
Caracterização Molecular da Via de Sinalização Mediada...
108
Universidade de São Paulo Faculdade de Medicina de Ribeirão Preto Caracterização Molecular da Via de Sinalização Mediada pelo Receptor Notch em S. mansoni Lizandra Guidi Magalhães Ribeirão Preto 2005
Transcript of Caracterização Molecular da Via de Sinalização Mediada...

Universidade de São Paulo
Faculdade de Medicina de Ribeirão Preto
Caracterização Molecular da Via de
Sinalização Mediada pelo Receptor Notch em
S. mansoni
Lizandra Guidi Magalhães
Ribeirão Preto
2005

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.


ÍNDICE
Abreviaturas ..........................................................................................................I
Resumo................................................................................................................ V
Summary ........................................................................................................... VII
1- Introdução ........................................................................................................1
1.1. Aspectos gerais da esquistossomose .........................................................2
1.2. Histórico ......................................................................................................3
1.3. Ciclo de vida de S. mansoni ........................................................................4
1.4. O genoma de S. mansoni............................................................................8
1.5. Vias de desenvolvimento celular ...............................................................11
1.5.1 Via de sinalização mediada pelo receptor Notch .................................15
2-Objetivos .........................................................................................................22
2.1. Objetivo geral ............................................................................................23
3- Materiais e Métodos ......................................................................................24
3.1. Análise in silico da via de desenvolvimento Notch ....................................25
3.1.1. Comparação da via Notch entre S. mansoni e S. japonicum..............26
3.2. Manutenção do ciclo biológico de S. mansoni...........................................26
3.3. Obtenção de ovos .....................................................................................27
3.4. Transformação mecânica de cercárias em esquistossômulos ..................27
3.5. Extração de RNA total...............................................................................28
3.6. Extração do DNA genômico ......................................................................30
3.7. Quantificação de DNA e RNA ...................................................................31
3.8. Síntese dos oligonucleotídeos iniciadores ................................................32
3.9. RT-PCR (Transcrição reversa e reação em cadeia da polimerase) ..........34
3.10. Purificação do produto de PCR...............................................................35

3.11. Clonagem do produto de PCR ................................................................36
3.12. Minipreparação de DNA plasmidial .........................................................37
3.13. Sequenciamento .....................................................................................38
3.14. Análise computacional das seqüências...................................................39
3.15. Northern Blot ...........................................................................................39
3.16. Preparo da sonda radioativa ...................................................................40
3.17. Reação de Hibridização ..........................................................................41
3.18. Autoradiografia ........................................................................................42
4- Resultados .....................................................................................................43
4.1. Análise in silico da via de sinalização Notch .............................................44
4.2. Amplificação dos fragmentos gênicos de Notch, Presenilina e Supressor
de Deltex a partir do DNA genômico do S. mansoni..............................................48
4.3. Sequenciamento e análise computacional dos fragmentos gênicos de
Notch, Presenilina e Supressor de Deltex .............................................................49
4.4. Perfil de expressão dos fragmentos gênicos de Notch, Presenilina e
Supressor de Deltex durante o ciclo de vida do parasita.......................................58
4.5. Estimativa por Northern Blot do tamanho da mensagem codificadora de
Notch e Presenilina................................................................................................62
5- Discussão.......................................................................................................64
6- Conclusões ....................................................................................................73
7- Referências Bibliográficas............................................................................76
8-Apêndice .........................................................................................................88

Abreviaturas

Abreviaturas
II
[αP32] DcTP Citosina trifosfato radiomarcada na posição α com 32P ºC Graus Celsius 32P Fosfato radiomarcado Ca 2+ Íon cálcio CaCl2 Cloreto de Cálcio cDNA DNA complementar cpm Contagem por minuto cm Centímetros CO2 Dióxido de Carbono CTAB Brometo de hexadeciltrimelilamônio DEPC Dietilpirocarbonato DNA Ácido dexorribonucleotídeo DNAg DNA genômico DNTP Deoxinucleosídeo trifosfato (N= A,C,G,T) D.O Densidade Óptica DTT Ditiotreitol EDTA Ácido etilenodiaminotetracético (sal dissódico) EST Expressed Sequence Tags g Grama HCl Ácido Clorídrico HEPES 2-4-(2-hidroxietil)-pperazinil-(1)-Ácido Etanosulfônico kDa Kilodaltons-1kDa equivale a 1000 Daltos Kb Kilobases- 1kb equivale a 1000 bases nucleotídicas KCl Cloreto de Potássio KH2PO4 Fosfato diácido de Potássio LB Lúria Bertani-Meio de cultura bacteriano LE Luis Evangelista – linhagem do parasita Schistosoma mansoni mV Milivolts M Mols/L MgCl2 Cloreto de magnésio MgSO4 Sulfato de magnésio mg Miligrama mL Mililitro mM Milimolar mm Milímetro mRNA Ácido Ribonucléico mensageiro MOPS [Ácido 3-(N-Morfolino)Propanosulfônico] NaCl Cloreto de Sódio NaHCO3 Bicarbonato de Sódio Na2HPO4 Fosfato de sódio dibásico NaOH Hidróxido de Sódio NaOAc Acetato de Sódio NEXT Domínio extracelular de Notch truncado NICD Domínio intracelular de Notch ng Nanograma nm Nanomols

Abreviaturas
III
Oligo(dt) Oligo desoxitimina ORF Open Reading Frame pb Pares de base PBS Solução salina tamponada com fosfato pmol Picomol RNAse Ribonuclease RNAseH Ribonuclease H RNA Ácido Ribonucléico rpm Rotações por minuto RT-PCR Transcriptase Reversa-Reação de Cadeia Polimerase SDS Dudecilsulfato de sódio SmNotch Fragmento gênico de Notch em S.mansoni SmPresenilina Fragmento gênico de Presenilina em S. mansoni SmSu(dx) Fragmento gênico de Supressor de deltex em S. mansoni Taq Thermos aquaticus TIGR The Institute for Genomic Research tris Tris-hidroximetilaminometano U Unidades v/v Volume/volume V Volts Xg Velocidade de sedimentação em unidade gravitacional WHO Organização Mundial da Saúde µCi Microcuries µg Micrograma µL Microlitros µM Micromolar
Abreviaturas
IV
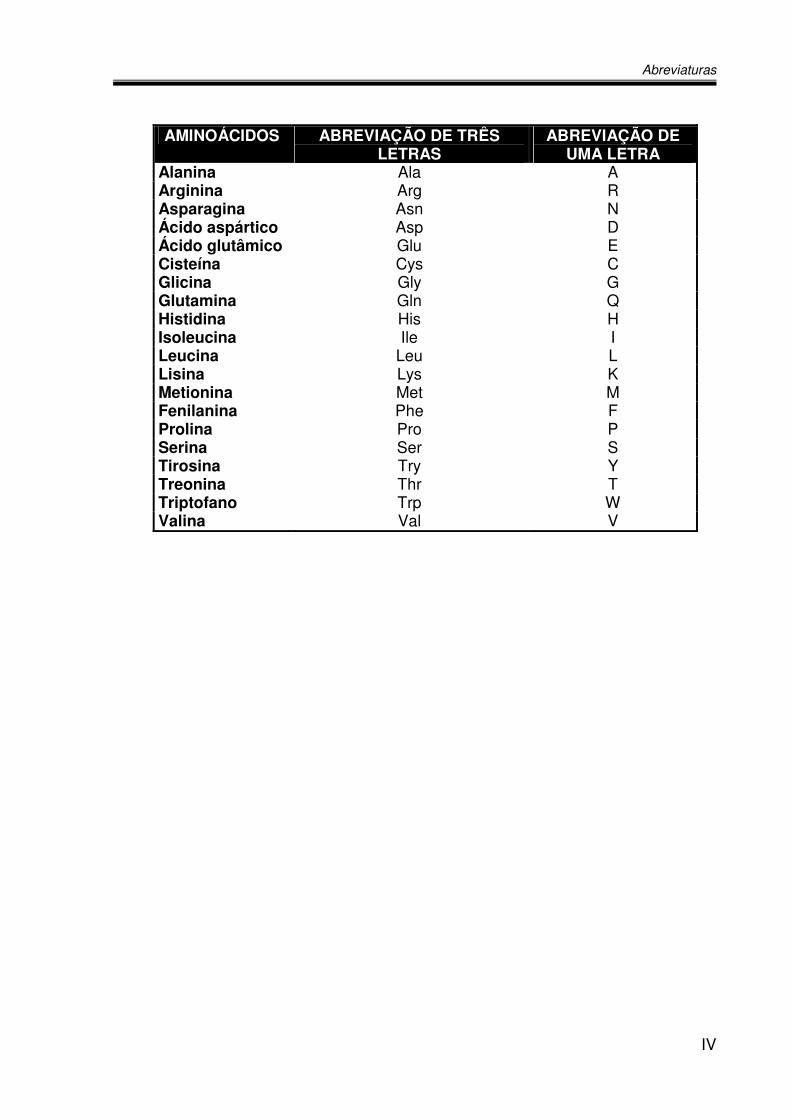
AMINOÁCIDOS ABREVIAÇÃO DE TRÊS LETRAS
ABREVIAÇÃO DE UMA LETRA
Alanina Ala A Arginina Arg R Asparagina Asn N Ácido aspártico Asp D Ácido glutâmico Glu E Cisteína Cys C Glicina Gly G Glutamina Gln Q Histidina His H Isoleucina Ile I Leucina Leu L Lisina Lys K Metionina Met M Fenilanina Phe F Prolina Pro P Serina Ser S Tirosina Try Y Treonina Thr T Triptofano Trp W Valina Val V

Resumo

Resumo
VI
Vias de transdução de sinal podem desempenhar um papel essencial para o
desenvolvimento e a homeostase de Schistosoma mansoni. Entre as diversas vias
de sinalização, a via mediada pelo receptor Notch se destaca por ser crítico na
diferenciação celular em muitos organismos. Alguns dos componentes dessa via,
como o receptor Notch, a protease Presenilina e a enzima ubiquitina ligase
Supressor de Deltex, desempenham papéis fundamentais durante a ativação e a
regulação da via Notch. O presente trabalho reconstituiu in silico essa via de
sinalização e analisou o perfil de expressão dos referidos genes durante o ciclo de
vida do parasita S. mansoni.
A reconstituição in silico demonstrou a presença de transcritos dos principais
componentes da via Notch. Através da técnica de RT-PCR foi possível demonstrar
que Notch é expresso em baixíssimos níveis durante o estágio de cercária,
entretanto, Presenilina e Supressor de Deltex são expressos nos principais estágios
evolutivos deste parasita. A análise dos fragmentos amplificados a partir do DNA
genômico mostrou a ausência de íntrons nas regiões flanqueadas pelos
oligonucleotídeos específicos.
Esta caracterização inicial constituiu uma abordagem pioneira no estudo dessa
via em S. mansoni. Desta forma, as análises subseqüentes de caráter estrutural e
funcional poderão ser empregadas para a elucidação dos mecanismos pelo qual a
via Notch coopera para garantir tanto a diferenciação como o desenvolvimento deste
parasita.

Summary

Summary
VIII
Signaling transducing pathways may have an essential role in the development
and the homeostasis of Schistosoma mansoni. Among the diverse signaling
pathways, the Notch pathway is emphasized by its critical role in cellular
differentiation in various organisms. Some components of this pathway, such as the
Notch receptor, the Presenilin protease and the Suppressor of Deltex ubiquitin ligase
enzyme, have an essential role during activation and regulation of the Notch
pathway. The present work highlights the in silico reconstitution of this pathway and
analyzes the expression profile of the mentioned genes during the life cycle of the
parasite S. mansoni.
The in silico reconstitution demonstrated the presence of transcripts coding for
several components of the Notch pathway. By using RT-PCR we were able to
demonstrate that Notch is lower expression during the cercariae stage whereas
Presenelin and Supressor of Deltex were expressed throughout different larval and
adult stage. In addition, the analyses of amplified fragments from genomic DNA
showed absence of introns concerning the regions flanked by each specific pair of
oligonucleotides.
This initial characterization constitutes the first evidence suggesting the
existence of the Notch pathway in S. mansoni. In this way, subsequently structural
and functional analysis towards the elucidation of the mechanisms by which the
Notch pathway cooperates to guarantee parasite development and differentiation.

1- Introdução

Introdução
2
1.1. Aspectos gerais da esquistossomose
Schistosoma mansoni é um trematódeo digenético, da família
Schistosomatidae, gênero Schistosoma, cuja principal característica é o seu
acentuado dimorfismo sexual em sua forma adulta. A infecção por S. mansoni
determina uma patologia denominada esquistossomose intestinal, popularmente
conhecida no Brasil como "Xistose", "Mal do caramujo" ou "Barriga d'água", devido à
ascite que acompanha as formas mais graves (MELO & COELHO, 2005). O sucesso
desse parasita está relacionado com a sua adaptação e na habilidade de se
transformar rapidamente durante os diferentes ambientes.
Existem 6 espécies de Schistosoma que podem infectar o ser humano, sendo:
S. mansoni, S haematobium, S. japonicum, S. intercalatum, S. mekongi e S.
malayensis. Contudo, no Brasil, somente a espécie S. mansoni se adaptou,
provavelmente devido à presença de moluscos do gênero Biomphalaria, que atuam
como hospedeiros intermediários.
Atualmente, estima –se existir pelo menos 300 milhões de pessoas infectadas
em todo mundo, e um total de 600 milhões de pessoas expostas ao risco de contrair
uma das esquistossomoses, em áreas tropicais e subtropicais do mundo (WHO,
1996). No Brasil, a área endêmica abrange 19 estados com aproximadamente 26
milhões de habitantes expostos ao risco, e cerca de 8 a 10 milhões de indivíduos
infectados pelo parasita, os quais em muitos casos apresentam severas deficiências
orgânicas, comprometendo o desenvolvimento de jovens e a produtividade de
adultos e fazendo desta doença um dos mais sérios problemas de saúde nos países

Introdução
3
em desenvolvimento (MELO & COELHO, 2005; LAMBERTUCCI & BARRAVIEIRA,
1994).
O tratamento com a droga praziquantel constitui uma importante arma no
controle da morbidade e na diminuição da prevalência e da incidência dessa doença
nas áreas endêmicas (KATZ, 1998), no entanto, a resistência do parasita contra essa
droga tem sido relatada em muitas localizações (ISMAIL et al, 1999). Desse modo,
vários métodos têm sido usados para combater a esquistossomose, incluindo
saneamento básico e educação sanitária, no entanto, esses métodos têm se
mostrado pouco eficiente no controle da esquistossomose.
1.2. Histórico
A esquistossomose é uma doença conhecida desde a antiguidade. O exame de
múmias do antigo Egito revelou a presença de granulomas hepáticos que são
característicos da infecção por S. mansoni. Em 1851, o médico alemão Theodoro
Bilharz, durante a realização de uma autópsia em um egípcio, observou a presença
de um parasita intravascular o qual deu o nome de Distoma haematobia.
Posteriormente, Winland denominou o mesmo helminto de Schistosoma e, em 1907
a denominação da espécie – Schistosoma mansoni foi dada por Sambon.
Acredita-se que as espécies do gênero Schistosoma chegaram no Brasil
durante o tráfico de escravos e com os imigrantes vindos da África. A descoberta dos
primeiros casos no Brasil aconteceu em 1951, pelo cientista brasileiro Manuel
Augusto Pirajá da Silva, na Bahia, quando estava fazendo numerosos exames de
fezes e necropsias. Os trabalhos realizados por Pirajá da Silva esclareceram todas

Introdução
4
as dúvidas em relação à espécie, mas a denominação coube a Sambon (MELO &
COELHO, 2005).
1.3. Ciclo de vida de S. mansoni
A biologia de S. mansoni apresenta um ciclo do tipo heteroxeno, o qual requer
dois hospedeiros para completar sua evolução, um hospedeiro invertebrado que são
os caramujos do gênero Biomplalaria e um hospedeiro definitivo vertebrado (Figura
1). Devido a esse complexo plano de desenvolvimento, o parasita S. mansoni sofre
profundas alterações morfológicas, como também alterações funcionais de diversas
vias metabólicas (HOCKLEY, 1973 ; RUMJANEK, 1987).
Assim que os ovos são liberados na água pelo hospedeiro definitivo, esses
eclodem devido a vários fatores como a temperatura, luz intensa e oxigenação da
água, liberando a forma larval denominada de miracídio. Uma vez liberado, esses
irão à procura do seu hospedeiro intermediário. Os mecanismos envolvidos no
reconhecimento do caramujo hospedeiro pelo miracídio têm sido alvo de muitos
estudos, no entanto, alguns relatos mostram que o miracídio é controlado por dois
tipos de respostas. Primeiramente, o miracídio responderia a estímulos físicos do
ambiente e num segundo momento, o miracídio responderia a estímulos químicos
originados pelo molusco (KAPP, et al; 2003). Após penetrar pelas partes moles dos
caramujos susceptíveis, os miracídios desenvolvem-se em esporocisto mães, por
poliembrionia, e posteriormente, diferenciam em esporocistos filhos, que migram
para as glândulas digestivas e ovoteste do molusco. Por reprodução assexuada,
originam as formas de vida livre conhecidas como cercárias, cerca de 20 a 30 dias

Introdução
5
após a infecção do hospedeiro intermediário. Um miracídio pode produzir de 100.000
a 300.000 cercárias, sendo que cada miracídio define o sexo das cercárias que serão
produzidas (ROLLINSON & SIMPSON, 1987).
Sob condições ideais de temperatura e luminosidade, as cercárias são liberadas
para o meio externo (água), podendo viver livremente por algumas horas até que
suas reservas energéticas, principalmente glicogênio, sejam consumidas. Neste
período, elas nadam ativamente na água, e como são fototrópicas positivas, elas se
concentram na superfície das águas, tornando o contato com o hospedeiro definitivo
mais provável.
Ao alcançarem a pele do hospedeiro, se fixam preferencialmente entre os
folículos pilosos com o auxilio de duas ventosas e de uma substância mucoprotéica
secretada por suas glândulas acetabulares. Através da ação lítica devido à presença
de proteases presentes nas glândulas pré-acetabulares e sub tegumentares, e por
ação mecânica, ocorre à penetração do corpo cercariano através da epiderme e
derme (GONZALES, 1989). Durante a penetração no hospedeiro definitivo, as
cercárias perdem a cauda e, algumas horas após, completam a metamorfose,
atingindo o estágio de esquistossômulo. Estes últimos migram pelo tecido
subcutâneo e, ao penetrarem num vaso, são levados da pele até os pulmões, onde
permanecem, por pelo menos 48 horas. Do pulmão, os esquistossômulos dirigem-se
para o sistema porta-hepático e, nesse ambiente ocorre à maturação sexual e o
acasalamento dos parasitas.
Num espaço de aproximadamente seis semanas pós-infecção, os vermes
encontram–se na sua forma adulta e migram para as veias mesentéricas, onde
ocorre a postura dos ovos na submucosa. Da submucosa, os ovos passam para a

Introdução
6
luz intestinal. Os casais de vermes adultos podem produzir de 300 a 1000
ovos/dia/casal, sendo que, parte desses ovos chegando à luz intestinal, é eliminada
para o exterior junto com o bolo fecal completando desse modo, o ciclo de vida do
parasita. Cerca de 50% desses ovos ficam retidos no hospedeiro, fato este
responsável pela patologia da doença, uma vez que, a presença de ovos induz
reações imuno–teciduais que desencadeiam a formação dos granulomas (WARREN,
1987). A formação desses granulomas é mediada por uma resposta celular,
constituída por subpopulações de linfócitos T e outros grupos celulares como
neutrófilos, eosinófilos, macrófagos e fibroblastos, os quais, migram para o local da
lesão devido aos componentes dos ovos que liberam determinados fatores solúveis
que induzem esta resposta imune (STADECKER, 1992).
Em relação ao sistema imune do hospedeiro, diversos grupos independentes
demonstraram que este desempenha um papel muito importante tanto na
modulação da carga parasitária como na capacidade de evitar ou minimizar re-
infecções, especialmente em áreas endêmicas. O grau da sintomatologia
apresentada pelo paciente como também a possibilidade de ocorrência de outras
afecções, devido à imunossupressão induzida pelo parasita, também está
intimamente relacionada ao sistema imune do hospedeiro (PEARCE &
MACDONALD, 2002.). Desta forma, no início da infecção ocorre um aumento de
IgE, decorrente da dermatite cercariana, e em seguida, uma elevação tanto de IgM
como de IgG2a, sendo que esta última imunoglobulina, encontra-se mais elevada
até o início da ovoposição. Nas fases mais tardias, observa-se uma diminuição do
título de IgG2a e a “modulação” ou diminuição do tamanho dos granulomas.

Introdução
7
Figura 1- Diagrama esquemático do ciclo de vida do parasita S. mansoni. Dentro do seu hospedeiro vertebrado, as formas larvais dão origem a parasitas sexualmente maduros os quais se acasalam e produzem os ovos que são liberados para o meio aquático. Os ovos eclodidos liberam os miracídios, e esses penetram nos caramujos dando origem a numerosos esporocistos, os quais culminam com a liberação de cercarias, a forma infectante deste parasita para o hospedeiro vertebrado. Adaptado de - Introdução à Parasitologia. Wilson, R.A,.p.13, 1980.

Introdução
8
Pouco se conhece sobre os aspectos da reprodução em S. mansoni. No
entanto, o parasita e seus hospedeiros têm uma relação intima que envolve a
exploração de sinais endócrinos e imunes dos hospedeiros, resultando na expressão
coordenada de um conjunto de genes distintos, necessários para o desenvolvimento,
diferenciação sexual e adaptação do parasita (SALZET, et al, 2000; DAVIES, et al.
2001). Desta forma, o conhecimento de genes e dos mecanismos que controlam as
suas expressões, permitirá uma melhor compreensão dos mecanismos envolvidos
com o desenvolvimento, diferenciação sexual, maturação e na adaptação deste
parasita em ambientes distintos. O conhecimento molecular também poderá ajudar
na escolha de novos alvos para drogas, bem como, de futuras vacinas, as quais
poderão desencadear uma resposta protetora, ou o bloqueio do desenvolvimento do
parasita no hospedeiro vertebrado.
1.4. O genoma de S. mansoni
S. mansoni apresenta um genoma estimado em 2.7X108 pb, organizado em oito
pares de cromossomos sendo sete pares autossômicos e um par de cromossomos
sexuais. Os machos são homogaméticos (ZZ) e as fêmeas heterogaméticas (ZW)
(TANAKA, 1995).
Considerando que este parasita apresenta diversas formas evolutivas para
completar um ciclo biológico, e sendo este processo acompanhado de profundas
alterações tanto morfológicas como bioquímicas em diversas vias metabólicas
durante este ciclo, provavelmente o genoma do parasita deve apresentar uma
grande plasticidade e conseqüentemente manter o padrão de expressão gênica

Introdução
9
diferencial. Desta forma, além dos genes constitutivos, pode–se esperar a existência
de dois outros grupos gênicos: um grupo, cujos genes são regulados por
mecanismos de estresse (como por exemplo: térmico, osmótico ou oxidativo) e um
outro, de genes regulados durante o desenvolvimento do parasita, o que sustentaria
um padrão de proteínas tanto estágio-específica como com níveis regulados durante
os distintos estágios evolutivos.
Nos últimos anos, através da iniciativa do Programa de Pesquisas em Doenças
Tropicais da Organização Mundial de Saúde e de outras agências de fomento
diversos grupos vêm estudando de uma forma sistemática o genoma desse parasita,
resultando numa rápida expansão do conhecimento (JOHNSTON, 1997).
A produção de ESTs (Expressed Sequence Tags), que são seqüências parciais
de cDNAs, com cerca de 150-600 pb, obtidas das extremidades 3’ e 5’ e a produção
de ORESTES (Open Reading Frame Expressed Sequence Tags), que são
seqüências parciais de cDNA com cerca de 150-800 pb, correspondentes à região
central dos transcritos, têm mostrado serem métodos relativamente simples e rápidos
tanto para descobrir novos genes (FRANCO, et al, 1995; HILLIER, et al, 1996; DIAS-
NETO, et al, 2000) como para examinar o padrão de expressão gênica, incluindo a
identificação dos genes diferencialmente expressos (LEE, et al, 1995 ; MANGER et
al, 1998).
Até 2002, aproximadamente 12.000 ESTs tinham sido geradas, o que
representa um número pouco significativo, se for levado em consideração o tamanho
do genoma do parasita, onde se estima a existência de 20.000 genes (FRANCO, et
al, 2000).

Introdução
10
Utilizando a metodologia ORESTES, Verjovski e colaboradores (2003), geraram
163,000 ESTs provenientes de bibliotecas representativas dos seis estágios de
desenvolvimento do parasita. Essas seqüências resultaram em 31,000 contigs e a
estimativa da existência de aproximadamente 14,000 genes no genoma do parasita,
sendo que 92% destes possuem seqüências depositadas nos bancos públicos
(VERJOVSKI, et al., 2003). Esses dados também abriram novas perspectivas para o
entendimento dos mecanismos moleculares envolvidos com desenvolvimento,
evasão do sistema imune, organização tecidual, sinalização, dimorfismo sexual e
interação com o hospedeiro, além da identificação de novos genes que codificam
para proteínas candidatas a vacinas e potenciais alvos para novas drogas
(VERJOVSKI, et al, 2003).
A Figura 2, mostra os genes diferencialmente expressos durante o
desenvolvimento, bem como, genes cuja expressão esta relacionada com a
diferenciação sexual tanto em S.mansoni como S. japonicum.

Introdução
11
1.5. Vias de desenvolvimento celular
Estudos indicam que o desenvolvimento de qualquer organismo baseia-se no
uso de pequenos sinais moleculares provenientes de dezessete vias de transdução
de sinal (Tabela 1). Cinco dessas vias agem durante o estágio inicial de
desenvolvimento em metazoários, vertebrados, artrópodes e nematódeos. Outras
cinco vias estão agindo no estágio tardio de desenvolvimento, que corresponde a
formação dos órgãos e a diferenciação celular além das vias que atuam no estágio
inicial. Para a manutenção do funcionamento fisiológico do feto, jovens e adultos, o
qual é acompanhado por diferenciação celular, outras sete vias estão atuando
(GERHART, 1999).
Figura 2: Resumo de alguns genes expressos o durante os estágios de desenvolvimento dos parasitas S. mansoni e S.japonicum. Os dados foram baseados dos transcriptomas de S.mansoni e S. japonicum. Adaptado de- Schistosome transcriptomes: new insights into the parasite and schistosomiasis. Wei HU, et al. TRENDS in Molecular Medicine, v. 10, p. 3491-3496, 2004.

Introdução
12
Vias de transdução de sinal atuantes durante o desenvolvimento
Estágio inicial e tardio do desenvolvimento
1- Via Wingless (Wg)
2- Via Hedgehog (Hh)
3- Via de Fator de Crescimento e Transformação β (TGF-β)
4- Receptor de Tirosina Quinase (RTK)
5- Via Notch
Estágio intermediário e tardio do desenvolvimento
6- Receptor de Citocina
7- Receptor de Hormônio Nuclear
8- Via NF-kB IL1/Toll
9- Via apoptótica
10- Via do Receptor Fosfotirosina Fosfatase
Manutenção do Funcionamento Fisiológico na Fase Larval/Adulto
11- Via do Receptor de Guanilato ciclase
12- Via do Receptor de Oxido Nítrico
13- Via do Receptor Acoplado a Proteína G
14- Via da Integrina
15- Via da Caderina
16- Vias de Junções do Tipo Gap
17- Via de Canal de cátion dependente de ligante
Tabela 1- Dezessete vias de sinalização intracelular que atuam durante o desenvolvimento. As cinco primeiras vias agem durante o estágio inicial do desenvolvimento. Durante o estágio intermediário e tardio do desenvolvimento além das cinco vias que agem durante o estágio inicial, outros cincos estão atuando. As outras sete vias atuariam na manutenção fisiológica nas fases larvais e adultos. Adaptado de- 1998 Warkany Lecture: Signaling Pathways in Development. Gerhart, J., Teratology,. v. 60 p. 226-239, 1999.

Introdução
13
A maioria das vias de transdução de sinal envolve um receptor
transmembrânico que interage a um ligante específico. Essa ligação faz com que o
receptor sofra uma mudança conformacional em seu domínio intracelular,
desencadeando posteriormente sucessivas modificações de proteínas
intermediárias. A maioria dessas proteínas intermediárias são proteínas quinases, e
a ativação ou inativação dessas proteínas são dadas por reações em cascata de
fosforilação/desfosforilação (Figura 3). Ao final, a proteína quinase ativada fosforila
uma proteína alvo, levando a ativação ou a inibição de um processo celular
particular.
Figura 3- Diagrama esquemático das vias de transdução de sinal mediadas por receptores transmembrânicos. A interação de ligantes na porção extracelular do receptor induz uma mudança conformacional no domínio intracelular. Essa mudança desencadeia alterações nas proteínas quinases através de reações de fosforilação/desfosforilação, resultando na ativação ou inativação de proteínas alvo. Adaptado de- 1998 Warkany Lecture: Signaling Pathways in Development. Gerhart, J., Teratology, v. 60 p. 226-239, 1999.

Introdução
14
Em S.mansoni as vias envolvidas com o desenvolvimento do parasita, melhores
caracterizadas até o momento são as seguintes: Fator de Crescimento e
Transformação β (TGF-β), via mediada por Receptor de Tirosina Quinase (RTK) e a
via mediada por Receptores de Hormônios Nucleares (BELL, et al, 2000;
SHOEMARKER, et al, 1992: VICOGNE, et al, 2003; MENONÇA, et al, 2000).
Com relação à via TGF-β, ancorado no tegumento do parasita foi identificado o
Receptor Quinase tipo I. Esse receptor denominado de SmRK1 contém um motivo
de glicina-serina conservado, com 58% de similaridade com domínio quinases de
ortólogos de receptores tipo I (DAVIES, et al, 1998). Além do receptor, foi identificado
duas R-Smads (SmSmad1 e SmSmad2), as quais mostram interagir com o SmRK1
in vitro (OSMAN, et al, 2001). Outras evidências demonstraram que o SMRK1 é
capaz de sinalizar via SmSmad2 em resposta ao TGF β (BEALL., et al, 2000).
Estudos relacionados com a via RTK em S mansoni identificaram três
receptores com essa atividade: um Receptor Fator de Crescimento Epidermal (EGF)
presente no músculo de parasitas adultos, um Receptor de Insulina e um terceiro
receptor não usual pertencente a uma nova classe de Receptor de Tirosina Quinase,
sendo que esses dois últimos estão presentes em esporocisto e ovocistos
(SHOEMARKER, et al, 1992; VICOGNE, et al, 2003).
Finalmente, dentre os receptores caracterizados em S. mansoni relacionados
com a transdução de sinal mediada por Receptor Nuclear estão os receptores para o
hormônio da tireóide, esteróides e para ácido retinóico, além de muitos receptores
órfãos, cujos ligantes ainda permanecem por ser identificados (MENDONÇA, et al,
2000).

Introdução
15
1.5.1 Via de sinalização mediada pelo receptor Notch
A via de sinalização Notch é evolutivamente conservada, estando presente em
todos metazoários (ARTAVANIS-TSAKONAS, et al, 1999). O nome Notch é derivado
de mutações em Drosophila melanogaster que afetam o crescimento da asa e causa
"notches" (rachaduras) nas mesmas. Posteriormente, o gene Notch ficou conhecido
como neurogênico, devido a sua capacidade de inibir o desenvolvimento neural em
D. melanogaster (POULSON, 1950).
Em vertebrados, por exemplo, esta via de sinalização esta envolvida com o
controle da diferenciação pancreática (MURTAUGH, et al., 2003), com a regulação
da hematopoiese (VIRAG, et al, 2005), no desenvolvimento e manutenção vascular
(KARSAN, 2005). Além disso, o comprometimento da função de qualquer um dos
componentes centrais desta via, tem sido associado com a formação de tumores
sólidos e linfáticos (EGAN, et al,1998 ; GRIDLEY, 1997). Em D. melanogaster, dentre
outros processos, esta via esta envolvida com o desenvolvido do sistema nervoso
central e periférico, da asa, do olho, na oogênese (PORTIN, 2002). Em C. elegans,
por exemplo, tanto a especificação inicial do blastômero como a indução da
assimetria direita-esquerda e indução da proliferação de células germinativa, são
reguladas pela via Notch (GREENWALD, 1998).
A Tabela 2 resume os componentes centrais da via Notch já caracterizados em
várias espécies. Nesse trabalho, a nomenclatura utilizada é a descrita para D.
melanogaster.

Introdução
16
Componentes Centrais
C.elegans D.melanogaster Mamíferos
Ligante LAG-2, APX-1, ARG-2, F16B12.2
Delta, Serrate Delta-like 1, Delta like-2,
Delta-like-3, Jagged 1, Jagged 2
Receptor LIN-12, GLP-1
Notch Notch1, Notch 2, Notch 3, Notch 4
Fator transcricional
(CSL)
LAG-1 Supressor de Hairless[Su(H)]
CBF1/RBJκ RBPL
Dados experimentais têm mostrado que a ativação de Notch envolve clivagens
proteolíticas em três sítios que são denominados de S1, S2 e S3, como demonstrado
na Figura 4 (MUMM, et al, 2000; BARON, et al, 2002). A clivagem S1 ocorre por
uma protease presente no complexo de Golgi denominada Convertase Furina Símile
(LOGEAT, et al, 1998). Essa clivagem tem como função promover o processo de
maturação do receptor e favorecer a montagem do heterodímero que será enviado
para a membrana plasmática. O ataque proteolítico, entretanto, não resulta em
dissociação e conseqüente separação das cadeias, mas permite a associação iônica
entre elas através da coordenação com átomos de Ca2+. O transporte do receptor
para a membrana culmina com a inserção do mesmo, orientado com sua porção C-
terminal para o meio intracelular, e a porção N-terminal, voltada para o meio
extracelular (RAND, et al, 2000).
A clivagem S2 é catalisada pela protease Desintegrina em resposta a interação
dos ligantes Delta ou Serrate com o receptor Notch; essa clivagem libera a maioria
do domínio extracelular do receptor, deixando somente o fragmento denominado
NEXT (domínio extracelular de notch truncado), o qual é o alvo para a clivagem S3.
Tabela 2- Componentes centrais da via Notch de sinalização em diferentes espécies. Adaptado de- Notch signaling: control of cell communication and cell fate .Eric, C. Lai. Development, v. 131, p. 965-973, 2004.

Introdução
17
A clivagem S3 por sua vez, irá liberar o NICD (domínio intracelular de Notch), o
qual é translocado para o núcleo para exercer sua função. Esta última clivagem é
realizada por um complexo γ - secretase, sendo a Presenilina, a componente chave
desse complexo proteolítico. Além disso, outra protease que faz parte desse
complexo é a Nicastrina. Estudos demonstram que essa proteína pode ser
imunopurificada do complexo γ - secretase juntamente com Presenilina (YU, et al,
2000). Aparentemente a função molecular da Nicastrina é regular a estabilidade da
Presenilina (CHUNG & STRUHL, 2001; HU, 2002). A Figura 5 ilustra a ativação de
Notch.
No núcleo, o alvo molecular do NICD é um complexo protéico denominado CSL
(CBF1, Su(H), Lag-1) que ligado ao DNA tem a propriedade de atuar como um
sistema repressor da transcrição gênica (na ausência de ativação da via), ou como
um ativador de genes responsivos, após a interação com NICD.
O estado de repressão é garantido através da ligação de vários elementos
repressores a CSL, como por exemplo, as histonas deacetilases e os SMRT
(Silencing Mediator of Retinoid and Thyroid hormone receptors). Entretanto, a
ativação do receptor intramembrana e conseqüente liberação para o núcleo de NICD
induzem que o mesmo recrute complexos desrepressores da transcrição gênica,
como as histonas acetilases. Em seguida, NICD liga-se ao complexo CSL
desestabilizando o estado de repressão. Finalmente, por meio do recrutamento de
complexos ativadores adicionais como Lag-3/ Mastermind, o conjunto CSL, NICD e
proteínas acessórias atuam como ativadores da transcrição gênica, dando início às
mudanças celulares associadas com a sinalização através da via Notch (MUMM &
KOPAM, 2000; WU, et al, 2002).

Introdução
18
ECN
Figura 4- Cascata proteolítica da ativação de Notch. Sítios de clivagens em Notch 1 de camundongo. A localização dos sítios de clivagem pode não ser conservada entre as espécies. Após a ligação do receptor com o ligante, o receptor sofre uma mudança conformacional expondo o sítio para a segunda clivagem (S2). O produto resultante é o NEXT e, esse será o substrato para a clivagem 3. Nessa última clivagem será formado o NICD e esse, sendo translocado para o núcleo, ativa o fator transcricional Supressor de Hairless. Abreviações: EGF, Fator de Crescimento Epidermal; LNR, domínio de repetições Lin-12/Notch; domínio RAM, RAM 23; nls, sinal de localização nuclear; ANK domínio de repetições de ankirin; PEST região rica em prolina, glutamina e serina. ECN, domínio extracelular de Notch; TMIC, domínio transmembrânico e Intracelular de Notch; NEXT, domínio extracelular de Notch truncado; NICD, domínio intracelular de Notch. Adaptado de: Notch Signaling: From the Outside In. Mumm, J.S & Kopam.R. Developmental Biology, v.228, p. 151-165, 2000.

Introdução
19
A via Notch é regulada por uma variedade de mecanismos moleculares, porém,
estudos têm demonstrado uma grande importância da via de ubiquitinação no
controle dessa sinalização. A via de modificação pós-traducional dependente de
ubiquitina inicia-se quando a ubiquitina é ativada pela enzima ativadora de ubiquitina
(E1) e posteriormente, transferida para a enzima conjugadora de ubiquitina (E2). A
enzima ubiquitina ligase (E3) confere a especificidade do substrato alvo, e se
associa com E2 e substrato para catalisar a transferência da ubiquitina para o
substrato (ERIC, 2003). Até o momento, quatro isoformas de E3 ligases parece
estarem diretamente envolvidas com a regulação dos níveis intracelulares do
receptor Notch e de outros componentes dessa via. O Supressor de Deltex promove
a ubiquitinação do domínio intracelular do Notch na membrana plasmática
(FOSTIER, et. al, 1998); Sel-10, no núcleo, conjuga ubiquitina no domínio
intracelular do Notch (WU, et al, 2001); "Neuralized", esta envolvida com a
ubiquitinação do domínio intracelular do ligante delta (DI) (YEH, et al, 2001; ERIC, et
al, 2001) e finalmente, a ubiquitina E3 ligase ligante de Numb (LNX) que medeia a
ubiquitinação de Numb (NIE, et al., 2002). Esta proteína é assimetricamente
localizada em células progenitoras em divisão, segregando preferencialmente para
uma das células filhas (SPANA & DOE, 1996; UEMURA, et al., 1989). Estudos
indicam que Numb regula negativamente a via de sinalização Notch, e desse modo,
é proposto que LNX pode aumentar a sinalização de Notch através a diminuição de
Numb.
Como qualquer via de transdução de sinal que finalmente resulta em
transcrição gênica, uma célula que tenha recebido um sinal da célula vizinha que
possui o ligante e o internaliza através do receptor Notch sofrerá profundas

Introdução
20
alterações em seu conteúdo protéico. Conseqüentemente, alterações metabólicas e
estruturais resultarão em uma célula diferenciada da população de células vizinhas.
Os mecanismos moleculares primordiais que definem o momento e o número de
células que devem se diferenciar em uma população, ainda são pouco
compreendidos, entretanto, parecem depender da distribuição diferencial e
quantitativa entre ligantes de superfície e receptores presentes em diferentes células
embrionárias (KIMBLE & SIMPSON, 1997).
Figura 5- Diagrama esquemático da via Notch. (a) O receptor Notch é sintetizado como uma proteína precursora de 300 kDA. (b) Através da ação da protease "Furin Convertase" o receptor é clivado no compartimento trans Golgi e o heterodímero maturo, liberado para a superfície celular (c) O receptor maturo presente na superfície celular, (d) sofre uma clivagem mediada por TACE após a sua ligação com o ligante. Essa clivagem libera o domínio extracelular do receptor. O fragmento que fica na membrana, denominado NEXT é clivado posteriormente pela Presenilina, (e) liberando desse modo, o domínio intracelular o qual transloca para o núcleo (f) e ativa a proteína CSL que induz a transcrição de genes alvo. Abreviaturas: NEXT, domínio extracelular truncado. Adaptado de- Notch and Presenilin: a proteolytic mechanism emerges. Fortini, M.A. Current Opinion in Cell Biology, v. 13, p.627-634, 2001.
indução de genes alvos

Introdução
21
Deste modo, o rastreamento molecular das etapas pelas quais S. mansoni sofre
um processo de desenvolvimento significa indiretamente, entender a fonte e a
natureza química dos estímulos recebidos de seus hospedeiros ou do ambiente
aquático. Neste sentido, este estudo poderá contribuir para a elucidação de alguns
mecanismos moleculares que podem permitir a complexa adaptação de S. mansoni
ao homem, à água, e ao caramujo, resultando no conseqüente sucesso evolutivo
deste parasita.

2-Objetivos

Objetivos
23
2.1. Objetivo geral
O objetivo geral deste trabalho foi à reconstituição in silico dos componentes da
via Notch e a identificação molecular da expressão dos fragmentos gênicos de
Notch, Presenilina e Supressor de Deltex que compõe a via de sinalização mediada
pelo receptor Notch durante o ciclo de via do parasita S. mansoni. Para isso foram
abordados os seguintes objetivos específicos:
1. Análise in silico dos componentes da via Notch utilizando dados do banco
originado pelo Projeto Transcriptoma de S. mansoni.
2. Comparação in silico da via Notch em S. mansoni e S. japonicum.
3. Obtenção de seqüências genômica.
4. Análise do perfil de expressão, por RT-PCR nas fases de vermes adultos,
ovos, esporocistos, cercárias e esquistossômulos.
5. Determinar a organização genômica evidenciando exóns e íntron.
6. Determinar o tamanho dos transcritos de Notch e Presenilina.

3- Materiais e Métodos

Materiais e Métodos
25
3.1. Análise in silico da via de desenvolvimento Notch
Os componentes da via de desenvolvimento Notch, que foram descritos para D.
melanogaster e C. elegans (ARTAVANIS-TSAKONAS, et al, 1999; GREENWALD,
1998) foram analisados nos 30,988 contigs gerados pelo Projeto Transcriptoma de
S.mansoni a partir do alinhamento de ORESTES, obtidos dos seis estágios de vida
do parasita (VERJOVISK et al, 2003), disponível no endereço
http://verjo18.iq.usp.br/schisto/. A seguir, as seqüências recuperadas foram
analisadas com auxílio do algoritmo BLASTp, disponível em:
http://www.ncbi.nlm.nih.gov/BLAST/. As seqüências foram consideradas confiáveis
quando apresentavam um e value < e –5.
As seqüências referentes aos clusters de Notch (608600.1) e Presenilina
(705353.1) foram extendidas com o auxílio do banco de dados da TIGR (The Institute
for Genomic Research), disponível no endereço http://www.tigr.org/tdb/e2K1/sma1/
(Tabela 3). Para certificarmos que as seqüências recuperadas relacionavam com os
contigs mencionados acima, as seqüências foram submetidas ao BLAST2 presente
no endereço http://www.ncbi.nlm.nih.gov/BLAST/ e ao programa BLASTx presente
no banco de dados do Projeto Transcriptoma de S. mansoni.
Cluster Tamanho (nt) ORF Tamanho após extensão (nt) ORF 608600.1 763 nt 231 aa 984 nt 327 aa 705353.1 427 nt 135 aa 486 nt 161 aa
Tabela 3- Tamanho das seqüências dos clusters 608600.1 e 705353.1 após a extensão. Na extensão, foram utilizadas seqüências do banco de dados genômico da TIGR (http://www.tigr.org/tdb/e2K1/sma1/)

Materiais e Métodos
26
3.1.1. Comparação da via Notch entre S. mansoni e S. japonicum
As seqüências do banco de dados de S. mansoni que deram similaridade com
os componentes da via Notch, foram comparados com os 13.131 genes gerados
pelo Transcriptoma de Schistosoma japonicum (WEI, et al, 2004). As análises foram
realizadas pelo programa BLASTn(base de dados dsEST) presente no endereço
http://www.ncbi.nlm.nih.gov/BLAST/. Os alinhamentos foram considerado confiável
quando apresentavam um e value < que e–5.
3.2. Manutenção do ciclo biológico de S. mansoni
O ciclo biológico de S.mansoni, linhagem LE é rotineiramente mantido no
Laboratório de Biologia Molecular de Parasitas, do Departamento de Bioquímica e
Imunologia da Faculdade de Medicina de Ribeirão Preto - USP.
Os ovos de S.mansoni presentes nas fezes de camundongos das linhagens
Swiss ou Balb/C previamente infectados com o parasita são recolhidos pelo método
de Hoffmann e expostos por aproximadamente 1 hora sob luz, para a liberação dos
miracídios. Os miracídios foram utilizados para infectar o hospedeiro intermediário,
os caramujos da espécie B. glabrata, que após 38 a 43 dias liberaram a forma
infectante do parasita, as cercárias, que por sua vez infectarão o hospedeiro
vertebrado. As cercárias são inoculadas nos camundongos via subcutânea e após
aproximadamente 45 dias os vermes adultos são recuperados do sistema porta-
hepático por perfusão (SMITHERS & TERRY, 1965).

Materiais e Métodos
27
Após a coleta, os parasitas foram mantidos em meio RPMI 1640 (Invitrogen) ou
em solução de NaCl a 0.9% e mantidos em gelo. Para a obtenção dos vermes
(macho ou fêmea), estes foram mecanicamente separados com o auxilio de um
pincel, sendo em seguida congelados e estocados à -70ºC, até o momento do uso.
3.3. Obtenção de ovos
Para a obtenção dos ovos, aproximadamente dez fígados de camundongos
infectados foram coletados durante perfusão e homogeneizados em 200 mL de
solução pH 8,3 contendo 1.79 g de Na2HPO4, 0.09 g de KH2PO4, e 20 mg de tripsina
(Invitrogen). O homogeneizado foi incubado em banho-maria a 37ºC durante 3 horas
e posteriormente, os ovos foram recuperados por tamização em malhas de 300 e
180 µm em solução de NaCl a 0.9%. (Castro-Borges, 2005).
3.4. Transformação mecânica de cercárias em esquistossômulos
Inicialmente 120.000 cercárias ficaram em repouso por 90 minutos dentro de
um béquer no gelo. Posteriormente, o sobrenadante foi aspirado e descartado
cuidadosamente com o auxílio de uma pipeta. As cercárias presentes no fundo do
béquer foram transferidas para tubos tipo falcon de 15 mL e submetidas a um pulso
de centrifugação cuja velocidade variou de 10.000 a 13.000 rpm. O sobrenadante foi
descartado e as cercárias foram ressuspendidas em 10 mL de água declorada
previamente filtrada em membranas com poros de 0.2 µm. O processo de
centrifugação e descarte do sobrenadante foi repetido por duas vezes.

Materiais e Métodos
28
Para a transformação mecânica das cercárias em esquistossômulos, as
cercárias presentes em tubo tipo falcon contendo 6 mL de meio RPMI 1640
(Invitrogen) foram agitadas em vortex sob velocidade máxima, durante 90 segundos
para a indução do rompimento da cercaria em corpo cercariano/cauda.
Em condições estéreis, os corpos e caudas cercarianas foram ressuspendidos
e transferidos para um frasco de cultura de 40 mL contendo 30 mL do meio RPMI
1640 suplementado com 100mg/mL de estreptomicina e 100mg/mL de ampicilina e
incubados em estufa a 37ºC/CO2 5%, durante um período de 4 horas.
Após o período de incubação, aproximadamente 28mL do sobrenadante foi
aspirado e descartado; cerca de 2 mL restantes foram distribuídos em dois tubos tipo
eppendorf de 2 mL. Posteriormente, para remoção das caudas, 1mL de meio RPMI
1640 foi adicionado em cada eppendorf e após 4 minutos, aproximadamente 1 mL
do sobrenadante foram descartados. Esse procedimento foi repetido de 3 a 5 vezes.
Após a remoção das caudas, os corpos cercarianos foram armazenados a –70 ºC
até o momento do uso (HARROP & WILSON, 1993).
3.5. Extração de RNA total
De posse do material biológico necessário (vermes adultos, ovos, cercárias,
esquistossômulos, hepatopâncreas de caramujos infectados (com 40 dias de
infecção como fonte de esporocistos) e hepatopâncreas de caramujos não
infectados), procedemos a extração do RNA total, como brevemente descrito a
seguir:

Materiais e Métodos
29
Cerca de 100mg de vermes adultos foram homogeneizados em 1mL de
TRIZOL LS (Invitrogen) com auxílio de um homogeneizador de tecidos (politron), até
completa solubilização. Em seguida, esse extrato foi incubado por 15 minutos a
temperatura ambiente.
Decorrido este intervalo de tempo, adicionamos a esse extrato, 200 µL de
clorofórmio, e as amostras agitadas em vortex vigorosamente por 1 minuto e
incubadas a temperatura ambiente durante 15 minutos, sendo posteriormente
centrifugadas a 10000 x g por 10 minutos a 4 °C.
Após esta etapa de centrifugação, a mistura separa-se em uma fase inferior de
cor vermelha (fase fenol-clorofórmio), interfase e fase aquosa superior incolor. O
DNA fica na interfase e as proteínas na fase orgânica, enquanto o RNA permanece
exclusivamente na fase aquosa. A fase aquosa foi transferida para um tubo estéril
(tipo eppendorf) e o RNA foi precipitado pela adição de 500 µL de isopropanol. Após
30 minutos de incubação à temperatura ambiente, o RNA foi então recuperado por
centrifugação a 10000 x g por 10 minutos a 4°C. O sobrenadante foi desprezado e o
precipitado lavado com 1,0 mL de etanol 70% em água previamente tratada com
dietilpirocarbonato (DEPC), sendo em seguida centrifugado a 10000 x g por 8
minutos a 4°C. O precipitado final foi seco a vácuo e ressuspendido em 30 µL de
água tratada com DEPC.
Para a extração do RNA total das formas larvais de S. mansoni foram utilizados
aproximadamente 120.000 cercárias, 80.000 esquistossômulos e 5.000 ovos. A
Figura 6 mostra um gel típico do procedimento de extração que foi conduzido como
descrito acima.

Materiais e Métodos
30
3.6. Extração do DNA genômico
Os parasitas adultos mantidos a –70 °C foram transferidos para um grau de
porcelana juntamente com gotas de nitrogênio líquido, com a finalidade de facilitar a
pulverização dos vermes. Em seguida, procedeu-se a extração do DNA genômico,
conforme o seguinte protocolo simplificado por COSTA PI (Tese de Doutorado,
1997).
Para cada 100 mg de vermes pulverizados, foram adicionados 250 µL do
tampão de lise (Tris 0,05 mM, pH 7,5, EDTA 1 mM, 1% de N-laurilsarcosina e 100
µg/mL de proteinase K) e mantidos a 37 °C durante 1 hora. A seguir, foi
acrescentado 100µL de NaCl 5M e a mistura incubada por 10 minutos a 65°C.
Posteriormente, foi adicionado 50µL de uma solução de CTAB/NaCl a 10%, seguido
por uma incubação de 20 minutos a 65°C.
Figura 6 - Análise da integridade do RNA total. O RNA foi extraído pelo método de Trizol a partir de macho (M), fêmea (F), vermes adultos (VA), esporocisto (EP), caramujo não infectado (C-),ovos (O) , cercárias (CE) e esquistossômulos (ES). Cerca de 3 µg de RNA de cada preparação foram analisadas em gel de agarose/formaldeído a 1% e visualizados com a coloração de brometo de etideo.
M F VA EP C- O CE ES

Materiais e Métodos
31
O DNA foi extraído com clorofórmio (v/v) e posteriormente precipitado pela
adição (v/v) de isopropanol e incubado à -20°C, durante 30 minutos. Posteriormente
o DNA foi centrifugado a 12000 x g, lavado com etanol a 70%, seco, ressuspendido
em TE e armazenados a 4°C.
A Figura 7 mostra uma preparação de DNA genômico realizada como descrito
acima e analisada em gel de agarose.
3.7. Quantificação de DNA e RNA
As concentrações tanto de DNA como de RNA foram estimadas a partir da
medida de absorbância a 260nm. Uma unidade de absorbância a 260nm
corresponde aproximadamente a 50µg/mL de DNA (fita dupla) e 40µg/mL para RNA
e DNA de fita simples. O grau de pureza da preparação foi estimado através da
relação entre as leituras a 260 e 280nm. As preparações foram consideradas boas,
quando o valor da razão A=260/280 variava entre 1,8-2,2.
Figura 7- Análise da integridade do DNA genômico. O DNA genômico foi extraído pelo método do CTAB a partir de vermes adultos. Cerca de 50 ng de DNA genômico foi analisado em gel de agarose 0.7% e visualizados com coloração de brometo de etídio.

Materiais e Métodos
32
3.8. Síntese dos oligonucleotídeos iniciadores
Para a idealização dos oligonucleotídeos iniciadores, a seleção dos clusters,
correspondentes aos fragmentos gênicos de Notch, Presenilina e Supressor de
Deltex foram realizadas baseadas em dois critérios: 1º presença do domínio que
caracteriza a proteína e 2º tamanho da seqüência.
Desse modo os oligonucleotideos correspondentes para aos fragmentos
gênicos de Notch e Presenilina foram idealizados com base nas seqüências dos
clusters 608600.1 e 705353.1, que foram posteriormente extendidas com o auxílio
do banco de dados da TIGR e descrito em 3.1. Os oligonucleotídeos para Supressor
de Deltex foram idealizados com base na seqüência do cluster 610048.1. Os
oligonucleotídeos iniciadores para os fragmentos gênicos em estudo foram
idealizados utilizando o programa Generunr. A Tabela 4 mostra as seqüências dos
oligonucleotideos utilizadas com as respectivas temperaturas de anelamento e os
tamanhos dos produtos esperados. Os oligonucleotideos idealizados estão indicados
nas seqüências dos referentes clusters, mostradas na Figura 8.
Gene Número do
cluster
Seqüência dos oligonucleotideos Temperatura de
anelamento
Produto esperado
(pb) Notch 608600.1 5'-CATGTGACAAAGGGTATCACC-3’
3'-GACATACACATTCTCCAGTCG-5'
47º C 850
Presenilina 705353.1 5’-CCGAACCGATGCAATTGTAG -3' 3'-GAACACACGTTCAATGGCTG- 5'
47º C 400
Supressor de Deltex
610048.1 5’-TATGGGAGTCCCATTGCAAT-3’ 3’-AATCTCTGGCATACCGGATA- 5’
47º C 800
Tabela 4-Seqüências dos oligonucleotideos específicos referentes a Notch, Presenilina e Supressor de Deltex. Os oligonucleotídeos de Notch e Presenilina foram idealizados com base nas seqüências dos clusters 608600.1 e 705353.1 que foram posteriormente extendidos com o auxílio do banco de dados da TIGR. Nota-se que a temperatura de anelamento para os três oligonucleotideos é de 47ºC e os produtos esperados são de 850 pb para Notch, 400pb para Presenilina e 800 pb para Supressor de Deltex.

Materiais e Métodos
33
Figura 8- Seqüência consenso obtida durante o Projeto Transcriptoma de S. mansoni: A)
Receptor Notch (608600.1), (B) Presenilina (705353.1), C) Supressor de Deltex (610048.1). Em negrito e sublinhado estão os oligonucleotídeos direto e reverso.
C
1 ATGGATGAAACACAACCTCTCCCGCCTGGATGGGAGAAACGTTATACGGCCCAGGGACAA 60
61 AGGTTTTTCATTGATCATAATACCCATACTACTACCCTTGTTGATCCACGTACTGGACAA 120
121 CATGCTGGTTCACTAGGTAGTATGGGAGTCCCATTGCAATATGAAAGGAATTTCCGATCC 180
181 AAAGTGAATTATTTTCGTGCTTGTTGTACAAATGCCATGTTAAATGGGCAAACTAAACTT 240
241 CTTATATCAAGAGACAATTTATTAGAGGACTCGTTCCAACTGGTATCTCAAATGTCTTCA 300
301 TCTGGTTTAAGAAGACGTTTATCAATTACATTTCTACATGAAGAAGGTTTGGATTACGGT 360
361 GGAGTAGCCAGAGAATGGTTCTATCGTCTTTCAAGAGAAATTTTAAACCCAATGTTTGGC 420
421 TTGTTTGAGTATACTGGAACAGATTATGCCTTACAAGTAAATCCAGCCTCTCATGTTAAT 480
481 CCAAATCATATGGCGTATTTTCGATTTGTTGGACGCTTTATTGGTATGGCACTTTTTCAT 540
541 GGTCGCTGTATTGACGGCGGGCTTACTCTTGCTTTTTATAAACAAATTTTGAAGCGGAAA 600
601 TTAACATTAGAAGATCTTGGTCATACGGATCACAGTTATTATCAATCGTTAATTTATGTC 660
661 AGAGACAATCCAGTTGATGAGTGTGATTTGGACTTGTACTTTATTGGAACCTATGACCTT 720
721 TTAGGCGTTTTGCATGAAGACGAACTTATTGAAGGTGGCAAAGATATTAAAGTGACTGAA 780
781 GAAAATAAATCCGATTATATCAGACTGATGGTTGATTGGCGGTTCAACAGAGGAGTAAAA 840
841 AAACAAACAGAAGAAATTCACAAAGGTATTTTTGAAGTACTAAATCCCGAATGGCTGGAA 900
901 TTATTCGATGAGCGTGAACTTGAACTCCTATTATCCGGTATGCCAGAGATTGATGTTGAT 960
961 GATTGGGAAAAGAACACGGTCTACATAAAGTACACACGATCAAGTAAACAGATTATTTGG 1020
1021 TTTTGGAAGTTGATTCAAAAACTTGATAATGAACATCGTGCTCGTCTTCTTCAATTCGTC 1080
1081 ACTGGTACCTGTCATTTACCATTAGGAGGCTTCTCTGAATTGACTGGCAGCGGTGGTCGT 1140
1141 CAATTATTTTGTATTGAGCGTACAGGTGATGAACAATGGCTTCCACGTAGTCACACATGT 1200
1201 TTCAACCGGTTAGATTTACCGCCATACAATTCGTACGATCAATTGTGCGAAAAACTGCAA 1260
1261 ATGGCAATTGAAGAAACTAAGGGATTTGGACAAGAATAA 1299
A
1 ATGGGTCATATTATTGTACATGTGACAAAGGGTATCACCAAGATTACAGAGGTCACTAGT 60
61 CTTCCAAACAAATGTAGTCCTGAATGTGTTCATGGACAAGGCGAATGTACATCTAATGGA 120
121 AAATGCTTGTGTAATCCAGGTTTTCGAGGGTTATCTTGTGAGACAGACGTAGATGAATGC 180
181 GCTGAAGGAAAACATAATTGTCAACAGGAATGTATCAATACTCCTGGGTCTTTTAAATGT 240
241 TCTTGTCATTCAGGTTACAAATTATCCACTGTAGATCCATCAAAATGTGAACCTATTTCT 300
301 TGTCCTTCTCAATGCAATTTAATAGATGGAGAATGTGACTGCGAATGCAAGCTTGGACAT 360
361 TTTGGACTTAATTGTAATCAAACAGTAGATTCATGTGCAATTAAACCATGTGACCAAATT 420
421 TGTGTGGATTTAGGTGATGGGAAGTATACATGCAAATGTAATGAAGGTTTTGAACCAAGT 480
481 CCAAATAATCCTAGACGATGTCAGCGTATATGTAAAGATGGTATAGACTGTATATATGGA 540
541 AGCTGTCGAACTGTGTTTTCAAATGACACAGATCAATCTTTATGTACTTGCCTAGAAGGA 600
601 TTTTACGGAACAAACTGTCAAAACGATGTAAACGAATGCTTGAATGAGAATGGTCAAAAA 660
661 CATTTTTGTGAACATCATTGCATAAATACATTAGGATCATACCAATGTTTTTGTGATCCG 720
721 GATTATGAATTACAATCTGATGGTCGAACGTGTAAAAAACAGTTTAAAGGTGAAATGAAA 780
781 TGTCCTGAATACTGCCTAAATGGTGCAACTTGTAAATCGACTGGAGAATGTGTATGTCAA 840
841 CCTGGTTATGAAGGAATATATTGTGAAAAAATAAAAGATATTTGTGCAATACTTAAGTTA 900
901 TGTGATCAACAATGTTTTCCTAAAGAAGTAAGTCGACTACATTATTTAATAACAGACAAC 960
961 ATTTGTGATACTGTACTTAAGTAG 984
B 1 ATGACAGGGAATTCAAGAAACATGTTAAATATTCCTAATAATACGTCCCGAACCGATGCA 60
61 ATTGTAGTTGATACTCCTGATGACAATACGATTAATAATGATGCCAGGAATAACAATGAA 120
121 ACTATATCACAGGCACCTCATGAAACAGATACACTTGTAGCAGGCCGACCCAATCGTTGG 180
181 CAAGAACTGCAAGCAGATTTAATTGATCGTGACTCAAAGGGTGTAAAACTTGGTTTAGGC 240
241 GATTTTGTATTTTATAGTTTACTGATTGGTCGTGCAACACTTGATGGCGATGCAGTCACT 300
301 GTGGTCACATGTTATGTGGCTATATTGGTTGGAATGTGTATTACTGTTATAGTATTGGGA 360
361 ATTACACGACAAGCATTACCAGCTTTACCAATATCCATAACATGTGGTATATTATTTTAT 420
421 TTTGTCAGTTCAGCGACGATATCACCGTTTCTTGAAGCAACAGCCATTGAACGTGTGTTC 480
481 TTCTGA 486

Materiais e Métodos
34
3.9. RT-PCR (Transcrição reversa e reação em cadeia da polimerase)
Para a obtenção dos cDNAs codificando para Notch, Presenilina e Supressor
de Deltex em S. mansoni e para analisar a presença desses transcritos nas várias
fases evolutivas do parasita, utilizamos a técnica de RT- PCR.
Nesse procedimento, inicialmente foi sintetizada a fita simples de cDNA,
utilizando o kit ThermoScript RT-PCR System (Invitrogen) e como iniciador oligo
(dT), como resumidamente descrito abaixo.
Foram utilizados 3µg de RNA total obtido a partir de parasitas e suas formas
intermediárias, 50pmol do iniciador oligo (dT) e água livre de RNAse, para um
volume final de 10µL. Esta mistura foi incubada por 5 minutos a 65ºC, sendo em
seguida adicionado dNTPs, tampão da enzima e enzima transcriptase reversa.
Posteriormente a amostra foi transferida para um termociclador, sendo incubada por
60 minutos a 50ºC. Após este intervalo de tempo, a reação foi interrompida,
incubando-se as amostras por 5 minutos a 85ºC. Em seguida foi adicionado 1,0 µL
de RNAse H e a amostra foi incubada durante 20 minutos a 37ºC e posteriormente
estocada a -20ºC.
Para a amplificação do cDNA de fita dupla e do DNA genômico, foi adotado o
seguinte procedimento. Em um tubo eppendorf de 200µL foi adicionada a mistura de
reação para um volume de 50µL. Os componentes desta mistura são: água (RNAse
livre), tampão para Taq DNA polimerase, MgCl2, iniciadores gene específicos direto e
reverso, dNTPs, cDNA ou DNA genômico e a enzima Taq DNA polimerase Platinum
(Invitrogen). Como controles negativos das reações de PCR, foram utilizados todos
os componentes descrito acima, exceto o cDNA ou DNA genômico.

Materiais e Métodos
35
A reação de amplificação de Notch, Presenilina e Supressor de Deltex
consistiram de uma etapa de desnaturação a 95ºC durante 3 minutos, seguido de 40
ciclos, de acordo com seguinte programa: desnaturação a 95º C durante 1 minuto; 1
minuto de anelamento a 47ºC e extensão a 72ºC durante 1:3 minutos. Ao término
dos 40 ciclos a reação foi mantida por um tempo adicional de 10 minutos a 72 ºC e
posteriormente a 4ºC.
Em seguida, 10µL desta reação foram analisadas em gel de agarose 1,5% e
visualizados com a coloração com brometo de etídeo (0,5 µg/mL).
3.10. Purificação do produto de PCR
Após a análise do PCR no gel de agarose, as bandas de interesse (850pb para
Notch, 400pb para Presenilina e 800pb e 850pb para Supressor de Deltex), foram
removidas do gel com o auxílio de um estilete. A purificação das bandas foram
realizadas utilizando o Kit In Concert Rapid Gel Extraction System (Invitrogen)
conforme o protocolo simplificado.
As bandas removidas foram transferidas para um tubo eppendorf de 2 mL. Em
cada eppendorf foram adicionados 500 µl do tampão de solubilização, e incubado
por 15 minutos a 50ºC. Posteriormente, o conteúdo do eppendorf foi transferido para
uma coluna de purificação e centrifugado a 12000 X g por 1 minuto. O eluído foi
descartado e após adicionar 500 µL da solução de eluição, a amostra ficou em
repouso durante 1 minuto e depois novamente centrifugado a 12000 X g por 1
minuto. Após esse procedimento o eluído foi descartado e adicionado 700 µL de
tampão de lavagem, novamente repouso por 1 minuto e depois centrifugado a

Materiais e Métodos
36
12000X g por 1 minuto. Por fim, cada coluna foi transferida para um tubo eppendorf e
adicionado aproximadamente 30 µl TE (TRIS-EDTA) previamente aquecido no
centro da coluna, e centrifugado 12000 x g. A eficiência da purificação foi analisada
em gel de agarose 1,5% e visualizados com a coloração com brometo de etídeo (0,5
µg/mL.
3.11. Clonagem do produto de PCR
Os produtos de PCR purificados foram posteriormente clonados no vetor
pGEM- T Easy (Promega), utilizando o kit conforme instruções do fabricante.
Cerca de 5,0 µL das reações de ligação foram utilizados nos procedimentos de
transformação. Nestes experimentos foram utilizadas células competentes
preparadas pelo método descrito por HANAHAN (1985).
O plasmídio foi introduzido em bactéria competente E. coli da linhagem DH5α,
através de choque térmico (90 segundos a 42°C; 3 minutos em banho de
gelo),crescimento sob agitação em meio SOC (SOB enriquecido com glicose) a 200
RPM e 37°C, por 1h e 30 min. Decorrido este intervalo, a suspensão bacteriana foi
plaqueada em meio LB/ampicilina contendo X-GAL(20µL/mL), para que ocorra a
seleção dos plasmídeos com inserto (colônias brancas) e sem inserto (colônias
azuis) e mantidas em estufa durante uma noite a 37°C.

Materiais e Métodos
37
3.12. Minipreparação de DNA plasmidial
Após a realização da clonagem no vetor pGEM-T Easy (Promega),
selecionamos dez colônias que poderiam conter inserto (seleção azul/branca) e
posteriormente, foram crescidas em LB/ampicilina durante uma noite e submetidas à
amplificação específica como descrito a seguir.
Os componentes da mistura de reação foram: água (livre RNAse), tampão para
enzima Taq DNA polimerase, MgCl2, oligonucleotideos direto e reverso, dNTPs, 1,0
µL da cultura bacteriana e enzima Taq DNA polimerase. Como iniciadores foi
utilizado o par de oligonucleotídeos M13 Universal (Invitrogen). A reação de
amplificação consistiu de 40 ciclos, segundo o programa: desnaturação a 95ºC
durante 45 segundos; 45 segundos de anelamento a 55ºC e extensão a 72ºC
durante 1 minuto. Ao término dos 40 ciclos a reação foi incubada por um tempo
adicional de 5 minutos a 72ºC e mantida posteriormente a 4ºC. Cerca de 10µL dessa
preparação foram analisados em gel de agarose 1,5%e visualizados com a
coloração com brometo de etídeo (0,5 µg/mL)
Após identificarmos quais colônias continham os insertos com os pesos
moleculares esperado, ou seja, em torno de 850 pb, 400 pb e 800pb, procedemos à
extração do DNA plasmidial segundo o método brevemente descrito.
Cerca de 1,5 mL da cultura bacteriana crescida em LB/ampicilina, durante uma
noite foram transferidos para tubos "eppendorf" e o precipitado bacteriano, foi obtido
por centrifugação a 10.000 x g.
Em seguida, este precipitado foi solubilizado em 300 µL de solução contendo
Tris-HCl 500 mM; EDTA 10mM e 100 µg de Rnase A; pH em 8,0. Após 15 minutos

Materiais e Métodos
38
de incubação à temperatura ambiente foram adicionados, 300 µL de solução
contendo NaOH 200mM e SDS 1%, e incubou-se 5 minutos à temperatura ambiente.
Logo após, adicionou-se 300 µL da solução de acetato de potássio 3.0 M pH 5,5 e
centrifugou-se durante 10 minutos a 14.000g. O sobrenadante foi transferido para um
novo tubo e em seguida, adicionados 400·µL de isopropanol. Posteriormente, as
amostras foram centrifugadas durante 10 minutos a 14.000 x g à temperatura
ambiente. Decorrido este intervalo de tempo, foi descartado o sobrenadante e o
precipitado resultante foi lavado com 700 µL de etanol 70%. As amostras foram
novamente centrifugadas a 4oC por 5 minutos.
Por fim, descartou-se o etanol e o precipitado seco foi ressuspendido em 23 µL
de tampão TE (EDTA 1.0 mM; Tris 10mM; pH em7,5). Alternativamente, para se
obter uma preparação de plasmídeos com elevado grau de pureza, utilizamos o kit
Concert Rapid Plasmid System (Invitrogen), conforme instruções do fabricante.
3.13. Sequenciamento
Para o sequenciamento dos DNAs genômico e do cDNAs, utilizamos a técnica
do término do crescimento da cadeia, inicialmente desenvolvida por SANGER et al,
1997. As reações de sequenciamento foram realizadas utilizando o kit Big-Dye
Terminator (Applied Biosystems), de acordo com as instruções do fabricante e as
reações, analisadas no sequenciador automático de DNA, ABI 3100 Genetic
Analyzer (Applied-Biosystems). Os plasmídeos foram sequenciados nas direções
“direta” e “reversa".
Materiais e Métodos
39
A Figura 9 mostra um cromatograma típico obtido utilizando o protocolo descrito
acima.
3.14. Análise computacional das seqüências
As seqüências obtidas foram submetidas à busca de similaridade com
nucleotídeos e aminoácidos utilizando os algoritmo BLASTn (base de dados dsEST)
e BLASTx, respectivamente disponível no site www.ncbi.nlm.nih.gov. Também foi
utilizado o algoritmo PFAM (www.sanger.ac.uk/Software/Pfam/) para identificar os
domínios conservados nas seqüências de aminoácidos preditas. Os alinhamentos
das seqüências de nucleotídeos e aminoácidos de S. mansoni obtidas com as
seqüências ortólogas, foram realizada com o auxílio do programa CLUSTALW
(http://www.ebi.ac.uk/clustalw/) e do algoritmo BLAST 2 www.ncbi.nlm.nih.gov.
3.15. Northern Blot
As confecções dos géis de agarose/formaldeído 1,2% foram realizadas de
acordo com SAMBROOK et al., 1989. Para cada amostra de RNA total de vermes
adultos (10 a 20µg), foi adicionado o tampão da amostra de RNA (formamida
deionizada 62,5%, formaldeído 1,14M, MOPS 1,25X, azul de bromofenol 0,25% e
Figura 9- Cromatograma obtido após a leitura em gel de seqüênciamento automático. O aparelho utilizado foi o sequenciador automático ABI 3100 Genetic Analyzer (Applied-Biosystems).

Materiais e Métodos
40
brometo de etídeo 0,5µg/mL de concentração). As amostras de RNA foram
desnaturadas a 65ºC por 15 minutos, seguido de banho de gelo por 3 minutos,
aplicadas nos poços de gel de agarose e a corrida eletroforética ajustada para 90
volts durante 1 hora e 30 minutos.
Em seguida, os géis foram tratados com SSC 10X durante 1 hora, e os RNAs
transferidos para uma membrana de nylon Hybond-N+ (Amersham - Biosciences)
durante 18 horas. Posteriormente, o sistema de transferência foi desmontado e as
membranas lavadas durante 5 minutos em SSC 6X, colocadas entre duas folhas de
papel de filtro 3mm, embrulhada em papel alumínio e mantidas em estufa a 80ºC por
duas horas para permitir a imobilização dos RNA na membrana. Decorrido esse
intervalo de tempo, as membranas foram armazenadas a temperatura ambiente até
o momento do uso.
3.16. Preparo da sonda radioativa
Para a obtenção das sondas radioativas, utilizamos a técnica de Random
Primer Extension (FEINBERG & VOGELSTEIN, 1983), e o kit Random Primer
Labelling System (Invitrogen). Esse procedimento permite obter fragmentos de DNA
marcados com 32P que podem ser empregados como sonda em experimentos de
hibridização. O método baseia-se na utilização de hexanucleotídeos sintéticos
aleatórios, como iniciadores, para a polimerização de qualquer segmento de DNA
molde. Para que ocorra a síntese de uma nova fita, são necessários uma mistura de
deoxinucleotídeos e o fragmento Klenow da DNA polimerase I. Neste caso, um dos
deoxinucleotídeos, o dCTP, possui um 32P na posição α.

Materiais e Métodos
41
Na reação de obtenção da sonda, cerca de 50-100ng do cDNA de Notch e
Presenilina foram desnaturadas por 5 minutos a 100°C, sendo em seguida
adicionados 2µL de cada um dos deoxinucleotídeos (dTTP,dGTP e dATP, na
concentração final de 0,5mM), 4µCi [α32P] dCTP, 1 unidade de DNA polimerase I
(fragmento Klenow) e água milli-Q estéril para completar um volume final de 50µL. A
reação foi incubada a temperatura ambiente por 1 hora, sendo posteriormente
interrompida pela adição de 2µL de EDTA 0,5M. Em seguida, para a remoção dos
nucleotídeos não incorporados, a sonda foi purificada em coluna de Sephadex G-50,
previamente equilibrada com tampão NT (Tris-HCl 10mM, NaCl 50mM, EDTA 0,1mM
pH em 8,0). A coluna foi montada em pipeta Pasteur, contendo lã de vidro na sua
extremidade e o empacotamento feito por sedimentação. Foi adicionado à sonda,
7µL do corante blue dextran e a mistura aplicada no topo da coluna. A sonda
marcada foi eluida no mesmo tampão (NT), sendo recolhida em um tubo eppendorf.
3.17. Reação de Hibridização
As membranas de náilon (contidas em saco plástico), inicialmente foram pré-
hibridizadas a 65°C, sob agitação moderada, no mínimo 4 horas, em uma solução
previamente aquecida composta por: 6X SSC, 2X reagente de Denhardt e 0,1%
SDS (100mg/mL) e 20µg de DNA de esperma de salmão para 1mL de solução de
pré-hibridização, por 4 a 6 horas sob agitação leve.
Posteriormente, foi adicionada a sonda previamente desnaturada, com uma
atividade específica de 2x108 cpm/µL. A incubação foi mantida por 18 horas. Em
seguida, para retirar o excesso de sonda não incorporada e remover os híbridos não

Materiais e Métodos
42
homólogos, as membranas foram lavadas seqüencialmente, uma vez, com as
seguintes soluções:
1) Solução 1: SSC 2X, SDS 0,1X, 15 minutos a temperatura ambiente;
2) Solução 2: SSC 0,5X, SDS 0,1%, 15 minutos, temperatura ambiente, com
agitação moderada;
3) Solução 3: SSC 0,1X, SDS 0,1%, 15 minutos, temperatura ambiente, com
agitação moderada;
4) Solução 4: SSC 0,1X, SDS 1% por 30 minutos a 50°C, com agitação
moderada.
3.18. Autoradiografia
Para verificar as regiões de homologia, as membranas obtidas após a reação
de hibridização, foram levemente secas, em papel de filtro, embrulhadas em filme de
PVC e acondicionada em cassetes contendo intensificador e expostas a filmes
Kodak (X-OMAT-XAR-5). O período de exposição foi variável e de acordo com a
intensidade do sinal, monitorada previamente com contador VICTOREEN, modelo
489-110C.
As revelações foram feitas de acordo com as instruções do fabricante,
utilizando como solução reveladora o Revelador e Reforçador GBX e como fixadora,
o Fixador e Revelador GBX (KODAK).

4- Resultados

Resultados
44
4.1. Análise in silico da via de sinalização Notch
A presença dos componentes da via de sinalização Notch, descritos para D.
melanogaster e C.elegans (ARTAVANIS-TSAKONAS, et al, 1999; GREENWALD,
1998) foram analisados nos 30,988 contigs gerados pelo Projeto Transcriptoma de
S.mansoni (VERJOVISKI et al, 2003). Posteriormente, as seqüências recuperadas
foram analisadas conforme descrito em Materiais e Métodos.
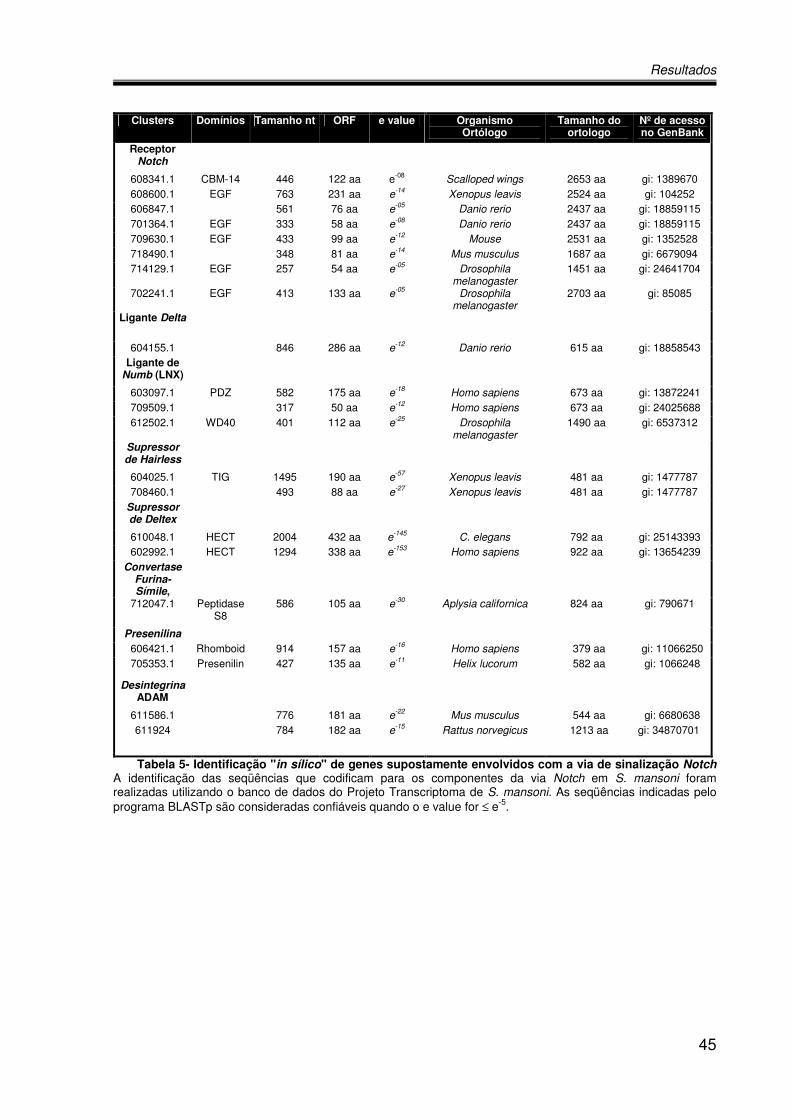
A Tabela 5 mostra os genes supostamente envolvidos com a via Notch de
sinalização em S.mansoni. Como podemos observar, essas análises permitiram a
identificação de oito clusters para o receptor Notch, um do ligante Delta, três do
ligante Numb, dois para o Supressor de Hairless, dois para Supressor de Deltex, dois
para Desintegrina, dois para Preselinina e um para a Convertase Furina-Símile,
sugerindo fortemente que todos os principais componentes dessa via apresentam
sequências depositadas no banco de dados.
Resultados
45
Clusters Domínios Tamanho nt ORF e value Organismo Ortólogo
Tamanho do ortologo
Nº de acesso no GenBank
Receptor Notch
608341.1 CBM-14 446 122 aa e-08 Scalloped wings 2653 aa gi: 1389670 608600.1 EGF 763 231 aa e
-14 Xenopus leavis 2524 aa gi: 104252
606847.1 561 76 aa e-05
Danio rerio 2437 aa gi: 18859115 701364.1 EGF 333 58 aa e
-08 Danio rerio 2437 aa gi: 18859115
709630.1 EGF 433 99 aa e-12
Mouse 2531 aa gi: 1352528 718490.1 348 81 aa e
-14 Mus musculus 1687 aa gi: 6679094
714129.1 EGF 257 54 aa e-05
Drosophila melanogaster
1451 aa gi: 24641704
702241.1 EGF 413 133 aa e-05
Drosophila melanogaster
2703 aa gi: 85085
Ligante Delta
604155.1 846 286 aa e-12
Danio rerio 615 aa gi: 18858543 Ligante de
Numb (LNX)
603097.1 PDZ 582 175 aa e-18
Homo sapiens 673 aa gi: 13872241 709509.1 317 50 aa e
-12 Homo sapiens 673 aa gi: 24025688
612502.1 WD40 401 112 aa e-25
Drosophila melanogaster
1490 aa gi: 6537312
Supressor de Hairless
604025.1 TIG 1495 190 aa e-57
Xenopus leavis 481 aa gi: 1477787 708460.1 493 88 aa e
-27 Xenopus leavis 481 aa gi: 1477787
Supressor de Deltex
610048.1 HECT 2004 432 aa e-145
C. elegans 792 aa gi: 25143393 602992.1 HECT 1294 338 aa e
-153 Homo sapiens 922 aa gi: 13654239
Convertase Furina-Símile,
712047.1 Peptidase S8
586 105 aa e-30
Aplysia californica
824 aa gi: 790671
Presenilina 606421.1 Rhomboid 914 157 aa e
-16 Homo sapiens 379 aa gi: 11066250
705353.1 Presenilin 427 135 aa e-11
Helix lucorum 582 aa gi: 1066248
Desintegrina ADAM
611586.1 776 181 aa e-22
Mus musculus 544 aa gi: 6680638 611924 784 182 aa e
-15 Rattus norvegicus
1213 aa gi: 34870701
Tabela 5- Identificação "in sílico" de genes supostamente envolvidos com a via de sinalização Notch A identificação das seqüências que codificam para os componentes da via Notch em S. mansoni foram realizadas utilizando o banco de dados do Projeto Transcriptoma de S. mansoni. As seqüências indicadas pelo programa BLASTp são consideradas confiáveis quando o e value for ≤ e-5.
Resultados
46
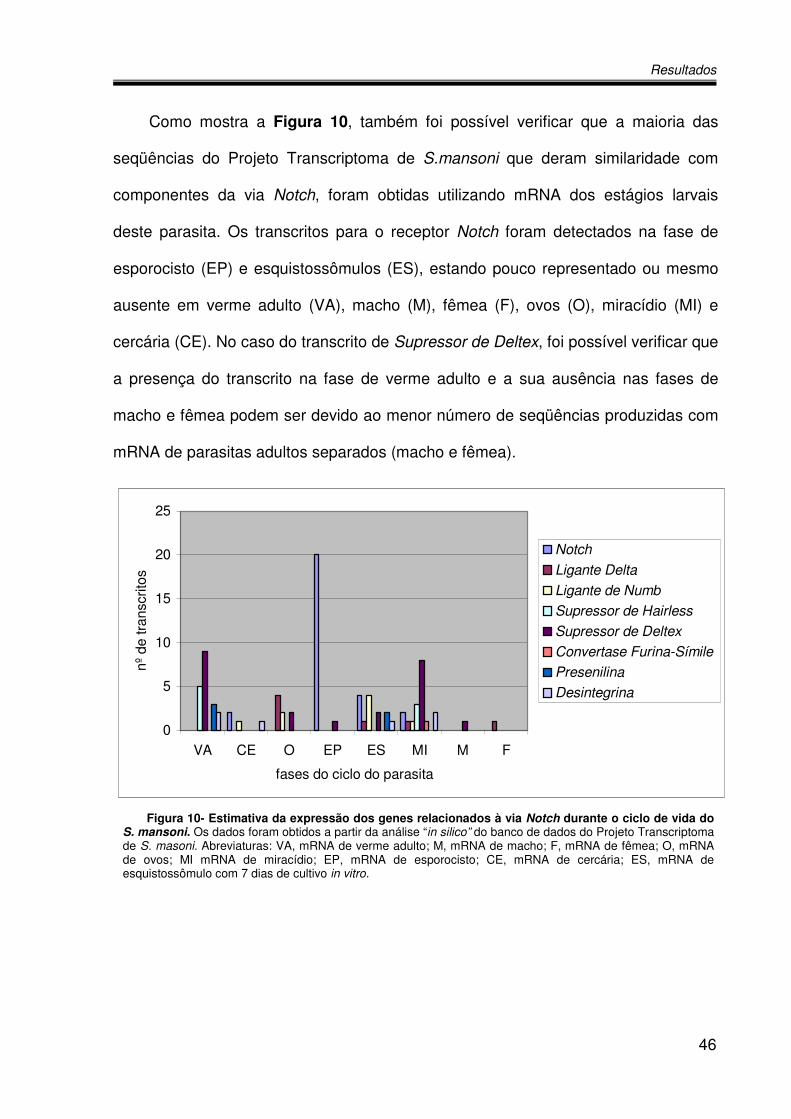
Como mostra a Figura 10, também foi possível verificar que a maioria das
seqüências do Projeto Transcriptoma de S.mansoni que deram similaridade com
componentes da via Notch, foram obtidas utilizando mRNA dos estágios larvais
deste parasita. Os transcritos para o receptor Notch foram detectados na fase de
esporocisto (EP) e esquistossômulos (ES), estando pouco representado ou mesmo
ausente em verme adulto (VA), macho (M), fêmea (F), ovos (O), miracídio (MI) e
cercária (CE). No caso do transcrito de Supressor de Deltex, foi possível verificar que
a presença do transcrito na fase de verme adulto e a sua ausência nas fases de
macho e fêmea podem ser devido ao menor número de seqüências produzidas com
mRNA de parasitas adultos separados (macho e fêmea).
Figura 10- Estimativa da expressão dos genes relacionados à via Notch durante o ciclo de vida do S. mansoni. Os dados foram obtidos a partir da análise “in silico” do banco de dados do Projeto Transcriptoma de S. masoni. Abreviaturas: VA, mRNA de verme adulto; M, mRNA de macho; F, mRNA de fêmea; O, mRNA de ovos; MI mRNA de miracídio; EP, mRNA de esporocisto; CE, mRNA de cercária; ES, mRNA de esquistossômulo com 7 dias de cultivo in vitro.
0
5
10
15
20
25
VA CE O EP ES MI M F
fases do ciclo do parasita
nº d
e tr
ansc
ritos
Notch
Ligante Delta
Ligante de Numb
Supressor de Hairless
Supressor de Deltex
Convertase Furina-Símile
Presenilina
Desintegrina

Resultados
47
Os transcritos de S. mansoni que deram similaridade com os componentes da
via Notch de outros organismos, foram analisados quanto a sua presença no banco
de dados gerado pelo Projeto Transcriptoma de S. japonicum (WEI, et al.2004)
conforme descrito em Materiais e Métodos.
Como mostra a Tabela 6 essa análise, ao contrário do que foi observado para o
S. mansoni, não identificou a maioria dos componentes da via de sinalização
mediada pelo receptor Notch em S. japonicum. Esse resultado pode ser justificado
pelo fato de que a maioria das seqüências de S. japonicum depositadas nos bancos
de dados públicos foram geradas a partir de bibliotecas de cDNA construídas a partir
de mRNA de ovos, verme adulto, macho e fêmea (WEI, et al, 2004). Indiretamente
esse resultado corrobora com os obtidos no banco de S. mansoni, sugerindo que os
transcritos relacionados com a via Notch são mais abundantes nos estágios larvais
relacionados tanto com o hospedeiro invertebrado como vertebrado,
respectivamente esporocistos e esquistossômulos.
Clusters do S. mansoni Nº de acesso do S. japonicum no GenBank
Estágio do desenvolvimento
e value
Ligante Delta 604155.1 gi: 28326430 M 1e-19 604025.1 gi: 28345625 O
Supressor de Deltex 610048.1 gi: 28354711 F 0.0
Presenilina 606421.1 gi: 28352486 F 8e-34 705353.1 gi:28332365 M 5e-45
Tabela 6- Comparação dos componentes da via Notch entre S. mansoni e S. japonicum. A comparação foi realizado utilizando o programa BLASTn. As seqüências indicadas pelo programa BLASTn são consideradas confiávies quando o e value for ≤ e5 . .Abreviaturas: M, mRNA de macho; F, mRNA de fêmea; O mRNA de ovos.

Resultados
48
4.2. Amplificação dos fragmentos gênicos de Notch, Presenilina e Supressor
de Deltex a partir do DNA genômico do S. mansoni
Com objetivo de confirmamos os resultados obtidos na análise in silico,
realizamos reações de amplificações de Notch, Presenilina e Supressor de Deltex a
partir de oligonucleotídeos gene-específico e DNA genômico, como descrito em
Materiais e Métodos.
Como podemos observar na Figura 11, os oligonucleotídeos para Notch
amplificaram um fragmento de aproximadamente 850 pb (A), para Presenilina,
utilizando os oligonucleotídeos específicos obtivemos um fragmento de
aproximadamente 400pb (B). Podemos observar que os oligonucleotídeos gene-
específico para Supressor de Deltex, amplificaram dois fragmentos, um de
aproximadamente 800 e outro com 850 pb (C).

Resultados
49
4.3. Sequenciamento e análise computacional dos fragmentos gênicos de
Notch, Presenilina e Supressor de Deltex
Para confirmarmos se os produtos amplificados estavam relacionados com os
genes em estudo, os produtos obtidos foram purificados, clonados no vetor pGEMT-
PM VA C
600pb 400pb
PM VA C
600pb
850 pb
PM VA C
600pb
850 pb 800 pb
A B
C
Figura 11- Amplificação dos fragmentos gênicos correspondentes ao gene Notch (A), Presenilina (B) e Supressor de Deltex.(C). Aproximadamente 50 ng de DNA genômico proveniente de vermes adultos foram utilizados nas reações de amplificações (PCR). Cerca de 10µl da preparação foram separados por eletroforese em gel de agarose 1,5% e corados com brometo de etídeo a 0,5 µg/ml. Abreviaturas: (PM) Peso Molecular 100pb, (VA) verme adulto, (C) controle negativo da reação de PCR.

Resultados
50
Easy (Promega) e seqüenciados em sequenciador automático ABI 3100 Genetic
Analyzer (Applied-Biosystems) como descrito em Materiais e Métodos.
As seqüências foram submetidas à busca de identidade com outras seqüências
de nucleotídeos e aminoácidos depositadas no GenBank (NCBI-NIH, Bethesda,
USA), utilizando-se o algoritmo BLASTn (base de dados dbEST) e BLASTx.
Podemos notar que, utilizando as seqüências de nucleotídeos e a base de
dados dbEST, foi possível verificar que as seqüências de Notch, Presenilina e
Supressor de Deltex apresentam identidade com diversas EST obtidas de S.
mansoni. No caso da Presenilina e do Supressor de Deltex as seqüências também
apresentam identidade com seqüências de S. japonicum, como pode ser observado
nas Tabela 07 A, B e C. Os resultados da análise computacionais também
mostraram que o fragmento de 850 pares de bases obtida com os oligonucleotídeos
específicos para Supressor de Deltex era inespecífico.
A
Nº de acesso no GenBank Organismo Identidade % e value
gi:34714368 S. mansoni 99% 0.0 gi:34707103 S. mansoini 99% 0.0
B
Nº de acesso no GenBank Organismo Identidade% e-value gi:12354025 S. mansoni 100% 0.0 gi:34712029 S. mansoni 97% 0.0
gi:5788730 S. mansoni 99% e-120 gi:5788642 S. mansoni 100% e-154 gi:60232785 S. japonicum 90% e-105 gi:60234653 S. japonicum 91% e-101

Resultados
51
C
Para a identificação das prováveis regiões codificadoras, as seqüências obtidas
após o sequenciamento foram submetidas à análise utilizando o algoritmo ORFfinder
disponível no endereço: http://www.ncbi.nlm.nih.gov/gorf/gorf.html.
Na Figura 12-A, B e C, podemos observar que os fragmentos amplificados
possuem uma seqüência predita com 273, 144 e 270 aminoácidos, para Notch,
Presenilina e Supressor de Deltex, respectivamente.
Nº de acesso no GenBank Organismo Identidade % e-value gi:34629380 S. mansoni 99% 0.0 gi:34689860 S. mansoni 99% 0.0 gi:34714442 S. mansoni 98% 0.0 gi:28354711 S. japonicum 88% e-161
Tabela 7- Análise das seqüências de Notch (A), Presenilina (B) e Supressor de Deltex (C) utilizando o programa BLASTn (base de dados dbEST).A identidade indica a % de resíduos idênticos observados na região de alinhamento. As seqüências indicadas pelo programa BLASTn são consideradas estatisticamente confiáveis quando o e value for ≤ e-5.

Resultados
52
A
1 catgtgacaaagggtatcaccaagattacagaggtcactagtctt
H V T K G I T K I T E V T S L
45 ccaaacaaatgtagtcctgaatgtgttcatggacaaggcgaatgt
P N K C S P E C V H G Q G E C
90 acatctaatggaaaatgcttgtgtaatccaggttttcgagggtta
T S N G K C L C N P G F R G L
135 tcttgtgagacagacgtagatgaatgcgctgaaggaaaacataat
S C E T D V D E C A E G K H N
180 tgtcaacaggaatgtatcaatactcctgggtcttttaaatgttct
C Q Q E C I N T P G S F K C S
225 tgtcattcaggttacaaattatccactgtagatccatcaaaatgt
C H S G Y K L S T V D P S K C
270 gaacctatttcttgtccttctcaatgcaatttaatagatggagaa
E P I S C P S Q C N L I D G E
315 tgtgactgcgaatgcaagcttggacattttggacttaattgtaat
C D C E C K L G H F G L N C N
360 caaacagtagattcatgtgcaattaaaccatgtgaccaaatttgt
Q T V D S C A I K P C D Q I C
405 gtggatttaggtgatgggaagtatacatgcaaatgtaatgaaggt
V D L G D G K Y T C K C N E G
450 tttgaaccaagtccaaataatcctagacgatgtcagcgtatatgt
F E P S P N N P R R C Q R I C
495 aaagatggtatagactgtatatatggaagctgtcgaactgtgttt
K D G I D C I Y G S C R T V F
540 tcaaatgacacagatcaatctttatgtacttgcctagaaggattt
S N D T D Q S L C T C L E G F
585 tacggaacaaactgtcaaaacgatgtaaacgaatgcttgaatgag
Y G T N C Q N D V N E C L N E
630 aatggtcaaaaacatttttgtgaacatcattgcataaatacatta
N G Q K H F C E H H C I N T L
675 ggatcataccaatgtttttgtgatccggattatgaattacaatct
G S Y Q C F C D P D Y E L Q S
720 gatggtcgaacgtgtaaaaaacagtttaaaggtgaaatgaaatgt
D G R T C K K Q F K G E M K C
765 cctgaatactgcctaaatggtgcaacttgtaaatcgactggagaa
P E Y C L N G A T C K S T G E
810 tgtgtatgtc
C V C
B
2 cgaaccgatgcaattgtagttgatactcctgatgacaatacg
R T D A I V V D T P D D N T
43 attaataatgatgccaggaataacaatgaaactatatcacag
I N N D A R N N N E T I S Q
85 gcacctcatgaaacagatacacttgtagcaggccgacccaat
A P H E T D T L V A G R P N
127 cgttggcaagaactgcaagcagatttaattgatcgtgactca
R W Q E L Q A D L I D R D S
169 aagggtgtaaaacttggtttaggcgattttgtattttatagt
K G V K L G L G D F V F Y S
211 ttactgattggtcgtgcaacacttgatggcgatgcagtcact
L L I G R A T L D G D A V T
253 gtggtcacatgttatgtggctatattggttggaatgtgtatt
V V T C Y V A I L V G M C I
295 actgttatagtattgggaattacacgacaagcattaccagct
T V I V L G I T R Q A L P A
337 ttaccaatatccataacatgtggtatattattttattttgtc
L P I S I T C G I L F Y F V
379 agttcagcgacgatatcaccgtttcttgaagcaacagccatt
S S A T I S P F L E A T A I
421 gaacgtgtgttc
E R V F

Resultados
53
Como observado na Tabela 08, a análise das seqüências de aminoácidos
preditas para os fragmentos gênicos de Notch, Presenilina e Supressor de Deltex
apresentaram identidade com diversas seqüências de aminoácidos de Notch,
Presenilina e Supressor de Deltex descritas para vários organismos.
C
2 atgggagtcccattgcaatatgaaaggaatttccgatccaaagtg M G V P L Q Y E R N F R S K V
46 aattattttcgtgcttgttgtacaaatgccatgttaaatgggcaa
N Y F R A C C T N A M L N G Q
91 actaaacttcttatatcaagagacaatttattagaggactcgttc
T K L L I S R D N L L E D S F
136 caactggtatctcaaatgtcttcatctggtttaagaagacgttta
Q L V S Q M S S S G L R R R L
181 tcaattacatttctacatgaagaaggtttggattacggtggagta
S I T F L H E E G L D Y G G V
226 gccagagaatggttctatcgtctttcaagagaaattttaaaccca
A R E W F Y R L S R E I L N P
271 atgtttggcttgtttgagtatactggaacagattatgccttacaa
M F G L F E Y T G T D Y A L Q
316 gtaaatccagcctctcatgttaatccaaatcatatggcgtatttt
V N P A S H V N P N H M A Y F
361 cgatttgttggacgctttattggtatggcactttttcatggtcgc
R F V G R F I G M A L F H G R
406 tgtattgacggcgggcttactcttgctttttataaacaaattttg
C I D G G L T L A F Y K Q I L
451 aagcggaaattaacattagaagatcttggtcatacggatcacagt
K R K L T L E D L G H T D H S
496 tattatcaatcgttaatttatgtcagagacaatccagttgatgag
Y Y Q S L I Y V R D N P V D E
541 tgtgatttggacttgtactttattggaacctatgaccttttaggc
C D L D L Y F I G T Y D L L G
586 gttttgcatgaagacgaacttattgaaggtggcaaagatattaaa
V L H E D E L I E G G K D I K
631 gtgactgaagaaaataaatccgattatatcagactgatggttgat
V T E E N K S D Y I R L M V D
676 tggcggttcaacagaggagtaaaaaaacaaacagaagaaattcac
W R F N R G V K K Q T E E I H
721 aaaggtatttttgaagtactaaatcccgaatggctggaattattc
K G I F E V L N P E W L E L F
766 gatgagcgtgaacttgaactcctattatccggtatgccagagatt
D E R E L E L L L S G M P E I
Figura 12- Seqüências dos nucleotídeos e aminoáciods preditas
referentes a Notch (A), Presenilina (B) e Supressor de Deltex (C) . As melhores sequências de aminoácidos correspondentes às possíveis regiões codificadoras foram obtidas com auxílio do programa ORF finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html). Os nucleotídeos sublinhados correspondem aos oligonucleotideos direto e reverso utilizados nas amplificações.

Resultados
54
A
B
C
Nº de acesso no Genbank
Organismo Identidade% e value
gi:25143391 C. elegans 51% 7e-72 gi:27477109 Homo sapiens 52% 9e-70 gi:40352723 Mus musculus 47% 6e-69 gi:50758591 Gallus gallus 47% 2e-69
Para confirmar a identidade das sequências de aminoácidos preditas dos
fragmentos gênicos de Notch, Presenilina e Supressor de Deltex, as seqüências
também foram analisadas em banco de dados que permite a identificação de
domínios conservados numa seqüência primária de aminoácidos. Neste caso,
utilizamos a base de dados do algoritmo Pfam disponível no endereço
(www.sanger.ac.uk/Software/Pfam/).
Nº de acesso no Genbank
Organismo Identidade% e value
gi: 104252 Xenopus leavis 32% 6e-30 gi: 22770986 Boophilus microplus 32% 8e-29 gi: 13242247 Rattus novergicus 29% 8e-25
gi: 85085 D. melanogaster 29% 8e-24
Nº de acesso no Genbank
Organismo Identidade% e value
gi: 2062395 D.melanogaster 52% 2e-24 gi: 17529699 Branchiostoma floridae 46% 2e-24 gi: 40950170 Canis familiaris 47% 3e-23 gi: 47575859 Rattus novergicus 47% 5e-23
gi: 4506163 Homo sapiens 47% 5e-23
Tabela 8- Análise das seqüências de Notch (A), Presenilina (B) e Supressor de Deltex (C) utilizando o programa BLASTx, com seqüências ortólogas de outros organismos. A identidade indica a % de resíduos idênticos observados na região de alinhamento. As seqüências indicadas pelo programa BLASTp são consideradas estatisticamente confiáveis quando o e value for ≤ e-5.

Resultados
55
A base de dados Pfam evidenciou a presença de cinco domínios EGF
(Epidermal Growth Factor) no fragmento gênico de Notch. Esse resultado sugere que
o fragmento amplificado provavelmente é a porção extracelular do receptor Notch.
No caso da Presenilina, a análise da seqüência predita, mostrou a presença de um
domínio Presenilina, que é característico dessa família de protease. Por fim, a
análise do Supressor de Deltex, demonstrou a presença do domínio Hect
(Homologous to E-6AP carboxy-terminus), que caracteriza a porção C-terminal da
família das proteínas ubiquitina ligase. Esses alinhamentos podem ser observados
no Apêndice1.
Como observado na Figura 13, as seqüências de aminoácidos preditas
referentes aos fragmentos gênicos de Notch, Presenilina e Supressor de Deltex em
S. mansoni, foram alinhadas com os respectivos ortólogos utilizando o algoritmo
CLUSTALW (http://www.ebi.ac.uk/clustalw/). Os resultados dos alinhamentos
confirmaram que as seqüências dos fragmentos gênicos de Notch, Presenilina e
Supressor de Deltex estão alinhando nas regiões de domínios das seqüências
ortólogas, como descrito anteriormente.

Resultados
56
A XlNotch NTYGGYNCVCVNGWTGEDCSENIDDCANAACHSGATCHDRVASFYCECPHGRTGLLCHLD 374 BmNotch NTIGGYSCICVNGWTGADCSENIDDCAVAACFNGATCHDRVGSFYCQCAPGKTGLLCHLD 383 DmNotch NTHGSYSCICVNGWAGLDCSNNTDDCKQAACFYGATCIDGVGSFYCQCTKGKTGLLCHLD 411 SmNotch -------------------------------------------------HVTKGITKITE 11
XlNotch NACISNPCNEGSNCDTNPVNGKAICTCPPGYTGPACNNDVDECSLG-ANPCERGGRCTNT 433
BmNotch DACASNPCHAGAICDTSPIDGTYLCSCPNGFQGVDCTEDVDECAQG--FPCEHQGLCVNT 441
DmNotch DACTSNPCHADAICDTSPINGSYACSCATGYKGVDCSEDIDECDQG--SPCEHNGICVNT 469
SmNotch VTSLPNKCSPECVHGQGECTSNGKCLCNPGFRGLSCETDVDECAEG--SKHNCQQECINT 69
XlNotch LGSFQCNCPQGYAGPRCEIDVNECLSNPCQNDSTCLDQIGEFQCICMPGYEGLYCETNID 553
BmNotch PGSFRCNCTRGFAGPRCEVNINECDSNPCQNEGTCLDERGGFRCVCMPGYRGVHCEVDID 561
DmNotch PGSYRCNCSQGFTGPRCETNINECESHPCQNEGSCLDDPGTFRCVCMPGFTGTQCEIDID 589
SmNotch PGSFKCSCHSGYK-----------------------------------------------
XlNotch ECASNPCLHNGKCIDKINEFRCDCPTGFSGNLCQHDFDECTSTPCKNGAKCLDGPNSYTC 493
BmNotch ECLPNPCLNGGICSDRINGFQCLCPVGFSGKKCETNEDDCSSFPCRNGGSCHDGIASYTC 501
DmNotch ECQSNPCLNDGTCHDKINGFKCSCALGFTGARCQINIDDCQSQPCRNRGICHDSIAGYSC 529
SmNotch -------------------------------LSTVDPSKCEPISCPSQCNLIDG—-ECDC 109
XlNotch QCTEGFTGRHCEQDINECIPDPCHYGTCKDGIATFTCLCRPGYTGRLCDNDINECLSKPC 613
BmNotch HCPPGFTGSNCETNMDDCQSSPCHHGECIDGTNAFTCNCHPGYTGLLCQNQINECLSSPC 621
DmNotch ECPPGYTGTSCEININDCDSNPCHRGKCIDDVNSFKCLCDPGYTGYICQKQINECESNPC 649
SmNotch ECKLGHFGLNCNQTVDSCAIKPCDQICVDLGDGKYTCKCNEGFE---------PSPNNPR 160
XlNotch LNGGQCTDRENGYICTCPKGTTGVNCETKIDDCASNLCDNGK-CIDKIDGYECTCEPGYT 672
BmNotch QHGGTCEDLVNGYQCRCRRGTSGPDCQFNVNECFSNPCRNGAKCVDGIDSYICECLPGYT 681
DmNotch QFDGHCQDRVGSYYCQCQAGTSGKNCEVNVNECHSNPCNNGATCIDGINSYKCQCVPGFT 709
SmNotch RCQRICKDGIDCIYGSCRTVFS-----------------------NDTDQSLCTCLEGFY 197
XlNotch GKLCNININECDSNPCRNGGTCKDQINGFTCVCPDGYHDHMCLSEVNECNSNPCIH-GAC 731
BmNotch GIHCETDINECASNPCANGGVCIDLINGFKCNCPRGYFDARCLSNVNECASNPCQHGGTC 741
DmNotch GQHCEKNVDECISSPCANNGVCIDQVNGYKCECPRGFYDAHCLSDVDECASNPCVNEGRC 769
SmNotch GTNCQNDVNECLNE------------NGQKHFCEH-----------------------HC 222
XlNotch HDGVNGYKCDCEAGWSGSNC—DINNNECESNPCMNGGTCKDMTGAYICTCKAGFSGPNC 789
BmNotch EDEVNHFVCHCPPGYGGHRC—EQDIDECQSNPCQHGGSCHDALNNYSCACIEGYSGRNC 799
DmNotch EDGINEFICHCPPGYTGKRC—ELDIDECSSNPCQHGGTCYDKLNAFSCQCMPGYTGQKC 827
SmNotch INTLGSYQCFCDPDYELQSD--------------------------GRTCKKQFKG--- 251
XlNotch QTNINECSSNPCLNHGTCIDDVAGYKCNCMLPYTGAICEAVLAPCAGSPCKNGGRCKESE 849
BmNotch ETNLDDCSPNPCRNGGSCIDLVGTFRCVCEVPFTGPTCEVELNPCSPNKCRNGAQCSPSS 859
DmNotch ETNIDDCVTNPCGNGGTCIDKVNGYKCVCKVPFTGRDCESKMDPCASNRCKNEAKCTPSS 887
SmNotch ------------------------------------------------------------
XlNotch DFETFSCECPPGWQGQTCEIDMNECVN-RPCRNGATCQNTNGSYKCNCKPGYTGRNCEMD 908
BmNotch NYLDFACSCKLGFTGRLCDEDIDECAVSSPCRNEATCINTNGSYECACTKGYEGRDCLIN 919
DmNotch NFLDFSCTCKLGYTGRYCDEDIDECSLSSPCRNGASCLNVPGSYRCLCTKGYEGRDCAIN 947
SmNotch -----------------------EMKCPEYCLNGATCKSTG---ECVC------------ 273

Resultados
57
B
HsPres HWKGPLVLQQAYLIMISALMALVFIKYLPEWSAWVILGAISVYDLVAVLCPKGPLRMLVE 279
CfPres HWKGPLRLQQAYLIMISALMALVFIKYLPEWTAWLILAVISVYDLVAVLCPKGPLRMLVE 268
RnPres HWKGPLRLQQAYLIMISALMALVFIKYLPEWTAWLILAVISVYDLVAVLCPKGPLRMLVE 273
DmPres HWQGPLRLQQGYLIFVAALMALVFIKYLPEWTAWAVLAAISIWDLIAVLSPRGPLRILVE 295
SmPres -----------------------------------------RTDAIVVDTPDD--NTINN 17
HsPres TAQERNEPIFPALIYSSAMVWTVG----------MAKLDPSSQG------ALQLPYDPEM 323
CfPres TAQERNETLFPALIYSSTMVWLVN----------MAEGDPEAQRKVSK--NSNYNAQSTE 316
RnPres TAQERNETLFPALIYSSTMVWLVN----------MAEGDPEAQRRVPK--NPKYSTQGTE 321
DmPres TAQERNEQIFPALIYSSTVVYALVNTVTPQQSQATASSSPSSSNSTTTTRATQNSLASPE 355
SmPres DARNNNETISQAPHETDTLV----------------AGRP-------------------- 41
HsPres EEDSYDS------------------FGE-------PSYPEV--FEPPLTGYPG------- 349
CfPres S-ETQDP------------------VTESDDGGFSEEWEAQR--DSRLGPHHSTTETRAA 355
RnPres REETQDT------------------GTGSDDGGFSEEWEAQR--DSHLGPHRSTPESRAA 361
DmPres AAAASGQRTGNSHPRQNQRDDGSVLATEAEAAGFTQEWSANLSERVARRQIEVQSTQSGN 415
SmPres -----------------------------------NRWQELQ------------------ 48
HsPres ------------------------------EELEEEEERGVKLGLGDFIFYSVLVGKAAA 379
CfPres VQELSS----------------------NILASEDPEERGVKLGLGDFIFYSVLVGKASA 393
RnPres VQELSG----------------------SILTSEDPEERGVKLGLGDFIFYSVLVGKASA 399
DmPres AQRSNEYRTVTAP---------------DQNHPDGQEERGIKLGLGDFIFYSVLVGKASS 460
SmPres ------------------------------ADLIDRDSKGVKLGLGDFVFYSLLIGRATL 78
HsPres NG--DWNTTLACFVAILIGLCLTLLLLAIFKKALPALPISITFGLIFNFATANLVTPFTD 439
CfPres TGSGDWNTTLACFVAILIGLCLTLLLLAVFKKALPALPISITFGLIFYFSTDNLVRPFMD 453
RnPres TASGDWNTTIACFVAILIGLCLTLLLLAIFKKALPALPISITFGLVFYFATDYLVQPFMD 459
DmPres TASGDWNTTIACFVAILIGLCLTLLLLAIFKKALPALPISITFGLIFYFATDYLVQPFMD 518
SmPres YG— DWTTTIACFVAILIGLCLTLLLLAIWRKALPALPISITFGLIFCFATSAVVKPFME 136
144
HsPres ALASQQVY 447
CfPres TLASHQLY 461
RnPres QLAFHQFY 467
DmPres QLAFHQFY 526
SmPres DLSAKQVF 144
C
MmSu(dx) DPRTGKSALDN---GPQIAYVRDFKAKVQYFRFWCQQLAMPQHIKITVTRKTLFEDSFQQ 525
CeSu(dx) DPRTGKPVGPLGVVGVQMAMEKSFRWKIAQFRYLCLSNSVPNHVKITVSRNNVFEDSFQE 453
SmSu(dx) -------------MGVPLQYERNFRSKVNYFRACCTNAMLNGQTKLLISRDNLLEDSFQL 47
MmSu(dx) IMSFSPQDLRRRLWVIFPGEEGLDYGGVAREWFFLLSHEVLNPMYCLFEYAGKDNYCLQI 585
CeSu(dx) IMRKNAVDLRRRLYIQFRGEEGLDYGGVAREWFFLLSHEVLNPMYCLFMYAGNNNYSLQI 513
SmSu(dx) VSQMSSSGLRRRLSITFLHEEGLDYGGVAREWFYRLSREILNPMFGLFEYTG-TDYALQV 106
MmSu(dx) NPASYINPDHLKYFRFIGRFIAMALFHGKFIDTGFSLPFYKRILNKPVGLKDLESIDPEF 645
CeSu(dx) NPASFVNPDHLKYFEYIGRFIAMALFHGKFIYSGFTMPFYKKMLNKKIVLKDIEQVDSEI 573
SmSu(dx) NPASHVNPNHMAYFRFVGRFIGMALFHGRCIDGGLTLAFYKQILKRKLTLEDLGHTDHSY 166
MmSu(dx)_YNSLIWVKENNIEECGLEMYFSVDKEILGEIKSHDLKPNGGNILVTEENKEEYIRMVAEW 705
CeSu(dx) YNSLMWIKDNNIDECDMELYFVADYELLGELKTYELKEGGTEIAVTEENKLEYIELLVEW 633
SmSu(dx) YQSLIYVRDNPVDECDLDLYFIGTYDLLGVLHEDELIEGGKDIKVTEENKSDYIRLMVDW 226
MmSu(dx) RLSRGVEEQTQAFFEGFNEILPQQYLQYFDAKELEVLLCGMQEIDLNDWQRHAIYRHYTR 765
CeSu(dx) RFNRGVEQQTKAFFTGFNSVFPLEWMQYFDERELELLLCGMQDVDVDDWQRNTVYRHYAP 693
SmSu(dx) RFNRGVKKQTEEIHKGIFEVLNPEWLELFDERELELLLSGMPEI---------------- 270
Figura 13- Alinhamento da região codificadora dos fragmentos gênicos de Notch (A), Presenilina (B) e Supressor de Deltex (C) de S. mansoni com as seqüências ortólogas. Os alinhamentos representam somente as regiões onde os fragmentos de S. mansoni estão alinhando. Na cor preta estão representados os aminoácidos idênticos e em cor cinza, os aminoácidos conservados. Abreviaturas: Sm, S. mansoni; Rn, R. novergicus; Xl, X. leavis; Bm, B. microphilus; Dm, D. melanogaster; Cf, C. familiaris; Hs, H. sapiens; Mm, M. musculus; e Ce, C. elegans. Nº de acesso das seqüências: notch, Xl 104252, Bm 22770986, Dm 85085; presenilina, Dm 2062395 Cf 40950170, Rn 47575859, Hs 4506163; supressor de deltex, Mm40352723, Ce 25713391.

Resultados
58
4.4. Perfil de expressão dos fragmentos gênicos de Notch, Presenilina e
Supressor de Deltex durante o ciclo de vida do parasita
Neste trabalho, a expressão de Notch, Presenilina e Supressor de Deltex
foram investigados em diferentes estágios de desenvolvimento do parasita, com a
finalidade de evidenciar a existência ou não de uma possível expressão diferencial
estágio específico durante o ciclo de vida do parasita.
Desse modo, a partir do RNA total de vermes adultos, hepatopâncreas de
caramujo infectado (fonte de esporocistos), hepatopâncreas de caramujo não
infectado (controle negativo), cercárias e esquistossômulos, e oligonucleotídeos
gene-específicos, os respectivos cDNAs foram sintetizados, utilizando a técnica de
RT-PCR, conforme descrito em Materiais e Métodos. Após a amplificação, os
produtos do PCR, foram analisados em gel de agarose 1,5%. Para confirmar se os
cDNAs amplificados eram correspondentes aos genes Notch, Presenilina e
Supressor de Deltex os cDNAs foram clonados no vetor pGEM-T easy (Promega),
e posteriormente seqüenciados. As seqüências obtidas foram idênticas a dos
correspondentes genômicos e podem ser analisadas no Apêndice 2 e 3.
Como pode ser observado na Figura 14 A, a análise do perfil de expressão
durante o ciclo de vida do parasita mostra que os baixíssimos níveis de expressão
do receptor Notch não foram detectados pela técnica de RT-PCR. Esse resultado
corrobora com os resultados obtidos durante a análise in silico, onde mostra que há
poucos transcritos para Notch nessa fase, por outro lado, pode observar a
amplificação do transcrito de Notch nas fases de esporocisto, esquistossômulo, ovo
e verme adulto.

Resultados
59
Em relação aos transcritos de Presenilina e Supressor de Deltex, esses se
mostraram expressos em todas as fases como podem ser observados nas Figuras
14B e 14C.
PM VA EP C- CE ES O C
600 pb
A
850 pb
PM VA EP C- CE ES O C
600 pb
B
400 pb

Resultados
60
Com o objetivo de avaliarmos se haveria expressão sexo específica dos
fragmentos gênicos de Notch, Presenilina e Supressor de Deltex, RNA total dos
parasitas machos e fêmeas foram separadamente submetidos à transcrição reversa
e os produtos gênicos amplificados exatamente como descrito em Materiais e
Métodos. Como mostra as Figuras 15 A, B e C, os genes em estudo foram
expressos em ambos sexo do parasita.
C PM VA EP C- CE ES O C
600 pb 800 pb
Figura 14- Perfil de expressão durante o ciclo de vida do parasita. RT- PCR do RNAm codificador para Notch (A), Presenilina (B) e Supressor de Deltex.(C). Cerca de 10µl de cada preparação foram aplicados em gel de agarose 1,5% e o gel corado com brometo de etídeo. Abreviaturas: Peso Molecular 100pb (PM); verme adulto (VA); esporocisto (EP), caramujo não infectado (C-), cercária (CE), esquistossômulo (ES) , ovo (O) e controle negativo da reação de PCR (C).

Resultados
61
A
600 pb
600 pb 850 pb
PM VA M F C
PM VA M F C
400 pb
B
PM VA M F C C
600 pb 800 pb
Figura 15- Perfil de expressão dos transcritos em relação ao sexo do parasita. RT- PCR do RNAm codificador para Notch (A), Presenilina (B) e Supressor de Deltex (C). Cerca de 10µl de cada preparação foram aplicados em gel de agarose 1,5% e o gel corado com brometo de etídeo. Peso Molecular 100pb (PM); verme adulto (VA); macho (M), fêmea (F) e controle negativo da reação de PCR (C).

Resultados
62
4.5. Estimativa por Northern Blot do tamanho da mensagem codificadora de
Notch e Presenilina
Com a finalidade de estimarmos o tamanho da mensagem codificadora de
Notch e Presenilina, e a presença de sub-populações de mRNAs utilizamos a técnica
de Northern Blot. Para isso, cerca de 15 µg de RNA total de vermes adultos foram
separados em gel de agarose formaldeído 1.2% e transferidos para membrana
Hybond N+ e posteriormente hibridizados, utilizando como sonda os cDNAs de Notch
e Presenilina conforme descrito em Materiais e Métodos.
Embora anteriormente tendo amplificado um produto de aproximadamente 850
pb codificando para Notch e um produto de aproximadamente 400 pb codificando
para Presenilina, a análise de Northern Blot evidenciou uma mensagem de
aproximadamente 2.7Kb para Notch; (Figura 16 A), e para Presenilina, uma
mensagem de aproximadamente 3.25 Kb, (Figura 16 B). Essa diferença entre os
cDNAs amplificados e o tamanho estimado da mensagem codificadora,
provavelmente esta relacionado com a utilização de oligonucleotídeos específicos
durante a técnica de RT-PCR que estão flanqueando parte da região codificadora do
gene e, também, até o momento não amplificamos as regiões 5’e 3’ não traduzidas
dos genes Notch e Presenilina.
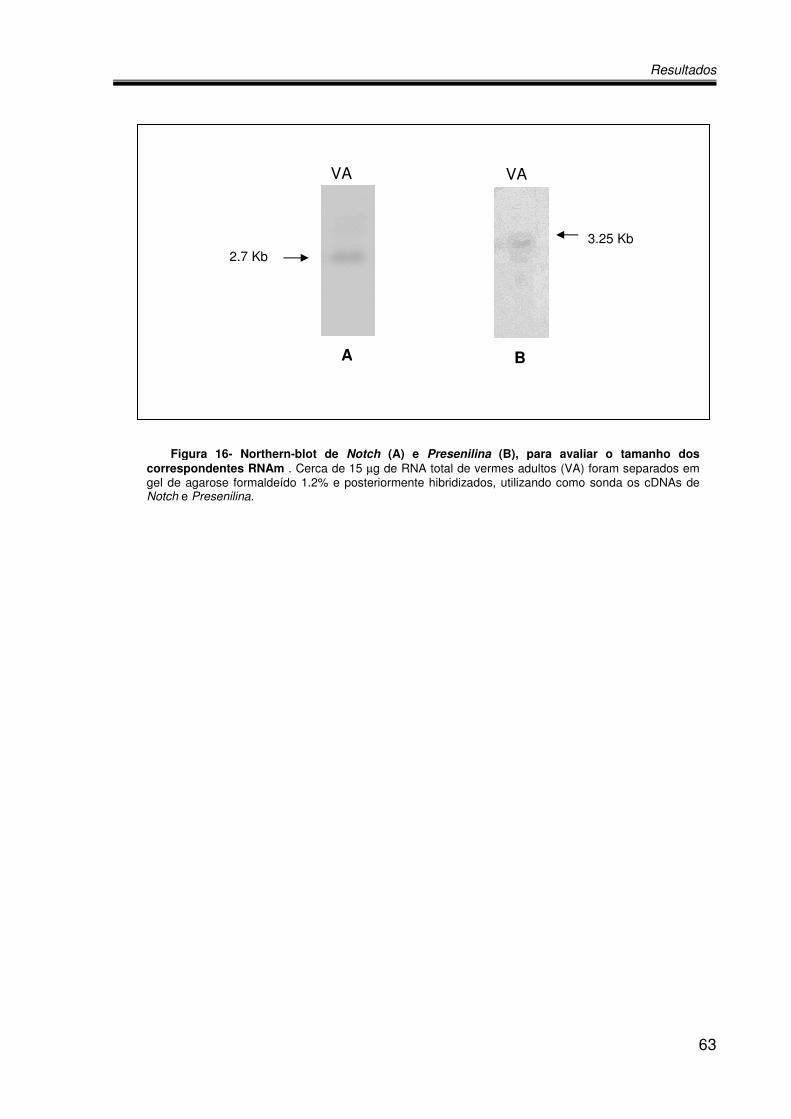
Resultados
63
2.7 Kb 3.25 Kb
VA VA
A B
Figura 16- Northern-blot de Notch (A) e Presenilina (B), para avaliar o tamanho dos correspondentes RNAm . Cerca de 15 µg de RNA total de vermes adultos (VA) foram separados em gel de agarose formaldeído 1.2% e posteriormente hibridizados, utilizando como sonda os cDNAs de Notch e Presenilina.

5- Discussão

Discussão
65
Para que ocorra o desenvolvimento de organismos multicelulares é necessário
que as células interpretem e transmitam sinais extrínsecos provenientes do meio ou
do hospedeiro em questão para o meio intracelular onde, culminará na ativação
transcricional de vários genes, resultando na diferenciação de um tipo celular. A
maioria desses sinais é liberada por um pequeno número de vias de sinalização, os
quais são altamente conversados entre as espécies. Dentre os mecanismos de
sinalização estão: a via Wingless, Fator de Crescimento e Transformação β,
Receptor de Tirosina Quinase, Hedgehog, Receptores Nucleares, JAK/STAT e Notch
(HANSSON, et al, 2004).
Estudos com S.mansoni, têm mostrado que as vias envolvidas com o
desenvolvimento do parasita, melhores caracterizadas até o momento são: Fator de
Crescimento e Transformação β (TGF-β), via mediada por Receptor de Tirosina
Quinase (RTK) e a via mediada por Receptores Nucleares (BELL & PEARCE, 2000;
SHOEMARKER, et al,1992, VICOGNE, et al, 2003; MENONÇA, et al 2000).
Nas últimas duas décadas, a via de transdução mediada pelo receptor Notch
vem sendo amplamente estudada em diversos modelos experimentais. Os
receptores Notch e seus ligantes foram primeiramente identificados em D.
melanogaster e C. elegans, onde mostraram regular tanto a proliferação como a
diferenciação celular. Estudos subseqüentes em camundongos e humanos
demonstraram que a sinalização mediada pelo receptor Notch era essencial em
múltiplos estágios da hematopoiese, como também na regulação do
desenvolvimento ou na homeostase celular em muitos tecidos (MCLNNES &
MICHAUD, 2003).

Discussão
66
Desse modo, a reconstituição in silico e a obtenção de seqüências dos
componentes essenciais dessa via, podem ser indicativos da presença da via Notch
durante o ciclo de vida de S. mansoni.
Neste contexto, a comparação de genes individuais de D. melanogaster e C.
elegans, por homologia de função, podem sugestivamente desempenhar papéis
semelhantes em parasitas como S. mansoni. Deste modo, a análise in silico da via
Notch pelo banco de dados do Projeto Transcriptoma de S. mansoni (VERSOVSKI,
et al, 2003), mostrou que este parasita apresenta várias seqüências codificando para
grande parte dos constituintes da via de sinalização mediada pelo receptor Notch. A
análise in silico também demonstrou que a maioria das seqüências para o receptor
Notch estavam presentes nas fases de esporocisto e esquistossômulos, no entanto,
a técnica de sequenciamento utilizada durante o Projeto Transcriptoma de S.
mansoni (VERSOVSKI, et al, 2003) somente nos permitiu estimar a expressão de
genes relacionados a via Notch, devido ao fato de as EST’s geradas terem sido
seqüenciadas aleatoriamente dos diversos estágios do parasita.
A análise comparativa dos componentes da via Notch com o Transcriptoma de
S. japonicum , mostrou que o parasita apresenta poucas seqüências codificando
para os constituintes da via Notch. Esses dados podem ser explicados devido a
utilização somente das fases de verme adulto, macho, fêmea e ovos na confecção
das bibliotecas de cDNA (WEI, et al, 2004).
Na caracterização molecular, o primeiro fragmento gênico investigado refere-se
ao próprio receptor Notch. Em S. mansoni a seqüência obtida durante o Projeto
Transcriptoma do parasita com similaridade significativa com este receptor em outros
organismos, contém apenas 763 pb (608600.1). A partir deste achado, houve a

Discussão
67
necessidade de extensão da seqüência consenso para o receptor utilizando-se de
ferramentas de bioinformática que permitiram a comparação entre a seqüência
proveniente do transcrito em S. mansoni com aquela depositada no banco genoma
do parasita, disponível no site: http://www.tigr.org. Com esta estratégia, foi possível
obtermos uma nova seqüência consenso para este receptor estendida em 221
nucleotídeos diferentemente distribuídos nas regiões 3' e 5' do transcrito original.
Uma nova idealização de oligonucleotideos permitiu a obtenção de um fragmento de
820 pares de bases, nas reações de amplificação utilizando o material genético do
parasita, e que corresponde a uma matriz de leitura aberta de 273 aminoácidos.
O segundo gene de interesse tem como produto um componente enzimático de
membrana denominado Presenilina. A função deste gene, já estabelecida, refere-se
a clivagem proteolítica da porção intracelular do receptor Notch de modo a liberar
esta última para atuação como fator de transcrição de genes alvos no núcleo celular
(CHUNG & STRUHL, 2001). Em S. mansoni, a seqüência obtida no Projeto
Transcriptoma deste parasita com similaridade significativa com a de outros
organismos, contém apenas 427 pb (705353.1). A partir deste achado, foi possível
de maneira análoga à utilizada para o receptor Notch, estender a seqüência
consenso para Presenilina em 59 nucleotídeos, diferentemente distribuídos nas
regiões 3' e 5' do transcrito original. Uma nova idealização de oligonucleotideos
permitiu a obtenção de um fragmento de 420 pares de bases nas reações de
amplificação utilizando material genético do parasita e que corresponde a uma matriz
de leitura aberta de 144 aminoácidos.
Um terceiro componente da via denominado Supressor de Deltex foi avaliado
neste estudo. Este gene codifica para uma enzima E3 ligase responsável pela

Discussão
68
ubiquitinação da porção intracelular do receptor Notch num evento mediado pelo
domínio HECT (Homologous to E-6AP carboxy-terminus) presente em sua estrutura.
Embora o mecanismo de ação desta enzima ainda permaneça por ser elucidado,
dados da literatura suportam a hipótese desta enzima atuar como um regulador
negativo da transdução de sinal mediada pela via Notch (ERIC,C.L, 2003). Utilizando
oligonucleotídeos específicos, foi possível a obtenção de um fragmento gênico de
811 pares de base, que corresponde a uma matriz de leitura aberta de 270
aminoácidos.
Estudos de organização estrutural dos genes Notch, Presenilina e Supressor de
Deltex em outros organismos demonstram a existência de íntrons (GREENWALD,
1985; MARTÍNEZ-MIR, et al, 2001; CORNELL, et al., 1999). Em nosso estudo, a
análise comparativa entre o DNA genômico e o cDNA dos fragmentos gênicos de
Notch, Presenilina e Supressor de Deltex através do alinhamento das seqüências e
do algoritmo BLAST 2 mostrou a ausência de introns no DNA genômico nas regiões
que estudamos.
Como descrito por WHARTON et al (1985). o receptor Notch é constituído por
uma porção extracelular, o qual é formada por repetições consecutivas de domínio
EGF e 3 repetições consecutivas Notch/Lin-12 rico em cisteína. Na porção
intracelular existem 6 repetições consecutivas de ankirin, um domínio rico em
glutamina (opa), e uma seqüência PEST (Região rica em prolina,glutamato, serina e
treonina) (Figura 17).
Com relação à Presenilina, dados da literatura demonstram que essa protease
é uma proteína com oito seções transmembrâna que apresenta o domínio
Presenilina (XIAJUN & GREENWALD, 1996; DOAN, et al. 1996). Para o Supressor

Discussão
69
de Deltex, estudos demonstraram que essa E3 ligase é constituída na região N-
termina de um domínio C2 (Protein kinase C conserved region 2 ), quatro domínios
WW (Two conserved tryptophans domain) e um domínio C-terminal HECT
(Homologous to E-6AP carboxy-terminus) (PERRY, et al., 1998). Desse modo.
através do algoritmo Pfam e do alinhamento com seqüências ortólogas, podemos
sugerir que o fragmento gênico Notch caracterizado faz parte da porção extracelular
do receptor, devido à presença de domínios de EGF.No caso do fragmento gênico
Presenilina, a análise pelo Pfam e o alinhamento demonstrou que o fragmento
caracterizado faz parte do domínio Presenilina. Por fim, o fragmento gênico de
Supressor de Deltex caracterizado faz parte da região C-terminal, devido a presença
do domínio HECT.
Neste trabalho, uma característica importante foi o perfil de expressão de
Notch, Presenilina e Supressor de Deltex durante o ciclo de vida do parasita. O
principal achado no contexto da biologia do S. mansoni, refere-se aos baixíssimos
Figura17: Similaridade estrutural entre os produtos de glp-1, lin-12, e Notch. Os 1295 aminoácidos de glp-1 são comparados com os 1429 aminoácios de lin-12 e os 2703 aminoácidos de Notch de D. melanogaster. Adaptado de - Yochem, J. and Greenwald, I. glp-1 and lin-12, Genes implicated in Distinct Cell-Cell Interations in C.elegans, encoded similar Transmembrane Proteins. Cell., v.58, p. 533-563, 1989.

Discussão
70
níveis de expressão do receptor, não detectados no estágio cercariano pela técnica
de RT-PCR mas, que foram observados durante a análise in silico poucos transcritos
para Notch nesse estágio. Esses baixíssimos níveis de expressão do receptor,
aliada à sua presença em níveis significativos, detectados nas fases de ovos,
esporocisto e esquistossômulo, sugerem que este gene poderia potencialmente
mediar o processo de diferenciação deste parasita, quando mantido no interior do
hospedeiro definitivo e intermediário. Sabendo que a via de sinalização Notch já foi
descrita estar envolvida com os processos de oogênese e espermatogênese em
outros organismos (PORTIN, 2002) e, considerando que o parasita S. mansoni
possui dimorfismo sexual e que no seu estágio de verme adulto a espermatogênese
e oogênese são constantes, (WERNER, 2001) sugerimos que a amplificação do
receptor Notch observada na fase de verme adulto poderia ser devido à presença de
células indiferenciadas relacionadas durante esses processos de oogênese e
espermatogênese. Experimentos futuros como de imunohistoquímica e de hibridição
em situ serão necessários para comprovarmos essa hipótese.
Conforme Wei, et al (2004), baseado nos dados dos Transcriptomas de S.
mansoni e S. japonicum, demonstrou a expressão de Notch somente no estágio de
esporocistos, esse fato pode ser devido a grande abundância de transcritos
observados nesse estágio de vida do parasita em relação aos outros estágios. Como
observamos nos nossos resultados, o receptor Notch se encontra expresso em
todos os estágios de vida do parasita S. mansoni.
A análise de expressão de Presenilina demonstrou a presença do transcrito em
todas as fases investigadas, inclusive no estágio de cercária. Este resultado reforça
a hipótese do envolvimento da Presenilina com outras vias de sinalização celular

Discussão
71
não diretamente relacionadas ao receptor Notch. (VIDAL, et al, 2005). Neste sentido,
a presença também em todas as fases do transcrito de Supressor de Deltex ao
longo do ciclo do parasita pode ser indicativa de funções celulares adicionais à
regulação desta via.
Outra possibilidade do controle da expressão que poderia ocorrer seria a
expressão sexo específico dos referidos fragmentos gênicos, no entanto, os
resultados, mostraram que os fragmentos gênicos foram expressos em ambos os
sexos do parasita.
Para estimarmos os tamanhos das mensagens de Notch e Presenilina, foram
realizados experimentos de Northern Blot. Dados da literatura demonstram que o
gene Notch em D. melanogaster codifica um transcrito único de aproximadamente
10kb (WHARTON, et al 1985). Em C. elegans os genes Lin-12 e Glp-1, os quais são
os homólogos de Notch codificam um transcrito único de aproximadamente 4.6 Kb
no caso de Lin-12 e um de 4.2 kb no caso de Glp-1 (AUSTIN. & KIMBLE., 1989). Em
S. mansoni, nossos resultados demonstraram um transcrito único de
aproximadamente 2.7 Kb. As diferenças, especificamente com relação ao tamanho
dos transcritos podem ser suportadas pelos dados da literatura que demonstra que
apesar do alto grau de conservação dos receptores Notch já caracterizados, as
proteínas de D. melanogaster e vertebrados diferem da proteína de C. elegans em
dois aspectos principais. Primeiro, o número e o espaçamento das repetições
semelhantes à EGF varia entre os organismos. O segundo fator que os distingue,
refere-se à presença de uma seqüência rica em poliglutamina denominada “opa”
localizada anteriormente à seqüência PEST C-terminal (Figura 17) (WHARTON et
al., 1985).

Discussão
72
Com relação a Presenilina, em nosso estudo foi obtido um transcrito único de
aproximadamente 3.25 Kb. Estudos realizados utilizando as fases de embrião, larva
e adulto de D. melanogaster demonstraram a presença de um transcrito único de
aproximadamente 2 Kb (CHANG-SOOK.& KOO, 1997). Contrapondo com esses
resultados, um outro estudo demonstrou a presença de transcritos de 2 a 5 Kb em
todas as fases de D.melanogaster, sugerindo a ocorrência de processamento
alternativo (BOULIANNE,et al, 1997)
Ao término da caracterização molecular de alguns componentes da via Notch
em S. mansoni, foi possível determinar que as fases de vida do parasita que são
mantidos no interior do hospedeiro definitivo e intermediário merecem ser
investigada com mais detalhes. Desta forma, os resultados desse trabalho
permitiram uma abordagem pioneira no estudo dessa via em S. mansoni e, para a
confirmação destas evidências tornam-se necessárias análises em caráter estrutural
e funcional, que contribuirão para um melhor entendimento da atuação tecido-
específica desta via.

6- Conclusões

Conclusões
74
Os resultados obtidos durante a realização desse trabalho nos permitiram
concluir que:
- A análise in silico da via Notch pelo banco de dados do Projeto Transcriptoma
de S. mansoni, mostrou que este parasita apresenta várias seqüências
codificando para grande parte dos constituintes da via de sinalização mediada
pelo receptor Notch.
- A análise comparativa dos componentes da via Notch com o Transcriptoma de
S. japonicum , mostrou que esse parasita apresenta poucas seqüências
codificando para os constituintes da via Notch. Esse dado pode ser explicado
devido à utilização somente das fases de verme adulto, macho, fêmea e ovos na
confecção das bibliotecas de cDNA.
- Os fragmentos de Presenilina e Supressor de Deltex apresentaram expressos
em todos os estágios do ciclo de vida do parasita S. mansoni analisados. Por
outro lado, os baixíssimos níveis de expressão de Notch no estágio de cercária
não detectados pela reação de RT-PCR, forneceu uma indicação de que esse
receptor pode ser essencial para a diferenciação celular das fases do parasita
dentro dos seus hospedeiros definitivo e intermediário.
- Não foi observadas íntrons nas seqüências de DNA genômico dos três
fragmentos gênicos analisados.

Conclusões
75
- A análise de Northern Blot demonstrou que o Notch possui um transcrito único
de aproximadamente 2.7 Kb enquanto que Presenilina possui um único transcrito
de aproximadamente 3.25 Kb.

7- Referências Bibliográficas

Referências Bibliográficas
77
ARTAVANIS-TSAKONAS, S.; RAND, M.D.; LAKE, R.J. Notch signaling: cell fate
control and signal integration in development. Science. 284:770-776, 1999.
AUSTIN, J.; KIMBLE, J. Transcript analysis of glp-1 and lin-12, homologous genes
required for cell interactions during development of C. elegans. Cell. 58:565-
571, 1989.
BARON, M.; ASLAM, H.; FLASZA, M.; FOSTIER, M.; HIGGS, J.E.; MAZALEYRAT
S.L.; WILKIN, M.B. Multiple levels of Notch signals regulation. Mol Membr
Biol.19:2269 – 2277, 2002.
BEALL, M. J.; MCGONIGLE, S.; PEARCE, E. J. Functional conservation of
Schistosoma mansoni Smads in TGF-beta signaling. Mol. Biochem. Parasitol.,
111 (1):131 – 142, 2000.
BOULIANNE, G.L..; LIVNE-BAR, I.; HUMPHREYS, J.M.; LIANG, Y.; LIN, C.;
ROGAEV, E.; GEORGE-HYSLOP, P.Cloning and characterization of the
Drosophila presenilin homologue. Neuroreport. 8:1025-1029, 1997.
CASTRO-BORGES, W. Diversidade do Proteassoma 26S e perfil de ubiquitinação
durante o desenvolvimento do parasita Schistosoma mansoni: Uma
abordagem proteômica. Tese de Doutorado, apresentada à Faculdade de
Medicina de Ribeirão Preto – Universidade de São Paulo, 2005.
CHANG-SOOK, H and KOO, E.H. Isolation and characterization of Drosophila
presenilin homolog. Neuroreport. 8:665-668, 1997.
CHUNG, H.M. & STRUHL, G. Nicastrin is required for Presenilin- mediated
transmembrane cleavage in Drosophila. Nat. Cell. Biol. 3:1129-1132, 2001.
CORNELL, M.; EVANS, D.; MANN, R.; FOSTIER, M.; FLASZA, M.;
MONTHATONG, M.; ARTAVANIS-TSAKONAS, S.; BARON, M. The Drosophila

Referências Bibliográficas
78
melanogaster Supressor of deltex gene, a regulator of the Notch receptor
signaling pathway, is an E3 Class Ubiquitin ligase. Genetics.152:567-576, 1999.
COSTA, P.I. Estudo da expressão de um gene codificando para o maior precursor
protéico do envoltório do ovo do trematoda Schistosoma mansoni . Tese de
Doutorado, apresentada à Faculdade de Mediciana de Ribeirão Preto -
Universidade de São Paulo, 1997.
DAVIES, S.J.; SHOEMAKER, C.B.; PEARCE, E.J. Divergent member of the
transforming growth factor beta receptor family from Schistosoma mansoni is
expressed on the parasite surface membrane. J Biol Chem. 273 (18):11234-40,
1998.
DAVIES, S.J; GROGAN, J.L; BLANK, R.B; LIM, K.C; LOCKSLEY, R.M;
MCKERROW, J.H. Modulation of blood fluke development in the liver by hepatic
CD4+ lymphocytes.Science. 294(5545):1358-1361, 2001.
DIAS - NETO, E; CORREA, R.G; VERJOVSKI-ALMEIDA, S; BRIONES, M.R;
NAGAI, M.A; DA SILVA, W. JR; ZAGO, M.A; BORDIN, S; COSTA, F.F;
GOLDMAN ,G.H; CARVALHO, A.F; MATSUKUMA, A; BAIA, G.S; SIMPSON,
D.H; BRUNSTEIN, A; DE OLIVEIRA, P.S; BUCHER, P; JONGENEEL, C.V;
O'HARE, M.J; SOARES, F; BRENTANI, R.R; REIS, L.F; DE SOUZA, S.J;
SIMPSON, A.J. Shotgun sequencing of the human transcriptome with ORF
expressed sequence tags. Proc Natl Acad Sci U S A. 97(7):3491-3496,2000.
DOAN, A.; THINAKARAN, G.; BORCHELT, R.D.; SLUNT, H.H.; RATOVITSKY, T.;
PODLISNY, M.; SLEKOE, J.D.; SEEGER, M.; GANDY, E.S.; PRICE, L.D.
Protein Topology of Presenilin 1. Neuron, 17: 1023-1030,1996.
EGAN, S.E.; ST – PIERRE, B.; LEOW, C.C. Notch receptors, partners and
regulators: from conserved domains to powerful functions. Curr. Top. Microbiol
Immun. 228: 273 – 324, 1998.

Referências Bibliográficas
79
ERIC, C.L..; DEBLANDRE, G.; KINTNER, C.; RUBIN, G.M. Drosophila Neutralized
is na ubiquitin ligase that promotes internalization and degradation of Deltex.
Dev Cell. 1:783 - 794, 2001.
ERIC, C.L. Protein Degradation: Four E3s for The Notch Pathway. Current Biology,
12: 74-78, 2003.
ERIC, C. L. Notch signaling: control of cell communication and cell fate.
Development. 131: 965-973, 2004.
FEINBERG, A.P.; VOGELSTEIN, B. A technique for radiolabeling DNA restriction
endonuclease fragments to high specific activity. Anal Biochem. 132: 06-13,
1993.
FORTINI, M.A. Notch and Presenilin: a proteolytic mechanism emerges. Current
Opinion in Cell Biology. 13: .627-634, 2001.
FOSTIER, M.; EVANS, D.; ARTAVANIS-TSAKONAS, S.; BARON ,M. Genetic
characterization of the Drosophila melanogaster Suppressor of deltex gene: a
regulator fo Notch signaling. Genetics. 150: 1477-1485, 1998.
FRANCO, G.R.; ADAMS, M.D.; BENTO SOARES, M.; SIMPSON, A.J.G.; VENTER,
J.C.; PENA, S.D.J. Identification of new Schistosoma mansoni genes by EST
strategy using a directional cDNA library. Gene. 152: 141 -147, 1995.
FRANCO, G.R.; VALADÃO, A.F.; AZEVEDO, V.; RABELO, E.M.L. The Schistosoma
mansoni gene Discovery program: State of Art. Intern. Journal for Parasitol. 30:
453-463, 2000.
GERHART, J. 1998 Warkany Lecture: Signaling Pathways in Development.
Teratology. 60: 226-239, 1999.

Referências Bibliográficas
80
GONZALEZ, E. Schistosomiasis, cercarial dermatitis, and marine dermatitis.
Dermatol Clin. 7(2):291-300, 1989.
GREENWALD, I. Lin-12, a nematode homeotic gene, is homologous to a set of
mammalian proteins that includes epidermal growth factor. Cell. 43: 583-590,
1985.
GREENWALD, I. Lin. 12/ Notch signaling: lessons from worms and flies. Genes &
Development.12: 1751 - 1762, 1998.
GRIDLEY, T. Notch signaling in vertebrate development and disease. Mol. Cell
Neurosci. 9(2): 103 – 108, 1997.
HANAHAN D., DNA cloning. A practical approach., v. 1 D. M. Glover, Edit. IRL,
Press Oxford, p. 109 - 135, 1985.
HANSSON, E. M; LENDAHL, U; CHAPMAN, G. Notch signaling in development and
disease. Seminars in Cancer Biology. 14(5): 320-328, 2004.
HARPER, J.A.; YUAN, J.S.; TAN, I.; VISAN, I.; GUIDOS, C.J. Developmental
Biology: Frontiers for Clinical Genetics. Clin Genet. 64: 461-472, 2003.
HARROP, R.; WILSON, R.A. Protein synthesis an release by cultures schistosomula
of Schistosoma mansoni. Parasitology. 107 (3): 265-274, 1993.
HILLIER, L.D.; LENNON, G.; BECKER, M. Generation and analysis of 280.000
human expressed sequence tags. Genome Res. 6: 807-828, 1996.
HOCKLEY, D.J. Ultrastructure of the Tegument of Schistosoma. In Advances in
Parasitology (ed. Dawes, B.), p.233-305, .Academic Press, London and New
York, 1973.

Referências Bibliográficas
81
HU, Y.; YE, Y.; FORTINI, M. Nicastrin is required for γ secretase cleavage of the
Drosophila Notch receptor. Dev. Cell. 2: 69-78, 2002.
ISMAIL, M; BOTROS, S; METWALLY, A; WILLIAM, S; FARGHALLY, A; TAO, L.F;
DAY, T.A; BENNETT, J.L. Resistance to praziquantel: direct evidence from
Schistosoma mansoni isolated from Egyptian villagers. Am J Trop Med Hyg.
60(6):932-935, 1999.
JOHNDTON, D.A.J. The WHO/UDP/World Bank Schistosoma Mansoni Genome
initiative: Current Status. Parasitol. Today.13:.45-46, 1997.
KAPP, K.;COUSTAU, C.; WIPPERSTEG, V.; JOURDANE, J.; KUNZ W,
GREVELDING, C.G. Transplantation of in vitro-generated Schistosoma mansoni
mother sporocysts into Biomphalaria glabrata. Parasitol Res. 91(6):482-485,
2003.
KARSAN, A. The role of notch in modeling and maintaining the vasculature. Can. J.
Physiol. Pharmacol..83 (1): 14-23, 2005.
KATZ, N. Schistosomiais control in Brazil. Mem. Inst. Oswaldo Cruz. 93: 33-35,
1998.
KIMBLE,J.; SIMPSON,P. The Lin-12/Notch Signaling pathway and its regulation.
Annu. Rev. 13:333-361, 1997.
LAMBERTUCCI, J.R.; BARRAVIERA, B. Equistossomose mansônica: estudo
clínico. Jornal Brasileiro Medicina. 67: 59-100, 1994.
LEE, N.H.; WEINSTOCK, K.G.; KIRKNESS, E.F. Comparative expressed-sequence
tag analysis of differential gene expression profiles in PC-12 cells before and

Referências Bibliográficas
82
after nerve growth factor treatment. Proc. Natl. Acad. Sci. USA. 92: 8303-8307,
1995.
LOGEAT, F.; BESSIA, C.; BROU, C.; LEBAIL, O.; JARRIAULT, S.; SEIDAH, N. G.;
SRAEL, A. The Notch 1 receptor is cleaved constitutively by a furin - like
convertase. Proc. Natl. Acad. Sci. USA. 95: 8108 - 8112, 1998.
MANGER, I.D.; HEHL, A.; PARMLEY, S. Expressed sequence tags analysis of the
bradzoite stage of Toxoplasmose gondii: Identification of developmentally
regulated genes. Infect. Immunol. 66:1632-1637, 1998.
MARTÍNEZ-MIR, A.; CAÑESTRO, C.; GONZÁLES- DUARTE, R.; ALBALAT, R.
Characterization of the amphioxus presenilin gene in high gene density genomic
region illustrates duplication during the vertebrate lineage. Gene. 279:157-164,
2001.
MCLNNES,R.R.& MICHAUD, J. Developmental Biology: Frontiers for Clinical
Genetics. Clin. Genet. 64: 461-472, 2003.
MELO, A.L & COELHO, P.M.Z. S. mansoni e a Doença. In Neves, D.P.
Parasitologia Humana, 11 ed, São Paulo: Atheneu, p. 193 - 212, 2005.
MENDONÇA, R.L.; ESCRIVÁ, H.; BOUTON, C.; LAUDET, V.; PIERCE, R.J.
Hormones and Nuclear Receptores in Schistosome Development. Parasitology
Today. 16: 233-240, 2000.
MUMM, J.S.; SCHROETER, M.M.; SAXENA MT, GRIESEMER A, TIAN X, PAN DJ,
RAY WJ, KOPAN R. A ligand-induced extracellular cleavage regulates gama-
secretase-like proteolytic activation of Notch1. Moll Cell. 5: 197-206, 2000.
MUMM, J.S.; KOPAN R. Notch Signaling: From the Outside In. Developmental
Biology. 228:151 - 165, 2000.

Referências Bibliográficas
83
MURTAUGH, L.G.; STANGER, B.Z.; KWAN, K.M.; MELTON, D.A. Notch signaling
controls multiple steps of pancreatic differentiation. PNAS. 100 (25):14920 –
14925, 2003.
NEVES, D.P. Parasitologia Humana 9 ed. São Paulo, Atheneu, . Cap. 20., p. 212-
237, 1998.
NIE, J.; MCGILL, M.; DERMER, M.; DHO, S.; WOLTING, C.; MCGLADE, C. J. LNX
functions as a RING type E3 ubiquitin ligase that targets the cell fate determinant
Numb for ubiquitin dependent degradation. EMBO. J. 21: 93 - 102, 2002.
OSMAN, A.; NILES, E.G.; LO VERDE, P.T. Identification and characterization of a
Smad2 homologue from Schistosoma mansoni, a transforming growth factor-
beta signal transducer. J. Biol. Chem. 276 (13): 10072-10082, 2001.
PEARCE, E.J.; MACDONALD, A.S. The immunobiology of schistosomiasis. Nat Rev
Immunol. 2(7):499-511,2002.
PERRY, W.L.; HUSTAND, C.M.; SWING, D.A.; O’SULLIVAN, T.N.;JENKINS, N.A.;
COPELAND, N.G. The itchy locus encodes a novel ubiquitin protein ligase that
is disrupted in a 18H mice. Nat Genet. 18: 143-146, 1998.
PORTIN, P. General outlines of the molecular genetics of the Notch signalling
pathway in Drosophila melanogaster: Hereditas. 136(2): 89-96, 2002.
POULSON, D.F. Histogenesis, organogenesis and differentiation in the embryo of
Drosophila melanogaster. In Biology of Drosophila, M. Demerec, ed (New York:
Wiley), 168-274, 1950.
RAND, M. D.; GRIMM, L. M.; ARTAVAINS –TSAKONAS, S.; PARTIUB, V.;
BLACKLOW, S.C.; SKLAR, J.; ASTER, J. Calcium depletion dissociates and
activates heterodimeric Notch receptor. Mol. Cell. Biol. 20: 1825 - 1835, 2000.

Referências Bibliográficas
84
ROLLINSON, D.; & SIMPSON, A.J.C.,eds. The Biology of Schistosoma mansoni:
from Genes to latrines. London: Academic Press, p 163 - 183, 1987.
RUMJANEK, F.D. Biochemistry and physiology. In Rollinson, D., Simpson,
A.J.C.,eds. The Biology of Schistosoma mansoni: from Genes to latrines.
London: Academic Press, p 163 - 183, 1987.
SALZET, M; CAPRON, A; STEFANO, G.B. Molecular crosstalk in host-parasite
relationships: schistosome- and leech-host interactions. Parasitol.Today. 16(12):
536-540, 2000.
SANGER, F.; NICKLEN, S..; COULSON, A.R. DNA sequencing with chain
terminating inhibitors. Proc. Natl. Acad. Sci. USA. 74: 5463 - 5467, 1977.
SHOEMAKER, C.B.; RAMACHANDRAN, H.; LANDA, A.; DOS REIS, M.G.; STEIN,
L.D. Alternative splicing of the Schistosoma mansoni gene encoding a
homologue of epidermal growth factor receptor. Mol Biochem Parasitol. 53(1-2):
17-32, 1992.
SMITHERS, S.R. ;& TERRY, R.J.; Infection of Laboratory hosts with cercariae of
Schistosoma mansoni: and the recovery of adult worms. Parasitololgy. 55: 695 -
700, 1965.
SPANA, E. P.; DOE, C. Q. Numb antagonizes Notch signaling to specify sibling
neuron cell fates. Neuron. 17: 21 - 26, 1996.
STADECKER, M.J. The role of T-cell anergy in the immunomudulation of
schistosomiasis. Parasitol. Today. 8: 199 - 204, 1992.
TANAKA, M.; HIRAI, H.; LO VERDE, P.T.; NAGAUCHI, S.; FRANCO, G.R.;
SIMPSON, A.J.G.; PENA, S.D.J. Yeast artificial chromosome (YAC) – based

Referências Bibliográficas
85
genome mapping of Schistosome mansoni. Mol. Biochem. Parasitol. 69:41-51,
1995.
UEMURA, T.; SHEPHERD, S.; ACKERMAN, L..; JAN, L .Y.; JAN, Y. N. Numb, a
gene required in determination of cell fate during sensory organ formation in
Drosophila embryos. .Cell. 58 (2): 349 -360, 1989.
VERJOVSKI-ALMEIDA S, DEMARCO R, MARTINS EA, GUIMARAES PE, OJOPI
EP, PAQUOLA AC, PIAZZA JP, NISHIYAMA MY JR, KITAJIMA JP, ADAMSON
RE, ASHTON PD, BONALDO MF, COULSON PS, DILLON GP, FARIAS LP,
GREGORIO SP, HO PL, LEITE RA, MALAQUIAS LC, MARQUES RC,
MIYASATO PA, NASCIMENTO AL, OHLWEILER FP, REIS EM, RIBEIRO MA,
SA RG, STUKART GC, SOARES MB, GARGIONI C, KAWANO T,
RODRIGUES V, MADEIRA AM, WILSON RA, MENCK CF, SETUBAL JC,
LEITE LC, DIAS-NETO E.Transcriptome analysis of the acoelomate human
parasite Schistosoma mansoni. Nat Genet.. 35(2): 148-157, 2003.
VICOGNE, J.; PIN, J.P.; LARDANS. V.; CAPRON, M.; NOE, C.; DISSOUS. C. An
unusual receptor tyrosine kinase of Schistosoma mansoni contains a Venus
Flytrap module. Mol. Biochem. Parasitol. 126 (1): 51-62, 2003.
VIDAL, G.A., NARESH, A., MARRERO, L. And JONES FE. Presenilin-dependent
gamma -secretase processing regulates multiple ERBB4/HER4 activities. J Biol
Chem. In press, 2005.
VIRAG, V.; ZSUZSANNA, K.; PALOCZI ,K.; UHER F. Notch signaling in the
regulation of hematopoiesis. Orv. Hetil.146 (7): 309-316,2005.
XIAJUN, L., GREENWALD, I. Membrane Topology of the C. elegans SEL-12
Presenilin. Neuron. 17: 1015-1021, 1996.

Referências Bibliográficas
86
WARREN, K.S., DOMINGO, E.O. AND COWAN, R.B.T. Granuloma formation
around schistosome eggs as a manifestation of delayed hypersensitivity.
American Journal of Pathology. 51: 735-756, 1987.
WEI, HU.; BRINDELY, J.P.; MCMANUS, P.D.; FENG, Z.; GUANG-HA, Z.
Schistosome transcriptome:new insights into the parasite and schistosomiasis.
TRENDS in Molecular Medicine.10: 218-225, 2004.
WERNER, K. Schistosome male - female interaction: induction pf germ cell
differentiation. TRENDS in Parasitology. 17(5): 227 - 231, 2001.
WHARTON, K.A.; JOHANSEN, K.M.; XU, T.; ARTAVANIS-TSAKONAS, S.
Nucleotide sequence from the neurogenic locus Notch implies a gene product
that shares homology with proteins containing EGF-like repeats. Cell. 43: 567-
581, 1985.
WILSON, R.A. Introdução à Parasitologia. Tradução: Cláudio Santos Ferreira e
Annete Silva Foronda. São Paulo; EPU; Ed. da Universidade de São Paulo,
1980.
WORLD HEATH ORGANIZATION. Division of Control of Tropical Disease.
Schistosomiasis Control. www.who.int/inf-fs/en/fact115.htlm, 1996.
WU ,G.; LYAPINA, S; DAS, I; LI, J; GURNEY, M; PAULEY, A; CHUI, I; DSHAIES,
R; KITAJEWSKI, J. Sel-10 is an inhibitor of Notch signaling that targets Notch
for ubiquitin- mediated protein degradation. Mol. Cell. Biol. 21: 7403-7415, 2001.
WU, L..; SUN, T.; KOBAYASHI, K.; GAO, P.; GRIFFIN, J.D. Identification of a
Family of Mastermind – Like Transcriptional Coactivators for Mammalian Notch
Receptors. Mol. Cell. Biol. 22(21): 7688 – 7700, 2002.

Referências Bibliográficas
87
YEH, E.; DERMER, M.; COMMISSO, C.; ZHOU, L.; MCGLADE, C.; BOULIANNE,
G. L.. Neutralized functions as na E3 ubiquitin ligase during Drosophila
devepolment. Curr. Biol. 11: 1675 - 1679, 2001.
YOCHEM, J.; GREENWALD. I. Glp-1 and lin-12, Genes implicated in distinct cell-
celll interactions in C. elegans, encode similar transmembrane proteins. Cell.
58: 55-563, 1989.
YU, G; NISHIMURA; M; ARAWAKA, S; LEVITAN, D; ZAHAN, L; TANDON, A;
SONG, Y.Q; ROGAEVA, E; CHEN, F; KAWARAI, T; SUPALA, A; LEVESQUE,
L; YU, H; YANG, D.S; HOLMES, E; MILMAN, P; LIANG, Y; ZHANG, D.M; XU,
D.H; SATO, C; ROGAEV, E; SMITH, M; JANUS, C; ZHANG, Y; AEBERSOLD,
R; FARRER, L.S; SORBI, S.; BRUNI, A; FRASER, P; ST, GEORGE-HYSLOP.
P. Nicastrin modulates presenilin-mediated Notch/GLP-1 signal transduction
and β-APP processing. Nature.407:48-54, 2000.

8-Apêndice

Apêndice
89
1- Representação esquemática da analise de domínios dos fragmentos gênicos
de Notch, Presenilina e Supressor de Deltex, utilizando o algoritmo Pfam.
1.1 Representação de Notch
1.2. Representação de Presenilina
1.3.Representação de Supressor de Deltex

Apêndice
90
2- Alinhamentos das seqüências dos cDNA dos fragmentos gênicos de Notch,
Presenilina e Supressor de Deltex com os respectivos DNAg.
2.1 Alinhamento de Notch
SmcDNA CATGTGACAAAGGGTATCACCAAGATTACAGAGGTCACTAGTCTTCCAAACAAATGTAGT 60
SmDNAg CATGTGACAAAGGGTATCACCAAGATTACAGAGGTCACTAGTCTTCCAAACAAATGTAGT 60
************************************************************
SmcDNA CCTGAATGTGTTCATGGACAAGGCGAATGTACATCTAATGGAAAATGCTTGTGTAATCCA 120
SmDNAg CCTGAATGTGTTCATGGACAAGGCGAATGTACATCTAATGGAAAATGCTTGTGTAATCCA 120
************************************************************
SmcDNA GGTTTTCGAGGGTTATCTTGTGAGACAGACGTAGATGAATGCGCTGAAGGAAAACATAAT 180
SmDNAg GGTTTTCGAGGGTTATCTTGTGAGACAGACGTAGATGAATGCGCTGAAGGAAAACATAAT 180
************************************************************
SmcDNA TGTCAACAGGAATGTATCAATACTCCTGGGTCTTTTAAATGTTCTTGTCATTCAGGTTAC 240
SmDNAg TGTCAACAGGAATGTATCAATACTCCTGGGTCTTTTAAATGTTCTTGTCATTCAGGTTAC 240
************************************************************
SmcDNA AAATTATCCACTGTAGATCCATCAAAATGTGAACCTATTTCTTGTCCTTCTCAATGCAAT 300
SmDNAg AAATTATCCACTGTAGATCCATCAAAATGTGAACCTATTTCTTGTCCTTCTCAATGCAAT 300
************************************************************
SmcDNA TTAATAGATGGAGAATGTGACTGCGAATGCAAGCTTGGACATTTTGGACTTAATTGTAAT 360
SmDNAg TTAATAGATGGAGAATGTGACTGCGAATGCAAGCTTGGACATTTTGGACTTAATTGTAAT 360
************************************************************
SmcDNA CAAACAGTAGATTCATGTGCAATTAAACCATGTGACCAAATTTGTGTGGATTTAGGTGAT 420
SmDNAg CAAACAGTAGATTCATGTGCAATTAAACCATGTGACCAAATTTGTGTGGATTTAGGTGAT 420
************************************************************
SmcDNA GGGAAGTATACATGCAAATGTAATGAAGGTTTTGAACCAAGTCCAAATAATCCTAGACGA 480
SmDNAg GGGAAGTATACATGCAAATGTAATGAAGGTTTTGAACCAAGTCCAAATAATCCTAGACGA 480
************************************************************
SmcDNA TGTCAGCGTATATGTAAAGATGGTATAGACTGTATATATGGAAGCTGTCGAACTGTGTTT 540
SmDNAg TGTCAGCGTATATGTAAAGATGGTATAGACTGTATATATGGAAGCTGTCGAACTGTGTTT 540
************************************************************
SmcDNA TCAAATGACACAGATCAATCTTTATGTACTTGCCTAGAAGGATTTTACGGAACAAACTGT 600
SmDNAg TCAAATGACACAGATCAATCTTTATGTACTTGCCTAGAAGGATTTTACGGAACAAACTGT 600
************************************************************
SmcDNA CAAAACGATGTAAACGAATGCTTGAATGAGAATGGTCAAAAACATTTTTGTGAACATCAT 660
SmDNAg CAAAACGATGTAAACGAATGCTTGAATGAGAATGGTCAAAAACATTTTTGTGAACATCAT 660
************************************************************
SmcDNA TGCATAAATACATTAGGATCATACCAATGTTTTTGTGATCCGGATTATGAATTACAATCT 720
SmDNAg TGCATAAATACATTAGGATCATACCAATGTTTTTGTGATCCGGATTATGAATTACAATCT 720
************************************************************
SmcDNA GATGGTCGAACGTGTAAAAAACAGTTTAAAGGTGAAATGAAATGTCCTGAATACTGCCTA 780
SmDNAg GATGGTCGAACGTGTAAAAAACAGTTTAAAGGTGAAATGAAATGTCCTGAATACTGCCTA 780
************************************************************
SmcDNA AATGGTGCAACTTGTAAATCGACTGGAGAATGTGTATGTC 820
SmDNAg AATGGTGCAACTTGTAAATCGACTGGAGAATGTGTATGTC 820
****************************************

Apêndice
91
2.2 Alinhamento de Presenilina
SmcDNA CCGAACCGATGCAATTGTAGTTGATACTCCTGATGACAATACGATTAATAATGATGCCAG 60
SmDNAg CCGAACCGATGCAATTGTAGTTGATACTCCTGATGACAATACGATTAATAATGATGCCAG 60
************************************************************
SmcDNA GAATAACAATGAAACTATATCACAGGCACCTCATGAAACAGATACACTTGTAGCAGGCCG 120
SmDNAg GAATAACAATGAAACTATATCACAGGCACCTCATGAAACAGATACACTTGTAGCAGGCCG 120
************************************************************
SmcDNA ACCCAATCGTTGGCAAGAACTGCAAGCAGATTTAATTGATCGTGACTCAAAGGGTGTAAA 180
SmDNAg ACCCAATCGTTGGCAAGAACTGCAAGCAGATTTAATTGATCGTGACTCAAAGGGTGTAAA 180
************************************************************
SmcDNA ACTTGGTTTAGGCGATTTTGTATTTTATAGTTTACTGATTGGTCGTGCAACACTTGATGG 240
SmDNAg ACTTGGTTTAGGCGATTTTGTATTTTATAGTTTACTGATTGGTCGTGCAACACTTGATGG 240
************************************************************
SmcDNA CGATGCAGTCACTGTGGTCACATGTTATGTGGCTATATTGGTTGGAATGTGTATTACTGT 300
SmDNAg CGATGCAGTCACTGTGGTCACATGTTATGTGGCTATATTGGTTGGAATGTGTATTACTGT 300
************************************************************
SmcDNA TATAGTATTGGGAATTACACGACAAGCATTACCAGCTTTACCAATATCCATAACATGTGG 360
SmDNAg TATAGTATTGGGAATTACACGACAAGCATTACCAGCTTTACCAATATCCATAACATGTGG 360
************************************************************
SmcDNA TATATTATTTTATTTTGTCAGTTCAGCGACGATATCACCGTTTCTTGAAGCAACAGCCAT 420
SmDNAg TATATTATTTTATTTTGTCAGTTCAGCGACGATATCACCGTTTCTTGAAGCAACAGCCAT 420
************************************************************
SmcDNA TGAACGTGTGTTC 433
SmDNAg TGAACGTGTGTTC 433
*************

Apêndice
92
2.3 Alinhamento de Supressor de Deltex SmcDNA TATGGGAGTCCCATTGCAATATGAAAGGAATTTCCGATCCAAAGTGAATTATTTTCGTGC 60
SmDNAg TATGGGAGTCCCATTGCAATATGAAAGGAATTTCCGATCCAAAGTGAATTATTTTCGTGC 60
************************************************************
SmcDNA TTGTTGTACAAATGCCATGTTAAATGGGCAAACTAAACTTCTTATATCAAGAGACAATTT 120
SmDNAg TTGTTGTACAAATGCCATGTTAAATGGGCAAACTAAACTTCTTATATCAAGAGACAATTT 120
************************************************************
SmcDNA ATTAGAGGACTCGTTCCAACTGGTATCTCAAATGTCTTCATCTGGTTTAAGAAGACGTTT 180
SmDNAg ATTAGAGGACTCGTTCCAACTGGTATCTCAAATGTCTTCATCTGGTTTAAGAAGACGTTT 180
************************************************************
SmcDNA ATCAATTACATTTCTACATGAAGAAGGTTTGGATTACGGTGGAGTAGCCAGAGAATGGTT 240
SmDNAg ATCAATTACATTTCTACATGAAGAAGGTTTGGATTACGGTGGAGTAGCCAGAGAATGGTT 240
************************************************************
SmcDNA CTATCGTCTTTCAAGAGAAATTTTAAACCCAATGTTTGGCTTGTTTGAGTATACTGGAAC 300
SmDNAg CTATCGTCTTTCAAGAGAAATTTTAAACCCAATGTTTGGCTTGTTTGAGTATACTGGAAC 300
************************************************************
SmcDNA AGATTATGCCTTACAAGTAAATCCAGCCTCTCATGTTAATCCAAATCATATGGCGTATTT 360
SmDNAg AGATTATGCCTTACAAGTAAATCCAGCCTCTCATGTTAATCCAAATCATATGGCGTATTT 360
************************************************************
SmcDNA TCGATTTGTTGGACGCTTTATTGGTATGGCACTTTTTCATGGTCGCTGTATTGACGGCGG 420
SmDNAg TCGATTTGTTGGACGCTTTATTGGTATGGCACTTTTTCATGGTCGCTGTATTGACGGCGG 420
************************************************************
SmcDNA GCTTACTCTTGCTTTTTATAAACAAATTTTGAAGCGGAAATTAACATTAGAAGATCTTGG 480
SmDNAg GCTTACTCTTGCTTTTTATAAACAAATTTTGAAGCGGAAATTAACATTAGAAGATCTTGG 480
************************************************************
SmcDNA TCATACGGATCACAGTTATTATCAATCGTTAATTTATGTCAGAGACAATCCAGTTGATGA 540
SmDNAg TCATACGGATCACAGTTATTATCAATCGTTAATTTATGTCAGAGACAATCCAGTTGATGA 540
************************************************************
SmcDNA GTGTGATTTGGACTTGTACTTTATTGGAACCTATGACCTTTTAGGCGTTTTGCATGAAGA 600
SmDNAg GTGTGATTTGGACTTGTACTTTATTGGAACCTATGACCTTTTAGGCGTTTTGCATGAAGA 600
************************************************************
SmcDNA CGAACTTATTGAAGGTGGCAAAGATATTAAAGTGACTGAAGAAAATAAATCCGATTATAT 660
SmDNAg CGAACTTATTGAAGGTGGCAAAGATATTAAAGTGACTGAAGAAAATAAATCCGATTATAT 660
************************************************************
SmcDNA CAGACTGATGGTTGATTGGCGGTTCAACAGAGGAGTAAAAAAACAAACAGAAGAAATTCA 720
SmDNAg CAGACTGATGGTTGATTGGCGGTTCAACAGAGGAGTAAAAAAACAAACAGAAGAAATTCA 720
************************************************************
SmcDNA CAAAGGTATTTTTGAAGTACTAAATCCCGAATGGCTGGAATTATTCGATGAGCGTGAACT 780
SmDNAg CAAAGGTATTTTTGAAGTACTAAATCCCGAATGGCTGGAATTATTCGATGAGCGTGAACT 780
************************************************************
SmcDNA TGAACTCCTATTATCCGGTATGCCAGAGATT 811
SmDNAg TGAACTCCTATTATCCGGTATGCCAGAGATT 811
*******************************
Apêndice
93
3. Representação esquemática da análise dos cDNA dos fragmentos gênicos
de Notch, Presenilina e Supressor de Deltex com os respectivos DNAg,
utilizando o algoritmo Blast 2
3.1. Representação esquemática de Notch
3.2. Representação esquemática de Presenilina
3.3. Representação esquemática de Supressor de Deltex
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo


















