
Joana Inês Pereira ECOLOGIA COMPORTAMENTAL EM FELINOS ...
120
i Universidade de Aveiro 2021 Joana Inês Pereira Ferreira Dias ECOLOGIA COMPORTAMENTAL EM FELINOS
Transcript of Joana Inês Pereira ECOLOGIA COMPORTAMENTAL EM FELINOS ...

i
Universidade de Aveiro
2021
Joana Inês Pereira
Ferreira Dias
ECOLOGIA COMPORTAMENTAL EM FELINOS

ii
Universidade de Aveiro
2021
Joana Inês Pereira
Ferreira Dias
ECOLOGIA COMPORTAMENTAL EM FELINOS
Dissertação apresentada à Universidade de Aveiro para cumprimento dos
requisitos necessários à obtenção do grau de Mestre em Biologia Aplicada,
realizada sob a orientação científica do Doutor Ulisses M. Azeiteiro, Professor
Associado com Agregação do Departamento de Biologia da Universidade de
Aveiro.

iii
o júri
presidente Prof. Doutora Maria Adelaide de Pinho Almeida
Professora Catedrática da Universidade de Aveiro
Arguente Doutor Mário Jorge Verde Pereira
Professor Auxiliar da Universidade de Aveiro
Orientador Doutor Ulisses Manuel de Miranda Azeiteiro
Professor Associado com Agregação da Universidade de Aveiro

iv
agradecimentos
À Universidade de Aveiro e ao Departamento de Biologia por me terem
aceite como estudante de licenciatura e de mestrado, e por tudo o que
tive a oportunidade de viver durante estes 6 anos como estudante
universitária.
Ao meu orientador, Prof. Doutor Ulisses M. Azeiteiro, por ter aceite este
desafio, e também pelo apoio, disponibilidade e pela imensa paciência
que demonstrou sempre.
A todo o staff do Centro Nacional de Reprodução do Lince Ibérico, em
especial às videovigilantes, por me terem aceite na sua equipa, pelo
apoio que me deram e por tudo o que me ensinaram.
Aos voluntários do CNRLI com quem tive o prazer de viver lado a lado
durante vários meses, especialmente às Voluncheerfuls Beatriz
Pessoa, Mónica Vidal e Diane Megias, pelas aventuras vividas em
grupo no Algarve e pelas fantásticas videochamadas durante os
confinamentos que me motivaram a terminar este projeto.
Aos meus amigos de sempre, Inês Ribas, Valéria Giménez, Daniel
Gomes, João Pinho, Eduardo Gomes, Pedro Costa, por me terem feito
perceber que é saudável afogar as mágoas em grupo, discutindo tudo o
que corre mal nas nossas teses, e fazer planos para quando
(finalmente!!) terminarmos o curso.
Aos meus pais, pelo fornecimento continuo de “chutos” motivacionais
para terminar este trabalho, de forma a ser livre de seguir com a minha
vida e concretizar os meus planos futuros.
Ao fotógrafo Sebastian Kennerknecht e ao Snow Leopard Trust pelas
fotografias que disponibilizaram para este trabalho.
Por fim, aos gatos dos vizinhos da frente, por me fazerem sorrir mesmo
nos piores dias, e por me fazerem desejar ter um gato só para mim.

v
palavras-chave
Etologia; Ecologia Comportamental; Personalidade Animal; Reintrodução
animal; Conservação; Leopardus jacobita; Lynx pardinus; Panthera uncia;
resumo
A diminuição da biodiversidade da Terra devido à atividade humana e, em
particular, o número de espécies em risco de extinção, está entre as
tendências ambientais críticas que marcam esta como uma era sem
precedentes na história do nosso planeta. Este estudo de revisão bibliográfica
visa determinar como a ecologia comportamental pode influenciar e auxiliar na
conservação de felinos, com foco em três espécies: Gato-Andino, Lince Ibérico
e Leopardo das neves. Esta investigação documental mostra que a integração
do comportamento animal, em escalas ecológicas maiores, torna mais
auspiciosa a aplicação bem-sucedida da pesquisa comportamental na
conservação. Além disso, demonstra que a restauração ecológica por meio da
reintrodução é uma das áreas mais promissoras para as contribuições de
pesquisas comportamentais. O trabalho conjunto de conservacionistas e
ecologistas comportamentais pode levar à criação de revisões e diretrizes
sistemáticas, completas e facilmente acessíveis, que permitirão o
desenvolvimento com base em evidências de diretórios de soluções
comportamentais reais e potenciais para problemas de conservação e gestão.

vi
keywords
Ethology; Behaviour Ecology; Animal Personality; Animal Reintroduction;
Conservation; Leopardus jacobita; Lynx pardinus; Panthera uncia;
abstract
The diminishing of the Earth's biodiversity due to human activity, and in
particular the number of species that are at risk of extinction, is among the
critical environmental trends that marks this as an unprecedented era in our
planet’s history. This bibliographic review study aims to determine how
behavioural ecology can influence and help in the conservation of felines, with
focus on three species: Andean Cat, Iberian Lynx, and the Snow Leopard. This
research shows that the integration of animal behaviour, at larger ecological
scales, makes the successful application of behavioural research in
conservation more auspicious. Additionally, it demonstrates that ecological
restoration through reintroduction is one of the most promising areas for
contributions from behavioural research. The joint work of conservationists and
behavioural ecologists can lead to the creation of systematic, complete, and
easily accessible reviews and guidelines that will allow evidence-based
development of directories of potential and real behavioural solutions to
conservation and management problems.

vii
Índice
Lista de Figuras viii
Lista de Tabelas ix
Introdução 1
1. Etologia 3
1.1. Ecologia Comportamental 6
1.1.1. Ecologia Comportamental em Felinos 11
1.2. Personalidade Animal 13
1.2.1. Personalidade em felinos 17
1.2.2. Avaliação da personalidade animal 22
1.2.2.1. Confiabilidade das classificações 23
1.3. Reintrodução 27
1.3.1. Reintrodução em Felinos 32
2. Família Felidae 36
2.1. Gato-Andino 40
2.1.1. Programas de Conservação 43
2.2. Lince Ibérico 45
2.2.1. Programas de Conservação 49
2.3. Leopardo das Neves 54
2.3.1. Programas de Conservação 58
3. O Futuro da Conservação Animal 61
Referências Bibliográficas 67
Anexo 1 – Proposta para estudo empírico de Personalidade Animal no Lince Ibérico 101
I. Classificação dos Traços de Personalidade 101
II. Codificação dos Traços de Personalidade 102
Notas importantes para este estudo: 107

viii
Lista de Figuras
Figura 1 - Gato-andino adulto. © Sebastian Kennerknecht ______________________________ 41
Figura 2 - Lince Ibérico, Espanha. © Sebastian Kennerknecht ____________________________ 46
Figura 3 - Leopardo das neves, Mongólia. © Snow Leopard Trust ________________________ 56

ix
Lista de Tabelas
Tabela 1 - Resumo dos trabalhos publicados sobre personalidade animal em felinos selvagens _ 20
Tabela 3 - Casos de estudo de reintrodução e translocação de felinos. ____________________ 32
Tabela 4 - Linhagens da Família Felidae (Adaptado de: Johnson & O’Brien, 1997; Johnson et al.,
2006; Werdelin et al., 2010) ______________________________________________________ 36
Tabela 5 - Estatuto de Conservação, Tendência Populacional e Distribuição Geográfica das
espécies de felinos (Retirado de IUCN, 2021). ________________________________________ 38
Tabela 6 - Projetos focados na conservação do lince ibérico na Península Ibérica (Retirado de LIFE
Program, 2021) _______________________________________________________________ 50

1
Introdução
A biodiversidade é uma questão central na biologia da conservação, pois é o motor que
impulsiona a evolução e permite que as espécies evoluam num planeta em constante mudança. A
biologia da conservação foca-se na prevenção da perda de diversidade devido às ações
antropogénicas. O comportamento de um animal é resultado da interação entre os seus genes e o
meio ambiente, e cumpre o papel de mediador entre estes dois elementos. Assim, é importante
que o comportamento animal seja um componente da biodiversidade e que deve ser considerado
na biologia da conservação. A capacidade dos animais de responder à atividade antropogénica
depende das suas capacidades de aprendizagem e da sua diversidade comportamental, em que a
incapacidade de resposta pode contribuir para a sua extinção. Como o comportamento é o
resultado das interações entre os genes e o meio ambiente, as mudanças no comportamento
podem ser usadas para avaliar os impactos antropogénicos no meio ambiente, e qualquer plano
de conservação deve ter isso em consideração.
Assim, a relevância do comportamento animal na conservação, foi um dos principais
fatores para a escolha desta área científica para a minha dissertação. O facto de a ONU ter
iniciado este ano a Década da Restauração dos Ecossistemas (2021-2030), também contribuiu
para esta decisão.
A escolha dos felinos para o foco deste trabalho foi muito fácil de fazer, pois desde criança
que sou fascinada por felinos, e também porque os felinos selvagens são o resultado de forças
evolutivas que trabalharam para os tornar num dos predadores mais eficientes do reino animal.
Pouco se sabe sobre muitas das espécies devido às dificuldades que os investigadores enfrentam
ao estudar esta família taxonómica. Além disso, muitas populações estão ameaçadas de extinção,
devido principalmente a ações antropogénicas, como caça e perda de habitat. A grande maioria
do trabalho científico em felinos é dedicado à compreensão da sua ecologia e comportamento, e
na aplicação desse conhecimento na recuperação e conservação das populações selvagens.
Focar este trabalho no Gato-Andino, no Lince Ibérico, e no Leopardo das neves, permitiu
estudar espécies de diferentes tamanhos e de 3 continentes diferentes, e comparar as suas
ecologias, ameaças e estratégias de conservação já implementadas. Estas espécies também
representam alguns dos desafios na investigação e conservação de felinos, em que o Gato-Andino
representa os pequenos felinos, espécies pouco estudadas e com lacunas de informação; o Lince
Ibérico, sendo um superespecialista trófico, com uma distribuição geográfica muito pequena, que

2
representa as espécies especialistas; e o Leopardo das neves a representar os grandes felinos,
com distribuições geográficas enormes que abrangem vários países e que, consequentemente
abrangem diferentes legislações e abordagens de conservação.
Assim, o grande objetivo desta dissertação é o estudo bibliográfico alargado da ecologia
comportamental e o papel que desempenha na conservação de felinos, com referência à ecologia,
e programas de conservação das 3 espécies escolhidas para casos de estudo, o Gato-Andino, o
Lince Ibérico, e o Leopardo das neves.
O segundo objetivo era o estudo empírico da Personalidade Animal no Lince Ibérico (Lynx
pardinus) no Centro Nacional de Reprodução do Lince Ibérico (CNRLI), situado em Vale de
Fuzeiros, em Silves. Até agora nunca foi realizado nesta espécie este tipo de estudo, pelo que este
pode ser o primeiro passo neste ramo de investigação científica. A replicação deste estudo em
todos os centros de reprodução permitirá obter informação a nível da espécie e não apenas das
populações. Se bem-sucedido, pode ser um fator a ter em conta na conservação deste felino.
Contudo, não foi possível realizar este estudo empírico devido à pandemia de Covid-19, pelo que
o Anexo 1 desta dissertação contém uma metodologia sugerida para a realização deste estudo, na
esperança de que o Anexo 1 seja útil no futuro para alguém com o mesmo objetivo.

3
1. Etologia
Charles Darwin (12 de fevereiro de 1809 – 19 de abril de 1882), durante os seus estudos
para compreender a evolução, seleção natural e o sucesso reprodutivo de diferentes animais,
incluindo o ser humano, chegou à conclusão de que o comportamento era uma componente
importante para os seus trabalhos em evolução biológica. No capítulo sobre “Instintos” de “On
the Origins of Species” (1859), Darwin questiona se, os traços comportamentais, tais como os
anatómicos, podem evoluir como resultado da seleção natural. Mais tarde, em 1872 publicou o
livro “The Expression of the Emotions in Man and Animal”, que se tornaria uma das maiores
influências para a etologia moderna. Tal como em muitas outras áreas da biologia, Charles Darwin
é considerado um dos fundadores deste ramo científico, a Etologia.
Contudo, foi no início do século XIX, que a etologia começou realmente a desenvolver-se.
Oskar Heinroth (1 de março de 1871 – 31 de maio de 1945), nos seus estudos com aves sobre
exibições comportamentais, sinais sociais e rituais entre membros da mesma espécie, afirmava
que o comportamento instintivo específico da espécie podia ser usado, como a morfologia, para
determinar as afinidades genéticas de diferentes táxon (Heinroth, 1910, 1924-1934 in, Taborsky,
2019). O seu trabalho viria a influenciar Konrad Lorenz (7 de novembro de 1903 – 27 de fevereiro
de 1989), que considerou o conceito de padrões de comportamento instintivo não como uma
hipótese a ser testada, mas como uma suposição fundamental. Tal como Heinroth, Konrad Lorenz
usou aves nos seus trabalhos, usando uma abordagem para realizar observações imparciais
desprovidas de qualquer hipótese, com as quais desenvolveu ideias e conceitos, tal como a
hipótese de “mecanismos inatos de libertação”, elaborando também um conceito introduzido por
Oskar Heinroth, à qual ele chamou imprinting (Lorenz, 1935).
Lorenz e outros etologistas da Europa trabalharam com questões que eram até então de
principal interesse para psicólogos, apesar das abordagens diferentes destes dois grupos. Ao
contrário dos psicólogos, os etologistas queriam saber mais sobre os comportamentos para além
da sua causa imediata (ver Lorenz, 1950), tendo como objetivo ensinar uma abordagem
comparativa e evolutiva, a Etologia Comparativa, e a necessidade de compreender o
comportamento de animais no seu habitat natural (Taborsky, 2019). Nikolaas Tinbergen (15 de
abril de 1907 – 21 de dezembro de 1988), um grande defensor desta abordagem, juntamente com

4
Lorenz, desenvolveu o conceito de instinto baseado numa organização hierárquica de ações,
assumindo como categorias chave de causalidade comportamental, a energia específica da ação,
os atos de consumo e os mecanismos inatos de libertação (Tinbergen, 1951).
Apesar dos interesses e opiniões em comum, Lorenz e Tinbergen tinham várias
divergências (Taborsky, 2019). Nomeadamente, Lorenz não realizava experiências sistemáticas,
possuindo um interesse filosófico nos mecanismos subjacentes ao comportamento,
principalmente nas causas do comportamento como resultado da interação de estímulos externos
e padrões de ações inatos e pré-programados, assim como no imprinting como um mecanismo
que provoca efeitos duradouros na origem e função das interações sociais. Em contraste,
Tinbergen preconizava uma abordagem experimental sistemática, principalmente em condições
de campo, formalizando o conceito de instinto, tentando estabelecer conexões mais próximas
entre a análise causal do comportamento e uma abordagem fisiológica. Foi Tinbergen que
percebeu que confinar a etologia ao estudo da causalidade, a tornaria uma disciplina demasiado
específica, pelo que ele queria que o valor da sobrevivência e a evolução fossem incorporados no
estudo sistemático do comportamento, em conjunto com a ontogenia e causalidade (Tinbergen,
1963).
Conhecidas por as quatro “questões de Tinbergen”, estas servem ainda hoje como uma
diretriz para o estudo do comportamento (Bateson & Laland, 2013; Taborsky, 2014). As “quatro
questões de Tinbergen” são: (1) Que função adaptativa, se houver, é que o comportamento
realiza? (2) Quais são os padrões evolutivos de comportamento? Como é que o comportamento
está relacionado com as filogenias? (3) Quais são os mecanismos hormonais e neurológicos que
medem o comportamento? e (4) Como é que o desenvolvimento ou a ontogenia afetam o
comportamento? Estas questões - profundamente enraizadas na biologia organizacional,
populacional e evolutiva - enfatizam a natureza abrangente e integrativa da disciplina de
comportamento animal.
Por várias décadas, Konrad Lorenz e Nikolaas Tinbergen, foram os responsáveis pelo
crescimento, desenvolvimento e fundação do estudo científico do comportamento. Em 1973,
estes etologistas, juntamente com Karl von Frisch (20 de novembro de 1886 – 12 de junho de
1982), receberam o Prémio Nobel em Fisiologia e Medicina por terem estabelecido Etologia como
uma disciplina científica. Foram responsáveis pela criação da primeira revista científica
“Ethology”1 em 1937, dedicada inteiramente a esta disciplina e, em 1952, organizaram a primeira
1 Edição de Junho 2021: https://onlinelibrary.wiley.com/toc/14390310/2021/127/6

5
“International Ethological Conference” que ainda se realiza a cada 2 anos. Devido ao seu trabalho
por todo mundo, surgiram grupos de investigação de etologia e de subdisciplinas, tal como a
Ecologia Comportamental.
A Ecologia Comportamental trouxe a unificação da investigação comportamental na
Europa e na América, apesar da história divergente desta ciência nos dois continentes (Taborsky,
2019). A consequente aplicação da teoria da evolução ao estudo do comportamento revolucionou
não só a etologia, mas toda a biologia. Um outro progresso importante no início da ecologia
comportamental foi o seu foco nos efeitos da aptidão dos traços biológicos, o que abriu caminho
para a análise de custo e benefício do comportamento (Taborsky, 2019). Apesar destes
desenvolvimentos positivos, o foco da ecologia comportamental sobre apenas o valor adaptativo
do comportamento, demonstrou a falta de um entendimento abrangente desta ciência, pois o
estudo da causa do comportamento passou a ser realizado por especialistas de outras áreas
científicas, como a neurofisiologia, endocrinologia, biologia do desenvolvimento e psicologia
experimental. Mesmo com o progresso científico nestas disciplinas, está ausente uma visão da
origem, causalidade, função e evolução do comportamento (Taborsky, 2019).
Recentemente, a etologia começou a recuperar o equilíbrio entre as várias abordagens à
disciplina (Kappeler, 2010; Rubenstein & Alcock, 2018) contudo, devido às conotações da palavra
“etologia”, a amplitude recuperada de investigação sobre o comportamento animal passou
recentemente a ser chamada de “Biologia Comportamental”, um termo na verdade já proposto
por Tinbergen (1963).
Etologia é definida como o estudo científico de comportamento animal, incluindo
comunicação animal, predação, defesa, agressão, acasalamento, imprinting, padrões de ação e
migrações, normalmente nas condições naturais dos indivíduos em estudo. Uma das principais
áreas da etologia é o estudo do instinto (comportamentos que os animais herdam e exibem sem
razão) versus aprendizagem (comportamentos que os animais desenvolvem durante a sua vida).
Existem várias subdisciplinas relacionadas com a etologia incluindo, neuro-etologia
(estudo de como o sistema nervoso de um animal produz comportamento), ecologia
comportamental (estudo de interações comportamentais entre indivíduos dentro de populações
e comunidades, geralmente num contexto evolutivo), sociobiologia (estudo dos aspetos
biológicos, especialmente ecológicos e evolutivos, do comportamento social em animais), biologia
da vida selvagem (abordagem multidisciplinar para o estudo de animais selvagens e dos seus
habitats), psicologia evolutiva (estudo do comportamento de indivíduos em relação às suas
origens evolutivas), psicologia comparada (estudo de semelhanças e diferenças na organização

6
comportamental entre os seres vivos), endocrinologia comportamental (estudo da interação
entre hormonas e comportamento), entre outras.
Existem três maneiras principais pelas quais o comportamento animal pode ser útil na
gestão da conservação (Berger-Tal et al., 2011). Primeiro, o comportamento pode ser usado como
um indicador de um problema de conservação, como o uso da diversidade do canto dos pássaros
num determinado fragmento de floresta para medir a diversidade de espécies. Em segundo lugar,
uma compreensão do comportamento pode ajudar a revelar a causa de um problema de
conservação. Por exemplo, compreender que as crias de tartarugas marinhas encontram o mar
naturalmente caminhando em direção às estrelas no horizonte, explica o porquê de elas se
desorientarem por causa das luzes artificiais na costa. Finalmente, o comportamento pode ser
usado como uma ferramenta de gestão da conservação. Por exemplo, evitar o contacto humano
direto durante períodos sensíveis do desenvolvimento, pode impedir que os animais criados em
cativeiro façam erradamente o imprinting nos humanos, permitindo que formem corretamente
preferências de acasalamento com a sua própria espécie. As potenciais aplicações para estes três
usos do comportamento variam consideravelmente e cobrem muitas das subdisciplinas do
comportamento animal.
A Etologia é importante para várias atividades e para o bem-estar animal como, por
exemplo, atividades de caça e pesca, investigação médica com animais, treino de animais,
veterinários e conservação animal. Ao compreender o comportamento natural das espécies,
podemos encorajar comportamentos desejáveis e desencorajar os indesejáveis e até manter um
animal selvagem calmo durante um encontro inesperado na natureza. Para proteger espécies
vulneráveis e em risco de extinção, é vital entender o comportamento natural destas espécies de
forma que, em ações de conservação de reprodução em cativeiro e reintrodução, os animais
mantenham o máximo de comportamentos selvagens para assegurar as melhores hipóteses de
sobrevivência na Natureza.
1.1. Ecologia Comportamental
Ecologia Comportamental e Sociobiologia são termos que designam abordagens para o
estudo do comportamento, na qual a investigação está normalmente centrada em como o
comportamento evoluiu por seleção natural, especialmente nas diferenças comportamentais
entre as espécies que surgiram em relação aos seus habitats naturais. Como a sociobiologia,
limitada apenas aos comportamentos sociais, está incluída no termo ecologia comportamental,

7
passou a ser comum substituir sociobiologia por este termo mais inclusivo, apesar de ambos
envolverem a integração de ideias de ecologia, evolução e etologia (Brown & Choe, 2019).
O autor do primeiro livro publicado com o termo “Ecologia Comportamental” no seu
título foi Klopfer (1962), o qual continha informações de assuntos selecionados, com ênfase em
aspetos comportamentais da ecologia de comunidades e, ainda, exemplos de ecologia
comportamental. Contudo, o primeiro livro focado apenas em ecologia comportamental foi
escrito por Brown (1975), seguido por uma série de livros com cobertura abrangente em dois
níveis, pós-licenciatura (Krebs & Davies, 1978) e licenciatura (Krebs & Davies, 1981). O
comportamento social dos pássaros recebeu muita atenção em todos estes livros, especialmente
porque a maioria dos autores e editores tinham alguma experiência em ornitologia.
Em 1976, foi publicada a primeira revista científica da disciplina, “Behavioral Ecology and
Sociobiology”2. Mais tarde, em Outubro de 1986, Jerram Brown, Thomas Caraco e Christopher
Barkan organizaram o primeiro International Behavioral Ecology Meeting, com palestras de 22
pessoas sobre uma variedade de tópicos de interesse para ecologistas comportamentais (Brown
& Choe, 2019). Na repetição deste encontro em 1988, esta nova sociedade, que ficaria conhecida
por International Society for Behavioral Ecology (ISBE), já tinha um nome, um boletim informativo
e planos para uma nova revista científica.
A Ecologia Comportamental surgiu na década de 1960, quando as análises de custo-
benefício do comportamento social se tornaram interessantes e quando as teorias de procura de
recursos e de parentesco foram introduzidas. Era, inicialmente, amplamente expressa em estudos
de biologia populacional do comportamento social, uma tendência refletida pelos livros de Alcock
(1975), Brown (1975) e Wilson (1975). Mais tarde, ao juntar o comportamento de procura de
alimento ao comportamento social, Krebs e Davies (1978) unificaram esta área científica sob a
estrutura geral da ecologia comportamental. Durante o mesmo período, ferramentas derivadas
da biologia molecular em rápido desenvolvimento começaram a ser usadas para responder a
questões sobre paternidade e evolução. Como resultado, a ecologia comportamental tornou-se
mais metódica, com ideias teóricas a serem demonstradas por conhecimentos detalhados das
relações genealógicas entre os animais nas populações (Brown & Choe, 2019).
Durante os últimos 35 anos, a Ecologia Comportamental tem organizado a maioria das
suas questões em torno do significado adaptativo do comportamento. Ecologistas
comportamentais fazem perguntas sobre a variação do fitness. Normalmente, estes fazem dois
2 Edição de Junho 2021: https://link.springer.com/journal/265/volumes-and-issues/75-6

8
tipos de perguntas: Qual é o significado adaptativo, se houver, desta ou daquela caraterística? E
Quais são as características que evoluem em resposta a esta ou àquela pressão de seleção?
Cientistas estão a tentar reinventar a forma como a investigação comportamental é
aplicada atualmente aos esforços de conservação. A forte estrutura teórica fornecida pela
ecologia comportamental fornece a base para uma abordagem hipotético-dedutiva para a ciência
da conservação. Integrar os quatro níveis de explicação do comportamento animal, causalidade,
desenvolvimento, utilidade adaptativa e história evolutiva, em escalas ecológicas maiores,
população, comunidades e ecossistemas, torna mais promissora a aplicação bem-sucedida da
pesquisa comportamental na conservação. O desafio para os cientistas do comportamento animal
tem sido passar da fase de implicações do comportamento de conservação para aplicações mais
ativas que resolvam problemas de conservação do mundo real.
O elemento-chave da ecologia comportamental é a natureza adaptativa do
comportamento. As estratégias comportamentais numa população são o resultado de processos
evolutivos que dependem do fitness de estratégias particulares sob condições ambientais
prevalecentes (Norris, 2004). Os comportamentos devem evoluir para maximizar a aptidão dos
indivíduos que apresentam esses comportamentos (Owens, 2006). No campo da ecologia
comportamental, são reconhecidos três domínios de comportamento chave que são
fundamentais para a obtenção de alta aptidão em indivíduos de todas as espécies e, portanto, são
de grande preocupação na conservação: (1) Padrões de movimento e uso do espaço, (2)
Comportamentos relacionados com interações predadores-presas e com procura de recursos, e
(3) Comportamento social e reprodução. Todos os diferentes comportamentos nestes domínios
afetam a sobrevivência e a reprodução das espécies, fornecendo assim informações valiosas
sobre a dinâmica populacional e comunitária.
Com estes três domínios, é possível identificar alguns ramos de investigação nos quais os
estudos de conservação orientados para o comportamento se devem concentrar. O primeiro é
“Impactos Antropogénicos no Comportamento Animal”, uma vez que estes podem ocorrer por
distúrbios humanos diretos, como superexploração de recursos e, por distúrbios indiretos, como a
introdução de espécies exóticas. Esses distúrbios podem afetar a aptidão animal dependente do
comportamento de duas formas, (1) quando o ser humano altera o ambiente, o valor de aptidão
das estratégias comportamentais existentes muda. Se o fitness for drasticamente reduzido e as
estratégias não forem suficientemente "plásticas" para responder à mudança ambiental, ou uma
resposta evolutiva ao ambiente alterado for lenta em relação à taxa de mudança ambiental, a
população irá decrescer (Norris, 2004). Ou (2), se o comportamento for plástico, ele pode ser

9
alterado pelas mudanças antropogénicas e, embora isso possa ser adaptativo a curto prazo, a
resposta comportamental pode alterar outros comportamentos relacionados com a aptidão,
como a estrutura social ou o sucesso de acasalamento, alterando assim a trajetória evolutiva das
espécies ou dos ecossistemas em questão, o que pode facilitar o desaparecimento de opções
evolutivas (Ehrlich, 2001), criando preocupações com a conservação numa escala de tempo
evolutiva mais longa (Manor & Saltz, 2003). Além disso, uma mudança no comportamento de
uma espécie pode alterar a dinâmica de toda uma comunidade ou ecossistema (Wright et al.,
2010).
Nos casos em que os impactos antropogénicos no comportamento animal levam a
preocupações com a conservação, as melhores soluções são, na maioria dos casos, esquemas de
gestão com base no comportamento, o que levam ao segundo ramo de investigação, “Gestão
Baseada em Comportamentos”. Aqui também, existem duas possibilidades de incorporar o
comportamento animal na gestão ativa da conservação. (1) O comportamento da espécie é
considerado na sua conservação nas decisões e nos protocolos desenvolvidos. As considerações
comportamentais podem desempenhar um papel crucial na criação e planeamento de áreas
protegidas e de corredores verdes (Schultz, 1998; Pe’er et al., 2004; Afonso et al., 2008),
epidemiologia da vida selvagem (Craft et al., 2009), e planeamento de reintroduções e
translocações (Saltz et al., 2000; Bar-David et al., 2005; Shier, 2006; Zidon et al., 2009).
Os objetivos imediatos da gestão baseada em comportamento têm, geralmente, uma
forte vertente demográfica, sejam eles de estabilizar ou aumentar o número de populações
pequenas ou em declínio, ou o de controlar populações de espécies invasoras. No entanto, nos
casos em que a mudança no comportamento do animal é a causa de preocupação para a
conservação, o objetivo imediato dos esforços de gestão pode ser mudar o comportamento da
população-alvo. Assim, em oposição à primeira possibilidade deste ramo, onde as decisões de
gestão são feitas com base no comportamento da espécie, na segunda via, (2) o gestor procura
mudar ou preservar o próprio comportamento, sendo uma abordagem comum usada no treino
de indivíduos em cativeiro, que serão reintroduzidos (McLean et al., 1996; Griffin et al., 2000;
Alberts, 2007).
Um outro ramo de investigação a considerar é “Indicadores Comportamentais”. Os vários
comportamentos adaptativos dos organismos fornecem uma grande quantidade de informações
sobre as forças evolutivas que moldam esses comportamentos, os ambientes em que os
organismos habitam e quaisquer mudanças recentes nas forças de seleção natural ou no
ambiente. Por isso, é possível usar o próprio comportamento como um indicador do estado do

10
organismo, bem como do estado do seu habitat (Kotler et al., 2007). Tais indicadores incluem
comportamentos de procura e uso de recursos (Whelan & Jedlicka, 2007), comportamentos de
mergulho (Mori et al., 2007), seleção de habitat (Heithaus et al., 2007) e uso de territórios (Owen-
Smith & Cain, 2007). Os indicadores comportamentais já foram usados na conservação para
fornecerem um alerta precoce para o declínio da população ou degradação do habitat antes dos
dados numéricos o evidenciarem (Searle et al., 2007; van Gils et al., 2009), e também para
monitorizar a eficácia dos programas de gestão, ou avaliar o seu sucesso nas fases iniciais, antes
que a população ou as respostas em nível de ecossistema sejam evidentes (Ikuta & Blumstein,
2003; Lindell, 2008).
Os três ramos de investigação aqui referidos estão fortemente ligados. Por exemplo, o
impacto antropogénico no comportamento animal pode ser detetado usando indicadores
comportamentais e pode sugerir a necessidade de uma gestão sensível do comportamento (Ikuta
& Blumstein, 2003; Zidon et al., 2009). Alternativamente, a gestão ativa sensível ao
comportamento pode ser avaliada usando indicadores comportamentais, e esse conhecimento
pode alterar o plano de gestão.
Estes ramos podem ser vistos como pontos de entrada para ecologistas comportamentais,
que têm o objetivo de usar o seu conhecimento e experiência do comportamento animal na
conservação. Em cada um dos temas, novos estudos devem ser baseados em conhecimentos
prévios do comportamento animal, bem como em questões urgentes de conservação. Desta
forma, o comportamento conservacionista pode servir como ligação entre o conhecimento de
ecologia comportamental e as necessidades mais práticas dos biólogos conservacionistas. Embora
ecologistas comportamentais possam, em muitos casos, abordar apenas um dos três ramos
propostos em qualquer estudo, o papel do biólogo conservacionista que enfrenta um problema
de conservação é considerar e integrar todos eles num esquema de gestão adaptativo.
Muitos tópicos de ecologia comportamental parecem promissores para aplicação em
conservação - incluindo dispersão e seleção de habitat, padrões de procura de recursos, escolha
de parceiros e respostas comportamentais à fragmentação de habitat e impactos antropogénicos.
No entanto, os esforços até agora têm se concentrado mais em comportamentos como dispersão
e procura de alimento do que em comportamentos anti-predadores, sociais ou de aprendizagem
(Berger-Tal et al., 2015). Além disso, o comportamento ainda é amplamente subutilizado, mesmo
em áreas de conservação onde deveria ser altamente aplicável. Por exemplo, embora mais de
30% dos artigos sobre reintroduções de animais publicados entre 1990 e 2005 relatassem
dificuldades decorrentes do comportamento, apenas 5% realmente mencionam comportamento

11
(Berger-Tal et al., 2015). Apesar da desconexão histórica entre conservacionistas e ecologistas
comportamentais, foram identificadas formas para melhorar a influência e a usabilidade do
conhecimento comportamental, tais como promover uma melhor comunicação com os gestores
da conservação, conduzindo investigações científicas com aplicações de gestão definidas, obtendo
uma maior compreensão das decisões de gestão da vida selvagem e oferecendo informações
facilmente acessíveis.
1.1.1. Ecologia Comportamental em Felinos
Em alguns aspetos, as 37 espécies da família Felidae abrangem uma grande diversidade,
por exemplo, em tamanho, coloração, vocalizações e nos habitats que ocupam. Ainda assim,
todos são felinos e as suas ecologias comportamentais são variações de um mesmo tema. Por
exemplo, geralmente vivem sozinhos, mas os leões (Panthera leo) e, sob algumas condições, as
chitas (Acinonyx jubatus) podem ser visivelmente sociais e cooperativos, contudo os indivíduos da
maioria das espécies de felinos são, geralmente, solitários.
Entre os carnívoros, há evidências de que as sociedades refletem as suas circunstâncias
ecológicas (Kruuk, 1976; Macdonald, 1983; Gittleman, 1989). Carbone et al. (1999) descobriram
que os membros desta ordem se enquadram em dois grandes grupos dietéticos: pequenos
carnívoros, que se alimentam de presas muito pequenas (invertebrados e pequenos vertebrados),
e grandes carnívoros que caçam vertebrados de grande porte, tendo em vista a maximização da
energia obtida pela presa caçada. Uma exploração dos tamanhos dos felinos e das suas presas
pode, portanto, fornecer informação para a compreensão das suas sociedades.
Não só o tamanho do corpo do felino afetará a presa que caça, mas também determinará
a quantidade de presa necessária para sustentá-lo. Uma vez que a distribuição geográfica e a
biomassa da presa serão determinadas pelos seus próprios tamanhos corporais, predadores
maiores geralmente requerem áreas geográficas maiores. Razões para isto podem ser a
correlação entre a produtividade ambiental negativa com tamanhos corporais maiores e a
partilha do seu território com outros membros da mesma espécie (Reiss, 1988; Jetz et al., 2004).
A ligação entre o tamanho da presa, do predador e da área geográfica pode ser parte de
um conjunto de fatores interativos que moldam as sociedades felinas. Não só
as diferentes necessidades energéticas de comer presas pequenas e grandes levam à
dicotomia mencionada, mas o tamanho da presa também é um aspeto da sua dispersão (em
áreas maiores ou menores) e este, entre outras medidas de dispersão de recursos, foi sugerido
como um determinante da organização social (Macdonald, 1983; Johnson et al., 2001).

12
A dispersão é o prelúdio necessário para a existência solitária que caracteriza o estilo de
vida felino. As crias são animais muito sociais até que ocorre o processo instintivo de dispersão.
Com este processo, os indivíduos jovens iniciam um distanciamento social da mãe e do resto da
sua ninhada, afastando-se do território materno à procura de território próprio, normalmente,
para territórios muito afastados da região natal (Macdonald et al., 2010b).
O processo de dispersão é muito perigoso, principalmente para os machos que costumam
viajar distâncias maiores do que as fêmeas, as quais se costumam estabelecer em áreas próximas
ao território materno (Macdonald et al., 2010b). Devido à procura ampla dos machos, estes
podem invadir inadvertidamente os territórios estabelecidos por indivíduos mais velhos e
envolverem-se em confrontos com o proprietário do território, que podem terminar em
ferimentos ou morte dos machos mais inexperientes.
O comportamento de caça requer um olhar fixo, demorado e concentrado. As técnicas de
caça são semelhantes em todas as espécies e envolvem espreitar através da vegetação, avanço
lento e, em seguida, um salto final para a presa, que pode ser horizontal ou vertical, capaz de
atingir distâncias notáveis (Macdonald et al., 2010b). O procedimento da caça felina pode ser
resumido numa série de ações: (1) caminhar, parar, observar, (2) avistamento, seguido por um
olhar fixo, (3) rastejar e perseguir, com movimentos lentos rentes ao solo, (4) parar e manter uma
posição perfeitamente imóvel e baixa, (5) movimentos leves do corpo e preparação física para
uma ação repentina, (6) movimento súbito para iniciar a corrida rapidamente, (7) atacar a presa,
por meio de um salto para cima desta e para a frente, (8) agarrar e apanhar a presa com as patas
dianteiras e a boca, (9) aperto fixo a segurar a presa enquanto está posicionado em cima dela, e
(10) matar, mordendo o pescoço, sufocando ou sacudindo a presa até ela ficar imóvel.
Depois da corrida ser iniciada, o felino não consegue parar ou mudar de direção
repentinamente. Com um limite energético para as corridas, a presa muitas vezes consegue
escapar se tiver a capacidade de se livrar do predador. Muitos ungulados, como veados e
antílopes, são capazes de se libertar devido à sua maturidade e aptidão, o que significa que
capturar estes animais pode ser difícil. Uma tentativa de captura bem-sucedida, pode resultar do
alvo ser mais fraco ou estar doente.
A predação é um meio de alimentação que apresenta um certo grau de incerteza, em que
cada alimentação requer uma busca pela presa e um grande esforço físico. O território abriga
recursos alimentares e é protegido como um lugar privado pelo felino que o adquiriu uma vez
que, as associações sociais significariam uma partilha de recursos restritos. Entre os felinos
selvagens, a fome é uma das causas de morte mais comuns (Fuller et al., 1985; Andrén et al.,

13
2006; Pereira et al., 2006, 2010). e episódios recorrentes de privação de comida podem servir
como motivação para preservar a posse exclusiva do seu território de sustento.
A posse de um território fixo é o fator de motivação dominante na vida de um gato.
Mesmo entre felinos com agrupamento social, como o leão, a defesa territorial é de grande
importância para todos os indivíduos. Um grupo coeso terá uma forte aversão a qualquer adulto
estranho que invada o território, evento que geralmente resulta em confrontos e numa nova
marcação territorial, especialmente por qualquer membro do sexo masculino do grupo
(Macdonald et al., 2010b). Embora a sobreposição territorial seja comum ocorrer entre um macho
estabelecido e algumas fêmeas individuais, a partilha territorial normalmente não se estende
além destas circunstâncias (Macdonald et al., 2010b).
Visto que felinos vivem geralmente vidas solitárias, a comunicação com outros indivíduos
é exclusivamente feita através da marcação de cheiros e sinalização das fronteiras do seu
território. A deposição de odores é feita por pulverização de urina, deposição fecal e fricção do
corpo. A função destes sinais territoriais é indicar o território de um indivíduo, em que os
invasores têm de determinar a sua resposta a estas marcações (Macdonald et al., 2010b). Uma
fêmea, por exemplo, não será dissuadida pelo cheiro de um macho, enquanto um macho adulto
reconhecerá que o território foi reivindicado por um indivíduo do sexo masculino e que a invasão
do território ocupado resultará numa resposta agressiva.
1.2. Personalidade Animal
O sucesso dos programas de reprodução em cativeiro não depende apenas da gestão da
população em cativeiro, mas também, do bem-estar dos indivíduos. O ambiente de cativeiro tem
um efeito profundo no comportamento de animais selvagens (Carlstead, 1996; Hosey, 2005;
Morgan & Tromborg, 2007) e os indivíduos variam nas suas respostas às variáveis ambientais
(Carlstead et al., 1999a; Jones & Gosling, 2005; Kuhar et al., 2006; Hill & Broom, 2009).
Vários trabalhos revelaram que os animais, tal como o ser humano, demonstram
diferenças comportamentais consistentes entre si e, por vezes, também entre populações (Powell
& Gartner, 2011). Considera-se que estas diferenças entre os indivíduos e populações são de
grande importância para entender como os animais tomam decisões, como é que eles interagem
com indivíduos intraespecíficos e interespecíficos e a evolução da população (Powell & Gartner,
2011). Também foi sugerido que estas diferenças podem ter influência na sobrevivência de
populações face a mudanças ambientais antropogénicas (McDougall et al., 2006) e na

14
probabilidade de que populações de algumas espécies possam ser restabelecidas na natureza
(Bremner-Harrison et al., 2004).
Estas diferenças têm várias denominações, tais como, Personalidade (Gosling & John,
1999) e Temperamento (Hansen & Møller, 2001), conceitos que tiveram origem numa variedade
de áreas tais como a psicologia, ecologia comportamental, biologia evolutiva, etologia e genética
de populações (Powell & Gartner, 2011).
Nos últimos vinte anos, surgiu uma área científica com trabalhos relacionados com o
estudo da personalidade animal e das suas implicações para a criação, gestão e bem-estar de
animais em cativeiro (Powell & Svoke, 2008). Este campo de investigação tem como objetivo
entender a variação entre os indivíduos e avaliar o bem-estar animal da perspetiva de um
indivíduo (Hill & Broom, 2009; Whitham & Wielebnowski, 2009; Whitham & Wielebnowski, 2013;
Watters & Powell, 2012).
Até agora foram descritas personalidades para uma grande variedade de animais,
incluindo mamíferos, pássaros, peixes, repteis, anfíbios, artrópodes e moluscos (Gosling, 2001;
Tetley & O'Hara, 2012).
Contudo, existe inconsistência na literatura em relação aos termos usados na descrição da
personalidade animal (Réale et al., 2007; Freeman & Gosling, 2010; Coleman, 2012). Muitos
investigadores referem-se a temperamento (Freeman et al., 2004), outros usam a expressão perfil
comportamental (Carlstead et al., 1999b; Carlstead et al., 2000) e outros ainda se referem a
diferenças individuais (Wielebnowski, 1999; Blumstein et al., 2006) ou distinção individual
(Carlstead et al., 1999a). Os diferentes termos são, por vezes, usados alternadamente no mesmo
trabalho (por exemplo, Blumstein et al., 2006).
Historicamente, foram feitas distinções na literatura de psicologia entre os termos
temperamento e personalidade com base na idade, em que o temperamento descreve diferenças
comportamentais em crianças e a personalidade descreve diferenças em adultos (Coleman, 2012;
Watters & Powell, 2012). Distinções adicionais entre os termos foram realizadas, com alguns
autores a argumentar que o temperamento tinha um elemento genético, ao contrário da
personalidade (Coleman, 2012). Além disso, alguns autores consideram o termo personalidade
muito antropomórfico para ser utilizado na literatura de comportamento animal (Gosling, 2008;
Weinstein et al., 2008; Meagher, 2009; Freeman & Gosling, 2010). No entanto, atualmente, há
pouca distinção entre os termos temperamento e personalidade (Coleman, 2012) e há poucas
evidências de que os resultados da pesquisa em personalidade animal sejam distorcidos pelo
antropomorfismo (Kwan et al., 2008; Weiss et al., 2012). Assim, apesar das divergências

15
anteriores na literatura, atualmente, o uso do termo personalidade parece ser geralmente mais
aceite (Gosling, 2008; Freeman & Gosling, 2010).
Assim, podemos definir o termo Personalidade como “diferenças individuais de
comportamento que são consideradas estáveis ao longo do tempo e das situações” (Freeman &
Gosling, 2010), e em que Traços de Personalidade são definidos como “comportamentos
envolvidos em diferenças de personalidade” (ex: Agressividade) (Sih et al., 2004).
A existência de diferenças de personalidade nas populações de animais tem implicações
numa ampla gama de aplicações (Conrad et al., 2011), tal como a reprodução e criação de animais
(ex: seleção de caraterísticas desejáveis de produção); bem-estar animal (ex: níveis de stress);
programas de reintrodução e biologia da conservação; gestão de espécies invasoras e controlo de
doenças (Wolf & Weissing, 2012). Em particular, ter em conta a combinação de tipos
comportamentais presentes numa população, pode ser crucial para prever como essa mesma
população responderá às mudanças ambientais antropogénicas (ex: mudança climática, perda ou
fragmentação de habitat e poluição) (Wolf & Weissing, 2012).
As diferenças de personalidade são importantes pois referem-se a variações altamente
estruturadas ao longo do tempo e em diferentes situações e contextos. Essa estrutura pode
desencadear consequências ecológicas e evolutivas que não seriam esperadas na presença de
variações menos estruturadas (Wolf & Weissing, 2012). Para além disso, vários traços de
personalidade estão ligados ao movimento no espaço e à resultante distribuição não aleatória e
estrutura de interação dos fenótipos, os quais podem ter consequências ecológicas e evolutivas
substanciais (Wolf & Weissing, 2012). As diferenças de personalidade também promovem o
surgimento de todos os tipos de outras diferenças ecologicamente e evolutivamente relevantes,
por exemplo, diferenças na história da vida, diferenças fisiológicas e neurobiológicas e diferenças
na disseminação de doenças (Wolf & Weissing, 2012). Por fim, as diferenças de personalidade são
uma dimensão importante, mas até agora, amplamente negligenciada da biodiversidade, que
afeta a estabilidade, resiliência e sobrevivência de populações, comunidades e ecossistemas
inteiros, tanto em escalas de tempo ecológicas como evolutivas (Wolf & Weissing, 2012).
Há um número crescente de evidências de que traços de personalidade são hereditários e
afetam o fitness e, portanto, têm consequências evolutivas (Weiss et al., 2000; Drent et al., 2003;
Réale & Festa-Bianchet, 2003; Dingemanse et al., 2004; Dingemanse & Réale, 2005; McDougall et
al., 2006; Réale et al., 2007; Smith & Blumstein, 2008). A seleção natural pode operar em traços
de personalidade (Réale & Festa-Bianchet, 2003; Dingemanse & Réale, 2005) e tem sido sugerido
que a variação na personalidade dos indivíduos leva a uma variedade de estratégias

16
comportamentais para lidar com estímulos desafiadores, o que resulta numa população viável
(Dingemanse & Réale, 2005; Réale et al., 2007).
Smith e Blumstein (2008) analisaram as consequências da personalidade no fitness e
encontraram 31 publicações que correlacionam personalidade com sucesso reprodutivo ou
sobrevivência. Os resultados indicaram que, em geral, indivíduos ousados tiveram melhor sucesso
reprodutivo que os indivíduos tímidos e que estes exibiram maior longevidade que os indivíduos
ousados. Tal pode ocorrer porque indivíduos ousados têm uma maior probabilidade de abordar
novos estímulos e menor probabilidade de evitar situações potencialmente perigosas (Bremner-
Harrison et al., 2004).
Os resultados de alguns estudos de personalidade em animais de zoos levaram os
investigadores a recomendar reduzir o medo, a fim de melhorar o sucesso reprodutivo (por
exemplo, Powell et al., 2008) e, uma vez que a personalidade é hereditária (Weiss et al., 2000;
Drent et al., 2003), existe o perigo de que a seleção artificial contribua para a adaptação a um
ambiente em cativeiro. A seleção destes comportamentos pode ser prejudicial à sobrevivência de
indivíduos em programas de reintrodução (McDougall et al., 2006). Existe, portanto, uma tensão
emergente relativa à relação entre personalidade e fitness em animais selvagens e em cativeiro.
Em cativeiro, o medo parece ser um preditor de sucesso reprodutivo reduzido (Wielebnowski,
1999; Powell et al., 2008; Smith & Blumstein, 2008) e stress crónico, caraterizado por
concentrações elevadas de corticoides fecais (Wielebnowski et al., 2002). Por outro lado,
indivíduos medrosos na natureza geralmente têm maior probabilidade de evitar predadores e,
portanto, as suas chances de sobrevivência são aumentadas (Bremner-Harrison et al., 2004;
Watters & Meehan, 2007). A monitorização sistemática da personalidade em cativeiro pode
fornecer informações importantes sobre os efeitos da criação e seleção de animais em cativeiro
(McDougall et al., 2006).
Os zoos são um excelente recurso para o estudo da personalidade animal, pois os
tratadores estão familiarizados com os seus animais e estas instituições oferecem aos
investigadores a oportunidade de conduzir investigações longitudinais sobre vários aspetos da
personalidade, incluindo a herança genética e os efeitos ambientais (Watters & Powell, 2012).
Existe, portanto, um grande potencial para que as avaliações de personalidade sejam
incorporadas nas práticas de gestão de zoos, a fim de melhorar o bem-estar e o sucesso
reprodutivo dos animais. No entanto, apenas recentemente, a avaliação quantitativa da
personalidade foi usada para investigar alguns dos desafios enfrentados na criação, gestão e bem-
estar de animais de cativeiro, o que forneceu novos caminhos de investigação sobre a avaliação

17
do bem-estar e a falha reprodutiva dos indivíduos. Além disso, vários autores identificaram a
necessidade de a avaliação da personalidade animal ser incorporada às práticas existentes de
gestão de zoos (McDougall et al., 2006; Whitham & Wielebnowski, 2009, 2013; Watters & Powell,
2012).
1.2.1. Personalidade em felinos
Na revisão de estudos de personalidade felina, Gartner e Weiss (2013a) encontraram
apenas 21 estudos publicados, dos quais 17 focados no gato doméstico (Felis silvestris catus).
Numa revisão semelhante de pesquisa de personalidade animal em zoos, mais de metade dos
artigos revistos (53%) tinham como foco primatas, sendo que, as espécies felinas foram objeto de
apenas cinco estudos (Tetley & O'Hara, 2012).
Um resumo dos trabalhos publicados sobre personalidade em felinos selvagens alojados
em cativeiro pode ser encontrado na Tabela 1, tendo sido, até ao momento, avaliada em nove
espécies: chitas (Wielebnowski, 1999; McKay, 2003; Baker & Pullen, 2013; Chadwick & O’Hara,
2014; Razal et al., 2016); leopardos-nublados (Wielebnowski et al., 2002; Fazio, 2010; DeCaluwe
et al., 2013; Gartner et al., 2014, 2016); gatos selvagens escoceses (Gartner & Weiss, 2013b;
Gartner et al., 2014); leopardos das neves (Gartner & Powell, 2012; Gartner et al., 2014, 2016);
tigres-de-bengala (Phillips & Peck, 2007); tigres Amur (Wang et al., 2019); tigres (Pastorino et al.,
2017a); Leão Africano (Gartner et al., 2014, 2016; Dunston et al., 2016) e Leão Asiático (Pastorino
et al., 2017b; Goswami et al., 2020). Todos estes estudos, à exceção de Dunston et al. (2016),
adotaram um método de classificação de traços de personalidade, através de questionários para
os tratadores. A validação dos resultados, quando realizada, foi feita a partir de observações
comportamentais e/ou testes com estímulos ou objetos novos para os animais, como espelhos ou
pessoas desconhecidas.
A personalidade felina foi descrita anteriormente como tendo até oito dimensões,
amplamente definidas como: “sociável”; “dominante”; “curioso”; “ativo”; “agressivo”; “calmo”;
"tímido" e "excitável" (Gartner & Weiss, 2013a). Embora diferentes investigadores usem termos
variados para descrever a personalidade felina, estas dimensões parecem replicar-se em
diferentes espécies felinas. Baker e Pullen (2013) encontraram uma dimensão de sociabilidade em
chitas, com altas cargas positivas na caraterística “brincalhão”. O medo tem sido descrito em
várias espécies, incluindo chitas (Wielebnowski, 1999) e leopardos da neve (Gartner & Powell,
2012). Wielebnowski (1999) encontrou uma dimensão denominada “tenso-medroso” em chitas.
Os indivíduos que obtiveram uma pontuação alta nesta dimensão foram classificados como mais

18
inseguros, tensos, com medo de outros indivíduos e de pessoas e menos seguros de si mesmos.
Da mesma forma, Gartner e Powell (2012) encontraram uma dimensão "tímida / ansiosa" nos
leopardos da neve, constituída pelas caraterísticas "ansioso", "medroso", "inseguro" e "tenso".
Trabalhos anteriores concentraram-se nas relações entre a personalidade e o sucesso
reprodutivo, atividade adrenal e bem-estar subjetivo. DeCaluwe et al. (2013) avaliaram as
personalidades dos leopardos-nublados do sexo masculino e correlacionaram os resultados com
observações comportamentais e níveis de corticoides fecais, a fim de avaliar a ansiedade e a
agressão. Os resultados mostraram que indivíduos classificados como mais ansiosos pelos seus
tratadores tinham concentrações mais altas de corticoides e exibiam mais comportamentos
indicativos de medo (por exemplo, comportamento de se esconder) do que indivíduos
classificados como calmos. Um estudo anterior descobriu ligações entre medo, comportamentos
estereotípicos, como andar de um lado para o outro, automutilação e concentrações de
corticoides fecais (Wielebnowski et al., 2002). Em conjunto, estes resultados indicaram ansiedade
crónica na população de leopardos-nublados em cativeiro, evidenciada pelas classificações dos
tratadores, observações comportamentais e análises hormonais. As recomendações resultantes
destes estudos para melhorar o bem-estar dos leopardos-nublados incluíram o fornecimento de
áreas elevadas e o aumento da altura do recinto (Wielebnowski et al., 2002), o uso da avaliação
da personalidade para prever pares apropriados de machos e fêmeas e, assim, a redução da
incidência de agressão intraespecífica (DeCaluwe et al., 2013). Este facto constitui uma barreira
para a reprodução em cativeiro bem-sucedida nesta espécie (Law & Tatner, 1998; Wielebnowski
et al., 2002; DeCaluwe et al., 2013).
No estudo sobre a personalidade dos gatos selvagens escoceses e o bem-estar subjetivo,
Gartner e Weiss (2013b) encontraram três dimensões da personalidade, denominadas
“dominância”, “agradabilidade” e “autocontrolo”. Os tratadores também foram capazes de
avaliar os indivíduos de maneira confiável num questionário subjetivo de bem-estar, o qual estava
relacionado à dimensão de personalidade de “autocontrolo”. Gartner e Weiss (2013b)
argumentaram que esta relação entre personalidade e bem-estar subjetivo poderia ter
implicações para a saúde e o bem-estar dos indivíduos.
Embora estudos diferentes usem métodos diferentes para avaliar a personalidade felina
(Tabela 1), incluindo adjetivos diferentes para descrever traços e nomes diferentes para os
componentes ou dimensões resultantes, parece que alguns traços de personalidade se repetem
em várias espécies felinas. Isto sugere que o desenvolvimento de um único questionário para
avaliar a personalidade felina no futuro pode ser útil. De facto, Watters e Powell (2012)

19
defendiam o uso de um questionário de personalidade única entre espécies semelhantes. O
desenvolvimento do Hominóide Personality Questionnaire (HPQ), que tem sido utilizado para
investigar a personalidade do chimpanzé e do orangotango (King & Figueredo, 1997; Weiss et al.,
2002; King et al., 2005; Pederson et al., 2005; Weiss et al., 2009), mostra que é possível usar o
mesmo questionário para avaliar a personalidade entre as espécies. Estes questionários devem
ser amplamente validados para garantir que avaliam, com precisão, os traços fundamentais de
personalidade de todas as espécies pretendidas (Tetley & O’Hara, 2012; Watters & Powell, 2012).

20
Tabela 1 - Resumo dos trabalhos publicados sobre personalidade animal em felinos selvagens
ARTIGOS ESPÉCIE AMOSTRA
SEXO
IDADE AMBIENTE
METODOLOGIA
M F D Classificações dos
tratadores
Observações
comportamentais
Testes com
estímulos/objetos novos
Baker e Pullen (2013) Chita 34 16 18 -
1,1-13,3 anos Zoo X
29 15 14 - X X
Chadwick e O’Hara
(2014) Chita 120 - - - - Zoo X
DeCaluwe et al.
(2013)
Leopardo-
nublado 16 16 - - 4-17 anos Zoo X X X
Dunston et al. (2016) Leão Africano 11 2 9 - Subadultos-
adultos
Reserva
cercada X X
Fazio (2010) Leopardo-
nublado 24 11 13 - 3-16 anos Zoo X X X
Gartner e Powell
(2012)
Leopardo das
neves 11 4 7 - 2-19 anos Zoo X X
Gartner e Weiss
(2013b)
Gato selvagem
escocês 25 9 14 2 1-15 anos Zoo X
Gartner et al. (2014)
Gato selvagem
escocês 25 9 14 2 1-15 anos Zoo
X
Gato doméstico 100 45 53 2 1 mês-19 anos Abrigo de
animais
Leopardo-
nublado 16 7 9 -
6 meses-9,58
anos Zoo
Leopardo das
neves 17 10 7 - 1-15 anos Zoo

21
Leão Africano 21 5 16 - 3-15 anos Zoo
Gartner et al. (2016)
Leopardo-
nublado 16 7 9 -
6 meses-9,58
anos Zoo
X Leopardo das
neves 17 10 7 - 1-15 anos Zoo
Leão Africano 21 5 16 - 3-15 anos Zoo
Goswami et al.
(2020) Leão Asiático 35 14 21 - 1,7-15,7 anos Zoo X X X
McKay (2003) Chita 41 - - - - Zoo X X X
Pastorino et al.
(2017a) Tigre 8 4 4 - 2-12 anos Zoo X X
Pastorino et al.
(2017b) Leão Asiático 3 0 3 - 5-7 anos Zoo X X
Phillips e Peck (2007) Tigre-de-bengala 7 6 1 - 2-9 anos Zoo X
Razal et al. (2016) Chita 17 8 9 - 1-12 anos Zoo X X
Wang et al. (2019) Tigre Amur 45 - - - 2-15 anos Zoo X X X
Wielebnowski (1999) Chita 44 19 25 - 3-13 anos Zoo X X
Wielebnowski et al.
(2002)
Leopardo-
nublado 72 36 36 - 3-18 anos Zoo X
Legenda: M - Masculino, F - Feminino, D- Desconhecido

22
1.2.2. Avaliação da personalidade animal
A personalidade animal pode ser avaliada usando dois métodos: classificação de traços
de personalidade por pessoas informadas (por exemplo, tratadores) e codificação do
comportamento dos animais (Gosling, 2001; Meagher, 2009; Freeman & Gosling, 2010; Highfill
et al., 2010; Watters & Powell, 2012). A personalidade animal em cativeiro é normalmente
avaliada através do uso de classificações dos observadores, em que as pessoas familiarizadas
com os animais são solicitadas a classificá-las em vários traços de personalidade (Tetley &
O'Hara, 2012). Isto geralmente envolve o uso de um questionário composto por uma lista de
adjetivos, por vezes acompanhados de uma definição para cada um, pedindo-se aos
avaliadores que classifiquem os indivíduos nesses adjetivos usando uma escala definida pelo
pesquisador (Dutton et al., 1997; Meagher, 2009).
A codificação consiste em observações de comportamento mais convencionais usando
etogramas e as observações são registadas e analisadas no contexto de traços de
personalidade (Gosling, 2001; Highfill et al., 2010). A maioria dos estudos que usam este
método regista o comportamento dos animais quando apresentados com novos objetos (Rouff
et al., 2005; Blumstein et al., 2006; Powell & Svoke, 2008) ou durante testes comportamentais
específicos (Uher et al., 2008). A codificação comportamental é considerada mais objetiva do
que as classificações dos tratadores, no entanto, os observadores que estão a codificar os
comportamentos tendem a usar a sua própria opinião para verificar se os comportamentos
que estão a observar são os definidos no etograma (Jones & Gosling, 2005; Meagher, 2009).
Isto é verdade para todos os trabalhos comportamentais mas, a confiabilidade e a
repetibilidade de estudos nos quais as ocorrências de comportamento são registadas,
raramente são questionadas (Jones & Gosling, 2005; Vazire et al., 2007; Meagher, 2009;
Highfill et al., 2010). Além disso, Vazire et al. (2007) descobriram que a classificação dos traços
era mais confiável do que a codificação comportamental numa avaliação da personalidade do
chimpanzé e sugeriram que as codificações comportamentais podem, de facto, ser difíceis de
medir com confiança.
Revisões anteriores de trabalhos sobre personalidade animal revelaram que a
codificação comportamental é o método mais comum de colheita de dados (Gosling, 2001;
Smith & Blumstein, 2008; Freeman & Gosling, 2010). De facto, Freeman e Gosling (2010)
descobriram que 89% dos estudos de personalidade de primatas usavam codificação
comportamental. No entanto, numa revisão da pesquisa de personalidade animal de zoos,
Tetley e O’Hara (2012) descobriram que 80% dos estudos usavam as classificações do tratador.

23
Assim, parece que a escolha do método na maioria das pesquisas sobre personalidade animal
em cativeiro, contrasta com as tendências metodológicas em outros estudos sobre
personalidade animal. O uso de questionários elimina a necessidade de os investigadores
visitarem todas as instituições participantes no estudo, aumentando o tamanho da amostra e
permitindo a recolha de dados em animais de várias coleções (Carlstead et al., 1999a;
Carlstead et al., 2000; Kuhar et al., 2006; Meagher, 2009). Por outro lado, a codificação
comportamental consome muito tempo e, na maioria das vezes, não é logisticamente possível
em estudos de cativeiro, dependendo do número de coleções participantes, pois requer
observações diretas do comportamento. Isso pode explicar a aparente tendência dos cientistas
em confiarem apenas nas classificações dos tratadores.
Como a personalidade é mais fortemente expressa quando os animais são
apresentados com novidades (Réale et al., 2007), observar e quantificar as reações dos animais
a ensaios de enriquecimento ambiental, podem fornecer informações sobre a personalidade
(Watters & Meehan, 2007; Powell & Svoke, 2008). Este método pode ser útil para avaliar
rapidamente traços de personalidade específicos com implicações na gestão e no bem-estar,
como o medo, em alguns indivíduos (Watters & Powell, 2012). No entanto, para que estas
avaliações sejam relevantes, é necessário primeiro identificar os componentes gerais da
personalidade de uma determinada espécie. Isso requer um estudo em larga escala com várias
instituições a participar, semelhante ao realizado por King e Figueredo (1997) em chimpanzés
e Wielebnowski (1999) em chitas, nos quais as classificações fornecidas por tratadores
experientes seriam essenciais para identificar caraterísticas complexas que podem não ser
facilmente distinguíveis usando apenas codificação comportamental.
1.2.2.1. Confiabilidade das classificações
Para que as informações fornecidas por um estudo de personalidade sejam úteis, a
avaliação da personalidade deve ser confiável e válida (Gosling, 2001; Gosling & Vazire, 2002;
Kuhar et al., 2006; Meagher, 2009; Freeman & Gosling, 2010). Os avaliadores que pontuam os
animais, ou codificadores que os observam, devem mostrar concordância nas suas avaliações
ou observações. Isto pode ser confirmado testando a confiabilidade dos observadores e entre
os observadores (Gosling, 2001; Gosling & Vazire, 2002; Martin & Bateson, 2007; Meagher,
2009). Portanto, é importante que o maior número possível de pessoas forneça classificações
para cada animal e que, aqueles que as fornecem, o façam independentemente e não
discutam as suas respostas com as restantes pessoas (Gosling, 2001).

24
O uso de classificações de tratadores para avaliar a personalidade tem sido criticado
por ser muito subjetivo, antropomórfico e pouco científico (Gosling & John, 1999; Gosling,
2001; Meagher, 2009; Weiss et al., 2012), pois exige que os tratadores usem o seu
discernimento para classificar os animais com base no seu próprio conhecimento dos mesmos
e do seu comportamento (Wemelsfelder, 1997; Gosling, 2001; Highfill et al., 2010). No
entanto, o aumento de evidências que sugerem que as avaliações dos observadores são
confiáveis e válidas, deu voz ao argumento de que este método é cientificamente credível
(Gosling, 2001; Meagher, 2009; Highfill et al., 2010). Além disso, existem poucas evidências
que sustentam a afirmação de que as classificações são contaminadas pelo antropomorfismo.
Kwan et al. (2008) encontraram pouca correlação entre classificações de auto-personalidade e
classificações de cães (Canis familiaris) fornecidas pela mesma pessoa, sugerindo que os
avaliadores não estavam a projetar as suas próprias caraterísticas nos seus animais de
estimação. Da mesma forma, Weiss et al. (2009) não encontraram diferenças transculturais
entre as classificações de personalidade do chimpanzé obtidas de observadores americanos e
japoneses, indicando que os antecedentes culturais e as experiências dos avaliadores não
influenciam as classificações. Assim, os preconceitos e expectativas dos avaliadores não
poderiam explicar as dimensões da personalidade provenientes das classificações (Weiss et al.,
2012).
As avaliações dos observadores têm sido usadas com grande efeito na avaliação do
bem-estar e da personalidade dos animais de criação (Hessing et al., 1994) e da personalidade
dos animais de companhia (Feaver et al., 1986; Hsu & Serpell, 2003). De facto, a avaliação da
personalidade em cães domésticos tem sido usada para indicar o quão adequados são para
papéis como cães-guia (Serpell & Hsu, 2001) e cães policiais (Slabbert & Odendaal, 1999) ou
como animais de estimação da família (Hennessy et al., 2001; Hsu & Serpell, 2003). Da mesma
forma, as classificações dos tratadores podem ser usadas para investigar o bem-estar e a
personalidade dos animais em cativeiro.
Para garantir que as dimensões da personalidade extraídas das classificações de traços
de personalidade sejam as mais precisas possíveis, os traços com baixa confiabilidade entre
avaliadores (ou seja, aqueles com os quais não conseguem concordar) devem ser excluídos.
Portanto, a confiabilidade entre avaliadores tem de ser examinada a priori. Testes comuns de
confiabilidade para este propósito incluem o Coeficiente de Concordância de Kendall (por
exemplo, Wielebnowski et al., 2002; Martin, 2005; DeCaluwe et al., 2013), o Coeficiente de
Correlação de Spearman (por exemplo, McKay, 2003; Dutton, 2008; DeCaluwe et al., 2013) e
correlações entre classes dentro de um Modelo Linear Geral ou GLM (por exemplo, King &

25
Figueredo, 1997; Weiss et al., 2007; Uher & Asendorpf, 2008). O teste de confiabilidade mais
apropriado dependerá do desenho do estudo, portanto, não é possível aos investigadores usar
uma única medida padrão de confiabilidade. Além disso, verificar se os testes de confiabilidade
são significativos é frequentemente problemático na pesquisa em zoos, onde as amostras
pequenas podem significar que os testes são insuficientes (Powell & Svoke, 2008) e propensos
a erros do Tipo II (Falsos-negativos). Estes erros ocorrem se o investigador não rejeitar uma
hipótese nula que é na realidade falsa na população (Martin & Bateson, 2007; Banerjee et al.,
2009). Apesar de não serem totalmente evitáveis, é possível reduzir a probabilidade de
ocorrerem aumentando o tamanho da amostra, em que quanto maior for a amostra, menor é
a probabilidade de que esta seja substancialmente diferente da população (Banerjee et al.,
2009). Os investigadores necessitam, por isso, de usar o seu próprio discernimento para
decidir se as classificações obtidas são confiáveis ou não.
Powell e Svoke (2008) argumentaram que pode levar muitos meses a trabalhar com
um animal em particular antes das classificações por um tratador serem confiáveis. Esta
questão foi abordada por King et al. (2005) no seu estudo comparativo da personalidade de
chimpanzés alojados em zoológicos e chimpanzés alojados num habitat naturalista num
santuário africano. Os tratadores do zoo conheciam os animais há 6,5 anos, enquanto os
funcionários do santuário tinham uma média de 6,9 meses de experiência com os animais. A
confiabilidade entre examinadores, avaliada por correlações entre classes (King & Figueredo,
1997), mostrou que a confiabilidade das classificações de um indivíduo era menor entre os
avaliadores do santuário (entre 0,17 e 0,51) do que os avaliadores do zoo (entre 0,43 e 0,76).
No entanto, cada chimpanzé do santuário foi classificado por uma média de 16,2 avaliadores,
de modo que a confiabilidade das classificações médias em todos os avaliadores foi maior
(entre 0,77 e 0,94). A experiência limitada dos avaliadores do santuário foi desta forma
mitigada pelo grande número de pessoas que forneceram as classificações (King et al., 2005).
A experiência dos avaliadores afeta claramente a sua capacidade de fornecer
classificações confiáveis (Dutton et al., 1997; King et al., 2005; Weiss et al., 2007). Os cientistas
devem, portanto, procurar obter informações sobre a experiência daqueles que fornecem
classificações e incluir a experiência do avaliador como fator nas suas análises (Carlstead et al.,
2000; Gosling, 2001; Meagher, 2009).
Estudos com várias instituições podem exigir que tratadores de diferentes países e
culturas forneçam classificações de personalidade animal, o que pode afetar a confiabilidade
dessas classificações. Dois estudos (King et al., 2005; Weiss et al., 2009) investigaram os efeitos
do idioma e da cultura dos avaliadores na confiabilidade das classificações. King et al. (2005)

26
traduziram o HPQ (Hominóide Personality Questionnaire) para francês de forma a permitir que
os tratadores de um santuário de língua francesa em África classificassem os seus animais. Isso
resultou em pequenas diferenças na estrutura de personalidade observada dos chimpanzés do
santuário pois dois fatores, Abertura e Emocionalidade, observados em chimpanzés alojados
em zoológicos, não se replicaram nos chimpanzés do santuário. Esta discrepância pode ter sido
causada pelo pequeno número de adjetivos que definem os fatores de abertura e
emocionalidade (King et al., 2005). No entanto, os autores também observaram que essas
diferenças podem ter sido causadas por inconsistências no questionário traduzido, uma vez
que nenhuma tradução no sentido inverso foi realizada para verificar se as definições dos
adjetivos fornecidos na versão em inglês do questionário eram as mesmas que as fornecidas
na versão francesa (King et al., 2005).
A classificação de personalidade de chimpanzé obtida por Weiss et al. (2009) num
santuário no Japão foram comparados com os obtidos por King e Figueredo (1997) na América
do Norte. Ao contrário da versão francesa do questionário (King et al., 2005), o questionário
japonês foi retraduzido para corrigir inconsistências. As classificações obtidas por Weiss et al.
(2009) eram tão confiáveis quanto as obtidas por King e Figueredo (1997) e também
resultaram em seis dimensões da personalidade, fornecendo evidências de que a
personalidade do chimpanzé pode ser avaliada com segurança por avaliadores de diferentes
culturas (Weiss et al., 2009). Assim, estas descobertas têm implicações importantes para
futuras avaliações de personalidade em zoológicos, que devem ser comparáveis e consistentes
quando as classificações são fornecidas por diferentes culturas de tratadores e em diferentes
idiomas.
O aumento de evidências em trabalhos sobre a personalidade animal em zoos
demonstra que os tratadores destas instituições são capazes de avaliar de maneira confiável os
traços de personalidade animal com base no seu conhecimento e observações de longo prazo
dos animais sob os seus cuidados (King & Figueredo, 1997; Carlstead et al., 1999a, b;
Wielebnowski, 1999; Wielebnowski et al., 2002; Kuhar et al., 2006; Whitham & Wielebnowski,
2009; Freeman et al., 2010; Horback et al., 2013). O uso de classificações de tratadores para
avaliar a personalidade facilita a colheita de dados de muitos indivíduos em diferentes
instituições, e as classificações de tratadores que trabalham em diferentes jardins zoológicos,
em diferentes países, com idiomas diferentes podem ser indicadores confiáveis da
personalidade animal (King & Figueredo, 1997; King et al., 2005; Weiss et al., 2009).

27
1.3. Reintrodução
Para combater e reverter as mudanças ambientais provocadas pelo ser humano, os
esforços de conservação envolvem-se em três tipos de ação: (1) Determinar qual o nível de
diversidade de espécies ou ecossistemas deve ser conservado, (2) Identificar e quantificar as
ameaças às referidas unidades de diversidade ou ecossistemas, e (3) Mitigação dessas ameaças
(Berger-Tal & Saltz, 2016).
Quantificar as unidades de biodiversidade para conservação pode envolver a
monitorização de populações de animais, o que requer recursos consideráveis, embora seja muito
auxiliada pelas tecnologias emergentes, como foto-armadilhagem e localizadores de GPS. Como o
comportamento afeta a detetabilidade, os etologistas desempenham um papel cada vez mais
importante nos esforços de investigação e monitorização. Enquanto isso, obter uma melhor
compreensão da capacidade de suporte, assim como, o tamanho da população que pode ser
sustentado por um determinado habitat, informa quais são os fatores ecológicos que podem
limitar o crescimento populacional de um animal, como comida, água, abrigo, refúgios de
predadores ou áreas para nascimento e desenvolvimento de crias, ao mesmo tempo que destaca
áreas onde fatores naturais podem ser controlados ou melhorados. Modificar os fatores que
limitam a capacidade de suporte pode alterar, de forma positiva ou não, o tamanho de uma
população (Swaisgood & Greggor, 2019).
Segundo Swaisgood e Greggor (2019), um método de último recurso para mitigar a perda
de espécies é resgatar indivíduos da natureza e colocá-los em programas de reprodução em zoos
e outras instalações apropriadas. Estas pequenas populações precisam de ser geridas devido à
diversidade genética, para evitar os efeitos adversos da endogamia e para preservar o máximo
possível do potencial evolutivo da espécie. Esta gestão, de acordo com os autores, combate a
deriva genética aleatória e, mais importante ainda, a seleção artificial que causa efeitos
semelhantes aos da domesticação em populações em cativeiro ou mesmo selvagens que vivem
em condições cada vez mais artificiais. O objetivo a longo prazo da reprodução para conservação
e outros programas de gestão de pequenas populações é manter um reservatório genético seguro
até que as condições na natureza possam ser melhoradas. Quando um habitat adequado é
encontrado ou criado, as populações podem ser restabelecidas através de programas de
reintrodução e translocação. A reintrodução de espécies-chave pode servir para restaurar a
integridade ecológica de áreas naturais, especialmente se as espécies reintroduzidas forem
indispensáveis ao ecossistema (Swaisgood & Greggor, 2019). Um exemplo disto é o corvo

28
havaiano, o único dispersor de sementes conhecido para muitas das plantas ameaçadas do Havaí
(Culliney et al., 2012), em que a sua reintrodução na natureza pode servir como uma forma de
incentivar o crescimento da floresta nativa, assim como proteger e aumentar as populações desta
espécie ameaçada.
Se uma espécie ameaçada de extinção for reproduzida com sucesso em cativeiro, isso não
significa necessariamente que a sua reintrodução na natureza será bem-sucedida. Apenas um
pequeno número de programas de reprodução em cativeiro e reintrodução foram considerados
bem-sucedidos, onde os animais soltos podem potencialmente formar populações sustentáveis
na natureza. Por esta razão, a reprodução em cativeiro e a reintrodução são frequentemente
consideradas como uma estratégia de último recurso, especialmente também porque pode ser
muito cara e demorada (Snyder et al., 1996). No entanto, várias espécies, como o lince ibérico,
foram resgatadas da beira da extinção devido a estes métodos. Os esquemas de reintrodução
bem-sucedidos requerem experiência de uma variedade de grupos, incluindo biólogos, decisores,
sociólogos e consultores organizacionais, podendo surgir problemas se a comunicação entre estes
grupos for insuficiente (Kelley & García, 2019).
É importante salientar que o sucesso limitado dos programas de reintrodução é
frequentemente atribuído a problemas comportamentais associados à reprodução em cativeiro.
Estes animais criados em cativeiro apresentam, frequentemente, carências associadas a
habilidades na procura de recursos, em evitar predadores e em interações sociais (Snyder et al.,
1996). Estas deficiências comportamentais podem surgir por meio da adaptação genética ao
ambiente cativo, domesticação, ou também como resultado de efeitos ontogenéticos de
desenvolvimento. Estes podem ser de particular interesse para gerir a criação dos animais, pois os
seus efeitos no comportamento animal podem ser mais facilmente revertidos (por exemplo,
através do enriquecimento ambiental e aprendizagem) do que aqueles resultantes da seleção não
intencional. Numa outra categoria está o comportamento de acasalamento, em particular a
escolha do parceiro, em que as preferências adaptativas os tornam mais exigentes quando se
trata de acasalar. Na maioria das espécies, é improvável que as fêmeas aceitem qualquer parceiro
potencial, pelo que a escolha de parceiros limita o sucesso dos programas de reprodução e
introdução de indivíduos em cativeiro, quando a disponibilidade de parceiros é limitada. A
perturbação dos mecanismos de escolha do parceiro pode explicar a razão da utilização de
estratégias de reprodução assistida, como a inseminação artificial, resultando, muitas vezes, em
menor sucesso reprodutivo (taxas de nascimento) do que eventos de acasalamento natural em
animais criados em cativeiro (Kelley & García, 2019).

29
A capacidade de manipular a dispersão de animais é particularmente útil em programas
de reintrodução e translocação, os quais forçam um evento de dispersão sobre os animais,
estejam ou não preparados. Uma vez que a maioria da mortalidade ocorre logo após a libertação
(Armstrong et al., 2017), a gestão comportamental da dispersão durante o período pós-libertação
pode ser crítica para determinar o sucesso ou o fracasso do programa.
A compreensão de como as experiências juvenis influenciam o uso posterior do habitat
também é uma área altamente relevante onde os etologistas podem contribuir para os esforços
de conservação. A “indução de preferência de habitat natal” ocorre quando os animais a dispersar
preferem novos habitats que contêm estímulos comparáveis aos do seu habitat natal. Tal
comportamento pode ajudar a explicar a razão pela qual os animais deslocados são
frequentemente propensos a viajar longas distâncias, apesar dos riscos (Stamps & Swaisgood,
2007). Compreender este fenómeno permite a implementação de novas abordagens em
programas de reintrodução, incluindo esforços para combinar o tipo de habitat nos locais de
captura e libertação, ou para manipular pistas conspícuas no habitat pós-libertação para coincidir
com aqueles do local de origem do animal.
Os mecanismos de escolha de parceiros também podem ser um obstáculo à reprodução
em pequenas populações, especialmente em programas de reprodução para conservação. Se os
animais em cativeiro não puderem exercer a escolha natural do parceiro, eles podem recusar-se
totalmente à reprodução. Por exemplo, no panda gigante, a taxa de cópulas foi quatro vezes
maior quando machos e fêmeas foram colocados com seus parceiros preferidos em comparação
com os pares com parceiros não preferenciais (Martin-Wintle et al., 2015). Acertar nestes pares
requer observações comportamentais cuidadosas para que se possa selecionar parceiros que
reflitam as preferências dos pandas. Na verdade, um dos mecanismos que regem a escolha do
companheiro de panda é a personalidade, em que certas combinações de personalidades são
mais propensas a procriar do que outras, sendo por isso possível melhorar o sucesso de
reprodução se forem recolhidas informações sobre as personalidades dos pandas (Martin-Wintle
et al., 2017).
Apesar de muitas outras organizações terem desenvolvido as suas próprias Diretrizes para
Reintroduções e outras Translocações para fins de Conservação, as diretrizes da IUCN são das
mais usadas mundialmente. As primeiras diretrizes da IUCN foram criadas em 1995 e publicadas
em 1998, tendo sido atualizadas nos anos seguintes (IUCN/SSC, 2013). A IUCN até agora, como
forma de reunir e estudar vários casos de estudos de reintrodução, publicou 7 edições de Global
Reintroduction Perspectives, a primeira em 2008 e a mais recente em 2021.

30
As diretrizes para reintrodução e translocações da IUCN abrangem vários tópicos a ter em
conta no planeamento, realização e pós-libertação dos indivíduos envolvidos (IUCN/SSC, 2013).
Primeiro, é necessário decidir se a reintrodução/translocação é uma opção viável, pois a
avaliação de qualquer proposta deve incluir a identificação de potenciais benefícios e impactos
negativos, incluindo aspetos ecológicos, sociais e económicos. Quando um alto grau de incerteza
permanece ou, não é possível estabelecer de forma confiável se uma proposta apresenta baixos
riscos, esta não deve prosseguir e soluções alternativas para conservação devem ser encontradas.
Segundo, ao planear uma reintrodução/translocação, são necessários objetivos bem
definidos, claros e específicos, garantindo que abordam todas as ameaças atuais identificadas ou
presumidas às espécies e indicando como é que as metas serão atingidas. Estas metas têm de
articular o benefício pretendido para a conservação e, muitas vezes, serão expressas em termos
do tamanho desejado e do número de populações que vão alcançar o benefício necessário para
conservação, localmente ou globalmente, tudo dentro de um prazo geral de tempo. O
planeamento deve seguir um processo lógico do conceito inicial ao design, viabilidade e avaliação
de riscos, tomada de decisão, implementação, monitorização, ajustes e avaliação.
Em terceiro lugar, o delineamento da translocação proposta estará sujeito tanto às
oportunidades como às limitações, e ambas irão influenciar a viabilidade da operação proposta,
sendo que a avaliação da viabilidade deve cobrir toda a gama de fatores biológicos e não
biológicos relevantes. A viabilidade biológica depende de fatores como o conhecimento biológico
da espécie, o habitat de introdução, os requisitos climáticos, os indivíduos fundadores (de fontes
de cativeiro ou selvagens), o bem-estar animal, os parasitas e as doenças potencialmente
associados. Para haver uma viabilidade social, é preciso ter em conta as circunstâncias
socioeconómicas, as atitudes e valores da comunidade, as motivações e expectativas, os
comportamentos e mudanças de comportamento e os custos e benefícios antecipados da
translocação. A compreensão destes é fundamental para o desenvolvimento de atividades de
relações públicas e para orientar o público em favor de uma translocação. Esta pode também
precisar de ter em consideração a disponibilidade de recursos financeiros, as exigências
regulamentares, a qualquer nível, internacional, nacional, regional ou sub-regional, assim como a
concordância dos proprietários dos terrenos, tanto em áreas propostas para libertação, como
áreas para as quais se podem deslocar posteriormente.
O quarto tópico a ter em conta é a avaliação dos riscos, em que a extensão da avaliação
de riscos deve ser proporcional ao nível de riscos identificados. Quando os dados são insuficientes
ou equívocos, a avaliação de riscos deverá ser apenas qualitativa, mas é necessária uma vez que a

31
falta de dados não indica ausência de riscos. Conclusões sobre a avaliação de riscos e estudos de
exequibilidade devem determinar se uma translocação deve ou não ocorrer. As principais
categorias de risco de uma translocação são: Risco às populações-fonte, Risco ecológico, Risco de
doenças, Risco associado à invasão por outras espécies, Risco de hibridização com espécies
aparentadas ou subespécies (desaparecimento de genes), Riscos socioeconómicos e Riscos
financeiros.
O quinto tópico é a implementação e libertação. A implementação de uma translocação
abrange mais do que a libertação dos organismos. Uma translocação, incluindo as realizadas em
áreas altamente adequadas, pode falhar devido a um fraco planeamento da libertação. A
implementação, portanto, deve considerar os aspetos que incluem os requerimentos legais,
participação do público, gestão do habitat, as fontes de obtenção e libertação de organismos,
intervenções e monitorização pós-libertação, selecionando locais e áreas de libertação, assim
como estratégias de libertação adequadas. À medida que os indivíduos libertados se estabelecem
na área de destino, uma ênfase deve ser dada à monitorização da população, ajustando a sua
gestão, tendo em conta os resultados de monitorização.
Por fim, o último fator a ter em conta é a monitorização e gestão contínuas a longo prazo,
em que a monitorização deve identificar novas ameaças à população inserida, que não faziam
parte do plano original de translocação. É essencial monitorizar a evolução da demografia dos
indivíduos libertados, os seus comportamentos, a ecologia das áreas de introdução, a genética, a
saúde e a mortalidade das populações libertadas, assim como a monitorização social, cultural e
económica das comunidades locais.
A restauração ecológica por meio da reintrodução é uma das áreas mais promissoras para
as contribuições de pesquisas comportamentais. As translocações de animais, sejam de animais
criados em cativeiro ou selvagens, requerem intensa pesquisa comportamental e gestão para
preparar os animais para a libertação num ambiente novo e, em muitos casos, monitorizar e
controlar o seu comportamento após a libertação. A maioria das reintroduções até ao momento
adotou uma abordagem bastante simplista para a gestão pós-libertação (Swaisgood & Greggor,
2019), baseando-se principalmente em práticas de libertação suave, como zonas de aclimatização
e fornecimento de alimentos e recursos hídricos a curto prazo. Os investigadores podem redefinir
o significado de “suave” nestes programas, melhorando estas práticas com outras mais relevantes
do ponto de vista ecológico, guiadas por uma forte estrutura teórica. Além disso, as ferramentas
do comportamento animal podem ser usadas para influenciar e treinar os indivíduos na pré-
libertação para melhorar e aumentar as suas hipóteses de sobrevivência pós-libertação.
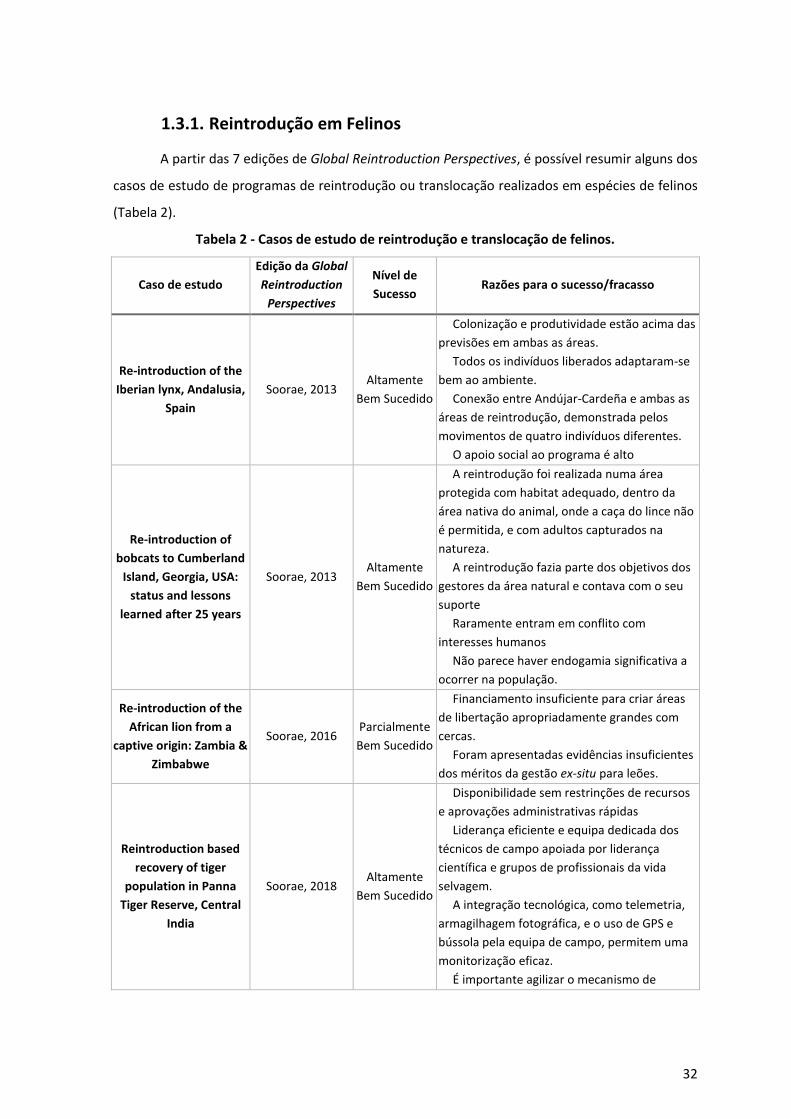
32
1.3.1. Reintrodução em Felinos
A partir das 7 edições de Global Reintroduction Perspectives, é possível resumir alguns dos
casos de estudo de programas de reintrodução ou translocação realizados em espécies de felinos
(Tabela 2).
Tabela 2 - Casos de estudo de reintrodução e translocação de felinos.
Caso de estudo
Edição da Global
Reintroduction
Perspectives
Nível de
Sucesso Razões para o sucesso/fracasso
Re-introduction of the
Iberian lynx, Andalusia,
Spain
Soorae, 2013 Altamente
Bem Sucedido
Colonização e produtividade estão acima das
previsões em ambas as áreas.
Todos os indivíduos liberados adaptaram-se
bem ao ambiente.
Conexão entre Andújar-Cardeña e ambas as
áreas de reintrodução, demonstrada pelos
movimentos de quatro indivíduos diferentes.
O apoio social ao programa é alto
Re-introduction of
bobcats to Cumberland
Island, Georgia, USA:
status and lessons
learned after 25 years
Soorae, 2013 Altamente
Bem Sucedido
A reintrodução foi realizada numa área
protegida com habitat adequado, dentro da
área nativa do animal, onde a caça do lince não
é permitida, e com adultos capturados na
natureza.
A reintrodução fazia parte dos objetivos dos
gestores da área natural e contava com o seu
suporte
Raramente entram em conflito com
interesses humanos
Não parece haver endogamia significativa a
ocorrer na população.
Re-introduction of the
African lion from a
captive origin: Zambia &
Zimbabwe
Soorae, 2016 Parcialmente
Bem Sucedido
Financiamento insuficiente para criar áreas
de libertação apropriadamente grandes com
cercas.
Foram apresentadas evidências insuficientes
dos méritos da gestão ex-situ para leões.
Reintroduction based
recovery of tiger
population in Panna
Tiger Reserve, Central
India
Soorae, 2018 Altamente
Bem Sucedido
Disponibilidade sem restrinções de recursos
e aprovações administrativas rápidas
Liderança eficiente e equipa dedicada dos
técnicos de campo apoiada por liderança
científica e grupos de profissionais da vida
selvagem.
A integração tecnológica, como telemetria,
armagilhagem fotográfica, e o uso de GPS e
bússola pela equipa de campo, permitem uma
monitorização eficaz.
É importante agilizar o mecanismo de
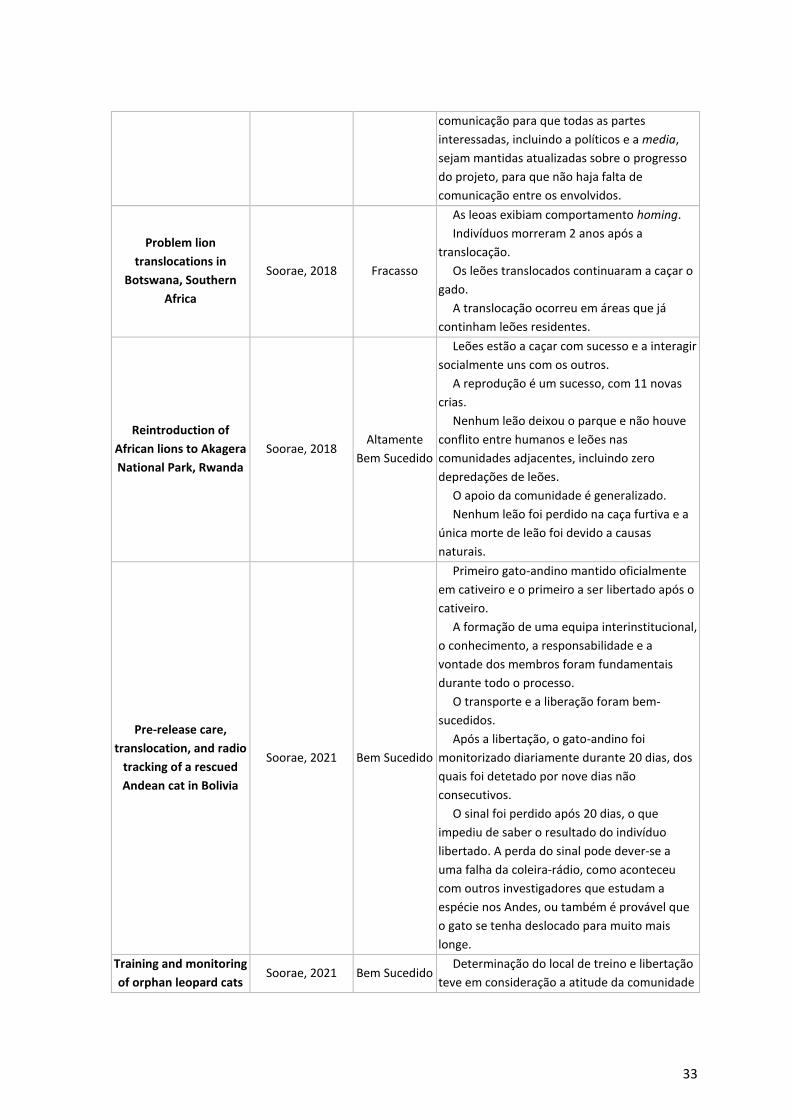
33
comunicação para que todas as partes
interessadas, incluindo a políticos e a media,
sejam mantidas atualizadas sobre o progresso
do projeto, para que não haja falta de
comunicação entre os envolvidos.
Problem lion
translocations in
Botswana, Southern
Africa
Soorae, 2018 Fracasso
As leoas exibiam comportamento homing.
Indivíduos morreram 2 anos após a
translocação.
Os leões translocados continuaram a caçar o
gado.
A translocação ocorreu em áreas que já
continham leões residentes.
Reintroduction of
African lions to Akagera
National Park, Rwanda
Soorae, 2018 Altamente
Bem Sucedido
Leões estão a caçar com sucesso e a interagir
socialmente uns com os outros.
A reprodução é um sucesso, com 11 novas
crias.
Nenhum leão deixou o parque e não houve
conflito entre humanos e leões nas
comunidades adjacentes, incluindo zero
depredações de leões.
O apoio da comunidade é generalizado.
Nenhum leão foi perdido na caça furtiva e a
única morte de leão foi devido a causas
naturais.
Pre-release care,
translocation, and radio
tracking of a rescued
Andean cat in Bolivia
Soorae, 2021 Bem Sucedido
Primeiro gato-andino mantido oficialmente
em cativeiro e o primeiro a ser libertado após o
cativeiro.
A formação de uma equipa interinstitucional,
o conhecimento, a responsabilidade e a
vontade dos membros foram fundamentais
durante todo o processo.
O transporte e a liberação foram bem-
sucedidos.
Após a libertação, o gato-andino foi
monitorizado diariamente durante 20 dias, dos
quais foi detetado por nove dias não
consecutivos.
O sinal foi perdido após 20 dias, o que
impediu de saber o resultado do indivíduo
libertado. A perda do sinal pode dever-se a
uma falha da coleira-rádio, como aconteceu
com outros investigadores que estudam a
espécie nos Andes, ou também é provável que
o gato se tenha deslocado para muito mais
longe.
Training and monitoring
of orphan leopard cats Soorae, 2021 Bem Sucedido
Determinação do local de treino e libertação
teve em consideração a atitude da comunidade

34
by soft release in Miaoli
County, north-western
Taiwan
local e a viabilidade do treino, localização via
rádio e monitorização.
Período suficiente de treino pré-libertação
na natureza, que poderia ser alcançado com
um cercado construído na natureza.
Adotadas as técnicas de libertação “suave” e
procedimentos detalhados.
Treinadores com conhecimento suficiente
em ecologia de gatos leopardo,
comportamento felino e experiência na
natureza.
Monitorização pós-libertação suficiente.
Todos estes casos de estudo tiveram níveis diferentes de sucesso, com apenas um
considerado como um caso de reintrodução falhado, nomeadamente, o da translocação de leões
para o Botswana. Contudo, todos enfrentaram problemas durante a preparação, realização e
monitorização destes programas de conservação, sendo o seu baixo financiamento e os elevados
custos para combater as ameaças a estas espécies (Soorae, 2013; Soorae, 2016; Soorae, 2018),
um dos problemas mais comuns. Também a perda de dados e informação sobre os indivíduos
envolvidos nas reintroduções, seja por mudanças nas equipas de investigação e monitorização, ou
devido à destruição inadvertida de amostras (Soorae, 2013), foi uma das dificuldades partilhadas
por alguns programas.
Para além destes, o apoio por parte das comunidades locais nem sempre foi favorável à
reintrodução destes predadores (Soorae, 2013; Soorae, 2018; Soorae, 2021), pelo que assegurar a
segurança dos locais de libertação e diminuir as ameaças como a caça ilegal, foram situações
difíceis de ultrapassar (Soorae, 2013; Soorae, 2016; Soorae, 2018).
Outros desafios que estes programas de reintrodução e translocação enfrentaram foram
de cariz biológico como, por exemplo, obter indivíduos fundadores (Soorae, 2018), a dispersão
dos animais para fora dos locais seguros de libertação (Soorae, 2018), permitir que os indivíduos
se familiarizem uns com os outros (leões) antes da sua libertação para assegurar a estabilidade
social (Soorae, 2016), manter uma população de presas capaz de sustentar os indivíduos
libertados, assim como, assegurar as futuras gerações (Soorae, 2018).
Por fim, os esforços de monitorização pós-libertação através de coleiras para
rastreamento e tecnologia de telemetria, foram dificultados devido à falta de experiência com
estas tecnologias, assim como pela dificuldade em obtê-las (Soorae, 2018; Soorae, 2021).
Contudo, a partir destes casos de estudo podemos retirar várias conclusões, que podem
ser vitais para futuros programas de reintrodução e translocação. A primeira, é que o apoio das

35
comunidades locais e das autoridades e governos envolvidos nos projetos são essenciais para
assegurar o seu sucesso, não só porque levarão à diminuição de ameaças como caça e comércio
ilegais, mas também, porque irá facilitar a obtenção de financiamento e aprovação dos programas
por parte das autoridades.
Em segundo lugar, apesar de as libertações “suaves” e “duras” terem funcionado para o
lince ibérico, é possível que, se tivesse sido feita uma libertação “suave”, através de um período
de cativeiro, para aclimatização, no local de libertação, teria sido possível prevenir a morte do
bobcat (Soorae, 2013). Por isso, é importante que os animais em cativeiro estejam em grandes
cercados que repliquem ao máximo o habitat onde serão libertados, com enriquecimentos
ambientais variados, de forma que estes não estejam sob stress e que não reajam negativamente
no momento de libertação. É fundamental manter os comportamentos naturais dos indivíduos
em cativeiro, assegurando que não desenvolvem comportamentos estereotípicos.
Por último, a monitorização dos indivíduos libertados é importante para determinar o que
acontece a cada um e garantir que o resto do ecossistema não sofre consequências negativas
como, por exemplo, a diminuição drástica das populações de presas, permitindo também verificar
o seu bem-estar e saúde.
Para além das espécies mencionadas na Tabela 2, outras espécies de felinos também já
foram alvos de programas de reintrodução ou translocação. Exemplos disso são o Leopardo
(Hayward et al., 2007; Kelly et al., 2013), o Lince-euroasiático (Breitenmoser & Haller, 1993;
Linnell et al., 2009), o Lince-canadiano (Wild et al., 2006; Devineau et al., 2010), e a Chita (van de
Meer et al., 2020).

36
2. Família Felidae
Os felinos fazem parte da Ordem Carnivora, mais especificamente da Família Felidae
(subfamília Felinae) (Johnson et al., 2006; Werdelin et al., 2010). Em termos paleogeológicos,
Felinae é relativamente recentemente e surgiu no final do Mioceno, há aproximadamente 13-14
milhões de anos atrás, com os géneros extintos e atualmente existentes da família Felidae
derivados de um ancestral comum, há cerca de 27 milhões de anos atrás (Werdelin et al., 2010).
Os primeiros esforços para organizar ou classificar os felinos em grupos basearam-se em registos
fósseis, semelhanças na aparência, nas caraterísticas dentárias, nas formas dos ossos dos ouvidos
e na sua distribuição geográfica. No entanto, com o avanço das tecnologias para estudar o DNA
das espécies, o conhecimento da evolução destes animais foi alterado, o que levou à criação da
primeira árvore genealógica da família Felidae (Johnson & O’Brien, 1997; Johnson et al., 2006;
Werdelin et al., 2010). Os geneticistas descobriram que as 37 espécies existentes se enquadram
em oito linhagens de espécies relacionadas, que evoluíram nos últimos 10 milhões de anos
(Tabela 3).
Tabela 3 - Linhagens da Família Felidae (Adaptado de: Johnson & O’Brien, 1997; Johnson et al.,
2006; Werdelin et al., 2010)
Lin
hag
ens
Panthera
ancestor
Lion Leopard Jaguar Tiger Snow
Leopard
Clouded
leopard
Sunda
clouded
leopard
Panthera leo Panthera pardus Panthera onca Panthera tigris Panthera
uncia
Neofelis
nebulosa
Neofelis
diardi
Bay cat
ancestor
Asiatic
golden cat
Bornean bay
cat Marbled cat
Catopuma
temminckii Catopuma badia
Pardofelis
marmorata
Caracal
ancestor
Caracal African golden
cat Serval
Caracal caracal Caracal aurata Leptailurus
serval
Ocelot
ancestor
Geoffroy’s
cat Guiña Oncilla Andean cat Pampas cat Margay Ocelot
Leopardus
geoffroyi
Leopardus
guigna
Leopardus
tigrinus
Leopardus
jacobita
Leopardus
colocolo
Leopardus
wiedii
Leopardus
pardalis
Lynx
ancestor
Iberian lynx Eurasian lynx Canada lynx Bobcat
Lynx pardinus Lynx lynx Lynx
canadensis Lynx rufus

37
Puma
ancestor
Puma Jaguarundi Cheetah
Puma concolor Herpailurus
yagouaroundi
Acinonyx
jubatus
Leopard Cat
ancestor
Leopard cat Fishing cat Flat-headed
cat
Rusty-
spotted cat Pallas’s cat
Prionailurus
bengalensis
Prionailurus
viverrinus
Prionailurus
planiceps
Prionailurus
rubiginosus
Otocolobus
manul
Domestic
ancestor
Domestic cat Wildcat Sand cat Black-footed
cat Jungle cat
Felis catus Felis silvestris Felis margarita Felis nigripes Felis chaus
Atualmente, de acordo com a Lista Vermelha de Espécies da IUCN (International Union for
Conservation of Nature and Natural Resources), 18 espécies de felinos estão nas categorias de
ameaçados, dos quais 5 estão classificadas como Em Perigo e 13 como Vulnerável. Das restantes
20 espécies, 6 apresentam-se como Quase Ameaçadas e as restantes 14 como Pouco
Preocupantes (Tabela 4). Foi apenas nesta última atualização dos estatutos que o felino mais
ameaçado do mundo até então, o Lince ibérico (Lynx pardinus), baixou na classificação de
Criticamente em Perigo para Em Perigo (Rodríguez & Calzada, 2015).
Com a exceção de dois continentes, Antártida e Oceânia, a família Felidae está presente
em todo o planeta, incluindo ilhas tais como Bornéu (no Oceano Pacífico) e Trinidad (no mar das
Caraíbas) (Macdonald et al., 2010a), vivendo numa grande variedade de habitats como florestas
boreais e tropicais, savanas, desertos, estepes e cadeias montanhosas. Algumas espécies de
felinos têm distribuições geográficas que incluem mais do que um continente (Tabela 4), como,
por exemplo, os leopardos (Panthera pardus) que estão presentes na Asia, Médio Oriente e
Africa, e o gato-selvagem (Felis silvestris) na Europa, Africa e Asia. No entanto, outras espécies são
altamente especializadas e, por isso, têm uma distribuição geográfica limitada a apenas alguns
países, como são os casos do gato-andino (Leopardus jacobita) e do lince ibérico, apresentados
mais à frente neste trabalho. As espécies mais especializadas, seja no habitat seja nas presas
consumidas, podem estar expostas a ameaças de origem antropogénica e/ou ambientais porém,
nem mesmo as espécies mais generalistas e com maior distribuição estão salvaguardadas destas
ameaças (Karanth & Chellam, 2009; Macdonald et al., 2010a).

38
Tabela 4 - Estatuto de Conservação, Tendência Populacional e Distribuição Geográfica das
espécies de felinos (Retirado de IUCN, 2021).
Espécie Estatuto de Conservação
(2021)
Tendência da
População
Distribuição Geográfica
(Continente)
African golden cat Vulnerável Decrescer África
Andean cat Em Perigo Decrescer América do Sul
Asiatic golden cat Vulnerável Decrescer Ásia
Black-footed cat Vulnerável Decrescer África
Bobcat Pouco Preocupante Estável América do Norte
Bornean bay cat Em Perigo Decrescer Ásia (Indonésia e Malásia)
Canada lynx Pouco Preocupante Estável América do Norte
Caracal Pouco Preocupante Desconhecida África, Ásia e Médio Oriente
Cheetah Vulnerável Decrescer África, Médio Oriente
Clouded leopard Vulnerável Decrescer Ásia
Eurasian lynx Pouco Preocupante Estável Ásia, Europa
Fishing cat Vulnerável Decrescer Ásia
Flat-headed cat Em Perigo Decrescer Ásia
Geoffroy’s cat Pouco Preocupante Estável América do Sul
Guiña Vulnerável Decrescer América do Sul
Iberian lynx Em Perigo Aumentar Europa (Península Ibérica)
Jaguar Quase Ameaçada Decrescer América Central e do Sul
Jaguarundi Pouco Preocupante Decrescer América Central e do Sul
Jungle cat Pouco Preocupante Decrescer África, Ásia, Médio Oriente
Leopard Vulnerável Decrescer África, Ásia, Médio Oriente
Leopard cat Pouco Preocupante Estável Ásia
Lion Vulnerável Decrescer África
Marbled cat Quase Ameaçada Decrescer Ásia
Margay Quase Ameaçada Decrescer América Central e do Sul
Ocelot Pouco Preocupante Decrescer América Central e do Sul
Oncilla Vulnerável Decrescer América do Sul
Pallas’s cat Pouco Preocupante Decrescer Ásia, Médio Oriente
Pampas cat Quase Ameaçada Decrescer América do Sul
Puma Pouco Preocupante Decrescer América do Norte, Centro e Sul
Rusty-spotted cat Quase Ameaçada Decrescer Ásia
Sand cat Pouco Preocupante Desconhecido África, Ásia, Médio Oriente
Serval Pouco Preocupante Estável África
Snow Leopard Vulnerável Decrescer Ásia
Sunda clouded
leopard Vulnerável Decrescer Ásia
Tiger Em Perigo Decrescer Ásia
Wildcat Pouco Preocupante Decrescer África, Ásia, Europa, Médio
Oriente
Apesar da variedade de tamanhos dos indivíduos da família Felidae, desde o tigre-
siberiano (Panthera tigris tigris) com 200-325 Kg até ao Rusty-Spotted Cat (Prionailurus
rubiginosus) com cerca de 1 Kg apenas (Macdonald et al., 2010a), a morfologia desta família

39
apresenta várias semelhanças entre as espécies. As cores e padrões da pelagem permitem aos
felinos esconderem-se e camuflarem-se com os habitats que os rodeiam, o que ajuda não só
durante a procura e caça das presas, mas também a evitar predadores e competidores (Kitchener
et al., 2010). Todos os sentidos destes animais são altamente desenvolvidos e adaptados ao seu
estilo de vida e para a localização e captura de presas, em que as vibrissas no focinho também são
importantes pois permitem navegar facilmente na escuridão noturna. A língua, com as suas
papilas em espinhos, é uma ferramenta para os felinos durante a sua alimentação porque é capaz
de arrancar a carne dos ossos das carcaças, além de servir como um pente durante a manutenção
e limpeza da pelagem (Kitchener et al., 2010). O género Panthera é o único que possui um
ligamento elástico em vez do osso hioide, que afasta a laringe da faringe permitindo a estes
felinos rugir (Kitchener et al., 2016).
Os felinos são carnívoros, pelo que os vertebrados são geralmente as presas escolhidas e
consumidas, sendo capturadas normalmente por emboscada. Possuem um corpo ágil, flexível e
com grande musculatura adaptado para os diferentes tipos de movimentos (saltos, corridas,
trepar), em que as caudas são 33-50% do comprimento total, salvo algumas exceções. As garras
curvas são retrateis, permitindo segurar as presas e os 28-30 dentes estão adaptados para matar
e rasgar a carne durante o consumo (Macdonald et al., 2010a).
À exceção dos leões, a maioria dos felinos são animais solitários que normalmente só
interagem com co-específicos durante a época de acasalamento e, para as fêmeas, durante a
criação e desenvolvimento das crias. Porém, a maioria das interações ocorre através das
marcações físicas e sensoriais deixadas como um aviso, dada à competição constante, por vezes
mortífera, por recursos, territórios e oportunidades reprodutivas (Macdonald et al., 2010a).
As ameaças e a mortalidade nos felinos estão associadas direta ou indiretamente a causas
antropogénicas. O comércio ilegal de peles e partes do corpo, em conjunto com a caça furtiva são
das maiores causas de mortalidade (Altrichter et al., 2006; Damania et al., 2008; Loveridge et al.,
2010). Situações de conflito com humanos, especialmente comunidades locais que partilham o
espaço com estes animais, são comuns (Inskip & Zimmerman, 2009; Karanth & Chellam, 2009)
devido, normalmente, à depredação de gado doméstico, em que as comunidades retaliam através
do envenenamento de engodos, armadilhas e caça furtiva (Breitenmoser et al., 2010; Loveridge et
al., 2010). No entanto, a maior ameaça é a perda e fragmentação dos habitats por todo o planeta
usados por estas espécies, em que todas as áreas protegidas e corredores verdes entre elas são
essenciais para a sua proteção e conservação (Weber & Rabinowitz, 1996)

40
Apesar da crescente bibliografia focada em felinos, existe ainda muito que não se sabe. As
grandes espécies, devido ao seu potencial como espécies bandeira e guarda-chuva na proteção de
ecossistemas, são normalmente as mais estudadas (Macdonald et al., 2010a). Ainda que a ênfase
nos grandes felinos contribua imenso para a sua conservação, as espécies mais pequenas e raras,
em especial da Asia e América do Sul, estão pouco estudadas pelo que muito se desconhece sobre
a sua ecologia (Macdonald et al., 2010a).
Neste trabalho são apresentadas 3 espécies de felinos (o Gato-andino, o Lince ibérico, e o
Leopardo das neves) como casos de estudo dos seus programas de conservação. Cada espécie é
ecologicamente caraterizada através da sua taxonomia, estatuto de conservação, distribuição
geográfica, morfologia corporal, dieta, habitat, estilo de vida, biologia reprodutiva, mortalidade e
ameaças. De seguida, todas as ações e planos de conservação, que foram até ao momento
realizados em cada uma destas 3 espécies, são descritas em pormenor.
2.1. Gato-Andino
O gato-andino (Leopardus jacobita (Cornalia, 1865)), também conhecido por gato-das-
montanhas-dos-andes, fazia até recentemente parte do género Oreailurus (Cabrera, 1940; Yensen
& Seymour, 2000) devido às diferenças de tamanho das cavidades auditivas encontradas numa
amostra pequena de indivíduos, apesar de acontecer o mesmo entre outras espécies de felinos
(Johnson et al., 1998; García-Perea, 2002). Contudo, estudos genéticos (Johnson et al., 2006)
colocam esta espécie no género Leopardus. Este é um dos únicos felinos para as quais não foram
descritas subespécies (Villalba et al., 2016).
Entre 1982 e 1990, o gato-andino estava classificado pela IUCN como Rara, sendo em
1996 alterado o seu estatuto de conservação para Vulnerável. Porém, desde 2002 que está
classificada com Em Perigo devido não só às baixas estimativas da quantidade de gatos-andinos
existentes em toda a sua distribuição geográfica (2755 indivíduos, em que 1378 têm maturidade
sexual), mas também por causa das várias ameaças a que esta espécie está sujeita (Villalba et al.,
2016).
Com uma distribuição geográfica irregular devido a uma especialização em habitats
rochosos naturalmente fragmentados, a maioria dos registos de presença do gato-andino são nos
Andes acima de 3.600 metros na Argentina (Perovic et al., 2003), Bolívia (Villalba et al., 2012),
Chile (Napolitano et al., 2008) e Peru (Cossíos et al., 2007). Existe, no entanto, registo desta
espécie a altitudes de 650m fora dos Andes, nas estepes da Patagónia e habitats de vegetação

41
rasteira na Argentina (Novaro et al., 2010; Martínez et al., 2008). Atualmente, o limite norte da
sua distribuição é no centro do Peru, enquanto o centro da Argentina é o limite sul (Cossíos et al.,
2007; Novaro et al., 2010). Um estudo da genética populacional desta espécie (Cossíos et al.,
2012) demonstrou que existem duas populações distintas, a população das grandes altitudes nos
Andes e a população nas estepes da Patagónia, tendo sido identificadas 10 possíveis
subpopulações (Villalba et al., 2016).
Esta espécie é pouco maior que um gato doméstico, com um peso entre os 3 e os 7 Kg
(Sunquist & Sunquist, 2014) e é caraterizada por uma pelagem densa de cor cinzento-claro ou
prateado, com riscas e manchas castanho-avermelhado e pretas no dorso, e uma pelagem clara
na zona ventral (Yensen & Seymour, 2000). Tem uma cauda longa e densa, cujo comprimento (41
a 48 cm) é cerca de 65% do comprimento total do felino, e tem 5 a 10 anéis escuros (García-
Perea, 2002; Sunquist & Sunquist, 2014). Os seus membros são marcados por 2 ou 3 barras pretas
e o seu rosto é pouco marcado, possuindo um nariz preto (Figura 1). A espécie exibe dimorfismo
sexual com os machos a serem ligeiramente maiores que as fêmeas (Tellaeche et al., 2018).
Figura 1 - Gato-andino adulto. © Sebastian Kennerknecht
A dieta do gato-andino é constituída principalmente por roedores sendo a chinchila-de-
cauda-curta (Chinchilla chinchilla) e a viscacha-da-montanha (Lagidium spp.) as suas principais
presas (Lucherini et al., 2009a; Napolitano et al., 2008), em que esta última é normalmente a mais

42
frequente. Outros animais, tais como ratos, roedores do género Phyllotis e indivíduos da família
Caviidae, assim como pássaros e carcaças de animais já foram identificados como parte da dieta
do gato-andino (Walker et al., 2007).
Tal como já foi referido, estes felinos estão presentes em locais a mais de 3000m de
altitude sendo por isso considerados especialistas de grandes altitudes. Os seus habitats
preferenciais são, geralmente, terrenos rochosos e ingremes e zonas áridas com pouca vegetação
nas áreas de grande altitude, enquanto na Patagónia são escolhidos habitats de estepes e com
vegetação rasteira (Napolitano et al., 2008; Novaro et al., 2010). Estes habitats são caraterizados
por temperaturas muito baixas, com uma grande variação térmica diária e pouca precipitação
(Villalba et al., 2016).
Lucherini et al. (2009b) mostraram, a partir de armadilhagem fotográfica, que o gato-
andino está mais ativo durante a noite (37,6%) e no crepúsculo (34,4%) do que durante o dia
(28%). Considera-se que este seja um felino solitário, que só se encontra com co-específicos para
reprodução (Villalba et al., 2004). Muito pouco se sabe sobre a biologia reprodutiva, o
desenvolvimento e crescimento das crias e a que idade atingem a maturidade sexual. Todavia, já
foram avistadas crias durante os meses da Primavera até aos de Outono (Villalba et al., 2016).
Atualmente, as principais ameaças a esta espécie são a perda e degradação de habitats,
caça dos felinos e das suas presas e a redução das populações das presas do gato-andino. As
alterações climáticas em conjunto com a expansão agrícola, gestão errada de gados e crescimento
das indústrias petrolíferas e mineiras estão a alterar o habitat ocupado por este felino (AGA,
2011; Villalba et al., 2016; Walker et al., 2014).
A caça ao gato-andino é um acontecimento frequente por ser considerado um predador
pelas populações locais (Lucherini & Merino, 2008; Novaro et al., 2010). Também existem registos
de caça deste felino para consumo e medicina tradicional no Peru (Cossíos et al., 2007) e, apesar
de ser considerado pelas tradições indígenas um animal sagrado na Bolívia, Peru e Chile,
espécimes embalsamados são guardados por estas populações por acreditarem que trazem maior
fertilidade aos seus gados e para usar nos festivais de colheitas (Cossíos et al., 2007; Villalba et al.,
2004, 2012).
A chinchila-de-cauda-curta foi provavelmente uma das principais presas do gato-andino,
tendo sido caçada quase até à extinção devido ao comércio das suas peles (Villalba et al., 2016).
Agora, a principal presa é a viscacha-da-montanha (Napolitano et al., 2008), que vive em
pequenas colónias e também está a sofrer pelo aumento da pressão de caça. A competição por

43
este roedor com outros predadores como o Pampas Cat pode afetar negativamente a
sobrevivência do gato-andino (Lucherini & Vidal, 2003; Reppucci, 2012).
2.1.1. Programas de Conservação
As espécies raras são de preocupação especial na conservação porque são
frequentemente mais propensas à extinção do que as espécies mais comuns (Dobson et al., 1995;
Yu & Dobson, 2000). A raridade também torna um desafio detetar estas espécies e estimar a sua
abundância e distribuição, pois constituem aspetos cruciais no desenvolvimento de estratégias de
conservação. Além disso, a falta de conhecimento de informações geográficas básicas dificulta a
gestão de programas de conservação, mesmo para espécies relativamente bem estudadas
(Anderson & Martínez-Meyer, 2004).
Primack et al. (2001) fez uma compilação das caraterísticas das espécies de carnívoros
que as tornam especialmente vulneráveis à extinção por causas antropogénicos sendo elas:
distribuições geográficas muito restritas, pequenas populações, diminuição do tamanho
populacional, baixa densidade populacional, grande tamanho do corpo que implica a necessidade
de grandes áreas para sobreviver, falta de dispersão efetiva, presença de migrações sazonais,
pouca variabilidade genética, requisitos de nicho especializado, associação com ecossistemas
antigos, distribuições geográficas agregadas, evolução isolada e estarem sujeitos à caça pelo ser
humano.
Os gatos-andinos podem ser enquadrados em algumas das categorias mencionadas
porém, também não existem informações sobre vários desses pontos sendo, por isso, importante
destacar a importância de continuar os estudos nesta espécie.
A espécie está listada como Criticamente Em Perigo na Bolívia, Em Perigo e Rara no Chile,
Em perigo no Peru e Vulnerável na Argentina (Villalba et al., 2016) contudo, ela tem proteção
total a nível nacional em toda a sua distribuição geográfica. Esta contradição deve-se ao facto de a
aplicação da lei ser problemática e os espécimes caçados serem, regularmente, observados no
campo e à venda, embora com menos frequência do que antes, em mercados especiais para
serem usados em cerimónias religiosas.
As legislações aplicáveis são (Villalba et al., 2016):
• Argentina: Lei Nacional 22421 de Conservação da Vida Selvagem e o seu Decreto
Estatutário 666/97, e também Resolução Nº 63/86 da Secretaria de Agricultura;

44
• Bolívia: Decreto Supremo Nº 22641, promulgado em 1990, que estabelece uma
proibição geral e indefinida de perseguição, captura, armazenamento e
acondicionamento de animais silvestres e derivados;
• Chile: Lei Nº 19473, de 1972, em que a caça ilegal de felinos no Chile é penalizada
com multas até US 6000$ e prisão até 3 anos;
• Peru: Decreto Supremo Nº 004-2014-MINAGRI, em que o comércio, caça, captura,
posse, transporte ou exportação para fins comerciais é proibido.
Após desenvolver as ações delineadas no Plano de Ação para a Conservação do Gato-
Andino (Villalba et al., 2004) e as informações obtidas posteriormente, a Aliança Gato-Andino
elaborou o Plano Estratégico para a Conservação do Gato-Andino (AGA, 2011), mantendo as
principais linhas de ação (Investigação, Educação Conservação e Gestão), com os seguintes
objetivos:
• Assegurar a conservação a longo prazo do gato-andino e do seu habitat natural,
incluindo a restauração ou reabilitação dos que estão degradados;
• Integrar a conservação do gato-andino e do seu habitat natural nas políticas locais
dos quatro países onde a espécie está presente, trabalhando localmente, mas
com uma abordagem global;
• Fortalecer as atividades de conservação e investigação em áreas protegidas com
populações de gatos-andinos e promover a criação ou extensão de áreas
protegidas existentes para fornecer conectividade e/ou proteção de habitats e
populações de gatos-andinos;
• Promover a investigação científica sobre as ameaças à conservação, os requisitos
ecológicos da espécie, das suas principais presas e outros carnívoros simpátricos;
• Padronizar as atividades dos grupos de trabalho sob objetivos comuns e mais
eficazes;
• Treinar equipas de áreas protegidas e comunidades locais em atividades de
investigação, educação e conservação, dentro e fora destas áreas.
Foram desenvolvidos, com a participação das comunidades e ONGs locais, e agências
governamentais dos quatro países, workshops que ajudaram a promover medidas de conservação
mais eficazes (AGA, 2011). Um grande avanço foi a assinatura de um Memorando de Acordo para
colaboração internacional entre as autoridades de Biodiversidade e Áreas Protegidas da
Argentina, Bolívia e Chile, patrocinado pela AGA e assinado em Tilcara, Argentina, em dezembro
de 2008. Sob este Memorando, as partes concordaram em trabalhar juntas para proteger o

45
ecossistema andino na região tripartida destes países. Os membros da AGA, em conjunto com o
Grupo para Conservação dos Flamingos Andinos, estão a liderar a criação de duas novas áreas
protegidas nesta região. Em colaboração com as áreas protegidas vizinhas, a AGA está a
desenvolver e a implementar o Projeto de "Estrada Única", que tem como objetivo a mitigação do
impacto que o turismo off-road está a causar nos habitats adequados para o gato andino e as
colônias migratórias de flamingo (AGA, 2011).
2.2. Lince Ibérico
O lince ibérico (Lynx pardinus (Temminck, 1827)) foi anteriormente considerado um
membro da mesma espécie do lince eurasiático (Lynx lynx (Linnaeus, 1758)). Contudo, atualmente
é considerado uma espécie distinta do lince eurasiático devido a dados morfológicos (Werdelin,
1981), paleontológicos, genéticos (Johnson et al., 2006).
Em 2002, o lince ibérico recebeu o estatuto de Criticamente em Perigo pela IUCN,
resultante do declínio constante da sua população selvagem, atingindo mínimos históricos com
números inferiores a 200 indivíduos selvagens em duas populações pequenas e isoladas em Sierra
Morena e Doñana, Espanha (Guzmán et al., 2004). Desde então, vários projetos de conservação,
ex situ e in situ, levaram a um aumento populacional com 686 indivíduos na Península Ibérica em
2018 (Simón, 2018), resultando na alteração do seu estatuto de conservação para Em Perigo em
2015 (Rodríguez & Calzada, 2015). Atualmente, tem uma distribuição geográfica com 8 áreas
diferentes espalhadas pelo sul e centro dos dois países (Andújar-Cardeña, Doñana-Alijarafe,
Guadalmellato, Guarrizas, Montes de Toledo, Sierra Morena Oriental, Vale do Guadiana, Valle del
Matachel; Iberlince, 2021).
O lince ibérico tem as caraterísticas comuns dos outros membros do gênero Lynx. O seu
tamanho corporal é semelhante ao das espécies americanas (lince canadiano (Lynx canadensis) e
lince pardo (Lynx rufus)), mas tem cerca de metade do tamanho do lince euroasiático (Lynx lynx)
(Delibes et al., 2000). A espécie possui uma cabeça relativamente pequena, pernas longas e cauda
muito curta com ponta preta. O seu rosto é curto e achatado, flanqueado por uma "barba"
caraterística, especialmente notável nos adultos e as orelhas triangulares terminam em tufos
pretos. As patas são largas com dedos que escondem garras retráteis. A cor de fundo dominante
da pelagem é castanho-avermelhada, salpicada de manchas escuras com tamanho, forma e
intensidade de cor variáveis (Figura 2). Em média, os machos adultos têm um peso de 13,64 Kg
enquanto as fêmeas adultas pesam 10,02 Kg (Rivas et al., 2016).

46
Figura 2 – Lince Ibérico, Espanha. © Sebastian Kennerknecht
O lince ibérico é considerado um superespecialista trófico (Ferrer & Negro, 2004) com o
coelho-bravo como presa fundamental e quase exclusiva deste carnívoro. Diferentes estudos
realizados no Parque Nacional de Doñana e na Serra Morena mostraram que esta espécie de
coelho é a sua principal presa, estando presente em 70-99% das amostras analisadas (Delibes,
1980; Palomares et al., 2001; Gil-Sánchez et al., 2006), percentagem que varia pouco entre áreas
geográficas e entre as estações do ano (Alfaya et al., 2020). As necessidades energéticas de um
lince adulto variam situando-se entre as 600-1000 kcal, o que corresponde aproximadamente à
energia contida num coelho adulto. O lince ibérico, no entanto, também consome outras espécies
de presas em proporções menores, mas sua importância relativa aumenta quando a
disponibilidade de coelhos diminui (Beltrán et al., 1985).
Outros vertebrados, incluindo roedores, lebre europeia (Lepus granatensis), perdiz-
vermelha (Alectoris rufa) (Delibes, 1980; Gil-Sánchez et al., 2006), patos e gansos (Anser anser)
podem ser regularmente mortos e comidos em algumas áreas e/ou estações do ano, mas
contribuem sempre pouco para a dieta do lince. Várias espécies de ungulados podem
ocasionalmente ser capturadas, especialmente no inverno. Presas unguladas incluem juvenis
vermelhos (Cervus elaphus), gamos (Dama dama) (Delibes, 1980; Beltrán et al., 1985; Gil-Sánchez
et al., 2006) e muflão (Ovis musimon) (Gil-Sánchez et al., 2006).

47
O lince ibérico também é um especialista em habitat, reproduzindo-se apenas em
matagais mediterrâneos e bosques que contêm densas populações de coelhos (Palomares et al.,
2000; Palomares, 2001). Esta escolha tem sido interpretada em termos da necessidade de abrigo
e reprodução fornecidos por estes habitats, pois favorecem as condições ecológicas para os
coelhos e uma estrutura adequada para a caça do lince (Palomares, 2001; Fernández et al., 2003).
Outros elementos essenciais do habitat incluem cavidades naturais (árvores ocas, cavernas
rochosas, pilhas de pedra, tocas de solo) que são usadas como tocas para as crias natais
(Fernández et al., 2002, 2006). Por outro lado, paisagens florestais, terrenos agrícolas ou outras
terras abertas desprovidas de arbustos nativos raramente são usados por linces residentes
(Palomares et al., 1991), mas ocasionalmente usados por adultos jovens durante a dispersão natal
(Palomares et al., 2000).
Como muitas outras espécies de felinos, o lince ibérico mantém um estilo de vida solitário
(Rivas et al., 2016). Os machos não ajudam a criar a ninhada e, normalmente, encontram as
fêmeas apenas na época de acasalamento. Os adultos defendem os seus territórios contra
intrusos do mesmo sexo, especialmente em áreas onde os coelhos são escassos e, também,
quando os territórios abrangem mais do que um território feminino (Sarmento et al., 2017).
Os padrões de atividade mostram uma variabilidade individual notável. Embora o lince se
possa mover a qualquer hora do dia, a atividade atinge o pico no crepúsculo, especialmente ao
pôr do sol. A atividade diurna ocorre principalmente no inverno (Penabad et al., 2012).
A biologia reprodutiva do lince ibérico ainda não é bem conhecida. Há indícios de que o
cio atinge o pico em janeiro, sendo que a maioria das ninhadas nasce entre março e abril (Rivas et
al., 2016). No entanto, quando as fêmeas não encontram um parceiro ou não engravidam,
quando a gestação é interrompida ou quando as ninhadas são perdidas numa fase inicial, as
fêmeas podem entrar no cio novamente. A gestação tem uma duração de dois meses, dando
origem a ninhadas que variam entre duas e quatro crias, sendo a média três (Rivas et al., 2016).
Estas ficam na toca de parto durante duas a quatro semanas, sendo em seguida movidas para
tocas auxiliares. Com quatro semanas de idade, começam a comer algumas presas trazidas pela
mãe para a toca e, aos quatro meses, as crias acompanham a mãe para desenvolverem as suas
habilidades de caça. Por volta dos três meses de idade, as crias desenvolvem uma agressividade
extrema nas brincadeiras com os restantes membros da ninhada, e que podem chegar a ser
mortais (Rivas et al., 2016). A maioria dos linces juvenis abandona definitivamente a área natal
entre os 8 e os 23 meses. Os machos dispersam-se mais cedo e para mais longe do que as fêmeas,
em que estas podem herdar o território materno ou parte dele, ou podem estabelecer-se num

48
território contíguo. Os machos juvenis não costumam ficar na área de origem natal ou próximos
desta (Delibes et al., 2000).
O lince adulto costuma matar outras espécies de carnívoros de menor tamanho que se
encontram dentro dos seus territórios, incluindo cães, gatos domésticos, raposa vermelha (Vulpes
vulpes), geneta (Genetta genetta), sacarrabo (Herpestes ichneumon) e lontra (Lutra lutra) (Delibes
et al., 2000). Como resultado da predação intraguilda, os pequenos carnívoros usam pouco os
territórios do lince, sendo que, as áreas habitadas pelo lince ibérico sustentam densidades mais
baixas de outras espécies carnívoras, comparando com as áreas sem lince. A baixa densidade de
carnívoros é refletida em algum relaxamento da pressão de predação o que, por sua vez, pode
produzir uma densidade de presa relativamente alta. Assim, através da supressão do
mesopredador, o lince diminui a competição e aumenta a densidade do coelho dentro do seu
território (Delibes et al., 2000; Ayanz, 2014).
Durante o século XX, a caça e as armadilhas foram as principais fontes de mortalidade
associadas ao controlo de predadores e à exploração de coelhos (Rodríguez & Delibes, 2004).
Apesar da diminuição da importância deste fator como ameaça para o lince ibérico (Ferreras et
al., 2010), nos últimos anos alguns linces foram alvejados ou capturados com armadilhas ilegais
(Iberlince LIFE Project 2014). Os acidentes rodoviários normalmente resultam em várias mortes
em cada ano (Simón et al., 2012; Iberlince LIFE Project 2014), uma vez que a extensão do número
de estradas pavimentadas, bem como uma maior quantidade de tráfego rodoviário, aumentaram
significativamente dentro e em redor das áreas de lince (Ferreras et al., 2010).
A homogeneização dos habitats em mosaico com matagais e áreas abertas devido à
intensificação da produção agrícola e florestal também contribuiu para o declínio do lince
(Rodríguez & Delibes, 2002; Ferreras et al., 2010). Mesmo em paisagens com estrutura adequada
e sujeitas a uma gestão de conservação intensiva, a abundância de coelho exibe grande
variabilidade temporal (Palomares et al., 2001; Fernández et al., 2007). Os incêndios florestais que
anualmente devastam áreas consideráveis da Península Ibérica são também um fator que tem
vindo a contribuir para a regressão do lince (Delibes et al., 2000).
Como uma manifestação da mudança global, a propagação de doenças virais com a ajuda
humana que afetou os coelhos-bravos, teve efeitos catastróficos nas populações do lince ibérico
no passado (Ferreras et al., 2010). No final dos anos 1950, a mixomatose entrou na Península
Ibérica e rapidamente se espalhou, provocando um declínio populacional que reduziu as
populações para menos de 5%. No final da década de 1980, uma nova patologia denominada
Doença Hemorrágica Viral (DHV) voltou a provocar um declínio dramático do coelho-bravo. Ao

49
mesmo tempo, mudanças ao nível do uso do solo acabaram por tornar as populações
extremamente vulneráveis e a sua recuperação bastante problemática. Embora os coelhos
possam eventualmente desenvolver resistência, as doenças virais continuam a ser uma ameaça
recorrente, visto que a chegada de novas estirpes pode causar novamente uma depressão
duradoura na disponibilidade de alimentos para o Lince Ibérico (Rodríguez & Calzada, 2015).
O pequeno tamanho populacional persistente ao longo de gerações de linces,
especialmente nas terras baixas da região de Doñana, produziu sinais de deterioração
demográfica e genética, incluindo endogamia, diminuição da idade de aquisição de território e
tamanho da ninhada e aumento da mortalidade devido a doenças e outras causas naturais
(Palomares et al., 2012). O desempenho demográfico e genético reduzido pode interagir
positivamente na forma de um vórtice de extinção (Palomares et al., 2012).
Finalmente, modelos detalhados combinando nicho ecológico e dinâmica de população
mostram que, sem uma intervenção intensiva, as mudanças climáticas diminuirão rapidamente as
populações de lince e, provavelmente, levarão à extinção do lince ibérico em 35 anos (Fordham et
al., 2013).
2.2.1. Programas de Conservação
Na década de 1960, a comunidade científica apercebeu-se do facto de que, tanto a
abundância como a distribuição das populações de lince ibérico, diminuíram drasticamente desde
o início do século XX na Península Ibérica (Valverde, 1963). Contudo, não foram implementados
planos de conservação até 1980, e muitos dos planos iniciais foram ineficazes (Palomares et al.,
2011), muito provavelmente devido ao financiamento insuficiente (Símon et al., 2012).
Em 1999, baseada no Plano de Ação Europeu para o Lince Ibérico, foi elaborada pela
Dirección General de Conservación de la Naturaleza a primeira “Estratégia para la Conservación
del Lince Ibérico” em território espanhol. O equivalente português “Plano de Ação para a
Conservação do Lince-ibérico” foi aprovado a 6 de Maio de 2008 através do despacho nº
12697/2008, publicado em Diário da República.
Várias iniciativas relativas à conservação do lince foram realizadas desde os anos 1990,
em que várias agências governamentais espanholas e portuguesas desenvolveram vários esforços
de conservação, a maioria deles no âmbito de projetos LIFE, cofinanciados em conjunto com a
Comissão Europeia (Símon et al., 2012) (Tabela 5).

50
Tabela 5 - Projetos focados na conservação do lince ibérico na Península Ibérica (Retirado de LIFE Program, 2021)
Projeto Duração Objetivos
Inventory and management of Portugal's natural heritage - 1st
phase
1994-
1996 Aumentar a Conscientização Pública; Recolha de Informação da espécie
Inventory and management of Portugal's natural heritage -
extension phase I
1994-
1996 Aumentar a Conscientização Pública; Recolha de Informação da espécie
Lince/Castilla Léon—Conservation of the Iberian lynx—Castilla y
Léon
1994-
1998
Diminuição da mortalidade; Gestão, Restauração e Conservação de Habitats; Redução das
Ameaças; Recolha de Informação da espécie
Conservation of the Iberian lynx (Comunidad de Madrid) 1994-
1998
Diminuição da mortalidade; Gestão, Restauração e Conservação de Habitats; Aumentar as
Populações de Presas; Gestão e Monitorização de Populações; Aumentar a
Conscientização Pública; Redução das Ameaças; Recolha de Informação da espécie
Conservation of the Iberian lynx (Madrid) 1994-
1998
Diminuição da mortalidade; Gestão, Restauração e Conservação de Habitats; Gestão e
Monitorização de Populações; Redução das Ameaças; Recolha de Informação da espécie;
Conservation of the Iberian lynx (Madrid) 1994-
1998
Diminuição da mortalidade; Gestão, Restauração e Conservação de Habitats; Aumentar a
Conscientização Pública; Redução das Ameaças; Recolha de Informação da espécie
Conservation of the Iberian lynx (Andalucia) 1994-
1998
Gestão, Restauração e Conservação de Habitats; Diminuição da mortalidade; Gestão e
Monitorização de Populações; Aumentar a Conscientização Pública; Redução das
Ameaças; Recolha de Informação da espécie
Conservation of the Iberian lynx (Castilla-la Mancha) 1994-
1998
Recolha de Informação da espécie; Redução das Ameaças; Gestão, Restauração e
Conservação de Habitats; Diminuição da mortalidade; Gestão e Monitorização de
Populações; Aumentar as Populações de Presas; Aumentar a Conscientização Pública
Conservation of the Iberian lynx (Extremadura) 1994-
1998
Recolha de Informação da espécie; Redução das Ameaças; Gestão, Restauração e
Conservação de Habitats; Diminuição da mortalidade; Gestão e Monitorização de
Populações; Aumentar as Populações de Presas; Aumentar a Conscientização Pública
Conservation of the Iberian lynx (Extremadura) 1994-
1998
Recolha de Informação da espécie; Redução das Ameaças; Gestão, Restauração e
Conservação de Habitats; Diminuição da mortalidade; Gestão e Monitorização de
Populações; Aumentar as Populações de Presas; Aumentar a Conscientização Pública
Conservation of the Iberian Lynx (Portugal) 1995-
1996
Recolha de Informação da espécie; Redução das Ameaças; Gestão, Restauração e
Conservação de Habitats; Aumentar as Populações de Presas; Aumentar a Conscientização
Pública

51
Conservation of the Iberian lynx (Castilla-la Mancha) 1995-
1998
Recolha de Informação da espécie; Redução das Ameaças; Gestão, Restauração e
Conservação de Habitats; Diminuição da mortalidade; Gestão e Monitorização de
Populações; Aumentar as Populações de Presas; Aumentar a Conscientização Pública
Conservation of the Iberian lynx (Castilla y Leon) 1995-
1998
Recolha de Informação da espécie; Redução das Ameaças; Gestão, Restauração e
Conservação de Habitats; Diminuição da mortalidade
Conservation of the Iberian lynx (Andalucia) 1995-
1998
Recolha de Informação da espécie; Redução das Ameaças; Gestão, Restauração e
Conservação de Habitats; Diminuição da mortalidade; Gestão e Monitorização de
Populações; Aumentar a Conscientização Pública
Conservation of the Iberian lynx (Madrid) 1995-
1998
Recolha de Informação da espécie; Redução das Ameaças; Gestão, Restauração e
Conservação de Habitats; Diminuição da mortalidade; Aumentar a Conscientização Pública
Conservation of the Iberian lynx (Madrid) 1995-
1998
Recolha de Informação da espécie; Redução das Ameaças; Gestão, Restauração e
Conservação de Habitats; Diminuição da mortalidade; Gestão e Monitorização de
Populações
Conservation of the Iberian lynx (Madrid) 1995-
1998
Recolha de Informação da espécie; Redução das Ameaças; Gestão, Restauração e
Conservação de Habitats; Diminuição da mortalidade; Gestão e Monitorização de
Populações; Aumentar as Populações de Presas; Aumentar a Conscientização Pública
CBD/especies - Conservation of the Imperial eagle, Black
vulture, Black stork and Iberian lynx on private protected land
in Extremadura and Castilla-La-Mancha
1999-
2002
Gestão e Monitorização de Populações; Gestão, Restauração e Conservação de Habitats;
Aumentar a Conscientização Pública
Lynx/Malcata - Recovery of habitats and preys of the Lynx
pardinus in Serra da Malcata
1999-
2003
Recolha de Informação da espécie; Gestão, Restauração e Conservação de Habitats;
Aumentar as Populações de Presas; Diminuição da mortalidade; Aumentar a
Conscientização Pública
Lince/Extremadura - Conservation of lynx pardinus in
Extremadura
1998-
2003
Gestão e Monitorização de Populações; Aumentar as Populações de Presas; Diminuição da
mortalidade; Aumentar a Conscientização Pública
Doñana - Land acquisition of strategic areas in Doñana district 1999-
2003 Gestão, Restauração e Conservação de Habitats
Cabañeros - Conservation of the threatened fauna and
vegetation in the Cabañeros National Park
1999-
2003 Gestão, Restauração e Conservação de Habitats; Aumentar as Populações de Presas
Lince Andalucía - Population recovery of Iberian Lynx in
Andalucia
2002-
2006
Gestão, Restauração e Conservação de Habitats; Aumentar as Populações de Presas;
Diminuição da mortalidade; Gestão e Monitorização de Populações; Avaliação de áreas

52
para Reintrodução; Aumentar a Conscientização Pública
Iberian lynx Conservation in Lugar Nuevo 2002-
2011 Gestão, Restauração e Conservação de Habitats; Gestão e Monitorização de Populações
Lince Toledo - Conservation of the Iberian Lynx in Montes de
Toledo - Guadalmena
2002-
2006
Gestão, Restauração e Conservação de Habitats Aumentar as Populações de Presas;
Gestão e Monitorização de Populações; Aumentar a Conscientização Pública
NORTENATUR - Management and conservation of the sites of S.
Mamede and Nisa/Laje de Prata
2004-
2009 Gestão, Restauração e Conservação de Habitats; Aumentar a Conscientização Pública
Lince Moura/Barrancos - Recovery of Iberian Lynx habitat in
Moura/Barrancos Site
2006-
2009
Gestão, Restauração e Conservação de Habitats; Aumentar a Conscientização Pública;
Avaliação de áreas para Reintrodução; Aumentar as Populações de Presas
Reintroducción Lince Andalucia - Conservation and
reintroduction of the Iberian lynx in Andalucia
2006-
2012
Gestão, Restauração e Conservação de Habitats; Gestão e Monitorização de Populações;
Avaliação de áreas para Reintrodução; Aumentar as Populações de Presas; Aumentar a
Conscientização Pública
Priorimancha - Conservation of Mediterranean priority species
in Castille-La Mancha
2009-
2014
Gestão, Restauração e Conservação de Habitats; Aumentar as Populações de Presas;
Redução das Ameaças; Aumentar a Conscientização Pública; Gestão e Monitorização de
Populações
Habitat Lince Abutre - Enhancing Habitat for the Iberian Lynx
and Black Vulture in the Southeast of Portugal
2010-
2014
Gestão, Restauração e Conservação de Habitats; Aumentar as Populações de Presas;
Aumentar a Conscientização Pública; Avaliação de áreas para Reintrodução
Iberlince - Recovering the historic distribution range of the
Iberian lynx (Lynx pardinus) in Spain and Portugal
2011-
2017
Gestão, Restauração e Conservação de Habitats; Gestão e Monitorização de Populações;
Diminuição da mortalidade; Avaliação de áreas para Reintrodução
LIFE SAFE-CROSSING - Preventing Animal-Vehicle Collisions –
Demonstration of Best Practices targeting priority species in SE
Europe
2018-
2023 Redução das Ameaças; Diminuição da mortalidade; Aumentar a Conscientização Pública
LIFE LYNXCONNECT - Creating a genetically and
demographically functional Iberian Lynx (Lynx pardinus)
metapopulation (Portugal and Spain)
2020-
2025
Avaliação de áreas para Reintrodução; Gestão e Monitorização de Populações; Aumentar
as Populações de Presas; Diminuição da mortalidade; Aumentar a Conscientização Pública

53
Na conservação do lince ibérico, os esforços devem ser direcionados em duas formas, a
conservação in situ e a conservação ex situ. A primeira diz respeito à monitorização, manutenção
e expansão das populações existentes; gestão das populações de coelho bravo; minimização das
causas não naturais de mortalidade; proteção, restauração e preparação de habitats para todas as
populações (incluindo as reintroduzidas); promoção de acordos com proprietários de terras
privadas e fazer a ligação genética entre as populações remanescentes (Vargas et al., 2008). No
caso da conservação ex situ, estão incluídas atividades como a criação em cativeiro de indivíduos;
preparação de animais para reintrodução; investigação científica sobre diferentes assuntos
relacionados com a espécie (ecologia, comportamento animal, etc.); gestão de um Banco de
Recursos Biológicos (BRB); gerir geneticamente todas as populações de lince (selvagens e em
cativeiro) como uma única meta população; educação da população e divulgação pública do
trabalho (Vargas et al., 2008).
Em 2003, começou um programa de reprodução em cativeiro com o objetivo de obter
indivíduos adequados para reintrodução ao nível fisiológico, psicológico e comportamental, assim
como, garantir um backup genético como garantia de uma potencial extinção da espécie na
natureza (Vargas et al., 2009). Quase uma década mais tarde, em 2011, foi iniciado um projeto
LIFE ibérico para identificar, preparar e estabelecer cinco áreas de reintrodução com capacidade
adequada para apoiar populações autossustentáveis de lince ibérico (Tabela 5). Seguindo as
diretrizes da IUCN para reintroduções (IUCN/SSC, 2013), as áreas de reintrodução foram
escolhidas devido ao facto de terem habitats, tamanho e alta densidade de coelhos adequados
para esta espécie. Assim, as libertações de lince criados em cativeiro foram iniciadas em Espanha
(primeiro na Andaluzia em 2011, depois na Extremadura e Castilla La Mancha em 2014) e em
Portugal (2015) (Sarmento & Carrapato, 2019).
Atualmente, de acordo com o Programa De Conservación Ex-Situ Del Lince Ibérico
(LYNXEXSITU, 2021) existem cinco centros de reprodução do lince ibérico, 4 em Espanha e 1 em
Portugal:
• Centro de Reprodução “El Acebuche” – inaugurado em 1992, localiza-se no
Parque Nacional de Doñana, Matalascañas (Huelva) e é constituído por 18
cercados. Foi nele que em 2005 nasceu a primeira ninhada em cativeiro.
• Zoobotanico de Jerez – abriu em 2005, localizado em Jerez de la Frontera (Cádiz),
com apenas 5 cercados;
• Centro Nacional de Reprodução de Lince Ibérico (CNRLI) – inaugurado em 2009,
localizado no Vale Fuzeiros, Silves, Portugal e é constituído por 16 cercados;

54
• Centro de Reprodução “Granadilla” – inaugurado em 2011, localiza-se em Zarza
de Granadilla (Cáceres) e possui 16 cercados;
• Centro de Reprodução “La Olivilla” – inaugurado em 2007, localiza-se nos
Montes de La Aliseda, Santa Elena (Jaén) e possui 23 cercados. Foi aqui que, em
2009, nasceu a primeira ninhada proveniente de progenitores também nascidos
em cativeiro.
Os esforços atuais de reprodução do lince ibérico concentram-se na criação em cativeiro
de indivíduos psicologicamente e fisicamente saudáveis e com comportamentos apropriados, que
sejam adequados para os programas de reintrodução. Para isso, são usadas técnicas de gestão e
investigação que dependem de subsídios e de conhecimentos multidisciplinares, provenientes da
história de vida da espécie, comportamento, nutrição, aspetos veterinários e de saúde, genética,
fisiologia reprodutiva, endocrinologia e ecologia. É particularmente importante destacar a
importância de adaptar os esquemas de criação com base em dados científicos para promover
comportamentos naturais em cativeiro (caça, territorialidade, interações sociais) e um ambiente
livre de stress que conduza à reprodução natural. Assim, é crucial minimizar o uso de métodos
invasivos, sendo para tal necessário mais informação de várias áreas de investigação relevantes
que incluem: a determinação de perfis de hormonas fecais para linces juvenis e adultos (Pelican et
al., 2009); o estudo do comportamento reprodutivo e desenvolvimento das crias (Antonevich et
al., 2009); a determinação da saúde reprodutiva dos indivíduos reprodutores (Roldan et al., 2009;
Göritz et al., 2009); o desenvolvimento de um teste de gravidez não invasivo (Jewgenow et al.,
2009); o estabelecimento de protocolos de biossegurança e biomédicos (Martínez et al., 2009);
estabelecer valores de referência para parâmetros sanguíneos (Pastor et al., 2009; García et al.,
2010); a genotipagem de todos os indivíduos fundadores e fazer recomendações de
emparelhamento de casais com base na sua distância genética (Godoy et al., 2009; Leus & Lacy,
2009).
Tal como já foi referido neste trabalho, o estudo da personalidade animal pode e deve ser
um fator em ter em conta na conservação ex situ de espécies ameaçadas.
2.3. Leopardo das Neves
Descrito pela primeira vez em 1775, o leopardo das neves (Panthera uncia (Schreber,
1775)) fez parte dos géneros Felis e, mais tarde Uncia (Pocock, 1916). Contudo, estudos genéticos
mostram que esta espécie pertence ao grupo Panthera (Johnson et al., 2006), sendo mais próxima

55
filogeneticamente do tigre (Panthera tigris), apesar destas espécies terem divergido há mais de 2
milhões de anos (Davis et al., 2010; O'Brien & Johnson, 2007).
Avaliada desde 1986 com o estatuto de conservação de “Em Perigo” pela IUCN, esta
espécie foi, em 2017, reclassificada como “Vulnerável” (McCarthy et al., 2017). A mudança de
estatuto deve-se, sobretudo, ao facto do tamanho populacional estimado ser entre 2500 e 10000
indivíduos, com um declino de 10% nos seus números devido a ameaças tais como a caça, a
exploração mineira e a perda de habitat (McCarthy et al., 2017). O leopardo das neves tem uma
larga distribuição geográfica no centro do continente asiático, com uma área estimada de 2.8
milhões km2 que atravessa 12 países, Afeganistão, Butão, China, Índia, Cazaquistão, Quirguistão,
Mongólia, Nepal, Paquistão, Rússia, Tajiquistão e Uzbequistão, desde os Himalaias a sul, passando
por cadeias montanhosas asiáticas e pelo Planalto de Qinghai-Tibet no centro, até à zona sul da
Sibéria a norte do continente (McCarthy et al., 2017).
O leopardo das neves é o felino mais pequeno do grupo dos grandes felinos do género
Panthera, com um peso médio entre os 20 e os 50 Kg, e um comprimento corporal entre 1 e
1,3m, com a cauda com 75% a 90 % do comprimento total do individuo (0,8 a 1,1m de
comprimento; Hemmer, 1972; Kitchener et al., 2016). O seu corpo está adaptado às condições
climatéricas adversas e aos habitats pela qual esta espécie tem preferência para viver e caçar. A
sua pelagem é a mais longa e densa de todos os felinos Panthera, com 4000 pelos/cm2, numa
proporção de 8 pelos inferiores, por cada pelo de guarda (Heptner & Sludskii, 1992), sendo estes
bastante eficazes no isolamento térmico do animal. A pelagem dorsal é em tons de cinzento e
castanho, com manchas escuras nas zonas da cabeça e membros e rosetas grandes no corpo e
cauda, enquanto a parte inferior do corpo tem pelagem branca (Figura 3). Estas colorações e
manchas da pelagem permitem aos leopardos das neves camuflarem-se nos ambientes rochosos
onde habitam e facilitar a perseguição e caça das suas presas. A cauda atua como um órgão de
equilíbrio, durante a mobilidade nas encostas ingremes, especialmente durante a caça, e também
é usada como um silenciador para isolar as patas e a cabeça do frio em grandes altitudes durante
o repouso (Rieger, 1984).

56
Figura 3 - Leopardo das neves, Mongólia. © Snow Leopard Trust
Apesar da falta de informação sobre adaptações fisiológicas às grandes altitudes, Marma
e Yunchis (1968) descobriram que os leopardos das neves têm uma grande quantidade de
glóbulos vermelhos pequenos (14.1-16.8 milhões/mm3; média de 5.5mm de diâmetro), elevada
concentração de hemoglobina (16.4 g%) e um hematócrito significativo (43% do volume total de
glóbulos vermelhos), que podem ajudar estes animais a uma hematose pulmonar mais eficiente a
grandes altitudes.
A dieta do leopardo das neves varia consoante a sua distribuição geográfica, consistindo
maioritariamente em ungulados de montanha e gado doméstico, suplementada por pequenos
mamíferos e aves. As duas espécies de presas mais frequentemente relatadas são a ovelha azul
(Pseudois nayaur) (Oli et al., 1993; Schaller et al., 1987, 1988; Wegge et al., 2012; Devkota et al.,
2013) e o íbex siberiano (Capra sibirica) (Schaller et al., 1988; Anwar et al., 2011; Shehzad et al.,
2012), em que as áreas de distribuição destas duas espécies cobrem praticamente toda a
extensão do leopardo da neve. Outras espécies de ungulados relevantes para a dieta deste felino
são: cabra-selvagem-paquistanesa (Capra falconeri) (Anwar et al., 2011), tahr-dos-himalaias
(Hemitragus jemlahicus) (Lovari et al., 2009; Ferretti et al., 2014), carneiro-da-montanha (Ovis

57
ammon) (Shehzad et al., 2012; Jumabay-Uulu et al., 2014), Urial (Ovis orientalis) (Mallon et al.,
2016), veado-vermelho (Cervus elaphus) (Lhagvasuren & Munkhtsog, 2000), veado-de-Thorold
(Cervus albirostris) (Mallon et al., 2016), corça-siberiana (Capreolus pygargus) (Lhagvasuren &
Munkhtsog, 2000), cervo-almiscarado (Moschus spp.) (Ferretti et al., 2014), javali (Sus scrofa) e
alce (Alces Alces) (Mallon et al., 2016). Pequenos mamíferos como as marmotas (Marmota spp.)
(Oli et al., 1993; Devkota et al., 2013; Jumabay-Uulu et al., 2014), lebres (Lepus spp.) (Schaller et
al., 1987), e aves como snowcock (Tetraogallus spp.) (Lhagvasuren & Munkhtsog, 2000), estão
também presentes na dieta do leopardo das neves. Como são felinos oportunistas, a predação de
gado consiste, frequentemente, em 15 a 30% da sua dieta (Schaller et al., 1988; Oli et al., 1993;
Bagchi & Mishra, 2006; Shehzad et al., 2012; Wegge et al., 2012). Estima-se que sejam necessárias
20 a 30 ovelhas azuis adultas anualmente, com predação a ocorrer a cada 10 a 15 dias, podendo
os leopardos das neves permanecer com as suas presas até uma semana após a matança
(Macdonald et al., 2010a).
Embora os leopardos-da-neve possam existir em áreas relativamente planas, como em
partes da Mongólia e do Planalto Tibetano (McCarthy et al., 2017), dentro do seu habitat, eles
preferem afloramentos rochosos e terrenos íngremes e acidentados (Jackson et al., 2010). Tal
como já foi referido, habitam em zonas montanhosas no centro da Ásia, a altitudes entre 3000 e
5000 metros nos Himalaias e no Planalto Tibetano, mas podem ocorrer em níveis tão baixos
quanto 500 m no Altai (Snow Leopard Network, 2014). A vegetação varia entre zonas alpinas sem
árvores na maioria dos sistemas montanhosos, densos ecótonos floresta-alpinos, florestas
abertas, zonas de pequenos arbustos e habitats de montanha desértica na porção central da
distribuição geográfica da espécie (Koshkarev, 1984; Mallon, 1984; Snow Leopard Network, 2014).
Os cumes montanhosos, bordas de penhascos e linhas de drenagem bem definidas servem como
rotas de viagem e locais comuns para a deposição de sinais, incluindo arranhões, fezes e marcas
de cheiro (Ahlborn & Jackson 1988).
De acordo com McCarthy et al. (2005), os padrões de atividade desta espécie mostram
que a maioria do seu tempo ativo ocorre durante a noite (51% do tempo) e não durante o dia
(35%), com uma tendência para atingir máximos no crepúsculo.
Apesar do leopardo das neves ser uma espécie normalmente solitária, nos meses de
Janeiro a Março, indivíduos adultos encontram-se para acasalar, altura em que a fêmea entra em
cio que pode durar até 8 dias (Fox & Chundawat, 2016), havendo uma intensificação nas
marcações e vocalizações do casal (Jackson & Ahlborn, 1988) durante este período. A gestação
dura 90 a 105 dias, com o nascimento das crias entre os meses de Abril a Junho, com ninhadas

58
geralmente com 2 ou 3 indivíduos, sendo raras as que têm diferente tamanho (Fox & Chundawat,
2016). Após uma semana, as crias abrem os olhos e na semana seguinte as orelhas, começando a
caminhar pouco depois. Com 8 semanas são capazes de comer sólidos e de brincar, e com 12
semanas já seguem a sua progenitora, sendo aos 5 meses totalmente desmamados (Fox &
Chundawat, 2016). A dispersão dos juvenis ocorre aos 18-22 meses de idade, em que indivíduos
da mesma ninhada podem permanecer temporariamente juntos (Jackson, 1996), atingindo a
maturidade sexual com 2 a 3 anos de idade (McCarthy et al., 2017).
Dentro dos países com leopardo das neves, existe uma variação no tipo e gravidade das
ameaças a que esta espécie está sujeita, principalmente em países como a China e a Mongólia,
em que a sua distribuição geográfica é extensa e abrange diferentes regiões administrativas.
Atualmente, as principais ameaças podem ser agrupadas de 3 formas: 1 – Competição com gado,
degradação de habitat e declínio na quantidade de presas; 2 – Depredação por leopardos das
neves no gado e matanças retaliatórias; e 3 – Comércio ilegal de peles, ossos e outras partes do
corpo (Snow Leopard Network, 2014; McCarthy et al., 2017).
O relatório da TRAFFIC por Nowell et al. (2016) mostra que 90% da caça furtiva do
leopardo das neves ocorre em apenas 5 dos 12 países abrangidos pela distribuição geográfica
desta espécie, China, Mongólia, Paquistão, India e Tajiquistão. Também é referido que, apesar da
China com uma das maiores áreas com leopardo das neves, ter os maiores níveis de caça furtiva e
comercio ilegal, os países que têm a menor área geográfica em que a espécie está presente
apresentam níveis elevadamente desproporcionais de caça furtiva (Nepal e Paquistão) e de
comércio ilegal (Afeganistão e Rússia) (Nowell et al., 2016).
Desde 2003 que as alterações climáticas, exploração mineira e grandes infraestruturas,
como estradas e linhas de comboio, são consideradas também ameaças para o leopardo das
neves, apesar de ser difícil quantificar o seu grau de ameaça na espécie e no seu habitat (Snow
Leopard Network, 2014; McCarthy et al., 2017). Outras ameaças incluem a falta de
consciencialização entre a população local e legisladores, falta de capacidade institucional,
armadilhagem e envenenamento acidentais, doenças, cães vadios atacarem estes felinos e as
suas presas e colaboração transfronteiriça limitada (Snow Leopard Network, 2014; McCarthy et
al., 2017).
2.3.1. Programas de Conservação
Conforme destacado acima, as populações existentes de leopardos das neves enfrentam
uma série de desafios de conservação. Isso inclui ameaças diretas das comunidades locais, guerras

59
ou falta de cooperação internacional e ameaças indiretas relacionadas com economias
subdesenvolvidas, comércio ilegal de peles e ossos ou falta de conscientização pública. A
importância de cada fator varia entre as paisagens e entre os países, necessitando assim de ações
de conservação apropriadas a cada região (McCarthy & Chapron, 2003).
Desde que a primeira versão da Snow Leopard Survival Strategy apareceu em 2003
(McCarthy & Chapron, 2003), inúmeros recursos e pessoas foram investidos na conservação dos
leopardos das neves, das suas presas e dos seus habitats. Esta resposta de conservação resultou
em acordos internacionais focados na proteção do leopardo das neves e restante biodiversidade,
proteção legal a nível nacional, novas e maiores áreas protegidas dentro da distribuição
geográfica da espécie, medidas para combater a caça furtiva (ex: remoção de armadilhas e o
recrutamento de ex-caçadores como guardas florestais das área protegida) (Paltsyn et al., 2016),
investigação científica, projetos de conservação de campo, suportes alternativos de subsistência
para as comunidades locais, iniciativas para reduzir o conflito com os pastores (por exemplo,
fortalecimento de currais de gado e zonas de pastagens), programas de ecoturismo, avaliações
socioeconómicas e programas de educação e sensibilização ambiental (Snow Leopard Network,
2014; McCarthy et al., 2017).
Projetos de conservação transfronteiriços fornecem benefícios importantes para esta
espécie (Rosen & Zahler, 2016). Exemplos disso são os projetos a decorrer na Ásia Central, (1) em
que o Parque de conservação da natureza Khan Tengri (3.257 Km²), no leste do Quirguistão, ligará
a Reserva Naryn, a Reserva Sarychat Ertash e duas outras no Quirguistão com a Reserva Natural
Tomur na China, e (2) um projeto transfronteiriço no norte de Tien Shan (Quirguistão e
Cazaquistão) com este felino como espécie focal, que tem como objetivo desenvolver a
cooperação e melhorar as áreas protegidas. O Projeto Altai-Sayan do PNUD / GEF (2007-2012)
também se focou no leopardo das neves, realizando vários projetos internacionais, incluindo
armadilhagem fotográfica (Rosen & Zahler, 2016).
É notória a ameaça constante da morte de indivíduos desta espécie por pastores em
retaliação à predação no gado, agravada por currais noturnos mal construídos ou mantidos que
facilitam a entrada de leopardos das neves. O fortalecimento dos currais para torná-los à prova de
predadores realizado no Afeganistão, Índia, Paquistão e Tajiquistão, mostrou que não houve
predação do gado após o reforço de mais de 260 currais (Mohammad et al., 2016). De forma
semelhante em Western Tyva, na Rússia, os currais fortalecidos em 46 campos de pastores não
sofreram nenhuma predação, pelo que as perdas anuais de gado caíram em 50-60% (Paltsyn et
al., 2016).

60
Programas comunitários inovadores foram estabelecidos no Afeganistão e no Paquistão,
com novas estruturas governamentais criadas para a gestão de recursos naturais locais, e com
várias dezenas de guardas a monitorizar a vida selvagem e a fazer cumprir os regulamentos locais
e nacionais contra a caça furtiva e outras atividades de extração de recursos (Zahler & Paley,
2016). O Snow Leopard Trust criou na Mongólia e no Quirguistão um programa pioneiro de
sucesso para aumentar os rendimentos das comunidades locais, através do fabrico de artesanato
e acesso destes produtos nos mercados internacionais, em troca de um compromisso para
prevenir a caça furtiva de leopardos das neves e das suas presas e de se envolverem em ações de
educação ambiental e outras atividades (Bayarjargal et al., 2016). Incentivos relacionados ao
ecoturismo levaram a um aumento de avistamentos de leopardos das neves no Parque Nacional
de Hemis, Índia, atribuídos a uma combinação da proteção dos currais contra predadores e
criação de rendimentos significativos associados a homestays administrados pela comunidade
(Jackson & Wangchuk, 2004; Jackson, 2015; Namgail & Dadul, 2016).
Li et al. (2014) demostraram que os 336 mosteiros budistas tibetanos da região de
Sanjiangyuan, na província chinesa Qinghai, estão dentro ou próximo das áreas geográficas
ocupadas pelo leopardo das neves, e estes são capazes de proteger, em média 75,1 Km2, para um
total de cerca de 8342 Km2, ultrapassando a área de habitat desta espécie na Reserva Natural de
Sanjiangyuan (7674 Km2). Os princípios básicos do budismo, nomeadamente, de amor, respeito e
compaixão por todos os seres vivos, levam a que estes mosteiros protejam espécies e locais
sagrados (Shen et al., 2012), e ensinem as comunidades locais sobre proteção ambiental,
afetando assim, de forma positiva, as suas atitudes e comportamentos em relação à vida
selvagem (Dudley et al., 2009).
Além desses programas, outras iniciativas de conservação incluem zonas apenas de
pastagens (Mishra et al., 2016), vacinação de gado (Nawaz et al., 2016), seguro para o gado
(Kunkel et al., 2016) e uma variedade de projetos de educação e de consciencialização (Hillard et
al., 2016). Iniciativas de conservação que combinam a ciência com o conhecimento tradicional,
que diminuem o conflito entre o ser humano e o leopardo das neves, que melhoram os
rendimentos familiares daqueles que dependem do gado e que destacam a importância cultural e
ecológica dos leopardos das neves, têm maior probabilidade de serem adotadas pelas
comunidades locais que partilham as remotas paisagens montanhosas com este felino (Jackson &
Lama, 2016).

61
3. O Futuro da Conservação Animal
Apesar dos bons resultados gerais dos esforços de conservação realizados em cada uma
das espécies mencionadas anteriormente, existem limitações que influenciam o sucesso dos
projetos de conservação, desde falta de conhecimentos científicos a legislações e políticas
insuficientemente implementadas. Assim, nesta secção serão abordadas as limitações e
problemas dos projetos de conservação do Gato-andino, Lince Ibérico e Leopardo das neves e
como podem ser resolvidos.
No caso do Gato-andino, é possível dizer que os fatores limitantes podem ser agrupados
em 4 grupos: Falta de Informação, Legislação e Políticas de Conservação, Violação das Leis, e
Áreas Protegidas e Corredores Verdes. A maioria dos estudos científicos está focada na
determinação da presença e distribuição geográfica desta espécie, enquanto aquilo que se sabe
da sua biologia e ecologia resultou de encontros inesperados e de estudos de presença (Perovic et
al., 2003). Baseada na informação disponível, é possível entender em que tipo de habitat é que
esta espécie ocorre e avaliar a qualidade de áreas com potencial para este felino. Contudo,
aspetos básicos da sua ecologia são ainda desconhecidos como, por exemplo, a sua biologia
reprodutiva, pelo que esta falta de informação restringe imenso o planeamento e realização de
ações de conservação para o gato-andino.
Existem iniciativas, principalmente a nível local, de autoridades e pessoas associadas com
áreas protegidas para avaliar prejuízos que animais carnívoros provocam nas comunidades, sendo
realizadas com o apoio de organizações ou instituições científicas (AGA, 2011).
Embora o gato-andino esteja protegido por leis nacionais nos quatro países onde existe e
tratados internacionais, estas leis não são específicas para esta espécie (Villalba et al., 2004). A
violação das leis dos quatro países ocorre principalmente porque as pessoas não estão
informadas sobre estas leis e também da falta de consciencialização das autoridades locais e do
público em geral. Nesse sentido, a aplicação de medidas de coerção para promover o
cumprimento da lei não é muito eficiente, sendo necessário desenvolver campanhas educativas
sobre a importância da conservação do gato-andino e da vida selvagem em geral (Villalba et al.,
2004).
Outro assunto importante relacionado com a falta de informação e gestão são as áreas
protegidas. A presença do gato-andino só foi verificada em 50% ou menos das áreas protegidas
onde a espécie poderia ocorrer em cada país (Villalba et al., 2004), não havendo informação sobre

62
algumas destas áreas. Com isto, é essencial perceber em quais áreas protegidas a espécie está
presente, em que medida essas áreas protegem a espécie e qual a eficiência da sua gestão.
De forma a ultrapassar estes problemas, é preciso insistir e continuar as investigações
científicas com esta espécie, para se poder ter uma melhor perceção da sua distribuição, ecologia,
comportamentos e ameaças. Assim, o envolvimento de ONG’s, instituições científicas e de ensino,
das autoridades e comunidades locais, é essencial para se obter novas informações sobre o gato-
andino. Ações de conservação como, desenvolver um projeto de treino para as pessoas das Áreas
Protegidas nos quatro países abrangidos, de forma coordenada e padronizada, e o
desenvolvimento em cada país de um módulo de treino e pesquisa em armadilhagem fotográfica
e outras técnicas de investigação de campo apropriadas ao estudo do gato-andino, dirigido a
estudantes universitários locais, permitirão não só adquirir novos dados mas, também, colocar
todos os países a níveis iguais de envolvimento e educação.
A investigação sobre a biologia e ecologia do lince ibérico tem desempenhado um papel
importante na orientação da conservação e implementação de planos de recuperação. Estudos
sobre a organização social e espacial, reprodução, padrões de atividade ou uso do habitat têm
sido importantes na compreensão dos requisitos do lince. No entanto, o foco da pesquisa
científica ganharia em focar-se em: (1) aprender como restaurar as condições ecológicas que
permitem o estabelecimento e reprodução do lince, (2) aprender mais sobre as novas ameaças,
como riscos associados à endogamia ou uma maior suscetibilidade a doenças infeciosas de
populações selvagens, e (3) avaliar a resposta de linces selvagens às medidas de gestão,
otimizando os procedimentos que conduzem à reprodução em cativeiro, repovoamento e
reintrodução.
Uma vez que as principais ameaças associadas ao lince ibérico são a caça furtiva ou
acidental e as armadilhas, derivadas de ações de controlo de predadores e caça de coelhos, e
também as mortes provocadas por acidentes rodoviários, é necessário melhorar as ações de
conservação focadas nestas áreas. A criação de legislações mais restritas para a exploração
cinegética e o aumento da severidade das penalizações para os caçadores furtivos podem ser a
solução para combater estas ameaças. Contudo, a proteção legal tem pouco efeito se não for
acompanhada por monitorização e aplicação de leis apropriadas, acompanhados por campanhas
de informação para aumentar a consciencialização das pessoas que vivem ou caçam em áreas de
lince (Delibes et al., 2000).
Para prevenir acidentes rodoviários são essenciais passagens subterrâneas que devem ser
construídas nas áreas onde existe lince, e a implementação de medidas para diminuir a

63
velocidade do tráfego, como a redução do limite máximo de velocidade, a instalação de lombas,
radares de velocidade e sinais de estrada que anunciam a entrada em áreas de lince. Estudos para
verificar os locais com maior número de mortes por este meio têm de ser realizados para
averiguar em que locais estas medidas devem ser implementadas, seguidos por estudos sobre a
sua eficácia e uso pelos linces.
Embora oficialmente o Leopardo das neves seja uma espécie totalmente protegida em
todos os 12 países, essa proteção raramente é aplicada devido à falta de sensibilização, à pouca
vontade de fazer cumprir as leis, à falta de fundos e de pessoal treinado e à baixa prioridade que
alguns governos dão à conservação da biodiversidade (McCarthy & Chapron, 2003; Theile, 2003).
Apenas cinco dos países envolvidos (Índia, Mongólia, Nepal, Paquistão e Rússia) desenvolveram
Planos de Ação Nacionais para o Leopardo da Neve, mas a sua implementação tem sido
frequentemente prejudicada pela insuficiente consciencialização ambiental dos políticos,
legislação inadequada e financiamento escasso (Jackson et al., 2010).
O verdadeiro desafio reside em mover as comunidades locais do seu modo de vida de
subsistência difícil e inseguro para atividades mais economicamente viáveis e ambientalmente
corretas. Em última análise, a população local deve ser encorajada a perceber que os leopardos
das neves e outros carnívoros valem "mais vivos do que mortos". Estas comunidades também
devem assumir uma maior responsabilidade pela proteção do seu gado contra predadores, o que
permitirá uma coexistência duradoura entre predadores e humanos em toda a distribuição
geográfica da espécie. A conservação eficaz do leopardo da neve depende de políticas nacionais
de conservação melhores e mais inclusivas, desenvolvendo maior consciencialização por meio de
programas de educação para a conservação direcionados aos níveis local e nacional e fomentando
a cooperação internacional e assistência financeira.
A conservação a longo prazo e em larga escala da vida selvagem em paisagens
humanizadas depende, maioritariamente, da nossa capacidade de identificar e lidar com os
impactos negativos das atividades humanas nas populações selvagens, bem como o impacto
negativo dessas mesmas populações nas atividades humanas. A recuperação de populações de
animais selvagens ainda enfrenta muita hostilidade devido aos seus impactos reais ou supostos
sobre, por exemplo, colheitas, gado ou até mesmo espécies de caça (Gomes & Sarrazin, 2016). Os
efeitos de e sobre as populações animais podem envolver processos comportamentais. Apesar do
facto de que a história da conservação e gestão populacional com base no comportamento ser
extensa, numerosos desafios conceptuais e técnicos ainda precisam ser resolvidos (Gomes &
Sarrazin, 2016). Ao nível individual e populacional, entender como o comportamento responde à

64
redução drástica na densidade populacional é essencial para evitar a fase proximal dos vórtices de
extinção (Henle et al., 2004). Da mesma forma, compreender como o comportamento responde à
alta densidade populacional continuará a ser crucial para regular as populações superabundantes
(Tavecchia et al., 2007).
Pesquisas contínuas sobre as bases genéticas e epigenéticas do comportamento, normas
de reação e plasticidade, bem como aprendizagem e transferência de informações por meio da
sociabilidade e da cultura, ainda são relevantes no contexto de crescentes mudanças globais. A
capacidade dos animais de manter os seus fenótipos adequados aos seus ecossistemas em
mudança dependerá amplamente da sua flexibilidade comportamental, por exemplo, na
capacidade de se alimentarem de novos recursos, de evitarem novos predadores, competidores
ou parasitas, ou de se reproduzirem em novos habitats (Rubenstein, 2016). O comportamento
terá um papel importante na capacidade da espécie de interagir com o seu ambiente e, portanto,
é um fator essencial da capacidade da espécie de se adaptar a ambientes em mudança (Gomes &
Sarrazin, 2016). Alternativamente, o comportamento de dispersão será uma resposta crucial para
vários organismos, mas pode também levar a que estes se instalem em habitats de baixa
qualidade, ou seja, aumenta o risco de armadilhas ecológicas (Gomes & Sarrazin, 2016).
Desde sempre houve uma dicotomia de ações humanas sobre os felinos selvagens, por
um lado eram caçados e perseguidos (Tuck, 2005) e, por outro lado, eram valorizados e
reverenciados (Callou et al., 2004). Esta dicotomia ainda persiste e é visível nas interações
contemporâneas entre as pessoas e estes animais. Por exemplo, o fascínio dos turistas com o
avistamento de grandes felinos em parques nacionais de todo o mundo, enquanto as populações
rurais locais fora das áreas protegidas sofrem depredações por estas mesmas espécies. Programas
e documentários sobre estes animais, apesar de mostrarem a biologia e ecologia destes felinos e
contribuírem para a educação da audiência, não mostram os desafios enfrentados pelas
comunidades locais com quem partilham territórios. Os valores culturais e estéticos que são
atribuídos às espécies carismáticas estão frequentemente em disputa com as pessoas que vivem
com elas. Os mecanismos para diminuir este conflito incluem delimitação e proteção de habitats
selvagens, compensação e proteção dos meios de subsistência, bem como a integração de
comunidades locais em atividades de conservação e ecoturismo para gerar receitas.
Embora estudos académicos possam não levar necessariamente a ações de conservação,
o aumento do número de estudos feitos em felinos selvagens mostra um crescente interesse por
estas espécies (Macdonald et al., 2010a). No entanto, ainda existem lacunas no conhecimento
destas mesmas espécies, em que as de maior porte tendem a receber mais atenção,

65
provavelmente devido ao seu carisma, maior facilidade de observação e maior potencial de atrair
financiamento para a realização de estudos sobre espécies que são conhecidas e atraentes para o
público. Embora a ênfase da investigação científica em grandes felinos ajude na sua conservação,
é necessário mais trabalho com os gatos pequenos, especialmente nas espécies mais raras. Os
esforços para promover o conhecimento e o interesse do público pelas espécies menores podem
corrigir este desequilíbrio de informação. Da mesma forma, é preciso reconhecer que há fortes
razões para a atenção dada às grandes espécies, devido ao seu potencial como espécie-bandeira e
guarda-chuva para a proteção dos ecossistemas naturais.
Contudo, Berger-Tal et al. (2015) mostraram que o número de artigos científicos que
tratam de problemas de conservação excede bastante o número de artigos que relatam soluções
de conservação. Esta tendência de relatar ameaças à biodiversidade com mais frequência do que
discutir as suas soluções pode impedir o sucesso da conservação (Lindenmayer et al., 2013).
Berger-Tal et al. (2015) demostraram também que esta tendência era menor quando o
comportamento dos animais era considerado, e que o comportamento era usado mais em
estudos de ações de conservação, o que sugere que a investigação de comportamento animal
pode ter um impacto positivo na conservação, fornecendo conhecimento comportamental que
pode ser usado na promoção de investigações baseadas em soluções.
O trabalho conjunto de conservacionistas e ecologistas comportamentais pode levar à
criação de revisões e diretrizes sistemáticas, completas e facilmente acessíveis (Dicks et al., 2014),
que permitirão o desenvolvimento baseado em provas de diretórios de soluções
comportamentais potenciais e reais para problemas de conservação e gestão. Porém, estar
consciente da importância do comportamento na conservação não se traduz necessariamente no
uso eficiente do comportamento na conservação. Os conservacionistas podem encontrar várias
restrições e problemas ao tentar aplicar princípios comportamentais, como a teoria de
forrageamento ideal ou aprendizagem associativa, a situações da vida real (Caro, 2007; Schakner
et al., 2014). Para que a ecologia comportamental se torne numa ferramenta eficaz, os ecologistas
comportamentais devem transformar os seus conhecimentos das implicações do comportamento
conservacionista em aplicações mais ativas, que resolvam problemas de conservação do mundo
real (Swaisgood & Greggor, 2019).
Finalmente, há amplo reconhecimento de que a conservação de espécies predatórias
precisa de ser conciliada com as necessidades humanas. Os felinos destacam-se como uma família
de espécies que envolve a nossa imaginação através do seu carisma e beleza. No entanto, estes
carnívoros também entram em conflito com as necessidades e atividades humanas, o que cria

66
aversões igualmente apaixonadas por aqueles cujos meios de subsistência sofrem com estes
animais. É vital que estes conflitos sejam resolvidos através da educação, que a conservação seja
planeada corretamente e bem gerida, o que requer uma ampla compreensão do conhecimento
relevante e um discernimento dos casos de estudos. Também exigirá uma abordagem radical à
conservação, com projetos novos de interdisciplinaridade e inovadores mecanismos de mercado
dado que, a sua posição precária no topo das pirâmides alimentares torna os felinos espécies
guarda-chuva importantes para a conservação para a biodiversidade em geral. A sua beleza,
significado cultural e carisma torna-os poderosas espécies-bandeira da natureza no cenário
mundial. Assim sendo, urge que todas as comunidades locais e internacionais se unam para
conservar as populações de felinos selvagens, sendo que, por vezes, basta um individuo ou uma
ação para colocar em risco todo o trabalho realizado. Assim, a perspetiva ampla da conservação
baseada no conhecimento, integrada com o desenvolvimento das comunidades, permitirá uma
conservação bem-sucedida dos felinos e de muitas outras espécies por todo o mundo.
Tendo em conta os objetivos estabelecidos pela ONU para a Década da Restauração dos
Ecossistemas, nomeadamente, a recuperação de ecossistemas degradados e, em especial, entre
outros, o combate à perda de biodiversidade, de mitigação e adaptação às alterações climáticas,
espero que este trabalho contribua para uma melhor divulgação deste assunto e aponte as
possíveis soluções para os problemas de conservação apresentados.

67
Referências Bibliográficas
Afonso, P., Fontes, J., Holland, K. N., Santos, R. S. (2008). Social status determines behavior and
habitat usage in a temperate parrotfish: implications for marine reserve design. Marine
Ecology Progress Series, 359, 215–227. https://doi.org/10.3354/meps07272
AGA. (2011). Strategic Plan for the Andean Cat Conservation. 2011-2016. Andean Cat Alliance –
Alianza Gato Andino.
Ahlborn, G. G., & Jackson, R. M. (1988). Marking in free ranging snow leopards in west Nepal: a
preliminar assessment. In H. Freeman (Ed), Proceedings of the Fifth International Snow
Leopard Symposium (pp. 25-49). International Snow Leopard Trust and Wildlife Institute
of India, Seattle.
Alberts, A. C. (2007). Behavioral considerations of headstarting as a conservation strategy for
endangered Caribbean rock iguanas. Applied Animal Behaviour Science, 102(3-4), 380–
391. https://doi.org/10.1016/j.applanim.2006.05.037
Alcock, J. (1975). Animal Behavior: An Evolutionary Approach. (1st Ed.). Sinauer Associates.
Alfaya, P., de Pablo, C. T. L., de Agar, P. M., Alonso, G. (2020). Assessing the influence of ecological
interaction patterns among habitat types on species distribution: studying the Iberian lynx
(Lynx pardinus Temminck 1827) in central Spain. Landscape Ecology, 35, 1923–1944.
https://doi.org/10.1007/s10980-020-01070-0
Altrichter, M., Boaglio, G., Perovic, P. (2006). The decline of jaguars Panthera onca in the
Argentine Chaco. Oryx, 40(3), 302–309. https://doi.org/10.1017/S0030605306000731
Anderson, R. P., & Martínez-Meyer, E. (2004). Modeling species' geographic distributions for
preliminary conservation assessments: an implementation with the spiny pocket mice
(Heteromys) of Ecuador. Biological Conservation, 116(2), 167–179.
https://doi.org/10.1016/S0006-3207(03)00187-3
Andrén, H., Linnell, J. D., Liberg, O., Andersen, R., Danell, A., Karlsson, J., Odden, J., Moa, P. F.,
Ahlqvist, P., Kvam, T., Franzén, R., Segerström, P. (2006). Survival rates and causes of
mortality in Eurasian lynx (Lynx lynx) in multi-use landscapes. Biological Conservation,
131(1), 23-32. https://doi.org/10.1016/j.biocon.2006.01.025

68
Antonevich, A. L., Sergey, V. N., Bergara, J., Vázquez, E., Vázquez, A., López, J., Pardo, A., Rivas, A.,
Martínez, F., Vargas, A. (2009). A comparative note on early sibling aggression in two
related species: the Iberian and the Eurasian lynx. In A. Vargas (Ed.), Iberian Lynx Ex situ
Conservation: An Interdisciplinary Approach (pp. 156-163). Fundación Biodiversidad,
Madrid.
Anwar, M. B., Jackson, R., Nadeem, M. S., Janecka, J. E., Hussain, S., Beg, M. A., Muhammad, G.,
Qayyum, M. (2011). Food habits of the snow leopard Panthera uncia (Schreber, 1775) in
Baltistan, Northern Pakistan. European Journal of Wildlife Research, 57, 1077–1083.
https://doi.org/10.1007/s10344-011-0521-2
Armstrong, D. P., Le Coeur, C., Thorne, J. M., Panfylova, J., Lovegrove, T. G., Frost, P. G. H., Ewen,
J. G. (2017). Using Bayesian mark-recapture modelling to quantify the strength and
duration of post-release effects in reintroduced populations. Biological Conservation 215,
39–45. https://doi.org/10.1016/j.biocon.2017.08.033
Ayanz, A. S. M. (Coord.). (2014). Manual para a gestão do habitat do lince ibérico (Lynx pardinus)
e da sua presa principal, o coelho-bravo (Oryctolagus cuniculus). (2ª Ed.). Fundação CBD-
Habitat. Madrid. Espanha.
Bagchi, S., & Mishra, C. (2006). Living with large carnivores: predation on livestock by the snow
leopard (Uncia uncia). Journal of Zoology, 268(3), 217–224.
https://doi.org/10.1111/j.1469-7998.2005.00030.x
Baker, K., & Pullen, K. (2013). The impact of housing and husbandry on the personality of cheetah
(Acinonyx jubatus). Journal of Zoo and Aquarium Research, 1(1), 35-40.
https://doi.org/10.19227/jzar.v1i1.11
Banerjee, A., Chitnis, U. B., Jadhav, S. L., Bhawalkar, J. S., Chaudhury, S. (2009). Hypothesis testing,
type I and type II errors. Industrial Psychiatry Journal, 18(2), 127–131.
https://doi.org/10.4103/0972-6748.62274
Bar-David, S., Saltz, D., Dayan, T. (2005). Predicting the spatial dynamics of a reintroduced
population: the Persian fallow deer. Ecological Applications, 15(5), 1833–1846.
https://doi.org/10.1890/04-0798

69
Bateson, P., & Laland, K. N. (2013). Tinbergen’s four questions: An appreciation and an update.
Trends in Ecology & Evolution, 28(12), 712–718.
https://doi.org/10.1016/j.tree.2013.09.013
Bayarjargal, A., Allen, P., Dashzeveg, U., Midgiddorj, Ts., Snell-Rullman, J. (2016). Handicrafts:
Snow Leopard Enterprises, Mongolia. In T. McCarthy & D. Mallon (Eds.), Snow Leopards,
Biodiversity of the World: Conservation from Genes to Landscapes (pp. 169-172).
Academic Press https://doi.org/10.1016/B978-0-12-802213-9.00013-4
Beltrán, J. F., San José, C., Delibes, M., Braza, F. (1985). An analysis of the Iberian lynx predation
upon fallow deer in the coto Doñana, SW Spain. XVIIth Congress of the International
Union of Game Biologists, (pp. 961-967).
Berger-Tal, O., & Saltz, D. (2016). Conservation Behavior: Applying Behavioral Ecology to Wildlife
Conservation and Management. Cambridge University Press.
Berger-Tal, O., Blumstein, D. T., Carroll, S., Fisher, R. N., Mesnick, S. L., Owen, M. A., Saltz, D.,
Claire, C. C. St., Swaisgood, R. R. (2015). A systematic survey of the integration of animal
behavior into conservation. Conservation Biology, 30(4), 744–753.
https://doi.org/10.1111/cobi.12654
Berger-Tal, O., Polak, T., Oron, A., Lubin, Y., Kotler, B. P., & Saltz, D. (2011). Integrating animal
behavior and conservation biology: a conceptual framework. Behavioral Ecology, 22(2),
236–239. https://doi.org/10.1093/beheco/arq224
Blumstein, D. T., Holland, B. D., Daniel, J. C. (2006). Predator discrimination and ‘personality’ in
captive Vancouver Island marmots (Marmota vancouverensis). Animal Conservation, 9(3),
274-282. https://doi.org/10.1111/j.1469-1795.2006.00033.x
Breitenmoser, U., & Haller, H. (1993). Patterns of predation by reintroduced European lynx in the
Swiss Alps. The Journal of Wildlife Management, 57(1), 135-144.
https://doi.org/10.2307/3809010
Breitenmoser, U., Ryser, A., Molinari-Jobin, A., Zimmermann, F., Haller, H., Molinari, P.,
Breitenmoser-Würsten, C. (2010). The changing impact of predation as a source of conflict
between hunters and reintroduced lynx in Switzerland. In D. W. Macdonald & A. J.
Loveridge (Eds.), Biology and Conservation of Wild Felids (pp. 493-505). Oxford University
Press.

70
Bremner-Harrison, S., Prodohl, P. A., Elwood, R. W. (2004). Behavioural trait assessment as a
release criterion: boldness predicts early death in a reintroduction programme of captive-
bred swift fox (Vulpes velox). Animal Conservation, 7(3), 313-320.
https://doi.org/10.1017/S1367943004001490
Brown, J. L. (1975). The evolution of behavior (No. 591.5 B7).
Brown, J. L., & Choe J. C. (2019). Behavioral Ecology and Sociobiology. In J. C. Choe (Ed.),
Encyclopedia of Animal Behavior (2nd Ed., Vol. 1, pp. 103-108). Elsevier.
https://doi.org/10.1016/B978-0-12-809633-8.20838-8
Cabrera, A. (1940). Notas sobre carnívoras sudamericanos. Notas del Museo de La Plata. Sección
Zoología, 5(29), 1-22.
Callou, C., Samzun, A., Zivie, A. (2004). Archaeology: a lion found in the Egyptian tomb of Maïa.
Nature, 427, 211–212. https://doi.org/10.1038/427211a
Carbone, C., Mace, G. M., Roberts, S. C., Macdonald, D. W. (1999). Energetic constraints on the
diet of terrestrial carnivores. Nature, 402, 286–288. https://doi.org/10.1038/46266
Carlstead, K. (1996). Effects of captivity on the behaviour of wild animals. In D. G. Kleiman, M. E.
Allen, K. V. Thompson & S. Lumpkin (Eds.), Wild Mammals in Captivity: Principles and
Techniques (1st Ed., pp. 317-333). Chicago: University of Chicago Press.
Carlstead, K., Fraser, J., Bennett, C., Kleiman, D. G. (1999b). Black rhinoceros (Diceros bicornis) in
U.S. zoos: II. Behaviour, breeding success and mortality in relation to housing facilities.
Zoo Biology, 18(1), 35-52. https://doi.org/10.1002/(SICI)1098-2361(1999)18:1<35::AID-
ZOO5>3.0.CO;2-L
Carlstead, K., Mellen, J., Kleiman, D.G. (1999a). Black rhinoceros (Diceros bicornis) in U.S. zoos: I.
individual behavior profiles and their relationship to breeding success. Zoo Biology, 18(1),
17-34. https://doi.org/10.1002/(SICI)1098-2361(1999)18:1<17::AID-ZOO4>3.0.CO;2-K
Carlstead, K., Shepherdson, D. J., Sheppard, C., Mellen, J., Ogden, J., Bennett, C. (2000).
Constructing Behaviour Profiles of Zoo Animals: Incorporating Behavioural Information
into Captive Population Management. Oregon Zoo: American Zoo and Aquarium
Association's Behaviour and Husbandry Advisory Group.

71
Caro, T. (2007). Behavior and conservation: a bridge too far?. Trends in ecology & evolution, 22(8),
394-400. https://doi.org/10.1016/j.tree.2007.06.003
Chadwick, C. L., & O’Hara, S. J. (2014). Social group cohesion and reproductive success: a survey of
captive cheetah (Acinonyx jubatus) personality. Poster abstract research showcase College
of Science and Technology, University of Salford, pp. 135.
Coleman, K. (2012). Individual differences in temperament and behavioral management practices
for nonhuman primates. Applied Animal Behaviour Science, 137(3–4), 106-113.
https://doi.org/10.1016/j.applanim.2011.08.002
Conrad, J. L., Weinersmith, K. L., Brodin, T., Saltz, J. B., Sih, A. (2011). Behavioural syndromes in
fishes: a review with implications for ecology and fisheries management. Journal of Fish
Biology, 78(2), 395-435. https://doi.org/10.1111/j.1095-8649.2010.02874.x
Cornalia, E. (1865). Descrizione di una nuova specie del genere: Felis, Felis jacobita. Memorie della
Societa Italiana di Scienze Naturalli (1st Ed., Vol. 1). Coi tipi di G. Bernardoni.
Cossíos, E. D., Madrid, A., Condori, J. L., Fajardo, U. (2007). Update on the distribution of the
Andean cat Oreailurus jacobita and the pampas cat Lynchailurus colocolo in Peru.
Endangered Species Research, 3(3), 313-320. https://doi.org/10.3354/esr00059
Cossíos, E. D., Walker, R. S., Lucherini, M., Ruiz-García, M., Angers, B. (2012). Population structure
and conservation of a high-altitude specialist, the Andean cat Leopardus jacobita.
Endangered Species Research, 16(3), 283-294. https://doi.org/10.3354/esr00402
Craft, M. E., Volz, E., Packer, C., Meyers, L. A. (2009). Distinguishing epidemic waves from disease
spillover in a wildlife population. Proceedings of the Royal Society B, 276(1663), 1777–
1785. https://doi.org/10.1098/rspb.2008.1636
Culliney, S., Pejchar, L., Switzer, R., Ruiz-Gutierrez, V. (2012). Seed dispersal by a captive corvid:
The role of the ‘Alala (Corvus hawaiiensis) in shaping Hawai‘i׳s plant communities.
Ecological Applications 22(6), 1718–1732. https://doi.org/10.1890/11-1613.1
Damania, R., Seidensticker, J., Whitten, T., Sethi, G., Mackinnon, K., Kiss, A., Kushlin, A. (2008). A
Future for Wild Tigers. World Bank.

72
Davis, B. W., Li, G., Murphy, W. J. (2010). Supermatrix and species tree methods resolve
phylogenetic relationships within the big cats, Panthera (Carnivora: Felidae). Molecular
Phylogenetics and Evolution, 56(1), 64–76. https://doi.org/10.1016/j.ympev.2010.01.036
DeCaluwe, H. B., Wielebnowski, N. C., Howard, J., Pelican, K. M., Ottinger, M. A. (2013). Behavioral
reactions relate to adrenal activity and temperament in male clouded leopards (Neofelis
nebulosa). Applied Animal Behaviour Science, 149(1-4), 63-71.
https://doi.org/10.1016/j.applanim.2013.09.008
Delibes, M. (1980). Feeding ecology of the Spanish lynx in the Coto Dońana. Acta Theriologica,
25(24), 309-324.
Delibes, M., Rodrıguez, A., Ferreras, P. (2000). Action Plan for the Conservation of the Iberian Lynx
(Lynx pardinus) in Europe, no. 111. Council of Europe Publishing. Nature and Environment,
Strasbourg, France.
Devineau, O., Shenk, T. M., White, G. C., Doherty Jr, P. F., Lukacs, P. M., Kahn, R. H. (2010).
Evaluating the Canada lynx reintroduction programme in Colorado: patterns in mortality.
Journal of applied Ecology, 47(3), 524-531. https://doi.org/10.1111/j.1365-
2664.2010.01805.x
Devkota, B. P., Thakur, S., Jaromir, K. (2013). Prey density and diet of Snow Leopard (Uncia uncia)
in Shey Phoksundo National Park, Nepal. Applied Ecology and Environmental Sciences,
1(4), 55–60. https://doi.org/10.12691/aees-1-4-4
Dicks, L. V., Walsh, J. C., Sutherland, W. J. (2014). Organising evidence for environmental
management decisions: a ‘4S’ hierarchy. Trends in Ecology & Evolution, 29(11), 607–613.
https://doi.org/10.1016/j.tree.2014.09.004
Dingemanse, N. J., & Réale, D. (2005). Natural selection and animal personality. Behaviour, 142(9-
10), 1159-1184. https://doi.org/10.1163/156853905774539445
Dingemanse, N. J., Both, C., Drent, P., J., Tinbergen, J. M. (2004). Fitness consequences of avian
personalities in a fluctuating environment. Proceedings of the Royal Society of London B,
271(1541), 847-852. https://doi.org/10.1098/rspb.2004.2680
Dobson, F. S., Yu, J., Smith, A. T. (1995). The importance of evaluating rarity. Conservation Biology,
9(6), 1648–1651. https://doi.org/10.1046/j.1523-1739.1995.09061648.x

73
Drent, P. J., van Oers, K., van Noordwijk, A. J. (2003). Realized heritability of personalities in the
great tit (Parus major). Proceedings of the Royal Society B, 270(1510), 45-51.
https://doi.org/10.1098/rspb.2002.2168
Dudley, N., Higgins-Zogib, L., Mansourian, S. (2009). The links between protected areas, faiths,
and sacred natural sites. Conservation Biology, 23(3), 568–577.
https://doi.org/10.1111/j.1523-1739.2009.01201.x
Dunston, E. J., Abell, J., Doyle, R. E., Evershed, M., Freire, R. (2016). Exploring African lion
(Panthera leo) behavioural phenotypes: individual differences and correlations between
sociality, boldness and behaviour. Journal of Ethology, 34, 277–290.
https://doi.org/10.1007/s10164-016-0473-9
Dutton, D. M. (2008). Subjective assessment of chimpanzee (Pan troglodytes) personality:
reliability and stability of trait ratings. Primates, 49(4), 253-259.
https://doi.org/10.1007/s10329-008-0094-1
Dutton, D. M., Clark, R. A., Dickins, D. W. (1997). Personality in captive chimpanzees: use of a
novel rating procedure. International Journal of Primatology, 18(4), 539-552.
https://doi.org/10.1023/A:1026311222491
Ehrlich, P. R. (2001). Intervening in evolution: ethics and actions. Proceedings of the National
Academy of Sciences of the United States of America, 98(10), 5477–5480.
https://doi.org/10.1073/pnas.091093898
Fazio, J. (2010). Behavioral assessment of the clouded leopard (Neofelis nebulosa); a comparative
study of reproductive success. Master Thesis, George Mason University.
Feaver, J., Mendl, M., Bateson, P. (1986). A method for rating the individual distinctiveness of
domestic cats. Animal Behaviour, 34(4), 1016-1025. https://doi.org/10.1016/S0003-
3472(86)80160-9
Fernández, N., Delibes, M., Palomares, F. (2006). Landscape evaluation in conservation: molecular
sampling and habitat modeling for the Iberian lynx. Ecological Applications, 16(3), 1037–
1049. https://doi.org/10.1890/1051-0761(2006)016[1037:LEICMS]2.0.CO;2
Fernández, N., Delibes, M., Palomares, F. (2007). Habitat-related heterogeneity in breeding in a
metapopulation of the Iberian lynx. Ecography, 30(3), 431-439.
https://doi.org/10.1111/j.0906-7590.2007.05005.x

74
Fernández, N., Delibes, M., Palomares, F., Mladenoff, D. J. (2003). Identifying breeding habitat for
the Iberian lynx: inferences from a fine-scale spatial analysis. Ecological Applications,
13(5), 1310–1324. https://doi.org/10.1890/02-5081
Fernández, N., Palomares, F., Delibes, M. (2002). The use of breeding dens and kitten
development in the Iberian lynx (Lynx pardinus). Journal of Zoology, 258(1), 1-5.
https://doi.org/10.1017/S0952836902001140
Ferrer, M., & Negro, J. J. (2004). The Near Extinction of Two Large European Predators: Super
Specialists Pay a Price. Conservation Biology, 18(2), 344-349.
https://doi.org/10.1111/j.1523-1739.2004.00096.x
Ferreras, P., Rodríguez, A., Palomares, F., Delibes, M. (2010). Iberian lynx: the difficult recovery of
a critically endangered cat. In D. W. Macdonald & A. J. Loveridge (Eds.), Biology and
Conservation of Wild Felids (pp. 507-520). Oxford University Press.
Ferretti, F., Lovari, S., Minder, I., Pellizzi, B. (2014). Recovery of the snow leopard in Sagarmatha
(Mt. Everest) National Park: effects on main prey. European Journal of Wildlife Research,
60, 559–562. https://doi.org/10.1007/s10344-014-0810-7
Fordham, D. A., Akçakaya, H. R., Brook, B. W., Rodríguez, A., Alves, P. C., Civantos, E., Triviño, M.,
Watts, M. J., Araújo, M. B. (2013). Adapted conservation measures are required to save
the Iberian lynx in a changing climate. Nature Climate Change, 3, 899-903.
https://doi.org/10.1038/nclimate1954
Fox, J. L., & Chundawat, R. S. (2016). What is a snow leopard? behavior and ecology. In T.
McCarthy & D. Mallon (Eds.), Snow Leopards, Biodiversity of the World: Conservation from
Genes to Landscapes (pp. 13-21). Academic Press. https://doi.org/10.1016/B978-0-12-
802213-9.00002-X
Freeman, E. W., Schulte, B. A., Brown, J. L. (2010). Using behavioral observations and keeper
questionnaires to assess social relationships among captive female African elephants. Zoo
Biology, 29(2), 140-153. https://doi.org/10.1002/zoo.20249
Freeman, E. W., Weiss, E., Brown, J. L. (2004). Examination of the interrelationships of behavior,
dominance status, and ovarian activity in captive Asian and African elephants. Zoo
Biology, 23(5), 431-448. https://doi.org/10.1002/zoo.20025

75
Freeman, H. D., & Gosling, S. D. (2010). Personality in nonhuman primates: a review and
evaluation of past research. American Journal of Primatology, 72(8), 653-671.
https://doi.org/10.1002/ajp.20833
Fuller, T. K., Berg, W. E., Kuehn, D. W. (1985). Survival rates and mortality factors of adult bobcats
in north-central Minnesota. The Journal of wildlife management, 49(2), 292-296.
https://doi.org/10.2307/3801518
García, I., Napp, S., Zorrilla, I., Vargas, A., Pastor, J., Muñoz, Á., & Martínez, F. (2010).
Determination of serum biochemical reference intervals for the Iberian lynx (Lynx
pardinus). The Veterinary Journal, 183(2), 201-204.
https://doi.org/10.1016/j.tvjl.2008.10.012
García-Perea, R. (2002). Andean mountain cat, Oreailurus jacobita: Morphological description and
comparison with other felines from the Altiplano. Journal of Mammalogy, 83(1), 110-124.
https://doi.org/10.1644/1545-1542(2002)083<0110:AMCOJM>2.0.CO;2
Gartner, M. C., & Powell, D. (2012). Personality assessment in snow leopards (Uncia uncia). Zoo
Biology, 31(2), 151-165. https://doi.org/10.1002/zoo.20385
Gartner, M. C., & Weiss, A. (2013a). Personality in felids: a review. Applied Animal Behaviour
Science, 144(1–2), 1-13. https://doi.org/10.1016/j.applanim.2012.11.010
Gartner, M. C., & Weiss, A. (2013b). Scottish wildcat (Felis silvestris grampia) personality and
subjective well-being: implications for captive management. Applied Animal Behaviour
Science, 147(3–4), 261-267. https://doi.org/10.1016/j.applanim.2012.11.002
Gartner, M. C., Powell, D. M., Weiss, A. (2014). Personality Structure in the Domestic Cat (Felis
silvestris catus), Scottish Wildcat (Felis silvestris grampia), Clouded Leopard (Neofelis
nebulosa), Snow Leopard (Panthera uncia), and African Lion (Panthera leo): A
Comparative Study. Journal of Comparative Psychology, 128(4), 414–426.
https://doi.org/10.1037/a0037104
Gartner, M. C., Powell, D. M., Weiss, A. (2016). Comparison of Subjective Well Being and
Personality Assessments in the Clouded Leopard (Neofelis nebulosa), Snow Leopard
(Panthera uncia), and African Lion (Panthera leo). Journal of Applied Animal Welfare
Science, 19(3), 294-302. https://doi.org/10.1080/10888705.2016.1141057

76
Gilbert-Norton, L. B., Wilson, R. R., Shivik, J. A. (2013). The effect of social hierarchy on captive
coyote (Canis lantrans) foraging behavior. Ethology, 119(4), 335-343.
https://doi.org/10.1111/eth.12070
Gil-Sánchez, J. M., Ballesteros-Duperón, E., Bueno-Segura, J. F. (2006). Feeding ecology of the
Iberian lynx Lynx pardinus in eastern Sierra Morena (Southern Spain). Acta Theriologica,
51, 85–90. https://doi.org/10.1007/BF03192659
Gittleman, J. L. (1989). Carnivore Group Living: Comparative Trends. In J. L. Gittleman (Ed.),
Carnivore Behavior, Ecology, and Evolution (pp. 183-207) Springer, Boston, MA.
https://doi.org/10.1007/978-1-4757-4716-4_8
Godoy, J. A., Casas-Marcé, M., Fernández, J. (2009). Genetic issues in the implementation of the
Iberian lynx ex situ conservation programme. In A. Vargas (Ed.), Iberian Lynx Ex situ
Conservation: An Interdisciplinary Approach (pp. 86-99). Fundación Biodiversidad, Madrid.
Gomes, C. B., & Sarrazin, F. (2016). From individual behavior to population viability: Implications
for conservation and management. In O. Berger-Tal & D. Saltz (Eds.), Conservation
Behavior: Applying Behavioral Ecology to Wildlife Conservation and Management (Vol. 21,
pp. 247-274). Cambridge University Press.
https://doi.org/10.1017/CBO9781139627078.013
Gosling, S. D. (2001). From mice to men: what can we learn about personality from animal
research?. Psychological Bulletin, 127(1), 45-86. https://doi.org/10.1037/0033-
2909.127.1.45
Gosling, S. D. (2008). Personality in non-human animals. Social and Personality Psychology
Compass, 2(2), 985-1001. https://doi.org/10.1111/j.1751-9004.2008.00087.x
Gosling, S. D., & John, O. P. (1999). Personality dimensions in nonhuman animals: a cross-species
review. Current Directions in Psychological Science, 8(3), 69-75.
https://doi.org/10.1111/1467-8721.00017
Gosling, S. D., & Vazire, S. (2002). Are we barking up the right tree? Evaluating a comparative
approach to personality. Journal of Research in Personality, 36(6), 607-614.
https://doi.org/10.1016/S0092-6566(02)00511-1

77
Goswami, S., Tyagi, P. C., Malik, P. K., Pandit, S. J., Kadivar, R. F., Fitzpatrick, M., Mondol, S. (2020).
Effects of personality and rearing-history on the welfare of captive Asiatic lions (Panthera
leo persica). PeerJ 8:e8425. https://doi.org/10.7717/peerj.8425
Griffin, A. S., Blumstein, D. T., Evans, C. S. (2000). Training captive-bred or translocated animals to
avoid predators. Conservation Biology, 14(5), 1317–1326. https://doi.org/10.1046/j.1523-
1739.2000.99326.x
Guzmán, J. N., García, F. J., Garrote, G., Pérez de Ayala, R., Iglesias, C. (2004). El lince ibérico (Lynx
pardinus) en España y Portugal. Censo-diagnóstico de sus poblaciones. Dirección General
para la Biodiversidad, Madrid.
Hansen, S. W., & Møller, S. H. (2001). The application of a temperament test to on-farm selection
of mink. Acta Agriculturae Scandinavica, Section A – Animal Science, 51(30), 93-98.
https://doi.org/10.1080/090647001316923144
Hayward, M. W., Adendorff, J., Moolman, L., Hayward, G. J., Kerley, G. I. (2007). The successful
reintroduction of leopard Panthera pardus to the Addo Elephant National Park. African
Journal of Ecology, 45(1), 103. https://doi.org/10.1111/j.1365-2028.2006.00673.x
Heinroth, O. (1910). Beiträge zur Biologie: Namentlich Ethologie und Psychologie der Anatiden.
Verhandlungen des 5. Internationalen Ornithologen-Kongresses in Berlin. Deutsche
Ornithologische Gesellschaft, Berlin.
Heinroth, O. (1924-1934). Die Vögel Mitteleuropas. (4 volumes). Bermühler, Berlin.
Heithaus, M. R., Wirsing, A. J., Frid, A., Dill, L. M. (2007). Behavioral indicators in marine
conservation: lessons from a pristine seagrass ecosystem. Israel Journal of Ecology and
Evolution, 53(3-4), 355–370. https://doi.org/10.1560/IJEE.53.3.355
Hemmer, H. (1972). Uncia uncia. Mammalian Species, 20, 1–5. https://doi.org/10.2307/3503882
Henle, K., Sarre, S., Wiegand, K. (2004). The role of density regulation in extinction processes and
population viability analysis. Biodiversity & Conservation, 13, 9–52.
https://doi.org/10.1023/B:BIOC.0000004312.41575.83
Hennessy, M. B., Voith, V. L., Mazzei, S. J., Buttram, J., Miller, D. D., Linden, F. (2001). Behavior
and cortisol levels of dogs in a public animal shelter, and an exploration of the ability of

78
these measures to predict problem behavior after adoption. Applied Animal Behaviour
Science, 73(3), 217-233. https://doi.org/10.1016/S0168-1591(01)00139-3
Heptner, V. G., & Sludskii, A. A. (1992). Mammals of the Soviet Union. Part 2 Carnivora (hyaenas
and cats) (Vol. II). E. J. Brill Publishing Company, Leiden, Netherlands.
Hessing, M. J. C., Schouten, W. G. P., Wiepkema, P. R., Tielen, M. J. M. (1994). Implications of
individual behavioural characteristics on performance in pigs. Livestock Production
Science, 40(2), 187-196. https://doi.org/10.1016/0301-6226(94)90048-5
Highfill, L., Hanbury, D., Kristiansen, R., Kuczaj, S., Watson, S. (2010). Rating vs. coding in animal
personality research. Zoo Biology, 29(4), 509-516. https://doi.org/10.1002/zoo.20279
Hill, S. P., & Broom, D. M. (2009). Measuring zoo animal welfare: theory and practice. Zoo Biology,
28(6), 531-544. https://doi.org/10.1002/zoo.20276
Hillard, D., Weddle, M., Padmanabhan, S., Ale, S., Khuukhenduu, T., Almashev, C. (2016).
Environmental education for snow leopard conservation. In T. McCarthy & D. Mallon
(Eds.), Snow Leopards, Biodiversity of the World: Conservation from Genes to Landscapes
(pp. 245-255). Academic Press. https://doi.org/10.1016/B978-0-12-802213-9.00017-1
Horback, K. M., Miller, L. J., Kuczaj II, S. A. (2013). Personality assessment in African elephants
(Loxodonta africana): comparing the temporal stability of ethological coding versus trait
rating. Applied Animal Behaviour Science, 149(1-4), 55-62.
https://doi.org/10.1016/j.applanim.2013.09.009
Hosey, G. (2005). How does the zoo environment affect the behaviour of captive primates?.
Applied Animal Behaviour Science, 90(2), 107-129.
https://doi.org/10.1016/j.applanim.2004.08.015
Hsu, Y., & Serpell, J. A. (2003). Development and validation of a questionnaire for measuring
behavior and temperament traits in pet dogs. Journal of the American Veterinary Medical
Association, 223(9), 1293-1300. https://doi.org/10.2460/javma.2003.223.1293
Iberlince LIFE Project. (2014). Recovering the historic distribution range of the Iberian lynx (Lynx
pardinus) in Spain and Portugal. (LIFE10 NAT/ES/000570).

79
Iberlince. (2021). Visor del Proyecto Life+IBERLINCE.
https://iberlince.maps.arcgis.com/apps/webappviewer/index.html?id=cabd0fcebcd947ac
8d0c8c6afcd7b07e
Ikuta, L. A., & Blumstein, D. T. (2003). Do fences protect birds from human disturbance?.
Biological Conservation, 112(3), 447–452. https://doi.org/10.1016/S0006-3207(02)00324-
5
Inskip, C. & Zimmermann, A. (2009) Human–felid conflict: a review of the patterns and priorities
worldwide. Oryx, 43(1), 18-34. https://doi.org/10.1017/S003060530899030X
IUCN (2021). The IUCN Red List of Threatened Species. (Version 2021-1).
https://www.iucnredlist.org
IUCN/SSC. (2013). Guidelines for Reintroductions and Other Conservation Translocations. (Version
1.0). Gland, Switzerland: IUCN Species Survival Commission, viiii + 57 pp.
Jackson, R. M. & Wangchuk, R. (2004). A community-based approach to mitigating livestock
depredation by snow leopards. Human Dimensions to Wildlife, 9(4), 1-16.
https://doi.org/10.1080/10871200490505756
Jackson, R. M. (1996). Home range, movements and habitat use of snow leopard (Uncia uncia) in
Nepal. Ph.D. Thesis, University of London.
Jackson, R. M. (2015). HWC Ten Years Later: Successes and Shortcomings of Approaches to Global
Snow Leopard Conservation. Human Dimensions of Wildlife, 20(4), 310-316.
https://doi.org/10.1080/10871209.2015.1005856
Jackson, R. M., & Ahlborn, G. (1988). Observations on the ecology of snow leopard in west Nepal.
In H. Freeman (Ed), Proceedings of the Fifth International Snow Leopard Symposium (pp.
65-97). International Snow Leopard Trust and Wildlife Institute of India, Seattle.
Jackson, R. M., & Lama, W. B. (2016). The role of mountain communities in snow leopard
conservation. In T. McCarthy & D. Mallon (Eds.), Snow Leopards, Biodiversity of the World:
Conservation from Genes to Landscapes (pp. 139-149). Academic Press.
https://doi.org/10.1016/B978-0-12-802213-9.00011-0

80
Jackson, R. M., Mishra, C., McCarthy, T. M., Ale, S. B. (2010). Snow leopards, conflict and
conservation. In D. W. Macdonald & A. J. Loveridge (Eds.), Biology and Conservation of
Wild Felids (pp. 417-430). Oxford University Press.
Jetz, W., Carbone, C., Fulford, J., Brown, J. H. (2004). The scaling of animal space use. Science,
306(5694), 266–268. https://doi.org/10.1126/science.1102138
Jewgenov, K., Braun, B. C., Göritz, F., Voigt, C. C., Martínez, F., Anaya, L., Vargas, A., Dehnhard, M.
(2009). Pregnancy diagnosis in Iberian lynx (Lynx pardinus) based on urinary and blood
plasma hormones. In A. Vargas (Ed.), Iberian Lynx Ex situ Conservation: An
Interdisciplinary Approach (pp. 376- 389). Fundación Biodiversidad, Madrid.
Johnson, D. D., Baker, S., Morecroft, M. D., Macdonald, D. W. (2001). Long-term resource
variation and group size: a large-sample field test of the resource dispersion hypothesis.
BMC Ecology, 1(2). https://doi.org/10.1186/1472-6785-1-2
Johnson, W. E., & O’Brien, S. J. (1997). Phylogenetic reconstruction of the Felidae using 16S rRNA
and NADH-5 mitochondrial genes. Journal of Molecular Evolution, 44(1), 98-116.
https://doi.org/10.1007/PL00000060
Johnson, W. E., Culver, M., Iriarte, J. A., Eizirik, E., Seymour, K. L. O'Brien, S. J. (1998). Tracking the
Evolution of the Elusive Andean Mountain Cat (Oreailurus jacobita) From Mitochondrial
DNA. Journal of Heredity, 89(3), 227-232. https://doi.org/10.1093/jhered/89.3.227
Johnson, W. E., Eizirik, E., Pecon-Slattery, J., Murphy, W. J., Antunes, A., Teeling, E., O'Brien, S. J.
(2006). The late Miocene radiation of modern Felidae: a genetic assessment. Science,
311(5757), 73-77. https://doi.org/10.1126/science.1122277
Jones, A. C., & Gosling, S. D. (2005). Temperament and personality in dogs (Canis familiaris): a
review and evaluation of past research. Applied Animal Behaviour Science, 95(1-2), 1-53.
https://doi.org/10.1016/j.applanim.2005.04.008
Jumabay-Uulu, K., Wegge, P., Mishra, C., Sharma, K. (2014). Large carnivores and low diversity of
optimal prey: a comparison of the diets of snow leopards Panthera uncia and wolves
Canis lupus in Sarychat-Ertash Reserve in Kyrgyzstan. Oryx 48(4), 529–535.
https://doi.org/10.1017/S0030605313000306
Kappeler, P. (2010). Animal Behavior: Evolution and Mechanisms. Springer, Berlin.
https://doi.org/10.1007/978-3-642-02624-9

81
Karanth, K. U., & Chellam, R. (2009). Carnivore conservation at the crossroads. Oryx, 43(1), 1-2.
https://doi.org/10.1017/S003060530843106X
Kelley, J. L., & García, C. M. (2019). Captive Breeding, Conservation and Welfare. In J. C. Choe
(Ed.), Encyclopedia of Animal Behavior (2nd Ed., Vol. 1, pp. 291-301). Elsevier.
https://doi.org/10.1016/B978-0-12-809633-8.01128-6
Kelly, P., Stack, D., Harley, J. (2013). A review of the proposed reintroduction program for the Far
Eastern leopard (Panthera pardus orientalis) and the role of conservation organizations,
veterinarians, and zoos. Topics in companion animal medicine, 28(4), 163-166.
https://doi.org/10.1053/j.tcam.2013.09.004
King, J. E., & Figueredo, A. J. (1997). The Five-Factor Model plus Dominance in chimpanzee
personality. Journal of Research in Personality, 31(2), 257-271.
https://doi.org/10.1006/jrpe.1997.2179
King, J. E., Weiss, A., Farmer, K. H. (2005). A chimpanzee (Pan troglodytes) analogue of cross-
national generalization of personality structure: zoological parks and an African sanctuary.
Journal of Personality, 73(2), 389-410. https://doi.org/10.1111/j.1467-6494.2005.00313.x
Kitchener, A. C., Driscoll, C. A., Yamaguchi, N. (2016). What is a snow leopard? Taxonomy,
morphology, and phylogeny. In T. McCarthy & D. Mallon (Eds.), Snow Leopards,
Biodiversity of the World: Conservation from Genes to Landscapes (pp. 3-11). Academic
Press. https://doi.org/10.1016/B978-0-12-802213-9.00001-8
Kitchener, A. C., van Valkenburgh, B., Yamaguchi, N., Macdonald, D. W., Loveridge, A. J. (2010).
Felid form and function. In D. W. Macdonald & A. J. Loveridge (Eds.), Biology and
Conservation of Wild Felids (pp. 83-106). Oxford University Press.
Klopfer, P. H. (1962). Behavioral Aspects of Ecology. Prentice-Hall, Inc, Englewood Cliffs, NJ.
Koshkarev, E. P. (1984). Characteristics of snow leopard (Uncia uncia) movements in the Tien
Shan. In L. Blomqvist (Ed.), International Pedigree Book of Snow Leopards, Panthera uncia
(Vol. 4, pp. 15–21). Helsinki Zoo, Helsinki.
Kotler, B. P., Morris, D. W., Brown, J. S. (2007). Behavioral indicators and conservation: wielding
‘‘The Biologist’s Tricorder’’. Israel Journal of Ecology and Evolution, 53(3-4), 237–244.
https://doi.org/10.1560/IJEE.53.3.237

82
Krebs, J. R., & Davies, N. B. (1978). Behavioural Ecology: An Evolutionary Approach. (1st Ed.).
Sinauer Associates.
Krebs, J. R., & Davies, N. B. (1981). An Introduction to Behavioural Ecology (1st Ed.). Sinauer
Associates.
Kruuk, H. (1976) Feeding and social behaviour of the striped hyena (Hyaena vulgaris Desmarest).
African Journal of Ecology, 14(2), 91–111. https://doi.org/10.1111/j.1365-
2028.1976.tb00155.x
Kuhar, C. W., Stoinski, T. S., Lukas, K. E., Maple, T. L. (2006). Gorilla Behaviour Index revisited: age,
housing and behaviour. Applied Animal Behaviour Science, 96(3-4), 315-326.
https://doi.org/10.1016/j.applanim.2005.06.004
Kunkel, K., Hussain, S., Khatiwada, A. (2016). A review of lessons, successes, and pitfalls of
livestock insurance schemes. In T. McCarthy & D. Mallon (Eds.), Snow Leopards,
Biodiversity of the World: Conservation from Genes to Landscapes (pp. 173-178).
Academic Press. https://doi.org/10.1016/B978-0-12-802213-9.00013-4
Kwan, V. S. Y., Gosling, S. D., John, O. P. (2008). Anthropomorphism as a special case of social
perception: a cross-species social relations model analysis of humans and dogs. Social
Cognition, 26(2), 129-142. https://doi.org/10.1521/soco.2008.26.2.129
Law, G., & Tatner, P. (1998). Behaviour of a captive pair of clouded leopards (Neofelis nebulosa):
introduction without injury. Animal Welfare, 7(1), 57-76.
Leus, K., & Lacy, R. C. (2009). Genetic and demographic management of conservation breeding
programmes oriented towards reintroduction. In A. Vargas (Ed.), Iberian Lynx Ex situ
Conservation: An Interdisciplinary Approach (pp. 75-84). Fundación Biodiversidad, Madrid.
Lhagvasuren, B., & Munkhtsog, B. (2000). The yak population in Mongolia and its relation with
snow leopards as a prey species. In H. Jianlin, C. Richard, O. Hanotte, C. McVeigh, J. Rege,
(Eds.), Proceedings of the Third International Congress on Yak production in Central Asian
Highlands (pp. 69–75). International Livestock Research Institute, Lhasa, China.
Li, J., Wang, D., Yin, H., Zhaxi, D., Jiagong, Z., Schaller, G. B., Mishra, C., McCarthy, T. M., Wang, H.,
Wu, L., Xiao, L., Basang, L., Zhang, Y., Zhou, Y., Lu, Z. (2014). Role of Tibetan Buddhist
monasteries in snow leopard conservation. Conservation Biology, 28(1), 87-94.
https://doi.org/10.1111/cobi.12135

83
LIFE Program. (2021). LIFE Project Database.
https://ec.europa.eu/environment/life/project/Projects/index.cfm
Lindell, C. A. (2008). The value of animal behavior in evaluations of restoration success.
Restoration Ecology, 16(2), 197–203. https://doi.org/10.1111/j.1526-100X.2008.00389.x
Lindenmayer, D. B., Piggott, M. P., Wintle, B. A. (2013). Counting the books while the library
burns: why conservation monitoring programs need a plan for action. Frontiers in Ecology
and the Environment, 11(10), 549–555. https://doi.org/10.1890/120220
Linnell, J. D., Breitenmoser, U., Breitenmoser-Würsten, C., Odden, J., von Arx, M. (2009). Recovery
of Eurasian lynx in Europe: What Part has Reintroduction Played?. In M. W. Hayward & M:
J. Somers (Eds.), Reintroduction of Top-Order Predators, (pp. 72-91). Blackwell Publishing.
https://doi.org/10.1002/9781444312034.ch4
Lorenz, K. (1935). Der Kumpan in der Umwelt des Vogels. Journal für Ornithologie, 83, 137–213.
https://doi.org/10.1007/BF01905355
Lorenz, K. (1950). The comparative method in studying innate behavior patterns. In Society for
Experimental Biology, Physiological mechanisms in animal behavior. (Society's Symposium
IV.) (p. 221–268). Academic Press.
Lovari, S., Boesi, R., Minder, I., Mucci, N., Randi, E., Dematteis, A., Ale, S. B. (2009). Restoring a
keystone predator may endanger a prey species in a human-altered ecosystem: the
return of the snow leopard to Sagarmatha National Park. Animal Conservation, 12(6),
559–570. https://doi.org/10.1111/j.1469-1795.2009.00285.x
Loveridge, A. J., Wang, S. W., Frank, L. G., & Seidensticker, J. (2010). People and wild felids:
conservation of cats and management of conflicts. In D. W. Macdonald & A. J. Loveridge
(Eds.), Biology and Conservation of Wild Felids (pp. 161-195). Oxford University Press.
Lucherini, M., & Merino, M. J. (2008). Perceptions of Human-Carnivore Conflicts in the High Andes
of Argentina. Mountain Research and Development, 28(1), 81-85.
https://doi.org/10.1659/mrd.0903
Lucherini, M., & Vidal, E. L. (2003). Intraguild Competition as a Potential Factor Affecting the
Conservation of Two Endangered Cats in Argentina. Endangered Species UPDATE, 20(6),
211-220.

84
Lucherini, M., Reppucci, J. I., Vidal, E. L. (2009a). A comparison of three methods to estimate
geografical variation in the relative abundance of Mountain vizcacha (Lagidium vizcacia)
in the High Andes ecosystem. Mastozoología Neotropical, 16(1), 223-228.
Lucherini, M., Reppucci, J. I., Walker, S., Villalba, L., Wurstten, A., Gallardo, G., Iriarte, A.,
Villalobos, R., Perovic, P. (2009b). Activity pattern segregation of carnivores in the High
Andes. Journal of Mammalogy, 90(6), 1404-1409. https://doi.org/10.1644/09-MAMM-A-
002R.1
LYNXEXSITU. (2021). Programa De Conservación Ex-Situ Del Lince Ibérico.
https://www.lynxexsitu.es
Macdonald, D. W. (1983). The ecology of carnivore social behaviour. Nature, 301, 379–384.
https://doi.org/10.1038/301379a0
Macdonald, D. W., Loveridge, A. J., & Nowell, K. (2010a). Dramatis personae: an introduction to
the wild felids. In D. W. Macdonald & A. J. Loveridge (Eds.), Biology and Conservation of
Wild Felids (pp. 3-58). Oxford University Press.
Macdonald, D. W., Mosser, A., & Gittleman, J. L. (2010b). Felid Society. In D. W. Macdonald & A. J.
Loveridge (Eds.), Biology and Conservation of Wild Felids (pp. 125-160). Oxford University
Press
Mallon, D. (1984). The snow leopard in Ladakh. In L. Blomqvist (Ed.), International Pedigree Book
of Snow Leopards, Panthera uncia (Vol. 4, pp. 23–37). Helsinki Zoo, Helsinki.
Mallon, D., Harris, R. B., Wegge, P. (2016). Snow leopard prey and diet. In T. McCarthy & D.
Mallon (Eds.), Snow Leopards, Biodiversity of the World: Conservation from Genes to
Landscapes (pp. 43-55). Academic Press. https://doi.org/10.1016/B978-0-12-802213-
9.00004-3
Manor, R., & Saltz, D. (2003). Impact of human nuisance disturbance on vigilance and group size
of a social ungulate. Ecological Applications, 13(6), 1830–1834.
https://doi.org/10.1890/01-5354
Marma, B. B., & Yunchis, V. V. (1968). Observation on the breeding, management and physiology
of snow leopards, Panthera u. uncia, at Kaunas Zoo from 1962 to 1967. International Zoo
Yearbook, 8(1), 66–74. https://doi.org/10.1111/j.1748-1090.1968.tb00440.x

85
Martin, J. E. (2005). The influence of rearing on personality ratings of captive chimpanzees (Pan
troglodytes). Applied Animal Behaviour Science, 90(2), 167-181.
https://doi.org/10.1016/j.applanim.2004.08.019
Martin, P., & Bateson, P. (2007). Measuring Behaviour: An Introductory Guide (3rd Ed.). Cambridge
University Press.
Martínez, F., Chebez, J. C., Berlanga, P., Yacante, R., Nigro, N. A. (2008). Nueva localidad para el
gato andino Oreailurus jacobita (Cornalia, 1865) en la provincia de Mendoza, Argentina.
Nótulas Faunísticas, 2(26),1-5.
Martínez, F., López, G., Pastor, J., Zorrilla, I., Muñoz, A., García, I., Peña, L., Jiménez, M. A., Pérez,
M. J., Molina, I., Aguilar, J. M., Quevedo, M. A., Meli, M. L., Lutz, H., Vargas, A. (2009).
Integrating health issues into the conservation of the Iberian lynx (Lynx pardinus). In A.
Vargas (Ed.), Iberian Lynx Ex situ Conservation: An Interdisciplinary Approach (pp. 167-
168). Fundación Biodiversidad, Madrid.
Martin-Wintle, M. S., Shepherdson, D., Zhang, G., Huang, Y., Luo, B., Swaisgood, R. R. (2017). Do
opposites attract? Effects of personality matching in breeding pairs of captive giant
pandas on reproductive success. Biological Conservation 207, 27–37.
https://doi.org/10.1016/j.biocon.2017.01.010
Martin-Wintle, M. S., Shepherdson, D., Zhang, G., Zhang, H., Li, D., Zhou, X., Li, R., Swaisgood, R. R.
(2015). Free mate choice enhances conservation breeding in the endangered giant panda.
Nature Communications, 6. https://doi.org/10.1038/ncomms10125
McCarthy, T. M., & Chapron, G. (2003). Snow Leopard Survival Strategy. International Snow
Leopard Trust and Snow Leopard Network, Seattle, USA.
McCarthy, T. M., Fuller, T. K., Munkhtsog, B. (2005). Movements and activities of snow leopards in
southwestern Mongolia. Biological Conservation, 124(4), 527–537.
https://doi.org/10.1016/j.biocon.2005.03.003
McCarthy, T. M., Mallon, D., Jackson, R., Zahler, P., McCarthy, K. (2017). Panthera uncia. The IUCN
Red List of Threatened Species 2017: e.T22732A50664030.
http://dx.doi.org/10.2305/IUCN.UK.2017-2.RLTS.T22732A50664030.en
McDougall, P. T., Réale, D., Sol, D., Reader, S. M. (2006). Wildlife conservation and animal
temperament: causes and consequences of evolutionary change for captive,

86
reintroduced, and wild populations. Animal Conservation, 9(1), 39-48.
https://doi.org/10.1111/j.1469-1795.2005.00004.x
McKay, S. (2003). Personality profiles of the cheetah in the UK and Ireland, in relation to
environmental factors and performance variables. In T. C. Gilbert (Ed.), Proceedings of the
Fifth Annual Symposium on Zoo Research (pp. 177-189). Federation of Zoological Gardens
of Great Britain and Ireland.
McLean, I. G., Lundie-Jenkins, G., Jarman, P. J. (1996). Teaching an endangered mammal to
recognize predators. Biological Conservation, 75(1), 51–62. https://doi.org/10.1016/0006-
3207(95)00038-0
Meagher, R. K. (2009). Observer ratings: validity and value as a tool for animal welfare research.
Applied Animal Behaviour Science, 119(1-2), 1-14.
https://doi.org/10.1016/j.applanim.2009.02.026
Mishra, C., Bhatnagar, Y. V., Trivedi, P., Timbadia, R., Bijoor, A., Sonam, K., Thinley, S., Namgail, T.,
Prins, H. T. (2016). The role of village reserves in revitalizing the natural prey base of the
snow leopard. In T. McCarthy & D. Mallon (Eds.), Snow Leopards, Biodiversity of the
World: Conservation from Genes to Landscapes (pp. 184-187). Academic Press.
https://doi.org/10.1016/B978-0-12-802213-9.00014-6
Mohammad, G., Mostafawi, S., Rosen, T., Dadul, J. (2016). Corral improvements. In T. McCarthy &
D. Mallon (Eds.), Snow Leopards, Biodiversity of the World: Conservation from Genes to
Landscapes (pp. 180-183). Academic Press. https://doi.org/10.1016/B978-0-12-802213-
9.00014-6
Morgan, K. N., & Tromborg, C. T. (2007). Sources of stress in captivity. Applied Animal Behaviour
Science, 102(3-4), 262-302. https://doi.org/10.1016/j.applanim.2006.05.032
Mori, Y., Mitani, Y., Watanabe, Y., Sato, K. (2007). A behavioral indicator of prey patch richness
derived from diving behavior: the proportion of residence time to the standard time.
Israel Journal of Ecology & Evolution, 53(3-4), 347–354.
https://doi.org/10.1560/IJEE.53.3.347
Namgail, T., & Dadul, J. (2016). Himalayan homestays: fostering human-snow leopard
conservation coexistence. In T. McCarthy & D. Mallon (Eds.), Snow Leopards, Biodiversity

87
of the World: Conservation from Genes to Landscapes (pp. 164-168). Academic Press.
https://doi.org/10.1016/B978-0-12-802213-9.00013-4
Napolitano, C., Bennett, M., Johnson, W. E., O’Brien, S. J., Marquet, P. A., Barria, I., Poulin, E.,
Iriarte, A. (2008). Ecological and biogeographical inferences on two sympatric and
enigmatic Andean cat species using genetic identification of faecal samples. Molecular
Ecology, 17(2), 678-690. https://doi.org/10.1111/j.1365-294X.2007.03606.x
Nawaz, M. A., Din, J. U., Buzdar, H. (2016). The Ecosystem Health Program: a tool to promote the
coexistence of livestock owners and snow leopards. In T. McCarthy & D. Mallon (Eds.),
Snow Leopards, Biodiversity of the World: Conservation from Genes to Landscapes (pp.
188-195). Academic Press. https://doi.org/10.1016/B978-0-12-802213-9.00014-6
Norris, K. (2004). Managing threatened species: the ecological toolbox, evolutionary theory and
declining-population paradigm. Journal of Applied Ecology, 41(3), 413–426.
https://doi.org/10.1111/j.0021-8901.2004.00910.x
Novaro, A. Waker, S., Palacios, R., Di Martino, S., Monteverde, M., Canadell, S., Rivas, L., Cossíos,
E. D. (2010). Endangered Andean cat distribution beyond the Andes in Patagonia. Cat
News, 53, 8-10.
Nowell, K., Li, J., Paltsyn, M., Sharma, R. K. (2016). An Ounce of Prevention: Snow Leopard Crime
Revisited. TRAFFIC International, Cambridge, UK.
O'Brien, S. J., & Johnson, W. E. (2007). The evolution of cats. Scientific American, 297(1), 68-75.
Oli, M. K., Taylor, I. R., Rogers, D. M. E. (1993). Diet of the snow leopard (Panthera uncia) in the
Annapurna Conservation Area, Nepal. Journal of Zoology, 231(3), 365–370.
https://doi.org/10.1111/j.1469-7998.1993.tb01924.x
Owens, I. P. F. (2006). Where is behavioural ecology going?. Trends in Ecology & Evolution, 21(7),
356–361. https://doi.org/10.1016/j.tree.2006.03.014
Owen-Smith, N., & Cain, J. W. (2007). Indicators of adaptive responses in home range utilization
and movement patterns by a large mammalian herbivore. Israel Journal of Ecology and
Evolution, 53(3-4), 423–438. https://doi.org/10.1560/IJEE.53.3.423

88
Palomares, F. (2001). Vegetation structure and prey abundance requirements of the Iberian lynx:
implications for the design of reserves and corridors. Journal of Applied Ecology, 38(1), 9-
18. https://doi.org/10.1046/j.1365-2664.2001.00565.x
Palomares, F., Delibes, M., Ferreras, P., Fedriani, J. M., Calzada, J., Revilla, E. (2000). Iberian lynx in
a fragmented landscape: pre-dispersal, dispersal, and post-dispersal habitats.
Conservation Biology, 14(3), 809–818. https://doi.org/10.1046/j.1523-1739.2000.98539.x
Palomares, F., Delibes, M., Revilla, E., Calzada, J., Fedriani, J. M. (2001). Spatial ecology of Iberian
lynx and abundance of European rabbits in southwestern Spain. Wildlife Monographs,
148, 1-36.
Palomares, F., Godoy, J. A., López-Bao, J. V., Rodríguez, A., Roques, S., Casas-Marcé, M., Revilla, E.,
Delibes, M. (2012). Possible extinction vortex for a population of Iberian lynx on the verge
of extirpation. Conservation Biology, 26(4), 689-697. https://doi.org/10.1111/j.1523-
1739.2012.01870.x
Palomares, F., Rodríguez, A., Laffitte, R., Delibes, M. (1991). The status and distribution of the
Iberian lynx, Felis pardina (Temminck), in the Coto Doñana area, SW Spain. Biological
Conservation, 57(2), 159-169. https://doi.org/10.1016/0006-3207(91)90136-W
Palomares, F., Rodríguez, A., Revilla, E., Lopez-Bao, J. V., Calzada, J. (2011). Assessment of the
conservation efforts to prevent extinction of the Iberian lynx. Conservation Biology, 25(1),
4–8. https://doi.org/10.1111/j.1523-1739.2010.01607.x
Paltsyn, M., Poyarkov, A., Spitsyn, S., Kuksin, A., Istomov, S., Gibbs, J. P., Jackson, R. M., Castner, J.,
Kozlova, S., Karnaukhov, A., Malykh, S., Korablev, M., Zvychainaya, E., Rozhnov, V. (2016).
Northern Range: Russia. In T. McCarthy & D. Mallon (Eds.), Snow Leopards, Biodiversity of
the World: Conservation from Genes to Landscapes (pp. 501-511). Academic Press.
https://doi.org/10.1016/B978-0-12-802213-9.00040-7
Pastor, J., Bach-Raich, E., Mesalles, M., García, I., Martínez, F., Vargas, A., Cuenca, R., Lavín, S.
(2009). Haematological reference values for the Iberian lynx. In A. Vargas (Ed.), Iberian
Lynx Ex situ Conservation: An Interdisciplinary Approach (pp. 185- 196). Fundación
Biodiversidad, Madrid.

89
Pastorino, G. Q., Paini, F., Williams, C. L., Faustini, M., Mazzola, S. M. (2017a). Personality and
Sociality in Captive Tigers (Panthera tigris). Annual Research & Review in Biology, 21(2), 1-
17. https://doi.org/10.9734/ARRB/2017/38122
Pastorino, G. Q., Viau, A., Curone, G., Pearce-Kelly, P., Faustini, M., Vigo, D, Mazzola, S. M.,
Preziosi, R. (2017b). Role of Personality in Behavioral Responses to New Environments in
Captive Asiatic Lions (Panthera leo persica). Veterinary Medicine International, 2017, pp.
17. https://doi.org/10.1155/2017/6585380
Pe’er, G., Saltz, D., Thulke, H., Motro, U. (2004). Response to topography in a hilltopping butterfly
and implications for modeling nonrandom dispersal. Animal Behavior, 68(4), 825–839.
https://doi.org/10.1016/j.anbehav.2004.02.006
Pederson, A. K., King, J. E., Landau, V. I. (2005). Chimpanzee (Pan troglodytes) personality predicts
behavior. Journal of Research in Personality, 39(5), 534-549.
https://doi.org/10.1016/j.jrp.2004.07.002
Pelican, K. M., Abáigar, T., Vargas, A., Rodriguez, J. M., Bergara, J., López, J., Vázquez, A.,
Chaparro, J. M., Brown, J. L., Wildt, D. E. (2009). Unusual gonadal hormone profiles in the
Iberian lynx as determined by fecal monitoring. In A. Vargas (Ed.), Iberian Lynx Ex situ
Conservation: An Interdisciplinary Approach (pp. 340-351). Fundación Biodiversidad,
Madrid.
Penabad, L., Rivas, A., Calzada, J. (2012). Ritmos de actividad del lince ibérico (Lynx pardinus) en
Cautividad. Galemys, 24, 57-63. https://doi.org/10.7325/Galemys.2012.A06
Pereira, J. A., Fracassi, N. G., Rago, V., Ferreyra, H., Marull, C. A., McAloose, D., Uhart, M. M.
(2010). Causes of mortality in a Geoffroy’s cat population—a long-term survey using
diverse recording methods. European Journal of Wildlife Research, 56(6), 939-942.
https://doi.org/10.1007/s10344-010-0423-8
Pereira, J. A., Fracassi, N. G., Uhart, M. M. (2006). Numerical and spatial responses of Geoffroy’s
cat (Oncifelis geoffroyi) to prey decline in Argentina. Journal of Mammalogy, 87(6), 1132–
1139. https://doi.org/10.1644/05-MAMM-A-333R2.1
Perovic, P., Walker, S., Novaro, A. (2003). New records of the Endangered Andean mountain cat in
northern Argentina. Oryx, 37(3), 374-377. https://doi.org/10.1017/S0030605303000644

90
Phillips, C., & Peck, D. (2007). The effects of personality of keepers and tigers (Panthera tigris
tigris) on their behaviour in an interactive zoo exhibit. Applied Animal Behaviour Science,
106(4), 244-258. https://doi.org/10.1016/j.applanim.2007.01.007
Pocock, R. I. (1916). XXXVI. – On the tooth-change, cranial characters and classification of the
snow-leopard or ounce (Felis uncia). Annals and Magazine of Natural History, 18(105),
306–316. https://doi.org/10.1080/00222931608693854
Powell, D. M., & Gartner, M. C. (2011). Applications of Personality to the Management and
Conservation of Nonhuman Animals. In M. Inoue-Murayama, S. Kawamura & A. Weiss
(Eds.), From Genes to Animal Behaviour (pp. 185-199). Primatology Monographs.
Springer, Tokyo. https://doi.org/10.1007/978-4-431-53892-9_8
Powell, D. M., & Svoke, J. T. (2008). Novel environmental enrichment may provide a tool for rapid
assessment of animal personality: A case study with giant pandas (Ailuropoda
melanoleuca). Journal of Applied Animal Welfare Science, 11(4), 301-318.
https://doi.org/10.1080/10888700802329673
Powell, D., Lin, H., Carlstead, K., Kleiman, D. G., Zhang, H-M., Zhang, G-Q., Yu, J-Q., Zhang, J-G., Lu,
Y-P., Timothy, S. K. N., Tang, J., Snyder, R. (2008). Relationships between temperament,
husbandry, management and socio-sexual behavior in captive male and female giant
pandas (Ailuropoda melanoleuca). Acta Zoologica Sinica, 54(1), 169-175.
Primack, R., Rozzi, R. C., Feisinger, P. C., (2001). Fundamentos de Conservación Biológica. Fondo
de Cultura Económica, México.
Razal, C. B., Pisacane, C. B., Miller, L. J. (2016). Multifaceted approach to personality assessment in
cheetahs (Acinonyx jubatus). Animal Behavior and Cognition, 3(1), 22–31.
https://doi.org/10.12966/abc.03.01.2016
Réale, D., & Festa-Bianchet, M. (2003). Predator-induced natural selection on temperament in
bighorn ewes. Animal Behaviour, 65(3), 463-470.
https://doi.org/10.1006/anbe.2003.2100
Réale, D., Reader, S. M., Sol, D., McDougall, P. T., Dingemanse, N. J. (2007). Integrating animal
temperament within ecology and evolution. Biological Reviews, 82(2), 291-318.
https://doi.org/10.1111/j.1469-185X.2007.00010.x

91
Reiss, M. (1988). Scaling of home range size: body size, metabolic needs and ecology. Trends in
Ecology & Evolution, 3(3), 85–86. https://doi.org/10.1016/0169-5347(88)90025-0
Reppucci, J. I. (2012). Ecología y abundancia poblacional del gato andino (Leopardus jacobita) y
gato del pajonal (L. colocolo) en los altos Andes Argentinos. Tesis Doctoral, Universidad
Nacional del Sur, Bahía Blanca, Argentina.
Rieger, I. (1984). Tail functions in ounces, Uncia uncia. In L. Blomqvist (Ed.), International Pedigree
Book of Snow Leopards, Panthera uncia (Vol. 4, pp. 85–97). Helsinki Zoo, Helsinki.
Rivas, A., Boixader, J., Vargas, A., Pérez, M. J., Serra, R., Asensio, V., Sánchez, Í., Lobato, V.,
Vázquez, A., Pardo, A. J., Chaparro, M., Penabad, L., Reeves, J., Jiménez, O. M., Ferreira,
C., Yerga, J., Gonçalves, N., Azevedo, A., Ferreira, L., Mendoza, J. L., … & Klink, L. (2016).
Manual de manejo del lince ibérico en Cautividad. Programa de Conservación Ex-Situ del
Lince Ibérico.
Rodríguez, A. & Calzada, J. (2015). Lynx pardinus (errata version published in 2020). The IUCN Red
List of Threatened Species 2015: e.T12520A174111773.
https://dx.doi.org/10.2305/IUCN.UK.2015-2.RLTS.T12520A174111773.en
Rodríguez, A., & Delibes, M. (2002). Internal structure and patterns of contraction in the
geographic range of the Iberian lynx. Ecography, 25(3), 314-328.
https://doi.org/10.1034/j.1600-0587.2002.250308.x
Rodríguez, A., & Delibes, M. (2004). Patterns and causes of non-natural mortality in the Iberian
lynx during a 40-year period of range contraction. Biological Conservation, 118(2), 151-
161. https://doi.org/10.1016/j.biocon.2003.07.018
Roldán, E. R. S., Gomendio, M., Garde, J. J., Gañán, N., González, R., Crespo, C., Arregui, L. (2009).
A genetic resource bank and assisted reproduction for the critically endangered Iberian
lynx. In A. Vargas (Ed.), Iberian Lynx Ex situ Conservation: An Interdisciplinary Approach
(pp. 304-314). Fundación Biodiversidad, Madrid.
Rosen, T., & Zahler, P. (2016). Transboundary initiatives and snow leopard conservation In T.
McCarthy & D. Mallon (Eds.), Snow Leopards, Biodiversity of the World: Conservation from
Genes to Landscapes (pp. 267-276). Academic Press. https://doi.org/10.1016/B978-0-12-
802213-9.00019-5

92
Rouff, J. H., Sussman, R. W., Strube, M. J. (2005). Personality traits in captive lion-tailed macaques
(Macaca silenus). American Journal of Primatology, 67(2), 177-198.
https://doi.org/10.1002/ajp.20176
Rubenstein, D. (2016). Anthropogenic impacts on behavior: the pros and cons of plasticity. In O.
Berger-Tal & D. Saltz (Eds.), Conservation Behavior: Applying Behavioral Ecology to
Wildlife Conservation and Management (Vol. 21, pp. 121-146). Cambridge University
Press. https://doi.org/10.1017/CBO9781139627078.008
Rubenstein, D., & Alcock, J. (2018). Animal Behavior. (11th Ed). Sinauer Associates.
Saltz, D., Rowen, M., Rubenstein, D. I. (2000). The effect of space-use patterns of reintroduced
Asiatic wild ass on effective population size. Conservation Biology, 14(6), 1852–1861.
https://doi.org/10.1111/j.1523-1739.2000.99227.x
Sarmento, P., & Carrapato, C. (2019). The use of spatially explicit capture-recapture models for
estimating Iberian lynx abundance in a newly reintroduced population. Mammalian
Biology, 98(1), 11-16. https://doi.org/10.1016/j.mambio.2019.07.004
Sarmento, P., Carrapato, C., Eira, C., Silva, J. P. (2017). Spatial organization and social relations in a
reintroduced population of Endangered Iberian lynx Lynx pardinus. Oryx, 53(2), 344-355.
https://doi.org/10.1017/S0030605317000370
Schakner, Z. A., Petelle, M. B., Berger-Tal, O., Owen, M. A., Blumstein, D. T. (2014). Developing
effective tools for conservation behaviorists: Reply to Greggor et al.. Trends in ecology &
evolution, 29(12), 651-652. https://doi.org/10.1016/j.tree.2014.09.008
Schaller, G. B., Hong, L., Talipu, Hua, L.., Jurang, R., Mingjiang, Q., Haibin, W. (1987). Status of
large mammals in the Taxkorgan Reserve, Xinjiang, China. Biological Conservation, 42(1),
53–71. https://doi.org/10.1016/0006-3207(87)90052-8
Schaller, G. B., Hong, L., Talipu, Junrang, R., Mingjiang, Q. (1988). The snow leopard in Xinjiang.
Oryx, 22(4), 197–204. https://doi.org/10.1017/S0030605300022328
Schultz, C. B. (1998). Dispersal behavior and its implications for reserve design in a rare Oregon
butterfly. Conservation Biology, 12(2), 284–292. https://doi.org/10.1111/j.1523-
1739.1998.96266.x

93
Searle, K. R., Hobbs, N. T., Gordon, I. J. (2007). It’s the ‘‘foodscape’’, not the landscape: using
foraging behavior to make functional assessments of landscape condition. Israel Journal
of Ecology and Evolution, 53(3-4), 297–316. https://doi.org/10.1560/IJEE.53.3.297
Serpell, J. A., & Hsu, Y. (2001). Development and validation of a novel method for evaluating
behavior and temperament in guide dogs. Applied Animal Behaviour Science, 72(4), 347-
364. https://doi.org/10.1016/S0168-1591(00)00210-0
Shehzad, W., McCarthy, T. M., Pompanon, F., Purevjav, L., Coissac, E., Riaz, T., Taberlet, P. (2012).
Prey preference of snow leopard (Panthera uncia) in South Gobi, Mongolia. PLoS ONE
7(2), 1-8. https://doi.org/10.1371/journal.pone.0032104
Shen, X., Lu, Z., Li, S., Chen, N. (2012). Tibetan sacred sites: understanding the traditional
management system and its role in modern conservation. Ecology and Society, 17(2), 1-
11. http://dx.doi.org/10.5751/ES-04785-170213
Shier, D. M. (2006). Effect of family support on the success of translocated black-tailed prairie
dogs. Conservation Biology, 20(6), 1780–1790. https://doi.org/10.1111/j.1523-
1739.2006.00512.x
Sih, A., Bell A., Johnson, J. C. (2004). Behavioral syndromes: an ecological and evolutionary
overview. Trends in Ecology & Evolution, 19(7), 372–378.
https://doi.org/10.1016/j.tree.2004.04.009
Simón, M. A. (2018). Censo De Las Poblaciones De Lince Ibérico Año 2018.
https://www.iberlince.eu
Símon, M. A., Gil-Sanchez, J. M., Ruiz, G., Garrote, G., Mccain, E. B., Fernandez, L., López‐Parra,
M., Rojas, E., Arenas‐Rojas, R., del Rey, T., Garcia-Tardio, M., López, G. (2012). Reverse of
the decline of the endangered Iberian lynx. Conservation Biology, 26(4), 731-736.
https://doi.org/10.1111/j.1523-1739.2012.01871.x
Slabbert, J. M., & Odendaal, J. S. J. (1999). Early prediction of adult police dog efficiency - a
longitudinal study. Applied Animal Behaviour Science, 64(4), 269-288.
https://doi.org/10.1016/S0168-1591(99)00038-6
Smith, B. R., & Blumstein, D. T. (2008). Fitness consequences of personality: a meta-analysis.
Behavioral Ecology, 19(2), 448-455. https://doi.org/10.1093/beheco/arm144

94
Snow Leopard Network. (2014). Snow Leopard Survival Strategy (Version 2014.1). Snow Leopard
Network, Seattle, Washington, USA.
Snyder, N. F., Derrickson, S. R., Beissinger, S. R., Wiley, J. W., Smith, T. B., Toone, W. D., Miller, B.
(1996). Limitations of captive breeding in endangered species recovery. Conservation
Biology, 10(2), 338–348. https://doi.org/10.1046/j.1523-1739.1996.10020338.x
Soorae, P. S. (Ed.) (2013). Global Re-introduction Perspectives: 2013. Further case studies from
around the globe. Gland, Switzerland: IUCN/ SSC Re-introduction Specialist Group and
Abu Dhabi, UAE: Environment Agency-Abu Dhabi. xiv + 282 pp.
Soorae, P. S. (Ed.) (2016). Global Re-introduction Perspectives: 2016. Case-studies from around the
globe. Gland, Switzerland: IUCN/SSC Reintroduction Specialist Group and Abu Dhabi, UAE:
Environment AgencyAbu Dhabi. xiv + 276 pp.
Soorae, P. S. (Ed.) (2018). Global Reintroduction Perspectives: 2018. Case studies from around the
globe. IUCN/SSC Reintroduction Specialist Group, Gland, Switzerland and Environment
Agency, Abu Dhabi, UAE. xiv + 286pp.
Soorae, P. S. (Ed.) (2021). Global conservation translocation perspectives: 2021. Case studies from
around the globe. Gland, Switzerland: IUCN SSC Conservation Translocation Specialist
Group, Environment Agency - Abu Dhabi and Calgary Zoo, Canada. xiv + 353pp.
Stamps, J. A., & Swaisgood, R. R. (2007). Someplace like home: Experience, habitat selection and
conservation biology. Applied Animal Behaviour Science, 102(3-4), 392–409.
https://doi.org/10.1016/j.applanim.2006.05.038
Sunquist, F., & Sunquist, M. (2014). Andean Cat. In F. Sunquist The Wild Cat Book (pp. 165-174).
University of Chicago Press. https://doi.org/10.7208/chicago/9780226145761.001.0001
Swaisgood, R. R., & Greggor, A. L. (2019). Applications of animal behavior to conservation. In J. C.
Choe (Ed.), Encyclopedia of Animal Behavior (2nd Ed., Vol. 1, pp. 220-229). Elsevier.
https://doi.org/10.1016/B978-0-12-809633-8.90027-X
Taborsky, M. (2014). Tribute to Tinbergen: The Four Problems of Biology. A Critical Appraisal.
Ethology, 120, 224–227. https://doi.org/10.1111/eth.12209

95
Taborsky, M. (2019). From Ethology to Behavioral Biology. In J. C. Choe (Ed.), Encyclopedia of
Animal Behavior (2nd Ed., Vol. 1, pp. 99-102). Elsevier. https://doi.org/10.1016/B978-0-12-
809633-8.20849-2
Tavecchia, G., Pradel, R., Genovart, M., Oro, D. (2007). Density-dependent parameters and
demographic equilibrium in open populations. Oikos, 116(9), 1481–1492.
https://doi.org/10.1111/j.0030-1299.2007.15791.x
Tellaeche, C. G., Reppucci, J. I., Morales, M. M., Luengos Vidal, E. M., Lucherini, M. (2018).
External and skull morphology of the Andean Cat and Pampas Cat: new data from the high
Andes of Argentina. Journal of Mammalogy, 99(4), 906-914.
https://doi.org/10.1093/jmammal/gyy065
Tetley, C. L., & O'Hara, S. J. (2012). Ratings of animal personality as a tool for improving the
breeding, management and welfare of zoo mammals. Animal Welfare, 21(4), 463-476.
https://doi.org/10.7120/09627286.21.4.463
Theile, S. (2003). Fading Footsteps: The Killing and Trade of Snow Leopards. TRAFFIC International,
Cambridge, UK.
Tinbergen, N. (1951). The Study of Instinct. Clarendon Press, Oxford.
Tinbergen, N. (1963). On aims and methods of ethology. Zeitschrift für Tierpsychologie, 20, 410–
433.
Tuck, S. L. (2005). The origins of Roman imperial hunting imagery: Domitian and the redefinition
of the virtus under the principate. Greece and Rome, 52(2), 221–245.
https://doi.org/10.1093/gromej/cxi024
Uher, J., & Asendorpf, J. B. (2008). Personality assessment in the Great Apes: comparing
ecologically valid behavior measures, behavior ratings, and adjective ratings. Journal of
Research in Personality, 42(4), 821-838. https://doi.org/10.1016/j.jrp.2007.10.004
Uher, J., Asendorpf, J. B., Call, J. (2008). Personality in the behaviour of great apes: temporal
stability, cross-situational consistency and coherence in response. Animal Behaviour,
75(1), 99-112. https://doi.org/10.1016/j.anbehav.2007.04.018
Valverde, J. A. (1963). Informacion sobre el lince ibérico en España. Boletín tecnico, serie
cinegetica, 1. Ministerio de Agricultura, Madrid.

96
van der Meer, E., Sousa, L. L., Loveridge, A. J. (2020). Reassessment of an introduced cheetah
Acinonyx jubatus population in Matusadona National Park, Zimbabwe. Oryx, 55(2) 294-
301. https://doi.org/10.1017/S0030605319000164
van Gils, J. A., Kraan, C., Dekinga, A., Koolhaas, A., Drent, J., de Goeij, P., Piersma, T. (2009).
Reversed optimality and predictive ecology: burrowing depth forecasts population change
in a bivalve. Biology Letters, 5(1), 5–8. https://doi.org/10.1098/rsbl.2008.0452
Vargas, A., Sánchez, I., Martínez, F., Rivas, A., Godoy, J. A., Roldán, E., Simón, M. A., Serra, R.,
Pérez, M., Enseñat, C., Delibes, M., Aymerich, M., Sliwa, A., Breitenmoser, U. (2008). The
Iberian lynx Lynx pardinus conservation breeding program. International Zoo Yearbook,
42(1), 190-198. https://doi.org/10.1111/j.1748-1090.2007.00036.x
Vargas, A., Sánchez, I., Martínez, F., Rivas, A., Godoy, J. A., Roldan, E., Símon, M. A., Serra, R.,
Pérez, M. J., Sliwa, A., Delibes, M, Aymerich, M., Breitenmoser, U. (2009) Interdisciplinary
methods in the Iberian lynx conservation breeding programme. In A. Vargas (Ed.), Iberian
Lynx Ex situ Conservation: An Interdisciplinary Approach (pp. 57- 71). Fundación
Biodiversidad, Madrid.
Vazire, S., Gosling, S. D., Dickey, A. S., Schapiro, S. J. (2007). Measuring personality in nonhuman
animals. In R. W. Robins, R. C. Fraley & R. F. Krueger (Eds.), Handbook of Research
Methods in Personality Psychology. New York: Guildford.
Villalba L., Bernal N., Nowell K., Macdonald, D. W. (2012). Distribution of two Andean small cats
(Leopardus jacobita and Leopardus colocolo) in Bolivia and the potential impacts of
traditional beliefs on their conservation. Endangered Species Research, 16(1), 85-94.
https://doi.org/10.3354/esr00389
Villalba, L., Lucherini, M., Walker, S., Cossíos, D., Iriarte, A., Sanderson, J., Gallardo, G., Alfaro, F.,
Napolitano, C., Sillero-Zubiri, C. (2004). The Andean cat: Conservation action plan. Andean
Cat Alliance – Alianza Gato Andino.
Villalba, L., Lucherini, M., Walker, S., Lagos, N., Cossios, D., Bennett, M., Huaranca, J. (2016).
Leopardus jacobita. The IUCN Red List of Threatened Species 2016: e.T15452A50657407.
http://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T15452A50657407.en

97
Walker, R. S., Funes, M., Heidel, L., Palacios, R., & Novaro, A. (2014). The Endangered Andean cat
and fracking in Patagonia. Oryx, 48(1), 14-15.
https://doi.org/10.1017/S0030605313001440
Walker, R. S., Novaro, A. J., Perovic, P., Palacios, R., Donadio, E., Lucherini, M., Pia, M., López, M.
S. (2007). Diets of three species of Andean carnivores in high-altitude deserts of
Argentina. Journal of Mammalogy, 88(2), 519-525. https://doi.org/10.1644/06-MAMM-A-
172R.1
Wang, Q., Liu, D., Holyoak, M., Jia, T., Yang, S., Liu, X., Kong, X., Jiang, G. (2019). Innate preference
for native prey and personality implications in captive amur tigers. Applied Animal
Behaviour Science, 210, 95-102. https://doi.org/10.1016/j.applanim.2018.10.006
Watters, J. V., & Meehan, C. L. (2007). Different strokes: can managing behavioural types increase
post-release success?. Applied Animal Behaviour Science, 102(3-4), 364-379.
https://doi.org/10.1016/j.applanim.2006.05.036
Watters, J. V., & Powell, D. M. (2012). Measuring animal personality for use in population
management in zoos: suggested methods and rationale. Zoo Biology, 31(1), 1-12.
https://doi.org/10.1002/zoo.20379
Webb, L. E., van Reenen, C. G., Jensen, M. B., Schmitt, O., Bokkers, E. A. M. (2015). Does
temperament affect learning in calves? Applied Animal Behaviour Science, 165, 33-39.
https://doi.org/10.1016/j.applanim.2015.01.013
Weber, W., & Rabinowitz, A. (1996). A global perspective on large carnivore conservation.
Conservation Biology, 10(4), 1046–1054. https://doi.org/10.1046/j.1523-
1739.1996.10041046.x
Wegge, P., Shrestha, R., Flagstad, Ø. (2012). Snow leopard Panthera uncia predation on livestock
and wild prey in a mountain valley in northern Nepal: implications for conservation
management. Wildlife Biology, 18(2), 131–141. https://doi.org/10.2981/11-049
Weinstein, T. A. R., Capitanio, J. P., Gosling, S. D. (2008). Personality in animals. In O. P. John, R. W.
Robins & L. A. Pervin (Eds.), Handbook of Personality: Theory and Research (3rd Ed., pp.
328-348). Guilford Press.
Weiss, A., Inoue-Murayama, M., Hong, K.-W., Inoue, E., Udono, T., Ochiai, T., Matsuzawa, T.,
Hirata, S., King, J. E. (2009). Assessing chimpanzee personality and subjective well-being in

98
Japan. American Journal of Primatology, 71(4), 283-292.
https://doi.org/10.1002/ajp.20649
Weiss, A., Inoue-Murayama, M., King, J. E., Adams, M. J., Matsuzawa, T. (2012). All too human?
Chimpanzee and orangutan personalities are not anthropomorphic projections. Animal
Behaviour, 83(6), 1355-1365. https://doi.org/10.1016/j.anbehav.2012.02.024
Weiss, A., King, J. E., Enns, R. M. (2002). Subjective well-being is heritable and genetically
correlated with Dominance in chimpanzees (Pan troglodytes). Journal of Personality and
Social Psychology, 83(5), 1141-1149. https://doi.org/10.1037/0022-3514.83.5.1141
Weiss, A., King, J. E., Figueredo, A. J. (2000). The heritability of personality factors in chimpanzees
(Pan troglodytes). Behavior Genetics, 30(3), 213-221.
https://doi.org/10.1023/A:1001966224914
Weiss, A., King, J. E., Hopkins, W. D. (2007). A cross-setting study of chimpanzee (Pan troglodytes)
personality structure and development: zoological parks and Yerkes National Primate
Research Centre. American Journal of Primatology, 69(11), 1264-1277.
https://doi.org/10.1002/ajp.20428
Wemelsfelder, F. (1997). The scientific validity of subjective concepts in models of animal welfare.
Applied Animal Behaviour Science, 53(1-2), 75-88. https://doi.org/10.1016/S0168-
1591(96)01152-5
Werdelin, L. (1981). The evolution of lynxes. Annales Zoologici Fennici, 18(1), 37-71
Werdelin, L., Yamaguchi, N., Johnson, W. E., & O'Brien, S. J. (2010). Phylogeny and evolution of
cats (Felidae). In D. W. Macdonald & A. J. Loveridge (Eds.), Biology and Conservation of
Wild Felids (pp. 59-82). Oxford University Press.
Whelan, C. J., & Jedlicka, D. M. (2007). Augmenting population monitoring programs with
behavioral indicators during ecological restorations. Israel Journal of Ecology and
Evolution, 53(3-4), 279–295. https://doi.org/10.1560/IJEE.53.3.279
Whitham, J. C., & Wielebnowski, N. (2009). Animal-based welfare monitoring: using keeper
ratings as an assessment tool. Zoo Biology, 28(6), 545-560.
https://doi.org/10.1002/zoo.20281

99
Whitham, J. C., & Wielebnowski, N. (2013). New directions for zoo animal welfare science. Applied
Animal Behaviour Science, 147(3–4), 247-260.
https://doi.org/10.1016/j.applanim.2013.02.004
Wielebnowski, N. C. (1999). Behavioral differences as predictors of breeding status in captive
cheetahs. Zoo Biology, 18(4), 335-349. https://doi.org/10.1002/(SICI)1098-
2361(1999)18:4<335::AID-ZOO8>3.0.CO;2-X
Wielebnowski, N. C., Fletchall, N., Carlstead, K., Busso, J. M., Brown, J. L. (2002). Noninvasive
assessment of adrenal activity associated with husbandry and behavioral factors in the
North American clouded leopard population. Zoo Biology, 21(1), 77-98.
https://doi.org/10.1002/zoo.10005
Wild, M. A., Shenk, T. M., Spraker, T. R. (2006). Plague as a mortality factor in Canada lynx (Lynx
canadensis) reintroduced to Colorado. Journal of wildlife Diseases, 42(3), 646-650.
https://doi.org/10.7589/0090-3558-42.3.646
Wilson, E. O. (1975). Sociobiology. (1st Ed.). Harvard University Press, Cambridge.
Wolf, M., & Weissing, F. J. (2012). Animal personalities: consequences for ecology and evolution.
Trends in Ecology and Evolution, 27(8), 452-461.
https://doi.org/10.1016/j.tree.2012.05.001
Wright, J. T., Byers, J. E., Koukoumaftsis, L. P., Ralph, P. J., Gribben, P. E. (2010). Native species
behavior mitigates the impact of habitat-forming invasive seaweed. Oecologia, 163, 527–
534. https://doi.org/10.1007/s00442-010-1608-2
Yensen, E., & Seymour, K. L. (2000). Oreailurus jacobita. Mammalian Species 2000(644), 1-6.
https://doi.org/10.1644/1545-1410(2000)644<0001:OJ>2.0.CO;2
Yu, J., & Dobson, F. S. (2000). Seven forms of rarity in mammals. Journal of Biogeography, 27(1),
131–139. https://doi.org/10.1046/j.1365-2699.2000.00366.x
Zahler, P., & Paley, R. (2016). Building community governance structures and institutions for snow
leopard conservation. In T. McCarthy & D. Mallon (Eds.), Snow Leopards, Biodiversity of
the World: Conservation from Genes to Landscapes (pp. 151-162). Academic Press.
https://doi.org/10.1016/B978-0-12-802213-9.00012-2

100
Zidon, R., Saltz, D., Shore, L. S., Motro, U. (2009). Behavioral changes, stress, and survival
following reintroduction of Persian fallow deer from two breeding facilities. Conservation
Biology, 23(4), 1026–1035. https://doi.org/10.1111/j.1523-1739.2008.01163.x

101
Anexo 1 – Proposta para estudo empírico de Personalidade
Animal no Lince Ibérico
O objetivo desde estudo era estudar a Personalidade Animal no Lince Ibérico (Lynx
pardinus) no Centro Nacional de Reprodução do Lince Ibérico (CNRLI), situado em Vale de
Fuzeiros, em Silves. Até agora nunca foi realizado nesta espécie este tipo de estudo, pelo que esta
proposta poderia ser o primeiro passo neste ramo de investigação científica. A replicação deste
estudo em todos os centros de reprodução permitiria ainda obter informação a nível da espécie e
não apenas das populações. Se bem-sucedido, pode ser um fator a ter em conta na conservação
deste felino.
Sugere-se que, numa primeira fase, este trabalho seja realizado a todos os indivíduos
adultos, com idades superiores a 24 meses e, numa fase posterior, aos indivíduos com idades até
aos 2 anos. São necessárias todas as informações genéricas dos indivíduos participantes para
termos de comparação e de influência nos resultados, tais como nome, idade, historial de criação,
genealogia e problemas de saúde associados.
A melhor forma de estudar a personalidade em animais é usar a combinação de duas
técnicas, Classificação de Traços de Personalidade e Codificação dos Traços de Personalidade. A
associação destes métodos gera mais informação e pode aumentar a validade e confiabilidade
dos resultados obtidos. Assim, a metodologia aqui sugerida está dividida em duas secções: I.
Classificação dos Traços de Personalidade e, II. Codificação dos Traços de Personalidade.
I. Classificação dos Traços de Personalidade
Usando uma versão adaptada do questionário desenvolvido por Wielebnowski (1999), foi
criado o questionário A, visto que o original demonstra ter alta confiabilidade entre avaliadores e
foi validado por um teste de estimulação de imagem em espelho.
No final deste anexo está o questionário A, que consiste numa tabela para avaliar 17
traços de personalidade, com uma escala de 1 a 7, em que 1 representa ‘Nada’, 4
‘Moderadamente’ e 7 ‘Extremamente’. Os observadores são solicitados para classificarem cada
animal com base no seu próprio conhecimento e impressão geral do comportamento do
indivíduo, indicando a sua classificação colocando uma cruz no número da escala que melhor o
descreve. A zona para comentários no final do questionário serve para os avaliadores adicionarem
quaisquer outros adjetivos que considerem que descrevam o comportamento dos linces. É

102
importante que todos os participantes preencham o questionário de forma independente, sem
conferirem com ninguém.
O propósito do questionário B (também no final do anexo) é saber a experiência com o
Lince Ibérico que cada observador participante possui, de forma a ser possível averiguar se terá
influência nos dados recolhidos por cada um e, consequentemente, nos resultados obtidos. Inclui
perguntas como, há quanto tempo trabalham com linces, há quanto tempo trabalham com
animais em geral e número de horas semanais em que trabalham com os linces.
Para responder aos questionários, sugerem-se todos os tratadores e videovigilantes que
trabalham no CNRLI porque, um grupo tem contacto direto com os animais diariamente (os
tratadores) e o outro tem contacto indireto através de videovigilância, também diariamente.
Assim, ambos os grupos têm conhecimento pessoal de como descreveriam cada lince, devido às
interações e observações constantes dos mesmos.
II. Codificação dos Traços de Personalidade
Para as observações comportamentais, propõe-se que sejam realizadas durante um
período contínuo de 24 horas, iniciado às 00:00h e a terminar às 23:59 do mesmo dia, a partir da
sala de videovigilância do CNRLI com as camaras de videovigilância, presentes em todos os
cercados com visão total dos mesmos e que estão em permanente funcionamento. Com a
capacidade de filmar e de gravação das camaras, as técnicas de amostragem focal e de gravação
contínua podem ser as mais adequadas para este estudo, pois permitem observar um único
indivíduo pelo período necessário e registar todos os comportamentos visualizados, fornecendo
um registo exato e fiel do comportamento, medindo as verdadeiras frequências e durações, e os
momentos em que os padrões de comportamento pararam e começaram.
Tendo em conta que os linces adultos estão sozinhos dentro de um cercado, com acesso a
toda a sua área, ou a partilhar o cercado com mais um lince adulto, sendo que um tem apenas
acesso à zona de campeio e o outro às zonas de maneio grande e pequeno e ao edifício de partos,
sugere-se restringir as observações apenas à zona de campeio (no caso de dois linces) e ao
cercado inteiro (no caso de apenas um lince). Visto que, periodicamente, os animais que
partilham o mesmo cercado, trocam um com o outro de zonas com acesso (com a ajuda dos
tratadores), é possível observar todos os indivíduos nas áreas escolhidas.
É importante obter informações sobre a temperatura e humidade da estação
meteorológica mais próxima obtidas a intervalos regulares, por exemplo a cada 30 minutos, a fim
de averiguar o efeito da temperatura e da humidade no comportamento e atividade dos linces.
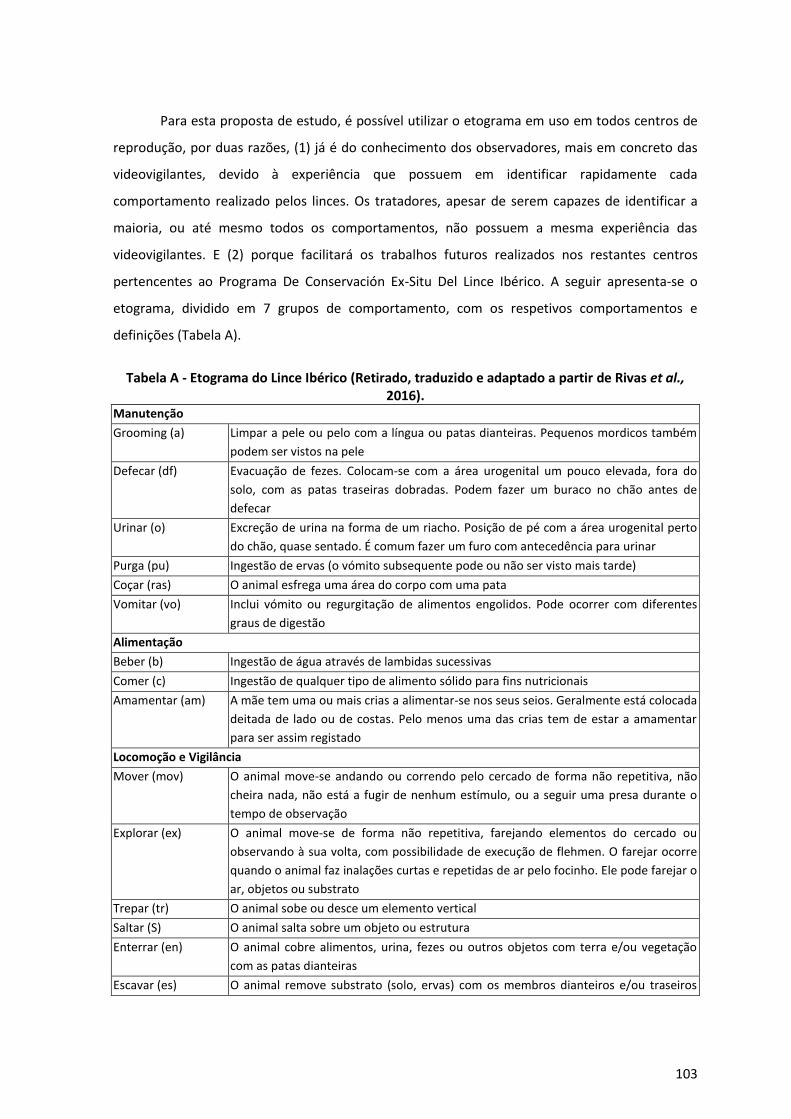
103
Para esta proposta de estudo, é possível utilizar o etograma em uso em todos centros de
reprodução, por duas razões, (1) já é do conhecimento dos observadores, mais em concreto das
videovigilantes, devido à experiência que possuem em identificar rapidamente cada
comportamento realizado pelos linces. Os tratadores, apesar de serem capazes de identificar a
maioria, ou até mesmo todos os comportamentos, não possuem a mesma experiência das
videovigilantes. E (2) porque facilitará os trabalhos futuros realizados nos restantes centros
pertencentes ao Programa De Conservación Ex-Situ Del Lince Ibérico. A seguir apresenta-se o
etograma, dividido em 7 grupos de comportamento, com os respetivos comportamentos e
definições (Tabela A).
Tabela A - Etograma do Lince Ibérico (Retirado, traduzido e adaptado a partir de Rivas et al., 2016).
Manutenção
Grooming (a)
Limpar a pele ou pelo com a língua ou patas dianteiras. Pequenos mordicos também
podem ser vistos na pele
Defecar (df) Evacuação de fezes. Colocam-se com a área urogenital um pouco elevada, fora do
solo, com as patas traseiras dobradas. Podem fazer um buraco no chão antes de
defecar
Urinar (o) Excreção de urina na forma de um riacho. Posição de pé com a área urogenital perto
do chão, quase sentado. É comum fazer um furo com antecedência para urinar
Purga (pu) Ingestão de ervas (o vómito subsequente pode ou não ser visto mais tarde)
Coçar (ras) O animal esfrega uma área do corpo com uma pata
Vomitar (vo) Inclui vómito ou regurgitação de alimentos engolidos. Pode ocorrer com diferentes
graus de digestão
Alimentação
Beber (b) Ingestão de água através de lambidas sucessivas
Comer (c) Ingestão de qualquer tipo de alimento sólido para fins nutricionais
Amamentar (am) A mãe tem uma ou mais crias a alimentar-se nos seus seios. Geralmente está colocada
deitada de lado ou de costas. Pelo menos uma das crias tem de estar a amamentar
para ser assim registado
Locomoção e Vigilância
Mover (mov) O animal move-se andando ou correndo pelo cercado de forma não repetitiva, não
cheira nada, não está a fugir de nenhum estímulo, ou a seguir uma presa durante o
tempo de observação
Explorar (ex) O animal move-se de forma não repetitiva, farejando elementos do cercado ou
observando à sua volta, com possibilidade de execução de flehmen. O farejar ocorre
quando o animal faz inalações curtas e repetidas de ar pelo focinho. Ele pode farejar o
ar, objetos ou substrato
Trepar (tr) O animal sobe ou desce um elemento vertical
Saltar (S) O animal salta sobre um objeto ou estrutura
Enterrar (en) O animal cobre alimentos, urina, fezes ou outros objetos com terra e/ou vegetação
com as patas dianteiras
Escavar (es) O animal remove substrato (solo, ervas) com os membros dianteiros e/ou traseiros

104
para fazer um buraco ou levantar pedaços do solo. Não se observa o animal a urinar
ou a defecar
Evitar (ev) O animal afasta-se de um estímulo negativo (pessoas, obras ou outro lince) e procura
refúgio em alguma área do cercado, escondido atrás de algum elemento ou longe
deste estímulo, com ou sem contacto visual. Ao se afastar do estímulo negativo, o
animal corre ou caminha com a barriga rente ao solo
Observar (obs)
O animal permanece imóvel (em pé sobre as 4 patas, sentado ou deitado), com os
olhos totalmente abertos. Às vezes, também pode mover as orelhas para explorar o
ambiente
Comportamento Predatório
Caçar (cz) O lince apresenta comportamentos (como stalking, perseguição) que fazem parte da
sequência direcionada à captura e morte da presa (coelho, perdiz, pássaros ou
qualquer animal que esteja no cercado que não seja outro lince)
Comportamento Sedentário
Descansar (des)
Numa posição relaxada. Se a cabeça estiver levantada, os olhos devem estar fechados
ou semifechados. Se a cabeça não estiver ereta e apoiada, os olhos devem estar
semifechados. O animal não está ciente de nada em particular. Exibe pequenos
movimentos, como mexer as orelhas ou mudar de posição.
Dormir (d) O animal está relaxado, permanece imóvel, os olhos fechados, nenhuma tensão é
exibida e a cabeça está apoiada, não ereta.
Social
Interação negativa
(i-)
Comportamento agressivo realizado por um lince em relação a outro lince. Observa-se
com um dos seguintes comportamentos
• Bufar: o animal sopra pela boca de forma breve e ameaçadora
• Rosnar: o animal faz um ruido grave
• Patada: o animal bate num outro lince com as patas dianteiras diretamente ou
através da cerca
• Pelo eriçado: o animal levanta os pelos do pescoço e/ou dorso
• Luta: dois animais envolvem-se num combate físico, mordendo-se e
arranhando-se
Interação positiva
(i+)
Comportamento amigável realizado por um animal em relação a outro indivíduo, sem
exibir comportamentos na categoria de interações negativas. Inclui os seguintes
comportamentos:
• Vocalizar: o animal emite um som semelhante ao de um miado e está próximo
ao animal com o qual interage. Pode ser acompanhada por uma ligeira elevação da
cabeça, com movimento para trás das orelhas e alongamento da garganta
• Mostrar genitais: o animal posiciona a região genital próxima ao rosto de outro
indivíduo com a cauda levantada.
• Marcação com urina: o animal marca o corpo de outro indivíduo com urina na
forma de spray
• Rebolar no solo: o animal faz voltas no solo apoiando-se no dorso, a menos de 5
metros de outro indivíduo. Mais frequente na época de acasalamento.
• Esfregar rosto/corpo: o animal roça com a sua barba ou com o corpo num outro
indivíduo
Interação neutra (i)
.
O animal apresenta um dos seguintes comportamentos, sem coincidir com a exibição
de comportamentos da categoria “i-” ou “i+”:
• Cabeceamento: os dois animais batem as cabeças um no outro. Ao contrário

105
dos "amigáveis", estes são fortes e bruscos
• Barbas abertas: o animal abre as barbas para os lados
• Andar paralelo: o animal move-se em sincronia com outro lince, seguindo-o ou
movendo-se em paralelo, no mesmo cercado ou em cercados adjacentes.
• Morder a cerca: o animal morde a rede que o separa de outro lince
• Farejar: o animal cheira o corpo de outro indivíduo
Allo-grooming (alo) O animal faz grooming num outro indivíduo. Só para indivíduos que realizam a ação
ou quando ambos realizam a ação mutuamente.
Jogo Social (js) Tipo de jogo disputado entre dois ou mais indivíduos, crias ou adultos. As formas mais
comuns são:
• Jogo de luta: os animais lutam entre si marcando com os dentes sem realmente
morder. Também usam as patas para bater suavemente no outro indivíduo, sem
estender as garras. O comportamento é semelhante ao de uma luta, mas com
movimentos mais lentos e menos intensos
• Jogo de perseguição: um animal corre atrás do outro até alcançá-lo. Os animais
são frequentemente separados novamente e ocorre uma nova perseguição, repetindo
este padrão várias vezes
• Jogo de montaria: um lince é posicionado na posição típica de cópula, com uma
mordida na nuca, por outro lince.
Jogo locomotor (jl) Jogo desenvolvido pelo animal por meio da interação com o meio ambiente, sem
envolver especificamente outro indivíduo ou a manipulação de objetos inanimados.
Jogos de corrida, salto e escalada.
Jogo com objetos
(jo)
O animal manipula um objeto inanimado com as patas. As formas mais frequentes
são: golpear com as patas (com as garras retraídas), atirar para o ar, saltar sobre o
objeto ou lutar com ele
Jogo com presa (jp)
O animal persegue ou luta com a presa, sem a intenção de a matar. O animal
manipula a presa ou exibe comportamentos que compõem a sequência dos
comportamentos de caça, estando a presa viva ou morta, e embora tenha a
oportunidade de capturá-la para matá-la ou comê-la, não o faz. É frequente pegar a
presa com a boca e soltá-la novamente para continuar o jogo
Marcação com urina
(xo)
O animal direciona a urina na forma de spray para um objeto, principalmente em
superfícies mais altas que o solo. Ao fazer isto, ele fica de pé com a cauda ereta e
pode vibrá-la
Marcação com patas
dianteiras (Xpd)
O animal arranha com as patas dianteiras a superfície, vertical ou horizontal, de um
tronco, cortiça ou estrutura
Marcação com patas
traseiras (Xpt)
O animal esfrega as patas traseiras no solo (areia, terra ou plantas)
Marcação com
barbas (xb)
O animal esfrega as barbas em objetos, vegetação ou solo
Marcação com
corpo (xc)
O animal gira o corpo esfregando-se contra o solo ou vegetação e não há outro
indivíduo a menos de 5 metros
Comportamentos Anormais
Mamar uma parte
do corpo (mc)
Ação de sucção, mas realizada numa alguma parte do corpo como a cauda, um
membro. Pode ser acompanhada por um movimento de “massagem”.
Movimento
repetitivo sem
estímulo de longa
Sem estímulo aparente, o animal move-se numa rota superior a 3 metros, que se
repete pelo menos 3 vezes num período de até 60 segundos. Pode parar para
observar, marcar, grooming ou outros, brevemente (<5 seg) interrompendo o
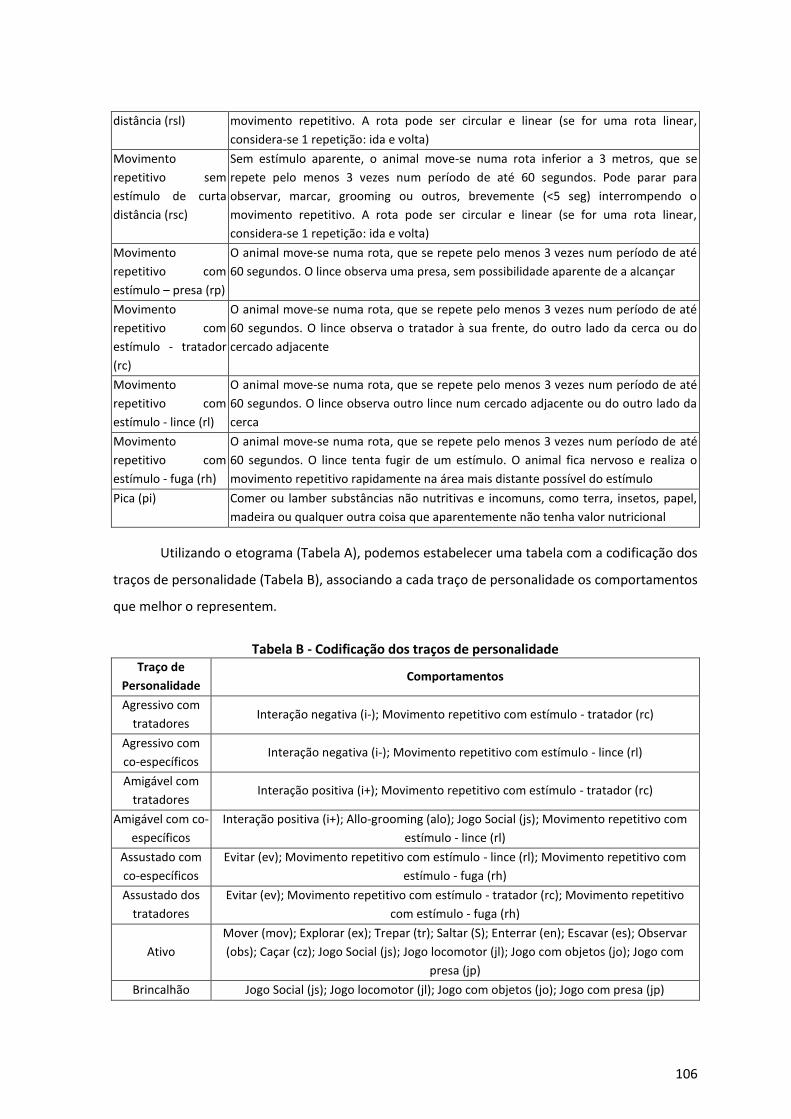
106
distância (rsl) movimento repetitivo. A rota pode ser circular e linear (se for uma rota linear,
considera-se 1 repetição: ida e volta)
Movimento
repetitivo sem
estímulo de curta
distância (rsc)
Sem estímulo aparente, o animal move-se numa rota inferior a 3 metros, que se
repete pelo menos 3 vezes num período de até 60 segundos. Pode parar para
observar, marcar, grooming ou outros, brevemente (<5 seg) interrompendo o
movimento repetitivo. A rota pode ser circular e linear (se for uma rota linear,
considera-se 1 repetição: ida e volta)
Movimento
repetitivo com
estímulo – presa (rp)
O animal move-se numa rota, que se repete pelo menos 3 vezes num período de até
60 segundos. O lince observa uma presa, sem possibilidade aparente de a alcançar
Movimento
repetitivo com
estímulo - tratador
(rc)
O animal move-se numa rota, que se repete pelo menos 3 vezes num período de até
60 segundos. O lince observa o tratador à sua frente, do outro lado da cerca ou do
cercado adjacente
Movimento
repetitivo com
estímulo - lince (rl)
O animal move-se numa rota, que se repete pelo menos 3 vezes num período de até
60 segundos. O lince observa outro lince num cercado adjacente ou do outro lado da
cerca
Movimento
repetitivo com
estímulo - fuga (rh)
O animal move-se numa rota, que se repete pelo menos 3 vezes num período de até
60 segundos. O lince tenta fugir de um estímulo. O animal fica nervoso e realiza o
movimento repetitivo rapidamente na área mais distante possível do estímulo
Pica (pi) Comer ou lamber substâncias não nutritivas e incomuns, como terra, insetos, papel,
madeira ou qualquer outra coisa que aparentemente não tenha valor nutricional
Utilizando o etograma (Tabela A), podemos estabelecer uma tabela com a codificação dos
traços de personalidade (Tabela B), associando a cada traço de personalidade os comportamentos
que melhor o representem.
Tabela B - Codificação dos traços de personalidade Traço de
Personalidade Comportamentos
Agressivo com
tratadores Interação negativa (i-); Movimento repetitivo com estímulo - tratador (rc)
Agressivo com
co-específicos Interação negativa (i-); Movimento repetitivo com estímulo - lince (rl)
Amigável com
tratadores Interação positiva (i+); Movimento repetitivo com estímulo - tratador (rc)
Amigável com co-
específicos
Interação positiva (i+); Allo-grooming (alo); Jogo Social (js); Movimento repetitivo com
estímulo - lince (rl)
Assustado com
co-específicos
Evitar (ev); Movimento repetitivo com estímulo - lince (rl); Movimento repetitivo com
estímulo - fuga (rh)
Assustado dos
tratadores
Evitar (ev); Movimento repetitivo com estímulo - tratador (rc); Movimento repetitivo
com estímulo - fuga (rh)
Ativo
Mover (mov); Explorar (ex); Trepar (tr); Saltar (S); Enterrar (en); Escavar (es); Observar
(obs); Caçar (cz); Jogo Social (js); Jogo locomotor (jl); Jogo com objetos (jo); Jogo com
presa (jp)
Brincalhão Jogo Social (js); Jogo locomotor (jl); Jogo com objetos (jo); Jogo com presa (jp)

107
Calmo Descansar (des); Observar (obs);
Curioso Explorar (ex);
Excêntrico
Mamar uma parte do corpo (mc); Movimento repetitivo sem estímulo de longa distância
(rsl); Movimento repetitivo sem estímulo de curta distância (rsc); Movimento repetitivo
com estímulo – presa (rp); Movimento repetitivo com estímulo - tratador (rc);
Movimento repetitivo com estímulo - lince (rl); Movimento repetitivo com estímulo -
fuga (rh); Pica (pi)
Excitável
Inseguro Movimento repetitivo com estímulo - fuga (rh);
Inteligente
Solitário Evitar (ev)
Tenso
Movimento repetitivo sem estímulo de longa distância (rsl); Movimento repetitivo sem
estímulo de curta distância (rsc); Movimento repetitivo com estímulo – presa (rp);
Movimento repetitivo com estímulo - tratador (rc); Movimento repetitivo com estímulo -
lince (rl); Movimento repetitivo com estímulo - fuga (rh)
Vocal Interação negativa (i-) (vocalizações); Interação positiva (i+) (vocalizações)
Notas importantes para este estudo:
No caso da classificação dos traços de personalidade recorrendo a questionários de
personalidade, os observadores podem ter histórias diferentes com os indivíduos, o que pode
representar um possível problema na escolha dos avaliadores (Vazire et al., 2007). Assim, o
comportamento dos animais na presença desses avaliadores pode ser mais um reflexo nas suas
expectativas de interação do que das caraterísticas de personalidade dos linces. Se os animais
reagem de maneira diferente a cada observador (tratador), uma medida de classificação pode não
descrever com precisão a personalidade do animal (Vazire et al., 2007). Consequentemente, a
escolha de avaliadores com diferentes histórias de interações com animais individuais pode
resultar numa baixa confiabilidade dos resultados. É, por isso, imperativo que a natureza da
relação entre cada observador e os animais em estudo seja especificada para permitir
comparações mais significativas das avaliações, sendo esta a razão para a especificação da função
do observador nos dois questionários (Anexos A e B).
A codificação dos traços de personalidade possui também fatores que são necessários ter
em conta aquando do planeamento e realização do estudo pois, facilmente, a validade dos
resultados pode ficar comprometida (Vazire et al., 2007). Tal pode acontecer se, por exemplo,
durante a criação da codificação, os comportamentos forem incorretamente interpretados ou, se
forem associados aos traços de personalidade comportamentos errados ou, ainda, não ter em
consideração o facto de que o mesmo comportamento pode ser indicativo de vários traços de
personalidade.

108
Uma vez que a presença de seres humanos próximos dos animais influencia o
comportamento e a reação dos mesmos, ainda que os observadores estejam fora de vista, os
animais ainda são capazes de cheirar e ouvir a presença humana (Vazire et al., 2007). Por esta
razão, a observação indireta dos animais através de videovigilância, tal como acontece no CNRLI,
é ideal para o estudo de comportamento e personalidade dos linces. Apesar dos problemas
óbvios que podem surgir com o uso de tecnologia, tal como falhas mecânicas e/ou software, falha
de eletricidade ou, a qualidade inferior de imagem, o uso de camaras de filmar tem também
grandes vantagens, como a gravação dos vídeos, permitindo visualizar eventos que não foram
visualizados em direto e analisar todos os comportamentos quando necessário, as vezes que
forem necessárias.
A utilização de tecnologias sofisticadas, como Noldus Observer XT, pode facilitar e
simplificar toda a recolha dos dados relativos às observações comportamentais e,
consequentemente, aumentar a validade e confiabilidade dos dados da codificação dos traços de
personalidade. Alguns estudos já publicados, tais como Gilbert-Norton et al. (2013) e Webb et al.
(2015), usaram esta tecnologia com bons resultados.

109
Questionário A – Avaliação dos traços de personalidade de cada
indivíduo em estudo
Procedimento: Avalie cada indivíduo na escala (de 1 a 7) fornecida para cada traço de
personalidade, em que 1 representa ‘Nada’, 4 ‘Moderadamente’ e 7 ‘Extremamente’ (ver
exemplo). O nome do indivíduo está colocado no topo da folha para permitir que avalie cada
indivíduo em relação aos outros. Por favor, não discuta as suas respostas com mais ninguém. É
fornecida uma zona para comentários, para cada indivíduo, para adicionar informações não
cobertas pelos adjetivos (pode incluir quaisquer adjetivos adicionais que entenda). Os
comentários também devem incluir o comportamento do animal na sua presença,
especificamente.
Exemplo: Indivíduo A
Adjetivo Caraterísticas 1 2 3 4 5 6 7
Ativo Move-se com frequência dentro do cercado (ex: passeios, corridas) X
Legenda: 1 – Nada; 2 – Muito pouco; 3 – Pouco; 4 – Moderadamente; 5 – Muito; 6 – Bastante; 7 – Extremamente.
*****
Nome: __________________________________ Tratador / Videovigilante Data: ___________
INDIVÍDUO: _________________
Traço de
personalidade Definição 1 2 3 4 5 6 7
Agressivo com
tratadores
Reage frequentemente de forma hostil e ameaçadora
para com as pessoas
Agressivo com co-
específicos
Reage de uma forma hostil (ex: ataques, rosnados) em
relação a outros linces
Amigável com
tratadores
Inicia a proximidade com os tratadores: aproxima-se da
cerca prontamente e de maneira amigável (ex: ronrona,
esfrega na cerca)
Amigável com co-
específicos Social; inicia e procura proximidade com outros linces
Assustado com co-
específicos Afasta-se e esconde-se rapidamente de outros linces
Assustado dos
tratadores Afasta-se imediatamente dos tratadores
Ativo Move-se com frequência dentro do cercado (ex: passeios,
corridas)
Brincalhão Inicia e envolve-se em comportamentos lúdicos

110
(comportamento aparentemente sem sentido e não
agressivo) com objetos e/ou outros linces
Calmo Não é facilmente perturbado por mudanças no seu
ambiente
Curioso Aborda e explora situações novas/mudanças no seu
ambiente
Excêntrico Mostra comportamentos estereotípicos ou incomuns
Excitável Reage exageradamente às mudanças no ambiente
Inseguro Parece facilmente assustado; “Nervoso” e com medo em
geral
Inteligente Aprende rapidamente a associar certos eventos (ex: sons
de coelhos associados com hora da alimentação)
Solitário Passa muito tempo sozinho; evita companhia
Tenso Mostra contenção de movimento e postura; altamente
sensível
Vocal Vocaliza com frequência e prontamente
Legenda: 1 – Nada; 2 – Muito pouco; 3 – Pouco; 4 – Moderadamente; 5 – Muito; 6 – Bastante; 7 – Extremamente.
Comentários:

111
Questionário B – Questionário Pessoal para Tratadores e
Videovigilantes
Por favor, preencha as seguintes questões sobre si e sobre a sua experiência profissional da forma
mais exata possível (anos e meses):
Nome:
Sexo: F M
Idade: Data:
Função: Tratador ou Videovigilante
No total, há quanto tempo é que trabalha com animais?
Em geral, há quanto tempo trabalha com linces ibéricos?
E, mais concretamente no Centro Nacional de Reprodução do Lince Ibérico?
Qual a média semanal de horas que trabalha com os linces?
Normalmente, entra dentro dos cercados com os linces aí presentes?
Acredita que existem personalidades distintas nos linces com quem trabalha?
Qual é a sua opinião sobre o facto de diferenças de personalidade estarem correlacionadas, por
exemplo, com o sucesso reprodutivo individual e com a forma como vários indivíduos lidam
com o stress?


















