
UNIVERSIDADE ESTADUAL DO NORTE …uenf.br/posgraduacao/ciencia-animal/wp-content/uploads/...Ele me...
157
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE DARCY RIBEIRO MARIA DE LOURDES AMARAL BERNARDINO VIABILIDADE DE ANTICORPOS IgY DE GALINHAS HIPERIMUNIZADAS CONTRA EXOANTÍGENOS DE Sporothrix spp. NO DIAGNÓSTICO DA ESPOROTRICOSE E INIBIÇÃO DO DESENVOLVIMENTO DO FUNGO CAMPOS DOS GOYTACAZES 2014
Transcript of UNIVERSIDADE ESTADUAL DO NORTE …uenf.br/posgraduacao/ciencia-animal/wp-content/uploads/...Ele me...

UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE DARCY RIBEIRO
MARIA DE LOURDES AMARAL BERNARDINO
VIABILIDADE DE ANTICORPOS IgY DE GALINHAS HIPERIMUNIZADAS
CONTRA EXOANTÍGENOS DE Sporothrix spp. NO DIAGNÓSTICO DA
ESPOROTRICOSE E INIBIÇÃO DO DESENVOLVIMENTO DO FUNGO
CAMPOS DOS GOYTACAZES 2014

MARIA DE LOURDES AMARAL BERNARDINO
VIABILIDADE DE ANTICORPOS IgY DE GALINHAS HIPERIMUNIZADAS CONTRA
EXOANTÍGENOS DE Sporothrix spp. NO DIAGNÓSTICO DA ESPOROTRICOSE E
INIBIÇÃO DO DESENVOLVIMENTO DO FUNGO
Tese de Doutorado apresentada ao Programa de
Pós-Graduação em Ciência Animal na área de
Doenças Infecto-contagiosas e Parasitárias dos
Animais, do Centro de Ciências e Tecnologias
Agropecuárias da Universidade Estadual do Norte
Fluminense Darcy Ribeiro como parte dos
requisitos para obtenção do título de Doutor em
Ciência Animal.
Orientador: Prof. Olney Vieira da Motta
CAMPOS DOS GOYTACAZES 2014

MARIA DE LOURDES AMARAL BERNARDINO
VIABILIDADE DE ANTICORPOS IgY DE GALINHAS HIPERIMUNIZADAS CONTRA
EXOANTÍGENOS DE Sporothrix spp. NO DIAGNÓSTICO DA ESPOROTRICOSE E
INIBIÇÃO DO DESENVOLVIMENTO DO FUNGO
Tese de Doutorado apresentada ao Programa de Pós-
Graduação em Ciência Animal na área de Doenças
Infecto-contagiosas e Parasitárias dos Animais, do Centro
de Ciências e Tecnologias Agropecuárias da Universidade
Estadual do Norte Fluminense Darcy Ribeiro como parte
dos requisitos para obtenção do título de Doutor em
Ciência Animal.
Aprovada em 18 de dezembro de 2014.
BANCA EXAMINADORA
Prof. Eulógio Carlos Queiroz de Carvalho (Doutor, Patologia – Anatomia Patológica) – LMPA/CCTA/UENF
Prof. Milton Masahiko Kanashiro (Doutor, Biociências e Biotecnologia) – LBR/CBB/UENF
Profa. Regina Célia de Souza Campos Fernandes (Doutora, Medicina) - FMC
Prof. Olney Vieira da Motta (Doutor, Biociências e Biotecnologia) – LSA/CCTA/UENF ORIENTADOR

Aos meus pais,
Dorival e Rosane, ao Martinho e ao nosso filho, Bernardo,
amor das nossas vidas.

AGRADECIMENTOS
Agradeço a Deus, por ter sido a minha força e a minha fortaleza, por ter
proporcionado todos os “encontros” de que necessitei para a execução deste
trabalho.
Este trabalho não seria possível sem a ajuda das pessoas que me
deram apoio técnico e pessoal no decorrer deste curso. Por isto, agradeço
especialmente:
Ao professor Olney Vieira da Motta, do Laboratório de Sanidade Animal
do Centro de Ciências Tecnológicas e Agropecuárias da Universidade Estadual do
Rio de Janeiro (UENF), pela oportunidade, orientação e compreensão dispensada
no decorrer do curso.
Aos professores Milton Masahiko Kanashiro, do Laboratório de
Biologia do Reconhecer, CBB/UENF e Eulógio Carlos Queiroz de Carvalho do
Laboratório de Morfologia e Patologia Animal do CCTA/UENF tanto pela participação
neste trabalho quanto pela contribuição em minha formação profissional. À
professora Regina Célia de Souza Campos Fernandes (Faculdade de Medicina de
Campos) pela participação na banca de avaliação e contribuições prestadas. Pelas
análises estatísticas, ao professor Cláudio Luiz Melo de Souza (LEAG/CCTA/UENF).
Ao apoio financeiro da FAPERJ (Fudação de Amparo à Pesquisa do
Estado do Rio de Janeiro), processo E-26/110.611/2012.
Ao David Gitirana (doutorando do LBR/CBB/UENF) por toda a ajuda
prestada desde a aquisição das aves para o experimento até os ensinamentos
técnicos de obtenção dos anticorpos IgY e testes de ELISA, muito obrigada pela
paciência e pela boa vontade com que sempre me atendeu.
Ao Arthur Rodrigues (LBCT/CBB/UENF) e Zila Sousa de Macedo
(LBT/CBB/UENF) pelos ensinamentos da técnica de eletroforese.
À colega Dra. Cláudia Costa de Almeida (LSA/CCTA/UENF), pela
amizade e ensinamentos da técnica de western blot.

Ao professor Carlos Jorge Logullo de Oliveira (LQFPP/CBB/UENF) e
colegas do setor, Josias Alves Machado (LCCA) e Newton Ruiz (doutorando
LQFPP), pelo apoio na aquisição e alojamento dos coelhos.
Às alunas da Escola Técnica Estadual João Barcelos Martins, Roberta
e Letícia Nascimento, pelo apoio nos cuidados com as aves e coleta dos ovos.
Ao doutorando, Anderson Barros, do Laboratório de Clínica e Cirurgia
Animal (LCCA/CCTA/UENF), pela doação dos soros felinos negativos. À Dra.
Mariane Távora e Dr. Valmir Laurentino Silva (FioCruz), pelos soros caninos de área
endêmica, positivos e negativos para leishmaniose. À médica veterinária Aline Lippi
(APA/Campos dos Goytacazes/RJ) pela permissão da coleta de sangue de cães da
APA e apoio técnico no Relato de Caso 2 deste experimento.
Ao apoio técnico nas análises histopatológicas, nas pessoas de
Luciano Grillo de Almeida e Luciana da Silva Lemos (TNS/LMPA/CCTA/UENF), nas
análises de imunohistoquímica: Maria Aparecida da Silva (doutoranda LMPA) e
Rachel Bittencourt Ribeiro (aluna de residência do LMPA); preparo das lâminas,
Elizabeth Gonçalves Pires (Auxiliar técnica LMPA) e fotografias, Hassan Jerdy
Leandro (mestrando LMPA).
Em momentos variados, ao apoio dos colegas Thiago da Silva Correa
(LMRGA/CCTA/UENF), professora Fernanda Antunes (LCCA/CCTA/UENF),
médicas veterinárias Maria Angélica Dutra Viestel (LCCA/CCTA/UENF), Fernanda
Ribeiro e Maria Clara (alunas de residência do LCCA), às colegas de doutorado
Luciana Mathias e Luize Néli (LSA/CCTA/UENF).
À colega Sueli Rosa Marins (LSA/CCTA/UENF) pelo apoio na limpeza
e esterilização de materiais.
À colega Gina Nunes Teixeira (TNS/CCTA/LSA/UENF) pela
contribuição em minha formação profissional, assim como, pela amizade e apoio
tanto técnico quanto no âmbito pessoal, a sua ajuda foi fundamental para este
empreendimento ir a termo. Agradecimento estendido à sua família, Prof. Leonardo
Serafim da Silveira (LMPA/CCTA/UENF), Laura e Luíza.

Ao meu marido, Martinho, agradeço pela paciência, pela preocupação
e ao apoio que me deu no decorrer do curso.
Ao meu filho, Bernardo, apesar de ainda criança, a compreensão da
minha ausência e, sem reclamar por atenção, pelas vezes que me deixou sozinha e
ficou quietinho para que eu pudesse estudar em casa. Te agradeço pelas muitas
vezes em que me acompanhou até a UENF tanto à noite, após as suas aulas,
quanto finais de semana. Os vídeo games facilitaram a nossa vida!
À Nete, a tranquilidade que me deu ao cuidar bem do meu filho e
contribuir para que tudo corresse bem no dia-a-dia da minha casa.
Aos meus pais, irmãos, Cida e tia Laia, pela torcida de que todas as batalhas
fossem vencidas e pela amizade e carinho que sempre deram. À Cida (Maria
Aparecida Fonseca do Amaral), agradeço a amizade de uma vida inteira e, por fim, o
presente de ter tido a sua presença no dia da minha defesa.

“O correr da vida embrulha tudo.
A vida é assim: esquenta e esfria,
aperta e daí afrouxa,
sossega e depois desinquieta.
O que ela quer da gente é coragem”
João Guimarães Rosa

O Senhor é o meu pastor;
nada me faltará.
Ele me faz repousar
em pastos verdejantes.
Leva-me para junto
das águas de descanso;
Refrigera-me a alma.
guia-me pelas veredas da justiça
por amor do seu nome.
Ainda que eu ande
pelo vale da sombra da morte,
Não temerei mal nenhum,
porque tu estás comigo;
o teu bordão e o teu cajado me consolam.
Preparas-me uma mesa
na presença dos meus adversários,
Unges-me a cabeça com óleo;
0 meu cálice transborda.
Bondade e misericórdia
certamente me seguirão
todos da minha vida;
e habitarei na Casa do Senhor
para todo o sempre.
(Salmo 23, 1-6)

RESUMO
A esporotricose é uma zoonose de grande importância no estado do Rio de Janeiro
e no país. Antígenos (AG) de leveduras e de micélio de Sporothrix schenckii ATCC
32285 e AG de levedura de cepa clínica foram submetidos a métodos de extração
física e química para estudos imunológicos através dos testes de ELISA, western
blot, imunohistoquímica e testes de inibição de crescimento. Animais experimentais,
galinhas poedeiras e coelhos foram imunizados, individualmente, com 200 μg de
proteína total de cada antígeno/imunização, a cada 21 dias de intervalo, totalizando
nove imunizações para as aves e cinco para os coelhos. A titulação de anticorpos
(AC) nestes modelos foi acompanhada por ELISA e foi observado que após a
terceira imunização já havia produção de anticorpos, tanto IgY quanto IgG, com
títulos satisfatórios para os testes imunológicos. Ensaios de ELISA, mostraram
reação cruzada entre AG (LSs, MSs e LSc) e AC (IgY anti-LSs, anti-MSs, anti-LSc;
IgG anti-LSs, anti-MSs, anti-LSc). Ensaios de western blot entre AC de animais
imunizados e AG de LSs, MSs e LSc mostraram que as bandas foram mais intensas
quando utilizado o AG LSc para sensibilização. Nas membranas sensibilizadas com
AG de leveduras houve o reconhecimento de várias bandas e nas sensibilizadas
com AG MSs poucas bandas proteicas reconhecidas. Quando confrontados o AG
MSs e anticorpos IgY, a banda de 64 – 49 kDa foi imunodominante e a única não
reconhecida pela IgY pré-imune. No ensaio de imunohistoquímica, os AC IgY
(5μg/mL) anti-LSs, anti-LMSs, anti-LSc ou IgG (1:500) anti-LSs, MSs e LSc testados
em pele de felinos positivos para esporotricose mostraram o reconhecimento
específico de leveduras de Sporothrix spp. pelos AC específicos e nenhum
reconhecimento por parte dos AC pré-imunes. Não foi observada nenhuma reação
cruzada entre AC específicos, IgY ou IgG, e formas amastigotas de Leishmania spp.
em seções de pulmão de cão positivo para leihsmaniose. Para avaliar o potencial
dos AC IgY na imunidade passiva da esporotricose, ensaios de inibição de
crescimento, in vitro, mostraram que houve inibição significativa (p<0,05) das
leveduras de S. schenckii ATCC 32285 e de Sporothrix spp., em caldo YPD, após
incubação 37oC com os AC imunes IgY anti-LSs, anti-LMSs ou anti-LSc (1mg/mL)
até 44 horas. Os AG produzidos também foram testados com soros de cão e gatos
positivos para esporotricose através de ensaios de ELISA e western blot. As bandas
de 64-49 kDa foram imunodominantes perante AC imunes e soros de animais

positivos nas membranas sensibilizadas com AG de LSs, MSs e LSc. A
diferenciação entre animais positivos e negativos pelo teste de western blot foi
melhor frente a AG de leveduras de S. sckenckii ATCC 32285. Foram relatados dois
casos clínicos, o de um canino com esporotricose de mucosa com diagnóstico
confirmado por ELISA e western blot utilizando os AG obtidos neste trabalho, devido
à impossibilidade de isolamento do agente. No outro caso relatou-se a transmissão
zoonótica onde houve isolamento do fungo por meio de cultivo tanto de material
extraído de felino quanto de biópsia da paciente humana, além de
imunohistoquímica positiva utilizando AC desenvolvidos neste trabalho.
Palavras-chave: DIAGNÓSTICO, ESPOROTRICOSE, ELISA, WESTERN BLOT,
ZOONOSE.

ABSTRACT
Sporothrichosis is a zoonotic disease with relevance in Rio de Janeiro state and
Brazil. Yeast and mycelium antigens (AG) from Sporothix schenckii ATCC 32285 and
yeast AG from clinical strain were submitted to physical and chemical extraction
methods for immunological studies by using ELISA, western blot,
immunohistochemistry and growth inhibition tests. Experimental animals (hens and
rabbits) were immunized with 200 μg of total protein of each antigen per
immunization, at 21 days interval, totalizing nine immunizations for hens and five for
rabbits. Antibodies (AB) titrations were followed by ELISA and after the third
immunization step the IgG and IgY titration was satisfactory. ELISA assays showed
cross reaction with AG (LSs, MSs and LSc) and AB (IgY anti-LSs, anti-MSs, anti-LSc;
IgG anti-LSs, anti-MSs, anti-LSc). By western blot assay among LSs, MSs and LSc
AG and immune AB showed that reactions were more intense when the sensitizing
AG was the LSc. There was a variety of bands recognized in sensitized strips with
yeast antigens and few protein bands recognized in the membranes coated with
MSs. For MSs AG x IgY pre and post-immune, the band profile between 64-49 kDa
were immunodominant because it was the only one not recognized by pre immune
IgY. By immunohistochemistry assay the IgY AB (5μg/mL) anti-LSs, anti-LMSs, anti-
LSc or IgG (1:500) anti-LSs, MSs and LSc tested in sporothrichosis-positive cat skin
showed specific reaction by the specific AB and negative towards pre-immunization
AB. No cross reaction between specific AB, IgY or IgG, was observed towards
leishmaniosis-positive tissues samples of dogs by immunohistochemistry assay. In
order to investigate passive immunity of sporothrichosis, an inhibition assay was
conducted in YPD broth, and showed a significant (p<0,05) growth inhibition of both
yeast strains (ATCC 32285 and clinical) at 37 ºC until 44 hours of incubation. The AG
produced in this work were contrasted towards sporothrichosis-positive serum
samples from dog and cats by ELISA and western blot. A band profile of 64-49 kDa
was immunodominant in the presence of immune AB and towards positive-serum in
the sensitized membranes with LSs, MSs and LSc AGs. Differentiation between
positive and negative animals by western blot assay was superior towards yeast AG
from S. sckenckii ATCC 32285. Two clinical cases were described, one from a dog
with mucosal sporothricosis confirmed by ELISA and western blot by using the AG
developed in the present work, due to the impossibility of isolation of the agent. The

second related case was a zoonotic transmission, confirmed by culture of both
samples, from cat and human biopses fragments, besides immunohistochemistry
assay utilizing AC developed in the present work.
Keywords: DIAGNOSTIC, SPOROTHRICOSIS, ELISA, WESTERN BLOT,
ZOONOSIS.

LISTA DE FIGURAS
Figura 1: Etapa de imunização das aves no músculo peitoral. 200 µg de
proteína do extrato antigênico/imunização em intervalos de 21
dias...............................................................................................................
70
Figura 2: Perfil eletroforético dos extratos proteicos de leveduras de S.
schenckii (LSs), Sporothrix spp. (LSc) e micélio de S. schenckii (MSs) (40
μg/raia). PM: padrão de peso molecular. .................................................
84
Figura 3: Perfil eletroforético de amostras das soluções de IgY extraídas
de gemas de ovos pré-imunes (raia 1, 20 μg) e pós-imunização: IgY anti-
LSs (raia 2, 30 μg), IgY anti-MSs (raia 3, 20 μg) e IgY anti-LSc (raia 4, 30
μg). PM: peso molecular..............................................................................
85
Figura 4: Titulação, por ELISA, dos anticorpos IgY séricos de galinhas
específicos para extratos proteicos de S. schenckii e Sporothrix spp. pré
e pós- imunizações. A: levedura de S. schenckii (IgY anti-LSs), B: micélio
de S. schenckii (IgY anti-MSs) e C: levedura de Sporothrix spp. (IgY anti-
LSc). Diluição do soro: 1:500.......................................................................
86
Figura 5: Titulação, por ELISA, de anticorpos IgY das soluções
purificadas das gemas específicos para extratos proteicos de S.
schenckii e Sporothrix spp. pré e pós- imunizações. A: levedura de S.
schenckii (IgY anti-LSs), B: micélio de S. schenckii (IgY anti-MSs) e C:
levedura de Sporothrix spp. (IgY anti-LSc). Diluições seriadas a partir de
um volume de 100 µl contendo 80 µg de solução de IgY/mL......................
87
Figura 6: Titulação, por ELISA, dos anticorpos IgG séricos de coelhos
específicos para extratos proteicos de S. schenckii e Sporothrix spp. pré
e pós- imunizações. A: levedura de S. schenckii (IgG anti-LSs), B:
micélio de S. schenckii (IgG anti-MSs) e C: levedura de Sporothrix spp.
(IgG anti-LSc). Diluição do soro: 1:500........................................................
88

Figura 7: Resultados médios de absorvância do ELISA indireto
cruzando anticorpos IgY (4 µg de proteína/poço) das gemas após a
terceira imunização x antígenos de S. schenckii ou Sporothrix spp. LSs:
antígeno de leveduras de S. schenckii; MSs: antígenos de micélio de S.
schenckii e LSc: antígenos de leveduras de Sporothrix spp......................
89
Figura 8: Resultados médios de absorvância do ELISA indireto
cruzando anticorpos IgG (1:500) após a terceira imunização x antígenos
de S. schenckii ou Sporothrix spp. LSs: antígeno de leveduras de S.
schenckii; MSs: antígenos de micélio de S. schenckii e LSc: antígenos
de leveduras de Sporothrix spp. ................................................................
90
Figura 9: Reconhecimento das moléculas antigênicas por anticorpos
das soluções de IgY das gemas (20 μg/mL) por meio da técnica de
western blot. (A) Tiras sensibilizadas com extratos proteicos de
leveduras de S. schenckii (LSs), (B) extratos proteicos de leveduras de
Sporothrix spp. (LSc) e (C) extratos proteicos de micélio de S. schenckii
(MSs). Linhas 1, 5 e 9: IgY pré-imunização. Linhas 2, 6 e 10: IgY anti-
MSs. Linhas 3, 7 e 11: IgY anti-LSs. Linhas 4, 8 e 12: IgY anti-LSc. PM:
peso molecular (kDa).................................................................................
91
Figura 10: Reconhecimento das moléculas antigênicas por anticorpos
IgG de coelhos (1:2000) por meio da técnica de western blot. (A) Tiras
sensibilizadas com extratos proteicos de leveduras de S. schenckii
(LSs), (B) extratos proteicos de leveduras de Sporothrix spp. (LSc) e (C)
extratos proteicos de micélio de S. schenckii (MSs). PI: IgG pré-imune.
Linhas 1, 4 e 7: IgG anti-MSs; linhas 2, 5 e 8: IgG anti-LSs; linhas 3, 6 e
9: IgG anti-LSc. PM: peso molecular. ........................................................
92
Figura 11: Reconhecimento das moléculas antigênicas por soros de
gatos negativos e positivos para esporotricose por meio da técnica de
western blot. Tiras sensibilizadas com antígenos de leveduras de S.
schenckii ATCC 32285. Diluição do soro: 1:300........................................
94

Figura 12: Reconhecimento das moléculas antigênicas por soros de
gatos negativos (linhas 1- 19) e positivos (linhas 20 – 26) para
esporotricose por meio da técnica de western blot. Tiras sensibilizadas
com antígenos de leveduras de Sporothrix spp. Diluição do soro: 1:300.
PM: peso molecular (kDa)..........................................................................
95
Figura 13: Reconhecimento das moléculas antigências por soros de
gatos negativos (linhas 1-20) e soros de gatos positivos (linhas 21 – 26)
para esporotricose por meio da técnica de western blot. Tiras
sensibilizadas com antígenos de micélio de S. schenkii ATCC 32285.
Diluição do soro: 1:300. PM: peso molecular (kDa)...................................
96
Figura 14: Reconhecimento das moléculas antigências por soros de
cães negativos (linhas 1-18) e soro de cão positivo (linhas 19) para
esporotricose versus antígeno de micélio de S. schenkii ATCC 32285.
PM: peso molecular (kDa). Diluição do soro: 1:300...................................
97
Figura 15: Reconhecimento das moléculas antigênicas por soros de
cães negativos e positivo para a esporotricose por meio da técnica de
western blot. (A) Versus antígeno de leveduras de S. schenckii:
amostras 1 – 10, soros de cães negativos, amostra 11: soro de cão
positivo. (B) Versus antígeno de leveduras de Sporothrix spp.: amostras
1 – 8, soros de cães negativos, amostra 9: soro de cão positivo. PM:
peso molecular. Diluição do soro: 1:300. ..................................................
98
Figura 16: Reconhecimento das moléculas antigênicas por soros de
animais positivos para esporotricose, antissoro de coelho e IgY imune
antiantígenos de leveduras de S. schenckii ATCC 32285 por meio da
técnica de western blot. Tiras sensibilizadas com antígenos de
leveduras de S. schenckii ATCC 32285. PM: peso molecular; linha 1:
IgG de coelho anti-LSs (1:2000); linha 2 IgY anti-LSs (20 μg/mL); linha
3: soro de cão (1:300) com esporotricose; linhas 4-10: soros de gatos
(1:300) positivos para esporotricose..........................................................
99

Figura 17: Fotomicrografia: imunohistoquímica para marcação de
leveduras de Sporothrix spp. em pele de gato. Formações
leveduriformes sem marcação (setas) nos tratamentos Controle e IgY
PI. Formações leveduriformes marcadas (setas) por anticorpos IgY
antiantígenos do Sporothrix spp. Controle= sem anticorpo; IgY PI= IgY
pré-imunização; IgY anti-LSs= IgY antiantígenos de leveduras de S.
schenckii, IgY anti-MSs= IgY antiantígenos de micélio de S. schenckii,
IgY anti-LSc= IgY antiantígenos de leveduras de Sporothrix spp. (IgY: 5
μg/mL). Barra: 50 µm.................................................................................
101
Figura 18: Fotomicrografia: imunohistoquímica para marcação de
leveduras de Sporothrix spp. em pele de gato. Formações
leveduriformes sem marcação (setas) no tratamento controle (IgG PI).
Formações leveduriformes marcadas (setas) por anticorpos IgG
antiantígenos do Sporothrix spp. IgG PI= IgG pré-imunização; IgG anti-
LSs= IgG antiantígenos de leveduras de S. schenckii, IgG anti-MSs=
IgG antiantígenos de micélio de S. schenckii, IgG anti-LSc= IgG
antiantígenos de leveduras de Sporothrix spp. Diluição do anticorpo:
1:500. Barra: 50 µm...................................................................................
102
Figura 19: Fotomicrografia: imunohistoquímica em fragmentos de
pulmões de cão positivo para leishmaniose visceral com anticorpos IgY
(5 μg/mL). Formas amastigotas de Leishmania spp. sem marcação em
todos os tratamentos (setas). Controle= sem IgY; IgY PI= IgY pré-
imune, IgY anti-LSs= IgY antiantígenos de leveduras de S. schenckii,
IgY anti-MSs= IgY antiantígenos de micélio de S. schenckii, IgY anti-
LSc= IgY antiantígenos de leveduras de Sporothrix spp. Barra: 10 µm....
104
Figura 20: Fotomicrografia: imunohistoquímica em fragmentos de
pulmões de cão positivo para leishmaniose visceral com anticorpos IgG
produzidos em coelhos. (A) IgG pré-imune; (B) IgG antiantígenos
miceliais de S. schenckii. Setas indicam formas amastigotas de
Leishmania spp. sem marcação. Barra: 10 µm..........................................
105

Figura 21: Regressão polinomial das curvas de crescimento do S.
schenckii (A) e Sporothrix spp. (B) frente a anticorpos IgY (1 mg/mL)
pré-imune (PI), anti-LSs, anti-MSs, IgY anti-LSc e Branco (sem solução
de IgY) às 0, 15, 18, 21, 24, 27, 38, 41 e 44 h de incubação a 37oC. ......
107
Figura 22: Dispersão das absorvâncias determinadas por ELISA de
sete amostras sorológicas de gatos com esporotricose comparados com
o grupo controle (gatos saudáveis). (A) Diluição dos soros: 1:900 versus
antígenos de extratos de leveduras de S. schenckii. Cutoff: 0,575 (B)
Diluição dos soros: 1:8100 versus antígenos de extratos de leveduras
de Sporothrix spp. Cutoff: 0,138................................................................
109
Figura 23: Cão (Amostra 1) com suspeita clínica de esporotricose de
mucosa antes e dois meses após o início do tratamento com solução
saturada de iodeto de potássio. ...............................................................
111
Figura 24 : Análise sorológica do cão com esporotricose pelo teste de
ELISA com o antígeno de levedura de S. schenckii. Curva de titulação
de duas amostras, uma amostra pré-tratamento (A) e outra após dois
meses de tratamento (B). ..........................................................................
115
Figura 25: (A) Resultados do teste de ELISA utilizando soros de cão
com histórico de esporotricose e soros de cães sadios versus o
antígeno de levedura de S. schenckii. (B) Versus o antígeno de
levedura de Sporothrix spp. Diluição dos soros: 1:100. ............................
116
Figura 26: Reconhecimento das moléculas antigênicas por soros de
cães negativos e positivo para a esporotricose por meio da técnica de
western blot. (A) Versus antígeno de levedura de S. schenckii:
amostras 1 – 10, soros de cães negativos, amostra 11: soro de cão
positivo. (B) Versus antígeno de levedura de Sporothrix spp.: amostras
1 – 8, soros de cães negativos, amostra 9: soro de cão positivo. PM:
peso molecular. Diluição dos soros: 1:300. ...............................................
117

Figura 27: Resultados do teste de ELISA utilizando soros de cães na
diluição 1:100 versus antígeno de levedura de S. schenckii . Amostra 1:
soro de cão positivo para esporotricose pré- tratamento e amostra: 2,
pós-tratamento. Amostra 3: soros de cães positivos para leishmaniose,
amostra 4: soros de cães negativos de zona endêmica de leishmaniose,
amostra 5: soro controle negativo para esporotricose. .............................
118
Figura 28: Face anterior da perna de mulher mordida por gato
positivo para esporotricose. Forma cutânea de esporotricose, antes
(setas) e após (setas) o tratamento com solução saturada de iodeto de
potássio. Setas indicam lesões residuais. ................................................
120
Figura 29: Fotomicrografia de pele de gato com esporotricose.
Infiltrado inflamatório composto por macrófagos repletos de leveduras
(setas) de cor púrpura (B). Hematoxilina-Eosina (A). Ácido Periódico de
Schiff (B). Barra 100 μm.............................................................................
122
Figura 30: Fotomicrografia de pele de humano com esporotricose.
Infiltrado inflamatório composto por macrófagos e neutrófilos. Ausência
de células fúngicas. (A) Hematoxilina-Eosina, (B) Ácido Periódico de
Schiff. Barra 50 μm ...................................................................................
123
Figura 31: Separação eletroforética por SDS-PAGE do extrato de
proteínas de leveduras de Sporothrix schenckii, após 7 dias de cultivo
em caldo cérebro-coração, 120 rpm, 37 ºC. Gel de poliacrilamida (10%).
Extratos proteicos brutos (20 μg de proteínas). P7: Tampão de lise Tris-
Cálcio e P8: Tampão de lise Ureia-Tioureia associados à lise física de
maceração em nitrogênio líquido. PM: Padrão de peso molecular -
Coloração por nitrato de prata. (Adaptada de Rodrigues, 2010). .............
125
Figura 32: Reconhecimento das moléculas antigênicas por soros de
gatos (n=21) por meio da técnica de western blot em tiras sensibilizadas
com antígenos de leveduras de S. schenckii ATCC 32285. Diluição do
soro: 1:300. PM: peso molecular (kDa)......................................................
153

Figura 33: Reconhecimento das moléculas antigênicas por 18 soros de
cães negativos (-) e um soro de cão positivo para esporotricose (+) por
meio da técnica de western blot em tiras sensibilizadas com antígenos
de leveduras de S. schenckii ATCC 32285. Diluição do soro: 1:300. PM:
peso molecular (kDa).................................................................................
153
Figura 34: Reconhecimento das moléculas antigênicas por 18 soros de
cães negativos (linhas 1- 8; 11- 20) e um soro de cão positivo para
esporotricose, em duplicata (linhas 9 e 10), versus antígeno de
leveduras de Sporothrix spp. Diluição do soro: 1:300. PM: Padrão de
peso molecular (kDa).................................................................................
154

LISTA DE TABELAS
Tabela 1: Esquema de imunização das galinhas para obtenção de IgY
antiantígenos de Sporothrix schenckii e Sporothrix spp. .......................
71
Tabela 2: Esquema de imunização dos coelhos para obtenção de IgG
antiantígenos de Sporothrix schenckii e Sporothrix spp. ........................
73
Tabela 3: Esquema de tratamentos para o teste de inibição de
crescimento de cepas de S. schenckii e Sporothrix spp. por anticorpos
IgY em meio líquido ..................................................................................
78
Tabela 4: Resultados médios de densidade óptica da curva de
crescimento do S. schenckii frente a anticorpos IgY com as respectivas
equações polinomiais, teste T e coeficiente de correlação (R2) ..............
106
Tabela 5: Resultados médios de densidade óptica da curva de
crescimento do Sporothrix spp. frente a anticorpos IgY com as
respectivas equações polinomiais, teste T e coeficiente de correlação
(R2) ...........................................................................................................
106
Tabela 6: Dados de identificação dos animais e densidades óticas
(D.O.) a 492 nm dos testes de ELISA utilizando soro de cão com
histórico de esporotricose e de cães aparentemente sadios (controles
normais) e incubados com os dois antígenos, levedura de S. schenckii e
levedura de Sporothrix spp.. Densidades óticas obtidas com soros
diluídos a 1:100, 1:300, 1:900, 1:2700 e 1:8100. DP = desvio padrão.
Cutoffs calculados como descrito acima ................................................
113
Tabela 7: Resultados de absorvância (492nm) do ensaio de ELISA
com soros de gatos saudáveis (animais controle) e infectados pelo
Sporothrix spp. versus antígenos de extratos de levedura de S.
schenckii. Diluições seriadas de 1:100 a 1:8100, médias, desvios
padrão e cutoffs ......................................................................................
155

Tabela 8: Resultados de absorvância (492nm) do ensaio de ELISA
com soros de gatos saudáveis (animais controle) e infectados pelo
Sporothrix spp. versus antígenos de extratos de levedura cepa clínica
de Sporothrix spp.. Diluições seriadas de 1:100 a 1:8100, médias,
desvios padrão e cutoffs ...........................................................................
156
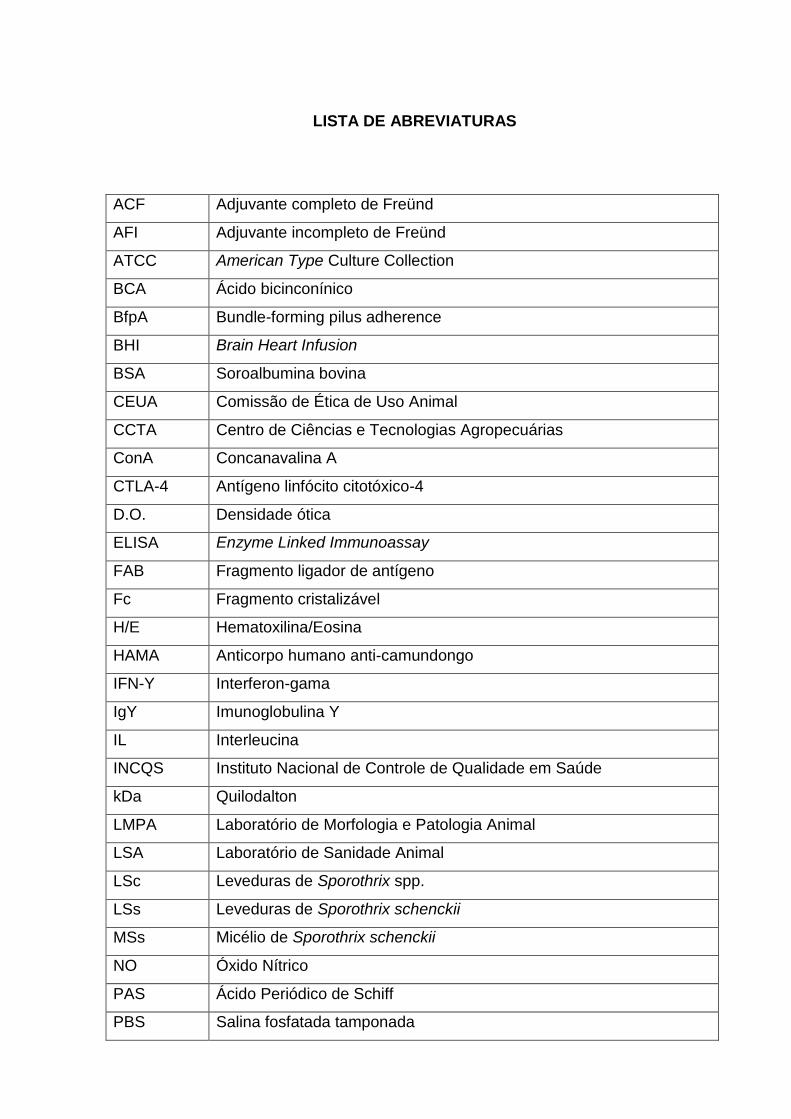
LISTA DE ABREVIATURAS
ACF Adjuvante completo de Freünd
AFI Adjuvante incompleto de Freünd
ATCC American Type Culture Collection
BCA Ácido bicinconínico
BfpA Bundle-forming pilus adherence
BHI Brain Heart Infusion
BSA Soroalbumina bovina
CEUA Comissão de Ética de Uso Animal
CCTA Centro de Ciências e Tecnologias Agropecuárias
ConA Concanavalina A
CTLA-4 Antígeno linfócito citotóxico-4
D.O. Densidade ótica
ELISA Enzyme Linked Immunoassay
FAB Fragmento ligador de antígeno
Fc Fragmento cristalizável
H/E Hematoxilina/Eosina
HAMA Anticorpo humano anti-camundongo
IFN-Y Interferon-gama
IgY Imunoglobulina Y
IL Interleucina
INCQS Instituto Nacional de Controle de Qualidade em Saúde
kDa Quilodalton
LMPA Laboratório de Morfologia e Patologia Animal
LSA Laboratório de Sanidade Animal
LSc Leveduras de Sporothrix spp.
LSs Leveduras de Sporothrix schenckii
MSs Micélio de Sporothrix schenckii
NO Óxido Nítrico
PAS Ácido Periódico de Schiff
PBS Salina fosfatada tamponada

PBST Salina fosfatada tamponada + Tween 20
PBST-L Salina fosfatada tamponada + Tween 20 + Leite em pó desnatado
PI Pré-imune
PM Padrão de Peso Molecular
ROS Espécies reativas de oxigênio
SC Sporothrix spp.
SDS-PAGE Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis
SRD Sem raça definida
Ss Sporothrix schenckii
SsCBF Sporothrix schenckii Con A Binding Fraction
Th Linfócitos T helper
TLR Tolll like receptors
TNF Fator de Necrose Tumoral
UENF Universidade Estadual do Norte Fluminense
UFC Unidades formadoras de colônia
YPD Levedura, peptona, dextrose

SUMÁRIO
PÁG.
1.0- INTRODUÇÃO 28
2.0 – REVISÃO DE LITERATURA 30
2.1 - AGENTE ETIOLÓGICO 30
2.1.1 - Classificação taxonômica do gênero Sporothrix 30
2.1.2 - Características de crescimento do agente 34
2.1.3 - Ultra-estruturas e dimorfismo celular 35
2.1.4 - Composição da parede celular 36
2.1.5 - Fatores de virulência 37
2.1.6 - Antígenos de Sporothrix schenckii 38
2.2 - HISTÓRICO DA ESPOROTRICOSE 40
2.3 - EPIDEMIOLOGIA 41
2.4 –ASPECTOS CLÍNICOS DA ESPOROTRICOSE 44
2.4.1 - Manifestações clínicas da esporotricose 44
2.4.2 - Reação imune à esporotricose 47
2.4.3 – Diagnóstico 48
2.4.4 - Citologia/ Histopatologia 49
2.4.5 - Métodos imunológicos 49
2.4.6 - Diagnóstico diferencial 51
2.4.7 - Prevenção e controle 51
2.4.8 – Tratamento 52
2.4.9 – Prognóstico 53
2.5 – IMUNOGLOBULINAS 53
2.5.1 - Transferência da IgY para o ovo 55
2.5.2 - Propriedades bioquímicas da IgY 57
2.5.3 - Vantagens da IgY 58
2.5.4 - Aplicações gerais da IGY 58
2.5.5 – Uso terapêutico da IgY 59
3.0 – JUSTIFICATIVA 64

4.0 – OBJETIVOS 65
5.0 - MATERIAL E MÉTODOS 66
5.1.0- Condições de cultivo das cepas de Sporothrix spp. para
extração de antígenos
66
5.1.1 - Extração dos antígenos 67
5.1.2 - Quantificação de proteínas 68
5.1.3 - Análise dos extratos proteicos antigênicos por SDS-PAGE 69
5.2.0 - Produção de anticorpos 69
5.2.1 - Produção de IgY - Imunização das aves 69
5.2.2 - Colheita de sangue e ovos 71
5.2.3- Purificação dos anticorpos IgY 72
5.2.4 - Análise em SDS-PAGE desnaturante 72
5.3.0 - IgG - Imunização dos coelhos 73
5.3.1 - Colheita de sangue 74
5.4 - Ensaio imunoenzimático (ELISA) dos soros e das preparações
de IgY purificadas
74
5.5 - Detecção de anticorpos antiantígenos de Sporothrix spp. por
western blotting
75
5.6 - Imunohistoquímica 76
5.7.0 - Avaliação da ação dos anticorpos IgY sobre o crescimento, in
vitro, das cepas de S. schenckii e Sporothrix spp.
77
5.7.1 - Análise estatística 79
5.8.0 - Colheita de sangue de cães e gatos 79
5.8.1 - Titulação por ELISA dos soros de gatos e cães versus
antígenos de Sporothrix spp.
80
5.8.2 - Detecção de anticorpos antiantígenos de Sporothrix spp. por
western blotting em soros de cães e gatos
80
5. 8.3 - Isolamento de Sporothrix spp. a partir de materiais clínicos 81
9.0- RESULTADOS 83
9.1 - SDS-PAGE DOS EXTRATOS ANTIGÊNICOS PRODUZIDOS 83
9.2.0 - ANÁLISE DOS ANTICORPOS IgY PRODUZIDOS POR SDS-
PAGE
84
9.2.1 - ENSAIOS DE ELISA DOS EXPERIMENTOS DE IMUNIZAÇÃO 85

9.2.1.1 - Titulação dos anticorpos IgY dos soros das galinhas e das
soluções de IgY purificadas das gemas
85
9.2.1.2 - Titulação por ELISA dos anticorpos IgG nos soros dos
Coelhos
88
9.2.1.3 - ELISA indireto para detecção do reconhecimento cruzado
entre anticorpos e antígenos de Sporothrix spp.
89
9.2.2 – WESTERN BLOT COM ANTISSOROS E SOLUÇÕES DE IgY
DOS ANIMAIS IMUNIZADOS
90
9.2.2.1- Western blot dos anticorpos IgY 90
9.2.2.2 - Western blot com anticorpos IgG de coelhos 92
9.3 – WESTERN BLOT COM SOROS DE ANIMAIS POSITIVOS E
NEGATIVOS PARA ESPOROTRICOSE
93
9.3.1 – Gatos 93
9.3.2 – Cães 96
9.4 - IMUNOHISTOQUÍMICA COM ANTICORPOS POLICLONAIS DE
GALINHAS E COELHOS
100
9.5 - AVALIAÇÃO DA AÇÃO DOS ANTICORPOS IgY SOBRE O
CRESCIMENTO, IN VITRO, DAS CEPAS DE S. schenckii e
Sporothrix spp.
105
9.6 - ANÁLISE SOROLÓGICA DE GATOS POSITIVOS PARA A
ESPOROTRICOSE
108
9.7 - RELATO DE CASO 1 110
9.8 - RELATO DE CASO 2 119
9.8.1 - Achados histopatológicos em seções tegumentares do gato
e da mulher
121
9.8.2 - Seções tegumentares do gato 121
9.8.3 - Seções tegumentares da mulher 122
10.0 – DISCUSSÃO 124
11.0 – CONCLUSÕES 134
12.0 – REFERENCIAS BIBLIOGRÁFICAS 136
APÊNDICE 1 153

28
1.0 - INTRODUÇÃO
A esporotricose é uma micose subcutânea subaguda ou crônica causada pelo
fungo dimórfico do complexo Sporothrix. Esta micose subcutânea é uma infecção de
implantação, que se desenvolve, na maioria dos casos, posteriormente a uma injúria
traumática seguida da inoculação do organismo, presente no ambiente, no
hospedeiro. Neste, ele afeta o tecido subcutâneo, a pele e outras estruturas
adjacentes (BARROS et al., 2001, SCHECHTMAN, 2010). Além do homem, a
esporotricose já foi descrita em várias espécies animais como cavalo, muar, cão,
gato, rato, camundongo, gambá, porco, camelo, chimpanzé, tatu e boi (LUTZ E
SPLENDORE, 1907; BARROS et al., 2001; CROTHERS et al., 2009).
A esporotricose tem distribuição mundial, porém é rara na Europa, mas é
comum na África, no Japão, na Austrália e nas Américas. No continente americano,
os países com maior endemicidade são El Salvador, Uruguai, Colômbia, Venezuela,
México e Brasil (PÉREZ et al., 2007). No Brasil, os casos têm sido relatados
principalmente nos estados do Rio de Janeiro, Rio Grande do Sul e São Paulo.
Atualmente, pertence à publicação brasileira, especificamente, no estado do Rio de
Janeiro, a maior coleção de casos de esporotricose em humanos, felinos e caninos e
a doença tem apresentado um caráter zoonótico.

29
No meio ambiente, o Sporothrix é isolado como saprófita, sob a forma
micelial, em associação com plantas vivas ou materiais orgânicos em decomposição
tais como: caules, folhas, palhas, madeiras, esterco (LACAZ et al., 1998). A
esporotricose era considerada como uma lesão adquirida através de atividades ao ar
livre ou como doença ocupacional para algumas classes de trabalhadores tais como:
fazendeiros, floristas, jardineiros, onde suas ocupações os obrigavam a estar em
frequente contato com materiais de plantas e solo (KOÇ et al., 2001; PÉREZ et al.,
2007, RAMÍREZ et al., 2010; VERMA et al., 2012). Após o surto epidêmico no Rio de
Janeiro, década de 80, a esporotricose passou a apresentar uma importância
zoonótica, principalmente, pela transmissão da doença do gato para o homem. A
partir daí passou a ser também uma doença ocupacional para os veterinários,
enfermeiros e auxiliares de consultórios veterinários (MARQUES et al., 1993,
BARROS et al., 2003, SILVA et al., 2012).

30
2.0 - REVISÃO DE LITERATURA
2.1 - AGENTE ETIOLÓGICO
2.1.1 - Classificação taxonômica do gênero Sporothrix
Reino - Fungi
Divisão - Ascomycota
Classe – Sordariomycetes
Ordem - Ophiostomatales
Família - Ophiostomataceae
Gênero - Sporothrix
Espécie – S. schenckii; S. brasiliensis; S. globosa; S. mexicana; S. albicans; S. luriei
Guarro et al. (1999); MARIMON et al. (2007); RODRIGUES (2010).
A heterogeneidade morfológica e genética de cepas de S. schenckii isoladas
a partir de material patológico foi registrada em diversos trabalhos ao longo dos
últimos anos (TRAVASSOS e LLOYD, 1980; De BEER et al., 2003). A discussão
sobre a filogenia do Sporothrix começou com Mariat et al. (1968), Mariat (1971ab),
Nicot and Mariat (1973), citados por De Beer et al. (2003), que sugeriram que o

31
Ophiostoma stenoceras poderia representar a forma teleomórfica do S. schenckii.
Desde então, a relação entre O. stenoceras e S. schenckii se tornou objeto de
muitas publicações. Em 2003, De Beer et al. Sequenciaram e compararam regiões
ITS (internal trancribed spacer) do gene ribossomal, inclusive o 5.8S do gene rRNA,
de isolados de O. stenoceras e O. nigrocarpum, colhidos de amostras de madeiras,
bezouros e solo de vários países; isolados de S. schenckii, obtidos de madeiras, do
solo e de pacientes humanos da África do Sul e de uma cepa ATCC (American Type
Culture Collection). Os autores confirmaram que O. stenoceras, O. nigrocarpum e S.
schenckii estão filogeneticamente relacionados, porém os dados de sequenciamento
do DNA separaram claramente estas três espécies e mostraram que o S. schenckii
pode ser classificado dentro do gênero teleomorfo Ophiostoma. Os isolados de S.
schenckii foram divididos em dois clados, os originários do solo e madeira e o
segundo com os isolados de tecidos humanos, sugerindo a existência de espécies
distintas dentro do grupo.
Marimon et al. (2006) sequenciaram o DNA de três diferentes loci (quitina
sintase, calmodulina e ß-tubulina) de 59 isolados clínicos de humanos e um isolado
ambiental, anteriormente identificados como S. schenckii. Os isolados humanos
foram originários do Brasil (Rio de Janeiro e São Paulo), Colômbia, Peru, Argentina,
Espanha e África do Sul. Os autores encontraram a existência de três grandes
clados: clado I com os isolados do Brasil, clado II com os isolados dos outros países
da América do Sul e da África e o clado III com os isolados da Espanha. Dentro dos
18 isolados do Rio de Janeiro, foram observados seis genótipos diferentes. Segundo
os autores, a população de S. schenckii estava em processo de diversificação com
possibilidade de haver uma especificidade geográfica para os grupos principais.
Em 2007, Marimon et al. estudaram 127 isolados, anteriormente identificados
como S. schenckii, através da amplificação, do sequenciamento e das análises
filogenéticas do gene nuclear calmodulina (CAL), das características macroscópicas,
da esporulação, das características microscópicas e dos estudos fisiológicos. Os
pesquisadores concluíram que os primers (CL1 e CL2A, O´Donnell et al., 2000)
utilizados foram capazes de amplificar e sequenciar o loci do gene CAL com 776 pb.
Os isolados foram subdivididos em cinco clados. Todos os isolados cresceram bem
entre 20 e 30oC e também apresentaram crescimento a 35oC, nenhum dos isolados
cresceu à temperatura de 40oC. A maioria dos isolados do clado III não apresentou
crescimento a 37oC. A mais importante variação na assimilação de carboidratos foi

32
observada para a sacarose, rafinose e adonitol. Todos os isolados foram capazes de
quebrar a ureia após oito dias de incubação, todos toleraram ciclohexamida a 0,25%
e a maior parte dos isolados fez a conversão para a fase leveduriforme, sem
diferenças significativas entre os clados. Diante dos resultados encontrados, os
pesquisadores propuseram a criação de três novas espécies para o complexo de
espécies do gênero Sporothrix: Sporothrix brasiliensis Marimon, Gené, Cano, et
Guarro, sp. nov. = Sporothrix schenckii, clade I sensu Marimon et al. (2006);
Sporothrix globosa Marimon, Gené, Cano, et Guarro, sp. nov. = Sporothrix schenckii,
clade III sensu Marimon et al. (2006) e Sporothrix mexicana Marimon, Gené, Cano,
et Guarro, sp. nov. = Sporothrix schenckii, clade IV sensu Marimon et al. (2007). As
espécies S. brasiliensis e S. globosa foram associadas com a infecção em humanos.
Posteriormente, foi proposto que a variante morfológica denominada S. schenckii
var. luriei fosse transformada em uma nova espécie com a denominação de
Sporothrix luriei.
Em São Paulo, Rodrigues (2010) estudou 161 cepas de Sporothrix spp.
provenientes de amostras clínicas e ambientais de diversas regiões do Brasil e de
outros países. Demonstrou que as espécies S. brasiliensis, S. globosa, S. mexicana
e S. schenckii têm ampla distribuição geográfica no país. Registraram, também, que
a espécie S. brasiliensis foi isolada com alta frequência de gatos nos estados do Rio
Grande do Sul e Rio de Janeiro, se mostrando um importante reservatório na
epidemiologia da esporotricose.
No Rio de Janeiro, Oliveira et al. (2011) utilizaram as características
fenotípicas propostas por Marimon et al. (2007) para caracterizar 246 isolados de
Sporothrix spp., sendo 245 isolados de pacientes humanos e uma amostra
ambiental. A maior parte (83,4%) foi caracterizada como sendo S. brasilensis, 6%
como S. schenckii e 0,5% como S. mexicana. Vinte e cinco isolados (10,1%) não
puderam ser classificados pelas características fenotípicas, seguindo-se então, o
sequenciamento do gene da calmodulina para a confirmação da identidade dos
isolados, 24 foram classificados como S. brasiliensis e um isolado como S. globosa,
os resultados mostraram a importância da análise, em nível de genótipo, para a
confirmação da classificação das espécies (OLIVEIRA et al., 2014).
Na Itália, Romeo et al. (2011) estudaram as relações filogenéticas entre
isolados de Sporothrix schenkii, 26 cepas ambientais e duas amostras clínicas de
humanos, através do gene da calmodulina. As cepas clínicas foram agrupadas

33
dentro do stricto sensu do Sporothrix schenckii e as cepas ambientais como S.
albicans, cujo nome foi recentemente substituído para S. pallida.
Segundo Oliveira et al. (2012), a chave de identificação baseada em testes
fenotípicos (MARIMON et al., 2007) foi descrita como fácil e suficiente para a
diferenciação de espécies sem a necessidade das técnicas moleculares. Entretanto,
os resultados são sempre inconclusivos ou ambíguos (OLIVEIRA et al., 2011; 2012;
ALMEIDA-PAES et al., 2014, RODRIGUES et al., 2014) e algumas espécies estão
tão proximamente relacionadas, que fenotipicamente não apresentam nenhuma
diferença. Por outro lado, o sequenciamento parcial do gene da calmodulina para
diferenciação de espécies pode se tornar bastante caro e dificultar a análise de um
grande número de amostras.
Buscando alternativas para se fazer um levantamento genotípico rápido, o
qual é importante para estudos epidemiológicos (RODRIGUES, 2010), visto que as
espécies diferem em distribuição, virulência (ARRILLAGA-MONCRIEFF, et al., 2009;
FERNANDES et al., 2013) e resistência aos antifúngicos (MARIMON et al., 2008),
Oliveira et al. (2012) propuseram a técnica da PCR fingerprinting para distinção das
espécies do complexo Sporothrix, utilizando o primer universal T3B. Para os autores
os perfis gerados por esta técnica foram altamente informativos, geraram padrões de
bandas específicas para cada espécie, permitiram a sua diferenciação e
apresentaram 100% de concordância com os resultados gerados pelo
sequenciamento do locus da calmodulina. Também foi sensível o suficiente para
classificar as espécies que apresentaram resultados inconclusivos nos testes
fenotípicos.
Rodrigues et al. (2014) sugeriram a PCR-RFLP do locus da calmodulina para
identificação das espécies do complexo Sporothrix de importância clínica (S.
schenckii, S. brasiliensis e S. globosa). Para as espécies S. mexicana e S. pallida, a
técnica não foi discriminatória, ambas apresentaram o mesmo sítio de clivagem
pelas enzimas de restrição, íntron 3 e exon 5 do locus da calmodulina.

34
2.1.2 - Características de crescimento do agente
O Sporothrix schenckii é um fungo dimórfico, ou seja, na sua forma saprofítica
ou in vitro, à temperatura inferior a 35 – 37oC, se desenvolve formando hifas e
conídios. A forma de levedura é obtida através do desenvolvimento, in vitro e in vivo,
a 35 – 37oC. A temperatura ótima para o crescimento micelial é de 25-27oC e a 39-
40oC o crescimento é inibido (LACAZ et al., 1998).
O isolamento do Sporothrix spp., em cultura, para o crescimento micelial,
pode ser feito utilizando o ágar Sabouraud dextrose a 2 ou 4% suplementado com
cloranfenicol e ciclohexamida. A princípio, as colônias filamentosas são lisas e
rugosas, de coloração branca a creme, tornando-se marrons, cinza-escuro a negras
após alguns dias. Algumas cepas, no entanto, podem apresentar a habilidade de
formar colônias escuras ao início do crescimento. Nos subcultivos, as colônias
podem perder, irreversivelmente, a cor escura, tornando-se branca-creme. As
colônias nunca se tornam flocosas ou cotonosas. As hifas aéreas são raras no
primeiro isolamento, mas são produzidas nos subcultivos. No cultivo em lâmina, à
temperatura de 25oC, as hifas são finas, hialinas, septadas e ramificadas, com
conidióforos delgados, em cujo ápice forma-se uma pequena vesícula com
dentículos simpodialmente dispostos. Em cada dentículo nasce um conídio, ficando
todos reunidos em disposição floral. Os conídios desprendem-se dos conidióforos,
ficando, por vezes, dispostos uns ao lado dos outros, enfileirados e em ambos os
lados do micélio. Os conídios podem ser hialinos com formato oval, elíptico a
piriforme (2 a 3 x 3 a 6 μm nas menores e maiores dimensões) ou escuros, de
parede celular espessa, esféricos a ovais, com 2 a 4 μm de diâmetro (LACAZ et al.,
1998).
As amostras de S. schenckii reproduzem-se por brotamento nos tecidos do
homem e dos animais. Considerando o dimorfismo, a forma de levedura pode ser
obtida in vitro, a partir de cultivos puros micelianos ou conidiais em ágar-infusão
cérebro-coração, ágar YPD (levedura/peptona/dextrose), dentre outros meios,
incubados a 37oC. Os blastoconídios produzidos in vitro podem ser ovais ou
fusiformes, com 2 - 6 μm de diâmetro, e normalmente apresentam brotações
alongadas em forma de charuto. Podem apresentar brotações únicas ou múltiplas.
Macroscopicamente, as colônias são lisas, de coloração creme a amarronzada
(LACAZ et al., 1998, BARROS et al., 2011).

35
O S. schenckii cresce em meios simples contendo sais inorgânicos,
asparagina, glicose e tiamina. Outros aminoácidos e vitaminas são estimulatórios,
mas não essenciais. Assimilam glicose, galactose, maltose, xilose e glicerol, ácidos
orgânicos não são utilizados. Outros sacarídeos são assimilados, mas não por todas
as cepas. Todas as cepas hidrolisaram amido (TRAVASSOS e LLOYD, 1980).
2.1.3- Ultraestruturas e dimorfismo celular
A estrutura celular do Sporothrix spp. é semelhante à dos outros eucariotos,
constituída basicamente por uma membrana, um citoplasma com as organelas
distribuídas aleatoriamente por todo interior celular e um compartimento especial, o
núcleo, que armazena o material genético. As células podem ser encontradas na
forma leveduriforme, ou então, formando conjuntos de hifas septadas, denominadas
de micélio. Tanto as células leveduriformes quanto o micélio estão envolvidos por
uma camada protetora externa denominada de parede celular (GARRISON et al.,
1975, FUKUDA et al., 2009).
A morfologia interna das estruturas do S. schenckii, assim como, a transição,
in vitro, da fase micelial para a leveduriforme, foram mostradas por microscopia
eletrônica por Garrison et al. (1975) e TRAVASSOS e LLOYD (1980). A parede
celular da levedura é mais espessa (100 a 300 nm) do que a do micélio (80 a 140
nm) e menos espessa do que a do conídio pigmentado (330 nm). Na levedura, são
observadas duas camadas eletro-densas, sendo a mais externa constituída de
microfibrilas, semelhante a um biofilme, presente somente na levedura e ausente na
fase micelial. As leveduras apresentaram grânulos de reserva e membrana
intracitoplasmática (GARRISON et al., 1975). Em hifas incubadas a 37oC/48 h,
observou-se grande número de protusões laterais na região do septo, ou próximo a
ele, e não se observou nenhuma alteração na fina estrutura do conteúdo
citoplasmático. Houve formação de brotações no ápice e por toda a hifa com
formação de células oidianas, que são as células iniciais da levedura. Não foi
observada brotação direta a partir de conídios. Nos conídios maduros foram
observados de um a vários corpos lipídicos de reserva que podem ocupar 50% ou
mais da área celular interna e que não estão presentes no micélio nem na levedura.
(GARRISON et al., 1975).

36
2.1.4 - Composição da parede celular
A parede celular fúngica pode ser caracterizada como uma estrutura
relativamente rígida, porém altamente dinâmica, que muda constantemente durante
a divisão celular, o crescimento e a morfogênese. Ela determina o formato da célula,
fornece suporte osmótico, proteção física, além de estar relacionada a eventos de
sinalização celular, adesão e reprodução, sendo por isso necessária para o
crescimento dos fungos nos ambientes onde são encontrados (FUKUDA et al.,
2009). É uma estrutura complexa composta tipicamente por quitina, homopolímero
linear, longo, formado por resíduos de N-acetilglicosamina com ligações β-(1→4);
1,3-β- e 1,6- β-glucanas, mananas e proteínas, entretanto estas composições variam
marcadamente entre as espécies de fungo (ADAMS, 2004).
Um dos componentes muito bem conhecidos da parede celular do S.
schenckii é a peptideoramnomanana. Este glicopeptídeo ou glicoconjugado é
formado por um complexo de moléculas, com uma larga variação de peso molecular
e de difícil purificação. Nos glicoconjugados da parede celular da forma do S.
schenckii foram identificadas glucanas solúveis e insolúveis unidas por ligações do
tipo β (1,3; 1,4 e 1,6) nas duas fases morfológicas do fungo, a existência de α
glucanas não foi identificada e não houve correlação entre a composição de β
glucanas e a morfologia transacional do fungo (PREVIATO et al., 1979). Segundo
Lloyd e Bitoon (1971), a peptideoramnomana da parede celular da levedura é
composta por 33,5% de ramnose, 57% de manose e 14,2% de proteína. Também
apresenta polissacarídeos contendo galactose na superfície do fungo, sugerindo a
presença de galactomananas na parede celular e resíduos de ácido glicurônico
foram descritos nas frações ácidas das ramnomananas. Componentes similares
foram detectados em frações isoladas de filtrados de cultura da fase micelial do S.
schenckii. Outro achado importante foi que estas peptideoramnomananas reagem
com a proteína concanavalina A, enquanto as ramnomananas extraídas por
tratamento alcalino a quente não reagem (LOPES-BEZERRA, 2011).

37
2.1.5 - Fatores de virulência
Pode-se definir fator de virulência como uma característica do micro-
organismo que permite ou estimula o seu crescimento no hospedeiro. Para estudar e
caracterizar os fatores de virulência é necessário comparar as interações micro-
organismo-hospedeiro de um isolado que expressa o fator suspeito e isolar um
mutante que perdeu a capacidade de expressá-la, o que pode ser conseguido por
mutagênese induzida através de estratégias moleculares. Se as diferenças nas
infecções causadas por estes diferentes isolados são notadas, é imperativo fazer o
mutante recuperar a capacidade de expressar o fator estudado e verificar se a
capacidade de causar a infecção é similar à da cepa selvagem parental (BARROS et
al, 2011).
A teoria mais aceita para a origem da virulência é a de que a interação
microbiana com outros organismos presentes no habitat natural do patógeno faz
com que os micro-organismos adquiram estratégias de sobrevivência e tenham uma
maior virulência do que quando encontram acidentalmente um animal hospedeiro.
Pouco se sabe sobre os fatores de virulência do S. schenckii devido aos poucos
estudos nesta área, em parte pelo fato do S. schenckii ser pouco responsivo às
análises genéticas. Mas, alguns fatores têm sido apontados: termotolerância,
melanina, adesinas, peróxido de ergosterol, enzimas e proteínas (BARROS et al.,
2011).
Quanto à termotolerância, sabe-se que isolados capazes de crescer a 35oC,
mas incapazes de crescer a 37oC são incapazes de causar a esporotricose linfática,
mas capazes de causar a esporotricose cutânea fixa. Ambas as fases morfológicas
do S. schenckii apresentam a habilidade de sintetizar a melanina que é um
composto insolúvel altamente relacionado com virulência em vários fungos
(BARROS et al., 2011). Romero-Martinez et al. (2000) demonstraram que a
melanina protege o S. schenckii contra certos compostos antimicrobianos oxidativos
e contra o ataque de macrófagos. Isolados pigmentados de Sporothrix spp. podem
apresentar uma maior capacidade de invasão do que o mutante albino em modelos
murinos. Em uma infecção, a cepa albina pode ficar restrita ao centro do granuloma
enquanto a cepa melanizada promove a formação de granulomas multifocais
(MADRID et al., 2010). O peróxido de ergosterol, formado em fungos patogênicos,
foi encontrado pela primeira vez no S. schenckii, é formado como um mecanismo de

38
proteção para fugir das espécies reativas de oxigênio durante a fagocitose pelas
células polimorfonucleares do hospedeiro (BARROS et al., 2011).
A adesão primária a células endoteliais e epiteliais, assim como aos
componentes da matriz extracelular é essencial para uma invasão efetiva dos
tecidos do hospedeiro pelos patógenos (LIMA et al., 1999). Tanto os conídios quanto
as leveduras do Sporothrix spp. são capazes de reconhecer e aderir a três
importantes glicoproteínas da matriz extracelular: fibronectina, laminina e colágeno
tipo II (LIMA et al., 1999). O efeito do fungo sobre monocamadas de células
epiteliais, in vitro, é citopático, há perda dos contatos celulares, da integridade da
monocamada e, consequentemente, da polaridade celular. O estudo bioquímico
mostrou que existem moléculas glicoproteicas na superfície dos epitélios que
interagem com as leveduras (SANDOVAL-BERNAL et al., 2009).
2.1.6 - Antígenos de Sporothrix schenckii
Os antígenos da parede celular do S. schenckii foram primeiramente
estudados por Gonzales-Ochoa e Figueroa (1947) através da análise de
precipitação e intradermorreação com a esporotriquina. Em 1967, de Bièvre e
Prevot, citados por Mendoza et al. (2002), descreveram um estudo eletroforético de
extratos proteicos do micélio e da levedura de S. schenckii. Em 1971, Lloyd e Bitoon
(1971) demonstraram que uma peptídeo-ramnomanana, glicopeptídeo predominante
na parede celular deste fungo, precipitava com soro de pacientes com esporotricose.
Scott e Muchmore (1989) produziram antígenos solúveis de leveduras do S.
schenckii, 15 bandas proteicas foram demonstradas pela SDS-PAGE, com variação
de peso molecular entre 22 e 70 kDa. À análise por western blot, os soros de
pacientes humanos com esporotricose reconheceram as proteínas de 40 e 70 kDa, a
banda de 40 kDa foi proeminente em todos os soros, enquanto a de 70 kDa mostrou
variabilidades de densidade. As bandas entre 22 e 36 kDa estiveram presentes
somente nos soros de pacientes humanos com esporotricose extracutânea. Os
autores também observaram que nos soros de pacientes com esporotricose
extracutânea, as bandas do western blot foram mais escuras do que nos soros de
pacientes com esporotricose cutânea, mesmo que estes apresentassem mesmo

39
título ao teste de ELISA. No ensaio de ELISA, 18% dos soros negativos e 20% dos
soros heterólogos apresentaram anticorpos detectáveis.
Mendoza et al. (2002) mostraram que a expressão de exoantígenos na fase
micelial do S. schenckii pode variar de acordo com o meio de cultura, o pH e a fase
de crescimento do fungo. Segundo os autores, o caldo Sabouraud apresenta um
padrão proteico complexo, com melhor expressão, na fase estacionária de
crescimento. Os autores constataram a expressão de dez bandas proteicas, com
pesos moleculares entre 29 e 200 kDa. A banda de 55 kDa foi a mais proeminente e
a de 40 kDa a mais estreita. O componente de 55 kDa foi correlacionado com a
peptideoramnomanana por ter sido caracterizada nas diversas preparações
antigênicas e por ter sido bastante reativa com os soros estudados. O antígeno de
90 kDa foi considerado como de grande valor diagnóstico, por ter reagido
especificamente com soros de pacientes com esporotricose e sem reação cruzada
com soros de pacientes com outras micoses.
Lima e Lopes Bezerra (1997) mostraram que a fração antigênica da parede
celular do S. schenckii (LLOYD e BITOON, 1971), SsCBF, apresenta três antígenos
principais, com peso molecular aproximado de 84, 70 e 58 kDa, que reagem com
soro de coelho anti-S. schenckii.
Nascimento e Almeida (2005) mostraram que antissoros de camundongos
reagiram somente contra a proteína de 70 kDa presente em exoantígenos
leveduriformes do S. schenckii. Em 2008, experimentos in vivo, mostraram uma
redução significativa do número de UFC nos órgãos de camundongos que
receberam anticorpos monoclonais, contra a glicoproteína de 70 kDa, antes e
durante a infecção (NASCIMENTO et al., 2008).
Em 2009, Ruiz-Baca et al. mostraram uma distribuição uniforme da proteína
de 70 kDa extraída da parede celular da levedura do S. schenckii sobre a superfície
de três diferentes isolados de S. schenckii. Ruiz-Baca et al. (2011) utilizaram o
ensaio de 2D-immunoblotting com anticorpos anti-S. schenckii produzidos em
coelhos para identificar proteínas antigênicas nas preparações da parede celular
obtidas das formas micelial e de levedura do fungo. Cinco antígenos de 48, 55, 66,
67 e 70 kDa foram imunodetectados na fase micelial do fungo. A glicoproteína de 70
kDa foi o principal antígeno detectado na parede celular das duas fases morfológicas
do fungo e a glicoproteina de 60 kDa esteve presente somente nas células
leveduriformes. As glicoproteínas de 70 e 60 kDa da parede celular de leveduras

40
também foram imunodetectadas com antissoros de coelhos em isolados de S.
brasiliensis e S. globosa (Ruiz-Baca et al., 2014).
Almeida-Paes et al. (2012) encontraram uma variedade de bandas
reconhecidas por antissoros de coelhos contra antígenos livres miceliais (220 – 40
kDa) e de levedura (220 – 20 kDa) de cepas do S. schenckii ATCC 23252 e S.
brasiliensis. Houve variação do número de bandas reconhecidas por cada cepa. O
extrato antigênico da levedura da cepa ATCC 23252 mostrou o maior número de
bandas reativas (22 bandas), entretanto, a intensidade das bandas foi menor que as
originadas pelos isolados de S. brasiliensis. A banda de 85 kDa foi reconhecida em
todos os extratos e nas duas fases morfológicas do fungo. Ao testarem soros
humanos positivos para a esporotricose, os autores identificaram sete proteínas
específicas do S. brasiliensis, que não apresentaram reações cruzadas com soros
heterólogos.
2.2 - HISTÓRICO DA ESPOROTRICOSE
O primeiro caso de esporotricose foi descrito por Benjamin Schenck , no
boletim do “Johns Hopkins Hospital”, em dezembro de 1898, intitulado de " On
Refractory Subcutaneous Abscesses caused by a Fungus possibly related to the
Sporotricha" (Abcesso subcutâneo refratário, causado por um fungo possivelmente
relacionado ao Sporotricha). O primeiro ponto da infecção foi no dedo indicador de
onde se estendeu para o braço, seguindo os vasos linfáticos e dando origem a
várias indurações circunscritas, algumas abriram e ulceraram. A infecção se mostrou
muito refratária ao tratamento. Schenck conseguiu isolar o agente das lesões e
descreveu o desenvolvimento do micro-organismo, as características da cultura e a
morfologia do agente. Enviou uma cepa para o micologista Erwin F. Smith, do
Departamento de Agricultura dos EUA em Washington, o qual concluiu que o micro-
organismo fazia parte do gênero Sporotrichum (HEKTOEN E PERKINS, 1900).
Poucos meses depois Hektoen e Perkins (1900) estudaram um caso que
apresentou características semelhantes às descritas por Schenck, com abcessos
subcutâneos refratários ao tratamento e o organismo isolado com características
idênticas às descritas por Schenck. Foi o caso de uma criança de cinco anos que
machucou o dedo indicador com um martelo e que 10 dias após apresentou-se

41
inchado e com uma ferida que não cicatrizava. O caso evoluiu para uma linfangite
nodular, supurada como no caso descrito por Schenck. Hektoen e Perkins
classificaram o agente etiológico como Sporothrix schenckii.
No Brasil, os primeiros casos de esporotricose foram descritos por Lutz e
Splendore (1907). Os pesquisadores tinham conhecimento de lesões que se
manifestavam espontaneamente no rato comum (Mus decumanus) na cidade de
São Paulo, iniciando principalmente na cauda. Os autores tiveram dificuldades em
identificar o agente e não encontraram na literatura nada parecido com o que
estavam vendo nos animais e “in vitro”. Inicialmente pensaram ser uma infecção
bacteriana e, posteriormente, devido à morfologia microscópica do agente, à
presença de hifas e às células semelhantes às vistas na doença do sapinho, os
autores passaram a empregar meios de cultura menos favoráveis para bactérias e
mais apropriados para tórulas e bolores. Desse modo, conseguiram facilmente isolar
um organismo que apresentava uma certa polimorfia, aparecendo, todavia, as
formas observadas sempre na mesma ordem e com os mesmos caracteres
microscópicos, acompanhando a transformação macroscópica da cultura.
Posteriormente, Lutz e Splendore tomaram conhecimento das publicações dos
americanos, Schench, Hektoen e Perkins, e constataram que os seus achados eram
bastante semelhantes aos encontrados por eles. Lutz e Splendore citaram a
ocorrência de cinco casos clínicos, sendo de um funcionário de laboratório, dois
açougueiros que tiveram lesão no dedo, um homem possivelmente mordido por rato
e outro que foi extensamente picado por insetos em zona rural, todos foram tratados
com solução de iodeto de potássio, curados, mas não se conseguiu isolar o agente
destas lesões.
2.3 - EPIDEMIOLOGIA
O primeiro grande surto de esporotricose foi descrito na África do Sul, entre
1941 e 1944, com 2825 mineiros contaminados devido à presença do fungo nas
madeiras que serviam de escoras nos corredores de uma mina de ouro no
Transvaal. Como a umidade do local atingia 100%, o ambiente era favorável à
multiplicação e esporulação do fungo, facilitando a contaminação de tantos

42
trabalhadores (Transvaal Mine Medical Officer´s Association, 1947) (FINDLAY, 1970,
citado por CRUZ, 2013).
O segundo grande surto foi descrito por DIXON et al. (1991), que relataram o
aparecimento de casos de esporotricose, no início do verão de 1988, em
trabalhadores que estavam participando do programa anual de reflorestamento em
Nova York e Illinois. Foram identificados 84 casos em 15 estados e constituiu a
maior epidemia de esporotricose nos EUA. Todos os casos foram associados com o
contato dos trabalhadores com o musgo esfagno que foi obtido de um único
distribuidor de Wisconsin. Nos viveiros, os musgos são desidratados e
armazenados, para, posteriormente, envolver as raízes das mudas e mantê-las
úmidas durante o embarque e transporte.
E o terceiro, com características bem diferentes, do que se sabia
anteriormente, é o que está ocorrendo no estado do Rio de Janeiro, há pelo menos
14 anos, onde a principal ocorrência tem sido relacionada à arranhadura e/ou
mordedura de gatos, com infecção dos familiares, além de casos em profissionais
que lidam com esses animais, como veterinários e auxiliares (SILVA et al., 2012).
Em vários países do mundo, os relatos de casos e a incidência da
esporotricose têm sido sobre a ocorrência da doença em humanos, sempre após
traumas relacionados com o exercício de atividades extradomiciliares, tais como:
cultivo da terra, recolhimento de lenha e forragens de florestas, criação de animais,
jardinagem, picadas de insetos, atividades de lazer em áreas rurais, como as
pescarias e caçadas (KOÇ et al., 2001;PÉREZ et al., 2007; RAMÍREZ et al., 2010;
VERMA et al., 2012). No Uruguai, a maior incidência da esporotricose foi
relacionada com a atividade de caça ao tatu (Dasipus septemcinctus), onde mais de
80% dos casos de esporotricose humana foram atribuídos a esta atividade (CONTÍ-
DIAS, 1980 citado por ALVES et al., 2010). Os tatus normalmente se escondem em
tocas, o caçador deve puxá-los pela cauda e não é incomum o caçador adquirir
traumas na pele durante esta operação. O solo das tocas dos tatus é rico em
matéria vegetal que pode ser um importante reservatório para o fungo (ALVES et al.,
2010).
No Brasil, no estado do Rio Grande do Sul, a ocorrência da doença em
humanos foi relacionada a atividades de lazer em zona rural, como pescarias e
caçadas, inclusive a caça ao tatu (LOPES et al., 1999; ALVES et al., 2010). No
estado do Rio de Janeiro, de 1987 a 1998, 13 casos de esporotricose humana foram

43
registrados pelo CPqHEC (Centro de pesquisa de dermatologias infecciosas do
Hospital Evandro Chagas -Fiocruz) e, em 2001, a doença foi caracterizada como
emergente neste estado (Barros et al., 2001). Entre 1997 e 2009, foram registrados
no Hospital Universitário Pedro Ernesto, Rio de Janeiro, 171 casos de esporotricose
em humanos (OROFINO-COSTA et al., 2011).
Posteriormente, os registros da Fiocruz mostraram que no período
compreendido entre 1997 e 2007 foram diagnosticados e tratados 1848 casos de
esporotricose humana, em residentes no Estado do Rio de Janeiro, dos quais 1289
foram registrados no último quadriênio. No detalhamento da fonte de infecção,
identificou-se que a maioria dos casos tinha registros de relatos de pacientes com
trauma envolvendo animais no ambiente doméstico. Um mil duzentos e vinte seis
casos relataram o trauma com gato como fonte de infecção (66,34%): gatos
domésticos, de rua ou sem especificação de origem, 16,61% trauma ao lidar com
plantas ou terra e 0,5% foram referentes à menção de picadas de insetos, aves ou
acidentes com cachorro (SILVA et al., 2012). Entre 2008 e 2011 foram confirmados
2.340 novos casos de esporotricose humana, segundo dados do Serviço de
Vigilância em Saúde da Fiocruz (SILVA et al., 2012).
Quanto ao atendimento de animais doentes, até 2010, foram atendidos,
aproximadamente 3.244 gatos e mais de 120 cães com esporotricose. Os cães não
desempenharam papel importante na cadeia epidemiológica da micose, pois não
houve nenhuma comprovação de transmissão ao ser humano por esses animais
(BARROS et al., 2010). Supõe-se que gatos adquirem a doença ao se arranharem
com vegetações e quando brigam com outros gatos de rua (CRUZ, 2013). Diante
dos relatos históricos da doença no Brasil e a relação na cadeia alimentar entre
gatos e ratos, seria interessante também, se avaliar o papel dos ratos na
epidemiologia desta doença.
Schubach et al. (2001; 2002) e Souza et al. (2006) mostraram que, além dos
gatos doentes, gatos sadios que têm acesso extradomiciliar ou em contato com
animais doentes podem ser portadores assintomáticos do Sporothrix spp. e carreá-lo
para o ambiente doméstico, onde poderá persistir por prolongados períodos de
tempo (REIS et al., 2009). A manutenção do Sporothrix spp. em clínicas e
consultórios veterinários, após atendimento de animal doente, também pode ser
uma fonte de infecção para os profissionais da área, assim como, para outros
animais que serão atendidos no recinto. Mattei et al. (2011) mostraram que o S.

44
schenckii pode ser isolado do ambiente hospitalar, após atendimento de gato com
lesões de esporotricose, mesmo após a desinfecção do local com álcool iodado.
Pelo levantamento dos dados epidemiológicos, a endemia da esporotricose
em gatos, no Rio de Janeiro, não atinge a Zona Sul, a região mais rica da cidade. Na
Zona Sul os gatos vivem em apartamentos, já nos bairros e nas cidades periféricas,
a população vive em casas, os gatos são mais livres, eles estão nos quintais e vão à
rua com frequência e sozinhos. Estes foram introduzidos como uma forma eficiente
de controle de ratos e passaram a ser vistos como animais tão úteis quanto os cães.
O aumento do número de casos de esporotricose nas regiões mais pobres do
Grande Rio pode ter aí a sua origem (ALVES, 2010; CRUZ, 2013).
2.4 – ASPECTOS CLÍNICOS DA ESPOROTRICOSE 2.4.1 - Manifestações clínicas da esporotricose
De acordo com Lopes-Bezerra et al. (2006), as apresentações clínicas da
esporotricose podem ser classificadas em:
Cutânea
◦ Linfocutânea
◦ Fixa
◦ Disseminada ou múltipla
Mucosa
◦ Ocular
◦ Nasal
◦ Outras
Extracutânea
◦ Pulmonar
◦ Osteoarticular
◦ Meningeal
◦ Generalizada
Residual (Sequela)
Formas especiais
◦ Regressão espontânea
◦ Hipersensibilidade (eritema nodoso, eritema multiforme)

45
Em humanos, a forma clínica mais frequente da esporotricose (em torno de
80%) é a forma linfocutânea que se inicia com uma única pápula no local da injúria,
mais comumente na mão, que aparece algumas semanas após a inoculação
(BARROS et al., 2001; ALVES et al., 2010; VERMA et al., 2012). A lesão se torna
ulcerada com um dreno purulento e geralmente não é dolorida. Semanas após o
desenvolvimento da lesão inicial, aparecem lesões adicionais, nódulos dérmicos e
subcutâneos típicos, ao longo dos vasos linfáticos, normalmente em direção ao
braço. Esta descrição clínica caracteriza a denominação da doença: “linfangite
nodular ascendente” (SCHECHTMAN, 2010). Em geral, a forma fixa cutânea é
caracterizada por lesões nodulares infiltradas, ulceradas ou eritematosas localizadas
na área exposta à inoculação fúngica. A forma cutânea disseminada ou múltipla
(BARROS et al., 2003) tem sido observada em pacientes imunosuprimidos, como os
positivos para o vírus HIV (SILVA-VERGARA et al., 2012), etilistas (SCHECHTMAN
et al., 2011; NASSIF et al., 2012), pessoas com idade avançada, que receberam
quimioterapia, diabetes, etc. A esporotricose disseminada foi responsável pelo
aumento do número de hospitalizações em 44% dos pacientes portadores do vírus
HIV contra 1% do grupo de pacientes somente com esporotricose, soronegativo para
o HIV (FREITAS et al., 2014). O acometimento de mucosas não é comum, mas pode
ocorrer e afeta preferencialmente a mucosa ocular (RIBEIRO et al., 2010). Entre as
formas extracutâneas, as mais comuns são a osteoarticular e a pulmonar, mas há
relatos de casos com severa disseminação hematogênica com acometimento de
múltiplos órgãos (SCHECHTMAN, 2010).
Nos cães, a via de entrada do fungo tem sido relacionada a perfurações com
espinhos, lascas de madeira, picadas de insetos, contato com gatos doentes. A
forma cutânea localizada ou fixa é a mais comumente relatada. Múltiplos nódulos
firmes, placas ulceradas com bordas elevadas e áreas alopécicas estão geralmente
presentes, em particular na região da cabeça e do tronco. Os nódulos podem ulcerar
e desenvolver tratos drenantes. As lesões normalmente não são doloridas ou
pruriginosas. Schubach et al. (2006) descreveram a ocorrência de esporotricose em
44 cães da região metropolitana do Rio de Janeiro. Todos os animais apresentaram
lesões fixas cutâneas únicas (40,9%) ou múltiplas (59,1%). A maior parte dos
animais apresentou lesões ulceradas no nariz e os sintomas respiratórios foram
considerados os sinais extracutâneos mais comuns da infecção, a transmissão por
gatos com esporotricose ocorreu em 84,1% dos animais.

46
A forma linfocutânea é considerada incomum nos cães, mas dois casos foram
relatados por Crothers et al. (2009), assim como, um caso de esporotricose
disseminada.
Madrid et al. (2007), em Pelotas/RS, descreveram o primeiro caso de
esporotricose óssea e cutânea em canino que apresentava lesões cutâneas
profundas caracterizadas por extensas úlceras e que evoluía há três anos, sem
diagnóstico.
Souza et al. (2009) relataram o caso clínico de esporotricose de um cão com
massa na fossa nasal e dificuldade respiratória, na cidade de Valença/RJ. O animal
estava sendo tratado para rinite alérgica com predinisolona. O diagnóstico da
esporotricose foi baseado no achado histopatológico de dois esporos diminutos na
porção central do piogranuloma.
Em 2012, Matos et al. relataram uma dermatite multifatorial causada pelo S.
schenckii, Demodex canis e bactérias dos gêneros Staphylococcus e Streptococcus
em canino da cidade de Pelotas/RS. No Canadá, Shany (2000) relatou o caso clínico
de co-infecção, de esporotricose e criptococose em canino, onde a apresentação
clínica da doença foi uma massa ulcerada protuberante nas duas narinas.
No gato, as lesões podem ocorrer no nariz (“nariz de palhaço”), na cabeça,
nas patas, na cauda e em outras partes do corpo. A inoculação do fungo ocorre por
lesões traumáticas, surgem pápulas e nódulos múltiplos que evoluem para úlceras
que podem chegar a extensas áreas de necrose com eliminação de exsudato
purulento, podendo haver também o envolvimento de tecido muscular e ósseo. Pela
auto-higienização com a língua, ocorre disseminação da infecção para outras áreas
do corpo. Do mesmo modo, por causa do prurido, o animal costuma esfregar as
patas sobre a lesão ulcerada, contaminando-a com as leveduras presentes em sua
superfície. Como acontece em outras espécies de animais, o S. schenckii invade a
circulação linfática e suas células são retidas nos linfonodos. Nos linfonodos, o fungo
continua a se multiplicar, logo ocorre a megalia, seguindo-se o seu rompimento e a
ulceração. Pelos vasos linfáticos eferentes, as células fúngicas chegam ao nódulo
seguinte e, desta maneira, os nódulos da cadeia ganglionar serão infectados
resultando, assim, em uma cadeia de nodulações perceptíveis visualmente ou pela
palpação (CRUZ, 2013). Barros et al. (2001) relataram que nos gatos, a maioria das
lesões cutâneas localizou-se na cabeça (46,6%) e nos membros (35,9%) e a maior
parte dos animais apresentou mais de uma lesão pelo corpo.

47
2.4.2 - Reação imune à esporotricose
A resposta imune contra o S. schenckii é estudada desde os anos 70 e, desde
então, ficou claro que pacientes com deficiência na resposta imune celular são mais
susceptíveis à infecção sistêmica (MARTÍNEZ-ÁLVAREZ et al., 2014). Scott et al.
(1986), através de estudos “in vitro”, levantaram a possibilidade da ativação da via
alternativa de complemento pelo S. schenckii como um dos mecanismos de defesa
do hospedeiro contra o fungo através da opsonização, fagocitose e pela produção
de uma inflamação aguda durante a infecção inicial. Os estudos “in vivo” de Kozel
(1996) mostraram que o sistema complemento é um importante componente na
resistência inata às infecções fúngicas, mas, os meios específicos pelos quais a
ativação de complemento medeia esta resistência não são claros.
Fernandes et al. (2008) observaram que camundongos infectados com
leveduras de S. schenckii apresentaram supressão de células T, alta carga fúngica
nos tecidos e sucumbiram à infecção. As leveduras se mostraram resistentes aos
mecanismos fungicidas intracelulares dos macrófagos e disseminaram pelos tecidos.
A infecção severa promoveu a liberação de mediadores pró-inflamatórios TNF-α, NO
(óxido nítrico) e subsequente indução de moléculas como a IL-10, FasL e CTLA-4
(antígeno linfócito citotóxico-4). A resistência foi atribuída principalmente à
proliferação de células T após desafio com ConA, equilíbrio de citocinas: IFN-γ e
IL10, e diminuição de células apoptóticas nos camundongos iNOS-/-. Os autores
mostraram que apesar do NO dos macrófagos ser um mediador essencial para a
morte, “in vitro”, do S. schenckii, a ativação do sistema NO, “in vivo”, contribui para a
imunossupressão e interfere no balanço de citocinas na fase inicial de infecção do S.
schenckii.
Para Carlos et al. (2009) não há dúvidas de que tanto as respostas Th1 e Th2
participam da esporotricose experimental, entretanto precisa-se investigar os
receptores presentes nas células que participam efetivamente do controle da
infecção, os primeiros resultados indicaram que TLR-4 (receptores semelhantes a
toll) participam da esporotricose murina, reconhecendo algumas moléculas lipídicas
extraídas da parede celular da levedura do S. schenckii. Negrini et al. (2013)
mostraram que também os TLR-2 são receptores importantes para o
reconhecimento do fungo, os camundongos TLR-2(-/-) apresentaram menores
porcentagens de macrófagos com leveduras internalizadas quando comparados aos

48
camundongos TLR-2 competentes. A ausência de TLR-2 levou a uma menor
produção de citocinas TNF-α, IL-1β, IL-12 e IL-10.
Nascimento e Almeida (2005) infectaram camundongos com leveduras de S.
schenckii por via intraperitoneal. Os resultados mostraram a participação de
anticorpos específicos no controle da infecção, foram observados altos níveis de
IgG1, indicando a ocorrência de neutralização, pois este isotipo está relacionado
com a opsonização, o que melhora a eficiência da fagocitose. De Lima Franco et al.
(2011) também mostraram que na esporotricose experimental, os anticorpos foram
importantes na proteção contra o S. schenckii e consideraram a opsonização um
processo importante para aumentar a produção de TNF-α e morte do fungo pelos
macrófagos. A ligação de diferentes opsoninas séricas na superfície de leveduras
(C3) e conídios (lectinas com resíduos de manose) guia o patógeno a receptores
específicos nos macrófagos, os quais levam a diferentes respostas celulares. Os
conídios, em culturas de macrófagos de células leucêmicas, induziram uma pobre
resposta pró-inflamatória e menores taxas de morte celular por ROS, quando
comparados às leveduras, aumentaram a sobrevivência do patógeno e favoreceram
a transição dimórfica de conídio para levedura (GUZMAN-BELTRAN et al., 2012).
Recentemente, Martínez-Álvarez et al. (2014) compilaram informações da
literatura sobre a resposta imune do hospedeiro frente ao S. schenckii. Segundo os
autores, as respostas imunes ainda não estão totalmente explicadas como as
descritas para a Candida e outros patógenos fúngicos, ainda sabe-se pouco sobre
os mecanismos moleculares de reconhecimento da resposta imune inata e os
receptores envolvidos. Além disto, não há informação sobre as outras espécies que
compõem o complexo Sporothrix.
2.4.3 - Diagnóstico
As amostras clínicas para análise laboratorial podem incluir fragmentos de
lesões ou swabs para análise microbiológica; e, amostras de tecido para biópsia em
formalina neutra a 10%, para estudos histopatológicos. O diagnóstico clínico da
esporotricose deve ser confirmado pela observação microscópica de estruturas
leveduriformes no material coletado das lesões e no isolamento e na identificação do
agente etiológico envolvido (QUINN et al., 1994).

49
2.4.4 - Citologia/ Histopatologia
A observação microscópica do fungo no material patológico pode ser
realizada em esfregaços submetidos às colorações convencionais como Gram,
Giemsa, panótico, azul de metileno ou em cortes histológicos corados pelo PAS,
Grocott-Gomori (GMS). Em todos os casos o Sporothrix spp. apresenta-se como
leveduras ovaladas, arredondadas ou em forma de charuto, livres ou no interior de
macrófagos. Sua parede celular é refrátil e o citoplasma pode se retrair, dando a
impressão de haver uma cápsula. Nas lesões ulceradas, as leveduras são
encontradas em grande número, especialmente em gatos, enquanto que no homem
e em outros animais como os cães e em lesões não ulceradas observa-se um
número menor ou, elas podem não ser encontradas (CRUZ, 2013).
Os achados histopatológicos predominantes em lesões piogranulomatosas da
pele de cães com esporotricose descritos por Miranda et al. (2010) foram :
granulomas mal definidos, de distribuição difusa com predomínio de células
epitelioides. Em 5% dos casos foi observada infiltração granulomatosa.
Os achados histopatológicos das lesões cutâneas de felinos com
esporotricose, descritas por Miranda et al. (2013), foram: granulomas supurativos,
com predomínio de granulomas mal definidos. A frequência de granulomas bem
definidos e de células epitelioides ficou em torno de 50 e 43%, respectivamente, nos
animais com mais de cinco anos de idade e com menor ocorrência nos animais com
menos de cinco anos de idade. Os autores também compararam a carga fúngica
das lesões e observaram que campos, com mais de 100 leveduras no aumento de
400x, foram encontradas nos granulomas mal delimitados (66,7% dos casos) e não
foi observada em granulomas bem delimitados e/ou com predominância de células
epitelioides. Os granulomas bem delimitados foram associados ao grupo de animais
com apenas uma lesão e as altas cargas fúngicas foram predominantes no grupo de
animais com mais de três lesões e granulomas mal delimitados.
2.4.5 - Métodos imunológicos
O primeiro teste imunológico sugerido para diagnóstico da esporotricose foi o
da intradermoreação, que utiliza um preparado com filtrados de cultura da fase

50
miceliana ou leveduriforme do S. schenckii, entretanto, em regiões endêmicas, um
número considerável de pessoas são reativas à esporotriquina sem apresentarem
sinais clínicos da doença (BARROS et al., 2011).
Nas formas não usuais de esporotricose os métodos de cultura são difíceis de
serem aplicados e, por isto, a detecção de anticorpos do paciente pode ser mais
uma opção no diagnóstico. Todavia, os métodos sorológicos ainda apresentam valor
limitado para o diagnóstico definitivo da esporotricose devido às reações cruzadas
com outras infecções ou aos resultados falsos positivos. A prova da aglutinação em
tubo, reação de fixação do complemento, aglutinação em partículas de látex,
imunodifusão em gel, imunoeletroforese, ELISA e imunofluorescência (BLUMER et
al., 1973; SCOTT et al., 1989, MENDONZA et al., 2002, FERNANDES et al., 2011,
ALMEIDA-PAES et al., 2007 e 2012, MIRANDA et al., 2010) fazem parte do rol de
metodologias já testadas para o Sporothrix spp. através de diferentes formas de
produção do antígeno.
Almeida-Paes et al. (2007b) utilizaram antígenos miceliais (Mendonza et al.,
2002) para o diagnóstico da esporotricose através do ensaio de ELISA. O ensaio
apresentou 97% de sensibilidade, 89% de especificidade e 92% de eficiência global,
todos os pacientes, com as diferentes apresentações clínicas da doença,
apresentaram valores de absorvância altos e similares, sem correlação com a
severidade da doença. Os experimentos mostraram que no decorrer do tratamento,
os títulos de anticorpos diminuíram, sendo este dado uma ferramenta importante
para o acompanhamento da resposta do paciente ao tratamento (BERNARDES-
ENGEMANN et al., 2005, ALMEIDA-PAES et al., 2007a).
Bernardes-Engemann et al. (2005) testaram uma fração antigênica da parede
celular da levedura, uma ramnomanana que se liga à concanavalina (SsCBF), para
o diagnóstico da esporotricose humana. O ensaio de ELISA mostrou 90% de
sensibilidade e 86% de eficiência global quando testado contra soros obtidos de 92
pacientes com diferentes apresentações clínicas da doença, juntamente com 117
soros controle obtidos de indivíduos saudáveis ou portadores de outras doenças. Em
2011, Fernandes et al. compararam os resultados de ELISA utilizando a fração
antigênica SsCBF ou exoantígenos brutos miceliais (MENDONZA et al., 2002) para
o diagnóstico da esporotricose em gatos. A fração antigênica SsCBF mostrou
73,33% de sensibilidade e 91,67% de especificidade enquanto o exoantígeno bruto
demonstrou 83,33% de sensibilidade e 91,67% de especificidade.

51
A imunohistoquímica também pode ser um método complementar que pode
ser aplicado para a detecção de antígenos intracelulares e extracelulares
aumentando a sensibilidade do diagnóstico em 80,5% e sem reações cruzadas com
lesões de leishmaniose (MIRANDA et al., 2011).
2.4.6 - Diagnóstico diferencial
Segundo Santos et al. (2007), a principal enfermidade a ser diferenciada da
esporotricose em humanos e caninos no Rio de Janeiro deve ser a leishmaniose
americana tegumentar, devido à similaridade dos sintomas clínicos nas
apresentações fixa cutânea da esporotricose e sobreposição de endemicidade
destas doenças no Rio de Janeiro.
Lopes-Bezerra et al. (2006) citam que a diversidade das formas clínicas da
esporotricose leva a um grande número de doenças que a esporotricose deve ser
diferenciada além da leishmaniose, nocardiose, cromomicoses, criptococose,
tuberculose, rosácea, doenças granulomatosas não infecciosas, psoríases,
carcinomas, dentre outras.
2.4.7 - Prevenção e controle
Para a esporotricose transmitida pelo contato com o solo e traumas, a
recomendação é a utilização de botas e luvas grossas nos trabalhos com a terra ou
plantas.
Para o controle da esporotricose transmitida por gatos, a prioridade deveria a
castração dos felinos, eutanásia dos casos sem possibilidade terapêutica, cremação
dos corpos que evoluíram para óbito e educação para a posse responsável de
animais domésticos. (BARROS et al., 2010).
Outro aspecto a ser pensado, considerando a dificuldade da realização do
tratamento nos gatos, é a oportunidade que essa epidemia representa para o
desenvolvimento de vacinas antifúngicas para animais, apesar dos fungos
representarem um desafio em pesquisas de vacinas (BARROS et al., 2010).

52
2.4.8 - Tratamento
O tratamento de escolha da esporotricose enquanto lesão cutânea é a
quimioterapia sistêmica com iodeto de potássio. Nas formas cutâneas disseminadas,
linfocutâneas recidivantes e extracutâneas, a anfotericina B é o fármaco mais
efetivo. Porém, a frequência de intolerância ao iodo e a alta toxicidade da
anfotericina B, muitas vezes, representam fatores que limitam o seu uso. Nas últimas
décadas, aumentou o emprego de derivados azólicos, dentre eles: o cetoconazol, o
itraconazol e o fluconazol, como alternativa terapêutica aos esquemas clássicos,
sendo o itraconazol o mais efetivo deles devido à boa eficácia e aos poucos efeitos
adversos tanto em adultos quanto em crianças: 100 mg/dia durante 120 dias
(KAUFFMAN et al., 2000, HEIDRICH et al., 2011). Por outro lado, a terbinafina, em
virtude da sua elevada atividade in vitro e in vivo, está sendo utilizada para diversas
infecções fúngicas. No entanto, a experiência clínica da terbinafina, no tratamento da
esporotricose, ainda é restrita (HEIDRICH et al., 2011).
Durante a gravidez, os derivados azólicos não devem ser administrados
devido ao seu potencial teratogênico e o iodeto de potássio, devido à sua toxicidade
para a tireoide fetal. A anfotericina B pode ser utilizada com segurança, mas é
indicada somente para a esporotricose disseminada ou pulmonar (KAUFFMAN et
al., 2000). Uma opção para a doença cutânea é a hipertermia local, banhos quentes,
com a temperatura entre 42 e 45°C, uma vez ao dia durante 30 minutos a uma hora,
ou duas ou três vezes ao dia durante 15 minutos por sessão (PÁVON et al., 2007).
Pode-se também esperar a gravidez ser completada para iniciar a terapia com
itraconazol, já que não existe risco de disseminação da infecção para o feto.
Nos animais, a esporotricose responde bem ao tratamento oral com soluções
saturadas de iodeto de potássio nas dosagens de 40mg/Kg para cães e de 20mg/Kg
para gatos a cada 12 a 24 horas e continuando por 30 dias após o desaparecimento
total dos sintomas. Em gatos, este tratamento tem sido evitado pelos veterinários
porque é comum a ocorrência de iodismo, caracterizado por vômitos, anorexia,
depressão, icterícia, tremores, corrimento nasal e ocular, hipotermia, insuficiência
cardíaca. Nestes casos, o tratamento deve ser interrompido por uma semana,
reiniciando-o com a mesma dosagem ou com dosagem mais baixa ou o iodeto pode
ser substituído por cetoconazol, itraconazol ou fluocitosina. Em gatos e cães, bons

53
resultados têm sido obtidos com dosagens de 100mg de itraconazol/dia. Em todos
os casos, o tratamento deve ser continuado por 30 dias depois de desaparecidos os
sinais. O tratamento de grandes animais pode ser feito com o iodeto de potássio
adicionado à ração e fornecido aos animais, inclusive de forma preventiva, quando
houver casos no plantel (CRUZ, 2013).
2.4.9 - Prognóstico
Embora a micose exiba muitas vezes um quadro clinico dermatológico
exuberante, costuma cursar sem gravidade e sem acometer outros órgãos além da
pele, mucosa e subcutâneo. O custo indireto é social — pelo absenteísmo ao
trabalho, pelo sofrimento durante a doença ativa e pelo aspecto desagradável das
lesões cicatriciais (BARROS et al., 2010).
Contrariamente nos gatos, a esporotricose tem curso muitas vezes longo,
frequentemente com acometimento sistêmico, levando a formas graves de difícil
tratamento e evolução para o óbito (BARROS et al., 2010).
2.5 - IMUNOGLOBULINAS
As Imunoglobulinas são glicoproteínas, produtos da diferenciação dos
linfócitos durante o desenvolvimento das células B, com atividade de anticorpo e são
encontradas no sangue, na linfa e nos tecidos vascularizados de todos os
vertebrados com mandíbula (LITMAN et al., 1993). A diversidade dos anticorpos é
especificada por uma molécula de imunoglobulina, heterodimérica, constituída de
duas cadeias pesadas e duas leves idênticas, que formam um monômero bivalente
e funcional. Variações nas sequências regionais das cadeias leves e pesadas
levaram à classificação de regiões variáveis (V) e constantes (C), que mais tarde
mostraram ser codificadas por exons separados (LITMAN et al., 1993).

54
Em aves, foi mostrada a existência de três classes de imunoglobulinas,
análogas às classes de imunoglobulinas dos mamíferos, IgA, IgM e IgY (IgG)
(LESLIE e CLEM, 1969, LEBACQ-VERHEYDEN et. al., 1974).
O peso molecular, a morfologia e a mobilidade imunoeletroforética da IgA e
IgM de galinhas são semelhantes aos da IgA e IgM de mamíferos (LESLIE e CLEM,
1969). A IgM de frangos é estruturalmente e funcionalmente homóloga à IgM de
mamíferos, apresenta um peso molécular médio de 890 kDa e é mais provável que
tenha uma configuração tetramérica (μ2L2)4 ao invés de pentamérica (μ2L2)5.
Invariavelmente, IgM é o isotipo produzido após a exposição inicial ao antígeno.
Assim como nos mamíferos, sua resposta é temporária (LEBACQ-VERHEYDEN et.
al., 1974, DAVISON et al., 2008).
IgA é o anticorpo predominante nas secreções corporais. Nos mamíferos, IgA
é um dímero (α2L2)2 unido pela cadeia J que se liga a um receptor de células
epiteliais nos tecidos. Este receptor torna-se integrado à molécula de imunoglobulina
como um componente de secreção e o complexo IgA é transportado através da
célula epitelial e secretada no lúmen do órgão em questão. A IgA está presente nas
secreções intestinais e biliares de galinhas (LEBACQ-VERHEYDEN et. al., 1972,
1974) . A IgA extraída da bile de frangos é normalmente maior do que a IgA
encontrada em mamíferos, sugerindo que a forma aviária é um trímero ou um
tetrâmero (α2L2)4 (LEBACQ-VERHEYDEN et. al., 1972, 1974, DAVISON et al.,
2008).
Em 1969, Leslie e Clem demonstraram que a imunoglobulina 7.1S de frangos
era o principal anticorpo do soro destes animais e baseado em suas características
físico-químicas e antigênicas, sem nenhuma similaridade com qualquer outra de
mamíferos, propuseram que esta molécula fosse chamada de IgY.
IgY é a imunoglobulina de baixo peso molecular, predominante no soro de
pássaros, répteis anfíbios e provavelmente peixes pulmonados (WARR et al., 1995).
Nos ovos, a IgY está presente predominantemente na gema, enquanto a IgA e IgM
estão presentes na clara (HAMAL et al., 2006) como resultado da secreção da
mucosa no oviduto (ROSE et al., 1974 citados por HAMAL et al., 2006). SCHADE et
al. (2005) relataram que, dependendo do método utilizado para a extração e da sua
sensibilidade, a concentração total de IgY pode ser de 100 a 200 mg/ovo e que esta
é prevalente na circulação sanguínea, com variações significativas entre indivíduos
(3 – 7 mg/ml) (HAMAL et al., 2006), também é encontrada no conteúdo duodenal, na

55
secreção traqueal, no fluido seminal, na bile, na saliva e nas lágrimas (LEBACQ-
VERHEYDEN et. al., 1974, DIAS DA SILVA, 2010).
A estrutura total da IgY é semelhante à da IgG dos mamíferos, com duas
cadeias leves e duas cadeias pesadas e massa molecular de 167,2 kDa, um pouco
maior do que a IgG (~160 kDa) (DIAS DA SILVA, 2010). A cadeia pesada apresenta
peso molecular de 65,105 kDa, é chamada de upsilon, υ, e possui um domínio
variável e quatro constantes (Cυ1, Cυ2, Cυ3 e Cυ4). A cadeia leve apresenta peso
molecular de 18,660 kDa, é composta de um domínio variável e um constante
(CARLANDER, 2002). Warr et al. (1995) citaram que as comparações das
sequências das regiões constantes da IgG e da IgY mostram que os domínios Cγ2 e
Cγ3 da IgG estão mais relacionados com os domínios Cυ3 e Cυ4 da IgY,
respectivamente, e que o equivalente ao domínio Cυ2 está ausente nas cadeias γ.
Supõe-se que o domínio Cυ2 foi condensado para formar a região de dobradiça da
IgG. A região de dobradiça é uma estrutura característica da Ig de mamíferos e que
não existe na IgY nem na IgA de aves. Na IgY, há regiões vizinhas aos domínios
Cυ1-Cυ2 e Cυ2-Cυ3 que contêm resíduos de prolina e glicina, estas regiões têm o
potencial para conferir flexibilidade limitada à molécula. Esta é provavelmente a
razão das diferentes propriedades da IgY em comparação com a IgG de mamíferos.
A região Fab (fragmento ligador de antígeno) é semelhante à das
imunoglobulinas de mamíferos, contém o sítio de ligação dos antígenos e a região
Fc (fragmento cristalizável) é o sítio efetor, responsável pela maioria das funções
biológicas, ele contém duas cadeias laterais de carboidratos, em contraste com
somente uma na IgG (WARR et al., 1995).
A IgY das aves é evidenciada como o progenitor imediato da IgG e IgE, a
evidência baseia-se em três pontos: as funções do anticorpos, a estrutura
polipeptídica das cadeias pesadas e a estrutura e expressão dos genes para a
cadeia pesada (WARR et al., 1995). Taylor et al. (2009) mostraram, pela análise
cristalográfica, que a estrutura da região Fc da IgY exibe características estruturais
da IgG e IgE.
2.5.1 - Transferência da IgY para o ovo
Dentro de um único ovário de uma ave existem numerosos e pequenos (<60
mg) oócitos imaturos. Em intervalos diários, um oócito desta população entra na fase

56
dois do processo de maturação. A primeira fase é caracterizada por um crescimento
relativamente lento e leva em torno de 15 dias. A segunda ocorre por um período de
6 dias e inclui o rápido sequestro de proteínas séricas selecionadas, resultando em
aumento de massa para 30 a 50 vezes. A concentração de IgY na gema é constante
durante a completa maturação do oócito, do pequeno (0,05 g) ao grande (20 g). A
massa do oócito cresce exponencialmente do dia -8 ao dia -2, e linearmente do dia -
2 até um pouco antes da entrada do oócito maduro no oviduto (dia zero). A
passagem de IgY segue este modelo, com taxas máximas de passagem ocorrendo
em torno do dia -3 e -2, quando quantidades tão grandes quanto 45 mg/dia são
acumuladas na gema. À maturidade, um oócito possuirá em torno de 100 – 200 mg
de IgY materna (KOWALCZYK et al., 1985).
Loeken e Roth (1983) analisaram amostras pareadas de IgY no soro e gema
de galinhas, os autores evidenciaram que todas as amostras de IgY apresentaram
pontos isoelétricos entre pH 6,5 – 9,0, sem nenhuma diferença qualitativa ou
quantitativa entre as IgYs do soro e da gema. As galinhas apresentaram
subpopulações de IgY que diferem na susceptibilidade à papaína e ao ponto
isolelétrico e todas as subpopulações são transferidas para a gema em proporção à
sua concentração no soro materno. Para a transferência, é necessária que a região
Fc e da dobradiça estejam intactas e as cadeias laterais de carboidratos associadas
às regiões Fc não interferem no processo. A passagem transovariana leva
aproximadamente cinco dias. O tempo médio de vida da IgY circulante de uma
galinha adulta é de aproximadamente 36 a 65 horas. Anticorpos específicos são
transportados para a gema com um atraso médio em torno de 5 dias (SCHADE et
al., 2005).
A transferência de anticorpos maternos para o feto ou neonato, é largamente
conhecida em mamíferos, pássaros e permite a aquisição passiva da imunidade
humoral. O saco vitelino do frango exibe ligadores de IgY semelhantes aos do Fc
neonatal para IgG com alta afinidade de ligação ao pH ácido e nenhuma ligação
observável ao pH neutro ou básico (TRESSLER e ROTH, 1987; TESAR et al.,
2008).
Em geral, os anticorpos policlonais IgY estão presentes no ovo como uma
emulsão de grânulos juntamente com lipoproteínas. A IgY é a fração γ-livetina da
gema que precisa ser separada de outras proteínas hidrossolúveis tais como α-
livetina, β-livetina e várias lipoproteínas (CARLANDER, 2002). Portanto, o

57
isolamento e a purificação da IgY aviária é essencial para um ótimo imunoensaio.
Segundo DIAS DA SILVA et al. (2010), o método de Bhanushali et al. (1994) é um
método simples e barato para tal procedimento.
Altos títulos de anticorpos IgY específicos no sangue são alcançados, em
média, após a terceira imunização. Almeida et al. (2003) utilizaram cinco
imunizações para a produção de IgY anti-BfpA, os anticorpos apareceram no sangue
após a segunda imunização, mas, ao ELISA, títulos altos de anticorpos foram
conseguidos somente 15 dias após a 4ª. imunização, diluição de 1:9000. Após a
terceira imunização há aumento dos títulos de anticorpos, portanto uma imunização
adicional deve ser feita dentro de, no máximo, três meses após as três primeiras
imunizações a fim de se manter títulos altos de anticorpos nos ovos por mais tempo
(CHALGHOUMI et al., 2008). Gassmann et al. (1990) conseguiram altos títulos de
anticorpos nas gemas após a terceira imunização e estes se mantiveram altos por
até 81 dias. Comparando adjuvantes, completo/incompleto de Freund com um
complexo de matriz imunoestimulante, Chalghoumi et al. (2008) observaram que os
títulos de anticorpos foram maiores nas gemas dos animais que receberam o
adjuvante de Freund e que este não causou danos nos tecidos ou anormalidades
nos animais que o receberam.
2.5.2 - Propriedades bioquímicas da IgY
A região das cadeias pesadas da IgY (Cυ2) apresenta mobilidade restrita,
torna o anticorpo mais rígido e afeta a capacidade do anticorpo precipitar ou
aglutinar antígenos. Somente parte dos anticorpos das galinhas são precipitados em
concentrações fisiológicas de sal e, aproximadamente, 25% dos anticorpos
permanecem no sobrenadante sob máxima precipitação. As condições que
permitem a precipitação podem afrouxar os movimentos restritos dos braços da
região Fab e fornecer independência funcional aos sítios de ligação (CARLANDER,
2002). Shimizu et al. (1992) demonstraram que a IgY é menos estável à temperatura
e às condições ácidas do que a IgG de coelho. IgY é bastante estável à digestão de
proteases internas tais como tripsina e quimiotripsina (CARLANDER, 2002).

58
2.5.3 - Vantagens da IgY
Os anticorpos IgG são as moléculas de escolha para uma variedade de
técnicas analíticas, bioquímicas e terapêuticas. Mas, infelizmente, o seu
envolvimento nas respostas imunes e patologias imunomediadas, juntamente com o
seu alto grau de conservação entre os mamíferos, podem torná-la susceptível a
interações indesejáveis com proteínas conservadas, dificultando o seu uso em
determinadas situações (SPILLNER et al., 2012).
Os anticorpos aviários podem reconhecer epitopos não reconhecidos pela IgG
dos mamíferos, dando acesso a um repertório diferente de anticorpos. A distância
filogenética entre mamíferos e aves aumenta a imunogenicidade das proteínas
conservadas dos mamíferos (GASSMANN et al., 1990) quando se utiliza anticorpos
IgY. Uma boa resposta imunológica pode ser alcançada se um anticorpo IgY anti-
IgG de mamífero é utilizado nos ensaios. IgY não se liga à proteína A estafilocóccica
ou proteína G estreptocóccica, não ativa o sistema complemento de mamíferos, IgY
não se liga ao fator reumatoide (FR) presente em pacientes com doenças
reumatoides ou não (LARSSON et al., 1991). Moléculas de IgG sempre causam
resultados falsos positivos devido à interação com FR nos imunoensaios. Um
número crescente de pacientes são tratados com anticorpos monoclonais de
camundongos, este tratamento leva a uma resposta imunológica no paciente
resultando na produção IgG anti- anticorpos monoclonais de camundongos (HAMA),
a vantagem da IgY está em não interagir com o fator HAMA (CARLANDER, 2002).
Além disto, o uso de aves para produção de anticorpos representa uma
redução do número de animais necessários para tal finalidade, pois as aves
produzem maiores quantidades de anticorpos do que os roedores, superior a 100
mg de IgY purificada de uma única gema (TRESSLER & ROTH, 1987) e também
eliminam a coleta de sangue que é dolorosa para o animal.
2.5.4 - Aplicações gerais da IgY
A imunização de galinhas é uma alternativa atrativa e elegante para a
produção de anticorpos. Este tipo de anticorpo possui propriedades distintivas que
podem ser exploradas em vários caminhos de pesquisa, diagnóstico e terapia.

59
Vários trabalhos relataram a viabilidade do uso dos anticorpos IgY nas
técnicas de imunodiagnóstico. Anticorpos IgY foram utilizados em testes de ELISA
para medir a concentração de biomarcadores sanguíneos como proteínas ou
peptídeos (LEE et al., 2004, RUAN et al., 2005; JINTARIDTH et al., 2006 e
SOTIROPOULOU et al., 2012), dosar princípio ativo de planta em produtos naturais
(FREIRE et al., 2004), diagnosticar vítimas humanas de veneno de cobra (BRUNDA
et al., 2006), diagnóstico de doenças virais (SILVA et al., 2012, PALANIYAPPAN et
al., 2012), de parasitos intestinais (LU et al., 2012, LEI et al., 2012, MANHANI et al.,
2012), de protozoários (SILVA, 1999; BERNARDO, 2009), enterotoxinas
bacterianas, seja em produtos alimentícios (JIN et al., 2013) ou em patologias
(PARMA et al. 2012), detecção de cepas de Staphylococcus aureus resistentes à
meticilina (YAMADA et al., 2013).
Pela técnica da imunohistoquímica, anticorpos IgY marcaram antígenos virais
no citoplasma de neurônios do glânglio trigeminal de camundongos infectados
experimentalmente pelo vírus da raiva (MOTOI et al., 2005) e distintas formas
antigênicas de Toxoplasma gondii em seções tissulares de cérebros de
camundongos cronicamente infectados (FERREIRA JÚNIOR et al., 2012).
Anticorpos IgY também foram testados para a produção de frações variáveis
de cadeia simples, em sistema bacteriano ou fagos, visando à produção de
anticorpos de alta afinidade e avidez sem a necessidade de novas imunizações
(NAKAMURA et al., 2004, ROCHA, 2010). Rocha (2010) iniciou a montagem de uma
biblioteca de cDNA de IgY visando à produção de frações variáveis de cadeia
simples com aplicabilidade na composição de soros antivenenos de cobra de alta
qualidade, ou mesmo em testes que exijam alto grau de sensibilidade.
2.5.5 - Uso terapêutico da IgY
A Escherichia coli enteropatogênica (EPEC), causadora de diarreia, utiliza o
homopolímero de BfpA, proteína do pili bacteriano, para se aderir aos enterócitos e
colonizar o intestino do hospedeiro. ALMEIDA et al. (2003) hiperimunizaram galinhas
com a proteína recombinante BfpA e concluíram que os anticorpos IgY foram
capazes de bloquear a colonização de células HeLa por EPEC, inibiram a
proliferação de EPEC em meio de cultura e foram capazes de diferenciar cepas de

60
E. coli produtoras de BfpA. Soluções de IgY podem ser produzidas para inibir o
crescimento específico de cepas homólogas (ALMEIDA et al., 2003; AMARAL et al.,
2008) ou, anticorpos podem ser produzidos simultaneamente, em um mesmo ovo,
para inibirem o crescimento de sorovares bacterianos diferentes (CHALGHOUMI et
al., 2008).
Estudos em ratos mostraram o potencial da IgY de aves imunizadas na
prevenção de cáries dentárias utilizando o Streptococcus mutans como agente
imunógeno (OTAKE et al., 1991) ou a sua proteína ligadora de glucanas (SMITH et
al., 2001). Os autores observaram uma redução significativa do número de cáries
nos molares de ratos que receberam gemas de aves imunizadas.
Vários autores estudaram a possibilidade de se utilizar a IgY de galinhas
imunizadas como fonte de anticorpos específicos para controle e prevenção da
diarreia bacteriana em suínos. A IgY mostrou ação efetiva, segura e protetiva,
principalmente contra infecções intestinais e apresentou atividade biológica
semelhante à do colostro em leitões recém-nascidos (YOKOYAMA et al., 1992).
Segundo Yokoyama et al. (1992), o valor terapêutico da IgY, administrada
oralmente, reside na sua capacidade em bloquear a colonização das células
epiteliais intestinais por cepas bacterianas enteropatogênicas. A IgY, assim como os
anticorpos colostrais, é absorvida somente nas primeiras horas pós-nascimento e
depois exerce um efeito terapêutico local, sendo o fornecimento contínuo preferível
ao somente ao nascer (RIBEIRO et al., 2007).
A imunidade passiva com IgY anti-Helicobacter pylori reduziu o crescimento
bacteriano, a atividade da urease, as injúrias gástricas em modelo animal (SHIN et
al. 2002). Quando a IgY anti-urease foi incorporada ao iogurte houve supressão da
infecção por H. pilori (HORIE et al., 2004).
Carlander (2002) demonstrou que o tratamento com IgY pode ser adjuvante
no tratamento convencional da fibrose cística. O autor observou que quando a
solução de IgY anti-Pseudomonas foi gargarejada e ingerida pelos pacientes, o
principal sítio de ação foi a cavidade oral, orofaringe e região linfática. Durante o
tratamento, nenhum paciente apresentou uma nova infecção e também houve
aumento do tempo de reincidência da infecção nos pacientes tratados.
A IgY específica, contra um coronavírus causador de diarreia em suínos,
levou à redução da taxa de mortalidade de leitões (KWEON et al. (2000). Na
parvovirose canina, os animais tratados com dois gramas de pó contendo IgY-

61
antiparvovírus canino, não apresentaram sintomatologia clínica da doença
(NGUYEN et al., 2006). A IgY específica preveniu a diarreia e diminuiu
significativamente a quantidade de vírus albergados nos intestinos de suínos
desafiados com uma cepa de rotavírus humano (VEGA et al., 2012).
Vários trabalhos na literatura relataram o potencial da imunização passiva
contra o Staphylococcus aureus ou suas enterotoxinas. Leclaire et al. (2002)
desafiaram macacos Rhesus com dose letal de SEB (enterotoxina B) e mostraram
que a taxa de sobrevivência foi de 100% para o grupo de animais tratados com a IgY
contra a enterotoxina B, enquanto os animais do grupo controle morreram. A IgY
anti-S. aureus foi mais eficiente na cura da mastite clínica e experimental por S.
aureus do que o tratamento com penicilina (ZHEN et al., 2009).
Vieira-da-Motta et al. (2001) mostraram que a IgY anti-RAP (RNA III ativador
de proteína do loci agr para produção de enterotoxinas) na concentração de 0,05
µg/ml suprimiu a produção, in vitro, de enterotoxinas por várias cepas de S. aureus.
GUIMARÃES et al. (2009) mostraram que a IgY anti-S. aureus na concentração de 5
µg/ml, inibe o crescimento do S. aureus em cultura. WANG et al. (2010) produziram
anticorpos IgY anti diferentes sorotipos de S. aureus, os anticorpos apresentaram
alta especificidade e foram capazes de bloquear a internalização de cepas
homólogas por células epiteliais da glândula mamária, entretanto mostraram baixa
atividade bacteriostática.
A efetividade da IgY na imunidade passiva também foi estudada na
aquacultura com resultados superiores para o grupo tratado com a IgY específica,
seja para a infecção viral (LU et al., 2008) ou bacteriana (LI et al., 2006), porém, a
sua utilização no campo ainda é limitada devido à falta de uma formulação
adequada para ser administrada juntamente com a ração.
Devido a vantagens como baixo custo, alta produtividade e menor risco de
choque anafilático, os anticorpos IgY antiveneno de cobras têm sido sugeridos como
alternativa ao uso da IgG de equinos no tratamento das vítimas de mordidas de
cobra. Experimentos, in vivo, mostraram que anticorpos IgY hiperimunes forneceram
100% de proteção a camundongos desafiados com doses letais de veneno de cobra
(PAUL et al., 2007, ALMEIDA et al., 2008, ROCHA, 2010), mas que a proteção está
relacionada com o aumento da afinidade do anticorpo hiperimune (ALMEIDA et al.,
2008, ROCHA, 2010).

62
Existem muitos trabalhos na literatura favoráveis à imunidade passiva via IgY
para diversos microrganismos, entretanto para os fungos tais trabalhos são
escassos, sendo que as infecções fúngicas são difícieis de serem debeladas, os
tratamentos são tóxicos e prolongados. Experimentos in vitro mostraram a inibição
do crescimento da alga Prototheca spp. (SILVA, 2005) e da C. albicans
(GONÇALVES, 2007) spp. por anticorpos IgY específicos contra os respectivos
micro-organismos. Ibrahim et al. (2008) estudaram a eficácia, in vitro e in vivo, dos
anticorpos IgY antiantígenos de C. albicans, os autores observaram que os
anticorpos reduziram a atividade de adesão da C. albicans às células faríngeas de
carcinoma humano, além disto, reduziram a disseminação sistêmica e a severidade
das lesões na língua de camundongos imunossuprimidos, que foram utilizados como
modelo para a infecção oral. Fujibayashi et al. (2009) mostraram que anticorpos IgY
produzidos anti-C. albicans inibiram a aderência da C. albicans e também de outras
espécies de Candida spp. às monocamadas de células epiteliais orais, entretanto,
nenhuma inibição foi observada quando houve formação do tubo germinativo.
Abdelnoor et al. (2006) imunizaram galinhas com uma combinação de tubos
germinativos e células leveduriformes de C. albicans inativadas pelo calor. No teste
in vivo, os autores desafiaram camundongos com doses letais de C. albicans e
observaram que todos os camundongos tratados com a IgY específica continuavam
vivos aos sete dias após desafio, não houve disseminação da C. albicans para os
rins dos animais e estes apresentaram morfologia normal. Em contraste, os animais
que receberam IgY pré-imune, permaneceram vivos por sete dias, mas
apresentaram rins com múltiplos focos de abscesso. No grupo que recebeu somente
os antígenos e nenhuma IgY, a sobrevivência foi de 20%.
O mecanismo pelo qual os anticorpos podem suprimir o crescimento
bacteriano não está claramente compreendido. À microscopia eletrônica, Lee et al.
(2002) observaram que a superfície bacteriana ligada à IgY específica apresentou-
se rugosa quando comparada à superfície de bactérias do grupo controle, IgY pré-
imunização. A IgY específica pode se ligar a componentes expostos na superfície
bacteriana e promover alterações estruturais desta superfície. Sabe-se que bactérias
com ligações cruzadas com anticorpos específicos são aglutinadas e apresentam
motilidade, oportunidade de capturar nutrientes e proliferação diminuídas quando
comparadas às células bacterianas livres. Por outro lado, Lee et al. (2002) citaram
que, em meios sólidos, já foi observada uma redução da contagem bacteriana e que

63
esta não foi atribuída à reação de aglutinação. Além disto, as propriedades de
aglutinação da IgY são exibidas somente em concentrações elevadas de sal ou
condições de baixo pH devido ao impedimento estérico dos braços Fab da IgY que
são alinhados de forma bastante próxima, portanto, fora a aglutinação, deve haver
outras reações entre IgY e bactéria para causar a inibição do crescimento.
Componentes específicos expressos na superfície bacteriana, os quais são fatores
cruciais para o crescimento bacteriano podem ser reconhecidos e ligados aos
anticorpos policlonais, IgY (LEE et al., 2002).

64
3.0 – JUSTIFICATIVA
O resultado do cultivo micológico positivo para a esporotricose é considerado
como diagnóstico definitivo, conhecido como “padrão ouro”, sendo um método de
identificação simples e de baixo custo, embora o resultado seja demorado, superior
a uma semana. No entanto, existem casos em que não é possível isolar o agente
etiológico e o diagnóstico imunológico pode ser uma alternativa para tais situações.
A esporotricose humana e animal estão descritas há pelo menos 14 anos no
estado do Rio de Janeiro e estes casos estão relacionados diretamente aos felinos
infectados pelo fungo. Na literatura existem vários trabalhos com estudos
sorológicos de pacientes humanos infectados, entretanto, para os animais estes são
escassos.
O tratamento da esporotricose com a administração da droga antifúngica
pode perdurar de semanas a meses até o restabelecimento do quadro clínico
favorável. A possibilidade de uma imunidade passiva via anticorpos IgY pode ser
uma alternativa para se diminuir o tempo de tratamento e o sucesso terapêutico,
principalmente na espécie felina.

65
4.0 - OBJETIVOS
1 . Produzir e desenvolver anticorpos IgY antiantígenos de Sporothrix schenckii e
Sporothrix spp. em galinhas para fins de diagnóstico;
2. Verificar a existência de reação cruzada entre os anticorpos produzidos e a
Leishmania spp. através da técnica de imunohistoquímica;
3. Estudar o perfil sorológico de animais positivos para esporotricose através dos
testes de ELISA e western blot;
4. Determinar moléculas imunodominantes nos soros de animais positivos para
esporotricose e animais imunizados experimentalmente.

66
5.0 - MATERIAL E MÉTODOS
5.1.0 - Condições de cultivo das cepas de Sporothrix spp. para extração de
antígenos
Para a produção de antígenos, foram cultivadas duas cepas: o Sporothrix
schenckii ATCC 32285 (Ss), gentilmente cedido pelo INCQS-Fiocruz, e uma cepa
clínica, Sporothrix spp., isolada pelo setor de micologia do LSA/CCTA/UENF, de um
felino com lesões cutâneas da cidade de Macaé/RJ.
Os isolados são mantidos em tubos de ensaio inclinados com 10-15 mL de
meio SDA (Sabouraud-dextrose-agar) (Himedia) à temperatura ambiente. A
manutenção dos isolados é realizada periodicamente através de repiques para
novos tubos.
A extração de antígenos para a cepa de Sporothrix schenckii ATCC 32285 foi
realizada para as duas fases morfológicas do fungo, micelial e leveduriforme. Na
cepa clínica, Sporothrix spp. (Sc), a extração de antígenos foi realizada somente
para a fase de levedura.

67
Para a extração de antígenos miceliais, fragmentos do fungo S. schenckii
foram passados pela extensão de ágars Sabouraud dextrose a 4% (Himedia) em
placas de petri 20mm x 100 mm e colocados para crescerem à temperatura
ambiente (25 a 30oC) por 14 dias. Após este período, os fungos foram retirados das
placas, evitando-se, ao máximo, a retirada do ágar, e caso este viesse junto, era
raspado com o auxílio de uma alça de platina. Os pellets recolhidos foram
transferidos para béqueres estéreis e cobertos com PBS, pH 7,2, e inativados com
timerosal (Sigma- Aldrich, EUA), 0,2 g/litro/48 horas. Após este período, a massa
fúngica foi lavada por duas vezes com o PBS, pH 7,2, e centrifugado a 10.000 xg
/10’ a 8oC. O material foi liofilizado e conservado a -20oC para posterior utilização.
Para a extração de antígenos do fungo na morfologia de levedura, as cepas
S. schenckii e Sporothrix spp. foram convertidas para a fase de levedura através do
repique da forma micelial em ágar Levedura(0,5%)-Peptona (0,5%)-dextrose (1%)
(YPD), pH 8,0, à temperatura de 37oC. Após a conversão, as leveduras de cada
cepa foram semeadas, em abundância, no ágar YPD e cultivadas a 37oC / seis dias.
Para a retirada das leveduras do meio de cultura, adicionou-se sobre o material
crescido 2 a 4 ml de PBS estéril, pH 7,2, executou-se uma raspagem superficial do
material para que houvesse uma dissolução das leveduras que, em seguida, foram
recolhidas com a ajuda de pipeta estéril e transferidas para béqueres estéreis para
inativação pelo timerosal, lavagem, liofilização e conservação conforme descrito
acima.
5.1.1 -Extração dos antígenos
A extração de antígenos deste experimento foi baseada no protocolo de
Rodrigues (2010), sendo que o passo de sucessivos descongelamentos em banho-
maria a 60oC e congelamentos com nitrogênio líquido com maceração, foi acrescido
no presente experimento.
Para a extração, as células liofilizadas foram ressuspensas em um almofariz
estéril e utilizou-se um tampão de lise de Tris-Cálcio em água Milli-Q, na proporção
de um grama de liófilo : 5,8 ml de tampão. Por um período de 60 minutos, o material
foi sucessivamente macerado com o auxílio de um pistilo sob congelamento em
nitrogênio líquido e descongelamento em banho-maria, 80oC. Em seguida, com o

68
auxílio de uma pipeta, adicionou-se o homogeneizado a um tubo do tipo Falcon de
50 ml, estéril, até a marca de 12 ml do tubo. Dentro do tubo havia, até a marca de 5
ml, areia fina, que foi previamente lavada com HCL e depois lavada sucessivamente
com água para retirar o resíduo do HCL, secada e esterilizada em estufa a 180oC/2
horas. Ditiotreitol (DTT) (Sigma-Aldrich, EUA) foi adicionado ao homogeneizado e,
em seguida, o tubo foi agitado vigorosamente em vórtex durante 40’ a 4oC. Após
esta etapa, o homogeneizado foi centrifugado a 15.000 g/30’, 4oC. Os
sobrenadantes foram homogeneizados em um tubo do tipo Falcon de 15 ml, estéril,
esterilizados por filtração (filtro Millipore, 0,22 μm de diâmetro, membrana de acetato
celulose), receberam um coquetel de inibidor de protease (GE Healthcare, USA)
(1X) e foram dialisados contra PBS, pH 7,2, em saco de diálise (porosidade 10 kDa,
Sigma-Aldrich, EUA), sob rotação constante, durante 24 horas a 4oC, contra PBS,
pH 7,2, com três trocas diárias. Após a diálise, o extrato proteico bruto foi submetido
à filtração estéril (filtro Millipore 0,22 μm) e armazenado imediatamente em freezer a
– 70 oC.
Tampão de lise Tris-Cálcio em água Milii-Q (Rodrigues, 2010) :
Tris-HCl, 2M, pH 8,8 1%
CaCl2, 1M 0,5%
Coquetel e inibidor de protease (1x) 10 mM
DTT 20 mM
Água Milli-Q , q.s.p 1000 ul
5.1.2 - Quantificação de proteínas
A quantificação de proteínas foi realizada, em duplicata, através de dosagem
pelo método do ácido bicinconínico (BCA; Sigma-Aldrich, EUA) (SMITH et al., 1985)
utilizando-se uma curva de calibração com a proteína padrão BSA (Soro albumina
bovina- Sigma-Aldrich, EUA) a 1mg/mL nas quantidades de 0, 2, 4, 6, 8 e 10 µl/
poço.

69
5.1.3 - Análise dos extratos proteicos antigênicos por SDS-PAGE
A eletroforese em gel de poliacrilamida (SDS-PAGE) foi realizada em sistema
vertical BioRad Mini Protean III, constituído de um gel de separação linear na
concentração de 10% de acrilamida/bisacrilamida e gel de empilhamento na
concentração de 4%, polimerizado entre 2 placas de vidro separadas por
espaçadores com 1,0 mm de espessura e tampão de corrida Tris-Glicina, segundo
metodologia descrita por Laemmli (1970).
Os extratos proteicos brutos foram preparados em tampão de amostra (5x),
equivalendo a concentrações proteicas finais de 1 µg/µl, fervidos por 5 minutos,
aplicados no gel em volumes finais de 40 µl por canaleta e submetidos a uma corrida
eletroforética de 25 mA até que o corante azul de bromofenol chegasse ao fim do
gel.
Foram utilizados padrões de peso molecular (BenchMarck TM Protein
Ladder/Gibco, BRL, USA) contendo proteínas com massas de 220, 160, 120, 100,
90, 80, 70, 60, 50, 40, 30, 25, 20, 15 e 10 kDa para calcular a massa das proteínas
reveladas no gel. Após a corrida o gel foi corado com o corante azul de coomassie
R-250 sob agitação lenta, por pelo menos uma hora, seguindo-se de banhos
repetidos em solução descorante, idêntica à corante, só que sem o azul de
coomassie, até que a visualização das bandas estivesse nítida. Após o término da
descoloração, o gel foi deixado em manutenção em solução contendo 5% de
metanol e 7% de ácido acético.
5.2.0 - Produção de anticorpos
5.2.1 - Produção de IgY - Imunização das aves
Foram utilizadas galinhas da linhagem Hysex Brown, com peso vivo médio de
1,7 kg, que foram divididas em três grupos de três animais e imunizadas,
separadamente, com os extratos proteicos produzidos: 1- Extrato proteico de micélio
- Sporothrix schenckii ATCC 32285 (MSs), 2- Extrato proteico de levedura -

70
Sporothrix schenckii ATCC 32285 (LSs) e 3- Extrato proteico de levedura -
Sporothrix spp. (LSc). Durante o experimento dois animais do grupo 1 morreram e
um animal do grupo 2, antes da terceira imunização, entretanto continuou-se as
imunizações com os animais restantes.
Injetou-se, por via intramuscular (músculo peitoral), 1mL de uma solução
contendo 200 μg/mL de proteína total, de acordo com o esquema de imunização
descrito na Tabela 1. A primeira injeção foi feita na forma de emulsão em adjuvante
completo de Freünd (ACF), a segunda com adjuvante incompleto de Freünd (AFI).
Da terceira à nona imunização, os adjuvantes foram substituídos por solução salina,
NaCl 0,15 M.
Figura 1: Etapa de imunização das aves no músculo peitoral. 200 µg de proteína
do extrato antigênico/imunização em intervalos de 21 dias.

71
Tabela 1- Esquema de imunização das galinhas para obtenção de IgY
antiantígenos de Sporothrix schenckii e Sporothrix spp.
Imuni-
zações
MSs- Extrato proteico
micelial-
Sporothrix schenckii
LSs - Extrato proteico
de levedura –
Sporothrix schenckii
LSc - Extrato proteico
de levedura –
Sporothrix spp.
Ordem Dose/ave/imunização Dose/ave/imunização Dose/ave/imunização
1ª. 200 µg do antígeno
em ACF
200 µg do antígeno em
ACF
200 µg do antígeno em
ACF
2ª. 200 µg do antígeno
em AIF
200 µg do antígeno em
AIF
200 µg do antígeno em
AIF
3ª.
200 µg do antígeno
em salina
200 µg do antígeno em
salina
200 µg do antígeno em
salina
4ª.
5ª.
6ª.
7ª.
8ª.
9ª.
5.2.2 - Colheita de sangue e ovos
Ovos controle, chamados de pré-imunes, foram recolhidos antes do início das
etapas de imunizações. Os ovos das aves imunizadas foram recolhidos a partir do
sexto dia após cada imunização, separados por período de imunização, por ave e
armazenados a 4oC.
As amostras de sangue foram recolhidas por punção da veia marginal da asa
antes do início da primeira imunização e 20 dias após cada imunização. Após a
coagulação, em temperatura ambiente, as amostras foram centrifugadas a 3000g/10
minutos, os soros separados foram aliquotados e armazenados a -20oC.

72
5.2.3 - Purificação dos anticorpos IgY
Os ovos recolhidos foram agrupados por ave e período de imunização. As
gemas de cada período de imunização foram previamente separadas da clara,
lavadas com água destilada e em seguida escorridas em béqueres estéreis,
homogeneizadas e uma alíquota de 20 mL foi retirada para a extração da IgY, o
restante do material foi armazenado a – 20oC.
A alíquota de 20mL foi transferida para béqueres estéreis, diluída com água
destilada na proporcão de 1:10 e acidificada (até atingir pH 5,0). A solução de gema
foi deixada em repouso por um período de 14-16h (overnight) a 4oC. O material
insolúvel foi removido por centrifugação a 10.000g por 30 min, a 4oC. O
sobrenadante foi recolhido em béqueres e neste foi adicionado sulfato de amônio,
sob agitação, até atingir 29% de saturação (p/v). A solução foi mantida em
temperatura ambiente, com pH reajustado para 7,4-7,5, sob agitacão por 2h, e
recentrifugada a 10.000g por 30 min (Almeida et al., 1998). O pellet resultante foi
dissolvido em solução salina 0,15M, passou por processo de diálise contra solução
salina 0,15M e, em seguida, foi submetido à filtração esterilizante (0,22 µm) . A
solução contendo IgY foi alíquotada e armazenada a -20oC.
A concentração proteica da solução foi determinada pelo método do ácido
bicinconínico (SMITH et al., 1985).
5.2.4 - Análise em SDS-PAGE desnaturante
Amostras da solução contendo IgY foram submetidas à SDS-PAGE com 10% de
acrilamida/bisacrilamida e gel de empilhamento na concentração de 4% (Laemmli,
1970). As amostras foram preparadas conforme descrito no item 5.4, aplicadas no
gel em volumes de 30 µl por canaleta e 30 µg de proteína total. As amostras foram
submetidas a uma corrente inicial de 100V/60mA por 50 a 60 minutos até que o
corante azul de bromofenol chegasse ao fim do gel.

73
5.3.0 - IgG - Imunização dos coelhos
Para a produção de IgG antiantígenos de Sporothrix spp. foram utilizados seis
coelhos da raça Nova Zelândia, do sexo masculino e com peso médio de 2,64 kg.
Os animais foram divididos em três grupos de dois animais.
O procedimento de imunização dos coelhos foi semelhante ao das aves, com
três grupos de coelhos imunizados com os respectivos extratos proteicos
produzidos: 1- Extrato proteico de micélio - Sporothrix schenckii ATCC 32285 (MSs),
2- Extrato proteico de levedura - Sporothrix schenckii ATCC 32285 (LSs) e 3- Extrato
proteico de levedura - Sporothrix spp. (LSc). Injetou-se, por via subcutânea, na
região da coxa, 1mL de uma solução contendo 200 μg/mL de proteína total, de
acordo com o esquema de imunização descrito na Tabela 2. A primeira injeção foi
feita na forma de emulsão em adjuvante completo de Freünd (ACF), sendo as duas
subsequentes em adjuvante incompleto de Freünd (AFI). Na quarta e quinta
imunização, os adjuvantes foram substituídos por solução salina, NaCl 0,15 M.
Tabela 2 - Esquema de imunização dos coelhos para obtenção de IgG
antiantígenos de Sporothrix schenckii e Sporothrix spp.
Imuni-
zações
MSs- Extrato proteico
micelial-
Sporothrix schenckii
LSs - Extrato proteico
de levedura –
Sporothrix schenckii
LSc - Extrato proteico
de levedura –
Sporothrix spp.
Ordem Dose/coelho/imunização Dose/coelho/imunização Dose/coelho/imunização
1ª. 200 µg do antígeno em
ACF
200 µg do antígeno em
ACF
200 µg do antígeno em
ACF
2ª. 200 µg do antígeno em
AIF
200 µg do antígeno em
AIF
200 µg do antígeno em
AIF
3ª.
200 µg do antígeno em
salina
200 µg do antígeno em
salina
200 µg do antígeno em
salina
4ª.
5ª.

74
5.3.1 - Colheita de sangue
As amostras de sangue foram recolhidas antes do início da primeira
imunização e 20 dias após cada imunização. Após a coagulação em temperatura
ambiente, as amostras foram centrifugadas a 3000g por 10 minutos e os soros
separados, foram aliquotados e armazenados a -20oC.
Todos os animais utilizados foram mantidos em condições adequadas
e dentro dos padrões exigidos pela Comissão de Ética para o Uso de Animais em
Experimentos da UENF (CEUA), protocolo número: 034.
5.4 - Ensaio imunoenzimático (ELISA) dos soros e das preparações de IgY
purificadas
Para a titulação dos anticorpos produzidos nos soros pré-imunes e imunes de
galinhas e coelhos utilizados no experimento, as placas de ELISA (96 poços, NUNC
MaxiSorp™) foram sensibilizadas com os extratos proteicos produzidos (MSs, LSs e
LSc), 0,2 μg/poço, em tampão carbonato/bicarbonato 0,05 M, pH 9,6 (50 μl/poço) por
uma hora a 37oC e, em seguida, por um período de 14-16h a 4oC. A seguir, os poços
foram lavados, uma vez, com (200 μl/poço) PBST 0,05% (PBS 1X + 0,05 % Tween
20, pH 7,2) e bloqueados com 150 μl/poço com PBST 0,05% acrescido de 10% de
soro fetal bovino (PBST 0,05% + 10% SFB : PBST-SFB) por uma hora a 37oC. Após
o bloqueio, as placas foram lavadas por 3x (200 μl/poço) com PBST 0,05%, em
seguida, os primeiros poços foram preenchidos com 100 μl de soros pré-imunes e
imunes na diluição 1:500, em PBST-SFB, e foram feitas diluições seriadas até a
diluição 1:32000, as placas foram incubadas em estufa a 37oC por uma hora. A
seguir, os poços foram lavados cinco vezes com PBST e incubados com 50 μl de
IgG de coelho anti-IgY (Sigma-Aldrich, EUA) conjugado com peroxidase na diluição
1:2000/uma hora a 37oC. Após esta incubação, os poços foram lavados por cinco
vezes com PBST 0,05%. A reação foi revelada pela adição de 50 μl de solução
ácida com substrato enzimático contendo H2O2 e ortofenildiamina – OPD (Sigma-

75
Aldrich, EUA). A reação foi interrompida após 2 minutos de incubação à temperatura
ambiente ao abrigo da luz, com a adição de 50 μl de solução de H2SO4 a 3N. As
placas foram lidas em espectrofotômetro (Multiskan GO, Thermo Scientific) a 492
nm.
Para a titulação dos anticorpos das preparações de IgY purificadas, o
protocolo foi semelhante ao descrito acima, com modificações somente no passo
referente à adição do anticorpo primário. Na titulação da solução de IgY, os
primeiros poços foram preenchidos 100 μl de PBST-SFB contendo 80 μg/mL de
proteína da solução de IgY, daí foram feitas diluições seriadas até a concentração
de 0,065 μg de proteína da solução de IgY/ poço.
Pelo teste de ELISA, os soros dos coelhos e as soluções de IgY, à terceira
imunização, foram contrastados com os diferentes extratos antigênicos para ver se
havia reconhecimento cruzado entre os diferentes antígenos e anticorpos
produzidos.
5.5 - Detecção de anticorpos antiantígenos de Sporothrix spp. por western
blotting
As soluções de cada antígeno produzido, 200 μl, com concentrações
proteicas de 1μg/μl, foram submetidas ao SDS-PAGE a 10% (Laemmli et al., 1970)
como descrito anteriormente.
Ao término da corrida, o gel foi submetido à etapa de eletrotransferência para
uma membrana de nitrocelulose com poros de 0,45 μm (Biorad), segundo
metodologia de Towbin (1979).
A membrana de nitrocelulose e outros componentes do sistema de
eletrotransferência foram imersos em tampão de transferência. Em seguida, o gel foi
montado segundo orientações do fabricante Bio-Rad – Mini Trans Blot Eletrophoretic
Transfer Cell (Bio-Rad Laboratories, Richmond, USA). Este sistema foi submetido à
corrente de 10V por duas horas. Para determinar a eficácia da eletrotransferência
das proteínas e comparação relativa dos pesos moleculares das bandas separadas
pela eletroforese, foi adicionado ao gel um marcador de peso molecular pré-corado
(BenchMark TM Pré-stainned Protein Ladder / Gibco, BRL) e visualizado na
membrana de nitrocelulose. Após a eletrotransferência, os géis também foram

76
corados e descorados conforme protocolo do item 5.1.3, para confirmação da
transferência.
Em seguida, a membrana de nitrocelulose foi imersa em uma solução
bloqueadora – PBST-L (PBS, pH 7,2, com 0,1% de Tween 20 e 3% de leite em pó
desnatado), por 14 a 16 horas, a 4oC. A parte da membrana que continha o antígeno
foi cortada em tiras de aproximadamente 5 mm, as quais foram utilizadas
individualmente nas reações imunoenzimáticas. As tiras de membranas foram
incubadas com os anticorpos primários diluídos em PBST-L, por 2 horas à
temperatura ambiente, sob agitação constante, que foram: soros dos coelhos pré-
imunes e após a terceira imunização na diluição 1:2000 ou com a solução de IgY
das aves pré-imunes e após a terceira imunização, na concentração de 20 μg/mL
utilizando-se tubos de vidro com rolha. Os soros, solução de IgY e extratos
antigênicos também foram contrastados entre si para ver se havia reconhecimento
cruzado entre os diferentes antígenos e anticorpos produzidos. Na etapa seguinte,
as membranas foram lavadas por três vezes com PBS, pH 7,2 com 0,1% de Tween
20 por cinco minutos a cada lavagem, e incubadas com o segundo anticorpo, IgG de
coelho anti-IgY de galinha conjugado a peroxidase (Sigma-Aldrich, EUA) diluído
1:2000 em PBST-L/ 4 horas à temperatura ambiente, sob agitação constante ou IgG
de cabra anticoelho na diluição 1:2000, a depender de qual anticorpo estava sendo
estudado. Em seguida foram feitas mais três lavagens e a reação foi revelada com
uma solução contendo substrato enzimático mais a substância cromógena 3,3’
diaminobenzidina (DAB-Sigma-Aldrich, EUA). Assim que as bandas tornaram-se
claramente visíveis, a reação foi interrompida pela adição de água.
5.6 - Imunohistoquímica
Amostras de pele de dois gatos, fixadas em formalina (10%) tamponada neutra,
positivas para esporotricose, contidas no arquivo do Setor de Morfologia e Patologia
Animal (LMPA) do Hospital Veterinário da UENF e uma amostra de pele e outra de
pulmão de cão positivo para leishmaniose tegumentar e visceral, respectivamente,
foram clivadas e submetidas ao processamento histológico de rotina. A técnica de
imunohistoquímica foi realizada sobre essas amostras. Os blocos de parafina foram
cortados (3 µm) e os cortes montados em lâminas silanizadas. A desparafinização

77
foi realizada com banhos de xilol e rehidratação em banhos de álcool e água
deionizada com posterior, incubação, em banho-maria, em tampão citrato a 96oC/30
minutos para a recuperação antigênica. Em seguida, foram esfriadas à temperatura
ambiente/20 minutos. Após cuidadosa secagem das lâminas, os cortes foram
circulados com Dako pen (Dako, CA, USA) e a atividade da peroxidase endógena
bloqueada com peróxido de hidrogênio, 10%, em solução de metanol (v/v) / 30
minutos. Logo depois, as lâminas foram incubadas em PBS com 1% de BSA e 10%
de leite em pó desnatado para o bloqueio de ligações inespecíficas/60 minutos e,
posteriormente, cada corte foi incubado em câmara úmida, em geladeira overnight,
com o anticorpos primários IgY pré-imune, IgY anti-LSs, IgY anti-MSs e IgY anti-LSc
(5 µg/mL) ou anticorpos IgG de coelhos pré-imune, anti-LSs, anti-MSs ou anti-LSc
(1:500) diluídos em PBS + 3% de leite em pó desnatado, no controle, não houve
adição de anticorpo. No dia seguinte, entre os anticorpos primário e secundário, fez-
se um segundo bloqueio/30 minutos com 2% de soro de coelho diluídos em PBS
para as lâminas contrastadas com os anticorpos IgY ou 2% de SFB diluídos em PBS
para as lâminas contrastadas com os anticorpos de coelho. Procedeu-se, então, a
incubação com o anticorpo secundário, por duas horas, IgG de coelho anti-IgY de
galinha conjugado a peroxidase (Sigma-Aldrich, EUA) diluído 1:3000 em PBS+ 3%
de leite em pó ou IgG de cabra anticoelho na diluição 1:1200. A reação foi revelada
com uma solução contendo substrato enzimático mais a substância cromógena 3,3’
diaminobenzidina (DAB-Sigma-Aldrich, EUA) e parada após três minutos. As seções
histológicas foram contracoradas com hematoxilina de Harris, desidratada com
álcool, clarificada com xilol e montadas com permount (Sigma-Aldrich, EUA).
5.7.0 - Avaliação da ação dos anticorpos IgY sobre o crescimento, in vitro, das
cepas de S. schenckii e Sporothrix spp.
Para a avaliação da atividade de anticorpos IgY sobre o crescimento in vitro
das cepas de S. schenckii e Sporothrix spp., os anticorpos IgY pré-imunes e pós-
imunização foram incubados com o S. schenckii ou Sporothrix spp., a 37oC/44 h,
conforme descrito a seguir e discriminado na Tabela 3. As cepas de S. schenckii e
Sporothrix spp., na fase morfológica de levedura, foram cultivadas em ágar YPD a
37oC/24h, em seguida foram feitos inóculos, em solução salina 0,15 M, com turbidez
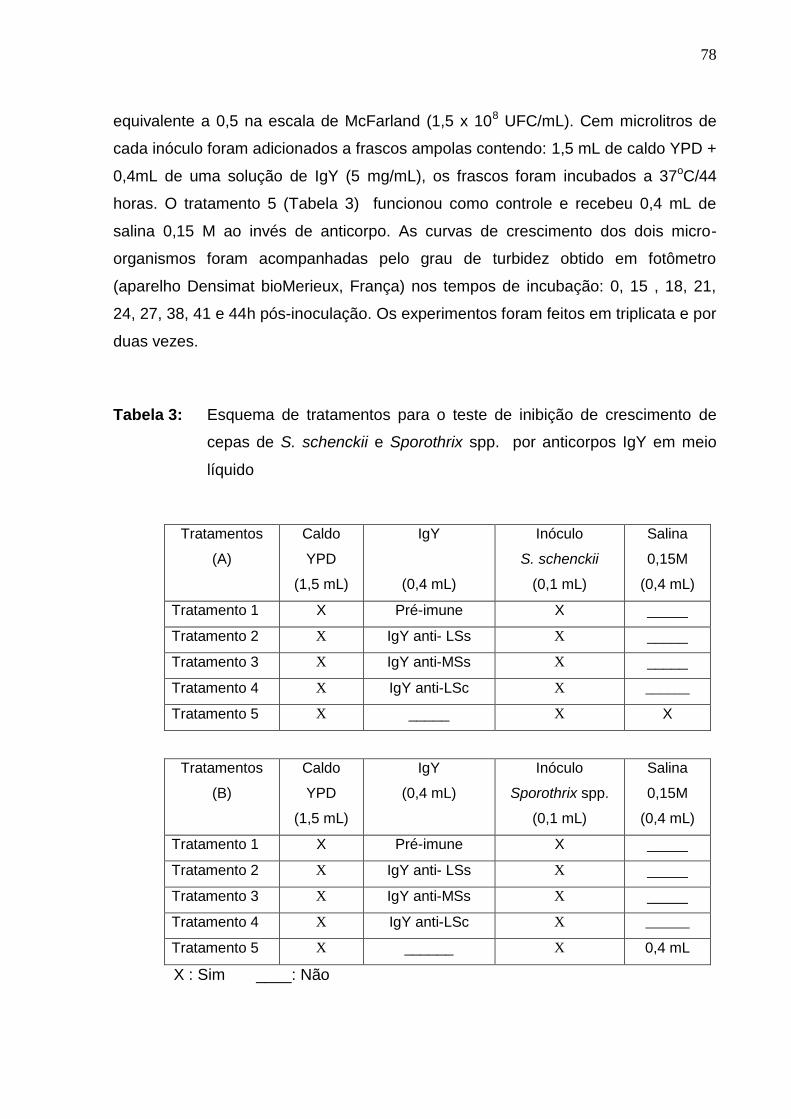
78
equivalente a 0,5 na escala de McFarland (1,5 x 108 UFC/mL). Cem microlitros de
cada inóculo foram adicionados a frascos ampolas contendo: 1,5 mL de caldo YPD +
0,4mL de uma solução de IgY (5 mg/mL), os frascos foram incubados a 37oC/44
horas. O tratamento 5 (Tabela 3) funcionou como controle e recebeu 0,4 mL de
salina 0,15 M ao invés de anticorpo. As curvas de crescimento dos dois micro-
organismos foram acompanhadas pelo grau de turbidez obtido em fotômetro
(aparelho Densimat bioMerieux, França) nos tempos de incubação: 0, 15 , 18, 21,
24, 27, 38, 41 e 44h pós-inoculação. Os experimentos foram feitos em triplicata e por
duas vezes.
Tabela 3: Esquema de tratamentos para o teste de inibição de crescimento de
cepas de S. schenckii e Sporothrix spp. por anticorpos IgY em meio
líquido
Tratamentos
(A)
Caldo
YPD
(1,5 mL)
IgY
(0,4 mL)
Inóculo
S. schenckii
(0,1 mL)
Salina
0,15M
(0,4 mL)
Tratamento 1 X Pré-imune X _____
Tratamento 2 X IgY anti- LSs X _____
Tratamento 3 X IgY anti-MSs X _____
Tratamento 4 X IgY anti-LSc X ______
Tratamento 5 X _____ X X
Tratamentos
(B)
Caldo
YPD
(1,5 mL)
IgY
(0,4 mL)
Inóculo
Sporothrix spp.
(0,1 mL)
Salina
0,15M
(0,4 mL)
Tratamento 1 X Pré-imune X _____
Tratamento 2 X IgY anti- LSs X _____
Tratamento 3 X IgY anti-MSs X _____
Tratamento 4 X IgY anti-LSc X ______
Tratamento 5 X ______ X 0,4 mL
X : Sim ____: Não

79
5.7.1 - Análise estatística
Realizou-se um experimento para cada cepa avaliada, neles utilizou-se o
delineamento inteiramente casualizado, com duas repetições em triplicatas, sob
esquema fatorial 5x8 caracterizado por 5 tratamentos avaliados em oito períodos. Os
tratamentos foram constituídos por (1) IgY pré-imune, (2) IgY anti-LSs, (3) IgY anti-
MSs, (4) IgY anti-LSc e (5) Branco, sem solução de IgY. Os tratamentos 1 e 5 foram
considerados controle. As leituras de densidade óptica (UFC mL-1) foram realizadas
nos períodos de 15, 18, 21, 24, 27, 38, 41 e 44 h pós-incubação do fungo. Os dados
obtidos foram submetidos aos testes (P≤0,05) de normalidade de Lilliefors e
homocedasticidade de Chrocan & Battlet, e não apresentaram restrições para a
análise da variância (P≤0,05) para efeito de regressão linear e múltipla em função do
período. Na análise descritva calculou-se as médias e os desvios padrão para os
tratamentos e na comparação utilizou-se o teste de Tukey (P≤0,05).
5.8.0 - Colheita de sangue de cães e gatos
Foram coletadas 21 amostras de sangue de gatos negativos e em bom
estado de saúde e de sete gatos, com lesões cutâneas, positivos para esporotricose,
confirmados pelo isolamento do fungo em cultivo (Apêndice 1, Tabela 7)
Foram coletadas 18 amostras de sangue de cães negativos e uma amostra
de um cão com histórico de esporotricose e com sintomatologia clínica da doença,
não houve isolamento fúngico a partir deste animal devido à ausência de lesões
externas. As amostras de sangue do cão positivo foram coletadas antes e 60 dias
após o tratamento com solução saturada de iodeto de potássio a 20%, 40 mg/kg,
duas vezes ao dia.
Após a coagulação em temperatura ambiente, as amostras de sangue foram
centrifugadas a 3000g por 10 minutos e os soros separados, foram aliquotados e
armazenados a -20oC.

80
5.8.1 - Titulação por ELISA dos soros de gatos e cães versus antígenos de
Sporothrix spp.
Para a titulação dos anticorpos nos soros de animais negativos e positivos, as
placas de ELISA (96 poços, NUNC MaxiSorp™) foram sensibilizadas com os
extratos proteicos produzidos (MSs, LSs e LSc), (0,2 μg/poço) em tampão
carbonato/bicarbonato 0,05 M, pH 9,6 (50 μl/poço) por uma hora a 37oC e, em
seguida, por um período de 14-16h a 4oC. A seguir, os poços foram lavados, uma
vez, com (200 μl/poço) PBST 0,05% (PBS 1X + 0,05 % Tween 20, pH 7,2) e
bloqueados com 150 μl/poço com PBST 0,05% acrescido de 3% de leite em pó
desnatado (PBST-L) por uma hora a 37oC. Após o bloqueio, as placas foram lavadas
por 3x (200 μl/poço) com PBST 0,05%, em seguida, os primeiros poços foram
preenchidos com 90 μl de soros na diluição 1:100, em PBST-L, foram feitas diluições
seriadas até a diluição 1:8100, as placas foram incubadas em estufa a 37oC por uma
hora. A seguir, os poços foram lavados 5 vezes com PBST e incubados com 50 μl
de IgG de coelho anti-IgG de gato (Abcam, EUA) conjugado com peroxidase na
diluição 1:1000/uma hora a 37oC ou com IgG de coelho anti-IgG de cão (Sigma-
Aldrich, EUA) conjugado com peroxidase na diluição 1:3000/uma hora a 37oC. Após
esta incubação, os poços foram lavados por cinco vezes com PBST 0,05%. A
reação foi revelada pela adição de 50 μl de solução ácida com substrato enzimático
contendo H2O2 e ortofenildiamina – OPD (Sigma-Aldrich, EUA). A reação foi
interrompida após 2 minutos de incubação à temperatura ambiente ao abrigo da luz,
com a adição de 50 μl de solução de H2SO4 a 3N. As placas foram lidas em
espectrofotômetro (Multiskan GO, Thermo Scientific) a 492 nm.
5.8.2 - Detecção de anticorpos antiantígenos de Sporothrix spp. por western
blotting em soros de cães e gatos
Para se testar os soros de cães e gatos frente aos antígenos produzidos pela
técnica do western blot, as tiras foram preparadas conforme descrito no item 5.5.
As tiras de membranas com antígenos dos extratos MSs, LSs ou LSc, foram
incubadas com os anticorpos primários, soros de gatos e cães positivos e negativos

81
para a esporotricose diluídos (1:300) em PBST-L, por 2 horas, à temperatura
ambiente, sob agitação constante. Na etapa seguinte, as membranas foram lavadas
por três vezes com PBS, pH 7,2 com 0,1% de Tween 20 por cinco minutos a cada
lavagem, e incubadas com o segundo anticorpo, IgG de coelho anti-IgG de gato
(Abcam, EUA) conjugado com peroxidase na diluição 1:1000/4 horas a 37oC ou com
IgG de coelho anti-IgG de cão (Sigma-Aldrich, EUA) conjugado com peroxidase na
diluição 1:3000. Em seguida foram feitas mais três lavagens e a reação foi revelada
com uma solução contendo substrato enzimático mais a substância cromógena 3,3’
diaminobenzidina (DAB-Sigma-Aldrich, EUA). Assim que as bandas tornaram-se
claramente visíveis, a reação foi interrompida pela adição de água.
5.8.3 - Isolamento de Sporothrix spp. a partir de materiais clínicos
Isolou-se o Sporothrix spp. a partir de fragmento para biópsia de uma
paciente humana, do sexo feminino, 48 anos, da cidade de Campos dos
Goytacazes/RJ, com lesão ulcerada na perna e histórico de mordida de gato com
suspeita clínica de esporotricose. A paciente foi atendida pelo Hospital Escola Álvaro
Alvim por médica dermatologista que encaminhou dois fragmentos para biópsia para
análise pelo setor de Micologia do LSA/UENF, sendo um em frasco com formol
tamponado a 10% para exame histopatológico e outro, em frasco estéril sem formol,
para a cultura fúngica, também foram encaminhados um suabe, uma lâmina com
imprint e soro sanguíneo da paciente.
Outro fragmento encaminhado foi do gato com mais de sete anos de idade,
que mordeu a referida paciente e que, após diagnóstico positivo para esporotricose,
foi eutanasiado devido à impossibilidade de tratamento do mesmo por parte da
paciente e gravidade da lesão no animal. Da lesão do gato, também foram feitos
imprints em lâminas de vidro para a coloração de Gram e visualização de leveduras
típicas de Sporothrix spp., assim como, a coleta de amostra dos exsudatos da úlcera
por suabe estéril.
Os materiais para o exame histopatológico foram encaminhados para o setor
de patologia do LMPA da UENF e processados de acordo com a rotina do setor.
Os materiais para isolamento fúngico, suabes e fragmentos de tecidos foram
processados no setor de micologia do Hospital Veterinário da UENF.

82
Para o isolamento da forma micelial do fungo, os suabes coletados foram
primeiramente passados em placas de petri contendo ágar Sabouraud Dextrose, a
4%, (Himedia) acrescido de cloranfenicol (50 mg/l) e cicloheximida (400 mg/l -
Sigma-Aldrich, EUA) com incubação a 25-30oC, e, em seguida, foram colocados em
cinco mL de caldo BHI (Brain Heart Infusion, Himedia) acrescido de cloranfenicol
(50 mg/l - Sigma-Aldrich, EUA) e incubados a 37oC/ 10 dias, depois os suabes foram
descartados, os tubos centrifugados a 3000 g/10 minutos, o pellet formado foi
transferido para o meio ágar Sabouraud Dextrose, a 4%, (Himedia) acrescido de
cloranfenicol (50 mg/l- Sigma-Aldrich, EUA) e cicloheximida (400 mg/l - Sigma-
Aldrich, EUA) e incubados à temperatura de 25-30oC, novamente. Para confirmação
do diagnóstico, as cepas isoladas na forma micelial foram convertidas para a forma
de levedura através do cultivo em ágar cérebro-coração a 37oC por 7 dias.

83
9.0 - RESULTADOS
9.1 - SDS-PAGE DOS EXTRATOS ANTIGÊNICOS PRODUZIDOS
O perfil eletroforético, em gel de poliacrilamida, dos extratos antigênicos
preparados do S. schenckii e Sporothrix spp. está apresentado na Fig. 2. O extrato
da amostra LSc apresentou em torno de 17 bandas, com PM na faixa de ≈120 a ≈15
kDa, enquanto o extrato da amostra LSs apresentou em torno de 14 bandas, com
PM na faixa de ≈95 a ≈27 kDa . Os extratos proteicos LSs e LSc apresentaram
diferenças principalmente para as proteínas expressas na faixa de PM entre ≈120 e
≈70 kDa, o Sporothrix spp. apresentou cinco bandas nesta faixa de peso, enquanto
o S. schenckii, somente duas bandas. Na linha da amostra LSc apareceu uma
banda bastante densa entre ≈80 e ≈70 kDa, a qual não ocorreu na LSs. Duas
bandas com pesos moleculares entre as faixas de ≈25 e ≈15 kDa foram mostradas
somente na linha da amostra LSc. A linha MSs apresenta o perfil eletroforético do
extrato micelial de S. schenckii. Houve concentração de bandas nas faixas de pesos
moleculares de aproximadamente, 120 - 80 kDa, e 60 - 50 kDa. Foi mostrada uma
banda com mais de ≈220 kDa e outras de aproximadamente, 40, 37, 35, 30 e 15
kDa .
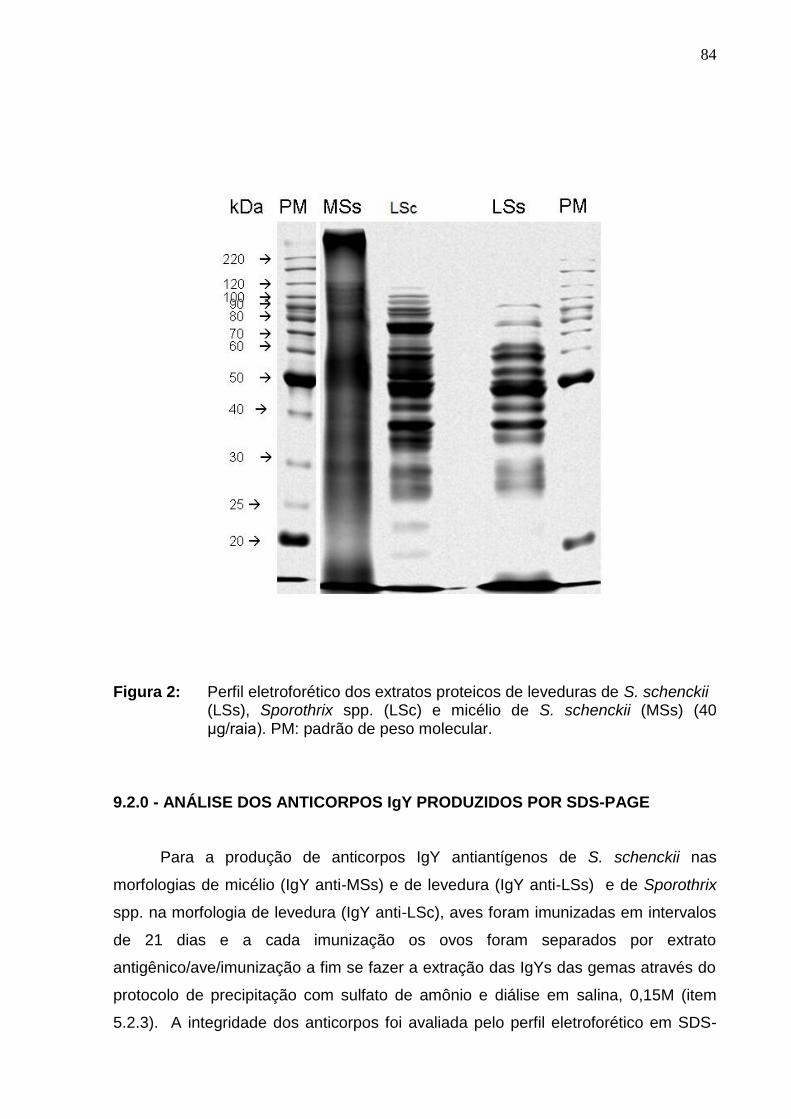
84
Figura 2: Perfil eletroforético dos extratos proteicos de leveduras de S. schenckii
(LSs), Sporothrix spp. (LSc) e micélio de S. schenckii (MSs) (40 μg/raia). PM: padrão de peso molecular.
9.2.0 - ANÁLISE DOS ANTICORPOS IgY PRODUZIDOS POR SDS-PAGE
Para a produção de anticorpos IgY antiantígenos de S. schenckii nas
morfologias de micélio (IgY anti-MSs) e de levedura (IgY anti-LSs) e de Sporothrix
spp. na morfologia de levedura (IgY anti-LSc), aves foram imunizadas em intervalos
de 21 dias e a cada imunização os ovos foram separados por extrato
antigênico/ave/imunização a fim se fazer a extração das IgYs das gemas através do
protocolo de precipitação com sulfato de amônio e diálise em salina, 0,15M (item
5.2.3). A integridade dos anticorpos foi avaliada pelo perfil eletroforético em SDS-

85
PAGE 10% e a Fig. 3 apresenta as duas subunidades das IgYs, cadeias leve (≈
18,66 kDa) e pesada (65,10 kDa) conforme apresentado.
Figura 3: Perfil eletroforético de amostras das soluções de IgY extraídas de
gemas de ovos pré-imunes (raia 1, 20 μg ) e pós-imunização: IgY anti-LSs (raia 2, 30 μg), IgY anti-MSs (raia 3, 20 μg) e IgY anti-LSc (raia 4, 30 μg). PM: peso molecular.
9.2.1 - ENSAIOS DE ELISA DOS EXPERIMENTOS DE IMUNIZAÇÃO
9.2.1.1 - Titulação dos anticorpos IgY dos soros das galinhas e das soluções
de IgY purificadas das gemas
O título de anticorpos IgY específicos foi monitorado pelo ensaio de ELISA
nos soros das aves e, posteriormente, nas soluções de IgY purificadas das gemas,

86
nos períodos pré-imunização e após a 1ª, 3ª, 5ª, 7ª e 9ª imunizações. A Fig. 4
mostra as titulações dos anticorpos nos soros das galinhas, diluições seriadas de
1:500 a 1:32000.
Para as soluções de IgY das gemas (Fig. 5), as diluições seriadas foram feitas
a partir de uma concentração de 4 µg de IgY/poço (80 µg de solução de IgY/mL) até
a concentração de 0,065 µg/poço (1,25 µg de solução de IgY/mL).
Figura 4: Titulação, por ELISA, dos anticorpos IgY séricos de galinhas
específicos para extratos proteicos de S. schenckii e Sporothrix spp. pré e pós- imunizações. A: levedura de S. schenckii (IgY anti-LSs), B: micélio de S. schenckii (IgY anti-MSs) e C: levedura de Sporothrix spp. (IgY anti-LSc). Diluição do soro: 1:500.

87
Os altos títulos de anticorpos obtidos, superiores a 1,2, a partir da terceira
imunização (Fig. 5), para uma solução de anticorpos com concentração de 80
μg/mL, indicaram a resposta imune humoral do animal frente aos antígenos
inoculados e habilitaram as soluções de IgY para os testes que seriam feitos
posteriormente (western blot, imunohistoquímica e inibição de crescimento).
Figura 5: Titulação, por ELISA, de anticorpos IgY das soluções purificadas das
gemas específicos para extratos proteicos de S. schenckii e Sporothrix spp. pré e pós- imunizações. A: levedura de S. schenckii (IgY anti-LSs), B: micélio de S. schenckii (IgY anti-MSs) e C: levedura de Sporothrix spp. (IgY anti-LSc). Diluições seriadas a partir de um volume
de 100 µl contendo 80 µg de solução de IgY/mL.
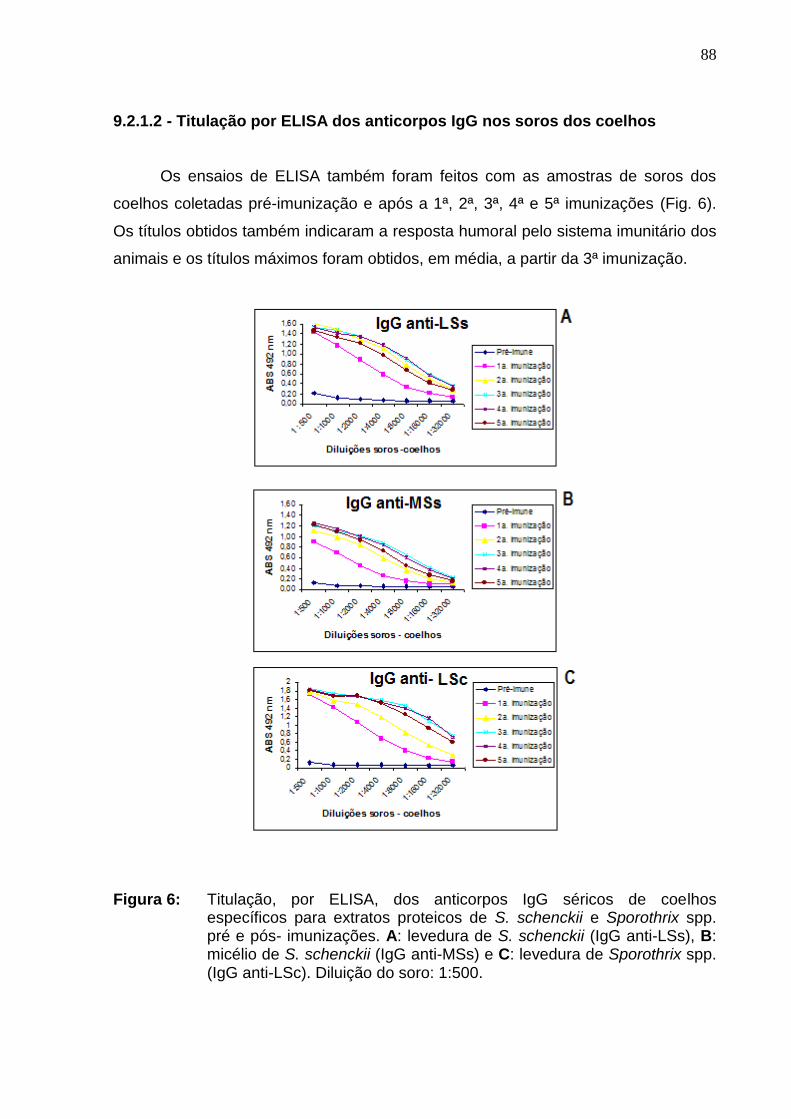
88
9.2.1.2 - Titulação por ELISA dos anticorpos IgG nos soros dos coelhos
Os ensaios de ELISA também foram feitos com as amostras de soros dos
coelhos coletadas pré-imunização e após a 1ª, 2ª, 3ª, 4ª e 5ª imunizações (Fig. 6).
Os títulos obtidos também indicaram a resposta humoral pelo sistema imunitário dos
animais e os títulos máximos foram obtidos, em média, a partir da 3ª imunização.
Figura 6: Titulação, por ELISA, dos anticorpos IgG séricos de coelhos específicos para extratos proteicos de S. schenckii e Sporothrix spp. pré e pós- imunizações. A: levedura de S. schenckii (IgG anti-LSs), B: micélio de S. schenckii (IgG anti-MSs) e C: levedura de Sporothrix spp. (IgG anti-LSc). Diluição do soro: 1:500.
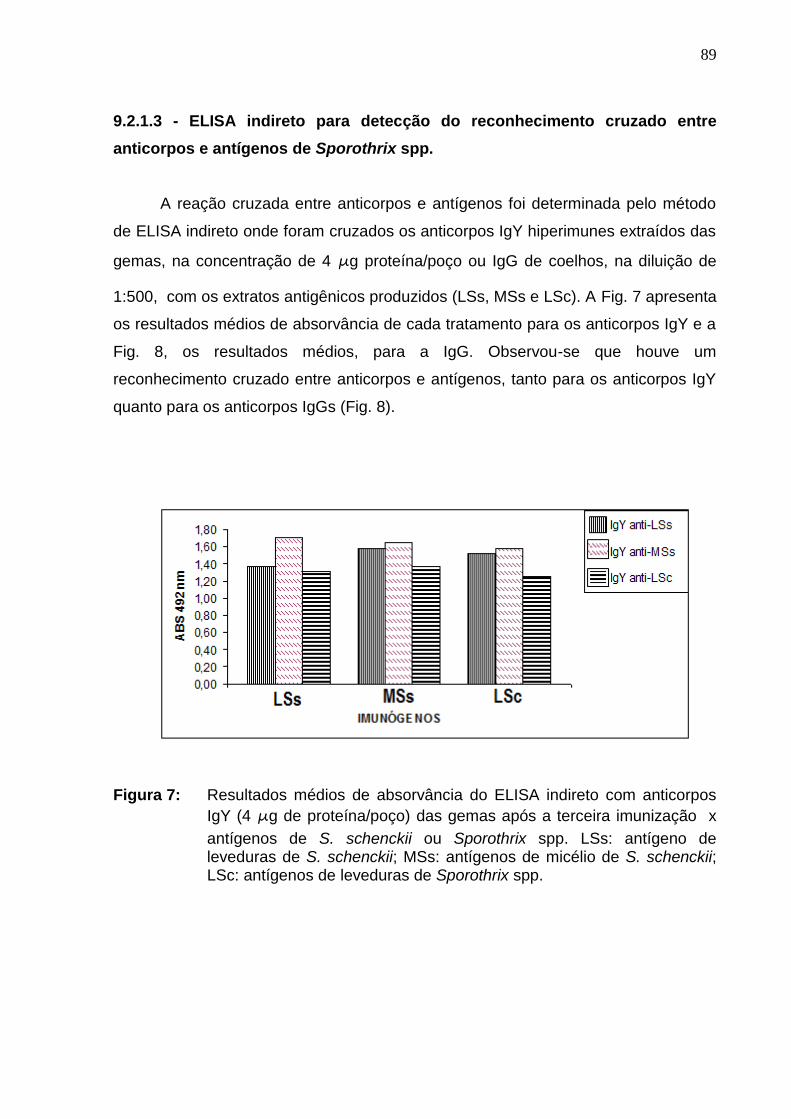
89
9.2.1.3 - ELISA indireto para detecção do reconhecimento cruzado entre
anticorpos e antígenos de Sporothrix spp.
A reação cruzada entre anticorpos e antígenos foi determinada pelo método
de ELISA indireto onde foram cruzados os anticorpos IgY hiperimunes extraídos das
gemas, na concentração de 4 µg proteína/poço ou IgG de coelhos, na diluição de
1:500, com os extratos antigênicos produzidos (LSs, MSs e LSc). A Fig. 7 apresenta
os resultados médios de absorvância de cada tratamento para os anticorpos IgY e a
Fig. 8, os resultados médios, para a IgG. Observou-se que houve um
reconhecimento cruzado entre anticorpos e antígenos, tanto para os anticorpos IgY
quanto para os anticorpos IgGs (Fig. 8).
Figura 7: Resultados médios de absorvância do ELISA indireto com anticorpos
IgY (4 µg de proteína/poço) das gemas após a terceira imunização x
antígenos de S. schenckii ou Sporothrix spp. LSs: antígeno de leveduras de S. schenckii; MSs: antígenos de micélio de S. schenckii; LSc: antígenos de leveduras de Sporothrix spp.

90
Figura 8: Resultados médios de absorvância do ELISA indireto cruzando
anticorpos IgG (1:500) após a terceira imunização x antígenos de S. schenckii ou Sporothrix spp. LSs: antígeno de leveduras de S. schenckii; MSs: antígenos de micélio de S. schenckii e LSc: antígenos de leveduras de Sporothrix spp.
9.2.2 – WESTERN BLOTS COM ANTISSOROS E SOLUÇÕES DE IgY DOS
ANIMAIS IMUNIZADOS
9.2.2.1 – Western blots dos anticorpos IgY
A Fig. 9 apresenta os resultados de western blot para as soluções de IgY.
Pode ser observado que um mesmo extrato é reconhecido de forma diferente por
cada anticorpo. Os anticorpos IgYs pré-imunes apresentaram um certo
reconhecimento antigênico, principalmente para o antígeno LSc (Fig. 9B, linha 5). As
reações de western blot foram mais fortes nas tiras sensibilizadas com o antígeno
LSc (Fig. 9B, linhas 6, 7 e 8). Nas tiras sensibilizadas com LSs foram reconhecidas
várias bandas com pesos moleculares entre ≈115 - 23 kDa. Houve reconhecimento
da banda de ≈ 23 kDa pelos três anticorpos utilizados para teste e esta não
apareceu nas tiras sensibilizadas com LSc. Nas tiras sensibilizadas com LSc (Fig.
9B) nas linhas 6, 7 e 8, houve forte reconhecimento de bandas entre ≈180 - 40 kDa,
a banda de ≈ 27 kDa foi marcada por todos os anticorpos e esta não apareceu nas
tiras sensibilizadas com LSs (FIG. 9A, linhas 2, 3 e 4).

91
O western blot com anticorpos IgY versus tiras sensibilizadas com antígeno
de micélio de cepa de S. schenckii (Fig. 9C) mostrou que o reconhecimento de
bandas na faixa de peso molecular de ≈115, 82 e 64 kDa foi comum para os três
anticorpos estudados (Fig. 9C, linhas 10, 11 e 12), sendo que as bandas de ≈115 e
82 kDa também foram reconhecidas pelos anticorpos pré-imunes (PI).
Figura 9: Reconhecimento das moléculas antigênicas por anticorpos das
soluções de IgY das gemas (20 μg/mL) por meio da técnica de western blot. (A) Tiras sensibilizadas com extratos proteicos de leveduras de S. schenckii (LSs), (B) extratos proteicos de leveduras de Sporothrix spp. (LSc) e (C) extratos proteicos de micélio de S. schenckii (MSs). Linhas 1, 5 e 9: IgY pré-imunização. Linhas 2, 6 e 10: IgY anti-MSs. Linhas 3, 7 e 11: IgY anti-LSs. Linhas 4, 8 e 12: IgY anti-LSc. PM: peso molecular (kDa).

92
9.2.2.2 – Western blots com anticorpos IgG de coelhos
A Fig. 10 apresenta o western blot com os anticorpos IgG de coelhos. Para as tiras
sensibilizadas com antígenos de leveduras (Fig. 10 A e B), houve um variado
reconhecimento de bandas, que coincidem com as diversas proteínas mostradas no
gel de eletroforese (Fig. 2). Para as tiras sensibilizadas com LSc, chama a atenção a
forte marcação de bandas na faixa de 64 a 49 kDa. Poucas proteínas foram
reconhecidas nas tiras sensibilizadas com antígeno micelial de S. schenckii (Fig. 10
C), houve reconhecimento de bandas na faixa de ≈115 e ≈82 kDa e as bandas na
faixa de ≈64 - 49 kDa foram fracamente marcadas.
Figura 10: Reconhecimento das moléculas antigênicas por anticorpos IgG de
coelhos (1:2000) por meio da técnica de western blot. (A) Tiras sensibilizadas com extratos proteicos de leveduras de S. schenckii (LSs), (B) extratos proteicos de leveduras de Sporothrix spp. (LSc) e (C) extratos proteicos de micélio de S. schenckii (MSs). PI: IgG pré-imune. Linhas 1, 4 e 7: IgG anti-MSs; linhas 2, 5 e 8: IgG anti-LSs; linhas 3, 6 e 9: IgG anti-LSc. PM: peso molecular.

93
9.3 – Western blot COM SOROS DE ANIMAIS POSITIVOS E NEGATIVOS PARA
ESPOROTRICOSE
9.3.1 - Gatos
A Fig. 11 mostra os resultados das reações de western blot com soros de
gatos saudáveis e soros de gatos positivos para esporotricose, confirmados pelo
isolamento do fungo em cultivo, versus o antígeno de levedura de S. schenckii. Os
testes foram feitos com 21 soros de animais saudáveis (Apêndice 1, Fig. 32), destes
retirou-se uma amostragem de sete soros para serem comparados mais
proximamente com os soros de animais positivos para esporotricose. Pode-se
observar que as bandas foram mais numerosas e mais fortes para os animais com
esporotricose do que nos animais saudáveis, comprovando a resposta humoral
frente aos antígenos do fungo. Dos sete animais positivos, cinco reconheceram
bandas próximas a ≈180 e ≈115 kDa, nenhuma destas foi marcada por soros de
animais negativos. A banda de ≈80 kDa foi marcada em soros de animais positivos e
negativos. A banda de ≈ 70 kDa foi reconhecida por cinco soros de animais
infectados, mas também foi identificada em pelo menos três soros de animais
saudáveis. Observa-se nos resultados dos animais positivos, uma banda larga na
faixa de ≈64 a 49 kDa.

94
Figura 11: Reconhecimento das moléculas antigênicas por soros de gatos
negativos e positivos para esporotricose por meio da técnica de western blot. Tiras sensibilizadas com antígenos de leveduras de S. schenckii ATCC 32285. Diluição do soro: 1:300.
Quando os soros dos animais negativos e positivos foram contrastados com
antígenos de levedura de Sporothrix spp. (Fig. 12), observou-se um forte
reconhecimento de antígenos por parte dos sete soros positivos e perfil de
reconhecimento antigênico semelhante (linhas 20 a 26). As proteínas na faixa de
peso molecular entre ≈ 64 e 49 kDa foram fortemente reconhecidas por todos os
soros positivos e por dois a três soros negativos (linhas 13, 14 e 16).

95
Figura 12: Reconhecimento das moléculas antigênicas por soros de gatos
negativos (linhas 1- 19) e positivos (linhas 20 – 26) para esporotricose por meio da técnica de western blot. Tiras sensibilizadas com antígenos de leveduras de Sporothrix spp. Diluição do soro: 1:300. PM: peso molecular (kDa).
Para os antígenos miceliais de S.schenckii, observou-se um forte
reconhecimento de bandas na faixa de ≈ 115 kDa por soros dos animais negativos
(Fig.13, linhas 1-10; 13; 14); já para os animais positivos para esporotricose, esta
banda não foi expressiva. Proteínas na faixa de ≈ 64 kDa foram reconhecidas
somente pelos soros dos animais positivos (Fig. 13, linhas 21 a 27).

96
Figura 13: Reconhecimento das moléculas antigências por soros de gatos
negativos (linhas 1-20) e soros de gatos positivos (linhas 21 – 26) para esporotricose por meio da técnica de western blot. Tiras sensibilizadas com antígenos de micélio de S. schenkii ATCC 32285. Diluição do soro: 1:300. PM: peso molecular (kDa).
9.3.2 - Cães
Dezoito soros de cães saudáveis e uma amostra de soro de cão com
esporotricose, na forma clínica de mucosa nasal, também foram contrastados com
antígenos miceliais de S. schenckii e antígenos de levedura de S. schenckii e
Sporothrix spp. pela técnica do western blot.
Para os antígenos miceliais, os resultados mostraram um forte
reconhecimento da banda de ≈ 115 kDa por parte dos soros de animais saudáveis
(Fig. 14, linhas 1-18) e pelo soro do animal positivo (Fig. 14, linha 19). Proteínas na
faixa de peso molecular de ≈64 kDa foram reconhecidas pelo soro do animal positivo
(Fig. 14, linha 19).

97
Figura 14: Reconhecimento das moléculas antigências por soros de cães negativos (linhas 1-18) e soro de cão positivo (linhas 19) para esporotricose versus antígeno de micélio de S. schenkii ATCC 32285. PM: peso molecular (kDa). Diluição do soro: 1:300.
As análises de western blot foram feitas para todas as amostras de soros dos
cães negativos (n=18) e positivo para esporotricose (n=1), na diluição 1:300.
Entretanto, na Fig. 15 apresenta-se somente os resultados dos soros dos animais
negativos (Fig. 15A, linhas 1-10 e 15B, linhas 1-8) que mostraram algum
reconhecimento antigênico para serem comparados com o soro do animal positivo
(Fig. 15A, linha 11 e 15B, linha 9) versus antígenos de leveduras de S. schenckii e
Sporothrix spp., Fig. 15A e 15B, respectivamente. Os resultados mostraram,
claramente, um perfil de reconhecimento antigênico diferenciado para o soro do
animal positivo. Houve um maior número de bandas reconhecidas e reação mais
intensa do que a observada para os soros dos animais negativos.

98
Figura 15: Reconhecimento das moléculas antigênicas por soros de cães negativos e positivo para a esporotricose por meio da técnica de western blot. (A) Antígeno de leveduras de S. schenckii: amostras 1 – 10, soros de cães negativos, amostra 11: soro de cão positivo. (B) Antígeno de leveduras de Sporothrix spp.: amostras 1 – 8, soros de cães negativos, amostra 9: soro de cão positivo. PM: peso molecular. Diluição do soro: 1:300.
Posteriormente realizou-se testes de western blot, utilizando-se tiras de uma
mesma eletrotransferência, para comparar os perfis de reconhecimento antigênico
por parte dos anticorpos dos animais imunizados (IgY-anti LSs e IgG de coelho anti-
LSs) e animais positivos para esporotricose (cães e gatos) versus o antígeno de
leveduras S. schenckii (Fig. 16). Observou-se em todas as linhas, um
reconhecimento variado de bandas, caracterizando a especificidade da resposta
humoral para os antígenos estudados. Houve o reconhecimento das bandas na faixa
de peso molecular de ≈180 e ≈115 kDa em todas as tiras e várias proteínas
reconhecidas nas faixas entre ≈64 e 37 kDa. O reconhecimento da banda de ≈26
kDa foi comum entre os anticorpos dos animais imunizados (linhas 1 e 2) e dos
animais que tiveram a doença, exceto pelos anticorpos do animal da linha 9.
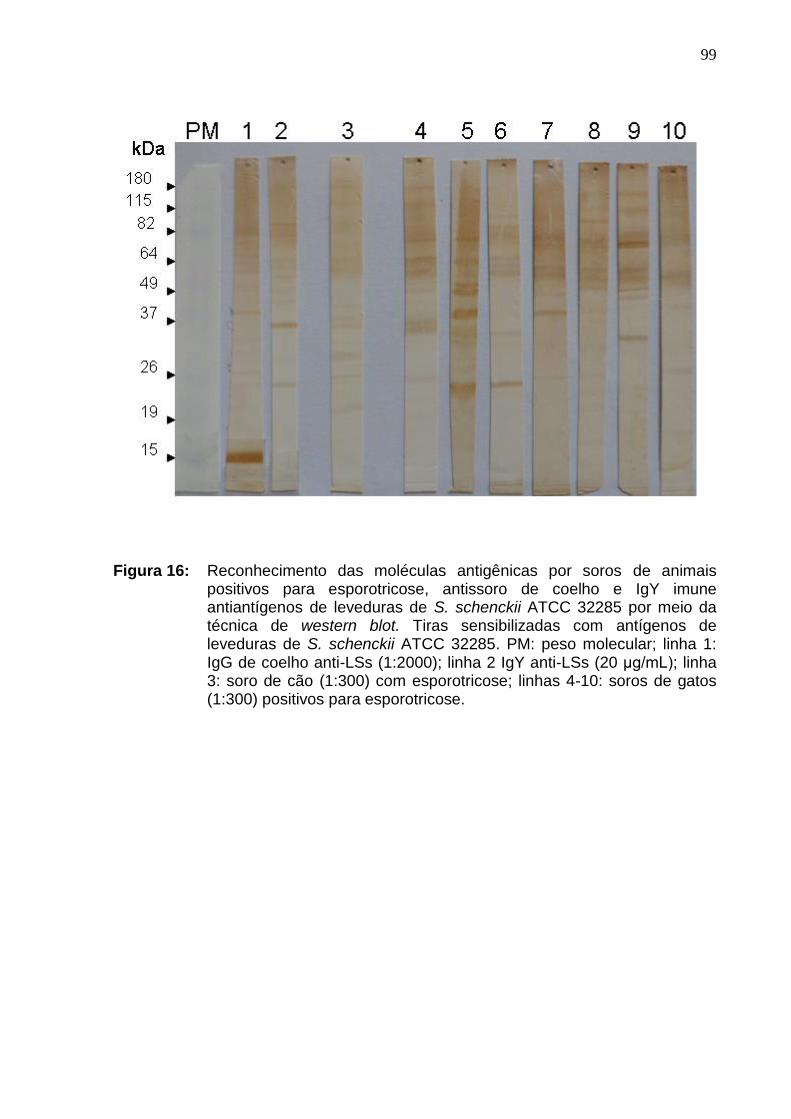
99
Figura 16: Reconhecimento das moléculas antigênicas por soros de animais
positivos para esporotricose, antissoro de coelho e IgY imune antiantígenos de leveduras de S. schenckii ATCC 32285 por meio da técnica de western blot. Tiras sensibilizadas com antígenos de leveduras de S. schenckii ATCC 32285. PM: peso molecular; linha 1: IgG de coelho anti-LSs (1:2000); linha 2 IgY anti-LSs (20 μg/mL); linha 3: soro de cão (1:300) com esporotricose; linhas 4-10: soros de gatos (1:300) positivos para esporotricose.

100
9.4 - IMUNOHISTOQUÍMICA COM ANTICORPOS POLICLONAIS DE GALINHAS E
COELHOS
Para saber se os anticorpos antiantígenos de S. schenckii e Sporothrix spp.
produzidos em galinhas e coelhos poderiam ser aplicados em imunohistoquímica
para o reconhecimento de leveduras de Sporothrix spp. em fragmentos para biópsia,
experimentos foram conduzidos em peles de gatos positivos para a esporotricose
utilizando-se os anticorpos policlonais de galinha (5 µg/mL) pré-imunização e pós-
imunização, assim como, soros de coelhos (1:500) pré e pós-imunização.
Os resultados mostraram que os anticorpos produzidos, nas duas espécies
animais, foram eficientes em reconhecer as leveduras do fungo em peles de gatos
positivos (Figuras 17 e 18).
Os anticorpos IgY marcaram leveduras dispersas no interstício e as
fagocitadas por macrófagos. Não houve marcação de leveduras pelo anticorpo IgY
pré-imune (Fig. 17).
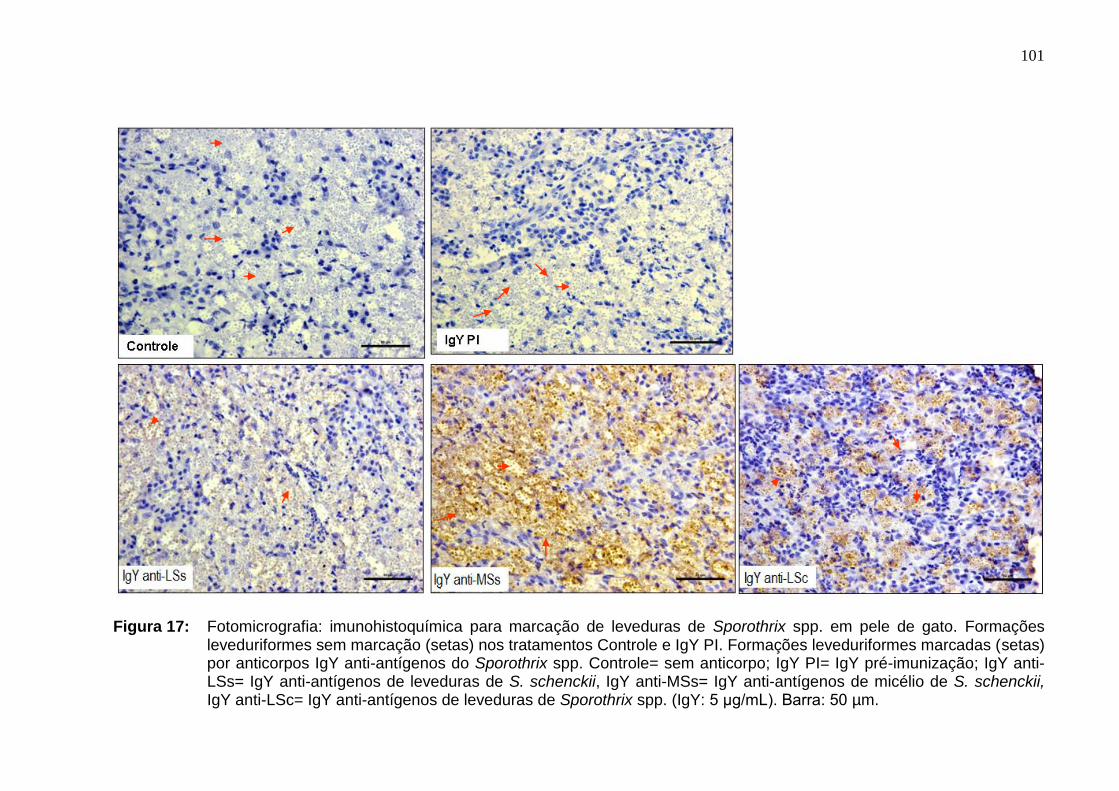
101
Figura 17: Fotomicrografia: imunohistoquímica para marcação de leveduras de Sporothrix spp. em pele de gato. Formações
leveduriformes sem marcação (setas) nos tratamentos Controle e IgY PI. Formações leveduriformes marcadas (setas) por anticorpos IgY anti-antígenos do Sporothrix spp. Controle= sem anticorpo; IgY PI= IgY pré-imunização; IgY anti-LSs= IgY anti-antígenos de leveduras de S. schenckii, IgY anti-MSs= IgY anti-antígenos de micélio de S. schenckii, IgY anti-LSc= IgY anti-antígenos de leveduras de Sporothrix spp. (IgY: 5 μg/mL). Barra: 50 µm.
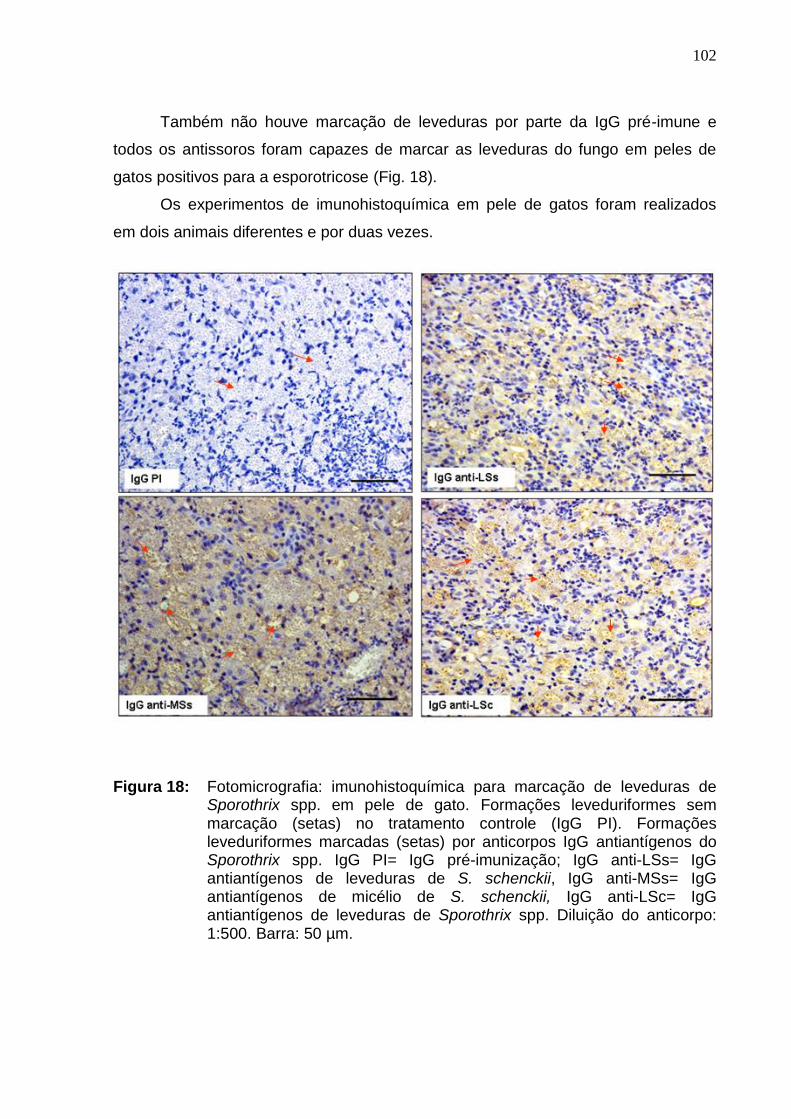
102
Também não houve marcação de leveduras por parte da IgG pré-imune e
todos os antissoros foram capazes de marcar as leveduras do fungo em peles de
gatos positivos para a esporotricose (Fig. 18).
Os experimentos de imunohistoquímica em pele de gatos foram realizados
em dois animais diferentes e por duas vezes.
Figura 18: Fotomicrografia: imunohistoquímica para marcação de leveduras de
Sporothrix spp. em pele de gato. Formações leveduriformes sem marcação (setas) no tratamento controle (IgG PI). Formações leveduriformes marcadas (setas) por anticorpos IgG antiantígenos do Sporothrix spp. IgG PI= IgG pré-imunização; IgG anti-LSs= IgG antiantígenos de leveduras de S. schenckii, IgG anti-MSs= IgG antiantígenos de micélio de S. schenckii, IgG anti-LSc= IgG antiantígenos de leveduras de Sporothrix spp. Diluição do anticorpo: 1:500. Barra: 50 µm.

103
Sabendo-se que cães com leishmaniose tegumentar apresentam lesões
ulceradas semelhantes às da esporotricose (SANTOS et al., 2007), procedeu-se,
também, a imunohistoquímica em fragmentos de biópsia de cão positivo para a
leishmaniose visceral. Os experimentos foram conduzidos em fragmentos de pulmão
utilizando-se os anticorpos policlonais de galinha (5 µg/mL) pré-imunização e pós-
imunização (anti-LSS, anti-MSs, anti-LSc) ou soros de coelhos (1:500) pré e pós-
imunização (anti-LSS, anti-MSs, anti-LSc). Os resultados mostraram que não houve
reação cruzada entre as IgYs específicas e formas amastigotas de Leishmania spp.
(Fig. 19).

104
Figura 19: Fotomicrografia: imunohistoquímica em fragmentos de pulmões de cão positivo para leishmaniose visceral com anticorpos IgY (5 μg/mL). Formas amastigotas de Leishmania spp. sem marcação em todos os tratamentos (setas). Controle= sem IgY; IgY PI= IgY pré-imune, IgY anti-LSs= IgY antiantígenos de leveduras de S. schenckii, IgY anti-MSs= IgY antiantígenos de micélio de S. schenckii, IgY anti-LSc= IgY antiantígenos de leveduras de Sporothrix spp. Barra: 10 µm.

105
Os antisoros de coelhos também não marcaram formas amastigotas (setas)
de Leishmania spp. (Fig. 20a e B). A Fig. 20B mostra marcações inespecíficas de
parênquima pulmonar, porém, a Leishmania spp.não foi marcada.
Figura 20: Fotomicrografia: imunohistoquímica em fragmentos de pulmões de cão
positivo para leishmaniose visceral com anticorpos IgG produzidos em coelhos. (A) IgG pré-imune; (B) IgG antiantígenos miceliais de S. schenckii. Setas indicam formas amastigotas de Leishmania spp. sem marcação. Barra: 10 µm.
9.5 - AVALIAÇÃO DA AÇÃO DOS ANTICORPOS IgY SOBRE O CRESCIMENTO,
IN VITRO, DAS CEPAS DE S. schenckii e Sporothrix spp.
O teste de inibição de crescimento foi conduzido para verificar o efeito dos
anticorpos IgY específicos sobre o crescimento, in vitro, das cepas de S. schenckii
(Tabela 4) e Sporothrix spp. (Tabela 5) estudadas. Como controle negativo de
inibição, foram utilizados os tratamentos Branco (sem solução de IgY) e o pré-imune
(PI), que continha anticorpos IgY pré-imunização. O crescimento das leveduras foi
acompanhado pelas leituras de densidade óptica (D.O) a 550 nm, medida nos
tempos zero, antes da incubação, e às 15, 18, 21, 24, 27, 38, 41 e 44h de
incubação, a 37oC. As Tabelas 4 e 5 apresentam os resultados médios de D.O para

106
cada observação e para cada cepa, sendo que no tempo zero, a D.O foi igual a zero
para todos os tratamentos.
Tabela 4 - Resultados médios de densidade óptica da curva de crescimento do S.
schenckii frente a anticorpos IgY com as respectivas equações polinomiais, o teste T e o coeficiente de correlação (R2)
PI : pré-imune, LSs: levedura de S. schenckii, MSs: micélio de S. schenckii, LSc: levedura de Sporothrix spp. Letras diferentes na mesma coluna diferem em P≤0,05 pelo teste de Tukey.
Tabela 5 - Resultados médios de densidade óptica da curva de crescimento do Sporothrix spp. frente a anticorpos IgY com as respectivas equações polinomiais, o teste T e o coeficiente de correlação (R2)
PI : pré-imune, LSs: levedura de S. schenckii, MSs: micélio de S. schenckii, LSc: levedura de Sporothrix spp. Letras diferentes na mesma coluna diferem em P≤0,05 pelo teste de Tukey.
A Fig. 21A e B apresenta as curvas de crescimento dos fungos S. schenckii e
Sporothrix spp. ajustadas para cada tratamento (Branco e anticorpos IgY) através de

107
regressões polinomiais (Tabelas 4 e 5). Nos dois experimentos, os resultados
mostraram a inibição significativa (P≤0,05) dos anticorpos específicos sobre o
crescimento do fungo, pois em ambos, as curvas de crescimento dos fungos perante
os anticorpos hiperimunes ficaram abaixo das curvas de crescimento dos
tratamentos com anticorpos PI e Branco. Neste experimento, foi mostrado mais uma
vez, o reconhecimento cruzado entre cepas e antígenos do Sporothrix spp., pois
cada inóculo de S. schenckii ou Sporothrix spp. teve o seu crescimento avaliado
perante os três anticorpos específicos produzidos: IgY anti-LSs, IgY anti-LMs e IgY
anti-LSc.
Figura 21: Regressão polinomial das curvas de crescimento do S. schenckii (A) e
Sporothrix spp. (B) frente a anticorpos IgY (1 mg/mL) pré-imune (PI), anti-LSs, anti-MSs, IgY anti-LSc e Branco (sem solução de IgY) às 0, 15, 18, 21, 24, 27, 38, 41 e 44 h de incubação a 37oC.

108
9.6- ANÁLISE SOROLÓGICA DE GATOS POSITIVOS PARA A ESPOROTRICOSE
A fim de se conhecer o perfil sorológico de gatos infectados, anticorpos IgG,
frente a antígenos de leveduras de S. schenckii e de cepa clínica de Sporothrix spp.
pelo ensaio de ELISA, coletou-se amostras de soros de gatos positivos (n=7) para
esporotricose cutânea, atendidos na clínica do Hospital Veterinário da UENF, antes
do início do tratamento e, como controle negativo, 21 amostras de soros de animais
saudáveis.
Todas as amostras de soro foram tituladas por ELISA em diluições seriadas
de 1:100 até 1:8100 versus os dois antígenos estudados, LSs (Tabela 7, Apêndice
1) e LSc (Tabela 8, Apêndice 1). Para a análise do ensaio, estabeleceu-se um ponto
de corte (cutoff), para cada antígeno testado e para cada diluição através da
seguinte fórmula: médias das absorvâncias dos soros dos gatos normais + 2 vezes o
desvio padrão destes soros (Média + 2x Desvio padrão) (Apêndice 1, Tabelas 7 e 8).
Para a análise dos resultados, considerou-se, os resultados de cutoff
(Absorvância492
= 0,575) obtidos com soros na diluição de 1:900 para a análise do
antígeno LSs (Tabela 7, Apêndice 1) e cutoff = 0,138, na diluição de 1:8100 para a
análise do antígeno de LSc (Tabela 8, Apêndice 1). Os resultados mostraram que
quando se avaliou o antígeno de leveduras de S. schenckii, 71,4% (n=5) das
amostras de soros positivos ficaram acima da linha de corte (cutoff=0,575), e duas
amostras de soro (28,6%), ficaram abaixo da linha de corte, falsos negativos (FIG.
22A). Com este antígeno, nenhuma das amostras de soros negativos foi dada como
falsa positiva, 100% das amostras ficaram abaixo da linha de corte.
Para o antígeno de leveduras de Sporothrix spp., considerando o
cutoff=0,138, 100% das amostras de soros positivos para a esporotricose ficaram
acima da linha de corte, verdadeiros positivos, e 95,2% das amostras de soros de
animais saudáveis ficaram abaixo da linha de corte estabelecida, verdadeiros
negativos (FIG. B ). Contudo, uma amostra de soro controle ficou acima do cutoff e
foi considerada como falsa positiva (4,8%).
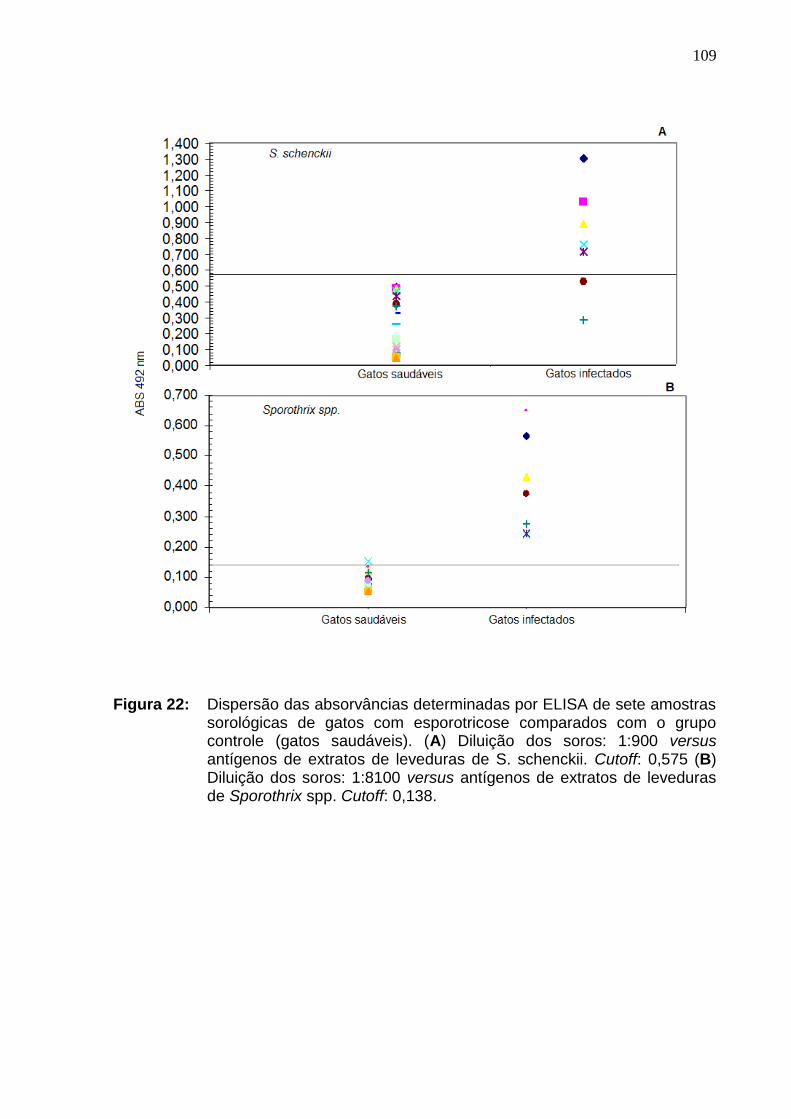
109
Figura 22: Dispersão das absorvâncias determinadas por ELISA de sete amostras sorológicas de gatos com esporotricose comparados com o grupo controle (gatos saudáveis). (A) Diluição dos soros: 1:900 versus antígenos de extratos de leveduras de S. schenckii. Cutoff: 0,575 (B) Diluição dos soros: 1:8100 versus antígenos de extratos de leveduras de Sporothrix spp. Cutoff: 0,138.

110
9.7 - RELATO DE CASO 1
Relata-se o caso clínico de um cão, SRD, do sexo feminino, com mais de sete
anos de idade. O animal era domiciliado na cidade do Rio de Janeiro, convivia com
dez gatos que tiveram esporotricose, sendo que sete destes felinos foram
sacrificados porque não responderam ao tratamento com itraconazol e os outros três
foram doados antes de adquirirem a doença. A proprietária relatou que, um ano
antes, o animal foi atendido por médico veterinário por apresentar uma massa
interna na narina direita que estava dificultando a respiração, com espirros e
escorria-lhe, pela narina, uma secreção fluída sanguinolenta. O animal não
apresentava nenhuma lesão externa e o diagnóstico foi de esporotricose baseado na
visualização de leveduras. Ainda durante a moradia na cidade do Rio de Janeiro, o
animal foi medicado com itraconazol, 100mg/dia, durante um ano. Entretanto,
durante todo o período de tratamento, a proprietária observou que a massa interna
persistia e que o animal passou a apresentar um inchaço no focinho. A proprietária
relatou que procurou várias ajudas médicas veterinárias e que lhe foi recomendada
a eutanásia do animal.
Ao levar o animal para domiciliar na cidade de Campos dos Goytacazes/RJ, a
proprietária recorreu ao atendimento do Hospital Veterinário da UENF. Ao exame
clínico, observou-se que o focinho e a região nasal direita do animal encontravam-se
edemaciadas, não havia lesões ulcerativas nas faces externa e interna da região
nasal, ausência de lesões na cavidade oral, secreção nasal fluída, transparente,
espumosa e abundante, principalmente na narina direita. O animal apresentava
dificuldade respiratória, devido ao aumento de volume interno na narina direita, ruído
na respiração inspiratória e sialorreia. Pelo histórico clínico, optou-se pelo início do
tratamento para esporotricose, com solução saturada 20% de iodeto de potássio
manipulada, na dosagem de 40 mg/kg, via oral, a cada 12 horas. Após 60 dias de
tratamento, o animal retornou ao atendimento, apresentando bom estado de saúde,
ausência de edema na face e no focinho, respiração normal, sem ruído e secreções.
Para garantir a cura clínica, optou-se por continuar com a medicação por mais três
meses (Fig. 1).
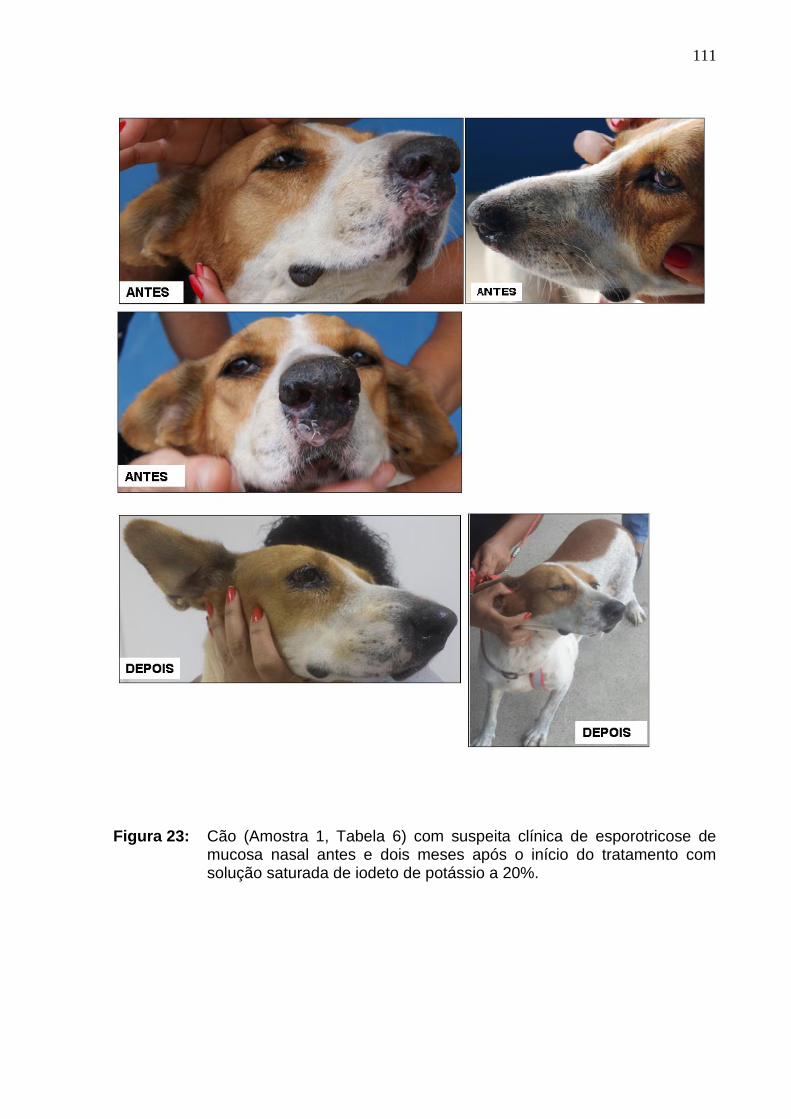
111
Figura 23: Cão (Amostra 1, Tabela 6) com suspeita clínica de esporotricose de
mucosa nasal antes e dois meses após o início do tratamento com solução saturada de iodeto de potássio a 20%.

112
Como não foi possível fazer uma punção aspirativa e não havia nenhuma
lesão externa para coleta de material por suabe e se tentar isolar o fungo por cultivo,
optou-se pela coleta de sangue, para obtenção do soro e realização da reação de
ELISA e western blot, utilizando como sensibilizadores, os antígenos de levedura de
S. schenckii e de cepa clínica de Sporothrix spp. estudados neste experimento (item
5.1.1). As coletas de sangue foram feitas na primeira e na segunda consulta. Os
resultados de D.O.s do ELISA e os obtidos por western blot foram contrastados com
os resultados de ELISA e western blot realizados com soros de cães clinicamente
negativos para esporotricose: cães de estimação e cães de uma associação
protetora dos animais (APA)/Campos dos Goytacazes/RJ (Tabela 6).
Os resultados de ELISA mostraram que, para todas as diluições dos soros, o
cão com histórico de esporotricose e apresentando os sintomas acima citados
(Tabela 6), apresentou valores de absorvância bem superiores aos dos animais
aparentemente saudáveis. Na diluição 1:100, a D.O. média para o antígeno LSs do
cão positivo (Tabela 6, amostra 1), antes do tratamento, foi de 1,62 e para o
antígeno LSc, o valor foi de 1,77. A média das D.O.s dos animais clinicamente
saudáveis foi de 0,452 (± 0,169) para o antígeno LSc e de 0,562 (± 0,217) para o
antígeno LSs. Se for considerado um ponto de cutoff (média + 2x desvio padrão das
absorvâncias dos soros de cães hígidos) pode-se dizer que todos os soros dos
animais saudáveis ficaram abaixo da linha de corte para o teste de ELISA com o
antígeno de levedura da cepa de Sporothrix spp. e podem ser considerados como
verdadeiros negativos. Já para o antígeno de levedura de S. schenckii, um animal
saudável (5,5%) apresentou valores de D.O. superiores ao ponto de corte sugerido
(Amostra 2, Tabela 6) e seria dado como um falso positivo.

113
Tabela 6 - Dados de identificação dos animais e densidades óticas (D.O.) a 492 nm dos testes de ELISA utilizando soro de cão com histórico de esporotricose e de cães aparentemente sadios (controles normais) e incubados com os dois antígenos, levedura de S. schenckii e levedura de Sporothrix spp.. Densidades óticas obtidas com soros diluídos a 1:100, 1:300, 1:900, 1:2700 e 1:8100. DP = desvio padrão. Cutoffs calculados como descrito acima.
Identificação dos animais
Antígeno Levedura de Sporothrix spp. Antígeno Levedura de S. schenckii
Diluição dos soros/D.O Diluição dos soros/D.O
Raça Idade Origem Amostra 1:100 1:300 1:900 1:2700 1:8100 1:100 1:300 1:900 1:2700 1:8100
SRD 7 anos Res. 1 1,771 1,326 0,738 0,329 0,151 1,617 1,469 1,167 0,752 0,358
SRD 5 anos APA 2 0,669 0,325 0,151 0,083 0,062 1,090 0,699 0,353 0,183 0,103
SRD 6 anos APA 3 0,662 0,295 0,139 0,083 0,061 0,869 0,440 0,221 0,121 0,079
SRD 2 anos Res. 4 0,597 0,280 0,128 0,073 0,057 0,802 0,358 0,165 0,094 0,067
SRD 5 anos Rua 5 0,470 0,244 0,123 0,073 0,058 0,592 0,293 0,143 0,083 0,065
SRD 5 anos SRD 6 0,412 0,179 0,107 0,068 0,055 0,552 0,237 0,115 0,074 0,065
SRD 1,5 anos Res. 7 0,588 0,273 0,152 0,086 0,064 0,695 0,331 0,182 0,096 0,071
SRD 3 anos APA 8 0,442 0,212 0,124 0,072 0,057 0,675 0,313 0,142 0,094 0,069
Boxer 3 anos Res. 9 0,289 0,132 0,076 0,061 0,058 0,636 0,308 0,137 0,079 0,062
Perdigueiro 7 anos Res. 10 0,383 0,171 0,088 0,063 0,053 0,596 0,258 0,126 0,074 0,060
SRD 2 anos APA 11 0,443 0,213 0,132 0,077 0,057 0,546 0,250 0,136 0,093 0,072
SRD 1,5 anos APA 12 0,397 0,189 0,098 0,068 0,058 0,545 0,270 0,133 0,094 0,068
SRD 6 anos APA 13 0,220 0,100 0,063 0,053 0,058 0,470 0,169 0,095 0,058 0,054
Labrador 8 anos Res. 14 0,314 0,135 0,079 0,057 0,050 0,420 0,194 0,102 0,071 0,064
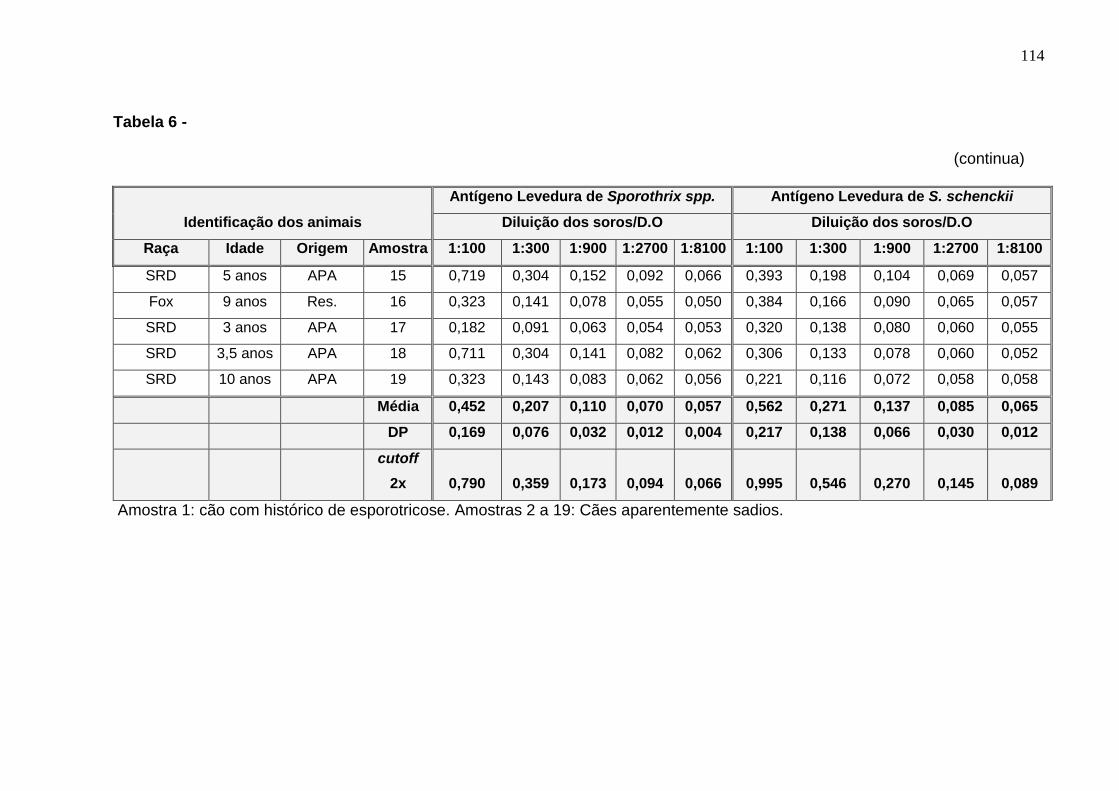
114
Tabela 6 - (continua)
Identificação dos animais
Antígeno Levedura de Sporothrix spp. Antígeno Levedura de S. schenckii
Diluição dos soros/D.O Diluição dos soros/D.O
Raça Idade Origem Amostra 1:100 1:300 1:900 1:2700 1:8100 1:100 1:300 1:900 1:2700 1:8100
SRD 5 anos APA 15 0,719 0,304 0,152 0,092 0,066 0,393 0,198 0,104 0,069 0,057
Fox 9 anos Res. 16 0,323 0,141 0,078 0,055 0,050 0,384 0,166 0,090 0,065 0,057
SRD 3 anos APA 17 0,182 0,091 0,063 0,054 0,053 0,320 0,138 0,080 0,060 0,055
SRD 3,5 anos APA 18 0,711 0,304 0,141 0,082 0,062 0,306 0,133 0,078 0,060 0,052
SRD 10 anos APA 19 0,323 0,143 0,083 0,062 0,056 0,221 0,116 0,072 0,058 0,058
Média 0,452 0,207 0,110 0,070 0,057 0,562 0,271 0,137 0,085 0,065
DP 0,169 0,076 0,032 0,012 0,004 0,217 0,138 0,066 0,030 0,012
cutoff
2x 0,790 0,359 0,173 0,094 0,066 0,995 0,546 0,270 0,145 0,089
Amostra 1: cão com histórico de esporotricose. Amostras 2 a 19: Cães aparentemente sadios.
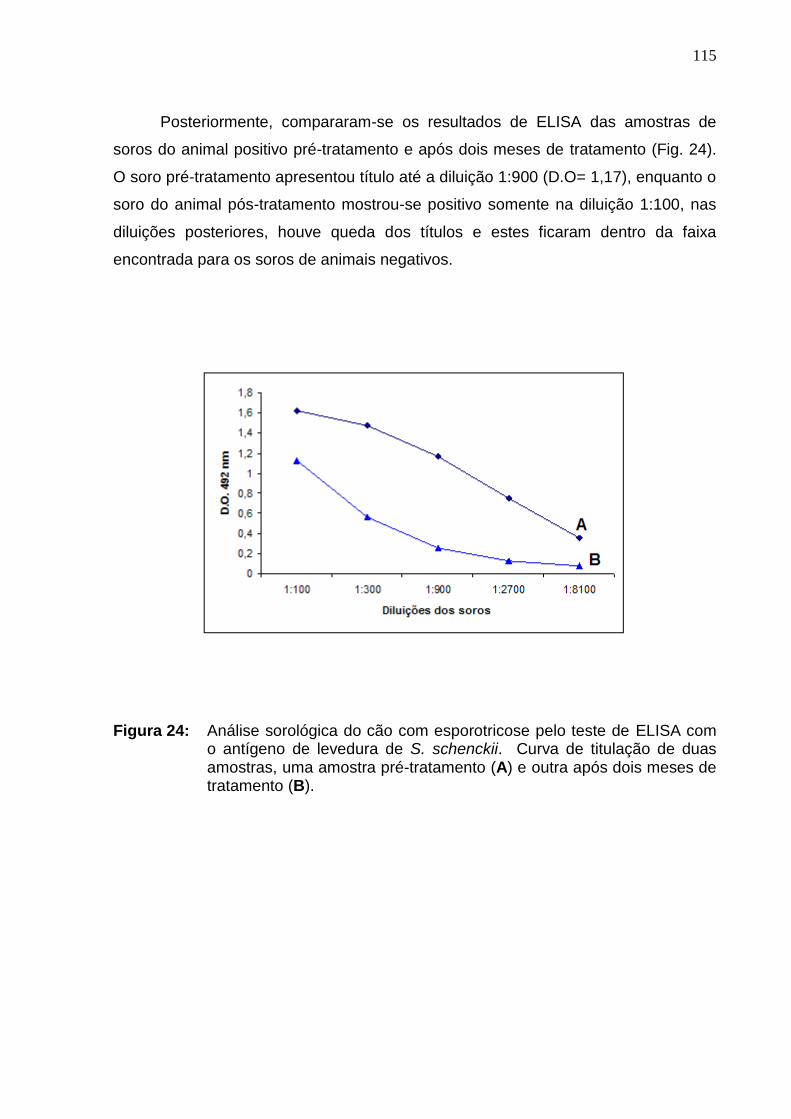
115
Posteriormente, compararam-se os resultados de ELISA das amostras de
soros do animal positivo pré-tratamento e após dois meses de tratamento (Fig. 24).
O soro pré-tratamento apresentou título até a diluição 1:900 (D.O= 1,17), enquanto o
soro do animal pós-tratamento mostrou-se positivo somente na diluição 1:100, nas
diluições posteriores, houve queda dos títulos e estes ficaram dentro da faixa
encontrada para os soros de animais negativos.
Figura 24: Análise sorológica do cão com esporotricose pelo teste de ELISA com
o antígeno de levedura de S. schenckii. Curva de titulação de duas amostras, uma amostra pré-tratamento (A) e outra após dois meses de tratamento (B).

116
A Fig. 25 ilustra a dispersão dos valores de D.O. para os soros dos animais
negativos e soro do animal com sintomas clínicos de esporotricose, na diluição de
1:100 versus os dois antígenos estudados: LSs e LSc.
Figura 25: (A) Resultados do teste de ELISA utilizando soros de cão com histórico
de esporotricose e soros de cães sadios versus o antígeno de levedura de S. schenckii. (B) Versus o antígeno de levedura de Sporothrix spp. Diluição dos soros: 1:100.
As análises por western blot mostraram uma resposta humoral mais intensa,
com maior número de bandas reconhecidas e maior intensidade da reação, para o
soro do animal positivo (Fig. 26A: linha 11 e Fig. 26B: linha 9) quando comparado
aos soros dos animais saudáveis (linhas 1-10: Fig. 26A e linhas 1-8, 10-11: Fig.
26B).

117
Figura 26: Reconhecimento das moléculas antigênicas por soros de cães negativos
e positivo para a esporotricose por meio da técnica de western blot. (A) Versus antígeno de levedura de S. schenckii: amostras 1 – 10, soros de cães negativos, amostra 11: soro de cão positivo. (B) Versus antígeno de levedura de Sporothrix spp.:: amostras 1 – 8, soros de cães negativos, amostra 9: soro de cão positivo. PM: peso molecular. Diluição dos soros: 1:300.
Objetivando um diagnóstico diferencial com a leishmaniose, foi realizado o
ensaio de ELISA com soros de animais positivos e negativos de área endêmica para
leishmaniose versus antígenos de leveduras S. schenckii (Fig. 27). Os resultados
mostraram que a D.O média para os animais positivos para leishmaniose (n= 9) foi
de 0,486 (± 0,32) e somente um animal (11,1 %) apresentou valor de D.O (1,06)
mais alto do que o valor de cutoff (0,995) estabelecido com a média dos soros
controle negativos para esporotricose (n= 18) e seria dado como falso positivo.

118
Figura 27: Resultados do teste de ELISA utilizando soros de cães na diluição 1:100
versus antígeno de levedura de S. schenckii. Amostra 1: soro de cão positivo para esporotricose pré- tratamento e amostra: 2, pós-tratamento. Amostra 3: soros de cães positivos para leishmaniose, amostra 4: soros de cães negativos de zona endêmica de leishmaniose, amostra 5: soro controle negativo para esporotricose.

119
9.8 - RELATO DE CASO 2 Relata-se o caso clínico de transmissão zoonótica da esporotricose,
após mordida de gato, para uma paciente humana, 48 anos, imunocompetente, com
histórico de paralisia infantil, na cidade de Campos dos Goytacazes/RJ.
O animal, do sexo masculino, com mais de sete anos de idade, apareceu com
uma lesão ulcerada na extremidade do membro anterior direito, foi submetido a uma
cirurgia, onde a médica veterinária optou pela amputação da falange e enviou
material para exame citológico. Enquanto aguardava o resultado, recomendou a
antibioticoterapia antibacteriana e o tratamento local da ferida cirúrgica. O animal
não apresentou nenhuma melhora clínica, o membro ficou edemaciado e
extremamente dolorido, impossibilitando o tratamento local e o animal se tornou
muito agressivo devido a dor. Ao tentar levar o animal para uma nova consulta, a
proprietária foi mordida pelo mesmo, na face anterior da perna direita e a partir daí
surgiram lesões ulceradas, granulomatosas, que aumentaram de tamanho. A mulher
procurou um atendimento médico no seu bairro e lhe foi receitada uma pomada para
tratamento local à base de antibiótico e antifúngico (cetoconazol), entretanto
nenhuma melhora clínica foi observada.
Em visita a APA (Associação Protetora dos Animais) de Campos dos
Goytacazes e após tomar conhecimento do caso, procedeu-se a visita à residência
da paciente, para exame do animal. O animal encontrava-se prostrado, apresentava
o membro anterior direito edemaciado, com lesão ulcerativa, que havia se estendido
de forma ascendente pelo membro e foram detectadas outras lesões ulceradas na
epiderme nos flancos esquerdo e direito do animal. Procedeu-se a coleta de material
das lesões ulceradas através de suabe estéril e fez-se um esfregaço em busca de
leveduras. Posteriormente, também foi coletado material para exame
histopatológico.
A mulher foi encaminhada para o Hospital Escola Álvaro Alvim/Campos dos
Goytacazes/RJ, setor de dermatologia, onde se procedeu a coleta de amostra da
lesão por suabe e retirada de dois fragmentos, sendo um para exame
histopatológico e outro para o cultivo de rotina laboratorial.
No material coletado do gato, procedeu-se a coloração de Gram da lâmina
com o esfregaço e foram visualizadas estruturas leveduriformes típicas de Sporothrix
spp., O diagnóstico de esporotricose foi confirmado pelo crescimento micelial do

120
fungo por meio de cultivo em ágar Sabouraud dextrose 4% (Himedia, EUA)
acrescido de cloranfenicol (50 mg/l) e cicloheximida (400 mg/l - Sigma-Aldrich, EUA)
à temperatura de 25oC após dez dias de incubação.
No material recolhido da mulher, isolou-se o fungo por meio de cultivo
do fragmento para biópsia. A mulher foi tratada com solução saturada de iodeto de
potássio e, dois meses após, logrou-se a cura clínica (Fig. 28).
Figura 28: Face anterior da perna de mulher mordida por gato positivo para
esporotricose. Forma cutânea de esporotricose, antes (setas) e após (setas) o tratamento com solução saturada de iodeto de potássio. Setas indicam lesões residuais.

121
9.8.1 - Achados histopatológicos em seções tegumentares do gato e da mulher
9.8.2 - Seções tegumentares do gato
A microscopia em seções de lesão tegumentar em felino, vista no H/E (Fig.
29A), mostrou reação granulomatosa, representada por abundantes macrófagos
epitelioides atulhados de estruturas fúgicas, leveduriformes (PAS) (Fig. 29B) e
dispersamente dispostos, caracterizando granulomas mal definidos.
Apresentou associação de exsudato misto com mononucleares
(linfoplasmocitário) e neutrófilos.
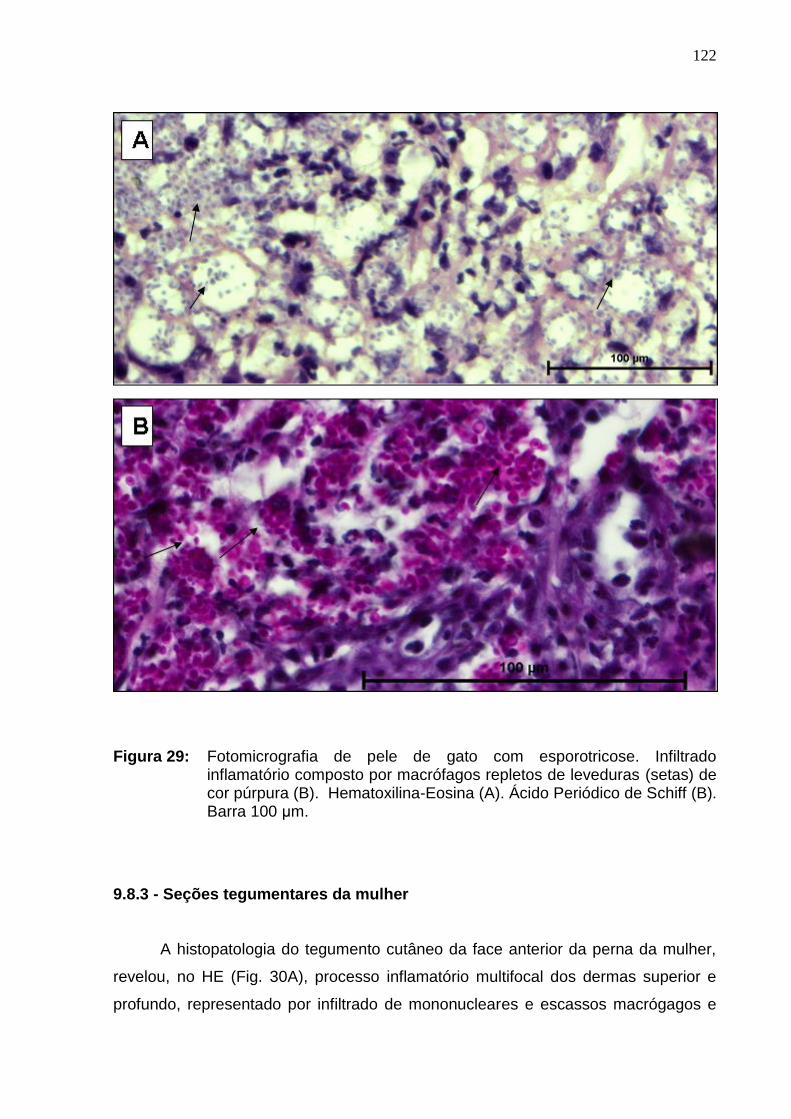
122
Figura 29: Fotomicrografia de pele de gato com esporotricose. Infiltrado
inflamatório composto por macrófagos repletos de leveduras (setas) de cor púrpura (B). Hematoxilina-Eosina (A). Ácido Periódico de Schiff (B). Barra 100 μm.
9.8.3 - Seções tegumentares da mulher
A histopatologia do tegumento cutâneo da face anterior da perna da mulher,
revelou, no HE (Fig. 30A), processo inflamatório multifocal dos dermas superior e
profundo, representado por infiltrado de mononucleares e escassos macrógagos e
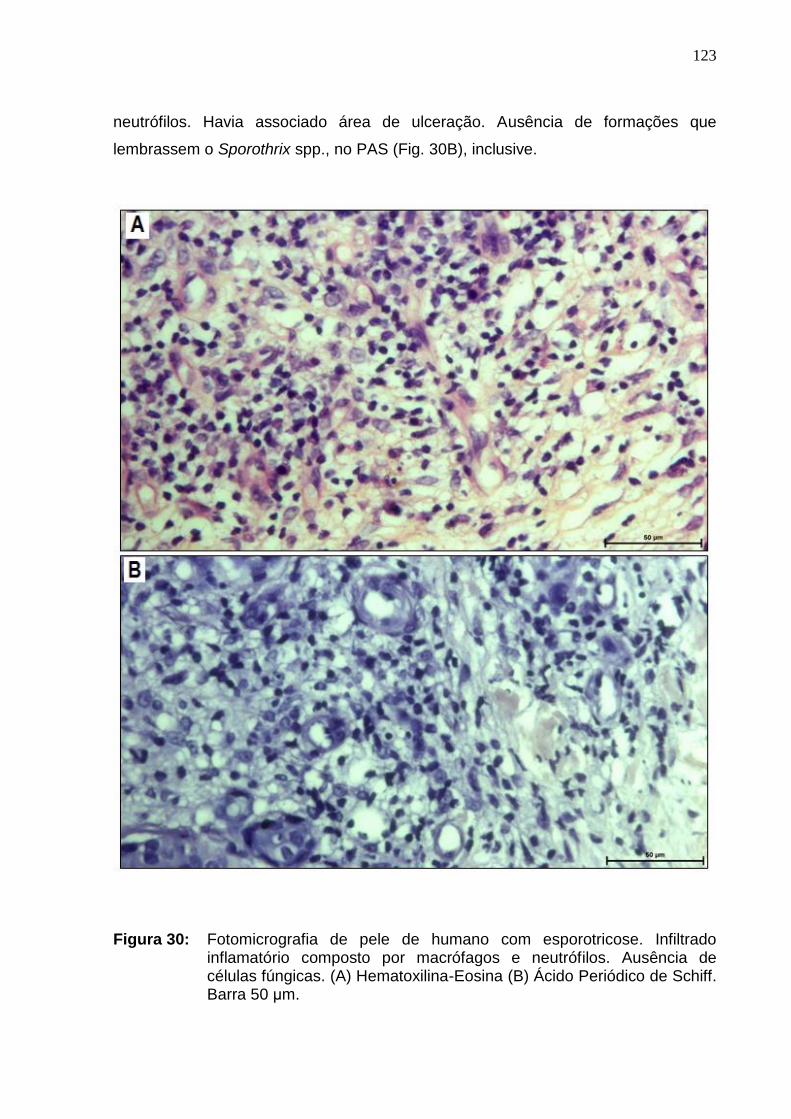
123
neutrófilos. Havia associado área de ulceração. Ausência de formações que
lembrassem o Sporothrix spp., no PAS (Fig. 30B), inclusive.
Figura 30: Fotomicrografia de pele de humano com esporotricose. Infiltrado
inflamatório composto por macrófagos e neutrófilos. Ausência de células fúngicas. (A) Hematoxilina-Eosina (B) Ácido Periódico de Schiff. Barra 50 μm.

124
10.0 - DISCUSSÃO
A secreção de proteínas/glicoproteínas é variável entre os isolados do
complexo S. schenckii e a sua composição pode alterar de acordo com o meio de
cultura utilizado para o cultivo do fungo (MENDONZA et al., 2002; FERNANDES,
2009). Para este experimento, as leveduras de S. schenckii e cepa clínica de
Sporothrix spp. foram cultivadas em ágar YPD, a 37oC, por seis dias e o micélio de
S. schenckii, em ágar Sabouraud dextrose 4%, a 25oC, por 14 dias. Optou-se pelo
protocolo de extração de proteínas com o tampão de lise tris-cálcio, associado à
maceração em nitrogênio líquido testado por Rodrigues (2010), acrescido da etapa
de descongelamento com maceração em banho-maria a 80oC. Rodrigues (2010)
testou oito metodologias de extração de antígenos e concluiu que, pela quantidade
de proteínas extraídas, diversidade de bandas, integridade das amostras, além da
reprodutibilidade das extrações, os tampões tris-cálcio e ureia-tioureia apresentaram
os melhores perfis de banda pela eletroforese unidimensional, sendo que, o tampão
tris-cálcio apresentou maior reprodutibilidade durante a eletroforese bidimensional.
O perfil proteico do extrato de leveduras de S. schenckii 32285 (Fig. 2) deste
trabalho apresentou semelhanças com o perfil do S. schenckii do trabalho de
Rodrigues (2010) (Fig. 31), as duas cepas apresentaram uma concentração de
bandas na faixa de 90 a 27 kDa. Várias bandas foram visualizadas no gel, fato

125
comumente visto em outras separações eletroforéticas de proteínas dos fungos do
complexo Sporothrix (MENDONZA et al., 2002; ALMEIDA-PAES et al., 2012,
FERNANDES et al., 2013).
Figura 31: Separação eletroforética por SDS-PAGE do extrato de proteínas de
leveduras de Sporothrix schenckii, após 7 dias de cultivo em caldo cérebro-coração, 120 rpm, 37 ºC. Gel de poliacrilamida (10%). Extratos proteicos brutos (20 μg de proteínas). P7: Tampão de lise Tris-Cálcio e P8: Tampão de lise Ureia-Tioureia associados à lise física de maceração em nitrogênio líquido. PM: Padrão de peso molecular - Coloração por nitrato de prata. (Adaptada de Rodrigues, 2010).
O extrato antigênico micelial do S. schenckii 32285 deste experimento
mostrou um perfil eletroforético bastante diferente dos perfis apresentados pelos
extratos de leveduras de S. schenckii 32285 e cepa clínica de Sporothrix spp. , os
quais apresentaram similaridades entre si. Para os dois extratos de leveduras, houve
maior concentração de bandas na faixa de peso molecular de, aproximadamente,
110 – 27 kDa.
Almeida-Paes et al. (2012) também consideraram os perfis eletroforéticos dos
extratos antigênicos, extratos livres das células, obtidos da fase de levedura de oito

126
isolados de Sporothrix spp. bastante similares entre si, com bandas variando de 220
– 20 kDa.
Fernandes et al. (2013) mostraram uma variedade de proteínas secretadas,
por leveduras, entre isolados do complexo Sporothrix, bandas com pesos
moleculares de 110, 90, 80, 60, 46, 38, 35, 28 kDa para o S. brasiliensis, de 160 a
38 kDa para o S. schenckii e de 160 a 28 kDa para o S. globosa. Os autores não
observaram associação entre perfil eletroforético de moléculas secretadas, espécies
e origens dos fungos.
Para se conhecer o perfil antigênico dos extratos proteicos produzidos foram
utilizados galinhas, para produção de anticorpos IgY, e coelhos, para a produção de
anticorpos IgG, antiantígenos miceliais de S. schenckii (anti-MSs) e de leveduras do
S. schenckii ATCC 32285 (anti-LSs) e Sporothrix spp. (anti-LSc). As titulações dos
anticorpos IgY específicos das gemas e dos soros dos coelhos (IgG) mostraram a
resposta humoral dos animais frente aos extratos antigênicos estudados. Os títulos
máximos foram obtidos, em média, após a terceira imunização.
Os resultados de western blot mostraram a resposta humoral dos animais
frente aos antígenos testados e evidenciaram um grande número de bandas
reconhecidas, principalmente nas tiras sensibilizadas com extratos de leveduras. As
bandas foram mais intensas nas tiras sensibilizadas com os extratos de leveduras de
Sporothrix spp., independente, do anticorpo primário utilizado na reação, IgG de
coelho ou IgY, anti-MSs, anti-LSs ou anti-LSc. Estes resultados corroboram os de
Almeida-Paes et al. (2012), que também relataram maior intensidade de bandas nos
extratos antigênicos de Sporothrix spp. do que nos extratos de S. schenckii.
Neste estudo, não foram observados perfis semelhantes de reconhecimento
antigênico quando antígenos de extratos miceliais e de levedura da mesma cepa de
S. schenckii 32285 foram analisados. Resultados que também corroboram os de
Almeida-Paes et al. (2012).
Poucas bandas foram reconhecidas pelos anticorpos versus antígenos
miceliais. Quando se contrastou antígenos miceliais com os anticorpos IgY, as
bandas de, aproximadamente, 115 e 82 foram reconhecidas pelos anticorpos pré-
imunes e pós-imunização (anti-MSs, anti-LSs e anti-LSc), em contrapartida, as
bandas de, aproximadamente, 64 e 49 kDa foram reconhecidas somente pelos
anticorpos pós-imunização. Os anticorpos policlonais, IgG, de coelhos, anti-MSs,
anti-LSs e anti-LSc versus antígenos miceliais reconheceram as bandas de 115 e 82

127
kDa e marcaram, fracamente, a banda de 64 kDa e não houve marcação de bandas
pelo soro pré-imune. Estes resultados são congruentes com os de Ruiz-Baca et al.
(2011) para antígenos miceliais da parede celular do S. schenckii que encontraram
cinco antígenos imunodetectados por antissoros de coelhos: 48, 55, 66, 67, e 70
kDa.
Para Nascimento e Almeida (2005), Nascimento et al. (2008) e Ruiz-Baca et
al., (2011) a glicoproteína de 70 kDa é o principal antígeno imunodetectado, em
animais experimentais, a partir de extratos da parede celular de leveduras e micélio
de S. schenckii (RUIZ-BACA et al., 2011) e a glicoproteína de 60 kDa é
imunodetectada somente nas células leveduriformes (RUIZ-BACA et al., 2011).
Almeida- Paes et al. (2012) encontraram padrões variados de
imunoreconhecimento entre cepas de Sporothrix. O antissoro policlonal de coelho
revelou 23 bandas glicoproteicas reativas com pesos moleculares aparentes de 240,
203, 186, 173, 152, 140, 130, 121, 112, 103, 97, 91, 85, 81, 72, 64, 60, 58, 55, 50,
47, 44 e 38 kDa para extratos livres de células de leveduras. Para os extratos
miceliais, os anticorpos policlonais de coelhos, reconheceram as bandas de 240,
230, 220, 203, 150, 143, 135, 90, 85, 81, 76, 70, 60 e 40 kDa, sendo que as bandas
de 230, 220, 135 e 85 kDa foram observadas em todas as cepas estudadas e a
banda de 40 kDa, em seis cepas.
Fernandes et al. (2013), encontraram um perfil heterogêneo de resposta
humoral, em camundongos infectados com S. brasiliensis, S. schenckii e S. globosa,
mas o reconhecimento da molécula de 60 kDa foi comum entre os animais
infectados.
Posteriormente, pela técnica do western blot, também se buscou conhecer o
reconhecimento antigênico dos extratos produzidos por parte dos anticorpos IgG de
cães (n=1) e de gatos (n=7) infectados, fato ainda não relatado na literatura, e
compará-los com soros de cães (n=18) e gatos (n=21) saudáveis.
Os resultados de western blot comprovaram a resposta humoral dos animais
infectados frente aos antígenos testados através do imunoreconhecimento de várias
bandas antigênicas, principalmente, nas tiras sensibilizadas com extratos de
levedura (LSs e LSc).
Assim como ocorreu nos soros dos animais experimentais, galinhas e
coelhos, as reações também foram mais intensas quando se contrastou soros de
animais infectados e extratos de levedura de Sporothrix spp.

128
As reações de western blot com soros de gatos infectados versus antígenos
de levedura de S. schenckii apresentaram maior intensidade de cor e maior número
de bandas reconhecidas do que as reações com soros de animais sadios. A banda
de, aproximadamente, 80 kDa foi reconhecida pelo soro de gatos saudáveis e
infectados, as bandas de ≈ 180 e 115 kDa não foram reconhecidas por soros de
animais saudáveis, somente por soros de animais infectados.
Em cães sadios, apesar de sete animais terem apresentado reconhecimento
de várias bandas, estas foram menos intensas do que as reconhecidas pelo soro do
animal positivo. O soro do animal positivo apresentou reconhecimento de bandas de
180 kDa até a faixa de 19 kDa.
Para os extratos de levedura de Sporothrix spp., observou-se uma forte
reação imunológica pelos soros de gatos infectados e por sete soros de animais
saudáveis (33,3%), o que indicou possível contato dos animais com antígenos do
fungo, sem, no entanto, terem adquirido a doença. A larga banda de 64 a 49 kDa foi
fortemente reconhecida por todos os soros positivos e foi mais estreita e fracamente
visualizada no western blot de dois soros de gatos saudáveis.
No western blot com soros de cães saudáveis versus antígenos de Sporothrix
spp. observou-se sete soros de animais saudáveis que apresentaram
reconhecimentos aleatórios de bandas com pesos moleculares na faixa de 110 a 80
kDa. O soro do animal positivo apresentou reações de forte intensidade, bem
diferente do observado nos animais negativos, a faixa compreendida por,
aproximadamente, 64 – 49 kDa apresentou as mesmas características de
intensidade de cor e espessura da apresentada pelos soros de gatos infectados. A
banda de, aproximadamente, 26 kDa pareceu ser uma característica para o
Sporothrix spp., pois foi reconhecida por anticorpos dos animais imunizados e
infectados.
Fernandes (2009) considerou as frações proteicas de 70 e 38 kDa de
exoantígenos de leveduras as principais frações reconhecidas, pela técnica de
western blot, por dois pools de soros humanos positivos para a esporotricose. O
reconhecimento foi independente da forma clínica avaliada (cutânea ou
linfocutânea). Contudo, quando a molécula de 38 kDa, purificada, foi utilizada em
ensaio de ELISA versus soros de gatos positivos e negativos para esporotricose, a
sensibilidade do teste foi de apenas 26,6% (FERNANDES, 2009).

129
Almeida-Paes et al. (2012) pesquisaram a aplicabilidade de antígenos
liberados das células obtidos de sete isolados de S. brasiliensis, fase de levedura,
no sorodiagnóstico da esporotricose humana, os autores encontraram uma
variedade de bandas reconhecidas por soros de pacientes infectados, de 131 a 40
kDa. O antígeno de 85 kDa foi o único reconhecido em todas as cepas e por todas
as amostras de soros, porém apresentou reação cruzada com amostras de soros de
pacientes infectados com outros patógenos (32,5%) e foi reconhecido por 40% dos
soros de indivíduos saudáveis. Os antígenos de 120, 112 e 103 kDa reagiram com a
maioria dos soros homólogos e sua reatividade com soros heterólogos foi menor do
que a observada para o antígeno de 85 kDa. Os antígenos de 72-, 55-, 50- e 40 kDa
foram reconhecidos por uma minoria dos indivíduos infectados.
Quanto à resposta humoral dos animais infectados e sadios frente ao extrato
de micélio, observou-se um reconhecimento antigênico semelhante ao dos animais
experimentais, bandas de, aproximadamente, 115 e 180 kDa, com
imunoreconhecimento comum em animais saudáveis e infectados e a de,
aproximadamente 64 – 49 kDa, mais característica dos animais infectados. Para
exoantígenos miceliais, as bandas de 90 e 55 kDa foram as que mais reagiram com
soros humanos com esporotricose (MENDONZA et al., 2002).
A banda, unanimemente reconhecida por anticorpos de animais
experimentais e infectados deste experimento, faixa de PM de 64-49 kDa, está
dentro ou bem próxima das glicoproteínas consideradas como possíveis
biomarcadores para o diagnóstico da esporotricose, moléculas de 60 para extratos
de leveduras (FERNANDES et al., 2013) e de 70 kDa para extratos de micélio,
(RUIZ-BACA et al., 2014).
O isolamento do Sporothrix em cultivo é considerado o padrão ouro para
diagnóstico da esporotricose, seguido pela biópsia da lesão com identificação do
agente. Entretanto, nem sempre é possível reconhecer as diferentes formas do
organismo nas seções tissulares e o desenvolvimento de técnicas específicas para a
identificação do Sporothrix spp. pode ser de grande valia, principalmente para a
diferenciação de doenças com apresentações clínicas semelhantes à da
esporotricose, como é o caso da leishmaniose tegumentar em cães.
Neste experimento, foi utilizada a técnica de imunohistoquímica com
anticorpos policlonais de galinhas, IgY, e coelhos, IgG, específicos para antígenos
de S. schenckii e Sporothrix spp. Tanto os anticorpos IgY de galinhas quanto os

130
anticorpos IgGs de coelhos antiextratos de levedura e antiextratos de micélio de
Sporothrix foram eficientes no reconhecimento do fungo em fragmentos de pele de
gatos considerados positivos para esporotricose, através do isolamento do agente
por cultivo. Os anticorpos foram eficientes em identificar as leveduras dentro de
macrófagos e no espaço intersticial, sem marcações inespecíficas de tecido.
Como a leishmaniose tegumentar é considerada o principal diagnóstico
diferencial da esporotricose em cães (SANTOS et al., 2006, MIRANDA et al., 2011),
pesquisou-se a existência de reações cruzadas entre anticorpos e formas
amastigotas de Leishmania spp. em fragmentos de biópsia de cães positivos para
leishmaniose. Os resultados mostraram a especificidade dos anticorpos produzidos,
tanto para a IgY de galinhas quanto para os IgGs de coelhos, pois nenhum
reconhecimento das formas amastigotas de Leishmania spp. em seções de biópsia
de pulmão e de pele de cão positivo para leishmaniose foi observado.
Segundo Marques et al. (1992), nem sempre é possível reconhecer diferentes
formas do Sporothrix spp. em seções tissulares e o desenvolvimento de ensaios
específicos para a identificação do agente pode ser bastante útil. Para estes autores,
a imunohistoquímica em seções de pele de humanos com anticorpos policlonais de
coelhos anti S. schenckii foi o método mais sensitivo (83%) e específico do que as
colorações de Gomori (37%), ácido periódico de Schiff (23%) ou hematoxilina-eosina
(23%) para a detecção do Sporothrix em seções histológicas de biópsia.
Para Miranda et al. (2011), a técnica de imunohistoquímica com antissoros de
coelhos aumentou a sensibilidade do diagnóstico histológico da esporotricose em
cães em 80,5% quando comparada às técnicas de PAS e Grocott.
Os resultados deste experimento corroboram os de Miranda et al. (2011), ao
mostrar que nenhuma reação cruzada foi encontrada entre anticorpos IgG de
coelhos antigênicos de Sporothrix e seções tissulares de cães com Leishmania spp.
Corroboram os resultados de Marques et al.(1992) e Miranda et al. (2011) ao
mostrar que anticorpos de coelhos antiextratos de Sporothrix schenckii e cepa clínica
de Sporothrix spp. reconhecem leveduras de Sporothrix spp. em fragmentos de
biópsia de animais positivos para esporotricose.
Pela primeira vez na literatura, descreve-se a utilização de anticorpos IgY (5
µg/ml) antiextratos antigênicos de S. schenckii e Sporothrix spp. no diagnóstico da
esporotricose através da técnica de imunohistoquímica e sem nenhuma reação

131
cruzada com seções tissulares de cães infectados com Leishmania spp. As
concentrações de anticorpos IgY utilizadas, neste experimento, foram semelhantes
às utilizadas por Inoue et al. (2005), 5 e 10 µg/ml, quando testaram IgY antiproteínas
recombinantes do vírus da raiva para detecção de antígenos virais no citoplasma de
neurônios de camundongos experimentalmente infectados.
Após os testes de reconhecimento das leveduras de Sporothrix spp. em
fragmentos de biópsia, os anticorpos IgY foram testados quanto à capacidade de
inibição do crescimento de leveduras de S. schenckii e Sporothrix spp. em caldo
YPD, na concentração final de 1mg/mL. Os resultados mostraram o potencial dos
anticorpos IgY antiantígenos de S. schenckii e Sporothrix spp. como possível
adjuvante no tratamento da esporotricose através do crescimento, significativamente
menor, das leveduras de S. schenckii e Sporothrix spp. no meio de cultura contendo
anticorpos IgY de animais imunizados quando comparados aos tratamentos controle
com anticorpos pré-imunes e branco, sem adição de anticorpos. Os testes, in vitro,
deste experimento foram concordantes com os experimentos, in vitro, de inibição de
crescimento realizados com anticorpos IgY hiperimunes anti-Prototheca spp. (SILVA,
2005), C. albicans spp. (GONÇALVES, 2007, VIEIRA et al., 2010), S. aureus
(GUIMARÃES et al., 2009) e antiprodução de enterotoxinas por S. aureus (Vieira-da-
Motta et al., 2001). Entretanto, é discordante de FUJIBAYASHI et al. (2009), onde
anticorpos IgY anti-C. albicans (2mg/mL) não inibiram o crescimento da C. albicans
em caldo YPD em comparação à IgY pré-imune. Pesquisas mostraram que os testes
de inibição de crescimento por anticorpos IgY, in vitro, são dependentes das
concentrações do inóculo e dos anticorpos (SUGITA-KONISHI et al., 1996, Vieira-
da-Motta et al., 2001, GUIMARÃES et al., 2009) no meio de cultivo.
As imunoglobulinas Y das gemas dos ovos de galinhas são
reconhecidamente uma fonte de anticorpos e o seu valor terapêutico contra vários
micro-organismos já foi comprovado em vários trabalhos. OTAKE et al. (1991),
YOKOYAMA et al. (1992), SMITH et al. (2001), CARLANDER (2002), LECLAIRE et
al. (2002), HORIE et al. (2004), RIBEIRO et al. (2005), LI et al. (2006), ZHEN et al.
(2009) demonstraram a eficiência da imunidade passiva com anticorpos IgY no
controle de infecções bacterianas, em infecções virais por coronavírus, rotavírus e
parvovírus (KWEON et al., 2000, NGUYEN et al., 2006, LU et al., 2008, VEGA et al.,
2012) e como alternativa à IgG de equinos no tratamento de vítimas de mordida de

132
cobra (PAUL et al., 2007, ALMEIDA et al., 2008) devido ao seu menor custo, à alta
produtividade e ao menor risco de choque anafilático.
A imunização passiva, por via oral, de anticorpos IgY anti-C. albicans reduziu
significativamente a disseminação sistêmica e a severidade das lesões na língua de
camundongos experimentalmente infectados (Ibrahim et al. (2008). Também
aumentou o tempo de sobrevivência e diminuiu a disseminação sistêmica da C.
albicans em camundongos desafiados com doses letais do agente por via parenteral
(ABDELNOOR et al., 2006).
Atualmente, não existem vacinas contra o Sporothrix e o desenvolvimento de
medidas terapêuticas, especialmente para indivíduos imunocomprometidos e casos
de resistência, como vem ocorrendo em gatos, aparece como medidas sinérgicas ao
tratamento. O efeito protetivo de anticorpos monoclonais antiglicoproteína de 70
kDa, em camundongos experimentalmente infectados pelo S. schenckii, foi mostrado
por Nascimento et al. (2008). Mas, sabe-se que o uso de anticorpos monoclonais e
policlonais, in vivo, aumenta o número de pacientes que contêm HAMA, a vantagem
da IgY está em não interagir com o fator HAMA (CARLANDER, 2002).
O efeito inibitório dos anticorpos IgY imunes sobre o crescimento in vitro do
fungo Sporothrix fortalece a ideia da sua utilização como adjuvante à terapia da
esporotricose. Entretanto, experimentos in vivo necessitam ser conduzidos, assim
como, a padronização da dose e frequência de administração do anticorpo.
Os resultados dos ensaios de ELISA deste experimento com soros de animais
(cães e gatos) positivos e negativos para a esporotricose mostraram que, a partir da
obtenção de uma amostragem mais representativa de soros de animais positivos, os
antígenos estudados poderão ser testados a fim de se obter a padronização de um
teste de ELISA para o diagnóstico sorológico nestas espécies animais. Apesar de
que em gatos, o diagnóstico sorológico é de menor importância devido à
apresentação clínica da doença nesta espécie, normalmente com lesões cutâneas e
abundância de leveduras nas lesões (MIRANDA et al., 2013), facilitando o
diagnóstico por isolamento do fungo em cultivo.
Em cães, conforme descrito no Caso clínico 1 e em conformidade com alguns
relatos da literatura (MADRID et al., 2007, SOUZA et al., 2009, Matos et al., 2012),
as lesões externas podem estar ausentes e o diagnóstico sorológico poderá ser de
grande valia nesta espécie. As técnicas de imunodiagnóstico adotadas no Caso
Clínico 1, foram importantes para o estabelecimento do diagnóstico, tal como

133
observado por outros autores (ALMEIDA-PAES et al., 2007, 2012, FERNANDES et
al., 2011) e o acompanhamento dos títulos de anticorpos no período pós-tratamento,
uma ferramenta importante para o acompanhamento da evolução clínica e/ou
resposta terapêutica do paciente (ALMEIDA-PAES et al., 2007). A cura clínica do
animal através do tratamento com solução saturada de iodeto de potássio a 20%
corroborou com Cruz (2013) e com os métodos de diagnóstico imunológico
adotados. O presente experimento é o primeiro trabalho a relatar uma avaliação
sorológica para antígenos de Sporothrix spp. em cães.
Ensaios de ELISA foram padronizados para o diagnóstico da esporotricose
humana (ALMEIDA-PAES et al., 2007ab, BERNARDES-ENGEMANN et al., 2005) e
felina (FERNANDES et al., 2011). No ensaio de ELISA para os felinos, Fernandes et
al. (2011) trabalharam soros na diluição de 1:800 e encontraram valores de
sensibilidade de 73,3 e 83,3% quando utilizaram, como sensibilizadores,
exoantígenos brutos miceliais de S. schenckii e a fração SsCBF (S. schenckii - Con A
Binding Factor) extraída da parede celular de leveduras. A especificidade do teste de
ELISA foi igual para os dois antígenos testados, 91,67%. Apesar da amostragem de
soros de gatos positivos deste experimento ser pequena (n=7), o resultado de
sensibilidade do ELISA com o antígeno LSs (71,4%), deste experimento, ficou
próximo do resultado obtido por Fernandes et al. (2011). Quando o ELISA foi testado
frente a antígenos de LSc, 100% dos soros positivos ficaram acima da linha de corte
estabelecida. A especificidade do teste com antígenos LSs foi de 100% e com os
antígenos LSc, de 95,2%.
Os achados histopatológicos dos fragmentos de biópsia do gato e do humano
do Relato de caso 2, estão de acordo com os achados da literatura (CRUZ, 2013,
MIRANDA et al., 2013), onde observa-se nos fragmentos de biópsia de gatos,
atulhamento de células fúngicas no interior de macrófagos e a presença de
granulomas mal definidos. E em fragmentos de biópsia de humanos, a ausência de
visualização de células fúngicas. A histopatologia do tegumento cutâneo de humano
revelou escassos macrófagos e neutrófilos, área de ulceração e células fúngicas não
foram visualizadas.

134
11.0 – CONCLUSÕES
- Bons títulos de anticorpos foram adquiridos após a terceira imunização dos
animais experimentais;
- Houve reconhecimento cruzado entre anticorpos hiperimunes, IgY ou IgG, e
antígenos de S. sckenckii e Sporothrix spp;
- A banda de 64-49 kDa foi imunodominante perante anticorpos hiperimunes e
soros de animais positivos para esporotricose, tanto em membranas
sensibilizadas com extratos antigênicos de leveduras de S. schenckii ATCC
32285 e Sporothrix spp. quanto de micélio de S. sckenckii ATCC 32285;
- As reações de western blot foram mais intensas nas membranas
sensibilizadas com o antígeno de leveduras de Sporothrix spp;
- A diferenciação entre animais positivos e negativos para esporotricose pela
técnica de western blot, foi melhor perante antígenos de leveduras de S.
sckenckii ATCC 32285;

135
- Os anticorpos IgY e IgG antiantígenos de LSs, LSc ou MSs reconheceram
leveduras de Sporothrix spp., pela técnica de imunohistoquímica, em seções
de pele de gatos infectados;
- Os anticorpos IgY e IgG antiantígenos de LSs, LSc ou MSs não
apresentaram reações cruzadas com formas amastigotas de Leishmania spp.
em seções de pulmão de cão infectado, pela técnina de imunohistoquímica;
- Os anticorpos IgY antiantígenos de LSs, LSc ou MSs inibiram o crescimento,
in vitro, de leveduras de S. sckenckii ATCC 32285 e Sporothrix spp.

136
12.0 - REFERÊNCIAS BIBLIOGRÁFICAS ABDELNOOR, A. M., RAHAL, E., ZEIDAN, J. A. et al. Preparation of anti-Candida albicans antibodies in an egg-laying hen and their protective efficacy in mice. Journal of Applied Research. Relato de caso. 2006. (Disponível em: http://www.thefreelibrary.com/Preparation+of+antiCandida+albicans+antibodies+in+an+egg-laying+hen...-a0159491750. Acesso em 01/04/2013). ADAMS, D. J. Fungal cell wall chitinases and glucanases. Microbiology. v. 150, n. 7, p. 2029 – 2035, 2004. ALMEIDA, C.M.C, SILVA, C. L., COUTO, H. P. et al. Development of process to produce polyvalent IgY antibodies anti-African snake venom. Toxicon. v. 52, p. 293–301, 2008. ALMEIDA, C.M.C., QUINTANA-FLORES, V.M., MEDINA-ACOSTA, E. et al. Egg yolk anti-BfpA antibodies as a tool for recognizing and identifying Enteropathogenic Escherichia coli. Scandinavian Journal of Immunology. v. 57, n. 6, 573–582, 2003. ALMEIDA-PAES, R., BAILÃO, A. M., PIZZINI, C. V. et al. Cell-free antigens of Sporothrix brasiliensis: antigenic diversity and application in an immunoblot assay. Mycoses. v. 55, n. 6, p. 467-475, 2012.

137
ALMEIDA-PAES, R., OLIVEIRA, M. M. E., FREITAS, D. F. S. et al. Sporotrichosis in Rio de Janeiro, Brazil: Sporothrix brasiliensis is associated with atypical clinical presentations. PLos Negleted Tropical Diseases. v. 8, n. 9, p. e3094, 2014. ALMEIDA-PAES, R. PIMENTA, M. A., MONTEIRO, P. C. F. Immunoglobulins G, M, and A against Sorothrix schenckii exoantigens in patients with sporothrichosis before and during treatment with itraconazole. Clinical and Vaccine Immunology. v. 14, n. 9, p. 1149- 1157, 2007a. ALMEIDA-PAES, R. PIMENTA, M. A., PIZZINI, C. V. et al. Use of mycelial-phase Sporothrix schenckii exoantigens in an enzyme-linked immunosorbent assay for diagnosis of sporotrichosis by antibody detection. Clinical and Vaccine Immunology. v. 14, n. 3, p. 244 – 249, 2007b. ALVES, S. H., BOETTCHER, C. S., OLIVEIRA, D. C. Sporothrix schenckii associated with armadillo hunting in Southern Brazil: epidemiological and antifungal susceptibility profiles. Revista da Sociedade Brasileira de Medicina Tropical. v. 43, n. 5, p. 523-525, 2010. AMARAL, J. A., TINO DE FRANCO, M., ZAPATA-QUINTANILLA, L. et al. In vitro reactivity and growth inhibition of EPEC serotype O111 and STEC serotypes O111 and O157 by homologous and heterologous chicken egg yolk antibody. Veterinary Research Communications. v. 32, p. 281–290, 2008. ARRILLAGA-MONCRIEFF, I., CAPILLA, J., MAYAYO, E. et al. Different virulence levels of the species of Sporothrix in a murine model. Clinical Microbiology and Infection. v.15, p. 651–655, 2009. BARROS, M. B. L., ALMEIDA-PAES, R., SCHUBACH, A. O. Sporothrix schenckii and sporotrichosis. Clinical Microbiology Reviews. v. 24, n. 4, p. 633–654, 2011. BARROS, M. B. L., SCHUBACH, T. M. P., GALHARDO, M. C. G. et al. Sporotrichosis: an Emergent Zoonosis in Rio de Janeiro. Memórias do Instituto Oswaldo Cruz. v. 96, n. 6, p. 777-779, 2001.e BARROS, M. B. L., SCHUBACH, T. M. P., GALHARDO, M. C. G. et al. Sporotrichosis with widespread cutaneous lesions: report of 24 cases related to transmission by domestic cats in Rio de Janeiro, Brazil. International Journal of Dermatology. v. 42, p. 677–681, 2003.

138
BARROS, M. B. L., SCHUBACH, T. P., COLL, J. O. et al. Esporotricose: a evolução e os desafios de uma epidemia. Revista Panamericana de Salud Publica. v. 27, n. 6, p. 455–60, 2010. BERNARDES-ENGEMANN, A. R., COSTA, R. C., MIGUENS, B. R. et al. Development of an enzyme-linked immunosorbent assay for the serodiagnosis of several clinical forms of sporotrichosis. Medical Mycology. v. 43, n. 6, p. 487–493, 2005. BERNARDO, A. R. Tecnologia IgY: produção de anticorpos aviários para Leishmania (Leishmania) amazonensis com o uso ético dos animais de experimentação. Seropédica:UFRRJ, 2009. 49 p., 2009. (Dissertação mestrado).
BHANUSHALI, J. K., GILBERT, J. M., MCDOUGALD, L. R. Simple method to purify chicken immunoglobulin G. Poultry Science. v. 73, n. 7, p. 1158-1161, 1994.
BLUMER, S. O., KAUFMAN, L., KAPLAN, W. Comparative valuation of five serological methods for the diagnosis of sporotrichosis. Applied Microbiology. v. 26, n. 1, p. 4-8, 1973. BRUNDA, G., SASHIDHAR, R.B., SARIN, R.K. Use of egg yolk antibody (IgY) as an immunoanalytical tool in the detection of Indian cobra (Naja naja naja) venom in biological samples of forensic origin. Toxicon. v.48, p. 183–194, 2006. CARLANDER, D. Avian IgY antibody. In vitro and in vivo. ACTA UNIVERSITATIS UPSALIENSIS: Uppsala, Suécia: 2002. 53p., 2002. (Dissertação doutorado). CHALGHOUMI, R., THÉWIS, A., PORTETELLE, D., BECKERS, Y. Production of hen egg yolk immunoglobulins simultaneously directed against Salmonella Enteritidis and Salmonella Typhimurium in the same egg yolk. Poultry Science. v. 87, p. 32–40. 2008. CROTHERS, S. L., WHITE, S. D., IHRKE, P. J., AFFOLTER, V. K. Sporotrichosis: a retrospective evaluation of 23 cases seen in northern California (1987–2007). Veterinay Dermatology. v. 20, n. 4, p. 249-259, 2009. CRUZ, L. C. H. Complexo Sporothrix schenckii. Revisão de parte da literatura e considerações sobre o diagnóstico e a epidemiologia. Veterinária e Zootecnia. v. 20, p. 08-28, 2013.

139
DAVISON, F., KASPERS, B., SCHAT, K. A. Avian immunology. London: Elsevier Ltda. 2008. 496 p. (Disponível em:books.elsevier.com, acesso em 01/03/2013). DE BEER, Z. W., HARRINGTON, T. C. VISMER, H. F. et al. Phylogeny of the Ophiostoma stenoceras-Sporothrix schenckii complex. Mycologia. v. 95, n. 3, p. 434 – 441, 2003. De LIMA FRANCO, D., NASCIMENTO, R. C., FERREIRA, K. S., ALMEIDA, S. R. Antibodies against Sporothrix schenckii enhance TNF-α production and killing by macrophages. Scandinavian Journal of Immunology. v. 75, p. 142–146, 2011. DIAS DA SILVA, W., TAMBOURGI, D. V. IgY: A promising antibody for use in immunodiagnostic and in immunotherapy. Veterinary Immunology and Immunopathology. V. 135, p. 173 -180, 2010. DIXON, D. M., SALKIN, I. F., DUNCAN, R. A. et al. Isolation and characterization of Sporothrix schenckii from clinical and environmental sources associated with the largest U.S. epidemic of sporotrichosis. Journal of Clinical Microbiology. v. 29, n. 6, p. 1106-1113, 1991. FERNANDES, G. F., LOPES-BEZERRA, L. M., BERNARDES-ENGEMANN, A. R. et al. Serodiagnosis of sporotrichosis infection in cats by enzyme-linked immunosorbent assay using a specific antigen, SsCBF, and crude exoantigens. Veterinary Microbiology. v. 147, p. 445-449, 2011. FERNANDES, G. F., SANTOS, P. O., RODRIGUES, A. M. et al. Characterization of virulence profile, protein secretion and immunogenicity of different Sporothrix schenckii sensu stricto isolates compared with S. globosa and S. brasiliensis species. Virulence. v.4, n. 3, p. 241–249, 2013. FERNANDES, K. S. S., HELAL NETO, E., BRITO, M. M. S. et al. Detrimental role of endogenous nitric oxide in host defence against Sporothrix schenckii. Immunology. v.123, p.469–479, 2008. FREIRE, R.B., FREIRE, M. F. I., BERBARA, L. R. Elisa de captura com IgY para quantificação de acetato de lupeol em Vernonia scorpioides lam. Pers (asteraceae). Ciência Rural. v.34, n.4, p.1069-1074, 2004. FREITAS, D. F. S., VALLE, A. C. F., da SILVA, M. B.T. et al. Sporotrichosis: an emerging neglected opportunistic infection in HIV-infected patients in Rio de Janeiro, Brazil. PLos Negleted Tropical Diseases. v. 8, n.8, p. e3110, 2014.

140
FUJIBAYASHI, T., NAKAMURA, M., TOMINAGA, A. et al. Effects of IgY against Candida albicans e Candida spp. adherence and biofilm formation. Japanese Journal of Infectious Diseases. v. 62, p. 337 – 432, 2009. FUKUDA, E. K., VASCONCELOS, A. F. D., MATIAS, A. C. et al. Polissacarídeos de parede celular fúngica: purificação e caracterização. Revisão. Semina: Ciências Agrárias. v. 30, n. 1, p. 117 – 134, 2009. GARRISON, R. G., BOYD, K. S., MARIAT, F. Ultrastructural studies of the mycelium-to-yeast transformation of Sporothrix schenckii. Journal of Bacteriology. v. 124, n. 2, p. 959-968, 1975. GARRO, J., GENÉ,J., STCHIGEL, A. M. Developments in fungal taxonomy. Clinical Microbiology Reviews. v. 12, n. 3, p. 454 – 500. 1999. GASSMANN, M.; THÖMMES, P.; WEISER, T.; HÜBSCHER, U. Efficient production of chicken egg yolk antibodies against a conserved mammalian protein. FASEB Journal. v.4, p. 2528-2532, 1990. GONÇALVES, D. S. Desenvolvimento de anticorpos contra o fungo Candida albicans e sua susceptibilidade a metabólitos secundários de Burkholderia sp. Campos dos Goytacazes: UENF/CCTA. 2007. 88 p., 2007. (Dissertação Mestrado). GONZALES-OCHOA, A., FIGUEROA, E. S. Polissacaridos del Sporothrix schenckii. Intradermoreacción en el diagnostico de la esporotricosis. Revista del Instituto de Salubridad y Enfermedades Tropicales. v. 8, p. 143-153, 1947. GUIMARÃES, M. C. C., AMARAL, L. G., RANGEL, L. B. A. et al. Growth inhibition of Staphylococcus aureus by chicken egg yolk antibodies. Archivum Immunologiae Therapiae Experimentalis. v. 57, p. 377–382, 2009. GUZMAN-BELTRAN, S., PEREZ-TORRES, A., CORONEL-CRUZ, C., TORRES-GUERRERO, H. Phagocytic receptors on macrophages distinguish between different Sporothrix schenckii morphotypes. Microbes and Infection. v. 14, p. 1093 - 1101, 2012. HAMAL, K. R., BURGESS, S. C., PEVZNER, I. Y., ERF, G. F. Maternal antibody transfer from dams to their egg yolks, egg whites, and chicks in meat lines of chickens. Poultry Science. v. 85, p.1364–1372, 2006.

141
HEIDRICH, D., STOPIGLIA, C. D. O., SENTER, L. et al. Sucesso terapêutico da terbinafina em um caso de esporotricose. Anais Brasileiros de Dermatologia. v.86, n. 4 (Supl.1), p. 182- 185, 2011. HEKTOEN, L. e PERKINS, C. F. Refractory subcutaneous caused by Sporothrix schenckii. A new pathogenic fungus. Journal of Experimental Medicine. v. 5, p. 77– 89, 1900. HORIE, K., HORIE, N., ABDOU, A. M. et al. Suppressive effect of functional drinking yogurt containing specific egg yolk immunoglobulin on Helicobacter pylori in humans. Journal of Dairy Science. v. 87, p. 4073–4079, 2004. IBRAHIM, E-S M, RAHMAN, A.K.M. S., ISODA, R. et al. In vitro and in vivo effectiveness of egg yolk antibody against Candida albicans (anti-CA IgY). Vaccine. v. 26, 2073—2080, 2008. JIN, W., KEIKO YAMADA, K., IKAMI, M. Application of IgY to sandwich enzyme-linked immunosorbent assays, lateral flow devices, and immunopillar chips for detecting staphylococcal enterotoxins in milk and dairy products. Journal of Microbiological Methods. v. 92, p. 323–331, 2013. JINTARIDTH, P., SRISOMSAP, C. VICHITTUMAROS, K. et al. Chicken egg yolk antibodies specific for the γ chain of human hemoglobin for diagnosis of thalassemia. International Journal of Hematology. v. 83, p. 408 – 414, 2006. KAUFFMAN, C. A., HAJJEH, R., CHAPMAN, S. W. Guidelines for the Management of Patients with Sporotrichosis. Clinical Infectious Diseases. v. 30, p. 684–687, 2000. KOÇ, A. N., ÜKSAL, OYMAK, O. Case Report. Successfully treated subcutaneous infection with Sporothrix schenckii in Turkey. Mycoses. v. 44,p. 330 – 333, 2001. KOWALCZYK, K., DAISS, J., HALPERN, J., ROTH, T. F. Quantitation of maternal-fetal IgG transport in the chicken. Immunology. v. 54, p. 755 – 762, 1985. KOZEL, T. R. Activation of the complement system by pathogenic fungi. Clinical Microbiology Reviews. v. 9, n. 1, p. 34-46, 1996.

142
KWEON, C., KWON, B., WOO, S. et al. Immunoprophylactic effect of chicken egg yolk immunoglobulin (IgY) agaist porcine epidemic diarrhea vírus (PEDV) in piglets. Journal of Veterinary Medical Science. v. 62, n. 9, p. 961-964,2000. LACAZ, C. S., PORTO, E., HEINS-VACCARI, E. M., MELO, N. T. Guia para identificação: fungos, actinomicetos, algas de interesse médico. São Paulo: SARVIER, 1988. 445 p. LARSSON, A., KARLSSON-PARRA, A., SJOQUIST, J. Use of chicken antibodies in enzyme immunoassays to avoid interference by rheumatoid factors. Clinical Chemistry. n. 37, v. 3, p. 411-414, 1991. LEBACQ-VERHEYDEN, A-M, VAERMAN, J. P., HEREMANS, J. F. A possible homologue of mammalian IgA in chicken serum and secretions. Immunology. v. 22, p. 165 – 175, 1972. LEBACQ-VERHEYDEN, A-M, VAERMAN, J. P., HEREMANS, J. F. Quantification and Distribution of Chicken Immunoglobulins IgA, IgM and IgG in Serum and Secretions. Immunology. v. 27, p. 683 – 692, 1974. LECLAIRE, R. D., HUNT, R. E., BAVARI, S. Protection against bacterial superantigen staphylococcal enterotoxin B by passive vaccination. Infection and Immunity. v. 70, n. 5, p. 2278–2281, 2002. LEE, D. Y, CHO, Y. W., KANG, S. G. et al. Development of a novel antigen capture-ELISA using IgY against porcine interleukin-6 and its application. Journal of Veterinary Science. v. 5, n. 4, p. 337–343, 2004. LEE, E. N., SUNWOO, H. H., MENNINEN, K., SIM, J. S. In vitro studies of chicken egg yolk antibody (IgY) against Salmonella enteritidis and Salmonella typhimurium. Poultry Science. v. 81, p. 632–641, 2002. LEI, J.-H., GUAN, F., XU, H. Application of an immunomagnetic bead ELISA based on IgY for detection of circulating antigen in urine of mice infected with Schistosoma japonicum. Veterinary Parasitology. v. 187, p. 196– 202, 2012.
LESLIE, G. A., CLEM, L. W. Phylogeny of immunoglobulin structure and function. III. Immunoglobulins of the chicken. Experimental Medicine. v.130, n. 6, p.1337–1352, 1969.

143
LI, X.–L., SHUAI, J.-B., FANG, W.-H. Protection of Carassius auratus Gibelio against infection by Aeromonas hydrophila using specific immunoglobulins from hen egg yolk. Journal of Zhejiang University Science B. v. 7, n. 11, p. 922-928, 2006. LIMA, O. C. & LOPES-BEZERRA, L. M. Identification of a concanavalin A-binding antigen of the cell surface of Sporothrix schenckii. Journal of Medical and Veterinary Mycology. v. 35, p. 167 – 172, 1997. LIMA, O. C., FIGUEIREDO, C. C., PEREIRA, B. A. S. Adhesion of the human pathogen Sporothrix schenckii to several extracellular matrix proteins. Brazilian Journal of Medical and Biological Research. v. 32, n. 5, p. 651-657, 1999. LITMAN, G. W., RAST, J. P., SHAMBLOTT, M. J., et al. Phylogenetic diversification of immunoglobulin genes and the antibody repertoire. Molecular Biology and Evolution. v.10, n. 1, p. 60-72, 1993. LLOYD, K. O., BITOON, M. A. Isolation and purification of a peptido-rhamnomannan from the yest form Sporothrix schenckii. Structural and Immunochemical studies. Journal of Immunology. v. 107, p. 160-171, 1971. LOEKEN, M. R., ROTH, T. F. Analysis of maternal IgY subpopulations which are transported into the chicken oocyte. Immunology. v. 49, p. 21 – 28, 1983. LOPES, J. O. , ALVES, S. H., MARI, C. R. et al. Epidemiologia da esporotricose na região central do Rio Grande do Sul. Revista da Sociedade Brasileira de Medicina Tropical. v. 32, n. 5, p. 541-545, 1999. LOPES-BEZERRA, L. M. Sporothrix schenckii cell wall peptidorhamnomannans. Mini review article. Frontiers in Microbiology. v. 2. 4 p. 2011. LOPES-BEZERRA, L. M., SCHUBACH, A., COSTA, R. O. Sporothrix schenckii and sporotrichosis. Anais da Academia Brasileira de Ciências. v.78, n. 2, p. 293-308, 2006. LU, Y., LIU, J., JIN, L. et al. Passive protection of shrimp against white spot syndrome virus (WSSV) using specific antibody from egg yolk of chickens immunized with inactivated virus or a WSSV-DNA vaccine. Fish & Shellfish Immunology. v. 25, p. 604–610, 2008.

144
LU, Y., XU, B., JU, C. et al. Identification and profiling of circulating antigens by screening with the sera from schistosomiasis japonica patients. Parasites & Vectors. v. 5, n. 115, 9 p., 2012. LUTZ, A. e SPLENDORE, A. Sobre uma micose observada em homens e ratos. Revista de Medicina de São Paulo. v. 21, p. 433- 450, 1907. MADRID, I. M., MATTEI, A. S., XAVIER, M. O. Esporotricose óssea e cutânea em canino. Brazilian Journal of Veterinary Research and Animal Science. v. 44, n. 6, p. 441-443, 2007. MADRID, I. M., XAVIER, M. O., MATTEI, A. S. Role of melanin in the pathogenesis of cutaneous sporotrichosis. Microbes and Infection. v. 12, p.162-165. 2010. MANHANI, M. N. Seleção de peptídeos reconhecidos por imunoglobulinas Y anti-metacestódeo de Taenia solium por phage display. Uberlândia: UFU/ICB, 2009. 98p. 2009. (Dissertação Mestrado). MARIMON R, GENÉ J, CANO J, GUARRO J. Sporothrix luriei: a rare fungus from clinical origin. Medical Mycology. v. 46, p. 621-625, 2008. MARIMON, R., GENÉ, J., CANO, J. et al. Molecular phylogeny of Sporothrix schenckii. Journal of Clinical Microbiology. v. 44, n. 9, p. 3251 – 3256, 2006. MARIMON, R., GENÉ, J., CANO, J. et al. Sporothrix brasiliensis, S. globosa, and S. mexicana, three new Sporothrix species of clinical interest. Journal of Clinical Microbiology. v. 45, n. 10, p. 3198 – 3206, 2007. MARQUES, M. E. A., COELHO, K. I. R., SOTTO, M. N., BACCKI, C. E. Comparison between histochemical and immunohistochemical methods for diagnosis of sporotrichosis. Journal of Clinical Pathology. v. 45, p. 1089-1093, 1992. MARQUES, S. A., FRANCO, S. R. V. S., CAMARGO, R. M. P. et al. Esporotricose do gato doméstico (Felis catus): transmissão humana. Revista do Instituto de Medicina Tropical de São Paulo. v. 35, n. 4, p. 327 – 330, 1993. MARTÍNEZ-ÁLVAREZ, J. A., PÉREZ-GARCÍA, L. A., FLORES-CARREÓN, MORA-MONTES, H. M. Mycologic Fórum: the immune response against Candida spp. and Sporothrix schenckii. Revista Iberoamericana de Micología. v. 31, n.1, p. 62 – 66, 2014.

145
MATOS, C. B., MADRID, I.M., SANTIN, R. Dermatite multifatorial em um canino. Arquivo Brasileiro de Medicina Veterinária e Zootecnia. v.64, n.6, p.1478-1482, 2012. MATTEI, A. S., MADRID, I. M., SANTIN, R. Sporothrix schenckii in a hospital and home environment in the city of PelotasRS – Brazil. Anais da Academia Brasileira de Ciências. v.83, n. 4, p. 1359-1362, 2011. MENDOZA, M., DIAZ, A. M., HUNG, M. B. et al. Production of culture filtrate of Sporothrix shcenckii in diverse culture media. Medical Mycology. v.40, p. 447-454,2002. MIRANDA, L. H. M., CONCEIÇÃO-SILVA, F., QUINTELLA, L. et al. Feline sporotrichosis: histopathological profile of cutaneous lesions and their correlation with clinical presentation. Comparative Immunology, Microbiology and Infectious Diseases. v.36, p. 425– 432, 2013. MIRANDA, L. H. M., QUINTELLA, L. P., SANTOS, I. B. et al. Comparative histopathological study of sporotrichosis and american tegumentar leishmaniasis in dogs from Rio de Janeiro. Journal of Comparative Pathology. v. 143, p. 1-7, 2010. MIRANDA, L.H.M., QUINTELLA, L. P., MENEZES, R. C. et al. Evaluation of immunohistochemistry for the diagnosis of sporotrichosis in dogs. The Veterinary Journal. v. 190, p. 408 - 411, 2011. MOTOI, Y., INOUE, S., HATTA, H. et al. Detection of rabies-specific antigens by egg yolk antibody (IgY) to the recombinant rabies virus proteins produced in Escherichia coli. Japanese Journal of Infectious Diseases. v. 58, p. 115 – 118, 2005. NAKAMURA, N., SHUYAMA, A., HOJYO, S. et al. Establishment of a chicken monoclonal antibody panel against mammalian prion protein. Journal of Veterinary Medical Science. v. 66, p. 807-814, 2004. NASCIMENTO, R. C., ALMEIDA, S. R. Humoral immune response against soluble and fractionate antigens in experimental sporotrichosis. FEMS Immunology and Medical Microbiology. v. 43, p.241–247, 2005.

146
NASCIMENTO, R. C., ESPÍNDOLA, N. M., CASTRO, R. A. et al. Passive immunization with monoclonal antibody against a 70-kDa putative adhesin of Sporothrix schenckii induces protection in murine sporotrichosis. European Journal of Immunology. v.38, p. 3080–3089, 2008. NASSIF, P. W., GRANADO, I. R. M. O., FERRAZ, J. S. et al. Esporotricose cutânea de apresentação atípica em paciente etilista. Dermatology Online Journal. v. 18, n. 6, 2012. NEGRINI, T. DE C. FERREIRA, L. S. , ALEGRANCI, P. et al. Role of TLR-2 and fungal surface antigens on innate immune response against Sporothrix schenckii. Immunological Investigations. v. 42 p. 36-48, 2013. NGUYEN, S. V., UMEDA, K., YOKOYAMA, H. et al. Passive protection of dogs against clinical disease due to Canine parvovirus-2 by specific antibody from chicken egg yolk. The Canadian Journal of Veterinary Research. v. 70, p. 62–64, 2006. O’DONNELL, K., NIRENBERG, H. I., AOKI, T., CIGELINK, E. A multigene phylogeny of the Gibberella fujikuroi species complex: detection of additional phylogenetically distinct species. Mycoscience. v. 41, n. 61-78, 2000. OLIVEIRA, M. M. E. SAMPAIO, P., ALMEIDA-PAES, R. et al. Rapid identification of Sporothrix species by T3B fingerprinting. Journal of Clinical Microbiology. v. 50, n. 6, p. 2159 – 2162, 2012. OLIVEIRA, M. M. E., VERÍSSIMO, C., SABINO, R. et al. First autochthone case of sporotrichosis by Sporothrix globosa in Portugal. Diagnostic Microbiology and Infectious Disease. v. 78, p. 388–390, 2014. OLIVEIRA, M.M.E., ALMEIDA-PAES, R., MUNIZ, M.M., GUTIERREZ-GALHARDO, M.C., ZANCOPÉ-OLIVEIRA, R. M. Phenotypic and molecular identification of Sporothrix isolates from a epidemic area of sporotrichosis of Brazil. Mycopathologia. v. 172, n. 4, p. 257- 267, 2011.
OROFINO-COSTA, R., BERNARDES-ENGEMANN, A. R., AZULAY-ABULAFIA, L. Esporotricose na gestação: relato de cinco casos numa epidemia zoonótica no Rio de Janeiro, Brasil. Anais Brasileiros de Dermatologia. v.86, n. 5, 2011. OTAKE, S., NISHIHARA, Y., MAKIMURA, M. et al. Protection of rats against dental caries by passive immunization with hen-egg-yolk antibody (IgY). Journal of Dental Research. v. 70, n.3, p. 162-166, 1991.

147
PALANIYAPPAN, A., DAS, D., KAMMILA, S. et al. Diagnostics of severe acute respiratory syndrome-associated coronavirus (SARS-CoV) nucleocapsid antigen using chicken immunoglobulin Y. Poultry Science. v.91, p. 636–642, 2012. PARMA, Y.R., CHACANA, P.A., LUCCHESI, P.M.A. Detection of Shigatoxin-producing Escherichia coli by sandwich enzyme-linked immunosorbent assay using chicken egg yolk IgY antibodies. Frontiers in Cellular and Infection Microbiology. v. 2, p. 1 – 8, 2012. PAUL, K., MANJULA, J., DEEPA, E.P. et al. Anti-Echis carinatus venom antibodies from chicken egg yolk: Isolation, purification and neutralization efficacy. Toxicon. v. 50, p. 893–900, 2007. PAVÓN, N., BONIFAZ, A., PONCE, R. M. Termoterapia en esporotricosis cutánea infantil. Comunicación de dos casos. Dermatología Rev. Mex. v. 51, n. 2, p. 68-72, 2007. PÉREZ, R. M., LOZANO, Y. R., ROLDÁN, E. C. et al. Epidemiología de la esporotricosis en el municipio de Huauchinango, Puebla. Enfermedades Infecciosas y Microbiología. v. 27, n. 2, p. 38 – 43, 2007. PREVIATO, J. O., GORIN, P. A. J., HASKINS, R. H., TRAVASSOS, L. R. Soluble and insoluble glucans from different cell types of the human pathogen Sporothrix schenckii. Experimental Mycology. v. 3, p. 92 – 105, 1979. QUINN, P. J., CARTER, M. E., MARKEY, B. K., CARTER, G. R. Clinical Veterinary Microbiology. London: WOLFE, 1994. 648 p. RAMÍREZ, F. A., HERRERA, A. V., BAHENA, M. T. Esporotricosis cutánea fija en un adolescente causada por Sporothrix schenckii (sensu stricto) y revisión comparativa de la bibliografía. Dermatología Revista Mexicana. v. 54, n.5, p.295-299, 2010. REIS, R. S., ALMEIDA-PAES, R., MUNIZ, M. M. et al. Molecular characterisation of Sporothrix schenckii isolates from humans and cats involved in the sporotrichosis epidemic in Rio de Janeiro, Brazil. Memórias do Instituto Oswaldo Cruz. v. 104, n. 5, p. 769-774, 2009. RIBEIRO, A. M., PINHEIRO, C. C., SILVA, I. M. Uso de gema de ovo de galinhas hiperimunizadas para vacinação de suínos. Acta Scientiae Veterinariae. v.35, p. 131 -137, 2007.

148
RIBEIRO, A. S. A., BISOL, T., MENEZES, M. S. Síndrome oculoglandular de Parinaud causada por esporotricose. Revista Brasileira de Oftalmologia. v.69, n. 5, p. 317-322, 2010. ROCHA, D. G. Isolamento transcriptômico das sequências variáveis de igy de galinhas poedeiras (Gallus gallus) antivenenos de Bitis arietans e Crotalus durissus terrificus. Campos dos Goytacazes: UENF/CBB, 2010. 90 p., 2010. (Dissertação Mestrado). RODRIGUES, A. M. Taxonomia polifásica e características proteômicas do complexo Sporothrix schenckii. São Paulo: UNIFESP. 265 p. 2010. (Dissertação Mestrado). RODRIGUES, A. M., HOOG, G. S., CAMARGO, Z. P. Genotyping species of the Sporothrix schenckii complex by PCR-RFLP of calmodulin. Diagnostic Microbiology and Infectious Disease. v.78, 383–387, 2014. ROMEO, O., SCORDINO, F., CRISEO, C. New Insight into Molecular Phylogeny and Epidemiology of Sporothrix schenckii Species Complex Based on Calmodulin-Encoding Gene Analysis of Italian Isolates. Mycopathologia. 2011. ROMERO-MARTINEZ, R., WHEELER, M., GUERRERO-PLATA, A. et al. Biosynthesis and functions of melanin in Sporothrix schenckii. Infection and Immunity. v. 68, n. 6, p. 3696–3703, 2000. RUAN, G-P., MA, L., HE, X-W. et al. Efficient production, purification, and application of egg yolk antibodies against human HLA-A*0201 heavy chain and light chain (β2m). Protein Expression and Purification. v. 44, p. 45–51, 2005. RUIZ-BACA, E., HERNÁNDEZ-MENDOZA, G., CUÉLLAR-CRUZ, M. et al. Detection of 2 immunoreactive antigens in the cell wall of Sporothrix brasiliensis and Sporothrix globosa. Diagnostic Microbiology and Infectious Disease. v. 79, p. 328 – 330, 2014. RUIZ-BACA, E., MORA-MONTES, H. M., LÓPES-HOMERO, E. et al. 2D-immunoblotting analysis of Sporothrix schenckii cell wall. Memórias do Instituto Oswaldo Cruz. v. 106, n. 2, p. 248 – 250, 2011.

149
RUIZ-BACA, E., TORIELLO, C. PEREZ-TORRES, A. et al. Isolation and some properties of a glycoprotein of 70 kDa (Gp70) from the cell wall of Sporothrix schenckii involved in fungal adherence to dermal extracellular matrix. Medical Mycology. v. 47, n. 2, p. 185 – 196, 2009.
SANDOVAL-BERNAL, G., SHIBAYAMA, M., TSUTSUMI, V. et al. Ultrastructural analysis of Sporothrix schenckii interaction with epithelium. Acta Microscópica. v. 18, n. 2, p. 89 – 97, 2009. SANTOS, I. B., SCHUBACH, T. M. P., LEME, L. R. P. et al. Sporotrichosis – The main differential diagnosis with tegumentary leishmaniosis in dogs from Rio de Janeiro, Brazil. Veterinary Parasitology. v.143, p. 1- 6, 2007. SCHADE, R., CALZADO, E. G., SARMIENTO, R. et al. Chicken egg yolk antibodies (IgY-technology): a review of progress in production and use in research and human and veterinary medicine. ATLA. v. 33, p. 1–26, 2005. SCHECHTMAN, R. C. Review – Sporotrichosis: Part I. Skinmed. v. 8, n. 4, p. 216 – 220, 2010. SCHECHTMAN, R. C., CRIGNIS, G. S. N., POCKSTALLER, M. P. Lesões molusco-símiles em paciente com esporotricose. Anais Brasileiros de Dermatologia. v.86, n. 6, p. 1217-1219, 2011.
SCHUBACH, T. M., de OLIVEIRA, S. A., dos REIS, R. S. et al. Sporothrix schenckii isolated from domestic cats with and without sporotrichosis in Rio de Janeiro, Brazil. Mycopathologia. v. 153, n. 2, p. 83-86, 2002.
SCHUBACH, T. M., VALLE, A.C., GUTIERREZ-GALHARDO, M. C. et al. Isolation of Sporothrix schenckii from the nails of domestic cats (Felis catus). Medical Mycology. v.39, n. 1, p.147- 149, 2001. SCHUBACH, T.M.P.; SCHUBACH, A.; OKAMOTO, T. et al. Canine sporotrichosis in Rio de Janeiro, Brazil: clinical presentation, laboratory diagnosis and therapeutic response in 44 cases (1998-2003). Medical Mycology. v.44, n. 1, p.87-92, 2006. SCOTT, E. N. e MUCHMORE, H. G. Immunoblot analysis of antibody responses to Sporothrix schenckii. Journal of Clinical Microbiology. v. 27, n. 2, p. 300 – 304, 1989.

150
SCOTT, E. N., MUCHMORE, H. G., FINE, D. P. Activation of the alternative complement pathway by Sporothrix schenckii. Infection and Immunity. v. 51, n. 1, p. 6-9, 1986.
SHANY, M. A mixed fungal infection in a dog: sporotrichosis and cryptococcosis. Canadian Veterinary Journal. v. 41, p. 799 – 800, 2000.
SHIMIZU, M. NAGASHIMA, H., SANO, K. et al. Molecular stability of chicken and rabbit immunoglobulin G. Bioscience Biotechnology, and Biochemistry. 56(2), p. 270-274, 1992. SHIN, JI-HYUN, YANG, M., NAM, S.W. et al. Use of egg yolk-derived immunoglobulin as an alternative to antibiotic treatment for control of Helicobacter pylori infection. Clinical and Diagnostic Laboratory Immunology. v. 9, n. 5, p. 1061–1066, 2002. SILVA, A. S., VASCONCELOS, G. A. L. B. M., KAPPEL, L. A. et al. An immunoenzymatic assay for the diagnosis of hepatitis A utilising immunoglobulin Y. Memórias do Instituto Oswaldo Cruz. v. 107, n. 7, p. 960-963, 2012. SILVA, L. G. Produção de anticorpos IgY contra o patógeno Prototheca sp. em aves hiperimunizadas. Campos dos Goytacazes: UENF/CCTA, 2005. 36 p. 2005. (Dissertação mestrado). SILVA, M. B. T., COSTA, M. M. M., TORRES, C. C. S. Esporotricose urbana: epidemia negligenciada no Rio de Janeiro, Brasil. Cadernos de Saúde Pública. v. 28, n. 10, p. 1867-1880, 2012. SILVA, V. L. Imunobiologia aviária e aplicação da imunoglobulina Y (IgY) na soroepidemiologia das leishmanioses caninas. Seropédica: UFRRJ, 1999. 167 p., 1999. (Tese Doutorado). SILVA-VERGARA, M. L., CAMARGO, Z. P., SILVA, P. F. et al. Case report: disseminated Sporothrix brasiliensis Infection with endocardial and ocular involvement in an HIV-infected patient. American Journal of Tropical Medicine and Hygiene. v.86, n. 3, p. 477–480, 2012.

151
SMITH, D. J., KING, W. F., GODISKA, R. Passive transfer of immunoglobulin y antibody to Streptococcus mutans glucan binding protein B can confer protection against experimental dental caries. Infection and Immunity. v. 69, n. 5, p. 3135–3142, 2001. SOTIROPOULOU, G., PAMPALAKIS, G., PROSNIKLI, E. et al. Development and immunochemical evaluation of a novel chicken IgY antibody specific for KLK6. Chemistry Central Journal. v. 6, n. 148, p. 1 – 8, 2012. SOUZA, L. L., NASCENTE, P. S., NOBRE, M. O. Isolation of Sporothrix schenkii from the nails of healthy cats. Brazilian Journal of Microbiology. v.37, p. 372-374, 2006. SOUZA, N. T., NASCIMENTO, A.C.B.M., SOUZA, J.O.T. Esporotricose canina: relato de caso. Arquivo Brasileiro de Medicina Veterinária e Zootecnia. v.61, n.3, p.572-576, 2009. SPILLNER, E., BRAREN, I., GREUNKE, K. et al. Review: avian IgY antibodies and their recombinant equivalents in research, diagnostics and therapy. Biologicals. v. 40, p. 313 – 322, 2012. SUGITA-KONISHI, Y., SHIBATA, K., YUN, S. S. et al. Immune functions of immunoglobulin Y isolated from egg yolk of hens immunized with various infectious bacteria. Bioscience Biotechnology, and Biochemistry. v. 60, n. 5, p. 886 – 888, 1996. TAYLOR, A. I., FABIANE, S. M., SUTTON, B. J., CALVERT, R. A. The crystal structure of an Avian IgY-Fc fragment reveals conservation with both mammalian IgG and IgE. Biochemistry. 2009.
TESAR, D. B., CHEUNG, E. J., BJORKMAN, P. J. The chicken yolk sac IgY receptor, a mammalian mannose receptor family member, transcytoses IgY across polarized epithelial cells. Molecular Biology of the Cell. v. 19, p. 1587–1593, 2008. TRAVASSOS, L. R. e LLOYD, K. O. Sporothrix schenckii and related species of Ceratocystis. Microbiological Reviews. v. 44, n. 4, p. 683-721, 1980. TRESSLER, R. L., ROTH, T. F. IgG receptors on the embryonic chick yolk sac. The Journal of the Biological Chemistry. V. 262, n. 32, p. 15406 – 15412, 1987.

152
VEGA, C.G, BOK, M., VLASOVA, A. N, CHATTHA, K. S. et al. IgY antibodies protect against human rotavirus induced diarrhea in the neonatal. Gnotobiotic Piglet Disease Model. Plos One. v. 7, n. 8, 16 p., 2012. VERMA, S., VERMA, G. K., SINGH, G. et al. Sporotrichosis in Sub-Himalayan India. PLos Negleted Tropical Diseases. v. 6, n. 6, 2012. VIEIRA-DA-MOTTA, O., RIBEIRO, P. D., SILVA, W. D., MEDINA-ACOSTA, E. RNAIII Inhibiting Peptide (RIP) inhibits agr-regulated toxin production. Peptides. v. 22, p. 1621–1627, 2001. WANG, L. -H, LI, X. -Y., JIN, L. -J. et al. Characterization of chicken egg yolk immunoglobulins (IgYs) specific for the most prevalent capsular serotypes of mastitis-causing Staphylococcus aureus. Veterinary Microbiology. 7 p., 2010. WARR, G. W., MAGOR, K. E., HIGGINS, D. A. IgY: clues to the origins of modern antibodies. Immunology Today. v. 16, n. 8, p. 392 -398, 1995. YAMADA, K., WANCHUN, J., OHKURA, T. Detection of methicillin-resistant Staphylococcus aureus using a specific anti-PBP2a chicken IgY antibody. Japanese Journal of Infectious Diseases. v. 66, p. 103-108, 2013. YOKOYAMA, H., PERALTA, R. C., DIAZ, R. et al. Passive protective effect of chicken egg yolk immunoglobulins against experimental enterotoxigenic Escherichia coli infection in neonatal piglets. Infection and Immunity. v. 60, n. 3, p. 998-1007, 1992. ZHEN, Y. –H., JIN, L. -J, LI, X. -Y. et al. Efficacy of specific egg yolk immunoglobulin (IgY) to bovine mastitis caused by Staphylococcus aureus. Veterinary Microbiology. v.133, p. 317–322, 2009.

153
APÊNDICE 1
Figura 32: Reconhecimento das moléculas antigênicas por soros de gatos (n=21)
por meio da técnica de western blot em tiras sensibilizadas com antígenos de leveduras de S. schenckii ATCC 32285. Diluição do soro: 1:300. PM: peso molecular (kDa).
Figura 33: Reconhecimento das moléculas antigênicas por 18 soros de cães
negativos (-) e um soro de cão positivo para esporotricose (+) por meio da técnica de western blot em tiras sensibilizadas com antígenos de leveduras de S. schenckii ATCC 32285. Diluição do soro: 1:300. PM: peso molecular (kDa).

154
Figura 34: Reconhecimento das moléculas antigênicas por 18 soros de cães
negativos (linhas 1- 8; 11- 20) e um soro de cão positivo para esporotricose, em duplicata (linhas 9 e 10), versus antígeno de leveduras de Sporothrix spp. Diluição do soro: 1:300. PM: Padrão de peso molecular (kDa).

155
Tabela 7- Resultados de absorvância (492nm) do ensaio de ELISA com soros de gatos saudáveis (animais controle) e infectados pelo Sporothrix spp. versus antígenos de extratos de levedura de S. schenckii. Diluições seriadas de 1:100 a 1:8100, médias, desvios padrão e cutoffs.
Amostras DILUIÇÕES DOS SOROS
Animais
Controle 1:100 1:300 1:900 1:2700 1:8100
1 1,438 0,893 0,492 0,253 0,111
2 1,133 0,873 0,483 0,252 0,116
3 1,306 0,936 0,482 0,408 0,189
4 1,354 0,868 0,469 0,241 0,115
5 1,261 0,923 0,436 0,197 0,105
6 1,193 0,755 0,384 0,181 0,098
7 1,079 0,812 0,373 0,168 0,090
8 1,016 0,647 0,326 0,172 0,098
9 1,055 0,551 0,258 0,120 0,071
10 0,850 0,376 0,184 0,090 0,063
11 0,717 0,369 0,158 0,077 0,055
12 0,517 0,230 0,118 0,072 0,056
13 0,609 0,247 0,118 0,064 0,052
14 0,514 0,219 0,103 0,065 0,054
15 0,328 0,147 0,081 0,061 0,052
16 0,140 0,101 0,074 0,061 0,053
17 0,261 0,128 0,071 0,054 0,049
18 0,158 0,081 0,061 0,054 0,048
19 0,080 0,064 0,056 0,049 0,046
20 0,057 0,049 0,050 0,050 0,048
21 0,056 0,050 0,046 0,055 0,052 Média 0,720 0,444 0,230 0,131 0,077 Desvio Padrão 0,717 0,369 0,158 0,077 0,056 Cutoff 1,689 1,131 0,575 0,324 0,149
DILUIÇÕES DOS SOROS
Animais
infectados 1:100 1:300 1:900 1:2700 1:8100
22 1,842 1,715 1,302 0,713 0,334
23 1,796 1,466 1,028 0,554 0,281
24 1,660 1,229 0,890 0,671 0,647
25 1,609 1,131 0,763 0,393 0,245
26 1,688 1,303 0,716 0,369 0,192
27 1,259 0,835 0,526 0,329 0,201
28 1,031 0,483 0,284 0,175 0,110

156
Tabela 8. Resultados de absorvância (492nm) do ensaio de ELISA com soros de gatos saudáveis (animais controle) e infectados pelo Sporothrix spp. versus antígenos de extratos de levedura cepa clínica de Sporothrix spp.. Diluições seriadas de 1:100 a 1:8100, médias, desvios padrão e cutoffs.
Amostras DILUIÇÕES DOS SOROS
Animais
Controle 1:100 1:300 1:900 1:2700 1:8100
1 0,855 0,396 0,245 0,123 0,080
2 1,473 1,066 0,594 0,290 0,135
3 1,437 0,944 0,503 0,233 0,119
4 1,593 1,315 0,789 0,353 0,155
5 0,703 0,371 0,175 0,102 0,066
6 1,202 0,588 0,320 0,146 0,094
7 1,114 0,706 0,413 0,226 0,114
8 1,090 0,540 0,246 0,130 0,090
9 0,993 0,357 0,184 0,099 0,067
10 0,411 0,173 0,114 0,075 0,062
11 0,606 0,345 0,219 0,114 0,077
12 0,481 0,232 0,123 0,078 0,059
13 0,753 0,355 0,159 0,090 0,060
14 0,142 0,079 0,064 0,057 0,055
15 0,635 0,376 0,203 0,115 0,087
16 0,106 0,069 0,052 0,051 0,052
17 0,120 0,090 0,063 0,057 0,053
18 0,097 0,067 0,054 0,054 0,054
19 0,069 0,057 0,054 0,054 0,054
20 0,056 0,056 0,054 0,054 0,052
21 0,067 0,060 0,054 0,054 0,054 Média 0,667 0,393 0,223 0,122 0,078 Desvio Padrão 0,516 0,359 0,201 0,085 0,030 Cutoff 1,699 1,111 0,625 0,291 0,138
DILUIÇÕES DOS SOROS
Animais
infectados 1:100 1:300 1:900 1:2700 1:8100
22 1,594 1,568 1,377 0,971 0,564
23 1,710 1,408 1,217 0,924 0,649
24 1,263 0,956 0,780 0,504 0,432
25 1,152 0,718 0,502 0,351 0,243
26 0,792 0,636 0,486 0,352 0,246
27 1,264 1,011 0,736 0,533 0,376
28 0,935 0,753 0,591 0,447 0,276

157


















