
metabolismo e exercício físico03 - FMUPruifonte/PDFs/PDFs_arquivados_anos... · A contração de...
22
1 Metabolismo dos carbohidratos e das gorduras no exercício físico Departamento de Bioquímica da Faculdade de Medicina do Porto [email protected]
-
Upload
nguyenthuan -
Category
Documents
-
view
221 -
download
1
Transcript of metabolismo e exercício físico03 - FMUPruifonte/PDFs/PDFs_arquivados_anos... · A contração de...

1
Metabolismo dos carbohidratos e das gorduras
no exercício físico
Departamento de Bioquímica da Faculdade de Medicina do Porto

A contração de uma fibra muscular é uma consequência da despolarização da
membrana sarcoplasmática. Esta despolarização origina-se na ligação da acetilcolina
libertada no terminal do axónio motor aos recetores nicotínicos da membrana pós-
sináptica da placa motora.
2
Com origem no retículo sarcoplasmático(e no meio extracelular)o Ca2+ move-se para o citoplasma.
A [Ca2+] citoplasmática ↑ 100 vezes (0,1 µM → 10 µM)
A contração termina quando o Ca2+ é bombeado para o interior do retículo sarcoplasmático e para o meio extracelular (ATPases do Ca2+) regressando às concentrações de repouso.
(A [Ca2+] também ↑ na mitocôndria)

Filamentos grossos (miosina)
Filamentos finos (actina, tropomiosina, troponina) Sarcómero distendido
Sarcómero contraído
Ca2+ ligado a troponina C⇒ ligação actina-miosina…
ATP + H2O
ADP + Pi
A contração muscular é o resultado do deslizamento entre os filamentos finos e grossos com aumento do grau de sobreposição entre estes filamentos ⇒encurtamento do sarcómero e das miofibrilas.
O processo de contração/relaxamento consome ATP: - ATPase da miosina- ATPase do Ca2+
Os músculos são “máquinas” que convertem energia química em movimento.
Ca2+
3
ATPase da miosina

4
0,28
18,9
46,2
1,12
21
49
0
10
20
30
40
50
60
descanço maratona sprint de 100 m
Despesa
energética
(kcal/min)
músculo esquelético
organismo
25%
90%
94%Despesa
energética nos
músculos/min Despesa
energética total/min
A razão…
Esta razão é, em repouso, de ≈≈≈≈ 25% e aumenta com o exercício. É maior nos exercícios de tipo anaeróbico (até ≈≈≈≈ 94%) que nos de tipo aeróbico (até ≈ 90%), aumentando com a intensidade do exercício.
A despesa energética do organismo como um todo é, maioritariamente, o somatório:
despesa energética basal (BMR) + despesa associada ao exercício físico.
0
5
10
15
20
25
30
35
40
45
50
BMR dormir barrer ochão
caminhar 5km/h
"jogging" maratona sprint 100m
Despesa
energética
total
(kcal/min)
Se considerarmos o organismo como um todo 1- um atleta durante a maratona tem uma despesa ≈ 20 xs superior à basal.2- um sprinter enquanto corre os 100 m tem uma despesa ≈ 50 xs superior à basal.
1,1 kcal/min

O ATP (5 mM) que está presente nas fibras musculares só seria capaz de manter a
contração muscular durante uma fração de segundo ou poucos segundos mas,
mesmo quando o exercício físico é intenso, a concentração de ATP praticamente não
varia. Cada molécula de ATP que sofre hidrólise é imediatamente reposta. Como?
Fosfo-creatina
Creatina
ADP
ATP
Glicogénio
G1P
lactato
ADP
ATP
VLDL no plasma
G6PADP
ATP
Glicogénio
G1P
CO2
TAG
das próprias
fibras
(triacilgliceróis
intramiocelulares)
ADP
ATP
CO2CO2
TAG do tecido adiposo
FFA
ADP
ATP
CO2
no plasma
Acil-CoA
Lípase de lipoproteínas
Acil-CoAADP
ATPCO2
FFA
no plasma
Acil-CoA
Os aminoácidos têm um contributo relativamente pequeno: talvez 1-5% da energia (?) 5

A maioria das atividades inclui exercícios de vários tipos (anaeróbico, aeróbico e
misto), mas é útil fazer a distinção entre exercício aeróbico e anaeróbico.
Tipo de exercício anaeróbico aeróbico
Exemplos halterofilismo, sprint de 100 m, saltos, rematar com força
caminhar, correr (por exemplo, a maratona)
Irrigação e consumo de O2
dos músculos implicados
Aumento nulo ou irrelevante Aumento muito marcado (pode ser de 100 xs)
Substituição do ATP
hidrolisado dependente
de...
Fosfocreatina, glicogénio armazenado nos músculos em contração e glicólise anaeróbica
Oxidação completa (a CO2) de nutrientes armazenadosno músculo, mas tb no tecido adiposo e no fígado
Potência (energia
gasta/unidade de tempo)
Muitíssimo elevada; nas fibras em contração, o gasto de ATP pode ser 300 xs superior ao de repouso
Menos elevada que no anaeróbico mas, quando a intensidade é elevada (taxa de consumo de O2 elevado),nas fibras em contração, o gasto de ATP pode ser 130 xssuperior ao de repouso
Duração Curta Pode ser longa
Tipo de fibras
predominantemente
implicadas
Tipo II (brancas, glicolíticas ou de contração rápida)
Tipo I (vermelhas, oxidativas, ou de contração lenta)
6

Nos exercícios de tipo anaeróbico, a ação da cínase da creatina é relevante, mas a
fosfocreatina esgota-se em alguns segundos.
ATPase da miosina
ADP ATP
Cínase da creatina
(junto da miofibrilas)
Cínase da creatina
(espaço intermembranar da mitocôndria)
matriz mitocondrialATPADP
ATPADP
creatinafosfocreatina
A reposição parcial ou total (durante o repouso) dos níveis de fosfocreatina ocorre por ação de moléculas da cínase da creatina situadas no espaço intermembranar da mitocôndria onde o quociente de reação favorece a reação inversa (a fosforilação da creatina pelo ATP).
descida da fosfocreatina em 10 s de exercício anaeróbico
10 s
H2OPi
7
ATP + H2O → ADP + Pifosfocreatina + ADP → ATP + creatina

No exercício anaeróbico, a fosforólise do glicogénio situado nas fibras musculares
que estão a contrair é muito estimulada sustentando um aumento da glicólise que
pode ficar 1000 vezes mais rápida que no repouso.
1º - O Ca2+ ↑↑↑↑ ⇒ ativação da cínase da fosforílase
2º- O aumento da hidrólise do ATP ⇒ [AMP] ↑↑↑↑
3º [Pi] ↑↑↑↑ resultante da ↓ de fosfocreatina
Fatores que contribuem para a ativação da glicogenólise:
4º - A adrenalina circulante (que aumenta durante o exercício) poderá ter algum papel facilitador via recetores β, ativação da PKA efosforilação (ativação) da cínase da
fosforílase.
Mas a adrenalina sem exercício muscular e ↑ da [Pi] NÃO provoca aumento da fosforólise.
⇒ ativação da fosforílase do
glicogénio
Glicogénio
Glicose-1-P
Pi
8
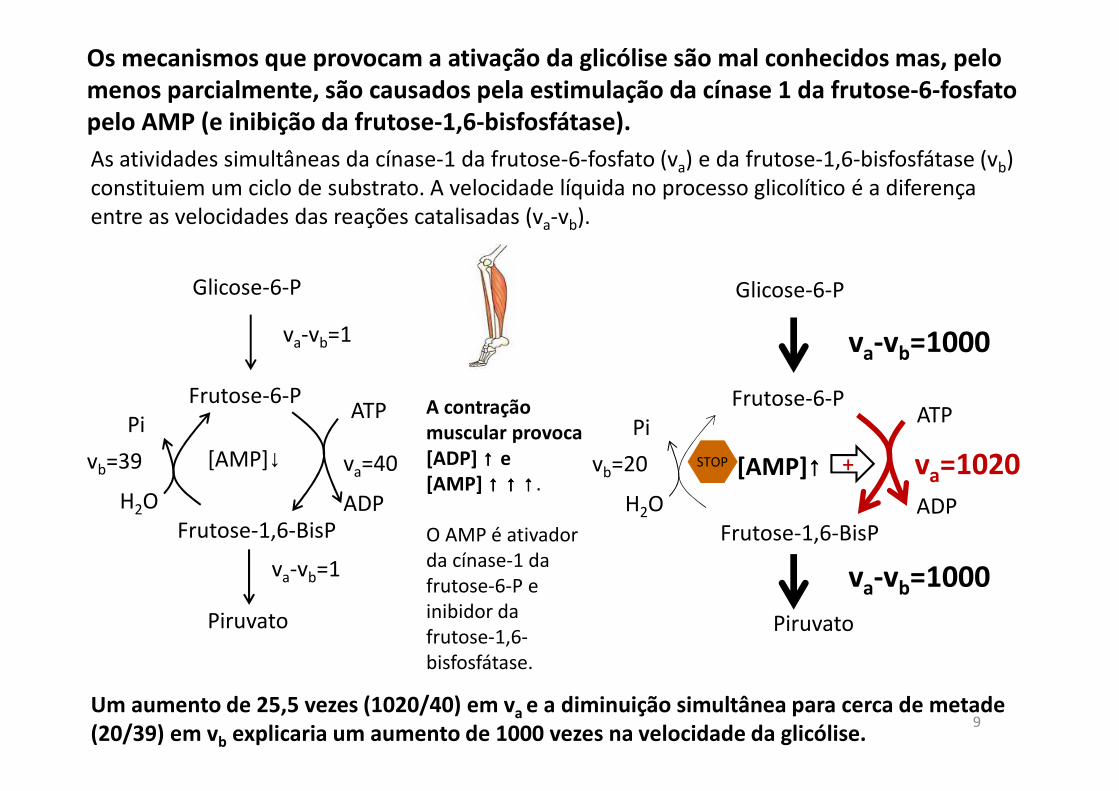
Os mecanismos que provocam a ativação da glicólise são mal conhecidos mas, pelo
menos parcialmente, são causados pela estimulação da cínase 1 da frutose-6-fosfato
pelo AMP (e inibição da frutose-1,6-bisfosfátase).
Glicose-6-P
Frutose-6-P
Frutose-1,6-BisP
Piruvato
va-vb=1
ATP
ADPH2O
Pi
va=40vb=39
va-vb=1
[AMP]↓
Glicose-6-P
Frutose-6-P
Frutose-1,6-BisP
Piruvato
va-vb=1000
ATP
ADPH2O
Pi
vb=20
va-vb=1000
[AMP]↑↑↑↑ va=1020
A contração
muscular provoca
[ADP] ↑↑↑↑ e
[AMP] ↑↑↑↑ ↑↑↑↑ ↑↑↑↑.
O AMP é ativador da cínase-1 da frutose-6-P e inibidor da frutose-1,6-bisfosfátase.
Um aumento de 25,5 vezes (1020/40) em va e a diminuição simultânea para cerca de metade
(20/39) em vb explicaria um aumento de 1000 vezes na velocidade da glicólise.9
STOP +
As atividades simultâneas da cínase-1 da frutose-6-fosfato (va) e da frutose-1,6-bisfosfátase (vb) constituiem um ciclo de substrato. A velocidade líquida no processo glicolítico é a diferença entre as velocidades das reações catalisadas (va-vb).
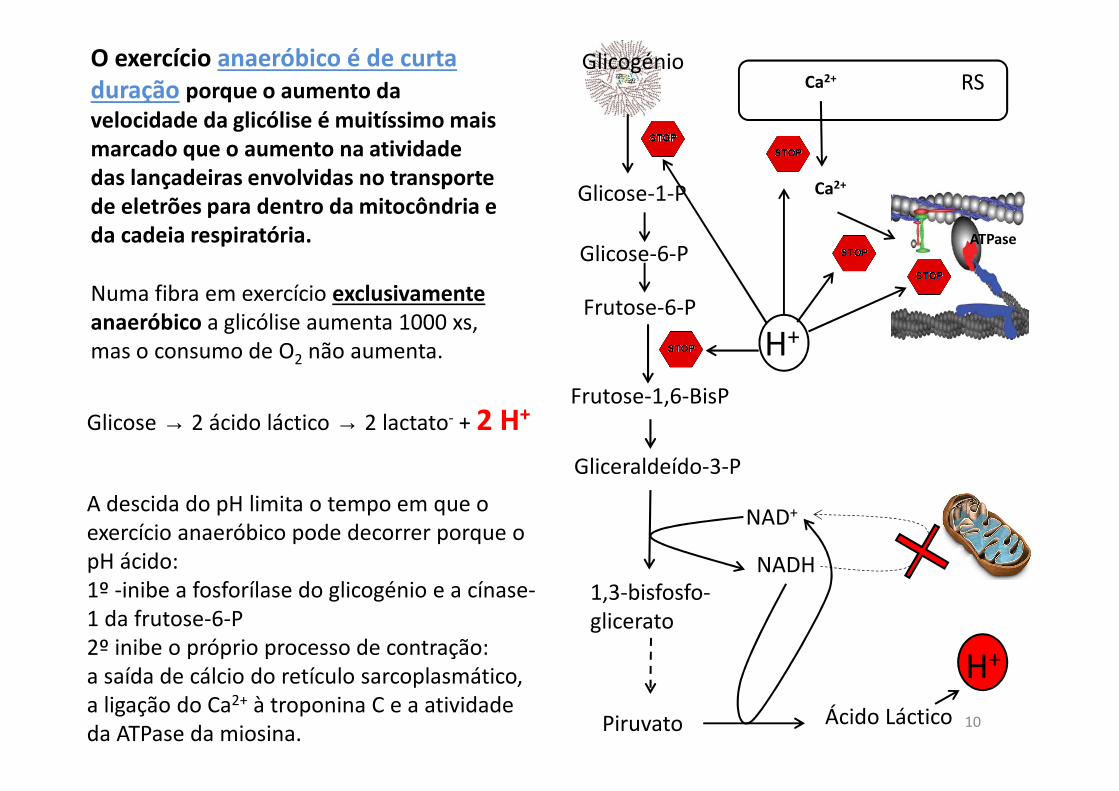
O exercício anaeróbico é de curta
duração porque o aumento da
velocidade da glicólise é muitíssimo mais
marcado que o aumento na atividade
das lançadeiras envolvidas no transporte
de eletrões para dentro da mitocôndria e
da cadeia respiratória. Glicose-6-P
Piruvato
Gliceraldeído-3-P
1,3-bisfosfo-glicerato
NAD+
NADH
Ácido Láctico
A descida do pH limita o tempo em que o exercício anaeróbico pode decorrer porque o pH ácido:1º -inibe a fosforílase do glicogénio e a cínase-1 da frutose-6-P2º inibe o próprio processo de contração: a saída de cálcio do retículo sarcoplasmático, a ligação do Ca2+ à troponina C e a atividade da ATPase da miosina.
Numa fibra em exercício exclusivamente
anaeróbico a glicólise aumenta 1000 xs, mas o consumo de O2 não aumenta.
Frutose-6-P
Frutose-1,6-BisP
H+
H+
10
Glicose-1-P
Glicogénio
Ca2+
ATPase
RSCa2+
Glicose → 2 ácido láctico → 2 lactato- + 2 H+
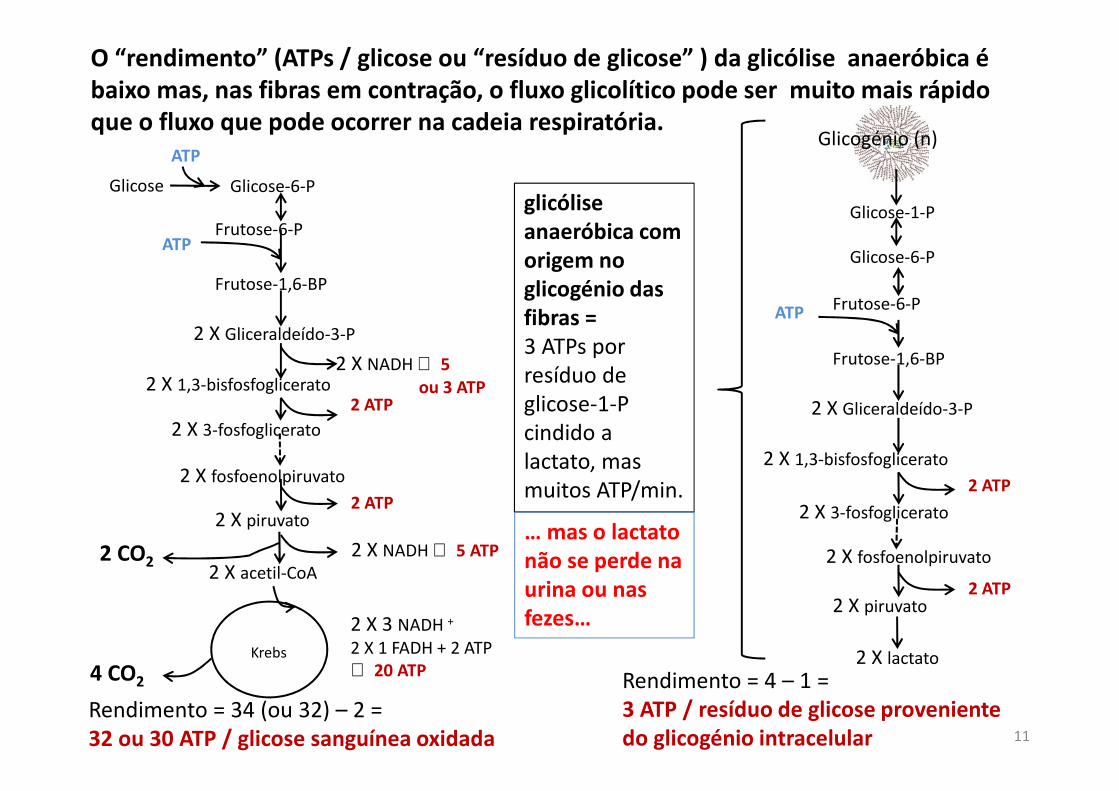
O “rendimento” (ATPs / glicose ou “resíduo de glicose” ) da glicólise anaeróbica é
baixo mas, nas fibras em contração, o fluxo glicolítico pode ser muito mais rápido
que o fluxo que pode ocorrer na cadeia respiratória.
Glicose Glicose-6-P
ATP
Frutose-6-P
Frutose-1,6-BP
ATP
2 X Gliceraldeído-3-P
2 X 1,3-bisfosfoglicerato 2 X NADH ⇔ 5
ou 3 ATP
2 X piruvato
2 X 3-fosfoglicerato
2 ATP
2 X fosfoenolpiruvato
2 ATP
2 X acetil-CoA 2 X NADH ⇔ 5 ATP
4 CO2
2 X 3 NADH +
2 X 1 FADH + 2 ATP⇔ 20 ATP
Rendimento = 34 (ou 32) – 2 =32 ou 30 ATP / glicose sanguínea oxidada
Glicose-6-P
Frutose-6-P
Frutose-1,6-BP
ATP
2 X Gliceraldeído-3-P
2 X 1,3-bisfosfoglicerato
2 X piruvato
2 X 3-fosfoglicerato
2 ATP
2 X fosfoenolpiruvato
2 ATP
2 X lactato
Rendimento = 4 – 1 =3 ATP / resíduo de glicose proveniente
do glicogénio intracelular
glicólise
anaeróbica com
origem no
glicogénio das
fibras =
3 ATPs por resíduo de glicose-1-P cindido a lactato, mas muitos ATP/min.
11
Glicose-1-P
Glicogénio (n)
2 CO2
Krebs
… mas o lactato
não se perde na
urina ou nas
fezes…

Embora o tempo em que o exercício de tipo aeróbico pode manter-se dependa da
intensidade (e do treino), em geral, pode prolongar-se por horas.
12
Num dado indivíduo a intensidade do exercício
aeróbico costuma medir-se como uma percentagem do VO2 máximo
Intensidade baixa ⇔≈ 25% do VO2 max
Intensidade intermédia ⇔geralmente de 45-60% do VO2 max
Intensidade alta ⇔> 80% do VO2 max
Exemplos: caminhar a 5 km/h ⇔ 4-5 kcal/min, correr a maratona ⇔ 20-25 kcal/min.A despesa energética em repouso ≈ 1,1 kcal/ min.
Exercício
anaeróbico
exercício aeróbico
Consumo de O2 mantém-se no máximo; 100 % VO2 max
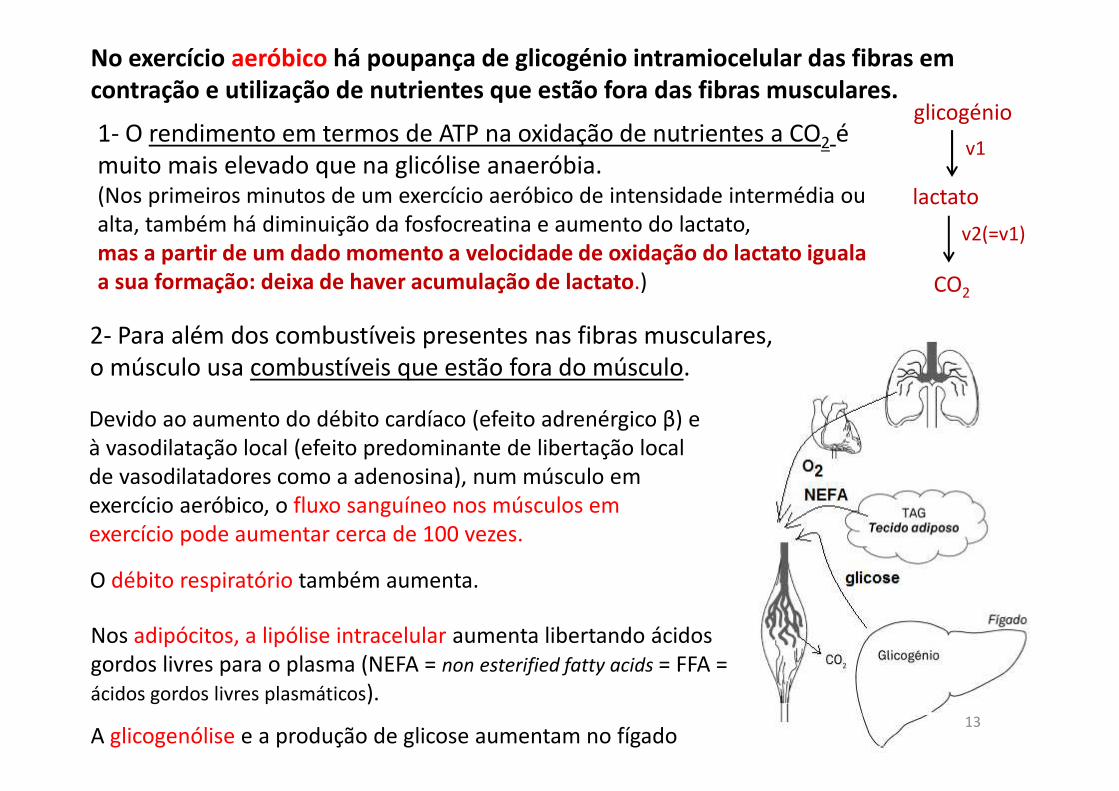
No exercício aeróbico há poupança de glicogénio intramiocelular das fibras em
contração e utilização de nutrientes que estão fora das fibras musculares.
1- O rendimento em termos de ATP na oxidação de nutrientes a CO2 é muito mais elevado que na glicólise anaeróbia. (Nos primeiros minutos de um exercício aeróbico de intensidade intermédia ou alta, também há diminuição da fosfocreatina e aumento do lactato,mas a partir de um dado momento a velocidade de oxidação do lactato iguala
a sua formação: deixa de haver acumulação de lactato.)
Devido ao aumento do débito cardíaco (efeito adrenérgico β) e à vasodilatação local (efeito predominante de libertação local de vasodilatadores como a adenosina), num músculo em exercício aeróbico, o fluxo sanguíneo nos músculos em exercício pode aumentar cerca de 100 vezes.
O débito respiratório também aumenta.
Nos adipócitos, a lipólise intracelular aumenta libertando ácidos gordos livres para o plasma (NEFA = non esterified fatty acids = FFA = ácidos gordos livres plasmáticos).
A glicogenólise e a produção de glicose aumentam no fígado
2- Para além dos combustíveis presentes nas fibras musculares, o músculo usa combustíveis que estão fora do músculo.
13
glicogénio
lactato
CO2
v1
v2(=v1)

As quantidades e o tipo de nutrientes oxidados durante os exercícios aeróbicos
variam com a intensidade do exercício.1- A oxidação da glicose sanguínea (= glicogénio hepático) aumenta com a intensidade do exercício.
2- O contributo do glicogénio das fibras musculares em atividade contratil aumenta muito para intensidades acima de 25% do VO2
max.
3- Entre o repouso e 25% do VO2max a oxidação dos ácidos gordos livres (FFA) aumenta, mas acima de 25% do VO2 max passa a haver diminuição.
4- A oxidação dos TAG intracelulares + TAG das VLDL (TAG=triacilgliceróis) aumenta com a intensidade, mas deixa de aumentar quando a intensidade do exercício fica acima de 50% do VO2 max.
14
0
5
10
15
20
25
0% 25% 50% 80%
kca
l/m
in
Intensidade do exercício (% VO2 max)
Glicogénio do músculo
Glicose sanguínea (glicogénio hepático)
TAG (VLDL e intramiocelular)
FFA (lipólise tecido adiposo)
5- O Quociente Respiratório aumenta quando a intensidade do exercício aumenta acima de 25% do VO2 max aproximando-se de 1 para exercícios de alta intensidade.
Repouso

Que fatores dentro dos músculos estimulam a captação e a oxidação
da glicose quando a intensidade do exercício aeróbico aumenta?
1-De forma independente da insulina, por um mecanismo que envolve a ativação da AMPK (pelo AMP ↑↑↑↑), o exercício provoca a mobilização para a membrana sarcoplasmática de moléculas de GLUT4.
3-O ↑↑↑↑ do [Ca2+] no sarcoplasma também difunde para as mitocôndrias onde a [Ca2+] também ↑.
piruvato
Acetil-CoA
H2O
PiFosfátase da desidrogénase
do piruvato
Ca2+
desidrogénase do
isocitrato
desidrogénase do
α-cetoglutarato
Ca2+NADH
Complexo I
Complexo IV
e-
O2
Síntase do ATP
ADP + Pi ATP
Ca2+
Ciclo de Krebs
5- O efeito ativador do ADP e do Ca2+ ajudam a explicar o aumento da oxidação de todos os
nutrientes (que, em todos os casos, é sempre maior durante o exercício que no repouso).
2- [AMP] ↑↑↑↑ ⇒ ↑ da cínase 1 da frutose-6-P e ↓ da fosfátase da frutose-1,6-BisP.
ADP ↑
4- O [ADP] ↑ e ativa a síntase do ATP.
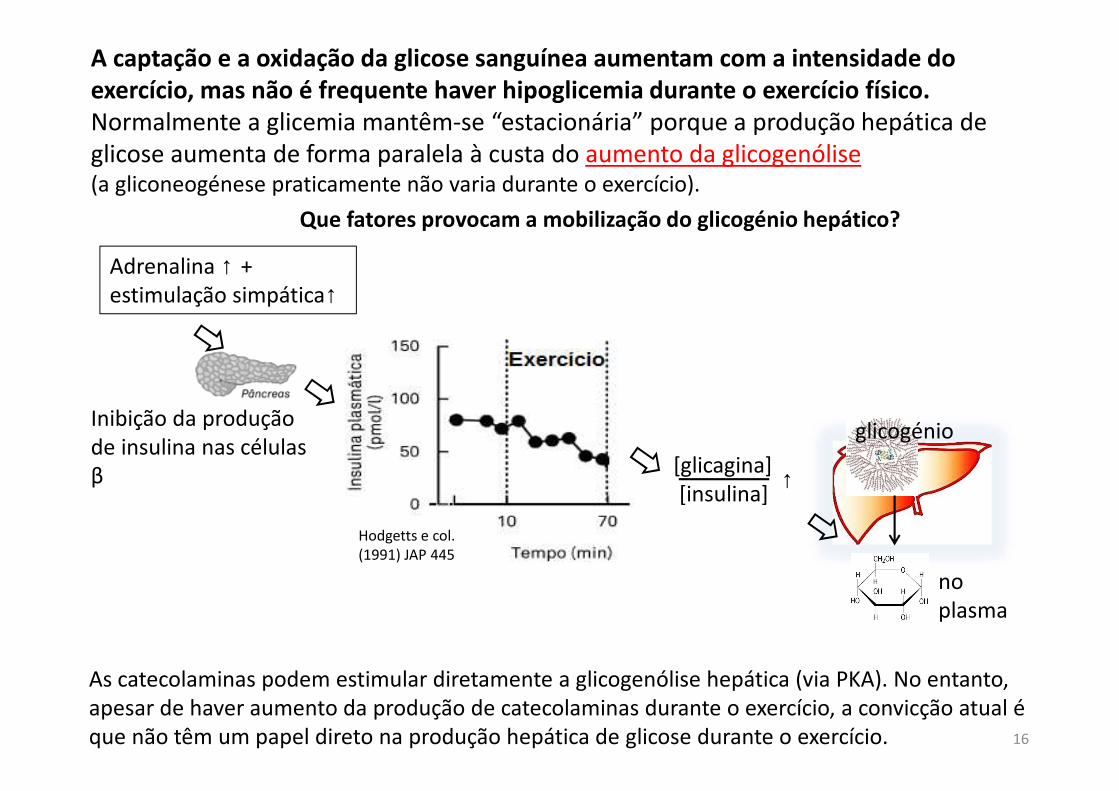
A captação e a oxidação da glicose sanguínea aumentam com a intensidade do
exercício, mas não é frequente haver hipoglicemia durante o exercício físico.
Normalmente a glicemia mantêm-se “estacionária” porque a produção hepática de glicose aumenta de forma paralela à custa do aumento da glicogenólise (a gliconeogénese praticamente não varia durante o exercício).
As catecolaminas podem estimular diretamente a glicogenólise hepática (via PKA). No entanto, apesar de haver aumento da produção de catecolaminas durante o exercício, a convicção atual é que não têm um papel direto na produção hepática de glicose durante o exercício.
Hodgetts e col. (1991) JAP 445
Que fatores provocam a mobilização do glicogénio hepático?
16
Adrenalina ↑ + estimulação simpática↑
[glicagina][insulina]
↑
no plasma
glicogénioInibição da produção de insulina nas células β

Em exercícios de baixa intensidade os
ácidos gordos plasmáticos (FFA) são o
principal combustível dos músculos em
exercício aeróbico.
Que fatores poderão ajudar a explicar este aumento de consumo de ácidos gordos livres plasmáticos?
A [FFA] no plasma mantém-se estacionária porque quer o ↑↑↑↑ da adrenalina e noradrenalina, quer a ↓↓↓↓ da insulina contribuem para o aumento da lipólise
Dentro dos músculos:
1- ↑↑↑↑ da irrigação sanguínea ⇒⇒⇒⇒ ↑ oferta de ácidos gordos livres2-A ativação pelo Ca2+ das desidrogénases do ciclo de Krebs, dos complexos da cadeia respiratória e síntase do ATP estimulam a oxidação dos ácidos gordos.3-O ADP estimula a síntase de ATP.
O malonil-CoA intramuscular não varia
durante o exercício e, por isso, pensa-se que não tem relevância na regulação da oxidação de ácidos gordos durante o exercício físico.
(via ↑ das formas fosforiladas da lípase hormono-sensível e da perilipina).

Quando a intensidade do exercício é
elevada, ↑↑↑↑ o contributo dos carbohidratos
(glicogénio muscular e glicose sanguínea),
mas ↓↓↓↓ o contributo dos ácidos gordos
livres (FFA).
Porque é que diminui a oxidação dos FFA?
Fatores intramusculares.
1- A par da estimulação da oxidação da glicose também há aumento da glicólise e aumento da produção de lactato com descida do pH
2- O aumento do fluxo na desidrogénase do piruvato leva à acumulação de acetil-CoA que reage com a carnitina (formando acetil-carnitina)limitando a quantidade de carnitina disponível.
glicose → piruvato → acetil-CoA ↑⇒ carnitina + acetil-CoA → acetil-carnitina + CoA⇒ [carnitina] intramiocelular ↓
18
↓ da oxidação em β
↓ atividade da carnitina-palmitil transférase 1
⇒
glicogénio das fibras
glicose
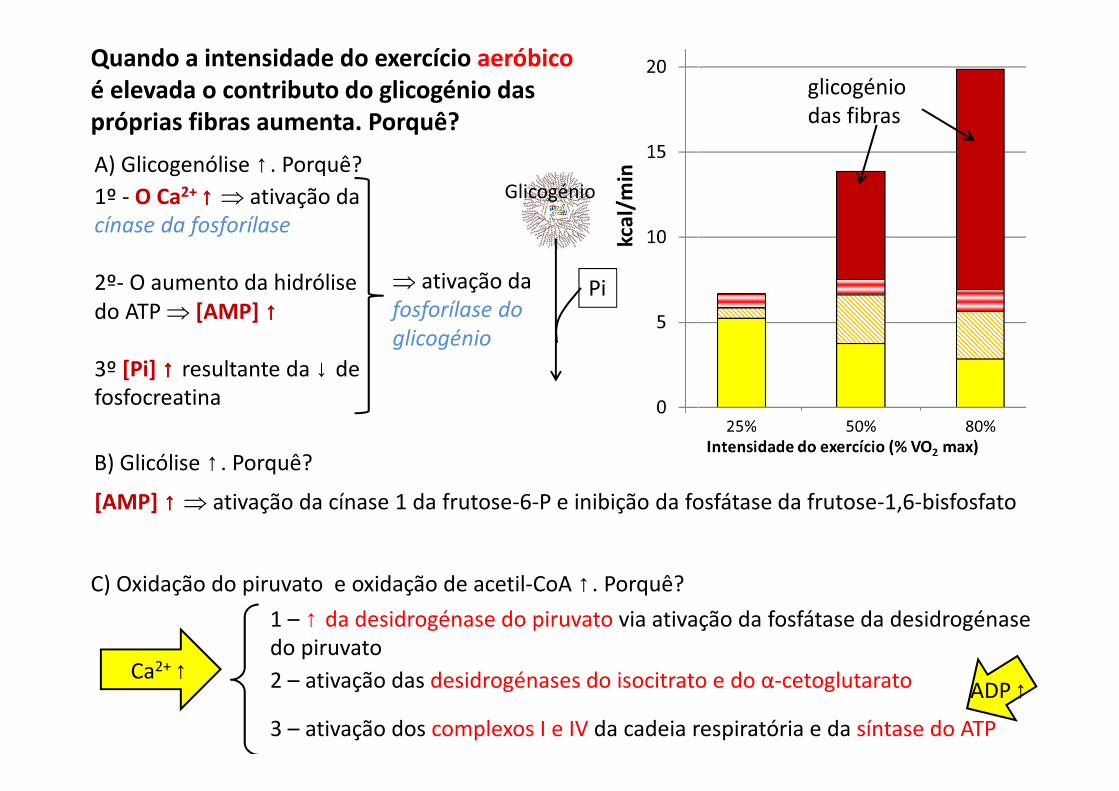
Quando a intensidade do exercício aeróbico
é elevada o contributo do glicogénio das
próprias fibras aumenta. Porquê?
glicogénio das fibras
A) Glicogenólise ↑. Porquê?
1º - O Ca2+ ↑↑↑↑ ⇒ ativação da cínase da fosforílase
2º- O aumento da hidrólise do ATP ⇒ [AMP] ↑↑↑↑
3º [Pi] ↑↑↑↑ resultante da ↓ de fosfocreatina
⇒ ativação da fosforílase do
glicogénio
Glicogénio
Pi
C) Oxidação do piruvato e oxidação de acetil-CoA ↑. Porquê?
Ca2+ ↑
1 – ↑ da desidrogénase do piruvato via ativação da fosfátase da desidrogénase do piruvato
2 – ativação das desidrogénases do isocitrato e do α-cetoglutarato
3 – ativação dos complexos I e IV da cadeia respiratória e da síntase do ATP
B) Glicólise ↑. Porquê?
[AMP] ↑↑↑↑ ⇒ ativação da cínase 1 da frutose-6-P e inibição da fosfátase da frutose-1,6-bisfosfato
ADP ↑

A hidrólise dos TAG acumulados nas
próprias fibras musculares (≈≈≈≈ 300 g no
total) e
a subsequente oxidação dos ácidos
gordos libertados
também contribuem para a substituição
do ATP durante o exercício aeróbico.
Pensa-se que o contributo dos TAG das VLDL para a despesa energética durante o
exercício muscular é quantitativamente pouco relevante.
O exercício físico aumenta a síntese da lípase de lipoproteínas do tecido muscular (efeito a longo prazo): este é um efeito com importância na descida das VLDL plasmáticas, mas o contributo dos TAG das VLDL para a despesa energética do músculo é irrelevante. A massa total de TAG plasmáticos num indivíduo saudável é de 3 g (1% da massa de TAG intramiocelulares). 20
Triacilgliceróis intramiocelulares (e das VLDL)
gotícula TAG
miofibrilas

Efeitos a longo prazo do exercício físico continuado no metabolismo.
1- O exercício físico aeróbico continuado aumenta a sensibilidade à insulina porque…… estimula a síntese de GLUT4 aumentado a capacidade da insulina para estimular a captação de glicose nas fibras musculares.
3- O exercício físico anaeróbico aumenta a síntese das proteínas que são componentes das miofibrilas: hipertrofia
das fibras musculares (quer as de tipo 1 quer as de tipo 2).
2- O exercício físico continuado estimula a síntese da
lípase de lipoproteínas muscular ⇒ ↓ [TAG do plasma] (das VLDL e dos quilomicra) porque acelera a sua degradação.
4- O exercício físico aeróbico não provoca hipertrofia muscular, mas aumenta a quantidade de mitocôndrias,
o transportador membranar FABPpm e
a densidade da rede capilar e
aumentando capacidade para exercícios aeróbicos. 21
FABPpmácidos gordos dentro da fibra muscular
ácidos gordos

Bibliografia usada:
Frayn, K. N. (2012) Regulação Metabólica. Uma perspetiva focada no organismo humano., U.P. Editorial, Porto.Stipanuk, M. H. (2012) Biochemical, Physiological and Molecular Aspects of Human Nutrition, 3rd edn, Elsevier, St. Louis.
van Loon, L. J., Greenhaff, P. L., Constantin-Teodosiu, D., Saris, W. H. & Wagenmakers, A. J. (2001) Theeffects of increasing exercise intensity on muscle fuel utilisation in humans, J Physiol. 536, 295-304.Romijn, J. A., Coyle, E. F., Sidossis, L. S., Gastaldelli, A., Horowitz, J. F., Endert, E. & Wolfe, R. R. (1993) Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity andduration, Am J Physiol. 265, E380-91.Murray, R. K., Granner, D. K. & Rodwell, V. W. (2012) Harper's Illustrated Biochemistry, 29th edn, Lange, Boston.Newsholme, E. A. & Leech, T. (2009) Functional Biochemistry in Health and disease, Wiley-Blackwell, Oxford.Stephens, F. B., Constantin-Teodosiu, D. & Greenhaff, P. L. (2007) New insights concerning the role ofcarnitine in the regulation of fuel metabolism in skeletal muscle, J Physiol. 581, 431-44.Wahren, J. & Ekberg, K. (2007) Splanchnic regulation of glucose production, Annu Rev Nutr. 27, 329-45.Van Hall, G., Jensen-Urstad, M., Rosdahl, H., Holmberg, H. C., Saltin, B. & Calbet, J. A. (2003) Leg andarm lactate and substrate kinetics during exercise, Am J Physiol Endocrinol Metab. 284, E193-205.Spriet, L. L., Howlett, R. A. & Heigenhauser, G. J. (2000) An enzymatic approach to lactate productionin human skeletal muscle during exercise, Med Sci Sports Exerc. 32, 756-63.Green, H. J., Duhamel, T. A., Holloway, G. P., Moule, J., Ouyang, J., Ranney, D. & Tupling, A. R. (2007) Muscle metabolic responses during 16 hours of intermittent heavy exercise, Can J Physiol Pharmacol. 85, 634-45.
22










![orgaos e tecidos03 - FMUP - Faculdade de Medicina da ...ruifonte/PDFs/PDFs_arquivados_anos... · a velocidade de entrada de glicose ↓⇒ [glicose] intracelular ↓ ⇒velocidade](https://static.fdocumentos.com/doc/165x107/5c441d0193f3c34c5a365c83/orgaos-e-tecidos03-fmup-faculdade-de-medicina-da-ruifontepdfspdfsarquivadosanos.jpg)






