
UNIVERSIDADE FEDERAL FLUMINENSE LEIDYANNE … · A massa de tecido adiposo aumenta durante a meia...
57
UNIVERSIDADE FEDERAL FLUMINENSE CAMPUS UNIVERSITÁRIO DE NOVA FRIBURGO CURSO DE GRADUÇÃO EM BIOMEDICINA LEIDYANNE FERREIRA GONÇALVES ESTUDO HISTOPATOLÓGICO DO TECIDO ADIPOSO BRANCO E MARROM DE CAMUNDONGOS C57Bl/6 FÊMEAS NO ENVELHECIMENTO NOVA FRIBURGO 2015
Transcript of UNIVERSIDADE FEDERAL FLUMINENSE LEIDYANNE … · A massa de tecido adiposo aumenta durante a meia...

UNIVERSIDADE FEDERAL FLUMINENSE
CAMPUS UNIVERSITÁRIO DE NOVA FRIBURGO
CURSO DE GRADUÇÃO EM BIOMEDICINA
LEIDYANNE FERREIRA GONÇALVES
ESTUDO HISTOPATOLÓGICO DO TECIDO ADIPOSO BRANCO E
MARROM DE CAMUNDONGOS C57Bl/6 FÊMEAS NO
ENVELHECIMENTO
NOVA FRIBURGO
2015

i
LEIDYANNE FERREIRA GONÇALVES
ESTUDO HISTOPATOLÓGICO DO TECIDO ADIPOSO BRANCO E
MARROM DE CAMUNDONGOS C57Bl/6 FÊMEAS NO
ENVELHECIMENTO
ORIENTADORA:
PROFa. DRª. CAROLINE FERNANDES DOS SANTOS BOTTINO
Nova Friburgo, RJ
2015
Monografia apresentada à Universidade
Federal Fluminense/Campus Universitário
de Nova Friburgo, como Trabalho de
Conclusão do Curso de graduação em
Biomedicina.

G635e Gonçalves, Leidyanne Ferreira
Estudo histopatológico do tecido adiposo branco e marrom de
camundongos C57BI/6 fêmeas no envelhecimento. / Leidyanne Ferreira
Gonçalves; Profᵃ. Drᵃ. Caroline Fernandes dos Santos Bottino, orientadora. --
Nova Friburgo, RJ: [s.n.], 2017.
46f. : il.
Trabalho de Conclusão de Curso (Graduação em Biomedicina) –
Universidade Federal Fluminense, Instituto de Saúde de Nova Friburgo,
2017.
1. Histologia. 2. Tecido adiposo. 3. Envelhecimento. I. Bottino, Caroline
Fernandes dos Santos, Orientadora. II. Título.
CDD M574.824

ii
LEIDYANNE FERREIRA GONÇALVES
ESTUDO HISTOPATOLÓGICO DO TECIDO ADIPOSO BRANCO E
MARROM DE CAMUNDONGOS C57Bl/6 FÊMEAS NO
ENVELHECIMENTO
Aprovado em: 21/12/ 2015
Banca Examinadora
_____________________________________ Profa. Dra. Caroline Fernandes dos Santos Bottino
Universidade Federal Fluminense - Campus Universitário de Nova Friburgo
_____________________________________ Profa. Dra. Elan Cardozo Paes de Almeida
Universidade Federal Fluminense - Campus Universitário de Nova Friburgo
_____________________________________ Profa. Dra. Bianca Martins Gregório
Universidade do Estado do Rio de Janeiro - Campus Maracanã
Nova Friburgo
2015
Monografia apresentada à Universidade
Federal Fluminense/Campus Universitário
de Nova Friburgo, como Trabalho de
Conclusão do Curso de graduação em
Biomedicina.

iii
A todos meus amigos, professores e familiares que estiveram ao meu
lado durante esses anos e me apoiaram.

iv
AGRADECIMENTOS
Agradeço primeiramente ao Senhor por ter me dado a oportunidade de tentar
me tornar a cada dia uma pessoa melhor, agradeço aos meus professores
por toda a instrução e sabedoria que foram passadas, aos meus amigos e
familiares por sempre me darem força nos momentos mais difíceis.

v
RESUMO
A massa de tecido adiposo aumenta durante a meia idade em humanos e declina em idosos, havendo redistribuição do depósito subcutâneo para o intra-abdominal. Os estudos sugerem que a deterioração do metabolismo do tecido adiposo branco (TAB) e marrom (TAM) com a idade contribua com a deterioração do metabolismo energético. O objetivo do presente estudo é avaliar as mudanças histopatológicas dos diferentes depósitos de gordura (branco [visceral e subcutâneo] e marrom) ao longo do envelhecimento em camundongos C57Bl/6 fêmeas e correlacionar estes achados com o perfil glicídico e lipídico. Os animais foram divididos em quatro grupos de acordo com a idade: 3, 6, 9 e 12 meses de idade. Antes da eutanásia foi avaliada a ingestão alimentar, massa corporal e a glicemia após jejum de 6 horas. Na eutanásia, os animais foram anestesiados e o sangue coletado para dosagem de triglicerídeo, colesterol total, HDL e insulina séricos. O TAB visceral e subcutâneo e o TAM foram coletados e pesados. Após processamento histológico, foi mensurado o diâmetro dos adipócitos (TAB e TAM), a densidade de gotículas lipídicas no TAM e o tamanho dessas gotículas por morfometria. A massa corporal teve aumento a partir dos 9 meses de idade, mesmo com a ingestão alimentar igual entre os grupos (3,7±0,1g/dia). Todos os depósitos de gordura aumentaram com a idade, mas mesmo com o ganho de adiposidade subcutânea (GI), a relação gordura subcutânea/visceral diminui progressivamente com a idade. Não houve diferença no peso da GM entre os grupos, porém a relação GM/gordura branca diminui progressivamente já a partir dos 6 meses de idade. Na quantificação morfológica, houve aumento do diâmetro dos adipócitos na GG, GI e GR. Em relação a GM, o grupo F3 apresentou 22,8% do tecido ocupado por gotículas lipídicas, e houve aumento dessa porcentagem em F6, F9 e F12, comparados ao grupo F3, assim como em F12 comparado a F6. Esta célula apresentou hipertrofia com a idade nos grupos F6, F9 e F12. Visto que há uma mudança fenotípica dos adipócitos de diferentes depósitos ao longo da vida, faz-se importante o estudo deste fenômeno e sua correlação com o metabolismo glicídico e lipídico. Esse entendimento auxiliará no desenvolvimento de novos medicamentos que tenham como objetivo o combate à obesidade e as doenças metabólicas.
Palavras chave: Tecido adiposo branco, tecido adiposo marrom, envelhecimento, fêmeas.

vi
ABSTRACT
The adipose tissue mass increases during the middle age in humans and declines in the elderly, when fat is redistributed from subcutaneous to intra-abdominal depots. Recent studies suggest that the deterioration in white (WAT) and brown (BAT) adipose tissue metabolism with age impairs energy metabolism. This study aimed to evaluate the histopathological changes of different fat depots (white [visceral and subcutaneous] and brown) during aging in C57Bl/6 female mice and to correlate these changes with glucose and lipid profile. Animals were allocated into four groups according to age: three, six, nine and 12 months old. Prior to euthanasia, we measured food intake, body weight and 6-hour fast glucose. At euthanasia, animals were anesthetized and blood was withdrawn to determine serum triglyceride, total cholesterol, HDL cholesterol and insulin. Visceral and subcutaneous WAT, and the subscapular BAT were harvested and weighed. After histological processing, we assessed adipocyte diameter (WAT and BAT), and lipid droplet area and size (BAT) by means of morphometry. Body weight had increased from 9 months of age, even with the same food intake between the groups (3.7 ± 0.1g / day). All deposits of fat increased with age, but even with subcutaneous fat gain (GI) the subcutaneous / visceral fat ratio decreases progressively with age. There was no difference in GM weight between the groups, but GM / white fat ratio decreases progressively from 6 months of age. In the morphological quantification, there was increase in diameter of adipocytes in GG, GI and GR. For GM, F3 group presented 22.8% of tissue occupied by lipid droplets, and this percentage was increased F6, F9 and F12, as compared to the group F3, and F12 compared to F6. This cell presented hypertrophy with aging in groups F6, F9 and F12. Since adipocytes from different depots exhibit phenotypic plasticity throughout life, it is important to study this phenomenon and to correlate it with glucose and lipid metabolism. This knowledge will guide the development of new drugs that aim to fight obesity and metabolic disorders.
Keywords: white adipose tissue, brown adipose tissue, aging, female.

vii
SUMÁRIO
1 INTRODUÇÃO ............................................................................... 1
2 REVISÂO DE LITERATURA............................................................2
2.1 TECIDO ADIPOSO ................................................................. .....2
2.1.1 Tecido adiposo branco...............................................................4
2.1.2 Tecido adiposo marrom..............................................................5
2.2 ENVELHECIMENTO.....................................................................7
2.3 METABOLISMO E HORMÔNIOS SEXUAIS.................................8
2.4 MODELO ANIMAL.......................................................................10
3 OBJETIVOS....................................................................................11
3.1 OBJETIVOS GERAIS...................................................................11
3.2 OBJETIVOS ESPECÍFICOS........................................................11
4 MATERIAIS E MÉTODOS .................................................. .........12
4.1 GRUPOS EXPERIMENTAIS.......................................................12
4.2 CITOLOGIA VAGINAL.................................................................12
4.3 TESTE ORAL DE TOLERÂNCIA À GLICOSE ........................... 14
4.4 BIOQUÍMICA SANGUÍNEA ....................................................... 14
4.5 EXTRAÇÃO DO TRIGLICERÍDEO HEPÁTICO ......................... 15
4.6 MORFOMETRIA E ESTEREOLOGIA ........................................ 15
4.7 ANÁLISE ESTATÍSTICA ......................................................... ..18
5 RESULTADOS ............................................................................. 19

viii
5.1 MASSA CORPORAL ................................................................. 19
5.2 INGESTÃO ALIMENTAR ........................................................... 20
5.3 ÚTERO E OVÁRIO .................................................................... 21
5.4 GLICEMIA DE JEJUM ............................................................... 22
5.5 LIPIDOGRAMA............................................................................23
5.6 FÍGADO ..................................................................................... 24
5.7 MASSA DE TECIDO ADIPOSO...................................................25
5.8 REMODELAMENTO DO TECIDO ADIPOSO BRANCO GENITAL
...........................................................................................................27
5.9 REMODELAMENTO DO TECIDO ADIPOSO BRANCO INGUINAL
...........................................................................................................28
5.10 REMODELAMENTO DO TECIDO ADIPOSO BRANCO
RETROPERITONEAL ......................................................................29
5.11 QUANTIFICAÇÃO DA GORDURA MARROM .........................30
5.12 INSULINA..................................................................................32
5.13 TESTE DE TOLERÂNCIA ORAL À GLICOSE..........................33
5.14 HOMA-IR e HOMA-B.................................................................34
6 DISCUSSÃO ............................................................................... .35
7 CONCLUSÕES ............................................................................ 39
8 REFERÊNCIAS ............................................................................ 40
ANEXO A – Aprovação CEUA ....................................................... 46

1
1 INTRODUÇÃO
O envelhecimento traz uma série de alterações no organismo, dentre elas
podemos observar a obesidade e o descontrole metabólico que vem ocorrendo cada
vez em maior número na população mundial. Um dos principais fatores envolvidos
no desenvolvimento dessas modificações fisiológicas é o tecido adiposo.
O tecido adiposo desempenha um papel fundamental no controle da
homeostase energética, com o passar dos anos vários estudos relataram a
importância crescente de uma maior investigação das características e
singularidades desse tecido, com isso vários aspectos importantes foram elucidados,
porém ainda há muito a ser descoberto.
Durante o envelhecimento são observadas várias alterações tanto na
morfologia desse tecido quanto na redistribuição entre os seus diferentes depósitos,
que podem ser correlacionadas com as alterações fisiológicas presentes durante
esse período. Os hormônios sexuais também têm grande influência no
envelhecimento e nas variações observadas no tecido adiposo.
Com isso, é importante um estudo mais aprofundado da relação existente
entre todos esses fatores, sendo muito comum a utilização de camundongos
C57BL/6, por serem excelentes modelos para o estudo da obesidade e do diabetes.

2
2 REVISÃO DA LITERATURA
2.1 TECIDO ADIPOSO
Durante muitos anos o tecido adiposo foi considerado apenas como um
importante órgão de armazenamento de energia, porém a identificação da leptina,
um hormônio secretado pelos adipócitos que tem efeito sobre o sistema nervoso e a
função endócrina, levou o tecido adiposo a ser visto também como um órgão
multifuncional, que produz e secreta peptídeos e proteínas bioativas denominadas
adipocitocinas ou adipocinas (Fonseca-Alaniz et al., 2007; Guimarães et al., 2007).
O tecido adiposo tem um papel essencial na sobrevivência de mamíferos, já
que consegue armazenar o excesso de calorias produzido na forma de lipídeos até
que se tornem necessários para o suprimento de energia, também proporcionam
isolamento térmico e proteção mecânica. Essas duas primeiras funções, de
estocagem e isolamento têm grande importância na regulação do metabolismo
energético. Os adipócitos que constituem o tecido adiposo têm a capacidade de
armazenar lipídeos sem que isso comprometa a integridade funcional das células.
Eles têm as enzimas necessárias para sintetizar os ácidos graxos (lipogênese) e
estocá-los em períodos de grande oferta de energia ou mobilizá-los quando há a
falta dela (lipólise). Essa capacidade de lipogênese ou lipólise pode variar de acordo
com a localização do tecido adiposo, sendo ele subcutâneo ou visceral, a secreção
das adipocinas também pode sofrer essa variação (Guimarães et al., 2007).
Vários sistemas como o sistema nervoso central, digestório e endócrino,
estão envolvidos na regulação dessa homeostase. O sistema nervoso central
participa desses dois processos (lipólise e lipogênese) tanto de forma direta quanto
indireta, levando por exemplo à busca pelo alimento ou a recusa do mesmo,
enquanto os sistemas digestório e endócrino realizam essa regulação através de
nutrientes e hormônios levando em consideração as necessidades do momento
(Fonseca-Alaniz et al., 2007). A capacidade de armazenamento é descrita em
alguns estudos como virtualmente ilimitada o que inicialmente é uma vantagem
adaptativa a curto prazo, já que permite a estocagem de grande quantidade de
energia, mas a longo prazo pode levar a uma disfunção metabólica (Costa e Duarte,

3
2006). Na prática sabe-se que os adipócitos não têm uma capacidade ilimitada de
armazenamento de triglicerídeos, quando seu crescimento atinge o grau máximo
eles se tornam mais lipolíticos podendo desencadear um aumento da concentração
de ácidos graxos livres no plasma e danificar a função de alguns órgãos pela
lipotoxicidade, assim que esse crescimento exacerbado ocorre, novas células são
lentamente recrutadas, porém essas células possuem uma capacidade de
armazenamento consideravelmente menor (Queiroz et al., 2009). Aproximadamente
um décimo do total de células adiposas é renovado a cada ano, seja pela
adipogênese que já está em curso ou pela morte de alguns adipócitos (Arner et al.,
2010).
O tecido adiposo está disperso por todo o organismo formando depósitos
que não possuem conexão física entre si, onde podem ser encontrados vários tipos
celulares, como macrófagos, fibroblastos, pré-adipócitos e adipócitos, além de uma
matriz de tecido conjuntivo (fibras colágenas e reticulares), fibras nervosas, estroma
vascular e nódulos linfáticos (Fonseca-Alaniz et al., 2007). Sua atividade secretória é
regulada tanto por mecanismos humorais quanto hormonais. As adipocinas
secretadas desempenham um papel importante não só na homeostase energética
como também na resposta imunológica, sensibilidade à insulina e doença vascular
(Costa e Duarte, 2006). Dentre essas adipocinas podemos citar: fator de necrose
tumoral-α (TNF-α) e interleucina-6 (IL-6) relacionadas ao sistema imunológico,
angiotensinogênio que influencia na regulação da pressão arterial e adiponectina,
resistina, visfatina e leptina que estão envolvidas na homeostase glicêmica
(Fonseca-Alaniz et al., 2007). A hipertrofia e/ou hiperplasia dos adipócitos leva a
alterações na secreção dessas adipocinas que podem influenciar no processo de
desenvolvimento da obesidade e suas complicações (Guimarães et al., 2007).
O tecido adiposo branco (TAB) e o tecido adiposo marrom (TAM) são
fundamentais no controle da massa corporal, pelo menos em pequenos mamíferos.
Enquanto o TAM é importante no gasto energético uma vez que ele é o principal
mediador da termogênese adaptativa, o tecido adiposo branco é importante no
armazenamento de energia, assim como na geração de sinais que informam o
encéfalo sobre o status da reserva energética corporal (p.ex., leptina) (Seydoux e
Girardier, 1978; Rothwell, 1989).
Estruturas chamadas “crown-like-stuctures” (CLS) foram identificadas em
grande número tanto em humanos quanto roedores no tecido adiposo em obesos,

4
são formadas por adipócitos mortos rodeados por macrófagos, na região visceral
esses CLS são encontrados em maior número que no tecido adiposo subcutâneo, o
número dessas estruturas diminui consideravelmente quando há grande perda de
peso, porém um estudo mais aprofundado é necessário para a avaliação do
envolvimento dessas estruturas com a obesidade e função dos adipócitos (Lee, Wu
e Fried, 2010).
2.1.1 Tecido Adiposo Branco
O tecido adiposo branco é formado por uma única e grande gotícula lipídica, o
citoplasma, núcleo e demais organelas ficam localizadas na periferia celular. Seu
volume é variável, de acordo com a quantidade de lipídeos armazenados, podendo
ser algumas centenas ou milhares de vezes maior que hemácias e células do
sistema imune. O tecido adiposo branco possui distribuição por todo o organismo
envolvendo ou se infiltrando em quase toda a região subcutânea, órgãos e
grupamentos musculares, oferecendo assim proteção mecânica e sendo essencial
na preservação da temperatura corporal.
Um fato curioso é que, inicialmente os adipócitos jovens apresentam
múltiplas gotículas lipídicas, que são características do adipócito marrom, porém
com o amadurecimento elas se unem para formar a tão representativa única gotícula
do adipócito branco (Fonseca-Alaniz et al., 2007). Para acomodar as grandes
variações na disponibilidade de nutrientes, o TAB (tecido adiposo branco) é capaz
de alterar de forma rápida e significativa o seu tamanho, em especial o tecido
adiposo subcutâneo (TASC), que não está submetido às limitações anatômicas ao
crescimento como ocorre com o tecido adiposo visceral (TAV). O crescimento do
TAB ocorre devido a alterações no tamanho e número das células adiposas, as
quais variam em magnitude entre os diferentes depósitos de gordura.

5
2.1.2 Tecido Adiposo Marrom
O tecido adiposo marrom é essencial para a sobrevivência de pequenos
mamíferos em ambientes frios, já que é ele o responsável pela termogênese
adaptativa através da produção de calor (Harms e Seale, 2013).
Comparado ao TAB, o TAM (tecido adiposo marrom) é mais vascularizado,
possui uma densa inervação adrenérgica e uma grande quantidade de mitocôndrias
e pequenas gotículas lipídicas. Até recentemente, acreditava-se que o TAM
desaparecia rapidamente após o nascimento, quando ele é essencial para que a
temperatura corporal seja mantida, já que a necessidade de ativação desses
adipócitos diminui depois de alguns meses de vida, e que ele desempenhava um
papel mínimo na fisiologia do adulto (Lean, 1989; Ravussin e Galgani, 2011).
Entretanto, o TAM foi consistentemente identificado em humanos adultos, os
adipócitos marrons estão localizados em dois tipos de depósito, depósito discreto ou
difuso. Os depósitos discretos se encontram na região cervical-supraclavicular (mais
comum), perirenal, adrenal, paravertebral e grandes vasos como a aorta e seus
principais ramos (aa. carótida, aa. subclávia, aa. intercostais e aa. renais), onde
provavelmente estão presentes para gerar calor e manter a temperatura central do
corpo. Já os depósitos difusos estão embebidos no próprio TAB e no músculo
esquelético (Nedergaard, Bengtsson e Cannon, 2007; Cypess et al., 2009; Ravussin
e Galgani, 2011).
O adipócito marrom tem formato poligonal e é formado por várias pequenas
gotículas lipídicas. Os adipócitos são menores do que os adipócitos brancos
(diâmetro médio de 60-100 μm), tendo em média 30-40 μm de diâmetro, e possuem
numerosas mitocôndrias onde se localiza a proteína desacopladora-1 (UCP-1 ou
termogenina) que leva a dissipação da energia em forma de calor. A coloração mais
escurecida do tecido ocorre pela alta concentração de citocromo oxidase dessas
mitocôndrias, a qual pode ser considerada um marcador desse tecido, assim como a
UCP-1. Alguns genes também são altamente expressos nessas células como, por
exemplo, o receptor β3 adrenérgico, citocromo C e o coativador 1 alfa do PPARγ
(PGC1-α), que estão relacionados a produção de calor, principal função do tecido
adiposo marrom (Fonseca-Alaniz et al., 2007; Ravussin e Galgani, 2011).

6
O gene PGC1-α tem um grande papel no desenvolvimento do tecido adiposo
marrom, reforçando a biogênese mitocondrial e a sinalização oxidativa metabólica. A
expressão ectópica desse gene nos adipócitos brancos tanto em humanos quanto
em ratos induziu genes mitocondriais e termogênicos incluindo UCP1 (Puigserver et
al., 1998; Tiraby et al., 2003).
Recentemente também foi descoberto um fator de transcrição que atua
diretamente na diferenciação desses adipócitos, o PRDM16 que é um potente
coativador da atividade transcricional do PGC1α e β, assim como do PPARα e γ.
Além disso, esse fator também suprime alguns genes específicos presentes no
tecido adiposo branco como a resistina e o angiotensinogênio (Seale et al., 2007;
Kajimura et al., 2008; Seale et al., 2008).
Em camundongos, e muito provavelmente em humanos também, ocorrem 2
tipos de população de adipócitos marrons, o clássico que se localiza na região
interescapular e profunda do pescoço e o segundo tipo, adipócitos que expressam
UCP-1 que podem ser encontrados interpostos no tecido adiposo branco e músculo
esquelético, esses adipócitos tem sido chamados de bege ou “brite” ("brite
adipocyte", junção dos termos em inglês "BRown adipocyte" e "whITE adipocyte").,
mesmo que ainda não se saiba qual o tipo é mais prevalente em humanos adultos, o
envelhecimento afeta negativamente a formação de ambos os tipos (Graja e Schulz,
2015). Sua presença está associada à proteção contra a obesidade induzida por
dieta e doenças metabólicas (Seale et al., 2011). Os adipócitos brite possuem
morfologia e bioquímica semelhantes ao adipócito marrom, incluindo a presença de
múltiplas gotículas lipídicas e a expressão da proteína desacopladora 1 (UCP-1),
porém eles não são originados da linhagem celular Myf-5, como ocorre normalmente
no desenvolvimento do TAM estimulado pela proteína PRDM16 (Seale et al., 2008).
Entre os genes compartilhados pelos adipócitos brite e adipócitos marrons clássicos
pode-se citar o PGC1-α e o CIDEA. Por outro lado, há genes que são expressos
predominantes dos adipócitos brite e não nos adipócitos marrons clássicos, como o
FGF21, CITED1 e HOX9a (Sharp et al., 2012).
O processo de termogênese desse tecido é ativado principalmente pela
noradrenalina que é secretada em resposta ao frio, ≤ 23ºC para humanos e ≤ 30ºC
para camundongos (Ravussin e Galgani, 2011), estimulado pelo sistema nervoso
simpático, sua atividade é maior em fêmeas, possivelmente hormônios sexuais
estão envolvidos na regulação, assim como a idade (Pfannenberg et al., 2010).

7
2.2 ENVELHECIMENTO
A massa de tecido adiposo aumenta durante a meia idade em humanos e
declina em idosos. A gordura é redistribuída entre os diferentes depósitos de
gordura, especialmente durante e após a meia idade, quando esta se redistribui do
depósito subcutâneo para o intra-abdominal (Visser et al., 2003; Raguso et al.,
2006). Como consequência, por exemplo, a circunferência abdominal aumenta
aproximadamente 4,0 centímetros a cada 9 anos nas mulheres (Hughes et al., 2004;
Koutsari et al., 2009). Em idades mais avançadas, a gordura é redistribuída para fora
dos depósitos de gordura (esteatose) e se acumula, por exemplo, na medula óssea,
músculo esquelético e fígado. Estas alterações estão associadas com o diabetes
tipo 2, hipertensão arterial, câncer, disfunção cognitiva e aterosclerose, podendo
levar ao infarto do miocárdio e acidente vascular encefálico (Guo et al., 1999; Lutz,
Sanderson e Scherbov, 2008).
Com o envelhecimento, há um declínio na atividade do TAM e esse
fenômeno está relacionado a diminuição da quantidade de depósitos pelo corpo (Au-
Yong et al., 2009). No idoso, o TAM passa a ser interposto por TAB no pescoço e
porção superior do tórax (Cypess et al., 2009). O autor Zingaretti (Zingaretti et al.,
2009) hipotetiza que em uma determinada idade os humanos passariam de um
fenótipo caracterizado por eutrofia e adipócitos brancos pequenos para um fenótipo
caracterizado por obesidade crescente, adipócitos brancos grandes e ausência de
adipócitos marrons. Entretanto os mecanismos biológicos que regulam a
manutenção ou o declínio da massa de TAM no envelhecimento são desconhecidos
no momento. Esta (re)descoberta do TAM tem incitado o interesse científico no
estudo do papel dele na regulação da massa corporal e das desordens metabólicas
que acometem os humanos, principalmente no que concerne à obesidade e doenças
associadas, assim como as doenças crônicas associadas ao envelhecimento (Lidell
e Enerback, 2010; Mattson, 2010).
Existe uma renovação das células adiposas ao longo da vida. Os pré-
adipócitos (células progenitoras dos adipócitos) são abundantes e estão intimamente
relacionados aos macrófagos. Eles se desdiferenciam em idades mais avançadas,
replicam em menor taxa, possuem menor capacidade adipogênica, maior
susceptibilidade a lipotoxicidade e adquirem um perfil pró-inflamatório e senescente

8
que favorece o remodelamento do TAB (Karagiannides et al., 2001; Guo et al., 2007;
Schipper et al., 2008). Outras células progenitoras mesenquimais também podem
adquirir um perfil pró-inflamatório e um fenótipo semelhante ao do adipócito com a
idade (Tchkonia et al., 2010). Também há aumento de macrófagos no TASC com a
idade em camundongos. Seu número já é grande em idades precoces no TAV intra-
abdominal, porém diferente do TASC esse número não se altera muito com a idade
(Jerschow e Al, 2007). O envelhecimento também está relacionado com o aumento
da hipóxia e dos níveis teciduais de espécies reativas de oxigênio no TAV de
camundongos C57Bl/6, onde o TAV apresentaria menor alteração na expressão de
genes envolvidos na regulação redox quando comparado ao TASC (Zhang et al.,
2011).
Enquanto os mecanismos que correlacionam a obesidade ao diabetes estão
começando a serem elucidados (Weisberg et al., 2003; Xu et al., 2003), o link entre
o envelhecimento e o diabetes tipo 2 continua obscuro. Um declínio na tolerância à
glicose como parte do envelhecimento humano foi primeiro notado em 1921
(Spence, 1921) e a redução da sensibilidade à insulina é agora reconhecida como a
causa primária da falência dependente da idade do metabolismo da glicose
(Silverstone et al., 1957; Defronzo, 1979). Um dos marcos do envelhecimento é a
perda da sensibilidade à insulina no TASC (Borkan et al., 1983), e acredita-se que
este declínio dependente da idade esteja relacionado a uma alteração na replicação
e diferenciação dos pré-adipócitos (Karagiannides et al., 2001; Miard et al., 2009).
Uma vez que a adipogênese reduzida e a deposição ectópica de lipídios estão
intimamente relacionados com a resistência à insulina (Muoio e Newgard, 2008), a
deterioração da função do tecido adiposo com a idade provavelmente contribui com
a deterioração do metabolismo da glicose. Assim, os mecanismos por trás da
disfunção do tecido adiposo durante o envelhecimento merecem investigação.
2.3 METABOLISMO E HORMÔNIOS SEXUAIS
As mulheres possuem um maior percentual de gordura corporal (Womersley,
1977; Jackson et al., 2002), apresentando uma quantidade menor de tecido adiposo

9
branco visceral e uma maior deposição de tecido adiposo subcutâneo, enquanto
homens acumulam mais gordura visceral (Schreiner et al., 1996; Després et al.,
2000; Demerath et al., 2007; Macotela et al., 2009; Camhi et al., 2011). A
distribuição da gordura corporal é um fator importante para a saúde metabólica, por
exemplo, uma menor quantidade de tecido subcutâneo é favorável, enquanto uma
grande quantidade de tecido adiposo visceral é desfavorável, podendo auxiliar nas
alterações da homeostase de lipídeos e da glicose.
Mesmo tendo um percentual de gordura corporal mais baixo, a maior
prevalência de diabetes do tipo 2 e de resistência à insulina ocorre nos homens, o
que pode ser explicado pela maior sensibilidade à insulina na mulher, que tem
também maior resistência a uma dieta hiperlipídica (high fat) (Macotela et al., 2009;
Varlamov, Bethea e Roberts, 2014).
O mecanismo responsável pelas diferenças sexuais na sensibilidade à
insulina ainda não é totalmente compreendido. Um estudo demonstrou que não há
diferença na secreção de insulina entre machos e fêmeas jovens (Basu et al., 2006),
mas outro já indicou que as fêmeas tem uma secreção maior de insulina de primeira
fase do que os machos (Flanagan et al., 2006).
A castração aumenta a resposta à insulina no tecido adiposo branco em
machos e a ovariectomia diminui essa resposta em fêmeas, as diferenças
relacionadas ao sexo em relação a sensibilidade a insulina e o desenvolvimento e
função do tecido adiposo podem ser atribuídas em parte ao estrogênio e a
testosterona, a diminuição do estrogênio e o aumento dos níveis de testosterona que
ocorrem na menopausa podem ser associados a perda de tecido adiposo
subcutâneo e ganho de gordura visceral, assim como aumento da resistência à
insulina (Macotela et al., 2009). Da mesma forma, foi visto que o uso de
contraceptivos orais está associado a redução da sensibilidade à insulina e baixo
nível de estrogênio, sugerindo que esse hormônio pode ser um fator protetor contra
a resistência à insulina. O estrogênio também pode suprimir a síntese de
triglicerídeos, reduzindo a lipogênese no fígado e aumentando a lipólise nos
adipócitos (Varlamov, Bethea e Roberts, 2014).

10
2.4 MODELO ANIMAL
O camundongo é amplamente utilizado como modelo experimental
principalmente por ser pequeno, muito prolífero, ter período de gestação curto, ser
de fácil domesticação e manutenção, além de ser capaz de mimetizar diversas
condições patológicas humanas (Chorilli e Al, 2007).
A linhagem de camundongos C57Bl/6 é a mais amplamente utilizada e foi a
primeira a ter o seu genoma sequenciado. Eles são altamente suscetíveis a
obesidade induzida por dieta, diabetes tipo 2 e aterosclerose, mimetizando o que é
visto em humanos e servindo assim como um bom modelo para o estudo destas
doenças (Fernandes-Santos et al., 2009; Fraulob et al., 2010). Mesmo sem um
estímulo externo (p.ex., dieta hiperlipídica), o camundongo C57Bl/6 apresenta
deterioração do metabolismo de carboidratos e lipídios com a idade, assim como da
função pancreática. Tanto em machos como em fêmeas, há aumento da glicemia de
jejum, triglicerídeos e colesterol total (Zhou e Hansson, 2004; Fernández et al.,
2010). O envelhecimento também resulta em hipertrofia das ilhotas pancreáticas e
aumento da secreção de insulina pelo pâncreas (Leiter et al., 1988), com redução da
expressão do transportador da glicose tipo 2 (GLUT-2) e aumento da expressão da
glicoquinase (Perfetti et al., 1996).

11
3 OBJETIVOS
3.1 OBJETIVOS GERAIS
A hipótese deste trabalho é a de que a deterioração do metabolismo com o
envelhecimento está correlacionada a alterações morfológicas do tecido adiposo.
Assim, o objetivo é avaliar se o envelhecimento influencia o remodelamento
morfológico do tecido adiposo branco (visceral e subcutâneo) e marrom de
camundongos e correlacionar estes dados com o perfil glicídico e lipídico.
3.2 OBJETIVOS ESPECÍFICOS
• Avaliar se o envelhecimento influencia o remodelamento morfológico do
tecido adiposo branco (visceral e subcutâneo);
• Avaliar o efeito do envelhecimento no tecido adiposo marrom;
• Correlacionar os achados morfológicos com os achados do perfil glicídico e
lipídico.

12
4 MATERIAIS E MÉTODOS
4.1 GRUPOS EXPERIMENTAIS
Os protocolos de manuseio e experimentação foram aprovados pelo Comitê
de Ética em experimentação animal da Universidade Federal Fluminense (CEUA:
Projeto nº 446/2014). A investigação respeitou os preceitos do “Guia para o uso e
cuidado de animais de laboratório” (Publicação do NIH Nº 85-23, revisada em 1996,
USA). Foram utilizados camundongos C57BL/6 fêmeas com 3, 6, 9 e 12 meses de
idade, obtidos do Biotério próprio da Universidade Federal Fluminense. Eles foram
alocados nos seguintes grupos de estudo:
▪ F3 – fêmeas eutanasiadas aos 3 meses de idade (n=15/grupo)
▪ F6 – fêmeas eutanasiadas aos 6 meses de idade (n=15/grupo)
▪ F9 – fêmeas eutanasiadas aos 9 meses de idade (n=13/grupo)
▪ F12 – fêmeas eutanasiadas aos 12 meses de idade (n=13/grupo)
Os animais foram alimentados com dieta própria para roedores (Nuvilab) e
tiveram a massa corporal e a ingestão alimentar aferidos antes da eutanásia. Água e
ração foram ofertadas ad libitum durante todo o experimento.
4.2 CITOLOGIA VAGINAL
Na semana anterior a eutanásia, foi realizado o exame de citologia vaginal
durante 5 dias, para observar se as fêmeas estavam ciclando ou não. Para isso, foi
utilizada uma pipeta de Pauster contendo aproximadamente 100 µl de água
destilada que foi posicionada na abertura do canal vaginal, gentilmente aperta-se o
bulbo da pipeta para que a água entre em contato com a abertura e então o bulbo
deve ser solto para que o líquido volte para a pipeta. Esse procedimento foi
realizado de 4 a 5 vezes. O líquido obtido foi desprezado em uma lâmina que
quando seca foi corada com cristal violeta (1 minuto) e lavada em água destilada (1

13
minuto). Após todo esse processo as lâminas ficaram prontas para análise, que foi
baseada na figura a seguir, retirada do mesmo estudo onde o protocolo acima foi
baseado (Mclean et al., 2012)
Adicionalmente, o útero e o ovário foram coletados para que juntamente com
a citologia vaginal pudessem indicar se os animais estavam na menopausa ou se
ainda estavam ciclando.
Figura 1 – Análise de Citologia Vaginal; A. Células epiteliais nucleadas; B. Células epiteliais
escamosas cornificadas; C. Leucócitos; D. Proestro; E. Estro; F. Metaestro; G Diestro; Cabeças de
seta preta em E, F e G apontam células epiteliais escamosas cornificadas representativas; setas
pretas em C, F e G indicam leucócitos; setas brancas em D e G sinalizam células epiteliais nucleadas
(Adaptado de Mclean 2012).

14
4.3 TESTE ORAL DE TOLERÂNCIA À GLICOSE
Para análise de um possível desenvolvimento de resistência à insulina, o
teste oral de tolerância à glicose (TOTG) foi realizado. Glicose a 50% diluída em
salina estéril (0,9% NaCl) foi administrada através de gavagem orogástrica após
jejum de 6 horas (dose de 1g/kg de massa corporal). Sangue foi coletado através de
uma incisão na ponta da cauda do animal para dosagem da glicose plasmática antes
da gavagem e após 15, 30, 45, 60, 90 e 120 minutos. A área sob a curva (ASC) foi
calculada usando a regra do trapézio para analisar a intolerância à glicose
(Fernandes-Santos et al., 2009).
4.4 BIOQUÍMICA SANGUÍNEA
Antes da eutanásia, os animais foram submetidos a jejum de 6 horas. A
glicemia sanguínea foi avaliada através de uma gota de sangue coletada por uma
pequena incisão na ponta da cauda do animal (glicosímetro One Touch Ultra,
Johnson& Johnson, SP, Brasil).
No momento da eutanásia, os animais foram anestesiados com ketamina
(100,0 mg/kg) e xilasina (10,0 mg/kg) intraperitoneal. Uma incisão foi realizada no
tórax e o coração foi exposto para que o sangue fosse coletado em seringa
previamente heparinizada diretamente do átrio direito. Após a centrifugação e
separação do plasma, foi feita a dosagem dos lipídios séricos (triglicerídeos,
colesterol total e HDL, ensaio colorimétrico, Bioclin, Belo Horizonte, MG, Brasil). A
insulina de jejum também foi verificada através de ensaio de Elisa (Cat#EZRMI-13K,
Millipore).
Os valores de glicemia e insulinemia de jejum foram utilizados para o cálculo
do índice de resistência à insulina HOMA-IR (homeostasis model assessment index)
e para a análise da função das células beta pelo índice HOMA-B, segundo as
equações: HOMA-IR = (insulina de jejum x glicemia de jejum)/ 22,5 e; HOMA-B = (20
x insulina de jejum)/(glicemia de jejum - 3,5)%, sendo a unidade da insulina mU/mL e
da glicose mmol/L (Matthews et al., 1985).

15
4.5 EXTRAÇÃO DO TRIGLICERÍDEO HEPÁTICO
Para a extração do triglicerídeo foi pesado aproximadamente 50 mg de
tecido hepático de cada animal. O fígado foi lisado em 1,0 ml de álcool isopropílico e
o lisado hepático foi centrifugado a 4ºC por 10 minutos a 5.500 rpm. Parte do
sobrenadante foi reservada em um novo microtubo (~400 µL) e 2,0 µL foram
utilizados em ensaio colorimétrico para detecção do triglicerídeo (Bioclin, Belo
Horizonte, MG, Brasil).
4.6 MORFOMETRIA E ESTEREOLOGIA
Após a punção cardíaca, o tecido adiposo branco visceral (gordura genital e
retroperitoneal), subcutâneo (gordura inguinal) e o tecido adiposo marrom
subescapular foram coletados, pesados em balança de precisão e então imersos em
formalina de Millonig (4% peso/volume em 0,1M tampão fosfato pH 7,2) por 48 horas
(Carson, Martin e Lynn, 1973). Fragmentos de cada tecido foram submetidos ao
processamento histológico de rotina, embebidos em parafina, seccionados a 3,0 μm
de espessura e corados com hematoxilina e eosina.
No tecido adiposo branco, imagens digitais foram obtidas de campos
aleatórios para a mensuração do diâmetro do adipócito (Mandarim-De-Lacerda,
Fernandes-Santos e Aguila, 2010) através de microscópio Diagtech acoplado a uma
videocâmera DCM510. A análise morfométrica foi realizada utilizando-se o software
Image-Pro® Plus versão 4.5 (Media Cybernetics, Silver Spring, USA). Pelo menos
50 adipócitos tiveram o seu maior e menor diâmetro mensurados por animal (Fig. 2).
A relação entre a massa de gordura visceral e subcutânea foi calculada como:
TASC:TAV = [gordura inguinal (g)]/ [gordura epididimal (g) + gordura retroperitoneal
(g)] (Fernandes-Santos et al., 2009).

16
Figura 2 - Mensuração do maior e do menor diâmetro de adipócitos pelo software Image-Pro® Plus 4.5.
O tecido adiposo marrom subescapular foi seccionado a 3,0 μm de
espessura e corado em hematoxilina e eosina. Uma vez que a célula adiposa
marrom é multilocular, imagens digitais foram obtidas para a quantificação que foi
feita em três abordagens diferentes:
▪ Porcentagem de gotículas lipídicas presentes no tecido, feita através
da análise por segmentação de imagem e posterior uso de
histograma, foram analisadas 8 imagens por animal;
▪ Foi mensurado o diâmetro das vesículas, sendo analisada a
circunferência das gotículas presentes dentro de um adipócito de
cada lâmina, sendo 8 lâminas usadas por animal;
▪ E por fim, o tamanho do adipócito marrom, foi medida a circunferência
de dois adipócitos por lâmina, num total de 8 lâminas por animal.
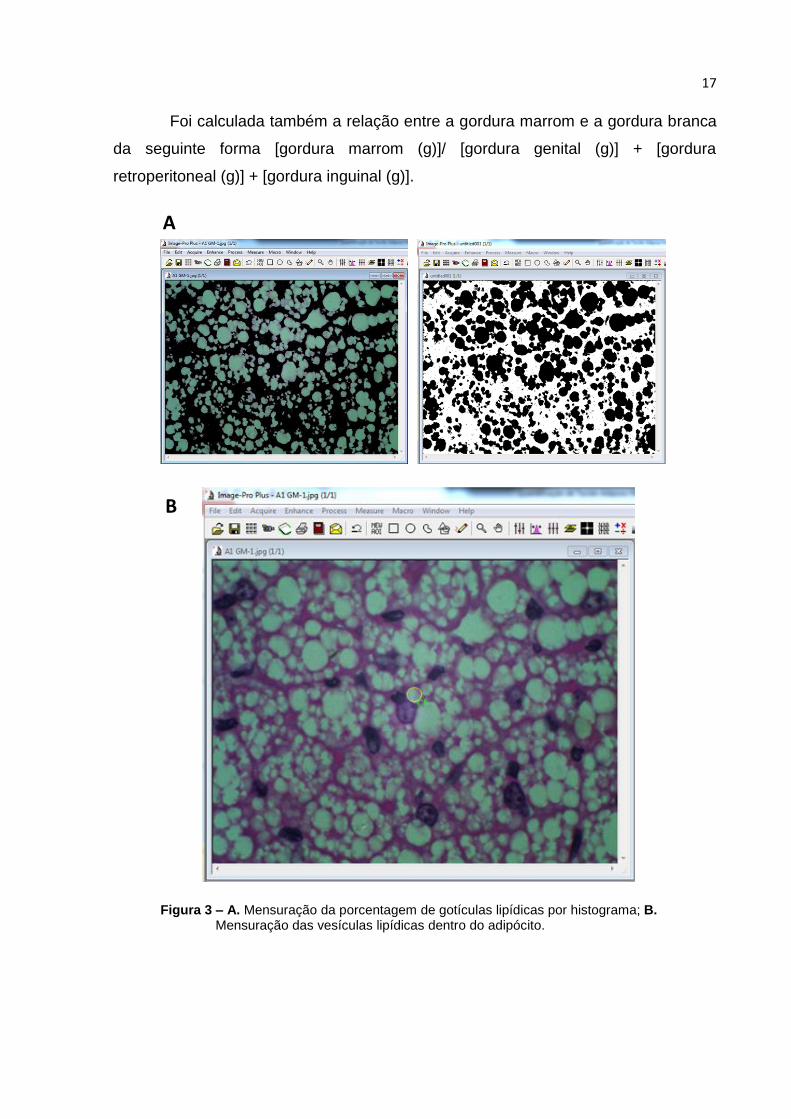
17
Foi calculada também a relação entre a gordura marrom e a gordura branca
da seguinte forma [gordura marrom (g)]/ [gordura genital (g)] + [gordura
retroperitoneal (g)] + [gordura inguinal (g)].
B
Figura 3 – A. Mensuração da porcentagem de gotículas lipídicas por histograma; B. Mensuração das vesículas lipídicas dentro do adipócito.
A

18
4.7 ANÁLISE ESTATÍSTICA
Os dados foram testados para distribuição normal e homocedasticidade das
variâncias e então analisados por one-way ANOVA com pós-teste de Tukey (dados
paramétricos) ou por ANOVA não paramétrico de Kruskal-Wallis com pós-teste de
Dunn (dados referentes a um mesmo gênero). Um P<0,05 foi considerado
estatisticamente significativo (GraphPad Prism versão 6).
A B

19
5. RESULTADOS
5.1 MASSA CORPORAL
A massa corporal das fêmeas foi aferida antes da eutanásia e é possível
observar que não houve um aumento significativo do peso entre as idades de 3 e 6
meses, mas foi visto aumento em relação aos animais de 9 meses quando
comparados aos de 3 e 6 meses, assim como também houve uma diferença entre
os de 3, 6 e 9 meses para os animais de 12 meses (Fig 4).
Figura 4 – Massa corporal (g) de camundongos fêmeas com 3 (n=10), 6 (n=9), 9 (n=7) e 12
(n=8) meses de idade. Dados expressos em média±EPM. Quando indicado, P<0,05 vs. [a] 3 meses;
[b] 6 meses e; [c] 9 meses.
20
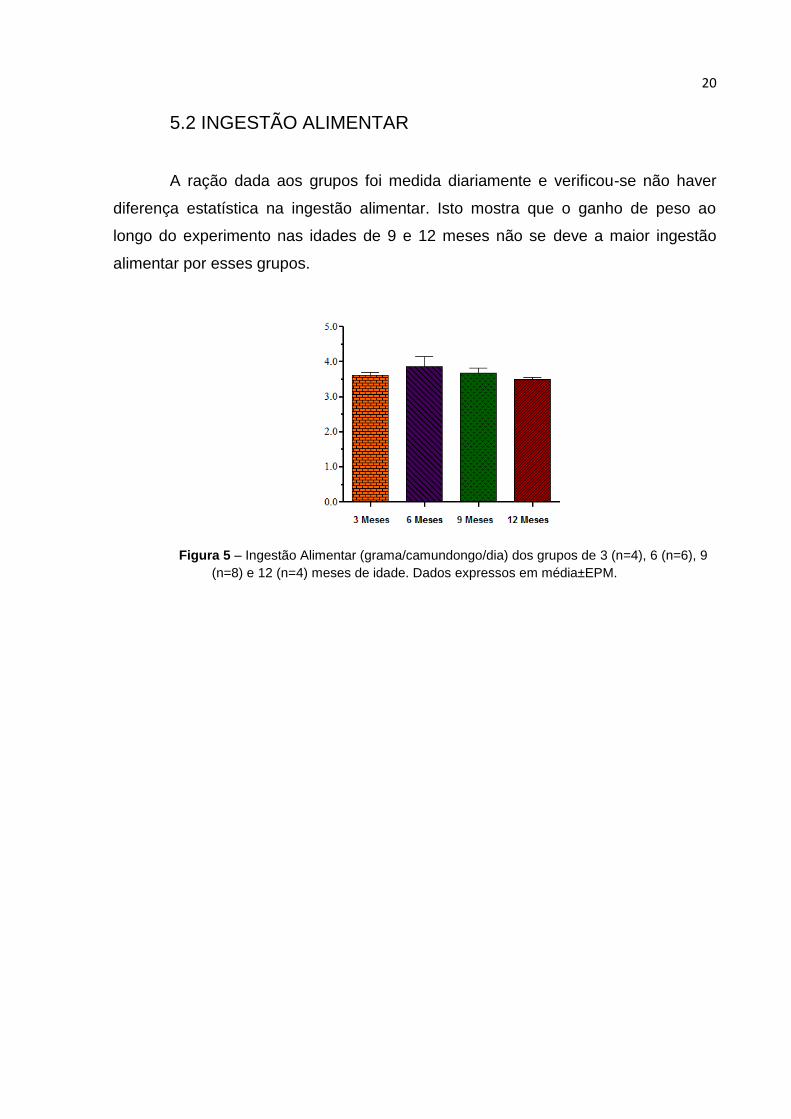
5.2 INGESTÃO ALIMENTAR
A ração dada aos grupos foi medida diariamente e verificou-se não haver
diferença estatística na ingestão alimentar. Isto mostra que o ganho de peso ao
longo do experimento nas idades de 9 e 12 meses não se deve a maior ingestão
alimentar por esses grupos.
Figura 5 – Ingestão Alimentar (grama/camundongo/dia) dos grupos de 3 (n=4), 6 (n=6), 9
(n=8) e 12 (n=4) meses de idade. Dados expressos em média±EPM.

21
5.3 ÚTERO E OVÁRIO
O útero e o ovário também foram coletados e pesados durante a eutanásia e
foi visto que não há nenhuma diferença estatística entre o peso desses órgãos com
o envelhecimento. Juntamente com o esfregaço vaginal que mostrou que as fêmeas
estavam ciclando, esses dados indicam que as fêmeas não entraram na
menopausa, sugerindo que as alterações observadas não ocorreram por esse
evento.
Figura 6 – A. Peso do útero (g) em camundongos fêmeas com 3 (n=5), 6 (n=6), 9 (n=6) e
12 (n=5) meses. Dados expressos em média±EPM; B. Peso do ovário (g) em camundongos fêmeas
com3 (n=5), 6 (n=6), 9 (n=6) e 12 (n=5) meses. Dados expressos em média±EPM.
B A
A B

22
5.4 GLICEMIA DE JEJUM
A glicemia foi aferida após 6 horas de jejum. Pode ser observado que não
houve variação significativa da glicemia até os 9 meses de idade, porém os animais
de 12 meses de idade apresentaram uma queda significativa desse parâmetro em
relação aos animais de 3, 6 e 9 meses. Esperava-se encontrar um aumento dos
níveis de glicemia nos últimos grupos devido a uma possível resistência à insulina
desenvolvida com a idade, mas o que ocorreu foi justamente o contrário.
Figura 7 – Glicemia de jejum (mg/dL) de camundongos fêmeas com 3 (n=10), 6 (n=9), 9
(n=7) e 12 (n=8) meses de idade. Dados expressos em média±EPM. Quando indicado, P<0,05 vs. [a]
3 meses; [b] 6 meses e; [c] 9 meses.

23
5.5 LIPIDOGRAMA
Houve variação dos níveis de triglicerídeo (Fig. 8A), em relação aos grupos
de 3, 6 e 9 meses para o grupo de 12 meses, que demonstrou uma queda em
relação aos mesmos. O colesterol HDL (Fig. 8C) sanguíneo diminuiu
significativamente nos grupos de 6, 9 e 12 meses em relação ao grupo de 3 meses,
enquanto o colesterol total (Fig. 8B) não apresentou diferença significativa entre os
grupos estudados.
Figura 8 – Resposta dos lipídios sanguíneos ao envelhecimento em camundongos fêmeas
com 3 (n=5), 6 (n=9), 9 (n=7) e 12 (n=8) meses de idade. A, Triglicerídeo (mg/dL); B, Colesterol total
(mg/dL); C, Colesterol HDL (mg/dL). Dados expressos em média±EPM. Quando indicado, P<0,05 vs.
[a] 3 meses e; [b] 6 meses.
A B A B
C

24
5.6 FÍGADO
É possível observar que não houve um aumento significativo do peso
hepático com a idade (Fig. 9A), mesmo tendo havido um aumento da massa
corporal. Entretanto, identificamos um maior armazenamento de triglicerídeo no
fígado, como evidenciado na Fig. 9B, especialmente nos grupos 9 e 12 meses de
idade quando comparados ao grupo de 3 meses.
Figura 9 – Resposta do fígado ao envelhecimento em camundongos fêmeas com 3 (n=10),
6 (n=9), 9 (n=7) e 12 (n=8) meses de idade. A. Peso do fígado (g); B. Triglicerídeo hepático
(mg/dL/mg). Dados expressos em média±EPM. Quando indicado, P<0,05 vs. [a] 3 meses.
A
A B
25
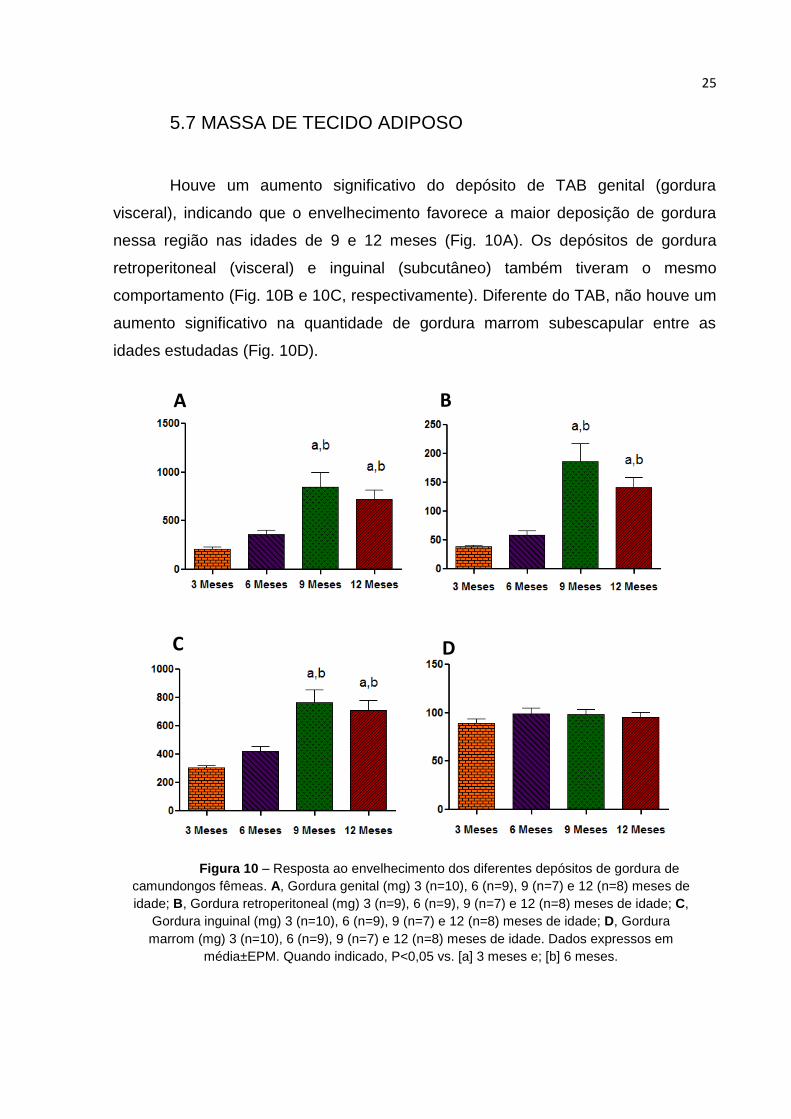
5.7 MASSA DE TECIDO ADIPOSO
Houve um aumento significativo do depósito de TAB genital (gordura
visceral), indicando que o envelhecimento favorece a maior deposição de gordura
nessa região nas idades de 9 e 12 meses (Fig. 10A). Os depósitos de gordura
retroperitoneal (visceral) e inguinal (subcutâneo) também tiveram o mesmo
comportamento (Fig. 10B e 10C, respectivamente). Diferente do TAB, não houve um
aumento significativo na quantidade de gordura marrom subescapular entre as
idades estudadas (Fig. 10D).
A B
C D
Figura 10 – Resposta ao envelhecimento dos diferentes depósitos de gordura de
camundongos fêmeas. A, Gordura genital (mg) 3 (n=10), 6 (n=9), 9 (n=7) e 12 (n=8) meses de
idade; B, Gordura retroperitoneal (mg) 3 (n=9), 6 (n=9), 9 (n=7) e 12 (n=8) meses de idade; C,
Gordura inguinal (mg) 3 (n=10), 6 (n=9), 9 (n=7) e 12 (n=8) meses de idade; D, Gordura
marrom (mg) 3 (n=10), 6 (n=9), 9 (n=7) e 12 (n=8) meses de idade. Dados expressos em
média±EPM. Quando indicado, P<0,05 vs. [a] 3 meses e; [b] 6 meses.
26
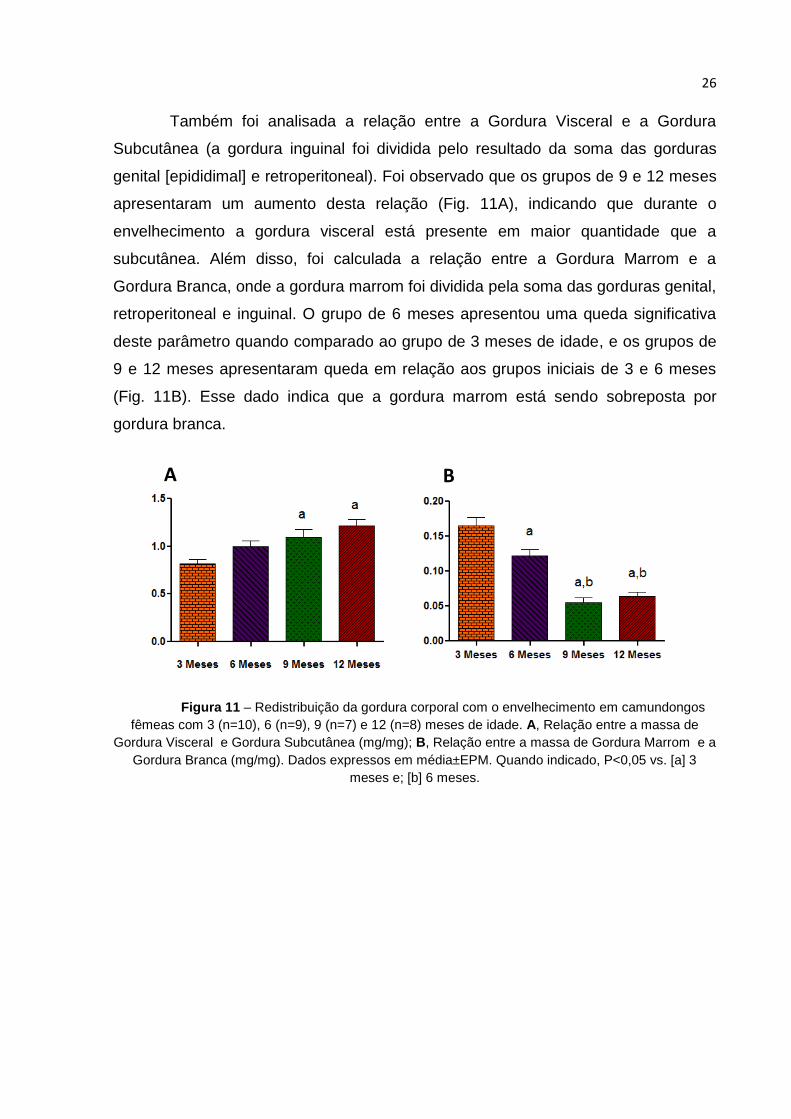
Também foi analisada a relação entre a Gordura Visceral e a Gordura
Subcutânea (a gordura inguinal foi dividida pelo resultado da soma das gorduras
genital [epididimal] e retroperitoneal). Foi observado que os grupos de 9 e 12 meses
apresentaram um aumento desta relação (Fig. 11A), indicando que durante o
envelhecimento a gordura visceral está presente em maior quantidade que a
subcutânea. Além disso, foi calculada a relação entre a Gordura Marrom e a
Gordura Branca, onde a gordura marrom foi dividida pela soma das gorduras genital,
retroperitoneal e inguinal. O grupo de 6 meses apresentou uma queda significativa
deste parâmetro quando comparado ao grupo de 3 meses de idade, e os grupos de
9 e 12 meses apresentaram queda em relação aos grupos iniciais de 3 e 6 meses
(Fig. 11B). Esse dado indica que a gordura marrom está sendo sobreposta por
gordura branca.
Figura 11 – Redistribuição da gordura corporal com o envelhecimento em camundongos
fêmeas com 3 (n=10), 6 (n=9), 9 (n=7) e 12 (n=8) meses de idade. A, Relação entre a massa de
Gordura Visceral e Gordura Subcutânea (mg/mg); B, Relação entre a massa de Gordura Marrom e a
Gordura Branca (mg/mg). Dados expressos em média±EPM. Quando indicado, P<0,05 vs. [a] 3
meses e; [b] 6 meses.
A B
27
5.8 REMODELAMENTO DO TECIDO ADIPOSO BRANCO
GENITAL
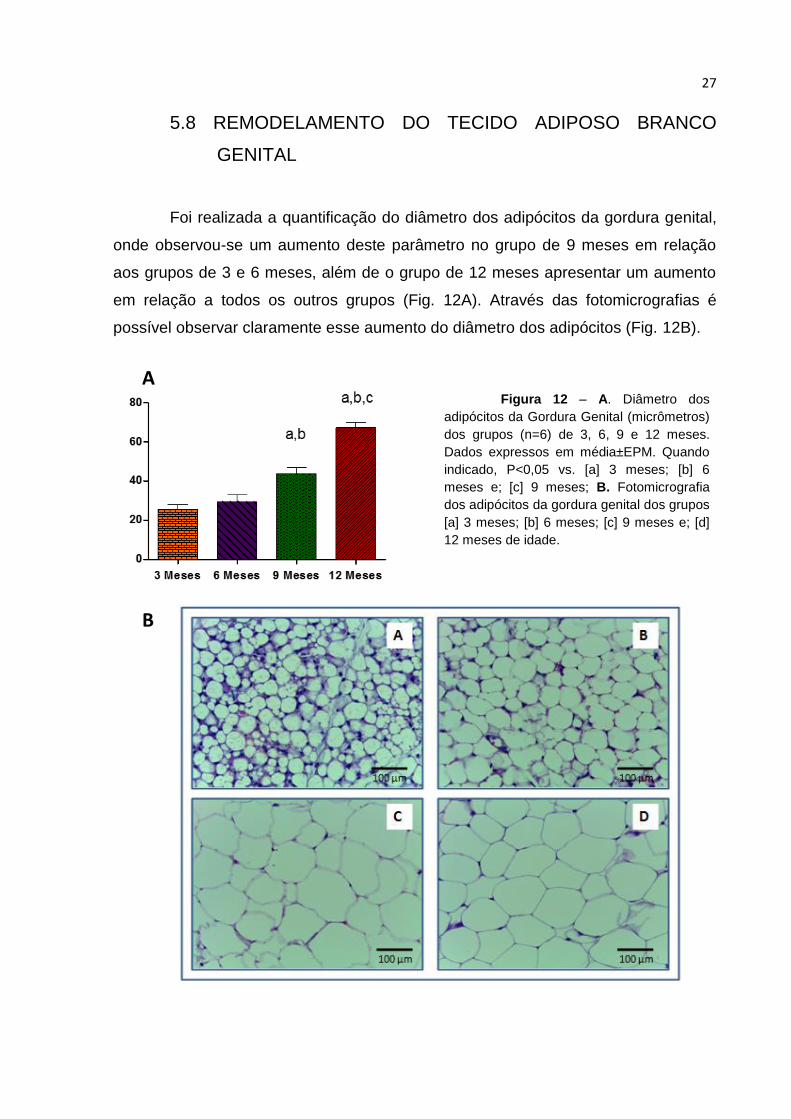
Foi realizada a quantificação do diâmetro dos adipócitos da gordura genital,
onde observou-se um aumento deste parâmetro no grupo de 9 meses em relação
aos grupos de 3 e 6 meses, além de o grupo de 12 meses apresentar um aumento
em relação a todos os outros grupos (Fig. 12A). Através das fotomicrografias é
possível observar claramente esse aumento do diâmetro dos adipócitos (Fig. 12B).
A
B
Figura 12 – A. Diâmetro dos
adipócitos da Gordura Genital (micrômetros)
dos grupos (n=6) de 3, 6, 9 e 12 meses.
Dados expressos em média±EPM. Quando
indicado, P<0,05 vs. [a] 3 meses; [b] 6
meses e; [c] 9 meses; B. Fotomicrografia
dos adipócitos da gordura genital dos grupos
[a] 3 meses; [b] 6 meses; [c] 9 meses e; [d]
12 meses de idade.
A
B
28
5.9 REMODELAMENTO DO TECIDO ADIPOSO BRANCO
INGUINAL
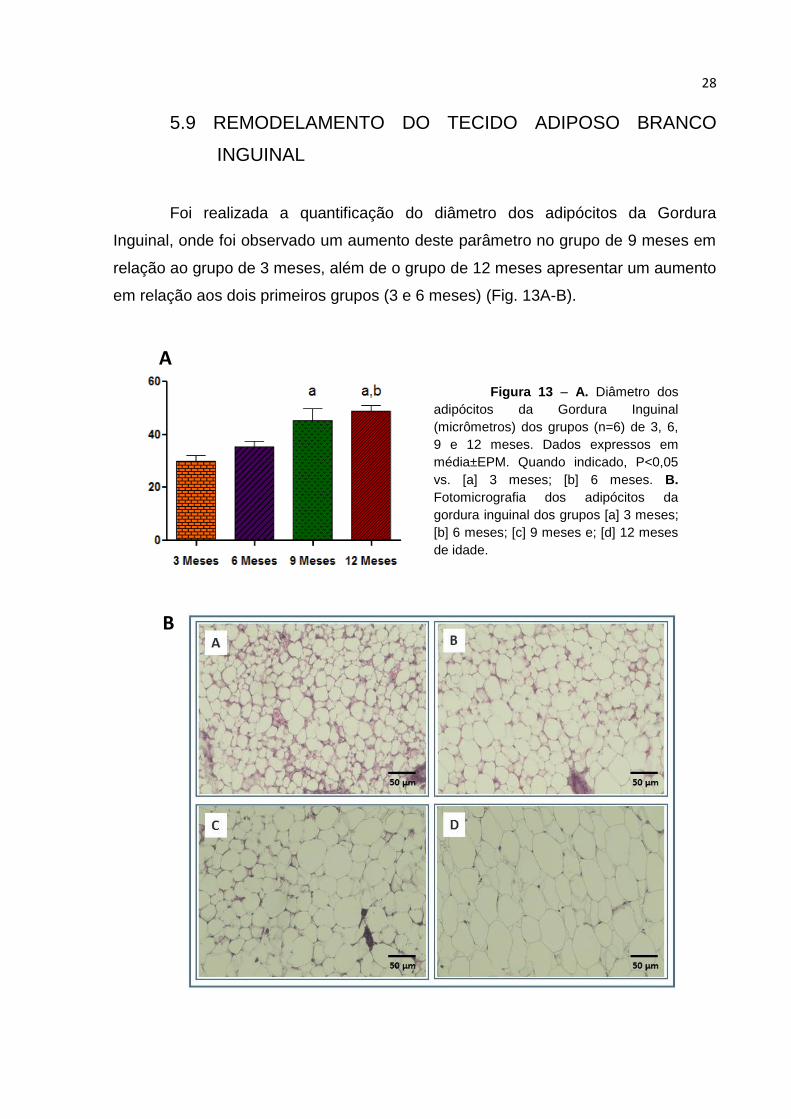
Foi realizada a quantificação do diâmetro dos adipócitos da Gordura
Inguinal, onde foi observado um aumento deste parâmetro no grupo de 9 meses em
relação ao grupo de 3 meses, além de o grupo de 12 meses apresentar um aumento
em relação aos dois primeiros grupos (3 e 6 meses) (Fig. 13A-B).
B
A
Figura 13 – A. Diâmetro dos
adipócitos da Gordura Inguinal
(micrômetros) dos grupos (n=6) de 3, 6,
9 e 12 meses. Dados expressos em
média±EPM. Quando indicado, P<0,05
vs. [a] 3 meses; [b] 6 meses. B.
Fotomicrografia dos adipócitos da
gordura inguinal dos grupos [a] 3 meses;
[b] 6 meses; [c] 9 meses e; [d] 12 meses
de idade.
A
B

29
5.10 REMODELAMENTO DO TECIDO ADIPOSO BRANCO
RETROPERITONEAL
A quantificação do diâmetro dos adipócitos da Gordura Retroperitoneal
demonstrou um aumento do diâmetro nos grupos de 9 e 12 meses em relação aos
grupos iniciais de 3 e 6 meses (Fig. 14A-B).
B
A Figura 14 – Diâmetro dos adipócitos
da Gordura Retroperitoneal (micrômetros) dos
grupos de 3 (n=4), 6 (n=6), 9 (n=6) e 12 (n=6)
meses. Dados expressos em média±EPM.
Quando indicado, P<0,05 vs. [a] 3 meses; [b] 6
meses. B. Fotomicrografia dos adipócitos da
gordura retroperitoneal dos grupos [a] 3
meses; [b] 6 meses; [c] 9 meses e; [d] 12
meses de idade.
A
B

30
5.11 QUANTIFICAÇÃO DA GORDURA MARROM
A quantificação da Gordura Marrom foi feita em duas etapas. Primeiramente,
a imagem foi segmentada em preto e branco e através do histograma da imagem foi
observada a porcentagem de área de tecido ocupada por gotículas lipídicas (Fig.
16A). Foi visto um aumento considerável nos grupos de 6, 9 e 12 meses em relação
ao grupo de 3 meses, e ainda do de 12 meses em relação ao de 6 meses.
A segunda etapa consistiu em medir o diâmetro do adipócito marrom (Fig.
16B), onde constatou-se um aumento nos grupos de 6, 9 e 12 meses comparando-
os ao grupo de 3 meses, e dos grupos de 9 e 12 meses em relação ao de 6 meses.
Adicionalmente, foi medido o diâmetro das gotículas lipídicas presentes no
citoplasma do adipócito marrom (Fig. 16C). Os grupos de 6, 9 e 12 meses tiveram
um aumento em relação ao grupo com 3 meses, mostrando assim tanto uma
hipertrofia do adipócito, quanto de suas gotículas lipídicas. Imagens representativas
das alterações morfológicas quantificadas são apresentadas nas fotomicrografias da
prancha da Fig.15.
Figura 15 – Fotomicrografia dos adipócitos da gordura marrom dos grupos [a] 3
meses; [b] 6 meses; [c] 9 meses e; [d] 12 meses de idade.

31
Figura 16 – A. Porcentagem de Gotículas lipídicas dos grupos de 3 (n=6), 6 (n=6), 9 (n=4) e
12 meses (n=6). Dados expressos em média±EPM. Quando indicado, P<0,05 vs. [a] 3 meses; [b] 6
meses; B. Diâmetro das vesículas (micrômetros) dos grupos de 3 (n=6), 6 (n=6), 9 (n=4) e 12 meses
(n=6). Dados expressos em média±EPM. Quando indicado, P<0,05 vs. [a] 3 meses; [b] 6 meses; C.
Diâmetro das gotículas (micrômetros) dos grupos de 3 (n=6), 6 (n=6), 9 (n=4) e 12 meses (n=6).
Dados expressos em média±EPM. Quando indicado, P<0,05 vs. [a] 3 meses.
A B
C C
A B
32
5.12 INSULINA
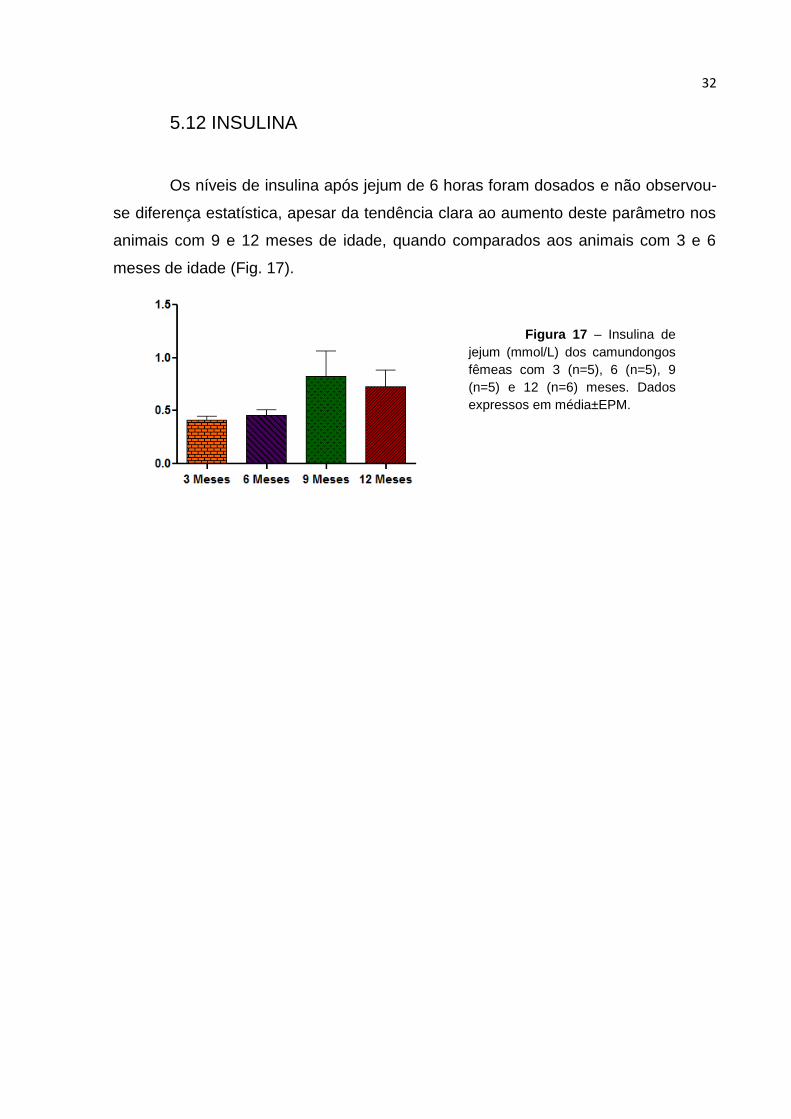
Os níveis de insulina após jejum de 6 horas foram dosados e não observou-
se diferença estatística, apesar da tendência clara ao aumento deste parâmetro nos
animais com 9 e 12 meses de idade, quando comparados aos animais com 3 e 6
meses de idade (Fig. 17).
Figura 17 – Insulina de
jejum (mmol/L) dos camundongos
fêmeas com 3 (n=5), 6 (n=5), 9
(n=5) e 12 (n=6) meses. Dados
expressos em média±EPM.

33
5.13 TESTE DE TOLERÂNCIA ORAL À GLICOSE
Após o teste ser feito, a análise estatística foi realizada e observou-se que
não há resistência à insulina, as curvas glicêmicas se comportaram de maneira
adequada, mesmo tendo uma diferença visual, não houve diferença estatística (Fig.
18A-B).
Figura 18 – A. Área sob a curva (ASC) do teste oral de tolerância à glicose (TOTG) dos
camundongos fêmeas com 3 (n=5), 6 (n=6), 9 (n=6) e 12 (n=5) meses (mg/dl). Dados expressos em
média±EPM; B. Curva do TOTG dos camundongos fêmeas de 3 (n=5), 6 (n=6), 9 (n=6) e 12 (n=5)
meses.
A
B
A
B
34
5.14 HOMA-IR e HOMA-B
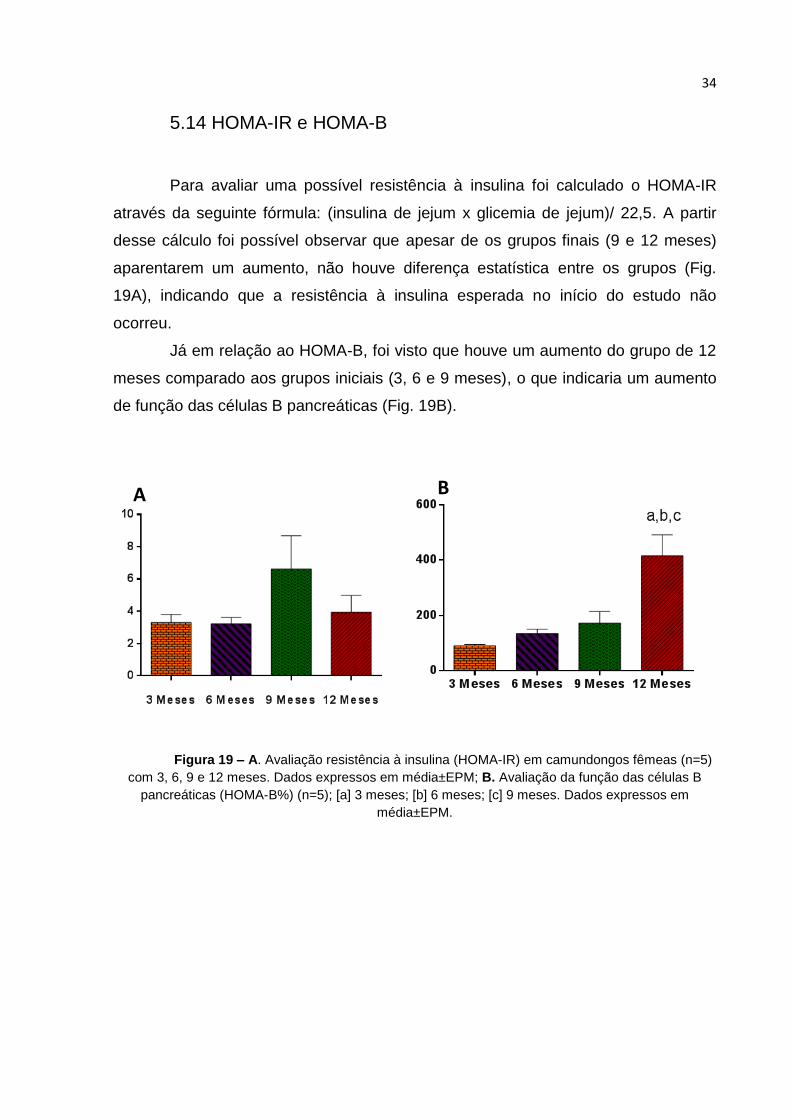
Para avaliar uma possível resistência à insulina foi calculado o HOMA-IR
através da seguinte fórmula: (insulina de jejum x glicemia de jejum)/ 22,5. A partir
desse cálculo foi possível observar que apesar de os grupos finais (9 e 12 meses)
aparentarem um aumento, não houve diferença estatística entre os grupos (Fig.
19A), indicando que a resistência à insulina esperada no início do estudo não
ocorreu.
Já em relação ao HOMA-B, foi visto que houve um aumento do grupo de 12
meses comparado aos grupos iniciais (3, 6 e 9 meses), o que indicaria um aumento
de função das células B pancreáticas (Fig. 19B).
Figura 19 – A. Avaliação resistência à insulina (HOMA-IR) em camundongos fêmeas (n=5)
com 3, 6, 9 e 12 meses. Dados expressos em média±EPM; B. Avaliação da função das células B
pancreáticas (HOMA-B%) (n=5); [a] 3 meses; [b] 6 meses; [c] 9 meses. Dados expressos em
média±EPM.
A B A B

35
6 DISCUSSÃO
Em humanos a massa corporal está associada com a idade, mostrando um
pico entre 40 e 70 anos dependendo da etnia e do gênero, após isso a massa
corporal permanece estável ou tem um pequeno declínio (Graja e Schulz, 2015).
Roedores se comportam da mesma forma também ganhando peso durante o
envelhecimento (Wu et al., 2007; Hemmeryckx et al., 2010; Sellayah e Sikder, 2014).
Muito se tem estudado sobre essa correlação entre envelhecimento e obesidade, e
como o tecido adiposo branco e marrom se comportam nessas diferentes situações,
nesse estudo visamos justamente a caracterização do remodelamento do tecido
adiposo tanto branco quanto marrom durante o envelhecimento em camundongos
C57BL/6 fêmeas.
Foi possível observar que com o aumento da idade não só a massa corporal
dos animais se tornou maior, como os depósitos de tecido adiposo branco também,
levando a uma redistribuição do depósito subcutâneo para o visceral, uma
característica do processo de envelhecimento (Rogers e Smith, 2012). Este
fenômeno pode representar um risco a saúde, já que a localização dos adipócitos
determina as diferentes características metabólicas que estes possuem. Além disso,
esta distribuição da gordura é um grande indicador da saúde metabólica, sendo a
adiposidade visceral a que apresenta maior impacto sobre a deterioração da
sensibilidade à insulina e do perfil lipídico, podendo resultar em uma síndrome
metabólica, e pode contribuir também para o desenvolvimento de doença arterial
coronariana em indivíduos não-obesos (Varlamov, Bethea e Roberts, 2014; Filho et
al., Abril 2006). O aumento da massa de tecido adiposo visceral também exerce um
fator mecânico, aumentando a pressão intra-abdominal, que sobre os rins ativariam
o sistema renina-angiotensina-aldosterona, contribuindo para a elevação da pressão
arterial, além disso a expressão de marcadores inflamatórios é maior na gordura
visceral do que na subcutânea (Filho et al., Abril 2006).
Todas essas mudanças não estão relacionadas a uma maior ingestão de
ração, nem a uma possível menopausa dos animais. A análise morfométrica também
evidenciou esse remodelamento dos depósitos mostrando que não só a massa de
tecido aumentou como também o diâmetro dos adipócitos.

36
O tecido adiposo marrom, tem um papel fundamental no controle da
termogênese, e é justamente por isso que tem sido um grande alvo para promover a
perda de peso (Harms e Seale, 2013). No presente estudo, foi visto que o peso do
depósito de gordura marrom não teve alteração ao longo dos meses, mas
interessantemente o remodelamento começou já aos 6 meses de idade. A
correlação feita com a gordura branca mostrou que com o envelhecimento ele vai
sendo interposto por gordura branca, fato também visto em um estudo onde animais
de 3 meses de idade apresentavam numerosos adipócitos marrons residindo no
tecido adiposo subcutâneo e alta expressão de UCP-1, no entanto com 6 meses já
foi observado uma menor quantidade desses adipócitos e de expressão de UCP-1.
Já aos 12 meses de idade, o depósito ganha uma aparência uniformemente branca
e com níveis de UCP-1 bem abaixo dos animais de 3 meses (Rogers e Smith, 2012),
Também foi visto na análise morfométrica do presente estudo que a
porcentagem de área ocupada por vesículas lipídicas, assim como o tamanho
destas aumentou no tecido marrom, da mesma forma que o tamanho do adipócito
marrom. Isto mostra que com o envelhecimento, o tecido adiposo marrom começa a
apresentar algumas características vistas no tecido adiposo branco. Foi mostrado
que grande parte da atividade do tecido marrom induzida pelo frio é perdida com o
envelhecimento, e essas características começam a serem vistas quando o tecido
adiposo marrom não é ativado adrenergicamente. Neste contexto, suas
propriedades morfológicas e moleculares se tornam similares as da gordura branca
e sua quantidade e ativação metabólica se tornam inversamente relacionadas ao
envelhecimento e a obesidade (Ravussin e Galgani, 2011).
A obesidade é um dos principais problemas de saúde pública do mundo,
caracterizada por um aumento da massa corporal resultante de um maior número
(hiperplasia) e/ou tamanho (hipertrofia) dos adipócitos. Isto se desenvolve por um
desbalanceamento entre o consumo energético e seu gasto (Graja e Schulz, 2015),
em ambos os tecidos, tanto branco quando marrom. Foi visto que com o
envelhecimento, os adipócitos tendem a se tornar maiores e esse aumento está
associado com várias mudanças nas funções metabólicas e no perfil lipídico. Além
disso, o aumento dos adipócitos no tecido subcutâneo é um grande fator de risco
para o desenvolvimento de diabetes do tipo 2 (Cartwright, Tchkonia e Kirkland, 2007;
Skurk et al., 2007). Um estudo in vitro identificou que a expressão e liberação de
vários fatores como a leptina, por exemplo, depende do tamanho dos adipócitos, não

37
só em humanos como em roedores (Jernås et al., 2006). Essa secreção diferenciada
pode promover a inflamação no tecido adiposo, que é uma característica da
obesidade, onde o tecido está infiltrado por macrófagos ativados que liberam
quantidades excessivas de citocinas pró-inflamatórias, como o fator-α de necrose
tumoral e interleucina-6. Recentemente foi visto que esses fatores secretados
prejudicam a diferenciação das células adiposas em humanos (Queiroz et al., 2009).
Arner sugere que a ocorrência de hiperplasia ou hipertrofia não é influenciada pelo
sexo ou peso corporal, levando a crer que pode ocorrer de forma igualmente comum
tanto em homens quanto mulheres, magros, com excesso de peso ou obesos da
população, porém foi visto que indivíduos obesos apresentam um maior número de
adipócitos adicionados por ano em relação a indivíduos magros (Arner et al., 2010) .
Observando o aspecto bioquímico do estudo, interessantemente foi visto que
os animais de 12 meses tiveram uma redução da glicemia, se aproximando mais dos
valores geralmente esperados, fato que tem maior chance de ocorrer aos 24 meses,
e se manter em níveis regulares até os 12 meses. Os níveis de insulina podem
apresentar uma queda a partir dos 12 meses, porém foi visto uma tendência ao
aumento desse parâmetro. A fim de explorar melhor esses resultados, também foi
realizado o HOMA-IR onde era esperada uma diminuição desse índice indicando
uma maior sensibilidade à insulina, que poderia explicar a redução da glicemia como
normalmente é visto aos 12 meses, atingindo a maior sensibilidade aos 24 meses,
mas na realidade foi apresentado uma tendência ao aumento desse índice que
talvez com um maior tempo de estudo poderia resultar numa diferença significativa,
o teste de tolerância oral à glicose ajudou a confirmar a ausência de resistência à
insulina até esse momento do estudo, já que a curva glicêmica se comportou da
maneira esperada. Ainda a fim de complementar esses dados, o HOMA-B foi
calculado e indicou que as células B pancreáticas tiveram a sua função aumentada,
podendo talvez explicar a tendência ao aumento da insulina (Hemmeryckx et al.,
2010; Spinelli et al., 2012).
Já ao observar o perfil lipídico, os níveis de triglicerídeo do grupo de 12
meses demonstraram uma queda, levando a crer que esse triglicerídeo estava
sendo estocado em grande quantidade, o que corrobora com o aumento dos
depósitos de gordura e a hipertrofia dos adipócitos. O colesterol HDL mostrou
diminuição nos grupos finais comparados ao de 3 meses, enquanto o colesterol total
não apresentou diferença significativa entre os grupos estudados. Os ácidos graxos

38
livres (vindos da lipólise dos adipócitos da gordura visceral) que são liberados em
grande quantidade na circulação apresentam um papel importante no
desenvolvimento da resistência tecidual à insulina, não só no fígado como em
tecidos periféricos (Filho et al., Abril 2006).
Outro fato interessante de ser notado é que o declínio associado à idade da
habilidade de armazenamento de lipídeos pode contribuir para a redistribuição de
gordura em outros tecidos (Cartwright, Tchkonia e Kirkland, 2007). Como foi visto no
presente estudo, o peso do fígado não teve alteração, porém o triglicerídeo hepático
aumentou nos dois grupos finais quando comparados ao grupo de 3 meses.
Esperava-se que com o aumento do triglicerídeo poderia também existir um
aumento do peso do fígado. Uma possível explicação para que isso não tenha
ocorrido é que pode ter havido morte celular, contrabalanceando o ganho de peso
que o triglicerídeo poderia ter acarretado. A gordura visceral vem sendo vista como o
principal fator para o depósito de gordura no fígado (Filho et al., Abril 2006),
podendo explicar o aumento observado.

39
7 CONCLUSÃO
O envelhecimento per si remodela os depósitos de gordura branca e marrom
em camundongos C57Bl/6 fêmeas, aumenta a adiposidade visceral e altera a
distribuição de gordura corporal. A partir dos dados obtidos, a hipótese inicial do
estudo de que o envelhecimento favorece uma maior deposição de tecido adiposo
branco é confirmada, uma vez que foi observado um aumento claro em todos os
depósitos de gordura branca estudados. Além disso, há acúmulo de triglicerídeos no
fígado, além de haver proporcionalmente menos gordura marrom. A quantificação da
gordura marrom demonstrou adipócitos marrom hipertrofiados e com maior
proporção de gotículas lipídicas em seu interior, mostrando assim que o tecido
adiposo sofre uma série de modificações durante o envelhecimento. Pela grande
capacidade que esse tecido possui de corrigir e influenciar desvios metabólicos, seu
estudo aprofundado se faz cada vez mais importante para auxiliar no combate a
obesidade e outras patologias associadas a degradação do metabolismo,
promovendo assim saúde e bem-estar a população.

40
8 REFERÊNCIAS
ARNER, E. et al. Adipocyte turnover: relevance to human adipose tissue morphology. Diabetes, v. 59, n. 1, p. 105-9, Jan 2010. ISSN 1939-327X. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/19846802 >. AU-YONG, I. T. et al. Brown adipose tissue and seasonal variation in humans. Diabetes, v. 58, n. 11, p. 2583-7, Nov 2009. ISSN 1939-327X. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/19696186 >. BASU, R. et al. Effects of age and sex on postprandial glucose metabolism: differences in glucose turnover, insulin secretion, insulin action, and hepatic insulin extraction. Diabetes, v. 55, n. 7, p. 2001-14, Jul 2006. ISSN 0012-1797. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/16804069 >. BORKAN, G. A. et al. Age changes in body composition revealed by computed tomography. J Gerontol, v. 38, n. 6, p. 673-7, Nov 1983. ISSN 0022-1422. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/6630900 >. CAMHI, S. M. et al. The relationship of waist circumference and BMI to visceral, subcutaneous, and total body fat: sex and race differences. Obesity (Silver Spring), v. 19, n. 2, p. 402-8, Feb 2011. ISSN 1930-739X. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/20948514 >. CARSON, F. L.; MARTIN, J. H.; LYNN, J. A. Formalin fixation for electron microscopy: a re-evaluation. Am J Clin Pathol, v. 59, n. 3, p. 365-73, Mar 1973. ISSN 0002-9173. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/4119901 >. CARTWRIGHT, M. J.; TCHKONIA, T.; KIRKLAND, J. L. Aging in adipocytes: potential impact of inherent, depot-specific mechanisms. Exp Gerontol, v. 42, n. 6, p. 463-71, Jun 2007. ISSN 0531-5565. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/17507194 >. CHORILLI, M.; AL, E. Animais de laboratório: o camundongo. Rev. Ciênc. Farm. Básica Apl. 28 n.1: 11-23 p. 2007. COSTA, J. V.; DUARTE, J. S. Tecido Adiposo e Adipocinas. Acta Med Port. 19: 251-256 p. 2006. CYPESS, A. M. et al. Identification and importance of brown adipose tissue in adult humans. N Engl J Med, v. 360, n. 15, p. 1509-17, Apr 2009. ISSN 1533-4406. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/19357406 >. DEFRONZO, R. A. Glucose intolerance and aging: evidence for tissue insensitivity to insulin. Diabetes, v. 28, n. 12, p. 1095-101, Dec 1979. ISSN 0012-1797. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/510806 >.

41
DEMERATH, E. W. et al. Anatomical patterning of visceral adipose tissue: race, sex, and age variation. Obesity (Silver Spring), v. 15, n. 12, p. 2984-93, Dec 2007. ISSN 1930-7381. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/18198307 >. DESPRÉS, J. P. et al. Race, visceral adipose tissue, plasma lipids, and lipoprotein lipase activity in men and women: the Health, Risk Factors, Exercise Training, and Genetics (HERITAGE) family study. Arterioscler Thromb Vasc Biol, v. 20, n. 8, p. 1932-8, Aug 2000. ISSN 1079-5642. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/10938014 >. FERNANDES-SANTOS, C. et al. Pan-PPAR agonist beneficial effects in overweight mice fed a high-fat high-sucrose diet. Nutrition, v. 25, n. 7-8, p. 818-27, 2009 Jul-Aug 2009. ISSN 1873-1244. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/19268533 >. FERNÁNDEZ, I. et al. Clinical biochemistry parameters in C57BL/6J mice after blood collection from the submandibular vein and retroorbital plexus. J Am Assoc Lab Anim Sci, v. 49, n. 2, p. 202-6, Mar 2010. ISSN 1559-6109. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/20353696 >. FILHO, F. F. R. et al. Gordura Visceral e Síndrome Metabólica: Mais Que Uma Simples Associação. Arq Bras Endocrinol Metab. 50 N2 Abril 2006. FLANAGAN, D. E. et al. Gender differences in the insulin-like growth factor axis response to a glucose load. Acta Physiol (Oxf), v. 187, n. 3, p. 371-8, Jul 2006. ISSN 1748-1708. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/16776662 >. FONSECA-ALANIZ, M. H. et al. O tecido adiposo como órgão endócrino: da teoria à prática. Jornal de Pediatria. Vol. 83, Nº5 2007. FRAULOB, J. C. et al. A Mouse Model of Metabolic Syndrome: Insulin Resistance, Fatty Liver and Non-Alcoholic Fatty Pancreas Disease (NAFPD) in C57BL/6 Mice Fed a High Fat Diet. J Clin Biochem Nutr, v. 46, n. 3, p. 212-23, May 2010. ISSN 1880-5086. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/20490316 >. GRAJA, A.; SCHULZ, T. J. Mechanisms of aging-related impairment of brown adipocyte development and function. Gerontology, v. 61, n. 3, p. 211-7, 2015. ISSN 1423-0003. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/25531079 >. GUIMARÃES, D. E. D. et al. Adipokines: a new view of adipose tissue. Revista Nutrição Campinas. 20(5): 549-559 p. 2007. GUO, S. S. et al. Aging, body composition, and lifestyle: the Fels Longitudinal Study. Am J Clin Nutr, v. 70, n. 3, p. 405-11, Sep 1999. ISSN 0002-9165. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/10479203 >. GUO, W. et al. Aging results in paradoxical susceptibility of fat cell progenitors to lipotoxicity. Am J Physiol Endocrinol Metab, v. 292, n. 4, p. E1041-51, Apr 2007. ISSN 0193-1849. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/17148751 >.

42
HARMS, M.; SEALE, P. Brown and beige fat: development, function and therapeutic potential. Nat Med, v. 19, n. 10, p. 1252-63, Oct 2013. ISSN 1546-170X. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/24100998 >. HEMMERYCKX, B. et al. Age-associated adaptations in murine adipose tissues. Endocr J, v. 57, n. 10, p. 925-30, 2010. ISSN 1348-4540. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/20686275 >. HUGHES, V. A. et al. Anthropometric assessment of 10-y changes in body composition in the elderly. Am J Clin Nutr, v. 80, n. 2, p. 475-82, Aug 2004. ISSN 0002-9165. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/15277173 >. JACKSON, A. S. et al. The effect of sex, age and race on estimating percentage body fat from body mass index: The Heritage Family Study. Int J Obes Relat Metab Disord, v. 26, n. 6, p. 789-96, Jun 2002. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/12037649 >. JERNÅS, M. et al. Separation of human adipocytes by size: hypertrophic fat cells display distinct gene expression. FASEB J, v. 20, n. 9, p. 1540-2, Jul 2006. ISSN 1530-6860. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/16754744 >. JERSCHOW, E.; AL, E. Macrophages accumulation in visceral and subcutaneous adipose tissue correlates with age. J Allergy Clin Immunol. 119 (Supply 1): p. S179 p. 2007. KAJIMURA, S. et al. Regulation of the brown and white fat gene programs through a PRDM16/CtBP transcriptional complex. Genes Dev, v. 22, n. 10, p. 1397-409, May 2008. ISSN 0890-9369. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/18483224 >. KARAGIANNIDES, I. et al. Altered expression of C/EBP family members results in decreased adipogenesis with aging. Am J Physiol Regul Integr Comp Physiol, v. 280, n. 6, p. R1772-80, Jun 2001. ISSN 0363-6119. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/11353682 >. KOUTSARI, C. et al. Fatty acid metabolism in the elderly: effects of dehydroepiandrosterone and testosterone replacement in hormonally deficient men and women. J Clin Endocrinol Metab, v. 94, n. 9, p. 3414-23, Sep 2009. ISSN 1945-7197. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/19567532 >. LEAN, M. E. Brown adipose tissue in humans. Proc Nutr Soc, v. 48, n. 2, p. 243-56, Jul 1989. ISSN 0029-6651 (Print) 0029-6651 (Linking). Disponível em: < http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=2678120 >. LEE, M. J.; WU, Y.; FRIED, S. K. Adipose tissue remodeling in pathophysiology of obesity. Curr Opin Clin Nutr Metab Care, v. 13, n. 4, p. 371-6, Jul 2010. ISSN 1473-6519. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/20531178 >.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=2678120

43
LEITER, E. H. et al. Aging and glucose homeostasis in C57BL/6J male mice. FASEB J, v. 2, n. 12, p. 2807-11, Sep 1988. ISSN 0892-6638. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/3044905 >. LIDELL, M. E.; ENERBACK, S. Brown adipose tissue--a new role in humans? Nat Rev Endocrinol, v. 6, n. 6, p. 319-25, Jun 2010. ISSN 1759-5037 (Electronic) 1759-5029 (Linking). Disponível em: < http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=20386559 >. LUTZ, W.; SANDERSON, W.; SCHERBOV, S. The coming acceleration of global population ageing. Nature, v. 451, n. 7179, p. 716-9, Feb 2008. ISSN 1476-4687. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/18204438 >. MACOTELA, Y. et al. Sex and depot differences in adipocyte insulin sensitivity and glucose metabolism. Diabetes, v. 58, n. 4, p. 803-12, Apr 2009. ISSN 1939-327X. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/19136652 >. MANDARIM-DE-LACERDA, C. A.; FERNANDES-SANTOS, C.; AGUILA, M. B. Image analysis and quantitative morphology. Methods Mol Biol, v. 611, p. 211-25, 2010. ISSN 1940-6029. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/19960334 >. MATTHEWS, D. R. et al. Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia, v. 28, n. 7, p. 412-9, Jul 1985. ISSN 0012-186X (Print). Disponível em: < http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=3899825 >. MATTSON, M. P. Perspective: Does brown fat protect against diseases of aging? Ageing Res Rev, v. 9, n. 1, p. 69-76, Jan 2010. ISSN 1872-9649. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/19969105 >. MCLEAN, A. C. et al. Performing vaginal lavage, crystal violet staining, and vaginal cytological evaluation for mouse estrous cycle staging identification. J Vis Exp, n. 67, p. e4389, 2012. ISSN 1940-087X. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/23007862 >. MIARD, S. et al. Aging alters PPARgamma in rodent and human adipose tissue by modulating the balance in steroid receptor coactivator-1. Aging Cell, v. 8, n. 4, p. 449-59, Aug 2009. ISSN 1474-9726. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/19485965 >. MUOIO, D. M.; NEWGARD, C. B. Mechanisms of disease:Molecular and metabolic mechanisms of insulin resistance and beta-cell failure in type 2 diabetes. Nat Rev Mol Cell Biol, v. 9, n. 3, p. 193-205, Mar 2008. ISSN 1471-0080. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/18200017 >.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=3899825

44
NEDERGAARD, J.; BENGTSSON, T.; CANNON, B. Unexpected evidence for active brown adipose tissue in adult humans. Am J Physiol Endocrinol Metab, v. 293, n. 2, p. E444-52, Aug 2007. ISSN 0193-1849. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/17473055 >. PERFETTI, R. et al. Molecular investigation of age-related changes in mouse endocrine pancreas. J Gerontol A Biol Sci Med Sci, v. 51, n. 5, p. B331-6, Sep 1996. ISSN 1079-5006. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/8808981 >. PFANNENBERG, C. et al. Impact of age on the relationships of brown adipose tissue with sex and adiposity in humans. Diabetes, v. 59, n. 7, p. 1789-93, Jul 2010. ISSN 1939-327X. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/20357363 >. PUIGSERVER, P. et al. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell, v. 92, n. 6, p. 829-39, Mar 1998. ISSN 0092-8674. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/9529258 >. QUEIROZ, J. C. F. D. et al. Controle da adipogênese por ácidos graxos Arq Bras Endocrinol Metab. 53/5 2009. RAGUSO, C. A. et al. A 3-year longitudinal study on body composition changes in the elderly: role of physical exercise. Clin Nutr, v. 25, n. 4, p. 573-80, Aug 2006. ISSN 0261-5614. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/16330136 >. RAVUSSIN, E.; GALGANI, J. E. The implication of brown adipose tissue for humans. Annu Rev Nutr, v. 31, p. 33-47, Aug 2011. ISSN 1545-4312. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/21548774 >. ROGERS, N. H.; SMITH, R. G. Brown-to-white transition in subcutaneous fat: linking aging and disease. Aging (Albany NY), v. 4, n. 11, p. 728-9, Nov 2012. ISSN 1945-4589. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/23241754 >. ROTHWELL, N. J. Central control of brown adipose tissue. Proc Nutr Soc, v. 48, n. 2, p. 197-206, Jul 1989. ISSN 0029-6651. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/2678115 >. SCHIPPER, B. M. et al. Regional anatomic and age effects on cell function of human adipose-derived stem cells. Ann Plast Surg, v. 60, n. 5, p. 538-44, May 2008. ISSN 1536-3708. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/18434829 >. SCHREINER, P. J. et al. Sex-specific associations of magnetic resonance imaging-derived intra-abdominal and subcutaneous fat areas with conventional anthropometric indices. The Atherosclerosis Risk in Communities Study. Am J Epidemiol, v. 144, n. 4, p. 335-45, Aug 1996. ISSN 0002-9262. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/8712190 >.

45
SEALE, P. et al. PRDM16 controls a brown fat/skeletal muscle switch. Nature, v. 454, n. 7207, p. 961-7, Aug 2008. ISSN 1476-4687. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/18719582 >. ______. Prdm16 determines the thermogenic program of subcutaneous white adipose tissue in mice. J Clin Invest, v. 121, n. 1, p. 96-105, Jan 2011. ISSN 1558-8238. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/21123942 >. ______. Transcriptional control of brown fat determination by PRDM16. Cell Metab, v. 6, n. 1, p. 38-54, Jul 2007. ISSN 1550-4131. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/17618855 >. SELLAYAH, D.; SIKDER, D. Orexin restores aging-related brown adipose tissue dysfunction in male mice. Endocrinology, v. 155, n. 2, p. 485-501, Feb 2014. ISSN 1945-7170. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/24248466 >. SEYDOUX, J.; GIRARDIER, L. Control of brown fat thermogenesis by the sympathetic nervous system. Experientia Suppl, v. 32, p. 153-67, 1978. ISSN 0071-335X. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/25781 >. SHARP, L. Z. et al. Human BAT possesses molecular signatures that resemble beige/brite cells. PLoS One, v. 7, n. 11, p. e49452, 2012. ISSN 1932-6203. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/23166672 >. SILVERSTONE, F. A. et al. Age differences in the intravenous glucose tolerance tests and the response to insulin. J Clin Invest, v. 36, n. 3, p. 504-14, Mar 1957. ISSN 0021-9738. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/13406065 >. SKURK, T. et al. Relationship between adipocyte size and adipokine expression and secretion. J Clin Endocrinol Metab, v. 92, n. 3, p. 1023-33, Mar 2007. ISSN 0021-972X. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/17164304 >. SPENCE, J. C. Some variation on sugar tolerance, with special reference to variations found at different ages. Quart J Med. 14 1921. SPINELLI, M. O. et al. Comparação dos parâmetros bioquímicos de camundongos criados em diferentes condições sanitárias Scientia Plena. 8, n1: 016101 p. 2012. TCHKONIA, T. et al. Fat tissue, aging, and cellular senescence. Aging Cell, v. 9, n. 5, p. 667-84, Oct 2010. ISSN 1474-9726. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/20701600 >. TIRABY, C. et al. Acquirement of brown fat cell features by human white adipocytes. J Biol Chem, v. 278, n. 35, p. 33370-6, Aug 2003. ISSN 0021-9258. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/12807871 >. VARLAMOV, O.; BETHEA, C. L.; ROBERTS, C. T. Sex-specific differences in lipid and glucose metabolism. Front Endocrinol (Lausanne), v. 5, p. 241, 2014. ISSN 1664-2392. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/25646091 >.

46
VISSER, M. et al. One- and two-year change in body composition as measured by DXA in a population-based cohort of older men and women. J Appl Physiol (1985), v. 94, n. 6, p. 2368-74, Jun 2003. ISSN 8750-7587. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/12598481 >. WEISBERG, S. P. et al. Obesity is associated with macrophage accumulation in adipose tissue. J Clin Invest, v. 112, n. 12, p. 1796-808, Dec 2003. ISSN 0021-9738. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/14679176 >. WOMERSLEY, J. A comparison of the skinfold method with extent of 'overweight' and various weight-height relationships in the assessment of obesity. Br J Nutr, v. 38, n. 2, p. 271-84, Sep 1977. ISSN 0007-1145. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/911746 >. WU, D. et al. Aging up-regulates expression of inflammatory mediators in mouse adipose tissue. J Immunol, v. 179, n. 7, p. 4829-39, Oct 2007. ISSN 0022-1767. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/17878382 >. XU, H. et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Invest, v. 112, n. 12, p. 1821-30, Dec 2003. ISSN 0021-9738. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/14679177 >. ZHANG, L. et al. Aging is associated with hypoxia and oxidative stress in adipose tissue: implications for adipose function. Am J Physiol Endocrinol Metab, v. 301, n. 4, p. E599-607, Oct 2011. ISSN 1522-1555. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/21586698 >. ZHOU, X.; HANSSON, G. K. Effect of sex and age on serum biochemical reference ranges in C57BL/6J mice. Comp Med, v. 54, n. 2, p. 176-8, Apr 2004. ISSN 1532-0820. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/15134363 >. ZINGARETTI, M. C. et al. The presence of UCP1 demonstrates that metabolically active adipose tissue in the neck of adult humans truly represents brown adipose tissue. FASEB J, v. 23, n. 9, p. 3113-20, Sep 2009. ISSN 1530-6860. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/19417078 >.

Serviço Público Federal
Universidade Federal Fluminense Pró-Reitoria de Pesquisa, Pós-Graduação e Inovação
Comissão de Ética no Uso de Animais
Certificamos que o projeto n° 446, intitulado “Alterações morfológicas e
moleculares do tecido adiposo branco e marrom em camundongos C57Bl/6 durante
o envelhecimento”, sob a orientação da Profª. Drª. Caroline Fernandes dos Santos
do Polo Universitário de Nova Friburgo, está de acordo com os Princípios Éticos na
Experimentação Animal da SBCAL e obteve a aprovação da Comissão de Ética no
Uso de Animais em 13 de março de 2014. A quantidade total de animais aprovada
pela CEUA para este projeto foi de 64 (sessenta e quatro) camundongos, e este
certificado é válido por três anos após sua aprovação.
Niterói, 13 de março de 2014.
_________________________________________
Prof. Dr. Fabio Otero Ascoli
Coordenador da CEUA








![A visão dos líderes industriais paranaenses - fiepr.org.br66939].pdf · responsabilidade primária dos entes públicos, como declina nosso Estatuto maior. Todavia, uma série de](https://static.fdocumentos.com/doc/165x107/5c4f02b993f3c3663b395eac/a-visao-dos-lideres-industriais-paranaenses-fieprorgbr-66939pdf-responsabilidade.jpg)









