





Línguas
Páginas
Legal


;ç
Universidade Federal do Pará
Campus Universitário de Bragança
Instituto de Estudos Costeiros
Programa de Pós-Graduação em Biologia Ambiental
LUZINETE LOURENÇO DA SILVA
RIQUEZA E ABUNDÂNCIA DE GALHAS EM MANGUES PLANTADOS
DE Avicennia germinans (L.) L.: TESTANDO A HIPÓTESE DA
ARQUITETURA DA PLANTA
Orientador: Dr. Marcus Emanuel Barroncas Fernandes
Co-orientadora: Dra. Rita de Cássia Oliveira dos Santos
BRAGANÇA, PARÁ
2015

i
Universidade Federal do Pará
Campus Universitário de Bragança
Instituto de Estudos Costeiros
Programa de Pós-Graduação em Biologia Ambiental
RIQUEZA E ABUNDÂNCIA DE GALHAS EM MANGUES PLANTADOS
DE Avicennia germinans (L.) L.: TESTANDO A HIPÓTESE DA
ARQUITETURA DA PLANTA
LUZINETE LOURENÇO DA SILVA
Dissertação apresentada ao Programa de Pós-graduação em
Biologia Ambiental (Ecologia de Ecossistemas Costeiros e
Estuarinos), da Universidade Federal do Pará, Campus de
Bragança, como parte dos requisitos necessários à obtenção
do título de Mestre em Biologia Ambiental.
Orientador: Dr. Marcus Emanuel Barroncas Fernandes
Co-orientadora: Dra. Rita de Cássia Oliveira dos Santos
BRAGANÇA, PARÁ
2015

ii
LUZINETE LOURENÇO DA SILVA
RIQUEZA E ABUNDÂNCIA DE GALHAS EM MANGUES PLANTADOS
DE Avicennia germinans (L.) L.: TESTANDO A HIPÓTESE DA
ARQUITETURA DA PLANTA
Dissertação apresentada ao Programa de Pós-graduação em
Biologia Ambiental (Ecologia de Ecossistemas Costeiros e
Estuarinos), da Universidade Federal do Pará, Campus de
Bragança, como parte dos requisitos necessários à obtenção
do título de Mestre em Biologia Ambiental.
Orientador: Dr. Marcus Emanuel Barroncas Fernandes
Co-orientadora: Dra. Rita de Cássia Oliveira dos Santos
BANCA EXAMINADORA
__________________________________________
Dr. Marcus Emanuel Barroncas Fernandes
Campus de Bragança, Universidade Federal do Pará
Orientador
___________________________________________
Titular 1: Dra. Moirah Paula Machado de Menezes
Campus de Bragança, Universidade Federal do Pará
___________________________________________
Titular 2: Dra. Erneida Coelho de Araujo
Campus de Bragança, Universidade Federal do Pará
___________________________________________
Titular 3: Dr. Cleidson Paiva Gomes
Campus de Bragança, Instituto Federal de Educação, Ciência e Tecnologia do Pará
___________________________________________
Suplente: Dra. Marlucia Bonifácio Martins
Museu Paraense Emílio Goeldi (MPEG)-Belém-PA
___________________________________________
Suplente: Dra. Marivana Borges
Campus de Bragança, Universidade Federal do Pará

iii
“A coisa mais indispensável a um homem é reconhecer o uso
que deve fazer do seu próprio conhecimento”.
Platão
Voltaire

iv
AGRADECIMENTOS
À UFPA pela oportunidade de mestrado em especial a todos os funcionários.
Ao Fundo Amazônia – BNDES pelo financiamento da pesquisa (Projeto No.3052) e a
FAPESPA – VALE (ICAAF No.068/2011) pelo apoio logístico.
Ao meus orientadores, prof. Dr. Marcus Fernandes e profª Dra. Rita Santos pela ajuda,
paciência, sugestões e pelo grande apoio profissional e pessoal.
Aos amigos do LAMA, pelo apoio pessoal, em especial ao Alex, Mayara Vieira, Pablo
Vilhena, Diego Ribeiro, Lanna Costa, Danilo Gardunho, Hudson, Cesar Braga e
Gustavo Moura.
Aos monitores das atividades de replantio de manguezal e produção de mudas: Moises,
Valdir, Roberto, Adelino e Valmir pela importante ajuda em campo.
Aos amigos Diego e Lenita pela ajuda com as análises estatísticas.
À amiga Sanae Hayashi pela elaboração do mapa.
A todos da minha família por todo apoio e incentivo, em especial minha avó Maria Diva
Ribeiro da Silva.
E à Deus pelo desafio deste trabalho e a coragem de tê-lo finalizado.

v
RESUMO
A hipótese da arquitetura da planta prediz que variações na complexidade estrutural da
planta hospedeira influenciam a comunidade de insetos herbívoros, sua dinâmica e
desempenho. Nós tentamos verificar o efeito da arquitetura da Avicennia germinans (L.)
L. sobre a riqueza e abundância dos morfotipos de galhas em sítios de mangue rplantado
na península de Ajuruteua, Bragança, costa amazônica brasileira. As amostragens foram
realizadas em cinco sítios usando 10 indivíduos e 20 ramos apicais por indivíduo. Os
atributos estruturais (variáveis explicativas) e a riqueza e abundância de galhas
(variáveis resposta) foram estimados e analisados com regressão múltipla através de
modelos lineares generalizados (GLM). Vinte e dois morfotipos de galha foram
registrados com a ocorrência de 16 em todos os sítios. Um total de 7.602 galhas com
média de 1,3±0,4 galhas por folha foi registrado. Os morfotipos MF7 e MF4 foram os
mais abundantes e representaram menos de 40% do total. Embora A. germinans seja
uma planta super-hospedeira, o parasitismo das espécies indutoras de galha não afetou o
seu desenvolvimento ou sua sobrevivência. Este fato somado à previsibilidade e
atratividade do recurso aos artrópodes galhadores podem explicar a alta riqueza e
abundância de morfotipos de galhas associadas a A. germinans nos manguezais.
Contudo, a complexidade arquitetônica dessa espécie arbórea do mangue não
influenciou a riqueza, apenas a abundância de morfotipos de galha, corroborando
parcialmente com a hipótese da complexidade estrutural em sítios de mangue plantados.
Palavras-chave: complexidade estrutural; herbivoria endofítica; mangue plantado; morfotipo de
galha.

vi
LEGENDAS DAS FIGURAS E TABELAS
FIGURAS
FIGURA 1. Mapa da área de estudo mostrando a localização dos cinco sítios de
trabalho. (A) Costa do Estado do Pará; (B) Península de Ajuruteua; (C) Sítios de
trabalho numerados de 1 a 5).
FIGURA 2. Curva de rarefação de morfotipos de galhas por indivíduo de Avicennia
germinans em sítios de replantio, na península de Ajuruteua, Bragança, costa amazônica
brasileira.
FIGURA 3. Curvas de rarefação de morfotipos de galhas por ramos coletados nos dez
primeiros indivíduos de Avicennia germinans em sítios de replantio, na península de
Ajuruteua, Bragança, costa amazônica brasileira.
FIGURA 4. Morfotipos (MF) de galhas associados a Avicennia germinans em sítios de
replantio, na península de Ajuruteua, Bragança, costa amazônica brasileira.
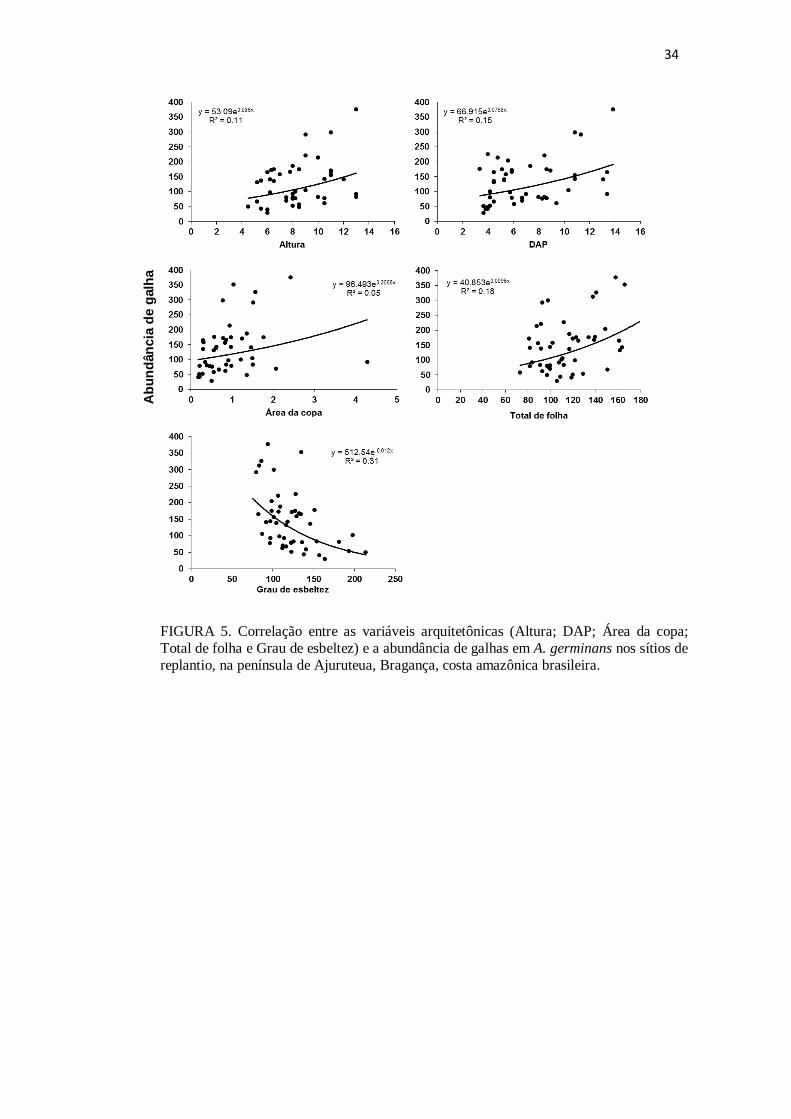
FIGURA 5. Correlação entre as variáveis arquitetônicas (Altura, Área da copa, DAP,
Total de folha e Grau de esbeltez) e a abundância de galhas em A. germinans nos sítios
de replantio, na península de Ajuruteua, Bragança, costa amazônica brasileira.
TABELAS
TABELA 1. Valores médios (± desvio padrão) das variáveis arquitetônicas e da
abundância e riqueza de galhas para cada sítio de replantio de Avicennia germinans, na
península de Ajuruteua, Bragança, costa amazônica brasileira. DAP = Diâmetro à
Altura do Peito.
TABELA 2. Análise de componentes principais com as variáveis arquitetônicas de
Avicennia germinans em sítios de replantio, na península de Ajuruteua, Bragança, costa
amazônica brasileira.
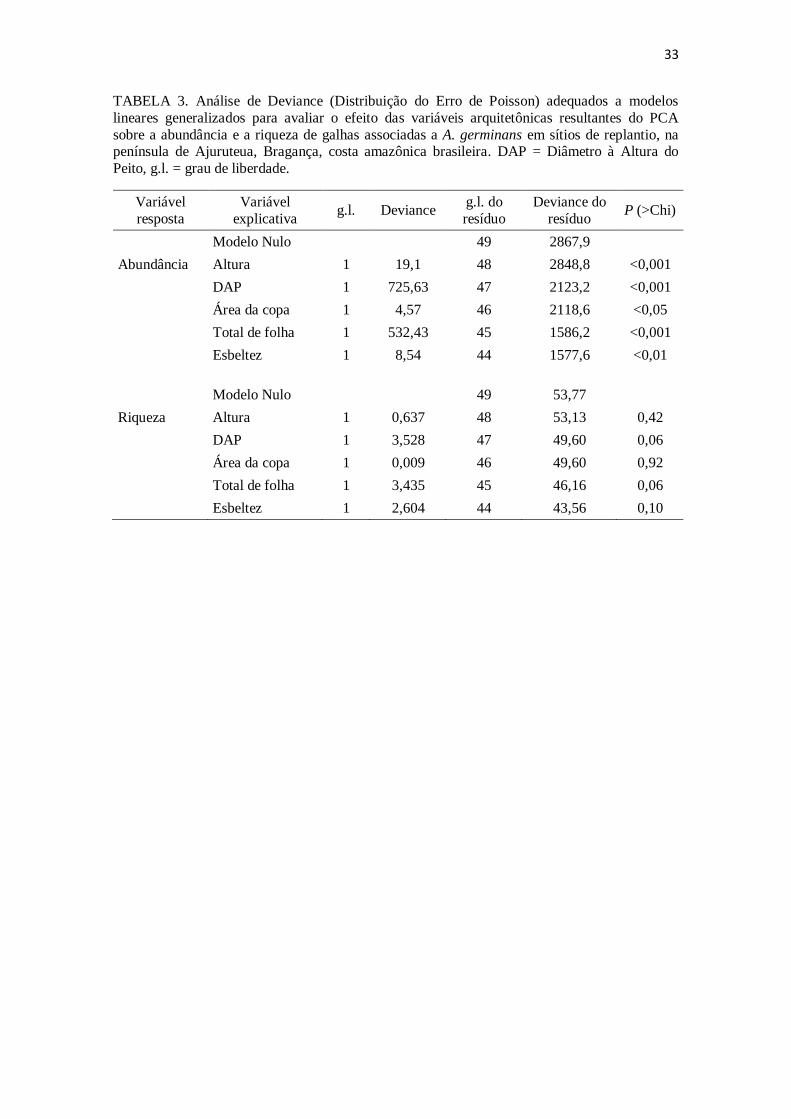
TABELA 3. Análise de Deviance (Distribuição do Erro de Poisson) adequados a
modelos lineares generalizados para avaliar o efeito das variáveis arquitetônicas
resultantes do PCA sobre a abundância e a riqueza de galhas associadas a A. germinans
em sítios de replantio, na península de Ajuruteua, Bragança, costa amazônica brasileira.
DAP = Diâmetro à Altura do Peito, g.l. = grau de liberdade.

vii
SUMÁRIO
RESUMO ..................................................................................................................... v
LEGENDAS DAS FIGURAS E TABELAS ................................................................ vi
CAPÍTULO 1 ............................................................................................................... 8
INTRODUÇÃO GERAL .............................................................................................. 9
O MANGUEZAL ...................................................................................................... 9
A HERBIVORIA NO MANGUEZAL .................................................................... 10
AS GALHAS .......................................................................................................... 11
AS GALHAS NO MANGUEZAL .......................................................................... 14
LITERATURA CITADA ............................................................................................ 15
CAPÍTULO 2 ............................................................................................................. 19
RIQUEZA E ABUNDÂNCIA DE GALHAS EM MANGUES PLANTADOS DE
Avicennia germinans (L.) L.: TESTANDO A HIPÓTESE DA ARQUITETURA DA
PLANTA .................................................................................................................... 19
ABSTRACT ............................................................................................................ 20
INTRODUCÃO ...................................................................................................... 20
MÉTODOS ............................................................................................................. 23
ÁREA DE ESTUDO............................................................................................ 23
ESPÉCIE HOSPEDEIRA .................................................................................... 24
AMOSTRAGEM ................................................................................................. 25
ANÁLISE DE DADOS ....................................................................................... 25
RESULTADOS ....................................................................................................... 27
DISCUSSÃO .......................................................................................................... 35
CONCLUSÃO ........................................................................................................ 38
LITERATURA CITADA ........................................................................................ 39
ANEXO .................................................................................................................. 44
REGRAS PARA SUBMISSÃO À REVISTA BIOTROPICA. ............................. 44

8
CAPÍTULO 1
INTRODUÇÃO GERAL

9
INTRODUÇÃO GERAL
O MANGUEZAL
O manguezal é um ecossistema costeiro submetido aos movimentos da maré, formado
por plantas típicas que se associam à uma fauna bem adaptada às condições de um solo
predominantemente lamoso de baixo teor de oxigênio e alto teor de enxofre, sendo
periodicamente inundado pela ação das marés que apresentam grande variação de
salinidade (Hogarth 1999).
Os manguezais são encontrados ao longo das costas tropicais e subtropicais no
Sul, Sudeste e Leste da Ásia, nas Américas, na África Oriental, Austral, Ocidental e
Central, na Austrália e na Nova Zelândia (Spalding et al. 2010). Na América do Sul,
especificamente no Brasil, encontra-se a segunda maior área de manguezal do mundo,
com 13.000 km2 (Spalding et al. 2010), ocorrendo desde o Oiapoque, no Estado do
Amapá, até Laguna em Santa Catarina (MMA 2009). A linha costeira que abrange
desde a baía de Marajó, no Pará, até a Ponta de Tubarão, baía de São José, no
Maranhão, é considerada a maior área contínua de manguezal do mundo, com 650 km
em linha reta (Souza Filho et al. 2005).
Esse ecossistema é de grande importância pois influencia no clima em âmbitos
local, regional e global. Participa de forma ativa nos fluxos de gases de efeito estufa
(GEE), funciona como um filtro biológico na retenção de poluentes, atua como uma
barreira para as águas das marés, ondas e ventos protegendo a zona costeira, tem grande
papel na manutenção dos estoques pesqueiros, apresenta alta produtividade primária e é
local de refúgio, alimentação, reprodução e desova de diversas espécies de animais
(Lugo & Snedaker 1974, Lacerda 1984, Vannucci 2001).
Apesar de toda importância ecológica dos manguezais, tem-se verificado que esse
ecossistema vem sofrendo forte pressão em todo o mundo. Nos últimos 30 anos,

10
grandes extensões de manguezais foram degradadas, algumas áreas em decorrência de
fenômenos naturais (Cahoon et al. 2003) e muitas áreas por ações antrópicas
(Suhogusoff & Piliackas 2007, Tsuji & Fernandes 2008).
Dentre as atividades humanas, as mais comuns são: superexploração dos recursos,
alteração de lagos para aquicultura, desenvolvimento de portos, áreas urbanas, turismo,
poluição, etc. No Brasil, todos esses efeitos antrópicos são observados e resultaram em
considerável degradação e perda dos manguezais existentes. Mas por outro lado, o
aumento do reconhecimento da importância desse ecossistema fez surgir a necessidade
de se elaborar e executar projetos que viabilizem a recuperação das áreas degradadas e a
conservação das remanescentes, bem como a elaboração de projetos de estudos que
buscam entender o funcionamento desse ecossistema.
A HERBIVORIA NO MANGUEZAL
A herbivoria é um processo ecológico que influencia o crescimento, a sobrevivência, a
reprodução dos indivíduos e, consequentemente, a abundância das populações vegetais
(Crawlei 1983). Os insetos fitófagos podem explorar as plantas de diferentes formas:
alimentando-se externamente, cortando e mastigando os tecidos vegetais, por sucção de
células individuais ou do sistema vascular, escavando o interior e ainda induzindo a
formação de galhas (Strong et al. 1984).
Em praticamente todos os ecossistemas florestais os insetos têm um relevante
impacto sobre a taxa de crescimento das árvores, forma, sobrevivência, reprodução e
produção (Schowalter et al. 1986, Crawley 1989). No manguezal, por exemplo, por
muito tempo, considerou-se que o impacto por insetos era de menor importância
comparados aos outros tipos de florestas (Macnae 1968). No entanto, pesquisas mais
recentes sobre a herbivoria no manguezal mostraram que a situação não é diferente de

11
outras florestas. O manguezal é dominado por espécies arbóreas, o que o faz semelhante
às florestas de terra firme em muitos aspectos. Burrows (2003), em uma revisão sobre o
tema “herbivoria no manguezal” não encontrou qualquer evidência de que as árvores de
mangue tenham menores níveis de herbivoria do que as espécies de árvores de outros
tipos de florestas. Para esse autor, as diferenças parecem estar relacionadas às
características das espécies de árvores, ao invés de uma característica relativa às
florestas de mangue.
As árvores de mangue são principalmente afetadas por folívoros, os quais podem
influenciar na produção de serapilheira e ciclagem de nutrientes do sistema (Fernandes
et al. 2009). Da mesma forma, a quantidade de material foliar perdido para a herbivoria
está relacionado às características das árvores e às condições ambientais locais (Feller
1995), resultando em taxas muito diferentes de consumo foliar (Anderson & Lee 1995).
De acordo com Cannicci et al. (2008), o verdadeiro papel ecológico desempenhado
pelos insetos herbívoros não é simplesmente a quantidade de material vegetal perdido
em função do consumo do material vegetal, mas também os efeitos de todo o
desempenho da planta e funcionamento do ecossistema.
Estudos sobre herbivoria são importantes não só para entender como a herbivoria
varia de acordo com a idade da folha, mas também para reconhecer que as folhas de
diferentes idades têm diferentes valores para a planta e diferentes papéis na
produtividade do ecossistema (Burrows 2003).
AS GALHAS
A herbivoria por insetos pode ser realizada pelo grupo dos i) endofíticos, que
consomem a planta internamente, como os galhadores, brocadores e minadores e ii)

12
pelos exofíticos, os quais se alimentam da parte externa da planta, como os folívoros e
nectarívoros.
As galhas, também conhecidas como tumores de plantas, são células, tecidos ou
órgãos de plantas induzidos por um crescimento no volume celular (hipertrofia celular)
e/ou no número de células (hiperplasia celular) provocados por parasitas ou patógenos,
que normalmente se desenvolvem no interior destas estruturas, ou seja, por insetos
endofíticos (Fernandes & Martins 1985, Raman et al. 2005, Santos et al. 2012). Do
ponto de vista evolutivo, as galhas podem ser vistas como fenótipos estendidos dos
insetos indutores, devido à influência que eles podem exercer sobre a diferenciação,
crescimento e desenvolvimento dos tecidos da planta. As galhas também podem ser
entendidas como adaptações induzidas dentro da planta hospedeira, permitindo a
alimentação com tecidos de alta qualidade e a proteção contra inimigos naturais e de
condições ambientais adversas dos seus insetos indutores (Stone & Schönrogge 2003).
As galhas são induzidas por uma ampla variedade de organismos como algas,
líquens, vírus, ácaros, nematóides e insetos. Mas os agentes formadores das galhas são
em sua grande maioria os insetos. Insetos indutores de galhas são encontrados nas
ordens Coleoptera, Diptera, Hemiptera, Hymenoptera, Lepidoptera e Thysanoptera,
sendo que aqueles pertencentes às ordens Diptera e Hymenoptera são os principais
indutores, apresentando uma grande riqueza de espécies galhadoras, especialmente nas
famílias Cecidomiiydae e Cynipidae, respectivamente (Santos et al. 2012).
O comportamento dos insetos de induzir galhas provavelmente foi selecionado
como um mecanismo de defesa contra adversidades climáticas e/ou refúgios contra
inimigos naturais (Price et al. 1986). Primeiramente, a fêmea do inseto galhador insere
seus ovos dentro de um tecido meristemático da planta. A larva do inseto galhador
manipula genética e quimicamente o tecido vegetal a sua volta, dando início ao processo

13
de formação da galha (Hartley 1998, Sopow et al. 2003). A diferenciação do tecido
vegetal para formar a galha pode envolver, entre outros, estímulos mecânicos e
químicos fornecidos pelo inseto, tais como: mastigação do tecido vegetal, fluidos
injetados durante a oviposição, secreções salivares, excretas e hormônios (Hori 1992).
Os sinais mais comumente propostos responsáveis pela formação de galhas são fatores
de crescimento vegetal, tais como: auxinas, citocininas, giberelinas, ácido abscísico,
zeatina ou seus agentes sinérgicos (Stone & Schönrogge 2003).
As galhas atuam como drenos fisiológicos de nutrientes nas plantas hospedeiras,
uma vez que nutrientes são deslocados dos tecidos sadios para o tecido da galha. Elas
também podem mudar a arquitetura e a fisiologia dos órgãos atacados ou vizinhos, bem
como de toda a planta hospedeira. Da mesma forma, nota-se um efeito na redução da
produção de flores, frutos, sementes e biomassa, causada pela formação da galha
(Santos et al. 2012). Além do mais, as galhas podem ser induzidas em todos os órgãos
da planta hospedeira, da extremidade da raiz até o cume do caule, tanto nas partes
vegetativas como nas reprodutivas (Fernandes & Martins 1985).
Buscando compreender os principais padrões do ciclo de vida, ataque, abundância
e riqueza dos insetos galhadores, algumas teorias já foram propostas. Na literatura estão
registradas várias hipóteses, como a “Hipótese do Vigor da Planta” Price (1991), a
“Hipótese do estresse ou estresse ambiental” (Fernandes & Price 1988), a “Hipótese da
sucessão ontogenética” (Medianero et al. 2003) e a “Hipótese da arquitetura da planta”
de Lawton (1983). Esta última hipótese sugere que plantas maiores suportam
populações mais numerosas de herbívoros, especialmente por apresentarem maior
complexidade estrutural. Assim, o tamanho da planta pode ter um efeito positivo sobre a
diversidade de insetos devido ao efeito da área per se (Strong et al. 1984).

14
Adicionalmente, plantas maiores disponibilizam maior quantidade de recursos (possuem
mais sítios de oviposição) e são mais aparentes para o inseto galhador (Lawton 1983).
AS GALHAS NO MANGUEZAL
Estudos sobre as interações inseto-planta e mais especificamente sobre a relação entre
insetos galhadores e suas plantas hospedeiras, no que se refere ao ecossistema
manguezal, são escassos. Os estudos existentes focam mais sobre as espécies de
mastigadores (Cannicci et al. 2008), do que acerca dos insetos endofíticos, como os
minadores e os galhadores (Gonçalves-Alvim et al. 2001, Burrows 2003, Menezes &
Peixoto 2009). No entanto, o trabalho de Santos et al. (2013) apresenta uma revisão da
associação entre as galhas e o ecossistema manguezal. De acordo com esses autores, das
nove espécies de artrópodes galhadores descritas para as espécies de plantas arbóreas de
mangue em diferentes partes do mundo, sete espécies foram registradas para o gênero
Avicennia L. (Acanthaceae). Além desses, já existem 22 morfotipos descritos,
totalizando 29 espécies de cecidomiídeos descritos em quatro espécies de Avicennia. As
espécies A. germinans (L.) L. e A. marina (Forssk.) Vierh. possuem a maior variedade
conhecida de artrópodes galhadores com 22 e 19 galhas, respectivamente, o que
caracteriza as duas espécies como plantas super-hospedeiras.
Assim, a presente dissertação trata da riqueza e abundância de morfotipos de
galhas associados à espécie arbórea de mangue super-hospedeira, A. germinans, em
cinco áreas plantadas de manguezal da península de Ajuruteua, usando como
pressuposto a hipótese da arquitetura da planta.

15
LITERATURA CITADA
ANDERSON, C., AND S.Y. LEE. 1995. Defoliation of the mangrove Avicennia marina
in Hong Kong: cause and consequences. Biotropica 27: 218–226.
BURROWS, D. W. 2003.The role of insect leaf herbivory on the mangroves Avicennia
marina e Rhizophora stylosa. Tese de Doutorado. School of Tropical
Ecology, James Cook University, Austrália.
CAHOON, D., P. HENSEL, J. RYBCZYK, K. MCKEE, E. PROFFITT, AND C.
PEREZ. 2003. Mass tree mortality leads to mangrove peat collapse at Bay
Islands, Honduras after Hurricane Mitch. J. Ecol. 91:1093–1105.
CANNICCI, S., D. BURROWS, S. FRATINI, T. J. SMITH, J. OFFENBERG, AND F.
DAHDOUH-GUEBAS. 2008. Faunal impact on vegetation structure and
ecosystem function in mangrove forests: A review. Aquatic Botany 89: 186–
200.
CRAWLEY, M. J. 1983. Herbivory: The dynamics of animal-plant interactions. pp.
437. Univ. California Press, Berkeley.
CRAWLEY, M. J. 1989. Insect herbivores and plant population dynamics. Ann. Rev.
Entomol. 34: 531–564.
FELLER, I. C. 1995. Effects of nutrients enrichment on growth and herbivory of dwarf
red mangrove (Rhizophora mangle). Ecol. Monographs. 65: 477–505.
FERNANDES, G.W., AND R. P MARTINS. 1985. Tumores de plantas: as galhas. Rev.
Ciên. Hj. 19: 59–64.
FERNANDES, G. W., AND P. W. PRICE. 1988. Biogeografical gradients in galling
species richness: tests of hypothesis. Oecologia 76: 161–167.
FERNANDES, M. E. B., A. A. M. NASCIMENTO AND M. L. CARVALHO. 2009.
Effects of herbivory by Hyblaea puera (Hyblaeidae: Lepidoptera) on litter

16
production in the mangrove on the coast of Brazilian Amazonia. J. Trop.
Ecol. 25: 337–339.
GONÇALVES-ALVIM, S. J., M. C. F. V. SANTOS, AND G. W FERNANDES. 2001.
Leaf gall abundance on Avicennia germinans (Avicenniaceae) along
interstitial salinity gradient. Biotropica 33: 69–77.
HARTLEY, S. E. 1998. The chemical composition of plant galls: are levels of nutrients
and secondary compounds controlled by the gall-former? Oecologia 113:
492–501.
HOGARTH, P. J. 1999. The biology of mangroves. Oxford University Press, Oxford.
HORI, K. 1992. Insect secretion and their effect on plant growth, with special reference
to hemipterans. In J. D. Shorthouse, and O. Rohfritsch (Eds.). Biology of
insect-induced galls, pp. 157–170. Oxford University Press, New York.
LACERDA, L. D. 1984. Manguezais: floresta de beira-mar. Rev. Ciên. Hj. 3: 63–70.
LAWTON, J. H. 1983. Plant architecture and the diversity of phytophagous insect.
Annu. Rev. Entomol. 28: 23–39.
LUGO, A. E., AND S. C. SNEDAKER. 1974. The ecology of mangroves. Annu. Rev.
Ecol. 5: 39–64.
MACNAE, W. 1968. Fauna and flora of mangrove swamps. Adv. Mar. Biol. 6: 73–
270.
MEDIANERO, E., A. VALDERRAMA, AND H. BARRIOS. 2003. Diversidad de
insectos minadores de hojas y formadores de agallas en el dosel y sotobosque
del bosque tropical. Acta Zoológica Mexicana 89: 153–168.
MMA-Ministério do Meio Ambiente. 2009. Mapeamento das áreas de manguezais do
Brasil. Disponível em:

17
<http://www.mma.gov.br/biodiversidade/biodiversidade-aquatica/zona-
costeira-e-marinha/manguezais> Acessado 20-Dec-2012.
MENEZES, L. F. T., AND A. L. PEIXOTO. 2009. Leaf damage in a mangrove swamp
at Sepetiba Bay, Rio de Janeiro, Brazil. Revista Brasileira Botânica. 32:
715724.
PRICE, P. W. 1991. Vigor hypothesis and herbivore attack. Oikos 62: 244–251.
PRICE, P. W.; G. WARING, AND G. W. FERNANDES. 1986. Hypothesis on the
adaptive nature galls. Proc. Entomol. Soc. Was. 88: 361–363.
RAMAN, A., C. W. SCHAEFER, AND T. H. WITHERS. 2005. Biology, ecology and
evolution of gall-inducing arthropods. pp. 817. Science Publishers, New
Hampshire.
SANTOS, J. C., M. A. A. CARNEIRO, AND G. W. FERNANDES. 2012. Insetos
galhadores neotropicais: diversidade e ecologia evolutiva dos herbívoros mais
sofisticados da natureza. In K. Del-Claro, and H. M. Torezan-Silingardi
(Eds.). Ecologia das interações plantas-animais: uma abordagem ecológico-
evolutiva, pp. 186–199. Technical Books, Rio de Janeiro.
SCHOWALTER, T. D., W. W. HARGROVE, AND D. A. CROSSLEY JR. 1986.
Herbivory in forested ecosystems. Annu. Rev. Entomol. 31: 177–196.
SPALDING, M., M. KAINUMA, AND L. COLLINS. 2010. World Atlas of
Mangroves. Earthscan, London, Washington, DC.
SOPOW, S. L., J. D. SHORTHOUSE, W. STRONG, AND D. T. QUIRING. 2003.
Evidence for long-distance, chemical gall induction by an insect. Ecology
Letters 6: 102–105.

18
SOUZA-FILHO, P. W. M. 2005. Costa de manguezais de macromaré da Amazônia:
cenários morfológicos, mapeamento e quantificação de áreas usando dados de
sensores remotos. Revista Brasileira de Geofísica 23: 427–435.
STONE, G. N., AND K. SCHONROGGE. 2003. The adaptive significance of insect
gall morphology. Trends in Ecology and Evolution 18: 512–521.
STRONG, D. R., J. H. LAWTON, AND S. R. SOUTHWOOD. 1984. Insects on plants:
community patterns and mechanisms. Harvard University Press, Cambridge,
Massachusetts, USA.
SUHOGUSOFF, V. G., AND J. M. PILIACKAS. 2007. Breve histórico da ação
antrópica sobre os ecossistemas costeiros do Brasil com ênfase nos
manguezais do estado de São Paulo. Revista Integração 51:343–352.
TSUJI, T., AND M. E. B. FERNANDES. 2008. Replantando os manguezais: técnicas
de reflorestamento. 1. Ed. Belém: Gráfica Alves.
VANNUCCI, M. 2001. What is so special about mangrove? Braz. J. Biol. 61: 599–603.

19
O presente artigo está no formato e será submetido ao periódico Biotropica, cujas regras
estão no ANEXO. Para a apresentação como capítulo da dissertação as figuras e tabelas
foram inseridas no corpo do texto para melhor visualização.
CAPÍTULO 2
RIQUEZA E ABUNDÂNCIA DE GALHAS EM MANGUES
PLANTADOS DE Avicennia germinans (L.) L.: TESTANDO
A HIPÓTESE DA ARQUITETURA DA PLANTA

20
Riqueza e Abundância de Galhas em Mangues Plantados de Avicennia germinans
(L.) L.: Testando a Hipótese da Arquitetura da Planta
ABSTRACT
The hypothesis of plant architecture predicts that variations in the structural complexity
of the host plant influences the insect herbivore community structure, dynamics, and
performance. We attempt to verify the effect of the architectural complexity of
Avicennia germinans (L.) L. on the richness and abundance of gall morphotypes in sites
of replanted mangrove on the Ajuruteua Peninsula, Bragança, Brazilian Amazon coast.
Sampling were carried out at five sites by using 10 individuals and 20 apical branches
by individual. The structural attributes (explanatory variables) and richness and
abundance of galls (response variables) were estimated and analyzed with multiple
regression by generalize linear models (GLM). Twenty-two gall morphotypes were
recorded with the occurrence of 16 at all sites. A total of 7602 galls with average of
1.3±0.4 galls by leaf was recorded. The mofophotypes MF7 e MF4 were the most
abundant and represent less than 40% of the total. Although A. germinans is a superhost
plant, gall-inducing species parasitism did not affected either its development or
survivorship. This fact added to predictibility and attractiveness of the resource to gall-
inducing arthropods may explain high richness and abundance of gall morphotypes
associated to A. germinans in the mangroves. However, the architectural complexity of
this mangrove tree species did not influenced richness but abundance of gall
morphotypes, corroborating partially the structural complexity hypothesis at replanted
mangrove sites.
Key words: endophytic herbivory; gall morphotype; replanted mangrove; structural complexity.

21
INTRODUCÃO
Diversos fatores interferem na riqueza e abundância de morfotipos de galhas
encontrados em plantas hospedeiras, principalmente aqueles relacionados às
características estruturais e fisiológicas dessas plantas. Além disso, algumas espécies
vegetais estão associadas a uma fauna de herbívoros muito mais rica do que outras. Esta
particularidade deve-se a alguns atributos intrínsecos a cada espécie, tais como: o
espectro da distribuição geográfica, o isolamento taxonômico e a fisionomia ou
arquitetura da planta (Cornell 1986). Diante dessas particularidades das interações
inseto-planta várias hipóteses têm sido propostas para explicar o padrão de distribuição
e abundância desses morfotipos e, consequentemente, dos insetos que os induzem em
suas respectivas plantas hospedeiras (Fleck & Fonseca 2007). Especificamente, a
hipótese da complexidade arquitetônica da planta, prediz que a estrutura física das
partes aéreas da planta hospedeira influenciam a estrutura da comunidade de insetos
herbívoros (Lawton 1983, Strong et al. 1984, Araújo et al. 2006). Tal complexidade da
planta é definida como a combinação do tamanho, forma de crescimento,
desenvolvimento sazonal, variedade das estruturas da planta acima do solo e
persistência dessas partes (Lawton 1983, Strong et al. 1984).
Segundo Araújo et al. (2006), as plantas estruturalmente mais complexas devem
apresentar maior riqueza de insetos herbívoros por i) promover maior diversidade de
microhábitats em uma determinada planta, com diferenças de temperatura e umidade
(Gonçalves-Alvim & Fernandes 2001); ii) abrigar populações maiores, reduzindo a
probabilidade de extinção e iii) permitir a existência de um número maior de locais de
refúgio, promovendo a persistência de espécies mais suscetíveis à predação.
Espírito-Santo et al. (2007), em sua revisão da literatura, salientam que a
arquitetura da planta é usada como um fator relevante e que influencia a distribuição e a

22
abundância dos herbívoros, mas que as características das plantas variam entre os
estudos dependendo da história de vida de cada espécie vegetal envolvida. É importante
ter em mente que a resposta de insetos indutores de galha à arquitetura da planta é muito
variável. Em resposta ao seu habito séssil, por exemplo, insetos indutores de galhas
dependem da disponibilidade de tecidos jovens indiferenciados para a indução e
desenvolvimento da galha (Rohfritsch 1992). Portanto, as características arquitetônicas
parecem ter um forte efeito sobre esta guilda quando comparadas aos herbívoros
mastigadores ou exofíticos.
Vários trabalhos sobre insetos galhadores e suas plantas hospedeiras têm
corroborado com a hipótese de que, de fato, há uma correlação positiva entre o aumento
da riqueza e abundância de insetos galhadores e o aumento da complexidade estrutural
da planta hospedeira (Lara et al. 2008, Araújo & Santos 2009, Gonçalves-Alvim et al.
2010). Outros trabalhos, porém, mostram que a complexidade estrutural parece não
influenciar significativamente na distribuição de insetos galhadores, sobretudo nos
ambientes florestais (Price et al. 1998, Lara et al. 2002, Costa et al. 2010, Araujo et al.
2013). Diante desta controvérsia, ainda há muito que se investigar sobre a relação entre
insetos galhadores e suas plantas hospedeira, principalmente no que se refere ao
ecossistema manguezal, onde os estudos sobre herbivoria e mais especificamente sobre
insetos indutores de galhas ainda são escassos. De acordo com Santos et al. (2013), as
espécies do gênero Avicennia L. (Acanthaceae) comportam-se como super-hospedeiras
nas florestas de mangue por apresentarem alta riqueza de insetos galhadores. Espécies
vegetais que se comportam como super-hospedeiras têm grande importância ecológica,
permitindo o teste de hipóteses de cunho ecológico e evolutivo, bem como os efeitos do
ambiente no próprio hospedeiro e na diversidade de galhadores associados (Ribeiro &
Basset 2007).

23
Nos manguezais da costa amazônica brasileira nós comparamos os diferentes
morfotipos de galhas associados às árvores de mangue preto, Avicennia germinans (L.)
L., nos cinco sítios de estudo. Especificamente, nós verificamos o efeito da estrutura das
árvores de A. germinans sobre a riqueza e abundância dos morfotipos de galhas. Os
resultados foram discutidos à luz da hipótese da complexidade estrutural, além de
ressaltar a suscetibilidade dessa planta super-hospedeira à indução de galha.
MÉTODOS
ÁREA DE ESTUDO.–O presente estudo foi realizado em cinco áreas de plantio de
manguezal, na comunidade de Tamatateua (0º57’12,5”S e 46º47’02,4”W), a 15 km do
município de Bragança, na península de Ajuruteua, costa amazônica brasileira (Fig. 1).
Segundo Moraes et al. (2005), o clima na costa amazônica brasileira é tropical,
quente e úmido, com precipitação anual de aproximadamente 2300 mm e temperatura
média de 26°C, devido à influência da Zona de Convergência Inter-Tropical. Os meses
mais secos compreendem o período de setembro a novembro, enquanto os mais
chuvosos de janeiro a abril. A península apresenta um regime de maré semidiurno
(DHN 2013) dominado por macromarés variando entre 4 e 6 m (Souza-Filho 2005).

24
FIGURA 1. Mapa de localização da área de estudo mostrando a
localização dos cinco sítios de trabalho. (A) Costa do Estado do Pará; (B) Península de Ajuruteua; (C) Sítios de trabalho numerados de 1 a
5).
ESPÉCIE HOSPEDEIRA.–O gênero Avicennia possui ampla distribuição geográfica
com 10 espécies válidas em toda a região tropical. No Brasil, ocorrem apenas duas
espécies: A. germinans e Avicennia schaueriana Stapf e Leechman ex Moldenke, ambas
encontradas ao longo do litoral, tanto na faixa tropical quanto subtropical do oceano
Atlântico (Tomlinson 1986). Essas espécies são caracterizadas por sua tolerância às
altas salinidades, em função da sua plasticidade fenotípica em resposta ao sal e às
adaptações fisiológicas e estruturais (Scholander 1968). A. germinans possui porte
arbóreo chegando a atingir mais de 30 m de altura, apresenta um pico de floração que
Sítio de trabalho

25
coincide com a estação seca, enquanto que o período de maior produção de frutos
ocorre na estação chuvosa (Silva & Fernandes 2011).
AMOSTRAGEM.–Foram escolhidas cinco áreas plantadas com idades variando entre 9
e 5 anos. Em cada uma das áreas foram sorteadas 10 árvores, totalizando 50 indivíduos
de A. germinans. As seguintes variáveis arquitetônicas foram mensuradas: Diâmetro à
Altura do Peito (DAP); Altura total (h); Grau de esbeltez; Comprimento do ramo; Total
de folha; Área da copa e Volume da planta.
Como A. germinans possui copa assimétrica, a área da copa foi obtida através da
fórmula, Área=π.D1.D2/4, sendo D1 e D2 os raios maior e menor respectivamente
(Cintrón & Schaeffer-Novelli 1984). O volume da planta foi calculado através da
fórmula do volume do cone, V=[(π.r².h)/3], sendo r = raio da copa e h = altura total da
árvore. A variável grau de esbeltez foi expressa como a razão entre a altura total e o
diâmetro da árvore (h/DAP) caracterizando a estabilidade das árvores, isto é, quanto
mais alto o grau de esbeltez mais instável é a árvore (Durlo & Denardi 1998).
A riqueza e abundância de galhas por planta foram determinadas através da
contagem das galhas presentes em 20 ramos terminais da copa de cada uma das 50
árvores selecionadas, totalizando 1.000 ramos amostrados. Estes ramos foram coletados
com o auxílio de um podão e levados para o laboratório para triagem e morfotipagem
das galhas. A morfotipagem foi realizada com base nas características morfológicas
externas das galhas, assumindo-se a premissa de que diferentes espécies de galhadores
produzem galhas com morfologia distinta (Stone & Schönrogge 2003).
ANÁLISE DE DADOS.–O esforço mínimo de coleta para amostragem representativa
da fauna de galhadores associada a A. germinans foi determinado através da curva de

26
rarefação de morfotipos de galhas em função dos indivíduos amostrados. Após a
determinação da quantidade necessária de indivíduos para a amostragem, esse número
foi utilizado para a elaboração das curvas de rarefação dos diferentes morfotipos em
função dos ramos coletados. A média dos valores correspondentes à saturação de cada
curva dos morfotipos representou o número aproximado de ramos que devem ser
coletados por planta para uma amostragem satisfatória.
Os pressupostos da normalidade (Teste de Shapiro-Wilk) e homocedasticidade
(Teste de Levene) foram testados. As variáveis arquitetônicas (DAP, h, Grau de
Esbeltez, Comprimento dos Ramos, Total de Folhas, Área da Copa e Volume da
planta), que expressam a complexidade dos diferentes bosques dos cinco sítios de
trabalho, foram comparadas entre os sítios através da análise de variância (ANOVA-um
fator). O teste post-hoc de Tukey HSD foi usado para detectar diferenças significativas
entre as médias das variáveis ao nível de α=0,05. Quando esses pressupostos foram
violados, os dados foram analisados pelo teste não-paramétrico de Kruskal-Wallis, com
o teste a posteriori de Dunn. O efeito das sete variáveis arquitetônicas sobre A.
germinans foi avaliado através da Análise de Componentes Principais (PCA),
permitindo a redução da multicolinearidade entre essas variáveis (Tabachnick & Fidell,
2007). Em seguida, foram realizados testes de regressão múltipla através da construção
de modelos lineares generalizados (GLM), utilizando uma distribuição de erros Poisson,
ajustados com a análise de resíduos adequada para cada variável resposta, de acordo
com a crítica ao modelo (Crawley 2002). Essas análises foram realizadas com o auxílio
do software (R-Project 2015).
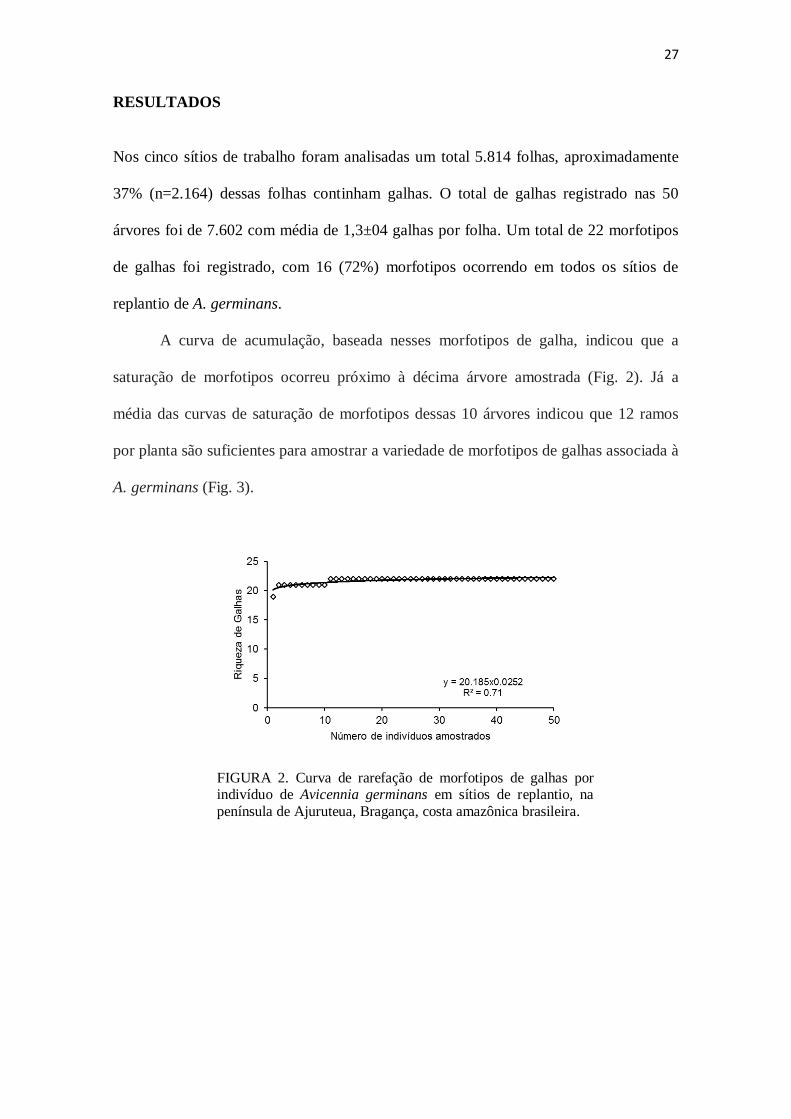
27
RESULTADOS
Nos cinco sítios de trabalho foram analisadas um total 5.814 folhas, aproximadamente
37% (n=2.164) dessas folhas continham galhas. O total de galhas registrado nas 50
árvores foi de 7.602 com média de 1,3±04 galhas por folha. Um total de 22 morfotipos
de galhas foi registrado, com 16 (72%) morfotipos ocorrendo em todos os sítios de
replantio de A. germinans.
A curva de acumulação, baseada nesses morfotipos de galha, indicou que a
saturação de morfotipos ocorreu próximo à décima árvore amostrada (Fig. 2). Já a
média das curvas de saturação de morfotipos dessas 10 árvores indicou que 12 ramos
por planta são suficientes para amostrar a variedade de morfotipos de galhas associada à
A. germinans (Fig. 3).
FIGURA 2. Curva de rarefação de morfotipos de galhas por indivíduo de Avicennia germinans em sítios de replantio, na
península de Ajuruteua, Bragança, costa amazônica brasileira.
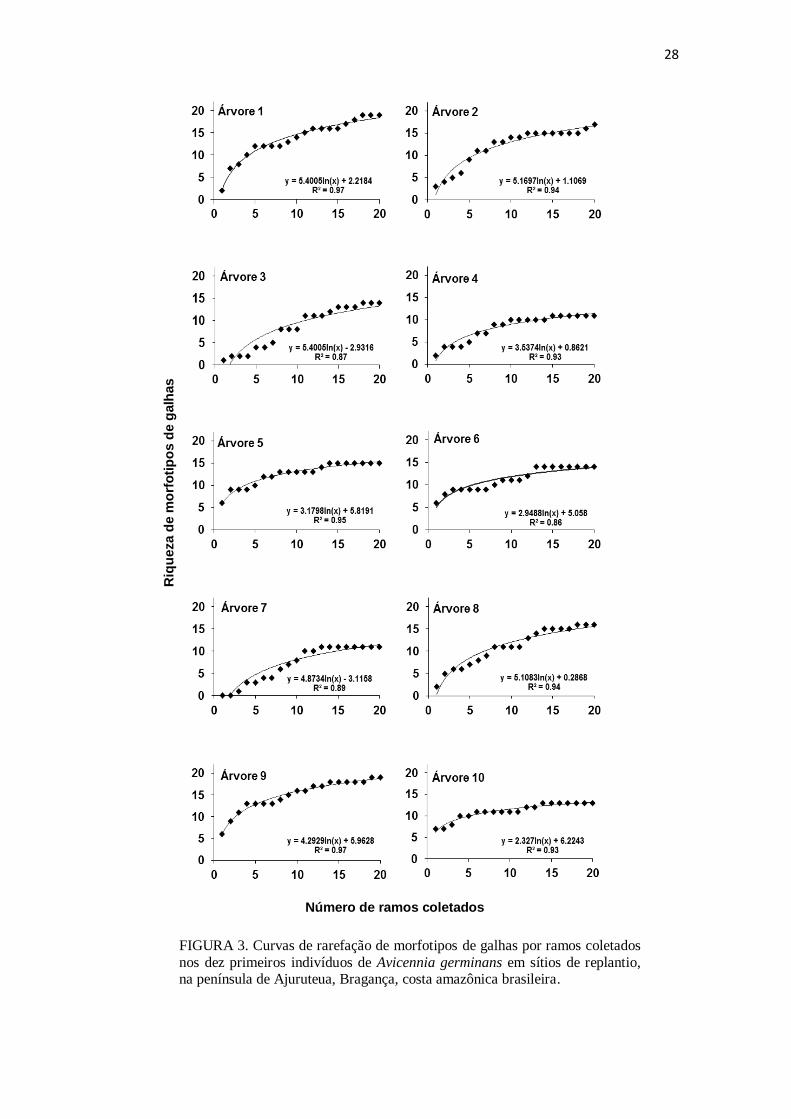
28
FIGURA 3. Curvas de rarefação de morfotipos de galhas por ramos coletados
nos dez primeiros indivíduos de Avicennia germinans em sítios de replantio, na península de Ajuruteua, Bragança, costa amazônica brasileira.
Número de ramos coletados
Riq
ueza d
e m
orf
oti
po
s d
e g
alh
as

29
Na análise comparativa das sete variáveis que expressam a complexidade
arquitetônica dos diferentes tamanhos de árvores de A. germinans representados pelos
cinco sítios de trabalho, somente o “Comprimento dos Ramos” não apresentou
diferença significante (Tabela 1). Por outro lado, os valores de abundância e riqueza de
galhas variaram significativamente entre os sítios (Kruskal-Wallis; P=0,0002 e Kruskal-
Wallis P=0,01, respectivamente), sendo que os sítios 1 e 4 apresentaram o maior e
menor valores, respectivamente, tanto para a abundância quanto para a riqueza de
galhas (Tabela 1).
Os morfotipos mais abundantes foram MF7 (23,6%) e MF4 (12,6%) (Fig 4).
Dentre as partes da folha avaliadas (lâmina, nervura e pecíolo) para ocorrência de
galhas, a lâmina foliar apresentou o maior percentual (90,2%), distribuídos tanto na face
adaxial quanto na abaxial, sendo também registradas no pecíolo (4%) e na nervura
central (5,8%) da folha.
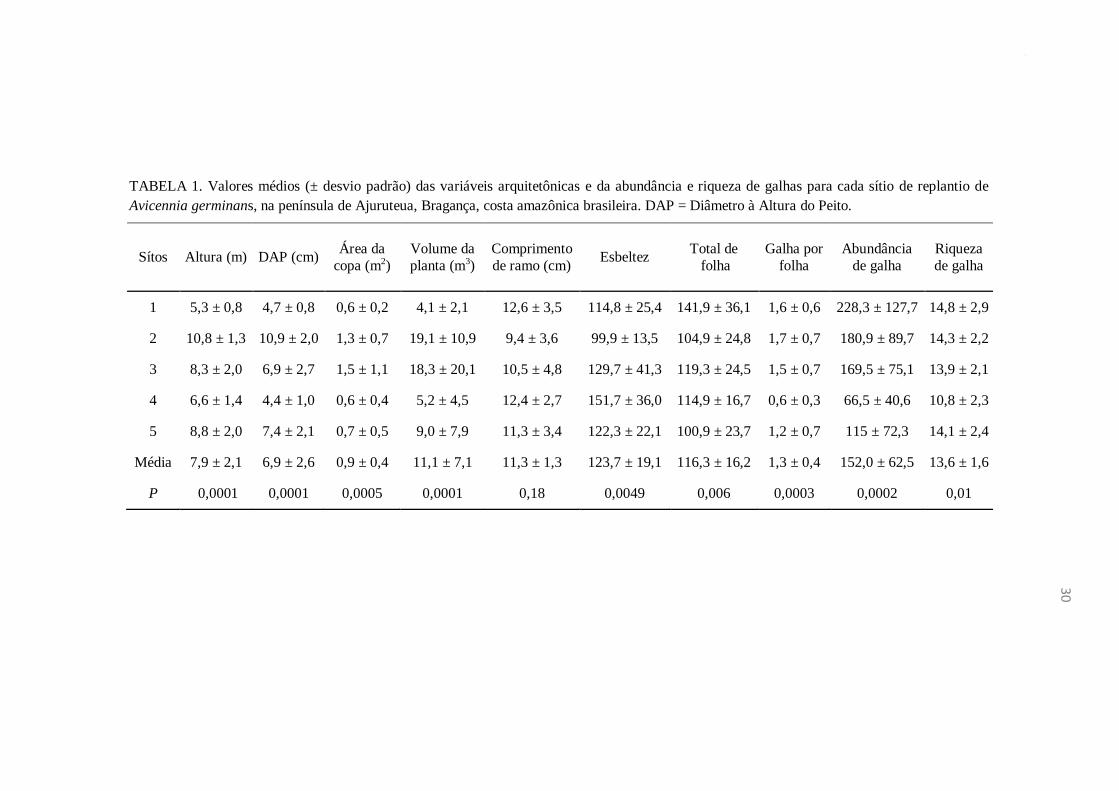
30
Sítos Altura (m) DAP (cm) Área da
copa (m2)
Volume da
planta (m3)
Comprimento
de ramo (cm) Esbeltez
Total de
folha
Galha por
folha
Abundância
de galha
Riqueza
de galha
1 5,3 ± 0,8 4,7 ± 0,8 0,6 ± 0,2 4,1 ± 2,1 12,6 ± 3,5 114,8 ± 25,4 141,9 ± 36,1 1,6 ± 0,6 228,3 ± 127,7 14,8 ± 2,9
2 10,8 ± 1,3 10,9 ± 2,0 1,3 ± 0,7 19,1 ± 10,9 9,4 ± 3,6 99,9 ± 13,5 104,9 ± 24,8 1,7 ± 0,7 180,9 ± 89,7 14,3 ± 2,2
3 8,3 ± 2,0 6,9 ± 2,7 1,5 ± 1,1 18,3 ± 20,1 10,5 ± 4,8 129,7 ± 41,3 119,3 ± 24,5 1,5 ± 0,7 169,5 ± 75,1 13,9 ± 2,1
4 6,6 ± 1,4 4,4 ± 1,0 0,6 ± 0,4 5,2 ± 4,5 12,4 ± 2,7 151,7 ± 36,0 114,9 ± 16,7 0,6 ± 0,3 66,5 ± 40,6 10,8 ± 2,3
5 8,8 ± 2,0 7,4 ± 2,1 0,7 ± 0,5 9,0 ± 7,9 11,3 ± 3,4 122,3 ± 22,1 100,9 ± 23,7 1,2 ± 0,7 115 ± 72,3 14,1 ± 2,4
Média 7,9 ± 2,1 6,9 ± 2,6 0,9 ± 0,4 11,1 ± 7,1 11,3 ± 1,3 123,7 ± 19,1 116,3 ± 16,2 1,3 ± 0,4 152,0 ± 62,5 13,6 ± 1,6
P 0,0001 0,0001 0,0005 0,0001 0,18 0,0049 0,006 0,0003 0,0002 0,01
TABELA 1. Valores médios (± desvio padrão) das variáveis arquitetônicas e da abundância e riqueza de galhas para cada sítio de replantio de
Avicennia germinans, na península de Ajuruteua, Bragança, costa amazônica brasileira. DAP = Diâmetro à Altura do Peito.
30

31
FIGURA 4. Morfotipos (MF) de galhas associados a Avicennia
germinans em sítios de replantio, na península de Ajuruteua,
Bragança, costa amazônica brasileira.
Abaxial Adaxial Abaxial Adaxial
MF1 (n=173) MF2 (n=321)
MF3 (n=218) MF4 (n=816)
MF5 (n=363) MF6 (n=235)
MF7 (n=1,534) MF8 (n=219)
MF9 (n=307) MF10 (n=163)
MF11 (n=34) MF12 (n=375)
MF13 (n=292) MF14 (n=28)
MF15 (n=317) MF16 (n=65)
MF17 (n=21) MF18 (n=38)
MF19 (n=146) MF20 (n=78)
MF21 (n=441) MF22 (n=303)

32
A análise de componentes principais (ACP), utilizando as sete variáveis
arquitetônicas, reduziu para apenas um componente principal (CP1) que explicou 95,1%
da variação das variáveis mensuradas. As variáveis Área da copa, DAP e Altura
apresentaram maior relevância caracterizando melhor a complexidade arquitetônica
entre os sítios de A. germinans (Tabela 2). Adicionalmente, as variáveis respostas
(abundância e riqueza) e as variáveis explicativas (Área da copa, DAP, Altura, Grau de
esbeltez e Total de folhas) foram correlacionadas. As variáveis que mais expressaram a
complexidade estrutural em A. germinans não explicaram a variação da riqueza de
morfotipos identificados, mas apenas a variação na abundância de galhas registrada
(Tabela 3). A abundância de galhas mostrou correlação positiva e significativa com
todas as variáveis, com exceção do “Grau de esbeltez”, cujo resultado foi uma
correlação negativa e significativa (Fig 5).
TABELA 2. Análise de componentes principais
com as variáveis arquitetônicas de Avicennia
germinans em sítios de replantio, na península de
Ajuruteua, Bragança, costa amazônica brasileira.
Variáveis arquitetônicas CP1 CP2
Área da copa 5.54 -4.86
DAP (cm) 4.82 2.56
Altura (m) 4.72 -0.10
Grau de esbeltez - 7.91 -2.14
Total de folha -7.17 2.27
Variação Cum. (%) 95.1 4,9
33
TABELA 3. Análise de Deviance (Distribuição do Erro de Poisson) adequados a modelos
lineares generalizados para avaliar o efeito das variáveis arquitetônicas resultantes do PCA
sobre a abundância e a riqueza de galhas associadas a A. germinans em sítios de replantio, na península de Ajuruteua, Bragança, costa amazônica brasileira. DAP = Diâmetro à Altura do
Peito, g.l. = grau de liberdade.
Variável
resposta
Variável
explicativa g.l. Deviance
g.l. do
resíduo
Deviance do
resíduo P (>Chi)
Modelo Nulo
49 2867,9
Abundância Altura 1 19,1 48 2848,8 <0,001
DAP 1 725,63 47 2123,2 <0,001
Área da copa 1 4,57 46 2118,6 <0,05
Total de folha 1 532,43 45 1586,2 <0,001
Esbeltez 1 8,54 44 1577,6 <0,01
Modelo Nulo
49 53,77
Riqueza Altura 1 0,637 48 53,13 0,42
DAP 1 3,528 47 49,60 0,06
Área da copa 1 0,009 46 49,60 0,92
Total de folha 1 3,435 45 46,16 0,06
Esbeltez 1 2,604 44 43,56 0,10
34
FIGURA 5. Correlação entre as variáveis arquitetônicas (Altura; DAP; Área da copa;
Total de folha e Grau de esbeltez) e a abundância de galhas em A. germinans nos sítios de
replantio, na península de Ajuruteua, Bragança, costa amazônica brasileira.
Ab
un
dân
cia
de g
alh
a

35
DISCUSSÃO
No presente trabalho, A. germinans apresentou um número elevado de morfotipos e de
número de galhas, sendo a maioria das ocorrências das galhas registradas na lâmina
foliar, tanto na parte adaxial quanto na abaxial e no pecíolo, corroborando com Santos et
al. (2013).
A oviposição em plantas com maior quantidade de folhas parece ser uma
estratégia de reprodução de insetos indutores de galha, a qual depende da
disponibilidade de tecidos meristemáticos ativos, pois são considerados locais de alta
qualidade para a oviposição (Vrcibradic et al. 2000). Da mesma forma, as plantas mais
complexas reiteram essa condição, pois possuem grande disponibilidade de sítios
meristemáticos favorecendo a colonização por herbívoros endofíticos (Larson &
Whitham 1997, Espírito-Santo et al. 2007).
Adicionalmente, quando comparadas às outras espécies de mangue dos gêneros
Rhizophora L. e Laguncularia L., A. germinans apresenta uma quantidade muito baixa
de compostos secundários, alto teor de nitrogênio, baixa razão C/N, menor quantidade
de lignina e de substâncias tóxicas (Bernini & Rezende 2010, Medina et al. 2001), além
de serem menos coriáceas oferecendo menor resistência aos ataques por herbívoros
(Tomlinson 1986). Tais características também propiciam o ataque de herbívoros em
geral, principalmente os endofíticos, transformando A. germinans na espécie arbórea
mais palatável e, portanto, uma das principais espécies super-hospedeiras dos
manguezais neotropicais (Santos et al. 2013). Segundo esses autores, A. schaueriana
também é considerada potencialmente uma super-hospedeira, pois já foram descritos
três artrópodes indutores de galha associados a esta espécie. Assim, as características
físico-químicas, acima descritas, somadas à previsibilidade do recurso e ao

36
comportamento especialista dos galhadores podem explicar a alta riqueza (n=22) de
morfotipos de galhas registrada no presente estudo.
No entanto, embora tenha sido registrada uma alta riqueza de galhas, apenas dois
morfotipos foram mais abundantes (MF7 e MF4), representando juntos menos de 40%
do total das espécies registradas. Dessa forma, assumimos que a maioria dos morfotipos
associados a A. germinans apresenta baixa frequência de ocorrência. O mesmo resultado
foi obtido por Santos et al. (2013), cujos resultados mostraram que dos 14 morfotipos
identificados os dois de maior frequência de ocorrência alcançou o percentual de 28,5%.
Assim, é importante ressaltar que 72% (n=16) desses morfotipos ocorreram em
todos os sítios de trabalho. Como consequência, houve uma distribuição mais uniforme
dos diferentes morfotipos entre as árvores de diferentes atributos estruturais, ou seja, a
riqueza de galhas não corroborou com a hipótese da complexidade arquitetônica de A.
germinans nos bosques de mangue plantados.
Contudo, nossos resultados corroboram apenas parcialmente com a hipótese da
arquitetura da planta hospedeira. Analisando pelo prisma da abundância dos diferentes
morfotipos de galhas identificados, é de se esperar que plantas maiores suportam uma
maior população de herbívoros, reduzindo, portanto, a probabilidade de sua extinção por
disponibilizar mais sítios de refúgio, permitindo uma maior persistência das espécies
mais suscetíveis à predação (Espirito-Santos & Fernandes 2002, Fernandes & Negreiro
2001). Esse é o caso de artrópodes galhadores que passam boa parte do seu ciclo de vida
como organismos sésseis em suas plantas hospedeiras. Estas vantagens,
consequentemente, são traduzidas em um aumento da sobrevivência para este grupo
(Gonçalves-Alvim & Fernandes 2001).
Nossos resultados também apontam para o fato de que a complexidade estrutural
influência positiva e significativamente a abundância de galhas em A. germinans,

37
corroborando em parte com a hipótese da arquitetura da planta hospedeira, já que a
riqueza de galhas não variou em função dos atributos estruturais da planta. De fato,
plantas com maior complexidade estrutural e, consequentemente, mais aparentes no
espaço, oferecem um número maior de microhabitats suscetíveis à colonização
(Gonçalves-Alvim & Fernandes 2001). O sucesso da sobrevivência de uma prole de
insetos indutores de galha também depende diretamente da habilidade das fêmeas
adultas em encontrar esses substratos adequados para o crescimento e o
desenvolvimento da sua prole.
A riqueza e abundância de morfotipos de galhas associados a A. germinans
caracterizam essa planta como um recurso-chave para os insetos indutores de galhas nas
florestas de mangue da região Neotropical, especialmente para a Amazônia brasileira,
onde esses morfotipos têm sido estudados (Santos et al. 2013). Da mesma forma, é
também relevante ter em mente que apesar da alta suscetibilidade de A. germinans à
infestação em grande quantidade de diferentes morfotipos de insetos indutores de
galhas, os resultados referentes aos atributos estruturais mostraram que as árvores
plantados estão bastante desenvolvidos. Isto implica dizer que diferentemente de
algumas espécies arbóreas, cuja presença desses insetos galhadores é capaz de reduzir o
desenvolvimento da sua planta hospedeira (Teas & McEwan 1982; Gonçalves-Alvim et
al. 1999, Fernandes et al. 2007, Pincebourde & Casas 2015), no caso de A. germinans,
embora seja uma planta super-hospedeira, o parasitismo de espécies endofíticas não
afeta o seu desenvolvimento, muito menos a sua sobrevivência.

38
CONCLUSÃO
Nossos resultados confirmam que A. germinans é uma planta super-hospedeira de
insetos galhadores e que a ampla ocorrência e distribuição desse gênero nos manguezais
torna esse sistema interessante para estudos ecológicos. Nossos dados corroboram
parcialmente com a hipótese da complexidade arquitetônica da planta, sendo a
abundância e não a riqueza influenciada pela complexidade da estrutura da planta
hospedeira. O fato de A. germinans ser uma planta super-hospedeira a torna um recurso-
chave para a sobrevivência da guilda dos indutores de galha, bem como dos herbívoros
em geral, fazendo com que aja uma colonização maciça de inúmeras espécies de
artrópodes indempendente dos atributos estruturais da planta. Apesar da grande
infestação de insetos indutores de galhas, os resultados também mostraram que as
árvores plantadas de A. germinans nas cinco áreas de estudo estão bastante
desenvolvidas e que o parasitismo não afeta o seu desenvolvimento, muito menos a sua
sobrevivência.

39
LITERATURA CITADA
ARAÚJO, A. P. A., J. D. PAULA, M. A. A. CARNEIRO, AND J. H. SCHOEREDER.
2006. Effects of host plant architecture on colonization by galling insects.
Aus. Ecol. 31: 343–348.
ARAÚJO, W. S., AND B. B. SANTOS. 2009. Efeitos da sazonalidade e do tamanho da
planta hospedeira na abundância de galhas de Cecidomyiidae (Diptera) em
Piper arboreum (Piperaceae). Rev. Bras. Entomol. 53: 300−303.
ARAÚJO, W. S., SCARELI-SANTOS. CLAUDIA, F. A. G. GUILHERME AND P.
CUEVAS-REYES. 2013. Comparing galling insect richness among
Neotropical savannas: effects of plant richness, vegetation structure and
super-host presence. Biodivers. Conserv. 22: 1083–1094.
BERNINI, E., AND C. E. REZENDE. 2010. Concentração de nutrientes em folhas e
sedimento em um manguezal do norte do estado do Rio de Janeiro. Revista da
Gestão Costeira Integrada. Número especial.
CORNELL, H. V. 1986. Oak species attributes and host size influence cynipine wasp
species richness. Ecology 67: 1582–1592.
COSTA, F. V., M. FAGUNDES, AND F. S. NEVES. 2010. Arquitetura da planta e
diversidade de galhas associadas à Copaifera langsdorffii (Fabaceae). Ecol.
Aus. 20: 9−17.
CRAWLEY, M. J. 2002. Statistical computing: An introduction to data analysis using
S-Plus. Wiley, England.
DURLO, M. A., AND L. DENARDI. 1998. Morfometria de Cabralea canjerana, em
mata secundária do Rio Grande do Sul. Ciência Florestal 8: 55−66.
DHN-Diretoria de Hidrografia e Navegação, Marinha do Brasil. Disponível em
<https://www,mar,mil,br/dhn/dhn/index,html>, Acessado em 20-May-2013.

40
ESPÍRITO-SANTO M. M., AND G. W FERNANDES. 2002. Host plant effects on the
development and survivorship of the galling insect Neopelma baccharidis
(Homoptera: Psyllidae). Austral. Ecol. 27:249–57.
ESPÍRITO-SANTO, M. M., F. S. NEVES, F. R. ANDRADE-NETO, AND G. W.
FERNANDES. 2007. Plant architecture and meristem dynamics as the
mechanisms determining the diversity of gall-inducing insects. Oecologia
153: 353−364.
FERNANDES G. W., AND D. NEGREIROS. 2001. The occurrence and effectiveness
of hypersensitive reaction against galling herbivores across host taxa. Ecol.
Entomol. 26: 46–55.
FERNANDES, G. W., J. C. SANTOS, F. M. C. CASTRO, AND A. CASTILHO. 2007.
Impact of a gall midge Parkiamyia paraensis (Diptera, Cecidomyiidae) on
the Amazonian plant Parkiapendula (Fabaceae). Rev. Bras. Entomol. 51:
471−475.
FLECK, T., AND C. R. FONSECA. 2007. Hipóteses sobre a riqueza de insetos
galhadores: uma revisão considerando os níveis intra-específico,
interespecífico e de comunidade. Neotropical Biology Conservation 2: 36−45.
GONÇALVES-ALVIM, S. J., M. L. FARIA, AND G. W. FERNANDES. 1999.
Relationships between four neotropical species of galling insects and shoot
vigor. An. Soc. Entomol. Bras. 28: 147−155.
GONÇALVES-ALVIM, S. J., AND G. W. FERNANDES. 2001. Comunidades de
insetos galhadores (Insecta) em diferentes fisionomias do cerrado em Minas
Gerais, Brasil. Rev. Bras. de Zool. 18: 289−305.
GONÇALVES-ALVIM, S. J., T. C. LANA, B. D. RANIERI, F. A. O. SILVEIRA, V.
RIBEIRO, AND G. W. FERNANDES. 2010. Growth, defense and herbivory

41
on young leaves of Qualeaparviflora (Vochysiaceae) in three different
Cerrado habitats. Neotropical Biology and Conservation 5: 86–92.
LARA, A. C. F., G. W. FERNANDES, AND S. J. GONÇALVES-ALVIM. 2002. Test
of hypotheses on patterns of gall distribution along an altitudinal gradient.
Trop. Zool. 15: 219–232.
LARA, D. P., L. A. OLIVEIRA, I. F. P. AZEVEDO, M. F. XAVIER, F. A. O.
SILVEIRA, M. A. A. CARNEIRO AND G. W. FERNANDES. 2008.
Relationships between host plant architecture and gall abundance and
survival. Rev. Bras. Entomol. 52: 78–81.
LAWTON, J. H., 1983. Plant architecture and the diversity of phytophagous insect.
Ann. Rev. Entomol. 28: 23−39.
LARSON, K. C., AND T. G. WHITHAM. 1997. Competition between gall aphids and
natural plant sinks: plant architecture affects resistance to galling. Oecologia
109:1432–1439.
MEDINA, E., T. GIARRIZO, M. MENEZES, M. L. CARVALHO, E. A.
CARVALHO, A. P. ESPIRITO-SANTOS, A. S. BRITO, R. E. S. VILHENA,
A. REISE, AND F. C. BRAGA. 2001. Mangal communities of the “Salgado
Paraense”: Ecological heterogeneity along the Bragança peninsula assessed
through soil and leaf analyses. Amazoniana 16: 397−416.
MORAES, B. C., J. M. N. COSTA, A. C. L. COSTA, AND M. H. COSTA. 2005.
Variação espacial e temporal da precipitação no estado do Pará. Acta
Amazonica 35: 207−214.
PINCEBOURDE, S. AND J. CASAS. 2015. Hypoxia and hypercarbia in endophagous
insects: larval position in the plant gas exchange network is key. Journal of
insect physiology. http://dx.doi.org/doi:10.1016/j.jinsphys.2015.07.006.

42
PRICE, P. W., H. ROININEN, AND A. ZINOVJEV. 1998. Adaptive radiation of gall-
inducing sawflies in relation to architecture and geographic range of willow
host plants. In G. Csóka, W. J. Mattson, G. N. Stone, and P. W. Price (Eds.).
The biology of gall-inducing arthropods, pp. 196–203. USDA. St. Paul,
Minnesota.
R-PROJECT. 2015. The R Project for Statistical Computing. – World Wide Web
electronic publication, accessible at http://www.r-project.org/ (Acessed
01/03/2015).
RIBEIRO, S. P., AND Y. BASSET. 2007. Gall-forming and free-feeding herbivory
along vertical gradients in a lowland tropical rainforest: the importance of leaf
sclerophylly. Ecography 30: 663−672.
ROHFRITSCH, O. 1992. Patterns in gall development. In J. D. Shorthouse, and O.
Rohfritsch (Eds.). Biology of insect-induced galls, pp. 60–86. University
Press, Oxford.
SANTOS, R. C. O., M. E. B. FERNANDES, AND M. B. MARTINS. 2013. Are the
species of the genus Avicennia L, (Acanthaceae) a “Superhost” plants of gall-
inducing arthropods in Mangrove forests?, In B. Barros, and M. E. B.
Fernandes (Eds.). Herbivory, pp. 17−32. http://dx.doi.org/10.5772/2718.
SCHOLANDER, P. F. 1968. How mangrove desalinate seawater. Physiol Plantarum 21:
258−268.
SILVA, L. L., AND M. E. B. FERNANDES. 2011. Relação entre os atributos
estruturais da árvores de Avicennia germinans (L.) Stearn e sua fenologia
reprodutiva. Bol. Lab. Hidro. 24: 51−57.

43
SOUZA-FILHO, P. W. M. 2005. Costa de manguezais de macromaré da Amazônia:
cenários morfológicos, mapeamento e quantificação de áreas usando dados de
sensores remotos. Revista Brasileira de Geofísica 23: 427−435.
STONE, G. N., AND K. SCHONROGGE. 2003. The adaptive significance of insect
gall morphology. Trends in Ecology e Evolution 18: 512–521.
STRONG, D. R., J. H. LAWTON AND T. R. E. SOUTHWOOD. 1984. Insects on
plants: community patterns and mechanisms. Blackwell Scientific
Publications, Oxford.
TABACHNICK, B. G., AND L. S. FIDELL. 2007. Using multivariate statistics.
Pearson international edition.
TEAS, H. J., AND R. J. McEWAN. 1982. An epidemic dieback gall disease of
Rhizophora mangroves in the Gambia, West África. Plant Disease 66: 522–
523.
TOMLINSON, P. B. 1986. The botany of mangroves. Cambridge University Press,
Cambridge.
VRCIBRADIC, D., C. F. D. ROCHA AND R. F. MONTEIRO. 2000. Patterns of
gallforming in Ossaea confertiflora (Melastomataceae) by Lopesia
brasiliensis (Diptera: Cecidomyiidae) in an area of Atlantic Rainforest in
Southeastern Brazil. Revista Brasileira de Biologia 60: 159–166

44
ANEXO
REGRAS PARA SUBMISSÃO À REVISTA BIOTROPICA.
BIOTROPICA – JOURNAL OF THE ASSOCIATION FOR TROPICAL BIOLOGY
AND CONSERVATION CHECKLIST FOR PREPARATION OF MANUSCRIPTS
AND ILLUSTRATIONS (updated February 2010)
I- General Instructions
Publication must be in English, but second abstract in other languages (such as Spanish,
French, Portuguese, Hindi, Arabic, Chinese etc.) may be published as online Supporting
Information. BIOTROPICA offers assistance in editing manuscripts if this is required
(see English Editorial Assistance below). Second abstracts will not be copy-edited and
the author(s) must take full responsibility for content and quality.
Manuscripts may be submitted in the following categories, based on these suggested
word limits: Paper (up to 5000 words); Insights (up to 2000 words); Review (up to 8000
words); Commentary (up to 2000 words).
Word counts exclude title page, abstract(s), literature cited, tables, figures, or
appendices.
Use 8.5" x 11" page size (letter size). Double space everything, including tables, figure
legends, abstract, and literature cited.
Use a 1" margin on all sides. Align left. Avoid hyphens or dashes at ends of lines; do
not divide a word at the end of a line.
Use standard 12 point type (Times New Roman).
Indent all but the first paragraph of each section.
Use italics instead of underline throughout. Italicize non-English words such as e.g., i.e.,
et al., cf., ca, n.b., post-hoc, and sensu (the exceptions being ‘vs.’ and ‘etc.’).
Include page number in the centre of all pages. Do use line numbering starting on each
page.
Cite each figure and table in the text. Tables and figures must be numbered in the order
in which they are cited in the text.
Use these abbreviations: yr (singular & plural), mo, wk, d, h, min, sec, diam, km, cm,
mm, ha, kg, g, L, g/m2

45
For units, avoid use of negative numbers as superscripts: use the notation /m2 rather
than m-2.
Write out other abbreviations the first time they are used in the text; abbreviate
thereafter: "El Niño Southern Oscillation (ENSO)…"
Numbers: Write out one to ten unless a measurement (e.g., four trees, 6 mm, 35 sites, 7
yr, 10 × 5 m, > 7 m, ± SE) or in combination with other numbers (e.g., 5 bees and 12
wasps). Use a comma as a separator in numbers with more than four digits (i.e., 1000,
but 10,000); use decimal points as in 0.13; 21°C (no spaces); use dashes to indicate a set
location of a given size (e.g., 1-ha plot).
Spell out ‘percent’ except when used in parentheses (20%) and for 95% CI.
Statistical abbreviations: Use italics for P, N, t, F, R2, r, G, U, N, χ2 (italics,
superscripts non-italics); but use roman for: df, SD, SE, SEM, CI, two-way ANOVA, ns
Dates: 10 December 1997; Times: 0930 h, 2130 h
Latitude and Longitude are expressed as: 10°34′21″ N, 14°26′12″ W
Above sea level is expressed as: asl
Regions: SE Asia, UK (no periods), but note that U.S.A. includes periods.
Geographical place names should use the English spelling in the text (Zurich, Florence,
Brazil), but authors may use their preferred spelling when listing their affiliation
(Zürich, Firenze, Brasil).
Lists in the text should follow the style: … : (1)… ; (2)…; and (3)…, as in, “The aims
of the study were to: (1) evaluate pollination success in Medusagyne oppositifolia; (2)
quantify gene flow between populations; and (3) score seed set.”
Each reference cited in text must be listed in the Literature Cited section, and vice versa.
Double check for consistency, spelling and details of publication, including city and
country of publisher.
For manuscripts ACCEPTED for publication but not yet published, cite as Yaz (in
press) or (Yaz, in press). Materials already published online can be cited using the
digital object identifier (doi)
Literature citations in the text are as follows: One author: Yaz (1992) or (Yaz 1992);
Two authors: Yaz and Ramirez (1992); (Yaz & Ramirez 1992) and Three or more
authors: Yaz et al. (1992), but include ALL authors in the literature cited section.
Cite unpublished materials or papers not in press as (J. Yaz, pers. obs.) or (J. Yaz,
unpubl. data). Initials and last name must be provided. ‘In prep’ or ‘submitted’ are NOT

46
acceptable, and we encourage authors not to use ‘pers. obs.’ or ‘unpubl. data’ unless
absolutely necessary. Personal communications are cited as (K. A. Liston, pers. comm.).
Use commas (Yaz & Taz 1981, Ramirez 1983) to separate citations, BUT use
semicolon for different types of citations (Fig. 4; Table 2) or with multiple dates per
author (Yaz et al. 1982a, b; Taz 1990, 1991). Order references by year, then
alphabetical (Azy 1980, Yaz 1980, Azy 1985).
Assemble manuscripts in this order: Title page; Abstract (s); Key words; Text;
Acknowledgments (spelled like this); Literature cited; Tables; Appendix (when
applicable); Figure legends (one page); Figures
For the review purpose, submit the entire manuscript, with Tables, Figure legends and
Figures embedded at the end of the manuscript text, as a Microsoft Word for Windows
document (*.doc), or equivalent for Mac or Linux. Do NOT submit papers as pdf files.
II- Title Page
(Do not number the title page)
Running heads two lines below top of page. LRH: Yaz, Pirozki, and Peigh (may not
exceed 50 characters or six author names; use Yaz et al.) RRH: Seed Dispersal by
Primates (use capitals; may not exceed 50 characters or six words).
Complete title, flush left, near middle of page, Bold Type and Initial Caps, usually no
more than 12 words.
Where species names are given in the title it should be clear to general readers what
type(s) of organism(s) are being referred to, either by using Family appellation or
common name. For example: ‘Invasion of African Savanna Woodlands by the Jellyfish
tree Medusagyne oppositifolia’, or ‘Invasion of African Savanna Woodlands by
Medusagyne oppositifolia (Medusagynaceae)’
Titles that include a geographic locality should make sure that this is clear to the general
reader. For example: ‘New Species of Hummingbird Discovered on Flores, Indonesia’,
and NOT ‘New Species of Hummingbird Discovered on Flores’.
Below title, include author(s) name(s), affiliation(s), and unabbreviated complete
address (es). Use superscript number(s) following author(s) name(s) to indicate current
location(s) if different than above. In multi-authored papers, additional footnote
superscripts may be used to indicate the corresponding author and e-mail address.
Abstract Page
(Page 1)

47
Abstracts should be concise (maximum of 250 words for papers and reviews; 50 words
for Insights; no abstract for Commentary). Include brief statements about the intent,
materials and methods, results, and significance of findings. The abstract of Insights
should emphasise the novelty and impact of the paper.
Do not use abbreviations in the abstract.
Authors are strongly encouraged to provide a second abstract in the language
relevant to the country in which the research was conducted, and which will be
published as online Supporting Information. This second abstract should be embedded
in the manuscript text following the first abstract.
Provide up to eight key words after the abstract, separated by a semi-colon (;). Key
words should be listed alphabetically. Include location, if not already mentioned in the
title. See style below. Key words should NOT repeat words used in the title. Authors
should aim to provide informative key words— avoid words that are too broad or too
specific.
Key words: Melastomataceae; Miconia argentea; seed dispersal; Panama; tropical wet
forest.— Alphabetized and key words in English only.
III- Text
(Page 2, etc) See General Instructions above, or recent issue of BIOTROPICA (Section
I).
No heading for Introduction. First line or phrase of Introduction should be SMALL
CAPS.
Main headings are METHODS, RESULTS, and DISCUSSION: All CAPITALS and
Bold. Flush left, one line.
One line space between main heading and text
Second level headings: SMALL CAPS, flush left, Capitalize first letter, begin sentence
with em-dash, same line (e.g., INVENTORY TECHNIQUE.—The ant inventory…).
Use no more than second level headings.
Do not use footnotes in this section.
References to figures are in the form of ‘Fig. 1’, and tables as ‘Table 1’. Reference to
online Supporting Information is as ‘Fig. S1’ or ‘Table S1’.
IV- Literature Cited
(Continue page numbering and double spacing)

48
No ‘in prep.’ or ‘submitted’ titles are acceptable; cite only articles published or ‘in
press’. ‘In press’ citations must be accepted for publication. Include journal or
publisher.
Verify all entries against original sources, especially journal titles, accents, diacritical
marks, and spelling in languages other than English.
Cite references in alphabetical order by first author's surname. References by a single
author precede multi-authored works by the same senior author, regardless of date.
List works by the same author chronologically, beginning with the earliest date of
publication.
Insert a period and space after each initial of an author's name; example: YAZ, A. B.,
AND B. AZY. 1980.
Authors Names: use SMALL CAPS.
Every reference should spell out author names as described above. BIOTROPICA no
longer uses ‘emdashes’ (—) to substitute previously mentioned authors.
Use journal name abbreviations (see
http://www.bioscience.org/atlases/jourabbr/list.htm). If in doubt provide full journal
name.
Double-space. Hanging indent of 0.5 inch.
Leave a space between volume and page numbers and do not include issue numbers. 27:
3–12
Article in books, use: AZY, B. 1982. Title of book chapter. In G. Yaz (Ed.). Book title,
pp. 24–36. Blackwell Publications, Oxford, UK.
Dissertations, use: ‘PhD Dissertation’ and ‘MSc Dissertation’.
V- Table
(Continue page numbering)
Each table must start on a separate page, double-spaced. The Table number should be in
Arabic numerals followed by a period. Capitalize first word of title, double space the
table caption. Caption should be italicized, except for words and species names that are
normally in italics.
Indicate footnotes by lowercase superscript letters (a, b, c, etc.).
Do not use vertical lines in tables.
Ensure correct alignment of numbers and headings in the table (see current issues)

49
Tables must be inserted as a Word table or copy and pasted from Excel in HTML
format.
VI- Figure Legends
(Continue page numbering)
Double-space legends. All legends on one page.
Type figure legends in paragraph form, starting with ‘FIGURE’ (uppercase) and
number.
Do not include ‘exotic symbols’ (lines, dots, triangles, etc.) in figure legends; either
label them in the figure or refer to them by name in the legend.
Label multiple plots/images within one figure as A, B, C etc, as in ‘FIGURE 1. Fitness
of Medusagyne oppositifolia as indicated by (A) seed set and (B) seed viability’,
making sure to include the labels in the relevant plot.
Top Related