
UNIVERSIDADE FEDERAL DA PARAÍBA CENTRO …€¦ · pela paciência recíproca que tivemos. ......
53
UNIVERSIDADE FEDERAL DA PARAÍBA CENTRO DE CIÊNCIAS EXATAS E DA NATUREZA CURSO DE BACHARELADO EM CIÊNCIAS BIOLÓGICAS Caracterização morfológica e da capacidade fagocítica de hemócitos da hemolinfa de ostras de mangue Crassostrea gasar Natanael Dantas Farias Orientadora: Profª Dra. Patricia Mirella da Silva Scardua João Pessoa – 2014
-
Upload
dinhnguyet -
Category
Documents
-
view
213 -
download
0
Transcript of UNIVERSIDADE FEDERAL DA PARAÍBA CENTRO …€¦ · pela paciência recíproca que tivemos. ......

UNIVERSIDADE FEDERAL DA PARAÍBA
CENTRO DE CIÊNCIAS EXATAS E DA NATUREZA
CURSO DE BACHARELADO EM CIÊNCIAS BIOLÓGICAS
Caracterização morfológica e da capacidade fagocítica de hemócitos da hemolinfa de
ostras de mangue Crassostrea gasar
Natanael Dantas Farias
Orientadora: Profª Dra. Patricia Mirella da Silva Scardua
João Pessoa – 2014

UNIVERSIDADE FEDERAL DA PARAÍBA
CENTRO DE CIÊNCIAS EXATAS E DA NATUREZA
CURSO DE BACHARELADO EM CIÊNCIAS BIOLÓGICAS
Caracterização morfológica e da capacidade fagocítica de hemócitos da hemolinfa de
ostras de mangue Crassostrea gasar
Natanael Dantas Farias
Orientadora: Profª. Dra. Patricia Mirella da Silva Scardua
Trabalho - Monografia apresentada ao
Curso de Ciências Biológicas (Trabalho
Acadêmico de conclusão de Curso), como
requisito parcial à obtenção do grau de
Bacharel em Ciências Biológicas.
João Pessoa – 2014

Catalogação na publicação
Universidade Federal da Paraíba
Biblioteca Setorial do CCEN
F224c Farias, Natanael Dantas.
Caracterização morfológica e da capacidade fagocítica de hemócitos da
hemolinfa de ostras de mangue Crassostrea gasar / Natanael Dantas Farias. -
João Pessoa, PB, 2014.
52 p. : il.
Monografia (Bacharelado em Ciências Biológicas) - Universidade
Federal da Paraíba.
Orientador: Profª Drª Patrícia Mirella da Silva Scardua.
1. Ostras. 2. Biologia da ostra. 3. Ostra de mangue. 4. Crassostrea gasar
I. Título.
UFPB/BS-CCEN CDU 564.1(043.2)

UNIVERSIDADE FEDERAL DA PARAÍBA
CENTRO DE CIÊNCIAS EXATAS E DA NATUREZA
CURSO DE BACHARELADO EM CIÊNCIAS BIOLÓGICAS
Natanael Dantas Farias
Caracterização morfológica e da capacidade fagocítica de hemócitos da hemolinfa de
ostras de mangue Crassostrea gasar
Trabalho – Monografia apresentada ao Curso de Ciências Biológicas, como requisito parcial à
obtenção do grau de Bacharel em Ciências Biológicas
Data:_________________________________
Resultado: ____________________________
BANCA EXAMINADORA:
Dra. Patrícia Mirella da Silva Scardua - Orientadora
UFPB/ Centro de Ciências Exatas e da Natureza/ Departamento de Biologia Molecular
Dra. Márcia Rosa de Oliveira- Avaliador
UFPB/ Centro de Ciências Exatas e da Natureza/ Departamento de Biologia Molecular
Dr. Luis Fernando Marques dos Santos - Avaliador
UFPB/ Centro de Ciências Exatas e da Natureza/ Departamento de Biologia Molecular

AGRADECIMENTOS
Primeiramente agradeço a minha orientadora Dra. Patrícia Mirella da Silva Scardua pela
confiança, pelo carinho e atenção em mim depositados. Por aceitar a minha entrada no
Laboratório de Imunologia e Patologia de Invertebrados (LABIPI), desde a época da monitoria.
Agradeço imensamente pela orientação de grande valor e também por ter conhecido essa pessoa
maravilhosa que és, desde o início da graduação. Agradeço também pela paciência ímpar que
teve e tem em me orientar.
Ao professor Dr. Luis Fernando Marques dos Santos (DBM), a professora Dra. Márcia
Regina Piuvezam (DFP) e a professora Dra. Márcia Rosa de Oliveira (DBM), por aceitarem o
convite de serem membros da minha banca, bem como por fazerem parte de maneira
construtiva, direta ou indiretamente, da minha graduação.
Aos professores Dr. Isac Almeida e Dr. Robson, por disponibilizarem o citômetro de
fluxo FACSCantoTM II, BD Biosciences.
Aos meus pais José Edmar Leal Farias e Nilza Maria Dantas Farias, bem como às
minhas irmãs Luciana Dantas e Marianne Farias, por todo apoio dedicado à minha formação.
Bem como pela atenção e dedicação neste importante período da minha vida.
Ao meu amigo e integrante do LABIPI, Msc. Fernando Ramos Queiroga, que me ajudou
bastante na construção desse trabalho e na obtenção e análise dos resultados. Agradeço também
pela paciência recíproca que tivemos.
Aos meus amigos Anna Carolina, Cairé Barreto e Raianna Boni, por estarem juntos
comigo desde o início da graduação e, juntos, atravessarmos os altos e baixos que ela nos trouxe.
À Sâmia Sousa Duarte, Lucas Nunes Santana e Carol Costa, também membros do
LABIPI pela companhia e pela amizade.
À Cristina Toledo, Jaíse Paiva, Matheus Ramos e Heytor Queiroz, pela dedicação nos
momentos que me senti para baixo e pela amizade.
Ao pessoal do Laboratório de Biologia do Desenvolvimento (LABID), Leonardo Lima,
Elis Torrezan, Jocelmo Cássio, Thaís Mangeon, Andrezza de Araújo, Thyago Fernandes e
Dalliane Macedo, pelos construtivos e divertidos momentos que passamos juntos.

RESUMO
Os hemócitos são as células de defesa de moluscos bivalves e estão presentes na
hemolinfa, líquido circulante análogo ao sangue de vertebrados. Estas células são capazes de
realizar a fagocitose, principal mecanismo de defesa, e produzir moléculas citotóxicas capazes
de destruir o agente patogênico. O objetivo de realizar este estudo foi caracterizar morfológica
e funcionalmente as células imunológicas da ostra Crassostrea gasar. As ostras (n = 60) foram
coletadas de um cultivo no estuário do Rio Mamanguape, na Paraíba, nos meses de março,
agosto e setembro de 2013 e agosto, setembro e outubro de 2014. A hemolinfa foi extraída do
músculo adutor. Para análise morfológica por microscopia óptica (n = 30) as células foram
deixadas aderir espontaneamente em uma lâmina e foram coradas com vermelho neutro ou
Giemsa. Para análise morfológica por citometria de fluxo, a mesma hemolinfa foi fixada com
formol 4% em água do mar filtrada estéril, na proporção de 1:1. Para análise da fagocitose por
citometria de fluxo (n = 30) as células foram adicionadas em contado com partículas
fluorescentes inertes (látex) ou de natureza biológica (zymosan e E. coli). Quanto a
caracterização morfológica, os resultados obtidos pelas duas técnicas mostram que a hemolinfa
da ostra é bastante diversa e apresenta cinco subpopulações hemocitárias: granulócitos grandes,
granulócitos pequenos, hialinócitos, hemócitos vesiculares e as células tipo-blásticas. As
maiores células observadas foram os granulócitos grandes e hemócitos vesiculares e as menores
foram as células do tipo-blásticas. As células mais abundantes na hemolinfa foram os hemócitos
vesiculares e hialinócitos. Quanto a fagocitose, os hemócitos mostraram capacidade diferencial
de fagocitar os três tipos de partículas com os quais foram desafiados; a maior taxa de fagocitose
foi do zymosan (15,1% 0,94), seguida do látex (9,7% 0,45) e da E. coli (5,0% 0,54). Os
diferentes tipos de hemócitos podem reconhecer e fagocitar diferencialmente as partículas; as
de látex foram as mais fagocitadas pelos granulócitos (40,1% 1,87), enquanto que as de E.
coli pelo hialinócitos e hemócitos vesiculares (7,2% 0,36). Os granulócitos foram as células
com maior capacidade fagocítica de látex, quando comparado aos hialinócitos e hemócitos
vesiculares juntos. Este é o primeiro estudo da caracterização morfológica de hemócitos de
ostras C. gasar, uma espécie de grande importância comercial no Nordeste brasileiro.

ABSTRACT
The hemocytes are the defense cells of bivalve mollusks and are present in the
hemolymph, analogous to the blood of vertebrates. These cells are able to perform
phagocytosis, the main defense mechanism, and produce cytotoxic molecules capable of
destroying the pathogen. The aim of this study was to characterize morphologically and
functionally immune cells of the oyster Crassostrea gasar. Oysters (n = 60) were sampled in a
oyster facility located at the estuary of the Mamanguape River, Paraíba, in March, August and
September 2013 and August, September and October 2014. The hemolymph was extracted from
the adductor muscle. For morphological analysis by light microscopy (LM; n = 30) cells were
allowed to adhere spontaneously on a slide and stained with Giemsa or Neutral Red. To
morphological analysis yet, the same hemolymph was fixed with formol 4% in filtered and
sterilized water of sea, in proportion of 1:1. To analyze the phagocytosis by flow cytometry
(FC; n = 30), the cells were added in contact with inert (latex) or biological (zymosan and E.
coli) fluorescent particles.The results obtained by the two techniques (LM and FC) showed that
oysters hemolymph is composed by five hemocytes subpopulations: large granulocytes, small
granulocytes, hyalinocytes, vesicular hemocytes and blast-like cells. The largest cells were
granulocytes and vesicular hemocytes and smallest cells were blast-like cells. The most
abundant cells in the hemolymph were vesicular hemocytes and hyalinocytes. Concerning
phagocytosis, C. gasar hemocytes showed differential ability to phagocyte the three types of
particles which were challenged; the highest rate of phagocytosis was with zymosan (15,1%
0,94), followed by latex (9,7% 0,45) and then E. coli (5,0% 0,54). The different types of
hemocytes can differentially recognize and phagocyte the particles; latex were preferentially
phagocytosed by granulocytes (40,1% 1,87), whilst E. coli by hyalinocytes and vesicular
hemocytes (0,36% 7,2). Granulocytes were more phagocytic than hyalinocytes and vesicular
hemocytes for latex particles. This is the first study on morphological characterization of
hemocytes of oysters C. gasar, a species with great commercial importance in the Northeast
Brazil.

LISTA DE ILUSTRAÇÕES
Figura 1. Sistema circulatório da ostra americana Crassostrea virginica 12
Figura 2. Diferenciação celular de hemócitos 14
Figura 3. Diferenciação celular proposta por análises morfológicas por citometria de
fluxo 16
Figura 4. Mecanismos de defesa hemocitários 18
Figura 5. Mecanismos líticos para degradação de patógenos 20
Figura 6. Modelos de ação de peptídeos antimicrobianos por ruptura de membrana 21
Figura 7. Modelo de funcionamento de um citômetro de fluxo 23
Figura 8. Citograma e histogramas representativos para fagocitose de partículas de
látex fluorescentes 28
Figura 9. Citogramas e histogramas representativos para fagocitose de Escherichia coli
e zymosan fluorescentes 29
Figura 10. Granulócitos grandes da hemolinfa de Crassostrea gasar 33
Figura 11. Granulócitos pequenos da hemolinfa de Crassostrea gasar 34
Figura 12. Hemócitos vesiculares da hemolinfa de Crassostrea gasar 34
Figura 13. Hialinócitos da hemolinfa de Crassostrea gasar 35
Figura 14. Células tipo-blásticas da hemolinfa de Crassostrea gasar 35
Figura 15. Populações de hemócitos da hemolinfa de Crassostrea gasar observadas
por citometria de fluxo 37
Figura 16. Fagocitose de diferentes partículas por hemócitos de Crassostrea
gasar 38
Figura 17. Fagocitose (%) de diferentes partículas pelos hemócitos da ostra
Crassostrea gasar 39
Figura 18. Fagocitose (%) diferencial de partículas de látex e Escherichia coli em ostras
Crassostrea gasar 40
Figura 19. Capacidade fagocítica (%) de partículas de látex pelos hemócitos de
Crassostrea gasar 40

LISTA DE TABELAS
Tabela 1. Média (± EP) do tamanho celular (µm e unidades arbitrárias) e da porcentagem (±
EP) das subpopulações de hemócitos da hemolinfa da ostra Crassostrea gasar.
Diferentes letras indicam diferenças significativas do tamanho entre as diferentes
populações celulares. Análises independentes para cada técnica 31

LISTA DE ABREVIATURAS E SIGLAS
BPI - Bactericidal /permeability-increasing protein (Proteína bactericida de aumento de
permeabilidade)
FSC - Forward Scattered (Dispersão Frontal)
LPS - Lipopolissacarídeo
NET - Neutrophil extracellular traps (Armadilhas extracelulares de neutrófilos)
NO - Nitric Oxide (Óxido Nítrico)
NOS - Nitric Oxide Synthase (Óxido Nítrico Sintase)
PAM - Peptídeo Antimicrobiano
PAMP - Pathogen-associated molecular patterns (Padrão Molecular Associado ao Patógeno)
PRP - Pattern Recognition Proteins (Proteínas de Reconhecimento Padrão)
PRR - Pattern Recognition Receptors (Receptores de Reconhecimento Padrão)
RNS - Reactive Nitrogen Species (Espécies Reativas de Nitrogênio)
ROS - Reactive Oxygen Species (Espécies Reativas de Oxigênio)
SOD - Superoxide Dismutase (Superóxido Dismutase)
SSC - Side Scattered (Dispersão Lateral)

SUMÁRIO
1. INTRODUÇÃO 11
1.1. Sistema de defesa de bivalve 11
1.2. Ontogenia e células imunoefetoras de bivalves 13
1.3. Mecanismos de defesa hemocitários 16
1.4. Mecanismos líticos na interação parasito-hospedeiro 19
1.4.1. Produção de espécies reativas de oxigênio 19
1.4.2. Produção de espécies reativas de nitrogênio 19
1.4.3. Produção de peptídeos anti-microbianos (PAMs) 20
1.4.4. Enzimas líticas 22
1.5. Outras funções dos hemócitos 22
1.6. Métodos de estudo dos hemócitos 22
1.7. Estudo de hemócitos em bivalves do Brasil 24
2. OBJETIVOS 25
2.1. Objetivo geral 25
2.2. Objetivos específicos 25
3. MATERIAL E MÉTODOS 26
3.1. Coleta dos animais 26
3.2. Extração de hemolinfa 26
3.3. Caracterização morfológica dos hemócitos por microscopia óptica 26
3.3.1. Monocamadas coradas com vermelho neutro 26
3.3.2. Monocamadas coradas com Giemsa 26
3.4. Caracterização morfológica dos hemócitos por citometria de fluxo 27
3.5. Caracterização da fagocitose dos hemócitos por citometria de fluxo 27
3.6. Análises estatísticas 30

4. RESULTADOS 31
4.1. Caracterização morfológica dos hemócitos por microscopia óptica 31
4.2. Caracterização morfológica dos hemócitos por citometria de fluxo 36
4.3. Caracterização funcional de hemócitos por citometria de fluxo 37
5. DISCUSSÃO 41
5.1. Caracterização morfológica dos hemócitos de Crassostrea gasar 41
5.2. Caracterização funcional dos hemócitos de Crassostrea gasar por
citometria de fluxo 43
6. CONCLUSÕES 45
7. REFERÊNCIAS 46

11
1. INTRODUÇÃO
1.1. Sistema de defesa de bivalves
O sistema imunológico de invertebrados, ao contrário dos vertebrados, é exclusivamente
inato ou natural, pois não produz anticorpos específicos da imunidade adaptativa, não possuindo
assim memória imunológica.
As células imunoefetoras de bivalves são denominadas hemócitos e estão presentes no
líquido circulante, a hemolinfa, análoga ao sangue de vertebrados, onde encontram-se também
moléculas solúveis no plasma, que atuam no reconhecimento e degradação de agentes não-
próprios (Vargas-Albores & Barracco, 2001; Song et al., 2010)
A hemolinfa de bivalves percorre um sistema circulatório aberto, que é bombeado por
um coração protegido por uma fina camada de epitélio, o pericárdio, e contém 3 câmaras, sendo
um ventrículo e dois átrios. Após ser bombeada pelo coração, a hemolinfa sai do ventrículo e
penetra na artéria aorta anterior, onde supre o sistema gástrico e a região anterior do corpo e
também penetra na artéria posterior. Essa artéria supre o músculo adutor e também o reto. A
hemolinfa percorre as artérias até desembocar em seios hemolinfáticos que banham os diversos
tecidos e órgãos, direcionando-a para as brânquias, retornando para o coração através de um
conjunto de veias e desembocando no átrio por três veias (Fig. 1) (Cheng, 1981; Eble, 2009).

12
Figura 1 - Sistema circulatório da ostra americana Crassostrea virginica
Sistema arterial (A) e venoso (B) observados lateralmente. A hemolinfa é bombeada pelo coração
(sistema arterial) do átrio (AT) para o ventrículo (V), seguindo para a artéria aorta anterior (AA), onde
supre o sistema gástrico e outros órgãos da região anterior e posterior (PA), suprindo o músculo adutor
(MA) e o reto. Ao atingir os seios hemolinfáticos que suprem esses órgãos, a hemolinfa retorna por um
conjunto de veias e chega ao átrio por três veias (não mostradas). Adaptado de Eble (2009).
Brânquias

13
1.2. Ontogenia e células imunoefetoras de bivalves
Embora existam algumas hipóteses para a origem dos hemócitos, o tecido
hematopoiético em bivalves permanece desconhecido. Cheng (1981) sugere que o tecido
conjuntivo seja o responsável pela origem destas células, que posteriormente migram para a
hemolinfa, onde sofrem maturação e diferenciação. Esta hipótese teve o apoio do trabalho de
Smolowitz; Miosky; Reinisch (1989) que identificaram, através da utilização de anticorpos,
antígenos de superfície de hemócitos semelhantes à antígenos de superfície de células do tecido
conjuntivo.
Com respeito aos tipos hemocitários da hemolinfa de bivalves, existem controvérsias
nas diferentes hipóteses de sua classificação. O primeiro trabalho de revisão sobre hemócitos
de várias espécies de bivalves propôs sua classificação simples baseada em características
morfológicas e citoquímicas e divide as células em duas grandes populações, os granulócitos e
os hialinócitos (Cheng, 1981).
Os granulócitos são geralmente as maiores células encontradas na hemolinfa, com a
maior capacidade fagocítica e de produção de espécies reativas de oxigênio, sendo, portanto as
células mais associadas à defesa (Fisher, 1988; Hine, 1999). Elas apresentam muitos filipódios
e são facilmente distinguidas das outras pela abundância de vesículas ou grânulos
citoplasmáticos que se dispõem geralmente na região endoplasmática, podendo ocupar também
a ectoplasmática. Além disso, essas vesículas podem ser basófilas ou acidófilas, dependendo
da espécie de bivalve analisada e do conteúdo enzimático presente em seu interior. O núcleo é
excêntrico, basófilo e apresenta uma baixa relação núcleo:citoplasma. Os hialinócitos são
células menores, com núcleo maior que o dos granulócitos e podem apresentar pouca ou
nenhuma vesícula. Seu citoplasma é geralmente basófilo e possui uma alta relação
núcleo:citoplasma. São células aptas a fagocitar, embora em menor proporção (Cheng, 1981).
A diferenciação de hemócitos também foi postulada pela primeira vez por Cheng (1981)
e hipotetiza duas linhagens celulares progenitoras: os granuloblastos e hialinoblastos. Essas
células seriam capazes de dar origem aos granulócitos e aos hialinócitos maduros (Fig. 2).
Como proposto por Cheng (1981), os granuloblastos se diferenciariam primariamente em
progranulócitos, células menores que os granulócitos, contendo poucos grânulos
citoplasmáticos, com núcleo e vesículas basófilas e capazes de fagocitar. Posteriormente, os
progranulócitos se diferenciariam em granulócitos do tipo I, com tamanho mediano, numerosas
vesículas citoplasmáticas basófilas ou acidófilas, com altos níveis de atividade de hidrolases e
capazes de fagocitar. Por fim, os granulócitos do tipo I maturariam para o tipo II, sendo as

14
maiores células observadas, com presença do complexo de Golgi e lisossomos maduros, bem
como com a maior atividade de hidrolases e maior atividade fagocítica. No caso de realizar uma
fagocitose, os granulócitos do tipo II se caracterizariam por possuir poucas projeções,
geralmente em polos opostos da célula, pouco ou nenhum grânulo, vacúolos de diversos
formatos e tamanhos e menor atividade de hidrolases. Essa alteração morfológica ocorreria
devido à ativação dos mecanismos de degradação intracelular, levando os granulócitos do tipo
II à um aspecto “gasto”. Entretanto, os granulócitos do tipo II poderiam ainda se fusionar,
formando células multinucleadas (Fig. 2).
Os hialinoblastos por sua vez, se diferenciariam em prohialinócitos, com núcleo
relativamente grande e pouco citoplasma, sem vesículas citoplasmáticas e essencialmente
basófilas. Em seguida ocorreria a maturação de prohialinócitos em hialinócitos, que seriam as
células maiores dessa linhagem, com poucos grânulos citoplasmáticos (Cheng, 1981) (Fig. 2).
Figura 2 - Diferenciação celular de hemócitos
Os hemócitos teriam sua origem em duas linhagens celulares, os granuloblastos e hialinoblastos. Os
granuloblastos se diferenciariam em progranulócitos, granulócitos tipo I e granulócitos tipo II,
aumentando gradativamente a capacidade fagocítica e amadurecimento de organelas. A fagocitose
acarretaria uma mudança dos granulócitos tipo II para granulócitos “gastos” ou, no caso de fusão para
células multinucleadas. Os hialinoblastos se diferenciariam em prohialinócitos e hialinócitos. Cheng
(1981).
Recentemente, trabalhos reunidos na revisão de Hine (1999) propuseram novas
classificações envolvendo as subpopulações de granulócitos e hialinócitos.
Os granulócitos com vesículas basófilas ou eosinófilas, bem como neutras, já foram
observados nos bivalves das famílias Ostreidae (Crassostrea virginica, Crassostrea gigas e
Ostrea edulis), Mytilidae (Mytilus edulis, Mytilus. galloprovincialis e Mytilus californianus),

15
Veneridae (Mercenaria mercenaria, Mercenaria campechiensis, Meretrix lusoria, Sunetta
scripta, Villorita cyprinoides, Ruditapes decussatus e Tapes semidecussata), Myidae (Mya
arenaria), Cardiidae (Cerastoderma edule), Arcidae (Scapharca inaequivalvis, Anadara
ovalis), Tridacnidae (Tridacna crocea, Tridacna maxima) e Pinnidae (Pinna nobilis) (vide
revisão de Hine, 1999).
Os hialinócitos, apesar de conterem pouco ou nenhum grânulo, são células muito
heterogêneas em nível de microscopia eletrônica. Por sua vez, podem ser subdivididos em
grandes e pequenos, sendo este último muitas vezes chamado de células tipo-blásticas. Os
hialinócitos grandes possuem núcleo grande, oval ou reniforme, com diversas organelas em seu
citoplasma, e já foi identificado em espécies da família Ostreidae, Mytilidae e Veneridae. As
células tipo-blásticas possuem núcleo central, relativamente pequeno e um citoplasma bem
escasso em volume e organelas, sendo identificadas em alguns bivalves das famílias Ostreidae,
Mytilidae, Dreissenidae, Veneridae, Cardiidae e Tridacnidae. Estas células também foram
descritas para as espécies de ostras C. virginica, C.gigas, O. edulis e Saccostrea glomerata
(Hine, 1999; Hégaret et al., 2003a; Lambert et al., 2003; Dang et al., 2012).
Estudos recentes sugerem que as células tipo-blásticas, para a maioria dos bivalves, são
precursoras dos hemócitos por apresentarem uma porção do citoplasma altamente basófila, o
que indica a presença de ribossomos livres e sugere a imaturidade celular, além de
características morfológicas semelhantes aos hialinócitos (Russell-Pinto et al., 1994; Wen et
al., 1994; Carballal et al., 1997; Rebelo et al., 2013).
Um novo grupo proposto por Hine (1999), os hemócitos vesiculares, encontrados apenas
em alguns bivalves das famílias Ostreidae, Cardiidae, Dreissenidae e Veneridae, possuem
características que são facilmente confundidas com os granulócitos, como a baixa relação
núcleo:citoplasma e com os hialinócitos, pela ausência ou pouca quantidade de grânulos. Nos
bivalves O. edulis, Dreissena polymorpha, M. mercenaria este grupo foi identificado como
hialinócitos, hialinócitos e hemócitos grandes basófilos e hialinócitos e fibrócitos,
respectivamente. Em M. lusoria, C. edule e S. inaequivalvis estes hemócitos foram classificados
como hialinócitos granulares, hemócitos tipo III e hemócitos do tipo II, respectivamente.
Rebelo et al. (2013) propõem ainda outro modelo de diferenciação celular de hemócitos
da ostra C. rhizophorae baseado em características morfológicas, no tamanho e granulosidade
interna, por análise de citometria-de-fluxo. Eles propuseram que as células tipo-blásticas
iniciam o processo diferenciando-se em hialinócitos maiores, pelo aumento da quantidade de
organelas. Em seguida, os hialinócitos podem desenvolver vesículas, adquirindo uma maior
granulosidade interna e maior tamanho, tornando-se granulócitos. Os granulócitos, por fim,

16
poderiam degranular por estímulos extracelulares, conservando seu tamanho, mas com uma
diminuição da granulosidade interna, sendo associados então aos hemócitos vesiculares (Fig.
3).
Figura 3 - Diferenciação celular proposta por análises morfológicas por citometria de
fluxo
Este modelo de diferenciação corrobora a hipótese das células do tipo-blásticas serem precursoras de
todos os hemócitos. O eixo X (FSC) representa o tamanho celular e o eixo Y (SSC) a granulosidade
celular. O amadurecimento intracelular das células tipo-blásticas (BL) levaria à diferenciação em
hialinócitos (HH) maiores, que posteriormente aumentariam a granulosidade e tamanho celular, se
diferenciando em granulócitos pequenos (GP) e grandes (GG), que por sua vez poderiam degranular e
tornarem-se hemócitos vesiculares (HV). Adaptado de Rebelo et al. (2013).
1.3. Mecanismos de defesa hemocitários
A defesa contra os agentes infecciosos, realizada por hemócitos, acontece
principalmente pelo mecanismo de fagocitose ou encapsulamento (Fig. 4). A fagocitose se
realiza em sucessivas etapas: quimiotaxia, reconhecimento, englobamento e degradação.
Primeiro ocorre a quimiotaxia, que é o movimento celular direcionado através de um gradiente
de concentração químico, com objetivo de chegar a um destino, geralmente acontece da
circulação para os tecidos. A quimiotaxia não acontece estritamente para a defesa do organismo,
ela pode ocorrer também no reparo de tecido ou concha, onde os hemócitos têm que chegar no

17
local afetado (Donaghy et al., 2009b). Posteriormente, ocorre o reconhecimento do
microrganismo por moléculas conservadas evolutivamente, chamadas de Padrões Moleculares
Associados aos Patógenos (PAMPs, do inglês Pathogen-Associated Molecular Patterns), como
por exemplo, os lipopolissacarídeos (LPS) presentes na parede celular de bactérias Gram-
negativas e β1,3-glicanas presentes nas paredes de fungos. O reconhecimento dos PAMPs se
dá por moléculas específicas denominadas Receptores ou Proteínas de Reconhecimento Padrão
(PRR, do inglês Pattern Recognition Receptors ou PRP, Pattern Recognition Proteins) que se
encontram na membrana dos hemócitos ou solúveis na hemolinfa (Song et al., 2010; Schmitt et
al., 2011).
Sete grupos de PRR são descritos em bivalves: receptores do tipo-Toll, scavengers,
proteínas de reconhecimento de peptideoglicanas, proteínas de ligação à bactérias Gram-
negativas, lectinas do tipo-C, galectinas e proteínas contendo thio-éster (Song et al., 2010).
Após o reconhecimento, finalmente ocorre o englobamento do microrganismo em um vacúolo
fagocítico para a sua destruição (Fig. 4). Várias vias de destruição podem ser ativadas, como a
da produção e liberação de espécies reativas de oxigênio e de nitrogênio, a produção de
peptídeos antimicrobianos (PAM) e produção de enzimas hidrolíticas, dentre elas a lisozima
(Song et al., 2010; Schmitt et al., 2011).
O encapsulamento, por sua vez, está relacionado com parasitas de maior tamanho, que
neste caso, não poderiam ser fagocitados (Fig. 4). Esta resposta envolve a interação hemócito-
parasita, onde os hemócitos se dispõem em várias camadas por volta do parasita, impedindo
sua proliferação (Barracco & da Silva, 2008; Vargas-Albores & Barracco, 2001). Estudos
sugerem que, posteriormente, os hemócitos secretam enzimas lisossomais e espécies reativas
de oxigênio para degradar o parasita encapsulado (Chagot et al., 1987; Montes et al., 1997).
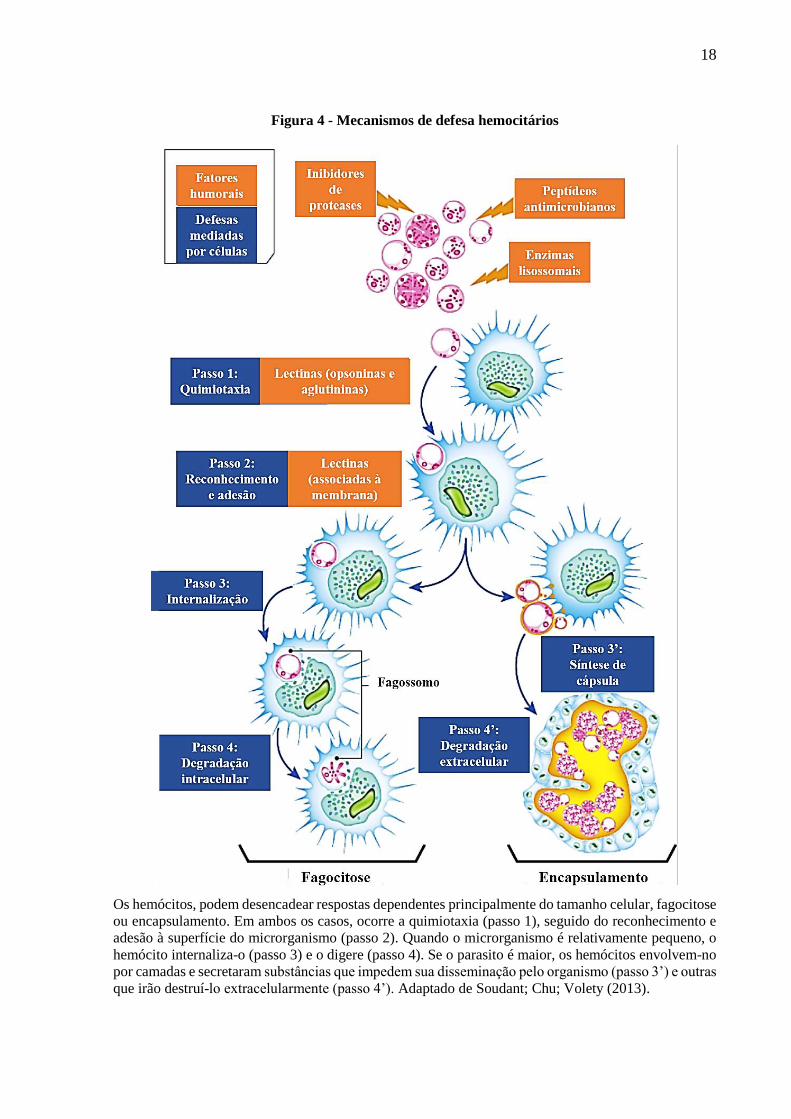
18
Figura 4 - Mecanismos de defesa hemocitários
Os hemócitos, podem desencadear respostas dependentes principalmente do tamanho celular, fagocitose
ou encapsulamento. Em ambos os casos, ocorre a quimiotaxia (passo 1), seguido do reconhecimento e
adesão à superfície do microrganismo (passo 2). Quando o microrganismo é relativamente pequeno, o
hemócito internaliza-o (passo 3) e o digere (passo 4). Se o parasito é maior, os hemócitos envolvem-no
por camadas e secretaram substâncias que impedem sua disseminação pelo organismo (passo 3’) e outras
que irão destruí-lo extracelularmente (passo 4’). Adaptado de Soudant; Chu; Volety (2013).

19
1.4. Mecanismos líticos na interação parasito-hospedeiro
1.4.1. Produção de espécies reativas de oxigênio
A produção de espécies reativas de oxigênio (ROS, do inglês Reactive Oxygen Species)
é um dos principais mecanismos de ação degradativa nas células de defesa de invertebrados, e
seu maior efeito direto consiste na oxidação de proteínas, carboidratos ou bases do DNA. Sua
produção pode ocorrer em grandes quantidades após o contato com os antígenos, processo
chamado de “explosão respiratória” devido ao alto consumo de oxigênio (do inglês, respiratory
burst). A catalisação da reação ocorre com o consumo de oxigênio molecular (O2) e é realizada
pela ação da enzima NADPH oxidase presente nas membranas da célula: NADPH + 2O2 →
NADP+ + 2O2- + H+ (Fang 2004, Robinson 2008). O produto desta reação, o ânion superóxido
(O2-), pode ainda ser dismutado por enzimas como a superóxido dismutase (SOD, do inglês,
Superoxide Dismutase) pela reação: SOD + O2- + O2
- + 2H+ → 2H2O2 + O2, produzindo o
peróxido de hidrogênio (H2O2), também um radical livre, tóxico para as membranas celulares.
Outros radicais livres podem ser produzidos a partir da interação do O2- e H2O2 com outras
moléculas intracelulares, como o radical hidroxila (OH∙), o oxigênio singlet (1O2) e o ácido
hipocloroso (HOCl) (Robinson, 2008) (Fig. 5).
Vários estudos têm verificado a produção de ROS pela detecção basal ou do respiratory
burst em diversas ostras como C. virginica (Larson et al., 1989; Hégaret et al., 2003b), C. gigas
(Lambert et al., 2003), Crassostrea ariakensis (Donaghy et al., 2009a) e Crassostrea gasar
(Queiroga et al., 2013), mexilhões M. edulis (Pipe, 1992) e mariscos Ruditapes philipinarum
(Cima et al., 2000) e M. mercenaria (Buggé et al., 2007).
1.4.2. Produção de espécies reativas de nitrogênio
A produção de espécies reativas de nitrogênio (RNS, do inglês Reactive Nitrogen
Species), principalmente o óxido nítrico (NO), parece ter uma importante função na defesa de
bivalves, embora não muito bem elucidada, e ocorre através da reação: Arginina + NADPH +
O2 → Citrulina + NO + NADP+, catalisada pela enzima óxido nítrico sintase (NOS, do inglês
Nitric Oxide Synthase) (Fang, 2004; Schmitt et al., 2011) (Fig. 5).
Os efeitos atribuídos às RNS envolvem passos bioquímicos muito mais complexos que
o das ROS e dependem da condição redox do meio celular. O NO pode inibir, o crescimento
celular de bactérias, através de mecanismos que sequestram o zinco de metaloproteínas
necessárias à replicação e pode bloquear diretamente o processo respiratório. A produção de
óxido nítrico pode ainda culminar no aumento da citotoxicidade das espécies reativas de

20
oxigênio, onde o NO pode interagir com O2- e produzir peroxinitrito (ONOO-) e outros
intermediários tóxicos (Fang, 2004) (Fig. 5).
Figura 2 - Mecanismos líticos para degradação de patógenos
Fagossomo e diversas enzimas que agem para degradação de um parasito englobado. Complexo da
NADPH oxidase que gera ânion superóxido (O2-) (1) a partir do oxigênio molecular (O2). O O2- pode
ser dismutado (2) pela superóxido dismutase (SOD), produzindo peróxido de hidrogênio (H2O2) ou
reagir com óxido nítrico (3) (NO) produzindo peroxinitrito (ONOO-). A mieloperoxidase (MPO) pode
converter H2O2 em HOCl (4) na presença de Cl-. Adaptado de Soudant et al. (2008).
1.4.3. Produção de peptídeos anti-microbianos (PAMs)
Os peptídeos antimicrobianos são moléculas pequenas (menores que 10 kDa) que
apresentam atividade microbicida e possuem natureza anfipática e catiônica. Atuam
principalmente pela formação de poros na membrana e desequilíbrio iônico do patógeno,
levando à lise celular (Schmitt et al., 2012).
Dois modelos principais para a ruptura da membrana por peptídeos antimicrobianos são
propostos, o modelo barril (do inglês “Barrel-stave” Model) e o modelo tapete (do inglês
“Carpet” Model). No modelo barril, a formação de poros se dá pela agregação dos peptídeos,
seguido da sua inserção na bicamada lipídica, onde os aminoácidos hidrofóbicos interagem com
os lipídeos de membrana e os hidrofílicos entre si, formando um canal (Fig. 6A). O modelo
tapete propõe que, os peptídeos em solução interagem primeiramente com os lipídeos e
grupamentos fosfatos da face externa da membrana e posteriormente entre si, criando poros

21
transitórios que, com o aumento da concentração de peptídeos na membrana, levará a formação
de micelas e a ruptura celular (Fig. 6B).
Figura 3 – Modelos de ação de peptídeos antimicrobianos por ruptura de membrana
(A) O modelo barril propõe a agregação dos peptídeos e sua interação posterior com a bicamada lipídica
formando poros que desestabilizarão ionicamente a célula. (B) O modelo tapete sugere que a interação
dos peptídeos primeiramente com a superfície externa da bicamada lipídica e posteriormente entre si,
formando micelas e destruindo a membrana plasmática. Li; Zhao; Song (2009).
Outros peptídeos antimicrobianos podem atuar sobre moléculas envolvidas em
processos vitais como enzimas ou moléculas menores relacionadas ao metabolismo celular, não
causando ruptura da membrana. Os PAMs atuam como barreira primária do sistema de defesa,
prevenindo o crescimento e a instalação de patógenos nos tecidos, sendo altamente expressos
nos epitélios e liberados no muco (Li et al., 2009; Casas et al., 2011).
A síntese de PAMs ocorre de maneira constitutiva e são armazenados em vesículas
dentro dos hemócitos para liberação após interação com o patógeno. Essa síntese ocorre
principalmente em órgãos que estão em contato frequente com o meio externo, como brânquias
e manto (Mitta et al., 2000; Zhao et al., 2007).
Diversos PAMs foram descritos em bivalves, como nos mexilhões M. galloprovincialis
(Hubert et al., 1996), M. edulis (Charlet et al., 1996) e nas ostras C. gigas (Gonzalez et al.,
2007). Em C. gigas, alguns peptídeos antimicrobianos foram caracterizados e incluídos nas
famílias: defensinas e PAM ricos em prolina. As defensinas foram identificadas primeiramente
a partir do manto (Cg-Defm) e posteriormente nos hemócitos (Cg-Defh1 e Cg-Defh2) e ambos
possuem a capacidade de impedir a síntese de peptídeoglicanos de bactérias Gram-positivas.

22
Um PAM rico em prolina foi identificado também nos hemócitos de C. gigas (Cg-Prp) e,
embora sua função seja pouco conhecida, observa-se uma atuação em sinergia com as
defensinas hemocitárias. Outro PAM foi identificado e caracterizado em C. gigas pela atividade
bactericida contra bactérias Gram-negativas e atua através da permeabilização da membrana
externa e interna (BPI, do inglês Bactericidal /permeability-increasing protein) (Schmitt et al.,
2011, 2012).
1.4.4. Enzimas líticas
Os lisossomos de hemócitos contém uma variedade de enzimas hidrolíticas, como a β-
glucuronidase, arilsulfatase, lisozima e catepsina B, no interior de vesículas que podem ter o
pH ácido ou básico e ocupam o citoplasma (Pipe, 1990).
As lisozimas desempenham duas funções necessárias para a sobrevivência de bivalves,
auxiliam na defesa contra patógenos e na digestão intracelular. Nos hemócitos, as lisozimas
secretadas atuam principalmente quebrando moléculas de peptideoglicanos, fundamentais na
estrutura da parece bacteriana (McHenery et al., 1986, Schmitt et al., 2011). Em relação à
digestão, as lisozimas já foram identificadas em células da glândula digestiva de M. edulis e de
C. gigas (Takahashi et al., 1986; Bachali et al., 2002).
1.5. Outras funções dos hemócitos
Atualmente, outras funções têm sido atribuídas aos hemócitos de ostras de acordo com
Marigómez et al. (2002). Como por exemplo, a remoção de metais tóxicos da circulação por
fagocitose (complexos maiores) ou pinocitose (íons livres) e seu posterior transporte para
órgãos de detoxificação, como brânquias e glândula digestiva. Elementos como Cu (Cobre), Zn
(Zinco) e Cd (Cádmio) podem então ser absorvidos pelo organismo dos bivalves, em seguida
serem associados com proteínas citosólicas ou internalizadas em lisossomos de hemócitos para
posterior transporte (vide revisão MARIGÓMEZ et al., 2002).
1.6. Métodos de estudo dos hemócitos
Diversos métodos são empregados nos estudos de caracterização morfológica e
funcional das células imunoefetoras de bivalves (Hine, 1999). Entre eles estão o uso de corantes
histológicos para ressaltar o caráter básico ou ácido do citosol ou dos seus componentes
intracelulares e a microscopia eletrônica de transmissão que revela características
ultraestruturais (Wootton & Pipe, 2003; Chang et al., 2005). Existe ainda a produção de

23
anticorpos monoclonais específicos para antígenos de superfície de hemócitos ou intracelulares,
utilizados tanto na tentativa de caracterizar ou localizar as células em estádios de
desenvolvimento (Renault et al., 2001; Xue & Renault, 2001), quanto de desvendar sua
ontogenia (Smolowitz et al., 1989).
Mais recentemente uma técnica muito utilizada em estudos de vertebrados, a citometria
de fluxo, vem despontando nos estudos da morfologia e funções de diferentes subpopulações
celulares da hemolinfa de bivalves. Esta técnica consiste na utilização de um equipamento que
contém lasers que irão atingir células em suspensão, unitariamente e por um fluxo contínuo,
difratando a luz ao atravessá-las e, posteriormente, atingindo detectores específicos, gerando
assim diversas medidas simultâneas (Fig. 7). Dentre os detectores, destacam-se o Forward
Scattered (FSC), que detecta o tamanho celular por receber a luz refratada frontalmente e o Side
Scattered (SSC), que detecta a complexidade interna celular, recebendo a luz difratada nas
laterais (Fig. 7). Existem ainda detectores de fluorescência, que serão atingidos por diferentes
comprimentos de onda provenientes da excitação de fluoróforos usados para marcar
componentes celulares (BD Biosciences, 2007) (Fig. 7). A utilização da citometria de fluxo traz
a vantagem de permitir a realização de análises simultâneas em uma mesma amostra (Allam et
al., 2002; Hégaret et al., 2003b; Donaghy et al.; 2009, Wang et al.; 2012).
Figura 4 - Modelo de funcionamento de um citômetro de fluxo
Vários parâmetros celulares são medidos em citometria-de-fluxo. O laser azul (488 nm) atinge as células
em suspensão e em fluxo contínuo. A célula refrata a luz e é captada por um sensor de tamanho
(Forward Scattered, FSC), localizado à frente do laser; por um sensor de complexidade interna (Side
Scattered, SSC), que capta a luz difratada, localizado nas laterais. BD Biosciences (2007).

24
1.7. Estudo de hemócitos em bivalves do Brasil
No Brasil, pouco se sabe ainda sobre os mecanismos de defesa de bivalves e poucos
trabalhos de caracterização morfológica e funcional dos tipos hemocitários foram realizados. O
primeiro estudo foi o da caracterização dos hemócitos do mexilhão Perna perna utilizando
diversos métodos de coloração, como o Giemsa para afinidade de grânulos, método de Gomori
para detecção de fosfatases ácidas e Azul de Sudan para marcação de lipídeos. Neste estudo foi
caracterizada a produção de ânion superóxido e capacidade fagocítica dos hemócitos (Barracco
et al., 1999). Posteriormente, os hemócitos da ostra C. rhizophorae foram caracterizados de
forma similar (Barth et al., 2005). Schleder et al. (2008) caracterizou morfologicamente os
hemócitos do pectinídeo Nodipecten nodosus utilizando a coloração de Giemsa e
funcionalmente, detectando a produção de PAMs, atividade de fenoloxidase e atividade de
aglutinação. E recentemente, dois estudos utilizaram a citometria de fluxo, um deles para
caracterizar a produção de ROS, capacidade fagocítica e de aglutinação de hemócitos da ostra
C. gasar (Queiroga et al., 2013) e outro para caracterizar morfológica e ultraestruturalmente os
hemócitos da ostra C. rhizophorae (Rebelo et al., 2013)
Até o presente momento não há estudos da caracterização morfológica de hemócitos de
ostras C. gasar, uma espécie de grande importância comercial no Nordeste brasileiro.

25
2. OBJETIVOS
2.1. Objetivo geral
Caracterizar morfológica e funcionalmente as células imunológicas da ostra Crassostrea
gasar.
2.2. Objetivos específicos
2.2.1. Quantificar e caracterizar morfologicamente as células da hemolinfa da ostra C.
gasar;
2.2.2. Caracterizar a capacidade fagocítica das células da hemolinfa da ostra C. gasar.

26
3. MATERIAL E MÉTODOS
3.1. Coleta dos animais
Foram realizadas seis coletas de ostras da espécie Crassostrea gasar adultas (> 60 mm)
nos meses de março (n = 10), agosto (n = 10) e setembro (n = 10) de 2013 e agosto (n = 10),
setembro (n = 10) e outubro (n = 10) de 2014, em um cultivo comercial localizado no estuário
do Rio Mamanguape (Marcação, PB - S06°47’08,2”; WO34°59’46,7”). As três primeiras
coletas foram direcionadas para a caracterização morfológica dos hemócitos enquanto que as
três últimas para a caracterização funcional.
Todos os animais foram identificados a nível de espécie por biologia molecular
(Queiroga et al. no prelo).
3.2. Extração de hemolinfa
A hemolinfa das ostras foi retirada do músculo adutor com uma seringa de 1 ml acoplada
a uma agulha (21 G) e foi imediatamente colocada em microtubos sobre gelo e em seguida
preparada para análises em microscopia óptica e citometria de fluxo, como descrito abaixo.
3.3. Caracterização morfológica dos hemócitos por microscopia óptica
3.3.1 Monocamadas coradas com vermelho neutro
A hemolinfa (225 µl) de cada ostra (n = 10) foi corada com uma solução do corante vital
vermelho neutro (concentração final de 0,005%) que penetra nos compartimentos ácidos das
células vivas. Após 5 min. as células em suspensão foram observadas ao microscópio com
contraste de fase (Olympus BX41) e fotografadas durante 20 min. Nesta preparação foram
observadas a emissão de pseudópodes e a presença ou ausência de vesículas ácidas nos
hemócitos.
3.3.2 Monocamadas coradas com Giemsa
A hemolinfa (100 µl) de cada ostra (n = 27) foi adicionada em uma lâmina posicionada
sobre papel umedecido, para formação de uma monocamada de hemócitos por adesão
espontânea, durante aproximadamente 5 min a 25 °C. Após este tempo, a monocamada foi

27
fixada com metanol (5 min.) e corada com Giemsa puro (Newprov) por 5 min. e após secas
foram montadas com Entellan (MERCK). As amostras foram observadas em campo claro e
fotografadas no microscópio Olympus BX41.
Cada célula (n 100 células / animal) foi caracterizada segundo o padrão de coloração
de seus grânulos (basófilos ou acidófilos), relação núcleo:citoplasma (baixa ou alta), formato
(circular ou estrelado) e tamanho celular (medida do maior eixo, sem considerar a projeção dos
pseudópodes). Os tipos celulares encontrados foram quantificados e a porcentagem
determinada sobre o total de células analisadas.
3.4 Caracterização morfológica dos hemócitos por citometria de fluxo
A hemolinfa (200 µl) de cada ostra (n = 10) foi imediatamente fixada em solução de
formaldeído à 4% em água do mar filtrada (0,22 µm) estéril na proporção de 1:1. Cada amostra
foi analisada em citômetro de fluxo pelo período de 30s, onde foi obtido o primeiro gráfico de
tamanho em função da complexidade interna (FSC vs SSC) e o experimento foi repetido três
vezes (FACSCalibur, BD Biosciences, San Jose, California, USA).
3.5 Caracterização da fagocitose dos hemócitos por citometria de fluxo
Para a avaliação da capacidade fagocítica dos hemócitos da ostra C. gasar, diferentes
partículas foram utilizadas nos ensaios: partículas inertes fluorescentes de látex (Polysciences,
2 µm), porções da parede de fungo, o zymosan (Life Technologies, 3µm) e bactérias Gram-
negativas Escherichia coli (Life Technologies, 1µm).
A hemolinfa de cada ostra (200 µl) (n = 10) foi subdividida em duas subamostras, em
uma foi adicionada a partícula teste (8 µl) na proporção de 1:10 (hemócito:partículas) e na outra,
foi adicionado o volume correspondente de água do mar filtrada estéril (amostra controle). As
suspensões celulares foram incubadas por 1 hora à 25 °C e analisadas em citômetro de fluxo
(FACSCantoTM II, BD Biosciences, San Jose, California, USA). O experimento foi repetido
três vezes.
Para a análise da fagocitose no gráfico de morfologia (FSC vs SSC), as populações de
hemócitos foram selecionadas para serem representadas em um histograma contendo o número
de hemócitos em função da fluorescência das partículas, verde (partículas de látex) ou vermelha
(zymosan e E. coli). As células-blásticas não apresentaram capacidade fagocítica e foram
retiradas desta análise.

28
Para a fagocitose de látex visualizou-se populações de hemócitos contendo diferentes
quantidades de partículas fagocitadas (1 a 2 partículas e 3 ou + partículas) (Fig. 8A e C).
Na fagocitose de zymosan e E. coli, não houve formação de picos definidos de
fluorescência. Com isto, para determinação da porcentagem de células fagocíticas o processo
foi distinto. Uma região no histograma da amostra sem adição de partículas (controle; Fig. 9B)
foi feita e a mesma região foi analisada na amostra com adição da partícula fluorescente
(tratado; Figs. 9C e 9D). A porcentagem de células com fluorescência da amostra tratada foi
diminuída da amostra controle (Fig. 9B).
Figura 5 - Citogramas e histogramas representativos para fagocitose de partículas de látex
fluorescentes
Citogramas representativos das populações analisadas, correspondendo aos hemócitos totais (A) e as
subpopulações hemocitárias (C). Histogramas (B e D) indicando a quantidade de hemócitos não
fagocíticos (M0) e fagocíticos (M1) que fagocitaram 1 a 2 partículas (M2) e mais que 3 (M3). O eixo X
(FSC) representa o tamanho celular e o Y (SSC) a granulosidade interna. Nos histogramas, o eixo Y
representa a quantidade de células e no eixo X o canal de fluorescência vermelho.

29
Figura 96 - Citogramas e histogramas representativos para fagocitose de Escherichia coli e
zymosan fluorescentes
Citogramas representativos da população total de hemócitos analisada (A). Histogramas de
fluorescência dos hemócitos sem adição de partículas (B; amostra controle) e com adição de E. coli (C)
e de zymosan (D). As regiões de fagocitose foram determinadas pelas linhas. No citograma, o eixo X
(FSC) representa o tamanho celular e o Y (SSC) a granulosidade interna. Nos histogramas, o eixo Y
representa a quantidade de células e no eixo X o canal de fluorescência vermelho.

30
3.6 Análises estatísticas
Os dados de porcentagem foram transformados para arco cosseno [acos(sqrt (X/100))]
para respeitar aos requerimentos da ANOVA. A normalidade de todos os grupos de dados foi
verificada com o teste D’Agostino & Person.
O teste de Kruskal-Wallis com post-hoc teste de separação de médias de Dunns foi
realizado para analisar dados não-paramétricos, como o tamanho celular das populações de
hemócitos obtidos por análise em microscopia óptica das monocamadas de hemócitos e por
citometria de fluxo. Para a comparação das proporções celulares obtidas com as duas técnicas
citadas anteriormente, foi realizado o teste de independência (teste G), utilizando uma tabela de
contingência e o teste χ² (Rohlf & Sokal, 1981).
Uma ANOVA Two-way foi realizada para comparar a fagocitose de diferentes partículas:
E. coli, Zymosan e látex. A réplica do experimento (coletas de agosto, setembro e outubro de
2014) foi considerada um segundo fator. Não havendo diferenças entre as réplicas, foi realizada
uma ANOVA one-way, sendo o tipo de partícula o fator principal, e um post-hoc teste de
separação de médias de Tukey.
O teste T não-pareado foi utilizado para 1) comparar a fagocitose de E. coli e látex
(separadamente); e 2) comparar a capacidade fagocítica de partículas de látex (separadamente)
entre as subpopulações de hemócitos (granulócitos e hemócitos hialinos + hemócitos
vesiculares).
Os dados estão representados como média ± erro padrão (EP). Todas as análises foram
feitas no software GraphPad versão 5.0.

31
4. RESULTADOS
Caracterização morfológica dos hemócitos por microscopia óptica
A hemolinfa das ostras da espécie C. gasar apresentou em sua composição diferentes
populações celulares que se mostraram mais ou menos definidas segundo a técnica utilizada.
Por microscopia óptica cinco populações puderam ser distinguidas por suas
características morfológicas e tintoriais, sendo assim denominadas: granulócitos grandes,
granulócitos pequenos, hemócitos vesiculares, hialinócitos e células tipo-blásticas.
Os granulócitos grandes apresentaram o maior tamanho entre as subpopulações
observadas (31,5 ± 0,31 µm; n = 27; Tab. 1). Este tipo celular apresentou muitos grânulos
basófilos (Figs. 10B e D) distribuídos por todo o citoplasma e se coraram também com o
vermelho neutro (Figs. 10A e C). A porcentagem observada deste tipo celular foi de 14,4% (±
1,58; Tab. 1). Em ambos os tipos de monocamadas, estas células projetaram filipódios (Fig.
10A) e lamelipódios (Fig. 10C). Observou-se o ectoplasma e o endoplasma contendo os
grânulos quando corados com Giemsa (Figs. 10B e D). O núcleo mostrou-se excêntrico na
maioria das células, redondo ou oval e com baixa relação núcleo:citoplasma.
Tabela 1. Média (± E. P.) do tamanho celular (µm e unidades arbitrárias) e da porcentagem (± E. P.)
das subpopulações de hemócitos da hemolinfa da ostra Crassostrea gasar. Diferentes letras indicam
diferenças significativas do tamanho entre as diferentes populações celulares. Análises independentes
para cada técnica.
Hemócitos
Giemsa* Citometria* Giemsa Citometria
Tamanho (µm) Tamanho (u.a.) Porcentagem (%) Porcentagem
(%) Nt Np
Granulócito
Grande 31,5 ± 0,31a 369,5 ± 12,96 a 14,4 ± 1,59 6,3 ± 0,79 694 27#
Granulócito
Pequeno 11,9 ± 0,22 c 160,3 ± 4,32 b 4,8 ± 1,10 1,3 ± 0,15 282 27#
Hemócito
Vesicular 23,1 ± 0,24 b 303,1 ± 2,34 a 31,7 ± 2,66 38,2 ± 2,13 528 27#
Hialinócito 10,7 ± 0,13 c 138,7 ± 2,3 b 33,0 ± 2,12 43,8 ± 1,85 529 27#
Células Blásticas 5,4 ± 0,06 d 43,55 ± 0,35 d 16,0 ± 1,53 10,6 ± 1,24 489 27#
* p < 0,0001 (Kruskal-Wallis com post-hoc teste de Dunns); Nt = Número de células medidas por
microscopia óptica; Np = Número de animais usados para cálculo da porcentagem. # 3 animais foram
descartados.

32
Os granulócitos pequenos apresentaram tamanho médio de 11,9 µm (± 0,21; n = 27;
Tab. 1) e foram as células menos abundantes (4,8% ± 1,09; Tab. 1). Os grânulos mostraram-
se abundantes e quando corados com vermelho neutro recobriam o núcleo (Figs. 11A e B), que
foi visualizado quando a célula foi corada com Giemsa. Seus grânulos eram basófilos (Figs.
11A e B). Estas células apresentaram baixa relação núcleo:citoplasma e raramente emitiam
pseudópodes.
Os hemócitos vesiculares apresentaram tamanho médio de 23,1 µm (± 0,24; n = 27;
Tab. 1) com grânulos refringentes (Fig. 12A) e raramente basófilos, com citoplasma levemente
acidófilo (Fig. 12B) e sua porcentagem na hemolinfa foi de 31,7% (± 2,65; Tab. 1). A célula
mostrou dois tipos de formatos após adesão espontânea, circular ou estrelado. O núcleo era
excêntrico ou central e a célula possuía uma baixa relação núcleo:citoplasma. A célula
apresentou projeções na forma de lamelipódios, sendo possível observar o ectoplasma e o
endoplasma (Fig. 12B).
Os hialinócitos apresentaram tamanho de 10,7 µm (± 0,12; n = 27; Tab. 1), sendo este
similar ao dos granulócitos pequenos. O citoplasma não apresentou grânulos, exceto em raras
exceções onde observou-se grânulos basófilos. Este tipo celular foi o mais abundante na
hemolinfa (33,0% ± 2,12; Tab. 1). As células eram geralmente arredondadas ou alongadas e
raramente emitiam projeções citoplasmáticas (Fig. 13A). O núcleo era grande, excêntrico ou
central e possuía escasso citoplasma basófilo (Fig. 13B). Esta célula apresentou alta relação
núcleo:citoplasma (Fig. 13).

33
Figura 10 - Granulócitos grandes da hemolinfa de Crassostrea gasar
Hemócitos corados com vermelho neutro (A e C, contraste de fase) e Giemsa (B e D). Note as projeções
citoplasmáticas do tipo filipódios e lamelipódios (setas largas), o ectoplasma (setas finas), o endoplasma
contendo numerosos grânulos (setas pontilhadas) e os núcleos (pontas de flecha).

34
Figura 11 - Granulócitos pequenos da hemolinfa de Crassostrea gasar
Hemócitos corados com vermelho neutro (A, contraste de fase) e Giemsa (B). Note as projeções
citoplasmáticas do tipo filipódios (seta larga), o ectoplasma (seta fina), o endoplasma contendo
numerosos grânulos (setas pontilhadas) e o núcleo (ponta de flecha).
Figura 7 - Hemócitos vesiculares da hemolinfa de Crassostrea gasar
Hemócitos corados com vermelho neutro (A, contraste de fase) e Giemsa (B). Note as projeções
citoplasmáticas do tipo filipódios e lamelipódios (setas largas), o ectoplasma (setas finas), o endoplasma
contendo alguns grânulos refringentes ou raramente basófilos (setas pontilhadas) e os núcleos (pontas
de flecha).

35
Figura 8 - Hialinócitos da hemolinfa de Crassostrea gasar
Hemócitos corados com vermelho neutro (A, contraste de fase) e Giemsa (B). Note as projeções
citoplasmáticas do tipo filipódios (setas largas), o endoplasma (seta pontilhada) e os núcleos (pontas de
flecha).
As células do tipo-blásticas foram as menores observadas, apresentando um tamanho
médio de 5,4 µm (± 0,05; n = 27; Tab. 1) e porcentagem de 16,0% (± 1,53; Tab. 1). Seu núcleo
era excêntrico, com citoplasma fortemente basófilo e possuia uma alta razão núcleo:citoplasma.
Raramente se observou filipódios em células do tipo-blásticas coradas com vermelho neutro
(Fig. 14).
Figura 9 - Células tipo-blásticas da hemolinfa de Crassostrea gasar
Células tipo-bláticas coradas com vermelho neutro (A, contraste de fase) e Giemsa (B). Observe a
pequena área citoplasmática (seta pontilhada) e o núcleo (ponta de flecha).

36
Caracterização morfológica dos hemócitos por citometria de fluxo
Os citogramas (tamanho vs compexidade interna, respectivamente, FSC vs SSC)
indicaram a presença de cinco subpopulações de hemócitos (Fig. 15; Tab. 1). Os hemócitos
com menor tamanho e granulosidade foram denominados de células tipo-blásticas (Fig. 13.
BL). Duas outras populações celulares com maior tamanho e granulosidade foram classificadas
como hialinócitos (Fig. 15. HH) e hemócitos vesiculares (Fig. 15. HV). As duas populações
com maior granulosidade, mas similares entre si, foram classificados como granulócitos, e estas
apresentaram diferenças em tamanho, correspondendo aos granulócitos grandes (Fig. 15. GG)
e pequenos (Fig. 15. GP).
As células mais frequentes na hemolinfa das ostras foram os hialinócitos e hemócitos
vesiculares e as menos frequentes foram os granulócitos pequenos (Tab. 1). Este resultado
ocorreu tanto para as monocamadas coradas com Giemsa quanto por citometria de fluxo. No
entanto, as proporções celulares obtidas com elas diferiram (G = 9,27; GL = 4) (Tab. 1).

37
Figura 105 - Populações de hemócitos da hemolinfa de Crassostrea gasar observadas por
citometria de fluxo
Citograma morfológico representativo onde o eixo X (FSC-Height) representa o tamanho celular e o
eixo Y (SSC-Height) a granulosidade interna. Destacam–se 5 subpopulações de hemócitos,
denominadas de células tipo-blásticas (BL); hialinócitos (HH); hemócitos vesiculares (HV);
granulócitos grandes (GG) e granulócitos pequenos (GP).
Caracterização da capacidade fagocítica de hemócitos por citometria de fluxo
Os hemócitos da ostra C. gasar foram capazes de fagocitar os três tipos diferentes de
partículas com os quais foram desafiados: látex (Fig. 16A e B), E. coli (Fig. 16C e D) e zymosan
(Fig. 16E e F). No entanto, os hemócitos mostraram uma atividade fagocítica diferencial, ou
seja, as partículas de zymosan (15,1% 0,94), seguidas das partículas de látex (9,7% 0,45)
foram as mais fagocitadas pelos hemócitos quando comparadas com as bactérias E. coli (5,0%
0,54) (p < 0.0001) (Fig. 17).

38
Figura 11 - Fagocitose de diferentes partículas por hemócitos de Crassostrea gasar
Fagocitose de partículas de látex (A e B), E. coli (C e D) e zymosan (E e F) observadas em microscopia
de contraste de fase e (coluna da direita) e em campo escuro (coluna da esquerda). As partículas ou
agregados de partículas fagocitados estão indicadas pelas flechas.

39
Figura 17 - Fagocitose (%) de diferentes partículas pelos hemócitos da ostra Crassostrea gasar
Lát
ex
Zym
osan
E. c
oli
0
5
10
15
20
b
a
c
% d
e h
emóci
tos
As diferentes letras indicam diferenças da fagocitose entre os três tipos de partículas testadas. (ANOVA
One-way; p < 0.0001).
No caso das partículas de látex e de E. coli foi possível analisar a fagocitose diferencial
entre duas populações de hemócitos: os granulócitos e os hialinócitos + hemócitos vesiculares
(Fig. 18). Para as partículas de látex, os granulócitos foram significativamente (teste t; p <
0.0001) mais fagocíticos (40,1% 1,87) que os hialinócitos + hemócitos vesiculares (7,2%
0,36) (Fig. 18A). Ao contrário, em relação as bactérias E. coli, os hialinócitos + hemócitos
vesiculares foram mais fagocíticos (teste t; p < 0.0001) (5,0% 0,80) que os granulócitos (0,3%
0,15) (Fig. 18B).
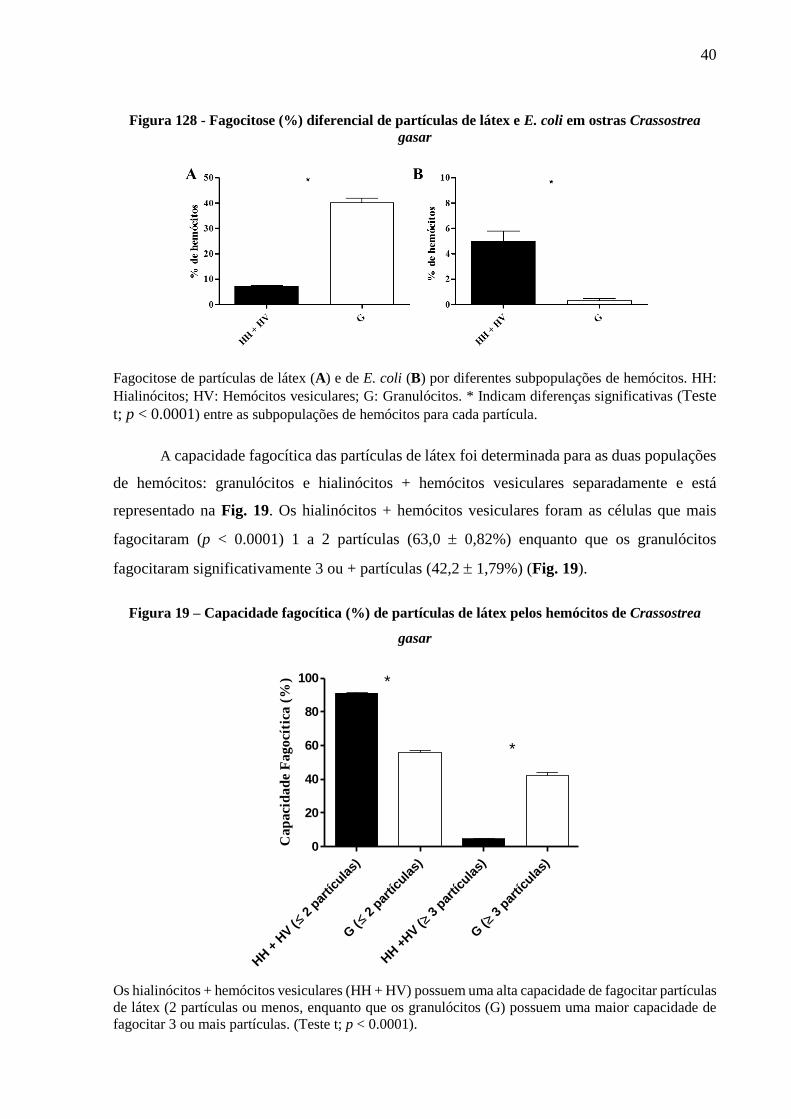
40
Figura 128 - Fagocitose (%) diferencial de partículas de látex e E. coli em ostras Crassostrea
gasar
Fagocitose de partículas de látex (A) e de E. coli (B) por diferentes subpopulações de hemócitos. HH:
Hialinócitos; HV: Hemócitos vesiculares; G: Granulócitos. * Indicam diferenças significativas (Teste
t; p < 0.0001) entre as subpopulações de hemócitos para cada partícula.
A capacidade fagocítica das partículas de látex foi determinada para as duas populações
de hemócitos: granulócitos e hialinócitos + hemócitos vesiculares separadamente e está
representado na Fig. 19. Os hialinócitos + hemócitos vesiculares foram as células que mais
fagocitaram (p < 0.0001) 1 a 2 partículas (63,0 0,82%) enquanto que os granulócitos
fagocitaram significativamente 3 ou + partículas (42,2 1,79%) (Fig. 19).
Figura 19 – Capacidade fagocítica (%) de partículas de látex pelos hemócitos de Crassostrea
gasar
2 par
tícula
s)
HH +
HV (
2 par
tícula
s)
G (
3 p
artíc
ulas)
HH +
HV (
3 p
artíc
ulas)
G (
0
20
40
60
80
100 *
*
Ca
pa
cid
ad
e F
ag
ocí
tica
(%
)
Os hialinócitos + hemócitos vesiculares (HH + HV) possuem uma alta capacidade de fagocitar partículas
de látex (2 partículas ou menos, enquanto que os granulócitos (G) possuem uma maior capacidade de
fagocitar 3 ou mais partículas. (Teste t; p < 0.0001).

41
5. DISCUSSÃO
5.1. Caracterização morfológica dos hemócitos de C. gasar
O presente trabalho representa a primeira caracterização morfológica e funcional dos
hemócitos da ostra C. gasar. Recentemente, nosso grupo de pesquisa fez o primeiro estudo em
células do sistema de defesa (hemócitos) desta espécie (Queiroga et al., 2013) e avaliou algumas
respostas celulares (fagocitose e produção de espécies reativas de oxigênio) frente à infecção
pelo protozoário do gênero Perkinsus.
Na hemolinfa das ostras C. gasar foram identificadas duas populações principais de
hemócitos, os granulócitos e os hialinócitos, já descritas e bem caracterizadas para os bivalves
(Cheng, 1981; Hine, 1999). Queiroga et al. (2013) não tiveram a pretensão de realizar um estudo
minucioso das células de defesa desta ostra, mas igualmente identificaram por citometria de
fluxo as duas principais populações (granulócitos e hialinócitos).
No presente estudo, os hemócitos diferenciaram-se por suas características citoquímicas
e morfológicas, incluindo o tamanho, granulosidade interna e relação núcleo:citoplasma, em
cinco subpopulações: os granulócitos grandes, os granulócitos pequenos, os hialinócitos, os
hemócitos vesiculares e as células do tipo-blásticas.
Subpopulações hemocitárias foram descritas para várias espécies de bivalves (Hine, 1999)
e foram correspondentes as observadas no presente estudo.
Os granulócitos em geral se diferenciam dos demais hemócitos por possuírem inúmeras
vesículas no seu citoplasma, facilmente distinguíveis no citosol por um aspecto granuloso,
quando coradas (associação com corantes) ou não (refringência da luz) e observadas ao
microscópio de luz. Esta característica não é observada em nenhum outro tipo de hemócito
(Hine, 1999). Um estudo com hemócitos do mexilhão M. edulis, mostrou que estas vesículas
comportam diversas enzimas, dentre elas hidrolases ácidas e a lisozima, responsáveis pela
destruição de partículas estranhas ao organismo (Pipe, 1990). Por este motivo, os granulócitos
são considerados o tipo celular mais apto à defesa imune (Pipe, 1990).
Neste estudo não foi feita nenhuma caracterização enzimática, mas foi possível confirmar
a presença das vesículas ácidas coradas no citosol e a maior granulosidade intracelular, que se
associam diretamente com a quantidade de vesículas e organelas celulares. As características
dos granulócitos da ostra C. gasar também foram observadas em outras espécies do gênero,
como em C. virginica (McCormich-Ray & Howarwt, 1991) e C. rhizophorae (Rebelo et al.,
2013) e em berbigões R. decussatus (Prado-Alvarez et al., 2012).

42
Os hialinócitos também foram encontrados na hemolinfa de C. gasar. Este tipo celular foi
caracterizado por possuir alta relação núcleo:citoplasma e pouco ou nenhum grânulo
intracelular. Tais características foram descritas para hemócitos de outros ostreídeos, como a
ostra nativa do Brasil C. rhizophorae (Rebelo et al., 2013), mas também em C. virginica
(Cheng, 2009), C. gigas e O. edulis (Auffret, 1989). Além de outros bivalves, como A. cygnea
(Soares-da-Silva et al., 2002; Salimi et al., 2009) e M. galloprovincialis (Carballal et al., 1997).
Os hemócitos vesiculares encontrados na hemolinfa de C. gasar são semelhantes
morfologicamente aos granulócitos, sendo grandes e apresentando baixa relação
núcleo:citoplasma. Entretanto, diferem destes por não possuírem nenhum ou poucos grânulos
citoplasmáticos, o que foi evidente pelo aspecto refringente mesmo após adição de corantes. Os
hemócitos vesiculares foram descritos também em ostras O. edulis e C. gigas (Auffret, 1989)
e C. rhizophorae (Rebelo et al., 2013).
As células do tipo-blásticas foram as menores células encontradas neste estudo. Possuem
alta relação núcleo:citoplasma e alta basofilia. Essa grande afinidade pelo corante básico é uma
característica que pode ser explicada pela ocorrência de muitos ribossomos livres (ricos em
RNA ribossomal de caráter ácido) (Auffret, 1989; Russell-Pinto et al., 1994; Wen et al., 1994;
Carballal et al., 1997). Estas características estruturais sugerem que as células tipo-blásticas
estão envolvidas no processo hematopoiético (Hine, 1999; Rebelo et al., 2013). Sugere-se que
estas células sejam capazes de maturar na hemolinfa, ou seja, transformar-se morfologicamente
e fisiologicamente em outros tipos hemocitários como os hialinócitos, granulócitos ou
hemócitos vesiculares (Rebelo et al., 2013).
Através da técnica de citometria de fluxo, foi possível identificar e quantificar com
sucesso as subpopulações hemocitárias da hemolinfa de C. gasar. Diversos estudos têm
empregado esta técnica para compreender e elucidar, principalmente, a função dos hemócitos
de outros bivalves, como nos ostreídeos O. edulis (Renault et al., 2001), C. gigas (Lambert et
al., 2003), C. ariakensis (Donaghy et al., 2009), C. rhizophorae (Rebelo et al., 2013), os
venerídeos, como R. decussatus (Prado-Alvarez et al., 2012) e R. philippinarum (Donaghy et
al., 2009b) e A. cygnea (Soares-da-Silva et al., 2002).
Os hialinócitos e hemócitos vesiculares constituíram as subpopulações hemocitárias mais
abundantes. Esta observação foi coincidente tanto por citometria de fluxo quanto por
microscopia óptica e este perfil foi descrito também para a hemolinfa de ostras C. ariakensis
(Donaghy et al., 2009) e C. rhizophorae (Rebelo et al., 2013) e de pectinídeos Argopecten
irradians (Zhang et al., 2006). Sabe-se que a alteração na proporção dos tipos hemocitários está
associada principalmente à susceptibilidade dos animais aos parasitos, como descrito na ostra

43
O. edulis infectada pelo protozoário Bonamia ostreae (Comesaña et al., 2012). Inclusive na
ostra C. gasar foi observado que na infecção avançada pelo protozoário Perkinsus sp., os
parasitos podem induzir ou causar uma alteração nas proporções de hialinócitos e células do
tipo-blásticas (Queiroga et al., 2013). Outros fatores ambientais podem levar à esta alteração,
como a presença de químicos no ambiente, como foi observado no gastrópode Bellamya
bengalensis e no bivalve Lamellidens marginalis (Ray et al., 2013).
Portanto, conhecer os tipos celulares da hemolinfa e as proporções das subpopulações de
hemócitos de uma espécie em condições normais é fundamental para análises futuras do
impacto de agentes biológicos, químicos ou de outra natureza.
5.2. Caracterização da capacidade fagocítica dos hemócitos de C. gasar por citometria
de fluxo
Independentemente do tipo de hemócito, o zymosan e o látex foram as partículas mais
fagocitadas quando comparadas às bactérias E. coli. Isto demonstra que estas células são
capazes de reconhecer diversas moléculas não-próprias. Dentre elas, as que contém β1,3-
glicanos (zymosan), presentes nas paredes de fungos, e algumas moléculas presentes na parede
da bactéria E. coli, como o LPS. Contudo, a baixa fagocitose de E. coli em relação a de outras
partículas pode estar relacionada à sua não-patogenicidade para os bivalves (Tubiash, 1971) ou
estas bactérias podem estar sendo mortas extracelularmente, através do uso de redes (NETs, do
inglês, Neutrophil extracellular traps), e não fagocitadas (Ng et al. 2013). Este fato, pode ainda
estar associado aos tipos de receptores expressos e o tempo de análise (Zhang et al. 2014). Nos
berbigões R. decussatus (Prado-Alvarez et al., 2012) e M. galloprovincialis (García-García et
al., 2008) foi observado resultado semelhante, sendo a fagocitose de bactérias a menor dentre
as partículas testadas.
As bactérias do gênero Vibrio são Gram-negativas e abundantes na água do mar, assim
como a E. coli, mas ao contrário destas, podem ser bastante patogênicas para bivalves (Tubiash,
1971). A infecção de diversas larvas de bivalves por bactérias deste gênero causa anormalidades
na formação da concha, problemas de crescimento e mortalidade (Figueras & Novoa, 2011).
Curiosamente, neste estudo, subpopulações de hemócitos mostraram uma diferente
capacidade fagocítica segundo o tipo de partícula. Os granulócitos foram as células que mais
fagocitaram as partículas de látex, enquanto que os hialinócitos e hemócitos vesiculares foram
as células que mais fagocitaram a E. coli. De forma semelhante ao observado no venerídeo R.
decussatus (Prado-Alvarez et al., 2012) e no ostreídeo C. ariakensis (Donaghy et al., 2009a),

44
os granulócitos foram as células que mais fagocitaram látex, demonstrando que essas células
são mais aptas a fagocitar partículas estranhas ao organismo, como o poliestireno presente nas
microesferas de látex.
Em relação a E. coli, apenas, os resultados em M. galloprovincialis (García-García et
al., 2008) e C. gigas (Terahara et al., 2006) contrapuseram o observado aqui, pois foi visto uma
maior fagocitose dessa bactéria pelos granulócitos. No presente estudo, o fato da ausência de
um número elevado de granulócitos que fagocitam E. coli pode ter ocorrido devido a
degranulação desta célula, como observado em C. gigas (Iizuka et al., 2014), visto que o
hemócito vesicular aparece em grande quantidade na hemolinfa, ou ainda pode ter ocorrido a
formação de armadilhas extracelulares (NETs), o que levaria a morte dos hemócitos (Ng et al.
2013).
A fagocitose é a principal resposta de defesa desencadeada por parasitas de pequeno
tamanho e pode ser inibida ou potencializada por um agente infeccioso. Como verificado em
ostras C. gasar infectadas pelo protozoário Perkinsus sp. (Queiroga et al., 2013) e em Anadara
trapezia infectados com trematódeos (Dang et al., 2013), este mecanismo de defesa pode ser
inibido.

45
6. CONCLUSÕES
A hemolinfa da ostra Crassostrea gasar é composta por cinco subpopulações de
hemócitos: os granulócitos grandes, granulócitos pequenos, hialinócitos, hemócitos
vesiculares e células do tipo-blásticas.
A citometria de fluxo mostrou-se uma técnica útil em estudos de aspectos morfológicos e
funcionais de hemócitos de bivalves.
A maior fagocitose de zymosan e partículas de látex mostra que os hemócitos da espécie
Crassostrea gasar são capazes de reconhecer partículas estranhas ao organismo, como o
β1,3-glucano presente na parede de fungos.
Mesmo sendo considerada não-patogênica para bivalves, a E. coli é fagocitada em grande
parte por hialinócitos e hemócitos vesiculares, indicando que há reconhecimento de
bactérias Gram-negativas pelos hemócitos.
A maior fagocitose de látex (≥ 3 partículas) pelos granulócitos indica que esta é célula
mais responsável pela defesa da ostra.
A maior fagocitose de látex (≤ 2 partículas) por hialinócitos e hemócitos vesiculares indica
que estas células também respondem à estímulos externos.

46
7. REFERÊNCIAS
Allam B., Ashton-Alcox KA, Ford SE (2002) Flow cytometric comparison of haemocytes
from three species of bivalve molluscs. Fish & Shellfish Immunology 13:141–158
Auffret M. (1989) Comparative Study of the hemocytes of two oyster species: the european
flat oyster, ostrea edulis, linnaeus, 1750 and the pacific oyster crassostrea gigas
(thunberg,1793). Journal of Shellfish Research 8:367–373
Bachali S, Jager M, Hassanin A, Schoentgen F, Jollès P, Fiala-Medioni A, Deutsch JS. (2002)
Phylogenetic analysis of invertebrate lysozymes and the evolution of lysozyme function.
Journal of molecular evolution 54:652–64
Barracco MA, Medeiros ID, Moreira FM (1999) Some haemato-immunological parameters in
the mussel Perna perna. Fish & Shellfish Immunology 9:387–404
Barracco MA., Silva PM. da (2008) HEMOLINFA E SISTEMA IMUNE NO MEXILHÃO
Perna perna.
Barth T, Moraes N, Barracco MA. (2005) Evaluation of some hemato-immunological
parameters in the mangrove oyster Crassostrea rhizophorae of different habitats of Santa
Catarina Island, Brazil. Aquatic Living Resources 186:179–186
BD Biosciences (2007) BD FACSCalibur Instructions For Use.
Buggé DM, Hégaret H, Wikfors GH, Allam B (2007) Oxidative burst in hard clam
(Mercenaria mercenaria) haemocytes. Fish & shellfish immunology 23:188–96
Carballal MJ., Lopez MC, Azevedo C, Villalba A (1997) Hemolymph cell types of the mussel
Mytilus galloprovincialis. Diseases of aquatic organisms 29:127–135
Casas SM., Comesaña P, Cao A, Villalba A (2011) Comparison of antibacterial activity in the
hemolymph of marine bivalves from Galicia (NW Spain). Journal of invertebrate
pathology 106:343–5
Chagot D., Comps M., Boulo V (1987) Histological study of a cellular reaction in Ruditpaes
decussatus infected by a protozoan. Aquaculture 67:3–4
Chang S, Tseng S, Chou H (2005) Morphological characterization via light and electron
microscopy of the hemocytes of two cultured bivalves: a comparison study between the
hard clam (Meretrix. Zool Stud 44:144–152
Charlet M, Chernysh S, Philippe H, Hetru C, Hoffman JA, Bulet P (1996) Innate Immunity.
ISOLATION OF SEVERAL CYSTEINE-RICH ANTIMICROBIAL PEPTIDES FROM
THE BLOOD OF A MOLLUSC, MYTILUS EDULIS. Journal of Biological Chemistry
271:21808–21813
Cheng TC (1981) Bivalves. In: Ratcliffe NA, Rowley AF (eds) Invertebrate Blood Cells, 1st
edn. Academic Press, London, p 233–300

47
Cima F, Matozzo V, Marin MG, Ballarin L (2000) Haemocytes of the clam Tapes
philippinarum (Adams & Reeve, 1850): morphofunctional characterisation. Fish &
shellfish immunology 10:677–93
Comesaña P, Casas SM, Cao A, Abollo E, Arzul I, Morga B, Villalba A (2012) Comparison
of haemocytic parameters among flat oyster Ostrea edulis stocks with different
susceptibility to bonamiosis and the Pacific oyster Crassostrea gigas. Journal of
invertebrate pathology 109:274–86
Dang C, Cribb TH., Osborne G, Kawasaki M, Bedin A-S, Barnes AC (2013) Effect of a
hemiuroid trematode on the hemocyte immune parameters of the cockle Anadara trapezia.
Fish & shellfish immunology 35:951–6
Dang C, Tan T, Moffit D, Deboutteville JD, Barnes AC (2012) Gender differences in
hemocyte immune parameters of bivalves: the Sydney rock oyster Saccostrea glomerata
and the pearl oyster Pinctada fucata. Fish & shellfish immunology 33:138–42
Donaghy L, Kim B-K, Hong H-K, Park H-S, Choi K-S (2009a) Flow cytometry studies on the
populations and immune parameters of the hemocytes of the Suminoe oyster, Crassostrea
ariakensis. Fish & shellfish immunology 27:296–301
Donaghy L, Kim B-K, Hong H-K, Park H-S, Choi K-S (2009b) Flow cytometry studies on
the populations and immune parameters of the hemocytes of the Suminoe oyster,
Crassostrea ariakensis. Fish & shellfish immunology 27:296–301
Donaghy L, Lambert C, Choi K-S, Soudant P (2009) Hemocytes of the carpet shell clam
(Ruditapes decussatus) and the Manila clam (Ruditapes philippinarum): Current
knowledge and future prospects. Aquaculture 297:10–24
Eble AF (2009) The circulatory system. In: Kennedy VS, Newell RIE, Eble AF (eds) The
Eastern Oyster Crassostrea virginica, 1st edn. Sea Grant, Maryland, p 271–298
Fang FC. (2004) Antimicrobial reactive oxygen and nitrogen species: concepts and
controversies. Nature reviews Microbiology 2:820–32
Figueras A, Novoa B (2011) Enfermedades de moluscos bivalvos de interés en Acuicultura
Enfermedades de moluscos bivalvos de interés en Acuicultura. Madrid
García-García E, Prado-Alvarez M, Novoa B, Figueras A, Rosales C (2008) Immune
responses of mussel hemocyte subpopulations are differentially regulated by enzymes of
the PI 3-K, PKC, and ERK kinase families. Developmental and comparative immunology
32:637–53
Gonzalez M, Gueguen Y, Desserre G, Lorgeril J de, Romestand B, Bachère E (2007)
Molecular characterization of two isoforms of defensin from hemocytes of the oyster
Crassostrea gigas. Developmental and comparative immunology 31:332–9
Hégaret H, Wikfors GH, Soudant P (2003a) Flow cytometric analysis of haemocytes from
eastern oysters, Crassostrea virginica, subjected to a sudden temperature elevation. Journal
of Experimental Marine Biology and Ecology 293:237–265

48
Hégaret H, Wikfors GH, Soudant P (2003b) Flow-cytometric analysis of haemocytes from
eastern oysters, Crassostrea virginica, subjected to a sudden temperature elevation. Journal
of Experimental Marine Biology and Ecology 293:237–248
Hine P (1999) The inter-relationships of bivalve haemocytes. Fish & Shellfish
Immunology:367–385
Hubert F, Noël T, Roch P (1996) A member of the arthropod defensin family from edible
Mediterranean mussels (Mytilus galloprovincialis). European Journal of Biochemistry
306:302–306
Iizuka M, Nagasaki T, Takahashi KG. (2014) Involvement of Pacific oyster CgPGRP-S1S in
bacterial recognition, agglutination and granulocyte degranulation. Developmental & …
43:30–4
Lambert C, Soudant P, Choquet G, Paillard C (2003) Measurement of Crassostrea gigas
hemocyte oxidative metabolism by flow cytometry and the inhibiting capacity of
pathogenic vibrios. Fish & Shellfish Immunology 15:225–240
Larson K, Roberson B, Hetrick F (1989) Effect of environmental pollutants on the
chemiluminescence of hemocytes from the American oyster Crassostrea virginica.
Diseases of aquatic organisms 6:131–136
Li C, Zhao J, Song L (2009) A review of advances in research on marine molluscan
antimicrobial peptides and. Molluscan Research 29:17–26
Marigómez I, Soto M, Cajaraville MP, Angulo E, Giamberini L (2002) Cellular and
subcellular distribution of metals in molluscs. Microscopy research and technique 56:358–
92
McCormich-Ray MG., Howarwt T. (1991) Morphology and Mobility of Oyster Hemocytes:
Evidence for Seasonal Variations. Journal of Invertebrate Pathology 58:219–230
McHenery J, Allen J, Birkbeck T (1986) Distribution of lysozyme-like activity in 30 bivalve
species. Comparative Biochemistry and … 85:581–584
Mitta G, Vandenbulcke F, Roch P (2000) Original involvement of antimicrobial peptides in
mussel innate immunity. FEBS Letters 486:185–190
Montes JF., Del-Río JA, Durfort M., García-Valero F. (1997) The protozoan parasite
Perkinsus atlanticus elicits a unique defensive response in the clam Tapes semidecussatus.
Parasitology 114:339–349
Ng TH, Chang S-H, Wu M-H, Wang H-C (2013) Shrimp hemocytes release extracellular
traps that kill bacteria. Developmental and comparative immunology 41:644–51
Pipe RK (1990) Hydrolytic enzymes associated with the granular haemocytes of the marine
musselMytilus edulis. The Histochemical Journal

49
Pipe RK (1992) Generation of reactive oxygen metabolites by the haemocytes of the mussel
Mytilus edulis. Developmental & Comparative Immunology 16:111–122
Prado-Alvarez M, Romero a, Balseiro P, Dios S, Novoa B, Figueras a (2012) Morphological
characterization and functional immune response of the carpet shell clam (Ruditapes
decussatus) haemocytes after bacterial stimulation. Fish & shellfish immunology 32:69–78
Queiroga FR, Marques-Santos LF, Hégaret H, Soudant P, Farias ND, Schlindwein AD,
Mirella da Silva P (2013) Immunological responses of the mangrove oysters Crassostrea
gasar naturally infected by Perkinsus sp. in the Mamanguape Estuary, Paraíba state
(Northeastern, Brazil). Fish & shellfish immunology 35:319–27
Queiroga FR, Vianna R, Vieira CB, Farias ND, Silva PM da (2014) Parasites infecting the
cultured oyster Crassostrea gasar (Adanson, 1757) in Northeast Brazil. Parasitology
Ray M, Bhunia AS, Bhunia NS, Ray S (2013) Density shift, morphological damage,
lysosomal fragility and apoptosis of hemocytes of Indian molluscs exposed to pyrethroid
pesticides. Fish & shellfish immunology 35:499–512
Rebelo MDF, Figueiredo E de S, Mariante RM, Nóbrega A, Barros M de, Allodi S (2013)
New insights from the oyster Crassostrea rhizophorae on bivalve circulating hemocytes.
PloS one 8:e57384
Renault T, Xue QG, Chilmonczyk S (2001) Flow cytometric analysis of European flat oyster,
Ostrea edulis, haemocytes using a monoclonal antibody specific for granulocytes. Fish &
shellfish immunology 11:269–74
Robinson JM (2008) Reactive oxygen species in phagocytic leukocytes. Histochemistry and
cell biology 130:281–97
Rohlf FJ., Sokal RR. (1981) Statistical tables. In: Statistical tables, 2nd edn. New York, p
737–738
Russell-Pinto F, Reimão R, Sousa M De (1994) Haemocytes in Cerastoderma edule
(Mollusca, Bivalvia)- distinct cell types engage in different responses to sheep
erythrocytes. Fish & Shellfish Immunology 4:383–397
Salimi L, Jamili S, Motalebi A, Eghtesadi-Araghi P, Rabbani M, Rostami-Beshman M (2009)
Morphological characterization and size of hemocytes in Anodonta cygnea. Journal of
invertebrate pathology 101:81–5
Schleder DD, Kayser M, Sühnel S, Ferreira JF, Rupp GS, Barracco M a. (2008) Evaluation of
hemato-immunological parameters during the reproductive cycle of the scallop Nodipecten
nodosus in association with a carotenoid-enriched diet. Aquaculture 280:256–263
Schmitt P, Duperthuy M, Montagnani C (2011) Immune responses in the Pacific oyster
Crassostrea gigas an overview with focus on summer mortalities. In: Qin JG (ed) Oysters.p
1–47

50
Schmitt P, Rosa RD, Duperthuy M, Lorgeril J de, Bachère E, Destoumieux-Garzón D (2012)
The Antimicrobial Defense of the Pacific Oyster, Crassostrea gigas. How Diversity may
Compensate for Scarcity in the Regulation of Resident/Pathogenic Microflora. Frontiers in
microbiology 3:160
Smolowitz RM, Miosky D, Reinisch CL (1989) Ontogeny of leukemic cells of the soft shell
clam. Journal of invertebrate pathology 53:41–51
Soares-da-Silva IM, Ribeiro J, Valongo C, Pinto R, Vilanova M, Bleher R, Machado J (2002)
Cytometric, morphologic and enzymatic characterisation of haemocytes in Anodonta
cygnea. Comparative biochemistry and physiology Part A, Molecular & integrative
physiology 132:541–53
Song L, Wang L, Qiu L, Zhang H (2010) Invertebrate immunity. In: Söderhäl K (ed)
Invertebrate Immunity, 1st edn. Springer Science+Business Media, Austin, p 44–65
Soudant P, Chu FLE, Villalba A, Cancela L (2008) Bivalves-Perkinsus spp. interactions. In:
Villalba A (ed) Workshop for the analysis of the impact of Perkinsosis to the European
Shellfish Industry. Vilanova de Arousa
Soudant P, E Chu F-L, Volety A (2013) Host-parasite interactions: Marine bivalve molluscs
and protozoan parasites, Perkinsus species. Journal of invertebrate pathology 114:196–216
Takahashi K, Mori K, Nomura T (1986) Occurrence and characterization of lysozyme in the
marine bivalves . Bulletin of the Japanese Society of …:2007
Terahara K, Takahashi KG, Nakamura A, Osada M, Yoda M, Hiroi T, Hirasawa M, Mori K
(2006) Differences in integrin-dependent phagocytosis among three hemocyte
subpopulations of the Pacific oyster “Crassostrea gigas.” Developmental and comparative
immunology 30:667–83
Tubiash HS (1971) Soft-Shell Clam, Mya arenaria, a Convenient Laboratory Animal for
Screening Pathogens of Bivalve Mollusks. Applied Microbiology 22:321–324
Vargas-Albores F, Barracco M (2001) Mecanismos de defensa de los moluscos bivalvos con
énfasis en pectínidos. Los Moluscos Pectínidos de Iberoamérica: Ciencia y
Acuicultura:127–146
Wang Y, Hu M, Chiang MWL, Shin PKS, Cheung SG (2012) Characterization of
subpopulations and immune-related parameters of hemocytes in the green-lipped mussel
Perna viridis. Fish & shellfish immunology 32:381–90
Wen C-M, Kou G-H, Chen S-N (1994) Light and electron microscopy of hemocyte of the
hard clam, Meretrix lusoria (Röding). Comparative biochemistry and physiology Part A,
Molecular & integrative physiology 108A:279–286
Wootton EC, Pipe RK (2003) Structural and functional characterisation of the blood cells of
the bivalve mollusc, Scrobicularia plana. Fish & Shellfish Immunology 15:249–262

51
Xue Q, Renault T (2001) Monoclonal antibodies to European flat oyster Ostrea edulis
hemocytes: characterization and tissue distribution of granulocytes in adult and developing
animals. Developmental and comparative immunology 25:187–94
Zhang T, Qiu L, Sun Z, Wang L, Zhou Z, Liu R, Yue F, Sun R, Song L (2014) The
specifically enhanced cellular immune responses in Pacific oyster (Crassostrea gigas)
against secondary challenge with Vibrio splendidus. Developmental and comparative
immunology 45:141–50
Zhang W, Wu X, Wang M (2006) Morphological, structural, and functional characterization
of the haemocytes of the scallop, Argopecten irradians. Aquaculture 251:19–32
Zhao J, Song L, Li C, Ni D, Wu L, Zhu L, Wang H, Xu W (2007) Molecular cloning,
expression of a big defensin gene from bay scallop Argopecten irradians and the
antimicrobial activity of its recombinant protein. Molecular immunology 44:360–8


















