
UNIVERSIDADE REGIONAL DE BLUMENAU - FURB CENTRO...
77
UNIVERSIDADE REGIONAL DE BLUMENAU - FURB CENTRO DE CIÊNCIAS TECNOLÓGICAS PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA FLORESTAL MANOELA DREWS DE AGUIAR ESTRUTURA DA PAISAGEM E COMPOSIÇÃO FUNCIONAL DE ESPÉCIES ARBÓREAS NA FLORESTA OMBRÓFILA DENSA DE SANTA CATARINA BLUMENAU, SC 2015
Transcript of UNIVERSIDADE REGIONAL DE BLUMENAU - FURB CENTRO...

1
UNIVERSIDADE REGIONAL DE BLUMENAU - FURB
CENTRO DE CIÊNCIAS TECNOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA FLORESTAL
MANOELA DREWS DE AGUIAR
ESTRUTURA DA PAISAGEM E COMPOSIÇÃO FUNCIONAL DE ESPÉCIES
ARBÓREAS NA FLORESTA OMBRÓFILA DENSA DE SANTA CATARINA
BLUMENAU, SC
2015

2
MANOELA DREWS DE AGUIAR
ESTRUTURA DA PAISAGEM E COMPOSIÇÃO FUNCIONAL DE ESPÉCIES
ARBÓREAS NA FLORESTA OMBRÓFILA DENSA DE SANTA CATARINA
Dissertação apresentada ao Programa de Pós-
Graduação em Engenharia Florestal do Centro
de Ciências Tecnológicas da Universidade
Regional de Blumenau, como requisito parcial
para a obtenção do grau de Mestre em
Engenharia Florestal.
Prof. Dr. Alexander Christian Vibrans - Orientador
Prof. Dr. Pedro Higuchi - Co-orientador
BLUMENAU, SC
2015

3
Ficha Catalográfica elaborada pela
Biblioteca Universitária da FURB
______________________________________________________________________
Aguiar, Manoela Drews de, 1990-
A282e Estrutura da paisagem e composição funcional de espécies arbóreas na Floresta
Ombrófila Densa de Santa Catarina / Manoela Drews de Aguiar. - 2015.
77 f. : il.
Orientador: Alexander C. Vibrans.
Co-orientador: Pedro Higuchi.
Dissertação (mestrado) - Universidade Regional de Blumenau,
Centro de Ciências Tecnológicas, Programa de Pós-Graduação em Engenharia Florestal.
Inclui bibliografia.
1. Florestas tropicais – Santa Catarina. 2. Florestas – Conservação – Santa Catarina. 3. Recursos
florestais – Santa Catarina. I. Vibrans, Alexander Christian. II. Higuchi, Pedro. III. Universidade
Regional de Blumenau. Programa de Pós-Graduação em Engenharia Florestal. III. Título.
CDD 634.9
______________________________________________________________________

4

5
Dedico este trabalho as duas pessoas mais importantes
da minha vida, meu pai Manoel Aguiar e minha mãe Traudi Aguiar.
Este trabalho só foi possível pela confiança deles em mim.

6
AGRADECIMENTOS
Aos meus pais, agradeço de forma especial, por todos os ensinamentos, por não me
deixarem desistir nos momentos ruins, por sempre me apoiarem e confiarem em mim.
Agradeço ao Professor Dr. Alexander Christian Vibrans, que é um grande pesquisador
e com toda a paciência me trouxe grandes aprendizados. Ao Professor Dr. Pedro Higuchi, que
já em minha graduação colaborou com o meu crescimento como aluna e estudiosa. Estou para
conhecer alguém com a inteligência, paciência e carinho pelo que faz, como o professor Pedro.
Serei eternamente grata pela grande ajuda que me deu!
Agradeço também imensamente ao Inventário Florístico Florestal por ter
disponibilizado sua base de dados, bem como à Professora Dra. Sandra C. Müller pela cessão
dos dados dos atributos funcionais das espécies estudadas. Katia Zaninni pela atenção prestada
a mim para que os dados fossem corretamente disponibilizados.
Agradeço à Capes, pela concessão da bolsa que possibilitou a realização deste trabalho.
De forma indireta, mas extremamente importante, meu parceiro de todas as horas, André
Schollemberg, fez que todo o fardo do mestrado se tornasse mais leve, me ouvindo, me
aconselhando, me ajudando, se necessário, e fazendo com que me sentisse bem sempre.
Aos meus eternos amigos, Fabiana, Diego e Sabrina (Parças), que estiveram presentes
todo o tempo ao meu lado, verdadeiros amigos! Só quem já experimentou de uma verdadeira
amizade, sabe da importância dela para a felicidade.
Aos amigos que fiz durante essa caminhada, não foram poucos e foram muito especiais,
talvez alguns nem saibam de sua importância nesse período, mas foram essenciais. Menciono
aqui alguns. Murel (muito obrigada por ouvir meus desabafos incessantes!), Patrícia, Joanna,
Daiane, Laio (peça raríssima) e João. Ao povo muito disciplinado do laboratório, em especial
à Débora que me ajudou muito (a eterna anja do inventário!). Karine Souza, aquela pessoa que
você sabe que pode contar. Mario José Marques, Danielle Ramos, Gabrielle Beca e Francielle
Missio, são de uma solidariedade imensa!
Esse é o fim de mais uma etapa, a partir daqui não sei qual será o desenrolar das coisas,
mas sei da importância do que tive até aqui e das pessoas que fizeram parte dessa história.
Muito obrigada a todos!

7
“Ali lhe parece que o céu é mais transparente e que o sol luz com claridade nova. Oferece-se-
lhe aos olhos uma aprazível floresta de tão verdes e frondosas árvores composta, que alegra
a vista sua verdura.”
O engenheiro fidalgo D. Quixote da Mancha - Miguel de Cervantes Saavedra

8
LISTA DE FIGURAS
Figura 1: Processo de fragmentação da paisagem em relação ao tempo em função de mudanças no uso
do solo. Áreas em preto representam o habitat original e as áreas em branco representam a matriz. ....13
Figura 2: Representação de dois pontos, escolhidos aleatoriamente em uma paisagem, não pertencerem
a um mesmo fragmento, fornecido em probabilidade pela métrica DIVISION (Índice de Divisão da
Paisagem). ..............................................................................................................................................16
Figura 3: Grau de subdivisão da paisagem. Imagem A representando alta subdivisão e B baixa
subdivisão. ..............................................................................................................................................17
Figura 4: Mapa com a localização das 202 unidades amostrais referentes a Floresta Ombrófila Densa
(FOD), alocadas pelo Projeto Inventário Florístico-Florestal de Santa Catarina (IFFSC). ....................28
Figura 5: Conglomerado utilizado para a amostragem da vegetação catarinense pelo Inventário Florístico
Florestal de Santa Catarina. ....................................................................................................................29
Figura 6: Representação do buffer de 2,8 km de raio, utilizado para o cálculo das métricas de paisagem,
estando representados, o remanescente-foco (mancha), no qual está localizada a Unidade Amostral do
IFFSC, remanescentes florestais inseridos na área buffer e que compõem a classe floresta, e as áreas não
florestais e de estágio sucessional inicial, não considerados no estudo. ................................................30
Figura 7: Histograma do Número de Manchas (NP) e porcentagem de floresta nas 202 áreas analisadas
de Floresta Ombrófila Densa no Estado de Santa Catarina. ...................................................................36
Figura 8:Histogramas da área do remanescente-foco e da porcentagem de área núcleo (CAI) para a classe
floresta das 202 áreas analisadas na Floresta Ombrófila Densa no Estado de Santa Catarina. ..............36
Figura 9: Histogramas do índice de forma do remanescente-foco (Shape_Idx) e do índice médio de
forma (MSI) da classe floresta para as 202 áreas analisadas na Floresta Ombrófila Densa de Santa
Catarina. .................................................................................................................................................37
Figura 10: Histogramas do índice de divisão (DIVISION) expresso em porcentagem (%) para as 202
áreas analisadas na Floresta Ombrófila Densa de Santa Catarina. .........................................................37
Figura 11: Histogramas da distância do vizinho mais próximo de mesma classe (NNDist) para as 202
áreas analisadas na Floresta Ombrófila Densa de Santa Catarina. .........................................................38
Figura 12: Mapa com a localização das unidades amostrais classificadas em mais e menos fragmentadas
na Floresta Ombrófila Densa de Santa Catarina ....................................................................................39
Figura 13: Boxplots dos valores de cada métrica de paisagem, a nível de classe e mancha, analisadas na
Floresta Ombrófila Densa de Santa Catarina nos grupos mais e menos fragmentado. 1: Métricas de área
florestal; 2: métricas de borda; 3: métricas de divisão; 4: distância do vizinho mais próximo; 5: métricas
de forma. n = 202, *P <0,05, ***P<0,001, ns = não significativo. ........................................................41
Figura 14: Dendrograma para as vinte espécies arbóreas mais abundantes de 202 unidades amostrais
alocados na Floresta Ombrófila Densa de Santa Catarina, a partir dos atributos funcionais altura máxima,
área foliar específica, densidade da madeira, síndrome de dispersão, fenologia foliar, conteúdo de
matéria seca, nitrogênio e fósforo foliar. ................................................................................................43
Figura 15: Análise de correspondência entre os atributos fenologia foliar (dec = decídua, semi =
semidecídua, per = perene) e síndrome de dispersão (anem = anemocoria, zoo = zoocoria) e a associação
com os grupos funcionais. ......................................................................................................................45
Figura 16: Análise de componentes principais para ordenação dos grupos funcionais por meio dos
atributos área foliar específica (SLA), conteúdo de matéria seca foliar (LDMC), altura máxima (Hmáx),
densidade da madeira (SSD), conteúdo de nitrogênio e fósforo foliar (LNC e LPC). ...........................46
Figura 17: Mapa das microrregiões do Estado e localização das 86 unidades amostrais alocadas pelo
Projeto Inventário Florístico Florestal de Santa Catarina (IFFSC) na Floresta Ombrófila Densa. Em

9
verde, a cobertura florestal remanescente de acordo com o mapa de uso do solo realizado pelo Projeto
de Proteção a Mata Atlântica em Santa Catarina (GEOAMBIENTE, 2008). ........................................58
Figura 18: Conglomerado utilizado para a amostragem da vegetação catarinense pelo Inventário
Florístico Florestal de Santa Catarina. ...................................................................................................59
Figura 19: Representação do buffer de 2,8 km de raio, utilizado para o cálculo das métricas de paisagem,
estando representados, o remanescente-foco, no qual está localizada a Unidade Amostral do IFSSC,
remanescentes florestais inseridos na área buffer e que compõem a classe floresta, e as áreas não
florestais e de estágio sucessional inicial, não considerados no estudo. ................................................61
Figura 20: Análise de Componentes Principais realizada a partir da matriz atributos médios por unidade
amostral (CWM), do conjunto de espécies mais abundantes da Floresta Ombrófila Densa de Santa
Catarina. SSD = densidade da madeira; LNC = conteúdo de nitrogênio foliar; SLA = área foliar
específica; LPC = conteúdo de fósforo foliar; LDMC = conteúdo de matéria seca foliar; hmáx = altura
máxima. ..................................................................................................................................................64
Figura 21: Gráfico do melhor modelo encontrado para explicação do Eixo 2 da Análise de Componentes
Principais (PCA), relacionado (p-value=0,0238) com a métrica DIVISION, que quantifica o grau de
subdivisão dos fragmentos florestais em uma dada paisagem, na Floresta Ombrófila Densa de Santa
Catarina. .................................................................................................................................................66
Figura 22: Estimativa de densidade de Kernel para a síndrome de dispersão zoocórica (1 – cinza claro)
e anemocórica (0 – cinza escuro) em relação à totalidade de bora –TE (A) e o índice médio de forma –
MSI (B). .................................................................................................................................................67

10
LISTA DE TABELAS
Tabela 1: Métricas de paisagem a nível de classe e a nível de mancha, com seus respectivos
significados e unidades de medida. .......................................................................................... 30
Tabela 2: Espécies arbóreas mais abundantes pertencentes à Floresta Ombrófila Densa do
Estado de Santa Catarina. ......................................................................................................... 31
Tabela 3: atributos funcionais observados com suas respectivas siglas, unidade de medida
(para variáveis quantitativas) e categorias (variáveis qualitativas) .......................................... 32
Tabela 4: Valores das métricas de paisagem a nível de classe e mancha, obtidos para 202
áreas analisadas na Floresta Ombrófila Densa de Santa Catarina. ........................................... 34
Tabela 5: Valores dos atributos funcionais obtidos das 20 espécies arbóreas mais abundantes
da Floresta Ombrófila Densa de Santa Catarina. ...................................................................... 42
Tabela 6: Análise de Variância para os atributos de área foliar específica (SLA), conteúdo de
matéria seca foliar (LDMC), altura máxima (Hmáx), densidade da madeira (SSD), conteúdo
de nitrogênio foliar (LNC) e conteúdo de fósforo foliar (LPC) por grupo funcional. ............. 44
Tabela 7: Frequências absolutas, com valores percentuais entre parênteses, por grupo
funcional para os atributos categóricos fenologia foliar e síndrome de dispersão, com os
resultados da análise da tabela de contingência........................................................................ 44
Tabela 8: Resultados dos modelos GLS para a abundância dos grupos funcionais. Somente o
melhor modelo de acordo com os valores de AIC são mostrados. São apresentados os
coeficientes estimados e os erros padrão. ................................................................................. 48
Tabela 9: Espécies arbóreas mais abundantes pertencentes à Floresta Ombrófila Densa do
Estado de Santa Catarina. ......................................................................................................... 59
Tabela 10: atributos funcionais observados com suas respectivas siglas, unidade de medida
(para variáveis quantitativas) e categorias (variáveis qualitativas) .......................................... 60
Tabela 11: Métricas de paisagem a nível de classe e a nível de mancha, com seus respectivos
significados e unidades de medida. .......................................................................................... 62
Tabela 12: Resultados da Análise de Componentes Principais (PCA) para os atributos
funcionais analisados, com a correlação das variáveis para os dois primeiros eixos da
ordenação. ................................................................................................................................. 64
Tabela 13: Resultados dos modelos lineares, usando o método dos mínimos quadrados
generalizados (GLS) para os dois primeiros eixos da Análise de Componentes Principais
(PCA) e para o índice de diversidade funcional RaoQ e modelos lineares generalizados
(GLM), com distribuição binomial, para a síndrome de dispersão. ......................................... 65

11
SUMÁRIO
RESUMO GERAL ___________________________________________________________________ 12
INTRODUÇÃO GERAL ______________________________________________________________ 13
REFERÊNCIAS BIBLIOGRÁFICAS ___________________________________________________ 19
CAPÍTULO I
FLORESTA OMBRÓFILA DENSA DE SANTA CATARINA: A ESTRUTURA ESPACIAL DOS
REMANESCENTES FLORESTAIS E OS ATRIBUTOS FUNCIONAIS DAS ESPÉCIES
ARBÓREAS ________________________________________________________________________ 23
RESUMO __________________________________________________________________________ 23
ABSTRACT ________________________________________________________________________ 24
INTRODUÇÃO _____________________________________________________________________ 25
MATERIAL E MÉTODOS ___________________________________________________________ 26 ÁREA DE ESTUDO _____________________________________________________________________ 26
ESTRUTURA ESPACIAL DOS REMANESCENTES FLORESTAIS _______________________________________ 29
SELEÇÃO E CARACTERIZAÇÃO FUNCIONAL DAS ESPÉCIES ARBÓREAS _______________________________ 31
ANÁLISES REALIZADAS PARA O 1º OBJETIVO: DESCREVER A CONDIÇÃO DE FRAGMENTAÇÃO, APONTANDO AS
REGIÕES ONDE A FRAGMENTAÇÃO OCORREU DE FORMA MAIS INTENSA _____________________________ 32
ANÁLISES REALIZADAS PARA O 2º OBJETIVO: AGRUPAMENTO FUNCIONAL E AVALIAÇÃO DA INFLUÊNCIA DE
MÉTRICAS DA PAISAGEM SOBRE A ABUNDÂNCIA DOS GRUPOS FUNCIONAIS IDENTIFICADOS. ______________ 33
RESULTADOS E DISCUSSÃO _______________________________________________________ 34 ESTRUTURA DA PAISAGEM _______________________________________________________________ 34
CLASSIFICAÇÃO DAS ÁREAS ANALISADAS EM MAIS E MENOS FRAGMENTADAS _________________________ 38
ATRIBUTOS FUNCIONAIS E ESTRATÉGIA DAS PLANTAS ___________________________________________ 41
RELAÇÃO ENTRE OS GRUPOS FUNCIONAIS E A ESTRUTURA DA PAISAGEM ____________________________ 47
CONCLUSÃO ______________________________________________________________________ 48
REFERÊNCIAS BIBLIOGRÁFICAS __________________________________________________ 49
CAPÍTULO II
A FRAGMENTAÇÃO COMO PREDITORA DE MUDANÇAS FUNCIONAIS DO COMPONENTE
ARBÓREO NA FLORESTA OMBRÓFILA DENSA DE SANTA CATARINA _________________ 53
RESUMO __________________________________________________________________________ 53
ABSTRACT ________________________________________________________________________ 54
INTRODUÇÃO _____________________________________________________________________ 54
MATERIAL E MÉTODOS ___________________________________________________________ 56 ÁREA DE ESTUDO _____________________________________________________________________ 56
SELEÇÃO DAS ESPÉCIES ________________________________________________________________ 57
CARACTERIZAÇÃO FUNCIONAL DAS ESPÉCIES ARBÓREAS ________________________________________ 60
ESTRUTURA DA PAISAGEM DOS REMANESCENTES FLORESTAIS ____________________________________ 60
ANÁLISES ESTATÍSTICAS_________________________________________________________________ 62
RESULTADOS _____________________________________________________________________ 63
DISCUSSÃO _______________________________________________________________________ 67
CONCLUSÃO ______________________________________________________________________ 70
REFERÊNCIAS BIBLIOGRÁFICAS __________________________________________________ 70
CONSIDERAÇÕES FINAIS ___________________________________________________________ 75
APÊNDICE_________________________________________________________________________76

12
RESUMO GERAL
A fragmentação florestal causa diversas modificações na estrutura e composição de
comunidades vegetais. Atualmente, de forma mais aprofundada, estudos têm avaliado a
influência da fragmentação com uma perspectiva funcional das plantas em metacomunidades.
Este aprofundamento é essencial em ecossistemas de complexa biodiversidade e em acelerada
degradação, como é o caso das florestas tropicais. Dessa forma, neste trabalho, avaliou-se a
influência da estrutura espacial de remanescentes florestais sobre um conjunto de atributos
funcionais das espécies arbóreas mais abundantes da Floresta Ombrófila Densa de Santa
Catarina. A dissertação foi dividida em dois capítulos. O primeiro capítulo teve dois objetivos:
(i) descrever a condição de fragmentação da Floresta Ombrófila Densa de Santa Catarina,
apontando as regiões onde a fragmentação ocorreu de forma mais intensa, classificando-as em
mais e menos fragmentadas segundo as métricas de paisagem mais explicativas dos dados
funcionais; (ii) descrever os atributos funcionais das espécies arbóreas mais abundantes,
agrupando-as funcionalmente e avaliando a influência das métricas da paisagem sobre a
abundância dos grupos funcionais identificados. Para isso, avaliaram-se, de forma exploratória,
os dados de métricas de paisagem e de atributos funcionais, realizaram-se análises de
agrupamento, Análise de Variância (ANOVA), ordenações e regressão. Os resultados
indicaram diferentes graus de fragmentação na Floresta Ombrófila Densa de Santa Catarina,
com maior intensidade nas regiões Sul, Litoral Norte e Alto Vale do Itajaí. Também
identificaram-se diferentes estratégias de vida das espécies avaliadas para a ocupação de
habitats e para o desenvolvimento e sobrevivência. O segundo capítulo teve como objetivo
investigar a influência da estrutura espacial dos remanescentes florestais sobre os atributos
funcionais e a diversidade funcional do conjunto de espécies mais abundantes da Floresta
Ombrófila Densa de Santa Catarina. Para isso, calculou-se o índice de diversidade de RaoQ e
realizaram-se Análise de Componentes Principais e regressões. Os resultados indicaram que as
áreas mais fragmentadas (aquelas que ocorreram em paisagens mais subdivididas, com maior
quantidade de borda e com maior complexidade de forma) apresentaram alterações nos padrões
de dispersão, na densidade da madeira e no conteúdo de matéria seca foliar. Em contrapartida,
a diversidade funcional foi mantida nas diferentes intensidades de fragmentação. De forma
geral, concluiu-se que parâmetros estruturais da paisagem demonstram-se importantes na
explicação de alterações da composição funcional da Floresta Ombrófila Densa; estes
resultados são importantes para a construção de uma base teórica no gerenciamento da
configuração de habitats e da composição da paisagem em áreas fragmentadas.
Palavras-chave: Fragmentação florestal. Atributos funcionais. Diversidade funcional.

13
INTRODUÇÃO GERAL
A fragmentação de habitats naturais é uma das maiores ameaças à biodiversidade global,
impulsionando mudanças na composição e configuração das paisagens. O alto grau de
fragmentação na Floresta Atlântica, por exemplo, contribuiu para que mais de 80% de seus
fragmentos florestais apresentassem menos de 50 hectares (RIBEIRO et al., 2009), e, devido
ao seu alto endemismo e grande número de espécies ameaçadas de extinção, foi classificada
como um hotspot terrestre no final do século XX (MYERS et al., 2000).
O bioma está presente em 17 Estados da costa brasileira, originalmente cobria todo o
Estado de Santa Catarina, esse com extensão de 95.346,18 km² (VIBRANS et al, 2013), possui
atualmente 26.337,8 km² de floresta em seu território (VIBRANS et. al, 2013) e ocupa a quinta
colocação no ranking de desmatamento da Floresta Atlântica (SOS MATA ATLÂNTICA e
INPE, 2014).
O desmatamento das florestas provoca redução e fragmentação de habitats, dois
conceitos distintos que foram confundidos por muito tempo (FAHRIG, 2003). Entende-se
fragmentação como processo de subdivisão, em que uma grande área contínua é convertida em
pequenas manchas isoladas por uma matriz diferente do habitat original (Figura 1) (WILCOVE
et al. 1986; FAHRIG, 2003). Na redução de habitat pode ou não ocorrer subdivisão, porém há
inevitável perda de espécies, sendo todas as envolvidas prejudicadas. Na fragmentação as
consequências dependem do grupo funcional das espécies considerado, algumas espécies se
beneficiam, outras nem tanto (VILLARD e METZGER, 2014).
A divisão de uma grande área em pequenos fragmentos faz com que estes adquiram
condições ambientais diferentes, causando a redução significativa do fluxo de animais, pólen e
sementes e aumento dos casos de invasão biológica. Uma paisagem fragmentada, em função de
sua configuração espacial, ainda pode manter diversas populações de organismos, porém, a
partir de um certo grau de fragmentação, os efeitos tornam-se intensos e, às vezes, irreversíveis
(METZGER, 2008).
Fonte: Fahrig, 2003.
Figura 1: Processo de fragmentação da paisagem em relação ao tempo em função de mudanças no uso
do solo. Áreas em preto representam o habitat original e as áreas em branco representam a matriz.

14
O efeito mais evidente da fragmentação é a redução do tamanho populacional das
espécies mais sensíveis às novas condições bióticas e abióticas impostas, porém os efeitos não
são homogêneos para todas elas. De acordo com Chan-Dzul et al. (2011), “quando um
ecossistema experimenta uma forte perturbação, a taxa de recuperação dependerá do tipo,
frequência e intensidade da mesma, assim como a capacidade de resiliência do ecossistema”.
Viana e Tabanez (1996) colocam ainda que a intensidade desses efeitos sobre as populações
depende da biologia das espécies e também das características do fragmento na qual a
comunidade está inserida. As variações na abundância, riqueza e composição de espécies em
paisagens fragmentadas ocorrem principalmente porque as espécies apresentam características
ecológicas próprias que as fazem responder de maneira particular as alterações de seu habitat
(OLIFIERS e CERQUEIRA, 2006).
Os efeitos da fragmentação florestal foram objeto de estudo para muitos pesquisadores
no Brasil (TABARELLI et al., 1999; NUNES et al., 2003; NASCIMENTO e LAURANCE,
2006; TABARELLI et al., 2006; MACHADO et al., 2008; LAURANCE e VASCONCELOS,
2009; UZÊDA et al., 2011). As evidências científicas colocam a fragmentação como uma
importante causadora de mudanças que afetam, de forma diferenciada, a estrutura e a dinâmica
de ecossistemas florestais. Em outros termos, a fragmentação gera comunidades empobrecidas,
biossimplificadas e com pouca representatividade de espécies raras e/ou ameaçadas
(OLIVEIRA et al., 2004).
Os mais discutidos e preocupantes efeitos da fragmentação são: o aumento do efeito de
borda, causado pela maior área de contato com o ambiente externo (matriz), principal
provocador de alterações ambientais nos remanescentes (NASCIMENTO e LAURANCE,
2006), redução da conectividade funcional (indicada pelo deslocamento da fauna polinizadora
e dispersora) e estrutural dos fragmentos (METZGER, 2008). E ainda, tamanho, forma, grau de
isolamento, tipo de vizinhança e histórico de perturbações, são os principais fatores
mencionados por Viana et al. (1992), que afetam a dinâmica de fragmentos florestais e se
relacionam com importantes fenômenos biológicos.
Diferenças nas taxas de colonização refletem-se principalmente nos modos de dispersão
(METZGER, 2000). Espécies dispersadas pelo vento (anemocóricas) têm maior capacidade de
mover-se a maiores distâncias entre fragmentos florestais. Espécies zoocóricas dependem do
deslocamento de animais dispersores que, devido a vários fatores, podem estar ausentes ou em
baixo número, ou impedidos de deslocar-se pela matriz do entorno (SCARIOT et al., 2003).

15
A ecologia da paisagem, como um novo programa de pesquisa, tornou-se proeminente
dentro da ecologia, na década de 1980 (TURNER et al., 2001). Concentra-se no estudo da
influência da estrutura espacial (heterogeneidade espacial) sobre processos ecológicos, um
campo altamente interdisciplinar, que tem ajudado a compreender a magnitude dos efeitos de
atividades humanas sob populações de espécies vegetais e animais.
Forman e Godron (1986) definem Ecologia de Paisagem como o “estudo da estrutura,
função e dinâmica de áreas heterogêneas compostas por ecossistemas interativos”. A estrutura,
é, segundo Soares Filho (1998), um arranjo ou padrão espacial da paisagem, descrito pelos
tamanhos, formas, números e tipos de configurações dos ecossistemas, e que governa a
distribuição de energia, materiais e organismos.
A paisagem é composta por três tipos de elementos: manchas, corredores e matriz. As
manchas (do inglês patch) em uma paisagem, normalmente, representam ecossistemas
compostos por comunidades de animais e plantas (FORMAN e GODRON, 1986) e são
definidas como uma superfície não linear e relativamente reduzida e homogênea, numa
determinada escala, que difere em aparência de seu entorno (METZGER, 2003a). No âmbito
do geoprocessamento, uma mancha é representada por uma unidade em um mapa, formada por
um único polígono (no caso da representação vetorial), e recebem atributos nominais definidos
pelos tipos de elementos da paisagem que representam, como as florestas, os campos agrícolas,
reflorestamentos, dentre outros (SOARES FILHO, 1998). Alguns dos aspectos que
caracterizam a mancha são seu tamanho, número e forma. As mudanças de uso do solo
modificam estes aspectos, diminuindo o tamanho, alterando a forma, aumentando o número das
manchas e a distância entre elas, caracterizado como processo de fragmentação da paisagem
(SOARES FILHO, 1998). Os corredores são resumidamente definidos como sendo, uma área
homogênea, discreta e linear da paisagem, relativamente estreita, que difere das áreas vizinhas
(METZGER, 2001; TURNER et al., 2001). A matriz é o tipo de cobertura mais extenso em
uma paisagem, mais conectado e funcionalmente mais dominante, que determina a dinâmica da
paisagem (FORMAN, 1995). A matriz não seria um ambiente totalmente inóspito e, em geral,
possui baixa similaridade florística e fisionômica em relação à mancha (METZGER, 2008).
Para o estudo da influência da estrutura espacial sobre processos ecológicos é necessário
quantificar os padrões espaciais. Uma das formas de quantificação é utilizar as chamadas
métricas da paisagem (METZGER, 2003b).
Para Metzger (2003b), as métricas de paisagem são agrupadas em duas categorias: as de
composição e as de disposição. Os parâmetros de composição dão uma ideia de quais unidades
ou elementos estão presentes na paisagem, da riqueza dessa unidade, bem como da área por

16
elas ocupadas. Já, segundo o mesmo autor, os parâmetros de disposição vão quantificar o
arranjo das unidades e são utilizados para caracterizar as unidades em termos de bordas,
diversidade de contatos, grau de isolamento, conectividade, área e formato.
A fragmentação condiciona a um maior comprimento de borda, criando um novo
habitat, ambientalmente mais limitante e propício para espécies menos sensíveis às condições
de borda, resultando em um aumento dessas espécies e diminuição do habitat de espécies de
interior, o que não é desejável do ponto de vista da conservação. A quantidade de borda
influencia também na quantidade de área núcleo, que é a área efetiva de uma mancha ou
conjunto de manchas na paisagem. As métricas de área núcleo são consideradas medidas de
qualidade de um dado habitat e são medidas pela relação da área de habitat descontada a borda
e a área total.
A forma dos remanescentes está diretamente relacionada ao efeito de borda, pois quanto
mais complexa a forma, maior o efeito externo sobre o fragmento (exposição à luz e ao vento,
por exemplo) e maior o contato com a matriz. Neste sentido, determinadas métricas de paisagem
que quantificam a forma dos remanescentes na paisagem avaliam “a complexidade da forma
por meio da comparação com uma feição padrão” (LANG e BLASCHKE, 2009). Segundo os
mesmos autores, quanto mais a forma do fragmento desviar do padrão redondo, maior será o
índice de forma e maior é a complexidade dos fragmentos na paisagem.
As métricas que quantificam a divisão da paisagem (DIVISION - LANG e TIEDE,
2003) é definido como a probabilidade (%) de que dois locais escolhidos aleatoriamente em
uma paisagem sob investigação não estejam situados num mesmo fragmento remanescente
(Figura 2), e é calculado a partir da função de distribuição do tamanho das manchas
remanescentes (JAEGER, 2000).
Disponível em: www.umass.edu/landeco/teaching/landscape_ecology/schedule/chapter9_metrics.pdf
Figura 2: Representação de dois pontos, escolhidos aleatoriamente em uma paisagem, não pertencerem
a um mesmo fragmento, fornecido em probabilidade pela métrica DIVISION (Índice de Divisão da
Paisagem).

17
A
B
A importância das métricas de subdivisão está relacionada com a subdivisão das
populações, com a interrupção de continuidade de processos ecológicos e com a conectividade
da paisagem (Figura 3), e é o conceito central no estudo da fragmentação de habitats
(METZGER, 1999).
Disponível em: www.umass.edu/landeco/teaching/landscape_ecology/schedule/chapter9_metrics.pdf
Figura 3: Grau de subdivisão da paisagem. Imagem A representando alta subdivisão e B baixa
subdivisão.
Pelo estudo de Lavorel et al. (1997), já foram reconhecidas as mudanças no uso da terra
como uns dos principais promotores da mudança global (bióticas e abióticas), e devido às
crescentes perturbações de ambientes naturais, constatou-se que haveria a necessidade de
identificar grupos de organismos classificados com base na sua resposta a essas perturbações.
Atualmente, propõe-se o estudo funcional das plantas como um diferencial às análises
tradicionais da vegetação. Estes estudos analisam a vegetação por meio de um conjunto de
características intrínsecas das plantas, características estas que podem ser morfológicas,
fisiológicas ou fenológicas mensuráveis a nível de indivíduo, conhecidas como atributos
funcionais (VIOLLE et al., 2007).
Os atributos funcionais são manifestações das estratégias das plantas para ultrapassarem
filtros ambientais (filtros de seleção impostos pelas condições ambientais - clima,
luminosidade, encharcamento do solo, stress hídrico e etc.) e também competirem com as
espécies coexistentes. Estão envolvidas nas maneiras de adquirir, processar e investir os
recursos disponíveis (DÍAZ et al., 2004). Espécies com uma mesma estratégia de vida
costumam apresentar atributos semelhantes, interpretados como adaptações a determinados
padrões de uso de recursos (LAVOREL et al., 1997).

18
Pillar (1999) observou que o estudo das respostas vegetais às mudanças ambientais em
uma escala global não pode basear-se na unidade taxonômica de espécies, uma vez que a
maioria delas apresenta distribuição geográfica limitada. O uso de grupos funcionais pode,
nesse caso, otimizar a percepção entre vegetação e mudanças ambientais, já que grupos de
plantas com estratégias de vida similares apresentam uma resposta similar às variações
ambientais em qualquer escala (local à global) (CARNEIRO, 2013).
De fato, observa-se relações dos atributos fucionais com diferentes condições
ambientais nas comunidades vegetais (CUNNINGHAM et al., 1999; MÜLLER et al., 2007;
CASTRO et al., 2010; LIENIN e KLEYER, 2011). Quando determinadas essas relações,
contribuem para a compreensão sobre a amplitude dos nichos ecológicos das espécies árbóreas
em diversas escalas (MISSIO, 2014).
De acordo com Carneiro (2013), as limitações de recursos e as condições abióticas nas
bordas, resultantes da fragmentação florestal, agem como filtros ambientais restringindo a
variação nos atributos das plantas. O que resultaria, ainda, em uma menor diversidade funcional
e levaria a um comprometimento do funcionamento e da manutenção de processos nas
comunidades (MAYFIELD et al., 2005).
Existe ainda, o que os pesquisadores chamam de trade-off em atributos funcionais,
ocorrendo, nesse caso, um tipo de espectro de variação de atributos (LAMBERS e POOTER,
1992; WESTOBY et al., 2002; DÍAZ et al. 2004; WRIGHT et al., 2004; FALSTER e
WESTOBY, 2005; CHAVE et al., 2009). Um exemplo seria o trade-off entre aquisição e
conservação de recursos por parte das plantas, em que, em um extremo do espectro estariam
espécies com atributos que facilitam a aquisição dos recursos, trazendo vantagens em ambientes
ricos em recursos e de alta competição interespecífica. E no outro extremo estariam espécies
que apresentam limitações na aquisição e, por isso, conservam por mais tempo os recursos
assimilados, apresentando, por outro lado, crescimento lento, maior estoque e defesa. Seus
atributos garantem potencial para a sobrevivência em ambientes pobres em recursos e/ou com
alta taxa de predação, por exemplo.
A preocupação dos efeitos da redução de florestas sobre os organismos vivos e a real
necessidade de reverter à situação de biossimplificação dos ecossistemas de forma acertada,
foram algumas questões que fizeram com que, atualmente, ocorresse uma maior
conscientização de empresas e pessoas da importância de manter as florestas. Porém, mesmo
com os olhares voltados à conservação e recuperação ambiental, faltam estudos que mostrem
como a intensidade da fragmentação exerce efeito sobre as características ecológicas das

19
comunidades vegetais e animais, a fim de elaborar estratégias para manter o equilíbrio entre
exploração e conservação.
Nesse sentido, a oportunidade de utilização de dados coletados de forma sistemática e
homogênea em um grande espaço geográfico, que é o caso do Inventário Florístico Florestal de
Santa Catarina, e a possibilidade de contribuir com estudos dos efeitos da fragmentação florestal
utilizando um novo enfoque (as características funcionais arbóreas e a estrutura de fragmentos
florestais), foram as principais motivações para este trabalho.
O objetivo geral deste trabalho é encontrar evidências da influência da estrutura da
paisagem sob a diversidade e os atributos funcionais de espécies arbóreas mais abundantes da
Floresta Ombrófila Densa de Santa Catarina. Espera-se verificar influência direta em
diversidade e em atributos específicos, quando analisado um gradiente estrutural da paisagem.
REFERÊNCIAS BIBLIOGRÁFICAS
CARNEIRO, M. S. Efeito da estrutura da paisagem sobre a riqueza e diversidade de
árvores em grupos funcionais reprodutivos. Universidade Federal de Alfenas. Dissertação
de Mestrado em Ecologia e Tecnologia Ambiental. Minas Gerais, 2013.
CASTRO, H.; LEHSTEN, V.; LAVOREL, S.; FREITAS, H. Functional response traits in
relation to land use change in the Montado. Agriculture, Ecossystems and Environment 137,
183-191, 2010.
CHAN-DZUL, A.; FINEGAN, B.; CASANOVES, F.; DELGADO, D. Aplicación del análisis
de varianza en el studio de la diversidad funcional. In: Validación y análisis de la diversidad
functional y su relación con los servicios ecossistémicos. CASANOVES, F.; PLA, L.; DI
RIENZO, J. A. (eds). Turrialba, Costa Rica: CATIE, 84 p., 2011.
CHAVE, J.; COOMES, D.; JANSEN, S.; LEWIS, S. L.; SWENSON, N. G; ZANNE, A. E.
Towards a worldwide wood economics spectrum. Ecology Letters, Montpellier, v. 12, p. 351-
366, 2009.
CUNNINGHAM, S. A.; SUMMERHAYES, B.; WESTOBY, M. Evolutionary divergences in
leaf structure and chemistry, comparing rainfall and soil nutrient gradients. Ecology 69: 569–
588, 1999.
DÍAZ, S. et al. The plant traits that drive ecosystems: Evidence from three continents. Journal
of Vegetation Science 15: 295-304, 2004.
FAHRIG, L. Effects of habitat fragmentation on biodiversity. Annual Reviews of Ecology and
Systematics, n. 34, p. 487-515, 2003.
FALSTER, D. S.; WESTOBY, M. Alternative height strategies among 45 dicot rain forest
species from tropical Queensland, Australia. Journal of Ecology, Oxford, v. 93, n. 3, p. 521-
535. 2005.

20
FORMAN, R. T. T. Land Mosaics. Cambridge University Press, Cambridge, UK, 1995.
FORMAN, R. T. T.; GODRON, M. Landscape ecology. New York: John Wiley & Sons. 619
p, 1986.
JAEGER, J. A. G. Landscape division, splitting index, and effective mesh size: new measures
of landscape fragmentation. Landscape Ecology, v. 15, p. 115 – 130, 2000.
LAMBERS, H.; POORTER, H. Inherent variation in growth rate between higher plants: a
search for physiological causes and ecological consequences. Advances in Ecological
Research, v. 23, p. 187 – 261, 1992.
LANG, S.; BLASCHKE, T. Análise da paisagem com SIG. Tradução Hermann Kux. São
Paulo: Oficina de Textos, 2009.
LAURANCE, W. F.; VASCONCELOS, H. L. Consequências ecológicas da fragmentação
florestal na Amazônia. Oecologia Brasiliensis. 13(3): 434-451, 2009.
LAVOREL, S.; McINTYRE, S.; LANDSBERG, J.; FORBES, T. D. A. Plant functional
classification: from general groups to specific groups based on response to disturbance. Tree,
v. 12, n. 12, 1997.
LIENIN, P., KLEYER, M. Plant leaf economics and reproductive investment are responsive to
gradients of land use intensity. Agric. Ecosyst. Environ. 145: 67–76, 2011.
MACHADO, E. L. M.; OLIVEIRA FILHO, A. T.; VAN DEN BERG, E.; CARVALHO, W.
A. C.; SOUZA, J. S.; MARQUES, J. J. G. S. M.; CALEGÁRIO, N. Efeitos do substrato, bordas
e proximidade espacial na estrutura da comunidade arbórea de um fragmento florestal em
Lavras, MG. Revista Brasil Bot., v.31, n. 2, p. 287-302, abr.-jun. 2008.
METZGER, J. P. Estrutura da Paisagem e Fragmentação: Análise Bibliográfica. Anais da
Academia Brasileira de Ciências, v. 71, n. 3-I, p.445-462, 1999.
METZGER, J. P. Tree Functional Group Richness and Landscape Structure in a Brazilian
Tropical Fragmented Landscape. Ecological Applications, 10(4), pp. 1147–1161, 2000.
METZGER, J. P. O que é ecologia de paisagens? Biota Neotropica. Vol.1. nº 1/2, 2001.
METZGER, J. P. Como restaurar a conectividade de paisagens fragmentadas? In: Restauração
Ecológica de Ecossistemas Naturais. KAGEYAMA, P. Y. et al. São Paulo: FEPAF, p. 49 –
76, 2008.
METZGER, J. P. Delineamento de Experimentos numa perspectiva de ecologia da paisagem.
In: CULLEN, Jr, RUDRAN, R.; VALLADARES-PADUA, C. Métodos de Estudos em
Biologia da Conservação e Manejo da Vida Silvestre. Curitiba: Editora UFPR, pp. 539-553,
2003a.
METZGER, J. P. Estrutura da Paisagem: o uso adequado de métricas. In: CULLEN, Jr,
RUDRAN, R.; VALLADARES-PADUA, C. Métodos de Estudos em Biologia da
Conservação e Manejo da Vida Silvestre. Curitiba: Editora UFPR, pp. 423-453, 2003b.
MISSIO, F. F. Traços funcionais e estratégias ecológicas de espécies arbóreas em Floresta
Ombrófila Mista no planalto catarinense. Universidade do Estado de Santa Catarina.
Dissertação de Mestrado em Engenharia Florestal. Santa Catarina, Lages, 2014.

21
MÜLLER, S. C.; OVERBECK, G. E.; PFADENHAUER, J.; PILLAR, V. D. Plant functional
types of woody species related to fire disturbance in forest–grassland ecotones. Plant Ecology,
Berlim, v. 189, n. 1, p. 1-14, 2007.
MURCIA C. Edge effects in fragmented forests: implications for conservation. Tree 10: 58-
62, 1995.
MYERS, A. N. M.; MITTERMEIER, C. G.; FONSECA, G. A. B. DA; KENT, J. Biodiversity
hotspots for conservation priorities. Nature, v. 403, p.853-858, 2000.
NASCIMENTO, H. E. M.; LAURANCE, W. F. Efeitos de área e de borda sobre a estrutura
florestal em fragmentos de floresta de terra firme após 13-17 anos de isolamento. Acta
Botanica. v. 36(2), p. 183-192, 2006.
NUNES, Y. R. F.; MENDONÇA, A. V. R.; BOTEZELLI, L.; MACHADO, E. L. M.;
OLIVEIRA-FILHO, A. T. Variações da fisionomia, diversidade e composição de guildas da
comunidade arbórea em um fragmento de floresta semidecidual em Lavras, MG. Acta bot.
bras. 17(2): 213-229, 2003.
OLIFIERS, N.; CERQUEIRA, R. Fragmentação de habitat: efeitos históricos e ecológicos. In:
Biologia da conservação: essências. ROCHA, C. F. D. et al. São Carlos: RiMa, p. 261-316,
2006.
OLIVEIRA, M. A.; GRILLO, S. A.; TABARELLI, M. Forest edge in the Brazilian Atlantic
Forest: drastic changes in tree species assemblages. Oryx, n. 38, p. 389-394, 2004.
PILLAR, V. D. On the identification of optimal plant functional types. Journal of Vegetation
Science, n. 10, p. 631-640, 1999.
RIBEIRO, M. C.; METZGER, J. P.; MARTENSEN, A. C.; PONZONI, F.; HIROTA, M. M.
The Brazilian Atlantic Forest: how much is left, and how is the remaining forest distributed?
Implications for conservation. Biological Conservation 142, 1141–1153, 2009.
SCARIOT, A.; FREITAS S. R.; NETO E. M. NASCIMENTO, M. T.; OLIVEIRA, L. C.;
SANAIOTTI T.; SERVILHA C.; VILLELA D. M. Vegetação e Flora. In RAMBALDI, D. M.;
OLIVEIRA, D. A. S. (orgs). Fragmentação de ecossistemas: causas, efeitos sobre a
diversidade e recomendações de políticas públicas. Brasilia: MMA/SBF, 508 p., 2003.
SOARES FILHO, B. S. Análise de Paisagem: fragmentação e mudanças. Instituto de
Geociências UFMG, 1998.
SOS MATA ATLÂNTICA; INPE. Atlas dos Remanescentes Florestais da Mata Atlântica,
Período 2012-2013. São Paulo: Fundação SOS Mata Atlântica / Instituto Nacional de Pesquisas
Espaciais, 61 p, 2014.
TABARELLI, M.; AGUIAR, A. V.; GRILLO, A. S. e SANTOS, A. M. M. Fragmentação e
perda de habitat na floresta Atlântica ao norte do Rio São Francisco. In: SIQUEIRA FILHO, J.
A.; LEME, E. M. C. (Org.). Fragmentos da Mata Atlântica do Nordeste: Biodiversidade,
Conservação e suas Bromélias. 1 ed. Rio de Janeiro: Andrea Jacobsson Estúdio Editorial Ltda,
p. 80 – 99, 2006.
TABARELLI, M.; MANTOVANI, W.; PERES, C. A. Effects of habitat fragmentation on plant
guild structure in the Montane Forest of Southeastern Brazil. Biological Conservation, 91:
119-127, 1999.

22
TURNER, M. G. G. GARDNER, R. H.; O’NEILL, R. V. Landscape and practice: pattern
and process. New York: Springer-Verlag, 401 p, 2001.
UZÊDA, M. C; FIDALGO, E. C. C.; IGUATEMY, M. A.; ALVES, R. C.; ROWS, J. R. C.
Explorando relações entre estrutura da paisagem e atributos de qualidade de fragmentos em
região de Mata Atlântica no Estado do Rio de Janeiro. Embrapa Solos, 2011.
VIANA, V. M.; TABANEZ, A. A. J. Biology and conservation of forest fragments in the
Brazilian atlantic moist forest. In: SCHELLAS, J.; GREENBERG, R. Forest patches in
tropical landscapes. Washington D.C.: Island Press, p. 151-167, 1996.
VIANA, V. M.; TABANEZ, A. A. J.; MARTINS, J. L. A. Restauração e manejo de fragmentos
florestais. In: Congresso Nacional Sobre Essências Nativas, 2, São Paulo, 1992. Anais. São
Paulo: Instituto Florestal de São Paulo. p. 400-407, 1992.
VIBRANS, A. C.; MCROBERTS, R. E.; MOSER, P.; NICOLETTI, A. L. Using satellite
image-based maps and ground inventory data to estimate the area of the remaining Atlantic
forest in the Brazilian state of Santa Catarina. Remote Sensing of Environment: 130: 87–95,
2013.
VILLARD, Marc-André; METZGER, J. P. Beyond the fragmentation debate: a conceptual
model to predict when habitat configuration really matters. Journal of Applied Ecology,
51:309-318, 2014.
VIOLLE, C.; NAVAS, M.; VILE, D.; KAZAKOU, E.; FORTUNEL, C.; HUMMEL, I.;
GARNIER, E. Let the concept of trait be functional! Oikos 116:882-892, 2007.
WESTOBY, M.; FALSTER, D. S.; MOLES, A. T.; VESK, P. A.; WRIGHT, I. J. Plant
Ecological Strategies: some leading dimensions of variation between species. Annu. Rev. Ecol.
Syst 33, 125-159, 2002.
WILCOVE, D. S.; MCLELLAN, C. H.; DOBSON, A. P. Habitat fragmentation in the
temperate zone. In Conservation Biology, ed. ME Soulé, pp. 237–56. Sunderland, MA:
Sinauer, 1986.
WRIGHT I. J. et al. The worldwide leaf economics spectrum. Nature 428(6985): 821–827,
2004.

23
CAPÍTULO I
Floresta Ombrófila Densa de Santa Catarina: a estrutura espacial dos remanescentes
florestais e os atributos funcionais das espécies arbóreas
RESUMO
A fragmentação florestal é, atualmente, a principal causa de perda de biodiversidade em
florestas tropicais. As espécies arbóreas de mesma estratégia de vida, que compõem
metacomunidades florestais, podem ser agrupadas funcionalmente, o que otimiza a percepção
entre vegetação e mudanças ambientais. Os objetivos deste trabalho são (i) descrever a condição
de fragmentação da Floresta Ombrófila Densa (FOD) de Santa Catarina, apontando as regiões
onde a fragmentação ocorreu de forma mais intensa, classificando-as em mais e menos
fragmentadas segundo as métricas de paisagem mais explicativas dos dados funcionais; (ii)
agrupar funcionalmente as espécies arbóreas mais abundantes da FOD e avaliar a influência das
métricas da paisagem sobre a abundância dos grupos funcionais identificados. Para isso,
analisou-se, de forma exploratória, a estrutura da paisagem utilizando métricas de 202
remanescentes florestais amostrados pelo Inventário Florístico Florestal de Santa Catarina
(IFFSC). Em seguida, foram determinadas as áreas mais e menos fragmentadas utilizando um
cluster construído com as métricas de paisagem mais explicativas dos dados funcionais. Para a
descrição dos atributos funcionais (altura máxima (hmáx), síndrome de dispersão, fenologia
foliar (FF), densidade da madeira (SSD), área foliar específica (SLA) e conteúdos de matéria
seca (LDMC), nitrogênio (LNC) e fósforo foliar (LPC)) foram selecionadas as 20 espécies
arbóreas mais abundantes da base de dados do IFFSC em toda área de estudo; para o
agrupamento funcional utilizou-se um dendrograma. Os modelos para estudar a influência das
métricas de paisagem na abundância dos grupos funcionais foram construídos por meio de GLS
e os melhores modelos selecionados pelo Critério de Informação de Akaike (AIC). Apesar dos
dados indicarem uma maior fragmentação nas microrregiões de Itajaí, Tubarão, Criciúma e
Ituporanga, os mesmos demonstram que, de forma geral, a área de FOD estudada ainda
apresenta uma paisagem com a presença de remanescentes grandes e conectados entre si. Por
meio da combinação dos atributos síndrome de dispersão, SLA, LDMC e FF foi possível
identificar quatro grupos funcionais com estratégias de vida distintas. A abundância do grupo
anemocóricas de intermediária eficiência na aquisição e conservação de recursos e de fenologia
foliar variada e do grupo zoocóricas aquisitivas perenifólias, foi significativamente afetada pela
estrutura da paisagem, ao contrário dos outros dois grupos (zoocóricas conservativas
perenifólias e zoocóricas de intermediária eficiência na aquisição e conservação de recursos e
de fenologia foliar variada) que mantiveram sua abundância independente das condições de
fragmentação.
Palavras-chave: Florestas tropicais. Fragmentação florestal. Métricas de paisagem.

24
ABSTRACT
Dense Ombrophylous Forest of Santa Catarina: the spatial structure of forest remnants
and functional traits of tree species. Forest fragmentation is currently the leading cause of
loss of biodiversity in tropical forests. Tree species of the same strategy of life, that make up
forest metacomunidades, can be grouped functionally, which optimizes the perception among
vegetation and environmental changes. Our objectives are twofold (i) to describe the
fragmentation of the Dense Ombrophylous Forest (FOD) of Santa Catarina, pointing out the
areas where fragmentation was more intense, classifying them into more and less fragmented
according to the landscape metrics that are explanatory of functional data; (ii) describe the
functional traits of the most abundant tree species on order to group them functionally and to
evaluate the influence of landscape metrics on the abundance of these functional groups.
Therefore, at an exploratory level, the landscape structure was described using metrics from
202 forest remnants sampled by Floristic and Forest Inventory of Santa Catarina. More and less
fragmented areas were identified by use of a cluster built with the most explanatory landscape
metrics of functional data. Functional traits like maximum height (hmax), dispersion syndrome,
leaf phenology (FF), wood density (SSD), specific leaf area (SLA), dry matter content (LDMC),
leaf nitrogen concentration (LNC) and leaf phosphorus concentration (LPC)) were extracted of
an existing database for the 20 most abundant tree species in the study area; the traits were
grouped by use of a dendrograma. The models used to study the influence of landscape metrics
on the abundance of functional groups were built by GLS, which allows the incorporation of
spatial structure; the best fitted models were selected by the Akaike Information Criterion
(AIC). Although the data indicate amore intense forest fragmentation in the regions of Itajaí,
Tubarão, Criciúma and Ituporanga, the data showed that, in general, the FOD study area still
presents a landscape with the presence of large and interconnected areas. Class metrics were
more significant for selecting traits than patch metrics, indicating that the structure of the set of
patches in a landscape is as important as the structure of a single patch for the functional
composition of communities. Combining the traits of dispersion syndrome, SLA, LDMC and
FF we identified four functional groups with different life strategies. The abundance of wind
dispersed of intermediate efficiency in the acquisition and conservation of resources and varied
leaf phenology group and zoochorous acquisitive evergreen group, was significantly affected
by landscape structure, unlike the other two groups (zoochorous conservative evergreen and
zoochorous intermediate efficiency acquisition and conservation of resources and varied leaf
phenology) that maintained their abundance independently of the fragmentation degree.
Keywords: Tropical forests. Forest fragmentation. Landscape metrics.

25
INTRODUÇÃO
O Estado de Santa Catarina com 95.346,18 km² de extensão está com todo o seu
território inserido no bioma Mata Atlântica. Sua cobertura florestal foi reduzida à 28,9% e está
em quinto lugar no ranking de desmatamento do bioma considerando os anos de 2012 e 2013,
segundo SOS Mata Atlântica e INPE (2014). No entanto, de acordo com os mesmos autores, é
o segundo Estado (que contempla o bioma) que possui maior área de remanescentes da Mata
Atlântica.
A mudança de uso do solo, com consequente fragmentação florestal, causa efeitos que
impedem o pleno desenvolvimento das florestas. Os mais discutidos e preocupantes efeitos da
fragmentação, de acordo com a literatura, são: a perda de diversidade genética (OLIFIERS e
CERQUEIRA, 2006); a redução do habitat e da interação com a fauna polinizadora e dispersora
(PARDINI et al., 2005; TABARELLI et al., 2006) e o efeito de borda, causado pela maior área
de contato com o ambiente externo (matriz), provocando o aumento da luminosidade e da
intensidade do vento, com aumento da temperatura e diminuição da umidade (MURCIA, 1995;
LAURANCE e VASCONCELOS, 2009). Logo, o tamanho, a forma, o número de fragmentos,
o tipo de vizinhança e demais características estruturais dos fragmentos em uma dada paisagem,
influenciarão no número de espécies capazes de sobreviver (VIANA et al., 1992).
O processamento e a interpretação de imagens de satélites e de fotografias aéreas são
técnicas que possibilitam uma análise mais criteriosa da paisagem e de seus componentes. Com
a utilização de métricas de paisagem é possível a quantificação das características estruturais
dos remanescentes florestais, por exemplo. Tornam-se então variáveis ambientais capazes de
explicar, por exemplo, variações na funcionalidade de metacomunidades vegetais.
No que diz respeito ao estudo da distribuição, diversidade e padrões de organização de
espécies arbóreas, atualmente propõe-se o estudo funcional das plantas como um diferencial às
medidas tradicionais de estudos da vegetação. Estes estudos analisam a vegetação por meio de
um conjunto de características intrínsecas das plantas (atributos funcionais), características
estas que podem ser morfológicas, fisiológicas ou fenológicas mensuráveis (VIOLLE et al.,
2007). As características estão envolvidas nas maneiras de adquirir, processar e investir os
recursos disponíveis (DÍAZ et al., 2004) e são manifestações das estratégias das plantas para
ultrapassarem filtros ambientais e competirem com outras espécies coexistentes.
Segundo Cianciaruso et al. (2009), informações de características funcionais das
espécies podem ser melhores que medidas tradicionais para muitas finalidades, permitem
comparações e generalizações significativas entre sítios com baixa afinidade taxonômica, já

26
que as espécies são analisadas por suas características e não por sua classificação taxonômica,
e ainda, identificar mais facilmente mudanças no ecossistema.
Em estudos recentes, focados principalmente em atributos reprodutivos e regenerativos
das plantas, encontrou-se a perda de riqueza e abundância de certos grupos funcionais, como
fator importante que influencia a funcionalidade dos ecossistemas (METZGER, 2000; GIRÃO
et al., 2007; LOPES et al., 2009; CARNEIRO, 2013; MAGNAGO et al., 2014).
Propõe-se no presente estudo uma análise de grupos funcionais que vão além destes
atributos, envolvendo também características foliares, de lenho e altura das plantas. Esperam-
se encontrar evidências de que a fragmentação florestal esteja atuando também sobre a seleção
de grupos que envolvam estratégias de captação e uso dos recursos (manifestos nas
características foliares e do lenho).
Ao considerar que (i) atributos funcionais estão associados às estratégias ecológicas,
que refletem o desempenho da planta às condições ambientais (REICH et al., 2003), (ii)
espécies podem ser agrupadas por compartilharem atributos similares (KEDDY, 1992), pois
apresentam respostas similares às mesmas variações ambientais, estabelecemos como hipótese
de trabalho que é possível detectar e quantificar os reflexos de diferentes graus de perturbação
do ambiente (na forma dos graus de fragmentação da paisagem) na abundância relativa dos
diferentes grupos na vegetação remanescente nestes ambientes. Considera-se que por meio
dessas análises criteriosas do papel dos atributos funcionais, seja possível entendermos melhor
as consequências das perturbações causadas pelo homem nas comunidades vegetais.
Os objetivos deste trabalho são (i) descrever a condição de fragmentação da Floresta
Ombrófila Densa (FOD) de Santa Catarina, apontando as regiões onde a fragmentação ocorreu
de forma mais intensa, classificando-as em mais e menos fragmentadas segundo as métricas de
paisagem mais explicativas dos dados funcionais; (ii) descrever os atributos funcionais das
espécies arbóreas mais abundantes, agrupando-as funcionalmente e avaliando a influência das
métricas da paisagem sobre a abundância dos grupos funcionais identificados.
MATERIAL E MÉTODOS
Área de estudo
O estudo foi realizado na área de ocorrência de Floresta Ombrófila Densa no Estado de
Santa Catarina, delimitada segundo o mapeamento de Klein (1978). A fitofisionomia está
situada entre as coordenadas 25º 57’ 40’’ e 29º 19’ 13’’ de latitude sul e 48º 24’ e 21’’ e 50º
14’ 14’’ de longitude oeste, em que sua área territorial original é de 29.282,00 km² (KLEIN,

27
1978), correspondendo a aproximadamente 31% do Estado, e que, atualmente, devido a um
histórico processo de fragmentação, restam apenas 12.632,7 km² (VIBRANS et al., 2013). A
altitude nas regiões de ocorrência da FOD, varia de alguns metros acima do nível do mar, a
pouco mais de 1.000 metros em alguns pontos.
Nessa região predomina o clima temperado úmido de verão quente (Cfa) (KOTTEK et
al., 2006), sem um período seco definido. As temperaturas médias anuais, dependendo da
altitude e da influência oceânica, chegam ao mínimo de 16ºC e ao máximo de 22ºC (NIMER,
1990). A precipitação apresenta grande variação, com registros de 1.250 a 2.000 mm, sendo
mais abundante ao norte (NIMER, 1990). Em consideração aos solos da região, predominam
os Cambissolos e Argissolos nas encostas e Gleissolos e Organossolos nas planícies
(EMBRAPA, 2006).
Este trabalho conta com informações da base de dados do Inventário Florístico Florestal
de Santa Catarina para a FOD. Este levantamento teve início em setembro de 2009 e foi
concluído em agosto de 2010. Foram ao todo 202 unidades amostrais analisadas, alocadas a
partir de uma rede de pontos sistematizada (grades) de dimensão 10 x 10 km. Foram compiladas
a lista florística com a abundância das espécies e as coordenadas geográficas das unidades
amostrais.
A distribuição das unidades amostrais no Estado e a cobertura florestal remanescente da
Floresta Ombrófila Densa em Santa Catarina (de acordo com o mapa de uso do solo realizado
pelo Projeto de Proteção a Mata Atlântica em Santa Catarina - PPMA/FATMA-SC –
GEOAMBIENTE, 2008) estão representadas na Figura 4, atentando para a divisão do Estado
em suas microrregiões, consideradas na discussão dos dados.

28
Figura 4: Mapa com a localização das 202 unidades amostrais referentes a Floresta Ombrófila Densa
(FOD), alocadas pelo Projeto Inventário Florístico-Florestal de Santa Catarina (IFFSC).
Cada unidade amostral possui 4.000 m², sendo implantada a partir de um ponto central,
quatro subunidades de 1.000 m² cada, orientadas na direção Norte, Sul, Leste e Oeste, distantes
30 m do ponto central. Em cada subunidade, de 20 m x 50 m, foi realizado o levantamento de
todos os indivíduos com mais de 1,5 metros de altura e com Diâmetro à Altura do Peito, (medido
a 1,30 m do solo) superior ou igual a 10 cm (DAP ≥ 10 cm) (Figura 5) (VIBRANS et al., 2012).

29
Fonte: Vibrans et al., 2012
Figura 5: Conglomerado utilizado para a amostragem da vegetação catarinense pelo Inventário Florístico
Florestal de Santa Catarina.
Estrutura espacial dos remanescentes florestais
Para a análise da estrutura dos remanescentes florestais, quantificada por meio de
métricas de paisagem, foi utilizado o mapa temático elaborado pelo Projeto de Proteção à Mata
Atlântica em Santa Catarina (PPMA/FATMA-SC), com base em 53 imagens multiespectrais
SPOT-4 do ano de 2005 na escala 1:50.000 (GEOAMBIENTE, 2008). A área mínima
considerada no mapeamento foi de 2,5 ha e, do total de usos do solo classificados (12), foi
considerado para este trabalho somente a classe ‘florestas em estágio médio ou avançado e/ou
primárias’. As métricas foram calculadas com auxílio do Software ArcGis 10.0, utilizando a
extensão V-LATE 1.1 (Vector-based Landscape Analysis Tools Extension) (LANG e TIEDE,
2003).
As métricas da paisagem foram subdivididas em métricas de classe e de mancha
(remanescente-foco). Os remanescentes-foco foram considerados aqueles onde estão inseridas
as unidades amostrais do IFFSC e as métricas medidas fazem referência a suas áreas, áreas
núcleo, formas e proximidades da mancha mais próxima.
A fim de obter métricas a nível de classe, ou seja, que envolvam outros fragmentos na
paisagem, demarcou-se para cada unidade amostral uma área circular a partir do ponto central
da unidade amostral do IFFSC, na qual foram identificadas todas as florestas em estágio médio
ou avançado e/ou primárias presentes. Para a demarcação da área circular no ArcGis utilizou-
se um buffer no entorno do ponto central do conglomerado do IFFSC. A distância determinada
(raio do buffer) foi de 2,8 km, seguindo a proposta do estudo multi-temporal de Schaadt (2012),
sendo a área do respectivo buffer possível de ser avaliada utilizando cinco fotografias aéreas.

30
Assim, foram determinadas ao todo 11 métricas (de classe e de mancha) em 202 recortes de
paisagem (buffer), com área aproximada de 2.462 ha, cada um (Tabela 1, Figura 6). Adotou-se
como borda a faixa marginal de 50 m de largura em cada fragmento avaliado (MURCIA, 1995).
Fonte: figura baseada na de Bernardo, 2012.
Figura 6: Representação do buffer de 2,8 km de raio, utilizado para o cálculo das métricas de paisagem,
estando representados, o remanescente-foco (mancha), no qual está localizada a Unidade Amostral do
IFFSC, remanescentes florestais inseridos na área buffer e que compõem a classe floresta, e as áreas não
florestais e de estágio sucessional inicial, não considerados no estudo.
Tabela 1: Métricas de paisagem a nível de classe e a nível de mancha, com seus respectivos significados
e unidades de medida.
CLASSE
Categoria Sigla Definição Unidade
ÁREA CA Área da classe ha
ÁREA NÚCLEO CAI Índice de área núcleo %
FORMA MSI Índice médio de forma ---
BORDA ED Densidade de borda m/ha
TE Total de borda m
SUBDIVISÃO DIVISION Índice de Divisão da Paisagem %
NP Número de manchas ---
MANCHA
Categoria Sigla Definição Unidade
ÁREA Area Área do fragmento ha

31
ÁREA NÚCLEO Área Core Área núcleo do fragmento ha
FORMA Shape_Idx Índice de Forma ---
PROXIMIDADE NNDist Distância do Vizinho mais próximo m
Seleção e caracterização funcional das espécies arbóreas
Da lista de 656 espécies da tabela fitossociológica do IFFSC, selecionou-se para a
caracterização dos atributos funcionais somente as 20 espécies lenhosas (DAP ≥ 10 cm) mais
abundantes (Tabela 2). Optou-se pela exclusão de samambaias e palmeiras da análise, pois
introduziriam um outro tipo de variabilidade aos resultados, já que utilizam de estratégias
específicas para a ocupação de ambientes e acabariam mascarando os padrões da grande
maioria das espécies.
Tabela 2: Espécies arbóreas mais abundantes pertencentes à Floresta Ombrófila Densa do Estado de
Santa Catarina.
FAMÍLIA ESPÉCIE
ANACARDIACEAE Tapirira guianensis Aubl.
ANNONACEAE Guatteria australis A.St.-Hil.
APOCYNACEAE Aspidosperma australe Müll.Arg.
ASTERACEAE Piptocarpha axillaris (Less.) Baker
CLETHRACEAE Clethra scabra Pers.
ELAEOCARPACEAE Sloanea guianensis (Aubl.) Benth.
EUPHORBIACEAE Alchornea triplinervia (Spreng.) Müll.Arg.
PERACEAE Pera glabrata (Schott) Poepp. ex Baill.
LAURACEAE Nectandra oppositifolia Nees
MELASTOMATACEAE Miconia cinnamomifolia (DC.) Naudin
MELIACEAE Cabralea canjerana (Vell.) Mart.
Cedrela fissilis Vell.
NYCTAGINACEAE Guapira opposita (Vell.) Reitz
PHYLLANTHACEAE Hieronyma alchorneoides Allemão
RUBIACEAE Bathysa australis (A.St.-Hil.) K.Schum.
Psychotria vellosiana Benth.
SALICACEAE Casearia sylvestris Sw.
SAPINDACEAE Matayba intermedia Radlk.
Cupania vernalis Cambess.
URTICACEAE Cecropia glaziovii Snethl.
Dos oito atributos funcionais analisados neste trabalho (Tabela 3), cinco foram
considerados por Cornelissen et al. (2003) como sendo relacionados com respostas a distúrbios,
que são: altura máxima da planta (hmáx), densidade da madeira (SSD), síndrome de dispersão
(Dispersão), fenologia foliar (FF) e conteúdo de nitrogênio e fósforo foliar (LNC e LPC). Os
outros, que são área foliar específica (SLA) e conteúdo de matéria seca foliar (LDMC),

32
envolvidos mais diretamente com variáveis climáticas e de disponibilidade de recursos no solo,
foram selecionados por entendermos que, de alguma forma, podem ser influenciados pelas
mudanças abióticas causados pelo efeito de borda no fragmento.
Os valores médios dos atributos por espécies foram disponibilizados pelo Departamento
de Ecologia da Universidade Federal do Rio Grande do Sul (UFRGS) (Apêndice – Tabela 1).
Tabela 3: atributos funcionais observados com suas respectivas siglas, unidade de medida (para variáveis
quantitativas) e categorias (variáveis qualitativas)
Atributo Sigla Unidade Variável Categórica
Altura Máxima Hmáx m -
Densidade da Madeira SSD g/cm³ -
Síndrome de Dispersão Dispersão categórica 0 - Anemocórica
1 - Zoocórica
Fenologia Foliar FF categórica
0 - Decídua
1 - Semidecídua
2 – Perenifólia
Concentração de Nitrogênio Foliar LNC mg/g -
Concentração de Fósforo Foliar LPC mg/g -
Atributo Sigla Unidade Variável Categórica
Área Foliar Específica SLA mm²/mg -
Conteúdo de Matéria Seca Foliar LDMC mg/g -
Análises realizadas para o 1º objetivo: descrever a condição de fragmentação, apontando as
regiões onde a fragmentação ocorreu de forma mais intensa
Foram confeccionados histogramas das frequências de cada uma das onze métricas, a
fim de observar, de forma geral, em que condições de fragmentação se encontram os
remanescentes florestais da Floresta Ombrófila Densa de Santa Catarina. Os resultados foram
comparados com os do estudo de Schaadt (2012), que analisou a estrutura da paisagem na
Floresta Ombrófila Mista de Santa Catarina.
Uma matriz de atributos médios por unidade amostral, ponderada pela densidade de
espécies (community-weighted mean trait matrix - CWM) (GARNIER et al., 2004) utilizada
para a o cruzamento com variáveis ambientais, foi obtida pela multiplicação da matriz de
abundância relativa de espécies por Unidade Amostral (UA x espécies) com a matriz funcional
(espécies x atributos), esse processo foi realizado utilizando a função dbFD do pacote FD no

33
programa estatístico R. Dessa forma, cada linha da matriz CWM possui médias de determinado
atributo em cada comunidade.
Um cluster com as métricas mais explicativas dos dados funcionais (matriz CWM),
selecionadas por meio da Seleção Progressiva de Variáveis Explicativas (Forward Selection of
Explanatory Variables, utilizando o pacote packfor do programa R) (BLANCHET et al., 2008),
foi utilizado para a separação das áreas analisadas em mais e menos fragmentadas. Boxplots
foram construídos para melhor visualização da distribuição dos valores de métricas em cada
um dos dois grupos, e um teste t para identificação de existência de diferenças significativas.
Análises realizadas para o 2º objetivo: agrupamento funcional e avaliação da influência de
métricas da paisagem sobre a abundância dos grupos funcionais identificados.
Foram definidos grupos funcionais a partir de um dendrograma construído por meio do
algoritmo de Ward, com base nos atributos avaliados. Para isto, aplicou-se o método de Gower
(GOWER, 1971), para a construção de uma matriz de dissimilaridades dos atributos funcionais
entre as espécies, que posteriormente foi transformada em distância Euclidiana pelo método de
Cailliez (CAILLIEZ, 1983).
Uma Análise de Variância para cada um dos atributos funcionais por grupo funcional
foi realizada, a fim de se identificar quais atributos estiveram significativamente envolvidos na
separação dos grupos, seguido pelo teste Tukey para averiguação dos grupos que se diferiam
em tal atributo. As análises, a partir da confecção do dendrograma até aqui foram realizadas
com o auxílio do Programa R (R DEVELOPMENT CORE TEAM, 2014).
Uma tabela de contingência, conjuntamente com uma análise de correspondência foi
utilizada para a classificação dos grupos funcionais, neste caso, para os dados categóricos de
síndrome de dispersão e fenologia foliar. Em seguida, foi realizada uma Análise de
Componentes Principais (PCA) para avaliar a correlação dos grupos formados com cada um
dos atributos analisados neste trabalho. Para essas últimas análises foi utilizado o programa
PAST (Paleontological Statistcs) (HAMMER et al., 2001).
Para analisar a relação das métricas de paisagem com a abundância das espécies em
cada grupo funcional foram empregados modelos lineares, usando o método dos mínimos
quadrados generalizados (GLS - Generalized Least Squares model), com a incorporação da
dependência espacial quando necessário. Foi construído um modelo global, com todas as
variáveis, após a remoção daquelas colineares (Fator de Inflação da Variância: VIF < 10), e
testada a autocorrelação espacial dos resíduos do modelo, utilizando a função “spline.correlog”

34
no pacote ncf do programa estatístico R com 1000 permutações (BJORNSTAD, 2008).
Identificada a autocorrelação no modelo global, incorporou-se a estrutura espacial no modelo
e, com base nisso, identificou-se o melhor modelo pelo o critério de informação de Akaike
(AIC), utilizando a biblioteca MASS e o comando stepAIC.
RESULTADOS E DISCUSSÃO
Estrutura da paisagem
As áreas dos remanescentes-foco estudados variaram de 11,1 a 2461,8 ha, a maioria com
uma área superior a 900 ha, distanciados da mancha florestal mais próxima em, no máximo,
262,8 m (Tabela 4).
Ao nível de classe, o menor coeficiente de variação foi obtido para índice de área núcleo
(CAI) e, a nível de mancha, para índice de forma (Shape_Idx), o que indica que para os buffers
analisados, as áreas efetivas de floresta, ou seja, livres do efeito de borda, se diferenciam pouco
entre si. Já para os remanescentes-foco avaliados, a dispersão dos dados foi menor para a forma
dos fragmentos.
Ao nível de mancha, a maior dispersão de dados foi observada para a distância do
vizinho mais próximo (NNDist). A maior variação desta métrica pode ser explicado pelo fato
de, em alguns casos, os remanescentes-foco terem sido os únicos representantes de floresta no
buffer, não existindo assim manchas vizinhas, sendo atribuído valor zero à esta métrica. Em
20% das áreas, a mancha foco representou toda a classe floresta, com a métrica NP (número de
manchas) assumindo o valor um e a métrica Índice de Divisão da Paisagem (DIVISION) o valor
zero.
Tabela 4: Valores das métricas de paisagem a nível de classe e mancha, obtidos para 202 áreas analisadas
na Floresta Ombrófila Densa de Santa Catarina.
CLASSE
Categoria Sigla Definição Unidade Mínimo Máximo Média CV%*
ÁREA CA Área da classe ha 170,1 2461,8 1597,8 101,46
ÁREA NÚCLEO CAI Índice de área
núcleo % 29,9 96,45 74,91 19
FORMA MSI Índice médio de
forma --- 1,002 7,366 2,426 41
BORDA ED
Densidade de
borda m/ha 7,16 176,04 54,84 61,05
TE Total de borda m 17628,83 134545,5 72214,42 37,3
SUBDIVISÃO
DIVISION Índice de divisão
da paisagem --- 0 92,14 17,48 148,89
NP Número de
manchas --- 1 34 7,51 101,46

35
MANCHA
Categoria Sigla Definição Unidade Mínimo Máximo Média CV%*
ÁREA Area Área do
fragmento ha 11,1 2461,8 1458,6 48,5
ÁREA NÚCLEO Área Core Área núcleo do
fragmento ha 0,23 2374,42 1045,75 69,84
FORMA Shape_Idx Índice de forma
do fragmento --- 1,002 9,387 4,271 43,53
PROXIMIDADE NNDist
Distância do
vizinho mais
próximo
m 0 262,88 48,82 89,47
*CV%: Coeficiente de Variação expresso em porcentagem.
Os resultados evidenciariam que ainda existe a predominância de grandes
remanescentes de Floresta Ombrófila Densa no Estado. A maior parte das áreas avaliadas (72%)
apresentou remanescentes maiores do que 1.000 ha. Os remanescentes considerados médios
(>100 ha e <1000 ha) representaram 22% dos casos, e os pequenos remanescentes (<100 ha),
somente 6%.
Em 75% das áreas avaliadas, o número de manchas (NP) apresentou valores de 1 a 10.
Neste caso, a ocorrência de poucas manchas não indica a escassez de floresta, mas a existência
de grandes remanescentes que cobrem quase a totalidade do buffer. Este fato é evidenciado
também quando analisado a porcentagem de floresta no buffer (CA/área total do buffer*100),
em que 70% dos casos apresentaram mais da metade da área analisada coberta por florestas
(Figura 7).
O número de manchas variou de 1 a 34, sendo que somente 8% das áreas apresentaram
NP>20, sugerindo baixa segmentação da classe. No estudo de Schaadt (2012), que analisou as
mesmas métricas para a Floresta Ombrófila Mista, no Estado de Santa Catarina, o número de
manchas na mesma área de buffer variou de 1 a 45, com maior segmentação quando comparado
à FOD, sendo que cerca de 50% das áreas eram compostas por 5 a 19 fragmentos. Na Floresta
Ombrófila Densa, 55% das áreas pertencem a classe de 2 a 10 fragmentos.

36
Figura 7: Histograma do Número de Manchas (NP) e porcentagem de floresta nas 202 áreas analisadas
de Floresta Ombrófila Densa no Estado de Santa Catarina.
Em relação à área núcleo da mancha, ou seja, a área efetiva de floresta retiradas as
bordas, foi também um indicador de grandes florestas, sendo que, 10% dos remanescentes-foco
avaliados possuem mais de 2000 hectares de área núcleo, área pouco menor a área total do
buffer de 2.462 ha.
Por consequência dos grandes e muitos remanescentes, 95% dos buffers analisados
apresentaram a porcentagem de área núcleo da classe floresta (CAI) maior que 50%, ou seja,
grande porcentagem de área de interior, que podem apresentar condições abióticas menos
severas, importantes para a manutenção da biodiversidade (Figura 8).
Figura 8:Histogramas da área do remanescente-foco e da porcentagem de área núcleo (CAI) para a classe
floresta das 202 áreas analisadas na Floresta Ombrófila Densa no Estado de Santa Catarina.
A classe floresta apresentou forma pouco complexa, com 44% das áreas com MSI entre
1 e 2, o que indica fragmentos menos irregulares. Porém, ao se analisar o índice de forma para
os remanescentes-foco (Shape_Idx), este variou de manchas mais circulares, quando os valores
estão próximos a um, até manchas bastantes complexas em forma, quando os valores estão mais
distantes de um (Figura 9).
0%
10%
20%
30%
40%
50%
60%
1 2 a 10 11 a 20 >20
Número de manchas - NP
0%
10%
20%
30%
40%
0-20 20-40 40-60 60-80 >80
Porcentagem de floresta
MANCHA CLASSE
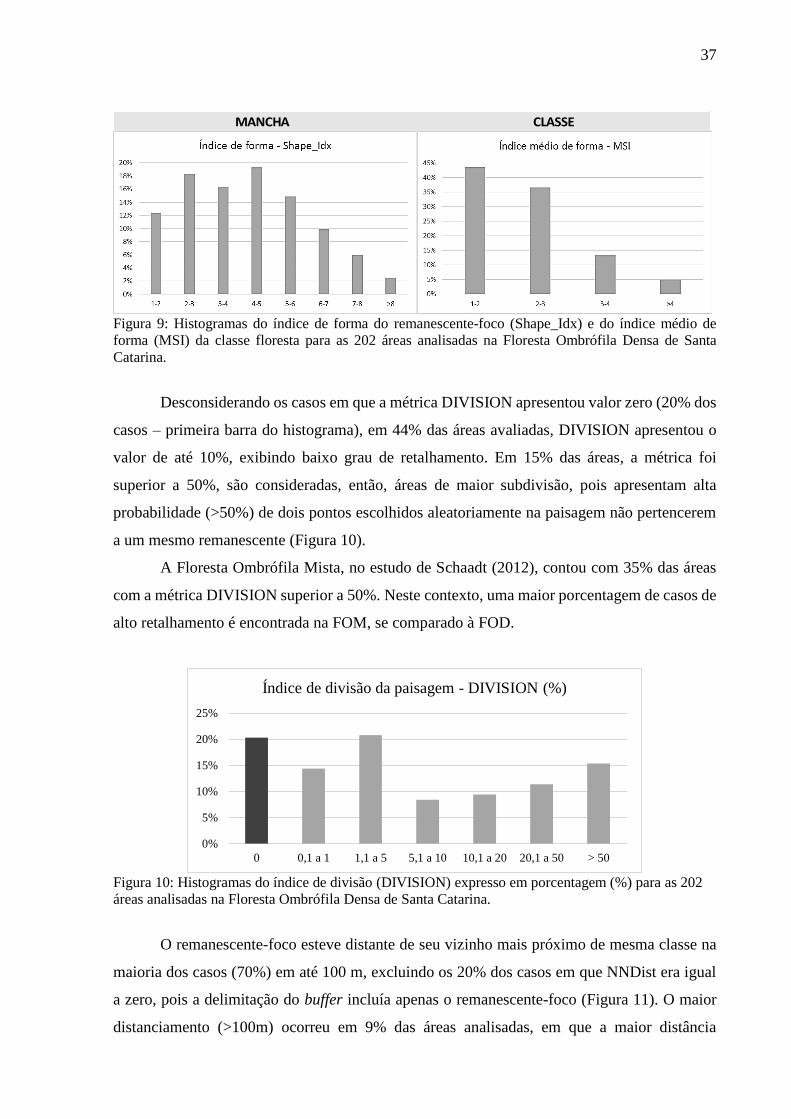
37
Figura 9: Histogramas do índice de forma do remanescente-foco (Shape_Idx) e do índice médio de
forma (MSI) da classe floresta para as 202 áreas analisadas na Floresta Ombrófila Densa de Santa
Catarina.
Desconsiderando os casos em que a métrica DIVISION apresentou valor zero (20% dos
casos – primeira barra do histograma), em 44% das áreas avaliadas, DIVISION apresentou o
valor de até 10%, exibindo baixo grau de retalhamento. Em 15% das áreas, a métrica foi
superior a 50%, são consideradas, então, áreas de maior subdivisão, pois apresentam alta
probabilidade (>50%) de dois pontos escolhidos aleatoriamente na paisagem não pertencerem
a um mesmo remanescente (Figura 10).
A Floresta Ombrófila Mista, no estudo de Schaadt (2012), contou com 35% das áreas
com a métrica DIVISION superior a 50%. Neste contexto, uma maior porcentagem de casos de
alto retalhamento é encontrada na FOM, se comparado à FOD.
Figura 10: Histogramas do índice de divisão (DIVISION) expresso em porcentagem (%) para as 202
áreas analisadas na Floresta Ombrófila Densa de Santa Catarina.
O remanescente-foco esteve distante de seu vizinho mais próximo de mesma classe na
maioria dos casos (70%) em até 100 m, excluindo os 20% dos casos em que NNDist era igual
a zero, pois a delimitação do buffer incluía apenas o remanescente-foco (Figura 11). O maior
distanciamento (>100m) ocorreu em 9% das áreas analisadas, em que a maior distância
MANCHA CLASSE
0%
5%
10%
15%
20%
25%
0 0,1 a 1 1,1 a 5 5,1 a 10 10,1 a 20 20,1 a 50 > 50
Índice de divisão da paisagem - DIVISION (%)
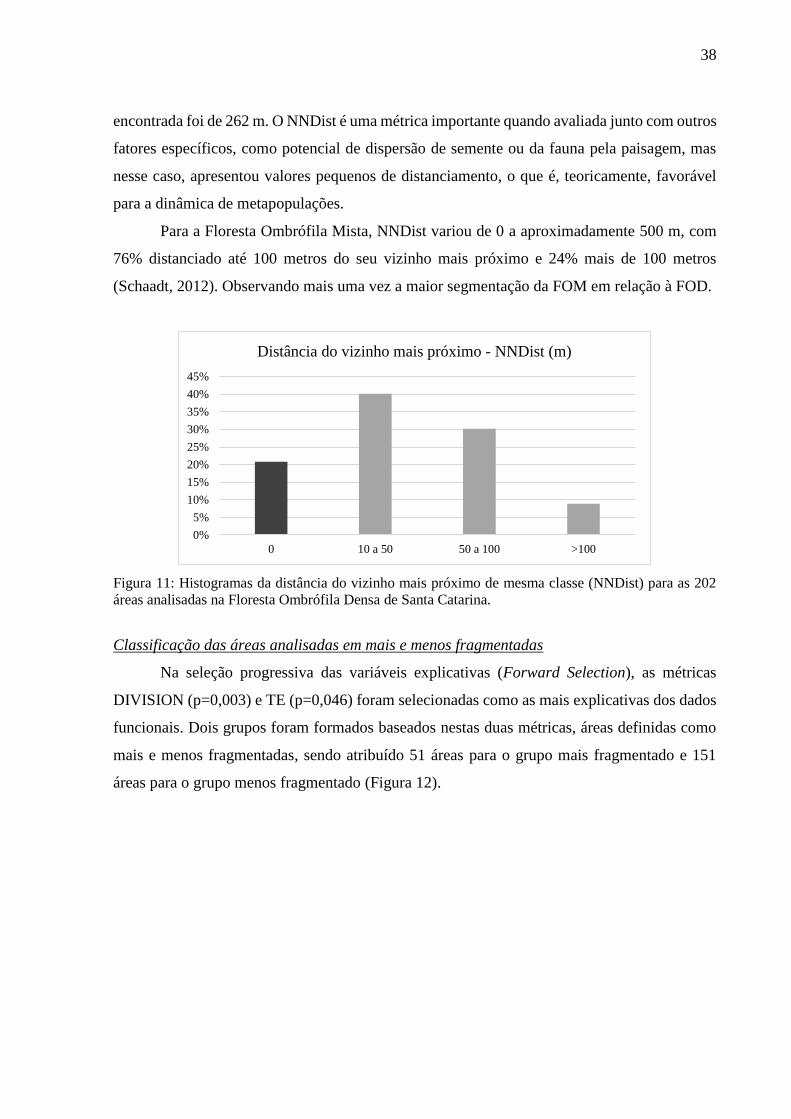
38
encontrada foi de 262 m. O NNDist é uma métrica importante quando avaliada junto com outros
fatores específicos, como potencial de dispersão de semente ou da fauna pela paisagem, mas
nesse caso, apresentou valores pequenos de distanciamento, o que é, teoricamente, favorável
para a dinâmica de metapopulações.
Para a Floresta Ombrófila Mista, NNDist variou de 0 a aproximadamente 500 m, com
76% distanciado até 100 metros do seu vizinho mais próximo e 24% mais de 100 metros
(Schaadt, 2012). Observando mais uma vez a maior segmentação da FOM em relação à FOD.
Figura 11: Histogramas da distância do vizinho mais próximo de mesma classe (NNDist) para as 202
áreas analisadas na Floresta Ombrófila Densa de Santa Catarina.
Classificação das áreas analisadas em mais e menos fragmentadas
Na seleção progressiva das variáveis explicativas (Forward Selection), as métricas
DIVISION (p=0,003) e TE (p=0,046) foram selecionadas como as mais explicativas dos dados
funcionais. Dois grupos foram formados baseados nestas duas métricas, áreas definidas como
mais e menos fragmentadas, sendo atribuído 51 áreas para o grupo mais fragmentado e 151
áreas para o grupo menos fragmentado (Figura 12).
0%
5%
10%
15%
20%
25%
30%
35%
40%
45%
0 10 a 50 50 a 100 >100
Distância do vizinho mais próximo - NNDist (m)

39
Figura 12: Mapa com a localização das unidades amostrais classificadas em mais e menos fragmentadas
na Floresta Ombrófila Densa de Santa Catarina
A seleção dessas duas métricas como as mais explicativas foi coerente, se pensarmos na
sensibilidade das florestas úmidas, neste caso a Floresta Ombrófila Densa, às mudanças
abióticas de borda (SAMPAIO e SCARIOT, 2011), apresentando espécies adaptadas a uma
condição de maior umidade. Mais precisamente, na borda observa-se um acréscimo na
temperatura do ar e do solo, na velocidade do vento e no déficit de pressão de vapor (relacionada
com a capacidade de absorção de água pelo ar) e um decréscimo na umidade do ar e do solo, se
comparado com o interior da floresta (POHLMAN et al., 2009). Estas alterações
microclimáticas condicionam efeitos na fisiologia das plantas (HERBST et al., 2007), que
possuem sensíveis estratégias para controle de água em seu sistema (RODRIGUES et al., 2011).
Se considerarmos estes fatos, a pressão pela seleção de espécies menos sensíveis a alterações
ambientais é possivelmente mais significativa na FOD, já que apresenta espécies com
características funcionais condizentes a ambientes mais úmidos.

40
As 202 áreas avaliadas estavam distribuídas em 13 microrregiões do Estado, e 11 delas
tiveram pelo menos uma área classificada como mais fragmentada, com maior destaque para
Criciúma e Itajaí, com 67% das áreas avaliadas em condição de maior fragmentação, além de
Tubarão com 53% e Ituporanga com 44%. Esse fato pode ser causado pelo desenvolvimento
urbano, principalmente no caso da microrregião de Itajaí, e às atividades econômicas, como
mineração e agricultura, para as outras regiões. A microrregião de Tijucas teve 16 áreas
avaliadas, mas nenhuma pertencendo ao grupo mais fragmentado.
As áreas classificadas como menos fragmentadas estiveram localizadas principalmente
próximas a regiões de maior relevo (como é o caso das áreas do limite Sul da Floresta Ombrófila
Densa com a Floresta Ombrófila Mista) e também próximas e dentro de Unidades de
Conservação. As áreas florestais em relevo mais acidentado, ainda são as áreas mais protegidas,
porém, devido as já saturadas áreas planas - pela urbanização e atividades econômicas -, fazem
com que o avanço das atividades humanas em áreas de maior relevo seja cada vez mais intenso,
se não tomadas medidas para a sua proteção. Mostra-se também, mais uma vez, a importância
de Unidades de Conservação para a preservação de ecossistemas, favorecendo, por
consequência, as pesquisas científicas e a educação ambiental, importantes para o
conhecimento e conscientização.
Algo importante a ser considerado é a forma da classe (MSI), que para as áreas “menos
fragmentadas” apresentou maiores valores do que para as áreas menos fragmentadas. Verificou-
se que grandes remanescentes, permeados por outros usos do solo (áreas de avanço de
atividades antrópicas), há maior complexidade de forma e, consequentemente, maior contato
com a matriz. Em áreas mais fragmentadas, com atividades antrópicas já consolidadas, os
remanescentes eram menores e de formato mais regular (Figura 13). É característico de
paisagens com maior interferência humana, que os componentes da paisagem apresentem
formas geométricas definidas, como círculos, retângulos e/ou quadrados.

41
Figura 13: Boxplots dos valores de cada métrica de paisagem, a nível de classe e mancha, analisadas na
Floresta Ombrófila Densa de Santa Catarina nos grupos mais e menos fragmentado. 1: Métricas de área
florestal; 2: métricas de borda; 3: métricas de divisão; 4: distância do vizinho mais próximo; 5: métricas
de forma. n = 202, t = valor do teste t, *P <0,05, ***P<0,001, ns = não significativo.
Atributos funcionais e estratégia das plantas
As espécies selecionadas pertencem a 18 famílias botânicas e 20 gêneros distintos,
compreendendo duas espécies tardias (10%), 14 espécies secundárias (70%) e quatro pioneiras
(20%) (Apêndice). A abundância variou de 5,87 a 20,83 indivíduos por hectare, com a menor
abundância para a Cedrela fissilis e a maior para Alchornea triplinervia.
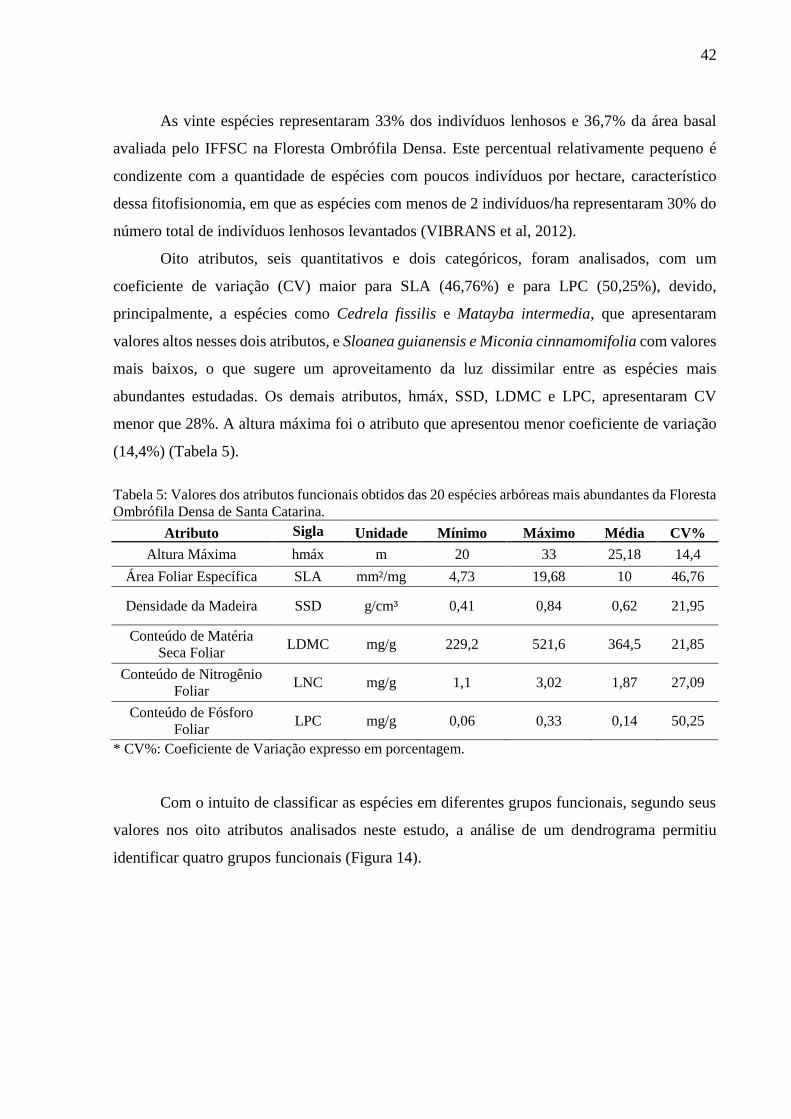
42
As vinte espécies representaram 33% dos indivíduos lenhosos e 36,7% da área basal
avaliada pelo IFFSC na Floresta Ombrófila Densa. Este percentual relativamente pequeno é
condizente com a quantidade de espécies com poucos indivíduos por hectare, característico
dessa fitofisionomia, em que as espécies com menos de 2 indivíduos/ha representaram 30% do
número total de indivíduos lenhosos levantados (VIBRANS et al, 2012).
Oito atributos, seis quantitativos e dois categóricos, foram analisados, com um
coeficiente de variação (CV) maior para SLA (46,76%) e para LPC (50,25%), devido,
principalmente, a espécies como Cedrela fissilis e Matayba intermedia, que apresentaram
valores altos nesses dois atributos, e Sloanea guianensis e Miconia cinnamomifolia com valores
mais baixos, o que sugere um aproveitamento da luz dissimilar entre as espécies mais
abundantes estudadas. Os demais atributos, hmáx, SSD, LDMC e LPC, apresentaram CV
menor que 28%. A altura máxima foi o atributo que apresentou menor coeficiente de variação
(14,4%) (Tabela 5).
Tabela 5: Valores dos atributos funcionais obtidos das 20 espécies arbóreas mais abundantes da Floresta
Ombrófila Densa de Santa Catarina.
Atributo Sigla Unidade Mínimo Máximo Média CV%
Altura Máxima hmáx m 20 33 25,18 14,4
Área Foliar Específica SLA mm²/mg 4,73 19,68 10 46,76
Densidade da Madeira SSD g/cm³ 0,41 0,84 0,62 21,95
Conteúdo de Matéria
Seca Foliar LDMC mg/g 229,2 521,6 364,5 21,85
Conteúdo de Nitrogênio
Foliar LNC mg/g 1,1 3,02 1,87 27,09
Conteúdo de Fósforo
Foliar LPC mg/g 0,06 0,33 0,14 50,25
* CV%: Coeficiente de Variação expresso em porcentagem.
Com o intuito de classificar as espécies em diferentes grupos funcionais, segundo seus
valores nos oito atributos analisados neste estudo, a análise de um dendrograma permitiu
identificar quatro grupos funcionais (Figura 14).
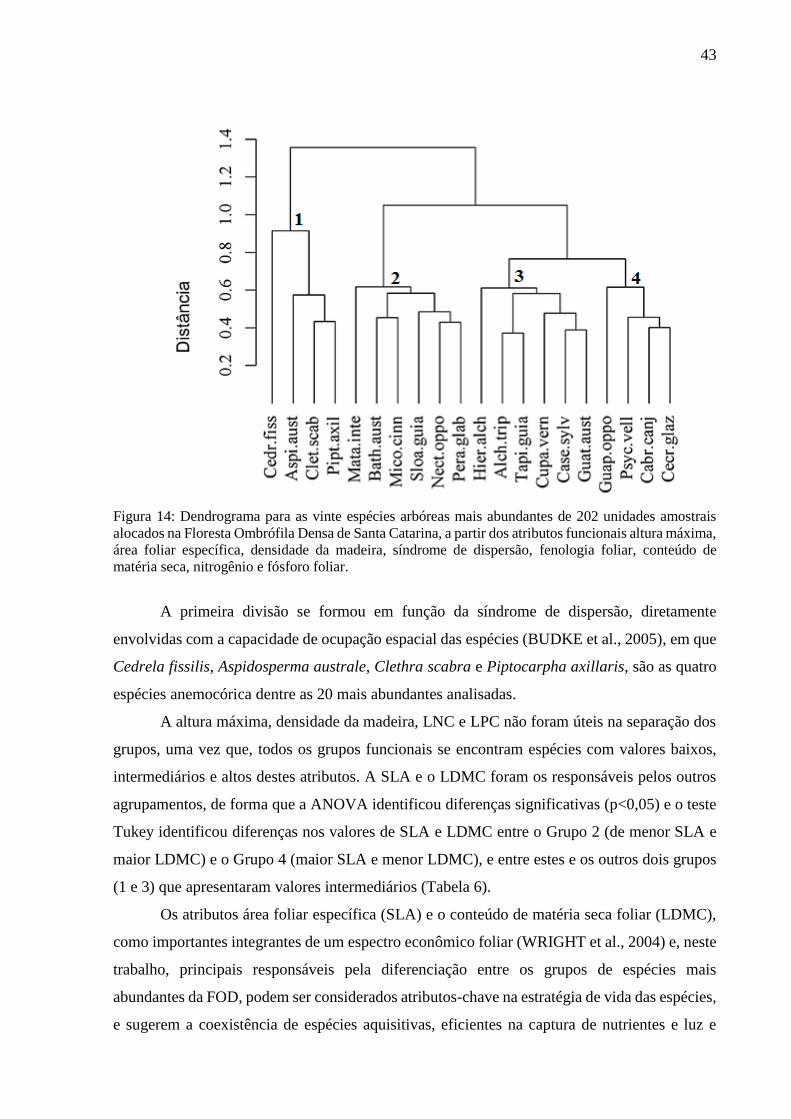
43
Figura 14: Dendrograma para as vinte espécies arbóreas mais abundantes de 202 unidades amostrais
alocados na Floresta Ombrófila Densa de Santa Catarina, a partir dos atributos funcionais altura máxima,
área foliar específica, densidade da madeira, síndrome de dispersão, fenologia foliar, conteúdo de
matéria seca, nitrogênio e fósforo foliar.
A primeira divisão se formou em função da síndrome de dispersão, diretamente
envolvidas com a capacidade de ocupação espacial das espécies (BUDKE et al., 2005), em que
Cedrela fissilis, Aspidosperma australe, Clethra scabra e Piptocarpha axillaris, são as quatro
espécies anemocórica dentre as 20 mais abundantes analisadas.
A altura máxima, densidade da madeira, LNC e LPC não foram úteis na separação dos
grupos, uma vez que, todos os grupos funcionais se encontram espécies com valores baixos,
intermediários e altos destes atributos. A SLA e o LDMC foram os responsáveis pelos outros
agrupamentos, de forma que a ANOVA identificou diferenças significativas (p<0,05) e o teste
Tukey identificou diferenças nos valores de SLA e LDMC entre o Grupo 2 (de menor SLA e
maior LDMC) e o Grupo 4 (maior SLA e menor LDMC), e entre estes e os outros dois grupos
(1 e 3) que apresentaram valores intermediários (Tabela 6).
Os atributos área foliar específica (SLA) e o conteúdo de matéria seca foliar (LDMC),
como importantes integrantes de um espectro econômico foliar (WRIGHT et al., 2004) e, neste
trabalho, principais responsáveis pela diferenciação entre os grupos de espécies mais
abundantes da FOD, podem ser considerados atributos-chave na estratégia de vida das espécies,
e sugerem a coexistência de espécies aquisitivas, eficientes na captura de nutrientes e luz e

44
espécies conservativas de baixa produtividade, com estratégias de conservação de água e
nutrientes (WRIGHT et al., 2004).
Tabela 6: Análise de Variância para os atributos de área foliar específica (SLA), conteúdo de matéria
seca foliar (LDMC), altura máxima (Hmáx), densidade da madeira (SSD), conteúdo de nitrogênio foliar
(LNC) e conteúdo de fósforo foliar (LPC) por grupo funcional.
Grupos
Funcionais SLA
(mm²/mg) LDMC (mg/g)
Hmáx (m)
SSD (g/cm³)
LNC (mg/g)
LPC (mg/g)
F=8,07
p=0,001
F=3,6
p=0,037
F=1,041
p=0,4
F=2,54
p=0,09
F=2,17
p=0,13
F=0,03
p=0,99
1 10,86 b 361,59 b 27,5 0,54 1,94 0,14
2 5,49 a 401,44 c 25 0,73 1,53 0,13
3 10,2 b 392,38 b 25,2 0,60 1,90 0,14
4 15,6 c 270,04 a 23 0,58 2,28 0,14
* Médias seguidas de mesma letra não diferem estatisticamente entre si, pelo teste de Tukey, α≤0,05.
As variáveis categóricas, fenologia foliar e agente dispersor, foram úteis ao mostrar
associações significativas (p<0,0001, nos dois casos) por meio da tabela de contingência
(Tabela 7).
Tabela 7: Frequências absolutas, com valores percentuais entre parênteses, por grupo funcional para os
atributos categóricos fenologia foliar e síndrome de dispersão, com os resultados da análise da tabela de
contingência.
Nº de
Espécies Grupo
1 Grupo
2 Grupo
3 Grupo
4 X² p
Fenologia
Foliar
Perenefólia 12 1 (8,3) 4 (33,3) 3 (25) 4 (33,3)
137,4 <0,0001 Semidecídua 6 2 (33,3) 2 (33,3) 2 (33,3) 0 (0)
Decídua 2 1 (50) 0 (0) 1 (50) 0 (0)
Nº de
Espécies Grupo
1 Grupo
2 Grupo
3 Grupo
4 X² p
Síndrome de
Dispersão
Anemocórica 4 4 (100) 0 (0) 0 (0) 0 (0) 199,0
<
0,0001 Zoocórica 16 0 (0) 6 (37,5) 6 (37,5) 4 (25)
Uma análise de correspondência permitiu visualizar as associações dos atributos
categóricos com os grupos funcionais (Figura 15), com uma explicação de 93,72% da variação
encontrada (eixo 1: 77,83% e eixo 2: 15,89%). Ao longo do eixo 1, que foi o mais explicativo,
observa-se, em um extremo, associado a valores negativos, os grupos 2 e 4, formados
predominantemente por espécies zoocóricas e perenifólias. No outro extremo, associado a
valores positivos, ocorreu o Grupo 1, formado por espécies anemocóricas. O Grupo 3 ordenou-
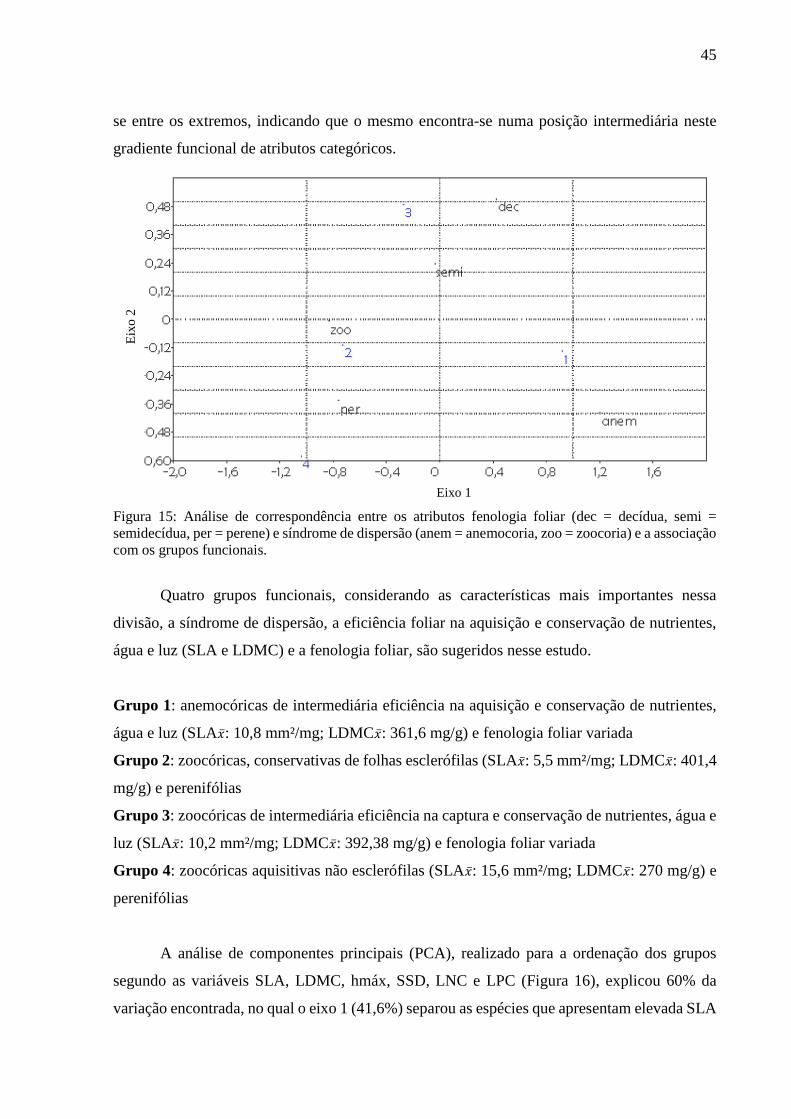
45
Eixo 1
Eix
o 2
se entre os extremos, indicando que o mesmo encontra-se numa posição intermediária neste
gradiente funcional de atributos categóricos.
Figura 15: Análise de correspondência entre os atributos fenologia foliar (dec = decídua, semi =
semidecídua, per = perene) e síndrome de dispersão (anem = anemocoria, zoo = zoocoria) e a associação
com os grupos funcionais.
Quatro grupos funcionais, considerando as características mais importantes nessa
divisão, a síndrome de dispersão, a eficiência foliar na aquisição e conservação de nutrientes,
água e luz (SLA e LDMC) e a fenologia foliar, são sugeridos nesse estudo.
Grupo 1: anemocóricas de intermediária eficiência na aquisição e conservação de nutrientes,
água e luz (SLA�̅�: 10,8 mm²/mg; LDMC�̅�: 361,6 mg/g) e fenologia foliar variada
Grupo 2: zoocóricas, conservativas de folhas esclerófilas (SLA�̅�: 5,5 mm²/mg; LDMC�̅�: 401,4
mg/g) e perenifólias
Grupo 3: zoocóricas de intermediária eficiência na captura e conservação de nutrientes, água e
luz (SLA�̅�: 10,2 mm²/mg; LDMC�̅�: 392,38 mg/g) e fenologia foliar variada
Grupo 4: zoocóricas aquisitivas não esclerófilas (SLA�̅�: 15,6 mm²/mg; LDMC�̅�: 270 mg/g) e
perenifólias
A análise de componentes principais (PCA), realizado para a ordenação dos grupos
segundo as variáveis SLA, LDMC, hmáx, SSD, LNC e LPC (Figura 16), explicou 60% da
variação encontrada, no qual o eixo 1 (41,6%) separou as espécies que apresentam elevada SLA

46
e LNC e baixo LDMC, isto é, espécies com folha de vida mais curta e maior capacidade
fotossintética (WESTOBY et al. 2002), daquelas que geralmente estão associadas a ambientes
oligotróficos e com stress hídrico (menor SLA e LNC e alto LDMC) (ACKERLY et al., 2002),
de concentração elevada de compostos secundários na parede celular, protegendo contra fatores
abióticos e bióticos nocivos, mas prejudicando a taxa fotossintética da planta (LAMBERS e
POORTER, 1992).
O Grupo 4 apresentou um média de altura inferior aos outros três grupos (23 m) e forte
correlação com os atributos SLA e LNC. Este grupo podendo ser também denominado como
grupo de árvores frutíferas de sub-bosque, importante fonte de alimento para animais de hábito
terrícola.
Figura 16: Análise de componentes principais para ordenação dos grupos funcionais por meio dos
atributos área foliar específica (SLA), conteúdo de matéria seca foliar (LDMC), altura máxima (Hmáx),
densidade da madeira (SSD), conteúdo de nitrogênio e fósforo foliar (LNC e LPC).
O Grupo 2 ordenou-se ao lado negativo do primeiro eixo, com correlações positivas
com o conteúdo de matéria seca foliar – LDMC e com a densidade da madeira - SSD, com uma
estratégia voltada para a longevidade das folhas e resistência a fatores bióticos e abióticos
nocivos e em menor grau voltada para a capacidade fotossintética.
hmáx
SLA
LDMCSSD
LNC
LPC Cedr.fiss
Aspi.aust
Clet.scab
Pipt.axil
Mata.inte
Bath.aust
Mico.cinn
Sloa.guia
Nect.oppo
Pera.glab
Hier.alch
Alch.trip
Tapi.guia
Cupa.vern
Case.sylv
Guat.aust
Guap.oppo
Psyc.vell
Cabr.canj
Cecr.glaz
-2,4 -1,6 -0,8 0,8 1,6 2,4 3,2 4,0
Component 1
-2,0
-1,5
-1,0
-0,5
0,5
1,0
1,5
2,0
2,5
Co
mp
one
nt
2

47
O Grupo 3, de forma geral, apresentou valores intermediários em todos os atributos em
relação aos três outros grupos, não se destacando em nenhum quesito. Tapirira guianensis e
Alchornea triplinervia, pertencentes a este grupo, são conhecidas por serem espécies
generalistas e que se sobressaem em condições mais severas de perturbação. No cenário atual
de fragmentação, as mesmas são frequentes e abundantes na floresta Atlântica, como observado
por Oliveira et al. (2004).
O cedro (Cedrela fissilis) foi a espécie que mais se diferenciou dentre as 20 analisadas,
por apresentar uma estratégia de alta capacidade fotossintética, diferenciando-se das outras
espécies, principalmente, por apresentar alta SLA (19,68 mm²/mg), que correspondeu ao valor
máximo encontrado, e o que fez com que o Grupo 1, ao qual pertence, apresentasse, em média,
valores intermediários nos atributos foliares analisados. Se não considerada a Cedrela fissilis,
o grupo apresentaria uma baixa altura potencial e alta relação com o atributo LDMC. Por esse
motivo, o grupo parece estar mais caracterizado pela síndrome de dispersão anemocórica à qual
pertence, que pelos outros atributos considerados.
Outra espécie que merece destaque neste sentido é a Sloanea guianensis que, ao
contrário de C. fissilis, apresentou o menor valor para SLA e ainda o maior para LDMC,
indicando uma espécie esclerófila característica, de maior resistência e longevidade foliar.
Relação entre os grupos funcionais e a estrutura da paisagem
Os melhores modelos GLS selecionados pelo Critério de Informação de Akaike (AIC)
(Tabela 8) mostram que o Grupo 1 e 4 tem sua abundância influenciada significativamente pela
estrutura da paisagem, ao contrário dos grupos 3 e 2, que não apresentaram relação de sua
abundância com o gradiente estrutural da paisagem.
O Grupo 1 foi influenciado de forma significativa, com relações positivas, pela
complexidade de forma e área florestal total da paisagem (MSI e CA) e, com relações negativas,
pelo tamanho da área núcleo do remanescente. Como observado anteriormente, na classificação
das áreas em mais e menos fragmentadas, as áreas menos fragmentadas foram as que
apresentaram maior complexidade de forma (>MSI), o que leva a interpretação de que estas
paisagens mais naturais, que sofrem mais com o avanço das atividades antrópicas do que
paisagens já mais alteradas, são as áreas em que o Grupo 1 é mais presente.
O Grupo 4 foi positivamente influenciado pela área total de floresta na paisagem e pela
forma da mancha (Shape_Idx), o que demostra que o grupo de espécies zoocóricas de
capacidade fotossintética alta e perenifólias são mais abundantes em paisagens mais florestais

48
e em remanescentes de forma mais complexa. Um resultado semelhante foi encontrado no
trabalho de Metzger (2000), que identificou a influência da complexidade da borda e do
tamanho do fragmento para o grupo de espécies classificadas como tolerantes intermediárias à
sombra.
Tabela 8: Resultados dos modelos GLS para a abundância dos grupos funcionais. Somente o melhor
modelo de acordo com os valores de AIC são mostrados. São apresentados os coeficientes estimados e
os erros padrão.
Métricas de
Paisagem G1 G2 G3 G4
NP -- -- -- --
CA 0,007(0,003)* -- -- 0,004(0,001)*
MSI 4,64(0,77)*** -1,8(1,2) ns -- - 1,9 (1,1) ns
DIVISION -- -- -- --
Area core -0,005(0,002)* -- -- --
Shape_Idx -- -- -- 1,45 (0,6)* NP: número de manchas na paisagem; CA: área da classe floresta; MSI: Índice médio de forma; DIVISION: Índice
de divisão da paisagem; Area core: área núcleo da mancha; Shape_Idx: forma da mancha; G1: Grupo funcional 1
(anemocóricas de capacidade fotossintética média e fenologia foliar variada; G2: zoocóricas de capacidade
fotossintética baixa e perenifólias; G3: zoocóricas de capacidade fotossintética média e fenologia foliar variada;
G4: zoocóricas de capacidade fotossintética alta e perenifólias. Parâmetros positivos indicam efeitos positivos e
parâmetros negativos indicam efeitos negativos. n = 202, *P <0,05, ** P <0,01, ***P<0,001, ns = não significativo.
Influências significativas das métricas de paisagem também podem ser observadas nos
trabalhos de Metzger (2000) e Carneiro (2013). O primeiro focou seu estudo na influência da
estrutura da paisagem na riqueza de espécies zoocóricas e de síndrome de dispersão abiótica e
espécies tolerantes a sombra. Carneiro (2013), por sua vez, estudou características reprodutivas
das espécies. Os dois trabalhos indicaram importante influência da estrutura da paisagem na
seleção de grupos de espécies.
CONCLUSÃO
Apesar do histórico de fragmentação florestal, os resultados demonstraram que
predominaram para a Floresta Ombrófila Densa a existência de remanescentes florestais com
grandes extensões e espacialmente próximos de outras áreas florestais. Porém, este padrão não
ocorreu de forma homogênea em todas as microrregiões estudadas, de forma que as
microrregiões de Itajaí, Criciúma, Tubarão e Ituporanga, apresentaram os maiores níveis de
fragmentação no Estado, que pode estar envolvido com atividades econômicas da região e/ou
ao desenvolvimento urbano.

49
A quantidade de borda e o grau de subdivisão da paisagem foram métricas importantes
para a classificação das áreas analisadas em mais e menos fragmentadas, e o que sugere sua
relevância em alterações funcionais nas comunidades vegetais da Floresta Ombrófila Densa.
A classificação das espécies em quatro grupos funcionais com relação à área foliar
específica, ao conteúdo de matéria seca foliar, à síndrome de dispersão e à fenologia foliar,
sugere que estes grupos apresentam variação significativas nestes atributos e apresentam
diferentes estratégias de vida para a ocupação de habitats e para seu desenvolvimento e
sobrevivência. Em que a área florestal, a complexidade de forma da paisagem e dos
remanescentes-foco, aparentemente, desempenham papel fundamental na seleção de certos
grupos funcionais.
A estrutura da paisagem teoricamente afeta a dinâmica e a sobrevivência de populações
vegetais, alterando a capacidade de colonização e sucessão de certos grupos funcionais. Mas
este não é a único e, em muitos casos, nem o mais importante fator influente em estruturação
de comunidades. Estão envolvidos outros fatores físicos e bióticos como tipo de matriz do
entorno, intensidade de interferência humana, condições climáticas, predação e extinção de
animais. Aqui, ateve-se somente a parâmetros estruturais da paisagem, que, mesmo assim,
demonstraram quesito importante para consideração em estudos ecológicos.
REFERÊNCIAS BIBLIOGRÁFICAS
ACKERLY, D.D., KNIGHT, C.A., WEISS, S.B., BARTON, K., STARMER, K.P. Leaf size,
specific leaf area and microhabitat distribution of chaparral woody plants: contrasting patterns
in species level and community level analyses. Oecologia 130, 449–457, 2002.
BERNARDO, P. V. S. Padrões de distribuição de mamíferos de médio e grande porte em
paisagens fragmentadas. Universidade Federal de Goiás. Dissertação de Mestrado em
Ecologia e Evolução. Goiânia – GO, 2012.
BJORNSTAD, O. N. ncf: Spatial nonparametric covariance functions. R package version 1.1-
1, 2008.
BLANCHET, F. G.; LEGENDRE P.; BORCARD, D. Forward Selection of Explanatory
Variables. Ecology, 89(9), p. 2623–2632, 2008.
BUDKE, J. C.; ATHAYDE, E. A.; GIEHL, E. L. H.; ZÁCHIA, R. A.; EISINGER, S. M.
Composição florística e estratégias de dispersão de espécies lenhosas em uma floresta
ribeirinha, arroio Passo das Tropas, Santa Maria, RS, Brasil. Iheringia, Série Bot., Porto
Alegre, v. 60, n. 1, p. 17-24, 2005.
CAILLIEZ, F. The analytical solution of the additive constant problem. Psychometrika,
Lisboa, v. 48, n. 2, p. 305-310. 1983.

50
CARNEIRO, M. S. Efeito da estrutura da paisagem sobre a riqueza e diversidade de
árvores em grupos funcionais reprodutivos. Universidade Federal de Alfenas. Dissertação
de Mestrado em Ecologia e Tecnologia Ambiental. Minas Gerais, 2013.
CIANCIARUSO, M. V.; SILVA, I. A.; BATALHA, M. A. Diversidade Filogenética e
Funcional: novas abordagens para a ecologia de comunidades. Biota Neotrop. 9 (3): 093 – 103,
2009.
CORNELISSEN, J. H. C. et al. Handbook of protocols for standardised and easy measurement
of plant functional traits worldwide. Australian Journal of Botany, Melbourne, v. 51, n. 4, p.
335-380. 2003.
DÍAZ, S. et al. The plant traits that drive ecosystems: Evidence from three continents. Journal
of Vegetation Science 15: 295-304, 2004.
EMBRAPA. Empresa Brasileira de Pesquisa Agropecuária. Centro Nacional de Pesquisa de
Solos. Sistema brasileiro de classificação de solos. 2. ed. Rio de Janeiro: EMBRAPA-SPI,
2006.
GARNIER, E., CORTEZ, J., BILLES, G., NAVAS, M.L., ROUMET, C., DEBUSSCHE,
M.,LAURENT, G., BLANCHARD, A., AUBRY, D., BELLMANN, A., NEILL, C.;
TOUSSAINT, J.P. Plant functional markers capture ecosystem properties during secondary
succession. Ecology, 85, 2630-2637, 2004.
GEOAMBIENTE. Projeto de Proteção da Mata Atlântica em Santa Catarina (PPMA-
SC/FATMA). Relatório Técnico Geral do Estado de Santa Catarina. São José dos Campos.
Geoambiente, 2008.
GIRÃO, L. C.; LOPES, A. V.; TABARELLI M.; BRUNA, E. Changes in tree reproductive
traits reduce functional diversity in a fragmented Atlantic Forest landscape. Plos One, 9:1-12,
2007.
GOWER, J. C. A general coefficient of similarity and some of its properties, Biometrics,
Washington, v. 27, n. 4, p. 857-874. 1971.
HAMMER, O.; HARPER, D. A. T.; RYAN, P. D. PAST: Paleontological Statistics software
package for education and data analysis. Paleontological Electronica 4 (1): 9 pp, 2001.
HERBST, M .J. M.; ROBERTS, J. M.; ROSIER, P. T. W.; TAYLOR, M. E.; GOWING, D. J.
Edge effects and forest water use: A field study in a mixed deciduous woodland. Forest
Ecology and Management, v.250, n.1, p.176-186, 2007.
KEDDY, P. A. Assembly and resume rules: two goals for predictive community ecology.
Journal of Vegetation Science, n. 3, p. 157-164, 1992.
KLEIN, R. M. Flora Ilustrada Catarinense: Mapa fitogeográfico do Estado de Santa Catarina.
Itajaí. Herbário Barbosa Rodrigues, 1978.
KOTTEK, M.; GRIESER, J.; BECK, C.; RUDOLF, B.; RUBEL, F. World Map of the Köppen-
Geiger climate classification updated. Meterologische Zeitschrift, v. 15, n. 3, p.259-263, 2006.
LAMBERS, H.; POORTER, H. Inherent variation in growth rate between higher plants: a
search for physiological causes and ecological consequences. Advances in Ecological
Research, v. 23, p. 187 – 261, 1992.

51
LANG, S.; TIEDE, D. V-LATE Extension für ArcGIS – vektorbasiertes Tool zur quantitativen
Landschaftsstrukturanalyse. ESRI Anwenderkonferenz 2003 Innsbruck. CD-ROM. 2003.
LAURANCE, W. F.; VASCONCELOS, H. L. Consequências ecológicas da fragmentação
florestal na Amazônia. Oecologia Brasiliensis. 13(3): 434-451, 2009.
LOPES, A. V.; GIRÃO, L. C.; SANTOS, B. A.; PERES, C. A.; TABARELLI, M. Long-term
erosion of tree reproductive trait diversity in edge-dominated Atlantic forest fragments. Biol
Conserv 142:1154–1165, 2009.
MAGNAGO, L. F. S.; EDWARDS, D. P.; EDWARDS, F. A.; MAGRACH, A.; MATINS, S.
V.; LAURANCE, W. F. Functional attributes change but functional richness is unchanged after
fragmentation of Brazilian Atlantic forests. Journal of Ecology, v. 102, p. 475–485, 2014.
METZGER, J. P. Tree Functional Group Richness and Landscape Structure in a Brazilian
Tropical Fragmented Landscape. Ecological Applications, 10(4), pp. 1147–1161, 2000.
MURCIA C. Edge effects in fragmented forests: implications for conservation. Tree 10: 58-
62, 1995.
NIMER, E. Clima. In: IBGE. Fundação Instituto Brasileiro de Geografia e
Estatística. Geografia do Brasil: Região Sul. Rio de Janeiro, p151-187, 1990.
OLIFIERS, N.; CERQUEIRA, R. Fragmentação de habitat: efeitos históricos e ecológicos. In:
Biologia da conservação: essências. ROCHA, C. F. D. et al. São Carlos: RiMa, p. 261-316,
2006.
PARDINI, R.; SOUZA, S. M.; BRAGA-NETO, R.; METZGER, J. P. The role of Forest
structure, fragment size and corridors in maintaining small mammal abundance and diversity
in an Atlantic forest landscape. Biological Conservation 124: 253 – 266, 2005.
POHLMAN, C. L.; TURTON, S. M.; GOOSEM M. Temporal variation in microclimatic edge
effects near powerlines, highways and streams in Australian tropical rainforest. Agricultural
and Forest Meteorology, v.149, n.4, p.84-95, 2009.
R DEVELOPMENT CORE TEAM. R: A Language and Environment for Statistical
Computing. R Foundation for Statistical Computing. Vienna, Áustria. Disponível em:
(http://www.R-project.org), 2014.
REICH, P. B.; WRIGHT, I. J.; CAVENDER-BARES, J.; CRAINE, J. M.; OLEKSYN, J.;
WESTOBY, M.; WALTERS, M. B. The evolution of plant functional variation: traits, spectra,
and strategies. Int. J. Plant Sci, 164:143-164, 2003.
RODRIGUES, H. J. B; COSTA, R. F.; RIBEIRO, J. B. M.; SOUZA FILHO, J. D. C.; RUIVO,
M. L. P.; SILVA JÚNIOR, J. A. Variabilidade sazonal da condutância estomática em um
ecossistema de manguezal amazônico e suas relações com variáveis meteorológicas. Rev. bras.
meteorol., vol.26, nº 2, 189 – 196, 2011.
SAMPAIO, A. B.; SCARIOT, A. Edge Effect on Tree Diversity, Composition and Structure in
a Deciduous Dry Forest in central Brazil. Revista Árvore, Viçosa – MG, v. 35, n.5, p. 1121-
1134, 2011.
SCHAADT, S. S. A fragmentação da Floresta Ombrófila Mista em Santa Catarina,
analisada a partir de dados do Inventário Florístico Florestal, fotografias aéreas e mapa

52
temático. Dissertação de Mestrado em Engenharia Ambiental. Universidade Regional de
Blumenau, Santa Catarina, 2012.
SOS MATA ATLÂNTICA; INPE. Atlas dos Remanescentes Florestais da Mata Atlântica,
Período 2012-2013. São Paulo: Fundação SOS Mata Atlântica / Instituto Nacional de Pesquisas
Espaciais, 61 p., 2014.
TABARELLI, M.; AGUIAR, A. V.; GRILLO, A. S. e SANTOS, A. M. M. Fragmentação e
perda de habitat na floresta Atlântica ao norte do Rio São Francisco. In: SIQUEIRA FILHO, J.
A.; LEME, E. M. C. (Org.). Fragmentos da Mata Atlântica do Nordeste: Biodiversidade,
Conservação e suas Bromélias. 1 ed. Rio de Janeiro: Andrea Jacobsson Estúdio Editorial Ltda,
p. 80 – 99, 2006.
VIANA, V. M.; TABANEZ, A. A. J.; MARTINS, J. L. A. Restauração e manejo de fragmentos
florestais. In: CONGRESSO NACIONAL SOBRE ESSÊNCIAS NATIVAS, 2, São Paulo,
1992. Anais. São Paulo: Instituto Florestal de São Paulo. p. 400-407, 1992.
VIBRANS, A. C.; MCROBERTS, R. E.; MOSER, P.; NICOLETTI, A. L. Using satellite
image-based maps and ground inventory data to estimate the area of the remaining Atlantic
forest in the Brazilian state of Santa Catarina. Remote Sensing of Environment: 130: 87–95,
2013.
VIBRANS, A. C.; SEVEGNANI, L.; GASPER, A. L. de; LINGNER, D. V. Inventário
Florístico Florestal de Santa Catarina: Diversidade e conservação dos remanescentes
florestais. Blumenau: Edifurb. Vol. 1, 344 p., 2012.
VIOLLE, C.; NAVAS, M.; VILE, D.; KAZAKOU, E.; FORTUNEL, C.; HUMMEL, I.;
GARNIER, E. Let the concept of trait be functional! Oikos 116:882-892, 2007.
WESTOBY, M.; FALSTER, D. S.; MOLES, A. T.; VESK, P. A.; WRIGHT, I. J. Plant
Ecological Strategies: some leading dimensions of variation between species. Annu. Rev. Ecol.
Syst 33, 125-159, 2002.
WRIGHT I. J. et al. The worldwide leaf economics spectrum. Nature 428(6985): 821–827,
2004.

53
CAPÍTULO II
A fragmentação como preditora de mudanças funcionais do componente arbóreo na
Floresta Ombrófila Densa de Santa Catarina
RESUMO
Os impactos aos ambientes naturais são consequências inevitáveis das atividades humanas,
sendo a fragmentação florestal um de seus resultados. A severidade da fragmentação é
dependente de características estruturais dos fragmentos, influenciando na estrutura e
composição funcional de comunidades vegetais. Dessa forma, o objetivo deste estudo foi
investigar a influência da estrutura de remanescentes florestais sobre os atributos funcionais e
a diversidade funcional do conjunto de espécies mais abundantes da Floresta Ombrófila Densa
de Santa Catarina. Para isso, a estrutura de 86 remanescentes florestais amostrados pelo
Inventário Florístico Florestal de Santa Catarina foi quantificada por meio de métricas de
paisagem. A descrição dos atributos funcionais (altura máxima (hmáx), síndrome de dispersão,
densidade da madeira (SSD), área foliar específica (SLA) e conteúdos de matéria seca (LDMC),
nitrogênio (LNC) e fósforo foliar (LPC)) foi realizada para as 20 espécies arbóreas mais
abundantes em toda área de estudo. Para cada uma das unidades amostrais foram determinados
os atributos funcionais médios, ponderados pela densidade das espécies (community-weighted
mean trait matrix - CWM). Em seguida, a matriz CWM, com exceção da variável binária
(síndrome de dispersão), foi ordenada por meio de uma Análise de Componentes Principais
(PCA). Para a análise da diversidade funcional, foi utilizado o índice de Rao (Rao’s Quadratic
Entropy- RaoQ). A influência das métricas de paisagem sobre os atributos funcionais médios
das áreas, sintetizados pelos eixos da PCA, e sobre a diversidade funcional (RaoQ) foi
determinada por meio modelos lineares, usando o método GLS (Generalized Least Squares
model). Modelos globais foram construídos a partir de todas as variáveis explicativas (exceto
as colineares - Fator de Inflação da Variância: VIF < 10) e selecionados os melhores modelos
pelo critério de informação de Akaike (AIC). Para a síndrome de dispersão, utilizou-se modelos
lineares generalizados (GLM), com distribuição binomial, para cada uma das variáveis
explicativas, cujo a qualidade de ajuste verificada por meio do teste de Hosmer-Lemeshow. A
estrutura da paisagem influenciou de forma significativa as variações de atributos de dispersão,
densidade da madeira (SSD) e no conteúdo de matéria seca foliar (LDMC) nas comunidades
arbóreas. Porém, a diversidade funcional não foi influenciada pelas métricas de paisagem
analisadas. A intensidade de fragmentação foi um importante impulsionador na seleção de
atributos funcionais, envolvidos principalmente na aquisição e conservação de recursos, porém
não teve influência na diversidade funcional, pelo menos quando considerado o conjunto de
espécies e de atributos avaliados neste estudo.
Palavras-chave: Métricas de paisagem. Fragmentação florestal. Atributos funcionais. Modelos
lineares. Diversidade funcional.

54
ABSTRACT
The fragmentation as a predictor of functional changes on the arboreous component in
the Dense Ombrophylous Forest of Santa Catarina. Impacts on natural environments are
inevitable consequences of human activities and the forest fragmentation is one of its results.
The severity of fragmentation depends on structural features of fragments, which may influence
the structure and functional composition of plant communities. Thus, the aim of this study is to
investigate the influence of the remaining forest structure on the functional traits and functional
diversity of the most abundant tree species of the Dense Ombrophylous Forest of Santa
Catarina. Therefore, the structure of 86 forest remnants sampled by Floristic and Forest
Inventory of Santa Catarina was quantified by landscape metrics. The description of the
functional traits (maximum height (hmax), dispersion syndrome, wood density (SSD), specific
leaf area (SLA), leaf dry matter content (LDMC), leaf nitrogen concentration (LNC), leaf
phosphorus concentration (LPC)) was performed for the 20 most abundant tree species. For
each sample units we determined the mean functional traits, weighted by the density of the
species (community-weighted mean trait matrix - CWM). Then the CWM matrix, except for
the binary variable (dispersion syndrome), was ordered by a Principal Component Analysis
(PCA). For the analysis of functional diversity, we used the Rao index (Rao's Quadratic
Entropy- RaoQ). The influence of landscape metrics on mean functional traits of each areas,
synthesized by the PCA axes, and on functional diversity (RaoQ) was determined by linear
models using the method GLS (Generalized Least Squares model), with the incorporation of
dependence space, if necessary, from geostatistical models. Global models were built based on
all the explanatory variables (except the collinear - Variance Inflation Factor: VIF <10) and the
best fitted models were selected by the Akaike information Criterion (AIC). For the dispersion
syndrome, we used generalized linear models (GLM), with binomial distribution, for each of
the explanatory variables, whose goodness of fit was verified by the Hosmer-Lemeshow test.
The landscape structure significantly influenced the changes in dispersion traits, wood density
(SSD) and leaf dry matter content (LDMC) in tree communities. However, the functional
diversity was not influenced by the analyzed landscape metrics. As a results we conclude that
fragmentation may act on the selection of more generalist species, while the edge effect acts on
the selection of anemochoric species.
Keywords: Landscape metrics. Forest fragmentation. Functional traits. Linear models.
Functional diversity.
INTRODUÇÃO
A fragmentação de habitats naturais é considerada, atualmente, um importante problema
ambiental da atualidade, pois representa uma das principais causas de extinção de espécies,
alterando a composição, estrutura e funcionalidade de comunidades vegetais (METZGER,
2000; OLIVEIRA et al., 2004; LAURANCE et al., 2006; MAYFIELD et al., 2006; GIRÃO et
al., 2007; LIENIN e KLEYER, 2011).
São diversos os ecossistemas ameaçados pela fragmentação. A Floresta Atlântica, por
exemplo, que se estende por quase toda a costa oriental brasileira, considerada um hotspot
terrestre desde o final do século XX (MYERS et al., 2000), tem seu processo de fragmentação
acelerado por abrigar a maior parte da população brasileira e por concentrar atividades de

55
importância econômica para o país (VIANA e TABANEZ, 1996). Na região Sul, a Floresta
Ombrófila Densa, região fitoecológica que compõe o domínio da Floresta Atlântica, possui
pouco mais de 100 anos de histórico de ocupação por núcleos coloniais (GOULARTI FILHO,
2002). Desde então, ela vem cedendo espaço à expansão agrossilvipastoril e urbano, com
consequente fragmentação. A região é caracterizada pela alta umidade e complexidade de
associações de espécies, e necessita de estudos mais específicos para esclarecer como a
fragmentação influencia esses ambientes.
Os efeitos da fragmentação sobre as populações de organismos dependem, sob vários
aspectos, da biologia das espécies e também das características estruturais da paisagem e do
fragmento na qual a comunidade está inserida (VIANA e TABANEZ, 1996; METZGER,
2000). A criação de bordas entre floresta e matriz circundante provoca, em um primeiro
momento, alterações no microclima da floresta, que envolvem aumento na temperatura e na
intensidade do vento e diminuição da umidade (MURCIA, 1995). As espécies pioneiras são
beneficiadas pelas novas condições, pois apresentam um conjunto de características que
possibilitam a ocupação de áreas abertas de condições ambientais mais severas, e passam a ter
uma maior participação na comunidade, anteriormente restritas às áreas naturalmente
perturbadas. As espécies “tolerantes à sombra”, adaptadas aos interiores úmidos e sombreados
das florestas, são as mais prejudicadas nesse novo contexto (LAURANCE et al., 2006;
OLIVEIRA et al., 2004). Essas diversas modificações, influenciam também em efeitos
biológicos indiretos (MURCIA, 1995), os quais envolvem mudanças nas interações ecológicas
como herbivoria, polinização e dispersão de sementes (MURCIA, 1995; CUNNINGHAM,
2000; METZGER, 2000; GIRÃO et al., 2007), e são agravados com o aumento do isolamento
entre os remanescentes.
A fragmentação pode ser quantificada e descrita pelas mudanças das chamadas
“métricas” de paisagem, que começaram a ser frequentemente empregadas na Ecologia da
Paisagem para o estudo da estrutura, função e dinâmica de áreas heterogêneas compostas por
ecossistemas interativos (FORMAN e GODRON, 1986). Elas tornaram-se, então, variáveis
ambientais importantes para o estudo da influência da estrutura da paisagem sobre aspectos
ecológicos de comunidades fragmentadas.
No que diz respeito aos estudos ecológicos, de distribuição, diversidade e padrões de
organização de espécies arbóreas, foi proposto o estudo funcional das plantas. Esta abordagem
permite analisar a vegetação por meio de um conjunto de características intrínsecas (atributos
funcionais), que podem ser morfológicas, fisiológicas ou fenológicas mensuráveis (VIOLLE et
al., 2007). Elas constituem um importante meio de otimizar a percepção entre vegetação e

56
mudanças ambientais trazidas pela fragmentação, por estarem envolvidas com as maneiras das
plantas adquirirem, processarem e investirem os recursos disponíveis (DÍAZ et al., 2004). Elas
também são responsáveis pela performance das plantas frente as mudanças no ambiente
(LEBRIJA-TREJOS et al., 2010).
Ainda são escassos os estudos que envolvam características estruturais dos fragmentos
e atributos funcionais de plantas arbóreas em florestas tropicais. Pouco se sabe sobre a atuação
da intensidade de fragmentação de florestas tropicais úmidas na seleção de atributos funcionais
específicos. Sabe-se que as limitações quanto a disponibilidade de recursos e outras condições
abióticas adversas agem como filtros ambientais em comunidades fragmentadas e favorecem
estratégias específicas, principalmente as de retenção de recursos (LAVOREL et al., 2007;
MASON et al., 2013; HOLDAWAY et al., 2011). Os efeitos da fragmentação interferem ainda
no sucesso da dispersão por espécies zoocóricas, devido aos vários fatores que impedem o
deslocamento de animais dispersores, provocando sua ausência ou menor ocorrência
(METZGER, 2000). A atuação mais intensa de filtros ambientais em ambientes fragmentados
(KEDDY, 1992) pode, nesse caso, acarretar em uma menor diversidade funcional, o que levaria
um comprometimento do funcionamento e da manutenção de processos nas comunidades
(MAYFIELD et al., 2005).
Diante disso, o presente trabalho tem como objetivo avaliar a influência da estrutura de
fragmentos florestais na paisagem sobre atributos funcionais e diversidade funcional do
conjunto de espécies mais abundantes da Floresta Ombrófila Densa de Santa Catarina, testando
as hipóteses de que (i) a intensidade da fragmentação, medida neste estudo, por meio de
métricas de paisagem, atua na seleção de diferentes estratégias, que são refletidas nos valores
dos atributos funcionais analisados, e (ii) na diminuição da diversidade funcional, em resposta
às alterações bióticas e abióticas causadas pela fragmentação de habitats.
MATERIAL E MÉTODOS
Área de estudo
O estudo foi realizado na área de ocorrência de Floresta Ombrófila Densa no Estado de
Santa Catarina, delimitada segundo o mapeamento de Klein (1978). A fitofisionomia está
situada entre as coordenadas 25º 57’ 40’’ e 29º 19’ 13’’ de latitude sul e 48º 24’ e 21’’ e 50º
14’ 14’’ de longitude oeste, em que sua área territorial original é de 29.282,00 km² (KLEIN,
1978), correspondendo a aproximadamente 31% da área territorial do Estado, e que, atualmente,
devido a um histórico processo de fragmentação, restam apenas 12.632,7 km² (VIBRANS et

57
al., 2013a). A altitude nas regiões de ocorrência da FOD, varia de alguns metros acima do nível
do mar, a pouco mais de 1.000 metros em alguns pontos.
Nessa região predomina o clima temperado úmido de verão quente (Cfa) (KOTTEK et
al., 2006), sem um período seco definido. As temperaturas médias anuais, dependendo da
altitude e da influência oceânica, chegam ao mínimo de 16ºC e ao máximo de 22ºC (NIMER,
1990). A precipitação apresenta grande variação, com registros de 1.250 a 2.000 mm, sendo
mais abundante ao norte (NIMER, 1990). Em consideração aos solos da região, predominam
os Cambissolos e Argissolos nas encostas e Gleissolos e Organossolos nas planícies
(EMBRAPA, 2006).
Seleção das espécies
Os dados de abundância das espécies foram extraídos da base de dados do Inventário
Florístico Florestal de Santa Catarina (IFFSC) (VIBRANS et al., 2013b), que possui 202
unidades amostrais levantadas em área de Floresta Ombrófila Densa, sendo utilizado para este
estudo 86 delas (Figura 17).
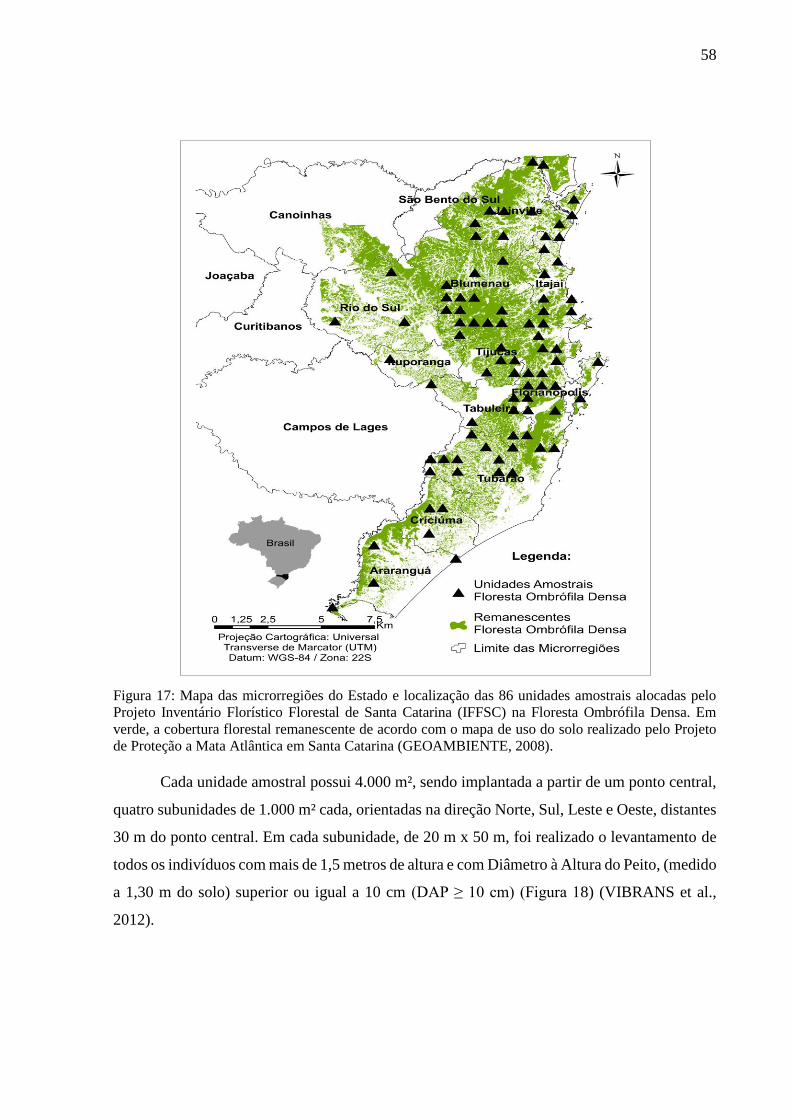
58
Figura 17: Mapa das microrregiões do Estado e localização das 86 unidades amostrais alocadas pelo
Projeto Inventário Florístico Florestal de Santa Catarina (IFFSC) na Floresta Ombrófila Densa. Em
verde, a cobertura florestal remanescente de acordo com o mapa de uso do solo realizado pelo Projeto
de Proteção a Mata Atlântica em Santa Catarina (GEOAMBIENTE, 2008).
Cada unidade amostral possui 4.000 m², sendo implantada a partir de um ponto central,
quatro subunidades de 1.000 m² cada, orientadas na direção Norte, Sul, Leste e Oeste, distantes
30 m do ponto central. Em cada subunidade, de 20 m x 50 m, foi realizado o levantamento de
todos os indivíduos com mais de 1,5 metros de altura e com Diâmetro à Altura do Peito, (medido
a 1,30 m do solo) superior ou igual a 10 cm (DAP ≥ 10 cm) (Figura 18) (VIBRANS et al.,
2012).

59
Fonte: Vibrans et al., 2012.
Figura 18: Conglomerado utilizado para a amostragem da vegetação catarinense pelo Inventário
Florístico Florestal de Santa Catarina.
Da lista de 656 espécies da tabela fitossociológica do IFFSC, selecionou-se para a
caracterização dos atributos funcionais somente as 20 espécies lenhosas (DAP ≥ 10 cm) mais
abundantes, representando parte considerável da estrutura da floresta (Tabela 9). Optou-se pela
exclusão de samambaias e palmeiras da análise, pois introduziriam um outro tipo de
variabilidade aos resultados, já que utilizam de estratégias específicas para a ocupação de
ambientes e acabariam mascarando os padrões da grande maioria das espécies. Também foram
removidas das análises as unidades amostrais onde as espécies avaliadas foram pouco
representativas da comunidade como um todo (quando somam menos de 30% da abundância
relativa), mantendo-se, dessa forma, as 86 unidades amostrais mencionadas acima.
Tabela 9: Espécies arbóreas mais abundantes pertencentes à Floresta Ombrófila Densa do Estado de
Santa Catarina.
FAMÍLIA ESPÉCIE
ANACARDIACEAE Tapirira guianensis Aubl.
ANNONACEAE Guatteria australis A.St.-Hil.
APOCYNACEAE Aspidosperma australe Müll.Arg.
ASTERACEAE Piptocarpha axillaris (Less.) Baker
CLETHRACEAE Clethra scabra Pers.
ELAEOCARPACEAE Sloanea guianensis (Aubl.) Benth.
EUPHORBIACEAE Alchornea triplinervia (Spreng.) Müll.Arg.
PERACEAE Pera glabrata (Schott) Poepp. ex Baill.
LAURACEAE Nectandra oppositifolia Nees
MELASTOMATACEAE Miconia cinnamomifolia (DC.) Naudin
MELIACEAE Cabralea canjerana (Vell.) Mart.
Cedrela fissilis Vell.
60
NYCTAGINACEAE Guapira opposita (Vell.) Reitz
PHYLLANTHACEAE Hieronyma alchorneoides Allemão
RUBIACEAE Bathysa australis (A.St.-Hil.) K.Schum.
Psychotria vellosiana Benth.
SALICACEAE Casearia sylvestris Sw.
SAPINDACEAE Matayba intermedia Radlk.
Cupania vernalis Cambess.
URTICACEAE Cecropia glaziovii Snethl.
Caracterização funcional das espécies arbóreas
Dos sete atributos funcionais analisados neste trabalho, cinco, segundo Cornelissen et
al. (2003), estão relacionados com respostas a distúrbios, que são: altura máxima da planta
(hmáx), densidade da madeira (SSD), síndrome de dispersão (Dispersão) e conteúdo de
nitrogênio e fósforo foliar (LNC e LPC). Os outros, que são área foliar específica (SLA) e
conteúdo de matéria seca foliar (LDMC), envolvidos mais diretamente com variáveis climáticas
e de disponibilidade de recursos no solo, foram selecionados por entendermos que, de alguma
forma, respondem as mudanças abióticas do efeito de borda no fragmento (Tabela 10). Os dados
de atributos médios por espécie foram disponibilizados pelo Departamento de Ecologia da
Universidade Federal do Rio Grande do Sul (UFRGS) (Apêndice – Tabela 1).
Tabela 10: atributos funcionais observados com suas respectivas siglas, unidade de medida (para
variáveis quantitativas) e categorias (variáveis qualitativas)
Atributos Sigla Unidade Variável Categórica
Altura Máxima Hmáx m -
Densidade da Madeira SSD g/cm³ -
Síndrome de Dispersão Dispersão Categórica 0 - Anemocórica
1 - Zoocórica
Concentração de Nitrogênio Foliar LNC mg/g -
Concentração de Fósforo Foliar LPC mg/g -
Área Foliar Específica SLA mm²/mg -
Conteúdo de Matéria Seca Foliar LDMC mg/g -
Estrutura da paisagem dos remanescentes florestais
Para a análise da estrutura dos remanescentes florestais, quantificada por meio de
métricas de paisagem, foi utilizado o mapa temático elaborado pelo Projeto de Proteção à Mata
Atlântica em Santa Catarina (PPMA/FATMA-SC), com base em imagens multiespectrais
SPOT-4 do ano de 2005 na escala 1:50.000 (GEOAMBIENTE, 2008). Foi considerado para

61
este trabalho somente a classificação ‘florestas em estágio médio ou avançado e/ou primárias’.
As métricas foram calculadas com auxílio do Software ArcGis 10.0, utilizando a extensão V-
LATE 1.1 (Vector-based Landscape Analysis Tools Extension) (LANG e TIEDE, 2003).
No ArcGis demarcou-se a partir do ponto central de cada unidade amostral (UA) uma
área circular (buffer) de 2,8 km de raio, seguindo a proposta de Schaadt (2012). Dentro da área
de aproximadamente 2.462 ha, delimitada pelo buffer no entorno das 86 unidades amostrais
todos os remanescentes mapeados das florestas em estágio médio ou avançado e ou primárias
foram identificados. As métricas de paisagem foram calculadas a nível de classe e de mancha,
em que a classe seria todo o conjunto de remanescentes presentes na paisagem (área do buffer)
e a mancha seria o remanescente-foco, onde está localizada a UA do IFFSC (Figura 19).
Fonte: figura baseada na de Bernardo, 2012.
Figura 19: Representação do buffer de 2,8 km de raio, utilizado para o cálculo das métricas de paisagem,
estando representados, o remanescente-foco, no qual está localizada a Unidade Amostral do IFSSC,
remanescentes florestais inseridos na área buffer e que compõem a classe floresta, e as áreas não
florestais e de estágio sucessional inicial, não considerados no estudo.
Todas as métricas consideradas neste trabalho estão descritas na Tabela 11,
categorizadas como métricas que se referem a área, área núcleo, forma, borda, subdivisão e
proximidade dos fragmentos; foram consideradas sete a nível de classe e quatro a nível de

62
mancha. Adotou-se como borda a faixa marginal de 50 m de largura em cada fragmento
avaliado (MURCIA, 1995).
Tabela 11: Métricas de paisagem a nível de classe e a nível de mancha, com seus respectivos significados
e unidades de medida.
CLASSE
Categoria Sigla Definição Unidade
ÁREA CA Área da classe ha
ÁREA NÚCLEO CAI Índice de área núcleo %
FORMA MSI Índice médio de forma ---
BORDA ED Densidade de borda m/ha
TE Total de borda m
SUBDIVISÃO DIVISION Índice de Divisão da Paisagem %
NP Número de manchas ---
MANCHA
Categoria Sigla Definição Unidade
ÁREA Area Área do fragmento ha
ÁREA NÚCLEO Área Core Área núcleo do fragmento ha
FORMA Shape_Idx Índice de Forma ---
PROXIMIDADE NNDist Distância do Vizinho mais próximo m
Análises estatísticas
Para cada uma das unidades amostrais foram determinados os atributos funcionais
médios, ponderados pela densidade das espécies (community-weighted mean trait matrix -
CWM) (GARNIER et al., 2004). Em seguida, com o propósito de identificar gradientes
associados às variações dos atributos funcionais entre as áreas, a matriz CWM, com exceção da
variável binária (síndrome de dispersão), foi ordenada por meio de uma Análise de
Componentes Principais (PCA). Para a análise da diversidade funcional, foi utilizado o índice
de Rao (BOTTA-DUKÁT, 2005) (Rao’s Quadratic Entropy- RaoQ). De acordo Weigelt et al.
(2008), o índice RaoQ é uma medida contínua da diversidade funcional, que inclui informações
sobre a regularidade da distribuição dos atributos funcionais dentro de uma comunidade, por
considerar a abundância das espécies em seu cálculo. Há um debate sobre a medida de
dissimilaridade apropriada para utilizar no cálculo de Rao (DE BELLO et al, 2013), para este
estudo foi utilizada, a distância de Gower (GOWER, 1971), recomendada quando utilizados
dados quantitativos e qualitativos simultaneamente (PAVOINE et al., 2009) e adequada para a
detecção de mudanças na diversidade funcional ao longo de gradientes ambientais (DE BELLO
et al., 2013).

63
A influência das métricas de paisagem sobre os atributos funcionais das áreas,
sintetizados pelos eixos da PCA, e sobre a diversidade funcional (RaoQ) foi determinada por
meio de modelos lineares, usando o método dos mínimos quadrados generalizados (GLS -
Generalized Least Squares model), com a incorporação da dependência espacial quando
necessário, a partir de modelos geoestatísticos. Para verificar a existência de autocorrelação
espacial nos resíduos foram utilizados correlogramas de Moran, com 1.000 permutações.
Modelos globais foram construídos a partir de todas as variáveis explicativas (exceto as
colineares - Fator de Inflação da Variância: VIF < 10) e selecionados os melhores modelos pelo
critério de informação de Akaike (AIC). Para a síndrome de dispersão, por ser representada por
dados binários, utilizou-se modelos lineares generalizados (GLM), com distribuição binomial,
para cada uma das variáveis explicativas, cujo a qualidade de ajuste dos melhores modelos
verificada por meio do teste de Hosmer-Lemeshow.
Todas as análises foram realizadas no R (R DEVELOPMENT CORE TEAM, 2014),
utilizando os pacotes FD (LALIBERTÉ e LEGENDRE, 2010; LALIBERTÉ e SHIPLEY,
2011), para a determinação da matriz CWM e do índice de RaoQ; Vegan (OKSANEN et al.,
2009) para a PCA; nlme (PINHEIRO et al., 2011) para a construção dos modelos, ncf
(BJORNSTAD, 2008), para a determinação da autocorrelação espacial dos resíduos dos
modelos; MASS, para seleção dos modelos por meio de AIC; e ResourceSelection (LELE,
2009; LELE e KEIM, 2006), para o teste de qualidade de ajuste de Hosmer-Lemeshow.
RESULTADOS
Os dois primeiros eixos da Análise de Componentes Principais realizada a partir da
matriz atributos médios por unidade amostral (CWM) (Tabela 12; Figura 20) explicaram
62,53% da variação encontrada (eixo 1: 39,26% e eixo 2: 23,27%). O primeiro eixo da
ordenação esteve negativamente correlacionado principalmente aos atributos área foliar
específica (SLA) e concentração de nitrogênio foliar (LNC), que por sua vez, estiveram
fortemente autocorrelacionadas. O eixo 1, interpretado como um gradiente associado sobretudo
a taxas fotossintéticas e de potencial de crescimento das espécies (WESTOBY et al. 2002),
segrega as áreas analisadas nas que apresentaram altos valores para os dois atributos no lado
negativo do eixo e nas que obtiveram os menores valores no lado positivo. O segundo eixo da
ordenação esteve fortemente correlacionado a densidade da madeira (SSD) e ao conteúdo de
matéria seca foliar (LDMC), que por sua vez estiveram negativamente correlacionadas. O eixo
2, interpretado como um gradiente de uso e capacidade de assimilação de recursos, divide o

64
quadrante superior com áreas com predominância de espécies de alta densidade da madeira e
baixa matéria seca foliar, e o quadrante inferior as áreas predominantemente ocupadas por
espécies de baixa densidade da madeira e alta matéria seca foliar.
Tabela 12: Resultados da Análise de Componentes Principais (PCA) para os atributos funcionais
analisados, com a correlação das variáveis para os dois primeiros eixos da ordenação.
VARIÁVEIS SIGLA EIXO PCA 1 EIXO PCA 2
Total de variação explicada --- 39,26% 23,27%
Altura Máxima hmáx 0,29 0,15
Área Foliar Específica SLA -0,92 0,01
Densidade da Madeira SSD 0,15 0,67
Conteúdo de Matéria Seca Foliar LDMC 0,4 -0,84
Conteúdo de Nitrogênio Foliar LNC -0,89 0,1
Conteúdo de Fósforo Foliar LPC -0,66 -0,45
Figura 20: Análise de Componentes Principais realizada a partir da matriz atributos médios por unidade
amostral (CWM), do conjunto de espécies mais abundantes da Floresta Ombrófila Densa de Santa
Catarina. SSD = densidade da madeira; LNC = conteúdo de nitrogênio foliar; SLA = área foliar
específica; LPC = conteúdo de fósforo foliar; LDMC = conteúdo de matéria seca foliar; hmáx = altura
máxima.

65
A relação inversa significativa (p=0,0008), porém fraca (r = -0,35) entre as variáveis
SLA e LDMC (por vezes referido como a densidade do tecido foliar), foi deparada também na
Análise de Componentes Principais. São atributos associados com um espectro econômico
foliar (leaf economics espectrum - DÍAZ et al. (2004)), partindo de uma estratégia de rápida
aquisição de recursos (alto SLA e baixo LDMC) até uma estratégia conservativa (baixo SLA e
alto LDMC) das plantas. Ao considerar a PCA realizada, SLA se relaciona fortemente com o
primeiro eixo de ordenação e o LDMC com o segundo eixo, e considerando os melhores
modelos para a explicação dos dois eixos (Tabela 13), o primeiro não foi significativamente
influenciado por nenhuma métrica de paisagem analisada. Por outro lado, o eixo 2, que diz
respeito a qualidade de fuste e estrutura foliar das espécies, apresentou correlação negativa com
o grau de divisão da paisagem (DIVISION) (Figura 21).
Tabela 13: Resultados dos modelos lineares, usando o método dos mínimos quadrados generalizados
(GLS) para os dois primeiros eixos da Análise de Componentes Principais (PCA) e para o índice de
diversidade funcional RaoQ e modelos lineares generalizados (GLM), com distribuição binomial, para
a síndrome de dispersão.
Variáveis Dependentes Variáveis
Explicativas Modelo Teste Coeficiente p
Eixo 1 PCA --- --- --- --- ---
Eixo 2 PCA DIVISION GLS t = -2,30 -0,001 0,0238
Diversidade Funcional
(RaoQ) --- --- --- --- ---
Síndrome de Dispersão TE GLM (binomial) z = -2,23 -0,12 0,03
Síndrome de Dispersão MSI GLM (binomial) z = -2,7 -0,002 0,007
DIVISION = índice de divisão da paisagem; TE= total de borda na paisagem; MSI = índice médio de forma da
classe
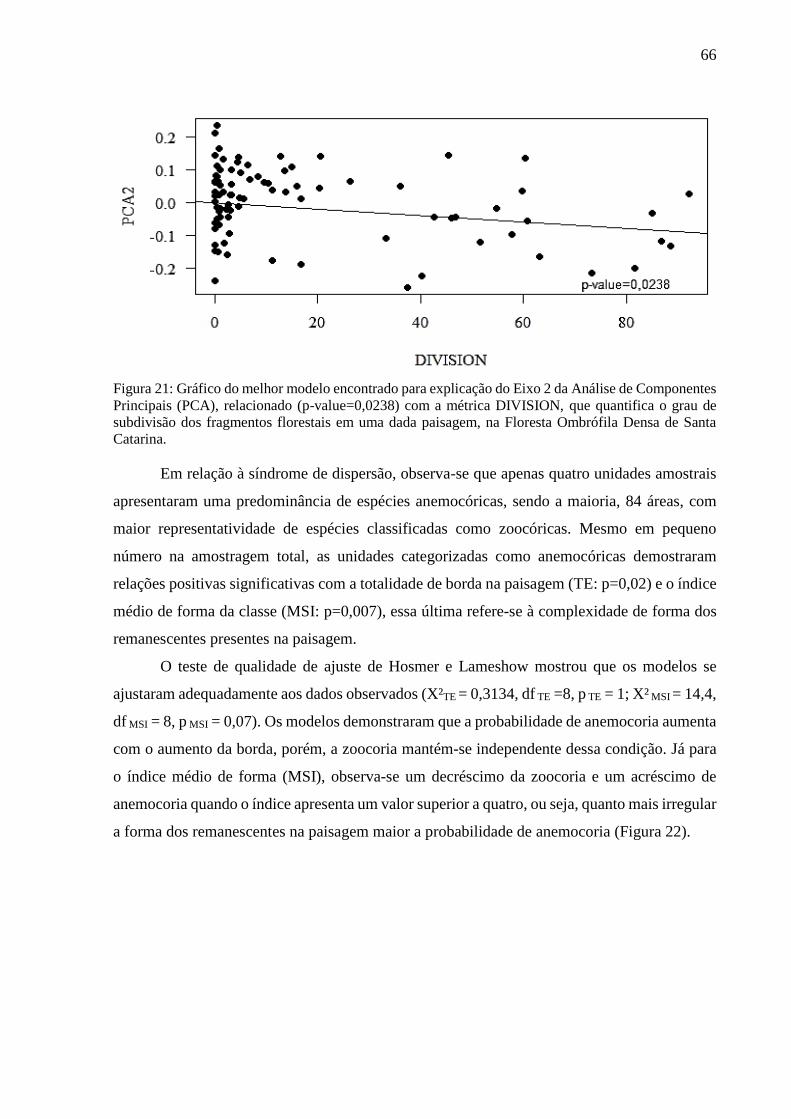
66
Figura 21: Gráfico do melhor modelo encontrado para explicação do Eixo 2 da Análise de Componentes
Principais (PCA), relacionado (p-value=0,0238) com a métrica DIVISION, que quantifica o grau de
subdivisão dos fragmentos florestais em uma dada paisagem, na Floresta Ombrófila Densa de Santa
Catarina.
Em relação à síndrome de dispersão, observa-se que apenas quatro unidades amostrais
apresentaram uma predominância de espécies anemocóricas, sendo a maioria, 84 áreas, com
maior representatividade de espécies classificadas como zoocóricas. Mesmo em pequeno
número na amostragem total, as unidades categorizadas como anemocóricas demostraram
relações positivas significativas com a totalidade de borda na paisagem (TE: p=0,02) e o índice
médio de forma da classe (MSI: p=0,007), essa última refere-se à complexidade de forma dos
remanescentes presentes na paisagem.
O teste de qualidade de ajuste de Hosmer e Lameshow mostrou que os modelos se
ajustaram adequadamente aos dados observados (X²TE = 0,3134, df TE =8, p TE = 1; X² MSI = 14,4,
df MSI = 8, p MSI = 0,07). Os modelos demonstraram que a probabilidade de anemocoria aumenta
com o aumento da borda, porém, a zoocoria mantém-se independente dessa condição. Já para
o índice médio de forma (MSI), observa-se um decréscimo da zoocoria e um acréscimo de
anemocoria quando o índice apresenta um valor superior a quatro, ou seja, quanto mais irregular
a forma dos remanescentes na paisagem maior a probabilidade de anemocoria (Figura 22).

67
Figura 22: Estimativa de densidade de Kernel para a síndrome de dispersão zoocórica (1 – cinza claro)
e anemocórica (0 – cinza escuro) em relação à totalidade de bora –TE (A) e o índice médio de forma –
MSI (B).
DISCUSSÃO
Os dois primeiros eixos da PCA sintetizaram dois aspectos relevantes das estratégias
ecológicas das espécies arbóreas mais abundantes da Floresta Ombrófila Densa em Santa
Catarina. Um primeiro aspecto, indicado pelo eixo 1 da ordenação, com explicação de 39,26%
da variação, esteve associado ao investimento em área foliar específica (SLA) e compostos
químicos foliares (LNC e LPC), que possibilitam maior taxa fotossintética e potencial de
crescimento à planta (LAMBERS e POORTER, 1992). A área foliar específica é considerada
um importante atributo que sintetiza um dos principais eixos de variação ecológica entre as
espécies (WESTOBY et al., 2002; DÍAZ et al., 2004), e está relacionada com a disponibilidade
de recursos (LIENIN e KLEYER, 2011). Na escala espacial considerada neste trabalho, não foi
encontrada nenhuma evidência significativa de associação dos atributos funcionais
representados pelo eixo 1 à estrutura da paisagem. Isto indica que estes estejam mais envolvidos
com outras variáveis não mensuradas nesse trabalho, como por exemplo, o tipo de matriz de
(A)
(B)

68
entorno (CARNEIRO, 2013), a intensidade de uso do solo e variáveis edáficas (CASTRO et
al., 2010; LIENIN e KLEYER, 2011), do que com o gradiente estrutural da paisagem.
O outro aspecto importante foi indicado pelo eixo 2 da ordenação, com explicação de
23,27% da variação; ele esteve associado à densidade da madeira (SSD) e ao conteúdo de
matéria seca foliar (LDMC). Variações destes atributos ao longo do eixo 2 foram
significativamente influenciados pela variável chave em estudos de fragmentação, o grau de
retalhamento da paisagem, que pode ser medido pelo índice de divisão da paisagem
(DIVISION). Dessa forma, os resultados encontrados indicam que a fragmentação, por si só,
represente um importante filtro na seleção de estratégias de aquisição e investimento dos
recursos pelas plantas.
Em locais de maior DIVISION ocorreu um favorecimento de um pequeno grupo de
espécies, porém de elevada abundância, com menor densidade da madeira e folhas de aspecto
mais coriáceo, como Tapirira guianensis e Alchornea triplinervia. Estas, por sua vez, são
espécies generalistas, amplamente distribuídas e diversos tipos de vegetação e comuns em
florestas mais jovens perturbadas pelo homem (OLIVEIRA et al., 2004; LIEBSCH et al., 2008).
Em áreas de menor DIVISION, por sua vez, as espécies Hieronyma alchorneoides (SSD: 0,7
g/cm³; LDMC: 274 mg/g) e Miconia cinnamomifolia (SSD: 0,73 g/cm³; LDMC: 316 mg/g)
foram as mais abundantes.
Dentre os efeitos da fragmentação florestal, o aumento da intensidade luminosa, a menor
umidade e a maior susceptibilidade ao ataque de herbívoros, fazem com que o ambiente
fragmentado torne-se mais “estressado” (CUNNINGHAM et al., 1999; LIENIN e KLEYER,
2011). Em consequência, as plantas que para mitigar os danos as suas células, protegem-nas
com alocação de compostos químicos de defesa, aumentando a esclerofilia das folhas (maior
LDMC); e aumentando também a resistência à perda de água e ataque de herbívoros (MONSON
e SMITH, 1982; TURNER, 1994; MUNGUÍA-ROSAS et al, 2014; ACKERLY et al., 2002;
SHIPLEY et al., 2006), têm maior aptidão em ambientes “estressados”.
No caso da densidade da madeira, observamos que espécies com menor densidade
mostraram-se importantes em locais de maior retalhamento da paisagem; isto sugere que o
menor investimento em lenho esteja envolvido com a maior capacidade das espécies na
ocupação de ambientes perturbados, facilitando seu desenvolvimento nestes locais com
alterações abióticas mais severas. No trabalho de Chave et al. (2009), os autores relacionam as
caraterísticas mecânicas da madeira com a resistência a distúrbios, sugerindo que o menor
investimento das plantas em densidade da madeira e o maior investimento em eficiência de
transporte de água e nutrientes (WRIGHT et al., 2007), garantem uma maior competitividade

69
para a planta, fazendo com que ela consiga ocupar mais rapidamente um determinado habitat.
O maior grau de subdivisão de paisagem parece causar, nesse caso, a seleção de um pequeno
conjunto dominante de espécies com menos carbono por unidade de biomassa (CHAVE et al.,
2009), fato observado também por Santos et al. (2008).
A pressão para a diferenciação de nicho entre as espécies coexistentes é menos intensa
em comunidades estressadas (MASON et al., 2013). Nesses casos, segundo o mesmo autor e
como observado também por Holdaway et al. (2011), são favorecidos os atributos que
aumentam o potencial para a aquisição e retenção de recursos limitantes.
A constatação de que paisagens mais fragmentadas selecionam espécies mais
conservadoras no quesito foliar, que investem em proteção foliar (aumento de compostos
orgânicos como fenóis, taninos e fibras – CUNNINGHAM et al., 1999), garantindo a
longevidade de suas folhas e que relativo ao seu lenho, a estratégia seja a eficiência no
transporte de água e nutrientes, nos leva a entender que nestes ambientes a escassez de água
seja um importante limiar para a seleção de estratégias (CUNNINGHAM et al., 1999; LIENIN
e KLEYER, 2011). No entanto, esta constatação necessitaria de estudos mais específicos.
O gradiente estrutural da paisagem observada, no entanto, parece não alterar a
diversidade funcional da vegetação, pelo menos quando são consideradas as 20 espécies mais
abundantes da Floresta Ombrófila Densa. Este resultado sugere que as diversas estratégias para
utilização dos recursos são mantidas, independentemente do grau de alteração da estrutura da
paisagem. Isto mostra que o conjunto de espécies analisado, possui estratégias de vida
adaptadas às diferentes configurações da paisagem, possuindo tais atributos que possibilitem
esse desempenho.
O incremento em borda e na complexidade de forma da paisagem (TE e MSI,
respectivamente) resultou em um incremento da abundância de espécies classificadas como
anemocóricas. No entanto, o incremento em borda não resultou em uma redução da importância
de espécies zoocóricas; o incremento na métrica MSI (complexidade de forma), por sua vez,
acarretou na diminuição da probabilidade de ocorrência de áreas ditas zoocóricas e no aumento
substancial da probabilidade de ocorrência de áreas ditas anemocóricas.
A maior ocorrência de espécies zoocóricas em florestas tropicais é resultado da forte
coevolução das plantas com os animais, porém, alterações nesse cenário foram percebidas com
a fragmentação da paisagem, como mostrado no estudo de Metzger (2000), que observou a falta
de conectividade entre fragmentos como fator que mais limitou a dispersão zoocórica.

70
Apesar de espécies anemocóricas serem mais comuns em fisionomias mais abertas,
como é o caso da Floresta Estacional Decidual no seu estado atual (LEYSER et al., 2009), a
fragmentação demonstrou-se uma importante força impulsionadora na seleção dessas espécies.
A impermeabilidade da matriz, a incapacidade de alguns animais de cruzarem áreas
abertas (VIANA e TABANEZ, 1996) e, ainda, a maior aptidão das espécies anemocóricas em
ambientes de maior intensidade de vento, resultam no favorecimento deste grupo de espécies e,
consequentemente, em implicações para a fauna e para a sucessão florestal. Já que, como
mencionado por Mayfield et al. (2005), as plantas são a principal fonte de alimento para muitos
animais, e os animais são importantes dispersores para muitas espécies de plantas nas florestas
tropicais.
CONCLUSÃO
Conclui-se que, na escala espacial considerada no presente trabalho, a quantidade de
borda, a intensidade de subdivisão da paisagem e a complexidade de forma do conjunto de
remanescentes em uma paisagem influenciaram de forma significativa as variações dos
atributos funcionas analisados nas diferentes áreas. Influenciando principalmente padrões de
dispersão, densidade da madeira (SSD) e conteúdo de matéria seca foliar (LDMC) nas
comunidades arbóreas. Por sua vez, a diversidade funcional não foi influenciada pelas métricas
da paisagem analisada.
Os processos de filtragem atuam em várias escalas espaciais, e os efeitos da
fragmentação florestal podem ser ainda pouco perceptíveis considerando o tempo de resposta
das espécies às alterações ambientais. Sem contar que a existência de uma espécie de maior
aptidão em ambientes naturais, ocorrendo em uma área degradada, não indica necessariamente
que ela seja tolerante aos efeitos da fragmentação, ela pode ser uma espécie remanescente da
fragmentação de seu habitat; mais sujeita ao desaparecimento do que as espécies de maior
aptidão nas condições bióticas e abióticas impostas pela fragmentação. Mesmo assim, foi
constatado prevalecimento de um grupo pequeno, mas abundante de espécies generalistas, com
menor densidade da madeira e folhas de aspecto coriáceo.
REFERÊNCIAS BIBLIOGRÁFICAS
ACKERLY, D. D., KNIGHT, C. A., WEISS, S. B., BARTON, K., STARMER, K. P. Leaf size,
specific leaf area and microhabitat distribution of chaparral woody plants: contrasting patterns
in species level and community level analyses. Oecologia 130, 449–457, 2002.

71
BERNARDO, P. V. S. Padrões de distribuição de mamíferos de médio e grande porte em
paisagens fragmentadas. Universidade Federal de Goiás. Dissertação de Mestrado em
Ecologia e Evolução. Goiânia – GO, 2012.
BJORNSTAD, O. N. ncf: Spatial nonparametric covariance functions. R package version 1.1-
1, 2008.
BOTTA-DUKÁT, Z. Rao’s quadratic entropy as a measure of functional diversity based on
multiple traits. Journal of Vegetation Science 16: 533-540, 2005.
CAILLIEZ, F. The analytical solution of the additive constant problem. Psychometrika,
Lisboa, v. 48, n. 2, p. 305-310. 1983.
CARNEIRO, M. S. Efeito da estrutura da paisagem sobre a riqueza e diversidade de
árvores em grupos funcionais reprodutivos. Universidade Federal de Alfenas. Dissertação
de Mestrado em Ecologia e Tecnologia Ambiental. Minas Gerais, 2013.
CASTRO, H.; LEHSTEN, V.; LAVOREL, S.; FREITAS, H. Functional response traits in
relation to land use change in the Montado. Agriculture, Ecossystems and Environment 137,
183-191, 2010.
CHAVE, J.; COOMES, D.; JANSEN, S.; LEWIS, S. L.; SWENSON, N. G; ZANNE, A. E.
Towards a worldwide wood economics spectrum. Ecology Letters, Montpellier, v. 12, p. 351-
366, 2009.
CORNELISSEN, J. H. C. et al. Handbook of protocols for standardised and easy measurement
of plant functional traits worldwide. Australian Journal of Botany, Melbourne, v. 51, n. 4, p.
335-380, 2003.
CUNNINGHAM, S. A.; SUMMERHAYES, B.; WESTOBY, M. Evolutionary divergences in
leaf structure and chemistry, comparing rainfall and soil nutrient gradients. Ecology 69: 569–
588, 1999.
DE BELLO, F., CARMONA, C. P., MASON, N. W. H., SEBASTIÀ, M. T.; LEPS, J. Which
trait dissimilarity for functional diversity: trait means or trait overlap? Journal of Vegetation
Science, 24: 807-819, 2013.
DÍAZ, S. et al. The plant traits that drive ecosystems: Evidence from three continents. Journal
of Vegetation Science 15: 295-304, 2004.
EMBRAPA. Empresa Brasileira de Pesquisa Agropecuária. Centro Nacional de Pesquisa de
Solos. Sistema brasileiro de classificação de solos. 2. ed. Rio de Janeiro: EMBRAPA-SPI,
2006.
FORMAN, R. T. T.; GODRON, M. Landscape ecology. New York: John Wiley & Sons. 619
p, 1986.
GARNIER, E.; CORTEZ, J.; BILLES, G.; NAVAS, M. L.; ROUMET, C.; DEBUSSCHE, M.
LAURENT, G.; BLANCHARD, A.; AUBRY, D.; BELLMANN, A.; NEILL, C.;
TOUSSAINT, J. P. Plant functional markers capture ecosystem properties during secondary
succession. Ecology, 85, 2630-2637, 2004.
GEOAMBIENTE. Projeto de Proteção da Mata Atlântica em Santa Catarina (PPMA-
SC/FATMA). Relatório Técnico Geral do Estado de Santa Catarina. São José dos Campos.
Geoambiente, 2008.

72
GIRÃO, L. C.; LOPES, A. V.; TABARELLI M.; BRUNA, E. Changes in tree reproductive
traits reduce functional diversity in a fragmented Atlantic Forest landscape. Plos One, 9:1-12,
2007.
GOULARTI FILHO, A. Formação econômica de Santa Catarina. Ensaios FEE, v.23, n.2,
p.977-1007, 2002.
GOWER, J. C. A general coefficient of similarity and some of its properties, Biometrics,
Washington, v. 27, n. 4, p. 857-874, 1971.
HOLDAWAY, R. J., RICHARDSON, S. J., DICKIE, I. A., PELTZER, D. A.; COOMES, D.
A. Species- and community-level patterns in fine root traits along a 120 000-year soil
chronosequence in temperate rain forest. Journal of Ecology 99: 954–963, 2011.
KEDDY, P. A. Assembly and resume rules: two goals for predictive community ecology.
Journal of Vegetation Science, n. 3, p. 157-164, 1992.
KLEIN, R. M. Flora Ilustrada Catarinense: Mapa fitogeográfico do Estado de Santa Catarina.
Itajaí. Herbário Barbosa Rodrigues, 1978.
KOTTEK, M.; GRIESER, J.; BECK, C.; RUDOLF, B.; RUBEL, F. World Map of the Köppen-
Geiger climate classification updated. Meterologische Zeitschrift, v. 15, n. 3, p.259-263, 2006.
LALIBERTÉ, E.; LEGENDRE, P. A distance-based framework for measuring functional
diversity from multiple traits. Ecology, Ithaca, v.91, p. 299-305, 2010.
LALIBERTÉ, E.; SHIPLEY, B. FD: measuring functional diversity from multiple traits, and
other tools for functional ecology. R package version 1.0-11, 2011.
LAMBERS, H.; POORTER, H. Inherent variation in growth rate between higher plants: a
search for physiological causes and ecological consequences. Advances in Ecological
Research, v. 23, p. 187 – 261, 1992.
LANG, S.; TIEDE, D. V-LATE Extension für ArcGIS – vektorbasiertes Tool zur
quantitativen Landschaftsstrukturanalyse. ESRI Anwenderkonferenz 2003 Innsbruck. CD-
ROM, 2003.
LAURANCE, W. F; NASCIMENTO, H. E. M.; LAURANCE, S.G.; ANDRADE, A.;
RIBEIRO, J; et al. Rapid decay of tree-community composition in Amazonian forest fragments.
Proc Natl Acad Sci 103: 19010–19014, 2006.
LAVOREL, S.; DÍAZ, S.; CORNELISSEN, H.; GARNIER, E.; HARRISON, S.; McINTYRE,
S.; PAUSAS, J. PÉREZ-HARGUINDEGUY, N.; ROUMET, C.; URCELAY, C.; Terrestrial
Ecosystem in a changing World. In chapter 13 Plant Functional Types: Are we Getting Any
Closer to the Holy Grail? The IGBP Series, Springer-Verlag, Berlin Heidelberg, 2007.
LEBRIJA-TREJOS, E.; PÉREZ-GARCIA, E. A.; MEAVE, J. A.; BONGERS, F.; POORTER,
L. Functional traits and environmental filtering drive community assembly in a species-rich
tropical system. Ecology, Washington, v. 91, n. 2, p. 386-398, 2010.
LELE, S. R. A new method for estimation of resource selection probability function. Journal
of Wildlife Management 73, 122–127, 2009.
LELE, S. R.; KEIM, J. L. Weighted distributions and estimation of resource selection
probability functions. Ecology 87, 3021–3028, 2006.

73
LEYSER, G.; VINISKI, M.; DONIDA, A. L.; ZANIN, E. M.; BUDKE, J. C. Espectro de
dispersão em um fragmento de transição entre Floresta Ombrófila Mista e Floresta Estacional
na região do Alto Uruguai, Rio Grande do Sul, Brasil. Pesquisas, série botânica, v. 60, p. 355-
366, 2009.
LIEBSCH, D., MARQUES, M. C. M., GOLDENBERG, R. How long does the Atlantic Rain
Forest take to recover after a disturbance? Changes in species composition and ecological
features during secondary succession. Biological Conservation, 141: 1717-1725, 2008.
LIENIN, P., KLEYER, M. Plant leaf economics and reproductive investment are responsive to
gradients of land use intensity. Agric. Ecosyst. Environ. 145: 67–76, 2011.
MASON N. W. H., DE BELLO F., MOUILLOT, D., PAVOINE S., DRAY S. A guide for
using functional diversity indices to reveal changes in assembly processes along ecological
gradients. J. Veg. Sci. 24: 794–806, 2013.
MAYFIELD, M. M.; BONI, M. F.; DAILY, G. C.; ACKERLY, D. Species and Functional
Diversity of Native and Human - Dominated Plant Communities. Ecology 86:2365 – 2372,
2005.
MAYFIELD, M. M.; ACKERLY, D.; DAILY, G. C. The diversity and conservation of plant
reproductive and dispersal functional traits in human-dominated tropical landscapes. Journal
of Ecology, London, v. 94, n. 3, p. 522-536, 2006.
METZGER, J. P. Tree Functional Group Richness and Landscape Structure in a Brazilian
Tropical Fragmented Landscape. Ecological Applications, 10(4), pp. 1147–1161, 2000.
MONSON, R. K., SMITH, S. D. Seasonal water potential components of sonoran desert plants.
Ecology 63: 113–123, 1982.
MUNGUÍA-ROSAS, M. A.; JURADO-DZIB, S. G.; MEZETA-COB, C. R.; MONTIEL, S.;
ROJAS, A.; PECH-CANCHÉ, J. M. Continuous forest has greater taxonomic, functional and
phylogenetic plant diversity than an adjacent naturally fragmented forest. Journal of Tropical
Ecology 30, 323 – 333, 2014.
MURCIA C. Edge effects in fragmented forests: implications for conservation. Tree 10: 58-
62, 1995.
MYERS, A. N. M.; MITTERMEIER, C. G.; FONSECA, G. A. B. DA; KENT, J. Biodiversity
hotspots for conservation priorities. Nature, v. 403, p.853-858, 2000.
NIMER, E. Clima. In: IBGE. Fundação Instituto Brasileiro de Geografia e Estatística.
Geografia do Brasil: Região Sul. Rio de Janeiro, p. 151-187, 1990.
OLIVEIRA, M. A.; GRILLO, S. A.; TABARELLI, M. Forest edge in the Brazilian Atlantic
Forest: drastic changes in tree species assemblages. Oryx, n. 38, p. 389-394, 2004.
OKSANEN, J. et al. Vegan: community ecology package. R package version, v. 1, p. 8-8. 2009.
PAVOINE, S.; VALLET, J.; DUFOUR, A. B.; GACHET, S.; DANIEL, H. On the challenge
of treating various types of variables: application for improving the measurement of functional
diversity. Oikos 118: 391–402, 2009.

74
PINHEIRO, J.; BATES, D.; DEBROY, S.; SARKAR, D.; R Development Core Team. nlme:
Linear and Nonlinear Mixed Models. R package version 3.1 – 100, Vienna, Austria, 2011.
R DEVELOPMENT CORE TEAM. R: A Language and Environment for Statistical
Computing. R Foundation for Statistical Computing. Vienna, Áustria. Disponível em:
<(http://www.R-project.org)> Acesso em: 2 de março de 2014.
SANTOS, B. A.; PERES, C. A.; OLIVEIRA, M. A.; GRILLO, A.; ALVES-COSTA, C. P.;
TABARELLI, M. Drastic erosion in functional attributes of tree assemblages in Atlantic forest
fragments of northeastern Brazil. Biol. Conserv. 141, 249–260, 2008.
SCHAADT, S. S. A fragmentação da Floresta Ombrófila Mista em Santa Catarina,
analisada a partir de dados do Inventário Florístico Florestal, fotografias aéreas e mapa
temático. Dissertação de Mestrado em Engenharia Ambiental. Universidade Regional de
Blumenau, Santa Catarina, 2012.
SHIPLEY, B.; LECHOWEICZ, M. J.; WRIGHT, I.; REICH, P. B. Fundamental trade-offs
generating the worldwide leaf economics spectrum. Ecology 87, 535–541, 2006.
TURNER, I. M. Sclerophylly: primarily protective? Functional Ecology 8:669–675, 1994.
VIANA, V. M.; TABANEZ, A. A. J. Biology and conservation of forest fragments in the
Brazilian Atlantic moist forest. In: SCHELLAS, J.; GREENBERG, R. Forest patches in
tropical landscapes. Washington D.C.: Island Press, p. 151-167, 1996.
VIBRANS, A. C.; SEVEGNANI, L.; GASPER, A. L.; LINGNER, D. V. Inventário Florístico
Florestal de Santa Catarina: Diversidade e conservação dos remanescentes florestais.
Blumenau: Edifurb. Vol. 1, 344 p., 2012.
VIBRANS, A. C.; MCROBERTS, R. E.; MOSER, P.; NICOLETTI, A. L. Using satellite
image-based maps and ground inventory data to estimate the area of the remaining Atlantic
forest in the Brazilian state of Santa Catarina. Remote Sensing of Environment: 130: 87–95,
2013a.
VIBRANS, A. C.; SEVEGNANI, L.; GASPER, A. L.; LINGNER, D. V. (Eds.). Inventário
Florístico Florestal de Santa Catarina: Floresta Ombrófila Densa. Blumenau: Edifurb,
2013b.
VIOLLE, C.; NAVAS, M.; VILE, D.; KAZAKOU, E.; FORTUNEL, C.; HUMMEL, I.;
GARNIER, E. Let the concept of trait be functional! Oikos 116:882-892, 2007.
WEIGELT, A.; SCHUMACHER, J.; ROSCHER, C.; SCHMID, B. Does biodiversity increase
spatial stability in plant community biomass? Ecology Letters, 11, 338–347, 2008.
WESTOBY, M.; FALSTER, D. S.; MOLES, A. T.; VESK, P. A.; WRIGHT, I. J. Plant
Ecological Strategies: some leading dimensions of variation between species. Annu. Rev. Ecol.
Syst 33, 125-159, 2002.
WRIGHT, I. J., ACKERLY, D. D., BONGERS, F., HARMS, K., IBARRA-MANRIQUEZ, G.
et al. Relationships among major dimensions of plant trait variation in 7 Neotropical forests.
Ann. Bot., 99, 1003–1015, 2007.

75
CONSIDERAÇÕES FINAIS
As análises realizadas, considerando a escala avaliada, demonstraram significativa
influência de alguns parâmetros estruturais da paisagem na seleção de estratégias das espécies,
aqueles envolvidos com efeito de borda e subdivisão. Isto leva a entender que estes fatores
sejam os principais impulsionadores das mudanças funcionais nas comunidades da Floresta
Ombrófila Densa, que por estarem interligados com alterações, como no microclima da floresta,
por exemplo, induzem a seleção de características que facilitam, principalmente, o acesso à
água e nutrientes.
Não foi identificada, no entanto, nenhuma alteração significativa na diversidade
funcional, que foi mantida nas diferentes intensidades de fragmentação. Este resultado fortalece
discussões sobre a importância dos pequenos fragmentos para a conservação da biodiversidade.
Estes podem garantir conexão funcional entre os remanescentes, que permite, por sua vez, o
fluxo gênico e, com isso, a manutenção das metacomunidades. Por outro lado, deve-se
considerar, que os efeitos da fragmentação florestal podem ser ainda pouco perceptíveis, tendo
em vista que o tempo de resposta das espécies às alterações ambientais pode ser mais longo do
que o período das nossas observações.
Teoricamente, a estrutura da paisagem afeta a dinâmica e a sobrevivência de populações
vegetais, alterando a capacidade de colonização e sucessão de certos grupos funcionais, mas
que não é a único e, em muitos casos, nem o mais importante fator responsável pela estruturação
de comunidades vegetais. Estão envolvidos outros fatores físicos, históricos e bióticos como o
tipo de matriz do entorno, a intensidade de interferência humana, as condições climáticas,
predação de propágulos e flutuação nas populações de plantas e animais.
É importante identificar os níveis mais críticos de fragmentação, aqueles em que há
alterações significativas, por exemplo, na funcionalidade e em processos nas comunidades
arbóreas, com aspectos estatísticos e teóricos concretos, para efetivamente coibir atividades
impactantes e recuperar áreas em estado crítico de fragmentação, melhorando, por exemplo, a
efetividade de corredores ecológicos na redução do efeito do isolamento entre manchas.
Correlacionar a estrutura espacial de remanescentes florestais com variáveis ecológicas
de metacomunidades, contribui para o desenvolvimento de uma base teórica para o
gerenciamento da configuração do habitat e da composição da paisagem em áreas
fragmentadas.
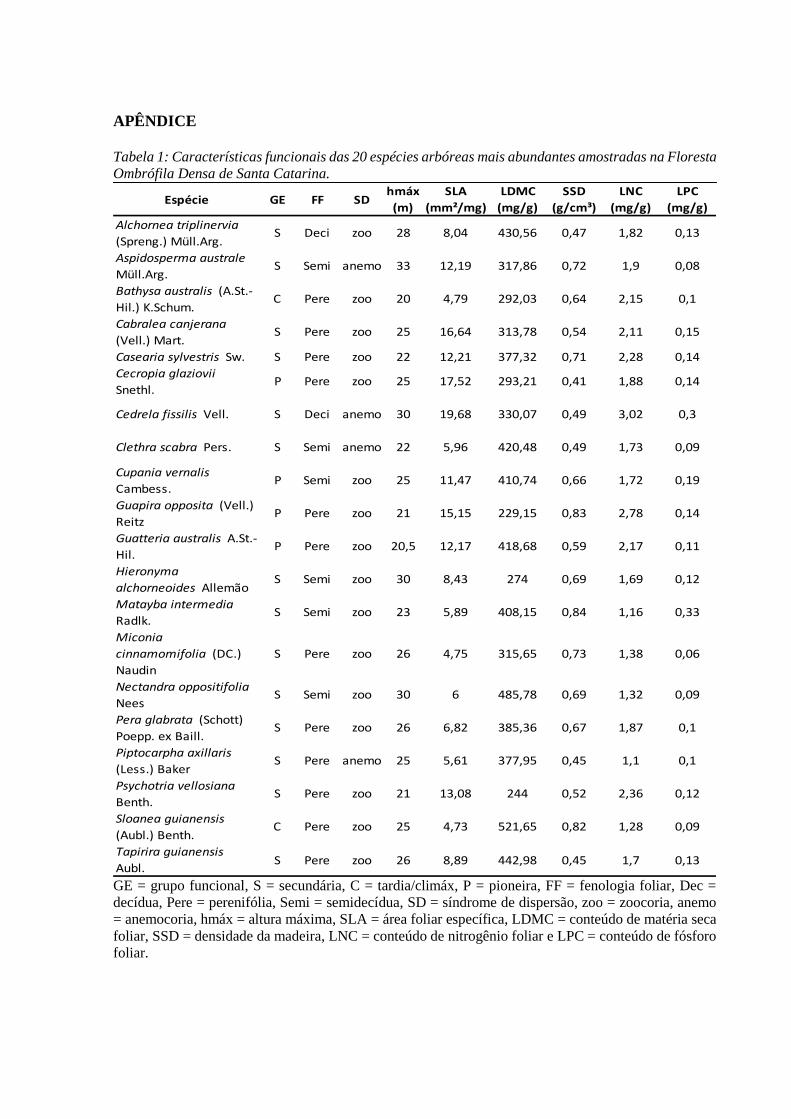
76
APÊNDICE
Tabela 1: Características funcionais das 20 espécies arbóreas mais abundantes amostradas na Floresta
Ombrófila Densa de Santa Catarina.
GE = grupo funcional, S = secundária, C = tardia/climáx, P = pioneira, FF = fenologia foliar, Dec =
decídua, Pere = perenifólia, Semi = semidecídua, SD = síndrome de dispersão, zoo = zoocoria, anemo
= anemocoria, hmáx = altura máxima, SLA = área foliar específica, LDMC = conteúdo de matéria seca
foliar, SSD = densidade da madeira, LNC = conteúdo de nitrogênio foliar e LPC = conteúdo de fósforo
foliar.
Espécie GE FF SDhmáx
(m)
SLA
(mm²/mg)
LDMC
(mg/g)
SSD
(g/cm³)
LNC
(mg/g)
LPC
(mg/g)
Alchornea triplinervia
(Spreng.) Müll.Arg.S Deci zoo 28 8,04 430,56 0,47 1,82 0,13
Aspidosperma australe
Müll.Arg.S Semi anemo 33 12,19 317,86 0,72 1,9 0,08
Bathysa australis (A.St.-
Hil.) K.Schum.C Pere zoo 20 4,79 292,03 0,64 2,15 0,1
Cabralea canjerana
(Vell.) Mart.S Pere zoo 25 16,64 313,78 0,54 2,11 0,15
Casearia sylvestris Sw. S Pere zoo 22 12,21 377,32 0,71 2,28 0,14
Cecropia glaziovii
Snethl.P Pere zoo 25 17,52 293,21 0,41 1,88 0,14
Cedrela fissilis Vell. S Deci anemo 30 19,68 330,07 0,49 3,02 0,3
Clethra scabra Pers. S Semi anemo 22 5,96 420,48 0,49 1,73 0,09
Cupania vernalis
Cambess.P Semi zoo 25 11,47 410,74 0,66 1,72 0,19
Guapira opposita (Vell.)
ReitzP Pere zoo 21 15,15 229,15 0,83 2,78 0,14
Guatteria australis A.St.-
Hil.P Pere zoo 20,5 12,17 418,68 0,59 2,17 0,11
Hieronyma
alchorneoides AllemãoS Semi zoo 30 8,43 274 0,69 1,69 0,12
Matayba intermedia
Radlk.S Semi zoo 23 5,89 408,15 0,84 1,16 0,33
Miconia
cinnamomifolia (DC.)
Naudin
S Pere zoo 26 4,75 315,65 0,73 1,38 0,06
Nectandra oppositifolia
NeesS Semi zoo 30 6 485,78 0,69 1,32 0,09
Pera glabrata (Schott)
Poepp. ex Baill.S Pere zoo 26 6,82 385,36 0,67 1,87 0,1
Piptocarpha axillaris
(Less.) BakerS Pere anemo 25 5,61 377,95 0,45 1,1 0,1
Psychotria vellosiana
Benth.S Pere zoo 21 13,08 244 0,52 2,36 0,12
Sloanea guianensis
(Aubl.) Benth.C Pere zoo 25 4,73 521,65 0,82 1,28 0,09
Tapirira guianensis
Aubl.S Pere zoo 26 8,89 442,98 0,45 1,7 0,13

77
Figura 1: Gráficos em ordem crescente dos valores dos atributos funcionais analisados para as vinte
espécies mais abundantes da Floresta Ombrófila Densa de Santa Catarina.


















