
NOVOS COMPONENTES PARA O MANEJO DA VASSOURA DE …
143
UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO PRÓ-REITORIA DE PESQUISA E PÓS-GRADUAÇÃO PROGRAMA DE PÓS-GRADUAÇÃO EM FITOPATOLOGIA Tese de Doutorado NOVOS COMPONENTES PARA O MANEJO DA VASSOURA DE BRUXA DO CACAUEIRO Antônio Alves Pimenta Neto Recife – PE 2016
Transcript of NOVOS COMPONENTES PARA O MANEJO DA VASSOURA DE …

UNIVERSIDADE FEDERAL RURAL
DE PERNAMBUCO
PRÓ-REITORIA DE PESQUISA E PÓS-GRADUAÇÃO
PROGRAMA DE PÓS-GRADUAÇÃO
EM FITOPATOLOGIA
Tese de Doutorado
NOVOS COMPONENTES PARA O MANEJO
DA VASSOURA DE BRUXA DO CACAUEIRO
Antônio Alves Pimenta Neto
Recife – PE
2016

ii
ANTÔNIO ALVES PIMENTA NETO
NOVOS COMPONENTES PARA O MANEJO DA VASSOURA DE BRUXA DO
CACAUEIRO
Tese apresentada ao Programa de Pós-Graduação em
Fitopatologia da Universidade Federal Rural de
Pernambuco, como parte dos requisitos para obtenção do
título de Doutor em Fitopatologia.
COMITÊ DE ORIENTAÇÃO:
Orientador(a): DELSON LARANJEIRA
Co-Orientador(a): EDNA DORA MARTINS NEWMAN LUZ
RECIFE-PE
FEVEREIRO – 2016

iii
Ficha Catalográfica
P644n Pimenta Neto, Antônio Alves.
Novos componentes para o manejo da vassoura de bruxa do
cacaueiro / Antônio Alves Pimenta Neto. – Recife, 2016.
142 f. : il.
Orientador: Delson Laranjeira.
Tese (Doutorado em Fitopatologia) – Universidade Federal
Rural de Pernambuco, Departamento de Agronomia, Recife, 2016.
Inclui referências.
1. Theobroma cação. 2. Manejo integrado. 3. Leveduras nativas.
4. Melhoramento genético. 5. Biocontrole. I. Laranjeira, Delson,
Orientador. II. Título
CDD 632

iv
NOVOS OMPONENTES PARA O MANEJO DA VASSOURA DE BRUXA DO
CACAUEIRO
ANTÔNIO ALVES PIMENTA NETO
Tese defendida e aprovada pela Banca Examinadora em: 26/02/2016
ORIENTADOR(A):
____________________________________________________
DELSON LARANJEIRA
EXAMINADORES:
_____________________________________________________
EDNA DORA MARTINS NEWMAN LUZ (CEPLAC)
_____________________________________________________
JOSÉ LUIZ BEZERRA (UFRB)
_____________________________________________________
ELINEIDE BARBOSA DE SOUZA (UFRPE)
_____________________________________________________
MARCO AURÉLIO SIQUEIRA DA GAMA (UFRPE)
RECFE-PE
FEVEREIRO – 2106

v
Ao Grande Arquiteto do Universo, por
me conceder a vida, e oportunidade diária de
crescimento e evolução. Aos meus amados pais
Joel e Islene, meus irmãos Felipe e Renata, e a
Thaís pelo companheirismo, apoio e amor
incondicional.
DEDICO

vi
A existência na Terra é um livro que estás escrevendo...
Cada dia é uma página...
Cada hora é uma afirmação de tua personalidade, através das pessoas e das situações que te
buscam.
Não menosprezes o ensejo de criar uma epopéia de amor em torno de teu nome.
As boas obras são frases de luz que endereças à Humanidade inteira.
Em cada resposta aos outros, em cada gesto para com os semelhantes, em cada manifestação
dos teus pontos de vista e em cada demonstração de tua alma, grafas com tinta perene, a
história de tua passagem...
CHICO XAVIER
A sabedoria não é um produto da escolaridade, mas
da tentativa, ao longo de uma vida, para obtê-la
ALBERT EINSTEIN
O rio atinge seus objetivos, porque
aprendeu a contornar obstáculos.
LAO-TSÉ

AGRADECIMENTOS
Ao Grande Arquiteto do Universo, por me conceder saúde, sabedoria e segurança,
para manter sempre de pé e a ordem diante das provas e desafios;
Aos meus pais por sempre me incentivarem e abrirem mão de seus sonhos para a
realização dos meus;
Aos meus irmãos, Renata e Felipe, pelo incentivo e companheirismo;
À Thaís, pela tolerância, cumplicidade, companheirismo, amor, paciência e incentivo;
À professora Dra. Edna Dora Martins Newman Luz, pela orientação durante todos
estes anos trabalhos, pela amizade e carisma;
Ao Professor Delson, pelo apoio e ensinamentos;
Aos professores Dra Ana Paula T. Uetanabaro, Dr Eduardo Gross, Dr Carlos Priminho
e Dra Andréa Costa, pela orientação e disponibilização dos Laboratórios de Microbiologia,
Microscopia Eletrônica e Proteômica da UESC;
Aos pesquisadores MSc Dilze A. Magalhães, Dr Givaldo Niella e Dr João de Cássia,
Dra Karina Gramacho, Dra Elizabeth Duarte, Dr Thiago Oliveira, pelo incentivo e instrução
nas tomadas de decisão;
Ao Dr José Luiz Pires pela colaboração científica desde o planejamento à execução e
discussão, além disponibilização de recursos para a realização dos experimentos;
Aos técnicos, operários de campo, bolsistas e a todos os funcionários da CEPLAC
(Cenilda, Joel, Maguinaldo, Orlando, Ananias, Elenízio, Orlando, Belmiro, Virgínia, Ana
Rosa, Catarino, Edarcy (Tita), Lourdes (Lurdinha), Denise, Laiane, João Marcos, William,
Francis, Tacila, Carolina, Elizângela, Gisele, Raquel, Carlisly e Vanessa), pela colaboração na
execução das atividades e manutenção de mudas em casa de vegetação;
À Universidade Federal Rural de Pernambuco pelo apoio institucional e a
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pela concessão de
bolsa de estudo;
À Comissão Executiva do Plano da Lavoura Cacaueira, pela estrutura e recurso
concedido;
Aos amigos e irmãos pelo incentivo e por suportarem as ausências e afastamento
durante esta fase tão importante na minha vida. Enfim, a todos aqueles que de forma direta ou
indireta participaram da minha trajetória me incentivando a seguir em frente e nunca desistir
principalmente nos momentos mais difíceis.

8
SUMÁRIO
RESUMO GERAL ................................................................................................................... 10
GENERAL ABSTRACT .......................................................................................................... 11
CAPÍTULO 1 - 1 INTRODUÇÃO GERAL ........................................................................... 13
1.1 O CACAUEIRO (Theobroma cacao L.) E A CULTURA DO CACAU.................... 13
1.2 A VASSOURA-DE-BRUXA DO CACAUEIRO ...................................................... 16
1.4 MANEJO INTEGRADO DA VASSOURA-DE-BRUXA DO CACAUEIRO .......... 20
2 REFERÊNCIAS BIBLIOGRÁFICAS .......................................................................... 23
CAPÍTULO 2 - SELEÇÃO DE PROGÊNIES DE CACAUEIRO QUANTO A
RESISTÊNCIA À VASSOURA DE BRUXA E MONILÍASE ............................................. 50
Resumo - ........................................................................................................................... 51
Introdução .......................................................................................................................... 52
Material e métodos ............................................................................................................ 53
Resultados e Discussão ..................................................................................................... 56
Referências ........................................................................................................................ 62
CAPÍTULO 3 - DIVERSIDADE E ANTAGONISMO DE LEVEDURAS NATIVAS
CONTRA O AGENTE DA VASSOURA DE BRUXA DO CACAUEIRO ........................... 71
Resumo .............................................................................................................................. 72
Abstract ............................................................................................................................. 73
Introdução .......................................................................................................................... 74
Materiais e Métodos .......................................................................................................... 75
Resultados ......................................................................................................................... 81
Discussão ........................................................................................................................... 87
Conclusões ........................................................................................................................ 91
Agradecimentos ................................................................................................................. 91
Referências ..................................................................... Erro! Indicador não definido.91
CAPÍTULO 4 - LEVEDURAS NATIVAS COMO POTENCIAIS AGENTES DE
BIOCONTROLE À VASSOURA DE BRUXA E PROMOTORES DE CRESCIMENTO EM
PLÂNTULAS DE CACAUEIRO .......................................................................................... 109
Resumo ............................................................................................................................ 110
Abstract ........................................................................................................................... 111
Introdução ........................................................................................................................ 111
Material e Métodos .......................................................................................................... 113

9
Resultados ....................................................................................................................... 119
Discussão ......................................................................................................................... 123
Conclusões ...................................................................................................................... 127
Agradecimentos ............................................................................................................... 128
Referências ...................................................................................................................... 128
CONCLUSÕES GERAIS....................................................................................................... 143

10
RESUMO GERAL
A vassoura de bruxa (VB) é considerada a principal doença do cacaueiro no Brasil. Os
objetivos deste trabalho de tese foram: 1. Gerar uma população de cacaueiros resistentes à VB
e que, preventivamente, também carreguem em sua estrutura genética, genes de clones que
tem indicativo, através de testes realizados em outros países, resistência a monilíase (MN)
(praga quarentenária A1); 2. Avaliar a diversidade de fungos leveduriformes presentes na
superfície de órgão vegetais de genótipos de cacaueiro, selecionados para a resistência à VB e
MN; 3. Estudar os mecanismos que permitem a colonização e o estabelecimento das
leveduras nos substratos vegetais em condições de cultivo, e seu efeito no crescimento e
esporulação de Moniliophthora perniciosa (Mp); 4. Avaliar a capacidade de leveduras nativas
no controle da VB e, na promoção de crescimento em plântulas seminais. Dos 59
cruzamentos avaliados, 58 geraram grupos de progênies que diferiram significativamente
(p<0,05) de catongo e 52 de SIC 23, padrões de suscetibilidade. Progênies de nove
cruzamentos, diferiram significativamente (p<0,05) do padrão de resistência SCA 6,
demonstrando alto potencial para seleção quanto a resistência à VB. Houve destaque para os
descendentes diretos de materiais oriundos da Amazônia Brasileira (CAB) e clones que foram
adicionados à população como portadores de resistência a monilíase, a exceção do clono ICS
95 que gerou grupos de indivíduos com índice de doença médio similar à testemunha
suscetível. Todas as leveduras caracterizadas molecularmente, foram pela primeira vez
identificadas na microbiota epifítica de órgãos vegetais do cacaueiro e, mesmo apresentando
comportamentos distintos para as variáveis analisadas, são potenciais agentes de biocontrole
da vassoura de bruxa do cacaueiro. As leveduras LEV 169 (Rhodosporidium sp.) e LEV 207
(Hannaella sp.) se destacaram nos testes in vitro por inibirem 87,5% dos basidióporos e 100%
o crescimento micelial de Mp em meio líquido, respectivamente. Os isolados LEV 207 e LEV
179 (Rhodosporidium sp.) foram responsáveis pelos maiores níveis de controle, tanto em
plântulas quanto na inibição da produção de basidiomas em vassouras secas. O trabalho de
tese apresenta dois novos grupos de integrantes para o manejo da vassoura de bruxa do
cacaueiro: 1. Progênies de cacaueiro portadoras de genes de resistência à VB obtidos de
diferentes origens; 2. Leveduras nativas com ação inibitória contra Mp, efetivas no controle
da VB e possibilidade de associação com utilização de fungicidas cúpricos.
Palavras-chaves: Theobroma cacao, manejo integrado, leveduras nativas, melhoramento de
plantas.

11
GENERAL ABSTRACT
The witches' broom (WB) is considered the main disease of cacao in Brazil. The objectives of
this thesis were: 1. Generate a population of cocoa trees resistant to WB and preventively also
carry in their genetic structure, clone genes has indicated, through tests conducted in other
countries, moniliasis resistance (MN ) (A1 quarantine pest); 2. Evaluate the diversity of yeast
fungi present in the plant body surface cacao genotypes, selected for resistance to WB and
MN; 3. To study the mechanisms that allow the colonization and the establishment of yeast in
plant substrates in cultivation conditions and their effect on growth and sporulation of
Moniliophthora perniciosa (Mp); 4. To assess the ability of native yeasts in controlling the
WB and the growth promotion in seminal seedlings. Of 59 crosses this, 58 groups generated
progeny that differed significantly (p <0.05) of catongo and 52 of SIC 23, susceptibility
patterns. Progeny nine intersections, differed significantly (p <0.05) resistance pattern SCA 6,
showing high potential for selection for resistance to WB. There was emphasis on the direct
descendants of materials from the Brazilian Amazon (CAB) and clones that were added to the
population as moniliasis resistance carriers, except for the clonus ICS 95 generating groups of
individuals with similar medium disease index to the susceptible control. All yeasts
molecularly characterized, were first identified in the epiphytic microflora of plant organs of
the cacao tree, and even with different behaviors for the variables are potential biocontrol
agents witch's broom cacao. The LEV 169 yeast (Rhodosporidium sp.) And LEV 207
(Hannaella sp.) Stood out in vitro in tests to inhibit 87.5% of basidióporos and 100% mycelial
growth in liquid medium Mp, respectively. Isolated LEV LEV 207 and 179 (Rhodosporidium
sp.) Were responsible for the highest levels of control, both in seedlings and in the inhibition
of the mushroom production in dry brooms. The thesis presents two new members of groups
for the management of witches' broom disease of cocoa: 1. Progeny of cocoa with WB
resistance genes obtained from different sources; 2. native yeasts with inhibitory activity
against Mp, effective in controlling the WB and possible association with the use of copper
fungicides.
Keywords: Theobroma cacao, integrated management, native yeasts, plant breeding.

12
CAPÍTULO 1
INTRODUÇÃO GERAL

13
NOVOS COMPONENTES PARA O MANEJO DA VASOURA DE BRUXA DO
CACAUEIRO
1 INTRODUÇÃO GERAL
1.1 O CACAUEIRO (Theobroma cacao L.) E A CULTURA DO CACAU
1.1.1 CLASSIFICAÇÃO E DESCRIÇÃO BOTÂNICA
A espécie Theobroma cacao L. foi descrita inicialmente por Charles de L'Êcluse e
citada pela primeira vez na literatura botânica como Cacao fructus (L’ÊCLUSE, 1601). Em
1737, a espécie foi classificada pelo botânico sueco Carolus Linneu (Linnaeus) como
Theobroma fructus, que é traduzido como “alimento dos deuses”, em alusão à crença indígena
na origem divina do cacaueiro. Mais tarde (1753), foi modificado pelo próprio Linnaeus, para
o binômio T. cacao, designação que permanece até hoje. A palavra cacau deriva de
cacauhuatl, diretamente do Nahuatl, a língua maia (DIAS, 2001a; CEPLAC, 2013).
O cacaueiro é uma planta umbrófila, arbórea, perene, que pertencia anteriormente a
família Sterculiaceae e atualmente integra a família Malvaceae, ordem Malvales, e gênero
Theobroma (ALVERSON et al., 1999; SOUNIGO et al., 2003). É considerada uma espécie
cauliflora, devido a formação das inflorescências ao longo do tronco, em estruturas
denominadas de almofadas florais, compostas por flores hermafroditas e pentâmeras,
apresentando pétalas, sépalas, estames e estaminoides (TOXOPEUS, 1972).
A espécie T. cacau pode atingir 5 a 8 m de altura e 4 a 6 m de diâmetro de copa,
quando oriunda de semente. Entretanto, pode ocorrer variações devido às condições de
cultivo, sendo também encontradas em habitat natural com porte de até 20 m. As folhas são
alternas simples, oblongas, acuminadas e glabras com nervura central proeminente e
secundárias filiformes, pecíolo cerca de um a três cm de comprimento, espessados nas duas
extremidades (SILVA NETO et al., 2001); as folhas apresentam lâmina hipoestomática, com
estômatos do tipo anomocítico, quatro tipos de tricomas em ambas as faces do limbo, sendo
dois tectores e dois glandulares, e presença de glândulas secretoras de mucilagem na epiderme
adaxial (NAKAYAMA et al., 1996); flores hermafroditas, completas, pentâmeras, cálice
formado de cinco pétalas, avermelhada e limbo branco-amarelado, inflorescência nos ramos
ou no tronco, sobre pequenas dilatações nodulares; fruto drupáceo, pericarpo carnoso-
consistente e amarelo quando maduro variando na forma e no tamanho; a semente é de cor
roxa e em número de 20 a 40 amêndoas, tem forma quase arredondada e dispõe-se em cinco
fileiras, achatado pela compressão mútua vertical; a testa é recoberta por uma polpa escassa,

14
branca e adocicada e encerra o embrião, composto de dois cotilédones plicado-enrugado
(CUATRECASAS, 1964; SILVA NETO et al., 2001).
Apresenta sistema reprodutivo misto que se caracteriza pela predominância de
fecundação por cruzamentos, ocorrendo autofecundações inferiores a 50% e superiores a 5%,
podendo a taxa de cruzamento alcançar até 100% (VELLO; NASCIMENTO, 1971).
A polinização do cacaueiro é essencialmente entomófila por possuir características
morfológicas das estruturas florais que impedem a autofecundação natural (ASCENSO,
1962), além da disposição em massa dos grãos de pólen devido a sua viscosidade, o que
dificulta o transporte pelo vento (VELLO; NASCIMENTO, 1971). Esta dificuldade de
transporte promove a deposição de grãos de pólen de flores da própria planta e de plantas
vizinhas sobre o estigma das flores, resultando assim em heterogeneidade na obtenção das
sementes (TOXOPEUS, 1972).
A polinização natural no cacaueiro é realizada principalmente por um pequeno grupo
de dípteros da família Ceratopogonidae e do gênero Forcipomyia, podendo também ter a
participação acidental de outros insetos como trips, formigas e afídeos. Tais insetos pelo
hábito de locomoção garantem, na maioria das vezes, a ocorrência da polinização cruzada
(SORIA et al., 1975; CHAPMAN; SORIA, 1983).
1.1.1 CENTRO DE ORIGEM E DISPERSÃO
O cacaueiro cresce de forma espontânea desde o sul do México até a Bolívia, e nas
margens dos rios da Floresta Amazônica, com provável centro de origem no Alto
Amazonas, na confluência dos rios Solimões, Putamaio e Caquetá (LACHENAUD; ZHANG,
2008). O cacaueiro encontrado sob condições silvestres, se desenvolve em bosques e
florestas úmidas, sombreado com árvores de grande porte e margeando os grandes rios,
onde predominam condições de temperatura e umidade elevadas, típicas das regiões tropicais
(CHEESMAN, 1944; CUATRECASAS, 1964; BARTLEY, 2005). Entretanto, em condições
de cultivo apresenta porte que varia de 3 a 5 metros.
Trabalhos com marcadores moleculares, isoenzimáticos, agronômicos e morfológicos,
indicam que a partir do centro de origem, provavelmente o cacaueiro tenha sido dispersado
em duas direções, sendo uma para o leste, sentido do curso do rio Amazonas e a outra para o
norte, rumo a Mesoamérica, dando origem aos dois grupos raciais, Crioulo e Forasteiro
Amazônico, respectivamente. A partir de cruzamentos naturais entre os referidos grupos
genéticos ocorridos em Trinidad, originou-se um terceiro tipo denominado Trinitário (DIAS,

15
2001b). Além disso, a população do grupo Forasteiro pode ser subdivida em Forasteiro do
Alto Amazonas e Forasteiro do baixo Amazonas, conforme a localização de sua ocorrência
(TOXOPEUS, 1969).
Genótipos do grupo Crioulo caracterizam por sementes redondas ou róseas, de sabor
adocicado e fermentação rápida, apresentam padrão de suscetibilidade à maioria das doenças
que acometem a cultura; os frutos são grandes, geralmente apresentam casca fina e rugosa,
coloração verde-escura quando imaturos, passando para amarelo ou alaranjado quando
amadurecem (HANCOCK, 1994; BART-PLANGE; BARYEH, 2003). As variedades
Forasteiro, cultivadas no Baixo Amazonas, principalmente no Brasil e na Venezuela
(SOUNIGO et al., 2003), são amplamente cultivados devido à alta produtividade e resistência
às doenças e a variabilidade genética (MARITA et al., 2001). O híbrido Trinitário produz
sementes de coloração que varia de amarelo pálido a roxo escuro, e no geral produzem cacau
de qualidade intermediária (HANCOCK, 1994; BART-PLANGE; BARYEH, 2003).
O crescente interesse e incentivo ao cultivo do cacaueiro proporcionou sua dispersão
para regiões de semelhantes aspectos edafoclimáticos, e suas sementes foram se
disseminando gradualmente pelo mundo, atravessando o oceano e chegando ao continente
africano na segunda metade do século XIX. As primeiras plantações africanas foram feitas
por volta de 1855, nas ilhas de São Tomé e Príncipe, colônias portuguesas ao largo da costa
ocidental africana (CEPLAC, 2013).
O cacaueiro foi introduzido na Bahia em meados de 1746, quando o colono francês
Luís Frederico Warneaux, plantou sementes de cacaueiro oriundas da população de
forasteiros baixo amazônicos, trazidas do Pará, no atual município de Canavieiras (VELLO;
GARCIA, 1971). Devido ao ambiente favorável ao cultivo, a região sul da Bahia veio a se
tornar a maior região produtora de amêndoas no país, e até a penúltima década do século
passado o cacau era o principal produto de exportação na Bahia (ALMEIDA, 2009). Apesar
do estado da Bahia ainda ser o maior produtor nacional, a cacauicultura brasileira está
distribuída nas regiões Nordeste (Bahia), Sudeste (Espírito Santo), Centro-Oeste (Mato
Grosso) e Norte (Pará, Rondônia e Amazonas) (IBGE, 2014).
1.1.2 IMPORTÂNCIA ECONÔMICA
O cacaueiro é um dos cultivos perenes mais importantes do mundo, com uma
produção mundial estimada em 4,2 milhões de toneladas de sementes secas (ICCO, 2015). A
exploração comercial desta cultura não se restringe apenas à produção de chocolate, mas para

16
outros ramos da indústria, como de cosméticos e farmacêuticos, geleias, sucos e licores
obtidos através do processamento da polpa do fruto de cacaueiro. A Costa do Marfim é
líder na produção mundial de cacau, com 42%, seguida de Gana, com 17,9%, e Indonésia
com 7,16%. Na sequência vem, Equador, República dos Camarões e Brasil. Os principais
países consumidores de cacau são os Estados Unidos, com 689 mil t, seguidos pela
Alemanha, França e Reino Unido (ICCO, 2015). O Brasil, que há três décadas se manteve como
um dos maiores produtores de cacau do planeta, produziu no ano de 2014, 273.793 t de
amêndoas secas, equivalente à R$ 1.589.535 reais (IBGE, 2014), O sul da Bahia é a principal
região produtora de cacau do Brasil, retomando o crescimento no ano de 2015 com a
participação no mercado internacional através da exportação de 6,6 mil toneladas de
amêndoas de cacau – avaliadas em R$ 15 milhões (BRASIL, 2015).
Durante muitos anos o cacau da Bahia contribuiu para que o Brasil fosse um dos
maiores produtores mundiais, entretanto, a competição nos países concorrentes, o aumento
das áreas de produção, atraídos pelos elevados preços no mercado internacional, a
descapitalização dos produtores brasileiros, problemas climáticos, com a ocorrência de seis
anos consecutivos de estresse hídrico na região e principalmente o surgimento da vassoura-
de-bruxa (VB), causada pelo fungo Moniliophthora perniciosa (Stahel) Aime & Phillips-
Mora, promoveram uma enorme crise socioeconômica, agronômica e ecológica na região,
levando o Brasil a alterar sua situação de exportador para importador de cacau (MIDLEJ;
SANTOS, 2012).
Na safra de 1984/1985 o Brasil foi o segundo maior produtor mundial de amêndoas,
produzindo aproximadamente 400.000 t, e no ano de 2000, a produção caiu para 210.000 t.,
passando de exportador para importador a partir de 1997 (FAO, 2002). O principal fator
responsável pela queda nas produções foi o surgimento da VB, a mais prejudicial enfermidade
do cacaueiro, constatada em 1989 na região Sul da Bahia (PEREIRA et al., 1989), que ao
encontrar condições favoráveis na região, disseminou-se rapidamente, atingindo níveis
epidêmicos e acarretando redução drástica na produção.
1.2 A VASSOURA-DE-BRUXA DO CACAUEIRO
O cacaueiro é ameaçado por diversas enfermidades fúngicas, sendo a VB, causada
pelo basidiomiceto M. perniciosa (=Crinipellis perniciosa) (AIME; PHILLIPS-MORA,
2005), a doença de maior impacto atualmente e, portanto, a mais preocupante no Brasil (LUZ
et al., 2006).

17
A doença foi mencionada pela primeira vez por Went em 1904, causando sérios
prejuízos no município de Saramacca, no Suriname (HOLLIDAY, 1952). No entanto o seu
agente etiológico só foi descrito por Stahel em 1915. Mesmo com esforços para impedir a
introdução do patógeno no estado da Bahia, a doença foi constatada em 1989, e apesar da
erradicação de 12 ha do cultivo na cidade de Uruçuca, novos focos foram identificados no
mesmo ano na cidade de Camacã, disseminando-se assim para todas as regiões produtoras do
estado (PEREIRA et al., 1989; ROCHA et al., 1993; ANDEBRHAN et al., 1999), sendo
atribuída a ela a devastação da lavoura cacaueira no Sul da Bahia no final do século passado.
A VB que já era um dos maiores entraves ao desenvolvimento do cultivo do cacaueiro na
América do Sul, incluindo a região amazônica brasileira, ao ser introduzida na Bahia, causou
sérios prejuízos à cacauicultura baiana, que utilizava principalmente variedades altamente
suscetíveis (PEREIRA et al., 1989).
A vassoura de bruxa distribui-se geograficamente por todos os países produtores da
América do Sul e Central como o Brasil, Bolívia, Colômbia, Equador, Guiana, Peru,
Venezúela, Panamá e nas ilhas do Caribe, Trinidad, Tobago, Granada, Santa Lucia e São
Vicente. Baker e Holliday (1957) afirmaram que desde 1920, a doença já estaria disseminada
por todo o vale do Amazonas e seus inúmeros afluentes, indo desde o leste no estado do Pará
até os planaltos aos pés da cordilheira dos Andes.
Atualmente essa doença ainda é um entrave para implantação de novas áreas de
cultivo e para a competitividade da cacauicultura nacional no mercado internacional
(BRASIL, 2015). Uma das consequências da doença para a sociedade regional foi o elevado
índice de desemprego e êxodo rural, que afetou todos os segmentos da cadeia produtiva do
cacau, particularmente os trabalhadores rurais e os pequenos produtores (TREVIZAN;
MARQUES, 2004). O fitopatógeno contribuiu também para grande índice de desmatamento
na região com a eliminação em várias propriedades dos remanescentes da Mata Atlântica,
sendo o cultivo de cacaueiro substituído pela atividade madeireira que foi estimulada como
alternativa para a então desempregada população de trabalhadores rurais (LUZ et al., 2006).
O fungo afeta principalmente os tecidos meristemáticos da planta (caules, almofadas
florais, flores e frutos), causando sintomas que variam de acordo com o órgão infectado e o
seu estádio de desenvolvimento (SILVA et al., 2003), levando à morte da planta, quando
afetada por sucessivos surtos de infecção e fatores abióticos em concomitância
(ANDEBRHAN, 1984; QUEIROZ et al., 2003).
As plântulas infectadas apresentam engrossamento do meristema apical, acompanhada
da proliferação de brotos inchados e lançamento foliar, folhas retorcidas, podendo apresentar

18
pecíolos e pulvinos inchados. Ainda visualiza-se em plântulas de alguns genótipos, uma
proliferação a partir das axilas (vassoura axilar) e cotilédones (vassoura cotiledonar), além do
fendilhamento da casca (cancro). Em plantas adultas, as gemas vegetativas infectadas dão
origem a ramos hipertrofiados, com internódios curtos, intensa brotação lateral (vassoura
vegetativa) e folhas grandes e retorcidas. Inicialmente as vassouras são verdes e depois
começam a secar da base para o ápice (SILVA et al., 2002).
Os basidiósporos são os únicos propágulos comprovadamente infectivos (SILVA et
al., 2002) e penetram no tecido hospedeiro por aberturas naturais (estômatos) e ferimentos
(FRIAS et al., 1991; SREENIVASAN; DABYDEEN, 1989) ou diretamente
(SREENIVASAN; DABYDEEN, 1989). Os frutos podem ser infectados a partir de flores
infectadas, e através da penetração dos basidiósporos através do epicarpo, apresentando uma
variedade de sintomas, dependentes do tipo de infecção e idade. Quando originado de flores
afetadas, os frutos são partenocárpicos, assumem forma de “morango” e paralisam o
crescimento sendo inicialmente verdes ou avermelhados, tornando-se negros e petrificados
quando secos. Quando infectados ainda jovens (± 1 cm de comprimento) e diretamente
através do pericarpo, assumem a forma de “cenoura”, e paralisam o crescimento com
aproximadamente 15 cm de comprimento. Os frutos que são infectados com 2 a 5 cm de
comprimento, tornam-se inchados e deformados, com amadurecimento precoce; já quando a
infecção ocorre frutos maduros, forma-se lesões externas com manchas negras, brilhantes, de
formato circular. Com o desenvolvimento da infecção, os frutos se tornam secos e
petrificados, com formação de basidiocarpos na superfície. O fungo coloniza inicialmente os
tecidos internos dos frutos e, quando é possível a visualização dos sintomas externos, as
sementes já estão impróprias para a comercialização, pois já ocorreu a invasão do micélio e as
sementes já se encontram fortemente aderidas entre si (ALBUQUERQUE et al., 2005).
O risco de infecção aumenta com a proximidade da fonte de inóculo e exposição de
regiões meristemáticas (LUZ et al., 2006), diminuindo gradualmente a medida em que se
distancia, pois a quantidade de basidiósporos no ar cai sensivelmente a distâncias superiores a
300 m da fonte de inóculo. Entretanto, quando as condições climáticas são favoráveis à
disseminação, poderá ocorrer o transporte de esporos e infecções em plantas situadas a alguns
quilômetros da fonte de inóculo (LUZ et al., 1997). Nas gemas dormentes a infecção torna-se
latente, assumindo o aspecto de pequenos cancros ou pontos necróticos que entram em
atividade quando a planta reinicia a brotação. Essas infecções têm importância
epidemiológica, pois permitem a sobrevivência do fungo entre os períodos sucessivos de
crescimento (lançamentos foliares) e de frutificação (safras) da planta. Após infecção e

19
colonização, ocorre uma intensa multiplicação celular (hipertrofia) que dura algumas semanas
e, em seguida as células morrem ocorrendo a necrose generalizada dos tecidos (SILVA et al.,
2002). O micélio saprofítico cresce nos tecidos necrosados (frutos e vassouras secas), e após
um período de dormência, se dá o aparecimento dos basidiomas ou frutificações do patógeno.
Este ciclo de basidioma a basidioma ocorre uma vez por ano na Amazônia, mas, na Bahia
pode ocorrer até duas vezes: uma na safra temporã (1° semestre) e outro na safra principal
(2°semestre) (LUZ et al., 2006).
Estas desordens fisiológicas são frequentemente causadas pela liberação intracelular
de grande quantidade de compostos fenólicos, etilênicos, açúcares solúveis e taninos, levando
a morte celular ou alterações no balanço hormonal (ODJAKOVA; HADJÜVANOVA, 2001;
SCARPARI et al., 2005).
A VB é considerada uma doença monocíclica, pois os tecidos infectados não
produzem esporos capazes de iniciar novas infecções na mesma estação (safra). Mesmo
havendo a contínua liberação dos basidiósporos durante toda a estação (LUZ et al., 1994),
estes provêm de basidiomas desenvolvidos em tecidos infectados em estações anteriores
(LUZ et al., 2006). Os períodos de duração das diferentes fases do ciclo vital de M. perniciosa
foram estudados na Bahia por LUZ et al. (1994), sendo período de incubação (4 semanas em
média), período entre o aparecimento de vassouras verdes e o seu secamento (7,5 semanas em
média), período de dormência das vassouras secas (13 semanas, a depender do tamanho e
localização da vassoura), período de produção de basidiomas em frutos mumificados (24
meses); e período de liberação de esporos (na Bahia ocorre o ano todo, com picos maiores nos
meses mais frios). Um basidioma se mantém ativo durante cinco dias em média, liberando
milhões de esporos nas primeiras horas do dia durante sua vida útil (LUZ et al., 1997).
As condições ambientais são preponderantes para o processo de esporulação,
disseminação e infecção de tecidos vegetais. Rocha e Wheeler (1985) descrevem como
condições favoráveis para a liberação dos basidiósporos, umidade próximo a saturação e
temperaturas entre 20 e 30°C. Quando os basidiósporos são expostos a radiação ultravioleta,
são facilmente desidratados, diminuindo drasticamente a porcentagem de germinação dos
esporos viáveis. Entretanto, quando os basidiósporos são liberados à noite, a probabilidade de
infecção aumenta, como ocorrido em Trinidad e Equador, onde foi verificado que a maior
liberação ocorreu entre 22 h e 4 h, com umidade relativa em torno de 95% e temperatura
entre 20 e 24 °C (BAKER; CROWDY, 1943; EVANS; SOLORZANO, 1982).
Na Amazônia Brasileira as vassouras localizadas na copa são as principais fontes
de inóculo enquanto aquelas na superfície do solo produzem poucos basidiomas e com

20
menor chance do basidiósporo atingir os órgãos suscetíveis (ANDEBRHAN, 1981). O vento
e altura em que o basidioma é produzido é preponderante para o mecanismo de dispersão e
progresso da doença (COSTA, 1993), entretanto, os casqueiros também se constituem
importantes fontes de inóculo, pois no estado da Bahia, vassoura e frutos infectados quando
dispostos sobre o solo, são importante fonte de inóculo para infecções futuras (NIELLA,
1997).
Mesmo o basidiósporo sendo o principal propágulo infectivo, o micélio saprotrófico
do fungo pode sobreviver por vários anos, em vassouras secas e frutos mumificados, bem
como no interior dos tecidos infectados como vassouras vegetativas, frutos e folhas, que ao
serem transportados indevidamente, quando expostos aos efeitos alternados de chuva e sol,
podem produzir basidiomas favorecendo a infecção pelo fungo (BAKER; CROWDY, 1943;
BASTOS, 1990; NIELLA et al., 1999).
1.3 BIOLOGIA E TAXONOMIA DO FUNGO Moniliophthora perniciosa
A vassoura de bruxa foi notificada inicialmente em 1700, mas somente em 1890 foi
esclarecido o agente causal através de Gregor Stahel, que isolou e nomeou o fungo causador
Marasmius perniciosa. Em 1942, foi transferido para o gênero Crinipellis por Singer e, na
revisão da sistemática do fungo, foi mantido o binômio Crinipellis perniciosa (Stahel) Singer
(SINGER, 1942). Por fim, em 2005, foi transferido para o gênero Moniliophthora por Aime e
Phillips-Mora (AIME; PHILLIPS-MORA, 2005) com base em características moleculares.
O fungo M. perniciosa pertence à divisão Basidiomycota, subdivisão
Agaricomycotina, classe Agaricomycetes, sub classe Agaricomycetidae, ordem Agaricales,
família Marasmiaceae (AIME; PHILLIPS-MORA, 2005), que é constituída de espécies com
basidiomas pileados, lignicolas ou habitantes da serrapilheira, e que possuem a capacidade
recuperar sua morfologia após o secamento quando novamente são umedecidos
(ALEXOPOULOS et al., 1996; EVANS, 1980).
Apesar de existirem outros patógenos causando doenças de impacto econômico à
cultura do cacau, o fungo M. perniciosa quando em condições favoráveis, é o principal
patógeno do cacaueiro na Bahia atualmente, sendo capaz de colonizar, além do cacaueiro,
várias outras plantas hospedeiras, como espécies do gênero Theobroma e Herrania
(Malvaceae) e alguns representantes das famílias Solanaceae e Bixaceae (BASTOS;
ANDEBRHAN, 1986), cuja adaptação nestas plantas podem contribuir para o aumento da
variabilidade genética desse microrganismo (LUZ et al., 2006).

21
O patógeno M. perniciosa é um fungo hemibiotrófico, pois apresenta ciclo de vida
com uma fase parasítica e outra saprofítica. O micélio primário formado após emissão do tubo
germinativo e que coloniza intercelular mente o tecido vegetal é monocariótico e representa a
fase biotrófica, já o micélio secundário é dicariótico e corresponde a fase necrotrófica.
(PEGUS, 1972). Os dois tipos de micélios são distintos e podem ser separados morfológica,
fisiológica e geneticamente em: micélio monocariótico, parasítico sem grampo de conexão,
encontrado em tecidos vivos infectados; e micélio dicariótico, saprofítico, com grampo de
conexão, encontrado em tecidos mortos e em meio de cultura (EVANS e BASTOS, 1979;
EVANS, 1980; CALLE et al., 1982).
Na fase biotrófica, o fungo se desenvolve a partir dos tecidos vivos do hospedeiro,
direcionando o metabolismo da planta para as suas necessidades básicas, como nutrição,
crescimento e desenvolvimento do micélio (SCARPARI et al., 2005; KILARU,
HASENSTEIN, 2005). A mudança da fase biotrófica para necrotrófica é estimulada pela
alteração na concentração carboidratos específicos, pois o fungo ao desviar o fluxo destes
carboidratos das partes saudáveis da planta para as regiões infectadas, leva à necrose rápida
dos tecidos, tanto por inanição como por resposta de defesa da planta, pela alteração de sua
fisiologia (ODJAKOVA, HADJÜVANOVA, 2001; SCARPARI et al., 2005; GALANTE,
2008).
O fungo M. perniciosa possui quatro biótipos, sendo o biótipo C específico para o
cacau, possuindo dois patótipos (A e B). O A está presente na Bolívia, Colômbia e Equador,
sendo considerado mais virulento e o B, com uma menor virulência está presente no Brasil,
Trinidade-Tobago e Venezuela (MOTILA; BUTLER, 2003). O conhecimento dos aspectos
biológicos do patógeno, hospedeiro e condições edafo-climáticas subsidiam a adoção de
diferentes métodos de controle, minimizando assim, os impactos causados pelo patógeno na
cultura (FALEIRO et al., 2004). Assim como Delgado e Cook (1976) relataram a severidade
da vassoura de bruxa em variedades selecionadas como resistentes no Equador, casos
semelhantes podem surgir, pois a anastomose de hifas provenientes da germinação de
basidiósporos de origens distintas, possibilitam o surgimento de heterocário no micélio
saprofítico, aumentando a variabilidade genética quando da formação de novos basidiósporos.
Os basidiósporos, produzidos no himenóforo localizado na face abaxial do píleo dos
basidiomas, são esporos sexuais unicelulares, hialinos, com parede celular delgada, e tamanho
médio de 12 x 6 µm (BAKER, CROWDY, 1943), que germinam rapidamente quando em
condições de saturação de umidade, com início da germinação aproximadamente 1 hora após
exposição a estas condições e término quatro horas depois (BASTOS, 1990; NIELLA et al.,

22
1999). Os basidiomas apresentam coloração avermelhadas a purpúrea, com píleo variando
entre 5 – 30 mm de diâmetro, radialmente sulcados, convexos à deprimidos no centro,
himenóforo branco com aproximadamente 15 lamelas, correspondente aos sulcos dos píleos;
o estipe apresenta coloração esbranquiçada, medindo de 5 a 10 mm de altura, com
engrossamento na base e coloração parda. Os basídios produzem quatro basidiósporos
unicelulares (BAKER, CROWDY, 1943).
1.4 MANEJO INTEGRADO DA VASSOURA-DE-BRUXA DO CACAUEIRO
O manejo integrado consiste na forma mais eficiente e econômica de controle,
associando todos os métodos de controle disponíveis (cultural, químico, biológico,
genético e outros), de maneira que o custo do controle seja reduzido, permitindo um
cultivo economicamente rentável. Os maiores benefícios do manejo integrado são seus
efeitos acumulativos, em que, através do emprego de combinações de diferentes métodos de
controle, cada um deles com diferentes modos de ação, reduzam progressivamente a
possibilidade do desenvolvimento da doença (PEREIRA; VALLE, 2012).
Apesar da utilização de defensivos químicos ainda ser a alternativa mais utilizada para
o controle de doenças de plantas, para o patossistema cacaueiro x M. perniciosa, existem
vários fatores que induzem a uma grande variação na eficiência desta técnica, principalmente
quando utilizada isoladamente. A alta variabilidade dentro da espécie tem sido detectada
quando isolados, de várias regiões de um país, de diferentes países ou de diferentes
hospedeiros são comparados. Tal variabilidade tem sido traduzida, por exemplo, em termos
de crescimento micelial em meios de cultura, tipos de reação em alguns testes bioquímicos,
compatibilidade somática e patogenicidade ao cacaueiro e a outros hospedeiros (BARTLEY,
1977; BASTOS, 1986; McGEARY; WHEELER, 1988). Em função desse comportamento,
pode ocorrer variação do patógeno até mesmo com referência à sensibilidade a fungicidas, o
que poderia eventualmente explicar o maior ou menor sucesso das recomendações de controle
da doença em determinadas situações (DELGADO; COOK, 1976).
A Comissão Executiva da Lavoura Cacaueira (CEPLAC) recomenda o manejo
integrado para o controle da VB composto por clones resistentes, aplicação de fungicidas,
poda fitossanitária, e agentes de biocontrole (OLIVEIRA; LUZ, 2012). Esforços têm sido
voltados para a busca de medidas alternativas que aumentem a eficiência do pacote de
manejo, racionalizando o uso de agroquímicos, e reduzindo a frequência de práticas agrícolas

23
como realização de podas e retiradas de vassouras (COSTA et al., 2009; MEDEIROS et al.,
2010).
Desde os primeiros estudos referentes ao controle desta enfermidade, foi recomendado
o uso do controle cultural associado ao químico (Sthael, 1915). No entanto, o controle
químico com fungicidas a base de cobre não protege os tecidos em crescimento (OLIVEIRA,
2004a) e as aplicações visam principalmente proteger os frutos, sendo necessárias aplicações
em períodos corretos e obedecendo rigorosamente doses, intervalos e forma de aplicação
adequada, evitando os períodos chuvosos que podem facilmente lixiviar os fungicidas
protetores. Em diversas pesquisas realizadas na região cacaueira da Bahia, constatou-se que o
fungicida sistêmico tebuconazole (OLIVEIRA, 2004a) e outros do grupo das estrobilurinas
(OLIVEIRA, 2004b), com destaque para a azoxystrobina, apresentaram os melhores
resultados para o controle da VB, tanto em viveiros quanto em campo (OLIVEIRA; LUZ,
2005).
Objetivando reduzir as fontes de inóculo, recomenda-se ainda a remoção de tecidos
infectados, seguido de queima, cobertura com folhas, e/ou aplicação de cal, para acelerar a
decomposição e evitar a esporulação do fungo (LUZ et al., 2006; STAHEL, 1915). Contudo,
a determinação do número, época de remoções e eficiência dessa prática depende de vários
fatores, principalmente, das condições climáticas da região (LUZ et al., 1997) e do nível de
infecção da planta (BASTOS, 1996). Na Bahia, são necessárias pelo menos quatro remoções
ao ano, realizadas normalmente nos meses de fevereiro, maio, agosto e novembro,
antecedendo os lançamentos foliares do cacaueiro. A eficácia deste método também é
influenciada pela presença de frutos mumificados ou vassouras que ficam no solo, ou ainda
através de inóculo proveniente de áreas onde o controle cultural não foi realizado (LUZ et al.,
2006). Tais riscos tornam o controle cultural menos eficiente, sendo necessária a aplicação de
fungicida para minimizar as perdas (ANDEBRHAN; BASTOS, 1985; OLIVEIRA; LUZ,
2005).
Além da aplicação de antifúngicos e poda fitossanitária, acrescenta-se às medidas de
controle da vassoura de bruxa, a busca por variedades resistentes, indução de resistência do
hospedeiro e a prospecção por micro-organismos antagônicos ao M. perniciosa, garantindo
desta forma a utilização de tecnologias com menor impacto ambiental e mais econômicas
(COSTA et al., 2009).
1.4.1 CONTROLE GENÉTICO DO CACAUEIRO QUANTO À RESISTÊNCIA A VASSOURA DE BRUXA

24
O emprego de variedades resistentes e de alta produtividade, desenvolvidas em
programas de melhoramento genético do cacaueiro é a alternativa mais recomendada para se
manejar a VB (PINTO; PIRES, 1998). Essa medida de controle é fundamental haja vista os
controles químico e cultural terem se mostrado onerosos e ineficazes quando não executados
rigorosamente de acordo com as recomendações técnicas da pesquisa, em se tratando de
lavouras formadas por variedades de alta suscetibilidade e de baixa produtividade (PINTO;
PIRES, 1998).
Alguns autores dividem didaticamente a história do melhoramento genético do
cacaueiro em antes e depois do desenvolvimento do cacaueiro híbrido, cuja implementação
ocorreu em Trinidad no início da década de 1930, realizando os primeiros estudos sobre a
frutificação, a propagação vegetativa e a herança da pigmentação dos cotilédones
(CHEESMAN; POUND, 1934). Também, foi a partir destes estudos que se detectou a
presença da autoincompatibilidade em cacaueiros do Alto Amazonas e foram definidas as
características para a seleção de cacaueiros superiores (DIAS, 2001b). O primeiro programa
de melhoramento do cacaueiro visando a obtenção de genótipos resistentes a M. perniciosa
foi iniciado no Equador em 1918, através da seleção massal de plântulas originadas de árvores
sem sintomas (BARTLEY, 1986).
Com a disseminação da VB nos cacauais do sul da Bahia a partir de 1989 (PEREIRA
et al., 1989) os métodos de melhoramento visando à obtenção de variedades clonais, bem
como, a utilização da clonagem para a rápida substituição de plantas suscetíveis, quer na
implantação de plantios novos ou por substituição de copas, tornou-se estratégia para a
recuperação da cacauicultura daquela região e consequentemente o resgate da cacauicultura
nacional (LOPES et al., 2011).
Uma das dificuldades encontradas no melhoramento genético do cacaueiro é a
limitada fonte de resistência utilizada até o início das pesquisas com a doença na Bahia. Os
clones e cultivares conhecidos como resistentes eram quase todos descendentes dos clones
Scavina-6 e 12 (SCA 6 e SCA 12), que foram selecionados no Peru e vem sendo utilizados
como fonte de resistência à VB desde 1940 (BARTLEY, 2005). No entanto, o clone SCA 6, o
mais utilizado, havia apresentado quebra de resistência à VB em algumas regiões. Wheeler e
Mepsted (1988) relatam a suplantação de resistência no Equador e Andebrhan et al. (1998)
constataram quebra da resistência em Rondônia, na Amazônia. Em meados da primeira
década deste século, descendentes deste clone, também começaram a apresentar
susceptibilidade na Bahia, o que justifica a busca de novas fontes de resistência (PAIM et al.,
2006).

25
Desde 1996, a CEPLAC disponibilizou aos produtores mais de 40 variedades clonais
de cacaueiro com diferentes níveis de resistência à VB entretanto, algumas foram retiradas e
não mais recomendadas e distribuídas, devido a suscetibilidade à outras doenças. Mesmo com
a atual recomendação de variedades clonais com características qualitativas e quantitativas de
frutos e sementes (FA-13, VEM-20, SJ-02, CEPEC 2002, CCN-51, PH-09, LP-06, PH-15,
BJ-11, CA-1.4, PS-13.19, CCN-10, PH-16) adequadas às exigências do mercado (PEREIRA;
VALLE, 2012), o programa de melhoramento genético do cacaueiro segue em busca do maior
número possível de fontes de resistência, combinando-as em cruzamentos de primeira e
segunda geração e proporcionando a obtenção de material genético com genes de resistência
de diferentes origens, aumentando assim a estabilidade, durabilidade e dificultando o aumento
da população e a evolução do fungo sobre tais materiais (PINTO; PIRES, 1998; PIRES,
2003).
Modelos experimentais de cruzamentos visando estimar parâmetros genético
quantitativos são desenhados de acordo com as especificidades de cada cultura. Comstock e
Robinson (1952), desenvolveram uma série de projetos experimentais para situações em que
um ou ambos os sexos podem proporcionar múltiplos cruzamentos. Tais protocolos
conhecidos como delineamento Carolina do Norte, receberam tal designação por serem muito
utilizados em programas de melhoramento naquele estado americano. Os delineamentos
Carolina do Norte I e II são utilizados quando pólen geralmente é produzido em abundância,
espécies com sexos separados, e o tamanho do experimento é geralmente limitado, pelo
número de flores produzidas por plantas fêmeas (LYNCH; WALSH, 1998).
Um sistema de cruzamentos dialélicos define-se como sendo aquele no qual é
escolhido um grupo de linhagens autofecundadas, as quais são cruzadas entre si, fornecendo
um máximo de p2 combinações (HAYMAN, 1954; GRIFFING, 1956). O termo dialelo,
porém, tem sido utilizado para expressar um conjunto de p (p – 1) /2 híbridos, resultantes do
acasalamento entre p genitores (linhagens, cultivares, clones etc.) (CRUZ et al., 2004).
As metodologias de análise dialélica têm por finalidade analisar o delineamento
genético, fornecendo estimativas de parâmetros úteis na seleção de parentais para a hibridação
e no entendimento dos efeitos gênicos envolvidos na herança dos caracteres (CRUZ et al.,
2004). De acordo com Geraldi e Miranda Filho (1988), os sistemas de cruzamentos dialélicos
são bastante utilizados no melhoramento de plantas, uma vez que possibilitam a avaliação da
capacidade combinatória e do potencial heterótico de variedades ou linhas em cruzamentos,
ou nos estudos básicos da estrutura genética das populações.

26
A seleção de parentais com base na avaliação “per se” nem sempre conduz a
resultados satisfatórios (ALLARD, 1971). Assim, a escolha de parentais deve ser
fundamentada na informação genética como um todo e no conhecimento do potencial da
capacidade combinatória dos parentais (AHMED et al., 1998).
Devido as características morfo-genéticas do cacaueiro, os dialelos formados são
desbalanceados, construídos de acordo com a disponibilidade de sementes obtidas dos
cruzamentos, seguindo o delineamento Carolina do Norte II (SILVA et al., 2010).
O emprego de variedades resistentes e de alta produtividade, desenvolvidas em
programas de melhoramento genético do cacaueiro (PMGC) é a base para o manejo de
doenças de plantas (PINTO; PIRES, 1998). Entretanto, alguns materiais mesmo apresentando
genes de resistência, têm, com o tempo e extensão do cultivo, demonstrado suscetibilidade,
devido às condições ambientais favoráveis à doença, à rápida mutação e grande variabilidade
do patógeno (BARTLEY, 2005). Sendo assim, o PMGC segue buscando ampliar a base
genética para resistência, combinando o maior número possível de genes através de
cruzamentos em primeira e segunda geração, no chamado processo de piramidação de genes,
para obter resistência mais estável, durável e efetiva, dificultando o aumento da população do
fungo e a sua evolução do sobre tais materiais (PINTO; PIRES, 1998; PIRES, 2003).
O uso de alguns clones, com diferentes níveis de resistência, em associação a outras
práticas do manejo integrado da vassoura-de-bruxa, tem possibilitado avanços na recuperação
da lavoura cacaueira sul baiana (LUZ et al., 2013). Nas últimas décadas, vários acessos foram
identificados como fontes de resistência à VB e monilíase, através de programas de
melhoramento em vários países produtores, que estão utilizando as coleções de germoplasma
que possuem em busca de fontes de resistência estáveis e duradouras. A CEPLAC possui duas
grandes coleções de germoplasma de cacau. A coleção localizada no município de Ilhéus, no
estado da Bahia, é considerada a mais diversificada por possuir 1.300 acessos, incluindo
germoplasmas selvagens e melhorados (LOPES et al., 2011), e também alguns clones
considerados na Costa Rica como resistentes à monilíase como: EET 75 e 233, ICS 95 e UF
273 (PHILLIPS-MORA et al., 2007). A outra coleção trata-se do maior banco de
germoplasma de cacau do mundo e está localizada no município de Marituba, no estado do
Pará (MONTEIRO; AHNERT, 2012).
O PMGC no Brasil, por outro lado, já gerou várias populações estruturadas e não
estruturadas visando a piramidação de genes para a resistência à VB, e destas vários clones
foram indicados para o cultivo na região (MONTEIRO; AHNERT, 2012). As populações
estruturadas foram geradas a partir de cruzamentos dialélicos realizados entre materiais

27
selecionados de três outras populações: A- alto amazônicos; B- Baixo amazônicos; e AB
cruzamentos entre alto e baixo amazônicos, distantes geneticamente, para exploração da
heterose, adição de características interessantes, além de resistência à VB e outras doenças, e
produção (PIRES et al., 1999). Uma dessas população estruturada foi construída a partir de
um ciclo de seleção recorrente e as plantas desta população avaliadas a campo por mais de
seis anos, para compor um segundo ciclo de seleção. Plantas elite destas populações foram
cruzadas entre si para a piramidação de genes de diferentes fontes e testadas neste trabalho em
um dialelo (10 x 8) pelo delineamento Carolina do Norte II (COMSTOCK; ROBINSON,
1952), incluindo também clones doadores de genes de resistência a monilíase.
Diversos trabalhos já foram também realizados visando associar marcadores
moleculares para acelerar o processo de melhoramento para resistência. Dantas Neto et al.
(2005) realizaram estudos em plantas com oito anos de idade, visando identificar oito genes
de resistência e Quantitative trait loci (QTL), dissimilares dos encontrados no clone Scavina-
6, em populações derivadas do cruzamento entre os clones SIC-864 e CCN-51. Faleiro et al
(2006) e Santos et al (2007) identificaram novos QTL relacionados a resistência à VB.
Populações estão sendo desenvolvidas e testadas gerando a possibilidade de futuros
materiais a serem distribuídos que apresentem além do caráter resistência, genes para outras
características agronomicamente desejáveis (MONTEIRO et al., 2006). Através de técnicas
de RAPD (Random amplified polymorphic DNA), Yamada et al. (2002) demonstraram
diversidade genética em estudo com 27 acessos de cacaueiro da série Cepec. Leal et al. (2008)
utilizando também técnicas de RAPD encontraram elevada diversidade genética entre as
seleções de cacaueiros em 17 fazendas de sete municípios do Sul da Bahia. Paim et al. (2006)
demonstraram o alto nível de resistência de progênies da série CAB (Cacau da Amazônia
Brasileira) na Bahia e que estas são distintas geneticamente de SCA 6 (PAIM, 2005).
Portanto, há variabilidade de resistência à VB tanto nos materiais de bancos de germoplasma
(BAG) como nas plantações de cacau no Sul da Bahia.
1.4.2 CONTROLE BIOLÓGICO DA VASSOURA-DE-BRUXA DO CACAUEIRO
Alternativamente, o controle biológico por meio de micro-organismos antagônicos a
M. perniciosa tem sido bastante utilizado, recebendo destaque o Tricovab, produto
desenvolvido pela CEPLAC e formulado a partir do fungo Trichoderma stromaticum Samuels
& Pardo-Schultheiss, que parasita M. perniciosa impedindo a produção de basidiomas

28
(BASTOS, 2000; LOGUERCIO et al., 2009; BEZERRA et al., 2000; SANTOS, 2005;
SOUZA et al., 2006).
As primeiras pesquisas com biocontrole da VB foram realizadas no final da década de
1970. Nesse período, foram testados vários micro-organismos em condições de laboratório,
havendo destaque para Cladobotryum amazonense C.N. Bastos, H.C. Evans & Samson, por
apresentar potencial para o controle da doença. Este fungo foi encontrado no Pará,
parasitando basidiomas de M. perniciosa (BASTOS et al., 1981). Em ensaios de laboratório e
de campo, verificou-se que este antagonista era capaz de colonizar basidiomas do patógeno,
impedindo a produção de inóculo, mas é de difícil esporulação. Optou-se então pelo uso do
sobrenadante obtido a partir do cultivo in vitro deste antagonista, que já havia demonstrado
ser ativo contra M. perniciosa e outros patógenos de outras culturas. O composto ativo foi
identificado e demonstrou ser absorvido e translocado por mudas de cacaueiro (BASTOS,
1988). Assim como C. amazonense, Verticillium lamellicola (F.E.V. Sm.) W. Gams também
foi detectado parasitando basidiomas de M. perniciosa, sendo capaz de produzir uma
substância altamente antagônica ao patógeno (ANDEBRHAN, 1981). Entretanto, mesmo
sendo realizadas várias pesquisas, por razões comerciais, biofungicidas baseados nestes
fungos não foram desenvolvidos LOGUERCIO et al., 2012).
Em 1987, um isolado fúngico do gênero Trichoderma com capacidade de colonizar
vassouras secas, foi identificado no Pará, inicialmente como T. viride Pers. Posteriormente, o
isolado foi classificado como T. polysporum (Link) Rifai e finalmente descrito como uma
nova espécie, recebendo o binômio, Trichoderma stromaticum (SAMUELS et al., 2000).
Os resultados das primeiras pesquisas com T. stromaticum, realizadas no Pará e
Rondônia, mostraram sua capacidade de reduzir a formação de basidiomas de M. perniciosa
em vassouras secas, culminando com a morte do patógeno (BASTOS, 1996a). Sua maior
vantagem sobre os outros antagonistas avaliados é a alta capacidade de sobrevivência em
vassouras secas, especificidade por M. perniciosa, alta esporulação no ambiente e fácil
cultivo. Ensaios enzimáticos demonstraram que o fungo é capaz de produzir enzimas
hidrolíticas como quitinase, endoglucanase e amilase (BASTOS, 1996 b), além da citação de
efeitos indiretos no aumento da produção de frutos de cacau após três anos em ensaios de
campo (COSTA et al., 2009; MEDEIROS et al., 2010).
Apesar dos estudos relacionados ao controle biológico da VB concentrar-se na
interação T. cacao x Trichoderma x M. perniciosa, a mistura de organismos benéficos,
buscando conjuntamente efeitos antagônicos, protetores e/ou antiesporulante do patógeno,
deve ser realizada visando promover um sistema integrado de biocontrole (ELAD, 2003).

29
As leveduras são promissores agentes de biocontrole de doenças de plantas, por
estarem tanto na filosfera, quanto na rizosfera de vários cultivos, inclusive o cacaueiro
(CABRAL et al., 2009), atuando de forma antagônica à diversos patógenos, através da
competição por nutrientes e espaço, parasitismo, antibiose, além de apresentarem potencial
indutor de resistência à doenças e promoção de crescimento de plantas (EL-TARABILY;
SIVASITHAPARAM, 2006; VALDEBENITO-SANHUEZA, 2000).
1.4.3 LEVEDURAS COMO AGENTES DE BIOCONTROLE E PROMOTORES DE
CRESCIMENTO
Leveduras são fungos unicelulares e não filamentosos, que são frequentemente
encontradas na natureza (TORTORA; FUNKE; CASE, 2005), apresentam geralmente células
ovais, esféricas, ou cilíndricas com dimensões maiores do que a de uma bactéria,
apresentando núcleo como estrutura interna diferencial, e sendo capaz de se dividir por fissão
binária ou brotamento celular, são encontradas abundantemente em habitats que contenham
açúcar como por exemplo flores, frutos e cascas de árvores (MADIGAN; MARTINKO;
PARKER, 2004), sendo também são encontrados em solos, sedimentos, água bem como sobre
as superfícies externas e tratos intestinais de animais, especialmente insetos (LACHANCE;
STARMER, 1998). As condições ideais para o crescimento da maioria das leveduras
encontram-se na faixa de pH entre 4 e 5, temperatura de crescimento entre 20 e 30ºC e sendo
o Carbono (C), o Fosforo (P) e o Nitrogênio (N) elementos químicos limitantes ao seu
crescimento (REHN, 1995).
As espécies de levedura integram os filos Ascomycota e Basidiomycota
(FUENTREFIA, 2007). São bastante utilizadas pelo homem para fins variados, a exemplo da
espécie Sacharomyces cerevisiae, microrganismo bastante utilizado pela indústria alimentícia
na produção de pães e massas, produção de álcool, vinhos, cerveja (SCHENBERG, 2010) na
alimentação humana e animal (COSTA, 2004) e no controle de doenças de plantas (LOPES et
al., 2015; SHALABY; EL-NADY, 2008; WACHOWSKA et al., 2013).
Leveduras, bactérias e fungos são constantemente isolados do filoplano de diversas
espécies vegetais, devido a riqueza de nutrientes disponíveis (ANDREWS; HARRIS, 2000).
Entretanto, rápidas alterações das condições ambientais consideradas ótimas para o
crescimento microbiano, promovem alterações na densidade populacional e na quantidade de
organismos epifíticos (INÁCIO et al., 2005). As bactérias iniciam o processo de colonização
foliar (BLAKEMAN, 1991), mas com o passar do tempo, as leveduras se tornam mais
abundantes devido a aumento na disponibilidade de nutrientes, oriundos da deposição de

30
grãos de polén e extravasamento de fotoassimilados com o processo de alimentação de afídios
e outros insetos (STADLER; MULLER, 1996). Após o estabelecimento na superfície foliar,
os micro-organismos epifíticos aderem fortemente à superfície, utilizando a matriz
extracelular como proteção (BEATTIE; LINDOW, 1999), e formação de biofilmes como
estratégia de sobrevivência em ambientes variáveis (CHEN et al., 2013).
As leveduras predominam nas gemas e nas folhas em termos de biomassa e, nesses
locais, sobressaem as leveduras com colônias vermelhas dos gêneros Sporobolomyces e
Rhodothorula e as brancas do gênero Cryptococcus. Na microbiota dos solos, mesmo em
quantidade menor que as bactérias, podem ser encontradas leveduras, e os principais gêneros
encontrados nesse ambiente são Cryptococcus, Torulaspora, Tremella e Trichosporon.
(VALDEBENITO-SANHUEZA, 2000). Dentre as leveduras citadas na literatura como
agentes de biocontrole, destacam-se os gêneros: Debaryomyces, Kloeckera, Saccharomyces,
Zygosaccharomyces, Sporobolomyces, Metschnikowia, Tilletiopsis, Rhodotorula,
Cryptococcus, Aureobasidium, Pichia e Candida. Espécies pertencentes a alguns destes
gêneros correspondem ao composto ativo de produtos registrados em muitos países como
biocontroladores de patologias pós-colheita, a exemplo de Aureobasidium pullulans (de Bary)
G. Arnaud, Cryptococcus albidus (Saito) C. E. Skinner, Metschnikowia fruticola (Kurtzman e
Droby), inibindo o desenvolvimento de patógenos como Botrytis, Penicillium, Rhizopus,
Monilia, Sclerotinia, Erwinia amylovora (Burril) Winslow (EL-TARABILY;
SIVASITHAPARAM, 2006).
A rápida capacidade de reprodução e assimilação de nutrientes, confere às leveduras
uma ótima ação competitiva no ambiente em que se encontram, sendo a competição por
nutrientes e espaço um dos principais mecanismos de biocontrole destes micro-organismos
(LAHLALI et al., 2011). Alguns representantes do gênero Candida e Rhodotorula também já
foram relatados como produtores de compostos quelantes de íons férricos (sideróforos) (EL-
TARABILY; SIVASITHAPARAM, 2006; ROSA et al., 2010). Estas características também
permitem o bloqueio da entrada de patógeno por meio de lesões na epiderme devido a
velocidade de colonização de ferimentos (ROBLIGLIO et al., 2011).
Pesquisas recentes têm revelado o potencial de leveduras no controle da VB do
cacaueiro, ao inibir o crescimento de M. perniciosa (CABRAL et al., 2009). A utilização de
tais micro-organismos como agentes biocontroladores se dá devido ao alto potencial
competidor (ZHANG et al., 2007) ao secretarem toxinas killer (WALKER et al., 1995),
produzirem compostos voláteis (BRUCE et al., 2004) e secretarem enzimas hidrolíticas
capazes de degradar a parede celular de fitopatógenos (MASIH; PAUL, 2002; URQUHART;

31
PUNJA, 2002), capacidade de suportar mudanças no ambiente e condições de estresse (SUI et
al., 2015), tolerância à poluentes, óleos essenciais dos vegetais, fitoalexinas e alguns
fungicidas (VALDEBENITO-SANHUEZA, 2000); sobreviverem saprofiticamente (AFSAH-
HEJRI, 2013); e tolerarem condições adversas (HU et al., 2015). Apesar destas
características, algumas práticas de manejo podem influenciar a microbiota epifítica das
leveduras, inclusive a utilização de alguns defensivos como os ditiocarbamatos e os
benzimidazóis, os quais apresentam efeito inibitório (VALDEBENITO-SANHUEZA, 2000).
A utilização de leveduras para o biocontrole de fitopatógenos ainda se concentra na
patologia pós-colheita (TIAN et al., 2004; HU et al., 2015). Entretanto, pesquisas recentes
têm demonstrado o potencial destes micro-organismos no controle de doenças ocasionadas
ainda no campo, promovendo também a germinação de sementes, o crescimento de plantas
(EL-MEHALAWY et al., 2004; EL-TARABILY, 2004; KAMAL et al., 2009; KHAN et al.,
2012) e a indução de resistência, através do próprio organismo, ou seus derivados como
carboidratos, proteínas, ácidos graxos, entre outros (SPADARO; GULLINO, 2004;
WALTERS et al., 2005), atuando como elicitores capazes de ativar respostas de defesa
localizada ou sistêmica em plantas (DI PIERO et al., 2005; ZANARDO et al., 2009).
Diversas pesquisas estão sendo realizadas no sentido de elucidar quais as substâncias
secretadas pelas leveduras que estão atuando na inibição do crescimento, germinação e/ou
esporulação do patógeno, ou como bioelicitores de respostas de defesa da planta (ZANARDO
et al., 2009) e secreção de fitohormônios (SUN et al., 2014). Dentre estas substâncias
destacam-se enzimas como peroxidases, glucanases e quitinase, que além de atuarem
diretamente sobe o patógeno, são consideradas bioelicitores pela indução à produção de
substâncias que estão relacionadas à resistência das plantas à patógenos como fitoalexinas, a
felilalamina amônio liase e o fitohomônio etileno (EL-TARABILY; SIVASITHAPARAM,
2006; HAÏSSAM, 2011). Diversos relatos sobre a produção de enzimas hidrolíticas por
leveduras são citados na literatura, a exemplo de isolados das espécies de A. pullulans, T.
albescen e C. saitoana, com ação sobre os patógenos P. expansum, P. xantii e B. cinerea,
repectivamente (EL-TARABILY; SIVASITHAPARAM, 2006; ROSA et al., 2010;
URQUHART; PUNJA, 2002)
Paredes das células fúngicas contêm α quitina, que difere da alomorfa β quitina em
relação ao empacotamento e à polaridade de cadeias adjacentes (AAM et al., 2010; CHEN et
al., 2010). As quitinases são enzimas sintetizadas por vários organismos como bactérias,
fungos filamentosos e leveduras, que possuem atividade quitinolítica responsável por clivar
ligações glicosídicas β-1,4 presentes nos polímeros de N-acetilglicosamina (GlcNac),

32
apresentando como principal função a remodelação da parede celular e outros, como
processos que envolvem a patogênese e a separação de células filhas, função utilizado pela S.
cerevisiae em reprodução (GOHEL et al.,2006). Além de quitina, a parede celular de
leveduras é formada por outros componentes principais como mananaproteínas e glucanas
(polímeros de β-1,3 e β-1-6 glicose), presentes na parede externa e interna respectivamente
(KIM; YUN, 2006). As glucanases também estão envolvidas em eventos morfogenéticos,
como brotamento celular, conjugação e esporulação, entretanto o aumento da atividade desta
enzima foi relatado quando a levedura S. cerevisae foi cultivada meios indutores com
concentrações de glicose entre 2 e 8 % m/v (DRAKE et al., 2012).
As leveduras têm sido relatadas como promissores agentes de biocontrole, pois além
de atuarem por diversos mecanismos, apresentam características que as diferem de alguns
fungos filamentosos utilizados em formulações comerciais, a exemplo da capacidade de
produzirem toxinas killer, uma glicoproteína que pode agir diretamente sobre o patógeno
(COELHO et al., 2011) e atuar em concomitância com outros mecanismos antagônicos, e por
não produzirem micotoxinas e esporos alergênicos como os fungos filamentosos (GOUVEA,
2007; VIANA et al., 2012).
Apesar da crescente busca por produtos e métodos sustentáveis, que minimizem os
impactos causados pela utilização de agroquímicos no controle de doenças de plantas, o
número de bioprodutos disponíveis para comercialização ainda é pequeno. Até o ano de 2014,
apenas sete produtos microbiológico possuíam registro no Ministério da Agricultura Pecuária
e Abastecimento (MAPA) para utilização na agricultura brasileira, sendo seis deles a base de
fungos filamentosos e um a base de bactéria (GIARETTA, 2014). Atualmente alguns
produtos cujo princípio ativo são fungos leveduriformes, estão disponíveis comercialização e
utilização em alguns países, a exemplo do Aspire® e Next® que são produzidos a base da
espécie Candida oleophila e atuam sobre diversas culturas em pós-colheita (DROB et al.,
1998; DROB et al., 2002; VERO et al., 2002).
Mesmo diante dos resultados obtidos até o momento, a bioprospecção por agentes de
biocontrole que co-habitem o mesmo nicho de espécies vegetais de grande importância
econômica como cacaueiro, associada a busca por genótipos que apresentem genes
responsáveis pela manutenção e durabilidade do caráter de resistência a vassoura de bruxa e
monilíase, é de suma importância para a mitigação dos impactos causados por estas
fitomoléstias.
Diante do exposto, o presente trabalho objetivou a seleção genótipos de cacaueiro com
resistência a vassoura de bruxa, ampliando-se a fonte de resistência a VB nos materiais já

33
anteriormente selecionados como resistentes a esta doença, e incorporar à população gerada,
genes de resistência à monilíase (M. roreri), além de determinar a diversidade e biologia de
leveduras que habitam a superfície de órgãos vegetais de cacaueiros, avaliar o potencial
antagônico destas leveduras contra M. perniciosa, investigar a sua ação no biocontrole da VB
e na ativação de crescimento de plântulas e mudas de cacaueiro.
2 REFERÊNCIAS BIBLIOGRÁFICAS
AFSAH-HEJRI, L. Saprophytic yeasts: effective biocontrol agents against Aspergillus flavus.
International Food Research Journal, Toronto, v. 20, n. 6, p. 3403-3409, 2013.
AHMED, N.; SHAH, F. A.; ZANGAR, G. H.; WANI, S. A. Line x tester analysis for the
study of combining ability in hot pepper (Capsicum annuum L.). Capsicum and Eggplant
Newsletter, Turin, n. 17, p. 38-41, 1998.
AIWE M. C.; PHILLIPS-MORA, W. The causal agents of witches´broom and frost pod rot of
cacao (chocolate, Theobroma cacao) form a new linage of Marasmiaceae. Mycologia, New
York, v. 97, p. 1012-1022, 2005.
ALBUQUERQUE, P. S. B.; BASTOS, C. N.; LUZ, E. D. M. N.; SILVA, S. D. V.
M. Doenças do cacaueiro. In: KIMATI, H.; AMORIM, L.; REZENDE, J. A. M.;
BERGAMIN FILHO, A.; CAMARGO, L. F. A. (Eds.). Manual de fitopatologia. 4.
ed., Piracicaba: Ceres, 2005. v. 2, cap. 18, p. 151-164.
ALEXOPOULOS, C. J.; MIMS, C. W.; BLACKWELL, M. Introductory Mycology. 4ª ed.
New York: John Wiley & Sons, 1996. 878 p.
ALLARD, R. W. Princípios do melhoramento genético das plantas. São Paulo: Edgard
Blucher, 1971. 381 p.
ALMEIDA, R. B. Traços da história econômica da Bahia no último século e meio. Revista de
desenvolvimento econômico, Salvador, n. 19, p. 82-101, 2009.
ALVERSON. W. S.; WHITLOCK, B. A.; NYFFELER, R.; BAYER, C.; BAUM, D. A.
Phylogeny of the core Malvales: evidence from ndhF sequence data. American Journal of
Botany, Lancaster, v. 89, p. 1474-1486, 1999.
ANDEBRHAN, T. Relação entre a idade do fruto do cacaueiro e suscetibilidade a
Crinipellis perniciosa. Belém: CEPLAC/ CEPEC, 1981. p. 315-317 (Informe Técnico).

34
ANDEBRHAN, T. Studies on the epidemiology and controlo f witches’ broom disease of
cocoa in the Brazilian Amazon. In: 9th Internacional Cocoa Research Conference.
Proceedings… Lome, 1984. p. 395-402.
ANDEBRHAN, T.; ALMEIDA, L. C.; NAKAYAMA, H. I. Resistência de Theobroma cacao
L. a Crinipellis perniciosa (Stahel) Singer: a experiencia da Amazônia Brasileira.
Agrotrópica, Itabuna, v. 10, p. 49-60, 1998.
ANDEBRHAN, T.; FIGUEIRA, A.; YAMADA, M. M.; CASCARDO, J.; FURTEK, D. B.
Molecular fingerprinting suggests two primary outbreaks of witches’ broom disease
(Crinipellis perniciosa) of Theobroma cacao in Bahia, Brazil. European Journal of Plant
Pathology, Dordrecht, v. 105, n. 2, p. 167-175, 1999.
AAM, B. B.; HEGGSET, E. B.; NORBERG, A. L.; SORLIE, M.; VARUM, K. M.; EIJSINK,
V. G. Production of chitooligosaccharides and their potential applications in medicine.
Marine Drugs, v. 8, p. 1482-1517, 2010.
CHEN, C. Y.; YANG, C. Y.; CHEN, A. H. Biosorption of Cu(II), Zn(II), Ni(II) and Pb(II)
ions by cross-linked metal-imprinted chitosans with epichlorohydrin. Journal of
Environmental Management, London, v. 92, p. 796-802, 2011.
ANDEBRHAN, T.; BASTOS, C. N. Variabilidade de isolados de Crinipellis perniciosa.
Belém: CEPLAC/ CEPEC, 1985. p. 51-56 (Informe Técnico).
ANDREWS, J. H.; HARRIS, R. F. The ecology and biogeography of microorganisms of plant
surfaces. Annual Review Phytopathology, Saint Paul, v. 38, p. 145-180, 2000.
ASCENSO, J. C. O seminário sobre seleção e melhoramento do cacaueiro da VIII conferência
internacional do cacau. Estudos Agronómicos, Lisboa, v. 3, n. 8, p. 135-141, 1962.
ASSIS, S. M. P.; MARIANO, R. L. R.; SILVA-HANLIN, D. M. W.; DUARTE, V. Mancha-
aquosa do melão causada por Acidovorax avenae subsp. citrulli no Estado do Rio Grande do
Norte. Fitopatologia Brasileira, Brasília, v. 24, n. 2, p. 191, 1999.
BAKER, R.E.D; CROWDY, S.H. Studies in the witches’ broom disease of cocoa caused by
Marasmius pernicious Stahel.: Introduction, symptons and a etiology. Mem Imperial
College Tropical Agriculture, Trinidad and Tobago, n. 7, p. 1-28, 1943.

35
BAKER, R. E. D.; HOLLIDAY, P. Witches' broom disease of cocoa (Marasmius perniciosus
Stahel). Phytopathological Papers, Oxon, n. 2, 1957.
BARTLEY, B. G. D. The genetic diversity of cacao and its utilization. Cambridge: CABI
Publishing, 2005. 341 p.
BARTLEY, B. G. D. Cacao, Theobroma cacao. FAO Plant Production and Protection
Paper, Rome, v. 70, p. 25-42, 1986.
BARTLEY, B. G. D. The status of genetic resistance in cocoa to Crinipellis perniciosa
(Stahel) Singer. In: INTERNATIONAL COCOA RESEARCH CONFERENCE, 6. 1977,
Caracas. Proceedings... Caracas: Alliance of Cocoa Producing Countries, 1977. p. 1-18.
BART-PLANGE, A.; BARYEH, E. A. The physical properties of Category B cocoa beans.
Journal of Food Engineering, Essex, v. 60, n. 3, p. 219-227, 2003
BASTOS, C. N. Resultados preliminares sobre a eficácia de Trichoderma viride no controle
da vassoura-de-bruxa (Crinipellis perniciosa) do cacaueiro. Fitopatologia Brasileira,
Brasília, v. 13, p. 340-342, 1988.
BASTOS, C. N. Comparação morfológica e patológica de isolados de Crinipellis perniciosa
(Stahel) Singer. Belém: CEPLAC/ CEPEC, 1986. p. 45-49 (Informe Técnico).
BASTOS, C. N. Patogenicidade e características do isolado de Crinipellis perniciosa de
Uruçuca, Bahia. Fitopatologia brasileira, Brasília, v. 15, p. 344-346, 1990.
BASTOS, C. N. Potential de Trichoderma viride no controle da vassoura-de-bruxa
(Crinipellis perniciosa) do cacaueiro. Fitopatologia Brasileira, Brasília, v. 21, p. 509-512,
1996.
BASTOS, C. N. Effect of water potential on growth and viability of conidia of Trichoderma
viride after storage at different temperatures. Summa Phytopathologica, Jaguariúna, v. 22, p.
258-261, 1996.
BASTOS, C. N. Mycoparasitic nature of the antagonism between Trichoderma viride and
Crinipellis perniciosa. Fitopatologia Brasileira, v. 21, n. 1, 1996.
BASTOS, C. N. Retrospectiva e avanços no controle da vassoura-de-bruxa do cacaueiro.
Fitopatologia Brasileira, v.25, p. 305-306, 2000.

36
BASTOS, C. N.; ANDEBRHAN, T. Urucu (Bixa orellana): nova espécie hospedeira da
vassoura-de-bruxa (Crinipellis perniciosa) do cacaueiro. Fitopatologia Brasileira, Brasília,
v. 11, p. 963-965, 1986.
BASTOS, C. N.; EVANS, H. C.; SAMSON, R. A. A new hiperparasitic fungus,
Cladobobryum amazonense, with potential for control of fungal pathogens of cocoa.
Transactions of the British Mycological Society, Cambridge, v. 77, p. 273-278, 1981.
BLAKEMAN, J. P. Foliar bacterial pathogens: epiphytic grown and interaction on leaves.
Journal of Applied Bacteriology, Oxford, v. 70, p. 49-50, 1991.
BEATTIE, G. A.; LINDOW, S. E. Bacterial colonization of leaves: a spectrum of strategies.
Phytopathology, Saint Paul, v. 89, n. 5, p, 353-359, 1999.
BEZERRA, J. L.; COSTA, J. C. B.; POMELA, A. W. V.; ALMEIDA, O. C. Como produzir
Tricovab para controlar a vassoura-de-bruxa do cacaueiro. Fitopatologia Brasileira, Brasília,
v.25, p.359, 2000.
BRUCE, A.; VERRALL, S.; HACKETT, C. A.; WHEATLEY, R. E. Identification of volatile
organic compounds (VOCs) from bacteria and yeast causing growth inhibition of sapstain
fungi. Holzforschung, Berlin, v.58, p.193-198, 2004.
BRASIL. Portal Brasil. Economia e emprego. Bahia retoma exportações de cacau
após vencer a vassoura-de-bruxa. Disponível em <http://www.brasil.gov.br/economia-e-
emprego/2015/11/bahia-retoma-exportacoes-de-cacau-apos-vencer-a-vassoura-de-
bruxa>. Acesso em: 26 jan. 2016.
CABRAL, A. S. Killer yeasts inhibit the growth of the phytopathogen Moniliophthora
perniciosa, the causal agent of witches’ broom disease. Brazilian Journal of Microbiology,
São Paulo, v. 40, p. 108-110, 2009.
CHAPMAN, R. K.; SORIA, S. J. Comparative Forcipomyia (Diptera, Ceratopogonidae)
pollination of cacao in Central America and Southern Mexico. Revista Theobroma, Itabuna,
v. 13, p. 129-139, 1983.
CHEESMAN, E. E. Notes on the nomenclature, classification and possible relationships of
cacao populations. Tropical Agriculture, Ibadan, v. 21, p. 144-159, 1944.
CHEESMAN, E. E.; POUND, F. J. Further notes on criterion of selection in cacao. Annual
Report on Cacao Research. Trinidad, n. 3, p. 21-24, 1934.

37
CHEN, Y.; YAN, F.; CHAI, Y.; LIU, H.; KOLTER, R.; LOSICK, R.; GUO, J. Biocontrol of
tomato wilt disease by Bacillus subtilis isolates from natural environments depends on
conserved genes mediating biofilm formation. Environmental microbiology, Oxford, v. 15,
n. 3, 2103.
COMISSÃO EXECUTIVA DO PLANO DA LAVOURA CACAUEIRA. Cacau:
História e Evolução. Disponível em: <http://www.ceplac.gov.br/radar/radar_cacau.htm>
Acesso em: 04 fev. 2016.
COSTA, J. C. B.; BEZERRA, J. L.; SANTOS FILHO, L. P.; ALVES, M. C.; MOURA, E. M.
Controle biológico da vassoura-de-bruxa do cacaueiro da Bahia, Brasil. In: W. BETTIOL; M.
A. B. MORANDI (Orgs.). Biocontrole de Doenças de Plantas: uso e perspectivas. Embrapa
Meio Ambiente, Jaguariúna, p. 245–266, 2009.
CALLE, H.; COOK, A. A.; FERNAND, S. Y. Histology of witches’ broom caused in cacao
by Crinipellis perniciosa. Phytopathology, Saint Paul, v. 72, p. 147-148, 1982.
CHEN, Y. C.; MILLIMAN, E. J.; GOULET, I.; COTE, J.; JACKSON, C. A.;
VOLLBRACHT, J. A.; YU, M. C. Protein Arginine Methylation Facilitates
Cotranscriptional Recruitment of Pre-mRNA Splicing Factors. Molecular and Cellular
Biology, Washington, v. 30, n. 21, p. 5245-5256, 2010.
COELHO, A. R.; NÓBREGA, G. M. A.; PAGNOCCA, F. C.; HOFFMANN, F. L.;
HARADA, K. I.; HIROOKA, E. Y. Avaliação do potencial antagônico de leveduras, visando
biocontrole de deterioração por Penicillium expansum. Semina Ciências Agrárias,
Londrina, v. 32, p. 1879-1892, 2011.
COMSTOCK, R. E.; ROBINSON, H. F. Estimation of overage dominance of genes. In:
GOWEN, J. W. Heterosis. Ames: Iowa State College, 1952. p. 494-516.
COSTA, J. C. B. Progresso da vassoura-de-bruxa em órgãos vegetativos do cacaueiro em
Altamira e Tomé- Açu, Pa. 1993. 52 f. Dissertação (Mestrado em Fitopatologia) -
Universidade Federal de Viçosa, Viçosa.
COSTA, J. C. B. et al. Controle biológico da vassoura-de-bruxa do cacaueiro da Bahia,
Brasil. In: BETTIOL, W. Biocontrole de doenças de plantas: uso e perspectivas. Jaguariúna:
Embrapa meio ambiente, 2009. p. 245-266.
CRUZ, C. D.; REGAZZI, A. J.; CARNEIRO, P. C. S. Modelos biométricos aplicados ao
melhoramento genético. Viçosa, MG: UFV, 2004. Cap. VII, p. 223-375.

38
CUATRECASAS, J. Cacao and its allies: a taxonomic revision of the genus Theobroma.
Contributions from the United States National Herbarium. Washington, n. 35, p. 379-
614, 1964.
DANTAS NETO, A. et al. Caracterização de uma população de cacaueiro para mapeamento
de genes de resistência à vassoura-de-bruxa e podridão-parda. Fitopatologia Brasileira,
Brasília, v. 30, p. 380-386, 2005.
DELGADO, J. C.; COOK, A. A. Nuclear condition of the basidia, basidiospores, and
mycelium of Marasmius perniciosus. Canadian. Journal.of Botany, Ottawa, v. 54, p. 66-72,
1976.
DROBY, S.; COHEN, L.; DAUS, A.; WEISS, B.; HOREV, E.; CHALUTZ, E.; KATZ, H.;
KEREN-TZOUR; M.; SHACHNAI, A. Commercial testing of Aspire: a biocontrol
preparation for the control of postharvest decay of citrus. Biological Control, Orlando, v.12,
p.97-101, 1998.
DROBY, S., VINOKUR, V., WEISS, B., COHEN, L., DAUS, A., GOLDSCHMIDT, E. E.,
PORAT, R. Induction of resistance to Penicillium digitatum in grapefruit by the yeast
biocontrol agent Candida oleophila. Phytopathology, Saint Paul, v. 92, p. 393-399, 2002.
DI PIERO, R. M.; GARCIA JUNIOR, D.; TONUCCI, N. M. Indutores bióticos. In:
ALVALCANTI, L. S.; DI PIERO, R. M.; CIA, P.; PASCHOLATI, S. F.; RESENDE, M.
L.V.; ROMEIRO, R. da S. (Eds.). Indução de resistência em plantas a patógenos e insetos.
Piracicaba: FEALQ, 2005. p. 29-50.
__________. Origem e dispersão de Theobroma cacao L.: um novo cenário. In: DIAS, L. A.
S (editor). Melhoramento genético do cacaueiro. Viçosa, MG: FUNAPE, UFG, 2001a, p.
81-127.
DIAS, L. A. S. Contribuições do melhoramento. In: DIAS, L. A. S. (editor) Melhoramento
genético do cacaueiro. Viçosa, MG: FUNAPE, UFG, 2001b, p. 493-529
ELAD Y. Biocontrol of foliar pathogens: mechanisms and application. Communications in
Agricultural and Applied Biological Sciences, v. 68, p. 17-24, 2003
EL-MEHALAWY, A. A. et al. Influence of maize root colonization by the rhizosphere
actinomycetes and yeast fungi on plant growth and on the biological control of late wilt
disease. International journal of agriculture and biology, Beijing, v. 6, n. 4, 599-605,
2004.

39
EL-TARABILY, K. A. Suppression of Rhizoctonia solani diseases of sugar beet by
antagonistic and plant growth-promoting yeasts. Journal of Applied Microbiology, Oxford,
v. 96, p. 69-75, 2004.
EL-TARABILY, K. A.; SIVASITHAMPARAM, K. Potential of yeasts as biocontrol agents
of soil-borne fungal plant pathogens and as plant growth promoters. Mycoscience, Tokyo, v.
47, p. 25-35, 2006.
EVANS, H. C. Pleomorphism in Crinipellis perniciosa causal agent witches’broom disease of
cocoa. Transactions of the British Mycological Society, Cambridge, v. 74 p. 515-523, 1980.
EVANS, H.C.; BASTOS, C.N. Uma reavaliação do ciclo de vida da vassoura de bruxa,
Crinipellis perniciosa. Fitopatologia Brasileira, Brasília, v.4, p.104, 1979.
EVANS, H. C.; SOLORZANO, G. R. Witches’s broom disease:wrong experimente right
results. In International Cocoa Research Conference, 8., Cartagena, Colombia, 1982.
Proceeedings...Lagos: Cocoa Producers Alliance, 1982. p. 415-318.
FALEIRO, F. G. et al. Mapping QTLs for witches’broom (Crinipellis perniciosa) resistance
in cacao. Euphytica, Wageningen, v. 149, p. 227-235, 2006.
FALEIRO, F. G.; JUNQUEIRA, N. T. V.; BELLON, G.; BORGES, T.A.; ANJOS, J. R. N.;
PEIXOTO, J. R.; BRAGA, M. F.; SANTOS, D. G. Diversidade genética de espécies silvestres
de maracujazeiro com resistência múltipla a doenças com base em marcadores RAPD.
Fitopatologia Brasileira, Brasília, v. 29, p. 325, 2004. Suplemento.
FAO. Prodution Yearbook. v. 54. Roma. 2002.
FRIAS, G. A.; PURDY, L. H.; SCIMIDT, R. A. Infection biology of Crinipellis perniciosa
on vegetable flushes of cacao. Plant Disease, Sain Paul, v. 75, n. 6, p.552-556, 1991.
FUENTEFRIA, A. M.; FAGANELLO, J.; PAZZINI, F.; SCHRANK, A.; VALENTE, P.;
VAINSTEIN, M. Typing and patterns of cellular morphological alteration in Cryptococcus
neoformans and Cryptococcus gatti isolates exposed to a panel of killer yeats. Medical
Mycology, Oxford, v. 45, p. 1-10, 2007.
GALANTE, R. S. Estudos de quitinases de Moniliophthora perniciosa (Stahel) Aime &
Phillips-Mora, fungo causador da vassoura-de-bruxa no cacaueiro: purificação,
caracterização e modelagem comparativa. 2008. 90 f. Dissertação (Mestrado em
Biotecnologia). Universidade Estadual de Feira de Santana, 2008.

40
GERALDI, I. O.; MIRANDA FILHO, J. B. Adapted models for the analysis of combining
ability of varieties in partial diallel crosses. Revista Brasileira de Genética, Ribeirão Preto,
v. 11, n. 2, p. 419-430, 1988.
GOHEL, V.; SINGH, A.; VIMAL, M.; ASHWINI, P.; CHHATPAR, H. S Bioprospecting
and antifungal potential of chitinolytic Microorganisms. African Journal Biotechnology,
Nairobi, v. 5, p. 54-72, 2006.
GOUVEIA, A. Controle em campo de pó-colheita de doenças e metabolismo do
morangueiro após tratamento com Saccharomyces cerevisiae. 2007. 85 f. Tese (Doutorado
em Agronomia) - Universidade Federal do Paraná, Curitiba.
GIARETTA R. D.; FREITAS, L. G.; XAVIER, D. M.; ZOOCA, R. J. F.; FERRAZ, S.;
LOPES, E. A. Incorporação ao solo de substrato contendo micélio e conídios de Pochonia
chlamydosporia para o manejo de Meloidogyne javanica. Ciência Rural, Santa Maria, v. 44,
n. 4, p. 629-633, 2014.
GRIFFING, B. A concept of general and specific combining ability in relation to diallel
crossing systems. Australian Journal of Biological Sciences, East Melbourn, v. 9, p. 463-
493, 1956.
HANCOCK, B. L. Producción y transporte del haba de cacao. In: BECKETT, S. T. (Ed.).
Fabricación y utilización industrial del chocolate. Zaragoza: Acribia, 1994. p. 9-32
HAYMAN, B. I. The theory and analysis of diallel crosses. Genetics, Bethesda, v. 39, p. 789-
809, 1954.
HOLLIDAY, P. Witches’s broom disease of cacao (Marasmius pernicious Stahel). London.
Her Majesty’s Stationery Office (HMSO). 1952.
HU, H.; XU, Y.; LU, H.; XIAO, R.; ZHENG, X.; YU, T. Evaluation of yeasts from Tibetan
fermented products as agents for biocontrol of blue mold of Nashi pear fruits. Journal of
Zhejiang University-SCIENCE B (Biomedicine & Biotechnology), Hangzhou, v. 16, n. 4,
275-285, 2015.
HAISSAM, J.M. Pichia anomala in biocontrol for apples: 20 years of fundamental research
and practical applications. Antonie Van Leeuwenhoek, Amsterdam, v. 99, n. 1, p. 93-105,
2011.

41
ICCO. International Cocoa Organization. Produção mundial de cacau. Disponível em:
<http://www.icco.org/>. Acesso em 16 fev. 2016.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA (IBGE). Sistema IBGE de
Recuperação Eletrônica (SIDRA). 2014. Disponível em:
http://www.sidra.ibge.gov.br/bda/agric/. Acesso em: 26 jan. 2016.
INACIO, J.; PORTUGAL, L.; MARTINS, I. S.; FONSECA, A. Phylloplane yeasts from
Portugal: seven novel anamorphic species in the Tremellales lineage of the Hymenomycetes
(Basidiomycota) producing orange-coloured colonies. FEMS Yeast Research, Oxford, v. 5,
p. 1167-1183, 2005.
KAMAL A. M. et al. Biological control of Fusarium wilt in tomato by plant growth
promoting yeasts and rhizobacteria. Plant Pathology Journal, v. 25, p. 199-204, 2009.
KILARU, A.; HASENSTEIN, K. H. Development and pathogenicity of the fungus Crinipellis
perniciosa on interaction with cacao leaves. Phytopathology, Saint Paul, v. 95, p. 101–107,
2005.
KANG, B. G.; KIM, W. T.; YUN, H. S.; CHANG, S. C. Use of plant growth-promoting
rhizobacteria to control stress responses of plant roots. Plant Biotechnology Reports, New
York, v. 4, p. 179-183, 2010.
KIM, K. S.; YUN, H. S. Production of soluble β-glucan from the cell wall of Saccharomyces
cerevisiae. Enzyme and Microbial Technology, New York, v. 39, n. 3, p. 496-500, 2006.
LACHANCE, M. A.; STARMER, W. T. Ecology and yeasts. In: KURTZMAN, C. P.; FELL,
W. F. (Eds.). The Yeast, a Taxonomic Study. 4ª. ed. Amsterdam: Elsevier, 1998, p. 21-30.
LACHENAUD, P.; ZHANG, D. Genetic diversity and population structure in wild stands of
cacao trees (Theobroma cacao L.) in French Guiana. EDP Sciences, Montpellier, v. 65, n.
310, p. 1-7, 2008.
L'ÊCLUSE, C. Rariorum plantarum historia. 1601. Disponível em: <http://caliban.mpiz-
koeln.mpg.de/~stueber/ecluse/index.html>. Acesso em: 14 jan. 2016.
LOGUERCIO, L. L. et al. Controle biológico das doenças do cacaueiro. In: VALLE, R. R.
(Ed.). Ciência, Tecnologia e Manejo do Cacaueiro. 2ª. ed. Brasília: CEPLAC, 2012. p. 11-
30.

42
LOGUERCIO, L. L.; CARVALHO, A. C.; NIELLA, G. R.; SOUZA J. T.; POMELLA, A.
W. V. Selection of Trichoderma stromaticum isolates for efficient biological control of
witches’ broom disease in cacao. Biological Control, Orlando, v. 51, p. 130-139, 2009.
LOPES, U. V.; et al. Cacao breeding in Bahia, Brazil - strategies and results. Crop breeding
and applied biotechnology, Londrina, p. 73-81, 2011.
LUZ, E. D. M. N. et al. Evaluating resistance/tolerance to Witches' Broom disease in
Bahia, Brazil. Final Report to ACRI. CEPLAC/CEPEC, p. 56, 1997.
LUZ, E. D. M. N.; ALMEIDA, H. A.; MACHADO, R. C. R. Períodos de incubação,
secamento de vassouras, produção de basidiomas e atividade de Crinipellis perniciosa em
ramos e frutos de cacaueiro na Bahia. In: CONGRESSO BRASILEIRO DE
FITOPATOLOGIA, 27. 1994. Itajaí. Anais... Itajaí: Sociedade Brasileira de Fitopatologia,
1994. p. 341-341.
LUZ, E. D. M. N. et al. Vassoura-de-bruxa do cacaueiro: novos enfoques sobre uma velha
doença. Revisão Anual de Patologia de Plantas, Passo Fundo, v. 14, p. 59-111, 2006.
LUZ, E. D. M. N. et al. Atualidades no manejo de doenças do cacaueiro. In: Patologia
Florestal: desafios e perspectivas. NEFIT: Suplema, p. 313-329, 2013.
LEAL, J. B.; SANTOS, L. M.; SANTOS, C. A. P.; PIRES, J. L.; AHNERT, D.; CORRÊA, R.
X. Diversidade genética entre acessos de cacau de fazendas e de banco de germoplasma na
Bahia. Pesquisa Agropecuária Brasileira, Brasília, v. 43, n. 7, 2008.
LOPES, R. B. M.; COSTA, L. E. O; VANETTI, M. C. D.; ARAÚJO, E. F.; QUEIROZ, M.
V. Endophytic bacteria isolated from common bean (Phaseolus vulgaris) exhibiting high
variability showed antimicrobial activity and quorum sensing inhibition. Current
microbiology, New York, v. 71, n. 4, p. 509-516, 2015.
LAHLALI, R.; HAMADI, Y.; EL GUILLI, M.; JIJAKLI, M. H. Efficacy assessment of
Pichia guilliermondii strain Z1, a new biocontrol agent, against citrus blue mould in Morocco
under the influence of temperature and relative humidity. Biological Control, Orlando, v. 56,
p.:217-242011
LYNCH, M.; WALSH, B. Genetics and analysis of quantitative traits. Sinauer Associates,
Inc. Sunderland 1998.
MADIGAN, M. T.; MARTINKO, J. M.; PARKER, J. Microbiologia. São Paulo: Pearson
Education, 2004. 608 p.

43
MASIH, E. I.; PAUL, B. Secretion of beta-1,3-glucanase by the yeast Pichia
membranifaciens and its possible role in the biocontrol of Botrytis cinerea causing mold
disease of the grapevine. Current Microbiology, New York, v. 44, p. 391-395, 2002.
McGEARY, F. M.; WHEELER, B. E. J. Growth rates of, and mycelial interactions, between
isolates of Crinipellis perniciosa from cocoa. Plant Pathology, Saint Paul, v. 37, p. 489-498,
1988.
MEDEIROS, F. H. V. et al. A novel, integrated method for management of witches’broom
disease in Cacao in Bahia, Brazil. Crop Protection. Guildford, n. 29, p. 704-711, 2010.
MIDLEJ, R. R.; SANTOS, A. M. Economia do Cacau. In: VALLE, R. R. (Ed.). Ciência,
Tecnologia e Manejo do Cacaueiro. 2ª. ed. Brasília: CEPLAC, 2012. p. 655-672.
MONTEIRO, W. R. et al. Population breeding activities in Brazil. In: ESKES, A. B.;
EFRON, Y. (Org.). Global approaches to cocoa germplasm utilization and conservation.
1 ed.: CFC/OCCO/IPGRI, v. 1, p. 28-34, 2006.
MONTEIRO, W. R.; AHNERT, D. Melhoramento Genético do Cacaueiro. In: VALLE, R.
R. (Ed.). Ciência, Tecnologia e Manejo do Cacaueiro. 2ª. ed. Brasília: CEPLAC, 2012. p.
11-30.
MOTILAL L; BUTLER D. Verification of identities in global cacao germplasm collections.
Genetic Resources and Crop Evolution, Dordrecht, v. 50, p. 799–807, 2003.
NAKAYAMA, L. H. I.; SOARES, M. K. M.; DA APPEZZATO-DA-GLÓRIA, B.
CONTRIBUIÇÃO AO ESTUDO ANATÔMICO DA FOLHA E DO CAULE DO
CACAUEIRO (Theobroma cacao L.) Scientia Agricola, Piracicaba, v. 53, n. 1, 1996.
NIELLA, G. R. Esporulação de Crinipellis perniciosa (Stahel) Singer em frutos de cacau
no Sudeste da Bahia e sensibilidade “in vitro” a quatro compostos sulfurados. 1997. 60
f. Dissertação (Mestrado em Fitopatologia) - Universidade Federal de Lavras, Lavras.
NIELLA, G. R.; RESENDE, M. L. V.; CASTRO, H. A.; CARVALHO, G. A.; SILVA, L. H.
C. P. Aperfeiçoamento da metodologia de produção artificial de basidiocarpos de Crinipellis
perniciosa. Fitopatologia Brasileira, Brasília, v. 24, p. 523-527, 1999.
ODJAKOVA M. E.; HADJÜVANOVA, C. The complexity of pathogens defense in plants.
Journal of plant physiology, Stuttgart, v. 27, n. 12, p. 101-109, 2001.

44
OLIVEIRA, M. L. Eficácia de fungicidas triazóis no controle da vassoura-de-bruxa do
cacaueiro causada por Crinipellis perniciosa. Fitopatologia Brasileira, Brasília, v. 29, p.
149, 2004a.
__________. Estrobilurinas: novo grupo de fungicidas com eficácia contra a vassoura-de-
bruxa do cacaueiro. Fitopatologia Brasileira, Brasília, v. 29, p. 285, 2004b.
OLIVEIRA, M. L.; LUZ, E. D. M. N. Vassoura-de-bruxa. In:__________. Identificação e
manejo das principais doenças do cacaueiro no Brasil. Ilhéus, CEPLAC/CEPEC/SEFIT,
2005, p. 1533.
PAIM, V. R. L. et al. Sources of resistance to Crinipellis perniciosa in progenies of cacao
accessions collected in the Brazilian Amazon. Scientia Agricola, Piracicaba, v. 63, n. 6, p.
572-578, 2006.
PAIM, V. R. L. M. Estudo da diversidade genética de novas fontes de resistência à
vassoura-de-bruxa do cacaueiro. 2005. Dissertação (Mestrado em Genética e Biologia
Molecular) - Universidade Estadual de Santa Cruz, Ilhéus.
PEGUS, J. E. Aspects of the host-parasite relationship in the Theobroma cacao
L./Marasimus perniciosus Stahel disease complex. 1972. Dissertação (Mestrado) -
University of West Indies, Trinidad.
PEREIRA, J. L.; VALLE, R. R. Manejo Integrado da vassoura de bruxa do cacaueiro. In:
VALLE, R.R. Ciência, tecnologia e manejo do cacaueiro. 2ª. ed. Brasília: CEPLAC, 2012
p. 11-30, 2012.
PEREIRA J. L.; RAM, A.; FIGUEIREDO J. M; ALMEIDA L. C. C. Primeira ocorrência de
vassoura-de-bruxa na principal região produtora de cacau do Brasil. Agrotrópica, Itabuna, v.
1, p.79-81, 1989.
PINTO, L. R. M.; PIRES, J. L. Seleção de plantas de cacau resistentes à vassoura-de
bruxa. Ilhéus: CEPLAC/ CEPEC, 1998. 181 p. (Boletim Técnico).
PIRES, J. L. Avaliação quantitativa e molecular de germoplasma para o melhoramento
do cacaueiro com ênfase na produtividade, qualidade do fruto e resistência a doenças.
2003. Tese (Doutorado em Melhoramento de Plantas) - Universidade Federal de Viçosa,
Viçosa.

45
PIRES, J. L.; MONTEIRO, W. R.; LUZ, E. D. M. N.; SILVA, S. D. V. M.; PINTO, L. R. M.;
FIGUEIRA, A.; GRAMACHO, K. P.; LOPES, U. V.; AlBUQUERQUE, P. S. B.;
YAMADA, M. M.; AHNERT, D. E.; BRUGNEROTTO, M. I. B.; Cocoa breeding for
Witches´ broom resistance at CEPEC, Bahia, Brazil. In: INTERNATIONAL WORKSHOP
ON THE CONTRIBUTION OF DISEASE RESISTANCE TO COCOA VARIETY
IMPROVEMENT, 1999. Salvador. Proceedings... Salvador: INGENIC, UK. p. 91-101.
PHILLIPS-MORA, W.; AIME, M. C.; WILKINSON, M. J. Biodiversity and biogeography of
the cacao (Theobroma cacao) pathogen Moniliophthora roreri. Plant Pathology, London, v.
56, p. 911-922, 2007.
QUEIROZ, V. T. et al. Identification of a major QTL in cocoa (Theobroma cacao L.)
associated with resistance to witches’ broom disease. Plant Breeding, Berlin, v. 122, n. 3, p.
268-272, 2003.
REHM, H. J. et al. Biotechnology a multi-volume comprehensive treatise, 2ª ed. vol.9.
Enzymes, biomass, food and feed. G. REED; T. W. NAGODAWITHANA. (Eds.). 1995. Cap.
5.
ROCHA, H. M. et al. Wiches’ broom in Bahia Brazil. In: RUDGARD, S. A. et al. (Eds.).
Disease Management in cocoa: comparative epidemiology of witches’ broom. London:
Chapman and Hall, p. 189-200, 1993.
ROCHA, H. M.; WHEELER, B. E. J. Factor influencing production of basidiocarps and the
deposition and germination of basidiospores of Crinipellis perniciosa, the causal fungus of
witches'broom on cocoa (Theobroma cacao). Plant Pathology, Oxford, v. 34, p. 319-328,
1985.
ROSA, M. M.; TAUK-TORNISIELO, S. M.; RAMPAZZO, P. E.; CECCATO-ANTONINI,
S. R. Evaluation of the biological control by the yeast Torulaspora globosa against
Colletotrichum sublineolum in sorghum. World Journal of Microbiology and
Biotechnology, Oxford, v. 26, n. 8, p. 1491-1502, 2010.
ROBIGLIO, A.; SOSA, M. C.; LUTZ, M. C.; LOPES, C. A. SANGORRÍN, M. P. Yeast
biocontrol of fungal spoilage of pears stored at low temperature. International Journal of
Food Microbiology, Amsterdam, v.1 47, n.3, p.211-216, 2011
SAMUELS, G. J. et al. Trichoderma stromaticum sp. nov., a parasite of the cacao witche’s
broom pathogen. Mycological research, Cambridge, v. 104, p. 760-764, 2000.
SANTOS, L. S. Interação antagônica Trichoderma stromaticum - Crinipellis perniciosa:
efeito do clima no biocontrole da vassoura de bruxa do cacaueiro e desenvolvimento de

46
marcadores microssatélites. 2005. Dissertação (Mestrado em Genética e Biologia
Molecular). Universidade Estadual de Santa Cruz, Ilhéus.
SANTOS, R. M. F. et al. Marcadores microssatélites relacionados com a resistência à
vassoura-de-bruxa do cacaueiro. Pesquisa Agropecuária Brasileira, Rio de Janeiro, v. 42, p.
1137-1142, 2007.
SAS Institute Inc (1989) SAS/STAT User´s Guide, version 6.4 ed. Cary, SAS Institute, North
Caroline State University. 846 p.
SCARPARI, L. M. et al. Biochemical changes during the development of witches’ broom: the
most important disease of cocoa in Brazil caused by Crinipellis perniciosa. Journal
Experimental Botany, Oxford, p. 113, 2005.
SCHENBERG, A. C. G. Biotecnologia e Desenvolvimento Sustentável. Estudos Avançados,
São Paulo, v. 24, n. 70, 2010.
SHALABY, M. E.; EL-NADY, M. F. Application of Saccharomyces cerevisiae as a
biocontrol agent against Fusarium infection of sugar beet plants. Acta Biologica
Szegediensis, Szeged, v. 52, n. 2, p. 271-275, 2008.
SILVA, S. D. V. M. et al. Parent selection for cocoa resistance to witche' s broom. Pesquisa
Agropecuária Brasileira, Rio de Janeiro, v. 45, p. 680-685, 2010.
SILVA, S. D. V. M. et al. Redescrição da sintomatologia causada por Crinipellis
perniciosa em cacaueiro. Agrotrópica, Itabuna, v. 14, n. 1, p. 1-28, Edição especial, 2002.
SILVA, S. D. V. M. Histologia e seleção de variáveis para avaliar resistência de
cacaueiro a Crinipellis perniciosa. 1997. Tese (Doutorado em Fitopatologia) - Universidade
Federal de Viçosa, Viçosa.
SILVA, S. D. V. M.; LUZ, E. D. M. N.; PIRES, J. L.; YAMADA, M. M.; SANTOS FILHO,
L. P. Cacao resistance to Crinipellis perniciosa: diallelic crosses increase genes for
resistance. In: 8th International Congress of Plant Pathology, Christchurch Addendum to
Abstracts of Offered Papers, Christchurch, p. 8, 2003.
SILVA NETO, P. J. et al. Sistema de produção de cacau para a Amazónia brasileira.
Belém, CEPLAC, 2001. 125 p. ISSN 0102-5511

47
SINGER, R. A monographic study of the genera “Crinipellis” and “Chaetocalathus”. Lilloa,
Tucuman, v. 8, p. 441-514, 1942.
SORIA, J. S. de; TONOSAKI, S.; MORENO, J. A polinização do cacaueiro ao vivo. Cacau
Atualidades, Itabuna, n. 3, v. 12, p. 14-18, 1975.
SOUNIGO, O.; LACHENAUD, P.; BASTIDE, P.; CILAS, C.; N'GORAN, J.; LANAUD, C.
Assessment of the value of doubled haploids as progenitors in cocoa (Theobroma cacao L.)
breeding. Journal Applied Genetics, v. 44, p. 339-353, 2003.
SREENIVASAN, T. N., DABYDEEN, S. Modes of penetration of young cocoa leaves by
Crinipellis perniciosa. Plant Disease, Saint Paul, v. 73, n. 6, p. 478-481, 1989.
SOUZA, C. A. S.; DIAS, L. A. S. Melhoramento ambiental e sócio-economia. In: DIAS, L.
A. S (editor). Melhoramento genético do cacaueiro. Viçosa: FUNAPE-UFG, 2001. p. 1-47.
SOUZA, J. T.; POMELLA, A. W. V.; BOWERS, J. H.; PIROVANI, C. P.; LOGUERCIO, L.
L.; HEBBAR, K. P. Genetic and biological diversity of Trichoderma stromaticum, a
mycoparasite of the cacao witches’-broom pathogen. Proceedings of the American
Phytopathological Society, Saint Paul, v. 96, n. 1, p. 61-67, 2006.
SPADARO, D.; GULLINO, M. L. State of the art and future prospects of the biological
control of postharvest fruit diseases. International Journal Food Microbiology, Amsterdan,
v. 91, p. 185-194, 2004.
STADLER, B.; MULLER, T. Aphid honeydew and its effect on the phyllosphere microflora
of Picea abies (L.) Karst. Oecologia, Berlin, v. 108, p. 771-776, 1996.
STAHEL, G. Marasmius perniciosus nov.spec.: the cause of the krulloten-disease of cacao in
Suriname. Bulletin, van den Landbouw, n. 33, p. 1-26, 1915.
SUI, Y.; WISNIEWSKI, M.; DROBY, S.; LIUA, J. Responses of yeast biocontrol agents to
environmental stress. Applied and environmental microbiology, Washington, v. 81, n. 9, p.
2968-2975 2015.
TIAN, S.; QIN, G.; XU, Y.; WANG, Y. Application of antagonistic yeasts under field
conditions and their biocontrol ability against postharvest diseases of sweet cherry. Acta
Botanica Sinica, Peiping, v. 46, n. 11, p. 1324-1330, 2004.
TORTORA, G. J.; FUNKE, B. R.; CASE, C. L. Microbiologia. 8. ed. Porto Alegre: Artmed,
2005.

48
TOXOPEUS, H. Cacao (Theobroma cacao L.). In: FERWERDA, F. P; WIT, F. Outlines of
perennial crop breeding in the tropics. Wageningen: Veenman and Zonen, 1969. p. 79-108.
TOXOPEUS, H. Cacao breeding: a consequence of mating system, heterosis and population
structure. In: International Cocoa Research Conference, 4th, 1972. Saint Augustine.
Proceedings…Port-of-Spain: Governnment of Trinidad and Tobago, 1972. p. 3-12.
TREVIZAN, S. D. P.; MARQUES, M. Impactos socioeconômicos da crise do cacau: um
estudo de comunidade-caso. Agrotrópica, Itabuna, v. 14, n. 3, p. 127-136, 2002.
URQUHART, E. J.; PUNJA, Z. K. Hydrolytic enzymes and antifungal compounds produced
by Tilletiopsis species, phyllosphere yeasts that are antagonists of powdery mildew fungi.
Canadian Journal of Microbiology, Ottawa, v.48, n.3, p.219-229, 2002.
VALDEBENITO-SANHUEZA, R. M. V. Leveduras para o biocontrole de fitopatógenos. In:
MELO, I. S; AZEVEDO, J. L. (Eds.). Controle biológico. Jaguariúna: Embrapa Meio
Ambiente. p. 41-56, 2000.
VELLO, F.; GARCIA, J. R. Características das principais variedades de cacau cultivadas na
Bahia. Revista Theobroma, Itabuna, v. 1, p. 3-10, 1971.
VELLO, F.; NASCIMENTO, I. F. Influência da origem do pólen na produção do cacaueiro.
Revista Theobroma, Itabuna, v. 1, p. 7-14, 1971.
VERO, S.; MONDINO, P.; BURGAENO, J.; SOUBES, M.; WISNIEWSKI, M.
Characterization of biological activity of two yeast strains from Uruguay against blue mold of
apple. Postharvest Biology and Technology, Amsterdam, v. 26, p. 91-98, 2002.
VIANA, P. T.; PIRES, A. J. V.; OLIVEIRA, L. B.; CARVALHO, G. G. P.; RIBEIRO, L. S.
O.; CHAGAS, D. M. T.; NASCIMENTO FILHO, C. S.; CARVALHO, A. O. Fracionamento
de carboidratos e de proteína das silagens de diferentes forrageiras. Revista Brasileira de
Zootecnia, Viçosa, v. 41, n. 2, p. 292-297, 2012.
WALTERS, D. et al. Induced resistance for plant disease control: maximizing the efficacy of
resistance elicitors. Phytopathology, Saint Paul, v. 95, p. 1368-1373, 2005.
WACHOWSKA, U.; KUCHARSKA, K.; JEDRYCZKA, M.; ŁOBIK, N. Microorganisms as
biological control agents against Fusarium pathogens in winter wheat. Polish Journal of
Environmental Studies, Olsztyn, v. 22, n. 2, p. 591-597, 2013.

49
WALKER, V. K.; RANCOURT, D. E.; DUNCKER, B. P. The transfer of fish antifreeze
genes to Drosophila: A model for the generation of transgenic beneficial insects. Proceedings
of the Entomological Society of Ontario, Ontario, v. 126, p. 3-13, 1995.
WHEELER, B. E. J.; MEPSTED, R. Pathogenic variability amongst isolates of Crinipellis
perniciosa from cocoa (Theobroma cacao). Plant Pathology, Saint Paul, v. 37, p. 475-488.
1988.
YAMADA, M. M. et al. Diversidade genética de acessos de cacaueiro da série CEPEC por
meio de marcadores RAPD. Agrotrópica, Itabuna, v. 14, n. 3, p. 137-140, 2002.
ZANARDO, N. M. T.; PASCHOLATI, S. F.; FIALHO, M. B. Resistência de plântulas de
pepineiro a Colletotrichum lagenarium induzida por frações de extrato de Saccharomyces
cerevisae. Pesquisa Agropecuária Brasileira, Rio de Janeiro, v. 44, n.11, p. 1499-1503,
2009.
ZHANG, H. Y. et al. Postharvest biological control of gray mold decay of strawberry with
Rhodotorula glutinis. Biological Control, Orlando, v. 40, p. 287-292, 2007.
ZHAO, K.; SUN, L.; MA, X.; LI, X.; WANG, X.; PING, W.; ZHOU, D. Improved taxol
production in Nodulisporium sylviforme derived from inactivated protoplast fusion. African
Journal of Biotechnology, v.10, n. 20, p. 4175-4182, 2011.

50
CAPÍTULO 2
Seleção de progênies de cacaueiro quanto a resistência à vassoura de bruxa e monilíase

51
Seleção de progênies de cacaueiro quanto a resistência à vassoura de bruxa 1
2
Antônio Alves Pimenta Neto(1), José Luiz Pires(2), Edna Dora Martins Newman Luz(2) 3
4
(1)Universidade Federal Rural de Pernambuco, Rua Dom Manuel Medeiros, s/n, Dois Irmãos, 5
CEP 52171-900, Recife, PE, Brasil. Email: [email protected] (4)Comissão 6
Executiva do Plano da Lavoura Cacaueira, Centro de Pesquisas do Cacau, Rodovia Ilhéus-7
Itabuna, Km 22, s/n, CEP 45660-000, Ilhéus, BA, Brasil. Email: [email protected], 8
10
Resumo - Objetivando a ampliação e a estabilidade do caráter resistência à vassoura de bruxa 11
em progênies de cacaueiro, e a incorporação de genes de resistência à monilíase, genótipos 12
pertencentes a uma população estruturada, foram selecionados em avaliação a campo, para 13
compor o segundo ciclo de seleção, e suplementados com doadores de genes de resistência a 14
monilíase pelos clones EET 233, EET 75, ICS 95 e UF 273. Cruzamentos dialélicos (10x8) 15
foram realizados com delineamento Carolina do Norte II. Vinte dias após a emergência, 56 16
plântulas de cada cruzamento (4 repetições de 14 plântulas) foram inoculadas com uma 17
suspensão de 2 x 105 basidiósporos/mL de Moniliophthora perniciosa depositadas no 18
meristema apical. Aos 60 dias da inoculação, as plântulas foram avaliadas quanto aos 19
sintomas da VB, calculado o índice de doença (ID) e os dados corrigidos para o efeito de 20
ensaio e de repetição por ensaio. A análise dos contrastes entre as progênies e os demais 21
procedimentos foram realizados utilizando o pacote estatístico LS Means SAS. Dos 59 22
cruzamentos avaliados, todas as progênies geradas, a exceção de uma, diferiram 23
significativamente (p<0.05) de Catongo e 52 delas de SIC 23, padrões de suscetibilidade. O 24
ID médio em progênies de nove cruzamentos diferiu significativamente (p<0,05) do padrão 25
de resistência SCA 6, demonstrando alta a resistência à VB. Houve destaque para os 26
descendentes diretos de materiais oriundos da Amazônia Brasileira (CAB) e clones portadores 27
de resistência a monilíase, a exceção do clone ICS 95 que gerou progênies com ID médio 28
similar à testemunha suscetível. São apresentadas progênies que carregam genes de 29
resistência à VB oriundos de fontes diversas, e seis oriundas de parentais portadores de genes 30
de resistência à monilíase. 31
32
Palavras Chave: Melhoramento genético, Theobroma cacao, Moniliophthora perniciosa, 33
Moniliophthora roreri, seleção recorrente. 34

52
35
Introdução 36
37
O cacaueiro, espécie arbórea, nativa das florestas equatoriais da América do Sul, pertencente 38
à ordem Malvales, família Malvaceae e gênero Theobroma (Alverson et al. 1999), é cultivada 39
e explorada comercialmente para produção de chocolate, sucos, geleia, licores, sorvete e 40
fortemente pela indústria de cosméticos nas Américas do Sul e Central, África e Ásia. O 41
cacaueiro é um dos cultivos perenes mais importantes do mundo, com uma produção mundial 42
estimada em 4,2 milhões de toneladas de sementes secas (ICCO 2015), com cerca de 70% das 43
sementes produzidas no mundo, oriundas do leste da África, com destaque para a Costa do 44
Marfim (40%), Gana (20%), Nigéria (5%) e Camarões (5%) (Lopes et al. 2011). O Brasil, já 45
foi o segundo maior produtor de cacau do mundo, mas após a introdução do fungo causador 46
da vassoura de bruxa, Moniliopththora (ex. Crinipellis) perniciosa na região sul da Bahia em 47
1989 (Pereira et al, 1989), maior região produtora do país, caiu para a quinta posição no 48
ranking dos maiores produtores de cacau (ICCO 2015). 49
A VB é considerada a principal doença do cacaueiro no Brasil. Entretanto, existem 50
outras doenças com igual ou maior poder devastador que ainda não foram relatadas no Brasil, 51
e representam uma ameaça potencial à cultura caso sejam introduzidas e encontrem condições 52
favoráveis ao seu estabelecimento. A monilíase (Moniliophthora roreri (Cif.) H.C. Evans, 53
Stalpers, Samson e Benny) é uma doença de grande importância nos países da América do Sul 54
e Central onde ocorre, sendo limitante à expansão da cacauicultura (Albuquerque et al. 2005). 55
No Brasil, embora ainda não tenha sido detectada, apresenta-se como uma séria ameaça uma 56
vez que levantamentos realizados na década atual constataram a doença na fronteira do Peru 57
com o Brasil (Moraes et al. 2012). 58
Nos últimos 50 anos, a monilíase se espalhou do Equador para o sul do Peru, 59
Colômbia, e países centro americanos, resultando em abandono das plantações, devido as 60
perdas que podem variar de 50 a 100% (Ram et al. 2004). Seu comportamento invasivo 61
representa uma séria ameaça para a cacauicultura do Brasil, que começa a soerguer-se do 62
impacto da VB, com reflexos na produção mundial. O fruto, em qualquer idade de seu 63
desenvolvimento, é a única parte do cacaueiro infectada por M. roreri, daí a importância da 64
doença, já que o fruto é a parte comercial do cacaueiro (Ram et al. 2004), de onde são 65
extraídas as amêndoas e todos os demais subprodutos que são utilizados na manufatura de 66
alimentos, cosméticos, etc (Efraim et al. 2011). 67

53
O emprego de variedades resistentes e de alta produtividade, desenvolvidas em 68
programas de melhoramento genético do cacaueiro (PMGC) é a base para o manejo de 69
doenças do cacaueiro e outras espécies arbóreas (Pinto e Pires 1998). Entretanto, alguns 70
materiais mesmo apresentando genes de resistência, têm demonstrado suscetibilidade, devido 71
às condições ambientais favoráveis, à rápida mutação e grande variabilidade de M. perniciosa 72
(Leung et al. 1993, McDonald e Linde 2002, Patrocínio et al. 2012) além de limitadas fontes 73
de resistência, predominando descendentes dos clones Scavina-6 e 12, selecionados no Peru, e 74
que indubitavelmente carregam genes de resistência a vários patógenos (Bartley 2005). Sendo 75
assim, o PMGC vem buscando ampliar a base genética para resistência à VB, combinando por 76
piramidação, o maior número possível de genes cruzando genótipos em primeira e segunda 77
geração, para obter resistência mais estável, durável e efetiva, dificultando o aumento da 78
população e a evolução do fungo sobre tais materiais (Lopes et al. 2012, Paim et al. 2006, 79
Pinto e Pires 1998, Pires 2003, Silva et al. 2010). 80
O PMGC no Brasil já gerou várias populações estruturadas e não estruturadas visando 81
a piramidação de genes para a resistência à VB, e destas, vários clones foram indicados para o 82
cultivo na região cacaueira do sul da Bahia (Monteiro e Ahnert 2012). As populações 83
estruturadas foram geradas a partir de cruzamentos dialélicos realizados entre materiais 84
selecionados de três populações: A- alto amazônicos; B- Baixo amazônicos; e AB- 85
cruzamentos entre alto e baixo amazônicos, distantes geneticamente, para exploração da 86
heterose, adição de características interessantes, além de resistência à VB e outras doenças, e 87
produção (Albuquerque et al. 2011, Paim et al. 2006, Pires et al. 1999). De uma dessas 88
populações foram selecionados clones com alta capacidade combinatória (Silva et al. 2011), 89
alguns dos quais serviram para formar o 2º ciclo de seleção de que trata o presente estudo. 90
Com isso, pretende-se ampliar as fontes de resistência a VB nos materiais já anteriormente 91
selecionados como resistentes a esta doença, e incorporar à população gerada, genes de 92
resistência à monilíase, através da inclusão dos clones ICS 95, EET 75, EET 233, e UF 273 93
como doadores de genes de resistência a monilíase (Phillips-Mora et al., 2007), no dialelo 94
(10x8), utilizando o delineamento Carolina do Norte II (Comstock e Robinson 1952). 95
96
Material e métodos 97
98
Os ensaios foram realizados em laboratórios, casas de vegetação e áreas experimentais da 99
Seção de Fitopatologia do Centro de Pesquisa do Cacau (CEPEC) na sede da Comissão 100
Executiva do Plano da Lavoura Cacaueira (CEPLAC), situada em Ilhéus, BA. 101

54
102
103
Material genético 104
105
Cruzamentos entre dois grupos de genótipos selecionados quanto a resistência a vassoura de 106
bruxa em avaliação a campo " per se " foram realizadas seguindo o delineamento Carolina do 107
Norte II (Comstock e Robinson 1952), e algumas progênies selecionadas dentro das 108
populações com o caráter resistência, para a composição do segundo ciclo de seleção 109
recorrente (Silva et al. 2010). A seleção de plantas dentro das progênies foi realizada com 110
base no número de vassouras e frutos infectados, número de frutos produzidos e aspectos 111
vegetativos das plantas em avaliações realizadas periodicamente durante no mínimo seis anos 112
de plantio em campo. Igualmente foram selecionadas plantas obtidas de progênies de 113
polinização livre dos clones CAB 270, CAB 208, CAB 214 e EET 392. Assim, 16 genótipos 114
oriundos desta seleção (CAB 270, CAB 208, CAB 214, EET 392, e dos cruzamentos: CSUL 115
3 x SCA 6, SGU 26 x SCA 6, NA 33 x RB 39, CEPEC 86 x SCA 6, RB 36 x SCA 6, CEPEC 116
90 x CHUAO 120, NA 33 x CSUL 7, MA 16 x SCA 6, CEPEC 86 x RB 39) foram utilizados 117
para compor o grupo de progenitores, que foi suplementado com genótipos selecionados 118
quanto a resistência à monilíase (EET 233, EET 75, ICS 95 e UF 233) (Phillips-Mora et al. 119
2007), visando a obtenção da agregação de características de interesse, e seleção de clones e 120
progênies para plantio comercial (Pires et al. 1999). 121
Cruzamentos entre os progenitores foram realizados e após a maturação dos frutos, as 122
sementes foram pré-germinadas por 48 horas em serragem esterilizada e umedecida e 123
plantadas em tubetes de 288 cm3 contendo aproximadamente 300 g de terriço esterilizado 124
(substrato comercial Plantmax® + solo, proporção 1:1). As plantas permaneceram em casa de 125
vegetação em condições de cultivo, submetidas à irrigação diária por um período de 20 126
minutos às 9:00 e às 14:00h, adicionando mais uma rega em dias com temperatura mais 127
elevada, por três a quatro semanas antecedendo a inoculação. As condições de temperatura, 128
umidade relativa e luminosidade foram monitoradas através de sensor HOBO® Data Logger 129
(U12-012). 130
Sementes dos clones SCA 6, SIC 23 e Catongo, sendo as do primeiro utilizadas como 131
padrão de resistência e as dos últimos como padrões de suscetibilidade, foram plantadas em 132
todos os ensaios. Devido às condições climáticas, às vezes adversas, e à disponibilidade de 133
flores nos genitores selecionados, os cruzamentos foram sendo realizados em diferentes 134
épocas e tiveram que ser avaliados quanto a resistência à VB em vários ensaios. Os ensaios 135

55
foram estabelecidos em delineamento inteiramente casualizado, com quatro repetições de 14 136
plântulas cada, e o número de cruzamentos variado, devido à dificuldade de obtenção de 137
frutos por polinização cruzada. Foram realizados ao todo 24 ensaios que totalizaram 59 138
progênies avaliadas, de 80 previstas. 139
140
Obtenção de suspensão de basidiósporos 141
142
Para obtenção de inóculo, ramos de cacaueiro apresentando sintoma de “vassoura vegetativa 143
seca” foram coletadas, desinfestados com lavagens rápidas em hipoclorito de sódio a 0,1%, 144
fixadas em arames esticados horizontalmente em casa de vegetação otimizada (vassoureiro), 145
coberta com sombrite de 50% de luminosidade, e então submetidas a um regime diário de 8 h 146
de molhamento e 16 h de secamento. Após o período de dormência, as vassouras foram 147
pulverizadas com solução de estreptomicina a 1%, e lavadas mais duas vezes em água 148
destilada. Com o surgimento dos basidiomas de M. perniciosa, estes foram então coletados e 149
secos em folhas de papel absorvente, seccionando os píleos que foram fixados com vaselina 150
pastosa (Silinol SG) em tampas de placas de Petri de vidro, com o himênio voltado para 151
baixo, sobre um béquer contendo uma solução de glicerina a 16%, e sob agitação constante, 152
permitindo a captação dos basidiósporos liberados. As suspensões de basidiósporos foram 153
quantificadas em câmara de Neubauer e estocadas em nitrogênio líquido, acondicionadas em 154
tubos criogênicos de 2 mL, até o momento de serem utilizadas, conforme metodologia 155
descrita por Dickstein et al. (1987). Antes da estocagem, uma alíquota de 25 µL de cada uma 156
das suspensões de basidiósporos obtidas foi retirada e espalhada com alça de Drigalski sobre a 157
superfície do meio ágar-água a 2%, em placas de Petri de 5 cm de diâmetro, para avaliar a 158
germinação dos esporos. Antes da estocagem e por ocasião da inoculação, procedia-se o teste 159
de germinação para determinar a viabilidade dos basidiósporos (Frias e Purdy 1995). Após 160
seis horas de incubação a 25°C, a percentagem de basidiósporos germinados foi mensurada 161
através da visualização em microscópio ótico e somente suspensões que apresentaram 162
percentual de germinação acima de 80% foram utilizadas. Estes foram repetidos antes da 163
inoculação para todos os ensaios. 164
165
Inoculação 166
167
No dia precedente à inoculação, o tamanho das folhas do lançamento apical das plântulas foi 168
reduzido a 1/3, para acelerar o crescimento apical e melhor expor a área da gema apical onde 169

56
seria depositado o inóculo. Em seguida as plântulas foram transportadas para câmara de 170
inoculação e mantidas sob condições de saturação de umidade e temperatura entre 25-27°C. 171
Vinte e quatro horas após, as plântulas foram inoculadas com uma gota de 20 µL da 172
suspensão do inóculo, na concentração de 2x105 basidiósporos/mL em meio ágar-água 0,2%, 173
depositada na gema apical das plântulas, permanecendo em câmara de inoculação com 174
saturação de umidade por mais 24h e em seguida transportadas e mantidas em casa de 175
vegetação sob as mesmas condições de cultivo, até a data da avaliação. 176
177
Avaliação dos sintomas 178
179
Aos 60 dias após a inoculação as plântulas foram avaliadas individualmente quanto a 180
incidência e severidade da doença, observando o tipo de vassoura formada: VT (vassoura 181
terminal), VA (vassoura axilar) e VC (vassoura cotiledonar), com tomada de dados binários, 182
além da quantificação de vassouras axilares maiores que 1 cm e mensuração do comprimento 183
da vassoura terminal. Para análise dos dados, foi calculado o índice de doença (calculado pela 184
seguinte fórmula: ID = VT + VA * (0,5 * CVT) + NVA, em que: VT = número de vassouras 185
terminais; VA = número de vassouras axilares; CVT = comprimento da vassoura terminal; 186
NVA = número de vassouras axilares maiores que 1cm (Frias e Purdy 1995) 187
188
Análise dos dados 189
190
As médias foram corrigidas por ensaio e ensaio dentro de repetição, obedecendo o princípio 191
de blocos incompletos, considerando progênie como única fonte de variação não casual. 192
Todas as análises foram conduzidas ao nível de 5% de significância utilizando-se o pacote 193
estatístico LSmeans SAS (SAS 1998), análise de variância e contrastes realizados entre as 194
médias das progênies dos cruzamentos e as das progênies dos controles, bem como entre si. 195
196
Resultados e Discussão 197
198
Foram avaliadas progênies de 59 cruzamentos e de três controles, dois suscetíveis 199
(CATONGO e SIC 23) e um resistente (SCA 6), totalizando 62 progênies. As progênies dos 200
59 cruzamentos apresentaram comportamento variáveis nos diferentes ensaios realizados (24), 201
bem como nas repetições dentro de cada ensaio e quando comparadas entre progênies obtidas 202
de cruzamentos diferentes (Tabela 1). Isto é fácil de entender, tendo em vista que em 203

57
condições diferentes nem sempre genes que estão presentes em um genótipo, nesse caso os de 204
resistência à VB, conseguem expressar-se, por não ser um caráter dominante (Silva et al. 205
2014). A capacidade de transferência de genes de um genótipo em combinação com outros é 206
conhecida como capacidade geral de combinação e é considerada um indicador da 207
superioridade do parental e de sua divergência relativa entre os demais parentais (Viana 208
2000). 209
Quando as progênies de cada cruzamento foram comparadas ao nível de p<0,05 com 210
testemunhas suscetíveis (SIC 23 e Catongo) e resistentes (Scavina 6 -SCA 6), inoculadas 211
conjuntamente em cada ensaio realizado, dos 59 cruzamentos realizados 52 geraram grupos 212
de progênies que diferiram da testemunha SIC 23, a qual apresentou ID médio de 5,47. 213
Apenas as plantas oriundas dos cruzamentos 11, 18, 39, 51, 54, 55 e 57 diferiram 214
estatisticamente da testemunha SIC 23, entretanto, dentre estes sete cruzamentos, o 215
cruzamento 39 não diferiu da testemunha resistente (Tabela 2). Apesar de ser observado 216
diferenças entre a suscetibilidade de plântulas seminais obtidas das testemunhas SIC 23 e 217
SCA 6, alguns materiais mesmo apresentando genes de resistência, têm demonstrado 218
suscetibilidade, devido às condições ambientais favoráveis, à rápida mutação e grande 219
variabilidade do patógeno, além de limitadas fontes de resistência, predominando 220
descendentes dos clones SCA-6 e 12, selecionados no Peru, e que indubitavelmente carregam 221
genes de resistência a vários patógenos (Bartley 2005). Apenas o cruzamento 26 não diferiu 222
da outra testemunha suscetível (Catongo) avaliada, destacando a importância da utilização 223
desta testemunha como padrão de suscetibilidade e avaliação do potencial do inóculo em 224
causar doença. 225
De modo geral pode-se avaliar a eficiência da seleção para resistência à VB 226
comparando as médias de ID obtidas nas progênies, quando contrastadas com as dos padrões 227
de suscetibilidade (Tabela 2), 58 das progênies foram estatisticamente diferentes das de 228
Catongo, cujo ID médio foi de 13,43, o mais alto entre as 61 progênies avaliadas (Tabela 2). 229
Apenas a progênie do cruzamento 26 [(SCA 6 x CSUL 3) x (ICS 95], com ID = 11,12, foi tão 230
suscetível quanto o padrão de suscetibilidade. Em relação ao SIC 23, cujo média do ID diferiu 231
significativamente do Catongo e, portanto, pode ser considerado moderadamente suscetível à 232
VB, 52 progênies foram distintas dele, apresentando médias inferiores a 5,48, o valor 233
corrigido para a média de ID obtida pelas progênies de SIC 23 testadas nestes ensaios. 234
Diferiram do padrão de resistência, SCA 6 (ID = 1,45), por apresentarem médias de ID 235
inferiores à daquele padrão, nove progênies dos seguintes cruzamentos: [CAB 270 livre x 236
(EET 75)]; [(CHUAO 120 x CEPEC 90) x (EET 75)]; [(NA 33 x CSUL 7) x (EET 75)]; 237

58
[(SCA 6 x P4 B) x (EET 233)]; [(SCA 6 x P4 B) x (UF 273)]; [(EET 233) x (CAB 214 livre)]; 238
[(UF 273) x (CHUAO 120 x CEPEC 90)], entre elas, seis portando como um dos genitores 239
EET 75 (3), EET 233 (1) e UF 273 (2), clones considerados como resistentes à monilíase (Ali 240
et al. 2015, Arciniega 2005; Evans 1981, Evans et al. 1998, Morera et al. 1991, Phillips et al. 241
2009). 242
Duas das progênies que se mostraram altamente resistentes à VB, têm como uma das 243
fontes de resistência os clones CAB 270 (cruzamento 1) e o CAB 214, considerados como 244
resistentes à VB e que já apresentaram outras progênies F1 resistentes em condições de 245
campo no Pará e na Bahia destacando-se por manterem-se resistentes por vários anos em 246
condições de campo (Albuquerque et al. 2010). De CAB 214 (progênie 232), 11 cruzamentos 247
foram avaliados (Tabela 2), demonstrando que a planta selecionada para o 2º ciclo de seleção 248
produziu flores suficientes para suportar todos os cruzamentos, indício de alta produção. 249
Dentre essas progênies de CAB 214, nove delas diferiram (p≤0,01) de SIC 23 e não diferiram 250
de SCA 6 [(CAB 214 livre) x (EET 75)]; [(SCA 6 x CEPEC 86) x (CAB 214 livre)]; [(SCA 251
6 x RB 36) x (CAB 214 livre)]; [(SCA 6 x SGU 26) x (CAB 214 livre)]; [(SCA 6 x CSUL 3) 252
x (CAB 214 livre)]; [(NA 33 x CSUL 7) x (CAB 214 livre)]; [(SCA 6 x MA 16) x (CAB 214 253
livre)]; [(SCA 6 x P4 B) x (CAB 214 livre)]; [(SCA 6 x SILECIA 1) x (CAB 214 livre)]; [(UF 254
273) x (CAB 214 livre)]; uma (cruzamento 50) foi mais resistente que o padrão e apenas 255
uma (cruzamento 11) teve ID médio similar ao de SIC 23. 256
O genótipo CAB 214 é, portanto, importante fonte de resistência à VB e segundo Leal 257
et al. (2007) foram identificados e caracterizados neste clone e também no CAB 208, genes 258
relacionados à defesa (Albuquerque et al. 2009). Estes dois clones são oriundos da bacia do 259
rio Purus, no Amazonas (Almeida et al. 1997, Barriga et al. 1985). Outro clone, CAB 270, 260
que também gerou uma das progênies altamente resistentes a VB é originário de Tefé, na 261
bacia do rio Solimões, também no Amazonas (Almeida 1982). Este clone participou como 262
genitor em sete cruzamentos avaliados neste trabalho, gerando além da progênie já 263
mencionada como altamente resistente (1), três resistentes por não diferirem de SCA 6 (21, 29 264
e 36), e três que foram distintos de SCA 6 e SIC 23 (15 e 39), portanto, consideradas 265
moderadamente resistentes. Famílias também dele originadas mostraram-se resistentes a VB 266
no Pará (Albuquerque et al. 2010) e na Bahia (Gramacho et al. 2016). 267
Outro clone da série CAB utilizado neste ciclo de seleção, foi o CAB 208 (progênie 268
225), que foi genitor de oito das progênies testadas (4, 10, 16, 22, 30, 37, 42, 52), sendo que 269
destes, seis não diferiram estatisticamente de SCA 6, comportando-se como resistentes e duas 270

59
(10 e 22), com valores médios de ID superiores a SCA 6 e inferiores, e distintos de SIC 23, 271
consideradas moderadamente resistentes. 272
Os cruzamentos que possuem descendentes diretos de genótipos obtidos de acessos da 273
Amazônia Brasileira (CAB) apresentaram resultados importantes para a piramidação de 274
genes/alelos para a ampliação da estabilidade e durabilidade da resistência. Dos 59 275
cruzamentos realizados, 26 possuem genitores descendentes diretos dos clones CAB 208, 276
CAB 214 e CAB 270, portanto, 46,06% das progênies avaliadas. Destes 26 cruzamentos, dois 277
estão entre os nove mais resistentes que a testemunha SCA 6, 84,61% diferiram (p<0,01) da 278
testemunha suscetível SIC 23 e comportaram-se similarmente ao SCA 6, sendo considerados 279
resistentes à VB. 280
Dentre os nove cruzamentos que diferiram da testemunha suscetível (SCA 6), o 281
cruzamento 50 gerou o grupo de progênies com menor índice de doença avaliado (-4.7273), 282
se destacando e diferenciando estatisticamente dos demais (Tabela 3). Não houve diferença 283
estatística entre os demais cruzamentos que compõem o grupo das interações que geraram as 284
melhores progênies para o caráter de resistência à VB. 285
O EET 233, usado como progenitor em apenas três cruzamentos avaliados, gerou duas 286
progênies altamente resistentes a VB (46 e 50) e outra (51) cujo ID médio corrigido foi de 287
4,7277, distinto de SIC 23, mas também de Catongo. Os clones CHUAO 120 e CEPEC 90, 288
outros parentais envolvidos neste cruzamento são considerados tolerantes a VB (Luz et al., 289
1997). SCA 6 e P4 B, são reconhecidos por apresentarem alta capacidade combinatória e 290
gerarem progênies no 1º ciclo de seleção com resistência a VB (Silva et al. 2010). 291
Embora a resistência dos clones SCA já tenha sido suplantada em campo (Pires et al., 292
2012), e cepas de M. perniciosa capazes de infecta-los já tenham sido selecionadas 293
(Patrocinio et al. 2012) e alguns dos seus descendentes já mostrarem no seu comportamento 294
ao longo dos anos este efeito (Benjamin et al. 2016, Paim et al. 2006), outras, mantem-se 295
resistentes quando o outro clone doador também é resistente (Benjamin et al. 2016, Pires et al. 296
2012; Paim et al. 2006). Ressalta-se que o comportamento das progênies de SCA 6 usadas 297
para inoculações artificiais em casa de vegetação, continua sendo de resistência (Albuquerque 298
et al. 2009, Kuhn et al. 2003, Yamada et al. 2005), o que ficou evidenciado também neste 299
trabalho, mesmo com a variação do inóculo do patógeno utilizado, pois, as vassouras são 300
coletadas a campo, nas áreas experimentais do CEPEC, e renovadas a cada seis meses para a 301
produção de basidiomas e estocagem dos basidiósporos usados nas inoculações. Portanto, 302
teoricamente, acompanha-se a possível evolução do patógeno. O comportamento do grupo de 303
progênies de SCA 6 que compõem estes cruzamentos, sugere que este clone possui genes de 304

60
resistência, de menor expressão, que não foram suplantados pelo patógeno. Além do locus 305
dominante para a resistência quantitativa (QRL), responsável por 51% da variação fenotípica 306
para a resistência identificada em SCA 6 através de mapeamento molecular (Brown et al. 307
2005, Faleiro et al. 2006), os clones originários do Peru, como Scavina, carregam também 308
genes de resistência à vários patógenos (Bahia et al. 2015, Bartley 2005). 309
Progenitores de elite dentre os testados em um dos dialelos (8 x 8) do 1º ciclo de 310
seleção recorrentes e envolvidos também no cruzamento que originou outra progênie destaque 311
(35), NA 33 e RB 39 são também clones conhecidos como resistentes a VB (Bemjamin et al. 312
2014). O clone CEPEC 90 também foi considerado como resistente a VB em avaliações em 313
campo e em casa de vegetação (Luz et al. 1997, Pires et al. 1996). CHUAO 120, um crioulo 314
venezuelano, tem se comportado como resistente a VB e tolerante em condições de campo no 315
Brasil (Pires et al. 1996) e foi considerado por Phillips-Mora et al. (2009), como 316
moderadamente resistente à monilíase, na Costa Rica. 317
Os outros dois clones resistentes a monilíase (Phillips-Mora et al. 2009) usados como 318
genitores das progênies testadas, EET 75 e UF 273, também geraram progênies que se 319
destacaram como altamente resistentes a VB, sendo três (1, 27, 35) descendentes de EET 75 e 320
uma (59) de UF 273. Não foram encontradas referências quanto ao comportamento em 321
relação a VB no banco internacional de dados de germoplasma do cacau (Turnbull e Hadley 322
2016), mas EET 75 é citado como resistente a Phytophthora palmivora na Costa Rica 323
(Arciniegas e Phillips-Mora 2006). As origens distintas destes clones e a distância genética já 324
determinada entre muitos deles (Marita et al. 2001, Montamayor et al. 2008, Sereno et al. 325
2006) atestam o sucesso da piramidação de genes. 326
Em relação aos clones utilizados no dialelo como padrão de resistência à M. roreri, 327
apenas o clone ICS 95, não combinou positivamente para o caráter resistência com o grupo de 328
progenitores selecionados quanto a resistência à VB, os demais EET 75 (3), EET 233 (2), UF 329
273 (1) integraram cruzamentos que apresentaram as menores médias de ID nos ensaios 330
avaliados (Tabela 3). Mesmo que não tenha sido identificada a transferência do caráter 331
resistência a M. perniciosa para as progênies oriundas de cruzamentos entre genótipos 332
selecionados como portadores deste caráter e o clone ICS 95, o PMGC segue na busca de 333
novas fontes de resistência que possam capacidade combinatória com este clone, que foi 334
citado por Argüello (2000), Phillips-Mora e colaboradores (2005) como um genótipo com 335
significante nível de resistência a isolados de M. roreri distintos geneticamente e avaliados 336
quanto ao índice de doença, severidade externa e severidade interna de sintomas de monilíase. 337

61
Destaca-se a importância da utilização do CATONGO como padrão de suscetibilidade 338
em experimentos realizados para avaliação a VB. No entanto deve ser observado o 339
procedimento de seleção das sementes brancas, tendo em vista que, embora auto compatível 340
(Yamada et al. 2005), pode ocorrer a fertilização cruzada em algumas flores, gerando algumas 341
sementes roxas, o que acontece com frequência. Estas sementes não têm homozigose para 342
suscetibilidade a VB. 343
Assim, demonstrou-se que está ocorrendo a piramidação de genes para a resistência a 344
M. perniciosa e a incorporação de genes de resistência a M. roreri, praga quarentenária A1, 345
nas progênies ora selecionadas e que estas constituem um importante material para o 346
programa de melhoramento genético visando resistência ás duas doenças. 347
Os resultados alcançados por este trabalho apresentam-se como satisfatórios quanto à 348
seleção de plantas para compor o segundo ciclo de seleção e apresenta não mais potenciais 349
progenitores, mas progênies que carregam genes de resistência à VB oriunda de fontes 350
diversas, além da incorporação de genes de resistência à monilíase, praga quarentenária A1 de 351
relevante impacto à cultura do cacaueiro. Todos as progênies obtidas dos cruzamentos e que 352
não apresentaram sintomas de VB foram transplantadas para sacos de polietileno (16 x 40 x 353
08 cm) e conduzidas ao campo para avaliação da estabilidade da resistência, produtividade e 354
autocompatibilidade em ensaios multiregionais. 355
O desenvolvimento de técnicas e produtos que visam mitigar os impactos causados por 356
doenças de plantas é de suma importância para o estabelecimento de cultivos agrícolas e 357
melhor exploração dos recursos ambientais em determinadas localidades. O presente trabalho 358
evidencia resultados de altíssima relevância para os países produtores de cacau afetados pela 359
VB e monilíase, gerando um grupo de indivíduos portadores de genes de resistência à essas 360
doenças, oriundos de fontes diversas. Além disso, demonstra a possibilidade de seleção de 361
progênies para a resistência à VB com base na resistência à monilíase, sendo sem dúvida, uma 362
valiosa contribuição para a viabilidade em bases sustentáveis da lavoura cacaueira em países 363
que já convivem com as duas doenças ou que trabalham de modo a conter a entrada do 364
patógeno em seus territórios. 365
366
Agradecimentos 367
368
Ao FINEP, FAPESB e MAPA através do orçamento da CEPLAC pelo apoio financeiro; À 369
Joel Feitosa, Magnaldo Nascimento, Layane Alves, Orlando Garcia, João Marcos Gouveia e 370
William Dias pela valiosa ajuda nas polinizações; À Virgínia Damaceno, Ananias, Orlando, 371

62
Elenísio e demais operários de campo pela ajuda nas inoculações e manutenção das plantas na 372
casa de vegetação. 373
374
Referências 375
376
Albuquerque PSB et al (2005) Doenças do cacaueiro. In: Kimati H et al Manual de 377
fitopatologia, 4th edn. v. 2. Ceres, Piracicaba, pp 151-164 378
Albuquerque PSB, Silva SDVM, Luz EDMN, Pires JL, Vieira AMC, Demétrio CGB, 379
Pascholatti SF, Figueira A (2009) Novel sources of witches’ broom resistance (causal agent 380
Moniliophthora perniciosa) from natural populations of Theobroma cacao from the Brazilian 381
Amazon. Euphytica. DOI 10.1007/s10681-009-0068-4 382
Ali SS, Shao J, Strem MD, Phillips-Mora W, Zhang D, Meinhardt LW, Bailey BA (2015) 383
Combination of RNAseq and SNP nanofluidic array reveals the center of genetic diversity of 384
cacao pathogen Moniliophthora roreri in the upper Magdalena Valley of Colombia and its 385
clonality. Frontiers in Microbiology 6. doi: 10.3389/fmicb.2015.00850 386
Almeida CMVC de (1982) Relatório da expedição botânica para coleta de cacau nativo na 387
região de Ariquemes, Rondônia, 1982. CEPLAC/DEPEA Belém, PA, Brasil. pp 23 388
Almeida CMVC, Barriga JP, Machado PFR, Bartley, BGD (1987) Evolução do programa de 389
conservação dos recursos genéticos de cacau na Amazônia Brasileira. CEPLAC/DEPEA, 390
Boletim Técnico, Belém, Brasil 5: pp.108. 391
Alverson WS et al (1999) Phylogeny of the core Malvales: evidence from ndhF sequence 392
data. Amer. Journal of Botany 89:1474-1486 393
Arciniegas A, Phillips-Mora W (2006) Caracterización de Genotipos Superiores de Cacao 394
Seleccionados por el Programa de Mejoramiento Genético del CATIE por Su Rendimiento 395
Y/O Resistencia a Moniliasis In: Proceedings of the 15th International Cocoa Research 396
Conference, San Jose, Costa Rica (2006). Cocoa Producers' Alliance, Lagos, Nigeria 397
Argüello O (2000) Variabilidad morfoagronómica de 59 árboles elite de cacao (Theobroma 398
cacao L.) en Santander. In: Mejía LA, Argüello CO (ed) Tecnología para el Mejoramiento del 399
Sistema de Producción de Cacao. Corpoica, Bucaramanga, pp 50-4 400
Bartley BG (2005) The genetic diversity of cacao and its utilization. CABI Publishing, 401
Massachusetts, 341p 402
Benjamin CS, Luz EDMN, Monteiro WR, Santos Filho LP, Santos WO (2014) Avaliação de 403
progênies de clones de cacaueiros (série CP) quanto à resistência a Moniliophthora 404
perniciosa. Agrotrópica 26:17 405

63
Comstock RE, Robinson HF (1952) Estimation of overage dominance of genes. In: Gowen J. 406
Heterosis. Iowa State: Iowa State College, pp 494-516. 407
Danquah OA (1986) Growth of Crinipellis perniciosa in cacao resistant and susceptible to 408
witches’ broom. PhD thesis, University of London. 409
Dickstein ER, Purdy LH, Frias GA (1987) Crinipellis perniciosa, the cacao witches’ broom 410
fungus: Inoculum production and storage. Phytopathology 77:1747 411
Efraim P, ALVES AB, Jardim DCP (2011) Polifenóis em cacau e derivados: teores, fatores de 412
variação e efeitos na saúde. Brazilian Journal Food Technology 14:181-201 413
Evans H, Krauss U, Rios RR, Zecevich AT, Arevalo GE (1998). Cocoa in Peru. Cocoa 414
Growers’. Bulletin 51:7–22 415
Evans, H. (1981). Pod rot of cacao caused by Moniliophthora (Monilia) roreri 416
Phytopathological papers 24:1–44 417
Frias GA, Purdy LH, Schmidt RA (1995) An Inoculation Method for Evaluating Resistance 418
of Cacao to Crinipellis perniciosa. Plant Disease 79:787-791. doi: 10.1094/PD-79-0787 419
Gramacho KP, Luz EDMN, Silva FS, Lopes UV, Pires JL, Pereira L (2016) Pathogenic 420
variability of Moniliophthora perniciosa in three agroecological zones of the cacao region of 421
Bahia, Brazil. Crop Breeding and Applied Biotechnology 16: 7-13. doi:10.1590/1984-422
70332016v16n1a2 423
ICCO (2015) Production of cocoa beans. International Cocoa Organization. http:// 424
http://www.icco.org/statistics/production-and-grindings/production.html. Acessado em 26 425
Janeiro 2016 426
Kuhn DN, Heath M, Wisser RJ, Meerow A, Brown JS, Lopes U, Schnell RJ (2003) 427
Resistance gene homologues in Theobroma cacao as useful genetic markers Theor Appl 428
Genet 107:191–202. DOI 10.1007/s00122-003-1239-7 429
Leung H, Nelson R.J, Leach JE (1993) Population structure of plant pathogenic fungi and 430
bacteria. Plant Pathology 10:157-205 431
Lopes UV, Monteiro WR, Pires JL, Clement D, Yamada, MM, Gramacho KP (2011) Cacao 432
breeding in Bahia, Brazil - strategies and results. Crop breeding and applied biotechnology 433
11:73-81.doi: 10.1590/S1984-70332011000500011 434
Marita JM, Nienhuis J, Pires JL, Aitken WM (2001) Analysis of Genetic Diversity in 435
Theobroma cacao with Emphasis on Witches’ Broom Disease Resistance. Crop Science. 436
41:1305-1316. 437
Mcdonald BA, Linde C (2002) Pathogen population genetics, evolutionary potential, and 438
durable resistance. Annual Review of Phytopathology 40:349-379 439

64
Monteiro WR, Ahnert D (2012) Melhoramento genético do cacaueiro. In: Valle RRM (ed) 440
Ciência, tecnologia e manejo do cacaueiro. 2ed. Brasilia, p.11-30, 2012. 441
Moraes WB, Jesus Júnior WC, Peixoto LA, Moraes WB, Furtado EL, Silva LG, Cecílio RA, 442
Alves FR (2012) An analysis of the risk of cocoa moniliasis occurrence in Brazil as the result 443
of climate change. Summa Phytopathologica 38:30-35. doi: 10.1590/S0100-444
4052012000100005 445
Morera J, Paredes A, Mora A (1991) Germoplasma de cacao en el CATIE entre 1947 y 1991, 446
Programa II: Generacion y transferencia de tecnologia. IICA, San Jose, Costa Rica, pp 49. 447
Paim VRLM, Luz EDMN, Pires JL, Silva SDVM, Souza JT, Albuquerque PSB, Santos Filho 448
LP (2006) Sources of resistance to Crinipellis perniciosa in progenies of cacao accessions 449
collected in the Brazilian Amazon. Scientia Agricola 63:572-578 450
Patrocinio NGRB, Moreira RFC, Serra WO, Pires JL, Gramacho KP (2012) Temporal genetic 451
characterization of isolates of Moniliophthora perniciosa in southeast of Bahia, Brazil 452
Agrotrópica 24(3) 453
Phillips W, Castillo J, Arciniegas A, Mata A, Sánchez A, Leandro M, Astorga C, Motamayor 454
J, Guyton B, Seguine E, Schnell R (2009) Overcoming the main limiting factors of cacao 455
production in central america through the use of improved clones developed at CATIE In: 456
Proceedings of the 16th International Cocoa Research Conference, Bali, Indonesia. Cocoa 457
Producers' Alliance, Lagos, Nigeria. 458
Phillips-Mora W, Aime MC, Wilkinson MJ (2007) Biodiversity and biogeography of the 459
cacao (Theobroma cacao) pathogen Moniliophthora roreri. Plant Pathology 56:911-922. doi: 460
10.1111/j.1365-3059.2007.01646.x 461
Phillips-Mora W, Castillo J, Krauss U, Rodríguez E, Wilkinson MJ (2005) Evaluation of 462
cacao (Theobroma cacao) clones against seven Colombian isolates of Moniliophthora roreri 463
from four pathogen genetic groups. Plant Pathology 54:483-490. doi:10.1111/j.1365-464
3059.2005.01210.x 465
Pinto LRM, Pires JL (1998) Seleção de plantas de cacau resistentes à vassoura-de-bruxa. 466
Ilhéus, CEPLAC/ CEPEC. Boletim Técnico. 181p 467
Pires JL (2003) Avaliação quantitativa e molecular de germoplasma para o melhoramento do 468
cacaueiro com ênfase na produtividade, qualidade do fruto e resistência a doenças. Tese, 469
Universidade Federal de Viçosa 470
Pires JL, De Melo GP, Yamada MM, Gramacho KP (2012) Association among sources of 471
resistance to witches’ broom disease for the increment of the level and durability of the 472
character. Agrotropica 24:27–30 473

65
Pires JL, Monteiro WR, Pinto LRM, Figueira A, Yamada MM, Ahnert D (1999) A proposal 474
for cacao breeding. In: Proceedings of the 12th International Cocoa Research Conference, 475
Cocoa Producers’ Alliance, Lagos, Nigeria. pp 287-292 476
Pires, JL., Monteiro, WR., Pinto, LRM; Luz, EDMN. (1996) Resistance to Witches Broom - 477
Evaluation of genotypes from different origins. In: Proceedings of the 12th International 478
Cocoa Research Conference, Salvador, Brasil 1996 Cocoa Producers' Alliance (in press). 479
Pokou ND, N’Goran JAK, Lachenaud Ph., Eskes AB, Montamayor JC, Schnell R, 480
Kolesnikova-Allen M, Clément D, Sangaré A (2009), Recurrent selection of cocoa 481
populations in Côte d’Ivoire: comparative genetic diversity between the first and second 482
cycles. Plant Breeding, 128: 514–520. doi: 10.1111/j.1439-0523.2008.01582.x 483
Ran A, Valle RR, Gardini EA (2004). Monília do Cacaueiro. Fundação Cargill, São Paulo, pp 484
36 485
RC Bahia, Aguilar-Vildoso CI, Luz EDMN, Lopes UV, Machado RCR, Corrêa RX (2015) 486
Resistance to black pod disease in a segregating cacao tree population Tropical plant 487
pathology. DOI 10.1007/s40858-014-0003-7 . 488
SAS INSTITUTE INC. SAS/STAT User’s Guide. Release 6.03. Cary, NC: SAS Institute 489
Inc.1028p. 1998. 490
Silva CRS, Albuquerque PSB, Ervedosa FR, Mota JWS, Figueira A, Sebbenn AM (2010) 491
Understanding the genetic diversity, spatial genetic structure and mating system at the 492
hierarchical levels of fruits and individuals of a continuous Theobroma cacao population from 493
the Brazilian Amazon. Heredity 106:973–985 494
Silva DV, Araújo IS, Branco SMJ, Aguilar‐Vildoso CI, Lopes UV, Marelli JP, Motamayor 495
JC, Royaert S, Rebouças RC, Corrêa RX (2014) Analysis of resistance to witches’ broom 496
disease (Moniliophthora perniciosa) in flower cushions of Theobroma cacao in a segregating 497
population Plant Pathology 63:1264-1271 498
Silva SDVM, Luz EDMN, Pires JL, Yamada MM, Santos Filho LP (2010) Parent selection 499
for cocoa resistance to witches’‐broomParent selection for cocoa resistance to witches’‐broom 500
Pesquisa Agropecuária Brasileira. doi: 10.1590/S0100-204X2010000700007 501
Turnbull CJ, Hadley P. International Cocoa Germplasm Database (ICGD). [Online Database]. 502
CRA Ltd./ICE Futures Europe/University of Reading, UK. http://www.icgd.reading.ac.uk. 503
Acessado 11 February 2016 504
Viana JMS (2000) The parametric restristions of the gardner and eberhart diallel analysis 505
model: heterosis analysis. Genetics and Molecular Biology 23:869-75 506

66
Yamada MM, Flores AB, Faleiro FG, Melo GRP, MM Macedo, Lopes UV, Corrêa RX, 507
Santos RF (2005) Identificação e variabilidade genética de acessos de cacaueiros 508
autocompatíveis selecionados para resistência à vassoura-de-bruxa em fazendas produtoras. 509
Agrotrópica 17:17-22. 510

67
Tabela 1. Análise de variância do índice de doença em plântulas de cacaueiro com sintomas
causados por Moniliophthora perniciosa, oriundas de cruzamentos (64) entre progênies
selecionadas quanto a resistência à M. perniciosa e M. roreri e características agronômicas
desejáveis, avaliadas em 24 ensaios e 4 repetições/ensaio.
FONTE DE VARIAÇÃO GL SQ TIPO 3 QM VALOR F P
ENSAIO 21 19059.1063 907.5765 21.09 <.0001
REP(ENSAIO) 60 4938.4887 79.6530 1.85 <.0001
PROG/CRUZAMENTO 60 59619.1705 961.5995 22.35 <.0001

68 Tabela 2. Índice de doença em plântulas de cacaueiro com sintomas causados por Moniliophthora perniciosa, oriundas de
cruzamentos entre progênies selecionadas quanto a resistência à M. perniciosa (213, 225, 232, 238, 243, 245, 247, 253, 258, 278,
282, 285, 300, 309, EET 392), M. roreri (ICS 95, EET 75, EET 233, UF 273) e características agronômicas desejáveis, e a
significância pelo teste F.
(CONTINUA)
Nº CRUZAMENTOS
(PROGENITORES)
CRUZAMENTOS
(PROGÊNIES)
ID1
LSMEAN
TESTEMUNHAS
SIC 23 CATONGO SCA 6
Pr > F2 Pr > F2 Pr > F2
1 (CAB 270 X LIVRE) X (EET 75) 213 X EET 75 -0.2470 <0.0001 <0.0001 0.0429
2 (CAB 214 X LIVRE) X (EET 75) 232 X EET 75 -0.1095 <0.0001 <0.0001 0.0899
3 (SCA 6 X CEPEC 86) X (CAB 270 X LIVRE) 238 X 213 3.4264 0.0078 <0.0001 0.0106
4 (SCA 6 X CEPEC 86) X (CAB 208 X LIVRE) 238 X 225 1.9929 <0.0001 <0.0001 0.4541
5 (SCA 6 X CEPEC 86) X (CAB 214 X LIVRE) 238 X 232 1.4130 0.0008 <0.0001 0.9751
6 (SCA6 X CEPEC 86) X (CHUAO 120 X CEPEC 90) 238 X 278 1.2815 <0.0001 <0.0001 0.8186
7 (SCA6 X CEPEC 86) X (NA 33 X CSUL 7) 238 X 282 1.9183 0.0054 <0.0001 0.7147
8 (SCA 6 X CEPEC 86) X (CEPEC 86 X RB 39) 238 X 309 2.1440 <0.0001 <0.0001 0.3304
9 (SCA 6 X RB 36) X (CAB 270 X LIVRE) 243 X 213 3.1460 0.0056 <0.0001 0.0444
10 (SCA 6 X RB 36) X (CAB 208 X LIVRE) 243 X 225 3.6561 0.0205 <0.0001 0.0051
11 (SCA 6 X RB 36) X (CAB 214 X LIVRE) 243 X 232 5.7655 0.8679 <0.0001 0.0131
12 (SCA 6 X RB 36) X (CHUAO 120 X CEPEC 90) 243 X 278 -0.2096 0.0479 <0.0001 0.5525
13 (SCA 6 X RB 36) X (CEPEC 86 X RB 39) 243 X 309 2.3941 0.0024 <0.0001 0.3525
14 (NA 33 X RB 39) X (EET 75) 245 X EET 75 -0.6965 <0.0001 <0.0001 0.1144
15 (SCA 6 X SGU 26) X (CAB 270 X LIVRE) 253 X 213 3.1039 0.0013 <0.0001 0.0252
16 (SCA 6 X SGU 26) X (CAB 208 X LIVRE) 253 X 225 1.8135 <0.0001 <0.0001 0.5936
17 (SCA 6 X SGU 26) X (CAB 214 X LIVRE) 253 X 232 1.7687 <0.0001 <0.0001 0.6936
18 (SCA 6 X SGU 26) X (CHUAO 120 X CEPEC 90) 253 X 278 5.2880 0.8723 <0.0001 0.0011
19 (SCA 6 X SGU 26) X (P4 B X OC 7) 253 X 285 1.5198 0.0263 <0.0001 0.9692
20 (SCA 6 X SGU 26) X (CEPEC 86 X RB 39) 253 X 309 2.8275 0.001 <0.0001 0.0883
21 (SCA 6 X CSUL 3) X (CAB 270 X LIVRE) 258 X 213 1.8234 <0.0001 <0.0001 0.5994
22 (SCA 6 X CSUL 3) X (CAB 208 X LIVRE) 258 X 225 2.9663 <0.0001 <0.0001 0.0033
23 (SCA 6 X CSUL 3) X (CAB 214 X LIVRE) 258 X 232 2.0542 <0.0001 <0.0001 0.3749
24 (SCA 6 X CSUL 3) X (NA 33 X RB 39) 258 X 245 0.9536 0.0003 <0.0001 0.6927
25 (SCA 6 X SGU 26) X (P4 B X OC 67) 258 X 285 1.5188 0.0005 <0.0001 0.9523
26 (SCA 6 X CSUL 3) X (ICS 95) 258 X ICS 95 11.1237 0.0008 0.1813 <0.0001
27 (CHUAO 120 X CEPEC 90) X (EET 75) 278 X EET 75 -0.1521 <0.0001 <0.0001 0.0041
28 (CHUAO 120 X CEPEC 90) X (ICS 95) 278 X ICS 95 3.4260 0.0210 <0.0001 0.0248
29 (NA 33 X CSUL 7) X (CAB 270 LIVRE) 282 X 213 2.3021 <0.0001 <0.0001 0.2428
30 (NA 33 X CSUL 7) X (CAB 208 X LIVRE) 282 X 225 1.5908 <0.0001 <0.0001 0.8544
31 (NA 33 X CSUL 7) X (CAB 214 X LIVRE) 282 X 232 1.9396 0.0101 <0.0001 0.7229
32 (NA 33 X CSUL 7) X (CHUAO 120 X CEPEC 90) 282 X 278 3.1630 <0.0001 <0.0001 <0.0001
33 (NA 33 X CSUL7) X (P4 B X OC 67) 282 X 285 -0.0751 <0.0001 <0.0001 0.0328
34 (NA 33 X CSUL 7) X (CEPEC 86 X RB 39) 282 X 309 1.0115 <0.0001 <0.0001 0.6927
35 (NA 33 X CSUL 7) X (EET 75) 282 X EET 75 -1.1474 <0.0001 <0.0001 0.0042
36 (NA 33 X CSUL 7) X (CAB 270 X LIVRE) 284 X 213 1.8534 0.0193 <0.0001 0.7949
37 (SCA 6 X MA 16) X (CAB 208 X LIVRE) 284 X 225 2.8301 0.0003 <0.0001 <0.0001
38 (SCA 6 X MA 16) X (CAB 214 X LIVRE) 284 X 232 -0.9195 0.0005 <0.0001 0.1950
39 (SCA 6 X MA 16) X (NA 33 X RB 39) 284 X 245 4.0529 0.2966 <0.0001 0.0568
40 (SCA 6 X MA 16) X (CHUAO 120 X CEPEC 90) 284 X 278 2.3228 <0.0001 <0.0001 0.2083
41 (SCA 6 X MA 16) X (CEPEC 86 X RB 39) 284 X 309 -0.1117 <0.0001 <0.0001 0.0254
42 (SCA 6 X P4 B) X (CAB 208 X LIVRE) 300 X 225 0.6585 <0.0001 <0.0001 0.1413
43 (SCA 6 X P4 B) X (CAB 214 X LIVRE) 300 X 232 0.7979 0.0002 <0.0001 0.6076
44 (SCA 6 X P4 B) X (NA 33 X RB 39) 300 X 245 0.6829 <0.0001 <0.0001 0.4683

69 Tabela 2. Índice de doença em plântulas de cacaueiro com sintomas causados por Moniliophthora perniciosa, oriundas de
cruzamentos entre progênies selecionadas quanto a resistência à M. perniciosa (213, 225, 232, 238, 243, 245, 247, 253, 258, 278,
282, 285, 300, 309, EET 392) e M. roreri (ICS 95, EET 75, EET 233, UF 273) e características agronômicas desejáveis, e a
significância pelo teste F.
(CONCLUSÃO)
CRUZAMENTOS
(PROGENITORES)
CRUZAMENTOS
(PROGÊNIES)
ID1
LSMEAN
TESTEMUNHAS
Nº SIC 23
Pr > F2
CATONGO
Pr > F2
SCA 6
Pr > F2
45 (SCA 6 X P4 B) X (CEPEC 86 X RB 39) 300 X 309 2.2035 <0.0001 <0.0001 0.2725
46 (SCA 6 X P4 B) X (EET 233) 300 X EET 233 -0.9673 <0.0001 <0.0001 <0.0001
47 (SCA 6 X P4 B) X (EET 75) 300 X EET 75 -0.0001 <0.0001 <0.0001 0.1197
48 (SCA 6 X P4 B) X (ICS 95) 300 X ICS 95 -1.6167 0.0001 <0.0001 0.0991
49 (SCA 6 X P4 B) X (UF 273) 300 X UF 273 -1.1336 <0.0001 <0.0001 <0.0001
50 (EET 233) X (CAB 214 X LIVRE) EET 233 X 232 -4.7243 <0.0001 <0.0001 <0.0001
51 (EET 233) X (CHUAO 120 X CEPEC 90) EET 233 X 278 4.7277 0.4258 <0.0001 0.0005
52 (SCA 6 X SILECIA1) X (CAB 208 X LIVRE) EET 392 X 225 0.3433 <0.0001 <0.0001 0.1193
53 (SCA 6 X SILECIA1) X (CAB 214 X LIVRE) EET 392 X 232 2.3448 0.0007 <0.0001 0.3326
54 (SCA 6 X SILECIA1) X (NA 33 X RB 39) EET 392 X 245 5.4786 0.9986 <0.0001 0.0025
55 (SCA 6 X SILECIA1) X (CHUAO 120 X CEPEC 90) EET 392 X 278 3.8655 0.2121 <0.0001 0.0615
56 (SCA 6 X SILECIA1) X (CEPEC 86 X RB 39) EET 392 X 309 2.9211 0.0031 <0.0001 0.0885
57 (SCA 6 X SILECIA1) X (ICS 95) EET 392 X ICS 95 5.2349 0.8264 <0.0001 0.0006
58 (UF 273) X (CAB 214 X LIVRE) UF 273 X 232 -1.1098 0.0007 <0.0001 0.1871
59 (UF 273) X (CHUAO 120 X CEPEC 90) UF 273 X 278 -1.3041 <0.0001 <0.0001 <0.0001
60 CATONGO - 13.4348 <0.0001 - <0.0001
61 SCA 6 - 1.4510 <0.001 <0.0001 -
62 SIC 23 - 5.4764 - <0.0001 <0.0001 1ID LSMEANS – Índice de doença obtido através da quantificação e mensuração de sintomas: VA – presença de vassoura axilar;
NVA – número de vassouras axilares maiores que 1cm; VT – presença de vassoura terminal; CVT – comprimento da vassoura
terminal, calculado pela fórmula VT + VA (0,5 * CVT) + NVA, avaliado pelo procedimento LSMEANS do software estatístico
SAS e corrigido para o efeito de ensaio e de repetição por ensaio. 2Probabilidade de cometer um erro ao aceitar a hipótese de
diferença entre as médias das progênies.

70
Tabela 3. Índice de doença em plântulas de cacaueiro com sintomas causados por Moniliophthora perniciosa, oriundas de cruzamentos entre progênies selecionadas
quanto a resistência à M. perniciosa (213, 232, 278, 282, 285, 300), M. roreri (EET 75, EET 233, UF 273) e características agronômicas desejáveis, e a significância pelo
teste F.
PROGÊNIES
ID
LSMEANS1 Nº Nº PROGÊNIES (CRUZAMENTOS E TESTEMUNHAS) Pr > F4
1 27 33 35 41 46 49 50 59 60 61 62
CRUZ.2
213 x EET 75 -0.2470 1 - 0.9682 0.8753 0.4096 0.9007 0.4063 0.3091 0.0032 0.2593 <0.0001 0.0444 <0.0001
278 X EET 75 -0.1521 27 - - 0.9318 0.291 0.9637 0.2007 0.1277 0.0046 0.1013 <0.0001 0.0041 <0.0001
282 X 285 -0.0751 33 - - - 0.3506 0.9636 0.3231 0.2454 0.0017 0.1925 <0.0001 0.0328 <0.0001
282 X EET 75 -1.1474 35 - - - - 0.3632 0.8477 0.9883 0.0218 0.8753 <0.0001 0.0042 <0.0001
284 X 309 -0.1117 41 - - - - - 0.3366 0.2556 0.0018 0.2003 <0.0001 0.00254 <0.0001
300 X EET 233 -0.9673 46 - - - - - 0.7809 0.0069 0.6435 <0.0001 <0.0001 <0.0001
300 X UF 273 -1.1336 49 - - - - - - <0.0001 0.8166 <0.0001 <0.0001 <0.0001
EET 233 X 232 -4.7243 50 - - - - - - - 0.0132 <0.0001 <0.0001 <0.0001
UF 273 X 278 -1.3041 59 - - - - - - - - <0.0001 <0.0001 <0.0001
TEST.3
CATONGO 13. 4348 60 - - - - - - - - - <0.0001 <0.0001
SCA 6 1.4510 61 - - - - - - - - - - <0.0001
SIC 23 5.4664 62 - - - - - - - - - - - 1ID LSMEANS – Índice de doença obtido através da quantificação e mensuração de sintomas: VA – presença de vassoura axilar; NVA – número de vassouras axilares
maiores que 1cm; VT – presença de vassoura terminal; CVT – comprimento da vassoura terminal, calculado pela fórmula VT + VA (0,5 * CVT) + NVA, avaliado pelo
procedimento LSMEANS do software estatístico SAS e corrigido para o efeito de ensaio e de repetição por ensaio; 2CRUZ. – Genótipos que compõem os cruzamentos; 3TEST. – Testemunhas com padrão de suscetibilidade e resistência, utilizadas em todos os ensaios realizados.4Probabilidade de cometer um erro ao aceitar a hipótese de
diferença entre as médias das progênies.

71
CAPÍTULO 3
Diversidade e antagonismo de leveduras nativas contra o agente da vassoura de bruxa
do cacaueiro

72
Diversidade e antagonismo de leveduras nativas contra o agente da vassoura de bruxa 1
do cacaueiro 2
3
Antônio Alves Pimenta Neto(1), Tacila Ribeiro Santos(2), Elizabeth Amélia Alves Duarte(3) 4
Francis Tocafundo(2), Thiago Alves Santos de Oliveira(3), Edson Mário de Andrade Silva(2), 5
Edna Dora Martins Newman Luz(4), Ana Paula Trovatti Uetanabaro(2), Delson Laranjeira(1) 6
7
(1)Universidade Federal Rural de Pernambuco, Rua Dom Manuel Medeiros, s/n, Dois Irmãos, 8
CEP 52171-900, Recife, PE, Brasil. Email: [email protected], 9
[email protected] (2)Universidade Estadual de Santa Cruz, Campus Soane Nazaré de 10
Andrade, Rod. Jorge Amado, Km 16, Salobrinho, Ilhéus, BA, Brasil. Email: 11
[email protected], [email protected], [email protected], 12
[email protected] (3) Universidade Federal do Recôncavo Baiano, Laboratório de 13
Biologia Molecular, Av. Rui Barbosa, 710, 44380-000, Cruz das Almas, BA, Brasil. Email: 14
[email protected], [email protected] (4)Comissão Executiva do Plano da 15
Lavoura Cacaueira, Centro de Pesquisas do Cacau, Rodovia Ilhéus-Itabuna, Km 22, s/n, CEP 16
45660-000, Ilhéus, BA, Brasil. Email: [email protected] 17
18
19
Resumo - Os objetivos deste trabalho foram avaliar a diversidade de leveduras na superfície 20
de órgão vegetais do cacaueiro e caracterizá-las quanto ao potencial antagônico à M. 21
perniciosa. Foi avaliado o comportamento de compostos difusíveis e voláteis de leveduras 22
nativas na inibição do crescimento micelial e germinação de basidiósporos de M. perniciosa. 23
Das 172 leveduras avaliadas, 18 apresentou inibição ≥50% o crescimento dos quatro isolados 24
do patógeno estudados. Oitenta isolados de leveduras foram identificados molecularmente 25
com amplificação da região D1/D2 da subunidade maior (26S) do rDNA. Os isolados (LEV 26
162, 194, 210 e 211), identificados como Hanseniaspora sp., se encontram no grupo de 27
isolados que apresentaram as maiores médias para as quatro variáveis analisadas. A levedura 28
LEV 169 (Rhodosporidium sp.) destacou-se por apresentar porcentagem média de 87,5% de 29
inibição da germinação de basidiósporos do patógeno. Quanto à inibição do crescimento de 30
M. perniciosa em meio líquido o isolado LEV 207 (Hannaella sp.) atingiu 100%, 31
demonstrando atuar na competição por nutrientes conforme os mecanismos avaliados e 32
supramencionados. Das leveduras identificadas, 18 do gênero Hannaella, quatro do gênero 33
Hanseniaspora, um do gênero Kwoniella, três do gênero Occultifur, 17 do gênero 34

73
Rhodosporidium, dois do gênero Pichia, um do gênero Sporobolomyces, um do gênero 35
Wicherhamomyces, um do gênero Yamadazyma e são relatadas pela primeira vez na 36
microbiota epifítica do cacaueiro e, mesmo apresentando comportamentos distintos para as 37
variáveis analisadas, são potenciais agentes de biocontrole da vassoura de bruxa do cacaueiro, 38
sendo recomendados para testes posteriores in vivo. 39
40
Termos para indexação: leveduras nativas, Moniliophthora perniciosa, Theobroma cacao, 41
bioprospecção, controle biológico. 42
43
44
Diversity and antagonism of native yeasts against the causal agent of the cocoa witch's 45
broom disease of cocoa 46
47
Abstract - The objectives of this study were to evaluate the diversity of epiphytic yeasts of the 48
cocoa and characterize them as potential antagonistic to Moniliophthora perniciosa. It 49
evaluated the behavior of diffusible and volatile compounds of native yeasts in inhibiting 50
mycelial growth and germination of basidiospores of M. perniciosa. Of the 172 yeast 51
evaluated, 18 showed ≤50% inhibition of growth of the four isolates studied pathogen. Eighty 52
yeast isolates were identified molecularly amplification of D1 / D2 region of the large subunit 53
(26S) rDNA. Isolates (LEV 162, 194, 210 and 211), identified as Hanseniaspora sp. Are in 54
the group of isolates showed the highest average for the four variables. The LEV 169 yeast 55
(Rhodosporidium sp.) stood out by presenting average percentage of 87.5% inhibition of the 56
pathogen basidiospores germination. As for the inhibition of M. perniciosa growth in liquid 57
isolated LEV 207 (Hannaella sp.) was 100%, demonstrating work in competition for nutrients 58
as the evaluated and above mechanisms. The identified yeast, 18 gender Hannaella, four 59
Hanseniaspora gender, one Kwoniella gender, three Occultifur gender, 17 of gender 60
Rhodosporidium, two Pichia genus, one Sporobolomyces gender, one Wicherhamomyces 61
gender, one Yamadazyma and gender are first reported in epiphytic microbiota cocoa and 62
even with different behaviors for the variables are potential biocontrol agents witch's broom 63
cacao, being recommended for further testing in vivo. 64
65
Index terms: Native yeast, Moniliophthora perniciosa, Theobroma cacao, bioprospecting, 66
biocontrol. 67
68

74
Introdução 69
70
O cacaueiro (Theobroma cacao L.) é considerado um dos cultivos perenes mais 71
importantes do mundo, com uma produção estimada em 4,2 milhões de toneladas de 72
amêndoas secas (ICCO, 2015). Esta espécie se desenvolve de forma espontânea em florestas 73
tropicais da América do Sul (Cuatrecasas, 1964) e é explorada comercialmente pela indústria 74
alimentícia para a produção de chocolate, geleias, polpas e outros derivados em países da 75
região equatorial das Américas, África e Ásia. 76
A Costa do Marfim é líder na produção mundial de cacau, com 42%, seguida de 77
Gana, com 17,9%, e Indonésia com 7,16%. Equador, Camarões e Brasil aparecem em 78
seguida dentre os maiores produtores. Os principais países consumidores de cacau são os 79
Estados Unidos que em 2014, apresentaram m o a g e m d e 418 mil toneladas de amêndoas 80
secas, seguidos pela Alemanha, França e Reino Unido (ICCO, 2015). O Brasil, que há três 81
décadas se manteve como um dos maiores produtores de cacau do planeta, produziu 273.793 82
toneladas de amêndoas secas no ano de 2014, o equivalente R$ 1.589.535 (IBGE, 2014), e 83
hoje ocupa a quinta posição (ICCO, 2015). 84
Durante muitos anos, a produção de cacau no sul do estado da Bahia contribuiu para 85
que o Brasil fosse um dos maiores produtores mundiais, entretanto, fatores econômicos, 86
problemas climáticos, e principalmente o surgimento da vassoura-de-bruxa (VB), causada 87
pelo fungo Moniliophthora perniciosa (Stahel) Aime & Phillips-Mora, ocasionaram uma 88
enorme crise sócio-econômica na região, levando o Brasil passar da situação de exportador 89
para importador de cacau (Midlej & Santos, 2012). 90
Apesar do impacto causado à lavoura cacaueira baiana, a vassoura de bruxa não está 91
restrita somente ao território brasileiro, mas ocorre em todos os países produtores da América 92
do Sul e Central como a Bolívia, Colômbia, Equador, Guiana, Peru, Venezuela, Panamá e nas 93
ilhas do Caribe, Trinidad, Tobago, Granada, Santa Lucia e São Vicente (Purdy & Schmidt, 94
1996). Em 2008, a doença foi relatada pela primeira vez em Union Vale, La Dauphine e Saint 95
Lucia, ilhas caribenhas (Kelly et al., 2009). 96
Mesmo com a diversificação através da exploração de outros cultivos, o cacaueiro 97
ainda é uma cultura de grande importância econômica, ecológica e social para os países 98
produtores. Grandes impactos na cultura do cacau foram provocados pela vassoura de bruxa e 99
esta permanece causando queda de produção, aumento dos custos e gastos diretos, em função 100
da necessidade do uso de medidas de controle, nem sempre aplicadas pelos produtores 101

75
descapitalizados. Sendo assim, torna-se de suma importância o estudo e a descoberta de 102
tecnologias que visem integrar o programa de manejo da cultura. 103
O controle biológico é um importante componente no manejo das doenças do 104
cacaueiro, entretanto o produto comercial Tricovab® desenvolvido e produzido pela 105
CEPLAC é o único produto biológico com registro no Ministério da Agricultura Pecuária e 106
Abastecimento (MAPA) para a comercialização e utilização nesta cultura. Mesmo com os 107
resultados promissores obtidos com o fungo Trichoderma stromaticum (princípio ativo do 108
Tricovab®) para o biocontrole do agente causal da VB, novos micro-organimos devem ser 109
investigados quanto ao potencial biocontrolador, principalmente os que co-habitam no mesmo 110
nicho, por serem ótimos competidores e competentes. As leveduras apresentam este potencial 111
por estarem tanto na filosfera quanto na rizosfera de vários cultivos, inclusive o cacaueiro 112
(Cabral et al., 2009). 113
Os micro-organismos são potenciais agentes biocontroladores por apresentarem 114
diversas características e mecanismos funcionais tais como o alto potencial competidor 115
(Zhang et al., 2007) ao secretarem toxinas “killer” (Platania et al., 2012), produzirem 116
compostos antifúngicos voláteis (Bruce et al., 2004) e secretarem enzimas hidrolíticas capazes 117
de degradar a parede celular de fitopatógenos (Lu et al., 2014; Masih & Paul, 2002; Urquhart 118
& Punja, 2002), capacidade de suportar mudanças no ambiente e condições de estresse (Sui et 119
al., 2015), tolerância a poluentes, óleos essenciais vegetais, fitoalexinas e alguns fungicidas 120
(Valdebenito-Sanhueza, 2000), além de sobreviverem saprofiticamente (Afsah-Hejri, 2013) e 121
tolerarem condições adversas (Hu et al., 2015). 122
Diante do exposto, buscamos neste estudo isolar, identificar e avaliar o potencial de 123
leveduras obtidas a partir de órgãos vegetais do cacaueiro como possíveis agentes de 124
biocontrole contra M. perniciosa. 125
126
Materiais e Métodos 127
128
Isolamento e preservação de fungos leveduriformes 129
Fungos leveduriformes foram obtidos a partir de diferentes órgãos vegetais (flor, folha 130
e fruto) de cacaueiro de acordo com o método descrito por Assis et al. (1999). As amostras 131
foram obtidas de plantas de cacaueiro pertencentes ao Programa de Melhoramento Genético 132
do Cacaueiro da CEPLAC e previamente selecionadas como padrão de resistência à 133
Moniliophthora perniciosa e M. roreri. Cinco repetições dos diferentes órgãos vegetais (flor, 134
folha e fruto) por genótipos foram coletadas e transportadas para isolamento em laboratório. 135

76
Um grama de cada amostra foi suspensa em 9 ml de solução salina (NaCl 0,85%) contendo 136
cloranfenicol (50 mg/L) e sonicados durante 15 min em ultrassonicador (Quimis 0335D). 137
Diluições em série da suspensão da amostra de levedura foram realizadas em solução salina + 138
cloranfenicol (50mg/L). Alíquotas de 300 µL de cada diluição foram distribuídas em placas 139
esterilizadas contendo o meio de cultura extrato de levedura peptona ágar - yeast extract 140
peptone dextrose agar (YEPD - 0,15% extrato de levedura, 1% neopeptona, 4% dextrose, 2% 141
ágar e 0,05% cloranfenicol). As placas foram incubadas a 28°C por 48 h e as colônias de 142
leveduras foram analisadas pelas características morfológicas de colônias e células. Uma 143
amostra representativa de cada colônia de levedura foi re-semeada em YEPD para obtenção 144
de culturas puras e posteriormente mantidos em tubos contendo meio batata dextrose ágar 145
(BDA) Himedia® à 4°C, e glicerol 2% a -80°C. 146
147
Organismos e condições de cultivo 148
Para cada experimento, os isolados de leveduras testados foram cultivados em meio 149
YEPD, em triplicata, por 72h a 28°C. Transcorrido o tempo de cultivo, as colônias de 150
leveduras foram raspadas e suspensas em solução salina (NaCl 0,85%). A concentração da 151
suspensão de leveduras foi ajustada a 108 ufc/mL com base na quantificação em câmara de 152
Neubauer e mensuração da densidade ótica com leitura (SpectraMax® Paradigm Multi-Mode 153
Detection Platform, Molecular-Devices Japan, Tóquio, Japão) a 595 nm. Quatro isolados de 154
M. perniciosa (Mp 4009, Mp 4010, Mp 4031 e Mp 4145) distintos molecularmente e 155
pertencentes à Coleção de Cultura Fúngica do Brazilian Culture Collection de Moniliophthora 156
(ex. Crinipellis) perniciosa (BCCCp), foram cultivados em meio BDA Himedia® e incubados 157
a 25°C por 15 dias. Basidiósporos foram obtidos a partir de vassouras coletadas na área da 158
Estação Experimental Arnaldo Medeiros, no CEPEC. As vassouras foram desinfestadas com 159
lavagens rápidas em hipoclorito de sódio a 0,1% e penduradas em arames esticados 160
horizontalmente sob um telado (vassoureiro), e então submetidas a um regime diário de 8 h de 161
molhamento e 16 h de secamento. Após o período de dormência das vassouras, foi aplicada 162
solução de estreptomicina a 1%, e lavadas mais duas vezes em água destilada. Os basidiomas 163
produzidos foram então coletados e secos em folhas de papel absorvente, seccionando os 164
píleos e fixando-os com vaselina pastosa (Silinol SG) em tampas de vidro, com himenóforos 165
voltados para baixo, sobre um béquer contendo uma solução de glicerina a 16%, e sob 166
agitação constante, permitindo a deposição dos basidiósporos liberados. As suspensões de 167
basidiósporos foram quantificadas em câmara de Neubauer e estocadas em nitrogênio líquido, 168
acondicionadas em tubos criogênicos de 2 mL, até o momento de serem utilizadas, conforme 169

77
metodologia descrita por Dickstein et al. (1987). Antes da estocagem, uma alíquota de 25 µL 170
de cada uma das suspensões de basidiósporos obtidas foi retirada e espalhada com alça de 171
Drigalski sobre a superfície do meio ágar-água a 2%, em placas de Petri de 5 cm de diâmetro, 172
para avaliar a germinação dos esporos. Após seis horas de incubação a 25°C, a percentagem 173
de basidiósporos germinados foi mensurada através da visualização em microscópio ótico. 174
Este procedimento foi realizado tanto no momento da estocagem como também vinte e quatro 175
horas antes da utilização do inóculo. Somente suspensões com mais de 80 % de basidiósporos 176
germinados foram utilizadas. 177
178
Detecção da atividade antifúngica por compostos difusíveis e voláteis 179
A atividade antifúngica dos isolados de levedura (172) foi avaliada através do 180
confronto em co-cultivo com quatro isolados de M. perniciosa distintos molecularmente (Mp 181
4009, Mp 4010, Mp 4031, Mp 4145), conforme Mariano et al. (2005). Discos (5mm) de meio 182
de cultura contendo estruturas fúngicas de M. perniciosa foram retirados da região periférica 183
do isolado em crescimento e repicados ao centro de placas de Petri (Ø 9,5 cm) contendo meio 184
BDA Himedia®. Os isolados de leveduras foram repicados em placas contendo isolados de 185
M. perniciosa, margeando os discos depositados ao centro da placa. As placas foram 186
incubadas por 10 dias à temperatura de 25°C em câmara de crescimento e avaliadas quanto à 187
inibição do crescimento micelial, através da mensuração diária do diâmetro da colônia a partir 188
do sexto dia. O diâmetro da colônia foi avaliado com paquímetro Mitutoyo® (Japão) em duas 189
medidas ortogonais. O delineamento experimental foi inteiramente casualizado, com cinco 190
repetições, sendo cada repetição constituída por uma placa de Petri. A testemunha consistiu 191
de placas contendo meio de cultura, semeado apenas com o patógeno. Quando o crescimento 192
do patógeno na testemunha alcançou os bordos das placas, as porcentagens de inibição 193
(PIN%) foram calculadas de acordo a fórmula de Topps e Wain (1957): I% = [(A - B) / A] x 194
100 onde, I% = percentagem de inibição de A = diâmetro médio do crescimento no controle, 195
B = crescimento médio de diâmetro num dado tratamento. 196
197
Determinação do potencial produtor de compostos voláteis com ação antifúngica 198
Para detecção de compostos voláteis com ação antifúngica, foi utilizada a técnica de 199
cultivo em placas de Petri bipartidas contendo meio BDA Himedia®. O confronto foi 200
realizado entre 66 isolados de leveduras previamente selecionados com atividade antifúngica 201
à M. perniciosa e os quatro isolados deste. Em um lado da placa foram realizadas estrias 202
contínuas com o isolado de levedura e no centro, do outro lado da placa, depositado um disco 203

78
contendo estruturas fúngicas do patógeno. As placas foram incubadas por 10 dias à 204
temperatura de 25°C em câmara de crescimento e avaliadas diariamente através da 205
mensuração com paquímetro, com precisão de abertura de 0,5 mm, em sentidos 206
diametralmente opostos. O delineamento experimental foi inteiramente casualizado, com 207
cinco repetições, sendo cada repetição constituída por uma placa de Petri. A testemunha 208
consistiu de placas contendo meio de cultura semeada apenas com o patógeno. A 209
porcentagem de inibição do crescimento foi calculada conforme Topps e Wain (1957). 210
211
Aplicação de bioinformática para avaliação dos compostos difusíveis e voláteis 212
Uma matriz gerada para os tratamentos (isolados de levedura) foi avaliada através do 213
software R 2.12, o que permitiu a construção de heatmaps a partir de distância Euclidiana, 214
para avaliar a ação de compostos difusíveis e voláteis na inibição do crescimento micelial de 215
4 isolados de M. perniciosa. 216
217
Determinação do potencial inibidor da germinação de basidiósporos 218
Os isolados de levedura a serem testados foram cultivados em meio YEPD líquido 219
incubados sob agitação de 150 e 28°C em incubadora com agitação orbital tipo Shaker 220
(ACBLABOR® AC-830) por 72h. A obtenção do secretoma ou extrato bruto foi baseado no 221
trabalho de Coelho et al. (2011) e Cabanas (2013) com otimizações em seus métodos. Foram 222
utilizados 45 ml de meio líquido YEPD mais 5 ml da suspensão de leveduras padronizadas à 223
108 ufc/mL através da mensuração de densidade ótica em espectrofotômetro a 595 nm. A 224
amostra foi centrifugada a 6.500 g por 15 minutos a 10ºC, com separação entre o secretoma e 225
as células das leveduras. O sobrenadante foi filtrado com seringa estéril e filtro para seringa 226
Millipore® 0,22 µm, separando células remanescentes e posteriormente armazenado em 227
ultrafreezer a -80ºC. Alíquotas do secretoma das leveduras testadas e suspensão de 228
basidiósporos (106) na proporção 1:1 foram homogeneizadas em microtubos eppendorf 2 mL 229
e vertidas em placas de Petri (60 x 15mm). O desenho experimental foi inteiramente 230
casualizado, com duas placas divididas em quadrantes e cada quadrante constituindo uma 231
repetição, totalizando oito repetições/tratamento. A avaliação do ensaio foi realizada 6 h após 232
o início da incubação, através da contagem do número de esporos totais e germinados, no 233
campo de visão da objetiva de 40X do microscópio óptico, em três pontos distintos de cada 234
quadrante. Foi considerado germinado o basidiósporo cujo tubo germinativo era maior ou 235
igual ao seu comprimento e calculada a porcentagem de esporos em cada repetição. 236
237

79
Determinação do fator “killer” 238
A atividade “killer” foi investigada através do co-cultivo dos isolados de levedura 239
selecionados e linhagens de referência como padrão de sensibilidade “killer”. Os testes foram 240
realizados utilizando a técnica de semeadura em meio ágar YEPD-MB (extrato de levedura 241
1%, glicose 2%, peptona 2%, ágar 2%, azul de metileno 0,003%, tamponado ao pH 4,0 com 242
tampão citrato fosfato 0,5 M) contido em placa de Petri conforme descrito por Sangorrín et al. 243
(2001). As leveduras foram ressuspensas em solução salina 0,85% até a concentração de 244
aproximadamente 5 x 105 ufc/mL. Em seguida, 100μL desta suspensão celular correspondente 245
a cada uma das linhagens sensíveis (Saccharomyces cerevisae NCYC1006 e Candida 246
glabrata Y55 ATCC90525) foram espalhados, com auxílio da alça de Drigalski, sobre a 247
superfície do meio ágar YEPD-MB contido em placas de Petri (90 x 15 mm). Após a secagem 248
da suspensão, os isolados de levedura foram inoculados em forma de ponto com auxílio da 249
alça de platina, sobre cada uma das placas correspondentes a uma linhagem sensível de 250
referência, de forma que, em cada placa de Petri, foi possível inocular 11 isolados teste mais a 251
linhagem produtora de toxina “killer”. O delineamento experimental foi inteiramente 252
casualizado, com três repetições, senda cada repetição constituída por uma placa de Petri. As 253
placas foram incubadas em estufa B.O.D. a 25 C por 72 horas e observadas diariamente. A 254
presença do fenótipo “killer” (K +) foi constatada nos isolados em que houve formação de 255
uma zona clara de inibição em volta da colônia, onde não houve crescimento da levedura 256
sensível, delimitada por um halo adjacente de células coradas em azul que evidencia morte 257
celular e é indicativo de atividade “killer” (Young, 1987). 258
259
Análise multivariada dos dados de atividade antifúngica, compostos voláteis, inibição da 260
germinação de basidiósporos e atividade “killer” 261
A análise dos dados binários e agrupamento por dissimilaridade dos perfis, utilizando 262
o algoritmo UPGMA (Unweighted Pair-Grou Method wiht Arithmetical Average) e distância 263
euclidiana, foi realizada através da utilização do software SAS® (SAS Corporation, Cary, 264
NC). Foi empregada a Análise de Componentes Principais (Principal Coordenates Analysis – 265
PCA) para avaliar a variação entre os grupos encontrados nas diferentes análises (Clarke, 266
1993). 267
268
Identificação molecular das leveduras 269
Para a identificação molecular das leveduras isoladas de diferentes órgãos do 270
cacaueiro, foi realizada a extração do DNA total utilizando o kit UltraClean® Microbial DNA 271

80
Isolation (MoBio, USA), seguindo recomendações do fabricante. A integridade e a quantidade 272
do DNA foram verificadas usando eletroforese em gel de agarose a 1,0% corado com brometo 273
de etídio e visualizado sobre luz ultravioleta, seguida da fotodocumentação. As amplificações 274
da região D1/D2 da subunidade maior (26S) do rDNA foram realizadas com os primers NL1 275
(5’- GCATATCAATAAGCGGAGGAAAAG - 3’) e NL4 (5’-276
GGTCCGTGTTTCAAGACGG-3’) (O’DONNELL, 1993). Para tanto, as reações foram 277
preparadas com os seguintes reagentes e concentrações: 60 ng de DNA de cada amostra; 1x 278
de tampão da enzima Taq DNA polimerase; 3,7 mM de MgCl2; 0,6 pmol/μL de dNTPs; 0,4 279
pmol/μL de cada primer; 2,5 U de Taq DNA polimerase, com volume final ajustado para 50 280
μL com água ultra pura. Os ciclos de amplificações foram realizados no Veriti Thermal 281
Cycler PCR (Appplied Biosystems) com temperatura inicial de desnaturação 95 °C por 5 282
minutos; 35 ciclos de amplificação a 95 °C por 30 segundos; 54 °C por 30 segundos; 72 °C 283
por 7 minutos e temperatura de extensão final de 72 °C por 10 minutos. Os produtos 284
amplificados foram visualizados em gel de agarose a 1%, corados com brometo de etídio e 285
visualizados sobre luz ultravioleta. Em seguida, os amplicons foram purificados utilizando o 286
kit PureLink™ PCR Purification (Invitrogen), quantificados com o Qubit® 2.0 Fluorometer 287
(Invitrogen) para posterior identificação nucleotídica, utilizando o sequenciador automático 288
ABI-Prism 3500 Genetic Analyzer (Applied Biosystems). A edição e montagem das 289
sequências foram realizadas com o programa Sequencher 4.1.4 (Gene Code Corporation). A 290
identidade taxonômica dos isolados foi verificada através do banco de dados GenBank, 291
usando o BLASTn “basic local alignment search tool” (BLAST) do NCBI 292
(http://www.ncbi.nlm.nih.gov). 293
294
Avaliação da interação levedura x Moniliophthora perniciosa em meio de cultivo líquido 295
Foi realizado um co-cultivo da LEV 207 com o patógeno M. perniciosa (Mp 4009) em 296
erlenmeyers de 125mL contendo 22,5 mL de meio batata dextrose (BD), adicionando a estes 297
alíquotas de 2,5 mL de suspensão de células do isolado de levedura e disco contendo 298
estruturas fúngicas do patógeno em diferentes tipos de tratamento e reinóculo da levedura, 299
com agitação de 150 rpm a 28ºC em termo shaker. Os tratamentos utilizados foram: T1 – 107 300
células da levedura 207; T2 – M. perniciosa; T3 - 107 células da levedura 207 + M. perniciosa 301
(simultâneo); T4 - 107 células da levedura 207 + M. perniciosa (simultâneo) + 1 reinóculo de 302
107 células da levedura 207 (120h após); T5 - 107 células da levedura 207 + M. perniciosa 303
(simultâneo) + 2 reinóculo de 107 células da levedura 207 (96 e 1920h após); T6 - 107 células 304
da levedura 207 + M. perniciosa (simultâneo) + 3 reinóculo de 107 células da levedura 207 305

81
(48, 144 e 216h após); T7 - 107 células da levedura 207 + M. perniciosa (simultâneo) + 1 306
reinóculo de 107 células da levedura 207 (48, 96, 144 e 192h após); T8 - 107 células da 307
levedura 207 + M. perniciosa (simultâneo) + 1 reinóculo de 107 células da levedura 207 (192h 308
após); T9 - M. perniciosa + 107 células da levedura 207 (168h após + 1 reinóculo 216h após). 309
O reinóculo foi realizado preservando a inocuidade da amostra, sendo retirada 310
momentaneamente das condições de cultivo e retornando em seguida. Dez dias após o início 311
do ensaio, as amostras foram coletadas e filtradas a vácuo em papel filtro Whatman® 110 mm 312
Ø, quantificando em seguida a biomassa fresca e seca em balança analítica. A amostra seca 313
foi obtida a partir da desidratação em estufa a 70°C por 48h. O consumo de glicose foi 314
inferido pela quantificação de açúcares redutores em ensaio de DNS (ácido dinitrosalicílico). 315
O experimento foi realizado em delineamento inteiramente casualizado em triplicata. 316
317
Resultados 318
319
Foram encontrados 172 isolados de leveduras na superfície de folhas (62,8%), frutos 320
(12,8%) e flores (24,4%) de genótipos de cacaueiro selecionados como portadores de genes 321
de resistência à vassoura de bruxa (VB) e monilíase. A maior quantidade de isolados oriundos 322
do limbo foliar se deve à maior disponibilidade deste órgão vegetal. Leveduras foram 323
encontradas em 100% das plantas amostradas. A densidade total de leveduras variou de 3,2 x 324
104 ufc/g a menos de 50 ufc em fruto; de 2,8 x 104 ufc/g a menos de 50 ufc em flor; e de 2,0 x 325
104 ufc/g a menos de 50 ufc em folha. Devido a maior disponibilidade de genótipos com 326
resistência a VB, 84% dos isolados de levedura foram obtidos destes materiais e apenas 16% 327
de genótipos utilizados como padrões de resistência a monilíase. 328
A análise de agrupamentos (heatmap) baseada na ação antifúngica de 172 isolados de 329
leveduras nativas do cacaueiro (eixo y) à quatro isolados distintos do patógeno M. perniciosa 330
em cinco dias de avaliação (6 – 9 dias após pareamento) (eixo x), permitiu inferir sobre a 331
separação de dois grupos (clados) (Figura 1). A maioria dos isolados de leveduras avaliados 332
foi capaz de inibir o crescimento micelial dos quatro isolados do patógeno com níveis acima 333
de 50%, à exceção dos isolados LEV 136, LEV 019 e LEV 110. Estes três isolados fazem 334
parte do clado que reúne as leveduras que menos inibiram o crescimento do patógeno (Figura 335
1). 336
Os isolados de levedura LEV 079, 139, 002, 047, 023, 034, 210, 194, 211, 030, 028, 337
097, 227, 204, 206, 189, 007, 021, 162 e LEV 212 destacaram-se por compor o clado de 338

82
segundo nível que se distingue das demais leveduras que mais inibiram o crescimento dos 339
quatro isolados do patógeno nos cinco dias de avaliação. 340
O padrão de inibição de cada levedura não foi uniforme para todos os isolados de M. 341
perniciosa, apresentando variações entre os isolados e dentre os dias de avaliação para cada 342
isolado (Figura 1). Houve variações na velocidade de crescimento e produção de compostos 343
antimicrobianos difusíveis e voláteis. O isolado Mp 4145 apresentou a menor velocidade de 344
crescimento dentre os demais isolados do patógeno testados e, devido a esta característica, 345
quando os pareamentos com as leveduras foram comparados com testemunhas somente com 346
plug (discos de 0,5 cm) do patógeno, geraram um falso positivo em relação à sensibilidade 347
deste isolado aos compostos antifúngicos secretados pelas leveduras, caracterizando-o como o 348
menos sensível. O potencial fungistático das leveduras foi semelhante para os isolados de Mp 349
4010 e Mp 4031, já para o isolado Mp 4009 foram identificados os maiores percentuais de 350
inibição (Figura 1). 351
Houve uma tendência crescente na evolução do percentual de inibição ao longo dos 352
dias para a maioria dos tratamentos avaliados, entretanto alguns tratamentos apresentaram 353
inibição constante durante todo o período de avaliação para alguns isolados: LEV 039, 168, 354
169, 18, 006, 164, 018, 166, 182, 067, 036, 039, 010, 084, 023, 047, 002, 139, 179, 085, 212, 355
013 e LEV 202 para o isolado Mp 4009; LEV 010, 067, 212, 021, 162, 097, 078, 194 e LEV 356
210 para o isolado Mp 4010; LEV 001, 018, 006, 212, 084, 002, 139, 162, 021, 097, 078, 194 357
e LEV 210 para o isolado Mp 4031; LEV 006, 018, 182, 036, 039, 010, 047, 162 e LEV 021 358
para o isolado Mp 4145. Também foi observado oscilação de inibição ao longo dos dias de 359
avaliação para as leveduras LEV 067, 202, 064, 069, 096, 074, 062, 110, 210 e LEV 211. As 360
leveduras LEV 039, 074, 212, 096 e LEV 069 apresentaram tendência decrescente na 361
evolução do percentual de inibição para alguns isolados de M. perniciosa (Figura 1). 362
A maioria dos tratamentos (leveduras) oscilaram em relação ao potencial inibidor do 363
crescimento micelial por compostos voláteis, entretanto alguns mantiveram constância 364
durante os dias de avaliação para alguns dos quatro isolados de M. perniciosa testados (Figura 365
2): Os isolados de levedura LEV 165, 002, 118, 084, 140, 111, 169, 082, 209, 059, 226 e LEV 366
088 para o isolado Mp 4009; LEV 002, 101, 140, 161, 178, 109, 090, 077, 185, 189 e LEV 367
225 para o isolado Mp 4010; LEV 162, 118, 166, 149, 143, 111, 178, 109, 100, 211 e LEV 368
080 para o isolado Mp 4031; e o somente LEV 162 para o isolado Mp 4145. 369
Para a avaliação de compostos voláteis, houve a formação de dois grupos (clados) de 370
levedura separados por distância euclidiana e visualizadas em heatmap (Figura 2). Vinte e 371
dois isolados (33,33 %), LEV 001, 014, 038, 139, 229, 088, 027, 078, 147, 023, 034, 115, 372

83
079, 063, 080, 226, 059, 209, 082, 169, 170 e LEV 225 compõem o clado que se distingue por 373
apresentar os menores percentuais de inibição, entretanto os isolados LEV 170, LEV 169 e 374
LEV 082, quando avaliados somente com os isolados Mp 4009, Mp 4010 e Mp 4031, 375
apresentam comportamento semelhante aos isolados de levedura que mais inibiram o 376
crescimento do patógeno. 377
Dentre os indivíduos que compõem o grupo que apresentou os maiores percentuais de 378
inibição por compostos voláteis, quatro isolados (LEV 225, LEV 206, LEV 166, LEV 092) se 379
distanciam em um subgrupo (clado de 2º nível) por se destacarem com percentuais acima de 380
50% de inibição, principalmente para os isolados Mp 4009 e Mp 4010. Leveduras 381
pertencentes ao segundo subgrupo formado (LEV 074, LEV 210, LEV 089, LEV 189 e LEV 382
211), mantiveram o padrão de inibição para os quatro isolados de M. perniciosa (Figura 2). 383
Todos os filtrados obtidos de cultivo de 66 isolados de levedura em meio YEPD, em 384
diluição 1:1 (filtrado:suspensão de basidiósporos), inibiram a germinação de basidiósporos 385
em meio ágar água após seis horas de incubação, quando comparados com a testemunha 386
(Figura 3). Três grupos foram formados a partir da comparação de médias pelo teste Scott-387
Knott. Dentre os isolados que mais inibiram, dezenove se destacaram (LEV 049, 063, 065, 388
093, 101, 105, 111, 147, 149, 166, 168, 169, 185, 204, 207, 210, 211, 219 e LEV 225) por 389
apresentarem percentuais de inibição de germinação de basidiósporos variando de 54,5 – 390
87,5%, produzidos por LEV 014 e LEV 169 respectivamente (Figura 3). 391
Apenas 13 isolados (19,69 %) não produziram halo de inibição quando cultivados com 392
padrões “killer” sensíveis: LEV 049, 077, 78, 88, 090, 092, 101, 106, 109, 118, 143, 161 e 393
LEV 229 (Figura 3). Apenas dois isolados (LEV 049 e LEV 101) foram do grupo com 394
maiores inibições. 395
Os resultados da análise multivariada mostraram haver diferença significativa para os 396
tratamentos (leveduras) e variáveis (antibiose, compostos voláteis, inibição de germinação e 397
atividade “killer”). 398
Na distribuição biplot (Figura 4), observa-se a dispersão dos isolados de levedura no 399
gráfico tridimensional das três primeiras componentes principais (variáveis sintéticas), que 400
juntas, correspondem a 79,78 % da variação total da análise. 401
O componente principal 1(PC1) representa 31,72 % da variação total e a variável 402
antibiose (AB) representa a maior variação desta componente por apresentar o maior 403
coeficiente (auto vetor: 0,62). As variáveis atividade “killer” (AK), compostos voláteis (CV) e 404
inibição da germinação (IG) apresentaram coeficientes de 0,60, 0,48 e 0,06, respectivamente. 405
A variável IG apresentou maior peso (0,85) na variação da componente principal 2 (PC2), já 406

84
para a componente principal 3 (PC3), a variável CV foi a que mais contribuiu para a variação 407
deste componente. As componentes CP2 e CP3 são responsáveis por 26 % e 22% da variação 408
total dos dados, respectivamente. 409
Quanto menor o ângulo entre os vetores das variáveis, maior a correlação. A variável 410
AB não apresentou alta correlação com as demais variáveis, porém se correlacionou 411
positivamente com as variáveis CV e AK, mais do que com a inibição da germinação (IG). A 412
correlação negativa entre as variáveis CV e IG é demonstrada através da vetorização oposta. 413
Os coeficientes dos autovetores explicam a correlação entre as variáveis reais e as variáveis 414
sintéticas, entretanto destaca-se a correlação negativa significativa da variável CV para PC2 e 415
variáveis AB e AK para PC3. A relação entre as variáveis (vetores) e distribuição dos 416
tratamentos (dispersão) são explicadas pela análise de componentes principais (PCA) 417
representada no biplot (Figura 4). 418
Os tratamentos que apresentaram as maiores médias em cada uma das variáveis são 419
representados na figura 4 próximos ao ápice do vetor, como os tratamentos LEV 169 e 219 420
para a variável IG, localizados no ápice do biplot. Quando os tratamentos apresentam as 421
menores médias para as variáveis, estes localizam-se em direção oposta ao vetor da variável 422
analisada, como os tratamentos LEV 49, 106, 88, 78, 109, 229, 74, 148, 90, 100, 179, 92, 161, 423
143, 118 que se distanciam inversamente do vetor PC1, indicando que esses isolados 424
apresentaram as menores médias para as variáveis que mais explicam a variação PC1, AB e 425
AK. 426
O LEV 210 está em posição central em relação aos vetores das quatro variáveis, por 427
ser o tratamento que mais se destacou com primeira, segunda e quarta maior média para as 428
variáveis AB, CV e IG, além de apresentar a característica qualitativa AK. Apesar do 429
tratamento LEV 169 apresentar a maior média para a variável IG, este se encontra próximo 430
apenas do vetor desta variável e do vetor CP2, componente que mais sofre influência desta 431
variável. O tratamento LEV 169 não proporcionou alta inibição do crescimento micelial por 432
compostos difusíveis e voláteis. 433
A divergência do comportamento dos tratamentos (leveduras) nas quatro variáveis 434
analisadas foi observada pelo dendograma de análise de agrupamento (Figura 5), formando 435
seis grupos distintos. No grupo 1, formado pelos tratamentos LEV 169 e 219, encontram-se os 436
dois isolados que mais inibiram a germinação (IG) dos basidiósporos de M. perniciosa, 437
entretanto sua atuação na inibição do crescimento micelial do patógeno foi mediana e 438
atribuída em maior parte aos compostos difusíveis e toxinas “killer”, pois apresentaram baixa 439
inibição do crescimento micelial por compostos voláteis (Tabela 2). O grupo 2 é formado por 440

85
nove leveduras com resultados medianos para todas as variáveis analisadas, sendo importante 441
ressaltar que apresentaram as maiores médias para a produção de compostos voláteis, mesmo 442
esta característica correspondendo com 50% para a inibição do crescimento micelial, a 443
exemplo do isolado LEV 174 que inibiu 50,28 % e 24, 58 % do crescimento micelial de 444
isolados de M. perniciosa por compostos difusíveis e voláteis (em conjunto) e voláteis 445
isoladamente (Figuras 2 e 3, respectivamente). Mesmo sendo agrupados separadamente, os 446
tratamentos do grupo 3 também apresentaram comportamento mediano. O maior número de 447
tratamentos (40%) foi reunido no grupo 4 por atuarem na inibição da germinação e 448
crescimento micelial principalmente por compostos difusíveis, pois apresentaram baixos 449
percentuais para a variável compostos voláteis. O grupo 5 se assemelha ao grupo 4 por reunir 450
tratamentos que atuam principalmente por compostos liberados no meio de cultivo, entretanto 451
difere por apresentar percentual de inibição do crescimento micelial maior do que o grupo 4. 452
Um sexto grupo foi formado pela proximidade de quatro tratamentos que apresentaram 453
atuação média-baixa para os parâmetros avaliados. 454
Sessenta isolados de levedura foram selecionados como amostras significativas do total de 455
leveduras isoladas por inibirem o crescimento micelial de M. perniciosa acima de 60% e 20 456
isolados selecionados aleatoriamente foram adicionados a este grupo, formando um conjunto 457
de 80 isolados com características representativas, de inibição do crescimento micelial e 458
germinação de basidiósporos, além de plasticidade fenotípica, correspondendo a 46,5% do 459
total de leveduras isoladas (Tabela 1). 460
Amostras do DNA genômico das 80 amostras foram sequenciadas e indexadas (Index 461
fungorum) como integrantes dos filos Ascomycota (25%) e Basidiomycota (67,5%) e seis 462
com identidade indeterminada (7,5%). Em relação ao filo Ascomycota, houve a 463
predominância da família Saccharomycetaceae e apenas dois integrantes da família 464
Pichiaceae. Integrantes do filo Basidiomycota foram identificados como membros de duas 465
famílias (Trichosporonaceae e Cystobasidiaceae) e isolados com incapacidade de estabelecer 466
posição exata de sua família dentro da classificação (Insertae sedis). Foram identificadas 14 467
espécies e 7 gêneros, levando em consideração as sinonímias. Os fungos leveduriformes 468
isolados com a maior frequência eram da ordem Tremellales, principalmente do gênero 469
Hannaella (18 isolados) e o gênero Rhodosporidium (17 isolados) dentre os isolados 470
sequenciados. Não houve predominância de gêneros representantes de um determinado filo 471
por genótipo ou órgão de cacaueiro, demonstrando haver grande diversidade na distribuição 472
de leveduras na planta (Tabela 1). 473

86
Houve variação em relação a prevalência de gêneros de leveduras por genótipo, 474
padrão de resistência (VB e monilíase) e órgão vegetal do cacaueiro. Os tratamentos LEV 169 475
e LEV 219 são formados por metabólitos de leveduras identificadas como Rhosdosporidium. 476
sp. e Candida sp. e isoladas de folha e fruto de genótipos de cacaueiro selecionados com 477
resistência a vassoura de bruxa (VB) e monilíase, respectivamente (Tabela 2). Os isolados 478
LEV 162, LEV 194, LEV 210 e LEV 211, foram identificados como Hanseniaspora sp., 479
sendo o primeiro obtido de folha e os três últimos de flores de diferentes genótipos de 480
cacaueiro, três dos quais selecionados para resistência à VB e um para monilíase (Tabela 2). 481
Os metabólitos obtidos destes micro-organismos mantiveram alto padrão de inibição para 482
todas as variáveis analisadas. 483
. O isolado 207 (Hannaella sp.) foi selecionado dentre os isolados pertencentes ao 484
gênero com maior prevalência, para avaliar a interação direta entre a levedura e o patógeno, 485
além de inferir sobre a competição por nutrientes através da determinação de açúcares 486
redutores. Quando cultivados em meio líquido, o isolado LEV 207 (Hannaella sp.) inibiu o 487
crescimento de M. perniciosa em todos os tratamentos avaliados, independentemente do 488
número de reinóculo e época de aplicação (Figura 6). A biomassa micelial do patógeno, 489
mensurada após 240h de cultivo, foi semelhante para as variáveis biomassa fresca (BF) e 490
biomassa seco (BS) em relação aos tratamentos avaliados (Figura 6A-B). Os pesos fresco e 491
seco do tratamento 1, correspondente ao cultivo apenas do fungo M. perniciosa, diferiram 492
significativamente dos demais tratamentos que receberam alíquotas de suspensão de células 493
(108 ufc/mL). Os tratamentos em que a levedura foi adicionada ao meio em concomitância 494
(T3, T4, T5, T6, T7) proporcionaram inibição mínima de 95,3% (Figura 6B), já os 495
tratamentos em que foi adicionado a levedura 192h (T8) e 168h com reinóculo após 216h 496
(T9), mesmo apresentando médias maiores, não diferiram estatisticamente (p<0.05) em 497
relação à variável PS. Quando a levedura foi adicionada ao meio de cultivo apenas uma vez, 498
em conjunto com M. perniciosa, no início do experimento, a inibição do crescimento micelial 499
de M perniciosa não diferiu dos tratamentos com 1 (T4), 2 (T5), 3 (T6), 4 (T7) reinóculos, 500
demonstrando que o isolado LEV 207 não necessita de reinóculos para aumentar a capacidade 501
de inibição do crescimento do patógeno in vitro. Mesmo não apresentando diferença 502
significativa (p<0.05) em relação aos tratamentos T3, T4, T5, o peso seco de co-cultivos do 503
T7 (4 reinóculos) possibilitou um maior crescimento do patógeno, indicando que a partir de 3 504
reinóculos, o aumento do número de células de levedura não influencia positivamente na 505
inibição do crescimento micelial do patógeno. 506

87
O consumo de carboidratos no meio de cultivo foi maior para tratamentos que 507
receberam alíquotas de suspensões de células de levedura, não havendo diferença significativa 508
(p<0.05), entre T2, tratamento com crescimento apenas de levedura, e os tratamentos T3, T4, 509
T5, T6 e T7, onde foram realizados co-cultivos de levedura e M. perniciosa (Figura 6C). Nos 510
tratamentos T8 e T9, que receberam células de levedura tardiamente, sem ou com um 511
reinóculo respectivamente, foram identificadas as maiores concentrações de açúcares 512
redutores. 513
Houve diferenças significativas (p<0,05) em relação à concentração de células de 514
levedura nos cultivos dos tratamentos avaliados (Figura 6D). O tratamento T3 (apenas uma 515
alíquota de células de levedura no início do co-cultivo) e os tratamentos T5 e T6 (alíquotas no 516
início do co-cultivo + 2 e 3 reinóculos respectivamente), diferiram do tratamento T2, onde 517
houve crescimento de levedura na ausência do patógeno, com média menor e maiores, 518
respectivamente. Foi identificado um decréscimo na concentração de células no meio de 519
cultivo após 3 reinóculos (T7) e baixo crescimento em tratamentos que receberam células de 520
leveduras após o crescimento micelial do patógeno por no mínimo 168h em meio caldo de 521
BD (Figura 6D). 522
523
Discussão 524
525
Vários trabalhos têm relatado a diversidade de leveduras associadas ao processo de 526
fermentação de amêndoas do cacaueiro (Daniel et al., 2009; Moreira et al., 2013), bem como 527
a utilização destes microorganismos como potencializadores de atributos da qualidade do 528
produto destinado a fabricação de chocolate (Leal Jr et al., 2008). Entretanto o potencial 529
biotecnológico destes microorganimos ainda se restringe ao desenvolvimento de tecnologias 530
para o beneficiamento do cacau. 531
Trabalhos a respeito da bioprospecção de microorganimos e metabólitos de interesse 532
são citados na literatura (Duarte et al., 2013; Gohel et al., 2006; McCormick et al., 2012), 533
entretanto evidencia-se o elevado número de organismos avaliados devido a diversidade de 534
mecanismos de ação e interação com outros organismos e meio ambiente. Rubini et al. (2005) 535
avaliaram a diversidade de fungos endofíticos em ramos sadios e infectados por M. perniciosa 536
de genótipos de cacaueiro selecionados como resistentes e suscetíveis à vassoura de bruxa, e 537
apesar de um total de 265 fungos endofíticos avaliados, apenas 16,22% dos isolados inibiram 538
o crescimento micelial do patógeno e apenas o isolado identificado como Gliocladium 539
catenulatum reduziu em 70,84% os sintomas da vassoura de bruxa em plântulas. Os 540

88
resultados alcançados na fenotipagem dos isolados de levedura obtidos quanto à inibição do 541
crescimento e germinação de esporos de M. perniciosa permitem inferir sobre a capacidade de 542
estabelecimento e colonização de sítios de infecção ocupados por estes micro-organimos e 543
consequente controle da vassoura de bruxa. 544
A maioria dos fungos conhecidos estão associados a outros organismos através de 545
relações parasíticas, comensalísticas ou mutualísticas (Barbieri & Carvalho, 2001). A 546
superfície de órgãos vegetais tem sido descrita como substrato colonizado por diversos micro-547
organimos como bactérias, fungos leveduriformes e filametosos (Andrews & Harris, 2000; 548
Leff & Fierer, 2013; Mautone, 2008), que desenvolvem estratégias de estabelecimento, 549
desenvolvimento e competição. O isolado 207 (Hannaella sp.) inibiu completamente o 550
crescimento de M. perniciosa quando cultivados concomitantemente em caldo BD devido a 551
rápida capacidade de reprodução e assimilação de nutrientes inerente à maioria das leveduras, 552
conferindo à estes micro-organimos uma ótima ação competitiva no ambiente em que se 553
encontram, sendo a competição por nutrientes e espaço um dos principais mecanismos de 554
biocontrole (Lahlali et al., 2011; Zhao et al., 2011). Alguns representantes do gênero Candida 555
e Rhodotorula já foram relatados como produtores de compostos quelantes de íons férricos 556
(sideróforos) (El-Tarabily & Sivasithaparam, 2006; Rosa et al., 2010). 557
Fungos leveduriformes preferencialmente colonizam ambientes ricos em carboidratos, 558
como a carposfera, espermosfera e antosfera, mas também estão presentes na superfície de 559
folhas, atuando por mecanismos diversos como produção de enzimas hidrolíticas, toxinas 560
“killer”, inibição da germinação de esporos fúngicos (Lutz et al., 2013), produção de 561
fitohormôios (Sun et al., 2014), indução de resistência (Lu et al., 2013) e formação de 562
biofilmes (Li et al., 2015), possibilitando a sua estabilização em ambientes hostis, expostos a 563
variações de temperatura, umidade relativa. A anatomia e morfologia dos órgãos vegetais, 564
como estômatos, nervuras, e apêndices na superfície, incluindo tricomas e hidatódios, podem 565
afetar a disponibilidade de nutrientes na filosfera, contribuindo assim para o estabelecimento 566
de microorganimos nestes substratos (Leveau & Lindow, 2001), sendo assim, muitos 567
genótipos selecionados para a resistência a doenças de plantas, podem ser favorecidos pela 568
interação com microorganimos benéficos capazes de potencializar características fenotípicas 569
desejáveis. 570
A prospecção de microorganismos que co-habitem o mesmo nicho do patógeno 571
aumenta a chance de eficiência na seleção de indivíduos com características relacionadas ao 572
biocontrole, pois a seleção do antagonista é relativa para cada patossistema (Bettiol, 1997). 573

89
Dentre os microorganismos citados na literatura como agentes de biocontrole, El-574
Tarabily e Sivasithamparan (2006) destacam leveduras pertencentes aos gêneros 575
Debaryomyces, Kloeckera, Saccharomyces, Zygosaccharomyces, Sporobolomyces, 576
Metschnikowia, Tilletiopsis, Rhodotorula, Cryptococcus, Aureobasidium, Pichia e Candida. 577
Sun et al. (2014) isolaram leveduras da filosfera de Drosera indica L. e identificaram o 578
potencial de leveduras pertencentes aos filos Ascomycota (Aureobasidium pullulans, Candida 579
apicola e Candida floricola) e Basidiomycota (Cryptococcus flavus, Hannaella 580
coprosmaensis, Pseudozyma aphidis, Sporisorium cruentum, Sporisorium reilianum, Ustilago 581
alcornii), para a produção de ácido indol-acético (AIA). 582
A espécie Hanseniaspora uvarum , pertencente à família Saccharomycetaceae, ordem 583
Saccharomycetales, classe Saccharomycetes, filo Ascomycota, é amplamente utilizada no 584
processo de fermentação de uvas para a produção de vinhos (Escalante et al., 2011; Moreira et 585
al., 2011) e produção de enzimas de interesse agroindustriais (López et al., 2016), além de ser 586
relatada como micro-organismo utilizado para o controle de patologias pós colheita 587
(Janisiewicz et al., 2010), atuando através da secreção de toxinas “killer” (Schmiti & 588
Neuhausen, 1994) compostos voláteis (Moreira et al., 2011), corroborando com os resultados 589
alcançados por este trabalho, onde todos os isolados deste gênero (LEV 162, LEV 194, LEV 590
210 e LEV 211) se destacaram com alto potencial antagônico à M. perniciosa para as 591
variáveis analisadas. Apesar da H. uvarum já ser citada como integrante da microbiota 592
presente na fermentação de amêndoas de três híbridos de cacaueiro, este é o primeiro relato 593
deste gênero habitando a superfície de folhas e flores de cacaueiro, além do seu potencial para 594
inibir o crescimento de M. perniciosa. 595
Outros gêneros da família Saccharomycetaceae, como Rhodosporidium, 596
Wicherhamomyces, Hansenula, Pichia, foram identificadas pela primeira vez na filosfera, 597
carposfera e antosfera do cacaueiro. Alguns destes gêneros já foram citadas na literatura como 598
portadoras de características de interesse para o biocontrole de doenças de plantas, a exemplo 599
de R. palludigenum (Lu et al., 2014; Lu et al., 2013), P. sporocuriosa (Péter et al., 2000), 600
além de Wickerhamomyces anomalus (Hashem et al., 2014). No entanto deve-se realizar um 601
estudo mais criterioso sobre a utilização de alguns representantes deste clado, como as 602
espécies do gênero Candida que devem ser estudadas cuidadosamente pois apesar de ser um 603
gênero muito utilizado no controle de doenças de plantas (Droby et al., 2003; Zhang et al., 604
2014), abriga espécies que podem ser consideradas patógenos humanos (Bertini et al., 2013). 605
O filo basidiomycota também reúne uma diversidade de fungos leveduriformes que 606
habitam a superfície de órgãos vegetais como Hannaella oryzae em arroz (Nakase & Suzuki, 607

90
1985) e em feijão (Phaseolus vugaris L.) (Gonzaga et al., 2015); H. pagnoccae em néctar de 608
Heliconia psittacorum (Heliconiaceae), flores de pimenta (Myrtaceae), raízes e folhas de 609
cana-de-açúcar (Saccharum spp.) no Brasil, folhas de Cratoxylum maingayi, Arundinaria 610
pusilla e Vitis vinifera na Tailândia, amostras de solo em Taiwan e USA (Landell et al., 611
2014). Estudos recentes têm revelado a diversidade de leveduras residentes de órgãos 612
vegetais, com identificação e descrição de novas espécies, a exemplo de Cryptococcus 613
foliicola e Cryptococcus taibaiensis (Wang et al., 2011). Representantes do clado Tremellales 614
são utilizadas como biocontroladores de doenças de plantas, a exemplo das espécies 615
Cryptococcus albidus (Tian et al., 2002), Cryptococcus laurentii (Tian et al., 2007; Wang et 616
al., 2008), Cryptococcus flavus, Hannaella sinensis, R. paludigenum, (Nutaratat et al., 2014). 617
Divergências em relação ao mecanismo intraespecífico de biocontrole também são 618
citadas em diversos trabalhos, por haver particularidades quanto ao modo de ação antagônica 619
em leveduras da mesma espécie. Portes et al. (2013) estudando o papel de leveduras “killer” 620
na inibição e atividade de biocontrole contra Penicillium expansum e Aspergillus orchraceus, 621
demonstraram que as leveduras avaliadas foram mais efetivas na inibição da germinação dos 622
conídios do que em relação à inibição do crescimento micelial dos patógenos. No presente 623
trabalho, isolados identificados como Rhosdosporidium sp. diferiram em relação a todos os 624
parâmetros avaliados, alguns apresentando atividade “killer”, mas não sendo identificada 625
secreção de tais toxinas para todos os isolados (LEV 161, LEV 101 e LEV 092). Em relação à 626
inibição da germinação de basidiósporos, o isolado LEV 169 se destaca com a maior 627
porcentagem de inibição e o LEV 161 se enquadra dentre os isolados que pouco inibiram a 628
germinação. 629
Apesar de receber pouca atenção quando comparados com compostos difusíveis, os 630
compostos voláteis orgânicos tem sido alvo de trabalhos que visam compreender os 631
mecanismos de síntese, reconhecimento e papel no ecossistema (Hung et al., 2015). Mesmo 632
não identificando a capacidade de produção de compostos antifúngicos voláteis em todos os 633
isolados testados, estes compostos podem ser responsáveis pela inibição do crescimento 634
micelial de M. perniciosa. As leveduras podem reduzir o crescimento micelial de 635
fitopatógenos por compostos difusíveis, mas não constatar semelhante ação inibitória por 636
compostos voláteis (Rosa et al., 2010). Isto pode significar na prática que se essas leveduras 637
que apresentam esta característica, se aplicadas nas vassouras secas, podem impedir o 638
desenvolvimento do micélio do patógeno e, portanto, a produção de basidiomas. 639
O presente trabalho contribui para o conhecimento de fungos leveduriformes, ainda 640
que genéricos, que habitam e colonizam a superfície de órgãos vegetais do cacaueiro, até 641

91
então pouco explorados (Cabral, 2009), além de caracterizá-los quanto ao potencial de 642
inibição do crescimento de M. perniciosa, agente causador da vassoura de bruxa do cacaueiro, 643
determinando mecanismos de ação direta às estruturas fúngicas do agente etiológico desta 644
doença. Os resultados alcançados apresentam leveduras nativas tais como Hannaella sp., 645
Cryptococcus sp., Hanseniasora sp., Rhodosporidium sp., Candida sp., que atuando na 646
inibição do crescimento micelial e germinação de basidiósporos, por compostos difusíveis e 647
voláteis, são potenciais agentes de biocontrole, sendo recomendados para testes de 648
estabilidade dos mecanismos in vivo e em condições de cultivo para posterior inserção como 649
novos componentes do manejo integrado da vassoura de bruxa do cacaueiro. 650
651
Conclusões 652
653
1. A microbiota epifítica residente em folhas, flores e frutos de cacaueiro abriga uma 654
diversidade de fungos leveduriformes, até então não citados na literatura. 655
2. Leveduras nativas do cacaueiro são produtoras de compostos difusíveis e voláteis 656
que atuam na inibição da germinação de basidiósporos e crescimento micelial de M. 657
perniciosa. 658
4. A maioria dos isolados de leveduras nativas do cacaueiro são produtoras de toxina 659
“killer”. 660
5. Na superfície de diferentes órgãos vegetais de cacaueiros selecionados como 661
resistentes à vassoura de bruxa e monilíase, existem uma diversidade de fungos 662
leveduriformes que podem estar atuando de forma positiva na inibição de M. perniciosa e 663
evolução do quadro sintomatológico. 664
665
Agradecimentos 666
667
À Financiadora de Estudos e Projetos (FINEP), através da Fundação Pau Brasil 668
(FUNPAB) e Comissão Executiva do Plano da Lavoura Cacaueira (CEPLAC), 669
à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Capes) e a Fundação de 670
Amparo à Pesquisa do Estado da Bahia (FAPESB) por concessão de bolsas e recursos. 671
672
Referências 673
674

92
ASSIS, S.M.P.; MARIANO, R.L.R.; MICHEREFF, S.J.; SILVA, G.; MARANHÃO, E.A.A. 675
Antagonism of yeasts to Xanthomonas campestris pv. Campestris on cabbage phylloplane in 676
field. Revista de Microbiologia, v. 30, p. 191-195, 1999. 677
AFSAH-HEJRI, L. Saprophytic yeasts: effective biocontrol agents against Aspergillus flavus. 678
International Food Research Journal, v.20, n.6, p.3403-3409, 2013. 679
BARBIERI, R.; CARVALHO, I. F. Coevolução de plantas e fungos fitopatogênicos. Revista 680
Brasileira de Agrociência, v.7, p.79-83, 2001. 681
BERTINI, A.; DE BERNARDISB, F.; HENSGENSA, L.A.M.; SANDINIB, S.; SENESIA, 682
S.; TAVANTIA, A. Comparison of Candida parapsilosis, Candida orthopsilosis, and 683
Candida metapsilosis adhesive properties and pathogenicity. International Journal of 684
Medical Microbiology, v.303, p.98-103, 2013. 685
BETTIOL, W. Biocontrole na filosfera: problemas e perspectivas. Revisão Anual de 686
Patologia de Plantas, v.5, p.59-97, 1997. 687
BRUCE, A.; STEWARTA, D.; VERRALLB, S.; WHEATLEY, R.E. Effect of volatiles from 688
bacteria and yeast on the growth and pigmentation of sapstain fungi. International 689
Biodeterioration & Biodegradation, v.51, n.2, p.101-108, 2003. 690
CABANAS, Andreia Sofia Pereira. Avaliação da atividade antifúngica de quitinases de 691
macieira. 2013. 64 f. Dissertação (Mestrado) - Curso de Engenharia Agronómica, Instituto 692
Superior de Agronomia Univercidade de Lisboa, Lisboa, 2013. 693
CABRAL, A.S. Killer yeasts inhibit the growth of the phytopathogen Moniliophthora 694
perniciosa, the causal agent of witches’ broom disease. Brazilian Journal of Microbiology, 695
v.40, p.108-110, 2009. 696
COELHO, A.R.; NÓBREGA, G.M.A.; PAGNOCCA, F.C.; HOFFMANN, F.L.; HARADA, 697
K.; HIROOKA, E.Y. Avaliação do potencial antagônico de leveduras, visando biocontrole de 698
deterioração por Penicillium expansum. Semina: Ciência Agrárias, v.32, p.1879-1892, 2011. 699
CLARKE, K.R. Non-parametric multivariate analyses of changes in community structure. 700
Australian Journal of Ecology, v.18, p.117-143, 1993. 701
CUATRECASAS, J. Cacao and its allies: a taxonomic revision of the genus Theobroma. 702
Contributions from the United States National Herbarium. n.35, p.379-614, 1964. 703
COSTA, J.C.B.; BEZERRA, J.L.; SANTOS FILHO, L.P.; ALVES, M.C.; MOURA, E.M. 704
Controle biológico da vassoura-de-bruxa do cacaueiro da Bahia, Brasil. In: BETTIOL, W.; 705
MORANDI, M.A.B. (Org.). Biocontrole de Doenças de Plantas: uso e perspectivas. 706
Jaguariúna: Embrapa Meio Ambiente, 2009, p.245–266. 707

93
DANIEL, H.; VRANCKEN, G.; TAKRAMA, J.F.; CAMU, N.; De Vos, P.; DE VUYS, L. 708
Yeast diversity of Ghanaian cocoa bean heap fermentations. FEMS Yeast Research, v.9, p. 709
774–783, 2009. 710
DICKSTEIN, E.R.; PURDY, L.H.; FRIAS, G.A. Crinipellis perniciosa, the cacao witches’ 711
broom fungus: Inoculum production and storage. Phytopathology, v.77, p.1747, 1987. 712
DROBY, S.; WISNIEWSKI, M.; EL GHAOUTH, A.; WILSON, C. Influence of food 713
additives on the control of postharvest rots of apple and peach and efficacy of the yeast-based 714
biocontrol product Aspire. Postharvest Biology and Technology, v.27, p.127-135, 2003. 715
DUARTE, S.H.; de ANDRADE, C.C.; GHISELLI, G.; MAUGERI, F. Exploration of 716
brazilian biodiversity and selection of a new oleaginous yeast strain cultivated in raw 717
glycerol. Bioresource Technology, v.138, p.377–381, 2013. 718
EL-TARABILY, K.A.; SIVASITHAMPARAM, K. Potential of yeasts as biocontro agents of 719
soil-borne fungal plant pathogens and as plant growth promoters. Mycoscience, v.47, p.25-720
35, 2006. 721
ESCALANTE, W.E.; RYCHTERAA, M.; MELZOCHA, K.; SAKODAB, B.H.; POLOC, 722
E.Q.; CERVANTESD, Z.L.; CASAVILCAD, V.S.; QUILCAD, G.C. Actividad fermentativa 723
de Hanseniaspora uvarum y su importancia en la producción de bebidas fermentadas. Revista 724
de la Sociedad Venezolana de Microbiología, v. 31, p.57-63, 2011. 725
GOHEL, V.; SINGH, A.; VIMAL, M.; ASHWINI, P.; CHHATPAR, H.S.Bioprospectingand 726
antifungal potential of chitinolytic microorganism. African Journal Biotecnology, v.5, n.3, 727
p.54-72, 2006. 728
GONZAGA, L.L.; COSTA, L.E.O.; SANTOS, T.T.; ARAUJO, E.F.; QUEIROZ, M.V. 729
Endophytic fungi from the genus Colletotrichum are abundant in the Phaseolus vulgaris and 730
have high genetic diversity. Journal Applied Microbiology, v.118, n.2, p.1-12, 2015. 731
HASHEM, M.; ALAMRI, S. A.; HESHAM, A.E.; AL-QAHTANI, F.M.H.; KILANY, M. 732
Biocontrol of apple blue mould by new yeast strains: Cryptococcus albidus KKUY0017 and 733
Wickerhamomyces anomalus KKUY0051 and their mode of action. Biocontrol Science and 734
Technology, v.24, v.10, 2014. 735
HU, H.; XU, Y.; LU, H.; XIAO, R.; ZHENG, X.; YU, T. Evaluation of yeasts from Tibetan 736
fermented products as agents for biocontrol of blue mold of Nashi pear fruits. Journal of 737
Zhejiang University-SCIENCE B (Biomedicine & Biotechnology), v.16, n.4, 275-285, 2015. 738
HUNG, R.; LEE, S.; BENNETT, J.W. Fungal volatile organic compounds and their role in 739
ecosystems. Applied Microbiology and Biotechnology, v.99, p. 3395-3405, 2015. 740
ICCO Quarterly Bulletin of Cocoa Statistics, Vol. XLI, No. 4, Cocoa year 2014/15 741

94
Published: 27-11-2015 742
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA (IBGE). Sistema IBGE de 743
Recuperação Eletrônica (SIDRA). 2014. Disponível em: 744
http://www.sidra.ibge.gov.br/bda/agric/. Acesso em: 26 jan. 2016. 745
JANISIEWICZ, W.J.; KURTZMAN, C P.; BUYER, J.S. Yeasts associated with nectarines 746
and their potential for biological control of brown rot. Yeast, v.27, p.389–398, 2010. 747
KELLY, P.L.; REEDER, R.; RHODES, S.; EDWARDS, N. First confirmed report of 748
witches’ broom caused by Moniliophthora perniciosa on cacao, Theobroma cacao, in Saint 749
Lucia. Plant Pathology, v.58, p.798-798, 2009. 750
LANDELL, M.F.; BRANDÃO, L.R.; BARBOSA, A.C.; RAMOS, J.P.; SAFAR, S.V.B.; 751
GOMES, F.C.O.; SOUSA, F.M. P.; MORAIS, P.B.; BROETTO, L.; LEONCINI, O.; 752
RIBEIRO, J.R.; FUNGSIN, B.; TAKASHIMA, M.; NAKASE, T.; LEE, C.; VAINSTEIN, 753
M.H.; FELL, J.W.; SCORZETTI, G.; VISHNIAC, H.S.; ROSA, C.A.; VALENTE, P. 754
Hannaella pagnoccae sp. nov., a tremellaceous yeast species isolated from plants and soil. 755
International Journal of Systematic and Evolutionary Microbiology, v.64, p.1970-1977, 756
2014. 757
LAHLALI, R.; HAMADI, Y.; EL GUILLI, M.; JIJAKLI, M.H. Efficacy assessment of Pichia 758
guilliermondii strain Z1, a new biocontrol agent, against citrus blue mould in Morocco under 759
the influence of temperature and relative humidity. Biological Control, v.56, n.3, p.217-224, 760
2011. 761
LEAL JR, G.A.; GOMES, L. H.; EFRAIM, P.; TAVARES, F. C. A.; FIGUEIRA, A. 762
Fermentation of cacao (Theobroma cacao L.) seedswith a hybrid Kluyveromyces marxianus 763
strain improved product quality attributes. FEMS Yeast Research, v.8, p.t88–798, 2008. 764
LEFF, J.W.; FIERE, N. Bacterial communities associated with the surfaces of fresh fruits and 765
vegetables. PLoS ONE, v.8, n.3, 2013. 766
LEVEAU, J.H.; LINDOW, S.E. Appetite of an epiphyte: quantitative monitoring of bacterial 767
sugar consumption in the phyllosphere. Proceedings of the National Academy of Science of 768
The United States of America. v.98, p.3446–3453, 2001. 769
LI, X.; YU, V.Y.; LIN, Y.; CHOMVONG, K.; ESTRELA, R.; PARK, A.; LIANG, J.M.; 770
ZNAMEROSKI, E.A.; FEEHAN, J.; KIM, S.R.; JIN, Y; GLASS, N.L.; CATE, J.H.D. 771
Expanding xylose metabolism in yeast for plant cell wall conversion to biofuels. Elife, v.4, 772
p.1-16, 2015. 773
LÓPEZ, S.; MATEO, J.J.; Maicas, S. Screening of Hanseniaspora strains for the production 774
of enzymes with potential interest for winemaking. Fermentation, v.1, p.1-16, 2016. 775

95
LU, L.; LU, H.; WUC, C.; FANGD, W.; YUA, C.; YEA, C.; SHIA, Y.; YUA, T.; ZHENGA, 776
X. Preharvest application of antagonistic yeast Rhodosporidium paludigenum induced 777
resistance against postharvest diseases in mandarin orange. Biological Control, v. 67, n. 2, p. 778
130–136, 2013. 779
LU, H.; LU, L.; ZENG, L.; FU, D.; XIANG, H.; YU, T.; ZHENG, X. EFFECT of chitin on 780
the antagonistic activity of Rhodosporidium paludigenum against Penicillium expansum in 781
apple fruit. Postharvest Biology and Technology, v.92, p.9-15, 2014. 782
MARIANO, R.L.R; GOMES, A. M. A.; ASSIS, S. M. P; SILVEIRA, E. B. Mecanismos de 783
ação de bactérias promotoras de crescimento de plantas. In: MARIANO, R. L. R.; 784
SILVEIRA, E. B. (Coord.). Manual de práticas em fitobacteriologia. 2.ed. Recife: UFRPE, 785
2005. p. 143-157. 786
MILLER, W.G.; BRAND, M.T.; QUIÑONES, B.; LINDOW, S.E. Biological sensor for 787
sucrose availability: relative sensitivities of various reporter genes. Applied and 788
Environment Microbiology, v.67, p.1308–1317, 2001. 789
MASIH, E.I.; PAUL, B. Secretion of beta-1,3-glucanase by the yeast Pichia membranifaciens 790
and its possible role in the biocontrol of Botrytis cinerea causing mold disease of the 791
grapevine. Current Microbiology, v.44, p.391-395, 2002. 792
MIDLEJ, R. R.; SANTOS, A. M. Economia do Cacau. In: VALLE, R. R. (Org). Ciência, 793
Tecnologia e Manejo do Cacaueiro. Brasília: Ceplac, 2012. p. 655-672. 794
MOREIRA, N.; PINA, C.; MENDES, F.; COUTO, J.A.; HOGG, T.; VASCONCELOS, I. 795
Volatile compounds contribution of Hanseniaspora guilliermondii and Hanseniaspora 796
uvarum during red wine vinifications. Food Control, v. 22, n. 5, p.662-667, 2011. 797
Moreira, N.; Pina, C.; Mendes, F.; Couto, J.A.; Hogg, T.; Vasconcelos, I. Volatile compounds 798
contribution of Hanseniaspora guilliermondii and Hanseniaspora uvarum during red wine 799
vinifications. Food Control, v 22, p.662-667, 2011. 800
McCORMICK, M.K.; WHIGHAM, D.F.; O’NEILL, J.P.; BECKER, J.J.; WERNER, S.; 801
RASMUSSEN, H.N.; et al. Abundance and distribution of Corallorhiza odontorhiza reflects 802
variations in climate and ectomycorrhizae. Ecological Monographs, v.79, n.4, p.619-635, 803
2009. 804
MAUTONE, J.N. Biodiversidade e potencial biotecnológico de leveduras e fungos 805
semelhantes a leveduras isoladas de folhas de figueiras do Parque de Itapuã-Viamão/RS. 806
2008. 109p. Dissertação (Mestrado em Microbiologia Agrícola e do Meio Ambiente) 807
Universidade Federal do Rio Grande do Sul, Porto Alegre. 808

96
NAKASE, L.; M. SUZUKI. Ballistospore-forming yeasts found on the surface of the 809
Japanese rice plant, Oryza sativa L. Journal of General and Applied Microbiology, v.31, 810
n.5, p.468, 1985. 811
NUTARATAT, P.; SRISUK, N.; ARUNRATTIYAKORN, P.; LIMTONG, S. Plant growth-812
promoting traits of epiphytic and endophytic yeasts isolated from rice and sugar cane leaves 813
in Thailand. Fungal biology, v.118, n.8, p.683-194, 2014. 814
OLIVEIRA, M. L.; LUZ, E. D. M. N. Principais doenças do cacaueiro e seu manejo. In: 815
VALLE, R. R. M. (Ed.). Ciência, tecnologia e manejo do cacaueiro. Brasília: 2012, 688 p. 816
O’DONNELL, K. (1993). Fusarium and its near relatives. In: REYNOLDS, D.R.; TAYLOR, 817
J.W. (Ed.). The Fungal Holomorph: Mitotic, Meiotic and Pleomorphic Speciation. Fungal 818
Systematics, pp. 225–233. 819
Wallingford, UK: CAB International.PÉTER, G.; TORNAI-LEHOCZKI, J.; DLAUCHY, 820
D.; VITÁNYI, G. Pichia sporocuriosa sp. nov., a new yeast isolated from rambutan. Antonie 821
van Leeuwenhoek, v.77, p.37-42, 2000. 822
PLATANIAA, A.; RESTUCCIAB, C.; MUCCILLIB, S.; CIRVILLERIA, G. Efficacy of 823
killer yeasts in the biological control of Penicillium digitatum on Tarocco orange fruits 824
(Citrus sinensis). Food Micrrobiology, v.30, n.1, p.219-225, 2012. 825
PORTES, C.S.; DE OLIVEIRA A.V.; SIMER, P.; LUNKES, A.M.; COELHO, A.R. Role of 826
killer factors in the inhibitory activity of bio-control yeasts against Penicillium expansum and 827
Aspergillus ochraceus. Brazilian Archives of biology and technology, v.56, n.4, p.619-627, 828
2013. 829
PURDY, L.H.; SCHMIDT, R A. Status of cacao witches’ broom, biology, epidemiology, and 830
management. Annual Review of Phytopathology, v.34, p.573-594, 1996 831
ROSA, M.M.; TAUK-TORNISIELO, S.M.; RAMPAZZO, P.E.; CECCATO-ANTONINI, 832
S.R. Evaluation of the biological control by the yeast Torulaspora globosa against 833
Colletotrichum sublineolum in sorghum. World Journal of Microbiology and 834
Biotechnology, v.26, n.8, p.1491-1502, 2010. 835
RUBINI, A.S. et al. Molecular identification and characterization of canine Hepatozoon 836
species from Brazil. Parasitology Research, v.97, n.2, p.91-93, 2005. 837
SANGORRÍN, M.P., ZAJONSKOVSKY, I.E., LOPES, C.A., RODRÍGUEZ, M.E., DE VAN 838
BROOCK, M.R.G., CABALLERO, A.C. Killer behaviour in wild wine yeasts associated with 839
Merlot and Malbec type musts spontaneously fermented from northwestern Patagonia 840
(Argentina). Journal of Basic Microbiology, v.41, p.105-113, 2001. 841

97
SCHMITI, M.J.; NEUHAUSEN, F. Killer Toxin-secreting double-stranded RNA 842
mycoviruses in the yeasts Hanseniaspora uvarum and Zygosaccharomyces bailii. Journal of 843
virology, p.1765-1772, 1994. 844
SUI, Y.; WISNIEWSKI, M.; DROBY, S.; LIU, J. Responses of Yeast Biocontrol Agents to 845
Environmental Stress. Applied and environmental microbiology, v.81, n.9, p.2968-2975 846
2015. 847
SUN, P.; FANG, W.; SHIN, L.; WEI, J.; FU, S.; CHOU, J. Indole-3-acetic acid-producing 848
yeasts in the phyllosphere of the carnivorous plant Drosera indica L. Plos one, p.1-22, 2014. 849
TIAN, S.P.; FAN, Q.; XU, Y.; LIU, H. B. Biocontrol efficacy of antagonist yeasts to gray 850
mold and blue mold on apples and pears in controlled atmospheres. Plant Disease, v.86, 851
p.848-853, 2002. 852
TIAN, S.P.; YAO, H.J; DENG, X.; XU, X.B.; QIN, G.Z.; CHAN, Z.L. Characterization and 853
expression of β-1,3-glucanase genes in jujube fruit induced by the microbial biocontrol agent 854
Cryptococcus laurentii. Phytopathology, v.97, p.260-268, 2007. 855
TOPPS, J.H.; WAIN, R.L.; Investigation on fungicides. III. The fungi toxicity of 3-and 5- 856
alkyl salicylanilide and P-chloronilines. Annals of Applied Biology, v.45, n.3, p.506-511, 857
1957. 858
URQUHART, E.J.; PUNJA, Z.K. Hydrolytic enzymes and antifungal compounds produced 859
by Tilletiopsis species, phyllosphere yeasts that are antagonists of powdery mildew fungi. 860
Canadian Journal of Microbiology, v.48, n.3, p.219-229, 2002. 861
VALDEBENITO-SANHUEZA, R. M. V. Leveduras para o biocontrole de fitopatógenos. In: 862
MELO, I. S; AZEVEDO, J. L. (Ed.). Controle biológico. Jaguariúna: Embrapa Meio 863
Ambiente. p. 41-56, 2000. 864
YOUNG, T.W. Killer yeasts. In: Rose, A.H.; Harrison, J.S.(Ed.). The yeasts. v.2. London: 865
Academic Press, 1987. 866
pp. 131 -164. WANG, Y.F., BAO, Y.H., SHEN, D.H., FENG, W., YU, T., ZHANG, J., 867
ZHENG, X.D., 2008. Biocontrol of Alternaria alternata on cherry tomato fruit by use of 868
marine yeast Rhodosporidium paludigenum. International. Journal of Food Microbiology, 869
Amsterdam, v.123, p.234-239, 2008. 870
WANG, Y.F.; TANG, F.; XIA, J.D.; YU, T.; WANG, J.; AZHATI, R.; ZHENG, X.D. A 871
combination of marine yeast and food additive enhances preventive effects on postharvest 872
decay of jujubes (Zizyphus jujuba). Food Chemistry, v.125, n.3, p.835-840, 2011. 873

98
ZHANG, H.Y.; WANG, L.; DONG, Y.; JIANG, S.; CAO, J.; MENG, R. Postharvest 874
biological control of gray mold decay of strawberry with Rhodotorula glutinis. Biological 875
Control, v 40, p.287-292, 2007. 876
ZHANG, D.; LU, C.; ZHANG, T.; SPADARO, D.; LIU, D.; LIU, W. Candida pruni sp. nov. 877
is a new yeast species with antagonistic potential against brown rot of peaches. Archives of 878
Microbiology, v.196, n.7, p.525-530, 2014. 879

99
Tabela 1. Classes e gêneros de isolados de leveduras nativas do cacaueiro,
verificados através do banco de dados GenBank, utilizando o BLASTn “basic
local alignment search tool” (BLAST) do NCBI (http://www.ncbi.nlm.nih.gov).
(continua)
CLASSE ISOL TAXONOMIA
ASCOMYCETES
LEV 002 Candida sp.
LEV 008 Yamadazyma sp.
LEV 030 Candida sp.
LEV 047 Wicherhamomyces sp.
LEV 049 Candida sp.
LEV 068 Candida sp.
LEV 074 Candida sp.
LEV 082 Candida sp.
LEV 084 Candida sp.
LEV 089 Candida sp.
LEV 098 Candida sp.
LEV 118 Candida sp.
LEV 162 Hanseniaspora sp.
LEV 189 Pichia sp.
LEV 194 Hanseniaspora sp.
LEV 204 Pichia sp.
LEV 210 Hanseniaspora sp.
LEV 211 Hanseniaspora sp.
LEV 219 Candida sp.
LEV 225 Candida sp.
BASIDIOMYCETES
LEV 001 Sporobolomyces sp.
LEV 007 Rhodosporidium sp.
LEV 014 Hannaella sp.
LEV 016 Cryptococcus sp.
LEV 023 Rhodosporidium sp.
LEV 027 Hannaella sp.
LEV 029 Hannaella sp.
LEV 034 Rhodosporidium sp.
LEV 038 Hannaella sp.
LEV 063 Hannaella sp.
LEV 065 Occultifur sp.
LEV 077 Hannaella sp.
LEV 078 Occultifur sp.
LEV 079 Rhodosporidium sp.
LEV 080 Hannaella sp.
LEV 088 Hannaella sp.
LEV 090 Cryptococcus sp.
LEV 092 Rhodosporidium sp.
LEV 093 Cryptococcus sp.
LEV 100 Cryptococcus sp.

100
Tabela 1. Classes e gêneros de isolados de leveduras nativas do cacaueiro,
verificados através do banco de dados GenBank, utilizando o BLASTn “basic
local alignment search tool” (BLAST) do NCBI (http://www.ncbi.nlm.nih.gov).
(conclusão)
CLASSE ISOL TAXONOMIA
BASIDIOMYCETES
LEV 101 Rhodosporidium sp.
LEV 105 Hannaella sp.
LEV 106 Cryptococcus sp.
LEV 109 Hannaella sp.
LEV 111 Cryptococcus sp.
LEV 115 Hannaella sp.
LEV 122 Cryptococcus sp.
LEV 140 Hannaella sp.
LEV 143 Hannaella sp.
LEV 147 Hannaella sp.
LEV 148 Occultifur sp.
LEV 149 Cryptococcus sp.
LEV 154 Cryptococcus sp.
LEV 161 Rhodosporidium sp.
LEV 164 Hannaella sp
LEV 165 Rhodosporidium sp.
LEV 166 Rhodosporidium sp.
LEV 168 Rhodosporidium sp.
LEV 169 Rhodosporidium sp.
LEV 170 Rhodosporidium sp.
LEV 174 Rhodosporidium sp.
LEV 178 Rhodosporidium sp.
LEV 179 Rhodosporidium sp.
LEV 185 Rhodosporidium sp.
LEV 187 Cryptococcus sp.
LEV 190 Cryptococcus sp.
LEV 192 Rhodosporidium sp.
LEV 193 Hannaella sp.
LEV 199 Cryptococcus sp.
LEV 206 Cryptococcus sp.
LEV 207 Hannaella sp.
LEV 216 Cryptococcus sp.
LEV 226 Hannaella sp
LEV 227 Kwoniella sp.
INDEFINIDO
LEV 059 .
LEV 097 .
LEV 137 .
LEV 146 .
LEV 209 .
LEV 229 .

101
Tabela 2. Grupos de 66 leveduras nativas do cacaueiro, formados a partir das medidas de dissimilaridade
das variáveis antibiose (AB), compostos voláteis (CV), inibição da germinação de basidiósporos (IG) e
toxina killer (TK) e análise de agrupamento obtido pelo método UPGMA (distância euclidiana). Os
isolados de levedura foram obtidos a partir do isolamento de diferentes genótipos (GEN), padrões de
resistência (RESIST) à vassoura de bruxa (VB) e monilíase (ML) e diferentes substratos (SUBST)
(continua)
GRUPO LEV TAXONOMIA
FENOTIPAGEM
GEN RESIT SUSBT AB CV IG TK SM
1 169 Rhodosporidium sp. 5 1 10 1 17 EET233 ML FOLHA
219 Candida sp. 5 1 9 1 16 EET 392 VB FRUTO
2
92 Rhodosporidium sp. 6 3 6 0 15 P253 VB FOLHA
165 Rhodosporidium sp. 6 3 6 1 16 EET 233 ML FOLHA
174 Rhodosporidium sp. 6 3 5 1 15 EET 392 VB FOLHA
162 Hanseniaspora sp. 7 3 6 1 17 P232 VB FOLHA
194 Hanseniaspora sp. 8 3 5 1 17 ICS 95 ML FLOR
211 Hanseniaspora sp. 7 3 7 1 18 P282 VB FLOR
206 Cryptococcus sp. 7 4 5 1 17 UF 273 ML FLOR
2 Candida sp. 7 3 4 1 15 P243 VB FOLHA
189 Pichia sp. 7 3 6 1 17 EET 75 ML FLOR
3
101 Rhodosporidium sp. 6 2 8 0 16 P213 VB FLOR
168 Rhodosporidium sp. 6 2 7 1 16 EET233 ML FOLHA
149 Cryptococcus sp. 6 3 8 1 18 P247 VB FLOR
166 Hannaella sp. 6 3 7 1 17 EET233 ML FOLHA
207 Hannaella sp. 6 2 8 1 17 UF 273 ML FLOR
210 Hanseniaspora sp. 8 4 8 1 21 P282 VB FLOR
225 Candida sp. 5 3 7 1 16 EET 233 ML FRUTO
4
49 Candida sp. 5 1 8 0 14 P245 VB FOLHA
185 Rhodosporidium sp. 6 1 8 1 16 ICS 95 ML FOLHA
63 Hannaella sp. 6 1 7 1 15 P282 VB FOLHA
147 Hannaella sp. 6 1 7 1 15 P247 VB FLOR
14 Hannaella sp. 6 1 7 1 15 P243 VB FOLHA
27 Hannaella sp. 6 1 7 1 15 P225 VB FOLHA
88 Hannaella sp. 5 1 5 0 11 P284 VB FOLHA
109 Hannaella sp. 5 1 5 0 11 P232 VB FLOR
77 Hannaella sp. 5 1 5 0 11 P309 VB FOLHA
65 Occultifur sp. 5 1 7 1 14 P284 VB FOLHA
78 Occultifur sp. 5 1 6 0 12 P285 VB FOLHA
111 Cryptococcus sp. 5 1 7 1 14 P253 VB FOLHA
16 Cryptococcus sp. 6 1 6 1 14 P213 VB FOLHA
106 Cryptococcus sp. 5 1 6 0 12 P232 VB FLOR
93 Cryptococcus sp. 4 1 7 1 13 P309 VB FLOR
79 Rhodosporidium sp. 7 1 5 1 14 P258 VB FOLHA
23 Rhodosporidium sp. 7 1 5 1 14 P253 VB FOLHA
34 Rhodosporidium sp. 6 1 6 1 14 P253 VB FOLHA
178 Rhodosporidium sp. 6 1 6 1 14 P258 VB FOLHA
7 Rhodosporidium sp. 7 1 4 1 13 P243 VB FOLHA
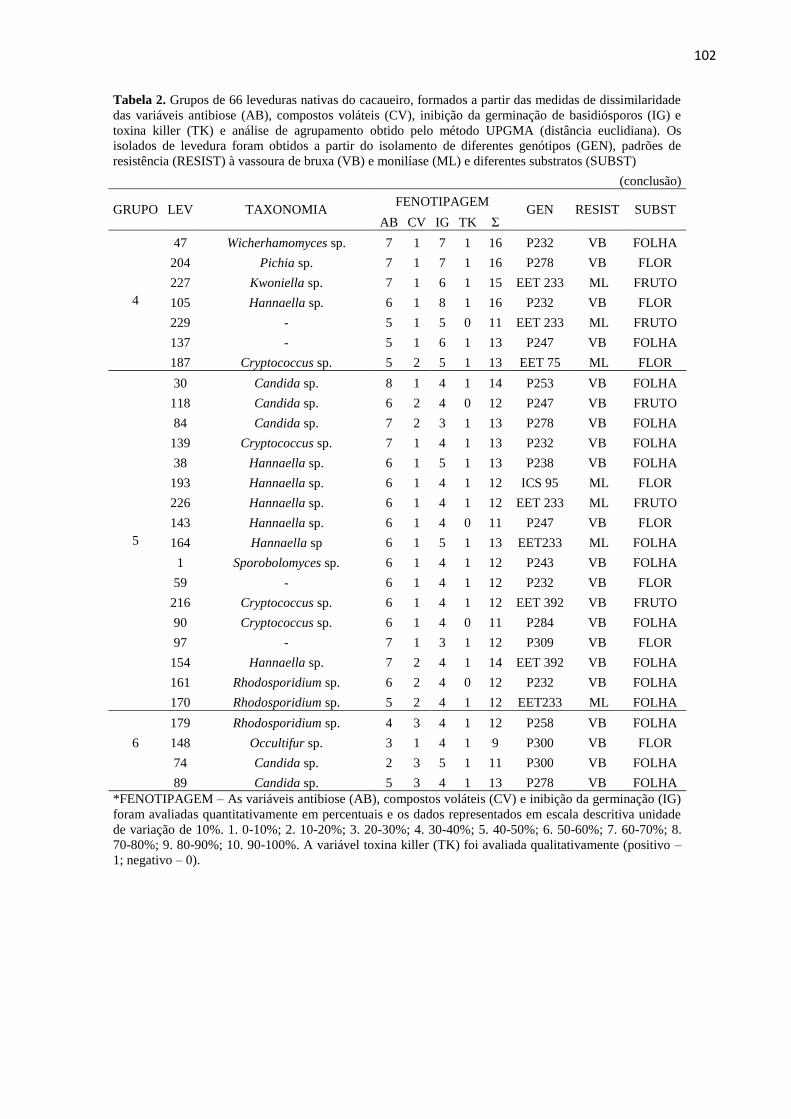
102
Tabela 2. Grupos de 66 leveduras nativas do cacaueiro, formados a partir das medidas de dissimilaridade
das variáveis antibiose (AB), compostos voláteis (CV), inibição da germinação de basidiósporos (IG) e
toxina killer (TK) e análise de agrupamento obtido pelo método UPGMA (distância euclidiana). Os
isolados de levedura foram obtidos a partir do isolamento de diferentes genótipos (GEN), padrões de
resistência (RESIST) à vassoura de bruxa (VB) e monilíase (ML) e diferentes substratos (SUBST)
(conclusão)
GRUPO LEV TAXONOMIA FENOTIPAGEM
GEN RESIST SUBST AB CV IG TK Σ
4
47 Wicherhamomyces sp. 7 1 7 1 16 P232 VB FOLHA
204 Pichia sp. 7 1 7 1 16 P278 VB FLOR
227 Kwoniella sp. 7 1 6 1 15 EET 233 ML FRUTO
105 Hannaella sp. 6 1 8 1 16 P232 VB FLOR
229 - 5 1 5 0 11 EET 233 ML FRUTO
137 - 5 1 6 1 13 P247 VB FOLHA
187 Cryptococcus sp. 5 2 5 1 13 EET 75 ML FLOR
5
30 Candida sp. 8 1 4 1 14 P253 VB FOLHA
118 Candida sp. 6 2 4 0 12 P247 VB FRUTO
84 Candida sp. 7 2 3 1 13 P278 VB FOLHA
139 Cryptococcus sp. 7 1 4 1 13 P232 VB FOLHA
38 Hannaella sp. 6 1 5 1 13 P238 VB FOLHA
193 Hannaella sp. 6 1 4 1 12 ICS 95 ML FLOR
226 Hannaella sp. 6 1 4 1 12 EET 233 ML FRUTO
143 Hannaella sp. 6 1 4 0 11 P247 VB FLOR
164 Hannaella sp 6 1 5 1 13 EET233 ML FOLHA
1 Sporobolomyces sp. 6 1 4 1 12 P243 VB FOLHA
59 - 6 1 4 1 12 P232 VB FLOR
216 Cryptococcus sp. 6 1 4 1 12 EET 392 VB FRUTO
90 Cryptococcus sp. 6 1 4 0 11 P284 VB FOLHA
97 - 7 1 3 1 12 P309 VB FLOR
154 Hannaella sp. 7 2 4 1 14 EET 392 VB FOLHA
161 Rhodosporidium sp. 6 2 4 0 12 P232 VB FOLHA
170 Rhodosporidium sp. 5 2 4 1 12 EET233 ML FOLHA
6
179 Rhodosporidium sp. 4 3 4 1 12 P258 VB FOLHA
148 Occultifur sp. 3 1 4 1 9 P300 VB FLOR
74 Candida sp. 2 3 5 1 11 P300 VB FOLHA
89 Candida sp. 5 3 4 1 13 P278 VB FOLHA
*FENOTIPAGEM – As variáveis antibiose (AB), compostos voláteis (CV) e inibição da germinação (IG)
foram avaliadas quantitativamente em percentuais e os dados representados em escala descritiva unidade
de variação de 10%. 1. 0-10%; 2. 10-20%; 3. 20-30%; 4. 30-40%; 5. 40-50%; 6. 50-60%; 7. 60-70%; 8.
70-80%; 9. 80-90%; 10. 90-100%. A variável toxina killer (TK) foi avaliada qualitativamente (positivo –
1; negativo – 0).

103
Figura 1. Heatmap do efeito in vitro de compostos difusíveis e voláteis, produzidos por leveduras nativas do
cacaueiro, no crescimento micelial de diferentes isolados de Moniliophthora perniciosa do sexto ao décimo dia
após pareamento dos microorganismo.

104
Figura 2. Heatmap do efeito in vitro de voláteis, produzidos por leveduras nativas do cacaueiro, no crescimento
micelial de diferentes isolados de Moniliophthora perniciosa do sexto ao décimo dia após pareamento dos
microorganismo.

105
Figura 3. Inibição da germinação de basidiósporos (%) e capacidade de produção de toxinas killer por
diferentes isolados de leveduras nativas do cacaueiro. *Leveduras não produtoras de toxinas killer. As médias
das porcentagens de inibição foram transformadas Sqrt (x + 0.5) e analisadas pelo teste de Scott-Knott (p
<0.05).

106
Figura 4. Diagrama Biplot (3D em cubo) da relação entre mecanismos de biocontrole
(variáveis reais), antibiose: compostos difusíveis e voláteis (AB), compostos voláteis
(CV), inibição da germinação (IG), atividade killer (AK), produzidos por leveduras
nativas do cacaueiro e avaliadas contra o patógeno Moniliophthora perniciosa.
1
2
7
14
16 23
34
27
47
30
49
38 59
63 65
74
77 78
79
84
88
89
90
92
93
97
100
101
106
109
111
118
137
139
143
47 147
148
149
154
161
164
165
166 168
169
170
174
178
179
185
187
189
193
194
204
206
207
210
211 216
219
225
226
227
229 162
1
AK
IG
AB
CV

107
Figura 5. Dendrograma de análise de agrupamento obtido pelo método de UPGMA
(distância Euclidiana), a partir das medidas de dissimilaridade entre 66 isolados de
leveduras nativas do cacaueiro avaliados in vitro para as variáveis antibiose, compostos
voláteis, inibição da germinação dos basidiósporos, capacidade de produção de toxina
killer. Grupos formados após corte na altura de θ = 25: I, II, III, IV, V, VI, VII.

108
Figura 6. Avaliação da interação levedura (Hanella sp.) x Moniliophthora perniciosa cultivados em meio líquido batata dextrose por 240h. A.
Biomassa Fresca – BF; B. Biomassa Seca - BS; C. Açúcares totais – AT; Concentração de células - CC (_x107; ufc); 1. Moniliophthora
perniciosa; 2. Hanella sp.; 3. M. perniciosa + Hanella sp. (mesmo dia); 4. M. perniciosa + Hanella sp. (mesmo dia + 1 reinóculo); 5. M.
perniciosa + Hanella sp. (mesmo dia + 2 reinóculos); 6. M. perniciosa + Hanella sp. (mesmo dia + 3 reinóculos); 7. M. perniciosa + Hanella
sp. (mesmo dia + 4 reinóculos); 8. M. perniciosa + Hanella sp. (192h após); 9. M. perniciosa + Hanella sp. (168h após + 1 reinóculo 216h
após). Médias seguidas com a mesma letra não diferem entre si, pelo teste de Tukey a 5% de probabilidade. Os dados de PS foram
transformados para √x+1.
A
D C
B
a
d d d d d
b c
e
c
e
b
d cd cd bcd d
bc
b
bc
c
a
b
c c c c
c
ab
d
e
c
e
bc
ab
a

109
CAPÍTULO 4
Leveduras nativas como potenciais agentes de biocontrole à vassoura de bruxa e
promotores de crescimento em plântulas de cacaueiro

110
Leveduras nativas como potenciais agentes de biocontrole à vassoura de bruxa e 1
promotores de crescimento em plântulas de cacaueiro 2
3
Antônio Alves Pimenta Neto(1), Tacila Ribeiro dos Santos(2), Carolina Benjamin (2), Eduardo 4
Gross(2), Edna Dora Martins Newman Luz(3), Delson Laranjeira(1) 5
6
(1)Universidade Federal Rural de Pernambuco, Rua Dom Manuel Medeiros, s/n, Dois Irmãos, 7
CEP 52171-900, Recife, PE, Brasil. Email: [email protected], 8
[email protected] (2)Universidade Estadual de Santa Cruz, Campus Soane Nazaré de 9
Andrade, Rod. Jorge Amado, Km 16, Salobrinho, Ilhéus, BA, Brasil. Email: 10
[email protected], [email protected], [email protected] (3)Comissão 11
Executiva do Plano da Lavoura Cacaueira, Centro de Pesquisas do Cacau, Rodovia Ilhéus-12
Itabuna, Km 22, s/n, CEP 45660-000, Ilhéus, BA, Brasil. Email: [email protected] 13
14
Resumo - O objetivo deste trabalho foi avaliar a eficiência de leveduras nativas no controle da 15
vassoura de bruxa (VB) e promoção de crescimento do cacaueiro (PC). Cem leveduras foram 16
avaliadas quanto ao controle da VB em plântulas, através da inoculação em sementes e 17
pulverização na parte aérea. A influência do período de aplicação do tratamento (1, 2, 7, 14 18
dias antes da inoculação do patógeno) foi avaliado pela pulverização de quatro leveduras e o 19
produto Agromós®. Leveduras foram analisadass isoladamente e em concomitância com 20
outras leveduras e produtos comerciais quanto ao controle em plântulas e inibição da 21
produção de basidiomas em vassouras secas (IPB). Foi investigado o efeito de aplicações (1, 22
2, 3) de leveduras quanto a PC, através da aplicação isolada e em consórcio. Houve diferenças 23
estatísticas (p<0,05) entre os 100 isolados de levedura, com distribuição em três grupos 24
(clusters) separados por dissimilaridade das médias dos sintomas de VB. O grupo II.1 reúne 25
os isolados que mais controlaram a VB. Tratamentos com isolados pertencentes aos gêneros 26
Rhodosporidium, Hanseniaspora, Candida e Hannaela foram responsáveis pelos maiores 27
níveis de controle e promoção de crescimento independente do experimento realizado. Houve 28
diferenças significativas entre período e número de aplicação da levedura. O Agromós® 29
proporcionou 100% de controle de VB independente da época de aplicação. Os fungicidas 30
quando aplicados isoladamente e associados a leveduras inibiram o aparecimento dos 31
sintomas em 100% das plântulas. Todas as leveduras avaliadas colonizaram os sítios de 32
infecção do patógeno e, permaneceram por 10 dias após a aplicação. Leveduras nativas 33

111
mostraram potencial para o biocontrole da vassoura de bruxa e promoção de crescimento do 34
cacaueiro. 35
Termos para indexação: controle biológico, leveduras, Rhodosporidium, Hanseniaspora, 36
Candida, Hannaela, Moniliotphora perniciosa, Theobroma cacao. 37
38
39
Native yeasts as potential biocontrol agent to witches' broom and growth promoters in 40
cocoa seedlings 41
42
Abstract - The objective of this study was to evaluate the efficiency of native yeasts to control 43
witches' broom (WB) and promote cacao growth (PG). One hundred yeasts were evaluated for 44
control of WB in seedlings by inoculating seeds and spraying aerial part of the plant. The 45
influence of the application of the treatment period (1, 2, 7, 14 days before pathogen 46
inoculation) was evaluated by spraying four yeasts and Agromós® product. Yeasts were 47
evaluated alone and associated with other yeasts and commercial products as to control in 48
seedlings and inhibition of the mushroom production in dry brooms (IMP). It investigated the 49
effect of applications (1, 2, 3) yeast as the PG with alone application and consortium. There 50
were statistical differences (p <0.05) among 100 isolates from yeast, with distribution in three 51
groups (clusters) separated by means of the dissimilarity of the symptoms of WB. The group 52
meets II.1 isolated more controlled the VB. Treatments with isolates from genre 53
Rhodosporidium, Hanseniaspora, Candida and Hannaela were responsible for the highest 54
levels of control and promotion of growth, independent of the experiment conducted. There 55
were significant differences between time and yeast application number. The Agromós® 56
provided 100% control, independent of the application time. The fungicides when applied 57
alone and in combination with yeast, inhibited the onset of the symptoms in 100% of the 58
seedlings. Native yeasts are potential agents of biocontrol of witches' broom disease of cocoa 59
and growth promoters. 60
61
Index terms: Biocontrol, yeast, Rhodosporidium, Hanseniaspora, Candida, Hannaela, 62
Moniliotphora perniciosa, Theobroma cacao. 63
64
65
66
67

112
Introdução 68
69
O fungo Moniliophthora perniciosa (Stahel) Aime & Phillips-Mora, anteriormente 70
descrito como Crinipellis perniciosa, é o agente etiológico da vassoura de bruxa (VB) do 71
cacaueiro e um dos patógenos mais importantes para os países produtores de cacau nas 72
Américas (Meinhardt et al. 2008). Esta doença foi responsável pela queda da produção de 73
cacau no sul da Bahia, principal região produtora do Brasil, de 400.000 toneladas em 1980, 74
para 12.000 após a entrada deste patógeno nos cacauais baianos (FAO, 2002), levando o 75
Brasil a alterar sua situação de exportador para importador de cacau (Midlej & Santos, 2012) 76
e permanece sendo a doença de maior relevância para a lavoura cacaueira baiana e o principal 77
entrave para a retomada do Brasil ao topo do ranking internacional dos maiores produtores de 78
cacau. 79
Este patógeno é um basidiomiceto hemibiotrófico que apresenta dois tipos de micélio, 80
biotrófico (parasítico) e necrotrófico (saprofítico) (Scarpari et al., 2005), afeta principalmente 81
os tecidos meristemáticos da planta (caules, almofadas florais, flores e frutos), causando uma 82
gama de sintomas, que varia de acordo com o órgão infectado e o seu estádio de 83
desenvolvimento (Silva et al., 2003), levando à morte da planta, quando afetada por 84
sucessivos surtos de infecção e fatores abióticos em concomitância (Andebrhan, 1984; 85
Queiroz et al., 2003). 86
Desde o estabelecimento da doença em regiões produtoras no Brasil e em países das 87
Américas do Sul e Central, diferentes estratégias de manejo da doença têm sido buscadas. 88
Atualmente é recomendado pela CEPLAC, a utilização de várias medidas de controle em 89
concomitância. Esforços têm sido voltados para a busca de medidas alternativas para o 90
controle efetivo da VB, podendo ir desde a racionalização do uso de agroquímicos, à redução 91
da frequência de realizações do saneamento vegetal (Costa et al., 2009; Medeiros et al.,2010). 92
O controle biológico, importante componente no manejo de doenças de plantas, utiliza 93
micro-organismos antagônicos e competidores, envolvido em diferentes mecanismos de ação: 94
competição, antibiose, hiperparasitismo, indução de resistência, e promoção de crescimento 95
(Valdebenito-Sanhueza, 2000). Resultados promissores estão sendo obtidos com o fungo 96
Trichoderma stromaticum para o biocontrole do agente causal da VB, entretanto novos micro-97
organimos devem ser testados, principalmente os que co-habitam no mesmo nicho, e que 98
possam ter ação sobre várias doenças, sendo ótimos competidores e competentes. As 99
leveduras apresentam este potencial por estarem tanto na filosfera, quanto na rizosfera de 100
vários cultivos, inclusive o cacaueiro (Cabral et al., 2009). 101

113
Apesar da utilização de fungos leveduriformes como agentes de biocontrole ainda se 102
concentrar na patologia pós colheita (Janisiewicz et al., 2010), estes micro-organismos 103
reúnem características de interesse que estimulam a investigação do potencial biocontrolador 104
em doenças pré colheita, bem como a ação antagônica a patógenos habitantes do solo, 105
rizosfera e filosfera. Diversos relatos descrevem o alto potencial competidor (Zhang et al., 106
2007) destes micro-organismos, que se reproduzem rapidamente em ambientes ricos em 107
açúcares (Lutz et al., 2013), secretam toxinas “killer” (Platania et al., 2012), compostos 108
voláteis (Bruce et al., 2004) e enzimas hidrolíticas capazes de degradar a parede celular de 109
fitopatógenos (Lu et al., 2014; Masih & Paul, 2002; Urquhart & Punja, 2002), apresentam a 110
capacidade de suportar mudanças no ambiente e condições de estresse (Sui et al., 2015), 111
tolerância à poluentes, óleos essenciais vegetais, fitoalexinas e alguns fungicidas 112
(Valdebenito-Sanhueza, 2000), além de sobreviverem saprofiticamente (Afsah-Hejri, 2013) e 113
tolerarem condições adversas (Hu et al., 2015). 114
O objetivo deste trabalho foi avaliar o potencial de leveduras nativas como possíveis 115
agentes de biocontrole à vassoura de bruxa e promotores de crescimento de plântulas do 116
cacaueiro. 117
118
Material e Métodos 119
120
Microrganismos e condições de cultivo 121
Os isolados de leveduras testados foram cultivados em meio de cultivo yeast extract peptone 122
dextrose (YEPD - 0,15% extrato de levedura, 1% neopeptona, 4% dextrose, 2% ágar e 0,05% 123
cloranfenicol), em triplicata, por 72h à 28°C. Transcorrido o tempo de cultivo as colônias de 124
leveduras separadas do meio com alça de Drigaslki previamente flambada e suspensas em 125
solução salina (NaCl 0,85%). A concentração da suspensão de leveduras foi ajustada à 108 126
ufc/mL com base na quantificação em câmara de Neubauer (marca) e mensuração da 127
densidade ótica em espectrofotômetro (SpectraMax® Paradigm Multi-Mode Detection 128
Platform, Molecular-Devices Japan, Tóquio, Japão) à 595 nm. Para obtenção de suspensão de 129
basidiósporos de M. perniciosa, vassouras vegetativas secas foram coletadas, desinfestadas 130
com lavagens rápidas em hipoclorito de sódio a 0,1%, fixadas em arames esticados 131
horizontalmente em casa de vegetação otimizada (vassoureiro), e então submetidas a um 132
regime diário de 8 h de molhamento e 16 h de secamento. Após o período de dormência das 133
vassouras, foi aplicada solução de estreptomicina a 1%, e lavadas mais duas vezes em água 134
destilada. Com o surgimento dos basidiomas, estes foram então coletados e secos em folhas 135

114
de papel absorvente, seccionando os píleos e fixando-os com vaselina pastosa (Silinol SG) em 136
tampas de vidro, com o himênio voltado para baixo, sobre um béquer contendo uma solução 137
de glicerina a 16%, e sob agitação constante, permitindo captação dos basidiósporos 138
liberados. As suspensões de basidiósporos foram quantificadas em câmara de Neubauer e 139
estocadas em nitrogênio líquido, acondicionadas em tubos criogênicos de 2 mL, até o 140
momento de serem utilizadas, conforme metodologia descrita por Dickstein et al. (1987). 141
Antes da estocagem, uma alíquota de 25 µL de cada uma das suspensões de basidiósporos 142
obtidas foi retirada e espalhada com alça de Drigalski sobre a superfície do meio ágar-água a 143
2%, em placas de Petri de 5 cm de diâmetro, para avaliar a germinação dos esporos. Após seis 144
horas de incubação a 25°C, a percentagem de basidiósporos germinados foi mensurada 145
através da visualização em microscópio ótico e somente suspensões que apresentaram 146
percentual de germinação acima de 80% foram utilizadas. Este procedimento foi realizado 147
tanto no momento da estocagem como também 24 h antes da utilização do inóculo. 148
149
Produção de plântulas 150
Plântulas de cacaueiro (Catongo), suscetíveis a M. perniciosa, foram obtidas a partir de 151
sementes pré-germinadas por 48h em serragem esterilizada e umedecida, e transferidas para 152
tubetes de 288 cm3 contendo terriço (solo + substrato) previamente esterilizados por 153
tindalização (autoclavados por 2x com intervalo de 24h) na proporção 1:1. As plântulas foram 154
mantidas em condições de casa-de-vegetação, submetidas à irrigação diária por um período de 155
20 minutos às 9:00 e às 14:00h, adicionando mais uma rega em dias com temperatura mais 156
elevada, por três a quatro semanas antecedendo a inoculação. As condições de temperatura, 157
umidade relativa e luminosidade foram monitoradas através de sensor HOBO® Data Logger 158
(U12-012). 159
160
Testes de biocontrole 161
Potencial biocontrolador em plântulas 162
O potencial biocontrolador foi avaliado através da inoculação dos isolados de leveduras em 163
sementes “nuas” e no meristema apical em genótipo suscetível de cacaueiro. Sementes de 164
cacau desprovidas de pericarpo e tegumento foram pré-germinadas durante 48 horas em água 165
corrente, desinfestadas por imersão em NaOCl a 0,5% durante 2 min, lavadas várias vezes em 166
água esterilizada e seca em câmara de fluxo laminar. As sementes foram imersas nas 167
suspensões dos isolados de levedura e mantidas nesta condição em over night, sendo em 168
seguida semeadas em tubetes contendo terriço e mantidas sob condição de cultivo em casa de 169

115
vegetação. Sementes não tratadas com leveduras foram utilizadas como controle, e imersas 170
em água. Vintes dias após a semeadura, as plântulas foram pulverizadas até o ponto de 171
escorrimento com suspensões de células de leveduras nos tratamentos correspondentes às 172
inoculações em sementes e posteriormente transportadas para a câmara de inoculação. Em 173
câmara de inoculação, as plântulas foram mantidas sob condições de saturação de umidade e 174
temperatura entre 25-27°C. 175
176
Influência do período de aplicação de suspensão de leveduras 177
Plântulas de cacaueiro (Catongo) mantidas em casa de vegetação sob condições de cultivo por 178
20 dias foram tratadas com a pulverização de suspensão de células de leveduras pré-179
selecionadas com potencial biocontrolador (LEV 074 e LEV 098 – Candida sp., LEV 162 e 180
LEV 210 – Hanseniaspora sp.) com 1, 2, 7, 14 dias antes da inoculação do patógeno. 181
182
Associação de leveduras e produtos comerciais 183
Dez isolados de levedura pré-selecionados com potencial biocontrolador (LEV 007 – 184
Rhodosporidium sp.; LEV 027 – Hannaella sp.; LEV 074; LEV 079 – Rhodosporidium sp.; 185
LEV 098; LEV 162; LEV 179 – Rhodosporidium sp.; LEV 193 – Hannaella sp.; LEV 207 – 186
Hannaella sp.; LEV 210) foram aplicados isoladamente, em consórcio com outra levedura e 187
todos em conjunto, em plântulas de cacaueiro suscetíveis, a fim de avaliar a interação entre os 188
mesmos para o biocontrole da vassoura de bruxa do cacaueiro. Também foram adicionados 189
como tratamentos, os fungicidas comerciais Folicur 200 EC® (tebuconazol, Bayer S.A.) e 190
Cobre Atar BR® (óxido cuproso, Atar do Brasil Defensivos Agrícolas LTDA) nas 191
concentrações de 0,3 mL/L e 0,9 g/L, respectivamente e a associação destes com a mistura de 192
células das 10 leveduras avaliadas. Paralelamente a este experimento, foi realizado um ensaio 193
in vitro para avaliar o efeito fungicida dos produtos comerciais utilizados. Alíquotas das 194
concentrações 1 (1,8g/L), 2 (0,9/L) e 3 (0,45/L) de Cobre Atar®, 1 (10 ml/L), 2 (5 mL/L) e 3 195
(1/L) de Agromós®, 1 (0,6 mL/L), 2 (0,3 mL/L) e 3 (0,1 mL/L) de Folicur®, foram 196
incorporadas em meio YEPD fundente, sendo em seguida semeado o isolado LEV 207 (108 197
ufc/mL) e espalhado com alça de Drigaslki. As placas foram incubadas à 28°C por 48h e 198
avaliadas através da visualização e contagem de colônias formadas. O experimento in vitro foi 199
realizado em triplicata. 200
201
Inoculação e avaliação dos sintomas 202

116
Vinte e quatro horas após a pulverização com leveduras, as plântulas de cada tratamento 203
foram inoculadas na gema apical com 30 µL da suspensão de basidiósporos na concentração 204
de 2x105 esporos/mL de meio ágar-água 0,2%. Plântulas tratadas apenas com leveduras e 205
apenas com o patógeno foram utilizadas como testemunhas relativas, recebendo apenas uma 206
gota de ágar-água a 0,2% ou com suspensão de basidiósporos respectivamente, enquanto que 207
testemunhas absolutas cresceram na ausência de leveduras e patógeno. Após a inoculação, 208
todas as plântulas permaneceram por 24 h em câmara de inoculação, sendo então transferidas 209
novamente para a casa de vegetação sob condições de cultivo. 210
As plantas foram avaliadas 30 dias após a inoculação quanto ao aparecimento dos 211
sintomas, incidência e severidade da doença, observando o tipo de vassoura formada, o 212
número de vassouras axilares, o diâmetro e a altura das vassouras terminais. Para análise dos 213
dados, foram analisadas: incidência (INC), percentagem de plantas apresentando qualquer 214
sintoma da doença, (vassoura terminal, axilar, cotiledonar e seca, intumecimento do caule, do 215
hipocótilo, dos pecíolos e dos pulvinos, cancro, superbrotamento ou hipertrofia); 216
comprimento da vassoura terminal; e índice de doença (calculado pela seguinte fórmula: ID= 217
VT+VA (0,5*CVT)+NVA, em que: VT = número de vassouras terminais; VA = número de 218
vassouras axilares; CVT = comprimento da vassoura terminal; NVA = número de vassouras 219
axilares maiores que 1cm. O desenho experimental foi em blocos casualizados, com quatro 220
repetições de 20 plantas cada. 221
222
Potencial biocontrolador em vassouras secas 223
A inibição da produção de basidiomas em vassouras secas foi avaliada pulverizando a 224
suspensão de células de leveduras, individualmente ou em conjunto. O experimento foi 225
realizado na Unidade de Biocontrole do Centro de Pesquisas do Cacau (CEPEC/CEPLAC) 226
em vassoureiros construídos para possibilitar a produção de basidiomas e em câmara de 227
inoculação. Vassouras secas foram coletadas em cacaueiros infectados nas áreas 228
experimentais do CEPEC, padronizadas, por diâmetro e comprimento, desinfestadas com 229
lavagens rápidas em hipoclorito de sódio a 0,1%, fixadas em arames esticados 230
horizontalmente em casa de vegetação otimizada (vassoureiro), e então submetidas a um 231
regime diário de 8 h de molhamento e 16 h de secamento. Com o surgimento de basidiomas 232
(quebra de dormência), vassouras produtivas foram selecionadas, transferidas para gabinetes 233
de vidro em condições ideais para a produção de basidioma, onde foram penduradas em 234
hastes de aço inox. O experimento foi realizado em delineamento inteiramente casualizado, 235
com 40 repetições (vassouras) por tratamento. Cada tratamento foi aplicado em vassouras 236

117
dispostas em um único gabinete para evitar transferência de inóculo e contaminação. Em cada 237
gabinete foi aplicado suspensão de células por pulverização das leveduras: LEV 007, LEV 238
027, LEV 074, LEV 079, LEV 098, LEV 162, LEV 179, LEV 193, LEV 207, LEV 210, além 239
da aplicação da mistura de todas as leveduras testadas. Foram incluídos como controle 240
positivo e negativo, o tratamento com o Trichovab, (Trichoderma stromaticum), biofungicida 241
registrado no Ministério da Agricultura Pecuária de Abastecimento (MAPA) para a cultura do 242
cacau, e testemunha absoluta apenas com aplicação de água. As vassouras foram submetidas a 243
um regime diário de 8 h de molhamento e 16 h de secamento, fotoperíodo de 12h e 244
temperatura de 25-27°C, durante 60 dias e avaliadas através da contagem e retirada dos 245
basidiomas produzidos, duas vezes por semana. As vassouras secas foram pulverizadas com 246
leveduras três vezes a cada 15 dias. Os basidiomas coletados foram armazenados em solução 247
salina com cloranfenicol em tubos de ensaio e submetidos a banho de ultrassom em 248
ultrassonicador (Quimis 0335D) para posterior isolamento de micro-organismos em placas de 249
Petri contendo meio YEPD. 250
251
Microscopia Eletrônica de Varredura 252
Pequenos fragmentos (3x3) do limbo, nervura e meristema apical de amostras foram 253
coletados em plântulas de cacaueiro (CATONGO) uma e 240 horas (10 dias) após a 254
pulverização da suspensão de células (1x 108 ufc/mL) das leveduras LEV 098, LEV 162 e 255
LEV 210 no meristema apical e limbo foliar. Alíquotas de 3µL da suspensão de basidiósporos 256
de M. perniciosa foram depositadas no meristema apical de plântulas inoculadas com 257
suspensão de leveduras, 168 h (7 dias) após a pulverização. As plântulas da testemunha 258
absoluta foram pulverizadas apenas com água destilada esterilizada, e plântulas inoculadas 259
apenas com M. perniciosa foram utilizadas como testemunha relativa. As amostras foram 260
selecionados em fragmentos de aproximadamente 3mmx3mm, fixados em gluteraldeído 2,5% 261
tamponado com cacodilato de sódio 0,1M em pH 6,9 e armazenados à 4°C. Após a fixação, os 262
espécimes foram lavados em tampão cacodilato de sódio 0,1M pH 6,9, duas vezes em 263
intervalo de 10 min. Os espécimes foram posteriormente desidratados em série gradual (50, 264
60, 70, 80, 90 %) de acetona e três vezes em acetona 100% com intervalos de 10 min cada. 265
Após a desidratação, os espécimes foram levados ao ponto crítico (BALZER, CPD020), 266
fixados em stub com fita dupla face de carbono e cobertos por pulverização catódica com um 267
filme de aproximadamente 20 a 30 nm de espessura de ouro, utilizando o aparelho SPUTTER 268
COATER, marca BALZERS. Posteriormente, foram observados no microscópio eletrônico de 269
varredura, modelo JSM T200, marca JEOL, à voltagem de aceleração a 10 ou 15 kV. 270

118
271
Testes de Promoção de Crescimento de Plantas 272
Avaliação do crescimento de plântulas seminais 273
O substrato utilizado para o cultivo das plântulas de cacaueiro nos experimentos 274
realizados neste trabalho apresentou pH 6,6, acidez total (Al + H) de 4,3 cmol/dm3, saturação 275
de bases (SB) de 19,6 cmol/dm3, potássio (K) – 0,34 cmol/dm3, magnésio (Mg) – 5,3 276
cmol/dm3, cálcio (Ca) – 14,0 cmol/dm3, capacidade de troca catiônica (CTC) de 23,9 277
cmol/dm3, além de fósforo (P) – 294 mg/Kg, Ferro (Fe) – 211,1 mg/Kg, zinco (Zn) – 0,7 278
mg/Kg, cobre (Cu) – 0,1 mg/Kg e manganês 1,7 mg/Kg. 279
Sementes de cacaueiro (clone PS 1319) foram selecionadas, escarificadas através da 280
retirada de pericarpo e tegumento, pré-germinadas durante 48 horas em água corrente, 281
desinfestadas por imersão em NaOCl a 0,5% durante 2 min, lavadas em água esterilizada e 282
imergidas em suspensão de células de leveduras [LEV 210, LEV 84 (Candida 283
nondorbophila), LEV 206 (Cryptococcus heimaeyensis), LEV 93 (Cryptococcus sp.), LEV 284
149 (Cryptococcus sp.), LEV 162, LEV 98, LEV 170 (Rhodosporidium paludigenum), LEV 285
79, LEV 168 (Rhodosporidium paludigenum)] por 24 h a 25-27°C. Foram adicionados como 286
tratamentos, uma mistura com as 10 leveduras que foram inoculadas individualmente, a 287
adição com fosfafato monoamônico (NH4H2PO4) (1g/ 200mL - 5mL/plântula) como 288
testemunha relativa e aplicação apenas de água destilada esterilizada como testemunha 289
absoluta. Após o período de incubação, as sementes foram plantadas em tubetes de 288 cm3 290
contendo terriço (substrato vegetal + solo 1:2) esterilizado e mantidas em casa de vegetação 291
sob condições de cultivo. O efeito da quantidade de aplicações foi avaliado através da injeção 292
de 5 mL de suspensão de levedura correspondente ao tratamento da semente, no terriço ao 293
redor da plântula, uma e duas vezes após a semeadura, correspondendo a um fatorial 11 x 3 + 294
2 (10 leveduras aplicadas isoladamente e em conjunto x 3 quantidade de aplicação do 295
tratamento + 2 testemunhas), totalizando 35 tratamentos. Sessenta dias após a semeadura, o 296
diâmetro da região do coleto foi mensurado com paquímetro Mitutoyo® (Japão), as plântulas 297
coletadas, separadas do terriço, as raízes lavadas em água corrente, transferidas para bancada 298
em laboratório para retirada de umidade e posterior mensuração da biomassa fresca da parte 299
aérea e sistema radicular (SR). Após a pesagem, as plântulas (parte aérea e SR) foram 300
transferidas para sacos de papel e secas em estufa de circulação forçada à 65°C por 48h (até a 301
estabilização do peso). O peso fresco da parte aérea e SR foi mensurado, assim como o peso 302
fresco, em balança semi-analítica centesimal (BEL M5202). 303
304

119
Análises estatísticas 305
Os dados foram submetidos a da análise de variância, testes de normalidade (Shapiro Wilk) e 306
homocedasticidade (Bartellet) e ajustados [Sqrt (x +0,5)] quando necessário, para posterior 307
análise de variância, seguida de teste Tukey a 5% de probabilidade, através do pacote 308
estatístico LSmeans SAS (SAS Institute, 1988). As médias das variáveis CVT, NVA e ID 309
foram avaliadas a partir do teste não paramétrico de Kruskal-Wallis. Uma matriz gerada para 310
os tratamentos (isolados de levedura) foi avaliada através do software PAST 3.1, o que 311
permitiu a construção de dendograma a partir de distância Euclidiana, para avaliar o 312
agrupamento dos isolados quanto ao potencial biocontrolador. 313
314
Resultados 315
316
Os tratamentos (leveduras) variaram significativamente (p<0,5) quanto ao potencial 317
bioncontrolador a vassoura de bruxa em mudas de cacaueiro (Tabela 1), para todas as 318
variáveis analisadas (incidência - INC, comprimento de vassoura terminal - CVT, número de 319
vassoura terminal maior que 1 cm – NVA e índice de doença (ID). 320
A análise de agrupamento (Figura 1), a partir da comparação de todas as variáveis 321
(sintomas) analisadas, permitiu inferir sobre a separação dos isolados de leveduras avaliados 322
em três grupos (clados), após corte na altura de θ = 12. O grupo 1, formado por tratamentos 323
com leveduras LEV 080, LEV 118, LEV 226, LEV 089, LEV 097, LEV 219, se distanciou 324
dos demais por proporcionarem as maiores médias de incidência e sintomas em plântulas de 325
cacaueiro, divergindo ainda da testemunha relativa, que foi inoculada apenas com 326
basidiósporos de M. perniciosa. Apesar da alocação, por dissimilaridade euclidiana, em um 327
terceiro grupo, as leveduras LEV 016, LEV 092, LEV 147, LEV 059, LEV 082, LEV 106, 328
LEV 206, LEV 140, LEV 201, além da testemunha inoculada, também não inibiram 329
significativamente o aparecimento e evolução do quadro sintomatológico da vassoura de 330
bruxa (VB). O grupo 2 reúne o maior número de tratamentos, entretanto se subdivide em 331
outros dois clados, um com características semelhantes ao grupo 3 (II.2), e outro se 332
destacando dentre todos os tratamentos avaliados, por proporcionarem os maiores níveis de 333
controle. Subdivisões abaixo do clado II.1 permitem selecionar leveduras ainda mais 334
promissoras (LEV 020, LEV 050, LEV 054, LEV 046, LEV 088, LEV 001, LEV 139, LEV 335
154, LEV 005, LEV 090, LEV 055, LEV 174, LEV 006, LEV 014, LEV 210, LEV 079). 336
Houve variações quanto ao comportamento das leveduras aplicadas em diferentes 337
tempos (Tabela 2). Plântulas tratadas com o isolado LEV 079 apresentaram menor incidência 338

120
de VB quando pulverizadas 7 e 14 dias antes da inoculação do patógeno (DAI), entretanto, 339
apenas o tratamento com aplicação 7 dias antes, diferiu estatisticamente da testemunha 340
inoculada quanto a INC. Para ID, o isolado LEV 079 proporcionou resultados semelhantes, 341
mas não se distanciaram estatisticamente (p<0.05) da testemunha. Todas as plântulas tratadas 342
com o isolado LEV 098 apresentaram menor incidência que a testemunha inoculada, 343
independente da época de aplicação, havendo destaque para aplicação do tratamento 7 DAI 344
(Tabela 2). O grupo de plântulas tratadas com LEV 098, diferiu significativamente (p<0,05) 345
da testemunha, para todas as variáveis com aplicação 1 DAI. O isolado LEV 162, quando 346
aplicado 1 DAI, proporcionou controle de 100% de controle, não diferindo (p<0,05) dos 347
resultados obtidos para o período de 14 DAI em relação à incidência. Todos os grupos de 348
plântulas tratadas com LEV 210 apresentaram incidência significativamente menor do que a 349
testemunha inoculada, independente da época de aplicação. Os melhores resultados para este 350
isolado foram obtidos quando as aplicações foram realizadas 14 DAI. O produto comercial 351
Agromós proporcionou 100% de controle independente do período de aplicação (Tabela 2). 352
Os tratamentos apresentaram diferenças significativas (p<0,05) quando comparados 353
dentro de cada período de aplicação (Tabela 2). O isolado LEV 162 apresentou controle 354
significativamente semelhante ao Agromós, quando aplicado somente 1 DAI, diminuindo a 355
incidência e o desenvolvimento de sintomas da vassoura de bruxa do cacaueiro, demonstrado 356
através do ID. O isolado LEV 098 apresentou o segundo menor ID para este período, 357
proporcionando resultados discrepantes dos isolados LEV 210 e LEV 079. Para o período de 358
2 DAI, apenas o isolado LEV 210 apresentou controle semelhante ao Agromós. Com 7 DAI, 359
os isolados de levedura testados não controlaram significativamente o patógeno. O tempo de 360
14 DAI permitiu que os isolados LEV 210 e LEV 162 proporcionassem os melhores controles 361
dentre os tratamentos e períodos avaliados. Mesmo o isolado 162 em 14 DAI não diferindo 362
estatisticamente (p<0,05) do LEV 210 para este período, este tratamento se destaca como 363
combinação que mais controlou a vassoura de bruxa em plântulas de cacaueiro seminais. 364
Os isolados de leveduras (LEV 079, LEV 162, LEV 027, LEV 007, LEV 210, LEV 365
193, LEV 207, LEV 098, LEV 074) quando avaliados separadamente e em conjunto com 366
outras leveduras e produtos comerciais, proporcionaram resultados discrepantes em relação à 367
incidência e desenvolvimento de sintomas da vassoura de bruxa do cacaueiro, com variação 368
de ID de 33,07 (LEV 207), à 83,27 (LEV 027 + LEV 207)), com exceção dos tratamentos 369
com fungicidas comerciais que controlaram 100% da doença (Tabela 3). 370
Plântulas tratadas com os fungicidas Folicur ® e Cobre Atar®, isoladamente e em 371
consórcio com leveduras, não apresentaram sintomas e, portanto, utilizados como padrão de 372

121
controle e comparação com demais tratamentos. Somente os isolados LEV 179 e LEV 207 373
apresentaram INC similares aos tratamentos com os fungicidas, mesmo não havendo 374
diferença significativamente em relação aos tratamentos LEV 162, LEV 210 e LEV 027 e a 375
combinação LEV 027 + LEV 207. Mesmo apresentando variação em relação ao número de 376
vassouras axilares menores que 1 cm (NVA), todos os tratamentos foram similares, não 377
diferindo estatisticamente. As variáveis CVT e ID também não proporcionaram diferenciação 378
estatística entre os tratamentos avaliados. 379
Quando os 10 isolados de levedura e combinações entre os mesmos foram avaliados 380
para a inibição da formação de basidiomas em “vassouras secas”, a maioria dos tratamentos 381
apresentaram resultados positivos, não diferindo da testemunha Tricovab® (Figura 2). Apenas 382
os tratamentos LEV 074 e LEV 193 diferiram significativamente (p<0,05) do Tricovab. O 383
número médio de basidiomas formados durante o período de avaliação variou de 0,37 (LEV 384
193) à 0,00 (LEV 079, LEV 162, LEV 179, LEV 210 e as combinações LEV 27 + LEV 207, 385
LEV 79 + LEV 179). 386
Os isolados LEV 179 e LEV 207 destacam-se positivamente dentre os demais 387
tratamentos por apresentarem eficiência no controle da VB, com possibilidade de atuação 388
tanto nos estágios iniciais do quadro epidemiológico desse patossistema, como germinação, 389
infecção e colonização, como no processo de esporulação e disseminação. O isolado LEV 390
207, quando combinado com o LEV 027, conseguiu manter comportamento positivo em 391
relação ao controle da vassoura de bruxa em plântulas de cacaueiro, entretanto o padrão de 392
controle do LEV 179 não foi evidenciado quando este foi aplicado em consórcio com o LEV 393
079. 394
As concentrações 1 (0,6 mL/L), 2 (0,3 mL/L) e 3 (0,1 mL/L) de Folicur®, inibiram 395
100% o crescimento do isolado LEV 207 in vitro (Figura 2). As concentrações utilizadas in 396
vivo para os produtos comerciais Cobre Atar® e Agromós® (0,9 g/L e 0,5 mL/L, 397
respectivamente) não influenciaram na reprodução de células da levedura analisada, 398
entretanto a maior concentração destes dois produtos, foi capaz de inibir o crescimento do 399
LEV 207 totalmente (Cobre Atar – 1,8g/L) e parcialmente (Agromós – 10mL/L). 400
Observações de eletromicrografias obtidas por microscopia eletrônica de varredura 401
(MEV) possibilitaram a investigação ultraestrutural da interação leveduras x cacaueiro x M. 402
perniciosa. Leveduras foram visualizadas em todas as amostras obtidas de folhas e 403
meristemas apicais de plântulas de cacaueiro, permanecendo nestes substratos vegetais desde 404
as primeiras horas à 10 dias após a pulverização de células de leveduras. Estas leveduras 405
permaneceram fortemente aderidas aos sulcos formados pela junção de uma célula epidérmica 406

122
e outra, e em irregularidades presentes nas nervuras abaxial e adaxial, pois mesmo as 407
plântulas estando sob turno de rega diário, as leveduras não foram lixiviadas (Figuras 3, 4 e 408
5). Mesmo visualizando maior densidade de células nas nervuras centrais (Figura 3A; Figura 409
4B, C, D, E; Figura 5D), as células se distribuem uniformemente pelo limbo foliar, havendo a 410
predominância na face abaxial, próximo a estômatos (Figura 3C; Figura 5C). 411
Na figura 3, observa-se a presença de células do isolado LEV 098 (C. saopaulonenis) 412
na face abaxial (B, C) e adaxial (A, F) do limbo foliar, bem como no meristema apical (D, E, 413
F) 10 dias após a pulverização da levedura. Nas mostras obtidas de plântulas pulverizadas 414
com o isolado LEV 162 (H. uvarum), é possível visualizar a presença de células de levedura 415
na base e aderidos à tricomas tectores (Figura 4B, C, D, E) e glandulares (Figura 4. B). 416
Células do LEV 210 também permaneceram aderidas ao limbo abaxial, adaxial, 417
meristema apical, base de tricomas e nervura (Figura 5), com formação de pseudomicélio 418
aderido a um tricoma tector (Figura 5B). 419
Os cortes transversais de folhas e meristema apical permitiram a observação de 420
possíveis canais de entrada e colonização de camadas epidérmicas e parenquimáticas (Figura 421
5A, B, E), além da provável formação de agregados citoplasmáticos de cálcio (Figura 5A). 422
Quando os isolados LEV 079, LEV 084, LEV 093, LEV 098, LEV 149, LEV 162, 423
LEV 168, LEV 170, LEV 206 e LEV 210 foram avaliados isoladamente e em conjunto 424
quanto a promoção do crescimento de plântulas seminais de cacaueiro (PS 1319), diferiram 425
para as variáveis analisadas, entre si e, para alguns isolados em relação ao número de 426
aplicações (1, 2 ou 3 vezes) da suspensão de células na parte aérea (Tabela 4). 427
Somente os isolados LEV 079, LEV 149, LEV 162, LEV 168, LEV 170 e 206 foram 428
influenciados pelo número de aplicações para manifestação da característica de promoção de 429
crescimento. Os isolados LEV 079 (variável DC), LEV 168 (variável CP e BRF) e LEV 170 430
(variáveis CP, DC e PSA) foram mais eficientes quando inoculados por três vezes, quantidade 431
de aplicação que diferiu significativamente (p<0,05) das demais para variáveis específicas. Os 432
isolados LEV 149 (variável DC), LEV 162 (variável CP) e LEV 168 (variável BRF) 433
proporcionaram melhores resultados quando inoculados uma ou duas vezes, diferindo do 434
tratamento com três aplicações. 435
O tratamento com LEV 093 proporcionou o maior comprimento da parte aérea (24,96 436
cm) dentre os demais isolados avaliados, quando inoculados apenas na semente (única 437
aplicação), entretanto só diferiu estatisticamente do LEV 170 (22,43 cm), tratamento que 438
apresentou comportamento semelhante à testemunha relativa (adubação mineral) e 439
testemunha absoluta, com 22,76 e 22,36 cm de comprimento, respectivamente. Quando foram 440

123
realizadas duas ou três aplicações, referentes a uma ou duas pulverizações de suspensões de 441
células na parte aérea, o LEV 210 se destaca dentre os demais em relação a variação CP, 442
gerando comprimento médio de 25,08 e 25,25 cm, respectivamente. Com estas doses reforço 443
(2 e 3 aplicações), as plântulas tratadas com LEV 149 e MIX apresentaram respectivamente 444
as menores médias. 445
O diâmetro médio do coleto das plântulas com apenas uma aplicação foi maior quando 446
estas foram inoculadas pelo LEV 149 (4,41 cm), diferindo apenas do LEV 079 (4,06 cm). O 447
comportamento destes dois isolados também foi evidenciado quando as plântulas foram 448
tratadas com duas aplicações de suspensão de células, com maior (4,63 cm) e menor (4,09 449
cm) média para DC. Dentre os tratamentos que receberam três aplicações, o LEV 210 450
destaca-se por apresentar o maior diâmetro médio (4,47 cm), entretanto não se diferencia 451
estatisticamente (p<0,05) dos demais. Todos os tratamentos (isolados de levedura) inoculados 452
uma, duas ou três vezes, influenciaram positivamente para a variável DC, pois todas 453
apresentaram médias maiores que as testemunhas, relativa (3,89 cm) e absoluta (3,85 cm). 454
A média de plântulas tratadas com LEV 210 foi maior para a variável PFA, com uma 455
(2,84 g) e 3 (2,94 g) aplicações, entretanto os resultados obtidos com este isolado nesta 456
variável não diferiu estatisticamente dos demais quando as plântulas receberam duas 457
aplicações. As menores médias para esta variável foram obtidas nos tratamentos LEV 170 458
(2,27 g) e MIX (2,32 g), com uma e três aplicações, respectivamente. 459
A variável PSA não foi responsável pela variação existente entre os tratamentos, pois 460
estes proporcionaram resultados semelhantes entre si e em comparação com a testemunha 461
relativa (0,61). Todos os tratamentos influenciaram positivamente no desenvolvimento da 462
parte aérea, pois o peso médio da parte aérea em plântulas da testemunha absoluta foi 463
aproximadamente 1/3 menor (0,20 g). 464
Os tratamentos não diferiram estatisticamente (p<0,05) entre si para PFR com apenas 465
uma aplicação. Entretanto, quando avaliados com duas ou três aplicações, os isolados LEV 466
093 (0,66 g) e LEV 084 (0,75 g) proporcionaram as maiores médias, respectivamente. Os 467
menores PFR foram observados nos tratamentos LEV 168 (0,52 g) quando as plântulas foram 468
tratadas com duas aplicações e o LEV 079 (0,54 g) quando houve três aplicações de células 469
da levedura. 470
Assim como para a variável PSA, o PSR não gerou variação entre os tratamentos à 5% 471
de probabilidade pelo teste de Tukey. 472
473
Discussão 474

124
475
O controle químico ainda é o recurso mais utilizado para o controle de doenças de 476
plantas, mas para o patossistema cacaueiro x M. perniciosa, existem vários fatores que 477
induzem à uma grande variabilidade na eficiência desta técnica, principalmente quando 478
utilizada isoladamente. A variabilidade genética do patógeno tem sido expressa através da 479
observação da plasticidade fisiológica no crescimento micelial em meios de cultura, tipos de 480
reação em alguns testes bioquímicos, compatibilidade somática e patogenicidade ao cacaueiro 481
e a outros hospedeiros (Bartley, 1977; Bastos, 1986; Mcgeary & Wheeler, 1988). Dessa 482
forma, a variação do patógeno até mesmo com referência à sensibilidade a fungicidas, pode 483
eventualmente explicar a eficácia das recomendações de controle da doença em determinadas 484
situações. 485
A combinação de diferentes métodos consiste na forma mais eficiente e econômica de 486
controle, cada um deles com diferentes modos de ação, reduzindo progressivamente o 487
desenvolvimento da doença (Pereira & Valle, 2012). Atualmente o Manejo Integrado de 488
Pragas (MIP) recomendado pela CEPLAC (Comissão Executiva do Plano da Lavoura 489
Cacaueira) aos agricultores brasileiros é constituído de quatro a seis aplicações de fungicidas 490
cúpricos, duas a quatro podas fitossanitárias das vassouras por ano e também aplicações do 491
fungo Trichoderma stromaticum (Tricovab) como medida de biocontrole (Rudgard & Buttle, 492
1987; Almeida et al., 2003; Medeiros et al, 2010). No entanto, pode-se constatar que tais 493
medidas não são amplamente adotadas pelos agricultores, visto o seu alto custo de 494
implementação e retorno a longo prazo (Midlej & Santos, 2007). 495
Esforços têm sido voltados para a busca de medidas alternativas para o controle 496
efetivo da VB, incorporando ao MIP, novos agentes e práticas que possam atuar desde a 497
racionalização do uso dos fungicidas cúpricos (ainda muito caros) a redução da frequência de 498
realizações do saneamento vegetal (Costa et al., 2009; Medeiros et al., 2010). Apesar dos 499
estudos relacionados ao controle biológico da VB concentrar-se na interação T. cacao x 500
Trichoderma x M. perniciosa, a mistura de organismos benéficos, buscando conjuntamente 501
efeitos antagônicos, protetores e/ou antiesporulante dos patógenos, deve ser realizada visando 502
promover um sistema integrado de biocontrole (Elad, 2003). 503
O controle biológico é um importante componente no manejo integrado de doença de 504
plantas, devido a possibilidade de associação a outros métodos de controle e uma alternativa à 505
diminuição do uso de defensivos agrícolas e práticas com menor impacto ambiental 506
(Valdebenito-Sanhueza, 2000). 507

125
A comunidade microbiana residente na superfície de órgãos vegetais tem sido 508
estudada, objetivando determinar biodiversidade existente e os mecanismos de interação entre 509
os micro-organismos presentes neste habitat e as plantas (Berghofer et al., 2003; Laca et al., 510
2006). Nestes substratos são identificados micro-organismos que interagem simbioticamente 511
com as plantas, utilizando nutrientes requeridos para seu crescimento e reprodução, e em 512
contrapartida controlando a densidade populacional de agentes patogênicos às plantas (Lima 513
et al., 2000). 514
Diversos relatos têm demonstrado o efeito de leveduras nativas na inibição do 515
crescimento micelial do patógeno de várias espécies vegetais (Cabral et al., 2009; Lutz et al., 516
2013; Nutaratat et al., 2014). Lutz et al. (2013) relataram o efeito antagônico de leveduras 517
nativas de Pyrus communis L. à Penicillium expansum e Botrytis cinerea, micro-organismos 518
responsáveis por podridões pós-colheita em frutos de várias espécies vegetais, destacando 519
assim, a importância da obtenção e seleção de micro-organismos biocontroladores que co-520
habitem o mesmo nicho ecológico. 521
Apesar da preferência por ambientes ricos em carboidratos, como a superfície de 522
frutos, florese sementes, as leveduras também estão presentes na superfície de folhas, atuando 523
por mecanismos diversos como produção de enzimas hidrolíticas, toxinas killer, inibição da 524
germinação de esporos fúngicos (Lutz et al., 2013), produção de fitohormônios (Sun et al., 525
2014), indução de resistência (Lu et al., 2013) e formação de biofilmes (Li et al., 2015), 526
possibilitando a sua estabilização em ambientes hostis, expostos a variações de temperatura, 527
umidade relativa. A distribuição espacial das leveduras aplicadas em folhas e meristema 528
apical de plântulas de cacaueiro, demonstraram que a anatomia e morfologia dos órgãos 529
vegetais, como estômatos, nervuras, e apêndices na superfície, incluindo tricomas e 530
hidatódios, podem afetar a disponibilidade de nutrientes na filosfera, contribuindo assim para 531
o estabelecimento de micro-organimos nestes substratos (Leveau & Lindow, 2001). 532
As variáveis utilizadas para comparação entre a severidade em mudas tratadas com 533
leveduras e produtos comerciais não proporcionaram resultados que permitissem a 534
diferenciação estatística (p<0,05) das médias, pois após a penetração e colonização do tecido 535
vegetal, os isolados de levedura utilizados não foram capazes de retardar ou impedir o 536
crescimento do patógeno e desenvolvimento dos sintomas. A penetração de M. perniciosa em 537
tecidos meristemáticos de cacaueiro acontece através de aberturas naturais e estômatos, além 538
de lesões na base de tricomas (Sena et al., 2014), desta forma, é essencial a colonização ou 539
presença de micro-organimos antagonistas nestes ambientes, como evidenciado nas 540
eletromicrografias presentes na Figuras 3, 4 e 5. A anatomia e morfologia dos órgãos 541

126
vegetais, podem afetar a disponibilidade de nutrientes na filosfera, contribuindo assim para o 542
estabelecimento de micro-organimos nestes substratos (Leveau & Lindow, 2001). 543
Fungos leveduriformes preferencialmente colonizam ambientes ricos em carboidratos, 544
como a carposfera, espermosfera e antosfera, mas também estão presentes na superfície de 545
folhas, atuando por mecanismos diversos como produção de enzimas hidrolíticas, toxinas 546
killer, inibição da germinação de esporos fúngicos (Lutz et al., 2013), produção de 547
fitohormôios (Sun et al., 2014), indução de resistência (Lu et a., 2013) e formação de 548
biofilmes (Li et al., 2015), possibilitando a sua estabilização em ambientes hostis, expostos a 549
variações de temperatura, umidade relativa. 550
Apesar do pequeno número de produtos comerciais biocontroladores no mercado, 551
quando comparados com a quantidade de produtos químicos sintéticos comercializados, 552
diversos produtos a base de leveduras têm demonstrado eficiência no controle de doenças em 553
várias culturas, a exemplo do Aspire®, cujo composto ativo são células da espécia Candida 554
oleophila (Droby et al., 2003). A eficiência do controle é dependente de variáveis como 555
época, doses, intervalo e formas de aplicação (Oliveira, 2004a), além da capacidade de tolerar 556
condições ambientais adversas, seja por variações climáticas ou plasticidade da concentração 557
de compostos ativos em determinados substratos. A velocidade de crescimento e estabilização 558
dos micro-organimos nos substratos colonizados é variável, sendo portanto, essencial o 559
conhecimento a cinética microbiana, bem como os mecanismos de biocontrole e tempo para 560
expressão de características de interesse envolvidas no biocontrole, como secreção de enzimas 561
hidrolíticas, toxinas, colonização de ferimentos e indução de resistência. 562
Em diversas pesquisas realizadas na região cacaueira da Bahia, constatou-se que o 563
fungicida sistêmico tebuconazole (Oliveira, 2004a) e outros do grupo estrubilurinas (Oliveira, 564
2004b), com destaque para a azoxystrobina, apresentaram os melhores resultados para o 565
controle da VB tanto em viveiros quanto em campo (Oliveira & Luz, 2005), entretanto esta 566
classe de fungicidas também impedem o crescimento de fungos leveduriformes benéficos que 567
habitam o mesmo ambiente, como as leveduras. A capacidade da associação de leveduras a 568
fungicidas protetores (Valdebenito-Sanhueza, 2000) foi reiterada por este trabalho (Figura 2), 569
entretanto novos testes devem ser realizados para determinação de dosagens, concentrações e 570
isolados de levedura tolerantes a tais condições. A concentração (0,9g/L) do fungicida cúprico 571
Cobre Atar® utilizado para os testes em plântulas, não inibiu o crescimento in vitro de um 572
representante do gênero Hannaella sp., mas os tratamentos da utilização de levedura e 573
fungicida em consórcio não diferiu do tratamento somente com o fungicida, proporcionando 574
100% de controle da VB. Destaca-se a importância da prospecção e associação de leveduras 575

127
com fungicidas a base de cobre que promovam níveis satisfatórios de controle, pois apesar 576
desta classe de fungicidas serem menos seletivos que os sistêmicos, apresentam menor 577
impacto ambiental. 578
Organismos envolvidos no controle de doenças de plantas podem atuar através de 579
características relacionadas diretamente aos processos de germinação, penetração, 580
colonização e esporulação, ou contribuírem para o melhor desenvolvimento vegetal, 581
promovendo o crescimento de plantas através da solubilização de fosfatos, fixação de 582
nitrogênio, produção de fitohormônios (Altomare et al., 1999; Oliveira et al., 2010; Oliveira et 583
al., 2012). Os resultados alcançados pelo presente trabalho, através da avaliação de leveduras 584
nativas do cacaueiro na promoção do crescimento de plântulas, corroboram com as 585
informações citadas por Nutaratat et al. (2014), que identificaram leveduras nativas epifíticas 586
e endofiticas (Hannaella sinensis, Cryptococcus flavus, Rhodosporidium paludigenum, 587
Torulaspora globosa), com alta capacidade de produção do fitohormônio ácido indol-acético 588
(AIA) e por consequência promotoras do crescimento de plantas. 589
Alguns isolados estudados neste trabalho (Hannaella sp. Hanseniaspora sp., Candida 590
sp. e Rhodosposporidium sp.) apresentaram resultados positivos para o controle da VB, 591
proporcionando as menores incidências em plântulas tratadas com suspensões de células 592
destas leveduras e inibição da formação de basidiomas em vassouras secas, além da promoção 593
do crescimento de plântulas seminais de cacaueiro. O presente trabalho ressalta a importância 594
da busca por micro-organismos que habitem e colonizem o sitio de infecção e tecidos vegetais 595
do hospedeiro, que podem atuar na manifestação do caráter de resistência a doenças de 596
plantas em condições de cultivo e ambientes naturais, e apresenta novos agentes de 597
biocontrole para o manejo integrado da VB. Ressalta-se ainda a necessidade do conhecimento 598
e elucidação dos mecanismos de biocontrole envolvidos, bem como as condições que mais 599
favorecem a estabilização destes micro-organismos no tecido hospedeiro. 600
601
Conclusões 602
603
1. Leveduras nativas são potenciais agentes de biocontrole da vassoura de bruxa do 604
cacaueiro, atuando na diminuição dos sintomas em plântulas seminais e inibição da produção 605
de basidiomas. 606
2. Há possibilidade de integração de Cobre Atar® com leveduras nativas para o 607
manejo da vassoura de bruxa do cacaueiro. 608

128
3. O período de aplicação da suspensão de células de leveduras é preponderante para a 609
manifestação do biocontrole da vassoura de bruxa do cacaueiro. 610
4. Leveduras nativas que apresentam potencial biocontrolador a vassoura de bruxa, 611
podem atuar como promotoras de crescimento de plântulas seminais do cacaueiro, havendo 612
variações intraespecíficas e em relação ao número de aplicações na manifestação desta 613
característica. 614
5. O produto comercial AgroMós pode atuar e agir em concomitância com leveduras 615
como indutores de resistência à vassoura de bruxa do cacaueiro 616
617
Agradecimentos 618
619
À Financiadora de Estudos e Projetos (FINEP), através da Fundação Pau Brasil 620
(FUNPAB) e Comissão Executiva do Plano da Lavoura Cacaueira (CEPLAC), 621
à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Capes) e a Fundação de 622
Amparo à Pesquisa do Estado da Bahia (FAPESB) por concessão de bolsas e recursos. 623
624
Referências 625
626
AFSAH-HEJRI, L. Saprophytic yeasts: effective biocontrol agents against Aspergillus flavus. 627
International Food Research Journal, v.20, n.6, p.3403-3409, 2013. 628
ALMEIDA, L.C.C.; COSTA, A.Z.M.; MENDONÇA, R.C.S. Efeito do número de remoções 629
de partes doentes do cacaueiro e número de aplicações de óxido cuproso no controle da 630
vassoura de bruxa na Bahia. Agrotrópica, Itabuna, n.15, p.79-84, 2003. 631
ALTOMARE, C.; NORVELL, W.A.; BJORKMAN, T.; HARMAN, G.E. Solubilization of 632
phosphates and micronutrients by the plant-growth-promoting and biocontrol fungus 633
Trichoderma harzianum Rifai 1295-22 C. Applied and Environmental Microbiology, 634
Washington, v.65, n.7, p.2926-2933, 1999. 635
ANDEBRHAN, T. et al. Molecular fingerprinting suggests two primary outbreaks of witches’ 636
broom disease (Crinipellis perniciosa) of Theobroma cacao in Bahia, Brazil. European 637
Journal of Plant Pathology, Dordrecht, v.105, n.2, p.167-175, 1999. 638
BARTLEY, B. G. D. The genetic diversity of cacao and its utilization. Cambridge, CABI 639
Publishing, 2005 341p. 640
BASTOS, C.N. Comparação morfológica e patológica de isolados de Crinipellis perniciosa 641
(Stahel) Singer. In Belém, CEPLAC/ DEPEA. Informe Técnico. p.45-49, 1986. 642

129
BERGHOFER, L.K.; HOCKING, A.D.; MISKELLY, D. JANSSON, E. Microbiology of 643
wheat and flour milling in Australia. International Journal of Food Microbiology, 644
Amsterdam, v.85, p.137-149, 2003. 645
BRUCE, A.; VERRALL, S.; HACKETT, C.A.; WHEATLEY, R.E. Identification of volatile 646
organic compounds (VOCs) from bacteria and yeast causing growth inhibition of sapstain 647
fungi. Holzforschung, Berlin, v.58, p.193-198, 2004. 648
CABRAL, A.S. Killer yeasts inhibit the growth of the phytopathogen Moniliophthora 649
perniciosa, the causal agent of witches’ broom disease. Brazilian Journal of Microbiology, 650
São Paulo, v.40, p.108-110, 2009. 651
COSTA, J.C.B. et al. Controle biológico da vassoura-de-bruxa do cacaueiro da Bahia, Brasil. 652
In: BETTIOL, W. Biocontrole de doenças de plantas: uso e perspectivas. Jaguariúna: 653
Embrapa meio ambiente, 2009, p.245-266. 654
COSTA, J. C. B. et al. Controle biológico da vassoura-de-bruxa do cacaueiro da Bahia, 655
Brasil. In: BETTIOL, W. Biocontrole de doenças de plantas: uso e perspectivas. Jaguariúna: 656
Embrapa meio ambiente, 2009, p.245-266. 657
DICKSTEIN, E.R.; PURDY, L.H.; FRIAS, G.A. Crinipellis perniciosa, the cacao witches’ 658
broom fungus: Inoculum production and storage. Phytopathology, Saint Paul, v.77, p.1747, 659
1987. 660
DROBY, S.; WISNIEWSKI, M.; EL GHAOUTH, A.; WILSON, C. Influence of food 661
additives on the control of postharvest rots of apple and peach and efficacy of the yeast-based 662
biocontrol product Aspire. Postharvest Biology and Technology, v.27, p.127-135, 2003. 663
ELAD Y. Biocontrol of foliar pathogens: mechanisms and application. Communications in 664
Agricultural and Applied Biological Sciences, v.68, p.17-24, 2003 665
FAO. Prodution Yearbook. v.54. Roma. 2002. 666
HU, H.; XU, Y.; LU, H.; XIAO, R.; ZHENG, X.; YU, T. Evaluation of yeasts from Tibetan 667
fermented products as agents for biocontrol of blue mold of Nashi pear fruits. Journal of 668
Zhejiang University-SCIENCE B (Biomedicine & Biotechnology), v.16, n.4, 275-285, 669
2015. 670
JANISIEWICZ, W.J.; KURTZMAN, C P.; BUYER, J.S. Yeasts associated with nectarines 671
and their potential for biological control of brown rot. Yeast, Davis, v.27, p.389–398, 2010. 672
LACA, A.; MOUSIA, Z.; DIAZ, M.; WEBB, C.; PANDIELLA, S.S. Distribution of 673
microbial contamination within cereal grains. Journal of food engineering, Essex, v.72, 674
p.332-338, 2006. 675

130
LEVEAU, J.H, LINDOW, S.E. Appetite of an epiphyte: quantitative monitoring of bacterial 676
sugar consumption in the phyllosphere. Proceedings of the national academy of sciences, 677
Washington, v.98, p.3446-3453, 2001. 678
LI, X.; YU, V.Y.; LIN, Y.; CHOMVONG, K.; ESTRELA, R.; PARK, A.; LIANG, J.M.; 679
ZNAMEROSKI, E.A.; FEEHAN, J.; KIM, S.R.; JIN, Y; GLASS, N.L.; CATE, J.H.D. 680
Expanding xylose metabolism in yeast for plant cell wall conversion to biofuels. Elife, v.4, 681
p.1-16, 2015. 682
LIMA, L.H.C.; DE MARCO, J.L.; FELIX, C.R. Enzimas hidrolíticas envolvidas no controle 683
biológico por micoparasitismo. In: Melo, I.S.; Azevedo, J.L (Ed.). Controle Biológico. 684
Jaguariúna: Embrapa Meio Ambiente, 2000. v.2, p.263-304. 685
LU, L.; LU, H.; WUC, C.; FANGD, W.; YUA, C.; YEA, C.; SHIA, Y.; YUA, T.; ZHENGA, 686
X. Preharvest application of antagonistic yeast Rhodosporidium paludigenum induced 687
resistance against postharvest diseases in mandarin orange. Biological Control, Maryland 688
Heights, v.67, n.2, p.130–136, 2013. 689
LU, H.; LU, L.; ZENG, L.; FU, D.; XIANG, H.; YU, T.; ZHENG, X. EFFECT of chitin on 690
the antagonistic activity of Rhodosporidium paludigenum against Penicillium expansum in 691
apple fruit Postharvest Biology and Technology, Amsterdam, v.92, p.9-15, 2014. 692
MASIH, E.I.; PAUL, B. Secretion of beta-1,3-glucanase by the yeast Pichia membranifaciens 693
and its possible role in the biocontrol of Botrytis cinerea causing mold disease of the 694
grapevine. Current Microbiology, New York, v.44, p.391-395, 2002. 695
MCGEARY, F.M.; WHEELER, B.E.J. Growth rates of and mycelial interaction betwenen 696
isolates of Crinipellis perniciosa from cocoa. Plant Pathology, London, v.37, p. 489-498, 697
1988. 698
MEDEIROS, F. H.V.; POMELLA, A.W..; SOUZA, J.T.; NIELLA, G.R.; VALLE, R.; 699
BATEMAN, R.P.; FRAVEL, D.; VINYARD, B.; HEBBAR, P.K. A novel, integrated method 700
for management of witches’broom disease in Cacao in Bahia, Brazil. Crop Protection. 701
Guildford, n.29, p.704-711, 2010. 702
MEINHARDT, L.W.; RINCONES J.; BAILEY, B.A.; AIME, M.C.; GRIFFITH, G.W.; 703
ZHANG, D.; PEREIRA, G.A. Moniliophthora perniciosa, the causal agent of witches’ broom 704
disease of cacao: what’s new from this old foe? Physiological and Molecular Plant 705
Pathology, London, v.9, n.5, p.577-588, 2008. 706
MIDLEJ, R. R.; SANTOS, A. M. Economia do Cacau. In: VALLE, R. R. (org). Ciência, 707
Tecnologia e Manejo do Cacaueiro. Brasília: Ceplac, 2012. p.655-672. 708
MIDLEJ, R.R.; SANTOS, A.M. Economia do cacau. In: VALLE, R. (Ed.). Ciência, 709

131
Tecnologia e manejo do cacaueiro. CEPLAC, Ilhéus, Brasil, 2007, p.422458. 710
MILLER, W.G.; BRANDL, M.T.; QUINONES. B.; LINDOW, S.E. Biological sensor for 711
sucrose availability: relative sensitivities of various reporter genes. Applied and 712
Environmental Microbiology, v.67, p.1308-1317, 2001. 713
NUTARATAT, P.; SRISUK, N.; ARUNRATTIYAKORN, P.; LIMTONG, S. Plant growth-714
promoting traits of epiphytic and endophytic yeasts isolated from rice and sugar cane leaves 715
in Thailand. Fungal biology, v.118, n.8, p.683-194, 2014. 716
OLIVEIRA, A.G.; CHAGAS JUNIOR, A.F.; SANTOS, G.R.; MILLER, L.O.; CHAGAS, 717
L.F.B. Potencial de solubilização de fosfato e produção de AIA por Trichoderma spp.. 718
Revista Verde, v.7, n.3, p.149-155, 2012. 719
OLIVEIRA, J.R.G.; MORAES, T.A.L.; MELO, N.F.; MELO, A.M.Y. Fungos micorrízicos 720
arbusculares e rizobactérias promotoras de crescimento na aclimatização de Zingiber 721
spectabile. Bragantia, v.69, n.3, p.687-694, 2010 722
OLIVEIRA, M. L. Eficácia de fungicidas triazóis no controle da vassoura-de-bruxa do 723
cacaueiro causada por Crinipellis perniciosa. Fitopatologia Brasileira, v.29, p.149, 2004a. 724
__________. Estrobilurinas: novo grupo de fungicidas com eficácia contra a vassoura-de- 725
bruxa do cacaueiro. Fitopatologia Brasileira, v.29, p.285, 2004b. 726
OLIVEIRA, M. L.; LUZ, E. D. M. N. Vassoura-de-bruxa. In:__________. Identificação e 727
manejo das principais doenças do cacaueiro no Brasil. Ilhéus, CEPLAC/CEPEC/SEFIT, 728
2005, p. 1533. 729
PEREIRA,J.L.; VALLE, R.R. Manejo Integrado da vassoura de bruxa do cacaueiro. In: 730
VALLE, R.R. Ciência, tecnologia e manejo do cacaueiro. Brasília: 731
CEPLAC/CEPEC/SEFIS. 2ed. p.11-30, 2012. 732
PLATANIAA, A.; RESTUCCIAB, C.; MUCCILLIB, S.; CIRVILLERIA, G. Efficacy of 733
killer yeasts in the biological control of Penicillium digitatum on Tarocco orange fruits 734
(Citrus sinensis). Food Micrrobiology, v.30, n.1, p.219-225, 2012. 735
QUEIROZ, V. T. et al. Identification of a major QTL in cocoa (Theobroma cacao L.) 736
associated with resistance to witches’ broom disease. Plant Breeding, v. 122, n. 3, p. 268-737
272, 2003. 738
RUDGARD, S.A.; BUTLER, D.R. Witches' broom disease on cocoa in Rondonia, Brazil: pod 739
infection in relation to pod susceptibility, wetness, inoculum, and phytosanitation. Plant 740
Patholology. n.36, p.512-522, 1987. 741
SAS Institute Inc (1989) SAS/STAT User´s Guide, version 6.4 ed. Cary, SAS Institute, North 742
Caroline State University. 846 p. 743

132
SCARPARI, L. M.; MEINHARD, L.M.; MAZZAFERA, P.; POMELLA, A.W.V.; 744
SCHIAVINATO, M.A.;CASCARDO, J.C.M.; PEREIRA, G.A.G. Biochemical changes 745
during the development of witches’ broom: the most important disease of cocoa in Brazil 746
caused by Crinipellis perniciosa. Journal Experimental Botany, p. 113, 2005. 747
SENA, K.; ALEMANNO, L.; GRAMACHO, K.P. The infection process of Moniliophthora 748
perniciosa in cacao. Plant Pathology, 2014. 749
SILVA, S.D.V.M.; LUZ, E.D.M.N.; PIRES, J.L.; YAMADA, M.M.; SANTOS FILHO, L.P. 750
Cacao resistance to Crinipellis perniciosa: diallelic crosses increase genes for resistance. 751
In: 8th International Congress of Plant Pathology, Christchurch Addendum to Abstracts of 752
Offered Papers, Christchurch, p.8, 2003. 753
SUI, Y.; WISNIEWSKI, M.; DROBY, S.; LIUA, J. Responses of yeast biocontrol agents to 754
environmental stress. Applied and environmental microbiology, v.81, n.9, p.2968-2975 755
2015. 756
SUN, P.; Fang, W.; SHIN, L.; WEI, J.; FU, S.; CHOU, J. Indole-3-acetic acid-producing 757
yeasts in the phyllosphere of the carnivorous plant Drosera indica L. Plos one, p.1-22, 2014. 758
URQUHART, E.J.; PUNJA, Z.K. Hydrolytic enzymes and antifungal compounds produced 759
by Tilletiopsis species, phyllosphere yeasts that are antagonists of powdery mildew fungi. 760
Canadian Journal of Microbiology, v.48, n.3, p.219-229, 2002. 761
VALDEBENITO-SANHUEZA, R.M.V. Leveduras para o biocontrole de fitopatógenos. In: 762
MELO, I.S; AZEVEDO, J.L. (Ed.). Controle biológico. Jaguariúna: Embrapa Meio 763
Ambiente. p.41-56, 2000. 764
ZHANG, H. Y. et al. Postharvest biological control of gray mold decay of strawberry with 765
Rhodotorula glutinis. Biological Control, v.40, p.287-292, 2007. 766
767
768
769
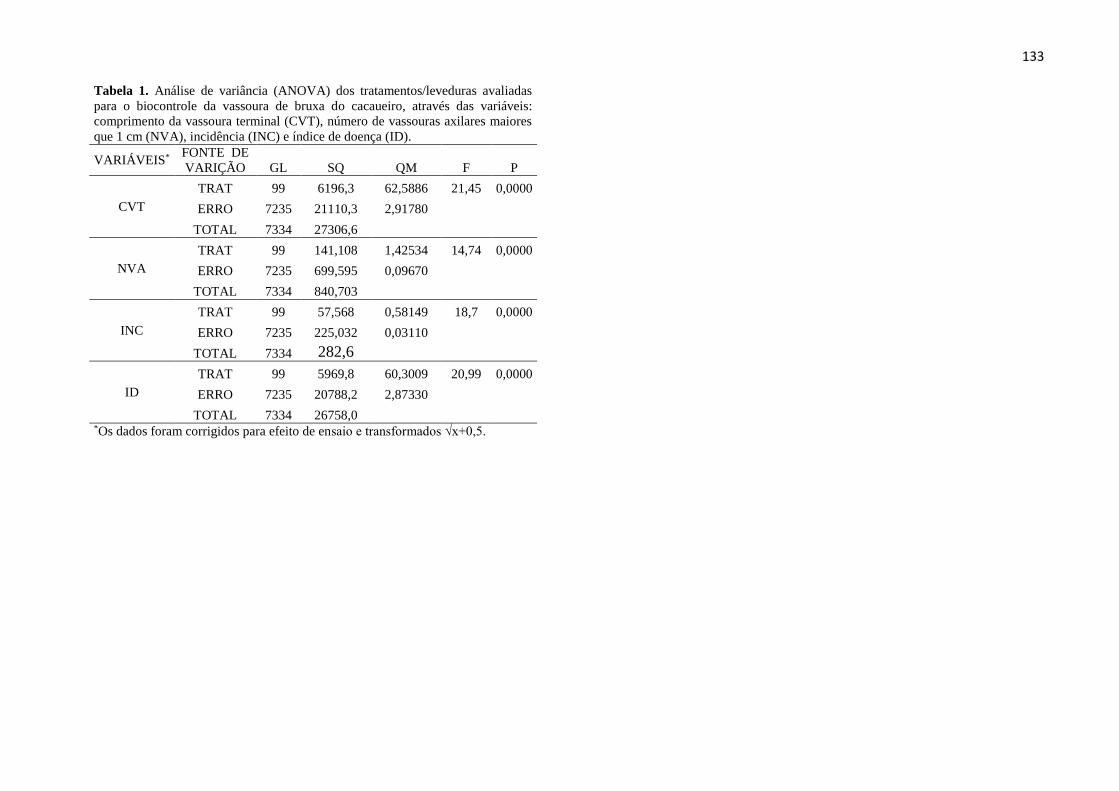
133 Tabela 1. Análise de variância (ANOVA) dos tratamentos/leveduras avaliadas
para o biocontrole da vassoura de bruxa do cacaueiro, através das variáveis:
comprimento da vassoura terminal (CVT), número de vassouras axilares maiores
que 1 cm (NVA), incidência (INC) e índice de doença (ID).
VARIÁVEIS* FONTE DE
VARIÇÃO GL SQ QM F P
CVT
TRAT 99 6196,3 62,5886 21,45 0,0000
ERRO 7235 21110,3 2,91780
TOTAL 7334 27306,6
NVA
TRAT 99 141,108 1,42534 14,74 0,0000
ERRO 7235 699,595 0,09670
TOTAL 7334 840,703
INC
TRAT 99 57,568 0,58149 18,7 0,0000
ERRO 7235 225,032 0,03110
TOTAL 7334 282,6
ID
TRAT 99 5969,8 60,3009 20,99 0,0000
ERRO 7235 20788,2 2,87330
TOTAL 7334 26758,0 *Os dados foram corrigidos para efeito de ensaio e transformados √x+0,5.
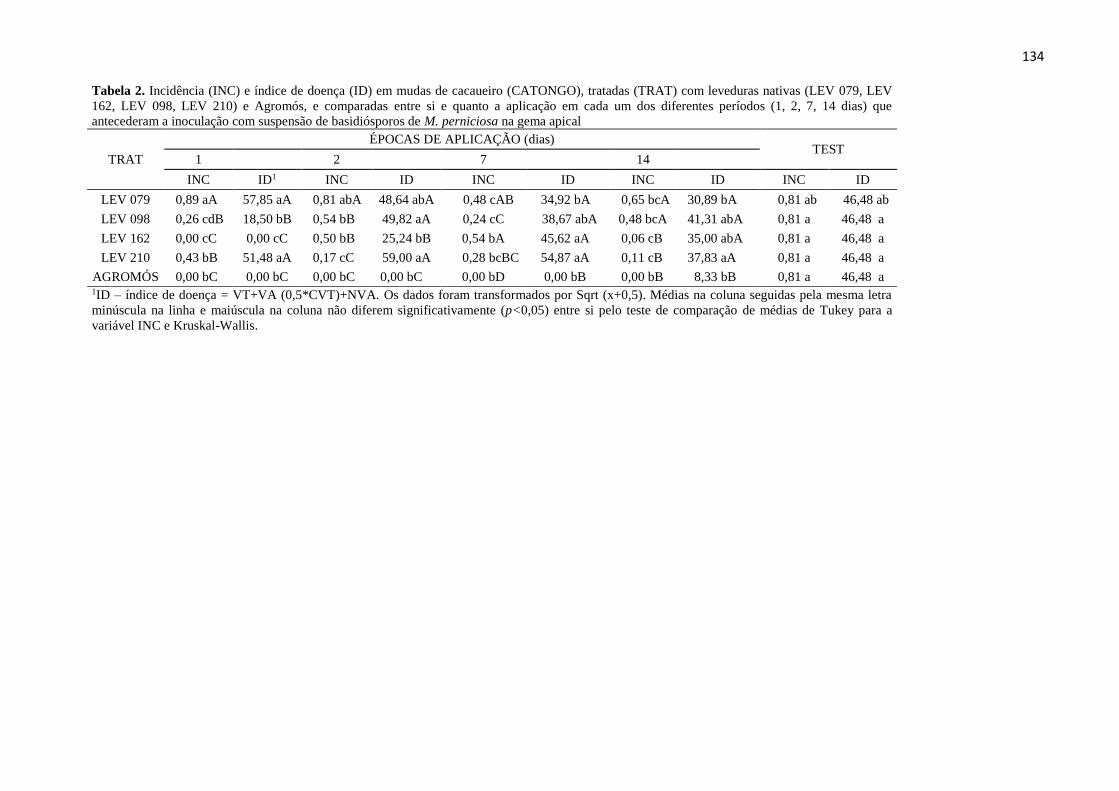
134 Tabela 2. Incidência (INC) e índice de doença (ID) em mudas de cacaueiro (CATONGO), tratadas (TRAT) com leveduras nativas (LEV 079, LEV
162, LEV 098, LEV 210) e Agromós, e comparadas entre si e quanto a aplicação em cada um dos diferentes períodos (1, 2, 7, 14 dias) que
antecederam a inoculação com suspensão de basidiósporos de M. perniciosa na gema apical
TRAT
ÉPOCAS DE APLICAÇÃO (dias) TEST
1 2 7 14
INC ID1 INC ID INC ID INC ID INC ID
LEV 079 0,89 aA 57,85 aA 0,81 abA 48,64 abA 0,48 cAB 34,92 bA 0,65 bcA 30,89 bA 0,81 ab 46,48 ab
LEV 098 0,26 cdB 18,50 bB 0,54 bB 49,82 aA 0,24 cC 38,67 abA 0,48 bcA 41,31 abA 0,81 a 46,48 a
LEV 162 0,00 cC 0,00 cC 0,50 bB 25,24 bB 0,54 bA 45,62 aA 0,06 cB 35,00 abA 0,81 a 46,48 a
LEV 210 0,43 bB 51,48 aA 0,17 cC 59,00 aA 0,28 bcBC 54,87 aA 0,11 cB 37,83 aA 0,81 a 46,48 a
AGROMÓS 0,00 bC 0,00 bC 0,00 bC 0,00 bC 0,00 bD 0,00 bB 0,00 bB 8,33 bB 0,81 a 46,48 a 1ID – índice de doença = VT+VA (0,5*CVT)+NVA. Os dados foram transformados por Sqrt (x+0,5). Médias na coluna seguidas pela mesma letra
minúscula na linha e maiúscula na coluna não diferem significativamente (p<0,05) entre si pelo teste de comparação de médias de Tukey para a
variável INC e Kruskal-Wallis.

135
Tabela 3. Incidência (INC), comprimento de vassouras terminais (CVT),
número de vassouras axilares maiores que 1 cm (NVA) e índice de doença
(ID) em plântulas de cacaueiro (CATONGO) tratadas com leveduras
nativas (pulverização da parte aérea), mistura de duas leveduras, mistura de
10 leveduras (MIX) e produtos comerciais, 24 h antes da inoculação com
suspensão de basidiósporos de M. perniciosa na gema apical, 30 dias após
a inoculação.
TRAT PLÂNTULAS
INC CVT (mm) NVA ID1
LEV 007 0,81 ab 45,48 a 2,72 a 46,34 a
LEV 027 0,41 defg 44,69 a 1,45 a 44,62 a
LEV 074 0,91 a 48,10 a 2,28 a 49,70 a
LEV 079 0,91 a 57,69 a 2,48 a 58,76 a
LEV 098 0,81 ab 40,00 a 2,00 a 40,56 a
LEV 162 0,47 cdefg 55,50 a 2,21 a 53,87 a
LEV 179 0,25 fgh 40,71 a 1,43 a 35,88 a
LEV 193 0,68 abcde 39,90 a 3,75 a 41,29 a
LEV 207 0,22 gh 39,83 a 1,40 a 33,07 a
LEV 210 0,56 bcdef 48,82 a 2,13 a 48,00 a
193+210 0,75 abc 42,00 a 2,32 a 41,24 a
27+207 0,34 efg 27,81 a 2,36 a 83,27 a
7+74 0,66 abcde 50,48 a 2,11 a 52,59 a
79+179 0,91 a 42,68 a 2,39 a 40,78 a
98+162 0,91 a 50,93 a 2,72 a 51,52 a
MIX 0,69 abcd 45,64 a 2,63 a 45,27 a
COBRE 0,00 h 0,00 0,00 0,00
FOLICUR 0,00 h 0,00 0,00 0,00
MIX/FOLICUR 0,00 h 0,00 0,00 0,00 1ID – índice de doença = VT+VA (0,5*CVT)+NVA. 2Os dados foram
transformados por Sqrt(x+0.5). Médias na coluna seguidas pela mesma letra
não diferem significativamente (P<0,05) entre si pelo teste de comparação
de médias de Tukey para a variável INC e Krukal-Wallis para as variáveis
CVT, NVA e ID.

136 Tabela 4. Crescimento de plântulas seminais de cacaueiro (PS 1319) submetidas a tratamentos com suspensões de células de levedura em diferentes
dosagens/aplicações (APL), e avaliadas quanto ao comprimento (CP), diâmetro do coleto (DC), peso fresco da parte aérea (PFA), peso seco da parte aérea (PSA),
peso fresco da raiz (PFR) e peso seco da raiz (PSR), 60 dias após a semeadura.
V1 APL TRATAMENTOS
TR2 TA3 CV4
MIX LEV 079 LEV 084 LEV 093 LEV 098 LEV 149 LEV 162 LEV 168 LEV 170 LEV 206 LEV 210
CP
1 22,98 abA 24,87 aA 24,16 abA 24,96 aA 24,04 abA 23,34 abA 24,80 aA 24,18 abAB 22,43 bB 23,71 abA 24,55 abA
22,76 22,36 6,08 2 24,04 aA 24,13 aA 23,96 aA 24,95 aA 24,16 aA 24,40 bA 23,07 aAB 23,35 aB 23,52 aAB 24,67 aA 25,08 aA
3 22,71 bA 24,32 abA 24,09 abA 24,84 abA 24,53 abA 24,70 abA 24,64 abB 24,88 abA 24,41 abA 23,22 abA 25,25 aA
DC
1 4,11 a abA 4,06 bB 4,29 abA 4,42 aA 4,21 abA 4,41 aAB 4,40 aA 4,38 abA 4,16 abAB 4,41 aA 4,27 abA
3,89 3,85 4,3 2 4,31 abcA 4,09 cB 4,42 abcA 4,35 abcA 4,20 bcA 4,63 aA 4,52 abA 4,41 abcA 4,11 cB 4,34 abcA 4,25 bcA
3 4,18 aA 4,39 aA 4,33 aA 4,42 aA 4,36 aA 4,36 aB 4,44 aA 4,33 aA 4,43 aA 4,47 aA 4,36 aA
PFA
1 2,48 abA 2,75 aA 2,63 abA 2,68 abA 2,68 abA 2,55 abA 2,66 abA 2,71 abA 2,27 bA 2,49 abA 2,84 aA
2,76 2,40 25,17 2 2,53 aA 2,65 aA 2,74 aA 2,85 aA 2,80 aA 3,25 aA 3,46 aA 2,67 aA 2,41 aA 2,78 aA 2,71 aA
3 2,32 bA 2,62 abA 2,69 abA 2,69 abA 2,75 aA 2,70 abA 2,73 abA 2,79 aA 2,63 abA 2,55 abA 2,94 aA
PSA
1 0,56 aA 0,58 aA 0,57 aA 0,59 aA 0,58 aA 0,54 aB 0,57 aA 0,61 aA 0,52 aB 0,55 aB 0,62 aA
0,61 0,20 18,40 2 0,58 aA 0,59 aA 0,61 aA 0,60 aA 0,60 aA 0,59 aAB 0,54 aA 0,61 aA 0,58 aAB 0,64 aA 0,61 aA
3 0,56 aA 0,56 aA 0,58 aA 0,56 aA 0,61 aA 0,61 aA 0,59 aA 0,59 aA 0,59 aA 0,54 aB 0,62 aA
PFR
1 0,63 abA 0,66 abA 0,69 aA 0,66 aA 0,60 abA 0,65 abA 0,67 abA 0,64 aA 0,53 aA 0,67 aA 0,59 aA
0,62 0,46 8,24 2 0,72 abA 0,58 abcA 0,63 abcA 0,75 aA 0,59 abcA 0,67 abcA 0,57 abcA 0,52 cB 0,53 bcA 0,68 abcA 0,55 bcA
3 0,57 abA 0,54 bA 0,75 aA 0,66 abA 0,61 abA 0,64 abA 0,63 abA 0,56 abAB 0,61 abA 0,64 abA 0,63 abA
PSR
1 0,15 aA 0,15 abAB 0,19 aA 0,17 abA 0,14 abA 0,17 aA 0,16 abA 0,15 abA 0,15 abA 0,19 aA 0,14 abA
0,17 0,13 3,36 2 0,17 aA 0,17 aA 0,16 aA 0,21 aA 0,17 aA 0,16 aA 0,14 aA 0,14 aA 0,19 aA 0,19 aA 0,17 aA
3 0,18 aA 0,14 aB 0,19 aA 0,16 aA 0,14 aA 0,18 aA 0,16 aA 0,15 aA 0,19 aA 0,14 aA 0,14 aA 1Variáveis analisadas; 2Testemunha relativa (adubo MAP 5mL/planta); 3Testemunha absoluta; 4Coeficiente de variação. Médias seguidas por letras minúsculas na linha e maiúsculas
na coluna, não diferem entre si pelo teste de Tukey a 5% de probabilidade.

137
Figura 1. Dendrograma de análise de agrupamento obtido pelo método de
UPGMA (distância Euclidiana), a partir das medidas de dissimilaridade entre 100
isolados de leveduras nativas do cacaueiro avaliadas quanto ao potencial
biocontrolador vassoura de bruxa do cacaueiro. Grupos formados após corte na
altura de θ = 12: I, II, III.

138
Figura 2. Número médio de basidiomas em vassouras secas (ramos de Theobroma cacao infectados com
Moniliophthora perniciosa) tratadas com leveduras nativas (pulverização), mistura de duas leveduras, mistura
de 10 leveduras e produtos comerciais, avaliadas durante 60 dias após incubação em câmaras climáticas
(gabinetes/vassoureiro). T1 – LEV 007; T2 – LEV 027; T3 – LEV 074; T4 – LEV 079; T5 – LEV 098; T6 –
LEV 162; T7 – LEV 179; T8 – LEV 193; T9 – LEV 207; T10 – LEV 210; T11 – MIX 193 + 210; T12 – MIX
027 + 207; T13 – MIX 007 + 074; T14 – MIX 098 + 162; T15 – MIX TOTAL; T16 – Cobre Atar®; T17 -
Tricovab®; T18 – Testemunha.

139
Figura 3. Microscopia eletrônica de varredura da superfície abaxial (LAb) e adaxial (LAd) do limbo foliar,
nervura central (NCAd e NCAb) e meristema apical (MA) de plântulas de cacaueiro, pulverizadas com a
levedura LEV 098 (Candida sp.), inoculadas com basidiósporos de Moniliophthora perniciosa., e coletadas 2 h
(2hA) e 10 dias (10dA) após a aplicação da levedura. Inoculação do patógeno 7 dias após a pulverização da
levedura. Detalhes estômato (Et), grãos de amido (Ad).
2hA
Et
br
Et
Et Et
Et
Et
NCAd
br
40 µm 20 µm
5 µm 50 µm
10 µm 5 µm
MA NCAd
MA NCAd
10dA
LAb
LAd
2hA
LAb
10hA

140
Figura 4. Microscopia eletrônica de varredura da superfície adaxial (LAd) do limbo foliar,
nervura central (NCAd e NCAb) e meristema apical (MA) de plântulas de cacaueiro,
pulverizadas com a levedura LEV 162 (Hanseniaspora sp.), inoculadas com basidiósporos
de Moniliophthora perniciosa e coletadas 2 h (2hA) e 10 dias (10dA) após a aplicação da
levedura. Inoculação do patógeno 7 dias após a pulverização da levedura. Tricomas tectores
(tt) e glandulares (tg)
tt
tg
br
10 µm 50 µm
20 µm 40 µm
20 µm 10 µm
10 µm 10 µm
br
NCAd NV
NCAd
LAd
NCAd
NCAd
MA NCAd
MA NCAd
MA
10dA NCAd
10dA NCAd
10dA NCAd
10dA NCAd
10dA NCAd
10dA NCAd
2hA
2hA
2hA 2hA
tt
tt
tt
tt

141
Figura 5. Microscopia eletrônica de varredura da superfície abaxial (LAb) e adaxial (LAd) do limbo foliar,
nervura central (NCAd e NCAb) e meristema apical (MA) de plântulas de cacaueiro, pulverizadas com a
levedura LEV 210 (Hanseniaspora sp.), inoculadas com basidiósporos de Moniliophthora perniciosa, e
coletadas 2 h (2hA) e 10 dias (10dA) após a aplicação da levedura. Inoculação do patógeno 7 dias após a
pulverização da levedura. Detalhes células epidérmicas (CEp), células parênquima paliçádico (CPp), estômato
(Et), grãos de amido (Ad), pseudohifa (PsH).
LAb
2hA
NCAd
2hA
LAd
2hA
LAd 2hA
LAd 2hA
NCAb
10dA NCAd
CEp
CPp
Et
Et
CEp
Ad
br
br
PsH
10 µm
10 µm
10 µm 10 µm
10 µm
10 µm

142
CAPÍTULO 5
Conclusões Gerais

143
CONCLUSÕES GERAIS
- São apresentadas progênies que carregam genes de resistência à VB oriunda de fontes
diversas, além da incorporação de genes de resistência à monilíase, praga quarentenária A1.
- A microbiota epifítica residente em folhas, flores e frutos de cacaueiro abriga uma
diversidade de fungos leveduriformes, até então não citados na literatura.
- Leveduras nativas do cacaueiro são produtoras de compostos difusíveis e voláteis que atuam
na inibição da germinação de basidiósporos e crescimento micelial de M. perniciosa.
- Na superfície de diferentes órgãos vegetais de cacaueiros selecionados como resistentes à
vassoura de bruxa e monilíase, existem uma diversidade de fungos leveduriformes que podem
estar atuando de forma positiva na inibição de M. perniciosa e evolução do quadro
sintomatológico.
- Leveduras nativas são potenciais agentes de biocontrole da vassoura de bruxa do cacaueiro,
atuando na diminuição dos sintomas em plântulas seminais e inibição da produção de
basidiomas.
- Há possibilidade de integração do controle químico com fungicidas cúpricos e controle
biológico com leveduras nativas para o manejo da vassoura de bruxa do cacaueiro.
- O período de aplicação da suspensão de células de leveduras é preponderante para a
manifestação do biocontrole da vassoura de bruxa do cacaueiro.
- Leveduras nativas colonização do cacaueiro colonizam prováveis sítios de infecção de
Moniliophthora perniciosa, como estômatos, bases de tricomas e nervuras foliares.
- Leveduras nativas que apresentam potencial biocontrolador a vassoura de bruxa, podem
atuar como promotoras de crescimento de plântulas seminais do cacaueiro, havendo variações
intraespecíficas e em relação ao número de aplicações na manifestação desta característica.


















