
Modelos matemáticos no estudo da malária · 2019-07-19 · a Modelos matemáticos no estudo da...
67
a Modelos matemáticos no estudo da malária Jorge Diquissone Ndacherenga Mestrado em Matemática para Professores Departamento de Matemática Ano 2019 Supervisora Prof. Doutora Helena Mena Matos Professora Auxiliar Faculdade de Ciências da Universidade do Porto a
Transcript of Modelos matemáticos no estudo da malária · 2019-07-19 · a Modelos matemáticos no estudo da...

a
Modelos matemáticos no estudo damalária
Jorge Diquissone NdacherengaMestrado em Matemática para ProfessoresDepartamento de MatemáticaAno 2019
SupervisoraProf. Doutora Helena Mena MatosProfessora AuxiliarFaculdade de Ciências da Universidade do Porto
a

a
a
Todas as correções de-terminadas pelo júri, e sóessas, foram efectuadas.
O presidente do Júri,
Porto, ___ /___ /_____

Às minhas filhas: Rosa e Chaquila.

Agradecimentos
Antes de mais, quero agradecer à Professora Doutora Helena Mena Matos pelaorientação que me deu na tese. Quero enaltecer a sua constante preocupação e de-dicação, mesmo com muitas outras atividades e supervisandos. A ela vai um sinceroobrigado.
Os meus agredecimentos são extensivos aos Professores que me deram aulasna parte curricular do curso, o que de certa forma me preparou para trabalhar comentusiasmo na Tese.
Agradeço igualmente a todos os meus colegas do curso de Matemática para Pro-fessores, especialmente a Christiane, o Bernardino e o Artálio, pelo companherismo eos momentos interessantes de estudo.

Resumo
Na presente dissertação foram estudados modelos matemáticos na transmissãode doenças infeciosas com especial enfoque na malária. Fez-se primeiramente umaabordagem geral dos modelos epidemiológicos simples SIR e SIS. No modelo SIR
destaca-se o facto de que uma epidemia só poderá ocorrer se a proporção da popula-ção suscetível for maior do que a taxa de remoção relativa, enquanto que no modeloSIS, uma epidemia só poderá ocorrer se a taxa de remoção for menor do que a taxade transmissão. Seguidamente foram analisados dois modelos de Ross-Macdonald, osimples proposto por Ross em 1911 e um outro mais recente (2008) levando em contaos períodos de latência do parasita no mosquito e no ser humano.

Abstract
In the present dissertation we study mathematical models for the transmission of in-fectious diseases with a special focus on malaria. A general approach was first madeto the simple epidemiological models SIR and SIS. In the SIR model, an epidemiccan only occur if the susceptible population’s proportion is greater than the relative re-moval rate, whereas in the SIS model, an epidemic can only occur if the removal rateis lower than the transmission rate. Two Ross-Macdonald models were then analyzed:the simple one proposed by Ross in 1911 and another more recent (2008) taking intoaccount the latency periods of the parasite in humans and in mosquitos.
i

Conteúdo
Conteúdo ii
Lista de Tabelas iv
Lista de Figuras v
1 Introdução 1
2 Doenças infeciosas 32.1 Definições básicas no estudo de doenças infeciosas . . . . . . . . . . . 42.2 Malária . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
2.2.1 Modelos matemáticos no estudo da malária . . . . . . . . . . . . 7
3 Modelo SIR simples 103.1 Modelo epidémico de Kermack-McKendrick . . . . . . . . . . . . . . . . 10
3.1.1 Propriedades matemáticas do modelo SIR . . . . . . . . . . . . 123.1.1.1 Prevalência máxima . . . . . . . . . . . . . . . . . . . . 153.1.1.2 Teorema do limiar da Epidemiologia . . . . . . . . . . . 153.1.1.3 Estimação de parâmetros . . . . . . . . . . . . . . . . . 17
3.1.1.3.1 Taxa de recuperação . . . . . . . . . . . . . . 173.1.1.3.2 Taxa de transmissão . . . . . . . . . . . . . . 18
3.2 Praga em Eyam . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183.3 Modelo SIR com vacinação . . . . . . . . . . . . . . . . . . . . . . . . . 22
3.3.1 Um modelo com vacinação à nascença . . . . . . . . . . . . . . 233.3.1.1 Pontos de equilíbrio e sua estabilidade . . . . . . . . . 23
4 Modelo epidémico SIS simples 274.1 Solução explícita . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 284.2 Modelo epidémico SIS com tratamento saturado . . . . . . . . . . . . . 30
4.2.1 Pontos de equilíbrio . . . . . . . . . . . . . . . . . . . . . . . . . 304.2.1.1 Estudo da estabilidade dos pontos de equilíbrio . . . . 33
ii

5 Modelo da malária de Ross-Macdonald 375.1 Modelo simples de Ross-Macdonald . . . . . . . . . . . . . . . . . . . . 38
5.1.1 Isóclinas e pontos de equilíbrio . . . . . . . . . . . . . . . . . . . 405.1.2 Número básico de reprodução da malária . . . . . . . . . . . . . 45
5.2 Modelo de Ross-Macdonald com atrasos . . . . . . . . . . . . . . . . . 475.2.1 Pontos de equilíbrio . . . . . . . . . . . . . . . . . . . . . . . . . 48
5.2.1.1 Estabilidade do ponto de equilíbrio livre de doença . . 485.2.1.2 Estabilidade do ponto de equilíbrio endémico . . . . . . 50
6 Conclusão 53
Bibliografia 55
iii

Lista de Tabelas
3.1 Número de indivíduos suscetíveis e infetados durante a grande Pestede Eyam [18]. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
3.2 Passos para calcular o coeficiente de determinação com os dados dapeste em Eyam em proporções. . . . . . . . . . . . . . . . . . . . . . . 21
3.3 Resolução da equação (3.6) pelo método de Newton para ρ = 0.61. . . 22
5.1 Parâmetros usados no modelo simples de Ross-Macdonald. . . . . . . 395.2 Valores numéricos de alguns parâmetros do modelo de Ross-Macdonald
([10], p.7). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
iv

Lista de Figuras
2.1 Evolução e agrupamento dos diferentes tipos de modelos de maláriaSEIR. Os subscritos h e m representam humano e mosquito respetiva-mente [10]. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
3.1 Fluxograma do modelo epidémico SIR de Kermack-McKendrick. . . . . 123.2 Órbitas do sistema (3.3), convergem para o ponto de equilíbrio livre de
doença da forma (s∗, 0). As curvas são determinadas pelas condiçõesiniciais i(0) = i0 e s(0) = s0. Uma situação epidémica existe se s0 > ρ,como pode ser verificado nas órbitas de (3.3). . . . . . . . . . . . . . . 14
3.3 Órbita da solução de (3.3) para ρ = 0.61, s0 = 254261
, i0 = 7261
e r0 = 0. . . . 203.4 Condição para a existência do ponto de equilíbrio endémico. . . . . . . 25
4.1 Fluxograma do modelo epidémico SIS simples. . . . . . . . . . . . . . . 274.2 Solução da equação logística (4.3), considerando alternativamente r <
0 e r > 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 294.3 Gráfico das funções g(i) e h(i) = α
γ. A figura ilustra a existência de uma
única intersecção para i ∈]0, 1[, dando um ponto de equilíbrio positivoúnico. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
4.4 Existência de equilíbrios endémicos para R0 < 1. . . . . . . . . . . . . . 324.5 Análise gráfica das soluções da equação (4.7) para R0 > 1. . . . . . . . 334.6 Análise gráfica da estabilidade dos pontos de equilíbrio para R0 < 1 e
∆ > 0. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 344.7 Análise gráfica das soluções da equação (4.7) para R0 < 1 e ∆ = 0. . . 354.8 Ocorrência de pontos críticos para R0 < 1 e ∆ < 0. . . . . . . . . . . . . 354.9 Gráficos da função f para R0 < 1 e ∆ < 0. . . . . . . . . . . . . . . . . 364.10 Análise gráfica das soluções da equação (4.7) para R0 < 1 e ∆ < 0. . . 36
5.1 Dinâmica da malária entre humanos e mosquitos no modelo simplesde Ross-Macdonald, sem considerarmos a inclusão de períodos de la-tência. Neste fluxograma assim como no modelo só se considera atransmissão da malária por picadas de mosquitos. . . . . . . . . . . . . 40
v

5.2 Esboço das curvas isóclinas para a = 0.3, p = q = 0.5, m = 3, α = 0.05
e µ = 0.35. Pontos de interseção: (0, 0) e (x∗, y∗) =(23, 29
). . . . . . . . . 41
5.3 Esboço das curvas isóclinas para a = 0.2, p = q = 0.5, m = 0.5, α = 0.05
e µ = 0.5. As curvas intersetam-se somente em (x, y) = (0, 0). . . . . . 445.4 Campo vetorial associado ao sistema (5.2). . . . . . . . . . . . . . . . . 455.5 Evolução das proporções de humanos e mosquitos infetados em função
do número básico de reprodução da doença para um índice de estabili-dade maior ou menor que 1. . . . . . . . . . . . . . . . . . . . . . . . . 46
5.6 Condição de existência do equilíbrio endémico. . . . . . . . . . . . . . . 51
vi

Capítulo 1
Introdução
Ao longo da História humana modelos matemáticos vêm sendo usados para aju-dar epidemiologistas na tomada de decisão acerca do melhor caminho a seguir nocombate às doenças infeciosas. Nesses modelos e em função da natureza da do-ença, a população é dividida em compartimentos epidemiológicos. Uma doença quese desenvolve numa comunidade, onde a população suscetível (S) se infeta por con-tacto com um indivíduo infetado (I) e o período de incubação do agente patogénicoé desprezável, e se recupera (R) com imunidade permanente, é modelada pelo mo-delo compartimental SIR. São exemplos de doenças modeladas por este modelo, osarampo, a parotidite e a rubéola. As doenças que se desenvolvem como o expostoacima, mas em que a recuperação não garante nenhuma imunidade, são modeladasgeralmente pelo modelo epidémico SIS. São exemplos de doenças modeladas pelomodelo epidémico SIS, a meningite e as doenças causadas por protozoários, como amalária e a doença do sono1.
A malária é uma doença infeciosa que é causada por parasitas protozoários dogénero Plasmodium. Em humanos ela é causada por Plasmodium falciparum, Plas-modium malariae, Plasmodium ovale e Plasmodium vivax . Os parasitas da maláriasão transmitidos para humanos por picadas infetadas da fêmea do mosquito do gé-nero Anopheles.
Desde a descoberta das propriedades inseticidas e tóxicas do DDT (dicloro-difenil-tricloroetano) em 1938-19392, muitas ações têm sido tomadas no combate a essa do-ença que está predominantemente presente em zonas de clima tropical e temperado.As ações de combate à malária têm se centrado em matar mosquitos que já tenhamvivido tempo suficiente para transmitirem a doença, através da pulverização intradomi-ciliária e do uso de redes mosquiteiras impregnadas com inseticidas, em campanhaspromovidas pela Organização Mundial de Saúde (OMS).
1http://webpages.fc.ul.pt/~mcgomes/aulas/biopop/Mod7/Text%20%20Model.pdf. Acesso:05.05.2019
2https://www.infopedia.pt/$ddt?uri=siglas-abreviaturas/DDT. Acesso: 05.05.2019.
1

O presente trabalho foi baseado essencialmente em livros e revistas científicas quefalam de modelos matemáticos epidemiológicos. Também se consultaram sites queestão devidamente referenciados nas notas de rodapé.
O trabalho está organizado em sete capítulos, sendo o primeiro a introdução.No capítulo 2 apresentam-se alguns conceitos relacionados com doenças infeciosas(úteis para a modelação matemática), o historial da descoberta do agente patogénicoe do vetor transmissor da malária assim como a hierarquia dos modelos matemáticosno estudo da malária. No capítulo 3 apresenta-se o modelo SIR simples, cuja princi-pal abordagem se concentra no estudo das condições de ocorrência duma epidemia ena percentagem da população a vacinar para a proteger duma epidemia. No capítulo 4apresenta-se o modelo SIS simples, onde se faz estudo das condições de ocorrênciaduma epidemia e dos efeitos duma taxa de recuperação decrescente (decrescendocom aumento do número de infetados) nos pontos de equilíbrio e sua estabilidade. Nocapítulo 5 faz-se uma análise qualitativa do modelo matemático de Ross-Macdonald,primeiro sem inclusão do período de latência do parasita e em seguida levando emconta os períodos de latência do parasita no mosquito e no ser humano. A análisedeste capítulo baseou-se no estudo da estabilidade dos pontos de equilíbrio e possí-veis intervenções a serem feitas para reduzir o número básico de reprodução da ma-lária, visto que matematicamente a existência da epidemia é explicada por um númerode reprodução da doença superior a um (R0 > 1). No capítulo 6 vêm as conclusões epor fim as referências bibliográficas.
2

Capítulo 2
Doenças infeciosas
Ao longo da História há registo de ocorrência de vários tipos de doenças e o Ho-mem tem procurado conhecer as causas e estudar meios de controlar ou até irradicaras doenças. A ciência que se ocupa do estudo dos padrões de saúde, doenças e fa-tores a elas associados ao nível da população denomina-se Epidemiologia. A palavraepidemiologia deriva dos termos gregos epi , que significa sobre, demos, que significapessoas, e logos, que significa estudo. A etimologia desta palavra mostra que ela dizrespeito somente a seres humanos ([11], p. 1).
Infelizmente ameaças de epidemias e pandemias1 existem continuamente, pois osvírus ou outros agentes infeciosos sofrem mutações muito rapidamente e podem saltarbarreiras de espécies e infetar seres humanos em grande escala.
Uma doença infeciosa é uma doença clinicamente evidente resultante da presençade um agente patogénico microbiano. Os agentes microbianos causadores da doençapodem ser bactérias, vírus, fungos, protozoários, ou podem ser proteínas tóxicas, cha-madas priões. Um conceito frequentemente usado no âmbito das doenças infeciosasé o de doenças transmissíveis, que são doenças que podem ser transmitidas de umapessoa infetada para outra, direta ou indiretamente. A transmissão de doenças infe-ciosas ocorre de várias maneiras. Na modelação matemática é habitual destinguir 4tipos de transmissão:
• Transmissão direta: o agente patogénico causador é transmitido de uma pessoainfetada para outra, por contacto direto ou de proximidade. Por exemplo: HIV,gonorreia e sífilis.
• Transmissão vetorial: o agente infecioso é transmitido de um vetor (veículo detransmissão) para um humano. São exemplos: malária, dengue e vírus do Nilo
1Quando uma doença existe apenas numa determinada região é considerada uma endemia.Quando a doença é transmitida para outras populações, atinge mais de uma cidade ou região, designa-se epidemia. Porém, quando uma epidemia se alastra de forma desequilibrada e se espalha peloscontinentes, ou pelo mundo, ela é considerada uma pandemia. [In: https://www.infoescola.com/
doencas/endemia-epidemia-e-pandemia/. Acesso: 30.10.2018.]
3

Ocidental2, que são transmitidas por mosquitos.
• Transmissão vertical: o agente patogénico é transmitido através da placenta demãe infetada para filho antes ou durante o nascimento. Por exemplo o HIV.
• Transmissão pelo meio ambiente: um humano é infetado pelo contacto com umagente infecioso presente no meio ambiente. A cólera é um exemplo de doençatransmitida pelo meio ambiente, que é provocada pelo consumo de água conta-minada.
O nicho ecológico onde um agente patogénico vive e se multiplica denomina-sereservatório. De acordo com o seu reservatório, os agentes microbianos são classifi-cados como humanos, animais e ambientais. Agentes patogénicos humanos circulamprincipalmente entre humanos e os seres humanos desempenham um papel funda-mental na sua transmissão. Os agentes patogénicos animais usam os animais verte-brados como reservatório e circulam principalmente entre os animais. Muitos dessesagentes infeciosos adaptam-se para infetar posteriormente humanos. Infeções que seespalham primeiramente em animais vertebrados e depois se transmitem para huma-nos são chamadas zoonoses. Agentes patogénicos ambientais multiplicam-se prin-cipalmente no meio ambiente (normalmente água e solo) e espalham-se a partir daípara populações animais e humanas ([11], p. 3).
Muitas doenças infeciosas têm mais de uma via de transmissão. Por exemplo, amalária para além da transmissão natural através da picada de uma fêmea do mos-quito Anopheles infetada, pode ser também transmitida por transfusão de sangue, usocompartilhado de agulhas e/ou seringas contaminadas, ou através de acidentes detrabalho em meio laboratorial ou hospitalar.
2.1 Definições básicas no estudo de doenças infecio-
sas
Existem vários conceitos em epidemiologia estritamente relacionados com doen-ças infeciosas e que são fundamentais na modelação matemática. Alguns dessesconceitos estão listados abaixo.
Indivíduos suscetíveis: indivíduos saudáveis mas não imunes a uma dada do-ença.
Indivíduo exposto: indivíduo suscetível que faz um contacto potencialmente trans-missor de uma doença. Indivíduos expostos podem ou não desenvolver a doença.
2O vírus do Nilo Ocidental (VNO) é transmitido por meio da picada de mosquitos infetados, prin-cipalmente do género Culex . [In: https://pt.wikipedia.org/wiki/V%C3%ADrus_do_oeste_do_Nilo.Acesso: 30.04.2019]
4

Indivíduos infetados e infeciosos: se o agente infecioso se estabelecer em umindivíduo exposto, então esse indivíduo é infetado. Indivíduos infetados que podemtransmitir a doença são chamados infeciosos. Indivíduos infetados podem não serinfeciosos durante todo o tempo da infeção.
Indivíduos latentes: são aqueles que estão infetados, mas ainda não são infeci-osos. O período de latência é o tempo que decorre desde o início da infeção até aomomento em que o hospedeiro é capaz de transmitir o agente infecioso para outroindivíduo.
Período de incubação: intervalo de tempo entre a exposição a um agente infe-cioso e o aparecimento dos sintomas da doença. O período de incubação tambémé definido como o tempo necessário para que o agente infecioso se multiplique atéum limiar necessário para produzir sintomas ou evidências laboratoriais de infeção. Operíodo de incubação não coincide necessariamente com o período de latência. Porexemplo indivíduos infetados com a gripe se tornam infeciosos aproximadamente umdia antes de exibirem sintomas visíveis da doença.
Incidência: número de indivíduos que ficam doentes por unidade de tempo. A inci-dência pode ser expressa em termos de proporção pelo quociente entre o número deindivíduos que ficam doentes por unidade de tempo e o tamanho total da população.
Prevalência: número de pessoas infetadas pela doença em um determinado mo-mento t. Em termos de proporção, a prevalência é o quociente entre o número depessoas que estão infetadas pela doença em um determinado momento t e o tama-nho total da população.
Força de infeção: incidência por suscetível ou probabilidade, por unidade detempo, de que um suscetível seja infetado.
Imunidade antiparasitária: imunidade ao parasita.
2.2 Malária
A malária é provavelmente uma das doenças mais antigas e tem sido um flagelonas populações do mundo desde a Antiguidade. Esta doença também é conhecidapelo seu termo francês, paludisme, embora o nome italiano, malária, descreva maisprecisamente a doença. O termo italiano se refere a mala aria, mau ar , isto é, osmiasmas dos pântanos e águas estagnadas, que os antigos povos acreditavam ser aorigem da doença ([3], p. 187).
Até à descoberta do agente causador e do vetor transmissor da malária um longocaminho foi percorrido. Vários cientistas e epidemiologistas se empenharam durantequase um século no estudo da malária. Em 1880 Charles Laveran, médico e militarfrancês na Argélia, descreveu parasitas da malária no sangue de doentes durante epi-
5

sódios de febre malárica. Ele chamou estes organismos microscópicos de Oscillariamalariae. Pela sua descoberta, Laveran é considerado o pai da parasitologia protis-tológica ([3], p. 187). Quase na mesma altura C. Gerhardt provou que uma pessoasaudável podia ficar doente pela inoculação de sangue de pessoas sofrendo com amalária ([15], p. 9).
Sir Patrick Manson, médico escocês, demonstrou em 1879 que a filariose de Ban-croft (Wuchereria bancrofti) era transmitida pela picada de um mosquito do géneroCulex . Ele pensava que um mosquito desse género também estivesse envolvido natransmissão da malária e que os parasitas eram soltos na água por mosquitos mortos,e em seguida transferidos para humanos quando bebiam a água. Em 1897 Sir RonaldRoss durante seus estudos sobre malária em aves, descreveu oocistos do parasita damalária nas paredes do estômago de um mosquito não classificado (mas chamadopor ele de: um mosquito cinzento, mosquito alado manchado), que ele acreditava sero Culex . Este foi o ponto de partida que acabou com o labirinto das origens da malária([3], p. 187,188). Pelas suas descobertas em malária, Ross recebeu o Prémio Nobelem 1902.
Atualmente sabe-se que o agente patogénico da malária é transmitido através dapicada da fêmea de um mosquito infetada do genéro Anopheles durante a sua ali-mentação de sangue humano. A malária é causada pelos parasitas protozoários dogénero Plasmodium. Nos seres humanos é causada por um dos ou uma combinaçãode Plasmodium falciparum, Plasmodium malariae, Plasmodium ovale e Plasmodiumvivax . A malária está predominantemente presente nos países tropicais e de climatemperado. O P. falciparum é a causa comum de infeção na África e no Sudeste daÁsia, e é responsável por cerca de 80% de todos os casos de malária e cerca de 90%das mortes. Na Índia, P. vivax , tem sido o agente patogénico primário, responsávelpela malária, embora os casos de P. falciparum estejam em ascensão nos últimostempos ([10], p. 1).
Na década de 50 do século passado George Macdonald seguindo as ideias deRoss, provou a validade dos dados epidemiológicos de campo, colhidos cerca demeio século antes e inferiu que para a irradicação da malária se devia diminuir otempo de vida do mosquito adulto. A proposta de Macdonald abriu caminho a umextenso programa de pulverização intra-domiciliária para matar os mosquitos adultoscom inseticidas DDT promovido pela Organização Mundial da Saúde (OMS) ([17], p.6).
Apesar dos esforços na sua erradicação, a malária ainda permanece um problemade saúde pública em 109 países, em que a doença foi declarada endémica em 2008.Sem nenhuma vacina eficaz à vista e muitos dos anti maláricos mais antigos e drogasperdendo eficácia devido à evolução da resistência do parasita aos medicamentos, aprevenção (utilização de redes mosquiteiras) é ainda o único conselho dado às pes-
6

soas que vivem em zonas endémicas ([10], p. 1).
2.2.1 Modelos matemáticos no estudo da malária
Um modelo matemático é uma descrição de um sistema usando ferramentas e lin-guagem matemática. Por outras palavras, um modelo matemático é uma aproximaçãoda realidade por meio de escrita e resolução de um problema (de biologia ou de ou-tra ciência) em linguagem matemática. Modelos matemáticos são desenvolvidos paraajudar a explicar um sistema, estudar os efeitos de seus vários componentes e fazerprevisões sobre o seu comportamento. Os modelos matemáticos são desenvolvidosa partir de um conjunto de suposições para simplificar a realidade ([11], p. 6).
Uma das ideias base para o desenvolvimento de modelos matemáticos epidemi-ológicos é a de que quando uma doença infeciosa invade uma população, os seusindivíduos podem ser classificados em categorias (ou compartimentos), que são re-presentados pela notação padrão S − E − I − R. O primeiro grupo S consiste nosindivíduos suscetíveis a contrair a doença. A classe E (exposta) consiste dos indi-víduos que estão infetados mas não são infeciosos (devido ao período de latência).A classe I é composta pelos indivíduos que se encontram infetados e capazes detransmitir a doença aos suscetíveis. Finalmente a classe R (recuperados/removidos)é constituída pelos indivíduos cujo período infecioso terminou não podendo transmitira doença (ou por imunidade adquirida ou por isolamento ou morte). Pode haver varia-ções na estrutura do compartimento dependendo do tipo de doença. Por exemplo, osindivíduos infetados (I) podem não recuperar e morrer; A classe R pode consistir emindivíduos que se recuperam com imunidade permanente ou temporária que se subdi-vide em mais compartimentos epidemiológicos. Usando essas notações, oito classesde modelos compartimentais são possíveis - SI, SIS, SEI, SEIS, SIR, SIRS, SEIR eSEIRS. Por exemplo, num modelo SEIRS, uma fração dos suscetíveis (S) da popu-lação fica exposta (E) à infeção, uma parte dos quais, em seguida, torna-se infeciosa(I). Alguns dos indivíduos infetados (I) recuperam da doença, e tornam-se parte daclasse recuperada (R) com imunidade temporária. Quando a imunidade é perdida,eles se tornam suscetíveis ao ataque de agentes patogénicos novamente e entram naclasse (S) dos suscetíveis ([10], p. 3).
Ronald Ross foi o primeiro a publicar um modelo matemático para o estudo quan-titativo da malária. O modelo de Ross, inclui a dinâmica do parasita no mosquito e noser humano. As equações para a dinâmica do parasita no ser humano são geradaspelo modelo compartimental SIS pois uma vez infetado, geralmente se recupera dadoença, mas sem imunidade, enquanto que a população de mosquitos é modeladapelo modelo compartimental SI ([13], p. 19-22). O seu modelo foi desenvolvido combase na suposição de que a população humana e de mosquitos era constante. Quer
7

dizer não considerou a migração nem nascimentos nem mortes em humanos e consi-derou a taxa de nascimento de mosquitos igual à taxa de mortalidade dos mosquitosadultos. Mas tarde Macdonald incluiu no modelo de Ross o período de latência doparasita no mosquito, e portanto, a população de mosquitos passou a ser modeladapelo modelo compartimental SEI.
Uma revisão histórica sistemática sugere que vários matemáticos e cientistas con-tribuíram para o desenvolvimento do modelo de Ross-Macdonald. Mais fatores foramsendo adicionados ao modelo simples de Ross, de forma a incluir a complexidade dadinâmica do parasita da malária. Na Figura 2.1 está representado um esquema daevolução dos modelos matemáticos no estudo da malária.
Nessa figura, os três modelos a partir dos quais todos os outros foram desenvolvi-dos estão na caixa cinzenta. A inclusão pela primeira vez de um novo compartimentoé mostrado em vermelho. O subscrito j = (1, 2, 3) indica subdivisão adicional do com-partimento correspondente. Setas pontilhadas mostram a incorporação de fatorescomplexos em diferentes modelos ou num compartimento específico (círculo verme-lho). O tamanho total da população é constante para todos os modelos, exceto os queestão dentro da caixa tracejada.
Modelos matemáticos vêm sendo usados para auxiliar os epidemiologistas na to-mada de decisões acerca do melhor caminho a seguir no combate da malária. Noentanto a integração de interações complexas entre parasitas e hospedeiros leva atermos não lineares e a modelos cada vez mais complexos.
Figura 2.1: Evolução e agrupamento dos diferentes tipos de modelos de malária SEIR.Os subscritos h e m representam humano e mosquito respetivamente [10].
8

Falando acerca dos modelos matemáticos complexos para a malária destacamosa seguinte observação de Jacob C. Koella ([8], p. 2):
Apesar do reconhecimento precoce por Ross e Macdonald da importân-cia de uma descrição quantitativa da transmissão da malária, a epidemiolo-gia matemática encontrou muitas dificuldades em obter aceitação por partede epidemiologistas e profissionais de saúde pública. Uma das razões paraessa falta de aceitação pode estar na crescente complexidade dos mode-los publicados nos anos 60 e 70 (revistos por Bailey, 1977). Esses modelosnão são difíceis apenas para os não-matemáticos, mas a interpretação dosseus resultados pode, na verdade, ser enganosa. Por um lado, quanto maisvariáveis incluídas num modelo, mais próximas das observaçoes se encon-tram geralmente as previsões do modelo, simplesmente porque mais grausde liberdade estão envolvidos. No entanto, uma proximidade entre previ-são e observação não implica necessariamente uma concordância entre aestrutura do modelo e os processos biológicos.
A inclusão de mais variáveis ou detalhes num modelo está relacionada sem dú-vida com o aumento de suposições para o modelo. E quanto maior for o número desuposições, maior é a probabilidade de se fazer uma suposição errada.
E fazendo uma comparação entre modelos simples e complexos Jacob C. Koellateceu as seguintes considerações ([8], p. 2):
As previsões qualitativas de modelos simples podem ser mais significa-tivas do ponto de vista biológico do que as previsões quantitativas precisasde modelos complexos envolvendo muitos parâmetros. Por outro lado, mo-delos mais detalhados não resultam necessariamente num maior poder deprevisão. De facto, os resultados de modelos mais complexos podem sermenos confiáveis do que os de modelos mais simples.
Neste trabalho, para além de se apresentar os modelos simples SIR e SIS noscapítulos 3 e 4 respetivamente, faz-se uma análise qualitativa dos modelos simplesde Ross-Macdonald sem levar em conta o período de latência do parasita da maláriana primeira parte do capítulo 5 e, na segunda parte faz-se uma análise qualitativada proposta de inclusão de dois períodos de latência, respetivamente a latência doparasita no ser humano e a latência do parasita no mosquito no modelo simples deRoss-Macdonald apresentado em [16].
A análise qualitativa feita, está ligada ao estudo da estabilidade dos pontos deequilíbrio e intervenções possíveis para reduzir o número básico de reprodução damalária.
9

Capítulo 3
Modelo SIR simples
Os resultados deste capítulo foram baseados essencialmente nos livros de Braun[2], Chasnov [5], Martcheva [11], Polya [14] e nos artigos científicos [7], [9], [18] e [19].
O modelo SIR para o estudo da propagação de uma doença foi publicado pelaprimeira vez por Kermack e McKendrick em 1927. Este modelo considera a populaçãodividida em três classes: suscetível (S), infetado (I) e removido (R). A classe dossuscetíveis é constituída por indivíduos saudáveis, mas que podem contrair a doença.A classe dos infetados é constituída pelos indivíduos que contraíram a doença e sãocapazes de a transmitir. A classe dos removidos / recuperados é constituída por in-divíduos que se recuperaram da doença e agora estão imunes, ou foram vacinados,ou foram isolados do resto da população, ou talvez tenham morrido da doença. Estesindivíduos não podem contrair novamente a doença.
O número de indivíduos S, I e R em cada uma dessas classes varia com o tempo,ou seja, S(t), I(t) e R(t) são funções do tempo t. O tamanho total da população N
é a soma dos tamanhos dessas três classes: N = S(t) + I(t) + R(t). Neste modelosupõe-se que o tamanho da população é constante. Hipoteticamente consideramosuma população fechada à migração e sem nascimentos nem mortes não relacionadascom a doença em estudo, ou por outras palavras consideramos o estudo num curtointervalo de tempo, de modo que a população não sofre grandes alterações no seutamanho.
3.1 Modelo epidémico de Kermack-McKendrick
O modelo SIR é baseado em equações diferenciais ordinárias (EDOs) e descrevea dinâmica ao longo do tempo em cada classe em que é dividida a população. Quandoum indivíduo suscetível entra em contacto com um indivíduo infetado, esse indivíduosuscetível fica infetado com uma certa probabilidade e se move da classe suscetívelpara a classe infetada. A população suscetível diminui por unidade de tempo de uma
10

quantidade igual à dos indivíduos que se infetaram nesse tempo. Ao mesmo tempo,a classe de infetados aumenta do mesmo número de indivíduos infetados. Em epide-miologia é designado por incidência o número de indivíduos infetados por unidade detempo. E a taxa de variação da classe suscetível é:
S ′(t) = −incidência.
Para determinarmos a incidência, consideremos um indivíduo infetado numa po-pulação de tamanho N . Ele realiza cN contactos por unidade de tempo (c taxa decontacto per capita), com probabilidade S
Nde que tal contacto seja com um indivíduo
suscetível. Mas nem todo contacto transmite infeção. Seja p a probabilidade de quetal contacto resulte em infeção, então pcS é o número de indivíduos suscetíveis quesão infetados por cada indivíduo infetado por unidade de tempo.
Se assumirmos que pc = β, então βSI é o número de indivíduos que são infetadospor unidade de tempo (incidência). βI é chamado de força de infeção e, o coeficienteβ é a constante de proporcionalidade chamada taxa de transmissão da doença.
Obtemos assim a seguinte equação diferencial para o número de indivíduos sus-cetíveis:
S ′(t) = −βIS.
Os indivíduos suscetíveis que se infetam passam para a classe dos infetados. Os indi-víduos que se recuperam ou morrem, deixam a classe dos infetados a uma probabili-dade per capita constante α por unidade de tempo, chamada de taxa de recuperação.Assim,
I ′(t) = βIS − αI.
Os indivíduos que se recuperam ou morrem, deixam a classe dos infetados e mudam-se para a classe dos recuperados:
R′(t) = αI.
Assim, o modelo é dado pelo seguinte sistema de EDOs:
S ′(t) = −βISI ′(t) = βIS − αIR′(t) = αI.
(3.1)
O sistema (3.1) pode ser formulado para proporções. Assim, tomando
s(t) =S(t)
N, i(t) =
I(t)
Ne r(t) =
R(t)
N,
11

temos:s′(t) = −γis
i′(t) = γis− αir′(t) = αi,
(3.2)
com γ = βN , também designada por taxa de transmissão. Daqui em diante a análisedeste modelo será feita com base no sistema (3.2).
Num modelo matemático há que ter em conta as unidades das quantidades envol-vidas. As unidades também são importantes na estimação dos parâmetros. Assim,as unidades de ambos os lados de (3.2) devem ser as mesmas. A partir da primeiraequação, vemos que i e s são proporções, de modo que são adimensionais. Assim asunidades de γ devem ser 1/[unidade de tempo]. Da mesma forma, a partir da terceiraequação, vemos que as unidades de α são 1/[unidade de tempo].
Se o tamanho N da população é constante, s(t) + i(t) + r(t) = 1 em qualquerinstante t. De facto, somando todas as três equações do sistema (3.2), obtemos
s′(t) + i′(t) + r′(t) = 0,
e no instante inicial tem-se: s(0) + i(0) + r(0) = S(0)+I(0)+R(0)N
= 1
Figura 3.1: Fluxograma do modelo epidémico SIR de Kermack-McKendrick.
Como está apresentado na Figura 3.1 cada indivíduo pode residir em exatamenteum compartimento e pode mover-se de um compartimento para outro. Setas indicama direção de movimento de indivíduos entre as classes. As setas do movimento sãotipicamente rotuladas pelas taxas de transição.
3.1.1 Propriedades matemáticas do modelo SIR
Pelo teorema de existência e unicidade de solução de equações diferenciais ordi-nárias ([2], p. 412), quaisquer que sejam s0, i0 e r0, o sistema (3.2) tem uma únicasolução que satisfaz, s(0) = s0, i(0) = i0 e r(0) = r0. Os pontos de equilíbrio do sis-tema (3.2) são claramente os pontos da família: (s∗, 0, 1 − s∗), pois r′(t) = 0 se e sóse i = 0, e da relação: s∗ + i∗︸︷︷︸
=0
+r∗ = 1, resulta r∗ = 1− s∗. Tais pontos de equilíbrio
apelidam-se livres da doença uma vez que i = 0.Uma epidemia ocorre quando uma pequena quantidade de infetados, introduzida
numa população totalmente suscetível, resultar no aumento da proporção de pessoas
12

infetadas. Para análise da ocorrência ou não de epidemia, consideremos apenas aequação de dinâmica da classe dos infetados em (3.2):
i′ = (γs− α)i
Uma epidemia só poderá ocorrer se γs0 − α > 0 ⇔ s0 >αγ. De facto se s0 < α
γ, então
s(t) < αγ
para todo t, pois s é monótona decrescente. Assim, i′(t) < 0 para qualquerinstante t e portanto não ocorre epidemia. No caso γs0 − α > 0, s(t) > α
γ∀t ∈ [0, t1[ e
nesse intervalo i′(t) > 0. Há portanto aumento da proporção de infetados até que s(t)passe a ser menor que
α
γe consequentemente i′(t) < 0.
A quantidade ρ =α
γé chamada em epidemiologia de limiar da epidemia, pois serve
de referência na dinâmica da doença. Também ρ é conhecido como taxa de remoçãorelativa, pois é razão entre a taxa de remoção (recuperação) e a taxa de transmissão.
A condição s0 > ρ para a existência de epidemia pode ser reescrita na forma R0 >
1, sendo R0 =s0γ
αdesignado por número básico de reprodução. Em Epidemiologia o
número básico de reprodução é o número médio de infeções secundárias causadaspor um único infetado numa população totalmente suscetível (à exceção do infetado)ao longo do tempo que demora ao infetado inicial a sair da classe. Quando R0 > 1 adoença consegue invadir uma população totalmente suscetível. Se R0 < 1 a doençaacaba por desaparecer.
As duas primeiras equações de (3.2) não dependem de r e portanto podem seranalisadas independentemente:
s′ = −γisi′ = (γs− α)i
(3.3)
Note-se que os pontos de equilíbrio de (3.3) são os pontos da forma (s∗, 0), com 0 ≤s∗ ≤ 1. Além disso,
di
ds= −1 +
ρ
s⇒ i = −s+ ρ ln(s) + C
onde C é uma constante arbitrária. A equação acima pode ser reescrita como:
i+ s− ρ ln(s) = C (3.4)
Usando a condição inicial (s0, i0) em (3.4), encontramos C = i0 + s0 − ρ ln(s0).Substituindo o valor de C em (3.4) resulta,
i+ s− ρ ln(s) = i0 + s0 − ρ ln(s0). (3.5)
É possível assim obter um esboço das órbitas do sistema (3.3) (Ver Figura 3.2).
13
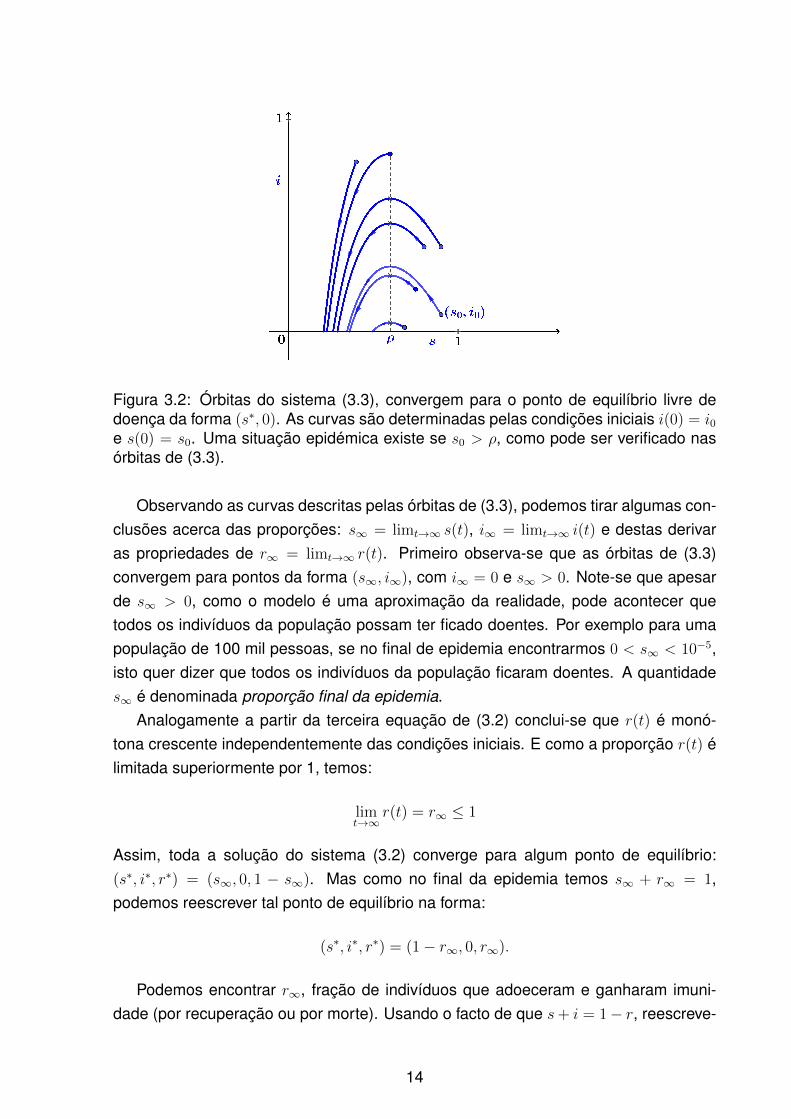
Figura 3.2: Órbitas do sistema (3.3), convergem para o ponto de equilíbrio livre dedoença da forma (s∗, 0). As curvas são determinadas pelas condições iniciais i(0) = i0e s(0) = s0. Uma situação epidémica existe se s0 > ρ, como pode ser verificado nasórbitas de (3.3).
Observando as curvas descritas pelas órbitas de (3.3), podemos tirar algumas con-clusões acerca das proporções: s∞ = limt→∞ s(t), i∞ = limt→∞ i(t) e destas derivaras propriedades de r∞ = limt→∞ r(t). Primeiro observa-se que as órbitas de (3.3)convergem para pontos da forma (s∞, i∞), com i∞ = 0 e s∞ > 0. Note-se que apesarde s∞ > 0, como o modelo é uma aproximação da realidade, pode acontecer quetodos os indivíduos da população possam ter ficado doentes. Por exemplo para umapopulação de 100 mil pessoas, se no final de epidemia encontrarmos 0 < s∞ < 10−5,isto quer dizer que todos os indivíduos da população ficaram doentes. A quantidades∞ é denominada proporção final da epidemia.
Analogamente a partir da terceira equação de (3.2) conclui-se que r(t) é monó-tona crescente independentemente das condições iniciais. E como a proporção r(t) élimitada superiormente por 1, temos:
limt→∞
r(t) = r∞ ≤ 1
Assim, toda a solução do sistema (3.2) converge para algum ponto de equilíbrio:(s∗, i∗, r∗) = (s∞, 0, 1 − s∞). Mas como no final da epidemia temos s∞ + r∞ = 1,podemos reescrever tal ponto de equilíbrio na forma:
(s∗, i∗, r∗) = (1− r∞, 0, r∞).
Podemos encontrar r∞, fração de indivíduos que adoeceram e ganharam imuni-dade (por recuperação ou por morte). Usando o facto de que s+ i = 1− r, reescreve-
14

mos (3.5) na formar + ρ ln(s) = r0 + ρ ln(s0).
Fazendo t tender para infinito e como s∞ = 1− r∞, vem:
r∞ + ρ ln(1− r∞) = r0 + ρ ln(s0)
que é equivalente à seguinte equação transcendental para r∞:
1− r∞ − s0e−1ρ(r∞−r0) = 0. (3.6)
A equação (3.6) pode ser resolvida numericamente pelo método de Newton1.
3.1.1.1 Prevalência máxima
Se ocorrer uma epidemia, então o número de infetados atinge seu valor máximono instante em que i′(t) = 0. Pela segunda equação de (3.2) temos:
i′ = (γs− α)i = 0⇔ i = 0 ∨ γs− α = 0⇒ s =α
γ= ρ
Da equação (3.5), isolando i e substituindo o valor de s para i máximo, temos:
imáx = s0 + i0 − ρ+ ρ ln
(ρ
s0
). (3.7)
Aqui imáx é a proporção máxima de indivíduos infetados atingidos pela epidemia,denominado prevalência máxima.
Se formos capazes de estimar a prevalência máxima para uma doença infeciosa,então se ela ocorrer novamente, saberemos quando o número de infeções começaráa decrescer.
3.1.1.2 Teorema do limiar da Epidemiologia
Como já se viu anteriormente, se a proporção inicial de suscetíveis s0 for maiorque o valor limiar ρ, então ocorrerá epidemia, isto é, a proporção de infetados irácrescer primeiro até um nível máximo decrescendo depois para zero. No caso de s0
ser maior do que ρ mas próximo desse valor, então é possível estimar a proporção deindivíduos que em última análise contrairão a doença: s0 − s∞. Mais especificamente,se ε = s0−ρ ρ então s0−s∞ ≈ 2ε, ou seja s∞ = ρ−ε. Por outras palavras a proporçãode suscetíveis é reduzido a um ponto tão abaixo do limiar como originalmente estavaacima dele.
1É um método de aproximação sucessiva que consiste em obter xn+1, a partir de xn, f(xn) e f ′(xn),tal que: xn+1 = xn − f(xn)
f ′(xn)[14].
15

Este é o famoso teorema do Limiar da Epidemiologia, que foi provado pela primeiravez em 1927 por Kermack e McKendrick.
Especificamente, o teorema do valor limiar diz que:
Teorema 3.1. ([2], p. 459) Seja s0 = ρ+ ε e suponha que ερ 1. Suponha, além disso,
que i0 é muito pequeno. Então, a proporção de indivíduos que finalmente contrairão adoença é aproximadamente 2ε.
Prova: Fazendo t tender para o infinito na equação (3.5) e desprezando i0 por sermuito pequeno:
0 = s0 − s∞ + ρ ln
(s∞s0
)= s0 − s∞ + ρ ln
(s0 − (s0 − s∞)
s0
)= s0 − s∞ + ρ ln
(1− s0 − s∞
s0
)Pretende-se obter s0 − s∞. Como s0 − ρ é pequeno comparado com ρ, s0 − s∞ é
pequeno comparado com s0. Aproxima-se então ln
(1− s0 − s∞
s0
)pelo polinómio de
Taylor de grau 2:
−(s0 − s∞s0
)− 1
2
(s0 − s∞s0
)2
.
Então,
0 = s0 − s∞ − ρ(s0 − s∞s0
)− ρ
2
(s0 − s∞s0
)2
,
colocando em evidência s0 − s∞, temos:
0 = (s0 − s∞)
[1− ρ
s0− ρ
2s20(s0 − s∞)
].
E portanto, tem-se:
ρ
2s20(s0 − s∞) = 1− ρ
s0⇔ s0 − s∞ =
s0−ρs0ρ2s20
(s0 − s∞) =s0 − ρs0
· 2s20ρ
= 2s0(s0 − ρ)
ρ= 2(ρ+ ε)
(ρ+ ε− ρ
ρ
),
s0 − s∞ = 2ρ
(1 +
ε
ρ
)ε
ρ,
mas a quantidade 1 + ερ≈ 1, pois ε
ρ 1, assim,
s0 − s∞ ≈ 2ε,
16

portanto a diminuição na população suscetível é 2 vezes a quantidade com que elasuperava o valor do limiar de epidemiologia.
3.1.1.3 Estimação de parâmetros
No modelo SIR simples, conhecendo-se alguns dados sobre a epidemia, pode-seestimar os parâmetros que estão envolvidos na dinâmica da doença, nomeadamenteα e γ. Vejamos os passos que podem ser seguidos abaixo.
3.1.1.3.1 Taxa de recuperação Muitas vezes é conhecido o tempo médio de infe-ção. Esse conhecimento permite estimar a taxa de recuperação α.
Para estimar a taxa de recuperação, supomos que não há influxo na classe infetadae que uma certa proporção de indivíduos (i0) está nesta classe no tempo zero. Entãoo problema de valor inicial para esta classe é:
i′(t) = −αi, i(0) = i0
e portanto,i(t)
i0= e−αt
A equação acima, para t ≥ 0 descreve a proporção de indivíduos que ainda estãoinfetados no momento t, ou em linguagem de probabilidade, é a probabilidade de umapessoa ainda estar na classe infetada no momento t. E a relação abaixo apresenta aproporção de indivíduos que deixaram a classe infetada no momento t.
1− e−αt
Assim, considerando a variável aleatória X representativa do tempo de permanênciana classe infetada, a função de distribuição de X é:
P (X ≤ t) =
1− e−αt, t ≥ 0
0, t < 0,
que é a probabilidade de recuperar ou deixar a classe infetada no intervalo [0, t[.A função densidade de probabilidade é dada por:
f(t) =
αe−αt, t ≥ 0
0, t < 0(3.8)
O tempo médio gasto na classe infetada é dado então pela média (valor esperado) davariável aleatória X.
17

E(X) =
∫ +∞
−∞tf(t)dt,
que é equivalente a:
E(X) =
∫ +∞
0
tαe−αtdt,
Usando a integração por partes, podemos escrever:
E(X) =(−te−αt
) ∣∣+∞0 +
∫ +∞
0
e−αtdt =
[−te−αt − 1
αe−αt
]+∞0
= limb→+∞
[(−be−bα − 1
αe−bα)−
(−0− 1
α
)]=
1
α.
Assim, a taxa de recuperação α é igual ao inverso do tempo médio E(X) gasto naclasse infetada:
α =1
E(X).
No caso da gripe o tempo médio gasto na classe infetada é de 5 dias. Assim, a taxade recuperação é de 1/5[dias]−1.
3.1.1.3.2 Taxa de transmissão Para o modelo SIR simples, conhecendo-se a taxade recuperação α, as condições iniciais (s0, i0) e finais (s∞, i∞), podemos determinarγ ( taxa de transmissão). Da equação (3.5) temos:
i0 + s0 −α
γln(s0) = i∞ + s∞ −
α
γln(s∞) (3.9)
Como i∞ = 0, rearranjando os termos, temos:
i0 + s0 − s∞ =α
γln
(s0s∞
)⇔ α
γ=i0 + s0 − s∞
ln( s0s∞
),
E isolando a taxa de transmissão (γ), resulta:
γ =α ln( s0
s∞)
i0 + s0 − s∞. (3.10)
3.2 Praga em Eyam
A vila de Eyam, na Inglaterra, em Derbyshire, sofreu um surto de peste bubónicaem 1665-1666. Acreditava-se que a fonte dessa praga era a Grande Peste de Londres.A vila é mais conhecida por ser a “vila de peste” que escolheu se isolar quando a pestefoi descoberta lá em Agosto de 1665. Registos detalhados das datas das mortes
18

devido à peste ( apenas 3 dos indivíduos infetados não morreram) foram preservados.Considerando o período infecioso da peste bubónica como sendo de 11 dias, essesregistos permitiram a construção da tabela abaixo em que os intervalos temporais sãode 15 dias e meio.
Data 1666 no de suscetíveis no de infetados18 de Junho 254 73/4 de Julho 235 14,519 de Julho 201 22
3/4 de Agosto 153,5 2919 de Agosto 121 21
3/4 de Setembro 108 819 de Setembro 97 8
3/4 Outubro Desconh. Desconh.20 de Outubro 83 0
Tabela 3.1: Número de indivíduos suscetíveis e infetados durante a grande Peste deEyam [18].
Em seguida aplicar-se-á o modelo SIR com início em 18 de Junho, data em que apopulação total era de 261 indivíduos dos quais 254 suscetíveis e 7 infetados.
Como o período infecioso é de cerca de 11 dias, então a taxa de recuperação (emdias−1) é dada por:
α =1
11⇒ α = 0.0909 ≈ 0.091
Para encontrar a taxa de transmissão (em dias−1), temos de considerar os dados da
Tabela 3.1. Em termos de proporções temos: s0 =254
261, i0 =
7
261e r0 = 0. E no final
da epidemia, temos s∞ =83
261e i∞ = 0, então de (3.10) temos:
γ =0.091 ln
(25483
)7+254−83
261
≈ 0.1492435
Assim,ρ =
0.091
0.1492435≈ 0.61.
A epidemia estabeleceu-se e dizimou muitas vidas humanas em Eyam, visto que aproporção inicial de suscetíveis é muito maior que o limiar da epidemia ou taxa deremoção relativa.
A Tabela 3.1 não apresenta detalhadamente o número de infetados todos os dias,mas podemos estimar através do modelo a proporção máxima de infetados atingida.
19
Para tal, por (3.7), temos:
imáx =254
261+
7
261− 0.61 + 0.61 ln
(0.61254261
)= 0.10506 ≈ 10.51%.
Portanto a proporção máxima de infetados em Eyam segundo o modelo seria de10.51% de sua população, ou seja, o número máximo de pessoas infetadas seriade cerca de 27 pessoas.
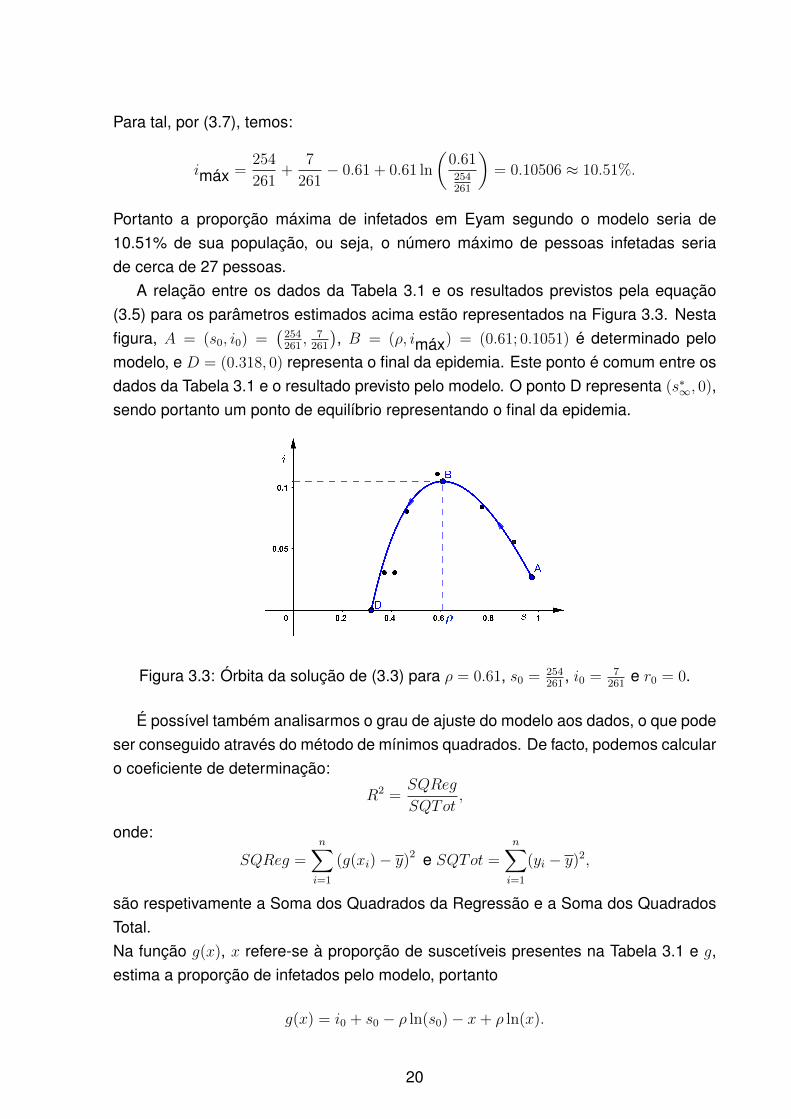
A relação entre os dados da Tabela 3.1 e os resultados previstos pela equação(3.5) para os parâmetros estimados acima estão representados na Figura 3.3. Nestafigura, A = (s0, i0) =
(254261, 7261
), B = (ρ, imáx) = (0.61; 0.1051) é determinado pelo
modelo, e D = (0.318, 0) representa o final da epidemia. Este ponto é comum entre osdados da Tabela 3.1 e o resultado previsto pelo modelo. O ponto D representa (s∗∞, 0),sendo portanto um ponto de equilíbrio representando o final da epidemia.
Figura 3.3: Órbita da solução de (3.3) para ρ = 0.61, s0 = 254261
, i0 = 7261
e r0 = 0.
É possível também analisarmos o grau de ajuste do modelo aos dados, o que podeser conseguido através do método de mínimos quadrados. De facto, podemos calcularo coeficiente de determinação:
R2 =SQReg
SQTot,
onde:
SQReg =n∑i=1
(g(xi)− y)2 e SQTot =n∑i=1
(yi − y)2,
são respetivamente a Soma dos Quadrados da Regressão e a Soma dos QuadradosTotal.Na função g(x), x refere-se à proporção de suscetíveis presentes na Tabela 3.1 e g,estima a proporção de infetados pelo modelo, portanto
g(x) = i0 + s0 − ρ ln(s0)− x+ ρ ln(x).
20

A designação yi refere-se aos dados da proporção de infetados na Tabela 3.1.Assim, podemos calcular o coeficiente de determinação e tirar conclusões acerca
do grau de precisão do modelo.
xi g(xi) yi (g(xi)− y)2 (yi − y)2
0.973 0.027 0.027 0.676× 10−3 0.676× 10−3
0.9 0.0524 0.06 0.00036× 10−3 0.049× 10−3
0.77 0.0873 0.084 1.17649× 10−3 0.961× 10−3
0.588 0.105 0.111 2.704× 10−3 3.364× 10−3
0.464 0.0843 0.08 0.97969× 10−3 0.729× 10−3
0.414 0.0647 0.031 0.13689× 10−3 0.484× 10−3
0.372 0.0415 0.031 0.13225× 10−3 0.484× 10−3
0.318 0 0 2.809× 10−3 2.809× 10−3∑− − 0.424 8.61468× 10−3 9.556× 10−3
Tabela 3.2: Passos para calcular o coeficiente de determinação com os dados dapeste em Eyam em proporções.
Pelos dados da Tabela 3.2,∑
(g(xi)− y)2 = 8.61468×10−3 e∑
(yi − y)2 = 9.556×10−3 e substituindo na fórmula do coeficiente de determinação, temos:
R2 =8.61468× 10−3
9.556× 10−3= 0.901494349,
isto quer dizer que cerca de 90,15% dos casos da Peste em Eyam é explicada pelomodelo, mostrando que ele é de facto apropriado para estudar esta doença.
Conhecendo-se a proporção final da epidemia, podemos encontrar r∞ pela relaçãos∞ + r∞ = 1. Mas geralmente há interesse em estimar a proporção de indivíduosque serão afetados por uma determinada doença infeciosa, que se desenvolve nascondições do modelo SIR em estudo neste capítulo. De facto, conhendo-se ρ, pode-seestimar r∞ se previamente definirmos uma margem de erro a estabelecer na resoluçãoda equação (3.6).
Definindo ε < 0.005, podemos estimar a proporção de indivíduos que serão afeta-dos pela doença. Usando os dados da Tabela 3.1, temos ρ ≈ 0.61.
Assim, tomandoF (r∞) = 1− r∞ − s0e−
r∞ρ ,
então
F ′(r∞) = −1 +s0e− r∞
ρ
ρ.
21

E o método de Newton para resolver a equação F (r∞) = 0, é dado pela fórmula:
r(n+1)∞ = r(n)∞ −
F (r(n)∞ )
F ′(r(n)∞ )
Tomando r(0)∞ = 1, podemos encontrar a fração aproximada de pessoas que adoe-ceram e ganharam imunidade até ao final da epidemia.
r(1)∞ = r(0)∞ −F (r
(0)∞ )
F ′(r(0)∞ )
= 1− −0.18887
−0.69038= 0.726426.
Verifica-se então se a diferença entre o valor obtido para r(1)∞ e r(0)∞ é menor do queo erro proposto, calculando |r(1)∞ − r(0)∞ |.
Como |r(1)∞ − r(0)∞ | = 0.273574 > ε, então continuando o processo de cálculo para
encontrar r(n+1)∞ , obtemos uma aproximação da solução de (3.6) conforme a Tabela
3.3.
n F(r(n)∞
)F ′(r(n)∞
)r(n+1)∞ |r(n+1)
∞ − r(n)∞ |n = 0 −0.18887 −0.69038 0.726426 0.273574 > εn = 1 −0.02218 −0.51516 0.68337 0.043056 > εn = 2 −0.000752 −0.4797 0.6818 0.0016 < ε
Tabela 3.3: Resolução da equação (3.6) pelo método de Newton para ρ = 0.61.
Na Tabela 3.3 vemos que r(3)∞ = 0.6818 e |r(3)∞ − r
(2)∞ | < ε, portanto temos uma
estimativa da proporção de indivíduos r∞ que em última instância irá se recuperar(esse número é r∞ − r0, mas neste caso r0 = 0). Assim, temos pelo modelo quecerca de 68, 18% da população seria infetada pela doença para uma taxa de remoçãorelativa ρ = 0.61.
Assim se conhecermos a taxa de remoção relativa de uma doença infeciosa, pode-mos estimar a proporção da população que será infetada pela doença e assim efetuara preparação em termos de logistíca para lidar com a epidemia.
3.3 Modelo SIR com vacinação
Perante uma situação de incerteza em relação a uma doença infeciosa, podemosprocurar controlar o seu desenvolvimento. Isto pode ser conseguido de diversos mo-dos. Por exemplo através da vacinação, reduzindo assim a proporção de suscetíveisou através de quarentenas e isolamento dos doentes de modo a diminuir a taxa decontacto e consequente força de infeção.
22

Analisando o caso da vacina, embora ela possa ter efeitos colaterais em algunsindivíduos, suponhamos que ela tenha 100% de eficácia na imunização em relação auma certa doença. Devido a limitações económicas, humanas e materiais, podemosestar interessados em saber qual a percentagem limiar da população que deve servacinada para protegê-la da doença. Para tal, consideremos o modelo SIR com vaci-nação a seguir, cuja abordagem se baseou nos livros de Braun [2], Martcheva [11] enos artigos científicos [4], [12] e [20].
3.3.1 Um modelo com vacinação à nascença
Consideremos o modelo SIR com demografia com taxa de nascimento per capitaµ (igual à taxa de mortalidade natural) e suponha-se que todos os recém-nascidos sãosuscetíveis à doença.
Seja p a fração de recém-nascidos vacinados. Portanto, se µN é o termo de influxoda classe suscetível por nascimentos, uma fração pµN vai diretamente para a classerecuperada, e proporção qµN , onde q = 1 − p, entra na classe suscetível. Assim, omodelo SIR com vacinação torna-se:
S ′(t) = (1− p)µN − βSI − µS,I ′(t) = βSI − (µ+ α)I,
R′(t) = pµN + αI − µR.(3.11)
Note-se que neste caso o tamanho da população N continua a ser constante, umavez que o modelo anterior foi alterado considerando uma taxa de natalidade igualà taxa de mortalidade. De facto, (S + I + R)′ = µ (N − (S + I +R)) e portanto seS(0) + I(0) +R(0) = N então S(t) + I(t) +R(t) = N para ∀t.
Passando para proporções, tomando: s(t) = S(t)N
, i(t) = I(t)N
e r(t) = R(t)N
, o sistema(3.11), torna-se:
s′(t) = (1− p)µ− γsi− µs,i′(t) = γsi− (µ+ α)i,
r′(t) = pµ+ αi− µr,(3.12)
com γ = βN .Como o tamanho da população N é constante, tem-se s(t) + i(t) + r(t) = 1 para
todo t.
3.3.1.1 Pontos de equilíbrio e sua estabilidade
Quanto a pontos de equilíbrio, há a destacar a existência de dois tipos de pontos:o ponto de equilíbrio livre de doença e o ponto de equilíbrio endémico (onde a doença
23

persiste na população).O ponto de equilíbrio livre da doença pode ser encontrado considerando i = 0 na
terceira equação de (3.12), donde resulta r = p e, da primeira equação temos s = 1−p.Portanto, o ponto de equilíbrio livre da doença é da forma: (1− p, 0, p).
Para estudar a estabilidade do mesmo, calculemos a matriz Jacobiana neste ponto.
J(1−p,0,p) =
−µ −γ(1− p) 0
0 γ(1− p)− (µ+ α) 0
0 α −µ
(3.13)
Se todos os valores próprios de J(1−p,0,p) têm parte real negativa, então (1− p, 0, p)é assintoticamente estável. Caso exista um valor próprio de J(1−p,0,p) com parte realpositiva, o ponto (1− p, 0, p) é instável ([2], p. 389).
Analisemos a equação característica dada por |J − λI| = 0. Se expandirmos pelaprimeira linha e primeira coluna, obtemos:
|J − λI| = (λ+ µ)2(λ− γ(1− p) + µ+ α) = 0.
Assim, os valores próprios de J(1−ρ,0,ρ) são: λ1,2 = −µ com multiplicidade 2 e
λ3 = γ(1− p)− (µ+ α) = (α + µ) ((1− p)R0 − 1) ,
com R0 = γµ+α
.
Como µ > 0 e α > 0 concluímos que, se p < 1 − 1
R0
, o ponto de equilíbrio (1 −
p, 0, p) é instável. No caso em que p > 1 − 1
R0
, o ponto de equilíbrio (1 − p, 0, p) é
assintoticamente estável.Assim, a percentagem limiar a vacinar é p = 1 − 1
R0
, pois para uma percentagem
de vacinação acima deste valor, a doença não persiste na população e abaixo dele adoença torna-se endémica.
Para a varíola por exemplo R0 ≈ 4. Assim para se prevenir esta doença numacomunidade deve-se vacinar mais de 75% de sua população ([20], p. 12). E para adoença de Chagas, R0 = 1.25 ([12]), então para proteger uma determinada comuni-dade da epidemia deve ser vacinada mais de 20% de sua população.
Para encontrar o ponto de equilíbrio endémico, igualamos a zero os segundosmembros do sistema (3.12) tendo em conta que i 6= 0.
Da segunda equação temos s∗ =α + µ
γe substituindo na primeira equação resulta,
i∗ =µ(γ(1− p)− α− µ)
γ(α + µ)e substituindo este valor na terceira equação encontramos:
r∗ =pγ(α + µ) + α(γ(1− p)− α− µ)
γ(α + µ).
24

Para determinar as condições de existência do ponto de equilíbrio endémico, con-sideremos as duas primeiras equações de (3.12). Igualando a zero os segundos mem-bros dessas equações, temos
i∗(s∗) =(1− p)µγs∗
− µ
γ, (3.14)
es∗ =
µ+ α
γ.
O ponto de equilíbrio endémico existe somente se o gráfico de i∗(s∗) interseta a
reta s∗ =µ+ α
γnum ponto do triângulo
i > 0, s ≥ 0, s+ i ≤ 1 .
Facilmente se verifica que tal acontece se e só se (1 − p) > µ+αγ. Esta condição é
equivalente a i∗ > 0 e já implica s∗ ≤ 1.Na Figura 3.4 é ilustrada a condição necessária de existência do ponto de equilíbrio
endémico.
Figura 3.4: Condição para a existência do ponto de equilíbrio endémico.
Por outras palavras, o ponto de equilíbrio endémico existe se e só se
(1− p) > µ+ α
γ⇔ (1− p)γ > µ+ α⇔ (1− p)γ
µ+ α> 1
Assim Rv = (1 − p)R0 é o novo número básico de reprodução da doença na pre-sença de vacinação. Nota: se p = 0 então Rv = R0.
Para estudar a estabilidade do ponto de equilíbrio endémico, consideremos a ma-triz Jacobiana no ponto (s∗, i∗, r∗) e calculemos a sua equação característica.
25

∣∣∣∣∣∣∣−µγ(1−p)
α+µ− λ −(α + µ) 0
µ(γ(1−p)−α−µ)α+µ
0− λ 0
0 α −µ− λ
∣∣∣∣∣∣∣ = 0
⇔ −(µ+ λ)
[λ2 +
µγ(1− p)α + µ
· λ+ µ (γ(1− p)− α− µ)
]= 0
⇔ λ1 = −µ < 0, ∨ λ2 + µRv · λ+ µ (γ(1− p)− α− µ) = 0.
O descriminante da equação do 2o grau é:
∆ = (µRv)2 − 4µ (γ(1− p)− (α + µ)) ,
eλ2,3 = −µRv ±
√(µRv)2 − 4µ (γ(1− p)− (α + µ)).
Assim, para analisar o sinal da parte real dos restantes valores próprios, consideremoso argumento da raiz acima, (µRv)
2−4µ (γ(1− p)− (α + µ))︸ ︷︷ ︸A
. Como µ (γ(1− p)− (α + µ))
é um número positivo, então podemos observar as seguintes situações:Se A > (µRv)
2, os valores próprios λ2,3 são complexos com parte real negativa.Entretanto, se A < (µRv)
2, então o argumento da raiz é um número positivo, massua raiz é menor do que µRv, assim, mais uma vez λ2,3 têm parte real negativa.
Portanto, os valores próprios de J(s∗,i∗,r∗) têm necessariamente partes reais nega-tivas. Desse modo o ponto de equilíbrio endémico é assintoticamente estável.
Concluindo: Se Rv < 1 o sistema (3.12) tem único ponto de equilíbrio que é as-sintoticamente estável. No entanto se Rv > 1 o sistema tem dois pontos de equilíbrio:o ponto de equilíbrio lívre de doença (1− p, 0, p) que é instável e o ponto de equilíbrioendémico (s∗, i∗, r∗) que é assintoticamente estável.
26

Capítulo 4
Modelo epidémico SIS simples
No modelo epidémico SIS, a população é dividida em duas classes, a classe dossuscetíveis e a classe dos infetados, pois a recuperação da doença não garante imu-nidade e o indivíduo torna-se imediatamente suscetível.
Para estudar a dinâmica da doença pelo modelo SIS simples, recorreu-se aos livrosde Chasnov [5] e Martcheva [11].
A dinâmica da doença pelo modelo epidémico SIS é ilustrada pelo fluxogramaabaixo.
Figura 4.1: Fluxograma do modelo epidémico SIS simples.
O sistema de equações que modela a dinâmica da doença pelo modelo SIS sim-ples é:
s′(t) = −γis+ αi,
i′(t) = γis− αi(4.1)
Aqui, s(t) e i(t) são as proporções de indivíduos suscetíveis e infetados respetiva-mente. Ainda no sistema acima, γ e α são respetivamente a taxa de transmissão e derecuperação da doença.
O modelo epidémico SIS, considera o tamanho da população constante, portanto,s(t) + i(t) = 1. Condições iniciais para (4.1) são s(0) = s0 e i(0) = i0 com s0 + i0 = 1.
27

4.1 Solução explícita
Como o tamanho total da população é constante, o sistema (4.1) pode ser reduzidoa uma única equação. De facto, substituindo s por 1− i na segunda equação, temos:
i′(t) = γi(1− i)− αi. (4.2)
Colocando em evidência i na equação (4.2) e fazendo algumas transformações algé-bricas temos:
i′(t) = i(γ − α− γi) = (γ − α)i
(1− γi
γ − α
)= (γ − α)i
(1− i
γ−αγ
)
Tomando r = γ − α e k =r
γtemos a equação logística a seguir:
i′(t) = ri(1− i
k). (4.3)
A equação logística é um dos modelos clássicos na dinâmica populacional e, podeser resolvida explicitamente.
Note-se que i(t) = 0 e i(t) = k são soluções constantes da equação (4.3). Asrestantes soluções satisfazem:
k
i(k − i)· didt
= r. (4.4)
Para integrar (4.4) começa-se por decompork
i(k − i)em frações simples:
k
i(k − i)=
1
i+
1
k − i.
Como∫
k
i(k − i)di = ln
∣∣∣∣ i
k − i
∣∣∣∣, a equação (4.4) pode ser reescrita como:
d
dt
[ln
∣∣∣∣ i
k − i
∣∣∣∣] = r.
E integrando ambos os membros entre 0 e t :
lni(t)
|k − i(t)|− ln
i(0)
|k − i(0)|= rt.
Assim,
lni(t)|k − i(0)|i(0)|k − i(t)|
= rt⇒ i(t)
|k − i(t)|=
i(0)
|k − i(0)|ert.
Uma vez que i(t) = k é solução da equação (4.3), k− i(t) e k− i(0) têm mesmo sinal.
28

Assim, isolando i(t) temos:
i(t) =kBert
1 +Bert, (4.5)
onde B =i(0)
k − i(0)e r = γ − α ( γ e α são as taxas de transmissão e de recuperação
da doença respetivamente).Assim r pode ser positivo ou negativo. Para r < 0 a doença por si só desaparece,
poislimt→+∞
i(t) = 0.
No entanto para r > 0,limt→+∞
i(t) = k =r
γ= 1− α
γ,
isto é, a doença permanece na população se não houver intervenções no seu com-bate.
A condição limite r > 0 pode ser reescrita como
γ − α > 0⇒ γ > α⇒ γ
α> 1 ou R0 > 1,
onde R0 = γα
é o número básico de reprodução da doença. Portanto, a doença persistena população se R0 > 1 e acaba por desaparecer se R0 < 1.
Na Figura 4.2 estão representados os comportamentos das proporções i(t) e s(t)para a condição limite r < 0 e r > 0.
Figura 4.2: Solução da equação logística (4.3), considerando alternativamente r < 0 er > 0.
(a) Solução da equação lo-gística (4.3) para r < 0.
(b) Solução da equação lo-gística (4.3) para r > 0 eγ < 2α.
(c) Solução da equaçãologística (4.3) para r > 0e γ > 2α.
Por outras palavras, a Figura 4.2 ilustra as condições para ocorrência de uma epi-demia e os valores esperados das proporções finais para cada sinal do parâmetro r,que constituem os pontos de equilíbrio do sistema.
Como visto anteriormente, i(t) tende para zero ou k conforme r < 0 ou r > 0
respetivamente. E para r > 0, pode-se fazer intervenção no sentido de mudar o sinal
29

de r. As intervenções passam necessariamente por diminuir a taxa de transmissão γ(diminuição da taxa de contacto colocando os doentes em quarentena até à cura final)ou aumentar a taxa de recuperação α (diminuição do período infecioso).
Os valores i∗1 = 0 e i∗2 = γ−αγ
= k são equilíbrios de (4.3) e o segundo existe apenasquando r > 0.
Assim, quando r < 0, a equação (4.3) tem um único ponto de equilíbrio i∗1 = 0 queé globalmente assintoticamente estável. De facto todas as soluções convergem parai∗1 = 0 quando t tende para infinito e nunca se afastando desse valor.
Quando r > 0, a equação (4.3) para além do equilíbrio livre de doença que éinstável, tem o equilíbrio endémico que é globalmente assintoticamente estável. Todasas soluções não nulas tendem para i∗2 = k quando t tende para infinito e nunca seafastando desse valor.
4.2 Modelo epidémico SIS com tratamento saturado
Suponhamos que no sistema (4.1) a taxa de recuperação per capita α depende dotratamento. Neste caso, podemos supor que os recursos de tratamento são escassose que a taxa de recuperação per capita α diminui com o aumento da proporção deinfetados. Assumindo que a taxa de recuperação é expressa pela função:
α(i) =α
1 + iN,
onde α é a taxa de tratamento / recuperação quando há poucos infetados e N > 1
é o tamanho total da população, o sistema de equações para o modelo SIS comtratamento saturado é:
s′(t) = −γis+ αi1+iN
i′(t) = γis− αi1+iN
(4.6)
Para o modelo SIS com tratamento saturado, o tamanho da população é constantee portanto s + i = 1. Condições iniciais para (4.6) são s(0) = s0 e i(0) = i0 coms0 + i0 = 1.
4.2.1 Pontos de equilíbrio
É possível reduzir a uma única equação o sistema de equações (4.6). Substituindos = 1− i na segunda equação de (4.6) temos:
i′(t) = γi(1− i)− αi
1 + iN(4.7)
Note-se que i∗ é ponto de equilíbrio da equação (4.7) se e só se (1− i∗, i∗) é ponto
30
de equilíbrio do sistema (4.6).Estudemos a possibilidade de ocorrência de pontos de equilíbrio de (4.7). Igua-
lando o lado direito de (4.7) a zero, vemos claramente que um dos pontos de equilíbrioé i∗1 = 0. Os pontos de equilíbrio endémicos satisfazem:
γ(1− i) =α
1 + iN,
que é equivalente a:(1− i)(1 + iN) =
α
γ. (4.8)
Na equação (4.8), tomemos g(i) = (1− i)(1 + iN) e h(i) =α
γ.
Como g(0) = 1 e g(1) = 0, se αγ< 1 então a equação (4.8) sempre tem somente
uma solução positiva i∗2. Então a equação (4.7) tem um único equilíbrio endémico (verFigura 4.3). A condição α
γ< 1, pode ser reescrita como γ
α> 1 ou R0 > 1.
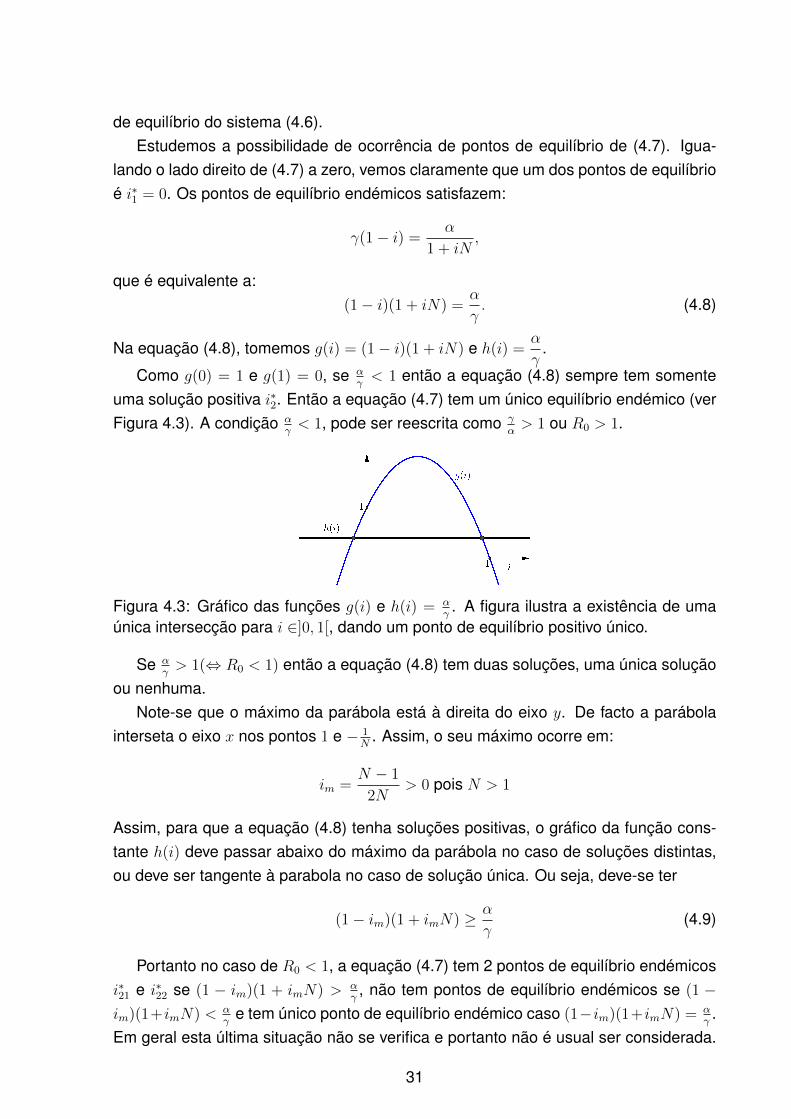
Figura 4.3: Gráfico das funções g(i) e h(i) = αγ. A figura ilustra a existência de uma
única intersecção para i ∈]0, 1[, dando um ponto de equilíbrio positivo único.
Se αγ> 1(⇔ R0 < 1) então a equação (4.8) tem duas soluções, uma única solução
ou nenhuma.Note-se que o máximo da parábola está à direita do eixo y. De facto a parábola
interseta o eixo x nos pontos 1 e − 1N
. Assim, o seu máximo ocorre em:
im =N − 1
2N> 0 pois N > 1
Assim, para que a equação (4.8) tenha soluções positivas, o gráfico da função cons-tante h(i) deve passar abaixo do máximo da parábola no caso de soluções distintas,ou deve ser tangente à parabola no caso de solução única. Ou seja, deve-se ter
(1− im)(1 + imN) ≥ α
γ(4.9)
Portanto no caso de R0 < 1, a equação (4.7) tem 2 pontos de equilíbrio endémicosi∗21 e i∗22 se (1 − im)(1 + imN) > α
γ, não tem pontos de equilíbrio endémicos se (1 −
im)(1+imN) < αγ
e tem único ponto de equilíbrio endémico caso (1−im)(1+imN) = αγ.
Em geral esta última situação não se verifica e portanto não é usual ser considerada.
31

A Figura 4.4, ilustra as possibilidades de ocorrência de pontos de equilíbrio endémicospara R0 < 1.
Figura 4.4: Existência de equilíbrios endémicos para R0 < 1.
(a) R0 < 1 e (1− im)(1 +imN) > α
γ .(b) R0 < 1 e (1− im)(1 +imN) < α
γ .(c) R0 < 1 e (1− im)(1+imN) = α
γ
Podemos encontrar os valores numéricos dos pontos de equilíbrio endémicos re-solvendo a equação
γ(1− i)(1 + iN)− α = 0. (4.10)
para as condições de ocorrência de interseção entre os gráficos de g(i) e h(i) parai ∈]0, 1[ discutidas anteriormente.
Desembaraçando parênteses e agrupando termos semelhantes temos:
−Ni2 + (N − 1)i+ 1− α
γ= 0,
e calculando o descriminante da equação do 20 grau:
∆ = (N − 1)2 + 4N
(1− α
γ
).
(A) Consideremos R0 > 1 (⇔ αγ< 1).
Neste caso ∆ > 0 e portanto, temos os seguintes pontos de equilíbrio para aequação (4.7):
i∗1 = 0 e i∗2 =1
2N
(N − 1 +
√∆).
(B) Consideremos R0 < 1 (⇔ αγ> 1) e ∆ > 0.
A equação (4.10) tem duas raizes positivas distintas. Neste caso a equação (4.7)para além do equilíbrio livre de doença i∗1 = 0, tem dois equilíbrios endémicos
i∗21 =1
2N
(N − 1 +
√∆)
e i∗22 =1
2N
(N − 1−
√∆).
(C) Consideremos R0 < 1 (⇔ αγ> 1) e ∆ = 0.
A equação (4.10) tem duas raizes positivas iguais. Neste caso a equação (4.7)
32

tem os seguintes pontos de equilíbrio:
i∗1 = 0 e i∗21 = i∗22 =1
2N(N − 1)
(D) Consideremos R0 < 1 (⇔ αγ> 1) e ∆ < 0.
Se ∆ < 0, a equação (4.10) não tem solução para i ∈]0, 1[, assim a equação(4.7) tem um único ponto de equilíbrio i∗1 = 0.
4.2.1.1 Estudo da estabilidade dos pontos de equilíbrio
Para decidir sobre a estabilidade dos pontos de equilíbrio discutidos na secçãoanterior, estudemos o comportamento das soluções positivas da equação (4.7):
i′(t) = f(i) com f(i) = γi(1− i)− αi
1 + iN
através de diagramas de fase construídos a partir do gráfico de f . Analisemos primei-ramente o comportamento das soluções no caso R0 > 1.
Figura 4.5: Análise gráfica das soluções da equação (4.7) para R0 > 1.
(a) Para R0 > 1 existem doispontos de equilíbrio (o livre dadoença e o endémico).
(b) Diagrama de fase das solu-ções da equação (4.7) para R0 >1 e as concavidades das respeti-vas soluções.
Pelo diagrama de fase (Figura 4.5 direita) para R0 > 1 vemos que o ponto deequilíbrio i∗1 é instável enquanto que o ponto de equilíbrio endémico i∗2 é globalmenteassintoticamente estável. E ainda estão representadas na Figura 4.5 direita as conca-vidades das soluções de i(t). As concavidades são obtidas a partir do sinal de
d2i
dt2= f ′(i) · f(i), (4.11)
para i ∈]0, 1[. As soluções da equação (4.7) têm concavidade voltada para baixo nointervalo em que f ′(i) · f(i) é negativo e, concavidade voltada para cima no intervaloem que f ′(i) · f(i) é positivo. As soluções da equação f ′(i) = 0 são os únicos pontos
33

que poderão ser pontos de inflexão das soluções da equação (4.7). No diagrama defase da Figura 4.5, c é o ponto de inflexão e é o ponto onde f(i) atinge o máximo (verFigura 4.5 esquerda).
E para estudar a estabilidade dos pontos de equilíbrio para R0 < 1 e ∆ > 0,consideremos a Figura 4.6.
Figura 4.6: Análise gráfica da estabilidade dos pontos de equilíbrio para R0 < 1 e∆ > 0.
(a) Para R0 < 1, ∆ >0 existem três pontos deequilíbrio i∗1, i∗21 e i∗22.
(b) Diagrama de fase e concavida-des das soluções de (4.7) paraR0 <1 e ∆ > 0.
Para R0 < 1 e ∆ > 0, a equação (4.7) tem três pontos de equilíbrio i∗1, i∗21 e i∗22,tal que i∗21 < i∗22. Pelo diagrama de fase da Figura 4.6 direita, vemos que as soluçõescom condição inicial i0 < i∗21 são atraídas para o ponto de equilíbrio livre de doençai∗1 = 0 quando t tende para infinito. Portanto o ponto de equilíbrio livre de doença éassintoticamente estável. Entretanto todas as soluções com condição inicial i0 > i∗21
são atraídas pelo ponto de equilíbrio i∗22 quando t tende para infinito. Neste caso, oequilíbrio endémico i∗22 é assintoticamente estável.
Os intervalos ]0, i∗21[ e ]i∗21, 1[ são os domínios de atração dos equilíbrios livre dedoença e endémico respetivamente. Esta situação é conhecida por biestabilidade.
Em relação ao ponto de equilíbrio i∗21 vemos que independentemente das condi-ções iniciais i0, as soluções da equação (4.7) se afastam dele. Portanto i∗21 é um pontode equilíbrio instável.
Ainda na Figura 4.6 direita, estão ilustradas as concavidades das soluções de (4.7)determinadas a partir do sinal de f ′(i)·f(i). Os pontos c1 e c2 são denominados pontosde inflexão, pois existe mudança de concavidade nas soluções.
Se para além de R0 < 1 se verificar ∆ = 0, a equação (4.7) tem 2 pontos deequilíbrio i∗1 e i∗21 = i∗22. Quanto à estabilidade dos pontos de equilíbrio, temos: i∗1 éassintoticamente estável, sendo seu domínio de atração ]0, i∗21[ enquanto que i∗21 = i∗22
é instável, sendo seu domínio de atração ]i∗21, 1[ (ver Figura 4.7).Para R0 < 1 e ∆ < 0 a equação (4.7) tem um único equilíbrio i∗1 = 0, que é global-
mente assintoticamente estável. Para decidir acerca das concavidades das soluções
34
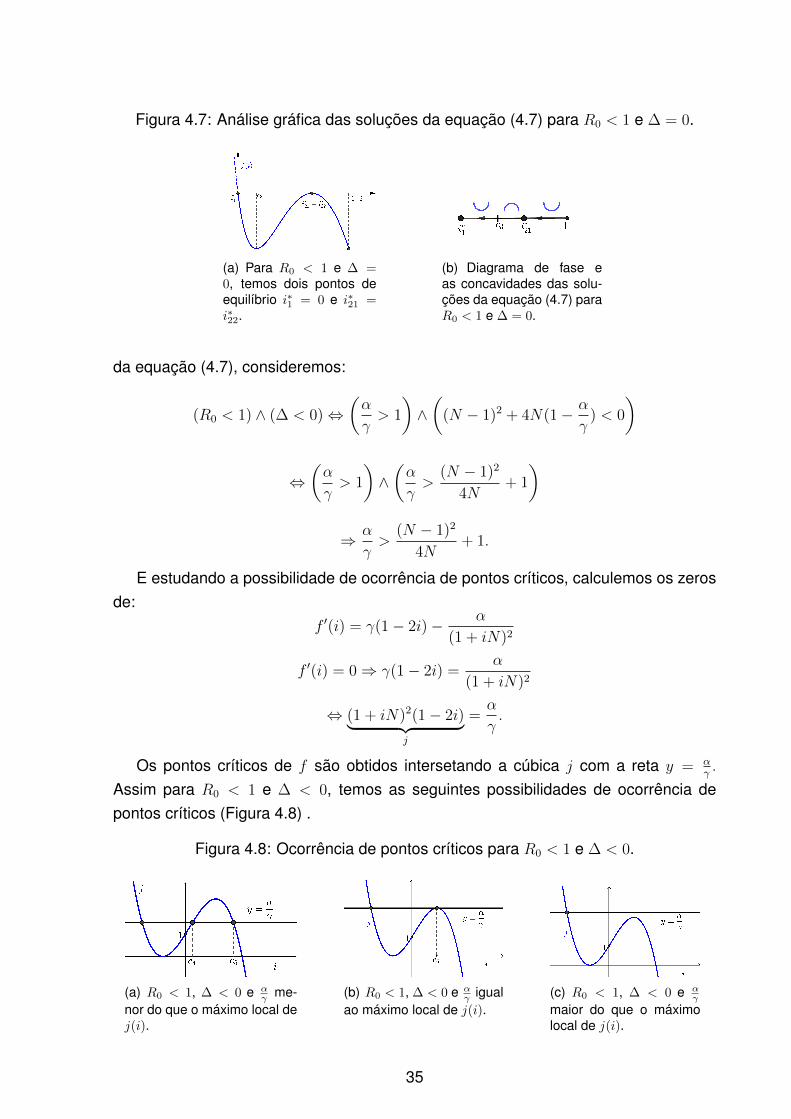
Figura 4.7: Análise gráfica das soluções da equação (4.7) para R0 < 1 e ∆ = 0.
(a) Para R0 < 1 e ∆ =0, temos dois pontos deequilíbrio i∗1 = 0 e i∗21 =i∗22.
(b) Diagrama de fase eas concavidades das solu-ções da equação (4.7) paraR0 < 1 e ∆ = 0.
da equação (4.7), consideremos:
(R0 < 1) ∧ (∆ < 0)⇔(α
γ> 1
)∧(
(N − 1)2 + 4N(1− α
γ) < 0
)
⇔(α
γ> 1
)∧(α
γ>
(N − 1)2
4N+ 1
)
⇒ α
γ>
(N − 1)2
4N+ 1.
E estudando a possibilidade de ocorrência de pontos críticos, calculemos os zerosde:
f ′(i) = γ(1− 2i)− α
(1 + iN)2
f ′(i) = 0⇒ γ(1− 2i) =α
(1 + iN)2
⇔ (1 + iN)2(1− 2i)︸ ︷︷ ︸j
=α
γ.
Os pontos críticos de f são obtidos intersetando a cúbica j com a reta y = αγ.
Assim para R0 < 1 e ∆ < 0, temos as seguintes possibilidades de ocorrência depontos críticos (Figura 4.8) .
Figura 4.8: Ocorrência de pontos críticos para R0 < 1 e ∆ < 0.
(a) R0 < 1, ∆ < 0 e αγ me-
nor do que o máximo local dej(i).
(b) R0 < 1, ∆ < 0 e αγ igual
ao máximo local de j(i).(c) R0 < 1, ∆ < 0 e α
γ
maior do que o máximolocal de j(i).
35

Assim, podemos obter os seguintes esboços da função f (ver Figura 4.9) levandoem conta as possibilidades de ocorrência de pontos críticos descritos na Figura 4.8acima.
Figura 4.9: Gráficos da função f para R0 < 1 e ∆ < 0.
(a) R0 < 1, ∆ < 0 e αγ
menor do que o máximolocal de j(i).
(b) R0 < 1, ∆ < 0e αγ igual ao máximo
local de j(i).
(c) R0 < 1, ∆ < 0 e αγ
maior do que o máximo localde j(i).
Assim, podemos ter os seguintes diagramas de fase (ver Figura 4.10) para as situ-ações discutidas na Figura 4.9, respetivamente (a), (b) e (c).
Figura 4.10: Análise gráfica das soluções da equação (4.7) para R0 < 1 e ∆ < 0.
(a) Concavidadespara R0 < 1, ∆ < 0e αγ menor do que o
máximo de j(i).
(b) Concavidades paraR0 < 1, ∆ < 0 e α
γ igualao máximo de j(i).
(c) Concavidadespara R0 < 1, ∆ < 0e α
γ maior do quemáximo de j(i).
Pelo diagrama de fase (ver Figura 4.10) vemos que todas as soluções independen-temente das condições iniciais convergem para o ponto de equilíbrio i∗1 = 0 quando t
tende para infinito. Portanto i∗1 = 0 é ponto de equilíbrio globalmente assintoticamenteestável.
36

Capítulo 5
Modelo da malária deRoss-Macdonald
O modelo de Ross-Macdonald, foi desenvolvido primeiramente por Ronald Ross(1857-1932). Ele desenvolveu sua pesquisa sobre malária na Índia quando pres-tava serviço militar ao Governo Britânico. Depois de ter descoberto que os mosquitoseram transmissores da malária aos humanos, ele associou à dinâmica das interaçõeshumano-mosquito equações diferenciais para o estudo quantitativo da doença e publi-cou o seu primeiro trabalho em 1908: Report on the Prevention of Malaria in Mauritiusonde constava o primeiro modelo matemático e em 1911 na segunda edição de ThePrevention of Malaria, propôs o segundo modelo para o estudo da doença ([17], p.3,4).
Em meados do século XX, George Macdonald (1903-1967) seguiu literalmente ospassos de Ross e testou a teoria de Ronald Ross com dados epidemiológicos e ento-mológicos de campo. Em 1947 Macdonald propôs a introdução do período de latênciado agente patogénico no mosquito na modelação das interações humano-mosquitono processo da transmissão da malária. A análise de Macdonald ajudou a explicarque a pulverização com inseticida funcionava porque reduzia bastante o número demosquitos que já tinham vivido tempo suficiente para se tornarem infeciosos e trans-mitirem a malária. Desde então muitas atividades de controlo da malária focaram-seno combate aos mosquitos adultos ([6], p. 5).
Ao longo do tempo foram desenvolvidas várias versões dos modelos de Ross-Macdonald. Sem uma formulação canónica, o modelo de Ross-Macdonald é maisproveitosamente descrito como um conjunto de modelos todos assentes num conjuntoconsensual de hipóteses simplificadoras.
Um modelo de Ross-Macdonald baseia-se num processo quantitativo simplificadoconsiderando a descrição do ciclo de vida do agente patogénico em quatro etapas:(1) o agente patogénico é passado de um mosquito infetado para um hospedeiro hu-mano durante a alimentação de sangue (pela picada de uma fêmea Anopheles infe-
37

tada, portadora de formas infetantes (esporozoítas) na glândula salivar); (2) o agentepatogénico infeta e depois multiplica-se no hospedeiro humano, atingindo densidadessuficientemente altas no sangue deste para infetar um novo mosquito; (3) um mosquitosuscetível absorve o agente patogénico do hospedeiro humano infetando-se durantea alimentação; e (4) o agente patogénico desenvolve-se no mosquito até ficar fixadonas glândulas salivares ou partes da boca e pronto para ser transmitido durante umamordida subsequente num hospedeiro humano ([17], p. 8).
Os modelos matemáticos Ross-Macdonald diferem uns dos outros na tipologia domodelo matemático e na maneira de implementar a latência no mosquito ou no hu-mano, mas há um conjunto consensual de hipóteses simplificadoras comuns sobre atransmissão e dinâmica da malária: picadas de mosquitos são distribuídas aleatoria-mente e uniformemente entre as populações de hospedeiros humanos, as populaçõesestão fechadas ao nascimento ou migração, os hospedeiros tornam-se suscetíveis àinfeção após a recuperação, a razão de mosquitos para humanos é constante e háapenas uma espécie de mosquito vetor.
Este capítulo está dividido em duas partes principais, a primeira parte está con-centrada no modelo simples de Ross-Macdonald (sem levar em conta a latência), cujaprincipal análise é observar a influência dos parâmetros, presentes nas equações, naprevalência da malária. E na segunda parte há inclusão dos períodos de latência nomosquito e no hospedeiro humano. A sua principal análise está ligada ao númerobásico de reprodução da doença, que serve como limiar de epidemia para doençastransmitidas por vetores. O estudo da dinâmica da malária levando em conta dois pe-ríodos de latência, isto é no mosquito e no ser humano foi incluído no modelo simplesde Ross-Macdonald em 2008 ([16]).
Para falar da dinâmica da malária, recorreu-se aos seguintes livros: Anderson [1],Braun [2], Martcheva [11] e aos artigos científicos: [6], [8], [10], [13], [16], [17] e [21]para suporte teórico.
5.1 Modelo simples de Ross-Macdonald
O modelo mais simples para a população de mosquitos é o modelo SI, pois umavez infetados, os mosquitos não se recuperam. Denotando o número de mosquitos naclasse suscetível por SM e os na classe infetada por IM , temos:
SM + IM = M,
onde M é o número total de mosquitos num determinado tempo t. Assume-se que onúmero total da população de mosquitos é constante.
E o modelo mais simples para humanos é da forma SIS sem demografia (isto é
38

não se considera migração, nascimentos nem mortes), pois uma vez infetados, geral-mente se recuperam mas não ganham imunidade. Assume-se que o tamanho total dapopulação humana é constante e dada por
SH + IH = N,
onde SH e IH são os números de humanos suscetíveis e infetados respetivamente.Assim, os tamanhos SH e SM podem ser escritos como: SH = N − IH e SM = M − IM ,respetivamente.
Apresentam-se na Tabela 5.1 alguns termos utilizados no modelo de Ross-Macdonald.
N número total de humanosp proporção de picadas em que um mosquito suscetível adquire infeçãoα taxa de recuperação de humanosa taxa de picada de mosquitos em humanosM número total de mosquitosq proporção de picadas que produzem infeção em humanosµ taxa de mortalidade de mosquitos
Tabela 5.1: Parâmetros usados no modelo simples de Ross-Macdonald.
O modelo de Ross-Macdonald assume que a transmissão do parasita da maláriado mosquito ao homem e vice-versa é proporcional ao produto do número de indiví-duos suscetíveis pelo número de infetados da espécie apropriada.
Se a transferência do parasita for do mosquito para o homem, o produto em ques-tão é IM×(N − IH). Por outro lado, a taxa de transferência do homem para o mosquitoé presumida proporcional a IH×(M − IM). Isso nos leva ao seguinte sistema de equa-ções
dIHdt
= ( aN
)qIM(N − IH)− αIH
dIMdt
= ( aN
)pIH(M − IM)− µIM(5.1)
onde os parâmetros, a, p, q, µ e α estão definidos na Tabela 5.1.A dinâmica da malária entre mosquitos e humanos pode ser ilustrada pelo fluxo-
grama a seguir (ver Figura 5.1).Considerando alternativamente, as proporções de humanos infetados x = IH
Ne de
mosquitos infetados y = IMM
, o sistema (5.1) fica:
x = aqym(1− x)− αxy = apx(1− y)− µy
(5.2)
onde m =M
N, é chamada densidade de mosquitos para humanos. Daqui em diante
39

Figura 5.1: Dinâmica da malária entre humanos e mosquitos no modelo simples deRoss-Macdonald, sem considerarmos a inclusão de períodos de latência. Neste fluxo-grama assim como no modelo só se considera a transmissão da malária por picadasde mosquitos.
nesta secção, a análise do modelo matemático de Ross-Macdonald para malária seráfeita com base no sistema (5.2).
5.1.1 Isóclinas e pontos de equilíbrio
Para encontrar os pontos de equilíbrio do sistema (5.2), igualamos os segundosmembros do sistema a zero. E facilmente vemos que uma das soluções do sistema éo ponto (0, 0). É facil provar que um ponto de equilíbrio em que x = 0, obriga y = 0, omesmo se passando com o y = 0. Para encontrar outra solução do mesmo sistema,consideramos x 6= 0 ou y 6= 0. De facto, resolvendo em relação a y a primeira equação,se x = 1, a primeira equação leva a uma equação impossível, logo não há ponto deequilíbrio com x = 1
y =αx
aqm(1− x), (1− x) 6= 0, (5.3)
e substituindo o valor encontrado na segunda equação do mesmo sistema, temos:
apx
(1− αx
aqm(1− x)
)− µ
(αx
aqm(1− x)
)= 0 x 6= 0, 1
⇔ ap(aqm(1− x)− αx)− µα = 0
⇔ a2pqm− a2pqmx− apαx− µα = 0
⇔ −(a2pqm+ apα)x = µα− a2pqm
⇒ x∗ =a2pqm− µαa2pqm+ apα
.
Substituindo x∗ em (5.3) e simplificando a expressão resultante,
y∗ =a2pqm− µαa2pqm+ aqmµ
.
40

Portanto o único outro ponto de equilíbrio do sistema (5.2) é o ponto:
(x∗, y∗) =
(a2pqm− µαa2pqm+ apα
,a2pqm− µαa2pqm+ aqmµ
)(5.4)
O ponto de equilíbrio (0, 0) é chamado de livre de doença pois x = y = 0. E o ponto(x∗, y∗) é chamado de ponto de equilíbrio endémico, e a sua existência fica garantidase x∗ e y∗ pertencem ao intervalo ]0, 1[. Ora
0 < x∗ < 1⇔ 0 < a2pqm− µα < a2pqm+ apα,
A desigualdade da direita é trivialmente satisfeita pois todos os parâmetros são positi-vos. A desigualdade da esquerda é equivalente a:
µα < a2pqm (5.5)
Do mesmo modo se conclui que 0 < y∗ < 1 se e só se µα < a2pqm. Assim a existênciado ponto de equilíbrio endémico fica garantida pela condição (5.5).
Suponha-se que se verifica (5.5) e consideremos o quadrado definido por 0 < x < 1
e 0 < y < 1 dividido em regiões em que x e y tenham sinais fixos. Igualando a zero ossegundos membros do sistema (5.2), obtém-se: x = 0⇔ y = f(x) e y = 0⇔ y = h(x),onde f(x) =
αx
aqm(1− x)e h(x) =
apx
apx+ µ.
Estas funções são funções crescentes de x e intersetam-se em (0, 0) e (x∗, y∗) e
limx→1
f(x) = +∞ e limx→+∞
h(x) = 1.
Facilmente se conclui que x é positivo para qualquer ponto (x, y) acima da curvay = f(x) e negativo para qualquer ponto abaixo da mesma curva. O mesmo acontececom y, que é positivo abaixo da curva y = h(x) e negativo acima dessa curva. Por-tanto, de acordo com a Figura 5.2 na região I, x > 0 e y > 0, na região II, x < 0 ey > 0, na região III, x < 0 e y < 0 e na região IV , x > 0 e y < 0.
Figura 5.2: Esboço das curvas isóclinas para a = 0.3, p = q = 0.5, m = 3, α = 0.05 eµ = 0.35. Pontos de interseção: (0, 0) e (x∗, y∗) =
(23, 29
).
41

Outra observação em relação às funções f e h é que na origem h cresce maisrapidamente do que f . De facto, por (5.5)
h′(0) =ap
µ>
α
aqm= f ′(0). (5.6)
Portanto, h(x) está acima de f(x) para 0 < x < x∗ e está abaixo para x∗ < x < 1.Estudemos o comportamento das soluções em cada uma das regiões I, II, III, IV.
Lema 5.1. Qualquer solução (x(t), y(t)) que começa na região I, no tempo t = t0 per-manece nesta região em qualquer instante t > t0 e aproxima-se do ponto de equilíbrio(x∗, y∗) quando t tende para infinito.
Prova: Suponhamos que (x(t), y(t)) deixa a região I no momento t = t1. Então,x(t1) = 0 ou y(t1) = 0, visto que a única maneira da solução sair da região I é cruzary = f(x) ou y = h(x). Assumamos que x(t1) = 0. Derivando a primeira equação de(5.2) em relação ao tempo e tomando t = t1 , temos:
x(t1) = aqmy(t1)(1− x(t1)).
Esta quantidade é positiva pois x(t1) < 1 e y(t1) é positivo na parte da curva y =
f(x) contida em 0 < x < x∗. Assim x(t) tem um mínimo em x(t1), o que é impossível,pois x(t) é crescente em I. Alternativamente, suponhamos que (x(t), y(t)) abandonaa região I cruzando a curva y = h(x) em t = t1, então y(t1) = 0 e
y(t1) = apx(t1)(1− y(t1)).
Esta quantidade é positiva, visto que y(t1) é menor que 1 e a quantidade x(t1) épositiva na parte da curva y = h(x) contida em 0 < x < x∗. Assim y(t) tem um mínimoem t = t1, o que é impossível, pois y(t) é crescente na região I.
Conclui-se assim que qualquer solução de (5.2) que começa na região I no tempot = t0, irá se manter nesta região para qualquer instante t > t0. Assim x(t) e y(t)
são funções monotonamente crescentes para t ≥ t0, com x(t) < x∗ e y(t) < y∗.Consequentemente ([2], p.426,427) (x(t), y(t)) tem limite (ε, η) quando t tende parainfinito, e (ε, η) é um ponto de equilíbrio de (5.2). Existem dois pontos de equilíbrio, oponto (0, 0) e o ponto (x∗, y∗), mas (ε, η) não pode ser o ponto (0, 0), visto que x(t) ey(t) são funções crescentes no tempo. Portanto (ε, η) = (x∗, y∗).
Usando argumentos semelhantes, mostra-se que:
Lema 5.2. Qualquer solução (x(t), y(t)) do sistema (5.2) que começa na região III,no tempo t = t0, permanece nesta região para t > t0 e tende para o ponto (x∗, y∗)
quando t tende para infinito.
42

Lema 5.3. Qualquer solução (x(t), y(t)) de (5.2) que começa na região II, no tempot = t0 e que permanece nesta região para todo o instante t, aproxima-se do ponto deequilíbrio (x∗, y∗) quando t tende para infinito.
Prova: Se a solução (x(t), y(t)) de (5.2) permanece na região II para t ≥ t0, entãox(t) é monótona decrescente e y(t) é monótona crescente para t ≥ t0. No entanto, x(t)
é positivo e y(t) é menor que 1 para t ≥ t0. Consequentemente, x(t) e y(t) têm limitesε e η respetivamente, quando t tende para infinito e (ε, η) é um ponto de equilíbrio de(5.2). Mas este ponto não pode ser (0, 0) porque y(t) é uma função crescente parat ≥ t0. Portanto (ε, η) = (x∗, y∗).
De modo análogo se mostra o resultado seguinte:
Lema 5.4. Qualquer solução (x(t), y(t)) de (5.2) que começa na região IV , no tempot = t0 e que permanece nesta região para todo o instante t, aproxima-se do ponto deequilíbrio (x∗, y∗) quando t tende para infinito.
Usando os resultados dos Lemas (5.1, 5.2, 5.3, 5.4 ), prova-se o seguinte teorema:
Teorema 5.1. ([2], p. 466)(a) Se µα < a2pqm, então todas as soluções (x(t), y(t)) do sistema (5.2) com condiçãoinicial em 0 < x < 1 e 0 < y < 1 tendem para o ponto (x∗, y∗) quando t tende parainfinito.(b) Se µα > a2pqm, então todas as soluções (x(t), y(t)) do sistema (5.2) com condiçãoinicial em 0 < x < 1 e 0 < y < 1 tendem para o ponto (0, 0) quando t tende para infinito.
Prova: Para provar a parte (a) deste teorema, observemos que pelos Lemas 5.1 e 5.2,toda a solução (x(t), y(t)) de (5.2) que se inicia na região I ou III, no tempo t = t0
permanece nestas regiões e converge para (x∗, y∗) quando t tende para infinito. Demodo similar, pelos Lemas 5.3 e 5.4, toda a solução que se inicia nas regiões II ouIV no tempo t = t0 e que permanece nessas regiões converge para (x∗, y∗) quando ttende para infinito. Agora, supondo que uma dada solução (x(t), y(t)) de (5.2), deixaas regiões II ou IV , então esta solução interseta as curvas y = f(x) ou y = h(x) eentra imediatamente nas regiões I ou III. Consequentemente as soluções (x(t), y(t))
de (5.2) que se iniciam nas regiões II ou IV ou nas curvas y = f(x) ou y = h(x) notempo t = t0 também convergem para (x∗, y∗).
Para provar a parte (b) do teorema temos de analisar as soluções de (5.2) paraµα > a2pqm. Neste caso as curvas y = f(x) e y = h(x) intersetam-se apenas em (0,0)e definem três regiões no quadrado 0 ≤ x ≤ 1 e 0 ≤ y ≤ 1, como na Figura 5.3.
De facto, como a2pqm < µα,
h′(0) =ap
µ<
α
aqm= f ′(0) (5.7)
43

e portanto na origem f cresce mais rapidamente do que h.Quanto ao sinal de x e y, temos o seguinte: na região I, x > 0 e y < 0, na região
II, x < 0 e y < 0 e na região III, x < 0 e y > 0.
Figura 5.3: Esboço das curvas isóclinas para a = 0.2, p = q = 0.5, m = 0.5, α = 0.05 eµ = 0.5. As curvas intersetam-se somente em (x, y) = (0, 0).
Assim como foi feito para provar a parte (a), podemos mostrar que qualquer solu-ção (x(t), y(t)) converge para o ponto de equilíbrio livre de doença (0, 0).
Consideremos a região II na Figura 5.3. Aqui, x < 0 e y < 0, e portanto, qualquersolução (x(t), y(t)) do sistema (5.2) que se inicia na região II permanece nesta regiãoe converge para (0, 0), quando t tende para infinito. E qualquer solução (x(t), y(t)) de(5.2) que se inicia na região I ou III abandona essas regiões intersetando a curvay = f(x) ou a curva y = h(x) e passa para a região II convergindo depois para(0, 0).
Na Figura 5.4 ilustra-se os resultados do Teorema 5.1 através da representação docampo vetorial associado ao sistema (5.2) para µα < a2pqm e µα > a2pqm respetiva-mente à esquerda e à direita.
Analisando a condição de ocorrência do ponto de equilíbrio endémico, baseado nainclinação das retas tangentes às curvas y = f(x) e y = h(x) em x = 0, temos: seh′(0) f ′(0) então o ponto de equilíbrio endémico existe e está muito próximo docanto superior direito do quadrado 0 < x < 1, 0 < y < 1. Se houver algum tipo deintervenção, por exemplo diminuição da taxa de picadas ou aumento da taxa de mor-talidade do mosquito, pode-se reduzir a diferença entre
ap
µe
α
aqm. Quanto menor for
a diferença entreap
µe
α
aqmmais perto de (0,0) estará o ponto de equilíbrio endémico.
A quantidadeap
µrepresenta o número médio de mordidas infeciosas
em hospedeiros humanos feitas por um mosquito em sua vida. Esta quan-tidade foi chamada por Macdonald de índice de estabilidade, pois em áreasonde os mosquitos picam os seres humanos com relativa frequência e têmesperança de vida relativamente longa, esse índice é alto e a malária tende
44
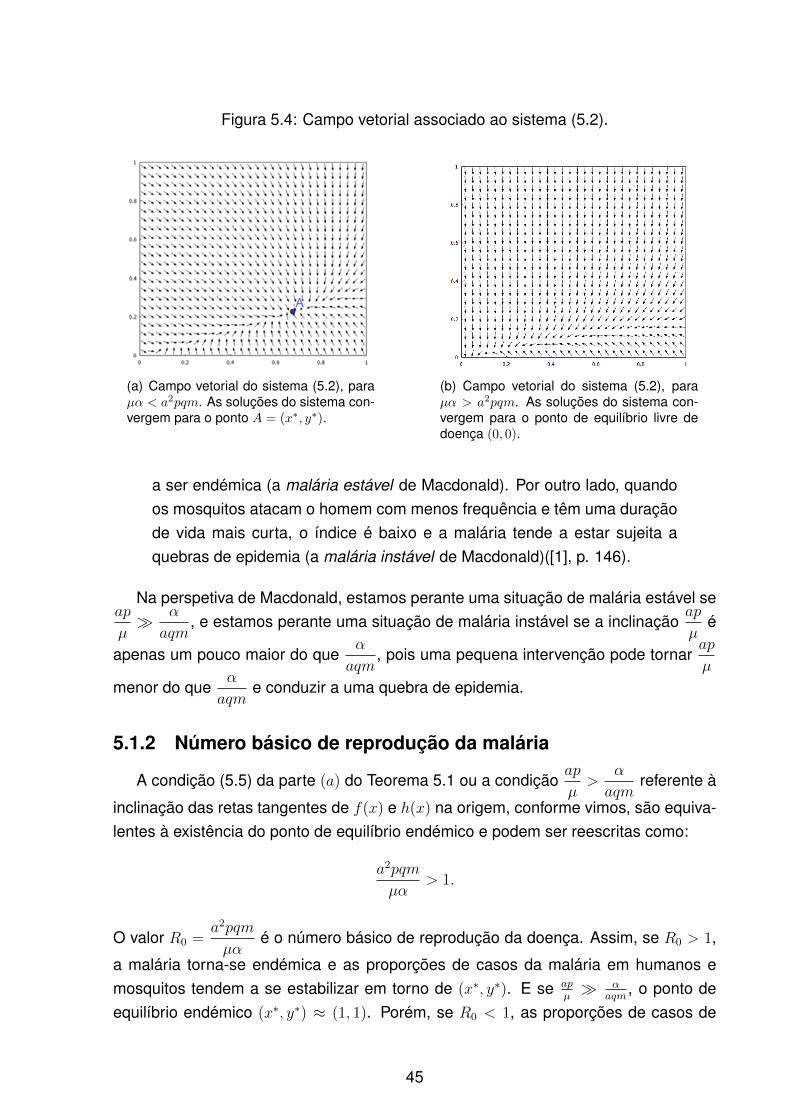
Figura 5.4: Campo vetorial associado ao sistema (5.2).
(a) Campo vetorial do sistema (5.2), paraµα < a2pqm. As soluções do sistema con-vergem para o ponto A = (x∗, y∗).
(b) Campo vetorial do sistema (5.2), paraµα > a2pqm. As soluções do sistema con-vergem para o ponto de equilíbrio livre dedoença (0, 0).
a ser endémica (a malária estável de Macdonald). Por outro lado, quandoos mosquitos atacam o homem com menos frequência e têm uma duraçãode vida mais curta, o índice é baixo e a malária tende a estar sujeita aquebras de epidemia (a malária instável de Macdonald)([1], p. 146).
Na perspetiva de Macdonald, estamos perante uma situação de malária estável seap
µ α
aqm, e estamos perante uma situação de malária instável se a inclinação
ap
µé
apenas um pouco maior do queα
aqm, pois uma pequena intervenção pode tornar
ap
µ
menor do queα
aqme conduzir a uma quebra de epidemia.
5.1.2 Número básico de reprodução da malária
A condição (5.5) da parte (a) do Teorema 5.1 ou a condiçãoap
µ>
α
aqmreferente à
inclinação das retas tangentes de f(x) e h(x) na origem, conforme vimos, são equiva-lentes à existência do ponto de equilíbrio endémico e podem ser reescritas como:
a2pqm
µα> 1.
O valor R0 =a2pqm
µαé o número básico de reprodução da doença. Assim, se R0 > 1,
a malária torna-se endémica e as proporções de casos da malária em humanos emosquitos tendem a se estabilizar em torno de (x∗, y∗). E se ap
µ α
aqm, o ponto de
equilíbrio endémico (x∗, y∗) ≈ (1, 1). Porém, se R0 < 1, as proporções de casos de
45

malária tenderão a se estabilizar em torno do equilíbrio livre de doença, ou seja doponto (0, 0).
O ponto de equilíbrio (x∗, y∗) pode ser reescrito na forma:
(x∗, y∗) =
(R0 − 1apµ
+R0
,(R0 − 1)ap
µ
R0(1 + apµ
)
)e podemos estudar a sua evolução em função de R0 fixando previamente o valor deh′(0) (ver Tabela 5.2).
Figura 5.5: Evolução das proporções de humanos e mosquitos infetados em funçãodo número básico de reprodução da doença para um índice de estabilidade maior oumenor que 1.
(a) Prevalência da málaria em humanos emosquitos para ap
µ = 3.(b) Prevalência da málaria em humanos emosquitos para ap
µ = 0.3.
Observando a Figura 5.5, vemos que à esquerda para o índice de estabilidadeigual a 3 tanto a proporção de humanos infetados assim como a proporção de mosqui-tos infetados aumentam grandemente e tendem para (1, 0.75) quando R0 tende parainfinito. Na Figura 5.5 à direita, a proporção de mosquitos infetados é reduzida pelofacto do índice de estabilidade ser menor que 1.
Assim é possível controlar a transmissão da malária para níveis baixos de pre-valência, fazendo intervenções no índice de estabilidade da malária ou em outrosparâmetros, tais como diminuir a densidade (m) ou aumentar a taxa de recuperação(diminuir o período infecioso).
Os vários estudos que já foram feitos acerca do desenvolvimento da malária condu-ziram a faixas numéricas a que pode pertencer cada parâmetro presente na expressãode R0. A Tabela 5.2 apresenta esta informação.
46

Parâmetro Valor ou intervalo a que pertencea (taxa de picada em humanos por dia) [0.01-0.5]
p (Proporção de picadas em queum mosquito suscetível adquire infeção) [0.5]
q (Proporção de picadas queproduzem infeção em humanos) [0.2-0.5]
m (Razão entre mosquitos fêmeas e humanos) [0.5-40]α (taxa média de recuperação de humanos por dia) [0.005-0.05]
µ (Taxa per capita de mortalidade de mosquitos por dia) [0.05-0.5]τM (Período latente no mosquito em dias) [5-15]
τH (Período de latência no ser humano em dias) [10-100]
Tabela 5.2: Valores numéricos de alguns parâmetros do modelo de Ross-Macdonald([10], p.7).
5.2 Modelo de Ross-Macdonald com atrasos
Em ([16]) foi proposto um sistema de equações diferenciais com atraso para oestudo da malária. Este modelo inclui dois atrasos: um que leva em conta o períodode latência do parasita no mosquito e o outro relacionado com o período de latência doparasita no corpo humano. Para deduzir tais equações, consideremos que o períodode latência do agente patogénico no mosquito é τ1. Portanto, dos mosquitos infetadosτ1 unidades de tempo anteriormente, apenas uma proporção pa[1 − y(t − τ1)]x(t −τ1)e
−µτ1 são infeciosos no momento atual t. A expressão e−µτ1 é a probabilidade domosquito sobreviver ao período de latência do parasita. Da mesma forma, seja τ2 operíodo de latência do agente patogénico no ser humano. Assim, daqueles humanosinfetados τ2 unidades de tempo atrás, apenas uma proporção qam[1 − x(t − τ2)]y(t −τ2)e
−ατ2 são infeciosos no momento atual t. A expressão e−ατ2 é a probabilidade deum ser humano permanecer infetado durante todo o período de latência do parasita.O modelo com atraso para a dinâmica de mosquitos e humanos infetados é
x = qame−ατ2(1− x(t− τ2))y(t− τ2)− αx
y = pae−µτ1(1− y(t− τ1))x(t− τ1)− µy.(5.8)
onde a, p, q,m, µ, α são como definidos na Tabela 5.1 apresentada na secção ante-rior e x(t), y(t) são as proporções de humanos e mosquitos infetados no instante t
respetivamente.
47

5.2.1 Pontos de equilíbrio
Para encontrar os pontos de equilíbrio do sistema (5.8), partimos do facto de que(x∗, y∗) é um ponto de equilíbrio do sistema se satisfaz
qame−ατ2(1− x)y − αx = 0
pae−µτ1(1− y)x− µy = 0.(5.9)
Uma das soluções do sistema (5.9) é claramente o ponto (x, y) = (0, 0). Para encontraro outro ponto de equilíbrio (x∗, y∗), resolvemos o sistema de equações (5.9) para x 6= 0,y 6= 0 e encontramos o ponto:
(x∗, y∗) =
(a2pqme−µτ1e−ατ2 − µα
a2pqme−µτ1e−ατ2 + αape−µτ1,
a2pqme−µτ1e−ατ2 − µαa2pqme−ατ2e−µτ1 + µqame−ατ2
)(5.10)
O ponto (0,0) é chamado de ponto de equilíbrio livre de doença e o ponto (x∗, y∗)
é chamado de ponto de equilíbrio endémico, cuja existência fica garantida de modoanálogo ao modelo simples estudado na secção anterior para µα < a2pqme−µτ1e−ατ2.Esta desigualdade pode ser reescrita como:
R0 =a2pqme−µτ1e−ατ2
µα> 1
Assim no quadrado 0 ≤ x ≤ 1 e 0 ≤ y ≤ 1, se R0 > 1 o sistema (5.8) tem doispontos de equilíbrio: o ponto (0, 0) e o ponto (x∗, y∗) definido em (5.10). Se R0 ≤ 1, osistema tem um único ponto de equilíbrio (0, 0).
5.2.1.1 Estabilidade do ponto de equilíbrio livre de doença
Estudemos a estabilidade do ponto de equilíbrio livre de doença, isto é o ponto(0, 0).
Teorema 5.2. ([21], p. 1879)Se R0 < 1 o ponto de equilíbrio (0, 0) é globalmente assintoticamente estável no qua-drado Ω = 0 ≤ x ≤ 1 e 0 ≤ y ≤ 1.
Prova: Consideremos
R0 =a2pqme−µτ1e−ατ2
µα=aqme−ατ2
α· ape
−µτ1
µ< 1.
E consideremos os seguintes casos:
10 Casoaqme−ατ2
α< 1,
ape−µτ1
µ< 1
48

20 Casoaqme−ατ2
α> 1,
ape−µτ1
µ< 1
30 Casoaqme−ατ2
α< 1,
ape−µτ1
µ> 1
Em seguida provamos que o ponto de equilíbrio livre de doença é globalmenteassintoticamente estável nos três casos acima construindo uma função de Lyapunovapropriada.
Assim para o 10 caso,
V1(t, x, y) =1
αx+
∫ t
t−τ2
aqme−ατ2
αydt+
1
µy +
∫ t
t−τ1
ape−µτ1
µxdt,
que é uma função definida positiva no quadrado Ω. E calculando a derivada deV1(t, x, y) ao longo da solução de (5.8) temos:
V1(t, x, y) = −x+aqme−ατ2
αy(t−τ2)−
aqme−ατ2
αx(t−τ2)y(t−τ2)+
aqme−ατ2
αy−aqme
−ατ2
αy(t−τ2)−y
+ape−µτ1
µx(t− τ1)−
ape−µτ1
µx(t− τ1)y(t− τ1) +
ape−µτ1
µx− ape−µτ1
µx(t− τ1)
= −x− aqme−ατ2
αx(t−τ2)y(t−τ2)+
aqme−ατ2
αy−y− ape
−µτ1
µx(t−τ1)y(t−τ1)+
ape−µτ1
µx
=
[aqme−ατ2
α− 1
]y+
[ape−µτ1
µ− 1
]x−aqme
−ατ2
αx(t−τ2)y(t−τ2)−
ape−µτ1
µx(t−τ1)y(t−τ1)
<
[aqme−ατ2
α− 1
]y +
[ape−µτ1
µ− 1
]x < 0
Como V1(t, x, y) < 0, então o equilíbrio livre de doença é globalmente assintotica-mente estável.
Para o 20 caso, construimos a seguinte função de Lyapunov:
V2(t, x, y) =1
aqme−ατ2·ape
−µτ1
µx+
1
µ· α
aqme−ατ2y+
∫ t
t−τ2
α
aqme−ατ2ydt+
∫ t
t−τ1
ape−µτ1
µ· α
aqme−ατ2xdt
E calculando a derivada ao longo da solução de (5.8), temos:
V2(t, x, y) = − α
aqme−ατ2· ape
−µτ1
µx+
ape−µτ1
µy(t− τ2)−
ape−µτ1
µx(t− τ2)y(t− τ2)
− α
aqme−ατ2y +
α
aqme−ατ2· ape
−µτ1
µx(t− τ1)−
α
aqme−ατ2· ape
−µτ1
µx(t− τ1)y(t− τ1)
+α
aqme−ατ2y − α
aqme−ατ2y(t− τ2) +
ape−µτ1
µ· α
aqme−ατ2x− ape−µτ1
µ· α
aqme−ατ2x(t− τ1)
49

=
[ape−µτ1
µ− α
aqme−ατ2
]y(t−τ2)−
ape−µτ1
µx(t−τ2)y(t−τ2)−
α
aqme−ατ2·ape
−µτ1
µx(t−τ1)y(t−τ1)
Assim, V2(t, x, y) < 0, pois para R0 =aqme−ατ2
α· ape
−µτ1
µ< 1⇒ ape−µτ1
µ<
α
aqme−ατ2.
Portanto, mais uma vez o ponto de equilíbrio livre de doença é globalmente assin-toticamente estável.
Para o 30 caso, consideremos a seguinte função de Lyapunov,
V3(t, x, y) =1
α· µ
ape−µτ1x+
1
ape−µτ1·aqme
−ατ2
αy+
∫ t
t−τ2
aqme−ατ2
α· µ
ape−µτ1ydt+
∫ t
t−τ1
µ
ape−µτ1xdt
E calculamos V3(t, x, y) ao longo da solução do sistema (5.8):
V3(t, x, y) = − µ
ape−µτ1x+
aqme−ατ2
α· µ
ape−µτ1y(t−τ2)−
aqme−ατ2
α· µ
apeµτ1x(t−τ2)y(t−τ2)
− µ
ape−µτ1· aqme
−ατ2
αy +
aqme−ατ2
αx(t− τ1)−
aqme−ατ2
αx(t− τ1)y(t− τ1) +
µ
ape−µτ1x
− µ
ape−µτ1x(t− τ1) +
aqme−ατ2
α· µ
ape−µτ1y − aqme−ατ2
α· µ
ape−µτ1y(t− τ2)
= −aqme−ατ2
α· µ
ape−µτ1x(t−τ2)y(t−τ2)−
aqme−ατ2
αx(t−τ1)y(t−τ1)+
[aqme−ατ2
α− µ
ape−µτ1
]x(t−τ1)
Note que R0 =aqme−ατ2
α· ape
−µτ1
µ< 1, portanto
aqme−ατ2
α<
µ
ape−µτ1, assim
V3(t, x, y) < 0. Mais uma vez o ponto de equilíbrio livre de doença (0, 0) é globalmenteassintoticamentre estável, o que conclui a prova do Teorema.
5.2.1.2 Estabilidade do ponto de equilíbrio endémico
Considere-se agora R0 > 1. Usando o processo de linearização do sistema (5.8),a estabilidade dos pontos de equilíbrio livre de doença e endémico foi estudada em[16] e as conclusões foram as seguintes:
(1) O ponto de equilíbrio livre de doença (0, 0) é instável, possuindo uma variedadeestável e uma variedade instável.
(2) O ponto de equilíbrio endémico (x∗, y∗) é estável no caso de
a2pqm < aαp+ 2µα.
50
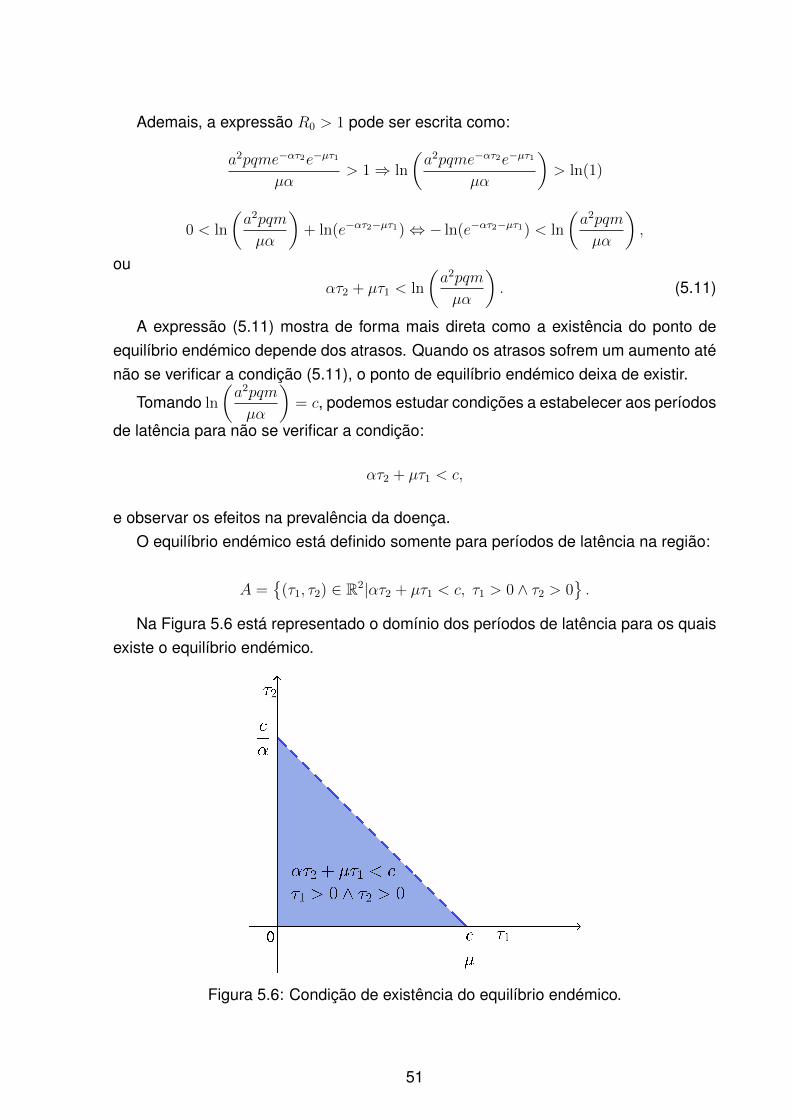
Ademais, a expressão R0 > 1 pode ser escrita como:
a2pqme−ατ2e−µτ1
µα> 1⇒ ln
(a2pqme−ατ2e−µτ1
µα
)> ln(1)
0 < ln
(a2pqm
µα
)+ ln(e−ατ2−µτ1)⇔ − ln(e−ατ2−µτ1) < ln
(a2pqm
µα
),
ouατ2 + µτ1 < ln
(a2pqm
µα
). (5.11)
A expressão (5.11) mostra de forma mais direta como a existência do ponto deequilíbrio endémico depende dos atrasos. Quando os atrasos sofrem um aumento aténão se verificar a condição (5.11), o ponto de equilíbrio endémico deixa de existir.
Tomando ln
(a2pqm
µα
)= c, podemos estudar condições a estabelecer aos períodos
de latência para não se verificar a condição:
ατ2 + µτ1 < c,
e observar os efeitos na prevalência da doença.O equilíbrio endémico está definido somente para períodos de latência na região:
A =
(τ1, τ2) ∈ R2|ατ2 + µτ1 < c, τ1 > 0 ∧ τ2 > 0.
Na Figura 5.6 está representado o domínio dos períodos de latência para os quaisexiste o equilíbrio endémico.
Figura 5.6: Condição de existência do equilíbrio endémico.
51

Podemos pois tirar a seguinte conclusão: para períodos de latência do parasitamaiores do que
c
µnos mosquitos ou
c
αnos humanos, o equilíbrio endémico deixa de
existir, e todas as soluções do sistema (5.8) convergem para o ponto de equilíbrio livrede doença. Neste caso, a doença por si só acabará por desaparecer.
O aumento dos períodos de latência do parasita em humanos ou mosquitos (viamedicamentos ou medidas de controlo) pode reduzir as proporções de humanos emosquitos infetados e, assim controlar a doença localmente ([16], p. 1111). No en-tanto, longos períodos de incubação podem ter um papel significativo na transmissãonão local da doença se levarmos em consideração a migração de humanos e mosqui-tos, já que humanos e mosquitos expostos podem certamente disseminar os parasitaspara outros locais.
52

Capítulo 6
Conclusão
Depois de percorrer alguma bibliografia relevante acerca dos modelos matemáti-cos epidemiológicos, eis algumas conclusões acerca dos modelos estudados nestetrabalho.
No modelo SIR simples se o tamanho da população suscetível for superior aolimiar de epidemia ou taxa de remoção relativa ρ = α
γ(sendo α a taxa de recuperação
da doença e γ a taxa de transmissão) ocorrerá epidemia, isto é, a introdução de umapequena quantidade de indivíduos infetados irá provocar o aumento da proporçãode infetados pela doença, portanto a prevalência da doença vai ser crescente até aquantidade de suscetíveis ser menor do que o limiar da epidemia. Daí o número decasos da doença irá decrescer tendendo para zero. No caso em que a população dossuscetíveis é menor do que ρ, então não ocorre epidemia.
De facto as epidemias tendem a se desenvolver mais rapidamente quando a densi-dade de suscetíveis é alta e a taxa de remoção é baixa por causa do desconhecimentoda doença e de cuidados médicos inadequados. Por outro lado, surtos tendem a serapenas de forma limitada quando as boas condições sociais acarretam menores den-sidades de suscetíveis, e quando as taxas de remoção são altas por causa de boavigilância e controlo por parte dos serviços de saúde pública ([2], p. 458).
Em 1912 Mckendrick escreveu acerca da influência do tamanho da população desuscetíveis na dinâmica de doenças infeciosas nos seguintes termos: se uma deter-minada comunidade que está a enfrentar uma doença infeciosa receber uma certaquantidade de pessoas totalmente suscetíveis em sua população, então a chegadados visitantes cria uma perturbação. Neste caso, a quantidade de suscetíveis vai su-perar por um valor ainda maior o limiar de epidemia, e sem controlo adequado, muitagente será afetada e demorará mais tempo até que a doença fique extinta. Quantomais tempo a epidemia se mantiver, maior será a catástrofe, se a densidade da popu-lação continuar a aumentar e o limiar de epidemia permanecer constante, esse atrasoprolongado pode levar à quase completa extinção da população ([7], p. 720).
Entretanto perante uma incerteza acerca da dimensão de uma doença infeciosa,
53

o Homem tem procurado controlá-la ou até irradicá-la através de campanhas de va-cinação. Devido à escassez de recursos (humanos e materiais) e com intenção deminimizar os custos, no modelo SIR com vacinação à nascença, viu-se que é possí-vel garantir uma imunidade vacinando apenas uma certa proporção da população. Eneste caso a percentagem limiar a vacinar é dada por ρ = 1− 1
R0, onde R0 é o número
básico de reprodução da doença, que é o número médio de casos secundários que umcaso primário de doença produz numa população totalmente suscetível. Assim parauma percentagem de vacinação acima de ρ, a população fica protegida à invasão dadoença.
Em relação ao modelo SIS simples foi possível observar que uma doença só po-derá invadir com sucesso uma população se a taxa de remoção/recuperação (α) formenor do que a taxa de transmissão (γ), caso contrário acabará por desaparecer.No caso da taxa de recuperação ser menor do que a taxa de transmissão, pode sefazer uma intervenção no combate à doença. As intervenções passam necessaria-mente por aumentar a taxa de recuperação (garantindo tratamentos médicos eficazesaos doentes, diminuindo assim o período infecioso) ou diminuir a taxa de transmissão(colocando os doentes em quarentena até ao final da cura ).
Falando acerca da malária, uma das doenças muito antigas que tem sido umgrande flagelo na população humana, a sua existência e persistência numa regiãoé explicada matematicamente pelo número básico de reprodução da doença, cuja in-terpretação está ligada a complexas interações entre a fêmea do mosquito do géneroAnopheles e o ser humano no processo de transmissão do parasita. Se o númerobásico de reprodução é maior do que um (R0 > 1), então a doença persistirá napopulação. Caso contrário (R0 < 1) não persistirá. Houve várias propostas para aerradicação da malária, a primeira delas foi proposta por Ronald Ross, e está ligadaà diminuição da densidade de mosquitos, eliminando mosquitos jovens através de lar-vicidas. Mas tarde, Macdonald argumentou que para a erradicação da malária, eranecessário matar os mosquitos adultos, cuja justificação está ligada à existência determos exponenciais na expressão de R0 (levando em conta o período de latênciado parasita no mosquito). A proposta de Macdonald ajudou a irradicar a malária emmuitos países, mas em 2008 ela foi considerada ainda endémica em 109 países. Atu-almente há várias medidas que são usadas para o controlo ou irradicação da maláriaentre elas a pulverização intra-domiciliária, o uso de redes mosquiteiras impregnadascom inseticidas e campanhas de sensibilização para as comunidades destruirem oscriadouros naturais de mosquitos. Entretanto a doença ainda persiste devido à com-plexidade de fatores ligados ao ciclo de vida do parasita e dos vetores e às migraçõesconstantes de indivíduos entre zonas endémicas e não endémicas da malária.
54

Bibliografia
[1] Anderson, R. M.(1982). Population Dynamics of Infectious Diseases: Theory andApplications[PDF]. DOI: 10.1007/978-1-4899-2901-3.
[2] Braun, M.(1983). Diferencial Equations and their Aplications, An Introduction toApplied Mathematics, United States of America: University of New York.
[3] Capanna, E.(2015), Battista Grassi: a zoologist for malaria. CONTRIBUTIONS toSCIENCE, 3 (2): 187–195. DOI: 10.2436/20.7010.01.5
[4] Chauhan, S., Misra, O., Dhar, J.(2014). Stability Analysis of SIR Model with Vacci-nation. American Journal of Computational and Applied Mathematics, 4(1): 17-23.doi: 10.5923/j.ajcam.20140401.03.
[5] Chasnov, J.(2009). Mathematical Biology: Lecture notes for MATH 4333 (S/N).Disponível em: https://www.math.ust.hk/~machas/mathematical-biology.
pdf.
[6] Chitnis, N.(2011). Introduction to mathematical epidemiology-further exem-ples (S/N). Disponível em: http://www.luchsinger-mathematics.ch/
ME-AdvancedModels.pdf. Acesso: 20.01.2019.
[7] Kermack, W. 0., McKendrick, A. G. (1927). A Contribution to the MathematicalTheory of Epidemics. Royal Society, 115(772), 700-721.
[8] Koella, J. C.(1990). On the use of mathematical models of malaria transmission.Acta Tropica, 49(1991)1-25. Doi: ACTROP 00141.
[9] Larson, R., Farber, B.(2010). Estatística Aplicada. São Paulo: Pearson PrinticeHall.
[10] Mandal, S., Sarkar, R. R., Sinha, S.(2011). Mathematical models of malaria.Malaria Journal2011,10:202. Disponível em: http://www.malariajournal.com/
content/10/1/202. Acesso: 22.06.2018.
[11] Martcheva, M.(2015). An Introduction to Mathematical Epidemiology [PDF]. doi:10.1007/978-1-4899-7612-3.
55

[12] MASSAD, E.(2008). The elimination of Chagas’ disease from Brazil. Epidemiol.Infect. (2008), 136, 1153–1164. doi: 10.1017/S0950268807009879.
[13] Plemmons, W. R.(2006). A Mathematical Study of Malaria Models of Ross andNgwa.(Dissertação de mestrado, University of Central Florida). Disponível em:http://etd.fcla.edu/CF/CFE0001406/Plemmons_William_R_200612_MS.pdf.
[14] Polya, G.(1977). Mathematicaly Methods in Science. United States of America:The Mathematicaly Association of America.
[15] Ross, R. (1911). The prevention of Malaria. 2nd edition. Disponívelem: https://archive.org/details/pr00eventionofmalarossrich/page/n23.Acesso: 22.04.2019.
[16] Ruan, S., Xiao, D., Beier, J.(2008). On the Delayed Ross–Macdonald Model forMalaria Transmission. Bulletin of Mathematical Biology (2008) 70: 1098–1114.DOI: 10.1007/s11538-007-9292-z.
[17] Smith D. L., Battle, K. E., Hay, S. I., Barker, C. M., Scott, T. W., Mc-Kenzie, F. E.(2012). Ross, Macdonald, and a Theory for the Dynamics andControl of Mosquito-Transmitted Pathogens.PLoS Pathog, 8(4), e1002588.doi:10.1371/journal.ppat.1002588.
[18] Sulsky, D.(2012). Using Real Data in an SIR Model (S/N). Disponível em: http://www.math.unm.edu/~sulsky/mathcamp/ApplyData.pdf. Acesso: 07.11.2018.
[19] Sulsky, D.(2012). SIR Model (S/N). Disponível em: https://www.math.unm.edu/~sulsky/mathcamp/ApplyData.pdf. Acesso: 07.01.2019.
[20] Tavares, J.(2017). Modelo SIR em epidemiologia, Rev. Ciência Elem., 5(02),1-12.doi: 10.24927/rce2017.020.
[21] Wu, C., Jiang, Z.(2012). Global Stability for the Disease Free Equilibriumof a Delayed Model for Malaria Transmission. Int. Journal of Math. Analysis,6(38), 1877 - 1881. Disponível em: http://www.m-hikari.com/ijma/ijma-2012/ijma-37-40-2012/wuchunqingIJMA37-40-2012.pdf. Acesso: 31.03.2019.
56


















