Bioacústica e sítio de vocalização em taxocenoses de anuros de ...

i
Maria Silva Cunha
SELEÇÃO DE SÍTIO DE VOCALIZAÇÃO PELO ANFÍBIO
BROMELÍGENO Phyllodytes melanomystax
(ANURA, HYLIDAE)
Programa de Pós-Graduação em Ecologia e Biomonitoramento Universidade Federal da Bahia
Salvador, 2011

ii
Salvador, 2011 UNIVERSIDADE FEDERAL DA BAHIA
INSTITUTO DE BIOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA E
BIOMONITORAMENTO
SELEÇÃO DE SÍTIO DE VOCALIZAÇÃO PELO ANFÍBIO
BROMELÍGENO Phyllodytes melanomystax (ANURA, HYLIDAE)
Maria Silva Cunha
Dissertação apresentada ao Programa
de Pós-Graduação em Ecologia e
Biomonitoramento do Instituto de
Biologia da Universidade Federal da
Bahia, sob orientação do Prof. Dr.
Marcelo Felgueiras Napoli, como pré-
requisito para a obtenção do título de
Mestre em Ecologia e
Biomonitoramento.
Salvador
2011

iii
Ficha Catalográfica
Cunha, Maria Silva. Seleção de sítio de vocalização pelo anfíbio bromelígeno Phyllodytes melanomystax (Anura, Hylidae) / Maria Silva Cunha. - 2011. 69 f. : il.
Orientador: Prof. Dr. Marcelo Felgueiras Napoli. Dissertação (mestrado) - Universidade Federal da Bahia, Instituto de Biologia, Salvador, 2011.
1. Anfíbio. 2. Perereca. 3. Bromélia. 4. Ecologia. I. Napoli, Marcelo Felgueiras. II. Universidade Federal da Bahia. Instituto de Biologia. III. Título.
CDD - 597.8 CDU - 597.8

iv
Maria Silva Cunha
Seleção de sítio de vocalização pelo anfíbio bromelígeno Phyllodytes
melanomystax (Anura, Hylidae)
Dissertação apresentada ao Programa de Pós-Graduação em Ecologia e
Biomonitoramentto do Instituto de Biologia da Universidade Federal da Bahia como
partes dos requisitos necessários para a obtenção do título de Mestre em Ecologia e
Biomonitoramento.
Aprovada por:
Prof. Dr. Marcelo Felgueiras Napoli Orientador
Prof.ª Dr.ª Flora Acuña Juncá Universidade Estadual de Feira de Santana
Prof. Dr. Mirco Solé Universidade Estadual de Santa Cruz

v
Dedicatória
À minha mamãe, sempre!

vi
Epígrafe
(...) de qualquer forma, eu gostaria de saber
se alguma vez aparecerei em histórias e
canções (...), quero dizer, ser posta em
palavras, sabe, contadas ao pé da lareira ou
lida de um grande livro grosso com letras
douradas, anos e anos depois. E as pessoas
dirão “Vamos ouvir a história de Maria e
dos Phyllodytes”! E elas dirão “Essa é uma
das minhas preferidas”.
J. R. R. Tolkien, O senhor dos anéis
(adaptado).

vii
Agradecimentos
Ao meu bom Deus, que me guarda a cada segundo, ilumina e protege os meus
passos por onde quer que eu passe. A ti, Senhor, toda a glória.
À, minha mamãe, Adenita, por ser esta figura única na minha vida, que tantas
vezes esqueceu-se de si para me proporcionar o melhor e que faz de tudo pela filhota
aqui. Ao meu pai, Antônio (in memorian), sou grata por ajudar tanto na formação do
que eu sou hoje, mesmo estando ausente. Às minhas tias “Anas” e ao meu tio/pai
Adelson, por mais uma vez acompanharem e apoiarem as minhas decisões e pela boa
educação que sempre me deram. Hoje compreendo a importância de vocês apagarem
toda a palavra quando eu errava apenas a última letra no dever de casa do primário! Aos
meus avós, às minhas primas Brunna e Mariana e ao meu afilhado Bruno, obrigada
por existirem na minha vida.
À Mariza, Marcely e toda sua família, por terem me acolhido em Salvador
como se eu fosse um deles, pela confiança e cuidado que só na minha própria família eu
encontraria. Muito obrigada!
Ao meu namorado lindo, Élton, por ser tão fofo! Por ser sempre uma companhia
maravilhosa, pelo carinho e por cuidar tão bem de mim. E a toda a família Milanês, em
especial à minha sogra Ozelita.
Ao meu orientador, Marcelo, pela oportunidade, confiança e experiência
passada. Obrigada por me ensinar tanto sobre o curioso mundo dos sapos e por ser um
exemplo de dedicação à ciência.
Ao Programa de Pós-Graduação em Ecologia e Biomonitoramento, por me
mostrar o que é Ecologia de verdade. Obrigada a cada professor que contribuiu tão
fortemente para minha formação profissional e que, através de suas disciplinas,
colaborou para a qualidade deste trabalho. E à Jussara, que facilitou a minha vida
burocrática na pós-graduação.
À Fundação de Amparo à Pesquisa do Estado da Bahia (FAPESB), que me
proporcionou a bolsa de mestrado durante os meus dois anos de curso.

viii
Ao Sr. Alberto Bolzico, por ter permitido a realização da coleta de dados na
área da Fazenda Praia do Forte. Ao Samuel, por ajudar alguns procedimentos logísticos
do campo e pela disponibilidade em cooperar conosco. E ao Seu Anísio! Que nos
conduziu tão bem pelas areias escaldantes da restinga de Praia do Forte e que, no final,
já estava até coletando os Phyllodytes conosco!
Aos meus super ajudantes de campo, que se revezaram durante as semanas de
coleta com toda a boa vontade do mundo! Obrigada a Rafa, Júnior, Lucas, Vanessa
Bonfim (VaiNessa!), Joice Ruggeri, Manu, Joice Herrera, Daniele, Jucilene,
Ariane, Thiago Filadelfo e Thaís (Kuen).
A Rafael e Fila pelas fotos maravilhosas e a Lucas pela ótima ilustração da
bromélia.
Aos meus super amigos da “Liga da UESC”, Euvaldo Júnior, Marlla (sem
alça) e Camila “Pitty”, por tornarem meus dias na UFBA mais familiares, por assim
dizer. Agradeço ao Júnior por me transmitir o vírus da herpetologia, à Marlla por ser tão
“Marlla” e à Pitty por ter se transformado de uma simples colega da graduação a uma
das minhas melhores amigas. Ao amigo Gilson Ximenes, por ser um exemplo de
professor e de pessoa, pela sua humildade e simplicidade e por não nos abandonar,
mesmo nos intitulando de “pragas”! E aos meus demais amigos que, mesmo não
estando presentes no meu dia-a-dia, torceram muito para que desse tudo certo. Amo
muito vocês!
Aos meus colegas da turma 2009.1 do PPGEcoBio, pelas tantas risadas e pelas
contribuições ao meu trabalho.
Agradeço imensamente ao povo do Laboratório de Taxonomia e História
Natural de Anfíbios (AMPHIBIA), por terem me recebido da melhor forma possível!
Eu que cheguei meio tímida e que aos poucos fui me deixando levar por essas pessoas
maravilhosas! São essas figuras pitorescas e tão especiais: Ariane Xavier (Aribio – a
alça), Thaís Dória (Kuen), Deise Cruz (“Valeu Neymar”!), Patrícia Fonseca (Pati –
Amuu!), Emanuela Petersen (Manuuuu!), Milena Camardelli (“Se eu pudesse eu
matava Mil, Mil Mil, ê iê!), Rafael Abreu (que gosta de biscoito de morango e faz
biscuit...), Joice Ruggeri (Cota), Lucas Menezes (Loucas), Laís Encarnação (Lai),

ix
Euvaldo Jr. (Juninho da Baêa, pai!), Joice Herrera (Joicebunda), Tamiris, (Têimiris)
e aos novatos Camila e Robson. E não posso esquecer do nosso mais ilustre anexo,
Tiago Porto (Tiko). Não dá para contar quantas resenhas surgiram, de quantas idiotices
de Rafa nós rimos, de quantas alucinações de Kuen, Pati e/ou Manu nós tivemos crises
de gargalhadas, mas dá pra saber que vocês foram essenciais nestes dois anos de
convivência.
E, finalmente, aos Phyllodytes (tão fofinhos), que permitiram que eu conhecesse
um pedacinho do curioso mundo em que vivem.

x
Sumário
Texto para divulgação 1
Introdução Geral 2
Referências 8
Figura 1 13
Figura 2 14
Artigo. Revista Journal of Animal Ecology 15
Título do Artigo: Calling site selection by the bromeliad-dwelling treefrog
Phyllodytes melanomystax (Amphibia: Anura: Hylidae) in a coastal sand
dune habitat
16
Summary 17
Introduction 19
Material and Methods
Study area 22
Research Design and Field Methods 22
Data Analysis 25
Results
Microhabitat variables and natural history notes 27
Statistical analyses 28
Discussion 29
Acknowledgements 34
References 35
Figure captions 40
Figure 1 41
Figure 2 42
Figure 3 43
Table 1 44
Table 2 45
Table 3 46

xi
Table 4 47
Regras para publicação no Journal of Animal Ecology 48

1
Texto De Divulgação
Bromélias-tanque são plantas capazes de acumular água da chuva dentro de suas
rosetas. Tal capacidade permite a formação de pequenos corpos d‟água conhecidos como
fitotelmos. Esse micro-hábitat é usado por uma grande variedade de vertebrados e
invertebrados. Além disso, algumas espécies chegam a passam todo o seu ciclo de vida dentro
das bromélias, como as atuais onze espécies de pererecas do gênero Phyllodytes.
É sabido que anfíbios anuros, que utilizam as bromélias-tanque, as selecionam de forma
não aleatória. Entretanto, os fatores ambientais que direcionam essa seleção ainda não estão
claros. Na presente dissertação, foram investigadas as variáveis associadas à seleção de
bromélias por machos vocalizantes do anuro bromelígeno Phyllodytes melanomystax.
Ademais, foram feitas observações acerca da história natural dessa espécie e um experimento de
marcação e recaptura.
O estudo foi conduzido em um ambiente de restinga arbóreo-arbustiva, localizada no
município de Mata de São João, litoral norte do estado da Bahia, Brasil. Duzentas e duas
bromélias terrestres foram amostradas, 101 com e 101 sem machos vocalizantes de P.
melanomystax. Foram medidas nove variáveis ambientais que, de acordo com a literatura,
possivelmente influenciam a seleção de bromélias por anuros. Os dados foram analisados
através da análise de regressão logística múltipla e do teste de Wald.
Foi encontrada uma relação significativa entre a presença/ausência de machos de P.
melanomystax e duas das variáveis medidas: quantidade de detrito acumulado no tanque da
bromélia e número de bromélias em um raio de 2m. O experimento de marcação e recaptura
indicou que os machos possuem alta fidelidade ao sítio de vocalização. As observações de
história natural sugeriram que as bromélias escolhidas para vocalização são também utilizadas
para reprodução, oviposição e crescimento dos girinos.
A partir dos resultados, entendemos que P. melanomystax segue um dos padrões
já descritos para alguns invertebrados (aranhas e opiliões) que utilizam as bromélias-tanque.
Para esses animais já foi demonstrado que o detrito acumulado pela bromélia os impede de
utilizar o tanque de modo eficiente. Ao conjugarmos as observações de história natural ao fato
de que P. melanomystax parece evitar bromélias que estão isoladas espacialmente, assumimos
que a área de vida dos machos é restrita, e que o número de bromélias com potencial de serem
usadas pode ser uma variável importante na escolha dos machos pelas fêmeas.

2
Introdução Geral 1
Na história de vida de espécies que habitam locais sob condições variáveis, um 2
dos processos decisivos para sobrevivência e sucesso reprodutivo dos indivíduos é a 3
seleção de hábitat (Huey 1991, Rudolf & Rödel 2005, Brown et al. 2008a, Lin et al. 4
2008). As características do hábitat selecionado influenciam desde disponibilidade de 5
alimento e vulnerabilidade do adulto ao predador, até condições do hábitat para o 6
desenvolvimento e sobrevivência da prole (Resetarits & Wilbur 1989, Romero & 7
Vasconcellos-Neto 2005, Rudolf & Rödel 2005). Os indivíduos são capazes de avaliar 8
previamente o hábitat em questão e tal avaliação é mais crítica durante o período 9
reprodutivo, pois a prole geralmente é menos tolerante a condições físico-químicas 10
extremas e normalmente é incapaz de mudar para outro local caso o ambiente não 11
apresente condições favoráveis à sobrevivência (Huey 1991, Rudolf & Rödel 2005). As 12
características do hábitat que influenciam tal processo de seleção dependerão, entre 13
outros fatores, do tipo de hábitat utilizado e das necessidades intrínsecas a cada espécie, 14
as quais estão associadas aos seus aspectos reprodutivos, ecológicos e comportamentais. 15
Entre os anfíbios anuros, características do hábitat que parecem influenciar fortemente a 16
seleção de tais locais durante o período reprodutivo são temperatura e pH da água, além 17
de presença de predadores e densidade de coespecíficos (Bursik 1988, Resetarits & 18
Wilbur 1989, Gascon 1992, Oliveira & Navas 2004, Brown et al. 2008a,b, Lin et al. 19
2008). Entretanto, anfíbios anuros que habitam regiões tropicais apresentam um grande 20
número de modos reprodutivos, o que leva ao uso de micro-hábitats diversos por estes 21
animais (Haddad & Prado 2005) e, por conseguinte, pode levar a mudanças nas 22
características associadas à seleção do hábitat. 23
Anfíbios anuros que habitam fitotelmos enfrentam riscos e situações incomuns a 24
outros hábitats. O termo fitotelmo (do grego phytos = planta, telm = poça) foi designado 25

3
em 1928 pelo alemão Ludwig Varga, que estudava a fauna aquática associada a plantas 1
carnívoras e é definido como corpos d‟água acumulados em plantas ou partes de plantas 2
(Kitching 2000). Entre os exemplos mais comuns de fitotelmos estão água acumulada 3
em bromélias-tanque, colmos de bambu, cavidades de árvores e brácteas florais 4
(Kitching 2001, Lehtinen et al. 2004). Anuros que habitam fitotelmos têm que lidar 5
frequentemente com situações como a exposição à dessecação, a imprevisível 6
disponibilidade de alimento e baixas concentrações de oxigênio, sendo tais situações 7
mais críticas para o indivíduo em estágio larval (Lehtinen et al. 2004). Anuros 8
habitantes de fitotelmos apresentam uma série de estratégias comportamentais que lhes 9
permitem sobreviver nestes ambientes, como oofagia e cuidado parental (Schiesari et al. 10
2003, Lehtinen et al. 2004, Brown et al. 2008 a,b), além de outras fisiológicas, como 11
redução do aparato branquial para minimizar a perda de oxigênio para o meio (Lanoo et 12
al. 1987). 13
Entre os tipos de fitotelmo supracitados, as bromélias são conhecidas como 14
plantas-chave em ambientes tropicais onde, em muitas situações, representam o único 15
reservatório de água disponível no ambiente (Neill 1951). Devido à sua alta 16
adaptabilidade, a família Bromeliaceae, atualmente composta por mais de 2800 17
espécies, se distribuiu amplamente nos diversos ecossistemas do Novo Mundo, sob uma 18
grande variedade de formas e tamanhos (Smith & Downs 1979). O micro-hábitat 19
formado pelas bromélias-tanque, aquelas cuja estrutura permite o armazenamento de 20
água da chuva, é utilizado por muitos grupos de animais (Neill 1951, Laessle 1961, 21
Armbruster et al. 2002) e até mesmo por outras plantas que utilizam tal ambiente para a 22
germinação de suas sementes (Sampaio et al. 2005). A relação entre as bromélias e sua 23
fauna associada possui graus de dependência entre animal e planta, podendo esta 24
relação ser espécie-específica, como entre a aranha Psecas chapoda Peckham & 25

4
Peckham, 1894 e Bromelia balansae Mez, 1891 (Romero & Vasconcelos-Neto 2005) e 1
entre a perereca dourada de Trinidad Phytotriades auratus (Boulenger, 1917) e a 2
bromélia-tanque Glomeropitcairnia erectiflora Mez, 1905 (Jowers et al. 2008). 3
Sob o propósito de investigar a relação entre os anfíbios anuros e as bromélias 4
da Mata Atlântica, Peixoto (1995) estudou os tipos de associação entre anuros e 5
bromélias neste ecossistema, fornecendo definições que nortearam os trabalhos 6
subsequentes que versaram sobre anuros em bromélias no Brasil. De acordo com o 7
autor, os anuros associados às bromélias podem ser classificados em: 8
1) Bromelícolas – espécies que utilizam as bromélias como refúgio, mas que se 9
reproduzem em outros locais, não havendo dependência do anuro para com a 10
bromélia. Os bromelícolas podem ainda ser divididos em (1) eventuais, 11
quando a espécie é encontrada frequentemente em outros ambientes e (2) 12
habituais, quando, apesar de se reproduzir em outros locais, a espécie é mais 13
frequentemente encontrada em bromélias. 14
2) Bromelígenas – espécies cujo ciclo reprodutivo é dependente do ambiente 15
proporcionado pelas bromélias. Quanto ao tipo de desenvolvimento larvar, as 16
espécies foram categorizadas em espécies de desenvolvimento direto e 17
espécies de fase larvar livre. 18
Dentre as espécies bromelígenas que ocorrem no Brasil, as mais conhecidas são 19
as espécies do grupo de Scinax perpusillus (sensu Peixoto 1987) [S. alcatraz (Lutz, 20
1973); S. atratus (Peixoto, 1989); S. littoreus (Peixoto, 1988); S. melloi (Peixoto, 1989); 21
S. perpusillus (Lutz & Lutz, 1939); S. v-signatus (Lutz, 1968); S. peixotoi Brasileiro 22
Haddad, Sawaya & Martins, 2007; S. faivovichi Brasileiro, Oyamaguchi & Haddad, 23
2007; S. tupinamba Silva & Alves-Silva, 2008; S. belloni Faivovich, Gasparini & 24
Haddad, 2010], as espécies do gênero Flectonotus [F. fissilis (Miranda-Ribeiro, 1920); 25

5
F. fitzgeraldi (Parker, 1934); F. goeldii (Boulenger, 1895); F. ohausi (Wandolleck, 1
1907); F. pygmaeus (Boettger, 1893)], Gastrotheca fissipes (Boulenger, 1888), 2
Crossodactylodes izecksohni Peixoto, 1983 e todas as espécies do gênero Phyllodytes 3
Wagler, 1830 (Peixoto 1995, Caramaschi et al. 2004, Silva & Britto-Pereira 2006, 4
Alves-Silva & Silva 2009). 5
O gênero Phyllodytes é composto por pequenas pererecas bromelígenas com 6
fase larvar livre, distribuídas em diferentes fisionomias da Mata Atlântica. São onze as 7
espécies atualmente descritas de Phyllodytes: P. acuminatus Bokermann, 1966; P. 8
brevirostris Peixoto & Cruz, 1988; P. edelmoi Peixoto, Caramaschi & Freire, 2003; P. 9
gyrinaethes Peixoto, Caramaschi & Freire, 2003; P. kautskyi Peixoto & Cruz, 1988; P. 10
luteolus (Wied-Neuwied, 1824); P. maculosus Cruz, Feio & Cardoso, 2007; P. 11
melanomystax Caramaschi, Silva & Britto-Pereira, 1992; P. punctatus Caramaschi & 12
Peixoto, 2004; P. tuberculosus Bokermann, 1966 e P. wuchereri (Peters, 1873). As 13
espécies de Phyllodytes apresentam como principais características a presença de 14
odontóides bastante evidentes e o ciclo de vida altamente relacionado às bromélias 15
(Caramaschi et al. 2004). 16
Pouco se sabe sobre ecologia e comportamento das espécies de Phyllodytes. 17
Eterovick (1999) avaliou a seleção de bromélias por P. luteolus em um ambiente 18
modificado e concluiu que esta espécie seleciona as bromélias de acordo com suas 19
características físico-químicas em apenas uma das quatro espécies de bromélias 20
avaliadas no estudo. Entretanto, problemas de delineamento amostral (p.ex., pseudo-21
replicação espacial) podem ser identificados neste estudo e o fato de ter sido feito em 22
“jardim” de bromélias pode ter influenciado o resultado encontrado, como sugerido pela 23
própria autora. Giaretta (1996) investigou os aspectos reprodutivos de P. luteolus e 24
atentou para a baixa fecundidade das fêmeas desta espécie, além do pequeno tamanho 25

6
da desova, o menor das espécies de Hylidae. Este autor discutiu ainda que a baixa 1
abundância de machos por bromélia pode ser um indício de territorialidade. Entretanto, 2
existem casos em que há mais de um macho vocalizante de Phyllodytes por bromélia, 3
como em P. luteolus (Eterovick 1999), P. melanomystax e P. tuberculosus (Marcelo F. 4
Napoli comunicação pessoal). As demais publicações sobre espécies do gênero 5
Phyllodytes são de caráter taxonômico (p.ex., Bokermann 1966, Caramaschi et al. 1992, 6
Peixoto et al. 1999, Caramaschi & Peixoto 2004, Caramaschi et al. 2004, Simon & 7
Gasparini 2003, Jowers et al. 2008, Vieira et al. 2009), ou são estudos com fins diversos 8
[p.ex., Weygoldt (1981) descreveu parte da biologia reprodutiva de P. luteolus; Papp & 9
Papp (2000) relataram o declínio em população de P. luteolus após queimada na área de 10
estudo] e que não objetivaram investigar a relação entre determinada espécie de 11
Phyllodytes e bromélias. 12
Entretanto, alguns trabalhos que investigaram a relação entre espécies 13
bromelígenas de outros gêneros de anfíbios anuros e as bromélias habitadas geraram 14
informações consistentes. Oliveira & Navas (2004) investigaram a seleção de bromélias 15
por Scinax perpusillus e sugeriram que o tamanho da bromélia e o nível de agrupamento 16
da planta (medido através do número de bromélias em um raio de 2 metros da planta 17
amostrada) são características influentes na seleção de bromélias por Scinax perpusillus. 18
Ainda sobre o grupo de S. perpusillus, Alves-Silva & Silva (2009) estudaram o 19
comportamento reprodutivo de seis das 11 espécies do grupo e revelaram um sistema 20
reprodutivo complexo nas espécies estudadas. Os machos das espécies do grupo de S. 21
perpusillus possuem um elaborado repertório acústico e as fêmeas apresentam um 22
sofisticado controle sobre a oviposição, que consiste em inspecionar previamente o sítio 23
reprodutivo e pôr geralmente apenas um ovo por axila foliar, característica considerada 24
sinapomorfia do grupo de S. perpusillus (Alves-Silva & Silva 2009). Sistemas 25

7
reprodutivos complexos também são observados em outros anuros bromelígenas, como 1
Anomaloglossus beebei (Noble, 1923) (Bourne et al. 2001) e anuros de outros tipos de 2
fitotelmo, como Kurixalus eiffingeri (Boettger, 1895) (Lin et al. 2008), Ranitomeya 3
variabilis (Zimmermann & Zimmermann, 1988), R. imitator (Schulte, 1986) (Brown et 4
al. 2008b) e Mantella laevigata Methuen & Hewitt, 1913 (Heying 2008). Nestas 5
espécies a seleção de hábitat já foi evidenciada e é possível que tal processo também 6
ocorra com as espécies de Phyllodytes, pois, como visto acima, talvez este seja um 7
processo crucial para a sobrevivência e sucesso reprodutivo das espécies que habitam 8
fitotelmos. 9
O local de estudo do presente trabalho está localizada numa área de restinga 10
arbóreo-arbustiva localizada na Área de Proteção Ambiental (APA) Litoral Norte (Fig. 11
1). A área é paralela à orla da praia e possui cerca de 1700 m de comprimento por 400 12
metros de largura. É delimitada ao sul por condomínios residenciais, ao norte por cerca 13
de arame de um grande resort, ao leste por moitas de arbustos próximos à orla da praia e 14
ao oeste por mata ciliar que margeia o Rio Timeantube (para mais detalhes, ver 15
Bastazini et al. 2007). A espécie escolhida como modelo para a realização deste 16
trabalho é Phyllodytes melanomystax (Fig. 2) que, assim como as demais espécies de 17
Phyllodytes (à exceção de P. luteolus, ver acima) possui muito pouco de sua história 18
natural descrita e carece de informações acerca de suas relações com as bromélias que 19
utilizam. 20
A presente dissertação tem por objetivos (1) investigar se há seleção de 21
bromélias por Phyllodytes melanomystax numa área de restinga arbóreo-arbustiva e, 22
caso haja, (2) determinar quais variáveis do microhabitat estão associadas à seleção das 23
bromélias por esta espécie. 24
25

8
Referências Bibliográficas 1
Alves-Silva, R. & Silva, H.R. (2009) Life in bromeliads: reproductive behaviour and the 2
monophyly of the Scinax perpusillus species group (Anura: Hylidae). Journal of 3
Natural History, 43, 205–217. 4
Armbruster, P., Hutchinson, R.A. & Cotgreave, P. (2002) Factors influencing 5
community structure in a South American tank bromeliad fauna. Oikos 96, 225–234. 6
Bastazini, C.V., Munduruca, J.F.V., Rocha, P.L.B. & Napoli, M.F. (2007) Which 7
environmental variables better explain changes in anuran community composition? A 8
case study in the Restinga of Mata de São João, Bahia, Brazil. Herpetologica, 63, 459–9
471. 10
Bokermann, W.C. (1966) O gênero Phyllodytes Wagler (1830). Anais da Academia 11
Brasileira de Ciências 38, 335–344. 12
Bourne, G.R., Collins, A.C., Holder, A.M. & McCarthy, C.L. (2001) Vocal 13
communication and reproductive behavior of the frog Colostethus beebei in Guyana. 14
Journal of Herpetology 35, 272–281. 15
Brown, J.L., Morales, V. & Summers, K. (2008a). Divergence in parental care, habitat 16
selection and larval life history between two species of Peruvian poison frogs: an 17
experimental analysis. Journal of Evolutionary Biology 21, 1534–1543. 18
Brown, J.L., Twomey, E., Morales, V. & Summers, K. (2008b) Phytotelm size in 19
relation to parental care and mating strategies in two Peruvian poison frogs. Behaviour 20
145, 1139–1165. 21

9
Bursik, J.V. (1988) Interactive effects of dragonfly predation in experimental pond 1
communities. Ecology 69, 857–867. 2
Caramaschi, U., Da Silva, H.R. & Britto-Pereira, M.C. (1992) A new species of 3
Phyllodytes (Anura, Hylidae) from southern Bahia, Brazil. Copeia 1, 187–191. 4
Caramaschi, U. & Peixoto, O.L. (2004) A new species of Phyllodytes (Anura: Hylidae) 5
from the State of Sergipe, northeastern Brazil. Amphibia-Reptilia 25, 1–7. 6
Caramaschi, U., Peixoto, O.L. & Rodrigues, M.T. (2004) Revalidation and redescription 7
of Phyllodytes wuchereri (Peters, 1873) (Amphibia, Anura, Hylidae). Arquivos do 8
Museu Nacional 62, 185–191. 9
Duellman, W.E. & Trueb, L. (1986) Biology of Amphibians. McGraw-Hill, New York. 10
Eterovick, P.C. (1999) Use and sharing of calling and retreat sites by Phyllodytes 11
luteolus in modified environment. Journal of Herpetology 33, 17–22. 12
Gascon, C. (1992) Predators and tadpole prey in central Amazonia: field data and 13
experimental manipulations. Ecology 73, 971–980. 14
Giaretta, A.A. (1996) Reproductive specializations of the bromeliad hylid frog 15
Phyllodytes luteolus. Journal of Herpetology 30, 96–97. 16
Haddad, C.F.B. & Prado, C.P.A. (2005) Reproductive modes in frogs and their 17
unexpected diversity in the Atlantic Forest of Brazil. BioScience 55, 207–217. 18
Heying, H.E. (2008) Social and reproductive behaviour in the Madagascan poison frog, 19
Mantella laevigata, with comparisons to the dendrobatids. Animal Behaviour 61, 567–20
577. 21
Huey, R.B. (1991) Physiological consequences of habitat selection. The American 22
Naturalist 137, 91–115. 23

10
Kitching, R.L. (2000) Food Webs and Container Habitats: The Natural History and 1
Ecology of Phytotelmata. Cambridge University Press, Cambridge, UK. 2
Kitching, R.L. (2001) Food webs in phytotelmata: “bottom-up” and “top-down” 3
explanations for community structure. Annual Review of Entomology 46, 729–60. 4
Jowers, M.J., Downie, J.R., & Cohen, B.L. (2008) The golden tree frog of Trinidad, 5
Phyllodytes auratus (Anura: Hylidae): systematic and conservation status. Studies on 6
Neotropical Fauna and Environment 43, 181–188. 7
Laessle, A.M. (1961) A micro-limnological study of Jamaican bromeliads. Ecology 42, 8
499–517. 9
Lannoo, M.J., Townsend, D.S. & Wassersug, R.I. (1987) Larval life in the leaves: 10
arboreal tadpole types, with special attention to the morphology, ecology and behavior 11
of the oophagous Osteopilus brunneus (Hylidae) larva. Fieldiana Zoology 38, 1–31. 12
Lehtinen, R.M. Lanoo, M.J. & Wassersug, R.J. (2004) Phytotelm-breeding anurans: 13
past, present and future research. Ecology and Evolution of Phytotelm Breeding 14
Anurans, (ed R.M. Lehtinen), pp. 1–9. Miscellaneous Publications of the Museum of 15
Zoology, University of Michigan. 16
Lin, Y.S., Lehtinen, R.M. & Kam, Y.C. (2008) Time-and context-dependent oviposition 17
site selection of a phytotelm-breeding frog in relation to habitat characteristics and 18
conspecific cues. Journal of Herpetology 64, 413–421. 19
Neill, W.T. (1951) A bromeliad herpetofauna in Florida. Ecology 32,140–143. 20
Oliveira, F.B. & Navas, C.A. (2004) Plant selection and seasonal patterns of vocal 21
activity in two populations of the bromeligen treefrog Scinax perpusillus (Anura, 22
Hylidae). Journal of Herpetology 38, 331–339. 23

11
Papp, M.G. & Papp, C.O.G. (2000) Decline in a population of the treefrog Phyllodytes 1
luteolus after fire. Herpetological Review 31, 93–95. 2
Peixoto, O.L. (1987) Caracterização do grupo „„perpusilla‟‟ e revalidação da posição 3
taxonômica de Ololygon perpusilla perpusilla e Ololygon perpusilla v-signata 4
(Amphibia, Anura, Hylidae). Arquivos da Universidade Federal Rural do Rio de 5
Janeiro 10, 37–49. 6
Peixoto, O.L. (1995) Associação de anuros a bromeliáceas na Mata Atlântica. Revista 7
Universidade Rural Série Ciencias da Vida 17, 75–83. 8
Peixoto, O.L., Caramaschi, U., Freire, E.M.X. (1999) Two new species of Phyllodytes 9
(Anura: Hylidae) from the state of Alagoas, northeastern Brazil. Herpetologica 59, 235–10
246. 11
Resetarits, W.J. & Willbur, H.M. (1989) Choice of oviposition site by Hyla 12
chrysoscelis: role of predators and competitors. Ecology 70, 220–228. 13
Romero, G.Q. & Vasconcellos-Neto, J. (2005) The effect of plant structure on the 14
spatial and microspatial distribution of a bromeliad-living jumping spider (Salticidae). 15
Journal of Animal Ecology 74, 12–21. 16
Rudolf, V.H.W. & Rödel, M.O. (2005) Oviposition site selection in a complex and 17
variable environment: the role of habitat quality and conspecific cues. Oecologia 142, 18
316–325. 19
Sampaio, M.C., Picó, F.X. & Scarano, F.R. (2005) Ramet demography of a nurse 20
bromeliad in Brazilian restingas. American Journal of Botany 92, 674–681. 21

12
Schiesari, L., Gordo, M. & Hodl, W. (2003) Treeholes as calling, breeding, and 1
developmental sites for the amazonian canopy frog, Phrynohyas resinifictrix (Hylidae). 2
Copeia 2, 263–272. 3
Schineider, J.A. & Teixeira, R.L. (2001) Relacionamento entre anfíbios anuros e 4
bromélias da restinga de Regência, Linhares, Espírito Santo, Brasil. Iheringia 91, 41–5
48. 6
Silva, H.R. & Britto-Pereira, M.C. (2006) How much fruit do fruit-eating frogs eat? An 7
investigation on the diet of Xenohyla truncata (Lissamphibia: Anura: Hylidae). Journal 8
of Zoology 270, 692–698. 9
Simon, J.E. & Gasparini, J.L. (2003) Descrição da vocalização de Phyllodytes kautskyi 10
Peixoto e Cruz, 1988 (Amphibia, Anura, Hylidae). Boletim do Museu de Biologia Mello 11
Leitão 16, 47–54. 12
Smith, L.B. & Downs, R.J. (1979) Bromeliaceae (Bromelioideae). Flora Neotropica 14, 13
1493–2142. 14
Vieira, W.S., Santana, G.G., Dos Santos, S.C.D.C., Alves, R.R.N. & Pereira-Filho, G.A. 15
(2009) Description of the tadpoles of Phyllodytes brevirostris (Anura: Hylidae). 16
Zootaxa 2119, 66–68. 17
Weygoldt, P. (1981). Beobachtungen zur fortpflanzungsbiologie von Phyllodytes 18
luteolus (Wied 1824) im terrarium (Amphibia: Salientia: Hylidae). Salamandra 17, 1–1. 19

13
Figura 1. Área de estudo na Praia do Forte, Município de Mata de São João, Estado da Bahia, Brasil. A – área de estudo: ambiente de restinga
arbóreo-arbustiva com moitas de bromélias terrestres, estas últimas indicadas por setas na cor vermelha. B – detalhe da bromélia terrestre
Aechmea cf. aquilega, espécie abundante na área de estudo e utilizada por Phyllodytes melanomystax como sítio de vocalização.
A B

14
Figura 2. O anuro Phyllodytes melanomystax em quatro diferentes momentos. A, girino
no tanque central da bromélia; B, macho vocalizante no tanque central da bromélia; C,
macho e fêmea juntos em uma mesma bromélia; D, vista frontal de P. melanomystax
destacando o “bigode” característico da espécie.
B A
C D

15
Artigo
Este capítulo é aqui apresentado sob a forma de artigo a ser submetido ao
periódico Journal of Animal Ecology, sob o título “Calling site selection by the
bromeliad-dwelling treefrog Phyllodytes melanomystax (Amphibia: Anura: Hylidae) in
a coastal sand dune habitat”. As normas de publicação no respectivo periódico se
encontram como anexo a esta dissertação.

16
Calling site selection by the bromeliad-dwelling treefrog Phyllodytes melanomystax 1
(Amphibia: Anura: Hylidae) in a coastal sand dune habitat 2
Maria Silva CUNHA¹,³ and Marcelo Felgueiras NAPOLI² 3
¹Programa de Pós-Graduação em Ecologia e Biomonitoramento, Instituto de Biologia, 4
Universidade Federal da Bahia, Rua Barão de Jeremoabo, Campus Universitário de 5
Ondina, 40170-115 Salvador, Bahia, Brazil 6
²Museu de Zoologia, Instituto de Biologia, Universidade Federal da Bahia, Rua Barão 7
de Jeremoabo, Campus Universitário de Ondina, 40170-115 Salvador, Bahia, Brazil. E-8
mail: [email protected] 9
3Corresponding author: [email protected] 10
Running headline: Calling site selection by a bromeliad-dwelling frog 11
12

17
Summary 1
1. Tank bromeliads are able to accumulate rainwater within their rosettes, forming small 2
bodies of water known as phytotelms. These bromeliads are used by a great variety of 3
vertebrates and invertebrates and some species spend their entire life cycle inside tank-4
bromeliads. It is known that for anurans there is a non-random use of bromeliads, but 5
the environmental factors that drive this selection are still poorly known. 6
2. In this study, variables associated to bromeliad selection by calling males of the 7
bromeliad-inhabiting frog Phyllodytes melanomystax were investigated. Novel natural 8
history observations of the focused species also were provided. 9
3. The study site was a sand-dune restinga environment, located in the Municipality of 10
Mata de São João, northern coast of Bahia State, northeastern Brazil. Two hundred and 11
two terrestrial bromeliads were sampled, 101 with and 101 without calling males of P. 12
melanomystax. Nine environmental variables (predictors) related in literature as 13
possibly associated to bromeliad selection by anurans were measured. Multiple logistic 14
regression analysis and Wald test were performed to test which environmental variables 15
could explain the occurrence of calling males of P. melanomystax within terrestrial 16
bromeliads. 17
4. The presence/absence of calling males of P. melanomystax in bromeliads was 18
influenced by number of bromeliads in a 2 meters radius and amount of debris inside 19
the rosettes, while physical and/or chemical variables of bromeliads and from stored 20
water inside the rosettes had no influence. The marking-recapture procedure of P. 21
melanomystax evidenced strong site fidelity for males. Yet, it was observed that males 22
calling sites were also used by the species as oviposition sites and for development of 23
tadpoles. 24

18
5. This is the first research to explain the pattern of bromeliad selection by a species of 1
the bromeliad-dwelling frog genus Phyllodytes. 2
3
Keywords: bromeligen species, microhabitat selection, phytotelm.4

19
Introduction 1
Selection of calling sites is a critical factor that affects the success of males of 2
many anuran species, as females may choose their mates not only by its acoustic and 3
physical characteristics, but also by the quality of the site for offspring (Duellman & 4
Trueb 1986). Anurans from tropical regions show a great variety of reproductive modes 5
(Haddad & Prado 2005), using an array of different microhabitats. By using different 6
microhabitats they face different situations, and the environmental factors that influence 7
the mechanisms of calling site selection also can change. 8
Phytotelm-dwelling anurans face risks and situations unusual to traditional 9
habitats like ponds and streams (Lehtinen et al. 2004). Phytotelms are small bodies of 10
water formed by accumulation of rainwater within plants or parts of plants, such as 11
treeholes, bromeliad rosettes, and bamboo stumps (Kitching 2001; Schiesari, Gordo & 12
Hödl 2003). Not surprisingly, phytotelm-inhabiting anurans have to deal with situations 13
as unpredictable food availability, low dissolved oxygen concentrations, and high 14
desiccation risk (Lehtinen et al. 2004). However, compared to pond and streams, 15
phytotelms are known as safe reproduction sites, because these sites do not have major 16
traditional predators for eggs and tadpoles, and consequently forces like predation and 17
competition are not so pronounced (Schiesari, Gordo & Hödl 2003), though predators 18
like larvae of Zygoptera and Anisoptera (Insecta, Odonata) and crabs are not rare in 19
terrestrial bromeliads of restinga habitats (MF Napoli, personal observation). Predation 20
pressure on eggs and tadpoles are referred as major forces that influence the evolution 21
of alternative reproductive strategies (Magnusson & Hero 1991), like use of phytotelms 22
as reproductive sites by anurans. 23
Among phytotelms, bromeliads are known as key plants for tropical regions 24
because sometimes they hold inside their tanks the unique source of available 25

20
freshwater (Neill 1951). Many species of vertebrates and invertebrates use bromeliads 1
for shelter or during foraging activity, and some species can spend their entire life cycle 2
inside tank-bromeliads (Neill 1951; Laessle 1961; Juncá & Borges 2002; Srivastava et 3
al. 2004; Lopez et al. 2005). Tank-bromeliad is a common name given to bromeliad 4
species that accumulate water because of the configuration of its leaves. These leaves 5
alternate and arrange in spiral and this arrangement permit them to accumulate water 6
and dry leaves from the canopy within the leaf axils and inside the central tank (Fig. 1). 7
Bromeliad-dwelling anurans usually have pronounced mechanisms of 8
microhabitat selection. Males of Scinax perpusillus (Lutz & Lutz, 1939), for example, 9
prefer clustered and terrestrial bromeliads, a preference associated to territorial behavior 10
(Oliveira & Navas 2004). Additionally, species of the S. perpusillus group have a 11
sophisticated reproductive system, which involves from an elaborated acoustic repertory 12
in males to a complex oviposition control in females (Alves-Silva & Silva 2009). 13
Complex reproductive systems and non-random bromeliad use were also reported for 14
other bromeliad frogs as Anomaloglossus beebei (Noble, 1923) (Bourne et al. 2001), 15
and for other phytotelm anurans like Kurixalus eiffingeri (Boettger, 1895) (Lin et al. 16
2008), Ranitomeya variabilis (Zimmermann & Zimmermann, 1988), R. imitator 17
(Schulte, 1986) (Brown et al. 2008), and Mantella laevigata Methuen & Hewitt, 1913 18
(Heying 2008). 19
The genus Phyllodytes Wagler, 1830 is entirely composed by small bromeliad-20
dwelling treefrogs distributed along the Tropical Atlantic Forest (Caramaschi, Peixoto 21
& Rodrigues 2004), but little is known about the nature of the relationship between 22
Phyllodytes and bromeliads. These frogs spend their entire life cycle inside the 23
bromeliads, using them as calling and retreat sites (Eterovick 1999), and local for 24
reproduction and deposition of eggs and tadpoles (Peixoto 1995). Few data on natural 25

21
history and ecology are available for P. luteolus (Eterovick 1999; Schineider & Teixeira 1
2001), and the remaining studies on Phyllodytes have a taxonomic approach (e.g.: 2
Peixoto, Caramaschi & Freire 1999, Caramaschi, Peixoto & Rodrigues 2004, Vieira et 3
al. 2009). In view of the high microhabitat specificity of Phyllodytes species, and of the 4
poorly known relationship between them and bromeliads, it is important to conduct 5
studies to clarify some aspects of microhabitat use by these species. 6
The treefrog Phyllodytes melanomystax Caramaschi, Silva & Britto-Pereira, 7
1992, is distributed from southern State of Sergipe to southern State of Bahia, Brazil 8
(Peixoto & Pimenta 2008). This species is very abundant in northern coast of the State 9
of Bahia, in a region known as Litoral Norte, which is composed in most part by coastal 10
sand dune habitats covered with herbaceous and shrubby vegetation, which is common 11
along the Brazilian coast and known as ‘restinga’ (Suguio & Tessler, 1984). The region 12
is suffering a process of habitat loss due to unregulated tourism and to conversion of the 13
restingas into resorts and residential areas. 14
In this paper, Phyllodytes melanomystax was used as a model species for 15
investigating the variables associated to site selection by the bromeliad-dwelling frog 16
genus Phyllodytes. Two questions were addressed: (1) is there any pattern in bromeliad 17
use by these frogs? (2) Which environmental variables influence the selection of 18
bromeliads by calling males of P. melanomystax? Calling males were used as a model 19
for selection system, and we also made some novel natural history observations that 20
could help to understand our results. 21
22

22
Material and Methods 1
Study Area 2
The field study was conducted from 9 April to 7 May 2010 in a private property 3
located in Praia do Forte, Municipality of Mata de São João, State of Bahia, Brazil (12º 4
34' 12'' S; 38º 00' 04'' W, 10 m above sea level). The property is located in the State 5
environmental protected area APA Litoral Norte (see Bastazini et al. 2007 for a location 6
map). The study area is a coastal sand dune habitat covered with herbaceous and 7
shrubby vegetation, known as ‘restinga’, a sandy coastal plain vegetation from the 8
Brazilian Atlantic Forest biome (Scarano et al. 2002). 9
10
Research Design and Field Methods 11
A sampling polygon of 1700 x 400 m was delimited within the study area, in 12
which 202 bromeliads (sample units—SU) were randomly sampled following a point 13
map. For the construction of the map, 202 values ranging from 0 to 1700 m 14
(representing the major axis of the polygon) and 202 values from 0 to 400 m 15
(representing the minor axis of the polygon) were obtained at a random, and then 16
randomly combined into ordered pairs. Each ordered pair corresponded to a SU. When a 17
bromeliad was not in the exact point previously chosen at random, the nearest 18
bromeliad was measured. A single species of tank-bromeliad was sampled to avoid 19
sampling biases due to species-specific structural distinctiveness, of which was elected 20
for sampling the most abundant tank-bromeliad in the study area, Aechmea aquilega 21
(Salisb.) Griseb., 1864. Bromeliads (SUs) were divided into two categories: unoccupied 22
(category 0) and occupied bromeliads (category 1) by calling males of P. melanomystax. 23
The same number of bromeliads was sampled for each category (101 bromeliads per 24

23
category), totalizing 202 SUs. Voucher specimens of P. melanomystax were deposited 1
in the Museu de Zoologia da Universidade Federal da Bahia (UFBA 10653–10712). 2
Bromeliads with or without calling males of Phyllodytes melanomystax were 3
determined by active search from 17h30min to 22h30min, time interval corresponding 4
to the vocal activity for this species (M.S. Cunha personal observation). This procedure 5
was adopted assuming that we only could determine whether a bromeliad was occupied 6
or not by males of P. melanomystax when searching at night, because during periods of 7
sunlight these treefrogs always drop into the leaf axils, which makes it quite impossible 8
to see and reach them without damage the bromeliad. After each night, 10 bromeliads 9
were marked with small plastic strips and then georeferenced. These procedures were 10
repeated until we reached the total of 202 sampled bromeliads (20 days of searching). 11
It was respected the minimal distance of 20 m between SUs and this value was 12
assumed as enough to assure independence of sample units. In order to test the 13
efficiency of this value, the home range of Phyllodytes melanomystax was investigated 14
with a capture-marking-recapture experiment. Thus, if the home range of P. 15
melanomystax calling males was smaller than 20 m, the samples were independent. 16
From the 101 males observed in calling activity in bromeliads from category 1, it was 17
captured the maximal number of males as possible throughout the 20 days of data 18
collection. Fifty-two males of P. melanomystax were marked using the toe-clipping 19
method (Martoff 1953) with a small modification: instead of clipping the toe as a whole, 20
only the disc was removed. Marked individuals were released soon after the marking in 21
the same bromeliad where they were captured. The recapture period was in the last 22
week of field research (03–07 May 2010). Some natural history observations (for 23
example, calling position inside the bromeliad, presence of another calling males etc.) 24
were also made during data collection. Before capturing a male, it was spent 25

24
approximately 5 minutes in silent to observe how many active males there were in that 1
clump. This observation was also made during the time spent for marking the males and 2
signalizing the sample unit, which took approximately 10–15 minutes of observations in 3
each point. 4
In the morning after marking the bromeliads, from 6h to 11h, nine predictor 5
variables from each marked bromeliad were measured: (1) volume, (2) temperature, and 6
(3) pH of stored water in bromeliad rosette, (4) amount of debris inside the bromeliad 7
tank, (5) number of leaf axils with water, (6) bromeliad tank height, (7) maximum 8
bromeliad diameter, (8) number of bromeliads in a 2 m radius, and (9) distance of the 9
referred bromeliad to the next bromeliad cluster. Variables 8 and 9 correspond to a 10
measure of bromeliad/cluster isolation. Bromeliad clumps varied in size and shape, but 11
they were elliptical-shaped by most times. The size of the clump varied from clumps 12
formed by just three bromeliads (± 1 m2) to clumps with more than one hundred 13
bromeliads (more than 20 m2). Water temperature was recorded with a digital thermo-14
hygrometer to the nearest 0.1ºC directly from inside the bromeliad. For variables 1 and 15
3 the entire volume of water was extracted inside the bromeliad rosette with a plastic 16
tube connected to a recipient, a device similar to that used by Guimarães-Souza et al. 17
(2006) and Jabiol, Corbara & Céréghino (2009). Water volume was measured in a 18
graduated cylinder to the nearest 0.1 ml, and pH with a digital pH meter to the nearest 19
0.1. The amount of debris was recorded following a similar procedure used by Osses, 20
Martins & Machado (2008), as an ordinal variable (1–4): 1—smallest amount of debris; 21
2—large amount of debris, but not enough to prevent a specimen of P. melanomystax 22
from passing through the bromeliad central tank or leaf axils; 3—very large amount of 23
debris, but still passage free; 4—largest amount of debris, blocking passage to central 24
tank and leaf axils. In order to avoid temporal biases during the period of the 25

25
measurement of variables (from 6h to 11h), as temperature increasing, we alternated the 1
sampling of bromeliads without (category 0) and bromeliads with P. melanomystax 2
(category 1). 3
4
Data Analysis 5
For the purpose of analysis, three data matrices were produced: (A) a matrix of 6
202 sample units (bromeliads, objects) vs. the presence (1) or absence (0) of Phyllodytes 7
melanomystax (attribute), representing the dependent variable of the analysis; (B) a 8
matrix of 202 sample units (bromeliads, objects) vs. 9 predictor variables, representing 9
the independent variables of the analysis; (C) similar to matrix B, but only with 10
uncorrelated predictor variables (n = 7). Five bromeliads lacked stored water in their 11
rosettes, and therefore no values referring to water related variables were entered in 12
matrices B and C. Descriptive statistics of predictor variables are in Table 1. 13
The multiple logistic regression was used to determine which predictor variables 14
(matrix C) were important for the selection of calling sites (bromeliads) by P. 15
melanomystax and to create a predictive habitat model based on presence/absence data 16
(matrix A). As in linear regressions, multiple logistic regressions have the assumption 17
of collinearity absence between independent variables (Gotelli & Ellison 2004). As 18
some variables did not present normal distribution, the existence of significant 19
correlations between predictor variables were tested using the Spearman rank 20
correlation (R), adjusting the α-level (P ≤ 0.05) by the Bonferroni correction (Bland 21
2000). Three significant correlations resulted from this analysis: number of leaf axils 22
with water vs. volume of stored water in bromeliad rosette, number of leaf axils with 23
water vs. amount of debris inside the bromeliad tank, and bromeliad tank height vs. 24

26
volume of stored water in bromeliad rosette (Table 2). The variables number of leaf 1
axils with water and bromeliad tank height were excluded from analysis because 2
volume of stored water in bromeliad rosette has a great biological meaning for 3
phytotelm-dwelling anurans (Bourne et al. 2001, Schiesari, Gordo & Hödl 2003), and 4
recent studies showed that amount of debris is a microhabitat selection criterion for 5
some bromeliad-inhabiting animals (Romero & Vasconcelos-Neto 2005; Osses, Martins 6
& Machado 2008). Therefore, the multiple logistic regression analysis was performed 7
with seven predictor variables. The Wald statistics (Quinn & Keough 2002) was used to 8
test the null hypothesis that any of the independent variables recorded influenced the 9
presence/absence of P. melanomystax in bromeliads. The Wald statistics tests the 10
contribution of each independent variable to the response variable individually when all 11
other variables are constant, revealing the effect of each variable independently. The α-12
level (P ≤ 0.05) was Bonferroni corrected to control Type I error rate when using 13
multiple tests (Bland 2004). For evaluating the goodness of fit of the model and its 14
predictive performance, the statistics of Hosmer-Lemeshow was conducted (Hosmer & 15
Lemeshow 2000). In this analysis, the null hypothesis is that there is no evidence for 16
lack of fit of the model, and therefore if null hypothesis is rejected the model is not well 17
adjusted. Specificity or true negative fraction (proportion of correct predictions for the 18
absence), sensitivity or true positive fraction (proportion of correct predictions for the 19
presence), and overall success rate of the model were calculated (Rudolf & Rödel 20
2005). 21
22

27
Results 1
Microhabitat variables and natural history notes 2
Calling specimens of Phyllodytes melanomystax always were obtained from 3
bromeliads filled with water. Out of 52 marked males, 27 (52%) were recaptured, all of 4
them during calling activity. The distance between release and recapture points was low 5
(0.27–11.5 m, X = 4.30 m ± 3.4 m), and three recaptured males were found in the same 6
bromeliads they were released. The amount of recaptured males from the first sampling 7
days was similar to that from the last sampling days. 8
Calling activity of Phyllodytes melanomystax took place from sunset to dawn, 9
with the activity peak ranging from 19h to 21h. Calling males were located 10
preferentially in the central tank (n = 50, 94%), with the head pointed to the external 11
side of the bromeliad (Fig. 2). Males calling from leaf axils were less frequent (n = 3, 12
6%). By three times we saw a female in the same bromeliad of a calling male, but not in 13
amplexus, and by one time we observed a female perched on the vegetation above the 14
bromeliad with a calling male. In two occasions we saw two calling males in a single 15
bromeliad, one on the central tank and the other in a leaf axil. Both pairs of calling 16
males were performing advertisement call. More frequently (not quantified), we 17
observed only one male per bromeliad cluster, but there were a few occasions where 18
there were more than one calling male in a single cluster. 19
We found tadpoles of Phyllodytes melanomystax in 25 (25%) of the 101 20
bromeliads with calling males, with 1 to 32 tadpoles per bromeliad ( X = 6.0±6.0). 21
Tadpoles or eggs were not observed in bromeliads without calling males of P. 22
melanomystax. The volume of water inside bromeliads rosettes with tadpoles varied 23

28
from 130 ml to 1237 ml ( X = 429.4±257.0 ml). We observed egg clutches of P. 1
melanomystax in two bromeliads that also had tadpoles, one with 12 eggs and the other 2
with 14 eggs. The 12-eggs clutch was found in a bromeliad of 480 ml with four 3
tadpoles; the 14-eggs clutch in a bromeliad of 270 ml with 32 tadpoles. All tadpoles and 4
egg clutches were found in the water stored inside the central tank of the bromeliad. It 5
was not possible to determine the stages (Gosner 1960) of the tadpoles found in the 6
same bromeliads, but it was visually possible to confirm that there were tadpoles two to 7
three times larger than others from the same site. 8
9
Statistical analysis 10
The Wald hypothesis test indicated that only predictor variables (1) number of 11
bromeliads in a 2 meter radius and (2) amount of debris inside the bromeliad tank 12
influenced the presence/absence of calling specimens of Phyllodytes melanomystax in 13
terrestrial bromeliads (Table 3). The number of bromeliads in a 2 meter radius 14
influenced positively the probability of occurrence of calling males in bromeliads, 15
indicating an avoidance of isolated bromeliads, and a preference for clustered 16
bromeliads (Fig. 3a). On the other hand, the amount of debris influenced negatively the 17
probability of occurrence of calling males in bromeliads, indicating an avoidance of 18
bromeliads with a great amount of debris, and a preference for calling sites with no or 19
with small amount of debris (Fig. 3b). The Hosmer-Lemeshow statistics accepted the 20
null-hypothesis that there is no evidence for lack of fit of the data (Chi-Square = 4.04, 21
DF = 8, P > 0.85), and therefore the model is well data adjusted. The high specificity 22
(71.6%), sensitivity (73.5%), and overall success rate (72.5%) of the model indicated 23
that it predicted most of the unused and used sites correctly (Table 4). 24

29
Discussion 1
The calling site of Phyllodytes melanomystax in terrestrial bromeliads of the 2
study area was similar to that published for P. luteolus (Wied-Neuwied, 1824) 3
(Eterovick 1999). Both species use the central tank of bromeliads and their leaf axils to 4
call, but P. melanomystax differs by using the central tank as the primary calling site, 5
while in P. luteolus there was no detected difference. The more frequent use of the 6
bromeliad central tank as the main calling site by P. melanomystax possibly is due to 7
the structure of the terrestrial bromeliads investigated, with close leafs and central tank 8
as the main bromeliad water reservoir. The morphological structure of these bromeliads 9
may be a response to the high sunlight incidence over these plants. It is known that 10
bromeliads can modify structural characteristics, as morphology of leaves, shape and 11
size of the tank, and photochemical efficiency in response to variations in light and 12
flooding regimes (Cogliatti-Carvalho, Almeida & Rocha 1998; Scarano et al. 2002). 13
Although up till now there is no study providing these responses for Aechmea cf. 14
aquilega, we assume that as phenotypic plasticity is a common characteristic of 15
Bromeliaceae, it is possible to be happening to this species in our study site. 16
The volume of stored water in terrestrial bromeliads was not able to explain the 17
calling site selection by adult males of Phyllodytes melanomystax. The results presented 18
here agree with Schineider & Teixeira (2001), which argued that the volume of water 19
stored in bromeliads is not important for P. luteolus. Nevertheless, Eterovick (1999) 20
observed for P. luteolus that volume and pH of water were statistically different 21
between occupied and unoccupied bromeliads. In fact, in the study area, calling males 22
of P. melanomystax did not occur in terrestrial bromeliads without stored water, which 23
points out to the presence of it as a primary condition for choice of the bromeliad by this 24

30
treefrog. A water reservoir is essential for the maintenance of hydric balance by the 1
animal, and for the development of tadpoles (Duellman & Trueb 1986). On the other 2
hand, bromeliads with small amounts of stored water (minimum of 85 ml) were used as 3
call sites by males of P. melanomystax, which led us to conclude that the presence of 4
stored water, even in low volumes, qualifies a terrestrial bromeliad of an open habitat to 5
host a calling male of P. melanomystax, but the increase of stored water does not 6
improve the chance of the bromeliad to be inhabited by a calling male. 7
In the present research, isolated or clustered terrestrial bromeliads were often 8
occupied by a single calling male of Phyllodytes melanomystax, which suggests a 9
territorial behavior for adult males, and the choice of calling sites as a valuable 10
parameter for the species reproductive success. In assuming the territoriality of adult 11
males, the preference for clustered bromeliads by a calling male also implies that its 12
territory was represented by the entire bromeliad cluster. Similar territoriality was 13
reported for Scinax perpusillus, where adult males defend not only a single bromeliad, 14
but also prefer isolated clusters of bromeliads, possibly to have access to several 15
potential oviposition sites that might confer reproductive advantages, and to reduce 16
intra-sexual competition (Oliveira & Navas 2004). As for P. melanomystax, variables 17
related to spatial distribution of bromeliads were more important to call site selection in 18
S. perpusillus than physical and chemical parameters of the bromeliad (Oliveira & 19
Navas 2004). 20
The presence of tadpoles and eggs of Phyllodytes melanomystax in the same 21
terrestrial bromeliads of conspecific calling males points out that the species possibly 22
uses the same sites for calling, egg-laying, and tadpole development, as observed for P. 23
luteolus by Eterovick (1999). Nevertheless, the reproductive characteristics of P. 24

31
melanomystax in the study site differs from that of P. luteolus because eggs and 1
tadpoles of P. melanomystax were found in the central tank of the bromeliads, whereas 2
that of P. luteolus were found not only in the central tank, but also in leaf axils. 3
Furthermore, we observed a higher amount of tadpoles and eggs per clutch in P. 4
melanomystax (1–32 tadpoles, 12–14 eggs) than in P. luteolus (1–3 tadpoles, 1–3 eggs, 5
Giaretta 1996, Eterovick 1999). In respect to the number of eggs per leaf axil, 6
Caramaschi, Silva & Britto-Pereira (1992) accounted a single egg for P. melanomystax 7
(and also only 1–2 tadpoles per bromeliad), similar number observed for Scinax 8
littoreus, S. perpusillus and S. v-signatus by Alves-Silva & Silva (2009), which greatly 9
differ from our results. As any data is provided on the bromeliads investigated by 10
Caramaschi, Silva & Britto-Pereira (1992), we can not relate this distinct number of 11
eggs and tadpoles (compared to the numbers presented here) to environmental 12
differences between the two sites. However, an important question that remains 13
unresolved is whether the eggs and tadpoles housed in a same bromeliad came from the 14
same couple, or from the same male with different females, as tadpoles were of distinct 15
body sizes and perhaps from different hatchings. This information is important, as it 16
implies that a same bromeliad, or bromeliad cluster, could house a male that is chosen 17
by distinct females and reproduce with them during the same breeding season. 18
The importance of amount of debris accumulated inside the rosettes regarding to 19
its influence on bromeliad selection was first reported for the bromeliad-living spider 20
Psecas chapoda Peckham & Peckham, 1894 (Romero & Vasconcellos-Neto 2005), and 21
later, to the bromeliad-dwelling harvestmen Bourguyia hamata Kury, 2003 (Osses, 22
Martins & Machado 2008). In both cases, the influence of amount of debris was 23
negatively related for bromeliad selection, as the dry leaves prevent these animals of 24

32
reaching the center of the rosettes, and thus they cannot use bromeliads as reproduction 1
and/or shelter sites. In the case of P. chapoda, the authors conclude that these 2
observations partially agree with their previous hypothesis that this species did not 3
occur in bromeliads in the forest because of the dry leaves that fall from the forest 4
canopy inside the bromeliads. Bastazini et al. (2007) reported the high abundance of P. 5
melanomystax in our study area, and devoted this to the great number of terrestrial 6
bromeliads. However, in an adjacent area of arboreal restinga (covered by arboreal 7
vegetation, containing springs, streams, and rivers), the authors also observed plots with 8
a great amount of terrestrial bromeliads, but where P. melanomystax occurred in a much 9
lower abundance. As Romero & Vascocellos-Neto (2005) observed, terrestrial 10
bromeliads from forested areas tend to accumulate more dry leaves inside their rosettes 11
than in open areas. Based on the results presented here that amount of debris 12
(represented by dry leaves) inside the bromeliads had a negative influence on 13
occurrence of P. melanomystax, we conclude that the low abundance of this species in 14
forested areas compared to open areas (Bastazini et al. 2007) is possibly due to the great 15
amount of debris inside bromeliads in forested areas. We understand that, as in P. 16
chapoda and B. hamata, debris accumulated inside bromeliads prevent individuals of P. 17
melanomystax to reach the center of the bromeliads during the day, when they use 18
bromeliads as shelter sites, and during the night, when they go out to forage or 19
reproduce, but in danger enter through the bromeliad tank. 20
The present research determined which variables were able to explain the pattern 21
of bromeliad selection by calling males of the bromeliad-dwelling frog Phyllodytes 22
melanomystax. The number of clustered bromeliads and the amount of debris 23
accumulated inside the rosette are the most important variables affecting the presence of 24

33
calling males. None of the physical and/or chemical variables of the bromeliads, and 1
from the stored water inside the rosettes, influenced the occurrence of calling males. 2
Mark-recapture experiments showed strong site fidelity for males, as the distance 3
between the release and recapture point was very low and, in some cases, males were 4
recaptured in the same bromeliad they were released. Natural history observations 5
suggested that the pattern of bromeliad use by calling males is similar to that for 6
reproduction, egg-laying, and tadpoles’ development, but these observations are not 7
conclusive, and future studies should focus on these unanswered questions to ameliorate 8
the understanding about the narrow relationship between bromeligen anurans and 9
bromeliads. 10
11

34
Acknowledgements 1
We are grateful to E.M.S. Silva Jr., L. Menezes, R.O. Abreu, J. Ruggeri, V. 2
Bonfim, E. Petersen, T. Dória, T. Filadelfo, D. Lima, J. Melo, J. Herrera, A.L. Xavier 3
and Mr. Anísio for the excellent field assistance; we acknowledge A. Bolzico for 4
allowing us to access research areas under his care; we are grateful to L. Menezes by 5
figure 1 and T. Filadelfo by figure 2; to F.A. Juncá and M. Solé for helpful comments 6
on the manuscript; the Conselho Nacional de Desenvolvimento Científico e 7
Tecnológico (CNPq) and the Fundação de Amparo à Pesquisa do Estado da Bahia 8
(FAPESB) for fellowships to M.F. Napoli (proc. 302542/2008-6) and M.S. Cunha (term 9
of grant 0213/2009), respectively; the Instituto Brasileiro do Meio Ambiente e dos 10
Recursos Naturais Renováveis (IBAMA) that permitted collection and mark-recapture 11
of specimens (license number 22354-1). 12
13

35
References 1
Alves-Silva, R. & Silva, H.R. (2009) Life in bromeliads: reproductive behaviour and the 2
monophyly of the Scinax perpusillus species group (Anura: Hylidae). Journal of 3
Natural History, 43, 205–217. 4
Bastazini, C.V., Munduruca, J.F.V., Rocha, P.L.B. & Napoli, M.F. (2007) Which 5
environmental variables better explain changes in anuran community composition? A 6
case study in the Restinga of Mata de São João, Bahia, Brazil. Herpetologica, 63, 459–7
471. 8
Bland, M. (2000) An introduction to medical statistics. Oxford University Press, USA. 9
Bourne, G.R., Collins, A.C., Holder, A.M. & McCarthy, C.L. (2001) Vocal 10
communication and reproductive behavior of the frog Colostethus beebei in Guyana. 11
Journal of Herpetology 35, 272–281. 12
Brown, J.L., Twomey, E., Morales, V. & Summers, K. (2008) Phytotelm size in relation 13
to parental care and mating strategies in two Peruvian poison frogs. Behaviour 145, 14
1139–1165. 15
Caramaschi, U., Da Silva, H.R. & Britto-Pereira, M.C. (1992) A new species of 16
Phyllodytes (Anura, Hylidae) from southern Bahia, Brazil. Copeia 1, 187–191. 17
Caramaschi, U., Peixoto, O.L. & Rodrigues, M.T. (2004) Revalidation and redescription 18
of Phyllodytes wuchereri (Peters, 1873) (Amphibia, Anura, Hylidae). Arquivos do 19
Museu Nacional 62, 185–191. 20
Cogliatti-Carvalho, L., Almeida, D.R. & Rocha, C.F.D. (1998) Phenotypic response of 21
Neoregelia johannis (Bromeliaceae) dependent on the amount of light reaching the 22
plant microhabitat. Selbyana 19, 240–244. 23
Duellman, W.E. & Trueb, L. (1986) Biology of Amphibians. McGraw-Hill, New York. 24

36
Eterovick, P.C. (1999) Use and sharing of calling and retreat sites by Phyllodytes 1
luteolus in modified environment. Journal of Herpetology 33, 17–22. 2
Gotelli, N.J. & Ellison, A.M. (2004) A primer of ecological statistics. Sinauer 3
Associates, Sunderland, Massachusetts, USA. 4
Giaretta, A.A. (1996) Reproductive specializations of the bromeliad hylid frog 5
Phyllodytes luteolus. Journal of Herpetology 30, 96–97. 6
Gosner, K.L. (1960) A simplified table for staging anuran embryos and larvae with 7
notes on identification. Herpetologica 16, 183–190. 8
Guimarães-Souza, B.A., Mendes, G.B., Bento. L., Marotta, H., Santoro, A.L., Esteves 9
F.A., Pinho L., Farjalla, V.F., & Enrich-Prast, A. (2006) Limnological parameters in the 10
water accumulated in tropical bromeliads. Acta Limnologica Brasiliensis 18, 47–53. 11
Haddad, C.F.B. & Prado, C.P.A. (2005) Reproductive modes in frogs and their 12
unexpected diversity in the Atlantic Forest of Brazil. BioScience 55, 207–217. 13
Heying, H.E. (2008) Social and reproductive behaviour in the Madagascan poison frog, 14
Mantella laevigata, with comparisons to the dendrobatids. Animal Behaviour 61, 567–15
577. 16
Hosmer, D.W. & Lemeshow, S. (2000) Applied Logistic Regression. Wiley, New York. 17
Jabiol, J., Corbara, B. & Céréguino, R. (2009) Structure of aquatic insect communities 18
in tank-bromeliads in an east-amazonian rainforest in French Guiana. Forest Ecology 19
and Management 257, 351–360. 20
Juncá, F.A. & Borges, C.L.S. (2002) Fauna associada a bromélias terrícolas da Serra da 21
Jibóia, Bahia. Sitientibus 2, 73–81. 22
Kitching, R.L. (2001) Food webs in phytotelmata: “bottom-up” and “top-down” 23
explanations for community structure. Annual Review of Entomology 46, 729–60. 24

37
Laessle, A.M. (1961) A micro-limnological study of Jamaican bromeliads. Ecology 42, 1
499–517. 2
Lehtinen, R.M. Lanoo, M.J. & Wassersug, R.J. (2004) Phytotelm-breeding anurans: 3
past, present and future research. Ecology and Evolution of Phytotelm Breeding 4
Anurans, (ed R.M. Lehtinen), pp. 1–9. Miscellaneous Publications of the Museum of 5
Zoology, University of Michigan. 6
Lopez, L.C.S., Filizola, B., Deiss, I. & Rios, R.I. (2005) Phoretic behaviour of 7
bromeliad annelids (Dero) and ostracods (Elpidium) using frogs and snakes as dispersal 8
vectors. Hydrobiologia 549, 15–22. 9
Lin, Y.S., Lehtinen, R.M. & Kam, Y.C. (2008) Time-and context-dependent oviposition 10
site selection of a phytotelm-breeding frog in relation to habitat characteristics and 11
conspecific cues. Journal of Herpetology 64, 413–421. 12
Magnusson, W.E. & Hero, J.M. (1991) Predation and the evolution of complex 13
oviposition behaviour in amazon rainforest frogs. Oecologia 86, 310–318. 14
Martof, B.S. (1953) Territoriality in the green frog, Rana clamitans. Ecology 34, 166–15
174. 16
Neill, W.T. (1951) A bromeliad herpetofauna in Florida. Ecology 32,140–143. 17
Oliveira, F.B. & Navas, C.A. (2004) Plant selection and seasonal patterns of vocal 18
activity in two populations of the bromeligen treefrog Scinax perpusillus (Anura, 19
Hylidae). Journal of Herpetology 38, 331–339. 20
Osses, F., Martins, E.G. & Machado, G. (2008) Oviposition site selection by the 21
bromeliad-dweller harvestman Bourguyia hamata (Arachnida: Opiliones). Journal of 22
Ethology 26, 233–241. 23
Peixoto, O.L. (1995) Associação de anuros a bromeliáceas na Mata Atlântica. Revista 24
Universidade Rural Série Ciências da Vida 17, 75–83. 25

38
Peixoto, O.L., Caramaschi, U., Freire, E.M.X. (1999) Two new species of Phyllodytes 1
(Anura: Hylidae) from the state of Alagoas, northeastern Brazil. Herpetologica 59, 235–2
246. 3
Peixoto, O.L. & Pimenta, B. (2008) Phyllodytes melanomystax. In: IUCN 2010. IUCN 4
Red List of Threatened Species. Version 2010.4. Available in www.iucnredlist.org. 5
Downloaded on 28 February 2011. 6
Quinn, G.P. & Keough, M.J. (2002) Experimental Design and Data Analysis for 7
Biologists. Cambridge University, Cambridge, USA. 8
Romero, G.Q. & Vasconcellos-Neto, J. (2005) The effect of plant structure on the 9
spatial and microspatial distribution of a bromeliad-living jumping spider (Salticidae). 10
Journal of Animal Ecology 74, 12–21. 11
Rudolf, V.H.W. & Rödel, M.O. (2005) Oviposition site selection in a complex and 12
variable environment: the role of habitat quality and conspecific cues. Oecologia 142, 13
316–325. 14
Schiesari, L., Gordo, M. & Hodl, W. (2003) Treeholes as calling, breeding, and 15
developmental sites for the amazonian canopy frog, Phrynohyas resinifictrix (Hylidae). 16
Copeia 2, 263–272. 17
Schineider, J.A. & Teixeira, R.L. (2001) Relacionamento entre anfíbios anuros e 18
bromélias da restinga de Regência, Linhares, Espírito Santo, Brasil. Iheringia Série 19
Zoologia 91, 41–48. 20
Scarano, F.R., Duarte, H.M., Rôças, G., Barreto, S.M.B, Amado, E.F., Reinert, F., 21
Wendt, T., Mantovani, A., Lima, H.R.P. & Barros, C. F. (2002) Acclimation or stress 22
symptom? An integrated study of intraspecific variation in the clonal plan Aechmea 23
bromeliifolia, a widespread CAM tank-bromeliad. Botanical Journal of the Linnean 24
Society 140, 391–401. 25

39
Srivastava, D.S., Kolasa, J., Bengtsson, J., Gonzalez, A., Lawler, S.P., Miller, T.E., 1
Munguia, P., Romanuk, T., Schneider, D.C. & Trzcinski, K. (2004) Are natural 2
microcosms useful model systems for ecology? Trends in Ecology and Evolution 19, 3
379–384. 4
Suguio, K. & Tessler, M.G. (1984) Planícies e cordões litorâneos quaternários do 5
Brasil: origem e nomenclatura. Restingas: origem, estrutura, processos, (eds L.D. 6
Lacerda, D.S.D. Araujo, R. Cerqueira & B. Turcq), pp. 15–26. Universidade Federal 7
Fluminense, Rio de Janeiro. 8
Vieira, W.S., Santana, G.G., Dos Santos, S.C.D.C., Alves, R.R.N. & Pereira-Filho, G.A. 9
(2009) Description of the tadpoles of Phyllodytes brevirostris (Anura: Hylidae). 10
Zootaxa 2119, 66–68. 11
12

40
Figure captions 1
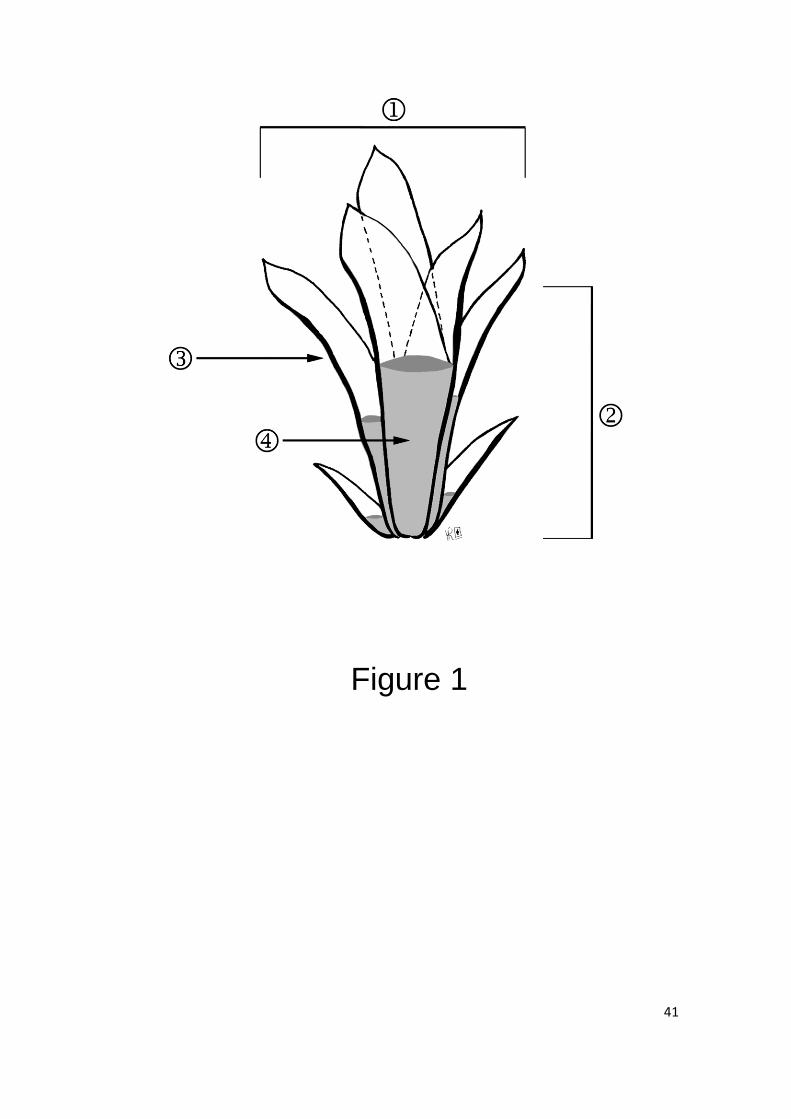
Figure 1. Schematic tank bromeliad similar to that used as sample unit in the present 2
research. 1, diameter of the bromeliad; 2, height of the central tank; 3, leaf axil; 4, 3
central tank. 4
Figure 2. Live calling male of Phyllodytes melanomystax in the central tank of a 5
terrestrial bromeliad. Males calling from central tank were more frequent than calling 6
from leaf axils. 7
Figure 3. A: presence and absence of Phyllodytes melanomystax according to the 8
number of bromeliads in a 2 meter radius. Circles of varying sizes (bubbles) represent 9
the relative frequencies of the number of points represented by a single plot position. B: 10
percentage of bromeliads used as oviposition site by the treefrog Phyllodytes 11
melanomystax (black bars) compared to the number of bromeliads not used (white bars) 12
according to the classes of amount of debris found inside the rosettes. 13
41
Figure 1

42
Figure 2

43
Figure 3

44
Table 1. Descriptive statistics of environmental variables in bromeliads with and
without calling males of Phyllodytes melanomystax. N, number of bromeliads; Mean,
arithmetic mean; SD, standard deviation; range, minimum and maximum values.
Abbreviations of environmental variables: temperature (TEMP), volume of stored water
in bromeliad rosettes (VOL), number of bromeliads in a 2 meter radius (NBRO),
distance of bromeliad to the next bromeliad cluster (DCL), maximum bromeliad
diameter (DIAM), amount of debris inside the bromeliad tank (DEB), number of leaf
axils with water (NAX); bromeliad tank height (HGH).
With P. melanomystax Without P. melanomystax
Variables N Mean SD Range N Mean SD Range
TEMP 99 28.9 2.5 (24.4–35.9) 95 29.4 2.7 (25.2–36.7)
PH 101 5.0 1.0 (3.4–8.0) 96 5.2 1.1 (3.5–8.1)
VOL 101 396.4 222.3 (85.0–1237.0) 101 269.4 243.2 (0.0–1275.0)
NBRO 101 30.0 14.3 (3.0–103.0) 101 20.0 12.3 (3.0–72.0)
DCL 101 5.07 3.4 (0.86–20.9) 101 5.1 3.6 (0.5–21.0)
DIAM 101 32.2 10.9 (10.0–55.0) 101 28.4 10.2 (8.0–54.0)
DEB 100 1.0 0.6 (1.0–3.0) 101 2.0 1.1 (1.0–4.0)
NAX 101 6.0 2.4 (1.0–14.0) 101 5.0 2.8 (0.0–13.0)
HGH 101 44.2 12.7 (18.0–79.0) 101 42.9 11.9 (20.0–77.0)

45
Table 2. Spearman rank correlation coefficients for predictor variables (see caption of
table 1 for variables abbreviations). The α-level (P ≤ 0.05) was Bonferroni corrected
(*P ≤ 0.001).
Variables TEMP PH VOL NBRO DCL DIAM DEB NAX HGH
TEMP 1.00
PH 0.04 1.00
VOL 0.02 -0.07 1.00
NBRO -0.12 -0.02 0.07 1.00
DCL -0.10 0.01 0.05 -0.13 1.00
DIAM 0.12 0.18 0.12 0.11 -0.02 1.00
DEB -0.13 0.18 -0.19 -0.02 0.20 0.12 1.00
NAX 0.08 -0.01 0.46* 0.09 -0.03 0.17 -0.38* 1.00
HGH -0.08 -0.21 0.24* 0.15 -0.05 -0.20 0.04 -0.08 1.00

46
Table 3. Variables in the equation of the multiple logistic regression analysis. See
caption of table 1 for variables abbreviations. β, regression coefficient; W, Wald value;
P, significance value (P ≤ 0.007).
Variables β Wald P
CONST 0.244 0.012 0.912
NBRO 0.065 18.665 0.000
DEB 0.750 9.930 0.002

47
Table 4. Percentage of correct classification of bromeliads in two categories of response
variables: without (0) and with (1) calling males of P. melanomystax. The percentage of
correct predictions of bromeliads without males is called specificity, while that from
bromeliads with males is called sensitivity of the logistic model.
Observed Predicted % Correct
Without (0) With (1)
Without (0) 68 27 71.6
With (1) 26 72 73.5
% Overall 72.5

48
Author Guidelines
INSTRUCTIONS FOR AUTHORS
The British Ecological Society publishes five international journals that between them
cover research in all aspects of the science of ecology. The subject matter and types of
paper accepted vary between journals, but all journals employ a uniform format. Please
follow these instructions carefully to ensure that your manuscript is dealt with speedily.
There are no pages charges for authors.
Journal of Animal Ecology only accepts submissions online. When your manuscript has
been prepared in accordance with the following instructions, please access the online
submission site at http://mc.manuscriptcentral.com/jae-besjournals. You must select the
Journal of Animal Ecology when you log on. Instructions (and a helpline) for the
submission web site are accessible from the 'Get Help Now' icon on the ScholarOne
Manuscripts submission site.
Editorial policy and process Journal of Animal Ecology publishes original papers on any aspect of animal ecology,
from molecular to community ecology. We aim to publish papers that are novel,
interesting and make a substantial contribution to our understanding of animal ecology.
They should also offer insights into issues of general interests to ecologists. Types of
paper published: Standard, Essay Review, Forum and In Focus (by invitation).
To achieve our aim of publishing top-quality papers, we have an editorial process
consisting of two stages; pre-review and peer review.
After submission, a paper enters the pre-review stage. The aim of pre-review is to
identify papers that have the potential to make novel, interesting and significant
contributions to broad ecological areas rather than being mainly taxon- or system-
specific in these respects. Rejecting papers at pre-review that lack the potential we are
looking for ultimately saves everyone time and reduces the burden on our referee
community and editorial system. The pre-review assessment is based on the following
criteria.
1. Does the paper fall within the broad remit of Journal of Animal Ecology?
2. Does the paper address a broad rather than narrow ecological subject area?
3. Does the paper have the potential to make a substantial contribution to the
development of a broad ecological subject area?
4. Is the subject area covered by the paper topical and, hence, potentially of interest
to a wide readership?
5. Is the paper appropriately formatted for Journal of Animal Ecology?
6. Is the paper novel and interesting?
A member of the editorial team undertakes the pre-review assessment, and they then
communicate a decision to the corresponding author if the paper is to be rejected. About
20-30% of papers submitted to the journal will be rejected at this stage.

49
Types of decisions If a paper is not rejected at the pre-review stage it then goes into peer-review. Each
paper is typically reviewed by two independent referees and a member of the editorial
team then makes a decision concerning publication, i.e:
Reject - the paper is not acceptable for publication and re-submission is not
possible,
Decline without prejudice - the paper is rejected but the option is open to submit
a completely new version of the paper that has undergone considerable changes,
Major Revision - the paper requires major changes and needs to go through the
review process again with no guarantee of acceptance, or
Minor Revision - the paper is provisionally accepted, subject to conditions that
need to be addressed in producing a final version of the manuscript.
Journal of Animal Ecology endeavours to maintain a high standard of reviewing
manuscripts, coupled with a prompt turnaround of getting decisions to authors. The
average time from submission to first decision is approximately 40 days.
The Decision of the Editors is final and will only be changed under exceptional
circumstances. Correspondence should be directed to the Managing Editor.
Welfare and legal policy Researchers must have proper regard for conservation and animal welfare
considerations. Attention is drawn to the 'Guidelines for the Treatment of Animals in
Research and Teaching'. Any possible adverse consequences of the work for
populations or individual organisms must be weighed against the possible gains in
knowledge and its practical applications. Authors are required to sign a declaration that
their work conforms to the legal requirements of the country in which it was carried out
(See below), but editors may seek advice from referees on ethical matters and the final
decision will rest with the editors.
This journal is a member of and subscribes to the principles of the Committee on
Publication Ethics.
Typescripts
Papers are expected to be no more than 10 published pages in length (8500 words
maximum), unless otherwise agreed or invited by the editor. There is no page charge to
authors.
Submission Manuscripts must be in English and spelling should conform to the Concise Oxford
Dictionary of Current English.
Lines must be numbered, preferably within pages.
Manuscripts should be in double-spacing.
Use the 'cover letter' section to highlight anything about your paper that needs to
be drawn to the attention of the Editors or the Editorial Office.

50
Editors reserve the right to modify manuscripts that do not conform to scientific,
technical, stylistic or grammatical standards.
During submission, all authors must confirm that:
the work as submitted has not been published or accepted for publication, nor is
being considered for publication elsewhere, either in whole or substantial part;
the work is original and all necessary acknowledgements have been made;
all authors and relevant institutions have read the submitted version of the
manuscript and approve its submission;
all persons entitled to authorship have been so included;
the work conforms to the legal requirements of the country in which it was
carried out, and to accepted international ethical standards, including those
relating to conservation and animal welfare, and to the journal's policy on these
matters (see 'Welfare and Legal Policy' above).
Pre-submission English-language editing
Authors for whom English is a second language may wish to consider having their
manuscript professionally edited before submission to improve the English. A list of
independent suppliers of editing services can be found at
www.blackwellpublishing.com/bauthor/english_language.asp. All services are paid for
and arranged by the author, and use of one of these services does not guarantee
acceptance or preference for publication.
Types of papers
STANDARD PAPERS
A standard paper should not normally be longer than 10 printed pages (8500 words). (A
page of printed text, without figures/tables carries c. 800 words.) The passive voice is
preferred in describing methods and results. The active voice may be used occasionally
to emphasize a personal opinion (typically in Introduction and Discussion sections). The
typescript should be arranged as follows, with each section starting on a separate page.
Title page. This should contain:
1. A concise and informative title (as short as possible). Do not include the
authorities for taxonomic names in the title. Titles with numerical series
designations (I, II, III, etc.) are acceptable provided the editors agreement is
sought and that at least Part II of the series has been submitted and accepted
before Part I is sent to the printer. Such series must begin in one of the journals
of the BES.
2. A list of authors' names with names and addresses of Institutions.
3. The name, address and e-mail address of the correspondence author to whom
proofs will be sent.
4. A running headline of not more than 45 characters.
Summary. This should summarize the main results and conclusions of the paper, using
simple, factual, numbered statements. It must not exceed 350 words.

51
Summaries should be understandable in isolation.
Summaries must start with point 1 describing, for example, the broad conceptual
question addressed by the study
Summaries should end with a final point highlighting the broader conceptual
implication of the results and conclusions of the current study.
Advice for optimizing your Summary (and Title) so that your paper is more
likely to be found in online searches is provided at
http://www.blackwellpublishing.com/bauthor/seo.asp
Key-words. Listed in alphabetical order, the key-words should not exceed 10 words or
short phrases.
Please pay attention to the keywords you select: they should not already appear in the
title or abstract. Rather, they should be selected to draw in readers from wider areas that
might not otherwise pick up your paper when they are using search engines.
Introduction. This should state the reason for doing the work, the nature of the
hypothesis or hypotheses under consideration, and should outline the essential
background.
Materials and methods. This should provide sufficient details of the techniques to
enable the work to be repeated. Do not describe or refer to commonplace statistical tests
in Methods but allude to them briefly in Results.
Results. This should state the results, drawing attention in the text to important details
shown in tables and figures.
Discussion. This should point out the significance of the results in relation to the
reasons for doing the work, and place them in the context of other work.
Acknowledgements. In addition to acknowledging collaborators and research assistants,
include relevant permit numbers (including institutional animal use permits),
acknowledgment of funding sources, and give recognition to nature reserves or other
organizations that made this work possible. Do not acknowledge Editors by name.
References (see Specifications).
Tables (see Specifications). These should be referred to in the text as Table 1, etc. Do
not present the same data in both figure and table form.
Figure legends (see Specifications). Figures should be referred to in the text as Fig. 1,
etc. (note Figs 1 and 2 with no period).
Illustrations (see Specifications) should be referred to as Figures.
REVIEWS
The editors welcome short review articles on topical subjects which are of general

52
interest to the readership. Intending authors are advised to consult with the editors
before preparing essays for submission.
FORUM
This section of the journal is designed to stimulate scientific debate. Contributions may
be short articles presenting new ideas (without data), opinions or responses to material
published in the journal. A cover letter is a requirement for all forum pieces.
IN FOCUS
In Focus articles are short pieces designed to draw attention to papers of high expected
interest in each issue of the journal. The In Focus articles act as mini-reviews that
expand the context of the paper they are featuring, and they are generally commissioned
by the Editors and Associate Editors. Guidelines for invited authors are available on the
ScholarOne Manuscripts site at http://mc.manuscriptcentral.com/jae-besjournals.
BOOK REVIEWS
Journal of Animal Ecology no longer publishes book reviews.
For general tips and guidance for authors or article preparation and submission visit the
Wiley-Blackwell Author Services site.
Specifications
Figures
The publishers would like to receive your artwork in electronic form. Please save vector
graphics (e.g. line artwork) in Encapsulated Postscript Format (EPS), and bitmap files
(e.g. half-tones) in Tagged Image File Format (TIFF). Ideally, vector graphics that have
been saved in a metafile (.WMF) or pict (.PCT) format should be embedded within the
body of the text file. Detailed information on the Wiley-Blackwell digital illustration
standards is available at: http://www.blackwellpublishing.com/bauthor/illustration.asp
Figures should not be boxed (superfluous bounding axes) and tick marks must be on the
inside of the axes. Where possible, figures should fit on a single page in the submitted
paper. In a final version they will generally be reduced in size by about 50% during
production. Wherever possible, they should be sized to fit into a single column width (c.
70mm final size). To make best use of space, you may need to rearrange parts of figures
(e.g. so that they appear side by side). Please ensure that symbols, labels, etc. are large
enough to allow reduction to a final size of c. 8 point, i.e. capital letters will be about 2
mm tall. Lettering should use a sans serif font (e.g. Helvetica and Arial) with capitals
used for the initial letter of the first word only. Bold lettering should not be used. Units
of axes should appear in parentheses after the axis name. Please note that line figures
should be at least 600 dpi and half-tones (photos) should be at least 300 dpi.
Images in the printed version of the Journal of Animal Ecology are in black and white
as it is the policy of the Journal of Animal Ecology for authors to pay the full cost
for colour paper print reproduction (currently £150 for the first figure, £50 thereafter).
Free colour reproduction is available for the on line version: if authors require this, they
should write their figure legend to accommodate both versions of the figure, and
indicate their colour requirements on the Colour Work Agreement Form. This form

53
should be completed in all instances where authors require colour, whether in print or
online. Therefore, at acceptance, please download the form and return it to the
Production Editor (Penny Baker, Wiley-Blackwell, John Wiley & Sons, 9600
Garsington Road, Oxford OX4 2DQ, UK. E-mail: [email protected]). Please
note that if you require colour content your paper cannot be published until this form is
received.
Figure legends Legends should be grouped on a separate sheet. Furnish enough detail so that the figure
can be understood without reference to the text. In the full-text online edition of the
journal, figure legends may be truncated in abbreviated links to the full screen version.
Therefore, the first 100 characters of any legend should inform the reader of key aspects
of the figure.
Tables
Each table should be on a separate page, numbered and accompanied by a title at the
top.
Supporting Information
Journal of Animal Ecology rarely publishes Appendices in the printed version.
However, Supporting Information that is referred to in the text may be made available
in the online version of the article. Guidelines for the preparation of Supporting
Information are available here.
For the printed version, any Appendices should be listed under 'Supporting
Information', and added after the References, with the opening statement: 'The
following Supporting Information is available for this article online' followed by
brief captions for the Appendices/Figs/Tables to be included. These should be
numbered Appendix S1, Fig. S1, Table S1, etc.
Any literature referred to in the Appendix or online Supporting Information should also
be referenced in the Appendix or online Supporting Information so that it is a self-
contained piece of work. This may mean duplicating references if any literature is cited
in both the main text and the Supporting Information.
All Supporting Information should be submitted online as part of the main manuscript.
Please name your online supporting files as „online supporting information' and upload
them with the main document. This allows the submission web site to combine all the
relevant files together but keep them separate when it comes to publication stage.
References References in the text to work by up to three authors should be in full, e.g. (Johnson,
Myers & James 2006). If there are more than three authors, they should always be
abbreviated thus: (Nilsen et al. 2009). When different groups of authors with the same
first author and date occur, they should be cited thus: (Jonsen, Myers & James 2006a;
Jonsen, James & Myers 2006b), then subsequently abbreviated to (Jonsen et al. 2006a,
b). The references in the list should be in alphabetical order with the journal name in
full. The format for papers, entire books, chapters in books, and PhD theses is as
follows.

54
Underwood, N. (2009) Effect of genetic variance in plant quality on the population
dynamics of a herbivorous insect. Journal of Animal Ecology, 78, 839–847.
Jonsen, I.D., Myers, R.A. & James, M.C. (2006) Robust hierarchical state–space models
reveal diel variation in travel rates of migrating leatherback turtles. Journal of Animal
Ecology, 75, 1046–1057.
Nilsen, E.B., Linnell, J.D.C., Odden, J. & Anderson, R. (2009) Climate, season, and
social status modulate the functional response of an efficient stalking predator: the
Eurasian lynx. Journal of Animal Ecology, 78, 741–751
Otto, S.P. & Day, T. (2007) A Biologist's Guide to Mathematical Modeling in Ecology
and Evolution. Princeton University Press, Princeton, New Jersey, USA.
Conway. G. (2007) A Doubly Green Revolution: ecology and food production.
Theoretical Ecology: Principles and Applications, 3rd edn (eds R. May & A. McLean),
pp. 158–171. Oxford University Press, Oxford.
Stevenson, I.R. (1994) Male-biased mortality in Soay sheep. PhD thesis, University of
Cambridge, Cambridge.
References should only be cited as 'in press' if the paper has been accepted for
publication. Other references should be cited as 'unpublished' and not included in the
list. Work not yet accepted for publication may be cited in the text and attributed to its
author as: author name (including initials), unpublished data. In press articles should be
uploaded with the manuscript as „supplementary files‟.
We recommend the use of a tool such as EndNote or Reference Manager for reference
management and formatting.
EndNote reference styles can be searched for here:
http://www.endnote.com/support/enstyles.asp
Reference Manager reference styles can be searched for here:
http://www.refman.com/support/rmstyles.asp
Citations from the World Wide Web
Citations from the world-wide-web are only allowed when alternative hard literature
sources do not exist for the cited information. Authors are asked to ensure that:
Fully authenticated addresses are included in the reference list, along with titles,
years and authors of the sources being cited.
The sites or information sources have sufficient longevity and ease of access for
others to follow up the citation.
The information is of a scientific quality at least equal to that of peer reviewed
information available in learned scientific journals.

55
Units, symbols and abbreviations Authors are requested to use the International System of Units (SI, Système
International d'Unités) where possible for all measurements (see Quantities, Units and
Symbols, 2nd edn (1975) The Royal Society, London). Note that mathematical
expressions should contain symbols not abbreviations. If the paper contains many
symbols, it is recommended that they should be defined as early in the text as possible,
or within a subsection of the Materials and methods section.
Scientific names Give the Latin names of each species in full, together with the authority for its name, at
first mention in the main text. If they appear in the Summary/Abstract, use the common
and Latin name only in the first instance, then the Latin or common name thereafter. If
there are many species, cite a Flora or checklist which may be consulted for authorities
instead of listing them in the text. Do not give authorities for species cited from
published references. Give priority to scientific names in the text (with colloquial names
in parentheses, if desired).
Makers' names When a special piece of equipment has been used it should be described so that the
reader can trace its specifications by writing to the manufacturer; thus: 'Data were
collected using a solid-state data logger (CR21X, Campbell Scientific, Utah, USA)'.
Mathematical material Mathematical expressions should be carefully represented. Suffixes and operators such
as d, log, ln and exp will be set in Roman type; matrices and vectors will be set in bold
type; other algebraic symbols will be set in italic. Make sure that there is no confusion
between similar characters like 'l' (ell) and '1' (one). Also make sure that expressions are
spaced as you would like them to appear, and if there are several equations they should
be identified by eqn 1, etc.
Numbers in tables Do not use an excessive number of digits when writing a decimal number to represent
the mean of a set of measurements (the number of digits should reflect the precision of
the measurement).
Numbers in text Numbers from one to nine should be spelled out except when used with units; e.g. two
eyes but 10 stomata and 5°C.
Accepted papers At the conditionally accepted stage all authors are asked to submit their final manuscript
text in word format with figures and tables conforming to the specifications above and
must include all required forms (nb Colour Artwork form for any colour figures and
Exclusive Licence form).
Exclusive Licence Form
Authors will be required to sign an Exclusive Licence Form (ELF) for all papers
accepted for publication. This can also be downloaded via the 'Instructions and forms'
icon located on the ScholarOne Manuscripts website. Please read carefully before

56
signing: conditions are changed from time to time and may not be the same as the last
time you completed one of these forms.
Signature of the ELF is a condition of publication, and papers will not be passed to the
publisher for production unless a signed form has been received. Please note that
signature of the Exclusive Licence Form does not affect ownership of copyright in the
material. After submission, authors will retain the right to publish their paper in various
media/circumstances (please see the form for further details).
Production tracking is available for your article through Author Services
Author Services enables authors to track their article, once it has been accepted, through
the production process to publication online and in print. Authors can check the status
of their articles online and choose to receive automated e-mails at key stages of
production. The author will receive an e-mail with a unique link that enables them to
register and have their article automatically added to the system. Authors can nominate
up to 10 colleagues to have complimentary access to the article once published on line.
Visit our Author Services site for more details on online production tracking, tips on
article preparation and submission, and more.
Please ensure that a complete e-mail address is provided when submitting the
manuscript. Visit our Author Services site for more details on online production
tracking, tips on article preparation and submission, and more.
Handling the proofs The correspondence author will receive an e-mail alert when the proof, an Acrobat PDF
(portable document format) file, can be downloaded from the e-proofing site. Acrobat
Reader will be required in order to read this file. This software can be downloaded (free
of charge) from the following web site:
www.adobe.com/products/acrobat/readstep2.html
This will enable the file to be opened, read on screen, and printed out for any
corrections to be added. Further instructions will be sent with the proofs. Proofs will be
posted if no e-mail address is available; in your absence, please arrange for a colleague
to access your e-mail to retrieve the proofs.
Alterations in the text, other than printers errors, may be charged to the author. Proofs
must be returned by e-mail, first-class mail, or fax within 3 days of receipt to:
Journal of Animal Ecology,
Wiley-Blackwell
John Wiley & Sons
9600 Garsington Road,
Oxford OX4 2DQ, UK.
Tel: +44 (0) 1865 476477
Fax: +44 (0) 1865 476772
E-mail: [email protected]
Foreign contributors might consider nominating someone in the UK to correct their
proofs for them; they should do this when submitting their revised manuscript. The
editors reserve the right to correct the proofs themselves, using the accepted version of

57
the manuscript, if the authors corrections are overdue and the journal would otherwise
be delayed.
Early View publication
Journal of Animal Ecology is covered by the Early View service. Early View articles
are complete, full-text articles published online in advance of their publication in a
printed issue. Articles are therefore available as soon as they are ready, rather than
having to wait for the next scheduled print issue. They have been fully reviewed,
revised and edited for publication, and the authors' final corrections have been
incorporated. To register to receive an e-mail alert when your Early View article is
published, click here and log-in to Wiley Online Library.
Because Early View articles are in their final, complete form, no changes can be made
after online publication. Articles are given a Digital Object Identifier (DOI), which
allows them to be cited and tracked before allocation to an issue. After print publication,
the DOI remains valid and can continue to be used to cite and access the article. More
information about DOIs can be found at http://www.doi.org/faq.html
OnlineOpen
OnlineOpen is available to authors of primary research articles who wish to make their
article available to non-subscribers on publication, or whose funding agency requires
grantees to archive the final version of their article. With OnlineOpen, the author, the
author's funding agency, or the author's institution pays a fee to ensure that the article is
made available to non-subscribers upon publication via Wiley Online Library, as well as
deposited in the funding agency's preferred archive. For the full list of terms and
conditions, click here.
Any authors wishing to send their paper OnlineOpen will be required to complete the
payment form.
Prior to acceptance there is no requirement to inform an Editorial Office that you intend
to publish your paper OnlineOpen if you do not wish to. All OnlineOpen articles are
treated in the same way as any other article. They go through the journal's standard
peer-review process and will be accepted or rejected based on their own merit.
Offprints
The corresponding author will receive a PDF file of their article free of charge at the
time of publication within an issue of the journal (i.e. once the article is paginated).
Printed offprints are normally dispatched by surface mail c.3 weeks after publication of
the issue in which the paper appears; however, please note that they are sent by surface
mail so delivery of overseas orders may take up to 6 weeks. Please contact the
publishers if they fail to arrive.
The PDF file is e-mailed to the correspondence author at the e-mail address given on the
title page of the paper (unless advised otherwise); therefore, please ensure that the
name, address and e-mail of the correspondence author are clearly indicated on the
manuscript title page if he/she is not the first author of the paper. A copy of the
Publisher's Terms and Conditions for the use of the PDF file will accompany it and the
file can only be distributed in accordance with these requirements. Authors can also

58
nominate up to 10 colleagues to receive complimentary access to their article by
registering for Author Services.