
UNIVERSIDADE FEDERAL DO AMAPÁ PRÓ-REITORIA DE … · O milagre da morte O vento, as cidades e as...
105
LEIDIANE LEÃO DE OLIVEIRA INTERAÇÕES DA ESTRUTURA DA VEGETAÇÃO COM A TOPOGRAFIA E SOLO NA FLORESTA NACIONAL DO AMAPÁ MACAPÁ-AP 2012 UNIVERSIDADE FEDERAL DO AMAPÁ PRÓ-REITORIA DE PESQUISA E PÓS-GRADUAÇÃO PROGRAMA DE PÓS-GRADUAÇÃO EM BIODIVERSIDADE TROPICAL UNIFAP/IEPA/EMBRAPA/CI
Transcript of UNIVERSIDADE FEDERAL DO AMAPÁ PRÓ-REITORIA DE … · O milagre da morte O vento, as cidades e as...

LEIDIANE LEÃO DE OLIVEIRA
INTERAÇÕES DA ESTRUTURA DA VEGETAÇÃO COM A TOPOGRAFIA E
SOLO NA FLORESTA NACIONAL DO AMAPÁ
MACAPÁ-AP
2012
UNIVERSIDADE FEDERAL DO AMAPÁ
PRÓ-REITORIA DE PESQUISA E PÓS-GRADUAÇÃO
PROGRAMA DE PÓS-GRADUAÇÃO EM BIODIVERSIDADE TROPICAL
UNIFAP/IEPA/EMBRAPA/CI

i
LEIDIANE LEÃO DE OLIVEIRA
INTERAÇÕES DA ESTRUTURA DA VEGETAÇÃO COM A TOPOGRAFIA E
SOLO NA FLORESTA NACIONAL DO AMAPÁ
Tese apresentada ao Programa de Pós-
graduação em Biodiversidade Tropical do
convênio Universidade Federal do Amapá
(UNIFAP), Instituto de Pesquisas Científicas
e Tecnológicas do Amapá (IEPA), Empresa
Brasileira de Pesquisa Agropecuária
(EMBRAPA-Amapá) e Conservação
Internacional do Brasil (CI-Brasil), como
parte dos requisitos para obtenção do título
de Doutor em Biodiversidade Tropical.
Área de Concentração: Biodiversidade
Linha de Pesquisa: Gestão e Conservação
da Biodiversidade
Orientador: Prof. Dr. Alan Cavalcanti da
Cunha
MACAPÁ-AP
2012

ii
Dados Internacionais de Publicação na Catalogação (CIP)
Biblioteca da Embrapa Amapá
Oliveira, Leidiane Leão de
Interações da estrutura da vegetação com a topografia e solo na Floresta Nacional
do Amapá / Leidiane Leão de Oliveira; Orientador Alan Cavalcanti da Cunha.
Macapá, 2012.
103 f.
Tese (Doutorado) - Fundação Universidade Federal do Amapá – Programa de
Pós-Graduação em Biodiversidade Tropical.
1. Vegetação. 2. Floresta Nativa. 3. Biomassa. 4. Biodiversidade Tropical. 5.
Amapá. 6. Amazônia brasileira. I. Cunha, Alan Cavalcanti da Cunha, Orient.. II.
Fundação Universidade Federal do Amapá. III. Título.
CDD 634.92098116

iii
LEIDIANE LEÃO DE OLIVEIRA
INTERAÇÕES DA ESTRUTURA DA VEGETAÇÃO COM A, TOPOGRAFIA,
SOLO E HIDROLOGIA NA FLORESTA NACIONAL DO AMAPÁ
__________________________________________
Orientador: Prof. Dr. Alan Cavalcanti da Cunha
Universidade Federal do Amapá – UNIFAP/CCA/PPGBIO
__________________________________________
Examinador: Prof. Dr. Antonio Carlos Lôla da Costa
Universidade Federal do Pará – UFPA/CG/UM
__________________________________________
Examinador: Prof. Dr. Cristoph Jaster
Instituto Chico Mendes de Biodiversidade - ICMBIO
__________________________________________
Examinador: Prof. Dr. Silas Mochiutti
Empresa Brasileira de Pesquisa Agropecuária - EMBRAPA
__________________________________________
Examinador: Profa. Dra. Helenilza F. Albuquerque Cunha
Universidade Federal do Amapá - UNIFAP/CCA/PPGBIO

v
Sempre…
Em algum lugar, uma voz chama
Do fundo do meu coração
Que eu possa sempre sonhar
Os sonhos que tocam meu coração
Tantas lágrimas de tristeza
Infinitas lágrimas rolaram
Mas sei que do outro lado
Encontrarei você
Toda vez que caímos no chão
Olhamos para o céu lá no alto
E acordamos para o seu azul
Como se fosse a primeira vez
Como o caminho é longo e solitário
E não enxergamos o fim
Posso abraçar a luz
Com meus dois braços
Quando digo adeus meu coração pára
Com ternura eu sinto
Que meu corpo silencioso
Passa a ouvir o que é verdadeiro
O milagre da vida
O milagre da morte
O vento, as cidades e as flores
Todos nós dançamos numa só unidade
Em algum lugar, uma voz chama
Do fundo do meu coração
Continue sonhando seus sonhos
Não os deixe morrer
Por que falar de sua melancolia
Ou dos tristes pesares da vida
Deixe seus lábios cantarem
Uma linda canção para você
Não esqueceremos a voz sussurrante
Em cada lembrança ela ficará
Para sempre, para guiar você
Quando um espelho quebra
Estilhaços se espalham pelo chão
Lampejos de uma vida nova
Refletem se por toda parte
Janela de um recomeço
Quietude, nova luz da aurora
Deixe que meu corpo vazio e silente
Seja preenchido e nasça outra vez
Não é preciso procurar lá fora
Nem velejar através do mar
Porque brilha aqui dentro de mim
Está bem aqui dentro de mim
Encontrei uma luz
Que estará sempre comigo
Yumi Kimura (compositora)
Canção tema da animação “A viagem de Chihiro”

vi
DEDICATÓRIA
A minha amada família: Minha filha Mariana e
meu marido Cledinaldo.
Em especial aos meus pais Terezinha de Jesus e
Valdo Mendes e as minhas irmãs, Erika e Luciane.

vii
AGRADECIMENTOS
A Deus, fonte de fé, energia e sabedoria, causa primária de todas as coisas.
Este trabalho não seria possível sem o apoio financeiro do Programa em
Pesquisa em Biodiversidade Tropical – PPBio/AP (2009-2010) “Protocolo do clima”.
Ao PROCAD/CAPES – Novas Fronteiras 2007. Programa de Pós-Graduação em
Biodiversidade Tropical - PPGBIO. Ao Conselho Nacional de Desenvolvimento
Científico e Tecnológico – CNPq pela bolsa concedida.
A EMBRAPA-AP, pelo empréstimo de material para as coletas de solo e pelas
análises feitas em seu Laboratório.
Aos analistas ambientais do ICMBIO/FLONA-AP pelo apoio logístico em
campo.
A Willian Magnusson “Bill” pelo apoio financeiro para a compra do trado para a
instalação dos poços piezométrico, e pelo incentivo para escolha do tema, área de
estudo e amostragem. Na reta final, também contribuiu no segundo capítulo em forma
de artigo, quando participei da oficina de artigos, proporcionada pelo PPGBIO, onde o
Bill foi palestrante.
A Claudia Keller pelo financiamento da última expedição para coleta de dados.
Flávia Regina Costa, que tem o “dom” de descomplicar as coisas, contribuiu de
maneira significativa para o avanço do segundo capítulo da tese, orientando melhor
minhas perguntas e gerando ajuda nas análises estatísticas e dos gráficos.
A todas as inúmeras pessoas (pesquisadores, bolsistas, voluntários, auxiliares de
campo) que se envolveram no trabalho de campo, desde a implantação das “polêmicas”
parcelas de curva de nível, a instalação dos poços piezométricos e coletores de
precipitação interna, coleta dos dados topográficos e de solo até a medição das árvores,
palmeiras e lianas. Formando assim uma super equipe de campo, com destaque
merecido a:
Graciliano Galdino que foi o responsável em coordenar a equipe para o
estabelecimento das parcelas de curva de nível. Também pela obtenção dos dados de
inclinação do terreno. Além de tudo contribuiu com inúmeras discussões positivas a
respeito dos rumos da tese. Durante o tempo de convivência no campo e fora do campo
aprendi a ter alguém que sempre pude contar. Nem que seja para escutar seus famosos
jargões.
Paulo Paiva responsável pelas coletas de amostras deformadas de solo. Pelas
discussões metodológicas e estatísticas na reta final. Principalmente pela amizade

viii
concedida pelo casal “Dani e Paulo” e pelos momentos de alegria e descontração ao
longo dos anos.
A minha segunda família, com quem passei mais tempo no começo do estudo:
Luiz Carlos, Seu Galvão, Dona Joana, Ocicleide, Cíntia, Paquinha, Seu Zé, Cledinaldo,
Clemilson Seu Conte (in memorian), que além do apoio concedido, vou levar
recordações desde o bolinho da Dona Joana, passando pelas piadas do seu Galvão, até
as estórias do Seu Conté (in memorian).
Ao meu orientador Alan Cunha que durante esses anos não desistiu pela
orientação, pelas oportunidades científicas, pela troca de conhecimento, pela total
liberdade e confiança em mim depositada e por final sua amizade.
A Coordenadora do PPGBIO Helenilza Cunha, pelo seu esforço a frente da
coordenação, pelos cursos e encontros oferecidos, principalmente pela oficina de artigos
e as ajudas para as viagens de campo e pelo apoio burocrático.
A Rejane Peixoto, eterna secretária do PPGBIO, que além dos tramites
burocráticos que fazem a vida acadêmica funcionar, me ofereceu sua amizade e alegria.
A Cledinaldo Marques, hoje meu marido que desde o começo não poupou
esforços em me ajudar, principalmente durante as minhas malárias e a gravidez, onde,
ficou responsável em manter as coletas de campo. Sempre acreditou em minha
capacidade, é meu maior incentivador. Obrigada pelo amor e dedicação oferecidos a nós
duas.
Aos meus amigos que de perto ou de longe sempre torcem por mim: Daniel,
Nete, Edmir, Jonathan, Marcia, Dani, Claudinha. Estes que em alguma fase da tese me
ajudaram sorrir.
Aos meus alunos da UEAP, Daniele, Edielem, Roney e Elielson que durante a
minha gravidez me deram o pique de campo que a Mariana já vinha me tirando. Eles
ajudaram nas últimas medições das árvores de DAP >30 cm e coleta de precipitação
interna.
A minha família, marido, pai, mãe, irmãs, sogro, sogra e cunhada, por serem o
esteio da minha vida, meus verdadeiros amigos, por estarem ao meu lado nas horas de
alegria e de tristeza, pelo incentivo à conclusão do doutorado, pelo auxílio nos momentos de
fraqueza, pelo companheirismo, pela confiança. Principalmente ao meu marido e cunhada
por cuidarem com amor da minha filha, para que eu pudesse estudar e por toda dedicação a
mim concedida.

ix
A minha filha Mariana, por entender minha ausência, falhas e sempre se
comportar e principalmente por me receber sempre com um lindo sorriso. Hoje, minha
maior fonte de inspiração, meu amor incondicional.
Eu sou eternamente agradecida por esta oportunidade, por ter passado tanto
tempo em campo (floresta), pelo que aprendi, pelas pessoas que conheci, pelos
problemas para ir a campo e do campo que aprendi a resolver. Finalmente aprendi com
cada pessoa e problema que convivi nestes últimos anos, tudo isso, em parte, serviu para
ser responsável pelo que sou hoje. Assim expresso os meus mais profundos
agradecimentos a todos que de alguma forma contribuíram direta ou indiretamente para
a realização deste trabalho.

x
LISTA DE FIGURAS
Capítulo I
Figura 1 Localização da Floresta Nacional do Amapá e da grade do PPBIO. ....................... 33
Figura 2 Grade do PPBIO em mapa de hidrografia com as parcelas de curva de
nível numeradas. .................................................................................................... 34
Figura 3 Distribuição das faixas de amostragem para a vegetação na parcela de
curva de nível......................................................................................................... 37
Figura 4 Distribuição da densidade de árvores por hectare em diferentes classes
diamétricas (A). Variação da densidade de árvores por hectare por
parcela (B) nas 30 parcelas de curva de nível do PPBIO na FLONA do
Amapá .................................................................................................................... 42
Figura 5 Distribuição da área basal de árvores por hectare em diferentes classes
diamétricas (A). Variação da área basal de árvores por hectare por
parcela (B) nas 30 parcelas de curva de nível do PPBio na FLONA do
Amapá. ................................................................................................................... 43
Figura 6 Distribuição da biomassa de árvores por hectare por parcela (A).
Distribuição de freqüência da biomassa arbórea (B) Variação da
biomassa de árvores por hectare em diferentes classes diamétricas (C)
nas 30 parcelas de curva de nível do PPBio na FLONA do Amapá. .................... 45
Figura 7 Distribuição da densidade (A) área basal (B) e biomassa (C) de palmeiras
por hectare em diferentes classes diamétricas nas 30 parcelas de curva de
nível do PPBio na FLONA do Amapá. ................................................................. 49
Figura 8 Distribuição da densidade (A) área basal (B) e biomassa (C) de lianas por
hectare em diferentes classes diamétricas nas 30 parcelas de curva de
nível do PPBio na FLONA do Amapá. A última classe representa a
densidade e a área basal total de árvores (DAP≥1). .............................................. 51
Capítulo II
Figura 1 - Localização da FLONA do Amapá e das parcelas (pontos) instaladas ao
longo das trilhas de acesso..................................................................................... 71
Figura 2. Distribuição de freqüência da biomassa total em 30 parcelas 1ha na
FLONA do Amapá. ............................................................................................... 82
Figura 3 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa árborea total (árvores, palmeiras e lianas) e os gradientes de (a)
Altitude, (b) Inclinação e (c) Soma de Bases – SB. .............................................. 85
Figura 4 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa árborea total (árvores, palmeiras e lianas) e os gradientes de (a)
Altitude, (b) inclinação. ......................................................................................... 86

xi
Figura 5 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa árborea total (árvores, palmeiras e lianas) e os gradientes de (a)
Argila, (b) Soma de Bases – SB. ........................................................................... 87
Figura 6 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa de árvores e os gradientes de (a) Altitude, (b) Inclinação e (c)
Soma de Bases – SB. ............................................................................................. 88
Figura 7 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa árvores e os gradientes de (a) Altitude, (b) inclinação. ......................... 89
Figura 8 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa árvores e os gradientes de (a) Argila, (b) Soma de Bases - SB. ............ 90
Figura 9 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa palmeiras e os gradientes de (a) altitude, (b) inclinação. ....................... 91
Figura 10 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa palmeiras e os gradientes de (a) Argila, (b) Soma de Bases -
SB. ......................................................................................................................... 92
Figura 11 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa de Palmeiras e os gradientes de (a) Altitude, (b) Inclinação e
(c) Soma de Bases – SB. ........................................................................................ 92

xii
SUMÁRIO
RESUMO GERAL ......................................................................................................... 14
GENERAL ABSTRACT ................................................................................................ 15
INTRODUÇÃO GERAL ............................................................................................... 16
REFERÊNCIAS ............................................................................................................. 21
CAPITULO I .................................................................................................................. 25
RESUMO ....................................................................................................................... 26
ABSTRACT ................................................................................................................... 27
1. INTRODUÇÃO .......................................................................................................... 28
2. MATERIAL E MÉTODOS ........................................................................................ 31
2.1. Área e período de estudo ..................................................................................... 31
2.2. Delineamento amostral ........................................................................................ 33
2.3. Dados de Estrutura da vegetação ......................................................................... 36
2.4. Cálculo da densidade, área basal e biomassa ...................................................... 39
3. RESULTADOS .......................................................................................................... 41
3.1. Densidade, área basal e biomassa de árvores ...................................................... 41
3.2. Densidade, área basal e biomassa de Palmeiras .................................................. 48
3.3. Densidade, área basal e biomassa de lianas ......................................................... 50
4. DISCUSSÃO .............................................................................................................. 52
4.1. Densidade, área basal – o que representam ......................................................... 52
4.2. Como a biomassa varia entre as parcelas na FLONA do Amapá e sua
contribuição no contexto local e regional ................................................................... 52
4.3. Contribuição de palmeiras para as estimativas de biomassa viva acima do solo 55
4.4. Contribuição das lianas para as estimativas de biomassa viva acima do solo ..... 55
4.5. A variação de biomassa depende do tamanho da árvore ..................................... 56
5. CONSIDERAÇÕES FINAIS ..................................................................................... 58
6. REFERÊNCIAS ......................................................................................................... 59
CAPITULO II ................................................................................................................. 64
RESUMO ....................................................................................................................... 65
ABSTRACT ................................................................................................................... 66
1. INTRODUÇÃO .......................................................................................................... 67
2. MATERIAL E MÉTODOS ........................................................................................ 70
2.1. Área de estudo, delineamento amostral e dados de estrutura da vegetação ........ 70
2.2. Dados de solo e topografia .................................................................................. 72
2.2.1. Solo ............................................................................................................... 72

xiii
2.2.2. Topografia .................................................................................................... 73
2.3. Estimativas de biomassa arbórea viva acima do solo .......................................... 73
2.4. Análise dos dados ................................................................................................ 75
3. RESULTADOS .......................................................................................................... 76
3.1. Topografia e gradientes do solo .......................................................................... 76
3.2. Gradientes do solo e sua relação com a topografia ............................................. 78
3.3. Estimativas de biomassa vegetal viva acima do solo .......................................... 80
3.4. Relações entre a biomassa vegetal viva acima do solo e as variáveis topográficas,
edáficas ....................................................................................................................... 83
3.4.1. Biomassa total .............................................................................................. 83
3.4.2. Árvores ......................................................................................................... 87
3.4.3. Palmeiras ...................................................................................................... 90
3.4.4. Lianas ........................................................................................................... 92
3.4.5. As diferentes classes de tamanhos de árvores e a relação entre suas estimativas
de biomassa e as características do solo e topografia ................................................. 93
4. DISCUSSÃO .............................................................................................................. 95
4.1. A variação de biomassa total acima do solo na FLONA do Amapá foi explicada
pela topografia e textura do solo ................................................................................ 95
4.2. Textura e a altitude do terreno e a sua relação com a variação espacial da
biomassa viva acima do solo ...................................................................................... 96
4.3. A relação da variação da biomassa com os gradientes do solo e topografia
dependem do tamanho das árvores ............................................................................. 97
4.4. A variação na quantidade de biomassa neste estudo é afetada pela textura do solo
e topografia comparando com os obtidos para Amazônia Central ............................. 98
5. CONSIDERAÇÕES FINAIS ..................................................................................... 99
2.8. REFERÊNCIAS .................................................................................................... 100

14
Resumo Geral
As variações de estrutura e biomassa vegetal são alvos de estudos nos últimos anos devido aos
temas de interesses globais, como mudanças climáticas, desenvolvimento sustentável e
estoque de carbono (crédito de carbono). A floresta Amazônica desempenha um papel
importante no ciclo global de carbono, porém ainda são incertas suas estimativas de
quantidade e distribuição estocado na floresta. A variação espacial das estimativas de
biomassa arbórea viva (árvores, palmeiras e lianas) acima do solo (AGLB) e suas relações
com o solo (química e física), topografia (altitude e inclinação) e hidrologia (precipitação
interna) foram investigadas em parcelas permanentes do Programa de Pesquisa em
Biodiversidade – PPBio na Floresta Nacional do Amapá – FLONA - AP. Foram instaladas 30
parcelas compridas (250 m) e estreitas (40 m), que seguiram as curvas de nível do terreno,
cobrindo toda variação topográfica existente na grade. Indivíduos com Diâmetro à Altura do
Peito (DAP) ≥ 30 cm foram amostrados em áreas de 1 ha e 0,5 ha para indivíduos com 1 ≤
DAP < 30 cm. Análises químicas e físicas do solo foram realizadas em amostras coletadas em
três pontos de cada parcela na profundidade de 0 – 40 cm. As medidas de altitude de cada
parcela foram obtidas por meio de georreferenciamento do ponto inicial de cada parcela. A
altitude foi calculada através de imagens do modelo digital de elevação do terreno derivados
do radar interferométrico SRTM (Shutter Radar Topographic mission). A inclinação de cada
parcela representa uma média de cinco medidas a cada 50 m e ao longo do eixo da parcela. O
peso seco das árvores foi obtido a partir de uma combinação de equações alométricas
utilizando o DAP ≥ 1 cm de árvores, palmeiras e lianas. A estimativa de AGLB variou de
194,6 a 968,4 Mg.ha-1
(média de 530,5 Mg ha-1
). Árvores com DAP > 100 cm contribuíram
com 21,7% da estimativa de biomassa de árvores por parcela. A textura do solo e topografia
explicam 30% da variação da AGLB FLONA-AP. Os resultados obtidos para FLONA-AP
indicam que o poder preditivo das variáveis edáficas e topográficas, na variação espacial da
biomassa arbórea, poderá ser utilizado para melhorar as estimativas de carbono estocado em
florestas Amazônicas.
Palavras-chave: Biomassa, solo, topografia, precipitação interna Amazônia Oriental.

15
General Abstract
Changes in structure and biomass are the target of studies in recent years due to global
concerns such as climate change, sustainable development and carbon (carbon credit). The
forest plays an important role in the carbon global cycle, but estimates are still uncertain of
the amount and distribution of carbon stored in forest. The spatial variation of aboveground
arboreal live biomass (tress, palms and lianas) and its relationship to soil (chemical and
physical) and topography (altitude and slope) were studied in permanent plots in Program
Biodiversity Research - PPBio in Amapá National Forest - FLONA – AP 30 plots that were
installed long (250 m) and narrow (40 m), and thin and followed isoclines, in order to sample
most of the variation in soil and topography found in FLONA – AP. The width of each plot
varied depending on the class of diameter at breast height (DBH) sampled. Individuals with
DBH ≥ 30 cm were sampled in 1 ha whereas subsamples of 0,5 and 0,1 ha were used for
individuals with 10 ≤ DBH < 30 cm and 1 ≤ DBH < 10 cm, respectively. Chemical and
physical analyses were undertaken using soil samples collected in the Ah horizon (0-40 cm
deep), in each plot. Measures altitude of each plot was obtained by means of georeferencing
of the starting point of each parcel as the parcel follow the contour, the altitude value is
constant throughout the length of the parcel. The altitude was calculated using images of the
digital elevation model of the terrain derived from radar interferometry SRTM (Shutter Radar
Topographic Mission). The slope of each plot was represented by the mean value of 5
consecutive measurements taken at intervals of 50 m along the length of the plot. The dry
weight of trees, palms e lianas was estimated by a combination of allometric equations using
individuals with DBH ≥ 1 cm. The AGLB estimates ranged from 194,6 a 968,4 Mg.ha-1
(mean 530,5 Mg ha-1
). Trees with DBH> 100 cm contributed 21,7% of the estimated biomass
of trees per plot. The soil texture and topography explain 30% of the variation of AGLB
FLONA-AP. The results obtained for FLONA-AP indicate that the predictive power of soil
characteristics and topography, the spatial variation in tree biomass, can be used to improve
estimates of carbon stored in Amazonian forests.
Keywords: Biomass, soil, topography, precipitation inside the eastern Amazon.

16
Introdução Geral
Um dos ecossistemas mais ricos em biodiversidade e mundialmente ameaçados são as
florestas tropicais. Alguns estudos em ecologia vegetal realizados em florestas tropicais, ou
em outras classes de vegetação no mundo, investigam as relações existentes entre a
distribuição das populações de espécies arbóreas e as variáveis ambientais que possam afetar
tal distribuição. (AUSTIN e GAYWOOD, 1994; CASTILHO et al., 2006; ENGELBRECHT
et al., 2007; RENNÓ et al., 2008).
A Amazônia é a maior floresta tropical do mundo, com uma área total de
aproximadamente 7 milhões de Km². Representa cerca de 60% do território brasileiro. Sua
importância ambiental, social e econômica é incontestável. Em termos quantitativos de água,
a região abriga cerca de 20% da disponibilidade mundial de água doce (MANZI, 2008,
BISPO, 2007). Quanto à sua dimensão, a Amazônia representa 56% das florestas tropicais do
planeta, sendo que somente a parcela brasileira contém cerca de 40% das florestas tropicais
restantes no mundo e dotadas de um papel vital na manutenção da biodiversidade, clima e
hidrologia regional, bem como na manutenção do estoque terrestre de carbono (HOUGHTON
et al., 2000; LAURANCE et al., 2001, MANZI, 2008).
É consenso que a biomassa florestal desempenha um papel importante no ciclo global
do carbono, principalmente nos dias atuais, cuja relevância é maior, devido aos debates acerca
do papel que as florestas desempenham sobre as mudanças climáticas globais. Biomassa
florestal ou fitomassa é a quantidade, expressa em unidade de massa, do material lenhoso
contido em uma unidade de área da floresta (ARAÚJO, HIGUCHI e CARVALHO, 1999).
Em florestas tropicais os reservatórios de biomassa viva acima do solo (AGBL)
desempenham um papel importante no ciclo global de carbono, sendo responsável por uma
porção significativa de armazenamento de carbono e estoque de nutrientes (PHILIPS et al.,
1998; BROWN et al, 1995).
A floresta Amazônica conta com um estoque de carbono de aproximadamente 100
bilhões de toneladas, equivalente a aproximadamente 14 vezes as emissões anuais globais
pela utilização de combustíveis fósseis (carvão mineral, petróleo e gás natural) e 60 vezes as
emissões anuais por desflorestamento em todo planeta. Além disso, é importante observar que
a capacidade que a vegetação apresenta em absorver carbono está muito intimamente
associada aos padrões espaço-temporais dos regimes de precipitação (ARAGÃO et al., 2008;
SOMBROCK, 2001, MALHI et al., 2006).

17
Nesse cenário, o Estado do Amapá tem contribuição significativa no esforço mundial,
devido aos serviços ambientais prestados, pois é o Estado (em termos percentuais) mais
protegido na faixa tropical, com 10.476.117 de hectares, ou 72% do seu território, em 12
unidades de conservação e cinco terras indígenas (SILVA et al., 2007). Os serviços
ambientais prestados pelas florestas têm valor econômico que precisa ser remunerado
(SOARES, 2007). Um dos maiores desafios do Estado do Amapá é o desenvolvimento
sustentável através do conhecimento de suas riquezas e a valoração dos serviços ambientais
prestados pela preservação de suas áreas.
As Florestas Nacionais são Unidades de Conservação (UC) pertencentes à categoria de
Uso Sustentável. Entretanto, uma condição preliminar ao estabelecimento de programas de
uso é a formulação e aprovação do Plano de Manejo da Unidade (BRASIL, 2000). Os
resultados deste trabalho também poderão colaborar com o Plano de Manejo da Floresta
Nacional do Amapá – FLONA-AP, uma vez que as informações abióticas associadas à
ecologia das florestas são importantes para a tomada de decisão no manejo florestal, pois
podem fortalecer as técnicas de exploração a apontar para estudos que garantam práticas
florestais sustentáveis.
O entendimento das funções que os ecossistemas florestais desempenham através da
interação água – solo – vegetação – atmosfera motivam diversas pesquisas na floresta
amazônica. Contudo, as informações ainda são escassas, especialmente na região equatorial.
A biodiversidade de espécies florestais e a riqueza mineral da região amazônica têm chamado
atenção do mundo inteiro. Mas a maior preocupação recai nos possíveis impactos que o
desmatamento nessa região causaria sobre o clima do planeta (PHILLIPS et al., 2009;
FERREIRA DA COSTA, 2008; MALHI, 2010). Por essa razão, a manutenção dos
ecossistemas amazônicos, como a floresta em pé e os ciclos hidrológicos, representam
importantes contribuições à mitigação do aquecimento global, principalmente nesta fase de
esforço mundial para redução de Gases do Efeito Estufa – GEE (MALHI, 2010).
Grande parte das incertezas nas estimativas de fluxo de carbono nas regiões tropicais é
conseqüência de inconsistências nas estimativas de biomassa da Floresta Amazônica
(KELLER, PALACE e HURTT, 2001; HOUGHTON et al., 2001). A principal dificuldade
para modelar a distribuição da biomassa florestal é que as parcelas onde a biomassa foi
medida geralmente são muito pequenas e não foram estabelecidas aleatoriamente através da
paisagem (KELLER, PALACE e HURTT, 2001). Alem disso, muitos estudos que relacionam

18
a variação de biomassa com as variáveis topográficas e do solo foram realizados em pequenas
escalas amostrais, que não abrangem a variabilidade da área (da SILVA et al., 2002;
VASCONCELOS et al., 2003). A implantação de parcelas permanentes, para estudos de
longo prazo em florestas tropicais, oferece um grande potencial para monitorar os estoques de
carbono acima do solo (MALHI e GRACE, 2000). A quantidade de carbono estocada na
vegetação pode ser obtida através de estimativas de biomassa.
Para entender o efeito das mudanças climáticas sobre a floresta e das perdas de floresta
sobre o clima são necessários estudos no nível de comunidades e em escala regional (FIELD,
2001), que levem em conta as variações espaciais de estrutura e estoque de biomassa da
floresta (QUESADA et al., 2009).
O Programa de Pesquisa em Biodiversidade (PPBio) foi um dos programas instituídos
pelo governo brasileiro como “[...] estratégia de investimento em Ciência, Tecnologia e
Informação que priorize e integre competências em diversos campos do conhecimento, gere,
integre e dissemine informações sobre biodiversidade que possam ser utilizadas para
diferentes finalidades” (PPBIO, 2006, p.2).
Protocolos padronizados para pesquisa de diversos grupos taxonômicos e variáveis
ambientais foram desenvolvidos com o objetivo de serem executados na forma de inventários
rápidos associados a estudos ecológicos de longa duração (RAPELD) e possibilitar um
diagnóstico dos ambientes estudados tanto em escalas locais como em escalas mais amplas
(MAGNUSSON et al., 2005). Unidades de Conservação (UC) e locais de relevância em
termos de biodiversidade têm sido escolhidas para o estabelecimento de grades de trilhas
constituindo um sítio de pesquisa. A Floresta Nacional (FLONA) do Amapá foi uma das áreas
escolhidas pelo PPBio na Amazônia Oriental.
O PPBio, núcleo Amapá é desenvolvido em uma área de 25 km² na FLONA do
Amapá, onde foram instaladas 30 parcelas de 1 ha para avaliar a variação de biomassa viva
acima do solo. As parcelas obedeceram ao método RAPELD (MAGNUSSON et al., 2005).
Este método consiste na distribuição sistemática de parcelas na paisagem, o qual permite
estimativas não tendenciosas da distribuição, abundância e biomassa das espécies em cada
sítio, além de comparações biogeográficas entre sítios. Este desenho minimiza a variação
interna de topografia e solo em cada parcela e permite o uso destas variáveis como preditoras
das distribuições de espécies (MAGNUSSON et al., 2005).

19
Na Amazônia Central a relação da biomassa de árvores e lianas com o solo e
topografia depende do tamanho do diâmetro (CASTILHO et al., 2006; NOGUEIRA, 2006).
Porém, ainda é necessário determinar grupos de tamanho de acordo com as relações entre a
biomassa com as características do solo. A textura do solo foi altamente associada com a
topografia na Amazônia central (LUIZÃO et al., 2004; CASTILHO et al., 2006). Nessa região
geralmente solos argilosos ocorrem em áreas mais altas e solos arenosos ocorrem em áreas
mais baixas (CHAVEAL et al., 1987).
Baseados nestes resultados a variação de biomassa viva acima do solo pode ser
explicada apenas com dados topográficos (altitude e inclinação) (CASTILHO et al., 2006).
Estes dados podem ser obtidos com facilidade através de mapas topográficos disponibilizados
para a Amazônia ou podem ser derivados do modelo digital de elevação (DEM). Estes
modelos são obtidos através do radar Shuttle Radio Topographic Mission (SRTM),
disponibilizados pela NASA. Na Amazônia oriental ainda são restritos os estudos que
relacionam a distribuição espacial de biomassa e a sua relação com o solo e topografia.
Segundo o INPE/MCT (2011) o Amapá é o Estado com o menor índice de
desmatamento da Amazônia Legal, nos último 7 anos, contando ainda com mais de 90% dos
seus ecossistemas naturais intactos, e considera a biodiversidade como um dos seus mais
importantes recursos naturais para a promoção do desenvolvimento social e econômico. O
Estado detém uma área de 143.537 km2, possuindo uma biodiversidade extraordinária,
estimada em mais de 400.000 espécies, muitas das quais são endêmicas (SILVA et al., 2007).
Diante deste cenário de preservação e das políticas públicas concernentes ao uso
sustentável da floresta no Estado do Amapá, tais como o manejo florestal sustentável, o uso
dos recursos florestais não madeireiros e turismo ecológico, é necessário realizar estimativas
de biomassa viva acima do solo. Além disso, também é necessário explicar sua relação
segundo as características ambientais quantificáveis e representadas por gradientes físicos e
químicos do solo. Este tipo de informação é de fundamental importância, pois pode ser
utilizada como base para realizar projeções da dinâmica da floresta e a sua relação espacial
com o solo e topografia e futuramente alimentar inventários de carbono (NASCIMENTO e
LAURANCE, 2002/ CASTILHO et al, 2006; QUESADA et al., 2009).
Esta tese foi dividida em dois capítulos com o objetivo de aperfeiçoar a análise e
exposição dos resultados derivados da pesquisa "interações da estrutura da vegetação com a
topografia e solo na FLONA do Amapá": O primeiro trata da distribuição espacial da

20
estrutura da vegetação incluindo árvores, palmeiras e lianas na FLONA do Amapá. Foi
definida e discutida a área basal, densidade e biomassa. No segundo capítulo se discute a
relação entre o padrão de distribuição de biomassa vegetal e as variáveis do solo (física e
química) e topografia (altitude e inclinação).

21
Referências
ARAGÃO, L.E.O.C. et al. Interactions between rainfall, deforestation and fires during recent
years in Brazilian Amazonia. Philosophical Transactions of the Royal Society – Biological
Sciences, n. 363, p. 1779-1985, 2008.
ARAÚJO, T.M.; HIGUCHI, N.; CARVALHO, J.A. Comparison of formulae for biomass
content determination in a tropical rain Forest site in the state of Pará, Brazil. Forest Ecology
and Management, n.117, p. 43-52, 1999.
AUSTIN, M.P.; GAYWOOD, M.J. Current problems of environmental gradients and species
response curves in relation to continuum theory. Journal of Vegetation Science, v. 5, n. 4, p.
473-482, 1994.
BISPO, C.J.C. Balanço de água em cultivo de soja no Leste da Amazônia. Dissertação
(mestrado em ciências ambientais). Universidade federal do Pará/Embrapa/Museu Emílio
Goeldi. Belém-PA. 148 p. 2007.
BRASIL. Lei nº 9.985, de 18 de julho de 2000. Diário Oficial República Federativa do
Brasil, Poder Executivo, Brasília, DF, 19 jul. 2000.
BROWN, I.F., et al. Uncertainty in the biomass of Amazonian forests: an example from
Rondonia, Brazil. Forest Ecol. Manag., n. 75, p. 175–189. 1995.
CASTILHO, C.V. et al. Variation in aboveground tree live biomass in a central Amazonian
forest: effects of soil and topography. Forest Ecology and Management, n. 234, p. 85-96,
2006.
CHAUVEL, A.; LUCAS, Y.; BOULET, R. On the genesis of the soil mantle of the region of
Manaus, Central Amazonia, Brazil. Experientia, n. 43, p. 234-241, 1987.
da SILVA, R.P.; SANTOS, J.; TRIBUZY, E.S.; CHAMBERS, J.Q.; NAKAMURA, S.,
HIGUCHI, N. Diameter increment and growth patterns for individual tree growing in Central
Amazon, Brazil. Forest Ecology and Management, v.166, p.295-301, 2002.
ENGELBRECHT BMJ, COMITA LS, CONDIT R, KURSAR TA, TYREE MT, TURNER
BL, HUBBELL SP. Drought sensitivity shapes species distribution patterns in tropical forests.
Nature 447: 80–82, 2007.
FERREIRA DA COSTA, R. Efeito da exclusão parcial da chuva na transpiração de
espécies vegetais da FLONA de Caxiuanã (Projeto ESECAFLOR-LBA), Pará. 2008.

22
96p. Tese (Doutorado em Meteorologia), Universidade Federal de Campina Grande, Campina
Grande.
FIELD, C. B.. Plant physiology of the ‘missing’ carbon sink. Plant Physiology, v. 125, n. 25–
28, 2001.
HOUGHTON, R. A.; LAWRENCE, K. T.; HACKLER, J. L.; BROWN, S. The spatial
distribution of forest biomass in the Brazilian Amazon: a comparison of estimates. Global
Change Biology, v. 7, p. 731-746, 2001.
HOUGHTON, R.A., SKOLE, D.L., NOBRE, C.A., HACKLER, J.L., LAWRENCE, K.T.,
CHOMENTOWSKI, W.R. Annual fluxes of carbon from deforestation and regrowth in the
Brazilian Amazon. Nature 403:301-304, 2000.
INPE/MCT. Estimativa da Taxa de Desmatamento da Amazônia no período de 2010-
2011. Brasília - DF, 2011. Disponível em www.obt.inpe.br/prodes/Prodes_Taxa2011.pdf.
Acessado em: 21 de jan. de 2012.
KELLER. M.; PALACE, M.; HURTT, G. Biomass estimation in the Tapajos National Forest,
Brazil – examination of sampling and allometric uncertainties. Forest Ecology and
Management, v. 154, p.371–382, 2001.
LAURANCE, W.F.; ALBERNAZ, A.K.M.; DA COSTA, C. Is deforestation accelerating in
the Brazilian Amazon? Environmental Conservation, 28(4):305-311, 2001.
LUIZÃO, R.C.C.; LUIZÃO, F.J.; PAIVA, R.Q.; MONTEIRO, T.F.; SOUSA, L.S.; KRUIJT,
B. Variation of carbon and nitrogen cycling processes along a topographic gradient in a
Central Amazonian forest. Global Change Biology, v. 10, p. 592-600, 2004.
MAGNUSSON, W.E. et al. A modification of the Gentry method for biodiversity surveys in
long-term ecological research sites. Biota Neotrop. v.2, n.5 , p. 1–6, 2005.
MALHI, Y. e GRACE, J.. Tropical forests and atmospheric carbon dioxide. Trends in
Ecology and Evolution, v. 15, n. 8, p. 332-337, 2000.
MALHI, Y. et al. The regional variation of aboveground live biomass in old-growth
Amazonian forests. Global Change Biology, n.12, p.1107–1138, 2006.
MALHI, Y. The carbon balance of tropical forest regions, 1990-2005. Current Opinion in
Sustainability Science, v. 2, n. 4, p. 237-244, 2010.

23
MANZI, A.O. Aquecimento global, Mudanças climáticas e o futuro da Amazônia.
GEAA: Grupo de Estudos estratégicos Amazônicos/ Mudanças Climáticas; Água no
mundo moderno; Biodiversidade Amazônica. Manaus: Editora INPA, 2008.
NASCIMENTO, H.E.M. e LAURANCE, W.F. Total aboveground biomass in central
Amazonian rainforest: a landscape-scale study. Forest Ecology and Management, n. 168: p.
311-321, 2002.
NOGUEIRA, A. Variação da densidade, área basal e biomassa de lianas em 64 km² de
floresta de terra firme na Amazônia Central. Dissertação de mestrado, INPA/UFAM.
Manaus, 2006.
PHILLIPS O.L.et al. Drought sensitivity of the Amazon rainforest. Science, n. 323, p. 1344-
1347, 2009.
PPBIO: Programa de Pesquisa em Biodiversidade/Amazônia. Documento Técnico do
Programa. Manaus: INPA/MPEG, 30 p. 2006.
QUESADA, C. A. et al. Effect of soils on forest structure and dynamics in Amazonia.
Biogeosciences Discuss., v. 6, p. 3993–4057, 2009.
QUESADA, C.A. et al. Effect of soils on forest structure and dynamics in Amazonia.
Biogeosciences Discuss., v. 6, p. 3993–4057, 2009.
RENNÓ, C.D., NOBRE, A.D., CUARTAS, L.A., SOARES, J.V., HODNETT, M.G.,
TOMASELLA, J., WATERLOO, M.J.. HAND, a new terrain descriptor using SRTM-DEM:
Mapping terra-firme rainforest environments in Amazonia. Remote Sensing of Environment
v. 112, p. 3469-3481, 2008.
SILVA, J.M.C. et al. O corredor de biodiversidade do Amapá. In:Corredor de
Biodiversidade do Amapá – Biodiversity Corridor. Belém, CI-Brasil, 54 p., 2007.
SOARES, T.J. O efeito estufa e o clima global. In: Efeito Estufa: a Amazônia e os aspectos
legais. Manaus: Editora INPA, 171p., 2007.
SOMBROCK, W. G. Spatial and temporal patterns of Amapá rainfall. Consequences for the
planning of agricultural occupation and the protection of primary forest. Ambio v. 30, n. 7, p.
388-396, 2001.

24
VASCONCELOS, H.L.; MACEDO, A.C.C.; VILHENA, J.M.S. Influence of topography on
the distribution of ground-dwelling ants in an Amazonian Forest. Studies on Neotropical
Fauna and environment, v. 38 n. 2, p. 115-124, 2003.

25
CAPITULO I
VARIAÇÃO ESPACIAL DA DENSIDADE, ÁREA BASAL E BIOMASSA VEGETAL EM
25 KM² DE FLORESTA DE TERRA - FIRME NA AMAZÔNIA ORIENTAL¹
Leidiane Leão de Oliveira
Alan Cavalcanti da Cunha
¹Scientia Plena ISSN: 1808-2793

26
Resumo
Na Amazônia Brasileira existem poucos trabalhos em meso-escala considerando a variação
espacial de estrutura e biomassa arbórea viva acima do solo (AGLB). O objetivo foi
quantificar a variabilidade espacial da biomassa na Floresta Nacional do Amapá (FLONA-
AP). Foram amostradas 30 parcelas de 1 ha, distribuídas por 25 km², em uma reserva de terra
- firme da Amazônia oriental. A densidade e área basal de árvores, palmeiras e lianas foram
quantificadas em cada uma das parcelas, e estimativas de biomassa foram realizadas com
diferentes equações alométricas. A área basal média de árvores, palmeiras e lianas,
respectivamente, foi de 256.518,6 cm².ha-1
; 6.837,2 cm².ha-1
e 5.914,3 cm².ha-1
. A densidade
média de árvores, palmeiras e lianas, respectivamente, foi de 389,4 ind.ha-1
; 91,3 ind.ha-1
e
470 ind.ha-1
. A biomassa média de árvores, palmeiras e lianas, respectivamente, foi de 523,9
Mg.ha-1
(variando de 177,6 a 966,4); 0,6 (variando de 0 a 14) e 6 Mg.ha-1
(variando de 1,9 a
11,7). A biomassa média total (árvores + palmeiras + lianas) foi de 530,5 Mg.ha-1
(variando de
194,6 a 968,4) e foi similar a outros estudos na região. Os maiores valores de biomassa de
árvores, palmeiras e lianas estão diretamente relacionados com os maiores valores de área
basal para a FLONA do Amapá. As últimas classes de diâmetro concentram os maiores
valores de biomassa de árvores e lianas. Por outro lado, as primeiras classes de diâmetro
concentram os maiores valores de biomassa de palmeiras. Na FLONA do Amapá a biomassa
de árvores representou 98,5%, enquanto palmeiras e lianas representaram, respectivamente,
0,3% e 1,3% da biomassa total. Este estudo mostrou que a variabilidade de biomassa, cerca de
80% de variação de valores extremos de biomassa, se deve à utilização da amostragem em
parcelas que incorporaram grandes áreas de variação do ambiente florestal-, o que detecta a
grande heterogeneidade ao longo da paisagem. Assim, podemos fazer extrapolações
confiáveis sobre uma área maior, sem superestimar a biomassa.
Palavras chave: Biomassa viva acima do solo, área basal, densidade, Amazônia Oriental

27
Abstract
There are few research on mesoscale considering the spatial variation of structure and
aboveground arboreal live biomass (AGBL). The objective of this investigation was to
quantify the spatial variability of biomass in the National Forest of Amapá (FLONA-AP). In a
25 km² of Terra-Firme forest Reserve in Oriental Amazon, 30 plots of 1 ha were distributed.
The density and basal area of trees, palms and lianas were quantified in each of the plots, and
biomass estimates were performed with different allometric equations. The average basal area
of trees, palms and lianas, respectively, was 256.518,6 cm².ha-1
, 6837,2 cm².ha-1
and 5914,3
cm².ha-1
. The average density of trees, palms and lianas, respectively, were 389,4 ind.ha-1
;
91.3 ind.ha-1
and 470 ind.ha-1
. The average biomass of trees, palms and lianas, respectively,
was 523,9 Mg.ha-1
(ranging from 177,6 to 966,4), 0,6 (ranging from 0 to 14) and 6 Mg.ha-1
(ranging from 1,9 to 11,7). The average total biomass (trees + palms + lianas) was 530,5
Mg.ha-1
(ranging from 194,6 to 968,4) and was similar to other studies in the region. Os
higher biomass values of trees, palms and lianas are directly related to the higher values of
basal area for the National Forest of Amapá. The final diameter classes concentrate the higher
biomass of trees and lianas. While the first diameter classes concentrate the higher biomass of
palms. In National Forest Amapá tree biomass accounted for 98.5%, while palm trees and
lianas represent respectively 0,3% and 1,3% of total biomass. This study showed great
variation of biomass, about 80% variation of extreme values of biomass, this is because the
use of the sampling parts incorporating major areas of variation of the environment forest,
which detects the heterogeneity along the landscape. Thus, we can make reliable
extrapolations over a larger area, without overestimating the biomass.
Key words: Aboveground tree live biomass, basal area, density, Amazonia oriental.

28
1. Introdução
A floresta Amazônia é diferenciada por sua extensa [área e principalmente por sua
megabiodiversidade e seu potencial econômico madeireiro (FRANCEZ et al., 2007).
Contudo, esta riqueza natural vem sofrendo pressão antrópica traduzida pela explora;’ao
desordenada de madeira e expansão dos sistemas agrícolas e outras formas de uso do solo
(FURTADO, 1997). Estas atividades antropog ênicas causam mudanças de cobertura e uso do
solo e representam a liberação dos estoques de carbono contidos na vegetação para a
atmosfera (FEARNSIDE, 1996).
Nos últimos 250 anos as taxas de aumento nos níveis de CO2 na atmosfera têm
crescido sem precedentes, partindo de 280 p.p.m. antes da revolução industrial, para 367
p.p.m. em 1999 (PRENTICE et al., 2001). Com isso, a taxa de aumento de temperatura na
Amazônia nas últimas décadas tem sido de aproximadamente 0,25o C por década, o que pode
ser considerada uma taxa elevada, já que no fim do último período glacial a atmosfera
amazônica esquentou apenas cerca de 0,1o C por século (MALHI e WRIGHT, 2004; BUSH et
al,. 2004). Além da alta taxa de aumento de temperatura, têm havido mudanças na
precipitação, particularmente na estação seca, que provavelmente será o determinante mais
importante para o destino da Amazônia frente às mudanças climáticas (MALHI et al., 2008).
Tais mudanças no clima podem diminuir a qualidade dos tecidos das plantas (modificando a
razão C:N nas folhas e galhos) e reduzir as taxas de produção de biomassa em florestas
tropicais. O estresse fisiológico causado por secas pode ainda levar a uma queda na
produtividade florestal e aumento da mortalidade de árvores (CLARK, 2004). Além dos
efeitos do clima sobre a floresta tropical. Por outro lado, a perda de florestas tropicais também
pode alterar o clima regional e global (MALHI et al., 2008; MALHI e WRIGHT, 2004).
Para entender o efeito das mudanças climáticas sobre a floresta e das perdas de floresta
sobre o clima são necessários estudos no nível de comunidades e em escala regional (FIELD,
2001), que levem em conta as variações espaciais de estrutura e estoque de biomassa da
floresta (QUESADA et al., 2009). Com efeito, estima-se que a principal conseqüência dessas
mudanças ocorra sobre o padrão de precipitação de chuvas, particularmente na estação seca,
que provavelmente serão o determinante mais importante para o destino da Amazônia frente
às mudanças climáticas (MALHI et al. 2008). O estresse fisiológico causado por secas pode
ainda levar a uma queda na produtividade florestal e aumento da mortalidade (CLARK et al.
2004). Além das secas mais rigorosas e prolongadas na Amazônia, a taxa de aumento de

29
temperatura na Amazônia nas últimas décadas tem sido de aproximadamente 0,25o C por
década, o que pode ser considerada uma elevada taxa, já que no fim do último período glacial
a atmosfera amazônica esquentou apenas cerca de 0,1o C por século (MALHI e WRIGHT,
2004; BUSH et al. 2004).
A importância de estimativas acuradas do estoque de biomassa florestal é que dão
origem às estimativas de emissão de carbono, resultantes das mudanças de cobertura e uso da
terra. Incertezas a respeito do estoque de biomassa florestal é a maior fonte de dúvidas nas
estimativas de emissão de carbono (FEARNSIDE 1996; BAKER et al. 2004; HOUGHTON,
2005; SAATCHI et al., 2007; SILVA, 2011).
Por seu turno, os inventários florestais podem contribuir efetivamente para
negociações do carbono no mercado internacional. Segundo o Intergovernmental Panel on
Climate Change - IPCC (2007) e a Global Forest Resources Assessments/ Food and
Agriculture Organization of the United Nations FRA/FAO o carbono poderá ser usado como
mercadoria a ser negociada sob o Protocolo de Quioto e transformar-se em importante valor
agregado para viabilizar o manejo florestal na Amazônia brasileira (SILVA, 2007; WUNDER
et al., 2008; SILVA, 2011).
Com base nesses argumentos, duas lacunas do conhecimento em estudos ecológicos
realizados na Amazônia têm sido consideradas como relevantes. Em primeiro lugar, apenas
um grupo vegetal (árvores) tem sido mais investigado. Isso se deve ao fato das árvores terem
maior importância econômica e praticidade de obtenção em inventários florestais
(FRANCEZ, CARVALHO e JARDIM, 2007). Talvez, por estes motivos, tende-se a desprezar
o fato de que a estrutura da floresta é formada também por outros grupos vegetais
importantes. As palmeiras e as lianas são bons exemplos. Portanto, é possível observar que
estes últimos são grupos importantes como fontes de observação da estrutura de vegetação e,
principalmente, para o correto manejo da floresta (SCHNITZER, DEWALT e CHAVE, 2006;
NASCIMENTO e LAURENCE, 2002; CASTILHO et al., 2006).
A segunda lacuna notada é com relação aos métodos adotados para estimar a biomassa
vegetal. Estes métodos abrangem áreas pequenas (< 1ha), muitas vezes selecionadas de
maneira não aleatória, ou seja, por métodos sistemáticos, e nem sempre são áreas
representativas de floresta tropical não perturbada. Já outros estudos abordam grandes áreas,
que não são replicáveis. Em síntese, este tipo de estudo não tem sido garantido o bastante,

30
pois generaliza atributos dos ecossistemas sem uma base confiável para sua extrapolação
(CASTILHO et al., 2006; LANDEIRO e MAGNUSSON, 2011).
As características de estrutura e biomassa vegetal foram alvos de estudos nos últimos
anos devido aos interesses globais, tais como, mudanças climáticas, desenvolvimento
sustentável e estoque de carbono (crédito de carbono). As estimativas de biomassa de um
ecossistema podem ser obtidas por métodos diretos (destrutivos) ou indiretos (não
destrutivos). O Método Direto é mais acurado e eficaz, mas metodologicamente apresenta
dificuldades na pesagem de todas as árvores de uma determinada área e, em geral, trabalhos
que o utilizam baseiam-se em poucas e pequenas parcelas, que não foram escolhidas de
maneira aleatória, normalmente não sendo representativas ou economicamente inviáveis
(SANTOS, MIRANDA e TOURINHO, 2004).
O Método indireto para estimativa de biomassa vegetal é mais rápido, não corta, não
pesa e nem seca nenhum indivíduo, além de permitir amostrar uma área maior e maior
número de indivíduos. As vantagens é que se utiliza de variáveis mais facilmente obtidas no
campo, como o diâmetro a 1,30 m acima do solo (diâmetro à altura do peito - DAP). O DAP é
inserido na equação alométrica representativa para aquele ambiente, no qual se deriva o valor
de biomassa. Contudo, como qualquer outro método indireto, está sujeito a erros de
quantificação que nem sempre são mencionados (HIGUCHI et al., 1998; HAIRIAH et al.,
2001; MALHI, 2010).
Considerando o processo de conhecimento da estrutura da vegetação em uma floresta
não perturbada e sua importante ciclagem de carbono, além do reduzido número de estudos
realizados sobre florestas nas unidades de conservação do Estado do Amapá, esta pesquisa
apresenta os seguintes objetivos: Estudar a estrutura espacial da vegetação e da biomassa
vegetal em uma área de 25 km². Neste aspecto, identificou-se padrões de distribuição da
vegetação devido aos mosaicos gerados pelos diferentes tipos de solo, topografia e
disponibilidade de água.
Além disso, apresentou-se a variação da estrutura da vegetação para três categorias ou
grupos de vegetal: árvores, palmeiras e lianas, todas na extensão de 25 km². Para tanto foram
estabelecidas os seguintes questionamentos:
(1) Ocorre variação espacial na estrutura da vegetação (árvore, palmeira e lianas) entre as 30
parcelas de curva de nível (25 km²)?
(2) Como se correlacionam biomassa e área basal de árvores, palmeiras e lianas?

31
(3) Como a biomassa se distribui ao longo das classes de diâmetro para árvores, palmeiras e
lianas ao longo das trinta parcelas?
(4) Qual é a contribuição percentual da estimativa de biomassa vegetal de árvores, palmeiras e
lianas?
Assim, de acordo com as perguntas levantadas, para a obtenção das respostas foram
elaborados as seguintes etapas metodológicas.
2. Material e Métodos
2.1. Área e período de estudo
O estudo experimental foi desenvolvido na Floresta Nacional (FLONA) do Amapá
durante o período de outubro de 2009 a agosto de 2010. O ambiente é definido como uma
floresta tropical úmida de terra firme e inserida totalmente na Bacia Hidrográfica do Rio
Araguari (OLIVEIRA et al., 2010).
A FLONA do Amapá foi criada pelo Decreto-Lei Federal nº 97.630, de 10 de abril de
1989. Está localizada no centro do Estado e vizinha ao Parque Nacional (PARNA) Montanhas
do Tumucumaque, com área estimada de 412.000 ha (Figura 1.1).
A área de estudo na FLONA do Amapá agrega importantes ecossistemas vegetais,
dentre eles a floresta densa de terra-firme e áreas alagadas (igapós). Fitofisionomicamente
constitui-se de uma formação luxuriante, densa, com árvores altas e emergentes, e ricas em
espécies de valor comercial como o acapu, maçaranduba e angelim. A altura máxima do
dossel encontrada foi em torno de 50 m e o diâmetro à altura do peito (DAP) máximo foi de
267,4 cm, enquanto Pereira et al. (2007) encontrou em expedições realizadas em áreas dentro
da FLONA do Amapá um DAP máximo 133,69 cm.
Inserida no Escudo das Guianas, a FLONA do Amapá apresenta vegetação bastante
diversa, predominante de florestas tropicais do tipo Ombrófila Densa, com a existência de
manchas de outros tipos de vegetação circundados por estas florestas. Dentre estas tipologias
podem ser mencionadas formações de floresta menos densa, manchas de tabocais, capoeiras
e de vegetações adaptadas aos afloramentos rochosos. A vegetação ao longo dos rios e
principais corpos d’água pode ser inundada durante os períodos de cheia (SIMONIAN et al.,
2010).

32
A FLONA possui áreas com ocupação humana. Estas se concentram na parte Sul, nos
rios Araguari e Falsino, constituindo onze ocupações típicas de ribeirinhos que desenvolvem
atividades características tais como agricultura de coivara, a pesca e a caça. Alguns produzem
farinha de mandioca para comércio em Porto Grande e vilarejos próximos (SIMONIAN et al.,
2010).
No destaque da Figura 1 observamos a grade do Projeto de Pesquisa em
Biodiversidade - PPBIO. Esta grade é padronizada e alocada em diferentes pontos da
Amazônia. Na FLONA do Amapá é constituída de um sistema de trilhas e a grade tem área de
25 km2. A unidade amostral empregada pelo PPBIO consiste em uma grade de 1x1 km,
formada por 12 trilhas de 5 km de comprimento, totalizando 25 km2 de área. As trilhas são
paralelas e se entrecruzam a cada 1 km. As trilhas são definidas e marcadas com um sistema
GPS geodésico. A cada 100 m de trilha é fixado um marco de PVC informando a coordenada
no sistema de trilhas. Nesses pontos se toma medidas topográficas de altitude. A cada km do
lado esquerdo da trilha de trilha, se estabelece uma parcela de amostragem, em curva de nível,
com 250 m de comprimento. A largura da parcela é variável, em função do grupo biológico a
ser amostrado.
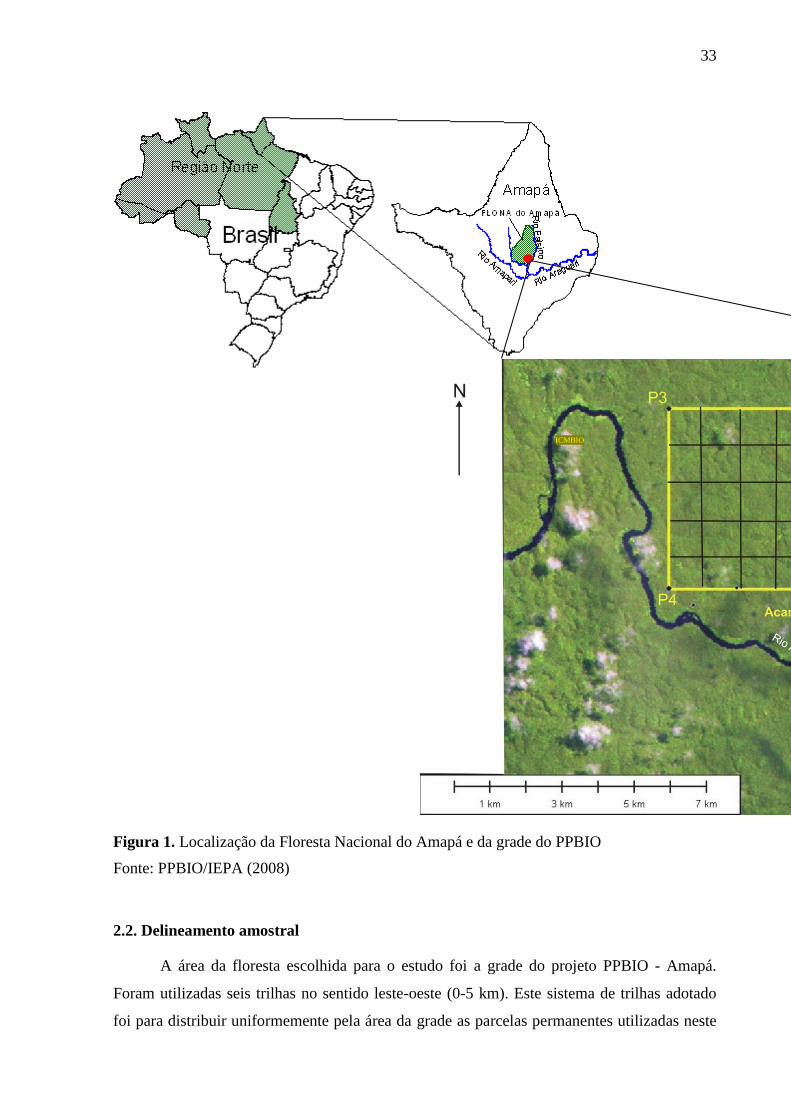
33
Figura 1. Localização da Floresta Nacional do Amapá e da grade do PPBIO
Fonte: PPBIO/IEPA (2008)
2.2. Delineamento amostral
A área da floresta escolhida para o estudo foi a grade do projeto PPBIO - Amapá.
Foram utilizadas seis trilhas no sentido leste-oeste (0-5 km). Este sistema de trilhas adotado
foi para distribuir uniformemente pela área da grade as parcelas permanentes utilizadas neste
ICMBIO

34
estudo. Foram estabelecidas cinco parcelas/trilha, ao longo de cada uma das seis trilhas que
correm no sentido leste-oeste (0-5 km), totalizando 30 parcelas. As parcelas foram
estabelecidas em intervalos de 1 km, sempre do lado esquerdo da trilha. A posição do início
da parcela foi definida arbitrariamente, seguindo a curva de nível do terreno, a partir de uma
distância mínima de 10 m da trilha, para evitar perturbações (ver detalhes da Figura 2).
Figura 2 Grade do PPBIO em mapa de hidrografia com as parcelas de curva de nível
numeradas
Fonte: Adaptada de PPBIO/IEPA (2008)
250 m

35
As parcelas utilizadas neste estudo têm 250 m de comprimento e seguiram uma curva
de nível do terreno (isoclina), determinando parcelas com formas variáveis. Algumas parcelas
curvaram-se e cruzaram a trilha principal, enquanto outras se curvaram e passaram em cima
de parte da parcela já demarcada. Nestes casos, foi aumentado a extensão da parcela para
compensar a área cortada pela trilha ou onde ocorreu sobreposição. A vantagem deste sistema
é que parcelas estreitas e orientadas ao longo de uma curva de nível tendem a manter as
condições de solo relativamente homogêneas (CASTILHO, 2004; MAGNUSSON et al.,
2005; LANDEIRO e MAGNUSSON, 2011). Nessas parcelas as variáveis ambientais podem
ser estimadas com maior precisão do que em parcelas quadradas que captam maior variação
interna de solos e topografia (TOLEDO, 2009).
A linha central das parcelas não é reta, mas segue a curva de nível do terreno na
altitude do ponto de intersecção da parcela com a trilha. Isso permite usar a altitude como
variável preditiva. O método de amostragem RAPELD (Programa de Avaliação Rápida de
Pesquisas Ecológicas de Longa-Duração). Baseia-se nessa premissa. Ele é a adaptação de dois
métodos já existentes, conforme descrito a seguir.
Segundo Magnusson et al. (2005) a filosofia por trás do método é a de maximizar a
probabilidade de amostrar adequadamente às comunidades biológicas. Para isso, são
necessárias áreas amostrais grandes, e ao mesmo tempo minimizar a variação nos fatores
abióticos que afetam estas comunidades, o que requer amostragem de áreas pequenas. Para
obter sucesso neste tipo de procedimento, as parcelas são longas e estreitas, com seu maior
eixo orientado ao longo das curvas de nível do terreno. Este desenho minimiza a variação
interna de topografia e solo em cada parcela, e permite o uso destas variáveis como preditores
das distribuições de espécies (LANDEIRO e MAGNUSSON, 2011).
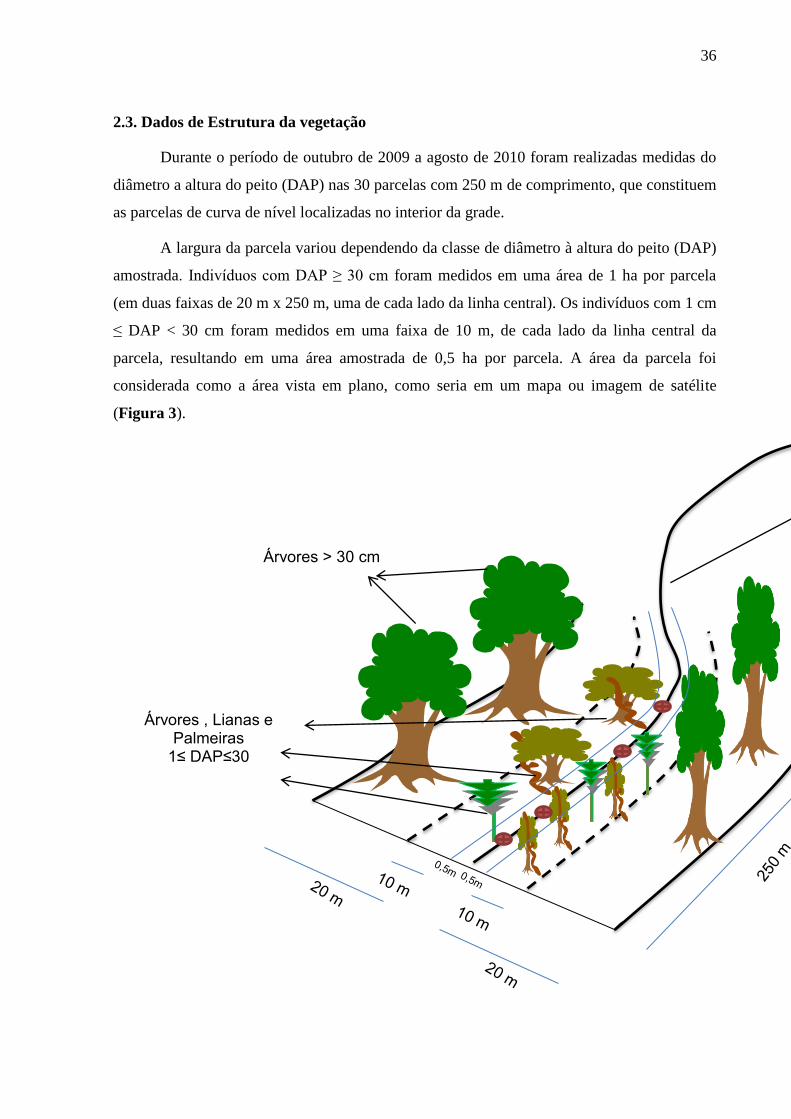
36
2.3. Dados de Estrutura da vegetação
Durante o período de outubro de 2009 a agosto de 2010 foram realizadas medidas do
diâmetro a altura do peito (DAP) nas 30 parcelas com 250 m de comprimento, que constituem
as parcelas de curva de nível localizadas no interior da grade.
A largura da parcela variou dependendo da classe de diâmetro à altura do peito (DAP)
amostrada. Indivíduos com DAP ≥ 30 cm foram medidos em uma área de 1 ha por parcela
(em duas faixas de 20 m x 250 m, uma de cada lado da linha central). Os indivíduos com 1 cm
≤ DAP < 30 cm foram medidos em uma faixa de 10 m, de cada lado da linha central da
parcela, resultando em uma área amostrada de 0,5 ha por parcela. A área da parcela foi
considerada como a área vista em plano, como seria em um mapa ou imagem de satélite
(Figura 3).
Parcela de Curva de nível
Árvores > 30 cm
Árvores , Lianas e Palmeiras
1≤ DAP≤30

37
Figura 3 Distribuição das faixas de amostragem para a vegetação na parcela de curva de nível
Fonte: Oliveira (2012)

38
As faixas de medição se iniciam a 0,5 m da linha central, para evitar a área de trânsito
de pessoas. Estão incluídos na vegetação lenhosa as árvores, palmeiras e lianas, conforme
classificação a seguir:
a) Árvores: O DAP das árvores foi medido a 1,30 m de altura. A altura para a medida do DAP
foi tomada na parte mais alta do terreno, quando ele não era plano.
b) Sapopemas: O diâmetro foi medido cerca de 50 cm acima do final da sapopema, com
auxílio de uma escada ou ajuda de forquilhas.
c) Palmeiras: foram incluídas as palmeiras acaules e os indivíduos jovens das palmeiras com
caule em seu estágio jovem. O DAP foi medido a 1,30 m de altura do solo, da mesma forma
que as árvores, para as palmeiras com caule. O Diâmetro dos indivíduos jovens das palmeiras
foi medido no nível do solo.
d) Lianas: Os indivíduos de lianas podem não ser fáceis de delimitar, pois os ramos se
espalham e se entrelaçam. Portanto, neste estudo, a presença de irregularidades no tronco
interfere no local do ponto de medida, sempre deslocado para secções mais cilíndricas do
tronco a ser medido.
Para determinar se o indivíduo está incluído na parcela, foi necessário medir a
distância perpendicular do indivíduo até a linha central da parcela. Esta medida foi feita com a
trena esticada horizontalmente e não acompanhando o terreno.
As medidas foram feitas com fita métrica, onde foi obtido o CAP (circunferência à
altura do peito) e não o DAP. Neste caso foi necessário dividir o valor medido por π para
obter o DAP. Para as medidas de diâmetro, foi utilizada uma fita métrica de costura ou uma
fita diamétrica (Forestry Suppliers, modelo 283D), ambas com precisão de ±1mm.
Os dados coletados foram identificados por parcela e divididos em 8 classes de DAP
para árvores e palmeiras (CASTILHO et al., 2006), já para lianas foi utilizado 9 classes de
DAP (NASCIMENTO e LAURENCE, 2002). As classes de DAP tiveram variação de 10 cm
(1-10, 10,1-20, 20,1-30, 30,1-40, 40,1-50, 50,1-60, 60,1-70 e >70 cm) para árvores e
palmeiras e de 2 cm para lianas (1-1,9, 2-2,9, 3-3,9, 4-4,9, 5-5,9, 6-6,9, 7-7,9, 8-8,9, 9-10 cm).
Depois foram calculados os totais de árvores, palmeiras e lianas por hectare. Todos os valores
foram extrapolados para indivíduos por hectare.

39
2.4. Cálculo da densidade, área basal e biomassa
A densidade de árvores, palmeiras e lianas é expressa em número de indivíduos por
hectare. A área basal corresponde a somatória das áreas transversais dos troncos por hectare.
Foram utilizadas diferentes equações alométricas disponíveis na literatura obtidas a
partir de diferentes metodologias. A utilidade dos modelos alométricos é estimar a biomassa
em função de parâmetros mensuráveis como o DAP.
Para o cálculo da biomassa foram utilizadas equações alométricas calibradas para o
ambiente de floresta Amazônica de acordo com cada forma de vida vegetal amostrada
(Tabela 1). No caso das árvores, ainda foram empregadas três equações, buscando o melhor
ajuste matemático, de acordo com a classe de diâmetro (OLIVEIRA, SOTTA e HIGUCHI,
2012; HIGUCHI et al.,1998; NASCIMENTO e LAURANCE, 2002). Para as estimativas de
biomassa de palmeiras, foi utilizada uma equação alométrica desenvolvida a partir de várias
espécies de palmeiras que ocorrem na região do alto Rio Negro na Venezuela e Colômbia
(SALDARRIAGA et al., 1988). Utilizamos uma equação alométrica para as estimativas de
biomassa de lianas (SCHNITZER et al., 2006).

40
Tabela 1 Equações alométricas utilizadas para estimar a biomassa seca acima do solo (kg. ha-
1) com base no diâmetro à altura do peito (DAP, cm) e altura (H, cm)
Indivíduos Equação usada no cálculo da Biomassa aérea
Árvores DAP≥ 1 cm e DAP< 5 cm e(-1,7689 + 2,3770*ln (DAP)) (a)
Árvores DAP≥ 5 cm e DAP< 10 cm e(-1,754 + 2,665* ln (DAP))*0,6 (b)
Árvores DAP≥ 10 cm e(-1,096367 + 2,437071*ln (DAP)) *0,6 (c)
Palmeiras DAP≥ 1 cm e(-6,3789 – 0.877*ln (1/DAP2)+2,151* ln (H) (d)
Lianas DAP ≥ 0,5 cm
e(-1,484 + 2,657*ln (DAP)) (e)
aNascimento e Laurance (2002)
bHiguchi et al. (1998)
cOliveira, Sotta e Higuchi, (2012)
dSaldarriaga et al. (1988)
eSchnitzer et al. (2006). As equações de
bHiguchi et al. (1998) e
cOliveira et al. (2010) fornecem estimativas de biomassa em peso fresco. Uma vez que o peso
seco do tronco corresponde acerca de 60% de seu peso fresco; e o da copa corresponde a 58%
de seu peso fresco, o resultado da equação foi multiplicado por 0,6 para ser expresso em peso
seco (Castilho, 2004).
A equação alométrica para estimativa de biomassa de palmeiras utiliza além do
diâmetro, a altura dos indivíduos. Foi definida uma altura padrão espécie-específica, com base
nos dados obtidos em campo.
Os cálculos de densidade, área basal e estimativa de biomassa de cada classe de DAP
foram corrigidos para expressar valores em termos de 1 ha. Devido à variação da largura da
parcela com as diferentes classes de diâmetro amostradas.

41
3. Resultados
3.1. Densidade, área basal e biomassa de árvores
A densidade de árvores (indivíduos.ha-¹) variou significativamente entre as diferentes
classes diamétricas (Figura 4A). As classes menores apresentaram um número maior de
indivíduos, contendo a maior amplitude de variação entre as parcelas. As estimativas de
densidade da primeira classe variaram de 77 a 419 indivíduos.ha-¹. A primeira classe de DAP
representou 90,9% da abundância de indivíduos por hectare nas 30 parcelas avaliadas A
primeira classe foi retirada e, assim, o gráfico evidenciou a verdadeira distribuição por classes
diamétricas. Foi possível observar a presença de 15 indivíduos de grande porte com DAP>70
cm.
O somatório da abundância dos indivíduos acima de 10,1 cm não superou o número de
indivíduos acumulados na primeira classe. A presença de indivíduos acima de 30,1 cm foi
menor, com média de 18,5 indivíduos por hectare entre as 30 parcelas avaliadas (Figura 4A).
Em relação à variação da abundância de árvores com dentro das parcelas foi possível
observar que entre as trinta totais apenas a parcela P25 apresentou densidade abaixo de 1600
indivíduos por hectare. Foi possível observar que a maioria das parcelas apresentou uma
abundância de indivíduos de grande porte, destacando-se as parcelas P6, P11, P26 (Figura
4B).
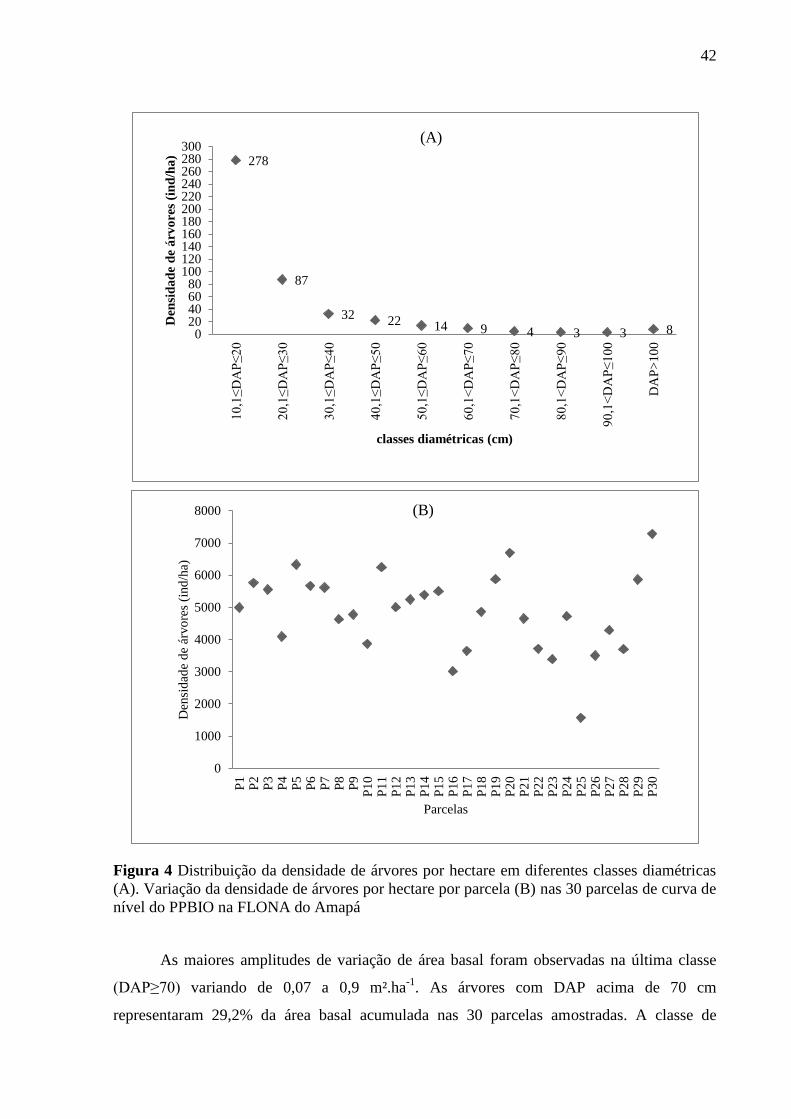
42
Figura 4 Distribuição da densidade de árvores por hectare em diferentes classes diamétricas
(A). Variação da densidade de árvores por hectare por parcela (B) nas 30 parcelas de curva de
nível do PPBIO na FLONA do Amapá
As maiores amplitudes de variação de área basal foram observadas na última classe
(DAP≥70) variando de 0,07 a 0,9 m².ha-1
. As árvores com DAP acima de 70 cm
representaram 29,2% da área basal acumulada nas 30 parcelas amostradas. A classe de
278
87
32 22 14 9 4 3 3 8 0
20406080
100120140160180200220240260280300
10
,1≤
DA
P≤
20
20
,1≤
DA
P≤
30
30
,1≤
DA
P≤
40
40
,1≤
DA
P≤
50
50
,1≤
DA
P≤
60
60
,1<
DA
P≤
70
70
,1<
DA
P≤
80
80
,1<
DA
P≤
90
90
,1<
DA
P≤
10
0
DA
P>
10
0
Den
sid
ad
e d
e á
rvo
res
(in
d/h
a)
classes diamétricas (cm)
0
1000
2000
3000
4000
5000
6000
7000
8000
P1
P2
P3
P4
P5
P6
P7
P8
P9
P1
0P
11
P1
2P
13
P1
4P
15
P1
6P
17
P1
8P
19
P2
0P
21
P2
2P
23
P2
4P
25
P2
6P
27
P2
8P
29
P3
0
Den
sid
ade
de
árvo
res
(ind
/ha)
Parcelas
(A)
(B)
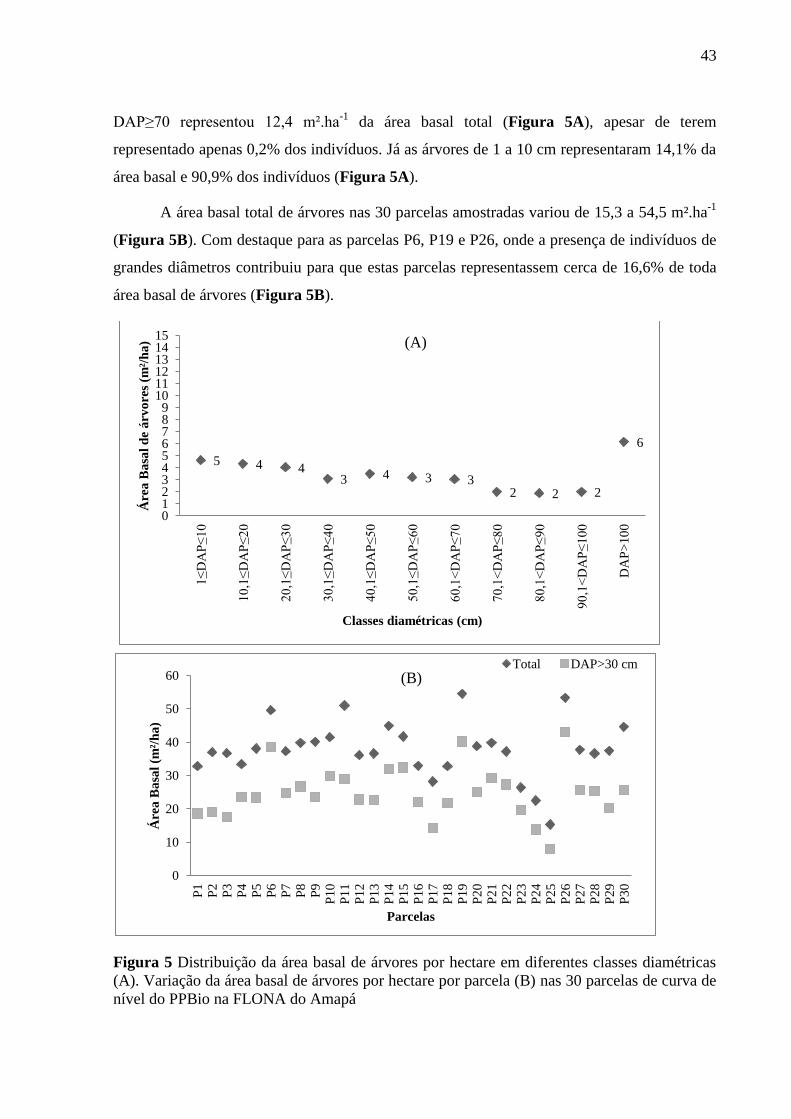
43
DAP≥70 representou 12,4 m².ha-1
da área basal total (Figura 5A), apesar de terem
representado apenas 0,2% dos indivíduos. Já as árvores de 1 a 10 cm representaram 14,1% da
área basal e 90,9% dos indivíduos (Figura 5A).
A área basal total de árvores nas 30 parcelas amostradas variou de 15,3 a 54,5 m².ha-1
(Figura 5B). Com destaque para as parcelas P6, P19 e P26, onde a presença de indivíduos de
grandes diâmetros contribuiu para que estas parcelas representassem cerca de 16,6% de toda
área basal de árvores (Figura 5B).
Figura 5 Distribuição da área basal de árvores por hectare em diferentes classes diamétricas
(A). Variação da área basal de árvores por hectare por parcela (B) nas 30 parcelas de curva de
nível do PPBio na FLONA do Amapá
5 4 4 3 4 3 3
2 2 2
6
0123456789
101112131415
1≤
DA
P≤
10
10
,1≤
DA
P≤
20
20
,1≤
DA
P≤
30
30
,1≤
DA
P≤
40
40
,1≤
DA
P≤
50
50
,1≤
DA
P≤
60
60
,1<
DA
P≤
70
70
,1<
DA
P≤
80
80
,1<
DA
P≤
90
90
,1<
DA
P≤
10
0
DA
P>
10
0
Áre
a B
asa
l d
e á
rvo
res
(m²/
ha
)
Classes diamétricas (cm)
0
10
20
30
40
50
60
P1
P2
P3
P4
P5
P6
P7
P8
P9
P1
0P
11
P1
2P
13
P1
4P
15
P1
6P
17
P1
8P
19
P2
0P
21
P2
2P
23
P2
4P
25
P2
6P
27
P2
8P
29
P3
0
Áre
a B
asa
l (m
²/h
a)
Parcelas
Total DAP>30 cm
(A)
(B)
44
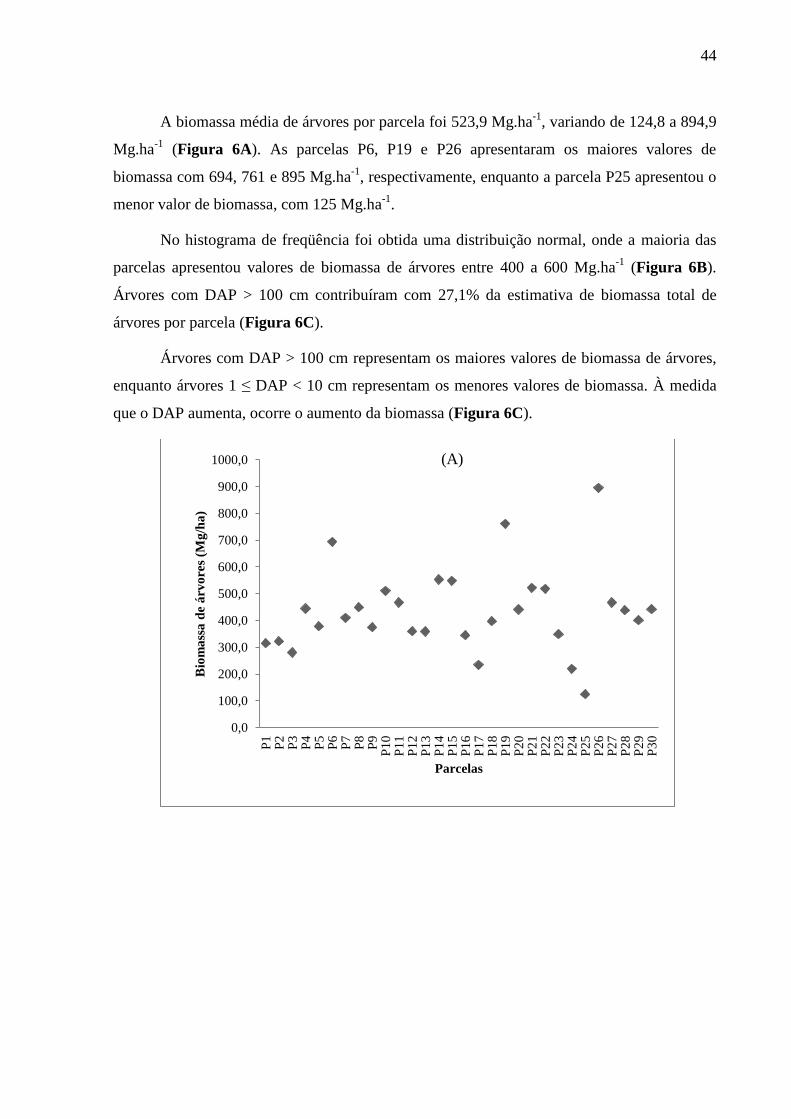
A biomassa média de árvores por parcela foi 523,9 Mg.ha-1
, variando de 124,8 a 894,9
Mg.ha-1
(Figura 6A). As parcelas P6, P19 e P26 apresentaram os maiores valores de
biomassa com 694, 761 e 895 Mg.ha-1
, respectivamente, enquanto a parcela P25 apresentou o
menor valor de biomassa, com 125 Mg.ha-1
.
No histograma de freqüência foi obtida uma distribuição normal, onde a maioria das
parcelas apresentou valores de biomassa de árvores entre 400 a 600 Mg.ha-1
(Figura 6B).
Árvores com DAP > 100 cm contribuíram com 27,1% da estimativa de biomassa total de
árvores por parcela (Figura 6C).
Árvores com DAP > 100 cm representam os maiores valores de biomassa de árvores,
enquanto árvores 1 ≤ DAP < 10 cm representam os menores valores de biomassa. À medida
que o DAP aumenta, ocorre o aumento da biomassa (Figura 6C).
0,0
100,0
200,0
300,0
400,0
500,0
600,0
700,0
800,0
900,0
1000,0
P1
P2
P3
P4
P5
P6
P7
P8
P9
P1
0P
11
P1
2P
13
P1
4P
15
P1
6P
17
P1
8P
19
P2
0P
21
P2
2P
23
P2
4P
25
P2
6P
27
P2
8P
29
P3
0
Bio
ma
ssa
de
árv
ore
s (M
g/h
a)
Parcelas
(A)
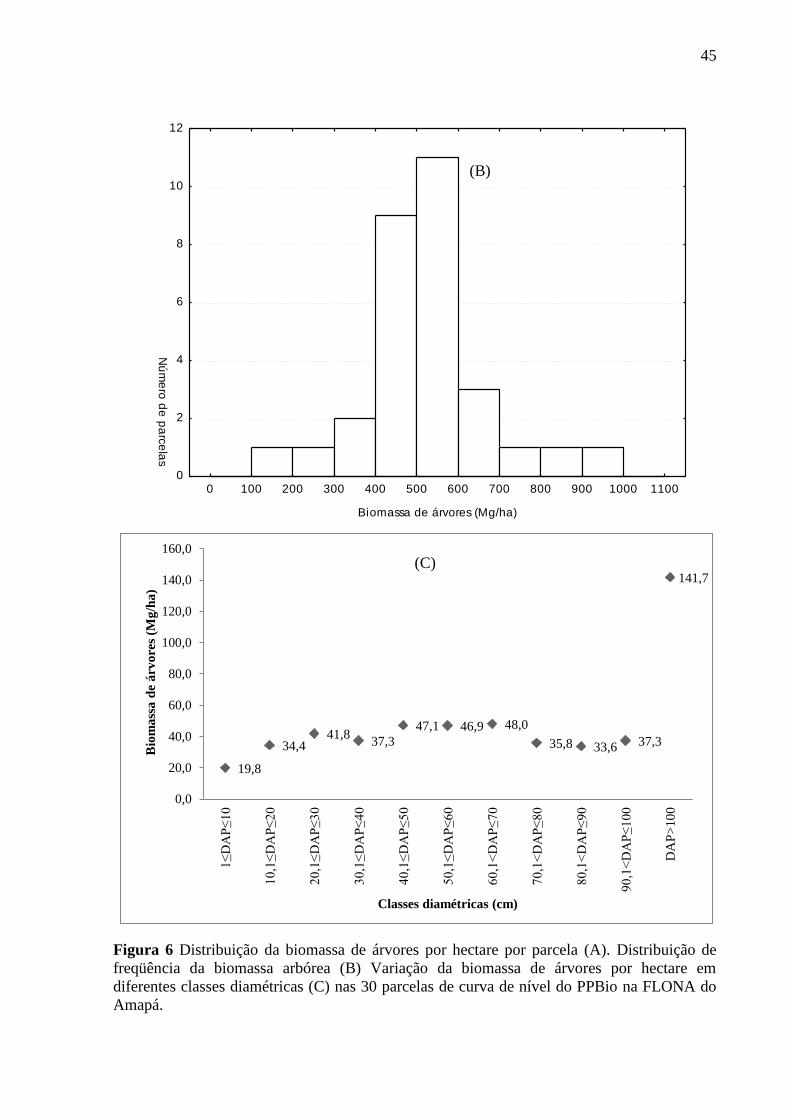
45
Figura 6 Distribuição da biomassa de árvores por hectare por parcela (A). Distribuição de
freqüência da biomassa arbórea (B) Variação da biomassa de árvores por hectare em
diferentes classes diamétricas (C) nas 30 parcelas de curva de nível do PPBio na FLONA do
Amapá.
0 100 200 300 400 500 600 700 800 900 1000 1100
Biomassa de árvores (Mg/ha)
0
2
4
6
8
10
12
Nú
me
ro d
e p
arc
ela
s
19,8
34,4 41,8
37,3
47,1 46,9 48,0
35,8 33,6 37,3
141,7
0,0
20,0
40,0
60,0
80,0
100,0
120,0
140,0
160,0
1≤
DA
P≤
10
10
,1≤
DA
P≤
20
20
,1≤
DA
P≤
30
30
,1≤
DA
P≤
40
40
,1≤
DA
P≤
50
50
,1≤
DA
P≤
60
60
,1<
DA
P≤
70
70
,1<
DA
P≤
80
80
,1<
DA
P≤
90
90
,1<
DA
P≤
10
0
DA
P>
10
0
Bio
ma
ssa
de
árv
ore
s (M
g/h
a)
Classes diamétricas (cm)
(B)
(C)

46
A biomassa de árvores também variou entre as diferentes classes de tamanho de
árvores. Árvores emergentes DAP > 100 cm ocorreram com densidade 8 indivíduos/ha em
todas as parcelas. Em toda a área amostrada, 15 parcelas apresentaram árvores gigantes (DAP
≥ 150 cm), apresentando variação 1 a 3 indivíduo/parcela. Quando presente, um único
indivíduo deste porte, em média, representou 16,6% da biomassa por parcela (79,0 Mg ha-1
,
com base em 22 indivíduos com DAP entre 159,2 e 318,3 cm) (Tabela 2).
A classe de DAP > 100 cm foi a que revelou o maior valor de biomassa, representando
27,1% do valor da biomassa total de árvores. O maior valor de área basal também foi
representado por esta última classe de DAP > 100, significa que cerca de 16,2% da área basal
é concentrada nesta classe de diâmetro. Porém, o menor número de indivíduos é encontrado
nesta última classe de DAP > 100 (0,2%) (Tabela 2).
Enquanto a menor classe 1≤DAP<10 representou o maior percentual de indivíduos,
cerca de 91% dos indivíduos encontram-se na primeira classe. Nesta mesma classe é notado o
menor valor e percentual de biomassa (3,8%). As classes 70,1≤DAP<80, 80,1≤DAP<90 e
90,1≤DAP<100 foram as que apresentaram menores valores em porcentagem de área basal
(15,5%) (Tabela 2).
De maneira geral árvores com diâmetros maiores, representam o maior percentual de
biomassa de árvores e área basal. Porém, estas árvores de diâmetros maiores são encontradas
em menor número na floresta. Às vezes um exemplar deste indivíduo em uma classe de
diâmetro grande, significa a maior concentração de biomassa da parcela.
Na FLONA do Amapá a biomassa está concentrada na maior classe de diâmetro DAP
> 100. A primeira classe 1≤DAP<10 apresentou o maior número de indivíduos mais não
representou a maior concentração de biomassa.
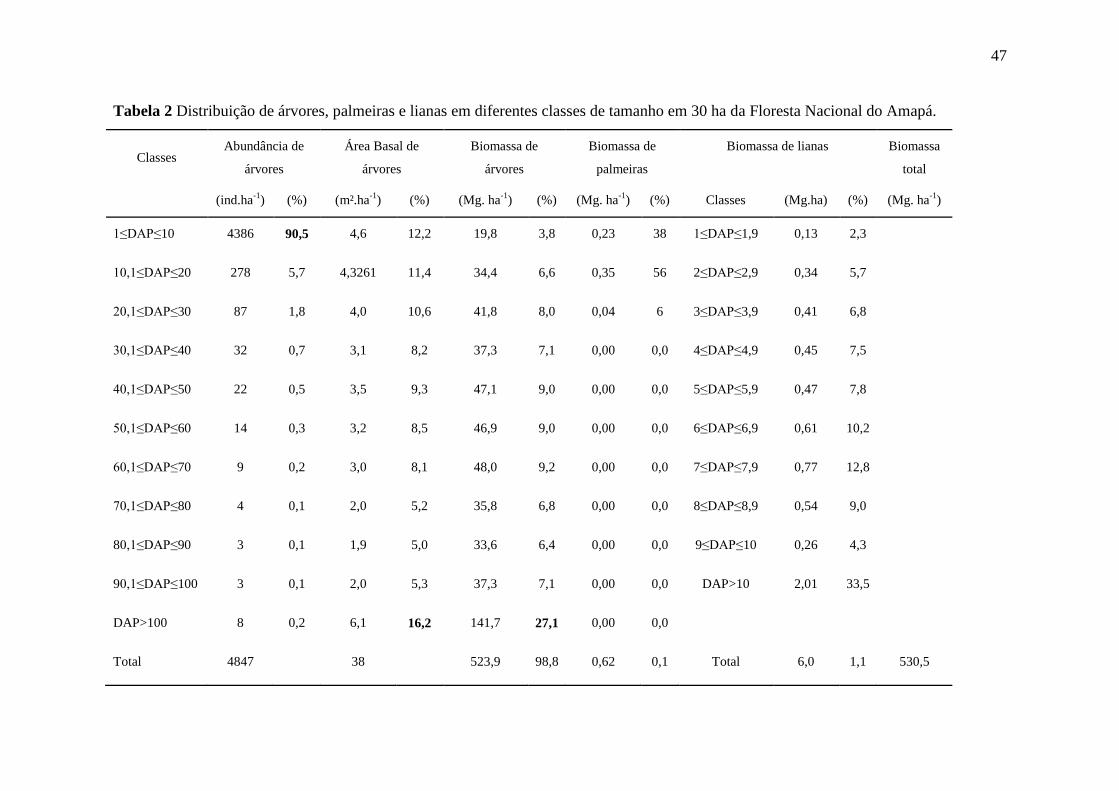
47
Tabela 2 Distribuição de árvores, palmeiras e lianas em diferentes classes de tamanho em 30 ha da Floresta Nacional do Amapá.
Classes Abundância de
árvores
Área Basal de
árvores
Biomassa de
árvores
Biomassa de
palmeiras
Biomassa de lianas Biomassa
total
(ind.ha-1
) (%) (m².ha-1
) (%) (Mg. ha-1
) (%) (Mg. ha-1
) (%) Classes (Mg.ha) (%) (Mg. ha-1
)
1≤DAP≤10 4386 90,5 4,6 12,2 19,8 3,8 0,23 38 1≤DAP≤1,9 0,13 2,3
10,1≤DAP≤20 278 5,7 4,3261 11,4 34,4 6,6 0,35 56 2≤DAP≤2,9 0,34 5,7
20,1≤DAP≤30 87 1,8 4,0 10,6 41,8 8,0 0,04 6 3≤DAP≤3,9 0,41 6,8
30,1≤DAP≤40 32 0,7 3,1 8,2 37,3 7,1 0,00 0,0 4≤DAP≤4,9 0,45 7,5
40,1≤DAP≤50 22 0,5 3,5 9,3 47,1 9,0 0,00 0,0 5≤DAP≤5,9 0,47 7,8
50,1≤DAP≤60 14 0,3 3,2 8,5 46,9 9,0 0,00 0,0 6≤DAP≤6,9 0,61 10,2
60,1≤DAP≤70 9 0,2 3,0 8,1 48,0 9,2 0,00 0,0 7≤DAP≤7,9 0,77 12,8
70,1≤DAP≤80 4 0,1 2,0 5,2 35,8 6,8 0,00 0,0 8≤DAP≤8,9 0,54 9,0
80,1≤DAP≤90 3 0,1 1,9 5,0 33,6 6,4 0,00 0,0 9≤DAP≤10 0,26 4,3
90,1≤DAP≤100 3 0,1 2,0 5,3 37,3 7,1 0,00 0,0 DAP>10 2,01 33,5
DAP>100 8 0,2 6,1 16,2 141,7 27,1 0,00 0,0
Total 4847 38 523,9 98,8 0,62 0,1 Total 6,0 1,1 530,5
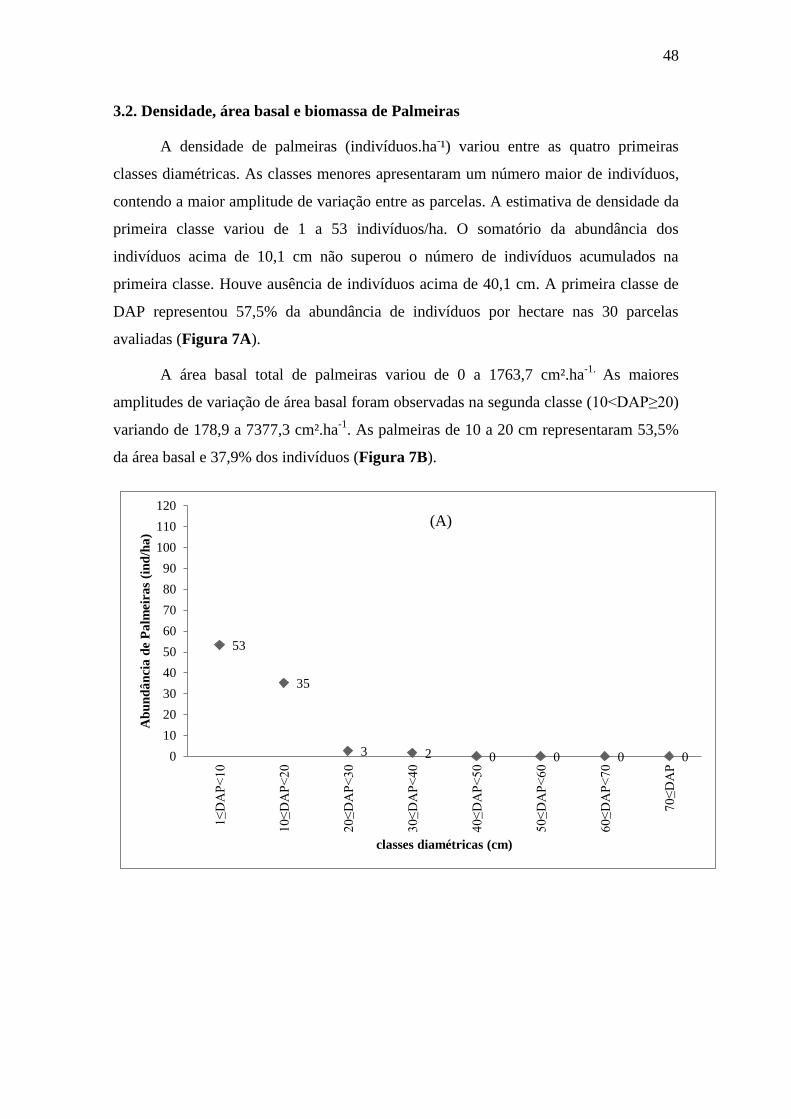
48
3.2. Densidade, área basal e biomassa de Palmeiras
A densidade de palmeiras (indivíduos.ha-¹) variou entre as quatro primeiras
classes diamétricas. As classes menores apresentaram um número maior de indivíduos,
contendo a maior amplitude de variação entre as parcelas. A estimativa de densidade da
primeira classe variou de 1 a 53 indivíduos/ha. O somatório da abundância dos
indivíduos acima de 10,1 cm não superou o número de indivíduos acumulados na
primeira classe. Houve ausência de indivíduos acima de 40,1 cm. A primeira classe de
DAP representou 57,5% da abundância de indivíduos por hectare nas 30 parcelas
avaliadas (Figura 7A).
A área basal total de palmeiras variou de 0 a 1763,7 cm².ha-1.
As maiores
amplitudes de variação de área basal foram observadas na segunda classe (10<DAP≥20)
variando de 178,9 a 7377,3 cm².ha-1
. As palmeiras de 10 a 20 cm representaram 53,5%
da área basal e 37,9% dos indivíduos (Figura 7B).
53
35
3 2 0 0 0 0 0
10
20
30
40
50
60
70
80
90
100
110
120
1≤
DA
P<
10
10
≤D
AP
<2
0
20
≤D
AP
<3
0
30
≤D
AP
<4
0
40
≤D
AP
<5
0
50
≤D
AP
<6
0
60
≤D
AP
<7
0
70
≤D
AP
Ab
un
dâ
nci
a d
e P
alm
eira
s (i
nd
/ha
)
classes diamétricas (cm)
(A)
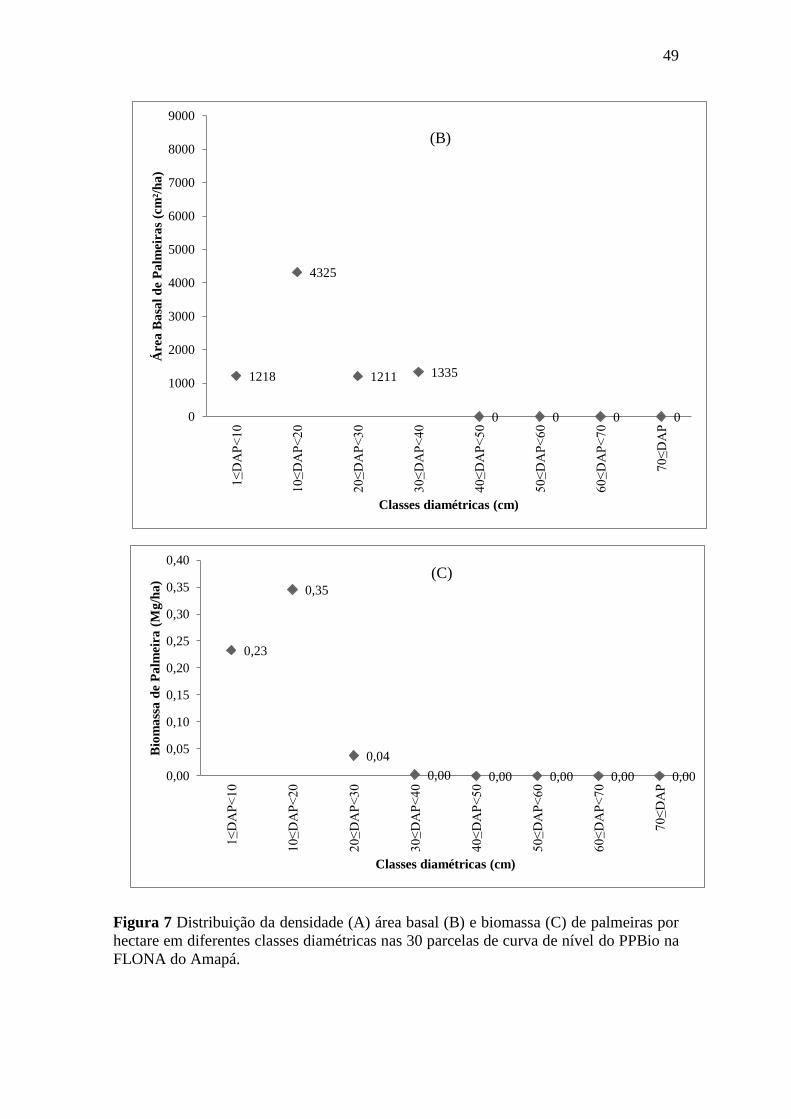
49
Figura 7 Distribuição da densidade (A) área basal (B) e biomassa (C) de palmeiras por
hectare em diferentes classes diamétricas nas 30 parcelas de curva de nível do PPBio na
FLONA do Amapá.
1218
4325
1211 1335
0 0 0 0 0
1000
2000
3000
4000
5000
6000
7000
8000
9000
1≤
DA
P<
10
10
≤D
AP
<2
0
20
≤D
AP
<3
0
30
≤D
AP
<4
0
40
≤D
AP
<5
0
50
≤D
AP
<6
0
60
≤D
AP
<7
0
70
≤D
AP
Áre
a B
asa
l d
e P
alm
eira
s (c
m²/
ha
)
Classes diamétricas (cm)
0,23
0,35
0,04
0,00 0,00 0,00 0,00 0,00 0,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
0,40
1≤
DA
P<
10
10
≤D
AP
<2
0
20
≤D
AP
<3
0
30
≤D
AP
<4
0
40
≤D
AP
<5
0
50
≤D
AP
<6
0
60
≤D
AP
<7
0
70
≤D
AP
Bio
ma
ssa
de
Pa
lmei
ra (
Mg
/ha
)
Classes diamétricas (cm)
(B)
(C)
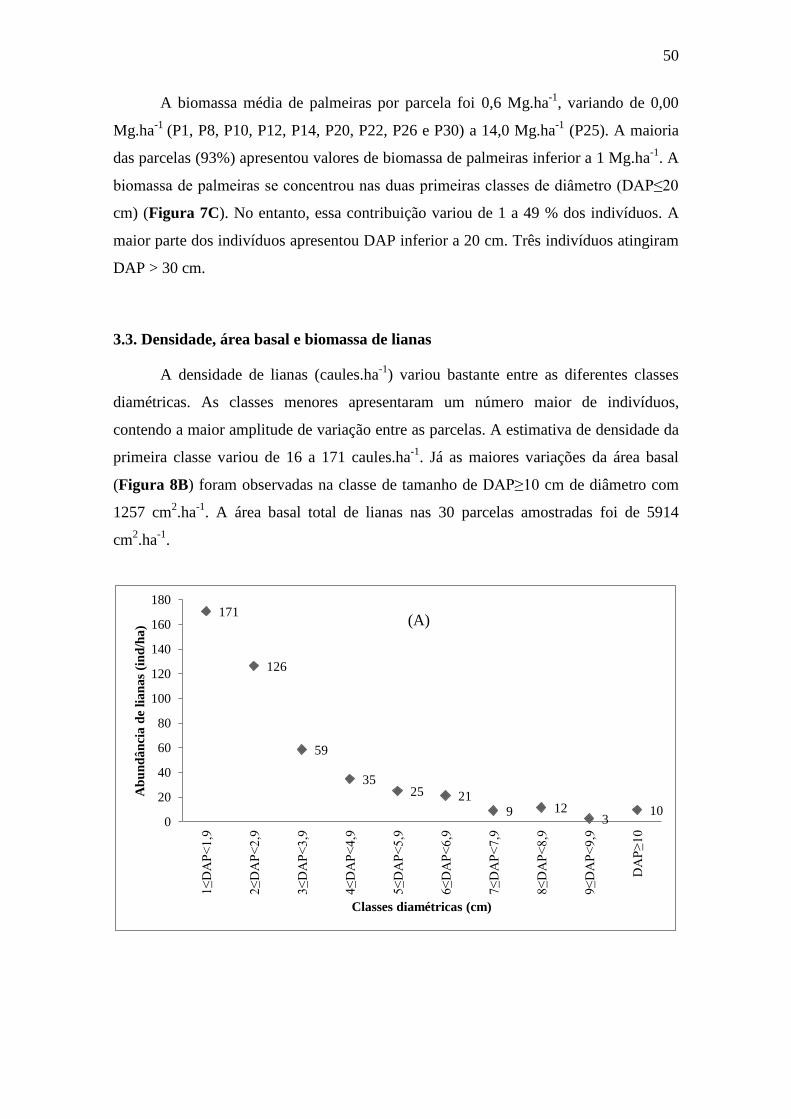
50
A biomassa média de palmeiras por parcela foi 0,6 Mg.ha-1
, variando de 0,00
Mg.ha-1
(P1, P8, P10, P12, P14, P20, P22, P26 e P30) a 14,0 Mg.ha-1
(P25). A maioria
das parcelas (93%) apresentou valores de biomassa de palmeiras inferior a 1 Mg.ha-1
. A
biomassa de palmeiras se concentrou nas duas primeiras classes de diâmetro (DAP≤20
cm) (Figura 7C). No entanto, essa contribuição variou de 1 a 49 % dos indivíduos. A
maior parte dos indivíduos apresentou DAP inferior a 20 cm. Três indivíduos atingiram
DAP > 30 cm.
3.3. Densidade, área basal e biomassa de lianas
A densidade de lianas (caules.ha-1
) variou bastante entre as diferentes classes
diamétricas. As classes menores apresentaram um número maior de indivíduos,
contendo a maior amplitude de variação entre as parcelas. A estimativa de densidade da
primeira classe variou de 16 a 171 caules.ha-1
. Já as maiores variações da área basal
(Figura 8B) foram observadas na classe de tamanho de DAP≥10 cm de diâmetro com
1257 cm2.ha
-1. A área basal total de lianas nas 30 parcelas amostradas foi de 5914
cm2.ha
-1.
171
126
59
35 25 21
9 12 3
10 0
20
40
60
80
100
120
140
160
180
1≤
DA
P<
1,9
2≤
DA
P<
2,9
3≤
DA
P<
3,9
4≤
DA
P<
4,9
5≤
DA
P<
5,9
6≤
DA
P<
6,9
7≤
DA
P<
7,9
8≤
DA
P<
8,9
9≤
DA
P<
9,9
DA
P≥
10
Ab
un
dâ
nci
a d
e li
an
as
(in
d/h
a)
Classes diamétricas (cm)
(A)
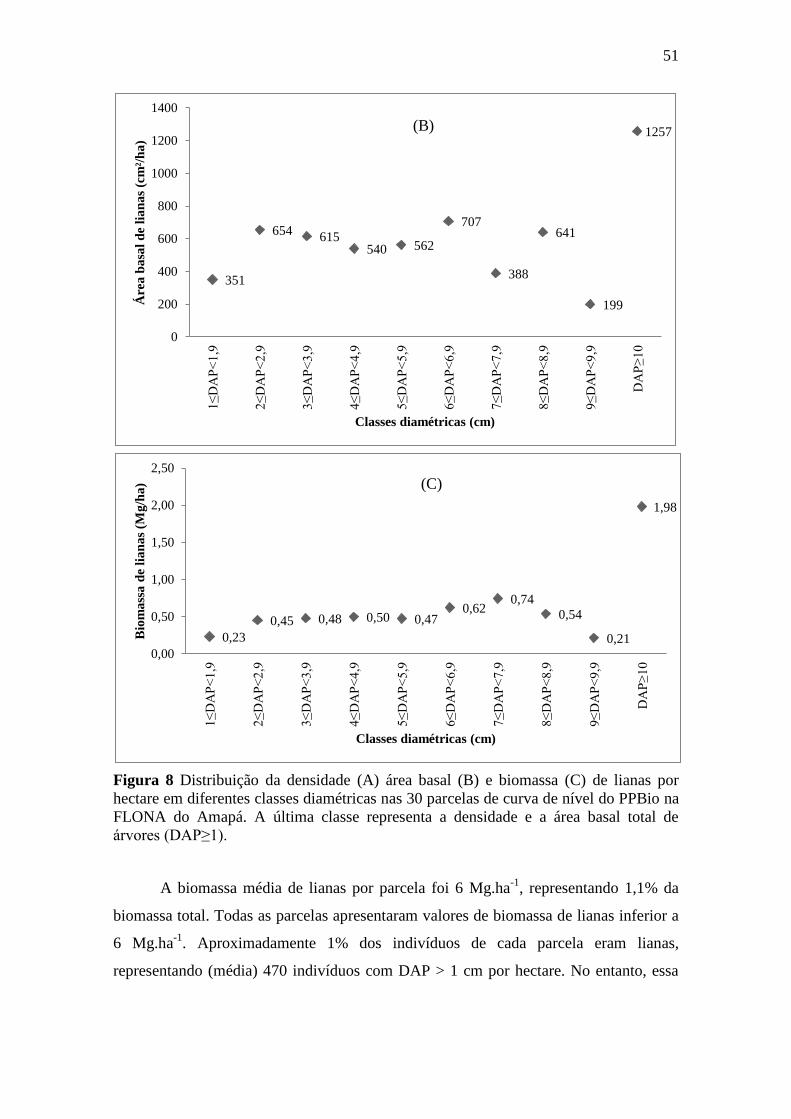
51
Figura 8 Distribuição da densidade (A) área basal (B) e biomassa (C) de lianas por
hectare em diferentes classes diamétricas nas 30 parcelas de curva de nível do PPBio na
FLONA do Amapá. A última classe representa a densidade e a área basal total de
árvores (DAP≥1).
A biomassa média de lianas por parcela foi 6 Mg.ha-1
, representando 1,1% da
biomassa total. Todas as parcelas apresentaram valores de biomassa de lianas inferior a
6 Mg.ha-1
. Aproximadamente 1% dos indivíduos de cada parcela eram lianas,
representando (média) 470 indivíduos com DAP > 1 cm por hectare. No entanto, essa
351
654 615
540 562
707
388
641
199
1257
0
200
400
600
800
1000
1200
1400
1≤
DA
P<
1,9
2≤
DA
P<
2,9
3≤
DA
P<
3,9
4≤
DA
P<
4,9
5≤
DA
P<
5,9
6≤
DA
P<
6,9
7≤
DA
P<
7,9
8≤
DA
P<
8,9
9≤
DA
P<
9,9
DA
P≥
10
Áre
a b
asa
l d
e li
an
as
(cm
²/h
a)
Classes diamétricas (cm)
0,23
0,45 0,48 0,50 0,47 0,62
0,74
0,54
0,21
1,98
0,00
0,50
1,00
1,50
2,00
2,50
1≤
DA
P<
1,9
2≤
DA
P<
2,9
3≤
DA
P<
3,9
4≤
DA
P<
4,9
5≤
DA
P<
5,9
6≤
DA
P<
6,9
7≤
DA
P<
7,9
8≤
DA
P<
8,9
9≤
DA
P<
9,9
DA
P≥
10
Bio
ma
ssa
de
lia
na
s (M
g/h
a)
Classes diamétricas (cm)
(B)
(C)

52
contribuição variou de 2,3 a 33,5% dos indivíduos. A última classe sozinha contribui
com 2 Mg ha-1
, representando 33,5% da biomassa total das classes (Figura 8C).
4. Discussão
4.1. Densidade, área basal – o que representam
De forma geral a densidade analisada por classes diamétricas representa a
distribuição de indivíduos jovens e adultos na área amostrada. Isto aborda um dos
aspectos da distribuição de estrutura arbórea (árvores, palmeiras e lianas). A variação do
número total de indivíduos por hectare representa principalmente a variação da
densidade dos indivíduos mais finos, representados pela primeira classe de diâmetro.
A distribuição da área basal representa de maneira geral a variação das maiores
classes. As últimas classes de diâmetros representam as maiores árvores. Já para
palmeiras a área basal representa a segunda menor classe, talvez pela presença de um
açaizal em uma parcela observada de forma diferenciada. Os dois fatores analisados,
densidade e área basal são expressões de condições prévias ambientais, ou seja, das
variáveis do solo, luz, nível piezométrico e topografia.
As estimativas de biomassa vegetal estão altamente correlacionadas às
distribuições de área basal, ou seja, as classes de diâmetro que representaram maior área
basal, também representaram maiores estimativas de biomassa vegetal. Estes resultados
foram notados para árvores, palmeiras e lianas.
4.2. Como a biomassa varia entre as parcelas na FLONA do Amapá e sua
contribuição no contexto local e regional
As estimativas de biomassa de árvores para a FLONA do Amapá variaram de
177,6 a 966,4 Mg.ha-1
, com biomassa média por parcela de 523,9 Mg.ha-1
(dp = ±157).
Uma área de estudo próxima à FLONA, dentro da Floresta Estadual do Amapá –
FLOTA, administrada pelo Instituto Estadual de Florestas – IEF, apresentou valores
médios de biomassa de árvores vivas acima do solo de 622,75 Mg.ha-1
(dp = ±251,91)
(OLIVEIRA, SOTTA e HIGUCHI, 2012), sendo obtido através de inventários e coleta
por método destrutivo, porém os resultados foram próximos, quando comparados o
desvios padrão. De maneira geral o resultado do presente trabalho mostrou-se
satisfatório, pois o inventário foi feito em uma área de 30 ha. Em seguida utilizamos

53
equações alométricas derivadas por método indireto obtido em parcelas próximas a área
de estudo para estimar a biomassa total de árvores. Assim, mostramos a distribuição da
biomassa de árvores em uma grande área.
Outros resultados na Amazônia Oriental mostraram valores de biomassa
próximos ao encontrado neste estudo. As estimativas de biomassa arbórea encontradas
na FLONA de Caxiuanã, foram em média de 430,33 Mg.ha-1
(área do projeto
RAINFOR apud MALHY et al., 2006) e 385,48 Mg.ha-1
(ALMEIDA et al., – dados não
publicados apud MALHY et al., 2006). A FLONA do Tapajós apresentou biomassa
variando entre 222 a 270 Mg.ha-1
, com valor médio de 253,5 Mg.ha-1
(KELLER,
PALACE e HURTT, 2001) (Tabela 1.1).
De maneira geral, os trabalhos aqui comparados obtiveram estimativas de
biomassa média muito semelhantes à deste estudo, mesmo utilizando metodologias de
amostragens diferentes (a maioria utilizou parcelas retas e amostrou áreas de 1 ha). A
maioria também apresentou pouca variação de biomassa entre as parcelas. Nossa
pesquisa mostrou grande variabilidade de biomassa entre 177,6 a 966,4 Mg.ha-1
. Outros
autores, entre eles, Clark e Clark (2000), Keller, Palace e Hurtt (2001); Houghton et al.
(2001) e Castilho et al. (2006), comentam que as estimativas de biomassa para grandes
áreas devem ser obtidas a partir de amostras aleatórias, replicáveis, de tamanho
adequado, dispostas de modo a cobrir toda a área em questão ou estratificadas de acordo
com a classificação espacial da vegetação. A presente pesquisa foi realizada
efetivamente em uma área de 30 ha, dividida em 30 parcelas de 1 ha, o que é
considerado um estudo de grande escala espacial, quando comparado com outros que
estimam a biomassa ou baseados em inventários feitos apenas em parcelas de 1 e 2 ha.
Um estudo semelhante a este foi realizado por Castilho (2004) em uma área na
Amazônia Central, utilizando 72 parcelas em curva de nível iguais às usadas aqui, e
distribuídas uniformemente sobre 64 km2. Este estudo obteve biomassa média de
árvores e palmeiras por parcela (média ± dp) foi 321,8±44,3, variando entre 215 a 421
Mg.ha-1
. Laurance et al. (1999) estimaram o estoque de biomassa entre 231 a 492
Mg.ha-1
, com média de 356±47 Mg.ha-1
. Brown et al. (1995) encontraram uma
amplitude de variação de biomassa entre 155 a 352 Mg.ha-1
. Contudo, Houghton et al.
(2000) obtiveram uma amplitude de variação de biomassa entre 290 a 464 Mg.ha-1
.
Ambos os trabalhos encontraram cerca de 50% de variação entre os valores extremos de
biomassa calculados, revelando uma distribuição heterogênea da biomassa arbórea ao
longo da paisagem nas florestas de terra-firme na Amazônia Central (Tabela 3).

54
Tabela 3 Comparativo dos valores de biomassa total (Mg.ha-1
) obtidos para estudos
realizados em diferentes sítios experimentais na Amazônia brasileira.
Na FLONA do Amapá a biomassa (árvores+palmeiras) por parcela (média ± dp)
foi (516,4 ± 89 Mg.ha-1
) a amplitude de variação da biomassa foi (177,6 a 966,4 Mg.ha-
1). Nos estudos realizados na Amazônia central a amplitude variação da biomassa foram
menores do que os obtidos na FLONA do Amapá. Em geral, o presente estudo mostrou
grande variabilidade de biomassa, cerca de 80% de variação de valores extremos de
biomassa. Isso se deve à utilização da amostragem em parcelas que incorporaram
grandes áreas de variação do ambiente floresta, o que captura a ampla heterogeneidade
ao longo da paisagem. Assim, é possível fazer extrapolações confiáveis sobre uma área
maior, sem superestimar a biomassa.
Biomassa
Local Média (±dp ) Mín. Máx. Autor
(Mg.ha-1
)
Floresta Estadual
do Amapá (AP)
622,75±251,91 - - Oliveira, Sotta e Higuchi
(2012)
Floresta Nacional
de Caxiuanã (PA)
430,3 - - Malhy et al. (2006)
Floresta Nacional
do Tapajós (PA)
253,5 222 270 Keller, Palace e Hurtt (2001)
Reserva Florestal
Ducke (AM)
321,8±44,3 215 421 Castilho (2004)
Reserva Florestal
Ducke (AM)
356±47 231 492 Laurance et al.(1999)
Reserva Florestal
Ducke (AM)
155 352 Brown et al. (1995)
Reserva Florestal
Ducke (AM)
290 464 Houghton et al. (2000)
Florestal Nacional
do Amapá (AP)
530,5±155 177,6 966,4 Este estudo

55
4.3. Contribuição de palmeiras para as estimativas de biomassa viva acima do solo
As palmeiras representaram 0,1% da biomassa arbórea total por parcela. As
palmeiras são excluídas nas estimativas de biomassa, ou consideradas como árvores. As
equações alométricas especificas para a estimativa de biomassa de palmeiras são
escassas. Na maioria das vezes, para estimar a biomassa de palmeiras são utilizadas as
equações alométricas específicas de árvores, o que superestima a biomassa de
palmeiras. Clark e Clark (2000) sugerem que estas superestimativas devem ter pouco
efeito nas estimativas de biomassa total viva acima do solo, visto que as palmeiras
representam menos de 1% da biomassa total (CASTILHO, 2004). Entretanto, o presente
estudo mostrou que nas áreas próximas a igarapés, as palmeiras representaram cerca de
7% da biomassa arbórea viva total.
Segundo o IBGE (1997) cerca de 20% da área original de floresta, localizada na
Amazônia Brasileira são cobertas por palmeiras. Sendo assim, assumir que as palmeiras
não contribuem para a biomassa florestal pode ser um erro. Mesmo fora da área
mapeada pelo IBGE como “floresta dominada por palmeiras” as palmeiras podem
atingir grande cobertura e contribuir significativamente para a biomassa florestal. Isto
ressalta a importância de uma amostragem com várias parcelas na escala média e local,
para cobrir toda a heterogeneidade associada à variação na dominância dos
componentes florestais. Alguns autores (BROWN e LUGO, 1992; CHAVE et al., 2001)
alertam a necessidade de melhorar nosso entendimento de como componentes não-
arbóreos em florestas perturbadas podem ser diferentes do que em florestas maduras.
Para uma melhor caracterização da estrutura da floresta é necessário considerar as
palmeiras como componente independente nas estimativas de biomassa, desta forma
teríamos referências para estudos comparativos de florestas perturbadas e não
perturbadas.
4.4. Contribuição das lianas para as estimativas de biomassa viva acima do solo
As lianas representaram apenas 1,3% de toda biomassa total viva acima do solo.
Em estudo realizado em média escala na Amazônia Central, Nogueira (2006) obteve
estimativas de 1 a 4%, dependendo da equação alométrica utilizada. Este estudo
concorda com os resultados obtidos para a Reserva Ducke, na Amazônia Central
(NOGUEIRA, 2006). A biomassa (média ±dp) variou entre 3,9±1,4 a 12,3±4,2 Mg.ha-1
,
a amplitude de variação da biomassa foi de 2 a 19 Mg.ha-1
, dependendo da equação
alométrica utilizada (NOGUEIRA, 2006). Na FLONA do Amapá a biomassa média

56
(média ±dp) foi 6 ±3,2 Mg.ha-1
e amplitude de variação de biomassa foi de 1,9 a 11,7
Mg.ha-1
. Em estudo realizado em 20 ha na Amazônia Central, Nascimento e Laurance
(2001) sugerem que as lianas representam 2,1% da biomassa total viva acima do solo,
com biomassa média de 8,3 Mg.ha-1
e amplitude de variação de 4,6 a 13,7 Mg.ha-1
.
Nestes estudos realizados na Amazônia central os valores médios e a amplitude de
variação da biomassa de lianas foram maiores do que os obtidos na FLONA do Amapá.
Resultados diferenciados foram encontrados por Gerwing e Farias (2000) em
estudo realizado em Paragominas – PA (Amazônia Oriental), onde a biomassa média foi
43 Mg.ha-1
representando 14% da biomassa total. Esta elevada proporção de biomassa
de lianas é atribuída a hipótese de que a floresta ou é muito propensa a distúrbios ou
está se recuperando de um distúrbio em larga escala ocorrido no passado. Em qualquer
caso, por causa do aumento na freqüência e intensidade de distúrbios por atividades
humanas, é provável que a biomassa de lianas aumente (GERWING e FARIAS, 2000).
Estes resultados de estudos realizados em vários estágios de floresta antropizada
diferem dos resultados obtidos no estudo realizado na FLONA do Amapá, onde a
biomassa de lianas representa apenas 1,3% da biomassa total, este resultado é
característica de floresta não perturbada. Segundo Hegarty e Caballé (1991) em florestas
não perturbadas a biomassa de lianas representa 5% da biomassa de uma floresta, as
lianas participam com mais de 30% da área foliar total e da produção de serrapilheira
Das 30 parcelas amostradas 4 apresentaram lianas acima de 14 cm de diâmetro
(17,5; 14,6; 18,1;15 cm), já em estudo realizado por Nogueira (2006) em 40 parcelas na
Reserva Florestal Ducke, 33 parcelas apresentaram indivíduos maiores que 14 cm.
4.5. A variação de biomassa depende do tamanho da árvore
Os maiores valores de biomassa foram dependentes dos maiores tamanhos de
árvores. As estimativas de biomassa de árvores evidenciaram a última classe (DAP >
100 cm) como a mais importante, alcançando em media 27,1% da biomassa total. A
classe de 1≤DAP<10 cm concentrou a maior parte dos indivíduos (90,6%), mas
representou apenas 3,8% da biomassa total de árvores.
Clark e Clark (1996) encontraram em algumas florestas tropicais árvores
emergentes com DAP ≥ 60 cm ocorreram em baixa densidade (aproximadamente 4
indivíduos/ha) e representaram cerca de 25 - 35% da biomassa total de árvores por

57
parcela. Diferente dos resultados encontrados para a Amazônia Central, onde árvores
com DAP entre 20 e 50 cm (classes intermediárias) contribuem com aproximadamente
50% da estimativa de biomassa de árvores por parcela (NASCIMENTO e LAURANCE,
2002; CASTILHO, 2004).
Na FLONA do Amapá mais de 50% da concentração de biomassa arbórea viva
total por hectare encontra-se na classe de tamanho de DAP≥60 cm. Árvores gigantes
(DAP>150) foram presentes na maioria das parcelas. Os dados evidenciam que a
presença de uma única árvore deste porte, contribui em média com 16,6% da biomassa
por parcela.
Os padrões obtidos com análise de dados por classes de tamanhos (pequeno,
médio e grande porte), mostram em detalhes como cada classe de tamanho responde
diferentemente as variáveis ambientais de cada região (NOGUEIRA et al., 2008).
Nossos resultados sugerem que a divisão das árvores em intervalos de tamanho menor,
pode ser mais apropriada para determinar padrões de variação espacial da biomassa em
escala regional, porque assim podemos investigar melhor as causas de variáveis que
influenciam sua distribuição.

58
5. Considerações Finais
A presença de grandes árvores com diâmetros acima de 70 cm junto com a
concentração de lianas que representam apenas 1,1% de toda biomassa lenhosa são
indicadores que a área de estudo dentro da Floresta Nacional do Amapá é uma área
pouco perturbada, caracterizada como uma floresta primária, também conhecida como
floresta de clímax. Trata-se da conhecida floresta intocada ou aquela que a ação do
homem não provocou significativas alterações das suas características originais de
estrutura e de espécies.
Os resultados apontam a área com grande potencial para o manejo florestal. Existem
muitas árvores de grande porte que estão presentes na maioria das 30 parcelas
estudadas. Assim este estudo contribui com dados indicativos para promover a
exploração sustentável de madeira na Floresta Nacional do Amapá.
Este estudo revelou a importância de uma amostragem com várias parcelas na escala
média e local, na qual pode cobrir toda a heterogeneidade associada à variação na
dominância dos componentes florestais. As parcelas em curva de nível contribuíram
para estes resultados, pois os dados foram representativos da variabilidade da
distribuição espacial, ao contrário de parcelas retas onde se tem heterogeneidade dentro
das parcelas. Este fato sugere que há um grande potencial da aplicação da metodologia
seguinte este tipo de delineamento amostral.
Nosso estudo mostrou a importância em considerar lianas e palmeiras nos
inventários florestais para o cálculo de biomassa. Dependendo do tipo de ambiente
específico dentro da floresta, pode ocorrer que estes grupos vegetais representam
grandes proporções. Com efeito, mostramos que a contribuição destes grupos são
significativos e não podem ser negligenciados.
Nossos resultados corroboram com outros autores da literatura que reforçando as
estimativas anteriores de que as florestas na Amazônia oriental estocam grande
quantidade de biomassa viva acima do solo. A liberação destes estoques, através de
mudanças no uso da terra, pode significar uma contribuição significativa para os
ecossistemas florestais e em consequência para o aquecimento global.

59
6. Referências
BAKER, T. R. et al. Increasing biomass in Amazonian forest plots. Phil. Trans. R. Soc.
Lond. B, n. 359, p. 353–365, 2004.
BROWN, I.F., et al. Uncertainty in the biomass of Amazonian forests: an example from
Rondonia, Brazil. Forest Ecol. Manag., n. 75, p. 175–189. 1995.
BROWN, S. E LUGO, A.E.. Aboveground biomass estimates for tropical moist forests
of the Brazilian Amazon. Interciencia, 17(1): 8-18, 1992.
BUSH, M.B.; SIMAN, M.R.; URREGO, D.H. 48,000 Years of Climate and Forest
Change in a Biodiversity Hot Spot. Science, n. 303, p. 827 – 829, 2004.
CASTILHO, C.V. et al. Variation in aboveground tree live biomass in a central
Amazonian forest: effects of soil and topography. Forest Ecology and Management,
234: 85-96. 2006.
CASTILHO, C.V. Variação espacial e temporal da biomassa arbórea viva em 64
Km2 de floresta de terra-firme na Amazônia Central. Tese de doutorado,
INPA/UFAM, Manaus, AM. 72p., 2004.
CHAVE, J.; RIERA, B.; DUBOIS, M.A. Estimation of biomass in a neotropical forest
of French Guiana: spatial and temporal variability. Journal of Tropical Ecology, 17,
79–96, 2001.
CLARCK, D. B. e CLARCK, D. A. Landscape-scale variation in forest structure and
biomass in a tropical rain forest. Forest Ecology and Management, v. 137, n. 1-3, p.
185-198, 2000.
CLARK, D. Sources or sinks? The responses of tropical forests to current and future
climate and atmospheric composition. Philosophical Transaction of the Royal Society
Biological Science, n. 359, p. 477-491, 2004.
FEARNSIDE, P.M.. Amazonian deforestation and global warming: carbon stocks in
vegetation replacing Brazil’s Amazon forest. Forest Ecology and Management, n. 80:
p. 21-34, 1996.
FEARSIDE, P.M. e LEAL FILHO, N.. Soil and development in Amazonia. In:
BIERREGAARD, R.O.; GASCON, C.; LOVEJOY, T.E.; MESQUITA, R.C.G. (Eds.),

60
Lessons from Amazonia: The Ecology and consevation of a fragmented forest. Yale
University Press, New Haven e London, p. 291-312, 2001.
FIELD, C. B. Plant physiology of the ‘missing’ carbon sink. Plant Physiology, n.125,
p. 25–28, 2001.
FRANCEZ, L.M.B.; CARVALHO, J.O.P.; JARDIM, F.C.S. Mudanças ocorridas na
composição florística em decorrência da exploração florestal em uma área de floresta de
Terra Firme na região de Paragominas, PA. Acta Amazonica, v.37, n.2, p.219-228.
2007.
FURTADO, L.G.. Amazônia: Desenvolvimento, sociodiversidade e qualidade de vida.
Belém: UFPA. NUMA. Universidade e meio ambiente, n. 9, 165p, 1997.
GERWING, J. J. e FARIAS, D. L. Integrating liana abundance and forest stature into na
estimate of total aboveground biomass for an eastern Amazonian forest. Journal of
Tropical Ecology, v. 16, p. 327-335, 2000.
GERWING, J. J. e FARIAS, D. L. Integrating liana abundance and forest stature into na
estimate of total aboveground biomass for an eastern Amazonian forest. Journal of
Tropical Ecology v. 16, p. 327-335, 2000.
HAIRIAH, K.; SITOMPULL, S.M.; NOORDWIJK, M.VAN.; PALM, C. Methods for
sampling carbon stocks above and below ground. In: NOORDWIJK, M.V.;
HEGARTY, E.E. Leaf life-span and leafing phenology of lianes and associated trees
during a rainforest sucession.. Journal of ecology, v. 78, p. 300-312, 1990.
HIGUCHI, N.; SANTOS, J.; RIBEIRO, R.J.; MINETTE, L.; BIOT, Y.. Biomassa da
parte aérea da vegetação de floresta tropical úmida de terra firme da Amazônia
Brasileira. Acta Amazonica, 28 (2): 153-165. 1998.
HOUGHTON, R. A.. Aboveground forest biomass and the global carbon balance.
Global Change Biology 11:945-958, 2005.
HOUGHTON, R. A.; LAWRENCE, K. T.; HACKLER, J. L.; BROWN, S. The spatial
distribution of forest biomass in the Brazilian Amazon: a comparison of estimates.
Global Change Biology, v. 7, p. 731-746, 2001.
IBGE. DIAGNÓSTICO AMBIENTAL DA AMAZÔNIA LEGAL. IBGE, Rio de
Janeiro. (CD-Rom), 1997.

61
INPE/MCT. Estimativa da Taxa de Desmatamento da Amazônia no período de
2010-2011. Brasília - DF, 2011. Disponível em
www.obt.inpe.br/prodes/Prodes_Taxa2011.pdf. Acessado em: 21 de jan. de 2012.
INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE (IPCC) (2007) Climate
Change 2007. Synthesis report, Valência, 2007. 52 p.
KELLER, M.; PALACE, M.; HURTT, G. Biomass estimation in the Tapajós National
Forest, Brazil. Examination of sampling and allometric uncertainties. Forest Ecology
and Management, v. 154, n. 3, p. 371-382, 2001.
LANDEIRO, V.L. e MAGNUSSON, E.M. The Geometry of Spatial Analyses:
Implications for Conservation Biologists. Natureza e Conservação, v. 1, n. 9, p. 7-20,
2011.
LAURANCE, W.F.; FEARNSIDE, P.M.; LAURANCE, S.G.; DELAMONICA, P.;
LOVEJOY, T.E.; RANKIN-DE-MERONA, J.M.; CHAMBERS, J.; GASCON, C..
Relationship between soils and Amazon forest biomass: a landscape-scale study. Forest
Ecology and Management, v. 118, p. 127-138, 1999.
MAGNUSSON, W.E. et al. A modification of the Gentry method for biodiversity
surveys in long-term ecological research sites. Biota Neotrop, v. 2, n. 5, p.1-6, 2005.
MALHI, Y. e WRIGHT, J.. Spatial patterns and recent trends in the climate of tropical
rainforest regions. Philosophical Transaction of the Royal Society B Biological
Science. v. 359, p. 311-329, 2004.
MALHI, Y. et al. The regional variation of aboveground live biomass in old-growth
Amazonian forests. Global Change Biology. V.12, pg. 1107–1138, 2008.
MALHI, Y. The carbon balance of tropical forest regions, 1990-2005. Current
Opinion in Sustainability Science, v.2, n.4, p.237-244, 2010.
NASCIMENTO, H.E.M. e LAURANCE, W.F.. Total aboveground biomass in central
Amazonian rainforest: a landscape-scale study. Forest Ecology and Management, v.
168, p. 311-32, 2002.
NOGUEIRA, A. Variação da densidade, área basal e biomassa de lianas em 64 km²
de floresta de terra firme na Amazônia Central. Dissertação de mestrado,
INPA/UFAM. Manaus, 2006.

62
NOGUEIRA, E.M. et al. Estimates of forest biomass inthe Brazilian Amazon: New
allometric equations and adjustments to biomass from wood-volume inventories. Forest
Ecology and Management, v. 256, p. 1853-1867, 2008.
OLIVEIRA, L.L. et al. Características Hidroclimáticas da bacia do rio Araguari. In:
CUNHA, A.C.; SOUZA, E.B.; CUNHA, H.F.A. Tempo, Clima e Recursos Hídricos:
resultados do Projeto REMETAP no Estado do Amapá. Macapá: IEPA, p. 83 – 96,
2010.
OLIVEIRA, L.P.S.; SOTTA, E.D.; HIGUCHI, N. Biomass Quantification in Amapá’s
State Forest: Carbon Stock Allometry and Estimate. 1st Edition. (Amapá’s State
Forest Institution-IEF. Technical Series), 52 p, 2012.
PEREIRA, L.A.; SENA, K.S; SANTOS, M.R.; COSTA NETO, S.V. Aspectos
florísticos da FLONA do Amapá e sua importância na conservação da biodiversidade.
Nota Cientifica da Revista Brasileira de Biociências, Porto Alegre, v. 5, n. 2, p. 693-
695, 2007.
PRENTICE, I. C. et al. The carbon cycle and atmospheric carbon dioxide. In: Climate
change 2001: the scientific basis, p. 183–237. Cambridge University Press, 2001.
QUESADA, C.A. et al. Effect of soils on forest structure and dynamics in Amazonia.
Biogeosciences Discuss., v. 6, p. 3993–4057, 2009.
SAATCHI, S.S. et al.. Distribution of aboveground live biomass in the Amazon
basin. Global Change Biology 13, 816-837. 2007.
SALDARRIAGA, J.G. et al. Long-term chronosequence of forest succession in the
upper Rio Negro of Colombia and Venezuela. Journal of Ecology, v. 76: p. 938-958.
1988.
SANTOS, S.R.M; MIRANDA, I.S.; TOURINHO, M.M. Estimativa de biomassa de
sistemas agroflorestais das várzeas do rio juba, Cametá, Pará. Acta Amaz. v.34, n.1,
Manaus, 2004.
SCHNITZER, S.A.; DeWALT, S.J. e CHAVE, J.. Censusing and Measuring Lianas: A
Quantitative Comparison of the Common Methods. Biotropica, v. 5, n. 38(): 581–591,
2006.
SILVA, J.M.C. et al. O corredor de biodiversidade do Amapá. In: Corredor de
Biodiversidade do Amapá – Biodiversity Corridor. Belém, CI-Brasil, 54 p. 2007.

63
SILVA, R. P. Alometria, estoque e dinâmica da biomassa de florestas primárias e
secundárias na região de Manaus (AM). 2007. 135 f. Tese (doutorado)--
INPA/UFAM, AM.
SILVA, W. da C.. Viabilidade econômica do pagamento por serviços ambientais no
Estado do Amapá utilizando Análise de Risco. 2011. Dissertação de Mestrado:
UNIFAP, Macapá- AP.
SIMONIAN, L. T. et al. Floresta Nacional do Amapá: um histórico breve, políticas
públicas e (IN) sustentabilidade. In: SIMONIAN, L.T.L. (Org.). Políticas públicas,
desenvolvimento, unidades de conservação e outras questões socioambientais no
Amapá. Belém: NAEA-UFPA/MPEAP, p. 115-180, 2010.
TOLEDO, J.J.. Influência do solo e topografia sobre a mortabilidade de árvores e
decomposição de madeira em uma floresta de terra-firme na Amazônia Central.
Tese de doutorado, 2009, 75p., INPA/UFAM, Manaus, AM.
WUNDER, S. et al. Pagamentos por Serviços Ambientais: perspectivas para a
Amazônia Legal. Brasília: Ministério do Meio Ambiente, 2008 (Série Estudos, 10).
Disponível em:
http://www.ekosbrasil.org/media/file/PagtoServAmbietaisAmzMMA.pdf. Acessado em:
05 jun. 2011.

64
CAPITULO II
INFLUÊNCIA DO SOLO E TOPOGRAFIA SOBRE A BIOMASSA VEGETAL EM
UMA FLORESTA DE TERRA - FIRME NA AMAZÔNIA ORIENTA¹
Leidiane Leão de Oliveira
Alan Cavalcanti da Cunha
Flavia Capelloto Costa
¹Acta Amazônica ISSN: 0044-5967

65
Resumo
Em escala Amazônica, diferenças na biomassa entre regiões são fortemente
influenciadas pelos nutrientes do solo. No entanto, em estudos de mesoescala na
Amazônia Central a variação na biomassa foi principalmente relacionada com a
topografia, com pouco efeito dos nutrientes. Este fato pode indicar que, nas áreas da
Amazônia Central onde foram realizados estes estudos, há diferenças de padrão onde as
variáveis que mais influenciam a biomassa dependem da escala. Neste trabalho,
mostramos que a variação de biomassa em mesoescala num sítio no Leste da Amazônia
é semelhante à área da Amazônia Central. Nós usamos inventários florestais conduzidos
em trinta parcelas permanentes de 1ha para avaliar a variação espacial nas estimativas
de biomassa arbórea viva acima do solo (AGLB), e a relação entre o solo (textura e
nutrientes) e topografia (altitude e inclinação). As parcelas, localizadas na Floresta
Nacional do Amapá, no Leste da Amazônia, distribuídas sistematicamente sobre 25
km². As parcelas apresentaram 250 m de comprimento e 40 m de largura que seguiram a
curva de nível. Análises químicas e físicas do solo foram realizadas utilizando amostras
superficiais do solo, coletadas em cada parcela.As parcelas representaram um gradiente
variando de 33,5% a 81,7% no teor de areia, a inclinação variou de 0,5º a 12,9º e a
diferença de altitude foi 50 m, variando de 99 a 149. A média total de AGLB
(arvores+palmeiras+lianas) para troncos acima de 1cm e diâmetro a altura do peito
(DAP =1,3 m), foi de 530,5 Mg.ha-1
(variando de 194,6 a 968,4) e foi similar a outros
estudos da região. Regressões múltiplas mostram que a AGLB foi positivamente
relacionada com a altitude do terreno e não esteve correlacionada com a inclinação,
enquanto os nutrientes não afetaram esta variação. A altitude do terreno explicou 30%
da variação na AGLB. Isto indica que as diferenças entre os estudos anteriores foram
devido à diferenças nas escalas de estudo e não a efeitos locais. Nós consideramos
árvores, palmeiras e lianas separadamente, e verificamos que a biomassa de árvores
tendeu a aumentar em solos argilosos e localizados em áreas mais elevadas. Enquanto a
biomassa de palmeiras tendeu a diminuir com o aumento da inclinação, as lianas não
apresentaram efeitos com as variáveis analisadas. A biomassa de árvores de tamanhos
grandes (DAP≥100 cm) concentrou-se principalmente nas áreas planas e altas. A
variação da biomassa foi influenciada pela heterogeneidade dos diferentes ambientes
gerados pelo solo e a topografia. Este fato pode ser explicado pela altitude que foi
correlacionada com as características do solo e a mesma pode sobrepor as características
do solo para prever a variação espacial de biomassa viva acima do solo na FLONA do
Amapá. Concluímos que os dados de altitude podem ser facilmente obtidos usando
imagem de satélite, os quais poderiam melhorar as estimativas atuais de estoques de
carbono sobre grandes áreas de floresta no Estado do Amapá.
Palavras-chave: Biomassa, solo, topografia, Amazônia Oriental.

66
Abstract
In Amazonian scale, differences in biomass between regions are strongly influenced by
soil nutrients. However, in a study of mesoscale variation in Central Amazonia biomass
was primarily related to topography and nutrients had little effect. This may indicate
that the area of the Central Amazon, where the study was conducted is different, or that
the variables that most influence the biomass depend on the scale of the study. We show
that the mesoscale variation of biomass at a site on the outskirts of the Amazon is
similar to the area of Central Amazonia. We used forest inventories, conducted in thirty
1-ha permanent plots to assess the variation in tree, palms e lianas aboveground live
biomass (AGLB), and its relation with soil gradients (texture and nutrients) and
topography (altitude and slope). Our plots, located at Forest National Amapá in
Amazonia oriental, were systematically spread over 25 km². The plots were long
(250m) and narrow (up to 40 m), following elevational contours. Chemical and physical
soil analyses were undertaken using topsoil samples, collected in each plot. The plots
covered a soil textural gradient ranging from 33,5 to 81,7% percent of sand, slopes
varying from 5º to 12,9º, and the maximum difference of altitude was 50 m, ranging
from 99 to 149. The mean total AGLB (Lianas + palm + trees) for stems over 1 cm
diameter breast height (dbh = 1,3 m), was 530,5 Mg.ha-1
(ranging from 149,6 to 968,4)
and was similar to other studies in the region. Multiple regressions show that AGLB
was positively related to the altitude of the terrain and was not correlated with the slope,
while the nutrients did not affect this variation. The terrain altitude explained 30% of
the variation in AGLB. This indicates that the difference between the earlier studies due
to the difference in scales of study and not the effect locations. When considering trees,
palms and lianas separately, we found that the trees biomass tended to increase in clay
soils and located in higher areas, while the palm biomass tended to decrease with
increasing slope, lianas showed no effects with the variables analyzed. The biomass of
trees of large sizes (DBH ≥ 100cm) are concentrated mainly in flat areas and high. The
variation of biomass was influenced by the heterogeneity of different environments
generated by the soil and topography. For the altitude was highly correlated with soil
characteristics. The altitude can replace soil characteristics to predict the spatial
variation in living biomass above the ground in the National Forest of Amapá. Elevation
data can be easily obtained using satellite image and could improve current estimates of
carbon stocks over large areas of forest in Amapá.
Key words: Biomass, soil, topography, Amazonia oriental.

67
1. Introdução
Ao longo da bacia Amazônica, variações de biomassa (MALHI et al., 2004) na
composição florística (ter STEEGE et al., 2006) e na densidade da madeira (BAKER et
al., 2004; CHAO et al.,2008) estão associadas à fertilidade do solo. Porém, o que
controla a produtividade da floresta Amazônica ou da bacia Amazônica em grande
escala ou regional deverá ser determinada com precisão (QUESADA et al., 2009).
Em escala Amazônica as diferenças na biomassa entre regiões são fortemente
influenciadas pelos nutrientes do solo. No entanto, em estudos de mesoescala na
Amazônia Central a variação na biomassa tem sido principalmente relacionada com a
topografia com pouco efeito dos nutrientes (CHAVEAL et al., 1987; LAURANCE et
al., 1999; NASCIMENTO e LAURANCE, 2001; LUIZÃO et al., 2004; CASTILHO et
al., 2006). Isto pode indicar que a área da Amazônia Central, onde foi feito o estudo é
diferente, ou que as variáveis que mais influenciam a biomassa dependem da escala do
estudo.
A importância desse tipo de abordagem metodológica é obter estimativas mais
acuradas do estoque de biomassa florestal. Em conseqüência, é possível quantificar ou
estimar emissões de carbono com base na biomassa. Isto facilita a compreensão dos
processos ecológicos resultantes das mudanças de cobertura e uso da terra (PHILLIPS
et al., 2009; MALHI, 2010).
Incertezas a respeito do estoque de biomassa florestal são a maior fonte de
dúvidas nas estimativas de emissão de carbono (FEARNSIDE 1996, BAKER et al.
2004; HOUGHTON, 2005; SAATCHI et al., 2007; MALHI, 2010). Para entender o
efeito das mudanças climáticas sobre a floresta e as perdas de floresta sobre o clima são
necessários estudos no nível de comunidades e em escala regional (FIELD, 2001,
NEVES et al., 2011), ou que considerem as variações espaciais de estrutura e estoque de
biomassa da floresta (QUESADA et al., 2009).
Estudos realizados na Amazônia Central sugerem que cerca de 30% da variação
espacial de biomassa viva acima do solo é determinada por características físicas do
solo e topografia (NASCIMENTO e LAURANCE, 2001; CASTILHO et al., 2006). A
distribuição de espécies de palmeiras (COSTA et al., 2009) e ervas de sub-bosque
(KINUPP e MAGNUSSON, 2005; COSTA et al., 2005), mortalidade de árvores

68
(TOLEDO, 2009) e densidade de lianas (NOGUEIRA, 2006) também são relacionados
com a textura do solo e a topografia da região.
Castilho et al. (2006) mostraram que um terço da variação espacial da biomassa
arbórea foi associada com a textura do solo, altitude e inclinação. Como os solos na
Amazônia não são mapeados em alta resolução, os autores sugeriram que a altitude e a
inclinação fossem utilizadas para prever a variação de biomassa em larga escala, pois
essas variáveis topográficas podem ser obtidas com maior facilidade através de mapas,
ou de imagens do modelo digital de elevação do terreno derivadas do radar
interferométrico SRTM (Shuttle Radar Topographic Mission), disponibilizadas pela
NASA.
A implantação de parcelas permanentes, para estudos de longo prazo em
florestas tropicais, oferece um grande potencial analítico para monitorar os estoques de
carbono acima do solo (LANDEIRO e MAGNUSSON, 2011; QUESADA et al., 2009;
MALHI E GRACE, 2000; NASCIMENTO e LAURANCE, 2001; CASTILHO et al.,
2006; MAGNUSSON et al., 2005). A quantidade de carbono estocada na vegetação
pode ser obtida através de estimativas de biomassa.
Incertezas a respeito de medidas confiáveis de biomassa e suas variações no
espaço, apesar da maioria dos estudos utilizar pequenas unidades amostrais, também se
apresentam com poucas repetições, sendo que estas parcelas muitas vezes não são
estabelecidas aleatoriamente através da paisagem (KELLER, PALACE E HURTT,
2001). Alem disso, muitos estudos que relacionam a variação de biomassa com as
variáveis topográficas e do solo foram realizados em pequenas escalas amostrais, que
não capturam a variabilidade efetiva da área (da SILVA et al., 2002; VASCONCELOS
et al., 2003).
O Projeto de Pesquisa em Biodiversidade (PPBio), núcleo Amapá, é
desenvolvido em uma área de 25 km² na Floresta Nacional (FLONA) do Amapá, na
qual foram instaladas 30 parcelas de 1 ha para avaliar a variação de biomassa viva
acima do solo. As parcelas obedeceram ao método RAPELD (MAGNUSSON et al.,
2005, LANDEIRO e MAGNUSSON, 2011), cujo método consiste na distribuição
sistemática de parcelas na paisagem e que permite estimativas não tendenciosas da
distribuição, obtenção da abundância e da biomassa das espécies em cada sítio, além de
comparações biogeográficas entre sítios. Este desenho amostral minimiza a variação
interna de topografia e do solo em cada parcela. Além disso, permite o uso destas

69
variáveis (ambientais) como preditoras das distribuições de espécies (MAGNUSSON et
al., 2005, LANDEIRO e MAGNUSSON, 2011).
Na Amazônia Central, a relação da biomassa de árvores e lianas com o solo
topografia depende do tamanho do Diâmetro à Altura do Peito (DAP) (CASTILHO et
al., 2006; NOGUEIRA, 2006). Porém, ainda falta determinar classes de DAP de acordo
com as relações entre a biomassa com as características do solo. A textura do solo foi
altamente associada com a topografia na Amazônia central (LUIZÃO et al., 2005;
CASTILHO et al., 2006). Nessa região geralmente solos argilosos ocorrem em áreas
mais altas e solos arenosos ocorrem em áreas mais baixas (CHAVEAL et al., 1987).
Com base nestes resultados a variação de biomassa viva acima do solo pode ser
explicada apenas com dados topográficos (altitude e inclinação) (CASTILHO et al.,
2006). E a vantagem deste tipo de observação é que estes dados podem ser obtidos
facilmente por intermédio de mapas topográficos disponibilizados para a Amazônia.
Outra vantagem é que podem ser obtidos do modelo digital de elevação do terreno
(DEM).
O DEM é obtido através do radar SRTM, disponibilizado pela NASA. No
presente estudo, demonstramos uma correlação entre as variáveis topográficas e
edáficas. De modo similar, é possível utilizar a mesma técnica aplicada na Amazônia
Central para melhor estimar as distribuições da biomassa na FLONA do Amapá. Deste
modo, a topografia poderia ser utilizada em substituição parcial às variáveis edáficas e,
assim, ser utilizada como variável preditora efetiva para as distribuições da biomassa
com menor esforço experimental e economia logística de campo.
Neste estudo, iremos mostrar que a variação de biomassa em mesoescala num
sítio da periferia da Amazônia é semelhante à área da Amazônia Central. Para tanto,
utilizamos dados de estimativa de biomassa viva acima do solo (AGBL), solos e
topografia de 30 parcelas de 1 ha na Floresta Nacional do Amapá para responder às
seguintes questões:
1) Solo e topografia explicam a variação das estimativas de biomassa de árvores,
palmeiras e lianas na FLONA do Amapá?
2) Dada a correlação positiva entre a textura do solo e a altitude, somente a altitude
pode substituir as características do solo para prever a variação espacial da biomassa
viva acima do solo?

70
3) Diferentes classes dimensionais de árvores mostram a mesma relação entre suas
estimativas de biomassa e as características do solo e topografia?
4) Os fatores que afetam a variação de biomassa vegetal na Amazônia central são os
mesmos encontrados neste estudo?
Para responder a estas perguntas, os seguintes métodos de investigação foram
aplicados.
2. Material e MétodosErro! Indicador não definido.
2.1. Área de estudo, delineamento amostral e dados de estrutura da vegetação
O levantamento dos dados de campo foi realizado na Floresta Nacional do
Amapá, Amapá, Brasil entre outubro de 2009 a julho de 2010. A Floresta Nacional do
Amapá é definida como uma floresta tropical úmida de terra firme, com área de 412.000
ha (ICMBIO) (Figura 1). A altura máxima do dossel encontrada foi 50 m e o diâmetro
a altura do peito (DAP) máximo foi de 133,69 cm (PEREIRA et al., 2007).
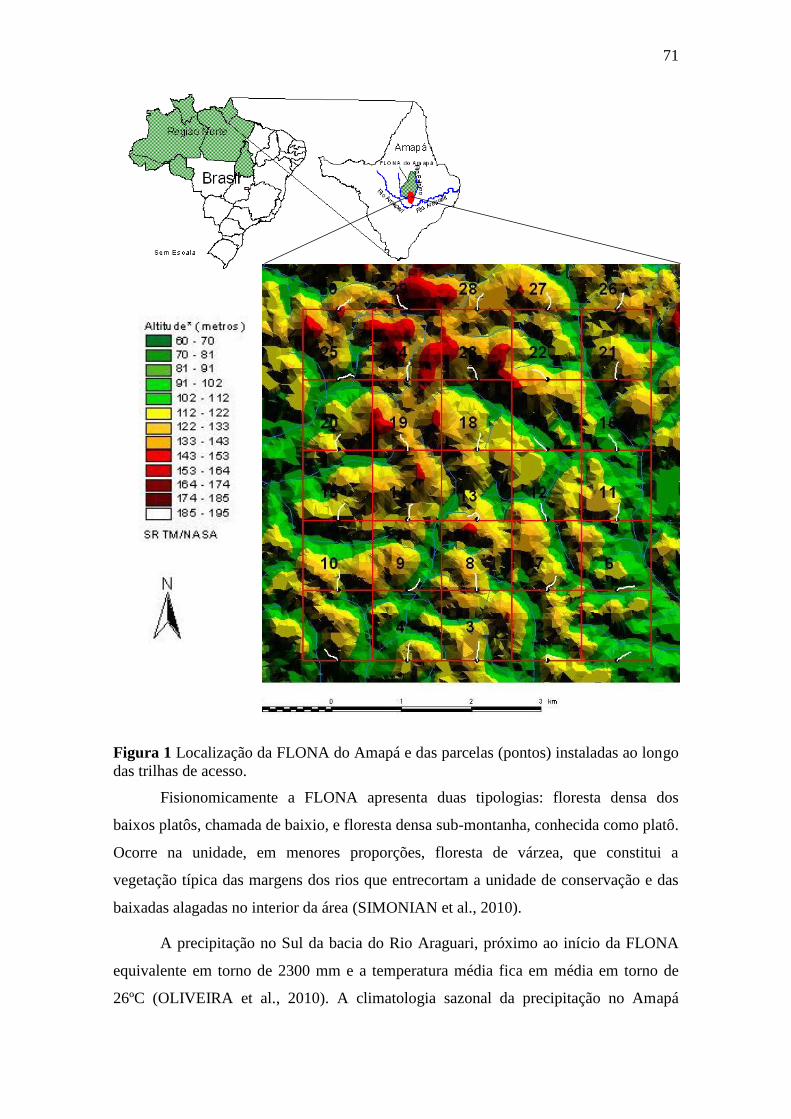
71
Figura 1 Localização da FLONA do Amapá e das parcelas (pontos) instaladas ao longo
das trilhas de acesso.
Fisionomicamente a FLONA apresenta duas tipologias: floresta densa dos
baixos platôs, chamada de baixio, e floresta densa sub-montanha, conhecida como platô.
Ocorre na unidade, em menores proporções, floresta de várzea, que constitui a
vegetação típica das margens dos rios que entrecortam a unidade de conservação e das
baixadas alagadas no interior da área (SIMONIAN et al., 2010).
A precipitação no Sul da bacia do Rio Araguari, próximo ao início da FLONA
equivalente em torno de 2300 mm e a temperatura média fica em média em torno de
26ºC (OLIVEIRA et al., 2010). A climatologia sazonal da precipitação no Amapá

72
(1978-2007) revelou que período mais seco ocorre de setembro a novembro e o mais
chuvoso ocorre de março a maio (SOUZA e CUNHA, 2010).
O relevo da FLONA do Amapá é predominantemente plano, com partes
suavemente onduladas e uma altitude média de 100 m acima do nível do mar. Existem
dentro da unidade vários afloramentos rochosos que se projetam tanto no interior da
floresta, quanto acima da copa das árvores. Os solos predominantes são os argissolos
(latossolo amarelo, latossolo vermelho-amarelo, latossolo vermelho-amarelo podzólico
e podzólico vermelho-amarelo) (SIMONIAN et al., 2010).
2.2. Dados de solo e topografia
2.2.1. Solo
Foram coletadas amostras de estrutura de solo deformadas (para análise da
composição química) e indeformadas (para análise da composição física) em três pontos
(10, 120 e 250m) ao longo do eixo mais longo de cada uma das 30 parcelas que seguem
a curva de nível do terreno, durante o período de janeiro a julho de 2010. O material
para análise da composição química do solo referente a pH, matéria orgânica (MO),
fósforo (P), potássio (K), cálcio-magnésio (Ca+Mg), cálcio (Ca), alumínio (Al), acidez
total (H+Al), soma de bases (SB), capacidade de troca catiônica (CTC), saturação por
bases (V) e saturação por alumínio (M), foi coletado com trado holandês, nas
profundidades de 0-5, 5-10, 10-20, e 20-40 cm, após remoção da camada de liteira. Uma
amostra composta foi obtida por ponto onde cada amostra era composta por quatro
profundidades. Esse procedimento reduz a quantidade de dados por ponto (VAN DE
BERG, 2001; VAN DE BERG e SANTOS, 2003).
As amostras foram homogeneizadas e armazenadas em sacos plásticos,
devidamente etiquetados, seguindo para análise no Laboratório de Solos da Embrapa
Amapá, conforme metodologia prescrita por Embrapa (1997).
As amostras de estrutura indeformada objetivaram a análise das propriedades
físicas (densidade aparente, densidade de partícula, porosidade, umidade e textura). As
coletas foram feitas com anéis volumétricos de 98cm3 com auxílio de um Trado Huland
na profundidade (0-5 cm). Apenas esta profundidade foi escolhida com base em Martins
et al. (2004), os quais relatam que nesta profundidade existe grande importância da
absorção de nutrientes pela camada de raízes finas presentes no perfil do solo. As

73
análises também foram realizadas no Laboratório de Solos da Embrapa Amapá, todas
conforme metodologia de Embrapa (1997).
Foram utilizadas 16 variáveis de solo: Argila (%), Silte (%), Areia (%),
Densidade aparente (DA), Densidade de Partícula (DP), Porosidade (PR), Umidade
(UM), pH, matéria orgânica (MO), fósforo (P), potássio (K), cálcio-magnésio (Ca+Mg),
alumínio (Al), acidez total (H+Al), soma de bases (SB) e capacidade de troca catiônica
(CTC) com o objetivo de relacionar estas variáveis com a biomassa viva acima do solo.
.
2.2.2. Topografia
As variáveis topográficas utilizadas neste estudo foram à altitude e a inclinação
do terreno. Todas as parcelas de curva de nível foram georreferenciadas com o auxílio
de um GPS (Global Positioning System) – Modelo Garmin GPSMAP 60CSx. Como as
parcelas seguem uma curva de nível, o valor da altitude é constante em toda extensão da
mesma, semelhante a metodologia utilizada por Castilho, 2004. A altitude foi calculada
através de imagens do modelo digital de elevação do terreno derivados do radar
interferométrico SRTM (Shutter Radar Topographic mission), disponibilizados através
de mapas pelo Programa de Pesquisa em Biodiversidade Tropical (PPBio – Núcleo
Amapá). Já as medidas de inclinação do terreno foram feitas com um clinômetro em
seis pontos distanciados 50 m um do outro ao longo da linha central de cada parcela (0,
50, 100, 150, 200, 250 m). Em cada ponto, a medida de inclinação foi obtida sobre uma
distância de 3 m, perpendicular à linha central da parcela (1,5 m de cada lado da linha).
A média das medidas foi utilizada para representar a inclinação da parcela (CASTILHO
et al., 2006).
2.3. Estimativas de biomassa arbórea viva acima do solo
Foi estimada a biomassa viva acima do solo de árvores, palmeiras e lianas nas 30
parcelas, distribuídas sistematicamente sobre 25 km². As parcelas mediam 250 m de
comprimento por 40 m de largura, e que seguiram com seu eixo maior a curva de nível
(MAGNUSSON et al., 2005; LANDEIRO e MAGNUSSON, 2011). Com base nas
medidas de Diâmetro à Altura do Peito (DAP), inicialmente para as duas primeiras
classes de DAP foram utilizadas equações alométricas “calibradas” para floresta
Amazônica (NASCIMENTO e LAURANCE, 2002; HIGUCHI et al., 1998). Para
árvores acima de 10 cm de DAP foi empregada a equação obtida próximo a área de

74
estudo (OLIVEIRA, SOTTA e HIGUCHI, 2012), com precisão de 97% para estimativa
de biomassa. Para as estimativas de biomassa de palmeiras utilizou-se uma equação
alométrica desenvolvida a partir de várias espécies de palmeiras que ocorrem na região
do alto Rio Negro na Venezuela e Colômbia (SALDARRIAGA et al., 1988). Para as
estimativas de biomassa de lianas utilizou-se uma equação alométrica desenvolvida por
Schnitzer et al. (2006) (Tabela 1).
Tabela 1 Equações alométricas utilizadas para estimar a biomassa seca acima do solo
(kg. ha-1
) com base no diâmetro à altura do peito (DAP, cm) e altura (H, cm).
Indivíduos Equação usada no cálculo da Biomassa aérea
Árvores DAP≥ 1 cm e DAP< 5 cm e(-1,7689 + 2,3770*ln (DAP)) (a)
Árvores DAP≥ 5 cm e DAP< 10 cm e(-1,754 + 2,665* ln (DAP))*0,6 (b)
Árvores DAP≥ 10 cm e(-1,096367 + 2,437071*ln (DAP)) *0,6 (c)
Palmeiras DAP≥ 1 cm e(-6,3789 – 0.877*ln (1/DAP2)+2,151* ln (H) (d)
Lianas DAP ≥ 0,5 cm
e(-1,484 + 2,657*ln (DAP)) (e)
aNascimento e Laurance (2002)
bHiguchi et al. (1998)
cOliveira, Sotta e Higuchi, (2012)
dSaldarriaga et al. (1988)
eSchnitzer et al. (2006). As equações de
bHiguchi et al. (1998)
e cOliveira et al. (2010) fornecem estimativas de biomassa em peso fresco. Uma vez que
o peso seco do tronco corresponde acerca de 60% de seu peso fresco; e o da copa
corresponde a 58% de seu peso fresco, o resultado da equação foi multiplicado por 0,6
para ser expresso em peso seco (Castilho, 2004).
A equação alométrica para estimativa de biomassa de palmeiras utiliza além do
diâmetro, a altura dos indivíduos. Uma vez que não medimos a altura dos indivíduos no
campo, definimos uma altura padrão espécie-específica, como 15 m. Para os testes de
hipóteses sobre as relações entre a estrutura da vegetação e as variáveis ambientais,
foram utilizadas as estimativas de biomassa como indicador categórico destas variações.

75
2.4. Análise dos dados
Para representar a variação nas condições edáficas em um número menor de
dimensões, foi feita uma Análise de Componentes Principais. Todas as variáveis
utilizadas foram padronizadas pela amplitude para que recebessem aproximadamente o
mesmo peso na análise. A ordenação resultante captou pouca variância nos dois
primeiros eixos (34 %), indicando que há pouca variação entre as variáveis edáficas na
área de estudo. Portanto, decidiu-se usar na regressão múltipla apenas as variáveis que
representam a textura e a fertilidade do solo.
Regressões múltiplas lineares foram utilizadas para relacionar as variáveis
ambientais (textura do solo, fertilidade do solo, altitude e inclinação) à biomassa
lenhosa total (biomassa de árvores + palmeiras + lianas) e à biomassa arbórea, de
palmeiras e de lianas. As variáveis edáficas e a altitude não foram utilizadas no mesmo
modelo porque foram fortemente correlacionadas (CASTILHO et al., 2006; TOLEDO,
2009). Foram testados 3 modelos para cada grupo biológico: 1) um modelo puramente
edáfico, incluindo textura e fertilidade do solo, 2) um modelo puramente topográfico,
incluindo altitude e inclinação; 3) um modelo completo, incluindo um preditor edáfico
(fertilidade) e os prediores topográficos (altitude e inclinação). A textura do solo não foi
incluída no modelo completo por estar correlacionada com a altitude e inclinação.
Visando determinar se a variação de biomassa com os diferentes gradientes do
solo e topografia é dependente do tamanho das árvores, as separamos em várias classes
de DAP (1≤DAP<10; 10,1≤DAP<20; 20,1≤DAP<30; 30,1≤DAP<40; 40,1≤DAP<50;
50,1≤DAP<60; 60,1<DAP≤70; 70<DAP) e calculamos a biomassa para cada uma delas.
Testamos o efeito do solo, altitude e inclinação sobre a biomassa arbórea total e depois
para árvores de cada classe de diâmetro. Todas as análises foram realizadas no
programa R 2.14.0 (R Development Core Team, 2011).

76
3. Resultados
3.1. Topografia e gradientes do solo
A inclinação do terreno variou desde terrenos planos (0,5º) até os mais íngremes
(12,9º), com uma inclinação média de 6,1º (dp = 3,6º). A diferença de altitude entre as
parcelas foi de 50 m (mínimo = 99 m; máximo = 149 m), e a altitude (média ± dp) das
parcelas amostradas foi 121 ± 13,4 m (Tabela 2).
As parcelas representaram uma variação de 33,5% a 81,7% no teor de areia. As
parcelas localizadas em solos arenosos (mais de 50% de areia) foram mais comuns em
toda grade totalizando 87% do total de parcelas, enquanto o teor de argila variou de
1,5% a 48,2% (Tabela 2).
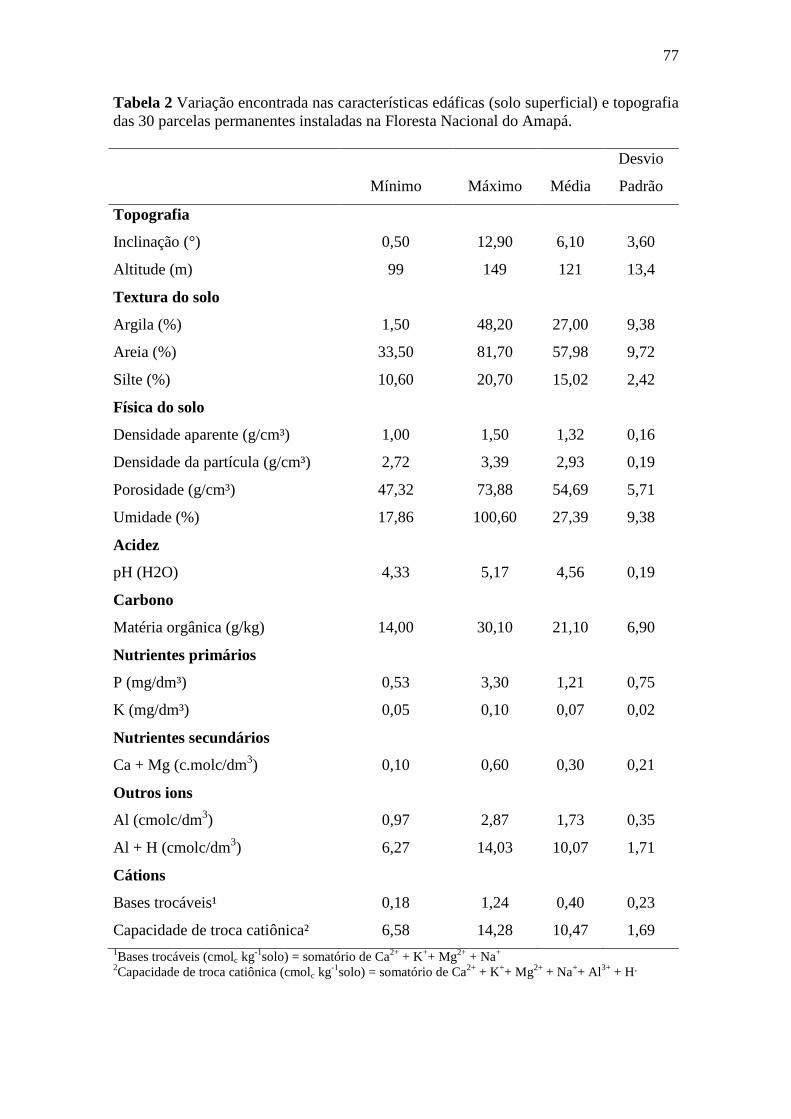
77
Tabela 2 Variação encontrada nas características edáficas (solo superficial) e topografia
das 30 parcelas permanentes instaladas na Floresta Nacional do Amapá.
Mínimo Máximo Média
Desvio
Padrão
Topografia
Inclinação (°) 0,50 12,90 6,10 3,60
Altitude (m) 99 149 121 13,4
Textura do solo
Argila (%) 1,50 48,20 27,00 9,38
Areia (%) 33,50 81,70 57,98 9,72
Silte (%) 10,60 20,70 15,02 2,42
Física do solo
Densidade aparente (g/cm³) 1,00 1,50 1,32 0,16
Densidade da partícula (g/cm³) 2,72 3,39 2,93 0,19
Porosidade (g/cm³) 47,32 73,88 54,69 5,71
Umidade (%) 17,86 100,60 27,39 9,38
Acidez
pH (H2O) 4,33 5,17 4,56 0,19
Carbono
Matéria orgânica (g/kg) 14,00 30,10 21,10 6,90
Nutrientes primários
P (mg/dm³) 0,53 3,30 1,21 0,75
K (mg/dm³) 0,05 0,10 0,07 0,02
Nutrientes secundários
Ca + Mg (c.molc/dm3) 0,10 0,60 0,30 0,21
Outros ions
Al (cmolc/dm3) 0,97 2,87 1,73 0,35
Al + H (cmolc/dm3) 6,27 14,03 10,07 1,71
Cátions
Bases trocáveis¹ 0,18 1,24 0,40 0,23
Capacidade de troca catiônica² 6,58 14,28 10,47 1,69
1Bases trocáveis (cmolc kg
-1solo) = somatório de Ca
2+ + K
++ Mg
2+ + Na
+
2Capacidade de troca catiônica (cmolc kg
-1solo) = somatório de Ca
2+ + K
++ Mg
2+ + Na
++ Al
3+ + H+

78
A densidade aparente das parcelas amostradas apresentou valores de 1,00 a 1,50
g/cm3, com (média ± dp) de 1,32 ± 0,16 g/cm
3 e densidade de partícula de 2,72 a 3,39
g/cm3, com (média ± dp) de 2,93 ± 0,19 g/cm
3. A porosidade apresentou valores
percentuais entre 47,32 e 73,88%, com (média ± dp) de 54,69 ± 5,71 g/cm3 e a umidade
variou de 17,86 a 100, 6 %. Com (média ± dp) de 27,39 ± 9,38% (Tabela 2).
Os valores de pH variaram de 4,3 a 5,2, com (média ± dp) de 4,56 ± 0,19. A
matéria orgânica apresentou valores de 14,0 a 30,1 g/Kg (média ± dp) de 21,1 ± 2,9. O
Fósforo apresentou pequena variação entre 0,53 (muito baixos) a 3,3 (baixos) com
(média ± dp) de 1,21 ± 0,75. Para os cátions trocáveis (K+, Ca
+2, Ca+Mg) estes
apresentaram valores baixos. A soma de bases (SB) e a capacidade de troca catiônica
(CTC) foram consideradas baixas, com valores de 0,18 a 1,24 (0,4 ± 0,23) para SB e de
6,6 a 14,3 (10,5± 1,7) para CTC (Tabela 2).
3.2. Gradientes do solo e sua relação com a topografia
O grau de correlação entre a maioria das variáveis edáficas e topográficas foi
baixo, com algumas exceções (Tabela 3). Dentre as variáveis correlacionadas, altitude e
graulometria do solo tiveram a relação mais forte. Quanto maior a altitude maior a
proporção de argila no solo e menor a de areia. A porosidade e a densidade aparente
apresentaram uma forte correlação inversa, ou seja, quanto maior a porosidade, menor a
densidade aparente, conforme esperado. Logo, a maior quantidade de partículas no
mesmo volume total só ocorrerá em detrimento de tal espaço, ou seja, quanto maior a
densidade aparente, menor a quantidade de poros ou porosidade (AMARO FILHO et
al., 2008; SOUSA, 2010).
Com relação às demais variáveis, que apresentaram correlações menores, a
altitude apresentou uma correlação positiva com a quantidade de alumínio no solo e
matéria orgânica. Por outro lado, o pH diminui com o aumento da quantidade de
alumínio.
Um dos fatores que contribuem com o aumento do pH é a rápida decomposição
da matéria orgânica, favorecida pelos ambientes de altitudes elevadas e terrenos planos,
uma vez que a umidade permanente em áreas baixas, dificulta o processo de
decomposição da matéria orgânica (LIMA et al. 2006).
Os nutrientes do solo (P, K, Ca+ Mg) tiveram uma relação positiva com o aumento da
quantidade de MO. A proporção de argila e alumínio aumentam com a inclinação do
terreno, enquanto a proporção de areia diminui.
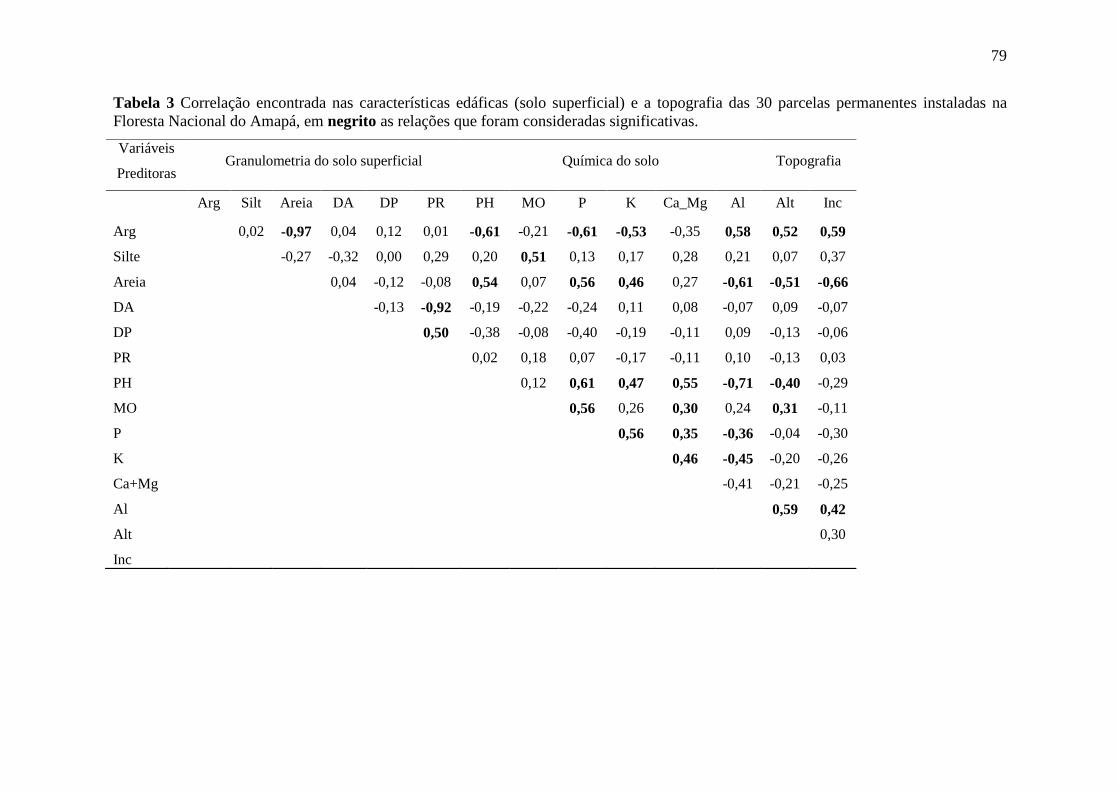
79
Tabela 3 Correlação encontrada nas características edáficas (solo superficial) e a topografia das 30 parcelas permanentes instaladas na
Floresta Nacional do Amapá, em negrito as relações que foram consideradas significativas.
Variáveis
Preditoras Granulometria do solo superficial Química do solo Topografia
Arg Silt Areia DA DP PR PH MO P K Ca_Mg Al Alt Inc
Arg 0,02 -0,97 0,04 0,12 0,01 -0,61 -0,21 -0,61 -0,53 -0,35 0,58 0,52 0,59
Silte -0,27 -0,32 0,00 0,29 0,20 0,51 0,13 0,17 0,28 0,21 0,07 0,37
Areia 0,04 -0,12 -0,08 0,54 0,07 0,56 0,46 0,27 -0,61 -0,51 -0,66
DA -0,13 -0,92 -0,19 -0,22 -0,24 0,11 0,08 -0,07 0,09 -0,07
DP 0,50 -0,38 -0,08 -0,40 -0,19 -0,11 0,09 -0,13 -0,06
PR 0,02 0,18 0,07 -0,17 -0,11 0,10 -0,13 0,03
PH 0,12 0,61 0,47 0,55 -0,71 -0,40 -0,29
MO 0,56 0,26 0,30 0,24 0,31 -0,11
P 0,56 0,35 -0,36 -0,04 -0,30
K 0,46 -0,45 -0,20 -0,26
Ca+Mg -0,41 -0,21 -0,25
Al 0,59 0,42
Alt 0,30
Inc

80
3.3. Estimativas de biomassa vegetal viva acima do solo
A biomassa total (árvores + palmeiras + lianas) média por parcela foi de 530,5
Mg.ha-1
, com mínimo de 194,6 e máximo de 968,4 Mg.ha-1
. A biomassa média de
árvores por parcela foi 515,8 Mg.ha-1
(98,5%), variando de 177,6 a 966,4 Mg.ha-1
. A
biomassa média de palmeiras foi de apenas 0,6 Mg/ha (0,3%), variando de 0 a 14
Mg.ha-1
. Já a biomassa média de lianas foi de 6 Mg.ha-1
(1,2%), variando de 1,9 a 11,7
Mg.ha-1
(Tabela 4). Analisando o histograma de freqüência notamos que a maioria das
parcelas apresentou valores de biomassa de árvores entre 400 e 600 Mg/ha (Figura 2).
A biomassa média de palmeiras por parcela foi de 0,6 Mg.ha-1
, variando de 0 a
14 Mg.ha-1
. A maioria das parcelas (93%) apresenta valores de biomassa de palmeiras
inferior a 1 Mg.ha-1
. Aproximadamente 2% dos indivíduos de cada parcela são
palmeiras, representando em média 93 indivíduos com DAP > 1 cm por hectare. No
entanto, essa contribuição varia de 1 a 49 % dos indivíduos. A maior parte dos
indivíduos apresenta DAP inferior a 20 cm, mas alguns indivíduos atingiram DAP > 30
cm (Tabela 4).
A biomassa média de lianas por parcela é 6 Mg.ha-1
, representando 1,2% da
biomassa total, e variando de 0,78 a 3 Mg.ha-1
. A maioria das parcelas apresenta valores
de biomassa de lianas inferior a 2 Mg.ha-1
(Tabela 4). Aproximadamente 9% dos
indivíduos de cada parcela são lianas, representando (média) 470 indivíduos com DAP
> 1 cm por hectare, comparados com 93 ind.ha-1
(palmeiras) e 4843 ind.ha-1
(árvores). A
biomassa de palmeiras representou, em média, menos de 1% da biomassa arbórea total
por parcela. No entanto, em uma parcela a biomassa de palmeiras contribuiu com
aproximadamente 7% da biomassa total. A biomassa de lianas foi pouco significativa,
contribuindo apenas com 0.34% da biomassa total por parcela.
A biomassa arbórea total (árvores, palmeiras e lianas) por parcela é de 530,5
Mg.ha-1
, variando de 194,6 a 968,4 Mg.ha-1
(Tabela 4). A biomassa de árvores
apresenta forte correlação com a biomassa arbórea total por parcela (r = 0,99).
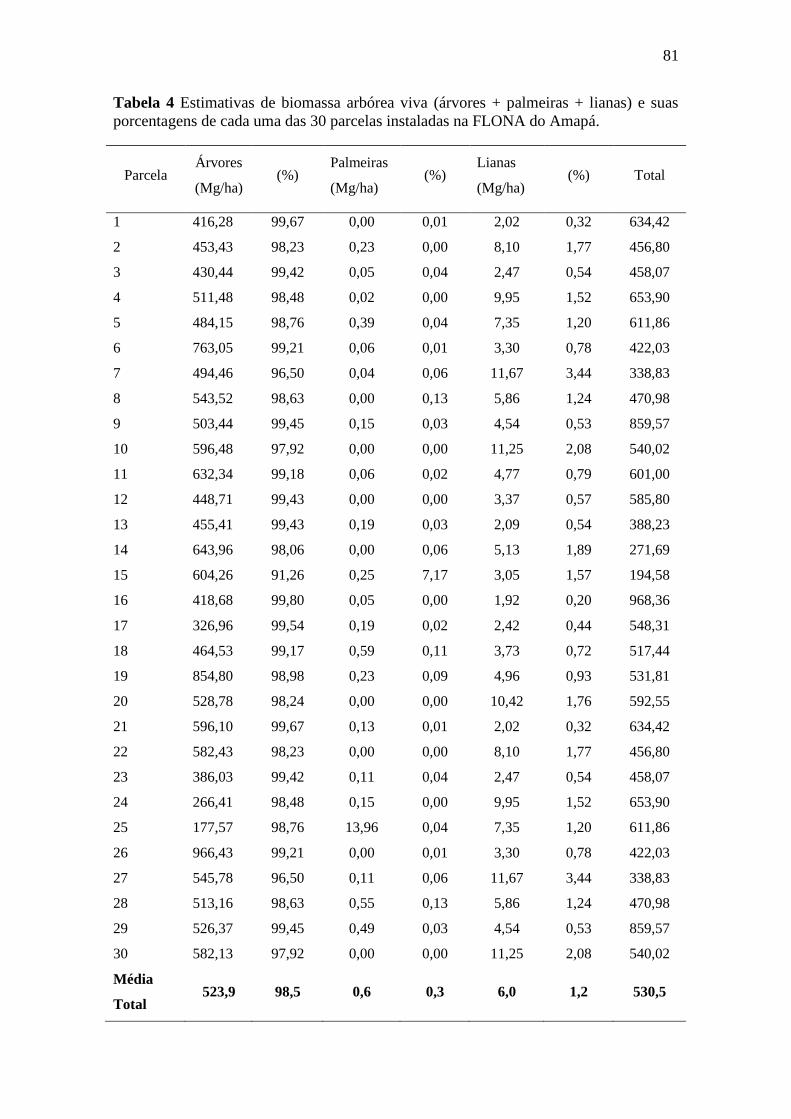
81
Tabela 4 Estimativas de biomassa arbórea viva (árvores + palmeiras + lianas) e suas
porcentagens de cada uma das 30 parcelas instaladas na FLONA do Amapá.
Parcela Árvores
(Mg/ha) (%)
Palmeiras
(Mg/ha) (%)
Lianas
(Mg/ha) (%) Total
1 416,28 99,67 0,00 0,01 2,02 0,32 634,42
2 453,43 98,23 0,23 0,00 8,10 1,77 456,80
3 430,44 99,42 0,05 0,04 2,47 0,54 458,07
4 511,48 98,48 0,02 0,00 9,95 1,52 653,90
5 484,15 98,76 0,39 0,04 7,35 1,20 611,86
6 763,05 99,21 0,06 0,01 3,30 0,78 422,03
7 494,46 96,50 0,04 0,06 11,67 3,44 338,83
8 543,52 98,63 0,00 0,13 5,86 1,24 470,98
9 503,44 99,45 0,15 0,03 4,54 0,53 859,57
10 596,48 97,92 0,00 0,00 11,25 2,08 540,02
11 632,34 99,18 0,06 0,02 4,77 0,79 601,00
12 448,71 99,43 0,00 0,00 3,37 0,57 585,80
13 455,41 99,43 0,19 0,03 2,09 0,54 388,23
14 643,96 98,06 0,00 0,06 5,13 1,89 271,69
15 604,26 91,26 0,25 7,17 3,05 1,57 194,58
16 418,68 99,80 0,05 0,00 1,92 0,20 968,36
17 326,96 99,54 0,19 0,02 2,42 0,44 548,31
18 464,53 99,17 0,59 0,11 3,73 0,72 517,44
19 854,80 98,98 0,23 0,09 4,96 0,93 531,81
20 528,78 98,24 0,00 0,00 10,42 1,76 592,55
21 596,10 99,67 0,13 0,01 2,02 0,32 634,42
22 582,43 98,23 0,00 0,00 8,10 1,77 456,80
23 386,03 99,42 0,11 0,04 2,47 0,54 458,07
24 266,41 98,48 0,15 0,00 9,95 1,52 653,90
25 177,57 98,76 13,96 0,04 7,35 1,20 611,86
26 966,43 99,21 0,00 0,01 3,30 0,78 422,03
27 545,78 96,50 0,11 0,06 11,67 3,44 338,83
28 513,16 98,63 0,55 0,13 5,86 1,24 470,98
29 526,37 99,45 0,49 0,03 4,54 0,53 859,57
30 582,13 97,92 0,00 0,00 11,25 2,08 540,02
Média
Total 523,9 98,5 0,6 0,3 6,0 1,2 530,5
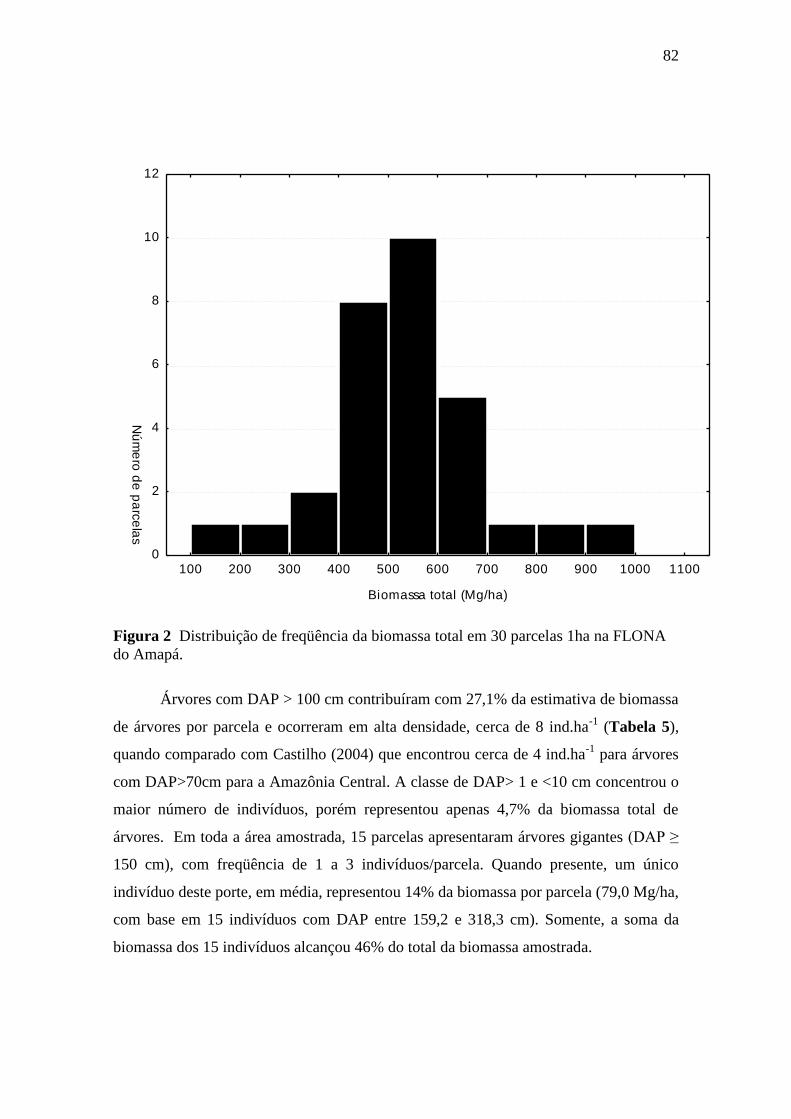
82
Figura 2 Distribuição de freqüência da biomassa total em 30 parcelas 1ha na FLONA
do Amapá.
Árvores com DAP > 100 cm contribuíram com 27,1% da estimativa de biomassa
de árvores por parcela e ocorreram em alta densidade, cerca de 8 ind.ha-1
(Tabela 5),
quando comparado com Castilho (2004) que encontrou cerca de 4 ind.ha-1
para árvores
com DAP>70cm para a Amazônia Central. A classe de DAP> 1 e <10 cm concentrou o
maior número de indivíduos, porém representou apenas 4,7% da biomassa total de
árvores. Em toda a área amostrada, 15 parcelas apresentaram árvores gigantes (DAP ≥
150 cm), com freqüência de 1 a 3 indivíduos/parcela. Quando presente, um único
indivíduo deste porte, em média, representou 14% da biomassa por parcela (79,0 Mg/ha,
com base em 15 indivíduos com DAP entre 159,2 e 318,3 cm). Somente, a soma da
biomassa dos 15 indivíduos alcançou 46% do total da biomassa amostrada.
100 200 300 400 500 600 700 800 900 1000 1100
Biomassa total (Mg/ha)
0
2
4
6
8
10
12
Nú
me
ro d
e p
arc
ela
s
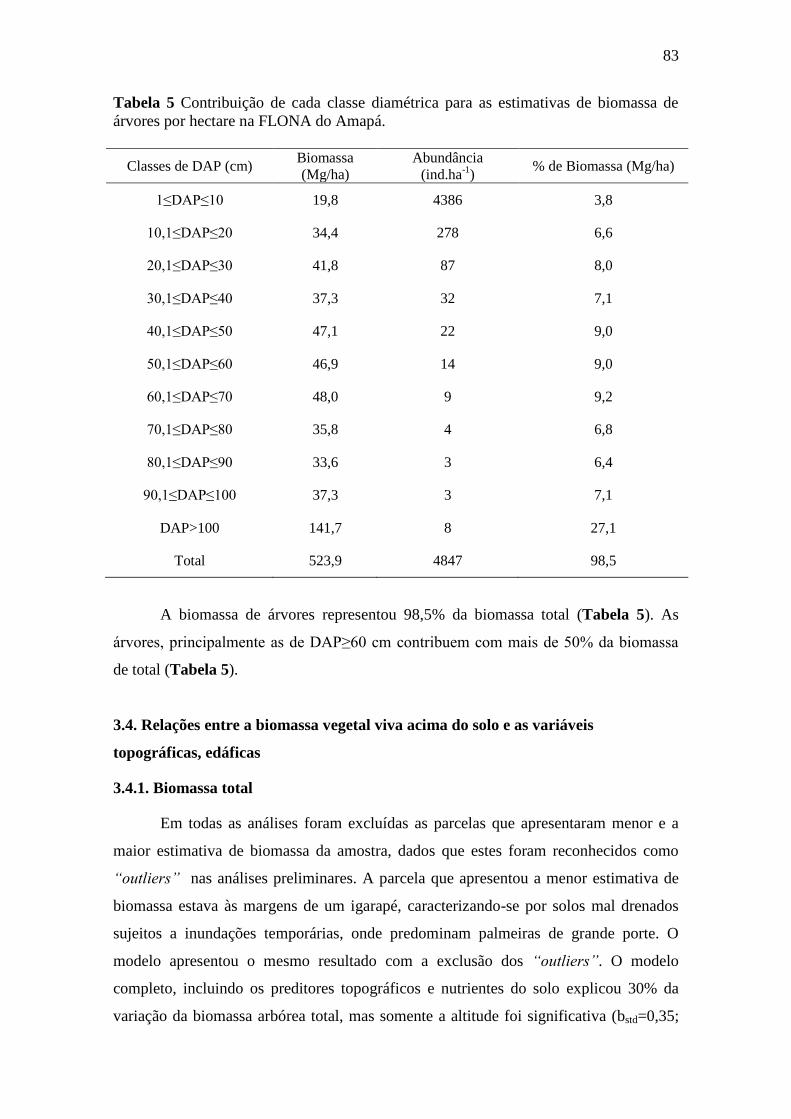
83
Tabela 5 Contribuição de cada classe diamétrica para as estimativas de biomassa de
árvores por hectare na FLONA do Amapá.
Classes de DAP (cm) Biomassa
(Mg/ha)
Abundância
(ind.ha-1
) % de Biomassa (Mg/ha)
1≤DAP≤10 19,8 4386 3,8
10,1≤DAP≤20 34,4 278 6,6
20,1≤DAP≤30 41,8 87 8,0
30,1≤DAP≤40 37,3 32 7,1
40,1≤DAP≤50 47,1 22 9,0
50,1≤DAP≤60 46,9 14 9,0
60,1≤DAP≤70 48,0 9 9,2
70,1≤DAP≤80 35,8 4 6,8
80,1≤DAP≤90 33,6 3 6,4
90,1≤DAP≤100 37,3 3 7,1
DAP>100 141,7 8 27,1
Total 523,9 4847 98,5
A biomassa de árvores representou 98,5% da biomassa total (Tabela 5). As
árvores, principalmente as de DAP≥60 cm contribuem com mais de 50% da biomassa
de total (Tabela 5).
3.4. Relações entre a biomassa vegetal viva acima do solo e as variáveis
topográficas, edáficas
3.4.1. Biomassa total
Em todas as análises foram excluídas as parcelas que apresentaram menor e a
maior estimativa de biomassa da amostra, dados que estes foram reconhecidos como
“outliers” nas análises preliminares. A parcela que apresentou a menor estimativa de
biomassa estava às margens de um igarapé, caracterizando-se por solos mal drenados
sujeitos a inundações temporárias, onde predominam palmeiras de grande porte. O
modelo apresentou o mesmo resultado com a exclusão dos “outliers”. O modelo
completo, incluindo os preditores topográficos e nutrientes do solo explicou 30% da
variação da biomassa arbórea total, mas somente a altitude foi significativa (bstd=0,35;
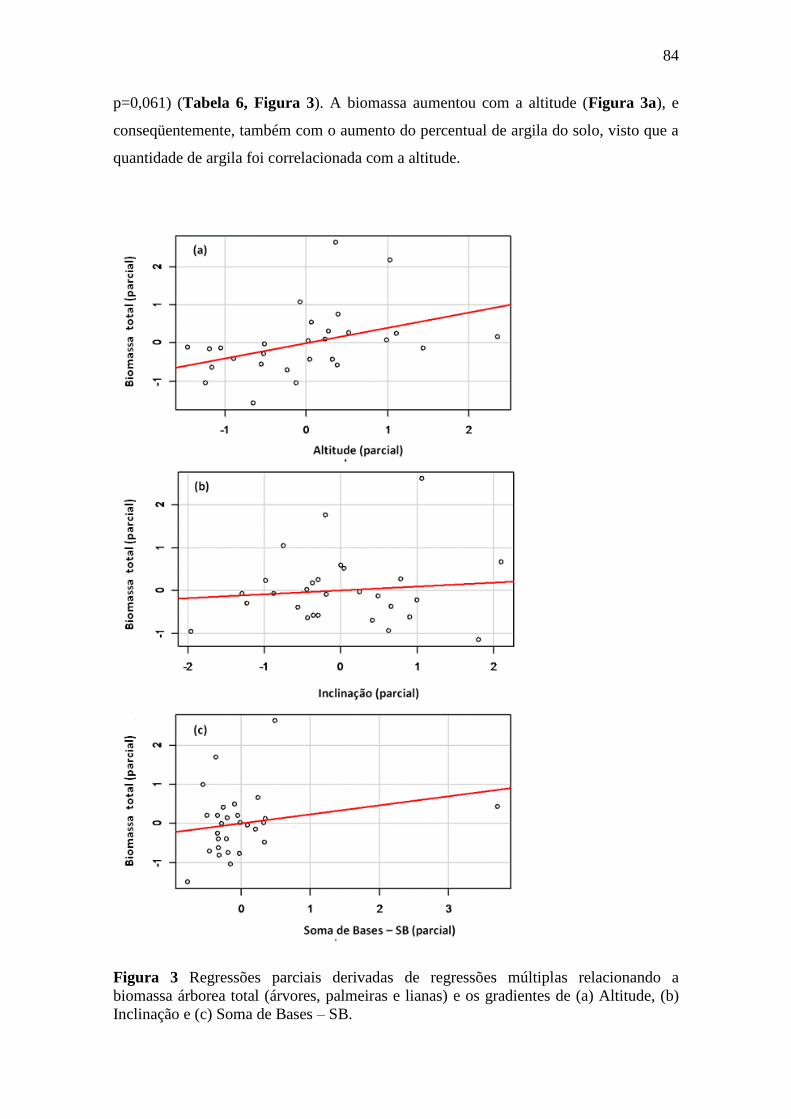
84
p=0,061) (Tabela 6, Figura 3). A biomassa aumentou com a altitude (Figura 3a), e
conseqüentemente, também com o aumento do percentual de argila do solo, visto que a
quantidade de argila foi correlacionada com a altitude.
Figura 3 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa árborea total (árvores, palmeiras e lianas) e os gradientes de (a) Altitude, (b)
Inclinação e (c) Soma de Bases – SB.
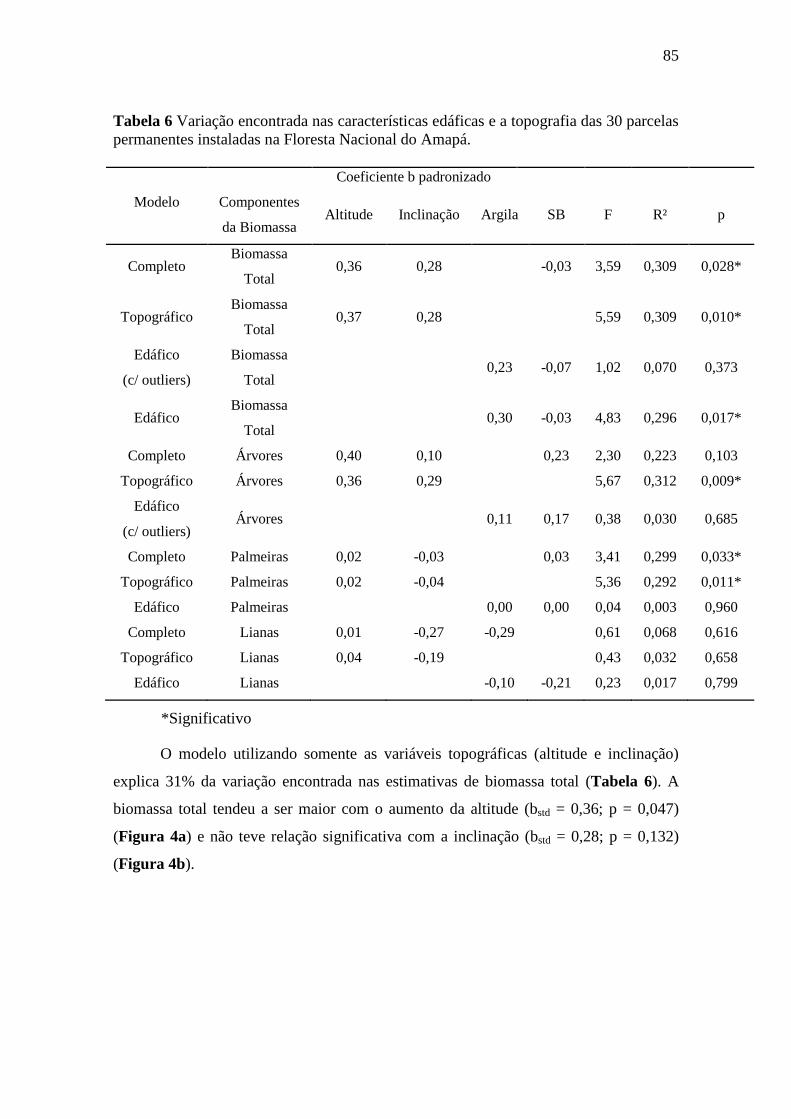
85
Tabela 6 Variação encontrada nas características edáficas e a topografia das 30 parcelas
permanentes instaladas na Floresta Nacional do Amapá.
Coeficiente b padronizado
Modelo Componentes
da Biomassa Altitude Inclinação Argila SB F R² p
Completo Biomassa
Total 0,36 0,28 -0,03 3,59 0,309 0,028*
Topográfico Biomassa
Total 0,37 0,28 5,59 0,309 0,010*
Edáfico
(c/ outliers)
Biomassa
Total 0,23 -0,07 1,02 0,070 0,373
Edáfico Biomassa
Total 0,30 -0,03 4,83 0,296 0,017*
Completo Árvores 0,40 0,10 0,23 2,30 0,223 0,103
Topográfico Árvores 0,36 0,29 5,67 0,312 0,009*
Edáfico
(c/ outliers) Árvores 0,11 0,17 0,38 0,030 0,685
Completo Palmeiras 0,02 -0,03 0,03 3,41 0,299 0,033*
Topográfico Palmeiras 0,02 -0,04 5,36 0,292 0,011*
Edáfico Palmeiras 0,00 0,00 0,04 0,003 0,960
Completo Lianas 0,01 -0,27 -0,29 0,61 0,068 0,616
Topográfico Lianas 0,04 -0,19 0,43 0,032 0,658
Edáfico Lianas -0,10 -0,21 0,23 0,017 0,799
*Significativo
O modelo utilizando somente as variáveis topográficas (altitude e inclinação)
explica 31% da variação encontrada nas estimativas de biomassa total (Tabela 6). A
biomassa total tendeu a ser maior com o aumento da altitude (bstd = 0,36; p = 0,047)
(Figura 4a) e não teve relação significativa com a inclinação (bstd = 0,28; p = 0,132)
(Figura 4b).
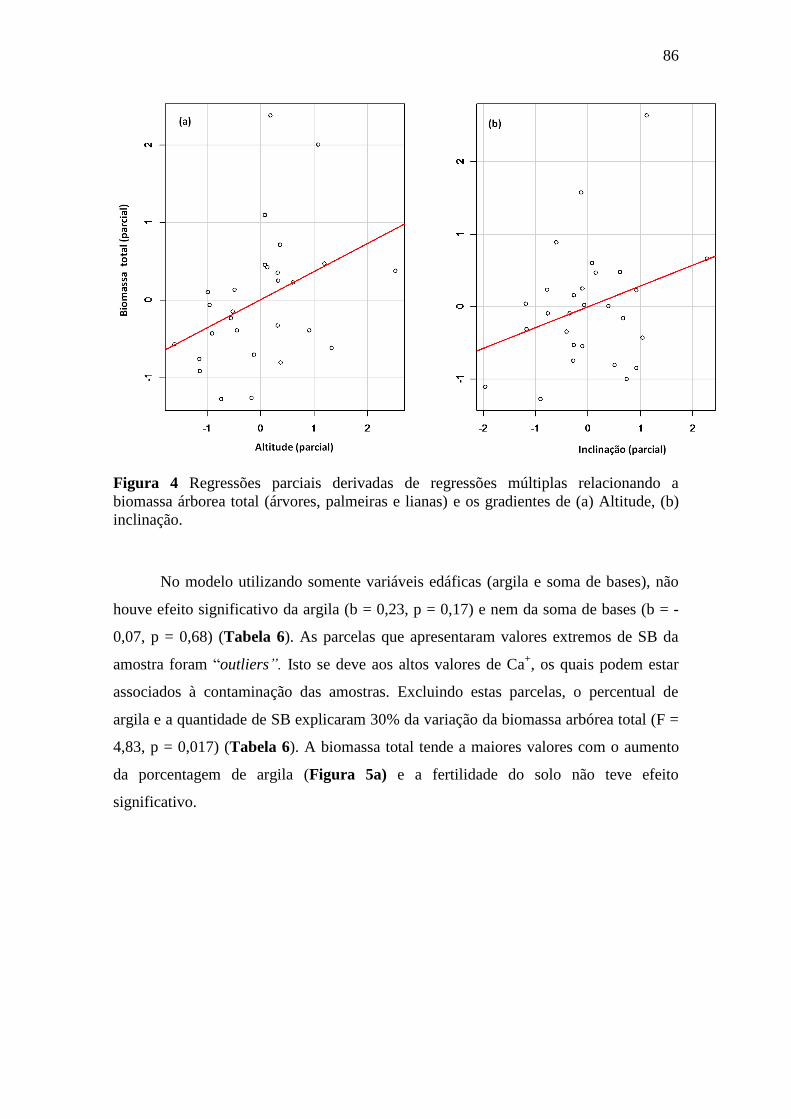
86
Figura 4 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa árborea total (árvores, palmeiras e lianas) e os gradientes de (a) Altitude, (b)
inclinação.
No modelo utilizando somente variáveis edáficas (argila e soma de bases), não
houve efeito significativo da argila (b = 0,23, p = 0,17) e nem da soma de bases (b = -
0,07, p = 0,68) (Tabela 6). As parcelas que apresentaram valores extremos de SB da
amostra foram “outliers”. Isto se deve aos altos valores de Ca+, os quais podem estar
associados à contaminação das amostras. Excluindo estas parcelas, o percentual de
argila e a quantidade de SB explicaram 30% da variação da biomassa arbórea total (F =
4,83, p = 0,017) (Tabela 6). A biomassa total tende a maiores valores com o aumento
da porcentagem de argila (Figura 5a) e a fertilidade do solo não teve efeito
significativo.
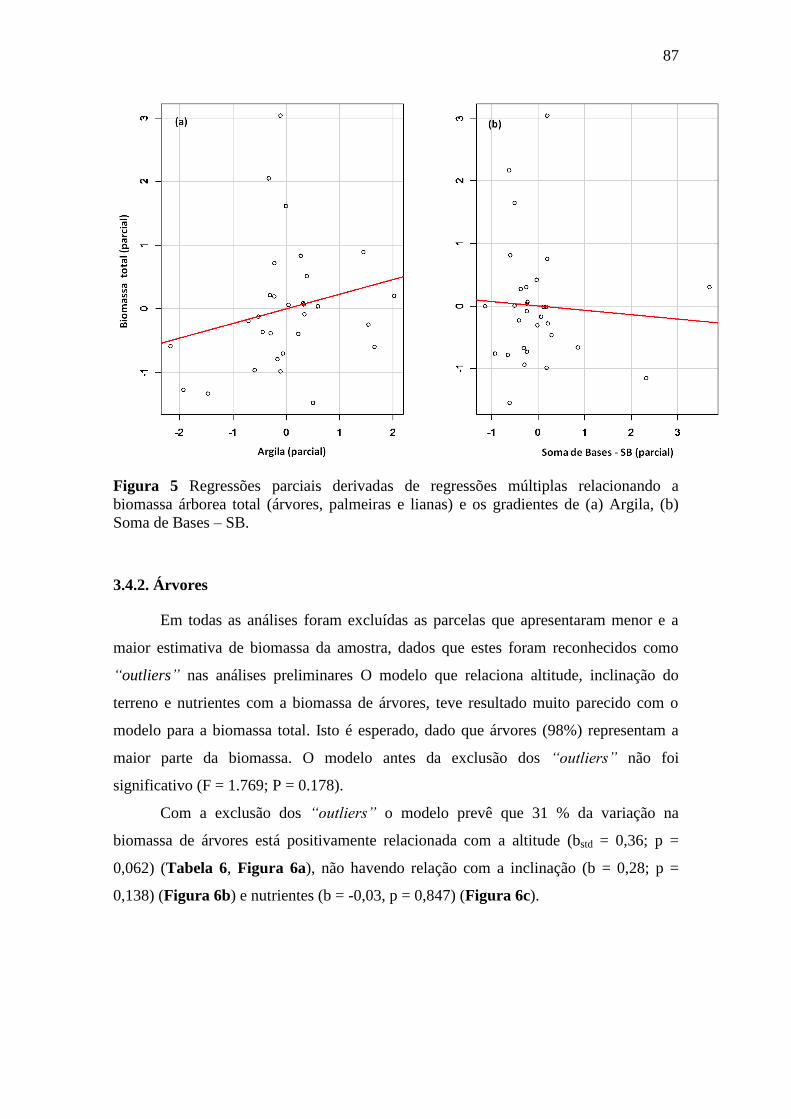
87
Figura 5 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa árborea total (árvores, palmeiras e lianas) e os gradientes de (a) Argila, (b)
Soma de Bases – SB.
3.4.2. Árvores
Em todas as análises foram excluídas as parcelas que apresentaram menor e a
maior estimativa de biomassa da amostra, dados que estes foram reconhecidos como
“outliers” nas análises preliminares O modelo que relaciona altitude, inclinação do
terreno e nutrientes com a biomassa de árvores, teve resultado muito parecido com o
modelo para a biomassa total. Isto é esperado, dado que árvores (98%) representam a
maior parte da biomassa. O modelo antes da exclusão dos “outliers” não foi
significativo (F = 1.769; P = 0.178).
Com a exclusão dos “outliers” o modelo prevê que 31 % da variação na
biomassa de árvores está positivamente relacionada com a altitude (bstd = 0,36; p =
0,062) (Tabela 6, Figura 6a), não havendo relação com a inclinação (b = 0,28; p =
0,138) (Figura 6b) e nutrientes (b = -0,03, p = 0,847) (Figura 6c).
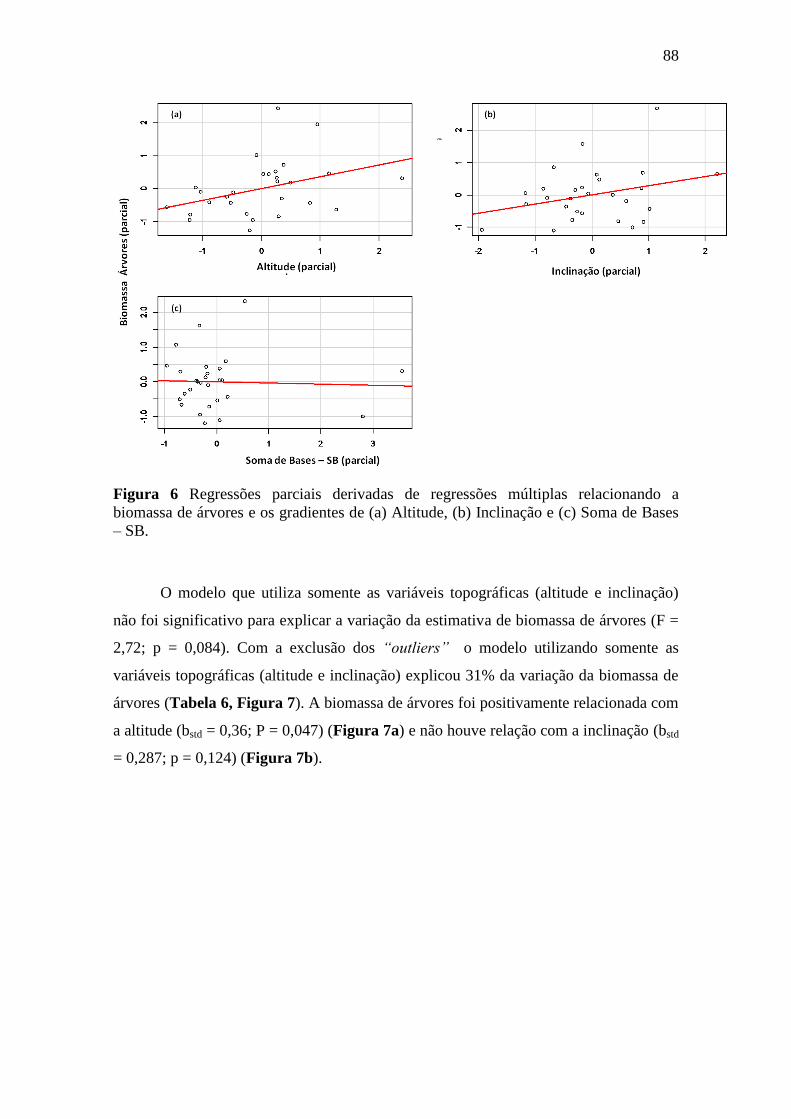
88
Figura 6 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa de árvores e os gradientes de (a) Altitude, (b) Inclinação e (c) Soma de Bases
– SB.
O modelo que utiliza somente as variáveis topográficas (altitude e inclinação)
não foi significativo para explicar a variação da estimativa de biomassa de árvores (F =
2,72; p = 0,084). Com a exclusão dos “outliers” o modelo utilizando somente as
variáveis topográficas (altitude e inclinação) explicou 31% da variação da biomassa de
árvores (Tabela 6, Figura 7). A biomassa de árvores foi positivamente relacionada com
a altitude (bstd = 0,36; P = 0,047) (Figura 7a) e não houve relação com a inclinação (bstd
= 0,287; p = 0,124) (Figura 7b).
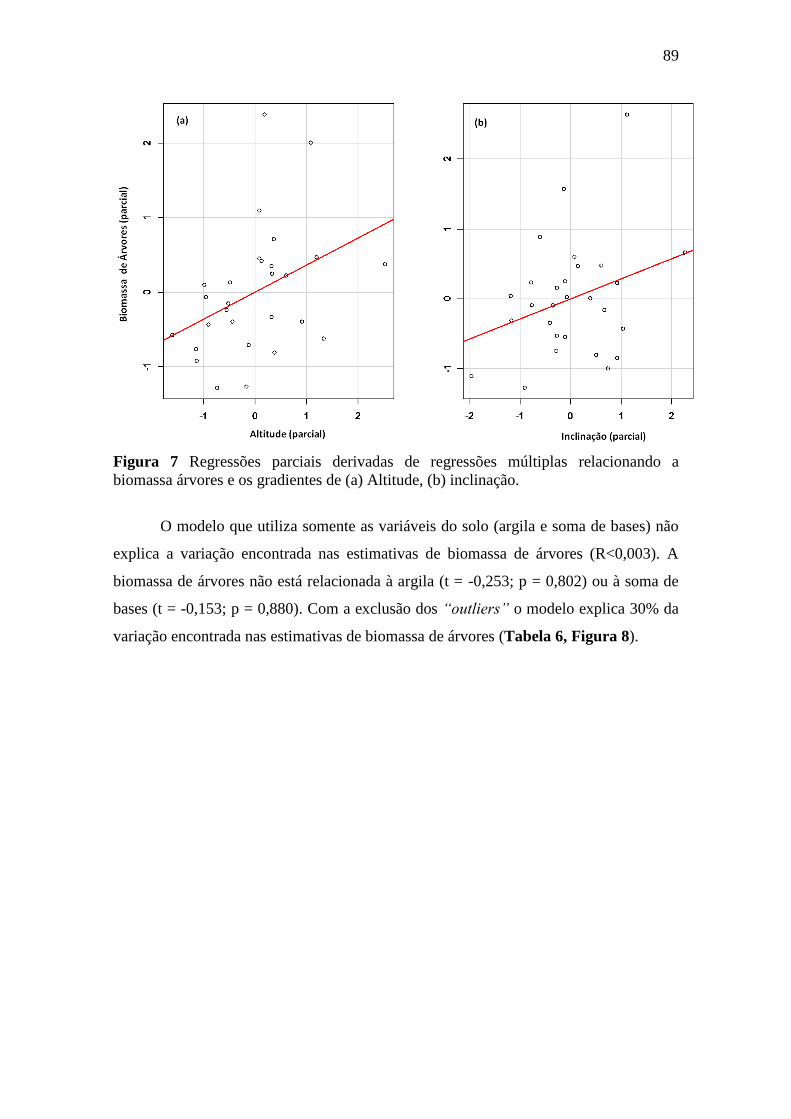
89
Figura 7 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa árvores e os gradientes de (a) Altitude, (b) inclinação.
O modelo que utiliza somente as variáveis do solo (argila e soma de bases) não
explica a variação encontrada nas estimativas de biomassa de árvores (R<0,003). A
biomassa de árvores não está relacionada à argila (t = -0,253; p = 0,802) ou à soma de
bases (t = -0,153; p = 0,880). Com a exclusão dos “outliers” o modelo explica 30% da
variação encontrada nas estimativas de biomassa de árvores (Tabela 6, Figura 8).
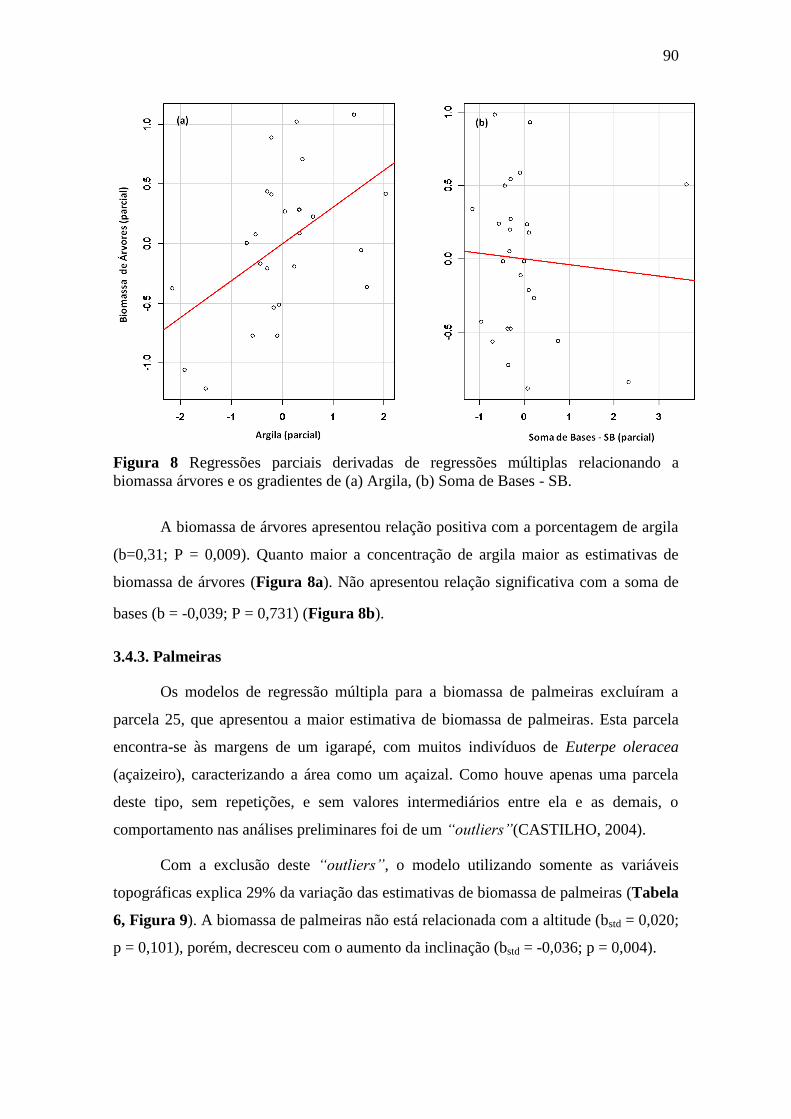
90
Figura 8 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa árvores e os gradientes de (a) Argila, (b) Soma de Bases - SB.
A biomassa de árvores apresentou relação positiva com a porcentagem de argila
(b=0,31; P = 0,009). Quanto maior a concentração de argila maior as estimativas de
biomassa de árvores (Figura 8a). Não apresentou relação significativa com a soma de
bases (b = -0,039; P = 0,731) (Figura 8b).
3.4.3. Palmeiras
Os modelos de regressão múltipla para a biomassa de palmeiras excluíram a
parcela 25, que apresentou a maior estimativa de biomassa de palmeiras. Esta parcela
encontra-se às margens de um igarapé, com muitos indivíduos de Euterpe oleracea
(açaizeiro), caracterizando a área como um açaizal. Como houve apenas uma parcela
deste tipo, sem repetições, e sem valores intermediários entre ela e as demais, o
comportamento nas análises preliminares foi de um “outliers”(CASTILHO, 2004).
Com a exclusão deste “outliers”, o modelo utilizando somente as variáveis
topográficas explica 29% da variação das estimativas de biomassa de palmeiras (Tabela
6, Figura 9). A biomassa de palmeiras não está relacionada com a altitude (bstd = 0,020;
p = 0,101), porém, decresceu com o aumento da inclinação (bstd = -0,036; p = 0,004).
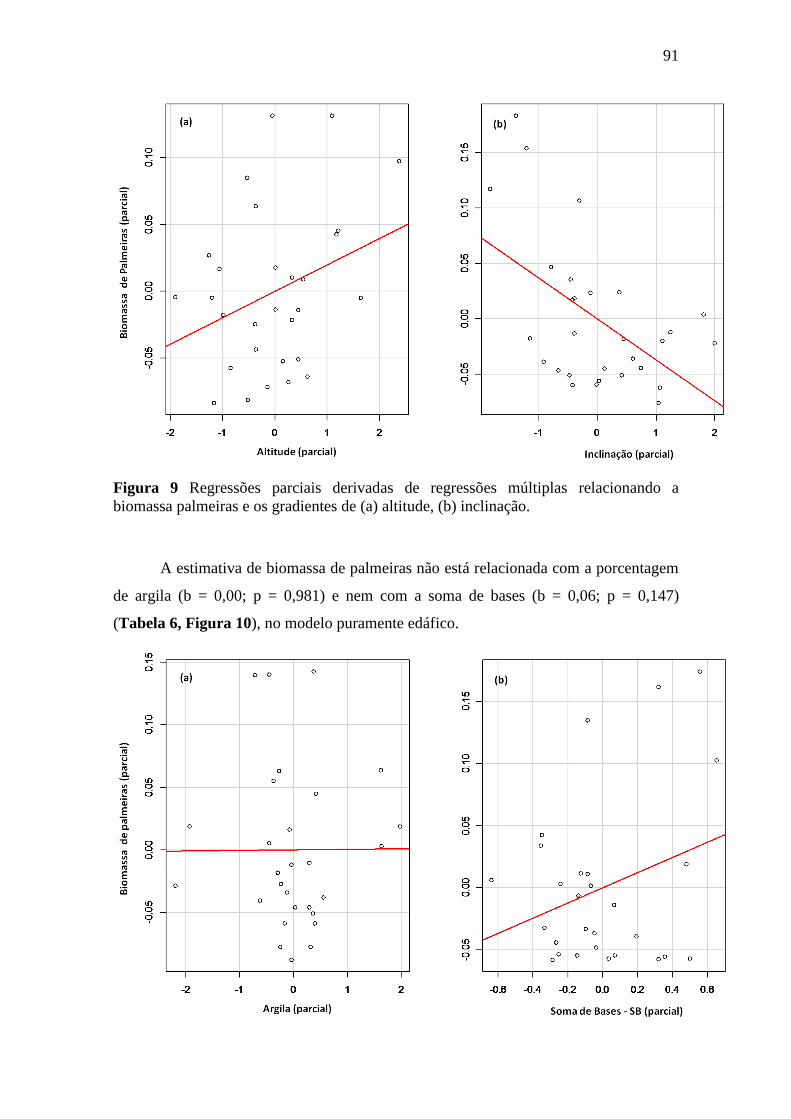
91
Figura 9 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa palmeiras e os gradientes de (a) altitude, (b) inclinação.
A estimativa de biomassa de palmeiras não está relacionada com a porcentagem
de argila (b = 0,00; p = 0,981) e nem com a soma de bases (b = 0,06; p = 0,147)
(Tabela 6, Figura 10), no modelo puramente edáfico.
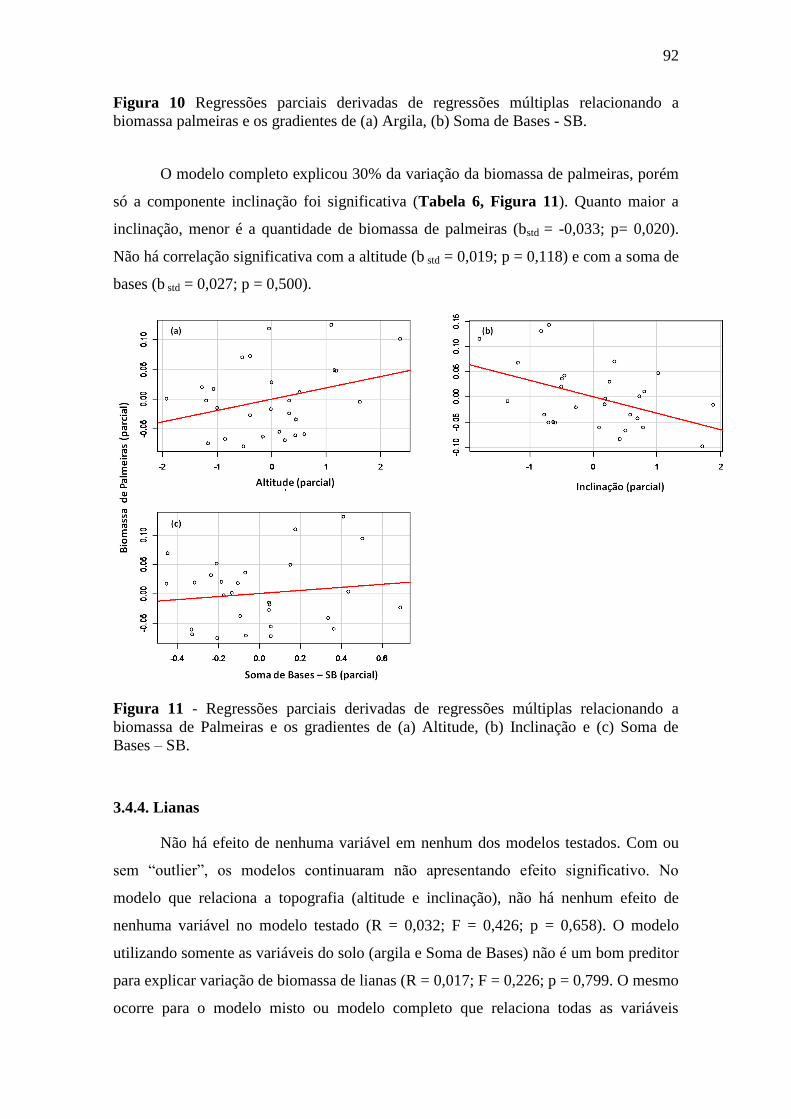
92
Figura 10 Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa palmeiras e os gradientes de (a) Argila, (b) Soma de Bases - SB.
O modelo completo explicou 30% da variação da biomassa de palmeiras, porém
só a componente inclinação foi significativa (Tabela 6, Figura 11). Quanto maior a
inclinação, menor é a quantidade de biomassa de palmeiras (bstd = -0,033; p= 0,020).
Não há correlação significativa com a altitude (b std = 0,019; p = 0,118) e com a soma de
bases (b std = 0,027; p = 0,500).
Figura 11 - Regressões parciais derivadas de regressões múltiplas relacionando a
biomassa de Palmeiras e os gradientes de (a) Altitude, (b) Inclinação e (c) Soma de
Bases – SB.
3.4.4. Lianas
Não há efeito de nenhuma variável em nenhum dos modelos testados. Com ou
sem “outlier”, os modelos continuaram não apresentando efeito significativo. No
modelo que relaciona a topografia (altitude e inclinação), não há nenhum efeito de
nenhuma variável no modelo testado (R = 0,032; F = 0,426; p = 0,658). O modelo
utilizando somente as variáveis do solo (argila e Soma de Bases) não é um bom preditor
para explicar variação de biomassa de lianas (R = 0,017; F = 0,226; p = 0,799. O mesmo
ocorre para o modelo misto ou modelo completo que relaciona todas as variáveis

93
(altitude, inclinação e Soma de Bases), nenhuma variável deste modelo apresenta efeito
significativo (R² = 0,068; F = 0,608; p = 0,616) (Tabela 6).
3.4.5. As diferentes classes de tamanhos de árvores e a relação entre suas
estimativas de biomassa e as características do solo e topografia
As estimativas de biomassa de árvores por classe diamétrica apresentam
respostas diferentes para a topografia (altitude e inclinação) e solo (argila e soma de
bases) (Tabela 2.8). De maneira geral, somente a biomassa de árvores com diâmetro
superior a 30 cm é afetada pela altitude, tanto na análise feita para o modelo
topográfico, quanto para o modelo completo. Não há efeito preditor de alguma variável
com relação à biomassa de árvores com diâmetro menor que 30 cm em nenhum modelo
analisado. Além disso, a biomassa por classe de diâmetro de árvores não respondeu a
nenhuma variável relacionada ao solo
Em geral, biomassa de árvores com classes de diâmetro de 20-30, 30-40 e 40-50
cm são todas positivamente relacionadas com a inclinação para todos os modelos
analisados (Tabela 2.8).
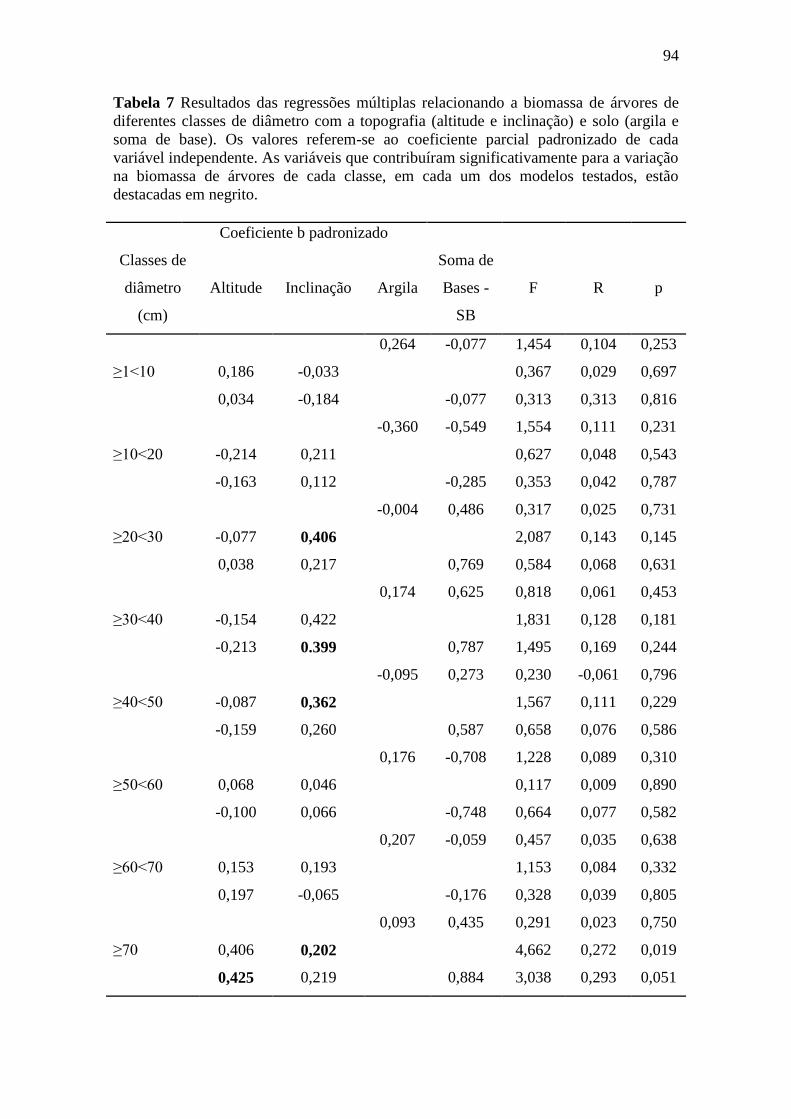
94
Tabela 7 Resultados das regressões múltiplas relacionando a biomassa de árvores de
diferentes classes de diâmetro com a topografia (altitude e inclinação) e solo (argila e
soma de base). Os valores referem-se ao coeficiente parcial padronizado de cada
variável independente. As variáveis que contribuíram significativamente para a variação
na biomassa de árvores de cada classe, em cada um dos modelos testados, estão
destacadas em negrito.
Coeficiente b padronizado
Classes de
diâmetro
(cm)
Altitude Inclinação Argila
Soma de
Bases -
SB
F R p
≥1<10
0,264 -0,077 1,454 0,104 0,253
0,186 -0,033 0,367 0,029 0,697
0,034 -0,184 -0,077 0,313 0,313 0,816
≥10<20
-0,360 -0,549 1,554 0,111 0,231
-0,214 0,211 0,627 0,048 0,543
-0,163 0,112 -0,285 0,353 0,042 0,787
≥20<30
-0,004 0,486 0,317 0,025 0,731
-0,077 0,406 2,087 0,143 0,145
0,038 0,217 0,769 0,584 0,068 0,631
≥30<40
0,174 0,625 0,818 0,061 0,453
-0,154 0,422 1,831 0,128 0,181
-0,213 0.399 0,787 1,495 0,169 0,244
≥40<50
-0,095 0,273 0,230 -0,061 0,796
-0,087 0,362 1,567 0,111 0,229
-0,159 0,260 0,587 0,658 0,076 0,586
≥50<60
0,176 -0,708 1,228 0,089 0,310
0,068 0,046 0,117 0,009 0,890
-0,100 0,066 -0,748 0,664 0,077 0,582
≥60<70
0,207 -0,059 0,457 0,035 0,638
0,153 0,193 1,153 0,084 0,332
0,197 -0,065 -0,176 0,328 0,039 0,805
≥70
0,093 0,435 0,291 0,023 0,750
0,406 0,202 4,662 0,272 0,019
0,425 0,219 0,884 3,038 0,293 0,051

95
O modelo utilizando apenas as variáveis topográficas explicou 27% da variação
encontrada nas estimativas de biomassa de árvores com DAP ≥ 70 cm. A biomassa de
árvores para esta classe foi relacionada positivamente apenas com a altitude. O modelo
completo (altitude. inclinação e soma de bases) explica 17% da variação de biomassa
para esta classe de diâmetro. A biomassa de árvores está relacionada somente com a
altitude (bstd = 0,26; p = 0,036). Com a exclusão dos “outliers”, o modelo explica 27%
da variação da biomassa de árvores para esta classe de diâmetro (F = 1,67; p = 0,069),
com efeitos significativos apenas da altitude (bstd = 0,42; p = 0,036).
4. Discussão
4.1. A variação de biomassa total acima do solo na FLONA do Amapá foi
explicada pela topografia e textura do solo
As características de solo e topografia explica cerca de um terço da variação de
biomassa arbórea total viva acima do solo na FLONA do Amapá. A biomassa total está
positivamente relacionada com a altitude. A textura do solo também afeta positivamente
a biomassa total viva acima do solo. Entretanto, altitude e porcentagem de argila são
correlacionadas na FLONA do Amapá. Portanto, não é possível separar seus efeitos
neste estudo.
Na FLONA do Amapá a variação de biomassa arbórea está relacionada com os
gradientes do solo e topografia. No Estado do Amapá a maior parte das florestas de terra
firme, localizadas no centro do Estado até o Parque Nacional Montanhas de
Tumucumaque tem grande influência da topografia apresentando áreas mais altas.
Nestas áreas podemos associar os gradientes edáficos à topografia.
Os resultados encontrados neste estudo estão de acordo com os encontrados por
Quesada et al. (2009); Castilho et al. (2006); Nascimento e Laurance (2001) que
mostraram que a textura e topografia são as componentes com melhor capacidade
preditora para a variação de biomassa arbórea viva acima do solo. Os gradientes de
nutrientes do solo (Soma de Bases – SB) não foram relacionado com a variação de
biomassa arbórea viva acima do solo.
Este estudo pode contribuir para alimentar modelos de variação espacial de
biomassa, que consideram que a topografia é um fator predominante para a variação de
biomassa. Nossas estimativas de biomassa total são realizadas em área considerada de

96
escala espacial grande, onde não trabalhamos com extrapolações, o que também conta
como um fator positivo para o mosaico real de variação de biomassa da região.
Estes resultados podem contribuir para o manejo florestal e com as políticas para
compensação de créditos de carbono, que ainda vêm sendo trabalhadas ao longo dos
anos no Estado. Entender a relação entre a variação da biomassa com as variáveis do
solo e topográficas ao longo da paisagem da floresta pode auxiliar na extrapolação dos
resultados para áreas maiores e de interesse à conservação ecológica.
4.2. Textura e a altitude do terreno e a sua relação com a variação espacial da
biomassa viva acima do solo
A área estudada da FLONA do Amapá a topografia e as variáveis do solo são
correlacionadas. A topografia e textura do solo influenciam diretamente a distribuição
de biomassa viva acima do solo, convergindo com os resultados de outros estudos
semelhantes realizados na Amazônia Central por Quesada et al. (2009); Castilho et al.
(2006); Nascimento e Laurance (2001), visto que os dados de solo são mais difíceis de
adquirir e quando disponíveis, em forma de mapas são imprecisos para a Amazônia.
Enquanto dados de altitude e inclinação são obtidos com boa resolução horizontal 0,8
ha, permitindo a extrapolação de resultados obtidos em parcelas de 1 ha para grandes
áreas (RENNÓ et al., 2008). Estas variáveis topográficas podem ser obtidas de imagens
do modelo digital de elevação do terreno, derivadas do radar interferométrico SRTM
(shuttle radar topographic mission).
Podemos utilizar apenas mapas com informações topográficas (altitude e
inclinação) para melhorar as estimativas de biomassa para toda a região da FLONA do
Amapá. Visto que a altitude é correlacionada positivamente com as características do
solo na região de estudo. Mostramos que a variação espacial da biomassa viva acima do
solo foi influenciada pela heterogeneidade do solo e da topografia, que para a região de
estudo podemos utilizar apenas dados topográficos. Outro ponto positivo é que estas
imagens são mais acessíveis que dados de solo, podendo ser parâmetro útil para a gestão
da conservação e estudos ecológicos.
Os resultados encontrados neste estudo com relação à correlação entre a
estrutura do solo e a topografia é uma alternativa analítica, onde o uso das variáveis
topográficas em substituição às variáveis do solo melhoram as estimativas de biomassa

97
viva acima do solo. Nossos resultados estão de acordo com os resultados obtidos por
Castilho (2004) e Laurance et al. (1999). Ambos os estudos apontam esta alternativa do
uso de variáveis topográficas para substituir as variáveis do solo para melhorar as
estimativas de biomassa viva acima do solo próxima às áreas de estudos.
Estes resultados facilitam as estimativas confiáveis de biomassa viva acima do
solo para grandes áreas utilizando apenas informações disponíveis em mapas
topográficos para a Amazônia ou obtendo estas variáveis topográficas (altitude e
inclinação) através de uma boa resolução da imagem de satélite obtida pelo radar SRTM
e disponibilizadas pela NASA (RENNÓ et al., 2008). Com base neste resultado as
contribuições científicas apresentam um ganho inédito para o Estado do Amapá,
especialmente quanto às suas aplicações nas políticas públicas de conservação da
floresta em pé. Futuramente estas significativas variações de estimativas de biomassa
viva acima do solo podem ser utilizadas como subsídios e informações para alimentar
os inventários de carbono. O carbono poderá ser usado como mercadoria a ser
negociada sob o Protocolo de Quioto (IPCC, 2007, SILVA, 2011), resultando
futuramente em uma potencial arrecadação financeira do Estado e contribuindo com a
conservação da Amazônia.
4.3. A relação da variação da biomassa com os gradientes do solo e topografia
dependem do tamanho das árvores.
O tamanho da árvore determina diferentes respostas ao solo e à topografia. A
topografia influencia de forma diferenciada, dependendo do tamanho da árvore. Em
geral,árvores com DAP≥30 cm são fortemente influenciadas pela altitude e inclinação,
porém esta influência não foi verificada em árvores com DAP inferior a 30 cm.
As árvores (menores) do sub-bosque, que possuem menores valores de
biomassa, devem ser influenciadas por outros fatores ambientais, tais como
disponibilidade de luz, enquanto as árvores consideradas de grande porte são
influenciadas pela altitude, porém podem ser influenciada por outros fatores não
testados neste estudo, tais como drenagem, lençol freático. A altitude tem uma relação
com solos argilosos. A textura do solo também determina a capacidade do solo em reter
cátions (Ca2+, Mg2+, K+ e Na+), água e matéria orgânica (FEARNSIDE e LEAL
FILHO, 2001; LUIZÃO et al., 2004). Os solos argilosos permitem melhor fixação de
raízes das árvores (DUPUY et al., 2005). Os processos de decomposição e ciclagem de

98
nutrientes são mais rápidos em solos argilosos do que em solos arenosos (LUIZÃO et
al., 2004). Assim, por essas razões, a argila encontrada nos platôs influencia a
quantidade de biomassa viva acima do solo (CASTILHO et al., 2006; LAURANCE et
al., 1999).
Neste estudo mais de 50% da concentração de biomassa arbórea viva total por
hectare encontra-se na classe de tamanho de DAP≥60 cm, diferente dos resultados
encontrados para Amazônia central, onde metade da biomassa total viva acima do solo
se concentrou em classes intermediárias (20-50 cm de DAP) (CASTILHO, 2004;
NASCIMENTO, 2002). Porém, em algumas áreas tropicais, de 25 –35% da biomassa
total está concentrada nas árvores com DAP>60 cm (CLARK e CLARK, 1996).
Árvores com DAP≥60 cm são altamente influenciadas pela altitude.
4.4. A variação na quantidade de biomassa neste estudo é afetada pela textura do
solo e topografia comparando com os obtidos para Amazônia Central
A quantidade de biomassa total está positivamente relacionada com a altitude. A
textura do solo também afeta positivamente a biomassa total viva acima do solo. Visto
que a altitude e porcentagem de argila são correlacionadas positivamente na FLONA do
Amapá, portanto não é possível separar seus efeitos neste estudo. Quesada et al. (2009);
Castilho et al. (2006); Nascimento e Laurance (2001); Laurance et al. (1999) mostraram
que a textura e topografia foram as componentes mais preditoras para a variação de
biomassa arbórea viva acima do solo. Os resultados encontrados nestes estudos
realizados na Amazônia Central são semelhantes aos resultados encontrados para no
leste da Amazônia. Segundo Aparício (2011) em estudo realizado na Reserva
Extrativista do Cajarí – Amapá não existiu relação entre a abundância de espécies de
árvores e a fertilidade do solo, pois o solo foi considerado pobre em nutrientes. O
gradiente de nutrientes do solo (Soma de Bases – SB) não foi relacionado com a
variação de biomassa arbórea viva acima do solo (este estudo).
Na FLONA do Amapá o padrão encontrado é que em solos argilosos dos platôs
se concentra a maior biomassa de árvores de que em solos arenosos nos baixios,
resultados estes também apontados por Castilho et al. (2006). A variável inclinação
apresenta uma relação negativa com a biomassa de palmeiras. Isto também corrobora
com os resultados encontrados por Castilho et al. (2006), mostrando que altitude e
inclinação tiveram uma relação negativa com a biomassa de palmeiras.

99
5. Considerações Finais
As características de solo e topografia explicam 30% da variação da biomassa na
FLONA do Amapá. Áreas altas com solos argilosos apresentaram maior biomassa que
em áreas baixas de solos arenosos.
A altitude pode substituir algumas características do solo para prever a variação
espacial de biomassa viva acima do solo na FLONA do Amapá. Pois a altitude é
altamente correlacionada com algumas características do solo. A variação da biomassa é
influenciada pela heterogeneidade dos diferentes ambientes gerados pelo solo e a
topografia.
A biomassa de árvores de diferentes classes de diâmetro respondeu de maneira
distinta às variações de altitude e inclinação. A maior parte da biomassa de árvores está
concentrada em árvores de grande porte (DAP≥70cm). Estas são encontradas em
terrenos planos e altos. Porém a biomassa de árvores com classe de diâmetro variando
de 20 a 50 cm são concentradas em áreas inclinadas.
Os fatores que afetam a variação de biomassa viva acima do solo na FLONA do
Amapá, região localizada no leste da Amazônia, são relacionados ao solo e à topografia.
Nestas condições, a variação de biomassa foi explicada pelas diferenças de altitude e
textura do solo. Além disso, a variação de biomassa não está relacionada à concentração
de nutrientes do solo, pois estes são considerados baixos nos solos da FLONA do
Amapá. Em geral estudos realizados na Amazônia Central apontam resultados
semelhantes.

100
6. Referências
AMARO FILHO, J.; ASSIS JUNIOR, R.N.; MOTA, J.C.A. Física do solo. Conceitos e
Aplicações. Fortaleza: Imprensa Universitária, 2008.
APARÍCIO, W. C.S.. Estrutura da vegetação em diferentes ambientes na RESEX
do Rio Cajari: interações solo-floresta e relações com a produção de castanha. Tese
em ciências florestais, UFRPE, Recife, Pernambuco. 150p, 2011.
BAKER, T. R. et al.. Increasing biomass in Amazonian forest plot.. Phil. Trans. R. Soc.
Lond. B, v. 359, p.353–365, 2004.
CASTILHO, C.V. et al. Variation in aboveground tree live biomass in a central
Amazonian forest: effects of soil and topography. Forest Ecology and Management, v.
234, p. 85-96. 2006.
CASTILHO, C.V.. Variação espacial e temporal da biomassa arbórea viva em 64
Km2 de floresta de terra-firme na Amazônia Central. Tese de doutorado,
INPA/UFAM, Manaus, AM. 72p. 2004.
CHAO, K.J. et al. Growth and wood density predict tree mortality in Amazon forests.
Journal of Ecology. v.96, p. 281-292, 2008.
CHAUVEL, A.; LUCAS, Y.; BOULET, R.. On the genesis of the soil mantle of the
region of Manaus, Central Amazonia, Brazil. Experientia, v. 43, p. 234-241, 1987.
CLARK, D.B. e CLARK, D.A. Abundance, growth and mortality of very large trees in
neotropical lowland rain forest. Forest Ecology and Management, v. 80, p. 235-244,
1996.
COSTA, F.R.C., GUILLAUMET, J.L., LIMA, A.P., PEREIRA, O.S. Gradients within
gradients: The mesoescale distribution patterns of palms in a central Amazonian forest.
Journal of Vegetation Science, v.20, p.69-78, 2009.
COSTA, F.R.C., MAGNUSSON, W.E, LUIZÃO, R.C. Mesoescale distribtion patterns
of Amazonian understorey herbs in relation to topography, soil and watersheds. Journal
of Ecology, v.93, p.863-878, 2005.
da SILVA, R.P.; SANTOS, J.; TRIBUZY, E.S.; CHAMBERS, J.Q.; NAKAMURA, S.,
HIGUCHI, N. Diameter increment and growth patterns for individual tree growing in
Central Amazon, Brazil. Forest Ecology and Management, v.166, p.295-301, 2002.

101
DUPUY, L.X., FOURCAUD, T., STOKES, A. A numerical investigation into the
influence of soil type and root architecture on tree anchorage. Palnt and Soil, v. 278, p.
119-134, 2005.
EMPRESA BRASILEIRA DE PESQUISA AGROPECUARIA — EMBRAPA.
Manual de métodos de análises de solo. 2.ed. Rio de Janeiro, Ministério da
Agricultura e do Abastecimento, 1997. 212p.
FEARNSIDE, P.M.. Amazonian deforestation and global warming: carbon stocks in
vegetation replacing Brazil’s Amazon forest. Forest Ecology and Management. V. 80,
p. 21-34, 1996.
FEARSIDE, P.M.; LEAL FILHO, N. Soil and development in Amazonia. In:
BIERREGAARD, R.O., GASCON, C., LOVEJOY, T.E., MESQUITA, R.C.G. (Eds.),
Lessons from Amazonia: The Ecology and consevation of a fragmented forest. Yale
University Press, New Haven & London, pp. 291-312, 2001.
FIELD, C. B. Plant physiology of the ‘missing’ carbon sink. Plant Physiology, v. 125,
p. 25–28, 2001.
HIGUCHI, N.; SANTOS, J.; RIBEIRO, R.J.; MINETTE, L.; BIOT, Y.. Biomassa da
parte aérea da vegetação de floresta tropical úmida de terra firme da Amazônia
Brasileira. Acta Amazonica, 28 (2): 153-165. 1998.
HOUGHTON, R. A. Aboveground forest biomass and the global carbon balance.
Global Change Biology, v.11, p.945-958, 2005.
ICMBIO: Instituto Chico Mendes de Biodiversidade. Disponível em:
www.icmbio.gov.br/portal/.../unidades.../1956-flona-do-amapa Acessado em: 21 de jan.
de 2012.
INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE (IPCC) (2007) Climate
Change 2007: the Physical Science Basis. Contribution of Working Group I to the
Fourth Assessment Report of the Intergovernmental Panel on Climate Change (eds
Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M, Miller
HL). Cambridge University Press, Cambridge, UK and New York. 2007.
KELLER, M.; PALACE, M.; HURTT, G. Biomass estimation in the Tapajos National
Forest, Brazil – examination of sampling and allometric uncertainties. Forest Ecology
and Management, v. 154, p.371–382, 2001.

102
KINUPP, V.F. e MAGNUSSON, W.E. Spatial patters in the understory shrub genus
Psychotria in Central Amazonia: Effects of distance and topography. Journal of
Tropical Ecology, v. 21, p.1-12, 2005.
LANDEIRO, V.L. e MAGNUSSON, E.M. The Geometry of Spatial Analyses:
Implications for Conservation Biologists. Natureza e Conservação, v. 1, n. 9, p. 7-20,
2011.
LAURANCE, W.F.; FEARNSIDE, P.M.; LAURANCE, S.G.; DELAMONICA, P.;
LOVEJOY, T.E.; RANKIN-DE-MERONA, J.M.; CHAMBERS, J.; GASCON, C..
Relationship between soils and Amazon forest biomass: a landscape-scale study. Forest
Ecology and Management, v. 118, p. 127-138, 1999.
LIMA, H.N. et al. Mineralogia e química de três solos de uma topossequência da bacia
sedimentar do Alto Solimões, Amazônia Ocidental. Revista brasileira de Ciência do
Solo, v. 30, p. 59-68, 2006.
LUIZÃO, R.C.C.; LUIZÃO, F.J.; PAIVA, R.Q.; MONTEIRO, T.F.; SOUSA, L.S.;
KRUIJT, B. Variation of carbon and nitrogen cycling processes along a topographic
gradient in a Central Amazonian forest. Global Change Biology, v. 10, p. 592-600,
2004.
MAGNUSSON, W.E., LIMA, A.P., LUIZÃO, R., LUIZÃO, F., COSTA, F.R.C.,
CASTILHO, C.V., KINUPP, V.F. RAPELD: A modification of the Gentry method for
biodiversity surveys in long-term ecological research sites. Biota Neotropica, v.2, n.5,
p.1–6, 2005.
MALHI, Y. e GRACE, J. Tropical forests and atmospheric carbon dioxide. Trends in
Ecology and Evolution, v. 15, n. 8, p. 332-337, 2000.
MALHI, Y. e WRIGHT, J. Spatial patterns and recent trends in the climate of tropical
rainforest regions. Philosophical Transaction of the Royal Society B Biological
Science, v. 359, p. 311-329, 2004.
MALHI, Y. The carbon balance of tropical forest regions, 1990-2005. Current
Opinion in Sustainability Science, v.2, n.4, p.237-244, 2010.
NASCIMENTO, H.E.M. e LAURANCE, W.F.. Total aboveground biomass in central
Amazonian rainforest: a landscape-scale study. Forest Ecology and Management, v.
168, p. 311-321, 2001.

103
NASCIMENTO, H.E.M.. Biomassa total acima do solo e a fragmentação de floresta
de terra-firme na Amazônia Central. Tese de doutorado, Instituto Nacional de
Pesquisas da Amazônia, Manaus, Amazonas. 98p, 2002.
NEVES, D.G. et al. Modelagem climática regional durante dois anos extremos de
precipitação sobre o Estado do Amapá. Revista Brasileira de Meteorologia
(Impresso), v. 26, p. 569-578, 2011.
NOGUEIRA, A. Variação da densidade, área basal e biomassa de lianas em 64 km²
de floresta de terra firme na Amazônia Central. Dissertação de mestrado,
INPA/UFAM. Manaus, 2006.
OLIVEIRA, L.L. et al. Características Hidroclimáticas da bacia do rio Araguari. In:
CUNHA, A.C.; SOUZA, E.B.; CUNHA, H.F.A. Tempo, Clima e Recursos Hídricos:
resultados do Projeto REMETAP no Estado do Amapá. Macapá: IEPA, p. 83 – 96,
2010.
OLIVEIRA, L.P.S.; SOTTA, E.D.; HIGUCHI, N. Biomass Quantification in Amapá’s
State Forest: Carbon Stock Allometry and Estimate. 1st Edition. (Amapá’s State
Forest Institution-IEF. Technical Series), 52 p, 2012.
PEREIRA, L.A.; SENA, K.S; SANTOS, M.R.; COSTA NETO, S.V. Aspectos
florísticos da FLONA do Amapá e sua importância na conservação da biodiversidade.
Nota Cientifica da Revista Brasileira de Biociências, Porto Alegre, v. 5, n. 2, p. 693-
695, 2007.
PHILLIPS O.L.et al. Drought sensitivity of the Amazon rainforest. Science, n. 323, p.
1344-1347, 2009.
QUESADA, C. A. et al. Effect of soils on forest structure and dynamics in Amazonia.
Biogeosciences Discuss., v. 6, p. 3993–4057, 2009.
R Development Core Team. R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, Austria. http//www.R-
project.or, 2011.
RENNÓ, C.D., NOBRE, A.D., CUARTAS, L.A., SOARES, J.V., HODNETT, M.G.,
TOMASELLA, J., WATERLOO, M.J.. HAND, a new terrain descriptor using SRTM-
DEM: Mapping terra-firme rainforest environments in Amazonia. Remote Sensing of
Environment v. 112, p. 3469-3481, 2008.

104
SAATH, S.S.; HOUGHTON, R.A.; ALVALLÁ, R.C.D.S.; SOARES, J.V.; YU, Y.
Distribution of aboveground live biomass in the Amazon basin. Global Change
Biology, v. 13, p. 816-387, 2007.
SALDARRIAGA, J.G.; WEST, D.C.; THARP, M.L.; UHL, C. Long-term
chronosequence of forest succession in the upper Rio Negro of Colombia and
Venezuela. Journal of Ecology, v. 76, p. 938-958, 1988.
SCHNITZER, S.A.; DeWALT, S.J. e CHAVE, J. Censusing and Measuring Lianas: A
Quantitative Comparison of the Common Methods. Biotropica, v. 38, n.5, p. 581–591,
2006.
SILVA, W. da C. Viabilidade econômica do pagamento por serviços ambientais no
Estado do Amapá utilizando Análise de Risco. Dissertação de Mestrado. Macapá:
UNIFAP, 2011.
SIMONIAN, L.T.L. (Org.). Políticas públicas, desenvolvimento, unidades de
conservação e outras questões socioambientais no Amapá. Belém: NAEA-
UFPA/MPEAP, p. 115-180, 2010.
SOUSA, M. A. R.. Anatomia ecológica do lenho de Mora Paraensis (Ducke) Ducke
em Várzea Estuarina no Estado do Amapá. Dissertação de Mestrado do Programa de
Pós-Graduação em Biodiversidade Tropical, Fundação Universidade Federal do Amapá.
Macapá, 2010.
SOUZA, E.B.; CUNHA, A.C. Climatologia da precipitação no Amapá e mecanismos
climáticos de grande escala. In: CUNHA, A.C.; SOUZA, E.B.; CUNHA, H.F.A.
Tempo, Clima e Recursos Hídricos: resultados do Projeto REMETAP no estado do
Amapá. Macapá: IEPA, 2010. p. 216.
TER STEEGE, H. et al. Cotinental-scale patterns of canopy tree composition and
function across Amazonia. Nature v. 443, p. 444-447, 2006.
TOLEDO, J.J.. Influência do solo e topografia sobre a mortabilidade de árvores e
decomposição de madeira em uma floresta de terra-firme na Amazônia Central.
Tese de doutorado, INPA/UFAM, Manaus, AM. 75p. 2009.
VAN DEN BERG, E. e SANTOS, F.A.M.. Aspectos da variação ambiental em uma
floresta de galeria em Itutinga, MG, Brasil. Revista Ciência Florestal, v. 13, n. 2, 2003.

105
VAN DEN BERG, E. Variáveis Ambientais e a dinâmica estrutural e populacional
de uma floresta de galeria em Utinga, MG. Tese de doutorado em Biologia vegetal -
UNICAMP, 2001.
VASCONCELOS, H.L.; MACEDO, A.C.C.; VILHENA, J.M.S. Influence of
topography on the distribution of ground-dwelling ants in an Amazonian Forest.
Studies on Neotropical Fauna and environment, v. 38, n. 2, p. 115-124, 2003.


















