
Fosforilação oxidativa - FMUPruifonte/PDFs/PDFs_arquivados_anos... · A desidrogénase do...
7
Fosforilação oxidativa Peter Mitchell, 1920 -1992 A fosforilação oxidativa é um processo metabólico em que as enzimas/transportadores envolvidos são proteínas da membrana interna da mitocôndria que, no seu conjunto, catalisam a formação de ATP (a partir de ADP + Pi) e a oxidação do NADH (e do FADH 2 ) pelo oxigénio molecular. NADH + ½ O 2 → H 2 O + NAD + (∆G ≈ -214 KJ; exergónico) 2,5 ADP + 2,5 Pi → 2,5 ATP + 2,5 H 2 O (∆G ≈ +125 KJ; endergónico) A “máquina de acoplagem” está na membrana interna da mitocôndria Porque o potencial de oxi-redução do par redox O 2 /H 2 O é muito positivo (Eº’ = + 0,815) e o do par NADH/NAD + tem um valor negativo elevado (Eº’ = - 0,315) os electrões têm tendência a moverem-se do NADH para o O 2 . A componente exergónica do processo é a oxidação do NADH pelo oxigénio. ∆Eº’= 1,13 V ⇔ Keq = 1,7 * 10 38 atm -1/2 ⇔ ∆Gº’ = -217 KJ NADH + ½ O 2 → H 2 O + NAD + NADH O processo de oxidação do NADH pelo oxigénio é catalisado por 3 complexos proteicos (I, III e IV) que usam a energia libertada no processo oxidativo para bombear (contra gradiente) protões da matriz da mitocôndria para o espaço intermembranar. A porina da membrana externa da mitocôndria permite a passagem de moléculas com massa ≈ 5000-10000 g/mol ⇒ existe equilíbrio (igual pH) entre o espaço intermembranar e o citosol.
Transcript of Fosforilação oxidativa - FMUPruifonte/PDFs/PDFs_arquivados_anos... · A desidrogénase do...

Fosforilação oxidativa
Peter Mitchell, 1920 -1992
A fosforilação oxidativa é um processo metabólico em que as enzimas/transportadores envolvidos são proteínas da membrana interna da mitocôndria que, no seu conjunto, catalisam a formação de ATP (a partir de ADP + Pi) e a oxidação do NADH (e do FADH2) pelo oxigénio molecular.
NADH + ½ O2 →H2O + NAD+
(∆G ≈ -214 KJ; exergónico)
2,5 ADP + 2,5 Pi →2,5 ATP + 2,5 H2O (∆G ≈ +125 KJ; endergónico)
A “máquina de acoplagem” está na membrana interna da mitocôndria
Porque o potencial de oxi-redução do par redox O2/H2O é muito positivo (Eº’ = + 0,815)
e o do par NADH/NAD+ tem um valor negativo elevado (Eº’ = - 0,315)
os electrões têm tendência a moverem-se do NADH para o O2.
A componente exergónica do processo é a oxidação do NADH pelo oxigénio.
∆Eº’= 1,13 V
⇔
Keq = 1,7 * 1038 atm-1/2
⇔
∆Gº’ = -217 KJ
NADH + ½ O2 →H2O + NAD+
NADH
O processo de oxidação do NADH pelo oxigénio é catalisado por 3 complexos proteicos (I, III e IV) que usam a energia libertada no processo oxidativo para bombear (contra gradiente) protões da matriz da mitocôndria para o espaço intermembranar.
A porina da membrana externa da mitocôndria permite a passagem de moléculas com massa ≈ 5000-10000 g/mol⇒ existe equilíbrio (igual pH) entre o espaço intermembranar e o citosol.
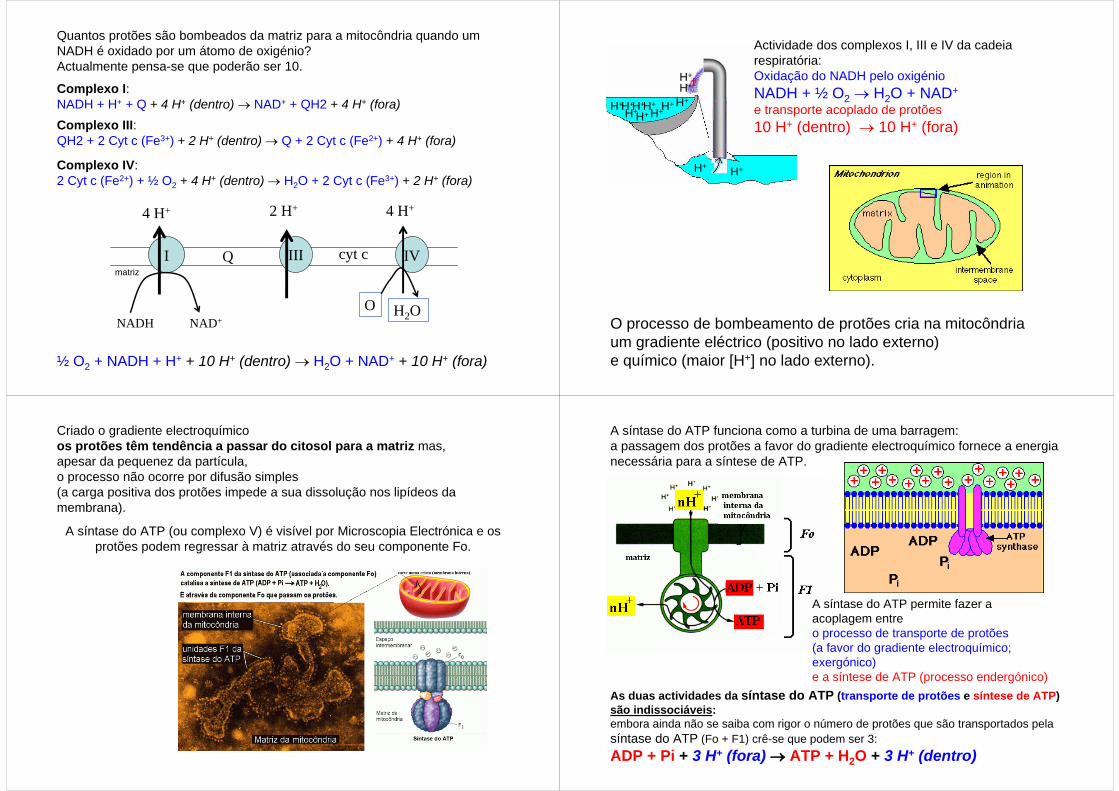
Quantos protões são bombeados da matriz para a mitocôndria quando um NADH é oxidado por um átomo de oxigénio? Actualmente pensa-se que poderão ser 10.
Complexo I: NADH + H+ + Q + 4 H+ (dentro) → NAD+ + QH2 + 4 H+ (fora)Complexo III: QH2 + 2 Cyt c (Fe3+) + 2 H+ (dentro) → Q + 2 Cyt c (Fe2+) + 4 H+ (fora)
Complexo IV: 2 Cyt c (Fe2+) + ½ O2 + 4 H+ (dentro) → H2O + 2 Cyt c (Fe3+) + 2 H+ (fora)
½ O2 + NADH + H+ + 10 H+ (dentro) → H2O + NAD+ + 10 H+ (fora)
I III IVQ cyt c
NADH NAD+
4 H+ 2 H+ 4 H+
O H2O
matriz
H+ H+
H+
H+H+H+H+
H+H+
H+ H+
H+
H+
Actividade dos complexos I, III e IV da cadeia respiratória:Oxidação do NADH pelo oxigénio NADH + ½ O2 → H2O + NAD+
e transporte acoplado de protões10 H+ (dentro) → 10 H+ (fora)
O processo de bombeamento de protões cria na mitocôndria um gradiente eléctrico (positivo no lado externo) e químico (maior [H+] no lado externo).
A síntase do ATP (ou complexo V) é visível por Microscopia Electrónica e os protões podem regressar à matriz através do seu componente Fo.
Criado o gradiente electroquímico os protões têm tendência a passar do citosol para a matriz mas, apesar da pequenez da partícula, o processo não ocorre por difusão simples (a carga positiva dos protões impede a sua dissolução nos lipídeos da membrana).
A síntase do ATP permite fazer a acoplagem entreo processo de transporte de protões (a favor do gradiente electroquímico; exergónico)e a síntese de ATP (processo endergónico)
A síntase do ATP funciona como a turbina de uma barragem: a passagem dos protões a favor do gradiente electroquímico fornece a energia necessária para a síntese de ATP.
As duas actividades da síntase do ATP (transporte de protões e síntese de ATP) são indissociáveis:embora ainda não se saiba com rigor o número de protões que são transportados pela síntase do ATP (Fo + F1) crê-se que podem ser 3: ADP + Pi + 3 H+ (fora) → ATP + H2O + 3 H+ (dentro)
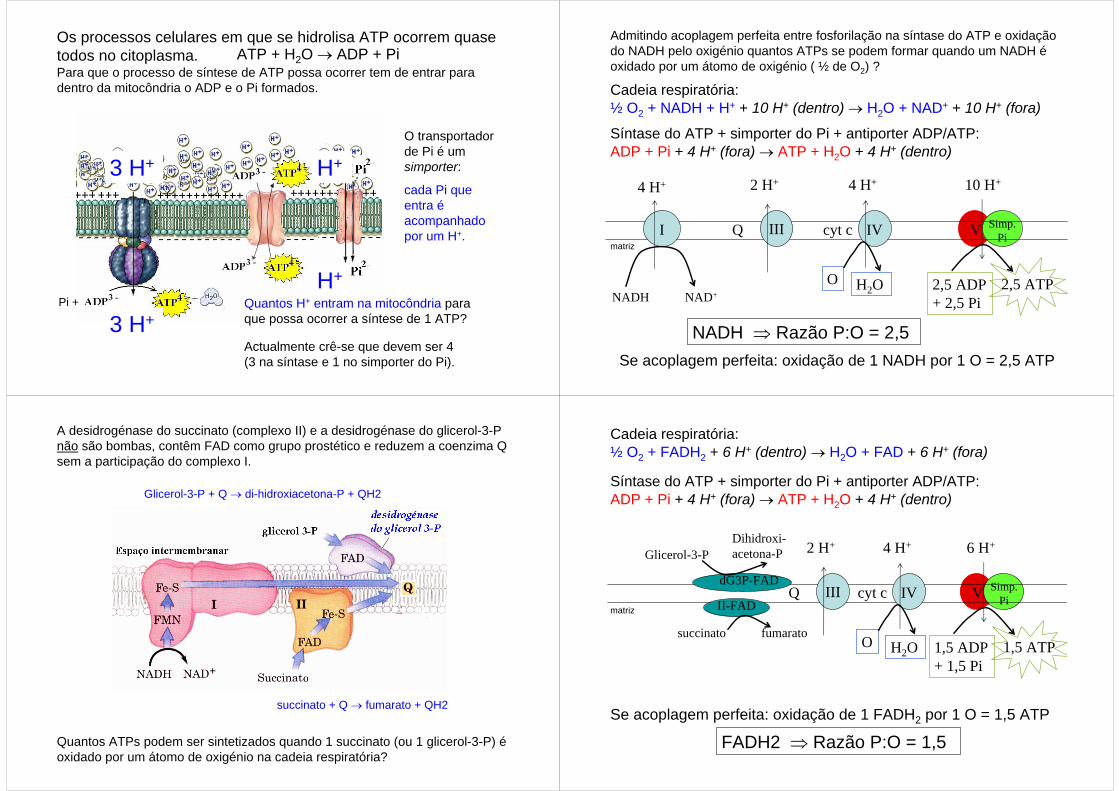
Os processos celulares em que se hidrolisa ATP ocorrem quase todos no citoplasma.Para que o processo de síntese de ATP possa ocorrer tem de entrar para dentro da mitocôndria o ADP e o Pi formados.
ATP + H2O → ADP + Pi
O transportador de Pi é um simporter:
cada Pi que entra é acompanhado por um H+.
Actualmente crê-se que devem ser 4 (3 na síntase e 1 no simporter do Pi).
3 H+
3 H+
H+
H+Pi + Quantos H+ entram na mitocôndria para
que possa ocorrer a síntese de 1 ATP?
Admitindo acoplagem perfeita entre fosforilação na síntase do ATP e oxidação do NADH pelo oxigénio quantos ATPs se podem formar quando um NADH é oxidado por um átomo de oxigénio ( ½ de O2) ?
Cadeia respiratória:½ O2 + NADH + H+ + 10 H+ (dentro) → H2O + NAD+ + 10 H+ (fora)
Síntase do ATP + simporter do Pi + antiporter ADP/ATP:ADP + Pi + 4 H+ (fora) → ATP + H2O + 4 H+ (dentro)
I III IVQ cyt c
NADH NAD+
4 H+ 2 H+ 4 H+
O H2O
V Simp. Pi
10 H+
2,5 ADP + 2,5 Pi
2,5 ATP
Se acoplagem perfeita: oxidação de 1 NADH por 1 O = 2,5 ATP
matriz
NADH ⇒ Razão P:O = 2,5
A desidrogénase do succinato (complexo II) e a desidrogénase do glicerol-3-P não são bombas, contêm FAD como grupo prostético e reduzem a coenzima Q sem a participação do complexo I.
Quantos ATPs podem ser sintetizados quando 1 succinato (ou 1 glicerol-3-P) é oxidado por um átomo de oxigénio na cadeia respiratória?
Glicerol-3-P + Q → di-hidroxiacetona-P + QH2
succinato + Q → fumarato + QH2
II-FADIII IV
dG3P-FADQ Vcyt c
fumarato
Simp. Pi
2 H+ 4 H+
succinato
6 H+
O H2O 1,5 ADP + 1,5 Pi
1,5 ATP
Glicerol-3-PDihidroxi-acetona-P
Se acoplagem perfeita: oxidação de 1 FADH2 por 1 O = 1,5 ATP
matriz
Cadeia respiratória:½ O2 + FADH2 + 6 H+ (dentro) → H2O + FAD + 6 H+ (fora)
Síntase do ATP + simporter do Pi + antiporter ADP/ATP:ADP + Pi + 4 H+ (fora) → ATP + H2O + 4 H+ (dentro)
FADH2 ⇒ Razão P:O = 1,5
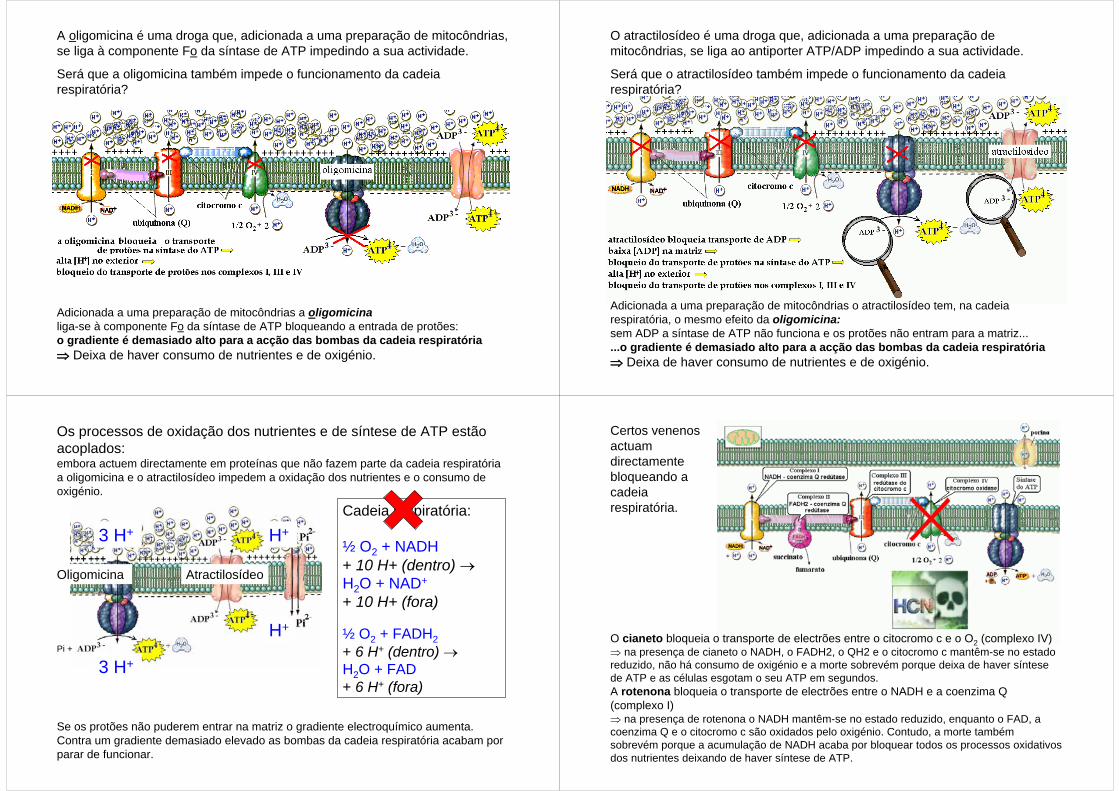
A oligomicina é uma droga que, adicionada a uma preparação de mitocôndrias, se liga à componente Fo da síntase de ATP impedindo a sua actividade.
Será que a oligomicina também impede o funcionamento da cadeia respiratória?
Adicionada a uma preparação de mitocôndrias a oligomicinaliga-se à componente Fo da síntase de ATP bloqueando a entrada de protões: o gradiente é demasiado alto para a acção das bombas da cadeia respiratória ⇒ Deixa de haver consumo de nutrientes e de oxigénio.
O atractilosídeo é uma droga que, adicionada a uma preparação de mitocôndrias, se liga ao antiporter ATP/ADP impedindo a sua actividade.
Será que o atractilosídeo também impede o funcionamento da cadeia respiratória?
Adicionada a uma preparação de mitocôndrias o atractilosídeo tem, na cadeia respiratória, o mesmo efeito da oligomicina:sem ADP a síntase de ATP não funciona e os protões não entram para a matriz... ...o gradiente é demasiado alto para a acção das bombas da cadeia respiratória ⇒ Deixa de haver consumo de nutrientes e de oxigénio.
3 H+
3 H+
H+
H+
Pi +
Oligomicina Atractilosídeo
Os processos de oxidação dos nutrientes e de síntese de ATP estão acoplados: embora actuem directamente em proteínas que não fazem parte da cadeia respiratória a oligomicina e o atractilosídeo impedem a oxidação dos nutrientes e o consumo de oxigénio.
Cadeia respiratória:
½ O2 + NADH+ 10 H+ (dentro) →H2O + NAD+
+ 10 H+ (fora)
½ O2 + FADH2+ 6 H+ (dentro) →H2O + FAD+ 6 H+ (fora)
Se os protões não puderem entrar na matriz o gradiente electroquímico aumenta. Contra um gradiente demasiado elevado as bombas da cadeia respiratória acabam por parar de funcionar.
Certos venenos actuam directamente bloqueando a cadeia respiratória.
O cianeto bloqueia o transporte de electrões entre o citocromo c e o O2 (complexo IV) ⇒ na presença de cianeto o NADH, o FADH2, o QH2 e o citocromo c mantêm-se no estado reduzido, não há consumo de oxigénio e a morte sobrevém porque deixa de haver síntese de ATP e as células esgotam o seu ATP em segundos.A rotenona bloqueia o transporte de electrões entre o NADH e a coenzima Q (complexo I) ⇒ na presença de rotenona o NADH mantêm-se no estado reduzido, enquanto o FAD, a coenzima Q e o citocromo c são oxidados pelo oxigénio. Contudo, a morte também sobrevém porque a acumulação de NADH acaba por bloquear todos os processos oxidativos dos nutrientes deixando de haver síntese de ATP.

Na glicólise aeróbia o NADH formado no citosol é oxidado na cadeia respiratória mas... não existe na membrana interna da mitocôndria um transportador para o NADH.O transporte dos equivalentes redutores do NADH formado no citosol para a matriz pode envolver a lançadeira do malato;nela cooperam (para além doutras enzimas e transportadores) a desidrogénase do malato e o “antiporter” do malato e do α-cetoglutarato.Neste caso, o NADH citosólico gera NADH na matriz da mitocôndria.
No músculo e no cérebro é mais importante alançadeira do glicerol-3-Pe nela cooperam 2 isoenzimas:
a) a desidrogénase do glicerol-3-P do citosol (dependente do NADH)b) a desidrogénase do glicerol-3-P da face externa da membrana interna da mitocôndria (que tem como grupo prostético o FAD)
Neste caso os equivalentes redutores fazem um “bypass” ao complexo I.
glicose
2 piruvato
2 acetil-CoA
4 CO2
2 piruvato
2 CO2
I IIIII IV
dG3PQ Vcyt c
2 ATP
2 ATP6 NAD+
6 NADH
2 NAD+
2 NADH
2 FAD
2 FADH2
2 NAD+
2 NADH
6 O2 H2OADP+Pi
26 ATP
28 ATP
Se houver acoplagem perfeita entre oxidação e fosforilação, na oxidação de um mole de glicose podem formar-se (1) admitindo a “lançadeira do malato” = 32 ATPs (28+4)
(2) admitindo a “lançadeira do glicerol-3-P” = 30 ATPs (26+4)
Simp. Pi
Chance e Williams(1955) JBC 217:383
Um automóvel aumenta de velocidade quando aumentamos a velocidade com que a gasolina é injectada no motor. E nos seres vivos como é que é regulada a velocidade de oxidação dos nutrientes?A 1ª resposta veio de estudos com mitocôndrias isoladas ainda antes de o modelo de Mitchell ter sido proposto (ADP = acelerador).
III-FAD
III IVdG3P-FAD
Q cyt c
NADH NAD+
H+ H+ H+
O H2O
V Simp. Pi
H+
↑ADP ATP ↓⇒ ↑ desidrogénases
Adaptando essa resposta à teoria de Mitchell.ADP ↑ e ATP ↓ ⇒ ↑ síntase do ATP ⇒↓ gradiente electroquímico da membrana mitocondrial ⇒↑ cadeia respiratória ⇒ ↓ [NADH] e ↑ [NAD+] ⇒↑ desidrogénases do ciclo de Krebs, glicólise e oxidação em β
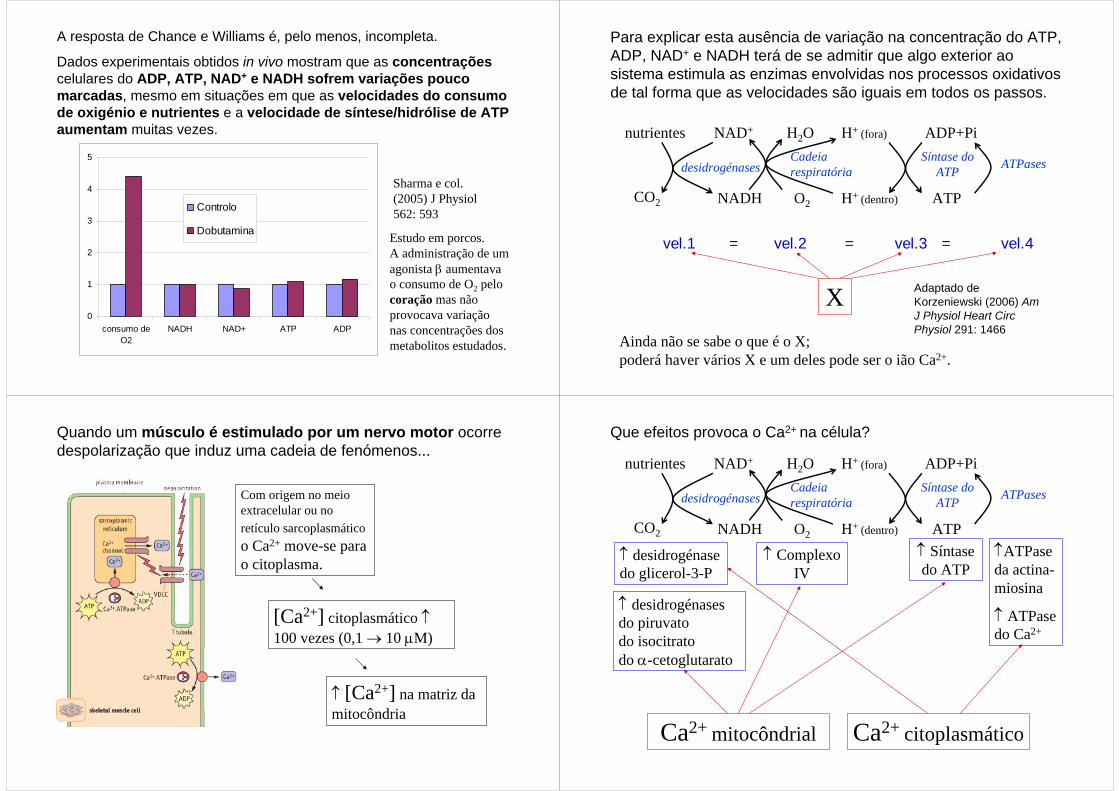
A resposta de Chance e Williams é, pelo menos, incompleta.
Dados experimentais obtidos in vivo mostram que as concentraçõescelulares do ADP, ATP, NAD+ e NADH sofrem variações pouco marcadas, mesmo em situações em que as velocidades do consumo de oxigénio e nutrientes e a velocidade de síntese/hidrólise de ATPaumentam muitas vezes.
0
1
2
3
4
5
consumo deO2
NADH NAD+ ATP ADP
Controlo
Dobutamina
Sharma e col. (2005) J Physiol562: 593
Estudo em porcos. A administração de um agonista β aumentava o consumo de O2 pelo coração mas não provocava variação nas concentrações dos metabolitos estudados.
Para explicar esta ausência de variação na concentração do ATP, ADP, NAD+ e NADH terá de se admitir que algo exterior ao sistema estimula as enzimas envolvidas nos processos oxidativos de tal forma que as velocidades são iguais em todos os passos.
NAD+
NADH
nutrientes
CO2 O2
H2O
H+ (dentro)
H+ (fora) ADP+Pi
ATP
ATPasesSíntase do ATP
Cadeia respiratóriadesidrogénases
vel.1 = vel.2 = vel.3 = vel.4
XAinda não se sabe o que é o X; poderá haver vários X e um deles pode ser o ião Ca2+.
Adaptado de Korzeniewski (2006) AmJ Physiol Heart CircPhysiol 291: 1466
Quando um músculo é estimulado por um nervo motor ocorre despolarização que induz uma cadeia de fenómenos...
Com origem no meio extracelular ou no retículo sarcoplasmáticoo Ca2+ move-se para o citoplasma.
[Ca2+] citoplasmático ↑100 vezes (0,1 → 10 µM)
↑ [Ca2+] na matriz da mitocôndria
Que efeitos provoca o Ca2+ na célula?
NAD+
NADH
nutrientes
CO2 O2
H2O
H+ (dentro)
H+ (fora) ADP+Pi
ATP
Síntase do ATP
Cadeia respiratóriadesidrogénases
Ca2+ citoplasmático
ATPases
↑ATPase da actina-miosina
↑ ATPase do Ca2+
↑ Síntasedo ATP
↑ Complexo IV
↑ desidrogénase do glicerol-3-P
↑ desidrogénases do piruvato do isocitratodo α-cetoglutarato
Ca2+ mitocôndrial
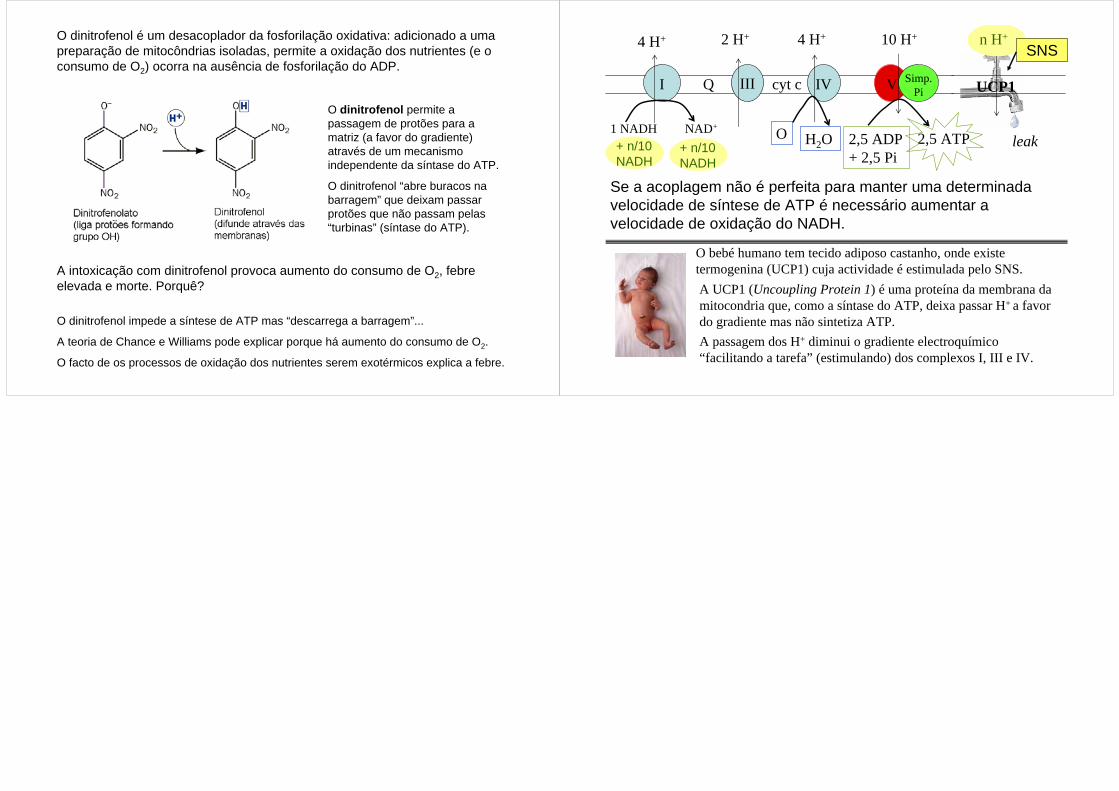
O dinitrofenol permite a passagem de protões para a matriz (a favor do gradiente) através de um mecanismo independente da síntase do ATP.
O dinitrofenol “abre buracos na barragem” que deixam passar protões que não passam pelas “turbinas” (síntase do ATP).
O dinitrofenol é um desacoplador da fosforilação oxidativa: adicionado a uma preparação de mitocôndrias isoladas, permite a oxidação dos nutrientes (e o consumo de O2) ocorra na ausência de fosforilação do ADP.
A intoxicação com dinitrofenol provoca aumento do consumo de O2, febre elevada e morte. Porquê?
O dinitrofenol impede a síntese de ATP mas “descarrega a barragem”...
A teoria de Chance e Williams pode explicar porque há aumento do consumo de O2.
O facto de os processos de oxidação dos nutrientes serem exotérmicos explica a febre.
I III IVQ cyt c
1 NADH NAD+
4 H+ 2 H+ 4 H+
O H2O
V Simp. Pi
10 H+
2,5 ADP + 2,5 Pi
2,5 ATP
Se a acoplagem não é perfeita para manter uma determinada velocidade de síntese de ATP é necessário aumentar a velocidade de oxidação do NADH.
O bebé humano tem tecido adiposo castanho, onde existe termogenina (UCP1) cuja actividade é estimulada pelo SNS.A UCP1 (Uncoupling Protein 1) é uma proteína da membrana da mitocondria que, como a síntase do ATP, deixa passar H+ a favor do gradiente mas não sintetiza ATP.A passagem dos H+ diminui o gradiente electroquímico “facilitando a tarefa” (estimulando) dos complexos I, III e IV.
UCP1
n H+
leak+ n/10 NADH
+ n/10 NADH
SNS


















