
Análise in vitro da expressão de conexinas e da comunicação...
88
Análise in vitro da expressão de conexinas e da comunicação celular em linhagens adeno-hipofisárias. Natalia Gonçalves Mação Dissertação de Mestrado apresentada ao Programa de Pós-graduação em Ciências Morfológicas da Universidade Federal do Rio de Janeiro. Orientadores: Prof. Dr. Luiz Eurico Nasciutti Prof. Dr. Leandro Miranda Alves Rio de Janeiro Setembro 2008
-
Upload
hoangnguyet -
Category
Documents
-
view
216 -
download
0
Transcript of Análise in vitro da expressão de conexinas e da comunicação...

Análise in vitro da expressão de conexinas e da
comunicação celular em linhagens adeno-hipofisárias..
Natalia Gonçalves Mação
Dissertação de Mestrado apresentada ao Programa de Pós-graduação em Ciências
Morfológicas da Universidade Federal do Rio de Janeiro.
Orientadores:
Prof. Dr. Luiz Eurico Nasciutti
Prof. Dr. Leandro Miranda Alves
Rio de Janeiro
Setembro 2008

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
Natalia Gonçalves Mação
Análise in vitro da expressão de conexinas e da comunicação celular em linhagens adeno-hipofisárias.
Dissertação de Mestrado apresentada ao Programa de Pós-Graduação em Ciências Morfológicas, Universidade Federal do Rio de Janeiro, como requisito para obtenção do título de Mestre em Ciências Morfológicas
Orientadores:
Prof. Dr. Luiz Eurico Nasciutti
Prof. Dr. Leandro Miranda Alves
Rio de Janeiro
2008

iii
Ficha Catalográfica
Mação, Natalia Gonçalves.
Análise in vitro da expressão de conexinas e da comunicação celular em linhagens adeno-hipofisárias. / Natalia Gonçalves Mação – 2008.
71f.
Dissertação de Mestrado em Ciências Morfológicas – Universidade Federal do Rio de Janeiro, Programa de Pós-Graduação em Ciências Morfológicas, Rio de Janeiro, 2008.
Orientador: Luiz Eurico Nasciutti
Leandro Miranda Alves
1. Glândula Hipófise. 2. Comunicação Intercelular. 3. Ciências Morfológicas – Dissertações.
I. Nasciutti, Luiz Eurico. Alves, Leandro Miranda. II. Programa de Pós-Graduação em Ciências Morfológicas. Universidade Federal do Rio de Janeiro.. III. Análise in vitro da expressão de conexinas e da comunicação celular em linhagens adeno-hipofisárias.

iv
Natalia Gonçalves Mação
Análise in vitro da expressão de conexinas e da comunicação celular em linhagens adeno-hipofisárias.
Dissertação de Mestrado apresentada ao Programa de Pós-Graduação em Ciências Morfológicas, Universidade Federal do Rio de Janeiro, como requisito para obtenção do título de Mestre em Ciências Morfológicas
Aprovada em ____________
___________________________________________________
Prof. Dr. Vânia Maria Correa da Costa – UFRJ
___________________________________________________
Prof. Dr. Valéria de Mello Coelho - UFRJ
__________________________________________________
Prof. Dr. Newton Gonçalves de Castro - UFRJ

v
Dedicatória
Ao meu grande e eterno companheiro Aquiles,
sol de todos os meus dias,
dedico esta dissertação.

vi
Agradecimentos
A Deus, acima de tudo, por todas as bênçãos, pelo amor, por me mostrar que existe e que está perto de mim nos meus momentos de desespero e de alegria. Aos meus pais, Sueli e Nélio, pelo amor, pelo apoio constante e pelos ensinamentos valiosos. Aos meus pais de coração, Wilka e Arakém, por ter me dado o privilégio de escutar seus conselhos, pelo carinho e apoio durante todos estes anos. Aos meus avós, por terem incentivado sempre meus estudos e me proporcionado a realização de um sonho. Às minhas irmãs, Marcele e Letícia, companheiras de uma vida, apoios de minha jornada. Ao meu “Lindo” marido, Aquiles Mação Júnior, meu companheiro, meu amigo, meu tudo, por estar sempre ao meu lado com um sorriso lindo, me incentivando e acreditando em mim, por ser meu maior suporte ao longo desta jornada, por “seus pés abrirem o caminho, aonde eu sigo e nunca me sinto só”. Lindo, te amo! Ao Fubá, meu filho canino, por toda alegria que nos tem proporcionado em tão pouco tempo de vida. A todos os meus amados amigos, irmãos de coração, por dividirmos problemas e multiplicarmos felicidades. Aos meus orientadores, Luiz Eurico Nasciutti e Leandro Miranda Alves, pelo seu precioso incentivo ao meu trabalho e à pesquisa científica, pela ajuda e apoio constantes, com sua experiência e orientações de enorme valor. À professora Regina Coeli dos Santos Goldenberg, pela valiosa colaboração, pela recepção em seu laboratório e por ceder material para a realização deste trabalho. A Leandro Vairo, pela paciência e dedicação dispensadas a todas as “injeções”. À professora Vânia Maria Corrêa da Costa, que gentilmente aceitou revisar este trabalho, pela grande contribuição. À professora Christina Maeda Takyia que de alguma forma contribuiu para a confecção dessa dissertação, sanando dúvidas e sempre dando sugestões. À professora Rosa Maria Marcos Mendes, por ter me apresentado aos campos da Histologia e Embriologia, por ter sempre me incentivado durante minha vida acadêmica.

vii
Aos amigos do laboratório Daniel, Michel, Antônio, Pedro, pelas ótimas horas de convivência dentro e fora do laboratório, pela amizade, pela atenção e pela ajuda quando precisei. À Célia, Elaine e Cida, por sermos componentes tão diferentes unidos por um envoltório tão bonito: a nossa amizade. Às secretárias Tânia, Graça e Simone, por sua indispensável ajuda. Aos membros da banca, professora Regina Coeli dos Santos Goldenberg, professora Valéria de Mello Coelho e professor Newton Gonçalves de Castro, por terem aceitado prontamente o convite para fazer parte desta.

viii
Epígrafe
“A utopia está lá no horizonte. Me aproximo dois passos, ela se afasta dois passos. Caminho dez passos e o horizonte corre dez passos.
Por mais que eu caminhe, jamais alcançarei. Para que serve a utopia?
Serve para isso: para que eu não deixe de caminhar.”
Eduardo Galeano

ix
Resumo
MAÇÃO, Natalia Gonçalves. Análise in vitro da expressão de conexinas e da comunicação
celular em linhagens adeno-hipofisárias. Rio de Janeiro, 2008. Dissertação (Mestrado em
Ciências Morfológicas) - Programa de Pós-Graduação em Ciências Morfológicas, Universidade
Federal do Rio de Janeiro, Rio de Janeiro, 2008.
Várias atividades da adeno-hipófise podem ser reguladas pela comunicação intercelular através de junções comunicantes (junções gap). Estas junções são canais intercelulares formados por proteínas denominadas conexinas, que se organizam formando conexons, por onde podem passar pequenas moléculas de até 1,2 kDa, coordenando diversos processos fisiológicos como proliferação, adesão, migração, diferenciação, sobrevivência celular e apoptose. No presente estudo, avaliamos a expressão in vitro das conexinas 26, 32 e 43 e a comunicação intercelular em linhagens celulares de adeno-hipófise de ratos, as células tumorais GH3B6 secretoras de prolactina e as células estromais APS9. Ensaios de RT-PCR realizados em cultura primária de adeno-hipófise de ratos revelaram a expressão de todas as conexinas estudadas. Quando cultivadas isoladamente, as células APS9 expressaram apenas a Cx43, enquanto que as células GH3B6 não expressaram nenhuma das isoformas. Para avaliar a comunicação entre as linhagens celulares foram realizadas microinjeções com o corante Lucifer Yellow, tendo sido observada a transferência deste apenas entre as células APS9, confirmando os dados obtidos nos ensaios de RT-PCR. No entanto, em sistemas de co-cultura, além do acoplamento entre os pares de células APS9, observamos também um acoplamento entre os pares de células GH3B6 e entre APS9/GH3B6. Por imunofluorescência indireta, confirmamos a presença da Cx43 nas regiões das membranas aposicionais de células APS9, de células GH3B6 e de pares de células GH3B6/APS9. Ao serem cultivadas com meio condicionado proveniente das células APS9, as células GH3B6 expressaram as conexinas 26 e 43, indicando que as interações existentes entre as células GH3B6 e as células APS9 levaram à formação de junções comunicantes funcionais. Desta forma, nossos estudos de co-cultura com linhagens celulares de adeno-hipófise revelaram a importância das células estromais na comunicação intercelular, levantando a questão do papel do estroma no microambiente glandular e, conseqüentemente, na fisiologia glandular. Esta influência pode estar relacionada com a secreção de diferentes fatores solúveis e/ou com o contato das células estromais com as células secretoras via elementos de matriz extracelular.
Palavras-chave: Hipófise. Conexina. Comunicação intercelular.

x
Abstract
MAÇÃO, Natalia Gonçalves. Análise in vitro da expressão de conexinas e da comunicação
celular em linhagens adeno-hipofisárias. Rio de Janeiro, 2008. Dissertação (Mestrado em
Ciências Morfológicas) - Programa de Pós-Graduação em Ciências Morfológicas, Universidade
Federal do Rio de Janeiro, Rio de Janeiro, 2008.
Many adenohypophysial activities can be regulated by gap junctions intercellular communication. These junctions are intercellular channels formed by proteins called connexins, which are organized forming a connexon, which is permeable to small molecules of up to 1.2 kDa, coordinating many physiological processes as proliferation, adhesion, migration, differentiation, cell survival and apoptosis. In the present study, we evaluated the in vitro expression of connexins 26, 32 and 43 in two rat adenohypophysis cell lines, the GH3B6 tumoral prolactin secreting cells and APS9 stromal cells. RT-PCR assays performed in adenohypophysis primary culture demonstrated the expression of all the connexins studied. When cultivated alone, APS9 cells only expressed Cx43, while GH3B6 didn’t express any connexin isoforms. To evaluate the communication between the cell lines, microinjections with Lucifer Yellow were performed, showing the dye transfer only between APS9 cells, confirming RT-PCR data. However, in co-culture systems, in addition to the coupling between APS9 cell pairs, we also observed coupling between GH3B6 cell pairs and APS9/GH3B6 cells. By indirect immunofluorescence, we have established the presence of Cx43 in appositional membranes regions of APS9 cells, GH3B6 cells and GH3B6/APS9 cells. When cultivated with conditioned medium from APS9 cells, GH3B6 cells expressed the connexins 26 and 43, indicating that interactions between GH3B6 and APS9 cells led to the formation of functional gap junctions. Thus, our co-culture studies with adenohypophysis cell lines shows the importance of the stromal cells in intercellular communication, raising the role of stroma in glandular microenvironment and, consequently, in glandular physiology. This influence could be related to the secretion of many soluble factors and/or the stromal cells contact between secreting cells through extracellular matrix elements.
Keywords: Pituitary. Connexin. Intercellular communication.

xi
Lista de Ilustrações
Figura 1: Suprimento sanguíneo da hipófise. Página 3. Figura 2: Corte histológico da adeno-hipófise. Página 5. Figura 3: Organização molecular e topologia esquemática de uma placa de junções gap. Página 18. Figura 4: Microscopia de contraste de fase das células GH3B6, APS9 e da cultura primária de adeno-hipófise . Página 40. Figura 5: Gel de integridade do RNA total extraído das linhagens GH3B6 e APS9, da cultura primária de adeno-hipófise, do fígado e do coração de ratos e o RT-PCR para GAPDH. Página 42. Figura 6: RT-PCR para as conexinas 26, 32 e 43 (C) na linhagem GH3B6. Página 43. Figura 7: RT-PCR para as conexinas 26, 32 e 43 (C) na linhagem APS9. Página 44. Figura 8: Perfil de acoplamento das células APS9 injetadas com corante LY. Página 47. Figura 9: Perfil de acoplamento das células GH3B6 injetadas com corante LY. Página 48. Figura 10: Perfil de acoplamento total da co-cultura das células APS9 e GH3B6 injetadas com corante LY. Página 49. Figura 11: Percentual de acoplamento entre os dois tipos celulares injetados com corante LY. Página 50. Figura 12: Imunofluorescência indireta para conexina 43. Página 52. Figura 13: RT-PCR das células GH3B6 cultivadas com meio condicionado das células APS9 para as conexinas 26, 32 e 43. Página 54.

xii
Lista de Abreviaturas
ACTH: hormônio adrenocorticotrófico
ADH: hormônio antidiurético
AMPc: adenosina monofosfato cíclico
APS9: linhagem celular derivada de estroma normal de hipófise de ratos
ATP: adenosina trifosfato
bFGF: fator de crescimento básico para fibroblasto
BMP-2: bone morphogenetic protein 2
cDNA: DNA complementar
CRH: hormônio liberador de corticotrofina
Cx: conexina
DAPI: 4,6-diamidino-2-fenilindol
DEPC: dietil-pirocarboneto
E2: 17β-estradiol
EGF: fator de crescimento epidérmico
FSH: hormônio folículo estimulante
GAPDH: gliceraldeído-3-fosfato-desidrogenase
GH: hormônio do crescimento
GH3B6: linhagem celular secretora de prolactina
GHRH: hormônio liberador do hormônio do crescimento
GnRH: hormônio liberador de gonadotrofina
HE: hematoxilina-eosina
HGF: fator de crescimento para hepatócito

xiii
ICSH: hormônio estimulador das células intersticiais
IL-1: interleucina-1
IL-2: interleucina-2
IL-6: interleucina-6
LH: hormônio luteinizante
LPH: hormônio lipotrófico
LY: Lucifer Yellow
MAPK: proteína quinase ativada por mitógeno,
MEC: matriz extracelular
NAD+: nicotinamida adenina dinucleotídeo
PDGF: fator de crescimento derivado de plaqueta
TPA: 12-O-tetradecanoil-forbol-13-acetato
PKC: proteína quinase C
PRL: prolactina
RER: retículo endoplasmático rugoso
RNA: ácido ribonucléico
RT-PCR: reação em cadeia da polimerase via transcriptase reversa
T3: triioditironina
T4: tetraiodotironina ou tiroxina
TGF beta: fator de crescimento transformante beta
TNF-alfa: fator de necrose tumoral alfa
TPS: tampão fosfato salina
TRH: hormônio liberador de tireotrofina
TSH: tireotrofina

SUMÁRIO
1. INTRODUÇÃO .................................................................................................................2
1.1. A hipófise e suas características histofisiológicas…...................................................... 2
1.2. As células secretoras de prolactina..................................................................................9
1.3. As células tumorais GH3B6............................................................................................10
1.4. As células estromais APS9.............................................................................................13
1.5. Comunicação intercelular e Junções Comunicantes......................................................14
1.6. Junções comunicantes em glândulas endócrinas e exócrinas........................................25
2. OBJETIVOS
2.1. Objetivo geral ................................................................................................................29
2.2 Objetivos específicos .....................................................................................................29
3. MATERIAIS E MÉTODOS
3.1. Cultivo das linhagens celulares GH3B6 e APS9.............................................................30
3.2. Cultura primária de hipófise de ratos.............................................................................30
3.3. Ensaios de co-cultura.....................................................................................................31
3.4. Análise por RT-PCR da expressão das conexinas 26, 32 e 43......................................31
3.5 Injeção iontoforética .....................................................................................................35
3.6. Imunocitoquímica indireta.............................................................................................36
3.7. Cultivo da linhagem celular GH3B6 com meio condicionado das células estromais
APS9.....................................................................................................................................38
4. RESULTADOS
4.1. Análise da morfologia das células GH3B6 e APS9........................................................39
4.2. Análise da morfologia da cultura primária de hipófise de ratos...................................39
4.3. Expressão do RNAm das conexinas 26, 32 e 43………………........................……....41
4.4. Análise da comunicação intercelular via junção gap.....................................................45
4.5. Imunodistribuição da conexina 43.................................................................................51

4.6. Expressão do RNAm das conexinas na linhagem celular GH3B6 cultivada na presença
de meio condicionado das células APS9...............................................................................53
5. DISCUSSÃO....................................................................................................................55
6. CONCLUSÃO.................................................................................................................62
7. REFERÊNCIAS .............................................................................................................63

2
1- INTRODUÇÃO
1.1- A hipófise e suas características histofisiológicas
A hipófise é um pequeno órgão que pesa em torno de 0,5 a 1g e se localiza na sela
túrcica do osso esfenóide (Comarck, 1987). Ela se liga por um pedículo ao hipotálamo na base
do cérebro, com quem guarda importantes relações anátomo-fisiológicas. A hipófise e o
hipotálamo juntos formam uma unidade que controla, de forma direta ou indireta, inúmeras
funções fisiológicas (Tirrell e cols., 1994).
A hipófise possui origem embriológica dupla (ectoderma neural e ectoderma
cutâneo), o que leva a uma divisão morfológica em duas regiões distintas (Weiss e Greep,
1981): a adeno-hipófise (porção glandular) ou lobo anterior, que origina-se do ectoderma oral
e é formada por três subdivisões – pars distalis (porção principal da glândula), pars tuberalis
(que envolve o infundíbulo) e a pars intermedia (que separa a pars distalis da neuro-hipófise)
e; a neuro-hipófise (porção nervosa) ou lobo posterior, que origina-se do neuroectoderma e é
formada por duas subdivisões – pars nervosa e o infundíbulo que, por sua vez, é constituído
por dois componentes: o processo infundibular e a eminência média.
O suprimento sanguíneo da hipófise é feito pelo sistema porta hipotálamo-hipofisário,
que exerce um importante papel na regulação das funções da adeno-hipófise. O sistema porta
hipotálamo-hipofisário permite o transporte de fatores de liberação e de inibição
hipotalâmicos para as células epiteliais produtoras de hormônios na adeno-hipófise. Além
disso, permite a secreção de hormônios da adeno-hipófise para a circulação geral e também a
integração funcional do hipotálamo com a adeno-hipófise. Através desse sistema, os
hormônios produzidos pelo hipotálamo são levados diretamente para a adeno-hipófise, sem
percorrer a circulação geral (Junqueira e Carneiro, 2004). O sistema porta origina-se nas

3
artérias carótidas internas, de onde parte a artéria hipofisária superior. Esta penetra na
eminência média e na parte superior do pedículo infundibular e forma o primeiro plexo
capilar sinusoidal (plexo capilar primário), que recebe a secreção das células neuroendócrinas
agrupadas no núcleo hipotalâmico. Os capilares oriundos do plexo capilar primário se
projetam para o infundíbulo e para a pars tuberalis para formar as veias porta. Os capilares
originados das veias porta formam um plexo capilar secundário, que supre a adeno-hipófise e
recebe secreções das células endócrinas da adeno-hipófise. Não existe fornecimento direto de
sangue arterial para a adeno-hipófise. Um terceiro plexo capilar, derivado da artéria
hipofisária inferior, supre a neuro-hipófise e coleta as secreções das células neuroendócrinas
presentes no hipotálamo. Os produtos de secreção são transportados ao longo dos axônios
para a neuro-hipófise (Kierszembaum, 2004).
Figura 1: Suprimento sanguíneo da hipófise. A artéria hipofisária superior forma o plexo
capilar primário no infundíbulo, que recebe fatores de inibição ou de liberação secretados pelo
hipotálamo. O plexo capilar primário drena para as veias porta hipofisárias que fornecem
sangue para o plexo capilar secundário, que encontra-se associado com as células presentes na
pars distalis. Um outro plexo capilar é formado pela artéria hipofisária inferior, responsável
por coletar hormônios secretados pelo hipotálamo (Adaptado de Kierszembaum, 2002).
Núcleo Hipotalamohipofisiotrófico
Núcleo Paraventricular
Núcleo Supra-óptico
Quiasma óptico
Corpo mamilar
Artéria hipofisária superior
Artéria hipofisária inferior
Lobo Posterior Plexo capilar da pars nervosa
Terminal axônico Lobo Anterior
Acidófilas Basófilas
Plexo capilar primário no infundíbulo superior
Veias porta
Plexo capilar secundário na pars distalis

4
Cerca de 80% da hipófise corresponde à adeno-hipófise, sendo a pars distalis a porção
mais desenvolvida desta, responsável pela secreção de três grupos distintos de hormônios: as
proteínas de cadeia simples, representadas pela prolactina (PRL) e pelo hormônio do
crescimento (GH); as glicoproteínas, destacando-se a tireotrofina (TSH) e os hormônios
folículo estimulante (FSH) e luteinizante (LH); e os peptídios da família das corticotrofinas, a
adrenocorticotrofina (ACTH) e os hormônios relacionados (True, 1990). Cada um dos
hormônios destacados acima possui regulação específica e funções fisiológicas primordiais
tanto no metabolismo, como no crescimento e reprodução (Bloom e Fawcett, 1994).
A pars distalis ou lobo anterior é revestida por uma cápsula fibrosa e constituída por
cordões celulares parenquimatosos envolvidos por uma delicada trama de fibras reticulares e
capilares sinusóides. Uma pequena quantidade de estroma glandular está localizada,
principalmente, ao redor das artérias hipofisárias e das veias porta. O revestimento endotelial
dos sinusóides, por formar uma camada descontínua, facilita a difusão de fatores de liberação
e/ou inibição para as células do parênquima glandular, desta forma proporcionando sítios de
penetração para as secreções liberadas. Tanto o epitélio glandular quanto o endotélio
fenestrado dos sinusóides estão apoiados numa delgada membrana basal, que apresenta vários
elementos de matriz extracelular como laminina, colágeno IV e heparan sulfato. Os elementos
celulares presentes neste estroma são representados principalmente por fibroblastos,
macrófagos e mastócitos, além de células ricas em prolongamentos que se relacionam por
junções comunicantes, denominadas de células folículo-estreladas, semelhantes às células de
origem glial modificadas. As células do parênquima da pars distalis, que possuem afinidade
por corantes, são chamadas de cromófilas e as células sem coloração citoplasmática são
denominadas cromófobas. As células cromófilas são classificadas em acidófilas e basófilas
(pela afinidade por corantes ácidos e básicos, respectivamente), constituindo a maior parte das
células secretoras da pars distalis (Gartner e Hiatt, 1999).
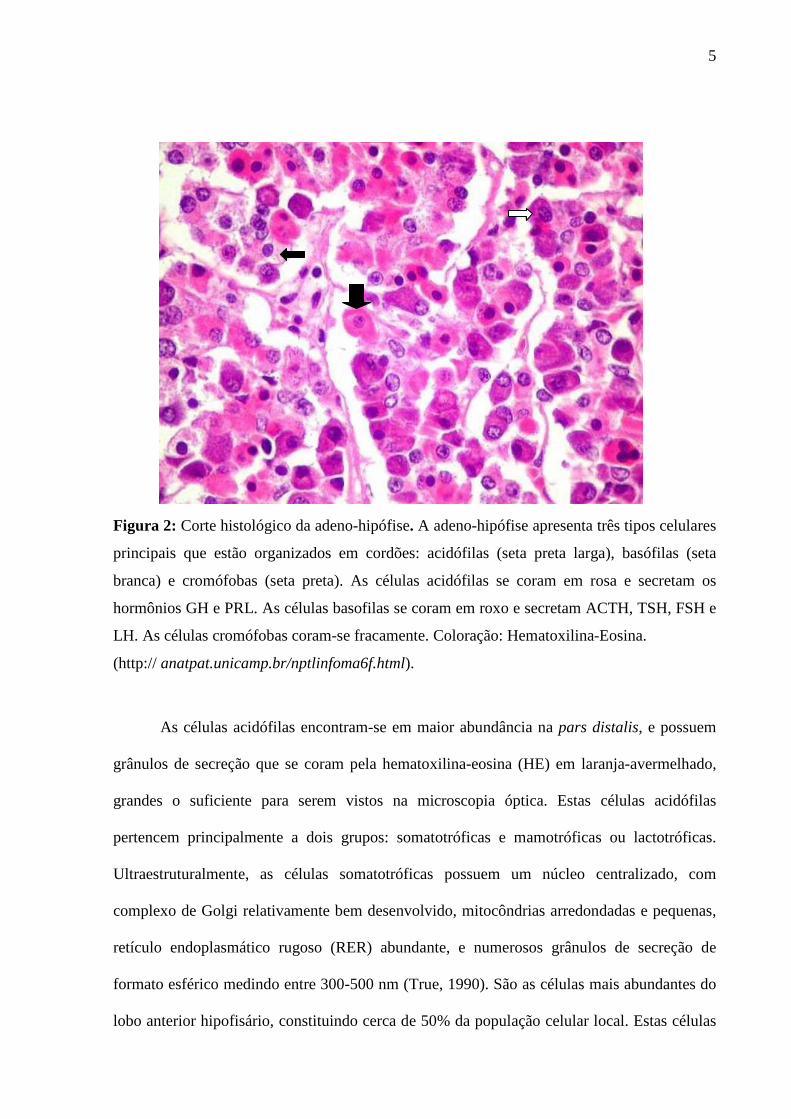
5
Figura 2: Corte histológico da adeno-hipófise. A adeno-hipófise apresenta três tipos celulares
principais que estão organizados em cordões: acidófilas (seta preta larga), basófilas (seta
branca) e cromófobas (seta preta). As células acidófilas se coram em rosa e secretam os
hormônios GH e PRL. As células basofilas se coram em roxo e secretam ACTH, TSH, FSH e
LH. As células cromófobas coram-se fracamente. Coloração: Hematoxilina-Eosina.
(http:// anatpat.unicamp.br/nptlinfoma6f.html).
As células acidófilas encontram-se em maior abundância na pars distalis, e possuem
grânulos de secreção que se coram pela hematoxilina-eosina (HE) em laranja-avermelhado,
grandes o suficiente para serem vistos na microscopia óptica. Estas células acidófilas
pertencem principalmente a dois grupos: somatotróficas e mamotróficas ou lactotróficas.
Ultraestruturalmente, as células somatotróficas possuem um núcleo centralizado, com
complexo de Golgi relativamente bem desenvolvido, mitocôndrias arredondadas e pequenas,
retículo endoplasmático rugoso (RER) abundante, e numerosos grânulos de secreção de
formato esférico medindo entre 300-500 nm (True, 1990). São as células mais abundantes do
lobo anterior hipofisário, constituindo cerca de 50% da população celular local. Estas células

6
secretam hormônio do crescimento (GH) ou somatotrofina, quando estimuladas pelo
hormônio liberador do hormônio do crescimento (GHRH); e são inibidas pela somatostatina
que, também como o GHRH, provém do hipotálamo (Bloom e Fawcett, 1994). O hormônio
do crescimento exerce uma ação global sobre o aumento da taxa do metabolismo celular; no
fígado, o GH induz à produção de somatomedinas, que vão estimular o índice mitótico dos
condrócitos da placa epifisária, promovendo alongamento dos ossos longos e,
consequentemente, o crescimento longitudinal do indivíduo (Gartner e Hiatt, 1999).
As células mamotróficas ou lactotróficas estão dispostas de maneira mais
individualizada, ou em grupos ou cordões localizados aleatoriamente na adeno-hipófise. Estas
pequenas células acidófilas constituem 10-15% das células da adeno-hipófise, possuem
grânulos de secreção com aproximadamente 200-600 nm, que contêm uma proteína
denominada de prolactina (True, 1990). Os grânulos e outras organelas típicas de células
secretoras aumentam muito em quantidade durante a gestação, devido principalmente aos
elevados níveis circulantes de estrogênio e progesterona (Tougard e Tixier-Vidal, 1994).
No homem as células basófilas predominam na região antero-central da pars distalis.
Em geral são maiores que as células acidófilas, possuindo forma ovóide ou arredondada
(Weiss e Greep, 1981). São descritos três tipos de células basófilas: as células tireotróficas, as
corticotróficas e as gonadotróficas. As células tireotróficas possuem como função principal a
secreção de um hormônio glicoprotéico, o hormônio estimulador da tireóide ou tireotrofina
(TSH). Este hormônio controla a atividade das células foliculares da tireóide que, por sua vez,
irão secretar a triioditironina (T3) e tetraiodotironina ou tiroxina (T4). Na microscopia
eletrônica, as células tireotróficas apresentam contornos angulares e pequenos grânulos de
secreção localizados próximos à membrana plasmática medindo entre 50 a 100 nm. Estas
células estão em geral localizadas na porção anteromedial e anterolateral da glândula. A
ativação destas células e conseqüente liberação do hormônio dependem do hormônio

7
liberador de tireotrofina (TRH), um tripeptídeo produzido no hipotálamo ( Bloom e Fawcett,
1994). As células corticotróficas encontram-se espalhadas pela pars distalis correspondendo a
15% das células da adeno-hipófise; têm formato arredondado ou ovóide, núcleo excêntrico e
contêm um número pequeno de organelas. Nos roedores podem ter uma forma estrelada, com
seus processos estendendo-se entre as células vizinhas, terminando próximo aos capilares. Os
seus grânulos secretores possuem cerca de 250-400 nm de diâmetro (True, 1990; Gartner e
Hiatt, 1999). As células corticotróficas, quando estimuladas pelo hormônio liberador de
corticotrofina (CRH) produzido no hipotálamo, secretam o hormônio adrenocorticotrófico
(ACTH) e o hormônio lipotrófico (LPH). O hormônio ACTH estimula as células do córtex
supra-renal a secretarem os hormônios corticóides (Bloom e Fawcett, 1994; Gartner e Hiatt,
1999). As células gonadotróficas estão distribuídas por toda a adeno-hipófise, constituem 10-
15% das células da adeno-hipófise, são arredondadas, possuem retículo endoplasmático
rugoso (RER) abundante, e complexo de Golgi bem desenvolvido. Os grânulos de secreção
têm um diâmetro que varia entre 200-400 nm. Secretam o hormônio folículo estimulante
(FSH) e o hormônio luteinizante (LH) ou hormônio estimulador das células intersticiais
(ICSH). Na verdade, não se sabe se existem sub-populações de células gonadotróficas, ou
seja, se existe um tipo celular secretor de FSH e outro de LH, ou se ambos os hormônios são
produzidos pela mesma célula em diferentes estágios do ciclo secretor. A secreção destes
hormônios é estimulada pelo hormônio liberador de gonadotrofina (GnRH) e inibida por
vários hormônios que são produzidos nos ovários e testículos (Comarck, 1987).
Além das células cromófilas, a pars distalis possui células pequenas que são pouco
coradas e dispostas em grupos, denominadas de células cromófobas. Estas células geralmente
possuem menos citoplasma do que as células cromófilas, podendo ser tanto células-fonte não
específicas quanto células cromófilas que já excretaram seu conteúdo hormonal e perderam
afinidade tintorial típica das células acidófilas ou basófilas (Kierszembaum, 2004). Sabe-se

8
que as células da adeno-hipófise apresentam uma atividade secretora cíclica: passando por
etapas de síntese, acúmulo, armazenamento e liberação de grânulos de secreção (Weiss e
Greep,1981).
Ainda na pars distalis, além das células cromófilas basófilas e acidófilas, as células
folículo-estreladas constituem uma grande população celular não secretora. Desde que as
células folículo-estreladas foram descritas como corticotrofos erroneamente por Farquhar em
1954, suas características histológicas foram continuamente investigadas por muitos
pesquisadores. A microscopia demonstrou que as células folículo-estreladas possuem longos e
complexos prolongamentos citoplasmáticos, formando uma rede anatômica tridimensional na
qual as células endócrinas residem (Fauquier e cols, 2001). As células folículo-estreladas
possuem um padrão epitelial de associação ao redor do lúmem de pequenos cistos ou
folículos, onde elas projetam microvilos e ocasionalmente cílios, formando junções
comunicantes com outras células folículo-estreladas. Sabe-se, que elas são capazes de sofrer
mitoses e que possuem filamentos de citoesqueleto do tipo gliofibrilar (Bloom e Fawcett,
1994; Gartner e Hiatt, 1999), são positivas para a proteína S-100 e são tidas por serem
importantes para a regulação das funções celulares endócrinas da hipófise. A importância do
sistema de rede das células folículo-estreladas assim como o sistema porta-hipofisário como
mecanismos envolvidos na secreção hormonal foi alvo de análises histológicas especialmente
envolvendo as junções comunicantes das células folículo-estreladas. (Sato e cols., 2005).
Além da função reguladora, essas células também foram especuladas como células de apoio,
células de limpeza e, células-tronco. Elas também secretam fatores como bFGF, EGF e IL-6.
Kabir e cols. (2005) sugeriram que a comunicação via junção comunicante seja importante
para as células folículo-estreladas.
A pars intermedia situa-se entre a pars distalis e a pars nervosa e representa menos de
0,8% do peso total da glândula. Ela caracteriza-se por possuir muitos cistos revestidos por

9
células cubóides (cistos de Rathke) contendo colóide, resquícios do ectoderma da bolsa de
Rathke. Circunda a haste hipofisária mas, frequentemente, está ausente na face posterior. É
ricamente vascularizada com cordões ou grupos de células cromófilas e cromófobas
(Kierszembaum, 2004).
A pars nervosa consiste em pituícitos (elementos gliais) e axônios amielínicos, cujos
corpos celulares encontram-se em núcleos localizados no hipotálamo (Bloom e Fawcett,
1994). As porções terminais destes axônios contêm inúmeros grânulos de secreção
concentrados, denominados corpos de Herring (Ross & Rowell, 1993). Estes grânulos
possuem os hormônios antidiurético (ADH) e ocitocina, liberados após estimulação do
hipotálamo (Weiss e Greep, 1981).
1.2 - As células secretoras de prolactina
As células secretoras de prolactina foram descritas pela primeira vez por Farquhar e
Rinehart (1954), como células que possuíam os maiores grânulos de secreção de todas as
células da hipófise. Possuem retículo endoplasmático rugoso e aparelho de Golgi bastante
desenvolvidos, e grânulos de secreção de forma e tamanhos variados (Tougard e Tixier-Vidal,
1994). O produto da secreção dessas células é a prolactina, armazenada sob a forma de
grânulos secretores que possuem aproximadamente 200-600 nm de diâmetro (True,1990).
A prolactina possui diversas funções na reprodução, na lactação, na osmorregulação e
na imuno-modulação, além de participar da manutenção do corpo lúteo em roedores (Weiss e
Greep,1981). A produção excessiva de prolactina (hiperprolactinemia) induz ao estado de
hipogonadismo em que há diminuição da libido ou impotência no homem e anovulação ou
amenorréia na mulher (True, 1990). As células lactotróficas têm sua proliferação aumentada
durante o período da gravidez devido à elevação dos níveis plasmáticos de estrogênio. A

10
síntese e a liberação de prolactina são processos dependentes de cálcio e podem ser
fortemente influenciados por manipulação do influxo deste íon através dos canais de cálcio da
membrana plasmática (Thorner, 1977).
A dopamina, que atua através de receptores do tipo D2 presentes nos lactotrofos, é o
fator fisiológico predominante de inibição da prolactina (Thorner, 1977; Bennett e Plum,
1996). Em células secretoras de prolactina GH3B6, há diminuição da secreção de prolactina
após 30 minutos de exposição à dopamina, efeito este dose-dependente. Alguns autores
defendem que a dopamina em baixas concentrações estimula a secreção de prolactina
(Melmed e cols., 1980; Faure e cols., 1980; Tixier-Vidal e cols., 1980; Gourdji e cols., 1972 e
1982). A somatostatina também diminui a liberação de prolactina, tendo sido demonstrada em
células GH3 uma inibição dose-dependente. (Gourdji e cols., 1982).
Em relação a fatores periféricos, o estradiol-17β (E2) possui um papel importante na
regulação total da prolactina. Seus efeitos são fortemente dependentes da dose administrada e
da duração de exposição. O E2 estimula a secreção, a síntese e armazenamento da prolactina,
além de aumentar o tamanho e o número de lactotrofos na adeno-hipófise. O E2 é um
importante regulador da expressão do gene da prolactina. Foi demonstrado que a expressão
dos genes de neuropeptídios, como o VIP e a galanina, também está implicada no controle da
secreção de prolactina (Croissandeau e cols., 1996).
1.3 - As células tumorais GH3B6
Em 1968, o grupo de Tashjian (Japão) isolou um tumor mamosomatotrófico de rato
Wistar Furth que, quando transplantado, secretava os hormônios adrenocorticotrófico
(ACTH), do crescimento (GH) e a prolactina (PRL). Por meio de várias passagens alternadas,
foi isolada uma linhagem de células que secretava GH e por isso foi denominada de células

11
GH. Posteriormente foram obtidos três clones dessa linhagem (GH1, GH1C1 e GH3). O clone
GH3 passou a ser utilizado para estudos de regulação neuroendrócrina. As células desse clone
se organizam em pequenos grupos de células menores que as células normais e secretam
prolactina e hormônio do crescimento (GH) (Tashjian e cols., 1968 e 1970; Gourdji e cols.,
1982).
As células GH3B6, um subclone obtido a partir das células GH3, secretam prolactina.
Foram caracterizadas pela presença de poucos pequenos grânulos de secreção, de um aparelho
de Golgi bem desenvolvido, de numerosos polissomas livres e de um retículo endoplasmático
rugoso formado de cisternas curtas e descontínuas (Gourdji e cols., 1972; Tixier-Vidal e cols.,
1980). A distribuição subcelular da prolactina é semelhante àquela observada nas células
adeno-hipofisárias normais, excetuando-se o fato de que as células GH3B6 possuem
compartimento reduzido de armazenamento do hormônio (Tougard e cols., 1980 e 1982).
Apesar deste fato, estas células liberam grandes quantidades de prolactina no meio de cultura.
Em 1984, Morin e cols. demonstraram que o turnover da prolactina nos compartimentos
membranosos envolvidos no processo secretor é mais rápido que em células normais. Além
disso, elas têm uma intensa atividade mitótica e renovam suas proteínas estruturais numa
velocidade mais rápida do que as células normais (Gourdji e cols., 1982). Nasciutti e cols.
(1992) demonstraram que o cultivo de células GH3B6 em baixas temperaturas e o tratamento
por drogas como a brefeldina A induzem uma modificação no transporte e na distribuição
intracelular de prolactina e de antígenos de membrana. Em 1997, Billis e White demonstraram
que a expressão gênica da prolactina basal é mantida através de atividades espontâneas dos
canais de cálcio dependentes de voltagem.
Uma maior adesão e proliferação celular ocorrem quando as células GH3B6 são
cultivadas sobre laminina. Prolongamentos celulares semelhantes a neuritos são formados,
onde se concentram grandes quantidades de grânulos de secreção de prolactina. Estas

12
mudanças se correlacionam com o aumento da secreção de prolactina, seja a nível basal, ou
em células estimuladas pelo TRH (Brunet de Carvalho e cols., 1989).
Em nosso laboratório (Carvalho e cols., 2000), foi realizada uma caracterização de
diferentes elementos de matriz extracelular constitutivamente sintetizados pelas células
GH3B6. Por citometria de fluxo, foi demonstrada uma concentração relevante dos receptores
de matriz α5β1 (receptor para fibronectina), α6β1 (receptor para laminina) e CD44 (receptor
para ácido hialurônico). Por procedimentos bioquímicos, foi demonstrada na superfície
celular a presença de grandes quantidades de heparan sulfato, condroitin sulfato e dermatan
sulfato. Ao contrário da secreção de prolactina, o TRH pareceu não influenciar a biossíntese
destes elementos de matriz extracelular nas células. Estes resultados sugerem uma
participação das células tumorais endócrinas na síntese dos componentes de MEC e seus
receptores in vivo. Conseqüentemente levantou-se a questão de um possível envolvimento
destes componentes de matriz como reguladores e/ou moduladores da secreção hormonal, em
particular da prolactina.
O emprego de células secretoras de prolactina em estudos do processo de secreção das
células da adeno-hipófise tem sido bastante utilizado (Tougard e cols., 1994). Os estudos em
culturas destas células têm revelado a grande importância na análise das etapas do processo
secretor, destacando-se particularmente as vias de liberação do hormônio e os fatores e as
interações celulares que participam da regulação da atividade secretora bem como os fatores
de crescimento/citocinas, metaloproteases e esteróides que regulam a secreção dos hormônios
adeno-hipofisários.

13
1.4 - As células estromais APS9
Em 2002, Alves e colaboradores isolaram, a partir da cultura primária de adeno-
hipófise de ratos Wistar, uma população estromal homogênea, que foi denominada APS9.
A cultura primária de adeno-hipófise inicialmente apresentou uma população celular
heterogênea, sem um arranjo definido. Com o desenvolvimento da cultura, em contraste ao
grande desenvolvimento das células estromais, as células de natureza epitelial foram se
tornando cada vez mais raras. A partir da nona passagem, uma única população celular foi
estabelecida. Esta população era do tipo fibroblastóide, formada por células alongadas, com
muitos prolongamentos citoplasmáticos, núcleo grande e proeminente e nucléolos numerosos
e evidentes. Similar a outras linhagens celulares estromais estabelecidas, as células APS9 são
miofibroblastos que expressam α-actina de músculo liso, produzindo uma membrana basal
complexa, formando contatos celulares específicos.
Já sabe-se que as células APS9, quando cultivadas juntamente com as células GH3B6,
com quem mantém uma interação funcional específica, podem promover uma organização
bastante eficiente destas em cordões característicos da organização da adeno-hipófise. Além
disso, as células APS9 foram claramente estimulatórias para o crescimento das células GH3B6
quando comparadas a fibroblastos e células derivadas da medula óssea e fígado fetal.
Já foi descrito que populações celulares podem se transformar em linhagens celulares
imortalizadas a partir das modificações induzidas e/ou espontâneas a nível de material
genético (Freshney e cols., 1975) e estas têm sido muito utilizadas em vários estudos de
fisiologia e fisiopatologia de vários tecidos e órgãos, constituindo modelos in vitro
extremamente importantes. Para estudo da fisiologia e fisiopatologia da adeno-hipófise, a
utilização dessa linhagem estabelecida de células estromais deverá ser de grande importância,

14
uma vez que pouco se sabe sobre a influência das interações celulares entre as células
secretoras e as células estromais na manutenção da integridade funcional glandular.
1.5 - Comunicação intercelular e Junções Comunicantes ou Gap
A homeostase tecidual é um fator importante para a manutenção da função de todos os
tecidos e, sua desordem geralmente resulta na disfunção dos órgãos e no desenvolvimento de
doenças (Momiyama e cols, 2003). Entre os aparatos que contribuem para a homeostase
tecidual está a comunicação intercelular (Kanczuga-Koda e cols, 2005). Usando este tipo de
comunicação, as células podem perceber diferenças nas condições ambientais e responder a
estas de maneira correta. Este conceito pode envolver tanto sinais enviados para células
vizinhas que geram uma resposta coordenada, quanto sinais enviados para células isoladas do
resto da comunidade, para que se mantenha a integridade tecidual. O caminho mais comum
deste tipo de comunicação é via junção comunicante ou junção gap Esse tipo de comunicação
é mediado via canais intercelulares que se localizam em regiões especializadas da membrana
plasmática, para formar essas junções (Mese e cols, 2007).
A comunicação intercelular através de canais de junções gap desempenha um papel
crucial não apenas na coordenação de sinais elétricos em células excitáveis, mas também
facilitando a sinalização intercelular em células não excitáveis durante o desenvolvimento,
influenciando também na regulação do crescimento e na proliferação e diferenciação celulares
(Ko e cols, 2000).
Muitos estudos envolvendo camundongos knock out revelaram a importância das
junções comunicantes no desenvolvimento normal e em processos fisiológicos. A ausência da
comunicação intercelular via junções gap em células humanas e de animais tem sido
associada à diminuição do controle do crescimento e aumento da tumorigênese (King e cols,

15
2005). Uma população de células acopladas por junções gap forma um tipo de sincício no
qual cada célula permanece autônoma em relação ao seu conteúdo macromolecular solúvel
intracelular, mas dependente ainda das células vizinhas a ponto de pequenos componentes
citoplasmáticos estarem relacionados (Morand e cols, 1996).
As junções gap são especializações de membrana, que permitem o movimento
intercitoplasmático de pequenas moléculas sinalizadoras de forma direta entre células
adjacentes (Ke e cols, 2005). Este tipo de canal intercelular permite que se estabeleça um
caminho direto e relativamente não específico para a difusão ou transporte de moléculas de
até 1,2 kDa. Entre estas moléculas encontramos água, íons, substratos metabólicos como
carboidratos, aminoácidos, nucleotídeos e segundos mensageiros, dentro dos quais estão o
cálcio, AMPc e o inositol 1,4,5-trifosfato (Decrouy, 2004). As junções gap são estruturas
dinâmicas; as trocas mediadas por elas podem ser interrompidas a qualquer momento em
resposta a sinais intracelulares específicos que induzem o fechamento reversível do canal da
junção gap (Morand e cols, 1996).
Nos cordados, as junções gap são formadas por proteínas de membrana chamadas de
conexinas. Estas proteínas são codificadas por uma família multigênica (Kanczuga-Koda e
cols, 2005). Estes genes podem ser categorizados em três grupos conhecidos como α,β e γ, de
acordo com sua estrutura genética, homologia genética e motivos seqüenciais específicos
(Mese e cols, 2007). Formam um grupo de pelo menos 20 proteínas altamente conservadas,
sendo diferentemente expressas em uma variedade de tecidos, as quais acredita-se que,
geralmente reflitam a regulação celular específica de acoplamento das junções gap e
demandas funcionais para estas junções em diferentes tipos celulares (Tanmahasamut e Sidell,
2005). São proteínas altamente homólogas, pois cerca de 50 e 80% dos aminoácidos são
iguais (Goodenough e cols., 1996). Existem duas convenções na literatura para a
nomenclatura das conexinas: uma depende da massa molecular da conexina (conexina 26

16
representa a proteína conexina de 26 kDa; a conexina 46 representa a isoforma de conexina de
46 kDa, etc), enquanto que outra utiliza símbolos gregos baseados nas considerações
evolutivas. A comunicação via junção gap em animais não cordados é, entretanto, mediada
via outra família de proteínas denominadas inexinas. As proteínas inexinas não são
homólogas às conexinas em termos de seqüência primária; no entanto, canais de junções gap
formados por inexinas dividem semelhanças funcionais com canais intercelulares formados
por conexinas. Recentemente, outro grupo de proteínas denominadas panexinas, as quais
podem estar distantemente relacionadas com as inexinas, foram identificadas em vertebrados
e foram demonstradas por estarem expressas em vários tecidos incluindo rim, olhos e
neurônios (Barbe e cols, 2006).
Cada conexina possui quatro domínios transmembranares que, em conjunto com
domínios das outras cinco conexinas, formam um poro simétrico. O arranjo de seis conexinas
forma um hemicanal na membrana plasmática conhecido como conexon. Os conexons são
transportados para a membrana plasmática e unem-se com conexons de células adjacentes
para formar os canais de junções gap (Laird e cols, 2005). Os domínios das conexinas estão
conectados por duas alças extracelulares que desempenham importante papel nos processo de
reconhecimento e adesão. Existem três resíduos cisteína preservados em cada alça, os quais
formam somente pontes dissulfeto intraconexina. Os domínios transmembrana e as alças
intracelulares são altamente conservados entre os membros da família. Além disso, as
proteínas conexinas apresentam domínios C terminal e N terminal citoplasmáticos e uma alça
citoplasmática ligando o segundo e o terceiro domínio transmembrana. Apesar do domínio N
terminal ser conservado, a alça citoplasmática e a porção C terminal demonstram grandes
variações em termos de seqüência e comprimento. Por exemplo, a conexina 26 apresenta uma
porção C-terminal menor, enquanto que a conexina 50 apresenta uma longa cauda C-terminal.
A alça e a cauda citoplasmática são suscetíveis a várias modificações pós-traducionais, como

17
por exemplo, a fosforilação. A maioria das conexinas são fosfoproteínas, e a fosforilação é
considerada importante para a regulação do arranjo e modulação das propriedades fisiológicas
dos canais (King e cols., 2005). As proteínas kinases que estão envolvidas na fosforilação das
conexinas são a MAPK, proteína kinase C, proteína kinase A, cdc2/ciclina B, a caseína kinase
1, v-src e x-scr. A fosforilação das conexinas regula vários passos do ciclo celular das junções
gap, incluindo o transporte da conexina para a membrana plasmática, arranjo dos conexons
em junções gap e passagem pelos canais de junção gap. Existe também uma forte evidência
que a fosforilação das conexinas seja importante na regulação da internalização e degradação
destas junções. A modulação da taxa de turnover da junção gap tem sido sugerida como um
importante mecanismo de regulação do nível de comunicação intercelular via junção gap. Há
uma crescente evidência que a ubiquitina é uma proteína chave na regulação da degradação da
junção comunicante (Leithe & Rivedal, 2007).

18
Figura 3: Organização molecular e topologia esquemática de uma placa de junções gap.
Hemicanais de membranas aposicionais de células adjacentes podem se acoplar e formar
junções de gap. Três tipos diferentes de junções de comunicação foram descritas: (1)
homomérica/homotípica; (2) heteromérica e (3) heterotípica. As conexinas são proteínas com
quatro domínios transmembrana que dividem três resíduos cisteína extracelulares
conservados, que são cruciais para o acoplamento. As subunidades de conexinas variam
principalmente na sua alça citoplasmática e na sua região carboxi-terminal. (Adaptado de
Willeck, 2005).
Existem pelo menos 21 isoformas de conexina no genoma humano e
aproximadamente todas as células no corpo expressam pelo menos um tipo desses genes em
algum ponto durante o desenvolvimento e na vida adulta. Por exemplo, a conexina 26 é
altamente expressa na cóclea, fígado, pele e placenta, enquanto que a conexina 46 e a
conexina 50 são encontradas exclusivamente no olho. Além disso, as conexinas mostram um
padrão de expressão sobreposto, onde uma única célula pode apresentar mais de um tipo de
Subunidade de conexina
Hemicanais
Alça extracelular
Membrana
Citoplasma
Conexina
Alça citoplasmática

19
isoforma. As conexinas 26, 30, 30.3, 31, 43 e outras, por exemplo, são encontradas nos
queratinócitos, enquanto que os cardiomiócitos utilizam as conexinas 31.9, 40, 43 e 45 para
comunicação intercelular. A co-expressão de múltiplos genes de conexinas em uma única
célula pode conseqüentemente afetar tanto a composição dos conexons quanto os canais
intercelulares formados e podem promover um mecanismo compensatório de perda de uma
isoforma (Segretain & Falk, 2004).
A biossíntese das junções gap e o seu arranjo são estritamente regulados e, estas
junções celulares possuem uma meia vida curta de apenas algumas horas. A maioria das
conexinas é co-traducionalmente integrada na membrana do retículo endoplasmático. A
oligomerização das seis conexinas em um hemicanal é conhecida por ocorrer de maneira a se
iniciar no retículo endoplasmático e terminar na região trans aparelho de Golgi (Laird, 2006).
Alguns estudos revelam que a conexina 26 e a conexina 43 podem também ser inseridas pós-
traducionalmente nas membranas do retículo endoplasmático e, uma outra pesquisa também
sugere que a conexina 26 pode também estar inserida nas membranas plasmáticas. Já foi
descrito que a conexina 43 e a conexina 46 se oligomerizam em conexons em compartimentos
incluindo a face trans do aparelho de Golgi (Laird e cols, 2005). Os conexons são então
levados à superfície celular via vesículas transportadas através dos microtúbulos, que se
fundem à membrana plasmática. Esse hemicanais podem também formar canais não
juncionais em áreas não opostas da membrana ou se difundir livremente a regiões de contato
célula-célula para encontrar um conexon “parceiro” de uma célula vizinha para completar a
formação de canais intercelulares. Os canais intercelulares então agrupam-se em placas de
junções gap, um evento altamente dinâmico envolvendo a remoção de velhos canais do centro
da placa enquanto há a adição de novas subunidade de junções de comunicação na periferia.
Os canais intercelulares do meio da placa são internalizados em estruturas vesiculares
denominadas “junções anulares” as quais se fundem com lisossomos para a degradação por

20
enzimas lisossomais ou são alvo da via proteosomal. A continua síntese e degradação de
conexinas através desse mecanismo pode promover uma rápida adaptação dos tecidos a
mudanças nas condições ambientais, incluindo estresse mecânico e isquêmico. Sob estas
circunstâncias, os hemicanais abertos são conhecidos por facilitar a liberação de vários fatores
como ATP, glutamato e NAD+ no espaço extracelular, gerando diferentes respostas
fisiológicas. Não se sabe se os hemicanais ativos se incorporam às junções gap antes da
degradação ou seguem um caminho de reciclagem (Evans e cols, 2006).
Se um conexon está composto de unicamente uma isoforma de conexina, é chamado
homomérico; se está composto por mais de uma isoforma de conexina, é denominado
heteromérico. Uma união homotípica pode ocorrer se dois conexons idênticos pareiam e uma
união heterotípica é composta por dois conexons diferentes (Bruzzone e cols, 1996). Por
exemplo, dois canais heteroméricos idênticos podem formar uma união homotípica. Também
existe a possibilidade da formação de canais homoméricos que constituiriam uma união
heterotípica; a união de um conexon heteromérico com um conexon homomérico formando
uma união mono-heteromérica, ou finalmente, a formação de um canal bi-heteromérico, que
seria composto por dois conexons heteroméricos (Wang e Peracchia, 1998). A formação
dessas estruturas depende da compatibilidade das conexinas formarem canais, apesar de nem
todas as conexinas poderem interagir entre si, como a conexina 26, que não forma canais
heteroméricos com a conexina 30 e 32, considerando que esta não pode formar canais
funcionais com a conexina 40. Estas interações complexas aumentam a diversidade estrutural
e funcional, permitindo várias possibilidades no tipo de moléculas a serem divididas entre
células (Segretain & Falk, 2004).
Modelos iniciais de comunicação via junção gap descreveram os canais como sendo
poros passivos não específicos, que permitiriam passagem livre de qualquer íon ou
metabólitos menores que 1.2 kDa. Entretanto, recentes desenvolvimentos nos campos da

21
permeabilidade seletiva dos canais de junção gap demonstraram que os canais formados por
diferentes conexinas são únicos em termos de condutância e permeabilidade a moléculas
específicas. Além disso, a associação de doenças a conexinas específicas também enfatiza a
individualidade de cada proteína, pois a perda de uma isoforma não pode ser compensada pela
presença de outras conexinas no mesmo tecido ou em um tipo celular, levando a efeitos
fisiopatológicos (Mese e cols, 2007). Altos graus de permeabilidade seletiva a íons e
moléculas foram observados para os canais homoméricos. Entretanto, a maioria das células
expressa mais de uma conexina, sendo canais heteroméricos de conexinas bastante comuns in
vivo. Uma mistura heteromérica de diferentes isoformas de conexinas, promovendo um
arranjo das isoformas nos hemicanais, pode permitir às células regularem dinamicamente suas
propriedades intercelulares de comunicação, incluindo a seletividade molecular (Ayad e cols,
2006).
Modelos de animais knock out para genes que codificam diferentes conexinas nos
mostra a importância destas proteínas na fisiologia. Sabe-se que animais knock out para
conexina 26 morrem 10-11 dias pós coitum (Simon e cols., 1998), os knock out para conexina
32 são viáveis, férteis porém apresentam mobilização de glicose hepática reduzida devido a
um defeito na transmissão do sinal disparado por estimulação do nervo simpático no fígado
(Nelles e cols., 1996). Da mesma forma animais knock out para o gene para conexina 37 são
viáveis porém, as fêmeas são inférteis apresentando perda dos folículos ovarianos maduros,
falhas no processo de ovulação e ainda desenvolvimento de numerosos e inapropriados corpos
lúteos (Simon e cols., 1997). Já o fenótipo apresentado por animais deficientes do gene da
Cx40 mostra animais viáveis, férteis e com anormalidades eletrocardiográficas (Simon e cols.,
1998), os animais knock out para conexina 43 apresentam letalidade na fase perinatal,
obstrução do fluxo do ventrículo direito e da artéria pulmonar e cianose (Reaume e cols.,
1995). Por outro lado animais deficientes para Cx46 são viáveis e férteis porém desenvolvem

22
catarata quatro semanas após o nascimento e os deficientes para Cx 50 também são viáveis e
também apresentam catarata (Gong e cols., 1997).
Uma abordagem alternativa para manipular a comunicação intercelular envolve a
super-expressão de genes de conexinas e isto tem sido realizado em animais transgênicos que
super-expressam conexina 43. Estes animais apresentam viabilidade baixa com defeitos no
tubo neural e no ventrículo direito do coração (Ewart e cols., 1997). Alguns estudos
demonstraram que, em certos tecidos, a comunicação intercelular pode estar mediada por
conexinas específicas, que asseguram sua função.
Por outro lado, um aspecto crítico da comunicação intercelular é que ela pode ser
modulada em resposta a eventos extra e intracelulares. A comunicação via junção gap pode
ser regulada ao nível da adesão célula-célula (Meyer e cols., 1992), transcrição do gene da
conexina, a tradução, a síntese, a degradação e/ou reutilização (Loewestein, 1987; Bruzzone e
cols., 1996). A determinação dos mecanismos pelos os quais estes efetores influenciam a
comunicação celular é essencial para entender o papel da comunicação através das junções
gap nas células. O mecanismo pelo qual um efetor particular influencia a comunicação via
junção gap pode ser extremamente complexo. Alguns agentes (AMPc, agonistas da PKC)
atuam em mais de um nível e o mesmo agente pode ter efeitos opostos sobre a comunicação
juncional em diferentes tipos celulares ou no mesmo tipo celular em diferentes concentrações
(Metha e cols., 1991,Rivedal e Leithe 2005).
Hormônios e fatores de crescimento também podem alterar a comunicação juncional o
que pode levar a efeitos diversos. Entre os fatores que podem modular estas junções gap
encontram-se o HGF (Moorby e cols., 1995; Ikejima e cols., 1995), TGF beta (Chandross e
cols., 1995, Chandross, 1998), IL-1 (Hu e cols., 1994), IL-2 (Wadamori e cols., 1996), EGF
(Díez e cols., 1998), TNF-alfa (van Rijen e cols., 1997), BMP-2 (Wyatt e cols., 2001), PDGF
(Pelletier e Boynton, 1994), hormônio paratireoideo (Schiller e cols., 2001) e os retinóides

23
(Metha, 1991; Tanmahasamut e Sidell, 2005). Estudos demonstram que o ácido retinóico e
outros retinóides aumentam a comunicação intercelular via junção gap e os níveis de conexina
43 em vários tipos celulares normais e malignos. Essa atividade, além das funções bem
caracterizadas das conexinas na implantação embrionária e desenvolvimento, sugerem uma
possível ligação entre os efeitos teratogênicos do ácido retinóico e sua modulação dos níveis e
do perfil de fosforilação da conexina 43 (Tanmahasamut e Sidell, 2005).
A capacidade de produtos químicos de inibir a comunicação via junções gap tem sido
associada com suas propriedades carcinogênicas. Alguns promotores de tumor como o TPA,
induzem a inibição temporária da comunicação intercelular via junção gap em muitos tipos
celulares diferentes. Não está claro em que medida a capacidade de induzir uma down-
regulation prolongada comparada como uma transitória esteja relacionada com a potência de
uma substância em induzir o câncer. Para melhor entendimento do papel da comunicação
intercelular via junção gap na carcinogênese, é importante entender os mecanismos
regulatórios envolvidos na inibição da comunicação por substâncias químicas causadoras de
câncer. (Rivedal & Leithe, 2005). Os álcoois e os derivados do ácido glicirretínico são os
bloquedores mais utilizados nos estudos de comunicação juncional. O mecanismo pelo qual
eles bloqueiam essas junções ainda é desconhecido, no entanto acredita-se que o heptanol
reduza a fluidez dos domínios de colesterol na membrana que por sua vez estão associados
com placas juncionais na mesma região, evitando a comunicação intercelular (Bastiaanse e
cols., 1993).
Além de produtos químicos, os hemicanais de conexinas se fecham em concentrações
fisiológicas milimolares de cálcio extracelular, mas abrem em resposta à diminuição de cálcio
extracelular (sozinho ou em combinação com outros cátions divalentes); forte despolarização
de membrana, estimulação mecânica; UTP extracelular; inibição metabólica; infecção por

24
Shighella e, mais recentemente, em reposta a um aumento citoplasmático de cálcio (De Vuyst
e cols, 2007).
Outro elemento importante na regulação das junções gap é a matriz extracelular,
aspecto ainda pouco estudado. Spray e cols. (1987) demonstraram a influência dos
glicosaminoglicanos sulfatados sobre a comunicação juncional. No entanto, o estudo não
relacionou diretamente o estado funcional dos canais com a matriz circundante. Alford e
Rannels (2001) mostraram a influência direta da matriz, sobre a formação de junções gap
compostas por conexina 43, mas da mesma maneira existem poucos estudos conclusivos
sobre a influência da matriz extracelular nas junções gap, apesar de parecer evidente a relação
existente para elas.
Estudos recentes de Kanczuga-Koda e cols. (2005) demonstraram que, em tecido
tumoral de cólon, a conexina 26 assume uma localização e distribuição distinta daquela
observada em tecido de cólon não patológico e que esta proteína está relacionada a proteínas
envolvidas com a apoptose Bcl-xL e Bax. Este estudo mostrou que a presença da conexina 26
e sua associação com marcadores de apoptose no citoplasma das células tumorais poderia
indicar um papel diferente daquele que classicamente se assume para as conexinas
(comunicação intercelular), o que sugeriria uma implicação desta proteína como modulador
da apoptose e possivelmente haveria uma implicação clínica nesta relação.
Recentemente foi observado que animais knock out para conexina 32 quando
submetidos a estresses químicos e/ou radioativos apresentam formação tumoral nos tecidos
hepático e pulmonar com decréscimo da expressão de gene supressor tumoral p27 (King e
cols., 2005).

25
1.6 - Junções comunicantes ou gap em glândulas endócrinas e exócrinas
As junções gap são visualizadas por microscopia eletrônica convencional e criofratura
em quase todos os tecidos animais adultos, incluindo as glândulas endócrinas. Existem dados
moleculares e funcionais disponíveis para ilhotas pancreáticas, córtex da adrenal, testículo,
ovário e tireóide (Morand e cols 1996).
Estudos demonstram que as células b do pâncreas endócrino produtoras de insulina
expressam conexina 43, a qual não é detectável nas células do pâncreas exócrino.
Mutuamente, as células acinares do pâncreas exógeno expressam as conexinas 32 e 26, as
quais não são encontradas nas ilhotas pancreáticas.
No ovário, acredita-se que as junções gap desempenhem um papel essencial na
organogênse e no controle da proliferação e diferenciação celulares. Essa comunicação entre
células ovarianas via junção gap pode também estar envolvida no controle do
desenvolvimento folicular, na maturação meiótica do ovócito e no crescimento e regressão
lúteos. As junções gap entre as células da granulosa ovariana contêm predominantemente
conexina 43, a qual está presente em todos os estágios do desenvolvimento folicular, com
folículos antrais grandes mostrando intensa marcação. Estudos recentes descreveram que, em
camundongos fêmeas deficientes de conexina 43, os folículos ovarianos ficavam presos em
estágios pré-antrais iniciais e que a comunicação celular entre as células da granulosa é
interrompida, sugerindo que a comunicação via conexina 43 da junção gap é crítica para a
foliculogênse ovariana. (Ke e cols, 2005)
No endométrio, as conexinas mais expressas são as isoformas 43 e 26, que constituem
a maioria das conexinas expressas no estroma e epitélio. Estudos recentes sugeriram que a
proteína da junção gap em células estromais do endométrio desempenha um papel regulatório
na manutenção dos níveis normais de comunicação intercelular via junção gap nas células

26
epiteliais. Desse modo, parece que a expressão da conexina 43 nas células estromais
endometriais faz a manutenção e a regulação das proteínas da junção gap em ambas as
populações celulares, sendo esta um processo dinâmico. Os níveis de conexina 43 no
endométrio aumentam durante a fase folicular sob influência do estrogênio e estão reduzidos
na fase luteínica do ciclo menstrual em resposta a progesterona. Esses padrões de expressão
da conexina 43 associados ao ciclo no endométrio apontam para um papel fisiológico da
conexina 43 no processo de implantação - a conexina 43 é tida por estar dramaticamente
reduzida durante a implantação, o que reduz a comunicação célula-célula. Esse processo pode
facilitar a invasão do trofoectoderma através das células estromais e, além disso, promover a
angiogênese. Esse papel da conexina 43 na reprodução é consistente com a função
demonstrada em progressão maligna, onde a expressão forçada de conexina 43 inibe o
crescimento da célula tumoral e o seu potencial invasivo e retarda a neo-vascularização
(Tanmahasamut e Sidell, 2005).
No testículo, as interações celulares são essenciais para o funcionamento adequado de
muitos tipos celulares diferentes, e 11 tipos de conexinas foram identificados nestes
constituintes celulares. As células de Leydig dissociadas mostram tanto acoplamento elétrico
como metabólico e a conexina 43 foi descrita como o principal componente funcional de suas
junções gap. Apesar das características elétricas das junções comunicantes nessas células
serem bem caracterizadas, os processos modulatórios ainda não foram bem estudados.
(Goldenberg e cols., 2003).
Em tireóides de ratos, estudos demonstram que três conexinas muito bem
estabelecidas são expressas - as conexinas 26, 32 e 43. Em folículos tireóideos organizados de
ratos, as três conexinas localizam-se nas diferentes superfícies celulares dos tireócitos. Por
exemplo, a conexina 43 está localizada na superfície basal, conectando as células de folículos
adjacentes; a conexina 32 está localizada na superfície lateral, unindo as células de um mesmo

27
folículo e a conexina 26 está localizada na porção apical. Existe um controle morfogenético
sobre a expressão da conexina 43 e da conexina 32, pois a expressão da conexina 43
predomina quando as células estão em monocamadas e a da conexina 32 predomina quando
as células começam a formar folículos. Já a localização da conexina 26 sugere que esta é
transportada para a superfície apical por transporte vesicular do complexo de Golgi, já que a
polaridade está estabelecida pelo arranjo lateral das junções de oclusão e de aderência. Essa
polaridade é um importante aspecto das glândulas endócrinas e exócrinas, o qual está
intimamente ligado às junções gap. Nos tireócitos, espera-se que essas conexinas contribuam
em diferentes aspectos na função glandular. A localização da conexina 32 na superfície lateral
estabelece cooperação metabólica entre as células acopladas. Isso as permite agir em conjunto
na resposta à sinalização pelo TSH/AMPc e subseqüente secreção do hormônio tireóideo.
Deste modo, a conexina 32 é a isoforma mais importante na coordenação do tireócitos na
formação dos folículos, sendo esse papel fundamental demonstrado pela up-regulation desta
proteína juntamente com o processo de foliculogênese e sua localização lateral (Green e cols,
1999). Em um estudo onde foi induzida uma tireoidite experimental em ratos, todas as três
isoformas de conexinas ficaram reduzidas. Além disso, as conexinas não estavam arranjadas
nas junções gap formando hemicanais, mas estavam quando comparados aos ratos controle
(Green e cols, 1999).
As junções gap têm sido demonstradas na adeno-hipófise por microscopia eletrônica
convencional. Entretanto, nessa glândula, a identidade das células conectadas por junções
comunicantes foi difícil de ser determinada devido a coexistência de diferentes tipos celulares
(5 tipos de células endócrinas e as células folículo-estreladas), as quais se conectam
intimamente umas às outras. (Morand e cols, 1996).
A promoção do crescimento na adeno-hipófise pode ser regulada pela comunicação
intercelular através de junções gap. A conexina 43 é o subtipo mais proeminente na adeno-

28
hipófise. As junções gap na adeno-hipófise são formadas principalmente entre células
folículo-estreladas ou entre as células folículo-estreladas e lactotrofos ou gonadotrofos. Há
alguns anos atrás, foi demonstrado que o crescimento celular está inversamente relacionado
ao grau de comunicação celular e a exposição aos mitógenos leva a uma rápida, mas
transitória inibição da comunicação celular antes das células entrarem em divisão. A
redistribuição da conexina 43 também foi associada com o controle do crescimento nas
células adrenais e o aumento da expressão leva a uma proliferação e diferenciação
aumentadas de células semelhantes a osteoblastos. Os hemicanais de conexina 43 também
parecem ser uns transdutores de sinais de sobrevivência em resposta sinais extracelulares,
pois, a adenosina, produzida na glândula hipófise, pode influenciar a expressão de conexina
43 e modificar a transmissão intercelular diretamente através das junções comunicantes e
indiretamente via hemicanais de conexina. (Lewis e cols, 2006).
Na verdade pouco se sabe sobre o papel das junções gap em glândulas e em especial
na hipófise. Até o presente momento poucos são os estudos que mostraram uma participação
das conexinas e dos canais por elas formados na manutenção da homeostasia glandular.

29
2 - OBJETIVOS
2.1 - Objetivo geral:
O objetivo desse trabalho é avaliar, in vitro, a expressão e a distribuição das conexinas
26, 32 e 43, e o acoplamento dos canais de junções comunicantes em linhagens de células
estromais (APS9) e secretoras de prolactina (GH3B6) derivadas de adeno-hipófise de rato.
2.2 - Objetivos específicos
• Analisar por RT-PCR, a expressão das conexinas 26, 32 e 43 nas linhagens celulares
GH3B6 e APS9;
• Analisar por métodos imunocitoquímicos, a distribuição das conexinas 26, 32 e 43 nas
linhagens celulares GH3B6 e APS9 ;
• Analisar por injeção iontoforética, a possibilidade de acoplamento celular através das
junções comunicantes nas linhagens celulares GH3B6 e APS9;
• Analisar por métodos imunocitoquímicos, a distribuição das conexinas 26, 32 e 43 em
co-culturas de células GH3B6 e APS9;
• Analisar por injeção iontoforética, a possibilidade de acoplamento celular através das
junções comunicantes em co-culturas de células GH3B6 e APS9.

30
3 - MATERIAIS E MÉTODOS
3.1 - Cultivo das linhagens celulares GH3B6 e APS9
A linhagem celular secretora de prolactina GH3B6 (Gourdji e cols., 1982) e a linhagem
celular estromal APS9 (Alves e cols., 2002) foram obtidas do Banco de Células do Estado do
Rio de Janeiro do Hospital Universitário Clementino Fraga Filho – UFRJ, sendo que as
células estromais APS9 foram originalmente obtidas em nosso laboratório por passagens
sucessivas de cultura primária de hipófise (Alves e cols., 2002). As células GH3B6 e APS9
foram rotineiramente mantidas em Ham F12 suplementado com 15% de soro de cavalo e
2,5% de soro fetal bovino, 100 U/mL de penicilina e 0,1mg/mL de estreptomicina e
Dulbecco´s Modified Medium (DMEM) suplementado com 10% de soro fetal bovino (SFB),
100 U/mL de penicilina e 0,1mg/mL de estreptomicina, respectivamente, em estufa a uma
temperatura de 37ºC e atmosfera de 5% de CO2. A cada 2 dias, o meio de cultura foi trocado e
o meio de cultura das células APS9 foi coletado e congelado. Quando entraram em
confluência, as células foram tripsinizadas com 0,25% de tripsina/EDTA e replaqueadas na
densidade de 5 x 104 células/garrafa, sob a mesma condição e cultivadas por mais sete dias.
3.2 - Cultura primária de hipófise de rato
As culturas primárias de hipófise de rato foram obtidas de 15 ratos Wistar jovens. Os
animais foram mortos por decapitação e suas hipófises removidas assepticamente. As adeno-
hipófises foram separadas e mecanicamente dispersadas e então incubadas três vezes por 20
minutos com agitação a 37ºC em meio de dispersão contendo 0,05% de tripsina e 0,05% de
albumina de soro bovino. As células dispersadas foram centrifugadas por 5 minutos a 12000 x

31
g. e ressuspensas em DMEM suplementado com 10% de SFB contendo penicilina e
estreptomicina. As células foram plaqueadas em garrafas e cultivadas em uma atmosfera
umidificada contendo 5% CO2 a 37ºC. Após seis dias de cultura, as células alcançaram a
confluência e então foi realizada a extração do RNA para análise por RT-PCR para as
conexinas 26, 32 e 43.
3.3 - Ensaios de co-cultura
Inicialmente, a linhagem celular APS9 foi plaqueada isoladamente em lamínulas de
vidro, em placas de cultura de 24 poços, em uma densidade de 3 x 104 células/poço. Após
dois dias de cultivo, a linhagem celular GH3B6 foi plaqueada nas lamínulas de vidro
previamente recobertas por uma monocamada de células APS9, em uma densidade de 5 x 104
células. As co-culturas foram mantidas como descrito anteriormente, por mais cinco dias,
somando um total de sete dias de cultivo. Após esse período, as co-culturas foram submetidas
à técnica de injeção iontoforética e imunocitoquímica indireta.
3.4 - Análise por RT-PCR da expressão das conexinas 26, 32 e 43
Para extração do RNA das linhagens celulares GH3B6 e APS9 e das culturas primárias
de hipófise de rato , foi utilizado o reagente TRIzol (GibcoBRL, NY, USA). As células
foram homogeneizadas em 1 mL de TRIzol. Aos lisados celulares foram adicionados 0,2
mL de clorofórmio, para separação do RNA total. Após centrifugação (12000 x g., 4°C por 15
minutos) foi possível observar uma fase superior aquosa, transparente e uma fase inferior
mais densa e rosada. A fase aquosa foi transferida para um novo tubo, onde foram
adicionados 0,5 mL de isopropanol, para precipitação do RNA total. Após centrifugação

32
(12000 x g., 4°C por 10 minutos), o sobrenadante foi descartado e 1 mL de etanol 75% foi
adicionado para lavagem do lisado remanescente. Após centrifugação (8000 x g., 4°C por 5
minutos), o etanol foi descartado e o precipitado depois de seco, foi ressuspenso em água
tratada com dietil-pirocarboneto (DEPC) (Sigma, MO, USA).
Os RNAs totais extraídos foram diluídos em água tratada com DEPC na proporção
1:500. As absorvâncias das amostras foram medidas nos comprimentos de onda de 260 e 280
nm em espectrofotômetro (Hitachi, U 3000). Através de cálculos (absorvância a 260 nm x
fator de diluição x 40)/1000 = RNA g/L) foi determinada a quantidade de RNA de cada
amostra e µg/µL. Para a verificação da pureza do RNA extraído, calculamos: absorvância a
260 nm/ absorvância a 280 nm. O resultado desta razão deve variar entre 1,5 a 2, indicando
que as amostras estão dentro dos padrões de pureza.
Depois da quantificação dos RNAs, as amostras foram submetidas a eletroforese em
gel de agarose (1% de agarose + tampão contendo Tris 40 mM, ácido acético 25 mM e EDTA
1 mM – TAE + 0,01% de brometo de etídio (intercalante de ácido nucléico) para a verificação
da integridade das amostras, através da observação das bandas correspondentes aos RNAs
ribossomais 28S e 18S. A visualização das bandas foi possível através da utilização de
transilumindor UV (Bio Rad).
Para nos assegurarmos de que não havia contaminação por DNA em nossas amostras
de RNA total, utilizamos a técnica de purificação do RNA total através do tratamento das
amostras com a enzima que degrada DNA, desoxiribonuclease I (Boehringer Mannheim,
Germany). Utilizamos 10 µg de RNA total e adicionamos 10 U da enzima DNase I,
juntamente com 1 µL do tampão DNase (25 mM Tris-HCl. pH 7,2, 5 mM MgCl2, 0,1 mM
EDTA) e água DEPC. Após 30 minutos a temperatura de 37ºC, adiciona-se 40 µL de água
DEPC e 50 µL de fenol : clorofórmio (1:1), para a retirada da enzima do meio. O tubo foi
submetido à centrifugação (12000 x g., 4°C por 2 minutos). Depois da transferência da fase

33
aquosa (fase superior) para um novo tubo, foram adicionados 2,5 volumes de etanol 100% e
0,3 volumes de acetato de amônia, para precipitação do RNA total. Depois de 18 horas em
freezer – 20°C, o tubo foi submetido à centrifugação (12000 x g., 4°C por 30 minutos) para a
obtenção do precipitado. Após a lavagem do precipitado com etanol 75%, este foi ressuspenso
em 15 µL de água DEPC. Uma alíquota foi retirada para quantificação do RNA total e outra
alíquota foi submetida à eletroforese em gel de agarose 1% para a verificação da integridade
do RNA total.
Na síntese de fitas simples de DNA complementar (cDNA) a partir do RNA total
purificado, foram utilizados 2 µg de RNA, 50 µM de oligonucleotídeo dT (Pharmacia
Biotech, NJ, USA), transcriptase reversa (Superscript RNase H- Reverse Transcriptase,
Gibco-BRL, NY, USA), tampão 5x first stand [250 mM Tris-HCl (pH 8,3), 375 mM KCl, 15
mM MgCl2], 100 mM DTT (Gibco-BRL, NY, USA), 10 unidade de RNAsin (Gibco-BRL,
NY, USA) (inibidor de RNase) e 10 µM dNTP (Gibco-BRL, NY, USA) (coquetel de
nucleotídeos). O RNA total foi purificado e incubado a 70°C, por 10 minutos, na presença de
água DEPC e oligonucleotídeo dT, para reconhecimento e ligação à cauda poli A dos RNA
mensageiros. Em seguida, adicionou-se ao tubo o restante dos reagentes e incubou-se a 42°C
por 1 hora. Terminado o tempo de incubação, a mistura foi submetida a 90°C por 5 minutos
para a inativação da enzima transcriptase reversa (RT). Foram adicionados ao tubo 0,3
volumes de 7,5 M de acetato de amônia, pH 8,0 e 2,5 volumes de etanol 100% e colocou-se
em freezer – 20°C, durante 18 horas, para precipitação do cDNA. A seguir, as amostras foram
submetidas à centrifugação (12000 x g., 4°C por 30 minutos) para precipitação do cDNA. O
sobrenadante foi descartado e acrescentou-se 50 µL de etanol 75% ao precipitado. O etanol
foi todo retirado, o cDNA foi ressuspenso em 11 µL de H2O MilliQ e armazenado em freezer
– 20°C.

34
Após a obtenção do DNA fita única (cDNA), através da etapa de transcrição reversa,
foi realizada a amplificação do fragmento do cDNA correspondente aos RNAs mensageiros
em estudo, utilizando-se oligonucleotídeos sense e antisense para uma região específica das
conexinas 26, 32 e 43. A contaminação por DNA genômico foi controlada pelo uso de
amostras de cDNA nas quais a transcriptase reversa foi omitida (RT-).
Para cada amplificação específica, foram utilizados 2 µL do cDNA ressuspenso em
água MilliQ, tampão para PCR na concentração final de 1 x 2,5 mM de MgCl2, 16 pmoles de
cada um dos primers, 0,5 mM de dNPT, 1,5 unidades de DNA polimerase e H2O MilliQ para
completar o volume final de 20 µL. O cDNA de cada amostra foi amplificado por PCR
utilizando-se os seguintes primers específicos para cada gene de conexina: conexina 26,
primer reverse: 5’ AGC ATA GTC ACA CCA GCA CG 3’, primer foward: 5’ GCT AAA
CAT CAC AGA GCT GT 3’ (produto final 322 pb, 30 ciclos); conexina 32, primer reverse:
5’CCT GCG TCC CAA TTC CAT C 3’, primer foward: 5’ CCT GTC TGG TGT TCT ATC
TCC C 3’ (produto final 283 pb, 37 ciclos); conexina 43, primer reverse: 5’ CTG CTG GCT
CTG CTG GAA GG 3’, primer foward: 5’ATC CAG TGG TAC ATC TAT GG 3’ (produto
final 593 pb, 31 ciclos).
As amostras foram levadas ao termociclador e desnaturadas a 94ºC por 30 segundos,
seguido pelo passo de anelamento a 58ºC (conexinas 26 e 43) e 60ºC (conexina 32) por 45
segundos, 45 segundos a 72ºC para extensão do primer. Cerca de 12 µL dos produtos finais
do RT-PCR foram separados por eletroforese em gel de agarose a 1,5% em 1x TAE (0,04
mol/L de Tris-acetato, 0,01% de ácido acético e 0,001 mol/L EDTA). O tamanho dos
produtos da reação foi determinado após 20 minutos de marcação com brometo de etídio (0,5
µg/mL em 1x TAE) seguido da visualização com um transiluminador UV. Como controle
positivo para a reação, estabelecemos uma cultura primária de adeno-hipófise de rato. Para
controle interno foi realizado o PCR para um gene constitutivo, o GAPDH, com a seguinte

35
seqüência: primer reverse 5’ CTG AGA ATG GGA AAC TTG TC 3’ e primer foward 5’
GTC ACT GTT AAA GTT GGA GG 3’ (produto final 377 pb, 31 ciclos). Para controles
positvos para as conexinas 26 e 32 foi utilizado fígado de rato para a conexina 43 foi utilizado
coração de rato. Todos os experimentos foram realizados 4 vezes. Os resultados foram
apresentados como média ± desvio-padrão. A análise estatística dos resultados foi realizada
por meio da aplicação da análise de variância – ANOVA. O valor de P< 0.05 foi considerado
significativo.
3.5 - Injeção Iontoforética
As linhagens celulares foram cultivadas sobre lamínulas de vidro, nas mesmas
condições descritas anteriormente, em placas de cultura de 35 mm, na densidade de 5x 104
células. Para a realização deste ensaio em sistema de co-cultivo, a linhagem celular APS9 foi
plaqueada isoladamente em lamínulas de vidro, em placas de cultura de 24 poços, em uma
densidade de 3 x 104. Após dois dias de cultivo, a linhagem celular GH3B6 foi plaqueada nas
lamínulas de vidro previamente recobertas por uma monocamada de células APS9, em uma
densidade de 5x 104 células, por mais cinco dias, somando um total de sete dias de cultivo.
Após esse período, as duas linhagens celulares, GH3B6 e APS9, foram injetadas com o
corante de baixo peso molecular (457 Da) Lucifer Yellow CH (5% em 150mM de LiCl) com
auxílio de microeletrodos de vidro (resistência entre 20-50 MΩ) por aplicação de pequenos
pulsos de corrente hiperpolarizante (9.1 nA, 100 ms), usando um amplificador WPI (modelo
7060). A transferência de corante foi observada no microscópio invertido Axiovert 100 (Carl
Zeiss, Oberkochen, Germany), equipado com arco de iluminação de xenônio. As fotografias
foram tiradas por sistema de captura digital acoplado ao microscópio, 5 minutos após cada
injeção. A cada injeção, as informações foram registradas e, posteriormente os dados foram

36
quantificados para produção de gráficos que revelaram a incidência do acoplamento e sua
extensão. Cada tipo celular foi injetado 90 vezes, em três placas diferentes, sendo 30
microinjeções em cada placa.
3.6 - Imunocitoquímica indireta
As linhagens celulares GH3B6 e APS9 foram plaqueadas de forma isolada na
densidade de 5 x 104 células em lamínulas de vidro, em placas de cultura de 24 poços por sete
dias. Para a realização deste ensaio em sistema de co-cultivo, a linhagem celular APS9 foi
plaqueada isoladamente em lamínulas de vidro, em placas de cultura de 24 poços, em uma
densidade de 3 x 104. Após dois dias de cultivo, a linhagem celular GH3B6 foi plaqueada nas
lamínulas de vidro previamente recobertas por uma monocamada de células APS9, em uma
densidade de 5x 104 células. As co-culturas foram mantidas como descrito anteriormente, por
mais cinco dias, somando um total de sete dias de cultivo. Todos os procedimentos
imunocitoquímicos foram efetuados diretamente nas células in vitro. Estas foram fixadas com
álcool 70%, durante 20 minutos a uma temperatura de -20°C. Após tal procedimento, as
células foram submetidas à técnica de imunofluorescência indireta, utilizando anticorpos anti-
conexina 43 e anti-α actina de músculo liso, conforme protocolo descrito a seguir:
1. Lavagem das células durante 30 minutos com tampão fosfato salina (TPS) 0,01M, pH
7,45, para retirada do fixador;
2. Incubação com Triton X-100 0,1% em TPS durante 5 minutos, para permeabilização
da membrana celular;
3. Lavagem das células durante 15 minutos com tampão fosfato salina (TPS) 0,01M, pH
7,45

37
4. Incubação com BSA 5% em TPS por 30 minutos, para bloqueio de sítios inespecíficos
5. Incubação com anticorpo primário overnight, conforme Tabela 1;
6. Lavagem das células durante 30 minutos com tampão fosfato salina (TPS) 0,01M, pH
7,45;
7. Incubação com anticorpo secundário biotinilado universal (Ig anti-mouse e Ig anti-
rabbit) (Dako) durante 120 minutos;
8. Lavagem das células durante 30 minutos com tampão fosfato salina (TPS) 0,01M, pH
7,45;
9. Incubar com 4,6-diamidino-2-phenylindole (DAPI), um corante nuclear, durante 5
minutos;
10. Lavagem com água destilada (4 vezes);
11. Montagem das lamínulas em lâmina com N-propilgalato (Sigma).
Tabela 1: Anticorpos Primários Utilizados nos Procedimentos Imunocitoquímicos
Anticorpos Procedência Diluição
Anticorpo policlonal anti-conexina 26 Zymed 1/100
Anticorpo policlonal anti-conexina 32 Zymed 1/100
Anticorpo policlonal anti-conexina 43 Zymed 1/100
Anticorpo monoclonal anti α-actina de
músculo liso
Sigma 1/100
OBS.: Para controle negativo dos anticorpos primários monoclonais, foi utilizado o TPS +
BSA 0,1%. Para os anticorpos policlonais, foi utilizado soro normal de coelho a 1/100.

38
3.7 - Cultivo da linhagem celular GH3B6 com meio condicionado das células estromais
APS9
Para a obtenção do meio condicionado das células APS9, cultivou-se esta linhagem
rotineiramente em Dulbecco´s Modified Medium (DMEM) por 2 dias.
A linhagem celular GH3B6 foi rotineiramente mantida em Ham F12 suplementado
com 15% de soro de cavalo e 2,5% de soro fetal bovino, 100 U/mL de penicilina e 0,1mg/mL
de estreptomicina por 5 dias. Após esse período, o meio foi retirado e adicionou-se o meio
condicionado obtido a partir da linhagem APS9, onde as células GH3B6 foram mantidas por
mais 2 dias. Ao final deste período de cultivo, foi realizada a extração do RNA para
realização da técnica de RT-PCR para as conexinas 26, 32 e 43. Os experimentos foram
realizados 2 vezes. Os resultados foram apresentados como média ± desvio-padrão. A análise
estatística dos resultados foi realizada por meio da aplicação da análise de variância –
ANOVA. O valor de P< 0.05 foi considerado significativo.

39
4 - RESULTADOS
4.1 – Análise da morfologia das células GH3B6 e APS9
As células GH3B6, quando cultivadas durante 7 dias em meio de cultura F12 de Ham
suplementado com 15% de soro de cavalo e 2,5% de soro fetal bovino e observadas à
microscopia de contraste de fase, apresentam-se com formato arredondado e com refringência
típica de células epiteliais secretoras. Elas crescem formando pequenos agregados em toda a
superfície da garrafa. As células APS9 cultivadas em meio de cultura DMEM suplementado
com 10% de soro fetal bovino à microscopia de contraste de fase apresentam-se com formato
alongado, com muitos prolongamentos citoplasmáticos, formando um tapete que recobre
quase toda superfície da garrafa de cultura e não apresentam inibição por contato (Figura 4A e
B).
4.2 - Análise da morfologia da cultura primária de hipófise de ratos
A cultura primária de hipófise de ratos machos adultos jovens, cultivada em meio de
cultura DMEM suplementado com 10% de soro fetal bovino por 6 dias e observadas a
microscopia de contraste de fase, mostrou uma população celular heterogênea (Figura 4C),
formada por células com características morfológicas de células epiteliais secretoras que
formavam agregados celulares refringentes e, circundando estes agregados, estavam presentes
células com características de células estromais com formato alongado apresentando
nucléolos bastante evidentes.

Figura 4: Microscopia de contraste de fase das células GH3B6 (A), APS9 (B) cultivadas por 7 dias, e da cultura primária de adeno-hipófise (C) cultivada por 6 dias. As células GH3B6 são arredondadas e organizam-se em pequenos agregados e as células APS9 são alongadas, com morfologia típica de células fibroblastóides. A cultura primária apresenta células refringentes (setas) com características de células epiteliais secretoras e células com características estromais. Barras: 100 µm.
A
B
C
40

41
4.3 – Expressão do RNAm das conexinas 26, 32 e 43
A Figura 5A mostra o gel de integridade do RNA total da cultura primária de adeno-
hipófise, das linhagens celulares APS9 e GH3B6, do fígado e do coração de rato, sendo os dois
últimos utilizados como controles positivos. A Figura 5B mostra o resultado do RT-PCR para
o controle interno da reação, o GAPDH.
A Figura 6 mostra o resultado do RT-PCR para a conexina 26 (A), para a conexina 32
(B) e para a conexina 43 (C) na linhagem celular GH3B6 secretora de prolactina. Nestas
células, não foi observada a expressão do RNAm de nenhuma das três conexinas estudadas.
Nas células estromais APS9 foi observado RNAm da conexina 43, entretanto, não
observamos a expressão do RNAm das conexinas 26 e 32 (Figura 7). Como controles
positivos, foram utilizados fígado de rato para as conexinas 26 e 32 e coração de rato para a
conexina 43.

GH
3B6
APS
9
377 pb
GH
3B6
APS
9 C
ultu
ra p
r im
ária
Fíga
doC
oraç
ãoR
T- C-
Cul
tura
pr i
már
iaFí
gado
Cor
ação
PM
←←←← 18 S←←←← 28 S
Figura 5: Gel de integridade do RNA total extraído das linhagens GH3B6 e APS9, da cultura primária de adeno-hipófise, do fígado e do coração de ratos (A), e o RT-PCR representativo para GAPDH (B). (RT-) corresponde à amostra onde a transcriptase reversa foi omitida e (C-) corresponde ao controle negativo.
A B
42
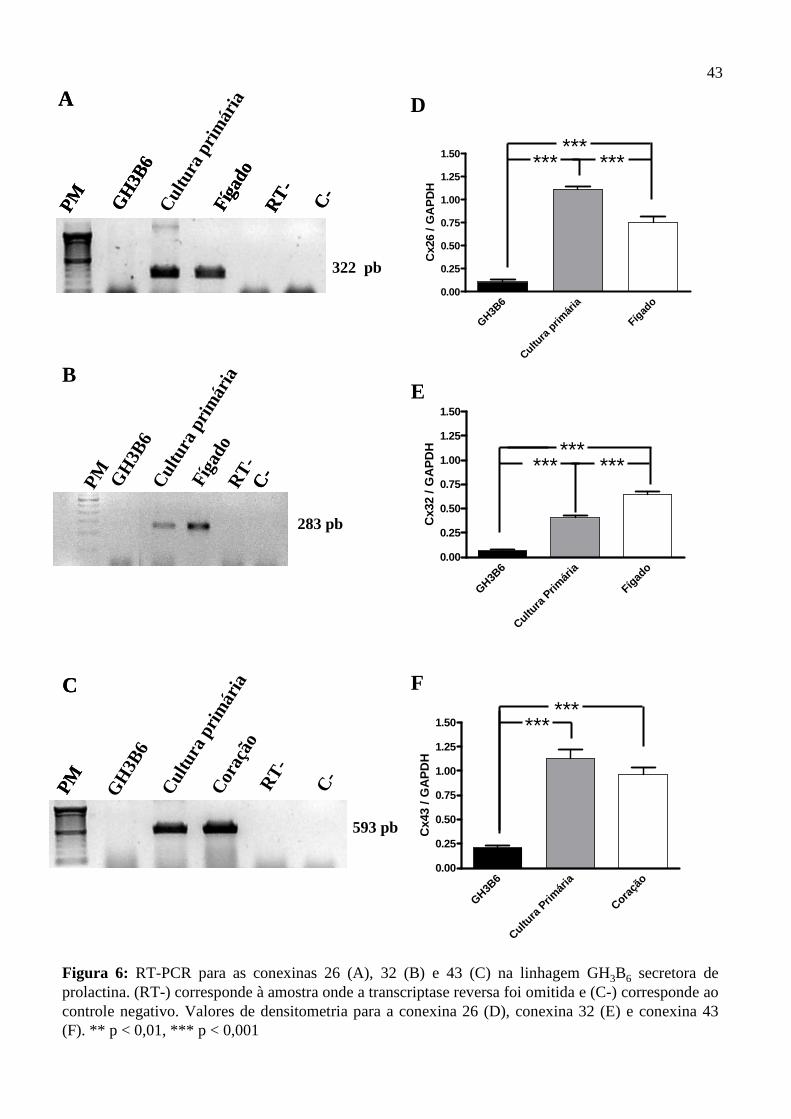
Figura 6: RT-PCR para as conexinas 26 (A), 32 (B) e 43 (C) na linhagem GH3B6 secretora de prolactina. (RT-) corresponde à amostra onde a transcriptase reversa foi omitida e (C-) corresponde ao controle negativo. Valores de densitometria para a conexina 26 (D), conexina 32 (E) e conexina 43 (F). ** p < 0,01, *** p < 0,001
GH
3B6
PM
C
Cor
ação
RT-
C-
PM
593 pb
C
Cul
tura
prim
ária
AG
H3B
6
Fíga
do
RT
-
C-
PM Cul
tura
prim
ária
322 pb
AG
H3B
6
Fíga
do
RT
-
C-
PM
B
Fíga
doR
T-C
-
PM
283 pb
GH
3B6
C-Cul
tura
prim
ária
D
F
E
43
GH3B6
Cultur
a prim
ária
Fígad
o0.00
0.25
0.50
0.75
1.00
1.25
1.50
Cx2
6 /
GA
PD
H
GH3B6
Cultur
a Prim
ária
Fígad
o0.00
0.25
0.50
0.75
1.00
1.25
1.50
Cx3
2 / G
AP
DH
GH3B6
Cultur
a Prim
ária
Coraç
ão0.00
0.25
0.50
0.75
1.00
1.25
1.50
Cx4
3 / G
AP
DH
******
****** ***
****** ***
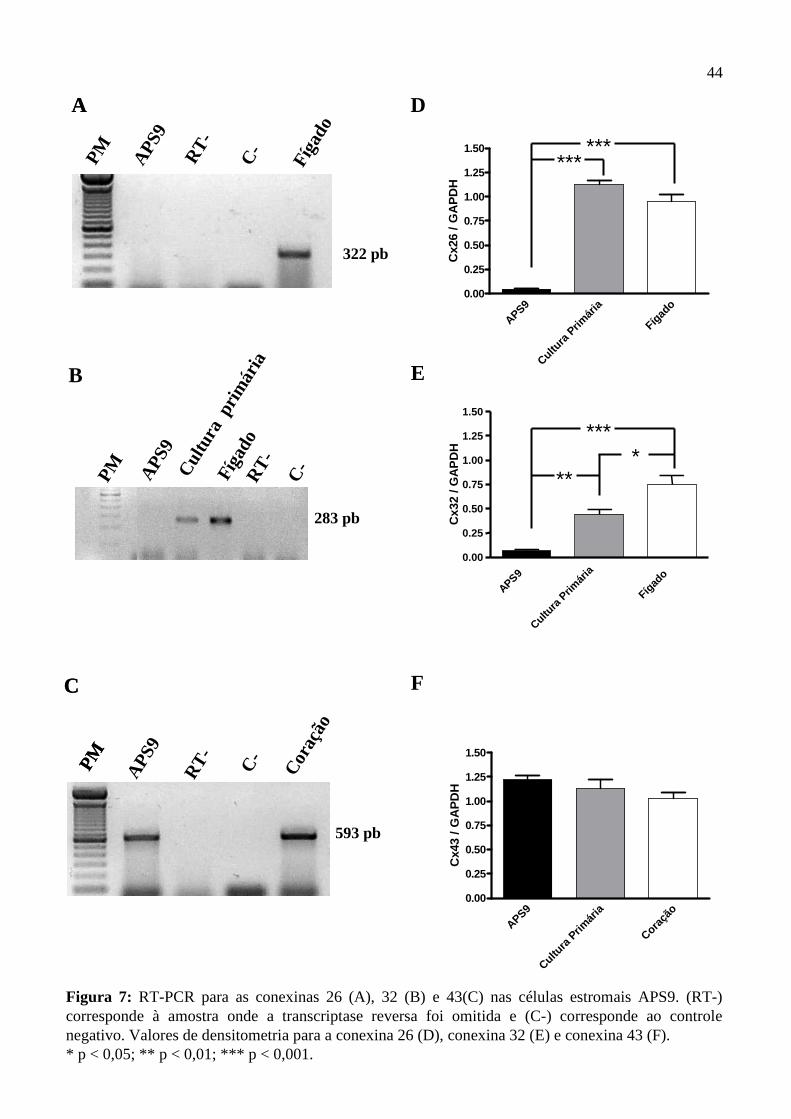
Figura 7: RT-PCR para as conexinas 26 (A), 32 (B) e 43(C) nas células estromais APS9. (RT-) corresponde à amostra onde a transcriptase reversa foi omitida e (C-) corresponde ao controle negativo. Valores de densitometria para a conexina 26 (D), conexina 32 (E) e conexina 43 (F). * p < 0,05; ** p < 0,01; *** p < 0,001.
B
Cul
tura
prim
ária
Fíga
do
C-
PM
283 pb
APS
9
RT-
AA
PS9
RT-
C-PM Fíga
do
322 pb
AA
PS9
RT-PM
C
C-PM
C
Cor
ação
593 pb
D
F
44
APS9
Cultur
a Prim
ária
Fígad
o0.00
0.25
0.50
0.75
1.00
1.25
1.50
Cx2
6 / G
AP
DH
APS9
Cultur
a Prim
ária
Coraç
ão0.00
0.25
0.50
0.75
1.00
1.25
1.50
Cx4
3 / G
AP
DH
E
APS9
Cultur
a Prim
ária
Fígad
o
0.00
0.25
0.50
0.75
1.00
1.25
1.50
Cx3
2 / G
AP
DH
***
***
******

45
4.4 – Análise da comunicação intercelular via junção gap
Três placas contendo células APS9 foram injetadas com o corante fluorescente LY e
apresentaram o seguinte perfil de acoplamento, com cerca de 70% de células não acopladas
(Figuras 8A e B). Das células que evidenciavam algum acoplamento, em cerca de 20% delas,
o corante foi observado passando para apenas uma célula vizinha; nos 10% restantes, a
presença do corante foi visualizada em duas ou mais células. As placas 1 e 3 apresentaram um
comportamento bastante semelhante no perfil de acoplamento.
As células GH3B6, quando injetadas com o corante fluorescente LY, apresentaram um
perfil de acoplamento diferenciado das células APS9, conforme mostrado na Figura 9A e B.
Aproximadamente 96% das células não apresentaram acoplamento. As placas 2 e 3
apresentaram um comportamento mais semelhante no perfil de acoplamento, diferentemente
da placa 1, onde nenhum acoplamento a células vizinhas foi observado.
As Figuras 10A e B mostram o perfil de acoplamento total na co-cultura das células
GH3B6 secretoras de prolactina e das células estromais APS9. Em cada uma das 3 placas da
co-cultura foram realizadas 30 injeções somando um total de 90 injeções. A primeira metade
destas injeções foi realizada na linhagem celular GH3B6 e a outra metade na linhagem APS9,
a fim de se observar o perfil de acoplamento entre o mesmo tipo celular e entre os dois
diferentes tipos. Quanto ao perfil de acoplamento total, pode-se observar que, quando em co-
cultura, apenas 44 % de células permaneceram não acopladas. Diferentemente das linhagens
cultivadas isoladamente, onde observou-se baixo perfil de acoplamento, em 50 % das células
foi possível observar a passagem do corante para pelo menos 1 célula vizinha. Nos 6 %
restantes, o corante foi observado em 2 ou mais células vizinhas. O perfil de acoplamento nas
três placas foi bastante semelhante, demonstrando apenas uma pequena diferença na placa 2.

46
A Figura 11A e B mostram o percentual de acoplamento entre o mesmo tipo celular e
entre os diferentes tipos. Cerca de 60% das células injetadas, que foram capazes de se
comunicar com pelo menos uma célula vizinha, demonstraram um perfil de acoplamento com
o mesmo tipo celular. Nos 40% restantes foi observada a passagem do corante para uma
linhagem celular diferente. As células APS9, quando cultivadas em sistema de co-cultura,
foram capazes transferir corante entre si, correspondendo a um percentual de 25,49% e
também para as células GH3B6. O mesmo comportamento foi observado na transferência de
corante entre as células GH3B6, onde esta transferência se deu em 35,29%, além da
comunicação com as células APS9. Este tipo de comportamento não foi observado
anteriormente, quando as células GH3B6 foram cultivadas de forma isolada.

10090303030Total
2,220115
1,110104
0,000003
5,652032
20,01841131
71,1642417230
Taxa de Acoplamento (%)
TotalPlaca 3Placa 2Placa 1Célulasacopladas
Células APS9
0 1 2 3 4 50
102030405060708090
Nº de células acopladas
Nº
de c
élul
as in
jeta
das
Figura 8: (A) Perfil de acoplamento das células APS9 injetadas com corante LY. (B) Incidência de acoplamento e sua extensão nas células APS9 de acordo com a Figura 8A. As células APS9 apresentaram algum grau de acoplamento com pelo menos umacélula vizinha.
B
A
47

Figura 9: (A) Perfil de acoplamento das células GH3B6 injetadas com corante LY. (B) Incidência de acoplamento e sua extensão nas células GH3B6 de acordo com a Figura 9A. As células secretoras de prolactina GH3B6 em sua maioria não apresentaram-se acopladas.
10090303030Total
0.000005
0,000004
0,000003
1,110102
3,332101
95,6862930270
Taxa de Acoplamento (%)
TotalPlaca 3Placa 2Placa 1Célulasacopladas
Células GH3B6
0 1 2 3 4 50
102030405060708090
Nº de células acopladas
Nº
de c
élul
as in
jeta
das
A
B
48
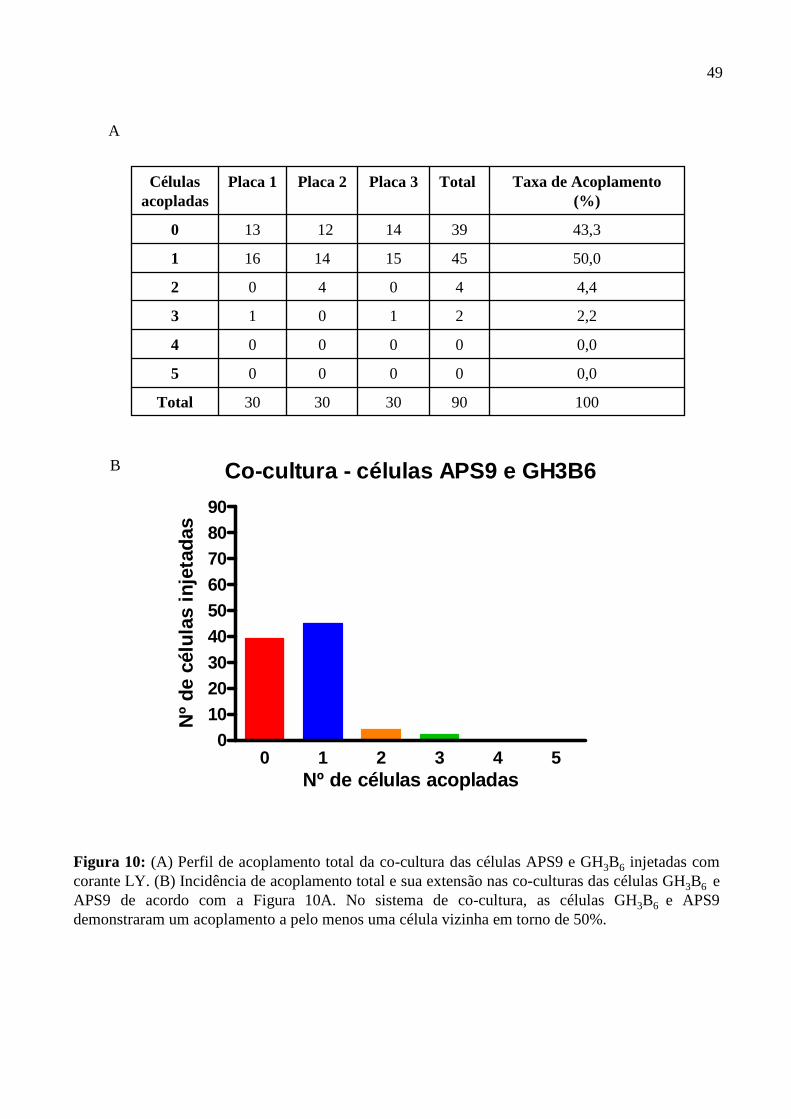
Figura 10: (A) Perfil de acoplamento total da co-cultura das células APS9 e GH3B6 injetadas com corante LY. (B) Incidência de acoplamento total e sua extensão nas co-culturas das células GH3B6 e APS9 de acordo com a Figura 10A. No sistema de co-cultura, as células GH3B6 e APS9 demonstraram um acoplamento a pelo menos uma célula vizinha em torno de 50%.
10090303030Total
0,000005
0,000004
2,221013
4,440402
50,0451514161
43,3391412130
Taxa de Acoplamento (%)
TotalPlaca 3Placa 2Placa 1Célulasacopladas
Co-cultura - células APS9 e GH3B6
0 1 2 3 4 50
102030405060708090
Nº de células acopladas
Nº
de c
élul
as in
jeta
das
A
B
49
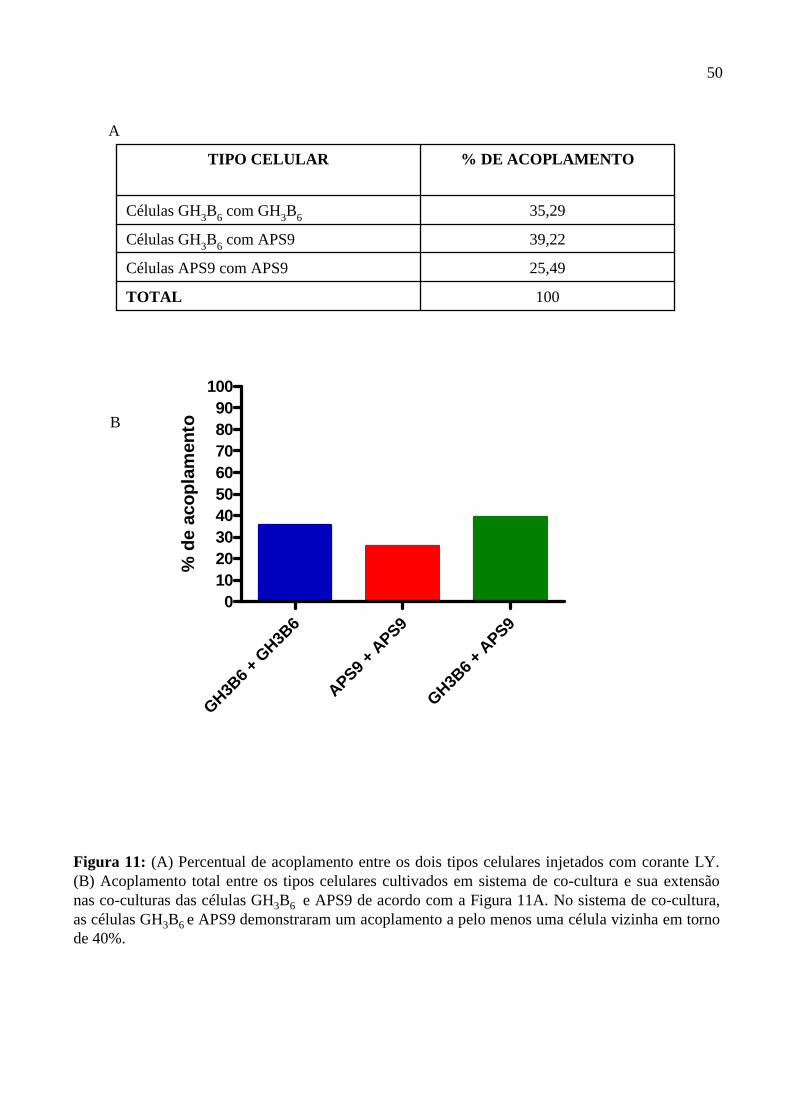
Figura 11: (A) Percentual de acoplamento entre os dois tipos celulares injetados com corante LY.(B) Acoplamento total entre os tipos celulares cultivados em sistema de co-cultura e sua extensão nas co-culturas das células GH3B6 e APS9 de acordo com a Figura 11A. No sistema de co-cultura, as células GH3B6 e APS9 demonstraram um acoplamento a pelo menos uma célula vizinha em torno de 40%.
GH3B6 +
GH3B
6
APS9 +
APS9
GH3B6 +
APS9
0102030405060708090
100
% d
e ac
opla
men
to
100TOTAL
25,49Células APS9 com APS9
39,22Células GH3B6 com APS9
35,29Células GH3B6 com GH3B6
% DE ACOPLAMENTOTIPO CELULAR
A
B
50

51
4.5 – Imunodistribuição da conexina 43
A imunofluorescência indireta foi utilizada para avaliar a localização celular da
conexina 43, já que foi a única isoforma de conexina expressa nos ensaios de RT-PCR. Nas
células APS9 cultivadas isoladamente, a imunorreatividade foi observada principalmente nas
regiões das membranas aposicionais (Figura 12A). O mesmo resultado não foi observado nas
células GH3B6, onde não foi detectada nenhuma imunomarcação (Figura 12C). Esses dados
corroboram os resultados obtidos nos ensaios de acoplamento celular.
Quando cultivadas em sistema de co-cultura, tanto nas células APS9 (dados não
mostrados) quanto nas células GH3B6 (Figura 12E) foi observada imunorreatividade intensa
nas regiões das membranas aposicionais, além de uma marcação difusa no citoplasma, o que
também corrobora os resultados obtidos nos ensaio de injeção do corante LY.
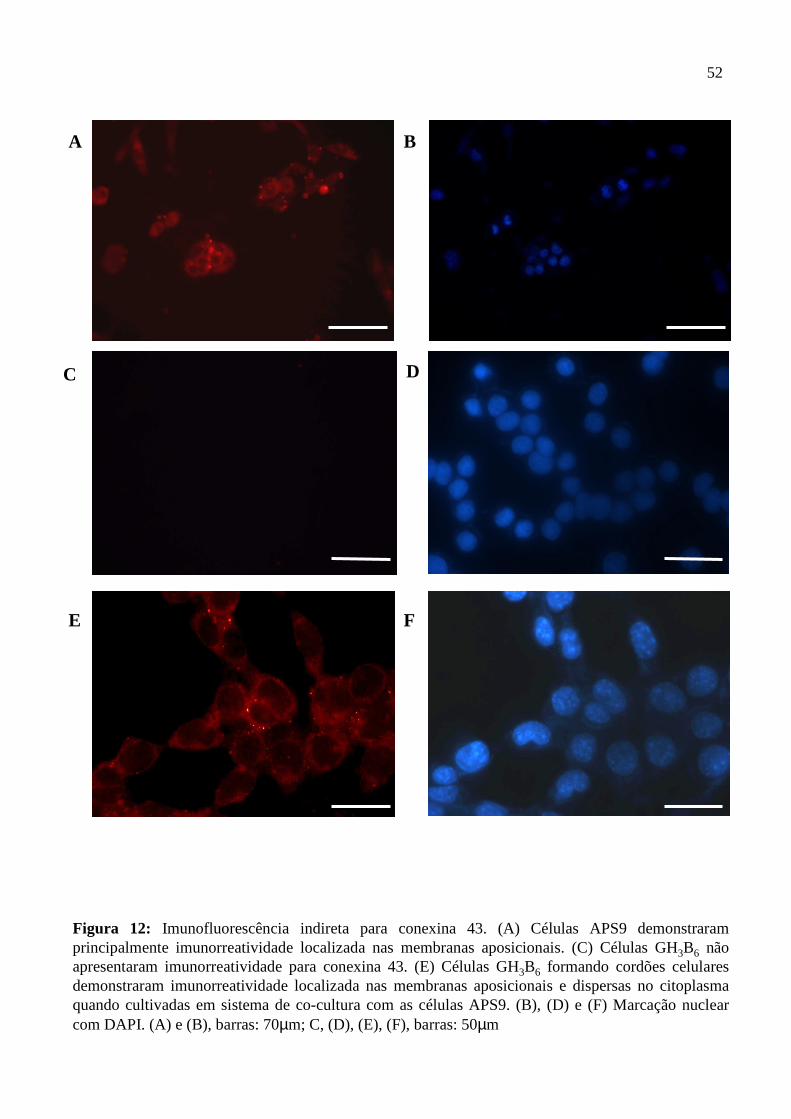
A
DC
B
Figura 12: Imunofluorescência indireta para conexina 43. (A) Células APS9 demonstraram principalmente imunorreatividade localizada nas membranas aposicionais. (C) Células GH3B6 não apresentaram imunorreatividade para conexina 43. (E) Células GH3B6 formando cordões celulares demonstraram imunorreatividade localizada nas membranas aposicionais e dispersas no citoplasma quando cultivadas em sistema de co-cultura com as células APS9. (B), (D) e (F) Marcação nuclear com DAPI. (A) e (B), barras: 70µm; C, (D), (E), (F), barras: 50µm
FE
52

53
4.6 – Expressão do RNAm das conexinas na linhagem celular GH3B6 cultivada na
presença de meio condicionado das células APS9
As células GH3B6 foram cultivadas por 2 dias com o meio condicionado obtido a partir
das células APS9. Logo após, o RNA total foi extraído e foi feita a análise da expressão do
RNAm das conexinas 26, 32 e 43, utilizando-se a técnica do RT-PCR. Quando as células
secretoras de prolactina foram cultivadas com o meio condicionado das células estromais,
observou-se a presença de RNAm para as conexinas 26 e 43 (Figura 13 A e C,
respectivamente). Entretanto, não foi observada a expressão do RNAm para a conexina 32
(Figura 13 B).

D
A
B
C
Figura 13: RT-PCR das células GH3B6 cultivadas com meio condicionado das células APS9. RT-PCR para, conexina 26 (A), conexina 32 (B) e conexina 43 (C) e GAPDH (D). (E) Valores de densitomentria para as conexinas 26, 32 e 43. * p < 0,05.
54
E
Cx26
Cx32
Cx43
0.00
0.25
0.50
0.75
1.00
1.25
Con
exin
as /
GA
PD
H
*

55
5- DISCUSSÃO
A comunicação direta célula-célula mediada por conexinas tem sido relacionada com a
coordenação de diversos processos fisiológicos como proliferação, adesão, migração,
diferenciação, sobrevivência celular e apoptose, provavelmente mediados por alterações
intracelulares envolvendo segundos mensageiros como o AMPc, 1,4,5-trifosfato de inositol e
cálcio. Por outro lado, a perda da comunicação celular pela expressão inadequada ou até mesmo
pela não expressão de conexinas pode ser responsável por processos de desdiferenciação e/ou
transdiferenciação, o que pode resultar em transformação neoplásica (Andrade-Rozental e cols.,
2000; Czyz, 2008).
Em hipófise o estudo pioneiro de Fletcher e cols., (1975) demonstrou, por microscopia
eletrônica, a presença de junções gap na adeno-hipófise de ratos. Utilizando parâmetros
morfológicos, este estudo revelou que as células acopladas via junções gap eram principalmente
as células secretoras de prolactina, além das células secretoras de GH, LH e TSH, que exibiam
um perfil de acoplamento de baixa freqüência.
Outros estudos demonstraram que as células da adeno-hipófise de ratos expressam
diferentes isoformas de conexinas - as conexinas 26 e 43, embora pouco se conheça sobre seus
efeitos na fisiologia e na fisiopatologia glandular (Meda e cols., 1993). Em nosso estudo,
demonstramos que duas linhagens celulares derivadas da adeno-hipófise de ratos, as células
GH3B6 secretoras de prolactina e as células estromais APS9, estão funcionalmente acopladas
através de junções gap.
A análise da expressão de conexinas em cultura primária de rato Wistar observada neste
trabalho, revelou a presença de RNAm para as conexinas 26, 32 e 43 (Figuras 6 e 7). Por outro
lado, em adeno-hipófises de ratos Sprague-Dawley foi observada, por imunofluorescência

56
indireta e por northern blot, apenas a presença das conexinas 26 e 43 (Meda e cols., 1993). É
importante ressaltar que na cultura primária, a análise da densitometria das bandas obtidas nos
ensaios de RT-PCR revelou uma quantidade diferencial na expressão dos RNAms das três
conexinas: Cx26 e Cx43 são as mais expressas quando comparadas com a Cx32, embora as
mesmas quantidades de cDNA tenham sido adicionadas para os ensaios. A ausência da expressão
de conexina 32 observada por Meda e cols., (1993) poderia então estar relacionada com as
diferentes abordagens metodológicas (ensaios in vitro e in vivo).
Na linhagem estromal APS9 encontramos a expressão de apenas uma isoforma, a
conexina 43. A conexina 43 é a conexina mais abundante e mais bem estuda nos mamíferos. A
comunicação intercelular envolvendo a conexina 43 tem despertado bastante interesse e tem sido
bem estudada, pois esta conexina é regulada por vários fatores fisiológicos e fisiopatológicos.
Nas células GH3B6 cultivadas isoladamente, não observamos a expressão de nenhuma
isoforma das conexinas estudas. Isto poderia ser explicado pelo fato de que células tumorais têm
sido relacionadas com uma baixa expressão de conexinas e, conseqüentemente, com um baixo
grau de acoplamento intercelular. Muitos trabalhos têm demonstrado que uma deficiência natural
ou provocada por agentes carcinogênicos na função dos genes de conexina está correlacionado
com um aumento da susceptibilidade das células para transformação neoplásica. Estudos in vitro
e in vivo têm demonstrado uma influência das junções gap no processo de tumorigênese, com
diminuição da expressão de conexinas e uma alteração na organização destas junções (Czyz,
2008).
Yamasaki e cols., (1999b) demonstraram que a transfecção de genes de conexinas em
células tumorais inibe a proliferação celular e afeta o contato intercelular, sugerindo que as
conexinas e as junções por elas formadas controlam o crescimento celular. Diversas linhagens
celulares têm a velocidade de transformação neoplásica diminuída quando é feita a transfecção

57
com genes de conexinas como: células SKHep1 de hepatoma, células de carcinoma de mama,
células de glioblastoma, células de carcinoma de pulmão, células HeLa de adenocarcinoma
cervical e células LNCaP de carcinoma prostático. Estes experimentos de transfecção têm
demonstrado, in vivo e in vitro, a inibição da proliferação destas células por diminuir o tempo de
crescimento celular, as células entram rapidamente na fase platô de crescimento, e da ancoragem
celular (Zhu e cols., 1991; Goldberg e cols., 2000). Além disto foi observada a reversão parcial
do fenótipo tumoral, pois houve uma diminuição da taxa de proliferação das células transfectadas
cultivadas em meio semi-sólido (Yamasaki e cols., 1999a; Chen e cols., 1995). A relação inversa
entre a expressão de conexina e proliferação das células tumorais poderia ser explicada como um
efeito da sincronização do acoplamento juncional (Loewenstein e cols., 1992).
Um dos principais efeitos da conexina sobre a fisiologia celular deve-se ao fato destas
proteínas formarem canais intercelulares. A formação destes canais afeta a proliferação celular de
maneira positiva e/ou negativa, pois, através destes, observamos a passagem de sinais (moléculas
e/ou íons) que podem reduzir ou estimular a proliferação, bem como causar a morte celular. No
entanto, estudos têm demonstrado que o papel da conexina em estimular ou inibir o crescimento
celular pode ser independente da formação de junções gap. Em linhagem de osteócitos tratadas
com AGA, agente que desorganiza as junções gap, foi demonstrado que a conexina 43 previne a
morte celular por apoptose causada por dexametasona e pela fosforilação de ERK induzida pelo
bifosfanato de alendronato (Plotkin e cols., 2002). Recentemente Prost e cols., (2008)
demonstraram que a Cx32 controla o crescimento normal da glândula tireóide, sendo que o
excesso de Cx32 é capaz de limitar o crescimento glandular mantendo as células foliculares no
estado basal de proliferação. Por outro lado, a perda da Cx32 confere aos tireócitos um aumento
na proliferação sugerindo que Cx32 e as junções gap por elas formadas são indispensáveis a
homeostasia glandular, mostrando-se fundamentais também em condições de hiperplasia das

58
células foliculares, que pode ter origem devido a hiperestimulação por altas e sustentada
concentrações de TSH sérico.
Considerando que a expressão de conexinas, a formação de conexons e de junções gap
podem não configurar em uma comunicação intercelular funcional, avaliamos a funcionalidade
das junções gap eventualmente formadas pelas isoformas Cx26, Cx32 e Cx43 expressas nas
linhagens celulares. Utilizando a técnica de microinjeção iontoforética com o corante Lucifer
yellow, observamos que as células APS9 estabelecem junções gap funcionais, provavelmente
com a participação da Cx43, revelada pelo RT-PCR. No entanto, não foi observada passagem de
corante entre as células GH3B6, o que poderia ser esperado uma vez que o RT-PCR não revelou a
expressão de nenhuma das conexinas estudadas nestas células. Munari-Silem e cols. (1996)
demonstraram em cultura primária de adeno-hipófise de ratos que a comunicação entre os
mesmos tipos celulares, comunicação homóloga, ocorria principalmente entre as células não
secretoras folículo-estreladas. Entre as células secretoras de prolactina, que se encontravam em
maior densidade, foi possível observar pouca comunicação entre as células. Estes autores
sugeriram que pequena comunicação entre as células secretoras poderia estar relacionada às
condições desfavoráveis de cultivo, que contribuem para a perda da organização histológica
observada na glândula in vivo. Estes resultados reforçam o fato das células GH3B6 secretoras de
prolactina não terem estabelecido junções funcionais quando cultivadas isoladamente. Por outro
lado, em estudo realizado por Soji e Herbert (1989) com tecido hipofisário de ratos, foi observada
a comunicação intercelular também apenas entre as células não secretoras, demonstrando que o
estabelecimento deste tipo de comunicação entre as células secretoras parece não depender
exclusivamente da organização estrutural da glândula in vivo.
A comunicação juncional entre tipos celulares diferentes, comunicação heteróloga, apesar
de ser menos estudada, já foi demonstrada em outros tecidos endócrinos, como por exemplo entre

59
as células α e β do pâncreas (Meda e cols., 1982). No sistema nervoso central, já foi observado
que há acoplamento juncional entre os neurônios e as células gliais (Bittman e cols., 2002), e o
mesmo tipo de comunicação intercelular também pode ser observado entre linfócitos e células
endoteliais (Oviedo-Orta e cols., 2002). Dados recentes de estudos desenvolvidos no nosso
laboratório, utilizando tecido hipofisário humano normal, revelaram imunomarcação positiva
para Cx43 nos cordões celulares, nas regiões de contato entre as células secretoras e não
secretoras, sugerindo a presença de comunicação heteróloga neste tecido (dados não publicados).
Nos experimentos de co-cultura com as células GH3B6 e APS9, observamos, como
esperado, comunicação homóloga entre as células estromais. No entanto, surpreendentemente,
nestas condições de cultivo, foi também observada a passagem de corante entre as células GH3B6,
que anteriormente não haviam estabelecido comunicação intercelular funcional. Estes resultados
levantam, por um lado, a questão da influência do estroma sobre fisiologia glandular, sugerindo
que as células estromais poderiam desempenhar um papel indutor da expressão de conexinas e da
formação de junções gap funcionais. Por outro lado, reforça a existência de comunicação
funcional entre células secretoras, diferentemente dos dados obtidos por Munari-Silem e cols.,
(1996).
O estroma pode influenciar a atividade glandular por contato célula-célula e pela secreção
de fatores que vão agir de forma parácrina nas células vizinhas. Nas co-culturas, houve
comunicação funcional entre as células GH3B6. Além disto, quando as células GH3B6 foram
cultivadas na presença de meio condicionado das células estromais APS9, observamos a
expressão do RNAm das conexinas 26 e 43. Estes resultados sugerem que algum fator solúvel
secretado pelas células APS9 estimula não só a expressão de conexinas, mas também a formação
de junções gap funcionais.

60
Estudos realizados por Oomizu e cols., (2004) utilizando linhagens celulares lactotróficas
provenientes de hipófise de ratos, estimuladas ou não pelo estrogênio, mostraram que estas
linhagens aumentavam suas atividades mitóticas quando estimuladas pelo estrogênio, apenas
quando co-cultivadas com células folículo-estreladas, sugerindo que a capacidade dos lactotrofos
de responder a este fator poderia estar sendo influenciada pela presença das células folículo-
estreladas. Hentges e cols., (2000) demonstraram que a ação mitogênica do estradiol em
lactotrofos envolvia a secreção de TGFβ-3 por estas células, e que este fator de crescimento
estimularia as células folículo-estreladas a produzir bFGF que, de maneira parácrina, agiria nos
lactotrofos aumentando a proliferação celular. Mais recentemente, Kabir e cols., (2005),
utilizando a mesma abordagem experimental e obtendo os mesmos resultados, verificaram que a
ação do TGFβ-3 nas células folículo-estreladas levaria a um aumento da síntese e da secreção de
bFGF e da ativação de seu receptor. Nesses estudos, os autores ainda verificaram um aumento da
expressão de conexina 43 e também um aumento da comunicação intercelular via junções gap
nas células folículo estreladas, o que justificaria ainda mais este aumento da secreção de bFGF.
Além do acoplamento entre o mesmo tipo celular, observamos também um acoplamento
entre as duas diferentes linhagens, sugerindo que as interações entre as células GH3B6 e as células
APS9 poderiam levar a formação de junções comunicantes funcionais pela indução da expressão
da conexina 43 nas células GH3B6, ou pela existência de outra isoforma de conexina, que poderia
estar sendo expressa nestas células.
Após análise da comunicação intercelular avaliamos, através da técnica de
imunofluorescência indireta, a distribuição da conexina 43 nas linhagens celulares cultivadas
isoladamente e em sistema de co-cultura. Embora a Cx43 tenha sido observada apenas nas células
APS9, confirmando os dados obtidos pela técnica de RT-PCR, nas co-culturas, houve

61
imunomarcação nas regiões das membranas aposicionais de pares de células GH3B6 e de células
GH3B6/APS9. Estudos anteriores de microscopia eletrônica de transmissão realizados por Alves e
cols. (2002) mostraram a presença de contatos focais entre células GH3B6, células APS9 e células
GH3B6/APS9. Em conjunto, estes resultados sugerem o aparecimento de junções gap nestas
células nestas condições de cultivo e reforçam o papel do estroma na fisiologia glandular. Em
adeno-hipófise de Mustela vison, foi demonstrado imunomarcação para a Cx43 nas regiões de
contato entre células folículo-estreladas, entre células secretoras e entre células secretoras/células
folículo-estreladas, sugerindo comunicação por junções gap homólogas e heterólogas (Vitale e
cols., 2001).

62
6- CONCLUSÃO
Nossos estudos de co-cultura com linhagens celulares de adeno-hipófise revelaram a
importância das células estromais na comunicação intercelular, a partir do estabelecimento de
junções comunicantes com as células secretoras. Por outro lado, levantam a questão do papel do
estroma no microambiente glandular e, conseqüentemente, na fisiologia glandular.
Em um modelo de co-cultura bidimensional, verificamos que estas células foram capazes
de mimetizar, em parte, este microambiente glandular, sugerindo que a influência das células
estromais possa estar relacionada com a secreção de diferentes fatores solúveis (fatores de
crescimento e citocinas) e/ou com o contato das células estromais com as células secretoras via
elementos de matriz extracelular.
Este estudo dá suporte para outros que serão necessários para uma melhor compreensão
do papel do estroma e das junções gap no tecido adeno-hipofisário.

63
7- REFERÊNCIAS
ALFORD, A.I., RANNELS, D.E. Extracellular matrix fibronectin alters connexin 43 expression by alveolar epithelial cells. Am J Physiol Lung Cell Mol Physiol, v. 280, p. 680-688. 2001.
ALVES. L.M., WERNECK, C.C., MARTINS, R.C.L., SILVA, L.C.F., TAFFAREL, M., BOROJEVIC, R., NASCIUTTI, L.E. A continuous lineage of rat adenohypophysis in stromal cells: characterisation and effects on GH3B6 prolactin-secreting cell behaviour. Biology of the cell, v. 94, p. 519-533. 2002.
ANDRADE-ROZENTAL, A.F., ROZENTAL, R., HOPPERSTAD, M.G., WU, J.K., VRIONIS, F.D., SPRAY, D.C. Gap junctions: the “kiss of death” and “the kiss of life”. Brain Research Reviews, v. 32, p. 308-315. 2000.
AYAD, W.A., LOCKE, D., KOREEN, I.V., HARRIS, A.L. Heteromeric, but not homomeric, connexin channels are selectively permeable to inositol phosphates. J Biol Chem, v. 281, p.16727-167239. 2006.
BARBE, M.T., MONYER, H., BRUZZONE, R. Cell-cell communication beyond connexins: the pannexin channels. Physiology, v. 21, p. 103-114. 2006.
BASTIAANSE, E.M., JONGSMA, H.J., VAN DER LAARSE, A., TAKENS-KWAK, B.R. Heptanol-induced decrease in cardiac gap junctional conductance is mediated by a decrease in the fluidity of membranous cholesterol-rich domains. J Membr Biol, v. 136, p. 135-45. 1993.
BILLIS W.M., WHITE B.A. Effect of the protein tyrosine kinase inhibitor, herbimycin A, on prolactin gene expression in GH3B6 and 235-1 pituitary tumor cells. Biochim Biophys Acta, v. 1358, p. 31-38. 1997.
BITTMAN K., BECKER D.L., CICIRATA F., PARNAVELAS J.G. Connexin expression in homotypic and heterotypic cell coupling in the developing cerebral cortex. J Comp Neurol, v. 443, p. 201-212. 2002.
BLOOM W., FAWCETT D.W. Hypohysis. In: A Textbook of Histology. 12. ed. New York: Chapman & Hall. cap. 17, p. 473-489.1994.
BRUNET-DE-CARVALHO, N., PICART, R., VAN DE MOORTELE, S., TOUGARD C., TIXIER-VIDAL, A. Laminin induces formation of neurite-like processes and potentiates prolactin secretion by GH3 rat pituitary cells. Differentiation, v. 40, p. 106-118. 1989.
BRUZZONE, R., WHITE, T., GOODENOUGH, D. The cellular internet: on-line with connexins. Bioessays, v. 18, p. 709-718. 1996.
CARVALHO, D.F., SILVA, K.L., DE OLIVEIRA, D.A.F., VILLA-VERDE, D.M.S., COELHO H.S., DA SILVA, L.C.F., NASCIUTTI L.E. Characterization and distribution of

64
extracellular matrix components and receptors in GH3B6 prolactin cells. Biology of the Cell, v. 92, p. 351-362. 2000.
CHANDROSS, K.J. Nerve injury and inflammatory cytokines modulate gap junctions in the peripheral nervous system. Glia. v. 24, p. 21-31. 1998.
CHANDROSS, K.J., CHANSON, M., SPRAY, D.C., KESSLER, J.A. Transforming growth factor-beta 1 and forskolin modulate gap junctional communication and cellular phenotype of cultured Schwann cells. J Neurosci, v. 15, p. 262-273. 1995.
CHEN, S.C., PELLETIER, D.B., AO, P., BOYNTON, A.L. Connexin43 reverses the phenotype of transformed cells and alters the expression of cyclin/cyclin-dependent kinases. Cell Growth Differ, v. 6, p. 681-690. 1995.
COMARCK, D. H.,. The Endocrine System. In: Ham’s Histology, 9. ed.. Philadelphia: J. B. Lippicontt Company. cap. 22, p. 591-602. 1987.
CROISSANDEAU, G., SCHUSSLER, N., GROUSELLE, D., PAGESY, P., RAUCH, C., BAYET, M.C., PEILLON, F., LE DAFNIET, M. Evidence of thyrotropin-releasing hormone (TRH) gene expression in rat anterior pituitaries and modulation by estrogens of TRH-like immunoreactivity and TRH-elongated peptide contents. J Endocrinol, v. 151, p. 86-87. 1996.
CZYZ, J. The stage-specific function of gap junctions during tumorigenesis. Cell Mol Biol Lett, v. 13, p. 92-102. 2008.
DECROUY, X., GASC, J.M., POINTIS, G., SEGRETAIN, D. Functional characterization of Cx43 based gap junctions during sepermatogenesis. J Cell Physiol, v. 200, p. 146-154. 2004.
DE VUYST, E., DECROCK, E., DE BOCK, M., YAMASAKI, H., NAUS, C.C., EVANS, W.H., LEYBAERT, L. Connexin hemichannels and gap junction channels are differentially influenced by lipopolysaccharide and basic fibroblast growth factor. Mol Biol Cell, v. 18, p. 34-46. 2007.
DÍEZ, J.A., ELVIRA, M., VILLALOBO, A. The epidermal growth factor receptor tyrosine kinase phosphorylates connexin 32. Mol Cell Biochem. v. 187, p. 201-210. 1998.
EVANS, W.H., DE VUYST, E., LEYBAERT, L. The gap junction cellular internet: connexin hemichannels enter the signalling limelight. Biochem J, v. 1, p. 1-14. 2006.
EWART, J.L., COHEN, M.F., MEYER, R.A., HUANG, G.Y., WESSELS, A., GOURDIE, R.G., CHIN, A.J., PARK, S.M., LAZATIN, B.O., BILLABON, S., LO, C.W, 1997. Heart and neural tube defects in transgenic mice overexpressing the Cx43 gap junction gene. Development, v. 124. p. 1281-1292. 1997.
FARQUHAR, M.G., RINEHART, J.F. Electron microscope studies of anterior pituitary of castrate rats. Endocrinology, v. 54, p. 516-541. 1954.

65
FAUQUIER, T., GUÉRINEAU, N.C., MCKINNEY, R.A., BAUER, K., MOLLARD, P. Folliculostellate cell network: a route for long-distance communication in the anterior pituitary. Proc Natl Acad Sci U S A, v. 98, p. 8891-8896. 2001.
FARQUHAR. M.G., WELLINGS, S.R. Electron microscopic evidence suggesting secretory granule formation within the Golgi apparatus. J Biophys Biochem Cytol, v. 3, p. 319-322. 1957.
FAURE, N., CRONIN, M., MARTIAL, J. A ., WEINER, R. I. Decreased responsiveness of GH3 cells to the dopaminergic inhibition of prolactin. Endocrinology, v. 107, p. 1022-1026. 1980.
FLETCHER, W.A., ANDERSON, N.C., EVERETT, J.W. Intercellular communication in the rat anterior pituitary gland: an in vivo and in vitro study. J Cell Biol, v. 67, p. 469-476. 1975.
FRESHNEY R.I., PAUL J., KANE I.M. Assay of anti-cancer drugs in tissue culture: conditions affecting their ability to incorporate 3H-leucine after drug treatment. Br J Cancer, v. 31, p. 89-99. 1975.
GARTNER , L.P., HIATT, J.L. Tratado de Histologia. 2. ed. Rio de Janeiro: Guanabara Koogan, 1999.
GOLDBERG, G.S., BECHBERGER, J.F., TAJIMA, Y., MARRITT, M., OMORI, Y., GAWINOVICZ, M.A., NARAYANAN, R., TAN, Y., SANAI, Y., YAMASAKI, H., NAUS, C.C., TSUDA, H., NICHOLSON, B.J. Connexin43 supresses MFG-E8 while inducing contact growth inhibition of glioma cells. Caner Res, v. 60, p. 6018-6026. 2000.
GOLDENBERG, R.C.S., FORTES, F.S.A., CRISTANCHO, J.M., MORALES, M.M., FRANCI, C.R., VARANDA, W.A., CAMPOS DE CARVALHO, A.C. Modulation of gap junction mediated intercellular communication in TM3 Leydig cells. J Endocrinol, v. 177, p. 327-335. 2003.
GONG, X., LI, E., KLIER, G, HUANG, Q., WU, Y., LEI, H., KUMAR, N.M., HORWITZ, J., GILULA, N.B. Disruption of alpha3 connexin gene leads to proteolysis and cataractogenesis in mice. Cell, v. 91, p. 833-843. 1997.
GOODENOUGH, D.A., GOLIJER, J.A., PAUL, D.J. Connexins, connexons and intercellular communication. Annu Rev Biochem, v. 65, p.475-502. 1996.
GOURDJI, D.; KERDELHUÉ, B., TIXIER-VIDAL, A. Ultrastructure d'un clone de cellules hypophysaires sécrétant de la prolactine (clone GH3). Modifications induites par l'hormone hypotalamique de libération de l'hormone thyréotrope (TRF), C. R. Acad Sc. Paris, v. 274, p. 437-440. 1972.
GOURDJI, D.; TOUGARD,C. & TIXIER-VIDAL, A. Clonal prolactin strains as a tool in neuroendocrinology. Frontiers. In: WF Ganong and L Martini, Neuroendocrinology. New York: Raven Press. 1982.

66
GREEN, L.M., MURRAY, D.K., TRAN. D.T., NELSON, G.A., SHAH, M.M., LUBEN, R.A. A spontaneously arising mutation in connexin 32 with repeated passage of FRTL-5 cells coincides with increased growth rate and reduced thyroxine release. J Mol Endocrinol, v. 27, p. 145-163. 1999.
GREENSPAN, F.S. & STREWLER, G.J. Basic and Clinical Endocrinology. London Prentice Hall International Inc. 1997.
HENTGES, S., BOYADJIEVA, N., SARKAR, D.K. Transforming growth factor-beta3 stimulates lactotrope cell growth by increasing basic fibroblast growth factor from folliculo-stellate cells. Endocrinology, v. 141, p. 859-67. 2000.
HU, V., XIE, H.Q. Interleukin1α suppresses gap junction-mediated intercellular communication in human endothelial cells. Exp Cell Res, v. 213, p. 2189-2223. 1994.
IKEJIMA, K., WATANABE, S., KITAMURA, T., HIROSE, M., MIYAZAKI, A., SATO, N. Hepatocyte growth factor inhibits intercellular communication via gap junctions in rat hepatocytes. Biochem Biophys Res Commun, v. 214, p. 440-446. 1995.
JUNQUEIRA, L.C., CARNEIRO, J. Glândulas Endócrinas. In: Junqueira, L.C., Carneiro, J. Histologia Básica. 10. ed. Rio de Janeiro: Guanabara Koogan. cap. 20, p. 332-354. 2004.
KABIR, N., CHATURVEDI, K., LIU, L.S., SARKAR, D.K. Transforming growth factor-beta3 increases gap-junctional communication among folliculostellate cells to release basic fibroblast growth factor. Endocrinology, v. 46, p. 4054-60. 2005.
KANCZUGA-KODA, L, SULKOWSKI, S., KODA, M., SKRZYDLEWSKA, E., SULKOWSKA, M. Connexin 26 correlates with Bcl-xL and Bax proteins expression in coloretal câncer. World J Gastroenterol, v. 11, p. 1544-1548. 2005.
KE, F.C., FANG, S.H., LEE, M.T., SHEU, S.Y., LAI, S.Y., CHEN, Y.J., HUANG, F.L., WANG, P.S., STOCCO, D.M., HWANG, J.J. Lindane, a gap junction blocker, supresses FSH and transforming growth factor β1-induced connexin43 gap junction formation and steroidogenesis in rat granulosa cells. J Endocrinol, v. 184, p. 555-566. 2005.
KIERSZEMBAUM, A.L. Neuroendocrine system. In: Kierszembaum, A.L Histology and cell biology: an introduction to pathology. Rio de Janeiro: Elsevier, cap. 18, p. 507-530. 2002.
KIERSZEMBAUM, A.L. Sistema Neuroendócrino. In: Kierszembaum, A.L Histologia e Biologia Celular: uma introdução à patologia. Rio de Janeiro: Guanabara Koogan, 2004. cap. 18, p. 507-530.
KING, T.J., GURLEY, K.E., PRUNTY, J., SHIN, J.L., KEMP, C.J., LAMPE, P.D. Deficiency in the gap protein connexin32 alters p27Kip1 tumor suppression and MAPK activation in a tissue-specific manner. Oncogene, v. 24, p. 1718-1726. 2005.

67
KO, K., ARORA, P., LEE, W., MCCULLOCH, C. Biochemical and functional characterization of intercellular adhesion and gap junctions in fibroblasts. Am J Physiol Cell Physiol, v. 279, p. 147-157. 2000.
LAIRD, D.W., THOMAS, T., JORDAN, K., SIMEK, J., SHAO, Q., JEDESZKO, C., WALTON, P. Mechanisms of Cx43 and Cx25 transport to the plasma membrane and gap junction regeneration. J Cell Science, v. 118, p. 4451-4462. 2005.
LAIRD, D.W. Life cycle of connexins in health and disease. Biochem J, v. 394, p. 527-43. 2006.
LEITHE, E., RIVEDAL, E. Ubiquitination of gap junction proteins. J Membr Biol, v. 217, p. 43-51. 2007.
LEWIS, B.M., PEXA, A., FRANCIS, K., VERMA, V., MCNICOL, A.M., SCANLON, M., DEUSSEN, A., EVANS, REES, D.A., HAM, J. Adenosine stimulates connexin 43 expression and gap junctional communication in pituitary folliculostellate cells. FASEB Journal, v. 20, p. E1-E9. 2006.
LOEWESTEIN, W. R. The cell to cell channel of gap junctions. Cell, v. 49, p. 725-726. 1987.
LOEWESTEIN, W.R., ROSE, B. The cell-cell channel in the control of growth. Semin Cell Biol, v. 3, p. 59-79. 1992.
MEDA P., KOHEN E., KOHEN C., RABINOVITCH A., ORCI L. Direct communication of homologous and heterologous endocrine islet cells in culture. J Cell Biol, v. 92, p. 221-226. 1982.
MEDA, P., PEPPER, M.S., TRAUB, O., WILLECKE, K., GROS, D., BEYER, E., NICHOLSON, B., PAUL, D., ORCI, L. Differential expression of gap junction connexins in endocrine and exocrine glands. Endocrinology, v. 133, p. 2371-2378. 1993.
MELMED, S.; CARLSON, H.E.; BRIGGS, J., HERSHMAN, J. M. Cell culture alters the hormonal response of rat pituitary tumors to dynamic stimulation. Endocrinology, v. 107, p. 789-793. 1980.
MESE, G., RICHARD, G., WHITE, T.W. Gap junction: basic structure and function. J Invest Dermat, v. 127, p. 2516-2524. 2007.
METHA, P, LOEWESTEIN, W. Differential regulation of communication by retinoid acid in homologous and heterologous junctions between normal and transformed cells. J Cell Biol, v. 113, p. 371-379. 1991.
MEYER, R., LAIRD, D.W., REVEL, J.P., JOHNSON, R. Inhibition of gap junction and adherens junction assembly by connexin and A-CAM antibodies. J Cell Biol, v. 119, v. 179-189. 1992.

68
MOMIYAMA, M., OMORI, Y., ISHIZAKI, Y., NISHIKAWA, Y ., TOKAIRIN, T., OGAWA, J., ENOMOTO, K. Connexin26-mediated gap junctional communication reverses the malignant phenotype of MCF-7 breast cancer cells. Cancer Sci, v. 94, p. 501-507. 2003.
MOORBY, C.D., STOKER, M., GHERARDI, E. HGF/SF inhibts junctional communication. Exp Cell Res, v. 219, p. 657-663. 1995.
MORAND, I.., FONLUPT, P., GUERRIER, A; TROUILLAS, J.; CALLE, A.; REMY, C.; ROUSSET, B.; MUNARI-SILEM, Y. Cell-to-cell communication in the anterior pituitary: evidence for gap junction-mediated exchanges between endocrine cells and folliculostellate cells. Endocrinology, v. 137, p. 3356-3367. 1996.
MORIN, A.; ROSENBAUM, E., TIXIER-VIDAL, A. Effects of thyrotropin-releasing hormone on prolactin compartments in clonal rat pituitary tumor cells. Endocrinology, v. 115, p. 2271-2277. 1984.
MUNARI-SILEM, Y., ROUSSET, B. Gap junctions-mediated cell-to-cell communication in endocrine glands – molecular and functional aspects: a review. Eur J Endocrinol, v. 135, p. 351-264. 1996.
NASCIUTTI, L.E.; PICART, R.; ROSENBAUM, E.; TIXIER-VIDAL, A., TOUGARD, C. Effect of reduced temperatures and brefeldin A on prolactin secretion and on subcellular distribution of the secretory product and membrane antigens in GH3 pituitary cells. Biol Cell, v. 75, p. 25-35. 1992.
NELLES, E., BÜTZLER, C., TEMME, A., GABRIEL, H.D., DAHL, U., TRAUB, O., STÜMPEL, F., JUNGERMANN, K., ZIELASEK, J. Defective propagation of signals generated by symphatetic nerve stimulation in the liver of connexin 32-deficient mice. Proc Natl Acad Sci, USA, v. 93, p. 9565-9570. 1996.
OOMIZU S. CHATURVEDI, K. , SARKAR, D.K. Folliculostellate cells determine the susceptibility of lactotropes to estradiol's mitogenic action. Endocrinology, v. 145, p. 1473-1480. 2004.
OVIEDO-ORTA E., ERRINGTON R.J., EVANS W.H. Gap junction intercellular communication during lymphocyte transendothelial migration.Cell Biol Int, v. 26, p. 253-63. 2002.
PELLETIER D.B., BOYNTON, A.L. Dissociation of PDGF receptor tyrosine kinase activity from PDGF-mediated inhibition of gap junction communication. J Cell Physiol, v. 158, p. 427-434. 1994.
PLOTKIN, L.I., MANOLAGAS, S.C., BELLIDO, T. Tranduction of cell survival signals by connexin43 hemichannels. J Biol Chem, v. 277, p. 8648-8657. 2002.

69
PROST, G., BERNIER-VALENTIN, F., MUNARI-SILEM, Y., SELMI-RUBY, S., ROUSSET, B. Connexin-32 acts as a downregulator of growth of thyroid gland. Am J Physiol Endocrinol Metab, v. 294, p. 291-299. 2008.
REAUMÉ, A.G., DE SOUZA, P.A., KULKARNI, S, LANGILLE, B.L., ZHU, D., DAVIES, T.C., JUNEJA, S.C., KIDDER, G.M., ROSSANT, L. Cardiac malformation in neonatal mice lacking connexin 43. Science. 267:1831-1834. 1995.
RIVEDAL, E., LEITHE, E. Connexin43 sysnthesis, phosphorilation and degradation in regulation of transient inhibition of gap junction intercellular communication by the phorbol ester TPA in rat liver epithelial cells. Experimental Cell Research, v. 302, p. 143-152. 2005.
ROSS, M. H., ROWRELL, L. J. Glândulas Endócrinas. In: ROSS, M. H., ROWRELL, L. J. Histologia: Texto e Atlas. São Paulo: Panamericana, cap. 20, p. 563-570. 1993.
SATO, Y., HASHITANI, H., SHIRASAWA, N., SAKUMA, E., NAITO, A., SUZUKI, H., ASAI, Y., SATO, G., WADA, I., HERBERT, D.C., SOJI, T. Interecellular communication within the rat anterior pituitary XII: immunohistochemical na physiological evidences for the gap junctional coupling of the folliculostellate cells in the rat abterior pituitary. Tissue cell, v. 37, p. 281-291. 2005.
SCHILLER P.C., D’IPPOLITO, G., BALKAN W., ROOS, B.A., HOWARD, G.A. Gap-junctional communication mediates parathyroid hormone stimulation of mineralization in osteoblastic cultures. Bone, v. 28, p. 38-44. 2001.
SEGRETAIN, D., FALK, M.M. Regulation of connexin biosynthesis, assembly, gap junction formation, and removal. Biochim Biophys Acta, v. 1662, p. 3-21. 2004.
SIMON, A., GOODENOUGH, D. Diverse functions of vertebrate gap junctions. Trends in Cell Biol, v. 8, p. 477-482. 1998.
SIMON, A.M., GOODENOUGH, D.A. Female infertility in mice lacking connexin 37. Nature, v. 385, p. 525-529. 1997.
SOJI, T., HERBERT, D.C. Intercellular communication between rat anterior pituitary cells. Anat Rec, v. 224, p. 523-533. 1989.
SPRAY, D.C., FUJITA, M., SAEZ, J.C., CHOI, H., WATANABE, T., HERTZBERG, E., ROSENBERG, L.C., REID, L.M. Proteoglycans and glycosaminoglycans induce gap junction synthesis and function in primary liver cultures. J Cell Biol, v. 105, p. 541-551. 1987.
TANMAHASAMUT, P., SIDELL, N. Up-regulation of gap junctional intercellular communication nd connexin43 expression by retinoic acid in human endometyrial stromal cells. J Clin Endocrinol Metabolism, v. 90, p. 4151-4156. 2005.

70
TASHJIAN, A.JR.; YASUMURA,Y.; LEVINEE; L.; SATO, G.H.; PARKER, M.L. Establishment of clonal strains of rat pituitary tumor cells that secrete growth hormone. Endocrinology, v. 82, p. 342-352. 1968.
TASHJIAN, A.Jr.; BANCROFT, F.C., LEVINE, L. Production of both prolactin and growth hormone by clonal strains of rat pituitary tumor cells. J. Cell Biol., v. 47, p. 61-70. 1970.
THORNER, M. Prolactin. Clinics in Endocrinology and Metabolism, v. 6, p. 201-221. 1977.
TIRRELL, J. B.; FINDING, J. W.; ARON, D.C. Hypothalamus and Pituitary. In GREENSPAN, F. S., BAXTER, J. D. Basic &Clinical Endocrinology, 4. ed. London: Prentice Hall, p. 64-127. 1994.
TIXIER-VIDAL, A.; BRUNET, N.; TOUGARD, C., GOURDJI, D. Morphological and molecular aspects of prolactin and growth hormone secretion by normal and tumoral pituitary cells in culture, In: FAGLIA, G., GIOVANELLI, M.A., MAC LEOD, R.M. Pituitary microadenomas. Academic Press, v. 29, p. 73-89. 1980.
TOUGARD, C.; PICART, R., TIXIER-VIDAL, A. Electron-microscopic cytochemical studies on the secretory process in rat prolactin cells in primary culture. Am. J. Anat., v. 158, p. 471-490. 1980.
TOUGARD ,C., PICART, R., TIXIER-VIDAL, A. Immunocytochemical localization of prolactina in the endoplasmatic reticulum of GH3 cells. Variations in response of thyroliberin. Biol Cell, v. 43, p. 89. 1982.
TOUGARD, C., TIXIER-VIDAL, A. Lactotropes and Gonadotropes, In: KNOBIL, E., NIEL, J. D. The Physiology of Reproduction. New York: Raven Press Ltda, p. 1711-1747. 1994
TRUE, L.D. Pituitary Hormones, In: Atlas of diagnostic Immunohistopathology. Philadelphia: J. B. Lippincott Company, 1990. p. 13.17.
VAN RIJEN H.V., VAN KEMPEN, M.J., ANALBERTS, L.J., ROOK, M.B., VAN GINNEKEN, A.C., GROS, D., JONGSMA, H.J. Gap junctions in human umbilical cord endothelial cells contain multiple connexins. Am J Physiol, v.272, p. 117-130. 1997.
VITALE, M.L., CARDIN, J., GILULA, N.B., CARBAJAL, M.E., PELLETIER, R.M. Dynamics of connexin 43 levels and distribution in the mink (Mustela vison) anterior pituitary are associated with seasonal changes in anterior pituitary prolactin content. Endocrinology, v. 64, p. 625-633. 2001.
WADAMORI, K., OKA, M., TOKUDA, M., FUJIKURA, Y., HAZAMA,S., FUKUMOTO, T., SUZUKI, T. Influence of continuous interleukin-2 administration via the portal vein on liver regeneration following partial hepatectomy in rats. Hepatology, v. 23, p. 1578-1583. 1996.

71
WANG X.G., PERACCHIA C. Molecular dissection of a basic COOH-terminal domain of Cx32 that inhibits gap junction gating sensitivity.Am J Physiol, v. 275, p. 1384-1390. 1998.
WEISS, L., GREEP, R., 1981. A Hipófise. In: WEISS, L., GREEP, Histologia. Rio de Janeiro: Guanabara Koogan, p. 875-897.1981
WILLECKE, K., SÖHL, G., MAXEINER, S. Expression and functions of neuronal gap junctions. Nature, v. 6, p. 191-200. 2005.
WYATT, T.A., CHUNG, C.Y., CARLSEN, B., IIDA-KLEIN, A., RUDKIN, G.H., ISHIDA, K., YAMAGUCHI, D.T., MILLER, T.A. Bone morphogenetic protein-2 (BMP-2) and transforming growth factor-beta1 (TGF-beta1) alter connexin 43 phosphorylation in MC3T3-E1 cells. BMC Cell Biol. 2:14-19. 2001.
YAMASAKI, H., KRUTOVSKIKH, V., MESNIL, M., TANAKA, T., ZAIDAN-DAGLI, M.L., OMORI, Y. Role of connexin (gap junction) genes in cell growth control and carcinogenesis. C R Acad Sci III, v. 322, p.151-159. 1999a.
YAMASAKI, H., OMORI, Y., ZAIDAN-DAGLI, M.L., MIRONO V, N., MESNIL, M., KRUTOVSKIKH, V. Genetic and epigenetic changes of intercellular communication genes during multistage carcinogenesis. Cancer Detect Prev, v. 23, p. 273-279. 1999b.
ZHU, D., CAVENEY, S., KIDDER, G.M., NAUSS, C.C. Transfection of C6 glioma cells with connexin 43 cDNA: analysis of expression, intercellular coupling and cell proliferation. Proc Natl Acad Sci USA, v. 88, p. 1883-1887. 1991.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo














![Microscopia Confocal Tem [Recuperado]](https://static.fdocumentos.com/doc/165x107/563dbbbb550346aa9aafc610/microscopia-confocal-tem-recuperado.jpg)



