
Aminoácidos, peptídeos e proteínas Prof. Dr. Francisco Prosdocimi.

\V
TOXINAS DO VENENO DE CROTALUS DURISSUS TERRIFICUS.
INTERAÇÃO PROTEÍNA - PROTEÍNA E CINÉTICA DE
TROCA ISOTÔPICA HIDROGÊNIO - TRICIO
José Roberto Rogero
DISSERTAÇÃO E TESE - IEA 119
IEADT-119ABRIL/1979

CONSELHO DELIBERATIVO
MEMBROS
Klaus Reinach — PresidenteRoberto D'Utr.i VazHelcio Modesto da CostaIvano Humbert MarchesiAdmar Cervellini
PARTICIPANTES
Regina Elisabete Azevedo Beretta
Flávio Gori
SUPERINTENDENTE
Rõmulo Ribeiro Pieroni

DISSERTAÇÃO E TESE-IEA 119 ABRIL/1979
IEA- DT- 119
TOXINAS DO VENENO DE CROTALUS DURISSUS TERRIFICUS.
INTERAÇÃO PROTEÍNA - PROTEÍNA E CIN£TtCA DE
TROCA ISOTOPICA HIDROGÊNIO - TRfCIO
Josá Roberto Rogero
Ta«a para obtançlo do Tftulo da "Doutor am CMnciai" -Araa da Bioquímica - Oriantador: Prof. Dr. Joié MouraGonçalvti Aprwantada a dafandida am 26 da agottoda 1978, no Instituto da Química da Univanidada da
Sao Paulo
INSTITUTO DE ENERGIA ATÔMICASAO PAULO-BRASIL

Série DISSERTAÇÃO E TESE IE A
INIS Categories and Ovscriptora
C45
Snakc*
Venoms
Toxins
Proteins
Interact k*v_
Hydrogen
Tritium
Isotopic «xehangt
Nota: A radaçfo, ortografia • ooncaltot ifo da raiponttWIklida dot autor*.

SUMARIO
Página
I - INTRODUÇÃO E PROPÓSITOS 1
|| _ MATERIAIS E MÉTODOS 3
11.1 - Purificação de Crotamina, Crotapotin e Fosfolipase A 3
11.2 — Eletroforese em Gel de Poliacrilamida 4
11.2.1 - Eletroforese de Crotapotin 4
11.2.2 - Eletroforese de Crotamina e Fosfolipase A 5
11.3 - Análise de Aminoácidos 5
11.3.1 - Análise do Padrão de Aminoácidos 511.3.2 - Análise das Proteínas 5
11.4 - Medida da Atividade Enzimática da Fosfolipase A 5
11.5 - Determinação da Presença de Fatores Coagulantes 6
11.6 — Dosagens das Proteínas 6
11.7 — Curvas de Absorbância das Pro temas 6
11.8 - Determinação de Grupos SH Livres 7
11.9 - Marcação de Crotapotin com 1 2 S I 7
11.9.1 - Controle Eletroforético do Pico de Crotapotin I 2 5 I 7
11.10 — Determinação da DLg Q das Proteínas 7
11.11 - Medida da Velocidade de Troca Hidrogênio-Trfcio 8
11.11.1 - Medida do "Exchange Out" da Crotamina 9
11.11.2 - Medida do "Exchange Out" da Fosfolipase A 10
11.11.3 - Medida do "Exchange Out" da Crotapotin 10
11.11.4 - Medida do "Exchange Out" da Crotoxina 10
11.11.5- Medida da Radioatividade 11
11.11.6 - Cálculo da Eficiência de Contagem 11
11.11.7 - Cálculo *> Número de Hidrogénios Teoricamente Trocáwii 11
11.11.8 - Ajustes aos Dados Obtidos p ela Troca Isotoplca Hidrogtnio-Trício 12
11.12 - Interaçffo das Proteínas 12
11.12.1 - Mídida da Area dos Picos 12
11.12.2 - Interação entre Fosfolipase A e Crotapotin 13
11.12.3 - Cromatografia de Crotoxina 13
11.12.4-InteracSo entre Fosfolipase A n Crotapotin Contendo Crotapotin111!
como Traçador 13
M.12.6 - InteracSo entre Crotamina t Crotapotin 13
11.12.6 - Interacfo entre Crotamina a Crotapotin Contando Crotapotin-1 a * I como
Traçador 13

Página
IM - RESULTADOS 14
111.1 - Purificação de Crotamina, Crotapotin e Fosfolipase A 14
111.2 - Ele^roforese em Gel de Poliacrilamida 20
1(1.3 - Presença de Fatores Coagulantes 20
\\\.A - Dosagem das Proteínas 20
111.5 - Análise de Aminoácidos 20
111.6 - Absorção das Proteínas no UV 20
111.7 - Medida da Atividade Enzimática da Fosfolipast A 20
II 1.8 - Determinação de Grupos SH Livres 24
If 1.9 - Interação entre Fosfolipase A e Crotapotin '. 24
111.10 - Interação entre Crotapotin e Crotamina 24
111.11 - Interação entre Fosfolipase A e Crotamina com Crotapotin Contando
Crotapotin-12SI como Traçador 24
111.12 - DeterminaçSo da DL g 0 30
111.13- Medida da Velocidade de Troca Hidrogftnio-Trício 30
111.13.1 - Troca H/T da Fosfolipa» A 32
111.13.2 - Troca H/T da Crotapotin 32
111.13.3 - Troca H/T da Crotoxina 37
111.13.4 - Troca H/T da Crotamina 37
111.14 - Medida da Radioatividade 37
IV - DISCUSSÃO 42
V - CONCLUSÕES 48
APÊNDICE 1 60
APÊNDICE 2 71
REFERÊNCIAS BIBLIOGRÁFICAS 78

TOXINAS DO VENENO DE CROTALUS DURISSUS TERRIFICUS.
INTERAÇÃO PROTEÍNA - PROTEllMA E CINÉTICA DE
TROCA ISOTÔPICA HIDROGÊNIO - TRICIO
José Roberto Roger o
RESUMO
A Fosfolipase A, Crotapotin e CroMirmu tuidm purificadas a part» do veneno de Crotalus duriaui MrrHkuf.Mediante análise de aminoácidos das proteínas ficou estabelecido um pero molecular 13400 para Fosfolipase A. 8300para Crotapotin e 4880 para Crotamina. Crotapotin interage com Fosfolipase A (DLgg = 0,55 mg/Kg peso) dando umcomplexo estável durante a filtraçSo em gel Sephadex G 75 fino, quando a proporção molar de interação é 1:1. O
complexo for mo do é cerca de 10 vezes mais tóxico do que a Fosfolipase A so2inha, quando injetado i.p. em
camundor.gos (DLJ-J-, = 0,05 mg/Kg peso). Estes fato;, levam a supor que a Crotoxina, toxina neurotôxica cristalizavel doveneno de Crotalus d. terrificus, resulta da interação Crotapotin-Fosfolipase A na proporção molar 1:1. A Crotapotin
interage com Crotamina (OL r , " 0,'' mg/Kg peso), formando um complexo dissociável durante a filtração emgtl
Sephadex G 75 fino. Quando o complexo formado " in vitro" na proporção molar de 1 1 é injetado em camundongosapresenta uma toxicidade cerca de 4 ve/cs maior do que a Crotamina sozinha ( D L , Q = 0,2 mg/Kg peso). O estudo d»cinética de troc? isotópica H/T foi feito em Fosfolipase A, Crotapotin, Crotamina e Crotoxina, utilizando o método d»filtraçSo em colunas de Sephadex G 20 "coarse". O melhor ajuste dos dados experimentais de troca H/T de Fosfolipase
A apresenta uma equação que é composta de duas expon*nciais, caracterizando duas classes de hidrogênios. Mostra
também que a Fosfolipase A tem 68% dos hidrogênios teoricamente trocáveis, muito rápidos e, nSo apresenta
hidrogênios com velocidades de troca próxima daqueles envolvidos em alfa-hélice. A Crotapotin pela mesma análise nSo
apresenta hidrogênios envolvidos em alfa-hélice e 83% dos teoricamente trocáveis, sá"o rapidamente trocados com o
solvente. A Crotamina pela medida da cinética de troca H/T, apôs um aquecimento inicial com H 2 0 a 70°C por 2
horas, mostra que dos hidrogênios teoricamente trocáveis, 31% sSo lentos. Quando o aquecimento inicial é prolongado
na mesma temperatura por 18 horas, os hidrogênios lentos diminuem para 11%. A aplicação da técnica da medida da -
cinética de troca H/T na Crotoxina, mostra o aparecimento de 26 hidrogênios que trocam com velocidades semelhante»
aqueles envolvido- em alfa-hélice. Este fato sugara que possivelmente ocorreria uma mudança conformacional após a
interação Fosfolipase A - Crotanitin, originando a Crotoxina.
I - INTRODUÇÃO E PROPÓSITOS
A Crotoxina, proteína neurotôxica do veneno Crotalus duriuus terrificus, foi pela primeira vezisolada e cristalizada por Stotta e í raenkel-Conrat , em 1 938. Estudos por eletroforese com aparelho
de Tíselius'601 e por ultraoentrifugaçlo137', mostraram ser a Crotoxina uma proteína homogênea depeso molecular aproximadamente 30000.
Durante algum tempo a Crotoxina permaneceu como uma proteína homogênea • cristalizavelapesar de apresentar além da neurotoxicidade, atividade fosfolipasica ' .
Em 1 955, Neumann e Habermanrv ' prcjrando retirar da Crotoxina a atividade
fosfolipásica, identificaram uma nova proteína que continha a mesma açlo tóxica da Crotoxina, porém
Aprovada para publicaclo em agosto/1978.

com uma atividade fosfolipásica menor.'A esta nova proteína eles denominaram Crotactina. Entretantoesta nova toxina não pode ser purificada e isolada em quantidade que possibilitasse sua caracterizaçãomais detalhada139'401
A primeira evidência definitiva da existência na Crotoxina de duas proteínas diferentes,apareceu com o tratamento da proteína pelo fluordinitrobenzeno, que em função da solubilidade emágua possibilitou o isolamento de duas dinitrofenil-proteínas que se diferenciavam em muito nas suascomposições de aminoácidos .
(43 81 82 83\Após um longo período onde so lemus trabalhos realizados por Vital brasil et aiií •'•"'•o*-°*'
sobre a farmacologia da Crotoxina, dois grupos de pesquisadores liderados por Hendon eFraenkel-Conrat 9 Rubsamen et alii publicam no mesmo ano de 1 971, o fracionamento daCrotoxina em coluna de troca iònica, isolando e caracterizando duas p. oteínas diferentes.
Estas proteínas se diferenciavam em primeiro lugar pela composição de aminoácidos, umaextremamente ácida e outra extremamente básica e, pela toxicidade, que mostrava que a proteína ácidanão era tóxica e a proteína básica tinha toxicidade baixa. A proteína ácida potenciava a básica e por issofoi chamada Crotapotin. A proteína básica foi identificada como sendo uma Fosfolipase A (EC 3.I.I.4.).As duas proteínas se recombinavam dando origem à Crotoxina.
As características das subunidades da Crotoxina e a interação entre elas, têm merecido umasérie de publicações no* últimos anos1 5 '4 2-4 8-4 9 7 0 1 .
A Crotamina, proteína tóxica de uma variedade de veneno de Crotalus d. terrificus, foi isolada 4caracterizada por eletroforese por Gonçalves e Vieira'361, em 1 950.
A Crotamina anteriormente, purificada por Amberlite IRC50 u 2 ) e SP Sephadex C25 ( 5 9 ) ,estudada quanto ao seu peso molecular'33 '34 ' e quanto a sua ação farmacológica'10 '31 '32 ' , teve suaseqüência de aminoácidos determinada recentemente mostrando um peso molecular de 4880* 5 8 ) .
É uma proteína muito básica de pi: 10,3 que apresenta, quase sempre, na purificação doveneno global por filtraçâo em gel Sephadex G 75 uma contaminação com uma fração ácida revelada porCheymol et alii após cromatografia em troca iònica com Carboximetil Sephadex C 25.
Os propósitos deste trabalho foram em primeiro lugar pela técnica de filtraçâo em gel, definirnas toxinas purificadas as estequiometrias das interações Crotapotin-Fosfolipase A eCrotapotin-Crotamina, bem como, a toxicidade dos complexos formados;em segundo lugar aplicando atécnica de troca isotópica hidrogênio-trício, estudar as possíveis mudanças conformacionais queocorreriam pela interação entre as proteínas citadas.
A técnica de filtraçâo em gel para determinar constantes de associação de proteínas, interaçõesproteínas-peptídeos, etc. tem sido muito usada com base no trabalho de Hummel e Dreyer1501 com ochamado "método do poço"* 1 ' 2 8 ' 5 6 1 .
No nosso caso a intenção primordial foi a de tentar isolar o complexo formado e daídeterminar as proporções molares de interações. As participações da Crotapotin nos complexos com aFosfolipase A e com Crotamina, foi melhor visualizada com o uso de Crotapotin marcada com I J 5 I ,como traçador.
O estudo do comportamento estrutural de uma proteína em solução pela técnica de trocaisoiópíca hidrogftnio-deutério, foi feito pela primeira vez na Insulina por Llnderstrom-Lang(61), am1955.
Vários trabalhos se seguiram, utilizando sempre o mesmo método inicial, aplicando-o amproteínas como Riboni-clea$e(61> e a Lisozima'5 2 '5 3 ' . Oestes trabalnos foi possível destacar qua

peptídeos simples ou polipeptídeos com conformação ao acaso em solução aquosa, trocavam seushidrogênios labels (aqueles ligados a O, N, S) cem o solvente, em poucos m>nutos. Por outro lado, umaproteína nativa troca os hidrogênios peptídicos com uma velocidade consideravelmente mais baixa. Esteshidrogênios da proteína que trocavam com o solvente com velocidades muito baixas, foram inicialmentedefinidos como participantes das pontes de hidrogênio de uma alfa hélice. Mais tarde porém, com o usoda técnica de RX para esclarecer as estruturas conformacionais de alçjmas proteínas, verificou-se quenão havia concordância entre as porcentagens de alfa hélice e as porcentagens de hidrogênios lentosobtidos pela troca H/D ou H/T.
A técnica original de Linderstrom-Lang que envolvia uma série muito grande de operações, taiscomo, I iofilização, aquecimento a 60°C, congelamento, etc. foi elegantemente modificada porEnglander1201, em 1 963.
O autor passou a utilizar trício e a retirar o excesso do isótopo por filtração rápida em
pequenas colunas de gel Sephadex G 25 "coarse". Pela quantidade de trício carregada pela proteína,
podia-se calcular o número de hidrogênios que foram trocados.
A técnica desenvolvida por Englarder passou a ser muito utilizada no estudo da conformação demuitos compostos, tais como polipeptídeos'2 4 '3 0 '6 9 '8 4 1 , proteínas'7 '1 4 '1 5 -8 8 1 , ácidos nucléicos'731 ehormônios de crescimento . As colunas de gel Sephadex foram algumas vezes substituídas por umamicrodiálise rápida, com resultados muito semelhantes .
No nosso trabalho aplicamos a técnica de medida da velocidade de troca isotópica
hidrogênio-trício, com a utilização de colunas de gel Sephadex G 25 "coarse".
A técnica foi inicialmente empregada às proteínas isoladas, para nos dar uma idéia daconformação destas em solução e, em seguida foi usada no estudo das possíveis mudançasconformacionais que ocorreriam pela interação Fosfolipase A-Crotapotin.
II - MATERIAIS E MÉTODOS
11.1 — Purificação de Crotamina, Crotapotin e Fosfolipase A
As proteínas usadas neste trabalho foram purificadas a partir do veneno bruto e seco decascavel brasileira (Crotalus durissus terrificus)
Foram trabalhados dois tipos de venenos, sendo que só um deles denominado por Gonçalves em
1 9 5 6 ( 3 2 ) variedade crotaminicus, continha crotamina. Em ambos os casos a tecnologia de purificação
foi a mesma, diferenciando-se apenas sobre qual proteína se desejava obter.
Os métodos utilizados para essa purificação foram os empregados em 1 971, por Hendon a
Fraenkel-Conrat'47' e Rubsamen et alii , com algumas modificações.
Cerca de 200 mg de veneno bruto seco, foram dissolvidos em 10 ml de ácido aoétioo 0,1 M,separando sobrenadanto por centrifugação a 10 000 rpm, e desprezando o precipitado. O sobrenadantefoi submetido â filtração em uma coluna de 3 cm x 80 cm de gel Sephadex G 75 fino. Com um fluxo de0,5 ml por minuto recolheu-se 10 ml por tubo de efluente num coletor de frações Gilson, acompanhandoo perfil cromatoyiatico com um >nonitor de UV equipado cum filtro para 280 nm e regisuaaut.
A maior quantidade em massa e consequentemente, o maior pico de absorbância a 280 nm doperfil cromatográfico, pertence a Crotoxina.
Os tubos correspondentes ao pico de Crotoxina foram separados, juntados e liofilizadot num

l iof i l izador V i t t i s , modelo 10-100. Quando o veneno contém Crotamina, aparece no perfil
cromatográfico mais um pico logo após a Crotoxina, ocasionando uma separação mais difícil, quase
sempre com contaminação cruzada, das duas proteínas.
A Crotoxina liofilizada foi algumas vezes recromatografada nas mesmas condições usadas para oveneno, sem porém mostrar melhoria no seu estado de pureza.
O segundo passo da purificação foi o fracionamento da Crotoxina em coluna de troca ibnica,
realizada a 4°C.
Cerca de 100 mg de Crotoxina foram dissolvidos em 5 ml de tampão ácido formico-formiato de
amônio 0,05 M, pH 3,5. A esta mistura levemente turva, acrescentou-se algumas gotas de ácido fórmico
diluído até obtenção de uma solução límpida.
A amostra assim trabalhada, foi colocada sobre uma coluna de 0,9 cm x 40 cm de SP Sephadex
C 25, equilibrado com tampão formiato 0,05 M, pH 3,5. A eluição foi feita com volume total de 400 ml
de um gradiente linear salino de 0 a 3 M de NaCI.
O perfil cromatográfico foi acompanhado através de um monitor UV Gilson equipado com
registrador e, frações de 4 ml do efluente foram coletadas num coletor de frações também Gilson, com
um fluxo de 0,2 ml/minuto.
Os picos correspondentes a Crotapotin, Fosfolipase A e Crotamina, foram recolhidos •
separadamente dialisados contra água deionizada até reação negativa para cloreto com nitrato do prata.
Os sacos de diálise utilizados foram previamente tratados com uma mistura de anidridoacéticc-piridina, na proporção de 1:1, por 12 horas, para redução do diâmetro dos poros, afim de evitarperda de proteínas no processo de diálise.
A Crotamina e a Fosfolipase A foram recromatografadas várias vezes em SP Sephadex C 25 nas
mesmas condições anteriores, até obter-se um pico único e homogêneo de cada proteína.
A Crotapotin foi recromatografada várias vezes em colunas de 0,9 cm x 30 cm de DEAE
Sephadex A 25, equilibrado com tampão formiato 0,05 M, pH 3,5 e aluída com 300 ml de um gradient»
linear salino de 0 a 3 M de NaCI, afim de se obter um pico único e homogêneo de Crotapotin.
As proteínas cromatograficamente puras foram dialisadas, liofilizadas e estocadas sob vácuo em
dessecadores mantidos na geladeira.
11.2 - Eletroforese em Gel de Poliacrilamida
O processo empregado foi o descrito por Davis112' em 1 964, usando-* equipamento específico
da Canalco, modelo 120, com tubos de vidro de 0,4 cm por 7,5 cm.
11.2.1 - EletrofomM d* Crotapotin
A eletroforese foi feita em tubos contendo gel de poliacrilamida a 7%. Em cada tubo colocou-st50 MI de amostra na concentração de 2 mg de Crotapotin por ml de tampfo Tris-gliclna pH9,6. Acorrida foi efetuada utilizando-se o mesmo tampfo nas cubas, o eletrodo colocado na posição normal(polo positivo na part* inferior) e uma corrente de 3 mA por tubo, dwani» UWÍ hora.
Após a corrida o gel fci retirado dos tubos e colocado por 30 minutos em ácido tricloroaoétkco

(TCA) a 12% para fixaçSo da proteína. Em seguida, o gel foi lavado varias vezes com água o corado comComassie Brilliant blue R 250. A descoloroçSo foi feita com lavagens seguidas com uma mistura demetanol :ácido acético:HaO na proporçSo em volume uc •50:75:875.
Após a descoloração os géis foram fotografados.
11.2.2 - Eletroforese de Crotamina e Fosfolipase A
A eletroforese foi realizada em tubos contendo gel de poliacrilamida a 15%. O tampSo utilizadofoi o de beta alanina - ácido acético, pH 4,3 e a concentração da amostra de 2 mg/ml do mesmotampão. Efetuou se a corrida com 3 mA por tubo, durante uma hora. Os géis foram fixados, cor ados edescorados como no item 11.2.1.
11.3 — Análise de Aminoácidos
A composição de aminoácidos foi determinada nas proteínas que mostraram apenas uma bandana eletroforese em gel de poliacrilamida. Utilizou-sf analisador de aminoácidos Beckman, modelo 120 C.
11.3.1 - Análise do Padrão de Aminoácidos
Uma solução estoque padronizada (Aminoacid Calibration Mixture Beckman' serviu de padrão
pa ra determinar as constantes relativas às áreas correspondentes a cada aminoácido.
Amostras contendo 0,1 /jmol de cada aminoácido foram diluídas em tampão citrato 0,2 N,
pH 2,2 e analisadas nas colunas contendo: resina PA 35 (Custom Resin type PA 35 - Beckman) eluídas
com tampão citrato 0,35 M, pH 5,8 para os aminoácidos básicos; resina AA 15 (Custom Resin type
AA 15 - Beckman) eluídas com tampão citrato 0,2 M, pH 3,28 e, em seguida com mesmo tampão com
pH 4,25, para os aminoácidos neutros e ácidos.
11.3.2 - Análise das Proteínas
Dois miligramas das proteínas (Cro tapo tin, Fosfolipase A e Crotamina) foram dissolvidos em1,0 ml de ácido clorídrico 6 N , em ampolas especiais que permitiram, após dissolução das proteínas, oseu fechamento sob vácuo. Em seguida, as amostras foram colocadas a 120° C por 10 h, 20 h e 24 h,para a hidrólise.
Após o tratamento as amostras foram lavadas três vezes com água bidestilada e secadas sobvácuo na presença de pentóxido de fósforo. Em seguida dissolvidas em 0,5 ml de tampão citrato •analisadas segundo procedimento usual.
11.4 - Medida da Atividade Enzimática da FosfolipaM A
O processo empregado foi o descrito por Kibermann et a l . ( 4 1 > onde uma suspensão de gema da
ovo 4 utilizada como substrato. A Fosfolipase A age na lecitina presente na gema, dando a lisolecitina
que na presença de glóbulos vermelhos provoca hemólise, que é quantificada pela absorblncia a 578 nm.
Foi utilizado para as leituras de absorbflncia um espactrofotometro Carl Zeiss, modelo PMQ I I .
A curva padrío de hemólise foi obtida com a Fosfolipase A pura da seguinte maneira: Q, 6, 20,
30, 40, 50 ^g de Fosfolipase A por 60^1 de solução fisiológica foram pipetados sobre 60pi de gema de

ovo fresca e, logo após agitação as amostras foram incubadas a 37°C por duas horas. Em seguida, 4 mlde suspensão de glóbulos vermelhos a 0,6% em soluçSo fisiológica foram adicionadas aos tubosprá-incubados de Fosfolipase A de gema de ovo. Os glóbulos vermelhos utilizados foram obtidos desangue de rato, lavados com soluçSo fisiológica e separados por centrifugação repetindo-se essa operaçãopor três ve/es.
Os tubos contendo Fosfolipase A, gema de ovo e glóbulos vermelhos foram agitados eincubados a 37°C por 40 minutos, ogo a seguir as amostras foram centrifugadas a 1500 rpm por 10minutos, e os sobrenadantes foram lidos a 578 nm.
Todos os outros testes de atividade enzimática de Fosfolipase A foram realizados usando-sesolução contendo 1 mg de Fosfolipase A/ml solução fisiológica, calculando-se porcentagem de hemôliseatravés da curva padrão.
11.5 — Determinação da Presença de Fatores Coagulantes
O veneno de Crotalus d. terrificus é muito rico em enzimas que tem a capacidade de coagular
outras proteínas. A estas enzimas ou complexos enzimáticos proteolfticos denomina-se "coagulase" ou
"fatores de coagulaçSo".
Para certificar-se de que a Crotamina, Crotapotin e Fosfolipase A não apresentavam
contaminação pela "coagulase", aplicou-se nas proteínas puras o teste descrito por Gonçalves'32' em
1 956, que usa a coagulação do fibrinogénio: a 0 , 100, 200, 500nq de Fosfolipase A, Crotamina e
Crotapotin, adicionou-se 1 ml de solução de fibrinogénio humano a 1 mg/ml de solução fisiológica.
Usou-se como padrão positivo 100/ig de veneno bruto de cascavel.
As amostras foram incubadas a temperatura ambiente por 30 minutos e observadas quanto ao
aparecimento do coágulo.
11.6 — Dosagem das Proteína»
A determinação do conteúdo protéico foi feito pelo • tétodo descrito por Lowry et a l i i ( 6 2 ) em
1 951, usando-se soro albumina bovina para curva padrão.
O coeficiente de extinção molar foi determinado pela absorbência a 280 nm de soluções de1 mg/ml, em cubas de quartzo com um cm de percurso óptico, no espectrofotômetro Carl Zeiss PMQ l i .
11.7 - Curvas de Absorbêneias das Proteína*
As curvas de absorbancfas da Fosfolipase A, Crotamina a Crotapotin foram realizadas noespectrofotômetro Carl Zeiss, modelo PMQ II utilizando-se cubas da quartzo com 1 cm da percursoóptico. Para Fosfolipase A e Crotapotin, absorbánda foi medida em tampão Trls 0,06 M, pH 8,06, na'concentração de t,36 mg/rm a 0,8 mg/ml respectivamente. Para a Crotamina, usou-se tampão tostato0,01 M, pH 7,3, na concentração de 1 mg/ml.
I I J - Detefmlnaçfo cte Grupos SH Livras
Para a constatação da existência da grupos SH livres nas proteínas, utlllzou-ss o mtftodo descritopor E l l m a n ' 1 * 1 ~ãm~ 1 9 6 9 , que usa a cistefna como padr fo • o reegente ácido6,6'dlt.obis-2-n!trobenz6ico-SJgma.

Inti(M)UZIU-SH na técnica as modificações piopostas nor Simizu em 1972, que seguem: a0,5 ml d d solução de pmteínas i:un tendo 0,1 mij, juntou-se 0,88 ml de tampão Tris 0,1 M, pH 8,0,0,03 ml d«í solução de EDTA 0,1 M no mesmo tampão e 3,0 ml de água destilada para completar ovolume, t m seguida, adicionou-se à mistura 0,02 ml do reativo de Ellrnan (20 mg de ácido5,S'ditiol>i$'2-mtrohKn7Óico em 5 ml de tampão Tris 0,1 M, pH 8,0).
Após 10 minutos fez-se a leitura de absorbância em 412 nm contra branco contendo somenteTris, EDTA, água e OTNB.
Para esclaiecer a existência cm yrupos SH livres, porém "escondidos" e não acessíveis aoreagente, adicionou-se ao meio 0,06 ml de uréia 0,01 M, na tentativa de expor à reação os SH livresporventura existentes.
11.9 - Marcação de Crotapotin com ''s I
O método para marcação de proteína com I 3 I I , foi publicado por Greenwood et alii , em1963.
A seqüência das reações foi a seguinte: a 5 M9 àe Crotapotin dissolvidos em 0,5 ml de tampãofosfato 0,25 M, de pH 7,4, juntou-se um mCi de N a 1 2 5 l contidos em aproximadamente 2/JI. A seguir,juntou-se 20 pI de urna solução (2,65 mg/ml de tampão fosfato) do oxidante cloraminaT.Imediatamente após, juntou-se 20pi de uma solução de metabissuifito de sódio (4,8 mg/ml de tampãofosfato), porá parar a reação.
A proteína marcada foi então purificada por filtraçâo em gel Sephadex G 50, empacotado numa
coluna de 35 cm:1 cm, previamente equilibrado com o mesmo tampão utilizado para marcação e que
serviu para eluir a coluna num fluxo de 1 ml/minuto.
Os tubos resultantes da eluicão, contendo aproximadamente 1 ml cada um, foram contados nocontador de radiação gama Tobor (Nuclear Chicago) para a identificação do pico de proteína radioativa.
11.9.1 - Controle Eletroforético do Pico de Crotapotin I 1 5 I
Pa.a controle da existência de iodeto livre ou de material radioativo degradado na amostracoletada, foi efetuada uma eletroforese da seguinte maneira; alíquotas das amostras foram colocadas emfitas (papel Whatman n° 1) de 2,5cm:33cm e submetidas a eletroforese por uma hora em tampiofosfato 0,25 M, pH 7,4, com 3 m A por fita. Utilizou-se Nal como carregador para localizar o 1 1 S I livraporventura existente.
Após a eletroforese, as fitas foram reveladas com solução de acetato da chumbo a 10%. Em
seguida, as fitas foram cortadas no sentido transversal, em pedaços de 1 cm e, contados no contador d *
radiação gama (Nuclear Chicago - série 1185).
Foram utilizados produtos marcados contendo no máximo 5% de iodo na forma de iodeto e
com ausência de frações protéicas degradadas.
11.10 - Determinação da DL g Q das Proteínas
A determinação da toxicidade da Crotapotin, Fosfolipase A • Crotamina foi feita pelo calculo
da dose letal 60% pelo método de Reed e Muench'7 4 ' , em 1 937.

A medkia da Dl S() foi tealtzada em camundongos suissos com peso aproximado de 25g cada
um, pela via intru|tt>Mtoneal, utilizando 6 grupos de 6 camundoiigos cada, mais um grupo controle.
De cada protefna foram feitas as seguintes diluições: 1/1, 1/2, 1/4, 1/8, 1/16, e 1/32, sendo adiluição inicial da proteína 1/1 de 500/ ig/ml de solução fisiológica.
A : seis diluições acima foram injetadas num volume de 0,2 ml usando para cada diluiçSo umgrupo de 6 animais.
As mortes e sobrevidas foram anotaoas 24 horas após as injeções. O cálculo foi feito da seguintemaneira:
X = % de mortalidade na diluiçSo logo acima do valor que provocou 50% de morte ou
mais.
Y = % de mortalidade na diluiçSo logo abaixo do valor que provocou 50% de morte ou
mais.
Z = diluição inferior ao valor que provocou 50% de morte,
d.p. = distância proporcional,
log 2 = logarítmo decimal do fator de diluição, no nosso caso 2.
50 - Yd.p. =
X - Y
C = logZ + (dp. x log 2)
O antilog de C forneceu a diluição exata em que ocorreu 50% de morte, e, consequentemente aconcentração em mg/Kg de peso.
Quando da medida da D L 5 0 dos complexos Crotapotin-Fosfolipase A e Crotapotin-Crotamina,realizou-se sempre a mistura das proteínas mantendo a primeira ailuíçáo 500pg de proteína total, obtidapela soma das massas das duas proteínas misturadas, mas em proporções molares de 1:1. 0 cálculo daD L 5 0 do complexo foi feito da mesma maneira descrita, porém deu-se maior importância è quantidadede proteína tóxica injetada (Crotamina ou Fosfolipase A), comparando-se o resultado obtido, com oencontrado pela injeçSo da proteína tóxica sozinha.
11.11 - Medida da Velocidade de Troca Isotopica Hidrogênio-Trlcio
A medida da velocidade de troca do trício incorporado a proteína com o solvente, "exchange
out", foi realizada usando-se a técnica descrita por Englander(20), em 1 963. A técnica foi aplicada a
Crotamina, Fosfolipase A, Crotapotin a Crotoxina cristalizada 3 vazas a gentilmente fornecida pela
equipe do professor Oswaldo Vital Brasil da UNICAMP-SP.
A técnica publicada por Englandar consta dos seguintes pastos: Inicialmente a proteína 4
Incubada am determinadas condições de pH e temperatura na presença de áflua triciada numa
concentraçlo aproximada de 10* cpm/ml (contagens por minuto), a seguir o excesso de trício tf retirado
pela filtreçfo rápida em coluna de gel Sephadex G 25 grosso de 3 cnr.8 cm, empacotada a equilibrada

o
com o tampão escolhido. Os tuhos que contém a proteína triciada sSo recolhidos e os seus volumesjuntados; a partir il«ste instante inicia-se a contagem do tempo de "exchange out". Retira-se alíquotas detempos em tempos e f iltta se cm outra coluna nas mesmas condições anteriores. Após a eluiçSo, nostubos que contém o pico de proteína determina se a absorbância a 280 nm e a quantidade deradioatividade; o cálculo do número de hidrogênios por mol de proteína que permaneceram nãotrocados, ou seja, cujo It feio nãn passou para o solvente decorrido determinado tempo, é feito daseguinte maneir.i:
C/An° Ha/mol = — x 111 x t
onde:
Ha = hidrogênios aparentes
C - contagem da radioatividade em cpm/ml carregada pela proteína
CQ = radioatividade inicial de incubaçlo em cpm/ml
A = absorbância das amostras a 280 nm
111 = concentração de átomos de hidrogênio na água
e = coeficiente de absorção molar
Na realidade, o número de hidrogênio por mol é calculado utilizando-se um valor médio dasatividades específicas (C/A) dos tubos que compõem o pico da proteína. £ possível obter na região demaior concentração de proteína, pelo menos 4 valores de C/A que variam entre si num máximo de 5%e, somente a média destes valores é que foi utilizada para calcular o n? Ha/mol, no tempo escolhido.
11.11.1 - Medida do "Exchange-Out" em Crotamina
As colunas para separação do excesso de 3 H2 0 foram empacotadas com gel Sephadex G 25grosso, previamente equilibrado com tampão formiato 0,1 M, pH 3,0. Contendo 3 cm x 8 cm de gel. ascolunas foram mantidas à temperatura de 3°C por meio de um banho de circulação Savant e eluídascom o mesmo tampão mantido também a 3°C.
0 fluxo das colunas foi sempre fixado em 1 ml/10 segundos e a troca dos tubos foi feitamanualmente de 10 em 10 segundos.
Foram utilizadas duas experiências com Crotamina, variando apenas o tempo de incubaçSo daproteína com água triciada.
Na primeira, 12 mg de Crotamina foram dissolvidos em. 1 ml da tampão formiato 0,1 M,pH 3,0, e em seguida adicionou-se gotas de 3H2O na concentração aproximada de 4 mCi/ml dando unwmedida da radioatividade inicial de aproximadamente 10* cpm/ml. Uma alíquota desta amostra foiretirada para determinação da radioatividade inicial. Em seguida a amostra foi incubada por 2 horas tmbanho termostatizado a 70°C. Apôs a incubacão e resfriamento, a amostra foi colocada em banho dtgelo.
Na segunda experiíncia, o procedimento foi o mesmo mudando» apenas o tampo dt Incubaçlopara 18 hora? a 70°C.

to
Os passos seguintes foram: filtração na primeira coluna de G 25, reunião dos volumes dos tubosque contém o pico de proteína, iniciando-se o tempo de contagem de saída do trício da proteína para osolvente; d«? tempos em tempos retirou se 1 ml deste "pool" para filtração numa segunda coluna e,final.netite, calculou-se a atividade específica C/A de cada tubo, a seguir usada para obtenção do n9 de
H/mol ile proteína.
O coeficiente de extinção inoiar usado tot de 11650 M~' x cm'*, calculado pela absorção a
280nrn de uma solução de 1 mg de Crotamina por ml de tampão formiato 0,1 M, pH 3,0, i.sondo-se um
peso molecular 48B01581 .
11.11.2 - Medida do "Exchange-Out" em Fosfolipase A
Cerca de 16 mg de Fosfolipase A foram dissolvidos em 1 ml de tampão formiato 0,1 M, pH 3,0.
Adicionou se gotas de água tricíada suficientes para dar uma radioatividade inicial próxima de 10s
cpm/ml e, incubou-se a 70°C por 2 horas.
Os passos seguintes foram os mesmos descritos nc item 11.11.1, utilizando-se as mesmas
condições para a fiitração.
O valor do coeficiente extinção molar usado no cálculo do n? H/mol, foi calculado pela
abscrçlo a 280 nm de uma solução 1 mg de Fosfolipase A/ml de tampão formiato 0,1 M, pH 3,0. Este
valor foi 22512 VT ' x c m ' ' , baseado num peso molecular 13400 calculado por análise de aminoácidos.
11.11.3 - Medida do "Exchange-Out" em Crotapotin
Cerca de 20 mg de Crotapotin foram dissolvidos em 1 ml de tampão formiato, 0,1 M pH 3,0.Adicionou-se a seguir gotas de 3 H 2 O para obter uma contagem de radioatividade final próxima de 10s
cpm/ml. A amostra foi então colocada para incubar em banho de água mantido a 60°C por 2 horas.
As colunas para filtração, bem como, os demais detalhes de procedimento, foram estabelecidosnos itens anteriores.
0 valor usado do coeficiente de extinção molar da Crotapotin foi 9074 M ' 1 x c m ' 1 , calculado .da mesma maneira descrita nos itens anteriores, usandose um peso molecular de 8300 calculado combase na análise de aminoácidos.
11.11.4 - Medida do "Exchange-Out" em Crotoxina
Cerca de 20 mg de Crotoxina foram dissolvidos em 1 ml de tampão formiato 0,1 M, pH 3,0,dando uma solução levemente turva, pois o valor do pH fica próximo do pi da Crotoxina: 4,7.Adicionou-se então gotas de ácido fórmico diluído para acertar o pH, tornando a solução límpida. Emseguida, colocou-se gotas de 3 H 2 O para dar ao meio uma radioatividade de aproximadamente 10*cpm/ml. A incubação foi feita em banho a 70°C por 2 horas.
Os passos seguintes de filtraçSo em gel, leitura de absorbflncía • medida de rad atividade, foramefetuados da maneira descrita nos itens anteriores.
0 valor do coeficiente de extinção molar da Crotoxina foi de 30597 M"1 x cm* ' calculado domesmo modo descrito, utilizando um valor para peso molecular de 21700, obtido pelas anáfiMS daaminoácidos de Fosfolipase A e Crotapotin, considerando a Crotoxina corto resultant» da inttraçlo da»duas proteínas, na proporção molar de 1:1.

11
11.11.5 — Medida da Radioatividade
Pata twins os rasos em que se necessitou medir a tadioatividade do trício presente na proteína,utilizou se o líquido de untilrição de Biay em 1 960.
Este líquido de cintilação é comporto drs secitiirii.fs renitentes:
Naftaleno
PPO
POPOP
Metanol
Etifenoglicol
Dioxano q.s.p.
60 g
4 9
0,2 g
100 ml
20 ml
10C0 ml
Nesta formulação temos: como soluto primário fluorescente o PPO (2,5 difenil-oxazol), comosoluto secundário fluorescente o POPOP {p-bis [ 2-(5-feniloxazolil) J benzeno} e o dioxano, etilenoglícol, metanol e naftaleno como solventes.
Para medida da radioatividade foi utilizado um contador de cintilação Nuclear Chicago, modeloISOCAP 300.
A radioatividade dos itens 11.1 a 11.4 foi medida da seguinte maneira: 5 /J I da amostraincubada foram adicionados a 5 ml do líquido de cintilaçJo, agitados e contados por 10 minutos.
Após a segunda filtracão na coluna de gel Sephadex G 25, dos tubos que continham a proteínaradioativa foram retirados 100pi adicionados a 5 ml de líquido de cintilaçSo, após a agitação, colocadopara contar por 10 minutos.
11.11.6 - Cálculo da Eficiência de Contagem
Para o .cálculo da eficiência de contagem, foram utilizados vários padrSes fornecidos pelaNuclear Chicago que continham valores de "quenching" diferentes. Com os valores obtidos nas contagens
destes padrões, estabeleceu-se a curva de eficiência do aparelho, usando o método de relação de canais.
Assim, com os yaiores de contagens no canal A/ contagens no canal B em abcissa e a % de eficiência em
ordenada, traçou-se uma curva de eficiência.
Quando da contagem das amostras, os valores da relação de canais A/B foram colocados na
curva padrSo e daí estabelecidas as eficiências de contagens, extrapolando-as para 100% da eficiência.
Tomou-se o cuidado, de tempos em tempos, da refazer a curva de eficiência padrlo paraverificar, as possíveis mudanças ocorridas nas condições de medida do cintilador líquido.
11,11.7 - Calculo do Número da Hidrogênio Teoricamente Trocaveis
Os hídrogtnios que teoricamente slo trocaveis com o solvente a uma velocidade mensurável sloaqueles ligados a ' xigênio, enxofre e nitrogênio. Os hidrogênios ligados a carbono têm uma velocidadetroca isotópica muito lenta e, nfo foram considerados no nosso trabalho.

12
O cálculo do número de hidrocjenios que teoricamente são troráveis pelo ti (cio foi feito com
base na análise de aminoáciiios, levando se em consideração os hidrogênios de cadeia lateral e os
peptftlicos.
Em todas as experiências de traça isotópica hidrogênio-trírio foram usadas condições próprias
de incubação, supondo-se que todos os hidrogênios teoricamente trocáveis foram substituídos pelo trírio.
11.11.8 - Ajuste dos Dados Obtidos pela Troca I so tópica H/T
Os dados obtidos pela medida da velocidade de troca de trfcio pelo hidrogênio, colocados emgráfico contra os respectivos tempos, delineavam uma curva que aparentemente representa uma funçãoexponential. Segundo Hvidt e Nielsen , a curva que representa a velocidade de troca isotópica H/T, éa resultante de urna soma de exponenciais do tipo de decaimento radioativo. Considerando este fato, foifeito para os pontos obtidos neste trabalho um ajuste para obtenção da melhor curva que passa pelospontos experimentais. Este ajuste de soma de exponenciais foi realizado com' auxílio do computadorIBM/370, modeio 155.
O programa utilizado foi desenvolvido pelo National Institute of Health, Laboratory ofTheoretical Biology, por Mones Berman e Marjory F. Weiss em 1 974. Este programa é denominadoSAAM (Simulation Analyses and Modeling) e foi utilizado na versão n? 25.
Todas as análises e ajustes foram realizadas no Centro de Processamentos de Dados do Institutode Energia Atômica.
11.12 — Interação das Proteínas
O estudo da interação entre as proteínas, Fosfolipase A + Crotapotin e Crotamina + Crotapotin,foi realizado com o intuito de definir as proporções mulares de interação. Para tanto foi utilizada acromatografia em coluna de gel Sephadex G 75 fino com a intenção de isolar os complexos formados. Acoluna usada para este estudo foi sempre a mesma, com as seguintes características: coluna refrigerada daPharmacia Uppsala de 100cm x 0,9 cm, e.iipacotada com Sephadex G 75 fino até altura de 80 cm,eluída com tampão formiato 0,1 M, pH 3,5 que previamente equilibrou o gel. O fluxo foi sempremantido em 0,2 ml/minuto. Os efluentes passavam por um monitor de UV LKB, modelo Uvicord.li 8300, sendo a absorbancia evidenciada com auxílio de registrador gráfico e recolhidos em coletor defrações LKB, modelo Ultrorac 7000.
Nos efluentes recolhidos foi doseada, pelo método de Lowry, a concentraçío das proteínas,utilizada depois no cálculo da proporção molar de interação.
11.12.1 - Medida das Areas dot Picos
Um outro método usado para o cálculo das proporções de interação foi o da medida das áreasdos picos obtidos pela tbsorçSo a 280 nm dos efluentes da coluna.
Mantendo-se sempre as mesmas condições de cromatografia, foi realizada, em primeiro lugar, amedida da área obtida por uma solução de concentraçío conhecida da Crotamina, Fosfolipase A aCrotapotin.
Nas experiências de interaefo utilizou-se excesso de uma proteína em relaçlo à outra obtendo-seportanto um pico que correspondia ao da proteína que nfo participou do complexo. Pela comparaçãocom os valores de áreas da quantidades da proteína conhecidas e, sabendo-se o total colocado na coluna,

13
pode sfi ':ali'liar (|ti.into «I» pmlfina p.'t tii inou do coinplttxo e i:uiiSH(|ueiitemente quanto constituiu
excesso.
As mudid.is daí ár«as foram efetuadas mm um planfmetro da KEU FEEL & ESSER Co. N. Y.,modelo 423R M.
11.12.2 — Interação entre Fosfolipase A e Crotapotin
O estudo da iníeração que ocorre entre a Fosfolipase A e Crotapotin foi realizado comodescrito nos itens 11.12 e 11.12.1. Foram realizadas uma série de experiências nas quais se procurou variara concentração de proteínas na coluna, modificando o excesso de uma ou de outra. Uma dasexperiências foi: 2,6 mg de Fosfolipase A foram dissolvidos em 0,5 ml de tampão formiato 0,05 M,pH 3,5; 0,8 mg de Crotapotin foram dissolvidos em 0,5 ml de tampão formiato 0,05 M, pH 3,5; a colunaa 4°C de ccl Sephadex G 75 fino, equilibrada e empacotada com o mesmo tampão, foi de 80 cm:0,9 cm; as soluções de Crotapotin e Fosfolipase A foram juntadas e colocadas na coluna; a eluição foifeita com o tampão citado mantendo-se um fluxo de 0,2 ml por minuto.
Os efluentes foram coletados, doseados, e a área do pico de Fosfolipase A em excesso foimedida. Os doseamentos e os valores de área foram utilizados p::ra a determinação da proporção molarde interação.
11.12.3 — Cromatografia de Crotoxina
Com o intuito de comparar o volume efluente obtido para o complexo Crotapotin-FosfolipaseA com o da Crotoxina, fez se a seguinte experiência: 2 mg de Crotoxina cristalizada três vezes, dandouma só banda na eletroforese em gel de poliacrílamid?. foram dissolvidos em 1 ml de tampão formiato0,05 M, pH 3,5 e colocados na mesma coluna descrita no item anterior. A eluição foi feita com o mesmotampão e o fluxo mantido a 0,2 ml por minuto.
11.12.4- Interact) entre Fosfolipase A e Crotapotin Utilizando Crotapotin - ' J 5 I como Tracador
Para confirmar a participação efetiva da Crotapotin na interação com Fosfolipase A. utilizou-se
Crotapotin - ' 2 51 misturada à proteína fria, como tracador.
A experiência foi a seguinte: 1,5 mg de Fosfolipase A foram dissolvidos em 0,5 ml de tamplo
formiato 0,05 M, pH3 ,5 ; 1,0 mg de Crotapotin dissolvidos em 0,5 ml do mesmo tampão e 60/ i l de
Crotapotin - I 2 5 I , foram juntados e a amostra resultante colocada na coluna de gel Sephadex nas
mesmas condições anteriores.
As absorbâncias dos efluentes foram lidas a 280 nm e as radioatividade» foram determinadas no
contador gama.
11.12.6 - Interaçfo entre Crotamina e Crotapotin
Para o estudo da interaçflo entre estas duas proteínas utilizou-se a mesma tecnologia empregada
nos itens 11.12.1., 11.12.2. e 11.12.3.

14
A cmmatoqiafia foi feita em roluna cte pel Seph.idex G 75 de 80 cm: 0,9 cm, equilibrada e
(•inp.HoiHfirt coin tmnpfo fnrmiato 0.0S M, pH 3,5, que tamtam serviu para eluir a coluna com um f'uxo
<le 0,? ml/mimito.
For am realizadas várias experiências na tentativa de isolar o complexo formado e elucidar a
proporçã.) molar (ie interação.
11.12 .6 - Interação entre Crotamina e Crotapotin Usando Crotapotin - l í s l como Traçador
Para demonstrar que realmente a Crotamina interape com Crotapotin, utilizou se Crotapotin —
' J * I como traçíirior.
As rondiçoes de cromatogiafia foram as mesmas descritas no item anterior. Uma das
experiências realizadas foi a seguinte: 3,0 mg de Crotamina foram dissolvidos em 0,5 ml de tamplo
fnrmiato 0,05 M pH 3,5; a 0,8 mg de Crotapotin dissolvidos em 0,5 ml do mesmo tampão, adicionou-se
20JJI d° Crotapotin - I 2 S I ; as soluções foram juntadas e colocadas na coluna. A concentração e
radioatividade -los cftientes foram determinadas.
I l l - RESULTADOS
•11.1 - Purificação da Crotamina, Fosfdipase A e Crotapotin
As proteínas utilizadas neste trabalho foram isoladas e purificadas a partir do veneno bruto •
seco de cascavel brasileira (Crotalus d. terrificui).
A individualização rias proteínas no veneno foi feita pela filtração em Sephadex G 75 como
mostra a Figura 1. Esta representa um perfil cromatográfico de amostra de veneno contendo crotamina
e, onde os picos III e IV correspondem à Crotoxina e Crotamina respectivamente.
Procurou se sempre reunir somente amostras dos tubos da região de maior concentração de cada
pico, para se evitar contaminações cruzadas. As proteínas assim recolhidas foram liofilizadas e
separadamente submetidas aos próximos passos de purificação.
A Figura 2 mostra os padrões de absorção a 280 nm obtidos pelo fracionamento da Crotoxina
em coluna de gel SP Sephadex C 25. Nesta figura podem ser visualizados, pelo menos seis picos onde os
I e VI correspondem respectivamente a Crotapotin e Fosfolipase A. Os picos I I , lit e IV são picos
intermediários de proteínas não identificadas. Os picos que correspondem à Crotapotin e Fosfolipase A
s3o recolhidos, dialisados contra água deionizada e liofilizados.
A Figura 3 mostra a recromatografia final da Fosfolipase A, apôs várias passagens em SP
Sephadex C 25, evidenciando apenas um pico que foi considerado cromatograficamente homogêneo.
A Crotapotin como não se liga ao SP Sephadex, foi recromatcçirafada várias vezes em DEAE
Sephadex A 25, até que se obteve um pico único e homogêneo, o que é nostrado na Figura 4.
A Crotamina isolada na filtração do veneno em Sephsdex G 76 foi também recromatografada
em coluna de gel SP Sephadex C 25 a seu perfil cromatográfico «stá mostrado na Figura 5. Pela
observação característica, após injeção, do afeito da Crotamina em cemundongos, ou seja, paralizaçfo das
patas posteriores, foi possível identificar no gráfico da Figura 5 os picos I I I , IV e V com atividade dt
Crotamina. O material do pico V, obtido com » cromatografia de vária» «mostras dt Crotamina em SP
Sephadex, foi recolhido, dialisado e liofilizado. A proteína foi seguidas vezes repassada no mesmo gel até
ohtençffo de pico único • homogêneo, como está mostrado com traço cheio na Figura 5.

' .9 ,
r ii ^ - -
*
Crotox in»
T i l
• » '
•"rotjr.ina
IV
1
1
l
t
30 40 5 0TUEC NO
Figurai — Filtraçio do Veneno Global em Coluna de Gel Sephadex G 75. Perfil Cromatográfico da Filtração de 300 mg de Veneno Total/10 ml de ÁcidoAoitioo 0.1 M. Fluxo de 0,5 ml/min oom Recolhimento de 10 ml/tubo. Coluna de Gel Sephadex G 75 Fino, de 3 cm: 80 cm. Equilibrada e Eluidacom Ácido Acético 0,1 M

o>
l . o
i i.o
Crotapot in
; \
I »
I I
h!
F r < s í o l i ; a s e A
VI
s
to TVIO M*
2 — Cromatoyafia de Crotoxina em SP Sephadex C 25. Perfil Cromatográfioo Obtido pelo Fracionamento de 100 mg Crotoxina/5 ml de TampãoFormiato 0,05 M. pH 3.5 em SP Sephadex C 25, Equilibrado com Tampão Formiato 0,05 NI, pH 3,5 e Elui'do com Gradiente Linear Salino de0a3MdeNeCI . Coluna de0,9 cm: 40 cm, Fluxo de 0,2 ml/min com Recolhimento de 4 ml/tubo.

2 , 0
gradiente
I10 20 40
TUBO
— Recromatografia de Fosfolipase A em SP Sephadex C 25. Recromatografia de 100 mg de Fosfolipase A/2 ml de Tampão Formiato 0,05 M oH 3,5em Coluna de SP Sephadex C 25 Equilibrado com Tampão Formiato 0,05 M, pH 3,5 e Elui'do com Gradiente Linear Salino de 0 a 3 M de NaCl.Flum de0.2 ml/mm com Recolhimento de 4 ml/tubo. Coluna de 0,9 cm: 40 cm

Í . O
n! I
(radian'.c
1»
TU80 NO
Figura* - Recromatografia de Crotapotin em DEAE Sephadex A 25. Coluna de DEAE Sephadex A 25, Equilibrada com Tampão Formiato 0,05 M, pH 3,5e Elufdo oom um Gradiente Linear Salino de 0 a 3 M de NaCI. Fluxo de 0,2 ml/min oom Recolhimento de 4 ml/tubo. Amostra Contendo 100 mgda Crotapotin/2 ml d* Tampèo Formiato 0,05 M, pH 3,5. Coluna de 0,9 cm: 30 cm

2 . 0
1 .0
. • *
S — Cromatografia d« Crotamina em SP Sephadex C 25. Com Linha Tracejada Ternos a Cromatograf ia de 200 mg de Crotamina/3 ml de Tampão For-miato 0.05 M. pH 3.5 • Elufda com um Gradiente Linear Salino de 0 a 3 de NaO. Com Traço Cheio Temos a Recromatografia Final de 100 mgdo Pico V/ml do Metmo Tampék» Após Varias Passagens. Os Picos I I I , IV e V têm Atividade de Crotimina tn

20
111.2 Eletroforese em Gel de Poliacrila'mida
Diante da possibilidade das proteínas cromatograficamente homogêneas nSo estarem realmentepuras, foram realizadas eletioforeses em gel de poliacrilamida para verificar a pureza ao níveleletrntorétiro. A Figura 6 mostra o aparecimento de uma única banda para Fosfolipay> A, Crotamina eCrotapotin indicando proteínas eletroforeticamente puras.
111.3 — Presença de Fatores Coagulants*
Nas proteínas consideradas puras realizou-se o teste de coagulação do fibrinogínio descrito porGonçalves132', em 1 956.
O teste revelou ausência completa de fatores coagulantes contaminando as proteínas. Comoreferência usou-se veneno bruto que apresenta alto poder de coaau lição sotre o fibrinogêniq.
f 11.4 - Oosagens das Proteínas
O doseamento das proteínas consideradas puras realizou-se pelo método convencional Lowry et
ali' usando soro albumina bovina, Sigma, pira se construir a curva padrSo.
Todos os pontos da curva padrSo bem como das amostras a serem doseadas, foram feitos em
triplicata. Os resultados mostraram que após a liofilizacio as proteínas ainda continham cerca de 10 a
15% de água.
111.5 - Análise de Aminoécidos
A Tabela I mostra os dados obtidos pela análise de aminoácidos num analisador Beckman
120 C. Os resultados evidenciam que s8o desprezíveis as diferenças encontradas com a variaçSo do tempo
de hidrôlise de 10 h, 20 h e 24 horas.
A soma dos valores médios para cada aminoácido, serviu para o cálculo do peso molecular das
proteínas.
Para cada partida nova das proteínas purificadas, realizou-se uma série de análises de
aminoácidos obtendo concordância com os dados mostrados na Tabela I.
Os pesos moleculares calculados com tose na análise de aminoácidos foram os seguinte*: 8300
para Crotapotin, 13400 para FosfoHpese A e 4880 par» a Crotamina.
111.6 - Absorçfo das Proteínas no UV
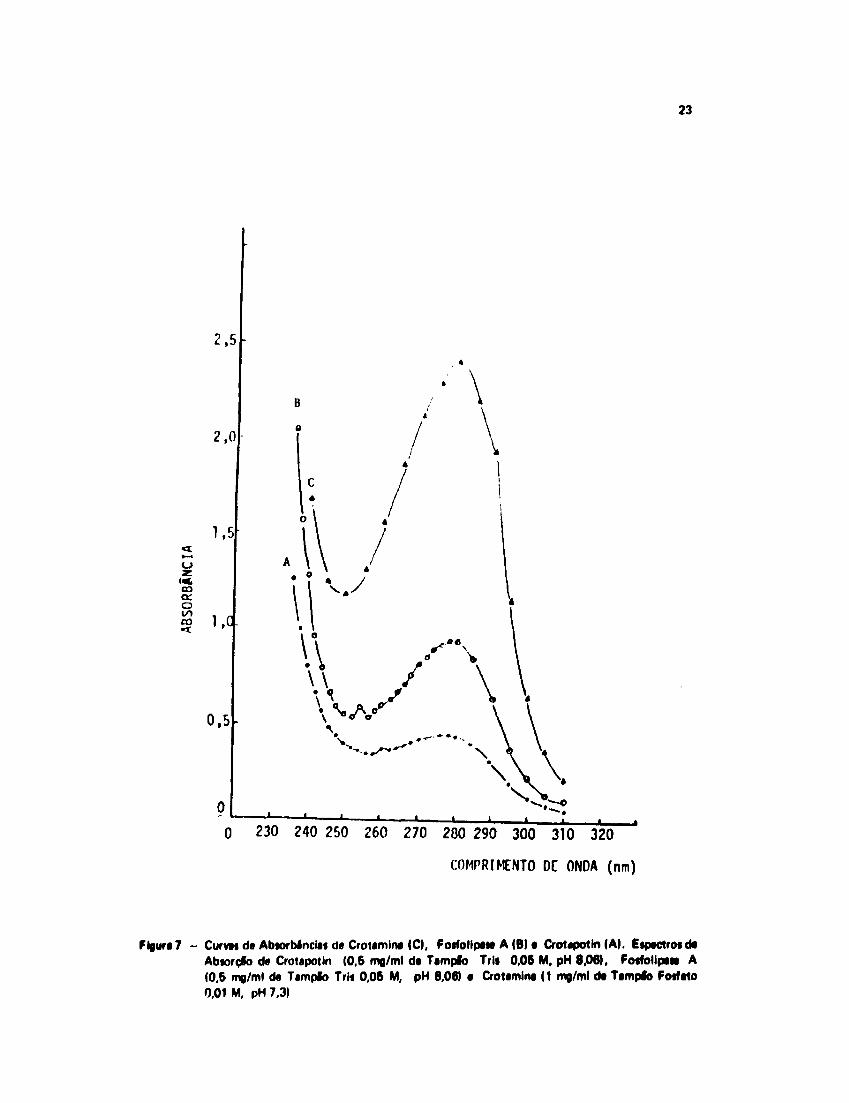
As curvas da absorção no UV da Fosfolipa» A, Crotapotin e Crotamina estlo mostradas na
Figura 7.
A Crotemina e • Fosfolipase A apresentam um máximo ds absorçfo em 280 nm, enquanto que
• Crotapotin tem um máximo em 278 nm. Estes máximos foram utilizados para o cálculo dos
coeficientes de absorçlo molar das três proteínas, ou seja, 9 O 4 7 M ' 1 x c m ' 1 para Crotapotin,
2 2 6 1 2 M ' 1 x c m * 1 para Fosfolipase A e 11660M"' x c m ' 1 para Crotamina.
111.7 - MedkJe da Atividade Enrimétiee da Fotfolipft» A

21
B•f
c+
'•V«#•'
Figura 6 - Eletroforesa em Gel de PoliacrilaminaA -Eletroforese de 50 JJI de Crotapotin (2 mg/ml) am Tampfo TRIS-glicina pH 9,5.
Corrida de 1,0 hora oom 3 m A/tubo, Utilizando o Meimo Tampfo nai Cubai a Gel daPoliacrilamida a 7%.
8 -Eletroforese de SOjil de Foifolipasa A (2 mg/ml) em Tampfo Bata-alamina pH 3,4.Corrida do 1,0 hora oom 3 m A/tubo, Utilizando o Mesmo Tampfo nas Cuba* a Gal d#Polu»crilamidae16%.
C -Eletroforese de 60/il de Crotamina (2 mg/ml) am Tampfo Beta-alamina pH 4,3. Corridada 1,0 hora oom 3 m A/tubo, Utilizando o Mesmo Tampfc nas Cubas a Gel daPoliacrilamida a 16%.Após as Corridas as Amostras Foram Fixadas oom TCA a 12% por 30 minuto*, Coradasoom Comaisie Brilliant Blue R 260 a Descoradas oom uma Mistura de Metanol: ÁcidoAoétioo: H,0

r?
Ta'iela I
Análise de Aminoácidos de Fosfolipase A, Crotapotine Crotamina*
pint\
a.a. \
Lys
His
Arg
Asp
Thr
Ser
G»u
Pro
Gly
Ala
Cys
Vai
Met
lie
Leu
Tyr
Phe
T r p "
10 h
9.3
1.8
9.3
9.0
6,5
5.5
9,0
4,3
11,0
5.8
11,8
2,0
2,3
4.0
5,8
9,5
6,5
—
Fosfolipase A
20 h
9,0
-
8,8
9,0
6,3
6,0
9,3
4,8
10,8
5,8
13,0
1,8
2,0
4,0
6,0
10,0
5,8
-
24 h
9,5
1.7
9.5
9,0
6,4
6,2
9.2
4.6
10,8
5,7
12.2
1.9
1.9
3.6
6.5
9.4
6.4
-
M
9.3
1.7
9.2
9,0
6.4
5,9
9,2
4,6
10,9
5,8
12,3
I.f
2.1
3.9
5,8
9,6
5.9
2.0
lOh
1,8
0,8
1.7
10.0
3.5
4.3
12,6
4,0
8,8
5.3
11.6
0,6
0,7
2.1
0,8
1,7
2.5
—
Crctapotin
20 h
1,7
0,7
1,4
10,0
3,6
4.3
12,4
4.3
8,3
5,0
12.0
0,5
0,7
1.7
1,0
2.4
2.1
-
24 h
1,7
1.0
1,7
10,0
3,6
4,3
12,6
4,3
8,8
5,0
11,9
0.5
0,7
1.7
1.0
2,4
2.1
-
M
1,7
0,8
1,6
10,0
3,6
4,3
12,5
4,2
8,6
5,1
11,8
0,5
0,7
1.8
0,9
2,2
2,2
1,0
10 h
8,8
2,0
1,7
1,9
-
2,5
1,8
3,1
4,9
-
5,4
-
0,8
1.2
0,7
0,8
1,7
-
Crotamina
20 h
9,0
2.0
2,0
2,1
-
2.7
2.0
3.1
5.0
—
5,6
—
1.0
0.9
0.9
0.9
2.0
—
24 h
9,1
2.2
1,9
1,9
-
2.6
1,8
3,2
6.0
-
5.5
-
0,8
1,1
0,9
0,7
1.9
-
M
8.9
2,0
1,8
1,9
-
2,6
1.8
3.1
4.9
-
5.5
-
0,8
1.0
0.8
0.8
1.8
2.0
* A tabela mostra os v&lores obtidos nas análises de aminoácidos pela hidrólise das protefnw a 120°C,
durante 10 h, 20 h e 24 horas num analisador de aminoácidos Beckman, modelo 120 C. Mostra
também os valores médios (M) para cada aminoáoido após as análises
' * Dados obtidos espectrofotometricamente.
23
2,5
2.0
o
03CgOiS)CO•X
1,5
..
0,5
-I L.
230 240 250 260 270 280 290 300 310 320
COMPRIMENTO DC ONDA (nm)
Figura 7 - Curvai da Abtorbinciai da Crotamina (O, Fotfolipata A (B) a CroUpottn (A). EipactrosdaAbiorçêo da Crotapotin (0,6 mg/ml da Tampio Trli 0,06 M, pH 8,06), Fo«folipa« A(0,5 mg/ml da Tampio Trii 0,06 M, pH 8,06) a Crotamina (1 mg/ml da Tamplò Fosfato0,01 M, pH 7.3)

24
A Figura 8 mostia a curva-padrão de atividade enzimálica da Fosfolipase A pura, tendo comosubstrato uma suspensão de gema (le ovo fresca. Em todas as experiências, tanto de purificação quantode interação, em que se suspeitou da presença de Fosfolipase A, mediu-se a atividade enzimática pelamesma técnica, usando a curva padrão para comparação.
111.8 - DeterminaçSo de Grupos SH Livres
Foram realizados para Fosfolipase A, Crotamir.a e Crotapotin puras, os testes de presença de SH
livres, utilizando o método colorimétrico de Eüman modificado por Simizu , que usa ácido
5,5'-ditiohis 2-nitrobenzóico, Sigma, como reagente e a cistefna como padrão.
Os resultados mostraram que mesmo em presença de uréia não aparecem SH livres, devendo
estar todos comprometidos nas pontes S-S das cadeias polipeptídicas.
111.9 - Interação entre a Fosfolipase A a Crotapotin
O estudo da interação entre as proteínas foi realizado pela filtração em gel Sephadex G 75,
mantendo-se sempre as mesmas condições de fluxo, temperatura, altura do gel, volume de amostras,
volume recolhido por tubo. etc. variando apenas as concentrações das proteínas colocadas na coluna.
Foram realizadas uma série muito grande de experiências, onde se procurou através o doseamento das
proteínas e a medida das áreas dos picos, evidenciar a proporção molar de interação.
Na Figura 9 tem-se os padrões de filtração na coluna de Sephadex G 75. O gráfico A representaa fíltraçjo de uma amostra de Crotoxina cristalizada e eletroforeticamente pura. O gráfico B representa acromatografia de uma amostra contendo excesso de Fosfolipase A e, no gráfico C uma amostra contendoexcesso de Crotapotin.
Na Tabela II tem-se dados obtidos para um grupo de experiências de interações entreFosfolipase A e Crotapotin, usando a técnica de filtração em gel Sephadex G 75. Os dados deconcentração da amostra, concentração do complexo formado e do excesso de proteína, foramcalculados pelo método Lowry e confirmados pela medida da área dos respectivos picos.
111.10 - Interação entre Crotapotin e Crotamina
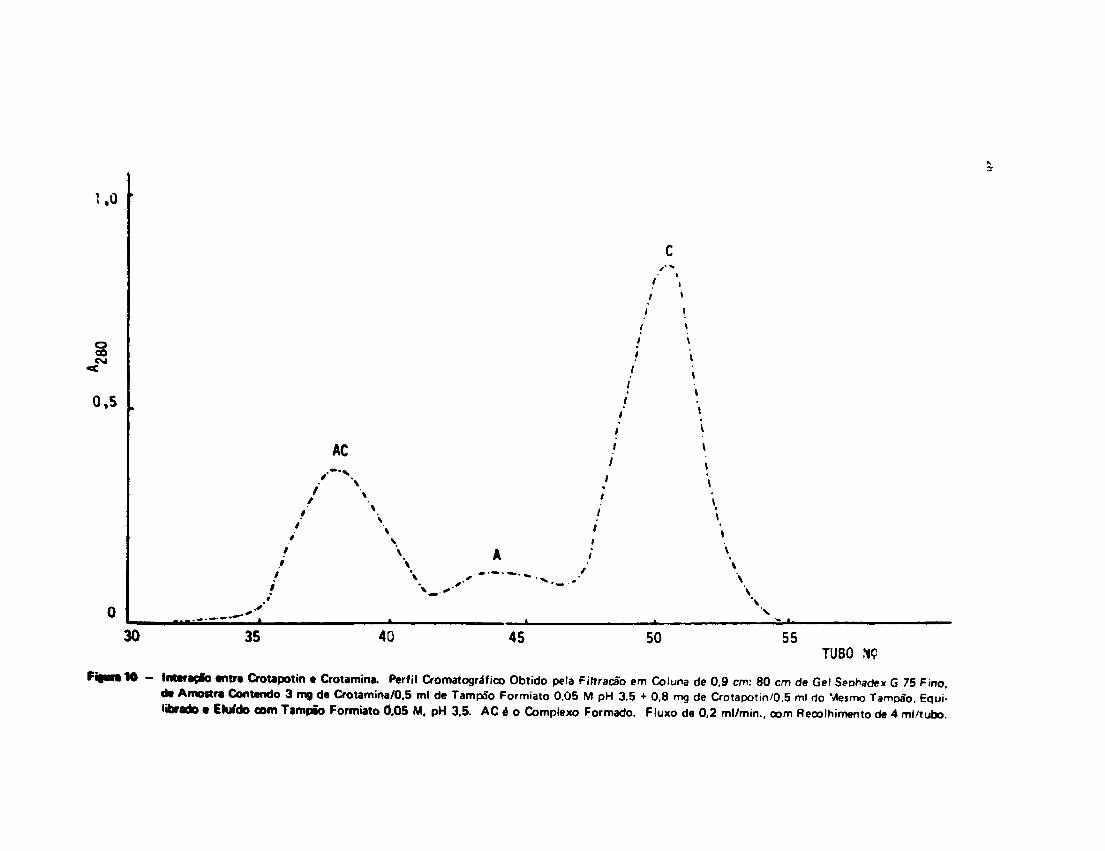
A Figura 10 mostra uma das experiências de interação Crotapotin-Crotamina realizada com •
técnica de filtração em gel Sephadex G 75.
A Tabela I I I mostra os dados obtidos para concentração de amostra, concentração de proteína*
dos picos, medidas pelo doseamento de Lowry e confirmadas pela medida das áreas dos picos.
A Tabela I I I mostra também os valore* da proporção molar de interação entre
Crotapotin-Crotamina nas experiências realizadas, evidenciando uma inconstância para os valore* obtido*
ds proporção molar de interação.
111.11 - Interação entre Fotfolipaw A a Crotamina com Crotapotin Contendo Crotapotin i a * l como
Treçadof
Para demonstrar a participação de Crotapotin no* complexo* Crotapotín-Fosfolipast A •
Crotapotin-Crotamina, realizamos experilncia* de interação em gel Sephadex G 76, utilizando na.
composição das amostra* colocada* na coluna, Crotapotin marcada com " * l . Neste» < «so» a proteína
marcada foi misturada ante* da interaçflo com a proteína fria.

25
0,9
0,7
0,5
0,3
0,1
010 20 30 40 50 60
uq de F0SF0LIPASE A
Figura 8 - Curva da Atividade Hemolftlca da Foifollpc*» * . Incubado Inicial da Foifollpe» A amSoluçio Fisiológica com Gama da Ovo Fraica por 2 horai a 37*C. A eita MitturaJ unta«auma Suipanilo a 0,8% da Glóbulot Vermelho» da Rato am Sotuçio fisiológica a Incube-tepor 40 minuto» a 38°C.Apói Centrifugaçlb, ai Amoitrti tio Lida» Quanto 4 Abaorblncla em 678 nm

26
I.i
n.S
0.!
0,5
A
AJ U.38 4S
TUBO NQ
Figura 9 - Interação entre Crotapotin e Fosfolipaie A . Perfi l Cromatográfico» Obtidos pala Fittraçfo e mGel Sephadex G 75 Fino, Coluna de 0,9 cm: 8 0 cm, Equilibrado e Elufdo com TamplòFormiato 0,05 M, pH 3,5.A -Cromatograf ia de 2 mg de Crotoxina/ml de Tampffo Formiato 0,05 M pH 3,6.B -Cromatografia de 3,25 mg de Foifolipaie AA>,5 ml de Tamplò Formiato 0,06 M, pH 3,5
+ 1,6 mg de Crotapotin/0,5 ml do Meimo Tamplò.C -Cromatografia de 1,6 mg de Crotapotin/0,5 ml de Tamplò Formiato > 1,3 mg de Fotfoli-
pe» A A), 5 ml do Mesmo Tampão.Para ai Experiência» A, B e C Foram Mantido* Fluxo de 0,2 ml/min, e 4 ml/tubo de VolumeF fluente.

27
Tabela II
Interação Crotapotin-Fosfolipase A na Filtraçío em Sephadex G 75*
Exper.
rr?
1
2
3
4
5
6
7
8
Prot. total
(mg)
2,9
3,4
4,8
2,5
2,5
2,5
2.5
2,5
Proporções
na amostra
A:B
(pmoles)
2
1
1
1,1
1.1
1.1
1.1
1.1
1
2
1.2
1
1
I
1
I
Complexo
(mg)
2,1
2.1
3,46
2.34
2,24
2,05
2,32
2,13
Proporções
de interação
B:A
(/imoles)
1
1
1
»
1
1
1
1
1
t
1.03
1.04
0,81
1,05
0.89
1.6
A tabela mostra os valores de proteína total na amostra, proporções molares de Crotapotin (A) para
Fosfolipase A (B) colocadas na coluna de gel Sephadex G 75 mg do complexo formado e proporções
de interação após a filtraçSo.
1.0
o00CM
c
0,5
0
>
t
i
i*.y
AC
/ V/ \
\' \\\\
A
i
ii
i
ii1
;
0 '
/
/
/
/
/Ê
1i
1
f
c
\í
\
\
\\
\
\
V
\
\
\vV
1
V
• \
• \
30
10
35 40 45 50 55TUBO
Intaraçfo antra Crotapotin a Crotamina. Perfil Cromatográfico Obtido pela Filtracão em Coluna de 0.9 cm: 80 cm de Gel SeDhadex G 75 Fino,de Amostra Contendo 3 mg de Crotamina/0,5 ml de Tampão Formiato 0,05 M pH 3,5 + 0,8 mg de Crotapotin/0,5 ml do Mesmo Tampão. Equi-librado « Elufdo com Tampão Formiato 0.05 M. pH 3.5. AC é o Complexo Formado. Fluxo de 0,2 ml/min., com Recolhimento de 4 ml/tubo.

29
Tabela I I I
Interação Crotapotin-Crotamina na Filtraçffo em Gel Sepnadex G 75*
Exper.
no
1
2
3
4
6
Prot. total
(mg)
6,68
3,80
3.06
2,81
3,62
Proporções
na amostra
C:A
(pmoles)
4:1
6:1
0,6:1
4:1
6:1
Complexo
(mg)
1.6
1.12
0,50
0.48
0,98
Proporções
de. interação
C:A
(^moles)
1:1
V«V1.11:1,1
1:2,1
A tabela mostra ot valores de proteína total na amostra, proporções molares de Crotamina (C) para
Crotapotin (A) colocadas na coluna de gel ãephadex G 76 mg do complexo formado t proporções
de interaçio após a filtraçfo.

30
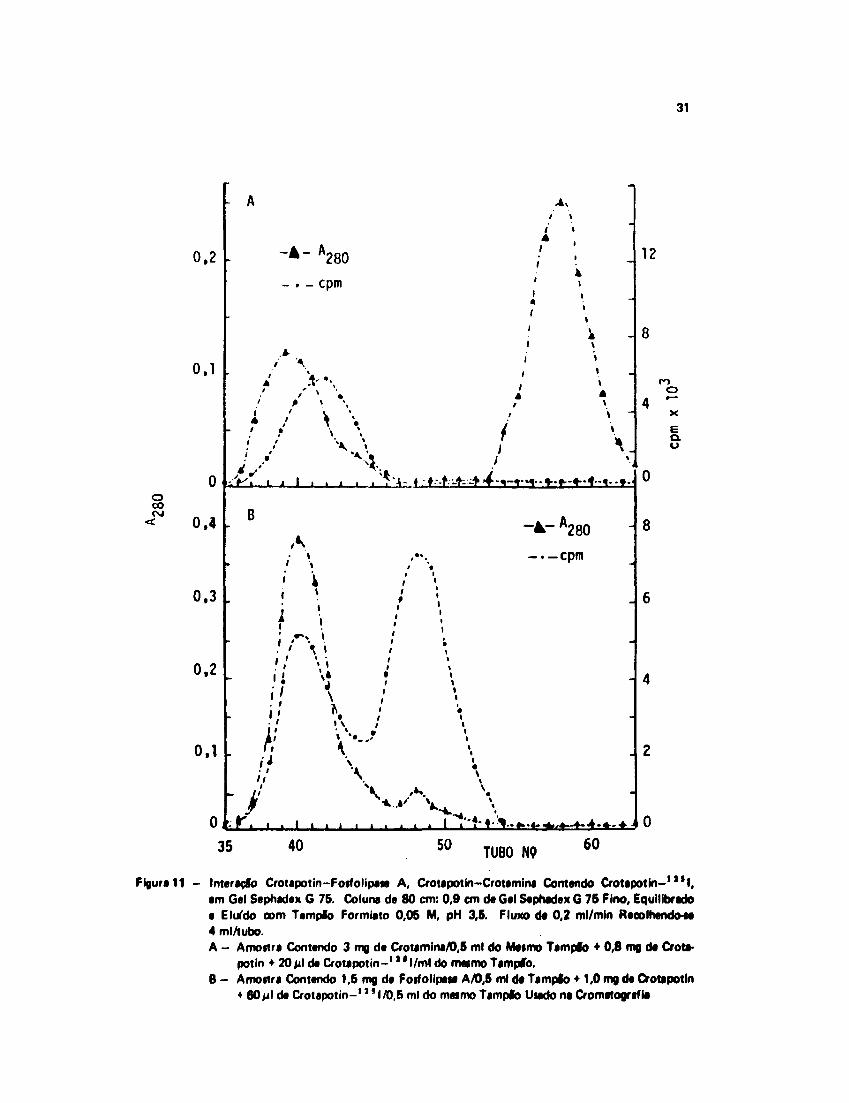
A Fiijura 11 mostra os jwif/s i.-romalotpáfiim « radioativo obtidos nas interações realizadas. Ográfico A representa o resultado da interação Ciotapotin Fosfolipase A e no B a interaçãoOotat'otin Ctotaniina.
ujiira evidencia claramente H participação da Ciutapotin nos dois complexos.
111.12 - Determinação da DL 5 Q
A determinação da D L 5 0 foi feita para Crotamina, Crotapotin, Fosfolipase A e para os
complexos Crotapotin Fosfolipase A, Crotapotin Crotamina na proporção molar de 1:1.
A Tabela IV apresenta as porcentagens de mortalidades para as várias diluições de cada proteínaou dos complexos, assim como os valores da Dl_ 5 0 obtidos.
Tabela IV
Toxicidade de Crotamina, Fosfolipase A e dos Complexos AB e AC"
Diluições
1: '
I . i
1:41:81:161:32
D L 60
% Mortalidade
Fosfolipase A
100
83,3
66,6
50,0
16,6
0
0,55 mg/Kg
Crotamina
10083,3
50,0
33,3
00
0,8 mg/Kg
%
AB
10083,3
83,3
66.6
33.3
0
0,05
mg/Kg
Mortalidade
AC
10010083,3
16,6
00
0,20
mg/Kg
* A tabela mostra as porcentagens de mortalidade para cada diluição obtidas pela injeçio i.p.
em camundongos, de Fosfolipase A, Crotamina e dos Complexos Crotapotin-Fosfolipaw A
(AB), Crotapotin-Crotamina (AC). Mos.ra também as D L 6 0 calculadas pelo método de
Reed e Muench, das proteínas isoladas e dos complexos AB, AC.
111.13- Medida da Velocidade de Troca HWroflenlo-Trícto
0 método utilizado foi descrito por Englander'20', em 1 963. Para os tempos pequenos como 1minuto, 2 minuto* e 3 minutos utilizou-se a técifca com ume coluna so, ou seja a amostra apôscolocada na coluna de G 26 era aluída até aproximadamente 2/3 da altura total para eliminar o excessode J H j O e, em seguida o fluxo da coluna era interrompido durante o tempo de 1 min., 2 min. e 3 min.,
logo apôs, iniciar se novamente a eluiçfo.
31
oco
<
0.2
. 2
TUBO N9
Figura 11 - Intersçfo Crotapotin-Fosfolipase A, Crotapotin-Crotamina Contendo Crotapotin-1 * * I,em Gel Sephadex G 75. Coluna de 80 cm: 0,9 cm de Gel Sephadex G 75 Fino, Equilibradoe Eluído com Tampfo Formiato 0,05 M, pH 3,5. Fluxo de 0,2 ml/min Recolhendo-M4 ml/tubo.A - Amostra Contendo 3 mg de Cfotimini/0,5 ml do Mesmo Tampfo • 0,8 mg de Crota-
potin + 20 jil de Crotapotin-1 l l l / m l do meimo Tampfo.8 - Amostra Contendo 1,5 mg de Fosfolipase A/0,6 ml de Tampfo • 1,0 mg de Crotapotin
• 80jil de Crotapotin-1 ' ' 1/0,6 ml do mesmo Tampfo Usado na Cromatografif

32
A Figura 12 mostra a separação que ocorre quando se utiliza a técnica descrita acima. Para ostempos mais longos de incubaçSo, utilizou-se a técnica de duas colunas, que sem dúvida é muito maiseficiente na separação.
III.13.1 - Troca d» Htdrogênio-Trlcio na Fotfolipaw A
A Figura 13 mostra a curva obtida através de um ajuste dos dados de n? H^/mol de proteína,contra tempo de saída do trício da proteína para o solvente. Os dados sobre os quais foi feito o melhorajuste são encontrados na Tabela V.
O tratamento dos dados pela computação, assim como os valores obtidos, encontram-se no
Apêndice 2.
A equação que representa a soma de duas exponenciais e que melhor ss ajusta nos dados
experimentais da troca H/T da Fosfolipase A foi a seguinte:
H ( t ) = 71 x e 0 - 9 * ' O ' * « + 59 x e " 0 0 5 5 t
onde 0,055 e 0,9 x IO* 4 representam as constantes de velocidade de trocas e, 59 e 71 os números de
hidrogênio* de cada classe.
Os dados de radioatividade inicial, radioatividade carregada pela proteína e absorbèncias a280 nm das amostras, que originaram os valores médios C/A e conseqüentemente o n? Hg/mol deproteína, para cada tempo considerado, podem ser vistos no Apêndice 1.
111.132 - Troca da Hidrogtnio-Trício na Crotapotin
Os dados experimentais obtidos para o n9 H /mol de Crotapotin com os respectivos tempos daincubaçSo, sSo mostrados na Tabela V.
O melhor ajuste obtido pela computação dos dados experimentais foi pala soma de duas
exponenciais, o que é mostrado na Figura 14.
0 modo como foi feito o ajuste é o mesmo do item anterior e os seus parâmetros podam ser
encontrados no Apêndice 2.
A equação da soma de duas exponenciais que originou a Figura 14 foi:
M ( | , = 22 x e o n * ' O ' 2 « • 84 x
Os valores 0,11 x 10~a • 0,22 slo as constantes da velocidade das d a n t t com 22 a 84
hidrogênio* respectivamente.
Os valores de n9 Ha/mol de Crotapotin, nos respectivos tampos òe incubaclo, originaram do*
resultados de radioatividade inicial, radioatividade carregada pala proteína a absorUnciM a 280 nm das
amostras que podem ser encontrados no Apêndice 1.

33
1,0
0.R
0.6
0.2
10
7 8 9 10 I I !,! I.) 14 IS 16
TIIÜO It?
Figura 12 - Separação da Radioatividade Carregada Pela Proteína do Excesso de *HjO na Filtraceoem Sephadex G 25 "Coarse". Amostra Contendo 3,42 mg de Fosfolipne A/ 0,8 ml daTampão Formiato 0,1 M, pH 3,0 • J H , 0 Dando uma Radioatividade Inicial da10" cpm/ml, foi Incubada por 2 h a 70°C e Resfriada. Filtracao em Coluna dt 8 cm: 3 cmde Gel Sephadex G 25 "Coarse" Resfriada a 3°C com Fluxo de 1 ml/10 segundos, Reco-lhendo-»» 1 ml/tubo. Após a Amostra ter Percorrido 2/3 da Coluna, o Fluxo foi Interrom-pido Durante 2 min para Logo em Seguida ser Restabelecido

34
1H0
f«0
SO
0 TOO SOO 1000 1500
TCMPO(mfn)
Figura 13 - Curva de Troca T/H da Fosfolipaw A. Ajuste Obtido pala Computação dos Dados Experi-mentai* de rrf> hidrogênio* Aparente* (H§) por mol de Fo*folipa*e A. A Curva Rapreientta* VelocidadM da Saída do Trício Incorporado é Proteína, para o Solvente (Exchange out)Obtidat pelo Método de Filtracfo em Coluna* de Sephadex G 25 "Coaria", Mantida*a 3°C, Equilibrada*« Eluída* com Templo Formiato 0,1 M, pH 3,0
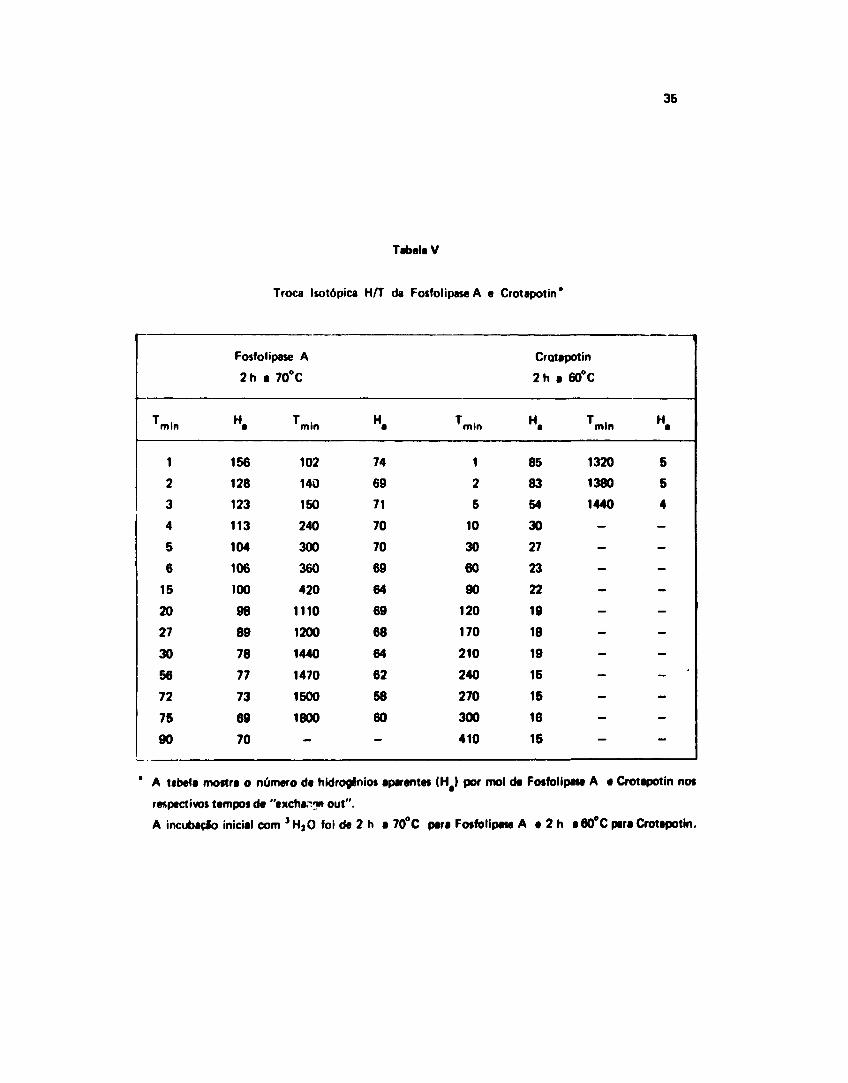
Tabela V
Troca Isotópica H/T da Fosfolipaie A e Crotapotin'
36
Tmln
1
2
3
4
5
6
15
20
27
30
56
72
75
90
Fosfol
2h a
H.
156
128
123
113
104
106
100
98
89
78
77
73
69
70
pase A
70°C
Tmin
102
140
150
240
300
360
420
1110
1200
1440
1470
1500
1800
-
H.
74
69
71
70
70
69
64
69
68
64
62
58
60
-
Tmln
1
2
5
10
30
60
90
120
170
210
240
270
300
410
Cratapotin
2h a
H.
85
83
54
30
27
23
22
19
18
19
15
15
16
15
60°C
Tmin
1320
1380
1440
-
-
—
—
-
-
-
-
-
-
—
1
H .
5
6
4
-
-
—
-
-
-
-
-
-
-
-
' A tabela mostra o número de hidrogênio» aparentes ( H § | por mol da Fosfolipaie A a Crotapotin nos
respectivos tempos de "txcbar.yi out".
A incubaclo inicial com J H , 0 foi de 2 h a 70°C para Fosfolipaie A a 2 h a 60°C para Crotapotin.

36
i no
so
10
f,00 1000 1400
TEWO(min)
Figura 14 - Curvt de Troca T/H da Crotapotin. A|u~te Obtido pela Computaçlò do* Dados Experi-mentai» de vfí Hidrogênio* Aparentei (H§)/mol de Crotapotin. A Curva Representa aiVelocidades de Saída do Trfcio Incorporado i Proteína, para o Solwite (Exchange out),Obtidas pelo Método de Filtracfo em Colunai de Gel Sephadex G 26 "Coaria", Mantida*a 3°C, Equilibradas e Elufdas com Tampão Formiato 0,1 M, pH 3,0

37
III.13.3 — Troce de Hidrogênio Trício na Crotoxina
Na Tabela VI t e m » os dados experimentais obtidos para o n° H§/mol de Crotoxina nosrespectivos termos de incubaçfo.
A Figura 16 mostra o ajuste feito pela computação dos dados experimentais da Tabela V I , cujosvalores estão no Apêndice 2.
O melhor ajuste para a Crotoxina foi obtido pela soma de três exponenciais fornecendo aseguinte equação:
H | t , = 9 6 x e ° - 1 3 x 1 0 ~ 3 ' + 26 x «"0.012 t + 3 2 x e0 1 6 >
Os valores de radioatividade inicial, radioatividade carregada pela proteína e absorbftncias queoriginaram os números da Tabela VI estão escritos no Apêndice 1.
A Figura 15 também mostra uma curva teórica obtida pela soma dos dados de troca H/T paraFosfolipase A + Crotapotin com intuito de comparar esta simples soma com os dados experimentais detroca H/T ria Crotoxina que é o produto da recombinaçao entre Fosfolipase A e Crotapotin.
III.13.4 - Troca da Hidrogênio-Trfcio da Crotamina
Os resultados da troca H/T da Crotamina estão escritos na Tabela VI . Foram realizadas paraCrotamina experiências com variação no tempo de incubação inicial: uma com 2 horas de incubaçfo daproteína com 3 H 2 O a 70°C e outra com 18 horas de incubação também a 70°C.
Os dados experimentais foram ajustados no computador e o resultado destas ajustes estfo na
Figura 16.
Os valores de radioatividade inicial, radioatividade carregada pela proteína • absorbtncias quepermitiram o cálculo do n? Hg/mol de Crotamina para cada tempo considerado, estfo no Apêndice 1.
O Apêndice 2 contém os parâmetros encontrados pela computação dos dados.
Os melhores ajustes aos dados experimentais surgiram da soma de duas exponenciais para ambas
as experiências. Para o caso da incubaçfo inicial de duas horas a 70°C, a equação obtida foi:
H m = 26 x e" 0 - 6 6 » 1O' J i + 38 „ , 0 . 2 6 «
Para incubaçfo de 18 horas a 70°C, a equação obtida foi:
H ( f ) - 16 x a " 0 3 3 « '«»"* « • a x e - 0 0 ' « •
I114 - Medida da Radioatividade
A Figura 17 mostra a curva da porcentagem de eficiências para os padrões de radioatividade

38
Tabela VI
Troca Isotópica H/T da Crotoxina e Crotamina*
min
1
2
3
5
12
15
30
35
55
60
70
75
go100
120
130
150
Crotoxina
2h s
H.
143
154
147
132
112
137
119
111
102
107
103
111
104
112
104
97
96
70°C
Tmin
150
240
280
300
310
360
420
480
930
960
1020
1260
1320
1430
1490
-
-
H.
94
94
98
91
93
93
92
87
87
85
88
85
78
81
76
-
-
Tmin
1
5
7
10
15
48
55
88
97
120
180
346
410
480
1410
1490
1650
Crotamina
2h.a
H.
58
47
29
39
29
28
26
30
25
23
22
18
20
20
12
9
10
60°C
Tm!n
5
20
40
55
60
150
165
180
210
220
240
260
270
290
1380
1440
-
H.
24
24
21
19
21
16
15
17
15
13
16
15
16
16
10
10
-
' A tabela mostra o número de hidrogênio» aparentei (Hf) por mol de Crotoxina e Crotamina not
respectivo* tempos de "exchange out".
A incubiçlo inicial comJ HjO foi de 2 h a 70°C para a Crotoxina e 2 h e 18 h a 70°C para Crotamina.

39
100
60
0 100 1000 1300
HMPO(min)
Figura IS - Cruvt da Troca T/H da Crotoxfna. Ajuste Obtido pala Computaçlo dos Dados Experi-mentai! da n<? Hidrogênio* Aparentes (H^l/mol de Crotoxina. A Cura Representa as Veloci-dades de Saída do Trício Incorporado i Proteína, para o Solvente (Exchange out), Obtidaspelo Método de Flttreçfo em Colunas da Gel Sephadex G 26 "Coarsa", Mantidas a 3°C,Equilibradas e Eluídas com Tamplo Formiato 0,1 M, pH 3,0. A Curvi A+B Representaa Soma Teórica da Equação Obtida pelo Ajuste aos Dados Experimentais da n9 H(/molde Crotapotin + a equação Obtida pelo Ajuste aos Dados da n9*Ha/mol da Fotfolipe» A

I'M
t« 50
1000 MOO
TLMPO(min)
Figura 10 - Curva de Troca T/H da Crotamina. Ajutte* Obtido* pala Computaçlo do* Dado* Expari-mantait da n? Hidrogênio* Aparente* (H^l/mol da Crotamina. At Curva* Representam atVelocidade* de Saída do Trício Incorporado è Proteína, para o Solvente (Exchange out),Obtida* pelo Método de Filtracfo em Coluna» da Gal Sephadex G 26 "Coana", Mantida*a 3oC, Equilibradas e Eluída* com Ttmpflb Formieto 0,1 M, pH 3,0. Na Curva I, Temo*os Valore* Obtido* Apót a Incubaçfo Inicial da Proteína com JHjO a 70°C por 2 hora*.Na curva II, Apói a IncubaçSo Inicial com JHjO a 70°C por 18 hora»

41
tjj•—*
O
u
Ui
o
50 .
0 0,2 0,6 1,0 1,2 1,4
CONTAGENS CANAL A/CONTAGENS CANAL B
17 - Curva da % Eficiência Obtida no Contador da Cintilaçfo ISOCAP/300. Curva Padrfo daEficiência Obtida no Contador da Cintilaçlò ISOCAP/300 (Nuclear Chicago) com Radio-atividade Conhecida de Amostra» Padrdei (Nuclear Chicago) Contendo "Quenching",Utilizando o Método da Relaçfò de Canaii para Corrigir a Eficiência dai Amoitrai Tridadat

42
cont»'i:iil,i. t:ontrd os V,IIOIHS da relação cfc? canais A/B. Esta curva foi utili/.-il.i I M I H O cádiilo daspiir .»•! i. i is •!.• cont.igeiis rias amostras i|iit? r.ontinhatn tricio.
IV - DISCUSSÃO
A .inálise dos resultados desle trabalho necessita em primeiro 'ugar que retiremos da literaturaalguns dados sohie as proteínas por nós purificadas.
A Crotapotin é uma proteína muito ácida (A) de ponto isoelétrico por volta de 3,4-3.7 ' ,desprovida de toxicidade, com peso molecular entre 8400-9400 ' .
A Fosfolipase A é uma protein muito básica (B) de ponto isoelétrico entre 8,4-9,7 ' , combaixa toxicidade e peso molecular de 13000 de acordo com Hendon e Fraenkel Conrat e 16294 deacordo com Breithaupt et alii151.
A Crotamina é uma proteína muito básica (C) de ponto isoelétrico 10,3 , altamente tóxica,com peso molecular 4 8 8 0 l 5 8 ) .
Os resultados da análise de aminoácidos da Fosfolipase A e Crotapotin descritos na Tabela I,
apresentam pequenas diferenças dos obtidos por Hendon e Fraenkel-Conrat . Estas diferenças
representam 4 a.a.* a mais na Fosfolipase A e 3 a.a. a mais na Crotapotin. 0 ponto principal discordante
é o número de cisteínas. onde temos 12 para A e para B enquanto que o referido autor encontra 11 .0$
resultados obtidos no item 111.8 mostram que não foi possível detectar grupos SH livres nas proteínas
mesmo em presença de uréia, o que nos leva a crer que o número de cisteínas deva ser par cr os SH
estejam comprometidos em pontes de enxofre.
Na Tabela I temos também dados da composição de aminoácidos da Crotamina que concordam
muito bem com os publicados por Laure . A Crotamina também não apresenta grupos íióis livres
reforçando a hipótese da existência de 3 pontes de enxofre envolvendo as 6 cisteínas.
Gonçalves e Poison ' em 1947. utilizando equipamento de Tiselius, analisaram ocomportamento eletroforético das proteínas do veneno de Crotalus d. terrificus. Usando pH 8,6 a 7,7puderam verificar o aparecimento de uma proteína muito básica mais tarde caracterizada comoCrotamina; nesses valores de pH era evidente a existência de complexos entre proteínas com cargaspositivas e negativas. Para minimizar a interação proteína proteína foi preciso aumentar a força iònica ourealizar a eletroforese a ^H 3,0 onde segundo os autores as proteínas possuíam a mesma carga 0forneciam diagramas eletroforéticos com imagens especulares nos ramos ascendente e descendente dacélula em U.
Com o isolamento e a caracterização da Fosfolipase A e Crotapotin em 1971 por Hendon •Fraenkel-Conrat e Rubsamen et alii'7 ficou evidenciado que a Crotoxina resulta jda interaçãoeletrostática entre as duas proteínas e que ocorre um fenômeno sinergístico quanto a ação neurotóxica.
A possibilidade de ocorrerem interações entre proteínas de cargas opostas que resultem numaumento de toxicidade, foi levantada por Jeng e Fraenkel-Conrat'66', em 1976. Os autores estudando •toxina de um cogumelo (Volvirtolla mrfvaoaa), a volvatoxina, que como na Crotoxina, tem uma aefotóxica resultante da interação de duas proteínas nlo tóxicas só que de mesmos pontos isoelétricot:pi 6,0, denominadas A t e A J ( verificaram que a volvatoxina A j interage com • Fosfolipase Aaumentando cerca da 6 vezes • toxicidade da lecitinase.
Como o complexo só se forma com • proteína A j da volvatoxina • como foi infrutífera •tentativa de se aumentar a toxicidade da Fosfolipase A, pela presença de outras proteínas ácidas, supflemos autores que o sinergisrno n lo pode ser explicado simplesmente por interações poliionicas.
• • ». = •mlnofcido

43
O surgimento destes complexos híbridos, mais o constante aparecimento de um contaminante
proteico uatregado negativamente, durante as purificações de Crotamina por nós realizadas, que mais
tarde identificamos como sendo a Crotapotin, levou-nos a estudar a possível formação de um complexo
Crotamina Crotapotin.
Na Figura 9, mostramos que A e B interagem formando um complexo estável, não dissociávelna filUdção de gel, com um volume efluente igual ao obtido para a Crotoxina pura e não fracionada. Asmudanças quantitativas de uma proteína em relação a outra, ou seja, excesso de B em relação a A ouvice-versa, não alteraram o volume efluente do complexo AB, como mostramos nas partes B e C daFigura 9.
A Tabela II mostra claramente que em qualquer que seja a proporção molar A : B colocada nacoluna, forma-se um complexo AB estável na proporção molar de 1 : 1.
Pela mesma técnica pudemos comprovar como mostramos na Figura 10, Crotamina (C) eCrotapotin (A) interagem formando um complexo que é entretanto dissociável pela filtração em gel, poiscom qualquer proporção molar A : C colocada na coluna, temos sempre como resultado o aparecimentode 3 picos AC, A e C na ordem crescente de volumes efluentes. Desta maneira pela técnica de filtraçãoem Sephadex G 75 foi difícil chegarmos à exata proporção molar de interação como constatamos nosdados da Tabela I I I . Os dados de medida de concentração de proteína e área dos picos forneceramproporções molares variadas, sugerindo que ocorre uma dissociação expontânea do complexo AC,durante a passagem pelo gel.
Este fato não causa surpresa, uma vez que, outros autores como por exemplo Hartley e
Smeaton'46 ' na interação entre Barnase e seu inibidor Barstar e Cuatrecasas e Hollemberg'11' numa
revisão sobre interação proteínas-membranas constataram a impossibilidade de em alguns casos definir as
proporções molares de interação pela filtração em gel Sephadex, devido a dissociação dos complexos
formados.
Entretanto, Nakazone et a | j j 1 6 5 . 6 6 - 6 7 . 7 5 ' e m nosso laboratório demonstraram de maneira
elegante, utilizando técnicas imunológicas, que o complexo AC se forma realmente na proporção molar
de 1:1.
O aumento da toxicidade da Crotamina e da Fosfolipase A, pela interação com Crotapotin podeser verificada na Tabela IV, onde temos as D L 5 0 obtidas para B e C isoladas e para os complexos AB eAC. Nota-se que a ação tóxica da Fosfolipase A é aumentada cerca de 10 vezes pela interação comCrotapotin e a toxicidade da Crotamina é aumentada cerca de 4 vezes, quando está complexada comCrotapotin.
Mais uma evidência da partic!paç3o de A na formação dos complexos AC e AB poda servisualizada na Figura 11 que resulta de experiências nas quais usamos Crotapotin marcada c o m l 2 5 l ,como traçador.
Como já dissemos o fenômeno de sinergismo que ocorre pela interaçlo entre as proteínas, nfo ésomente produzido pela aproximação eletrostática, alguns fatores estruturais devem estar envolvidos paraque a formação dos complexos seja possível.
Do ponto de vista conformacional o que w conhece sobre Crotamina, Crotapotin • Fosfolipase
A, é realmente muito pouco.
Brelthaupt et al í i '8 1 em 1 974, após a reduçio e alquilaçlo da Crotapotin, «pararam por
filtraçlo em gel 3 cadeias peptídicas, sugerindo que a proteína na tua estrutura original contém as 3
cadeias unidas por pontes de enxofre. Sugerem os autores ainda que a Fosfolipase A é uma cadeia
polipsptídica intramolecularmente ligada por pontes de enxofre.

44
Em 1 975, Paradies e Breithàupt' ' ü l atiavés das medidas de sedimentação, viscosidade edifusão, sugerem que a Crotoxina seria uma esfera com taio de Stokes 22,5 A e uma relação axial 1: 3.enquanto que a Fosfolipase A seria assimétrica, com uma relação axial muito alta e a Crotapotinassumiria uma formação de um elipsóide oblatn de revolução.
Em 1 976, Hampe e Gonçalves trabalhando sobre a dispersão óptica rotatória da Crotamina
encontraram um espectro totalmente atípico. Os mesmos autores em 1 978 analisando a dispersão óptica
rotatória em vários valores dp pH, definem a presença de 3 contôrmeros diferentes, com pH de transição2,6 do confòrmero I para o II e 9,7 do confôrmero II para o III .
A análise dos confòrmeros pelos autores sugerem que a Crotamina seria uma esfera com grau de
compactação decrescente com o aumento do pH.
0 estudo do comportamento estrutural de proteínas em solução, pela medida da velocidade de
troca hidrogênio/trício, tem merecido um certo destaque nos últimos anos. Assim podemos citar
importantes trabalhos com mioglobina e hemoglobina ' ' , conformação'do virus de Mosaico do
Tabaco , ácidos nuclêicos , anidrase carbônica e outras, que utilizam a tecnologia da filtração
em Sephadex G 25 e a interpretação matemática originalmente aplicada por Linderstrom-Lang ' , em
1 955. Esta análise considera que a velocidade de troca hidrogênio-trício (ou deutério) é representada,
para a lisozima, como uma soma de 4 exponenciais do tipo de decaimento radioativo.
0 número de exponenciais encontrado representa o número de classes de hidrogênios
caracterizadas pelos parâmetros destas exponenciais.
0 mesmo tipo de análise tem sido utilizada por outros autores como por exemplo:Englander , em 1 963, estudando a ribonuclease encontra o melhor ajuste na soma de 4 expanenciaispara a curva de velocidade troca H/T definindo 4 classes de hidrogênios, ou seja, uma classe I que seriarapidamente expostos ao solvente com ty, da ordem de segundos; uma classe II que seria hidrogêniospeptídicos em "random-chain" com ty} entre 4-10 minutos; uma classe I I I seria composta porhidrogênios peptídicos envolvidos em alfa-hélice com ty2 por volta de 160 min. e a classe IV hidrogêniospeptídicos ou de cadeias laterais, escondidos na estrutura protéica.
Duas críticas principais surgiram IOS trabalhos mais recentes, a primeira se refere i relaçãodireta da porcentagem de alfa-hélice ao número de hidrogênios da classe I I I , uma vez que, as medidasrealizadas por diversas técnicas não forneciam os mesmos resultados1541. Hoje já se procura associar avelocidade de troca H/T de uma proteína à localização dos seus hidrogênios, se estão mais ou menosexpostos ao solvente.
A segunda crítica está muito bem fundamentada no trabalho de Laiken e Printz'6 7 ' . Os autoras
ss referem è obtenção do melhor ajuste dos pontos experimentais pela computação dos dados,
constatando que é possível, a partir dos mesmos dados, obter-se ajustes razoáveis contendo duas ou três
exponenciais. Fica evidente que, a escolha entre a existência de duas ou de três exponenciais, leva •
erros na interpretação dos resultados.
Entretanto consideramos qua as críticas nío invalidam os bons resultados obtidos por trabalhos
mais recentes, principalmente aqueles que procuram analisar as curvas exponenciais do ponto da vista
comparativo.
Outro fato que devemos considerar é a existência dos afeitos isotópico químico a cinético nas
experiências de troca isotópíca H/T e H/D. A existência do efeito isotópico cinético já havia lido
demonstrada por Englander'201 «m 1 963 na ribonucleest, ou seja, a protafna dauterada pardia dautério
para o solvent» mais lentamente do que a tríciada.
Em 1 969, Englander e Poulsen'241 trabalhando com poliaminoácidos confirmaram a existência
de um efeito itotópico de equilíbrio para os hidrogênios das ligações peptídlcas ou seja, o equilíbrio

45
entre T/H no polímero é 21% maior rio que T/H no solvente. A partir deste feito os autores passaram aincluir na equação utilizada para o cálculo do número de hidrogfnios por mol, o valor 1,21.
H = (111e/1,21Co) (C/A)
Nos cálculos destes trabalhos não consideramos os possíveis efeitos ísotópicos, passando a nosreferir ao número H/mol como número de hidrogênios aparentes por mol(H /mol).
Considerando também que se a computação dos resultados não fornecem números exatos paraos hidrogênios ou constantes de velocidades, com certeza ela nos dá uma imagem do comportamentoestrutural da proteína em solução e essa imagem é que nós utilizamos para analisar os resultados destetrabalho.
Duas linhas de pensamentos diferentes sobre a interpretação do aparecimento de hidrogênioslentos nas proteínas, tem sido austeramente discutidas nos últimos anos.
A primeira linha admite que ocorre na proteína em solução o que é chamado de "respiração"ou flutuações da estrutura protéica, ou ainda desnaturações reversíveis e localizadas, acompanhadas pordestruiçOes de pontes de hidrogênios e conseqüentemente exposição ao solvente para at roca'2 5 '2 7 '5 4 '6 3».
Desta linha destacamos o trabalho de Nakanishi e Tsuboi164' sobre a flutuação da estrutura dalisozima. Após experiências com troca H/D com ou sem a presença de LiBr como desnaturante, osautores sugerem que existe na lisozima uma conformação N e uma conformação D. Na conformação Nocorre 44 pontes de hidrogênios peptídeo-peptídeo, as mesmas encontradas no estado cristalino. Naconformação D as pontes são quebradas e os 44 hidrogênios estão livres. Em solução, a conformação Nestá presente, mas quando é adicionado LiBr a quantidade da conformação D aumenta. Os autoresimaginam ainda um número de diferentes conformações D,, D2, etc. nas quais algumas das 44 pontes dehidrogênio da lisozima estariam quebradas e outras intactas.
A segunda linha de pensamento interpreta o aparecimento de hidrogênios lentos, pelasdiferentes acessibilidades ao solvente dos vários hidrogênios, ou seja, o número de hidrogênios lentosnuma proteína nativa é igual ao número de hidrogênios com acessibilidade ao solvente restrita, e estespodem ou nSo estar envolvidos em pontes de hidrogênios intra-moleculares. Dentro ainda desta linha, omecanismo de exposição ao solvente pode e provavelmente deve, envolver flutuações da proteína, masestas flutuações nSo envolvem necessariamente quebra de pontes de hidro^ín •
Desta linha destacamos os trabalhos de Browne et a l l 6 ) , Wickett et a l l i ' 8 7 ' * principalmente aspublicações de Ellis e Woodward" 8 ' 9 0 ' 9 " que aplicam a técnica de troca isotopica H/T no inibidor datripsina de soja, na tripsina e no complexo formado pela interação das duas proteínas.
A idéia fundamental do trabalho reside na obtençio das curvas cinéticas das proteínas isoladas,para a seguir após a Interação das proteínas, analisar o comportamento cinético da troca H/T nocomplexo. Os autores observam que ocorre uma grande diferença entre a cinética do complexo » •simples soma das cinéticas das proteínas isoladas. A cristalografia da RX descartou a possibilidade doretardamento da velocidade de troca, ser explicado pelo aparecimento de pontes de hidrogênio.
Numa discusslo multo bem feita os autores propõem que a atanuecfo da cinética de troca apósa formeçlo do complexo é devido primariamente as mudanças na distrlbuiçlo das velocidades de trocaem funçlo das mudanças da acessibilidade ao solvente, mas uma pequena contribuiçfo por formeçlo depontes de hidrogênio nío é descartada pelos autores.

46
Um fato importante que é aceito pelos dois grupos de pesquisadores é de que o transporte dehidrogênio <*> far segundo a teoria de E i g e n 1 1 6 ' 1 " , que determina três fases para a reação detramfeiência: a pumeira fase seria a colisão por difusão do doador de hidrogênio com o aceptor para aformação de uma ponte de hidrogênio no complexo; a segunda seria o equilíbrio de redistribuição dopróton entre os dois membros; e a terceira seria a dissociação do complexo com o proton, se a reaçãoteve sucesso, indo para o aceptor.
A análise dos resultados de cinética de troca H/T, aplicada a Crotapotin, Crotamina, FosfolipaseA e Crotoxina, pode ser feita considerando os fatos citados acima, mas não pode decidir sobre uma ououtra explicação das cinéticas encontradas, pela falta de maiores informações sobre as referidasproteínas.
Pela análise de aminoácidos temos que o número de hidrogênios teoricamente trocáveis da
Fosfolipase A é 223, sendo que destes, 122 são hidrogênios peptídicos e, 101 são hidrogênios de cadeias
laterais e de grupos terminais. A Figura 13 nos mostra o ajuste feito sobre os. dados experimentais de
troca H/T da Tabela V, cujo desenvolvimento matemático pode ser visto no Apêndice 2.
A equação que originou este ajuste nos fornece a existência de duas classes de hidrogênios.
sendo que aqueles mais rápidos, impossíveis de serem determinados pela técnica, são obtidos por simples
subtrações do total teórico. Assim teríamos
Classe I = 93 H muito rápidos
Classe II = 59 H com t,/a = 12,6 minutos
Classe I I I - 71 H com t% = 128 horas
Não aparece na Fosfolipase A a ciasse de hidrogênios envolvidos em alfa-hélice cujo t ^ seria de2 a 2,5 horas. Observa-se também que 68% dos hidrogênios teoricamente trocáveis slo rápidos, mesmoconsiderando o fato de trabalharmos a pH 3,0 que está próximo do pH mínimo ou do pH onde •velocidade de troca isotópica é mínima, que está entre pH 2 e pH 4 < 2 7 ' . Por outro lado sabemos que •Fosfolipase A i uma enzima que produz » hidrólist de ésteres de ácidos graxos ligados è posição 2 de1,2 díacilsn-fosfoglicfdeos > 4 < 7 6 > . Sabemos também que estes substratos sfo insolúveis e se encontramna forma micelar.
Estudos sobre mecanismos de ação da Fosfoüpast A de Naja'1 3 1 , de veneno de Crotatut
adamanteui'8 6 '8 6 1 e principalmente sobre a Fosfolipase A de pancreas de porco'71 > 7 2 > 8 9 1 , tem
mostrado que a enzima tem preferência sobre substratos na forma micelar e, que neste caso necessita
penetrar na micela para hidrolisar a ligação ésteres.
O número grande de hidrogênios rápidos encontrados na troca H/T da Fosfolipase A, portanto
acessíveis ao solvente e possivelmente solvatados. facilitariam a interação com o substrato na forma
micelar também solvatado.
Na Figuia 14 temos o melhor ajuste feito para a Crotapotin pela computação dm dados
experimentais da troca H/T da Tabela V, cuios parlmetros slo encontrados no APÊNDICE 2.
0 número de hidrogênios teoricamente trocáveis calculados pela análise de aminoácidos é 130,sendo que destes, 81 slo hidrogênios peptídicos e, 49 slo das cadeias laterais • d* grupos terminais. Daequação que originou o ajuste temos:
Classe I = 24 H muito rápidos
Classe II = 84 H com t)/4 = 3,5 minutos

47
Classe III = 22 H com t,- 10 5 horas
Não aparece na Crotapotin a exponenc.ial cai;icti»c(stica da alfa-hélice com ty de 2,5 horas e83'.Y dos hidrogênios teóticos são rapidamente trocados com o solvente.
Paradies e Breithaupt mostraram qun a Crotapotin é composta de três cadeias polipeptídicasunida? por pontes de enxofre e que a proteína em solução apresenta-se na forma de um elipsóide ofcdatode revolução com relação a/b = 3. Este fato mostra que a Crotapotin é mais esférica que a Fosfolipase Ao que poderia explicar o aparecimento de um maior número de hidrogênios rápidos na Crotapotin.
Por outro lado, do ponto de vista de acessibilidade ao solvente segundo Welch e Fasmann ,proteínas pequenas devem trocar seus hidrogênios com velocidades maiores do que proteínas de altopeso molecular e sem dúvida tanto a Crotapotin quanto a Fosfolipase A são consideradas proteínaspequenas.
O fenômeno de sinergismo que ocorre na interação não é inespecífico ou somente umaaproximação entre proteínas de cargas opostas; devem ocorrer algumas mudanças estruturaispossibilitando a formação de um complexo altamente tóxico que é a Crotoxina.
Na Figura 15 temos o melhor ajuste aos dados experimentais (Tabela VI) da troca H/T daCrotoxina cujos parâmetros podem ser encontrados no Apêndice 2.
O número de hidrogênios teoricamente trocáveis, calculados pela análise de aminoácidos é 353,sendo que destes, 203 são hidrogênios peptídicos e, 150 são de cadeias laterais e grupos terminais. Daequação que originou o ajuste temos:
Classe I = 199 H muito rápidos
Classe II = 32 H com Xy} = 4,3 minutos
Classe III - 26 H com \y} = 58 minutos
Classe IV = 96 H com t,/a = 90 horas
Na Figura 15 temos também a curva que representa a soma teórica das exponenciais da
Crotapotin + as da Fosfolipase A, considerando a interação com proporção molar 1 : 1 .
Em primeiro lugar devemos ressaltar que pela simples comparação das curvas da Figura 15,vemos que a Crotoxina apresenta uma curva mais lenta do que a da soma das duas proteínas que acompõem. Em segundo lugar pelos dados obtidos peio ajuste, temos o aparecimento de uma nova classede hidrogênios com tya = 58 min. podendo ser identificados como participantes de pontes de hidrogêniosque surgiriam da interação Fosfolipase A-Crotapotín.
No geral podemos dizer que pela interação Fosfolipase A - Crotapotin aparecem ra Crotoxina
cerca de 13% a mais de hidrogênios lentos que a simples soma das proteínas isoladas.
Segundo Paradies e Breithaupt'70' pela interação da Fosfolipase A-Crotapotin, a liberação daágua é acompanhada por uma liberação de ions H* da Fosfolipase A resultando num decréscimo na cargapositiva, pelo estabelecimento de pontes de hidrogênios entre grupos carboxil da Crotapotin por volta depH 6,0. Por outro lado os 26 hidrogênios que trocam com velocidade próxima daqueles participantes daalfa-hélice poderiam sugerir a presença de aproximadamente 8 voltas de uma espiral constituindo-senuma razoável alteração estrutural e que seria, quem sabe, o ponto de lígaçSo aos receptores econseqüentemente responsável pelo grande aumento de toxicidade.

48
Na Figura 16 temos os ajustes aos dados experimentais (Tabela VI) da troca H/T da Crotamina
em duas condições iniciais de incubação: uma com 2 horas a 79° C e outra com 18 horas a 70° C.
Na Crotamina temos 86 hidrogénios teoricamente trocáveis, sendo que destes, 44 sfo peptídicose 42 sa"o hidrogênio sm cadeias laterais e de grupos terminais. Das equações que originaram os ajustesteremos:
Classe I = 22 H muito rápidos
2 h a 70°C Classe II = 38 H com ty2 = 3 minutos
Classe I I I = 26 H com t % = 18 horas
Classe I = 61 H muito rápidos
18 h a 70°C Classe II = 9 H com t % = 43 minutos
Classe I I I = 16 H com t,/} = 35 horas
A Crotamina é uma proteína compacta e altamente resistente à temperatura ' . Com
aquecimento de 70°C por 18 horas, a Crotamina manteve sua toxicidade plenamente.
Os valores de troca H/T nos mostram que a Crotamina nSo apresenta hidrogénios envolvidos em
pontes, concordando com os dados já publicados de ORD ' . Os 3 1 % de hidrogénios lentos que
aparecem após a incubaçSo de duas horas a 70°C, diminuem para 18% se a incubaçlo prossegue até 18
horas e, há o aparecimento de um grupo de 9 hidrogénios com ti^ = 43 minutos, quê tf intermediário
entre os encontrados para hidrogSnios peptfdicos livres ou aqueles comprometidos em pontes de
hidrogénios'20'.
Estes fatos podem sugerir uma "abertura" da proteína sem no entanto levar a uma
desnaturaçfo.
Por outro lado, do ponto de vista de acessibilidade ao solvente podemos dizer que a Crotamina,como uma proteína pequena de peso molecular 4880, troca seus hidrogénios rapidamente mantendo umnúdeo contendo 26 H lentos, que diminuem para 16 H quando o aquecimento passa de 2 horas a 70°Ca 18 horas na mesma temperatura.
V - CONCLUSÕES
1) A Crotapotin interage com Fosfolipase A, formando um complexo estável, na proporclo
molar de 1 : 1 , aumentando a toxicidade da Fosfolipase A cerca d* 10 vazas.
2) A Crotapotin interage com Crotamina, formando um complexo dissociávtl pala filtraçlò
am gel, cuja toxicidade quando a proporclo molar do complexo é 1 : 1 , ê cerca da 4
vazes maior que a Crotamina sozinha.
3) A Crotapotin, Fosfolipase A a Crotamina quando estudadas pala medida da velocidade da
troca H/T, apresentam comportamento cinético qua pode sar representado pala soma da
duas exponenciais.

49
4) A cinética da troca isotòpira H/T da Crotamina submetida a um aquecimento com 3 H 2 O
a 70uC por duas horas e 18 horas, mostra que para um maior tempo de incubacSo ocorre
um aumento do número de hidroyênios rápidos com conspquente diminuição dos lentos.
5) A cinética de troca H/T da Crotoxina difere da cinética obtida pela simples soma
Fosfolipase A + Crotapotin, evidenciando o aparecimento de uma terceira exponencial
cujos parâmetros se assemelham aos característicos de hidrogênios envolvidos em
alfa-hélice.

50
APÊNDICE 1
O Apêndice 1, contém os valores de Absorbância (A), contagens por minuto (cpm/ml) e
contagens por minuto/Absorção (C/A) para cada tubo após a filtraclo das amostras contendo proteína +
'H2O.
Contém também, os valores de radioatividade inicial da incubação proteína + 3 H 2 O(C ) e os
tempos de "exchange out".
Finalmente o Apêndice 1 contém o n9 de hidrogênios calculados para cada amostra (H)
submetida à filtraçãV), com base na equação:
H/mol = ( E x 1 1 1 / C 0 ) (C/A)
Foram utilizados para o cálculo do n? H/mol, valores médios de C/A qua tinham uma variaçSo
entre si de no máximo 5%.

CROTOXINA
tubo
A
cpm/ml
C/A
H m
tubo
A
cpm/ml
C/A
co
20
0.60
20016
33360
C o
19
0.17
6617
38923
= 7 , 8 6 K
21
0,78
22516
28866
= 9.29x
20
0.1 S
6553
43686
10* cpm/ml tempo = 1
22 23 24
0.70 0.56 0.37
23319 18561 11834
33312 33144 31983
143
min
25
0.23
8117
34540
10* cpm/ml tempo = 5 min
21 22 23
0.15 0.11 0.08
5184 4008 2665
34560 36436 32812
132
24
0.05
1608
32160
co
17
0,56
19392
34628
C o
14
0.06
1890
31«»00
= 7.18x
18
0,72
22948
31872
= 9,29x
15
0.13
4252
32707
10* cpm/ml tempo = 2 min
19 19
0.69 0.55
21479 18323
31128 33314
154
20
0,39
13742
35235
10* cpm/ml tempo = 12
16 17
0,19 0.20
6193 6341
32594 31705
112
10
0,15
5786
38573
21
0,27
9227
3*174
min
19
0,10
3052
30520
Co
19
0,35
11390
32542
Co
16
0,12
5473
45623
= 7,24 >
20
0.50
14655
29310
= 9,5x
17
0,18
7387
41038
t 10* cpm/ml tempo = 3
21 22
0,48 0.42
15247 13368
31764 31828
147
23
0,33
11620
35212
min
24
0.25
9126
36504
10* cpm/ml tempo =15 min
18 19
0.22 0,17
8576 6396
38981 37623
137
20
0,11
4476
40690
21
0,08
3220
40250
OI

CROTOXINA
tubo
A
cpm/ml
C/A
Hm
tubo
A
cpm/ml
C/A
"m
Co
15
0.15
4879
32526
co
18
0.19
6210
32684
»9.5x
16
0.22
7499
34016
= 9.5x
19
0.22
6634
30154
10a cpm/ml tempo = 30 min
17 18
0.22 0.18
7543 5968
34286 33155
119
19
0.14
4820
34428
20
0.10
3903
39030
10a cpm/ml tempo = 60 min
20 21
0.21 0.17
6240 5543
29714 32605
107
22
0.12
3621
30175
23
0.08
2848
35600
18
0.18
5738
31877
Co
18
0,09
2431
27011
= 9.29x
19
0.17
5404
31788
= 9,29
19
0,15
4381
29206
10a cpm/ml tempo = 35
20 21 22
0,15 0,15 0,11
5738 4311 3192
38253 28740 29018
111
x 108cpm/ml tempo =
20 21 22
0.20 0,18 0,16
5662 5207 4995
28310 28927 31218
103
min
23
0,09
2737
30411
70 min
23
0,14
3802
27157
V17
0,14
4207
30050
Co
16
0,24
7445
31020
= 9,29 x
18
0,15
3923
26153
= 9,5x
17
0,24
7602
31675
10* cpm/ml tempo = 55 min
19 20
0,12 0,11
3208 2919
26733 26536
102
21
0,09
2523
28033
10* cpm/ml tempo =75
18 19
0,19 0,14
6009 4945
31626 35321
111
20
0,11
3519
30600
22
0,06
1752
29200
min
21
0,07
3106
44471

CROTOXINA
tubo
A
j|i
Co
14
0.12
= 9,29x
15
0.13
10a cpm/ml tempo =
16 17
0.11 0.10
18
0,09
90 min
19
0.09
C o
18
0,19
= 9.5x
19
0.19
10* cpm/ml tempo = 100
20 21
0.19 0,17
22
0.13
min
23
0,09
C o
17
0,06
= 9,29 x
18
0,13
108 cpm/ml tempo
19
0,18
20
0,20 0,
= 120 min j
21 22 !
19 0,14
cpm/ml • 3380 3622 2980 2980 2665 1722 6445 5823 6163 5356 4092 3002 I 1643 4036 5322 5735 5088 4294
C A 28166 27861 27090 29800 29611 24600 33921 30805 32436 31505 31476 33355|27383 31046 29566 27975 28266 30671
104 112 104
tubo
A
cpm/ml
C/A
C o
16
0.15
4036
26906
= 9,5 x 10a cpm/ml tempo =130 min
17
0.122
5183
23559
18 19
0.24 0.22
8003 6129
33345 27859
07
20
0.16
4418
27612
21
0.11
3121
26008
C o
18
0.09
2537
26989
= 9,5 x 10* cpm/ml tempo =150
19
0,16
5090
31812
20
0.23
6207
26986
í
21
0.23
6266
27243
)6
22
0,19
5060
26631
miii
23
0,10
4291
42910
Co =
15
0,14
3740
26714
9,29 x
16
0,19
4848
25515
10* cpm/ml tempo = 190
17 18
0,20 0,17
5191 4476
25955 26329
94
19
0,11
2876
25008
min
20
0,07
2162
26145
s
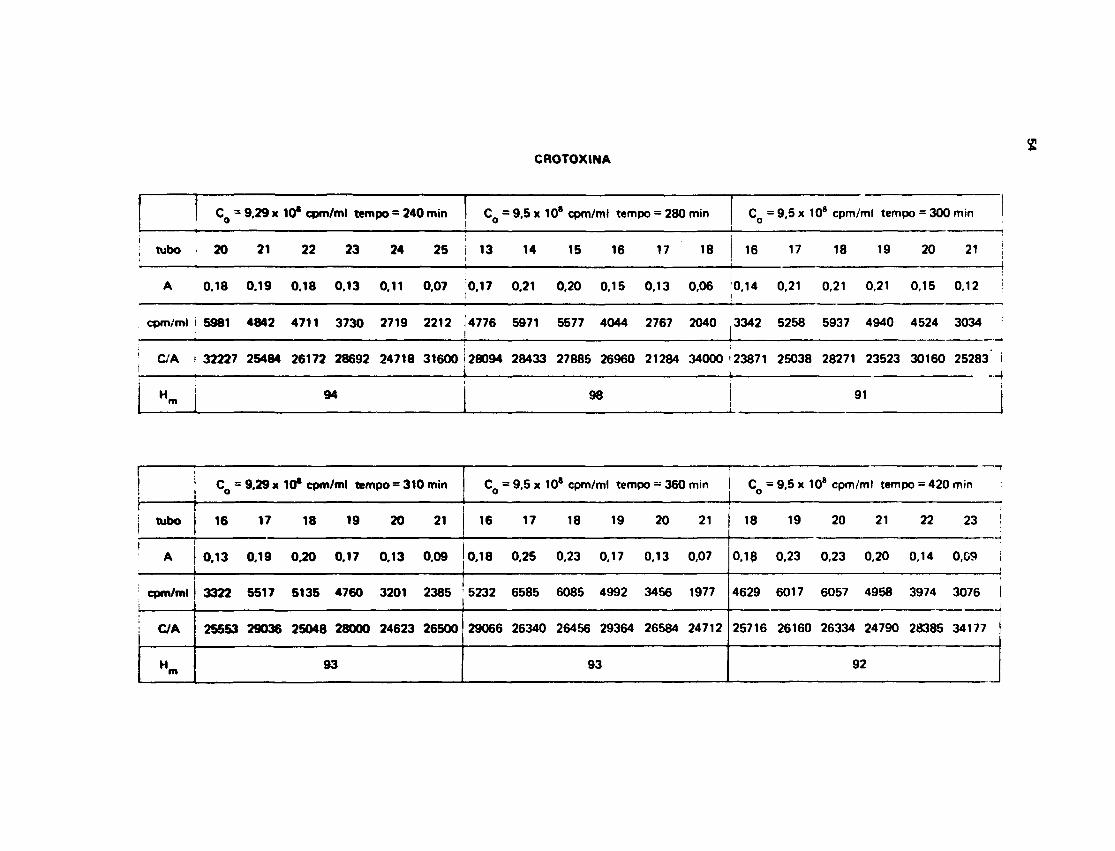
CROTOXINA
; tubo
A
cpm/ml
! aA
tubo
' A
; cpm/ml
| C/A
C o =
20
0.18
5981
32227
16
0.13
3322
25553
9.29 x
21
0.19
4842
25484
9.29 x
17
0,19
5517
29036
10* cpm/ml tempo - 240 min
22 23
0.18 0.13
4711 3730
26172 28692
94
24
0.11
2719
24718
25
0.07
2212
31600
10* cpm/ml tempo = 310 min
18 19
0.20 0.17
5135 4760
25048 28000
93
20
0,13
3201
24623
21
0.09
2385
26500
Co
I 13
0.17
4776
28094
Co
16
0.18
5232
29066
= 9,5 x 1
14
0.21
5971
28433
= 9,5 x 1
17
0.25
6585
26340
0s cpm/mt tempo = 280
15 16
0,20 0,15
5577 4044
27885 26960
98
17
0,13
2767
21284
0* cpm/ml tempo = 360
18 19
0,23 0,17
6085 4992
26456 29364
93
20
0,13
3456
26584
min
18
0,06
2040
34000
min
21
0,07
1977
24712
Co =
16
Í0.14
3342
23871
Co =
18
0,18
4629
25716
9,5 x
17
0,21
5258
108 cpm/ml tempo = 300
18 19
0,21 0,21
5937 4940
25038 28271 23523
9,5 x
19
0,23
6017
91
20
0,15
4524
30160
10* cpm/ml tempo = 420
20 21
0,23 0,20
6057 4958
26160 26334 24790
92
22
0,14
3974
28385
min
21I
0,12
3034 !
25283
min !
23
0,09 i
3076
34177
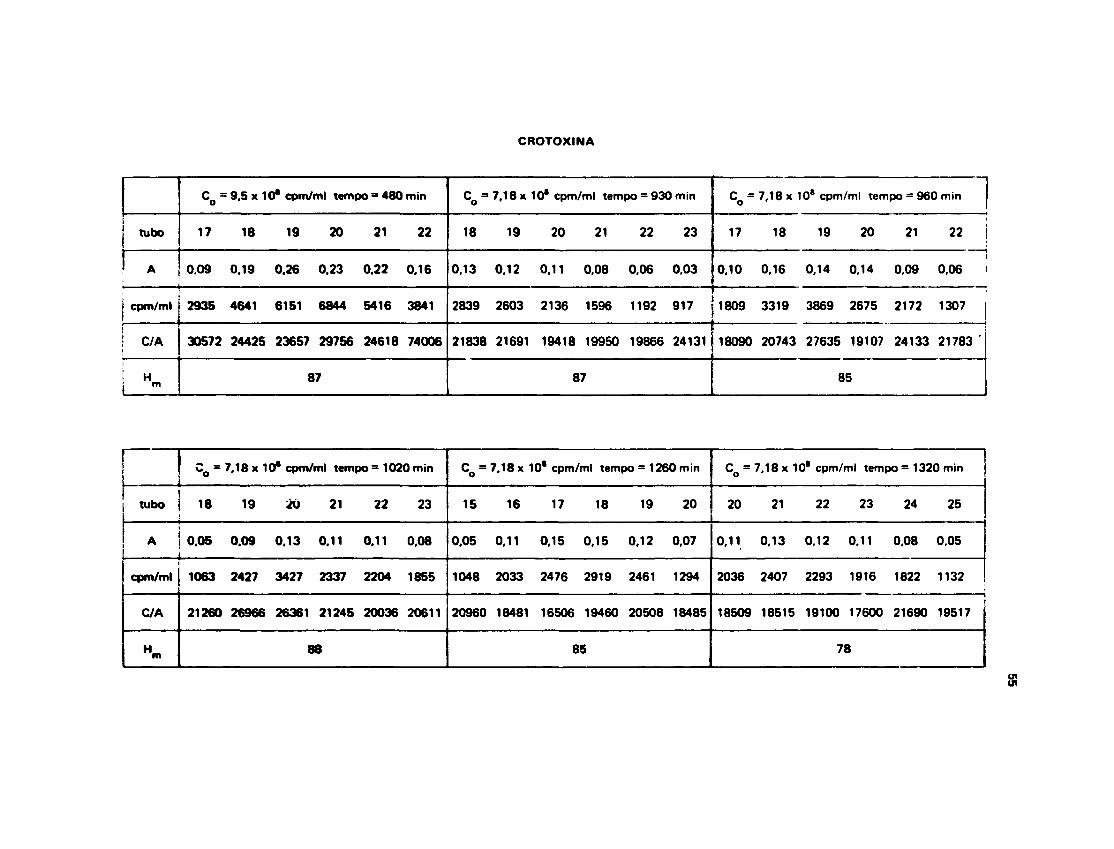
CROTOXINA
tubo
A
cpm/ml
i C/A
i
Hm
tubo
A
cpm/ml
C/A
V17
0.09
2935
30572
=.-
18
0.05
1063
21260
= 9.5x
18
0.19
4641
10* cpm/ml tempo = 480 min
19 20
0.26 0.23
6151 6844
2442S 23657 29756
7.18 x
19
0.09
2427
87
21
0.22
5416
24618
22
0.16
3841
74006
10* cpm/ml tempo = 1020 min
20 21
0,13 0.11
3427 2337
26966 26361 21245
88
22
0.11
2204
20036
23
0,08
1855
20611
Co =
18
0.13
2839
21838
Co =
15
0.05
1048
20960
7,18 x
19
0.12
2603
21691
7,18 x
16
0.11
2033
18481
10* cpm/ml tempo = 930 min
20 21
0.11 0.08
2136 1596
19418 19950
87
22
0,06
1192
19866
23
0,03
917
24131
10s cpm/ml tempo = 1260 min
17 18
0.15 0,15
2476 2919
16506 19460
85
19
0,12
2461
20508
20
0,07
1294
18485
Co =
17
0,10
1809
18090
Co =
20
0,11
2036
18509
7,18 x 1
18
0,16
3319
20743
0* cpm/ml tempo = 960 min
19 20
0,14 0.14
3869 2675
27635 19107
85
21
0,09
2172
24133
22
0,06
1307
21783 '
7,18 x 108 cpm/ml tempo = 1320 min
21
0,13
2407
18515
22 23
0,12 0,11
2293 1916
19100 17600
78
24
0,08
1822
21690
25
0,05
1132
19517

CROTOXINA
tubo
A
cpm/ml
C7A
C
16
0.11
2440
22181
:o = 9.29 x
17
0.18
3897
21650
10* cpm/ml tempo =
18
0.20
4533
22765
19
0.19
4343
22857
81
1430 min
20
0.14
3068
21914
21
0.08
2011
25137
C
15
0.10
2199
20942
o = 9' 2 9 x
16
0.18
4133
22961
10* cpm/ml tempo
17
0,22
4840
22000
18
0,20
4083
20415
75
=1490 min
19
0,16
3271
20443
20
0,11
2134
19400

FOSFOLIPASE A
C = 5,9 x 10s cpm/ml tempo = 1 min Co = 5,3 x 108 cpm/ml tempo = 2 min C = 4,05 x 108 cpm/ml tempo = 3 min
tubo 12 13 14 15 16 17 8 9 10 11 12 13 16 17 18 19 20 21
cpm/ml
C/A
0.12
5735
45880
0.18
6772
37622
0.19
7094
37366
0.18
6715
37305
156
0.18
7458
41433
0.18
-
-
0.06
2526
42100
0.43
12072
28074
0.84
21955
26136
0,76
21080
27736
128
0,45
11894
26431
0.28
9423
33653
0,37
8620
22986
0,74
15344
20735
0,89
17572
19743
0,82
15744
19200
123
0,61
12058
19767
0,41
8515
20768
1
tubo
A
cpm/ml
C/A
i
co
17
0.73
14256
19528
= 3.16x
18
1.0
19142
19142
10* cpm/ml tempo = 4 min
19 20
1.2 0.96
21513 17656
17927 18391
113
21
0.65
12064
18560
22
0.38
7570
19921
10
0.16
4048
25300
= 5.9€x
11
0.44
11342
25777
10* cpm/ml tempo = 5 min
12 13
0,69 0,75
16554 18251
23991 24384
104
14
0,64
17097
26714
15
0,40
14545
36362
co
7
0,13
3143
24176
= 5,99 x
8
0,48
15560
32416
IO8 cpm/ml tempo = 6
9 10
0,65 0.55
16542 14992
25449 27258
106
11
0,48
16228
33808
min
12
0,39
i
i

FOSFOLIPASE A
!
tubo
A
cpm/ml
C/A
tubo
A
cpm/ml
C/A
H m
Co
11
0.22
4503
20468
Co =
19
0.08
1988
24850
= 5 x 10* cpm/ml tempo =15 min
12
0.50
10010
20020
9.04 x
20
0.08
2404
30050
13 14
0.49 0.46
9863 9568
20128 20865
100
15 16
0.41 0.30
- -
- -
10* cpm/ml tempo = 30 min
21 22
0.09 0.07
2516 2066
27995 29514
78
23 24
0.05 0.03
1988
39760 -
Co
17
0.04
1932
48300
Co
18
0,06
1463
24383
= 7x
18
0.15
4296
10* cpm/ml tempo = 20 min
19 20
0.21 0.23
6038 5855
28640 28752 25456
= 7x
19
0.14
3051
98
21
0,16
4583
28643
10* cpm/ml tempo = 56 i
20 21
0.30 0,26
6527 5404
21792 21756 20784
77
22
0.21
3934
18733
22
0.09
2862
31800
nin
23
0,14
3051
21792
Co
17
0,07
1777
25385
Co
16
0,08
1700
21250
= 7 x 10* cpm/ml tempo = 27
18
0,16
4037
25231
19 20
0,22 0,24
5527 5800
25122 24166
89
21
0,25
4406
17624
min
22
0,21
2739
13042
= 7 x 108 cpm/ml tempo = 72 min
17
0.16
3877
24231
18 19
0,22 0,22
4334 4864
19700 22109
73
20
0,18
4048
21881
21
0,13
3335
25653
1
i
í1
|
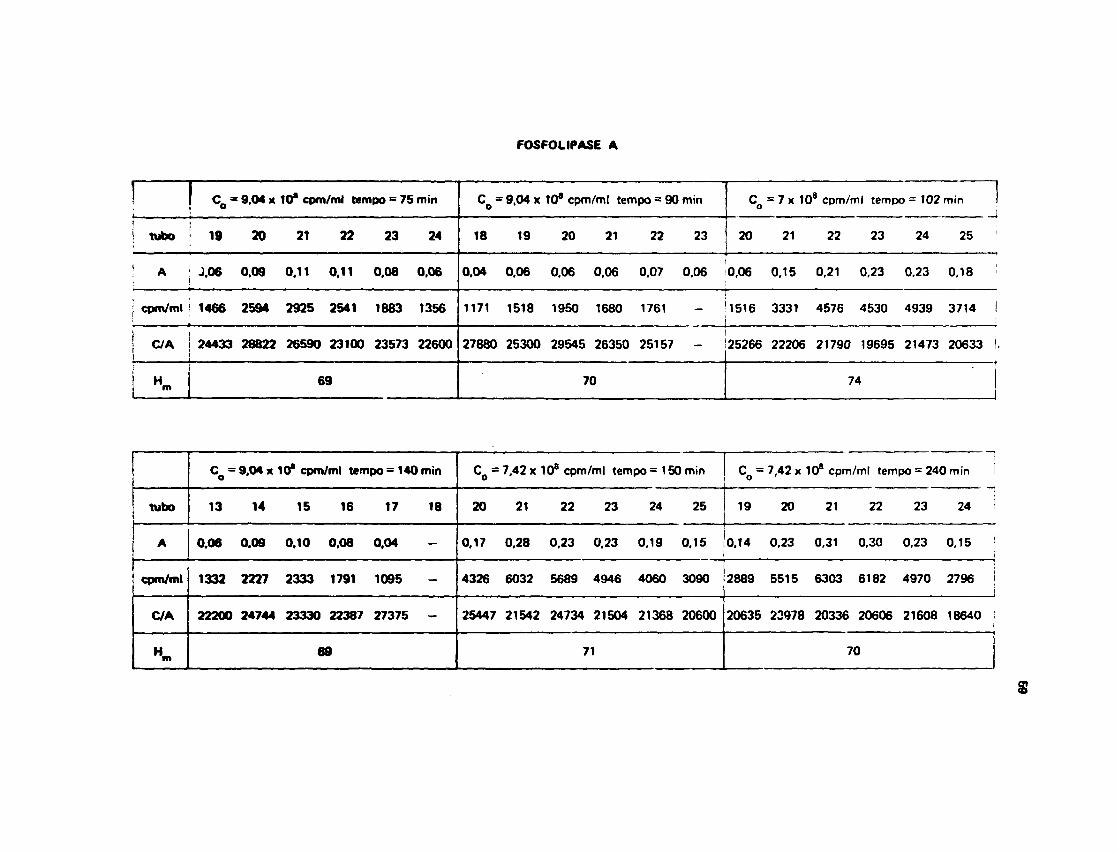
FOSFOLIPASE A
Itubo
! A
cpm/ml
C/A
tubo
A
cpm/ml
C/A
! <=•
• 19
; J.06
1466
= 9,04x
20
0.09
2594
24433 28822
Co
13
0.06
1332
= 9.04x
14
0.09
2227
22200 24744
10* cpm/ml tempo = 75 min
21 22
0.11 0.11
2925 2541
26590 23100
69
23
0,08
1883
23573
24
0.06
1356
22600
10* cpm/ml tempo = 140 min
15 16
0.10 0.08
2333 1791
23330 22387
69
17
0.04
1095
27375
18
—
—
-
Co
18
0,04
1171
= 9.04x
19
0.06
1518
27880 25300
Co
20
0.17
4326
= 7.42 x
21
0.28
6032
25447 21542
10* cpm/ml tempo = 90
20 21
0.06 0,06
1950 1680
29545 26350
70
22
0.07
1761
25157
108 cpm/ml tempo = 150
22 23
0,23 0.23
5689 4946
24734 21504
71
24
0,19
4060
21368
min
23
0.06
-
-
min
25
0.15
3090
20600
Co
20
0,06
1516
25266
Co =
19
0,14
2889
20635
= 7 x 108 cpm/ml tempo = 102 min
21
0,15
3331
22206
7,42 x
20
0,23
5515
23978
22 23
0,21 0,23
4576 4530
21790 19695
74
24
0.23
4939
21473
10s cpm/ml tempo = 240
21 22
0,31 0,30
6303 6182
20336 20606
70
23
0,23
4970
21608
25
0,18
3714
20633
min
24
0,15
2796
18640

FOSFOLIPASE A8
tubo
í A
cpm/ml
C/A
H m
C o =
7
0.06
1287
21450
7.42 x
8
0.57
11859
20805
10* cpm/ml tempo = 300 min
9 10
0.47 0.14
9716 2828
20672 20200
70
11
0.04
-
-
12
0.03
-
-
Co =
11
0.14
2701
19292
7,42 x
12
0,22
4587
20850
108 cpm/ml tempo = 360 min
13 14
0,25 0,19
5416 3542
21664 18642
69
15
0,14
3016
21542
16
0,10
-
-
Co =
20
0,15
10051
67006
7,42 x
21
0,25
4614
18456
10* cpm/ml tempo = 420
22 23
0,28 0.29
5477 5078
19560 18135
64
24
0,21
3884
18495
min
25
0,18 •
2857
15443
tubo
A
cpm/ml
C/A
18
0.04
821
20625
5,25x1
19
0.06
960
16000
IO* cpm/ml tempo = 1110 min
20 21
0.05 0,05
798 831
15960 16620
69
22 23
0.03
473
15766 -
C o =
17
0,06
855
14250
5,25 x 10* cpm/ml tempo = 1200 min
18
0,08
1256
15700
19 20
0,07 0,07
1037 1013
14814 14471
68
21 22
0,03 0,02
- -
- -
Co
20
0.19
2852
15010
= 7x 10'
21
0,20
3653
18265
cpm/ml tempo
22 23
0,23 0,17
4204 3016
18278 17976
64
= 1440
24
0,12
2325
19375
min
25
0,08
1683
21037

FOSFOLIPASE A
tubo
A
"t
19
0.06
• 5.52 x
20
0.08
10* cpm/ml tempo = 1470 min
21 22
0.08 0.06
23
0.04
24
0.03
C o
19
0.11
= 7x 10"
20
0,20
cpm/ml
21
0.23
tempo
22
0,25
= 1500 min
23 24
0,18 0,14
C o
19
0,03
*7x1Q*
20
0,06
cpm/m
21
0,07
tempo
22
0,07
= 1800
23
0,06
min
24
0,05
cpm/ml i
C/A
- . !
1127
18783
1059
13237
1098 927
13725 15450
62
595
14875
422
14066
1093
15390.
3879
19395
3464 4093
15060 16372
58
2790
15500
1990
14214
709
23633
1116
18600
1411 1337
20157 ^9100
60
1128
18800
1049
20980
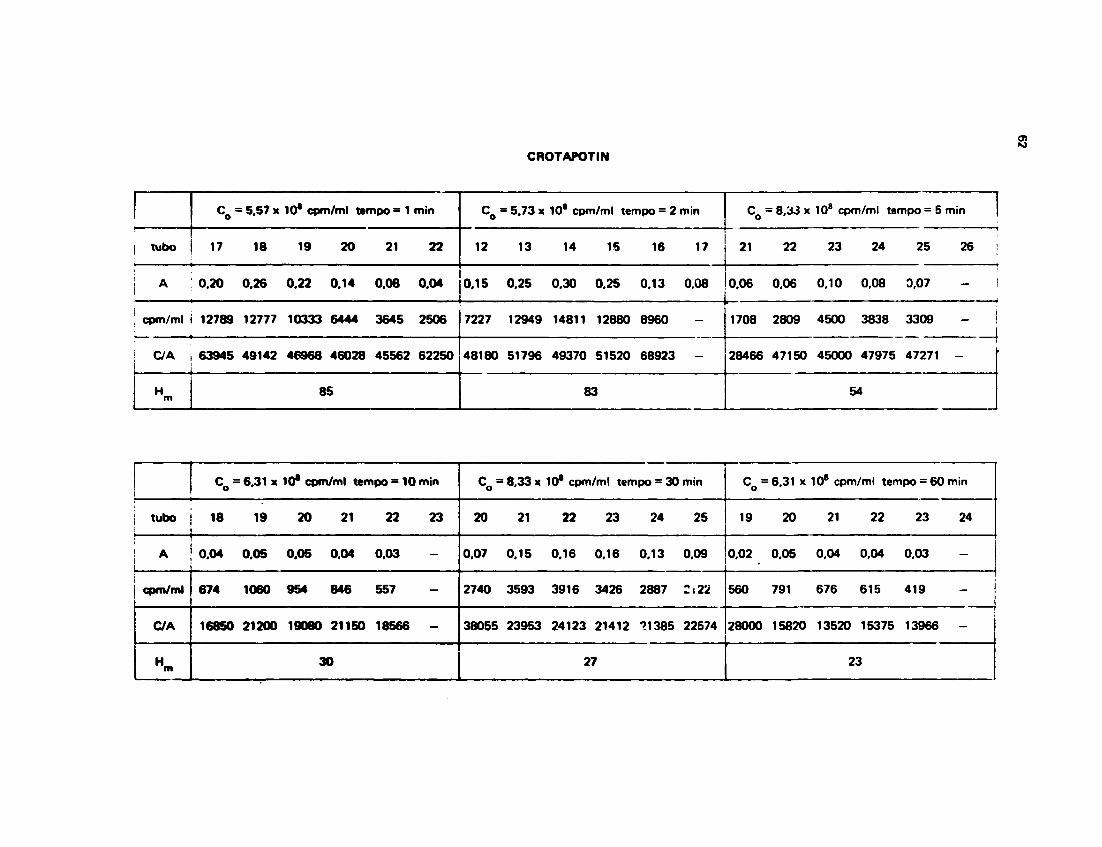
CROTAPOTIN
1tubo
í A
cpm/ml
1 C/A
co
17
0.20
12789
63945
= 5.57 x
18
0.26
12777
49142
10* cpm/ml tempo = 1
19 20
0.22 0.14
10333 6444
46968 46028
85
21
0.08
3645
45562
min
22
0.04
2506
62250
co
12
0.15
7227
48180
= 5.73 x
13
0,25
12949
51796
10* cpm/ml tempo = 2
14 15
0.30 0.25
14811 12880
49370 51520
83
16
0.13
8960
68923
min
17
0,08
-
-
co
21
0.06
1708
28466
= 8,3Jx
22
0,06
2809
47150
10* cpm/ml tempo = 5
23 24
0,10 0,08
4500 3838
45000 47975
54
25
0,07
3309
47271
min
26
-
-
-
; tubo
A
cpm/ml
C/A
co
18
0.04
674
16850
= 6.31 x
19
0.05
1060
21200
10* cpm/ml tempo = 10 min
20 21
0.05 0.04
954 846
19080 21150
30
22 23
0.03
557
18566 -
co
20
0,07
2740
38055
= 8.33x
21
0.15
3593
23953
10* cpm/ml tempo = 30 min
22 23
0.16 0.16
3916 3426
24123 21412
27
24
0.13
2887
?1385
25
0,09
2.22
22574
co
19
0,02
560
28000
= 6,31 x
20
0,05
791
15820
10* cpm/ml tempo = 60
21 22
0,04 0,04
676 615
13520 15375
23
23
0,03
419
13966
min
24
-
-
-

CROTAPOTIN
tub»
!
cpm/ml
C/A
co =
17
0.08
1617
20212
«.33 x
18
0.16
3228
20175
10* cpm/ml tempo = 90 min
19 20
0.23 0.19
4320 3261
18782 17163
22
21
0.10
1875
18750
22
0.04
1006
25150
co =
20
0.10
1726
17260
8.33 x
21
0,12
2132
17766
10s cpm/ml tempo = 120
22 23
0.13 0.12
1834 2101
14107 17508
20
24
0.11
1709
17354
min
25
0,10
1770
17700
Co =
16
0.09
1527
16966
8.33 x
17
0,10
1494
14940
108 cpm/ml tempo =170 min
18 19
0,08 0.07
1257 1230
15712 15375
18
20 21
0,06
937
15616 - f
tubo
A
cpm/ml
C/A
M m
Co
20
0.08
411
5137
= 6.85x
21
0.12
1275
10BI'S
10* cpm/ml tempo-210 min
22 23
0,11 0.10
1315 1122
11954 11220
16
24
0.09
878
9755
25
0.05
565
11300
Co
17
0.06
537
8950
= 6.85x
18
0.08
749
936?
10s cpm/ml tempo = 240
19 20
0,09 0,09
922 997
10244 11077
15
21
0,08
776
9700
min
22
0,07
503
7185
Co =
17
0,09 ,
1461
16233
8,33 x
18
0,14
1752
1k514
10s cpm/ml tempo = 270 mini
19 20
0,15 0,14
1969 2078
13126 14842
15
21
0,12
1496
12466
22
0,07
918i
13114

2CROTAPOTIN
I' tubo
A
cpm/ml
C/A
H m
1
c.-
14
0.11
1548
14072
8.33 x
15
0.17
2394
14082
10* cpm/ml tempo = 300 min
16 17
0.19 0.09
2619 1342
13784 14911
16
18
0.07
410
5857
19
0.05
342
6840
Co =
16
0,03
239
7966
6.31 x
17
0.05
447
8940
10* cpm/ml tempo = 410
18
0.04
372
9300
19
0.05
446
8920
15
20
0,04
203
5075
min
21
0,04
193
4825
Co =
17
0.04
249
6225
6,85 x
18
0.08
253
3162
10* cpm/ml tempo = 1320 min
19
0.10
281
2810
20
0,12
223
1858
5
21
0,11
351
3190
22
0.08
255
3187
: tubo
A
cpm/ml
C/A
C o • 6.85 x
20 21
0.08 0.10
294 381
3675 3810
10* cpm/ml tempo3 1380min
22
0.11
406
3690
23
0.10
259
2590
5
24 25
0.07
258
3685
Co =
16
0.09
222
2466
6.85 x
17
0.10
267
2670
10* cpm/ml tempo
18
0,11
288
2618
19
0.09
185
2055
4
= 1440 min
20 21
0.06
149
2483
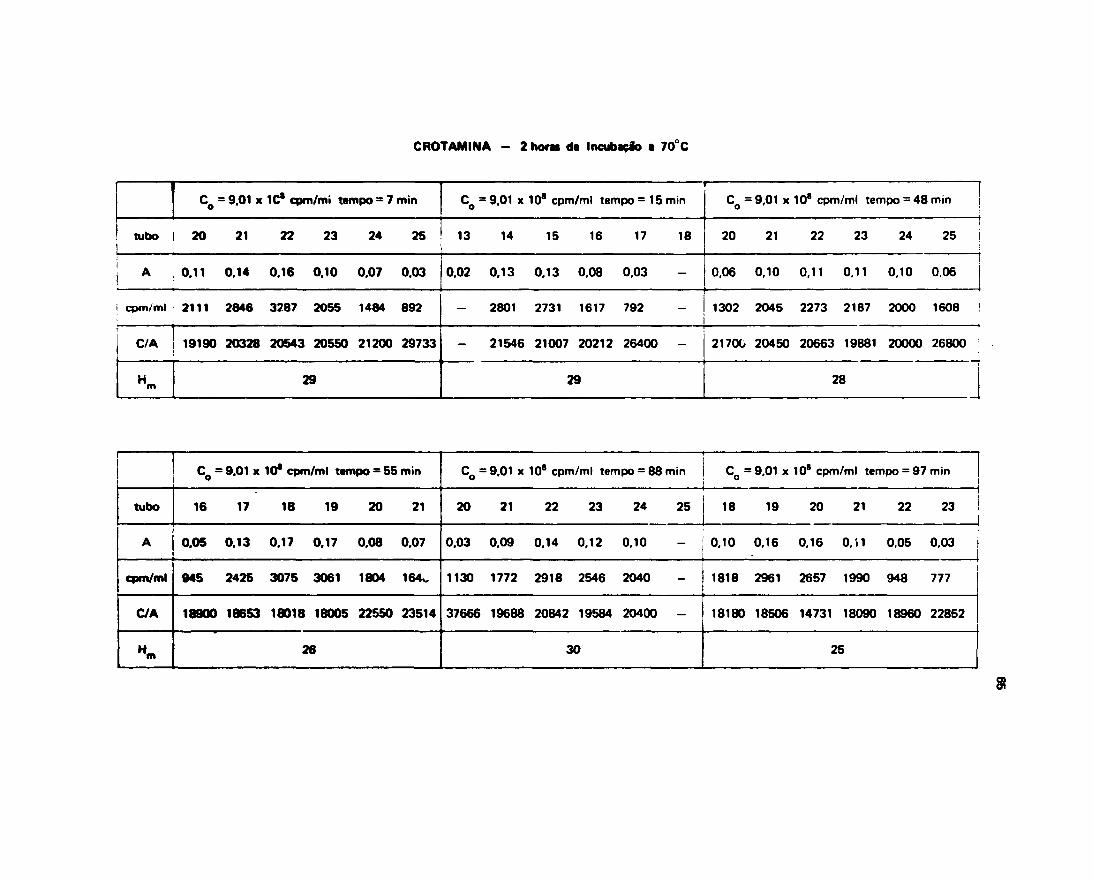
CROTAMINA - 2 horn d* Incubaçio a 70°C
tubo
A
i cpm/ml
C/A
1
tubo
A
cpm/ml
C/A
C = 9.01 xO
20
0.11
2111
19190
21
0.14
2846
20328
C o = 9.01x
16
O.OS
945
18900
17
0.13
2425
18653
1C* cpm/mi tempo = 7
22 23
0.16 0.10
3287 2055
20543 20550
29
24
0.07
1484
21200
min
25
0.03
892
29733
10s cpm/ml tempo = 55 min
18 19
0.17 0.17
3075 3061
18018 18005
26
20
0,08
1804
22550
21
0.07
164c
23514
Co
13
0.02
-
-
Co
20
0.03
1130
37666
= 9.01 x
14
0.13
2801
21546
= 9.01 x
21
0.09
1772
19688
10s cpm/ml tempo = 15 min
15 16
0.13 0.08
2731 1617
21007 20212
29
17 18
0.03
792
26400 -
10s cpm/ml tempo = 88 min
22 23
0,14 0.12
2918 2546
20842 19584
30
24 25
0,10
2040
20400 -
20
0,06
1302
21700
C =o
18
0,10
1818
18180
= 9,01 x
21
0,10
2045
20450
= 9,01 x
19
0,16
2961
18506
10" cpm/ml tempo = 48
22 23
0,11 0.11
2273 2187
20663 19881
28
24
0,10
2000
20000
10* cpm/ml tempo = 97
20 21
0,16 0.Í1
2657 1990
14731 18090
25
22
0,05
948
18960
min
25
0,06
1608
26800
min
23
0,03
777
22852

CROTAMINA - 2 hom d* Incutucio • 70°C
' tubo
A
cpm/ml
C/A
Co
13
0.11
1998
18163
= 3.93x
14
0.18
3316
18422
10* cpm/ml tempo = 1
15 16
0.30 0.36
5210 6090
17366 16916
58
17
0.40
6907
17267
min
18
-
—
—
Co
20
0.12
2163
18025
= 5,O4x
22
0.46
8589
18671
10* cpm/ml tempo = 5 min
23 24
0.57 0.59
10173 10806
17487 18315
47
26
0.35
6479
18511
27
0,22
3917
17804
V19
0,46
8261
17958
= 5,76 x
20
0,59
10243
17361
10* cpm/ml tempo = 10
21 22
0,61 0,45
10210 7786
16737 17302
39
23
0,34
5625
16544
min
24
0,25
4110
16440
tubo
A
cpm/ml
C/A
Hm
24
0.10
1222
12220
9.01 x
25
0.12
786
6550
10* cpm/ml tempo = 1490 min
26
0.13
593
7561
27
0.11
683
6209
S
28
0.07
471
6728
29
0.04
272
6800
19
0.07
486
6942
9.01 x
20
0.12
915
7625
10* cpm/ml tempo = 1550 min
21
0,15
975
6500
22
0,15
1117
7446
10
23
0,11
803
7300
24
I0,07
611
8728

CROTAMINA - 2 horas dt Incubaçio • 70°C
tubo
A
cpm/ml
C/A
Co = 9 ,0 ix
23 24
0.11 0.12
1720 1962
15636 16350
10a cpm/ml tempo = 120 min
25 26
0.11 0.09
2305 1517
20954 16855
23
27
0.07
1138
16257
28
0.04
770
18333
co =
19
0.05
1021
20420
= 9.01 x
20
0,10
1534
15340
10* cpm/ml tempo = 180
21 22
0.11 0,10
1604 1671
14581 16710
22
23
0,08
1557
19462
min
24
0.05
768
15360
co =
19
0.05
615
12300
= 9,01 x
20
0,10
1343
13430
10* cpm/ml tempo = 345
21 22
0,20 0.10
2425 1487
12125 14870
18
23
0,09
1040
11555
min
24
0,04
-
-
•
tubo
A
cpm/ml
C/A
C o =
21
0.06
816
13600
9.01 x
22
0.08
1069
13237
10* cpm/ml tempo = 410 min
23 24
0.10 0.11
1355 1897
13550 172*5
20
25 26
0.09
1302
14466 -
Co =
20
0.07
942
13457
9.01 x
21
0.09
1256
13955
10* cpm/ml tempo = 480
22 23
0.09 0.10
1285 1342
14277 13420
20
24
0.08
1091
13637
min
25
0.05
999
19980
Co
21
0,07
593
8471
= 9,01 x
22
0.10
873
8730
108 cpm/ml tempo = 1410 min
23
0.11
937
8518
24
0,10
1236
12360
12
25
0,08
679
8487

CROTAMINA - 18 horas d* Incubacão • 70°Cs
Co = 8.36 x 10* cpm/ml tempo = 5 min | CQ = 8.36 x 10a cpm/ml tempo = 20 min C =8,36 x 108 cpm/ml tempo = 40 min i
tubo 18 19 20 21 22 23 15 16 17 18 19 20 19 20 21 22 23 24
0.04 0.09 0.14 0,12 0.09 0.07 10.06 0.12 0,18 0,16 0.13 0.08 0,09 0,12 0,17 0,12 0,11 0,08 j
cpm/ml
C/A
"n.
670
16730
1470
16333
2225 1858
15892 15816
24
2388
25866
1795
25642
909
15150
1715
14291
2351 2502
13061 15637
24
1957
15053
872
10900
1981
22011
1S06
13383
2314
13670
21
1634
13616
1575
14318
1060
13250
tubo
A
cpm/ml
C/A
Co
17
0.12
1180
9833
= 8.39 x
18
0.27
3445
12759
10s cpm/ml tempo = 55 min
19 20
0.20 0.15
2570 1851
12850 12340
19
21
0.08
829
10362
22
0.05
552
11040
co
17
0,08
1660
20750
= 8,36 x
18
0,14
1783
12735
108 cpm/ml tempo = 60
19 20
0,17 0,16
2238 2693
13164 16831
21
21
0,12
1778
14816
min
22
0,07
1026
14657
co =
21
0,06 .
644
10733
8,39 x
22
0,11
1104
10036
108 cpm/ml tempo = 150
23 24
0,13 0,13
1406 1321
10815 10161
16
25
0,14
1492
10657
min
26
0,11
-
-
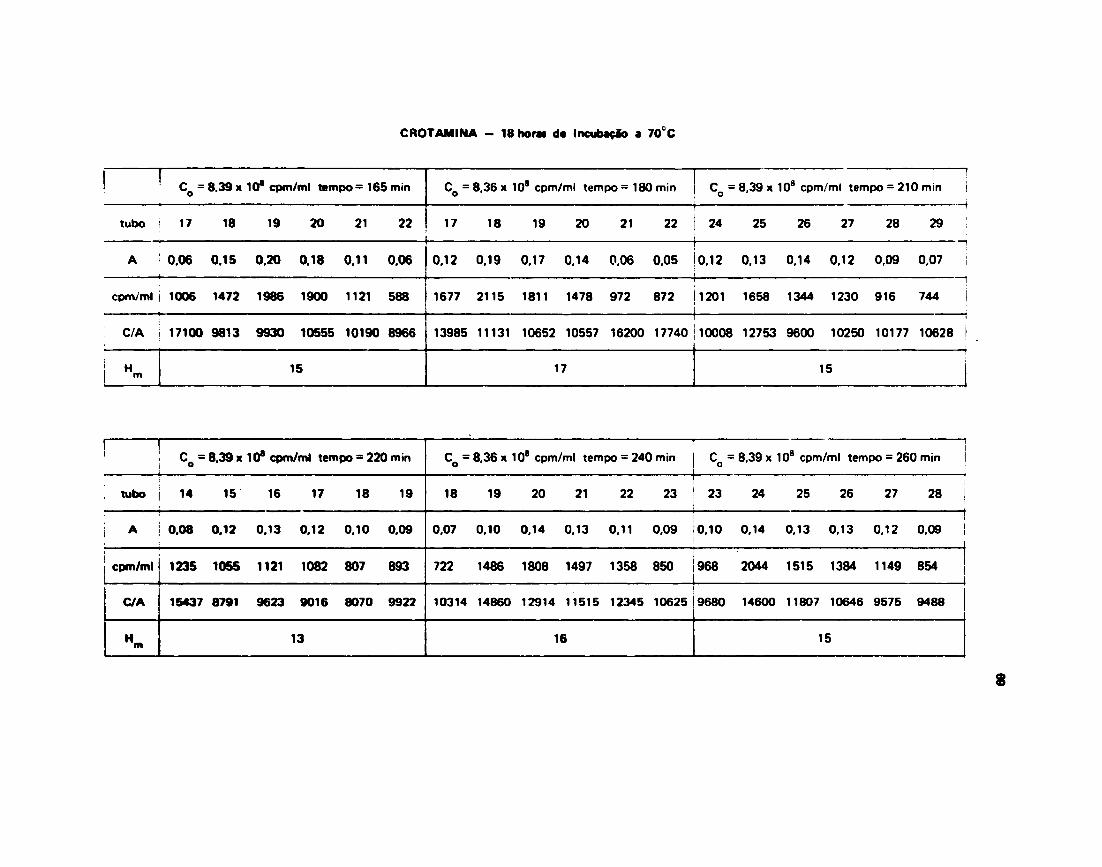
CROTAMINA - 18 hora de Incubarão a 70°C
tubo
A
cpm/ml
C/A
H m
tubo
cpm/ml
C/A
Co =
17
0.06
1006
17100
14
0.08
1235
15437
8.39 x
18
0.15
1472
9813
8.39 x
15
0.12
1055
8791
10* cpm/ml tempo =165 min
19
0.20
1986
9930
20
0.18
1900
10555
15
21
0,11
1121
10190
22
0.06
588
8966
10* cpm/ml tempo = 220 min
16
0.13
1121
9623
17
0.12
1082
9016
13
18
0.10
807
8070
19
0.09
893
9922
Co =
17
0,12
1677
13985
Co =
18
0.07
722
10314
8,36 x
18
0.19
2115
11131
8.36 x
19
0,10
1486
14860
10* cpm/ml tempo = 180
19 20
0,17 0,14
1811 1478
10652 10557
17
21
0,06
972
16200
10* cpm/ml tempo = 240
20 21
0.14 0,13
1808 1497
12914 11515
16
22
0,11
1358
12345
min
22
0.05
872
17740
min
23
0,09
850
10625
Co =
24
0,12
1201
10008
Co =
23
0,10
968
9680
= 8,39 x
25
0,13
1658
12753
= 8.39 x
24
0.14
2044
14600
10* cpm/ml tempo = 210 min
26
0,14
1344
9600
1
27
0,12
1230
10250
5
28
0,09
916
10177
29
0,07
744
10628
IO8 cpm/ml tempo = 260 min
25
0,13
1515
11807
26
0,13
1384
10646
15
27
0,12
1149
9575
28
0,09
854
9488
I
j
j
I

CROTAMINA - 18 hom de Incubaçfo • 70 C
1 1
tubo í
! A :
cpm/ml '
; C/A i
i H- í
Co = 8.36 x
19 20
0.11 0.18
1151 1917
10463 10650
10* cpm/ml tempo = 270 min
21 22
0.20 0.15
2092 1819
10460 12126
16
23 24
0.10
1005
10050 -
V15
0,06
668
1113J
: 8,36 X
16
0.14
1538
10985
IO8 cpm/ml tempo = 290
17 18
0,15 0,16
1611 1773
10740 11081
16
19
0,12
1730
14416
min
20
0,09
1611
17500
co =
20
0,11
739
6718
8,36 x
21
0,12
726
6050
10* cpm/ml tempo = 1380 min
22
0,12
636
5300
23
0,09
608
6755
10
24
0,08
483
6037
25
0,05
263 !
5260 I
tubo
í A
cpm/ffil
C/A
Co =
10
0.06
395
6500
3.36 x
11
0.18
1274
7077
10* cpm/ml tern
12
0.24
1544
6433
13
0.19
1220
6421
10
po- 1440 min
14
0,10
1138
11380
15
0.04
707
1767S

71
APÊNDICE 2
O Apêndice 2, contém os parâmetros obtidos pela computação dos dados experimentais do n?de H/mol de cada proteína.
Os ajustes foram alcançados num Computador IBM/370, modelo 155 do Centro deProcessamento de Dados do IEA, atrave's um programa denominado SAAM (Simulation, Analysis andModeling) na versão n? 25, desenvolvido pelo National Institutes of Health no Laboratory of TheoreticalBiology por Mones Berman e Marjory F. Weiss.

72
ftwF M o v * l Ht S
• JI:MF DF ^ f i
M * • t' » t " • » '
*»l •-. •11 " . 1t 1 ~ . .1 1 > . '
. 1r,. |
' " . 1
ft 1 -i <.<»«.', ' * ' . T *•.!
,* *. ,* ) *. * '
r ruin
• ' í•r I-r \-" 1
0 fI 1y *i «
*> *
* '
11;
i *\%i*
ni *rny?' i
; f
( r
(1
- ( i f
«
T< • » . ! • > * ' • » | » " | •> t
• * . . " M * ^ • « • - • ! * * ,
"" ' ^ 1 ^ - ' *"' P*r # i r» | »n/ fi .n . (i* " t • f ? * .r> . %<./"* f tnf •*.
'!v^-Í!o7 *,*
* " • 9 " - " ( • ' / ' ••» r< t\t\rt f # ' t - #
* ^ . I?*1* • »r % ' .'-.li"-» * • ' • ^ .
^ ^ . ; « ' " ^ T i r.
r. »l'l»»f.-i ,O4 r
tut Ik h KFF . I gr 1». I " " f
- . ir»' -f •f . \ r " f •
' . iff,
^,|r"r-F tft, |rr ,• I o
" . 117"«F»r «
ir *»«»f I
".mii*f«'7r.M777F»f7
f)!-t>.t»M 0.
,!M11H0l
uc/ooI.K440.9»*O.«*2! . « •l.inoB. Ml1.11* n,r«iit<(>0
' s o0.7IJ?f-010.64 «If-07
" . H6IÍ-0I0. 7?)lf-010.II47E-OIO.I
1.00»1.01)0.9*2
(I.TT70f»00
1.007
1.0*0
O.IO. IM71-0I0. l4Mf-<ll«•I11»!-01O.IHII -01l>. I4ME-01(l.t?07t-OI
?. 11771 «01CII2TF>n|(l.U77F»01
MIHU'I
0. W06Í-OI
O.l»nif-O»P.I4IH-010. I»»M-OTo.ii:7f-ri
o.wniioi
1.0*2
O.I7«4|-07O.l»n»f-^l0. H7lf-«1e.ii«ri-«in. 20IH-OI
«Ml» 4 ITO4IIIINS IS " . M M t| * • ! . >^• ' ,4 »/'if i r» .*•»4 l'"«âl|"s€, n - .? l» ;m«t» r 7 •'«Í--
HO»1** INtTIM
f «dl
f O»' « I 4 I K N •«?•!<f ' l ' j » . I 7 I
I i . r - r . * 4 n.74
« ' . 7 » I . » ' l.*<- n . l l ' . M - ' . 1 44 ' . n ' . " í , • i i . " • . K - r . >4
o - - . / « - r . < "

ç ç I- c c < f > c < t c <. c c c t f • c ç t < f t c c c • eU' U U' U ' U Uh U' U U U' Ü ' O U UJ U U 111 U * U U U U U
O ( C C ( i & C (Ui UI Ui Ui Ui UJ Ui Ui i< Ui Ui U' U> Ul Ui UJ
r c eoo cooC C COC C CC/
73
*-*^*OO r • - — - -
2r v*BO
e e" c> c c* c! c c c c o c c c o e c c © c* o c- c c c* c o
c c c o o o c o r - o o e & o o o o c C C O C J O C O O OC CC>CCC C C C v C C C'C C C O C C O C ' C C C C O O
j « 0 *t if *9 C O & O1 C ^ O C C' Ç C' & C Ç O C' Q C Cv OIIP N N ^ C CJ C' * ^ • ^ ' ^ f r o ^ * ' — Ç C v1 f V V « N B ^
c c < c c c r c r c c c c c c c c t c e c c c r c o c
1""">'"*'?nvu^'v?vvH??vi?v*'?iiUJ Ui Ui I
ui*> # •-*p « ci *-e
r c c c r t
• • C C I I C V < C t C < C < *"» « C t C ' < C « ( . C O C C Ç C *• « * # * * » • * • • • • • • • • • • • • • • • • • * • uy
u j i c. * c c c r . C ' o c c c i i c cc c c c c c ' c c c r*_ c t c c i * c t c
e coc• • • •o oe o
rs S|ei— — -CCI
o oooCC >"•<c c> c o• •II
I ' C C C ' l K O C t C C 6 <
'oecoc'i'oerci Coc r c c c c t c / c r c c c
c e c c c c e c c c e c t ' C c c e c ' C ' O b o c C ' C ' e c " < g .• « • • • » « • • « « • « * * • * * * « * * * • • * • »>r»i4
« í c c rc e c Ç L Cc C C r c c
;^2I a •' * C
•» ** U» J J
o r c c c c c r c c o c m c c c ^ ' C C c - c ^ r c c L C C
i** r c c <• c f c c c c b C b o c c r e o e e o o o c 1
3 2
n c oc« I I I
SKÍ;-Í58S
i • • •
m ™ t • . . i
s . . . . s —• ,°i^ • » • • ** • • • »
S:::". lissss

C0«9C0CCfO
1C">30CCC1'0-
tO»3CPCCl*O
o*eo*c
tc»?coctfc-tt»3ooeoi*o-
00*1 10*0- li'O10*0- 00M ?Z'OZl'O tl'O 00*110*0- «»*0 M*0» t i
X1C1VN
10*6- » "0»
^c•^c^rcc^^•p ic-3f>rcolC-3it«lOCf *0»0-3tilft9»*0
• 1001MO
t ?11f yI t
• 0• 1
*1 1 *
•s• <• 0•0
^ «Oil1 •Da
13W-O3
1 S
k l h C5
^o•^^•>*•let^•o
O S I
?c -hl-ft
K--Ç" n -c
T * 0l * C
K--C1---C
t t i t *c
' f c ? * c
* T 1 " I*C1 *1
f (f *C
»?f -C
e-ic «STfc-c
c to»3rcocí*c3*citfC ?c»ifrctfr361/t I'C-?C»ircC-«fC
c-?r«3rccsT*r
re»cc»
'C•r
t c » 3 ? c z t f t ^c•3^ccn•c ?o»3ctt i i *o•o-?c» :
e f 1
•c•c
•c ?o»3cccr»*c•c 2c«?rrc??"0•C 7C»3rfC»?T•0 ?C»'tCCi?'t•c-^c•3^ctct•c•C ?C»3ffC*CT•c ^c«3ccct^•c't-ec»aCCC4»*C
or
• 3?t"»C?'O• aricifo
•c
0 10»3CC001'C tC'JCOOCI'
3C00C10 10»300001n to»3rcoot'
t 0» =T00 t'1P»3CC001ie»3rcccftO»5CCOCttc«="crcfu -ncccrtc«i-coctp«-rrrNcite»?ccrrt
•o io»;rceot•c te»3r•c to»?(
10<?CCOCt'Cd93l«
COo-ec-eo*e0"Co-c0"CCTp'Co-c(.•tC'C0*0
c*c
c-c
»c•=^c^n•c
fO»3CCt<?"CrO»5CCC*-?*C
er«?cc:c'*c
tr• • H O C : ? ' !; - : : r - - c
S f t«. I t<. «1
( 1
11C!
CC»3-.tt.1t??"C
Ci»U

75
t i i i t i i t r i iC C t C C cI I t I I I
C C C O C O C C t
*€ C O O O« C C O O* C C O O
oo r — c c" c c c c -• — — -*o<-—o
ClCOC
1 f h N C r<
o c c c c oce cec cr. coce o
— C », ' C C C C f l C C t - C C C t C *• * • • • • • • • • • • • • • • • • I V
d if tu u> te ui li1 b UIU^IIIICUIWUJUIUI 9
• • — I* <
u. * * •
- . c tI I •
Ui Ul Ut
v* • W —
V V
• • 'l »
c r e r r e o t e í . c c t c c f
[ f ' 9 4 c t- C
r r. r i c r <-•<•. r t c c c r r c i
W tV M' M' U> U tb l$f U' U lir W tl' lU Uf HI Ui
c r t c r c c c c c c c c c r c c• - f « r r f r < - f . C C t * » < C — V C
C
<_ t c r t u < c c c e c• C Ml
I C C C C>
• • • *
* ! I/» v w w • t - » « * ^ l * r 1 * ^ • iff
tc c * cI I I t
• «fl c c <"- cL. • • I • * *
c ^ c u• • • •& C O * *| t
• t * • c c —<
^ • • • •« O O O
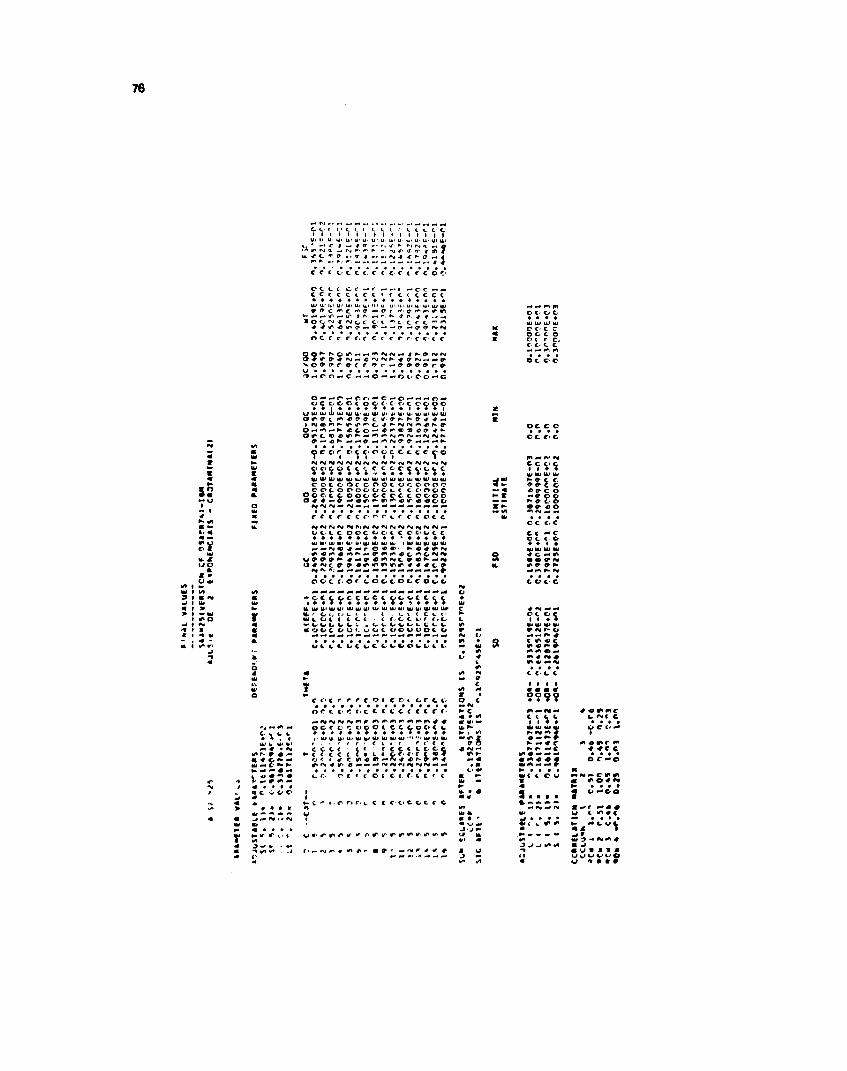
76
C C• r c» t i. i t. t t *. t c C CI I I I I I I I I I I I t I | t
c c> c e c c t c e o t-c c i
Ci ^ IT M IT I Al H '
* (T - . <T> fy f \ i #M
8?:> -• c **-* o-^-*
o c o c• • • •UUIUJIUe c> c co c c oO t r e .c o c c
o c c- c c c o c <• c c« c o o o o• • I * * * * * * * * I • * * I
UlfUJIUlulUUitUU U' >il Ui UI UJ UI U Üatra ( («•off-tfcw-^r-^-»-***-
? c o r C ' C c - o o c c c c c c - oi i i i i i
oc c o• • • •o c c c
C O C C O f C O O C O C C O O CCOCOOOOOCl-C C O O O C
o c c c c o o o & c c c c o & o c
UI UI Ui 111K» C C» *• c o< » c o
V4
C C C f O '
^ C C C C C C C C C C s C d C C Ç L *• • • • • • • • • • • • • • • • • • UI
U.UIU<UlU'UIUIUfUlU<UJUIUlU'UlUIU> Cu, r c-1. o t • i t '. i. i < c «. «'• ^>
c c — cÇ C ( O
• I •
m cr tr f• • • •
C O C' C
C C b CI » • •
• r» •>m « *«
c c c c t r c -
• • • f 4' UI U1 W U' Ul Ul W U* tát tM " ' '•*' UU Wurfawv f , i ' Í r ' C i i < r r £
, ? J i 2 t - ' i f t r i l i i f , ' ! - - - '
t - r . r < f . c c i c t c c t ' c
c c c cI l l I
C l C CI I * •
UI Ut UI UI
2S?
t; C c -
u c

77
\ •
o.irrrrrrnr) niC.-mcmccn C2o . e n r r r r r n 02O. 121 r n r r c r ) C30. L C K c c c c n C3n . 2 C i r r n r o n ciO.tMrrrrn CJ0.2 u r r c c m C3Ü. i ? K ( r r n i i C3fl.)',ifcrn rjO . ^ C l f : c r c n C30. « A i r o r r r o C3o . 4 ° i r r r r c n C3U.'J; if crren C3o.v-.irrrrrn nO.üT i r r r rc ' ) T3O.üMf.XCCn C3O . t P i r n r . f O l P3n . 7 2 i r r o r c i ) C3c. 7'>iccccon c iü .bCi rc r rnn C3o . á ^ i r r n r c n CJ0. j R i c c c c c n ci
C3C3
0 . ICC1CCC 3 0 C^o. i f * i r rcrn o««O.IOUKCCCO c<»o. i i í inoccn C4o.iuirrron 04
0.i?flif crn c
o.UÉiccccn C40. Hn i r c f Of) 0'*
O.l'jfllCCCCO C«
0.1561CCC0O C40.l6ClC0fCn C4o.i6«irnccn c
0 . 1 7 / 1 0 C C C ) C<»
0 .
O.21H41717H 0 )c.9fMoflsr>'?n 02o.s^n^nn^n 02
02020202020?
n . t i ' ^ S i q in 02"C . n n i ' H V ' n 02p.»'? »i.'<ivin 02O.C13JA7-'» »n 020. dn/.r ' i" n n 02n. 7r*'.7??'• n o?^ . 7nHr,3b-jsn 02C . 7 « l l y 5 ^ O 020 , 7 7'<qr? MO 0?0. jtoo^T'n 02
0202
0.7^647'??^n 02C.740l6?q5O 020.n«o"<«7io 02C. 7JflPü5lr>D 02C.7?23C7'56O 02owifioçsnio 0?0.711,239190 02C.70b93?690 0?0.70077t99n 020.695/^78«?O 02C.69C655410 02
0202
C.676912^70 020.672^92«>OD 020.669176420 02C.663965550 020.t»9949700 020.655326790 02C.651892910 0?0.64H044130 0?0.644277510 020.64Oi»8*C3O 0?0.63697:-. ?!? 0?0.633434320 02
0?

78
ABSTRACT
Phosphohpase A, Crotapotin and Crotamine mere purified from the venom uf Crotalus duritsut terrificui Tfw
aminoacid ramposttton» oi (Hottms shovwd rrtolecular weigths. 13400 ft» Phfupliiilip/w A, 8300 fer GreMpotin andAnnr>it%K rtntnniinr I M W ) Seph.idex gel filtration (G 1ft tine) Experiment», stable complex is formed by interaction
_ o - . l t - A - •.-.-. *f - • *v
(1:1 molar i.niul of Phospholip.i» A ILDj-f, 0,5b mg/Kg), in mice) and Crotapotin, Heoomoiimlion of Crotapotin
with Phospholip.K» A in 1:1 molar ratio, increase»», ttie toxicity of Phospholipase A appuMtimatety 10 fold to
LDe- - 0 , 0 5 mg/Kg. These facts indicate Crotoxin as a product of 1:1 molar ratio interaction of Crotapotin and
Phospholip.ise A, Ffemf Sephadex gel filtration (fr?6 f?rre) •xpemnems, unstable complex is formed by interaction of
Crotapotin and Crotamine ILD^-j ~0 ,8 mg/Kg, in mice), The-"m vitro" combination of Crotapotln and Crotamine tiTim " ' a r rf l l '"4 inueasi«^the toxiritv of Crotamine approximately 4 fold to L D c r i
= 0,2 mg/Kg. In pro f in
tritium-hydrogen exchange experiments, the back exchange kinetic of^tritium labelled o^"Mphff"p^y *w rririapntitr.
Cretamirra and C»oto»tn were measured in gel filtration columns of Sephadex G 25 "coarse'V Thy- best fit o f
experimental data of H/T exchange of Phospholipase A tbet>» two clearly distinguishable kinetic classes of
exchangeable hydrogens. From the exchangeable hydrogens only 68% were rapidly exchanged and the occurrence of
hydrogens envolver! in alpha-helix was practically absent. The Crotapotin has no hydrogens of alpna-helix and 83% of
exchangeable hydrogens were rapidly exchanged with solvitnt. The H/T exchange ki íetics of Crotanine^after a initial
heating in trmau'd water at 70.C for 1 imun.showedjthat 31% qf exchangeable hydrogens were slowly exchanged with
mlvent, \neteaimg trte intitai hratinij-W^ 18 hours, ma rawwbet of iloi»ly ewehangina pcotont diminished for 11%.
Fitting- corves of mperimentsi dattt o4 H/T exerianrjo o) Crotoxin showed three exponential classes of exchangeable
hydrogens and about 26 protons have alpha-helix characteristic exchange rate. These tactl suggest £ possibl*
conformational change after Phospholipas* A-Crotapotin interaction^-1 . ' . v / < ; <J.*_ : •( .
REFERÊNCIAS BIBLIOGRÁFICAS'
1. ACKLRS, G. K. & THOMPSON, T. E. Determination of stoichiomctry and equilibrium constantsfor revcrsibly association systems by molecular sieve chromatography. Proc. natn. Acad. Sci.USA, 53J42, 1965.
2. BRAY, G. A. A simple efficient liquid scintillation method for counting aqueous solutions in aliquid scintillation counter. Analyt. Biochem., 1279-85, 1960.
3. BREITHAUPT, H. Enzymatic characteristics of ootalus phospholipase A} and the crotoxincomplex. Toxicon, ]4:221, 1976.
4. BREITHAUPT, H.; 0M0RI-SAT0H, T.; LANG, J. Isolation and characterization of threephospholipases A from the crotoxin complex. Biochim. biophys. Acta, 403355-69, 1974.
5. BREITHAUPT, H.; RUBSAMEN, K.; HABERMANN, E. Biochemistry and pharmacology ofCTOtoxin complex. Bur. J. Biochem., 49333-45, 1974.
6. BROWNE, C. A. & WALEY, S. G. Studie* of those phosphate isomerase by hydrogen exchange.Biochem. J., 14J:753-60, 1974.
7. BUDZYNSKI, A. Z. Difference in conformation of fribrinogen degradation products as revealed byhydrogen exchange and spcctropolarimetry. Biochim. biophys. Acta, 229:663-71, 1971.
«. BUDZINSKI, A. Z. & FRAINKIL C0NRA1, H. Hydrogen exchange as a probe ofconformation to t obacco mosaic virus and its coat protein. Biochemistry (New York),9(J 7)3301-9. Aug. 1970.
(') Ai ref«r#nciM bibli^rific* relatival a df)cumento» Iwili/ado» pelo IEA foram revista» a enquadrada* n* NB-66d* ABNT.

79
9. CAMBIASO, C. L ; RETEGUI, L. A.; DELLACHA, J. M.; SANTOMÊ, J. A.; PAJLADINI, A. C.H-excliange behaviour and extent of reversible conformation charges in human, bovine, ovine,porcine and equine growth hormones. Biochim. biophys. Acta, 221:290-6, 1970.
10. CHEYMOL, J.; GONÇALVES, J. M.; BOURILLET, F.; ROUCH - ARVEILLER, M. Actionneuromusculaire comparée de la crotamine et du venin de "Crotalus durissus terrificus" var.crotaniinicus. Toxicon, 9279, 1971.
11. CUATREÍ ASAS, P. & HOLLKMBERG, M. D. Membrane receptors and hormone action. Adv.Protein Chem., 3J:251, 1976.
12. DAVIS, B. J. Disc eletrophoresis. II. Method and application to human serum proteins. Ann. N. Y.Acad. Set., m:404, 1964.
13. DAWSON, R. M. C. On the mechanism of action of phospholipase A. Biochem. J., 8 8 4 1 4 , 1963.
14. DOWNIiR, N. W. & FiNGLANDER, S. W. Hydrogen exchange study of membra, c-bound rhodopsin.I. Protein structure. J. biol. Chem., 252(22):8O92-1OO, Nov. 1977.
15. DOWNLR, N. W. & ENGLANDER, S. W. Hydrogen exchange study of membrane-bound rhodopsin.II. Light-induced protein strucutre change. J. biol. Chem., 252(22)«1014, Nov. 1977.
16. EIGEN, M. Proton transfer, acid-basic catalysis, and enzymatic hydrolysis. Angew. Chem., 3:1,1964.
17. EIGEN, M.; KUSE, W.; MAASS, G.; DEMAEYER, L. Rate constants of protolytic reactions inaqueous solution. Prog. React. Kinet.,2-2%5, 1963.
I?. ELLIS, L. M.; BLOCMFIELD, V. A.; WOODWARD, C. K. Hydrogen - tritium exchange kinetics ofsoybean try pun inhibitor (Kunitz). Solvent accessibility in the folded conformation.Biochemistry (New York), J4(15)J413-9, Jul. 1975.
19. ELLMAN, G. L. Tissue sulphydryl groups. Archs Biochem. Biophys., 82:70-7, 1959.
20. ENGLANDER, S. W. A hydrogen exchange method using tritium and sephadex: its application toribonuclease. Biochemistry (New York), 2:798-803, 1963.
21. ENGLANDER, S. W. & CROWE, D. Rapid microdialysis and Lydrogen exchange. Analyt. Biochem.,|2:579-84, 1965.
22. ENGLANDER, S. W. & ENGLANDER, J. J. Hydrogen exchange studies of sRNA. Proc. ntn Asad.Sei. USA, 5JJ70, 1975.
23. ENGLANDER, S. W. & MAUEL, C. Hydrogen exchange studies of respiratory proteins. II.Detection of discrete, ligand - induced changes in hemoglobin./ biol. Chem., 247(8):2387-94,Apr. 1972.
24. ENGLANDER, S. W. & POULSEM, A. Hydrogen-tritium exchange of the random chainpolypcplidc. Biopolyrmrs, 1319, 1969.
25. ENGLANDER, S. W. & ROLI E, A. Hydrogen exchange studies of respiratory proteins. III.Stiuctural and free cncifiy changes in hemoglobin by use of a difference. J. biol. Chem.,248(I3):4852-6I, Jul. 1973.
26. I-NGLANDER, S. W. A SIAII.Y, R. Measurement of ihc free and the ll-bondcd amides ofin. J. rrwlec. Biol.. 45:277 (>5, I<><V>.

80
27. ENGLANDER, S. W ; DOWNER, N. W.; TFITELBAUM, H. Hydrogen exchange. A. Rev. Biochem.,41 <>03. 1972.
28. FAIRCLOUGH JR., G. F. & FRUTON, J. S. Peptide protein interaction as studied by gel filtration.Biochemistry (New York), 5(2):673-83, Feb. 1966.
29. FRAENKEL-CONRAT, H. & SINGER, B. Fractionation and composition of crotoxin. ArchsBiochem. Biophys., 6J:64-73, 1956.
30. GALARDY, R. E.; CRAIG L. C; PRINTZ, M. P. Tritium - hydrogen exchange of the cyclic
peptide polymyxin B,. Biochemistry (New York), B(8): 1674-7, Apr. 1974.
31. GONÇALVES, J. M. Estudos sobre crotamina. RibeirJo Preto, 1961. (Tese de doutoramento).
32. CONÇALVES, J. M. Purification and properties of crotaminc. In: BUCKLEY, E.E. & PORGES, N.,eds. Venoms. Washington, D. C , A. A. A. S., 1956. p.261.
33. GONÇALVES J. M. & DEUTSCH, H. F. Ultracentrifugal and zone eletrophoresis studies of someCi.'iaiidac venoms. Archs Biochem. Biopphys., 60:402-11, 1956.
34. GONÇALVES, J. M. & GIGLIO, J. R. Amino acid composition and terminal group analysis ofcrotamine. In: PROCEEDINGS 6th int. congress Biochemistry, New York, 1964, v.2. I s.n.t. I.p. 170.
35. GONÇALVES, J. M. & POLSON, A. The electrophoretic analysis of snake venoms. Archs Biochem,43:253-9, 1947.
36. GONÇALVES, J. M. & VIEIRA, L. G. Estudos sobre veneno de serpentes brasileiras. I. Análise
clctroforética. Anais Acad. bras. Gene, 22(1 ):141-9, mar. 1950.
37. GRALÊN, N. & SVEDEBERG, T. The molecular weight crotoxin. Biochem. J., |2:1375, 1938.
38. GREENWOOD, F. C ; HUNTER, W. H.; GLOVER, J. S. The preparation of 1 3 I I labelled humangrowth hormone of high specific radioactivity. Biochem. J., 89:114-23, 1963.
39. HABERMANN, E. Gewinnung und Eingenshaften von Crotactin, Phospholipase A, Crotamin undToxin III aus dem Gift der Brasilianhischen Klapperschlange. Biochem. Z, 329:405, 1957.
40. HABERMANN, E. & NEUMANN, W. P. Crotactin, ein neues Pharmakologishes Wirkp.inzip aus demGift von "Crotalus terrificus". Naunyn-Schmiedebergs Arch. exp. Path. Pharmak., 228:217,1956.
41. HABERMANN, E. & NEUMANN. W. P. Die Hemmung der Hit/ekoagulation von Eigelb durchBicnengift - ein Phospholipasc - Effect. Hoppe - Seyler's. Z. physiol. Chem., 2%:I79, 1954.
42. HABERMANN, E.; WALSCH, P.; BREITHAUPT, H. Biochemistry and pharmacology of thecrotoxin complex. Naunyn-Schmiedebergs. Naunyn-Schmiedebergs Arch. exp. Path. Pharmak.,273:313, 1972.
43. HADLKR. W. A. At VITAL BRAZIL, O. Pharmacology of crystalline crotoxin IV. Nephrotoxicity.Mems Inst. Butantan, 3^3:1001, 1966.
44 MAMPE, O. (',. & v.ONÇAI VIS, J. M. Optical rotatory dispersion of crotamine: effect ofdenaturants. Polynwr, 17:638, 1976.

81
45. HAMPE, O. G. & GONÇALVES, J. M. Optical rotatory dispersion of crotamine: effect ofdenaturants. Toxicon, 1978.
46. HARTLEY, R W. & SMHATON, J. R. On the reaction between the extracellular ribonuciease ofBacillus amyloliquefaciens (Barnase) and its intracellular inhibitor (Barstar). J. biol. Chem.,248(16):5624-6, Aug. 1973.
47. HENIX)N, R. A. & I RAINKIL-CONRAT, H. Biological roles of the two components of crotoxin.Proc. natn. Acad. Sci. U.S.A., 68:1560, 1971.
48. HENDON, R. A. & FRAENKEL-CONRAT, H. The roles of complex formation in the neurotoxicityof crotoxin components A and B. Toxicon, |4:283, 1976.
49. HORST, J.; HENDON, R. A.; FRAENKEL-CONRAT, H. The active components of crotoxin.Biochem. biophys. Res. Commun., 46(3):1042-7, 1972.
•1. HUMMKL, J. P. & DREYER, W. J. Measurement of protein - binding phenomena by gel nitration.Biochim. biophys. Ada, 63:530, 1962.
51. HVIDT, A. Deuterium exchange between ribonuciease and water. Biochim. biophys. Acta, 18:307,1955. ~
52. HVIDT, A. The deuterium exchange of lysozyme as followed by an infrared spectrophotometricmethod. C. r. Trav. Carlsberg, 33:475, 1963.
53. HVIDT, A. 6L KANAREK, L. The deuterium exchange of lysozyme as followed by the gradient
method. C. r. Trav. Carlsberg, 33:463, 1963.
54. HVIDT, A. Ac NIELSEN, S. Hydrogen ex ;hange in proteins. Adv. Protein Chem., 2J:2£7, 1966.
55. JENG, T. W. &. FRAENKEL-CONRAT, H. Activation of crotoxin B by volvation A, . Biochem.biophys. Res. Commun., 70(4): 132s-9, Jun. 1976.
56. KELLET, G. L. The study of interacting protein systems by molecular sieve chromatogiaphy. Lab.Pract, 16(7):857-62, 1967.
57. LA1KEN, S. L. & PRINTZ, M. P. Kinetic class analysis of hydrogen - exchange data. Biochemistry(Nr« York), 9(7): 1547-53, Mar. 1970.
58 LAURE, C J. Di? Pi;>r.orstruktur des Crotamins. Hoppe - Seyler's Z. physiol. Chem., 356313.1975.
59. LAURE, C. J.; COUTINHO NETO, J.; GONÇALVES, J. M. ObtençSo de crotaminas e suaquantificação. Cienc. Cult. fS. Paulo), 23(Supl.):3l3-4, jun. 1971.
60. U , C. H. it FRAENKEL-CONRAT, H. Electrophoresis of crotoxin. J. Am. chem. Soe.,64(7): 1586-8, Jul. 1942.
61. LINDERSTROM-LANG, K. The ph dependence of deuterium exchange of insulin. Biochim.biophys. Actã, j|:3O8, 1955
62. LOWRY, 0 . H.; ROSEBOOGH. N. J.; FARR, L; RANDALL, R. J. Protein measurement with thefolin phenol reagent. J. biol. Chem., 193:265, 1951.
63. MARCIANI, D. J. & TOLBI.RT, B. M. Structural damage in r'fradiated lysozyme. Biochim.biophys. Acta, 351:387-05, 1974.

82
64. NAKANISHI, M.; TSUBOI, M.; IKEGAM, A. Fluctuation of the lysozyme stiuctuie. J. molec. Biol.,
65. NAKA/ONE, A. K. Aspectos imunoquímicos da crotoxina e suas subunidades. Sa"o Paulo, 1978.(Tese de doutoramento).
66. NAKAZONE, A. K.; ROWRO, J. R.; GONÇALVES, J. M. Caráter imunológico e potenciador dacrotapoim. In: II? REUNION regional de Ia Associación de Sociedades Americanas deBioquímica (PAAKS), Córdoba, Argentina, 2 a 4 de novembro de 1977. (Res., n? 179).
67. NAKAZONE, A. K.; ROGERO, J. R.; GONÇALVES, J. M. Potenciação da crotamina pela interaçãocom crotapotin. In: 6? ENCONTRO anual da Sociedade Brasileira de Bioquímica, Caxambú, 20a 23 abril, 1977. (Res., n? 79).
68. NEUMANN, W. P. & IIABERMANN, E. Uber Crotactin, das Haupttoxin des Giftes derBrasilianishen Klapperschlangcr (crolalus duriss"s terrificus). Biochim. Z., 327:170. 1955.
69. PAIVA, T. B.; PAIVA, A. C. M.; SCHERAGA, A. H. The conformation of angiotensin II jn aqueoussolution. Biochemistry (New York), 2(6):!327-34, Nov. 1963.
70. PARADIES, H. H. & BREITHAUPT, H. On the subunit structure of crotoxin: hydrodynamic andshape properties of crotoxin, phospholipase A and crotapotin. Biochim. biophys. Res.Commun., 66.496, 1975.
71. PIETERSON, W. A.; VIDAL, J. C ; VOLWERK, J. J.; HAAS, G. H. de Zymogen-catalyzedhydrolysis of monomeric substrates and the presence of a recognition site for |ipid-waterinterfaces in phospholipase A2- Biochemistry (New York). 13(7):1455-60, Mar. 1974.
72. PIETERSON, W. A.; VOLWERK, J. J.; HAAS, G. H. de Interaction of phospholipase A2 and itszymogen with divalent metal ions. Biochemistry (New York), ]3(7):143945, Mar. 1974.
73. PRINTZ, M. P. & HIPPEL, P. H. von Hydrogen exchange studies of DNA structure. Proc natn.Acad. Sei. U.S.A., 5J363, 1965.
74. REED, L. J. & MUENCH, H. A simple method of stimating fifty per cent endpoints. Am. J. Hyg.,27:493, 1937.
75. ROGERO, J. R.; NAKAZONE, A. K.; GONÇALVES, J. M. Interação de crotapotin com fosfolipaseA e crotamina. In: II* REUNION regional de la Associación de Sociedades Americanas deBioquímica (PAABS), Córdoba, Argentina, 2 a 4 de novembro de 1977. (Res., n? 178).
76. ROIIOLT, O. A. & SCHLAMOWITZ, M. Studies of the use of dihexanoyllecithins as substrates forphospholipasc A. with addendum on aspects of micelle properties of dihexanoy(lecithin. ArchtBiochem. Biophys., 94:364 79, 1961.
77. ROSENBERG, A. & CHAKRAVARTI, K. Studies of hydrogen exchange in proteins. I. Theexchange kinetics of bovine carbonic anhydrasc. J. biol. Chem., 24J(19):5193-201, Oct. 1968.
78. RUBSAMEN, K.; BREITHAUPT, H.; IIABERMANN, E. Biochemistry and pharmacology of t! .crotoxin complex. NaunynSchmiedebergs Arch. txp. Path. Pharmak., 270274, 1971.
79. SIMIZU, K. Ação do ácido frbromopirúvico sobr» aldolase da frutose 1,6 difotfato da múiculoesfriado da coelho. Suo Paulo, 1972. (Tese de doutoramento).
80. SLOTTA. C. H. & TRENKEL-CONRAT. If. Purificação e cristalízaçJo do veneno de cobra cascavel.Mems Inst. Butantan, 12:505, 19381939.

83
81. VITAL BRAZIL, U. Pharmacology of crystalline crotoxin. II. Neuromuscular blocking action. MemsInst. Butantan, 33:981, 1966.
82. VITAL BRAZIL, 0 . ; FARINA, R.; YOSHIDA, L.; OLIVEIRA, V. A. Pharmacology of crystallinecrotoxin. III. Cardiovascular and respiratory effects of crotoxin and "Crotalus durissusterrificus" venom. Mems Inst. Butantan, 33993 , 1966.
83. VITAL BRAZIL, 0. ; FRANCESCHI, J. P ; WAISBICH, E. Pharmacology of crystalline crotoxin. I.Toxicity. Mems Inst. Butantan, 33973, 1966.
84. WELCH JR., W. H. & FASMAN, C. D. Hydrogen-tritium exchange in polypeptides. Models ofa-helical and conformations. Biochemistry (New York), 13(12):2455-66, Jun. 1974.
85. WELLS, M. A. Effect of pH on the kinetic and spectral properties of "Crotalus adamantus"phospholipase A2 in H 2 0 and D 2 0 . Biochemistry (New York), 13(ll):2265-8, May 1974.
86. WELLS, M. A. A kinetic study of the phospholipase A] (Crotalus adamanteus) catalyzed hydrolysisof l,2-Dibutyryl-sn-glycero-3-phosphorylcholine. Biochemistry (New York), ]2(6):1030-41>
Mar. 1972. ~
87. WICKETT, R. R.; IDE, G. J.; ROSENBERG, A. A hydrogen-exchange study of lysozymeconformation changes induced by inhibitor binding. Biochemistry (New York), 13(16):3273-7,Jul. 1974. ~
8 8 . WILLUMSEN, L. Hydrogen-deuterium exchange studies on chymotrypsinogen A, alpha -chymotrypsin, and diisopropylphosphoryl-alpha-chymotrypsin. C. r. Trav. Lab. Carlsberg,36227, 1968.
89. WOLWERK, J. J.; PIETERSON, W. A.; HAAS, G. H. de Histidine at the active site of phospholipaieA2 . Biochemistry (New York), 13(7):1446-54, Mar. 1974.
90. WOODWARD, C. K. & ELLIS, L. M Hydrogen exchange kinetics changes upon formation of thesoybean trypsin inhibitor - trypsin complex. Biochemistry (New York), 14(15)3419-23, Jul.1975.
91. WOODWARD, C. K. ELLIS, L. M.; ROSENBERG, A. Solvent accessibility in folded protein*:studies of hydrogen exchange in trypsin. j , bio/. Chem., 250(2):432-9, Jan. 1975.

INSTITUTO DE ENERGIA ATÔMICACaix? Postal, 11049 - PinheirosCEP 0550801000 - São Pauto - S?
Telefone: 211-6011Endereço Telegráfico - IEATOMICATelex - 011-23592 IENA BR


















