
I. INTRODUÇÃO 1. Família Trypanosomatidae cristina de faria moreira.pdf · receptores para a...
33
1 I. INTRODUÇÃO 1. Família Trypanosomatidae A Família Trypanosomatidae é de grande interesse médico e econômico, sendo composta por protozoários uniflagelados parasitas, que podem ter seus ciclos de vida estabelecidos em um único hospedeiro (parasitas monoxênicos), ou em mais de um hospedeiro (parasitas heteroxênicos). Os gêneros heteroxênicos estão divididos entre os que parasitam invertebrados (em especial insetos) e vertebrados: Leishmania, Trypanosoma e Endotrypanum e o constituído de parasitas de insetos e vegetais: Phytomonas (Hoare & Wallace, 1966; Vickerman, 1994). 2. Leishmania Parasitos do gênero Leishmania foram descritos em 1903 por James Wright. As síndromes atribuídas a estes protozoários são chamadas de leishmanioses, de grande importância médica, essas doenças variam de lesões que se curam espontaneamente à formas viscerais fatais. (Vannier-Santos Martiny & De Souza, 2002). As leishmanioses são as mais importantes doenças emergentes, devido a sua incidência, alta mortalidade em indivíduos não tratados e crianças desnutridas, e estão em segundo lugar no número de mortes, entre as doenças parasitárias, logo depois da malária. Há cerca de 350 milhões de pessoas vivendo em áreas endêmicas. A leishmaniose cutânea afeta a pele, causando úlceras e a mucocutânea ulceração, seguida da destruição de membranas mucosas e do tecido do nariz, da boca e da garganta. É uma doença que pode levar à morte por infecção secundária das vias respiratórias. A Bolívia, o Brasil e o Peru contabilizam 90% de todos os casos mundiais. A forma mais perigosa da doença, no entanto, é a leishmaniose visceral (LV), que está entre as seis doenças mais importantes causadas por protozoários no mundo, devido a sua incidência, alta mortalidade em indivíduos não tratados e crianças desnutridas, e emergência em indivíduos portadores da infecção por HIV. Tem ampla distribuição geográfica, ocorrendo na Ásia, Europa, Oriente Médio, África e
Transcript of I. INTRODUÇÃO 1. Família Trypanosomatidae cristina de faria moreira.pdf · receptores para a...

1
I. INTRODUÇÃO
1. Família Trypanosomatidae
A Família Trypanosomatidae é de grande interesse médico e econômico,
sendo composta por protozoários uniflagelados parasitas, que podem ter seus
ciclos de vida estabelecidos em um único hospedeiro (parasitas monoxênicos),
ou em mais de um hospedeiro (parasitas heteroxênicos). Os gêneros
heteroxênicos estão divididos entre os que parasitam invertebrados (em
especial insetos) e vertebrados: Leishmania, Trypanosoma e Endotrypanum e
o constituído de parasitas de insetos e vegetais: Phytomonas (Hoare &
Wallace, 1966; Vickerman, 1994).
2. Leishmania
Parasitos do gênero Leishmania foram descritos em 1903 por James Wright.
As síndromes atribuídas a estes protozoários são chamadas de leishmanioses,
de grande importância médica, essas doenças variam de lesões que se curam
espontaneamente à formas viscerais fatais. (Vannier-Santos Martiny & De
Souza, 2002). As leishmanioses são as mais importantes doenças emergentes,
devido a sua incidência, alta mortalidade em indivíduos não tratados e crianças
desnutridas, e estão em segundo lugar no número de mortes, entre as doenças
parasitárias, logo depois da malária. Há cerca de 350 milhões de pessoas
vivendo em áreas endêmicas. A leishmaniose cutânea afeta a pele, causando
úlceras e a mucocutânea ulceração, seguida da destruição de membranas
mucosas e do tecido do nariz, da boca e da garganta. É uma doença que pode
levar à morte por infecção secundária das vias respiratórias. A Bolívia, o Brasil
e o Peru contabilizam 90% de todos os casos mundiais. A forma mais perigosa
da doença, no entanto, é a leishmaniose visceral (LV), que está entre as seis
doenças mais importantes causadas por protozoários no mundo, devido a sua
incidência, alta mortalidade em indivíduos não tratados e crianças desnutridas,
e emergência em indivíduos portadores da infecção por HIV. Tem ampla
distribuição geográfica, ocorrendo na Ásia, Europa, Oriente Médio, África e

2
Américas. Estima-se a ocorrência de 500 mil novos casos por ano, sendo que
90% deles em Bangladesh, Brasil, Índia e Sudão. Na América Latina, a doença
já foi descrita em 12 países (cerca de 90% dos casos no Brasil)
(http://www.who.int/en/).
A reprodução destes protozoários ocorre por divisão binária simples em
ambos os hospedeiros. Os hospedeiros vertebrados incluem uma grande
variedade de mamíferos. Como hospedeiro invertebrado são identificados,
exclusivamente, fêmeas de inseto hematófagos conhecidos como
flebotomíneos dos gêneros Lutzomyia (Novo Mundo) e Phlebotomus (Velho
Mundo) (Neves et al., 2005).
São reconhecidos dois estágios de desenvolvimento distintos durante o ciclo
de vida deste parasito (Fig. 1). As formas promastigotas (Fig. 1A) que estão
presentes dentro do flebotomíneo (hospedeiro invertebrado) são alongadas e
possuem flagelo longo e as formas amastigotas (Fig. 1B) que estão presentes
no hospedeiro vertebrado, são arredondadas, com flagelo curto e intracelulares
(Olivier, Gregory & Forget, 2005).
Figura 1: Formas evolutivas: A- Amastigota B- Promastigota
(adaptação de Brener & Andrade, 2002)
O ciclo biológico da Leishmania começa quando a fêmea pica um
hospedeiro vertebrado infectado (Figura 2), ingerindo o sangue com os
macrófagos e monócitos contaminados com as formas amastigotas do parasito,
que no interior do aparelho digestivo do inseto sofrem mudanças bioquímicas e
morfológicas e se diferenciam em promastigotas procíclicas (forma replicativa
no inseto). O hospedeiro vertebrado saudável é infectado quando as formas
B

3
promastígotas metaciclicas (infectante no hospedeiro vertebrado) são
inoculadas pelas fêmeas dos insetos vetores, durante o repasto sanguíneo.
(Neves et al., 2005).
Os macrófagos são as primeiras células a serem infectadas por estes
parasitos. Depois da ligação ao macrófago, as formas promastigotas são
internalizadas e começam a diferenciar para amastigotas que são resistentes
ao ambiente acídico e hidrolítico do fagolisossoma, por isso a maturação do
endossoma e a fusão endossoma-fagossoma são retardadas até que as
formas promastigotas possam diferenciar em amastigotas mais resistentes,
esse processo ainda não está bem entendido (Olivier, Gregory & Forget, 2005).
Para tentativa de controle da infecção, os macrófagos produzem um oxidante
chamado óxido nítrico (NO). O NO é gerado após ativação dos macrófagos, por
INF-γ e TNF-α, e é o mais importante no estabelecimento da morte de
amastigotas intracelulares. O NO foi descrito como atuante na eliminação de L.
major por macrófagos humanos, estimulados através do receptor de baixa
afinidade Fce, CD23 e o IFN-γ (Vouldoukis et al., 1995). Entretanto, os
mecanismos de subversão do sistema imune são extremamente eficientes e
estão relacionados com fatores de virulência destes protozoários, como LPG
(camada de lipofosfoglicana “protetora” presente nas espécies de Leishmania)
e a glicoproteína 63 (gp63), uma metalopeptidase, também conhecida como
leishmaniolisina, que é descrita como um importante fator de virulência, por
conferir resistência à lise mediada por complemento, clivando e convertendo
C3b para sua forma inativa iC3b. A gp63 ainda é descrita como um ligante de
proteínas de superfície do hospedeiro, como o receptor para fibronectina
(McKerrow et al., 2006). Dentro das células do sistema fagocítico e
mononuclear os parasitos se multiplicam e rompem as células, alcançando, via
vasos sanguíneos e linfáticos, muitos órgãos, especialmente baço, fígado,
medula óssea e tecidos linfóides, no caso da leishmaniose visceral (Vannier-
Santos, Martiny & De Souza, 2002).

4
Figura 2: Ciclo biológico
(adaptado CDC, www.dpd.cdc.gov/dpdx)

5
2.1. Leishmania chagasi
Os protozoários do complexo Leishmania donovani são os causadores da
leishmaniose visceral na África, Ásia, Europa e nas Américas, e incluem três
espécies de Leishmania: L. donovani, L. infantum, L. chagasi. Nas Américas a
Leishmania chagasi é a espécie responsável pelas formas clínicas da
leishmaniose visceral e tem como vetor a espécie de flebotomíneo Lutzomyia
longipalpis (Neves et al., 2005). Outras espécies estão relacionadas com o
calazar ou leishmaniose visceral do Velho Mundo como, L. donovani na Índia e
L. infantum na região do Mar Mediterrâneo, Europa, África e China (Sundar &
Chatterjee, 2006). Estes protozoários apresentam um grande tropismo por
células do sistema fagocítico e mononuclear (Vouldoukis et al., 1995). O
primeiro relato de infecção por Leishmania chagasi no Brasil foi feito em 1934,
quando encontraram formas amastigotas de Leishmania em cortes histológicos
do fígado de pessoas que morreram com suspeita de febre amarela. Somente
20 anos depois é que se registrou o primeiro surto da doença em Sobral, no
Ceará (Gontijo e Melo, 2004).
Devido à gravidade desta doença, são registradas cerca de 59.000 mortes
por ano, com incidência de 500.000 novos casos. O índice de mortalidade da
leishmaniose visceral é de quase 100%, se não tratada (Dantas-Torres &
Brandão-Filho, 2006). No Brasil, são registrados cerca de 3.500 casos anuais,
cerca de 90% do total de casos de toda América Latina (Queiroz, Alves &
Correia, 2004). Grande parte dos casos acomete crianças menores de cinco
anos, moradoras de áreas rurais com baixa renda familiar (Jerônimo et al,
2004; Rey et al, 2005).
3. Tratamento
O arsenal terapêutico contra a leishmaniose viceral é delimitado. Os
antimoniais pentavalentes (Sb5+), antimoniato de N-metilglucamina
(Glucantime®) e o estibogliconato sódico (Pentostam®) são na maioria dos
países, a primeira opção terapêutica. No Brasil, a droga de escolha é a
Glucantime®, que é de distribuição gratuita na rede de saúde publica. Estas

6
drogas são tóxicas, nem sempre efetivas, e na LV são usadas em esquemas
prolongados (Gontijo e Melo, 2004).
No Brasil, segundo o Ministério da Saúde, não existe documentações de
presença de cepas de L. chagasi resistentes aos antimoniais, in vitro. Apesar
de países do sudeste da África e Índia apresentarem índices preocupantes de
resistência aos antimoniais (Frezard et al., 2009).
Em caso de recidiva é recomendado um segundo tratamento com o
desoxicolato de sódio de anfotericina B e suas formulações lipossomais, além
de pentamidinas e imunomoduladores (Neves et al., 2005). As pentamidinas
(sulfato e mesilato) e os imunomoduladores (interferon gama e GM-CSF) ainda
se encontram em fase de investigação. A utilização destas drogas só deve ser
realizada em hospitais de referência (Gontijo e Melo, 2004).
O desenvolvimento de anfotericina B encapsulada em lipossomas
(AmBisome) tem mostrado bons resultados, com cura de 90-95% na Índia
(Gontijo e Melo, 2004). Em 2000 foi disponibilizada uma nova droga de uso
oral, o Miltefosine, um hexadecylphosphoscoline, originalmente desenvolvido
para o câncer. Este foi o mais significativo avanço no tratamento de
leishmaniose viceral desde 1940. Esse tipo de medicação é ainda de alto
custo, portanto o seu uso é restrito (Neves et al., 2005).
4. Fator de Ativação de Plaquetas (PAF)
Apesar de ainda serem pouco estudadas em tripanossomatídeos as vias de
sinalização celular são um potencial alvo para o desenvolvimento de novas
drogas, pois regulam virtualmente todos os aspectos do comportamento celular
(Cuatrecasas, 1986; Naula & Seebeck, 2000; Hammarton, 2007). A fosforilação
e a defosforilação de proteínas têm papel fundamental no controle de
fenômenos biológicos na maioria dos organismos (Hunter, 1995), incluindo
protozoários da família Trypanosomatidae (Parsons et al., 1993).
A ligação de muitos hormônios ou polipeptídios aos receptores na
membrana celular, transmitem seus sinais via (1) elevação de adenosina
monofosfato cíclica (AMPc) e transmissão, através de sinalização, via proteína
cinase dependente de AMP cíclico (PKA); (2) estimulando a hidrólise de
fosfatidilinositol 4,5-bisfosfato e ativando uma combinação de mensageiros

7
secundários, incluindo o inositol trifosfato (IP3), íons Ca+2 e o diacilglicerol
(DAG), que por sua vez ativam a proteína cinase C (PKC); ou (3) estimulando
níveis intracelulares de GMPc e, consequentemente, ativando uma sinalização
via proteína cinase G (Derynck & Wagner, 1995; Robbins & Hollenberg, 1999;
Siegel et al., 1991).
Nas três vias, é o receptor que transmite a informação para dentro da
célula, através da membrana plasmática, por intermédio das proteínas G, que
são ativadas quando se ligam a guanosina trifosfato (GTP) (Derynck & Wagner,
1995). Diferentemente dos receptores ligados às proteínas G, outros
receptores de superfície celular são diretamente ligados a enzimas
intracelulares, é a família dos receptores de proteínas tirosina cinase, que
fosforilam suas proteínas substratos em resíduos de tirosina. Esta família inclui
receptores para a maioria dos fatores de crescimento; por isso, a fosforilação
por tirosina cinase tem sido bem estudada, como um mecanismo sinalizador
envolvido no controle do crescimento e diferenciação celular animal, inclusive
de tripanossomatídeos (Hubbard et al., 1998; Chuenkova & Pereira-Perrin,
2004).
Em 1970, Henson descobriu um fosfolipídio que tinha a capacidade de
promover a agregação plaquetária em coelhos imunizados que ele chamou de
Fator de Ativação de Plaquetas (PAF). O PAF (1-O-alquil-2-acil-glicero-3-
fosfocolina) é um potente sinalizador molecular lipídico que apresenta uma
grande variedade de efeitos fisiológicos e fisiopatológicos em diversas células e
tecidos, incluindo a diferenciação celular, na inflamação e alergia (Rosa et al,
2001). Antes pensavam que as funções do PAF eram exclusivamente
estruturais ou de reserva energética, mas estudos comprovaram que este fator
solúvel liberado por leucócitos estimula a agregação de plaquetas e a liberação
de serotonina (Lenhinger, 2006 ).

8
Figura 3: Estrutura do PAF
Em mamíferos, o PAF é sintetizado e liberado por uma grande variedade de
células, incluindo plaquetas, macrófagos, neutrófilos, basófilos, eosinófilos e
células endoteliais, sendo mediador de diversos processos biológicos e
patofisiológicos, como a inflamação, alergia, bronquite asmática, doenças de
pele, infecção por Streptococcus e desordens do sistema nervoso central
(Benveniste, 1974; Bozza et al., 1994; Campbell & Halushka, 1996; Ishii,
Nagase, Shimizu, 2002). A presença de PAF também já foi observada em
outros organismos, como peixes (Turner & Lumb, 1989), vários invertebrados
(Sugiura et al., 1991, 1992), em Dictyostelium (Bussolino et al., 1991), plantas
(Mangold, Apte & Weber, 1991; Calligerou et al., 1996) e até mesmo em
microorganismos, como cianobactérias (Antonopoulou et al., 2002),
Tetrahymena (Tsoukatos Tselepis & Lekka, 1993) e Saccharomyces cerevisae
(Nakayama et al., 1994 a, b).
PAF age por meio da ativação de um receptor específico, presente na
superfície de suas células alvo. Este receptor, quando exposto ao seu
respectivo agonista, ativa uma proteína G, que por sua vez ativa e regula várias
respostas celulares (Hwang, Lam & Pong 1986; Zhu et al, 2006). Esta proteína
parece funcionar como um mediador, ligando o receptor do PAF a vários
sistemas celulares, como por exemplo as fosfolipases C e A2 (Chao & Olson,
1993). Além disso, através da ligação com seu receptor, o PAF também pode
estimular a reciclagem de fosfatidilinositol, fluxo de cálcio, ativação de proteína
cinase importantes como a proteína cinase C (PKC), produção de eicosanóides
e fosforilação de diversas proteínas (Gouill et al., 1997). Esses eventos levam à
ativação de diferentes fatores de transcrição, que controlam a expressão de

9
genes, que estão relacionados aos diferentes processos de ativação e
diferenciação celular (Bazan & Allan, 1996; Liu et al., 1997).
Figura 4: Estrutura do receptor de PAF
O WEB 2086 (3-[4-(2-clorofenil)-9-metil-6H-tieno-[3,2-f] [1,2,4]triazolo-[4,3,·]
[1,4] diazepina-2-·1-1-(4-morfolina-·1)-1-propanona), é um importante
antagonista do PAF, e que tem se mostrado uma poderosa ferramenta para o
estudo de receptores de membrana (Rodrigues et al., 1996; Lopes et al., 1997;
Dutra et al., 1998; Rodrigues et al., 1999)]. Em relação aos
tripanossomatídeos, em estudos anteriores, foi demonstrado que o PAF é
capaz de induzir um aumento de 40% na diferenciação celular dos
tripanossomatídeos T. cruzi (Rodrigues et al., 1996) e H. muscarum muscarum
(Lopes et al., 1997), além de modular uma atividade fosfatásica na superfície
destes protozoários (Dutra et al., 1998; Rodrigues et al., 1999; Dutra et al.,
2000). Também foi mostrado que o PAF é capaz de estimular a atividade da
enzima CK2 de H. m. muscarum, através de complexa via de transdução de
sinais, com participação de proteínas serina/treonina e tirosina fosfatases, PLC,
PKC e MAP cinases (Silva-Neto et al., 2002). Todos esses efeitos parecem

10
ocorrer via receptor de superfície, uma vez que foram revertidos na presença
do WEB 2086.
Recentemente, nosso grupo identificou uma molécula com característica
semelhante ao PAF em T. cruzi (Gomes et al., 2006). Além disso, a presença
de atividade de PAF acetil –hidrolase (enzimas capazes de hidrolisar a
molécula de PAF) em Rhodnius prolixus pode ser um indicativo da ação de
fosfolipídio no inseto vetor do T. cruzi (Figueiredo et al, 2008).
A cascata de eventos de transdução de sinais, disparada pela ligação do
PAF ao seu receptor, ativa diferentes fatores de transcrição, que controlam a
expressão de genes, que estão relacionados aos diferentes processos de
ativação e diferenciação celular. Estes estudos demostram como é complexa a
sinalização intracelular promovida por PAF (Rosa et al, 2001).
Uma droga promissora para o tratamento das leishmanioses, em especial a
leishmaniose visceral, é a Miltefosine. Curiosamente, a Miltefosine é uma
alquil-glicero-fosfocolina (um éter lipídio), que apresenta uma grande
semelhança estrutural com o PAF, além de se ligar ao receptor do PAF em
células de mamíferos (Croft & Engel, 2006).
Todos os estudos envolvendo as ações do PAF sobre L. chagasi, além dos
aspectos acadêmicos, tem o interesse de possibilitar a elucidação de vias de
sinalização celular, o que é de extrema importância para a descoberta de
novas drogas e estratégias quimioterápicas para a leishmaniose visceral.

11
II.OBJETIVOS
1. Objetivo Geral
Determinar o efeito que do Fator de Ativação de Plaquetas (PAF) comercial
e extraído de L. chagasi na proliferação e interação de Leishmania chagasi
com células do hospedeiro vertebrado.
2. Objetivos específicos
2.1. Obter o lipídio com atividade semelhante a PAF em L. chagasi;
2.2. Verificar a influência do PAF de Leishmania e comercial na proliferação de
L. chagasi;
2.3. Verificar a influência do PAF de Leishmania e comercial na infecção de
macrófagos peritoneais de camundongos por L. chagasi;
2.4. Analisar as possíveis mudanças, no perfil de expressão de proteínas, após
o tratamento com PAF de Leishmania e comercial, em Leishmania chagasi;

12
III.METODOLOGIA
1- Microorganismos
O parasita foi obtido no Centro de Referência em Leishmaniose do Instituto
Oswaldo Cruz. Leishmania (Leishmania) chagasi (MHO/BR/1974/PP75).
Os promastigotas foram mantidos em meio Schneider contendo 20% de
soro fetal bovino a 26oC .
2- Extração e purificação do lipídio semelhante a PAF de Leishmania
chagasi
Protozoários da espécie L. chagasi foram crescidos por 5 dias. Estes
parasitos (109células/mL) foram, então, centrifugados, lavados duas vezes com
solução salina. A extração ocorreu com a adição de uma solução de ácido
acético (50mM) em metanol, também foi adicionado clorofórmio e acetato de
sódio 0.1M, vortexar e centrifugar por 5 minutos a 5000 rpm. Foram formadas 3
fases, a fase superior foi descartada, a fase inferior foi seca em nitrogênio, à
fase mediana foi adicionada a mesma proporção de ácido acético (50mM) em
metanol, clorofórmio e acetato de sódio, esse procedimento foi repetido por
três vezes (adaptado de Bligh & Dryer, 1959). A parte inferior corrrespondente
aos lipídeos totais dos parasitos, foi ressuspensa em etanol (50 L) e
posteriormente, separada por cromatografia de camada fina (HPTLC)
preparativa, em placas de sílica (Si 60 F254, Merk, Darmstadt, Alemanha),
usando sistema clorofórmio: metanol: água (65:35:6, v/v). As bandas foram
visualizadas por vapores de iodo, H2SO4. A banda correspondente ao autêntico
PAF (PAF comercial foi utilizado como padrão) e ao lipídio semelhante a PAF
de L. chagasi foram extraídas da sílica. A concentração do lipídio semelhante a
PAF de L. chagasi foi estimada, com base em resultados de experimentos de
agregação plaquetária (principal função do PAF) e ajustada para os
experimentos posteriores.

13
3- Avaliação da proliferação celular
Protozoários da espécie L. chagasi foram crescidos por 1-7 dias na
ausência ou na presença de PAF-like extraído de Leishmania (10–5 M) ou PAF
comercial (10–5 M). Cerca de 5 x 105 parasitos (de uma cultura de 5 dias) foram
inoculados em 5 ml de meio de cultura. A contagem das células foi
determinada diariamente através de Câmara de Neubauer (Santos & Oliveira,
1988).
4- Macrófagos peritoneais de camundongos BALB/c
Os camundongos BALB/c fêmeas foram anestesiados com éter etílico e
sacrificados por deslocamento da coluna cervical e, então, a cavidade
peritoneal foi lavada com 5 a 6 mL de PBS estéril. O volume final obtido,
retirado da cavidade abdominal, foi distribuído em placas (24 poços) para
fixação e manutenção dos macrófagos em meio RPMI, em atmosfera contendo
4 % de CO2, por 16 horas.
5- Interação dos parasitos com macrófagos peritoneais de camundongos
Os macrófagos peritoneais, tratados ou não com PAF-like isolado de
Leishmania chagasi (10–5 M) e PAF comercial (10–5 M), foram infectados com
Leishmania chagasi tratadas ou não tratadas com PAF-like de Leishmania
chagasi (10–5 M) e PAF comercial (10–5 M) que foram adicionadas às culturas,
na proporção de 10 parasitos para cada macrófago. Após a infecção, as placas
foram incubadas durante 4 horas a 37 ºC, em atmosfera contendo 4 % de CO2.
Em seguida, as lamínulas foram lavadas em PBS, fixadas e coradas com
reagente de Giemsa. Depois de secas, as lamínulas foram montadas sobre
lâminas de vidro (24x76mm), utilizando-se Permount.
6- Sobrevivência dos parasitos em macrófagos peritoneais de
camundongos
Os macrófagos peritoneais, tratados ou não com PAF-like isolado de
Leishmania chagasi (10–5 M) e PAF comercial (10–5 M), serão infectados com
Leishmania chagasi tratadas ou não tratadas com PAF-like isolado de
Leishmania chagasi (10–5 M) e PAF comercial (10–5 M). Os parasitos serão
adicionados às culturas, na proporção de 10 parasitos para cada macrófago.

14
Após a infecção, as placas foram incubadas durante 24 horas a 37 ºC, em
atmosfera contendo 4 % de CO2. Em seguida, as lamínulas serão lavadas em
PBS, fixadas e coradas com reagente de Giemsa. Depois de secas, as
lamínulas serão montadas sobre lâminas de vidro (24x76mm), utilizando-se
para isso o Permount.
7- Aprovação para experimentação utilizando células de animais
Os macrófagos peritoneais de camundongos foram obtidos de acordo
com as normas para experimentação animal, aprovadas pela Comissão de
Ética para Experimentação Animal do Centro de Ciências da Saúde da
Universidade Federal do Rio de Janeiro.
8 - Perfil de expressão de proteínas
L. chagasi foram tratadas e/ou não tratadas com PAF-like isolado de
Leishmania chagasi (10–5 M) e PAF comercial (10–5 M), as células foram então
centrifugadas, lavadas duas vezes com solução salina. O extrato celular foi
ressuspenso em tampão de lise (Tris-HCl 20 mM, NaCl 15 mM, ortovanadato
de sódio 1 mM, EDTA 1 mM, EGTA 1 mM, NaF 10 mM, azida sódica 0,02 %;
pH 8,0). Os parasitos foram, então, congelados em nitrogênio líquido e
descongelados por 3 vezes. O material referente homogenato da célula foi
usado no gel. As proteínas foram separadas por SDS-PAGE, a 10%. A
eletroforese foi realizada a 150 volts e 60 mA. O gel foi corado com o corante
Coomassie Blue.
9. Análise estatística
Todos os experimentos foram feitos em triplicata. Os resultados foram
analisados estatisticamente pelo teste de Fisher e pelo teste de análise de
variância (ANOVA). Valores de P iguais ou menores que 0,05 foram
considerados significativos.

15
IV. RESULTADOS
1. Extração de lipídio com atividade semelhante a PAF em L. chagasi
De modo a verificar se o parasita Leishmania chagasi produz o lipídeo
com atividade semelhante ao PAF produzido pelos mamíferos foi feita a
extração total de lipídeos de uma cultura de L. chagasi. Estes lipídios foram
analisados por cromatografia em camada fina de alta resolução (HPTLC),
usando placas de sílica gel e, como sistema de separação, clorofórmio:
metanol: água (65: 35: 6, v/v). Lipídeos com fator de corrida semelhante ao
autêntico PAF foram observados (Figura 1). Neste ensaio foram isolados
lipídeos com atividade semelhante ao PAF comercial, que foram extraídos da
sílica pra serem utilizados nos experimentos seguintes.
Fig. 5. Cromatografia de camada fina de alta resolução (HPTLC) de lipídios
extraídos de Leishmania chagasi em sistema solvente composto por
clorofórmio:metanol:água (65:35:6, v/v/v). As placas foram secas e expostas a
vapor de iodo. Os lipídios foram identificados por comparação com PAF
comercial, usado como padrão.
PAF comercial PAF Leishmania chagasi

16
2. Efeito do PAF-like extraído de Leishmania chagasi e PAF comercial na
proliferação de L. chagasi
Também foi avaliado se o PAF-like extraído de Leishmania e PAF comercial
teria efeito na proliferação celular de L. chagasi. A figura 6 mostra a curva de
crescimento de parasitos crescidos na ausência (controle) e na presença das
seguintes drogas: PAF comercial (10-5M) e PAF-like isolado de L. chagasi (10-
5M). Podemos observar que nos sistemas onde os parasitos foram crescidos
na presença de PAF-like isolado de L. chagasi e PAF comercial houve uma
diminuição (30%) do número de células, em relação ao sistema controle. Este
resultado mostra que o PAF-like isolado de L. chagasi e o PAF comercial têm
efeito semelhante na modulação do crescimento de L. chagasi.
Fig. 6. Índice de proliferação de Leishmania chagasi. Parasitos tratados com
PAF comercial 10-5 M e PAF-like extraído de Leishmania chagasi 10-5 M (PAF
Lc).
24h 48h 72h 96h 120h 144h 168h
x 1
06
ce
ls
0
2
4
6
8
10
Controle
PAF comercial
PAF Lc

17
3. Efeito do PAF-like extraído de Leishmania e do PAF comercial na
infecção de macrófagos peritoneais de camundongos por L. chagasi
De maneira a investigar o efeito do PAF na infecção de macrófagos
peritoneais de camundongos balb/c por L. chagasi foram feitas interações na
presença e ausência de PAF comercial (10-5M), PAF-like extraído de L. chagasi
(10-5M) e WEB 2086 (antagonista do receptor do PAF) (10-5M). Observamos
na figura 7 que quando tratamos somente os parasitas tanto com o PAF
comercial quanto com o PAF-like isolado de L. chagasi há um aumento de
100% do índice de associação comparado ao controle. Também observamos
que quando tratamos apenas os macrófagos tanto com o PAF comercial
quanto com o PAF-like extraído de L. chagasi o índice de associação diminui
em 50% em relação ao controle e quando tratamos os parasitos e os
macrófagos tanto com PAF comercial quanto com o PAF-like isolado de L.
chagasi o aumento da interação promovido pelo PAF comercial e pelo PAF
isolado de L. chagasi é revertido em 85%.
O WEB 2086 é um antagonista sintético do PAF, que tem sido utilizado como
ferramenta de estudo de ligações do PAF com o seu receptor para averiguar se
o PAF age via receptor na superfície da celular. A figura 8 mostra a interação
de macrófagos peritoneais de camundongos com L. chagasi utilizando o PAF
comercial, o PAF-like extraído de L. chagasi e o WEB 2086, para verificarmos
se a ação do PAF acontece via receptor. Observamos que quando o parasito é
tratado com o WEB + PAF (comercial e isolado de Leishmania), há a
diminuição do índice de associação quando comparamos com os parasitos
apenas tratados com o PAF (comercial e isolado de Leishmania) , indicando a
participação do receptor no efeito do PAF no parasito. Além disso, quando
tratamos os macrófagos com WEB + PAF (comercial e isolado de Leishmania)
também há a reversão do efeito promovido pelo PAF indicando também a
participação do receptor na sinalização promovida pelo PAF nos macrófagos.
Fizemos também a avaliação da sobrevivência de L. chagasi nos
macrófagos de camundongos por 24 horas. Podemos observar na figura 9, que
da mesma maneira que acontece na interação (4 horas), quando tratamos os
parasitos tanto com PAF comercial, quanto com PAF-like isolado de
Leishmania chagasi há um aumento na infecção, em relação ao controle.

18
Também observamos que da mesma forma quando apenas os macrófagos
foram tratados com PAF comercial e PAF-like isolado de Leishmania o índice
de infecção diminuiu em relação ao controle, igualmente como ocorre na
interação, e quando tratamos os macrófagos e os parasitos com PAF comercial
e com PAF-like isolado de Leishmania o aumento do índice de infecção
promovido tanto pelo PAF comercial quanto pelo PAF-like isolado de
Leishmania é revertido, também corroborando com os resultados da interação.
Estes resultados demonstram similaridade nos efeitos do PAF comercial e
do PAF-like extraído de L. chagasi em relação à infectividade de L. chagasi.

19
Fig. 7. Índice de associação de Leishmania chagasi com macrófagos de
camundongos. Parasitos e/ou macrófagos tratados com PAF comercial 10-5 M
e PAF-like extraído de Leishmania chagasi 10-5 M.
MCPC – macrófagos controle/ parasitos controle
MCPPc – macrófagos controle/ parasitos PAF comercial 10-5M
MPcPC – macrófagos PAF comercial 10-5M/ parasitos controle
MPcPPc- macrófagos PAF comercial 10 -5M/ parasitos PAF comercial 10-5M
MCPPl – macrófagos controle/ parasitos PAF de Leishmania 10-5M
MPlPC – macrófagos PAF de Leishmania 10-5M/ parasitos controle
MPlPPl- macrófagos PAF de Leishmania 10-5M/ parasitos PAF de Leishmania 10-5M
Interação de Leishmania chagasi x macrófagos
(PAF comercial 10-5
M e PAF de Leishmania 10-5
M)
MCPC MCPPc MPcPC MPcPPc MCPPl MPlPC MPlPPl
Índ
ice
de
As
so
cia
çã
o
0
20
40
60
80
100
120
140

20
Fig. 8. Índice de infecção na sobrevivência por 24 horas de Leishmania chagasi
com macrófagos de camundongos. Parasitos e/ou macrófagos tratados com
PAF comercial 10-5 M e PAF-like extraído de Leishmania chagasi 10-5 M.
MCPC – macrófagos controle/ parasitos controle
MCPPc – macrófagos controle/ parasitos PAF comercial 10-5M
MPcPC – macrófagos PAF comercial 10-5M/ parasitos controle
MPcPPc- macrófagos PAF comercial 10 -5M/ parasitos PAF comercial 10-5M
MCPPl – macrófagos controle/ parasitos PAF de Leishmania 10-5M
MPlPC – macrófagos PAF de Leishmania 10-5M/ parasitos controle
MPlPPl- macrófagos PAF de Leishmania 10-5M/ parasitos PAF de Leishmania 10-5M
Sobrevivência de Leishmania chagasi x macrófagos
(PAF comercial 10-5
M e PAF de Leishmania 10-5
M)
MCPC MCPPc MPcPC MPcPPc MCPPl MPlPC MPlPPl
Índ
ice d
e I
nfe
cção
0
20
40
60
80
100
120
140
160
21
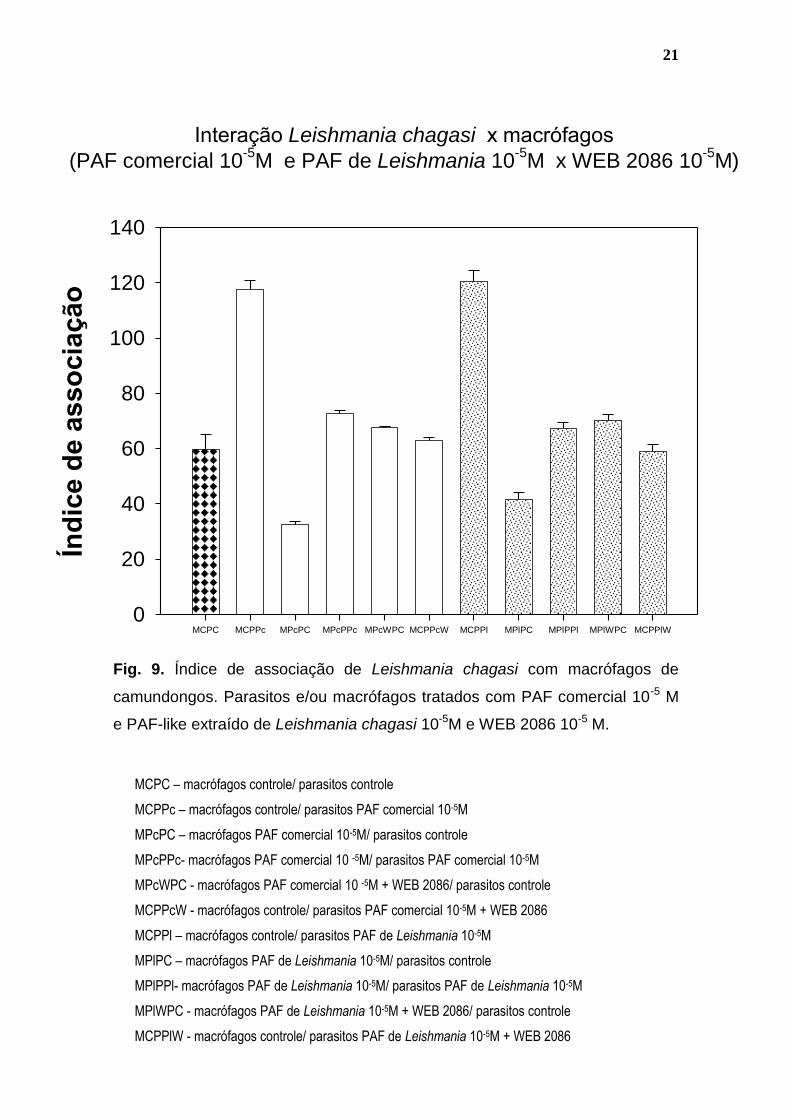
Fig. 9. Índice de associação de Leishmania chagasi com macrófagos de
camundongos. Parasitos e/ou macrófagos tratados com PAF comercial 10-5 M
e PAF-like extraído de Leishmania chagasi 10-5M e WEB 2086 10-5 M.
MCPC – macrófagos controle/ parasitos controle
MCPPc – macrófagos controle/ parasitos PAF comercial 10-5M
MPcPC – macrófagos PAF comercial 10-5M/ parasitos controle
MPcPPc- macrófagos PAF comercial 10 -5M/ parasitos PAF comercial 10-5M
MPcWPC - macrófagos PAF comercial 10 -5M + WEB 2086/ parasitos controle
MCPPcW - macrófagos controle/ parasitos PAF comercial 10-5M + WEB 2086
MCPPl – macrófagos controle/ parasitos PAF de Leishmania 10-5M
MPlPC – macrófagos PAF de Leishmania 10-5M/ parasitos controle
MPlPPl- macrófagos PAF de Leishmania 10-5M/ parasitos PAF de Leishmania 10-5M
MPlWPC - macrófagos PAF de Leishmania 10-5M + WEB 2086/ parasitos controle
MCPPlW - macrófagos controle/ parasitos PAF de Leishmania 10-5M + WEB 2086
Interação Leishmania chagasi x macrófagos
(PAF comercial 10-5
M e PAF de Leishmania 10-5
M x WEB 2086 10-5
M)
MCPC MCPPc MPcPC MPcPPc MPcWPC MCPPcW MCPPl MPlPC MPlPPl MPlWPC MCPPlW
Índ
ice d
e a
sso
cia
ção
0
20
40
60
80
100
120
140

22
4. Efeito do PAF no perfil de expressão de proteínas de L. chagasi
Foi determinado também o perfil de expressão de proteínas com o PAF
comercial 10-5 M e PAF-like extraído de Leishmania chagasi 10-5 M para
verificar o efeito do PAF nas vias de sinalização celular deste protozoário. A
figura 10 mostra o perfil de expressão de proteínas de L. chagasi crescidos na
ausência ou na presença de PAF comercial ou PAF-like extraído de
Leishmania chagasi. O perfil de expressão apresenta mudanças nas proteínas
expressas principalmente entre o sistema controle e os sistemas PAF
comercial ou PAF-like extraído de Leishmania chagasi (setas).
Fig. 10. Perfil de expressão de proteínas de L. chagasi tratadas ou não com PAF comercial 10-5M (PAFc) e/ou PAF- like extraído de L. chagasi 10-5M (PAFLc). Cont- controle.
PPM Cont PAFc PAFLc
40 kDa -
25 kDa -
15 kDa -

23
V. DISCUSSÃO
Os tripanossomatídeos são excelentes modelos para estudos sobre vários
aspectos de biologia celular, incluindo ultraestrutura e morfologia,
posicionamento de organelas, tráfego de vesículas e de macromoléculas,
divisão e diferenciação celular. A organização da estrutura celular dos
tripanossomatídeos é fundamental para seu ciclo celular. Assim, compreender
a biologia celular do desenvolvimento dos tripanossomatídeos significa dar
subsídios à elucidação tanto de processos fundamentalmente conservados,
como de outros totalmente diferentes, em relação aos aspectos organizacionais
e fisiológicos da célula eucariótica (Matthews, 2005). Os tripanossomatídeos
assemelham-se aos eucariontes superiores em vários aspectos, incluindo o
fato de suas funções celulares serem mediadas por vias de sinalização,
envolvendo receptores de superfície, proteínas cinases e fosfatases, e
mensageiros secundários. Embora alguns gêneros de tripanossomatídeos
sejam patogênicos para humanos, animais ou plantas, as vias de transdução
de sinais destes parasitos ainda são pouco conhecidas (Parsons & Ruben,
2000).
O fator de ativação de plaquetas (PAF) é um potente fosfolipídio mediador
de diversas funções celulares em vários processos biológicos e patofisiológicos
de mamíferos, como diferenciação celular, inflamação e alergia. Considerando
as vias de sinalização celular, o fator de ativação de plaquetas (PAF) tem
recentemente assumido um papel cada vez mais importante como um segundo
mensageiro lipídico. Uma variedade de bioatividades de PAF têm sido
elucidadas, incluindo embriogênese, diferenciação celular, choque e respostas
imunes (Deo et al., 2004). Vários tipos de células e tecidos, quando
estimulados, são capazes de sintetizar e liberar PAF, além de apresentar
diversas respostas biológicas a este composto (Chao & Olson, 1993).
PAF é tão potente que pode elicitar respostas biológicas importantes em
concentrações nanomolares, tanto in vivo, quanto in vitro. As atividades
mediadas por PAF ocorrem através de uma proteína G ligada ao receptor, que
é expressa na superfície de muitas células (Le Gouill et al, 1997; Prescott et al,
2000; Ishii, Nagase & Shimizu, 2002).

24
Apesar de PAF estar bem estabelecido como mediador de vias de
sinalização em sistemas de mamíferos, pouco é conhecido sobre suas funções
em tripanossomatídeos (Izumi & Shimizu, 1995; Kulikov & Muzya, 1997;
Rodrigues et al., 1999, Silva-Neto et al., 2002). Já foi demonstrado que PAF
promove um aumento de 40% na diferenciação celular de Herpetomonas
muscarum muscarum e T. cruzi. Além disso, este lipídeo também tem
capacidade de modular a atividade ecto-fosfostásica e a interação de
Leishmania com células de mamíferos (Rodrigues et al., 1996, 1999; Lopes et
al., 1997; Dutra et al., 1998, 2000; Rosa et al., 2001).
Em 2006, um estudo publicado por Gomes e colaboradores identificou uma
molécula com características semelhantes ao PAF em T. cruzi, com efeitos
biológicos importantes como diferenciação celular (Gomes et al., 2006). De
modo a verificar se a Leishmania chagasi também produz o lipídeo com perfil
semelhante ao PAF foi feita uma extração de lipídeos seguida por comparação
com o PAF comercial (utilizado como padrão). Na figura 5 podemos observar
que a L. chagasi produz lipídeos com perfil análogo ao PAF comercial.
Estudos anteriores demostraram que o PAF não tem efeito significativo na
proliferação celular de Herpetomonas muscarum muscarum (Lopes et al.,
1997). Curiosamente, na figura 6 mostramos que houve uma diminuição de
30% na proliferação das células crescidas na presença tanto de PAF comercial
quanto de PAF extraído de L. chagasi em relação ao sistema controle. Isto
sugere que de maneira diferente do que acontece com o tripanossomatídeo H.
m. muscarum há uma sinalização para a diminuição do crescimento celular, o
que poderia indicar uma indução da diferenciação celular quando os parasitos
entram em contato com o PAF (Alberts et al, 2004).
Em 2001, Rosa e colaboradores demonstraram que o PAF comercial é
capaz de modular a infecção de macrófagos peritoneais de camundongos por
Leishmania amazonensis, onde a infecção foi inibida quando os macrófagos ou
ambos macrófagos e parasitos foram tratados com PAF comercial, porém
estimulada quando somente os parasitos foram tratados com PAF comercial.
Estes resultados estão de acordo com os desse estudo como podemos
observar na figura 7 onde quando tratamos L. chagasi tanto com o PAF
comercial quanto com o PAF-like isolado de L. chagasi há um aumento de
100% de infecção em relação ao controle. Podemos observar também que

25
quando tratamos os macrófagos tanto com o PAF comercial quanto com o
PAF-like isolado de L. chagasi o índice de infecção diminui em relação ao
controle, e quando tratamos os macrófagos e os parasitos tanto com o PAF
comercial quanto com o PAF-like isolado de L. chagasi o índice de infecção
promovido é reduzido, tanto com o PAF comercial quanto com o PAF isolado
de L. chagasi, em comparação com o sistema controle. Também analisamos a
sobrevivência (Figura 8) onde observamos um perfil semelhante de infecção,
mostrando que esses efeitos podem ser duradouros possivelmente
influenciando também em uma infecção mais longa.
O WEB 2086, antagonista do receptor do PAF, têm sido utilizado como
ferramenta em estudos de ligação do PAF com seu receptor. O estudo
publicado por nosso grupo onde foi isolada e identificada uma molécula com
características semelhantes ao PAF em T. cruzi também mostrou que os
efeitos promovidos pelo PAF parecem ocorrer via receptor do PAF, já que
foram revertidos na presença do WEB 2086 (Gomes et al., 2006). Além disso,
outros estudos também demontraram que o WEB 2086 reverte efeitos
promovidos pelo PAF em tripanossomatídeos, como diferenciação celular
(Lopes et al., 1997; Rodrigues et al., 2001). A figura 9 demonstra que quando
tratamos os parasitos com o WEB 2086 mais o PAF comercial ou o PAF-like
isolado de L. chagasi, há uma diminuição do índice de infecção, quando
comparamos com os parasitos tratados apenas com PAF comercial ou o PAF-
like isolado de L. chagasi, indicando a participação do receptor na ação do PAF
sobre o parasito. E quando tratamos os macrófagos com o PAF comercial ou o
PAF-like isolado de L. chagasi e mais o WEB 2086, há a reversão de todo o
efeito promovido pelo PAF comercial ou o PAF-like isolado de L. chagasi,
indicando também uma participação do receptor na sinalização promovida pelo
PAF nos macrófagos.
A fosforilação e a defosforilação de proteínas têm papel fundamental no
controle de fenômenos biológicos na maioria dos organismos (Hunter, 1995),
incluindo protozoários da família Trypanosomatidae (Parsons et al., 1993).
Nas vias de sinalização é o receptor que transmite a informação para dentro
da célula, através da membrana plasmática, por intermédio das proteínas G,
que são ativadas quando se ligam a guanosina trifosfato (GTP) (Derynck &
Wagner, 1995). É intrigante o fato de que o sequenciamento dos genomas de

26
T.brucei, T.cruzi e L. major demonstrou ate o momento a ausência de qualquer
isótopo da proteína G. Apesar de não podermos explicar ainda como temos a
ação do PAF em tripassomatideos, na ausência da proteina G, sugerimos a
expressão de diferentes proteínas para atuar junto ao estimulo do PAF. Como
podemos observar na figura 10, o perfil de expressão de proteínas apresenta
mudanças nas proteínas expressas principalmente entre o sistema controle e
os sistemas tratados com PAF comercial ou com PAF-like extraído de L.
chagasi.
VI. CONCLUSÃO
O papel fisiológico de PAF e lipídios semelhantes à PAF precisa ser melhor
compreendido. De acordo com nossos resultados até agora, podemos
hipotetizar que a molécula lipídica produzida por L. chagasi, como o PAF
comercial, de funcionar como reguladora da infectividade de L. chagasi na
célula hospedeira, efeito que parece ocorrer via receptor, além disso, este
lipídeo isolado de Leishmania também age como modulador da proliferação
celular deste parasita e potencial modulador de vias de sinalização.
Obviamente esta molécula não é a única responsável por estes eventos,
contudo, o envolvimento de um composto intrínseco com efeitos importantes na
infectividade desses parasitas é de grande contribuição para o conhecimento
da biologia celular deste parasito.
O estudo dos efeitos do PAF produzido por Leishmania chagasi em
processos como proliferação, interação com células de mamífero e sinalização
celular pode ser bastante interessante para um maior entendimento do ciclo de
vida destes parasito. Além disso, o estudo das vias de sinalização induzidas
por PAF, assim como o mapeamento das proteínas envolvidas, parecem ser
bastante promissores como potencial alvo para drogas contra leishmaniose
visceral.

27
VII. REFERÊNCIAS BIBLIOGRÁFICAS
1. ALBERTS, B., JOHNSON, A., LEWIS, J., RAFF, M., ROBERTS, K., &
WALTER, P. Capítulo 15: Comunicação Celular. Sinalização por meio de
receptores de superfície associados a proteína G. Título: Biologia
Molecular da Célula. Páginas: 852-870. Quarta edição. Editora Artmed.,
2004.
2. ANTONOUPOU, S., OIKONOMOU, A., KARANTONIS, C. H.,
FRAGOPOULOU, E. & PANTAZIDOU, A. Isolation and elucidation of
biologically active phospholipids from Scytonema julianum
(cyanobacteria). Biochem. J. 367(Pt1): 287-293, 2002.
3. BAZAN, N.G., & ALLAN, G. Platelet-activating factor in the modulation of
excitatory amino acid neurotransmitter release and of gene expression.
J. Lipid. Mediat. Cell. Signal 14: 321-30, 1996.
4. BENVENISTE, J. Platelet-activating factor, a new mediator of
anaphylaxis and imune complex deposition from rabbit and human
basophils. Nature 249: 581-590, 1994.
5. BLIGH E. G. & DYER W. J. A rapid method of total lipid extraction and
purification. Can J Biochem Physiol. 37(8): 911-7, 1959.
6. BOZZA, P.T., CASTRO FARIA NETO, H.C., SILVA, A.R., LARANJEIRA,
A.P., SILVA, P.M.R., MARTINS, N.A. & CORDEIRO, R.S.B.
Lipopolysaccharide-induced pleural neutrophils and platelet-activating
factor. Eur. J. Pharmacol. 270: 143-149, 1994.
7. BUSSOLINO, F., SORDANO, C., BENFENATI, E. & BAZZARO, S.
Dictyostelium cells produce platelet-activating factor in response to
cAMP. Eur. J. Biochem. 196: 609-615, 1991.
8. CALLIGEROU, M., SIFAKA-KAPADAI, A., GALANOPOULOU, D.,
WEINTRAUB, S.T. & MAVRI-VAVAYANNI, M. Platelet-activating factor
and lyso-phosphotidylcholine from strawberry. Phytochemistry 41(1): 89-
92, 1996.
9. CAMPBELL, W.B. & HALUSHUKA, P.V. In: Goodman and Gilman’s, the
Pharmacological Basis of therapeutics, 9th edition, pp. 611-617, 1996.
10. CHAO, W. & OLSON, M. Platelet-activating factor: receptors and signal
transduction. Biochem. J., 292 : 617-629, 1993.

28
11. CROFT, S., ENGEL, J. Miltefosine discovery of the antileishmanial
activity of phospholipid derivatesTrans. Royal Society. Trop. Med. Hyg.
100S: 54-58, 2006.
12. CUATRECASAS, P. Hormones receptors, membrane phospholipids, and
protein kinases. The Harvey Lectures Series. 80-89, 1986.
13. DANTAS-TORRES, F. & BRANDÃO-FILHO, S. P. Visceral leishmaniasis
in Brazil: revisiting paradigms of epidemiology and control. Rev. Inst.
Med. Trop. S. Paulo. 48 (3): 151-156, 2006.
14. DEO, D. D., BAZAN, N. G. & HUNT, J. D. Activation of platelet-activating
factor receptor-coupled Gq leads to stimulation of Src and focal
adhesion kinase via two separate pathways in human umbilical vein
endothelial cells. J. Biol. Chem. 279 (5): 3497-3508, 2004.
15. DERYNCK, R. & WAGNER, E. F. Cell differentiation. Curr. Opin. Cell.
Biol. 7: 843-844, 1995.
16. DUTRA, P.M.L., RODRIGUES, C.O., JESUS, J.B., LOPES, A.H.C.S.,
SOUTO-PADRÓN, T. & MEYER-FERNANDES, J.R. A novel ecto-
phosphatase activity of Herpetomonas muscarum muscarum inhibited by
platelet-activating factor. Biochem. Biophys. Res. Commun, 253: 164-
169, 1998.
17. DUTRA, P. M. L., DIAS, F. A. RODRIGUES, C. O., ROMEIRO, ª,
ATTIAS, M., DE SOUZA, W., LOPES, A. H. C. S. & MEYER-
FERNANDES, J. R. Platelet-activating factor modulates a secreted
phosphatase activity of the trypanosomatid parasite Herpetomonas
muscarum muscarum Cur. Microbiol. 43: 288-292, 2001.
18. FIGUEIREDO, M, B, GENTA F. A., GARCIA, E. S., AZAMBUJA, P. Lipid
mediators and vector infection: Trypanosoma rangeli inhibits Rhodnius
prolixus hemocyte phagocytosis by modulation of phospholipase A2 and
PAF-acetylhydrolase activities. Journal of Insect Physiology 54 1528–
1537, 2008.
19. FRÉZARD, F, DEMICHELI, C. RIBEIRO, R. R. Pentavalent antimonials:
new perspectives for old drugs. Molecules: Jun 30;14 (7):2317-36, 2009
20. GOMES, M.T., MONTEIRO, R.Q., GRILLO, L.A., LEITE-LOPES, F.,
STROEDER, H., FERREIRA-PEREIRA, A., ALVIANO, C.S., BARRETO-
BERGTER, E., CASTRO-FARIA-NETO, H., CUNHA-E-SILVA, N.,

29
ALMEIDA, I.C., SOARES, R.M.A. & LOPES, A.H. Platelet-activating
factor-like isolated from Trypanosoma cruzi. Intern. J. Parasitol. 36: 165-
173, 2006.
21. GONTIJO, C. M. F. & MELO,M.N,Rev.bras.epidemiol. vol.7 no.3 São
Paulo Sept. 2004.
22. HAMMARTON, T.C. Cell cycle regulation in Trypanosoma brucei. Mol
Biochem Parasitol. 153 (1): 1-8, 2007, Review.
23. HUBBARD, S. R., MOHAMMADI, M. & SCHLESSINGER, J. J.
Autoregulatory mechanisms in protein-tyrosine kinases. Biol. Chem. 273:
11987-11990, 1998.
24. HUNTER, T. Epidermal growth factor induces rapid tyrosine
phosphorylation of proteins in A431 human tumor. Cell 80: 225-36, 1995.
25. HWANG, S.B., LAM, M H. & PONG, S.S. Ionic and GTP regulation of
binding of platelet-activating factor to receptor and platelet-activating
factor induced activation of GTPase in rabbit platelet membrane. J. Biol.
Chem. 261: 532-7, 1986.
26. ISHII, S., NAGASE, T. & SHIMIZU, T. Prostaglandins Other Lip. Med. 68-
69: 599-609, 2002.
27. IZUMI, T. & SHIMIZU T. Platelet-activating factor receptor: gene
expression and signal transduction. Biochim. Biophys. Acta. 1259: 317-
333, 1995.
28. JERONIMO, S. M.; DUGGAL, P.; BRAZ, R. F.; CHENG, C.; MONTEIRO,
G. R.; NASCIMENTO, E. T.; MARTINS, D.R.; KARPLUS, T. M.;
XIMENES, M. F.; OLIVEIRA, C. C.; PINHEIRO, V. G.; PEREIRA, W.;
PERALTA, J. M.; SOUSA, J.; MEDEIROS, I. M.; PEARSONI, R. D.;
BURNS, T. L.; PUGH, E. W. & WILSON, M. E. An emerging peri-urban
pattern of infection with Leishmania chagasi, the protozoan causing
visceral leishmaniasis in northeast Brazil. Scand. J. Infect. Dis. 36 (6-7):
443-449, 2004.
29. KULIKOV, V.I. & MUZYA, G.I. Ether lipids and platelet-activating factor:
evolution and cellular function. Biochemistry. (Moscow) 62: 1103-1108,
1997.
30. LE GOUILL, C., PARENT, J.L., ROLA-PLESZEZYNSKI & STANKOVA,
J. Structural and functional requirements for agonist-induced

30
internalization of the human platelet-activating factor receptor. J. Biol.
Chem. 272: 21289-21295, 1997.
31. LENHINGER, A. L., NELSON, D. L. & COX, M. M. Título: Principles of
Biochemistry. Páginas: 736-787. Quarta edição Editora: Worth
Publishers, Inc. New York, 2004.
32. LIU, B., NAKASHIMA, S., ADACHI, T., ITO, Y., TAKANO, T., SHIMIZU,
T. & NOZAWA, Y. Prolonged activation of phospholipase D in Chinese
hamster ovary cells expression platelet-activating factor receptor lacking
cytoplasmic C-terminal tail. Biochem. J. 327: 239-44, 1997.
33. LOPES, A.H.C.S., DUTRA, P.M.L., RODRIGUES, C.O., SOARES, M.J.,
ANGLUSTER, J. & CORDEIRO, R.S.B. Effect of platelet-activating
factor on the process of cellular differentiation of Herpetomonas
muscarum muscarum. J. Eukariot. Microbiol. 44: 321-325, 1997.
34. MANGOLD, H.K., APTE, S.S. & WEBER, N. Biotransformation of
alkylglycerols in plant cell cultures: production of platelet-activating factor
and other biologically active ether lipids. Lipids 26(12): 1086-1092, 1991.
35. MATTHEWS, K. R. The developmental cell biology of Trypanosoma
brucei. J. Cell Sci. 118: 283-290, 2005
36. MCKERROW J. H., Caffrey C., Kelly B., Loke P., Sajid M. Proteases in
parasitic diseases. Annu Rev Pathol. (1): 497-536. Review, 2006
37. NAKAYAMA, R., KUMAGAI, H. & SAITO, K. Evidence for production of
platelet-activating factor by yeast Saccharomyces cerevisiae cells.
Biochem. Biophys. Acta 1199: 137-142, 1994a.
38. NAKAYAMA, R., UDAGAWA, H., MITSUI, S & KUMAGAI, H.
Saccharomyces cerevisiae Cells Produce Platelet-activating Factor in
Response to Calcium Ionophore A23187. Biosci. Biotech. Biochem. 58
(6): 1115-1119, 1994b.
39. NAULA, C. & SEEBECK, T. Cyclic AMP Signaling in Trypanosomatids.
Parasitol. Today I: 35-38, 2000.
40. NEVES, D. P. Capítulo Leishmanioses. Parasitologia Humana. Editora
Atheneu, 2005.
41. OLIVIER, M., GREGORY, D. J. & FORGET, G. Subversion mechanisms
by which Leishmania parasites can escape the host immune response: a
signaling point of view. Clin. Microbiol. Rev. 18 (2): 293-305, 2005.

31
42. PARSONS, M., VALENTINE, M., & CARTER, V. Protein kinases in
divergent eukaryotes: identification of protein kinase activities regulated
during trypanosome development. Proc. Natl. Acad. Sci. USA 90: 2656-
60, 1993.
43. PARSONS, M. & RUBEN, L. Pathways involved in environmental
sensing in trypanosomatids. Parasitol. Today 16: 56-62, 2000.
44. PRESCOTT, S. M., ZIMMERMAN, G. A., STAFFORINI, D. M. & MC
INTYRE, T. M. Platelet-activating factor and related lipid mediators.
Annu. Rev. Biochem. 69: 419-445, 2000.
45. QUEIROZ, M. J., ALVES, J. G. & CORREIA, J. B. Visceral leishmaniasis:
clinical and epidemiological features of children in an endemic area. J.
Pediat. 80: 141-146, 2004.
46. REY, L. C., MARTINS, C. V., RIBEIRO, H. B. & LIMA, A. A. . American
visceral leishmaniasis (kala-azar) in hospitalized children from an
endemic area. J. Pediat. 81: 73-78, 2005.
47. ROBBINS, S.M. & HOLLENBERG, M.D. Plasma membrane-localized
signal transduction. In: Current topics in membranes-membrane
permeability London, Academic Press 48: 351-95, 1999.
48. RODRIGUES, C.O., DUTRA, P.M.L., SOUTO-PADRÓN, T., CORDEIRO
R.S.B. & LOPES, A.H.C.S. Effect of platelet-activating factor on cell
differentiation of Trypanosoma cruzi. Arch. Biochem. Biophys. 223: 735-
740, 1996.
49. RODRIGUES, C.O., DUTRA, P.M.L., BARROS, F.S., SOUTO-PADRÓN,
T., MEYER-FERNANDES, J.R. & LOPES, A.H.C.S. Platelet-activating
factor induction of secreted phosphatase activity in Trypanosoma cruzi.
Biochem. Biophys. Res. Commun. 266: 36-42, 1999.
50. ROSA, M. S. S, VIEIRA, R. B., PEREIRA A. F., DUTRA, P. M. L. &
LOPES, A. H. C. S. Platelet- activating factor (PAF) modulates peritoneal
mouse macrophage infectiom by Leishmania amazonensis. Curr.
Microbiol. 43: 33-37, 2001.
51. SANTOS, D. O. & OLIVEIRA, M. M. Effect of cAMP on macromolecule
synthesis in the pathogenic protozoa Trypanosoma cruzi. Mem. Inst.
Oswaldo Cruz. 83 (3): 287- 92, 1988.

32
52. SIEGEL, J. N., EGERTON, M., PHILLIPS, A. F. & SAMELSON, L. E.
Multiple signal transduction pathways activated through the T cell
receptor for antigen. Semin. Immunol. 3 (5): 325-34, 1991.
53. SILVA-NETO, M. A. C., CARNEIRO, A. B., VIEIRA, D. P., MESQUITA,
R. D. & LOPES, A. H. C. S. Platelet-activating factor (PAF) activates
casein kinase 2 in the protozoan parasite Herpetomonas muscarum
muscarum. Bioch. Biophys. Res. Commun. 293: 1358-1363, 2002.
54. STAFFORINI, D.M., PRESCOTT, S.M., ZIMMERMAN, G.A. &
MCINTYRE, T.M. Mammalian platelet-activating factor acetylhydrolases.
Biochem. Byophys. Acta 1301: 161-173, 1996.
55. SUGIURA, T., OJIMA, T., FUKUDA, T., SATOUCHI, K., SAITO, K. &
WAKU, K. J. Production of platelet-activating factor in slugs. Lipid Res.
32(11): 1795-1803, 1991.
56. SUGIURA, T., YAMASHIRA, A., KUDO, N., FUKUDA, T., MIYAMOTO,
T., CHENG, N.N., KISHIMOTO, S., WAKU, K., TANAKA, T., TSUKAKI,
H. & TOKOMURA, A. Distribution of alkyl and alkenyl ether-linked
phospholipids and platelet-activating factor-like lipid in various species of
invertebrates. Biochim. Biophys. Acta. 1126: 298-308, 1992.
57. SUNDAR, S. & CHATTERJEE, M. Visceral leishmaniasis - current
therapeutic modalities. Indian J. Med. Res. 123: 345-352, 2006.
58. TSOUKATOS, D.C., TSELEPIS, A.D. & LEKKA, M.E. Studies on the
subcellular distribution of 1-O-alkyl-2-aceytyl-sn-glycero phosphocholine
(PAF) and on the enzymic activities involved in its biosynthesis within the
ciliate Tetrahymena pyriformis. Biochim. Biophys. Acta 1170: 258-264,
1993.
59. TURNER, M. R. & LUMB, R.H. Synthesis of platelet activating factor by
tissues from the rainbow trout, Salmo gairgneri. Biochem. Biophys. Acta.
1004: 49-52, 1989.
60. VANNIER-SANTOS, M. A., MARTINY, A. & DE SOUZA, W. Cell biology
of Leishmania spp.: Invading and Evading. Curr. Pharmacol. Design. 8:
297-318, 2002.
61. VOULDOUKIS, I.; RIVEROS-MORENO, V.; DUGAS, B.; OUAAZ, F.;
BECHEREL, P.; DEBRE, P.; MONCADA, S.; MOSSALAYI, M. D. The
killing of Leishmania major by human macrophages is mediated by nitric

33
oxide induced after ligation of the Fc epsilon RII/CD23 surface
antigen.Proc. Natl. Acad. Sci. USA, v.92: 7804,1995.
62. ZHU, T., GOBEIL J. F., VASQUEZ-TELLO, A., LEDUC, M., RIHAKOVA,
L., BOSSOLASCO, M., BKAILY, G., PERI, K., VARMA, D. R.,
ORVOINE, R. & CHEMTOB, S. Intracrine signaling through lipid
mediators and their cognate nuclear G-protein-coupled receptors: a
paradigm based on PGE2, PAF, and LPA1 receptors. Can. J. Physiol.
Pharmacol. 84: 377-391, 2006.


















