
INFLUÊNCIA DE BACTÉRIAS ENDOFÍTICAS NA … · NA FISIOLOGIA DE PLANTAS DE CANA-DE-AÇÚCAR SOB...
81
INSTITUTO AGRONÔMICO CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA TROPICAL E SUBTROPICAL INFLUÊNCIA DE BACTÉRIAS ENDOFÍTICAS NA FISIOLOGIA DE PLANTAS DE CANA-DE-AÇÚCAR SOB RESTRIÇÃO HÍDRICA FERNANDA CASTRO CORREIA MARCOS Orientadora: Ana Maria Magalhães Andrade Lagôa Co-orientadora: Adriana Parada Dias da Silveira Dissertação submetida como requisito parcial para obtenção do grau de Mestre em Agricultura Tropical e Subtropical, Área de Concentração em Tecnologia de Produção Agrícola Campinas, SP Abril 2012
Transcript of INFLUÊNCIA DE BACTÉRIAS ENDOFÍTICAS NA … · NA FISIOLOGIA DE PLANTAS DE CANA-DE-AÇÚCAR SOB...

INSTITUTO AGRONÔMICO CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA
TROPICAL E SUBTROPICAL
INFLUÊNCIA DE BACTÉRIAS ENDOFÍTICAS NA FISIOLOGIA DE PLANTAS
DE CANA-DE-AÇÚCAR SOB RESTRIÇÃO HÍDRICA
FERNANDA CASTRO CORREIA MARCOS
Orientadora: Ana Maria Magalhães Andrade Lagôa Co-orientadora: Adriana Parada Dias da Silveira
Dissertação submetida como requisito parcial para obtenção do grau de Mestre em Agricultura Tropical e Subtropical, Área de Concentração em Tecnologia de Produção Agrícola
Campinas, SP Abril 2012

ii
Ficha elaborada pela bibliotecária do Núcleo de Informação e Documentação do Instituto Agronômico de Campinas M625 i Marcos, Fernanda Castro Correia Influência de bactérias endofíticas na fisiologia de plantas de cana-de-açúcar sobre restrição hídrica / Fernanda Castro Correia Marcos Campinas, 2012. 64fls Orientadora: Ana Maria Magalhães Andrade Lagôa Co-orientadora: Adriana Parada Dias da Silveira Dissertação (Mestrado) em Agricultura Tropical e Subtropical
1. Cana-de-açúcar – planta-bactéria I. Lagôa Ana Maria Magalhães Andrade II. Silveira, Adriana Parada Dias da III. Título
CDD 633.61

iii

iv
Aos meus pais Maria José e César,
Aos meus avós Maria da Graça e Antônio (in memorian)
Ao meu irmão Gabriel, e à Lilian
Pelo incentivo, compreensão e amor, e
A mais nova integrante da família a Beatriz
DEDICO

v
AGRADECIMENTOS
Aos meus pais, pela educação, valores e princípios que me fizeram aproveitar as
oportunidades que surgiram durante minha trajetória de vida e também pelo amor e carinho.
Em especial a minha mãe que me ajudou a cuidar do experimento em casa de vegetação;
À minha avó, por todos os mimos e ensinamentos;
Ao meu irmão Gabriel e esposa Lilian, pelo amor, amizade e conversas que sempre
tivemos, e eventual ajuda na coleta de dados;
À Dra. Ana Maria Magalhães Andrade Lagôa e à Dra. Adriana Parada Dias da
Silveira pela oportunidade que me deram e pela amizade e orientação;
A todos os pesquisadores do Laboratório de Fisiologia Vegetal Coaracy de Moraes
Franco em especial ao Dr. Eduardo Caruso Machado e Dr. Rafael Vasconcelos Ribeiro pelos
conselhos e ensinamentos;
À Dra. Sueli dos Santos Freitas pela amizade, ensinamentos e pelos momentos
gastronômicos;
Ao Laboratório de Microbiologia do Solo pela disponibilidade de uso da infra-
estrutura e a todos os amigos Matheus Cipriano, Júlia Talazzo, Elaine Rodrigues, Ana Olívia
Fernandes, Régia a técnica Rosana Gonçalves Giertz, ao Sr. Lazinho, e em especial à Raquel
de Paula Freitas por todos os momentos de trabalho e muita diversão;
À todos os colegas do laboratório de Fisiologia Vegetal Coaracy de Moraes Franco
Daniela Machado, Ricardo Machado, José Rodrigues, Paulo Marchiori, Karina Silva, Pedro
Rampazzo, Fernanda Vilela e Rafaely Lameira pela ajuda no experimento de campo e no
laboratório;
À amiga Verónica Lorena Dovis por todos os ensinamentos e ajuda durante grande
parte do mestrado e também pelos bons momentos de convívio;
À amiga Cristina Sales pela ajuda constante em todas as etapas de desenvolvimento do
trabalho;
Às minhas queridas amigas Flávia Maria, que sempre me incentivou a trabalhar com
pesquisa e esteve ao meu lado em momentos difíceis, e Andréa Borges, pela ajuda sempre que
necessária e pelo carinho e compreensão;
Aos amigos da academia que foram muitas vezes essenciais para a manutenção do
bem estar físico e mental durante esses dois anos;
Ao Jefferson, por ter me proporcionado carinho e tranqüilidade na fase final do
trabalho;

vi
À todos os amigos da PG/IAC;
À CAPES e à FAPESP/BIOEN pelo suporte financeiro;
A todos os professores do Programa de Pós Graduação do IAC, pelos ensinamos;
À Pós Graduação e ao Instituto Agronômico pela oportunidade;

vii
“Quanto maior é o obstáculo,
maior será a glória de tê-lo superado”
Molière

viii
SUMÁRIO
LISTA DE ABREVIAÇÕES ..................................................................................................... xLISTA DE TABELAS .............................................................................................................. xiLISTA DE FIGURAS .............................................................................................................. xiiRESUMO ................................................................................................................................ xviABSTRACT ........................................................................................................................... xvii1 INTRODUÇÃO ....................................................................................................................... 12 REVISÃO BIBLIOGRÁFICA ................................................................................................ 32.1 Aspectos gerais da cultura da cana-de-açúcar ...................................................................... 32.2 Efeitos da deficiência hídrica na fisiologia vegetal .............................................................. 42.3 Interações benéficas entre planta e bactérias endofíticas diazotróficas ................................ 93 MATERIAL E MÉTODOS ................................................................................................... 153.1 Material vegetal e isolados bacterianos .............................................................................. 153.2 Delineamento experimental ................................................................................................ 173.2.1 Primeiro experimento: desenvolvimento de plantas cana-de-açúcar tratadas com inóculos bacterianos e sob disponibilidade hídrica adequada. ................................................. 173.2.2 Segundo experimento: desenvolvimento de plantas de cana-de-açúcar tratadas com inóculos bacterianos e sob condição de restrição hídrica. ........................................................ 173.3 Medidas de crescimento ..................................................................................................... 183.4 Potencial hídrico e conteúdo relativo de água na folha ...................................................... 183.5 Trocas gasosas .................................................................................................................... 183.6 Atividade fotoquímica ........................................................................................................ 193.7 Conteúdo de clorofilas ........................................................................................................ 193.8 Extração de compostos por MCW ...................................................................................... 193.9 Teor de carboidratos foliares .............................................................................................. 203.10 Teor de prolina .................................................................................................................. 213.11 Teor de aminoácidos livres totais ..................................................................................... 213.12 Teor de nitrogênio total em folhas .................................................................................... 223.13 Extração enzimática .......................................................................................................... 223.14 Atividade enzimática in vitro da redutase do nitrato ........................................................ 233.15 Atividade enzimática in vitro da sintetase da glutamina .................................................. 233.16 Quantificação de bactérias presentes nos colmos e nas raízes ......................................... 243.17 Delineamento estatístico e análise de dados ..................................................................... 244 RESULTADOS ..................................................................................................................... 254.1 Efeitos da interação planta-bactéria na fisiologia de cana-de-açúcar sob disponibilidade hídrica adequada. ...................................................................................................................... 254.1.1 Contagem de bactérias endofíticas .................................................................................. 254.1.2 Aspectos biométricos ....................................................................................................... 254.1.3 Potencial hídrico e conteúdo relativo de água ................................................................. 264.1.4 Teores de prolina e aminoácidos livres totais .................................................................. 274.1.5 Atividade fotoquímica ..................................................................................................... 284.1.6 Clorofila a, b e total ......................................................................................................... 284.1.7 Trocas gasosas ................................................................................................................. 294.1.8 Carboidratos foliares ........................................................................................................ 314.1.9 Nitrogênio ........................................................................................................................ 314.1.10 Enzimas do metabolismo do nitrogênio ........................................................................ 324.2 Atenuação dos efeitos da restrição hídrica pela interação com bactérias diazotróficas endofíticas ................................................................................................................................. 334.2.1 Contagem de bactérias diazotróficas endofíticas ............................................................. 33

ix
4.2.2 Altura e número de folhas ............................................................................................... 344.2.3 Massa de matéria seca da parte aérea e das raízes ........................................................... 344.2.4 Potencial hídrico e conteúdo relativo de água ................................................................. 354.2.5 Teores de prolina e de aminoácidos livres totais (ALT) .................................................. 364.2.6 Atividade fotoquímica ..................................................................................................... 384.2.7 Clorofila total ................................................................................................................... 384.2.8 Trocas gasosas ................................................................................................................. 394.2.9 Carboidratos .................................................................................................................... 424.2.10 Nitrogênio ...................................................................................................................... 444.2.11 Atividade de enzimas do metabolismo do nitrogênio ................................................... 455 DISCUSSÃO ......................................................................................................................... 475.1 Efeitos da interação planta-bactéria na fisiologia de cana-de-açúcar ................................. 475.2 Atenuações dos efeitos do estresse hídrico por bactérias endofíticas diazotróficas ........... 516 CONCLUSÕES ..................................................................................................................... 557 REFERÊNCIAS BIBIOGRÁFICAS ..................................................................................... 56

x
LISTA DE ABREVIAÇÕES
A Assimilação líquida de CO2 ABA Ácido abscísico AMI Amido AST Açúcares solúveis totais ATP Adenosina trifosfato BSA Albumina de soro bovino Ci Concentração intercelular de CO2 CRA Conteúdo relativo de água DAP Dias após o plantio DTT Ditiotreitol EDTA Ácido etilenodiamino tetra-acético E Transpiração ETR Transporte aparente de elétrons FAD Flavina adenina dinucleotídeo FM Fluorescência máxima em tecidos adaptados ao escuro FV Fluorescência variável no escuro FV/FM Eficiência quântica potencial do fotossistema II em tecidos adaptados ao escuro GGH γ-glutamil-hidroximato Glu Glicose gs Condutância estomática JMV Meio de cultura semi-específico para o gênero Burkholderia JNFb Meio de cultura semi-específico para o gênero Herbaspirillum LGI-P Meio de cultura semi-específico para G. diazotrophius MCW Metanol, clorofórmio e água MF Massa fresca MS Massa Seca MT Massa Túrgida NADH Nicotinamida Adenina Dinucleotídeo NPQ Coeficiente de extinção não fotoquímica da fluorescência PMSF Fluoreto de fenilmetilsultonil PVPP Polivinilpolipirrolidona Q Radiação fotossinteticamente ativa (µmol m-2 s-1) R Respiração no escuro RN Redutase do nitrato SAC Sacarose SG Sintetase da glutamina TCA Ácido tricloroacético Ψw Potencial hídrico

xi
LISTA DE TABELAS Tabela 1 - Número mais provável (NMP) de bactérias, no início do experimento, nos colmos
dos genótipos IACSP94-2094 e IACSP95-5000, utilizados para produção de mudas, em três meios de cultura (JNFb, LGIP, JMV)...........................................25
Tabela 2 – Índice de eficiência do uso de nitrogênio (g biomassa2/g N-1) nos genótipos de
cana-de-açúcar IACSP94-2094 e IAC95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle)...................................................32
Tabela 3 - Número de bactérias (UFC 105 g-1) nas raízes dos genótipos IACSP94-2094 e
IACSP95-5000 mantidos em condição hidratada (H) ou restrição hídrica (RH), tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), em três meios de cultura (JNFb, LGIP, JMV). Contagem realizada no final do segundo experimento................................................................................34
Tabela 4 – Clorofila total nos dois genótipos de cana-de-açúcar mantidos hidratados (H) ou
sob restrição hídrica (RH) tratados com bactérias endofíticas diazotróficas (inóculo A e B) ou não (controle)..........................................................................39
Tabela 5 – Respiração nos dois genótipos de cana-de-açúcar mantidos hidratados (H) ou sob
restrição hídrica (RH) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle)............................................................................................42
Tabela 6 – Teores de nitrogênio (g kg -1) nos dois genótipos de cana-de-açúcar IACSP94-
2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou sob restrição hídrica (RH)........................................................................................................................44
Tabela 7 – Quantidade de nitrogênio acumulado (mg N planta-1) nos dois genótipos de cana-
de-açúcar IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou sob restrição hídrica (RH).............................................................................................46
Tabela 8 – Índice de eficiência de utilização de nitrogênio (g biomassa2/g nutriente-1) nos
dois genótipos de cana-de-açúcar IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou sob restrição hídrica (RH).........................................................46

xii
LISTA DE FIGURAS Figura 1 – Massa de matéria seca da parte aérea (a) e de raiz (b) dos genótipos IACSP94-
2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle). Cada histograma representa o valor médio (n=5) ± desvio-padrão. Letras iguais indicam ausência de diferenças estatísticas significativas entre os tratamentos bacterianos dentro do mesmo genótipo pelo teste de Tukey (p>0,05)..........................................................................................26
Figura 2 – Potencial da água na folha (Ψw, MPa) nos genótipos de cana-de-açúcar IACSP94-
2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculo A e B) ou não (controle), medido às 12h30min. Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras distintas indicam diferenças estatísticas significativas entre os tratamentos bacterianos dentro do mesmo genótipo pelo teste de Tukey (p<0,05)..........................................................................................27
Figura 3 – Teor de prolina em dois genótipos de cana-de-açúcar IACSP94-2094 e IACSP95-
5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle). Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras distintas indicam diferenças estatísticas significativas entre os tratamentos bacterianos dentro do mesmo genótipo pelo teste de Tukey (p<0,05)...................28
Figura 4 – Transporte aparente de elétrons (ETR) dos genótipos de cana-de-açúcar IACSP94-
2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) medido às 13 horas. Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras distintas indicam diferenças estatísticas significativas entre os tratamentos bacterianos dentro de genótipo pelo teste de Tukey (p<0,05).......................................................................................................29
Figura 5 – Assimilação de CO2
Figura 8 – Massa de matéria seca da parte aérea dos genótipos IACSP94-2094 (a,c) e IACSP95-5000 (b,d) tratados com bactérias endofíticas diazotróficas (inóculos A
(A-a), condutância estomática (gs-b) e transpiração (E-d) dos genótipos IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) medidos às 13 horas. Cada histograma representa o valor médio (n=4) ± desvio-padrão. Letras distintas indicam diferenças estatísticas significativas pelo teste de Tukey (p<0,05)..........30
Figura 6 – Teor de carboidratos: sacarose (SAC) (a), açúcares solúveis totais (AST) (b) nos
genótipos IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle). Cada histograma representa o valor médio (n=4) ± desvio-padrão. Letras distintas indicam diferenças significativas entre os tratamentos bacterianos dentro do genótipo pelo teste de Tukey (p<0,05).......................................................................................................31
Figura 7 – Atividade das enzimas redutase do nitrato (a) e sintetase da gutamina (b) dos
genótipos IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle). Letras distintas indicam diferenças significativas entre os tratamentos bacterianos dentro do genótipo pelo teste de Tukey (p<0,05)..........................................................................................33

xiii
e B) ou não (controle), mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=5) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro do mesmo genótipo pelo teste de Tukey (p<0,05)...................................................................35
Figura 9 – Potencial da água na folha (MPa) medido às 12h 30 min. nos genótipos de cana-
de-açúcar IACSP94-2094 (a) e IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou em restrição hídrica (RH). Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).......36
Figura 10 – Conteúdo relativo de água nos genótipos de cana-de-açúcar IACSP94-2094 (a) e
IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=5) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas iguais indicam ausência de diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p>0,05)......................................................37
Figura 11 – Teores de prolina nos genótipos de cana-de-açúcar IACSP94-2094 (a) e
IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05)..................................................................................37
Figura 12 – Teores de aminoácidos livres totais (ALT) nos genótipos de cana-de-açúcar
IACSP94-2094 (a) e IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=4) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).....................................38
Figura 13 – Assimilação de CO2 (A-a,b), condutância estomática (gs-c,d), concentração
intercelular de CO2 (Ci-e,f) de plantas dos genótipos IACSP94-2094 (a,c,e) e IACSP95-5000 (b,d,f) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantido hidratados (H) ou sob restrição hídrica (RH) medidos às 13 horas. Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano e genótipo. Letras

xiv
minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica e genótipo pelo teste de Tukey (p<0,05)..................................................................................................................40
Figura 14 – Transpiração (E) de plantas dos genótipos IACSP94-2094 (a) e IACSP95-5000
(b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH) medida às 13 horas. Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano e genótipo. Letras minúsculas iguais indicam ausência de diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica e genótipo pelo teste de Tukey (p<0,05)........41
Figura 15 – Teores de açúcares solúveis totais (AST) nos genótipos de cana-de-açúcar
IACSP94-2094 (a) e IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=4) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).....................................42
Figura 16 – Teor de sacarose (SAC) nos genótipos de cana-de-açúcar IACSP94-2094 (a) e
IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=4) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05)..................................................................................43
Figura 17 – Teores de amido (AMI) nos genótipos de cana-de-açúcar IACSP94-2094 (a) e
IAC95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=4) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05)..................................................................................44
Figura 18 – Atividade da redutase do nitrato (RN) nos genótipos de cana-de-açúcar
IACSP94-2094 (a) e IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).....................................45

xv
Figura 19 – Atividade da sintetase da glutamina (SG) nos genótipos de cana-de-açúcar IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05)..........................................48
Figura 20 – Representação das respostas fisiológicas dos genótipos IACSP94-2094 (a) e
IACSP95-5000 (b) com boa disponibilidade hídrica, tratados com inóculo A (quadrado) e inóculo B (triangulo) em relação aos respectivos controles (linha pontilhada). Os valores referem-se a mudanças relativas das seguintes variáveis: massa de matéria seca da parte aérea (MSPA), massa seca de raiz (MSR), fotossíntese (A), condutância estomática (gs), transpiração (E), transporte aparente de elétrons (ETR), coeficiente de extinção fotoquímica (NPQ), potencial hídrico (Ψw), teor de sacarose (Sac), teor de açúcares solúveis totais (AST), teor de nitrogênio (N), atividade da redutase do nitrato (RN), atividade da sintetase da glutamina (SG), teor de prolina (Prolina), teor de aminoácidos livres totais (ALT). * indicam diferença estatística em relação ao controle..........................................51

xvi
Influência de bactérias endofíticas na fisiologia de plantas de cana-de-açúcar sob restrição hídrica
RESUMO
O estresse hídrico é a principal causa de perdas agrícolas e o uso de novas tecnologias
pode melhorar o desenvolvimento de plantas submetidas a esse estresse. A utilização de
bactérias endofíticas capazes de promover alterações fisiológicas nas plantas é uma
possibilidade a ser examinada como nova tecnologia no cultivo da cana-de-açúcar. Desta
forma, buscou-se estudar se a interação planta-bactérias diazotróficas endofíticas seria
benéfica em condições de disponibilidade hídrica adequada, e se esta interação poderia
reduzir a suscetibilidade das plantas de cana-de-açúcar à restrição hídrica. Foram utilizados
dois genótipos um tolerante IACSP94-2094 e outro sensível IACSP95-5000 tratados com
inóculos bacterianos (A ou B) ou não (controle) em dois experimentos. Os inóculos
bacterianos foram formulados a partir de seleção de isolados bacterianos em meio semi-
seletivo para Herbaspirillum (A) e para Glucanoacetobacter (B). No primeiro experimento,
realizado em condição hídrica adequada, houve aumento da fotossíntese em plantas tolerantes
tratadas com os inóculo B, por terem aumento no transporte de elétrons, e em plantas
sensíveis o aumento da fotossíntese ocorreu no tratamento com inóculo A, por terem maior
abertura estomática. Ainda no primeiro experimento ocorreu aumento na atividade da
redutase do nitrato em IACSP95-5000 com o emprego do inóculo B. A interação planta-
bactérias endofíticas influenciou positivamente as trocas gasosas e o metabolismo do
nitrogênio, sem, entretanto, refletir em promoção de crescimento. No segundo experimento,
as plantas foram cultivadas sob restrição hídrica e plantas do genótipo sensível não tiveram
diminuição da massa de matéria seca quando tratadas com inóculo B. O tratamento com esse
inóculo ainda garantiu maior eficiência do uso do nitrogênio sob condição de restrição
hídrica. A influência da inoculação de bactérias endofíticas na atenuação dos efeitos da seca
foi mais evidente no genótipo sensível (IACSP95-5000).
Palavras chave: estresse abiótico, interação planta-bacteria, atenuação de efeitos da seca,
fotossíntese, nitrogênio.

xvii
Influence of endophytic bacteria in the physiology of sugar cane
ABSTRACT
plants under water restriction
Water stress is the main cause of agricultural losses, and the use of new technologies
can improve the development of plants under such stress. The use of endophytic bacteria that
is capable of promoting physiological changes in plants is a possibility to be examined as new
technology in the cultivation of cane sugar. Thus, we sought to study if the interaction plant-
bacteria would be beneficial in conditions of adequate water availability, and whether this
interaction could promote more benefits to plants under water stress and drought sensitive
genotype. For this we used tolerant genotype IACSP94-2094 and sensitive IACSP95-5000
treated with bacterial inocula (A or B) or not (control) in two experiments. The bacterial
inocula were formulated from the selection of bacterial isolates in semi-selective for
Herbaspirillum (A) and Glucanoacetobacter (B). In the first experiment, conducted in proper
water conditions, photosynthesis increased in tolerant genotype plants treated with inoculum
B, because of the increase in electron transport; and in sensitive plants the increase of
photosynthesis occurred in the treatment with the inoculums A, by a more open stomata.
Also in the first experiment there was an increase in the activity of nitrate reductase in
IACSP95-5000 with the use of inoculums B. The plant-endophytic bacteria positively
influenced gas exchange and nitrogen metabolism, without however, reflected in growth
promotion. In the second experiment, plants were grown under water stress and plant
genotype had no appreciable decrease in dry weight when inoculated with B. Treatment with
this inoculum has ensured greater nitrogen use efficiency under water stress condition. The
influence of inoculation of endophytic bacteria in mitigating the effects of drought was most
evident in the sensitive genotype (IACSP95-5000).
Keywords: abiotic stress, plant bacteria interaction, attenuation effects of drought
, photosynthesis, nitrogen.

1
1 INTRODUÇÃO
A importância da cana-de-açúcar para o Brasil é inquestionável, desde os tempos
coloniais até o presente. Atualmente o país se destaca pela produção não só de açúcar, mas
também de etanol, combustível que ganhou relevância na década de 70 com o Proálcool e que
atualmente tem grande espaço no mercado devido ao sucesso dos carros bi-combustíveis. Para
abastecer o mercado de forma adequada, a produção e a produtividade da cultura precisam
aumentar rapidamente. O aumento da produtividade pode ser atingido por meio de melhorias
das questões fitotécnicas ou da ocupação de novas áreas, muitas vezes localizadas em
ambientes restritivos à produção canavieira. No Estado de São Paulo, a expansão do cultivo
da cana-de-açúcar está ocorrendo sobre áreas que apresentam restrições hídricas moderadas
(BRUNINI, 2007) e que podem prejudicar o cultivo.
O estresse hídrico é a principal causa de perdas agrícolas e entender como as plantas
respondem a essa alteração ambiental é importante para traçar estratégias e evitar prejuízos.
A falta de água causa uma série de alterações morfo-fisiológicas como: diminuição da área
foliar, do turgor celular, do conteúdo relativo de água (CRA), do potencial hídrico foliar, a
absorção de nutrientes é alterada, além de grandes alterações no metabolismo do nitrogênio e
do carbono (FAROOQ et al., 2009).
Uma das primeiras respostas das plantas à seca é o fechamento estomático (CHAVES,
1991), causando uma limitação de substrato para a fotossíntese. A diminuição de CO2
acarreta menor assimilação de carbono, mas este não é o único fator limitante, já que pode
ocorrer também alteração metabólica do processo fotossintético (LAWLOR, 2002; FLEXAS
& MEDRANO, 2002). Em cana-de-açúcar, essas alterações metabólicas na fotossíntese
parecem acontecer a partir de um potencial hídrico foliar de -0,37 MPa (DU et al., 1996).
Estudos que considerem as respostas fisiológicas dos genótipos a condições restritivas
podem auxiliar programas de melhoramento genético. A melhoria dos cultivos em condições
de estresse também pode ocorrer com a utilização de tecnologias como a aplicação de
bactérias capazes de promover crescimento vegetal ou alterações fisiológicas benéficas às
plantas. Um exemplo interessante ligado à melhoria da capacidade fotossintética foi
apresentado por SHI et al. (2010), no qual o tratamento com bactérias endofíticas causou às
plantas de beterraba maior eficiência fotossintética, maior conteúdo de clorofila, além de
aumento no teor de carboidratos.

2
A interação entre plantas e bactérias promotoras de crescimento vegetal é bastante
conhecida. Essas bactérias podem influenciar diretamente o metabolismo das plantas,
fornecendo substâncias que normalmente estariam em menor quantidade, pela absorção de
nutrientes ou ainda pelo biocontrole de patógenos como, por exemplo, pela indução de
resistência sistêmica (BASHAN & DE-BASHAN, 2005).
A fixação biológica de nitrogênio (FBN) é uma das vantagens da associação entre
bactérias e plantas. Variedades brasileiras de cana podem obter pelo menos 40 kg N ha-1 ano-
1
pela FBN (URQUIAGA et al., 2011). Cerca de 60% a 70% do nitrogênio em cana-de-açúcar
é proveniente da FBN segundo URQUIAGA et al. (1992). No entanto, não está claro de que
forma ocorreria essa interação. Algumas hipóteses surgem como alternativas considerando
que as bactérias são responsáveis por uma melhoria na absorção de nitrogênio (MANTELIN
& TOURAINE, 2004).
Por auxiliar no desenvolvimento vegetal, promover o crescimento radicular
(OLIVEIRA et al., 2002) e melhorar a absorção de nutrientes (BARRETTI et al., 2008 a,b), o
uso de inoculantes contendo bactérias poderia beneficiar plantas sob restrição hídrica. A
presença de bactérias pode também levar à produção de substâncias osmorreguladoras pela
planta e assim atuar de forma sinérgica, colaborando na tolerância à seca (DIMKPA et al.
2009).
Trabalhos com inoculação de bactérias mostraram que plantas tratadas possuíam
potencial hídrico mais elevado do que as plantas controle sob estresse hídrico (CREUS et al.,
1998; 2004), garantindo assim vantagens na regulação do balanço hídrico. A inoculação
dessas bactérias em plantas pode ainda aliviar os efeitos negativos causados por estresse,
garantindo maior fotossíntese e abertura estomática (HAN & LEE, 2005). O tratamento com
bactérias parece ser mais favorável em situação de restrição hídrica. Plantas de arroz tratadas
com bactérias apresentaram maior diferença de massa de matéria seca entre raiz de plantas
tratadas em relação ao controle sem bactérias em condições de menor disponibilidade de
água, enquanto que em situação de boa disponibilidade hídrica essa diferença não foi
registrada (YUWONO et al., 2005)
Desta forma, buscou-se estudar se a interação planta-bactérias diazotróficas
endofíticas seria benéfica em condições de disponibilidade hídrica adequada, e se esta
interação poderia reduzir a suscetibilidade das plantas de cana-de-açúcar à restrição hídrica.

3
2 REVISÃO BIBLIOGRÁFICA 2.1 Aspectos gerais da cultura da cana-de-açúcar
Planta originária da Nova Guiné, a cana-de-açúcar (Saccharum spp.) foi introduzida
no Brasil no século XVI e parte da história do país confunde-se com a desta cultura. O Brasil
é o maior produtor mundial de açúcar e álcool, com produção de cana-de-açúcar estimada de
623 milhões de toneladas para a safra 2011/2012 (CONAB, 2011). A região Centro-Sul do
país concentra cerca de 90% da produção nacional, da qual cerca de 60% está no Estado de
São Paulo (CONAB, 2011). Mesmo com essa grande produção, na safra de 2009/2010 houve
queda e o principal motivo foi climático. A seca que atingiu a região centro-sul prejudicou o
desenvolvimento e a renovação dos canaviais (CONAB, 2011). Assim, a importância
econômica do cultivo de cana-de-açúcar revela a necessidade de um contínuo
aperfeiçoamento para aumentar sua produtividade, garantir melhores condições de cultivo e
reduzir impactos ambientais.
A cana-de-açúcar é uma cultura tropical e subtropical que produz grande quantidade
de biomassa e requer quantidades substanciais de água e nitrogênio para obter rendimento
máximo (WIEDENFELD, 2000). Plantas de metabolismo C4 possuem alta eficiência no uso
da radiação (ROBERTSON et al., 1996), alcançando altas taxas fotossintéticas e grande
acúmulo de carboidratos (WELBAUM & MEINZER, 1990). O destaque desse grupo de
plantas é o sistema de concentração de CO2
nas folhas que consiste em modificações
bioquímicas e estruturais garantindo melhor desempenho fotossintético (GHANNOUM,
2009). Outra característica inerente as plantas C4 é que a fotossíntese é mais competitiva em
situações que poderiam favorecer perda de carbono pela fotorrespiração tais como altas
temperaturas ou diminuição da disponibilidade de água (LOPES et al., 2011). O grande
acúmulo de sacarose em genótipos de cana pode ser alcançado pelo aumento da conversão da
energia solar em carboidratos, além da distribuição dos fotoassimilados para os colmos
(SINGELS et al., 2005). Colmos maduros são capazes de acumular altas concentrações de
sacarose chegando a 650 mM em algumas variedades (WELBAUM & MEINZER, 1990).
É sabido que o teor de sacarose dos colmos varia com o tempo de colheita. Ele
também varia com a idade da cultura, com os estresses abióticos e com o genótipo. O teor de
sacarose é maior no inverno, quando há aumento no armazenamento devido à redução na
força do dreno, regulada por temperatura, estresse hídrico e nitrogênio (SINGELS et al.,
2005).

4
Dentre as variáveis ambientais que podem afetar os processos de bioconversão de
energia na cana-de-açúcar estão a luz (intensidade e quantidade), a concentração de CO2
2.2 Efeitos da deficiência hídrica na fisiologia vegetal
, a
disponibilidade de água, os nutrientes e a temperatura (RODRIGUES, 1995), sendo o déficit
hídrico um dos principais fatores limitantes da produção da cana-de-açúcar. O estudo dos
efeitos do estresse hídrico sobre os componentes de produção e sua relação com a
produtividade pode ajudar os programas de melhoramento a selecionar genótipos produtivos
sob essa condição (SILVA et al., 2008).
Segundo a UNICA (União das Indústrias de Cana-de-açúcar), a área cultivada com
cana-de-açúcar no Brasil passou de 8 milhões em 2011 (CONAB, 2011), e deve dobrar até
2020 (UNICA, 2008). Esse crescimento é estimulado pela demanda do mercado por etanol,
principalmente para abastecer veículos bicombustíveis. O mapeamento da expansão da cultura
de cana-de-açúcar (AGUIAR et al., 2009) indicou crescimento de 73% na área disponível
para colheita entre as safras 2003/04 e 2008/09. Além disso, as áreas com maior expansão
estão localizadas na região oeste do Estado de São Paulo, que também apresentam as menores
proporções de extensão territorial ocupada pelo cultivo da cana-de-açúcar. Essas áreas do
Estado apresentam restrição hídrica de moderada a alta (BRUNINI, 2007), o que pode
desfavorecer o cultivo.
Sendo a restrição hídrica uma das principais causas de perdas agrícolas, e diante de um
cenário que preconiza o uso eficiente da água, estudos que auxiliem o entendimento das
respostas de genótipos em condições restritivas de água são essenciais, em especial os que
usam técnicas que possam melhorar o desempenho das plantas e aumentar produtividade em
condições adversas.
A seca como fenômeno meteorológico ocorre quando o balanço hídrico da cultura se
torna negativo, quando a entrada de água por precipitação ou irrigação é inferior ao potencial
de evapotranspiração (PIMENTEL, 2004). O tempo para a insuficiência de suprimento de
água causar reduções de crescimento depende do tipo de planta, da capacidade de retenção de
água no solo, das condições atmosféricas, principalmente da diferença de pressão de vapor
entre a folha e o ar (DPV) e da temperatura e de fatores que controlam a evapotranspiração
(PIMENTEL, 2004).
As respostas das plantas à seca são caracterizadas por alterações morfológicas como
diminuição da área foliar, reduzida pelo enrolamento das folhas e diminuição do crescimento

5
(PIMENTEL, 2004). Segundo FAROOQ et al. (2009), o estresse hídrico ainda impede a
divisão celular, leva à perda de turgor, inibindo o alongamento celular, e há uma série de
alterações fisiológicas que impedem o desenvolvimento normal da planta. Estas alterações
são: mudanças na razão raiz/parte aérea, nas relações hídricas com diminuição do potencial
hídrico (Ψ), no conteúdo relativo de água (CRA) e na absorção de nutrientes; diminuição da
fotossíntese, da abertura estomática, da respiração, dos teores de carboidratos, alterações no
metabolismo do nitrogênio e aumento da atividade de enzimas ligadas ao estresse oxidativo
(CHAVES et al., 2002; PIMENTEL, 2004; FAROOQ et al., 2009). Obviamente, esse
impedimento do crescimento irá afetar a produtividade das culturas, fato que torna o estudo
das conseqüências e das formas de evitar o déficit hídrico os objetivos de muitos trabalhos.
Para PIMENTEL (2004) existem três estádios de resposta à deficiência hídrica. No
primeiro (ligeira deficiência hídrica) não ocorre diminuição da transpiração, havendo aumento
de fitormônio ácido abscísico (ABA), diminuição da expansão celular e da turgescência. Em
seguida, no segundo estádio (deficiência hídrica moderada), a condutância estomática diminui
levando à redução das taxas de transpiração e de fotossíntese, além da redução da atividade da
redutase do nitrato, da sacarose fosfato sintase e de enzimas ligadas à assimilação
fotossintética, aumento do teor de carboidratos solúveis e aminoácidos livres. Já no terceiro
estádio (deficiência hídrica severa), a transpiração é mínima, há aumento da atividade de
enzimas hidrolíticas, diminuição do teor de amido, de proteínas solúveis e clorofila e aumento
de espécies reativas de oxigênio.
Dentre essas alterações, o fechamento estomático tem sido identificado como um dos
primeiros eventos na resposta das plantas à seca moderada, por controlar a perda de água
(CHAVES, 1991). Essa resposta pode estar ligada com a comunicação das raízes com a parte
aérea, pois ocorre sinalização da disponibilidade hídrica entre raiz e parte aérea levando a
regulação do desenvolvimento da planta quando em restrição hídrica (DAVIES & ZHANG,
1991). A principal mudança que pode indicar essa comunicação ocorre na seiva do xilema
com diminuição de cátions, de ânions, do pH, de aminoácidos e aumento de hormônios
vegetais, principalmente o ABA. Existe uma relação bastante significativa entre a
concentração de ABA e fechamento dos estômatos, sendo que quanto maior a quantidade
desse hormônio maior é a limitação da condutância e menor a taxa de transpiração das
plantas. Considera-se, então, que o ABA seja um sinal químico relevante no desenvolvimento
vegetal em restrição hídrica (DAVIES & ZHANG, 1991). Além disso, o fechamento
estomático pode estar ligado com a diminuição do turgor e do potencial hídrico foliar
(LAWLOR, 2002), com variações na umidade do ar (FRANKS et al., 1997), ou ainda com o

6
ritmo circadiano da planta (CHAVES et al., 2002). Para YORDANOV (2003), a seca
moderada induz as plantas a uma regulação do consumo de água permitindo a manutenção do
conteúdo relativo de água dentro de limites em que a capacidade fotossintética e a
produtividade sejam pouco afetadas. Se por um lado o fechamento estomático restringe a
perda de água, ele também diminui a entrada de CO2
Segundo LAWLOR (2002), a limitação da fotossíntese potencial acontece mais por
consequência da menor síntese de ATP pela ATPsintase com o decaimento do conteúdo
relativo de água (CRA), do que pela menor concentração de CO
, substrato para a assimilação de carbono
pela fotossíntese.
Encontram-se aqui duas formas de se tratar a influência da seca na limitação da
fotossíntese. Ocorreria, primeiramente, uma limitação ligada ao fechamento estomático que
pode levar a mudanças no metabolismo fotossintético logo no início do processo de
desidratação, mas também existem mudanças no metabolismo que acontecem devido ao
aumento da intensidade do estresse hídrico.
Plantas tolerantes à seca podem controlar a abertura dos estômatos para permitir a
fixação de carbono no estresse, assim melhorando a eficiência do uso da água, ou abrindo
estômatos rapidamente quando o déficit hídrico acaba (YORDANOV, 2003). Em estudo com
genótipos de cana-de-açúcar, MACHADO (2009) demonstrou que um genótipo de cana-de-
açúcar em condições de déficit hídrico apresentou fechamento estomático antecipado, o que
manteve um melhor balanço hídrico, garantindo posteriormente aos estômatos uma abertura
capaz de não afetar tanto a fixação de carbono e, desta forma, manter o crescimento.
A limitação metabólica é um processo mais complexo e menos esclarecido, o que tem
levado a divergências sobre o assunto. Para CHAVES (1991) e CHAVES et al. (2002) a
limitação metabólica só ocorreria após um déficit hídrico prolongado que causaria danos mais
severos na planta (CORNIC, 2000); para outros autores, os danos metabólicos iniciariam logo
no início do processo de resposta da planta à seca (GHANNOUM et al., 2003).
2. O autor não nega a
influência da diminuição de CO2, mas os efeitos metabólicos dessa redução seriam menos
marcantes. O ATP possui papel vital no metabolismo, desta forma os danos metabólicos
causados pela diminuição da quantidade desta molécula energética influenciariam uma série
de processos. Uma das moléculas que tem sua quantidade reduzida em condições de estresse é
a ribulose-1,5-bisfosfato (RuBP), aceptor primário de CO2. Essa diminuição de síntese ou
regeneração de RuBP está ligada a falta de ATP, cuja síntese parece ser afetada por
inativação progressiva ou perda do fator de acoplamento da enzima ATPsintase, ou pela
redução da capacidade do transporte de prótons e elétrons, ou ainda por um gradiente de

7
prótons inadequado no tilacóide (LAWLOR, 2002). A síntese de aminoácidos, proteínas e
mesmo a atividade de muitas enzimas nas células dependem diretamente da molécula
energética para seu funcionamento adequado.
A diminuição da entrada de CO2 associada a altos níveis de radiação solar,
comumente registrados em condições de seca, pode levar ao aumento da suscetibilidade a
danos no aparato fotoquímico (YORDANOV, 2003). Em condições ideais de irrigação, a
energia luminosa interceptada pela folha é transformada em energia química através da cadeia
de transporte de elétrons formando NADPH e ATP, moléculas utilizadas no ciclo de Calvin.
A redução na assimilação de carbono pode afetar a via fotoquímica, de modo que durante
períodos desfavoráveis para a atividade fotossintética, as plantas têm de lidar com um
excedente de energia de excitação. Nesta situação, uma série de eventos é desencadeada no
aparato fotossintético em resposta ao estresse luminoso, que pode ser definido como a
redução na eficiência de conversão da energia luminosa em energia química, devido ao
excesso de energia nos fotossistemas II (MAXWELL & JOHNSON, 2000; MATTOS, 2006).
Em condições de deficiência hídrica, pode ocorrer um efeito fotoinibitório devido ao excesso
de energia luminosa não convertida em energia química. SILVA et al. (2007) encontraram
redução nos valores da eficiência quântica do fotossistema II em tecidos adaptados ao escuro
(FV/FM) em todos os genótipos de cana-de-açúcar estudados sob estresse hídrico, de forma
mais acentuada nos genótipos suscetíveis à seca. A redução de FV/FM indica fotoinibição
(MAXWELL & JOHNSON, 2000) sendo a habilidade de manter valores altos de FV/FM sob
estresse hídrico um indicativo da eficiência do uso da radiação para a atividade fotoquímica e
assimilação de carbono.
Quanto às causas metabólicas de impedimentos na fotossíntese, por exemplo, a
atividade enzimática da ribulose-bisfosfato carboxilase oxigenase
Apesar da grande parte das respostas de plantas C3 serem semelhantes às de plantas
C4, existem algumas diferenças relevantes. Assim como em C3, ocorre em C4 o fechamento
estomático devido à diminuição do potencial hídrico foliar, que leva a uma diminuição da
concentração intercelular de CO
(Rubisco) diminui muito
em condições de seca (GHANNOUM, 2009). A atividade enzimática decai linearmente com a
redução do potencial hídrico, alterações na viscosidade e volume celular alteram a agregação
e a desnaturação de proteínas e aumentam a concentração de solutos que podem se tornar
tóxicos e deletérios às enzimas (FAROOQ et al., 2009)
2 (Ci) e, consequentemente, da fotossíntese. No entanto, essa
resposta acontece em estágios iniciais do estresse hídrico (DU et al., 1996). Em estudo com
cana-de-açúcar, a limitação estomática aparece primeiramente quando o estresse ainda é

8
moderado (-0,37 MPa). Com o aumento da severidade, o que se observa é uma diminuição
das trocas gasosas sendo o Ci encontrado alto. Essa resposta indica a limitação não-estomática
da fotossíntese, que é corroborada pela redução no teor de clorofila, de proteínas solúveis
totais e na atividade enzimática de fosfoenopiruvato carboxilase (PEPcase) e Rubisco (DU et
al. 1996).
Segundo PIMENTEL (2004) a desidratação causa alterações nos conteúdos de
carboidratos, ocorrendo diminuição de amido, quando em estádio moderado de desidratação,
além de aumento de açúcares solúveis pela hidrólise de amido que são então usados na
respiração de manutenção. Segundo INMAN-BAMBER & SMITH (2005), a extensibilidade
celular é afetada pelo status hídrico foliar e pelos processos bioquímicos influenciados pela
falta de água. Plantas de cana parecem ter a extensão foliar afetada a partir de -0,4 MPa
(LOPES et al., 2011) e reduzida a praticamente zero quando Ψ for de -1,3 MPa. No entanto,
a taxa de extensão foliar é rapidamente recuperada com a reidratação, inclusive podendo
superar em crescimento aquelas plantas que não passaram por estresse (INMAN-BAMBER &
SMITH, 2005). A diminuição da área foliar é uma estratégia para limitar a transpiração.
Folhas jovens de cana-de-açúcar são capazes de se enrolar e reduzir a área foliar projetada e
com isso reduzir a radiação recebida. INMAN-BAMBER & SMITH (2005) relatam o
enrolamento a partir de um potencial de -1,0 MPa ao meio dia, podendo ser esse considerado
com suscetibilidade do genótipo à seca, como também uma estratégia de evitar o estresse.
Pela diminuição de crescimento, a relação fonte-dreno é alterada. LOPES et al. (2011)
sugerem que os efeitos adversos da seca podem ser evitados quando ocorre uma mudança no
padrão de alocação de carbono para permitir a formação de um sistema radicular mais
profundo antes de uma limitação de crescimento pela escassez de água. Sob seca pode ocorrer
acúmulo de amido nas raízes como estratégia de resistência (FAROOQ et al., 2009). INMAN-
BAMBER & SMITH (2005) sugerem que, em cana-de-açúcar, a síntese de sacarose seria
afetada pelo estresse a partir de Ψ -0,9 MPa. Outra alteração citada pelos autores é a alocação
da sacarose que, no período de seca, seria armazenada nos colmos ao invés de ser usada no
alongamento de folhas e colmos.
O estresse hídrico pode ainda causar diminuição na atividade da redutase do nitrato
(RN), enzima que catalisa a etapa da redução de nitrato (NO3−)R a nitrito (NO2
−) na assimilação
do nitrogênio. O metabolismo do nitrogênio, assim como o do carbono, tem grande relevância
sobre a produção de fitomassa e na produtividade final das culturas. FOYER et al. (1998),
trabalhando com milho, verificaram que a atividade enzimática máxima da RN diminuiu
como resultado do estresse hídrico, e que existe uma correlação entre a queda da taxa de

9
assimilação de CO2 com a da atividade enzimática. A atividade dessa enzima é regulada pela
fosforilação/desfosforilação, processo que diminui com a redução de CO2.
Além disso, sob
estresse, o substrato NO3− diminuiu na célula, o que pode modular a transcrição da enzima,
levando à redução de sua atividade (FOYER et al., 1998). Já para CHAVES et al. (2002),
algumas mudanças reversíveis no metabolismo como a diminuição da atividade da RN podem
contribuir para a manutenção da pressão osmótica em células fotossintéticas pelo aumento do
nitrato e diminuição da exportação de carboidratos.
Açúcares solúveis, além de serem fontes de carbono para manutenção e recuperação
de crescimento (CHAVES, 2002), são também tidos como moléculas osmoticamente ativas,
assim como a prolina, a glicina-betaina, os ácidos orgânicos, o cálcio, o potássio e os cloretos.
O aumento na concentração dessas moléculas é uma estratégia de proteção, já que esses
solutos contribuem para ajuste osmótico, desintoxicação de espécies reativas de oxigênio,
estabilização de membranas, e das estruturas de enzimas e de proteínas (FAROOQ et al.,
2009).
Dentre as moléculas osmorreguladoras, a prolina recebe bastante atenção. Aminoácido
que costuma aparecer em maiores concentrações em plantas sob estresse hídrico (MAIA et al.,
2007), seu acúmulo ocorre em função de maior síntese, diminuição da sua oxidação, da
diminuição na incorporação do aminoácido em proteínas, além do aumento da proteólise
(PIMENTEL, 2004). Segundo MOLINARI et al. (2007), o papel como osmorregulador dessa
molécula é controverso e a prolina em plantas de cana-de-açúcar se mostrou eficiente na
proteção contra estresse oxidativo e não foi mediador de ajustamento osmótico. Os autores
verificaram que a prolina contribuiu para eliminar radicais livres e tamponar o potencial redox
sob condições de estresse e que as plantas com maior teor de prolina tiveram maior produção
de biomassa, indicando que essa molécula pode estar ligada à tolerância ao estresse hídrico
com outro papel que não o ajustamento osmótico em cana-de-açúcar.
Em cana-de-açúcar ocorre um aumento na concentração de hexoses, sacarose,
potássio e aminoácidos, mas aparentemente sem alteração na osmorregulação celular. Esses
metabólitos podem aumentar em 30% em folhas durante o estresse. O ajuste osmótico deve
ocorrer apenas depois de um estresse hídrico severo (INMAN- BAMBER & SMITH, 2005).
2.3 Interações benéficas entre planta e bactérias endofíticas diazotróficas

10
No solo, existe uma infinidade de espécies de bactérias capazes de viver livremente ou
em associação com plantas. Esses e outros organismos são responsáveis por uma série de
interferências no solo que garantem a qualidade do mesmo, sendo fundamental para a
sustentabilidade agrícola. Desta forma, a relação planta-solo-microrganismos é importante
para a produção agrícola. É na rizosfera, definida como região do solo que recebe influência
direta das raízes possibilitando proliferação microbiana (CARDOSO & NOGUEIRA, 2007),
que se encontra o maior número de rizobactérias promotoras do crescimento de plantas
(RPCPs). Segundo GRAY & SMITH (2005), as RPCPs poderiam ser divididas dependendo
do grau de proximidade com a raiz e a intimidade da associação, classificando-as como
rizobactérias extracelulares, que vivem na rizosfera ou no rizoplano (superfície entre raiz e o
solo), as que vivem entre os espaços intercelulares do córtex radicular (endofíticas), e as
rizobactérias intracelulares as quais existem dentro das células da raiz, geralmente em
estruturas especializadas como os nódulos.
Dentro dessa classificação, as bactérias endofíticas são aquelas que vivem no interior
da planta, não apenas das raízes, e que podem ser isoladas de tecidos cuja superfície foi
desinfestada, e que não causam danos visíveis às plantas (HALLMANN et al., 1997). As
bactérias endofíticas podem ainda ser chamadas de “endofíticas promotoras do crescimento de
plantas” (TAULÉ et al. 2011). As bactérias endofíticas colonizam as plantas primeiramente se
aderindo à superfície das raízes, em seguida com a colonização das raíses laterais com
penetração pela epiderme, envolvendo lipopolisacarideos, exopolissacarideos bacterianos,
posteriormente ocupando os espaços intercelulares, colonizando também o xilema e parte
aérea das plantas (MONTEIRO et al., 2012), ainda podem usar aberturas naturais (estômatos)
ou artificiais (ferimentos) para sua entrada (BASHAN & DE-BASHAN, 2005). JAMES et al.
(2002) comprovaram esse tipo de colonização de endofíticos com a espécie Herbaspirillum
seropedicae que se encontra em coleóptilos, raízes laterais e junções destas com a principal,
em espaços intercelulares da raiz, aerênquima e células corticais, com pequena entrada no
estelo e no tecido vascular, sendo encontradas colônias no xilema, folhas e caules de arroz.
Em experimento com cana-de-açúcar micropropagada, OLIVEIRA et al. (2006) observaram o
sucesso na colonização de plantas por inóculos contendo até cinco espécies de bactérias
endofíticas, indicando ainda que existe uma dinâmica de competição entre as espécies para
dominar o ambiente vegetal.
As bactérias promotoras de crescimento podem influenciar as plantas de duas formas,
segundo BASHAN & DE-BASHAN (2005). Uma delas, afetando diretamente o metabolismo
das plantas por fornecerem substâncias que normalmente estariam pouco disponíveis. Elas

11
podem fixar nitrogênio, solubilizar fósforo e ferro ou produzir hormônios como auxinas,
giberelinas, citocininas e etileno. A outra forma de atuação dessas bactérias é o biocontrole de
patógenos, promovendo o crescimento de forma indireta por permitir que a planta expresse
todo o seu potencial de crescimento, que poderia ser limitado, caso um patógeno estivesse
presente (BASHAN & DE-BASHAN, 2005).
Bactérias endofíticas podem desencadear um fenômeno conhecido como indução de
resistência sistêmica (IRS) fazendo com que a planta evite a entrada de patógenos (RYAN et
al., 2007). DIMPKA et al. (2009) fazem referência à proteção cruzada, sendo que genes
ativados pelo reconhecimento de rizobactérias pelas plantas, além de ativarem a IRS, podem
estar envolvidos nas respostas a estresses abióticos como seca, frio, salinidade. Além dessa
proteção, existe o controle de insetos por organismos endofíticos sendo que plantas tratadas
com esses microrganismos têm menores índices de herbivoria (AZEVEDO et al., 2000).
A maioria dos trabalhos, no entanto, está relacionada aos efeitos diretos das bactérias
no metabolismo de plantas. Uma característica importante das bactérias endofíticas é a
fixação de nitrogênio atmosférico, diminuindo aplicação de fertilizantes nitrogenados, o que
propicia menor utilização desses insumos, cujo uso comumente causa poluição ambiental
além de proporcionar economia na aquisição destes adubos. A disponibilidade deste nutriente
reflete diretamente no desenvolvimento e no rendimento da cultura da cana-de-açúcar
(TRIVELIN & VITTI, 2009). Essas bactérias são capazes de realizar fixação biológica de
nitrogênio (FBN) (BARBOSA et al., 2006), o que as denomina diazotróficas, e de viver no
interior dos tecidos vegetais (endofítico) sem induzirem uma resposta de defesa à sua
presença.
O processo de FBN é muito importante no Brasil onde existem áreas plantadas de
cana-de-açúcar produzindo razoavelmente com aplicações mínimas de fertilizante
nitrogenado (BODDEY et al., 1992). OLIVEIRA et al. (2003) relataram que os melhores
resultados quanto à contribuição da FBN foram observados em solo de menor fertilidade,
indicando que as bactérias contribuem mais para o aumento no conteúdo de nitrogênio quando
em condições restritivas. Essas bactérias não são capazes de suprir totalmente a demanda de N
das plantas somente pela FBN, porém, podem influenciar fortemente a nutrição nitrogenada
das culturas as quais estão associadas, aumentando a capacidade de assimilação de N,
indiretamente, com o aumento do sistema radicular, ou diretamente, estimulando o sistema de
transporte de N nas plantas (MANTELIN & TOURAINE, 2004).
Em relação à classificação quanto à localização, BALDANI et al. (1997) sugeriram
três categorias para bactérias fixadoras de nitrogênio que colonizam gramíneas: 1- os

12
organismos rizosféricos; 2- os endofíticos facultativos (capazes de viver na rizosfera ou em
vida livre); 3- os endofíticos obrigatórios (incapazes de viver no solo). Estudos com
inoculação de microrganismos diazotróficos mostram que bactérias endofíticas contribuem
mais com a FBN, além de haver influência de genótipos na relação planta/bactéria
(BALDANI & BALDANI, 2005).
Os melhores resultados na FBN por organismos endofíticos estão ligados,
provavelmente, à sua localização, por estarem próximos à fonte de energia disponibilizada
pelas plantas, como a sacarose (BALDANI & BALDANI, 2005), e porque o interior vegetal
garante uma baixa pressão de oxigênio que é ideal para a atividade da nitrogenase
(BALDANI & BALDANI, 2005). A enzima nitrogenase (responsável pelo processo) funciona
em sistemas anaeróbios, aeróbios facultativos e microaeróbios, sendo que a concentração de
O2 deve estar em um nível aceitável para que não cause inibição da atividade enzimática, mas
que garanta a respiração celular e a produção de ATP. A FBN necessita de muita energia,
cerca de 16 a 30 moléculas de ATP para cada molécula de NH3
O nitrogênio (N
produzida (BURRIS, 1991).
2
Os gêneros de bactérias endofíticas freqüentemente encontrados associados à cultura
da cana-de-açúcar são Herbaspirillum, Burkholderia, Gluconocetobacter, Azospirillum
(CAVALCANTE & DOBEREINER, 1988), mas não são os únicos, e nem exclusivos desta
cultura. O estudo da interação entre esses gêneros de bactérias e os genótipos de cana pode
fornecer subsídios para o aumento de produção e manejo desta cultura. OLIVEIRA et al.
(2002) obtiveram resultados que indicaram um aumento nos teores de N com contribuição da
FBN em até 30% em cana-de-açúcar inoculada com mistura de cinco espécies de bactérias
dos gêneros citados acima. MOREIRA et al. (2010) relataram uma contribuição de 25 a 50 kg
) é reduzido pela nitrogenase em amônia e esta é considerada como a
provável forma de fornecimento de nitrogênio às plantas (DONATO et al., 2004). A amônia é
então metabolizada pela via GS-GOGAT. A enzima sintetase da glutamina (SG) catalisa a
reação transformando glutamato e amônia em glutamina com uso de ATP. Posteriormente, a
glutamina é reduzida pela glutamato sintase (GOGAT) a glutamato, aminoácido que também
já pode ser usado pela célula (REIS & TEIXEIRA, 2005). No entanto, ainda existem
especulações sobre a forma como as bactérias auxiliam a garantir o suprimento de nitrogênio
para plantas. A RN é uma enzima que catalisa a reação de nitrato (NO3−) para nitrito (NO2
−) e
esse, por sua vez, é transformado em amônio que entra então nas reações do ciclo GS-
GOGAT. Existe ainda a idéia de que a contribuição das bactérias endofíticas para o aumento
do conteúdo de nitrogênio seria pelo aumento da absorção de nitrato pelas raízes, o que
aumentaria a atividade da RN (BALDANI & BALDANI, 2005).

13
de N/ha/ano fixado em gramíneas, equivalente a 17% da demanda dessas culturas.
URQUIAGA et al. (1992) demonstraram que 60% a 70% do N acumulado em algumas
variedades de cana-de-açúcar foram provenientes da FBN.
Em relação à nutrição de plantas tratadas com bactérias endofíticas, BARRETTI et al.
(2008a) em trabalho com tomateiros, relataram o aumento na eficiência nutricional com
plantas apresentando maiores teores de nutrientes na parte aérea e raiz. A melhoria na
qualidade nutricional acaba por proporcionar maior crescimento. Em outro trabalho do
mesmo grupo é relatado aumento de crescimento, área foliar, número de folhas e massa da
parte aérea e raiz (BARRETTI et al., 2008b). Em cana-de-açúcar já foram registrados
aumentos na massa fresca de colmos, na massa de matéria seca de raiz (OLIVEIRA et al.,
2002) e até aumento na produtividade dessa cultura (OLIVEIRA et al., 2006) em plantas
tratadas com bactérias endofíticas.
A maior produção de matéria seca em plantas tratadas com bactérias indica um
possível estímulo da fotossíntese. Apesar dessa relação, não são muitos os trabalhos que
abordam esse aspecto da interação planta-bactéria, menor ainda é o número de trabalhos com
bactérias endofíticas. SHI et al. (2010) relatam que plantas de beterraba tratadas com bactérias
endofíticas apresentaram maiores valores de eficiência quântica potencial do fotossistema II
(FV/FM
Hormônios como etileno e ABA estão relacionados a estresses ambientais e podem ser
regulados ou produzidos por bactérias em associação com plantas. O etileno produzido em
), maior fotossíntese e, esses resultados seriam devido ao melhor transporte de elétrons
na membrana do tilacóide, além do aumento do conteúdo de clorofila nas folhas de plantas
tratadas, levando a um aumento no teor de carboidratos e uma possível correlação com
produção de fitormônios pelas bactérias.
A produção de fitormônios como ácido indol acético (AIA), citocininas, giberelinas e
ABA por bactérias podem alterar o padrão e o crescimento das plantas, interferindo no seu
desenvolvimento (TSAVKELOVA et al., 2006a). A maioria das associações entre bactéria e
raízes que mostra efeitos promotores do crescimento em plantas tem sido relacionada com a
produção de indóis (AIA), e a inoculação de várias espécies de bactérias resultou no
crescimento da raiz e/ou no aumento da formação de raízes laterais. A promoção do
crescimento resulta em um aumento da superfície de absorção e pode, por conseqüência, ter
efeitos positivos na aquisição de água e nutrientes (DIMKPA et al., 2009). BOIERO et al.
(2007) consideram que estirpes produtoras de AIA em grandes quantidades podem ser fontes
naturais de hormônios promotores de crescimento, eliminando a necessidade de fontes
sintéticas.

14
situação de estresse afeta o crescimento radicular e, consequentemente, da parte aérea.
Bactérias que possuem a enzima aminociclopropano-1-carboxilase deaminase (ACC
deaminase) podem regular a produção desse hormônio sendo vantajoso para o crescimento
sob condições de estresse (SALEEM et al., 2007). O
CREUS et al. (2004), em trabalho com plantas de trigo inoculadas com Azospirillum,
relataram que plantas tratadas possuem maior hidratação, potencial hídrico e fração de água
apoplástica maiores do que plantas controle em condições de estresse, podendo ocorrer o
ABA, também relacionado a sinais anti-
estresses, principalmente estresse hídrico, estimula o fechamento dos estômatos (HARTUNG
& SLOVIK 1991).
Bactérias podem induzir ou ser mediadoras de tolerância a estresses abióticos como
salinidade, seca, inundações, temperaturas muito altas ou baixas, deficiência de nutrientes e
metais tóxicos (DIMKPA et al., 2009). Bactérias podem ainda produzir substâncias
osmotolerantes, como, por exemplo, glicina-betaína que pode atuar sinergicamente com os
outros compostos vegetais na redução do potencial hídrico das células, ajudando na tolerância
à seca (DIMKPA et al. 2009). A produção de prolina pode ser estimulada nas plantas em
presença de bactérias em resposta a estresses bióticos e abióticos, pode mediar o ajuste
osmótico, e proteger membranas e proteínas contra efeitos adversos do aumento da
concentração de íons inorgânicos (GROVER et al., 2011). Interessante falar que a percepção
de um estresse abiótico pela planta pode levar a uma proteção cruzada de outro estresse. Por
exemplo, o aumento da síntese de aminas quaternárias como glicina-betaína, aumenta a
resistência a deficiência hídrica, mas também protege a planta contra frio e salinidade
(DIMKPA et al., 2009).
BARKA et al. (2006) mostraram que plantas tratadas com uma estirpe de
Burkholderia phytofimans tiveram aumento nos teores de prolina, amido e compostos
fenólicos e aumento de resistência ao frio. Usando plantas transgênicas carregando o gene
derivado de Bacillus subtillis em Arabidopsis thaliana, verificou-se que houve um aumento
nos níveis de prolina levando a um aumento da tolerância a estresses osmóticos (CHEN et al.,
2007). Bactérias osmotolerantes, capazes de sintetizar solutos ou osmólitos que aumentam a
osmolaridade citoplasmática e conferem estabilidade osmótica às células, são usadas como
inoculantes em plantas e parecem também influenciar positivamente as plantas em condições
de estresse hídrico (YUWONO et al., 2005). Essas bactérias são capazes de promover
crescimento em massa de matéria seca de raiz e parte aérea em plantas de arroz submetidas a
estresse hídrico com efeitos mais significativos entre plantas tratadas e controle em condições
hídricas mais severas (YUWONO et al., 2005).

15
ajustamento elástico da parede para manutenção de status hídrico favorável em plantas
tratadas. A produtividade de trigo, quando em condições de restrição hídrica, foi menos
afetada e houve maior conteúdo de Mg, K e Ca nos grãos em plantas tratadas com as
bactérias do que plantas controle (CREUS et al., 2004). Para DIMKPA et al. (2009), bactérias
podem intermediar mudanças na elasticidade das paredes das células radiculares, o que seria
um dos primeiros passos para melhorar a tolerância ao estresse hídrico. Bactérias também
podem auxiliar na recuperação após o estresse hídrico, mesmo não evitando perda de água
durante o estresse, podendo fazer com que os níveis de água na célula retornem mais
rapidamente ao normal (MAYAK et al., 2004).
Em plantas tratadas com bactérias em situação de estresse, MAYAK et al. (2004)
encontraram estimulo de crescimento. Já LIDDYCOAT et al. (2009) observaram crescimento
diferencial dependendo do genótipo de aspargo, para um deles o crescimento foi favorecido
pela presença de bactérias apenas sob condições ideais de irrigação, enquanto que para outro
genótipo o favorecimento ocorreu em situação de estresse hídrico. Os autores demonstraram
assim uma resposta diferencial da interação entre bactéria e planta, associada à condição
hídrica.
Baseada neste contexto, a interação entre bactérias endofíticas diazotróficas e plantas
de cana-de-açúcar submetidas ao déficit hídrico deve ser compreendida para elucidar os
aspectos fisiológicos envolvidos nesse complexo sistema biológico.
3 MATERIAL E MÉTODOS 3.1 Material vegetal e isolados bacterianos
Os experimentos foram realizados com dois genótipos de Saccharum spp., o
IACSP94-2094 e o IACSP95-5000, tidos como tolerante (LANDELL et al., 2005a,
MACHADO et al., 2009) e suscetível (LANDELL et al., 2005b) a condições de restrição
hídrica, respectivamente. Foram geradas mudas pela brotação de discos dos colmos contendo
gemas em substrato comercial a base de turfa de Sphagno e vermiculita expandida,
esterilizado em autoclave a 121°C por uma hora, a 1,5 kgf/cm2
Na ocasião do plantio, os discos de colmos, provenientes de plantas cultivadas em
campo no Centro Experimental Central (CEC) do Instituto Agronômico (IAC), contendo
gemas maduras foram cortados e colocados em sacos plásticos para realização da inoculação
.

16
do material vegetal. Para isso, foram selecionados 10 isolados de bactérias endofíticas
diazotróficas da coleção da Seção de Microbiologia do Solo, do Centro de Solos e Recursos
Ambientais – Instituto Agronômico, Campinas, SP. A origem dos isolados foi a partir de
raízes e colmos de plantas cultivadas em solos do Estado de São Paulo. Esses isolados foram
selecionados pelos melhores resultados dentre os obtidos em experimento de isolamento e
caracterização bacteriana por métodos in vitro, de produção de indóis, redução de acetileno e
por testes de desempenho de plantas de cana-de-açúcar em casa de vegetação, que avaliaram a
massa seca de raízes e da parte aérea (FREITAS, 2011).
Os 10 isolados foram crescidos separadamente em frascos com meio de cultivo líquido
Dygs (DÖBEREINER et al.,1995) até a obtenção da concentração de 108
Durante o período de crescimento, as plantas permaneceram em ambiente de telado e
foram irrigadas, três vezes por semana, com solução nutritiva com baixo nível de nitrogênio
(57 mg L
células. No dia do
plantio, foram preparados dois inóculos com a mistura de 5 isolados bacterianos cada. O
primeiro, denominado inóculo A, composto por 5 isolados provenientes do isolamento em
meio de cultivo JNFb (semi–seletivo para Herbaspirillum) (DÖBEREINER et al.,1995), e o
segundo, denominado inóculo B, composto pelos isolados provenientes do isolamento em
meio LGI-P (semi–seletivo para Gluconoacetobacter diazotrophius) (DÖBEREINER et
al.,1995).
Amostras de cada material vegetal foram coletadas para quantificação de bactérias
endofíticas nativas presentes nos colmos utilizados no plantio (DÖBEREINER et al.,1995). A
inoculação foi realizada com a imersão dos discos dos colmos em meio de cultivo com os
inóculos (A ou B) ou sem inóculo (Controle), por um período de uma hora antes do plantio.
Em seguida, os discos dos colmos foram colocados em copos plásticos de 200 mL com
substrato esterilizado.
-1) (solução estoque A composta por: 200 g L-1 de Ca(NO3)2, 250 g L-1 de CaCl2, 20
g L-1 de ConMicros Standart (produto comercial); e solução estoque B composta por 200 g L-1
de KNO3, 150 g L-1 de KH2PO4, 300 g L-1 de MgSO4.7H2O e 100 g L-1 de KCl por litro: para
cada litro de água, a ser usada na irrigação, adicionou-se 2 mL da solução A e 3 mL da
solução B). Aos 32 dias após o plantio (DAP), as mudas foram transplantadas para vasos com
capacidade de 5 L contendo uma mistura de areia, terra e substrato na proporção 1:1:1 (v/v/v),
esterilizada em autoclave a 121°C, por uma hora a 1,5 kgf/cm2. No dia seguinte, foi realizada
nova aplicação de inóculos nas plantas, com 10 mL de cada inoculo (A ou B) com
concentração de 108 células ou meio de cultura Dygs estéril (controle). Aos 60 DAP, foi
realizada uma terceira inoculação da mesma forma como anteriormente descrita.

17
As plantas permaneceram em ambiente telado, com valores médios de temperatura
máxima de 37,4 ± 2,8°C, mínima de 18,2 ± 1,5°C e umidade relativa de 73,1 ± 6,8%. Durante
os experimentos, foi feita a retirada dos perfilhos, mantendo-se apenas um colmo principal.
Foram realizados dois experimentos com as mudas cultivadas: 3.2 Delineamento experimental 3.2.1 Primeiro experimento: desenvolvimento de plantas cana-de-açúcar tratadas com
inóculos bacterianos e sob disponibilidade hídrica adequada.
As plantas foram mantidas sob boa disponibilidade hídrica em vasos com 80% da
capacidade de retenção de água. Aos 71 DAP, foi feita a coleta de 30 plantas, sendo 5 plantas
de cada tratamento (inóculo A ou B e controle).
O delineamento do primeiro experimento foi inteiramente ao acaso em esquema
fatorial simples, sendo as causas de variação: inóculos bacterianos A ou B ou sem inoculação
(controle).
3.2.2 Segundo experimento: desenvolvimento de plantas de cana-de-açúcar tratadas com
inóculos bacterianos e sob condição de restrição hídrica.
As plantas foram submetidas a duas condições hídricas: vasos com 80% (hidratada -
H) e com 50% (restrição hídrica - RH) da capacidade de retenção, controlado pela massa dos
mesmos. As avaliações fisiológicas foram realizadas no 5º dia em que os vasos com
suspensão parcial da rega estavam a 50% da capacidade de retenção de água (78 DAP). No
dia da coleta as plantas apresentaram folhas amareladas e retorcidas indicando os efeitos do
tratamento hídrico. Nessa etapa, foram coletadas 60 plantas para análises laboratoriais
posteriores, sendo 5 plantas de cada tratamento bacteriano (inóculo A ou B e controle) em
condição hidratada (H) ou restrição hídrica (RH), totalizando 30 plantas de cada genótipo.
O delineamento experimental do segundo experimento foi inteiramente ao acaso em
esquema fatorial duplo, sendo as causas de variação: as condições hídricas e a inoculação.
Nos dois experimentos foram realizadas as avaliações descritas a seguir.

18
3.3 Medidas de crescimento
Foram avaliados os números de folhas, a altura das plantas utilizando-se uma trena, e a
massa de matéria seca da parte aérea e raízes pela secagem em estufa a 60°C.
3.4 Potencial hídrico e conteúdo relativo de água na folha
O potencial hídrico (Ψw) nas folhas + 3 de cana-de-açúcar foi determinado com o
auxílio da câmara de pressão tipo Scholander.
Na folha +2, foram extraídos cinco discos para a determinação do conteúdo relativo de
água (CRA) (SILVEIRA et al., 2003). Discos foliares de 1 cm de diâmetro foram pesados
imediatamente após a coleta, obtendo-se sua massa fresca (MF). Em seguida, foram dispostos
em placas de Petri e imersos em água destilada, por sete horas, em temperatura ambiente sob
radiação fotossinteticamente ativa de 40 µmol m-2 s-1
3.5 Trocas gasosas
. Então, os discos foram secos em papel
filtro e pesados novamente, obtendo-se a massa túrgida (MT). O material foi seco em estufa a
80°C por 48 horas e pesados novamente, para a obtenção da massa seca (MS). O CRA foi
calculado pela equação abaixo:
CRA =MF − MSMT − MS
× 100
Foi avaliada a assimilação líquida de CO2 (A, µmol m-2 s-1), a transpiração (E, mmol
m-2 s-1), a condutância estomática (gs, mol m-2 s-1) e a concentração intercelular de CO2 (Ci,
µmol mol-1) nas folhas +1. As medidas foram realizadas com concentração de CO2 constante
(380 µmol mol-1) e sob Q de 1600 µmol m-2 s-1 (semelhante ao do ambiente), às 14 horas. A
respiração no escuro (R, µmol m-2 s-1
) foi avaliada às 6 horas. As medidas foram realizadas
com um analisador portátil de gases a infravermelho, IRGA (Infra Red Gas Analyser) modelo
Li-6400 (Licor, EUA), na primeira folha totalmente expandida e com lígula aparente (folha
+1), no terço médio do limbo foliar e as medidas foram registradas quando o coeficiente de
variação (CV) total foi inferior a 0,5%.

19
3.6 Atividade fotoquímica
As medidas da emissão de fluorescência da clorofila a foram realizadas por meio de
um fluorômetro modulado (6400-40 LFC) integrado ao Li-6400F. As folhas +1, usadas para
medidas de trocas gasosas, foram previamente adaptadas ao escuro por 30 minutos com
auxílio de papel alumínio. Em seguida, um pulso de luz saturante foi aplicado e então foi
medida a eficiência quântica potencial do fotossistema II (FV/FM), o coeficiente de extinção
não fotoquímica da fluorescência [NPQ=(FM-FM’)/FM’] e o transporte aparente de elétrons
[ETR=Q x ΔF/FM
3.7 Conteúdo de clorofilas
’ x 0,5 x 0,84].
Os conteúdos foliares de clorofilas a e b e total foram avaliados com um medidor
eletrônico de teor de clorofila (ClorofiLOG CFL 1030, Faker- Brasil), nos mesmos dias e
folhas avaliadas para as trocas gasosas nas folhas +1.
3.8 Extração de compostos por MCW
As amostras de folhas +3 coletadas foram armazenadas em ultrafreezer (-80°C) e
posteriormente secas em liofilizador (Labconco – Freezone 4.5) a -50°C e 133x10-3 mbar.
Depois de secas, as amostras vegetais foram moídas em moinho marca IKA modelo A 11
basic, e armazenadas em frascos plásticos hermeticamente fechados até o momento das
análises.
Para extração de açúcares solúveis totais, sacarose, aminoácidos livres totais e prolina,
aproximadamente 75 mg de amostra vegetal juntamente com 3 mL de solução de
metanol:clorofórmio:água (MCW), na proporção 12:5:3 (v/v/v), segundo BIELESK &
TURNER (1966), foram colocados em tubos tipo rosca. A solução foi mantida em geladeira a
5°C, por três dias. Então, acrescentou-se água e clorofórmio na proporção 1:1,5 (v/v),
agitando-se os tubos e voltando-os à geladeira por mais 24 horas. Após a separação das fases,
coletou-se o sobrenadante (fração aquosa-metanólica) e foi feita a concentração das amostras
em banho-maria a 50°C. A solução foi quantificada em proveta (mL) e armazenada em
microtubos, em freezer (-20°C).

20
3.9 Teor de carboidratos foliares
As análises de carboidratos incluíram açúcares solúveis totais (AST), sacarose (Sac) e
amido (AMI).
Os AST foram determinados pelo método do fenol-sulfúrico (DUBOIS et al., 1956).
Em tubos de ensaio, foram adicionados 10 µL de extrato foliar e 490 µL de água mili-Q, 500
µL de solução de fenol 5% e 2 mL de ácido sulfúrico (H2SO4) concentrado. Depois de
agitados os tubos até a homogeneidade da solução e posterior resfriamento, foram feitas as
leituras de absorbância em espectrofotômetro (Micronal – B342II), no comprimento de onda
de 490 nm. As concentrações de sacarose foram calculadas a partir de uma curva-padrão
obtida com leituras de solução de glicose contendo concentrações de 0, 5, 10, 20, 30, 40, 50,
60, 70, 80 e 90 µg/mL.
A concentração de Sac foi determinada pelo método descrito por VAN HANDEL
(1968) e DUBOIS et al. (1956). Em tubos de ensaio, foram adicionados 10 µL de extrato, 490
µL de água mili-Q e 500 µL de solução de hidróxido de potássio (KOH) 30%. Os tubos foram
vedados e levados ao banho-maria a 100 °C, por 10 minutos. Em seguida, adicionaram-se 500
µL de solução de fenol 5% e 2 mL de ácido sulfúrico (H2SO4) concentrado. As leituras de
absorbância foram feitas em espectrofotômetro no comprimento de onda de 490 nm. As
concentrações de sacarose foram calculadas a partir de uma curva-padrão obtida com leituras
de solução de sacarose contendo concentrações de 0, 5, 10, 20, 30, 40 e 50 µg/mL.
Para a quantificação de AMI, foi utilizado o método descrito por AMARAL et al.
(2007). Em microtubos contendo 10 mg de material vegetal seco, foram realizadas quatro
extrações com 500 µL de etanol 80%, a 80 °C em banho-maria, por 20 minutos cada, para
solubilização de açúcares, pigmentos, fenóis e outras substâncias. O precipitado foi então seco
a temperatura ambiente, até a completa evaporação do resíduo de etanol. Em seguida, foram
adicionados 500 µL (120 U mL-1) d e α-amilase (EC 3.2.1.1) termoestável de Bacillus
licheniformis (cód. E-ANAAM, MEGAZYME, Irlanda), diluída em tampão MOPS 10 mM e
pH 6,5. As amostras foram incubadas em banho-maria a 75°C, por 30 minutos. Em seguida,
foram adicionados 500 µL (30 U mL-1) de amiloglucosidase (EC 3.2.1.3) de Aspergillus niger
(cód. E-AMGPU, MEGAZYME, Irlanda) em tampão acetato de sódio 100 m mol L-1 e pH
4,5. As amostras foram incubadas em banho-maria a 50°C, por 30 minutos. Foram
acrescentados 100 µL de ácido perclórico (HClO4) 0,8 mol L-1 para parar a reação e precipitar
proteínas. As amostras permaneceram em freezer (-20°C) até o momento da quantificação.

21
Para a quantificação, em microtubos de 1,5 mL, foram colocados 50 µL de amostra e
750 µL de Glicose PAP Liquiform (Labtest Diagnóstica S.A. Brasil). A solução foi encubada
em banho-maria a 37°C, por 15 minutos. O conteúdo de glicose foi determinado em leitor de
microplacas modelo EL307 (Bio-Tek Instruments, EUA), em comprimento de onda de 490
nm. As concentrações de amido foram calculadas a partir de uma reta-padrão obtida com
leituras de solução de glicose contendo concentrações de 0, 5, 10, 15, 25 e 30 µg/mL.
Os teor de AST, Sac e AMI foram expressos em mg g-1
3.10 Teor de prolina
de massa seca.
A quantificação de prolina foi realizada segundo RENA & MASCIOTTI (1976). Em
tubos de ensaio, foram adicionados 100 µL de glicina 0,1M, 300 µL de extrato de MCW, 2,6
mL de água mili-Q, e 2 mL de ácido acético concentrado. Após serem agitados para
homogeneizar a solução, foram acrescentados 2 mL de reagente de ninhidrina (600 mg de
ninhidrina + 15 mL de ácido acético concentrado + 10 mL de ácido fosfórico (H3PO4) 6 mol
L-1). Após agitação, a mistura foi encubada em banho-maria a 100°C, por 35 minutos. Em
seguida, os tubos foram colocados em banho de gelo para rápido resfriamento da reação e
adicionados 4 mL de tolueno concentrado. Agitou-se vigorosamente e a absorbância do
sobrenadante (tolueno) foi determinada em espectrofotômetro com o comprimento de onda de
515 nm. A reta-padrão foi obtida com solução de prolina em concentrações de 0, 0,5, 1, 2, 3 e
4 µg/mL. O conteúdo de prolina foi expresso em µmol g-1
3.11 Teor de aminoácidos livres totais
de massa seca.
Segundo YEMM & COCKING (1954), foi determinado o conteúdo de aminoácidos
solúveis totais nas folhas. Em tubos de ensaio, foram dispensados 50 µL de extrado de MCW,
950 µL de água mili-Q, em seguida 500 µL de tampão citrato 0,2 M (20,5 mL de ácido cítrico
0,2 M e 29,5 mL citrato de sódio 0,2 M misturados até o pH 5), 200 µL de ninhidrina em
metilglicol (2,5 g de ninhidrina em 50 mL de etilenoglicol monometiléter a 99%) e 1 mL de
solução de KCN (2 mL de KCN 0,01 N em 98 mL de metilglicol). Os tubos foram agitados,
vedados e encubados em banho-maria a 100°C, por 20 minutos. Posteriormente, foram
resfriados a temperatura ambiente e adicionado 1 mL de etanol 60%. Os tubos foram agitados
e as leituras foram feitas em espectrofotômetro com comprimento de onda de 570 nm. A

22
curva padrão foi feita utilizando-se solução de leucina em diferentes concentrações de 0, 20,
40, 60, 80 e 100 nmol/mL. O teor de aminoácidos solúveis totais foi expresso em mg g-1
de
massa seca.
3.12 Teor de nitrogênio total em folhas
Foi transferido 0,1 g de material vegetal seco e moido para tubos de digestão (tipo
Folin-Wu), contendo 1 g de mistura digestora (1000 g de K2SO4 + 100 g de CuSO4.5H2O +
10 g de Se), 3 mL de ácido sulfúrico concentrado e 1 mL de peróxido de hidrogênio (H2O2).
Os tubos foram então colocados em um bloco digestor e aquecidos lentamente até a
temperatura de 360 ºC, sendo as amostras digeridas ao ponto de ebulição da mistura, até a
obtenção de um extrato esverdeado. Após o resfriamento até temperatura ambiente, foram
adicionados 5 mL de água mili-Q. O conteúdo de cada tubo foi usado na dosagem de
nitrogênio, segundo o método de Kjeldahl (BREMNER, 1965).
Para a dosagem, as amostras passaram por processo de destilação. O tubo contendo a
amostra foi conectado a destilador e 15 mL de NaOH 10 N foram dispensados para reagir
com o conteúdo do tubo. Foi realizada a destilação, a fim de que o nitrogênio contido na
amostra fosse volatilizado em forma de amônia e recolhido em erlenmeyer contendo 20 mL
de solução de ácido bórico-indicador. A destilação foi interrompida após a produção de 20
mL de destilado. O conteúdo do erlenmeyer foi então titulado, usando solução padronizada de
ácido sulfúrico 0,0714 N. O volume de ácido foi utilizado para calcular a quantidade de
nitrogênio na amostra.
O teor de nitrogênio foi utilizado para calculador a quantidade acumulada (mg de N
planta-1) e o índice de eficiência de utilização (g biomassa2/ g nutriente-1). O teor de
nitrogênio foi expresso em g kg-1
de massa seca.
3.13 Extração enzimática
Para a extração enzimática, seguiu-se o método descrito por SILVEIRA et al. (2010)
com adaptações nas concentrações de inibidores de proteases. As amostras de folhas +1
preservadas em freezer -80°C foram pesadas (2,0 g), rapidamente transferidas para almofariz
e maceradas com auxílio de nitrogênio líquido e PVPP (Polivinilpolipirrolidona).
Acrescentaram-se ao macerado 5 mL de tampão de extração Tris-HCl 100 mM pH 7,5

23
contendo FAD 10 µM + EDTA 20 mM + DTT 5 mM + BSA 0,5% + mistura de inibidores
(PMSF 0,1 mM + leuptina 10 µM + benzidina 1 mM). O macerado foi então filtrado em duas
camadas de gaze e centrifugado a 10.000 rpm a 4°C, por 20 minutos. O sobrenadante foi
recolhido em microtubos de 2,0 mL para posterior quantificação da atividade enzimática.
3.14 Atividade enzimática in vitro da redutase do nitrato Para determinação da atividade da redutase do nitrato, 200 µL de extrato enzimático
foram adicionados a 500 µL de tampão de quantificação (Tris-HCl 100 mM pH 7,5 + EDTA
10 mM + KNO3 5 mM +DTT 5 mM + FAD 10 µM) e 15 µL de NADH 1 mM em
microtubos. A reação foi incubada em banho-maria a 30°C, por 30 minutos. Logo após esse
período, a reação foi paralisada colocando os microtubos em banho-maria a 100°C, por 10
minutos e posteriormente foi acrescentados 750 µL de sulfanilamida [sulfanilamida 1% +
dicloridrato de N-(1-naftil) etilenodiamina] dissolvida em HCl (2,4 N). Para eliminar a
turvação, os microtubos foram centrifugados a 13.000 rpm e o sobrenadante contendo o nitrito
formado foi quantificado por meio de leitura em espectrofotômetro, com comprimento de
onda de 540 nm. Uma curva padrão foi feita utilizando solução de NO2− em diferentes
concentrações: 15, 37,5, 60, 75, 150 e 225 ng/mL. A atividade foi expressa em nmol NO2− g-
1 MF h-1
3.15 Atividade enzimática in vitro da sintetase da glutamina
. O procedimento utilizado seguiu o descrito por SILVEIRA et al.(2010).
Para determinação da atividade da glutamina sintetase, 100 µL de extrato enzimático
foram acrescentados a 300 µL de tampão de quantificação (Tris-HCL 50 mM pH 7,0 +
MgSO4 5 mM + hidroxilamina NaOH 100mM + glutamato de Na 50 mM), 100µL de ATP
100 mM. A reação foi incubada em banho-maria a 30°C, por 30 minutos. Logo em seguida,
foram adicionados 500 µL de solução férrica (FeCl3 0,37M + TCA 0,2 M dissolvidos em
HCl 0,67M) para paralizar da reação. Para eliminar a turvação, os microtubos foram
centrifugados a 13.000 rpm e o sobrenadante foi quantificado por meio de leitura em
espectrofotômetro com comprimento de onda de 535 nm. Uma curva padrão foi feita
utilizando solução de γ -glutamil-hidroximato (GGH) em diferentes concentrações 0,05, 0,1,
0,25, 0,5, 1,0, 1,5, 2,0 µmol/mL. A atividade foi expressa em µmol GGH g-1 MF h-1. O
procedimento utilizado seguiu o descrito por SILVEIRA et al. (2010), com adaptações:

24
diminuição na concentração de MgSO4
3.16 Quantificação de bactérias presentes nos colmos e nas raízes
e hidroxilamina NaOH e aumento na concentração de
ATP.
No início do primeiro experimento, foi realizada a quantificação de bactérias
diazotróficas endofíticas nos colmos utilizados para a produção de mudas. A técnica utilizada
foi a descrita em DÖBEREINER et al. (1995). Para isso, foram coletados 10 g de cinco
colmos de cada material (IACSP94-2094 e IACSP95-5000), submetidos à desinfecção
superficial com álcool 70%, por 30 segundos, seguido de duas lavagens com água destilada
esterilizada, e Cloramina T 1% por 30 segundos, seguido de três lavagens com água destilada
esterilizada. Os colmos desinfestados foram triturados juntamente com 90 mL de solução de
sacarose 4%, em liquidificador, por 1 minuto. A seguir, foram feitas diluições.
Com o objetivo de verificar se a inoculação foi eficiente, no final do experimento foi
realizada a quantificação de bactérias diazotróficas endofíticas presentes nas raízes das
plantas. Foram coletados 10 g de raiz que foram submetidas à desinfecção superficial com
Cloramina T 1% por 15 minutos, seguido de três lavagens com água destilada esterilizada e
transferidos para uma solução tampão fosfato 0,05 M (pH 7,0) por 15 minutos, seguido de três
lavagens com água destilada esterilizada. As raízes desinfestadas foram trituradas juntamente
com 90 mL de solução de sacarose 4%, no liquidificador, por 1 minuto e feitas as diluições e
as inoculações.
As diluições seriadas das amostras foram feitas em solução de sacarose, de 10-2 a 10-6
3.17 Delineamento estatístico e análise de dados
.
Utilizou-se 0,1 mL de cada diluição para inoculação no meio semi-sólido para o diazotrófico
desejado, ou seja, JMV semi-específico para o gênero Burkholderia (BALDANI, 1996), JNFb
para o gênero Herbaspirillum e LGI-P para G. diazotrophius (DÖBEREINER et al.,1995).
O crescimento bacteriano foi evidenciado pela presença da película típica logo abaixo
da superfície do meio, depois de 7 dias de incubação a 30°C. A comunidade bacteriana foi
estimada pela técnica do número mais provável (MPN), consultando a tabela de McCrady
para 3 repetições por diluição.
Os dados foram submetidos à análise de variância e quando as médias apresentaram
diferenças significativas, foram comparadas pelo teste de Tukey a 5% de probabilidade.

25
4 RESULTADOS 4.1 Efeitos da interação planta-bactéria na fisiologia de cana-de-açúcar sob disponibilidade hídrica adequada. 4.1.1 Contagem de bactérias endofíticas A contagem de bactérias nos colmos, usados para produção das mudas, indicou
número médio, aproximadamente, sete vezes maior de bactérias nativas no genótipo
IACSP95-5000, quando comparado com o IACSP94-2094 (Tabela 1).
Tabela 1 - Número mais provável (NMP) de bactérias, no início do experimento, nos colmos dos genótipos IACSP94-2094 e IACSP95-5000, utilizados para produção de mudas, em três meios de cultura (JNFb, LGIP, JMV).
Meios de cultura Genótipos JMV LGI-P JNFb Média
UFC.105 g -1 IACSP94-2094 2,50 2,0 5,4 3,3 IACSP95-5000 6,00 18,8 41,3 22,0
Os valores são descritivos e não aplicado teste estatístico.
4.1.2 Aspectos biométricos
Plantas tratadas não diferiram das do controle em relação à altura nos genótipos
avaliados, variando de 37 a 39 cm para IACSP94-2094, e 25 a 27 cm para IACSP95-5000.
Quanto ao número de folhas, também não houve diferença entre os tratamentos
bacterianos e plantas controle, sendo 7,0 o número médio de folhas em cada planta.
As massas de matéria seca da parte aérea e das raízes não apresentaram diferenças
significativas entre os tratamentos controle, inóculo A ou B (Figura 1).
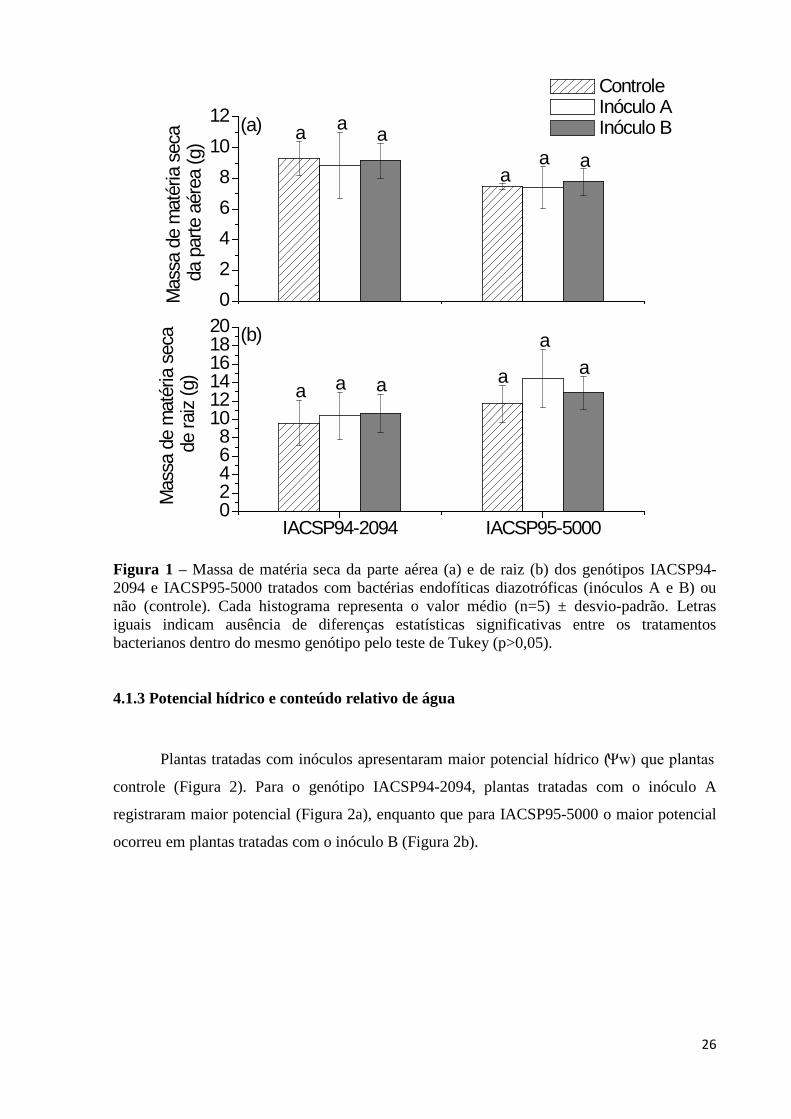
26
Figura 1 – Massa de matéria seca da parte aérea (a) e de raiz (b) dos genótipos IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle). Cada histograma representa o valor médio (n=5) ± desvio-padrão. Letras iguais indicam ausência de diferenças estatísticas significativas entre os tratamentos bacterianos dentro do mesmo genótipo pelo teste de Tukey (p>0,05). 4.1.3 Potencial hídrico e conteúdo relativo de água
Plantas tratadas com inóculos apresentaram maior potencial hídrico (Ψw) que plantas
controle (Figura 2). Para o genótipo IACSP94-2094, plantas tratadas com o inóculo A
registraram maior potencial (Figura 2a), enquanto que para IACSP95-5000 o maior potencial
ocorreu em plantas tratadas com o inóculo B (Figura 2b).
IACSP94-2094 IACSP95-500002468
101214161820 (b)
(a)
aa
aaaa
Mas
sa d
e m
atér
ia s
eca
de ra
iz (g
)
02468
1012
aaa
aaa
Mas
sa d
e m
atér
ia s
eca
da p
arte
aér
ea (g
)
Controle Inóculo A Inóculo B
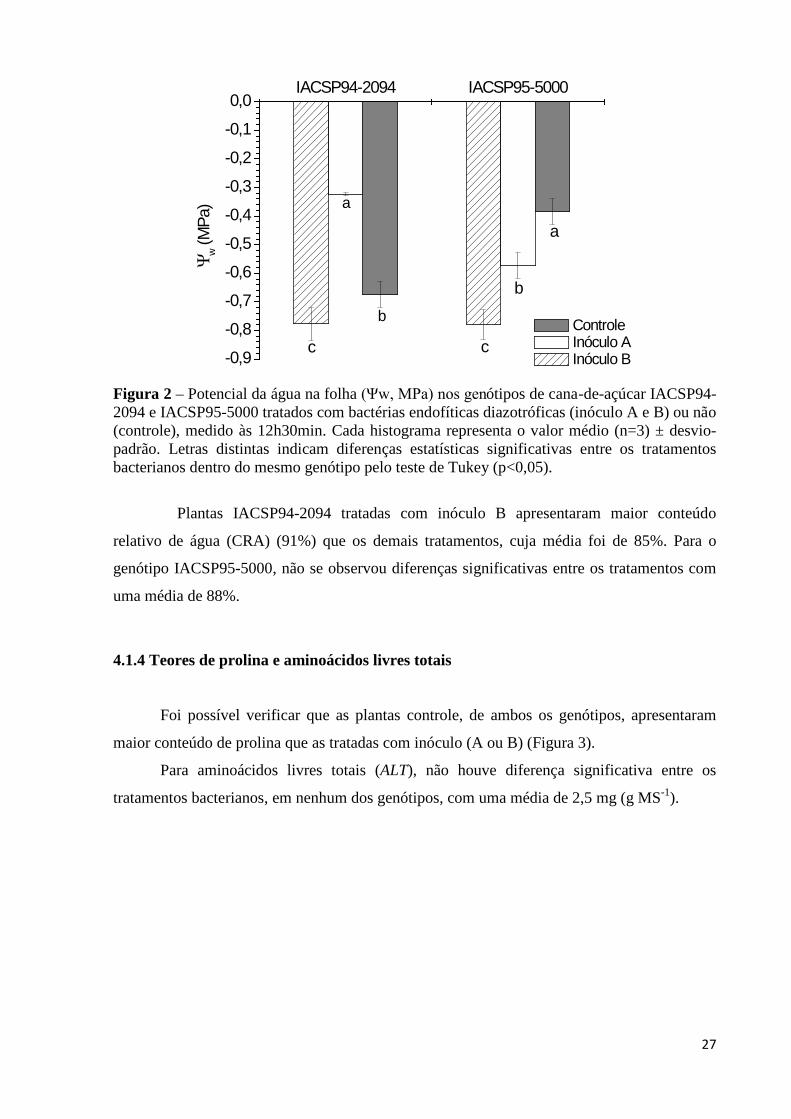
27
Figura 2 – Potencial da água na folha (Ψw, MPa) nos genótipos de cana-de-açúcar IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculo A e B) ou não (controle), medido às 12h30min. Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras distintas indicam diferenças estatísticas significativas entre os tratamentos bacterianos dentro do mesmo genótipo pelo teste de Tukey (p<0,05).
Plantas IACSP94-2094 tratadas com inóculo B apresentaram maior conteúdo
relativo de água (CRA) (91%) que os demais tratamentos, cuja média foi de 85%. Para o
genótipo IACSP95-5000, não se observou diferenças significativas entre os tratamentos com
uma média de 88%.
4.1.4 Teores de prolina e aminoácidos livres totais Foi possível verificar que as plantas controle, de ambos os genótipos, apresentaram
maior conteúdo de prolina que as tratadas com inóculo (A ou B) (Figura 3).
Para aminoácidos livres totais (ALT), não houve diferença significativa entre os
tratamentos bacterianos, em nenhum dos genótipos, com uma média de 2,5 mg (g MS-1
-0,9
-0,8
-0,7
-0,6
-0,5
-0,4
-0,3
-0,2
-0,1
0,0IACSP94-2094 IACSP95-5000
b
a
c
a
b
c
Ψw (
MPa
)
Controle Inóculo A Inóculo B
).

28
Figura 3 – Teor de prolina em dois genótipos de cana-de-açúcar IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle). Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras distintas indicam diferenças estatísticas significativas entre os tratamentos bacterianos dentro do mesmo genótipo pelo teste de Tukey (p<0,05). 4.1.5 Atividade fotoquímica Quanto à eficiência quântica potencial do fotossistema II (FV/FM) e coeficiente de
extinção não fotoquímica da fluorescência (NPQ) não foram observadas diferenças
significavas entre plantas tratadas e controle em ambos os genótipos. FV/FM
4.1.6 Clorofila a, b e total
variou entre
0,753 e 0,777 na medida realizada às 13h. Para NPQ o valor médio para o genótipo IACSP94-
2094 foi de 1,63, e 1,98 para IACSP95-5000.
O transporte aparente de elétrons (ETR) em plantas IACSP94-2094 tratadas com
inóculo B foi maior do que as plantas controle (Figura 4a), enquanto que para as plantas
IACSP95-5000 o ETR foi maior nas plantas controle (Figura 4b).
Quanto às medidas dos teores relativos clorofila, não foram observadas diferenças
significativas entre os tratamentos, sendo as médias de 35 para clorofila a, 13 para clorofila b
e 50 para clorofila total no genótipo IACSP94-2094. No genótipo IACSP95-5000, os valores
médios foram de 36 para clorofila a, 17 para clorofila b e 53 para clorofila total.
IACSP94-2094 IACSP95-50000,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
bb
b
ab
a
a
Prol
ina
[µm
ol (g
MS)
-1]
Controle Inóculo A Inóculo B

29
Figura 4 – Transporte aparente de elétrons (ETR) dos genótipos de cana-de-açúcar IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) medido às 13 horas. Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras distintas indicam diferenças estatísticas significativas entre os tratamentos bacterianos dentro de genótipo pelo teste de Tukey (p<0,05). 4.1.7 Trocas gasosas
Plantas do genótipo IACSP94-2094 tratadas com o inóculo B mostraram maiores
valores de fotossíntese, enquanto que o tratamento com inóculo A no genótipo IACSP95-
5000 foi o que promoveu maior estímulo da assimilação de CO2
IACSP94-2094 IACSP95-50000
20406080
100120140160180200
bab
aa
bbET
R (µ
mol
m-2 s
-1)
Controle Inóculo A Inóculo B
(Figura 5a) e maior
condutância estomática (Figura 5b). No genótipo IACSP94-2094, não houve diferença entre
os tratamentos quanto à condutância estomática (Figura 5b).
Os valores de transpiração (E) nas plantas IACSP95-5000 tratadas com inóculo B
foram reduzidos em relação ao controle. O genótipo IACSP94-2094 não apresentou diferença
significativa (Figura 5d).
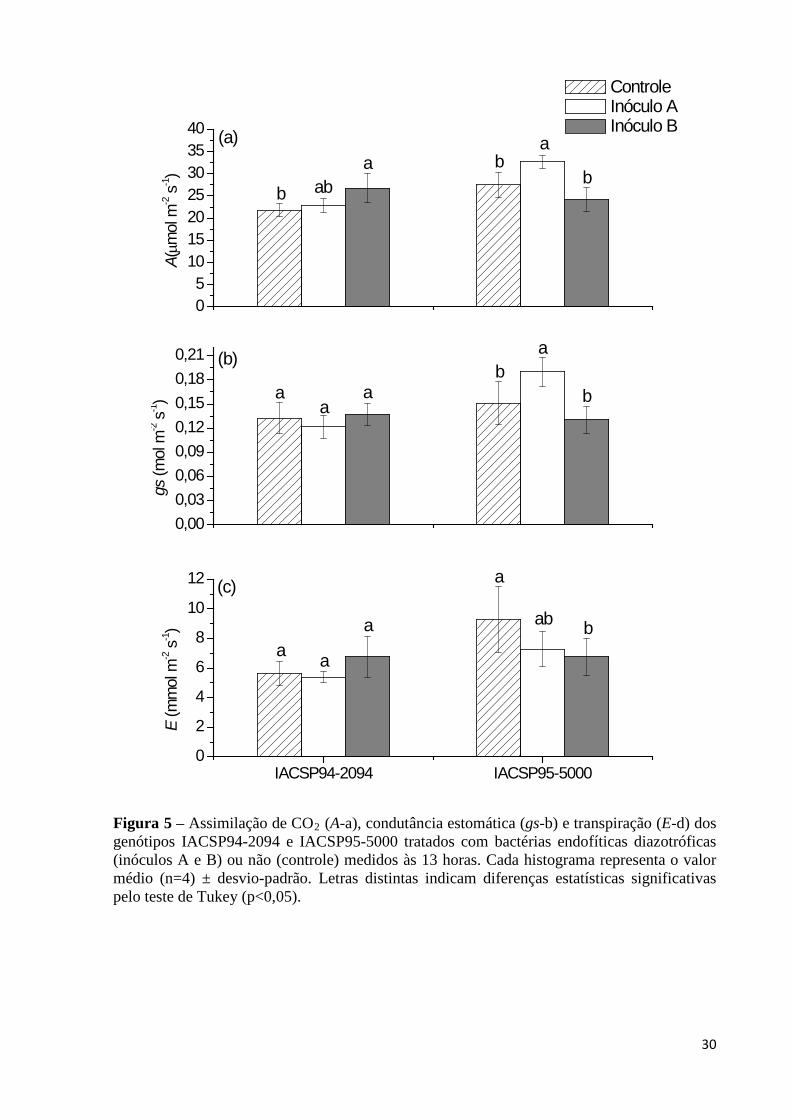
30
Figura 5 – Assimilação de CO2
05
10152025303540
(c)
(b)
(a)
Controle Inóculo A Inóculo B
bb
a
b aba
A(µm
ol m
-2 s
-1)
0,000,030,060,090,120,150,180,21
bb
a
aa
a
gs (m
ol m
-2 s
-1)
IACSP94-2094 IACSP95-50000
2
4
6
8
10
12
a
aabab
a
E (m
mol
m-2 s
-1)
(A-a), condutância estomática (gs-b) e transpiração (E-d) dos genótipos IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) medidos às 13 horas. Cada histograma representa o valor médio (n=4) ± desvio-padrão. Letras distintas indicam diferenças estatísticas significativas pelo teste de Tukey (p<0,05).
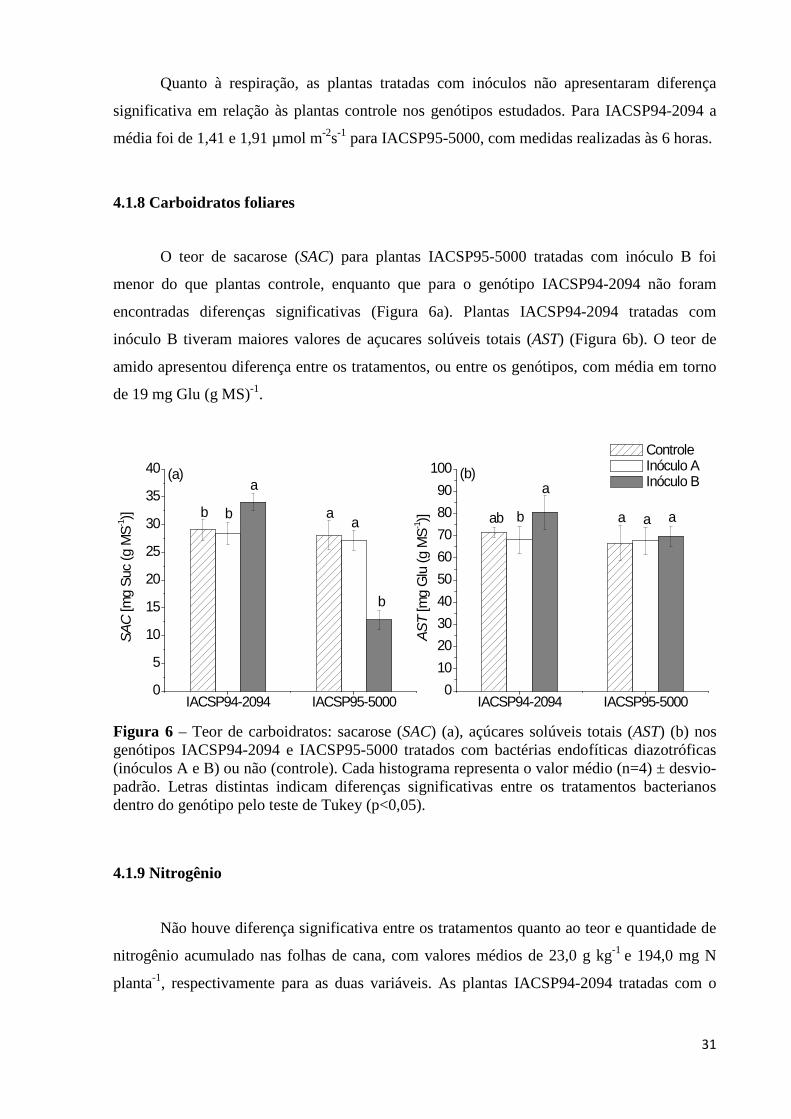
31
Quanto à respiração, as plantas tratadas com inóculos não apresentaram diferença
significativa em relação às plantas controle nos genótipos estudados. Para IACSP94-2094 a
média foi de 1,41 e 1,91 µmol m-2s-1
4.1.8 Carboidratos foliares
para IACSP95-5000, com medidas realizadas às 6 horas.
O teor de sacarose (SAC) para plantas IACSP95-5000 tratadas com inóculo B foi
menor do que plantas controle, enquanto que para o genótipo IACSP94-2094 não foram
encontradas diferenças significativas (Figura 6a). Plantas IACSP94-2094 tratadas com
inóculo B tiveram maiores valores de açucares solúveis totais (AST) (Figura 6b). O teor de
amido apresentou diferença entre os tratamentos, ou entre os genótipos, com média em torno
de 19 mg Glu (g MS)-1
Figura 6 – Teor de carboidratos: sacarose (SAC) (a), açúcares solúveis totais (AST) (b) nos genótipos IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle). Cada histograma representa o valor médio (n=4) ± desvio-padrão. Letras distintas indicam diferenças significativas entre os tratamentos bacterianos dentro do genótipo pelo teste de Tukey (p<0,05).
.
4.1.9 Nitrogênio
Não houve diferença significativa entre os tratamentos quanto ao teor e quantidade de
nitrogênio acumulado nas folhas de cana, com valores médios de 23,0 g kg-1 e 194,0 mg N
planta-1
IACSP94-2094 IACSP95-50000
5
10
15
20
25
30
35
40 (a)
b
aa
a
bb
SAC
[mg
Suc
(g M
S-1)]
IACSP94-2094 IACSP95-50000
102030405060708090
100 (b)
aaa
a
bab
AST
[mg
Glu
(g M
S-1)]
Controle Inóculo A Inóculo B
, respectivamente para as duas variáveis. As plantas IACSP94-2094 tratadas com o

32
inóculo A apresentaram menor eficiência na utilização do nitrogênio que os demais
tratamentos (Tabela 2).
Tabela 2 – Índice de eficiência do uso de nitrogênio (g biomassa2/g N-1
Valores referem-se às médias (n=3) ± desvios. Letras distintas indicam diferenças significativas entre tratamentos bacterianos pelo teste de Tukey (p<0,05).
) nos genótipos de cana-de-açúcar IACSP94-2094 e IAC95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle).
4.1.10 Enzimas do metabolismo do nitrogênio A atividade da enzima redutase do nitrato em plantas IACSP95-5000 tratadas com
inóculo B foi maior do que nas plantas do controle, sendo que o tratamento com inóculo B
promove maior atividade da enzima que com o inóculo A (Figura 7a). A atividade da sintetase
da glutamina nas plantas desse genótipo não foi influenciada pelos tratamentos bacterianos
(Figura 7b). Nas plantas do genótipo IACSP94-2094, os tratamentos bacterianos não
causaram diferenças significativas nas atividades da redutase do nitrato e sintetase da
glutamina (Figura 9a, b).
Tratamento IACSP94-2094 IACSP95-5000 Controle 0,438 ± 0,014 a 0,339 ± 0,019 a
Inóculo A 0,357 ± 0,034 b 0,352 ± 0,004 a Inóculo B 0,427 ± 0,031 a 0,329 ± 0,016 a
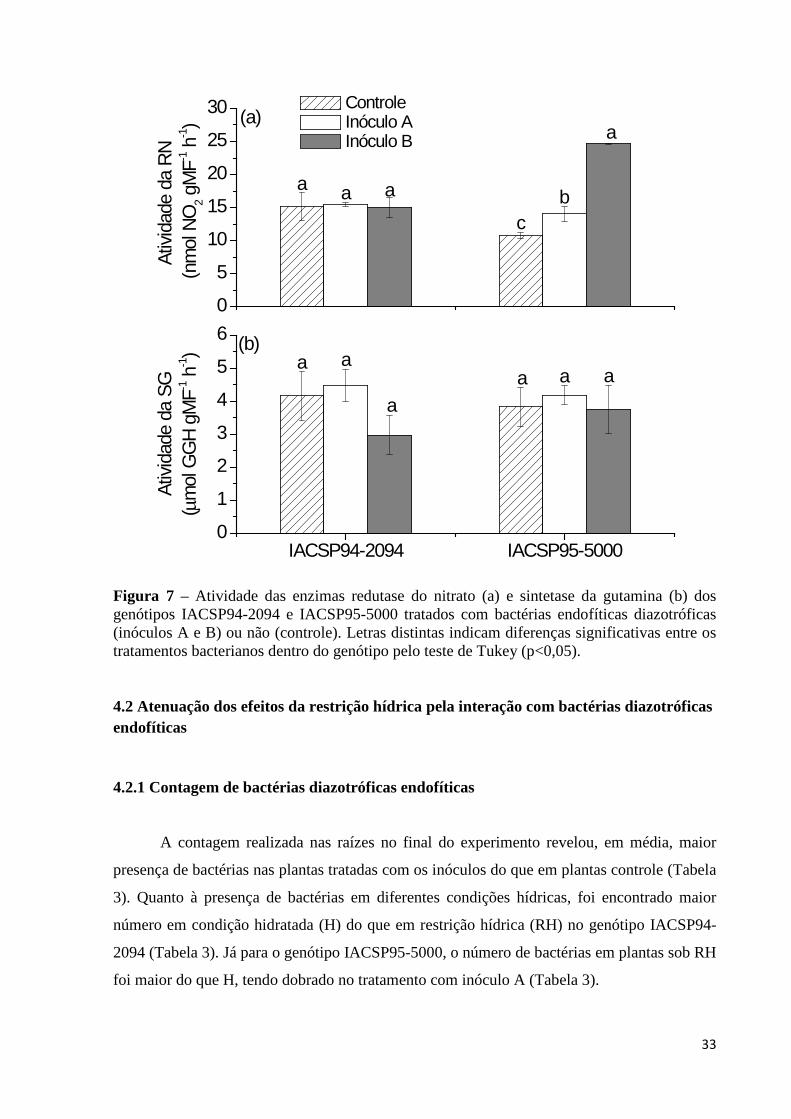
33
Figura 7 – Atividade das enzimas redutase do nitrato (a) e sintetase da gutamina (b) dos genótipos IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle). Letras distintas indicam diferenças significativas entre os tratamentos bacterianos dentro do genótipo pelo teste de Tukey (p<0,05). 4.2 Atenuação dos efeitos da restrição hídrica pela interação com bactérias diazotróficas endofíticas
4.2.1 Contagem de bactérias diazotróficas endofíticas A contagem realizada nas raízes no final do experimento revelou, em média, maior
presença de bactérias nas plantas tratadas com os inóculos do que em plantas controle (Tabela
3). Quanto à presença de bactérias em diferentes condições hídricas, foi encontrado maior
número em condição hidratada (H) do que em restrição hídrica (RH) no genótipo IACSP94-
2094 (Tabela 3). Já para o genótipo IACSP95-5000, o número de bactérias em plantas sob RH
foi maior do que H, tendo dobrado no tratamento com inóculo A (Tabela 3).
0
5
10
15
20
25
30
a
bc
aaa
(a)
Ativi
dade
da
RN
(nm
ol N
O2 g
MF-1
h-1)
Controle Inóculo A Inóculo B
IACSP94-2094 IACSP95-50000
1
2
3
4
5
6 (b)
aaaa
aa
Ativi
dade
da
SG(µ
mol
GG
H gM
F-1 h
-1)
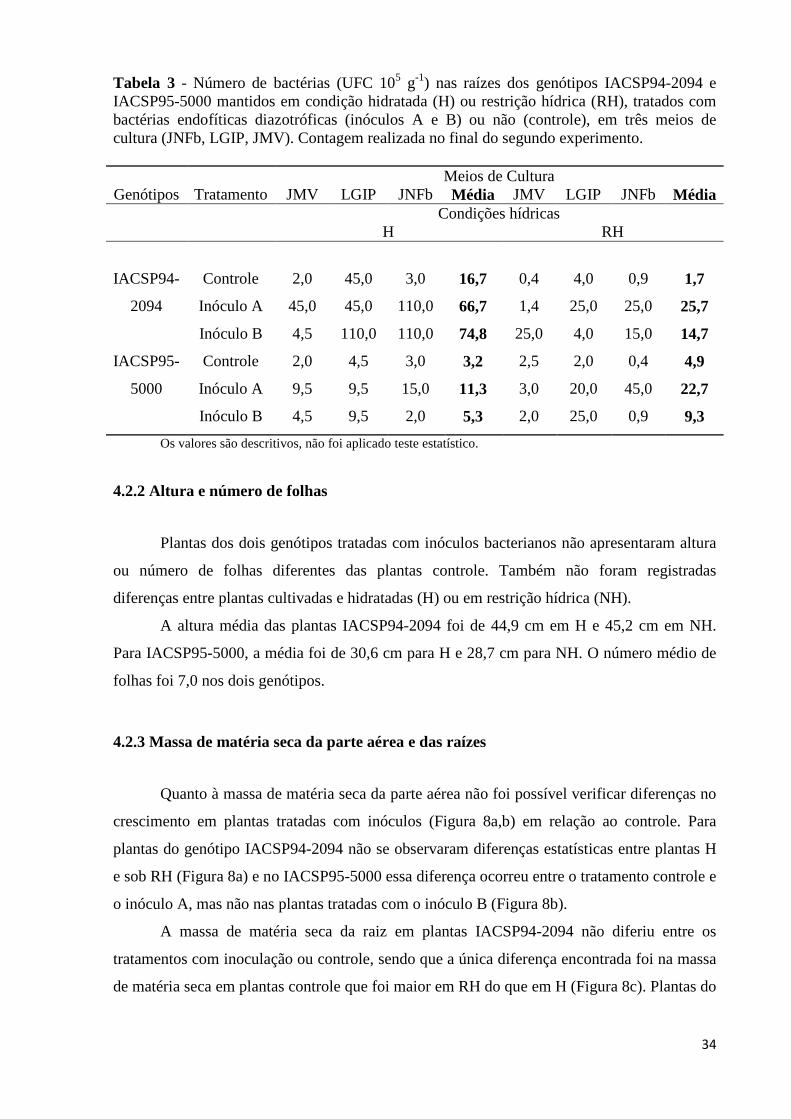
34
Tabela 3 - Número de bactérias (UFC 105 g-1
) nas raízes dos genótipos IACSP94-2094 e IACSP95-5000 mantidos em condição hidratada (H) ou restrição hídrica (RH), tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), em três meios de cultura (JNFb, LGIP, JMV). Contagem realizada no final do segundo experimento.
Meios de Cultura Genótipos Tratamento JMV LGIP JNFb Média JMV LGIP JNFb Média
Condições hídricas H RH
IACSP94-
2094
Controle 2,0 45,0 3,0 16,7 0,4 4,0 0,9 1,7
Inóculo A 45,0 45,0 110,0 66,7 1,4 25,0 25,0 25,7
Inóculo B 4,5 110,0 110,0 74,8 25,0 4,0 15,0 14,7
IACSP95-
5000
Controle 2,0 4,5 3,0 3,2 2,5 2,0 0,4 4,9
Inóculo A 9,5 9,5 15,0 11,3 3,0 20,0 45,0 22,7
Inóculo B 4,5 9,5 2,0 5,3 2,0 25,0 0,9 9,3 Os valores são descritivos, não foi aplicado teste estatístico.
4.2.2 Altura e número de folhas Plantas dos dois genótipos tratadas com inóculos bacterianos não apresentaram altura
ou número de folhas diferentes das plantas controle. Também não foram registradas
diferenças entre plantas cultivadas e hidratadas (H) ou em restrição hídrica (NH).
A altura média das plantas IACSP94-2094 foi de 44,9 cm em H e 45,2 cm em NH.
Para IACSP95-5000, a média foi de 30,6 cm para H e 28,7 cm para NH. O número médio de
folhas foi 7,0 nos dois genótipos.
4.2.3 Massa de matéria seca da parte aérea e das raízes
Quanto à massa de matéria seca da parte aérea não foi possível verificar diferenças no
crescimento em plantas tratadas com inóculos (Figura 8a,b) em relação ao controle. Para
plantas do genótipo IACSP94-2094 não se observaram diferenças estatísticas entre plantas H
e sob RH (Figura 8a) e no IACSP95-5000 essa diferença ocorreu entre o tratamento controle e
o inóculo A, mas não nas plantas tratadas com o inóculo B (Figura 8b).
A massa de matéria seca da raiz em plantas IACSP94-2094 não diferiu entre os
tratamentos com inoculação ou controle, sendo que a única diferença encontrada foi na massa
de matéria seca em plantas controle que foi maior em RH do que em H (Figura 8c). Plantas do
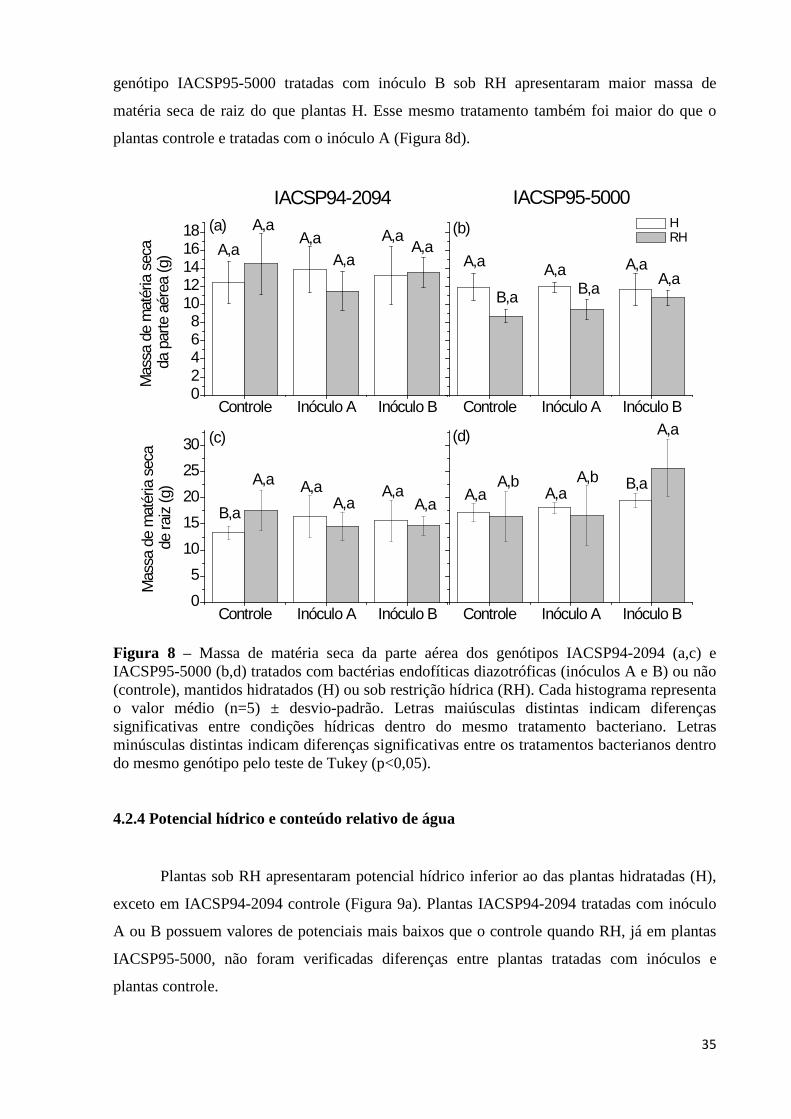
35
genótipo IACSP95-5000 tratadas com inóculo B sob RH apresentaram maior massa de
matéria seca de raiz do que plantas H. Esse mesmo tratamento também foi maior do que o
plantas controle e tratadas com o inóculo A (Figura 8d).
Figura 8 – Massa de matéria seca da parte aérea dos genótipos IACSP94-2094 (a,c) e IACSP95-5000 (b,d) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=5) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro do mesmo genótipo pelo teste de Tukey (p<0,05).
4.2.4 Potencial hídrico e conteúdo relativo de água Plantas sob RH apresentaram potencial hídrico inferior ao das plantas hidratadas (H),
exceto em IACSP94-2094 controle (Figura 9a). Plantas IACSP94-2094 tratadas com inóculo
A ou B possuem valores de potenciais mais baixos que o controle quando RH, já em plantas
IACSP95-5000, não foram verificadas diferenças entre plantas tratadas com inóculos e
plantas controle.
Controle Inóculo A Inóculo B02468
1012141618
A,aA,a
A,a
Mas
sa d
e m
atér
ia s
eca
da p
arte
aér
ea (g
)
Controle Inóculo A Inóculo B05
1015202530 (d)(c)
A,aA,a
A,aA,aA,a
B,a
Mas
sa d
e m
atér
ia s
eca
de
raiz
(g)
H RH
Controle Inóculo A Inóculo B
A,a
B,aA,bA,a
A,bA,a
A,aA,a
A,a
IACSP94-2094 IACSP95-5000
Controle Inóculo A Inóculo B
(b)(a)
A,aA,a
B,aA,a
B,a
A,a

36
Figura 9 – Potencial da água na folha (MPa) medido às 12h 30 min. nos genótipos de cana-de-açúcar IACSP94-2094 (a) e IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou em restrição hídrica (RH). Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).
O conteúdo relativo de água (CRA) não apresentou diferenças estatísticas entre
plantas tratadas com inóculos (A ou B) e plantas controle em condição H ou sob RH. Plantas
hidratadas apresentaram maior CRA do que plantas sob restrição em ambos os genótipos,
exceto para plantas IACSP94-2094 controle (Figura 10).
4.2.5 Teores de prolina e de aminoácidos livres totais (ALT)
Plantas IACSP95-5000 tratadas com o inóculo A apresentaram maior teor de prolina
do que o controle em RH (Figura 11b). Plantas IACSP94-2094 tratadas com inóculo A ou B
tiveram maiores teores de prolina quando em RH enquanto essa diferença não foi notada em
plantas controle.
-1,6
-1,4
-1,2
-1,0
-0,8
-0,6
-0,4
-0,2
0,0Controle Inóculo A Inóculo B
IACSP95-5000IACSP94-2094
(a)B,b
H RH B,b
A,a A,a
A,aA,a
Ψw (
MPa
)
Controle Inóculo A Inóculo B
(b) B,aB,a
B,a
A,a
A,aA,a
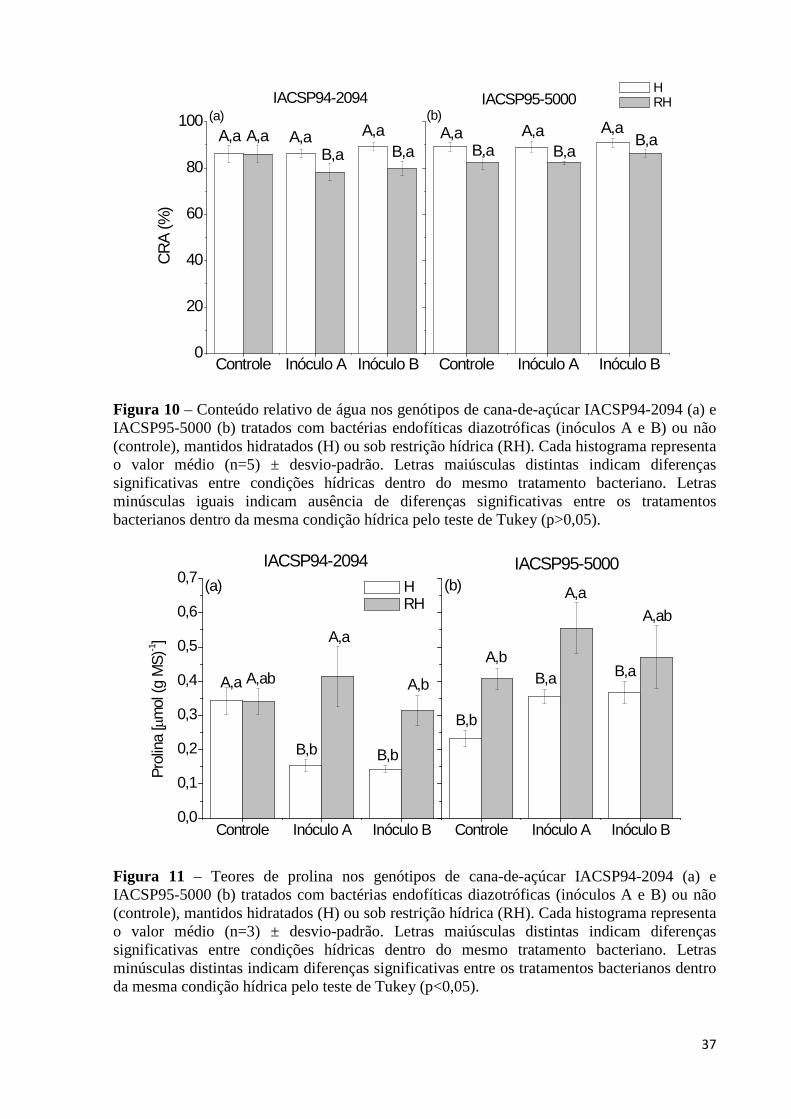
37
Figura 10 – Conteúdo relativo de água nos genótipos de cana-de-açúcar IACSP94-2094 (a) e IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=5) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas iguais indicam ausência de diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p>0,05).
Figura 11 – Teores de prolina nos genótipos de cana-de-açúcar IACSP94-2094 (a) e IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).
Controle Inóculo A Inóculo B0
20
40
60
80
100B,a
B,aB,aB,a B,aA,aA,aA,aA,a
CRA
(%)
Controle Inóculo A Inóculo B
(b)(a)A,aA,aA,a
IACSP95-5000IACSP94-2094 H RH
Controle Inóculo A Inóculo B0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7 (b)(a)
A,a
IACSP95-5000IACSP94-2094
A,b
B,bB,b
A,abA,a
Prol
ina
[µm
ol (g
MS)
-1]
Controle Inóculo A Inóculo B
A,ab
B,a
A,a
B,aA,b
B,b
H RH

38
As plantas de IACSP94-2094 sob restrição hídrica e tratadas com inóculo A
apresentaram maior quantidade ALT nas folhas que o controle (Figura 14a), enquanto que
para IACSP95-5000, as plantas tratadas com inóculo B foram as que apresentaram maiores
valores de ALT sob restrição hídrica (Figura 14b).
Figura 12 – Teores de aminoácidos livres totais (ALT) nos genótipos de cana-de-açúcar IACSP94-2094 (a) e IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=4) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05). 4.2.6 Atividade fotoquímica
Plantas IACSP95-5000 em RH apresentaram diminuição da eficiência quântica
potencial (FV/FM), sendo a média para plantas hidratadas 0,766 ± 0,016 e para plantas sob RH
de 0,682 ± 0,028. Os valores de transporte aparente de elétrons (ETR) também foram menores
em plantas sob RH, 71,11 ± 12,79 µmol m-2 s-1 em relação a plantas hidratadas 117,86 ± 9,31
µmol m-2 s-1
4.2.7 Clorofila total
. O coeficiente de extinção não fotoquímica da fluorescência (NPQ) aumentou em
plantas sob RH (1,95±0,29) em relação a plantas hidratadas (1,44 ± 0,13).
Controle Inóculo A Inóculo B0,00,51,01,52,02,53,03,54,04,55,0
A,abA,a
A,a
B,aA,b
B,a
IACSP95-5000IACSP94-2094(b)(a)
ALT
[mg(
g M
S-1)]
Controle Inóculo A Inóculo B
A,a
B,abA,b
B,b
A,bA,a
H RH

39
Plantas IACSP94-2094 tratadas com inóculo A tiveram diminuição da clorofila total
quando em RH. Para a condição H, o tratamento com inóculo B também apresentou menos
clorofila total que os demais tratamentos (Tabela 4). As plantas IACSP95-5000 foram mais
influenciadas pelo tratamento com os inóculos, principalmente sob restrição hídrica, tendo
valores de clorofila superiores às plantas controle. Em condição H, plantas tratadas com
inóculo B apresentaram os maiores valores dentre os tratamentos (Tabela 4).
Tabela 4 – Clorofila total nos dois genótipos de cana-de-açúcar mantidos hidratados (H) ou sob restrição hídrica (RH) tratados com bactérias endofíticas diazotróficas (inóculo A e B) ou não (controle). IACSP94-2094 IACSP95-5000 Tratamentos H RH H RH Controle 49,9 ± 1,1 A,a 49,9 ± 1,7 A,a 50,3 ± 2,5 A,b 45,5 ± 2,3 B,b Inóculo A 50,3 ± 0,7 A,a 45,9 ± 2,1 B,a 52,6 ± 2,5 A,ab 52,4 ± 1,5 A,a Inóculo B 44,6 ± 1,0 A,b 46,6 ± 1,8 A,a 54,8 ± 1,7 A,a 50,8 ± 3,5 B,a Letras maiúsculas distintas indicam diferenças entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças entre os tratamentos bacterianos dentro da mesma condição hídrica e período. Tukey (p<0,05). Valor médio (n=4) ± desvio-padrão. 4.2.8 Trocas gasosas
As plantas hidratadas tratadas com inóculo A apresentaram maior fotossíntese do que
os demais tratamentos (controle, inóculo B) para os dois genótipos, em condições H (Figura
13a,b). Já em condições restritivas (RH), plantas IACSP94-2094 tratadas com inóculo A
apresentaram os menores valores de fotossíntese (Figura 13a), e para IACSP95-5000 não
foram encontradas diferenças estatísticas (Figura 13b).
Assim como na fotossíntese, plantas dos dois genótipos tratadas com inóculo A
apresentaram maiores valores de condutância (gs) do que os outros tratamentos em condição
H (Figura 13c,d). Em condições restritivas (RH) não foram encontradas diferenças entre os
tratamentos bacterianos (inóculo A, B ou controle), mas as plantas apresentaram menor
condutância do que plantas H (Figura 13c,d).
Quanto à concentração intercelular de CO2, plantas IACSP94-2094 em RH tratadas
com inóculo A ou B apresentaram maiores valores de Ci em relação ao controle e em relação
a plantas tratadas com inóculos e hidratadas (H) (Figura 13e). Plantas IACSP95-5000 em RH
apresentaram maiores valores de Ci quando tratadas com inóculo A (Figura 13f).

40
Figura 13 – Assimilação de CO2 (A-a,b), condutância estomática (gs-c,d), concentração intercelular de CO2
Controle Inóculo A Inóculo B05
10152025303540
(b)(a)
IACSP94-2094 IACSP95-5000
B,aA,b
B,b
A,a
B,aA,b
A (µ
mol
m-2 s
-1)
H RH
Controle Inóculo A Inóculo B0,00
0,05
0,10
0,15
0,20
0,25
0,30(c)
B,aA,b
B,a
A,a
B,aA,ab
g s (m
ol m
-2 s
-1)
Controle Inóculo A Inóculo B
(d)
B,a
A,b
B,a
A,a
B,aA,b
Controle Inóculo A Inóculo B0
50
100
150
200
250
300 (f)(e)A,a
B,a
A,a
B,bA,b
A,a
C i (µm
ol m
ol-1)
Controle Inóculo A Inóculo B
B,cA,a
A,a
B,a
A,a
B,b
Controle Inóculo A Inóculo B
B,a
A,b
B,a
A,a
B,a
A,b
(Ci-e,f) de plantas dos genótipos IACSP94-2094 (a,c,e) e IACSP95-5000 (b,d,f) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantido hidratados (H) ou sob restrição hídrica (RH) medidos às 13 horas. Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano e genótipo. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica e genótipo pelo teste de Tukey (p<0,05).
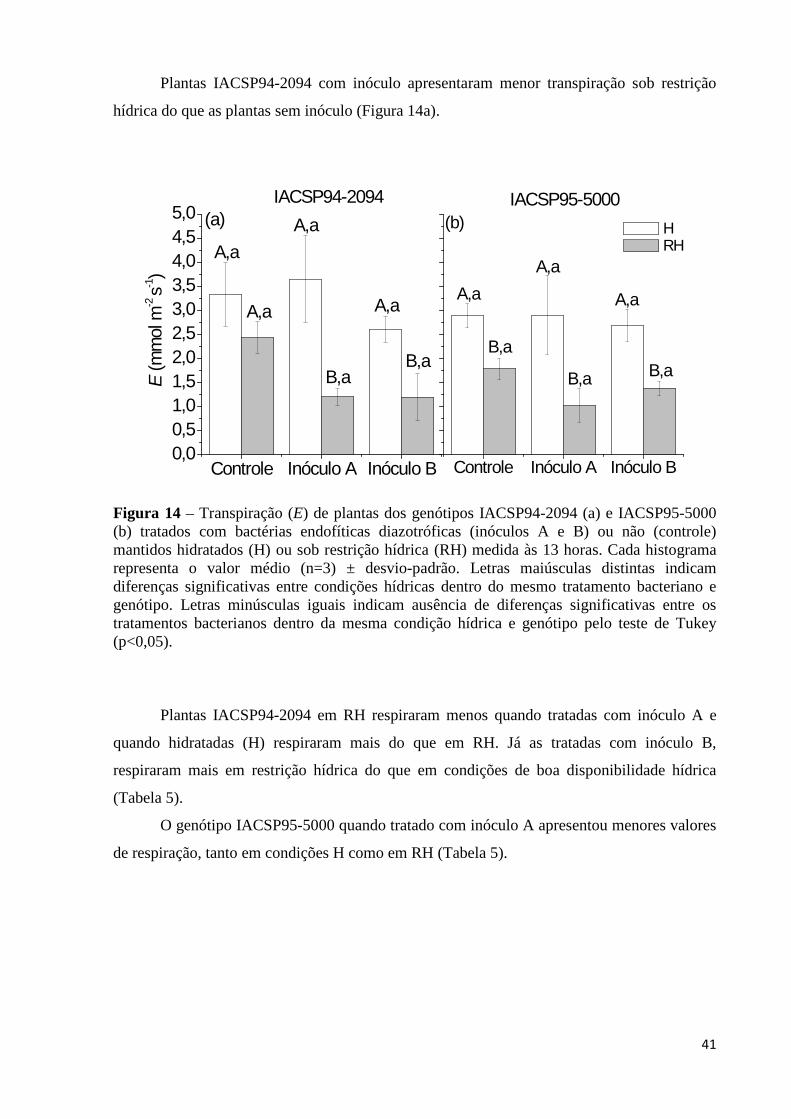
41
Plantas IACSP94-2094 com inóculo apresentaram menor transpiração sob restrição
hídrica do que as plantas sem inóculo (Figura 14a).
Figura 14 – Transpiração (E) de plantas dos genótipos IACSP94-2094 (a) e IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH) medida às 13 horas. Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano e genótipo. Letras minúsculas iguais indicam ausência de diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica e genótipo pelo teste de Tukey (p<0,05).
Plantas IACSP94-2094 em RH respiraram menos quando tratadas com inóculo A e
quando hidratadas (H) respiraram mais do que em RH. Já as tratadas com inóculo B,
respiraram mais em restrição hídrica do que em condições de boa disponibilidade hídrica
(Tabela 5).
O genótipo IACSP95-5000 quando tratado com inóculo A apresentou menores valores
de respiração, tanto em condições H como em RH (Tabela 5).
Controle Inóculo A Inóculo B0,00,51,01,52,02,53,03,54,04,55,0 (a)
IACSP95-5000IACSP94-2094
B,a
A,a
B,a
A,a
A,a
A,a
E (m
mol
m-2
s-1)
Controle Inóculo A Inóculo B
(b)
B,a
A,a
B,a
A,a
B,a
A,a
H RH
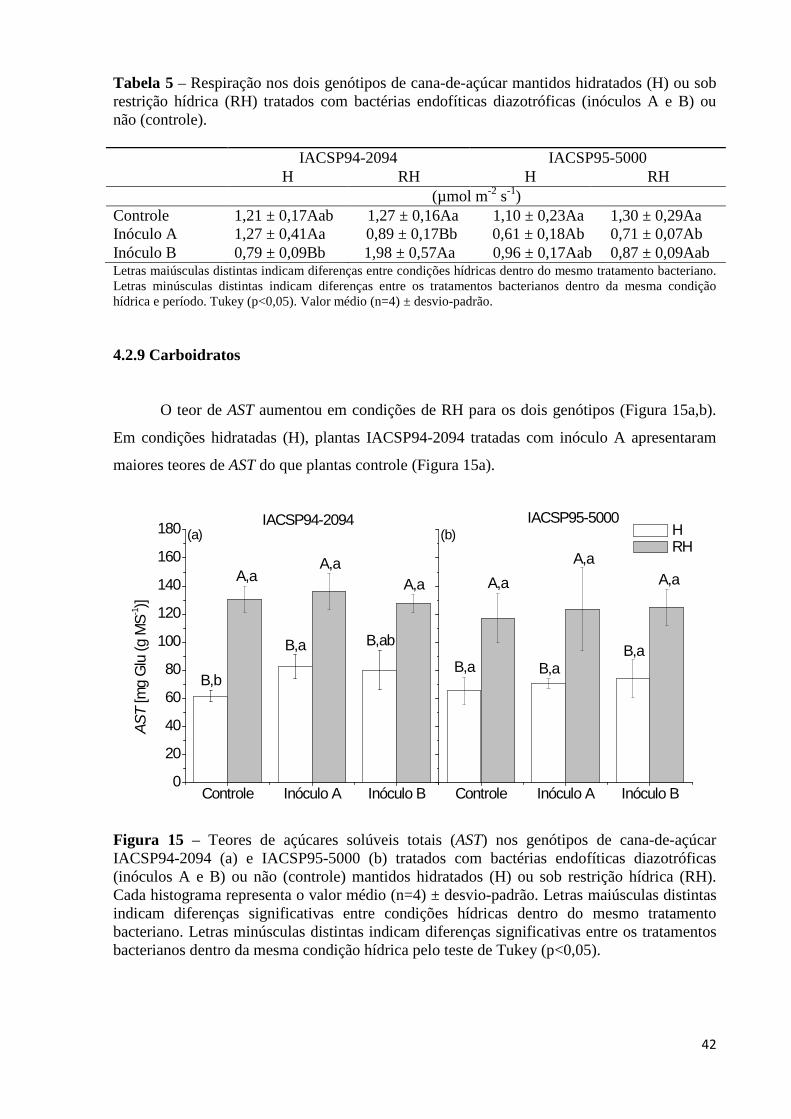
42
Tabela 5 – Respiração nos dois genótipos de cana-de-açúcar mantidos hidratados (H) ou sob restrição hídrica (RH) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle). IACSP94-2094 IACSP95-5000 H RH H RH (µmol m-2 s-1) Controle 1,21 ± 0,17Aab 1,27 ± 0,16Aa 1,10 ± 0,23Aa 1,30 ± 0,29Aa Inóculo A 1,27 ± 0,41Aa 0,89 ± 0,17Bb 0,61 ± 0,18Ab 0,71 ± 0,07Ab Inóculo B 0,79 ± 0,09Bb 1,98 ± 0,57Aa 0,96 ± 0,17Aab 0,87 ± 0,09Aab Letras maiúsculas distintas indicam diferenças entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças entre os tratamentos bacterianos dentro da mesma condição hídrica e período. Tukey (p<0,05). Valor médio (n=4) ± desvio-padrão. 4.2.9 Carboidratos O teor de AST aumentou em condições de RH para os dois genótipos (Figura 15a,b).
Em condições hidratadas (H), plantas IACSP94-2094 tratadas com inóculo A apresentaram
maiores teores de AST do que plantas controle (Figura 15a).
Figura 15 – Teores de açúcares solúveis totais (AST) nos genótipos de cana-de-açúcar IACSP94-2094 (a) e IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=4) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).
Controle Inóculo A Inóculo B0
20
40
60
80
100
120
140
160
180 (a)
A,aA,a
A,a
B,abB,a
B,b
AST
[mg
Glu
(g M
S-1)]
Controle Inóculo A Inóculo B
(b)
A,aA,a
A,a
B,aB,aB,a
IACSP95-5000IACSP94-2094 H RH
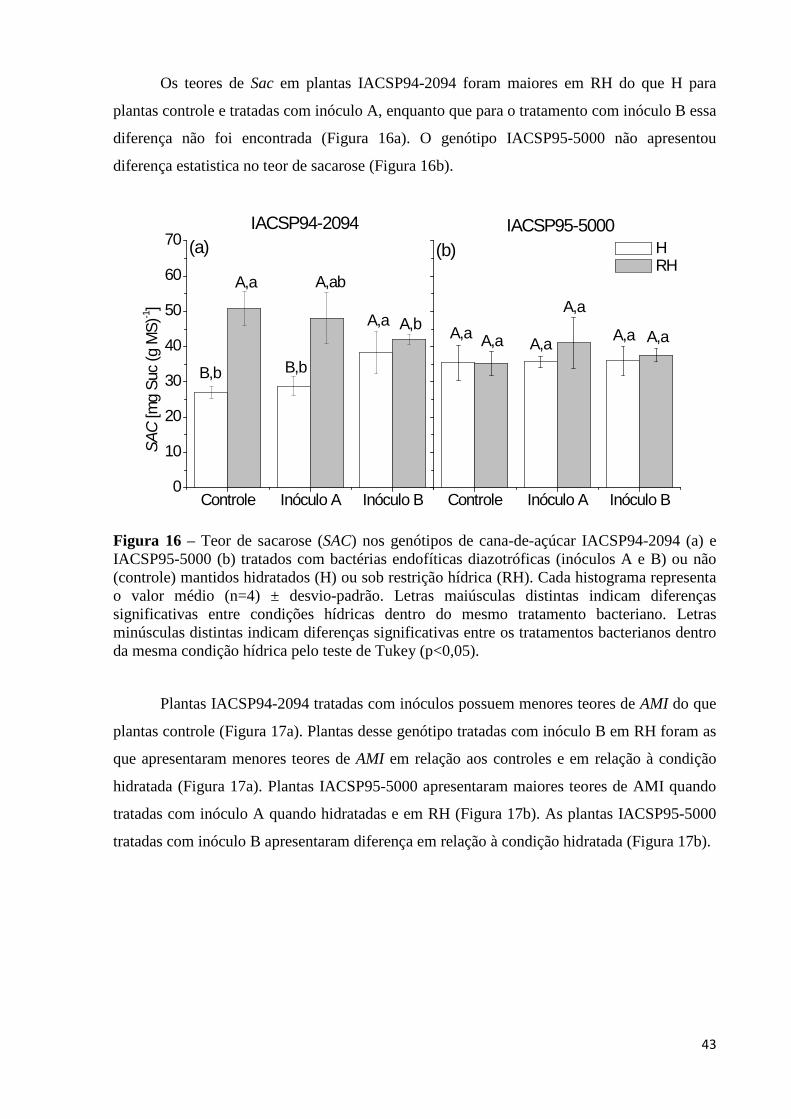
43
Os teores de Sac em plantas IACSP94-2094 foram maiores em RH do que H para
plantas controle e tratadas com inóculo A, enquanto que para o tratamento com inóculo B essa
diferença não foi encontrada (Figura 16a). O genótipo IACSP95-5000 não apresentou
diferença estatistica no teor de sacarose (Figura 16b).
Figura 16 – Teor de sacarose (SAC) nos genótipos de cana-de-açúcar IACSP94-2094 (a) e IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=4) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).
Plantas IACSP94-2094 tratadas com inóculos possuem menores teores de AMI do que
plantas controle (Figura 17a). Plantas desse genótipo tratadas com inóculo B em RH foram as
que apresentaram menores teores de AMI em relação aos controles e em relação à condição
hidratada (Figura 17a). Plantas IACSP95-5000 apresentaram maiores teores de AMI quando
tratadas com inóculo A quando hidratadas e em RH (Figura 17b). As plantas IACSP95-5000
tratadas com inóculo B apresentaram diferença em relação à condição hidratada (Figura 17b).
Controle Inóculo A Inóculo B
(b)(a)IACSP95-5000IACSP94-2094
SAC
[mg
Suc
(g M
S)-1]
A,aA,a
A,a
A,aA,aA,a
H RH
Controle Inóculo A Inóculo B0
10
20
30
40
50
60
70
A,b
A,abA,a
A,a
B,bB,b

44
Figura 17 – Teores de amido (AMI) nos genótipos de cana-de-açúcar IACSP94-2094 (a) e IAC95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=4) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).
4.2.10 Nitrogênio Não houve diferenças estatísticas entre os tratamentos com inóculos e controle, nem entre as condições hídricas (H e RH) em relação aos teores de nitrogênio (Tabela 6). Tabela 6 – Teores de nitrogênio (g kg -1
) nos dois genótipos de cana-de-açúcar IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou sob restrição hídrica (RH).
IACSP94-2094 IACSP95-5000 Condições hídricas
H RH H RH Controle 21,3 ± 0,9 19,1 ± 1,9ns 21,6 ± 1,5ns 20,4 ± 0,9ns
Inóculo A
ns
22,9 ± 1,8 20,7 ± 1,6 20,6 ± 0,6 19,5 ± 0,7 Inóculo B 21,1 ± 0,9 21,0 ± 0,8 20,4 ± 1,3 19,2 ± 0,3
ns indica diferenças não significativas pelo teste de Tukey (p<0,05). Valores referem-se às médias (n=5) ± desvios-padrão.
Controle Inóculo A Inóculo B
(b)(a)IASP95-5000IACSP94-2094
B,abA,a
A,bA,abA,a
A,b
H RH
Controle Inóculo A Inóculo B0
5
10
15
20
25
30
35
B,c
A,bA,a
A,bA,abB,a
AMI [
mg
Glu
(g M
S)-1]

45
O genótipo IACSP94-2094 tratado com inóculo B apresentou maior acúmulo de
nitrogênio em RH do que em H (Tabela 7). Plantas IACSP95-5000 tratadas com inóculo B
apresentaram maiores valores de nitrogênio acumulado do que plantas controle ou tratadas
com inóculo A quando em RH (Tabela7).
Quanto ao índice de eficiência de utilização de nitrogênio, plantas IACSP95-5000
tratadas com inóculo B em RH tiveram maior eficiência na conversão do N fixado ou
absorvido (Tabela 8 – destaque).
4.2.11 Atividade de enzimas do metabolismo do nitrogênio A atividade da RN em plantas IACSP94-2094 sob RH foi maior em plantas tratadas
com o inóculo B (Figura 19a). Em plantas IACSP95-5000 sob RH a maior atividade foi
verificada no tratamento com inóculo A e a menor com inóculo B (Figura 18b). Plantas
hidratadas (H) tratadas com inóculo B apresentaram maior atividade da RN (Figura 18b).
Figura 18 – Atividade da redutase do nitrato (RN) nos genótipos de cana-de-açúcar IACSP94-2094 (a) e IACSP95-5000 (b) tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=3) ± desvio-padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).
controle Inóculo A Inóculo B0
5
10
15
20
25
A,a
B,b
B,b
A,a
A,bA,b
IACSP95-5000IACSP94-2094
(b)(a)
Ativi
dade
da
RN
(nm
ol d
e NO
2 g M
F-1 h
-1)
controle Inóculo A Inóculo B
B,b
A,a
A,a
B,c
A,abA,b
H RH

46
Tabela 7 – Quantidade de nitrogênio acumulado (mg N planta-1
) nos dois genótipos de cana-de-açúcar IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou sob restrição hídrica (RH).
IACSP94-2094 IACSP95-5000 Condições hídricas
H RH H RH Controle 250,45 ±14,68 A b 262,53 ± 8,78 A a 262,10 ± 13,59 A a 182,65 ± 9,70 B b
Inóculo A 335,16 ± 30,85 A a 277,00 ± 10,90 A a 252,01 ± 5,43 A ab 194,37 ± 17,10 B ab Inóculo B 249,31 ± 4,70 B b 308,72 ± 9,57 A a 231,80 ± 1,09 A b 216,31 ± 10,12 A a
Valores médios (n=3) ± desvios-padrão. Letras maiúsculas distintas indicam diferenças entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças entre tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).
Tabela 8 – Índice de eficiência de utilização de nitrogênio (g biomassa2/g nutriente-1
) nos dois genótipos de cana-de-açúcar IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle), mantidos hidratados (H) ou sob restrição hídrica (RH).
IACSP94-2094 IACSP95-5000 Condições hídricas H RH H RH
Controle 0,508 ± 0,070 B a 0,697 ± 0,096 A a 0,596 ± 0,027 A a 0,459 ± 0,036 B b Inóculo A 0,481 ± 0,066 A a 0,480 ± 0,010 A b 0,605 ± 0,015 A a 0,538 ± 0,046 A ab Inóculo B 0,575 ± 0,025 B a 0,700 ± 0,031 A a 0,581 ± 0,067 A a 0,587 ± 0,027 A a
Valores médios (n=3) ± desvios-padrão. Letras maiúsculas distintas indicam diferenças entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças entre tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).
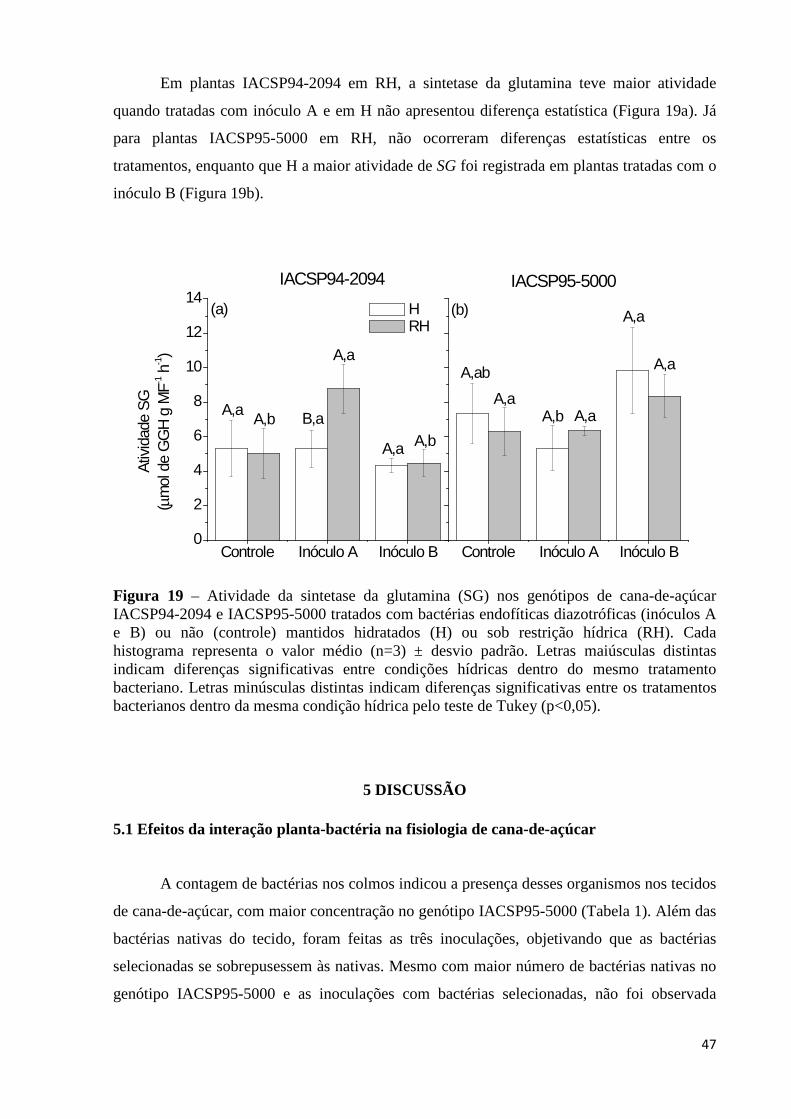
47
Em plantas IACSP94-2094 em RH, a sintetase da glutamina teve maior atividade
quando tratadas com inóculo A e em H não apresentou diferença estatística (Figura 19a). Já
para plantas IACSP95-5000 em RH, não ocorreram diferenças estatísticas entre os
tratamentos, enquanto que H a maior atividade de SG foi registrada em plantas tratadas com o
inóculo B (Figura 19b).
Figura 19 – Atividade da sintetase da glutamina (SG) nos genótipos de cana-de-açúcar IACSP94-2094 e IACSP95-5000 tratados com bactérias endofíticas diazotróficas (inóculos A e B) ou não (controle) mantidos hidratados (H) ou sob restrição hídrica (RH). Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças significativas entre condições hídricas dentro do mesmo tratamento bacteriano. Letras minúsculas distintas indicam diferenças significativas entre os tratamentos bacterianos dentro da mesma condição hídrica pelo teste de Tukey (p<0,05).
5 DISCUSSÃO 5.1 Efeitos da interação planta-bactéria na fisiologia de cana-de-açúcar
A contagem de bactérias nos colmos indicou a presença desses organismos nos tecidos
de cana-de-açúcar, com maior concentração no genótipo IACSP95-5000 (Tabela 1). Além das
bactérias nativas do tecido, foram feitas as três inoculações, objetivando que as bactérias
selecionadas se sobrepusessem às nativas. Mesmo com maior número de bactérias nativas no
genótipo IACSP95-5000 e as inoculações com bactérias selecionadas, não foi observada
Controle Inóculo A Inóculo B0
2
4
6
8
10
12
14
A,bB,aA,b
A,a
A,a
A,a
A,a
(b)(a)
IACSP95-5000IACSP94-2094
Ativi
dade
SG
(µm
ol d
e G
GH
g M
F-1 h
-1)
Controle Inóculo A Inóculo B
A,ab
A,b
A,a
A,a
A,a
H RH

48
promoção de crescimento. Plantas tratadas com os inóculos não tiveram maior massa seca de
parte aérea ou raiz (Figura 1) do que as plantas controle, sendo que a altura e o número de
folhas também não diferiram. Um dos resultados mais enfatizados na literatura em relação ao
uso de bactérias como inoculante é a promoção de crescimento (RYAN et al., 2008), com
estímulo no desenvolvimento tanto da parte aérea como das raízes (OLIVEIRA et al., 2002).
Tanto pelo referencial teórico do uso desse grupo de bactérias como pelos resultados obtidos
por FREITAS (2011), do qual foram selecionados os isolados para composição dos inoculos
usados, o resultado esperado não foi observado, ou seja, maior crescimento das plantas
tratadas com os inóculos em relação ao controle.
Apesar disto, a interação entre planta e bactérias causou um estímulo na assimilação
de CO2 (Figura 5a). O aumento da fotossíntese no genótipo IACSP95-5000 tratado com
inóculo A pode estar ligado ao aumento da abertura estomática (Figura 5b). O aumento da
condutância estomática pode ter sido estimulado por sinais hormonais enviados pelas
bactérias, tendo como conseqüência a maior disponibilidade de CO2
Segundo FUENTES-RAMÍREZ et al. (1999), uma diminuição da sacarose pode estar
relacionada ao seu uso pelas bactérias fixadoras de nitrogênio que colonizam a planta. A fonte
para plantas tratadas com
o inóculo A. Já para IACSP94-2094 tratado com inóculo B, o estímulo da fotossíntese pode
estar ligada ao aumento do ETR (Figura 4a), pois o transporte aparente de elétrons gera
moléculas energéticas, ATP e NADPH, que são usadas na assimilação de carbono. No
entanto, a maior fotossíntese não resultou no aumento da matéria seca das plantas tratadas.
O estímulo da fotossíntese proporcionou aumento na disponibilidade de carboidratos
nas plantas IACSP94-2094 quando tratadas com inóculo B (Figura 6). O maior teor de
sacarose nessas plantas provavelmente é resultado da maior fotossíntese, indicando um efeito
direto da presença das bactérias endofíticas na fisiologia. No entanto, para plantas IACSP95-
5000 tratadas com inóculo A, condição que estimulou a fotossíntese, não houve maiores
teores de carboidratos e as tratadas com inóculo B apresentam valores de SAC menores que o
controle (Figura 6a). A diminuição de SAC, um açúcar solúvel, não resultou em menores
teores AST nesse caso, podendo-se pensar na hipótese de que deve haver outro açúcar solúvel
sendo produzido, por exemplo, a frutose, como o encontrado em maiores concentrações em
plantas tratadas com bactérias endofíticas no trabalho de SHI et al. (2010). A SAC poderia
estar sendo consumida pela planta com sua quebra e utilização de açúcar para a respiração,
mas os valores de respiração não apresentam diferenças entre os tratamentos. Se as plantas do
genótipo produtivo não se utilizam do carboidrato produzido pelo estímulo da fotossíntese,
algum desvio deve ocorrer.

49
de carbono forneceria energia para o estabelecimento dessa relação entre planta e bactéria.
Essa poderia ser uma explicação para os baixos níveis de sacarose em plantas IACSP95-5000
(Figura 6a). Pode-se traçar um paralelo com outra forma de interação: planta-rizóbio. Nessa
interação, os bacteróides presentes nos nódulos, por necessitarem de muita energia para a
fixação de nitrogênio, acabam por ser um dreno de carboidratos, o que estimula a fotossíntese,
ocorrendo uma maior conversão de triose-P para sacarose, que é transportada para os drenos
(KASCHUK et al., 2009).
Outra alteração fisiológica registrada foi o aumento da atividade da redutase do nitrato
(RN) em plantas IACSP95-5000 tratadas com inóculo A e B, este último com maior atividade
(Figura 7b). A RN está ligada com a redução de nitrato a nitrito, que é transformado
posteriormente em amônio, e em sequência assimilado em glutamina pela sintetase da
glutamina (SG). Neste experimento, SG não teve aumento de atividade, não sendo então o
nitrito produzido pela RN das plantas IACSP95-5000 tratadas com inóculos A e B utilizado
pela SG na folha. Os teores de nitrogênio e de ALT também não foram alterados, indicando
que o nitrato poderia não estar sendo utilizado pelas plantas. DONATO et al. (2004) indicam
que a influência de bactérias no metabolismo de nitrogênio estaria no aumento da atividade da
RN, aumentando a entrada de nitrato e, consequentemente, favorecendo aumento nos teores
de nitrogênio na planta. Então, as plantas deste trabalho que apresentaram maior atividade de
RN deveriam ser favorecidas, fato que não ocorreu.
O teor de prolina também não aumentou em plantas tratadas com os inóculos, pelo
contrário, os maiores teores foram observados em plantas controle (Figura 3). Esse
aminoácido tem sido relacionado à proteção das plantas a diferentes estresses (ASHRAF &
FOOLAD, 2007), mas neste experimento, as plantas encontravam-se em condições de
adequada disponibilidade hídrica, o que não justificaria um aumento da produção de prolina
em plantas controle, mas talvez um consumo desse aminoácido em plantas tratadas. Além do
mais, os teores de prolina encontrados são muito baixos para terem função de proteção. Em
SARAVANAKUMAR et al. (2011), os valores de prolina de plantas de Vignia radiata
tratadas com bactérias aumentou 3 vezes da condição bem irrigada para condição de estresse
hídrico.
Quanto à caracterização hídrica das plantas, foi observado que plantas tratadas com
inóculos tiveram potencial hídrico maior do que plantas controle (Figura 2). Como todas as
plantas estavam sob adequada disponibilidade hídrica, as alterações registradas devem ter sido
provocadas pela interação com os inóculos. CREUS et al. (2004) relatam que, independente

50
de estresse hídrico, a fração de água no apoplasto aumenta em resposta a inoculação com
Azospirillum, o que pode justificar esse aumento no potencial hídrico de plantas tratadas.
A análise dos dados pelo gráfico radar (Figura 21) indica os principais parâmetros que
são influenciados pela inoculação de bactérias endofíticas em relação às plantas controle. Para
o genótipo IACSP94-2094, o tratamento com o inóculo B tem um maior número de
parâmetros influenciados, sendo A, ETR e prolina os principais (Figura 21a). Para o genótipo
IACSP95-5000, os dois inóculos influenciaram os parâmetros medidos. Plantas tratadas com
o inóculo A se diferenciaram estatisticamente do controle em A e gs, Ψw, RN, e prolina. Para
plantas tratadas com inóculo B, os parâmetros que mais se diferenciam do controle são Ψw,
Sac e RN (Figura 21b).
Figura 20 – Representação das respostas fisiológicas dos genótipos IACSP94-2094 (a) e IACSP95-5000 (b) com boa disponibilidade hídrica, tratados com inóculo A (quadrado) e inóculo B (triangulo) em relação aos respectivos controles (linha pontilhada). Os valores referem-se a mudanças relativas das seguintes variáveis: massa de matéria seca da parte aérea (MSPA), massa seca de raiz (MSR), fotossíntese (A), condutância estomática (gs), transpiração (E), transporte aparente de elétrons (ETR), coeficiente de extinção fotoquímica (NPQ), potencial hídrico (Ψw), teor de sacarose (Sac), teor de açúcares solúveis totais (AST), teor de nitrogênio (N), atividade da redutase do nitrato (RN), atividade da sintetase da glutamina (SG), teor de prolina (Prolina), teor de aminoácidos livres totais (ALT). * indicam diferença estatística em relação ao controle.
HARDOIM et al. (2008) sugerem um novo conceito: endofíticos competentes. Seriam
microrganismos que colonizam a planta com sucesso, entrando nos seus tecidos, e apresentam
0,0
0,5
1,0
1,5
2,0
2,5MSPA
MSR
A
gs
E
ETR
NPQ
ψwSac
AST
Ntotal
RN
GS
Prolina
ALT
IACSP94-2094
0,0
0,5
1,0
1,5
2,0
2,5MSPA
MSR
A
gs
E
ETR
NPQ
ψwSac
AST
Ntotal
RN
GS
Prolina
ALT
IACSP95-5000
Controle Inóculo A Inóculo B
(a) (b)
*
*
* *
*
*
*
*
*
*
*
*
*

51
capacidade de modular a fisiologia da planta a ser favorecida, levando à manutenção dos
benefícios da associação. Foram observadas mudanças benéficas na fisiologia dos dois
genótipos, no entanto, não houve alterações no crescimento vegetal. O estímulo à fotossíntese
e à atividade da redutase do nitrato deveria garantir maiores valores de matéria seca ou
maiores teores de nitrogênio. Se a influência na fisiologia da planta não é benéfica, seria a
bactéria a mais beneficiada nessa interação? O carboidrato fixado e o nitrogênio (nitrito –
produto da RN e a prolina) poderiam ser utilizados para o consumo da comunidade bacteriana
no interior dos tecidos vegetais. Como as plantas não estavam sob estresse, o excedente
desses compostos não poderiam estar sendo usados para manutenção do metabolismo vegetal,
fato corroborado pela não alteração dos valores de respiração. Além disso, não há explicação
plausível que justifique um aumento do metabolismo do C e do N sem que os produtos
fossem utilizados. Talvez a interação da planta com as bactérias inoculadas não tenha sido
suficientemente eficiente para se encaixar no conceito de endofíticos competentes, visto que a
promoção de crescimento não ocorreu (Figuras 1), apesar de outras alterações fisiológicas
observadas. As alterações fisiológicas poderiam ser destinadas à manutenção dos organismos
endofíticos.
5.2 Atenuações dos efeitos do estresse hídrico por bactérias endofíticas diazotróficas Pela contagem de bactérias nas raízes (Tabela 3) pode-se afirmar que ocorreu
colonização efetiva, já que as plantas que receberam inóculo (inóculo A ou B - bactérias
introduzidas) mostraram um número superior ao das plantas que não receberam (controle –
presença de bactérias nativas). Nas inoculações, usaram-se concentrações bacterianas
comumente recomendadas (OLIVEIRA et al., 2002; CREUS et al., 2004), objetivando que as
bactérias selecionadas se sobrepusessem às nativas.
O número de bactérias nas raízes sofreu alteração dependente da condição hídrica
(Tabela 3). Plantas do genótipo tolerante IACSP94-2094 apresentaram redução do número de
bactérias em RH. Para COMPANT et al. (2010), as condições ambientais podem influenciar o
número de bactérias presentes em condições de restrição hídrica, sendo a comunidade afetada
pela seca. No entanto, essa influência negativa não ocorreu para plantas do genótipo sensível
(IACSP95-5000). Pelo contrário, o número médio de bactérias presentes nas raízes das
plantas em RH foi maior do que plantas hidratadas (H). Aparentemente, as plantas desse

52
genótipo favorecem as bactérias apenas em RH e resta saber se essa interação também traria
benefícios para a planta que está sob condição hídrica restritiva.
Outro aspecto a ser considerado é a reestruturação de características do solo
pela introdução de microrganismos. O substrato autoclavado perdeu as características de um
solo com microrganismos vivos, a introdução do inóculo pode ter favorecido a reestruturação
de características como, por exemplo, solubilização de fosfatos e disponibilização de
nutrientes para plantas, ou, até mesmo, o estabelecimento da comunidade rizosférica. Nos
dois experimentos foram utilizados colmos não desinfestados que mantiveram os
microrganismos nativos. Não se conhecia o hábito das bactérias nativas podendo ser
endofíticas facultativas (podem viver fora de tecidos vegetais) ou rizosféricas, que também
podem influenciar a fisiologia do vegetal. Pela complexidade da interação e presença de
microrganismos nativos as respostas fisiológicas das plantas são muitas vezes difíceis de
serem esclarecidas, já que o material controle também possui bactérias, mesmo que em menor
quantidade, mas possíveis de influenciar a planta .
Sob restrição hídrica, plantas do genótipo IACSP95-5000, sensíveis à seca
(LANDELL et al., 2005b), quando tratadas com inóculo B não sofreram redução de massa de
matéria seca da parte aérea e apresentaram maior massa de matéria seca de raiz (Figura 8b).
Esses resultados mostram que o tratamento com inóculo B pode impedir a restrição de
crescimento da parte aérea e ainda promover o crescimento na raiz em condições hídricas
restritivas. Esta resposta pode ser respaldada pelo maior número de bactérias em plantas
IACSP95-5000 em RH. Bactérias podem alterar o padrão de crescimento das plantas pela
produção de fitormônios (TSAVKELOVA et al., 2006a), como o ácido 3,indol-acético (AIA)
aumentar a produção de massa seca (SHI et al., 2010). Para o genótipo IACSP94-2094,
tolerante à seca (LANDELL et al., 2005a; MACHADO et al., 2009), não ocorreu diminuição
da matéria seca (Figura 8a), mas também não ocorreu promoção de crescimento pelas
bactérias. Talvez, a interação com as bactérias seja mais eficiente em genótipos sensíveis à
seca do que nos tolerantes. LIDDYCOAT et al. (2009) também observaram respostas
diferentes dos genótipos de aspargos tratados com bactérias; um genótipo foi favorecido em
condição hídrica ideal e o outro em restrição hídrica. Em trabalho com arroz usando
rizobactérias osmotolerantes, YUWONO et al. (2005) observaram que os resultados mais
pronunciados dos inóculos ocorreram em condições de restrição hídrica.
A diminuição do crescimento das plantas IACSP95-5000 sob RH foi provavelmente
resultado das modificações fisiológicas que ocorrem sob deficiência hídrica. Foram
registrados menores potenciais hídricos foliares (Ψw) em condição de RH (Figura 9), que

53
podem causar alterações bioquímicas. Segundo PIMENTEL (2004), em estágio de deficiência
moderada, em que a planta tem a transpiração reduzida (Figura 15), pode haver redução da
atividade enzimática da redutase do nitrato, aumento de carboidratos solúveis e aminoácidos
livres totais e prolina. No presente trabalho, ocorreu aumento de carboidratos (Figura 16 e
17), prolina (Figura 11) e ALT (Figura 12).
O potencial hídrico sob RH em IACSP94-2094 foi menor em plantas tratadas com os
inóculos do que no controle (Figura 9a), assim como o conteúdo relativo de água (CRA) que
foi menor em plantas tratadas do que no controle (Figura 10a). O aumento da prolina poderia
estar ligado a um aumento de compostos osmorreguladores, importantes em situações de
estresse. Sugere-se que a prolina tenha relação com maior tolerância de cultivares de cana-de-
açúcar à seca (INMAN-BAMBER & SMITH, 2005; MOLINARI et al., 2007). Os teores de
prolina foram maiores em plantas sensíveis a seca, IACSP95-5000, tratadas com inóculo,
principalmente inóculo A, confirmando a hipótese de que esse genótipo foi mais favorecido
pelo tratamento bacteriano. No entanto, os valores de prolina encontrados são baixos e podem
ser apenas indicativos de alterações metabólicas devido ao estresse e não a uma resposta
adaptativa com efeitos protetores para planta (SOUZA et al., 2004). Outros compostos como a
glicina-betaina poderiam ter sido produzidos pelas bactérias em resposta ao estresse
aumentando a tolerância das plantas com menor potencial hídrico (DIMKPA et al., 2009).
Neste trabalho, o teor de AST aumentou em plantas sob RH (Figura 16). O acúmulo de
carboidratos também tem função osmoprotetora e de regulação osmótica (CHAVES et al.,
2002 e NIEDZWIEDZ-SIEGIEN et al., 2004). Carboidratos devem ser acumulados de forma
rápida e em quantidade suficiente para cumprir o seu papel de proteção, como por exemplo, a
sacarose que protege membranas de cloroplastos e tonoplastos da dissecação (DINAKAR et
al., 2012). Foi possível verificar aumento de SAC em plantas IACSP94-2094 (tolerantes à
seca) tanto tratadas com inóculo A quanto no controle (Figura 17a). A quantidade de amido
variou em função da condição hídrica apenas em IACSP94-2094 controle e inóculo B (Figura
18a). O amido, carboidrato de reserva, seria consumido e convertido em açúcares solúveis em
condições de restrição hídrica como forma de proteção celular (LEE et al., 2008).
Observaram-se maiores valores de amido em plantas IACSP94-2094 controle que nas plantas
tratadas com inóculos, indicando que plantas tratadas necessitariam de mais energia para sua
manutenção em condição de estresse, o que pode ter levado à maior degradação de AMI. Os
teores de AMI em IACSP95-5000 tratados com inóculo B e em RH diminuem em relação à
condição irrigada (Figura 18b), fato que pode estar relacionado a não diminuição de

54
crescimento. Plantas desse tratamento recorreriam à degradação de AMI para manutenção do
metabolismo em situação de estresse.
As alterações nos teores de carboidratos estão relacionadas à fotossíntese. A restrição
hídrica leva à diminuição do potencial hídrico (Figura 9) reduzindo a assimilação de CO2
(Figura 14a,b), que é intimamente associada ao fechamento estomático (Figura 14c,d)
(CHAVES, 1991). A redução da assimilação de carbono em plantas sob RH acaba por levar a
uma degradação de AMI elevando os teores de açúcares solúveis. A resposta das plantas à
falta de água com o fechamento estomático foi verificada neste trabalho (Figura 14c,d) e
apesar de não ter havido diferenças entre plantas tratadas com inóculos e controle, ainda assim
a diminuição da condutância estomática revelou um padrão de resposta interessante. Isto é,
nas plantas controle, a mudança da condição hidratada (H) para a restritiva (RH) ocasionou
uma redução da condutância estomática de 35%, em média, para ambos os genótipos.
Entretanto, para plantas IACSP94-2094, a redução na condutância foi de 69% nas plantas
tratadas com inóculo A e de 39% nas com inóculo B, enquanto que nas plantas IACSP95-
5000, a redução ficou em 78% e 60% para os inóculos A e B, respectivamente. Isso mostra
que os tratamentos com inóculos influenciaram o fechamento estomático e de forma mais
intensa no genótipo sensível (IACSP95-5000). O maior fechamento estomático pode ser um
indicativo de envio de sinais químicos da raiz para a parte aérea (DAVIES & ZHANG, 1991)
ou mesmo de substâncias osmorreguladoras que podem influenciar no fechamento estomático.
Observou-se também que em condição hidratada (H) plantas de ambos os genótipos,
tratadas com inóculo A, possuem maiores valores de fotossíntese (Figura 14a,b). Sugere-se
que a fotossíntese de plantas tratadas com bactérias poderia ser estimulada pela melhor
nutrição da planta ou ainda pela eliminação de limitações enzimáticas ou maior transporte de
elétrons, além de estimular o consumo de triose fosfato da célula, garantindo fluxo de saída
para os drenos (KASCHUK et al., 2009). SHI et al. (2010) registraram estímulo da
fotossíntese por bactérias endofíticas em plantas com maior conteúdo de clorofila, maior
FV/FM
O estímulo à fotossíntese não garantiu promoção de crescimento às plantas tratadas
com inóculo A e não explica o impedimento da restrição de crescimento em plantas
IACSP95-5000 em RH tratadas com inóculo B. Nenhum demais parâmetros discutidos até
agora parecem explicar esse favorecimento do genótipo sensível em condição restritiva.
. Neste trabalho, os dados de clorofila total em plantas IACSP95-5000 tratadas com
inóculo A corroboram essa hipótese de estímulo da fotossíntese por ter mais clorofila do que
plantas controle (Tabela 4). No entanto, a maior quantidade de clorofila total não causou
aumento na atividade fotoquímica.

55
Observou-se que as plantas do genótipo sensível apresentaram maiores teores de ALT quando
em RH (Figura 12b) e a diferença ainda é maior quando sob o tratamento com inóculo B. A
princípio, o aumento de ALT poderia estar ligado à proteólise devido à suscetibilidade à seca
(ROY-MACAULEY et al., 1992). No entanto, o genótipo sensível tratado com inóculo B
apresenta manutenção de crescimento sugerindo que o aumento de ALT poderia estar ligado à
síntese de aminoácidos, fazendo com que o crescimento se mantenha.
A atividade das enzimas redutase do nitrato e sintetase da glutamina não foram
maiores quando o genótipo sensível tratado com inóculo B e, portanto, não explica a maior
concentração de ALT. Mas, com o tratamento de bactérias endofíticas diazotróficas, o
aumento de nitrogênio na planta pode ocorrer por maior estimulo na absorção de nitrogênio,
inclusive aumentando a quantidade de proteínas nas plantas (SAUBIDET et al., 2002), o que
poderia explicar a maior concentração de ALT. A eficiência do uso do nitrogênio de plantas
do genótipo sensível tratadas com inóculo B é maior quando em condição de restrição hídrica
(Tabela 8), o que coincide com o de ALT, indicando favorecimento dessas plantas sob esse
tratamento bacteriano.
6 CONCLUSÕES
Em condições de disponibilidade hídrica adequada, a interação planta-bactérias
endofíticas influenciou positivamente as trocas gasosas e o metabolismo do nitrogênio dos
genótipos de cana-de-açúcar IACSP94-2094 e IACSP95-5000, sem, entretanto, refletir em
promoção de crescimento das plantas.
O tratamento do genótipo de cana-de-açúcar sensível à seca com os isolados
bacterianos que compuseram o inóculo B favoreceu a manutenção do crescimento das plantas,
além de garantir melhor eficiência do uso do nitrogênio sob condições hídricas restritivas.

56
7 REFERÊNCIAS BIBIOGRÁFICAS
AGUIAR, D. A.; SILVA, W. F.; RUDORFFI, B. F. T.; SUGAWARA, L. M.; CARVALHO, M.A.; Expansão da cana-de-açúcar no Estado de São Paulo: safras 2003/2004 a 2008/2009. Anais do XIV Simpósio Brasileiro de Sensoriamento Remoto, Natal, Brasil, INPE, 25-30 de abril 2009. p. 9-16. AMARAL, L.I.V.; GASPAR, M.; COSTA, P.M.F.; AIDAR, M.P.M.; BUCKERIDGE, M.S. Novo método enzimático rápido e sensível de extração e dosagem de amido em materiais vegetais. Hoehnea, v.34, p. 425-431, 2007. ASHRAF, M.; FOOLAD, M. R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environmental and Expirimental Botany, v.59, p. 206-216, 2007 AZEVEDO, J. L.; Jr MACCHERONI, W.; PEREIRA, J. O.; ARAÚJO, W. L. Endophytic microorganisms: a review on insect control and recent advances on tropical plants. Electronic Journal of Biotechnology, v.3, n.1, 2000. Disponível em: <http://www.ejb.org/content/vol3/issue1/full/4> BALDANI, V.L.D. Efeito da inoculação de Herbaspirillum spp. no processo de colonização e infecção de plantas de arroz e ocorrência e caracterização parcial de uma nova bactéria diazotrófica. 1996. 262p. Tese (Doutorado) - Universidade Federal Rural do Rio de Janeiro, Seropédica. BALDANI, J. I.; CARUSO, L.; BALDANI, V. L. D.; GOI, S. R.; DÖBEREINER, J. Recent advances in BNF with non-legume plants. Soil Biology and Biochemistry, v. 29, p.911-922, 1997. BALDANI, J. I.; BALDANI, V. L. D. History on the biological nitrogen fixation research in graminaceous plants: special emphasis on the Brazilian experience. Anais da Academia Brasileira de Ciências, v.77, p. 549-579, 2005. BARBOSA, E.A.; PERIN, L.; REIS, V.M. Uso de diferentes fontes de carbono por estirpes de Gluconacetobacter diazotrophicus isoladas de cana-de-açúcar. Pesquisa Agropecuária Brasileira, v.41, p. 827-833, 2006. BARKA, E. A.; NOWAK, J.; CLÉMENT, C. Enhancement of chilling resistance of inoculated grapevine plantlets with a plant growth-promoting rhizobacterium, Burkholderia phytofirmans strain PsJN. Applied and Environmental Microbiology, v.72, p. 7246-7252, 2006. BARRETTI, P. B.; SOUZA, R. M.; POZZA, A. A. A.; POZZA, E. A.; CARVALHO, J. G.; SOUZA, J. T. Aumento da eficiência nutricional de tomateiros inoculados com bactérias endofíticas promotoras de crescimento. Revista Brasileira de Ciência do Solo, v. 32, p. 1541-1548, 2008. BARRETTI, P. B.; SOUZA, R. M.; POZZA, E. A. Bactérias endofíticas como agentes promotores de crescimento de plantas de tomateiros e de inibição in vitro de Ralstonia solanacearum. Ciência e Agrotecnologia, v. 32, p. 731-739, 2008.

57
BASHAN, Y.; de-BASHAN, L. E. Plant Growth-Promoting In: HILLEL, D., In Encyclopedia of soils in the environment. 1.ed, Oxford, 2005.v. 1, pp. 103-115. BIELESKI, R.L.; TURNER, A. Separation and estimation of amino acids in crude plant extracts by thin-layer electrophoresis and chromatography. Analytical Biochemistry, v.17, p. 278-293, 1966.
BODDEY, R.M.; URQUIAGA, S.; ASSIS, R.L.P.; DÖBEREINER, J. Fixação Biológica de Nitrogênio por bactérias associadas a cana-de-açúcar. Comunicado Técnico Embrapa/CNPBS, n.6, p. 1-5, 1992.
BOIERO, L.; PERRIG, D.; MASCIARELLI, O.; PENNA, C.; CASSÁN, F.; LUNA, V.; Phytohormone production by three strains of Bradyrhizobium japonicum and possible physiological and technological implications. Applied Microbiology Biotechnology, v.74, p. 874 - 880, 2007. BREMNER, J.M. Total nitrogen. In: BLACK, C.A., ed. Methods of soil analysis. Madison American Society of Agronomy, p. 1149-1178, 1965. BRUNINI, O. Zoneamento de culturas bioenergéticas no estado de São Paulo, Disponível em: <http://www.infobibos.com/Artigos/2007_3/zoneamento/index.htm>, Acesso em 09 dezembro 2011. BURRIS, R. H. Nitrogenases. The Journal of Biological Chemistry. v.266, p. 9339-9342, 1991. CARDOSO, E. J. B. N.; NOGUUEIRA. M. A. A Rizosfera e seus Efeitos na Comunidade Microbiana e na Nutrição de Plantas. In: SILVEIRA, A.P.D., FREITAS, S.D.S. Bactérias Diazotróficas Associadas a Plantas Não-Leguminosas. In: Microbiota do Solo e Qualidade Ambiental. 1.ed. Campinas:Instituto Agronômico, 2007. p.79-96. CAVALCANTE V A ; DÖBEREINER J A new acid tolerant nitrogen-fixing bacterium associated with sugarcane. Plant and Soil v.108,p. 23–31, 1988. CHAVES M. M. Effects of water déficits on carbon assimilation. Journal of Experimental Botany, v.42, p.1-16, 1991 CHAVES, M. M.; PEREIRA, J. S.; MAROCO, J.; RODRIGUES, M. L.; RICARDO, C. P. P.; OSÓRIO, M. L.; CARVALHO, I.; FARIA, T.; PINHEIRO, C. How plants cope with water stress in the Field? Photosynthesis and growth. Annals of Botany, v.89, p. 907-916, 2002. CHEN, M.; WEI, H.; CAO, J.; LIU, R.; WANG, Y.; ZHENG, C. Expression of Bacillus subtilis proAB genes and reduction of feedback inhibition of proline synthesis increases proline reduction and confers osmotolerance in transgenic Arabdopsis. Journal of Biochemistry and Molecular Biology, v.40, p. 396-403, 2007. CONAB, 2011. Companhia Nacional de Abastecimento. Ministério da Agricultura, Pecuária e Abastecimento. Disponível em: <http://www.conab.gov.br/conabweb/download/safra/cana> Acesso em 10 de dezembro de 2011.

58
COMPANT, S.; VAN DER HEIJIDEN, M. G. A.; SESSITSCH, A. Climate change effects on beneficial plant-microorganism interaction. FEMS Microbiology Ecology, v.73, p. 197-214, 2010. CORNIC, G. Drought stress inhibits photosynthesis by decreasing stomatal aperture – not by affecting ATP synthesis. Trends in Plant Science, v.5, p. 187-188, 2000. CREUS, C. M; SUELDO, R. J.; BARASSI, C. A. Water relations in Azospirillum inoculated wheat seedlings under osmotic stress. Canadian Journal of Botany, v. 76, p.238-244, 1998. CREUS, C. M; SUELDO, R. J.; BARASSI, C. A. Water relations and yield in Azospirillum-inoculated wheat exposed to drought in the field. Canadian Journal of Botany, v.82, p.273-281, 2004. DAVIES, W. J.; ZHANG, J. Root signals and the regulation of growth and development of plants in drying soil. Annual Review of Plant Physiology and Plant Molecular Biology, v.42, p.55-76, 1991. DIMKPA, C.; WEINAND,T.; ASCH, F. Plant–rhizobacteria interactions alleviate abiotic stress conditions. Plant, Cell and Environment, v.32, p.1682–1694, 2009. DINAKAR, C.; DJILIANOV, D.; BARTELS, D. Photosynthesis in desiccation tolerant plants: Energy metabolism and antioxidative stress defense. Plant Science, v. 182, p. 29-41, 2012. DÖBEREINER, J.; BALDANI, J.I.; BALDANI, V.L.D. Como isolar e identificar bactérias diazotróficas de plantas não leguminosas. Brasília: EMBRAPA, SPI.; Itaguaí: EMBRAPA, CNPAB, 60p. 1995. DONATO, V. M. T. S.; ANDRADE, A. G. de; SOUZA, E. S. de; FRANÇA, J. G. E. de; MACIEL, G. A. Atividade enzimática em variedades de cana-de-açúcar cultivadas in vitro sob diferentes níveis de nitrogênio. Pesquisa Agropecuária Brasileira, v.39, p.1087-1093, 2004. DU, Y.C.; KAWAMITSU, Y.; NOSE, A. ; HIYANE, S.; MURAYAMA, S. ; WASANO, K. UCHIDA,Y. Effects of Water Stress on Carbon Exchange Rate and Activities of Photosynthetic Enzymes in Leaves of Sugarcane (Saccharum sp.). Australian Journal of Plant Physiology, v. 23, p.719-726, 1996. DUBOIS, M.; GILLES, K.A.; HAMILTON, J.K.; REBERS, P.A.; SMITH, F. Colorimetric method for determination of sugars and related substances. Analytical Chemistry, v. 28, n.30, p. 350-356, 1956. FAROOQ, M.; WAHID, A.; KOBAYASHI, D.; FUJITA, D.; BASRA, S. M. A. Plant drought stress effects, mechanisms and management. Agronomy for Sustainable Development, v. 29, p.185-212, 2009. FLEXAS, J.; MEDRANO, H. Drougt-inhibition of photosynthesis in C3 plants: stomatal and non-estomatal limitations revisited. Annals of Botany, v. 89, p.183-189, 2002.

59
FOYER, C. H.; VALADIER, M.H.; MIGGE, A. ; BECKER, T. W. Drought-induced effects on nitrate reductase activity an mRNA and on the coordination of nitrogen and carbon metabolism in maize leaves. Plant Physiology, v.117, p. 283-292, 1998. FRANKS, P. J.; COWAN, I. R.; FARQUHAR, G. D. The apparent feedforward response of stomata to air vapor pressure deficit: information revealed by different experimental procedures with two rainforest trees. Plant, Cell and Environment, v. 20, p.142-145, 1997. FREITAS, R. de P. Bactérias diazotróficas endofíticas associadas à cana-de-açúcar. 2011. 77f. Dissertação (Mestrado em Agricultura Tropical e Subtropical) – Instituto Agronômico, Campinas. FUENTES-RAMÍRES, L. E.; CABALLERO-MELLADO, J.; SEPÚLVEDA, J.; MARTÍNEZ-ROMERO, E. Colonization of sugarcane by Acetobacter diazotrophicus is inhibited by high N-fertilization. FEMS Microbiology Ecology, v.29, p.117-128, 1999. GHANNOUM, O.; CONROY, J. P.; DRISCOLL, S. P.; PAUL, M. J. FOYER, C. H.; LAWLOR, D. W. Nonstomatal limitations are responsible for droughtinduced photosynthetic inhibition in four C4 grasses. New Phytologist, v. 159, p. 599-608, 2003. GHANNOUM, O. C4 photosynthesis and water stress. Annals of Botany, v. 103, p.635 – 644, 2009. GRAY, E. J.; SMITH, D. L. Intracellular and extracellular PGPR: commonalities and distinctions in the plant–bacterium signaling processes. Soil Biology and Biochemistry, v. 37, p.395–412, 2005. GROVER, M.; ALI, S. Z.; SANDHYA, V.; RASUL, A.; VENKATESWARLU, B. Role of microorganisms in adaptation of agriculture crops to abiotic stress. World Journal of Microbiology Biotechnology, v. 27, p1231-1240, 2011. HALLMANN, J.; QUADT-HALLMANN, A.; MAHAFFEE, W. F., KLOEPPER J.W. Bacterial endophytes in agricultural crops. Canadian Journal of Microbiology, v.43, p.895-914. 1997. HAN, H. S.; LEE, K. D. Plant growth promoting rhizobacteria effect on antioxidant status, photosynthesis mineral uptake and growth of lettuce under soil salinity. Research Journal of Agriculture and Biological Sciences, v.1, p. 210-215, 2005 HARDOIM, P. R.; VAN OVERBEEK, L. S.; VAN ELSAS, J. D. Properties of bacterial endophytes and their proposed role in plant growth. Trends in Microbiology, v.16, p. 463-471, 2008. HARTUNG, W.; SLOVIK, S. Physicochemical properties of plant growth regulators and plant tissues determine their distribution and redistribution: stomatal regulation by abscisic acid in leaves. New Phytologist, v. 119, p. 361-382, 1991. INMAN-BAMBER, N., & SMITH, D. Water relations in sugarcane and response to water deficits. Field Crops Research, v.92, p.185-202, 2005.

60
JAMES, E. K.’; GYANESHWAR, P.; MATHAN, N.; BARRAQUIO, W. L.; REDDY, P. M.; IANNETTA, P. P. M.; OLIVARES, F. L.; LADHA, J.K. Infection and Colonization of Rice Seedlings by the Plant Growth-Promoting Bacterium Herbaspirillum seropedicae Z67. The American Phytopathological Society, v. 15, p. 894-906, 2002. KASCHUK, G.; KUYPER, T.W.; LEFFELAAR, P.A.; HUNGRIA, M.; GILLER, K. E. Are the rates of photosynthesis stimulated by the carbon sink strength of rhizobial and arbuscular mycorrhizal symbioses? Soil Biology & Biochemistry, v. 41, p. 1233-1244. LANDELL, M.G. de A.; CAMPANA, M.P.; FIGUEIREDO, P.; VASCONCELOS, A.C.M. de; XAVIER, M.A.; BIDOIA, M.A.P.; PRADO, H. do; SILVA, M. de A.; DINARDO-MIRANDA, L.L.; SANTOS, A. da S. dos; PERECIN, D.; ROSSETTO, R.; SILVA, D.N. da; MARTINS, A.L.M.; GALLO, P.B.; KANTACK, R.A.D.; CAVICHIOLl, J.C.; VEIGA FILHO, A. de A.; ANJOS, I.A. dos; AZANIA, C.A.A.M.; PINTO, L.R.; SOUZA, S.A.C.D. Variedades de cana-de-açúcar para o Centro-Sul do Brasil: 15° liberação do programa Cana IAC (1959-2005).Campinas, IAC, 2005a. 33p.
LANDELL, M.G. de A.; CAMPANA, M.P.; FIGUEIREDO, P.; VASCONCELOS, A.C.M. de; XAVIER, M.A.; BIDOIA, M.A.P.; PRADO, H. do; SILVA, M. de A.; DINARDO-MIRANDA, L.L.; SANTOS, A. da S. dos; PERECIN, D.; ROSSETTO, R.; SILVA, D.N. da; MARTINS, A.L.M.; GALLO, P.B.; KANTACK, R.A.D.; CAVICHIOLl, J.C.; VEIGA FILHO, A. de A.; ANJOS, I.A. dos; AZANIA, C.A.A.M.; PINTO, L.R.; SOUZA, S.A.C.D. Variedades de cana-de-açúcar para o Centro-Sul do Brasil: 16° liberação do programa Cana IAC (1959-2007).Campinas, IAC, 2005b. 33p. LAWLOR, D. W. Limitation to Photosynthesis in Water-stressed Leaves : Stomata vs . Metabolism and the Role of ATP. Annals of Botany, v.89, p.871-885, 2002. LEE, B. R.; JIN, Y. L.; JUNG, W. J.; AVICE, J. C.; MORVANPBERTRAND, A.; OURRY, A.; PARK, C. W.; KIM, T. H. Water-deficit accumulates sugars by starch degradation – not by de novo synthesis – in white clover leaves (Trifolium repens). Physiologia Plantarum, v.134; p. 403-411, 2008. LIDDYCOAT, S.; GREENBERG, B. M.; WOLYN, D. J. The effect of plant growth-promoting rhizobacteria on asparagus seedlings and germinating seeds subjected to water stress under greenhouse conditions. Canadian Journal of Microbiology, v.55, p. 388-394, 2009. LOPES, M. S.; ARAUS, J. L.; VAN HEERDEN, P. D. R.; FOYER, C. H. Enhancing drought tolerance in C4 crops. Journal of Experimental Botany, v. 62, p. 3235- 3153, 2011.
MACHADO, R. S. Respostas fisiológicas de genótipos de cana-de-açúcar ao déficit hídrico imposto na fase inicial de desenvolvimento. 2009. 77p. Dissertação (Mestrado) – Instituto Agronômico de Campinas, Campinas.
MACHADO, R. S.; RIBEIRO, R. V.; MARCHIORI, P. E. R.; MACHADO, D. F. S. P.; MACHADO, E. C.; LANDELL, M. G. DE A. Respostas biométricas e fisiológicas ao deficit hídrico em cana-de-açúcar em diferentes fases fenológicas. Pesquisa Agropecuária Brasileira, v.44, p.1575-1582, 2009.

61
MAIA, P. S. P. ; OLIVEIRA NETO, C. F. de; CASTRO, D. da S.; FREITAS, J. M. N. de; LOBATO, A. K. da S.; COSTA, R. C. L. da. Conteúdo relativo de água, teor de prolina e carboidratos solúveis totais em folhas de duas cultivares de milho submetidas a estresse hídrico. Revista Brasileira de Biociências, v. 5, p. 918-920, 2007. MANTELIN, S.; TOURAINE, B. Plant growth-promoting bacteria and nitrate availability: impacts on root development and nitrate uptake. Journal of Experimental Botany, v. 55, p. 27-34, 2004. MATTOS, E. A. De. O estudo da fluorescência da clorofila e a detecção de estresses ambientais em plantas. In: PRADO, C. H. B. de A. & CASALI, C. A. (Ed.). Fisiologia Vegetal – Praticas em Relações Hídricas Fotossíntese e Nutrição Mineral. 1.ed. Barueri: São Paulo, 2006. 255-266. MAXWELL, K.; JOHNSON, G. N. Chlorophyll fluorescence – a practical guide. Journal of Experimental Botany, v. 51, n. 345, p. 659-668, 2000. MAYAK, S.; TIROSH, T.; GLICK, B. Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiology and Biochemistry, v.42, p. 565-572, 2004. MOLINARI, H. B. C.; MARUR, C. J.; DAROS, E.; CAMPOS, M.K.F.DE; CARVALHO, J. F. R. P.; BESPALHOK FILHO, J. C.; PEREIRA, L. F. R.; VIEIRA, L. G. E.. Evaluation of the stress-inducible production of proline in transgenic sugarcane (Saccharum spp.): osmotic adjustment, chlorophyll fluorescence and oxidative stress. Physiologia Plantarum. v. 130, p. 218-229, 2007. MONTEIRO, R. A.; BALSANELLI, E.; WASSEM, R.; MARIN, A. M.; BRUSAMARELLO-SANTOS; L. C. C.; SCHIMIDT, M. A.; TADRA-SFEIR, M. Z.; PANKIEVICZ, V. C. S.; CRUZ, L.M.; CHUBATSU, L. S.; PEDROSA, F. O.; SOUZA, E. M. Herbaspirillum- plant interactions: microscopical, histological and molecular aspects. Plant Soil DOI 10.1007/s11104-012-1125-7, 2012 MOREIRA, F. M. S.; SILVA, K.; NÓBREGA, R. S. A.; CARVALHO, F. Bactérias diazotróficas associativas: diversidade, ecologia e potencial de aplicações. Comunicata scientiae, v.1, p. 74-99, 2010. NIEDZWIWDZ-SIEGIEN, I.; BOGATEK-LESZCZYNSKY, R.; COME, D.; CORBINEAU, F. Effects of drying rate on dehydration sensitivity of excised wheat seedlings shoots as related to sucrose metabolism and antioxidant enzyme activities. Plant Science, v.167, p.879-888, 2004. OLIVEIRA, A. L. M.; URQUIAGA, S.; DÖBEREINER, J.; BALDANI, J. I. The effect of inoculating endophytic N2-fixing bactéria on micropropagated sugarcane plants. Plant and Soil. v. 242, p. 205-215, 2002. OLIVEIRA, A. L. M.; CANUTO, E. L.; REIS, V. M.; BALDANI, J. I. Response of micropropagated sugarcane varieties to inoculation with endophytic diazotrophic bacteria. Brazilian Journal of Microbiology. v. 34, p.59-61, 2003.

62
OLIVEIRA, A. L. M.; CANUTO, E. L.; URQUIAGA, S.; REIS, V. M.; BALDANI, J. I. Yield of micropropagated sugarcane varieties in diferent soil types following inoculation with diazotrophic bacteria. Plant and Soil, v. 284, p.23-32, 2006. PIMENTEL, C. 2004. A relação da planta com a água. EDUR - Editora Universidade Federal Rural do Rio de Janeiro. Seropédica, RJ. 191 páginas 1ª edição REIS, V. M.; TEIXEIRA, K. Fixação biológica de nitrogênio – Estado da arte. In. AQUINO, A. M.; LINHARES, A. Processos biológicos no sistema solo-planta, Brasília:Embrapa Agrobiologia, 2005, p.151-180. RENA, A.B.; MASCIOTTI, G.Z. The effect of dehydration on nitrogen metabolism and growth of bean cultivars (Phaseolus vulgaris L.). Revista Ceres, v.23, p. 288-301, 1976. ROBERTSON, M. J.; WOOD, A. W.; MUCHOW, R. C. Growth of sugarcane under high input conditions in tropical Australia. I. Radiation use, biomass accumulation and partitioning. Field Crops Research v.48, p. 11-25, 1996. RODRIGUES, J.D. Fisiologia da cana-de-açúcar. 1995. 99 p. Universidade Estadual Paulista- Instituto de Biociências- Botucatu. ROHÁCEK, K.; Chlorophyll fluorescence parameters: the definitions, photosynthetic meaning, and mutual relationships. Photosynthetica, v.40, p. 13-29, 2002. ROY-MACAULEY, H.; ZUILY-FODIL, Y.; KIDRIC, M.; PHAM THI, A.T.; VIEIRA DA SILVA, J. Effects of drought stress on proteolytic activities in Phaseolus and Vigna leaves from sensitive and resistant plants. Physiologia Plantarum, v. 85, p.90-96, 1992. RYAN, R. P.; GERMAINE, K.; FRANKS, A.; RYAN, D. J.;.DOWLING D. N. Bacterial endophytes: recent developments and applications. FEMS – Microbiology Letters. v. 278, p. 1-9, 2007. SALEEM, M.; ARSHAD, M.; HUSSAIN, S.; BHATTI, A. S. Perspective of plant growth promoting rhizobacteria (PGPR) containing ACC deaminase in stress agriculture. Journal of Industrial Microbiology Biotechnology, v.34, p.635–648, 2007. SARAVANAKUMAR, D; KAVINO, M.; RAGUCHANDER, T.; SYBBUAN, P.; SAMIYAPPAN, R. Plant growth promoting bactéria enhance water stress resistence in Green gram plants. Acta Physiologiae Plantarum, v. 33, p. 203-209, 2011. SAUBIDET, M. I.; FATA, N.; BARNEIX, A. J. The effect of inoculation with Azospirillum brasilense on growth and nitrogen utilization by wheat plants. Plant Soil, v. 245, p. 215-222, 2002. SHI, Y.; LOU, K.; LI, C. Growth and photosynthetic efficiency promotion of sugar beet (Beta vulgaris L.) by endophytic bacteria. Photosynthesis Research. v. 105, p.5-13, 2010. SILVA, M. de A.; JIFON, J. L.; SILVA, J. A. G. da; SHARMA, V. Use of physiological as fast tools to screen for drought tolerance in sugarcane. Brazilian Journal of Plant Physiology, v. 19, p. 193 – 201, 2007.

63
SILVA, M.A.; SILVA, J.A.G.; ENCISO, J.; SHARMA, V.; JIFON, J. Yield components as indicators of drought tolerance of sugarcane. Scientia Agricola, v.65, p.620-627, 2008. SINGELS, A.; DONALDSON, R.A.; SMIT, M.A. Improving biomass production and partitioning in sugarcane: theory and practice. Field Crops Research, v. 92, p. 291-303, 2005. SILVEIRA, J.A.G.; VIÉGAS, R.A.; ROCHA, I.M.A.; MOREIRA, R.A.; OLIVEIRA, J.T.A. Proline accumulation and glutamine synthetase activity are increased by salt-induced proteolysis in cashew leaves. Journal of Plant Physiology, v.160, p. 115-123, 2003. SILVEIRA, J.A.G.; MARTINS, M. de O.; LIMA A.B.; FERREIRA-SILVA, S. Redução de nitrato e assimilação da amônia em sistemas vegetais: mensuração de atividade enzimática e metabólitos. In: Figueiredo, M.V.B. (Ed.). Biotecnologia Aplicada à Agricultura: Textos de Apoio e Protocolos. 1ª ed. Brasília, 2010. p. 93-124. SOUZA, R. P.; MACHADO, E. C.; SILVA, J. A. B.; LAGÔA, A. M. M. A.; SILVEIRA, J. A. G. Photosynthetic gas exchange , chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environmental and Experimental Botany, v. 51, p.45-56, 2004. TAULÉ, C.; MAREQUE, C. ; BARLOCCO, C.; HACKEMBRUCH, F.; REIS, V. M.; SACARDI, M.; BATTISTONI, F. The contribution of nitrogen fixation to sugarcane (Saccharum officiarum L.), and the identification and characterization of part of the associated diazotrophic bacterial community. Plant Soil DOI 10.1007/s11104-011-1023-4, 2012 TRIVELIN, P.C.O. & VITTI, A.C. Manejo do Nitrogênio e enxofre na nutrição e adubação da cana-de-açúcar. CENA/USP, Piracicaba, SP.p.46.2009 TSAVKELOVA,E. A.; KLIMOVA, S.YU.; CHEDYNTSEVA, T.A.; NETRUSOV, A.I.; Microbial Producers of Plant Growth Stimulators and Their Practical Use: A Review Applied Biochemistry and Microbiology, v. 42, p. 117–126,2006a.
TSAVKELOVA,E. A.; KLIMOVA, S.YU.; CHEDYNTSEVA, T.A.; NETRUSOV, A.I. Hormones and Hormone-Like Substances of Microorganisms: A Review, Applied Biochemistry and Microbiology, v. 42, p. 229–235, 2006b.
UNIÃO DA INDÚSTRIA DE CANA-DE-AÇÚCAR (UNICA). Dados e Cotações – Estatísticas, Produção Brasil. 2008. Disponível em <http://www.unica.com.br/dadosCotacao/ estatistica/>. Acesso em: 08.dezembro 2011. URGUIAGA, S.; CRUZ, K.H.S. BODDEY, R.M. Contribution of nitrogen fixation to sugarcane: nitrogen-15 and nitrogen balance estimates. Soil Science Society of America Journal, v.56, p.105-114, 1992. URQUIAGA, S.; XAVIER, R. P.; DE MORAIS, R. F.; BATISTA, R. B.; SCHULTZ, N.; LEITE, J. M.; SÁ, J. M.; BARBOSA, K. P.; RESENDE, A. D.; ALVES, B. J. R.; BODDEY, R. M. Evidence from field nitrogen balance and 15N natural abundance data for the contribution of biological N2 fixation to Brazilian sugarcane varieties. Plant Soil DOI 10.1007/s11104-011-1016-3, 2012

64
VAN HANDEL, E. Direct microdetermination of sucrose. Analytical Biochemistry, Amsterdam, v. 22, p. 280-283, 1968. WELBAUM, G.E., MEINZER, F.C. Compartmentation of solutes and water in developing sugarcane stalk tissue. Plant Physiology, v 93, 1147–1153, 1990. WIEDENFELD, R.P. Water stress during different sugarcane growth periods on yield and response to N fertilization. Agricultural Water Management, v. 43, p. 173-182, 2000. YEMM, E. W.; COCKING, A. C. The determination of amino-acids winth ninhydrin. Analyst, v. 80, p. 209-214, 1955. YORDANOV, I., VELIKOVA, V., & TSONEV, T. Plant responses to drought and stress tolerance. Bulgarian Journal of Plant Physiology, Special Issue, p.187-206, 2003 YUWONO, T.; HANDAYANI, D.; SOEDARSONO, J. The role of osmotolerant rhizobacteria in rice growth under different drought conditions. Australian Journal of Agricultural Research, n. 56, p. 715-721, 2005.


















