
Universidade de São Paulo Faculdade de Saúde Pública ... fileobtidos de amostras de queijo Minas...
74
Universidade de São Paulo Faculdade de Saúde Pública Determinação de grupos filogenéticos e pesquisa de genes de virulência em isolados de Escherichia coli obtidos de amostras de queijo Minas Suzete Contrera de Moura Pedro Tese apresentada ao Programa de Pós-Graduação em Saúde Pública para obtenção do título de Doutor em Saúde Pública. Área de concentração: Serviços de Saúde Pública. Orientadora: Professora Associada Maria Helena Matté. São Paulo 2009
Transcript of Universidade de São Paulo Faculdade de Saúde Pública ... fileobtidos de amostras de queijo Minas...

Universidade de São Paulo Faculdade de Saúde Pública
Determinação de grupos filogenéticos e pesquisa de genes de virulência em isolados de Escherichia coli
obtidos de amostras de queijo Minas
Suzete Contrera de Moura Pedro
Tese apresentada ao Programa de Pós-Graduação em Saúde Pública para obtenção do título de Doutor em Saúde Pública.
Área de concentração: Serviços de Saúde Pública.
Orientadora: Professora Associada Maria Helena Matté.
São Paulo 2009

2
Determinação de grupos filogenéticos e pesquisa de genes de virulência em isolados de Escherichia coli
obtidos de amostras de queijo Minas
Suzete Contrera de Moura Pedro
Tese apresentada ao Programa de Pós-Graduação em Saúde Pública para obtenção do título de Doutor em Saúde Pública.
Área de concentração: Serviços de Saúde Pública.
Orientadora: Professora Associada Maria Helena Matté.
São Paulo 2009

3
É expressamente proibida a comercialização deste documento, tanto na sua forma impressa como eletrônica. Sua reprodução total ou parcial é permitida exclusivamente para fins acadêmicos e científicos, desde que na reprodução figure a identificação do autor, título, instituição e ano da tese.

4
Dedico este trabalho a minha amada família,
ternos companheiros de vida.

5
AGRADECIMENTOS
Cada etapa da vida vem repleta de sentimentos e emoções. Nos
decepcionamos, sentimos tristeza, há momentos de alegria e animação, as
descobertas nos surpreendem, as dúvidas causam angústias, fazemos inúmeros
sacrifícios e também obtemos prazer e satisfação. Viver intensamente cada momento,
extrair o melhor de cada aprendizado torna a vida muito especial. Agradeço a Deus a
oportunidade de mudar com cada experiência. Agradeço aos meus companheiros de
jornada que vibram comigo nos contentamentos e me amparam nas consternações.
Agradeço ao meu marido Cláudio, infatigável co-participante, meu maior
incentivador, aos nossos filhos, Felipe e Frederico, pelo constante apoio e carinhoso
estímulo.
Aos meus pais, Zinho e Flora, ao meu irmão Silvio, aos meus saudosos sogro
e sogra, cunhados e cunhadas, sobrinhos e sobrinhas, agradeço a torcida e entusiasmo
nas vitórias alcançadas.
Meus agradecimentos aos meus saudosos, vovó Maria e Angel, que mesmo
sem entenderem muito, vibravam ao ouvir as novidades.
A Professora Associada Maria Helena Matté meus sinceros agradecimentos
pela orientação prestada, pelo incentivo em transpor barreiras e obstáculos.
Ao Professor Titular Pedro Manuel Leal Germano agradeço imensamente a
confiança e a oportunidade ímpar de crescimento profissional.
Aos Professores Associados Maria Helena e Glavur Rogério Matté agradeço
o amistoso acolhimento no Laboratório de Saúde Pública possibilitando de todas as
formas minha atualização e aprimoramento.
A Professora Associada Maria Cecília e Américo Pelicioni eternamente
agradecida pela generosa amizade e confiança que foram fundamentais para meu
progresso.
A Professora Doutora Nicolina Silvana Romano Lieber agradeço o constante
incentivo, nossa afetuosa convivência e sempre simpática amizade.

6
A Miriam Lopes da Silva agradeço a terna amizade, o firme apoio, o reiterado
auxílio e a vibração em todas as conquistas.
A Milena Dropa jovial amizade, agradeço a torcida e a freqüente ajuda no
laboratório.
Agradeço a agradável convivência e a todos os colegas que direta ou
indiretamente auxiliaram na execução deste trabalho: Ana Maria, Andréa, Angélica,
Agnes, Cláudia, Gillian, Lívia, Lícia, Lúcia, Elaine, Fernanda, Elenice, Karla,
Magali, Marisa, Maria do Carmo, Maria Tereza, Martha, Mônica, Rita, Rosinha,
Ronalda, Silvia, Silvana, Solange, Suzi, Therezinha, Thiago, Vandréa, Vera Cecília e
Viraneide.
Agradeço aos funcionários da Biblioteca, do Departamento de Prática de
Saúde Pública e do Departamento de Pós Graduação da Faculdade de Saúde Pública
a importante colaboração e o simpático convívio.
Agradeço o carinhoso apoio dos queridos amigos do Laboratório de Saúde
Pública da Lapa e da Supervisão de Saúde Lapa-Pinheiros.
Agradeço a paciência, a torcida e o incentivo na correção final do trabalho
dos amigos da Vigilância Ambiental, em especial a Teresa, Giuliano, Stela, Edna,
Valquíria, Celinha, Regina, Léo, Dirceu e Angélica.
A cada membro da banca Professora Doutora Maria Inês Zanoli Sato,
Professor Doutor Waldir Pereira Elias Júnior, Professora Doutora Maria Tereza Pepe
Razzolini, Professora Doutora Lúcia Baldassi e a Professora Doutora Márcia Regina
Franzolin, agradeço a atenção e a consideração em dispor de seu valioso tempo para
prestar uma inestimável e fundamental colaboração na correção do trabalho.

7
“Quem mexeu no meu queijo?”
Spencer Johnson

8
RESUMO
Pedro SCM. Determinação de grupos filogenéticos e pesquisa de genes de
virulência em isolados de Escherichia coli obtidos de amostras de queijo Minas
[Tese Doutorado]. São Paulo. Faculdade de Saúde Pública da USP. 2009.
Introdução. A pesquisa de Escherichia coli em alimentos é relevante para a Saúde
Pública porque indica a contaminação fecal e a qualidade do produto oferecido ao
consumidor. A determinação do grupo filogenético e de fatores de virulência de E.
coli permite identificar a existência de cepas patogênicas que poderiam causar
doença. Objetivos: Determinar o grupo filogenético e fatores de virulência de
isolados de E. coli pertencentes aos patotipos ETEC, EIEC, EPEC, STEC e EAEC
usando métodos moleculares, obtidos de amostras de queijo Minas, discutir a
presença dos patotipos nas amostras e o significado para Saúde Pública. Métodos.
250 isolados de Escherichia coli provenientes de 10 amostras de queijo Minas foram
utilizados para a realização do estudo. O DNA genômico foi extraído e neste foi
realizado a reação de PCR multiplex. Resultados. Os resultados demonstraram que
dos 250 isolados, 93,2% foi classificado como grupo filogenético A, 3,2% como B1,
2,8% como B2 e 0,8% como D. Dos 250 isolados estudados, em 96,8% (242) foram
encontrados fatores de virulência, sendo 91,6% de marcadores para ETEC, 0,4% para
EPEC e 4,8% para EAEC. Conclusões. Houve predominância de fatores de
virulência do patotipo ETEC e do grupo filogenético A. A presença de fatores de
virulência indica que as amostras de queijo estavam contaminadas e poderiam causar
doença, evidenciando a necessidade de medidas de controle efetivas por parte das
autoridades sanitárias, bem como campanhas educativas.

9
Descritores: Saúde Pública, queijo Minas, Escherichia coli enteropatogênica, grupos
filogenéticos, fatores de virulência.

10
ABSTRACT
Pedro SCM. Determinação de grupos filogenéticos e pesquisa de genes de virulência
em isolados de Escherichia coli obtidos de amostras de queijo Minas. [Determination
of phylogenetic groups and search of virulence genes in isolated Escherichia coli
from Minas cheese samples [Tese de doutorado]. São Paulo. Faculdade de Saúde
Pública da USP. 2009.
Introduction. The search of Escherichia coli in foods is relevant for the Public
Health because it indicates the fecal contamination and the product quality offered to
the consumer. The determination of phylogenetic group and virulence factors of E.
coli, allows the identification of the existence of pathogenic strains that could cause
disease. Objectives: Determine the phylogenetic group and the pathogenic of
Escherichia coli belonging to the patotipos isolated occurrence ETEC, EIEC, EPEC,
STEC and EAEC using molecular methods obtained from Minas cheese samples, to
discuss the presence of the patotipos in the samples and the meaning for Public
Health. Methods. 250 isolates of Escherichia coli from 10 Minas cheese samples
was used for the accomplishment of the study. Genomic DNA was extracted and in
this the multiplex reaction PCR was accomplished. Results. The results
demonstrated that the isolates 250, 93.2% were classified as phylogenetic group A,
3.2% as B1, 2.8% as B2 and 0.8% as D. Of the isolates 250 studied, in 96.8% (242)
were found virulence factors, being 91.6% of markers for ETEC, 0.4% for EPEC and
4.8% for EAEC. Conclusions. There was predominance to virulence factors of the
patotipo ETEC and the phylogenetic group A. The presence of virulence factors
indicates that the cheese samples were contaminated and they could cause disease,

11
put in evidence the need of effective control measures on the part of the sanitary
authorities, as well as educational campaigns.
Descriptors: Public Health, Minas cheese, enteropathogenic Escherichia coli,
phylogenetic group, virulence factors.

12
ÍNDICE
1.INTRODUÇÃO 16
1.1. JUSTIFICATIVA 16
1.2. ANÁLISE DA LITERATURA 17
1.2.1. Legislação de alimentos no Brasil 17
1.2.2. Coliformes em alimentos- Escherichia coli 19
1.2.3. Escherichia coli enterotoxigênica (ETEC) 22
1.2.4. Escherichia coli enteropatogênica (EPEC) 24
1.2.5. Escherichia coli enteroinvasiva (EIEC) 26
1.2.6. Escherichia coli produtora de toxina Shiga (STEC) 27
1.2.7. Escherichia coli enteroagregativa (EAEC) 31
1.2.8. Grupos Filogenéticos 34
1.2.9. Reação de polimerização em cadeia (PCR) 37
2.OBJETIVOS 38
3.MATERIAL E MÉTODOS 39
3.1. ISOLADOS BACTERIANOS- Escherichia coli 39
3.2.REISOLAMENTO E CONFIRMAÇÃO DA VIABILIDADE E PUREZA DAS CEPAS
40
3.3.MÉTODOS MOLECULARES 40
3.3.1. Extração do material genético 40
3.3.2. Reação em cadeia pela polimerase (PCR) 41
3.3.3. Eletroforese em gel de agarose 44
4.RESULTADOS 45
4.1. GRUPOS FILOGENÉTICOS 45
4.2. DETECÇÃO DE MARCADORES DE VIRULÊNCIA 48

13
5. DISCUSSÃO 56
6. CONCLUSÕES 62
7. REFERÊNCIAS 63
Lista de Quadros
Quadro 1. Tipo de queijo Minas e local de aquisição das amostras
utilizadas para obtenção dos isolados de E. coli. São Paulo, 2003
(PEDRO, 2003).
39
Quadro 2. Seqüência dos iniciadores da PCR e tamanho esperado dos
produtos que foram utilizados na determinação filogenética e de fatores
de virulência dos isolados de E. coli.
44
Lista de Figuras
Figura 1. Diagrama de Venn ilustrando patotipos de E. coli, EHEC/
VTEC/ EPEC (baseado no estudo de NAYLOR e col., 2005). 30
Figura 2. Esquema representando os grupos filogenéticos utilizando
iniciadores para os genes chuA, yjaA e o fragmento de DNA TSPE4.C2
(CLERMONT e col., 2000).
35
Figura 3. Gel de agarose contendo os fragmentos obtidos para isolados
de Escherichia coli representativos dos perfis filogenéticos. M-
Marcador de peso molecular 100pb, 1- isolado 298 Grupo A, 2- isolado
299 Grupo A, 3 - isolado 300 Grupo B1, 4- isolado 302 Grupo D, 5 -
isolado 303 Grupo A, 6 - isolado 309 Grupo D. São Paulo, 2009.
46

14
Figura 4. Proporção de E. coli do total dos isolados estudados por
grupo filogenético nas 10 amostras de queijo Minas. São Paulo, 2009.
47
Figura 5. Proporção de E. coli do total dos isolados estudados por
fatores de virulência nas 10 amostras de queijo Minas. São Paulo, 2009.
48
Figura 6. Diferentes perfis obtidos de isolados de Escherichia coli
utilizadas como controle positivos. M- Marcador de Peso molecular, 1 –
E. coli Verotoxigenica, 2 – IAL339, 3 – IAL399, 4 – IAL 1893, 5 –
IAL1895, 6 – IAL1896, 7 – IAL1848, 8 - E. coli enteroinvasiva
(isolado clínico), 9 - E. coli enteroagregativa (isolado clínico).
50
Figura 7. Proporção de E. coli do total dos isolados estudados por
patotipos nas 10 amostras de queijo Minas. São Paulo, 2009.
51
Lista de Tabelas
Tabela 1. Número e proporção dos isolados de E. coli nas amostras de
queijo Minas. São Paulo, 2009. 45
Tabela 2. Número dos isolados de E. coli por grupos filogenéticos nas
10 amostras de queijo Minas. São Paulo. 2009. 47
Tabela 3. Número dos isolados de E. coli por fatores de virulência nas
10 amostras de queijo Minas. São Paulo, 2009. 49
Tabela 4. Número e proporção dos isolados de E. coli por patotipo nas
10 amostras de queijo Minas. São Paulo, 2009. 51
Tabela 5. Número de isolados de E. coli por patotipo e por filogrupo na
amostra 1 de queijo Minas. São Paulo, 2009. 52
Tabela 6. Número de isolados de E. coli por patotipo e por filogrupo na
amostra 2 de queijo Minas. São Paulo, 2009. 52
Tabela 7. Número de isolados de E. coli por patotipo e por filogrupo na
amostra 3 de queijo Minas. São Paulo, 2009. 53

15
Tabela 8. Número de isolados de E. coli por patotipo e por filogrupo na
amostra 4 de queijo Minas. São Paulo, 2009. 53
Tabela 9. Número de isolados de E. coli por patotipo e por filogrupo na
amostra 5 de queijo Minas. São Paulo, 2009. 53
Tabela 10. Número de isolados de E. coli por patotipo e por filogrupo
na amostra 6 de queijo Minas. São Paulo, 2009. 54
Tabela 11. Número de isolados de E. coli por patotipo e por filogrupo
na amostra 7 de queijo Minas. São Paulo, 2009. 54
Tabela 12. Número de isolados de E. coli por patotipo e por filogrupo
na amostra 8 de queijo Minas. São Paulo, 2009. 54
Tabela 13. Número de isolados de E. coli por patotipo e por filogrupo
na amostra 9 de queijo Minas. São Paulo, 2009. 55
Tabela 14. Número de isolados de E. coli por patotipo e por filogrupo
na amostra 10 de queijo Minas. São Paulo, 2009. 55

16
1. INTRODUÇÃO
1.1. JUSTIFICATIVA
O queijo Minas é um produto de paladar suave que agrada desde crianças até
idosos, e é muito utilizado como componente de dietas, de preço acessível, alimento
facilmente encontrado em diversos pontos de venda, desde o comércio informal até
em grandes redes de hipermercados.
É um produto de alto valor nutritivo e que possui condições de pH e umidade
que favorecem o crescimento microbiano. Esses fatores quando associados às
deficientes condições da produção artesanal, de transporte, que na maioria das vezes
não é refrigerado, ao armazenamento em locais inapropriados e também à
comercialização do produto fracionado, podem representar um risco ainda maior de
contaminação microbiana.
Na dissertação de Mestrado (PEDRO, 2003) os resultados indicaram um
número expressivo de amostras de queijo Minas contaminadas por coliformes fecais
acima do permitido pela legislação em vigor, e desses foram isolados Escherichia
coli.
A presença de Escherichia coli em um alimento é um indicador da falta de
qualidade higiênico sanitária, pesquisando a literatura observa-se que há poucos
trabalhos publicados sobre E. coli enteropatogênica em alimentos.
A E. coli é habitante natural da microbiota humana e de animais
homeotérmicos, e quando presente no alimento sugere que outros patógenos

17
intestinais podem estar presentes e que o mesmo teve em algum momento contato
com fezes.
Os isolados de E. coli podem ser pesquisados por meio de métodos clássicos
(fenotípicos) e por meio de métodos moleculares (genotípicos).
O método molecular permite a identificação de genes de virulência e de
grupos filogenéticos presentes em microrganismos isolados, inclusive obtidos de
amostras de queijo, indicando a possibilidade da ocorrência de doença a partir da
ingestão desse alimento.
Os fatores descritos justificam a importância de se realizar um estudo na área
da Saúde Pública, visando avaliar a contaminação por E. coli enteropatogênica do
queijo Minas oferecido ao consumidor.
1.2. ANÁLISE DA LITERATURA
1.2.1. Legislação de alimentos no Brasil
Alguns estudos têm revelado uma deficiente qualidade higiênico sanitária dos
queijos comercializados. Todas as etapas de fabricação do queijo devem obedecer a
normas operacionais pré-estabelecidas, desde a recepção do leite utilizado como
matéria prima até o produto final, de modo a impedir a contaminação do produto ou
alterações em sua composição normal.
No Brasil, há uma legislação vigente que estabelece limites de tolerância para
microrganismos em alimentos, a Resolução da Diretoria Colegiada da Agência

18
Nacional de Vigilância Sanitária 12 de 2 de janeiro de 2001 (RDC 12/ 2001)
(BRASIL, 2001).
Pesquisas realizadas no Brasil, em queijo Minas e água demonstraram que
muitos produtos comercializados estavam fora dos padrões da legislação citada.
PERESI e col. (2001) realizaram análise microbiológica em 30 amostras
artesanais de queijo Minas frescal em feiras livres de São José do Rio Preto (SP) e
em 30 amostras industrializadas coletadas em supermercados revelando que 23,3%
das amostras industrializadas e 76,7% das amostras de produção artesanal estavam
em desacordo com a legislação vigente devido à presença de coliformes fecais e
cepas de estafilococos coagulase positiva acima do permitido.
SILVA e col. (2001) observaram que 95,7% do queijo Minas frescal artesanal
produzido em São José do Rio Preto (SP) e comercializado em feiras livres estavam
em desacordo com a legislação em vigor.
RABELO e col. (2001) analisaram 160 amostras de queijo Minas frescal no
Estado de Goiás, revelando que 43% das amostras apresentaram contaminação por
coliformes fecais e por estafilococos coagulase positiva em desacordo com a
legislação em vigor.
AMARAL e col. (2003) em um estudo realizado na região Nordeste do
Estado de São Paulo, determinaram que o número mais provável de coliformes
estava fora dos padrões microbiológicos de potabilidade para água destinada ao
consumo humano. Os autores concluíram que a água utilizada nas propriedades
rurais, que também pode ser utilizada na fabricação do queijo, é um importante fator
de risco à saúde dos seres humanos que a utilizam sendo necessário à adoção de
medidas preventivas.

19
BORGES e col. (2003) pesquisaram queijo coalho produzido em diferentes
regiões do Ceará e verificaram que cerca de 90% das amostras estavam acima do
permitido pela legislação para contagem de coliformes fecais.
Em um estudo realizado em São Paulo foi observado que do total de amostras
estudadas, estavam em desacordo com a legislação em vigor, 26,7% devido à
presença de estafilococos coagulase positiva e foi identificado Staphylococcus
aureus, 73,3% pela presença de coliformes fecais (PEDRO, 2003).
SALOTTI e col. (2006), em estudo realizado no município de Jaboticabal
(SP) com amostras de queijo Minas frescal, encontraram 83,4% das amostras
artesanais e 66,7% das amostras industriais em desacordo com a legislação para
coliformes, demonstrando risco potencial para a saúde da população consumidora.
QUINTANA e CARNEIRO (2007), pesquisando o queijo Minas
comercializado na cidade de Morrinhos (GO), detectaram amostras impróprias para
consumo, com contagens para coliformes fecais acima do permitido pela legislação.
Todos os estudos descritos são importantes para a Saúde Pública, pois visam
demonstrar a contaminação, do queijo comercializado, por coliformes e sugerir a
veiculação de agentes que possam causar doença.
1.2.2. Coliformes em alimentos – Escherichia coli
A contagem dos coliformes nos alimentos, sobretudo os de origem fecal,
indica as condições de higiene em que os queijos foram processados, o encontro
desse grupo implica em deficiências ou falhas no processamento industrial dos

20
alimentos, matéria prima inadequada ou uma contaminação pós-processo
(GRÜNSPAN e col., 1996; PEDRO, 2003; PEDRO e col., 2005).
As bactérias do grupo coliforme estão constituídas por gêneros classificados
na família Enterobacteriaceae dentre os quais se destaca o gênero Escherichia que
são bastonetes Gram-negativos, não esporulados, anaeróbios facultativos,
fermentadores da glicose com produção ou não de gás (TORTORA e col., 2005).
O habitat é o intestino do homem e estudos relatam que Escherichia coli
coloniza o trato intestinal do recém nascido a partir de 40 horas de vida localizando-
se na camada de muco do intestino grosso (FORBES e col., 1998; TORTORA e col.,
2005).
Do gênero Escherichia, E. coli é utilizada como indicador higiênico sanitário
de contaminação fecal, indicando a incidência de contato direto ou indireto dos
alimentos com matéria fecal. A presença de uma bactéria de origem fecal em
alimentos sinaliza também a possibilidade da presença de outras bactérias
patogênicas entéricas como Salmonella spp, Shigella spp (LECLERC e col., 2001;
TODAR, 2002; TORTORA e col., 2005).
Embora E. coli faça parte da biota intestinal humana e de animais
homeotérmicos, alguns organismos desta espécie podem ser patogênicos para ambos
(TODAR, 2002; KAPER e col., 2004).
Escherichia coli comensal pode adquirir específicos atributos de virulência
que são codificados em elementos genéticos, e tornar-se patogênica, as cepas
patogênicas de Escherichia coli são denominadas de patotipos (KAPER e col.,
2004).

21
Didaticamente, o resultado da infecção com um patotipo de E.coli pode ser
dividida em três síndromes gerais: entérica/diarréia, urinária e séptica/meningite
(RODRIGUEZ, 2002; TRABULSI e col., 2004; KAPER e col., 2004). Todas as
síndromes são devido a patotipos de E. coli oriundos do intestino.
A síndrome entérica é ocasionada por seis patotipos: E. coli enteropatogênica
(EPEC), E. coli enterotoxigênica (ETEC), E. coli enteroinvasiva (EIEC), E. coli
produtora de Shiga toxina (STEC), E. coli enteroagregativa (EAEC) e E. coli
difusamente aderente (DAEC) (GUTH, 2000; KAPER e col., 2004, KARCH e col.,
2005).
Para ocorrer à diarréia pelos enteropatotipos de E. coli, três paradigmas são
descritos: produção de enterotoxina (ETEC e EAEC) ou invasão (EIEC) ou aderência
íntima com a membrana (EPEC e EHEC) (KAPER e col 2004).
Os patotipos entéricos apresentam fatores de virulência que proporcionam
que a bactéria cause a infecção no hospedeiro como, por exemplo, as adesinas que
permitem que as bactérias colonizem a mucosa ileal como a intimina de EPEC e
EHEC; as enterotoxinas como a ST1 de ETEC e fatores de invasão que permitem a
invasão da mucosa, semelhante a pInv de EIEC (KAPER e col. 2004).
Os patotipos entéricos apresentam características diferentes, descritas a
seguir, relacionadas a patogenicidade, síndrome clínica e epidemiológica.

22
1.2.3. Escherichia coli enterotoxigênica (ETEC)
É geralmente associada à “diarréia dos viajantes”. Afeta crianças de países
em desenvolvimento e é uma importante causa de diarréia também nos animais
homeotérmicos. O reservatório é o homem. Os sintomas são similares aos da cólera,
afetando o intestino delgado provocando cólicas, vômitos e febre baixa, diarréia
profusa aquosa, tipo água de arroz, sem sangue, podendo ocorrer a desidratação e o
choque (NATARO e KAPER, 1998).
A dose infectante é alta estando entre 108 e 1010 microrganismos/g ou ml. O
período de incubação varia entre 8 e 44 horas, com média de 26 horas e a duração da
doença é curta, aproximadamente de 24 a 30 horas (NATARO e KAPER, 1998).
A contaminação fecal de água e alimentos é a principal razão da alta
incidência de infecção por ETEC em países em desenvolvimento (NATARO e
KAPER, 1998).
O patotipo ETEC coloniza o intestino delgado por meio de fatores de
colonização que são adesinas e produz uma ou mais toxinas, induzindo a liberação de
fluído intestinal (KAPER e col., 2004).
As toxinas produzidas são a termo estável (ST) e termo lábil (LT),
codificadas por plasmídios (KAPER e col., 2004).
A toxina termo-estável (ST) é uma proteína de 5kDa, tem esse nome porque
pode resistir à temperatura de 100°C por 30 minutos. Há dois tipos que diferem
biológica e quimicamente a ST I e ST II (NATARO e KAPER, 1998; KAPER e col.,
2004).

23
A STI é solúvel no metanol, seu mecanismo de ação aciona a guanilato
ciclase alterando o metabolismo celular e aumentando a secreção (GUTH, 2000;
TRABULSI e col., 2004; BRITO e col., 2004).
Há duas variantes genéticas com diferenças nas seqüências de aminoácidos:
STIh que ocorre em humanos e STIp observada em porcos (GUTH, 2000;
TRABULSI e col., 2004; BRITO e col., 2004).
A STII é insolúvel em metanol. Estudos divergem quanto à ocorrência de
casos em humanos. Atuam elevando as concentrações de cálcio estimulando a
prostaglandina e serotonina que aumentam a secreção de íons (GUTH, 2000;
TRABULSI e col., 2004; BRITO e col., 2004).
A toxina termo-lábil LT é inativada por aquecimento a 100°C por 30
minutos. É muito semelhante à toxina colérica, com atividade de adenilato ciclase. É
denominada de citotonina porque causa um bloqueio das funções. É uma proteína de
cerca de 80 kDa composta por 5 subunidades B arranjadas em forma de anel e uma
subunidade A que é responsável pela atividade enzimática da toxina, que estimula a
excreção de cloro e bicarbonato de sódio com acúmulo de líquido e diarréia
(NATARO e KAPER, 1998).

24
1.2.4. Escherichia coli enteropatogênica (EPEC)
É conhecida como agente de diarréia infantil ou diarréia neonatal causando
surtos em berçários hospitalares por acometer neonatos de 0 a 6 meses e crianças que
não se alimentam do leite materno é freqüente nos países em desenvolvimento
(NATARO e KAPER, 1998).
Muitos adultos possuem EPEC no trato intestinal, porém não expressam os
sintomas da doença o que sugere a ocorrência de imunidade a este microrganismo. O
reservatório é o homem, ressalta-se o papel das mães e enfermeiras como portadoras
assintomáticas (NATARO e KAPER, 1998; TORRES e col., 2005).
A transmissão ocorre por contato pessoal, água e alimentos contaminados. O
sítio de infecção é o intestino delgado ocasionando má absorção dos nutrientes e
conseqüente diarréia, outros sintomas são febre baixa e vômitos (TORRES e col.,
2005).
A lesão histológica que EPEC causa no epitélio intestinal recebe a designação
de Attachment/ Effacement ou lesão A/E, que ocorre também no patotipo EHEC. O
diferencial é que EPEC não produz Shiga toxina (STx) ou também chamada de
Verotoxina (VT) (NATARO e KAPER, 1998).
A lesão A/E envolve o gene eae e de outros que estão localizados na ilha de
patogenicidade denominada de LEE (locus of enterocyte effacement) (NATARO e
KAPER, 1998).
O LEE é organizado em 5 operons policistrônicos LEE 1 a LEE 5. Os
produtos codificados por ele são: o sistema de secreção tipo III (LEE 1 A LEE 3); o

25
sistema de translocação de proteínas (LEE 4) e os genes do operon eae e tir no
operon LEE 5 (BARBA e col., 2005; GYLES, 2007).
A adesina intimina, codificada pelo gene eae é uma proteína de 94-kDa, da
membrana externa da bactéria, responsável pela aderência íntima às células
hospedeiras (NATARO e KAPER, 1998).
Na lesão A/E, o receptor da intimina é denominado de Tir ("translocated
intimin receptor") e é um produto bacteriano que é inserido na célula hospedeira
durante a infecção, a descoberta de diferentes tipos de intimina demonstrou que há
um tropismo pelo tecido do hospedeiro (TORRES e col., 2005).
A EPEC adere intimamente à membrana das células intestinais e formam
pedestais com a condensação de actina (TRABULSI e col., 2004; KAPER e col.,
2004; SCHIMDT e HENSEL, 2004).
KAPER (1996) descreveu no Segundo Simpósio Internacional sobre EPEC
(1995) as denominações:
• EPEC típica para as cepas que:
o produzem lesão A/E (gene eae),
o não produzem a toxina Stx,
o contém o plasmídio EAF (EPEC adherence factor plasmid) o
qual codifica a produção do pili BFP (bundle-forming pili)
que produz a característica de aderência a célula epitelial
chamada de aderência localizada (LA) (Figura 1)
• EPEC atípica foi atribuída para as cepas:
o produzem lesão A/E (gene eae),
o não produzem a toxina Stx,

26
o não contém o plasmídio EAF.
1.2.5. Escherichia coli enteroinvasiva (EIEC)
Como o próprio nome sugere e a característica que diferencia Escherichia
coli enteroinvasiva dos outros patotipos é a invasão da célula sendo, portanto um
patógeno intracelular (NATARO e KAPER, 1998).
A EIEC apresenta uma patologia muito semelhante à Shigella spp, sendo que
a disenteria causada por ambas é clinicamente indistinguível.
Vários biossorotipos são responsáveis por infecções de EIEC, caracterizados
geralmente por serem imóveis, não descarboxilizarem a lisina e não fermentarem a
lactose diferente dos outros patotipos (NATARO e KAPER, 1998).
Acometem crianças maiores de 2 anos de idade e adultos. Os sintomas da
doença causada por EIEC são calafrios, febre, dores abdominais e diarréia que pode
ou não ser sanguinolenta com presença de leucócitos e muco, devido à inflamação e
necrose do cólon (NATARO e KAPER, 1998).
O reservatório é o homem, e a transmissão se dá por ingestão de alimentos e
águas contaminados (NATARO e KAPER, 1998).
Outro diferencial é a dose infectante que para Shigella spp é em torno de
5.000 microrganismos/g ou ml, e para EIEC é geralmente entre 106 a 108
microrganismos/g ou ml (NATARO e KAPER, 1998; KAPER e col., 2004).
O período de incubação varia de 8 a 24 horas com média de 11 horas e a
duração da doença é de vários dias (NATARO e KAPER, 1998; KAPER e col.,
2004).

27
O microrganismo invade células do epitélio intestinal, multiplica-se dentro
destas células e invadem células epiteliais adjacentes provocando ulcerações do
cólon, resultando em diarréia sanguinolenta. Os genes responsáveis pela penetração
localizam-se num plasmídio pInv de 120-140 MDa (megadaltons), que desencadeiam
a penetração na célula epitelial, ocorrendo a lise do vacúolo endocítico, multiplicação
intracelular, atingindo a célula epitelial adjacente (NATARO e KAPER, 1998;
KAPER e col., 2004).
Na infecção severa estes eventos causam uma forte reação inflamatória com
ulcerações. A invasão e disseminação induzem a apoptose, durante a infecção, do
macrófago, embora seja invasiva é raro encontrá-la na submucosa intestinal
(NATARO e KAPER, 1998; TRABULSI e col., 2004).
1.2.6. Escherichia coli produtora de toxina Shiga (STEC)
O patotipo EHEC é considerado como subgrupo de STEC (KARCH e col.,
2005; NAYLOR e col., 2005).
Ocorre em países desenvolvidos (sorotipo 0157:H7) e em desenvolvimento.
O sítio de infecção é o intestino grosso (NATARO e KAPER, 1998; TRABULSI e
col., 2004).
A transmissão ocorre através da ingestão de alimentos mal cozidos, tais
como hambúrguer. A dose infectante é baixa, de 10 a 100 microrganismos já podem
causar a doença. O período de incubação é de 3 a 4 dias (NATARO e KAPER, 1998;
TRABULSI e col., 2004).

28
Os sintomas são de dor abdominal, vômitos, diarréia que inicialmente é
aquosa podendo progredir para colite hemorrágica e síndrome urêmico hemolítico e
morte (NATARO e KAPER, 1998; TRABULSI e col., 2004).
A virulência se dá pela produção de uma ou duas citotoxinas semelhantes a
Shiga toxina produzida pela Shigella dysenteriae tipo 2 a (toxina de Shiga ou Stx ou
também chamada de Verotoxina-VT) (NATARO e KAPER, 1998; TRABULSI e
col., 2004).
São toxinas termolábeis, cujo peso molecular varia de 70.000 a 88.500
daltons. Similarmente à LT do patotipo ETEC, elas são constituídas por uma
subunidade A, enzimaticamente ativa, e cinco subunidades B, com função de
acoplamento a receptores celulares (NATARO e KAPER, 1998; KAPER e col.,
2004).
O mecanismo de ação da Vero toxina consiste na subunidade B se ligar ao
receptor Gb3, a subunidade A é internalizada na célula, inibindo desta forma a
síntese protéica e ocorrendo a morte celular, o efeito citotóxico caracteriza-se por
arredondamento celular, morte e descolamento do tapete de células (NATARO e
KAPER, 1998; KAPER e col., 2004).
Estas toxinas se classificam em dois tipos: Stx I e II, que são sorologicamente
distintas e apresentam efeito citotóxico em células de rim de macaco verde africano
(VERO) e células de adenoma de útero humano (HeLa) (NATARO e KAPER, 1998;
KAPER e col., 2004).
Os genes que codificam as Stx I e II estão relacionados a bacteriófagos
(NATARO e KAPER, 1998; KAPER e col., 2004).

29
O subgrupo EHEC também contém a ilha de patogenicidade LEE e aderência
da bactéria à célula intestinal é devido ao gene eae (NATARO e KAPER, 1998;
KAPER e col., 2004).
Utiliza-se o termo STEC ou VTEC para as cepas de E. coli que:
• produzem a toxina VT e
• não possuem a ilha de patogenicidade LEE (referente a produção da
adesina intimina).
O termo EHEC é utilizado para as cepas de E. coli que:
• produzem toxina VT e
• possuem a ilha de patogenicidade LEE (lesão A/E).
Também são denominadas de: (Figura 1)
• EHEC típica as cepas que:
o causam colite hemorrágica ,
o causam síndrome urêmico hemolítica ,
o contém a ilha de patogenicidade LEE causam a lesão A/E,
o produzem toxina VT e
o contém o plasmídio de 60MDa que codifica uma hemolisina.
• EHEC atípica as cepas que:
o causam colite hemorrágica ,
o causam síndrome urêmico hemolítica ,
o não contém a ilha de patogenicidade LEE,
o produzem toxina VT e

30
o e/ou não contém o plasmídio de 60MDa que codifica uma
hemolisina (NATARO e KAPER, 1998; KAPER e col., 2004,
NAYLOR e col., 2005; KARCH e col., 2005).
AEEC- E. coli com lesão A/E VTEC- E. coli produtora de verotoxina
EPEC típica- E. coli enteropatogênica típica VT- verotoxina
EHEC atípica- E. coli enterohemorrágica atípica pEAF- plasmídio EAF
LEE- ilha de patogenicidade LEE CH-SUH-colite hemorrágica/ síndrome urêmica
hemolítica
pEHEC- plasmidio de E. coli enterohemorrágica EHEC típica- E. coli enterohemorrágica típica
Figura 1. Diagrama de Venn ilustrando os patotipos de E. coli, EHEC/ VTEC/
EPEC (baseado no estudo de NAYLOR e col., 2005).

31
1.2.7. Escherichia coli enteroagregativa (EAEC)
EAEC é um grupo de bactérias que abrange sorogrupos de E. coli que aderem a
linhagens celulares "in vitro", tais como células HeLa e HEp-2 (células de carcinoma
laríngeo humano) formando agregados bacterianos, localizados na superfície celular
e em regiões da lamínula livres de células, em um arranjo semelhante a tijolos
empilhados (denominado de aderência agregativa) (NATARO e col., 1987).
A configuração em “pilha de tijolos” é considerada condição obrigatória para o
padrão típico de aderência agregativa (AA). Anteriormente denominada com a sigla
EaggEC, atualmente EAEC. Esse microrganismo causa uma diarréia aquosa e que
dura até 14 dias e que está relacionada com a formação de um biofilme (NATARO e
KAPER, 1998; OKEKE e NATARO, 2001).
Escherichia coli enteroagregativa acomete crianças e adultos e está implicada
em surtos causados por ingestão de alimentos (NATARO e col., 1998).
Além de diarréia aguda EAEC está fortemente associada como agente de diarréia
persistente (HARRINGTON e col., 2006).
A persistente diarréia causada pela EAEC pode ser explicada devido a sua
sobrevivência intracelular por até 72hs, esse nicho intracelular protege as cepas dos
mecanismos de defesa do hospedeiro e tratamento por antibióticos (PEREIRA e col.,
2008).
Por ser muito heterogênea em relação aos fatores de virulência e embora estes
fatores sejam freqüentes na categoria, não há uma prova sensível para identificação

32
genética (NATARO e col., 1998; ELIAS e col., 2002), porém CERNA e col. (2003)
afirmam em seu estudo terem desenvolvido uma reação de PCR multiplex rápida,
fácil e sensível para três genes do plasmídio de virulência pAA.
KAPER e col. (2004) sugerem o uso da denominação EAEC típica para as cepas
que contém o gene regulador aggR e denominação EAEC atípica para as que não
contém.
Como meio diagnóstico é utilizada a pesquisa do gene aatA do plasmídio pAA,
que está relacionado com a aderência agregativa (AA) (BAUDRY e col., 1990;
NISHI e col., 2003).
Muitos pesquisadores têm utilizado métodos moleculares para verificarem a
presença de fatores de virulência dos enteropatotipos de Escherichia coli em
amostras de alimentos, água e fezes de doentes, com intenção de determinar o
patotipo, a fonte de infecção e associar a doença.
JACKSON e col. (1998) estudaram um caso de diarréia com sangue ocorrido em
uma fazenda da cidade de Ontário, Canadá. Os autores analisaram as fezes da criança
doente, do gado e da água da fazenda, através de eletroforese de campo pulsado e
demonstraram que tanto o gado como a água eram fontes potenciais de E. coli
O157:H7.
E. coli O157:H7 foi encontrado em espinafres para consumo em 26 estados
dos Estados Unidos, resultando em 199 pessoas doentes e 3 mortes. Segundo relato
de MAKI (2006), mesmo utilizando a técnica de enzimas de restrição não foi
possível determinar o modo de contaminação do vegetal se foi por contato com os
animais homeotérmicos das fazendas produtoras, pela manipulação ou pela água.

33
PERELLE e col (2007) realizaram um estudo visando à prevenção de surtos
de intoxicação gastrintestinal por meio da reação de PCR multiplex para detecção da
presença de STEC a partir de matérias-primas de origem bovina.
PANETO e col. (2007) utilizando a reação de PCR multiplex, determinaram a
ocorrência de ETEC e VTEC em amostras de queijo Minas frescal colhidas na região
Centro Oeste do Brasil.
Os resultados obtidos no trabalho realizado por STEPHAN e col. (2008)
utilizando a reação de PCR multiplex em cepas de amostras de queijo Suíço
fabricado com leite cru, sugerem que o queijo pode ser um potencial veiculo de
transmissão de STEC para o homem.
No estudo, realizado por OUNDO e col. (2008), utilizando eletroforese de
campo pulsado em cepas obtidas de amostras de alimentos, água e de manipuladores
de alimentos dos hotéis no Quênia, África, foi possível determinar que os
manipuladores são importantes na transmissão da EAEC.
SCAVIA e col. (2008), estudando surtos de gastroenterite causados por
alimentos, concluíram que os casos por EAEC que ocorrem nos países europeus são
subnotificados devido à falta de treinamento do pessoal de laboratório e que as boas
práticas de higiene e manipulação dos alimentos são importantes para a prevenção
dos surtos. Sugerem também a possibilidade do leite não pasteurizado utilizado na
fabricação de queijo ser a origem da contaminação do alimento.
Com o objetivo de determinar a ocorrência de Escherichia coli produtora de
toxina Shiga (STEC) em carne moída e leite cru no sul do Brasil, TIMM e col.
(2009), utilizando PCR multiplex concluíram que a baixa prevalência de STEC em
alimentos e as características das cepas encontradas em bovinos podem estar

34
relacionadas, e explicaria a baixa prevalência de enfermidades de origem alimentar
causadas por STEC no Brasil.
Outra forma de pesquisa de E. coli é agrupar os isolados em filogrupos.
1.2.8. Grupos Filogenéticos
Utilizando a técnica de multilocus enzimático e ribotipagem SELANDER e
col. (1986) e HERZER e col. (1990) caracterizaram quatro grupos filogenéticos de
Escherichia coli: A, B1, B2 e D.
Os autores concluíram que cepas pertencentes aos grupos B2 são relacionadas
à doença extra intestinal, em menor frequência as do grupo D, e possuem mais
fatores de virulência. As cepas do grupo A e B1 são consideradas como linhagens
intestinais comensais e também foram descritas como patogênicas (PUPO e col.,
1997; CLERMONT e col., 2000; GIRARDEOU e col., 2005; TIBA e col., 2009).
Para se determinar o filogrupo utiliza-se à reação de PCR multiplex
empregando três marcadores: o gene chuA que é relacionado com o transporte de
ferro na EHEC O157:H7 (TORRES e PAYNE, 1997); o gene yjaA de função
desconhecida mas identificado na E. coli K-12 e um fragmento anônimo de DNA
designado de TSPE4.C2 (CLERMONT e col., 2000)
Os perfis para os grupos filogenéticos estão apresentados na Figura 2.

35
Figura 2. Esquema representando os grupos filogenéticos utilizando iniciadores para
os genes chuA, yjaA e o fragmento de DNA TSPE4.C2 (CLERMONT e col., 2000).
Várias pesquisas têm agrupado as cepas de E. coli em filogrupos, porém a
maioria dos trabalhos encontrados na literatura é de isolados clínicos e de ambiente.
Utilizando a reação de PCR triplex, DURIEZ e col. (2001), estudaram cepas
de E. coli isoladas de fezes de três populações humanas distintas geograficamente e
os dados obtidos mostraram que os grupos filogenéticos A e B1 são mais freqüentes,

36
seguidos do grupo filogenético D. Os autores concluíram que as linhagens do grupo
filogenético B2 são raras.
KOTLOWSKI e col. (2006), relacionam em seu estudo a ocorrência dos
grupos filogenéticos B2 e D de E. coli em pacientes com doença intestinal
inflamatória e colite ulcerativa.
O estudo filogenético de cepas de E. coli provenientes de amostras de esgoto
de origem humana e de origem animal, indicou que o grupo B2 é associado ao esgoto
de origem humana, e os grupos A, B1 e D estão em ambos os esgotos, SABATÉ e
col. (2008), sugerem também que o uso abusivo de antibiótico utilizado na produção
animal pode promover a resistência nos filogrupos de E. coli e que pode ser
transmitido depois para a população humana através do alimento de origem animal.
Num estudo epidemiológico com crianças saudáveis e com diarréia, AFSET e
col. (2008) relatam que os grupos filogenéticos A, B1 e D não apresentaram
associação significativa com diarréia. E que cepas pertencentes ao filogrupo B2
foram mais comuns em crianças saudáveis do que em crianças com diarréia.
NOWROUZIAN e col. (2009), compararam cepas do filogrupo B2 obtidas de
crianças suecas e paquistanesas e observaram que as cepas encontradas nas crianças
paquistanesas carregam um número significativamente menor de genes de virulência.
Concluem os autores que cepas de E. coli isoladas de diferentes populações humanas
podem diferir na distribuição filogenética.
HANNAH e col. (2009), pesquisando amostras de carne e amostras clínicas
de humanos de uma comunidade rural nos Estados Unidos, encontraram grupos
filogenéticos de E. coli similares, os autores sugerem que alimento pode ser o veículo
de transmissão da E. coli.

37
1.2.9. Reação de polimerização em cadeia (PCR)
A reação de PCR é um método que permite a amplificação de um fragmento de
DNA em poucas horas, teve início na década de 80, foi desenvolvida pelo
pesquisador Kary Mullis e sua equipe, tem mostrado grande aplicabilidade,
principalmente no diagnóstico de doenças (SCHUTZBANK e STERN, 1993;
RUPPENTHAL, 1999).
É considerada uma reação de grande sensibilidade para detectar os fatores de
virulência (NATARO e KAPER, 1998; RUPPENTHAL, 1999).
A reação da PCR consiste em um processo de replicação de DNA, utiliza
alterações de temperatura que contribuem para a separação da molécula de DNA,
ocorrendo à associação dos iniciadores com a atuação da DNA polimerase e
finalmente a síntese da cadeia complementar (RUPPENTHAL, 1999).
Vários autores a utilizaram por meio do multiplex (múltiplos iniciadores) para
determinarem a presença dos patotipos de E. coli (PASS e col., 2000; TOMA e col.,
2003; SARANTUYA e col., 2004; ARANDA e col., 2004; NGUYEN e col., 2005;
FRANZOLIN e col., 2005; VIDAL e col., 2005; ARANDA e col., 2007).
Em isolados de amostras de alimentos a PCR multiplex é muito utilizada nas
pesquisas de fatores de virulência (PERELLE e col., 2007; PANETO e col., 2007;
STEPHAN e col., 2008; TIMM e col., 2009).
VICENTE e col. (2005), confirmando a importância da reação da PCR
multiplex, demonstraram que um surto clinicamente diagnosticado como cólera em
vilarejos da floresta Amazônica, era na realidade causado por cepas de ETEC.

38
2.OBJETIVOS
Caracterização molecular dos isolados de E. coli obtidos de amostras de
queijo Minas:
• Verificar a freqüência com que ocorrem os grupos
filogenéticos: A, B1, B2 e D.
• Determinar a ocorrência dos fatores de virulência nos
isolados de E. coli pertencentes aos patotipos: ETEC,
EIEC, EPEC, STEC e EAEC.
• Discutir a presença dos referidos patotipos de E. coli nas
amostras de queijo Minas e o significado para a Saúde
Pública e Vigilância Sanitária.

39
3. MATERIAL E MÉTODOS
3.1. ISOLADOS BACTERIANOS – Escherichia coli
Foram utilizadas para o estudo, 250 isolados de Escherichia coli oriundas de
10 amostras de queijo Minas frescal e meia cura, adquiridas em supermercados,
feiras livres, vendedores ambulantes e mercados municipais da cidade de São Paulo
isoladas durante o período de 2001 a 2002 (PEDRO, 2003), e mantidas a -70oC em
caldo Luria Bertani (LB) contendo 15% de glicerol (Quadro 1).
Quadro 1. Tipo de queijo Minas e local de aquisição das amostras utilizadas
para obtenção dos isolados de E. coli. São Paulo, 2003 (PEDRO, 2003).
Amostra queijo Minas frescal Queijo Minas meia cura 1 supermercado 2 supermercado 3 supermercado 4 supermercado 5 supermercado 6 feira livre 7 ambulante 8 ambulante 9 mercado municipal
10 mercado municipal

40
3.2. REISOLAMENTO E CONFIRMAÇÃO DA VIABILIDADE E
PUREZA DOS ISOLADOS
Uma alíquota de cada uma dos isolados proveniente da coleção de cultura foi
cultivada em caldo LB, incubado 35oC ±0,5 por 24 horas. Os tubos que apresentaram
turvação foram subcultivados em ágar McConkey e incubados por um período de 24
horas a 37oC ±0,5 para isolamento das colônias.
Após o período de incubação, as colônias características de E. coli que
apresentaram coloração rosa, pela utilização da lactose, foram selecionadas e
submetidas à confirmação do posicionamento taxonômico com emprego de métodos
fenotípicos convencionais.
As provas bioquímicas empregadas foram as seguintes: produção de Indol e
prova de Vermelho de Metila (VM) que são positivas para E. coli e utilização de
Citrato segundo Simmons, prova de Voges – Proskauer (IMVC) também a prova da
citocromo oxidase que são negativas para E. coli (SILVA e col., 2001).
3.3.MÉTODOS MOLECULARES
3.3.1. Extração do material genético
O material genético (DNA genômico e plasmidial) foi extraído por meio de
choque térmico segundo a técnica descrita por CHAPMAN e col. (2001), os isolados

41
de Escherichia coli foram cultivadas em 10 mL de caldo LB por 24 horas a 35oC. As
culturas foram centrifugadas por 10 minutos em velocidade de 5.000 rpm à
temperatura ambiente, os sobrenadantes descartados e o precipitado correspondente
às células bacterianas foi lavado duas vezes com 1 mL de água MilliQ® estéril,
ressuspenso em 200 µL de água MilliQ® e transferido para microtubos do tipo
eppendorf estéreis. Os tubos contendo as suspensões bacterianas foram levados por
10 minutos em banho-maria fervente (95oC) e em seguida mantidos por 30 minutos a
-20oC. Após esse período, os tubos foram mantidos em temperatura ambiente por
aproximadamente 5 minutos e então foram submetidos à centrifugação por 10
minutos em velocidade de 12.000 rpm. O sobrenadante contendo o material genético
foi transferido para um novo microtubo e conservado até o momento do uso a -20oC.
3.3.2.Reação em cadeia pela polimerase (PCR)
Foram realizadas três reações de PCR multiplex:
• PCR para determinação do grupo filogenético de E. coli foi realizada baseada no
estudo realizado por CLERMONT e col. (2000), com a utilização simultaneamente
(multiplex) de três iniciadores específicos, para a identificação dos grupos
filogenéticos A e B1(comensais) e B2 e D (patogênicos). Os iniciadores
denominados chuA, para detecção do gene relacionado com o transporte ferro
(enterohemorrágicas), yjaA que detecta o gene fortemente relacionado com cepas
responsáveis por casos de meningite neonatal e TSPE4C gene de função

42
desconhecida observado em cepas de K12 representantes comensais de E. coli. As
seqüências dos iniciadores que foram utilizados para este estudo estão descritas no
Quadro 2.
As condições de ciclo e concentração de reagentes para a detecção de genes
relacionados à filogenia de E. coli foram os seguintes: para cada reação de 25 µL
foram utilizados 2,5 µL de tampão de reação 10x; 1,5 mM de cloreto de magnésio;
200µM de solução de DNTP; 200µM de cada um dos iniciadores; 1,25 U de Taq
DNA polimerase e 5 µL de DNA. Foram empregados para amplificação um ciclo de
desnaturação inicial a 94oC por 4 minutos, 30 ciclos compostos de desnaturação a
94oC por 5 segundos, anelamento a 59oC por 10 segundos e extensão a 72oC por 2
minutos, e ainda um ciclo único final a 72oC por 5 minutos, em um termociclador
Mastercycler gradient Eppendorf.
Os produtos de reação para estes iniciadores são fragmentos de 279, 211, e
152 pares de base (bp) para os genes chuA, yjaA e TSPE4C respectivamente.
• PCR para determinação da presença de fatores de virulência de E. coli foi
realizada com base no estudo realizado por KONG e col. (1999). Utilizou-se o
sistema multiplex com iniciadores específicos para amplificação de fragmentos
correspondentes a genes responsáveis pela produção de toxina termolábil 2 (LT2),
toxina termolábil 1 (LT1), toxina termoestável (ST1), verotoxina (VT1 e VT2) e
ainda o gene que codifica a adesina intimina (eae).
As condições de ciclo e concentração de reagentes para a detecção de genes
relacionados à virulência de E. coli foram os seguintes: para cada reação de 25 µL
foram utilizados 2,5 µL de tampão de reação 10x; 1,5 mM de cloreto de magnésio;
200µM de solução de DNTP; 200µM de cada um dos iniciadores; 1,25 U de Taq

43
DNA polimerase; e 5 µL de DNA. A reação composta por um ciclo inicial de
desnaturação a 94oC por 2 minutos, seguida por 40 ciclos composto de desnaturação
a 94oC por 1 minuto, anelamento a 52oC por 1 minuto e extensão a 72oC por 1
minuto, e ainda uma extensão final por 5 minutos a 72oC.
Os produtos de reação esperados para estes iniciadores são fragmentos de
720, 523, 360, 275 e 175 pares de base (bp) para os iniciadores LT2, VT (VT1 e
VT2), eae, LT1 e ST1 respectivamente (Quadro 2).
• A pesquisa de fatores virulência em E. coli foi complementada com a utilização
dos iniciadores Einv que detectam o gene ipaH do plasmídio de invasão (pInv) e
Eagg que detecta o gene aaTa do plasmídio pAA da aderência agregativa (AA),
presentes nos patotipos de E. coli enteroinvasiva e enteroagregativa de acordo com o
trabalho de ORLANDI e col. (2006).
As condições de ciclo e concentração de reagentes para a detecção de genes
relacionados à virulência de E. coli foram os seguintes: para cada reação de 25 µL
foram utilizados 2,5 µL de tampão de reação 10x; 2 mM de cloreto de magnésio;
200µM de solução de DNTP; 200µM de cada um dos iniciadores; 1,25 U de Taq
DNA polimerase; e 5 µL de DNA. A reação constou de 30 ciclos composto de
desnaturação a 92oC por 30 segundos, anelamento a 57oC por 1 minuto e extensão a
72oC por 1 minuto, e ainda uma extensão final por 5 minutos a 72oC.
Os produtos de reação esperados para estes iniciadores são fragmentos de 140
e 194 pares de base (bp) para os iniciadores Einv e Eagg, respectivamente (Quadro
2).

44
Quadro 2. Seqüência dos iniciadores da PCR e tamanho esperado dos produtos que
foram utilizados na determinação filogenética e de fatores de virulência dos isolados
de E. coli.
Patotipo/
Filogrupo
Iniciador Seqüência Tamanho do produto (bp)
Referência
chuAF 5` GAC GAA CCA ACG GTC AGG AT 3’ B2 e D chuAR 5` TGC CGC CAG TAC CAA AGA CA 3’ 279
yjaAF 5` TGA AGT GTC AGG AGA CGC TG 3’ A e B2 yjaAR 5` ATG GAG AAT GCG TTC CTC AAC 3’ 211
TSPE4CF 5` GAG TAA TGT CGG GGC ATT CA 3’ B1, D e B2 TSPE4CR 5` CGC GCC AAC AAA GTA TTA CG 3’ 152
CLERMONT e col. (2000).
LT2 F 5` ATA TCA TTT TCT GTT TCA GCA AA 3’ ETEC LT2R 5` CAA TAA AAT CAT CTT CGC TCA TG 3’ 720
VTF 5` GAA CGA AAT AAT TTA TAT GTG 3’ STEC VTR 5` CCT GAT GAT GGC AAT TCA GTA 3’ 523
eaeF 5` GGA ACG GCA GAG GTT AAT CTG CAG 3’ EPEC/ EHEC eaeR 5` CGA AGC CAT TTG CTG GGC GCT C 3’ 360
LT1F 5` TTA CGG CGT TAC TAT CCT CTC TA 3’ ETEC LT1R 5` GGT CTC GGT CAG ATA TGT GAT TC 3’ 275
ST1F 5` CTT TCC CCT CTT TTA GTC AG 3’ ETEC ST1R 5` TAA CAT GGA GCA CAG GCA GG 3’ 175
KONG e col. (1999).
Einv F* 5' TGG AAA AAC TCA GTG CCT CTG CGG 3' EIEC Einv R 5' TTC TGA TGC CTG ATG GAC CAG GAG 3' 140
Eagg F** 5' AGA CTC TGG CGA AAG ACT GTA TC 3' EAEC Eagg R 5' ATG GCT GTC TGT AAT AGA TGA GAA C 3' 194
ORLANDI e col. (2006).
*Einv=gene ipaH do pInv. **Eagg= gene aatA do pAA.
3.3.3. Eletroforese em gel de agarose
Os produtos da PCR foram visualizados em eletroforese em gel de agarose
3% em tampão TAE 1x (Tris-ácido acetico-EDTA, pH 8.0), corado com brometo de
etídeo, em uma intensidade de corrente de 6V/cm por 1 hora. O peso molecular de
cada fragmento foi calculado com base na comparação com um marcador incluso em
cada gel (100bp MassRuler™ DNA Ladder, Fermentas). As imagens foram
capturadas sob iluminação Ultra Violeta em equipamento Chemi II Darkroom (UVP)
e gravadas utilizando o programa Labworks (UVP, Inc Upland, CA, USA).

45
4. RESULTADOS
Foram avaliados quanto aos grupos filogenéticos e seu potencial patogênico
utilizando métodos moleculares, 250 isolados de Escherichia coli, provenientes de
10 amostras de queijo Minas (PEDRO, 2003), a distribuição dos isolados nas
amostras estão demonstrados na Tabela 1.
Tabela 1. Número e proporção dos isolados de E. coli nas amostras de queijo
Minas. São Paulo, 2009.
Amostra nº isolados(%) 1 42(16,8%) 2 21(8,4%) 3 46(18,4%) 4 24(9,6%) 5 110(44%) 6 1(0,4%) 7 2(0,8%) 8 2(0,8%) 9 1(0,4%)
10 1(0,4%) Total 250(100%)
4.1. GRUPOS FILOGENÉTICOS
Os isolados foram agrupados em filogrupos utilizando a reação de PCR
multiplex tendo como base a presença de fragmentos correspondentes aos genes
chuA, yjaA e o fragmento de DNA TSPE4.C2 (CLERMONT e col., 2000).
Na Figura 3 estão apresentados os perfis obtidos para cada grupo filogenético
em gel de agarose.

46
Figura 3. Gel de agarose contendo os fragmentos obtidos para isolados de
Escherichia coli representativos dos perfis filogenéticos. M- Marcador de peso
molecular 100pb, 1- isolado 298 Grupo A, 2- isolado 299 Grupo A, 3 - isolado 300
Grupo B1, 4- isolado 302 Grupo D, 5 - isolado 303 Grupo A, 6 - isolado 309 Grupo
D. São Paulo, 2009.
Das 10 amostras de queijo Minas estudadas foi possível classificar os isolados
em pelo menos um dos grupos filogenéticos, embora se tenha observado
concomitantemente 2 e 3 grupos presentes numa mesma amostra de queijo, conforme
a Tabela 2.

47
Tabela 2. Número dos isolados de E. coli por grupos filogenéticos nas 10 amostras
de queijo Minas. São Paulo, 2009.
Amostra/
filogrupo
1 2 3 4 5 6 7 8 9 10 Total
A 41 18 42 23 105 1 - 1 1 1 233
B1 1 - - 1 5 - - 1 - - 8
B2 - 3 3 - - - 1 - - - 7
D - - 1 - - - 1 - - - 2
Total 42 21 46 24 110 1 2 2 1 1 250
Os resultados demonstraram que o grupo filogenético A foi o mais freqüente
nas amostras estudadas onde 93,2% dos 250 isolados foram classificados como
grupo filogenético A, 3,2% como B1, 2,8% como B2 e 0,8% como D (Figura 4).
Figura 4. Proporção de E. coli do total dos isolados estudados por grupo
filogenético nas 10 amostras de queijo Minas. São Paulo, 2009.
93%
3%1%2,8%
A
B1
B2
D

48
4.2. DETECÇÃO DE MARCADORES DE VIRULÊNCIA
Os resultados do estudo dos 250 isolados de E. coli para os marcadores de
virulência demonstraram a presença de marcadores em 242 isolados (96,8%) (Tabela
3), as porcentagens dos isolados encontrados foram: 5,8% para ST1, 1,2% para LT1
+ ST1, o marcador para intimina (eae) em 0,4%, ST1+ aatA em 57,9% dos isolados,
ST1+ aatA +LT1+LT2 em 0,8%, ST1+ aatA +eae em 2,1% dos isolados, ST1+eae,
ST1+LT1+ aatA +eae e ST1+LT1+eae ambos foram detectados em 0,4% dos
isolados, ST1+LT1+ aatA em 25,6%, o marcador para o patotipo EAEC (aatA) em
5,0%, enquanto que para Verotoxina (VT) e o mecanismo enteroinvasivo (ipaH) não
foram detectados em nenhum dos isolados estudados (Figura 5).
.
Figura 5. Proporção de E. coli do total dos isolados estudados por fatores de
virulência nas 10 amostras de queijo Minas. São Paulo, 2009
0,4% 5,0%
5,8% 1,2%
57,9% 0,8%
2,1%
0,4%
0,4%
0,4%
25,6%
eae aatA
ST1
LT1+ST1
ST1+aatA ST1+aatA+LT1+LT2
ST1+aatA+ eae
ST1+eae
ST1+LT1+aatA+eae
ST1+LT1+eae
ST1+LT1+aatA

49
Tabela 3. Número dos isolados de E. coli por fatores de virulência nas 10 amostras
de queijo Minas. São Paulo, 2009.
Marcador/ Amostra 1 2 3 4 5 6 7 8 9 10 Total
ST1 2 - - 5 5 - 2 - - - 14
LT1+ST1 - 2 - - 1 - - - - - 3
LT1+ST1+LT2 - - - - - - - - - - -
eae 1 - - - - - - - - - 1
aatA 9 1 1 - 1 - - - - - 12
VT - - - - - - - - - - -
ipaH - - - - - - - - - - -
ST1+ aatA 18 13 18 14 73 1 - 1 1 1 140
ST1+ aatA +LT1+LT2 1 - - 1 - - - - - - 2
ST1+ aatA + eae 4 - - - 1 - - - - - 5
ST1+eae 1 - - - - - - - - - 1
ST1+LT1+ aatA +eae 1 - - - - - - - - - 1
ST1+LT1+eae 1 - - - - - - - - - 1
ST1+LT1+ aatA 1 3 27 4 26 - - 1 - - 62
Total 39 19 46 24 107 1 2 2 1 1 242

50
Na Figura 6 é possível visualizar os perfis de bandas obtidos em gel de
agarose para os diferentes marcadores de virulência.
Figura 6. Diferentes perfis obtidos de isolados de Escherichia coli utilizadas como
controle positivos. M- Marcador de Peso molecular, 1 – E. coli Verotoxigenica, 2 –
IAL339, 3 – IAL399, 4 – IAL 1893, 5 – IAL1895, 6 – IAL1896, 7 – IAL1848, 8 - E.
coli enteroinvasiva (isolado clínico), 9 - E. coli enteroagregativa (isolado clínico).
Com base nos resultados obtidos foi possível classificar os isolados de
Escherichia coli estudados de acordo com os diferentes patotipos, a saber: EHEC
(enterohemorrágica), ETEC (enterotoxigênica), EPEC (enteropatogênica), e EAEC
(enteroagregativa) e EIEC (enteroinvasiva) (Tabela 4).
Na Figura 7 é possível visualizar que os marcadores de virulência estavam
presentes em 242 isolados (96,8%) e mostra que o patotipo ETEC foi o mais
freqüente (91,6%) seguido por EAEC (4,8%) e por EPEC (0,4%). Não foram
observados marcadores indicativos da presença dos patotipos EIEC e EHEC.

51
Figura 7. Proporção de E. coli do total dos isolados estudados por patotipos nas 10
amostras de queijo Minas. São Paulo, 2009.
Tabela 4. Número e proporção dos isolados de E. coli por patotipo nas 10 amostras
de queijo Minas. São Paulo, 2009.
Amostra/
Patotipo 1 2 3 4 5 6 7 8 9 10 Total %
ETEC 29 18 45 24 106 1 2 2 1 1 229 94,6%
EPEC 1 - - - - - - - - - 1 0,4%
EHEC - - - - - - - - - - - -
EIEC - - - - - - - - - - - -
EAEC 9 1 1 - 1 - - - - - 12 5%
Total 39 19 46 24 107 1 2 2 1 1 242 100,0%
0%10%20%30%40%50%
60%70%80%90%
100%
ETEC EPEC EHEC EIEC EAEC Total
patotipos
% isolados

52
As tabelas seguintes expressam o que foi detectado em cada amostra de
queijo Minas relativo aos grupos filogenéticos e patotipos (Tabela 5, 6, 7, 8, 9, 10,
11, 12,13 e 14).
Tabela 5. Número de isolados de E. coli por patotipo e por filogrupo na amostra 1 de
queijo Minas. São Paulo, 2009.
Amostra 1 A B1 B2 D Total ETEC 29 - - - 29 EPEC 1 - - - 1 EAEC 9 - - - 9 EHEC - - - - 0 EIEC - - - - 0 Total 39 - - - 39
Tabela 6. Número de isolados de E. coli por patotipo e por filogrupo na amostra 2 de
queijo Minas. São Paulo, 2009.
Amostra 2 A B1 B2 D Total ETEC 15 - 3 - 18 EPEC - - - - - EAEC 1 - - - 1 EHEC - - - - - EIEC - - - - - Total 16 - 3 - 19
53
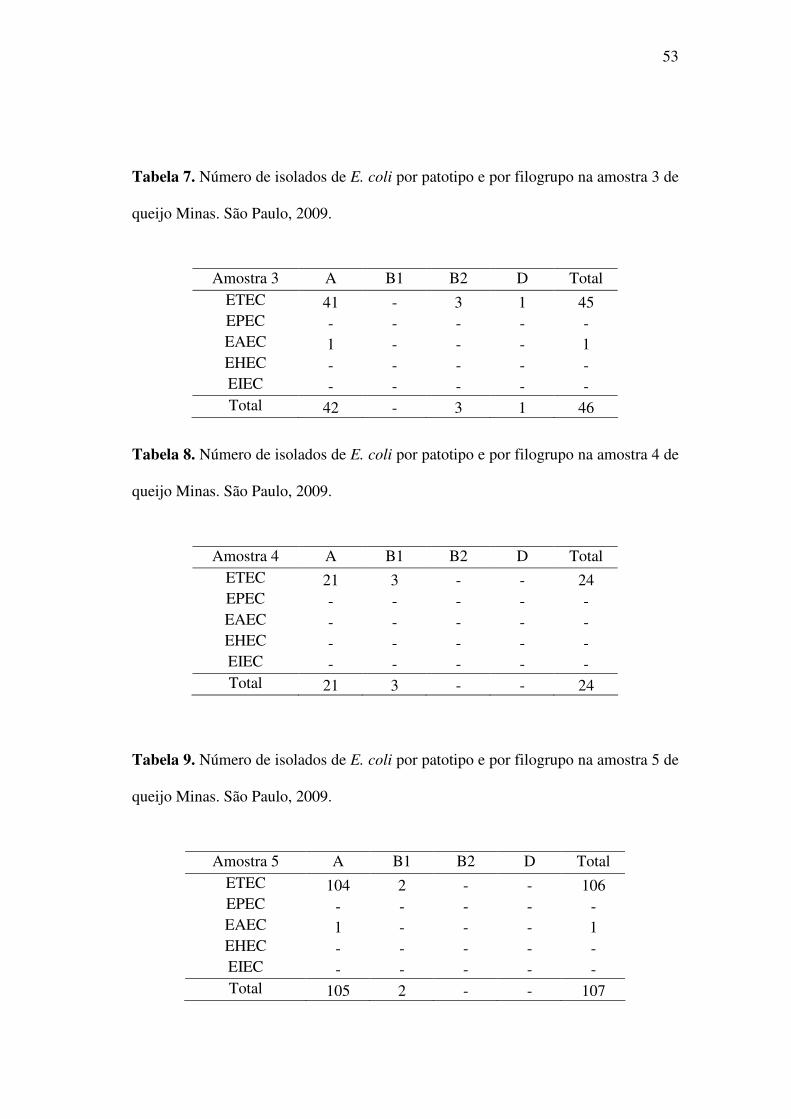
Tabela 7. Número de isolados de E. coli por patotipo e por filogrupo na amostra 3 de
queijo Minas. São Paulo, 2009.
Amostra 3 A B1 B2 D Total ETEC 41 - 3 1 45 EPEC - - - - - EAEC 1 - - - 1 EHEC - - - - - EIEC - - - - - Total 42 - 3 1 46
Tabela 8. Número de isolados de E. coli por patotipo e por filogrupo na amostra 4 de
queijo Minas. São Paulo, 2009.
Amostra 4 A B1 B2 D Total ETEC 21 3 - - 24 EPEC - - - - - EAEC - - - - - EHEC - - - - - EIEC - - - - - Total 21 3 - - 24
Tabela 9. Número de isolados de E. coli por patotipo e por filogrupo na amostra 5 de
queijo Minas. São Paulo, 2009.
Amostra 5 A B1 B2 D Total ETEC 104 2 - - 106 EPEC - - - - - EAEC 1 - - - 1 EHEC - - - - - EIEC - - - - - Total 105 2 - - 107

54
Tabela 10. Número de isolados de E. coli por patotipo e por filogrupo na amostra 6
de queijo Minas. São Paulo, 2009.
Amostra 6 A B1 B2 D Total ETEC - 1 - - 1 EPEC - - - - - EAEC - - - - - EHEC - - - - - EIEC - - - - - Total - 1 - - 1
Tabela 11. Número de isolados de E. coli por patotipo e por filogrupo na amostra 7
de queijo Minas. São Paulo, 2009.
Amostra 7 A B1 B2 D Total ETEC - - 1 1 2 EPEC - - - - - EAEC - - - - - EHEC - - - - - EIEC - - - - - Total - - 1 1 2
Tabela 12. Número de isolados de E. coli por patotipo e por filogrupo na amostra 8
de queijo Minas. São Paulo, 2009.
Amostra 8 A B1 B2 D Total ETEC 1 1 - - 2 EPEC - - - - - EAEC - - - - - EHEC - - - - - EIEC - - - - - Total 1 1 - - 2

55
Tabela 13. Número de isolados de E. coli por patotipo e por filogrupo na amostra 9
de queijo Minas. São Paulo, 2009.
Amostra 9 A B1 B2 D Total
ETEC 1 - - - 1 EPEC - - - - - EAEC - - - - - EHEC - - - - - EIEC - - - - - Total 1 - - - 1
Tabela 14. Número de isolados de E. coli por patotipo e por filogrupo na amostra 10
de queijo Minas. São Paulo, 2009.
Amostra 10 A B1 B2 D Total
ETEC 1 - - - 1 EPEC - - - - - EAEC - - - - - EHEC - - - - - EIEC - - - - - Total 1 - - - 1

56
5. DISCUSSÃO
A detecção de agentes patogênicos em alimentos faz-se necessária para que
possam ser definidas estratégias de prevenção e controle mais efetivas
(HANASHIRO e col., 1999; ARAUJO e col., 2002; BALLAL e RAMAMURTY,
2005; SABATÉ e col., 2008; HANNAH e col., 2009).
Escherichia coli é um importante agente causador de diarréia e infecções
extra-intestinais em humanos em todo o mundo (SABATÉ e col., 2008; HANNAH e
col., 2009). Esses organismos pertencem a diferentes classes de patotipos e são
classificados com base em características observadas na clínica, mecanismos de
virulência e sorotipos (MAREK e col., 2004; BALLAL e RAMAMURTY, 2005;
SCAVIA e col., 2008).
Diferentes formas de classificar Escherichia coli, provenientes de amostras
clínicas, de alimentos ou ambientais, foram descritas a fim de determinar uma
melhor forma de diferenciar as cepas patogênicas de não patogênicas (KONG e col.,
1999; CLERMONT e col., 2000; ORLANDI e col., 2006; SKYBERG e col., 2007;
AFSET e col., 2008; SCAVIA e col., 2008; HANNAH e col., 2009; GRAZIANI e
col., 2009).
SELANDER e col. (1986) utilizaram a técnica de multilocus enzimático para
caracterizar quatro grupos filogenéticos de Eschericha coli, A, B1, B2 e D (HERZER
e col., 1990).

57
CLERMONT e col. (2000) utilizaram a técnica de PCR multiplex para
caracterizarem isolados de Escherichia coli nesses quatro grupos. Diversos estudos
vêm aplicando este tipo de disposição para isolados de Escherichia coli, de amostras
clínicas e ambientais, com a finalidade de elucidar a distribuição dos diferentes
grupos (CLERMONT e col., 2000; WALK e col., 2007; GRAZIANI e col., 2009).
Estudos demonstraram que os grupos filogenéticos de Escherichia coli mais
freqüentemente relacionados com doenças extra-intestinais são do grupo B2 e com
menor freqüência o grupo D e o grupo filogenético A e B1 são mais habituais em
isolados intestinais (CLERMONT e col., 2000; GIRARDEOU e col., 2005;
KOTLOWSKI e col., 2006; SABATÉ e col., 2008; HANNAH e col., 2009;
MORENO e col., 2009; TIBA e col., 2009).
Os limites de coliformes termotolerantes em amostras de alimentos são
definidos pela legislação sanitária vigente (BRASIL, 2001). Contagens elevadas de
coliformes em amostras de alimento indicam que o produto não é adequado para o
consumo (HANASHIRO e col., 1999; ARAUJO e col., 2002; PEDRO e col., 2005).
No presente estudo, 250 isolados de Escherichia coli (Tabela 1) foram
caracterizados nos diferentes grupos filogenéticos de acordo com a metodologia
descrita por CLERMONT e col. (2000). Os resultados demonstraram que o grupo A
foi o mais freqüente (93,2%) (Figura 4), que corroboram com a literatura pesquisada
que demonstra que esse grupo é o mais freqüentemente isolado de amostras não
clínicas (CLERMONT e col., 2000; KOTLOWSKI e col., 2006; SABATÉ e col.,
2008; HANNAH e col., 2009; MORENO e col., 2009; TIBA e col., 2009). As
linhagens patogênicas intestinais pertencem geralmente aos grupos A, B1 e D

58
(Tabela 5, 6, 7, 8, 9, 10, 11, 12, 13, 14) (PUPO e col., 1997; GIRARDEOU e col.,
2005).
Estudos mostram, a presença dos grupos filogenéticos B2 e D, como
principais responsáveis por doenças extra-intestinais em amostras derivadas de
esgoto (SABATÉ e col., 2008), amostras de carne (HANNAH e col., 2009) e água
doce (WALK e col., 2007) mesmo que em baixos números (Tabela 2).
No presente estudo foi possível observar que o grupo B2 estava presente em
três amostras e o grupo D em duas amostras (Tabela 2, 6, 7 e 11).
Não foi possível observar na literatura pesquisada estudos que agrupassem,
em grupos filogenéticos, isolados de Escherichia coli provenientes de amostras de
queijo, sendo este o primeiro estudo demonstrando a distribuição dos grupos
filogenéticos de Escherichia coli em amostras de queijo Minas.
O estudo de marcadores de virulência demonstrou que os fatores de virulência
para ETEC foi o mais freqüente correspondendo a 91,6% (Figura 7) dos isolados
seguido dos fatores de virulência para os patotipos EPEC e EAEC. Escherichia coli
enterotoxigênica representa um importante patógeno responsável por diarréia
principalmente em países em desenvolvimento (MULLER e col., 2007; SUZUKI,
2007).
A detecção de isolados de ETEC portadores de marcador sugerindo a
produção de enterotoxina termoestável ST1 em maior número (Figura 5), como
pode ser observado na Tabela 3, indica que os produtos de onde foram obtidos os
isolados representam um perigo aos consumidores uma vez que cepas com essas
características podem produzir diarréia principalmente em crianças, sendo

59
responsáveis pela maioria dos casos endêmicos (NATARO e KAPER, 1998;
RODAS e col., 2009).
É importante ressaltar a presença de isolados classificados como Escherichia
coli enteroagregativa (EAEC) (Tabela 4). Conforme orientação da literatura
consultada, foram considerados como EAEC aqueles isolados que apresentaram o
gene aatA indicando a presença do plasmídio de virulência de EAEC e que não
apresentavam outro fator como ST ou LT onde foi observada uma baixa freqüência
desse patotipo (4,8%) (Figura 7) (NATARO e col., 1998)
Não há relatos na literatura sobre a ocorrência do gene aatA vinculado com o
marcador de ST1.
Pesquisas mostram que marcadores de EAEC podem ser também obtidos no
patotipo ETEC, pode conter em um plasmidio o gene homólogo ao da EAEC
produzindo reação positiva para os isolados de ETEC (YAMAMOTO e
ECHEVERRIA, 1996).
Esse fato foi observado no presente estudo, porém com outro marcador de
EAEC, foi utilizado o iniciador desenhado com base em seqüências descritas por
PASS e col. (2000).
Vários estudos com amostras clínicas apresentaram resultados com
freqüência de Escherichia coli enteroagregativa (ARANDA e col., 2004;
RODRIGUES e col., 2004; VAZ e col., 2004; SARANTUYA e col., 2004; REGUA-
MANGIA e col., 2004; NGUYEN e col., 2005, FRANZOLIN e col., 2005;
ORLANDI e col., 2006; MORENO e col., 2009).
Os resultados obtidos no estudo de NALÉRIO e col.(2004) demonstraram que
a técnica de PCR, com a utilização de seqüência do gene eae, apresentou maior

60
capacidade discriminatória que a técnica de cultivo celular em células HEp-2 para a
identificação do sorogrupo Escherichia coli enteropatogênica clássica. No presente
estudo foi detectado o marcador para intimina em um isolado, e associado a outros
fatores de virulência com ST1, LT1 e aatA em 8 isolados, não havendo relato
semelhante na literatura (Tabela 3).
Esses estudos, em sua maioria, buscam a etiologia da doença diarréica em
crianças, porém do ponto de vista da vigilância sanitária, conhecer a etiologia das
doenças diarréicas em adultos é primordial, pois estes são considerados os potenciais
manipuladores de alimento. Um estudo buscando a etiologia das diarréias em adulto
foi realizado por SUZUKI (2007) onde o autor observou que na maioria dos casos de
doença diarréica aguda, atendidas nos serviços de emergência na cidade de Itapevi
(SP) de agosto a setembro de 2006, E. coli enterotoxigênica foi o único organismo
isolado das fezes dos pacientes.
Alguns dos queijos utilizados no presente estudo não tinham procedência, não
sendo possível identificar, nos produtos, dados sobre a origem e data de validade.
Outro fato a ser considerado é a exposição do produto nos pontos de venda e a
inadequação da manipulação principalmente devido à falta de condições higiênico
sanitárias nos pontos de comercialização. A prevenção depende de medidas sanitárias
que evitem a transmissão fecal-oral, através da lavagem das mãos antes de manipular
os alimentos, cloração de água para abastecimento público e tratamento e disposição
de esgoto (NATARO e KAPER, 1998; KAPER e col., 2004).
Neste estudo muitos isolados apresentaram a toxina termo-estável ST1
(Figura 5) e são associadas a cepas que causam os casos endêmicos (NATARO e
KAPER, 1998). Com menor freqüência foram observados isolados com a toxina

61
termo-lábil LT1 ambas podem causar diarréia aquosa ao consumidor do queijo
Minas. Em poucos isolados foram encontrados a toxina termolábil LT2 que também
causa diarréia aquosa e é associada a cepas isoladas de animais homeotérmicos e em
alimentos (STELLA e col., 2008). O veículo mais comum de infecção por ETEC é a
água e alimento contaminado tanto para os casos endêmicos quanto para a diarréia do
viajante (NATARO e KAPER, 1998).
Considerando a alta freqüência de isolados com fatores de virulência
observados no presente estudo que pertençam ao grupo filogenético A que pode ser
de linhagens patogênicas intestinais (PUPO e col., 1997) e ainda que Escherichia
coli enterotoxigênica é responsável por surtos endêmicos no Brasil, também que a
transmissão de EPEC e EAEC ocorre pela manipulação de alimentos por portadores
sintomáticos ou assintomáticos (NATARO e KAPER, 1998), os resultados obtidos
demonstram que grande parte dos isolados tem potencial para causar doença.
Estudos adicionais com mais fatores de virulência dos patotipos encontrados
são necessários para melhor caracterizar os isolados de ETEC, EPEC e EAEC, pois
os marcadores utilizados só revelam que estão presentes nos isolados e não
necessariamente que esses organismos conseguem causar doença no consumidor.
De qualquer forma, independente do patotipo caracterizado, o consumo de
queijo Minas pode representar um risco à saúde do consumidor final, sendo
necessário maior rigor na fiscalização realizada pela vigilância sanitária visando
coibir a venda de produtos sem procedência e uma higiênica manipulação do queijo e
também campanhas educativas que alcancem tanto consumidores como
comerciantes, para que tenham consciência dos perigos envolvidos desde a produção
até o consumo desse tipo de alimento.

62
6. CONCLUSÕES
• A caracterização dos isolados de E. coli provenientes do queijo Minas
revelou maior freqüência do grupo filogenético A, sendo que os grupos B2 e
D também foram identificados.
• Foi possível observar que o fator de virulência para o patotipo ETEC foi o
mais freqüente nas amostras analisadas.
• É possível concluir que o queijo Minas comercializado pode representar um
risco à saúde do consumidor, devido à presença dos diferentes grupos
filogenéticos e fatores de virulência dos enteropatotipos de Escherichia coli
observados.
• São necessárias ações de vigilância sanitária que definam estratégias de
prevenção e controle efetivas na disseminação de doença diarreica causada
pelos queijos Minas comercializados e principalmente os de origem
desconhecida.
• Divulgar por meio de campanhas educativas a população, aos produtores e
vendedores de queijo os riscos envolvidos pelo consumo de alimentos
produzidos e comercializados fora das normas vigentes que garantam a
inocuidade do produto.

63
7. REFERÊNCIAS
1. Afset JE, Andersen E, Bruant G, Harel J, Wieler L, Bergh K. Phylogenetic
background and virulence profile of atypical enteropathogenic Escherichia coli from
a case control study using multilocus sequence typing and DNA microarray. J Clin
Microbiol. 2008. 46: 2280-2290.
2. Amaral LA, Nader Filho A, Rossi Junior OD, Ferreira FLA, Barros, LSS. Água
de consumo humano como fator de risco à saúde em propriedades rurais. Rev Saude
Publica. 2003. 37: 510-514.
3. Aranda KRS, Fabbricotti SH, Fagundes Neto U, Scaletsky ICA. Single multiplex
assay to identify simultaneously enteropathogenic, enteroaggregative,
enterotoxigenic, enteroinvasive and Shiga toxin-producing Escherichia coli strains in
brazilian children. FEMS Microbiol Lett. 2007. 267: 145-150.
4. Aranda KRS, Fagundes Neto U, Scaletsky CA. Evaluation of multiplex PCR for
diagnosis of infection with diarrheagenic Escherichia coli and Shigella spp. J Clin
Microbiol. 2004. 42: 5849-5853.
5. Araujo VS, Pagliares VA, Queiroz MLP, Freitas-Almeida AC. Occurence of
Staphylococcus and enteropathogens in soft cheese commercialized in the city of Rio
de Janeiro, Brazil. J Appl Microbiol. 2002. 92: 1172-1177.
6. Ballal M; Ramamurty T. Enteroaggregative Escherichia coli diarrhea in Manipal.
Indian Pedriatr. 2005. 42: 722-723.

64
7. Barba J, Bustamante VH, Flores-Valdez MA, Deng W, Finlay BB, Puente JL. A
positive regulatory loop controls expression of the locus of enterocyte effacement-
encoded regulators Ler and GrlA. J. Bacteriol. 2005. 187: 7918-7930.
8. Baudry B. Savarino SJ, Vial P, Kaper JB, Levine MM. A sensitive and specific
DNA probe to identify enteroaggregative Escherichia coli, a recently discovered
diarrheal pathogen. J Infect Dis. 1990. 161: 1249-1251.
9. Borges MF, Feitosa T, Nassu RT, Muniz CR, Azevedo EHF, Figueiredo EAT.
Microrganismos patogênicos e indicadores em queijo de coalho produzido no estado
do Ceará, Brasil. Bol Centro Pesquis Process Alimen. 2003. 21: 31-40.
10. Brasil. Ministério da Saúde. Resolução 12 de 02/01/2001. Aprova o regulamento
técnico sobre os padrões microbiológicos para alimentos. Diário Oficial da União.
Brasília. 10 de janeiro de 2001. Seção I: 45-53.
11. Brito BG, Vidotto MC, Berbel MM, Tagliari KC. Fatores de virulência presentes
em amostras de Escherichia coli uropatogênicas – UPEC para suínos. Cienc Rural.
2004. 34: 645-652.
12. Cerna JF, Nataro JP, Garcia TE. Multiplex PCR for detection of three plasmid-
borne genes of enteroaggregative Escherichia coli strains. J Clin Microbiol. 2003.
41: 2138-2140.
13. Chapman PA, Ellin M, Ashton R, Shafique W. Comparison of culture, PCR and
immunoassays for detecting Escherichia coli O157 following enrichment culture and
immunomagnetic separation perfomed on naturally contaminated raw meat products.
Int J Food Microbiol. 2001. 68: 11-20.
14. Clermont O, Bonacorsi S, Bingen E. Rapid and Simple determination of the
Escherichia coli phylogenetic group. Appl Environ Microbiol. 2000. 10: 4555-4558.

65
15. Duriez P, Clermont O, Bonacorsi, Bingen E, Chaventre A, Elion J, Picard B,
Denamur E. Commensal Escherichia coli isolates are phylogenetically distributed
among geographically distinct human populations. Microbiology. 2001.147: 1671-
1676.
16. Elias Jr WP, Barros SF, Moreira CG, Trabulsi, Gomes TAT. Enteroaggregative
Escherichia coli strains among classical enteropathogenic Escherichia coli O
serogroups. J Clin Microbiol. 2002. 40: 3540-3541.
17. Forbes BA, Sahm DF, Weissfeld AS. Diagnostic Microbiology. Ed Mosby. 1998.
509-526.
18. Franzolin MR, Alves RCB, Keller R, Gomes TAT, Beutin L, Barreto ML, Milroy
C, Strina A, Ribeiro H, Trabulsi LR. Prevalence of diarrheagenic Escherichia coli in
children with diarrhea in Salvador, Bahia, Brazil. Mem Inst Oswaldo Cruz. 2005.
100: 359-363.
19. Girardeou JP, Dalmasso A, Bertin Y, Ducrot C, Bord S, Livrelli V, Vernozy-
Rozand C, Martin C. Association of virulence genotype with phylogenetic
background in comparasion to different seropathotypes of Shiga toxin-producing
Escherichia coli isolates. J Clin Microbiol. 2005. 43: 6098-6107.
20. Graziani C, Luzzi I, Corrò M, Tomai F, Parisi G, Giufrè M, Morabito S, Caprioli
A, Cerquetti M. Phylogenetic background and virulence genotype of ciprofloxacin-
susceptible and ciprofloxacin-resistant Escherichia coli strains of human and avian
origin. J Infect Dis. 2009. 199: 1209-1217.
21. Guth BEC. Enterotoxigenic Escherichia coli- An Overview. Mem Inst Oswaldo
Cruz. 2000. 95: 95-97.

66
22. Grünspan ED, Ulon SN, Santos AF, Herrmann GP, Shirmer VR.Contaminação
microbiana em carne moída de açougues da cidade de Santa Maria, RS, Brasil. Cienc
Rural. 1996. 26: 263-267.
23. Gyles CL. Shiga toxinproducing Escherichia coli: an overview. J Anim Sci.
2007. 85: 45-62.
24. Hanashiro A, Torres EAFS, Germano MIS, Germano PML. Avaliação da
comercialização de refeições prontas- bentôs- no bairro da Liberdade, São Paulo.
Revista Higiene Alimentar. 1999. 13: 19-31.
25. Hannah EL, Johnson JR, Angulo F, Haddadim B, Willamson J, Samore MH.
Molecular analysis of antimicrobial-Susceptible and –Resistant Escherichia coli
from retail meats and human stool and clinical specimens in a rural community
setting. Foodborn Pathog Dis. 2009. 6: 1-11.
26. Harrington SM, Dudley EG, Nataro JP. Pathogenesis of enteroaggregative
Escherichia coli infection. FEMS Microbiol Lett. 2006. 254: 12-18.
27. Herzer PJ, Inouye S, Inouye S, Whittam TS. Phylogenetic distribution of
branched RNA-linked multicopy single-stranded DNA among natural isolates of
Escherichia coli. J Bacteriol. 1990. 172: 6175-6181.
28. Holko I, Bisova T, Holkova Z, Kmet V. Virulence markers of Escherichia coli
strains isolated from traditional cheeses made from unpasteurised sheep milk in
Slovakia . Food Control. 2006.17: 393-396.
29. Jackson SG; Goodbrand RB; Johnson RP; Odorico VG; Alves D; Rahn K;
Wilson JB; Welch MK; Khakhria R. Escherichia coli O157:H7 diarrhoea associated
with well water and infected cattle on an Ontario farm. Epidemiol Infect.1998.120:
17-20.

67
30. Kaper JB. Defining EPEC. Rev Microbiol . 1996. 27: 130-133.
31. Kaper JB, Nataro JP e Mobley HLT. Pathogenic Escherichia coli. Microbiology.
2004. 2: 122-140.
32. Karch H, Tarr PI, Bielaszewska M. Enterohaemorragic Escherichia coli in
human medicine. Int J Med Microbiol. 2005. 295: 405-18.
33. Kong RYC, SO CL, Law WF, Wu RSS. A Sensitive and Versatile Multiplex
PCR System for the Rapid Detection of Enterotoxigenic (ETEC),
Enterohaemorrhagic (EHEC) and Enteropathogenic (EPEC) Strains of Escherichia
coli. Mar Pollut Bull. 1999. 38: 1207-1215.
34. Kotlowski R, Bernstein CN, Sepehri S, Krause DO. High prevalence of
Escherichia coli belonging to the B2+D phylogenetic group in inflammatory bowel
disease. Gut. 2006.(Acesso em 28/03/2009) Disponível http://gut.bmj.com
35. Leclerc H, Mossel DAA, Edberg SC, Strijk CB. Advances in the bacteriology of
the coliform group: their suitability as markers of microbial water safety. Annu Rev
Microbiol. 2001. 55: 201-234.
36. Maki DG. Don’t eat the spinach- controlling foodborne infectious disease. N
Engl J Med. 2006. 355: 1952-1955.
37. Marek P, Nair MKM, Hoagland T, Venkitanarayanan K. Survival and growth
characteristcs of Escherichia coli O157:H7 in pasteurized and unpasteurized Cheddar
cheese whey. Int J Food Microbiol. 2004. 94: 1-7.
38. Moreno E, Johnson JR, Perez T, Prats G, Kuskowski MA, Andreu A. Structure
and urovirulence characteristcs of the fecal Escherichia coli population among
healthy women. Microbes Infect. 2009. 11: 274-280.

68
39. Müller D, Greune L, Heusipp G, Karch H, Fruth A, Tschäpe H, Schimdt MA.
Identification of unconventional intestinal pathogenic Escherichia coli isolates
expressing intermediate virulence factor profiles by using a novel single step
multiplex PCR. App Environ Microbiol. 2007. 73: 3380-3390.
40. Nalério ES, Araújo MR, Vidor T, Fischer G, Silva WP. Identificação de
Escherichia coli enteropatogênica clássica através de padrão de adesão células HEp 2
e PCR. 2004. (Acesso 10 março 2009). Disponível em:
http://www.ufpel.edu.br/cic/2004/arquivos/CA_01222.doc
41. Nataro JP, Kaper JB. Diarrheagenic Escherichia coli. Clin Microbiol Rev. 1998.
1: 142-201.
42. Nataro JP, Kaper JB, Robins-Browne R, Prado V, Vial PA, Levine MM. Patterns
of adherence of diarrheagenic Escherichia coli to HEp-2 cells. Pediatr Infect Dis
J.1987. 6: 829-831.
43. Nataro JP, Steiner T, Guerrant RL. Enteroaggregative Escherichia coli. Emerg
Infect Dis.1998. 4: 251- 261.
44. Naylor SW, Gally DL, Low JC. Enterohaemorrhagic E. coli in veterinary
medicine. Int J Med Microbiol. 2005. 295: 419-441.
45. Nguyen TV, Van PL, Huy CL, Gia KN, Weintraub A. Detection and
characterization of diarrheagenic Escherichia coli from young children in Hanoi,
Vietnam. J Clin Microbiol. 2005. 43: 755-760.
46. Nishi J, Sheikh J, Mizuguchi K, Luisi B, Burland V, Boutin A, Rose DJ, Blattner
FR, Nataro JP. The export of coat protein from enteroaggregative Escherichia coli by
a specific ATP-binding cassete transporter system. J Biol Chem. 2003. 278: 45680-
45689.

69
47. Nowrouzian FL, Östblom AE, Wold AE, Adlerberth I. Phylogenetic group B2
Escherichia coli strains from the bowel microbiota of Pakistani infants carry few
virulence genes and lack the capacity for long-term persistence. Clin Microb Infec.
2009. (Acesso 8 março 2009) Disponível em:
http://www.ncbi.nlm.nih.gov/pubmed/19260873
48. Okeke IN, Nataro JP. Enteroaggregative Escherichia coli. Lancet Infect Dis.
2001. 1: 304- 311.
49. Orlandi PP, Magalhães GF, Matos NB, Silva T, Penatti M, Nogueira PA, Silva
LHP. Etiology of diarrheal infections in children of Porto Velho (Rondonia, Weatern
Amazon region, Brazil). Braz J Med Biol Res. 2006. 39: 507-517.
50. Oundo JO, Kariuki SM, Boga HI, Muli FW, Iijima Y. High Incidence of
enteroaggregative Escherichia coli among food handlers in three areas of Kenya: a
possible transmission route of travelers diarrhea. J Travel Med. 2008. 15: 31-38.
51. Paneto BR, Schocken I, Macedo C, Santo E, Marin JM. Ocorrência de
Escherichia coli toxigênica em queijo-de-minas frescal no Brasil. Arq Bras Med Vet
Zootec. 2007. 59: 508-512.
52. Pass MA, Odedra R, Batt RM. Multiplex PCR for identification of Escherichia
coli virulence genes. J Clin Microbiol. 2000. 38: 2001-2004.
53. Pedro SCM. Avaliação da qualidade higiênico sanitária dos locais de venda e de
amostras de queijo Minas meia cura, comercializado na cidade de São Paulo.
[Dissertação de Mestrado]. São Paulo. Faculdade de Saúde Pública da Universidade
de São Paulo. 2003.

70
54. Pedro SCM, Matté GR, Germano PML, Matté MH. Aspectos sanitários da
comercialização de queijo minas meia cura no Município de São Paulo. Revista
Brasileira de Vigilância Sanitária. 2005.1: 246-253.
55. Pereira ACM, Brito-Filho JD, Carvalho JJ, Luna MG, Rosa ACP.
Enteroaggregative Escherichia coli (EAEC) strains enter and survive within cultured
intestinal epithelial cells. Microb Pathog. 2008. 45: 310-314.
56. Perelle S, Dilasser F, Grout J, Fach P. Screening food raw materials for the
presence of the world's most frequent clinical cases of Shiga toxin-encoding
Escherichia coli O26, O103, O111, O145 and O157. Int J Food Microbiol. 2007. 15:
284-288.
57. Peresi JTM, Graciano RAS, Almeida IAZC, Lima SI, Ribeiro AK, Carvalho IS.
Queijo Minas tipo Frescal artesanal e industrial: qualidade microscópica,
microbiológica e teste de sensibilidade aos agentes antimicrobianos. Revista Higiene
Alimentar. 2001. 15: 63-70.
58. Pupo GM, Karaolis DK, Lan R, Reeves PR. Evolutionary relationships among
pathogenic and nonpatogenic Escherichia coli strains inferred from multilocus
enzyme electroforesis and mdh sequence studies. Infect Immun. 1997. 65: 2685-
2692.
59. Quintana RC, Carneiro LC. Avaliação das condições higiênico sanitárias dos
queijos Minas frescal e mussarela produzidos na cidade de Morrinhos, GO. Rev
Bras Saúde Prod An. 2007. 8 :205-211.
60. Rabelo JA, Moulin MRI, Squilassi KMBS, Souza WM, Vieira MCM, Souza CM.
Qualidade microbiológica de queijo frescal comercializado no Estado de Goiás no
período de junho a dezembro de 1999. Revista Higiene Alimentar. 2001. 15: 119.

71
61. Regua- Mangia AH, Gomes TAT, Vieira MAM, Andrade JRC, Irino K, Teixeira
LM. Frequency and characteristics of diarrhoeagenic Escherichia coli strains isolated
from children with and without diarrhoea in Rio de Janeiro, Brazil. J Infect. 2004.
48:161-167.
62. Rodas C, Iniguez V, Qadri F, Wiklund G, Svennerholm AM, Sjöling.
Development of multiplex PCR assays for detection of enterotoxigenic Escherichia
coli (ETEC) colonization factor and toxins. J Clin Microbiol. 2009. 47: 1218-1220.
63. Rodrigues J, Thomazini CM, Morelli A, Batista GCM. Reduced etiological role
enteropathogenic Escherichia coli in case of diarrhea in brazilian infants. J Clin
Microbiol. 2004. 42: 398-400.
64. Rodriguez GA. Principales caracteristicas y diagnosticos de los grupos patogenos
de Escherichia coli. Salud Publica Mex. 2002. 44: 464-475.
65. Ruppenthal RD. Aplicação da reação em cadeia da polimerase (PCR) no
diagnóstico clínico. (Monografia). UFRGS. 1999. (acesso: 27 junho 2009)
Disponível : http://www6.ufrgs.br/bioquimica/
66. Sabaté M, Prats G, Moreno E, Ballesté E, Blanch AR, Andreu A .Virulence and
antimicrobial resistence profiles among Escherichia coli strains isolated from human
and animal wastewater. Res Microbiol. 2008. 159: 288-293.
67. Salotti BM, Carvalho ACFB, Amaral LA, Vidal-Martins AMC, Cortez AI.
Qualidade microbiológica do queijo Minas frescal comercializado no município de
Jaboticabal, SP, Brasil. Arq Inst Biol. 2006. 73: 171-175.
68. Sarantuya J, Nishi J, Wakimoto N, Erdene S, Nataro JP, Sheikh J, Iwashita M,
Manago K, Tokuda K, Yoshinaga M, Miyata K, Kawano Y. Typical

72
Enteroaggregative Escherichia coli is most prevalent pathotipe among E. coli strain
causing diarrhea in Mongolian children. J Clin Microbiol. 2004. 42: 133-139.
69. Scavia G, Staffolani M, Fisichella S, Striano G, Colletta S, Ferri G, Escher M,
Minelli F e Caprioli A. Enteroaggregative Escherichia coli associated with a
foodborne outbreak of gastroenteritis. J Med Microbiol. 2008. 57: 1141-1146.
70. Schimdt H, Hensel M. Pathogenicity island in bacterial pathogenesis. Clin
Microbiol Rev. 2004. 17: 14-56.
71. Schutzbank TE, Stern HJ. Princípios e aplicações da reação da cadeia de
polimerase. Rev Bras Anal Clin. 1993. 25: 112-118.
72. Selander RK, Caugant DA, Ochman H, Musser M, Gilmour MN , Whittam TS.
Methods of multilocus enzyme eletroforesis for bacterial population genetics and
systematics. Appl Environ Microbiol. 1986. 51: 873-884.
73. Silva N, Junqueira VCA, Silveira NFA. Manual de métodos de análise
microbiológica de alimentos. 2. ed. São Paulo. Editora Varela. 2001.
74. Silva JV, Hoffmann FL, Mansor AP, Coelho ARV, Maria T. Estudo higiênico
sanitário de queijos tipo Minas frescal fabricados artesanalmente. Revista Higiene
Alimentar. 2001. 15: 97.
75. Skyberg JA, Siek KE, Doetkot C e Nolan LK. Biofilm formation by avian
Escherichia coli in relation to media, source and phylogeny. J App Microbiol. 2007.
102: 548-554.
76. Stella AE, Rigobelo EC, Oliveira AC, Maluta RP, Marin JM, Ávila FA.
Características genotípicas de Escherichia coli shigatoxigênicas isoladas de água,
leite e fezes de bovinos da região de Ribeirão Preto-SP. 2008. (acesso 20 junho
2009).Disponível: www.sovergs.com.br/conbravet2008/anais/cd/.../R0200-1.pdf

73
77. Stephan R, Schumacher S, Corti S, Krause G, Danuser J e Beutin L. Prevalence
and characteristics of Shiga toxin- producing Escherichia coli in Swiss raw milk
cheeses collected at producer level. J Dairy Sci. 2008. 91: 2561-2565.
78. Suzuki E. Etiologia das diarréias dos adultos atendidos nos pronto-socorros do
município de Itapevi. [Dissertação de Mestrado]. São Paulo. Faculdade de Saúde
Pública da USP. 2007.
79. Tauschek M, Gorell RJ, Strugnell RA, Browne RMR. Identification of a protein
secretory pathway for the secretion of heat-labile enterotoxin by an enterotoxigenic
strain of Escherichia coli. Proc Nat Acad Sci USA. 2002. 99: 7066-7071.
80. Tiba MR, Nogueira GP, Leite DS. Estudo dos fatores de virulência associados à
formação de biofilme e agrupamento filogenético em Escherichia coli isoladas de
pacientes com cistite. Rev Soc Med Trop. 2009. 42: 58-62.
81. Todar K. Pathogenic E. coli. Todar's Online Textbook of Bacteriology.
University of Wisconsin-Madison Department of Bacteriology. 2002. (Acesso 16
outubro 2008). Disponível em: http://www.textbookofbacteriology.net/
82. Toma C, Lu Y, Higa N, Nakasone N, Chinen I, Baschkier A, Rivas M, Iwanaga
M. Multiplex PCR assay for identification of human diarrheagenic Escherichia coli.
J Clin Microbiol. 2003. 41: 2669-2671.
83. Torres AG, Payne SM. Haem iron- transport system in enterohaemorrhagic
Escherichia coli 0157:H7. Mol Microbiol. 1997. 23: 825-833.
84. Torres AG, Zhou X, Kaper JB, Adherence of diarrheaagenic Escherichia coli
strains to epithelial cells. Infect Immun. 2005. 73: 18-29.
85. Tortora GJ, Funke BR, Case CL. Microbiologia. 8. ed. Rio de Janeiro. Artmed,
2005.

74
86. Trabulsi LR, Altherthum F, Goompertz OF, Candeias JAN. Microbiologia. 4º ed.
Rio de Janeiro. Editora Atheneu. 2004.
87. Timm CD, Conceição FR, Menin A, Conceição RCS, Dellagostin AO, Aleixo
JAG. Prevalence of Shiga –toxin producing Escherichia coli in Southern Brazil
isolated from ground beef and raw milk. Ciência Animal Brasileira. 2009. 10: 641-
649.
88. Vaz TMI, Irino K, Kato MAMF, Dias AMG, Gomes TAT, Medeiros MIC,
Rocha MMM, Guth BEC. Virulence properties and characteristics of Shiga toxin-
producing Escherichia coli in São Paulo, Brazil, from 1976 throug 1999. J Clin
Microbiol. 2004. 42: 903-905.
89. Vicente ACP, Teixeira LFM, Iniguez Rojas L, Luna MG, Silva L, Andrade JRC,
Guth BEC. Outbreaks of cholera-like diarrhea caused by enterotoxigenic Esche-
richia coli in the Brazilian Amazon rainforest. J Trop Med Hyg. 2005. 99: 669-674.
90. Vidal M, Kruger E, Duran C, Lagos R, Levine M, Prado V, Toro C, Vidal R.
Single multiplex PCR assay to identify simultaneously the six categories of
diarrheagenic Escherichia coli associated with enterics infections. J Clin Microbiol.
2005. 43: 5362-5365.
91. Walk ST, Alm EW, Calhoun LM, Mladonicky JM, Whittam TS. Genetic
diversity and population structure of Escherichia coli isolated from freshwater
beaches. Environ Microbiol. 2007. 9: 2274-2288.
92. Yamamoto T, Echeverria P. Detection of the enteroaggregative Escherichia coli
heat-stable enterotoxin 1 gene sequences in enterotoxigenic E. coli strain pathogenic
for humans. Infect Immun. 1996. 64: 1441-1445.


















