
RODRIGO DE SOUZA AMARAL - Biblioteca Digital de Teses e ... · conjutos comerciais para dosagem de...
86
RODRIGO DE SOUZA AMARAL Uso de diferentes matrizes biológicas na dosagem de andrógenos em peixes-bois da Amazônia machos (Trichechus inunguis) mantidos em cativeiro São Paulo 2008
Transcript of RODRIGO DE SOUZA AMARAL - Biblioteca Digital de Teses e ... · conjutos comerciais para dosagem de...

RODRIGO DE SOUZA AMARAL
Uso de diferentes matrizes biológicas na dosagem de
andrógenos em peixes-bois da Amazônia machos
(Trichechus inunguis) mantidos em cativeiro
São Paulo
2008

RODRIGO DE SOUZA AMARAL
Uso de diferentes matrizes biológicas na dosagem de
andrógenos em peixes-bois da Amazônia machos
(Trichechus inunguis) mantidos em cativeiro
Dissertação apresentada ao Programa de Pós-Graduação em Reprodução Animal da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para obtenção do título de Mestre em Medicina Veterinária
Departamento:
Reprodução Animal
Área de concentração:
Reprodução Animal
Orientador:
Prof. Dr. Cláudio Alvarenga de Oliveira
São Paulo
2008





FOLHA DE AVALIAÇÃO
Nome: AMARAL, Rodrigo de Souza
Título: Uso de diferentes matrizes biológicas na dosagem de andrógenos em peixes-
bois da Amazônia machos (Trichechus inunguis) mantidos em cativeiro
Dissertação apresentada ao Programa de Pós-Graduação em Reprodução Animal da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para obtenção do título de Mestre em Medicina Veterinária
Data:____/____/____
Banca Examinadora
Prof. Dr. ___________________________ Instituição: ___________________
Assinatura: ___________________________ Julgamento: __________________
Prof. Dr. ___________________________ Instituição: ___________________
Assinatura: ___________________________ Julgamento: __________________
Prof. Dr. ___________________________ Instituição: ___________________
Assinatura: ___________________________ Julgamento: __________________

“Brindo à casa, brindo à vida, meus amores, minha família...”
Mar de Gente
O RAPPA

AGRADECIMENTOS
Ao Prof. Dr. Cláudio Alvarenga de Oliveira, meu orientador, pela amizade e pela
confiança depositada ao me aceitar como seu orientado, realizando meu sonho de
ingressar na pós-graduação do Departamento de Reprodução Animal –
VRA/FMVZ/USP;
Ao Dr. Fernando C. W. Rosas, pela amizade, pela confiança e incentivo na
realização deste trabalho, por abrir as portas do Laboratório de Mamíferos Aquáticos
– LMA/INPA para mim, e por todas as conversas e ensinamentos passados;
Ao Prof. Dr. Marcelo A. B. V. Guimarães pelo incentivo na realização deste
trabalho e pelo fornecimento de um dos conjutos comerciais para dosagem de
testosterona, impressindíveis para a realização deste trabalho;
À Dra. Vera M. F. da Silva, pelo apoio e confiança permitindo a realização deste
trabalho;
Ao Médico Veterinário José Anselmo D’Affonsêca Neto e aos tratadores Daniel,
Jeová, Marcelo, Nazaré e Raimundo, por toda a ajuda nas coletas das amostras, e
por todas as conversas e risadas;
Aos peixes-bois do LMA/INPA, em especial Tupy e Yanomama, por me aturarem
todos os dias durante o período de coleta;
A todos os pós-graduandos e funcionários do LMA/INPA, à Bernadete e ao
Javier, por terem feito o período de coleta mais divertido;
Aos meus avós e tias, por sempe me receberem muito bem, sempre fazendo de
tudo para me agradar, toda vez que vou a Manaus;
À Tecnopec Laboratórios, pelo fornecimento do hormônio Lecirelina,
impressindível para a realização deste trabalho;

À Médica Veterinária Josiane Almeida Sales pela ajuda no envio do hormônio
Lecirelina. Valeu Josi!!!;
À Técnica de Laboratório Maria do Socorro Pontes Silva e os demias técnicos da
Fundação de Medicina Tropical do Amazonas pelas dosagens de creatinina urinária;
À amiga Dra. Priscila Viau, grande companheira de laboratório, pela paciência, por
todos os ensinamentos passados, por todas as conversas e risadas, e pela
disposição em me aturar por mais quatro anos!;
À companheira de mestrado Renata Marino Romano pelo fornecimento dos
conjutos comerciais para dosagem de testosterona, impressindíveis para a
realização deste trabalho;
A todos os professores do VRA, pela amizade e pelos conhecimentos passados;
À secretária Harumi Dói Shiraishi, por toda ajuda nos trâmites burocráticos e por
sentir minha falta quando o corredor do VRA está silencioso;
Aos funcionários do VRA, por me ajudarem quando foi preciso, sempre com boa
vontade;
Aos pós-graduandos do VRA, grandes amigos, pelas conversas no corredor e
principalmente pelos churrascos;
A todos do LDH, grandes pesquisadores, por todas as discussões científicas,
conversas e risadas. Mulheres que levam a reprodução a sério!;
Aos agregados e visitantes constantes do LDH, por tornarem o local de trabalho
mais descontraído;
À minha irmã Fabiane, pela capa deste trabalho, sempre provando que seu diploma
serve para aguma coisa!;

Ao CNPq pela bolsa de mestrado concedida;
À Fundação Amaral, por sempre financiar meus projetos;
Ao Zé e à Ana Paula, por todo suporte na temporada paulistana;
Ao “núcleo candango da novela”, por fazerem me sentir em Brasília mesmo
estando em São Paulo;
Aos FF, por serem os FF...;
Aos grandes amigos “sirenólogos”, por reavivarem em mim a paixão pelos
peixes-bois, e por sempre me insentivarem a continuar trilhando este caminho;
A todos os meus grandes amigos espalhados pelo mundo, que se mostram muito
mais que “simples amigos”, me dando todo o apoio necessário e sempre se
esforçam para manter contato;
Aos meus pais e minhas irmãs, por sempre me apoiarem e permitirem que eu
seguisse em busca dos meus sonhos;
Ao Santo Expedito, por ouvir as nossas súplicas;
A Deus, sempre!

“O pescador, Ele sai bem cedinho, ele sai bem cedinho, ele vai pescar. O pescador, Ele ama o rio, ele ama o rio, o rio Amazonas. E você sabe por quê? Por que de baixo das águas Não existe só boto, Existe também o peixe-boi, o pirarucu e o tambaqui.”
(Autor desconhecido)

RESUMO
AMARAL, R. S. Uso de diferentes matrizes biológicas na dosagem de andrógenos em peixes-bois da Amazônia machos (Trichechus inunguis) mantidos em cativeiro. [Use of different biological matrices on androgens measurement in captive male Amazonian manatees (Trichechus inunguis)]. 2008. 85 f. Dissertação (Mestrado em Medicina Veterinária) - Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2008.
O objetivo deste estudo foi verificar a viabilidade da dosagem de andrógenos em
amostras de saliva, secreção lacrimal, urina e fezes de peixe-boi da Amazônia
realizando um desafio hormonal. Dois peixes-bois amazônicos adultos machos (A-1
e A-2) foram submetidos a um protocolo de experimentação de doze dias (D-1 a
D10). No D0 os animais receberam uma injeção intramuscular de GnRH exógeno.
Amostras de saliva, secreção lacrimal, urina e fezes foram coletadas diariamente
(entre 08h00 e 09h00) e mantidas a -20°C até o ensaio. As amostras de fezes foram
liofilizadas, extraídas com metanol 80% e diluídas em tampão antes do
radioimunoensaio (RIE). As amostras de urina sofreram hidrólise ácida e foram
diluídas em soro bovino depletado. As amostras de saliva e secreção lacrimal foram
dosadas sem etapa de extração, porém, o ensaio foi adaptado para aumentar a
sensibilidade do teste. Os ensaios hormonais foram realizados utilizando um
conjunto comercial de RIE para testosterona total. Um pico de andrógenos
(mediana+2DI) somente foi observado nas amostras de saliva, urina e fezes de
ambos os animais. Porém, os picos de andrógenos fecais ocorreram depois (cinco
dias) dos picos de andrógenos urinários e salivares. Este intervalo está
correlacionado com o longo tempo de passagem da digesta pelo trato
gastrointestinal na espécie. Os picos salivares e urinários ocorreram muito próximos,
provavelmente com poucas horas de intervalo. Esses resultados demonstram que as
concentrações de andrógenos em amostras de saliva, urina ou fezes refletem
consistentemente os eventos fisiológicos e são ferramentas de grande utilidade no
monitoramento reprodutivo de peixes-bois da Amazônia.
Palavras-chave: Peixe-boi Amazônico. Radioimuniensaio. Reprodução. Sirênio.
Testosterona.

ABSTRACT
AMARAL, R. S. Use of different biological matrices on androgens measurement in captive male Amazonian manatees (Trichechus inunguis). [Uso de diferentes matrizes biológicas na dosagem de andrógenos em peixes-bois da Amazônia machos (Trichechus inunguis) mantidos em cativeiro]. 2008. 85 f. Dissertação (Mestrado em Medicina Veterinária) - Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2008.
The aim of this study was to verify the viability of androgens measurement in saliva,
lacrimal secretion, urine and fecal samples of Amazonian manatee by carrying out
hormonal challenge. Two adult male manatees (A-1 and A-2) were submitted to an
experimentation protocol of twelve days (D-1 to D10). On D0 the animals received an
intramuscular injection of GnRH-analogue. Salivary, lacrimal, urinary and fecal
samples were collected daily (between 08h00 and 09h00) and frozen at -20°C until
assayed. Fecal samples were lyophilized, extracted with 80% methanol and diluted in
buffer before the radioimmunoassay (RIA). Urine samples underwent acid hydrolysis
and diluted in depleted bovine serum. Salivary and lacrimal samples were assayed
without extraction step, but, the assay was adapted to improve the sensibility.
Hormonal assays were carried out with a commercial testosterone RIA kit. An
androgen peak (>median+2IQR) was observed only in salivary, urinary and fecal
samples of both animals. However, the fecal androgens peaks occurred later than
urinary and salivary androgens peaks. These intervals are correlated with the long
digesta passage time in this species. The salivary and urinary peaks were very close,
probably with few hours of interval. These results show that androgens
concentrations in saliva, urine or feces samples reflect reliably physiological events
and are powerful tool for reproductive monitoring of Amazonian manatees.
Key words: Amazonian manatee. Radioimmunoassay. Reproduction. Sirenian.
Testosterone.

LISTA DE FIGURAS
Figura 1 – Peixe-boi da Amazônia (Trichechus inunguis).................................. 24
Figura 2 – Estrutura da molécula de testosterona.............................................. 29
Figura 3 – Figura esquemática das rotas de metabolismo e excreção da testosterona....................................................................................... 32
Figura 4 – Tanques dos peixes-bois da Amazônia no Laboratório de Mamíferos Aquáticos do Instituto Nacional de Pesquisas da Amazônia – LMA/INPA...................................................................... 42
Figura 5 – Aplicação do análogo de GnRH na região do pedúnculo caudal de um T. inunguis................................................................................... 43
Figura 6 – Coleta de fezes de T. inunguis após drenagem do tanque de cambiamento..................................................................................... 44
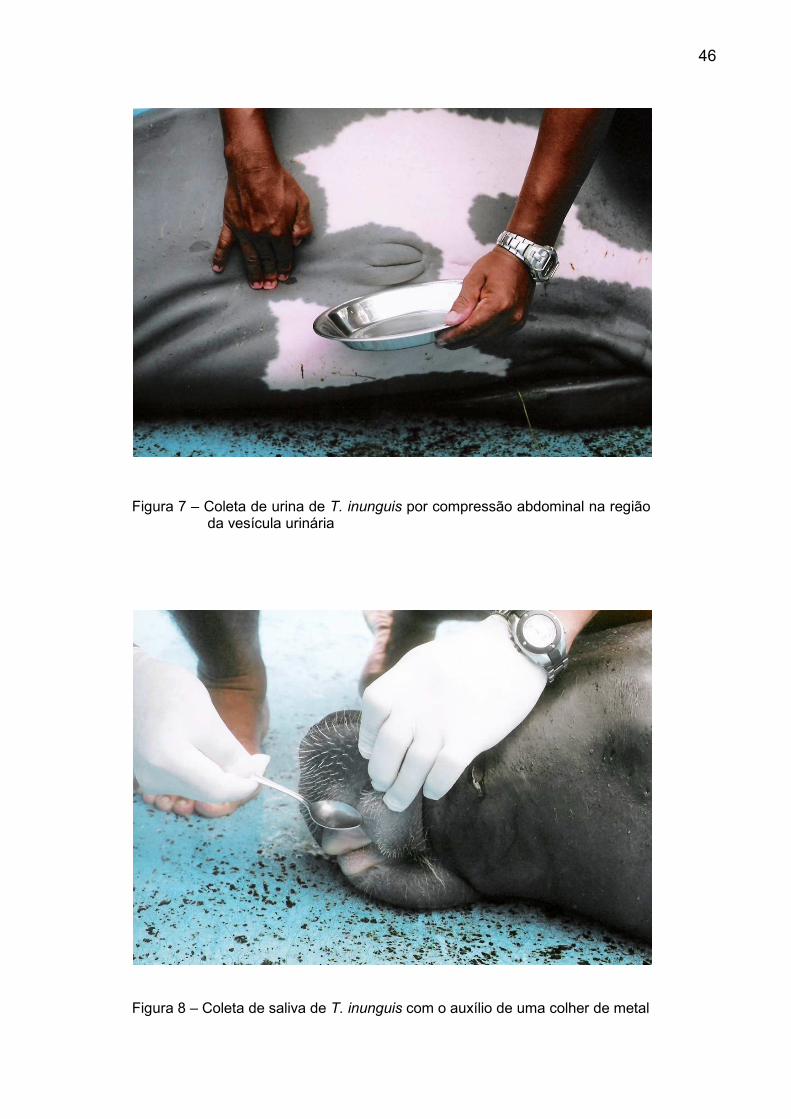
Figura 7 – Coleta de urina de T. inunguis por compressão abdominal na região da vesícula urinária................................................................ 46
Figura 8 – Coleta de saliva de T. inunguis com o auxílio de uma colher de metal.................................................................................................. 46

LISTA DE GRÁFICOS
Gráfico 1 – Representação gráfica da regressão linear entre a matriz salivar de T. inunguis e a curva padrão do conjunto diagnóstico comercial Testosterona Total Coat-A-Count® (Siemens).................................. 53
Gráfico 2 – Representação gráfica da regressão linear entre a matriz urinária de T. inunguis e a curva padrão do conjunto diagnóstico comercial Testosterona Total Coat-A-Count® (Siemens).................................. 54
Gráfico 3 – Representação gráfica da regressão linear entre a matriz fecal de T. inunguis e a curva padrão do conjunto diagnóstico comercial Testosterona Total Coat-A-Count® (Siemens).................................. 54
Gráfico 4 – Mudança na excreção de andrógenos salivares, lacrimais, urinários e fecais em resposta ao desafio hormonal com GnRH em dois peixes-bois da Amazônia machos mantidos em cativeiro (A-1 e A-2)................................................................................................. 58

LISTA DE QUADROS E TABELAS
Quadro 1 – Características dos peixes-bois da Amazônia utilizados neste estudo................................................................................................ 41
Tabela 1 – Controles de qualidade obtidos nos ensaios hormonais de andrógenos presentes na saliva, secreção lacrimal, urina e fezes de dois peixes-bois amazônicos machos adultos............................. 52
Tabela 2 – Resultados da regressão linear para o teste de paralelismo entre as matrizes salivar, urinária e fecal de peixes-bois da Amazônia e a curva do conjunto diagnóstico comercial Testosterona Total Coat-A-Count® (Siemens)................................................................. 53
Tabela 3 – Resultados das dosagens de andrógenos salivares, lacrimais, fecais e urinários, e de creatinina urinária (Cr) de um peixe-boi amazônico macho (A-1) durante o desafio hormonal........................ 55
Tabela 4 – Resultados das dosagens de andrógenos salivares, lacrimais, fecais e urinários, e de creatinina urinária (Cr) de um peixe-boi amazônico macho (A-2) durante o desafio hormonal........................ 56
Tabela 5 – Análise dos andrógenos salivares, lacrimais, urinários e fecais de dois peixes-bois da Amazônia machos em resposta ao desafio hormonal com GnRH......................................................................... 57
Tabela 6 – Resultados do teste de correlação linear de Pearson para as matrizes salivar, lacrimal, urinária e fecal de dois peixes-bois da Amazônia machos (A-1 e A-2) submetidos a um desafio hormonal com GnRH......................................................................................... 59

LISTA DE ABREVIATURAS E SIGLAS
A-1 Animal – 1
A-2 Animal – 2
cm Centímetro
Cr Creatinina
DI desvio interquartílico
dL Decilitro
G Giros
g Grama
GnRH hormônio liberador de gonadotrofinas
IUCN União Internacional para a Conservação da Natureza e dos Recursos Naturais
Kg Quilograma
LH hormônio luteinizante
M Molar
m Metro
m³ metro cúbico
mg Miligrama
mL Mililitro
mm Milímetro
µg micrograma
ng nanograma
pg Picograma
RIE radioimunoensaio

SUMÁRIO
1 INTRODUÇÃO.......................................................................................... 21
2 REVISÃO DA LITERATURA.................................................................... 23
2.1 O PEIXE-BOI DA AMAZÔNIA................................................................... 23
2.1.1 Exploração do peixe-boi da Amazônia no Brasil....................................... 24
2.1.2 Aspectos alimentares e fisiologia digestiva............................................... 27
2.1.3 Metabolismo.............................................................................................. 27
2.1.4 Aspectos reprodutivos............................................................................... 28
2.2 TESTOSTERONA – SÍNTESE, METABOLISMO E EXCREÇÃO............. 29
2.3 UTILIZAÇÃO DE DIFERENTES MATRIZES BIOLÓGICAS NA DOSAGEM HORMONAL........................................................................... 33
2.3.1 Utilização de diferentes matrizes biológicas na dosagem hormonal em sirênios....................................................................................................... 36
3 OBJETIVOS.............................................................................................. 39
4 MATERIAL E MÉTODOS.......................................................................... 41
4.1 ANIMAIS.................................................................................................... 41
4.2 DESAFIO HORMONAL............................................................................. 43
4.3 COLHEITA DO MATERIAL BIOLÓGICO.................................................. 44
4.3.1 Fezes......................................................................................................... 44
4.3.2 Urina.......................................................................................................... 45
4.3.3 Saliva......................................................................................................... 45
4.3.4 Secreção lacrimal...................................................................................... 45
4.4 ANÁLISE HORMONAL.............................................................................. 47
4.4.1 Fezes......................................................................................................... 47
4.4.2 Urina.......................................................................................................... 48
4.4.3 Saliva......................................................................................................... 48
4.4.4 Secreção lacrimal...................................................................................... 49
4.4.5 Paralelismo................................................................................................ 49
4.5 ANÁLISE DOS RESULTADOS................................................................. 50
5 RESULTADOS ......................................................................................... 52
5.1 PARÂMETROS DE QUALIDADE DOS ENSAIOS HORMONAIS............. 52
5.2 PARALELISMO......................................................................................... 53

5.3 DESAFIO HORMONAL............................................................................. 55
6 DISCUSSÃO............................................................................................. 61
6.1 DESAFIO HORMONAL X AMOSTRAS SALIVARES................................ 61
6.2 DESAFIO HORMONAL X AMOSTRAS LACRIMAIS................................ 62
6.3 DESAFIO HORMONAL X AMOSTRAS URINÁRIAS................................ 63
6.4 DESAFIO HORMONAL X AMOSTRAS FECAIS....................................... 64
6.5 COMPARAÇÃO ENTRE AS MATRIZES ANALISADAS........................... 65
7 CONCLUSÕES......................................................................................... 69
REFERÊNCIAS......................................................................................... 71

INTRODUÇÃO

21
1 INTRODUÇÃO
O peixe-boi amazônico (Trichechus inunguis, MAMALIA: SIRENIA) é o menor
do gênero medindo até três metros de comprimento e pesando no máximo 450kg
(AYRES; BEST, 1979; CALDWELL; CALDWELL, 1985). Ocorre na Bacia
Amazônica, sendo o único sirênio exclusivamente de água doce (CALDWELL;
CALDWELL, 1985). Nos séculos passados, foi intensamente caçado para o
comércio de sua carne, gordura e couro (DOMNING, 1982). Atualmente, a espécie é
protegida por lei no Brasil, Peru e Colômbia e está classificada como “Vulnerável”,
tendo alto risco de extinção na natureza em médio prazo, pela Lista Vermelha das
Espécies Ameaçadas da IUCN (IUCN, 2006) e pelo Plano de Ação para os
Mamíferos Aquáticos do Brasil (IBAMA, 2001). Porém, a caça de subsistência ainda
persiste com certo grau comercial (ROSAS, 1994).
Alguns estudos sobre os aspectos reprodutivos do peixe-boi amazônico têm
sido realizados nas ultimas décadas (BEST, 1982b; PIMENTEL, 1998;
NASCIMENTO et al., 2002; RODRIGUES, 2002; RODRIGUES et al., 2003;
NASCIMENTO, 2004), porém as informações sobre a biologia reprodutiva da
espécie continuam escassas.
Trabalhos sobre endocrinologia reprodutiva em animais domésticos
normalmente são realizados com amostras sanguíneas. Porém, como alternativa na
coleta de sangue, o uso de diferentes matrizes, como fezes, urina ou saliva, tem se
mostrado como uma excelente ferramenta no monitoramento endócrino-reprodutivo
de animais selvagens, minimizando o estresse causado pela captura e contenção, e
possibilitando um acompanhamento endócrino diário dos animais. Entretanto, é de
suma importância avaliar se a técnica empregada na dosagem hormonal nessas
matrizes é capaz de demonstrar os eventos fisiológicos ocorridos na espécie
estudada.
Sendo assim, foi proposto, neste estudo, avaliar a viabilidade da utilização de
diferentes matrizes biológicas como ferramentas no monitoramento reprodutivo de
peixes-bois da Amazônia machos.

22
REVISÃO DE LITERATURA

23
2 REVISÃO DE LITERATURA
A revisão de literatura foi redigida em três principais tópicos, compreendendo:
1 – os aspectos biológicos e históricos do peixe-boi da Amazônia; 2 – a síntese,
metabolismo e excreção da testosterona; e 3 – a utilização de diferentes matrizes
biológicas na dosagem hormonal.
2.1 O PEIXE-BOI DA AMAZÔNIA
A ordem Sirenia compreende três gêneros recentes, Trichechus (peixe-boi),
Dugong (dugongo) e Hydrodamalis (vaca marinha de Steller), sendo este último
extinto em 1768 pela caça predatória. O gênero Trichechus apresenta três espécies
viventes: Trichechus inunguis (peixe-boi amazônico), Trichechus senegalensis
(peixe-boi africano) e Trichechus manatus (peixe-boi marinho) (DOMNING; HAYEK,
1986).
Sendo o menor dos sirênios, o peixe-boi amazônico (Figura 1) nasce medindo
entre 85-105 cm e pesando entre 10 e 15kg (BEST, 1984), e pode atingir até três
metros de comprimento e pesar até 450kg quando adulto (AYRES; BEST, 1979;
CALDWELL; CALDWELL, 1985; ROSAS, 1994).
Apresenta uma coloração de pele variando de cinza escuro a preto com a
presença de manchas brancas na região ventral, sendo estas características da
espécie (HUSAR, 1977; CALDWELL; CALDWELL, 1985). Porém, alguns indivíduos
podem não apresentar essas manchas, tendo a região ventral só levemente mais
clara que o dorso (ROSAS, 1994). Sua pele tem em média dois centímetros de
espessura, apresentando uma camada de tecido adiposo subcutâneo
aproximadamente do mesmo tamanho (MENDES, 1958; VERÍSSIMO, 1970).

24
Figura 1 – Peixe-boi da Amazônia (Trichechus inunguis)
Possui pêlos esparsos pelo corpo, os quais especula-se terem a função de
sinalizar variações na velocidade e direção das correntes aquáticas (CALDWELL;
CALDWELL, 1985; REEP; MARSHALL; STOLL, 2002). Há uma maior concentração
de pêlos na região perioral de grande importância no contato entre os indivíduos, na
exploração do ambiente e também na manipulação dos alimentos (MARSHALL et
al., 2003). As nadadeiras peitorais não apresentam unhas, diferentemente dos
outros sirênios (CALDWELL; CALDWELL, 1985).
Sendo o único sirênio exclusivamente de água doce, o T. inunguis é
endêmico da Bacia Amazônica, ocorrendo desde os rios da Colômbia, Peru e
Equador até a ilha de Marajó no estado do Pará (BEST, 1984).
2.1.1 Exploração do peixe-boi da Amazônia no Brasil
A caça do peixe-boi amazônico no Brasil existe desde antes do período de
colonização, porém, seus registros são quase inexistentes. Sabe-se que a caça
tradicional era realizada pelos índios para o consumo da carne e gordura, sendo
estes considerados itens comuns na dieta local, e também para a utilização de seu

25
couro como escudo (HEATON, 1970) e de seus ossos na fabricação de gaponga
(isca na pesca de tambaqui), ponteira de lança e arpão, anzol e no artesanato
(PEREIRA, 1944; FERREIRA, 1972).
No final do século XVI a atividade comercial utilizando a carne fresca, a
manteiga (a gordura extraída do animal), a mixira (carne frita e conservada na
própria gordura do animal) e a carne seca (utilizando as técnicas portuguesas de
salga e secagem) já havia começado. Porém, por volta do meio do século XVII
houve um crescente aumento na exportação tanto para as grandes capitais (Manaus
e Belém) quanto para os países europeus (PEREIRA, 1944). Ferreira (1903) relata
que somente em dois anos da década de 1780 o Pesqueiro Real da Villa Franca
(próximo a Santarém – PA) produziu o equivalente a mais de 1500 peixes-bois na
forma de carne salgada e manteiga.
Com a caça indiscriminada, no ano de 1786 o naturalista Alexandre
Rodrigues Ferreira, em um de seus relatos para a Corte Real Portuguesa sobre sua
expedição pelo Brasil (FERREIRA, 1903, p. 171), já demonstrava certa preocupação
com a conservação da espécie quando relatou:
Sem embargo de tantas utilidades, quantas são as que deste animal se tirão, nenhuma Policia tem até agora a sua pesca. Hum peixe-boy para chegar ao seu devido crescimento deve gastar annos; e em todos elles se harpoão a elto os que aparecem, não se distingue o tempo, em que as femeas andão prenhes, porque ou prenhes ou não, as perseguem; ellas não parem mais de 1 até 2 filhos por anno, e os filhos tirados do ventre das Mãys assim mortas para nada servem. Não se distingue o tempo da criação, porque antes hé felicidade para o harpoador, surpreender o filho para arpoar a Mãy; não se destingue a idade, porque pequenos, e grandes todos são harpoados. A vista do que nenhum espanto deve causar a sua raridade em alguns Lagos, onde não ha muitos annos, que se observão bastantes.
Porém, essa preocupação não foi levada a diante já que, mesmo assim a caça
continuou se intensificando cada vez mais.
Com o passar dos séculos a mixira foi se tornando o principal produto
comercial do peixe-boi, provavelmente por ter um maior tempo de conservação (três
a quatro meses) e por ser mais apreciada (PEREIRA, 1944; DOMNING, 1982).
Durante todo este período, o couro sempre foi sub-aproveitado por causa da

26
dificuldade na curtição sendo assim, na maioria das vezes, descartado pelos
caçadores (FERREIRA, 1903). Porém, por volta de 1934 curtumes no sudeste do
Brasil aprimoraram as técnicas de curtição do couro dando um pico na exploração
do peixe-boi (MENDES, 1958; DOMNING, 1982). Seu couro, por ser grosso,
resistente e durável, era utilizado na confecção de artigos para a indústria como
polias, correias de transmissão, mangueiras, juntas, entre outros (PEREIRA, 1944;
MENDES, 1958; DOMNING, 1982; BEST, 1984). Durante 20 anos estima-se que
entre 80.000 a 140.000 peixes-bois foram mortos nos lagos do estado do Amazonas
tendo o couro exportado para todo Brasil, Estados Unidos e países da Europa. Mas
no ano de 1954 sua exportação cessou abruptamente por razões não muito claras.
Possivelmente o comércio do couro terminou como resultado do aumento do uso de
borracha e materiais sintéticos (DOMNING, 1982; BEST, 1984).
Apesar do fim da indústria do couro, o comércio de carne continuou na
década de 50, sendo ampliado e tendo como principal produto a carne fresca devido
à disponibilidade de refrigeração. A caça se tornou tão intensa nesse novo mercado
promissor que voltou a apresentar picos de quase 7.000 animais caçados por ano na
década de 60 (tendo isso ocorrido anteriormente no início da década de 40). Porém
esse pico teve uma queda contínua no final da década de 60 até chegar ao ano de
1973 (ano em que a caça foi teoricamente banida no Brasil). Este declínio não pode
ser justificado pela disponibilidade de substitutos sintéticos como na indústria do
couro, podendo assim ser reflexo da diminuição populacional dos animais
(DOMNING, 1982).
Desde 1973 o T. inunguis está citado no Apêndice I da CITES (como animal
ameaçado de extinção). A espécie é protegida no Brasil desde 1967 pela Lei de
Proteção à Fauna (nº 5.197, alterada para 7.653 em 1988) e a partir de 1998 pela
Lei de Crimes Ambientais (nº 9.605). Atualmente, além do Brasil, a espécie também
é protegida por lei na Colômbia e no Peru, e está classificada pela Lista Vermelha
das Espécies Ameaçadas da IUCN (IUCN, 2006) e pelo Plano de Ação para os
Mamíferos Aquáticos do Brasil (IBAMA, 2001) como “Vulnerável”, tendo alto risco de
extinção na natureza em médio prazo. Entretanto, a caça de subsistência ainda
persiste, com certo grau comercial (BEST, 1982a; ROSAS; PIMENTEL, 2001;
AGUILAR et al., 2006).

27
2.1.2 Aspectos alimentares e fisiologia digestiva
Os sirênios são os únicos mamíferos aquáticos estritamente herbívoros,
sendo animais monogástricos e apresentando fermentação cecal (BEST, 1981). Os
peixes-bois amazônicos passam de seis a oito horas por dia se alimentando sem
apresentar um ritmo circadiano aparente, podendo ser observados comendo a
qualquer hora do dia ou da noite (PEREIRA, 1944; BEST, 1981). Peixes-bois da
Amazônia de vida-livre comem exclusivamente plantas aquáticas e semi-aquáticas,
consumindo no mínimo 8% de seu peso vivo por dia com uma eficiência digestiva
entre 45-70%, dependendo do teor de fibras e minerais do alimento (BEST, 1981;
ROSAS, 1994).
Os sirênios apresentam um longo tempo de passagem do alimento pelo trato
gastrointestinal quando comparado com outros mamíferos (de um a três dias para
mamíferos de médio e grande porte), sendo esses tempos somente próximos aos
relatados para preguiça-de-três-dedos (Bradypus tridactylus) e coala (Phascolarctos
cinereus) (WARNER, 1981; CORK; WARNER, 1983; FOLEY; ENGELHARDT;
CHARLES-DOMINIQUE, 1995). O peixe-boi amazônico apresenta um tempo de
passagem entre 5-7 dias (ITAVO; ROSAS; CAVALLANTE, 1996), o peixe-boi da
Flórida entre 5-10 dias (LOMOLINO; EWEL, 1984; LARKIN; FOWLER; REEP, 2007)
e o dugongo entre 6-7 dias (LANYON; MARSH, 1995), variando de acordo com o
teor de fibra dos alimentos ingeridos.
2.1.3 Metabolismo
O peixe-boi amazônico apresenta uma baixa taxa metabólica, por volta de
64% menor do que a de mamíferos terrestres de tamanho similar. Este fator,
juntamente com a baixa freqüência respiratória e a tolerância a baixos níveis de
oxigênio e altos níveis de gás carbônico, são os que provavelmente permitem a
espécie realizar mergulhos de até mais de dez minutos de duração (GALLIVAN;
BEST, 1980).

28
Segundo Best (1983), o grande acúmulo de tecido adiposo durante os
períodos de maior disponibilidade de alimento (período de enchente e cheia dos
rios) e a baixa taxa metabólica são fatores essenciais para a sobrevivência do
animal durante as estações de secas prolongadas (época de baixíssima
disponibilidade de alimento). A combinação destes dois aspectos poderia fornecer
aproximadamente 200 dias de sobrevivência somente utilizando suas reservas de
gordura como fonte de energia.
2.1.4 Aspectos reprodutivos
Mesmo com a realização de alguns estudos sobre os aspectos reprodutivos
do peixe-boi amazônico nas ultimas décadas (BEST, 1982b; PIMENTEL, 1998;
NASCIMENTO et al., 2002; RODRIGUES, 2002; RODRIGUES et al., 2003;
NASCIMENTO, 2004), as informações sobre a biologia reprodutiva da espécie
continuam escassas.
Os testículos do peixe-boi da Amazônia são intra-abdominais e o dimorfismo
sexual mais aparente para a espécie é o posicionamento da abertura genital, onde
nos machos encontra-se mais próxima da cicatriz umbilical e nas fêmeas, mais
próxima ao ânus. Ambos os sexos possuem duas mamas localizadas na inserção
ventral das nadadeiras, sendo que as das fêmeas são mais desenvolvidas
(MARMONTEL; ODELL; REYNOLDS III, 1992).
De acordo com Best (1982b), o T. inunguis apresenta sazonalidade
reprodutiva, com os acasalamentos e nascimentos ocorrendo durante o período de
enchentes e cheias dos rios, coincidindo com o período de maior disponibilidade de
alimento (como relatado anteriormente). O ciclo estral é de aproximadamente 22
dias (NASCIMENTO, 2004), a gestação dura entre 11 a 12 meses (NASCIMENTO et
al., 2002) e em cada gestação nasce um filhote (BEST, 1984).
A idade de maturidade sexual é desconhecida, mas acredita-se que seja entre
5 e 10 anos baseado em observações anatômicas feitas por Rodrigues et al. (2003)
e em estimativas a partir dos dados para peixes-bois da Flórida (T. manatus
latirostris) (MARMONTEL; ODELL; REYNOLDS III, 1992; ROSAS, 1994).

29
2.2 TESTOSTERONA – SÍNTESE, METABOLISMO E EXCREÇÃO
A testosterona (17β-hidroxi-4-androsten-3-ona) (Figura 2) é um hormônio
andrógeno tendo, como todo hormônio esteróide, a molécula de colesterol como seu
precursor. Nos machos, a testosterona é produzida e secretada pelas células de
Leydig nos testículos. Nas fêmeas, pequenas quantidades desse andrógeno são
produzidas nas células da teca interna pela conversão de androstenediona e de
desidroepiandrosterona (DHEA) a testosterona, sendo essa, importante na síntese
de estradiol. A zona reticularis do córtex da adrenal também tem a capacidade de
sintetizar alguns andrógenos, como androsterona, 4-androstene-3,17-diona, DHEA e
3β,11β-diidroxi-4-androsten-17-diona, porém não consegue converter estes
andrógenos a testosterona por falta da enzima 17-cetoesteróide redutase
(NORMAN; LITWACK, 1997).
Figura 2 – Estrutura da molécula de testosterona
A síntese de testosterona está diretamente relacionada com a síntese de
GnRH (hormônio liberador de gonadotrofinas – gonadotropin releasing hormone)
pelo hipotálamo, o qual irá estimular a liberação de LH (hormônio luteinizante –
luteinizing hormone) pelo lóbulo anterior da hipófise. O LH, por sua vez, é o
responsável pela estimulação da síntese de testosterona nas células de Leydig
(NORMAN; LITWACK, 1997).
Parte da testosterona produzida se liga à ABP (proteína de ligação a
andrógeno - androgen binding protein), produzida nas células de Sertoli, mantendo,

30
assim, altas concentrações de testosterona nos túbulos seminíferos, fato este,
importante na espermatogênense. A outra parte é transportada para a corrente
sanguínea para atuação em outras partes do organismo (NORMAN; LITWACK,
1997).
Devido às suas propriedades lipofílicas, as moléculas de andrógenos livres
não são altamente solúveis em soluções aquosas como o sangue, podendo assim,
passar rapidamente pelas membranas celulares atingindo os órgãos-alvo ou ser
metabolizadas pelo fígado ou rim. Dessa forma, grande parte da testosterona
presente no sangue (por volta de 91%) está ligada a globulinas ligadoras de
hormônios sexuais (SHBG - sex hormone binding globulin) possibilitando um maior
tempo de circulação do hormônio na corrente sanguínea, o restante está na forma
livre (forma considerada biologicamente ativa – aproximadamente 3%) ou ligada à
albumina (uma ligação fraca, facilmente dissociada para a disponibilização da
testosterona – apoximadamente 6%) (NORRIS, 1997; NOZAKI, 2001).
A atuação dos andrógenos nos machos pode ser dividida em quatro
categorias: 1 – promover o desenvolvimento do trato reprodutivo masculino (pênis,
testículos, epidídimos, próstata, vesículas seminais, vasos deferentes e glândulas
bulbouretrais); 2 – desenvolver os caracteres sexuais secundários masculinos
(variando de acordo com cada espécie); 3 – promover um efeito estimulatório ou
anabólico no desenvolvimento do corpo (crescimento dos ossos, desenvolvimento
muscular, distribuição do tecido adiposo subcutâneo, crescimento de órgãos
acessórios); e 4 – atuação no sistema nervoso central (diferenciação de
determinadas áreas como hipotálamo e córtex cerebral e desenvolvimento da libido)
(NORMAN; LITWACK, 1997).
Os andrógenos também apresentam um papel importante nas glândulas
lacrimais, estimulando a secreção de lágrima, bem como, de seus constituintes
químicos, como sódio, potássio, proteínas e anticorpos. Estudos demonstram que
animais castrados apresentam uma redução no tamanho das glândulas lacrimais e
em suas funções secretórias, sendo estas, reestabelecidas após a suplementação
com andrógenos (SULLIVAN; BLOCH; ALLANSMITH, 1984; AZZAROLO et al.,
1997; MATHERS et al., 1998).
Os esteróides circulantes no sangue passam pelas células acinares e ductos
das glândulas salivares por difusão, devido sua solubilidade em membranas
lipídicas, e atingem a saliva. Esse mecanismo parece ser possível somente para os

31
esteróides não-conjugados, pois os esteróides conjugados não são suficientemente
lipossolúveis. A concentração dos esteróides salivares não é afetada por diferenças
na intensidade de produção de saliva (RIAD-FAHMY et al., 1982; VINING;
MCGINLEY; SYMONS, 1983). Em humanos, a testosterona salivar apresenta
valores próximos dos encontrados para a testosterona livre no sangue e por volta de
2% da concentração de testosterona total sanguínea (RIAD-FAHMY et al., 1982;
RILLING et al., 1996; FORDE et al., 2006).
Os hormônios esteróides presentes na circulação sanguínea são, em sua
grande maioria, metabolizados no fígado, porém alguma atividade catabólica
também ocorre nos rins (PALME et al., 1996; NORRIS, 1997; GRAHAM, 2004).
Inicialmente os andrógenos têm as suas duplas ligações reduzidas, inativando as
moléculas, e posteriormente são conjugados com glucoronídeos ou sulfatos
tornando-os hidrossolúveis. Os metabólitos podem retornar à circulação sanguínea
ou serem eliminados no duodeno através da bile. No sangue, os metabólitos são
filtrados pelos rins e eliminados na urina (SENGER, 2005). Os metabólitos
excretados no intestino podem sofrer ação bacteriana causando sua desconjugação.
Após isto, podem ser reabsorvidos pela circulação entero-hepática e transportados
para o fígado para serem conjugados novamente ou para os rins para serem
excretados. Os metabólitos não reabsorvidos (conjugados e não-conjugados) são
excretados pelas fezes (WHITTEN; BROCKMAN; STAVISKY, 1998; GRAHAM,
2004; TOUMA; PALME, 2005) (Figura 3).

32
Figura 3 – Figura esquemática das rotas de metabolismo e excreção da testosterona

33
2.3 UTILIZAÇÃO DE DIFERENTES MATRIZES BIOLÓGICAS NA DOSAGEM
HORMONAL
O estudo da endocrinologia reprodutiva, em animais domésticos, normalmnte
é realizado com amostras sangüíneas, necessitando, em estudos mais profundos,
de coletas seriadas com intevalos curtos. Em animais selvagens e animais de
pequeno porte, essas coletas podem ficar impossibilitadas devido ao estresse
causado pela coleta (contenção física e/ou química do animal) (GUIMARÃES, 2003),
risco de flebite (animais que não apresentam vaso periférico de fácil puncionamento)
e/ou por volume sanguíneo insuficiente (CHELINI et al., 2005).
A partir dos conhecimentos sobre a distribuição dos esteróides pela corrente
sanguínea, a sua fácil passagem pelas membranas celulares, e seu metabolismo e
vias de excreção, vários trabalhos vêm sendo realizados sobre a fisiologia
reprodutiva de diferentes espécies de animais tanto de laboratório quanto
domésticos e selvagens utilizando diferentes matrizes biológicas.
Atkinson et al (1999) mensuraram progesterona em amostras de secreção
lacrimal de falsas-orcas (Pseudorca crassidens), porém não obtiveram correlação
com amostas plasmáticas. Até o presente momento não existe relato da utilizaçào
de secreção lacrimal na dosagem de andrógenos em humanos ou animais,
possivelmente devido ao baixo volume de amostra que se consegue obter na grande
maioria dos animais. Entretanto, a secreção lacrimal em peixes-bois da Amazônia,
como nos outros mamíferos aquáticos, é de consistência viscosa, espessa,
transparente, e de produção constante, principalmente quando o animal se encontra
fora d’água. Possivelmente esta secreção tem a função bacteriostática, lubrificante,
de proteção corneal e de redução da resistência hidrodinâmica, como sugerido para
golfinhos (YOUNG; DAWSON, 1992).
Amostras de saliva têm sido utilizadas na mensuração de esteróides
reprodutivos femininos (estrógenos e progesterona) e de glucocorticóides em
animais (VINCENT; MICHELL, 1992; PIETRASZEK; ATKINSON, 1994; CZEKALA;
CALLISON, 1996; CROSS; PINES; ROGERS, 2004; CROSS; ROGERS, 2004;
PEDERNERA-ROMANO et al., 2006). Já o uso de testosterona salivar tem sido
muito relatado em estudos comportamentais e de puberdade em humanos, devido a
sua grande facilidade de coleta (DABBS JR, 1993; HALPERN; UDRY;

34
SUCHINDRAN, 1998; CAMPBELL; SCHULTHEISS; MCCLELLAND, 1999;
GRANGER et al., 1999; GRANGER et al., 2004; SCHULTHEISS; WIRTH;
STANTON, 2004), entretanto, já existem relatos se sua utilização em estudos
reprodutivos e/ou comportamentais de lobos (Canis lupus) (HORVÁTH; ÚJVÁRY;
MIKLÓSI, 2007) e alguns mamíferos aquáticos mantidos em cativeiro como as
focas-monges-do-Havaí (Monachus schauinslandi) (THEODOROU; ATKINSON,
1998) e os golfinhos-nariz-de-garrafa (Tursiops truncatus) (HOGG; VICKERS;
ROGERS, 2005).
Os andrógenos urinários vêm sendo muito utilizados em estudos sobre
biologia reprodutiva e/ou comportamental de animais de laboratório, domésticos e
selvagens (em cativeiro ou vida-livre) devido a sua facilidade de coleta, podendo até
ser coletado diretamente do chão logo após a micção, e sua estreita correlação com
os andrógenos sangüíneos (LASLEY, 1985; DRAISCI et al., 2000; WILLIAMS et al.,
2000; GANSWINDT et al., 2002; MÖHLE et al., 2002; HAGEY; CZEKALA, 2003;
DECATANZARO et al., 2004; BUSSO et al., 2005; MARSHALL; HOHMANN, 2005).
Entretanto, o uso de andrógenos fecais em estudos reprodutivos é o de maior
difusão, dentre as matrizes anteriormente citadas, por ser a de mais fácil obtenção e
por necessitar de menor contato com o animal. Os andrógenos fecais já foram
mensurados em várias espécies de mamíferos (domésticos, de laboratório e
selvagens) e aves (LASLEY, 1985; BROWN; TERIO; GRAHAM, 1996; PALME et al.,
1996; HIRSCHENHAUSER et al., 2000; MÖHLE et al., 2002; GUIMARÃES, 2003;
DLONIAK et al., 2004; MORATO et al., 2004a; MORATO et al., 2004b; DIAS;
OLIVEIRA, 2006; LIMA, 2006; SCHWARZENBERGER, 2007).
A principal rota de excreção (fezes ou urina), o principal metabólito excretado
e sua forma (se conjugado ou livre) e o tempo entre a síntese do esteróide e a
excreção de seu metabólito nas fezes ou urina podem variar consideravelmente
entre as espécies e entre cada esteróide em uma mesma espécie animal (PALME et
al., 1996; SCHWARZENBERGER et al., 1996; WHITTEN; BROCKMAN; STAVISKY,
1998; GRAHAM, 2004). Estas informações são extremamente importantes para
correlacionar os hormônios estudados com os eventos fisiológicos e/ou
comportamentais observados, influenciando assim, em todo o delineamento
experimental a ser feito.
O tempo para metabolização e excreção dos esteróides pela urina está
relacionado com a taxa metabólica do animal, já a excreção pelas fezes, além da

35
taxa metabólica, está também relacionada com a taxa de passagem da digesta pelo
trato intestinal (do duodeno – onde o metabólito foi excretado pela vesícula biliar –
ao reto) (PALME et al., 1996; SCHWARZENBERGER et al., 1996; WHITTEN;
BROCKMAN; STAVISKY, 1998; MÖHLE et al., 2002; KRETZSCHMAR;
GANSLOßER; DEHNHARD, 2004; PALME, 2005)
Uma forma para a determinação das vias e tempo de excreção dos
esteróides e seus principais metabólitos de excreção é a aplicação de hormônios
radiomarcados na corrente sanguínea e posterior realização de coletas seriadas,
com intervalos curtos, de fezes e urina para a mensuração dos metabólitos
radiomarcados. Porém, deve-se lembrar sempre dos riscos inerentes de utilização
de substâncias radioativas em animais (GRAHAM, 2004). Este procedimento tem
sido realizado em várias espécies de animais domésticos e selvagens como forma
de validação fisiológica da metodologia empregada para a mensuração hormonal,
além da determinação do tempo e rotas de excreção. Para a grande maioria dos
animais o tempo de aparecimento dos metabólitos na urina é curto (menos de um
dia), já nas fezes pode variar de 12 horas a três dias (HINDLE; HODGES, 1990;
BROWN et al., 1994; MONFORT et al., 1995; BROWN; TERIO; GRAHAM, 1996;
PALME et al., 1996; SCHWARZENBERGER et al., 1996; WASSER et al., 1996;
MONFORT et al., 1997; BILLITTI; LASLEY; WILSON, 1998; HEISTERMANN et al.,
1998; VELLOSO et al., 1998; MOORMAN et al., 2002; BUSSO et al., 2005; TOUMA;
PALME, 2005).
Outra forma de garantir que as metodologias utilizadas nas análises
hormonais são capazes de expressar os eventos fisiológicos ocorridos, além
fornecer dados sobre o tempo para a excreção do metabólito, é a realização de
desafios hormonais (PALME et al., 2005). Trata-se da administração de uma droga
conhecidamente estimulante ou inibitória da produção hormonal para demonstrar a
relação de causa-efeito entre a administração exógena e a subseqüente produção
hormonal. Para a avaliação dos hormônios andrógenos pode-se utilizar uma
aplicação intramuscular de GnRH (que causará um pico de LH e posterior elevação
nos níveis de testosterona) ou de hCG (gonadotrofina coriônica humana – human
chorionic gonadotropin) que mimetizará a atuação do LH estimulando a produção de
testosterona.
O pico de testosterona pode ser observado tanto no sangue quanto nas
outras matrizes biológicas, sendo na forma de metabólito ou não. No sangue, já foi

36
observado em vários mamíferos que uma única dose intramuscular de GnRH
exógeno causa um aumento nos níveis de LH em aproximadamente 30 minutos e
um pico na testosterona plasmática entre uma a três horas após a aplicação (POST;
REICH; BINDON, 1987; GILMORE et al., 1991; BENNETT et al., 1993; BENNETT;
FAULKES; SPINKS, 1997; JUHÁSZ et al., 2000; SPINKS et al., 2000; HERBERT et
al., 2004; BROWN-DOUGLAS et al., 2005; ALLEN et al., 2006; BEIJERINK et al.,
2007; OOSTHUIZEN; BENNETT, 2007). Porém, nas outras matrizes biológicas, o
tempo para o aparecimento dos picos de LH e andrógenos irá depender de outros
fatores como a taxa metabólica, as vias de excreção do hormônio e fisiologia
digestiva da espécie, conforme já descrito anteriormente.
2.3.1 Utilização de diferentes matrizes biológicas na dosagem hormonal em sirênios
Alguns estudos já foram realizados utilizando esteróides urinários ou fecais de
sirênios. Com dugongos, Lanyon et al. (2005) mensuraram estrógenos e andrógenos
fecais de animais de vida-livre para verificar a existência de diferenças nas
concentrações hormonais entre os sexos e entre as faixas etárias, e Wakai et al.
(2002) dosaram progestinas e estrógenos urinários em uma fêmea cativa
monitorando suas variações durante dois anos. Com peixe-boi marinho da Flórida,
foram utilizadas amostras fecais, tanto de animais cativos quanto de vida-livre, para
analisar o ciclo estral, sazonalidade reprodutiva e diferenças nas concentrações
hormonais entre os sexos e entre as faixas etárias, dosando progestinas, estrógenos
e andrógenos (LARKIN, 2000; LARKIN; GROSS; REEP, 2005). Já para o peixe-boi
da Amazônia, foram realizados trabalhos comparando andrógenos fecais de machos
cativos com diferentes períodos do ciclo hidrológico dos rios da Amazônia e com a
precipitação pluviométrica (PIMENTEL, 1998), e utilizando estrógenos e progestinas
fecais para a verificação de sazonalidade e ciclo estral em fêmeas mantidas em
cativeiro (NASCIMENTO, 2004). Até o presente momento, nenhum trabalho foi
realizado com sirênios utilizando amostras de saliva ou secreção lacrimal na
dosagem hormonal.
Porém, tendo em vista que os sirênios são animais que apresentam algumas
características fisiológicas peculiares (como a taxa metabólica e a fisiologia

37
digestiva), todos os trabalhos relatam a necessidade de maior entendimento sobre o
tempo entre a síntese e a excreção dos hormônios esteróides nas espécies
estudadas para uma melhor compreensão dos resultados encontrados.

38
OBJETIVOS

39
3 OBJETIVOS
- Determinar o tempo necessário para que um pico de testosterona sérica
seja observado em outras matrizes biológicas tais como: fezes, urina,
saliva e secreção lacrimal.
- Validar biologicamente a utilização de fezes, urina, saliva e secreção
lacrimal como meios não-invasivos de mensuração de andrógenos por
meio de radioimunoensaio.
- Determinar a eficiência e a aplicabilidade da dosagem de andrógenos por
diferentes matrizes biológicas na espécie.

40
MATERIAL E MÉTODOS

41
4 MATERIAL E MÉTODOS
As descrições sobre os animais e a metodologia utilizada seguem nos tópicos
abaixo.
4.1 ANIMAIS
Foram utilizados dois machos adultos de peixe-boi da Amazônia (Quadro 1)
mantidos em cativeiro no Instituto de Pesquisas da Amazônia (INPA), Manaus - AM,
alojados juntamente com mais 20 animais em três piscinas circulares (10m X 2,5m
de profundidade; 196m3) conectadas entre si por tanques de cambiamento (3,5m X
2,5m X 1,5m de profundidade; 13m3) (Figura 4). Cada tanque possui fonte de água e
drenagem independente. A água é retirada de um poço artesiano e circula pelos
tanques 24 horas por dia, tendo seus componentes químicos e físicos
periodicamente analisados. A temperatura da água variou entre 27 e 29ºC. Durante
o período de experimentação, cada animal foi mantido no tanque de cambiamento
para facilitar o manejo.
Os animais foram alimentados com capim colônia (Brachiaria mutica), além de
verduras e legumes diversos, em porções variadas, com o total de alimentos não
sendo inferior a 8% do peso de cada animal.
Animal Idade (anos) Peso (kg) Tamanho (cm)
A-1 > 28 325 251
A-2 > 27 216 244
Quadro 1 – Características dos peixes-bois da Amazônia utilizados neste estudo

42
Figura 4 – Tanques dos peixes-bois da Amazônia no Laboratório de Mamíferos Aquáticos do Instituto Nacional de Pesquisas da Amazônia – LMA/INPA

43
4.2 DESAFIO HORMONAL
Os animais foram submetidos a um protocolo de experimentação de 12 dias
(D-1 a D10). No D0, foi aplicado o análogo de GnRH (Lecirelina, Gestran Plus -
Tecnopec - São Paulo/SP) para promover a produção de um pico fisiológico de
testosterona. A aplicação foi por via intramuscular, na região do pedúnculo caudal
utilizando agulha 40X0,8mm, na dosagem de 0,7µg/Kg (Figura 5).
Figura 5 – Aplicação do análogo de GnRH na região do pedúnculo caudal de um T. inunguis

44
4.3 COLHEITA DO MATERIAL BIOLÓGICO
As coletas foram realizadas diariamente (entre 08:00 e 09:00 horas) após a
contenção física dos animais causada pela drenagem do tanque de cambiamento.
Todo o material coletado era identificado com o nome do animal, data e hora da
coleta e congelado a -20ºC até a análise.
4.3.1 Fezes
A compressão do abdômen do animal no fundo do tanque devido à drenagem
do cambiamento sempre resultava em defecação. As fezes eram coletadas
imediatamente, com o soerguimento da nadadeira caudal, e condicionadas em
sacos plásticos (Figura 6).
Figura 6 – Coleta de fezes de T. inunguis após drenagem do tanque de cambiamento. Seta: amostra de fezes

45
4.3.2 Urina
A urina foi obtida com a lateralização do animal e compressão abdominal na
região da vesícula urinária seguindo a metodologia descrita por Pantoja (2004) e
armazenadas em frascos plásticos (Figura 7). Para compensar as variações no
consumo de água e no clearance renal, uma alíquota de cada amostra foi separada
para dosagem de creatinina urinária (Cr).
4.3.3 Saliva
A coleta da saliva foi realizada com o auxílio de uma colher de metal,
raspando levemente a mucosa bucal, e armazenada em frascos plásticos (Figura 8).
4.3.4 Secreção lacrimal
As amostras de secreção lacrimal foram coletadas, por gravidade,
diretamente da face do animal, utilizando tubos plásticos, sendo estas armazenadas
nos próprios tubos.
46
Figura 7 – Coleta de urina de T. inunguis por compressão abdominal na região da vesícula urinária
Figura 8 – Coleta de saliva de T. inunguis com o auxílio de uma colher de metal

47
4.4 ANÁLISE HORMONAL
Todo o material coletado foi dosado técnica de radioimunoensaio (RIE) em
fase sólida para testosterona total utilizando o conjunto diagnóstico comercial Coat-
A-Count® (Siemens, Los Angeles, CA, USA; antiga DPC Medlab) desenvolvido para
dosagem hormonal em soro humano.
Este conjunto diagnóstico comercial é um teste de competição entre
testosterona marcada (125I) e testosterona não-marcada (testosterona da amostra),
durante um tempo fixo, por sítios de ligação dos anticorpos específicos que se
encontram imobilizados nas paredes dos tubos de polipropileno.
O anticorpo utilizado apresenta as seguintes reações cruzadas (fornecidas
pelo fabricante): 100% testosterona, 20% 19-nortestosterona, 20% 4-estren-17-ol-3-
ona, 16% 11-cetotestosterona, 3.3% 5α-diidrotestosterona, 2.0% 19-
hidroxiandrostenediona, 1.7% metiltestosterona, 1.1% 4-estren-7α-metil-17β-ol-3-
ona, e <1% com (lista parcial) aldosterona, androstenediona, androsterona, 5-
androsten-3β,17β-diol, corticosterona, cortisol, cortisona, desidroepiandrosterona,
estradiol, estrona, progesterona e 11β-hidroxitestosterona.
A etapa de análise hormonal foi realizada no Laboratório de Dosagens
Hormonais do Departamento de Reprodução Animal da Faculdade de Medicina
Veterinária e Zootecnia da Universidade de São Paulo (LDH-VRA-FMVZ/USP), São
Paulo – SP.
4.4.1 Fezes
As amostras de fezes foram secas em um liofilizador a vácuo (Savant
Instrument Speedvac Rotatory Evaporator, Forma Scientific Inc., OH, EUA) por 12
horas e peneiradas para remoção de alimentos não digeridos (pedaços de capim e
sementes). O material foi submetido à extração hormonal utilizando o protocolo
descrito por Graham et al. (2001).
Foi colocado 0,25g de fezes liofilizada e peneirada em um tubo de ensaio
juntamente com 5mL de metanol 80%. Os tubos foram tampados e suavemente

48
agitados por 15 horas utilizando um homogeneizador de sangue (AP22, Phoenix,
SP, Brasil). As amostras foram centrifugadas (500G, 15 minutos) e o sobrenadante
separado e armazenado a -20ºC até a análise.
Os extratos fecais obtidos foram diluídos (1:10) em tampão gelatina (0,061M
H2NaPO4.12H2O; 0,041M H2NaPO4.H2O; 0,015M NaN3; 0,154M NaCl; 1,0g gelatina;
pH 7,0) antes do RIE. A dosagem hormonal foi realizada de acordo com as
especificações do fabricante do conjunto diagnóstico comercial de RIE.
4.4.2 Urina
Antes do RIE, as amostras de urina foram submetidas a um processo de
hidrólise ácida para quebra das ligações com os conjugados. Foi pipetado 1mL de
urina para um tubo de ensaio e adicionado 200µL de ácido clorídrico (HCl, 12N). Os
tubos foram levemente tampados e incubados em banho-maria fervente por 15
minutos. Após isso, cada amostra de urina hidrolisada foi diluída (1:25) em soro
bovino depletado (carvão-dextran) e em seguida analisada hormonalmente.
Toda etapa laboratorial foi relizada de acordo com a metodologia proposta
pelo fabricante do conjunto diagnóstico comercial de RIE, com exceção do uso de
soro bovino depletado na diluição das amostras no lugar do calibrador “0” (soro
humano limpo de hormônio) por ser uma opção de baixo custo e com resultados
similares.
A creatinina urinária foi dosada colorimetricamente utilizando a reação de
Jaffé (TAUSSKY, 1954). Esta análise foi realizada na Fundação de Medicina
Tropical do Amazonas, Manaus – AM, usando o analisador químico Dade Behring
Dimension AR (Diamond Diagnostics, MA, EUA).
4.4.3 Saliva
As amostras de saliva passaram por dois ciclos de congelamento e
descongelamento, e posterior centrifugação para remoção dos mucopolissacarídeos

49
e outros resíduos. Após isso, foram submetidas a um protocolo de RIE adaptado das
metodologias propostas por Schultheiss et al. (2004) e Campbell, Schultheiss e
McClelland (1999) para aumentar a sensibilidade do teste.
Foram pipetados 200µL de saliva nos tubos de propileno e posteriormente
foram incubados por 24 horas em temperatura ambiente. Após a primeira incubação,
adicionou-se 1mL da testosterona radiomarcada e, então, os tubos foram para a
segunda incubação por 16-24 horas em temperatura ambiente. O material foi
decantado e os tubos foram medidos em contador gama por 2 minutos. Os
calibradores utilizados na construção da curva padrão foram diluídos 20 vezes em
água destilada.
4.4.4 Secreção lacrimal
Devido à caraterística peculiar das amostras de secreção lacrimal (formação
de uma gelatina consistente após o congelamento), essas foram submetidas a vários
ciclos de congelamento e descongelamento, e posterior centrifugação, até que aa
amostras se apresentassem liqüefeitas. O ensaio hormonal foi realizado seguindo o
protocolo adotado para as amostras de saliva.
4.4.5 Paralelismo
Foi realizado o teste de paralelismo utilizando matriz íntegra para avaliar se
as matrizes interferiram na ligação antígeno-anticorpo. Para tal, adicionou-se uma
concentração conhecida de testosterona a um “pool” de cada matriz com baixas
concentrações hormonais, o qual posteriormente foi diluído seriadamente utilizando
a própria matriz e dosado por RIE, comparando os resultados com a curva padrão. O
paralelismo não foi realizado para as amostras de secreção lacrimal por não ter
volume suficiente de amostras para a formação do “pool”.

50
4.5 ANÁLISE DOS RESULTADOS
Os resultados nas amostras urinárias foram corrigidos com o valor de
creatinina e expressos como ng/mg Cr, enquanto os andrógenos fecais foram
expressos como ng/g de fezes secas. Já a testosterona salivar e a testosterona
lacrimal foram expressas como pg/mL.
Devido ao número amostral ser pequeno, não foi possível realizar análises
estatísticas mais refinadas. Por não se ter um número grande de amostras do
período antes da aplicação hormonal, o qual possibilitaria o cálculo da média e do
desvio padrão, sendo estes os rotineiramente utilizados para a definição de pico
hormonal, optou-se por trabalhar com a mediana e o desvio interquartílico (DI) dos
dados, onde, assim, poderia se diluir os efeitos pós tratamento hormonal. A mediana
e o DI para cada matriz de cada animal foram calculados. Foi definido como pico
para cada animal como qualquer valor que ultrapassasse o dobro do DI acima da
mediana (pico > mediana+2DI) para cada matriz. O intervalo de tempo entre a
aplicação hormonal e o pico de andrógeno para cada matriz foi determinado e
comparado entre eles. Foram calculados os coeficientes de correlação de Pearson
entre as matrizes para cada animal e os valores foram considerados significantes
quando p<0,05.
A análise do paralelismo foi realizada comparando os resultados obtidos com
a curva padrão por regressão linear simples.
As análises estatísticas e os gráficos foram realizados utilizando os
programas estatísticos SPSS 12.0 (© SPSS, Inc.) e SigmaPlot 10.0 (© Systat
Software, Inc.).

51
RESULTADOS

52
5 RESULTADOS
Os resultados obtidos são apresentados a seguir.
5.1 PARÂMETROS DE QUALIDADE DOS ENSAIOS HORMONAIS
O controle de qualidade dos ensaios de RIE foi realizado através da análise
dos coeficientes de variação intra-ensaio, o qual foi inferior a 13,30% para todas as
matrizes. As doses mínimas detectadas foram de 0,34 ng/dL, 0,08 ng/dL, 2,73 ng/dL
e 3,18 ng/dL de testosterona para os ensaios de saliva, secreção lacrimal, urina e
fezes, respectivamente (Tabela 1).
Tabela 1 – Controles de qualidade obtidos nos ensaios hormonais de andrógenos presentes na saliva, secreção lacrimal, urina e fezes de dois peixes-bois amazônicos machos adultos
CPM Cap Lig. L.N.E Sensibilidade CV Intra CV Intra Ensaio
total B/B0 (%) % (dose ng/dL) Baixo Alto
Saliva 34152,5 48% 0,59% 98,9 (0,3448) 7,70% 12,49%
Secreção lacrimal
23336,3 56% 0,55% 99,5 (0,0784) 13,30% 1,44%
Urina 22778,5 46% 0,91% 90,8 (2,7253) 6,54% 8,02%
Fezes 36834,5 45% 0,30% 92,5 (3,1844) 1,63% 4,89%
53
5.2 PARALELISMO
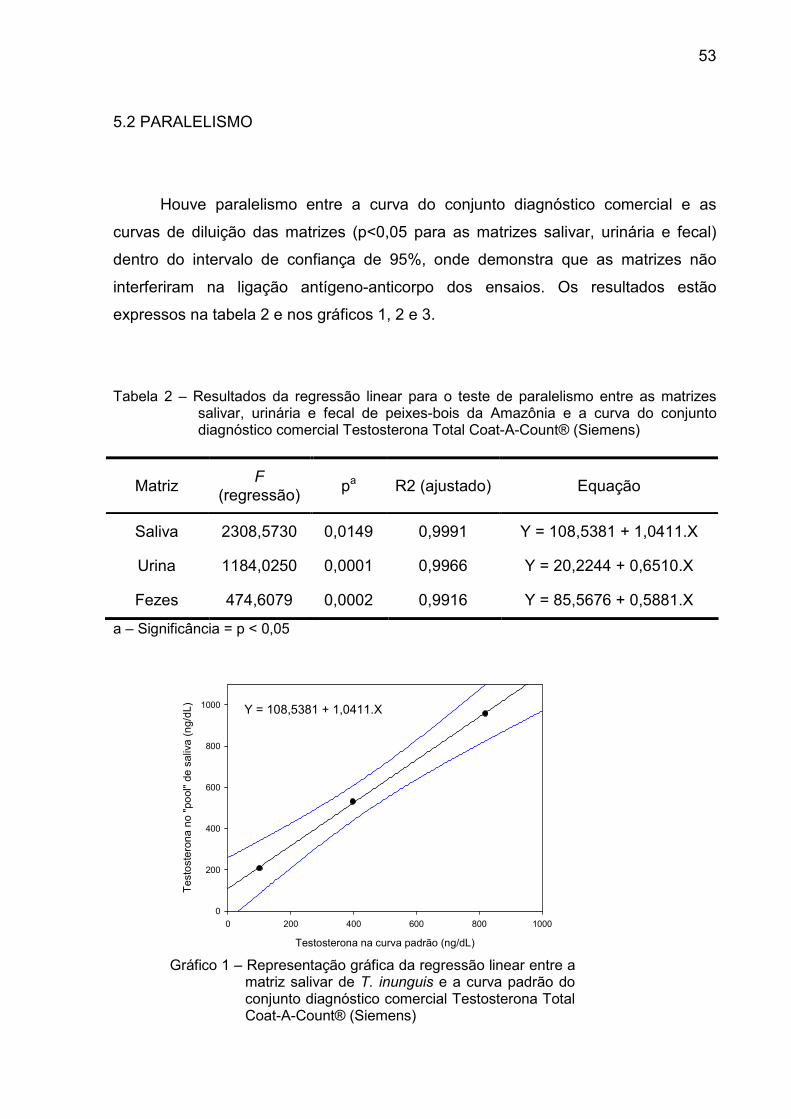
Houve paralelismo entre a curva do conjunto diagnóstico comercial e as
curvas de diluição das matrizes (p<0,05 para as matrizes salivar, urinária e fecal)
dentro do intervalo de confiança de 95%, onde demonstra que as matrizes não
interferiram na ligação antígeno-anticorpo dos ensaios. Os resultados estão
expressos na tabela 2 e nos gráficos 1, 2 e 3.
Tabela 2 – Resultados da regressão linear para o teste de paralelismo entre as matrizes salivar, urinária e fecal de peixes-bois da Amazônia e a curva do conjunto diagnóstico comercial Testosterona Total Coat-A-Count® (Siemens)
Matriz F
(regressão) pa R2 (ajustado) Equação
Saliva 2308,5730 0,0149 0,9991 Y = 108,5381 + 1,0411.X
Urina 1184,0250 0,0001 0,9966 Y = 20,2244 + 0,6510.X
Fezes 474,6079 0,0002 0,9916 Y = 85,5676 + 0,5881.X
a – Significância = p < 0,05
Gráfico 1 – Representação gráfica da regressão linear entre a matriz salivar de T. inunguis e a curva padrão do conjunto diagnóstico comercial Testosterona Total Coat-A-Count® (Siemens)
Y = 108,5381 + 1,0411.X
Testosterona na curva padrão (ng/dL)
0 200 400 600 800 1000
Testosterona no "pool" de
saliva (ng/dL)
0
200
400
600
800
1000
54
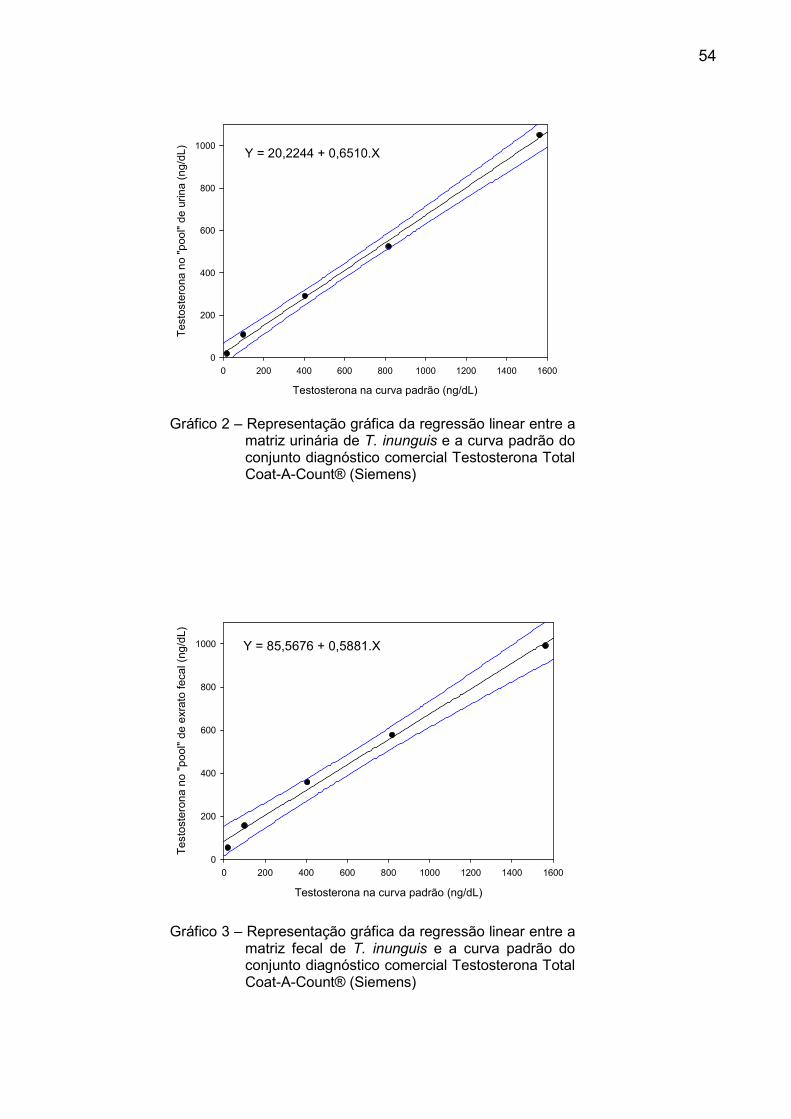
Gráfico 2 – Representação gráfica da regressão linear entre a matriz urinária de T. inunguis e a curva padrão do conjunto diagnóstico comercial Testosterona Total Coat-A-Count® (Siemens)
Gráfico 3 – Representação gráfica da regressão linear entre a matriz fecal de T. inunguis e a curva padrão do conjunto diagnóstico comercial Testosterona Total Coat-A-Count® (Siemens)
Y = 20,2244 + 0,6510.X
Testosterona na curva padrão (ng/dL)
0 200 400 600 800 1000 1200 1400 1600
Testosterona no "poo
l" de urin
a (ng/dL
)
0
200
400
600
800
1000
Y = 85,5676 + 0,5881.X
Testosterona na curva padrão (ng/dL)
0 200 400 600 800 1000 1200 1400 1600
Testosterona no "pool" de exrato fecal (ng/dL)
0
200
400
600
800
1000
55
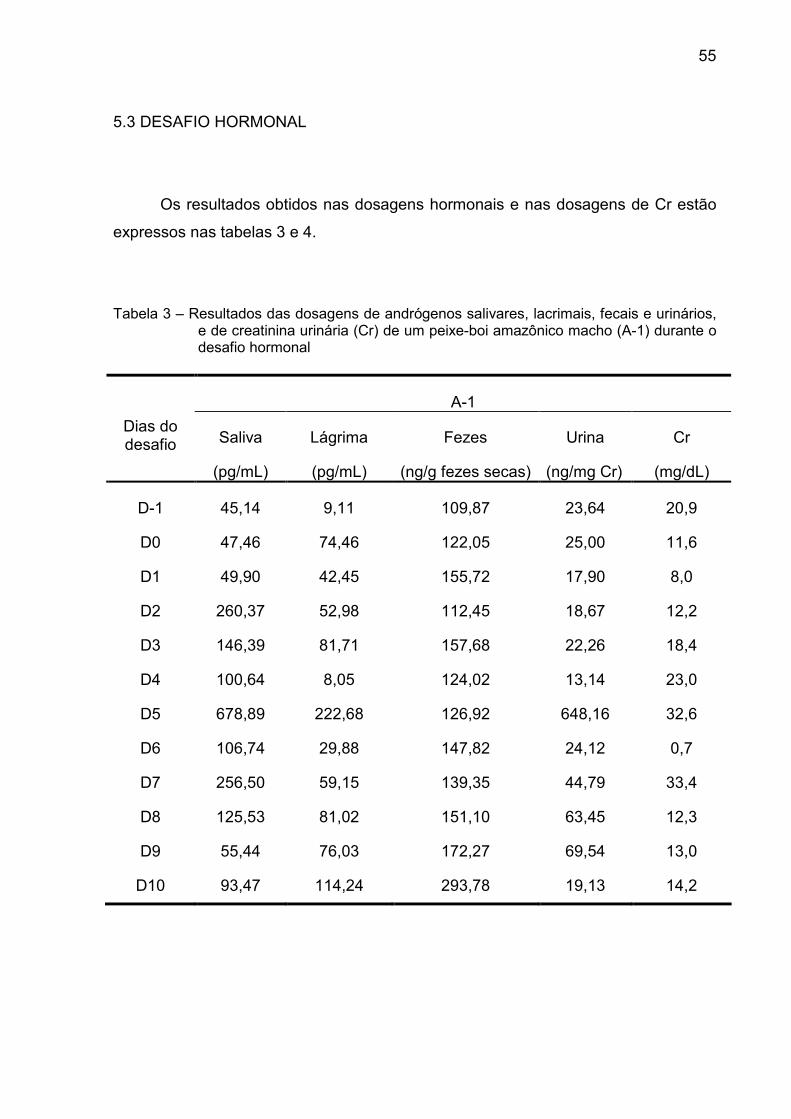
5.3 DESAFIO HORMONAL
Os resultados obtidos nas dosagens hormonais e nas dosagens de Cr estão
expressos nas tabelas 3 e 4.
Tabela 3 – Resultados das dosagens de andrógenos salivares, lacrimais, fecais e urinários, e de creatinina urinária (Cr) de um peixe-boi amazônico macho (A-1) durante o desafio hormonal
A-1
Saliva Lágrima Fezes Urina Cr Dias do desafio
(pg/mL) (pg/mL) (ng/g fezes secas) (ng/mg Cr) (mg/dL)
D-1 45,14 9,11 109,87 23,64 20,9
D0 47,46 74,46 122,05 25,00 11,6
D1 49,90 42,45 155,72 17,90 8,0
D2 260,37 52,98 112,45 18,67 12,2
D3 146,39 81,71 157,68 22,26 18,4
D4 100,64 8,05 124,02 13,14 23,0
D5 678,89 222,68 126,92 648,16 32,6
D6 106,74 29,88 147,82 24,12 0,7
D7 256,50 59,15 139,35 44,79 33,4
D8 125,53 81,02 151,10 63,45 12,3
D9 55,44 76,03 172,27 69,54 13,0
D10 93,47 114,24 293,78 19,13 14,2
56
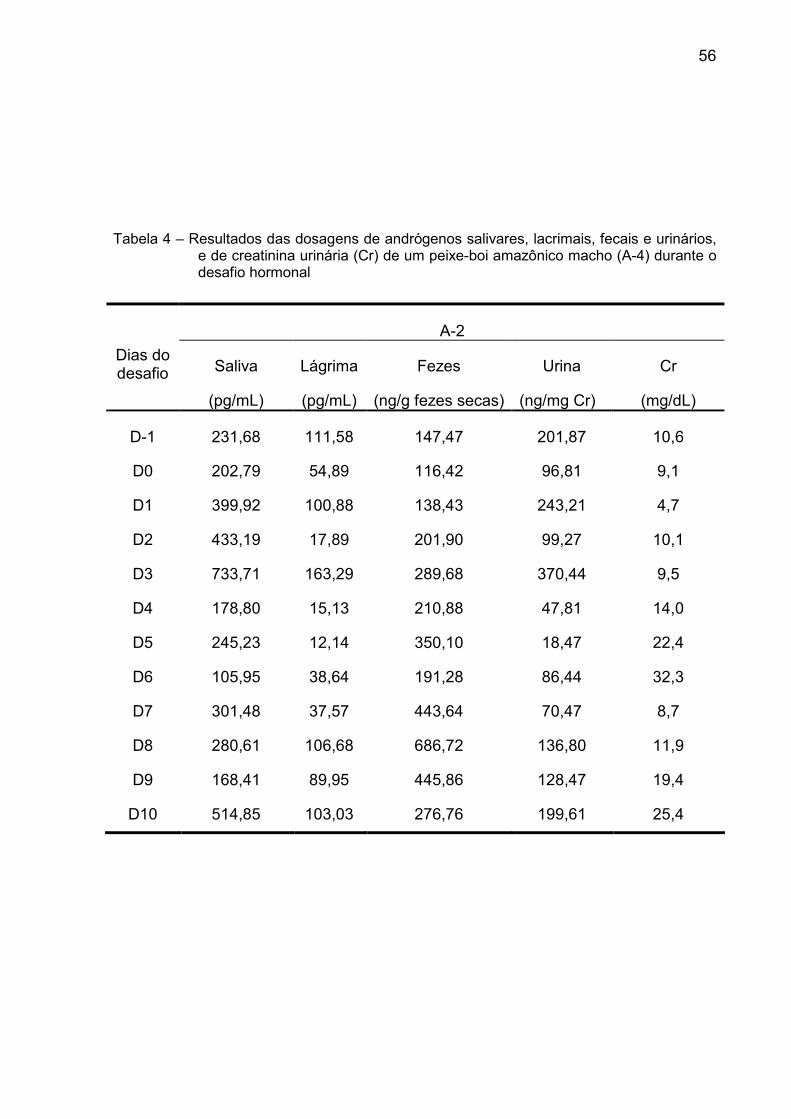
Tabela 4 – Resultados das dosagens de andrógenos salivares, lacrimais, fecais e urinários,
e de creatinina urinária (Cr) de um peixe-boi amazônico macho (A-4) durante o desafio hormonal
A-2
Saliva Lágrima Fezes Urina Cr Dias do desafio
(pg/mL) (pg/mL) (ng/g fezes secas) (ng/mg Cr) (mg/dL)
D-1 231,68 111,58 147,47 201,87 10,6
D0 202,79 54,89 116,42 96,81 9,1
D1 399,92 100,88 138,43 243,21 4,7
D2 433,19 17,89 201,90 99,27 10,1
D3 733,71 163,29 289,68 370,44 9,5
D4 178,80 15,13 210,88 47,81 14,0
D5 245,23 12,14 350,10 18,47 22,4
D6 105,95 38,64 191,28 86,44 32,3
D7 301,48 37,57 443,64 70,47 8,7
D8 280,61 106,68 686,72 136,80 11,9
D9 168,41 89,95 445,86 128,47 19,4
D10 514,85 103,03 276,76 199,61 25,4
57
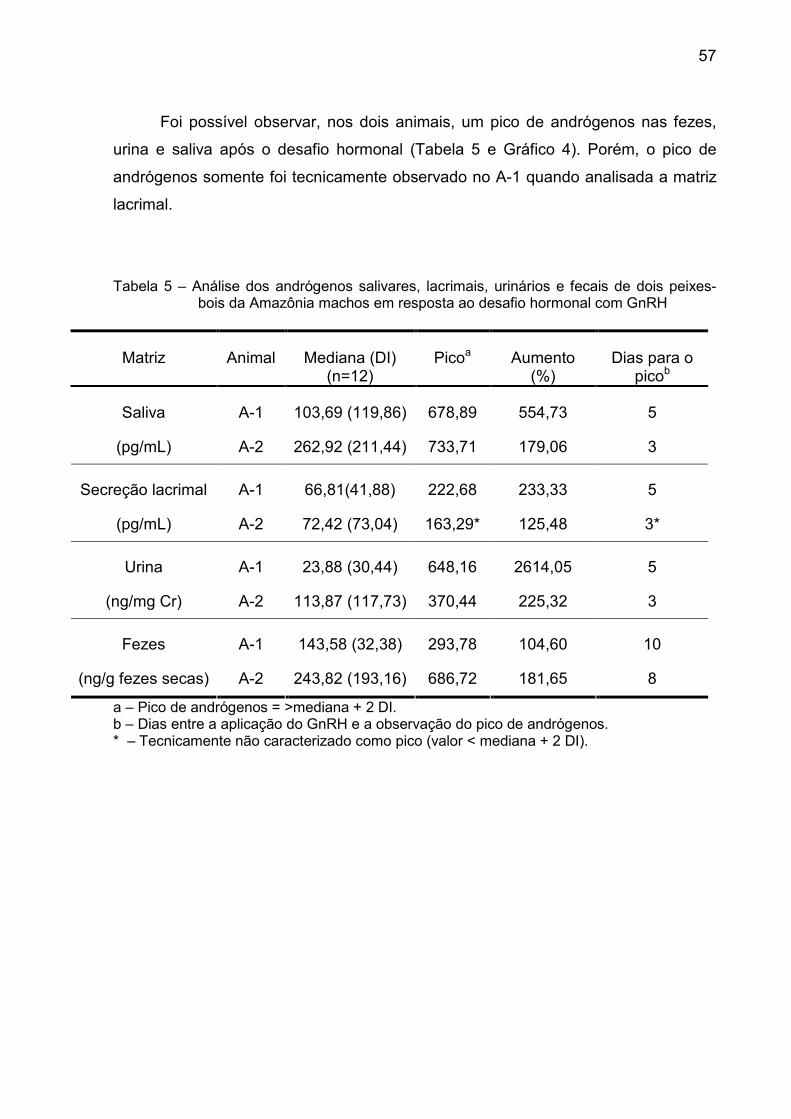
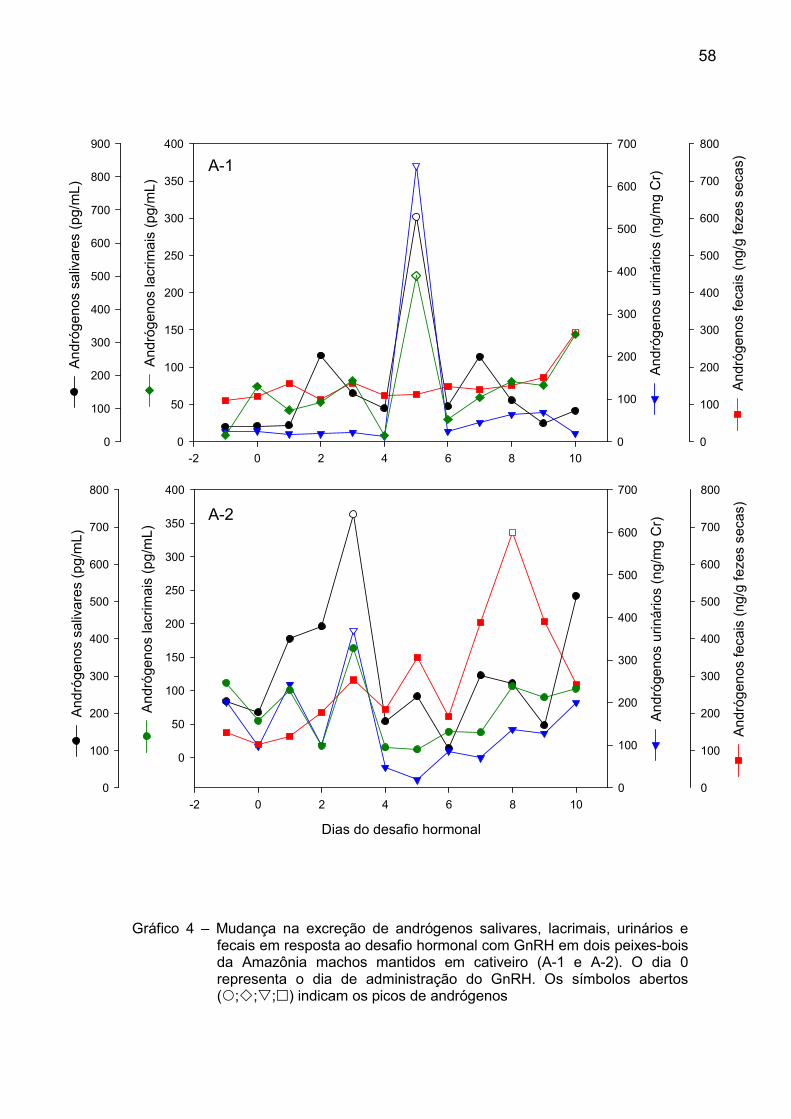
Foi possível observar, nos dois animais, um pico de andrógenos nas fezes,
urina e saliva após o desafio hormonal (Tabela 5 e Gráfico 4). Porém, o pico de
andrógenos somente foi tecnicamente observado no A-1 quando analisada a matriz
lacrimal.
Tabela 5 – Análise dos andrógenos salivares, lacrimais, urinários e fecais de dois peixes-bois da Amazônia machos em resposta ao desafio hormonal com GnRH
Matriz Animal Mediana (DI) (n=12)
Picoa Aumento (%)
Dias para o picob
A-1 103,69 (119,86) 678,89 554,73 5 Saliva
(pg/mL) A-2 262,92 (211,44) 733,71 179,06 3
A-1 66,81(41,88) 222,68 233,33 5 Secreção lacrimal
(pg/mL) A-2 72,42 (73,04) 163,29* 125,48 3*
A-1 23,88 (30,44) 648,16 2614,05 5 Urina
(ng/mg Cr) A-2 113,87 (117,73) 370,44 225,32 3
A-1 143,58 (32,38) 293,78 104,60 10 Fezes
(ng/g fezes secas) A-2 243,82 (193,16) 686,72 181,65 8
a – Pico de andrógenos = >mediana + 2 DI. b – Dias entre a aplicação do GnRH e a observação do pico de andrógenos. * – Tecnicamente não caracterizado como pico (valor < mediana + 2 DI).
58
Gráfico 4 – Mudança na excreção de andrógenos salivares, lacrimais, urinários e
fecais em resposta ao desafio hormonal com GnRH em dois peixes-bois da Amazônia machos mantidos em cativeiro (A-1 e A-2). O dia 0 representa o dia de administração do GnRH. Os símbolos abertos (�;�;�;�) indicam os picos de andrógenos
A-1
-2 0 2 4 6 8 10
Andrógenos salivares (pg/m
L)
0
100
200
300
400
500
600
700
800
900
Andrógenos urinários (ng/m
g Cr)
0
100
200
300
400
500
600
700
Andrógenos fecais (ng/g fezes secas)
0
100
200
300
400
500
600
700
800Andrógenos lacrimais (pg/m
L)
0
50
100
150
200
250
300
350
400
A-2
Dias do desafio hormonal
-2 0 2 4 6 8 10
Andrógenos salivares (pg/mL)
0
100
200
300
400
500
600
700
800
Andrógenos urinários (ng/mg Cr)
0
100
200
300
400
500
600
700
Andrógenos fecais (ng/g fezes secas)
0
100
200
300
400
500
600
700
800
Andrógenos lacrimais (pg/m
L)
0
50
100
150
200
250
300
350
400

59
Observou-se uma correlação significante entre as matrizes salivar, lacrimal e
urinária para ambos os animais, fato esse não observado entre a matriz fecal e as
outras matrizes (Tabela 6).
Tabela 6 – Resultados do teste de correlação linear de Pearson para as matrizes salivar,
lacrimal, urinária e fecal de dois peixes-bois da Amazônia machos (A-1 e A-2) submetidos a um desafio hormonal com GnRH
A-1 A-2 Matrizes
r pa r pa
Saliva X Lágrima 0,7825 0,0026 0,5815 0,0473
Saliva X Urina 0,9058 < 0,0001 0,7685 0,0035
Saliva X Fezes -0,2091 0,5142 -0,0187 0,9540
Lágrima X Urina 0,8514 0,0004 0,9170 < 0,0001
Lágrima X Fezes 0,2494 0,4344 0,1443 0,6547
Urina X Fezes -0,1515 0,6384 -0,1264 0,6955
a – Significância = p < 0,05

60
DISCUSSÃO

1 Informação fornecida por D’Affonsêca Neto em Manaus, em 2005.
61
6 DISCUSSÃO
O manejo populacional de uma espécie, tanto cativa quanto de vida-livre,
necessita, como base, de conhecimentos prévios sobre a biologia da espécie em
questão, sendo um dos pontos fundamentais as informações sobre a biologia
reprodutiva. Alguns estudos foram realizados sobre a endocrinologia reprodutiva de
sirênios (FRANCIS-FLOYD et al., 1991; PIMENTEL, 1998; LARKIN, 2000;
NASCIMENTO et al., 2002; WAKAI et al., 2002; NASCIMENTO, 2004; LANYON;
SMITH; CARRICK, 2005; LARKIN; GROSS; REEP, 2005). Entretanto, este é o
primeiro trabalho realizando um desafio hormonal.
6.1 DESAFIO HORMONAL X AMOSTRAS SALIVARES
Como previsto, um pico de andrógenos foi observado nas amostras de saliva
dos dois peixes-bois da Amazônia utilizados neste estudo. Porém, estes picos
aconteceram depois do período esperado (poucas horas após a administração
hormonal). Vários trabalhos têm demonstrado que as concentrações de testosterona
salivar estão intimamente correlacionadas com as concentrações plasmáticas em
humanos devido à grande facilidade que a molécula de testosterona é transportada
do sangue para a saliva (RIAD-FAHMY et al., 1982; VINING; MCGINLEY; SYMONS,
1983; DABBS JR, 1993; NEAVE et al., 2003; SCHULTHEISS; WIRTH; STANTON,
2004; WHEELER, 2006). Com isso, descartando a possibilidade de uma demora no
transporte da testosterona plasmática para a saliva, o atraso observado neste
estudo (5 e 3 dias para A-1 e A-2, respectivamente) pode ter ocorrido em
conseqüência de uma ação retardada do GnRH exógeno no eixo hipófise-gonadal.
A escolha da agulha utilizada a aplicação intramuscular (40X0,8mm) foi
baseada na rotina de manejo veterinário dos animais alojados no LMA – INPA
(informação verbal)1. Porém, possivelmente a agulha utilizada é curta para a
administração de fármacos pela via intramuscular em peixe-boi da Amazônia (devido
a sua grande espessura de pele e de camada de gordura

62
subcutânea (MENDES, 1958; VERÍSSIMO, 1970), fazendo com que a aplicação
tenha ocorrido na camada de tecido adiposo subcutânea, levando, com isso, a um
retardamento na absorção hormonal.
A absorção de fármacos pela via intramuscular ocorre tanto
hematogenicamente como por via linfática e geralmente é rápida. Uma das
desvantagens desta via é a possibilidade de deposição inadequada nos nervos,
vasos sanguíneos, gordura ou entre as fibras musculares dos revestimentos de
tecido conjuntivo. A taxa de absorção pela via subcutânea pode ser imprevisível e
depende de vários fatores, sendo o mais importante o fluxo sanguíneo na região,
porém, geralmente a taxa de absorção tende a ser mais lenta que pela via
intramuscular. Alguns fármacos são absorvidos tão rapidamente nos tecidos
subcutâneos como nos músculos, embora a absorção nos locais de aplicações em
regiões com concentração de gordura subcutânea fique sempre significantemente
retardada (FRASER et al., 1996; BROWN, 2001; SPINOSA; GORNIAK; BERNARDI,
2006).
Outro fator agravante para o atraso na absorção hormônio exógeno poderia
ser a taxa metabólica extremamente baixa apresentada pelo T. inunguis, sendo
equivalente a 36% da taxa de um mamífero terrestre de tamanho similar
(GALLIVAN; BEST, 1980; BEST, 1981).
Entretanto, não existem trabalhos sobre farmacocinética na espécie para
possibilitar o descarte de alguma dessas hipóteses (atraso na absorção pela
aplicação no tecido adiposo e/ou pela baixa taxa metabólica). Baseado nisso,
acredita-se que o GnRH exógeno tenha tido seu efeito (ativando o eixo hipófise-
gonadal) entre os dias 4 e 5 para o animal A-1 e entre os dias 2 e 3 para o animal A-
2.
6.2 DESAFIO HORMONAL X AMOSTRAS LACRIMAIS
Foi observado um aumento de testosterona nas amostras de secreção
lacrimal em ambos os animais (D5 e D3 para A-1 e A-2, respectivamente), porém,
somente no animal A-1 este aumento foi considerado tecnicamente como pico (<
mediana + 2 DI). Os baixos valores observados durante todo o desafio hormonal nas

63
amostras lacrimais e as baixas porcentagens de aumento observadas (179,06% e
125,48% para A-1 e A-2, respectivamente) sugerem que, apesar de ser possível
dosar testoserona em amostras de secreção lacrimal, a matriz, nas condições
propostas, não é capaz de expressar todas as possíveis variações hormonais que
possam ocorrer no animal. Entretanto, como a matriz não foi validada
laboratorialmente, realizando a teste de paralelismo, não se pode descartar uma
possível interferência da matriz nas reações antígeno-anticorpo do ensaio.
O único trabalho realizado avaliando hormônios esteróides em secreção
lacrimal foi realizado por Atkinson et al. (1999) mensurando progesterona em falsas-
orcas. Entretanto, os autores não observaram correlação entre as amostras de
secreção lacrimal e de plasma sanguíneo. Porém, para testosterona em peixe-boi
amazônico, a matriz lacrimal apresentou uma correlação significante com a matriz
salivar (p < 0,05) e principalmente com a matriz urinária (p < 0,0004) nos dois
animais, sugerindo uma possível correlação com a matriz sérica.
6.3 DESAFIO HORMONAL X AMOSTRAS URINÁRIAS
Os picos de andrógenos foram observados na urina após 5 e 3 dias, para A-1
e A-2 respectivamente. Apesar do peixe-boi amazônico apresentar uma baixa taxa
metabólica, e do tempo de metabolização e excreção dos esteróides pela urina estar
relacionado com a taxa metabólica do animal, os resultados obtidos não devem
expressar o tempo real de metabolização e excreção dos andrógenos.
Considerando o atraso na ação do GnRH exógeno e a correlação significante
observada entre as amostras de saliva e urina em ambos os animais (p < 0,01), o
pico observado nas amostras de urina provavelmente ocorreu poucas horas após o
pico aparecer na corrente sanguínea, corroborando com o tempo de excreção dos
andrógenos pela urina observado em outros mamíferos (menos de 24 horas)
(MONFORT et al., 1995; PALME et al., 1996; BILLITTI; LASLEY; WILSON, 1998;
WILLIAMS et al., 2000; MÖHLE et al., 2002; GRAHAM, 2004; BUSSO et al., 2005).

64
6.4 DESAFIO HORMONAL X AMOSTRAS FECAIS
O tratamento com GnRH também causou um aumento significativo dos
andrógenos fecais nos dois machos utilizados nesse estudo (A-1 no D-8 e A-2 no D-
10). Adotando o atraso na ação do GnRH exógeno, o intervalo entre o possível pico
de testosterona no sangue (entre os dias 4 e 5 para o animal A-1 e entre os dias 2 e
3 para o animal A-2) e o pico de andrógenos observado nas fezes (entre cinco e seis
dias para ambos os animais) foi maior do que os relatados para outras espécies de
mamíferos herbívoros. O intervalo de tempo para o aparecimento dos andrógenos
fecais é por volta de 12-24 horas em ruminantes e entre 24-50 horas em animais
com fermentação cecal (eqüinos, suínos, rinocerontes, elefantes e primatas)
(PALME et al., 1996; SCHWARZENBERGER et al., 1996; BROWN, 2000; MÖHLE et
al., 2002; KRETZSCHMAR; GANSLOßER; DEHNHARD, 2004;
SCHWARZENBERGER, 2007)
Os metabólitos de esteróides são excretados via bile dentro do duodeno e são
transportados junto com o bolo fecal. Com isso, em espécies não-ruminantes, a
excreção dos esteróides fecais está intimamente relacionada com a taxa de
passagem do alimento pelo trato gastrointestinal (PALME et al., 1996;
SCHWARZENBERGER et al., 1996; SCHWARZENBERGER, 2007). Os sirênios
apresentam um longo tempo de passagem em comparação com o tempo de transito
gastrointestinal de outros mamíferos (WARNER, 1981; LOMOLINO; EWEL, 1984;
LANYON; MARSH, 1995; ITAVO; ROSAS; CAVALLANTE, 1996; LARKIN; FOWLER;
REEP, 2007). Itavo, Rosas e Cavallante (1996) observaram um tempo de passagem
gastrointestinal entre cinco e sete dias para peixes-boi da Amazônia, e este
resultado corrobora com o tempo de excreção dos andrógenos pelas fezes
observado no desafio hormonal (entre cinco e seis dias).
Os resultados não demonstraram correlação entre os andrógenos fecais e os
andrógenos urinários, salivares ou lacrimais, sendo claramente explicado pelo curto
período experimental e o grande intervalo entre os picos observados (cinco dias),
sendo este influenciado pelas vias de excreção.

65
6.5 COMPARAÇÃO ENTRE AS MATRIZES ANALISADAS
Como o pico de andrógenos foi observado tanto na urina quanto nas fezes,
não foi possível definir uma principal rota de excreção de andrógenos para a
espécie, sendo possível que o T. inunguis utilize as duas vias de excreção em
proporções próximas. Uma maneira de provar esta hipótese seria a administração de
andrógenos radiomarcados.
A administração de hormônios radioativamente marcados em animais
domésticos e selvagens tem sido usada para determinar a rota de excreção, o tempo
para a excreção e o tipo de metabólito excretado em maior quantidade na urina e
nas fezes (HINDLE; HODGES, 1990; BROWN et al., 1994; MONFORT et al., 1995;
BROWN; TERIO; GRAHAM, 1996; PALME et al., 1996; SCHWARZENBERGER et
al., 1996; WASSER et al., 1996; MONFORT et al., 1997; BILLITTI; LASLEY;
WILSON, 1998; HEISTERMANN et al., 1998; VELLOSO et al., 1998; MOORMAN et
al., 2002; TOUMA et al., 2003; BUSSO et al., 2005). Entretanto, este procedimento é
muito trabalhoso, necessitando coletas de amostras seriadas com curtos intervalos
de tempo (30 minutos a três horas) durante duas semanas, impossibilitando assim a
sua realização em peixes-bois amazônicos. Outro fator agravante é o risco inerente
no uso de substâncias radioativas em animais (GRAHAM, 2004).
Os resultados demonstraram a viabilidade da utilização de amostras de
saliva, urina e fezes para o monitoramento reprodutivo de peixes-bois da Amazônia
machos. Porém, a escolha de qual matriz biológica será usada no manejo
reprodutivo deve considerar a facilidade de coleta das amostras, as etapas
laboratoriais necessárias e se a matriz escolhida será capaz de responder a questão
levantada.
As amostras de saliva apresentam como desvantagens a necessidade de
contato íntimo com o animal para a realização de sua coleta (drenagem do tanque e
manipulação do animal), o baixo volume da amostra (em média 500µL) e a
obrigatoriedade de um ajuste no protocolo de RIE (aumentando o tempo para a
obtenção dos resultados). Sendo que, a grande vantagem é a proximidade com o
evento fisiológico o qual está sendo demonstrado, sendo este fator de grande
importância no manejo reprodutivo de espécies em cativeiro.

66
Do mesmo modo, as amostras de urina também necessitam de manipulação
do animal para a sua coleta e de etapas laboratoriais mais laboriosas como a
hidrólise para a desconjugação dos metabólitos e a dosagem de creatinina urinária
para a correção dos resultados. Porém, a vantagem em sua utilização, assim como a
saliva, é o fornecimento de resultados mais próximos do evento fisiológico.
Tanto para saliva quanto para urina, o problema da necessidade de drenagem
do tanque para sua obtenção poderia ser resolvido com o condicionamento dos
animais para colaborarem com as coletas, como já foi demonstrado com falsas-
orcas (ATKINSON et al., 1999) e focas-monge-do-Havaí (Monachus schauinsland)
(THEODOROU; ATKINSON, 1998) para coleta de saliva, e com orcas (Orcinus orca)
(WALKER et al., 1988; ROBECK et al., 2004), golfinhos-nariz-de-garrafa (Tursiops
truncatus) (ROBECK et al., 2005), dugongos (WAKAI et al., 2002) e peixes-bois
marinhos (COLBERT et al., 2001; LIMA et al., 2005) para coleta de urina.
Já as amostras de fezes, são as mais fáceis de serem obtidas, podendo ser
coletadas boiando na água logo após a defecação. Porém, necessitam de liofilização
e extração hormonal tornado sua manipulação laboratorial mais trabalhosa. Outro
importante fator desfavorável à matriz fecal é o longo intervalo entre o resultado
obtido e o evento fisiológico. Apesar destas desvantagens, é a única matriz de fácil
trabalho em animais de vida-livre, por não necessitar de nenhum contato com o
animal (nem mesmo visual) para a sua coleta. Os animais podem ser rastreados por
radio-telemetria e as amostras coletadas nas proximidades de onde se encontra o
animal podem ser aliadas a técnicas de análises genéticas para a identificação do
indivíduo, como tem sido realizado com baleias-francas-do-Norte (Eubalaena
glacialis) (ROLLAND et al., 2005; HUNT et al., 2006; ROLLAND et al., 2006).
Há uma necessidade de mais estudos sobre o uso de secreção lacrimal no
monitoramento reprodutivo de peixes-bois da Amazônia machos, porém, nas
condições aqui apresentadas, sua utilização é inviável por não ser capaz de
demonstrar as variações hormonais ocorridas. Outros fatores que devem ser levado
em consideração são a forma de coleta (necessidade de manipulação do animal) e a
manipulação da amostra no laboratório (necessidade de vários ciclos de
congelamento e descongelamento para liqüefazer a amostra e uso do protocolo
ajustado como nas amostras salivares). Com isso, comparando as matrizes salivar e
lacrimal, onde as desvantagens de uso são as mesmas, a priimeira se apresenta
como uma melhor opção já é capaz de expressar os eventos fisiológicos.

67
Esses resultados, apesar de serem provenientes de somente dois animais e
do sexo masculino, podem ser utilizados como referência para estudos futuros sobre
a fisiologia reprodutiva de peixes-bois da Amazônia fêmeas, além de outros sirênios.
No entanto, é necessário sempre lembrar que a principal rota de excreção dos
esteróides pode variar consideravelmente entre as espécies, bem como entre os
esteróides em uma mesma espécie (SCHWARZENBERGER et al., 1996).

68
CONCLUSÕES

69
7 CONCLUSÕES
Nas condições em que foi realizado o experimento, é possível concluir que:
- O conjunto diagnóstico comercial Testosterona Total Coat-A-Count® (Siemens) foi
validado biologicamente para a quantificação de andrógenos salivares, urinários e
fecais de peixes-bois da Amazônia machos.
- Os andrógenos lacrimais, apesar de serem possíveis de ser mensurados pelo
conjunto diagnóstico comercial Testosterona Total Coat-A-Count® (Siemens),
aparentemente não são capazes de demonstrar os eventos fisiológicos em T.
inunguis de forma confiável.
- As concentrações de andrógenos presentes na saliva, urina ou fezes são capazes
de refletir consistentemente eventos fisiológicos, sendo uma importante ferramenta
no monitoramento reprodutivo de T. inunguis.
- Apesar dos andrógenos fecais apresentarem um longo intervalo entre o evento
fisiológico e sua observação, eles podem ser utilizados no monitoramento
reprodutivo de peixes-bois da Amazônia de vida-livre devido à facilidade na coleta
de amostras de fezes.
- Andrógenos salivares e urinários são capazes de refletir os eventos fisiológicos
mais próximos de sua ocorrência, se tornando ferramentas mais acuradas para o
manejo reprodutivo de peixes-bois amazônicos em cativeiro.

70
REFERÊNCIAS

71
REFERÊNCIAS
AGUILAR, C. V. C.; LUNA, F. O.; GUIMARÃES, D. A.; OLIVEIRA, J. M.; DUQUE, V. M. Caça e utilização do peixe-boi amazônico (Trichechus inunguis) no Rio Arapiuns. In: CONGRESSO INTERNACIONAL SOBRE MANEJO DE FAUNA SILVESTRE NA AMAZÔNIA E AMÉRICA LATINA, 7., 2006, Ilhéus. Resumos... Ilhéus: UESC, 2006. p. 393. ALLEN, C. D.; MCKINNON, A. J.; LISLE, A. T.; D'OCCHIO, M. J.; JOHNSTON, S. D. Use of a GnRH agonist and hCG to obtain an index of testosterone secretory capacity in the koala (Phascolarctos cinereus). Journal of Andrology, v. 27, n. 6, p. 720-724, 2006. ATKINSON, S.; COMBELLES, C.; VINCENT, D.; NACHTIGALL, P.; PAWLOSKI, J.; BREESE, M. Monitoring of progesterone in captive female false killer whales, Pseudorca crassidens. General and Comparative Endocrinology, v. 115, n. 3, p. 323-332, 1999. AYRES, J. M.; BEST, R. Estratégias para a conservação da fauna amazônica. Acta Amazonica, v. 9, n. 7, p. 81-101, 1979. Suplemento. AZZAROLO, A. M.; MIRCHEFF, A. K.; KASWAN, R. L.; STANCZYK, F. Z.; CTENSCHEIN, E.; BECKER, L.; NASSIR, B.; WARREN, D. W. Androgen support of lacrimal gland function. Endocrine, v. 6, n. 1, p. 39-45, 1997. BEIJERINK, N. J.; BUIJTELS, J. J. C. W. M.; OKKENS, A. C.; KOOISTRA, H. S.; DIELEMAN, S. J. Basal and GnRH-induced secretion of FSH and LH in anestrous versus ovariectomized bitches. Theriogenology, v. 67, n. 5, p. 1039-1045, 2007. BENNETT, N. C.; FAULKES, C. G.; SPINKS, A. C. LH responses to single doses of exogenous GnRH by social Mashona mole-rats: A continuum of socially induced infertility in the family Bathyergidae. Proceedings of the Royal Society of London - Series B, Biological Sciences, v. 264, n. 1384, p. 1001-1006, 1997. BENNETT, N. C.; JARVIS, J. U. M.; FAULKES, C. G.; MILLAR, R. P. LH responses to single doses of exogenous GnRH by freshly captured Damaraland mole-rats, Cryptomys damarensis. Journal of Reproduction and Fertility, v. 99, n. 1, p. 81-86, 1993.

72
BEST, R. C. Foods and feeding habits of wild and captive Sirenia. Mammal Review, v. 11, n. 1, p. 3-29, 1981. ______ A salvação de uma espécie: novas perspectivas para o peixe-boi da Amazônia. Revista IBM, v. 14, n. 1, p. 1-11, 1982a. ______ Seasonal breeding in the Amazonian manatee, Trichechus inunguis (Mammalia: Sirenia). Biotropica, v. 14, n. 1, p. 76-78, 1982b. ______ Apparent dry-season fasting in Amazonian manatees (Mammalia: Sirenia). Biotropica, v. 15, n. 1, p. 61-64, 1983. ______ The aquatic mammals and reptiles of the Amazon. In: SIOLI, H. The Amazon, limnology and landscape: ecology of a mighty tropical river and its basin. 1. ed. Dordhecht: Dr. W. Junk Publishers, 1984. p. 371-412. BILLITTI, J. E.; LASLEY, B. L.; WILSON, B. W. Development and validation of a fecal testosterone biomarker in Mus musculus and Peromyscus maniculatus. Biology of Reproduction, v. 59, n. 5, p. 1023-1028, 1998. BROWN-DOUGLAS, C. G.; FIRTH, E. C.; PARKINSON, T. J.; FENNESSY, P. F. The pituitary and testicular responses to GnRH challenge between 4 and 14 months of age in thoroughbred colts born in spring and autumn. Animal Reproduction Science, v. 88, n. 3-4, p. 287-298, 2005. BROWN, J. L. Reproductive endocrine monitoring of elephants: an essential tool for assisting captive management. Zoo Biology, v. 19, n. 5, p. 347-367, 2000. BROWN, J. L.; TERIO, K. A.; GRAHAM, L. H. Fecal androgen metabolite analysis for noninvasive monitoring of testicular steroidogenic activity in felids. Zoo Biology, v. 15, n. 4, p. 425-434, 1996. BROWN, J. L.; WASSER, S. K.; WILDT, D. E.; GRAHAM, L. H. Comparative aspects of steroid hormone metabolism and ovarian activity in felids, measured noninvasively in feces. Biology of Reproduction, v. 51, n. 4, p. 776-786, 1994. BROWN, S. A. Pharmacokinetics: disposition and fate of drugs in the body. In: ADAMS, H. R. Veterinary pharmacology and therapeutics. 8. ed. Ames: Iowa State University Press, 2001. p. 15-56.

73
BUSSO, J. M.; PONZIO, M. F.; DABBENE, V.; DE CUNEO, M. F.; RUIZ, R. D. Assessment of urine and fecal testosterone metabolite excretion in Chinchilla lanigera males. Animal Reproduction Science, v. 86, n. 3-4, p. 339-351, 2005. CALDWELL, D. K.; CALDWELL, M. C. Manatees: Trichechus manatus, Trichechus senegalensis and Trichechus inunguis. In: RIDGWAY, S. H.; HARRISON, R. J. Handbook of marine mammals: the sirenians and baleen whales. 1. ed. New York: Academic Press, 1985. p. 33-66. CAMPBELL, K. L.; SCHULTHEISS, O. C.; MCCLELLAND, D. C. A necessary adjustment of protocol for use of DPC Coat-a-Count Total Testosterone assay with saliva. Clinical Biochemistry, v. 32, n. 1, p. 83-85, 1999. CHELINI, M. O.; SOUZA, N. L.; ROCHA, A. M.; FELIPPE, E. C.; OLIVEIRA, C. A. Quantification of fecal estradiol and progesterone metabolites in Syrian hamsters (Mesocricetus auratus). Brazilian Journal of Medical and Biological Research, v. 38, n. 11, p. 1711-1717, 2005. COLBERT, D. E.; FELLNER, W.; BAUER, G. B.; MANIRE, C. A.; RHINEHART, H. L. Husbandry and research training of two Florida manatees (Trichechus manatus latirostris). Aquatic Mammals, v. 27, n. 1, p. 16-23, 2001. CORK, S. J.; WARNER, A. C. I. The passage of digesta markers through the gut of a folivorous marsupial, the koala Phascolarctos cinereus. Journal of Comparative Physiology - B Biochemical, Systemic, and Environmental Physiology, v. 152, n. 1, p. 43-51, 1983. CROSS, N.; PINES, M. K.; ROGERS, L. J. Saliva sampling to assess cortisol levels in unrestrained common marmosets and the effect of behavioral stress. American Journal of Primatology, v. 62, n. 2, p. 107-114, 2004. CROSS, N.; ROGERS, L. J. Diurnal cycle in salivary cortisol levels in common marmosets. Developmental Psychobiology, v. 45, n. 3, p. 134-139, 2004. CZEKALA, N. M.; CALLISON, L. Pregnancy diagnosis in the black rhinoceros (Diceros bicornis) by salivary hormone analysis. Zoo Biology, v. 15, n. 1, p. 37-44, 1996. DABBS JR, J. M. Salivary testosterone measurements in behavioral studies. Annals of the New York Academy of Sciences, v. 694, n. 1, p. 177-183, 1993.

74
DECATANZARO, D.; MUIR, C.; BEATON, E. A.; JETHA, M. Non-invasive repeated measurement of urinary progesterone, 17b-estradiol, and testosterone in developing, cycling, pregnant, and postpartum female mice. Steroids, v. 69, n. 10, p. 687-696, 2004. DIAS, E. A.; OLIVEIRA, C. A. Determinação do sexo de psitacídeos por radioimunoensaio (RIA) de esteróides sexuais a partir de excretas cloacais. Brazilian Journal of Veterinary Research and Animal Science, v. 43, n. 4, p. 5-11, 2006. Suplemento. DLONIAK, S. M.; FRENCH, J. A.; PLACE, N. J.; WELDELE, M. L.; GLICKMAN, S. E.; HOLEKAMP, K. E. Non-invasive monitoring of fecal androgens in spotted hyenas (Crocuta crocuta). General and Comparative Endocrinology, v. 135, n. 1, p. 51-61, 2004. DOMNING, D. P. Commercial exploitation of Manatee Trichechus in Brazil, c. 1785-1973. Biological Conservation, v. 22, n. 2, p. 101-126, 1982. DOMNING, D. P.; HAYEK, L.-A. C. Interspecific and intraspecific morphological variation in manatees (Sirenia: Trichechus). Marine Mammal Science, v. 2, n. 2, p. 87-144, 1986. DRAISCI, R.; PALLESCHI, L.; FERRETTI, E.; LUCENTINI, L.; CAMMARATA, P. Quantitation of anabolic hormones and their metabolites in bovine serum and urine by liquid chromatography-tandem mass spectrometry. Journal of Chromatography A, v. 870, n. 1-2, p. 511-522, 2000. FERREIRA, A. R. Memoria sobre o peixe-boy e do uso que lhe dão no estado do Grão Pará. Archivos do Museu Nacional do Rio de Janeiro, v. 12, n. 1, p. 169-174, 1903. ______ Viagem filosófica pelas capitanias do Grão Pará, Rio Negro, Mato Grosso e Cuiabá: memórias, zoologia e botânica. 2. ed. Rio de Janeiro: Conselho Federal de Cultura, 1972. 246 p. FOLEY, W. J.; ENGELHARDT, W. V.; CHARLES-DOMINIQUE, P. The digesta passage, particle size, and in vitro fermentation rate in the three-toed sloth Bradypus tridactylus (Edentata: Bradypodidae). Journal of Zoology, v. 236, n. 1, p. 681-696, 1995.

75
FORDE, M. D.; KOKA, S.; ECKERT, S. E.; CARR, A. B.; WONG, D. T. Systemic assessments utilizing saliva: part 1 - general considerations and current assessments. International Journal of Prosthodontics, v. 19, n. 1, p. 43-52, 2006. FRANCIS-FLOYD, R.; WHITE, J. R.; CHEN, C. L.; CARDEILHAC, P. T.; CICHRA, C. E. Serum progesterone and estradiol concentrations in captive manatees Trichechus manatus. Journal of Aquatic Animal Health, v. 3, n. p. 70-73, 1991. FRASER, C. M.; BERGERON, J. A.; MAYS, A.; AIELLO, S. E. Manual merk de veterinária: um manual de diagnóstico, tratamento, prevenção e controle de doenças para o veterinário. 7. ed. São Paulo: Roca, 1996. 2169 p. GALLIVAN, G. J.; BEST, R. C. Metabolism and respiration of the Amazonian manatee (Trichechus inunguis). Physiological Zoology, v. 53, n. 3, p. 245-253, 1980. GANSWINDT, A.; HEISTERMANN, M.; BORRAGAN, S.; HODGES, J. K. Assessment of testicular endocrine function in captive African elephants by measurement of urinary and fecal androgens. Zoo Biology, v. 21, n. 1, p. 27-36, 2002. GILMORE, D. P.; PERES DA COSTA, C.; VALENÇA, M.; DUARTE, D. P. F.; WILSON, C. A.; GRAY, C. E. Effects of exogenous LHRH on plasma LH and sex steroid levels in the three-toed sloth Bradypus tridactylus. Medical Science Research, v. 19, n. 1, p. 333-335, 1991. GRAHAM, L. H. Non-invasive monitoring of reproduction in zoo and wildlife species. Annual Review of Biomedical Sciences, v. 6, n. 1, p. 91-98, 2004. GRAHAM, L. H.; SCHWARZENBERGER, F.; MÖSTL, E.; GALAMA, W.; SAVAGE, A. A versatile enzyme immunoassay for the determination of progestogens in feces and serum. Zoo Biology, v. 20, n. 3, p. 227-236, 2001. GRANGER, D. A.; SCHWARTZ, E. B.; BOOTH, A.; ARENTZ, M. Salivary testosterone determination in studies of child health and development. Hormones and Behavior, v. 35, n. 1, p. 18-27, 1999. GRANGER, D. A.; SHIRTCLIFF, E. A.; BOOTH, A.; KIVLIGHAN, K. T.; SCHWARTZ, E. B. The "trouble" with salivary testosterone. Psychoneuroendocrinology, v. 29, n. 10, p. 1229-1240, 2004.

76
GUIMARÃES, M. A. B. V. The importance of the application of non-invasive techniques in the study of wild animal reproduction. Annual Review of Biomedical Sciences, v. 5, n. 1, p. 29-37, 2003. HAGEY, L. R.; CZEKALA, N. M. Comparative urinary androstanes in the great apes. General and Comparative Endocrinology, v. 130, n. 1, p. 64-69, 2003. HALPERN, C. T.; UDRY, J. R.; SUCHINDRAN, C. Monthly measures of salivary testosterone predict sexual activity in adolescent males. Archives of Sexual Behavior, v. 27, n. 5, p. 445-465, 1998. HEATON, H. C. The discovery of the Amazon according to the account of Friar Gaspar de Carvajal and others documents. New York: AMS Press, 1970. 467 p. HEISTERMANN, M.; AGIL, M.; BÜTHE, A.; HODGES, J. K. Metabolism and excretion of oestradiol-17b and progesterone in the Sumatran rhinoceros (Dicerorhinus sumatrensis). Animal Reproduction Science, v. 53, n. 1-4, p. 157-172, 1998. HERBERT, C. A.; TRIGG, T. E.; RENFREE, M. B.; SHAW, G.; ECKERY, D. C.; COOPER, D. W. Effects of a gonadotropin-releasing hormone agonist implant on reproduction in a male marsupial, Macropus eugenii. Biology of Reproduction, v. 70, n. 6, p. 1836-1842, 2004. HINDLE, J. E.; HODGES, J. K. Metabolism of oestradiol-17b and progesterone in the white rhinoceros (Ceratotherium simum simum). Journal of Reproduction and Fertility, v. 90, n. 2, p. 571-580, 1990. HIRSCHENHAUSER, K.; MÖSTL, E.; PÉCZELY, P.; WALLNER, B.; DITTAMI, J.; KOTRSCHAL, K. Seasonal relationships between plasma and fecal testosterone in response to GnRH in domestic ganders. General and Comparative Endocrinology, v. 118, n. 2, p. 262-272, 2000. HOGG, C. J.; VICKERS, E. R.; ROGERS, T. L. Determination of testosterone in saliva and blow of bottlenose dolphins (Tursiops truncatus) using liquid chromatography-mass spectrometry. Journal of Chromatography B: Analytical Technologies in the Biomedical and Life Sciences, v. 814, n. 2, p. 339-346, 2005.

77
HORVÁTH, Z.; ÚJVÁRY, D.; MIKLÓSI, Á. Dominance and social stress in the gray wolf (Canis lupus): an analysis of testosterone and cortisol levels in saliva from a captive pack. In: INTERNATIONAL ZOO AND WILDLIFE RESEARCH CONFERENCE ON BEHAVIOUR, PHYSIOLOGY AND GENETICS, 6, 2007, Berlin. Contributions... Berlin: Leibniz Institute for Zoo and Wildlife Research - IZW, 2007. p. 109. HUNT, K. E.; ROLLAND, R. M.; KRAUS, S. D.; WASSER, S. K. Analysis of fecal glucocorticoids in the North Atlantic right whale (Eubalaena glacialis). General and Comparative Endocrinology, v. 148, n. 2, p. 260-272, 2006. HUSAR, S. L. Trichechus inunguis. Mammalian Species, v. 72, n. 1, p. 1-4, 1977. IBAMA Mamíferos aquáticos do Brasil: plano de ação, versão II. 2. ed. Brasília: Instituto Brasileiro do Meio ambiente e dos Recursos Naturais Renováveis, 2001. 102 p. ITAVO, R. V.; ROSAS, F. C. W.; CAVALLANTE, A. P. Tempo de passagem do alimento no trato digestivo do peixe-boi da Amazônia, Trichechus inunguis, em cativeiro. In: REUNIÓN DE TRABAJO DE ESPECIALISTAS EN MAMÍFEROS ACUÁTICOS DE AMÉRICA DEL SUR & CONGRESSO SOLAMAC, 7., 1996, Viña del Mar. Resumenes... Viña del Mar: Sociedad Latinoamericana de Especialistas en Mamíferos Acuáticos - SOLAMAC, 1996. p. 105. IUCN. 2006 IUCN Red List of Threatened Species. Gland: The World Conservation Union, 2006. Disponível em: < http://www.iucn.org >. Acesso em: 15 ago. 2007. JUHÁSZ, J.; NAGY, P.; KULCSÁR, M.; HUSZENICZA, G. Methods for semen and endocrinological evaluation of the stallion: A review. Acta Veterinaria Brno, v. 69, n. 4, p. 247-259, 2000. KRETZSCHMAR, P.; GANSLOßER, U.; DEHNHARD, M. Relationship between androgens, environmental factors and reproductive behavior in male white rhinoceros (Ceratotherium simum simum). Hormones and Behavior, v. 45, n. 1, p. 1-9, 2004. LANYON, J. M.; MARSH, H. Digesta passage times in the dugong. Australian Journal of Zoology, v. 43, n. 2, p. 119-127, 1995. LANYON, J. M.; SMITH, K. M.; CARRICK, F. N. Reproductive steroids are detectable in the faeces of dugongs. The Australian Zoologist, v. 33, n. 2, p. 247-250, 2005.

78
LARKIN, I. L. V. Reproductive endocrinology of the Florida manatee (Trichechus manatus latirostris): estrous cycles, seasonal patterns and behavior. 2000. 354 f. Dissertation (Philosophy Doctor) - Department of Physiological Sciences, University of Florida, Gainesville, 2000. LARKIN, I. L. V.; FOWLER, V. F.; REEP, R. L. Digesta passage rates in the Florida manatee (Trichechus manatus latirostris). Zoo Biology, v. 26, n. 6, p. 503-515, 2007. LARKIN, I. L. V.; GROSS, T. S.; REEP, R. L. Use of faecal testosterone concentrations to monitor male Florida manatee (Trichechus manatus latirostris) reproductive status. Aquatic Mammals, v. 31, n. 1, p. 52-61, 2005. LASLEY, B. L. Methods for evaluating reproductive function in exotic species. Advances in Veterinary Science and Comparative Medicine, v. 30, n. 1, p. 209-228, 1985. LIMA, A. B. F. Avaliação endocrinológica da reprodução de muriquis do sul em cativeiro (Brachyteles arachnoides - E. GEOFFROY, 19806) por meio de dosagem de metabólitos de esteróides fecais. 2006. 104 f. Dissertação (Mestrado em Medicina Veterinária (Reprodução Animal)) - Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2006. LIMA, D. S.; VERGARA-PARENTE, J.-E.; YOUNG, R.-J.; PASZKIEWICZ, E. Training of Antillean manatee Trichechus manatus manatus (Linnaeus, 1758) as a management technique for individual welfare. The Latin American Journal of Aquatic Mammals, v. 4, n. 1, p. 61-68, 2005. LOMOLINO, M. V.; EWEL, K. C. Digestive efficiencies of the West Indian manatee (Trichechus manatus). Biological Sciences, v. 3, n. 1, p. 176-179, 1984. MARMONTEL, M.; ODELL, D. K.; REYNOLDS III, J. E. Reproductive biology of South American manatees. In: HAMLETT, W. C. Reproductive biology of South American vertebrates. 1. ed. New York: Springer-Verlag, 1992. p. 295-312. MARSHALL, A. J.; HOHMANN, G. Urinary testosterone levels of wild male bonobos (Pan paniscus) in the Lomako Forest, Democratic Republic of Congo. American Journal of Primatology, v. 65, n. 1, p. 87-92, 2005. MARSHALL, C. D.; MAEDA, H.; IWATA, M.; FURUTA, M.; ASANO, S.; ROSAS, F.; REEP, R. L. Orofacial morphology and feeding behaviour of the dugong, Amazonian,

79
West African and Antillean manatees (Mammalia: Sirenia): Functional morphology of the muscular-vibrissal complex. Journal of Zoology, v. 259, n. 3, p. 245-260, 2003. MATHERS, W. D.; STOVALL, D.; LANE, J. A.; ZIMMERMAN, M. B.; JOHNSON, S. Menopause and tear function: the influence of prolactin and sex hormones on human tear production. Cornea, v. 17, n. 4, p. 353-358, 1998. MENDES, A. As pescarías amazônicas e a piscicultura no Brasil: notas e sugestões. 1. ed. São Paulo: Livraria e Editora Record, 1958. 181 p. MÖHLE, U.; HEISTERMANN, M.; PALME, R.; HODGES, J. K. Characterization of urinary and fecal metabolites of testosterone and their measurement for assessing gonadal endocrine function in male nonhuman primates. General and Comparative Endocrinology, v. 129, n. 3, p. 135-145, 2002. MONFORT, S. L.; HARVEY, E.; GEURTS, L.; PADILLA, L.; SIMMONS, H. A.; WILLIAMSON, L. R.; WILDT, D. E. Urinary 3a,17b-androstanediol glucuronide is a measure of androgenic status in Eld's deer stags (Cervus eldi thamin). Biology of Reproduction, v. 53, n. 3, p. 700-706, 1995. MONFORT, S. L.; WASSER, S. K.; MASHBURN, K. L.; BURKE, M.; BREWER, B. A.; CREEL, S. R. Steroid metabolism and validation of noninvasive endocrine monitoring in the African wild dog (Lycaon pictus). Zoo Biology, v. 16, n. 6, p. 533-548, 1997. MOORMAN, E. A.; MENDOZA, S. P.; SHIDELER, S. E.; LASLEY, B. L. Excretion and measurement of estradiol and progesterone metabolites in the feces and urine of female squirrel monkeys (Saimiri sciureus). American Journal of Primatology, v. 57, n. 2, p. 79-90, 2002. MORATO, R. G.; BUENO, M. G.; MALMHEISTER, P.; VERRESCHI, I. T. N.; BARNABE, R. C. Changes in the fecal concentrations of cortisol and androgen metabolites in captive male jaguars (Panthera onca) in response to stress. Brazilian Journal of Medical and Biological Research, v. 37, n. 12, p. 1903-1907, 2004a. MORATO, R. G.; VERRESCHI, I. T. N.; GUIMARÃES, M. A. B. V.; CASSARO, K.; PESSUTI, C.; BARNABE, R. C. Seasonal variation in the endocrine-testicular function of captive jaguars (Panthera onca). Theriogenology, v. 61, n. 7-8, p. 1273-1281, 2004b. NASCIMENTO, C. C. Avaliação da função reprodutiva de fêmeas de peixe-boi da Amazônia (Trichechus inunguis, Natterer, 1883), mantidas em cativeiro, por

80
meio da extração e dosagem de esteróides fecais. 2004. 113 f. Dissertação (Mestrado em Medicina Veterinária (Reprodução Animal)) - Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2004. NASCIMENTO, C. C.; OLIVEIRA, C. A.; DA SILVA, V. M. F.; D'AFFONSÊCA NETO, J. A. Estimativa do período de gestação em fêmeas de peixe-boi da Amazônia (Trichechus inunguis), mantidas em cativeiro, com base nos níveis plasmáticos de progesterona. In: REUNIÓN DE TRABAJO DE ESPECIALISTAS EN MAMÍFEROS ACUÁTICOS DE AMÉRICA DEL SUR & CONGRESSO SOLAMAC, 10., 2002, Valdivia. Libro de Resúmenes... Valdivia: Sociedad Latinoamericana de Especialistas en Mamíferos Acuáticos - SOLAMAC, 2002. p. 41-42. NEAVE, N.; LAING, S.; FINK, B.; MANNING, J. T. Second to fourth digit ratio, testosterone and perceived male dominance. Proceedings of the Royal Society of London - Series B, Biological Sciences, v. 270, n. 1529, p. 2167-2172, 2003. NORMAN, A. W.; LITWACK, G. Hormones. 2. ed. San Diego: Academic Press, 1997. 558 p. NORRIS, D. O. Vertebrate endocrinology. 3. ed. San Diego: Academic Press, 1997. 634 p. NOZAKI, O. Steroid analysis for medical diagnosis. Journal of Chromatography A, v. 935, n. 1-2, p. 267-278, 2001. OOSTHUIZEN, M. K.; BENNETT, N. C. LH responses to single doses of exogenous GnRH in the Cape mole rat (Georychus capensis): the pituitary potential for opportunistic breeding. Journal of Zoology, v. 271, n. 1, p. 198-202, 2007. PALME, R. Measuring fecal steroids: Guidelines for practical application. Annals of the New York Academy of Sciences, v. 1046, n. 1, p. 75-80, 2005. PALME, R.; FISCHER, P.; SCHILDORFER, H.; ISMAIL, M. N. Excretion of infused 14C-steroid hormones via faeces and urine in domestic livestock. Animal Reproduction Science, v. 43, n. 1, p. 43-63, 1996. PALME, R.; RETTENBACHER, S.; TOUMA, C.; EL-BAHR, S. M.; MÖSTL, E. Stress hormones in mammals and birds: Comparative aspects regarding metabolism, excretion, and noninvasive measurement in fecal samples. Annals of the New York Academy of Sciences, v. 1040, n. 1, p. 162-171, 2005.

81
PANTOJA, T. M. A. Análise física e química da urina de Trichechus inunguis (Mammalia, Sirenia): valores-referência para o peixe-boi da Amazônia. 2004. 125 f. Dissertação (Mestrado em Ciências Biológicas (Biologia de Água Doce e Pesca Interior)) - Instituto Nacional de Pesquisas da Amazônia / Universidade Federal do Amazonas, Manaus, 2004. PEDERNERA-ROMANO, C.; VALDEZ, R. A.; SINGH, S.; CHIAPPA, X.; ROMANO, M. C.; GALINDO, F. Salivary cortisol in captive dolphins (Tursiops truncatus): A non-invasive technique. Animal Welfare, v. 15, n. 4, p. 359-362, 2006. PEREIRA, M. N. O peixe-boi da Amazônia. Boletim do Ministério da Agricultura, v. 33, n. 5, p. 21-95, 1944. PIETRASZEK, J.; ATKINSON, S. Concentrations of estrone sulfate and progesterone in plasma and saliva, vaginal cytology, and bioelectric impedance during the estrous cycle of the Hawaiian monk seal (Monachus schauinslandi). Marine Mammal Science, v. 10, n. 4, p. 430-441, 1994. PIMENTEL, G. P. Determinação da testosterona presente nas fezes do peixe-boi da Amazônia Trichechus inunguis (Sirenia: Trichechidae), utilizando a técnica de radioimunoensaio. 1998. 93 f. Dissertação (Mestrado em Ciências (Oceanorafia Biológica)) - Departamento de Oceanografia, Universidade Federal de Pernambuco, Recife, 1998. POST, T. B.; REICH, M. M.; BINDON, B. M. Characterization of LH and testosterone responses to intramuscular injection of GnRH in tropical postpubertal bulls. Theriogenology, v. 27, n. 2, p. 305-315, 1987. REEP, R. L.; MARSHALL, C. D.; STOLL, M. L. Tactile hairs on the postcranial body in Florida manatees: a Mammalian lateral line? Brain Behavior and Evolution, v. 59, n. 3, p. 141-154, 2002. RIAD-FAHMY, D.; READ, G. F.; WALKER, R. F.; GRIFFITHS, K. Steroids in saliva for assessing endocrine function. Endocrine Reviews, v. 3, n. 4, p. 367-395, 1982. RILLING, J. K.; WORTHMAN, C. M.; CAMPBELL, B. C.; STALLINGS, J. F.; MBIZVA, M. Ratios of plasma and salivary testosterone throughout puberty: Production versus bioavailability. Steroids, v. 61, n. 6, p. 374-378, 1996.

82
ROBECK, T. R.; STEINMAN, K. J.; GEARHART, S.; REIDARSON, T. R.; MCBAIN, J. F.; MONFORT, S. L. Reproductive physiology and development of artificial insemination technology in killer whales (Orcinus orca). Biology of Reproduction, v. 71, n. 2, p. 650-660, 2004. ROBECK, T. R.; STEINMAN, K. J.; YOSHIOKA, M.; JENSEN, E.; O'BRIEN, J. K.; KATSUMATA, E.; GILI, C.; MCBAIN, J. F.; SWEENEY, J.; MONFORT, S. L. Estrous cycle characterisation and artificial insemination using frozen-thawed spermatozoa in the bottlenose dolphin (Tursiops truncatus). Reproduction, v. 129, n. 5, p. 659-674, 2005. RODRIGUES, F. R. Características anatômicas e histológicas do aparelho reprodutor feminino de Trichechus inunguis (Natterer, 1883) (Mammalia: Sirenia). 2002. 114 f. Dissertação (Mestrado em Ciências Biológicas (Biologia de Água Doce e Pesca Interior)) - Instituto Nacional de Pesquisas da Amazônia / Universidade do Amazonas, Manaus, 2002. RODRIGUES, F. R.; DA SILVA, V. M. F.; MARQUES, J. F.; LAZZARINI, S. M. Evidence of infertile estrus cycles before the first conception on a female Amazonian manatee (Trichechus inunguis) kept in captivity at the Aquatic Mammals Laboratory, Amazon, Brazil. In: BIENNIAL CONFERENCE ON THE BIOLOGY OF MARINE MAMMALS, 15., 2003, Greensboro. Abstracts... Greensboro: Society of Marine Mammalogy, 2003. p. 138. ROLLAND, R. M.; HAMILTON, P. K.; KRAUS, S. D.; DAVENPORT, B.; GILLETT, R. M.; WASSER, S. K. Faecal sampling using detection dogs to study reproduction and health in North Atlantic right whales (Eubalaena glacialis). Journal of Cetacean Research and Management, v. 8, n. 2, p. 121-125, 2006. ROLLAND, R. M.; HUNT, K. E.; KRAUS, S. D.; WASSER, S. K. Assessing reproductive status of right whales (Eubalaena glacialis) using fecal hormone metabolites. General and Comparative Endocrinology, v. 142, n. 3, p. 308-317, 2005. ROSAS, F. C. W. Biology, conservation and status of the Amazonian manatee Trichechus inunguis. Mammal Review, v. 24, n. 2, p. 49-59, 1994. ROSAS, F. C. W.; PIMENTEL, T. L. Order Sirenia (manatees, dugongs and sea cows). In: FOWLER, M. E.; CUBAS, Z. S. Biology, medicine, and surgery of South American wild animals. 1. ed. Ames: Iowa State University Press, 2001. p. 352-362.

83
SCHULTHEISS, O. C.; WIRTH, M. M.; STANTON, S. J. Effects of affiliation and power motivation arousal on salivary progesterone and testosterone. Hormones and Behavior, v. 46, n. 5, p. 592-599, 2004. SCHWARZENBERGER, F. The many uses of non-invasive faecal steroid monitoring in zoo and wildlife species. International Zoo Yearbook, v. 41, n. 1, p. 52-74, 2007. SCHWARZENBERGER, F.; MÖSTL, E.; PALME, R.; BAMBERG, E. Faecal steroid analysis for non-invasive monitoring of reproductive status in farm, wild and zoo animals. Animal Reproduction Science, v. 42, n. 1-4, p. 515-526, 1996. SENGER, P. L. Pathways to pregnancy and parturition. 2. ed. Pullman: Current Conceptions, 2005. 368 p. SPINKS, A. C.; BENNETT, N. C.; FAULKES, C. G.; JARVIS, J. U. M. Circulating LH levels and the response to exogenous GnRH in the common mole-rat: Implications for reproductive regulation in this social, seasonal breeding species. Hormones and Behavior, v. 37, n. 3, p. 221-228, 2000. SPINOSA, H. S.; GORNIAK, S. L.; BERNARDI, M. M. Farmacologia aplicada à medicina veterinária 4. ed. Rio de Janeiro: Guanabara Koogan, 2006. 897 p. SULLIVAN, D. A.; BLOCH, K. J.; ALLANSMITH, M. R. Hormonal influence on the secretory immune system of the eye: androgen regulation of secretory component levels in rat tears. The Journal of Immunology, v. 132, n. 3, p. 1130-1135, 1984. TAUSSKY, H. H. A microcolorimetric determination of creatinine in urine by the Jaffe reaction. The Journal of Biological Chemistry, v. 208, n. 1, p. 853-851, 1954. THEODOROU, J.; ATKINSON, S. Monitoring total androgen concentrations in saliva from captive Hawaiian monk seals (Monachus Schauinslandi). Marine Mammal Science, v. 14, n. 2, p. 304-310, 1998. TOUMA, C.; PALME, R. Measuring fecal glucocorticoid metabolites in mammals and birds: The importance of validation. Annals of the New York Academy of Sciences, v. 1046, n. 1, p. 54-74, 2005. TOUMA, C.; SACHSER, N.; MÖSTL, E.; PALME, R. Effects of sex and time of day on metabolism and excretion of corticosterone in urine and feces of mice. General and Comparative Endocrinology, v. 130, n. 3, p. 267-278, 2003.

84
VELLOSO, A. L.; WASSER, S. K.; MONFORT, S. L.; DIETZ, J. M. Longitudinal fecal steroid excretion in maned wolves (Chrysocyon brachyurus). General and Comparative Endocrinology, v. 112, n. 1, p. 96-107, 1998. VERÍSSIMO, J. A pesca na Amazônia. 1. ed. Belém: Universidade Federal do Pará, 1970. 130 p. VINCENT, I. C.; MICHELL, A. R. Comparison of cortisol concentrations in saliva and plasma of dogs. Research in Veterinary Science, v. 53, n. 3, p. 342-345, 1992. VINING, R. F.; MCGINLEY, R. A.; SYMONS, R. G. Hormones in saliva: Mode of entry and consequent implications for clinical interpretation. Clinical Chemistry, v. 29, n. 10, p. 1752-1756, 1983. WAKAI, Y.; HASEGAWA, K.; SAKAMOTO, S.; ASANO, S.; WATANABE, G.; TAYA, K. Annual changes of urinary progesterone and estradiol-17b of the dugong (Dugong dugon) in captivity. Zoological Science, v. 19, n. 6, p. 679-682, 2002. WALKER, L. A.; CORNELL, L.; DAHL, K. D.; CZEKALA, N. M.; DARGEN, C. M.; JOSEPH, B.; HSUEH, A. J.; LASLEY, B. L. Urinary concentrations of ovarian steroid hormone metabolites and bioactive follicle-stimulating hormone in killer whales (Orcinus orchus) during ovarian cycles and pregnancy. Biology of Reproduction, v. 39, n. 5, p. 1013-1020, 1988. WARNER, A. C. I. Rate of passage digesta through the gut of mammals and birds. Nutritional Abstracts Review Series B, v. 51, n. 1, p. 789-820, 1981. WASSER, S. K.; PAPAGEORGE, S.; FOLEY, C.; BROWN, J. L. Excretory fate of estradiol and progesterone in the African elephant (Loxodonta africana) and patterns of fecal steroid concentrations throughout the estrous cycle. General and Comparative Endocrinology, v. 102, n. 2, p. 255-262, 1996. WHEELER, M. J. Measurement of androgens. Methods in molecular biology (Clifton, N.J.), v. 324, n. 1, p. 197-211, 2006. WHITTEN, P. L.; BROCKMAN, D. K.; STAVISKY, R. C. Recent advances in noninvasive techniques to monitor hormone-behavior interactions. Yearbook of Physical Anthropology, v. 41, n. 1, p. 1-23, 1998.

85
WILLIAMS, T. M.; KIND, A. J.; HYDE, W. G.; HILL, D. W. Characterization of urinary metabolites of testosterone, methyltestosterone, mibolerone and boldenone in greyhound dogs. Journal of Veterinary Pharmacology and Therapeutics, v. 23, n. 3, p. 121-129, 2000. YOUNG, N. M.; DAWSON, W. W. The ocular secretions of the bottlenose dolphin Tursiops truncatus. Marine Mammal Science, v. 8, n. 1, p. 57-68, 1992.


















