
UNIVERSIDADE FEDERAL DA PARAÍBA UNIVERSIDADE...
140
UNIVERSIDADE FEDERAL DA PARAÍBA UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO UNIVERSIDADE FEDERAL DO CEARÁ PROGRAMA DE DOUTORADO INTEGRADO EM ZOOTECNIA DISTRIBUIÇÃO ESPACIAL DE SERAPILHEIRA E CÍBALAS EM ÁREAS DE CAATINGA PASTEJADAS POR CAPRINOS MARIAH TENORIO DE CARVALHO SOUZA AREIA - PB 2015
-
Upload
nguyentram -
Category
Documents
-
view
214 -
download
0
Transcript of UNIVERSIDADE FEDERAL DA PARAÍBA UNIVERSIDADE...

UNIVERSIDADE FEDERAL DA PARAÍBA
UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO
UNIVERSIDADE FEDERAL DO CEARÁ
PROGRAMA DE DOUTORADO INTEGRADO EM ZOOTECNIA
DISTRIBUIÇÃO ESPACIAL DE SERAPILHEIRA E CÍBALAS EM
ÁREAS DE CAATINGA PASTEJADAS POR CAPRINOS
MARIAH TENORIO DE CARVALHO SOUZA
AREIA - PB
2015

UNIVERSIDADE FEDERAL DA PARAÍBA
UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO
UNIVERSIDADE FEDERAL DO CEARÁ
PROGRAMA DE DOUTORADO INTEGRADO EM ZOOTECNIA
DISTRIBUIÇÃO ESPACIAL DE SERAPILHEIRA E CÍBALAS EM
ÁREAS DE CAATINGA PASTEJADAS POR CAPRINOS
MARIAH TENORIO DE CARVALHO SOUZA
Zootecnista
AREIA - PB
2015

MARIAH TENORIO DE CARVALHO SOUZA
DISTRIBUIÇÃO ESPACIAL DE SERAPILHEIRA E
CÍBALAS EM ÁREAS DE CAATINGA PASTEJADAS POR
CAPRINOS
Tese apresentada ao Programa de
Doutorado Integrado em Zootecnia, da
Universidade Federal da Paraíba, do qual
participa a Universidade Federal Rural
de Pernambuco e a Universidade Federal
do Ceará, como requisito para obtenção
do título de Doutor em Zootecnia.
Área de Concentração: Forragicultura
Comitê de Orientação:
Prof. Dr. Alberício Pereira de Andrade - Orientador Principal
Prof. Dr. Divan Soares da Silva
AREIA - PB
2015

ADE FEDERAL DA PARAÍBA
PROGRAMA DE PÓS-GRADUAÇÃO EM ZOOTECNIA
PARECER DE FEDESA DO TRABALHO DE TESE
TÍTULO: “Distribuição espacial da serapilheira e cíbalas em áreas de caatinga
pastejada por caprinos”
AUTOR: MARIAH TENORIO DE CARVALHO SOUZA
ORIENTADOR: ALBERICIO PEREIRA DE ANDRADE
JULGAMENTO
CONCEITO:
EXAMINADORES:

5
DADOS CURRICULARES DO AUTOR
Mariah Tenório de Carvalho Souza, nasceu em Maceió, Alagoas em 23 de
fevereiro de 1987. Em dezembro de 2004 concluiu o 2º grau no Colégio Contato em
Maceió/AL. Colou grau do curso de Zootecnia em fevereiro de 2009 pelo Centro de
Ciências Agrárias da Universidade Federal de Alagoas, desenvolvendo pesquisa na área
de conservação de forragem com apoio da Embrapa CPATC – Tabuleiros Costeiros. Em
Março do mesmo ano ingressou no curso de pós-graduação em nível de Mestrado na
área de Produção Animal e subárea de forragicultura, especializando-se em cultivares
forrageiras tropicais, com apoio da Fundação de Amparo à Pesquisa do Estado de Alagoas -
FAPEAL, submetendo-se a defesa em dezembro de 2010. Ingressou no Programa de
Doutorado Integrado em Zootecnia em Março de 2011 e desenvolveu pesquisa em áreas
de caatinga, especializando-se mais uma vez em forragicultura com apoio financeiro do
Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq.

6
Epígrafe
"A vida é feita de capítulos e viver é acalentar sonhos e esperanças, fazendo da fé a nossa inspiração
maior. É buscar nas pequenas coisas, um grande motivo para ser feliz!"
(Adaptado de Mário Quintana)

7
Dedico.
Aos meus pais,
Pelo conforto, carinho, compreensão, pela
maravilhosa educação que recebi e recebo. Pelo amor
incondicional, pela atenção e dedicação de toda uma vida,
mesmo nos momentos de dificuldades. Pelo exemplo de
coragem, simplicidade e persistência em suas metas. É com
muito amor que dedico esta conquista.
Ao meu marido,
Márcio Eduardo Freire Silva.
Nem todas as palavras do mundo poderiam expressar a minha gratidão e o meu amor!
Por muitas vezes ter sentido na pele as minhas
adversidades. Por ser a certeza nos meus momentos de
dúvida; por ser meu alicerce, meu suporte. Por mostrar que
juntos, conseguimos ser um só coração e por fazer de um
grande amor uma linda realidade.
Que Deus retribua em dobro todo o carinho e
toda a dedicação. E que o fruto do nosso amor seja
concebido brevemente!
Com muito amor e gratidão,
Dedico.
Ofereço
Aos queridos colegas Zootecnistas e a essa
profissão tão árdua e nobre que tanto enriquece o nosso país.
Em especial à amiga Laíza Acioli (In memorian).

8
AGRADECIMENTOS
A Deus.
Por ter me proporcionado esta oportunidade incrível e inesquecível. Por me
proporcionar forças nas adversidades da vida.
A Universidade Federal da Paraíba, por ter me concedido a oportunidade de
ingressar no curso de Doutorado Integrado em Zootecnia.
A Capes, por ter auxiliado com bolsa de pós-graduação.
Aos meus Professores queridos, a quem tenho grande admiração e por servirem
de exemplo profissional, nos nomes: Prof. Ariosvaldo Nunes de Medeiros, Prof. Divan
Soares da Silva, Prof. Edilson Paiva, Prof. Edson Mauro, Prof. Ivandro de França,
Profa. Riselane Alcântara, Prof. Severino Gonzaga Neto, Prof. Walter Esfrain, Prof.
José Morais Pereira Filho, Prof. Jacob Silva Souto, Profa. Dulciene Karla de Andrade
Silva, Prof. Andé Luiz Rodrigues Magalhães e todos os que de alguma forma
participaram para com o meu aprendizado. Aos meus colegas e Professores José
Teodorico de Araújo e Prof. Elton Lima Santos. Agradeço pela compreensão, orientação
e dedicação; principalmente ao meu orientador Prof. Dr. Alberício Pereira de Andrade,
por ter acreditado na minha capacidade intelectual e por ter me proporcionado um
grande crescimento profissional; agradeço, além de tudo, pela amizade.
Aos servidores/trabalhadores da UFPB e da Estação Experimental de São João
do Cariri, que sempre estiveram presentes, servindo aos alunos e à instituição, em
especial a Marciane, Netinho, Paulo Henrique, Alessandro, Graça Medeiros, Sra.
Carmen e Sr. Damião.
Aos trabalhadores de campo, que me ajudaram e trabalharam incansavelmente
em tempo hábil e com toda a dedicação nas áreas de caatinga; meus agradecimentos,
respeito e admiração: Rogério, Cláudio, Neguinho, Darlan, entre outros.
Aos colegas que me ajudaram na execução deste projeto. Sem eles eu não teria
conseguido enfrentar nem a metade do caminho; em especial a Michel Lopes, Dinnara
Laiza, Laiza Sofia (in memorian), Paula Frassinetti, Meiry Cassuce, Adelílian Baracho,
Adeilson Melo, Paulo Sérgio, Mariana Maciel, entre outros.

9
Ao meu marido, Eduardo Freire, que além de tudo segurou a enchada na mão e
entrou bravamente nas áreas de caatinga para me ajudar quando mais precisei.
A minha mãe e melhor amiga, Vera Lúcia Tenorio de Carvalho pela positividade
mesmo depois de tantas dificuldades e depois de passar por um grave problema de
saúde. Por sempre ter depositado esperanças em mim; pela fé de que um dia vou
conseguir realizar o meu sonho profissional. Enfim, por todo amor e dedicação.
Ao meu pai e amigo Onaldo Souza, doutor e zootecnista; que apesar de
aposentado, ainda me veste de conhecimentos. Agradeço por estar sempre presente.
Enfim, a todos que de alguma forma me apoiaram na execução desse projeto e
contribuíram para a finalização deste trabalho.

10
SUMÁRIO
Página
Considerações iniciais..................................................................................... 1
CAPÍTULO 1 - Referencial teórico................................................................ 3
Introdução........................................................................................................ 4
1. CARACTERIZAÇÃO DO SEMIÁRIDO NORDESTINO: CLIMA E
SOLO..............................................................................................................
6
2. A IMPORTÂNCIA DA SERAPILHEIRA PARA REGIÕES
SEMIÁRIDAS: PRODUÇÃO, ACÚMULO E DECOMPOSIÇÃO..............
8
3. SAZONALIDADE DA DISPONIBILIDADE DE FORRAGEM
NO SEMIÁRIDO............................................................................................
13
4. COMPOSIÇÃO QUÍMICA DA FORRAGEM DISPONÍVEL
PARA PEQUENOS RUMINANTES EM ÁREAS DE
CAATINGA...................................................................................................
16
5. ENTRADA E SAÍDA DE NUTRIENTES NO SISTEMA................ 18
6. DISTRIBUIÇÃO ESPACIAL E ESTATÍSTICA
GEORREFERENCIADA: ESTUDOS E APLICAÇÃO PRÁTICA NAS
CIÊNCIAS AGRÁRIAS.................................................................................
20
7. REFERÊNCIAS BIBLIOGRÁFICAS................................................ 26
CAPÍTULO 2 - Distribuição espacial da serapilheira em áreas de caatinga
sob diferentes lotação de caprinos...................................................................
33
1 Introdução..................................................................................................... 35
2 Material e métodos....................................................................................... 36
3 Resultados e discussão................................................................................. 47
4 Conclusões................................................................................................... 73
5 Referências................................................................................................... 74
CAPÍTULO 3 - Acúmulo da serapilheira disponível para o pastejo caprino
em áreas de caatinga
76
1 Introdução..................................................................................................... 79
2 Material e métodos....................................................................................... 80
3 Resultados e discussão................................................................................. 89
4 Conclusões................................................................................................... 118
5 Referências................................................................................................... 118
CONSIDERAÇÕES FINAIS.......................................................................... 121

11
LISTA DE TABELAS
Página
Tabela 1 – Disponibilidade dos estratos arbóreo, arbustivo e sub-arbustivo em
áreas de caatinga, em diferentes Estados e os respectivos
autores................................................................................................................
14
Tabela 2 - Composição bromatológica, em percentagem, de herbáceas
dicotiledôneas e do capim panasco encontrada por diversos autores em áreas
de caatinga.........................................................................................................
17
Tabela 3. Composição química do pool da serapilheira em áreas de caatinga
no cariri paraibano............................................................................................
67
Tabela 4. Composição química do pool da serapiheira e das cíbalas de
caprinos SPRD em sistema de criação semiestensivo em áreas de
caatinga...............................................................................................................
70
Tabela 5. Relação C:N e C:P da serapilheira e cíbalas em áreas de
caatinga................................................................................................................
71
Tabela 6. Composição química e fertilidade do solo em áreas de
caatinga.................................................................................................................
72
Tabela 7. Estoque de serapilheira em nove áreas de caatinga no Cariri
paraibano para o pastejo caprino..........................................................................
91
Tabela 8. Espécies de inclusão por parcela em diferentes áreas de
caatinga................................................................................................................
93
Tabela 9. Número de famílias, gêneros, espécies e indivíduos ocorrentes nas
áreas 1, 2, 3, 4 e 5 em São João do Cariri – PB...................................................
94
Tabela 10. Relação das famílias encontradas e frequências de indivíduos nas
cinco áreas de estudo...........................................................................................
96
Tabela 11. Matriz presença/ausência, das espécies amostradas.......................... 97
Tabela 12. Espécies e frequências de indivíduos encontrados nas cinco áreas
de caatinga............................................................................................................
98
Tabela 13. Índice de equabilidade de Pielou (J’)................................................. 99
Tabela 14. Número de parcela (NP), área total amostrada em hectare (ATA),
Número de indivíduos encontrados (NIE) e Número de indivíduos por hectare
(NIH)....................................................................................................................
99
Tabela 15. Parâmetros fitossociológicos das espécies amostradas em cinco
áreas de caatinga...................................................................................................
101
Tabela 16. Abrangência de P. pyramidalis nos mapas em isolinhas avaliados
nas áreas de caatinga Índice de diversidade de Shannon H’................................
103
Tabela 17. Índice de dominância de Simson (C’) ............................................... 104
Tabela 18. Matriz de similaridade florística (Jaccard) em cinco áreas de
caatinga................................................................................................................
104
Tabela 19.Classificação do padrão de distribuição das espécies, segundo o
Índice de MacGuinnes (IGA)...............................................................................
106
Tabela 20. Abrangência de P. pyramidalis nos mapas em isolinhas avaliados
nas áreas de caatinga............................................................................................
117

12
LISTA DE FIGURAS
Página
Figura 1 - Sistema de resposta aos pulsos de entrada: (a) amplamente
separados, (b) agrupados, (c), com espaçamento intermediário..........................
7
Figura 2. Variável aleatória regionalizada Z(X)............................................................................ 21
Figura 3. Semivariograma experimental e modelo matemático
ajustado.................................................................................................................
23
Figura 4. Modelos e componentes do variograma: a) Esférico; b) Linear; c)
Exponencioal; d) Gausiano...................................................................................
24
Figura 5. Representação das cinco áreas experimentais. Em destaque, na cor
vermelha, a subdivisão das parcelas contíguas.....................................................
37
Figura 6. Fotos representativas das áreas experimentais com presença de
lajedo (a); fragmentos de caatinga hiperxerófila e solo parcialmente descoberto
(b); presença de animais em local de repouso, evidenciando a presença de
cíbalas no solo; e folhas compondo a serapilheira (d)..........................................
38
Figura 7. Croqui de distribuição das parcelas nos fragmentos de caatinga
estudados para levantamento dos dados, coleta de dejetos caprinos e
serapilheira............................................................................................................
39
Figura 8. Esquematização feita em programa de GPS dos pontos centrais
marcados nas áreas experimentais........................................................................
40
Figura 9. Fotos demonstrativas, identificando subparcelas da área 1 com
presença de lajedo (a), vegetação hiperxerófila (b) e solo descoberto (c)...........
40
Figura 10. Demonstração do molde de ferro quadrado........................................ 42
Figura 11. Balança de precisão 1g utilizada no experimento............................... 42
Figura 12. Pesagem da serapilheira alocada em sacos plásticos.......................... 43
Figura 13. Fracionamento dos constituintes da serapilheira em material
lignificado (a), folhas (b), miscelânea (c), estrutura reprodutiva (d) e cíbalas
(e)..........................................................................................................................
44
Figura 14. Amostras em laboratório da serapilheira............................................ 46
Figura 15. Demonstração das cíbalas depositadas em área experimental (a), do
estádio de conservação na época de coleta (b) e demonstração das baias com
caprinos (c)...........................................................................................................
47
Figura16. Dados da precipitação (mm) e temperatura (ºC) de 2011 a 2014........ 48
Figura 17. Gráfico em boxplot demonstrando médias e desvios da produção da
serapilheira nas quatro áreas de caatinga no ano base de 2012............................
49
Figura 18. Gráfico em boxplot com medianas e quartis do fracionamento da
serapilheira em miscelânea, folhas, pool do material lenhoso e cíbalas em
áreas de caatinga...................................................................................................
50
Figura 19. Gráfico em boxplot com médias e desvios do fracionamento da
serapilheira em miscelânea, folhas, pool do material lenhoso e cíbalas em
áreas de caatinga...................................................................................................
51
Figura 20. Fracionamento dos constituintes da serapilheira em cinco áreas de
caatinga. As linhas acima das barras representam o desvio padrão da média......
52
Figura 21. Gráfico em boxplot demonstrando a variação da produção da
serapilheira nas quatro áreas de caatinga no ano base 2014...............................
53
Figura 22. Semivariograma do peso da serapilheira total (2012)......................... 54

13
Figura 23. Isolinhas da distribuição do pool da serapilheira (g.m-²) em quatro
áreas de caatinga no ano de
2012.......................................................................................
55
Figura 24. Isolinhas de folhas (g.m-2
) no período seco em áreas de caatinga no
ano de 2012...........................................................................................................
58
Figura 25. Isolinhas da miscelânea (g.m-2
) no período seco em áreas de
caatinga no ano de 2012.......................................................................................
60
Figura 26. Isolinhas do pool do material lenhoso (g.m-2
) no período seco em
áreas de caatinga no ano de 2012........................................................................
61
Figura 27. Isolinhas das cíbalas (g.m-2
) no período seco na área 2 no ano base
de 2012. ....................................... ....................................... ...............................
62
Figura 28. Semivariograma do peso da serapilheira total no ano base de 2014... 63
Figura 29. Isolinhas do pool da serapilheira em áreas de caatinga no período
chuvoso (ano base 2014). ...................................................................................
65
Figura 30. Conteúdo de água presente na serapilheira e nas cíbalas em áreas de
caatinga no cariri paraibano................................................................................
69
Figura 31. Conteúdo de água das amostras de serapilheira em cinco áreas de
caatinga...............................................................................................................
70
Figura 32. Representação das nove áreas experimentais. Em destaque, na cor
azul, a subdivisão das parcelas de 1m² em três transectos por área.....................
82
Figura 33. Demonstração da demarcação das parcelas em área alagada (a);
com solo descoberto(b); da luva utilizada para colheita da serapilheira (c); e
pesagem do material (d).......................................................................................
83
Figura 34. Dados da precipitação pluvial e temperatura no ano base de 2013..... 89
Figura 35. Gráfico boxplot demonstrando a disponibilidade da serapilheira em
áreas de caatinga no ano base 2013. ....................................................................
90
Figura 36. Curva do coletor para as áreas cinco áreas em estudo, com o
número de espécies registradas em uma área acumulada de 15.000 m².............
92
Figura 37. Número das famílias amostradas nas áreas 1,2,3,4 e 5 em São João
do Cariri - PB. .....................................................................................................
95
Figura 38. Distribuição em classes de altura das espécies amostradas em cinco
áreas de caatinga. ................................................................................................
107
Figura 39. Distribuição em classes de diâmetro das espécies amostradas em
cinco áreas de caatinga. .......................................................................................
109
Figura 40. Gráficos de semivariograma das áreas interpoladas para o acúmulo
de serapilheira......................................................................................................
111
Figura 41. Isolinhas da distribuição de serapilheira em parcelas contíguas no
ano base de 2013. ..............................................................................................
113
Figura 42. Represenntação dos semivariogramas para a espécie P. pyramidalis. 115
Figura 43. Variabilidade espacial de P. pyramidalis em fragmentos de
caatinga. ...............................................................................................................
116

14
LISTA DE QUADROS
Página
Quadro 1. Comparação entre métodos de interpolação. Fonte: Krajewski e
Gibbs (1966)........................................................................................................
25
Quadro 2. Valores de efeito pepita, patamar, alcance e dependência espacial
dos modelos de semivariograma..........................................................................
55
Quadro 3. Valores de efeito pepita, patamar, alcance e dependência espacial
dos modelos de semivariograma..........................................................................
64
Quadro 4. Valores de efeito pepita, patamar, alcance e dependência espacial
segundo os modelos dos semivariogramas..........................................................
112
Quadro 5. Valores de efeito pepita, patamar, alcance e dependência espacial
segundo os modelos dos semivariogramas..........................................................
116

15
SOUZA, M. T. C. Distribuição espacial da serapilheira e cíbalas em áreas de
caatinga pastejada por caprinos. 2015. f. Tese (Doutorado em Zootecnia).
Universidade Federal da Paraíba, Centro de Ciências Agrárias, Areia – PB.
RESUMO GERAL
Objetivou-se avaliar a distribuição espacial da deposição da serapilheira e de
cíbalas em áreas de caatinga pastajadas por caprinos em período prolongado de seca e
período chuvoso com diferentes taxas de lotação animal. O experimento foi conduzido
na Estação Experimental pertencente a UFPB, localizada no município de São João do
Cariri-PB, Brasil. A área experimental representou, aproximadamente, 5 hectares, dos
quais foram subdivididos em cinco áreas (1, 2, 3, 4 e 5), de 3,3 hectares cada, com
diferentes lotações de animais e diferentes estados de antropização. O método utilizado
para a análise dos dados foi o de parcelas contíguas em áreas de um hectare e áreas de
3,3ha plotadas em transectos de 1m². A serapilheira acumulada na superfície do solo foi
estimada através de uma colheita na estação seca. Para a quantificação desse material
foi utilizado um molde de estrutura férrea, vazado, medindo 0,25m², lançado
aleatoriamente em cada subparcela cinco vezes, cujo material foi colhido e pesado para
obtenção da disponibilidade em g. m-². Os mapas de contorno das variáveis estudadas
foram construídos por interpolação utilizando o método da função da krigagem, por ser
considerado o melhor estimador linear não tendencioso. A amplitude da variação da
produção de serapilheira foi alta por tratar-se de um experimento a campo. Assim, o
valor máximo da área 4 foi de pouco mais de 400 g.m-2
. As áreas com maior taxa de
lotação animal demonstram maior desaparecimento da serapilheira. É provável que os
animais passem a maior parte do tempo alimentando-se de folhas do estrato arbustivo e
arbóreo no período chuvoso. Ainda, a deposição das cíbalas se deve ao acúmulo desse
material no período das águas.
Palavras-chave: Brasil, fezes, fonte alimentar, liteira, pequenos ruminantes, semiárido

16
SOUZA, M. T. C. . Distribuição espacial da serapilheira e cíbalas em áreas de
caatinga pastejada por caprinos. 2015. f. Tese (Doutorado em Zootecnia).
Universidade Federal da Paraíba, Centro de Ciências Agrárias, Areia – PB.
ABSTRACT
Aimed to evaluate the spatial distribution of deposition of litter and cíbalas in
areas of savanna pastajadas by goats in prolonged period of dry and rainy season with
different stocking rates. The experiment was conducted at the Experimental Station
belonging to UFPB, located in São João do Cariri-PB, Brazil. The experimental area
accounted for approximately 5 hectares of which were divided into five areas (1, 2, 3, 4
and 5) 3.3 hectares each, with different manning of animals and different states of
human disturbance. The method used for the analysis was the contiguous plots in areas
of one hectare and areas of 3,3ha plotted on transects of 1m². The accumulated litter on
the soil surface was estimated from a harvest in the dry season. To quantify this material
we used a cast iron structure, hollow, measuring 0,25m², released at random on each
subplot five times, whose material was collected and weighed to obtain the availability
in g. m ². The contour maps of the studied variables were constructed by interpolation
using the method of kriging function, being considered the best non-biased linear
estimator. The magnitude of the variation of litter production was high because it is an
experiment in the field. Thus, the maximum value of the area 4 was just over 400 gm-2.
The areas with higher stocking rate showed higher disappearance of litter. It is likely
that the animals spend most of their time feeding on leaves of the shrub and tree layer in
the rainy season. Moreover, the deposition of cíbalas is due to the accumulation of this
material in the rainy season.
Keywords: Brazil, feces, food source, litter, small ruminants, semiarid

1
CONSIDERAÇÕES INICIAIS
É fato que, há décadas, muitos pesquisadores e cientistas vêm interpretando
erroneamente sobre questões ambientais em áreas de caatinga, repercutindo muitas
vezes sobre um “clima adverso” e sobre uma “pobreza em recursos ambientais” nestas
áreas.
Sabe-se que o clima na região semiárida apresenta como característica marcante
a irregularidade do regime pluviométrico, com duas estações definidas: a estação
chuvosa, que corresponde a períodos com ocorrência de chuvas, distribuídas de forma
irregular, e a estação seca, correspondendo a períodos críticos de estiagem com baixa
oferta de volumosos para os animais criados a pasto. Mas, isto não impede que a
produção animal seja bem sucedida em ambientes semiáridos com áreas de caatinga
nativa.
Além disso, muitos produtores rurais sobrevivem e sustentam os seus familiares
com a produção de pequenos ruminantes criados de forma extensiva nestas áreas,
muitas estas de sucessão, antropizadas ou desmatadas para algum tipo de produção
agrícola em um passado próximo. Vale ressaltar que a vegetação predominante dessas
áreas são resilientes e oportunistas; plantas caducifólias espinhosas com uma ótima
capacidade de rebrota. Ainda, a produção da serapilheira proveniente dessa vegetação
pode contribuir com a ciclagem dos nutrientes do solo.
Porém, poucos estudos são realizados com a serapilheira proveniente destas
plantas e das cíbalas provenientes da criação de caprinos em áreas de caatinga nos
períodos de escassez de volumosos.
Portanto, estudos que visam a avaliação da distribuição e composição química
dos resíduos gerados por caprinos, assim como a distribuição espacial e composição
química da serapilheira, justificam-se pela possibilidade de obter novas informações
para otimizar a produção em sistemas extensivos nestas áreas.
Por conseguinte, deve-se a importância aos estudos mais aprofundados sobre
como ocorre a entrada e saída de nutrientes entre a serapilheira, o solo e as cíbalas de
caprinos.
Sabendo isso, surgem questionamentos tais quais: i) qual a variabilidade espacial
do estoque da serapilheira e arelação entre a presença de caprinos com o seu
desaparecimento? ; ii) quais as espécies vegetais que mais contribuem para a produção

2
da serapilheira?; iii) qual a composição química das cíbalas produzidas por caprinos e a
sua distribuição espacial em áreas de caatinga?; iv) qual a disponibilidade e o acúmulo
de serapilheira para esses animais e neste bioma?
É presumível que a presença de animais em áreas antropizadas, com uma leve
lotação, possa contribuir com a ciclagem dos nutrientes na caatinga, sem causar grandes
perturbações ao meio ambiente; e que a interação solo-planta-animal possa equilibrar-se
contribuindo assim, com a sustentabilidade do sistema.
Infelizmente, informações errôneas sobre este bioma são muitas vezes impedem
o interesse de muitos pesquisadores que deveriam estar envolvidos na conservação, no
estudo e na busca pelo uso sustentável deste bioma.
Baseado nisso, o objetivo geral deste trabalho foi avaliar o impacto do pastejo
caprino sobre o acúmulo da serapilheira e consequentemente, a composição química das
cíbalas e da serapilheira, assim como a distribuição espacial destes em áreas de
caatinga.
Para compreender melhor esses questionamentos, este trabalho foi dividido em
três capítulos, abordando os seguintes temas: Referencial teórico (Capítulo 1);
Distribuição espacial da serapilheira e cíbalas em áreas de caatinga pastejada por
caprinos (Capítulo 2); Acúmulo da serapilheira disponível para o pastejo caprino em
áreas de caatinga (Capítulo 3) e; Composição química das cíbalas, serapilheira e
constituintes do solo em áreas de caatinga (Capítulo 4).

3
CAPÍTULO 1
Referencial Teórico
Distribuição espacial da serapilheira e cíbalas em áreas de caatinga
pastejada por caprinos

4
INTRODUÇÃO
O clima na região semiárida apresenta como característica marcante à
irregularidade do regime pluviométrico, com duas estações definidas: a estação
chuvosa, que corresponde a períodos com ocorrência de chuvas e a estação seca,
correspondendo a períodos críticos de estiagem com baixa oferta de volumosos para os
animais criados a pasto.
Andrade et al. (2005), afirmaram que esses ambientes semiáridos são
caracterizados pela alta variabilidade das chuvas, onde os eventos são constituídos por
pulsos de precipitação. Desse modo, o conhecimento do sincronismo e da amplitude das
chuvas vem a ser fundamental para o estudo da dinâmica do ecossistema. A hidrologia
desta região é totalmente dependente do ritmo climático e as secas são caracterizadas
pela ausência e escassez quanto pela alta variação espacial e temporal das chuvas. A
limitação hídrica anual se verifica em função do longo período seco que leva a não
perenização dos rios e riachos endógenos. Além disso, a reduzida capacidade de
absorção de água da chuva pelo solo é dificultada pelas alterações do relevo e dos solos
rasos e pedregosos.
Nos ecossistemas florestais, o solo constitui-se no principal coletor dos detritos
vegetais, onde esse material orgânico acumulado sofre um processo de transformação
realizado pelos microrganismos, culminando com a transferência de nutrientes para o
solo e, posteriormente, parte deles para as plantas (Souto, 2006).
Parte desse processo de retorno de matéria orgânica e de nutrientes para o solo
florestal se dá através da produção de serapilheira, sendo este considerado o meio mais
importante de transferência de elementos essenciais da vegetação para o solo (Vital et
al., 2004).
A serapilheira pode ser caracterizada como sendo uma camada de resíduos
orgânicos constituída por folhas, gravetos, ramos, caules, cascas, frutos, flores, partes
vegetais não identificadas, corpos e dejetos de animais (Souto, 2006). Sendo assim,

5
diferentes quantidades de serapilheira, que também podem apresentar diferentes
proporções de frações constituintes, são depositadas em diferentes ecossistemas
florestais (Figueiredo Filho et al., 2003). Além disso, produz sombra e retém umidade,
criando condições microclimáticas que influem na germinação de sementes e
estabelecimento de plântulas (Moraes et al., 1998).
Os vários trabalhos que abordam a ciclagem de nutrientes nos mais diversos
ambientes constataram que diversos fatores interferem na deposição da serapilheira no
solo. Kolm e Poggiani (2003) enumeraram alguns desses fatores, como, por exemplo, o
clima, o solo, as características genéticas da espécie, a idade e a densidade de plantio.
Ademais, a quantidade de nutrientes na serapilheira depende da espécie, do
tamanho e tipo das folhas em relação aos demais componentes, da capacidade de
translocação do nutriente antes da senescência, bem como do tipo de solo (Schumacher,
1992).
Devido à sua importância para o equilíbrio dos ecossistemas florestais, tais
processos têm sido amplamente adotados como indicadores da integridade ecológica
desses ecossistemas, uma vez que além do seu papel na ciclagem de nutrientes, a
serapilheira confere heterogeneidade espacial e temporal ao ambiente, além de servir
como recurso para uma grande diversidade de microrganismos e invertebrados (Arato et
al. 2003).
Além da serapilheira, as cíbalas dos caprinos também contribuem para a
ciclagem de nutrientes de um ecossistema. Segundo Kiehl (1985) e Malavolta et al.
(1991), os animais devolvem ao solo de 40 a 50 % das rações que lhes é oferecida como
alimento. Não se deve esquecer que os animais jovens não devolvem os nutrientes
recebidos nas mesmas proporções. Além disso, a permanência de fezes na pastagem
também tem aspecto negativo, uma vez que de imediato trazem prejuízos ao
crescimento da forragem em função do bloqueio de nutrientes essenciais para o
desenvolvimento das plantas (Hirata et al., 1990). Segundo Malavolta et al. (1991) a
cada 1.000 kg do peso vivo de caprinos, estes, produzem em torno de 15 toneladas por
ano de cíbalas.
Assim, os nutrientes excretados nas fezes variam significativamente de acordo
com a ingestão de alimento (Van Horn, 1994). Além disso, outros fatores podem estar
envolvidos como a quantidade excretada de fezes pelos animais, entre eles pode- se

6
citar: peso, idade, nível de produção, estado fisiológico (animais prenhes, secas, em
lactação, em acabamento), quantidade e qualidade do alimento fornecido aos animais,
sistema de produção (pasto, confinamento ou semi-confinamento), e até mesmo com a
estação do ano.
Dessa forma, é presumível que a presença de animais em áreas antropizadas,
com uma baixa taxa de lotação, possa contribuir com a ciclagem dos nutrientes na
Caatinga, sem causar grandes perturbações ao meio ambiente; e que a interação solo-
planta-animal possa se equilibrar contribuindo assim, com a sustentabilidade do
sistema.
8. CARACTERIZAÇÃO DO SEMIÁRIDO NORDESTINO: CLIMA E SOLO
O clima predominante na região semiárida nordestina é do tipo BSw’h’,
conforme a classificação de Köppen, ou seja, tropical seco com a evaporação excedendo
a precipitação, com ocorrência de pequenos períodos de chuvas sazonais. Outra
característica importante da região é a imprevisibilidade das estações chuvosas, de
maneira que a época em que são elevados os índices pluviométricos varia ano a ano,
tornando-se difíceis às tomadas de decisão sobre o uso dos recursos desse ecossistema.
Andrade et al. (2010) afirmam que enquanto a temperatura, a radiação solar e os aportes
de nutrientes nos ecossistemas do semiárido variam relativamente pouco no ano, a
precipitação comumente ocorre em eventos descontínuos, em forma de pulsos de curta
duração.
Os pulsos de precipitação pluvial nas regiões semiáridas podem acontecer de três
formas distintas como mostra na Figura 1: (a) amplamente separados, (b) agrupados, (c)
com espaçamento intermediário. Entretanto, nos períodos chuvosos, os intervalos entre
cada evento podem estimular os pulsos de crescimento da planta (Noy-Meir, 1973), e
como conseqüência a recuperação dessas reservas.
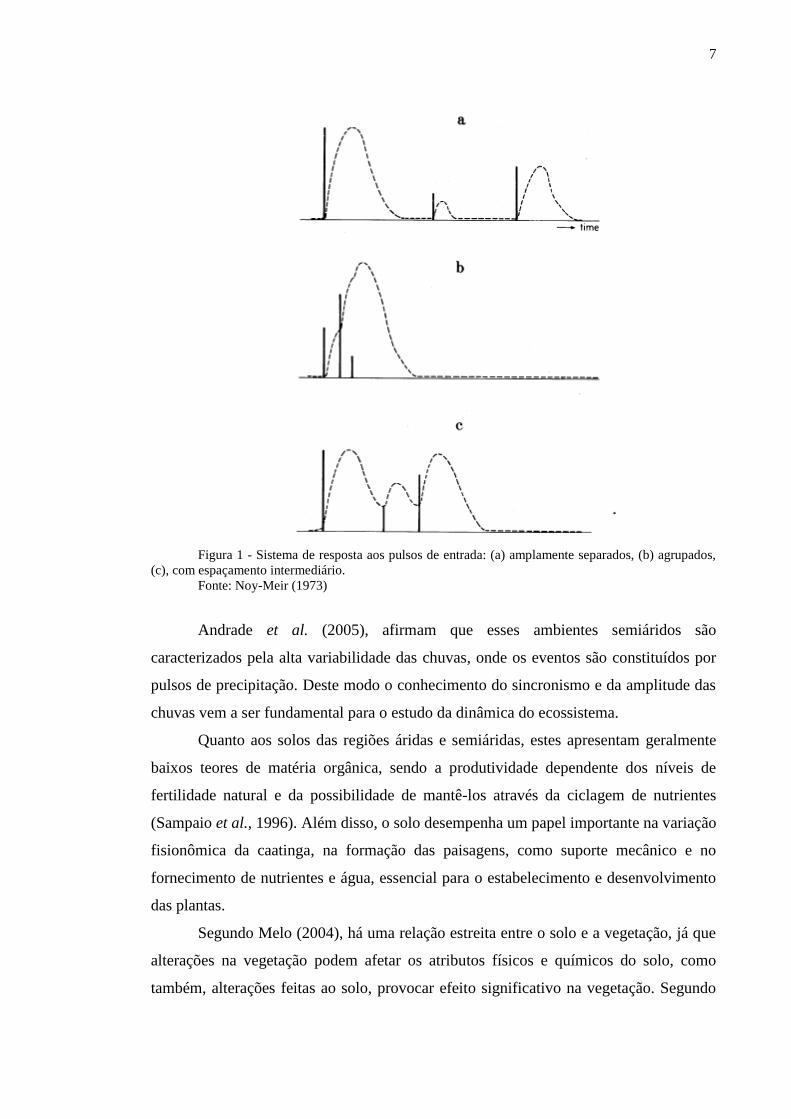
7
Figura 1 - Sistema de resposta aos pulsos de entrada: (a) amplamente separados, (b) agrupados,
(c), com espaçamento intermediário.
Fonte: Noy-Meir (1973)
Andrade et al. (2005), afirmam que esses ambientes semiáridos são
caracterizados pela alta variabilidade das chuvas, onde os eventos são constituídos por
pulsos de precipitação. Deste modo o conhecimento do sincronismo e da amplitude das
chuvas vem a ser fundamental para o estudo da dinâmica do ecossistema.
Quanto aos solos das regiões áridas e semiáridas, estes apresentam geralmente
baixos teores de matéria orgânica, sendo a produtividade dependente dos níveis de
fertilidade natural e da possibilidade de mantê-los através da ciclagem de nutrientes
(Sampaio et al., 1996). Além disso, o solo desempenha um papel importante na variação
fisionômica da caatinga, na formação das paisagens, como suporte mecânico e no
fornecimento de nutrientes e água, essencial para o estabelecimento e desenvolvimento
das plantas.
Segundo Melo (2004), há uma relação estreita entre o solo e a vegetação, já que
alterações na vegetação podem afetar os atributos físicos e químicos do solo, como
também, alterações feitas ao solo, provocar efeito significativo na vegetação. Segundo

8
Sampaio (2010) a variabilidade dos solos advém, principalmente, do efeito diferencial
da erosão geológica, descobrindo camadas distintas, até o limite da exposição das
rochas, formando lajedos e os pavimentos recobertos por rochas.
Sá et al., (2004) ressaltaram que os Estados do Ceará e da Paraíba tem as
maiores áreas, em termos percentuais, com problemas de degradação no nível severo,
seguidos de perto pelos Estados de PE e Bahia. Segundo o mesmo autor, o nível de
degradação ambiental severo aparece principalmente nas áreas dos Estados onde se
encontram os solos do tipo Luvissolos (antigo Bruno-não-cálcico). Ainda, o nível de
degradação ambiental acentuado está mais relacionado às áreas de solos Litólicos, ou
seja, solos mais recentes e em fase de desagregação da rocha que lhe deu origem (Sá et
al., 2004).
Por isso, deve-se dar importância a estudos conjugados de fitossociologia,
serapilheira e análise de solos, pois somente assim pode-se compreender a
complexidade desses sistemas de produção evitando-se uma maior degradação das áreas
de sucessão em sistemas de produção de regiões semiáridas.
9. A IMPORTÂNCIA DA SERAPILHEIRA PARA REGIÕES SEMIÁRIDAS:
PRODUÇÃO, ACÚMULO E DECOMPOSIÇÃO
O estudo do fluxo dos nutrientes no ecossistema, isto é, a produção e
decomposição da serapilheira com a conseqüente transferência desses para o ambiente,
é essencial para a caracterização dos padrões de ciclagem, pois representa a principal
via de retorno de nutrientes e matéria orgânica à superfície do solo (Pagano e Durigan,
2000).
A produção de serapilheira e a devolução de nutrientes em ecossistemas
florestais constituem a via mais importante no sistema solo-planta (Fernandes et al.
2003).
A ciclagem dos nutrientes no sistema solo-planta caracteriza-se, no primeiro
estádio, pela absorção de nutrientes pelas raízes e por sua distribuição pelas diferentes
partes da planta, sendo a taxa de absorção maior no período em que as plantas se
encontram em estádio juvenil, o que corresponde ao período de maior produtividade
dentro do processo de sucessão (Kimmins, 1993). Após esse período, os nutrientes são
transferidos novamente para o solo, pela deposição de serapilheira, lixiviação pelas

9
folhas, ramos e troncos e pela ação da chuva, além do trabalho da fauna herbívora e da
dispersão de frutos e sementes (Poggiani e Schumacher, 2000).
Parente (2009) estudando áreas de caatinga no Cariri paraibano verificou que a
serapilheira, composta por grande parte das folhas das espécies arbustivas e arbóreas
(e.x. catingueira, pereiro, marmeleiro etc), é responsável pela alimentação de pequenos
ruminantes no período seco, e exerce função importante na manutenção da cobertura do
solo e no incremento do teor de matéria orgânica que mantém as condições químicas,
físicas e biológicas do mesmo, devendo, portanto, fazem parte das premissas do manejo
a ser adotado.
O mesmo autor afirma ainda que o pastejo caprino interfere na vegetação da
caatinga de forma acentuada, sendo necessário ser monitorado constantemente a área
pastejada. Além disso, o solo e a vegetação da caatinga apresentam alta resiliência ao
pastejo, quando oferecido uma mínima cobertura vegetal e oportunidade de rebrota,
respectivamente, confirmando que a vegetação da caatinga apresenta potencial para a
produção animal.
Uma série de outros fatores (bióticos e abióticos) também influenciam na
deposição de serapilheira. Dentre eles destacam-se: latitude, altitude, temperatura,
precipitação, estágio sucessional, herbivoria, disponibilidade hídrica e estoque de
nutrientes do solo (Portes et al., 1996), umidade do solo (Burghouts et al., 1994) e vento
(Dias e Oliveira Filho, 1997).
Quanto à contribuição de cada fração na formação da serapilheira, verificou-se
nos diversos estudos (Andrade et al., 2000; Corrêa Neto et al., 2001; Vital, 2002;
Figueiredo Filho et al., 2003; Schumacher et al., 2003; Arato et al., 2003; Souto, 2006)
que a fração folhas contribui com valores que variam entre 60 a 70% ou mais.
Carpanezzi (1990) afirma que as folhas constituem a mais importante fração do
material orgânico decíduo. Essa assertiva confirma os valores reportados no trabalho de
Bray e Ghoran (1964), onde a participação das folhas em diferentes zonas macro-
ecológicas situa-se entre 58 e 79% do material orgânico depositado no solo.
Segundo Dias e Oliveira-Filho (1997) na estação seca, o pico de deposição de
serapilheira seria ocasionado pela grande queda de folhas, provocada como alternativa
ao estresse hídrico causado pela baixa precipitação pluviométrica e pela alta radiação
solar em áreas de caatinga.

10
Além disso, a quantidade de serapilheira depositada pode variar dentro de um
mesmo tipo de vegetação, dependendo dos graus de antropização das áreas. Áreas com
um grau de antropização maior possuem um número elevado de espécies pioneiras de
crescimento rápido, que investem muito em produção de biomassa e acabam produzindo
maior quantidade de serapilheira. Situação diferente das áreas menos antropizadas, pois
possuem menor número de espécies de crescimento rápido e, portanto, apresentam
menor produção de biomassa. Ademais, diferentes ecossistemas florestais depositam
diferentes quantidades de serapilheira que também podem apresentar diferentes
proporções de frações constituintes (Figueiredo Filho et al., 2003).
O desaparecimento acelerado da serapilheira foi observado por pesquisadores
em áreas de caatinga de sucessão secundária, pastejadas por caprinos sem padrão racial
definido (SPRD) ao longo dos meses. Segundo Parente (2009), o pastejo caprino
promove redução na cobertura do solo em função do consumo das espécies constituintes
do estrato herbáceo no período chuvoso e do consumo da serapilheira no período seco.
O mesmo autor verificou ainda a importância da deposição da serapilheira em
áreas de caatinga no período de escassez, pois com o avançar do período seco as folhas
das espécies arbóreas são rapidamente consumidas em função da extinção do estrato
herbáceo bem como dos ramos e das espécies arbustivas.
Formiga (2013), estudando o comportamento ingestivo de caprinos SPRD em
áreas de caatinga, observou que no período de transição (chuvoso-seco) a gramínea, a
serapilheira e o estrato arbóreo foram selecionados de forma semelhante, não diferindo
estatisticamente. A autora afirmou ainda, que a vegetação disponível nessa época do ano
pode ter se apresentado de forma homogênea. Dessa forma, pode-se considerar que a há
uma variabilidade da forragem disponível nas áreas de estudo. Por isso, deve-se ter um
conhecimento mais aprofundado sobre tal variabilidade.
Ainda, em áreas de caatinga, quando chegam as primeiras chuvas, a serapilheira
encontrada na superfície do solo é degradada por ação dos microrganismos
decompositores (fauna edáfica), não ocorrendo grande acúmulo desse material na
superfície do solo.
A decomposição dos resíduos orgânicos que formam a serapilheira é o principal
processo para a ciclagem de nutrientes em ecossistemas florestais (Montagnini e Jordan,
2002). Estes resíduos, ao serem depositados, sofrem inicialmente decomposição parcial

11
pela mesofauna e, posteriormente, ação decompositora dos microrganismos. Parte do
carbono presente nos resíduos é liberado para a atmosfera como CO2 e o restante passa
a fazer parte da matéria orgânica como componente do solo (Bayer e Mielniczuk, 1999).
Segundo Souto (2009), fatores como alta radiação e baixa precipitação em áreas
de caatinga, interagindo simultaneamente, limitam a atividade dos organismos
decompositores, propiciando decomposição mais lenta da serapilheira, quando
comparados com outros ecossistemas mais úmidos. No entanto, esse é um mecanismo
de sustentabilidade desse ecossistema, onde parte da serapilheira seca e mais
lignificada, não é decomposta, servindo como manta protetora para o solo contra os
fortes raios solares, já que sua vegetação na época seca fica quase que totalmente
desfolhada e com isso, ciclagem de nutrientes é mais lenta. Além disso, essa cobertura
de resíduos orgânicos protege o solo do impacto das gotas, por ocasião das primeiras
chuvas. Segundo dados da mesma autora, a serapilheira da caatinga apresentou uma
lenta taxa de decomposição, necessitando de mais de 14 meses para que ocorresse 50%
do seu desaparecimento.
Maior relevância do que a simples acumulação quantitativa é o processo de
mineralização do material orgânico, responsável pela liberação de nutrientes para o
solo, pois a dinâmica da decomposição de resíduos orgânicos disponibiliza
(mineralização) ou não (imobilização) os nutrientes para o sistema, regulado pela
qualidade dos resíduos, principalmente das relações C/N, C/P e C/S (Gonçalves, 1995).
Taylor et al. (1989) afirmaram que a relação C/N é considerada como o melhor
parâmetro para estimar as taxas de decomposição da serapilheira. Quando a relação fica
na faixa de 20-30:1 os processos de imobilização e mineralização se igualam e abaixo
de 20:1 ocorre a mineralização com a maior disponibilidade de compostos nitrogenados.
Souto (2006), trabalhando em áreas de caatinga, observou que o N na
serapilheira apresentou maiores concentrações no período chuvoso, principalmente
devido à contribuição de estruturas reprodutivas das plantas e excretas de animais que,
como as folhas, apresentam teores elevados desse elemento. A mesma autora verificou
relações C/N e C/P da serapilheira elevadas evidenciando uma taxa de decomposição
lenta, quando comparada a outros biomas nacionais.
Em termos de ecossistema, a ciclagem de nutrientes é determinada pela
quantidade de nutrientes que entra, pela quantidade retida na fitomassa, pelas taxas de

12
decomposição dos diferentes componentes da serapilheira e da matéria orgânica do solo,
pelas taxas de imobilização e mineralização de nutrientes e, pela absorção dos nutrientes
(Cuevas e Medina, 1996).
Diversos estudos comprovam que a manutenção dos ecossistemas florestal
depende da ciclagem de nutrientes via produção e decomposição da serapilheira, sendo
este o mais importante processo de transferência de nutrientes proveniente do material
vegetal e/ou animal presente na superfície do solo. Portanto, a velocidade com que esses
nutrientes presentes no solo vão ser reciclados vai influenciar diretamente na
produtividade primária da floresta (Souto, 2006).
Vários pesquisadores têm estudado a produção, qualidade e a contínua
decomposição da serapilheira em diversos ecossistemas no Brasil. No entanto, em áreas
de caatinga esses estudos são escassos. Sampaio (1996) afirma que estudos
desenvolvidos na caatinga nordestina trazem informações da biomassa da vegetação, de
produtividade de herbáceas e de relação solo e vegetação, mas por outro lado, trabalhos
visando a queda do folhedo, massa de serapilheira, decomposição e circulação de
nutrientes são escassos.
Camargo et al. (1999), citando vários autores, explicaram as fases da
decomposição nos resíduos orgânicos. A fase inicial da biodegradação microbiana é
caracterizada pela perda rápida dos compostos orgânicos prontamente decomponíveis
(açúcares, proteínas, amido, celulose, etc.). Na fase subseqüente, produtos orgânicos
intermediários e protoplasma microbiano, recentemente formado, são biodegradados por
vários microrganismos com produção de nova biomassa e liberação de CO2. O estágio
final é caracterizado pela decomposição gradual de compostos mais resistentes,
exercidas pela atividade de actinomicetes e fungos.
Assim, é importante salientar que a velocidade da decomposição da serapilheira
é influenciada pela fauna edáfica, por fatores abióticos (chuvas, temperatura, vento etc)
e pela composição química da serapilheira.
Além disso, é possível afirmar que a variação na disponibilidade dos estratos
vegetais em áreas de caatinga interferem na quantificação e na decomposição da
serapilheira.

13
10. SAZONALIDADE DA DISPONIBILIDADE DE FORRAGEM NO
SEMIÁRIDO
As condições edafoclimáticas da caatinga favoreceram ao desenvolvimento de
espécies vegetais bem adaptadas à região. O bioma é caracterizado por um ecossistema
de plantas xrófitas, caducifólias, espinhosas, muitas vezes esgalhadas e esparsas, mas
composta por diversidade considerável de espécies arbóreas, arbustivas, subarbustivas e
herbáceas, sendo muitas das espécies consideradas endêmicas (Giulietti et al., 2004).
Gonzaga Neto et al. (2001) afirmam que a caatinga constitui-se na mais
importante fonte de alimentação para os rebanhos desta região, chegando a participar
em até 90% da dieta de caprinos e ovinos durante o ano todo, principalmente para
aqueles rebanhos criados extensiva ou semi-intensivamente. Porém a caatinga apresenta
uma dinâmica de produção de fitomassa variável de acordo com as épocas do ano,
presença do estrato vegetativo, composição química do solo, entre outros.
Assim, no período chuvoso há um excesso de matéria seca, enquanto no período
seco há escassez de volumosos, porém a necessidade da ingestão de matéria seca pelos
pequenos ruminantes é constante durante todo o ano.
Na literatura, é possível encontrar que a disponibilidade de fitomassa varia de
2,0 a 4,0 t/ha e que a contribuição do estrato arbustivo-arbóreo na produção total de
fitomassa é bem significativa, e se situa entre 1.000 a 3.000 kg/ha, dependendo do local
e da época do ano (Oliveira, 1996).
Porém, é importante ressaltar que nem toda forragem contabilizada está
prontamente disponível para o consumo, visto que há um limite de alcance máximo para
a apreensão do alimento pelos caprinos (em ramoneio) e que algumas plantas do estrato
arbóreo possuem fatores antinutricionais.
Dessa forma, os animais perdem peso na época de escassez, prejudicando o
sistema de produção local. Esses aspectos reforçam a importância do uso de estratégias
alimentares, tais como: o plantio e manejo adequado de forrageiras xerófitas; a
conservação da forragem excedente e o uso de coprodutos da agroindústria, visando
melhorar os índices zootécnicos e econômicos, tornando viável a ovinocaprinocultura
na região.

14
Porém, existem situações em que a seca permanece constante com pouca
incidência de chuvas (anos de sequeiro), de forma que a vegetação não consegue
rebrotar. Por isso, o produtor deve se precaver para esses tempos, como o qual
presencia-se atualmente. Assim, o uso de silagens, produção de feno ou até mesmo o
cultivo de plantas xerófitas servem como alternativas alimentares.
Em épocas mais chuvosas, é possível observar a presença de folhas verdes e
suculentas dos três estratos vegetativos (herbáceo, arbóreo e sub-arbustivo). Assim,
torna-se relativamente fácil controlar a vegetação nativa para aumentar a
disponibilidade de forragem através do manejo da caatinga, como o raleamento ou
rebaixamento, porém com o avanço da estação seca os caprinos aumentam a
porcentagem de utilização de folhas de arbustos e árvores, passando a consumir troncos,
tubérculos e as folhas caídas no solo após a perda das espécies decíduas (Mesquita et
al., 1989). No entanto, Parente et al. (2012) afirmam que houve uma forte influência da
precipitação sobre o brotamento, floração e frutificação, mostrando o efeito do pulso de
precipitação sobre os aspectos fenológicos das espécies malva, catingueira e pereiro.
Dessa forma, a fenologia de algumas espécies arbóreas não são influenciadas pelo
ramoneio dos caprinos e sim pelo aparecimento do período seco.
Abaixo, segue uma tabela com as principais plantas do estrato herbáceo, sub-
arbustivo e arbóreo encontradas em áreas de caatinga no semiárido Paraibano e
Pernambucano por diversos autores.
Tabela 1 – Disponibilidade dos estratos arbóreo, arbustivo e sub-arbustivo em áreas de
caatinga, em diferentes Estados e os respectivos autores.
Estrato herbáceo e sub-arbustivo
Espécie Estado Autores
Nome comum Nome científico
Capim-búfel Cenchrus ciliaris L. PE, PB Cassuce, 2012
Capim-corrente Urochloa trichopus
Stapf
PE Santos et al.2010
Erva-de-ovelha Diodia sp PB Cassuce, 2012
Pimenta d’agua Phyllanthus sp PE Cassuce, 2012
Orelha-de-onça Macroptium martii
Benth
PE Santos et al.2010
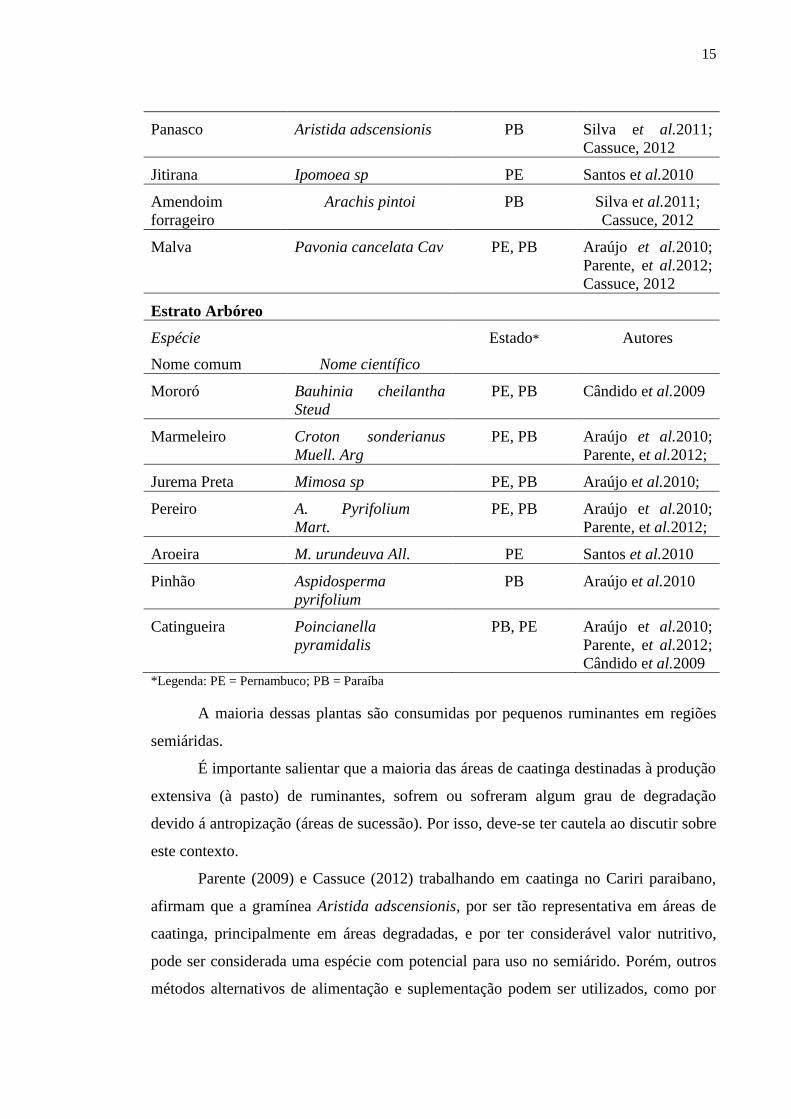
15
Panasco Aristida adscensionis PB Silva et al.2011;
Cassuce, 2012
Jitirana Ipomoea sp PE Santos et al.2010
Amendoim
forrageiro
Arachis pintoi PB Silva et al.2011;
Cassuce, 2012
Malva Pavonia cancelata Cav PE, PB Araújo et al.2010;
Parente, et al.2012;
Cassuce, 2012
Estrato Arbóreo
Espécie Estado* Autores
Nome comum Nome científico
Mororó Bauhinia cheilantha
Steud
PE, PB Cândido et al.2009
Marmeleiro Croton sonderianus
Muell. Arg
PE, PB Araújo et al.2010;
Parente, et al.2012;
Jurema Preta Mimosa sp PE, PB Araújo et al.2010;
Pereiro A. Pyrifolium
Mart.
PE, PB Araújo et al.2010;
Parente, et al.2012;
Aroeira M. urundeuva All. PE Santos et al.2010
Pinhão Aspidosperma
pyrifolium
PB Araújo et al.2010
Catingueira Poincianella
pyramidalis
PB, PE Araújo et al.2010;
Parente, et al.2012;
Cândido et al.2009 *Legenda: PE = Pernambuco; PB = Paraíba
A maioria dessas plantas são consumidas por pequenos ruminantes em regiões
semiáridas.
É importante salientar que a maioria das áreas de caatinga destinadas à produção
extensiva (à pasto) de ruminantes, sofrem ou sofreram algum grau de degradação
devido á antropização (áreas de sucessão). Por isso, deve-se ter cautela ao discutir sobre
este contexto.
Parente (2009) e Cassuce (2012) trabalhando em caatinga no Cariri paraibano,
afirmam que a gramínea Aristida adscensionis, por ser tão representativa em áreas de
caatinga, principalmente em áreas degradadas, e por ter considerável valor nutritivo,
pode ser considerada uma espécie com potencial para uso no semiárido. Porém, outros
métodos alternativos de alimentação e suplementação podem ser utilizados, como por

16
exemplo, a utilização de pastagens cultivadas ou até mesmo o cultivo de plantas xrófitas
(Andrade et al., 2010) para não sobrecarregar as pastagens nativas.
É importante ressaltar que a maioria das criações semiextensivas com caprinos e
ovinos é feita em áreas de sucessão, ou seja, áreas que apresentam algum estádio de
degradação. Dessa forma, áreas que se apresentam com algum tipo de sucessão
ecológica associada à altas taxas de lotação sofrem um processo acelerado de
degradação, tanto das pastagens como do solo. Sendo assim, é imprescindível um
manejo correto de caprinos e ovinos em áreas de caatinga com sucessão, uma vez que
tornar-se-á um fator problemático quando mal manejada, causando um aumento na
degradação da caatinga, diminuindo assim, a disponibilidade de forragem e
consequentemente, afetando a produção como um todo.
4 COMPOSIÇÃO QUÍMICA DA FORRAGEM DISPONÍVEL PARA PEQUENOS
RUMINANTES EM ÁREAS DE CAATINGA
A eficiência da utilização das plantas forrageiras pelos animais está na
dependência de vários fatores, entre os quais podem ser citados como mais
significativos: a qualidade e a quantidade de forragem disponível na pastagem e o
potencial do animal. Por isso, quando a disponibilidade de forragem e o potencial
animal não são limitantes, a qualidade da pastagem é definida pela produção animal,
estando diretamente relacionada com o consumo voluntário e a disponibilidade dos
nutrientes contidos na mesma (Silva e Medeiros, 2003). Assim, o grande desafio da
pecuária no semiárido é utilizar os recursos da caatinga preservando sua
sustentabilidade.
A avaliação da composição bromatológica é importante para verificar a
qualidade dessa forragem nativa, porém poucos estudos são realizados para verificar a
qualidade da forragem nativa disponível para a alimentação de pequenos ruminantes em
áreas de sucessão.
Silva et al. (2010) afirmam que a composição bromatológica das espécies
herbáceas na caatinga variam segundo a época do ano (seca ou chuvosa) e a
conservação das áreas nativas. Além disso, estas plantas são consideradas adequadas ao
consumo de animais adaptados à região do cariri paraibano.
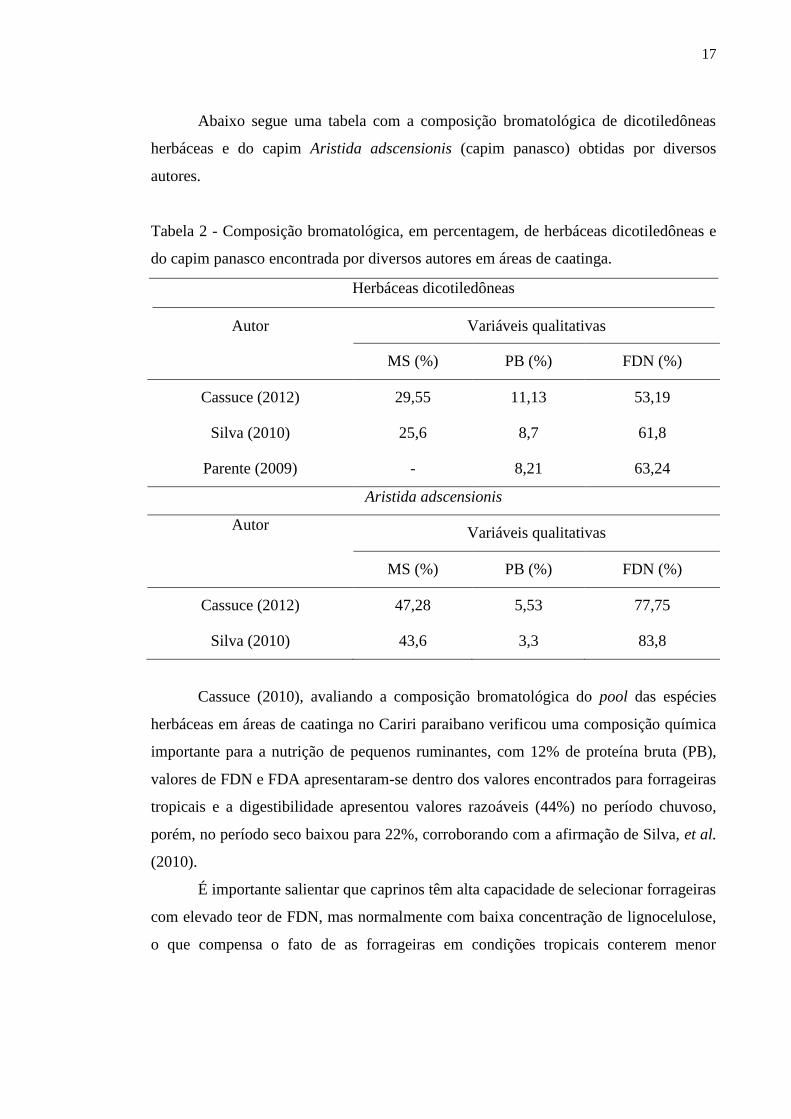
17
Abaixo segue uma tabela com a composição bromatológica de dicotiledôneas
herbáceas e do capim Aristida adscensionis (capim panasco) obtidas por diversos
autores.
Tabela 2 - Composição bromatológica, em percentagem, de herbáceas dicotiledôneas e
do capim panasco encontrada por diversos autores em áreas de caatinga.
Herbáceas dicotiledôneas
Autor Variáveis qualitativas
MS (%) PB (%) FDN (%)
Cassuce (2012) 29,55 11,13 53,19
Silva (2010) 25,6 8,7 61,8
Parente (2009) - 8,21 63,24
Aristida adscensionis
Autor Variáveis qualitativas
MS (%) PB (%) FDN (%)
Cassuce (2012) 47,28 5,53 77,75
Silva (2010) 43,6 3,3 83,8
Cassuce (2010), avaliando a composição bromatológica do pool das espécies
herbáceas em áreas de caatinga no Cariri paraibano verificou uma composição química
importante para a nutrição de pequenos ruminantes, com 12% de proteína bruta (PB),
valores de FDN e FDA apresentaram-se dentro dos valores encontrados para forrageiras
tropicais e a digestibilidade apresentou valores razoáveis (44%) no período chuvoso,
porém, no período seco baixou para 22%, corroborando com a afirmação de Silva, et al.
(2010).
É importante salientar que caprinos têm alta capacidade de selecionar forrageiras
com elevado teor de FDN, mas normalmente com baixa concentração de lignocelulose,
o que compensa o fato de as forrageiras em condições tropicais conterem menor

18
conteúdo de energia,o que faz com que o animal necessite de maiores quantidades de
alimento para atender suas exigências (Araújo et al., 2009).
Além disso, a mesma autora em conjunto com Parente (2009), ao avaliarem a
qualidade do capim panasco, afirmam que este é uma forrageira em potencial para o
semiárido do cariri paraibano.
No entanto, Silva (2010) ao avaliar a composição bromatológica de
dicotiledôneas herbáceas, verificou que a quantidade de PB caiu drasticamente ao
decorrer do tempo (de 8,65% para 4,0%), devido a fatores de baixa precipitação e pelo
avanço da idade fenológica deste estrato, pois com o passar do tempo estas plantas à
medida que se desenvolvem, passam a produzir maior quantidade de caules, ricos em
tecido de sustentação, mas pobres em proteína. Além disso, ao avaliar o capim panasco
a autora afirma que este capim tem baixo valor nutricional.
Assim, discutir o valor nutritivo da pastagem da caatinga exige muito cuidado
pela sua complexidade, visto que inúmeros aspectos devem ser considerados: (i) a
elevada biodiversidade vegetal (plantas lenhosas e herbáceas) dificulta o conhecimento
de seus valores nutricionais de forma individualizada; (ii) embora o potencial de
produção de matéria seca esteja em torno de quatro toneladas, existe grande variação na
quantidade e qualidade da matéria seca disponível para o pastejo/ramoneio dos animais,
em função da quantidade e distribuição das chuvas; (iii) a maioria das propriedades cria
caprinos, ovinos e bovinos sem padrão racial definido e muito adaptados à região, que
apresentam diferenças significativas na preferência e seletividade de plantas e/ou partes
destas ao longo do ano; (iv) a presença de substâncias antinutricionais/tóxicas em
algumas espécies vegetais que se consumidas podem influenciar na resposta nutricional
dos animais; (v) que a maioria das propriedades não respeita a capacidade de suporte de
sua pastagem, dificultando qualquer avaliação nutricional das plantas forrageiras
(Gariglio et al., 2010)
É claro que estas condições são variáveis de região para região. Por isso, é
importante o estudo mais aprofundado desses componentes qualitativos na avaliação de
forragens nativas em áreas de caatinga.
5. ENTRADA E SAÍDA DE NUTRIENTES NO SISTEMA

19
Para uma melhor compreensão da entrada e saída dos nutrientes no sistema é
necessário compreender de que forma os nutrientes são retirados e devolvidos ao
solo.
Os vegetais transformam a energia (em forma de carbono) através da
fotossíntese em nutrientes para a sua manutenção, crescimento e desenvolvimento. É
através das relações hídricas entre o solo e a planta (água carregada de sais - N, P,
K, Ca e Mg) que as estas conseguem gerar essa energia e assim, produzir
substâncias proteicas, ricas em carboidratos, lipídicas etc.
Além da serapilheira, os dejetos de animais também contribuem para a ciclagem de
nutrientes de um ecossistema. Segundo Kiehl (1985) e Malavolta et al. (1991), os animais
devolvem ao solo de 40 a 50 % das rações que lhes é oferecida como alimento. Não se deve
esquecer que os animais jovens não devolvem os nutrientes recebidos nas mesmas
proporções. Além disso, a permanência de fezes na pastagem também tem aspecto negativo,
uma vez que de imediato trazem prejuízos ao crescimento da forragem em função do
bloqueio de nutrientes essenciais para o desenvolvimento das plantas (Hirata et al., 1990).
Segundo Malavolta et al. (1991) fezes de caprinos, a cada 1.000 kg de peso vivo, produzem
em torno de 15 toneladas por ano.
Assim, os nutrientes excretados no nas fezes variam significativamente de acordo
com a ingestão de alimento (Van Horn, 1994). Além disso, outros fatores podem estar
envolvidos como a quantidade excretada de fezes pelos animais, entre eles pode- se citar:
peso, idade, nível de produção, estado fisiológico (animais prenhes, secas, em lactação, em
acabamento), quantidade e qualidade do alimento fornecido aos animais, sistema de
produção (pasto, confinamento ou semi-confinamento), e até mesmo com a estação do ano.
De forma sucinta, o solo comporta todos os nutrientes necessários para a
sobrevivência dos vegetais; a água é responsável pela hidratação e pelo transporte dessas
substâncias em todo o seu sistema vascular; e a luz solar é responsável pela geração de
energia (em ATP’s) através da fotossíntese. Dessa forma, todo o sistema fica comprometido
caso algum desses componentes venha a falhar.
No entanto, quando o vegetal alcança uma idade fenológica avançada, ou quando há
privação de água no sistema, forma-se a serapilheira (composta por folhas caducas, galhos e
estruturas reprodutivas), que carrega parte dos nutrientes das plantas, devolvendo ao solo os
nutrientes e assim, mantendo a sustentabilidade do sistema.
Por isso é importante a compreensão de como acontece a entrada e a saída desses
componentes no ecossistema.

20
Alguns autores avaliam a qualidade química dos resíduos orgânicos através do
conteúdo de N; P; relação C/N; hemiceluloses; celuloses; ligninas e polifenóis; juntamente
com suas interações (Mendonça e Sttot, 2003; Esse et al. 2001). Em função da maior ou
menor quantidade de certos componentes, o resíduo degrada mais lentamente ou mais
rapidamente (Moreira e Siqueira, 2006). Segundo Freitas, et al. (2012), materiais pobres em
nitrogênio e com alta relação C/N, C/P, C/S possuem maior tempo de resiliência no solo.
Já, segundo Thönnissen et al. (2000), a decomposição e liberação de
nutrientes varia também com o local de disposições dos resíduos. Resíduos
incorporados geralmente são decompostos mais rapidamente, devido ao aumento da
superfície de ataque microbiano. De acordo com esses autores, no interior do solo
geralmente ocorrem temperaturas mais amenas e uma maior umidade que propicia o
rápido crescimento da população de microrganismos, acelerando os processos de
decomposição, dados confirmados por Souto et al. (2005).
6. DISTRIBUIÇÃO ESPACIAL E ESTATÍSTICA GEORREFERENCIADA:
ESTUDOS E APLICAÇÃO PRÁTICA NAS CIÊNCIAS AGRÁRIAS
O avanço do conhecimento nas diversas áreas que compõem as ciências agrárias
tem evidenciado as limitações dos métodos tradicionais da estatística, no tratamento da
variabilidade espacial de variáveis do sistema solo – planta – água – atmosfera.
As técnicas da estatística clássica assumem os princípios da casualização,
repetição e do controle local, onde todas as amostras são aleatórias e independentes de
uma distribuição de probabilidade simples, sem continuidade espacial. Assim, a sua
aplicação não envolve qualquer conhecimento da posição atual das amostras ou do
relacionamento entre amostras. Já a estatística espacial é o estudo quantitativo de
fenômenos posicionados no espaço, e assume que os valores estão associados à
localização no espaço, com distribuição contínua dos valores e admite processos de
estimativas para valores não amostrados.
Na geoestatística, se assume que a distribuição das diferenças de variáveis entre
dois pontos amostrados é a mesma para toda a área, e que isto depende somente da
distância entre eles e da orientação dos pontos (Clark, 1979). Assim, é possível admitir
que a geoestatística multivariada é a aplicação de métodos matemáticos e estatísticos a

21
problemas das Ciências da Terra, com o objetivo principal de estimar simultaneamente
um conjunto de variáveis espacialmente correlacionadas (variáveis corregionalizadas).
Desta forma, a estimativa da dependência entre amostras vizinhas no espaço
pode ser realizada através da auto-correlação que é de grande utilidade quando se está
fazendo amostragem em uma direção. Quando a amostragem envolve duas direções
(x,y) o instrumento mais indicado na estimativa da dependência entre amostras é o
semivariograma (Silva, 1988).
A figura 2 demonstra o conjunto de variáveis Z(x) medidas em toda a área S
pode ser considerada uma função aleatória Z(x) uma vez que, segundo Isaaks e
Srivastava (1989), são variáveis aleatórias, regionalizadas e assume-se que a
dependência entre elas é especificada por algum mecanismo probabilístico.
Figura 2. Variável aleatória regionalizada Z(X).
O semivariograma analisa o grau de dependência espacial entre amostras dentro
de um campo experimental, além de definir parâmetros necessários para a estimativa de
valores para locais não amostrados, através da técnica de krigagem (Salviano, 1996).
Sendo assim, o variograma é a ferramenta básica, que permite descrever
quantitativamente a variação no espaço de um fenômeno regionalizado (Huijbregts,
1975).
De forma geral, a geoestatística calcula estimativas dentro de um contexto
regido por um fenômeno natural com distribuição no espaço e, desse modo, supõe que
os valores das variáveis, consideradas como regionalizadas, sejam espacialmente
correlacionados. Devido a essa característica tem sido grande a sua aplicação

22
principalmente para efetuar estimativas e/ou simulações de variáveis em locais não
amostrados.
Assim, a metodologia geoestatística procura extrair, de uma aparente
aleatoriedade dos dados coletados, as características estruturais probabilísticas do
fenômeno regionalizado, ou seja, uma função de correlação entre os valores situados
numa determinada vizinhança e direção no espaço amostrado.
Conhecido o variograma da variável, e havendo dependência espacial entre as
amostras, podem-se interpolar valores em qualquer posição no campo de estudo, sem
tendência e com variância mínima (Vieira, 2000). O método de interpolação chama-se
krigagem e tem como base os dados amostrais da variável regionalizada e as
propriedades estruturais do semivariograma obtido a partir destes dados, o que permite
visualizar o comportamento da variável na região através de um mapa de isolinhas ou de
superfície. Segundo Salviano (1996) os valores estimados das variáveis são
selecionados de forma que a estimativa obtida seja não tendenciosa, assim o estimador
de krigagem se torna o melhor estimador linear não tendencioso (BLUE = Best Linear
Unbiased Estimator), pois apresenta variância mínima e não tendencioso por assegurar
que o somatório dos pesos é igual à unidade.
Landim (2002) explica que, se uma variável regionalizada n(i) for coletada em
diversos pontos i, o valor de cada ponto estará relacionado com valores obtidos a partir
de pontos situados a uma certa distância ∆h e a influência será tanto maior quanto
menor for a distância entre os pontos. Sendo n(1), n(2), .... n(i), .... n(n), realizações de
uma variável regionalizada, a estimativa não tendenciosa da semivariância é dada por:
γ(h) = 1/2n ∑{v(i+h) – v (i)}²
Tais relações são mostradas quando a função g(h) é colocada em gráfico contra
∆h para originar o semivariograma e este não apresenta tendência. Este expressa o
comportamento espacial da variável regionalizada:
a) a amplitude (range) (a), que indica a distância a partir da qual as amostras
passam a não possuir correlação espacial e a relação entre elas torna-se aleatória; toda
amostra cuja distância ao ponto a ser estimado for menor ou igual à amplitude fornece
informações sobre o ponto;
b) o patamar ou silo (sill) (C + Co), que indica o valor segundo o qual a função
estabiliza-se no campo aleatório, correspondente à distância “a”; mostra a variabilidade

23
máxima entre pares de valores, isto é, a variância dos dados e, consequentemente,
covariância nula;
c) a continuidade, pela forma do variograma, em que para h=0, g(h) já apresenta
algum valor. Esta situação é conhecida como efeito pepita e é representada por Co; o
efeito pepita (nugget) pode ser atribuído a erros de medição ou ao fato de que os dados
não foram coletados a intervalos suficientemente, para mostrar o comportamento
espacial subjacente do fenômeno em estudo;
d) a anisotropia, quando os semivariogramas mostram parâmetros diferentes
para diferentes direções de amostragem.
Figura 3. Semivariograma experimental e modelo matemático ajustado. (Genú, 2004)
Seguem alguns modelos de variografia, segundo Lembo e Magri (2002):

24
Figura 4. Modelos e componentes do variograma: a) Esférico; b) Linear; c) Exponencioal;
d) Gausiano.
A krigagem usa informações a partir do variograma para encontrar os pesos
ótimos a serem associados às amostras com valores conhecidos que irão estimar pontos
desconhecidos. Nessa situação o método fornece, além dos valores estimados, o erro
associado à tal estimação, o que o distingue dos demais algoritmos de interpolação. É
entendida como uma série de técnicas de análise de regressão que procura minimizar a
variância estimada a partir de um modelo prévio, que leva em conta a dependência
estocástica entre os dados distribuídos no espaço (Landim e Sturaro, 2002). Por isso,
dentre os métodos de estimativas, comumente empregados, o método geoestatístico da
krigagem pode ser considerado como o melhor estimador linear sem viés (como citado
acima), cujo objetivo é a minimização da variância da estimativa.
Krajewski e Gibbs (1966), mostram a seguinte comparação entre métodos de
interpolação:
Quadro 1. Comparação entre métodos de interpolação. Fonte: Krajewski e Gibbs
(1966).

25
Assim, a krigagem, mostra-se mais uma vez, como o método de maior precisão.
Os estudos sobre a produtividade e acúmulo de serapilheira que são realizados
utilizam-se, na sua maioria, de métodos clássicos de análise estatística de dados, os
quais, em geral, supõem que as realizações das variáveis aleatórias são independentes
entre si (Snedecor e Cochran, 1967). No entanto, fenômenos naturais comumente
apresentam-se com certa estruturação nas variações entre vizinhos podendo apresentar
algum grau de dependência espacial (Guimarães, 1993). Logo, a distribuição da
vegetação de determinado local pode apresentar continuidade espacial (Miller et al.,
2007) e, consequentemente, os métodos estatísticos clássicos podem não ser são bons
estimadores para estudos de produtividade e acúmulo de serapilheira.
Aquino (2013), ao estudar a análise espacial da serapilheira (fração folhas e
total) pelo método da krigagem em região de mata, verificou que foi possível visualizar
regiões a partir das quais pode-se definir como zonas de manejo, que podem ajudar na
tomada de decisão com relação a transposição de serapilheira, coleta de banco de
sementes para recuperação de áreas, entre outros. Tal resultado é muito importante para
lidar com um trabalho sustentável em áreas nativas e de preservação ambiental.
Barbosa (2011), ao avaliar a distribuição espacial de espécies de cactáceas em
áreas de caatinga, observou que algumas espécies nativas se comportam de forma
agregada, enquanto que outras apresentam distribuição uniforme dentro das áreas de
caatinga. Dessa forma, o manejo pode ser diferenciado quanto as espécies nativas
presente nas áreas.
Assim, a elaboração dos mapas de contorno tem sido muito utilizados para a
avaliação dos atributos físicos dos solos de uma determinada região e, muitas vezes,
empregados para caracterizar os padrões espaciais dos atributos do solo (Zhu e Shao,
2008; Brocca et al., 2007; Cichota et al., 2006; Hébrard et al., 2006; Souza et al., 2004;
2005, entre outros).

26
Mas, recentemente, estes mapas têm sido utilizados para avaliação de outras
variáveis que permitem ser regionalizadas, como a distribuição dos estratos vegetativos
em determinadas áreas degradadas, distribuição do CO2 liberado pelo solo em áreas de
caatinga, distribuição da serapilheira, entre outros.
Assim, cada vez mais a estatística georreferenciada aplica-se a várias áreas de
estudo com o intuito de obter novas informações técnicas e contribuir ainda mais para
alcançar níveis ótimos de produtividade aliados à sustentabilidade.
Dadas as observações anteriores, torna-se mais fácil o entendimento da
importância da aplicação prática da estatística georreferenciada com a avaliação da
distribuição espacial de amostras populacionais para uma melhor compreensão dos
sistemas de produção e da caracterização ambiental, mantendo assim uma produção
sustentável sem causar danos ao ambiente.
7 REFERÊNCIAS BIBLIOGRÁFICAS
ANDRADE, A. P.; COSTA, R. G.; SANTOS, E. M.; SILVA, D. S. Produção animal no
semiárido: o desafio de disponibilizar forragem, em quantidade e com qualidade, na
estação seca. Revista Tecnologia & Ciência Agropecuária, João Pessoa, v.4, n.4,
p.01-14, dez. 2010
ANDRADE, A.P.; SOUZA, E.S.; SILVA, D.S.; et al. 2006. Produção animal no bioma
caatinga: paradigmas dos “pulsos-reservas”. In: REUNIÃO ANUAL DA SOCIEDADE
BRASILEIRA DE ZOOTECNIA, 43. Anais de Simpósios..., 2006. João Pessoa.
ANDRADE, L. A.; PEREIRA, I. M.; LEITE, U. T.; BARBOSA, M. R. V. Análise da
cobertura de duas fitofisionomias de caatinga, com diferentes históricos de uso, no
município de São João do Cariri, Estado da Paraíba. Cerne, v.11, n.3, p.253-262, 2005.
AQUINO, P. S. R. Análise espacial da produtividade e acúmulo de serapilheira em mata
de galeria. Dissertação (Mestrado em Ciências Florestais).Departamento de Engenharia
Florestal, Universidade de Brasília,DF,76p. 2013.
ARATO, H.D.; MARTINS, S.V. & FERRARI, S.H.S. 2003. Produção e decomposição
de serapilheira em um sistema agroflorestal implantado para recuperação de área
degradada em Viçosa-MG. Revista Árvore 27: 715-721.
ARAÚJO, K. D. Análise da vegetação e organismos edáficos em áreas de caatinga
sob pastejo e aspectos socioeconômicos e ambientais de São João do Cariri - PB.
Campina Grande - PB, Centro de Tecnologia e Recursos Naturais, UFCG, fevereiro de
2010. 151 p.il. (Tese de Doutorado). Programa de Pós-Graduação em Recursos
Naturais.

27
ARAÚJO, K. D.; DANTAS, R. T.; ANDRADE, A. P.; PARENTE, H. N.; SILVA, É. É.
Uso de espécies da caatinga na alimentação de rebanhos no município de São João do
Cariri. R. RA´E GA, Curitiba, n. 20, p. 157-171, 2010. Editora UFPR.
BARBOSA, A. S. Estrutura da vegetação e distribuição espacial de Cactaceae em
áreas de caatinga do semiárido paraibano 166f. (Dissertação de Mestrado em
Agronomia) Programa de Pós-Graduação em Agronomia. Centro de Ciências Agrárias –
Universidade Federal da Paraíba, 2011.
BAYER, C; MIELNICZUK, J. Dinâmica e função da matéria orgânica. In: SANTOS,
G. A.; CAMARGO, F. A. O. Fundamentos da Matéria Orgânica do Solo: ecossistemas
tropicais e subtropicais. Porto Alegre: Gênesis, 1999. Cap. 2. p.9-26.
BRAY, J.R.; GORHAM, E. Litter production in forests of the world. Advances in
Ecological Research, v.2, n.1, p.101-157,1964.
BROCCA, L.; MORBIDELLI, R.; MELONE, F.; MORAMARCO, T. Soil moisture
spatial variability in experimental areas of central Italy. Journal of Hydrology, v.333,
n.1, p.356-373, 2007.
BURGHOUTS, T. B. A.; CAMPBELL, E. J.F.; KODERMAN, P. J. Effects of tree
species heterogeneity on leaf fall in primary an logged dipterocarp forest in the Ulu
Segana Forest Reserve, Sabah, Malasia. Jornal of Tropical Ecology, v. 10, p. 1-26,
1994.
CÂNDIDO, M.J.D.; DE ARAÚJO, G.G.L, CAVALCANTE, M.A.B. Pastagens no
Ecossistema Semi-árido brasileiro: atualização e perspectivas Futuras. Publicado no
Núcleo de Ensino e Estudos em Forragicultura (NEEF), Universidade Federal do
Ceará/UFC, 2009.
CASSUCE, M. R. Fitossociologia e Composição Bromatológica de Espécies Herbáceas
e Subarbustivas em Áreas de Caatinga Sob Pastejo. 2012. 88 f. Dissertação (Mestrado
em Zootecnia). Universidade Federal da Paraíba, Centro de Ciências Agrárias, Areia-
PB.
CAMARGO, F.A.O.; SANTOS, G.A.; GUERRA, J.G.M. Macromoléculas e
substâncias húmicas. In: SANTOS, G.A.; CAMARGO, F.A.O. (Ed). Fundamentos da
matéria orgânica do solo: ecossistemas tropicais e subtropicais. Porto Alegre:
GENESIS, 1999, p.27-40.
CARPANEZZI, A. A.; COSTA, L. G. S.; KAGEYAMA, P. Y.; CASTRO, C. F. A.
Espécies pioneiras para recuperação de áreas degradadas: a observação de laboratórios
naturais. In: CONGRESSO FLORESTAL BRASILEIRO, 6., 1990, Campos do Jordão.
Anais...Campos do Jordão: SBS/SBEF, 1990. p. 216-221.
CICHOTA, R.; HURTADO, A. L. B.; LIER, Q. J. VAN. Spatio-temporal variability of
soil water tension in a tropical soil in Brazil. Geoderma, v.133, n.8, p.231-243, 2006
CLARK, I. The semivariogram - Part I. Eng. & Min. J., 180(7):90-94, 1979.

28
CORRÊA NETO, T. de A.; PEREIRA, M.G.; CORREA, M.E.F.; ANJOS, L.H.C. dos.
Deposição de serapilheira e mesofauna edáfica em áreas de eucalipto e floresta
secundária. Floresta e Ambiente, v.8, n.1, p.70-75, 2001.
CUEVAS, E.; MEDINA, E. Nutrient cycling in the conservation of soil fertility in the
Tropics. In: Congresso Latino Americano de Ciência do Solo, 13.; Reunião Brasileira
de Biologia do solo, 1.; Simpósio Brasileiro Sobre Microbiologia do Solo, 4.; Reunião
Brasileiro Sobre Micorrizas, 6; Reunião Brasileira de Manejo e Conservação do Solo e
da Água, 11., 1996, Águas de Lindóia, SP. Solo Suelo 96. Águas de
Lindóia:USP/SLCS/SBCS, 1996. 1 CD-ROM.Comissão 10: Solos Florestais.
DIAS, H. C. T.; OLIVEIRA FILHO, A. T. Variação temporal e espacial da produção de
serapilheira em uma área de Floresta Estacional Semidecídua Montana em Lavras-MG.
Revista Árvore, v. 21, n. 1, p. 11-26, 1997.
FIGUEIREDO FILHO, A.; MORAES, G.F.; LUCIANO BUDANT SCHAAF, L.B.;
FIGUEIREDO, D.J. de. Avaliação estacional da deposição de serapilheira em uma
floresta ombrófila mista localizada no sul do Estado do Paraná. Ciência Florestal, v.
13, n. 1, p. 11-18, 2003.
FERNANDES, M. M.; PEREIRA, M. G.; MAGALHÃES, L. M. S.; CRUZ, A. R.;
GIÁCOMO, R. G.; 2003. Aporte e decomposição de serapilheira em áreas de floresta
secundária, Plantio de sabiá (Mimosa caesalpiniaefolia benth.) e andiroba (Carapa
guianensis aubl.) na Flona mário xavier, RJ. Rev. Ciência Florestal, Santa Maria, v.
16, n. 2, p. 163-175.
FORMIGA, L., D., A., S. Evolução da taxa de CO2, organismos edáficos e
herbivoria em áreas de caatinga sob pastejo caprino. 2013. Tese (Doutorado em
Zootecnia) Centro de Ciências Agrárias, Universidade Federal da Paraíba, Areia.
GIULIETTI, A.M., BOCAGE NETA, A.L., CASTRO, A.A.J.F. Diagnóstico da
vegetação nativa do bioma da caatinga In: Biodiversidade da Caatinga: áreas e ações
prioritárias para a conservação. Brasilia: MMA-UFPE; Brasília, DF: 2004. p.47-90.
GENÚ, A. M. Geoestatística multivariada. Texto didárico.Universidade de São Paulo.
p. 2-17, 2004
GONZAGA NETO, S.; BATISTA, A. M. V.; CARVALHO, F. F. r. de; MARTÍNEZ, r.
L. V.; BARBOSA, J. E. A. S.; SILVA, E. O. Composição bromatológica, consumo e
digestibilidade In Vivo de dietas com diferentes níveis de feno de catingueira
(Caesalpinea bracteosa), fornecidas para ovinos Morada Nova. Revista Brasileira de
Zootecnia,v. 30, n. 2, p. 553-562, 2001.
GONÇALVES, J.L. de M. Efeito do cultivo mínimo sobre a fertilidade do solo e
ciclagem de nutrientes. In: SEMINÁRIO SOBRE CULTIVO MÍNIMO DO SOLO EM
FLORESTAS, 1, 1995, Curitiba. Anais ... Curitiba –PR, 1995, p. 43-60.
GUIMARÃES, E. C. Variabilidade espacial da umidade e da densidade do solo em
um Latossolo Roxo. Campinas: 1993. 135 f. Dissertação (Mestrado em Engenharia

29
Agrícola) - Faculdade de Engenharia Agrícola, Universidade Estadual de Campinas,
Campinas, 1993.
HÉBRARD, O.; VOLTZ, M.; ANDRIEUX, P.; MOUSSA, R. Spatio-temporal
distribution of soil surface moisture in a heterogeneously farmed Mediterranean
catchment. Journal of Hydrology, v.329, p.110-121, 2006.
HIRATA, M. et al. Return of dung to be hiagrass (Paspalum notatum Fliigge) pasture
by dairy catlle. Journal Japanese Grassland Society Science, Nishinasuno, v. 35, n. 4,
p.350-357, 1990.
HUIJBREGTS, C.J. Regionalized variables and quantitative analysis of spatial data.
In: Davis, J.C. & McCullagh, M.J. (ed) Displayand anaysis of spatial data. New York,
John Wiley. p.38-53. 1975.
ISAAKS, E.H.; SRIVASTAVA, R.M. An Introduction to Applied Geostatistics. New
York: Oxford University Press, 1989. 561 p.
KIEHL, E. J. Fertilizantes orgânicos. São Paulo: Agronômica Ceres, 1985. 492p.
KIMMINS, J. P. Forest ecology. New York: Collier Macmillan Canada, 1987; São
Paulo: Ed. UNESP, 1993.184 p.
KOLM, L; POGGIANI, F. Ciclagem de nutrientes em povoamentos de Eucalyptus
grandis submetidos à prática de desbastes progressivos. Scientia Forestalis, n.63, p.
79-93, 2003.
KRAJEWSKI, S. A. & GIBBS, B. L. Understanding Contouring: A pratical Guide to
Spatial Estimation and Contouring Using a Computer and Basics of Using Variograms:
Gibbs Associates, 1966.
LANDIM, P.M.B. & STURARO, J.R. Krigagem indicativa aplicada à elaboração de
mapas probabilísticos de riscos. DGA, IGCE, UNESP/Rio Claro, Lab.
Geomatemática,Texto Didático 06, 19 pp. 2002. Disponível em
<http://www.rc.unesp.br/igce/aplicada/textodi.html>. Acesso em: julho de 2013.
LEMBO, A. J.; MAGRI, A. (2002) Geostatistics. Paper apresentado durante a
ministração do curso Spatial Modeling and Analysis – CSS 620, do Department of Crop
and Soil Sciences, Cornell University, Ithaca, NY.
MALAVOLTA, E. et al. Micronutrientes, uma visão geral. In: FERREIRA, M.E.;
CRUZ, M. C. Micronutrientes na Agricultura. Piracicaba: POTAFOS / CNPq, 1991. p.
1-33.
MELO, M.S. Florística, fitossociologia e dinâmica de duas florestas secundárias
antigas com histórias de usos diferentes no Nordeste do Pará-Brasil. 2004, 116f.
Dissertação (Mestrado em Recursos Florestais). Escola Superior de Agricultura Luiz de
Queiroz. Universidade de São Paulo. Piracicaba.
MESQUITA, R. C. M., E. R. LEITE, & J. A. ARAÚJO-FILHO. 1989. Estacionalidade
da dieta de pequenos ruminantes em ecossistema de caatinga. Pp 59-82 in: EMBRAPA
(org.) Curso de Melhoramento e manejo de pastagem nativa no trópico semiárido.
EMBRAPA - CPAMN/SPI, Teresina.

30
MILLER, J., FRANKLIN, J., ASPINALL, R. Incorporating spatial dependence in
predictive vegetation models. Ecological Modelling, v.202, p.225-242, 2007.
PARENTE, H. N. Avaliação da vegetação e do solo em áreas de caatinga sob pastejo
caprino no Cariri da Paraíba. Tese (Doutorado em Zootecnia). Areia: UFPB/CCA, 2009.
35 p.
MONTAGNINI, F.; JORDAN, C.F. Reciclaje de nutrientes. In: GUARIGUATA, M.R.;
KATTAN, G.H. (Eds.). Ecologia y conservación de bosques neotropicais.Cartago:
Ediciones LUR, 2002. p.167-191.
MORAES, R. M.; DELITTI, W. B. C.; RINALDI, M. C. S.; REBELO, C. F. Ciclagem
mineral em Matas Atlênticas de encosta e mata sobre restinga, Ilha do Cardoso, SP:
nutrientes na serapilheira acumulada. In: SIMPÓSIO DE ECOSSISTEMAS
BRASILEIROS, 4., 1998, Águas de Lindóia. Anais... Águas de Lindóia: ACIESP,
1998. p. 71-77.
NOY-MEIR, I. 1973. Desert ecosystems: environment and producers. Annual Reviews
Ecology. Annual Review of Ecology and Systematics, v. 4 25-51p.
OLIVEIRA, A.N. Desempenho e características da carcaça de caprinos mestiços Anglo-
nubiano, Boer e caprinos sem padrão racial definido em pastagem e em confinamento.
Tese (Doutorado em Zootecnia). Fortaleza: Universidade Federal do Ceará. 2006, 65p.
OLIVEIRA, E.R. Alternativas de alimentação para a pecuária do Semi-Árido
nordestino. In: SIMPÓSIO NORDESTINO DE ALIMENTAÇÃO DE RUMINANTES.
Natal, 1996. Anais... Natal:SNPA, 1996. p. 127-147.
PAGANO, S.N. ; DURIGAN, G. 2000. Aspectos da ciclagem de nutrientes em matas
ciliares do oeste do Estado de São Paulo, Brasil. In:Rodrigues, R.R; Leitão Filho H.F.
(eds). São Paulo: matas ciliares : conservação e recuperação. São Paulo : Editora da
USP : Fapesp, p. 109-123.
PARENTE, H. N.; ANDRADE, A. P; SILVA, D. S.; SANTOS, E. M; ARAÚJO, K. D.;
PARENTE, M. O. M. INFLUÊNCIA DO PASTEJO E DA PRECIPITAÇÃO SOBRE
A FENOLOGIA DEQUATRO ESPÉCIES EM ÁREA DE CAATINGA. Revista
Árvore, Viçosa-MG, v.36, n.3, p.411-421, 2012
PARENTE, H. N. Avaliação da vegetação e do solo em áreas de caatinga sob
pastejo caprino no Cariri da Paraíba. Tese (Doutorado em Zootecnia). Areia:
UFPB/CCA, 2009.
PORTES, M. C. G. O.; KOEHLER, A.; GALVÃO, F. Variação sazonal de deposição
de serapilheira em uma Floresta Ombrófila Densa Altomontana no morro do Anhangava
– PR. Floresta, v. 26, n. 1/2, p. 3-10, 1996.
POGGIANI, F.; SCHUMACHER, M. V. Ciclagem de nutrientes em Florestas
Nativas. In: GONÇALVES, J. L. M.; BENEDETTI, V. (Eds.). Nutrição e fertilização
florestal. Piracicaba: IPEF/Escola Superior de Agricultura “Luiz de Queiroz”, 2000.
427 p.
RODAL, M. J. N. Fitossociologia da vegetação arbustivo-árborea em quatro áreas
de caatinga em PE. 1992. 198f. Tese (Doutorado em Biologia Vegetal) - Universidade
Estadual de Campinas, Campinas.

31
SÁ, I.B., RICHÉ, G.R., FOTIUS, G.A.As paisagens e o processo de degradação do
semi-árido nordestino In: Biodiversidade da Caatinga: áreas e ações prioritárias para a
conservação. Brasilia: MMA-UFPE; Brasília, DF: 2004. p.17-36.
SAMPAIO, E.V.S.B. Características e potencialidades. In: Uso sustentável e
conservação dos recursos florestais da caatinga. Brasília: Serviço Florestal
Brasileiro, p.29-42, 2010.
SAMPAIO, E.V.S.B. Ciclagem de nutrientes. In: SAMPAIO, E.V.S.B.; MAYO, S.J.;
BARBOSA, M.R.V. (eds) Pesquisa Botânica Nordestina: progresso e
perspectivas.Sociedade Botânica do Brasil, Recife, 1996. p.191-201.
SANTOS, M. V. F.; LIRA, M. A.; DUBEUX JR., J. C. B.; GUIM, A.; MELLO, A. C.
L.; CUNHA, M. V. Potential of Caatinga forage plants in ruminant feeding. R. Bras.
Zootec., v.39, p.204-215, 2010.
SALVIANO, A.A.C. Variabilidade de atributos de solo e de Crotalaria juncea em
solo degradado do município de Piracicaba-SP. Piracicaba, 1996. 91p. Tese
(Doutorado) - Escola Superior de Agricultura "Luiz de Queiroz", Universidade de São
Paulo.
SILVA, A.P. Variabilidade espacial de atributos físicos do solo. Piracicaba, 1988,
105p. Tese (Doutorado) - Escola Superior de Agricultura "Luiz de Queiroz",
Universidade de São Paulo.
SILVA, L. D. A. Ovinos e caprinos terminados em Caatinga raleada e enriquecida com
capim buffel (Cenchrus ciliarisL.). Patos, PB: UFCG, 2010, 86p. (Dissertação –
Mestrado em Zootecnia – Sistemas Agrossilvipastoris no Semiárido).
SILVA, D. S.; ANDRADE, M. V. M.; ANDRADE, A. P.; CARNEIRO, M. S. S.;
OLIVEIRA, J. S. Bromatologic composition of the herbaceous species of the
Northeastern Brazil Caatinga. R. Bras. Zootec., v.40, n.4, p.756-764, 2011.
SILVA D. S.; MEDEIROS A. N. Eficiência do uso dos recursos da caatinga: produção
e conservação. In: SEGUNDO SIMPÓSIO INETRNACIONAL DE CAPRINOS E
OVINOS DE CORTE. Anais... João Pessoa, PB, p.571-582, 2003.
SCHUMACHER, M.V.; BRUN, E.J.; RODRIGUES, L.M.; SANTOS, E.M. dos.
Retorno de nutrientes via deposição de serapilheira em um povoamento de acácia-negra
(Acacia mearnsiiDe Wild.) no Estado do Rio Grande do Sul. Revista Árvore, v.27, n.6,
p. 791-798, 2003.
SCHUMACHER, V. M. Aspectos da ciclagem de nutrientes e do microclima em
talhãoes de Eucalyptuscamaldulensis Dehnh, Eucalyptus grandis Hill ex Maiden e
Eucalyptus toreliana F. Muell. 1992. 87 f. Dissertação (Mestrado) – Escola Superior
de Agricultura de Luiz de Queiroz, Piracicaba, 1992.
SNEDECOR, G. W.; COCHRAN, W. G. Statistical methods. 6. ed., Ames, Iowa State
University Press, 1967. 593 p.
SOUTO, P. C. Acumulação e decomposição da serapilheira e distribuição de
organismos edáficos em área de caatinga na Paraíba, Brasil. 2006. 150f. Tese

32
(Doutorado em Agronomia) - Centro de Ciências Agrárias, Universidade Federal da
Paraíba, Areia.
SOUTO, P. C.; SOUTO, J. S.; SANTOS, R. V.; BAKKE, I. V. Características químicas
da serapilheira depositada em área de caatinga. Revista Caatinga (Mossoró,Brasil),
v.22, n.1, p.264-272, 2009.
SOUTO, P.C.; SOUTO, J.S.; SANTOS, R.V.;ARAÚJO, G.T.& SOUTO, L.S.
Decomposição de estercos dispostos em diferentes profundidades em área degradada no
semi-árido da Paraíba. R. Bras. Ci. Solo, 29:125-130, 2005.
SOUZA, Z. M.; MARQUES JUNIOR, J.; PEREIRA, G. T.; BARBIERI, D. M.
Variabilidade espacial da textura de um Latossolo Vermelho eutroférrico sob cultivo de
cana-de-açúcar. Engenharia Agrícola, v.24, n.2, p.309-319, 2004.
TAYLOR, B.R.; PARKINSON, D.; PARSONS, W.F.J. Nitrogen and lignin content as
predictors of litter decay rates: a microcosm test. Ecology, v.70, n.2, p.97-104, 1989.
VAN HORN, H.H.; WILKIE, A. C.; POWERS, W.J.; NORDSTEDT, R.A.
Components of dairy manure management systems. Journal Dairy Science, Savoy, v.
77, n.7, p. 2008 -2030, 1994.
VIEIRA, S.R. Geoestatística em estudos de variabilidade espacial do solo. In:
NOVAIS, R. F. de; ALVAREZ V., V. H.; SCHAEFER, C. E. G. R. (Ed.). Tópicos em
ciência do solo. Viçosa: Sociedade Brasileira de Ciência do Solo, 2000.v. 1, p. 1-54.
VITAL, A.R.T. Caracterização hidrológica e ciclagem de nutrientes em fragmento
de mata ciliar em Botucatu, SP. 2002. 117 f. Tese (Doutorado em Agronomia) –
Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Botucatu, SP.
VITAL, A.R.T.; GUERRINI, I.A.; FRANKEN, W.K.; FONSECA, R.C.B. Produção de
serapilheira e ciclagem de nutrientes de uma Floresta Estacional Semidecidual em Zona
Ripária. Revista Árvore, v. 28, n. 6, p. 793-800, 2004.
VAN HORN, H.H.; WILKIE, A. C.; POWERS, W.J.; NORDSTEDT, R.A.
Components of dairy manure management systems. Journal Dairy Science, Savoy, v.
77, n.7, p. 2008 -2030, 1994.
ZHU, Y.; SHAO, M. Variability and pattern of surface moisture on a small-scale
hillslope in Liudaogou catchment on the northern Loess Plateau of China. Geoderma,
v.147, p.185-191, 2008.

33
CAPÍTULO 2
Distribuição espacial e estacional da serapilheira em áreas de caatinga
sob diferentes lotação de caprino
________________________________________________________
RESUMO
O estudo da variabilidade espacial da serapilheira em áreas de caatinga de
vegetação nativa é importante para a compreensão da trajetória cíclica dos elementos
minerais essenciais à vida dentro desse ecossistema. Desta forma, o presente estudo
objetivou avaliar a distribuição espacial da deposição da serapilheira em áreas de

34
caatinga em período prolongado de seca com diferentes lotação animal. O experimento
foi conduzido na Estação Experimental pertencente a UFPB, localizada no município de
São João do Cariri-PB. A área experimental representou, aproximadamente, 5 hectares,
dos quais foram subdivididos em cinco áreas (1, 2, 3, 4 e 5), com diferentes lotação
animal e diferentes estados de antropização. Os animais pertencentes às áreas eram
caprinos machos, adultos, sem idade padrão e sem padrão racial definido. O método
utilizado para a análise dos dados foi o de parcelas contíguas em áreas de um hectare,
delimitadas com auxílio de GPS. Para a quantificação desse material foi utilizado um
molde de estrutura férrea, vazado, medindo 0,25m² e balança de precisão de 1g. Da
serapilheira amostrada, foram separados os seguintes constituintes: folhas (incluindo
folíolos + pecíolo), pool do material lenhoso (correspondente às partes lenhosas
arbóreas, arbustivas e subarbustivas de todas as dimensões + cascas), estrutura
reprodutiva (flores, frutos e sementes), miscelânea (material vegetal que não pôde ser
identificado) e cíbalas (dejetos caprinos). Duas áreas apresentaram maior acúmulo de
serapilheira e do pool do material lenhoso, enquanto outras apresentaram maior
produção de miscelânea, quando comparado os demais constituintes da serapilheira.
Além disso, a deposição das cíbalas está diretamente ligada ao acúmulo de folhas
produzidas e consequentemente, a presença do estrato arbóreo/arbustivo.
Palavras-chave: cariri, galhos, folhas, pequenos ruminantes, semiárido
ABSTRACT
The spatial variability of litter at Caatinga native vegetation is important for
understanding the cyclical trajectory of mineral elements essential to life within that
ecosystem. Thus, the present study aimed to evaluate the spatial distribution of litter
deposition in areas of savanna in prolonged dry period with different stocking. The
experiment was conducted at the Experimental Station belonging to UFPB, localized in
São João do Cariri. The experimental area represented approximately 5 hectares of
which were subdivided into five areas (1, 2, 3, 4 and 5), with different stocking.
Animals belonging to the areas were male goats, adults, no standard age and without

35
defined breed. The method used for data analysis was the contiguous plots in areas of
one hectare, delineated using GPS. The method used for data analysis was the
contiguous plots in areas of one hectare. To quantify this material a mold of iron
structure was used, leaked, 0,25m² measuring and precision balance 1g. Sampled the
litter, the following constituents were separated: leaves (including leaflets + petiole),
pool of woody material (corresponding to trees, shrubs and subshrubs woody parts of all
sizes + shells), reproductive structures (flowers, fruits and seeds), miscellaneous (plant
material that could not be identified) and cíbalas (goat droppings). Two areas showed
higher accumulation of leaf litter and woody material from the pool, while others
produced more miscellaneous compared the other constituents of the litter. Furthermore,
the deposition of cíbalas is directly linked to the accumulation of leaves produced and
hence the presence of the tree / shrub.
Key words: cariri, twigs, leaves, small ruminants, semiarid
1 INTRODUÇÃO
Segundo Souza (2009) a serapilheira depositada sob os solos florestados
compreende a camada mais superficial, sendo formados por folhas, galhos, órgãos
reprodutivos e miscelânea, que exercem inúmeras funções para o equilíbrio e dinâmica
destes ecossistemas. Este material protege o solo contra as elevadas temperaturas,
armazena grande quantidade de sementes e abriga uma abundante diversidade de
microrganismos que atuam diretamente nos processos de decomposição e incorporação
do material fornecendo nutrientes ao solo.
Para Moreira e Siqueira (2002), o tipo de vegetação e as condições ambientais são
os fatores que mais influem na quantidade e qualidade do material que cai no solo.
Além disso, o estágio sucessional da formação vegetal influi diretamente na produção
do folhedo, de tal modo que florestas secundárias caracterizam-se por apresentar menor
produção de serapilheira que florestas em início de regeneração, já que estas últimas
apresentam dominância de espécies pioneiras (Leitão Filho et al. 1993).
O estudo da variabilidade espacial da serapilheira em áreas de caatinga de
vegetação nativa é importante para a compreensão da trajetória cíclica dos elementos
minerais essenciais à vida dentro desse ecossistema.

36
Segundo Delitti (1995) a partir da avaliação da queda da serapilheira é possível
estimar de forma indireta, a via de absorção de nutrientes pelas plantas, uma vez que,
quando os ecossistemas estão em equilíbrio, a quantidade de nutrientes transferida pela
queda do folhedo é equivalente a absorvida pelas plantas. Além disso, a quantidade dos
macronutrientes que chegam ao solo via serapilheira, apresenta um padrão sazonal
semelhante àquele da produção da serapilheira (Cesar, 1993).
Em áreas de caatinga, a quantificação da deposição da serapilheira é variável não
só pela vegetação nativa presente nesse ecossistema; a precipitação pluvial (por estação)
influencia na serapilheira acumulada sobre a superfície do solo e consequentemente na
quantidade de matéria orgânica que volta para o sistema em forma de nutrientes para a
planta. Ainda, em períodos de seca prolongada com sistemas de produção que possuem
carga animal também interferem na quantificação desse material, uma vez que estes
consomem as folhas e galhos presentes na serapilheira.
A vegetação xerófita, presente em ambientes semiáridos possui uma eficiente
resiliência a esse habitat, com uma capacidade incrível de recuperação e rápida rebrota,
principalmente do estrato herbáceo. Segundo Souza (2009) o xerofilismo expressa uma
condição de sobrevivência ligada a um ambiente seco, ecologicamente com deficiência
hídrica, cuja água disponível às plantas procede unicamente do curto período da estação
chuvosa.
Assim, a importância de se avaliar a produção de serapilheira está na compreensão
dos reservatórios e fluxos de nutrientes, nestes ecossistemas, os quais se constituem na
principal via de fornecimento de nutrientes, por meio da mineralização dos restos
vegetais (Souza e Davide, 2001).
Por isso, a cada dia fica perceptível a necessidade de se realizarem pesquisas a
curto, médio e longo prazo, que possam dar subsídios ao maior entendimento de como
ocorrem os processos de ciclagem de nutrientes em áreas de caatinga. Por esta razão
faz-se necessário estudar a deposição da serapilheira para melhor subsidiar ações
conservacionistas e orientar práticas de manejo, associando a preservação do meio
ambiente ao desenvolvimento sustentável.
Desta forma, o presente estudo objetivou avaliar a distribuição espacial da
deposição da serapilheira em áreas de caatinga em período prolongado de seca e em
época chuvosa, com diferentes lotação animal.

37
2 MATERIAL E MÉTODOS
2.1 Área de estudo
O experimento foi conduzido na Estação Experimental pertencente a UFPB,
localizada no município de São João do Cariri-PB, nas coordenadas 7o23’30”S e
36o31’59”W, numa altitude de 458 m. A figura 5 demonstra a área delimitada para o
estudo.
Figura 5. Representação das cinco áreas experimentais. Em destaque, na cor vermelha, a subdivisão das
parcelas contíguas.
O município está inserido na zona fisiográfica do Planalto da Borborema,
fazendo parte da microrregião do Cariri Oriental.
Apresenta relevo suave ondulado sobre o embasamento cristalino. Com relação
ao sistema de drenagem, a área faz parte da bacia hidrográfica do rio Paraíba e sub-
bacia do rio Taperoá, riacho Namorado e demais cursos d’água tributários desse canal,
cujas nascentes encontram-se inseridas no território da UFPB. Os solos predominantes
na Estação, de acordo com Chaves e Chaves (2000), são: LUVISSOLO Crômico
vértico, que ocupa a maior parte das terras contidas na área da bacia, apresentando-se
severamente erodido à medida que o relevo torna-se mais acentuado; VERTISSOLO,

38
ocupando as posições mais baixas da bacia, nas circunvizinhanças do açude Namorado
e relevo mais plano no interior da bacia hidrográfica; E NEOSSOLO Lítico, encontrado
nas encostas mais íngremes e em posição de topo associado a afloramentos de rochas,
em relevo ondulado a forte ondulado (Embrapa, 1999). Na área experimental a textura
predominante é franco arenosa e franco argilo-arenosa.
Na área predomina o clima Bsh - semiárido quente com chuvas de verão e o
bioclima 2b apresentando um total de meses secos, variando de 9 a 11, denominado de
subdesértico quente de tendência tropical (Governo do Estado da Paraíba, 1985). A
temperatura média mensal varia de 27,2 ºC no período novembro-março a 23,1 ºC em
julho, sendo estes os valores máximos e mínimos, respectivamente, com precipitações
de 400 mm/ano e umidade relativa de 70%. A vegetação que recobre a região estudada é
a Caatinga hiperxrófita (vegetação caducifólia espinhosa), como demonstra a figura 6.
Figura 6. Fotos representativas das áreas experimentais com presença de lajedo (a); fragmentos de
caatinga hiperxrófita e solo parcialmente descoberto (b); presença de animais em local de repouso,
evidenciando a presença de cíbalas no solo; e folhas compondo a serapilheira (d).
39
A área experimental representou, aproximadamente, 5 hectares, dos quais foram
subdivididos em cinco áreas (1, 2, 3, 4 e 5), tal qual demonstra a figura 7, de 3,3
hectares com diferentes lotação animal e estados de antropização, da seguinte forma:
Área 1 (10 animais – 1 animal/3.200 m2), Área 2 (5 animais – 1 animal/6.400 m
2), Área
3, 4 e 5 (sem animais). Os animais pertencentes às áreas eram caprinos machos, adultos,
sem padrão racial definido.
2.2 Coleta e avaliação dos dados
O método utilizado para a análise dos dados foi o de parcelas contíguas (Muller-
Dombois e Ellemberg, 1974; Rodal et al., 1992) em áreas de um hectare. Esse método
vem sendo utilizado para o estudo da caatinga e permite inferências mais apropriadas
para determinados aspectos da avaliação em estudo. Cinco áreas de caatinga foram
delimitadas em um piquete de um hectare (10.000m²) cada, os quais foram subdivididos
em 100 subparcelas de 10mx10m (Figura 9) com auxílio de um GPS (Global
Positioning System), totalizando uma área amostral de 50.000m2.
100m
10
100m
9
8
7
6
5
4
3
2
1
A B C D E F G H I J

40
Figura 7. Croqui de distribuição das parcelas nos fragmentos de caatinga estudados para levantamento dos
dados, coleta de dejetos caprinos e serapilheira.
Cada subparcela foi identificada na área experimental com estacas de madeira
devidamente alocadas na parte central de cada parcela contígua, com o auxílio de um
profissional equipado com GPS, segundo as figuras 8 e 9.
Figura 8. Esquematização feita em programa de GPS dos pontos centrais marcados nas áreas
experimentais.

41
Figura 9. Fotos demonstrativas, identificando subparcelas da área 1 com presença de lajedo (a), vegetação
hiperxrófita (b) e solo descoberto (c).
2.3 Coleta do solo e dos dados meteorológicos
As coletas de solo tiveram o intuito de caracterizar os atributos físico-químicos
das áreas experimentais. Para tanto, foram realizadas três coletas de forma aleatória nas
cinco áreas, próximo ao local dos dejetos caprinos na profundidade de 0-20 cm,
descartando-se a serapilheira, e formada uma amostra composta de cada área. Foram
retiradas cinco amostras compostas de solo, no total, e levadas ao Laboratório de
Análises Físicas e Químicas do CCA/UFPB, colocadas para secar ao ar, destorroadas,
passadas em peneira de malha de 2 mm (Terra Seca ao Ar - TFSA) e enviadas para
análises físicas e químicas, segundo procedimentos metodológicos contidos no manual
de análise do solo da Embrapa (1997), conforme resultados apresentados no capítulo 4
deste trabalho.
Os dados meteorológicos (pluviometria e temperaturas diárias) foram obtidos na
Estação Meteorológica, classe A, localizada na Estação Experimental de São João do
Cariri.

42
2.4 Metodologia para a obtenção da Serapilheira
2.4.1 Estimativa do estoque de serapilheira acumulada sobre o solo
A serapilheira acumulada na superfície do solo foi estimada através de uma
coleta na estação seca (novembro e dezembro de 2012), e durante a estação das águas
(julho de 2014) em 24 meses de condução do experimento, visando detectar variações
sazonais de forma espacial, através de avaliações em isolinhas. Foram coletas amostras
em cada parcela contígua, como demonstrado na figura 8, em cada hectare demarcado
nas áreas de estudo. Para a quantificação desse material foi utilizado um molde de
estrutura férrea, vazado, medindo 0,5 m x 0,5 m (0,25m²) como demonstra a figura 10,
lançado aleatoriamente em cada subparcela cinco vezes.
Figura 10. Demonstração do molde de ferro quadrado.
A pesagem da serapilheira foi feita com a balança Balmak Actlife de precisão 1g
(própria para campo – capacidade até 5kg) como demonstra a figura 11.

43
Figura 11. Balança de precisão 1g utilizada no experimento.
Toda a serapilheira contida na moldura férrea foi alocada em sacolas plásticas
transparentes e foi feita a pesagem desse material, conforme mostra a figura 12, nos
cinco lances em cada subparcela.
Figura 12. Pesagem da serapilheira alocada em sacos plásticos.
Em seguida foi retirada uma amostra composta representativa de cada
subparcela e logo após, devolvido para o solo o excedente desse material, evitando
assim a degradação do solo.

44
Posteriormente tais amostras foram transferidas para a sede da Estação
experimental para a separação e identificação dos componentes da serapilheira. A
quantidade de serapilheira acumulada foi estimada para g. m-².
2.4.2 Identificação dos componentes da serapilheira
Da serapilheira amostrada, foram separados os seguintes constituintes: folhas
(incluindo folíolos + pecíolo), pool do material lenhoso (correspondente às partes
lenhosas arbóreas, arbustivas e subarbustivas de todas as dimensões + cascas), estrutura
reprodutiva (flores, frutos e sementes), miscelânea (material vegetal que não pôde ser
identificado) e cíbalas (dejetos caprinos) – figura 13. Logo após foram pesados cada
constituinte na mesma balança utilizada para a pesagem a campo, e em seguida,
alocados em sacos de papel devidamente identificados.
Figura 13. Fracionamento dos constituintes da serapilheira em material lignificado (a), folhas (b),
miscelânea (c), estrutura reprodutiva (d) e cíbalas (e).
2.4.3 Plotagem dos Mapas de Isolinhas de Distribuição Espacial
Os mapas de isolinhas de distribuição espacial para a quantificação da
serapilheira foram elaborados com o auxílio dos programas Microsoft Office ® – Excel

45
2010, Surfer® v. 9 (Golden software, Colorado, EUA) e Corel DRAW® v. X4. Com
auxílio de um GPS Garmin® e técnico responsável, foram obtidas as coordenadas de
cada área. O equipamento foi configurado para SAD-69 (Datum Sul Americano) e o
modo de coordenadas adotado foi o UTM (Universal Transverse Mercator). As
coordenadas foram transferidas para o computador com auxílio do aplicativo GPS
TrackMaker® v. 13.8.
A interpolação linear das parcelas perdidas das variáveis foram determinadas
para melhor ajustar os dados nas análises estatísticas, baseados na equação a seguir:
( )
( )
( )
Onde ƒ(a) e ƒ(b) são conhecidos como valores de ƒ(x) em x = a e x = b,
respectivamente.
Após a verificação das variáveis consideradas neste trabalho, observou-se nas
análises de semivariograma, ausência da correlação a curta distância, havendo
dependência espacial entre as amostras. As análises de semivariância foram feitas com
auxílio do programa GS+ (Geostatistics for the Environmental Sciences) v. 10. Assim, a
heterogeneidade espacial das variáveis foi caracterizada através da estatística clássica
com as médias de cada parcela contígua, representadas por mapas na forma de isolinhas
utilizando o programa Surfer® v. 9.0. Cada variável foi avaliada de forma
independente. Os mapas de contorno das variáveis estudadas foram construídos por
interpolação utilizando o método da função da krigagem, por ser considerado o melhor
estimador linear não tendencioso (BLUE = Best Linear Unbiased Estimator) (Ramalho,
2008).
O valor a ser interpolado foi estimado por:
∑
Onde:
Gj =Valor do estimador para o ponto j
NPT =Número de pontos usados para a interpolação
Zi =Valor estimado no ponto i com valor conhecido

46
wij =Peso associado ao valor estimado i
Os mapas com a distribuição espacial dos constituintes da serapilheira foram
elaborados na perspectiva de comparar a distribuição destes em cada área de caatinga
analisada.
A medida de dependência espacial (DE), foi calculada segundo a equação de
Biondi et al. (1994), conforme a fração abaixo:
(
)
Onde, C0 é o efeito pepita; e C1, a contribuição.
2.5 Avaliação da umidade e características químicas e bromatológicas das cíbalas e
da serapilheira
Para esta avaliação foram colhidas quatro amostras de forma aleatória da
serapilheira nas cinco áreas delimitadas neste experimento no período seco (janeiro de
2013). O material foi pesado e em seguida, seco em estufa com circulação e renovação
forçada de ar (± 55°C) até atingir peso constante, triturado em moinho tipo willey e
posteriormente analisado quanto ao conteúdo de água, determinadas as quantidades de
matéria seca (MS), matéria orgânica (MO), cinzas (CZ), proteína bruta (PB), fibra em
detergente neutro (FDN), fibra em detergente ácido (FDA), segundo a metodologia
descrita por Silva e Queiroz (2002). As análises foram realizadas no Laboratório de
Nutrição Animal (LANA) do CCA/UFPB.
Para a determinação da umidade da serapilheira foi feita a pesagem das alíquotas
antes e após a pré-secagem e, posteriormente, determinada a percentagem do conteúdo
de água através do programa Microsoft Excel 2007®.

47
Figura 14. Amostras em laboratório da serapilheira.
Para a análise das cíbalas, foram feitas amostras composta através de coletas das
áreas 1 e 2, onde predominava a aparição destas, conforme demonstra a figura 15.

48
Figura 15. Demonstração das cíbalas depositadas em área experimental (a), do estádio de conservação na
época de coleta (b) e demonstração das baias com caprinos (c).
Em seguida, foram secas em estufa com circulação e renovação forçada de ar (±
55°C) até atingir peso constante, triturado em moinho tipo willey e posteriormente
analisada quanto aos teores de macronutrientes (MO, C, N, K, P e Na) segundo a
metodologia da Embrapa (1997). As análises foram realizadas no Laboratório de Solos
da UFPB, Campus II.
3 RESULTADOS E DISCUSSÃO
3.1 Produção da Serapilheira Total e Fracionada e Relevo do solo
O experimento foi conduzido na época seca, porém com uma importante
observação, pois este período representou a pior seca dos últimos cinquenta anos (desde
1962) com mais de 1.400 municípios afetados (ONU, 2013).
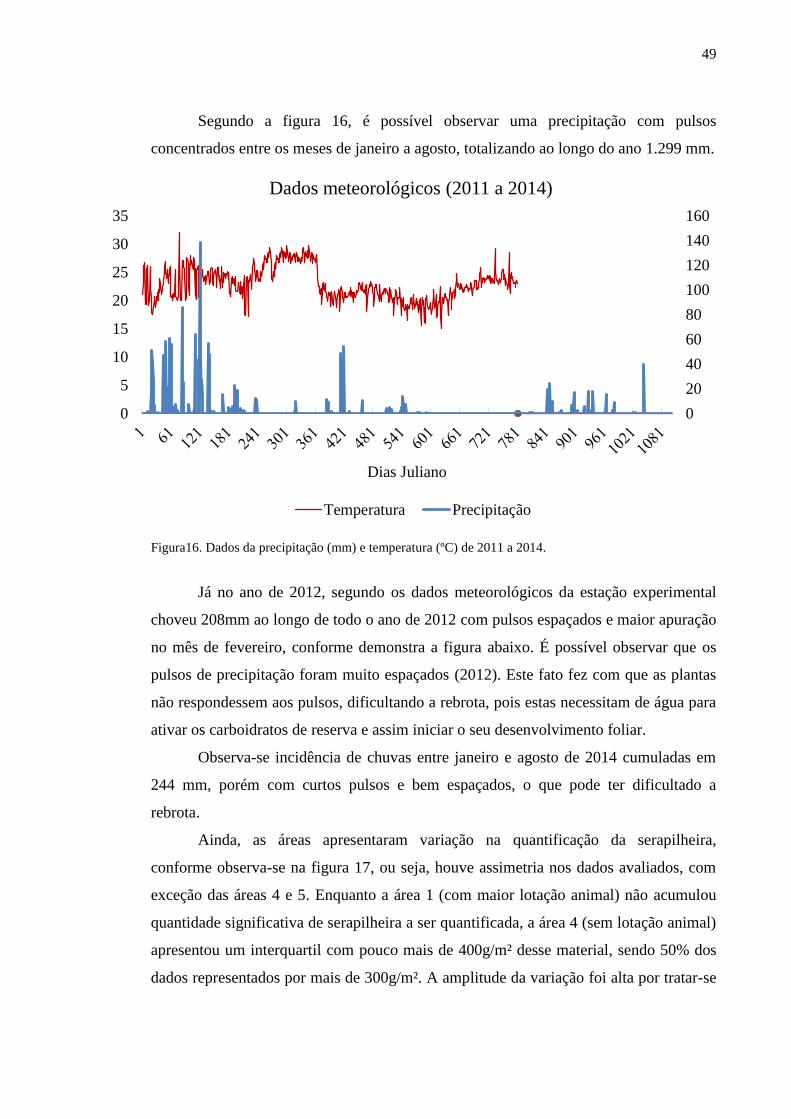
49
Segundo a figura 16, é possível observar uma precipitação com pulsos
concentrados entre os meses de janeiro a agosto, totalizando ao longo do ano 1.299 mm.
Figura16. Dados da precipitação (mm) e temperatura (ºC) de 2011 a 2014.
Já no ano de 2012, segundo os dados meteorológicos da estação experimental
choveu 208mm ao longo de todo o ano de 2012 com pulsos espaçados e maior apuração
no mês de fevereiro, conforme demonstra a figura abaixo. É possível observar que os
pulsos de precipitação foram muito espaçados (2012). Este fato fez com que as plantas
não respondessem aos pulsos, dificultando a rebrota, pois estas necessitam de água para
ativar os carboidratos de reserva e assim iniciar o seu desenvolvimento foliar.
Observa-se incidência de chuvas entre janeiro e agosto de 2014 cumuladas em
244 mm, porém com curtos pulsos e bem espaçados, o que pode ter dificultado a
rebrota.
Ainda, as áreas apresentaram variação na quantificação da serapilheira,
conforme observa-se na figura 17, ou seja, houve assimetria nos dados avaliados, com
exceção das áreas 4 e 5. Enquanto a área 1 (com maior lotação animal) não acumulou
quantidade significativa de serapilheira a ser quantificada, a área 4 (sem lotação animal)
apresentou um interquartil com pouco mais de 400g/m² desse material, sendo 50% dos
dados representados por mais de 300g/m². A amplitude da variação foi alta por tratar-se
0
20
40
60
80
100
120
140
160
0
5
10
15
20
25
30
35
Dias Juliano
Dados meteorológicos (2011 a 2014)
Temperatura Precipitação
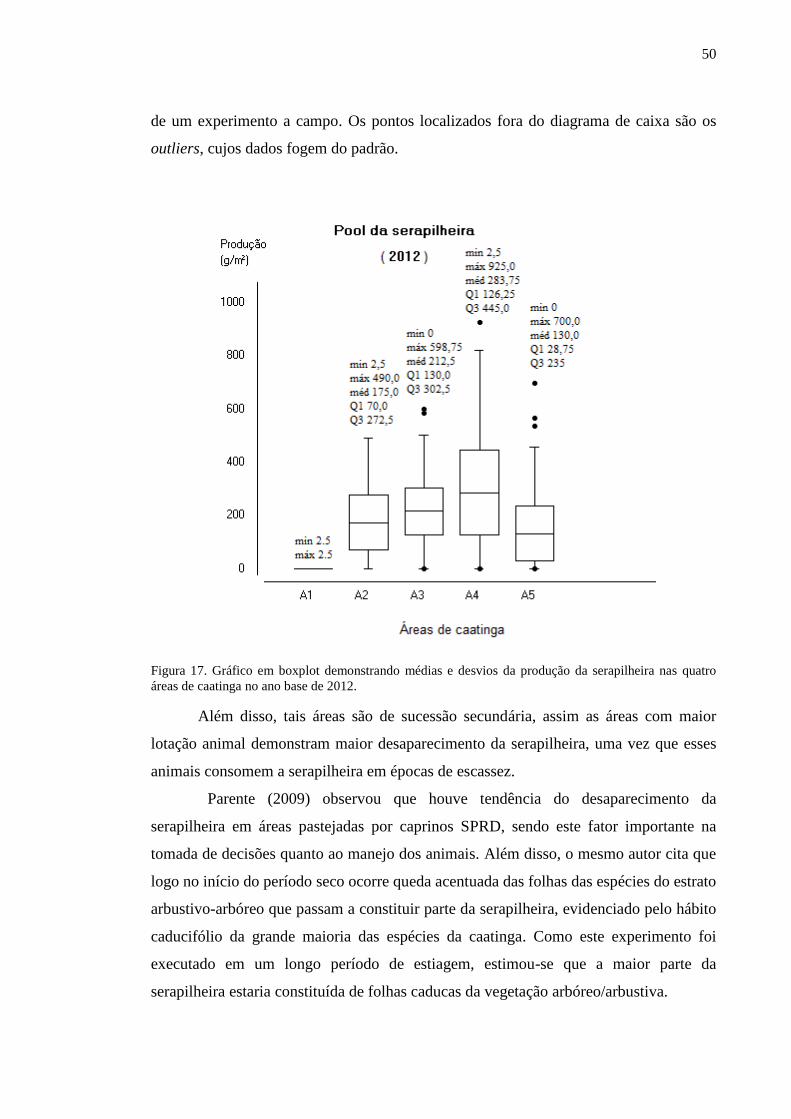
50
de um experimento a campo. Os pontos localizados fora do diagrama de caixa são os
outliers, cujos dados fogem do padrão.
Figura 17. Gráfico em boxplot demonstrando médias e desvios da produção da serapilheira nas quatro
áreas de caatinga no ano base de 2012.
Além disso, tais áreas são de sucessão secundária, assim as áreas com maior
lotação animal demonstram maior desaparecimento da serapilheira, uma vez que esses
animais consomem a serapilheira em épocas de escassez.
Parente (2009) observou que houve tendência do desaparecimento da
serapilheira em áreas pastejadas por caprinos SPRD, sendo este fator importante na
tomada de decisões quanto ao manejo dos animais. Além disso, o mesmo autor cita que
logo no início do período seco ocorre queda acentuada das folhas das espécies do estrato
arbustivo-arbóreo que passam a constituir parte da serapilheira, evidenciado pelo hábito
caducifólio da grande maioria das espécies da caatinga. Como este experimento foi
executado em um longo período de estiagem, estimou-se que a maior parte da
serapilheira estaria constituída de folhas caducas da vegetação arbóreo/arbustiva.
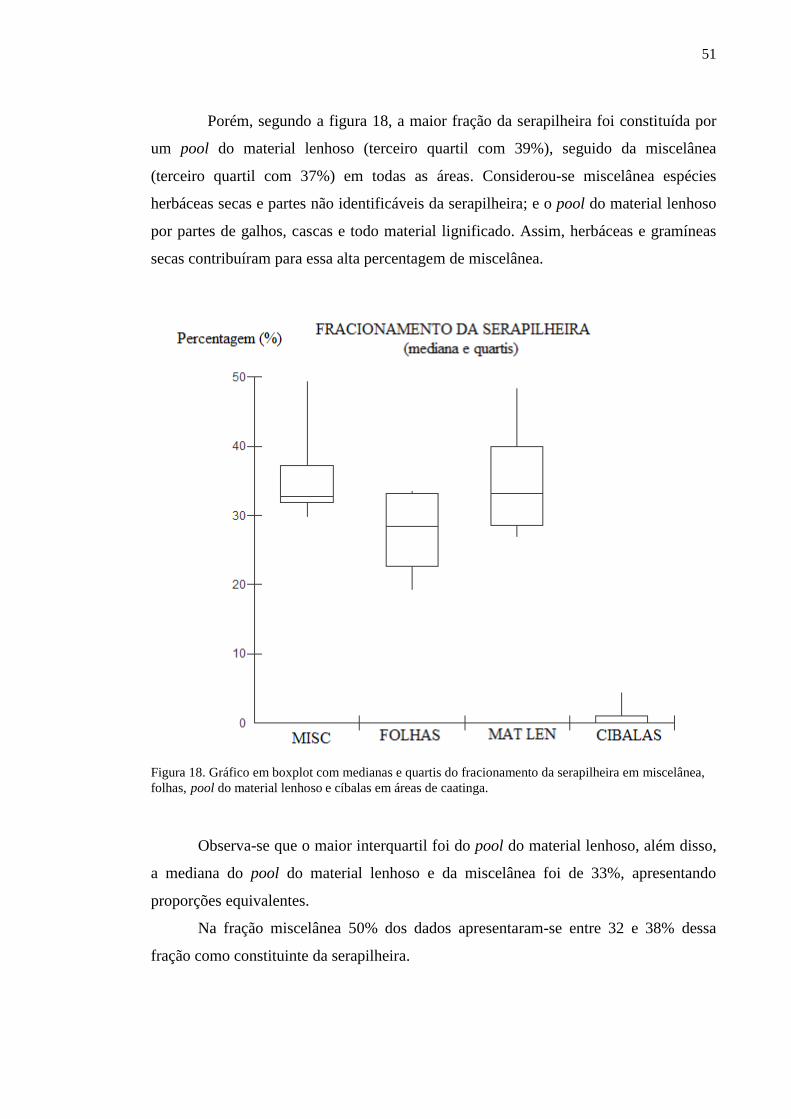
51
Porém, segundo a figura 18, a maior fração da serapilheira foi constituída por
um pool do material lenhoso (terceiro quartil com 39%), seguido da miscelânea
(terceiro quartil com 37%) em todas as áreas. Considerou-se miscelânea espécies
herbáceas secas e partes não identificáveis da serapilheira; e o pool do material lenhoso
por partes de galhos, cascas e todo material lignificado. Assim, herbáceas e gramíneas
secas contribuíram para essa alta percentagem de miscelânea.
Figura 18. Gráfico em boxplot com medianas e quartis do fracionamento da serapilheira em miscelânea,
folhas, pool do material lenhoso e cíbalas em áreas de caatinga.
Observa-se que o maior interquartil foi do pool do material lenhoso, além disso,
a mediana do pool do material lenhoso e da miscelânea foi de 33%, apresentando
proporções equivalentes.
Na fração miscelânea 50% dos dados apresentaram-se entre 32 e 38% dessa
fração como constituinte da serapilheira.
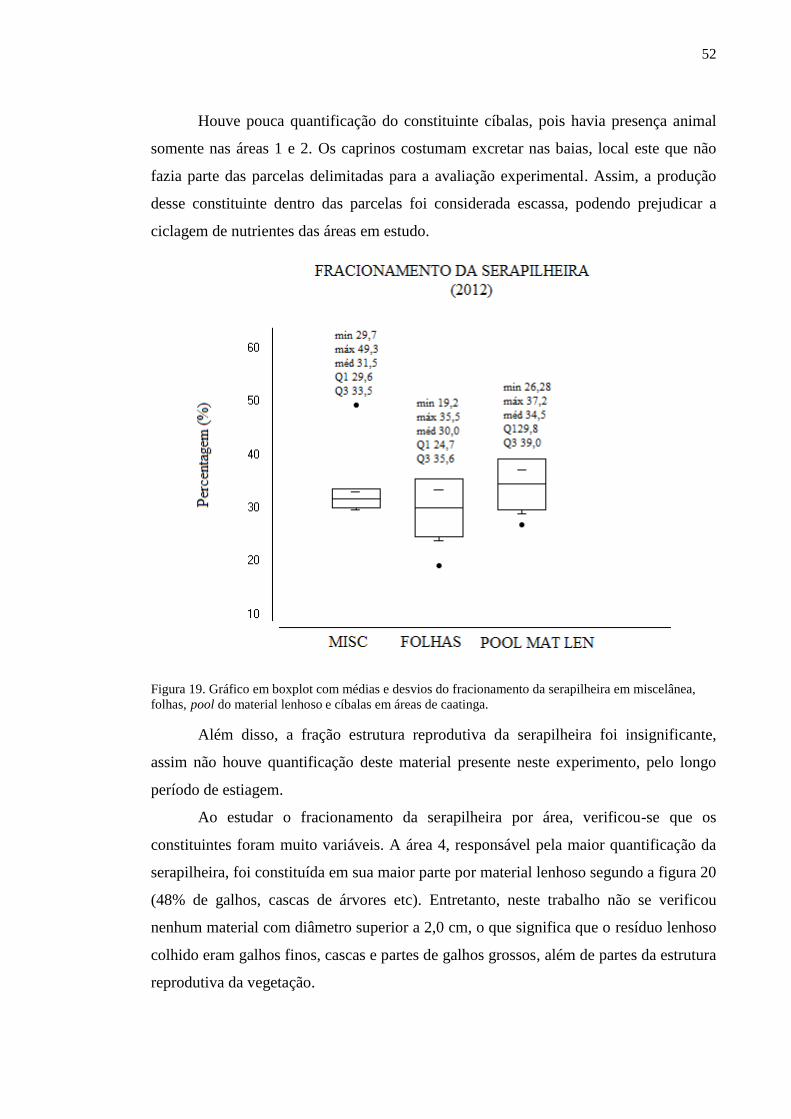
52
Houve pouca quantificação do constituinte cíbalas, pois havia presença animal
somente nas áreas 1 e 2. Os caprinos costumam excretar nas baias, local este que não
fazia parte das parcelas delimitadas para a avaliação experimental. Assim, a produção
desse constituinte dentro das parcelas foi considerada escassa, podendo prejudicar a
ciclagem de nutrientes das áreas em estudo.
Figura 19. Gráfico em boxplot com médias e desvios do fracionamento da serapilheira em miscelânea,
folhas, pool do material lenhoso e cíbalas em áreas de caatinga.
Além disso, a fração estrutura reprodutiva da serapilheira foi insignificante,
assim não houve quantificação deste material presente neste experimento, pelo longo
período de estiagem.
Ao estudar o fracionamento da serapilheira por área, verificou-se que os
constituintes foram muito variáveis. A área 4, responsável pela maior quantificação da
serapilheira, foi constituída em sua maior parte por material lenhoso segundo a figura 20
(48% de galhos, cascas de árvores etc). Entretanto, neste trabalho não se verificou
nenhum material com diâmetro superior a 2,0 cm, o que significa que o resíduo lenhoso
colhido eram galhos finos, cascas e partes de galhos grossos, além de partes da estrutura
reprodutiva da vegetação.
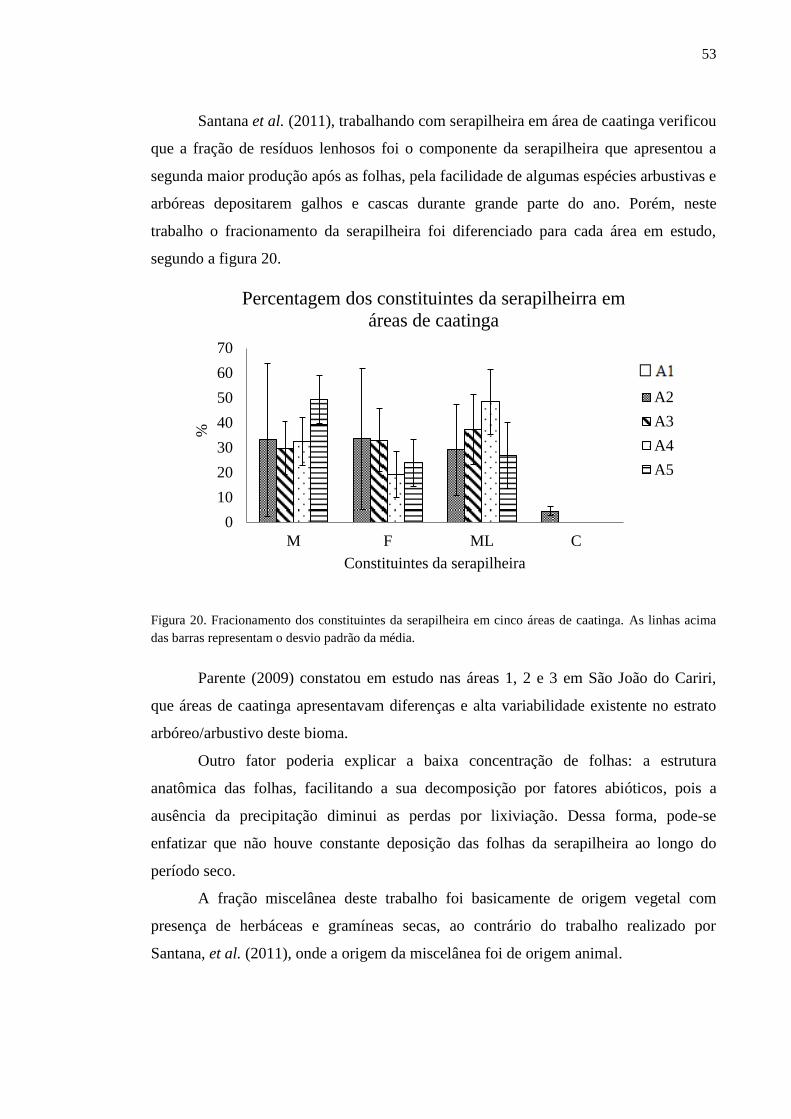
53
Santana et al. (2011), trabalhando com serapilheira em área de caatinga verificou
que a fração de resíduos lenhosos foi o componente da serapilheira que apresentou a
segunda maior produção após as folhas, pela facilidade de algumas espécies arbustivas e
arbóreas depositarem galhos e cascas durante grande parte do ano. Porém, neste
trabalho o fracionamento da serapilheira foi diferenciado para cada área em estudo,
segundo a figura 20.
Figura 20. Fracionamento dos constituintes da serapilheira em cinco áreas de caatinga. As linhas acima
das barras representam o desvio padrão da média.
Parente (2009) constatou em estudo nas áreas 1, 2 e 3 em São João do Cariri,
que áreas de caatinga apresentavam diferenças e alta variabilidade existente no estrato
arbóreo/arbustivo deste bioma.
Outro fator poderia explicar a baixa concentração de folhas: a estrutura
anatômica das folhas, facilitando a sua decomposição por fatores abióticos, pois a
ausência da precipitação diminui as perdas por lixiviação. Dessa forma, pode-se
enfatizar que não houve constante deposição das folhas da serapilheira ao longo do
período seco.
A fração miscelânea deste trabalho foi basicamente de origem vegetal com
presença de herbáceas e gramíneas secas, ao contrário do trabalho realizado por
Santana, et al. (2011), onde a origem da miscelânea foi de origem animal.
0
10
20
30
40
50
60
70
M F ML C
%
Constituintes da serapilheira
Percentagem dos constituintes da serapilheirra em
áreas de caatinga
A2
A3
A4
A5
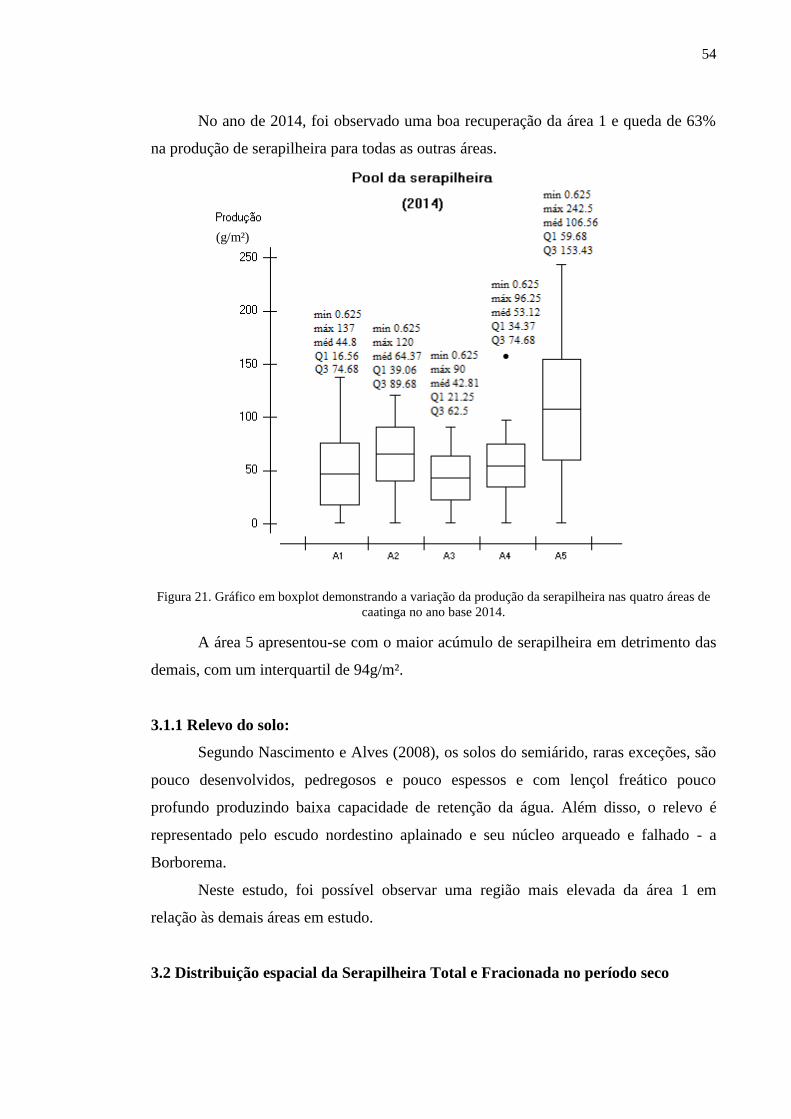
54
No ano de 2014, foi observado uma boa recuperação da área 1 e queda de 63%
na produção de serapilheira para todas as outras áreas.
Figura 21. Gráfico em boxplot demonstrando a variação da produção da serapilheira nas quatro áreas de
caatinga no ano base 2014.
A área 5 apresentou-se com o maior acúmulo de serapilheira em detrimento das
demais, com um interquartil de 94g/m².
3.1.1 Relevo do solo:
Segundo Nascimento e Alves (2008), os solos do semiárido, raras exceções, são
pouco desenvolvidos, pedregosos e pouco espessos e com lençol freático pouco
profundo produzindo baixa capacidade de retenção da água. Além disso, o relevo é
representado pelo escudo nordestino aplainado e seu núcleo arqueado e falhado - a
Borborema.
Neste estudo, foi possível observar uma região mais elevada da área 1 em
relação às demais áreas em estudo.
3.2 Distribuição espacial da Serapilheira Total e Fracionada no período seco
(g/m²)
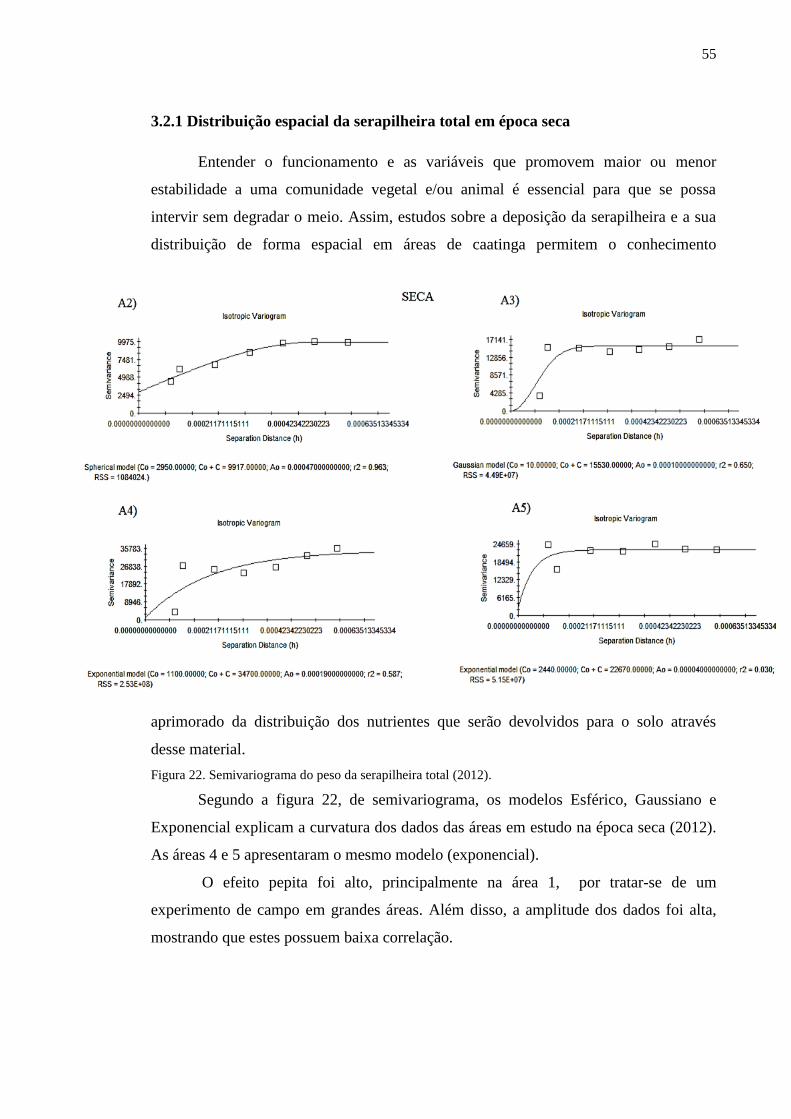
55
3.2.1 Distribuição espacial da serapilheira total em época seca
Entender o funcionamento e as variáveis que promovem maior ou menor
estabilidade a uma comunidade vegetal e/ou animal é essencial para que se possa
intervir sem degradar o meio. Assim, estudos sobre a deposição da serapilheira e a sua
distribuição de forma espacial em áreas de caatinga permitem o conhecimento
aprimorado da distribuição dos nutrientes que serão devolvidos para o solo através
desse material.
Figura 22. Semivariograma do peso da serapilheira total (2012).
Segundo a figura 22, de semivariograma, os modelos Esférico, Gaussiano e
Exponencial explicam a curvatura dos dados das áreas em estudo na época seca (2012).
As áreas 4 e 5 apresentaram o mesmo modelo (exponencial).
O efeito pepita foi alto, principalmente na área 1, por tratar-se de um
experimento de campo em grandes áreas. Além disso, a amplitude dos dados foi alta,
mostrando que estes possuem baixa correlação.
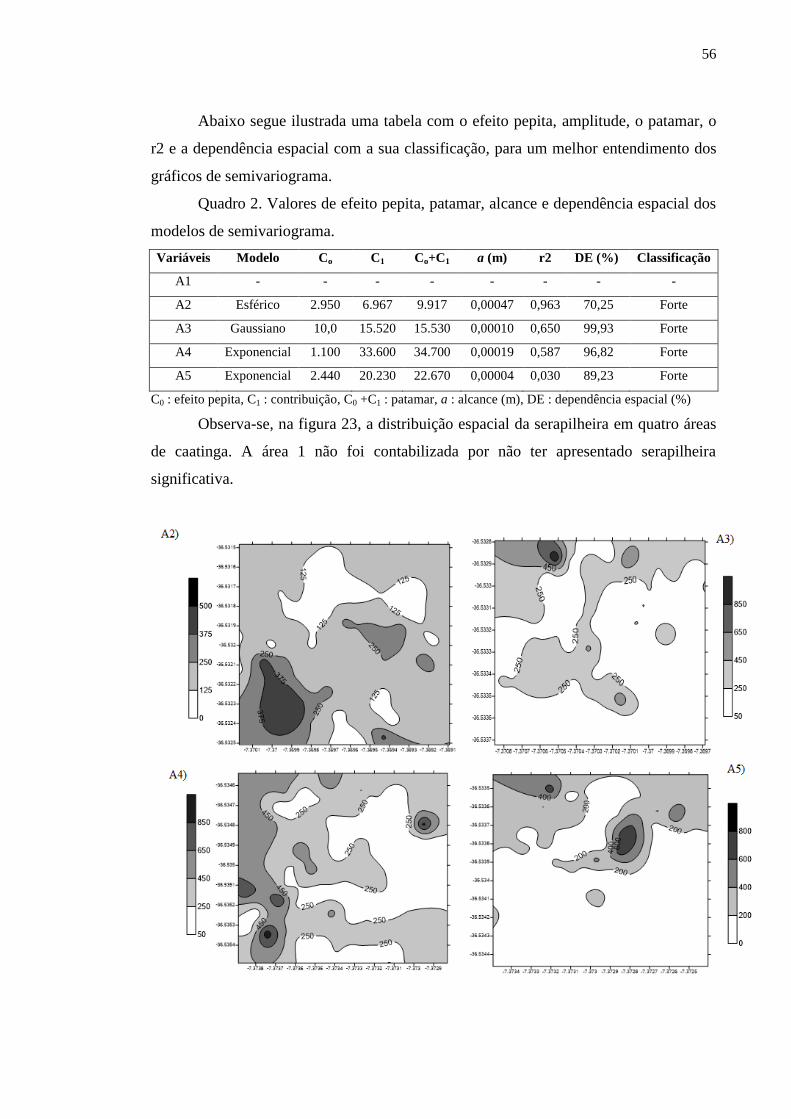
56
Abaixo segue ilustrada uma tabela com o efeito pepita, amplitude, o patamar, o
r2 e a dependência espacial com a sua classificação, para um melhor entendimento dos
gráficos de semivariograma.
Quadro 2. Valores de efeito pepita, patamar, alcance e dependência espacial dos
modelos de semivariograma.
Variáveis Modelo Co C1 Co+C1 a (m) r2 DE (%) Classificação
A1 - - - - - - - -
A2 Esférico 2.950 6.967 9.917 0,00047 0,963 70,25 Forte
A3 Gaussiano 10,0 15.520 15.530 0,00010 0,650 99,93 Forte
A4 Exponencial 1.100 33.600 34.700 0,00019 0,587 96,82 Forte
A5 Exponencial 2.440 20.230 22.670 0,00004 0,030 89,23 Forte
C0 : efeito pepita, C1 : contribuição, C0 +C1 : patamar, a : alcance (m), DE : dependência espacial (%)
Observa-se, na figura 23, a distribuição espacial da serapilheira em quatro áreas
de caatinga. A área 1 não foi contabilizada por não ter apresentado serapilheira
significativa.

57
Figura 23. Isolinhas da distribuição do pool da serapilheira (g.m-²) em quatro áreas de caatinga no ano de
2012.
Pode-se observar que as áreas 4 e 3 apresentaram maior acúmulo de serapilheira,
seguidas das áreas 5 e 2, respectivamente.
As áreas 2 e 3 são constituídas em parte, por uma área alagada na época
chuvosa, onde predomina o capim Aristida adscensionis, na época seca. Na época
experimental este capim fez parte da miscelânea por não ter sido classificado como
material lenhoso, folhas ou estrutura reprodutiva da serapilheira.
Segundo Araújo (2010), em estudo semelhante nas mesmas áreas de caatinga,
afirma que as espécies mais frequentes na área 1 foram Croton sonderianus
(Marmeleiro) representando 54,05%, Pilosocereus gounellei (Xique-xique) com
13,05% e Poincianella pyramidalis Tul. (Catingueira) com 11,22% no ano base de
2010. Neste estudo, asespécies mais encontradas na área 1 foram: P. pyramidalis
(47.7%), A. pyrifolium (Pereiro) (23.49%) e P. gounellei (17.92%) no ano base de 2014,
mostrando-se diferenciada das demais.
Os animais presentes nesta área têm preferência a consumir o Marmeleiro em
época chuvosa. Assim, é possível afirmar que a quantidade da serapilheira depositada
no solo foi considerada insuficiente pela presença de cactáceas nesta área, aliado ao
fator do consumo dos animais. Ademais, estes animais têm o hábito comportamental de
ingerir folhas de catingueira secas (que fazem parte da serapilheira acumulada no solo),
o que também pode ter contribuído para o desaparecimento da serapilheira.
Já na área 2, as espécies verificadas com maior frequência, foram Croton
sonderianus apresentando 51,70%, seguidas de Poincianella pyramidalis, Jatropha
mollisssima (Pinhão) e Pilosocereus gounellei com 15,30, 12,40 e 11,50% ,
respectivamente (Araújo, 2010) no ano base de 2010. Já em 2014, verificou-se a
predominância de P. pyramidalis (42.64%), C. sonderianus (15.32%) e A. pyrifolium
(14.71%). Assim, verificou-se uma semelhança entre as áreas 1 e 2.
Logo, a serapilheira será diferenciada em detrimento da lotação animal ficando
claro que a presença de uma alta lotação pode prejudicar o acúmulo da serapilheira e,
consequentemente a ciclagem dos nutrientes para o solo.
É possível afirmar ainda, com base na figura 23 e conforme o trabalho de Araújo
(2010), que nos locais onde ocorreu uma maior deposição da serapilheira estão

58
localizados os estratos arbóreo/arbustivos de maior frequência (citados acima). O local
com maior deposição da serapilheira apresentou apenas 1,24% da área amostral da
serapilheira e corresponde acima de 500g.m-2
. A maior parte representativa da área foi
de 55,80% de 100 a 200g.m-2
de serapilheira.
A área 3 (sem animais), mostrou-se diferenciada das demais em 2010. As
espécies que se destacaram foram o Croton sonderianus, seguido de Sida sp. (Malva),
Jatropha mollisssima e Poincianella pyramidalis, representando 42,73, 13,45, 12,08 e
11,89% (Araújo, 2010) no ano base de 2010. Já em 2014, as espécies predominantes
foram P. pyramidalis (29.52%), A. pyrifolium (17.78%) e C. sonderianus (16.51%), ou
seja, semelhantes à área 2. Esta área apresentou-se com a segunda maior deposição da
serapilheira acumulada sobre o solo. Tal fato pode ser comprovado pela ausência dos
animais na área em estudo. Observou-se nas análises das isoietas que 93,67% da área
correspondeu entre 0 a 600 g.m-2
de serapilheira.
Apesar dos resultados, não é possível afirmar que o pastejo caprino degrada as
áreas, pois a área 5 apresentou-se semelhante a área 2, apesar de não haver presença
animal. Da mesma forma, a área 4 mostrou-se semelhante a área 3, quando se trata da
deposição da serapilheira. Como todas estas áreas são de sucessão secundária, é
plausível afirmar que a antropização contribuiu significativamente para a degradação
deste bioma.
Além disso, é importante salientar a grande proporção do solo descoberto em
estádio avançado de degradação (erosão) na área 5. Observou-se ao avaliar as isolinhas,
que 68,47% do mapa apresentou uma produção de serapilheira entre 0 a 200g.m-2
.
Assim, é possível considerar que a baixa produção de serapilheira na maior parte na
área é devido a grande parte do solo descoberto.
Nas áreas 4 e 5, foi observado uma grande presença de Sida sp. (malva) e
Aristida adscensionis (capim panasco). Já foi comprovado que o surgimento do capim
panasco é uma prova circunstancial de áreas degradadas ou em estádio sucessional de
degradação, (Guedes et al. 2008) inferindo uma precedente antropização das áreas
atualmente estudadas.
Além disso, as espécies arbóreas predominantes da área 4 foram: C. sonderianus
(31,34%), P. pyramidalis (26,27%) e P. gounellei (17,78%), diferenciando-se das

59
demais. Já na área 5 foram: P. pyramidalis (24,16%) , J. molíssima (20.22%) e A.
pyrifolium (19,1%) no ano base 2014.
Segundo Pereira Júnior e Araújo (2012), um maior valor de importância da
espécie C. sonderianus pode indicar área de sucessão, evidenciando o aumento de
degradação da área.
3.2.2 Distribuição Espacial do fracionamento da serapilheira em época seca
É necessária a compreensão de como se distribuem os componentes da
serapilheira nas áreas de estudo, uma vez que tais elementos sofrem interferência do
vento e da lixiviação. Além disso, a estrutura vegetativa (herbáceo, arbóreo ou
arbustivo) também infere na variação espacial desse material.
Foi observado em estudos na mesma área (Araújo, 2010; Guedes et al. 2008;
Andrade, 2008) que há uma grande variabilidade de indivíduos da vegetação com
distribuição heterogênea nas áreas de sucessão secundária. Dessa forma, o estudo da
distribuição espacial dos constituintes da serapilheira pode explicar a contribuição da
ação do vento na deposição desse material sobre o solo.
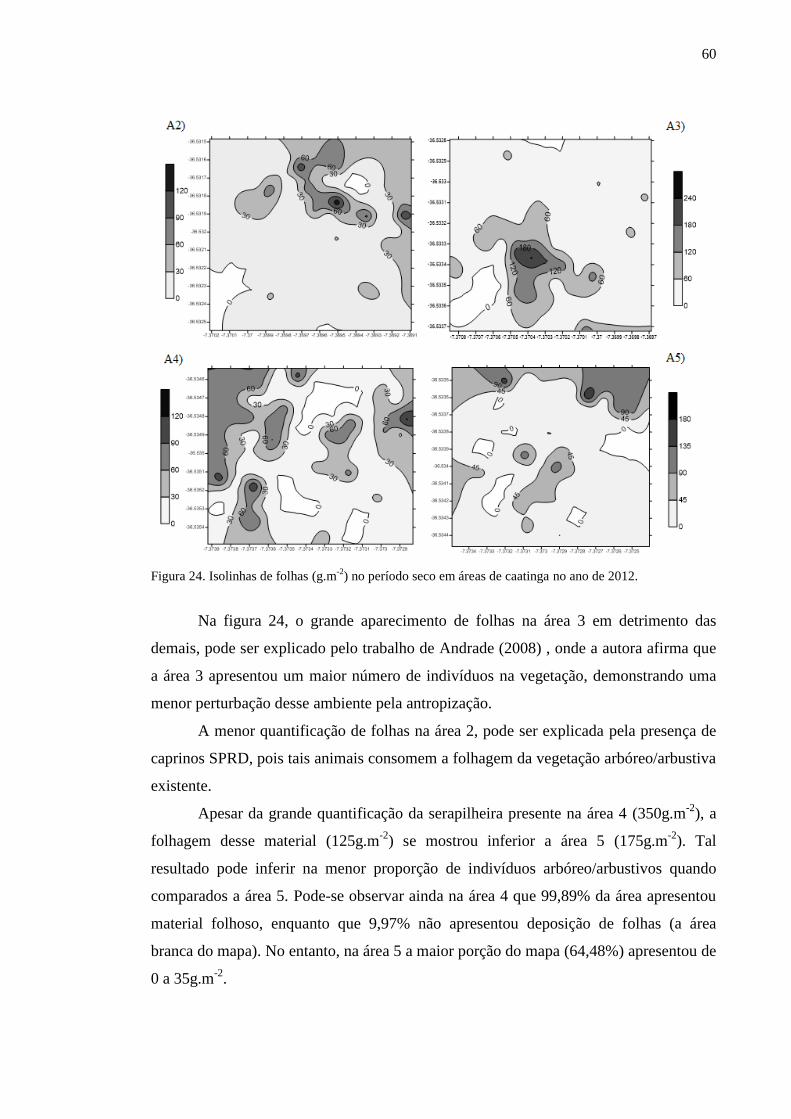
60
Figura 24. Isolinhas de folhas (g.m-2
) no período seco em áreas de caatinga no ano de 2012.
Na figura 24, o grande aparecimento de folhas na área 3 em detrimento das
demais, pode ser explicado pelo trabalho de Andrade (2008) , onde a autora afirma que
a área 3 apresentou um maior número de indivíduos na vegetação, demonstrando uma
menor perturbação desse ambiente pela antropização.
A menor quantificação de folhas na área 2, pode ser explicada pela presença de
caprinos SPRD, pois tais animais consomem a folhagem da vegetação arbóreo/arbustiva
existente.
Apesar da grande quantificação da serapilheira presente na área 4 (350g.m-2
), a
folhagem desse material (125g.m-2
) se mostrou inferior a área 5 (175g.m-2
). Tal
resultado pode inferir na menor proporção de indivíduos arbóreo/arbustivos quando
comparados a área 5. Pode-se observar ainda na área 4 que 99,89% da área apresentou
material folhoso, enquanto que 9,97% não apresentou deposição de folhas (a área
branca do mapa). No entanto, na área 5 a maior porção do mapa (64,48%) apresentou de
0 a 35g.m-2
.
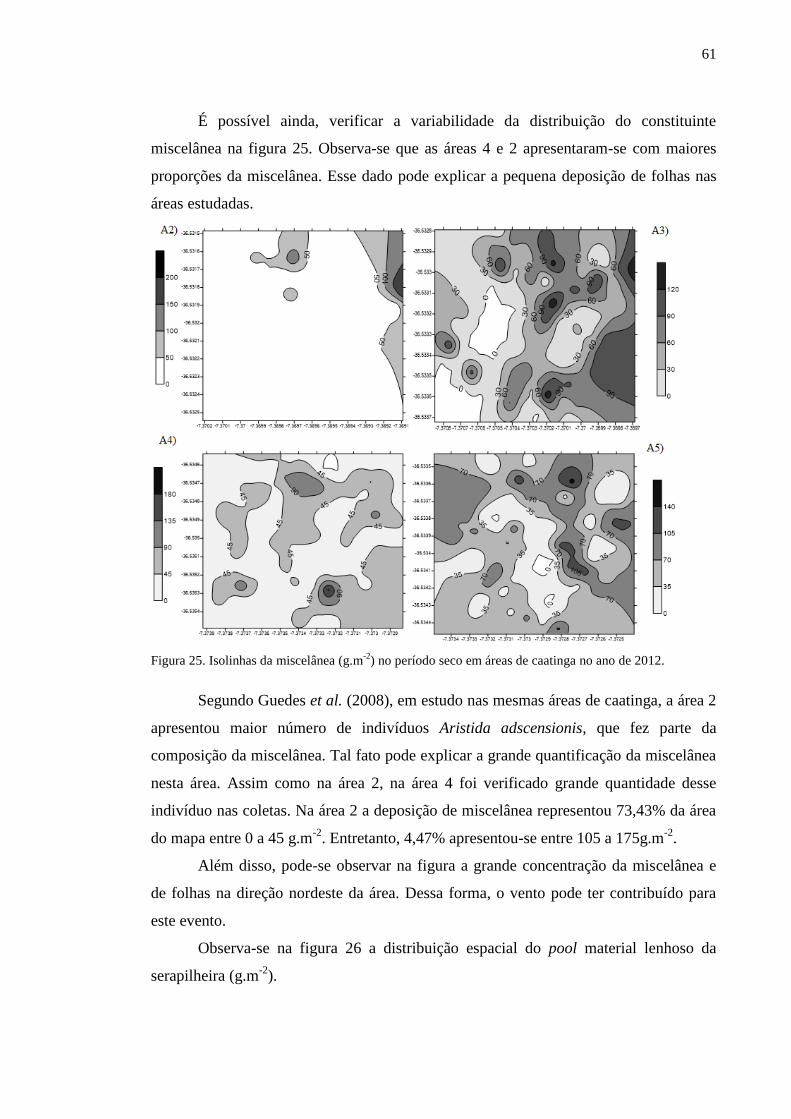
61
É possível ainda, verificar a variabilidade da distribuição do constituinte
miscelânea na figura 25. Observa-se que as áreas 4 e 2 apresentaram-se com maiores
proporções da miscelânea. Esse dado pode explicar a pequena deposição de folhas nas
áreas estudadas.
Figura 25. Isolinhas da miscelânea (g.m-2
) no período seco em áreas de caatinga no ano de 2012.
Segundo Guedes et al. (2008), em estudo nas mesmas áreas de caatinga, a área 2
apresentou maior número de indivíduos Aristida adscensionis, que fez parte da
composição da miscelânea. Tal fato pode explicar a grande quantificação da miscelânea
nesta área. Assim como na área 2, na área 4 foi verificado grande quantidade desse
indivíduo nas coletas. Na área 2 a deposição de miscelânea representou 73,43% da área
do mapa entre 0 a 45 g.m-2
. Entretanto, 4,47% apresentou-se entre 105 a 175g.m-2
.
Além disso, pode-se observar na figura a grande concentração da miscelânea e
de folhas na direção nordeste da área. Dessa forma, o vento pode ter contribuído para
este evento.
Observa-se na figura 26 a distribuição espacial do pool material lenhoso da
serapilheira (g.m-2
).
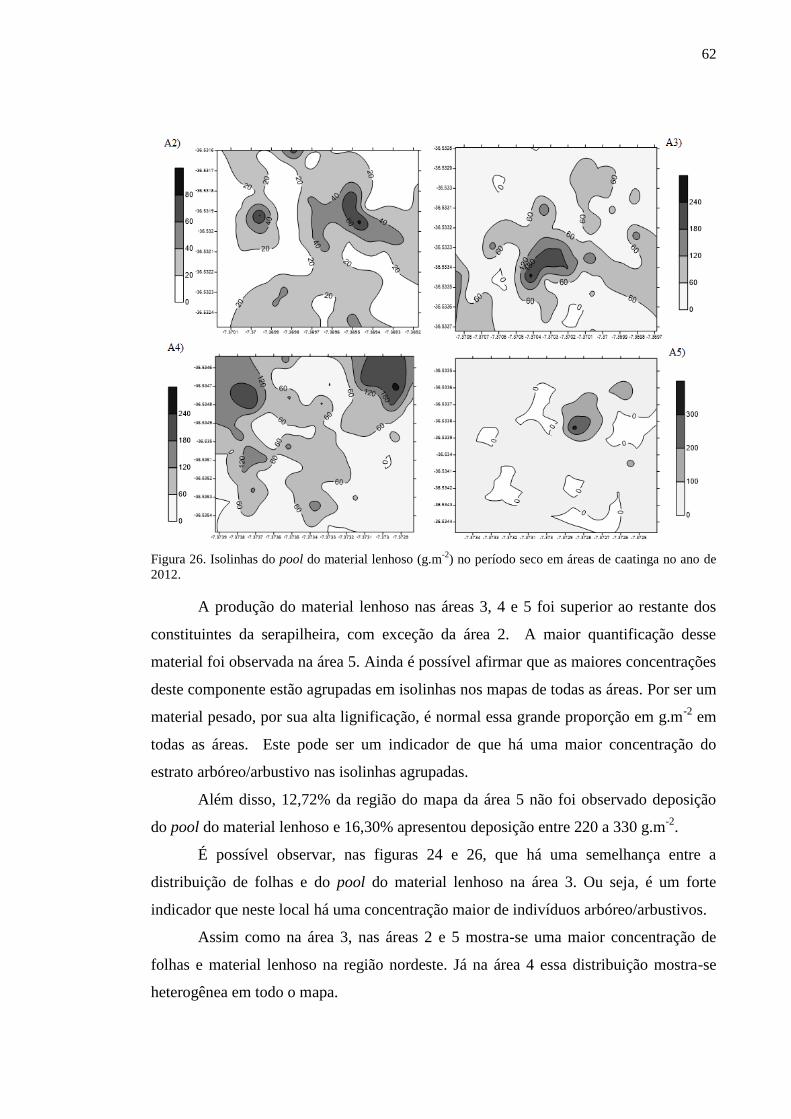
62
Figura 26. Isolinhas do pool do material lenhoso (g.m-2
) no período seco em áreas de caatinga no ano de
2012.
A produção do material lenhoso nas áreas 3, 4 e 5 foi superior ao restante dos
constituintes da serapilheira, com exceção da área 2. A maior quantificação desse
material foi observada na área 5. Ainda é possível afirmar que as maiores concentrações
deste componente estão agrupadas em isolinhas nos mapas de todas as áreas. Por ser um
material pesado, por sua alta lignificação, é normal essa grande proporção em g.m-2
em
todas as áreas. Este pode ser um indicador de que há uma maior concentração do
estrato arbóreo/arbustivo nas isolinhas agrupadas.
Além disso, 12,72% da região do mapa da área 5 não foi observado deposição
do pool do material lenhoso e 16,30% apresentou deposição entre 220 a 330 g.m-2
.
É possível observar, nas figuras 24 e 26, que há uma semelhança entre a
distribuição de folhas e do pool do material lenhoso na área 3. Ou seja, é um forte
indicador que neste local há uma concentração maior de indivíduos arbóreo/arbustivos.
Assim como na área 3, nas áreas 2 e 5 mostra-se uma maior concentração de
folhas e material lenhoso na região nordeste. Já na área 4 essa distribuição mostra-se
heterogênea em todo o mapa.

63
Segundo tais informações, pode-se afirmar que quanto maior a produção do do
pool do material lenhoso e de folhas da serapilheira nas áreas, maior é a probabilidade
da presença do estrato arbóreo/arbustivo.
Na figura 27, observa-se a distribuição espacial das cíbalas na área 2. É possível
estabelecer uma ligação entre o aparecimento de folhas e de cíbalas na região nordeste
do mapa. É provável que os animais passem a maior parte do tempo nesta região se
alimentando de folhas do estrato arbustivo e arbóreo no período chuvoso. Assim, a
deposição das cíbalas nesta região deve-se ao acúmulo desse material no período das
águas.
Figura 27. Isolinhas das cíbalas (g.m
-2) no período seco na área 2 no ano base de 2012.
Aproximadamente, 10,25% da região do mapa apresentou uma produção de 6 a
8 g.m-2
de cíbalas, enquanto que 54,95% da área do mapa representou de 0 a 4 g.m-2
.
Sendo assim, pode-se afirmar que há uma concentração de cíbalas na parte
norte/nordeste da área do mapa. Além disso, pode-se afirmar que a produção de cíbalas
na maior parte da área foi mínima.
3.3 Distribuição espacial da Serapilheira Total e Fracionada no período chuvoso
3.3.1 Distribuição espacial da serapilheira total em época chuvosa
Pode-se observar na figura 28, os semivariogramas da distribuição espacial na época
chuvosa (2014).
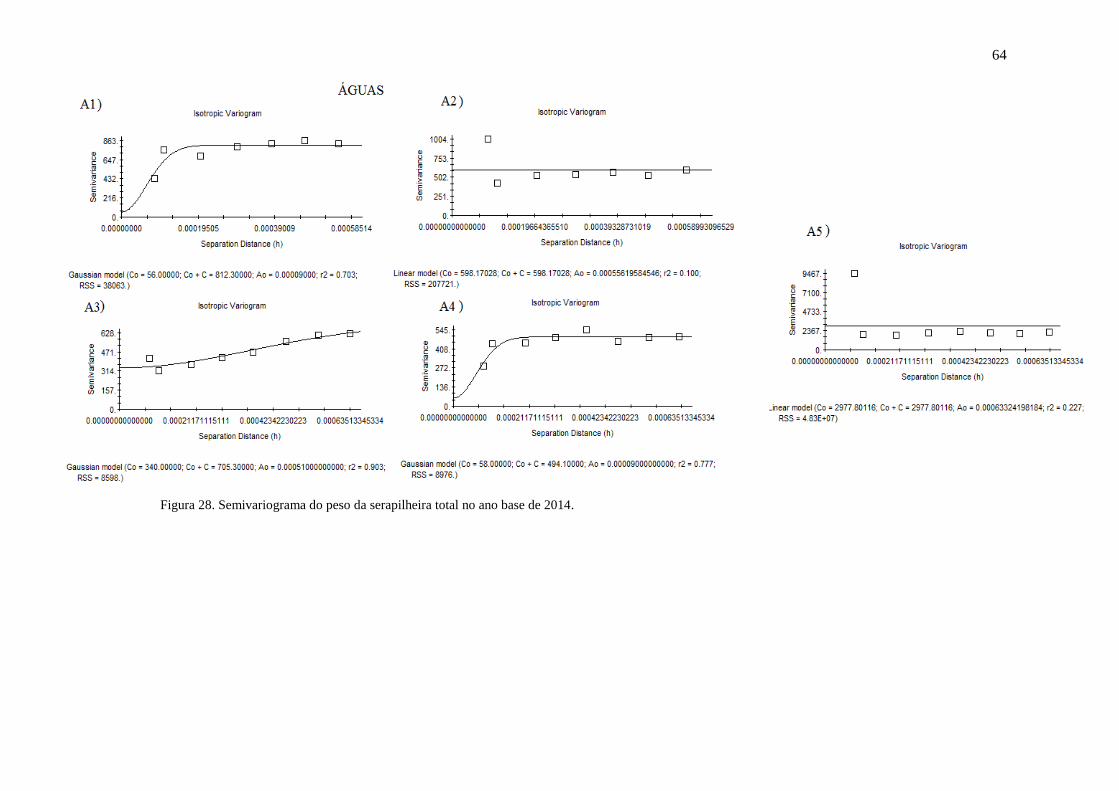
64
Figura 28. Semivariograma do peso da serapilheira total no ano base de 2014.
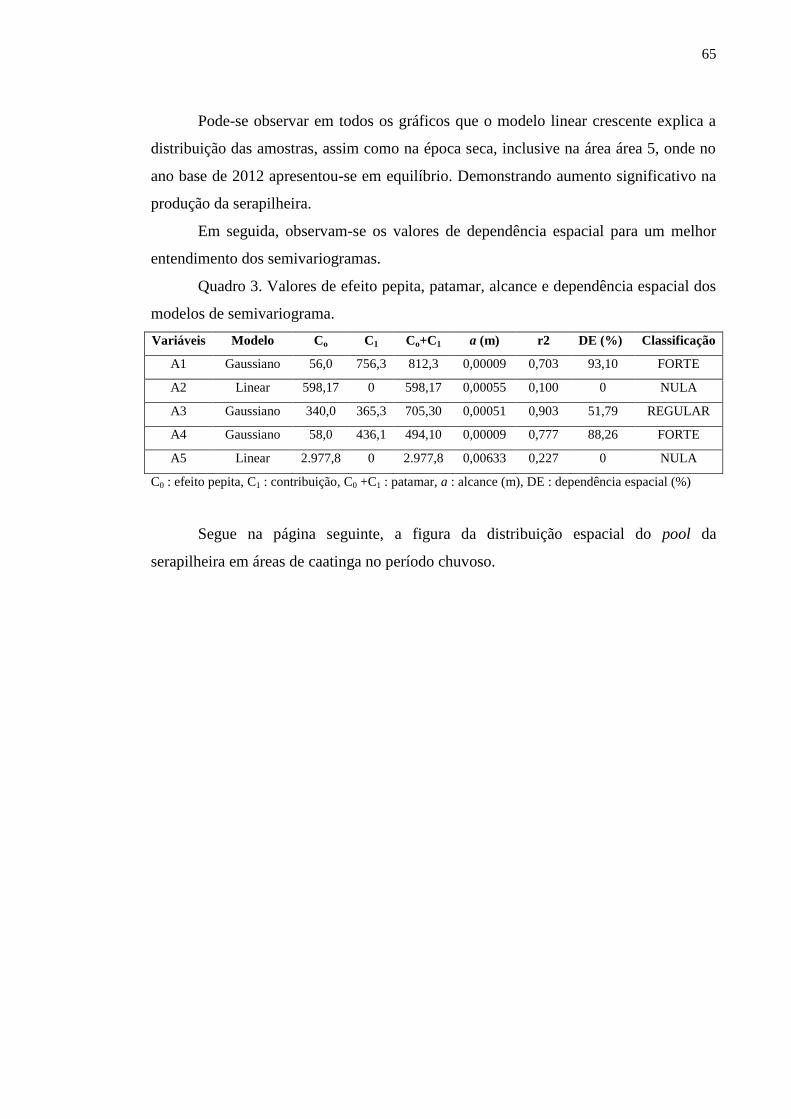
65
Pode-se observar em todos os gráficos que o modelo linear crescente explica a
distribuição das amostras, assim como na época seca, inclusive na área área 5, onde no
ano base de 2012 apresentou-se em equilíbrio. Demonstrando aumento significativo na
produção da serapilheira.
Em seguida, observam-se os valores de dependência espacial para um melhor
entendimento dos semivariogramas.
Quadro 3. Valores de efeito pepita, patamar, alcance e dependência espacial dos
modelos de semivariograma.
Variáveis Modelo Co C1 Co+C1 a (m) r2 DE (%) Classificação
A1 Gaussiano 56,0 756,3 812,3 0,00009 0,703 93,10 FORTE
A2 Linear 598,17 0 598,17 0,00055 0,100 0 NULA
A3 Gaussiano 340,0 365,3 705,30 0,00051 0,903 51,79 REGULAR
A4 Gaussiano 58,0 436,1 494,10 0,00009 0,777 88,26 FORTE
A5 Linear 2.977,8 0 2.977,8 0,00633 0,227 0 NULA
C0 : efeito pepita, C1 : contribuição, C0 +C1 : patamar, a : alcance (m), DE : dependência espacial (%)
Segue na página seguinte, a figura da distribuição espacial do pool da
serapilheira em áreas de caatinga no período chuvoso.
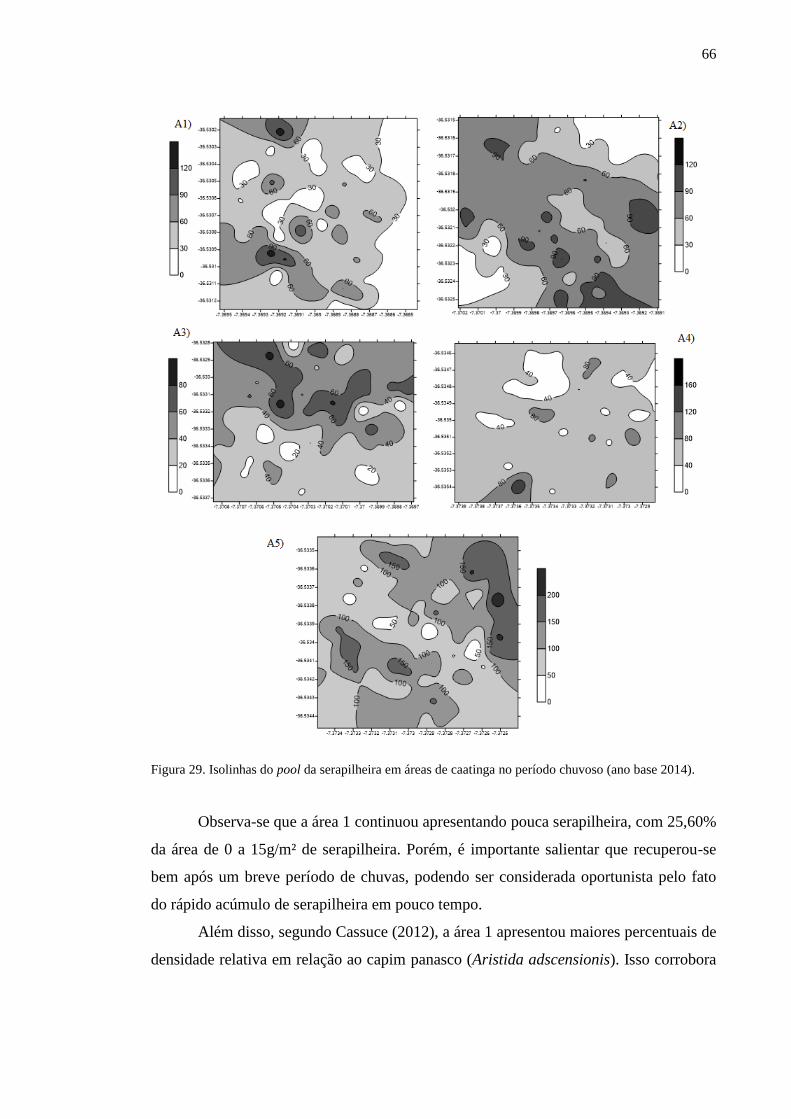
66
Figura 29. Isolinhas do pool da serapilheira em áreas de caatinga no período chuvoso (ano base 2014).
Observa-se que a área 1 continuou apresentando pouca serapilheira, com 25,60%
da área de 0 a 15g/m² de serapilheira. Porém, é importante salientar que recuperou-se
bem após um breve período de chuvas, podendo ser considerada oportunista pelo fato
do rápido acúmulo de serapilheira em pouco tempo.
Além disso, segundo Cassuce (2012), a área 1 apresentou maiores percentuais de
densidade relativa em relação ao capim panasco (Aristida adscensionis). Isso corrobora

67
com o fato de que a resposta de rebrota do capim em época chuvosa é muito rápida, e
isso fez com que esse acúmulo de serapilheira aumentasse em 2014 nesta área.
É notável também que a área 2 apresentou-se com baixo acúmulo de
serapilheira, sendo 12,61% da área representada com 0 a 30g/m². Sendo assim, houve
uma lenta recuperação do estrato vegetal da área.
Porém, a área 3, que representava o maior acúmulo desse material no período
seco, representou uma queda de 63% do acúmulo da serapilheira. Isso pode ser
explicado pela grande riqueza florística presente na área enfatizado por Cassuce (2012),
pois na época chuvosa esse material encontra-se verde.
A área 5 apresentou-se com maior acúmulo de serapilheira em detrimento das
demais, sendo 8,03% da área do mapa representada com 75 a 90 g/m² de serapilheira,
podendo ser considerada oportunista, pois na época das chuvas responde rapidamente ao
acúmulo de serapilheira, corroborando com os dados apresentados nos gráficos de
semivariograma.
3.4 Avaliação de qualidade da serapilheira
Alguns estudos feitos em regiões semiáridas do nordeste brasileiro avaliaram a
composição química da serapilheira para verificar a entrada e saída dos nutrientes (N, P,
K, S, Ca e Mg) do sistema de produção, verificando assim, a taxa de decomposição,
porém em sistemas extensivos com a criação de caprinos SPRD em áreas de caatinga a
serapilheira serve para a alimentação desses animais, hábito alimentar comprovado em
estudos de herbivoria.
Assim, o conhecimento da composição bromatológica é o ponto de partida para
o discernimento da concentração e disponibilidade dos nutrientes, o que contribui para
predizer a resposta animal em diferentes situações de pastejo. Por isso a importância de
avaliar a qualidade bromatológica desse material em áreas de caatinga, pois como visto
nos resultados do estoque da serapilheira, áreas com animais apresentaram menor
produção desse componente.
Pode-se observar na tabela 3 a composição química do pool da serapilheira
(folhas, material lignificado, miscelânea e estrutura reprodutiva vegetal) em áreas de
caatinga no cariri paraibano.
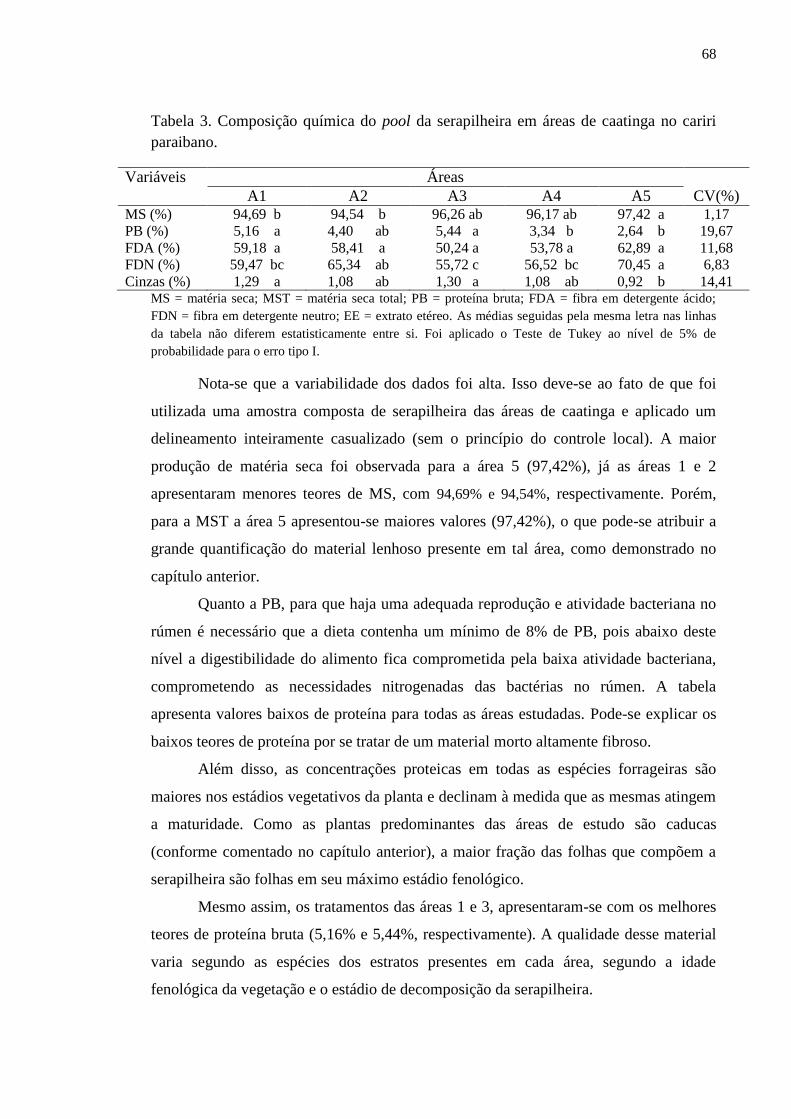
68
Tabela 3. Composição química do pool da serapilheira em áreas de caatinga no cariri
paraibano.
Variáveis Áreas
A1 A2 A3 A4 A5 CV(%) MS (%) 94,69 b 94,54 b 96,26 ab 96,17 ab 97,42 a 1,17
PB (%) 5,16 a 4,40 ab 5,44 a 3,34 b 2,64 b 19,67
FDA (%) 59,18 a 58,41 a 50,24 a 53,78 a 62,89 a 11,68
FDN (%) 59,47 bc 65,34 ab 55,72 c 56,52 bc 70,45 a 6,83
Cinzas (%) 1,29 a 1,08 ab 1,30 a 1,08 ab 0,92 b 14,41 MS = matéria seca; MST = matéria seca total; PB = proteína bruta; FDA = fibra em detergente ácido;
FDN = fibra em detergente neutro; EE = extrato etéreo. As médias seguidas pela mesma letra nas linhas
da tabela não diferem estatisticamente entre si. Foi aplicado o Teste de Tukey ao nível de 5% de
probabilidade para o erro tipo I.
Nota-se que a variabilidade dos dados foi alta. Isso deve-se ao fato de que foi
utilizada uma amostra composta de serapilheira das áreas de caatinga e aplicado um
delineamento inteiramente casualizado (sem o princípio do controle local). A maior
produção de matéria seca foi observada para a área 5 (97,42%), já as áreas 1 e 2
apresentaram menores teores de MS, com 94,69% e 94,54%, respectivamente. Porém,
para a MST a área 5 apresentou-se maiores valores (97,42%), o que pode-se atribuir a
grande quantificação do material lenhoso presente em tal área, como demonstrado no
capítulo anterior.
Quanto a PB, para que haja uma adequada reprodução e atividade bacteriana no
rúmen é necessário que a dieta contenha um mínimo de 8% de PB, pois abaixo deste
nível a digestibilidade do alimento fica comprometida pela baixa atividade bacteriana,
comprometendo as necessidades nitrogenadas das bactérias no rúmen. A tabela
apresenta valores baixos de proteína para todas as áreas estudadas. Pode-se explicar os
baixos teores de proteína por se tratar de um material morto altamente fibroso.
Além disso, as concentrações proteicas em todas as espécies forrageiras são
maiores nos estádios vegetativos da planta e declinam à medida que as mesmas atingem
a maturidade. Como as plantas predominantes das áreas de estudo são caducas
(conforme comentado no capítulo anterior), a maior fração das folhas que compõem a
serapilheira são folhas em seu máximo estádio fenológico.
Mesmo assim, os tratamentos das áreas 1 e 3, apresentaram-se com os melhores
teores de proteína bruta (5,16% e 5,44%, respectivamente). A qualidade desse material
varia segundo as espécies dos estratos presentes em cada área, segundo a idade
fenológica da vegetação e o estádio de decomposição da serapilheira.

69
Segundo Minson (1976), as gramíneas de clima tropical possuem níveis de PB
inferiores ao das espécies de clima temperado. O baixo nível de proteína observado nas
gramíneas tropicais se deve ao metabolismo fotossintético C4 apresentado pelas
mesmas, devido a anatomia foliar e o modo de fixação do CO2, assim como a alta
proporção de caule em relação às folhas.
Como esperado, os valores de FDN e FDA apresentaram-se altos, variando de
50 a 70%. Quanto aos valores de FDA não houve diferença significativa para todas as
áreas em estudo. Porém, os valores de FDN apresentaram diferença significativa
(P<0,05). A área 5 apresentou maior teor de fibra com 70,45%. Pode-se dizer que a
serapilheira da área 5, apesar de ter apresentado um maior teor de MST, apresentou
pouca proteína (2,64%) e muita fibra (70,45%). Sendo assim, a qualidade do material
torna-se baixa para o consumo de pequenos ruminantes.
Já a serapilheira da área 1, que apresentou menor teor de MST (94,69%),
apresentou-se com maior teor proteico (5,16%) e uma fração fibrosa intermediária entre
as demais com 59,47% de FDN, apesar de ter apresentado mais FDA (59,18%).
3.5 Conteúdo de água das cíbalas e da serapilheira
A água é um componente essencial para as células da planta e para todos os
processos metabólicos que dependem da sua presença. Assim, a deficiência de água no
solo reduz o crescimento das pastagens e, como consequência, reduz a quantidade de
forragem disponível e o desempenho dos animais em pastejo.
Embora alguns autores citem que a seca possa melhorar o valor nutritivo das
forragens, proporcionando maior teor de N nos tecidos das plantas e aumentando o
consumo de MS, o seu efeito no desempenho animal é negativo tendo em vista a
redução da produção das espécies forrageiras e, como consequência, a diminuição da
capacidade suporte das pastagens. Tais resultados são divergentes em várias autorias,
mas o certo é que estes resultados dependem do grau do estresse hídrico, que ocasionará
a senescência de folhas e mudanças na relação folha/caule.
O estudo do conteúdo de água presente na fração do material morto sobre o solo
é de extrema importância, pois está diretamente relacionado á decomposição desse
material e consequentemente, interfere na ciclagem dos nutrientes da planta para o solo.
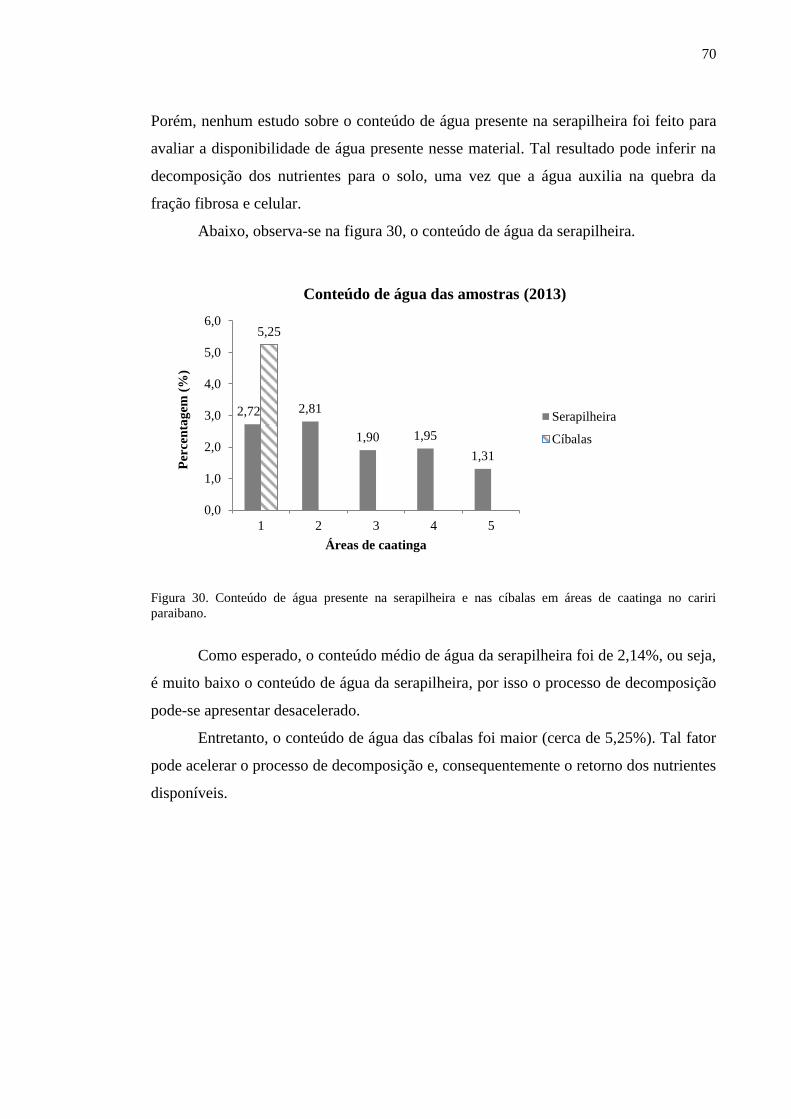
70
Porém, nenhum estudo sobre o conteúdo de água presente na serapilheira foi feito para
avaliar a disponibilidade de água presente nesse material. Tal resultado pode inferir na
decomposição dos nutrientes para o solo, uma vez que a água auxilia na quebra da
fração fibrosa e celular.
Abaixo, observa-se na figura 30, o conteúdo de água da serapilheira.
Figura 30. Conteúdo de água presente na serapilheira e nas cíbalas em áreas de caatinga no cariri
paraibano.
Como esperado, o conteúdo médio de água da serapilheira foi de 2,14%, ou seja,
é muito baixo o conteúdo de água da serapilheira, por isso o processo de decomposição
pode-se apresentar desacelerado.
Entretanto, o conteúdo de água das cíbalas foi maior (cerca de 5,25%). Tal fator
pode acelerar o processo de decomposição e, consequentemente o retorno dos nutrientes
disponíveis.
2,72 2,81
1,90 1,95
1,31
5,25
0,0
1,0
2,0
3,0
4,0
5,0
6,0
1 2 3 4 5
Per
cen
tag
em (
%)
Áreas de caatinga
Conteúdo de água das amostras (2013)
Serapilheira
Cíbalas
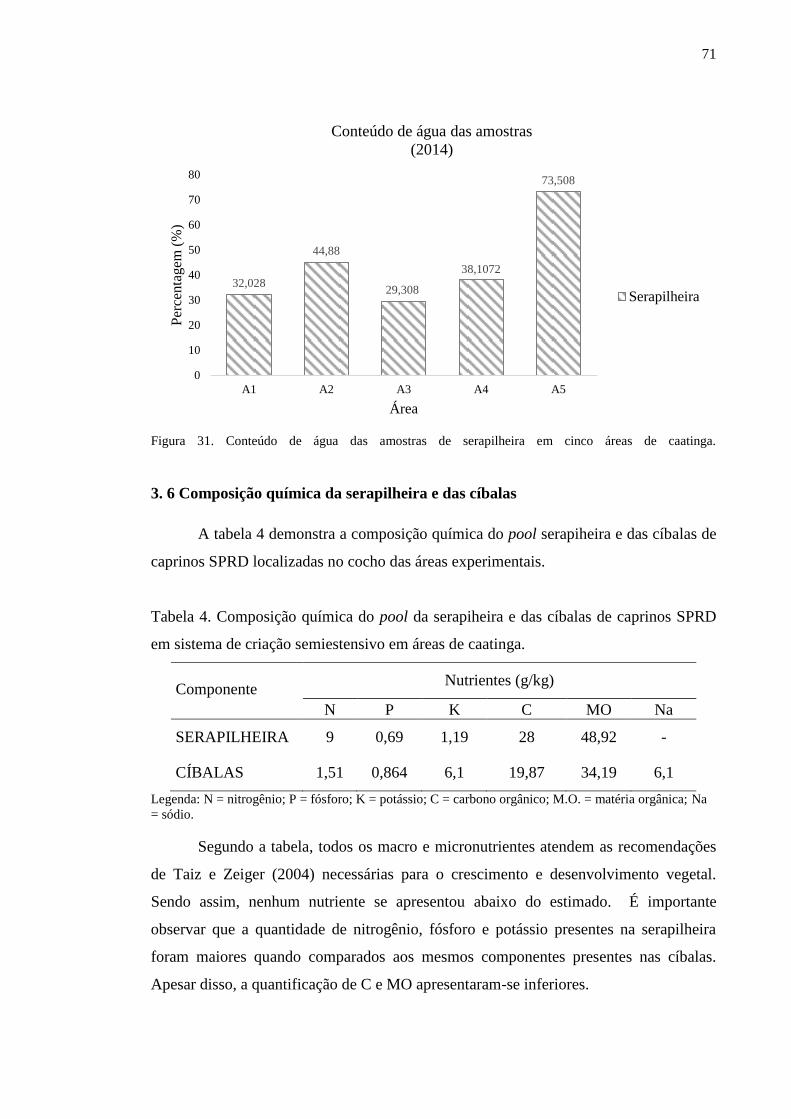
71
Figura 31. Conteúdo de água das amostras de serapilheira em cinco áreas de caatinga.
3. 6 Composição química da serapilheira e das cíbalas
A tabela 4 demonstra a composição química do pool serapiheira e das cíbalas de
caprinos SPRD localizadas no cocho das áreas experimentais.
Tabela 4. Composição química do pool da serapiheira e das cíbalas de caprinos SPRD
em sistema de criação semiestensivo em áreas de caatinga.
Componente Nutrientes (g/kg)
N P K C MO Na
SERAPILHEIRA 9 0,69 1,19 28 48,92 -
CÍBALAS 1,51 0,864 6,1 19,87 34,19 6,1
Legenda: N = nitrogênio; P = fósforo; K = potássio; C = carbono orgânico; M.O. = matéria orgânica; Na
= sódio.
Segundo a tabela, todos os macro e micronutrientes atendem as recomendações
de Taiz e Zeiger (2004) necessárias para o crescimento e desenvolvimento vegetal.
Sendo assim, nenhum nutriente se apresentou abaixo do estimado. É importante
observar que a quantidade de nitrogênio, fósforo e potássio presentes na serapilheira
foram maiores quando comparados aos mesmos componentes presentes nas cíbalas.
Apesar disso, a quantificação de C e MO apresentaram-se inferiores.
32,028
44,88
29,308
38,1072
73,508
0
10
20
30
40
50
60
70
80
A1 A2 A3 A4 A5
Per
cen
tagem
(%
)
Área
Conteúdo de água das amostras
(2014)
Serapilheira

72
A relação entre esses nutrientes é necessária para compreender o processo de
imobilização ou mineralização do componente estudado. Assim, pode-se inferir na
velocidade da decomposição para averiguar se a ciclagem desses nutrientes essenciais
para o crescimento e desenvolvimento dos vegetais estão sendo rapidamente
disponibilizados na sua forma orgânica.
Observou-se que as maiores quantidades totais dos nutrientes analisados na
biomassa seguem a seguinte ordem: MO > C > N > K > P para os dois componentes. Os
dados corroboram com os de Souto et al. (2009). É importante observar que a
quantidade de nitrogênio, fósforo e potássio presentes na serapilheira foram maiores
quando comparados aos mesmos componentes presentes nas cíbalas. Apesar disso, a
quantificação de C e MO apresentaram-se inferiores.
Ademais, Meguro et al. (1980) afirma que as diferenças entre elementos são
resultantes das características funcionais de cada nutriente no metabolismo da planta, na
diversidade do controle de fluxo e refluxo nos compartimentos de rápida ciclagem e
lenta ciclagem, antes da abscisão dos órgãos em cada espécie e, mesmo das
características do compartimento solo.
A relação entre esses nutrientes é necessária para compreender o processo de
imobilização ou mineralização do componente estudado. Assim, pode-se inferir na
velocidade da decomposição para averiguar se a ciclagem desses nutrientes essenciais
para o crescimento e desenvolvimento dos vegetais estão sendo rapidamente
disponibilizados na sua forma orgânica.
Tabela 5. Relação C:N e C:P da serapilheira e cíbalas em áreas de caatinga.
Componente Relação
C/N C/P
Serapilheira 3:1 41:1
Cíbalas 12:1 23:1
Observa-se na tabela 5 que a relação C:N de ambos os componentes foi muito
inferior ao esperado (30:1). Isso implica afirmar a imobilização desses componentes, ou
seja, o processo de decomposição é muito lento, principalmente da serapilheira. Apesar

73
disso, as cíbalas apresentaram uma relação C:N maior quando comparado à serapilheira,
ou seja, a decomposição dos dejetos caprinos é mais rápida.
Segundo Freire et al. (2010) há uma grande variação anual na decomposição da
serapilheira em região da zona da mata, porém pela baixa precipitação em áreas de
caatinga, essa variabilidade anual é mínima, uma vez que a vegetação só favorece o seu
crescimento em detrimento dos nutrientes disponíveis e da disponibilidade de água.
Além disso, a mesma autora afirma que a relação carbono/nitrogênio diminuiu na
medida em que os dias de incubação da serapilheira aumentaram. Tal fato pode explicar
a baixa relação C:N observada neste trabalho, uma vez que a serapilheira remanescente
presente nas áreas não são facilmente decompostas, pois a precipitação pluvial naquela
época foi baixa.
Segundo trabalhos de fitossociologia na mesma área de estudo indicam a
predominância de espécies altamente lignificadas, o que pode ser uma estratégia de
sobrevivência dessas plantas nativas, pois a lenta decomposição da serapilheira faz com
que os nutrientes continuem entrando no sistema por um período mais prolongado.
A relação C:P foi alta para a serapilheira (41:1). Tal fato pode ser explicado por
ser constituída por partes vegetativas, ricas em fósforo (P), uma vez que os vegetais
precisam de “P” para produzir ATP e assim gerar energia através do processo
fotossintético. Por esse motivo a relação C:P das cíbalas apresentou-se mais baixa em
comparação à serapilheira.
Assim, pode-se afirmar que a relação C:N é maior nas cíbalas, porém a C:P é
maior na serapilheira. Em geral, as cíbalas apresentam maiores teores de N, P, K e C do
que a serapilheira.
3.7 Composição química e fertilidade do solo
Para compreender melhor como as cíbalas contribuem para um maior retorno
dos nutrientes do solo, segue abaixo a tabela 6 com a composição química do solo em
áreas de caatinga.
Tabela 6. Composição química e fertilidade do solo em áreas de caatinga.
Amostra
Composição química
pH P K+ Na
+ H
+Al
+3 Ca
+2 Mg
+2 SB CTC M.O.
(mg/dm³) (cmolc/dm³) (g/kg)

74
1* 6,83 8,21 229,14 0,71 0,99 6,45 3,45 10,49 11,48 5,16
2*
5,92 27,77 120,12 0,34 2,64 9,5 6,7 16,85 19,49 14,62
*Legenda: 1 = amostra composta homogênea, correspondente ao solo de todas as áreas em estudo; 2 =
amostra composta do solo coberto por cíbalas.
É possível observar que o solo coberto por cíbalas possui maiores quantidades
de fósforo (P), H+Al
+, Ca
+2, Mg, soma de bases (SB), capacidade de troca catiônica
(CTC) e matéria orgânica (M.O.), e menores quantidades de potássio (K+), sódio (Na
+)
e pH mais ácido. Tal resultado deixa claro a importância que esse material tem para o
retorno dos nutrientes ao solo, uma vez que os nutrientes mais importantes para a
manutenção da vegetação são a MO, P, CTC e em menor proporção o Ca+2
e o Mg+2
.
Por isso, pode-se afirmar que os animais não somente retiram os nutrientes da
vegetação, uma vez que devolvem na forma de cíbalas esses nutrientes para o solo,
renovando o ciclo de nutrientes presentes no bioma.
É importante salientar que a presença de abrigos e do cocho dentro das áreas, faz
com que as cíbalas fiquem concentradas nesses ambientes, uma vez que os animais
permanecem a maior parte do tempo nos abrigos (na época seca), pois em épocas de
escassez não há muito suprimento volumoso dentro das áreas. Por isso, é imprescindível
a coleta e utilização dessas cíbalas como adubo orgânico para as plantas da caatinga,
assim os estratos podem se recuperar mais rapidamente nas primeiras chuvas e ao
mesmo tempo evita uma maior degradação do solo.
4 CONCLUSÕES
A distribuição espacial da serapilheira apresentou alta variabilidade em todas as
áreas em estudo.
Na época chuvosa, não foi observado aumento na deposição de serapilheira.
Duas áreas apresentaram maior acúmulo de serapilheira e do pool do material
lenhoso, enquanto outras apresentaram maior produção de miscelânea, quando
comparado os demais constituintes da serapilheira.
A deposição das cíbalas está diretamente ligada ao acúmulo de folhas produzidas
e consequentemente, a presença do estrato arbóreo/arbustivo;

75
A serapilheira apresentou baixa qualidade nutricional para ser considerada como
única fonte nutricional para caprinos em áreas de caatinga;
A decomposição da serapilheira apresentou-se muito lenta, fazendo com que o
retorno dos nutrientes para o solo torne-se um processo demorado e dependente da
precipitação pluvial.
5 REFERÊNCIAS
ANDRADE, M. V. M. DE. Dinâmica e qualidade da vegetação do estrato herbáceo
subarbustivo da caatinga no Cariri paraibano. Tese (Doutorado em Zootecnia) -
Universidade Federal da Paraíba - Centro de Ciências Agrárias, Areia, 2008.
ARAÚJO, K. D. Análise da vegetação e organismos edáficos em áreas de caatinga
sob pastejo e aspectos socioeconômicos e ambientais de São João do Cariri - PB.
Campina Grande - PB, Centro de Tecnologia e Recursos Naturais, UFCG, fevereiro de
2010. 151 p.il. (Tese de Doutorado). Programa de Pós-Graduação em Recursos
Naturais.
BIONDI, F.; MYERS, D.E. & AVERY, C.C. Geostatistically modeling stem size and
increment in an old-growth forest. Can. J. For. Res., 24:1354-1368, 1994.
CASSUCE, M. R. Fitossociologia e Composição Bromatológica de Espécies Herbáceas
e Subarbustivas em Áreas de Caatinga Sob Pastejo. 2012. 88 f. Dissertação (Mestrado
em Zootecnia). Universidade Federal da Paraíba, Centro de Ciências Agrárias, Areia-
PB.
CESAR, O. Produção de serapilheira na mata mesófila semidecídua da Fazenda
Barreiro Rico, município de Anhembi, SP. Revista Brasileira de Biologia, São Paulo,
v. 53, n. 4, p. 671-681, 1993.
CHAVES, L. H. G.; CHAVES, I. B.; VASCONCELOS, A. C. F. Salinidade das águas
superficiais e suas relações com a natureza dos solos na Bacia Escola do açude
Namorados. Campina Grande: BNB/UFPB, 54p, 2000. (Boletim Técnico).
DELITTI, W. B. C. Estudos de ciclagem de nutrientes: instrumentos para a análise
funcional de ecossistemas terrestres. Oecologia Brasiliensis, Rio de Janeiro, v. 1, p.
469-486, 1995.
EMBRAPA. Centro Nacional de Pesquisa de Solos. Manual de métodos de análise de
solos. 2 ed. Revista atual. Rio de Janeiro, 1997. 212p. (EMBRAPA - CNPS.
EMBRAPA. Centro Nacional de Pesquisa de Solos. Sistema brasileiro de
classificação de solos. Brasília: EMBRAPA Produção de Informação; Rio de Janeiro:
EMBRAPA Solos, 1999, 421p.
GOVERNO DO ESTADO DA PARAÍBA. Secretaria da Educação. Universidade
Federal da Paraíba. Atlas Geográfico da Paraíba. João Pessoa: Grafset, 1985, 100p.

76
GUEDES, D. .S; ANDRADE, M. V. M.; SILVA, D. S. da; ANDRADE, A. P.
Ocorrência e Qualidade do Capim Panasco (Aristida Adscensionis L.) em Áreas de
Caatinga no Cariri Paraibano. In: Zootec. Anais... João Pessoa, 2008.
GS+ - GEOSTATISTICS FOR THE ENVIRONMENTAL SCIENCES. P. O. Box 201,
Plainwell, Michigan 49080. Leeland Stanford Junior Univerity, USA. V. 10.0, 1989-
2014.
LEITÃO FILHO, H.F.; TIMONI, R.; PAGANO, S.N.; CESAR, O. Ecologia da Mata
Atlântica em Cubatão. São Paulo: Editora UNESP, Editora UNICAMP, 1993. 184p.
MINSON, P. J., STOBLES, T. H., HEGARY, M. P., et al. Measuring the nutritive value
of pasture plants. In: SHAW, N. H. e BRYAN, W. W. Tropical pasture research.
Principles and methods. common wealth Agricultural Bureaux, 1976. p.308-337
MEGURO, M.; VINUEZA, G.N.; DELITTI, W.B.C. Ciclagem de nutrientes na mata
mesófila secundária – São Paulo. I – Produção e conteúdo de nutrientes minerais no
folhedo. Bol. Botânica, Univ. S. Paulo, v.7, n.11, p. 11-21, 1980.
MOREIRA, F. M. S.; SIQUEIRA, J. O. Microbiologia e bioquímica do solo. Lavras:
Ed. UFLA, 2002. 626p.
MULLER-DOMBOIS; ELLENBERG, H. Aims and methods of vegetation ecology.
New York: John Willey e Sons, 1974. 525p.
NOY-MEIR, I. 1973. Desert ecosystems: environment and producers. Annual Reviews
Ecology. Annual Review of Ecology and Systematics, v. 4 25-51p.
ORGANIZAÇÃO DAS NAÇÕES UNIDAS. Pior seca dos últimos 50 anos no
nordeste brasileiro confirma estatísticas da ONU sobre escassez. Disponível em:
<http://www.onu.org.br/>. Acesso em: 17 de julho de 2013.
PARENTE, H. N. Avaliação da vegetação e do solo em áreas de caatinga sob pastejo
caprino no Cariri da Paraíba. Tese (Doutorado em Zootecnia). Areia: UFPB/CCA, 2009.
35 p.
PEREIRA JÚNIOR, L. R.; ANDRADE, A P. ARAÚJO, K. D. Composição florística e
fitossociológica de um fragmento de caatinga em Monteiro, PB. Rev. HOLOS, n 28, v.
6. 2013.
RAMALHO, C. I. Estrutura da vegetação e distribuição espacial do licuri (Syagrus
coronata (Mart) Becc.) em dois municípios do centro norte da Bahia, Brasil. 2008.
168f. (Tese de Doutorado em Agronomia) – Centro de Ciências Agrárias,
Universidade Federal da Paraíba, Areia, PB, 2008.
RODAL, M. J. N. Fitossociologia da vegetação arbustivo-árborea em quatro áreas de
caatinga em PE. 1992. 198 f. Tese (Doutorado) - Universidade Estadual de Campinas,
Campinas.
SILVA, D.J.; QUEIROZ, A.C. Análises de alimentos (métodos químicos e biológicos).
3.ed. Viçosa, MG: Editora UFV, 2002. 235p.
SOUZA, Bruna Vieira de. Avaliação da sazonalidade da deposição de serapilheira
em RPPN no semi-árido da Paraíba - PB 2009. Monografia (Graduação) Curso
Engenharia Florestal. CSTR/UFCG, Patos-PB, 2009.

77
SOUZA, J. A.; DAVIDE, A. C. Deposição de serapilheira e nutrientes em uma mata
não minerada e em plantações de bracatinga (mimosa scabrella) e de eucalipto
(eucalyptus saligna) em áreas de mineração de bauxita. Cerne, v. 7, n. 1, p.101-113,
2001.
SANTANA J. A. S; SOUTO, J. S. Produção de serapilheira na Caatinga da região semi-
árida do Rio Grande do Norte, Brasil. IDESIA (Chile) v. 29, nº 2, Mayo-Agosto, 2011.
TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 3ed. Porto Alegre: Artmed, 2004

78
CAPÍTULO 3
Acúmulo da serapilheira disponível para o pastejo caprino em áreas de
caatinga
________________________________________________________
RESUMO
A determinação da disponibilidade de forragem em pastagem é de fundamental
importância, tanto para a pesquisa científica quanto para o planejamento da exploração
racional de áreas manejadas comercialmente. Sabendo disso, o objetivo deste trabalho
foi avaliar o estoque da serapilheira disponível para o pastejo de caprinos, em diferentes
áreas de caatinga de sucessão secundária. O experimento foi conduzido na Estação
Experimental pertencente a UFPB, localizada no município de São João do Cariri-PB.
A área experimental representou, aproximadamente, 90m² que foram delimitados dentro
de nove áreas de caatinga em três transectos de cada área, com diferentes lotação de
animal. Os animais pertencentes às áreas eram caprinos machos, adultos, sem idade
padrão e sem padrão racial definido. O método utilizado para a análise dos dados foi o
de parcelas contíguas em transectos. A serapilheira acumulada na superfície do solo foi
estimada através de uma coleta após um longo período de seca (janeiro de 2013). Para a
quantificação da serapilheira foi feita a coleta todo o material que estava contido nas
parcelas, alocado em sacolas plásticas e posteriormente pesado. Foi realizado
levantamento florístico e fitossociológico em cinco áreas de caatinga pelo método de
parcelas (plot sampling), com distribuição sistemática dispostas de forma equidistante
(10 m x 10 m). Em cada área, foram mensuradas 30 parcelas de 100m², resultando em
um total de 15.000m² de área amostrada, e estimados índices de equabilidade,

79
similaridade, diversidade e agregação. Para avaliação estatística foram utilizados os
softwares Surfer v12.0, Mata Nativa v3 e Microsoft Excel v2013. A serapilheira pode
ser considerada como uma fonte alimentar alternativa em épocas de estiagem para o
consumo de pequenos ruminantes em sistemas de produção extensivos com acúmulo
médio de 190g.m-2
. A espécie arbórea predominante nas áreas de estudo foi a P.
pyramidalis (catingueira). A distribuição da serapilheira varia conforme os pulsos de
precipitação e à distribuição da catingueira nas áreas, tendo influência ao agrupamento
devido à fatores abióticos.
Palavras-chave: cariri, fitossociológico, levantamento florístico, liteira, pequenos
ruminantes, semiárido
ABSTRACT
Determination of forage availability in pastures is of fundamental importance
both for scientific research and for planning the rational exploitation of commercially
managed areas. Knowing this, the objective of this study was to evaluate the stock of
litter available for grazing goats in different areas of caatinga of secondary succession.
The experiment was conducted at the Experimental Station belonging to UFPB,
localized in São João do Cariri. The experimental area represented approximately
90sqm which were delimited within nine areas of caatinga in three transects in each
area, with different stocking animal. Animals belonging to the areas were male goats,
adults, no standard age and without defined breed. The method used for data analysis
was the contiguous plots in transects. The accumulated litter on the soil surface was
estimated through a collection after a long dry period (January 2013). quantification of
litter was collected all the material that was contained in the plots, allocated in plastic
bags and then weighed. Floristic and phytosociological survey was conducted in five
areas of the savanna plot method (plot sampling), with systematic distribution arranged
equidistantly (10 mx 10 m). In each area, 30 plots of 100 m were measured, resulting in
a total of 15.000 sampled area, and estimated indices of evenness, similarity, diversity
and aggregation. For statistical evaluation the Surfer v12.0, Native Forest v3 and

80
Microsoft Excel software were used V2013. The litter can be considered as an
alternative food source in times of drought for the consumption of small ruminant
production systems in extensivos with average accumulation 190g.m-2. The
predominant tree species in the study areas was P. pyramidalis (catingueira). The
distribution of litter varies pulses of precipitation and distribution of catingueira areas,
and influence the grouping due to abiotic factors.
Key words: cariri phytossociological, floristic survey, litter, small ruminants,
semiarid

81
1 INTRODUÇÃO
A determinação da disponibilidade de forragem em pastagem é de fundamental
importância, tanto para a pesquisa científica quanto para o planejamento da exploração
racional de áreas manejadas comercialmente. Por isso, há necessidade de se estimar de
forma simples e precisa a taxa de acúmulo de matéria seca (MS), permitindo assim, os
cálculos da taxa de lotação, capacidade de suporte, e o desempenho animal, com o
objetivo final de tornar o sistema produtivo e sustentável.
Entretanto, a grande variabilidade das características morfológicas, teores de MS
das diferentes espécies forrageiras, além da influência edafoclimática e das limitações
de recursos humanos e materiais, há uma enorme dificuldade na escolha do método
mais apropriado para se estimar a massa forrageira.
A literatura nacional e internacional relata vários métodos de determinação de
massa forrageira, uns com boa aplicabilidade para as gramíneas de clima tropical e
outros restritos às gramíneas temperadas; outros métodos ainda carentes de
metodologias mais precisas são aplicados para o estrato arbóreo/arbustivo e herbáceas.
Porém, a maior dificuldade se encontra em métodos precisos para a determinação da
disponibilidade de forragem em períodos críticos ou de seca em áreas de caatinga de
regiões semiáridas.
O bioma caatinga possui um complexo vegetacional de porte arbóreo, em sua
maioria composta por leguminosas arbóreo/arbustivas que são utilizadas como suporte
forrageiro em sistemas de criação extensiva de pequenos ruminantes. Além disso, a
serapilheira (camada de material morto sobre o solo) contribui significativamente para a
alimentação de pequenos ruminantes no período seco. Estes animais são explorados de
forma semiextensiva na região nordeste neste bioma, e no período seco há uma grande
escassez de volumosos.
A formação do rebanho caprino na região Nordeste é constituída, basicamente,
de animais sem padrão racial definido (SPRD), devido a cruzamentos desordenados
excessivos entre espécies nativas e exóticas (Melo e Oliveira, 2006) e, em maioria, de

82
raças nativas que, em geral, apresentam baixa exigência nutricional e baixa
produtividade (Silva, 2004). Apesar disso, a região Nordeste detém 94% do rebanho
caprino nacional (IBGE, 2001) e 54% do rebanho ovino, isto devido à rusticidade e
adaptabilidade destas espécies às condições edafoclimáticas da região, favorecendo
assim a exploração destas espécies em microrregiões como a do sertão paraibano.
Como esses animais são seletivos, na época seca uma das únicas alternativas
alimentares para estes animais a pasto é a serapilheira, tornando-se a principal fonte de
alimento em grandes épocas de estiagem, tendo assim, fundamental importância para a
produção desta região.
Então surge o questionamento: qual é o estoque de serapilheira disponível para a
alimentação de pequenos ruminantes em áreas de caatinga?
A hipótese foi que a produção da serapilheira em áreas de caatinga serve de base
para a alimentação de pequenos ruminantes em sistemas de criação extensivos no bioma
caatinga em regiões semiáridas.
Sabendo disso, o objetivo deste trabalho foi avaliar o acúmulo da serapilheira
disponível para o pastejo de caprinos, em diferentes áreas de caatinga de sucessão
secundária.
2 MATERIAL E MÉTODOS
2.1 Área de estudo
O experimento foi conduzido na Estação Experimental pertencente a UFPB,
localizada no município de São João do Cariri-PB, nas coordenadas 7o23’30”S e
36o31’59”W, numa altitude de 458 m. O município está inserido na zona fisiográfica do
Planalto da Borborema, fazendo parte da microrregião do Cariri Oriental.
Apresenta relevo suave ondulado sobre o embasamento cristalino. Com relação
ao sistema de drenagem, a área faz parte da bacia hidrográfica do rio Paraíba e sub-
bacia do rio Taperoá, riacho Namorado e demais cursos d’água tributários desse canal,
cujas nascentes encontram-se inseridas no território da UFPB. Os solos predominantes
na Estação, de acordo com Chaves e Chaves (2000), são: LUVISSOLO Crômico
vértico, que ocupa a maior parte das terras contidas na área da bacia, apresentando-se
severamente erodido à medida que o relevo torna-se mais acentuado; VERTISSOLO,

83
ocupando as posições mais baixas da bacia, nas circunvizinhanças do açude Namorado
e relevo mais plano no interior da bacia hidrográfica; E NEOSSOLO Lítico, encontrado
nas encostas mais íngremes e em posição de topo associado a afloramentos de rochas,
em relevo ondulado a forte ondulado (Embrapa, 1999). Na área experimental a textura
predominante é franco arenosa e franco argilo-arenosa.
Na área predomina o clima Bsh - semiárido quente com chuvas de verão e o
bioclima 2b apresentando um total de meses secos, variando de 9 a 11, denominado de
subdesértico quente de tendência tropical (Governo do Estado da Paraíba, 1985). A
temperatura média mensal varia de 27,2ºC no período novembro-março a 23,1ºC em
julho, sendo estes os valores máximos e mínimos, respectivamente, com precipitações
de 400 mm/ano e umidade relativa de 70%. A vegetação que recobre a região estudada é
a Caatinga hiperxrófita (vegetação caducifólia espinhosa) e arbustiva.
A área experimental representou, aproximadamente, 90m² que foram
delimitados dentro de nove áreas de caatinga em três transectos de cada área. As áreas 1
e 2 possuíam carga animal da seguinte forma: Área 1 - 10 animais (1 animal/3.200 m2),
Área 2 - 5 animais (1 animal/6.400 m2), e áreas 3, 4, 5, 6, 7, 8 e 9 - sem animais, com
diferentes estados de antropização. Os animais pertencentes às áreas eram caprinos
machos, adultos, sem idade padrão e sem padrão racial definido.
2.2 Coleta e análise dos dados
O método utilizado para a análise dos dados foi o de parcelas contíguas em
transectos. Foram utilizadas nove áreas de caatinga e em cada área foram estabelecidos
três transectos, distando-se aproximadamente 20m em largura. Em cada transecto,
foram delimitadas dez unidades experimentais equidistantes de 10m x 10m (em
comprimento) com sub-parcelas de 1m x 1m, totalizando 30 unidades, em cada área.
Todos os pontos foram identificados com auxílio de um aparelho GPS.

84
Figura 32. Representação das nove áreas experimentais. Em destaque, na cor azul, a subdivisão das
parcelas de 1m² em três transectos por área.
2.3 Metodologia para a obtenção da Serapilheira
2.3.1 Estimativa do estoque de serapilheira acumulada sobre o solo
A serapilheira acumulada na superfície do solo foi estimada através de uma
coleta após um longo período de seca (janeiro de 2013). Foram feitas coletas em 270
parcelas contíguas de 1m² cada, como demonstrado na figura 33.
Para a quantificação da serapilheira foi colhido todo o material que estava
contido nas parcelas, alocado em sacolas plásticas e posteriormente pesado. Em seguida
todo o material foi devolvido, evitando a degradação das áreas.

85
Figura 33. Demonstração da demarcação das parcelas em área alagada (a); com solo descoberto(b); da
luva utilizada para coleta da serapilheira (c); e pesagem do material (d).
2.4 Levantamento florístico e fitossociológico:
O levantamento florístico e fitossociológico foi realizado em cinco áreas de
caatinga pelo método de parcelas (plot sampling), com distribuição sistemática
dispostas de forma equidistante (10 m x 10 m). Em cada área foram mensuradas 30
parcelas de 10m², resultando em um total de 1.500m² de área amostrada.
2.4.1 Levantamento florístico:
O levantamento florístico foi realizado por meio de caminhadas nas parcelas,
onde foram coletados partes botânicas das plantas (folha, flor e fruto) amostradas com
presença aparente de descritores. O material foi acondicionado em estufa por 48 horas,
em seguida foram preparadas as exsicatas.
Para identificação dos indivíduos, as exsicatas foram enviadas ao Herbário
Lauro Pires Xavier no Departamento de Sistemática e Ecologia da Universidade Federal
da Paraíba (DSE-UFPB) para comparação com exsicatas que fazem parte da coleção do
herbário e lá permanecem para consultas futuras.

86
2.4.2 Levantamento fitossociológico:
Foram mensurados em cada parcela: a espécie, a altura (≥1,0m) e a
circunferência ao nível do solo de todos os indivíduos vivos (≥9cm) do estrato arbóreo
(Rodrigues, 1989). As espécies foram organizadas por família no sistema de Cronquist
(1988), incluindo-se informação sobre o hábito.
Para verificar a suficiência amostral das áreas foram confeccionadas curvas de
coletores para cada área de estudo (Rodal et al., 1992), plotando-se no eixo das
abscissas o número de parcelas adotadas. Para a identificação da similaridade florística
entre as áreas de estudo foi elaborada uma matriz de presença-ausência de espécies.
Para quantificar a diversidade do ecossistema com o intuito de se comparar os
diferentes tipos de vegetação, foi utilizado o Índice de diversidade de Shannon-Weaver
que considera igual o peso entre as espécies raras e as abundantes (Magurran, 1988).
.ln( ) ln( )
'
S
i lN N ni ni
HN
em que:
H’ = Índice de diversidade de Shannon-Weaver;
N = número total de indivíduos amostrados;
n i = número de indivíduos amostrados da i-ésima espécie;
S = número de espécies amostradas;
ln = logaritmo de base neperiana (e).
Quanto maior for o valor de H', maior será a diversidade florística da população
em estudo (Ramalho, 2008). Este índice pode expressar riqueza e uniformidade.
Para verificar se as espécies são igualmente abundantes, foi utilizado o índice de
equabilidade de Pielou (J’). É derivado do índice de diversidade de Shannon e permite
representar a uniformidade da distribuição dos indivíduos entre as espécies existentes
(Pielou, 1966). Seu valor apresenta uma amplitude de 0 (uniformidade mínima) a 1
(uniformidade máxima).

87
Onde:
J’= Equabilidade
H’= Índice de diversidade de Shannon-Weaver
ln(S)= Logarítmo neperiano do número total de espécies amostradas
Para quantificar a similaridade das comunidades foi realizada uma análise de
agrupamento usando o índice de similaridade de Jaccard (Muller-dombois e Ellenberg,
1974).
cSJij
a b c
em que:
a = número de espécies ocorrentes na parcela 1 ou comunidade 1;
b = número de espécies ocorrentes na parcela 2 ou comunidade 2;
c = número de espécies comuns às duas parcelas ou comunidade s.
O índice de dominância de Simpson (C’) mede a probabilidade de 2 (dois)
indivíduos, selecionados ao acaso na amostra, pertencer à mesma espécie (Brower e
Zarr, 1984) e foi utilizado para verificar a dominância de comunidades florísticas. Uma
comunidade de espécies com maior diversidade terá uma menor dominância. O valor
estimado de C varia de 0 (zero) a 1 (um), sendo que para valores próximos de um, a
diversidade é considerada maior.
C = índice de dominância de Simpson;
n i = número de indivíduos amostrados da i-ésima espécie;
N = número total de indivíduos amostrados;
Para a avaliação da distribuição espacial das espécies (agregação) foi aplicado o
Índice de MacGuinnes (IGA) (McGuinnes, 1934), a partir da equação:
ii
i
DIGA
d

88
Sendo:
ii
nD
uT
ln 1i id f i
i
uf
uT
em que:
IGA i = "Índice de MacGuinnes" para a i-ésima espécie;
Di = densidade observada da i-ésima espécie;
di = densidade esperada da i-ésima espécie;
fi = freqüência absoluta da i-ésima espécie;
ln = logaritmo neperiano;
ni = número de indivíduos da i-ésima espécie;
ui = número de unidades amostrais em que a i-ésima espécie ocorre;
uT = número total de unidades amostrais.
Classif. IGA = Classificação do padrão de distribuição dos indivíduos das espécies, que
obedeceu a seguinte escala:
IGAi < 1: distribuição uniforme;
IGAi = 1: distribuição aleatória;
1 < IGAi <= 2: tendência ao agrupamento;
IGAi > 2: distribuição agregada ou agrupada.
Para determinação dos parâmetros fitossociológicos, foram considerados todos
os indivíduos arbóreo-arbustivos vivos com Circunferência ao nível do solo (CNS) ≥ a 9
cm e altura (h) mínima de 1 m (Rodal, 1992). Em casos de indivíduos ramificados, a
área basal individual resultou da soma de áreas basais de cada ramificação (Rodrigues,
1989).
As medidas de altura foram realizadas com régua de mira de 5m de altura em
alumínio. Para medir a circunferência dos indivíduos foi utilizada suta digital e fita
métrica.
Em seguida foi calculado o diâmetro pela equação:

89
em que:
D = diâmetro;
CNS = Circunferência ao Nível do Solo;
Para caracterizar a estrutura da comunidade arbórea, foram calculados, para cada
espécie, os parâmetros fitossociológicos de acordo com Rodrigues (1989).
Foram determinados os seguintes parâmetros:
- Número de indivíduos (N);
- Número de parcelas de ocorrência (NPARC);
- Frequência Absoluta (FA) - relação entre o número de parcelas ou pontos que ocorre
uma dada espécie e o número total de amostras, expressa em percentagem.
% 100%Pi
FA xP
em que:
Pi = número de ocorrência da espécie i;
P = número total de amostras.
- Frequência Relativa (FR) - relação entre a frequência absoluta de uma dada espécie
com as frequências absolutas de todas as espécies, expressa em percentagem.
% 100%i
i
FAFR x
FA
- Densidade Absoluta - medida que expressa o número de indivíduos de uma dada
espécie (ni) por unidade de área (A).
niDA
A ha
em que:
ni = número de indivíduos da espécie i;

90
A = área total amostrada (ha).
- Densidade Relativa - relação entre o número de indivíduos de uma determinada
espécie (ni) e o número de indivíduos amostrados de todas as espécies (N), expressa em
percentagem.
% 100%ni
DR xN
- Dominância Absoluta (DoA) - dada a partir da somatória da área basal dos indivíduos
de cada espécie (ABi), dividido pela área total amostrada (A).
ABiDoA
A ha
- Dominância Relativa (DoR) - relação entre a área basal total de uma determinada
espécie e a área basal total de todas as espécies amostradas, expressa em percentagem.
% 100%ABi
DoR xABi
- Índice de Valor de Importância (IVI) - representa a soma dos valores relativos de
densidade, de frequência e de dominância de cada espécie.
IVI %= DR + FR + DoR
- Índice de Valor de Cobertura (IVC) - representa a soma dos valores relativos de
densidade e dominância de cada espécie.
IVC %= DR + DoR
2.5 Avaliação Estatística
Para a análise de variância foi utilizado o programa Assistat Beta v.7.6, onde as
áreas foram consideradas tratamentos e as parcelas as repetições. Dessa forma foram
analisados 9 tratamentos com 30 repetições em um delineamento totalmente
casualizado. As médias foram avaliadas através do teste de Tukey a 0,05 de
probabilidade para o erro tipo I.
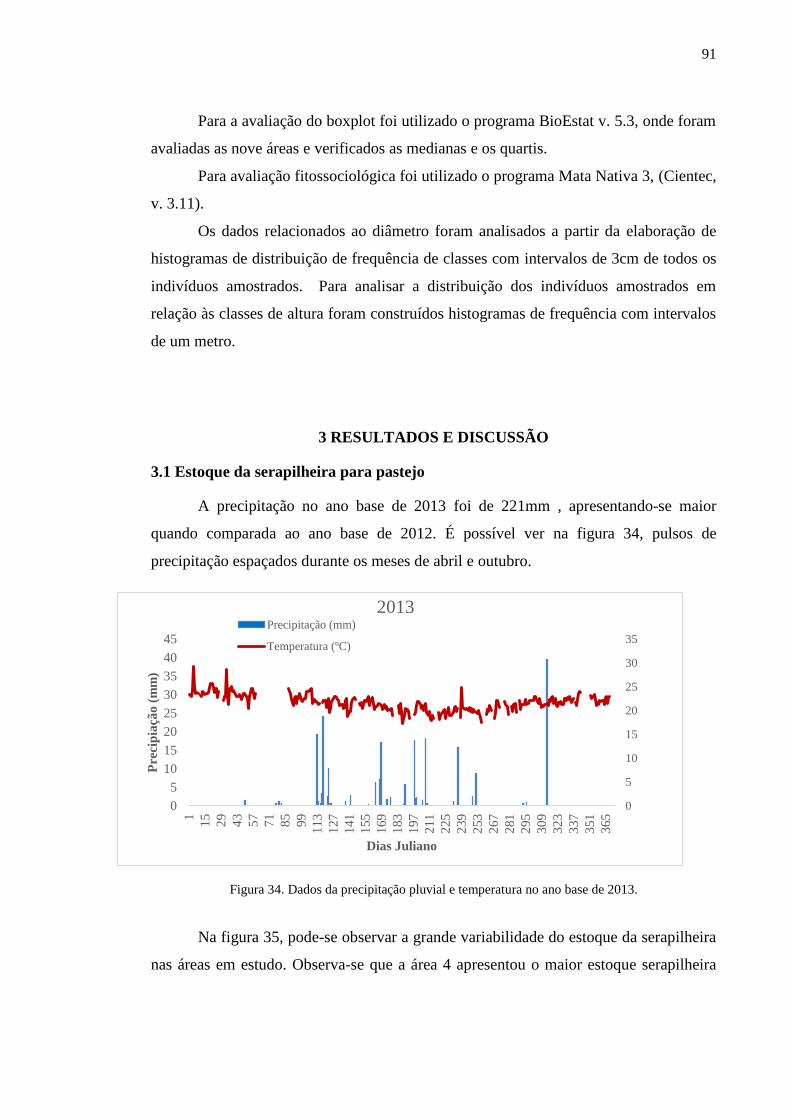
91
Para a avaliação do boxplot foi utilizado o programa BioEstat v. 5.3, onde foram
avaliadas as nove áreas e verificados as medianas e os quartis.
Para avaliação fitossociológica foi utilizado o programa Mata Nativa 3, (Cientec,
v. 3.11).
Os dados relacionados ao diâmetro foram analisados a partir da elaboração de
histogramas de distribuição de frequência de classes com intervalos de 3cm de todos os
indivíduos amostrados. Para analisar a distribuição dos indivíduos amostrados em
relação às classes de altura foram construídos histogramas de frequência com intervalos
de um metro.
3 RESULTADOS E DISCUSSÃO
3.1 Estoque da serapilheira para pastejo
A precipitação no ano base de 2013 foi de 221mm , apresentando-se maior
quando comparada ao ano base de 2012. É possível ver na figura 34, pulsos de
precipitação espaçados durante os meses de abril e outubro.
Figura 34. Dados da precipitação pluvial e temperatura no ano base de 2013.
Na figura 35, pode-se observar a grande variabilidade do estoque da serapilheira
nas áreas em estudo. Observa-se que a área 4 apresentou o maior estoque serapilheira
0
5
10
15
20
25
30
35
0
5
10
15
20
25
30
35
40
45
1
15
29
43
57
71
85
99
113
127
141
155
169
183
197
211
225
239
253
267
281
295
309
323
337
351
365
Pre
cip
iaçã
o (
mm
)
Dias Juliano
2013 Precipitação (mm)
Temperatura (ºC)
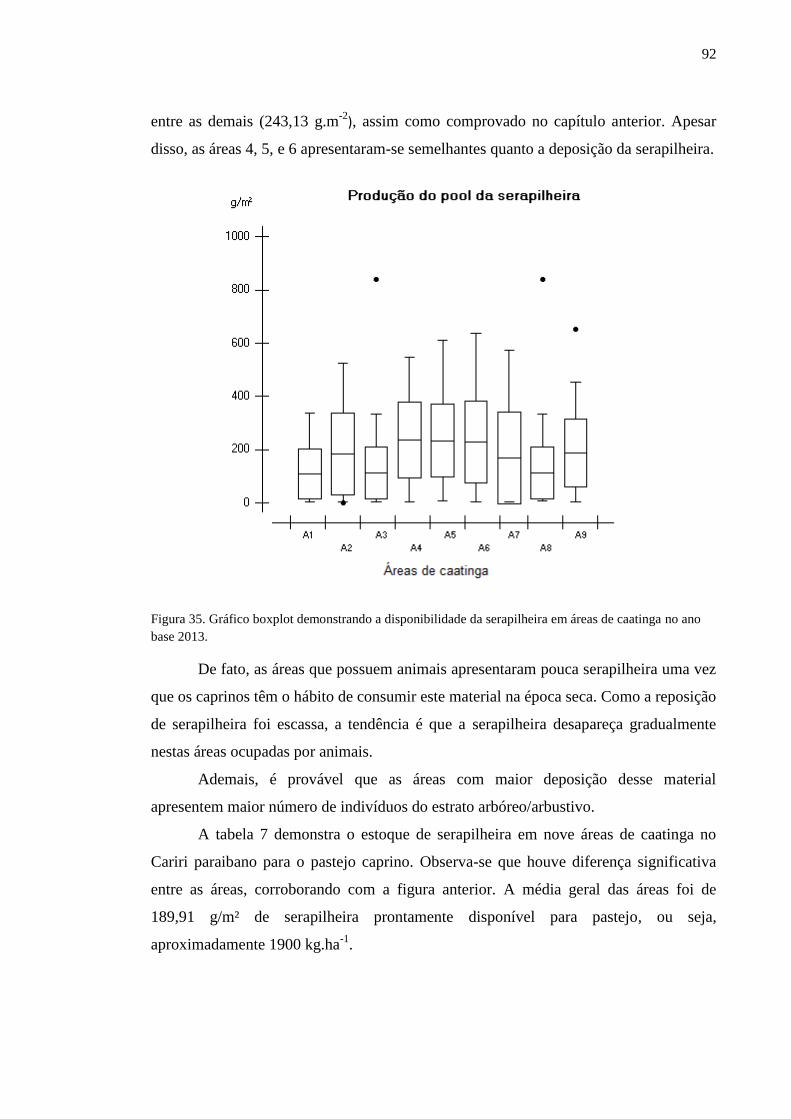
92
entre as demais (243,13 g.m-2
), assim como comprovado no capítulo anterior. Apesar
disso, as áreas 4, 5, e 6 apresentaram-se semelhantes quanto a deposição da serapilheira.
Figura 35. Gráfico boxplot demonstrando a disponibilidade da serapilheira em áreas de caatinga no ano
base 2013.
De fato, as áreas que possuem animais apresentaram pouca serapilheira uma vez
que os caprinos têm o hábito de consumir este material na época seca. Como a reposição
de serapilheira foi escassa, a tendência é que a serapilheira desapareça gradualmente
nestas áreas ocupadas por animais.
Ademais, é provável que as áreas com maior deposição desse material
apresentem maior número de indivíduos do estrato arbóreo/arbustivo.
A tabela 7 demonstra o estoque de serapilheira em nove áreas de caatinga no
Cariri paraibano para o pastejo caprino. Observa-se que houve diferença significativa
entre as áreas, corroborando com a figura anterior. A média geral das áreas foi de
189,91 g/m² de serapilheira prontamente disponível para pastejo, ou seja,
aproximadamente 1900 kg.ha-1
.

93
Tabela 7. Estoque de serapilheira em nove áreas de caatinga no Cariri paraibano para o
pastejo caprino.
Área Produção de serapilheira (g/m²)
1 110,86 b
2 194,40 ab
3 166,06 ab
4 243,13 a
5 240,00 a
6 235,90 a
7 173,86 ab
8 138,60 ab
9 206,43 ab
CV(%) 80,60
Média geral 189,91
As médias seguidas pela mesma letra não diferem estatisticamente entre si. Foi aplicado o
teste de Tukey ao nível de 5% de probabilidade para o erro tipo I.
Pode-se observar ainda que a área 1 foi a que apresentou menor deposição de
serapilheira (110,86g.m-2
).
A variação do erro foi alta (80,60%) devido a variabilidade dos estratos vegetais
das áreas e por se tratar de um experimento a campo, apesar de serem todas áreas de
caatinga. Como o delineamento foi considerado inteiramente ao acaso (sem o princípio
do controle local) obteve-se essa alta variabilidade no coeficiente de variação.
Observa-se que houve uma grande variabilidade nos dados, ou seja, os dados
foram assimétricos, com exceção das áreas 3, 6, 8 e 7, pelo fato de que estas
apresentaram menor quantificação da serapilheira. O máximo valor observado foi
837g/m² da área 3. O maior interquartil observado foi da área 6, com 75% dos dados
entre 200 a 400g/m² de serapilheira. Pode-se dizer, que a área 6 apresentou maior
estoque de serapilheira, podendo ser considerada a área mais preservada. É provável que
esta área possua maior número de indivíduos arbóreo/arbustivos, aumentando assim a
deposição da serapilheira.
3.2 Levantamento florístico e fitossociológico:
3.2.1 Curva do coletor:
Foi determinada a curva do coletor para cinco áreas amostradas, visando
representar as espécies em abundância, verificando o erro experimental devido a
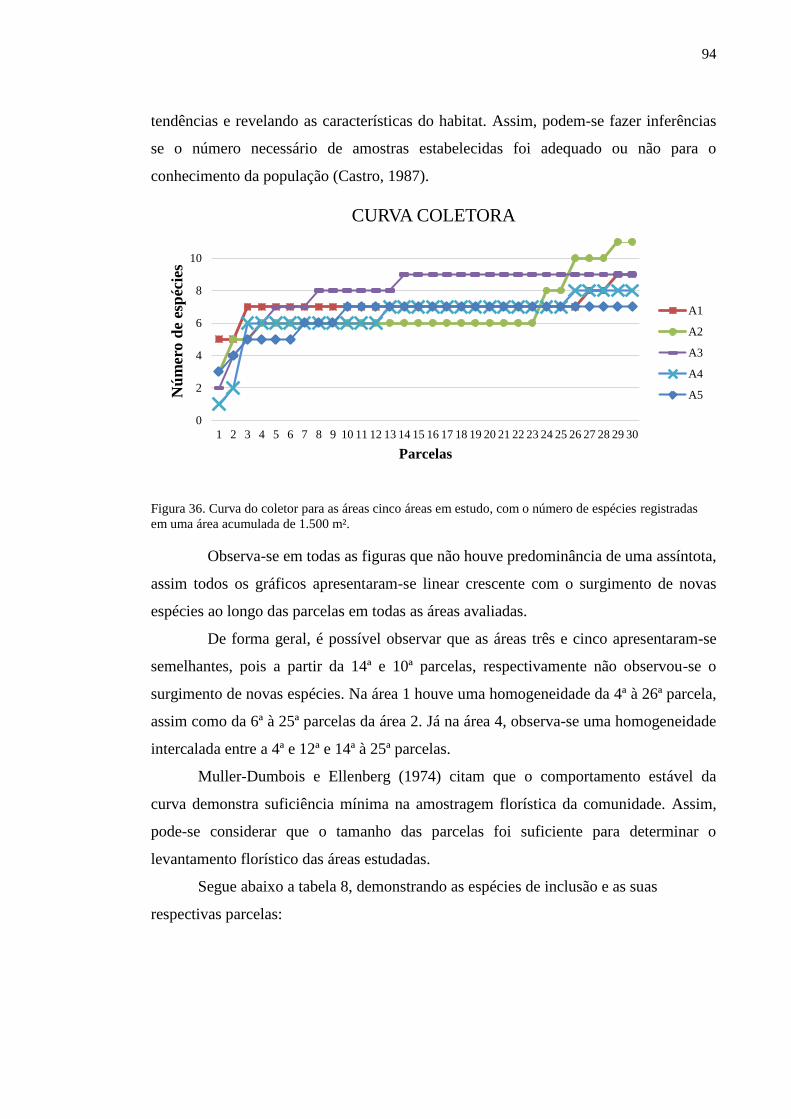
94
tendências e revelando as características do habitat. Assim, podem-se fazer inferências
se o número necessário de amostras estabelecidas foi adequado ou não para o
conhecimento da população (Castro, 1987).
Figura 36. Curva do coletor para as áreas cinco áreas em estudo, com o número de espécies registradas
em uma área acumulada de 1.500 m².
Observa-se em todas as figuras que não houve predominância de uma assíntota,
assim todos os gráficos apresentaram-se linear crescente com o surgimento de novas
espécies ao longo das parcelas em todas as áreas avaliadas.
De forma geral, é possível observar que as áreas três e cinco apresentaram-se
semelhantes, pois a partir da 14ª e 10ª parcelas, respectivamente não observou-se o
surgimento de novas espécies. Na área 1 houve uma homogeneidade da 4ª à 26ª parcela,
assim como da 6ª à 25ª parcelas da área 2. Já na área 4, observa-se uma homogeneidade
intercalada entre a 4ª e 12ª e 14ª à 25ª parcelas.
Muller-Dumbois e Ellenberg (1974) citam que o comportamento estável da
curva demonstra suficiência mínima na amostragem florística da comunidade. Assim,
pode-se considerar que o tamanho das parcelas foi suficiente para determinar o
levantamento florístico das áreas estudadas.
Segue abaixo a tabela 8, demonstrando as espécies de inclusão e as suas
respectivas parcelas:
0
2
4
6
8
10
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
Nú
mer
o d
e es
péc
ies
Parcelas
CURVA COLETORA
A1
A2
A3
A4
A5

95
Tabela 8. Espécies de inclusão por parcela em diferentes áreas de caatinga
PARCELA ESPÉCIE DE INCLUSÃO
AREA 1
3 J. molíssima
3 C. sonderianus
27 C. decídua
29 S. tuberosa
AREA 2
2 P. gounellei
2 J. molíssima
5 O. palmadora
26 C. flexuosa
26 C. decídua
29 L. microphylla
AREA 3
2 M. ophthalmocentra
2 C. sonderianus
3 O. palmadora
4 P. gounellei
5 J. molíssima
8 C. felxuosa
14 S. brasiliensis
AREA 4
2 PARCELA PERDIDA
3 C. sonderianus
3 J. molíssima
3 P. gounellei
3 A. pyrifolium
13 O. palmadora
26 M. ophthalmocentra
AREA 5
2 P. gounellei
3 A. pyrifolium
7 O. palmadora
10 M. ophthalmocentra
É possível observar que em todas as áreas, a partir da 2ª e 3ª parcelas há uma
tendência de inclusão de espécies.
3.2.2 Diversidade florística:

96
Segundo o levantamento florístico do estrato arbóreo realizado em cinco áreas
de caatinga, foi observado o aparecimento de sete famílias, nove gêneros e nove
espécies. Araújo (2010), em estudo realizado nas mesmas áreas de caatinga, observou
um maior número de gêneros e espécies no ano de 2010 ao avaliar os estratos arbóreo-
arbustivo.
Conforme a tabela 9, observa-se uma homogeneidade na diversidade de
famílias, gênero e espécies nas áreas 1, 2 e 3, enquanto que as áreas 4 e 5 apresentaram-
se com uma menor diversidade.
Tabela 9. Número de famílias, gêneros, espécies e indivíduos ocorrentes nas áreas 1, 2,
3, 4 e 5 em São João do Cariri – PB.
AREA 1 2 3 4 5
FAMILIAS 6 6 6 5 4
GENEROS 9 8 9 7 7
ESPÉCIES 9 8 9 7 7
INDIVÍDUOS 413 332 315 215 178
No total foram encontrados 1.288 indivíduos, 8 famílias,12 gêneros e 12
espécies. Os resultados deste estudo foram muito abaixo quando comparado ao trabalho
de Araújo (2010), uma vez que de 2010 para 2014, ou seja, em quatro anos, o número
de indivíduos diminuiu significativamente. Tal fato, pode ser explicado pelo longo
período de sequeiro que ocorreu de 2011 a 2013.
Todavia, considerando o fato que Bessa et al. 2011 encontraram 392 indivíduos,
distribuídos em 10 famílias 19 gêneros e 21 espécies, no total de três áreas de caatinga
no Rio Grande do Norte, neste estudo as áreas apresentaram-se com maior riqueza de
indivíduos.
Abaixo, segue a figura 37 demonstrando as famílias encontradas nas áreas em
estudo.
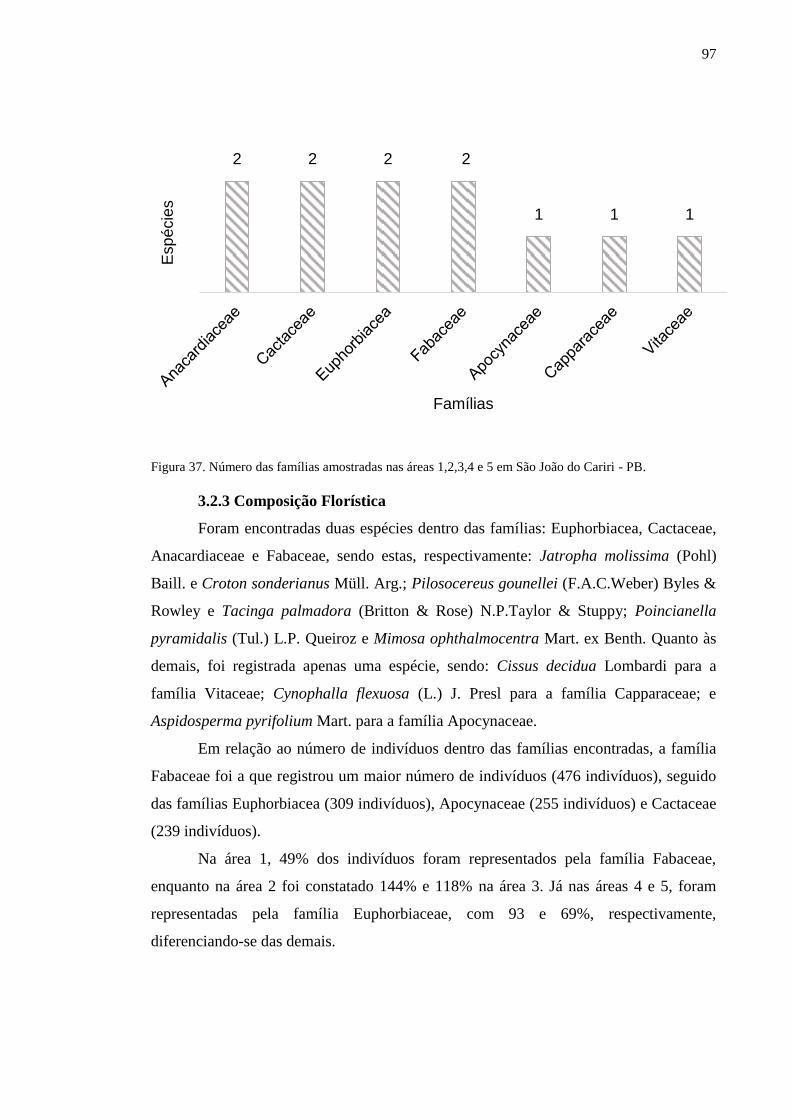
97
Figura 37. Número das famílias amostradas nas áreas 1,2,3,4 e 5 em São João do Cariri - PB.
3.2.3 Composição Florística
Foram encontradas duas espécies dentro das famílias: Euphorbiacea, Cactaceae,
Anacardiaceae e Fabaceae, sendo estas, respectivamente: Jatropha molissima (Pohl)
Baill. e Croton sonderianus Müll. Arg.; Pilosocereus gounellei (F.A.C.Weber) Byles &
Rowley e Tacinga palmadora (Britton & Rose) N.P.Taylor & Stuppy; Poincianella
pyramidalis (Tul.) L.P. Queiroz e Mimosa ophthalmocentra Mart. ex Benth. Quanto às
demais, foi registrada apenas uma espécie, sendo: Cissus decidua Lombardi para a
família Vitaceae; Cynophalla flexuosa (L.) J. Presl para a família Capparaceae; e
Aspidosperma pyrifolium Mart. para a família Apocynaceae.
Em relação ao número de indivíduos dentro das famílias encontradas, a família
Fabaceae foi a que registrou um maior número de indivíduos (476 indivíduos), seguido
das famílias Euphorbiacea (309 indivíduos), Apocynaceae (255 indivíduos) e Cactaceae
(239 indivíduos).
Na área 1, 49% dos indivíduos foram representados pela família Fabaceae,
enquanto na área 2 foi constatado 144% e 118% na área 3. Já nas áreas 4 e 5, foram
representadas pela família Euphorbiaceae, com 93 e 69%, respectivamente,
diferenciando-se das demais.
2 2 2 2
1 1 1
Esp
écie
s
Famílias
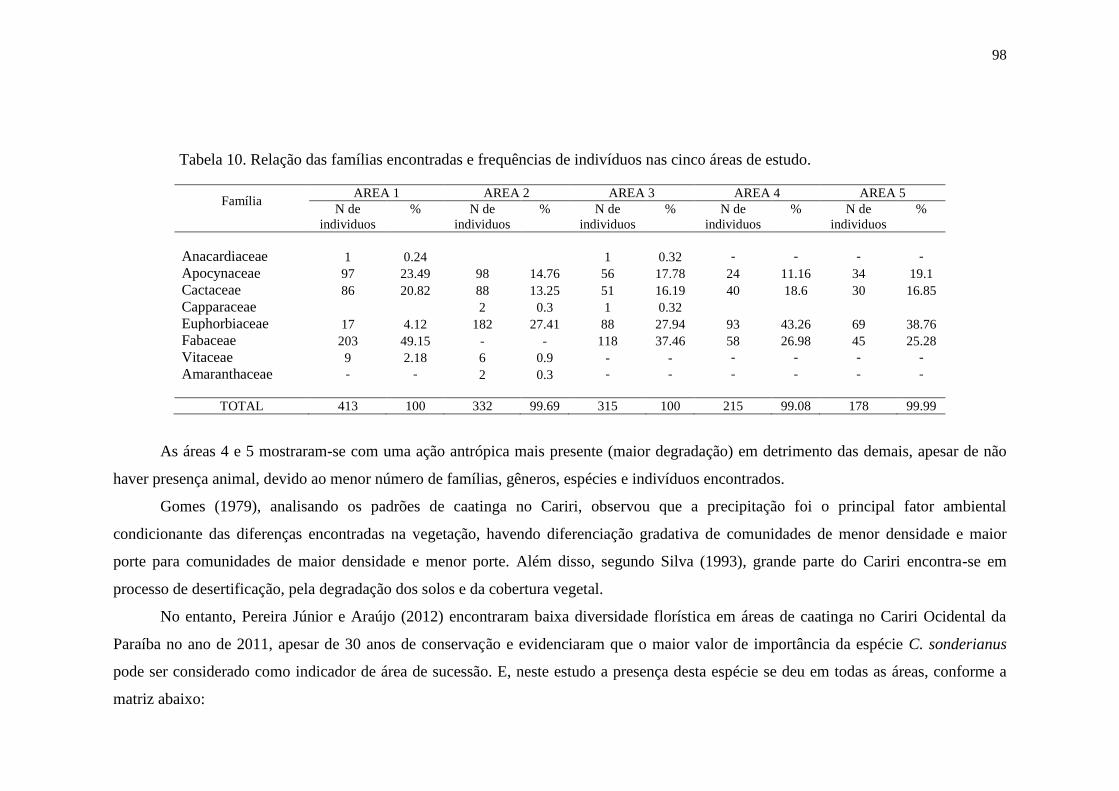
98
Tabela 10. Relação das famílias encontradas e frequências de indivíduos nas cinco áreas de estudo.
Família
AREA 1 AREA 2 AREA 3 AREA 4 AREA 5
N de
individuos
% N de
individuos
% N de
individuos
% N de
individuos
% N de
individuos
%
Anacardiaceae 1 0.24 1 0.32 - - - -
Apocynaceae 97 23.49 98 14.76 56 17.78 24 11.16 34 19.1
Cactaceae 86 20.82 88 13.25 51 16.19 40 18.6 30 16.85
Capparaceae 2 0.3 1 0.32
Euphorbiaceae 17 4.12 182 27.41 88 27.94 93 43.26 69 38.76
Fabaceae 203 49.15 - - 118 37.46 58 26.98 45 25.28
Vitaceae 9 2.18 6 0.9 - - - - - -
Amaranthaceae - - 2 0.3 - - - - - -
TOTAL 413 100 332 99.69 315 100 215 99.08 178 99.99
As áreas 4 e 5 mostraram-se com uma ação antrópica mais presente (maior degradação) em detrimento das demais, apesar de não
haver presença animal, devido ao menor número de famílias, gêneros, espécies e indivíduos encontrados.
Gomes (1979), analisando os padrões de caatinga no Cariri, observou que a precipitação foi o principal fator ambiental
condicionante das diferenças encontradas na vegetação, havendo diferenciação gradativa de comunidades de menor densidade e maior
porte para comunidades de maior densidade e menor porte. Além disso, segundo Silva (1993), grande parte do Cariri encontra-se em
processo de desertificação, pela degradação dos solos e da cobertura vegetal.
No entanto, Pereira Júnior e Araújo (2012) encontraram baixa diversidade florística em áreas de caatinga no Cariri Ocidental da
Paraíba no ano de 2011, apesar de 30 anos de conservação e evidenciaram que o maior valor de importância da espécie C. sonderianus
pode ser considerado como indicador de área de sucessão. E, neste estudo a presença desta espécie se deu em todas as áreas, conforme a
matriz abaixo:
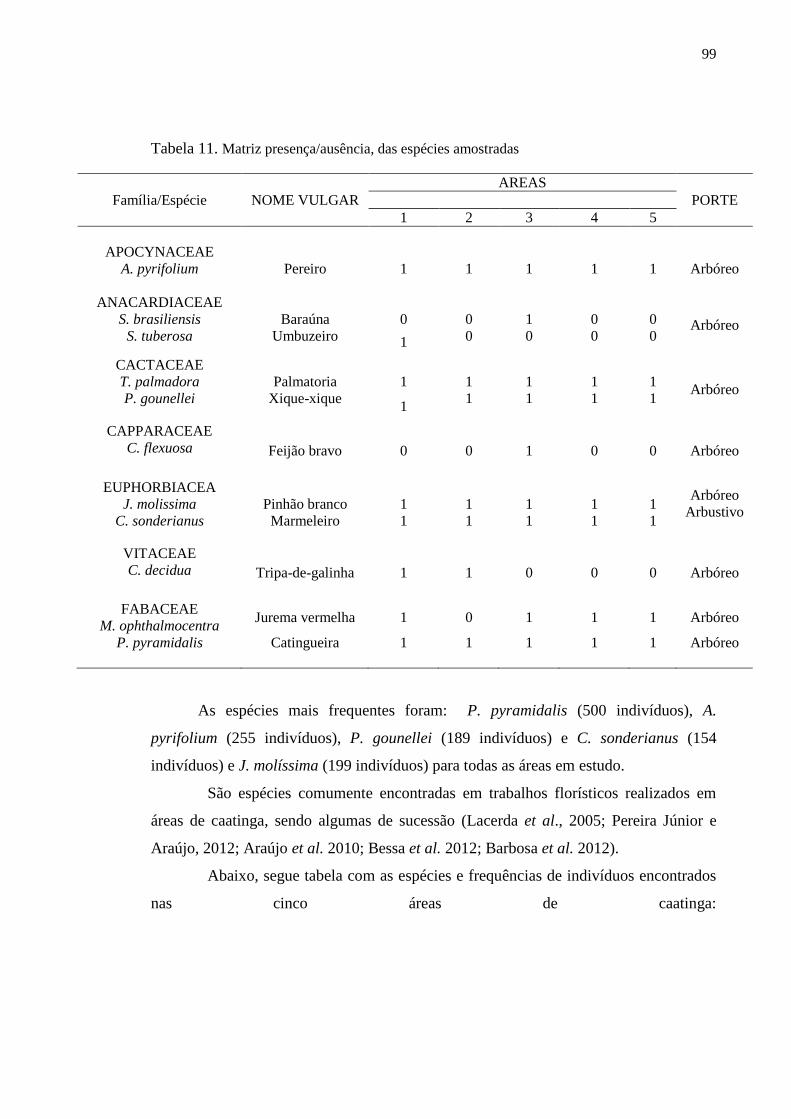
99
Tabela 11. Matriz presença/ausência, das espécies amostradas
Família/Espécie NOME VULGAR
AREAS
PORTE
1 2 3 4 5
APOCYNACEAE
A. pyrifolium Pereiro 1 1 1 1 1 Arbóreo
ANACARDIACEAE
Arbóreo S. brasiliensis Baraúna 0 0 1 0 0
S. tuberosa Umbuzeiro 1 0 0 0 0
CACTACEAE
Arbóreo T. palmadora Palmatoria 1 1 1 1 1
P. gounellei Xique-xique 1
1 1 1 1
CAPPARACEAE
Feijão bravo 0 0 1 0 0 Arbóreo C. flexuosa
EUPHORBIACEA Arbóreo
Arbustivo
J. molissima Pinhão branco 1 1 1 1 1
C. sonderianus Marmeleiro 1 1 1 1 1
VITACEAE
Tripa-de-galinha 1 1 0 0 0 Arbóreo C. decidua
FABACEAE Jurema vermelha 1 0 1 1 1 Arbóreo
M. ophthalmocentra
P. pyramidalis Catingueira 1 1 1 1 1 Arbóreo
As espécies mais frequentes foram: P. pyramidalis (500 indivíduos), A.
pyrifolium (255 indivíduos), P. gounellei (189 indivíduos) e C. sonderianus (154
indivíduos) e J. molíssima (199 indivíduos) para todas as áreas em estudo.
São espécies comumente encontradas em trabalhos florísticos realizados em
áreas de caatinga, sendo algumas de sucessão (Lacerda et al., 2005; Pereira Júnior e
Araújo, 2012; Araújo et al. 2010; Bessa et al. 2012; Barbosa et al. 2012).
Abaixo, segue tabela com as espécies e frequências de indivíduos encontrados
nas cinco áreas de caatinga:
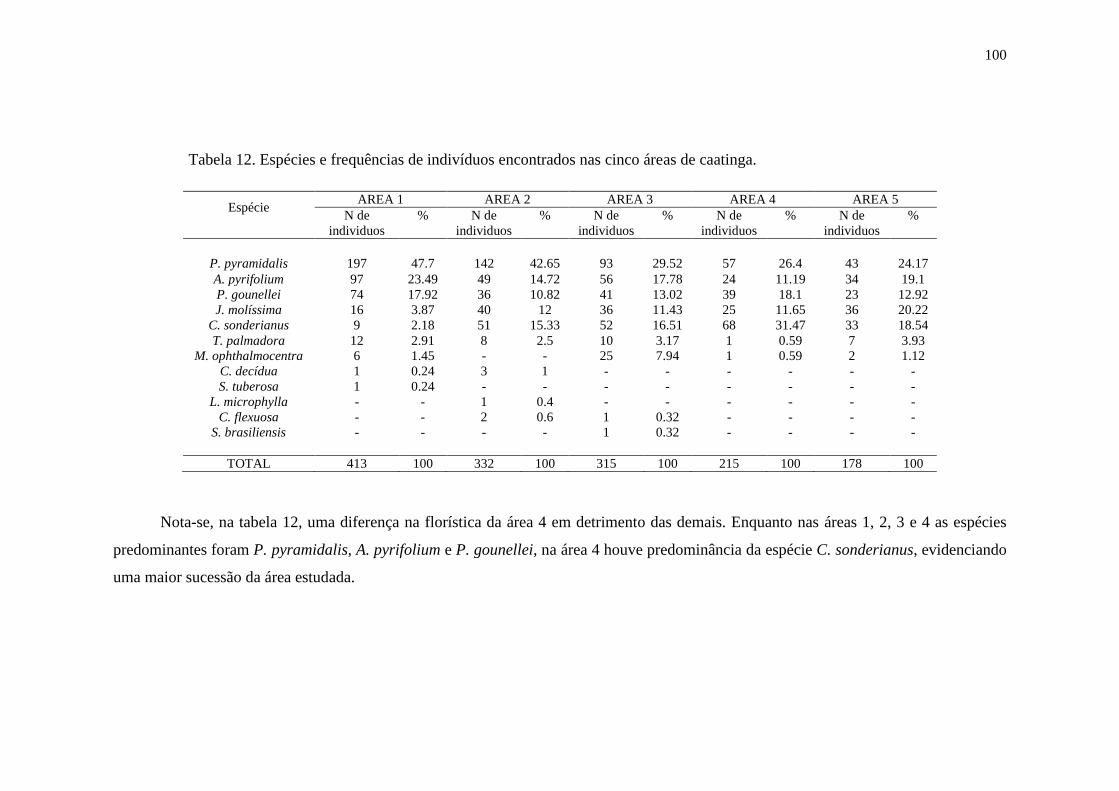
100
Tabela 12. Espécies e frequências de indivíduos encontrados nas cinco áreas de caatinga.
Espécie
AREA 1 AREA 2 AREA 3 AREA 4 AREA 5
N de
individuos
% N de
individuos
% N de
individuos
% N de
individuos
% N de
individuos
%
P. pyramidalis 197 47.7 142 42.65 93 29.52 57 26.4 43 24.17
A. pyrifolium 97 23.49 49 14.72 56 17.78 24 11.19 34 19.1
P. gounellei 74 17.92 36 10.82 41 13.02 39 18.1 23 12.92
J. molíssima 16 3.87 40 12 36 11.43 25 11.65 36 20.22
C. sonderianus 9 2.18 51 15.33 52 16.51 68 31.47 33 18.54
T. palmadora 12 2.91 8 2.5 10 3.17 1 0.59 7 3.93
M. ophthalmocentra 6 1.45 - - 25 7.94 1 0.59 2 1.12
C. decídua 1 0.24 3 1 - - - - - -
S. tuberosa 1 0.24 - - - - - - - -
L. microphylla - - 1 0.4 - - - - - -
C. flexuosa - - 2 0.6 1 0.32 - - - -
S. brasiliensis - - - - 1 0.32 - - - -
TOTAL 413 100 332 100 315 100 215 100 178 100
Nota-se, na tabela 12, uma diferença na florística da área 4 em detrimento das demais. Enquanto nas áreas 1, 2, 3 e 4 as espécies
predominantes foram P. pyramidalis, A. pyrifolium e P. gounellei, na área 4 houve predominância da espécie C. sonderianus, evidenciando
uma maior sucessão da área estudada.

101
Para avaliar a diversidade da área foram utilizados o índice de diversidade de
Shannon-Weaver (H’) e de equitabilidade de Pielou (J’), permitindo representar a
uniformidade de distribuição dos indivíduos entre todas as espécies existentes
(Magurran, 1988).
3.2.3 Índice de equabilidade
O índice de equabilidade pertence ao intervalo de 0 a 1, onde 1 representa a
máxima diversidade, ou seja, todas as espécies são igualmente abundantes e zero o
contrário.
Este valor é representativo ao encontrado para áreas de caatinga com algum grau
de antropização, outros trabalhos demonstram valores superiores quando a caatinga é
mais preservada (Pereira et al., 2002; Alcoforado Filho et al., 2003; Ramalho, 2008).
Tabela 13. Índice de equabilidade de Pielou (J’).
Área 1 Área 2 Área 3 Área 4 Área 5
0.64 0.67 0.83 0.77 0.89
Os índices apresentados mostram uma alta diversidade das áreas em estudo,
especialmente as áreas 3 e 5, segundo a tabela 13.
Segundo Araújo (2010), o conjunto de fatores responsáveis pelos níveis de
similaridade entre as áreas de caatinga analisadas podem ser provavelmente explicado
pelo nível de antropização das áreas e pela proximidade geográfica das mesmas.
3.2.4 Parâmetros fitossociológicos das espécies amostradas
A tabela 14, demonstra um maior número de indivíduos na área 1 (1.380),
enquanto que a área 5 registrou um número mais baixo (593).
Tabela 14. Número de parcela (NP), área total amostrada em hectare (ATA),
Número de indivíduos encontrados (NIE) e Número de indivíduos por hectare (NIH).
Áreas NP ATA (ha) NIE NIH
1 30 0,30 414 1.380
2 30 0,30 332 1.106
3 30 0,30 315 1.050
4 30 0,30 217 723
5 30 0,30 178 593

102
Observa-se que o tamanho da área basal do estrato arbóreo foi diferenciado em
todas as áreas. A maior abrangência para uma área basal com CAB ≥ 3cm foi de 8.067
m² para a área 2, seguidos de 4.506m² para a área 1; 3.217m² para a área 3; 1.662m²
para a área 4 e 1.466m² para a área 5. Observa-se assim, uma sequência decrescente
para a área basal: A2 >A1 > A3 > A4 > A5.
O maior número de indivíduos por hectare foi observado na área 2, com a
presença marcante da espécie P. pyramidalis em uma densidade absoluta (DA) de 4.866
indivíduos/ha.
A área 1 apresentou uma DA de 3.233 indivíduos/ha para a espécies A.
pyrifolium. Enquanto que na área 5 foi observado uma DA de 1.100 indivíduos/ha da
espécie C. sonderianus. Todas as espécies citadas, especialmente o marmeleiro,
descendem de áreas de sucessão conforme autores (Bessa, 2001; Pereira Júnior e
Araújo, 2012), confirmando a hipótese de que as áreas em estudo de sucessão em
regeneração natural.
Sampaio (1996), afirmou que P. pyramidalis é a espécie que aparece com mais
frequência no topo da maior parte das listas de estudos sobre a vegetação de caatinga, o
que foi corroborado por Silva et al. (2012), que comentaram que a espécie é uma das
que mostraram as maiores densidades e regeneração natural em área abandonada de
caatinga.
Além disso, todas as áreas apresentaram altos valores de DR e valor de
importância (VI) para a espécie P. pyramidalis, evidenciando a dominância da
catingueira em áreas de caatinga com sucessão.
Em estudo semelhante, Araújo (2010) verificou no ano de 2010 maiores VI e
DR para a espécie Croton sonderianus em três das áreas deste mesmo estudo. Tal fato,
evidencia a alta modificação e adaptação da vegetação nestas áreas de caatinga em
apenas quatro anos corridos.
Assim como os valores de importância e densidade, os de dominância também
apresentaram-se altos em todas as áreas para a espécie P. pyramidalis, chegando a
67,12% de dominância relativa (DoR) na a área 2.
Abaixo encontram-se os parâmetros fitossociológicos das espécies estudadas nas
áreas de caatinga avaliadas.

103
Tabela 15. Parâmetros fitossociológicos das espécies amostradas em cinco áreas de
caatinga.

104
Nome Científico Nome Comum AB
(m²)
DA
(N/ha)
DR
(%)
FA
(%)
FR
(%)
DoA
(m²/ha)
DoR
(%)
VC
(%)
VI
(%)
AREA 1
A, pyrifolium Pereiro 0,99 323,33 23,49 93,33 26,42 33,13 22,05 22,77 23,99
T, palmadora Palmatória 0,04 40 2,91 20 5,66 1,44 0,95 1,94 3,17
P, pyramidalis Catingueira 2,68 656,66 47,7 96,67 27,36 89,45 59,55 53,62 44,87
M, ophthalmocentra Jurema vermelha 0,05 200 1,45 20 5,66 1,73 1,15 1,3 2,76
P, gounellei Xique-xique 0,5 246,66 17,92 60 16,98 16,73 11,14 14,53 15,34
J, molíssima Pinhão Branco 0,02 53,33 3,87 30 8,49 0,65 0,43 2,15 4,27
C, sonderianus Marmeleiro 0,04 30 2,18 26,67 7,55 1,3 0,86 1,52 3,52
C, decídua Tripa-de-galinha 0,01 3,33 0,24 3,33 0,94 0,07 0,05 0,14 0,41
S, tuberosa Umbuzeiro 0,17 3,33 0,24 3,33 0,94 5,73 3,82 2,03 1,67
TOTAL 4,5 1.556,64 100 353,33 100 150,23 100 100 100
AREA 2
P. pyramidalis Catingueira 5,42 486,67 43,20 83,33 24,04 180,49 67,12 55,19 45,04
A. pyrifolium Pereiro 1,27 163,33 14,50 60,00 17,31 42,39 15,76 15,16 16,11
C. sonderianus Marmeleiro 0,55 176,67 15,68 56,67 16,35 18,35 6,82 11,28 13,21
P. gounellei Xique-xique 0,54 120,00 10,65 40,00 11,54 17,87 6,65 8,73 9,86
J. molíssima Pinhão Branco 0,16 133,33 11,83 60,00 17,31 5,40 2,01 6,94 9,24
T. palmadora Palmatória 0,06 26,67 2,37 20,00 5,77 2,10 0,78 1,60 3,24
C. flexuosa Feijão Bravo 0,03 0,00 0,00 6,67 1,92 1,15 0,43 0,24 1,03
C. decídua Tripa-de-galinha 0,02 10 0,89 10 2,88 0,62 0,23 0,59 1,58
L. microphylla Alecrim da Serra 0,02 3,33 0,30 3,33 0,96 0,54 0,20 0,28 0,74
TOTAL 8,07 1.120 99,42 340 98,08 268,91 100 100 100
AREA 3
P. pyramidalis Catingueira 1,67 310 29,52 90 22,31 55,73 51,98 40,75 34,60
A. pyrifolium Pereiro 0,79 186,67 17,78 66,67 16,53 26,17 24,40 21,09 19,57
M. ophthalmocentra Jurema vermelha 0,10 83,33 7,94 56,67 14,05 3,21 2,99 5,46 8,33
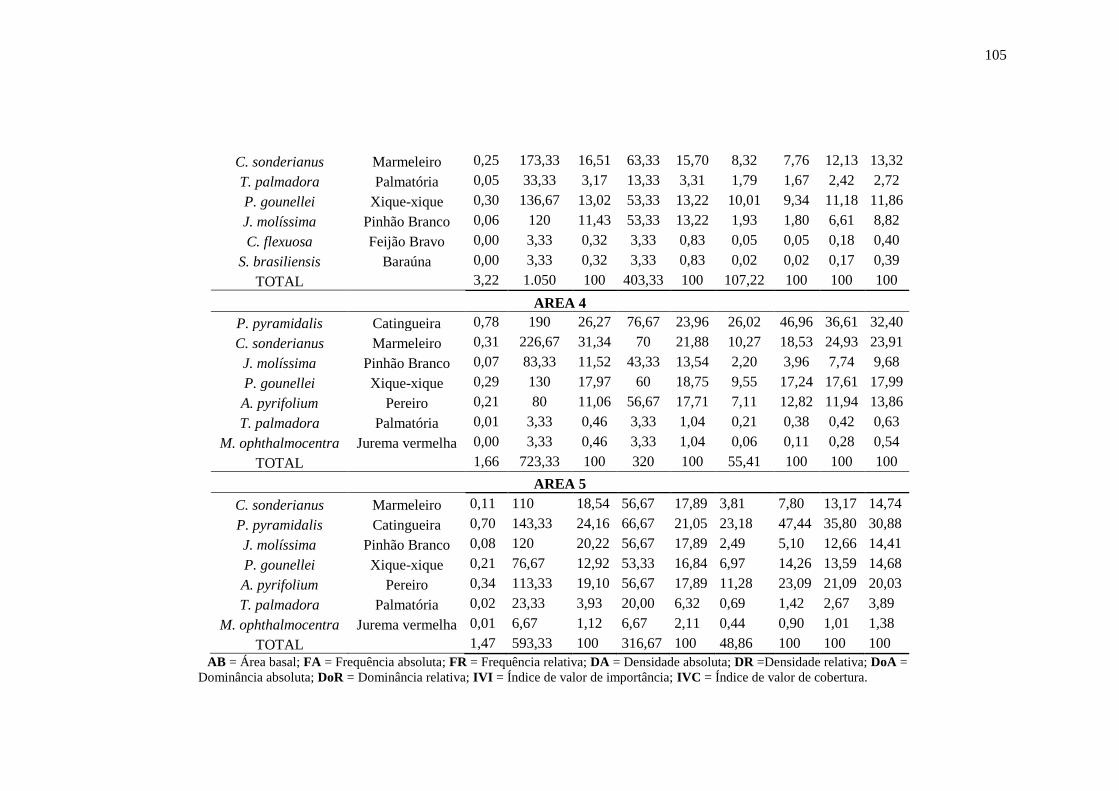
105
C. sonderianus Marmeleiro 0,25 173,33 16,51 63,33 15,70 8,32 7,76 12,13 13,32
T. palmadora Palmatória 0,05 33,33 3,17 13,33 3,31 1,79 1,67 2,42 2,72
P. gounellei Xique-xique 0,30 136,67 13,02 53,33 13,22 10,01 9,34 11,18 11,86
J. molíssima Pinhão Branco 0,06 120 11,43 53,33 13,22 1,93 1,80 6,61 8,82
C. flexuosa Feijão Bravo 0,00 3,33 0,32 3,33 0,83 0,05 0,05 0,18 0,40
S. brasiliensis Baraúna 0,00 3,33 0,32 3,33 0,83 0,02 0,02 0,17 0,39
TOTAL 3,22 1.050 100 403,33 100 107,22 100 100 100
AREA 4
P. pyramidalis Catingueira 0,78 190 26,27 76,67 23,96 26,02 46,96 36,61 32,40
C. sonderianus Marmeleiro 0,31 226,67 31,34 70 21,88 10,27 18,53 24,93 23,91
J. molíssima Pinhão Branco 0,07 83,33 11,52 43,33 13,54 2,20 3,96 7,74 9,68
P. gounellei Xique-xique 0,29 130 17,97 60 18,75 9,55 17,24 17,61 17,99
A. pyrifolium Pereiro 0,21 80 11,06 56,67 17,71 7,11 12,82 11,94 13,86
T. palmadora Palmatória 0,01 3,33 0,46 3,33 1,04 0,21 0,38 0,42 0,63
M. ophthalmocentra Jurema vermelha 0,00 3,33 0,46 3,33 1,04 0,06 0,11 0,28 0,54
TOTAL 1,66 723,33 100 320 100 55,41 100 100 100
AREA 5
C. sonderianus Marmeleiro 0,11 110 18,54 56,67 17,89 3,81 7,80 13,17 14,74
P. pyramidalis Catingueira 0,70 143,33 24,16 66,67 21,05 23,18 47,44 35,80 30,88
J. molíssima Pinhão Branco 0,08 120 20,22 56,67 17,89 2,49 5,10 12,66 14,41
P. gounellei Xique-xique 0,21 76,67 12,92 53,33 16,84 6,97 14,26 13,59 14,68
A. pyrifolium Pereiro 0,34 113,33 19,10 56,67 17,89 11,28 23,09 21,09 20,03
T. palmadora Palmatória 0,02 23,33 3,93 20,00 6,32 0,69 1,42 2,67 3,89
M. ophthalmocentra Jurema vermelha 0,01 6,67 1,12 6,67 2,11 0,44 0,90 1,01 1,38
TOTAL 1,47 593,33 100 316,67 100 48,86 100 100 100
AB = Área basal; FA = Frequência absoluta; FR = Frequência relativa; DA = Densidade absoluta; DR =Densidade relativa; DoA =
Dominância absoluta; DoR = Dominância relativa; IVI = Índice de valor de importância; IVC = Índice de valor de cobertura.

106
Assim, a P. pyramidalis pode ser considerada como um forte indicador do nível
de perturbação antrópica, característica que se expressa pelo comportamento de suas
populações nos ambientes avaliados, já que apresenta grande poder invasor, com
tendência a formar pontos densos, dominando frequentemente pastos limpos ou áreas
perturbadas.
3.2.5 Índice de diversidade de Shannon-Weaver (H’)
Para expressar a heterogeneidade florística da área foi utilizado o índice de
diversidade de Shannon-Weaver (H’).
O índice de diversidade de Shannon (H’) é um indicador da diversidade local e
pode ser utilizado na comparação de diferentes tipologias numa mesma área ou
diferentes áreas com a mesma tipologia (Martins, 1991). Este índice pode expressar
riqueza (número de espécies presentes na flora) e uniformidade (distribuição de
indivíduos entre as espécies).
De acordo com Ramalho (2008) o valor de H’ é maior quanto maior for à
diversidade florística da população estudada.
Tabela 16. Índice de diversidade de Shannon H’
Área 1 Área 2 Área 3 Área 4 Área 5
1.40 1.61 1.83 1.61 1.74
Nota-se, segundo a tabela 16, similaridade entre as áreas 2 e 4, quanto à
diversidade florística, enquanto que as demais mostram-se heterogêneas. O maior índice
de similaridade foi o da área 3, com valor de 1.83, enquanto a que mais apresentou
homogeneidade foi a área 1 (valor de 1.40).
3.2.6 Índice de dominância de Simpson (C’):
O Índice de dominância de Simpson mede a probabilidade de 2 (dois)
indivíduos, selecionados ao acaso na amostra, pertencer à mesma espécie (Brower e
Zarr, 1984). Uma comunidade de espécies com maior diversidade terá uma menor
dominância. O valor estimado de C varia de 0 (zero) a 1 (um), sendo que para valores
próximos de um, a diversidade é considerada maior.

107
Tabela 17. Índice de dominância de Simson (C’)
Área 1 Área 2 Área 3 Área 4 Área 5
0.68 0.74 0.82 0.78 0.82
Logo, os valores apresentaram-se próximos a um (tabela 17), o que representa
uma alta diversidade, ou seja, há pouca dominância da comunidade de espécies
presentes nas áreas. Observou-se uma maior dominância de comunidades na área 1, em
relação às demais.
Desta forma, os altos valores deste índice indicam a antropização das áreas, onde
evidenciam um maior número de indivíduos distribuídos em poucas espécies,
corroborando com Andrade et al. (2011)
3.2.6 Índice de similaridade de Jaccard (J’):
Segundo Mueller-Dombois e Ellemberg (1974), as áreas consideradas
floristicamente similares são as que apresentam índice de Jaccard superior a 0,25.
Tabela 18. Matriz de similaridade florística (Jaccard) em cinco áreas de caatinga.
A1 A2 A3 A4 A5
A1 1 0.73 1 0.89 0.78
A2 0.73 1 0.73 0.64 0.7
A3 1 0.73 1 0.89 0.78
A4 0.89 0.64 0.89 1 0.88
A5 0.78 0.7 0.78 0.88 1
Segundo a tabela 18, pode-se considerar que todas as áreas avaliadas apresentam
similaridade, corroborando com os resultados de Araújo (2010), em pesquisa nas
mesmas áreas de caatinga. Sendo assim, a autora afirma que a proximidade geográfica
pode ter colaborado para tal resultado.
3.2.7 Índice de Agregação ou Índice de MacGuinnes (IGA)

108
A análise do padrão de distribuição espacial dos indivíduos das espécies foi feita
por meio da estimativa de índices de agregação ou índice de MacGuinnes (IGA), que se
refere à distribuição espacial de uma espécie.
Conforme demonstra a tabela abaixo, pode-se afirmar uma tendência ao
agrupamento da espécie P. pyramidalis, com exceção da área 2.Dado este corroborado
pela variabilidade espacial deste espécie, conforme demonstrado nos próximos tópicos.
A espécie J. molíssima foi a única que apresentou tendência de agrupamento
para todas as áreas estudadas.
As espécies M. ophthalmocentra, C. decídua, S. tuberosa, L. microphylla, C.
flexuosa e S. brasiliensis mostraram-se distribuídos de forma uniforme nas áreas. Tal
fato pode ter ocorrido devido ao baixo número de indivíduos registrados nas áreas.
A espécie P. gounellei, possui maior hábito de agregação pelo próprio hábito de
crescimento e propagação da espécie por meio dos cladódios.
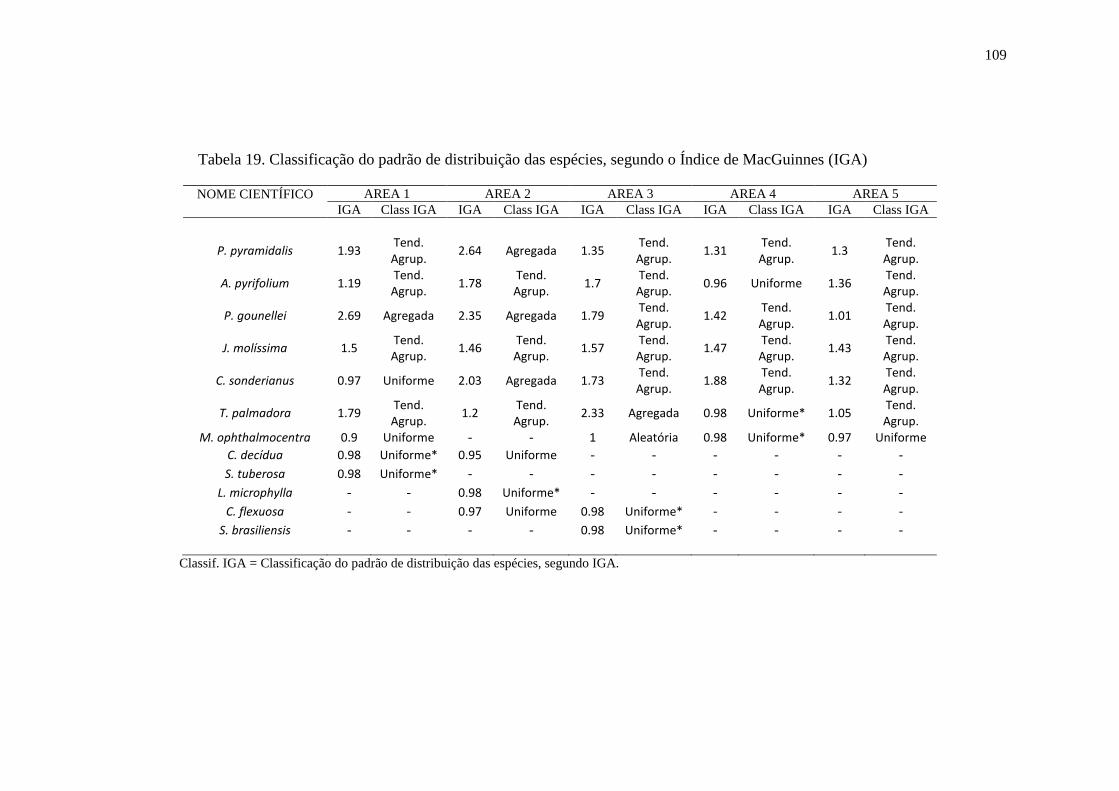
109
Tabela 19. Classificação do padrão de distribuição das espécies, segundo o Índice de MacGuinnes (IGA) NOME CIENTÍFICO
AREA 1 AREA 2 AREA 3 AREA 4 AREA 5
IGA Class IGA IGA Class IGA IGA Class IGA IGA Class IGA IGA Class IGA
P. pyramidalis 1.93 Tend. Agrup.
2.64 Agregada 1.35 Tend. Agrup.
1.31 Tend. Agrup.
1.3 Tend. Agrup.
A. pyrifolium 1.19 Tend. Agrup.
1.78 Tend. Agrup.
1.7 Tend. Agrup.
0.96 Uniforme 1.36 Tend. Agrup.
P. gounellei 2.69 Agregada 2.35 Agregada 1.79 Tend. Agrup.
1.42 Tend. Agrup.
1.01 Tend. Agrup.
J. molíssima 1.5 Tend. Agrup.
1.46 Tend. Agrup.
1.57 Tend. Agrup.
1.47 Tend. Agrup.
1.43 Tend. Agrup.
C. sonderianus 0.97 Uniforme 2.03 Agregada 1.73 Tend. Agrup.
1.88 Tend. Agrup.
1.32 Tend. Agrup.
T. palmadora 1.79 Tend. Agrup.
1.2 Tend. Agrup.
2.33 Agregada 0.98 Uniforme* 1.05 Tend. Agrup.
M. ophthalmocentra 0.9 Uniforme - - 1 Aleatória 0.98 Uniforme* 0.97 Uniforme
C. decídua 0.98 Uniforme* 0.95 Uniforme - - - - - -
S. tuberosa 0.98 Uniforme* - - - - - - - -
L. microphylla - - 0.98 Uniforme* - - - - - -
C. flexuosa - - 0.97 Uniforme 0.98 Uniforme* - - - -
S. brasiliensis - - - - 0.98 Uniforme* - - - -
Classif. IGA = Classificação do padrão de distribuição das espécies, segundo IGA.
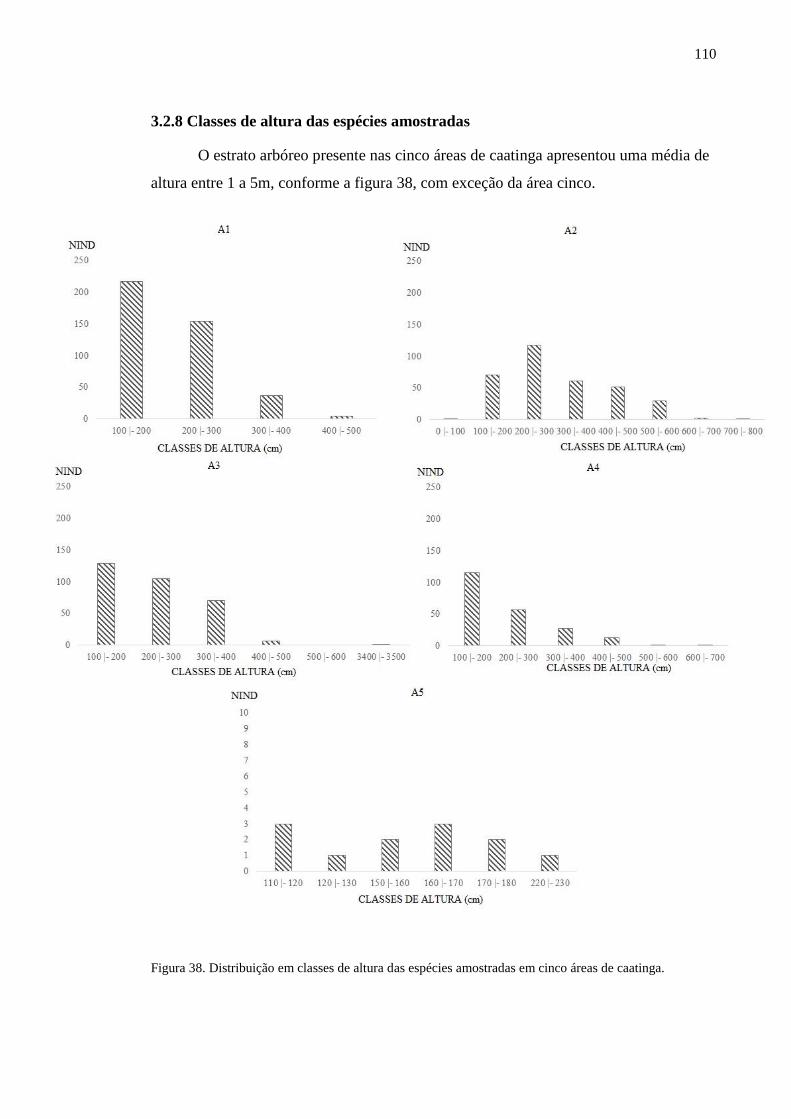
110
3.2.8 Classes de altura das espécies amostradas
O estrato arbóreo presente nas cinco áreas de caatinga apresentou uma média de
altura entre 1 a 5m, conforme a figura 38, com exceção da área cinco.
Figura 38. Distribuição em classes de altura das espécies amostradas em cinco áreas de caatinga.

111
A área 1 apresentou o maior número de indivíduos (pouco mais de 200)
concentrados em uma altura entre 100 a 200cm. Logo, observa-se um comportamento
semelhante nas áreas 3 e 4, porém com um menor número de indivíduos (pouco mais de
100) registrados com altura no intervalo de 100 e 200cm, indicando que nas áreas 3 e 4
há predominância de indivíduos de menor porte arbóreo. Tal fato pode ser explicado por
uma forte ação antrópica que contribuiu em mudanças no hábito de crescimento deste
estrato, com tendência de crescimento a partir de ramos laterais e um maior número de
perfilhos basais.
Segundo o levantamento florístico, estas áreas registraram predominância de
diferentes espécies, sendo a predominante da área 3 P. pyramidalis e da área 4 C.
sonderianus, confirmando a existência de sucessão por atividade antrópica.
Araújo (2010) encontrou as mesmas características para a área 1 na mesma área
em pesquisa, no entanto foram encontrados indivíduos com até 7m de altura e em maior
número. A autora encontrou ainda, valores semelhantes a este trabalho para a área 3 no
ano de 2010 para a variável classes de altura.
Amorim et al. (2005) registraram porte arbóreo com até 7m de altura em áreas
de caatinga no Rio Grande do Norte, enquanto Pereira Júnior e Araújo (2012)
registraram indivíduos com altura até 17m.
Logo, a presença de um menor porte arbóreo registrado neste estudo pode ser
explicado por tratarem-se de áreas de caatinga de sucessão secundária com regeneração
natural, em preservação há um curto período de tempo com indicadores meteorológicos
de baixa precipitação e distribuição irregular nos pulsos de precipitação pluvial. Sendo
Assim, pode-se considerar que os espécimes registrados se tratam de indivíduos jovens
em pleno estádio de desenvolvimento.
A área 5 mostrou-se diferenciada das demais, devido ao seu baixo número de
indivíduos de forma geral. Os maiores registros de altura foram encontrados nas classes
entre 110-120cm e 160-170cm, com pouco mais de 3 indivíduos. Tal fato pode ser
explicado pela distribuição uniforme da altura entre todas as classes avaliadas.
3.2.9 Classes de diâmetro de caule das espécies amostradas
Neste trabalho foram registrados espécimes com diâmetros entre 3 e 223cm, no
entanto o maior número de indivíduos foi observado com intervalo entre 3 e 60cm.
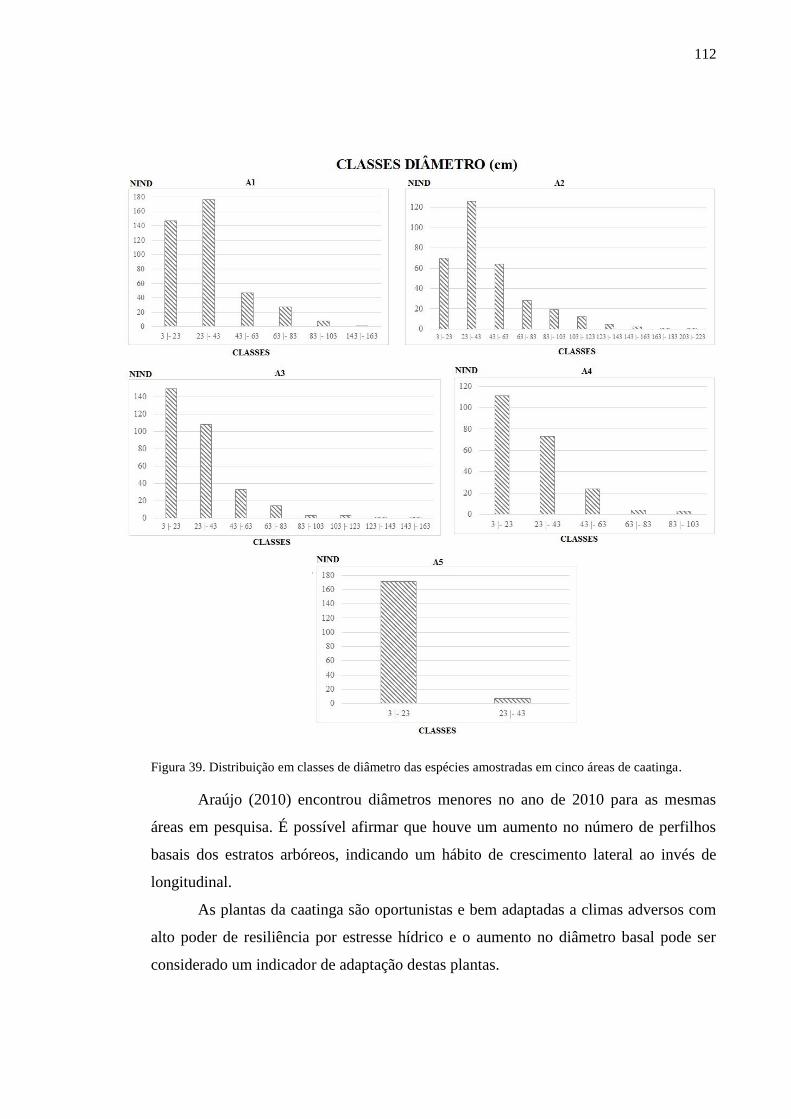
112
Figura 39. Distribuição em classes de diâmetro das espécies amostradas em cinco áreas de caatinga.
Araújo (2010) encontrou diâmetros menores no ano de 2010 para as mesmas
áreas em pesquisa. É possível afirmar que houve um aumento no número de perfilhos
basais dos estratos arbóreos, indicando um hábito de crescimento lateral ao invés de
longitudinal.
As plantas da caatinga são oportunistas e bem adaptadas a climas adversos com
alto poder de resiliência por estresse hídrico e o aumento no diâmetro basal pode ser
considerado um indicador de adaptação destas plantas.

113
Amorim et al. (2005) constataram diâmetros médios de 4,0cm, o que pode ser
considerado baixo ao comparar com os dados registrados neste estudo. Isso demonstra
alta adaptabilidade da vegetação nestas áreas de caatinga.
Além disso, pode-se observar uma alta incidência de indivíduos com caules de
diâmetro entre 3 e 23cm, indicando serem árvores jovens em estádio de
desenvolvimento. Segundo Pereira Júnior e Araújo (2012), a presença de muitos
indivíduos com o diâmetro do caule nas classes de diâmetro iniciais demonstra uma
característica de estágio secundário inicial por parte da vegetação estudada. Os autores
indicam que caso ocorra alguma perturbação na vegetação (estresse hídrico, por
exemplo) e os indivíduos mais velhos (menor número de indivíduos) venham a morrer,
os demais indivíduos jovens, juntamente com os regenerantes, rapidamente repovoarão
a área afetada.
3.3 Variabilidade espacial da serapilheira acumulada e do estrato vegetal em
fragmentos de áreas de caatinga:
Com o interesse de saber a representatividade dos dados avaliados para a
execução da krigagem (interpolação dos dados), foi elaborado o semivariograma de
todas as áreas, conforme demonstra a figura 40.
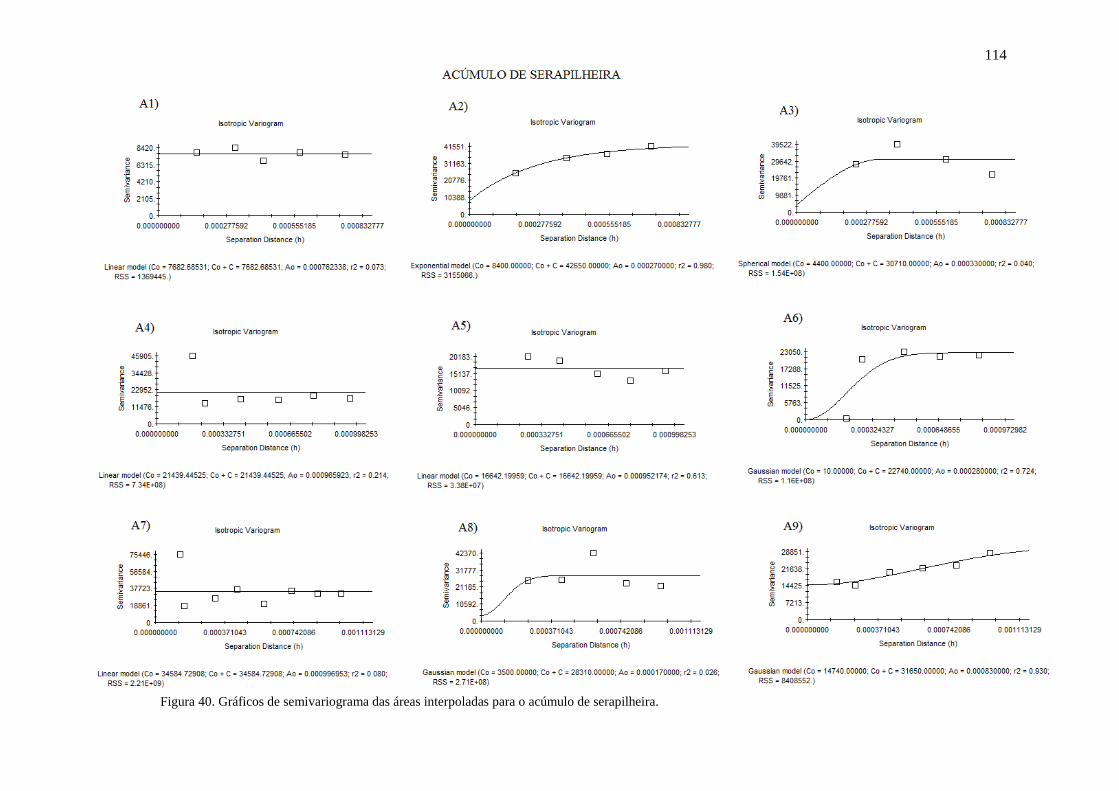
114
Figura 40. Gráficos de semivariograma das áreas interpoladas para o acúmulo de serapilheira.
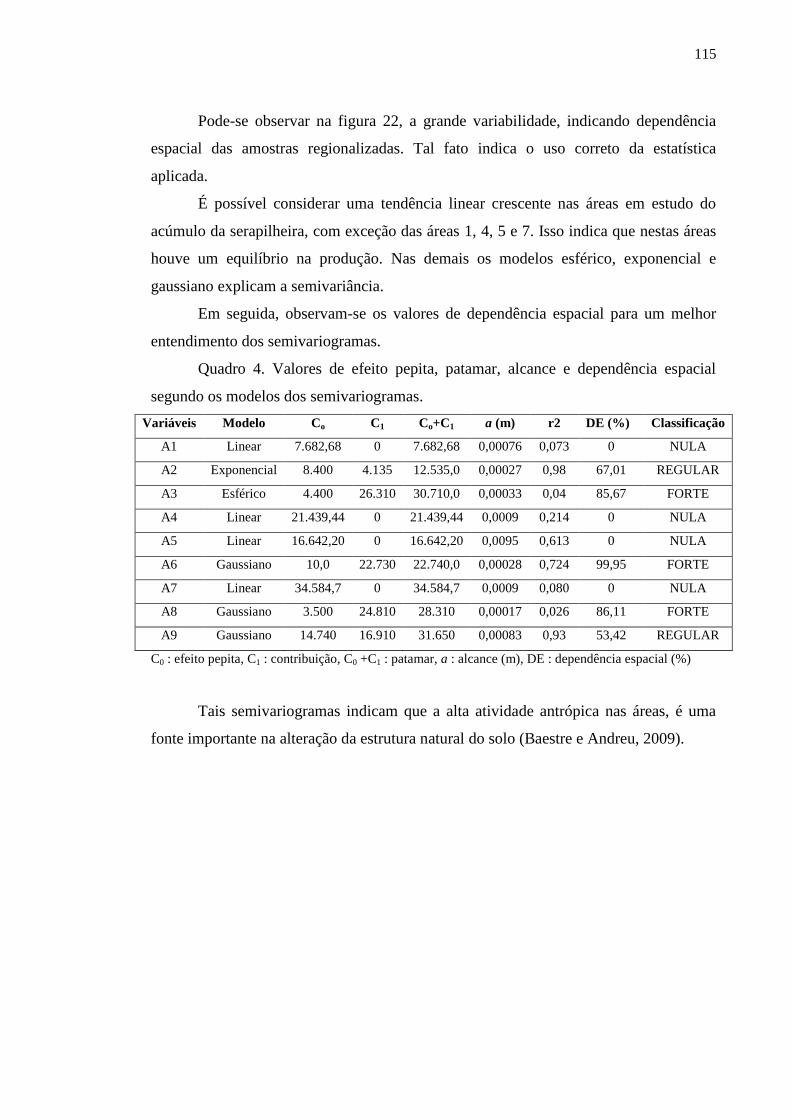
115
Pode-se observar na figura 22, a grande variabilidade, indicando dependência
espacial das amostras regionalizadas. Tal fato indica o uso correto da estatística
aplicada.
É possível considerar uma tendência linear crescente nas áreas em estudo do
acúmulo da serapilheira, com exceção das áreas 1, 4, 5 e 7. Isso indica que nestas áreas
houve um equilíbrio na produção. Nas demais os modelos esférico, exponencial e
gaussiano explicam a semivariância.
Em seguida, observam-se os valores de dependência espacial para um melhor
entendimento dos semivariogramas.
Quadro 4. Valores de efeito pepita, patamar, alcance e dependência espacial
segundo os modelos dos semivariogramas.
Variáveis Modelo Co C1 Co+C1 a (m) r2 DE (%) Classificação
A1 Linear 7.682,68 0 7.682,68 0,00076 0,073 0 NULA
A2 Exponencial 8.400 4.135 12.535,0 0,00027 0,98 67,01 REGULAR
A3 Esférico 4.400 26.310 30.710,0 0,00033 0,04 85,67 FORTE
A4 Linear 21.439,44 0 21.439,44 0,0009 0,214 0 NULA
A5 Linear 16.642,20 0 16.642,20 0,0095 0,613 0 NULA
A6 Gaussiano 10,0 22.730 22.740,0 0,00028 0,724 99,95 FORTE
A7 Linear 34.584,7 0 34.584,7 0,0009 0,080 0 NULA
A8 Gaussiano 3.500 24.810 28.310 0,00017 0,026 86,11 FORTE
A9 Gaussiano 14.740 16.910 31.650 0,00083 0,93 53,42 REGULAR
C0 : efeito pepita, C1 : contribuição, C0 +C1 : patamar, a : alcance (m), DE : dependência espacial (%)
Tais semivariogramas indicam que a alta atividade antrópica nas áreas, é uma
fonte importante na alteração da estrutura natural do solo (Baestre e Andreu, 2009).
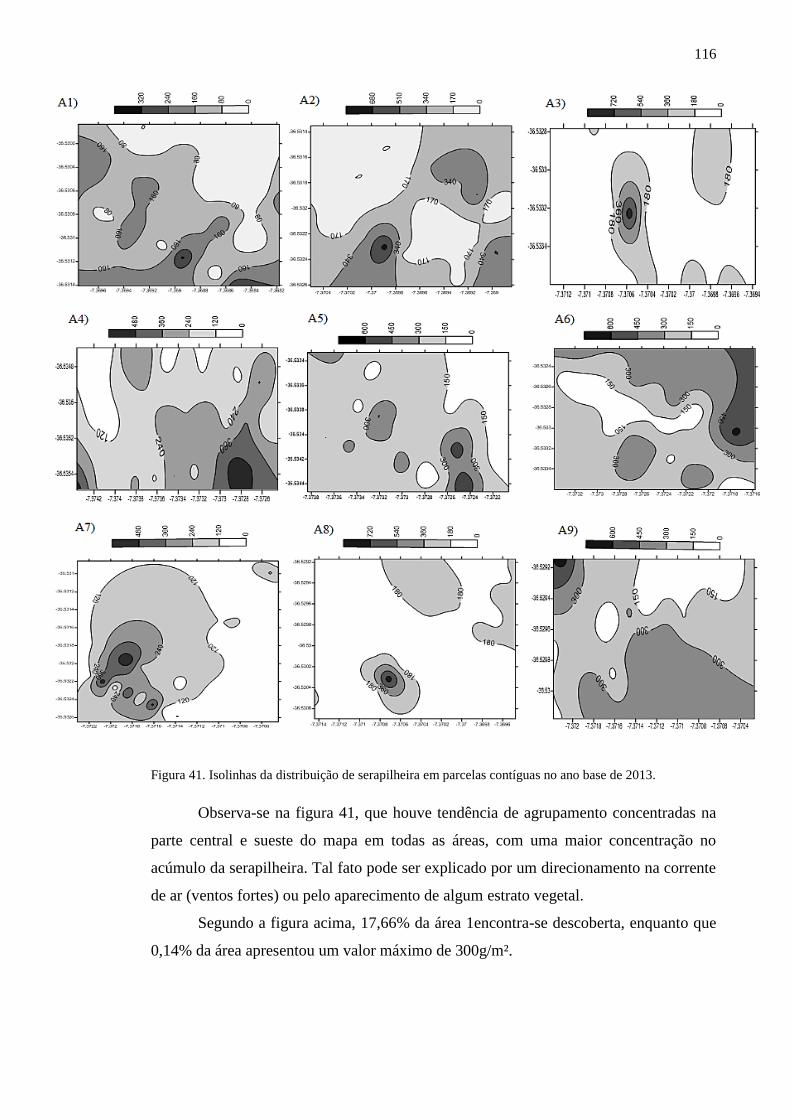
116
Figura 41. Isolinhas da distribuição de serapilheira em parcelas contíguas no ano base de 2013.
Observa-se na figura 41, que houve tendência de agrupamento concentradas na
parte central e sueste do mapa em todas as áreas, com uma maior concentração no
acúmulo da serapilheira. Tal fato pode ser explicado por um direcionamento na corrente
de ar (ventos fortes) ou pelo aparecimento de algum estrato vegetal.
Segundo a figura acima, 17,66% da área 1encontra-se descoberta, enquanto que
0,14% da área apresentou um valor máximo de 300g/m².

117
Ao mesmo tempo, a área 3 mostrou-se com um agravante de 66% de área
descoberta por serapilheira e apenas 34,74% da área coberta com 150 a 700g/m² de
serapilheira.
Cerca de 7,18% da área 4 apresentou pouco mais de 500g/m² de serapilheira,
tendendo ao agrupamento.
A figura dos semivariogramas, representados na página seguinte (figura 42),
para a variabilidade espacial da espécie P. pyramidalis, demonstram uma total
dependência espacial, indicando o uso correto da krigagem para a avaliação estatística.
Os modelos linear, exponencial e esférico representam a variabilidade dos dados.
Além disso, observa-se uma tendência muito forte ao equilíbrio no aparecimento
desta espécie vegetal nas áreas em estudo, corroborando coma avaliação
fitossociológica:
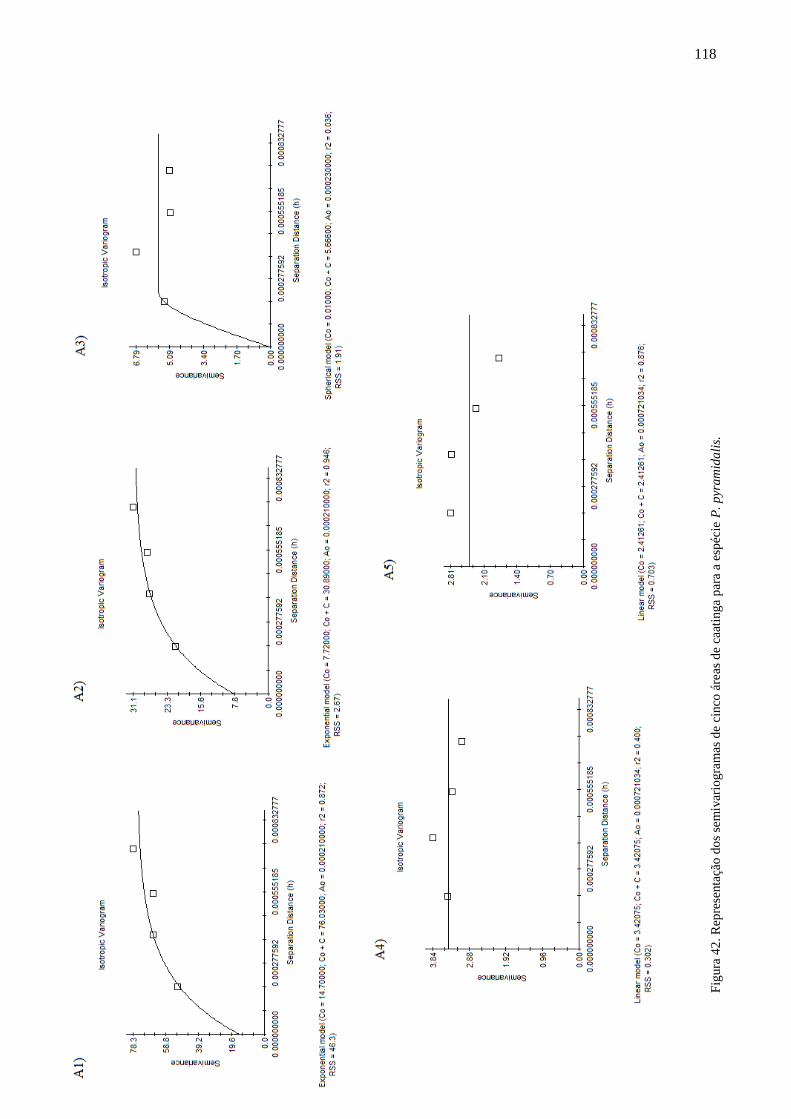
118
Fig
ura
42
. R
epre
senta
ção
do
s se
miv
ario
gra
mas
de
cinco
áre
as d
e ca
atin
ga
par
a a
esp
écie
P.
pyr
am
ida
lis.
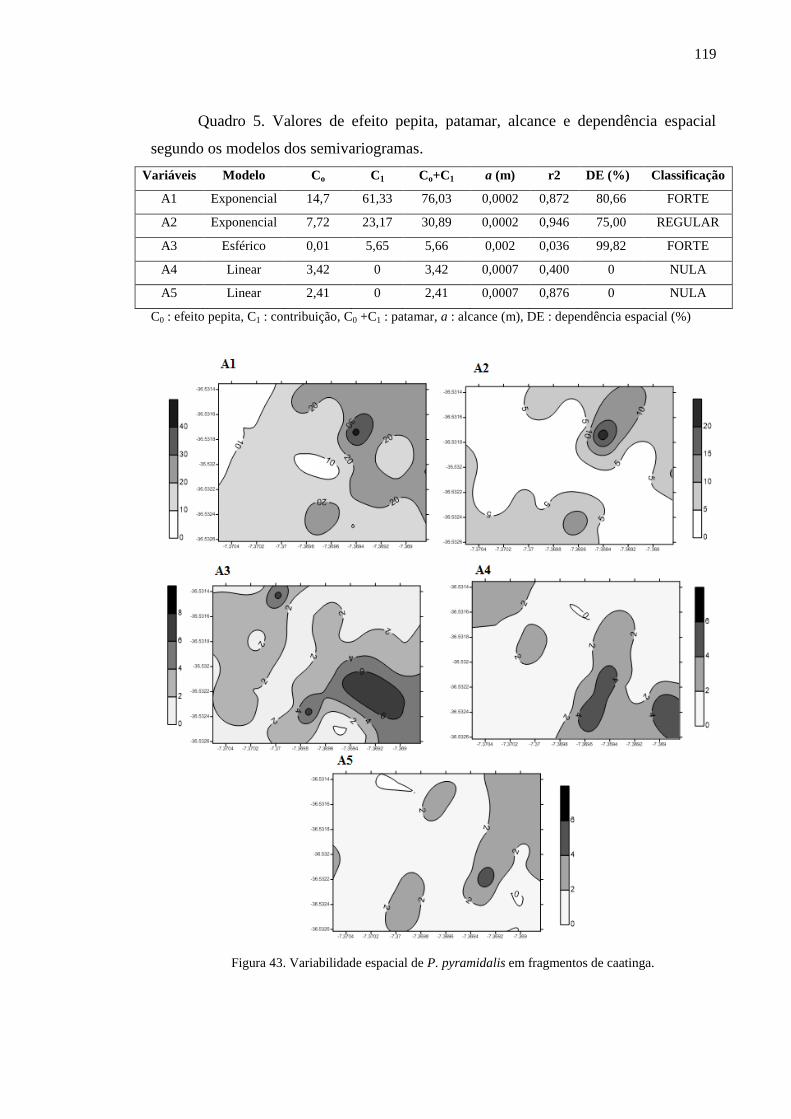
119
Quadro 5. Valores de efeito pepita, patamar, alcance e dependência espacial
segundo os modelos dos semivariogramas.
Variáveis Modelo Co C1 Co+C1 a (m) r2 DE (%) Classificação
A1 Exponencial 14,7 61,33 76,03 0,0002 0,872 80,66 FORTE
A2 Exponencial 7,72 23,17 30,89 0,0002 0,946 75,00 REGULAR
A3 Esférico 0,01 5,65 5,66 0,002 0,036 99,82 FORTE
A4 Linear 3,42 0 3,42 0,0007 0,400 0 NULA
A5 Linear 2,41 0 2,41 0,0007 0,876 0 NULA
C0 : efeito pepita, C1 : contribuição, C0 +C1 : patamar, a : alcance (m), DE : dependência espacial (%)
Figura 43. Variabilidade espacial de P. pyramidalis em fragmentos de caatinga.

120
É possível observar tendência de agrupamento em todas as áreas avaliadas para a
espécie P. pyramidalis, segundo a figura 43. As áreas 1 e 5 apresentaram o maior
número de indivíduos na posição leste do mapa, enquanto que as demais
apresentaram-se dispersas por toda a área.
Ao estudar a variabilidade espacial de duas espécies de cactáceas (Cereus
jamacaru e Pilosocereus pachycladus), Barbosa (2011) verificou tendência no
agrupamento das mesmas, o que pode indicar tendência no agrupamento de espécies
nativas da vegetação na caatinga.
Segue em sequência, o percentual de ocupação da espécie P. pyramidalis nas
áreas em estudo:
Tabela 20. Abrangência de P. pyramidalis nos mapas em isolinhas avaliados nas
áreas de caatinga.
Área Ocupação na área (%)
1 84,60
2 27,11
3 61,62
4 34,55
5 26,48
A tabela acima corrobora com os estudos de levantamento florístico e
fitossociológico, onde a área 1 registrou maior número de indivíduos de P.
pyramidalis, enquanto que a área 5 registrou um menor número. Da mesma forma,
ocorreu a margem de presença nas áreas, com exceção da área 3, onde a presença
desta espécie prevaleceu em 61,62% da área.
Pode-se notar que não houve relação direta entre o acúmulo da serapilheira com
a distribuição de P. pyramidalis em todas s áreas avaliadas, indicando que o maior
fator de ocorrência para o agrupamento da serapilheira podem ter sido proveniente de
correntes de ventos.
Observou-se alguma semelhança nas isolinhas das áreas 2, 4 e 5, apenas. Nestas
áreas, é possível inferir relações entre o acúmulo de serapilheira e a presença de P.
pyramidalis, no entanto mais estudos precisam ser realizados para confirmar tal afirmação.

121
4 CONCLUSÕES
A serapilheira pode ser considerada como uma fonte alimentar alternativa em
épocas de estiagem para o consumo de pequenos ruminantes em sistemas de produção
extensivos.
Não houve aumento no acúmulo de serapilheira devido ao longo período de
estiagem, indicando a presença de espécimes adaptados à situações de adversidade
climática.
As áreas de caatinga avaliadas apresentam-se em regeneração natural, com
presença de espécimes jovens e oportunistas.
A distribuição da serapilheira varia conforme os pulsos de precipitação e a
distribuição da catingueira nas áreas, tendo influência ao agrupamento devido à fatores
abióticos.
5 REFERENCIAS
ALCOFORADO FILHO, F. G.; SAMPAIO, E. V. S. B.; RODAL, M. J. N. Florística e
fitossociologia de um remanescente de vegetação caducifólia arbórea em Caruaru, PE.
Acta Botanica Brasilica, São Paulo , v. 17,n. 2, p. 287-303, 2003.
AMORIM, I. L. de; SAMPAIO, E. V. S. B.; ARAÚJO, E. de L. Flora e estrutura da
vegetação arbustivo-arbórea de uma área de caatinga do Seridó, RN, Brasil. Acta
Botânica Brasilica, v. 19, n. 3, p. 615- HOLOS, Ano 28, Vol 6 84 623, 2005.
ANDRADE, L. A.; FABRICANTE, J. R.; ARAÚJO, E. L. Fitossociologia no Brasil:
Métodos e estudos de casos. Estudos de Fitossociologia em vegetação de caatinga.
Universidade Federal de Viçosa, MG, v1, p 239-371, 2011.
ARAÚJO, K. D. Análise da vegetação e organismos edáficos em áreas de caatinga
sob pastejo e aspectos socioeconômicos e ambientais de São João do Cariri - PB.
Campina Grande - PB, Centro de Tecnologia e Recursos Naturais, UFCG, fevereiro de
2010. 151 p.il. (Tese de Doutorado). Programa de Pós-Graduação em Recursos
Naturais.
ARAÚJO, K. D.; DANTAS, R. T.; ANDRADE, A. P.; PARENTE, H. N.; SILVA, É. É.
Uso de espécies da caatinga na alimentação de rebanhos no município de São João do
Cariri. R. RA´E GA, Curitiba, n. 20, p. 157-171, 2010. Editora UFPR.

122
BAESTRE, R. B. e ANDREU, A. F. Variabilidad espacial de las propiedades físicas de
dos suelos cultivados con palma aceitera en el estado Monagas, Venezuela. Revista
Científica UDO Agrícola, Vol. 9, No. 4, October -December, 2009, pp. 912-924
BARBOSA, M. D.; MARANGON, L. C.; FELICIANO, A. L. P.; FREIRE, F. J.;
DUARTE, G. M. T. Florística e fitossociologia de espécies arbóreas e arbustivas em
uma área de caatinga em Arcoverde, PE, Brasil. Revista Árvore, Viçosa-MG, v.36, n.5,
p.851-858, 2012.
BESSA, M. A. P; MEDEIROS, J. F. Levantamento florístico e fitossociológico em
fragmentos de Caatinga no município de Taboleiro Grande-RN. Rev. Geo Temas. Pau
dos Ferros, Rio Grande do Norte, Brasil. v 1, n. 2, p. 69-83, 2011.
BROWER, J.E. & ZAR, J.H. 1984. Field & laboratory methods for general ecology.
W.C. Brown Publishers, Boston.
CASTRO, A. A. J. F. Florística e fitossociologia de um cerrado marginal brasileiro,
Parque Estadual de Vaçununga, Santa Rita do Passa Quatro - SP. 1987. 238f.
Dissertação (Mestrado em Ciências Biológicas) – Universidade Estadual de Campinas,
Instituto de Biologia.
CHAVES, L. H. G.; CHAVES, I. B.; VASCONCELOS, A. C. F. Salinidade das águas
superficiais e suas relações com a natureza dos solos na Bacia Escola do açude
Namorados. Campina Grande: BNB/UFPB, 54p, 2000. (Boletim Técnico).
CRONQUIST, A.The evolution and classification of flowering plants. 2. ed. New
York: New York Botanical Garden, 1988.
EMBRAPA. Centro Nacional de Pesquisa de Solos. Sistema brasileiro de
classificação de solos. Brasília: EMBRAPA Produção de Informação; Rio de Janeiro:
EMBRAPA Solos, 1999, 421p.
FREIRE, J. L; JÚNIOR, J. C. D. B; LIRA, M. A.; FERREIRA, L. C.; SANTOS, M. V.
F; FREITAS, E. V. Decomposição de serrapilheira em bosque de sabiá na Zona da Mata
de Pernambuco. R. Bras. Zootec., v.39, n.8, p.1659-1665, 2010.
GOVERNO DO ESTADO DA PARAÍBA. Secretaria da Educação. Universidade
Federal da Paraíba. Atlas Geográfico da Paraíba. João Pessoa: Grafset, 1985, 100p.
GOMES, M. A. F. Padrões de caatinga nos Cariris Velhos. Paraíba. 1979. Dissertação
de Mestrado. Universidade Federal de PE, Recife. 88p.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA - IBGE. Rebanho
nacional de caprinos e ovinos. Rio de Janeiro: 2001.
LACERDA, A. V.; NORDI, N. BARBOSA, F.M.; WATANABE,T. Levantamento
florístico do componente arbustivo-arbóreo da vegetação ciliar na bacia do rio Taperoá,
PB, Brasil. Acta bot. bras. 19(3):647-656p. 2005
PEREIRA, I. M.; ANDRADE, L. A.; BARBOSA, M. R. V.; SAMPAIO, E. V. S. B.
Composição florística e análise fitossociológica do componente arbustivo-arbóreo de
um remanescente florestal no agreste paraibano. Acta Botanica Brasilica 2002.

123
PEREIRA JÚNIOR, L. R.; ANDRADE, A P. ARAÚJO, K. D. Composição florística e
fitossociológica de um fragmento de caatinga em Monteiro, PB. Rev. HOLOS, Ano 28,
Vol 6. 2013.
MAGURRAN, A. Ecological diversity and its measurement. New Jersey: Princeton University Press, 179 p. 1988.
MARTINS, F. R. Estrutura de uma floresta mesófila. Campinas: Editora da UNICAMP,
1991. 246 p.
MELO, A. M.; OLIVEIRA, A. B. R. Caprinocultura na Bahia. Companhia Nacional de
Abastecimento-CONAB. 2006
McGUINNES, W. G. The relationship between frequency index and abundance as
applied to plant populations in a semi-arid region. Ecology, v.16, p.263-282, 1934.
MUELLER-DOMBOIS; ELLENBERG, H. Aims and methods of vegetation ecology.
New York: John Willey e Sons, 1974. 525p.
PEREIRA, I. M.; ANDRADE, L. A. de; BARBOSA, M. R. de V.; SAMPAIO, E. V. S.
B. Composição florística e análise fitossociológica do componente arbustivo-arbóreo de
um remanescente florestal no Agreste paraibano. Acta bot. bras. v.16 n.3, p.357-369,
2002.
RAMALHO, C. I. Estrutura da vegetação e distribuição espacial do licuri (Syagrus
Coronata (Mart) Becc.) em dois municípios do Centro Norte da Bahia, Brasil.
2008.131f. Tese (Doutorado em Agronomia) – Centro de Ciências Agrárias,
UniversidadeFederal da Paraíba, Areia.
RODRIGUES, R. R. Análise estrutural das formações florestais ripárias. In:BARBOSA,
L. M. (Coordenador). Simpósio sobre mata ciliar. Campinas. Anais... Campinas.
Fundação Cargill, 1989, p.99-119.
RODAL, M. J. N. Fitossociologia da vegetação arbustivo-árborea em quatro áreas
de caatinga em PE. 1992. 198f. Tese (Doutorado em Biologia Vegetal) - Universidade
Estadual de Campinas, Campinas.
SAMPAIO, E. V. S. B. Fitossociologia. In: SAMPAIO, E. V. S. B.; MAYO, S. J.; BARBOSA,
M. R. V. (Eds.). Pesquisa botânica nordestina: progressos e perspectivas. Recife: Sociedade
Botânica do Brasil/Seção Regional de PE. 1996. p. 203-230.
SILVA, G. G. A problemática da desertificação no ecossistema da caatinga do
município de São João do Cariri. Monografia de Especialização. Universidade
Federal do Piauí. 1993.93p.
SILVA, S. O.; FERREIRA, R.L. C.; SILVA, J. A. A.;LIRA, M. A.;ALVES JÚNIOR, F.
T.; CANO,M. O. O; TORRES, D. L.. Regeneração natural em um remanescente de
caatinga com diferentes históricos de uso no agreste pernambucano. Rev.
Árvore [online]. 2012, vol.36, n.3, pp. 441-450. ISSN 0100-6762.
SILVA, D. F.; SILVA, A. M. A.; LIMA, A. B.; MELO, J. R. M. Exploração da
Caatinga no Manejo Alimentar Sustentável de Pequenos Ruminantes. In: II Congresso
Brasileiro de Extensão Universitária. 2004. Anais... Belo Horizonte, MG.

124
CONSIDERAÇÕES FINAIS
Pode-se considerar que os resultados deste estudo contribuem para um melhor
entendimento de áreas de caatinga de sucessão secundária com criação de pequenos
ruminantes em sistemas extensivos de criação.
Foi observado que a serapilheira é utilizada pelos caprinos como fonte
alternativa de volumoso na época de escassez, em sistemas extensivos de criação em
áreas de caatinga. Tal observação serve para melhorias da alimentação alternativa e de
manejo em épocas de caatinga.
Além disso, foi verificado que a distribuição espacial da serapilheira é variável
segundo as intempéries e a presença dos estratos vegetais nas áreas de caatinga. No
entanto, este material possui baixa qualidade nutricional para a manutenção de caprinos
como forma exclusiva de fonte alimentar, pois apresentou baixos valores proteicos e
altos valores fibrosos. Dessa forma, serão necessários estudos sobre a melhoria da
qualidade nutricional desse volumoso e o acompanhamento do acúmulo deste material
durante um longo período, devido as variações climáticas da região.
É possível considerar que, uma leve lotação de caprinos em áreas de caatinga
para sistemas de criação semiextensiva, não prejudicam a biodiversidade do bioma,
auxiliando na entrada e saída dos nutrientes do solo e consequentemente na
sustentabilidade do sistema, uma vez que as áreas de caatinga do Cariri paraibano são
de sucessão secundária, com regeneração natural e presença de indivíduos jovens e bem
adaptados à situações de adversidade climática.


















