UNIVERSIDADE FEDERAL DO PARÁ NÚCLEO DE MEDICINA...
126
UNIVERSIDADE FEDERAL DO PARÁ NÚCLEO DE MEDICINA TROPICAL PROGRAMA DE PÓS-GRADUAÇÃO EM DOENÇAS TROPICAIS CARACTERIZAÇÃO DAS INFECÇÕES POR NOROVÍRUS NAS HOSPITALIZAÇÕES PEDIÁTRICAS POR GASTRENTERITE NA CIDADE DE BELÉM, PARÁ. JONES ANDERSON MONTEIRO SIQUEIRA Belém – Pará 2012
Transcript of UNIVERSIDADE FEDERAL DO PARÁ NÚCLEO DE MEDICINA...
UNIVERSIDADE FEDERAL DO PAR
NCLEO DE MEDICINA TROPICAL
PROGRAMA DE PS-GRADUAO EM DOENAS TROPICAIS
CARACTERIZAO DAS INFECES POR NOROVRUS NAS
HOSPITALIZAES PEDITRICAS POR GASTRENTERITE NA CIDADE DE
BELM, PAR.
JONES ANDERSON MONTEIRO SIQUEIRA
Belm Par
2012
JONES ANDERSON MONTEIRO SIQUEIRA
CARACTERIZAO DAS INFECES POR NOROVRUS NAS HOSPITALIZAES
PEDITRICAS POR GASTRENTERITE NA CIDADE DE BELM, PAR.
Belm Par
2012
Dissertao apresentada ao Programa de Ps-
graduao em Doenas Tropicais (rea de
concentrao de Patologia das Doenas
Tropicais), Ncleo de Medicina Tropical,
Universidade Federal do Par, como requisito
parcial para a obteno do grau de Mestre em
Doenas Tropicais.
Orientadora: Dr Yvone Gabbay Mendes
Dados Internacionais de Catalogao na Publicao (CIP)
Biblioteca do Ncleo de Medicina Tropical/UFPA, Belm-PA
____________________________________________________________
Siqueira, Jones Anderson Monteiro.
Caracterizao das infeces por norovrus nas hospitalizaes
peditricas por gastrenterite na cidade de Belm, Par / Jones Anderson
Monteiro Siqueira; orientadora: Yvone Gabbay Mendes 2012.
Dissertao (Mestrado) Universidade Federal do
Par. Ncleo de Medicina Tropical. Programa de Ps-Graduao em
Doenas Tropicais. BelmPar, 125p, 2012.
1. Gastrenterite Crianas Belm. 2. Norovrus Belm (PA). 3.
Infeces por norovrus Belm (PA). I. Mendes, Yvone Gabbay,
orient. II. Ttulo.
CDD: 22. ed. 618.923427 ___________________________________________________________
Dedico esta obra aos meus pais, Vera e Mrio, a
quem dedico todas as vitrias de minha vida e sem os
quais, nenhuma teria sentido.
AGRADECIMENTOS
A toda minha famlia, me, pai, irmos, tias, tios, primas e primos, que sempre festejaram
comigo as vitrias da minha vida.
minha orientadora Dr Yvone Gabbay Mendes por todos os conhecimentos compartilhados,
ensinamentos, confiana, incentivo e horas disponibilizadas. Obrigado pela oportunidade e
satisfao em trabalhar sob sua orientao por todos esses quase 6 anos, por ter acreditado em
meu potencial, mesmo eu no tendo muita experincia em laboratrio de pesquisa quando
comecei a atuar como seu estagirio. Obrigado pelo exemplo vvido de profissional
apaixonada, tica, dedicada e acima de tudo humana e humilde, a quem sempre terei como
referncia em minha vida e carreira.
Ao Dr. Alexandre da Costa Linhares e toda equipe do Projeto Efetividade, pelos conselhos e
por permitir o uso das amostras e dados provenientes deste projeto, o que ajudou muito no
desenvolvimento do presente estudo.
A todos os amigos da Seo de Virologia do IEC, principalmente ao grupo de gastrenterites
virais, em especial ao Darivaldo Neri, Luana Soares, Jane Kaiano e Patrcia Lobo.
Aos amigos de laboratrio: Dielle Teixeira, Silvia Lucena, Juliana Hernandez, Joo Felipe e
Maryelle Gonalves, pela ajuda na bancada, preparao de reagentes, suporte informtico e,
sobretudo, pela amizade e bons momentos de convivncia no laboratrio.
Aos companheiros mais recentes no convvio do laboratrio, porm no menos importantes,
Smya Costa, Thayara Morais, Tammy Amaral, Tallyta Barros, Evandro Rodrigues, Luciana
Damascena e Tlio Fumian pela prazerosa convivncia e certeza de amizade cada vez maior,
o que j facilmente observado.
s amigas Gliclia Arago e Paula Spada no s pela amizade e convivncia diria, mas,
sobretudo, por terem compartilhado comigo essa experincia, cheia de dvidas, descobertas e
dificuldades. Tenho certeza que crescemos juntos ao longo destes dois anos e posso dizer que
foi um grande prazer t-las ao meu lado.
Aos queridos amigos Ian Lima, Sylvia Guerra e Alessilva do Socorro pelo apoio profissional,
conselhos, advertncias, momentos de descontrao sempre constantes e, especialmente, pela
preciosa amizade.
Um agradecimento mais que especial a Thas Carvalho, amiga na vida pessoal e a qual tive o
prazer de conviver dentro do laboratrio, onde trabalhou comigo intensamente neste projeto.
Sem sua ajuda no seria possvel terminar meus testes laboratoriais com tanta antecedncia e
nem me dedicar a outras atividades no laboratrio. Muito obrigado.
s colegas, Milla Santos, Luana Soares e Luana Barbagelatta pela ajuda na montagem e
execuo do aparelho de sequenciamento nucleotdico muito utilizado neste trabalho.
Aos amigos Tlio Fumian e Darleise Oliveira pela ajuda na anlise dos cromatogramas e
criao dos dendogramas.
s colegas Anna Alice Nunes e Milene Silveira pelo apoio na realizao do grfico de
Controle de Sries Temporais.
Ao Ncleo de Medicina Tropical da Universidade Federal do Par (UFPA) pelo curso de ps-
graduao que me permitiu obter o grau de mestre em Doenas Tropicais.
Ao Instituto Evandro Chagas (IEC) por todos os recursos humanos e materiais
disponibilizados, os quais foram imprescindveis para a realizao deste e de outros projetos
anteriormente conduzidos por mim.
Coordenao de Aperfeioamento de Pessoal de Nvel Superior (CAPES) pelo suporte
financeiro fundamental para que pudesse desenvolver esse trabalho integralmente.
Ao Instituto Nacional de Metereologia (INMET) por ter gentilmente disponibilizado os dados
climticos utilizados neste estudo.
Ao corpo tcnico da biblioteca do IEC pelo auxlio na busca de peridicos.
dona Valdenira de Jesus Moreira, bibliotecria do Ncleo de Medicina Tropical/UFPA,
pela ajuda na construo da ficha catalogrfica deste trabalho.
Ao professor Esmaelino Neves de Farias por ter gentilmente realizado a reviso ortogrfica do
referencial terico desta dissertao.
dona Graa Santos da Assessoria de Comunicao (ASCOM) do IEC, pelo auxlio na
montagem do grfico de sazonalidade relacionado aos dados climticos e pela construo dos
inmeros psteres enviados a eventos cientficos.
s pesquisadoras Maisa Silva de Sousa, Rosa Helena Porto Gusmo, Ana Ceclia Ribeiro
Cruz e Luana da Silva Soares por aceitarem participar da banca examinadora desta dissertao
de mestrado, tendo muito contribudo para o aperfeioamento da mesma.
Por fim, a todos queles que direta ou indiretamente contriburam para a realizao do
trabalho aqui apresentado, meus sinceros agradecimentos.
s vezes, quando considero as tremendas consequncias
advindas das pequenas coisas ... sou tentado a pensar ... no
existem pequenas coisas.
Bruce Barton
...caminante no hay camino, se hace camino al andar....
Antnio Machado
Resumo
O Norovirus (NoVs), pertencente a famlia Caliciviridae, ocasiona quadro de gastrenterite
aguda (GEA) em pessoas de todas as faixas etrias. Sua relevncia como causa de surtos j
est confirmada, os quais ocorrem principalmente em locais fechados. Sua transmisso ,
sobretudo, pela via fecal-oral, por meio da ingesto de gua/alimentos contaminados e contato
pessoa-a-pessoa. A doena em geral se manifesta de forma sbita, caracterizada por diarreia,
vmito, nusea e clica abdominal. O objetivo deste estudo foi demonstrar a importncia dos
NoVs como patgenos associados s internaes de crianas com quadro de GEA em Belm,
Par. A coleta dos espcimes fecais ocorreu de maio/2008 a abril/2011, sendo examinados
somente aqueles com resultados negativos para rotavrus. Para a deteco dos NoVs foi
utilizado o ensaio imunoenzimtico (EIA) e a reao em cadeia mediada pela enzima
polimerase e precedida de transcrio reversa (RT-PCR). As amostras com resultado positivo
no EIA e negativo na RT-PCR foram submetidas a semi-nested RT-PCR, e aquelas que
permaneceram negativas foram testadas pela PCR em tempo real. Foram analisadas 483
amostras, sendo observada uma positividade de 35,4% (171/483). Adotando-se a RT-PCR
como o mtodo de referncia, o EIA apresentou sensibilidade de 85,9% e especificidade de
93,4%, com excelente reprodutibilidade entre ambas (Kappa= 0.8, p
Abstract
The Norovirus (NoVs), Caliciviridae family, is related with acute gastroenteritis (AGE) in
people of all ages groups. Its importance as cause of outbreaks have been confirmed, which
occur mainly indoors. Its transmission is mainly by the fecal-oral route through contaminated
water and food, and person to person contact. The disease usually is characterized by diarrhea,
vomiting, nausea and abdominal cramps. The aim of this study was to demonstrate the
importance of NoVs as pathogen associated with hospital admissions of children with AGE in
Belm, Par. The collection of fecal specimens occurred from May 2008 to April 2011, being
only tested the samples with negative results for rotavirus. The enzyme immunoassay (EIA)
and the reverse transcription-polymerase chain reaction (RT-PCR) were used for NoVs
detection. The samples with positive results by EIA and negative by RT-PCR, were submitted
to the semi-nested RT-PCR, and the ones with remain negative, were tested by the real-time
PCR. A total of 483 samples were analyzed with a positivity of 35.4% (171/483). Adopting
the RT-PCR as the reference method, the EIA had a sensitivity of 85.9% and a specificity of
93.4% with excellent reproducibility between them (kappa = 0.8, p
SUMRIO
LISTA DE ABREVIATURAS
1 INTRODUO
2 JUSTIFICATIVA
3 REFERENCIAL TERICO
3.1 GASTRENTERITES
3.1.1 Gastrenterite viral
3.2 HISTRICO
3.3 CLASSIFICAO
3.3.1 Famlia Caliciviridae
3.3.2 Gnero Norovirus
3.4 ESTRUTURA MORFOLGICA, PROTICA E GENMICA
3.5 PROPRIEDADES FSICO-QUMICAS E VIABILIDADE NO AMBIENTE
3.6 REPLICAO VIRAL
3.7 MODELOS DE INFECO IN VITRO
3.8 VARIABILIDADE E RECOMBINAO
3.9 PATOFISIOLOGIA DA INFECO VIRAL
3.10 ASPECTOS IMUNOLGICOS
3.11 NOROVRUS E O GRUPO ABO
3.12 SUSCEPTIBILIDADE E RESISTNCIA
3.13 PERSISTNCIA DAS NOROVIROSES EM POPULAES HUMANAS
3.14 DIAGNSTICO LABORATORIAL
3.15 EPIDEMIOLOGIA
3.15.1 Epidemiologia no mundo
3.15.1.1 Surtos
3.15.1.2 Hospitalizaes
3.15.2 Epidemiologia no Brasil
3.16 SAZONALIDADE
3.17 ASPECTOS CLNICOS
3.18 TRANSMISSO
3.18.1 Transmisso zoontica
3.19 TRATAMENTO, PREVENO E CONTROLE
3.20 VACINA
12
13
15
17
17
19
20
22
22
24
26
28
29
30
31
35
37
39
41
42
43
45
45
45
46
48
49
51
53
53
54
56
4 OBJETIVOS
4.1 OBJETIVO GERAL
4.2 OBJETIVOS ESPECFICOS
5 PROCEDIMENTOS METODOLGICOS
5.1 PACIENTES E ESPCIMES CLNICOS
5.2 CARACTERIZAO DO ESTUDO
5.3 ASPECTOS TICOS E DE BIOSSEGURANA
5.4 PROCESSAMENTO DAS AMOSTRAS
5.4.1 Ensaio imunoenzimtico (EIA)
5.4.2 Suspenso fecal a 10%
5.4.3 Extrao do RNA viral
5.4.4 Sntese do DNA complementar (cDNA)
5.4.5 Reao em cadeia da Polimerase (PCR Convencional)
5.4.6 Semi-nested RT-PCR
5.4.7 PCR em tempo real (Real Time)
5.4.8 Anlise dos produtos amplificados
5.4.9 Sequenciamento parcial do genoma viral
5.4.9.1 Purificao do DNA
5.4.9.2 Reao de sequenciamento
5.4.9.3 Dendograma de similaridade
5.5 ANLISE ESTATSTICA
6 RESULTADOS
6.1 DETECO DE NOROVRUS
6.2 AVALIAO DA EFICCIA DOS MTODOS DE DETECO (EIA E RT-PCR)
6.3 GENOTIPAGEM
6.4 DADOS CLNICOS
6.5 SAZONALIDADE
7 DISCUSSO
8 CONCLUSES
9 PERSPECTIVAS
REFERNCIAS
APNDICES
ANEXOS
58
58
58
59
59
60
60
61
62
63
63
64
65
65
68
69
70
70
71
73
73
75
75
76
77
81
83
87
96
97
98
12
LISTA DE ABREVIATURAS
Aminocidos
Centros de Controle de Doenas e Preveno dos EUA
DNA Complementar
Comit de tica em Pesquisa em Humanos do Instituto Evandro Chagas
Cloreto de Csio
Dimetil Sulfxido
Deoxinucleotdeos Trifosfato
cido Etileno Diamino Tetractico
Ensaio Imuno-Enzimtico
Estados Unidos
Genogrupo
cido Clordrico
Calicivrus Humano
Instituto Evandro Chagas
Imuno-Microscopia Eletrnica
Microscopia Eletrnica
Cloreto de Magnsio
Mistura
Hidrxido de Sdio
Nmero Necessrio para Causar um Evento Desfavorvel
Norovrus
Matriz de Leitura Aberta
Pares de Bases
Quantidade Suficiente Para
Gene que codifica a enzima RNA polimerase RNA dependente
Rotavrus
Rotaes por Minuto
Reao em Cadeia mediada pela Polimerase e precedida de Transcrio Reversa
Unidades
Unidade de Terapia Intensiva
Radiao Ultravioleta
Vacina Oral contra Rotavrus Humano
aa
CDC
cDNA
CEP/IEC
CsCl
DMSO
DNTPs
EDTA
EIA
EUA
G
HCl
HuCVs
IEC
IME
ME
MgCl
Mix
NaOH
NNH
NoVs
ORF
pb
q.s.p.
RdRp
RoVs
rpm
RT-PCR
U
UTI
UV
VORH
13
1 INTRODUO
A gastrenterite, tanto epidmica quanto espordica, uma das principais causas de
morbidade e mortalidade entre pessoas de todas as faixas etrias, sendo responsvel,
anualmente, por mais de 1,8 milhes de mortes de crianas menores de 5 anos de idade em
todo o mundo (BRYCE et al., 2005). A causa da doena inclui uma ampla variedade de
patgenos como bactrias, parasitas e vrus; contudo, em muitos locais, a relativa contribuio
desses agentes desconhecida. Estudos a respeito da doena raramente avaliam todos os
agentes etiolgicos possveis e, mesmo naqueles que o fazem, aproximadamente 40>50% dos
casos da doena permanecem sem uma definio etiolgica (PATEL et al., 2009). Estratgias
de preveno e tratamento so necessrias para que haja reduo nas taxas de incidncia das
diarreias graves causadas por diversos agentes (PATEL et al., 2008).
Dos principais enteropatgenos associados gastrenterite, os vrus apresentam grande
importncia epidemiolgica, dentre os quais se destacam os rotavrus (RoVs) do grupo A,
calicivrus humanos (HuCVs) (gneros Norovirus [NoVs] e Sapovirus), astrovrus humanos e
adenovrus entricos (tipos 40 e 41). Outros vrus menos conhecidos tambm esto
relacionados a casos de gastrenterite tais como aichivrus, bocavrus, picobirnavrus,
parechovrus humano, torovrus, coronavrus, pestivrus e enterovrus 22 (GLASS et al., 2001;
WILHELMI et al., 2003; USHIJIMA, 2009). No entanto, quando considerados os surtos de
diarreia em populaes humanas, os NoVs assumem maior importncia (CASTILHO et al.,
2006).
O NoVs categorizado como um patgeno de categoria B pelo National Institute of
Allergy and Infectious Diseases, agencia federal dos Estados Unidos (EUA) que promove
pesquisa mdica e investiga as causas e tratamentos para doenas comuns e raras,
classificando os patgenos em categorias A, B ou C (em ordem decrescente de importncia)
conforme a necessidade de biodefesa contra eles (NIAID, 2007).
14
Esse vrus merece essa classificao devido a sua importante capacidade de provocar
surtos de gastrenterite em populaes expostas, especialmente em locais fechados como
restaurantes, asilos, hospitais, acampamentos militares, navios, dentre outros. A ocorrncia
desses surtos pode assumir grandes propores afetando centenas de pessoas e exigindo
atendimento mdico imediato e medidas de conteno da doena, como fechamento de
estabelecimentos e descontaminao ambiental, o que nem sempre de fcil procedimento. O
NoVs de interesse para pediatras por causar gastrenterite aguda grave em crianas, estando
relacionado no raramente a surtos nosocomiais e em creches (MATSON E SZCS, 2003).
Estudos empregando mtodos moleculares de diagnstico tm demonstrado que o
NoVs responsvel por >90% dos surtos de gastrenterite no-bacteriana e 50% de todas as
causas de gastrenterite epidmica ocorridas no mundo (WIDDOWSON et al., 2005). Dados
recentes sugerem que o impacto do NoVs muito maior do que se previa antigamente,
especialmente em pesquisas realizadas antes da dcada de 90, em que as tcnicas de
identificao (microscopia eletrnica [ME] e ensaios imunoenzimticos [EIA]) eram restritas
e pouco sensveis, subestimando a verdadeira taxa de infeco desse agente na populao
(GLASS et al., 2000; AMAR et al., 2007).
Apesar da prevalncia global desse vrus, seu impacto econmico permanece
desconhecido, sendo que o uso generalizado de mtodos diagnsticos mais sensveis poder
melhorar a vigilncia e controle deste patgeno (LOPMAN, 2006; PATEL et al., 2008).
15
2 JUSTIFICATIVA
Estudos estimam que, anualmente, os NoVs sejam responsveis por 900.000
episdios de gastrenterite que requerem visita clnica e 64.000 hospitalizaes de crianas
menores de 5 anos de idade, em pases economicamente desenvolvidos. Nos pases em
desenvolvimento, a situao mais alarmante. Acredita-se que ocorram mais de 1,1 milhes
de hospitalizaes e 128.000 mortes por ano (PATEL et al., 2008).
Em todo o mundo, estudos de prevalncia demonstram a presena desse agente
infectando as mais diversas populaes. No Brasil, os dados sobre a circulao do NoVs ainda
so escassos, especialmente em pacientes hospitalizados ou vtimas de surtos, podendo-se
citar alguns estudos realizados no Distrito Federal, Par, Rio de Janeiro, Esprito Santo e em
So Paulo, dentre outros, com positividades que variam de 1,9% a 39,7% (BORGES et al.,
2006; CASTILHO et al., 2006; RIBEIRO et al., 2008; VICTORIA et al., 2007; ARAGO et
al., 2010).
A possibilidade de aplicao da tcnica de reao em cadeia mediada pela enzima
polimerase, precedida de transcrio reversa (RT-PCR), no incio da dcada de 1990, facilitou
o diagnstico do NoVs e permitiu um avano considervel na pesquisa deste agente. Ressalta-
se que j bem definida a importncia deste vrus como responsvel por ocasionar
gastrenterite grave em pessoas de todas as faixas etrias, bem como sua associao com surtos
(MEAD et al., 1999).
Contudo, tem sido observado um aumento nas taxas de deteco deste agente (por
exemplo, de 12,5% em 1990-1991 para 44,4% em 2010, nas cidades de Fortaleza e Belm,
respectivamente) (PARKS et al., 1999; SIQUEIRA, et al., 2010a). Este aumento pode estar
relacionado melhora dos mtodos diagnsticos ou possibilidade de as noroviroses serem
uma doena emergente na atualidade. Recentes pesquisas discutem se novas linhagens de
16
NoVs podem periodicamente emergir em nvel local ou global, aumentando assim a
incidncia da doena (HALE et al., 2000; NOEL et al., 1999).
Entretanto, um estudo de reviso sistemtica que avaliou pesquisas conduzidas em
diferentes localidades por um perodo de 18 anos, demonstrou que 75>100% dos casos de
gastrenterite associados ao NoVs foram ocasionados por cepas pertencentes ao genogrupo (G)
II, gentipo 4 (PATEL et al., 2008). No est totalmente esclarecido o porqu desta linhagem
ser a mais prevalente em todo o planeta, como tambm, se est relacionada a uma maior
gravidade do quadro clnico, maior facilidade de escapar da resposta imune do hospedeiro
e/ou maior transmissibilidade (WIDDOWSON et al., 2005). Assim sendo, pesquisas que
visem definir a variabilidade genotpica deste agente, nas mais diversas localidades do globo,
so importantes para estabelecer um panorama epidemiolgico-molecular deste vrus, que
pode embasar estudos futuros que visem concepo de uma vacina.
O aumento nas taxas de deteco do NoVs em diversas localidades e o grande prejuzo
econmico causado pelo mesmo, tornam-no um problema de sade pblica, requerendo
maiores esforos da comunidade cientfica mundial para melhorar as medidas de
tratamento/preveno e a vigilncia desse agente. No entanto, as particularidades de cada
regio devem ser levadas em considerao, uma vez que nem sempre procedimentos
previamente estabelecidos de logstica hospitalar/ambulatorial e laboratorial podem ser
aplicados em todas as localidades do mundo, mesmo que sejam de fcil execuo.
Diante deste panorama, de fundamental importncia a melhor caracterizao desse
vrus a fim de esclarecer seus aspectos epidemiolgicos e moleculares, especialmente na
Amaznia, onde em virtude das peculiaridades ambientais da regio, muitos dados de
sazonalidade e clima no se aplicam ao resto do mundo.
17
3 REFERENCIAL TERICO
3.1 GASTRENTERITES
Gastrenterite um termo usado para definir a inflamao do trato gastrintestinal,
inespecfico quanto aos possveis agentes etiolgicos envolvidos. Corresponde a vrios
estados patolgicos do estmago e intestino, cuja principal manifestao a diarreia, que
pode ser associada a nuseas, vmitos e dor abdominal. No existe um conceito universal de
diarreia e muitas definies sobre a frequncia, consistncia e teor de gua das fezes so
relatados por diferentes autores, contudo, simplificadamente, a diarreia pode ser caracterizada
como fezes que tomam a forma do recipiente em que esto armazenadas (DISKIN, 2009).
Segundo a Organizao Mundial de Sade, a diarreia consiste em trs ou mais evacuaes
lquidas ou semilquidas por um perodo de 24 horas, ou mais evacuaes do que o habitual
dirio para cada indivduo (WHO, 2011).
Fisiologicamente, a diarreia de natureza osmtica um tipo de fisiopatologia de
diarreia aguda, que compreende um movimento alterado de ons e gua que segue uma
diferena de gradiente eletroqumico. Sob condies normais, o trato gastrintestinal tem
grande capacidade de absorver lquidos e eletrlitos. O intestino recebe diariamente 8>9 litros
de fluidos, contudo, somente 100>200 ml so excretados nas fezes. Quando patgenos
entricos acometem o organismo, pode haver alterao no balano inico e lquido, podendo
gerar doena diarreica (HODGES E GILL, 2010).
Essa doena pode rapidamente atingir propores de uma epidemia dependendo do
enteropatgeno causativo, gerando superlotao dos sistemas de sade pblica, mesmo nas
sociedades economicamente favorecidas. Quando transmitida por gua e alimentos
contaminados que resultam em surtos em embarcaes e aeronaves, sua identificao passa a
ser de notificao compulsria conforme preconiza a Portaria n 104 de 25 de janeiro de 2011
do Ministrio da Sade, que define as terminologias adotadas em legislao nacional com
18
relao a doenas, agravos e eventos em sade pblica em todo o territrio nacional e
estabelece fluxo, critrios, responsabilidades e atribuies aos profissionais e servios de
sade (BRASIL, 2011).
Trata-se de um problema de sade pblica de preocupao mundial. Informaes
precisas sobre esta enfermidade em nvel global so subestimadas, especialmente nos pases
em desenvolvimento socioeconmico, nos quais se verifica constante inobservncia na
obrigatoriedade da notificao desta doena e a insuficincia de dados nas declaraes de
bito quanto causa bsica (OLIVEIRA et al., 1998).
Estima-se que ocorram anualmente cerca de 2>4 bilhes de episdios de diarreia,
apresentando 4>6 milhes de bitos em crianas, sendo os lactentes no primeiro ano de vida o
principal grupo de risco, tanto no que diz respeito incidncia, como no risco para
complicaes e morte (ALMEIDA et al., 1998; CLARK E MCKENDRICK, 2004; HODGES
E GILL, 2010). A prevalncia da doena varia com diversos fatores, tais como a condio
socioeconmica, a localizao geogrfica, a faixa etria da populao estudada e as estaes
do ano (ALMEIDA et al., 1998). Essa frequncia tambm varia de acordo com o
enteropatgeno responsvel (bactria [51%], vrus [40%], Giardia lamblia [7-15%],
Cryptosporidium parvum [7%]). Em pases de baixa renda, as condies socioeconmicas da
populao favorecem a ocorrncia de desnutrio infantil, o que se constitui um fator de
agravo para os episdios de gastrenterite (LEE E GREIG, 2010; TERLETSKAIA-LADWIG
et al., 2011).
Estimativas sobre mortalidade infantil em 193 pases demonstraram que 68% das
mortes de crianas menores de 5 anos de idade foram causadas por doenas infecciosas, sendo
15% em decorrncia de diarreia. No Brasil, 5,3% das mortes de crianas nessa faixa etria
esto associadas a essa enfermidade (BLACK et al., 2010).
19
No estado do Par, os casos de diarreia so mais registrados em crianas com idade
menor de 1 ano (mdia de 20 para 100 nascidos vivos); e nesta faixa etria que se observa o
maior nmero de internaes, o maior coeficiente de letalidade hospitalar (2,1 bitos em cada
100 internaes) e o maior coeficiente de mortalidade infantil (0,7 bitos para 1000 nascidos
vivos) em decorrncia de gastrenterite. Acredita-se que esses dados sejam subestimados
devido ao registro ineficiente das informaes, especialmente em virtude da falta de
notificao dos casos e da banalizao que as doenas diarreicas assumem entre a populao e
os profissionais de sade em geral (DIAS et al., 2010).
3.1.1 Gastrenterite viral
Vrios enteropatgenos virais esto associados gastrenterite que acomete pessoas de
todas as idades, dentre os quais podemos citar como principais, os RoVs, NoVs, astrovrus
humano e adenovrus entrico (OLIVEIRA et al., 1998).
Os RoVs so responsveis por 2 milhes de hospitalizaes e 600-875 mil mortes por
ano, apresentando infeco mais grave em crianas menores de 5 anos residentes em pases
em desenvolvimento. A maioria das infeces ocasionada pelos RoVs do grupo A, porm os
grupos B e C tambm podem causar diarreia em humanos. Os NoVs assumem maior
importncia quando considerados surtos de gastrenterite no-bacteriana, sendo responsveis
por >90% dos mesmos. Os astrovrus possuem um gnero que acomete o homem
(Mamastrovrus), o qual se subdivide em 8 sorotipos, sendo o tipo 1 o mais prevalente em
todo o mundo. J os adenovrus apresentam at 51 sorotipos e seis subgneros que infectam
seres humanos; contudo, os principais relacionados a quadros de gastrenterite so os sorotipos
40 e 41, membros da famlia F (considerados fastidiosos), que esto associados a at 15% das
diarreias (CLARK E MCKENDRICK, 2004).
20
3.2 HISTRICO
Nas dcadas de 1940 e 1950, e principalmente a partir dos anos 1970, iniciou-se, no
campo da virologia, a procura por um agente infeccioso que pudesse estar implicado nos
casos de gastrenterite, dos quais a etiologia bacteriana havia sido excluda. A doena ento
chamada de sndrome era caracterizada por um conjunto de sinais/sintomas clnicos que
envolviam vmito, diarreia e dores abdominais que podiam se manifestar isoladamente ou em
conjunto. Esta enfermidade acometia uma grande parte da populao e estudos envolvendo
famlias demonstraram que, por um perodo de 10 anos, esta foi a segunda mais frequente
molstia. Desde a dcada de 1940, tentativas de cultivar e caracterizar in vitro o patgeno
responsvel pela doena foram realizadas em clulas de vrios animais, incluindo macacos,
contudo sem sucesso (KAPIKIAN, 2000).
Em 1971, Kapikian et al. (1972) utilizaram um filtrado proveniente de uma amostra de
swab retal pertencente a um adulto acometido por diarreia durante um surto ocorrido em uma
escola primria na cidade de Norwalk, Ohio, EUA, para infectar voluntrios via oral. Dos 3
voluntrios inoculados, dois apresentaram sintomas tpicos da sndrome, os quais persistiram
mesmo aps duas inoculaes seriadas. Estudos de caracterizao revelaram que o agente
causador possua menos de 36 nm de dimetro e era resistente a ter, cido e aquecimento a
60C por 30 minutos.
Na tentativa de se identificar esse patgeno, foi empregada a tcnica de Imuno-
microscopia eletrnica (IME), utilizando fezes e soro de um desses voluntrios, em que foi
observado um agregado de partculas no envelopadas com cerca de 27 nm de dimetro
similares aos rinovrus (figura 1) (KAPIKIAN, 2000).
21
Figura 1. Aglomerado de partculas de norovrus observado, pela primeira vez por IME, no
estudo realizado por Kapikian et al. (1972), a partir de filtrado fecal de amostra de voluntrio.
Desta forma, foi ento identificado o fastidioso patgeno responsvel pela sndrome
que acometia, e ainda hoje acomete, grande parte da populao mundial, denominado
inicialmente de agente de Norwalk devido ao local onde ocorreu o surto. Amostras coletadas
em outras localidades e que apresentavam as mesmas caractersticas quando visualizadas
IME eram denominadas de Norwalk like virus ou nomeadas com o nome do local de deteco.
Genericamente, eram chamados de pequenos vrus de estrutura arredondada (Small Round
Structure Virus) (KAPIKIAN et al., 1972).
Apesar de terem sido descobertos oficialmente em 1972, o conhecimento acerca desse
patgeno ficou muito limitado em virtude de o mesmo no se propagar em cultura de clulas e
no existir, at ento, um modelo animal eficiente para os estudos. Em 1990, Jiang e
colaboradores realizaram os primeiros estudos envolvendo tcnicas mais avanadas como a
clonagem do genoma viral. Foram construdas bibliotecas de cDNA a partir da extrao do
cido nuclico de vrions, provenientes de amostras de fezes de voluntrios, que aps
clonadas, permitiram saber que o vrus possui genoma de RNA de fita simples, senso
positivo, poliadenilado na extremidade 3end, com cerca de 7,5 kilobases (kb) e com uma
enzima polimerase RNA dependente (RdRp).
22
Ainda no incio da dcada de 1990, Jiang et al. (1992) produziram a protena do
capsdeo utilizando um sistema de traduo em clulas livre e baculovrus recombinantes. Os
produtos expressos demonstraram que foi possvel produzir artificialmente protenas
similares, em tamanho e aparncia, quelas vistas nos NoVs nativos, cuja antigenicidade foi
testada por imunoprecipitao e EIA. Este estudo foi de grande importncia para a pesquisa
dos NoVs, pois permitiu trabalhar com um grande nmero de partculas virais recombinantes,
possibilitando o desenvolvimento de testes seguros, rpidos e sensveis. Pode ser considerado
um marco na pesquisa deste patgeno, pois iniciou os estudos do genoma viral em nvel
estrutural.
Em 1993, ainda Jiang e colaboradores fizeram mais uma importante contribuio para
a pesquisa dos NoVs. Dando continuidade ao estudo de clonagem desenvolvido em 1990, foi
realizada sua completa caracterizao molecular, que permitiu saber que o vrus composto
por um genoma com 7642 nucleotdeos (excluindo sequencias de cauda poli-A), apresenta
cerca de 48% de bases G+C e dividido em 3 matrizes de leitura. Com o conhecimento mais
detalhado da organizao genmica deste patgeno, somado aos aspectos morfolgicos, de
tamanho e polaridade, foi ento possvel observar que o mesmo se assemelhava muito a
outros dois calicivrus de interesse veterinrio, indicando que os ento chamados Norwalk
vrus, pertenciam a esta famlia, o que foi posteriormente aceito pela comunidade acadmica.
3.3 CLASSIFICAO
3.3.1 Famlia Caliciviridae
A famlia Caliciviridae apresenta cinco gneros, dois importantes na sade humana
(Norovirus e Sapovirus) e trs de interesse veterinrio (Lagovirus, Vesivirus e Nebovirus)
cada um representando um distinto clado gentico dentro da famlia. Embora a maioria dos
estudos diagnsticos tenham sido concebidos visando a deteco dos NoVs, o que
23
provavelmente levou a uma subvalorizao da relevncia dos sapovrus, inegvel a
importncia epidemiolgica dos NoVs como o principal membro dos HuCVs nos quadros de
diarreia grave (MATSON E SZCS, 2003; CLARK et al., 2012).
Em cada gnero, uma ou mais espcies tm sido definidas baseadas em relaes
genticas. Abaixo do nvel de espcie, sistemas de tipagem gentica provisria determinam os
genogrupos, que so divididos em gentipos (GREEN, 2007). O Nebovirus, gnero no
patognico aos humanos, foi includo recentemente na famlia (CARSTENS, 2010). Estudos
sugerem a introduo de outros potenciais gneros como os Tulane vrus detectados em
macacos reshus, Bavaria vrus encontrados em frangos e os Valovrus em porcos adultos,
contudo, os mesmos ainda no foram reconhecidos pelo Comit Internacional de Taxonomia
em Viroses (FARKAS et al., 2008; LHOMME et al., 2009; WOLF et al., 2011). O quadro 1
apresenta a estrutura taxonmica dos gneros de calicivrus.
Gnero Vrus representativo Linhagem representativa
Norovirus Norwalk virus Hu/NoV/GI.1/Norwalk/1968/US
Sapovirus Sapporo virus Hu/SaV/GI.1/Sapporo/1982/JP
Lagovirus
Vrus da doena hemorrgica de
coelhos Ra/LaV/RHDV/GH/1988/DE
Vrus da sndrome da lebre marrom
europeia Ha/LaV/EBHSV/GD/1989/FR
Vesivirus
Vrus do exantema vesicular de sunos Sw/VeV/VESV/A48/1948/US
Calicivrus felino Fe/VeV/FCV/F9/1958/US
Nebovirus Newbury-1 virus Bo/Newbury1/1976/UK
Quadro 1. Estrutura taxonmica dos calicivrus. Os criptogramas que simbolizam as
linhagens representativas so organizados da seguinte maneira: Hospedeiro de onde o vrus
foi obtido / Gnero / Vrus representativo ou Genogrupo / Nome da linhagem / Ano de
ocorrncia / Pas de origem. Quadro adaptado de Oliver et al. (2006) e Green et al. (2007).
24
3.3.2 Gnero Norovirus
No h consenso acerca de critrios para a padronizao da classificao genotpica do
NoVs. estabelecido que existam 5 genogrupos dentro do gnero (GI, GII, GIII, GIV e GV),
no entanto, poucos estudos filogenticos consideram a sequncia genmica completa do
capsdeo e raros incluem sequncias dos 5 genogrupos na determinao dos gentipos
(GREEN et al., 2000) (figura 2).
Figura 2. Classificao dos norovrus em 5 genogrupos (GI-GV) e 35 gentipos baseados na
diversidade gentica da protena completa do capsdeo (VP1). rvore filogentica adaptada
do CDC (2011).
GII (Humano e suno)
GV (Murino)
GI (Humano)
GIII (Bovino)
GIV (Humano e canino)
25
Com o objetivo de simplificar e de certo modo uniformizar os difceis critrios de
classificao dos NoVs, Zheng et al. (2006) propuseram uma nova classificao da
nomenclatura deste gnero, baseada em um esquema filogentico bem definido, envolvendo a
anlise de linhagens por mltiplos mtodos de clculo de distncia e avaliao das sequncias
genmicas da protena do capsdeo dos 5 genogrupos aceitos, determinando 29 gentipos
circulantes. Contudo, dados mais recentes publicados pelos Centros de Controle de Doenas e
Preveno dos EUA apresentam outra classificao genotpica (quadro 2).
Genogrupo Hospedeiro Gentipos N de Gentipos
GI Humano 1-8 8
GII Humano 1-10, 12-17, 20, 21
21 Suno 11, 18, 19
GIII Bovino 1-3 3
GIV Humano 1
2 Canino 2
GV Murino 1 1
Total 5 35
Quadro 2. Genogrupos e gentipos pertencentes ao gnero Norovirus. Quadro adaptado do
CDC (2011).
Em contrapartida, Phan et al. (2007) propuseram uma nova nomenclatura baseada na
anlise gentica do domnio S e N-terminal de sequncias do gene do capsdeo de linhagens
de referncia caracterizadas ao redor do mundo e disponveis no GenBank. Nesta
classificao, os autores dividem o NoVs em sete genogrupos (GI-GVII), sendo GI, GIV, GVI
e GVII encontrados exclusivamente em humanos, GII incluindo cepas humana e suna, GIII
encontrado apenas em bovinos e GV em murinos.
Outros estudos sugerem a presena de mais 3 gentipos: um em GII, que alm de
humanos e sunos, tambm infectaria bovinos assim como o GIII; outro gentipo em GIII que
26
estaria relacionado infeco em ovelhas; e um terceiro em GIV que foi descrito infectando
um leo (MARTELLA et al., 2007; MATTISON et al., 2007; WOLF et al., 2009).
.
3.4 ESTRUTURA MORFOLGICA, PROTICA E GENMICA
Os calicivrus possuem capsdeo icosadrico composto por mltiplas cpias da maior
protena estrutural. Cerca de 180 molculas da protena do capsdeo so codificadas pela
segunda matriz de leitura aberta do genoma e organizadas em 90 capsmeros dimricos que
formam 32 depresses em forma de clice. composto por duas protenas estruturais (VP1
[protena maior] e VP2 [protena menor]), na qual a VP1 assume maior importncia, levando
muitos autores a considerar a presena de apenas uma protena estrutural nos NoVs. Capsdeo
composto por uma nica protena estrutural caracterstica comum aos vrus que infectam
plantas, porm raro nos que infectam animais. Alm dos calicivrus, o nico exemplo
conhecido a famlia Nodaviridae que infecta insetos (HOSUR et al., 1987). Assim sendo, os
determinantes funcionais da integridade estrutural, imunogenicidade e infectividade so
definidos a partir dessa nica protena. Dessa maneira, os calicivrus possuem todas as
caractersticas dos vrus que infectam animais, contudo, tm a simplicidade estrutural
daqueles que infectam plantas, tornando-se nicos dentre os vrus (PRASAD et al., 2000).
O NoVs caracterizado como um vrus de RNA fita simples, senso-positivo, no
envelopado, medindo de 27 a 40 nm de dimetro e com 7,5 kb. Apresentam superfcie externa
de aspecto rugoso e com espculas pouco pronunciadas (GREEN et al., 2007).
Seu genoma contm trs matrizes de leitura aberta (ORF): ORF 1, ORF 2 e ORF 3
(figura 3). A ORF 1 codifica uma poliprotena no estrutural de 1738 aminocidos (aa) que
pode ser clivada em 6 protenas, incluindo a RdRp. A ORF 2 codifica a VP1 de 530 a 555 aa e
que possui dois domnios: S e P. O domnio S contm elementos essenciais para a formao
do icosadro. O domnio P est dividido em dois subdomnios: P1 e P2, os quais interagem
27
em contatos dimricos aumentando a estabilidade do capsdeo e formando as protruses no
vrion. A ORF 3 codifica a protena menor do capsdeo (VP2) de 208 a 268 aa que atua na
encapsidao do RNA genmico e na estabilizao dos blocos capsdicos da VP1 para
produzir partculas que sejam resistentes degradao proteoltica (ATMAR E ESTES, 2001;
HARDY, 2005).
Figura 3. Organizao genmica e protica dos norovrus baseada na linhagem representativa
(Hu/NoV/GI.1/Norwalk/1968/US), cdigo M87661 de acesso no GenBank. Faixa amarela
corresponde regio do genoma que codifica as protenas. p48: Protena p48; NTPase:
Protena Nucleosdeo Trifosfatase; p22: Protena p22; VPg: Protena de unio ao genoma;
3CLpro: Proteinase; RdRp: Polimerase; VP1: Protena principal do capsdeo; VP2: Protena
menor do capsdeo. Faixa azul: apresenta as matrizes de leitura aberta. Crculo laranja:
Protena VPg. (A)n: Cauda Poli (A). Esquema adaptado de Vinj et al. (2004), Hardy (2005),
Green et al. (2007) e Siebenga et al. (2009).
Os diversos estudos de filogenia realizados com o NoVs concentraram-se nas regies
da RdRp e da protena do capsdeo. As regies A e B (partes do gene da RdRp localizado na
ORF1) compreendem a regio mais conservada do genoma viral, sendo portanto, mais
adequada para a deteco dos NoVs (JIANG et al., 1999; FANKHAUSER et al., 2002). As
28
regies C (uma pequena extenso da regio 5end da ORF2) e D (regio 3end da ORF2)
codificam parcialmente a protena VP1 do capsdeo e so mais indicadas para a caracterizao
molecular de diferentes gentipos, sendo a regio D a mais recomendada para a
caracterizao de variantes GII.4 e utilizada como uma alternativa eficiente ao
sequenciamento completo da VP1 que apresenta cerca de 1600 nucleotdeos (VINJ et al.,
2004; MATTISON et al., 2009; SIEBENGA et al., 2009). Existe ainda a regio E localizada
entre as regies C e D.
3.5 PROPRIEDADES FSICO-QUMICAS E VIABILIDADE NO AMBIENTE
A densidade de sedimentao dos NoVs em CsCl de 1,33 a 1,41 g/cm. Os vrions
preservam sua infectividade ao serem expostos a agentes fsico-qumicos como solues
cloradas (concentraes de 3,75 a 6,25 mg/L), aps tratamento com ter a 20%, ao calor de
at 60C por 30 minutos ou a baixas temperaturas. Tambm so resistentes a pH cido. No
entanto, concentraes de cloro superiores a 10 mg/L so capazes de inativar esses agentes,
bem como a exposio a etanol a 70% por 30 minutos (DUIZER et al., 2004a; GREEN et al.,
2007).
O fato dos NoVs humanos serem incultivveis em sistema de cultura celular
rotineiramente utilizados nos laboratrios, dificulta a realizao de estudos que estimem a
infectividade destes vrus no ambiente, contudo, algumas pesquisas envolvendo possveis
substitutos aos NoVs permitem mensurar a viabilidade da partcula viral fora do hospedeiro
(MARSHALL E BRUGGINK, 2011).
Doultree e colaboradores (1999) utilizaram os calicivrus de felinos como substitutos
aos NoVs humanos em um estudo que verificou a sobrevivncia deste patgeno a uma
variedade de condies ambientais. Foi observado que esse vrus sobreviveu por at 60 dias a
29
4C (com perda mnima de infectividade) e permaneceu estvel temperatura ambiente por
14 a 21 dias quando suspensos em meio de cultura e 21 a 28 dias, em ambiente seco.
Bae e Schwab (2008) compararam a sobrevivncia dos calicivrus de felinos com o
NoVs murino em gua, e concluram que o ltimo apresentou maiores taxas de sobrevivncia,
com menos perda de infectividade e de integridade do cido nuclico a 25C. Outro estudo
conduzido por Lamhoujeb e colaboradores (2009) verificou a infectividade dos NoVs em
superfcies de materiais que comumente entram em contato com alimentos, como ao
inoxidvel (usado na fabricao de talheres) e policloreto de polivinila, conhecido por PVC
(usado em recipientes que armazenam alimentos). Neste estudo, foi observado que os NoVs
humanos permanecem infecciosos nestas superfcies por at 28 dias a 20C.
3.6 REPLICAO VIRAL
Os calicivrus compartilham muitas estratgias de replicao observadas em outros
vrus de polaridade positiva. Primeiramente, o vrus interage com a clula do hospedeiro via
receptor vrus-especfico, permitindo a entrada da partcula no citoplasma da clula.
Posteriormente, o RNA genmico do vrus liberado e ocorrem interaes com a clula que,
at o momento so pouco conhecidas. O incio da traduo mediada pela protena VPg e
ocorre pela interao do genoma viral com os mecanismos de traduo celular. A ORF 1
traduzida para produzir a poliprotena no estrutural. O incio da sntese de uma fita simples
de RNA antisense, a partir do template, ocorre envolvendo interaes com as protenas
celulares. Da mesma forma que todos os outros vrus de RNA senso-positivos, a replicao
est associada com as membranas da clula do hospedeiro. Os mecanismos de
empacotamento do RNA viral, maturao e liberao celular so pouco compreendidos,
contudo, a apoptose celular pode facilitar a liberao dos vrus (GREEN et al., 2007).
30
3.7 MODELOS DE INFECO IN VITRO
O tropismo celular dos NoVs humanos ainda desconhecido. Diversas tentativas de se
identificar um sistema de cultura eficaz para este patgeno j foram feitas, porm sem
sucesso. A inabilidade destes vrus em se propagarem em meios de cultivo artificiais o
maior obstculo para estud-los (DUIZER et al., 2004b; KARST, 2010). Bipsias intestinais
de voluntrios infectados com NoVs GI.1 e GII.1 demonstraram pela ME, no haver presena
de partculas virais neste tipo de material (AGUS et al., 1973; DOLIN et al., 1975). Contudo,
recentes estudos em modelos animais sugerem que mltiplos tipos de clulas intestinais,
incluindo entercitos, macrfagos e clulas dendrticas podem apresentar, mesmo que em
baixo nvel, alguma replicao in vitro (KARST, 2010).
Na literatura, j foi reportada a infeco de NoVs em um modelo tridimensional do
epitlio intestinal humano, contudo com resultados questionveis (STRAUB et al., 2007).
Pesquisas demonstram a infeco de sunos e bezerros gnotobiticos pela linhagem HS66,
pertencente ao NoVs GII.4. Esses animais desenvolveram diarreia e os bezerros apresentaram
mudanas histopatolgicas evidentes. Nos sunos, observaram-se partculas virais e capsdeo
viral nas vesculas citoplasmticas dos entercitos localizados na extremidade apical e ao
longo das vilosidades, porm ausentes nas criptas do duodeno e jejuno (CHEETHAM et al.,
2006; SOUZA et al., 2008).
O NoVs que infecta murino vem sendo alvo de muitas pesquisas envolvendo o cultivo
celular deste patgeno. Por ser naturalmente entrico, muito til neste campo de pesquisa,
haja vista ser oralmente infeccioso, podendo ser encontrado na mucosa do intestino e nos
ndulos linfticos mesentricos durante a infeco, bem como ser expelido pelas fezes, o que
permite seu isolamento. conhecido por se replicar em clulas dendrticas e macrfagos
derivados de moncitos isolados de clulas mononucleares do sangue perifrico, porm, no
cultivvel em outros tipos celulares (WOBUS et al., 2004, 2006; HSU et al., 2006;
31
MUMPHREY et al., 2007; PERRY et al., 2009; KIM et al., 2010). J foi descrito que este
NoVs provoca diminuio da consistncia das fezes e inflamao intestinal moderada em
camundongos selvagens infectados (MUMPHREY et al., 2007; LIU et al., 2009).
Baseado no tropismo observado no NoVs murino, tentou-se cultivar o vrus Norwalk
G1.1 em clulas dendrticas e macrfagos, porm sem evidencia de replicao viral (KARST,
2010). Tambm j foi relatado que ratos imunodeficientes naturalmente infectados com NoVs
murino, apresentaram antgeno viral no citoplasma de clulas inflamatrias no fgado, bao,
lmina prpria intestinal, folculos linfides do intestino, pulmo, cavidades pleural e
peritoneal, e ndulos linfticos mesentricos (WARD et al., 2006, PERDUE et al., 2007).
Acredita-se que o interferon possa restringir o tropismo celular de NoVs, como j foi
demonstrado para outros vrus (RYMAN et al., 2000). Se este for o caso, o diferente tropismo
celular apresentado pelos NoVs pode ser definido conforme a diferente sensibilidade dos
vrus s respostas de interferon (KARST, 2010).
3.8 VARIABILIDADE E RECOMBINAO
A recombinao de RNA uma das foras motrizes da evoluo dos vrus. Pode afetar
os agrupamentos filogenticos, aumentar a virulncia e confundir estudos epidemiolgicos e
moleculares, tendo grandes implicaes nas estratgias de concepo de vacinas (BULL et al.,
2007).
Pode-se definir um NoVs recombinante a partir da anlise filogentica de duas regies
diferentes do genoma, geralmente o capsdeo e a RdRp, de clusters de dois grupos distintos
(BULL et al., 2005). Os eventos de recombinao j foram descritos ocorrendo entre as
regies ORF 1/ORF 2, RdRp/ORF 2 e ORF2/ORF3 (WATERS et al., 2007; ROHAYEM et
al., 2005; CHHABRA et al., 2010). Existem trs tipos de recombinao: intergenogrupo
(entre genogrupos distintos), intergenotipo (entre gentipos distintos) e intragenotipo (entre
32
mesmos gentipos), sendo a recombinao intergenotpica a mais frequentemente observada.
Alguns pesquisadores ainda dividem as cepas em intersubgenotipos recombinantes (PHAN et
al., 2007). Somente uma recombinao intergenogrupo j foi reportada na literatura ocorrendo
entre GI.3/GII.4 e nove recombinaes intragenotpicas entre GII j foram relatadas
(ROHAYEM et al., 2005; ETHERINGTON et al., 2006; PHAN et al., 2006).
A recombinao mais observada em 3 dos 5 genogrupos (GI, GII e GIII)
(KATAYAMA et al., 2002; HAN et al., 2004). Contudo, GII apresenta o maior nmero de
gentipos variantes do tipo 4 que causam a maioria dos surtos e casos espordicos no mundo
(CASTILHO et al., 2006). No entanto, um recombinante GII.b/GII.3 tem sido descrito como
o principal responsvel por surtos na Europa, Austrlia e sia (AMBERT-BALAY et al.,
2005; BULL et al., 2005; PHAN et al., 2006, 2007).
No existe uma classificao especfica para linhagens recombinantes e um sistema de
genotipagem amplamente aceito ainda necessrio, gerando dificuldades em definir o nmero
e tipos de recombinantes em circulao. Apesar dessas dificuldades, pesquisas demonstram a
maior circulao de 20 diferentes tipos recombinantes de NoVs no mundo (um GI, 17 GII e
dois GIII) (BULL et al., 2007).
Existem vrions pertencentes ao genogrupo II, que ao terem a regio da RdRp de seus
genomas sequenciada, no apresentam classificao conhecida. Entretanto, quando
sequenciada a regio do capsdeo, podem apresentar alguma classificao, porm diferente
daquela observada pela RdRp, sendo ento denominadas de variantes. Para essas cepas, h a
necessidade de um sistema de classificao que as diferencie daquelas que apresentam igual
classificao quando caracterizadas por ambas as regies do genoma (RdRp e capsdeo),
sendo comumente caracterizadas como GII.a, GII.b, GII.c, GII.d ou GII.e (BUESA et al.,
2002; BULL et al., 2007).
33
A identificao de novos clusters no incomum. Diferentes grupos de pesquisa tm
caracterizado cepas circulantes de NoVs e constantemente tem relatado novos gentipos
(VIDAL et al., 2006).
Na Austrlia foram detectados diversos gentipos (GII.1, GII.3, GII.4, GII.6, GII.7 e
GII.10) de NoVs por Nested RT-PCR/sequenciamento em amostras fecais coletadas de surtos
e casos espordicos de diarreia entre 1997 e 2004. Neste estudo se observou a presena de um
GII.4 variante que foi responsvel por 18 surtos, sendo tambm descrito como agente
etiolgico de epidemias na Holanda, Japo e Taiwan entre 2004 e 2005 (BULL et al., 2006).
No Chile, a anlise parcial do genoma do capsdeo e da RdRp de amostras
provenientes de 13 surtos ocorridos entre 2001 e 2003 demonstrou que os mesmos foram
ocasionados por cepas geneticamente diversas, agrupadas principalmente dentro de GII e por
5 possveis linhagens recombinantes (VIDAL et al., 2006).
No Japo, entre 2004 e 2005, foram testadas por RT-PCR convencional amostras de
casos espordicos de gastrenterite em que se detectaram vrios gentipos (GII.1, GII.2, GII.3,
GII.4, GII.6 e GII.7). Neste estudo se observou quatro tipos recombinantes co-circulando e a
linhagem GII.b surgiu como a principal responsvel pelos quadros de diarreia no Japo
(PHAN et al., 2007).
Outras linhagens tambm foram observadas no Japo em uma pesquisa envolvendo
100 surtos ocorridos em hospitais, asilos, escolas e instalaes sociais em 2006. Foram
detectados os tipos GI.4, GI.8, GII.2, GII.3, GII.4 e GII.12. Com relao ao gentipo GII.4,
trs diferentes subtipos foram observados (GII.4d, GII.4e e GII.4f) sendo o ltimo o mais
prevalente (OKADA et al., 2007).
Na Sucia, amostras de 115 surtos nosocomiais ocorridos em hospitais, asilos e outras
instituies de sade entre 1997 e 2005 foram testadas e demonstraram que as duas principais
34
epidemias ocorridas nos invernos de 2002/2003 e 2004/2005 foram ocasionadas por duas
cepas variantes fortemente relacionadas com GII.4 (JOHANSEN et al., 2008).
Na ndia, um estudo da anlise parcial do gene da RdRp e do capsdeo, envolvendo
crianas diarreicas positivas para NoVs demonstrou uma nova variante de GII.4, denominada
GII.4/2007 e outros 3 novos intergenotipos recombinantes de GII, indicando a marcante
variabilidade gentica dos NoVs nesse pas (NAYAKA et al., 2009). Tambm na ndia,
espcimes fecais de crianas menores de 5 anos hospitalizadas e atendidas ambulatorialmente
foram testadas por RT-PCR convencional, observando-se a co-circulao de GII.1, GII.2,
GII.3 e da variante GII.b, detectada pela primeira vez nesse pas (CHHABRA E
CHITAMBAR, 2008). Esta ltima tambm foi a mais prevalente em crianas menores de 2
anos e meio de idade atendidas em um hospital holands durante 5 invernos consecutivos
(2002-2007) (BEERSMA et al., 2009).
Em 2006 foi registrado, na Austrlia e Nova Zelndia, o surgimento de duas novas
variantes GII.4 (2006a e 2006b) que foram implicadas como as responsveis pelo aumento da
gastrenterite em toda a Europa naquele ano (TU et al., 2008). Ainda em 2006, a cepa 2006a
foi a principal responsvel por um surto de gastrenterite ocorrido na Alemanha (HOFFMANN
et al., 2010). Em 2007 e 2008 foi descrita, na Coria do Sul, a ocorrncia de GII.4 em
amostras provenientes de crianas com gastrenterite aguda, estando as mesmas intimamente
relacionadas com a cepa 2006b tambm descrita na China, Hong Kong, e Japo (LE et al.,
2010).
Um estudo realizado no Canad analisou 707 surtos ocorridos na provncia de Alberta
e territrios do norte por um perodo de 9 anos (2000-2008). A caracterizao molecular das
amostras demonstrou uma grande variedade de cepas GII.4 consideradas variantes (GII.4-
1996, GII.4-2002, GII.4-2004, GII.4-2006a, GII.4-2006b, GII.4-2008a e GII.4-2008b). As
duas variantes que surgiram em 2006 tiveram ampla distribuio geogrfica e ocasionaram a
35
maioria dos surtos em comparao com as demais. As linhagens variantes foram mais
comumente associadas com surtos em hospitais, enquanto as no variantes foram mais
prevalentes em escolas e eventos sociais (PANG et al., 2010).
Um estudo analisou surtos ocorridos em vrios pases nos cinco continentes entre os
anos de 2001 e 2007 e observou oito variantes de GII.4, sendo quatro de distribuio global
(linhagens 1996, 2002, 2004 e 2006b). As variantes 2001Japan e 2001Henry tambm foram
encontradas em todo o mundo, porm em baixa frequncia; e a 2003Asia e 2006a causaram
epidemias geograficamente limitadas (SIEBENGA et al., 2009).
Na Alemanha, durante surto de gastrenterite ocorrido em 2006, duas cepas foram
detectadas, a GII.4-2006a que foi a predominante e a GII.7 que acometeu dois indivduos
imunocomprometidos. Nestes pacientes, o gentipo observado foi similar ao isolado em 1990
e 1994 na sia, sugerindo uma lenta evoluo viral, com evidencia de recombinao
(HOFFMANN et al., 2010).
3.9 PATOFISIOLOGIA DA INFECO VIRAL
A infeco por NoVs autolimitada, com durao em geral de 24-48 horas, que pode
ou no requerer hospitalizao e terapia de reidratao. Por muito tempo, o conhecimento
acerca da patognese da infeco viral foi habitualmente obtido de estudos de inoculao em
voluntrios (GRAHAM et al., 1994).
O vrus provoca leses histopatolgicas transitrias na mucosa do intestino delgado,
que inclui alargamento das vilosidades e o encurtamento dos microvilos. Infiltrao de clulas
mononucleares na lmina prpria e vacuolizao do citoplasma so observadas em pessoas
infectadas. Ainda no foi detectado o vrus ou o antgeno viral nas clulas intestinais, porm
esto presentes nas fezes e no vmito. A excreo viral atinge seu pico no primeiro e no
segundo dia aps a infeco, podendo continuar por mais duas semanas (alguns autores
36
afirmam at 100 dias), mesmo aps os sintomas clnicos terem cessado. Podem, tambm, ser
excretados por indivduos assintomticos (infeces subclnicas). Na fase aguda da doena, as
mucosas gstrica e retal no so afetadas histologicamente e os nveis de adenilato ciclase na
mucosa jejunal no so alterados, ou seja, sem o aumento da presena dessa enzima, no h a
produo de adenosina monofosfato cclico (AMPc), que resultaria em hipersecreo de
eletrlitos e gua, gerando diarreia do tipo secretora, frequentemente de grave intensidade
(KAPIKIAN et al., 1996; ESTES et al., 2000; KIRKWOOD E STREITBERG, 2008; LOPES
E PINTO, 2010).
Todavia, vrios fatores virais podem interferir com as funes celulares bsicas, sem
causar morte celular, porm gerando quadro diarreico. Uma delas a proteinase 3C-like
(3CLpro), que semelhante a uma enzima previamente descrita em rinovrus chamado 3Cpro.
Esta enzima interfere com traduo eucaritica pela clivagem da cauda poli (A) do material
gentico da clula infectada. A inibio da traduo na clula do hospedeiro pode limitar a
abundncia de reguladores importantes da homeostase intestinal. Outro mecanismo que tem o
potencial de interferir com protenas do hospedeiro envolve a protena no-estrutural p48 do
vrus, a qual interage com outras que so reguladoras de trfego de vescula. Tal como
acontece com a inibio da traduo, a inibio do trfego de protena susceptvel de
interferir com processos celulares crticos para manter a funo de absoro (HODGES E
GILL, 2010).
Recentemente foi realizado um estudo com bipsias intestinais de pacientes que
sofreram de infeco por NoVs. Em 123 observaes iniciais, verificou-se que a altura das
vilosidades foi reduzida em 25% dos casos, enquanto a profundidade das criptas permaneceu
inalterada, reduzindo a superfcie total de absoro do epitlio (figura 4). Os nveis de Na+
no foram alterados, no entanto houve um aumento da secreo de ons Cl- alm de uma
acentuada diminuio na resistncia transepitelial. Pelo fato de s recentemente as pesquisas
37
em cultura de clulas terem avanado, o entendimento dos mecanismos patognicos do NoVs
ainda muito limitado, necessitando de extensas pesquisas. O conhecimento dos mecanismos
fisiopatolgicos desse patgeno pode ajudar a identificar novos alvos teraputicos, no s
para a diarreia associada com infeces entricas, mas tambm para uma variedade de outras
doenas diarreicas (HODGES E GILL, 2010).
Figura 4. Reduo da altura das vilosidades do intestino em resposta infeco viral por
norovrus. Imagem adaptada de Hodges e Gill (2010).
3.10 ASPECTOS IMUNOLGICOS
Nos primeiros estudos realizados em voluntrios sobre a resposta imune do hospedeiro
infeco pelo NoVs, constatou-se que a maioria ficou doente aps exposio ao vrus. Isto
reforou a idia de que grande parte da populao no possua imunidade natural ao patgeno
ou este apresentava mecanismos que facilitavam sua evaso resposta imune do hospedeiro.
Estudos posteriores demonstraram que indivduos que apresentaram a doena no
desenvolveram novamente a enfermidade at 14 semanas aps reinfeco, sugerindo uma
imunidade de curto prazo (WYATT et al., 1974).
Normal Infectado
38
Essas observaes iniciais foram confirmadas nos anos posteriores e os conhecimentos
acerca da imunologia do NoVs foram ampliados quando testes sorolgicos mais sensveis se
tornaram comercialmente disponveis. Pode-se citar, como a principal ferramenta de
investigao, o imunoensaio em fase slida, que contou com antgenos virais nativos e soro
pareado de infeco humana (GREENBERG et al., 1978).
Muitos estudos soroepidemiolgicos visando a deteco de anticorpos contra NoVs
foram realizados envolvendo, principalmente, soro de crianas em vrias localidades. Em
geral, nos pases desenvolvidos, a prevalncia de anticorpos mais baixa em crianas de 0>5
anos, os quais aumentam com a idade (GREENBERG et al., 1979).
As diferenas observadas em termos de taxa de anticorpos envolvendo
crianas/adultos e crianas provenientes de distintas populaes podem ser justificadas por
fatores scio-econmico-culturais que influenciam na precoce ou tardia obteno de
anticorpos. Por exemplo, em adultos de pases economicamente desenvolvidos, o nvel de
anticorpos no soro pode ser um indicador de exposio passada, em vez de infeco recente.
Enquanto que em populaes de pases em desenvolvimento, a frequncia com que as
crianas so expostas ao vrus pode ser to habitual, devido viverem em aparente
contaminao ambiental (condies sanitrias inadequadas) e aglomeraes, que a imunidade
em longo prazo pode no ser observada (PARKER et al., 1994; MATSUI E GREENBERG,
2000).
So comuns os estudos imunolgicos com a utilizao de partculas virais
recombinantes produzidas em sistemas de expresso em baculovrus, comparveis em
tamanho, estrutura e propriedades antignicas aos vrions nativos. A possibilidade de possuir
uma fonte facilmente renovvel de partculas recombinantes uniformes ampliou o
desenvolvimento de imunoensaios mais sensveis e altamente especficos, como os NoVs
39
recombinantes e EIA, que at hoje esto amplamente disponveis para uma variedade de
aplicaes (GREEN et al., 1993).
Atualmente, sabe-se que a infeco causada por este patgeno induz uma resposta
imune pela produo de anticorpos da classe IgG, IgA e IgM, os quais proporcionam uma
proteo contra reinfeces por um perodo de 4 a 6 meses (LINDESMITH et al., 2005).
No Brasil, um estudo envolvendo 8 comunidades indgenas da Regio Amaznica
demonstrou que as tribos apresentaram de 39% a 100% de soropositividade para o NoVs, no
se observando uma diferena significativa na prevalncia de anticorpos entre as diferentes
faixas etrias, evidenciando a presena deste patgeno tambm em comunidades isoladas do
Pas (GABBAY et al., 1994). Em nossa regio, estudos sorolgicos realizados em 37 crianas
demonstraram que aos trs anos de idade, 94,6% j possuam anticorpos contra NoVs
(MENDES, 1997).
3.11 NOROVRUS E O GRUPO ABO
Diversos estudos tm demonstrado uma correlao entre o grupo sanguneo ABO e
infeco intestinal por vrios patgenos. Por exemplo, o fentipo O parece ter influncia na
gravidade dos sintomas de pacientes com clera, bem como no desenvolvimento de lceras
duodenais causadas por Helicobacter pylori (MENTIS et al., 1991; ALBERT, 1996).
Estudos demonstram a associao entre o fentipo ABO e o risco de
infeco/desenvolvimento de doena sintomtica gerada pelo NoVs. De acordo com Hutson
et al. (2002), indivduos do grupo sanguneo B tm menor chance de se infectarem com o
vrus do que aqueles pertencentes ao grupo O.
Autores sugerem que a presena da D-galactose terminal do antgeno B possa de
alguma forma alterar a partcula de NoVs e impedir sua ligao clula hospedeira, porm
isto no consenso entre diferentes pesquisadores (MEYER et al., 2004).
40
Outros, no entanto, atribuem ao gene FUT2 (codificador da enzima 1,2
fucosiltransferase) responsvel por expressar o carboidrato H tipo 1 (encontrado na superfcie
das clulas epiteliais do intestino delgado e em secrees como a saliva) a responsabilidade
por esta relao (infeco x sistema ABO). Acredita-se que este carboidrato seja um receptor
para o NoVs. Desta maneira, a proteo contra a infeco viral ocorre quando outras
glicosiltransferases (agora quelas que determinam o fentipo do sistema ABO sanguneo)
encontradas tambm na superfcie das clulas epiteliais do intestino, tm a capacidade de
modificar a estrutura do carboidrato H tipo 1, tornando-o inativo e prejudicando sua funo
como receptor dos NoVs. Supe-se que este processo ocorra com pessoas do grupo sanguneo
B, conferindo maior proteo queles que apresentam este fentipo (HARRINGTON et al.,
2002; MARIONNEAU et al., 2005).
Estudos indicam que as diversas linhagens de NoVs podem atuar de maneiras
diferentes na associao da partcula viral aos antgenos do grupo sanguneo ABO. O NoVs
tipo GII.4 conhecido globalmente por causar epidemias, tem mais facilidade de se ligar a
esses antgenos que os outros gentipos, o que pode explicar sua prevalncia mundial
(SHIRATO et al., 2008). Alm disso, existem evidncias de que componentes extracelulares
encontrados nas fezes possam ter papel importante na ligao do vrus ao antgeno do grupo
sanguneo, potencializando o ataque e a infectividade in vivo (HARRINGTON et al., 2004).
A pesquisa a respeito da relao do NoVs com o grupo ABO importante, pois este
vrus apresenta grupos antigenicamente diversos e a possvel identificao de um eptopo
comum de ligao com a clula hospedeira fundamental no desenvolvimento de futuras
vacinas (SHIRATO et al., 2008).
41
3.12 SUSCEPTIBILIDADE E RESISTNCIA
Embora o NoVs seja altamente infeccioso, experimentos em voluntrios demonstram
que alguns sujeitos permanecem sadios, sem desenvolver a doena, mesmo aps a inoculao
de grandes doses virais. Este mecanismo ainda no est bem esclarecido, porm poderia estar
relacionado a uma resistncia inata ou a uma imunidade pr-existente infeco viral
(MATSUI E GREENBERG, 2000).
Um estudo com amostras de saliva evidenciou que indivduo com mucosa secretora do
carboidrato H tipo 1 um importante preditor de susceptibilidade, enquanto a resposta imune
gravada na memria da mucosa pode estar associada proteo contra o patgeno, apesar de
nunca ter sido identificado nenhum componente imunolgico especfico de proteo.
(LINDESMITH et al., 2003). So descritas at 3 respostas em indivduos inoculados com
NoVs: (1) resistncia infeco apesar de repetidas inoculaes; (2) desenvolvimento de
imunidade protetora e (3) susceptibilidade apesar de vrias inoculaes (JOHNSON et al.,
1990).
No que diz respeito resistncia, autores sugerem que at dois mecanismos podem ser
observados: (1) resistncia gentica inata mediada pela inativao do alelo FUT2 (gene
responsvel por expressar o fentipo secretor) e (2) imunidade adquirida caracterizada pela
rpida produo de IgA NoVs-especfica na mucosa em indivduos secretores. Seja qual for o
mecanismo, observa-se alto percentual de indivduos resistentes (LINDESMITH et al., 2003).
Em relao susceptibilidade infeco pelo NoVs, as diferenas genticas no
explicam, totalmente, os diversos padres de respostas inoculao, indicando que essa
susceptibilidade multifatorial (GRAHAM et al., 1994).
42
3.13 PERSISTNCIA DAS NOROVIROSES EM POPULAES HUMANAS
As pesquisas sobre a persistncia do NoVs na populao humana tem como principal
alvo de estudo a cepa epidmica, globalmente conhecida, GII.4. Esta linhagem evoluiu na
populao durante aproximadamente 20 anos, uma vez que no h evidncias de sua
existncia antes de meados da dcada de 1980, enquanto que outros genogrupos tm sido
isolados desde a dcada de 1960. Sua emergncia foi marcada por uma evoluo em que os
clusters iniciais foram submetidos a perodos de hibernao, seguido de surgimento de
epidemias que posteriormente evoluram de forma mais rpida e linear (SIEBENGA et al.,
2007; LINDESMITH et al., 2008).
Atualmente, dados indicam que diferenas estruturais permitem que esta cepa escape
da resposta imune pr-existente. Novas linhagens emergem a partir da seleo do sistema
imune, resultando em variantes que so competentes para infectar os mesmos indivduos
infectados anteriormente. Como consequncia dessa seleo, essa variante pode evoluir e se
especializar na associao com determinado grupo sanguneo ABO, fazendo com que
indivduos resistentes a uma cepa sejam sensveis outra (DONALDSON et al., 2008).
Mudanas de aminocidos no subdomnio P1 ou P2 (responsveis pela estabilidade do
capsdeo e pelas protruses do vrion) podem resultar em grandes alteraes no uso do
receptor e na antigenicidade, gerando a persistncia do vrus por dois mecanismos: (1)
adaptao, conduzida pela imunidade, resultando na capacidade do vrus em se ligar a
diferentes receptores, inclusive queles relacionados ao grupo sanguneo ABO, permitindo
expanso para uma gama de hospedeiros e penetrao na populao previamente virgem. (2)
variao, que permite escapar da imunidade predominante, resultando em um vrus
competente em acometer uma mesma populao anteriormente infectada (DONALDSON et
al., 2008) (figura 5).
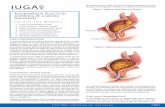
43
Figura 5. Ilustrao simplificando os dois mecanismos de persistncia da linhagem GII.4 na
populao humana. Adaptado de Donaldson et al. (2008).
Entender o porqu da linhagem GII.4 ter evoludo para cepas pandmicas, to bem
sucedidas em escapar do sistema imunolgico humano, enquanto outras linhagens estavam
co-circulando na poca de seu surgimento, uma questo que s pode ser respondida quando
comparados os perfis evolucionrios e os modelos estruturais dos diferentes tipos de NoVs
circulantes na populao (LINDESMITH et al., 2008).
3.14 DIAGNSTICO LABORATORIAL
A anlise laboratorial para a deteco do NoVs pode ser realizada em vrios espcimes
clnicos como fezes, vmito, saliva e soro sanguneo (MOE et al., 2004; TAKANASHI et al.,
2009; ATMAR et al., 2011; KIRBY et al., 2011).
A ausncia de um mtodo de diagnstico rpido, sensvel e de baixo custo financeiro
o maior obstculo na deteco de infeces por NoVs em laboratrios de sade pblica e
Mesma cepa
Populaes com diferentes
receptores
Variabilidade de cepas
Mesma populao
ADAPTAO VARIAO
44
hospitais, especialmente devido diversidade gnica e antignica deste grupo de vrus
(GONZLEZ et al., 2006).
Durante muitos anos, esse vrus foi diagnosticado pela ME, porm nas ltimas dcadas
a introduo da tcnica de RT-PCR convencional melhorou consideravelmente a sensibilidade
de deteco, tornando-se o teste padro para o diagnstico destes patgenos. Apesar da sua
sensibilidade, a RT-PCR um mtodo de alto custo, demorado, que requer pessoas bem
treinadas e equipamentos sofisticados, no sendo disponvel na rotina da maioria dos
laboratrios de diagnstico (KHAMRIN et al., 2008).
Estudos utilizando a RT-PCR apresentam grande variao no que diz respeito aos
iniciadores de cadeia utilizados. Trabalhos envolvendo a amplificao nucleotdica parcial das
regies A, B, C, D e E do genoma do vrus so comumente reportados na literatura (JIANG et
al., 1999; ANDERSON et al., 2001; KOJIMA et al., 2002; VINJ et al., 2004; SIEBENGA et
al., 2009).
Geralmente, h a necessidade do uso de metodologias complementares aps a
realizao da RT-PCR convencional, como o sequenciamento nucleotdico ou a hibridizao
por blot, a fim de caracterizar os genogrupos e gentipos de NoVs (BRUIN et al., 2006).
Alternativamente RT-PCR, tcnicas mais sensveis de amplificao do genoma viral como a
semi-nested RT-PCR tambm j foram descritas (BOXMAN et al., 2006).
O advento da tcnica de PCR em tempo real revolucionou o processo de
deteco/quantificao de fragmentos de RNA e DNA para diagnstico microbiolgico. Esta
tcnica tem como vantagem ser muito mais sensvel que todas as outras j descritas, ser de
fcil quantificao, rpida anlise, alta reprodutibilidade e preciso, melhor controle de
qualidade e baixo risco de contaminao. Sua desvantagem em relao s outras metodologias
de amplificao o alto custo para sua manuteno (TRUJILLO et al., 2006; VALONES et
al., 2009).
45
Recentemente, tm sido divulgados inmeros EIA comerciais e de
imunocromatografia, bem como tcnicas de amplificao isotrmica e DNA Microarray para
deteco e caracterizao simultnea de NoVs (ITURRIZA-GMARA et al., 2008;
KHAMRIN et al., 2008; PAGOTTO et al., 2008; SIQUEIRA et al., 2011). Estas tcnicas tm
sido descritas como metodologias complementares RT-PCR, podendo ser valiosa na
investigao de surtos de gastrenterite (DIMITRIADIS et al., 2006).
3.15 EPIDEMIOLOGIA
3.15.1 Epidemiologia no mundo
3.15.1.1 Surtos
O NoVs mundialmente conhecido como o principal responsvel por surtos de
gastrenterite viral em pessoas de todas as idades. Critrios epidemiolgicos descritos por
Kaplan et al. (1982) definem como surto de gastrenterite por NoVs, a presena de vmito em
mais da metade das pessoas afetadas, tempo mdio de incubao da doena de 24-48 horas,
durao mdia de sinais e sintomas clnicos de 12-60 horas e a ausncia de patgenos
bacterianos em cultura de fezes. Estudo realizado por Turcios et al. (2006) demonstrou que o
conjunto de critrios de Kaplan altamente especfico (99%) e moderadamente sensvel
(68%) para diferenciar surtos ocasionados pelo NoVs daqueles causados por bactrias, sendo
considerados, at hoje, muito teis para esta distino.
Novos mtodos moleculares permitiram rastrear a origem de surtos, levando a
identificao do vrus em guas/alimentos contaminados (especialmente mariscos) e
manipuladores de alimentos (GLASS et al., 2000).
Acredita-se que mais de 80% dos surtos e 18% dos casos espordicos de diarreia
registrados na Holanda entre 1998 e 1999 foram causados por NoVs (KOOPMANS et al.,
46
2000). Na Espanha, 60 surtos afetaram 1791 pessoas entre 2004 e 2005, apresentando uma
incidncia global de 24,6 por 100 pessoas/ano (ARIAS et al., 2010).
Durante a ocorrncia de um surto em uma escola primria do Reino Unido em 2001,
foi demonstrada a implicao de aerossis gerados por vmitos como veculo de infeco
area dos NoVs (MARKS et al., 2003). Na Austrlia, entre 1997 e 2000, observou-se uma
grande atividade da linhagem epidmica GII.4, com diminuio de sua ao entre 2001 e
2004 quando outra linhagem GII.4 (variante) surgiu no mundo e gerou surtos
predominantemente em hospitais e asilos. Acredita-se que s em 2004, mais de 400 surtos
ocorreram e 15.000 pessoas apresentaram a doena nesse pas (TU et al., 2007).
Nos EUA, em 2004, um surto nosocomial acometeu pacientes e profissionais de sade
de duas unidades (psiquitrica e coronariana) de um hospital. O custo estimado dos prejuzos
com apenas esse surto foi de $657.644 dlares entre fechamento das unidades, licena mdica
e despesas com limpeza (JOHNSTON et al., 2007). Um total de 38 surtos nosocomiais foi
confirmado em Hong Kong, no ano de 2006. A maioria dos pacientes acometidos foram
idosos com mobilidade reduzida e que apresentaram sintomas prolongados da doena causada
por NoVs (TSANG et al., 2008).
Um estudo realizado no Canad demonstrou que pacientes com quadro de gastrenterite
aps um surto provavelmente causado por NoVs, desenvolveram a Sndrome do Intestino
Irritvel ps-infeco, afeco crnica do trato gastrintestinal, cujos sintomas podem ser
secundrios a uma leso aguda na mucosa aps uma gastrenterite viral (MARSHALL et al.,
2007).
3.15.1.2 Hospitalizaes
O NoVs tambm tm grande importncia em casos de gastrenterite aguda espordica
que requerem hospitalizaes. Um estudo realizado com amostras coletadas entre 2005 e 2008
47
na Alemanha, investigou os principais patgenos responsveis por ocasionar gastrenterite
grave em crianas hospitalizadas, demonstrando ser o NoVs o segundo patgeno mais
prevalente, apresentando um percentual menor apenas que o do RoVs (WIEGERING et al.,
2011). Outro estudo semelhante desenvolvido na Tanznia, detectou 13,7% de NoVs
acometendo crianas hospitalizadas em um dos 4 principais hospitais peditricos de Dar es
Salaam (MOYO et al., 2011).
Nos EUA, a anlise de um conjunto representativo de dados nacionais a respeito de
todas as internaes por gastrenterite entre os anos de 1996 e 2007, revelou que, neste
perodo, houve aproximadamente 71.000 hospitalizaes associadas infeco por NoVs,
podendo ter chegado a 110.000 internaes em temporadas de epidemia, gerando um prejuzo
de $493 milhes (LOPMAN et al., 2011).
Pesquisa conduzida em Taiwan envolvendo crianas menores de 5 anos internadas em
decorrncia de diarreia aguda, demonstrou que 14,6% das hospitalizaes foram em virtude
de infeco por NoVs, sendo que alm da diarreia, 74,8% apresentaram vmito e 71,3% febre
branda (YANG et al., 2010).
Os NoVs foram implicados em 8,3% dos casos de gastrenterite aguda ocorridos em
crianas internadas no perodo de 2001 a 2004 em um hospital universitrio de Paris.
Contudo, apesar da necessidade de hospitalizao, essas crianas apresentaram quadro clnico
com menor gravidade que aqueles com diarreia associada infeco por RoVs, especialmente
quando comparados o tempo de hospitalizao e a necessidade do uso de reidratao
intravenosa (LORROT et al., 2011).
Estudo conduzido na Espanha envolvendo crianas menores de 5 anos hospitalizadas
com gastrenterite entre os anos de 2006 e 2007, demonstrou que o NoVs apresentou uma
prevalncia de 27%, sendo maior do que para RoVs (21%). Contudo, o quadro clnico das
48
crianas acometidas por NoVs foi mais brando do que o observado pelas infectadas por RoVs
(GONZALEZ-GALAN et al., 2011).
3.15.2 Epidemiologia no Brasil
No Brasil, algumas pesquisas j foram realizados envolvendo os aspectos
epidemiolgicos e moleculares desse vrus. A diversidade genmica do NoVs foi investigada
em um estudo longitudinal conduzido em favelas urbanas de Fortaleza envolvendo 120
crianas entre 1990 e 1991 cuja positividade foi de 12,5% (PARKS et al., 1999). Entre 2004 e
2005, esse patgeno tambm foi detectado em 15% das crianas hospitalizadas de Recife
(NAKAGOMI et al., 2008).
Em Salvador, um estudo longitudinal com amostras coletadas entre 2001 e 2002
envolvendo crianas moradoras de 21 reas de baixa-renda, demonstrou a circulao dos
NoVs GII em 5% dos casos (XAVIER et al., 2009). Em 2006, um surto acometeu jovens
adultos na mesma cidade, sendo identificado o genogrupo II, tipos 3 (8,8%), 4 (72,5%) e 9
(10%) (CAMPOS et al., 2008).
Em 1996, foi registrado na Baixada Santista (SP) o primeiro surto causado por esse
grupo de vrus no Brasil (UEDA et al., 1996), sendo tambm responsvel por outros ocorridos
na capital paulista nos anos de 1995, 2004, 2005 e 2006 com positividade de 15,7%
(MORILLO et al., 2008). Tambm em So Paulo, foi detectado NoVs GII.4 em 33,3% das
crianas atendidas ambulatorialmente, inclusive com infeco mista entre diferentes gentipos
e possveis linhagens recombinantes (CASTILHO et al., 2006).
O NoVs tambm j foi associado a diversos surtos de gastrenterite aguda descritos em
creches do Rio de Janeiro (capital [37,6%] e outros municpios [66%]) (GALLIMORE et al.,
2004; FERREIRA et al., 2008), bem como j foi encontrado em 14,5% de amostras clnicas
de crianas hospitalizadas e atendidas ambulatorialmente na capital carioca (SOARES et al.,
49
2007). Na regio Centro-Oeste, a positividade para NoVs observada entre crianas
hospitalizadas foi de 8,6% e no Esprito Santo de 39,7% (BORGES et al., 2006; RIBEIRO et
al., 2008).
Estudo sobre a diversidade gentica do NoVs no Brasil demonstrou a circulao de
vrios gentipos de GII (71%) em todas as regies do pas, tanto em casos espordicos de
gastrenterite aguda quanto em surtos. Foi observado o gentipo 2 em So Paulo; tipo 3
(Esprito Santo e So Paulo); tipo 6 (Rio de Janeiro e Pernambuco) em 2006; tipo 7 no
Distrito Federal em 2005; tipos 8 e 14 (Rio de Janeiro e Esprito Santo); tipo 12 (Sergipe,
Minas Gerais e Rio Grande do Sul) em 2009; e tipos 16 e 17 no Acre em 2005. Tambm
foram registradas as variantes 2006a circulando no Maranho, Rio Grande do Norte, Alagoas,
Cear, Bahia e Sergipe; e 2006b no Mato Grosso do Sul e em Minas Gerais, entre os anos de
2007 e 2009 (CASTILHO et al., 2006; BARREIRA et al., 2010; FERREIRA et al., 2010;
FIORETTI, et al., 2011).
Testes realizados em amostras fecais provenientes de crianas internadas em um
hospital pblico do Estado do Par demonstraram uma positividade de 15% pela RT-PCR em
amostras coletadas entre 1992-1994 (NAKAMURA et al., 2006; SILVA et al., 2006).
Posteriormente, estudos de biologia molecular envolvendo crianas internadas ou
atendidas ambulatorialmente, detectaram NoVs circulando em Belm nos anos de 1998-2000,
2003 e 2008-2009 com positividades que variaram de 7,1% a 44,4% (ARAGO et al., 2010;
SIQUEIRA et al., 2010a; 2010b).
3.16 SAZONALIDADE
A sazonalidade dos patgenos pode ser definida pelo aparecimento de recorrentes
epidemias da doena em definidos perodos do ano e est associada s mudanas ambientais e
ao comportamento do hospedeiro (DOWEL, 2001).
50
Devido ao pouco conhecimento a respeito da sazonalidade dos NoVs, foi realizada
uma anlise envolvendo 12 pesquisas conduzidas em 8 pases em um perodo de 21 anos
(1978 a 1998), a qual constatou que a maior incidncia de infeco ocorreu nos meses de
inverno e menor nos meses de vero, independentemente do mtodo de deteco usado, idade
dos acometidos pela doena ou suspeita do modo de transmisso. Contudo, os picos de
incidncia da doena no aconteceram obrigatoriamente nos mesmos meses durante o ano e
nem nas mesmas regies, sugerindo que essa sazonalidade possa ser diferente dependendo do
local estudado (MOUNTS et al., 2000).
Outros vrus apresentam um padro de transmisso sazonal no inverno, como o
influenza, vrus respiratrio sincicial, vrus do sarampo e RoVs, nos quais tambm se
atribuem a transmisso pelo ar, facilitada nos meses frios devido a maior aglomerao de
pessoas em locais fechados (PRINCE et al., 1986; MOUNTS et al., 2000).
Mudanas climticas em nvel local, regional ou global geram variaes no padro
pluviomtrico, temperatura do ar, ventos e correntes do oceano, que resultam em mudana no
padro de incidncia e transmisso do patgeno e alteram os conhecimentos epidemiolgicos
e de morbi-mortalidade (CHAN et al., 1999). Esses fatores, especialmente umidade,
contribuem para o aumento do risco de contaminao por RoVs e poliovrus pela manuteno
da viabilidade desses vrus nas superfcies (fmites) (NATHANSON E MARTIN, 1979;
PRINCE et al., 1986) e podem influenciar na resistncia, transmisso e/ou virulncia do
patgeno. O fato do NoVs e RoVs apresentarem padres similares de transmisso e tropismo,
faz presumir que ambos compartilham similaridade em termos de sazonalidade (ROHAYEM,
2009).
Independentemente da estao do ano, aglomerados de indivduos facilitam a
transmisso da doena. Pesquisas demonstram que o padro de transmisso depende da taxa
51
de contato entre as pessoas; por exemplo, verificada uma maior ocorrncia de infeces por
NoVs em crianas durante no perodo escolar do que nas frias (FINE E CLARKSON, 1982).
Estudos revelam que a vitamina D um importante regulador da funo fagocitria
das clulas imunes e est associada com a resposta antiviral infeco pelo vrus influenza.
Durante o inverno, a radiao ultravioleta (UV) diminui e consequentemente ocorre a menor
produo de vitamina D, levando ao comprometimento do sistema imune e possvel aumento
da susceptibilidade s infeces virais. No claro se este mesmo processo atribudo
infeco pelo NoVs, contudo, um campo de pesquisa que deve ser considerado para se
entender melhor a relao da sazonalidade do patgeno com o organismo do hospedeiro
(CANNELL et al., 2006; ROHAYEM, 2009).
Para o entendimento completo de como as mudanas ambientais podem afetar a
epidemiologia do NoVs, necessrio o conhecimento de todos os fatores que determinam sua
sazonalidade (comportamento e susceptibilidade do hospedeiro, transmisso viral e
competncia do vrus em seu ambiente natural). Vigilncia epidemiolgica, monitoramento
climtico, pesquisas multidisciplinares envolvendo virologistas, climatologistas, ecologistas e
epidemiologistas so medidas fundamentais para controlar a doena e diminuir os custos com
servios de sade pblica (ROHAYEM, 2009).
3.17 ASPECTOS CLNICOS
Diversos sintomas clnicos so observados em pessoas acometidas por NoVs, tais
como sbito incio de nuseas, meses, diarreia no sanguinolenta, hiperemia moderada, dores
abdominais/epigstricas, mialgia, cefalia, astenia, calafrios e mal-estar geral (CDC, 2011).
Autores sugerem que exista relao entre o gentipo do NoVs e os sinais/sintomas
apresentados durante a infeco. Em um estudo envolvendo 28 surtos em asilos da Holanda,
o NoVs GII.4 foi responsvel por 21 deles. Nos surtos causados por este gentipo, os
52
principais sinais/sintomas manifestados foram vmito, nuseas, clicas abdominais, febre e
muco nas fezes. Nos surtos causados pela variante GII.4-2004, houve maior predominncia
dos sintomas nuseas, dores de estmago e febre quando comparados linhagem GII.4-
2006a, demonstrando que conforme a cepa, pode ocorrer variaes nos sinais/sintomas
(FRIESEMA et al., 2009).
reconhecido que o NoVs seja um enteropatgeno viral que afeta indistintamente
todas as faixas etrias. Contudo, as faixas extremas de idade (infncia e velhice) so mais
gravemente acometidas em virtude de possurem sistema imunolgico mais debilitado. Um
estudo com recm nascidos prematuros de uma Unidade de Terapia Intensiva (UTI) neonatal
demonstrou que esses menores apresentaram um padro diferente de sintomas durante a
infeco por NoVs com predominncia de abdome distendido e apnia. Vmitos no foram
observados nestes casos (ARMBRUST et al., 2009). Outra pesquisa tambm realizada em
UTI neonatal revelou a associao do NoVs com a enterocolite necrosante em crianas
expostas (TURCIOS et al., 2008).
Um estudo analisou a infeco por NoVs como sendo fator de risco para complicaes
clnicas de outras enfermidades. A investigao de 5 surtos em enfermarias de um hospital
universitrio na Alemanha demonstrou que a infeco por NoVs gerou falncia renal aguda,
sinais de rejeio aps transplante de rins, arritmia cardaca, imunossupresso e aumento da
durao da diarreia em pacientes idosos (MATTNER et al., 2006).
Desde 1982, vrios trabalhos j demonstraram a associao de quadros clnicos de
convulses benignas com gastrenterite leve (MOROOKA, 1982). Atualmente, esta condio
reconhecida como uma entidade clnica distinta, caracterizada por critrios bem definidos,
relacionada geralmente s infeces gastrintestinais causadas por RoVs e NoVs, cuja terapia
envolve inclusive a administrao de anticonvulsivantes (KAWANO et al., 2007).
53
Estudos com pacientes peditricos demonstram que existe associao da infeco por
NoVs com a exacerbao de doena inflamatria intestinal


















