







Línguas
Páginas
Legal


EIX
O B
IOLÓ
GIC
O

I. Introdução
II. Carboidratos
III. Lipídeos
IV. Proteínas e reações enzimáticas
V. Referências
Introdução às biomoléculas: estrutura e função
Unidade 10
Autor: Professor Carlos Eduardo Anunciação

386 Módulo II — Processos biológicos na captação e na transformação da matéria e da energia
Num organismo, os alimentos podem ser utilizados de diferentes maneiras e, de acordo com as necessidades celulares, podem ser reciclados na sua forma funda-mental, ou catabolizados de modo a serem transformados em moléculas menos
complexas que serão usadas como precursores no anabolismo ou para prosseguir sua que-bra que cursa juntamente com a conservação de energia utilizada nos processos celulares.
Ao ingerir alimentos, o organismo inicia, por meio da mastigação, um processo de desagregação dos componentes desses alimentos. Esse processo físico continua quimica-mente até sua completa separação, nos intestinos, nas diversas biomoléculas, por ação das enzimas e demais componentes digestórios do organismo, possibilitando sua absorção seletiva pelas membranas celulares.
Nesta unidade, vamos discorrer sobre o papel das biomoléculas na nossa organi-zação celular, conhecer suas propriedades e entender como sua estrutura pode ser orga-nizada para desempenhar tantas funções diferentes, como reserva energética, proteção celular, lubrificação, etc.
Ao final desta unidade você deverá ser capaz de:
Reconhecer e analisar as propriedades dos carboidratos, lipídeos, aminoácidos e proteínas (aproveite para revisar as propriedades dos ácidos nucléicos na uni-dade 5 do módulo 1).
Avaliar o papel desempenhado por estas biomoléculas no organismo.
Compreender a estrutura e o mecanismo funcional das proteínas e enzimas.
#M2U10 I. Introdução

Consórcio Setentrional de Ensino a Distância 387
Biológico
P
BSC
B
Saiba mais
Metabolismo – conjunto de reações bioquímicas de transformações nas biomoléculas no nosso organismo, objetivando a obtenção, armazenamento e utilização de energia, no sentido de construção (anabolismo) ou quebra (catabolismo) de biomoléculas.
Catabolismo Anabolismo
CO2 + H2O + energia
Glicose
OCH2OH
OH
OH
OH
OH
Também chamados de glúcides ou glicídios, os carboidratos são, de longe, as bio-moléculas mais abundantes na natureza e têm funções biológicas muito importantes. São sintetizados pelos seres vivos e se apresentam de diversas maneiras, tais como a quitina nos insetos, ou como componente estrutural das paredes celulares, na forma de grandes polímeros como a celulose nos vegetais. Também são usados como um importante com-bustível para os seres vivos, sendo armazenados na forma de amido nas plantas e como glicogênio nas células dos animais. É interessante notar que tanto nas plantas como nos animais a D-glicose é o carboidrato mais utilizado pelas células e o único a compor a re-serva energética destes diferentes seres.
Eles são de fundamental importância, sendo usados como fonte energética em uma cadeia de reações, chamada glicólise, capaz de conservar grande parte da sua energia. Essa via central do nosso catabolismo, além de retirar energia para a produção de ATP, permite usar parte da molécula para biossíntese de novas outras como os aminoácidos e lipídeos. Observamos ainda a produção pela ação fermentativa de bactérias e leveduras, de produtos lácteos a partir da lactose do leite, outro tipo de açúcar e de etanol, utilizando o açúcar sacarose da cana-de-açúcar.
Quimicamente falando, os carboidratos são poliidroxialdeídos ou poliidroxicetonas (Aproveite para revisar as funções orgânicas aldeído e cetona na unidade 4 do módulo 1), compostos a base de “hidratos de carbono” na proporção de CnH2nOn, cujo número “n” vai de 3 a 10 car-bonos aproximadamente. A D-glicose (Figura 1), por exemplo, possui 6 carbonos e, portanto, tem a fórmula de C6H12O6, denominada de hexose, um poliidroxialdeído cuja fórmula é:
Na natureza podemos encontrar os carboidratos em três formas:
Como unidades simples em que são chamados de monossacarídeos, onde te-mos como exemplos glicose, frutose, galactose, glicerol, eritrose entre outros.
Como pequenas unidades de 2 ou mais monossacarídeos ligados covalentemen-te, ou oligossacarídeos, como os dissacarídeos, por exemplo, sacarose (glicose e frutose), lactose (glicose e galactose) e pequenas cadeias de açúcares nas mem-branas celulares.
Como longas cadeias constituídas de centenas a milhares de unidades monos-sacarídicas ligadas covalentemente entre si, puras ou mistas, os chamados po-lissacarídeos, como o amido, o glicogênio, a celulose e os peptideoglicanos nas paredes celulares das bactérias.
Cabe aqui uma observação – Esse padrão estrutural é comum em alguns grupos de biomoléculas e é chamado de oligomerização.
#M2U10 II. Carboidratos
Figura 1:alfa-D-glicose.

#M2U10
388 Módulo II — Processos biológicos na captação e na transformação da matéria e da energia
Introdução às biomoléculas: estrutura e função
Curiosidade
Importante: os átomos que formam uma biomolécula, não são seus monômeros. Os monômeros são moléculas. Cuidado para não confundir!
CHO
CH2OH
OH
OH
CH2OH
CH2OH
O
OH
D-eritrose (aldose) D-eritrulose (cetose)
Num determinado grupo de biomoléculas, encontramos unidades menores e me-nos complexas que são denominadas monômeros. Da ligação covalente entre monômeros são originados oligômeros e polímeros, que podem ser diferenciados entre si pelo núme-ro de monômeros que os constituem ou pela sua origem, dependendo do contexto em que são analisados.
Para o caso dos carboidratos, como acabamos de ver, os monômeros são chamados de monossacarídeos (ex: glicose), os oligômeros são os oligossacarídeos (ex: sacaro-se, formada por uma glicose ligada a uma frutose) e os polímeros são formados por centenas de monômeros (ex: amido, formado por diversas unidades de glicose).
Quando ingerimos a mandioca, o milho, a batata ou seus derivados, no ato da mas-tigação misturamos a enzima alfa-amilase, secretada pelas glândulas salivares, que ini-ciam o processo de quebra enzimática dos polissacarídeos em oligômeros e nas unidades monoméricas de glicose. Esse processo, interrompido pelo ácido que se encontra no estô-mago, se reinicia nos intestinos, onde o pH alcalino é restaurado e mais amilase é secreta-da, até sua completa clivagem, permitindo a rápida absorção pelas células intestinais.
Os açúcares possuem em sua fórmula um átomo de carbono que pode se apresentar como uma carbonila (contendo uma dupla ligação com o oxigênio) ou pode ser chamado de carbono anomérico, caso o açúcar esteja na forma cíclica. Na forma linear, quando esta dupla está no carbono 1 corresponde à função aldeído (sendo o carboidrato classificado como aldose) e, na sua outra única posição possível, no carbono 2, a função cetona (sendo o carboidrato classificado como cetose). (Figura 2)
Figura 2:A função cetona e aldeído;a) D-eritrose (aldose);b) D-eritrulose (cetose).

Consórcio Setentrional de Ensino a Distância 389
Biológico
P
BSC
B
C
Y
Y
C
B
X
A
A
X
B
CBX
A
Y
Y
A
B
Imagem no espelhode uma molécula
original
Moléculaoriginal
Molécula Quiral:mesmo sofrendo rotação a moléculanão pode ser superposta à imagem do espelho
C
C
B
X
A
A
X
B
CBX
A
Imagem do espelhode uma molécula
original
Moléculaoriginal
Molécula Aquiral:após rotação adequada a molécula pode sersuperposta à imagem do espelho
X
X
X
A)
B)
C
Y
Y
C
B
X
A
A
X
B
CBX
A
Y
Imagem no espelhode uma molécula
original
Moléculaoriginal
Molécula Quiral:mesmo sofrendo rotação a moléculanão pode ser superposta à imagem do espelho
C
C
B
X
A
A
X
B
CBX
A
B
Imagem do espelhode uma molécula
original
Moléculaoriginal
Molécula Aquiral:após rotação adequada a molécula pode sersuperposta à imagem do espelho
X
X
A
X
X
A)
B)
109.5º
109.5º
C
Os carboidratos mais comuns na natureza possuem 6, 5, 4 e 3 carbonos (Figura 3), sendo o gliceraldeído, o açúcar de 3 carbonos, a única triose a possuir isomeria óptica. Isto se dá porque os três átomos de carbono se ligam ao O e H formando funções químicas diferentes, resultando em uma propriedade já conhecida no carbono 2, o carbono quiral.
O carbono 2 do gliceraldeído, ao se ligar a 4 grupos diferentes, coloca seus quatro li-gantes em geometria tetraédrica, resultando em duas formas espaciais possíveis, D e L.
Na outra triose (cetose) possível, a dihidroxicetona, o carbono 2, ao fazer a dupla ligação com o oxigênio, se liga a apenas 3 átomos ou funções químicas diferentes, resul-tando em uma forma geométrica linear que não apresenta variações espaciais.
Este carbono com 4 ligantes diferentes, chamado de carbono assimétrico ou centro quiral, é importante para a determinação do tipo de estereoisômero (açúcar D ou L), e os açúcares que possuem mais carbonos apresentam mais carbonos quirais, entretanto o car-bono quiral mais distante da carbonila (o carbono da dupla ligação) é quem determina a forma isomérica D ou L.
Figura 3:Assimetria molecular:a) moléculas quiraisb) aquirais.
(b)
(a)

#M2U10
390 Módulo II — Processos biológicos na captação e na transformação da matéria e da energia
Introdução às biomoléculas: estrutura e função
CHO
C
CH2OH
OHH
CHO
C
CH2OH
HO H
D-Gliceraldeído L-Gliceraldeído
Fórmulas de projeção de Fischer
Fórmulas em perspectiva
CHO
C
CH2OH
OHH
CHO
C
CH2OH
HO H
D-Gliceraldeído L-Gliceraldeído
H O
H OH
H OH
C
C
C
H
H
H OH
O
H OH
C
C
C
HGliceraldeído Dihidroxicetona
CHO
C
CH2OH
OHH
CHO
C
CH2OH
HO H
D-Gliceraldeído L-Gliceraldeído
Fórmulas de projeção de Fischer
Fórmulas em perspectiva
CHO
C
CH2OH
OHH
CHO
C
CH2OH
HO H
D-Gliceraldeído L-Gliceraldeído
H O
H OH
H OH
C
C
C
H
H
H OH
O
H OH
C
C
C
HGliceraldeído Dihidroxicetona
CHO
C
CH2OH
OHH
CHO
C
CH2OH
HO H
D-Gliceraldeído L-Gliceraldeído
Fórmulas de projeção de Fischer
Fórmulas em perspectiva
CHO
C
CH2OH
OHH
CHO
C
CH2OH
HO H
D-Gliceraldeído L-Gliceraldeído
H O
H OH
H OH
C
C
C
H
H
H OH
O
H OH
C
C
C
HGliceraldeído Dihidroxicetona
A presença de carbono(s) quiral(is) no carboidrato gera uma forma geométrica sin-gular, de característica não plana, resultando, quando da formação de polímeros, em macromoléculas com formas e características especiais tão diferentes como o amido e a celulose.
Atividade complementar 11. Desenhe a molécula da glicose e calcule quantos carbonos assimétricos ela possui.
2. Vivencie esta diferença química espetando em duas bolinhas de isopor ou em dois limões, palitos de dente em ângulos de 109 graus.
3. Una as duas bolas ou mais através dos palitos e verifique se é possível assumir a forma plana? Use palitos partidos ao meio para representar os grupos –OH.
Na natureza e principalmente no meio aquoso, os carboidratos adquirem a for-ma cíclica, pois se tornam moléculas mais estáveis. A ciclização dos açúcares ocorre nas moléculas com 5 ou mais carbonos, através da carbonila e o carbono assimétrico mais distante desta.
Na molécula da glicose ela ocorre entre o carbono 1 com o carbono 5, resultando numa estrutura contendo 6 átomos na formação do anel. Esta forma assumida é seme-lhante à da estrutura química do pirano e a glicose é chamada de D-glicopiranose.
Outro monossacarídeo, chamado frutose, apresenta a dupla ligação com o oxigênio no carbono 2 (cetose), sendo que a ciclização ocorre entre o carbono 2 e 5, resultando em 5 átomos no anel, semelhante a molécula química furano e é chamada de D-frutofuranose.
Observe que na frutose tanto o carbono 1 quanto o 6 ficam de fora do anel. Observe, também, que na ciclização ocorre a atração do par de elétrons livre do oxigênio do carbo-no assimétrico pelo carbono quiral mais distante, resultando no deslocamento temporá-rio da dupla ligação com o oxigênio, surgindo uma função –OH (hidroxila).
Lembrando que os carbonos internos do açúcar são também assimétricos e, portan-to, não possibilitam ao carbono assumir qualquer posição plana, que torna inviável a for-mação do anel entre carbonos vizinhos. O grupo –OH pode surgir acima ou abaixo do anel, uma vez que o carbono que apresentava a função carbonila agora tornou-se assimétrico. Esse carbono assimétrico exclusivo da forma cíclica é chamado de carbono anomérico.
Este grupo –OH quando fica acima do anel é chamado de tipo alfa ou ά D-glicopiranose e, quando para baixo, é chamado de ß D-glicopiranose (figura 4).
Figura 4:Formas isoméricas do gliceraldeído e a dihidroxicetona.

Consórcio Setentrional de Ensino a Distância 391
Biológico
P
BSC
B
Saiba mais
Você sabia que esta descoberta permitiu que os cientistas desenvolvessem os adoçantes? Os açúcares sintéticos utilizados como adoçantes não são metabolizados pelo nosso organismo e foram desenvolvidos com a intenção de simular a forma geométrica do açúcar que mais nos desencadeia a sensação de doce, o dissacarídeo sacarose (figura 6). Eles estimulam os receptores de uma forma mais eficiente que a sacarose, portanto nos dão maior sensação de doçura.
Saiba Mais
As papilas gustativas estão localizadas na mucosa que recobre a língua. Em cada papila estão localizados vários receptores nervosos que, ao serem estimulados, transmitem impulsos que são decodificados no cérebro. Um fator que prejudica o paladar é o acúmulo de resíduos alimentares na mucosa da língua, pois estes bloqueiam o estimulo às papilas.
O
OH
H
H
HO
CH2OH
HOCH2 H1
2
3 4
5
6
β
O
CH2OH
HOOH
HHHOH H
H
α
23
4
5
6
1
O
Sacaroseα-D-glicopiranosil β-D-frutofuranosídeo
Glc(α1 2β)Fru
Atividade complementar 2A partir do modelo molecular que você fez com as bolas de isopor, tente repre-sentar a estrutura química da glicose, agora com seis bolas. Você vai verificar que apenas duas formas ou configurações de anel são possíveis de se obter, a de barco e de cadeira.
Voltando ao princípio da termodinâmica que você aprendeu, onde diz que quanto mais espaçados ou distantes estão os átomos de uma molécula, mais estável ela será, você verá que a forma de cadeira é a única que representa esta alternativa (Figura 5). Aproveite para verificar a posição do grupo –OH em alfa e beta.
Atividade complementar 3Tente representar a molécula da frutose e verifique a diferença estrutural das for-mas piranosídicas e furanosídicas.
Agora você está pronto para entender os reflexos nas nossas vidas das formas quí-micas que aprendemos. Os diferentes açúcares nos dão sensação de doçuras diferentes. As nossas papilas gustativas possuem moléculas de proteínas capazes de reconhecer os açúcares pela sua fora geométrica. Como os açúcares, mesmos entre as hexoses possuem a disposição dos grupos –OH distribuídos ao redor do anel diferentes, eles se encaixam com intensidades diferentes nos receptores das papilas, com maior e menor intensidade, nos impingindo maior ou menor sensação de doce.
O
CH2OH
HOOH
OH
H HHOH H
H
1
23
4
5
6
-D-Glicopiranose
O
CH2OH
HOOH
OHH
H
HOH H
Hß-D-Glicopiranose
ß-D-Frutofuranose
Piran
O
H2C
HC CH
CH
HC
Furan
O
C
HC CH
C
O
OH
H
H
HO
OHH
HOCH2 CH2OH1
2
34
5
6
-D-Frutofuranose
O
OH
H
H
HO
OH
H
HOCH2
CH2OHH H
Figura 5: d-glicopiranose e d-frutofuranose.

#M2U10
392 Módulo II — Processos biológicos na captação e na transformação da matéria e da energia
Introdução às biomoléculas: estrutura e função
O
CH2OH
HOOH
OH
H HHOH H
H
O
CH2OH
HOOH
OHH
H
HOH H
H
-D-Glicose ß-D-Glicose
Hemiacetal
Álcool
O
CH2OH
HOOH
H HHOH H
H
O
CH2OH
OH
OHH
H
HOH H
H
-D-Glicose
-D-Glicopiranosil-(1 4)-D-glicopiranose
ß- -D-Glicose
hidrólise condensação
H2O H2O
1
23
4
5
6
1
23
4
5
6
O
Acetal Hemiacetal
Maltose
Atividade complementar 4Vamos explorar mais nossa visão geométrica da glicose. Experimente construir duas moléculas cíclicas de glicose – uma na forma alfa e outra na forma beta.
Tente fazer a sobreposição das duas moléculas sem romper nenhuma ligação. Você vai verificar que, apesar de serem muito semelhantes e de conterem exatamente os mesmos átomos e os mesmos tipos de ligação, a disposição espacial do carbono 1 é diferente, de forma que somente poderia existir sobreposição entre as moléculas, caso as ligações covalentes fossem modificadas.
Indo mais adiante, os açúcares podem se unir para formar os oligo e polissacarídeos através da ligação entre os grupos –OH do carbono anomérico de um monossacarídeo. Esta ligação é chamada de ligação glicosídica e ocorre com a saída de uma molécula de água (Figura 7).
Atividade complementar 5Aproveitando o modelo molecular construído por você e observando o grupo –OH do carbono 1 (anomérico) na posição beta, verifique a possível união de dois anéis pelos carbonos 1 e 4 (β1 → 4).
Considerando a estrutura na configuração de cadeira do anel e a ligação β1 → 4, ve-rifique que o grupo –OH em alfa do carbono 1 do primeiro anel permite uma boa aproxi-mação do segundo açúcar sem torcê-lo, ou seja, mantendo-o no mesmo plano do primeiro anel. A figura resultante é como uma escada, onde os degraus representam o assento e o encosto da cadeira do nosso açúcar. Daí é possível construir uma “fita” em linha reta, embora na forma de escada, de um polissacarídeo. Esta forma linear permite que possa-mos justapor duas ou mais linhas de polissacarídeo, formando “folhas”, como acontece no caso da celulose (Figura 8).
Figura 7:Ligação glicosídica.

Consórcio Setentrional de Ensino a Distância 393
Biológico
P
BSC
B
O
OO
O O
OH OH
HO
OH
Unidades de D-glicose ligadas (β1 → 4)
OH
HO
O
OO
O O
OH OH
HO
OH
Unidades de D-glicose ligadas (β1 → 4)
OH
HO
Atividade complementar 6Agora experimente trocar o grupo –OH de beta para alfa no carbono 1 e aproximá-lo do grupo –OH do carbono 4 do outro anel. Verifique que isto só é possível com a torção da molécula de um dos anéis.
Ao ligar mais monossacarídeos por ligação glicosídica, a forma assumida pela es-trutura resultante é uma estrutura helicoidal, como uma verdadeira mola. Esta forma glicosídica resultante da união alfa1 → 4, forma uma grossa fi ta helicoidal contendo uma fenda central, permitindo que muitas moléculas de água se agreguem no seu interior.
Esta é a estrutura de um dos polímeros do amido, a amilose que tem a forma linear. A amilopectina é outra estrutura encontrada no amido e ocorre quando na cadeia linear fazemos ligações do tipo alfa1 → 6 (além das alfa1 → 4) criando ramifi cações na cadeia central. O carbono 4 do açúcar inserido na posição 6 permanece livre e é utilizado para criar uma nova fi ta helicoidal linear do tipo alfa1 → 4. (Figura 9)
Quanto mais ramifi cações acontecem na cadeia central, mais globuloso e espon-joso fi ca o polissacarídeo, formando grânulos na batata e, no caso da mandioca, menos ramifi cada e mais dura ela fi ca. Por isso, quando fritamos ou congelamos uma batatinha ou mandioca elas murcham, porque a água que se instala dentro da estrutura helicoidal dando consistência ao amido se desloca, deixando os espaços internos vazios.
Vale ressaltar que os grupos –OH do açúcar atraem moléculas de água via pontes de hidrogênio para o interior da estrutura da mola, dando o aspecto de fi rmeza (dureza) ao amido encontrado na batata e na mandioca.
Figura 8: Estrutura da celulose.
HO OH
CH2OHO
O
O
HO
OH
CH2OH
O
O
Unidades de D-Glicose ligadas ( l 4)
A)
B)
HO OH
CH2OHO
O
O
HO
OH
CH2OH
O
O
Unidades de D-Glicose ligadas ( l 4)
A)
B)
Figura 9: a) estrutura linear da amilose; b) estrutura helicoidal da amilopectina (cada hexágono é um resíduo de glicose).

#M2U10
394 Módulo II — Processos biológicos na captação e na transformação da matéria e da energia
Introdução às biomoléculas: estrutura e função
Outra diferença interessante, o amido, por formar grânulos, não induz agregação forte entre os polímeros, não possui forma definida e pode ter sua estrutura desfeita pelo aquecimento. A união β1 → 4 da celulose, por formar fitas ou folhas, permite um contato muito íntimo entre as fibras e pode ser desfibrilada pela moagem, mas volta a adquirir consistência de folha ao se juntar às fibras com uma peneira ou moldá-la com as mãos, como o papier-mâché.
A essa altura você deve ter se perguntado: como eu posso dissolver a sacarose no café e não posso dissolver o amido e nem a celulose?
Bem, isto acontece porque a massa molecular de um monossacarídeo e até de al-guns dissacarídeos, em relação às pontes de hidrogênio com as moléculas de água, são favoráveis para manter a molécula miscível na água. Quando o número de monossacarí-deos no polímero alcança um número muito grande, fica impossível manter o polissaca-rídeo miscível na água, porque a razão entre pontes de hidrogênio e massa molecular fica muito desfavorável, fazendo o polímero precipitar.
Esta propriedade é muito utilizada pelos trabalhadores da mandioca, que sepa-ram os polímeros de amido em duas frações, a totalmente insolúvel de grande massa molecular para fazer a farinha e a de massa molecular inferior, mas ainda insolúvel em água, o polvilho.
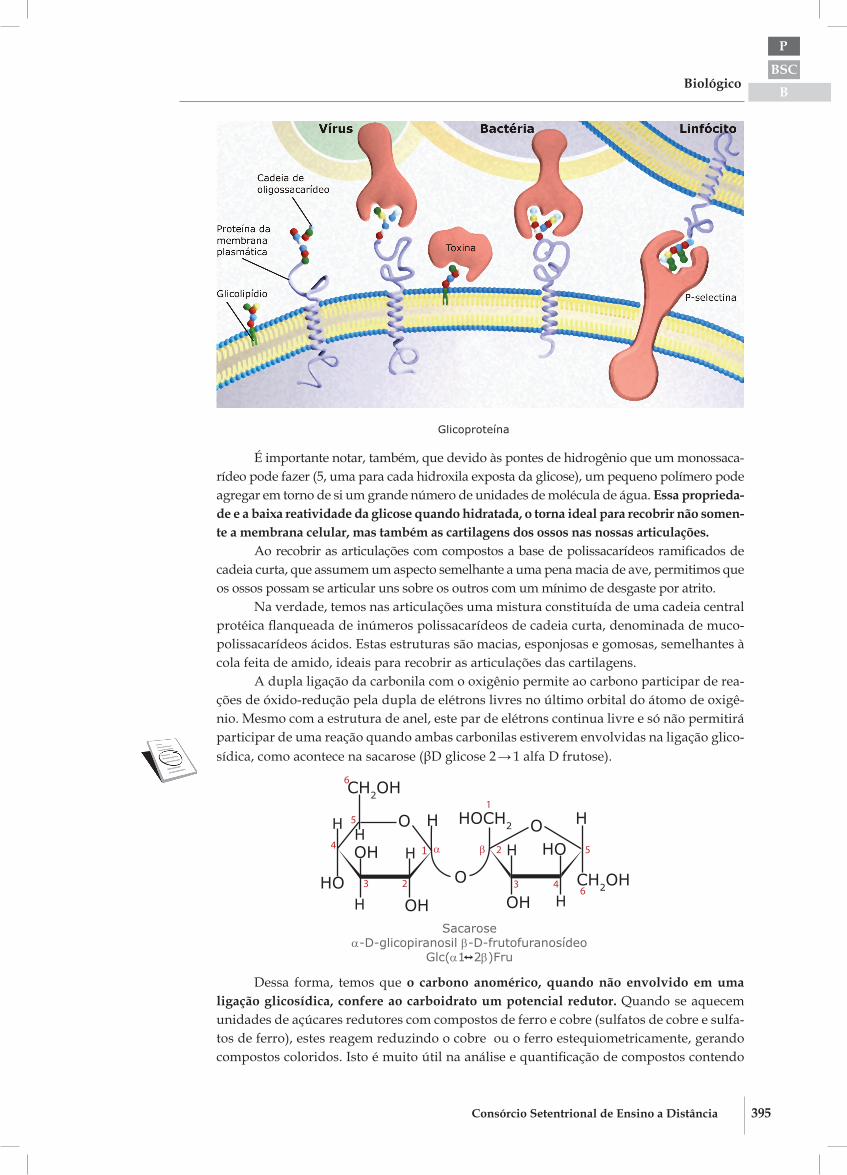
Nas membranas das nossas células a estrutura helicoidal resultante das ligações alfa1 → 4 e alfa1 → 6 são muito úteis. Ligadas aos lipídeos e também às proteínas da superfície externa da célula, encontramos uma camada de lipopolissacarídeos e glico-polissacarídeos ramificados (glicocálice), delgada ou espessa como no glicogênio e a zona pelúcida do óvulo.
As cadeias alfa1 → 4 sofrem diversas ramificações do tipo alfa1 → 6, que originam inúmeras novas pequenas cadeias polissacarídicas alfa1 → 4. A estrutura resultante se as-semelha a uma frondosa árvore, cuja copa esconde e protege a membrana citoplasmática e, pelo pequeno número de unidades de monossacarídeos, não são insolúveis.
Glicocálix
Consórcio Setentrional de Ensino a Distância 395
Biológico
P
BSC
B
O
OH
H
H
HO
CH2OH
HOCH2 H1
2
3 4
5
6
β
O
CH2OH
HOOH
HHHOH H
H
α
23
4
5
6
1
O
Sacaroseα-D-glicopiranosil β-D-frutofuranosídeo
Glc(α1 2β)Fru
É importante notar, também, que devido às pontes de hidrogênio que um monossaca-rídeo pode fazer (5, uma para cada hidroxila exposta da glicose), um pequeno polímero pode agregar em torno de si um grande número de unidades de molécula de água. Essa proprieda-de e a baixa reatividade da glicose quando hidratada, o torna ideal para recobrir não somen-te a membrana celular, mas também as cartilagens dos ossos nas nossas articulações.
Ao recobrir as articulações com compostos a base de polissacarídeos ramificados de cadeia curta, que assumem um aspecto semelhante a uma pena macia de ave, permitimos que os ossos possam se articular uns sobre os outros com um mínimo de desgaste por atrito.
Na verdade, temos nas articulações uma mistura constituída de uma cadeia central protéica flanqueada de inúmeros polissacarídeos de cadeia curta, denominada de muco-polissacarídeos ácidos. Estas estruturas são macias, esponjosas e gomosas, semelhantes à cola feita de amido, ideais para recobrir as articulações das cartilagens.
A dupla ligação da carbonila com o oxigênio permite ao carbono participar de rea-ções de óxido-redução pela dupla de elétrons livres no último orbital do átomo de oxigê-nio. Mesmo com a estrutura de anel, este par de elétrons continua livre e só não permitirá participar de uma reação quando ambas carbonilas estiverem envolvidas na ligação glico-sídica, como acontece na sacarose (βD glicose 2 → 1 alfa D frutose).
Dessa forma, temos que o carbono anomérico, quando não envolvido em uma ligação glicosídica, confere ao carboidrato um potencial redutor. Quando se aquecem unidades de açúcares redutores com compostos de ferro e cobre (sulfatos de cobre e sulfa-tos de ferro), estes reagem reduzindo o cobre ou o ferro estequiometricamente, gerando compostos coloridos. Isto é muito útil na análise e quantificação de compostos contendo
Glicoproteína

#M2U10
396 Módulo II — Processos biológicos na captação e na transformação da matéria e da energia
Introdução às biomoléculas: estrutura e função
OC C
CH2
CH
O
O C
CH2
O
O O1
2
3
açúcares, como o plasma ou soro e a urina. A base das reações químicas, utilizadas por sistemas e aparelhos simples utilizados para checar o nível de glicose no sangue, é muito útil na determinação do nível de glicemia e controle do diabetes.
Atualmente são utilizados métodos enzimáticos, que apresentam maior sensibi-lidade e maior especificidade, porém a simplicidade dos métodos que usam a proprie-dade redutora dos açúcares ainda faz desse método um recurso útil em situações de recursos limitados.
Na unidade 3 você viu a estrutura e alguns aspectos sobre as moléculas dos lipíde-os, suas interações com as moléculas da água e alguns aspectos na formação de micelas e das membranas celulares. Agora vamos conhecer um pouco mais destas moléculas tão conhecidas genericamente por nós e que exercem tantas funções importantes e diferentes no nosso organismo, muitas delas ainda desconhecidas.
Devido à natureza apolar e à baixa reatividade, apesar da alta concentração de energia nas suas ligações, os lipídeos são considerados moléculas neutras no nosso or-ganismo. Mas alguns tipos dessas moléculas no nosso organismo apresentam uma parti-ção seletiva com a água, originada pela presença de um grupo polar na molécula (cabeça polar), por isso são moléculas chamadas de anfipáticas. Esta propriedade permite a for-mação das membranas e manutenção de algumas funções celulares e, dada a estabili-dade destas, são muito exploradas para diferenciar a composição das membranas e sua superfície externa.
Atividade complementar 7Procure pesquisar quais são os tipos de lipídeos presente no organismo humano ou animal, sua estrutura química e funções.
A maioria dos tipos de lipídeos no nosso organismo é derivada dos ácidos graxos e em segundo plano dos colesteróis. Estes ácidos gra-xos geralmente estão ligados à molécula do gli-cerol via ligação éster, contendo de duas a três cadeias de ácidos graxos. Quando a ligação se
dá com três cadeias de ácido graxo, formando um triacilglicerol, to-dos os três grupos –OH da molécula do glicerol estão esterificados, não existindo nenhum gru-pamento iônico livre, assim, a molécula torna-se totalmente apolar e é utilizada como armazenamento de energia pelo organismo.
Quando apenas duas cadeias de ácido graxo estão ligadas ao glicerol, temos o diacilglicerol, que possui uma estrutura ci-líndrica, estrutura ideal para a formação de membranas, como você viu nas unidades anteriores.
#M2U10 III. Lipídeos
Triacilglicerol

Consórcio Setentrional de Ensino a Distância 397
Biológico
P
BSC
B
*Todos os ácidos têm suas fórmulas apresentadas na forma não-ionizada. Em pH 7, todos os ácidos graxos livres têm um carboxilatoionizado. Note que a numeração dos átomos de carbono se inicia no carbono carboxílico.
†O prefixo “n-” indica uma estrutura “normal”, não-ramificada. Por exemplo, “dodecanóico” simplesmente indica 12 átomos de carbono, quepoderiam ser dispostos em uma variedade de formas ramificadas; “n-dodecanóico” especifica a forma linear, não-ramificada. Para ácidosgraxos insaturados, a configuração de cada dupla ligação está indicada; em ácidos graxos biológicos, a configuração é quase sempre cis.
Esqueletocarbônico
12:0
14:0
16:0
18:0
20:0
24:0
16:1(Δ9)
18:1(Δ9)
18:2(Δ9,12)
18:3(Δ9,12,15)
20:4(Δ5,8,11,14)
Ponto defusão (ºC)
44,2
53,9
63,1
69,6
76,5
86,0
-0,5
13,4
-5
-11
-49,5
Água
0,063
0,024
0,0083
0,0034
Benzeno
2,600
874
348
124
Nome sistemático†
Ácido n-dodecanóico
Ácido n-tetradecanóico
Ácido n-hexadecanóico
Ácido n-octadecanóico
Ácido n-eicosanóico
Ácido n-tetracosanóico
Ácido cis-9-hexadecenóico
Ácido cis-9-octadecenóico
Ácido cis-,cis-9,12-octadecadienóico
Ácido cis-,cis-,cis-9,12,15-octadecatrienóico
Ácido cis-,cis-,cis-,cis-5,8,11,14-icosatetraenóico
Nome comum(derivação)
Ácido láurico(do latim, laurus,“árvore do louro”)
Ácido mirístico(do latim, Myristica,gênero de noz moscada)
Ácido palmítico(do latim, palma,“palmeira”)
Ácido esteárico(do grego, stear,“gordura dura”)
Ácido araquídico(do latim, Arachis,“gênero dos legumes”)
Ácido lignocérico(do latim, lignum,“madeira” + “cera”)
Ácido palmitoléico
Ácido oléico(do latim, oleum, “óleo”)
Ácido linoléico(do grego, linon, “linho”)
Ácido α-linolênico
Ácido araquidônico
Estrutura*
CH3(CH2)10COOH
CH3(CH2)12COOH
CH3(CH2)14COOH
CH3(CH2)16COOH
CH3(CH2)18COOH
CH3(CH2)22COOH
CH3(CH2)5CHCH(CH2)7COOH
CH3(CH2)7CHCH(CH2)7COOH
CH3(CH2)4CH CHCH2CHCH(CH2)7COOH
CH3CH2CHCHCH2CH CHCH2CHCH(CH2)7COOH
CH3(CH2)4CH CHCH2CHCHCH2CH CHCH2CHCH(CH2)3COOH
Solubilidade a 30ºC(mg/g solvente)
Quanto à cadeia carbônica, estes ácidos graxos podem apresentar somente ligações simples entre os átomos de carbono, ou apresentar uma ou mais duplas ligações espaça-das geralmente por 3 carbonos. Os sufixos –oléico, -linoléico e –linolênico são acrescenta-dos no nome da molécula quando ela possui 1, 2 ou 3 insaturações (Tabela 1).
Tabela 1: Nomenclatura dos ácidos graxos
É importante notar que os mamíferos não sintetizam ácidos graxos insaturados con-tendo uma dupla ligação após o carbono 9 (C9). Nossa maquinaria metabólica (mamíferos) apresenta limites na capacidade da síntese relativa ao número de carbonos e ao número de duplas ligações, ou seja, não possuímos enzimas para executar esses tipos de anabolismo.
Atividade complementar 8Pesquise sobre o que são e qual a importância dos ácidos graxos essenciais.
Você também viu na unidade 5 do módulo 1 que o número de cadeias instauradas nos diacilgliceróis da membrana citoplasmática é um fator crucial na adaptação das nos-sas células às mudanças climáticas.

#M2U10
398 Módulo II — Processos biológicos na captação e na transformação da matéria e da energia
Introdução às biomoléculas: estrutura e função
H OH
H
C
C
H
C
CHC
C9
O O
2
Lipídios dearmazenamento
(neutro)
Triacilgliceróis
Fosfolipídios
Ácido graxo
Ácido graxo
Ácido graxo
Glic
erol
Ácido graxo
Ácido graxo
Glic
erol
PO4 Álcool
Ácido graxo
PO4 Colina
Glicerofosfolipídios Esfingolipídios Esfingolipídios
Lipídios de membrana (polares)
Glicolipídios
Esf
ingolip
ídio
s
Ácido graxo
Mono ouoligossacarídeoE
sfin
golip
ídio
s
Ao contrário do princípio da termodinâmica, os dois átomos de carbono ao redor da dupla ligação do carbono C9 ocupam a posição Cis e não trans, como era de se esperar (Figura 10). Isso impõe uma torção na cadeia hidrocarbonatada, dificultando a interação inter-molecular desses compostos, conferindo característica de tendência ao estado líqui-do em temperatura ambiente, ideal para períodos de temperatura mais baixas.
Nas membranas, o terceiro grupo -OH livre da molécula do glicerol pode ser ligado a grupos polares, tendo como conseqüência um aumento na polaridade desses lipídeos, através da inserção de um fosfato (um fosfolipídeo) ou um carboidrato, mais especifica-mente a glicose (glicolipídeo). Além de aumentar a polaridade e a coesão destes compo-nentes das membranas, eles servem como elo para a adição de diferentes tipos de novas biomoléculas, originando classes especiais de lipídeos (Figura 11), resultando em ativida-des específicas ao tipo celular, conforme você verá a seguir.
Triacilgliceróis
Os triacilgliceróis são as gorduras de reserva comuns nos animais e vegetais. Nos animais, as cadeias de ácidos graxos apresentam carbonos saturados, flexíveis, capazes de formar uma estrutura cristalina organizada, portanto são sólidos a temperatura ambiente. Nos vegetais, estes ácidos graxos são insaturados, portanto líquidos a temperatura am-biente. Estes ácidos graxos são muito procurados pelas indústrias alimentícias e farma-cêuticas para a produção de margarinas, batons, sorvetes, etc.
Esfingolipídeos
As membranas de algumas células especializadas, como no caso das células nervo-sas, apresentam um tipo diferenciado dos diacilgliceróis que contêm um composto cha-
Figura 10:Diacilglicerol – linoléico. Ácido graxo insaturado com dupla ligação no carbono 9 da cadeia.
Figura 11: classes de lipídios de armazenamento e de membrana.

Consórcio Setentrional de Ensino a Distância 399
Biológico
P
BSC
B
HO 3CH
2CH N C
O
Esfingosina
Ácido graxo
Esfingolipídio(estrutura geral)
N
1CH2
CH CH3CH (CH2)12
O X
GalGlc
H
Neu5Ac
Gal GalNacGlc
+O CH2 CH2 N(CH3)3
O
P
O-
HH
H
H
H
OH
OH
CH2OH
OH
O
Fórmula de XNome de X
Fosfocolina
Oligossacarídeocomplexo
Glicose
Di, tri outetrassacarídeo
Nome de esfingolipídio
Esfingomielina
Lactosilceramida(um globosídio)
Gangliosídio GM2
Glicolipídios neutros
Glicosilcerebrosídio
Ceramida
mado de esfingosina (Figura 12). Nos esfingolipídeos, temos, substituindo o glicerol e um ácido graxo, um aminoálcool de cadeia longa ligado à molécula do ácido graxo denomi-nado ceramida.
Este tipo lipídico apresenta a porção polar com estrutura física muito semelhante aos diacilgliceróis, e a cabeça polar pode se ligar ao fosfato ou a uma molécula de açúcar, que serve como elo para a adição de variados tipos de moléculas, muito semelhante aos vistos nos diacilgliceróis (Figura 13). Esses tipos de lipídeos são comuns na bainha de mielina dos axônios das células nervosas.
Figura 12: Esfingosina.
Figura 13: Grupos R dos tipos de ligantes as esfingosinas.

#M2U10
400 Módulo II — Processos biológicos na captação e na transformação da matéria e da energia
Introdução às biomoléculas: estrutura e função
Saiba Mais
Os gangliosídeos e os cerebrosídeos são lipídios especiais (GM1 e GM2) encontrados no tecido nervoso, principalmente no cérebro, e seu catabolismo é complexo. Nos indivíduos afetados pela doença genético-bioquímica de Tay-Sachs observa-se o acúmulo de tais compostos dentro das células.
CH3
CH
CH2
CH2
CH2
CH CH3
CH3
A B
CH3
CH3
HO
C D
Cadeiaalquilalateral
Núcleo esteróideCabeçapolar
CH3
CH3
HOÁcido taurocólico(un ácido biliar)
CH3
C NH
O
CH2 CH2 SO3
OH
OH
CH2
HO
H3C
H3C
CH3
CH3
Colecalciferol(vitamina D3)
Quando a ceramida se liga a moléculas de carboidratos diferentes como a glicose e galactose e, a partir daí formam-se cadeias longas e até ramificadas, forma uma classe diferente de lipídeos denominados gangliosídeos e cerebrosídeos.
Colesterol e seus derivados
O colesterol é uma molécula derivada do esterol, que apresenta 4 anéis cíclicos fun-didos, representando um núcleo tetracíclico. O álcool de esterol mais abundante da natureza é o colesterol que serve, por sua vez, como base da biossíntese de várias moléculas do nosso organismo, como os sais biliares, vitamina D e alguns hormônios esferóides. Não são saponificáveis, uma vez que não apresentam ácido graxo na sua estrutura, mas apresentam uma forma tridimensional cilíndrica permitindo seu ajus-tamento na camada lipídica. Imerso na estrutura das membranas, o colesterol in-terfere no nível de fluidez destas. Embora o organismo animal sintetize o colesterol (Figura 14), ele é um alimento importante na dieta, devido ao seu elevado consumo.
Figura 14: Colesterol, Ácido taurocólico, Colecalciferol

Consórcio Setentrional de Ensino a Distância 401
Biológico
P
BSC
B
Figura 15: triacontanilpalmitato, componente da cera de abelhas.
Ceras
As ceras (Figura 15) são ésteres de ácidos graxos de cadeia lon-ga, com álcoois de cadeia longa. As cadeias carbônicas podem ser sa-turadas ou insaturadas e apresentam de 14 a 36 átomos de carbono.
São comuns nas secreções glandulares da pele, dos cabelos e cavidade auricular dos animais, recobrindo folhas das plantas e em pássaros, como impermeabilizante para as penas.
Devido à longa cadeia carbônica flexível, as ceras podem for-mar estruturas cristalinas estáveis, no estado sólido a temperatura ambiente, com elevado ponto de fusão.
Uma enzima é um catalisador biológico que trabalha em condições moderadas de temperatura e pH para efetuar uma catálise química, sem produzir produtos intermediá-rios ou produtos alterados. Ela é na realidade uma proteína que apresenta uma atividade catalítica. Esta fantástica biomolécula é capaz de assumir as mais variadas atividades na célula que vão desde o catabolismo/anabolismo, ao transporte pelas membranas como vimos anteriormente, proteção imunológica, etc. Antes de entendermos como ela pode processar estas atividades, vamos entender brevemente como é sua estrutura molecular.
As proteínas/enzimas são compostas por aminoácidos ligados covalentemente (Fi-gura 16), formando um longo cordão protéico, enovelado (“como um novelo de lã”) no caso das enzimas ou assumindo outras formas variadas como as fibras musculares (Figura 17).
#M2U10 IV. Proteínas e reações enzimáticas
Figura 16: estrutura química de um aminoácido.
Figura 17: Níveis estruturais de uma proteína.

#M2U10
402 Módulo II — Processos biológicos na captação e na transformação da matéria e da energia
Introdução às biomoléculas: estrutura e função
A coesão da estrutura do novelo é mantida por interações químicas e físicas fracas, mas que por serem numerosas se tornam fortes, como as pontes de hidrogênio, interações iônicas e hidrofóbicas. Assim, a estrutura da proteína não é rígida, isto é, ela é flexível, influenciável pelas condições do meio, já que estas interações são muito influenciadas pelo calor, teor de sais e íons no meio.
Os responsáveis por estas interações são os grupos “R” dos aminoácidos que com-põem a estrutura das proteínas. Grupos R dos aminoácidos com carga positiva atraem os de carga negativa, os grupos hidrofóbicos se atraem na água e o oxigênio, com seu par de elétrons livres, realiza pontes de hidrogênio (H-H) com o hidrogênio de outro. Quanto mais destes aminoácidos uma proteína possui, mais força de coesão terá. Mas verifique que todos os aminoácidos diferem na sua estrutura química apenas pelo grupo R e, que o grupo carboxila de um pode realizar uma ligação covalente com o grupo amino do outro, chamada de ligação peptídica.
Assim, dois aminoácidos unidos sempre estarão com um grupo amino e outro car-boxílico livre nas extremidades. Eles podem formar, então, longos cordões de aminoáci-dos, sempre ficando expostos nesta cadeia, chamada de cadeia polipeptídica, os grupos R, a dupla do oxigênio do ácido carboxílico e o hidrogênio do grupo amino.
Além das interações entre os grupos R, o oxigênio da carboxila e o hidrogênio do grupo amino podem se atrair formando inúmeras pontes de hidrogênio. Estas pontes de hidrogênio, quando ocorrem repetidamente, formam padrões estruturais, classifica-dos como estrutura secundária, por exemplo, uma estrutura espiralada chamada de alfa hélice, que se alterna com trechos aonde esta interação não existe, camada de trechos de beta-forma (lineares).
O resultado destas interações é a formação da estrutura tridimensional enovelada (Figura 17), que pode apresentar formas muito variadas e possuir fendas ou cavidades aonde acontecem as reações catalíticas e de regulação.
Durante o processo evolutivo, estas fendas passaram a apresentar a forma de um contramolde que permite a entrada do substrato e, como sua estrutura é flexível, elas po-dem se ajustar perfeitamente a eles durante o processo da reação, através da exposição de seus grupos R que podem atrair os grupos químicos da molécula do substrato.
Esta atração pode ser tão intensa que pode provocar reações químicas transitórias entre a enzima e o substrato, modificando-o, ou seja, realizando a catálise. Quando estes grupos R não podem interagir com o substrato, as enzimas adsorvem dentro de seu sítio catalítico, moléculas capazes de fazê-lo, moléculas chamadas de coenzimas, que intera-gem de modo transitório também. Estas coenzimas são fundamentais para a catálise e as enzimas fornecem o ambiente para que a reação molecular aconteça.
Alguns destes grupos alteram a estrutura da proteína, sendo que sua forma va-ria de acordo com a seqüência dos aminoácidos, ou seja, com as dobraduras da ca-deia. As interações que “unem” as cadeias ou dobras das proteínas são produtos dos grupos R dos aminoácidos que possuem atração. Elas são fracas, pontes de hidrogênio, interações iônicas e hidrofóbicas, e fortes, as pontes dissulfeto que ocorrem entre dois grupos R de aminoácidos cisteína próximos, sendo esse padrão estrutural que depende de interações entre grupos R de aminoácidos da mesma cadeia, chamado de estrutura terciária. (figura 18)
As enzimas geralmente possuem massa molecular muito grande, podendo ser cons-tituídas por mais de uma cadeia polipeptídica (estrutura quaternária) e ainda apresentar componentes químicos diferentes dos aminoácidos, chamados de grupos prostéticos ou coenzimas (vitaminas). Exemplos destes componentes são metais, moléculas orgânicas complexas como as hemoglobinas, vitaminas, etc. (Figura 19).

Consórcio Setentrional de Ensino a Distância 403
Biológico
P
BSC
B
Grupo R não-polares e alifáticos Grupo R aromáticos
Grupo R carregados positivamente
Grupo R carregados negativamente
Grupo R não-carregados,mas polares
AlaninaValina
Leucina
MetioninaIsoleucina
Serina
Treonina Cisteína
Asparagina
Prolina
Glutamina
Lisina
Arginina
Histidina
Aspartato
Glutamato
C HH3N+
COO-
HGlicina
C HH3N+
COO-
CH3
C HH3N+
COO-
CH3 CH3
CH C HH3N+
COO-
C HH3N+
COO-
CH3 CH3
CH2
CH
C H
H
H3N+
COO-
CH2
C HH3N+
COO-
C
CH2 CH2
S
CH3
CH3
CH3
C HH3N+
NH3+
COO-
CH2
CH2
CH2
C HH3N+
COO-
CH2
CH2
C HH3N+
COO-
CH2
C
CCH2
NH2
CH3H
C HH3N+
COO-
C
CH3
C HH3N+
COO-
CH2OH
C HH2N
+
COO-
CH2
CH2H2C
C HH3N+
COO-
CH2
C HH3N+
COO-
CH2
CH2C
C HH3N+
COO-
CH2
SH
H2N O C
H2N O
Triptofano
CH2
C HH3N+
COO-
CH2
OH
C HH3N+
COO-
CH2
C CHNH
NH
C NH2
+
NH
H
CHN
C HH3N+
COO-
COO-
CH2
C HH3N+
COO-
COO-
CH2
CH2
TirosinaFenilalanina
OH
Fe
N+
C
C
C
O O-O O
CCH3
CH2
CH3
CH2
CH3
CH
CH
CH
CH
CH
CH
CH2 CH2
CH2
C
C C
C
C
C
C C
C C CC
N
-NN
CH3
C
C
CH2
-
N
N
NH2
N
N
O
O
O
O
P
P
O
O
O
O
O
FMN
CH2 O
HH
OH
H
OH
H
NHCH3
CH3
CH2
CH2
HCOH
HCOH
HCOH
NN
N
FAD
Flavina adenina dinucleotídeo (FAD)e flavina mononucleotídeo (FMN)
-
-
CH2O O
O
P
P
O
O
O
O
O
HH
OH
NAD+
Adenina
H
OH
H
CH2 O
HH
OH
H
OH
H
NNH2
HC
O
N
N
NH2
N
N
-
-
+
Figura 18: Classificação dos aminoácidos baseado nas características do seu grupo R.
Figura 19: grupos prostéticos e coenzimas; a) grupo Heme das hemoglobinas e mioglobinas; b) NAD (nicotinamida Adenina Dinucleotídeo);
(a) (b)

#M2U10
404 Módulo II — Processos biológicos na captação e na transformação da matéria e da energia
Introdução às biomoléculas: estrutura e função
Fe
N+
C
C
C
O O-O O
CCH3
CH2
CH3
CH2
CH3
CH
CH
CH
CH
CH
CH
CH2 CH2
CH2
C
C C
C
C
C
C C
C C CC
N
-NN
CH3
C
C
CH2
-
N
N
NH2
N
N
O
O
O
O
P
P
O
O
O
O
O
FMN
CH2 O
HH
OH
H
OH
H
NHCH3
CH3
CH2
CH2
HCOH
HCOH
HCOH
NN
N
FAD
Flavina adenina dinucleotídeo (FAD)e flavina mononucleotídeo (FMN)
-
-
CH2O O
O
P
P
O
O
O
O
O
HH
OH
NAD+
Adenina
H
OH
H
CH2 O
HH
OH
H
OH
H
NNH2
HC
O
N
N
NH2
N
N
-
-
+
Substrato(bastão de metal)
Estado de transição(bastão dobrado)
Produtos(bastão partido)
Ener
gia
liv
re,
G
G
SP
Enzima complementar ao substrato
Ímãs
Ener
gia
liv
re,
G
G G
SP
GM
catnão-cat
ES
A
B
A forma tridimensional da proteína-enzima é fundamental para a sua atividade e nas dobraduras, ela apresenta fendas ou cavidades que simulam o contramolde do subs-trato (molde – substrato / enzima – contramolde), onde podem apresentar papel cata-lítico, este é o chamado sítio catalítico. Quanto mais esse contramolde é semelhante ao molde, mais específi ca é a enzima e maior poder catalítico ela apresentará (Figura 20). Esse modelo fi cou conhecido como chave-fechadura nos livros didáticos, pois para cada molde (substrato ou chave) existe uma enzima específi ca (contramolde ou fechadura).
Figura 19:Grupos prostéticos e coenzimas;c) FAD (Flavina Adenina Dinucleotídeo).
(c)
Consórcio Setentrional de Ensino a Distância 405
Biológico
P
BSC
B
Substrato(bastão de metal)
Estado de transição(bastão dobrado)
Produtos(bastão partido)
Ener
gia
liv
re,
G
G
SP
Enzima complementar ao substrato
Ímãs
Ener
gia
liv
re,
G
G G
SP
GM
catnão-cat
ES
A
B
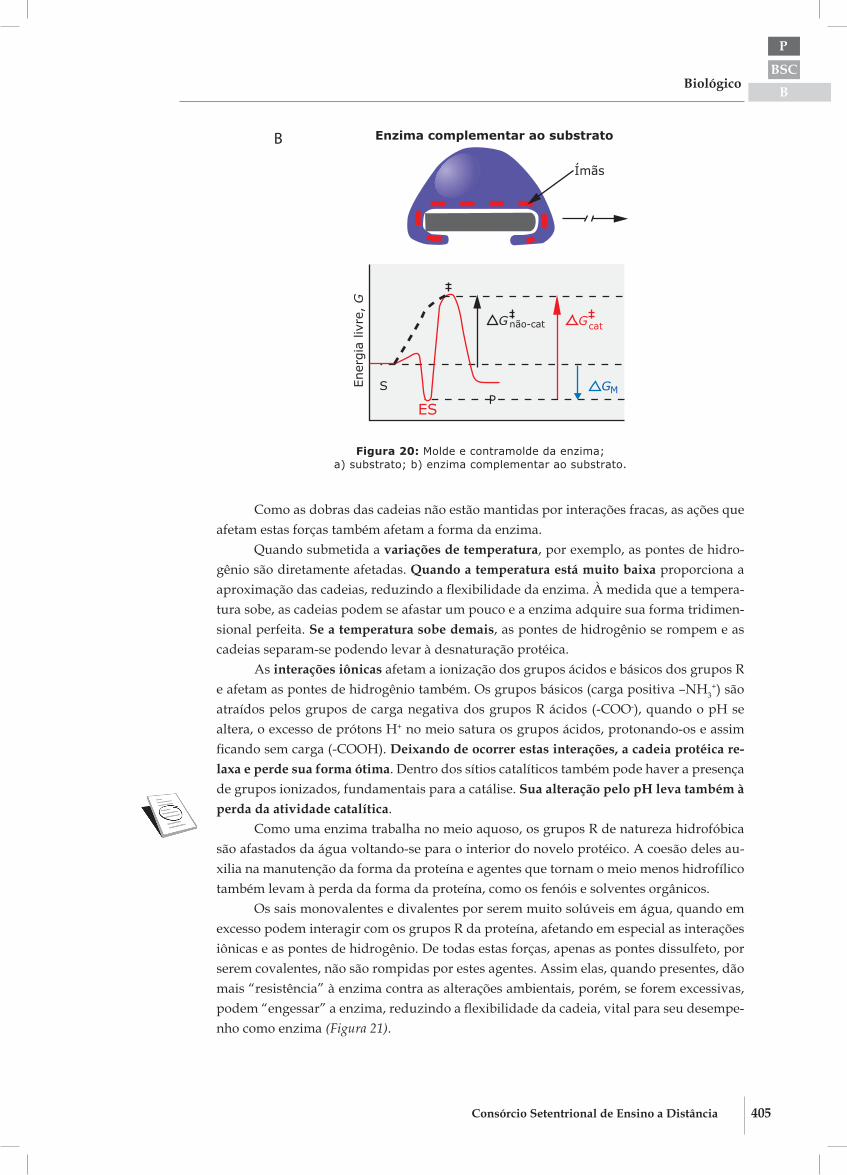
Como as dobras das cadeias não estão mantidas por interações fracas, as ações que afetam estas forças também afetam a forma da enzima.
Quando submetida a variações de temperatura, por exemplo, as pontes de hidro-gênio são diretamente afetadas. Quando a temperatura está muito baixa proporciona a aproximação das cadeias, reduzindo a fl exibilidade da enzima. À medida que a tempera-tura sobe, as cadeias podem se afastar um pouco e a enzima adquire sua forma tridimen-sional perfeita. Se a temperatura sobe demais, as pontes de hidrogênio se rompem e as cadeias separam-se podendo levar à desnaturação protéica.
As interações iônicas afetam a ionização dos grupos ácidos e básicos dos grupos R e afetam as pontes de hidrogênio também. Os grupos básicos (carga positiva –NH3
+) são atraídos pelos grupos de carga negativa dos grupos R ácidos (-COO-), quando o pH se altera, o excesso de prótons H+ no meio satura os grupos ácidos, protonando-os e assim fi cando sem carga (-COOH). Deixando de ocorrer estas interações, a cadeia protéica re-laxa e perde sua forma ótima. Dentro dos sítios catalíticos também pode haver a presença de grupos ionizados, fundamentais para a catálise. Sua alteração pelo pH leva também à perda da atividade catalítica.
Como uma enzima trabalha no meio aquoso, os grupos R de natureza hidrofóbica são afastados da água voltando-se para o interior do novelo protéico. A coesão deles au-xilia na manutenção da forma da proteína e agentes que tornam o meio menos hidrofílico também levam à perda da forma da proteína, como os fenóis e solventes orgânicos.
Os sais monovalentes e divalentes por serem muito solúveis em água, quando em excesso podem interagir com os grupos R da proteína, afetando em especial as interações iônicas e as pontes de hidrogênio. De todas estas forças, apenas as pontes dissulfeto, por serem covalentes, não são rompidas por estes agentes. Assim elas, quando presentes, dão mais “resistência” à enzima contra as alterações ambientais, porém, se forem excessivas, podem “engessar” a enzima, reduzindo a fl exibilidade da cadeia, vital para seu desempe-nho como enzima (Figura 21).
Figura 20: Molde e contramolde da enzima;a) substrato; b) enzima complementar ao substrato.

#M2U10
406 Módulo II — Processos biológicos na captação e na transformação da matéria e da energia
Introdução às biomoléculas: estrutura e função
R CO R
O OHH
Cisteína
CH
OH
O
PE = 100,5ºC
Ácidos carboxílicos
Pontes dissulfeto
Alto PE (formação de duas pontes de hidrogênio)
CH3CH2OH PE = 78ºC
COO-
CH
CH2
SH
SH
CH2
CH
COO-
NH3
H3N+
+
Cisteína
Cisteína
COO-
CH
CH2
S
S
CH2
CH
COO-
NH3
H3N+
+
CR
O H
O H O
OC R Dímero
2H+ + 2e-
2H+ + 2e-
‘
Pontes de hidrogênioEntre um grupo R ácido, ácidos carboxílicos e a H2O
Tendo essa idéia de como é uma estrutura protéica de uma enzima, vamos ver como ela realiza sua função.
Para que uma reação catalítica ocorra, é necessário haver empenho de energia e, ao final do processo, a enzima deve estar novamente pronta para um novo trabalho, ou seja, não pode sofrer alterações químicas permanentes.
Em outras palavras, uma enzima participa do processo catalítico, proporcionando um “local” (sitio catalítico) adequado para a reação ocorrer, reduzindo a energia que seria necessária para a realização do processo químico. (Figura 22)
Figura 21: Interações entre os grupos R.

Consórcio Setentrional de Ensino a Distância 407
Biológico
P
BSC
B
O C OH + H
O
HH
R1 R2 R1
:A-
O
C
O. .. .
OR2 OR2
H
HH
R1
O
C
O+
H A
H A
:B
:BHH
R1 R2
O
C
O. .. .
OR2
H BH
R1
C OH + H
O
R1 OR2
O
C
O +
Catálise ácido geral
Catálise básica geral
V0 =Vmáx[S]
Km + [S]
V0 =Vmáx[S]
Km + [S]
Vel
oci
dad
e in
icia
l, V
0(μ
M/m
in)
Concentração do substrato, [S] (mM)
Vmáx
1/2Vmáx
Km
figura 24
figura 22
Mas e a energia de onde vem? Vem do próprio sistema onde ela está quando a re-ação é exergônica e, quando ela é endergônica (a energia do produto é maior que energia do substrato) é necessário haver doação, que pode vir do ATP, de uma reação exergônica doadora acoplada ou outra molécula com muita energia.
Um exemplo de reação:Numa reação exergônica, a energia para “disparar” o processo catalítico vem
da própria energia livre do substrato ou do sistema da solução. Devido à cadeia po-lipeptídica ser flexível e ter alta massa molecular, muito maior que o substrato, ao se chocar com o substrato no sítio catalítico, ela aproveita a energia cinética do substrato para executar trabalho.
Pelo princípio da termodinâmica, a energia cinética pode ser transformada em tra-balho, e é isto que a enzima faz neste caso. No choque com o substrato, a enzima sofre uma distorção local, fazendo com que o sitio “abrace” o substrato, englobando-o todo ou no local onde vai haver a transformação.
A energia cinética nesse instante foi transformada em movimento mecânico sobre o substrato, a energia necessária para que a catálise ocorra. Por isso que em baixa tem-peratura as reações catalíticas desta natureza são muito lentas, difíceis de ocorrer, pois é necessária uma energia cinética do substrato que o meio não proporciona.
Então, desse ”abraço” da enzima com o substrato, a enzima mecanicamente aproxi-ma seus grupos químicos ionizados, o cofator ou coenzima, permitindo uma ação atrativa forte ao substrato, alterando-o (Figura 23). Essa ação é muito rápida e como resultado do efeito elástico sofrido com o impacto com o substrato, ela “empurra” o material para fora do sitio catalítico, que, transformado, já é o produto.
Figura 22: Gráfico representativo de uma reação enzimática simples.

#M2U10
408 Módulo II — Processos biológicos na captação e na transformação da matéria e da energia
Introdução às biomoléculas: estrutura e função
O C OH + H
O
HH
R1 R2 R1
:A-
O
C
O. .. .
OR2 OR2
H
HH
R1
O
C
O+
H A
H A
:B
:BHH
R1 R2
O
C
O. .. .
OR2
H BH
R1
C OH + H
O
R1 OR2
O
C
O +
Catálise ácido geral
Catálise básica geral
V0 =Vmáx[S]
Km + [S]
V0 =Vmáx[S]
Km + [S]
Vel
oci
dad
e in
icia
l, V
0(μ
M/m
in)
Concentração do substrato, [S] (mM)
Vmáx
1/2Vmáx
Km
figura 24
figura 22
Assim, quanto maior o número de moléculas de substrato na solução contendo a energia cinética apropriada para que no impacto ocorra o “abraço” efetivo da enzi-ma com o substrato, maior a quantidade de produto formado.
Assim, tudo que afetar a forma da proteína de forma a tornar o sítio catalítico mais próximo da forma do molde (substrato) afetará a catálise. Quanto mais relaxado esse sitio, maior será a energia cinética necessária do substrato para provocar esse “abraço” protéico.
Ou seja, menor será o número de moléculas da solução contendo esse alto nível de energia cinética, portanto mais difícil e lenta será a quantidade de produto (veloci-dade) da reação.
Uma outra característica da ação das enzimas em conseguir energia para a catálise é pela atração do substrato. A arquitetura do sitio catalítico é mantida por interações entre os grupos R dos aminoácidos. Ao entrar no sitio catalítico, o substrato atrai parte destas interações, provocando uma reação em cadeia, com deslocamentos sucessivos destas interações, o que resulta na mudança conformacional das estruturas tanto da proteína quanto do substrato.
Isto faz com que a estrutura do sítio (cadeias protéicas) se feche como um “abraço” sobre o substrato. O substrato sofre, então, mudanças químicas, e desfaz sua atração pelos grupos R da cadeia polipeptídica da enzima. Esta então refaz suas interações, voltando para sua forma original.
Esse modelo deixa claro a necessidade de uma complementaridade parcial entre substrato e sítio catalítico para que o processo possa se iniciar, mas o maior nível de com-plementaridade somente deve ser atingido durante o processo, quando enzima e substra-to já tiverem sofrido alterações estruturais, numa situação chamada estado de transição.
Essa especificidade do molde-contramolde é traduzida num símbolo estudado ma-tematicamente por duas pesquisadoras (Michaelis e Menten) que ganharam um prêmio Nobel por terem conseguido, pela primeira vez, explicar matematicamente como uma reação enzimática ocorre e seus parâmetros (Figura 24).
Figura 23: Catálise ácido base geral.
Figura 24: Equação de Michaelis e Menten (Equação de MM). Km = constante de MM = concentração de substrato específico, onde a enzima está na metade de sua velocidade máxima.

Consórcio Setentrional de Ensino a Distância 409
Biológico
P
BSC
B
1
V0
1
Vmáx
Km
Vmáx[S]= +
1
Km
1
[S]
Km
Vmáx
Inclinação=
1
V0
1
Vmáx
-
1
V0
1
Vmáx
Km
Vmáx[S]= +
1
Km
1
[S]
Km
Vmáx
Inclinação=
1
V0
1
Vmáx
-
Equaçao de Lineweaver-Burk
De acordo com o princípio da trigonometria, uma curva pode ser representada por uma equação de reta. Lineweaver-Burk fez esse tratamento com a equação (Figura 25) de MM e com o gráfico, multiplicando a equação por -1, assim:
Elas chamaram esta especificidade como a afinidade que a enzima tem pelo seu substrato específico ou Km.
O Km é um valor de concentração de substrato e pode ser medido baseado na ve-locidade máxima da reação.
Observe no gráfico que o valor de Km corresponde ao valor de concentração de substrato em que a enzima trabalha na metade de sua velocidade máxima (Vmáx). Ou seja, é a projeção da metade do valor de Vmáx na curva, sob o eixo da concentração.
Em termos práticos, significa que quanto menor o valor de Km maior o poder ca-talítico da enzima, pois menor concentração de substrato específico ela precisa para alcan-çar a metade da Vmáx. Falando molecularmente, quanto mais eficiente é a interação com o contramolde (sítio catalítico), menor deformação a enzima precisará sofrer para “abraçar” o substrato e, consequentemente, de menor energia cinética precisará o substrato.
FUTUYMA, Douglas. J., Biologia Evolutiva, Riberão Preto: Soc. Bras. Genet. 1997.
LEHNINGER , A.; NELSON, D.I.; COX, M.M. Princípios de Bioquímica. 2. ed. São Paulo: Savier, 1995.
MARZZOCO, A. & TORRES, B.B. Bioquímica Básica. 2. ed., Rio de Janeiro: Ed. Guanabara, 1999.
MURRAY, R. K. et al. Harper’s Biochemistry. 25. ed. LONDON: Ed. Appleton & Lange, 2000.
ROSKOSKI. Bioquímica. Rio de Janeiro: Ed. Guanabara Koogan, 1997
STRYER, L. Biochemistry. 4. ed. New York: Ed. W.E. Freeman and Company, 1995.
#M2U10 V. Referências
Figura 25: Equação e representação gráfica de Lineweaver-Burk.
Top Related