc catcttgagt taatttttgt 8401 ataaggtgta aggaaggggt ccagtttcaa ttttctgcat atggctaggc agtttcacca 8461 acaccattta ttaaatagga aatcttttcc ccattgcttt tgtgtgtcag gtttgtcaaa 8521 catcagatgg tagtagatgc atggtgttat ttctgaggcc tctgttctgt tccattgatc 8581 tatatttctg ttttggtacc tgtaccatgc tgttttggtt actgtagcct tttagtataa 8641 tttgaagtca ggtagcgtga tgcctccagt tttgttcttt ttgcttagga ttgtcttgtc 8701 tatgtgggct cttttttggt tccatatgaa ctttaaagta gttttttcca attctatgag 8761 gaaagtcagt ggtagcttga tggaaatagc attgaatcta taaattacct tgggcagtat 8821 ggccattttc atgatatgga gtcttcctac ccatgagcat ggaatgttct tccatttgtt 8881 tgtgtcctct tttatttcat tgagcagcgg tttgtagttc tccttgaaga ggagaacttc 8941 acatcccttg taagctggat tcctagatat tttattctct ttgtagtaat tgtgaatggg 9001 agttcactca tgatttggct ctctgtttgt ctattattgg tgtgtaggaa tgcttgtgat 9061 ttttgtaaat tgattttgta tcctgaggct ttcctgaagt tgcttatcag cttaaggagt 9121 tttggggctg agacgatggg gttttctaaa tatacaatca tgtcatctgc aaacagagac 9181 aatttgactt cctcttttcc taattgaata tccatttctt tctcttgcct gattgcccta 9241 ttcagaactt ccgacactat gttgaatagg agtggtgaga gaggacatcc ttgtcttgtg 9301 ccggttttca aagggaatgc ttctagtttt tgcccattca gtatgatatt ggctgtaggt 9361 ttgtcataaa tagctcttac tattttgaga tacgttccat tgatacctag tttattcaga 9421 gtttttagca tgaaaggctg ttgaattttg tcaaaggcct tttctgcatc tattgaggta 9481 attatgtggt ttttgtcatt ggttctgttt atgtgatgga ttacatttat tgatttggta 9541 ttttgaaccc agccttgcat cccagggatg atgctgactt gatcctggtg gataagtttt

9601 ttgatgtttt gctggatttg atttgccagt attttattga ggattttcgc atcgatattc 9661 attagggata ttggactaaa attctctttt tttgttgtgt ctctgtcagg ctttggtatc 9721 aggagatact ggcctcataa aatgagttag ggaggattcc ctccttttct attgttcaga 9781 aaaatttcag aaggaatgat aacagctcct ctttgtatct ctggtagaat tcagctgtga 9841 atccatttgg tcctggactt tttttggttg gtagcctatt aataattgcc tcaattcaaa 9901 acctattatt ggtctattca gagattcaac ttcttcctgg tttagtcttg ggagggggca 9961 tgtgtccagg aatttatcta tttcttctag attttctagt ttatttgcat agaggtgttt 10021 atagtattct ctgatggtaa tttgcatttc tgtgggatca gtggtgatat cctttttatc 10081 attatttatc gcatctattt gattctgctt ttttctttat tagtcttgct agcagtctat 10141 catattttgt tgaccttttt caaaaaacca gctcctggct tctctcattt tttgaagggt 10201 ttgttgtgtc tcaatctcct tctgttctgc tctgatctta gggctcagtc aacgtagcat 10261 agccacagta actcccgcct ctcgatggta aggggaacgt tacagagtca ggaatcgacc 10321 tatcctgcat tctctggggg gcaatatctt agttatttct tgtcttctgc tagcttttga 10381 atttgtttac tcttgcttct ctagttctgt taattgtgat gttactgtgt caattttaga 10441 tctttcctgc tttctctagt gggcatttaa tgctataaat ttccctctac acactgcttt 10501 aaatgtgtcc cagagattct ggtacattgt gcctttgttc tcattggttt catagaacat 10561 ctttatttct gccttcactt ccttatttac ccggtagtca ttcaggagca agttgttcag 10621 tttccatgta gttgtgtgat tttgagtcac tttcttaatc atgagttgta atttgatttc 10681 actgtggtct gagagacagt ttgttgtgat ttcttttctt ttacatttgc tgaggagtat 10741 attacttcca attatgtggt caattttaga ataagtgcga tgtggtgctg agaagaatgt 10801 atattctgtt aatttggggt ggagatttct gtagatgtct attaggtcct cttggtccag 10861 agctgagttc aagttctgaa tatccttgtt aattttctgt cttgttgatc tgtctaatat 10921 tgacagtggg gtgttaaaat ctcccattat tattgtgtgg aaatcttagt ctctttgtag 10981 gtctctaaga acttgcttta tgaatctggg tgctcctgta tttgggtgca tatatattta 11041 ggatatttag ctcttcctgt tgaattgatc ccatcaccat tatgtaatgc ccatctttgt 11101 ctcttttgat ctttgttggt ttaatattct ttttaaaata aaatatttta aacatatgaa 11161 acattatgga gaatggcatg ggaaatatcc acgtatgcac cacccagctt aacaaatgct 11221 ctactgtcat ttctaaccat ggtctctttg aagagctctt ttgtctttca atgtctcttc 11281 cttgtttggc ccacattatc cttcatcata tgaagacttg ggtggctcct gtgtcagact 11341 cttgctgtgt gtcataccta atgaactaga acctaagatt actgtgtatt gtacaactaa 11401 gggattatgt aaagtcagga tcaaagtctg gcttcctggg ttgggctcca gctgtagaat 11461 aaggctgttg atgtttaatc aactctgttt ttttcacaca gcttttatga tggtcaacag 11521 cctgtgctgg ctatcacaga tcctgacatg atcaaaacag tgctagtgaa agaatgttat 11581 tctgtcttca caaaccggag ggtaagcatt catgtgttga aattaaaata ctgattgatt 11641 aaatttatat tttgaaattc ttatatattc atagacagtt gcctaaaaaa tgtccaggaa 11701 ggttccacgt ccacttcatc ctgtcccccc cgaatggtaa catcttgcaa tcttgcataa 11761 ctataaatac agtatattca tgttactata tagtacagga aattgacatt gatacagttc 11821 acagagctta ttcagatttc accagcttta tgtgccctca tttctgttct atgcaaattt 11881 atcacaatca tagatttgtg caatgaccag catgatcaaa acgcagaacc catttgtgtt 11941 ctatacaaat ttatcacatc attagtagat ttgtgcaatg accagcatga tcaagatgca 12001 gatccactct gtctccatgt ggctccctcc tgctatccta cagtcacaag tctttttctc 12061 tgacagggtc catatcagag caaactattt tttattttga ggtgatcaat gtattaatat 12121 ttccttttct ggattgtact tttggtgcca tgtttgaaag ttctttgcct agccctcctt 12181 cctatattgc tgtttttttc taaaagtcat aaagtttaac gctttactaa atgtagctct 12241 ataatcaatt ttgtgttatt ttttgtataa ggtctgagat ttagatcaag gttcattttt 12301 tttgtggctt atggctgtcc aattcctcca acgccatttt tggaaagatg ggtatattgt 12361 taaaaatcaa ctgggcatac ttctttgcat ctctcacgta ctactgtgtc ccattgatca 12421 ctgtgtttat tattccacca ataccacact gtgttgaccc tagtagctgt acactaactc 12481 ttaacctcgt gtagagtgat tcttcccact tttattgact tattttttca gatttgtttt 12541 aactcttcct ggtcctctgc ttttccataa aaattcagaa tgagtttctc agtgtctaca 12601 aataaacgtg ctgttatttt gacaagaatt gaattaaata tatagaccag tttatggaaa 12661 atgtgtatct ttactgtttg atctttcaat tcatgaacat agtatgcctc tccatttcct 12721 tagattttct ttgattgctt ttaacatctt gtagttttca gcatagagat cctgtacatg 12781 ttttgttaga tttacagcta aactatttca tttttgttgg aatgaccgta aatgatatta 12841 tgtttttctt gtattttcag atattgattg ttgttacata gaaatgtgct ttgattttgt 12901 gtgttgatct ggtatcctag aaccttgcag aactgacata gtagttctag aagtctcttt 12961 gtatatgcct tgagatttta cacattgtca gttatgtcac ttgaaaatag agacaattct 13021 tttatttcct ttccaatctg tatgcctttt atttcttttc ttttgtctat tgcactagga 13081 cttcccagta taatgttgag taaatgtggt gaatttttga attgttccta atcttaggag 13141 ggaagccttc agttctttca tcattaagca tcatttagac gaagtgggct gtttttttgt 13201 agattctctt tatcaaattg aggaattttc tctccctagt ttgctgagtt tttatcataa 13261 atgaatgctg gaagtactca ttataaaaaa aatggctatg gaagatgaaa gaaagtttca 13321 agcatgttgg cttgataggc cagttccaag ttggcaaaaa taattatctc ttttttcttt 13381 ctatccatga aataaaaaat taagagccaa gaatgtttat ggaattgcat tatttcttca 13441 aaatatgttc ctagttttaa aggtattacc tactatttct tttaaaacca tcacattgag

13501 gcacttcttt ttcacgttgc ccatgctgca ggagaacata aagacagctt gtctgaggca 13561 acatacaatc caccaaagtc acctgcttgt ctgtctccac tccgtctcta cactgcagaa 13621 gtgctaggtc ttgattctgt ttattgtact ggaagaacac attctctacc acgtggataa 13681 tttgcatgta aaagaagact gggatataga ggctggagac gacatcaggg tctcctgaac 13741 actgccgcca cccccccgcc ccgccccaca caaatacatc ccaggacact gggcatctgg 13801 gataaatctc tattgagcat ctagcatagg gcccatcacc cagtagacag tcactaaata 13861 gttgttgaat aagtgttcct gtttaacaca ttttctacaa ccatggagac ctccacaact 13921 gatgtaggac aaaatgtttc tgctttgaac tctagccttt tggtccagtg ggatttatga 13981 aaagtgccat ctctatagct gaggatgaag aatggaagag attacgatca ttgctgtctc 14041 caaccttcac cagtggaaaa ctcaaggagg tatgaaaata acatgagttt taataagaaa 14101 cttaaagaat gaatctggtg gggacaggta taaaataaga tcacagtccc tttccaaggg 14161 gtagtccact gaatttgagc tgcctaaaaa tggtctttta tctttatgta cagaaaacac 14221 atcacaaaat tcattataaa atgtcactta ctgctccatg ctggggaaag ccatgtcctt 14281 ctgggactag agtctgcaca tttaactatg ggtggtgttg tgttttgtgc ttagatggtc 14341 cctatcattg cccagtatgg agatgtgttg gtgagaaatc tgaggcggga agcagagaca 14401 ggcaagcctg tcaccttgaa agagtaagta gaagcgcagc catggggttc tgagctgtca 14461 tgaacccctc cagctgcctg ccatggagct gatattcctg ctgttgggtt attccagtga 14521 ccagacaaaa ggagggctgt ggtaatgcaa cttcaatggg tctcccaaga tggggcagct 14581 ccgatgagga ggtggggcag ctggaggaaa aggatcttct cccctgtgca caggggccag 14641 ggtttacata tccattaaat tgtcaccttg gatattctag aagactaaat atatccttta 14701 gggggaaaaa gtgtgattgt accaaagttt taagcatgga gtgtatggga tggtggaagg 14761 ggaaggcact tggtatctgt tggttggcag tgagtaggtt gggagagtta taatggagaa 14821 cttagaataa ctttgatcat ttcatgtttt tttctgagga tatcagtaga atactaaata 14881 ttaaaattcc taccatttct ttttcctcca gtctcaaaga gagagggtgg taaaaacact 14941 ataggtaggg caagcctatt atttgctatc tacacttatg cagtaaaaac aggtgtaatc 15001 tgagtttgtc ctgggcagac cagggatatg tggtcactca ctatagaaat ttccaaatca 15061 aattttgaga gatttttttt taaccaggac attattggtc attatatttt acaaaaataa 15121 ttctgctgtc agggcaacct cagctcacca cagctgggga tagtggaatt ttccaaagct 15181 tgagcaggga gtatagagaa taaggatgat atttctagga gctcagaaca gggtactgtt 15241 gctttgtaaa gtgctgaaga ggaatcggct ctgggcatag agtctgcagt caggcaatat 15301 cacctgtctt gagcccctta ggaagagtta attattctac tcttgttctg ctgaagcaca 15361 gtgcttaccc atcttgtatc atccacaatc aatacatgct actgtagttg tctgatagtg 15421 ggtctctgtc ttcctatgat gggctccttg atctcagagg taggtctaat tcagttcagt 15481 gtctccatca cacccagcgt agggccagct gcatcactgg cacctgataa caccttctga 15541 tggagtgtga tagaaggtga tctagtagat ctgaaagtct gtggctgttt gtctgtcttg 15601 actggacatg tgggtttcct gttgcatgca tagaggaagg atggtaaaaa ggtgctgatt 15661 ttaattttcc acatctttct ccactcagcg tctttggggc ctacagcatg gatgtgatca 15721 ctagcacatc atttggagtg aacatcgact ctctcaacaa tccacaagac ccctttgtgg 15781 aaaacaccaa gaagctttta agatttgatt ttttggatcc attctttctc tcaataagta 15841 tgtggactac tatttccttt aatttatctt tctctcttaa aaataactgc tttattgaga 15901 tataaatcac catgtaattc atccacttaa aatatacagt tcagtgattt gtagtacatt 15961 tgaagatatg tgtgaccatc atcattttaa actttaaaac tttttttgtc aatctagaga 16021 cctcatacat ttttagctat cagccccctg tcacaaaccc tgtcatcata tgcaaccact 16081 aatcaacttt ctgcttctat ggatttgcct attctggaca cttcatagaa atgatattaa 16141 ttcatcaggg ttttttattc tctagttcat gaatttgtac tttagtctgt atcattttct 16201 ttcttctgct ggcttcaggc ttagtttgcc cttcttcgtt tactatgttg tggcatgaac 16261 atagattact gatttgtgat ttttttgttc ctctaaattt agacattaca gctgtaactt 16321 tccctctgag cacttccttt gctaaatccc atgagattgt ggcctatcac atcttagttt 16381 tgttcacctc aaaacagttt ctatttgccc tttgggtttc tactttgact cattgggtac 16441 ttaaatgttt attatttaac ttccacatat gtgtgagttt ctcaattttc tttcccttat 16501 tgattttatc tttattccat gataggtgac agagatatgc tgtgttattt ctatcttgac 16561 tacctactat ttcttgaaca gcaagattaa ttttgagctt cagattatga tttgggttat 16621 tctaggagac tgtagtccaa tagataaagg caaagagatt agggcattga attttgttcc 16681 ttttatcctt caaaagatgc acaaggggct gctgatctca ctgctgtagc ggtgctcctt 16741 atgcatagac ctgcccttgc tcagccactg gcctgaaaga ggggcaaaag tcatagaagg 16801 aatggcttcc agttgagaac cttgatgtct tttactcttc tggttggtag agaaaactag 16861 aattgctcca ggtaaatttt gcacattcac aatgaatttc tttttctgtt tttgttttgt 16921 ttttcctaca gcagtctttc cattcctcat cccaattctt gaagtattaa atatctgtgt 16981 gtttccaaga gaagttacaa attttttaag aaaatctgta aaaaggatga aagaaagtcg 17041 cctcgaagat acacaaaagg taaaatgtgg tggtagttat aggaggatgt ttagtttttc 17101 ataatttttt agataatata catatgatca gtgcagttac ctgtatgttt ttaaagaatg 17161 cttttaacat gaagactgct catgtttaga gcaagagaat tcatttggta gaaatgcaga 17221 aagtggggtt tggggaagga gatatgagaa tgagtcagag acagcacatg aaatttgata 17281 tcagacacaa caattagtat gccacggcat aaattttatt gagataaaac tttaccactt 17341 tacttccctt caataaattg tcagaggata aacattactg tttggaaata tattttactg

17401 atattatgct ttccccagat cataaattgg taaagactca tttcaagtac aaacctgtta 17461 ttttacacat tctcaaatga aagtccctat caggccacct gctgaagtgt acgtgtgtta 17521 aatttgagcc atctcacatg atagccagat ttgtttcagg aaaacatcct gctttccaag 17581 gatttagaag ggtatgtttt tcactggtga ttcaggcaac atgcatcaga tttctggtct 17641 tcaaggagcc atattctcag aagggagatc aaggaccacg ctgtgattac ttctgacttc 17701 aggagccact ttctgtcagt gaaatttctc tttttgcttc tagcaccgag tggatttcct 17761 tcagctgatg attgactctc agaattcaaa agaaactgag tcccacaaag gtaaccagag 17821 tgtttctgag ggctacttgt ggggcactca gagggaaggc cttgttctga aaatgtgcag 17881 gaagtattcc aggatgatga gaatttctgc cacatagcag aacgacacat gtttgaatgt 17941 tataagtggt agttggaggc actttctaga ggcatgcagg catagatagc catgttctaa 18001 gagtaaaggg caaccctaag caaacctggc atgctagaaa gtcagtctgc ggtctgtgga 18061 tcacctacat cagatcaaat gccaattctc agcctccttc agatccactg aataaaaatt 18121 tctgcctaga aatttattag gttgctgcaa aattaattgt gggtttttcc attactttta 18181 attgcaaaaa aatgcaatta aaagtaatgg caaaaaccac aataactttt gcagctacct 18241 aatacatcta acatccaatc aggagccacg ctgttccaaa ggggtgattt taactggcag 18301 tacttgtgaa agtgtgttca ccaggtaatc tactgcaagt attttcactt tggaatggat 18361 agtaactgtg gggaaaacct ctgagaccgt gtaaatatcc tctttgtctt caatgttcca 18421 cctacgagtt ttacaaccca tgtatgtttt ttactgaagt cactattact atgacagtgg 18481 caaaatgatg accatggtca attacaagcc accaagactt ggcaaccatc tcacaaaatt 18541 cctgaatatt taactattgg ttctagagag caggactggg cttactccag catactgctt 18601 taaatatatc catgtctaca tccacttttg tctgtatgtc tatgtatcta tctatgtatc 18661 tatctagcta tgtatctatc tatctatcta tcatctatct atctatcatc tatccatcta 18721 tcatctatca tttatccatc tatcatctat ctacttatat attatctatc tatcatctaa 18781 ctattcatct atctatccca atataacttg ctgtgataaa ggaaatagtc tattgtttta 18841 ctgtttcata tagaaatcac tagacacata tggctattga gcactggata tgtggctagt 18901 gccattgaag atcaatttta aatggtattt cagtttaatt taataaaatt tgattttaaa 18961 tagccactag tgggtagtgg ctaccatatt ggacagcaga gctctaaact ttgagattat 19021 agttcaattt cacatcagta tcatcaggtt catgataact gaatactatg attaggtagt 19081 tgaatttact tataactgca tcacagaagt cttcactggt aaatcacagc tctgtcgacc 19141 tttctcacac tcctttcata ttggttttgg ttgtgaatta catggttgga gcaggcatta 19201 tatttatttc tatgttccag gtctctaaag gtcctaatcc agtcctgatc aaacagacca 19261 gtgatggacc atcctgagct tctctcagga gagaaatcaa gaggggccca acttgtaatc 19321 ataggagctt atgctatttt aatgccatcc atcagactac aatcaattac cactcatcta 19381 gctttttgtc catctctcat tcttgtacat cctgagatag tcaattctga gaactgtagc 19441 ctagatctat cacctgatgc ctctcaaaga tataatccgt gcttctcaag ctaggctatg 19501 cacacaaatc actgcatctt gtgaaagttc agattttgaa tcagtagttc aagggtgggg 19561 tttgagattt tgcatttcta aatgagctct caagatgctt ctgacccatg gaccacactt 19621 tgaataccaa gaagtggtct gtagaccaat attggtccct taagttccct caaacatatc 19681 ttcgggaaac gtcctttgat tttccctaca tttaaccatt agtgttgcaa attctctcaa 19741 agtttgtcaa gatatattgt agctaaaata aattacattt ttcttggggg agagtactac 19801 ctcatattaa cttacaataa agtactttta ggatcattca aggaacacac ccataacact 19861 gagtatgtta tgcggaaatg ctctctctgg aaattacaca gctgtgcagg tggcgggggt 19921 ggcatgagga ggagtggatg gcccacattc tcgaagacct tggggaaaac tggattaaaa 19981 tgatttgcct tattctggtt ctgtaagata cacatcagaa tgaaaccacc cccagtgtac 20041 ctctgaattg cttttctatt cttttccctt agggatttga gggcttcact tagatttctc 20101 ttcatctaaa ctgtgatgcc ctacattgat ctgatttacc taaaatgtct ttcctctcct 20161 ttcagctctg tccgatctgg agctcgtggc ccaatcaatt atctttattt ttgctggcta 20221 tgaaaccacg agcagtgttc tctccttcat tatgtatgaa ctggccactc accctgatgt 20281 ccagcagaaa ctgcaggagg aaattgatgc agttttaccc aataaggtga gtggatggta 20341 catggagaag gagggaggag gtgaaacctt agcaaaaatg cctcctcacc acttcccagg 20401 agaattttta taaaaagcat aatcactgat tctttcactg actctatgta ggaaggctct 20461 gaaaagaaaa agaaagaaac atagcaaatg gttgctactg gcagaagcgt aagatctttg 20521 taaaacgtgc tggctctggt tcatctgctt tctattacta caataatgct aagtaaaaaa 20581 cctccaaaaa cctcagtggc atctaacaat aagcatttgt tgctcacact catttcaatt 20641 ggttttggtt gtgaattaca tgtttgcagc aggcaccata gtggtgtgtg atgtcccctt 20701 agctgtatcc acatatggac acaggaattg gctcttttta tctcttttta ttttcttggt 20761 tacagacatg tgactttttt ttttgaaagg taacaatcac tttctcatat gttatttgat 20821 gctagtggtc atagcctata gtcacatttg tttcaatgag aaagaaaaac cagtacacgg 20881 ttatgctaag gatttcagtc cctggggtga gagccgtctc gaatgtctcc ccacttcata 20941 actcctccac acatcatagt tggatagtga gctctgctga tattggcagg acttgctctg 21001 gtctggctgt agtctgacgg agcctggccc tgggtgtgct gtgcaggctg actcagctct 21061 ccccacacct atctcatgtt ccagtcaggc agtaactggt gaagaagcca agctaggaac 21121 caggatatct ggctcctgag ctaaagtctt aaaacactat catattgcct tccaaatata 21181 acaccaaata ctaggtgcat atcaccctca ctgttttcag acctctgcca aaattgggat 21241 tctttgtggt atgaagagac acggctttgg ggctggcccg gctgtgacag tgaggtgaac

21301 acaaagggat gttcttcaga gattacagtc cagccctgaa gcaacaacta ggagactgat 21361 tgagcaagtg aggacagggc tgtgtggggt tctatcctct ttataacttc cactagcact 21421 gaaatcgtgt tctctggtta catccaacca gagccttctt tgtcattttt gggatagaaa 21481 gggactagtt tatcctcaaa ttatttatgg agattttata taatatagtg tttctctcca 21541 cattttctgt atataacaaa agtcctcctt ttagtgtgtg tatacacata tatatacaca 21601 tatatatgtg tgtgtgtatg tgtgtgtgta tatatatata cacatatata cacggttatg 21661 ctaaggattt cagtccctgg ggtgagagcc ttcccgaatg cttcccacct tcataactcc 21721 tccacacatc tcagtgggcc actgagcaca gcaatgggca tgacagttat taaaacactt 21781 tataaatgct tcgatccttt accagtatga gttagtctct ggagctccta atacttcatt 21841 agtactgcat ggactgagtt aaaagttaat tcaaaatctc aatttatcca aatctgtttc 21901 gttctttcca ggcaccaccc acctatgata ctgtgctaca gatggagtat cttgacatgg 21961 tggtgaatga aacgctcaga ttattcccaa ttgctatgag acttgagagg gtctgcaaaa 22021 aagatgttga gatcaatggg atgttcattc ccaaaggggt ggtggtgatg attccaagct 22081 atgctcttca ccgtgaccca aagtactgga cagagcctga gaagttcctc cctgaaaggt 22141 acaaggcccc tgggaaggga gccctccctg aaccagcctg gttcaagcat attctgcctc 22201 tcttaatcta caggacagtc atgtggttgt ataattattt gcttgtattt ttatatttag 22261 agattttttt aatcatcaaa ttgattattg tcacacttta caaaccatag actagaaaaa 22321 agaaaactac agtcatccac aattccaaca acttacgatg aaggtcatca gttatgtcct 22381 tatgggtcat cagtgtccaa aatgtaagga ctcttttaaa aacacatgat cacaatgcta 22441 ttattatgtc ccccaaatga atattttttt cataaatata atcaaatatt taggaataac 22501 attttaataa aaaacatgca tctaatcttc aaagaatttt atagcttact ggaacagata 22561 gacaaagaaa gcagggatga tactgcatta catcggtaca gtagcatcat atgcctgtgt 22621 aaattatctg acttcaacta ttctatggag gtgtggggga gaaagaggga gagatggaga 22681 gaagaagaag gaggagaagg agggagaaga caaggtaagg aggagaagga ggagaattag 22741 aaaaacaaga gaggagatga gaaggaaagt gcaaaataac aattttgaaa tagtacaaga 22801 caatttcccc ttctccttcc tcatgaccaa tgtaagtgtg acttgaggca ggaacctact 22861 tttccatcag tcagtcccat cacttatgtg ccttttatag tgtggacaca tcaccaccct 22921 gaatataatt tcagtgttta gaaataagta ttctttgcaa cactatttat ctcatctcaa 22981 caagactgaa agctcctata gtgtcaggag agtagaaagg atctgtagct tacaattctc 23041 atagcaaaat aagcatagca ggatttcaat gaccagccca caaaagtatc ctgtgtacta 23101 ctagttgagg ggtggcccct aagtaagaaa ccctaacatg taactcttag gggtattatg 23161 tcattaactt tttaaaaatc taccaacgtg gaaccagatt cagcaagaag aacaaggaca 23221 acatagatcc ttacatatac acaccctttg gaagtggacc cagaaactgc attggcatga 23281 ggtttgctct catgaacatg aaacttgctc taatcagagt ccttcagaac ttctccttca 23341 aaccttgtaa agaaacacag gttagtcaat tttctataaa aataatgttg tattaataat 23401 tcttttaact gagtggtctg tattttttaa aaagaatatg cttgtttaat cttttactaa 23461 tttgttctct gggccaaaga atcaattagg cccatctgtg atctttaagg gtgcttcagt 23521 tctggagttc aaaagctgta gcatcaaaaa catcatgtaa agcccatgta gattagcatg 23581 gcataattat ctgcagtctc cttgaacttg agcaaagtta cattcatttc aagtcgattg 23641 gaaagatggg gaagtatttt tgcacagtca tgaagtgtaa tgattacctt gttgtgactt 23701 ttgaatgctg ctcttcccaa ccagaacttg gagaagcttt ctcatgagag tggctcccaa 23761 ccactagctg tacattggaa tcaccaggga gctttaaaaa ttcatgatgt ctgggatatc 23821 acagaaattc taaactaatt tgcccagagt gtggctttaa aagcttcccc attgcttctc 23881 atgtgaagcc aaggttgaga atgactaatt taaggcattt ctggtggata taaaggacta 23941 ccacagtcca aggccatcct gactgacctc accttccagg tgcctagctc catccagctg 24001 ggctcctttt caacccaatt ataactctat taatgttgtt cccagccagg catggtggct 24061 catgcctgta atcccagcac tttgggaggc cgaagcaggc ggatcatgag gtcaggagat 24121 cgagaccatc ctggctaaca cggtgaaacc ccgtctctac taaaaataca aaaaattacc 24181 ctggagaggt ggcaggcacc tgtagtccca gctactctgg aggctgaggc aggagaatgg 24241 catgaacccc agggagcgga gcctgcagtg agccgagatt gggccactgc actccagcct 24301 gggtgacagc gagactctgt ctcaaaaaaa aaaaaatgtt gttccctttc tcctcatttt 24361 gttcttatct ttcaagtcct agttcaatcc ccaagcccct ccaaagtgtc ttctcctcct 24421 agtccagggc ccatttactt ctctgctctg ttattggata ctggaggcta ttatcataaa 24481 tttgacaatt tgccattaaa tcattgagtt ttattctcta tattttcttt gtatctaaaa 24541 tgtcttcccc cctccattaa caatatcctc tcattttatt cctttttaaa atatcccagt 24601 ggtgccttgc aagggactgt atctaatgca agcatttggt aaatgtttaa ggagtgatgt 24661 gcagttgatg gcttgcatac atatattaag ctatttaatg tgaaccttta aacaaatgcc 24721 attcgtgcat atgcatgtgt gtgtgtgtgt gtgcacatgt gggcatgcat gtctgtctgc 24781 agtgaaaata tattcagggt ttttgaaaat ttttaaataa taaggtatta tatttataga 24841 aagatttgaa atattttctc tgaagaagtt aaagaacaga cgtcattgat tcatattaaa 24901 caatacccta taaatcttat ttctaggtct catgtattta ttattcaatt ccacccctta 24961 agtgggcttt ctatatagga gaggaagaag acagaaatag tttccatatt atttccatat 25021 tccatattat ttgtggcttt aggccagcag tgtagctgta ttatatgtgc ccagacaggg 25081 gactcagccc tgaataaaag tggtcctctg gcacacctgg gatggggaag gtactccttg 25141 gtaagctccc aacctggcac ttcttgatct ccctggcaat tttcttgccc attactccat

25201 ggagatcaga atatcactct gttgtgtccc ctcaacacgg aaggagtgtc tcaataagaa 25261 tggggctaaa agttgagtcc aaacactgta ggaattgaga ggttccccac cttgcactac 25321 ccttggaagc caagagaaga tgttaaaaat aaaatggtaa tgcttcctga aggtgtcttc 25381 ccatctttac actggatggg ttcaattggg aggaattact ggactctgga agttgaagac 25441 tgtccatata attaaaatgt acaataacta cccaggttta ccttgcaagt ttcaacatac 25501 acaaaattaa ctttatatga ctcttcaaaa acagtttgcc atcataccta ataatctggt 25561 ttaaatttta aaaactcatc cattttactt aaaatttaaa tcaaaaaaga acacaggttt 25621 ccatgaattt gtctcaggcc tggcacagaa tagtactcca taaatatttt gttaaatgat 25681 agatgatgaa tgctctcact gtccaatctt cacacatctt atagactaag tataaagaat 25741 ccaagattta tagtgctgaa agtagttttt atatgtttac aaagcattat tgtcattact 25801 gcattttttt tgcccattac tccatagaga tcagaatatc actctgttgt gtcccctcaa 25861 cactgaagga gtgtctcact cactttgatg ctatactttc tacttttgtt tatttaatgc 25921 ttctcaatat gcttgtttaa ctgttgcaga tccccctgaa attaagctta ggaggacttc 25981 ttcaaccaga aaaacccgtt gttctaaagg ttgagtcaag ggatggcacc gtaagtggag 26041 cctgaatttt cctaaggact tctgctttgc tcttcaagaa atctgtgcct gagaacacca 26101 gagacctcaa attactttgt gaatagaact ctgaaatgaa gatgggcttc atccaatgga 26161 ctgcataaat aaccggggat tctgtacatg cattgagctc tctcattgtc tgtgtagagt 26221 gttatacttg ggaatataaa ggaggtgacc aaatcagtgt gaggaggtag atttggctcc 26281 tttgcttctc acgggactat ttccaccacc cccagttagc accattaact cctcctgagc 26341 tctgataaga gaatcaacat ttctcaataa tttcctccac aaattattaa tgaaaataag 26401 aattattttg atggctctaa caatgacatt tatatcacat gttttctctg gagtattcta 26461 taagttttat gttaaatcaa taaagaccac tttacaaaag ta //
Primers 2fw e 2rv Primers 3fw e 3rv

Anexo II Trabalho apresentado no âmbito do estudo desenvolvido.
Participação em Congressos
- Julho, 2000
13th International Symposium on Microsomes and Drug Oxidations, Stresa, Italy.
- Setembro, 2000
XII Congresso Nacional de Bioquímica, Póvoa do Varzim, Portugal.
- Junho - Julho, 2001
27th Meeting of the Federation of European Biochemical Societies in collaboration with the PAN-
American Association for Biochemical Molecular Biology, Lisboa, Portugal.
- Setembro, 2001
12th International Conference on Cytochrome P450, La Grande Motte, France.
Comunicações
Cavaco I, Gil P, Ribeiro V. (2000) Analysis of CYP3A Gene Polymorphisms in a Portuguese
Caucasian Population. 13th International Symposium on Microsomes and Drug Oxidations, abst. p.
112, 10, Stresa, Italy.
Cavaco I, Gil P, Ribeiro V. (2000) CYP3A Genetic Polymorphism in a Portuguese Caucasian
Population. XII Congresso Nacional de Bioquímica, abst. p. 53, P2.4, Póvoa do Varzim, Portugal.
Cavaco I, Gil P, Ribeiro V. (2001) Incidence of cytochrome P450 strutural variants, CYP3A4*1B
and CYP3A4*2, in the Portuguese population. 27th Meeting of the Federation of European
Biochemical Societies in collaboration with the PAN-American Association for Biochemical
Molecular Biology, abst. p. 65, PS6-027, Lisboa, Portugal.

Cavaco I, Gil P, Ribeiro V. (2001) PCR-RFLP analysis of cytochrome P450 3A4 polymorphism in
Portuguese. 12th International Conference on Cytochrome P450, abst. p.110, P 326, La Grande Motte,
France.
Cavaco I, Gil P, Ribeiro V. (2001) Cytochrome P450 3A4 and MDR1 genotyping in the
Portuguese population. 6th International Meeting of the International Society for the Study of
Xenobiotics (ISSX), abst. p. 219, 436 Munich, Germany.
Comunicações Orais
- Setembro, 2001
PCR-RFLP analysis of cytochrome P450 3A4 polymorphism in Portuguese. 12th International
Conference on Cytochrome P450, La Grande Motte, France.
Publicações
Cavaco I, Gil JP, Ribeiro V. PCR-RFLP identification of Cytochrome P450 3A4 polymorphism in the
Portuguese population. Submitted.
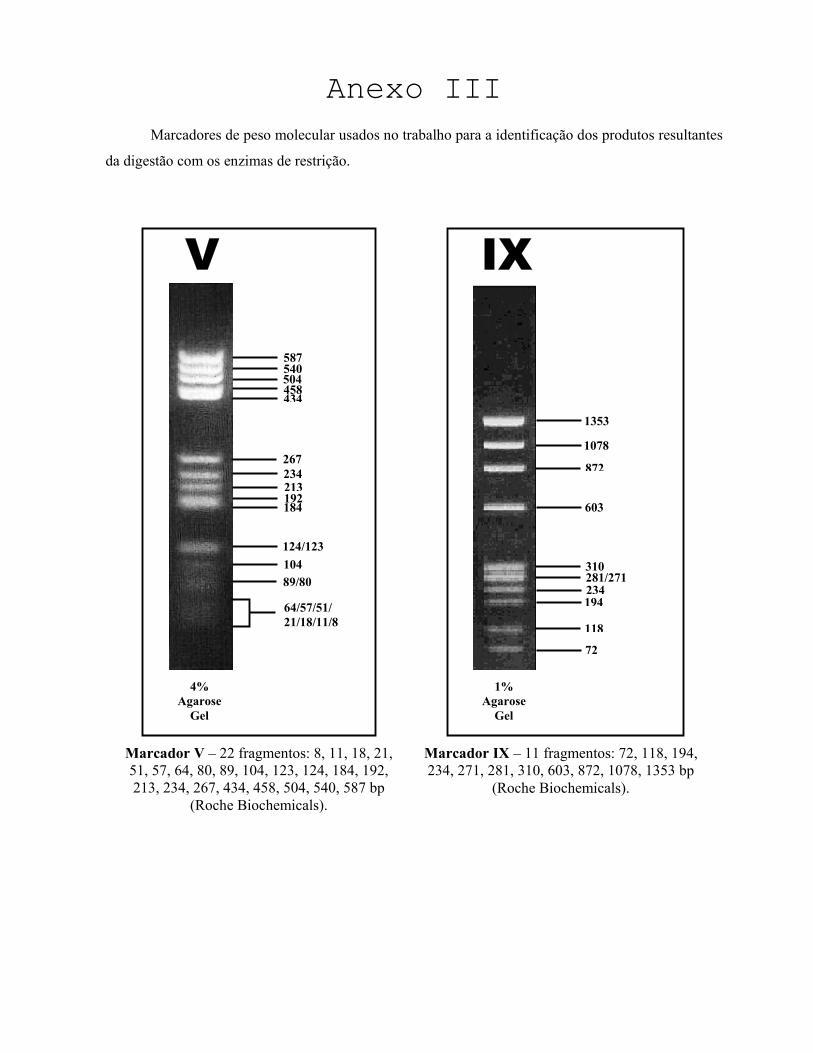
Anexo III Marcadores de peso molecular usados no trabalho para a identificação dos produtos resultantes
da digestão com os enzimas de restrição.
Marcador IX – 11 fragmentos: 72, 118, 194, 234, 271, 281, 310, 603, 872, 1078, 1353 bp
(Roche Biochemicals).
V
587 540 504 458 434
267 234 213192 184
124/123 104 89/80
64/57/51/ 21/18/11/8
4% Agarose
Gel
IX
1353
1078
872
603
310 281/271
72
1% Agarose
Gel
118
194234
Marcador V – 22 fragmentos: 8, 11, 18, 21, 51, 57, 64, 80, 89, 104, 123, 124, 184, 192, 213, 234, 267, 434, 458, 504, 540, 587 bp
(Roche Biochemicals).






![PDF - [Selma] Aula 3 - Metabolismo Dos Xenobióticos](https://static.fdocumentos.com/doc/165x107/55cf94cb550346f57ba470e0/pdf-selma-aula-3-metabolismo-dos-xenobioticos.jpg)











