UNIVERSIDADE FEDERAL DO ESPÍRITO SANTO NÚCLEO DE …livros01.livrosgratis.com.br/cp028926.pdf ·...
117
UNIVERSIDADE FEDERAL DO ESPÍRITO SANTO NÚCLEO DE DOENÇAS INFECCIOSAS Carla Couzi Marques Strongyloides stercoralis e Alcoolismo crônico Vitória 2005
Transcript of UNIVERSIDADE FEDERAL DO ESPÍRITO SANTO NÚCLEO DE …livros01.livrosgratis.com.br/cp028926.pdf ·...

UNIVERSIDADE FEDERAL DO ESPÍRITO SANTO NÚCLEO DE DOENÇAS INFECCIOSAS
Carla Couzi Marques
Strongyloides stercoralis e Alcoolismo crônico
Vitória 2005

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

Carla Couzi Marques
Strongyloides stercoralis e Alcoolismo crônico
Dissertação apresentada ao Programa de Pós-graduação em Doenças Infecciosas da Universidade Federal do Espírito Santo, como requisito parcial para obtenção do título de Mestre em Medicina - Doenças Infecciosas. Orientador: Prof. Dr. Fausto Edmundo Lima Pereira.
VITÓRIA 2005


Aos meus pais, que me deram a vida e aos meus irmãos pelo carinho e apoio.

LISTA DE TABELAS
Tabela 1: Idade e sexo de pacientes alcoolistas crônicos e respectivos controles, atendidos no Programa de Atendimento ao Alcoolista do HUCAM, no ano de 2003, nos quais os exames parasitológicos de fezes foram feitos com o método de Baermann............................................................................... Tabela 2: Idade e sexo em 156 alcoolistas crônicos atendidos no Programa de Atendimento ao Alcoolista do HUCAM e não alcoolistas tomados aleatoriamente pelo levantamento de prontuários do mesmo hospital, nos quais o exame parasitológico de fezes foi feito pelo método de sedimentação...................................................................................................... Tabela 3: Idade e sexo em 504 alcoolistas crônicos atendidos no Programa de Atendimento ao Alcoolista do HUCAM, nos anos de 2001 e 2002 e de 591 não alcoolistas tomados aleatoriamente pelo levantamento de prontuários do mesmo hospital, no mesmo período................................................................... Tabela 4: Idade e sexo de 49 pacientes alcoolistas crônicos e 129 controles não alcoolistas atendidos na Unidade Regional de Saúde Feu Rosa (município de Serra), no ano de 2003, nos quais os exames parasitológicos de fezes foram realizados pelo método de sedimentação................................. Tabela 5: Prevalência de parasitas intestinais em 7112 exames parasitológicos de fezes, realizados pelo método de sedimentação, no laboratório de Análises Clínicas do HUCAM, no ano de 2003...........................
68 69 69 70 71

LISTA DE FIGURAS Figura 1: Ciclo evolutivo do S. stercoralis........................................................... Figura 2: Prevalência de S. stercoralis em alcoolistas crônicos e não alcoolistas nas quatro diferentes amostras......................................................... Figura 3: Prevalência de S. stercoralis em alcoolistas crônicos e não alcoolistas nas quatro amostras, nos dois sexos................................................ Figura 4: Relação entre a quantidade de etanol ingerida e a prevalência de exame de fezes positivo para S. stercoralis........................................................ Figura 5: Prevalência de exame positivo para S. stercoralis em alcoolistas crônicos com e sem cirrose hepática................................................................................ Figura 6: Níveis de cortisol plasmático em 23 alcoolistas com exame de fezes positivo para S. stercoralis e em 23 pacientes alcoolistas, com exame de fezes negativo para o parasita. Os resultados mostram as medianas, os quartis e os percentis 5 e 95 (Teste de Mann-Whitney, p=0,602).......................
27 73 74 76 77 77

LISTA DE SIGLAS
ACTH: hormônio adreno-corticotrófico AIDS: Síndrome da imuno-deficiência adquirida CDC: Centers for disease control DP: desvio padrão ELISA: enzima imuno ensaio EPF: exame parasitológico das fezes HIV: vírus da imunodeficiência humana HTLV-1: vírus linfotrófico das células T de adultos HUCAM: Hospital Universitário Cassiano Antônio de Moraes IC: imunocompetente IFI: imunofluorescência IFN gama: interferon gama IgE: imunoglobulina E IgG: imunoglobulina G IgG 4: imunoglobulina G4 IL-4: interleucina 4 IL-5: interleucina 5 IL-12: interleucina 12 IL-13: interleucina 13 N= número ND: não determinado NS: não significativa OR: razão de chance SPSS= Statistical Package for the Social Sciences t= teste de Student URSFR= Unidade Regional de Saúde Feu Rosa X: média χ²= Qui quadrado

SUMÁRIO 1-Introdução.................................................................................................
2-Revisão de literatura.................................................................................
2.1-Strongyloides stercoralis e estrongiloidíase........................................
2.1.1-Histórico...........................................................................................
2.1.2-Morfologia........................................................................................
2.1.3-Ciclo Biológico.................................................................................
2.1.4-Auto-infecção (interna e externa) e hiperinfecção...........................
2.1.5-Modelos experimentais....................................................................
2.1.6-A resposta imunitária na infecção pelo Strongyloides
stercoralis.....................................................................................................
2.1.7-Os mecanismos envolvidos nas formas disseminadas da
estrongiloidíase..............................................................................................
2.1.8-Strongyloides stercoralis e infecção com o vírus HTLV-1................
2.1.9-Anatomia patológica e patogênese das lesões na infecção pelo S.
stercoralis.......................................................................................................
2.1.10-Quadro clínico..................................................................................
2.1.11- Diagnóstico da infecção..................................................................
2.1.12-Tratamento......................................................................................
2.1.13-Profilaxia.........................................................................................
2.2-Efeitos do uso abusivo do etanol sobre o sistema imunitário...............
2.3-Uso abusivo de etanol e prevalência de Strongyloides stercoralis......
3-Objetivos...................................................................................................
3.1-Objetivos gerais..................................................................................
3.2-Objetivos específicos..........................................................................
4-Pacientes e métodos..................................................................................
4.1-Amostras estudadas.............................................................................
4.1.1-Amostras para estudo da prevalência do S.stercoralis em
alcoolistas crônicos........................................................................................
4.1.2-Amostras para avaliação da prevalência de S. stercoralis nos
exames parasitológicos do HUCAM..............................................................
16
20
21
21
24
26
30
32
35
37
42
43
45
46
51
53
53
56
59
60
60
62
63
63
64

4.1.3-Amostra utilizada para avaliação dos níveis séricos de cortisol........
4.2-Exames parasitológicos de fezes.........................................................
4.3-Avaliação do cortisol plasmático...........................................................
4.4-Critérios para diagnóstico de alcoolismo..............................................
4.5- Critérios de exclusão de pacientes e controles...................................
4.6-Informações aos pacientes e controles................................................
4.7-Análise estatística dos resultados........................................................
5-Resultados..................................................................................................
5.1-Idade, sexo e procedência das amostras estudadas...........................
5.2-Prevalência de S. stercoralis nos exames parasitológicos de fezes
realizados no laboratório de rotina do HUCAM.............................................
5.3-Prevalência de S. stercoralis nas diferentes amostras de alcoolistas
e controles.....................................................................................................
6-Discussão...................................................................................................
6.1-Análise crítica das amostras.................................................................
6.1.1-Justificativa para a utilização de diferentes amostras.......................
6.1.2-Análise crítica de cada amostra.........................................................
6.1.3-Análise crítica da utilização de amostras onde o exame
parasitológico de fezes foi realizado com métodos de diferentes
sensibilidades para identificar o S. stercoralis...............................................
6.2-Prevalência do S. stercoralis nos alcoolistas........................................
6.3-Possíveis mecanismos envolvidos na relação entre uso crônico do
etanol e aumento da freqüência de exame parasitológico positivo para S.
stercoralis.......................................................................................................
7-Conclusões.................................................................................................
7.1-A análise dos resultados permite concluir que....................................
8-Referências bibliográficas..........................................................................
Anexo I .......................................................................................................
Anexo II......................................................................................................
Anexo III.....................................................................................................
Anexo IV.....................................................................................................
64
64
65
65
65
66
66
67
68
71
72
78
79
79
79
81
82
84
88
89
90
106
107
108
109

Anexo V......................................................................................................
Anexo VI.....................................................................................................
Anexo VII....................................................................................................
Anexo VIII......................................................................................................................
110
111
112
113

RESUMO


stercoralis. Conclusões: Os resultados demonstraram que: (a) é significativa a
prevalência de exame coprológico positivo para S. stercoralis em alcoolistas
crônicos de ambos os sexos; (b) houve relação direta entre a quantidade média de
etanol ingerida e a freqüência do S. stercoralis no exame coprológico; (c)
alcoolismo crônico foi um fator independente, associado à presença de S.
stercoralis no exame coprológico; (d) as variáveis analisadas não permitiram
identificar se a maior freqüência do exame positivo está relacionada à maior
prevalência do parasita ou se a uma facilitação na produção e eliminação de
larvas; (e) não houve relação entre cortisol plasmático e prevalência do S.
stercoralis nos alcoolistas crônicos onde o hormônio foi avaliado; (f) a prevalência
significativa de exame positivo para S. stercoralis no laboratório de rotina do
HUCAM, em pacientes do sexo masculino foi possivelmente relacionada com a
alta freqüência de pacientes alcoolistas (a maioria do sexo masculino) atendidos
no HUCAM.
Palavras-chaves: alcoolismo; Strongyloides stercoralis; cortisol

ABSTRACT

Introduction: High prevalence of S. stercoralis has been reported in chronic
alcoholic abusers. Objectives: To study (a) the prevalence of S. stercoralis in
chronic alcoholic patients and in non-alcoholic, using the Baermann and
sedimentation methods; (b) the prevalence of alcoholism in patients with S.
stercoralis detected in stools in a routine laboratory of a General Hospital;(c) the
plasma level of cortisol in alcoholic patients with and without S. stercoralis.
Patients and Methods: (1)190 alcoholic patients and 111 nonalcoholic (Baermann
method for detection of S. stercoralis, three stool samples ); (2) 156 alcoholic
patients and 591 non-alcoholic (sedimentation method for identification of S.
stercoralis, three stool samples); all alcoholic and non-alcoholic patients (controls)
attended at the University Hospital Cassiano Antônio de Moraes (HUCAM), belong
to the same socioeconomic class and lived in the same neighborhoods of the
Metropolitan Vitória. Evaluation of plasma cortisol was done in 46 alcoholic
patients, 23 with S. stercoralis. (3) 49 alcoholic patients and 129 non-alcoholic from
the out patient unit Feu Rosa in the municipal district of Serra; (4) file review of
504 alcoholic patients attended at a special unit for alcoholism in the out patients
unit of the HUCAM (during 2001 and 2002) and 591 non alcoholic patients
attended at the same Hospital. Both, the alcoholic and control groups, belong to
the same socioeconomic class and lived in similar neighborhoods of Metropolitan
Vitória; (5) review of records of patients with S. stercoralis diagnosed by stool
examination at the routine laboratory of HUCAM from January to December 2003.
Results: The prevalence of S. stercoralis was significantly higher in alcoholic
patients than in controls in all samples studied (OR from 3,41 to 4,92 with 95% CI
from 1,58 to 8,36). In the sample obtained by file review of 504 alcoholic patients,
the difference was significant for males and females, that was not observed in
other samples from HUCAM and in sample from Serra, because the small
number of alcoholic women in these groups. Among 188 patients (129 males and
59 females; p<0,05) with S. stercoralis detected in the routine laboratory of
HUCAM during 2003, chronic alcoholism was diagnosed in 41,17% of 51 patients
whose records presented a sure diagnosis of alcohol abuse. There was not

difference in the plasma cortisol levels in alcoholic patients with or without S.
stercoralis. Conclusion: Results demonstrated: (a) there was a significant
prevalence of S. stercoralis in male or female alcoholic patients; (b) there was a
significant correlation between the daily amount of ingested alcohol and the
prevalence of S. stercoralis; (c) chronic alcoholism was an independent factor
associated to the detection of S. stercoralis in stool examination; (d) the higher
prevalence of S. stercoralis in males, observed in stool examinations at the
HUCAM’s routine laboratory, probably was due to the high prevalence of alcoholic
patients (most of them males) that had medical treatment at this Hospital; (e) there
was not relation between the plasma cortisol levels and the prevalence of S.
stercoralis in the alcoholic patients in which the hormone was evaluated.
Key words: alcoholism; Strongyloides stercoralis; cortisol

INTRODUÇÃO

A estrongiloidíase é uma helmintíase primordialmente intestinal causada
pelo Strongyloides stercoralis e pelo Strongyloides fulleborni, nematódeos da
família Rhabdiasidae. Desta superfamília, um único gênero interessa à medicina:
gênero Strongyloides, e, no Brasil, a única espécie diagnosticada é o S.
stercoralis.
Segundo alguns cálculos devem existir 200 milhões de pessoas infectadas
com o S. stercoralis em todo o mundo (WHO, 1992).
O S. stercoralis é um parasita que tem como habitat o intestino delgado
mas pode disseminar-se por todo o organismo, como conseqüência da
imunossupressão natural ou adquirida, quando então pode tornar-se
potencialmente fatal. Acomete indivíduos de qualquer idade, predominando em
zonas de clima tropical ou sub-tropical (CUNHA, 1988 e GENTA, 1989).
Condições precárias de higiene, tratamento inadequado das fezes e poluição do
solo favorecem a infecção. É necessário que o terreno seja poroso, que tenha
certo grau de umidade e temperaturas favoráveis entre 25º a 30º C. Isso explica o
predomínio desta helmintíase nos trópicos (CUNHA , 1988).
No Brasil, a estrongiloidíase distribui-se por todo o país, sendo mais
freqüente nas áreas mais quentes. Os dados brasileiros sobre a prevalência desta
helmintíase são variáveis de acordo com a região, em função das condições
sócio-econômicas. A prevalência do S. stercoralis em crianças com idade variando
de 4 meses a 7 anos foi de 13%, na cidade de Uberlândia - estado de Minas
Gerais, e de 5,5% em população de adultos não alcoolistas (MACHADO e
COSTA-CRUZ, 1988; OLIVEIRA et al, 2002). No Espírito Santo, existe pouca
informação sobre a prevalência desse nematóide. Em uma amostra de 190
crianças internadas no Hospital Nossa Senhora da Glória, tomada aleatoriamente,
nas quais exames parasitológicos das fezes foram realizados em 4 amostras pelo
método de Baerman-Morais, 16 (8,4%) foram positivas para o S. stercoralis
(MOREIRA-SILVA, 1998).
A dependência do álcool, além de ser um problema médico e social,
representa um sério problema de saúde pública acometendo em torno de 11,2%
da população adulta brasileira (CARLINI et al, 2002). O uso crônico e até mesmo

agudo, pode aumentar a susceptibilidade do indivíduo a infecções (MAC
GREGOR, 1986; ROSELLE et al,1995; SZABO, 1999 ). De fato, observações em
pacientes alcoolistas crônicos ou com intoxicação aguda e em modelos
experimentais de intoxicação etílica mostram que o álcool altera a resposta
imunológica, tanto nos seus mecanismos inespecíficos ou inatos como nos seus
mecanismos específicos ou adaptativos (MAC GREGOR, 1986).
Existem poucos relatos na literatura sobre os possíveis efeitos do etanol na
evolução de parasitas intestinais, sendo todos baseados em observações
experimentais. STEVEN et al (1990) mostraram que a eliminação de Trichinella
spiralis foi retardada em ratos tratados com etanol. NA et al (1997) demonstraram
que ratos infectados com T. spiralis e tratados com etanol têm redução da
resposta inflamatória mediada por células com aumento da produção de citocinas
do tipo TH2. A baixa produção de IFN gama e o subseqüente aumento na
produção de citocinas TH2 se correlacionou com a maior sobrevivência e
fecundidade do parasita. Avaliaram os níveis séricos de corticosterona e eles
foram semelhantes nos dois grupos. Os autores admitiram que a redução nas
citocinas do tipo TH1, produzidas no início da infecção, possa ter sido o fator
fundamental na facilitação da proliferação e no retardo da eliminação do parasita.
GABURRI et al (1997) estudando pacientes cirróticos em Juiz de Fora -
Minas Gerais observaram prevalência significativamente maior de S. stercoralis
em pacientes cirróticos, especialmente nos de etiologia alcoólica. No entanto, não
teceram considerações sobre os possíveis mecanismos envolvidos, admitindo
inclusive que a doença hepática e não o etanol seria o fator mais importante na
facilitação da helmintíase.
OLIVEIRA et al (2002) avaliaram a freqüência de S. stercoralis em
alcoolistas. Foram avaliados 145 indivíduos, dos quais 45 eram dependentes do
álcool (27 alcoolistas sem cirrose hepática e sem pancreatite, 9 com cirrose
hepática e 9 com pancreatite crônica), 10 eram portadores de cirrose hepática
não-alcoólica e 90 eram indivíduos não alcoolistas assintomáticos. Foram feitos
três exames parasitológicos das fezes em dias alternados pelos métodos de
Baermann-Moraes e Lutz. A freqüência da estrongiloidíase no grupo total de

alcoolistas foi 33,3% e no subgrupo de alcoolistas com cirrose hepática, 44,4%.
Com pancreatite, 33,3% e naqueles sem cirrose hepática e sem pancreatite,
29,6%, o que foi estatisticamente mais alto que o encontrado nos controles (5,5%).
Nenhum dos indivíduos com cirrose hepática não alcoólica tinha infecção pelo S.
stercoralis. Estes resultados mostram que o álcool é um importante fator que
predispõe à estrongiloidíase.
Observação semelhante foi feita no Serviço de Gastroenterologia do
HUCAM, onde a freqüência de S. stercoralis em alcoolistas crônicos foi
significativamente maior do que em não alcoolistas atendidos no mesmo hospital
(ZAGO-GOMES, et al 2002).
Há portanto evidências de que o consumo excessivo de etanol possa estar
relacionado com um exame parasitológico positivo para o S. stercoralis, razão pela
qual planejamos a presente investigação com o objetivo de verificar a prevalência
de S. stercoralis em alcoolistas crônicos atendidos em dois diferentes serviços da
Região Metropolitana de Vitória e comparar com amostras de pacientes não
alcoolistas atendidos nos mesmos serviços.

REVISÃO DE LITERATURA

2.1- Strongyloides stercoralis e estrongiloidíase
2.1.1- Histórico
Este helminto foi descoberto em julho de 1876 por Louis Normand, um
médico do Hospital Naval em Toulon, França, nas fezes de soldados franceses
que retornavam da Cochinchina (Vietnam) razão pela qual, no passado, a doença
humana ter sido conhecida como diarréia da Cochinchina (PESSOA, 1978;
GROVE, 1996; NOLAN, GENTA, e SCHAD, 1998; CAMPOS e FERREIRA, 1999).
Os vermes foram enviados a Bavay, um colega de Normand e professor de
farmácia no Hospital Naval em Toulon, que os descreveu com o nome de
Anguillula stercoralis, no mesmo ano. Ele também reconheceu que quando as
larvas eram mantidas nas fezes por poucos dias sob condições favoráveis, estas
se desenvolviam em vermes adultos de vida livre (machos e fêmeas). Na
necropsia de alguns destes soldados que retornavam da Cochinchina e que
faleceram devido à diarréia, Normand encontrou larvas por todo o intestino, canais
biliares e pancreático e fêmeas adultas nos intestinos que, aparentemente,
diferiam dos primeiros vermes adultos de vida livre observados. Pensando tratar-
se de uma nova espécie, descreveu-os com o nome de Anguillula intestinalis.
Durante muito tempo foram assim conhecidos e até hoje os clínicos dão o nome
de “anguilulose” à moléstia determinada pelo Strongyloides, e de “anguílulas” às
larvas rabditóides encontradas nas fezes. Isso se justifica pelo fato de que fêmeas
parasitas diferem marcadamente dos adultos de vida livre e os primeiros foram
considerados uma espécie diferente. Fortaleceu esta dedução, a descoberta de
um segundo tipo de larva, a larva filarióide (posteriormente identificada como a
forma infectante) que, na história da estrongiloidíase, foi considerada a larva da
segunda espécie, A. intestinalis. Houve muita confusão com essas descobertas
nos anos que se seguiram, pois parecia que havia duas espécies com diferentes
ciclos de vida chamados de A. stercoralis, no qual a larva rabditóide aparecia nas
fezes e os vermes adultos no meio externo, e A. intestinalis, no qual os vermes
adultos eram parasitas intestinais e que geravam as larvas filarióides (NOLAN,
GENTA e SCHAD, 1998 )

Grassi e Parona (1878) mostraram que a fêmea parasita chamada de A.
intestinalis põe ovos que se rompem rapidamente, aumentando o número de
larvas rabditóides que foram conhecidas como A. stercoralis. Aparentemente, eles
tinham uma cepa homogônica do parasita porque todas as larvas rabditóides se
desenvolviam em larvas filarióides infectantes tal como foi descrito para A.
intestinalis. Isto permaneceu até que Perroncito (1881) mostrou que a larva
rabditóide se transforma em machos e fêmeas de vida livre e que estes produzem
a larva filarióide, esta última representando o estágio infectante do parasita.
Perroncito não se deu conta de que os vários estágios eram partes de um
complexo ciclo de vida alternando vida parasitária e gerações de vida-livre
(GROVE, 1996).
A correta relação entre esses estágios foi resolvida por Grassi e Parona na
Itália em 1878 (GROVE, 1996). Grassi, em 1879, criou uma nova classe que ele
chamou de Strongyloides (de strongylos, redondo, e eidos, semelhante) e
chamou o parasita de Strongyloides intestinalis. Perroncito, em 1881, cultivou o
verme adulto de vida-livre (que ele chamou de Pseudorhabditis stercoralis) a partir
da larva (agora chamada rabditóide) idêntica à do A. stercoralis, descrita por
Normand. Leuckart (1883) demonstrou que todas essas formas eram diferentes
fases no ciclo de vida de uma único parasita. Finalmente, em 1902, Stiles e Hassal
demonstraram que o parasita deveria ser chamado de Strongyloides stercoralis
(GROVE,1996 ).
Os franceses, Normand e Bavay, descobriram a doença e descreveram o
parasita; os italianos, Perroncito, Grassi e Parona elucidaram o ciclo de vida livre
e subseqüentemente, os parasitologistas alemães Looss, em 1905, e Fulleborn,
em 1914, mostraram, respectivamente, que a infecção ocorria pela penetração
pela pele e que a larva poderia migrar da pele para intestinos, via circulação,
pulmões e traquéia. Em 1911, Gage mostrou a ocorrência da auto-infecção
(NOLAN, GENTA e SCHAD, 1998).
Durme (1902), Looss (1905), Ranson (1907) e Fulleborn (1914) verificaram
que a larva de Strongyloides faz migração pulmonar e que as formas infectantes
penetram pela pele. Até recentemente, só se tinha conhecimento da existência de

formas femininas parasitas e assim eram elas consideradas partenogenéticas,
porém como Sandgroun (1926) encontrou nestas formas, espermatozóides,
considerou-as hermafroditas. Kreis (1932) descreveu o que considerou machos
parasita do gênero Strongyloides e Faust (1933-1935) assinalou a presença de
machos adolescentes no pulmão os quais inseminavam as fêmeas antes delas
alcançarem o intestino. Cram (1936-1939), trabalhando com S. ratti, concluiu que
as fêmeas do S. ratti são partenogenéticas (PESSOA, 1978).
Para alguns autores, os machos parasitas vivem no intestino delgado
profundamente encravados na mucosa. Para outros, a fêmea adolescente é
fertilizada pelo macho nos brônquios ou na traquéia antes que penetre no epitélio
intestinal. Outros pesquisadores não consideram o macho um parasita do tecido,
sendo expulso nas fezes após curto estágio no corpo. Finalmente, a maioria
considera a fêmea parasita como partenogenética, devido à impossibilidade de se
encontrar formas masculinas nas infecções experimentais ou espermatozóides
nas fêmeas, bem como pelo estudo dos cromossomos do Strongyloides. De
acordo com Chang e Grahm (1957), os cromossomos da fêmea parasita são
triplóides, sendo derivados de um ovo diplóide da fêmea de vida livre (devido à
falta de redução na divisão) que é fertilizado por um espermatozóide haplóide do
macho de vida livre (PESSOA, 1978).
Atualmente, considera-se uma forma feminina partenogenética parasita e
formas masculinas e femininas de vida livre.
Espécies do gênero Strongyloides têm sido descritas em aves, répteis,
anfíbios e mamíferos. São conhecidas pelo menos 52 espécies (GROVE, 1996;
CAMPOS e FERREIRA, 1999), entre elas o S. canis, que parasita o cão,
descoberto por Fulleborn em 1914. O rato pode ser parasitado pelo S. ratti e os
primatas pelo S. fulleborni. O S. fulleborni foi encontrado no homem por Blackie
(1932), na Rodésia, e por Wallace (1948), nas Filipinas. Pampligione e Riccard
(1972), demonstraram que esta espécie não é parasita acidental do homem,
porém parasita os habitantes de uma região da África. No Brasil esta espécie foi
estudada por Rego (1972) em macacos Rhesus naturalmente infectados.
Verificou-se que o S. fulleborni é incapaz de infectar macacos do Novo Mundo,

não conseguindo infectar rato, camundongo e cobaia. S. fulleborni “símile” tem
sido encontrado infectando crianças na Ilha de Nova Guiné (GROVE, 1996;
CAMPOS e FERREIRA, 1999; COSTA-CRUZ, 2000).
Uma característica das espécies do gênero Strongyloides é a uniformidade
morfológica, considerada como possível marcador da natureza primitiva do
parasita (Sandground, 1926).
Em resumo, a família Strongyloididae (Classe Secernentasida, Ordem
Rhabditida, Superfamília Rhabditoidea) é formada somente por três gêneros,
Strongyloides Grassi, 1879; Parastrongyloides, Morgan, 1928; e Leipernema, Sing,
1976. Os membros desta família, também chamados vermes filiformes, são
heterogônicos, com geração parasítica, de vida-livre e compreende pelo menos 52
espécies. A grande maioria das espécies não infecta o homem. Infecções patentes
não foram encontradas após infecção experimental de humanos com S. canis, S.
cebus, S. felis, S. myopotami, S. planiceps, e S. simiae (GROVE, 1996).
2.1.2- Morfologia
A fêmea parasita partenogenética é semitransparente, filiforme, medindo
aproximadamente 2,2 mm de comprimento por 0,04mm de diâmetro; parede do
corpo delicadamente estriada; extremidade anterior dotada de pequena abertura
oral que se comunica com o esôfago longo, cilíndrico, e em seguida ao intestino,
reto e ânus. Tem o aparelho genital constituído por ovário, oviduto, útero, vagina e
vulva; esta se localiza no terço posterior do corpo do verme, diferencia-se em uma
pequena vagina que se comunica com o útero; este, dirige-se tanto para a região
anterior como para a região posterior; no útero, observa-se uma fileira única de
ovos transparentes de casca fina; as alças uterinas, anterior e posterior,
diferenciam-se em ovidutos e ovários. O ovário duplo caracteriza o nematóide
como anfidelfo; ovário anterior dirige-se até próximo ao esôfago. Não há
receptáculo seminal. A fêmea parasita de S. stercoralis é considerada ovovivípara,
pois os ovos expulsos contêm uma larva no seu interior. A oviposição ocorre nas
criptas da mucosa intestinal.

Os ovos apresentam uma casca fina. Medem cerca de 50 a 58 µm de
comprimento por 30 a 34 µm de largura, em sua maioria, embrionados no
momento da postura. No interior dos ovos, encontram-se as larvas de primeiro
estágio (rabditóides). Raramente, os ovos são encontrados nas fezes do indivíduo
infectado, uma vez que as larvas eclodem muito rapidamente, nas criptas de
Lieberkuhn.
As larvas de primeiro estágio (rabditóides) eclodem dos ovos e se
insinuam no epitélio glandular e caem na luz intestinal, sendo assim encontradas
nas fezes nos infectados com S. stercoralis. Medem entre 200 a 300 µm de
comprimento por 14 a 16 µm de diâmetro. A forma em bastão do esôfago, neste
estágio, deu origem ao nome da larva. Esta larva tem o esôfago dividido em três
porções: corpo, istmo e bulbo. Ao esôfago seguem-se: intestino, reto e ânus.
Apresenta o vestíbulo bucal curto (2 a 3 µm) e o primórdio genital conspícuo. O
vestíbulo bucal corresponde à região que se inicia na cutícula da extremidade
anterior da larva até o início do esôfago; mede aproximadamente 2 µm
de comprimento. O primórdio genital corresponde a um conjunto de células
situado ao lado do intestino e no terço posterior do corpo da larva. A extremidade
posterior (cauda) termina bruscamente.
Em laboratório clínico há necessidade de se estabelecer o diagnóstico
diferencial entre larvas rabditóides de S. stercoralis com larvas de
ancilostomídeos. As larvas de ancilostomídeos apresentam um vestíbulo bucal
longo (10 µm de comprimento), o esôfago é menos nitidamente dividido em três
porções e o primórdio genital é menor do que nas larvas de Strongyloides
stercoralis. A extremidade posterior afila-se lentamente.
Antes de alcançar a fase infectante, as larvas de primeiro estágio de S.
stercoralis passam por uma fase intermediária que corresponde ao segundo
estágio ou fase pré-infectante: o esôfago perde a sua forma rabditóide, torna-se
alongado e surge uma cutícula que caracteriza a ocorrência da primeira muda
larval.
As larvas filarióides são encontradas no meio externo (fezes e solo); são
oriundas de um processo de diferenciação e segunda muda larval, a partir das

larvas rabditóides pré-infectantes. Medem aproximadamente 500 µm de
comprimento por 10 µm de largura. O tubo digestivo e constituído de esôfago,
intestino, reto e ânus. O esôfago é longo, filiforme e ocupa 40% do comprimento
do corpo da larva. A boca é provavelmente fechada e impermeável a pequenas
partículas e talvez a líquidos. A extremidade posterior termina sob forma de um
entalhe, uma estrutura típica deste estágio: a de vida livre é uma larva auto-
infectante, surgindo no hospedeiro infectado, tornando-se maior em diâmetro,
menor em comprimento e tendo um esôfago mais estrongiliforme que a larva
infectante de vida livre, conforme descreveram SCHAD et al (1993).
As fêmeas do ciclo de vida livre são encontradas no meio externo.
Medem cerca de 1mm de comprimento por 50 a 75 µm de largura. Apresentam o
esôfago do tipo rabditóide, semelhante ao da larva de primeiro estágio. Possuem o
aparelho genital do tipo anfidelfo. Os ovários, anterior e posterior, situam-se do
lado aposto à vulva. Ambos os ovários, após um certo trajeto, diferenciam-se em
oviduto, receptáculo seminal e útero. A fêmea madura apresenta o útero repleto
de ovos.
Os machos do ciclo de vida livre medem cerca de 0,7 mm de
comprimento por 40 µm de largura. Os vermes machos tem a cauda pontiaguda
que se curva anteriormente e dá ao verme uma forma de J. Possuem esôfago
rabditóide. O aparelho genital consiste de testículos, vesícula seminal e vaso
deferente que desemboca na cloaca. Quando mortos, os vermes machos
apresentam a extremidade posterior recurvada ventralmente. Nesta região, há
dois espículos iguais sustentados por uma pequena estrutura conhecida como
gubernáculo, que é uma estrutura na parede dorsal da cloaca que guia os
espículos durante sua extrusão.
2.1.3- Ciclo Biológico
O habitat das fêmeas partenogenéticas são vilosidades do duodeno e
porção posterior do jejuno, local em que depositam seus ovos e encontram
alimento. Em infecções maciças podem ser encontradas no piloro, íleo, intestino
grosso, ductos biliares e pancreáticos. Os ovos já contém as larvas no momento

da postura ao nível das criptas mucosas, onde a fêmea parasita vive. Os ovos
podem ser vistos nas mucosidades que acompanham o líquido obtido por
intubação duodenal. Como as larvas são muito ativas, elas saem dentro de
poucos instantes rompendo a fina casca do ovo.
O ciclo evolutivo do S. stercoralis é complexo. Apresenta uma fase no
hospedeiro humano (ciclo parasitário, ciclo direto ou homogônico) e uma fase no
meio externo (ciclo indireto ou heterogônico ou ciclo de vida livre).
Figura 1- Ciclo evolutivo do S. stercoralis (Copiado do CDC).
O ciclo direto ou desenvolvimento homogônico representa a
diferenciação de larvas filarióides a partir de larvas rabditóides eclodidas no
duodeno. Isso pode acontecer no meio externo ou no intestino. No meio externo,
as larvas rabditóides, alimentando-se de bactérias, sofrem duas ecdises e, após
24 a 36 horas, dão origem a larvas filarióides infectantes. Estas não se alimentam,
sobrevivem do glicogênio armazenado sob forma de reserva; permanecem na

superfície do solo ou em vegetações que lhes forneçam umidade por uma ou duas
semanas, a menos que encontrem um hospedeiro susceptível. As larvas filarióides
infectantes após atravessar a pele, circulação venosa e linfática do hospedeiro,
chegam nos pulmões via coração direito. Rompem os alvéolos pulmonares,
ascendem por via brônquica até a faringe, podendo ser expulsas com as
secreções pulmonares ou deglutidas. Ao serem deglutidas, chegam no intestino
delgado onde se transformam em fêmeas adultas, entre 17 e 21 dias após a
penetração através da pele do hospedeiro. Em seguida, inicia-se a oviposição pela
fêmea parasita. Segundo GROVE (1996), as duas ecdises do ciclo homogônico
ocorrem no intestino delgado.
Neste ciclo direto há quatro mudas ou ecdises, sendo que as duas
primeiras (L1→L2→L3) ocorrem no solo ou no intestino e as duas últimas
(L3→L4→fêmea parasita) no intestino delgado do hospedeiro, o habitat definitivo
da fêmea partenogenética.
No ciclo indireto ou heterogônico, as larvas rabditóides, oriundas da
fêmea parasita, sofrem quatro mudas dentro de 36 horas e se diferenciam em
vermes adultos machos e fêmeas sexualmente maduros. Tanto as larvas como os
vermes adultos apresentam o esôfago do tipo rabditóide. No meio ambiente, os
vermes adultos podem dar uma ou mais gerações de vida livre, antes de haver a
transformação da larva rabditóide em larva filarióide infectante. Estas podem
permanecer no solo durante muitas semanas e só continuarão o seu
desenvolvimento após alcançar o hospedeiro favorável. A larva infectante
depende de alimento armazenado para sobreviver no solo. Como não possui
cápsula envolvente, naturalmente resiste menos às agressões do meio externo do
que as larvas infectantes dos ancilostomídeos. Segundo Watson (1960), no
laboratório, a larva vive durante cinco semanas, mas no meio externo supõe-se
sobreviver menos tempo (PESSOA, 1978).
A infecção do homem ocorre com a penetração das larvas filarióides
através da pele, nas regiões menos espessas, como os espaços interdigitais ou,
menos freqüentemente, segundo de Langen (1936), através da mucosa bucal,
esofágica ou gástrica, com alimentos ou água contaminados com larvas

infectantes. A penetração da pele é rápida e, segundo Brumpt (1936), em uma
hora as larvas estariam na espessura da derme, atingindo os linfáticos ou vênulas
ou, então, na impossibilidade de atingirem estas estruturas, sucumbiriam. Através
dos vasos atingem o coração direito e daí até os capilares pulmonares. Após isso,
rompem os capilares septais e chegam à luz alveolar e bronquiolar. Ascendem por
via brônquica até a faringe, onde, deglutidas, caem no trato digestivo (PESSOA,
1978).
No homem, parece que o período pré-patente é em torno de 17 dias
(Brumpt, 1949). Admite-se em média um período entre 15 a 25 dias, tempo
necessário para a fêmea começar a expelir os ovos embrionados e larvas
rabditóides serem observadas nas fezes (PESSOA, 1978).
As larvas filarióides maduras do gênero Strongyloides, como as do
Ancylostoma e Necator, podem ocasionalmente ser ingeridas e, após
atravessarem o estômago, vão ao intestino em cuja mucosa podem penetrar,
desenvolvendo-se diretamente para dar os vermes adultos (PESSOA, 1978).
Os fatores que determinam se as larvas rabditóides do S. stercoralis vão se
desenvolver direta ou indiretamente são pobremente entendidos. Possíveis fatores
incluem diferentes cepas geneticamente determinadas, condições do organismo
do hospedeiro e a fase de vida-livre. O potencial para controle genético do
desenvolvimento é talvez melhor visto no S. ratti, no qual cepas têm sido bem
selecionadas para predizer o desenvolvimento homogônico ou heterogônico.
Segundo Berezhnaia, citado por Grove (1996, p.263), coproculturas isoladas de
diferentes áreas geográficas mostraram que as cepas tropicais desenvolvem
predominantemente de maneira indireta, em todas temperaturas, enquanto cepas
isoladas de áreas temperadas tendem a se desenvolver diretamente,
particularmente em temperaturas baixas.
Alguns autores têm estudado o papel de fatores ambientais influenciando o
S. stercoralis. Nem a temperatura nem o grau de diluição das fezes tem qualquer
efeito significativo no número de vermes adultos machos que se desenvolvem. O
número de vermes adultos fêmeas foi máximo a 20 a 30° C e o número de larvas
filarióides foi grande em temperaturas acima de 30° C. Os números de vermes

adultos fêmea caiu enquanto as larvas filarióides aumentaram quando as fezes
foram diluídas progressivamente. Estes resultados podem indicar que o sexo
masculino é estabelecido no estágio de ovo mas que o potencial de ovos com
embriões fêmeas se desenvolverem em vermes adultos fêmea ou larva filarióide
depende de circunstâncias ambientais (SHIWAKU et al, 1988).
Dos fatores do hospedeiro parece que a resposta imunitária, especialmente
anticorpos, pode influenciar no direcionamento da larva rabditóide para o ciclo
direto ou indireto: maiores taxas de IgG anti-epitopos da larva favorecem o ciclo
indireto (HARVEY et al, 2000).
Tanto no ciclo direto como no indireto as larvas filarióides migram nos
tecidos até chegar ao duodeno. Sabe-se que a larva infectante do S. stercoralis
secreta uma metaloprotease que pode facilitar a penetração na pele e a migração
através dos tecidos. Esta protease tem atividade de elastase e catalisa a
degradação da matriz extra-celular. Tem peso molecular de 40KD e é
imunogênica (BRINDLEY et al, 1995). A invasão da pele pela larva é prevenida
pelos inibidores de metaloproteases, enfatizando a importância desta enzima
como um fator de virulência do S. stercoralis (MCKERROW et al,1990).
Entretanto, não há razão para se admitir que infecção patente não se
desenvolverá se a larva infectante for ingerida; isso de fato já foi mostrado por
Wilms (1897).
Um ponto importante nessa migração é a rota, ou as rotas, que a larva
infectante segue. Os primeiros estudos foram feitos em cães traqueostomizados,
tendo sido demonstrado que a maioria das larvas passam da corrente sangüínea
para os pulmões, ascendendo para a árvore respiratória, onde são deglutidas,
chegando no intestino delgado onde elas completam o seu desenvolvimento
(Fulleborn,1914).
2.1.4- Auto-infecção (interna e externa) e hiperinfecção
A auto-infecção é uma característica que separa o S. stercoralis de quase
todos os outros vermes que infectam os humanos, possibilitando a replicação do
verme dentro do hospedeiro. Devido a isso, a infecção pode persistir por muitas

décadas; a persistência mais longa registrada é de 65 anos (GILL e BELL, 1979;
GROVE, 1980; PELLETIER et al, 1988; LEIGHTON e MACSWEEN, 1990). A
primeira suspeita da existência da auto-infecção foi em 1911, quando Gage
publicou um caso de infecção protraída e postulou que auto-infecção poderia
ocorrer por duas rotas, interna e externa.
Observações posteriores à de Gage têm reforçado a idéia de auto-
infecção. Representa forte argumento a favor da auto-infecção a comprovação da
parasitose em pacientes por longo prazo, durante o qual eles não tiveram acesso
a fontes externas de infecção. É também pouco provável a sobrevivência da
fêmea parasita por longo período. Os casos de parasitismo muito intenso podem
ser explicados por constantes auto-infecções (GENTA, 1989; LIU E WELLER,
1993; GROVE, 1996; ADEDAYO, GRELL e BELLOT, 2002).
É antiga a discussão sobre as rotas da auto-infecção na estrongiloidíase.
Fulleborn (1916) admitiu a auto-infecção, a qual se faria através da pele da região
perianal de indivíduos infectados, em cujos pêlos das margens do ânus seriam
abrigadas as larvas rabditóides, posteriormente transformadas em larvas
filarióides infectantes. Faust (1930) admitiu ainda a auto-infecção interna ou
hiper-infecção, quando as larvas rabditóides, na luz do intestino, se
metamorfoseiam em larvas filarióides durante seu trânsito intestinal. Tais
organismos infectantes podem produzir re-infecções pela invasão da mucosa do
íleo ou do cólon e alcançar o pulmão, via veia porta. Esse autor admitiu que, em
indivíduos com resistência diminuída, as larvas rabditóides, mesmo sem a
metamorfose em filarióides, poderiam invadir a parede intestinal, cair nas veias
mesentéricas e iniciar uma re-infecção. Alguns autores têm argumentado que a
rota pulmonar é justamente uma das muitas rotas possíveis para a larva atingir o
duodeno (SCHAD, AIKENS, SMITH, 1989).
Baseado em estudo de cães infectados, SCHAD et al (1993) descreveram o
que chamaram de “explosão auto-infectante”. Confirmaram que em infecções
crônicas, a maioria das larvas intestinais não atingem a infectividade mas são
eliminadas nas fezes como larva rabditóide. Em infecções primárias, em animais
imunologicamente virgens, o desenvolvimento segue adiante sem prejudicar o

hospedeiro e muitas larvas atingem o estágio infectante. Há um espectro de taxa
de desenvolvimento da larva e aquelas que se tornam infectantes podem ser uma
das que se desenvolveriam mais rapidamente. Por um curto período, há uma
hiper-infecção durante a qual ocorre aumento da população de vermes adultos no
intestino até o nível no qual o intestino suporta e o desenvolvimento é retardado.
Postula-se que, durante a auto-infecção, as larvas atravessam os linfáticos
intestinais para o ducto torácico, daí passam, via corrente sangüínea, para os
pulmões, penetrando nos alvéolos e ascendendo para vias respiratórias.
A exacerbação dos mecanismos de auto-infecção é de fundamental
importância na patogenia da estrongiloidíase humana, especialmente nas formas
graves ou fatais. Este fenômeno originou os termos estrongiloidíase sistêmica,
disseminada ou hiperinfecção. Nestas circunstâncias há disseminação de larvas
para as circulações pulmonar e sistêmica. Nas formas graves, fêmeas
partenogenéticas podem alcançar a maturidade sexual e realizar oviposição nos
pulmões. A exacerbação do ciclo auto-infectante pode resultar no aumento da
carga parasitária com agravamento do quadro intestinal, às vezes seguido de
óbito, sem disseminação de larvas pela circulação sistêmica.
2.1.5-Modelos Experimentais
Atualmente, S. stercoralis infectando cães (GROVE, 1980; SCHAD,
HELMMAN e MUNCEY, 1984) e macacos (Erythrocebus patas) (GENTA, 1984;
BARRET et al, 1988 ) são usados como modelos para estudo da estrongiloidíase
humana. Embora estas espécies sejam hospedeiros naturais para o S. stercoralis,
eles não são ideais como hospedeiros de laboratório pois são animais
relativamente grandes e caros para se comprar e manter. Outra possibilidade de
modelo laboratorial são os gatos, que toleram apenas infecções transitórias e o
furão (Mustela putorius furo), que desenvolve infecções importantes somente após
imunossupressão (DAVIDSON, 1988). Roedores investigados (camundongo, rato,
cobaios) e coelhos não desenvolvem infecções sustentadas pelo S. stercoralis.

A incapacidade do S. stercoralis desenvolver-se na maioria dos animais
pequenos tem sido o principal impedimento na obtenção de informações sobre a
patogênese dessa parasitose.
Foi mostrado que os gerbilos (Meriones unguiculatus) podem ser infectados
com S. stercoralis (NOLAN et al, 1993), mas os mesmos são resistentes a re-
infecção (NOLAN et al, 1995). Quando são infectados com quantidade suficientes
de larva infectante ( L3i ) e em idade adequada, estas irão em curto espaço de
tempo amplificar a população de vermes adultos, como ocorre em cães. Isso faz
dos gerbilos o modelo de escolha para investigações laboratoriais de infecções
pelo S. stercoralis (KERLIN, NOLAN e SCHAD, 1995; NOLAN, BHOPALE e
SCHAD, 1999).
Larva rabditóide e vermes adultos foram recuperados de gerbilos machos
por pelo menos 131 dias após a infecção. No caso de gerbilos fêmeas, vermes
intestinais não foram vistos após 70 dias da infecção. Os autores interpretaram
que em gerbilos machos a duração da vida de vermes adultos é 131 dias, pois
eles não encontraram nenhuma larva auto-infectante circulando. Pode ser que a
auto-infecção ocorra em baixos níveis, abaixo da sensibilidade da necropsia em
encontrar larvas migrando (NOLAN et al, 1993).
Gerbilos tratados com acetato de metilpredinosolona tiveram auto-infecção,
mimetizando uma característica da infecção em humanos. Auto-infecção nunca
ocorreu em gerbilos não tratados com corticóides (NOLAN et al, 1993).
KERLIN, NOLAN e SCHAD (1995) estudaram a histopatologia na
estrongiloidíase não complicada e na hiper-infecção em gerbilos. Gerbilos com
hiper-infecção desenvolveram grave hemorragia pulmonar, com vários graus de
pneumonia eosinofílica intersticial sub-aguda, associada com numerosas larvas
nos alvéolos, no interstício e no compartimento vascular. Hiper-infecção induzida
por corticosteróides, dado antes da inoculação de larvas do S. stercoralis ou após
a infecção crônica pelo S. stercoralis, produziu lesões semelhantes. Vermes
adultos não estavam associados com inflamação e não foram mais comuns em
gerbilos tratados com corticosteróides. De modo diverso, pulmões de gerbilos com

estrongiloidíase não-complicada tinham grave vasculite e peri-vasculite
eosinofílica com pouca hemorragia, sem pneumonia e sem larvas.
Transmissão trans-mamária em gerbilos, lactentes de fêmeas parasitadas
foi demonstrada (NOLAN et al, 1993).
Os ovos raramente são encontrados em fezes frescas, lembrando a
infecção humana com S. stercoralis, onde as larvas rabditóides (L1) são
caracteristicamente expelidas.
Furão (Mustela putorius furo) não parece ser suscetível à infecção sob
condições normais. Entretanto, após tratamento com metilprednisolona, tem
infecção importante, mas disseminação da infecção não foi vista
(DAVIDSON,1988). Pode ser infectado com cepas humanas de S. stercoralis e ser
usado como fonte para larvas.
Sabe-se que os cães são susceptíveis à infecção com S. stercoralis e,
durante algum tempo, foram utilizados como modelo experimental, tendo SCHAD,
HELLMAN e MUNCEY (1984) re-introduzido o seu uso nos últimos 20 anos. Eles
estudaram hiperinfecção em cães imunossuprimidos e observaram que auto-
infecção ocorre em cães infectados com S. stercoralis e que, se esta infecção
persiste por um longo tempo em hospedeiros imunossuprimidos, hiperinfecção
maciça e mesmo infecção disseminada, podem ocorrer, demonstrando que a
infecção do cão pode ser um excelente modelo para hiper-infecção humana e
estrongiloidíase disseminada (GROVE, HEENAN e NORTHERN, 1983; SCHAD,
HELLMAN e MUNCKEY, 1984; GENTA, SCHAD e HELLMAN, 1986).
Deficiência de IgA em cães, aparentemente, não afeta o curso ou gravidade
da infecção com S. stercoralis. Resultados de estudos em cães mostraram que
elevações da IgA no soro e nas fezes não estão correlacionadas com a resistência
ao parasita. Níveis aumentados de IgA, publicados em alguns estudos, pode ser
meramente um resultado da estimulação policlonal pelo parasita e não ter papel
funcional na resistência (MANSFIELD e SCHAD, 1992).
Transmissão trans-mamária e trans-placentária do S. stercoralis não foi
observada em uma cadela infectada (MANSFIELD e SCHAD, 1995). Como este
experimento envolveu apenas um animal, os resultados são meramente

especulativos, mas servem como preliminar para estudos adicionais sobre
transmissão vertical do parasita em modelos animais.
O macaco (Erythrocebus patas) é um excelente modelo para se estudar
estrongiloidíase. Doença disseminada pode ser produzida agudamente durante a
infecção primária em animal virgem, ou pelo uso de esteróides para converter
infecção crônica assintomática em doença fulminante letal. O espectro patológico
mimetiza o visto em humanos, incluindo graves lesões pulmonares e colônicas
pela hiper-infecção larval (HAPER et al 1984).
2.1.6- A resposta imunitária na infecção pelo Strongyloides stercoralis
O problema central na estrongiloidíase humana é a capacidade do parasita
se reproduzir e persistir no hospedeiro, indefinidamente (BARRET et al,1988).
Essa infecção crônica é usualmente silenciosa, mas, se o sistema imunitário é
alterado, o nematóide pode se multiplicar rapidamente e disseminar para órgãos
distantes, geralmente causando morte (GENTA, 1984) ou estrongiloidíase grave,
como citada na publicação de dois casos de estrongiloidíase grave em pacientes
portadores de hepatite C, após início da terapia com interferon e ribavirina, onde
postulou-se que a ação imunossupressiva ou imunomoduladora da ribavirina
poderia estar associada à estrongiloidíase grave pela alteração da resposta
imunitária celular e/ou humoral (PARANA et al, 2000).
Os pontos nos quais os mecanismos efetores de defesas atuariam para
controlar o número de vermes incluem: (1) diminuição da duração da vida de
vermes adultos no intestino; (2) diminuição da fecundidade de vermes adultos no
intestino; (3) prejuízo da transformação de larva rabditóide em larva infectante in
vivo; (4) diminuição da sobrevida da larva infectante no lúmen intestinal; (5)
destruição da larva filarióide migrando através dos tecidos; (6) indução de falha no
desenvolvimento da larva de terceiro-estágio para vermes parasitas adultos
fêmeas.
O mecanismo imunitário que confere proteção à disseminação não é bem
entendido, mas existem evidências de que a resposta Th2, através da síntese de
IL-4, IL-5, e conseqüente produção de IgE, eosinofilia e mastocitose está

envolvida na destruição do parasita. Em modelos experimentais há associação
entre a resposta Th2 e a proteção contra helmintos. A produção de IL-4 é
importante para essa proteção, limita a gravidade da infecção e age na fisiologia
do intestino, aumentando o conteúdo de fluidos no trato digestivo, além de ativar
mastócitos. O acúmulo de fluidos decorre do aumento da permeabilidade intestinal
e redução da absorção de líquidos. IL-12 e IFN-γ inibem a imunidade protetora
contra estes parasitos (FINKELMAN et al, 1994 e 1997).
A avaliação da resposta imunitária no indivíduo infectado mostra resultados
variáveis. A resposta “in vitro” de células mononucleares de sangue periférico a
antígenos do parasita pode ser pequena ou ausente. A avaliação de anticorpos
IgG, IgE total e específica contra antígenos do S. stercoralis não mostrou
correlação entre essas imunoglobulinas e as formas clínicas na estrongiloidíase
humana. É possível que mecanismos de resposta imunitária na mucosa e não os
que se manifestam perifericamente, sejam responsáveis pela proteção contra a
doença disseminada (BADARÓ et al, 1987 ; ROSSI et al 1993; NAWA et al, 1994).
Estudo em macacos demonstrou que os mastócitos podem degranular em
resposta aos antígenos dos parasitas e que este evento tem um papel na proteção
do hospedeiro. BARRET et al (1988) avaliaram o número de mastócitos e a
reatividade destas células aos antígenos do parasita em macacos (Erythrocebus
patas) infectados com cepas humanas de S. stercoralis. Tanto a infecção inicial
como a re-infecção estavam associadas com aumento de histamina e aumento do
número de mastócitos no jejuno e que estas células liberavam histamina em
resposta aos antígenos parasitários. Durante a fase crônica da infecção, o número
de mastócitos retornava ao normal. Após tratamento com esteróides, as células
perdiam a capacidade de responder à estimulação antigênica. Curso mais
prolongado de tratamento com esteróides foi associado com reativação da
infecção crônica para doença disseminada fatal e com marcada diminuição do
número de mastócitos e histamina no jejuno. Se os mastócitos se tornam
seletivamente dessensibilizados aos antígenos do parasita, isso pode representar
um mecanismo permissivo para que o parasita permaneça no intestino. Estes
dados são consistentes com a hipótese de que os mastócitos ativados podem

representar um importante mecanismo efetor na contenção da infecção inicial e re-
infecções por S. stercoralis em macacos e uma proteção contra doença
disseminada. Os mecanismos mediados pelos mastócitos podem ser diretos,
devido a capacidade dos mediadores destas células em lesar diretamente os
helmintos, ou indiretos, via capacidade desses mediadores em atrair e modular a
função de outras células “helmintocidas”, como os eosinófilos.
O mecanismo efetuador da resistência contra o S. stercoralis pode se
manifestar através de destruição do verme adulto ou das larvas durante a auto-
infecção. Em torno das larvas observa-se infiltrado de eosinófilos e tem sido
demonstrado que os grânulos liberados dos eosinófilos são tóxicos para as larvas
infectantes (L3) de S. stercoralis . Devido a isso, tem sido aventada a possibilidade
de que o mecanismo de citotoxicidade celular dependente de anticorpo (ADCC)
seja uma forma de defesa contra este helminto. Além do aumento do número de
eosinófilos, tem sido demonstrado, em pacientes infectados por helmintos, um
aumento da sua capacidade helmintotóxica (PORTO et al, 2002). “In vitro”, tem
sido mostrado que o parasita pode ativar a cascata do complemento, resultando
na produção da C5a e C3a que ativariam os eosinófilos e neutrófilos, causando
sua degranulação (TAKAFUJI et al, 1994).
Pacientes com formas leves e assintomáticas de estrongiloidíase
apresentam níveis elevados de IgE total e IgE específica contra antígenos do
parasita e pacientes com forma grave da doença apresentam-se com níveis
baixos destas imunoglobulinas (ROSSI et al 1993; PORTO et al, 2002).
2.1.7- Os mecanismos envolvidos nas formas disseminadas da
estrongiloidíase
A estrongiloidíase pode cronificar-se, devido à auto-infecção interna ou
externa, mas o indivíduo pode perder esta capacidade de limitar o tamanho
populacional do parasita quando se encontra imunossuprimido, em particular
quando usa corticosteróides. Sendo assim, ocorre um grande aumento da carga
parasitária, podendo as larvas filarióides ser encontradas em diversos órgãos do
organismo (CAMPOS e FERREIRA, 1999). Em estados de imunossupressão, até

as fêmeas partenogenéticas podem disseminar-se para diferentes órgãos (p.ex:
pulmões) e neles realizar a oviposição. A disseminação ocorre mais
freqüentemente em indivíduos com depressão da imunidade celular por drogas ou
doenças. Os corticóides, usados em transplantes de órgãos, tratamento de
neoplasias hematológicas e doenças auto-imunes, são as drogas mais
freqüentemente associadas à síndrome de hiperinfecção (IGRA-SIEGMAN et al,
1981). Enquanto linfoma, leucemia crônica, tumores sólidos, infecção por
micobactérias, a desnutrição e o alcoolismo são as doenças mais freqüentemente
a ela associadas. Em torno de 13% dos casos de disseminação e hiperinfecção,
não é possível identificar depressão da imunidade (IGRA-SIEGMAN et al, 1981).
Os mecanismos exatos envolvidos na disseminação da infecção pelo S.
stercoralis não são totalmente compreendidos (GENTA et al, 1983; GENTA, 1984;
NEVA, 1986; BARRET et al, 1988; LIU e WELLER, 1993). A existência de casos
de estrongiloidíase disseminada sem evidência de imunossupressão (GENTA et
al, 1983) e a baixa prevalência da disseminação do S. stercoralis em pacientes
com AIDS (Síndrome da imuno-deficiência adquirida) mostram serem muito
complexos os fatores envolvidos no processo. Seria esperado que indivíduos
infectados pelo HIV apresentassem freqüente disseminação do parasito. Porém
isso não ocorre, havendo poucos relatos de casos de disseminação em pacientes
HIV positivos. A prevalência de S. stercoralis em pacientes HIV positivos, avaliada
em diferentes regiões, não diferiu da observada, em pacientes pareados, HIV
negativos (VIEIRA et al, 1985; COURA, 1987; PETITHORY e DEROUIN, 1987;
DIAS et al, 1992; COSTA-CRUZ, FERREIRA, ROSSIN, 1996; TRIONE et al, 2001;
KIM e LUPATKIN, 2004). No Brasil, um estudo realizado por DIAS et al (1992), em
São Paulo, mostrou que a prevalência de larvas nas fezes de pacientes com AIDS
foi semelhante a de pacientes não-infectados pelo HIV (respectivamente 9,75% e
10,56%). No entanto, FEITOSA et al (2001), na Bahia, verificaram que a
prevalência S. stercoralis e G. lamblia foi maior em indivíduos HIV positivos. A
presença de infecção parasitária não foi associada a uma progressão mais rápida
da doença. Outras observações feitas em regiões endêmicas para parasitoses
intestinais e HIV, como na África, mostraram que a associação do S. stercoralis e

HIV parece não ser freqüente, e como não há evidências clínicas e
epidemiológicas que sustentem a hipótese que a infecção pelo HIV predispõe à
estrongiloidíase extra-intestinal, foi sugerido que esta parasitose não deveria ser
incluída na definição clínica de AIDS (PETITHORY e DEROUIN, 1987).
Uma explicação do motivo pelo qual a infecção pelo HIV não leva à
estrongiloidíase disseminada foi aventada recentemente, com base em estudo
realizado em africanos HIV positivos. Nos indivíduos HIV positivos, com função
imunitária deficiente, identificada pela contagem baixa de linfócitos CD4+ e pelo
estadiamento clínico, o desenvolvimento do S. stercoralis pelo ciclo indireto foi
mais freqüente, quando comparado com indivíduos HIV positivos com função
imunitária preservada. Como a auto-infecção é facilitada pelo ciclo direto do
desenvolvimento da larva, aquela seria menos freqüente nos pacientes com AIDS
(VINEY et al, 2004).
Estudos experimentais em ratos infectados com S. ratti, têm sugerido
importante papel da IgG na resposta protetora contra a migração larvar. A
ausência ou baixos títulos de anticorpos anti-S. ratti favorece o desenvolvimento
da larva no ciclo direto e a presença dos anticorpos favorecem o desenvolvimento
do ciclo indireto, em machos e fêmeas de vida livre (HARVEY et al, 2000).
A ausência de relação entre resposta IgG anti-S. stercoralis e contagem
de células CD4 e CD8 ou relação CD4:CD8, observada no estudo de VINEY et al
(2004), sugere que no estadio avançado da doença, a resposta IgG ao S.
stercoralis não estaria alterada, apesar da profunda depressão da imunidade
celular. De fato, na infecção pelo HIV a resposta Th1 está prejudicada e há
evidência que a resposta Th2 predomina. Portanto os níveis de IgG anti-
Strongyloides existentes mesmo nas formas avançadas da AIDS ainda mantêm o
estímulo para a diferenciação das larvas para machos e fêmeas e não para larvas
filarióides infectantes (VINEY et al, 2004).
Argumenta-se que a imunodepressão iatrogênica, responsável pela maior
parte dos casos de disseminação, é diferente daquela induzida pelo HIV. Isso
reforça a teoria de que a limitação da infecção está ligada, ao menos em parte, a
mecanismos de defesa de mucosa, como foi observado em estudo realizado em

macacos tratados com prednisona (BARRET et al, 1988). Nesses animais, a
disseminação ocorria independente da resposta sistêmica, tanto humoral quanto
celular, que se encontravam adequadas, porém estava fortemente ligada com a
dessensibilização de mastócitos e diminuição da produção de histamina à nível de
mucosa. Isso levanta a hipótese de que a defesa local mediada por mastócitos
pode ser responsável pelo controle da intensidade da infecção, tanto diretamente,
pela capacidade dessas células de lesar os vermes, quanto indiretamente, através
da habilidade de substâncias liberadas pelos mastócitos em atrair e modular a
função de células “helmintocidas” como os eosinófilos.
Embora falha imunitária tenha sido admitida como a base das infecções
graves, complicadas, com S. stercoralis (IGRA-SIEGMAN et al, 1981; GENTA,
1986; GROVE, 1989), recentemente, GENTA (1992) tem argumentado contra a
hipótese de a imunidade ser importante nessa disseminação, pelos seguintes
motivos: (1) tem havido poucas publicações de infecção disseminada em má
nutrição protéico-calórica (uma causa admitida como importante de falha de
imunidade); (2) infecção disseminada não é comum em lepra lepromatosa,
condição com depressão de resposta celular, a menos que os pacientes tenham
recebido corticosteróides; (3) infecção disseminada tem sido incomum em
transplante renal desde a introdução da ciclosporina A; (4) infecção disseminada
não é proeminente em AIDS ou em infecção pelo HTLV.
Parece que há razões para estas aparentes contradições. Estrongiloidíase
grave, complicada no contexto de má nutrição protéico-calórica é provável ocorrer
em áreas onde serviços médicos são precários e publicações são raras;
ciclosporina utilizada nos transplantados renais pode ter efeito anti-Strongyloides
(SCHAD, 1986) e disseminação do parasita pode ocorrer em infecções retro-
virais, mas pode ser pouco relatada em áreas endêmicas para HIV e S.
stercoralis, por razões já citadas acima.
NEVA (1986) levantou a possibilidade de um efeito direto dos
corticosteróides sobre as fêmeas partenogenéticas, levando a um aumento da
oviposição, ou sobre as larvas rabditóides, acelerando a sua transformação em

larvas filarióides como uma tentativa de justificar a forte ligação entre esse grupo
de drogas, em particular, e as formas graves de infecção pelo S. stercoralis.
GENTA (1992) propôs que a administração de corticosteróides e seu
subseqüente metabolismo pode resultar em aumento na produção de moléculas
semelhantes a ecdisteróides. Esses são hormônios de muda, relacionados ao 20-
hidroxi-ecdisona, que controlam a muda em insetos e possivelmente em
helmintos. Podem ser encontrados no soro e urina de pacientes infectados com
helmintos. Presume-se que essas substâncias são produzidas pelos parasitas e
podem ser usadas como marcador de infecção parasitária e na avaliação pós-
tratamento com anti-parasitários. Quantidades aumentadas destas substâncias
podem aumentar a taxa de muda levando ao ciclo direto e disseminação da
infecção. Esta é uma teoria que merece ser investigada, pois até o momento
permanece como especulativa. LANSOUD-SOUKATE et al (1990), pesquisaram
compostos semelhante a ecdisteróides no soro e urina de pacientes africanos
infectados com dois tipos de microfilárias Loa loa e Mansonella perstans e viram
que os títulos de ecdisteróides foram estatisticamente maiores em pacientes
microfilariêmicos, mas eles não se correlacionaram com a concentração da
microfilária no sangue. Pode ser que esta substância não tenha sido produzida
pela microfilária. Uma outra possibilidade é que níveis anormalmente altos de
ecdisteróides possa ser um efeito indireto da infecção, ou derivar do
desenvolvimento da larva, como ocorre com Schistosoma mansoni em que uma
larva de 11 dias tem maiores níveis de ecdisteróides que uma schistosomulo
infectante, ou ser devido a uma doença indetectável associada com os parasitas.
Devemos também considerar a possível contribuição da fonte dietética de
ecdisteróides.
Além disso, NEVA (1993) observou muitos casos de estrongiloidíase grave
no Caribe em pacientes que não receberam corticosteróides e especulou que a
hiperinfecção foi provavelmente devido a co-infecção com HTLV-1, naqueles que
tinham imunidade suprimida. Estas duas teorias não são mutuamente exclusivas e
é possível que ambas, imunidade e ação de ecdisteróides, sejam importantes.

2.1.8- Strongyloides stercoralis e infecção com o vírus HTLV-1
O HTLV (vírus linfotrópico para células T humanas tipos 1 e 2, sub-família
Oncovirinae, família Retroviridae) é retro-vírus que tem relação causal com
leucemia de células T em adultos e paraparesia espástica tropical (também
chamada de mielopatia associada ao HTLV-1)(MARSH, 1996). Infectam células T
CD4+, com integração do genoma viral ao genoma da célula hospedeira. Os
modos de transmissão são sexual, parenteral (via transfusão de sangue e
compartilhamento de seringa) e, vertical (transplacentária e via aleitamento
materno). As áreas de maior prevalência são o Caribe (4 a 9%) e ilhas do
sudoeste do Japão (37%). No Brasil é variável de 0,08 a 1,35%, sendo a maior
prevalência na cidade de Salvador (GABBAI et al, 1993; BROUTET et al, 1996;
CHIEFFI et al, 2000; GALVÃO LOURES E RODRIGUES,1997).
Foi estabelecido que HTLV-1 causa imunossupressão em pacientes com
leucemia de células T de adultos ou linfoma (ATL), mas imunossupressão sub-
clínica tem sido postulada para infecções não malignas. Infecção com HTLV-1 tem
sido associada com alteração da função dos linfócitos “in vitro” e com diminuição
significativa dos níveis séricos de IgE em pessoas saudáveis (MATSUMOTO et al,
1990).
A co-infecção S. stercoralis e HTLV-1 parece ser freqüente embora haja
resultados discrepantes nos relatos de diferentes regiões. Uma associação
significativa tem sido descrita em Okinawa e nas Ilhas do Caribe (NAKADA et al,
1984 e 1987; SATO et al, 1994; TERRY et al, 1989; ROBINSON et al, 1994). Às
vezes existe forte associação quando se avalia a infecção com S. stercoralis
através da detecção de larvas nas fezes, fato não observado quando se usa teste
sorológico (ROBINSON et al, 1994). A não associação entre as duas infecções foi

não co-infectados com HTLV-1 (TERRY et al, 1989; TOMA et al, 2000; SATO et
al, 1994; TERASHIMA et al 2002).
Trabalho realizado em doadores de sangue no estado de São Paulo –
Brasil (CHIEFFI et al, 2000), estudando doadores de sangue co-infectados com
HTLV-1 e S. stercoralis verificou que a freqüência do S. stercoralis foi
significativamente maior no grupo infectado com HTLV-1 (12,1% x 1,6%). Estes
resultados sugerem que pacientes infectados com HTLV-1 mesmo quando
assintomáticos, devem ser considerados como um grupo de alto risco para
infecção com S. stercoralis na municipalidade de São Paulo.
A associação entre HTLV-1 e S. stercoralis pode ser explicada pelo
estímulo da célula Th1 pelo vírus, com a redução de resposta Th2 favorecendo
assim a instalação da parasitose. De fato, indivíduos infectados pelo HTLV-1
apresentam redução na produção de IL-4, IL-5, IL-13 e IgE, componentes
participantes dos mecanismos de defesa contra S. stercoralis. Ocorre também
aumento do IFN-gama e IL-10. Estas alterações constituem a base para maior
freqüência e maior gravidade da estrongiloidíase em pacientes infectados pelo
HTLV-1 (NEVA et al, 1998; PORTO et al, 2001; PORTO et al, 2002; CARVALHO e
PORTO, 2004).
2.1.9- Anatomia patológica e patogênese das lesões na infecção pelo S.
stercoralis
Este helminto estimula resposta imune vigorosa, a qual pode, em muitos
indivíduos, erradicar por completo a infecção na ausência de qualquer tratamento
específico. No entanto, a maioria dos infectados não consegue eliminar o parasita,
tornando-se portador de uma forma crônica da infecção, na maior parte das vezes
assintomática ou oligossintomática.
As lesões devidas ao S. stercoralis relacionam-se com a penetração da
larva no hospedeiro, com sua migração durante o ciclo pulmonar e com sua
permanência e multiplicação na mucosa intestinal ou em locais ectópicos (REY,
1992).

Lesões cutâneas: em geral são discretas e podem ser vistas como pontos
ou placas eritematosas nos locais de penetração das larvas infectantes, tanto na
pele como na mucosa bucal, que desaparecem dentro de uma a duas semanas
(PESSOA, 1978). Quando ocorre auto-infecção externa, surgem em torno do ânus
ou regiões adjacentes lesões urticariformes transitórias, recorrentes (REY, 1992).
Lesões pulmonares: as larvas produzem hemorragias petequiais ou
profusas, quando passam dos capilares para os alvéolos pulmonares, onde
realizam suas mudas e aumentam de tamanho. As lesões inflamatórias são as de
uma pneumonite difusa que podem complicar-se com fenômenos alérgicos,
determinando infiltrados pulmonares transitórios, configurando a síndrome de
Loeffler, ou então broncopneumonia por invasão bacteriana secundária. As larvas
podem ser encontradas no escarro e em derrames pleurais (REY, 1992).
Lesões intestinais: no duodeno e no jejuno, a presença e atividade das
fêmeas, sua oviposição, bem como a eclosão e migração das larvas, na
espessura da mucosa, produzem lesões mecânicas, histolíticas e irritativas que
levam a uma inflamação catarral. As fêmeas raramente ultrapassam a muscular
mucosa; alojam-se, via de regra, no interior das glândulas de Lieberkuhn onde é
feita a postura dos ovos e posterior liberação das larvas. As larvas exercem ação
traumática e provavelmente tóxica na mucosa intestinal que mostra uma
inflamação catarral crônica. Pontos hemorrágicos e ulcerações de vários
tamanhos podem ser vistos em quantidade dependente da carga parasitária.
Congestão e edema, que tornam as paredes do duodeno e jejuno espessas, as
pregas mucosas tumefeitas e as vilosidades alargadas e achatadas, completam o
quadro da duodeno-jejunite catarral. O edema pode atingir a submucosa. Nas
infecções maciças, pode ocorrer espessamento da parede do intestino,
transformando-o num tubo rijo. Assim ocorre estreitamento da luz do intestino,
simulando uma obstrução do segmento atingido, que pode ser confundido com
neoplasia ou outras afecções do delgado, ao exame radiológico. A invasão
bacteriana secundária determina a formação de úlceras grosseiras e organização
fibrosa, caracterizando as formas graves da doença. Nesta forma ocorre também
a obstrução linfática devida à migração de larvas que determina um quadro de

endo e perilinfangite granulomatosa. Devido a isto, ocorre a linfangiectasia e o
edema da mucosa e submucosa (de PAOLA, 1962).
2.1.10- Quadro clínico Na estrongiloidíase não complicada, a maioria dos pacientes são
assintomáticos ou apresentam discretos sinais e sintomas cutâneos, pulmonares
e/ou abdominais de forma intermitente (GENTA, 1987; LIU e WELLER 1988;
PELLETIER et al, 1988; LIU e WELLER, 1993; GROVE, 1996).
A pequena freqüência de detecção da fase aguda da infecção pelo S.
stercoralis em áreas endêmicas sugere que essa fase é assintomática ou mal
diagnosticada (CARVALHO FILHO, 1978).
A penetração cutânea é geralmente assintomática, mas pode acompanhar-
se de eritema, prurido, edema local e manifestações urticariformes. Estas são
intensas em pacientes que desenvolvem hipersensibilidade aos produtos
parasitários.
Urticária recorrente, com freqüência envolvendo as nádegas, tronco e os
punhos, é a manifestação cutânea mais comum. As larvas que estão migrando
podem evocar um erupção urticariforme, migratória e serpiginosa, patognomônica,
denominada larva currens – uma lesão que avança rapidamente, 5 a 10 cm/hora,
o que pode ser perceptível durante o exame físico (LIU e WELLER, 1993;
GROVE, 1996).
O quadro pulmonar inicia-se poucos dias depois da penetração das larvas.
Os sintomas pulmonares são raros na estrongiloidíase não complicada. Pode se
manifestar como tosse, expectoração, febre e mal-estar. Por vezes, os sintomas
são os de uma broncopneumonia ou de uma pneumonia atípica (LIU e WELLER,
1993; GROVE, 1996).
A sintomatologia mais importante é a relacionada com o aparelho digestivo,
que pode variar desde quadros mais benignos ou assintomáticos até formas
graves e dramáticas. Pode ocorrer dor abdominal, principalmente mesogástrica,
que simula a dor da úlcera péptica, exceto por ser agravada pela ingestão
alimentar.

Surtos de diarréia com três a seis evacuações/dia, com fezes ora pastosas
ora líquidas, sem muco ou sangue, que intercalam-se com períodos de
constipação intestinal. As crises de disenteria com fezes muco-sanguinolentas
devem corresponder às formas agudas da doença. Os pacientes são
assintomáticos entre os surtos (CARVALHO FILHO, 1978).
Os pacientes queixam-se de desconforto abdominal ou dores vagas, tipo
cólica, em crises de 1 a 2 dias, com intervalos variáveis de mais ou menos uma
semana, além de náuseas e anorexia (LIU e WELLER, 1993; GROVE, 1996).
Podem ocorrer sintomas gerais como febre, emagrecimento, astenia,
desidratação, palpitações, sonolência, tonturas, irritabilidade e depressão.
2.1.11- Diagnóstico da infecção
O diagnóstico definitivo da estrongiloidíase é usualmente feito com base na
detecção das larvas nas fezes (GROVE, 1994; SIDDIQUI e BERK, 2001). Na
estrongiloidíase não complicada, a carga parasitária no intestino é geralmente
pequena e a eliminação das larvas é mínima e irregular (GENTA, 1986; NEVA,
1986; LIU e WELLER, 1993; SILVA et al, 2003).
Larvas rabditóides nas fezes são geralmente esparsas, com a maioria dos
pacientes eliminando 100-2000 larvas/dia, sendo o equivalente a 0-20 larvas por
grama de fezes. Cada verme adulto tem, em média, uma dúzia de ovos mas a
taxa de eliminação não é conhecida.
Pesquisa de Larvas em Amostras Fecais. Em fezes frescas são
encontradas as larvas rabditóides de S. stercoralis mas não os ovos, que eclodem
logo depois da oviposição, ainda na mucosa intestinal (LIU e WELLER, 1988). Em
fezes envelhecidas (eliminadas há mais de 24 horas) ou quando o trânsito
intestinal é muito demorado ou ainda, no caso de hiper-infecção, larvas filarióides
podem ser detectadas nas fezes (FERREIRA, 1991).
Infelizmente, as larvas rabditóides são difíceis de serem vistas nas fezes,
pois são excretadas em pequena quantidade e de forma intermitente (VAN DER
FELTZ et al, 1999).

O diagnóstico pode ser feito pelo exame microscópico direto, pelos métodos
de concentração com formol-éter (LIU e WELLER, 1993), mas o método de
Baermann é o mais sensível (PESSOA, 1978; FERREIRA, 1991; LIU e WELLER,
1993; CAMPOS e FERREIRA, 1999).
Para extração de larvas, recomenda-se o método de Baermann-Moraes

sete dias. No ciclo biológico do S. stercoralis exacerbações agudas são
intercaladas com períodos de remissão de 4 a 6 meses. Nesse período, é difícil
demonstrar a excreção larvar. Mesmo um tratamento ineficaz pode inativar
temporariamente o parasita e então, após o mesmo, o parasita pode ganhar
vitalidade e as larvas reaparecerem nas fezes.
Técnicas de coprocultura aumentam a possibilidade de se diagnosticar
infecções por S. stercoralis. É feita pelo método de Harada-Mori (em tubos de
ensaio) ou pelo método de Baermann de cultivo sobre carvão ativado em placas
fechadas, que requerem menores quantidades de fezes e se adaptam melhor aos
inquéritos parasitológicos de massa (REY, 1992).
SATO et al (1995) compararam a eficácia de quatro métodos de exames
coprológicos em diagnosticar S. stercoralis. A maior taxa de detecção foi obtida
pela cultura em placa de Agar (+96%) quando comparada com o exame direto,
concentração éter-formalina e método de cultura Harada-Mori, que são métodos
convencionais para detecção das lavras nas fezes. Esses métodos convencionais
não são sensíveis o suficiente para diagnosticar os casos crônicos pois, nestes
casos, as larvas estão presentes em pequena quantidade. A cultura em placa de
Agar usa mais fezes que os métodos convencionais e é duas a três vezes mais
efetiva que tais métodos. Embora a cultura em placa de Agar seja mais sensível
que o método de Baermann, esse tem vantagens em termos custo-efetividade e
tempo para se obter os resultados.
KAMINSKY (1993) avaliou três métodos (método direto, Baermann
modificado e cultura em placa de Agar) para o diagnóstico laboratorial da infecção
pelo S. stercoralis e mostrou que a cultura em placa de Agar é 4,4 vezes mais
eficiente que o método direto e 0,8% mais eficiente que o método de Baermann.
Os médicos devem ficar alertas quando o laboratório libera um resultado de
exame parasitológico positivo para larva de ancilostomídeo pois pode haver a
possibilidade de enganar-se e ser a larva de S. stercoralis (LIU e WELLER, 1993).
É importante dar-se conta de que resultados negativos não
necessariamente indicam ausência de infecção de forma inequívoca (SATO et al,

1995; SATO, KOBAYASHI e SHIROMA, 1995; VAN DER FELTZ et al, 1999;
CAMPOS e FERREIRA, 1999).
Pesquisa de larvas em material de conteúdo duodenal. Temos o

Os testes sorológicos apresentam os seguintes problemas: (1) falta de
antígenos padronizados, já que se usam extratos brutos de larvas ou de vermes
adultos; (2) necessidade de se ter antígenos de outros helmintos para adsorver os
soros antes do teste para minimizar as reações cruzadas; (c) os métodos
sorológicos de rotina identificam os anticorpos mas não indicam se resultam de
infecção presente ou passada (LIU e WELLER, 1993; GROVE 1996).
Os exames sorológicos têm utilidade nos inquéritos epidemiológicos para
separar os que tem reação positiva e neles fazer repetidos exames parasitológicos
(SATO, KOBAYASHI e SHIROMA, 1995). De modo semelhante, em casos com
suspeita clínica, mas com exame parasitológico negativo, um teste ELISA positivo
indica necessidade de repetir a pesquisa da larva e o teste ELISA negativo,
nesses casos, quase que elimina a possibilidade da infecção.
A sorologia pode ser útil na monitorização do tratamento. A permanência
dos títulos de anticorpos observados antes do tratamento é fortemente sugestivo
de insucesso terapêutico e requer repetidas pesquisas da larva nas fezes para
confirmação.
Uma outra ferramenta útil na definição do diagnóstico da estrongiloidíase,
em casos nos quais a sorologia deixa dúvida, é o Western blotting, utilizando
antígeno do S. ratti (SILVA et al, 2003).
Testes cutâneos de hipersensibilidade imediata com extratos da larva
infectante do parasita têm sido testados experimentalmente. Co-infecção com
HTLV-1 diminuiu a sensibilidade dos testes cutâneos (NEVA et al, 2001; PORTO
et al, 2001).
O diagnóstico presuntivo de estrongiloidíase pode ser suspeitado através
da eosinofilia que é uma alteração laboratorial não-específica, geralmente pouco
intensa (5 a 15%), encontrada em 50 a 80% dos indivíduos infectados,
especialmente se são sintomáticos (BERK et al, 1987; GENTA, 1989; LIU e
WELLER, 1993; GROVE, 1996). Apesar disso, a eosinofilia é um indicador útil de
estrongiloidíase, particularmente em pacientes recentemente infectados. Nas
infecções crônicas, como de prisioneiros de guerra (GIL e BELL, 1979) ou em

pacientes recebendo terapia imunossupressora (IGRA-SIEGMAN et al, 1981), a
contagem de eosinófilos é baixa ou normal.
2.1.12- Tratamento
A estrongiloidíase, mesmo quando assintomática, deve ser tratada por
causa do potencial de hiperinfecção fatal. Somente a completa erradicação do
parasita remove o perigo potencial da doença. Devido a isso, o anti-helmíntico
ideal deve matar todas larvas auto-infectivas L3, que são relativamente resistentes
aos agentes químicos (LIU e WELLER, 1993; GROVE, 1996). Dentre as
medicações disponíveis temos:
Tiabendazol: derivado imidazólico introduzido na terapêutica da estrongiloidíase
nos anos 60, constituiu a droga de escolha na erradicação desta helmintíase,
apesar dos efeitos colaterais e da alta taxa de recidiva (GROVE, 1996; ZAHA et al,
2000; KEISER e NUTMAN, 2004) . Administrado via oral (VO), é rapidamente
absorvido. A dose indicada é de 50mg/Kg/dia, dividida em duas tomadas, de
preferência às refeições, por dois ou três dias. Não se deve ultrapassar 3g/dia.
Nas formas disseminadas, o tempo de tratamento deve ser prolongado para 10 ou
mais dias ou até que os parasitos sejam erradicados. É eficaz contra as formas
adultas, mas não é larvicida. Por isso, deve-se levar em conta a possibilidade da
auto-infecção (PESSOA, 1978). Os efeitos colaterais mais freqüentes são
tonturas, cefaléia, sonolência, náuseas, vômitos e dor abdominal. Também pode
ocorrer hepatoxicidade e leucopenia. Seu uso é contra-indicado em mulheres
grávidas e durante a lactação. Os índices de cura ultrapassam 90% (CAMPOS e
FERREIRA, 1999).
Cambendazol: derivado imidazólico, com eficácia semelhante ao tiabendazol
(>90%). É administrado em dose única, na dose de 5mg/Kg, após a refeição.
Raramente observa-se dor abdominal, náuseas, vômitos e diarréia (CAMPOS e
FERREIRA, 1999).
Albendazol: imidazólico de amplo espectro, ativo inclusive na forma larvária,
porém não demonstra bons resultados na terapêutica da estrongiloidíase. A dose
preconizada é de 400mg/dia, VO, por três dias; nessa posologia, o índice de cura

é de apenas 40 a 50% (CAMPOS e FERREIRA, 1999). A dose recomendada é
de 800mg/dia, dividida em duas tomadas, durante três dias, e a cura
parasitológica pode chegar a 80 a 90% (VAN DER FELTZ et al, 1999). Não é
recomendado o uso desta medicação em casos graves ou disseminados desta
parasitose (CAMPOS e FERREIRA, 1999). Efeitos colaterais são cefaléia, tonturas
e desconforto gastro-intestinal. É contra-indicado na gestação.
Ivermectina: utilizada na dose de 150 a 200 µg/kg. É uma droga bem tolerada,
com poucas reações clínicas e bioquímicas (ADENUSI, 1997). Estudos recentes
têm mostrado que a ivermectina é a melhor droga para o tratamento da
estrongiloidíase não complicada (SALAZAR et al, 1994; SIDDIQUI e BERK, 2001;
KEISER e NUTMAN, 2004). Seu uso parece ser seguro na gravidez (VAN DER
FELTZ et al, 1999). ASHRAF, GUE e BADDOUR (1996) publicaram relato de caso
de estrongiloidíase refratária ao tratamento com ivermectina em treze cursos de
dois dias, em paciente com hipogamaglobulinemia. Isso reforça a necessidade de
pesquisa de outros agentes anti- S. stercoralis.
Em caso de estrongiloidíase disseminada ou grave, em especial em
pacientes imunodeprimidos, a ivermectina tem sido empregada em esquemas de
multidoses, ou seja, 200µg/kg nos dias 1, 2, 15 e 16, com altos índices de cura
clínica e parasitológica (CAMPOS e FERREIRA, 1999).
A eficácia terapêutica pode ser avaliada através de exames parasitológicos
de fezes realizados nos dias 7º, 14º e 21º após o tratamento. O método de
Baermann-Moraes deve ser o utilizado nestes exames para controle de cura desta
helmintíase (CAMPOS e FERREIRA, 1999).
Recentemente, a ivermectina foi registrada como a droga de escolha na
lista da Organização Mundial de Saúde (OMS) para o tratamento de S. stercoralis
(ALBONICO, CROMPTON e SAVIOLI, 1999).
Nas formas graves da doença, com disseminação larvária a múltiplos
órgãos, o uso de antimicrobianos com cobertura para bactérias Gram-negativas é
mandatório. Neste caso, deve-se usar as cefalosporinas de terceira ou quarta-
geração, os aminoglicosídeos, o aztreonam ou quinolonas.

2.1.13- Profilaxia
Atenção especial deve ser dada aos hábitos higiênicos. O uso de calçados,
a limpeza e a higiene adequada dos alimentos crus a serem ingeridos são pontos
cruciais na prevenção desta helmintíase. Em pacientes imunodeprimidos, exames
parasitológicos devem ser realizados periodicamente utilizando-se o método de
Baerman-Moraes. Em pacientes com AIDS, está indicado o uso profilático
secundário do tiabendazol por dois ou três dias, mensalmente, para evitar
recidivas da doença (CAMPOS e FERREIRA, 1999).
2.2- Efeitos do uso abusivo do etanol sobre o sistema imunitário
Devido a um aumento da freqüência e gravidade das infecções em
alcoolistas crônicos, existe uma convicção que o etanol inibe diretamente os
mecanismos imunitários inatos e adaptativos do organismo (MAC GREGOR,
1986).
Granulocitopenia tem sido observada em 4 a 8% dos alcoolistas admitidos
em hospital, especialmente naqueles com infecção (LIU, 1980). Exame da medula
óssea tem mostrado uma marcada diminuição no número de granulócitos
maduros, com vacuolização de mielócitos precursores. A injeção de endotoxicina
nos pacientes alcoolistas não causa um aumento na contagem de
polimorfonucleares, indicando uma diminuição da reserva medular. A diminuição
da produção parece ser a base para a granulocitopenia (MACGREGOR, 1986).
Isso pode ser devido à diminuição da produção de fatores estimuladores de
colônia, mais do que ao efeito tóxico direto do etanol em precursores mielóides.
Ainda não está bem definida a causa da diminuição do “pool” de reserva medular
de granulócitos em alcoolistas, embora os possíveis responsáveis sejam a má
nutrição, a doença hepática e a toxicidade pelo etanol (ADAMS e COLIN, 1984).
O mecanismo responsável pela diminuição da mobilização de
polimorfonucleares em indivíduos agudamente intoxicados é a diminuição da
aderência, impedindo a célula de sair do compartimento vascular (MACGREGOR,
GLUCKMAN e SENIOR, 1978).

Para avaliar o efeito imunossupressor do etanol na ausência de fatores
complicadores como cirrose hepática e má nutrição, foi estudada a ingestão
crônica do etanol, em um ambiente controlado, demonstrando-se inibição da
quimiotaxia de granulócitos que melhorou com a abstinência. A resposta de
anticorpos a novos antígenos não se alterou e o número de linfócitos no sangue
periférico não foi afetado pelo consumo de etanol em alcoolistas voluntários não
cirróticos (GLUCKMAN, DVORAK e MACGREGOR, 1977).
O etanol prejudica a resposta Th1 e aumenta a resposta Th2
(WALTENBAUGH, VASQUEZ e PETERSON, 1998). Esses autores, em modelo
animal, demonstraram que o etanol afeta diretamente as células apresentadoras
de antígeno, que têm papel central em determinar se o padrão predominante da
resposta será Th1 ou Th2.
O efeito do etanol na resposta imunitária é dose dependente . Em ratos
demonstrou-se que ingestão de etanol em baixa dose (0,5 a 2g/kg) estimulou a
resposta imunitária celular e doses maiores (6g/kg) suprimiram a resposta
imunitária basal (DEHNE et al 1989). Resultado semelhante foi observado in vivo:
pequenas doses de etanol aumentaram e doses mais elevadas de etanol
diminuíram significativamente a resposta intradérmica à fitohemoglutinina
(MENDENHALL et al 1977).
No entanto, outros tem demonstrado efeito deletério na resposta imunitária
com a ingestão de pequenas quantidades de etanol, tanto em animais de
laboratório (ALDO-BENSON, 1989) como em humanos (BOUNDS et al, 1994) .
Ainda que haja relatos de que o consumo moderado de etanol possa ter
impacto positivo sobre o sistema imunitário (DIAZ et al, 2002), a definição de
consumo moderado de álcool é muito variável. Segundo GONZÁLES-GROSS,
LEBRÓN e MARCOS (2000), consumo moderado pode ser definido em termos de
10-12g dia para mulheres e 20-24g dia para homens.
COOK (1998) mostrou que o consumo crônico do etanol altera a função
imunitária humoral. Embora o número de células B tenha sido normal ou
discretamente diminuído em alcoolistas sem doença hepática, elas são
significantemente diminuídas em pacientes com doença hepática alcoólica.

Alcoolistas crônicos em síndrome de abstinência têm aumento do nível
sérico de IgE, além do aumento de citocinas tipo TH2 (GONZÁLES-QUINTELA, et
al, 1999). Foi hipotetizado que o aumento dos níveis séricos de IgE em alcoolistas
seria devido ao etanol e seus metabólitos, e que o etanol atuaria como um
alergeno ou antígeno (DOMÍNGUEZ-SANTALLA et al, 2001).
O alcoolismo crônico é considerado como a causa mais comum de má
nutrição em países ricos ocidentais, havendo alto risco de imunossupressão
nutricional em alcoolistas crônicos. A ingestão crônica do etanol é associada com
uma alta ingestão de energia derivada do mesmo e uma inadequada ingestão de
proteínas, vitaminas e minerais (WATZL e WATSON, 1992). No entanto, fica difícil
incluir ou excluir o papel da nutrição na alteração imunitária induzida pelo etanol,
porque essas são observadas em animais de laboratório ou humanos, submetidas
ao etanol, mas com bom estado nutricional.
Além das numerosas doenças associadas com o consumo excessivo de
etanol, também ocorre aumento da atividade do eixo hipotálamo-pituitário-adrenal,
com aumento dos níveis plasmáticos de ACTH (hormônio adreno-corticotrófico) e
corticosterona. Em trabalho experimental com ratos, OGILVIE et al (1998)
introduziram etanol intragástrico por 3 dias, e, re-expuseram esses animais à
droga 3 a 12 dias mais tarde. Ratos pré-tratados com diluente do etanol e
injetados com etanol, 3 a 12 dias mais tarde, mostraram um marcado aumento
nas concentrações plasmáticas de ACTH e corticosterona. Ao contrário, animais
pré-tratados com etanol exibiram uma diminuição da resposta hormonal e
hipotalâmica durante a segunda exposição à droga. Esses animais mantiveram
uma resposta endócrina normal a outros sinais como eletro-choque ou injeção de
citocinas. Fica portanto difícil interpretar a participação do eixo hipotálamo-
hipofisário-adrenal nas alterações imunitárias induzidas pelo etanol.
Em resumo, os dados da literatura indicam que: (1) consumo baixo e
moderado de etanol, ou não altera ou até pode potencializar a resposta imunitária;
(2) consumo excessivo, de modo geral, altera quase todos os mecanismos
imunitários inatos e adaptativos, especialmente em alcoolistas com doença

hepática associada; (3) quanto aos mecanismos da interferência do etanol no
sistema imunitário, se diretos ou indiretos , há ainda pouca informação.
2.3- Uso abusivo de etanol e prevalência de Strongyloides stercoralis
Além de alterações nos mecanismos imunitários causados pelo alcoolismo
(MACGREGOR, 1986; COOK, 1996), as infecções são mais freqüentes e graves
em alcoolistas também devido ao prejuízo da função mental, que leva à menor
cuidado com a higiene pessoal e maior exposição aos patógenos, à quebra das
barreiras protetoras nas mucosas, à maior facilidade de aspiração e à má nutrição
(MACGREGOR, 1986).
Em trabalho prospectivo realizado em 35 pacientes cirróticos e 45 pacientes
não-cirróticos submetidos a três amostras de exames parasitológicos das fezes
pelos métodos de Hoffman, Pons-Janner, Baermann e Willis, GABURRI et al,
(1997) observaram que 14/35 cirróticos tinham exame parasitológico positivo para
S. stercoralis, sendo que nenhum caso de estrongiloidíase foi diagnosticado nos
45 pacientes (grupo controle 1). Quanto à etiologia da cirrose hepática temos:
álcool em 19 (54,3%), hepatite por vírus B (HBV) em 3 (8,6%), hepatite por vírus C
(HCV) em 5 (14,3%), HBV + HCV em 2 (5,7%), outras causas 6 (17,2%). No grupo
controle 2, representado por 1411 indivíduos que fizeram exames parasitológicos
das fezes no mesmo hospital, a prevalência de S. stercoralis foi de 27/1411
(1,91%). O S. stercoralis foi diagnosticado em 10/19 cirróticos alcoólicos, o que
comparado com portadores de cirrose hepática não alcoólica, não houve diferença
estatisticamente significante. Segundo os autores, “tal observação se reveste de
suma importância, sugerindo que estes resultados estariam relacionados a
alterações geradas pela própria cirrose hepática e não apenas ao alcoolismo”. Os
mesmos ainda citam que “à primeira vista, parece-nos não ser o álcool o fator
diretamente implicado em tal ocorrência, mas talvez razões de ordem imunológica,
geradas pela própria doença hepática, favoreçam tal associação”.
AVEDAÑO et al (1999) estudaram a freqüência da estrongiloidíase em 106
pacientes alcoolistas crônicos da Costa Rica internados em um Instituto Nacional
para Dependência de Fármacos e Alcoolismo que foi feita pelos métodos cultura

em placa de Agar, Baermann e exame direto das fezes, em apenas uma amostra
de fezes e S. stercoralis foi diagnosticado em 5,7%. Essa prevalência foi muito
maior do que a observada em inquérito nacional (0,1%) feito com exame através
de sedimentação. No entanto as amostras, como observou o autor, não são
comparáveis.
OLIVEIRA et al (2002) avaliaram a freqüência de S. stercoralis em
alcoolistas. Foram avaliados 145 indivíduos, dos quais 45 eram dependentes do
álcool, 10 eram portadores de cirrose hepática não alcoólica, e 90 eram indivíduos
não alcoolistas assintomáticos, que serviram como controle. Foram feitos três
exames parasitológicos das fezes em dias alternados pelos métodos de
Baermann-Moraes e Lutz. A freqüência da estrongiloidíase, no grupo total de
alcoolistas, foi de 15/45 (33,3%) e no subgrupo de alcoolistas com cirrose hepática
foi de 4/9 (44,4%), com pancreatite foi de 3/9 (33,3%) e naqueles sem cirrose
hepática e sem pancreatite foi de 8/27 (29,6%), que foi significativamente
(p<0,0001) mais alta que a encontrada nos controles 5/90 (5,5%). Nenhum dos
indivíduos com cirrose hepática não alcoólica tinham infecção pelo S. stercoralis.
Embora o número de pacientes alcoolistas seja pequeno, para conferir força
estatística dos resultados. Há indicação de que a prevalência de S. stercoralis seja
maior em alcoolistas.
ZAGO-GOMES et al (2002) realizaram um estudo retrospectivo da
freqüência de nematóides intestinais em 198 alcoolistas e em 440 controles, não
alcoolistas, atendidos no HUCAM em Vitória - Brasil. O grupo controle foi formado
por 144 pacientes não alcoolistas, pareados por procedência com o grupo de
alcoolistas e 296 pacientes atendidos no Hospital, com diferentes procedências.
Todos os pacientes foram submetidos a três exames parasitológicos de fezes pelo
método de sedimentação. Houve uma freqüência significativamente maior de
nematóides intestinais no grupo de alcoolistas do que no grupo controle (35,3% e
18,7%, respectivamente), devido à maior freqüência de S. stercoralis (21,7 e 4,1%,
respectivamente). Não houve diferença significante na freqüência de nematóides
entre alcoolistas com ou sem cirrose hepática (37,9% e 34,2%, respectivamente
para pelo menos um nematóide e 25,8% e 20% para S. stercoralis,

respectivamente). A freqüência dos outros nematóides foi semelhante entre os
alcoolistas quando comparado com o grupo controle. Para os autores, essas
observações indicam que o etanol está relacionado com o aumento do risco para
infecção pelo S. stercoralis e não com cirrose hepática, como admitida por
GABURRI et al (1997).
ADEDAYO, GRELL e BELLOT (2002) relataram 27 casos de
estrongiloidíase hiper-infectiva diagnosticados e internados em enfermaria médica
durante 5 anos (18 homens e 9 mulheres, de 26 a 91 anos de idade). Vinte e
quatro pacientes foram testados para HTLV-1, e 17 (71%) tiveram resultado
positivo. É interessante observar que 12/27 (44%) tinham história de alcoolismo
crônico. Desses 12 alcoolistas três eram HTLV-1 positivos. Com a freqüência de
alcoolistas nessa série de casos, sem infecção com o HTLV-1, os autores
chamaram a atenção para o possível papel do alcoolismo como indicador de pior
prognóstico nas formas graves de estrongiloidíase.
Quanto aos possíveis mecanismos dessa maior prevalência, nada se
conhece e admitindo-se que seja a redução dos mecanismos imunitários e/ou um
possível aumento de corticóides circulantes induzidos pelo uso do etanol, os
fatores mais importantes. No entanto, os alcoolistas, devido aos distúrbios do
comportamento, estão mais sujeitos à infecção com o parasita, fato difícil de ser
descartado.
Conclui-se, pelos dados da literatura, que o alcoolismo crônico parece ser
um fator de risco para ocorrência de um exame parasitológico de fezes positivo
para S. stercoralis. Nenhum dos autores, que relataram a maior prevalência do S.
stercoralis em alcoolistas, apresentaram dados sugerindo possíveis mecanismos
para explicar essa associação. Só apresentaram algumas especulações sobre o
assunto.

OBJETIVOS

3- Objetivos:
Tendo em vista dados da literatura apontando para uma associação entre
alcoolismo crônico e parasitismo pelo S. stercoralis e considerando alguns
mecanismos propostos para explicar a maior prevalência do S. stercoralis em
alcoolistas crônicos planejamos a presente investigação com os seguintes
objetivos:
3.1- Objetivos gerais
Estudar a prevalência de S. stercoralis em três diferentes amostras
de alcoolistas crônicos, atendidos em dois diferentes serviços da Região
Metropolitana de Vitória e comparar com amostras de pacientes não alcoolistas,
atendidos nos mesmos serviços, nas mesmas condições.
3.2- Objetivos específicos
1- Avaliar a prevalência do S. stercoralis em alcoolistas crônicos e
não alcoolistas, através do método de sedimentação ou método de Baermann,
atendidos no Hospital Universitário Cassiano Antônio de Moraes, em 2003.
2- Avaliar a prevalência de S. stercoralis através da revisão de
prontuários de pacientes alcoolistas crônicos atendidos no PAA (Programa de
Atendimento ao Alcoolista) do HUCAM nos anos de 2001 e 2002 e em controles
não alcoolistas atendidos no mesmo Hospital, no mesmo período.
3- Avaliar a prevalência de S. stercoralis em alcoolistas crônicos e
não alcoolistas, atendidos em uma Unidade de Saúde do Município de Serra,
através do método de sedimentação realizado no mesmo laboratório, no ano de
2003.
4- Avaliar os níveis séricos de glicocorticóides em pacientes
alcoolistas crônicos com e sem infecção com o S. stercoralis, para identificar

possível participação do eixo hipotálamo-hipófise-adrenal na maior prevalência da
infecção pelo S. stercoralis em alcoolistas crônicos, atendidos no HUCAM.
5-Verificar a freqüência de S. stercoralis nos exames parasitológicos
do Hospital Universitário Cassiano Antônio de Moraes, que atende alcoolistas
crônicos em serviço especializado, e avaliar, naqueles pacientes com S.
stercoralis, a prevalência de alcoolismo crônico.

PACIENTES E MÉTODOS

4.1-Amostras estudadas
Para atingir os objetivos propostos foram estudadas diferentes amostras no
sentido de fazer avaliação da prevalência com utilização de método mais sensível
para identificar o S. stercoralis, como também em amostras estudadas
rotineiramente, onde os exames parasitológicos são realizados pelo método de
sedimentação. Embora utilizando métodos com sensibilidade diferente, os
controles para cada amostra eram submetidos ao mesmo método de exame
parasitológico.
4.1.1- Amostras para estudo da prevalência do S. stercoralis em alcoolistas
crônicos
Foram estudadas três amostras de pacientes alcoolistas crônicos nos quais
foi pesquisada a prevalência de S. stercoralis:
A- Amostra constituída de 346 alcoolistas crônicos, atendidos no Programa
de Atendimento ao Alcoolista do HUCAM, nos quais foi feita a pesquisa do S.
stercoralis, pelo método de Baermann, em 190 pacientes. Em 156 pacientes o
exame parasitológico foi feito pelo método de sedimentação. Para controle, foram
usados 111 pacientes, não alcoolistas, procedentes das mesmas regiões e
atendidos no mesmo Hospital, submetidos a três exames parasitológicos de fezes
também pelo método de Baermann. A amostra controle, para os 156 alcoolistas
com exame parasitológico de fezes por método de sedimentação, foi tomada
aleatoriamente por revisão de prontuários, de 591 pacientes não alcoolistas,
atendidos no HUCAM, com exame parasitológico de fezes realizados no mesmo
laboratório, nos anos de 2001 e 2002.
B- Amostra obtida pela revisão dos prontuários de pacientes alcoolistas
crônicos, formada por 504 pacientes atendidos no Programa de Atendimento ao
Alcoolista no HUCAM, durante os anos de 2001 e 2002, e que tinham sido
submetidos a exame parasitológico de fezes. Para controle, foi tomada uma
amostra de pacientes atendidos no mesmo Hospital, formada por 591 pacientes
não alcoolistas, atendidos nos anos de 2001 e 2002, com procedência

semelhante, tomada aleatoriamente pela revisão de prontuários de pacientes
atendidos no HUCAM e que tenham o resultado do exame parasitológico anotado
no prontuário. Nos dois grupos, alcoolistas e controles, o exame parasitológico de
fezes foi realizado no Laboratório de Análises Clínicas do HUCAM, no mesmo
período, pelo método de sedimentação, em três amostras.
C- Amostra tomada na Unidade Regional de Saúde Feu Rosa, município de
Serra, formada por 49 alcoolistas crônicos e 129 não alcoolistas, atendidos no
mesmo serviço, no ano de 2003. Nos dois grupos os exames parasitológicos
foram realizados no mesmo laboratório, em três amostras de fezes, pelo método
de sedimentação.
4.1.2- Amostra para avaliação da prevalência de S. stercoralis nos exames
parasitológicos do Hospital Universitário Cassiano Antônio de Moraes
Foi feita a avaliação da prevalência de S. stercoralis em 7112 exames
parasitológicos realizados no ano de 2003, no laboratório de Análises Clínicas do
HUCAM. Dos casos positivos para S. stercoralis, tentou-se levantar os prontuários
e verificar a informação sobre história pregressa de alcoolismo crônico.
4.1.3- Amostra utilizada para avaliação dos níveis séricos de cortisol
Em 46 alcoolistas crônicos, sendo 23 com exame parasitológico de fezes
positivo, para S. stercoralis e 23 negativos, foi feita a dosagem do cortisol
plasmático. Essa amostra foi obtida dos pacientes atendidos no ano de 2003 no
HUCAM.
4.2- Exames parasitológicos de fezes
Os exames parasitológicos de fezes foram realizados pelos métodos de
Baermann e sedimentação conforme descrito resumidamente a seguir:
O método de Baermann (1917), modificado por Moraes (1948) foi realizado
conforme técnica padrão. Cerca de 8 a 10 g de fezes sobre uma gaze eram
colocadas em um funil de vidro de 10 a 12 cm ligado a um tubo de látex fechado,
com uma pinça de Mohr. O funil era parcialmente preenchido com água de

torneira, aquecida a 40 a 44°C, de modo que a água aquecida entrasse em
contacto com a gaze. Após 60 minutos, a pinça era aberta e parte do líquido era
coletado em tubo cônico, centrifugado rapidamente e o sedimento examinado
entre lâmina e lamínula.
O método de Sedimentação (Lutz, 1919) foi realizado conforme técnica
tradicional. As fezes eram diluídas com água de torneira, bem misturadas e, em
seguida, filtradas em gaze e transferidas para cálices cônicos de sedimentação.
Entre duas e quatro horas após, o sedimento era examinado entre lâmina e
lamínula.
Sempre foram examinadas três amostras de cada paciente, em dias
alternados.
4.3- Avaliação do cortisol plasmático
A dosagem do cortisol foi feita no soro pelo método de imunoensaio
competitivo por quimioluminescência direta, utilizando o sistema ADVIACentaur
(Bayer Health Care LLC). Nesse método, o cortisol da amostra compete com o
cortisol marcado com éster de acridina pela ligação ao anticorpo policlonal de
coelho anti-cortisol preso na fase sólida. O anticorpo policlonal de coelho está
ligado ao anticorpo anti Ig de coelho, monoclonal, obtido em rato, o qual está
ligado covalentemente a partículas paramagnéticas. O sangue de todos os
pacientes foi coletado pela manhã (7:30 às 8:00 hs), após um repouso de trinta
minutos. Para esse método os valores de referência são 5 a 25 µg/dL.
4.4- Critérios para o diagnóstico de alcoolismo crônico
O diagnóstico de Síndrome de Dependência do Álcool, foi feito de acordo com
a Classificação Internacional de Doenças (CID -10).
4.5- Critérios de exclusão de pacientes e controles
Foram excluídos todos os indivíduos HIV positivos.

4.6- Informações aos pacientes e controles
Todos os pacientes e controles foram convidados a participar do estudo em
caráter voluntário. Os que concordaram em participar, assinaram um termo de
consentimento (ANEXO I), após receberem as informações referentes ao
protocolo de pesquisa e responderem um questionário (ANEXO II).
4.7- Análise estatística dos resultados
Os resultados foram analisados utilizando-se o programa SPSS, versão 10.0
(Statistical Package for the Social Sciences). As comparações de variáveis
quantitativas foram feitas pelo teste t de Student ou pelo teste de Mann-Whitney.
As comparações de variáveis qualitativas foram feitas pelo teste χ² ou teste exato
de Fisher. A associação entre a presença de alcoolismo e o exame parasitológico
de fezes positivo para S. stercoralis foi feita pelo cálculo da razão de chances
(Odds-Ratio, indicado como OR nas tabelas de resultados). A força da
associação, foi calculada pelo teste do χ². Análise de regressão logística binária foi
utilizada para verificar a interferência de diferentes variáveis com o alcoolismo, na
ocorrência de exame parasitológico positivo para S. stercoralis. Foram
considerados significantes os valores de p menores do que 0,05, admitindo-se
todos como testes bi-caudal.

RESULTADOS

5.1- Idade, sexo e procedência das amostras estudadas
Os principais dados sobre idade e sexo das amostras estudadas para
verificação da prevalência do S. stercoralis estão resumidos nas Tabelas 1 a 4.
Como esperado a maior prevalência do alcoolismo foi no sexo masculino, o
que foi observado em todas as amostras. Houve uma diferença estatisticamente
significativa na média de idade dos alcoolistas em relação aos controles não
alcoolistas, tanto no sexo masculino como no sexo feminino na amostra do
HUCAM onde a pesquisa do S. stercoralis foi feita pelo método de Baermann
(Tabela 1) e no sexo feminino na amostra estudada na Serra (Tabela 4) .
Tabela 1- Idade e sexo de pacientes alcoolistas crônicos e respectivos controles, atendidos no
Programa de Atendimento ao Alcoolista do HUCAM, no ano de 2003, nos quais os exames
parasitológicos de fezes foram feitos com o método de Baermann.
Variáveis Alcoolistas (N=190) Controles (N=111) p
Sexo
Masculino
Feminino
Idade
Sexo masculino
Média ± DP
Mediana
Sexo feminino
Média ± DP
Mediana
171 (90%)
19 (10%)
42,1 ± 9,8
44,5
39,5 ± 7,8
41,0
50 (45%)
61 (55%)
48,3 ± 13,1
41,0
45,15 ± 12,15
37,5
0,000*
<0,001**
<0,001**
*χ² ; **Teste t de Student

Tabela 2- Idade e sexo em 156 alcoolistas crônicos atendidos no Programa de Atendimento ao
Alcoolista do HUCAM e não alcoolistas tomados aleatoriamente pelo levantamento de prontuários
do mesmo hospital, nos quais o exame parasitológico de fezes foi feito pelo método de
sedimentação.
Variáveis Alcoolistas (N=156) Controles (N=591) p
Sexo
Masculino
Feminino
Idade
Sexo masculino
Média ± DP
Mediana
Sexo feminino
Média ± DP
Mediana
141 (92,1%)
15 (7,9%)
46,4 ± 9,1
46
41,3 ± 7,3
42,5
465 (78,6%)
126 (21,4%)
44,8±16,1
45
41,6±13,2
43
0,020*
N.S.**
N.S.**
*χ ²; ** Teste t de Student; N.S. (não significativa)
Tabela 3- Idade e sexo em 504 alcoolistas crônicos atendidos no Programa de Atendimento ao
Alcoolista do HUCAM, nos anos de 2001 e 2002, e de 591 não alcoolistas tomados aleatoriamente
pelo levantamento de prontuários do mesmo Hospital, no mesmo período.
Variáveis Alcoolistas (N=504) Controles (N=591) p
Sexo
Masculino
Feminino
Idade
Sexo masculino
Média ± DP
Mediana
Sexo feminino
Média ± DP
Mediana
447 (88,6%)
57 (11,4%)
44,6 ± 9,3
44
43,2 ± 8,9
42
465 (78,6%)
126 (21,4%)
44,8 ± 16,1
41,6 ± 13,2
43
0,000*
N.S.**
N.S.**
* χ²; ** Teste t de Student; N.S. (não significativa)

A procedência de alcoolistas crônicos e de controles atendidos no Hospital
Universitário foi semelhante. Entre os alcoolistas, 86% eram provenientes da
periferia urbana que formam a Região Metropolitana da Grande Vitória (Vitória,
Cariacica, Serra, Vila Velha e Viana) e o restante, de outros municípios do estado.
Entre os controles a procedência era bem semelhante: 73,8% eram provenientes
da Região Metropolitana da Grande Vitória. A grande maioria dos pacientes
alcoolistas, atendidos no Programa de Atendimento ao Alcoolista do HUCAM, é
residente nos Municípios da Região Metropolitana de Vitória. Os controles
escolhidos aleatoriamente têm procedência semelhante.
Tabela 4- Idade e sexo de 49 pacientes alcoolistas crônicos e 129 controles não alcoolistas
atendidos na Unidade Regional de Saúde Feu Rosa (município de Serra), no ano de 2003, nos
quais os exames parasitológicos de fezes foram realizados pelo método de sedimentação.
Variáveis Alcoolistas (N=49) Controles (N=129) p
Sexo
Masculino
Feminino
Idade
Sexo masculino
Média ± DP
Mediana
Sexo feminino
Média ± DP
Mediana
46 (93,8%)
3 (6,2%)
45,5 ± 10,1
42
36,0 ± 6,2
38,0
98 (75,9%)
31 (24%)
44,5 ± 16,0
45,5
41,6 ± 14,4
42,0
0,012*
N.S.**
0,002**
*χ²; ** Teste t de Student; N.S. (não significativa)

5.2- Prevalência de S. stercoralis nos exames parasitológicos de fezes
realizados no laboratório de rotina do Hospital Universitário Cassiano
Antônio de Moraes
A prevalência dos parasitas intestinais diagnosticados em 7112 exames,
realizados no ano de 2003, no Laboratório de Análises Clínicas do HUCAM, está
resumida na tabela 5, separada por sexo.
Tabela 5- Prevalência de parasitas intestinais em 7112 exames parasitológicos de fezes, realizados pelo método de sedimentação, no laboratório de Análises Clínicas do HUCAM, no ano de 2003.
Parasitas Sexo
Masculino (N=3147) Feminino (N=3965)
Nematóides
Ascaris lumbricoides
Trichuris trichiura
Ancilostomídeos
Enterobius vermicularis
Strongyloides stercoralis
Schistosoma mansoni
Taenia sp
384 (12,2%)
161 (5,11%)
41 (1,30%)
20 (0,63%)
55 (1,74%)
129 (4,09%)
7 (0,22%)
1 (0,03)
372 (9,38%)
193 (4,86%)
44 (1,10%)
17 (0,42%)
59 (1,48%)
59 (1,48%)
6 (0,15%)
2 (0,05)
A tabela nos mostra que o único parasita para o qual houve uma diferença
significante entre os sexos é o S. stercoralis: respectivamente para o sexo
masculino e feminino: 4,09%, IC a 95% 3,33 a 4,84% e 1,48%, IC a 95%: 1,11 a
1,85%. Dos 188 casos (129 do sexo masculino e 59 do sexo feminino) de infecção
com S. stercoralis, no Laboratório do HUCAM, foram levantados os prontuários de
76 pacientes, dos quais, em 51 havia anotação confiável sobre o uso do etanol.
Desses, 21 eram alcoolistas crônicos, representando 41,17% dos 51 exames. Se
considerarmos a amostra válida para os 188 casos positivos, aproximadamente 77

casos positivos para S. stercoralis eram alcoolistas crônicos. Se subtrairmos os
alcoolistas do sexo masculino, a prevalência do parasita fica semelhante nos dois
sexos.
5.3- Prevalência de S. stercoralis nas diferentes amostras de alcoolistas e
controles
As prevalências da infecção com S. stercoralis e de outros nematóides nas
diferentes amostras estudadas estão resumidas nas Figuras 2 e 3 e nas Tabelas
1, 2, 3,4 dos ANEXOS (III,IV,V,VI).
Em todas as amostras a prevalência do S. stercoralis é maior no grupo de
alcoolistas, quando comparado com controles não alcoolistas. A chance de o
alcoolista crônico ter um exame de fezes positivo para S. stercoralis foi no mínimo
3,4 vezes maior do que os controles não alcoolistas, exceto na amostra da
Unidade Regional de Saúde Feu Rosa (URSFR) onde o número de casos não
permitiu o cálculo adequado da razão de chances, pelo ausência de casos
positivos para S. stercoralis no grupo controle (Figura 2). Mas nessa última
amostra o teste exato de Fisher mostra ser a prevalência do parasita
significativamente maior nos alcoolistas crônicos (Tabela 4 do ANEXO VI).
Quando separamos as amostras pelo sexo (Figura 3) observa-se que
apenas na amostra maior, dos casos analisados pela revisão dos prontuários dos
pacientes alcoolistas do Programa de Atendimento ao Alcoolista e controles
atendidos no HUCAM, a diferença é significativa nas mulheres. Provavelmente,
isso se deveu ao fato de ter nessa amostra um número de mulheres alcoolistas
muito maior do que nas demais amostras estudadas.

Figura 3- Prevalência de S. stercoralis em alcoolistas crônicos e não alcoolistas nas quatro
amostras, nos dois sexos. NS (não significativa); URSFR (Unidade Regional de Saúde Feu Rosa)
Nas amostras estudadas durante o ano de 2003 foram anotados os dados
sobre condições sanitárias (rede de esgoto, água encanada e tratada e hábito de
andar descalço) dos pacientes alcoolistas e controles. A tabela 6 mostra uma
análise de regressão logística, para avaliar o impacto do uso crônico do etanol
como fator de risco para infecção com S. stercoralis, levando em consideração as
condições sanitárias e hábitos dos pacientes do sexo masculino. Verifica-se que o
álcool é fator de risco significativo, mesmo quando ajustado para os co-fatores
citados. Foi feita a análise no sexo masculino porque a prevalência de alcoolismo
é significativamente maior nos homens e o sexo aparece como co-fator que
interfere no alcoolismo. Não foi feita nas mulheres devido ao pequeno número de
casos.
sexoAlcoolistas crônicos Nâo alcoolistas
0
5
10
15
20
25
M F M F M F M F
% d
e ca
sos
BaermannHUCAM
HUCAM2003
HUCAM2001/02
USFRSerra
S e d i m e n t a ç ã o
ns ns
ns
*
* *
**
* =p<0.005
sexoAlcoolistas crônicos Nâo alcoolistas
0
5
10
15
20
25
M F M F M F M F
% d
e ca
sos
BaermannHUCAM
HUCAM2003
HUCAM2001/02
USFRSerra
S e d i m e n t a ç ã o
ns ns
ns
*
* *
**
* =p<0.005

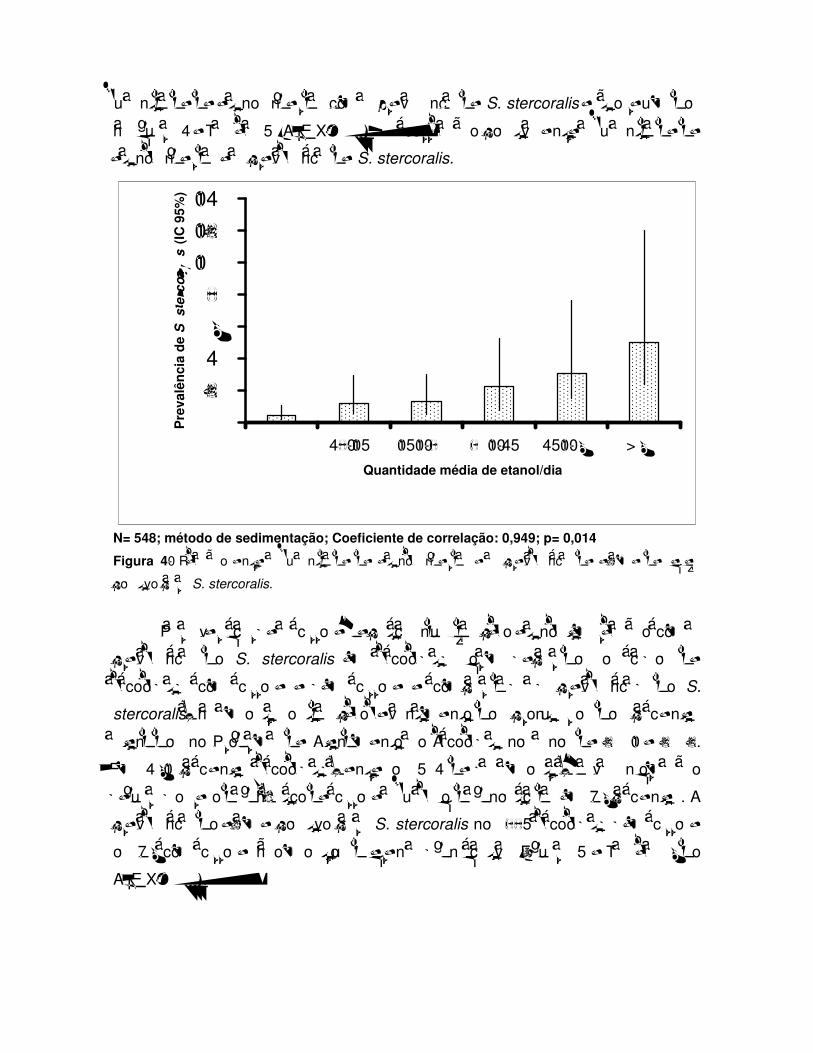
quantidade de etanol ingerida com a prevalência de S. stercoralis estão resumidos
na figura 4 e Tabela 5 (ANEXO VII). Há correlação positiva entre a quantidade de
etanol ingerida e a prevalência de S. stercoralis.
0
20
40
60
80
100
120
140
0 48-150 151-300 301-450 451-600 >600
Quantidade média de etanol/dia
Pre
valê
nci
a d
e S
. ste
rco
ralis (
IC 9
5%)
N= 548; método de sedimentação; Coeficiente de correlação: 0,949; p= 0,014 Figura 4- Relação entre a quantidade de etanol ingerida e a prevalência de exame de fezes
positivo para S. stercoralis.
Para verificar se a cirrose hepática induzida pelo etanol tem relação com a
prevalência do S. stercoralis em alcoolistas foram separados os casos de
alcoolistas com cirrose e sem cirrose e comparadas as prevalências do S.
stercoralis, na amostra obtida pelo levantamento dos prontuários dos pacientes
atendidos no Programa de Atendimento ao Alcoolista nos anos de 2001 e 2002.
Em 461 pacientes alcoolistas, entre os 504 dessa amostra, havia informação
segura sobre o diagnóstico de cirrose a qual foi diagnosticada em 76 pacientes. A
prevalência do exame positivo para S. stercoralis nos 385 alcoolistas sem cirrose
os 76 com cirrose não mostrou diferença significativa (Figura 5 e Tabela 6 do
ANEXO VIII).

Figura 5- Prevalência de exame positivo para S. stercoralis em alcoolistas crônicos com e sem cirrose hepática.
Os resultados da dosagem do cortisol plasmático nos 46 pacientes
alcoolistas crônicos, 23 com S. stercoralis positivo no exame parasitológico de
fezes e 23 com exame parasitológico de fezes negativo para o parasita, estão na
figura 6. Não há diferença estatisticamente significativa entre as medianas do
cortisol plasmático nos alcoolistas com ou sem S. stercoralis detectado no exame
parasitológico de fezes.
S. stercoralis
negativo positivo
CO
RT
ISO
L ug
/ dL
40
30
20
10
0
Figura 6- Níveis de cortisol plasmático em 23 pacientes alcoolistas, com exame de fezes positivo
para S. stercoralis, e em 23 pacientes alcoolistas, com exame de fezes negativo para o parasita.
Os resultados mostram as medianas, os quartis e os percentis 5 e 95 (Teste de Mann-Whitney,
p=0,602)
0
5
10
15
20
25
Sem cirrose Com cirrose
% d
e ca
sos
76/385 19,5%
9/76 11,8%
p= 0,144

DISCUSSÃO

6.1- ANÁLISE CRÍTICA DAS AMOSTRAS
6.1.1- Justificativa para a utilização de diferentes amostras
Nessa pesquisa foram utilizadas diferentes amostras de pacientes,
alcoolistas e não alcoolistas, para comparação entre os grupos, além de
comparações de médias de variáveis quantitativas ou ainda para avaliação de
prevalência de um ou mais desfechos de interesse. A utilização de diferentes
amostras foi decidida tendo em vista o objetivo principal da pesquisa: o de
verificar a força da associação entre etilismo crônico e parasitismo pelo S.
stercoralis. Desse modo, foram utilizadas amostras coletadas prospectivamente,
sendo que, em uma amostra, o diagnóstico da parasitose foi feito por método mais
sensível (Baermann), e, em duas amostras, o diagnóstico foi feito pelo método de
sedimentação. O uso dessas duas últimas amostras, ainda que com pesquisa do
parasita por método menos sensível, foi escolhido porque eram de dois serviços
diferentes (em dois municípios distintos da Região Metropolitana de Vitória) o que
possibilitaria o aumento do número de casos e a validação das observações. Uma
amostra tomada pela revisão de prontuários do Programa de Atendimento ao
Alcoolista foi utilizada porque representava um grande número de alcoolistas que
poderiam ser comparados com um grande número de controles, atendidos no
mesmo Hospital. A análise da prevalência do S. stercoralis nos exames
parasitológicos realizados no HUCAM, no ano de 2003, foi feita no sentido de se
tentar verificar a prevalência de etilismo crônico em pessoas com o exame
parasitológico positivo para o helminto, ou seja, uma confirmação da associação
entre alcoolismo crônico e o parasita a partir da identificação do uso crônico do
etanol em pessoas com exame parasitológico positivo para S. stercoralis.
6.1.2- Análise crítica de cada amostra
A amostra com a pesquisa do parasita pelo método Baermann, realizada
em 2003, tentou ser pareada, especialmente pelo local de residência. A
dificuldade de se ter um controle do mesmo sexo, levou a um grande número de
controles do sexo feminino e fez com que o número de controles ficasse reduzido:

111 controles para 190 casos, relação próxima de 2 casos para 1 controle. Se
considerarmos a prevalência de 8,75% de S. stercoralis nos exames realizados
pelo método de Baermann, em amostra de 190 crianças do Hospital Infantil Nossa
Senhora da Glória em Vitória, com procedência muito semelhante a dos pacientes
atendidos no HUCAM (73% provenientes dos municípios da Região Metropolitana
de Vitória), e considerando que observamos prevalência média de S. stercoralis
nos alcoolistas próxima de 25%, o número ideal de pacientes para comparação
do tipo caso controle, na relação de 2 casos para 1 controle, com erro beta de
20% e erro alfa de 5%, seria de 223 casos e 112 controles. Na amostra
estudada, os controles estão em número adequado (111) e o número de casos é
ligeiramente inferior, mas não tão inferior a ponto de reduzir muito a força
estatística da associação observada. A diferença entre a proporção de homens e
mulheres e as diferenças nas médias de idade não são relevantes, já que a
prevalência do parasitismo pelo S. stercoralis parece não ser influenciada pela
idade em adultos, e que tal prevalência tende a se estabilizar entre a terceira e
quarta décadas da vida.
Quanto ao sexo, alguns autores relatam uma predominância da parasitose
no sexo masculino, relacionada possivelmente a hábitos e condições de vida que
aumentam o risco de contacto com as larvas infectantes (MILDER et al, 1981;
WALZER et al, 1982; DAVIDSON, FLETCHER e CHAPMAN, 1984;
VANNACHONE et al, 1988). Alguns modelos experimentais sugerem influência
dos hormônios sexuais na resistência aos helmintos. Em trabalho experimental em
S. ra

maturação sexual ela era semelhante mas aumentava nos machos e diminuía nas
fêmeas, na medida em que os níveis séricos dos hormônios sexuais aumentavam
(RIVERO et al, 2002). No entanto, WIDJANA e SUTISNA (2000) não se referem a
diferenças importantes na prevalência desta parasitose entre homens e mulheres.
Na amostra controle de não alcoolistas, obtida da revisão de prontuários no
Hospital Universitário, a prevalência de S. stercoralis em homens foi maior do que
em mulheres, mas a diferença não foi significativa. Como será discutido a seguir,
maior prevalência do parasita em homens pode estar associada ao fato de que o
alcoolismo crônico é significativamente mais prevalente em homens.
Das duas amostras obtidas no ano de 2003, onde o exame parasitológico
foi feito por sedimentação, a da Unidade Feu Rosa é pequenas em relação aos
casos, mas suficiente se considerarmos as prevalências de S. stercoralis
observadas. Por outro lado, têm controles bem pareados pela procedência. A
outra, do HUCAM é numericamente satisfatória e foi comparada com controles
atendidos no mesmo Hospital.
A amostra oriunda da revisão dos prontuários tem a vantagem do número
de casos e controles, tendo inclusive um número razoável de mulheres nos dois
grupos, o que permitiu comparações da prevalência do S. stercoralis em
alcoolistas dos dois sexos. O grupo controle dessa amostra é adequado porque
tem procedência semelhante e foi tomado no mesmo Hospital onde foram
atendidos os alcoolistas.
A análise dos resultados dos exames de fezes no Laboratório de Análises
Clínicas do HUCAM, durante um ano, permitiu a identificação de um bom número
de casos com resultados positivo para S. stercoralis. Embora não tenhamos
conseguido informações completas nos prontuários sobre o uso do etanol em
todos os casos, em 51 isso foi possível, número que nos permitiu algumas
considerações.
6.1.3- Análise crítica da utilização de amostras onde o exame parasitológico
de fezes foi realizado com métodos de diferentes sensibilidades para
identificar o S. stercoralis

O fato de terem sido utilizadas amostras nas quais a pesquisa do S.
stercoralis foi feita pelo método de sedimentação não invalida os resultados, tendo
em vista que para cada amostra o método utilizado foi o mesmo nos alcoolistas e
respectivos controles e não se tomaram inferências comparando amostras em que
o parasita nos casos (alcoolistas) e controles foram investigados com metodologia
diferente. Apenas na análise de regressão logística para avaliar o alcoolismo como
fator independente associado a um exame positivo para S. stercoralis foram
utilizados pacientes e controles que tiveram o diagnóstico feito ou pelo método de
Baermann ou pelo método de sedimentação.
No que se refere à sensibilidade dos métodos, confirmou-se o que é muito
conhecido: o método de Baermann é muito mais sensível do que um exame por
sedimentação, detectando de duas a três vezes mais casos positivos do parasita.
6.2- Prevalência do S. stercoralis nos alcoolistas
A prevalência de S. stercoralis nos alcoolistas crônicos em todas as
amostras estudadas foi significativamente maior do que nos controles,
independente do método utilizado para identificar o parasita.
Quando analisamos os grupos separados por sexo a prevalência mantém-
se significativa em todas as amostras, no sexo masculino. No entanto, nas
amostras coletadas no ano de 2003 a diferença não foi observada nas mulheres.
Isso se explica, pelo pequeno número de casos de mulheres alcoolistas. O
etilismo é muito mais prevalente no sexo masculino, e no PAA do HUCAM a
relação masculino/feminino é de 11:1 (MACIERA, ZAGO-GOMES e GARCIA,
1993). No entanto, quando analisamos a amostra obtida pelo levantamento de
prontuários do Programa de Atendimento ao Alcoolista, que tem um número maior
de mulheres, permitindo avaliação estatística mais segura, a prevalência do S.
stercoralis é significativamente maior nas mulheres alcoolistas do que no grupo
controle.
Podemos concluir, ainda que levando em consideração possíveis falhas de
amostragem, que a prevalência do S. stercoralis em alcoolistas de ambos os
sexos é significativamente maior do que em não alcoolistas, confirmando e

ampliando observações anteriores de ZAGO-GOMES et al (2000) e OLIVEIRA et
al (2002).
A análise da relação entre a quantidade de etanol ingerida diariamente e a
presença do exame de fezes positivo para S. stercoralis mostrou correlação
significativa. Isso demonstra que quanto maior a ingestão do etanol maior o risco
de se albergar o parasita, o que indica uma possível participação direta do etanol
nos mecanismos envolvidos na relação do parasita com o hospedeiro.
Os dados sobre a prevalência do etilismo crônico em pacientes com S.
stercoralis identificados nos exames parasitológicos de fezes no laboratório do
HUCAM, ainda que incompletos, confirmam que o etilismo crônico é importante
fator associado à existência de exame positivo para S. stercoralis. Se
considerarmos os 51 pacientes nos quais houve possibilidade de identificar com
segurança o diagnóstico de etilismo crônico, 41,17% eram alcoolistas. Como o
HUCAM atende grande número de alcoolistas, a maioria do sexo masculino, se
explica a significativa maior prevalência de exame positivo para S. stercoralis nos
homens em relação às mulheres, no laboratório de rotina daquele hospital. Esta
observação é reforçada pelo fato de que nos controles não etilistas, com exame
feito no mesmo laboratório, a diferença de prevalência de S. stercoralis entre os
dois sexos não é significativa, ainda que maior no sexo masculino: 4,8% (IC a
95%: 2,94 a 6,86%) nos homens e 3,30% (IC a 95%:1,4 a 6,20%) nas mulheres
(p=0,620).
Outro dado importante que mostra a relação entre ingestão crônica de
etanol e presença de exame parasitológico positivo para S. stercoralis é a
demonstração de que o uso crônico do etanol é fator independente, associado à
existência de exame positivo para S. stercoralis mesmo eliminando a influência de
co-fatores como idade, andar descalço, usar água tratada e ter esgoto sanitário.
Nas amostras estudadas, não observamos diferença significativa na
prevalência do S. stercoralis em pacientes etilistas com ou sem cirrose hepática.
Essa observação difere da relatada por OLIVEIRA et al (2002) que encontraram
prevalência significativamente maior em cirróticos, mas confirma a de ZAGO-
GOMES et al (2000) que também não encontraram tal diferença. É possível que a

observação de OLIVEIRA et al (2002) seja resultante do tamanho da amostra (45
casos de alcoolistas sendo 9 cirróticos alcoólicos e 10 pacientes com cirrose não
alcoólica, onde nenhum dos pacientes tiveram resultado positivo para o helminto).
Embora GABURRI et al (1997) tenham admitido que a cirrose seja fator de risco
para exame positivo para S. stercoralis, nos seus casos, a maioria dos cirróticos
eram alcoolistas crônicos.
De modo semelhante ao observado por ZAGO-GOMES et al (2000), não
houve diferença significativa na prevalência de outros nematóides, considerados
em conjunto.
6.3- Possíveis mecanismos envolvidos na relação entre uso crônico do
etanol e aumento da freqüência de exame parasitológico positivo para S.
stercoralis
Quanto aos possíveis mecanismos que possam relacionar o etilismo
crônico com a maior prevalência de S. stercoralis, há muita discussão. Primeiro é
necessário saber se (i) a prevalência é maior ou seja, o alcoolista tem maior
chance de se infectar ou se uma vez infectado, a sobrevivência do verme é
facilitada; (ii) se a carga parasitária é maior porque há facilitação da auto-
infecção ou se a fecundidade das fêmeas em produzir larvas é maior, o que
aumentaria o número de larvas nas fezes facilitando e aumentando o diagnóstico
de casos positivos.
Se a prevalência é realmente maior ela poderia ser independente de efeitos
do etanol nos mecanismos que regulam a relação parasita hospedeiro mas, seria
dependente de alterações do comportamento que favoreceriam uma maior
exposição ao parasita. De fato, os alcoolistas tendem a ter hábitos de higiene mais
precários, e nesse caso, a larva rabditóide presente na região perianal, de
indivíduos infectados transforma-se em larva filarióide infectante penetrando
através da pele dessa região (GROVE, 1996). Isso pode ser favorecido devido a
higiene deficiente acompanhando a evacuação. Por outro lado, os indivíduos
alcoolizados podem cair com freqüência, entrando em contacto com o solo, e às
vezes aí permanecer por maior tempo, ficando assim mais expostos às larvas

infectantes. Nas amostras aqui estudadas, não foi possível avaliar esses hábitos
de higiene. No entanto, quando analisamos alguns hábitos de higiene (andar
descalço, disponibilidade de esgoto e água tratada) como co-fatores em relação
ao etilismo, como fator associado à um exame parasitológico positivo para S.
stercoralis, embora a inexistência de rede de esgoto seja significante, a
significância para o uso do etanol persiste. Essa observação indica que os hábitos
de higiene avaliados não explicariam isoladamente a maior prevalência do exame
positivo para S. stercoralis nos alcoolistas.
Quanto ao uso abusivo do etanol poder alterar os mecanismos de
resistência à infecção, as possibilidades existem podendo ser múltiplas e
complexas.
Relata-se que o uso do etanol reduza a resposta Th1 e aumenta ou altera
pouco a resposta Th2 e a produção das citocinas IL-3, IL-4, IL-5, IL-6, IL-10 e IL-
13 (WALTENBAUGH, VASQUEZ, PETERSON, 1998; SZABO, 1999). A resposta
Th1 nas helmintíases parece importante nos mecanismos de indução da reação
inflamatória na mucosa envolvida nos mecanismos de expulsão, como
demonstram alguns trabalhos experimentais (NA et al, 1997). É possível portanto,
que a resposta Th2 esteja presente, mas os mecanismos efetuadores iniciais que
envolvem as duas respostas podem estar comprometidos prejudicando assim a
eliminação da infecção.
Por outro lado, pouco conhecemos sobre os efeitos do etanol nos
mecanismo inatos de defesa, importantes na montagem da resposta imunitária,
nos primeiros contatos com o parasita. Pode ser que o reconhecimento de
antígenos pelas células dendríticas e a ação de células NK possam estar
comprometidas, impedindo uma resposta TH2 adequada, favorecendo a infecção.
Pode ainda haver alteração dos mecanismos executores da resposta Th2
na mucosa, por efeitos crônicos do etanol sobre os mastócitos, por exemplo,
importantes células executoras da resistência aos helmintos intestinais e na
infecção com o S. stercoralis (FINKELMANN et al, 1994 e 1997; BARRET et al,
1988; NAWA et al, 1994).

Outra possibilidade seria uma alteração, induzida pelo etanol, na produção
de anticorpos, com mudança quantitativa na produção dos diferentes isotipos
necessários para a resistência. Os anticorpos das classes IgE , IgG4 e IgA anti-S.
stercoralis têm papel importante na resistência ao parasita: IgA é importante no
controle da fecundidade das fêmeas parasitas, IgE é importante na ativação dos
mastócitos e IgG4 atua como contra-reguladora da IgE (COSTA-CRUZ, 2000;
ATKINS et al, 1999).
Nas circunstâncias descritas nos quatro parágrafos anteriores pode haver
facilitação da infecção e da sobrevivência do parasita o que justificaria um real


CONCLUSÕES

7.1- A análise dos resultados permite concluir que:
1- É significativa a maior prevalência de um exame de fezes positivo para
S. stercoralis em alcoolistas crônicos de ambos os sexos.
2- Há uma correlação significativa entre a quantidade média diária ingerida
de etanol e a prevalência do exame parasitológico de fezes positivo
para S. stercoralis.
3- O uso crônico de etanol é fator significativamente associado, de modo
independente, a um exame de fezes positivo para S. stercoralis no sexo
masculino, devendo ser considerado um fator de risco importante para
se albergar o parasita.
4- Nas amostras estudadas as variáveis analisadas não permitiram
identificar se a maior freqüência do exame positivo está relacionada à
maior prevalência do parasita ou se a uma facilitação na produção e
eliminação de larvas, aumentando a chance de seu achado nas fezes.
5- Não houve diferença nos níveis de cortisol plasmático em alcoolistas
com ou sem S. stercoralis, o que demonstra ser pouco provável a
participação do eixo hipotálamo-hipófise-adrenal na gênese das
modificações imunitárias ou outros efeitos relacionados à facilitação da
infecção, ao aumento da carga parasitária e /ou à eliminação de larvas
nas fezes.
6- A prevalência significativa de exame positivo para S. stercoralis no
laboratório de rotina do HUCAM, em pacientes do sexo masculino foi
possivelmente relacionada com alta freqüência de pacientes alcoolistas
(a maioria do sexo masculino) atendidos no HUCAM.

REFERÊNCIAS BIBLIOGRÁFICAS

Adams HG, Colin J. “Infections in the alcoholic”. Med Clin North Am.
1984;28:179- 200.
Adedayo O, Grell G, Bellot P. “Hyperinfective strongyloidiasis in the medical
ward: review of 27 cases in 5 years”. South Med J. 2002;95(7):711-6.
Adenusi AA. “Cure by ivermectin of a chronic, persistent, intestinal
strongyloidosis”. Acta Trop. 1997;66(3):163-7.
Albonico M, Crompton DW, Savioli L. “Control strategies for human
intestinal nematode infections”. Adv Parasitol. 1999;42:277-341.
Aldo-Benson M. “Mechanisms of alcohol-induced suppression of B-cell
response”. Alcohol Clin Exp Res. 1989;13(4):469-75.
Arakaki T, Asato R, Ikeshiro T, Sakiyama K, Iwanaga M. “Is the prevalence
of HTLV-1 infection higher in Strongyloides carriers than in non-carriers?” Trop
Med Parasitol. 1992;43(3):199-200.
Arakaki T, Iwanaga M, Asato R, Ikeshiro T. “Age-related prevalence of
Strongyloides stercoralis infection in Okinawa, Japan”. Trop Geogr Med. 1992; 44
(4 ) 299-303.
Ashraf M, Gue C, Baddour L. “Case report: Strongyloidiasis refractary to
treatment with ivermectin”. Am J Med Sci. 1996;311(4):178-179.
Atkins NS, Conway DJ, Lindo JF, Bailey JW, Bundy DA. “L3 antigen-specific
antibody isotype responses in human strongyloidiasis: correlations with larval
output”. Parasite Immunol. 1999;21(10):517-26.
Avedaño L, Hernandez F, Jimenez F, Avila A, Castro D. “Strongyloides
stercoralis em pacientes alcohólicos”. Parasitol. Dia. 1999;23:1-6.
Badaró R, Carvalho EM, Santos RB, Gam AA, Genta RM. “Parasite-
specific humoral responses in different clinical forms of strongyloidiasis”. Trans R
Soc Trop Med Hyg. 1987;81(1):149-50.
Barrett KE, Neva FA, Gam AA, Cicmanec J, London WT, Phillips JM,
Metcalfe DD. “The immune response to nematode parasites: modulation of mast
cell numbers and function during Strongyloides stercoralis infections in nonhuman
primates”. Am J Trop Med Hyg. 1988;38(3):574-81.


Costa-Cruz JM, Bullamah CB, Goncalves-Pires Mdo R, Campos DM, Vieira
MA. “Cryo-microtome sections of coproculture larvae of Strongyloides stercoralis
and Strongyloides ratti as antigen sources for the immunodiagnosis of human
strongyloidiasis”. Rev Inst Med Trop Sao Paulo. 1997;39(6):313-7.
Costa-Cruz JM. “Strongyloides stercoralis”. In: Neves, Melo, Genaro,
Linardi. Parasitologia Humana. 10ed. Rio de Janeiro: Atheneu, 2000.p.247-256.
Costa-Cruz JM, Madalena J, Silva DA, Sopelete MC, Campos DM,
Taketomi EA. “Heterologous antigen extract in ELISA for the detection of human
IgE anti-Strongyloides stercoralis”. Rev Inst Med Trop Sao Paulo.
2003;45(5):265-8.
Coura JR. “Parasitoses nos Portadores de AIDS”. 1987. JBM; 53(40):42-53.
Cunha AS. “Parasitoses Intestinais”. In: Dani/Paula Castro.
Gastroenterologia Clínica. 2.ed. Rio de Janeiro: Guanabara Koogan S.A.,
1988.p.830-44.
Davidson RA, Fletcher RH e Chapman LE. “Risk factors for strongyloidiasis.
A case-control study”. Arch Intern Med. 1984;144:321-324.
Davidson RA. “Strongyloides stercoralis infection in the ferret”. 1988. J
Parasitol;74(1):177-79.
de Paola, Dias LB, da Silva J. “Enteritis due to Strongyloides stercoralis. A
report of 5 fatal cases”. Am J Dig Dis. 1962;7:1086-98.
de Paula FM, de Castro E, Goncalves-Pires M, Marcal M, Campos DM,
Costa-Cruz JM. “Parasitological and immunological diagnoses of strongyloidiasis
in immunocompromised and non-immunocompromised children at Uberlandia,
State of Minas Gerais, Brazil”. Rev Inst Med Trop Sao Paulo. 2000;42(1):51-5.
Dehne NE, Mendenhall CL, Roselle GA, Grossman CJ. “Cell-mediated
immune responses associated with short term alcohol intake: time course and
dose dependency”. Alcohol Clin Exp Res. 1989;13(2):201-5.
Dias RMDS, Mangini ACS, Torres DMAGV, Vellosa SAG, da Silva MIPG, da
Silva RM, Corrêa MOA, Coletti C. “Ocorrência de Strongyloides stercoralis em

pacientes portadores da síndrome de imunodeficiência adquirida (AIDS)”. 1992.
Inst Med trop São Paulo; 4(1):15-7.
Diaz LE, Montero A, Gonzalez-Gross M, Vallejo AI, Romeo J, Marcos A.
“Influence of alcohol consumption on immunological status: a review”. Eur J Clin
Nutr. 2002;56 Suppl 3:S50-3.
Dominguez-Santalla MJ, Vidal C, Vinuela J, Perez LF, Gonzalez-Quintela A.
“Increased serum IgE in alcoholics: relationship with Th1/Th2 cytokine production
by stimulated blood mononuclear cells”. Alcohol Clin Exp Res. 2001;25(8):1198-
205.
Feitosa G, Bandeira AC, Sampaio DP, Badaro R, Brites C. “High prevalence
of giardiasis and stronglyloidiasis among HIV-infected patients in Bahia, Brazil”.
Braz J Infect Dis. 2001;5(6):339-44.
Ferreira MS. Estrongiloidíase. In: Veronesi R, Focaccia R, Dietze R.
Doenças Infecciosas e parasitárias. 8ed. Rio de Janeiro: Guanabara Koogan,
1991. p.856-865.
Ferrioli Filho F. “Condições que influem na extração de larvas do
Strongyloides stercoralis das fezes pelo método de Loss-Baerman Modificado
(Técnica do Pires)”. Rev Inst Med Trop São Paulo. 1961; 3(2):51-60.
Finkelman FD, Madden KB, Cheever AW, Katona IM, Morris SC, Gately MK,
Hubbard BR, Gause WC, Urban JF Jr. “Effects of interleukin 12 on immune
responses and host protection in mice infected with intestinal nematode parasites”.
J Exp Med. 1994 May 1;179(5):1563-72.
Finkelman FD, Shea-Donohue T, Goldhill J, Sullivan CA, Morris SC,
Madden KB, Gause WC, Urban JF Jr. ‘Cytokine regulation of host defense against
parasitic gastrointestinal nematodes: lessons from studies with rodent models”.
Annu Rev Immunol. 1997;15:505-33.
Gabbai AA, Bordin JO, Vieira-Filho JP, Kuroda A, Oliveira AS, Cruz MV,
Ribeiro AA, Delaney SR, Henrard DR, Rosario J, et al. “Selectivity of human T
lymphotropic virus type-1 (HTLV-1) and HTLV-2 infection among different
populations in Brazil”. Am J Trop Med Hyg. 1993 Dec;49(6):664-71.

Gaburri D, Gaburri AK, Hubner E, Lopes MH, Ribeiro AM, de Paulo GA,
Pace FH, Gaburri PD, Ornellas AT, Ferreira JO, Chebli JM, Ferreira LE, de Souza
AF. “Intestinal parasitosis and hepatic cirrhosis”. Arq Gastroenterol. 1997;34(1):7-
12.
Galvão CB, Loures L, Rodrigues LGM. “Geographic distribuition of human
T-lymphotropic virus type-I among blood donors: a Brazilian nationwide study”.
Transfusion. 1997; 37:242.
Gam AA, Neva FA, Krotoski WA. “Comparative sensitivity and specificity os
ELISA and IHA for serodiagnosis of strongyloidiasis with larval antigens”. Am J
Trop Med Hyg. 1987; 37(1):157-61.
Genta RM, Ottesen EA, Neva FA, Walzer PD, Tanowitz HB, Wittner M.
“Cellular responses in human strongyloidiasis”. Am J Trop Med Hyg.
1983;32(5):990-4.
Genta RM, Harper JS 3rd, Gam AA, London WI, Neva FA. “Experimental
disseminated strongyloidiasis in Erythrocebus patas. II”. Immunology. Am J Trop
Med Hyg. 1984;33(3):444-50.
Genta RM. “Immunobiology of strongyloidiasis”. Trop Geogr Med.
1984;36(3):223-9.
Genta RM, Schad GA, Hellman ME. “Strongyloides stercoralis:
parasitological, immunological and pathological observations in
immunosuppressed dogs”. Trans R Soc Trop Med Hyg. 1986;80(1):34-41.
Genta RM. “Strongyloides stercoralis: Immunobiological considerations on
an unusual worm”. Parasitol Today. 1986;2(9):241-6.
Genta RM. “Strongyloidiasis”. Baillière’s Clin Trop Med Communicable
Dis. 1987;2(3):645-665.
Genta RM. “Global prevalence of strongyloidiasis: critical review with
epidemiologic insights into the prevention of disseminated disease”. Rev Infect
Dis. 1989; 11(5):755-67.
Genta RM. “Dysregulation of strongyloidiasis: a new hypothesis”. Clin
Microbiol Rev. 1992;5(4):345-55.

Gill GV, Bell DR: “Strongyloides stercoralis infection in former Far East
prisoners of war”. Br Med J [Clin Res] 1979;279: 572-574.
Gluckman SJ, Dvorak VC, MacGregor RR. “Host defenses during prolonged
alcohol consumption in a controlled environment”. Arch Intern Med.
1977;137:1539-1543.
Gonzalez-Quintela A, Vidal C, Lojo S, Perez LF, Otero-Anton E, Gude F,
Barrio E. “Serum cytokines and increased total serum IgE in alcoholics”. Ann
Allergy Asthma Immunol. 1999;83(1):61-7.
Gonzalez-Gross M, Lebron M & Marcos A (2000): ”Revisión bibliográfica
sobre los efectos del consumo moderado de cerveza sobre la salud”.
Gotuzzo E, Terashima A, Alvarez H, Tello R, Infante R, Watts DM,
Freedman DO. “Strongyloides stercoralis hyperinfection associated with human T
cell lymphotropic virus type-1 infection in Peru”. Am J Trop Med Hyg.
1999;60(1):146-9.
Grove DI. “Strongyloidiasis in allied ex-prisioners of war in southeast Asia”.
Br Med J. 1980; 280:598-601.
Grove DI, Heenan PJ, Northern C. “Persistent and disseminated infections
with Strongyloides stercoralis in immunosupressed dogs”. Int J Parasitol. 1983;
13(5): 483-90.
Grove DI. Diagnosis. In: Grove, DI,ed. “Strongyloidiais: a major roundworm
infection in ma”n, London: Taylor & Francis,1989.p.175-97.
Grove DI. “Strongyloidiasis: a conundrum for gastroenterologists”. Gut.
1994; 35(4):437-40.
Grove DI. “Human strongyloidiasis”. Adv Parasitol. 1996;38:251-309.
Harper JS, Genta RM, Gam A, London WT, Neva FA. “Experimental
disseminated strongyloidiasis in Erythrocebus patas”. AM J Trop Med Hyg. 1984;
33(3):431-43.
Harvey SC, Gemmill AW, Read AF, Viney ME. “The control of morph
development in the parasitic nematode Strongyloides ratti”. Proc R Soc Lond B
Biol Sci. 2000;267(1457):2057-63.

Kaminsky RG. “Evaluation of three methods for laboratory dianosis of
Strongyloides stercoralis infection”. J Parasitol. 1993; 79(2):277-80.
Kerlin RL, Nolan TJ, Schad GA. “Strongyloides stercoralis: histopathology of
uncomplicated and hyperinfective strongyloidiasis in the Mongolian gerbil, a rodent
model for human strongyloidiasis [corrected]”. Int J Parasitol. 1995;25(4):411-20.
Erratum in: Int J Parasitol 1995 ;25(8):1007.
Keiser PB, Nutman TB. “Strongyloides stercoralis in the
Immunocompromised Population”. Clin Microbiol Rev. 2004;17(1):208-17.
Kim AC, Lupatkin HC. “Strongyloides stercoralis infection as a manifestation
of immune restoration syndrome”. Clin Infect Dis. 2004.1;39(3):439-40.
Igra-Siegman Y, Kapila R, Sen P, Kaminski ZC, Louria DB. “Syndrome of
hyperinfection with Strongyloides stercoralis”. Rev Infect Dis. 1981 ;3(3):397-407.
Lansoud-Soukate J, Gharib B, Baswaid S, Capron A, de Reggi M.
“Ecdysteroid-like compounds in the serum and urine of African patients infected
with Loa loa and Mansonella perstans microfilariae”. Trans R Soc Trop Med Hyg.
1990;84(2):269-71.
Leighton PM, MacSween HM. “Strongyloides stercoralis. The cause of an
urticarial-like eruption of 65 years' duration”. Arch Intern Med. 1990;150(8):1747-
8.
Liu YK. “Effects of alcohol on granylocytes and lymphocytes”. Semin
Hematol. 1980:17:130-136.
Liu LX and Weller PF. “Helmintíases”. In: HARRISON. Medicina Interna.
14.ed.Rio de Janeiro: Mc Graw Hill, 1988.p.1289-1296.
Liu LX and Weller PF. “Strongyloidiasis and other intestinal nematode
infections”. Infect Dis Clin North Am. 1993;7(3):655-683.
MacGregor RR, Gluckman SJ, Senior JR. “Granulocyte function and levels
of immunoglobulins and complement in patients admitted for withdrawal from
alcohol”. J Infect Dis. 1978;138(6):747-55
MacGregor RR. “Alcohol and immune defense”. JAMA. 1986; 256(11):1474-
9.

Machado ER, Costa-Cruz JM. “Strongyloides stercoralis and other
enteroparasites in children at Uberlandia city, state of Minas Gerais, Brazil”. Mem
Inst Oswaldo Cruz. 1998;93(2):161-4.
Machado ER, Ueta MT, Goncalves-Pires M do R, de Oliveira JB, Faccioli
LH, Costa-Cruz JM. “Diagnosis of human strongyloidiasis using particulate antigen
of two strains of Strongyloides venezuelensis in indirect immunofluorescence
antibody test”. Exp Parasitol. 2001;99(1):52-5.
Machado ER, Ueta MT, de Fatima Goncalves-Pires Mdo R, Alves de
Oliveira JB, Faccioli LH, Costa-Cruz JM. “Strongyloides venezuelensis alkaline
extract for the diagnosis of human strongyloidiasis by enzyme-linked
immunosorbent assay”. Mem Inst Oswaldo Cruz. 2003;98(6):849-51.
Macieira MS, Zago-Gomes MP, Garcia MLT. “Programa de atendimento ao
alcoolista do Hospital Universitário Cassiano Antônio de Moraes da Universidade
Federal do Espírito Santo (PAA-HUCAM-UFES)”. J Bras Psiq. 1993;42(2):97-109.
McKerrow JH, Brindley P, Brown M, Gam AA, Staunton C, Neva FA.
“Strongyloides stercoralis: identification of a protease that facilitates penetration of
skin by the infective larvae”. Exp Parasitol. 1990;70(2):134-43.
Mansfield LS, Schad GA. “Strongyloides stercoralis infection in IgA-deficient
dogs”. Am J Trop Med Hyg. 1992;47(6):830-6.
Mansfield LS, Schad GA. “Lack of transmammary transmission of
Strongyloides stercoralis from a preciously hyperinfected bitch to her pups”. J
Helminthol. Soc. Wash. 1995; 62(1):80-3.
Marsh BJ. “Infectious complications of human T cell leukemia/lymphoma
virus type I infection”. Clin Infect Dis. 1996;23(1):138-45.
Matsumoto T, Miike T, Mizoguchi K, Yamaguchi K, Takatsuki K, Hosoda M,
Kawabe T, Yodoi J. “Decreased serum levels of IgE and IgE-binding factors in
individuals infected with HTLV-I”. Clin Exp Immunol. 1990;81(2):207-11.
Mendenhall CL, Theus SA, Roselle GA, Grossman CJ, Rouster SD.
”Biphasic in vivo immune function after low- versus high-dose alcohol
consumption”. Alcohol. 1997;14(3):255-60.
Milder JE, Walzer PD, Kilgore G, Rutherford I, Klein M. Clinical features of

Strongyloides stercoralis infection in an endemic area of the United States.
Gastroenterology. 1981 ;80(6):1481-8.
Moreira-Silva, Sandra Fagundes. “Nematóides intestinais, infecção por
Toxocara e infecção estafilocóccica em crianças: uma possível associação”. 1998.
116f. Dissertação (Mestrado em Doenças Infecciosas)- Núcleo de Doenças
Infecciosas, Universidade Federal do Espírito Santo, Vitória, 1998.
Na HR, Zhu X, Stewart GL, Seelig LL Jr. “Ethanol consumption suppresses
cell-mediated inflammatory responses and increases T-helper type 2 cytokine
secretion in Trichinella spiralis-infected rats”. Alcoholism: Clinical and
Experimental Research. 1997; 21(7):1179-85.
Nakada K, Kohakura M, Komoda H, Hinuma Y. “High incidence of HTLV
antibody in carriers of Strongyloides stercoralis”. Lancet. 1984;17;1(8377):633.
Nakada K, Yamaguchi K, Furugen S, Nakasone T, Nakasone K, Oshiro Y,
Kohakura M, Hinuma Y, Seiki M, Yoshida M, et al. “Monoclonal integration of
HTLV-I proviral DNA in patients with strongyloidiasis”. Int J Cancer.
1987;40(2):145-8.
Nawa Y, Ishikawa N, Tsuchiya K, Horii Y, Abe T, Khan AI, Bing-Shi, Itoh H,
Ide H, Uchiyama F. “Selective effector mechanisms for the expulsion of intestinal
helminths”. Parasite Immunol. 1994;16(7):333-8.
Neva FA. “Biology and immunology of human strongyloidiasis”. J Infect
Dis. 1986; 153(3):397-406.
Neva FA. “Letter to the editor: a reply to Storngyloides hyperinfection in
patients coinfected with HTLV-a and S. stercoralis”. Am J Med. 1993;94:448-9.
Neva FA, Filho JO, Gam AA, Thompson R, Freitas V, Melo A, Carvalho EM.
“Interferon-gamma and interleukin-4 responses in relation to serum IgE levels in
persons infected with human T lymphotropic virus type I and Strongyloides
stercoralis”. J Infect Dis. 1998;178(6):1856-9.
Neva FA, Gam AA, Maxwell C, Pelletier LL. “Skin test antigens for
immediate hypersensitivity prepared from infective larvae of Strongyloides
stercoralis”. Am J Trop Med Hyg. 2001;65(5):567-72.

Newton RC, Limpuangthip P, Greenberg S, Gam A, Neva FA.
“Strongyloides stercoralis hyperinfection in a carrier of HTLV-I virus with evidence
of selective immunosuppression”. Am J Med. 1992;92(2):202-8.
Nielsen PB, Mojon M. “Improved diagnosis of strongyloides stercoralis by
seven consecutive stool specimens”. Zentralbl Bakteriol Mikrobiol Hyg [A].
1987; 263(4):616-8.
Nolan TJ, Megyeri Z, Bhopale VM, Schd GA. “Strongyloides stercoralis: the
first rodent model for uncomplicated and hyperinfective strongyloidiasis, the
Mongolian Gerbil (Meriones unguiculatus)”. J Infec Dis. 1993; 168:1479-84.
Nolan TJ, Rotman HL, Bhopale VM, Schad GA, Abraham D. “Immunity to a
challenge infection of Strongyloides stercoralis third-stage larvae in the jird”.
Parasite Immunol. 1995;17(11):599-604.
Nolan TJ, Genta RM e Schad GA. “Strongyloidosis”. In:Palmer SR.
Zoonoses. Biology, Clinical Practices, and Public Health Control. Oxford:
Oxford University Press, 1998.p.747-56.
Nolan TJ, Bhopale VM, Schad GA. “Hyperinfective strongyloidiasis:
Strongyloides stercoralis undergoes an autoinfective burst in neonatal gerbils”. J
Parasitol. 1999;85(2):286-9.
Ogilvie K, Lee Soon, Weiss Burt, Rivier Catherine. “Mechanisms mediating
the influence of alcohol on the hypothalamic-pituitary-adrenal axis responses to
immune and nonimmune signals”. Alcohol Clin Exp Res.1998; 22(5):243S-247S.
Oliveira LCM, Ribeiro CT, Mendes DM, Oliveira TC, Costa-Cruz JM.
“Frequency of Strongyloides stercoralis infection in alcoholics”. Mem Inst Oswaldo
Cruz. 2002;97(1):119-21.
Parana R, Portugal M, Vitvitski L, Cotrim H, Lyra L, Trepo C. “Severe
strongyloidiasis during interferon plus ribavirin therapy for chronic HCV infection”.
Eur J Gastroenterol Hepatol. 2000;12(2):245-6.
Pelletier LL Jr, Baker CB, Gam AA, Nutman TB, Neva FA. “Diagnosis and
evaluation of treatment of chronic strongyloidiasis in ex-prisoners of war”. J Infect
Dis. 1988;157(3):573-6.

Pessoa, SB. “Superfamília Rhabdiasoidea-Strongyloides stercoralis e
estrongiloidose”. In: Pessoa. Parasitologia Médica. 10.ed. Rio de Janeiro:
Guanabara Koogan, 1978. cap. 40, p. 569-80.
Petithory JC, Derouin F. “AIDS and strongyloidiasis in Africa”. Lancet. 1987
18;1(8538):921.
Porto AF, Neva FA, Bittencourt H, Lisboa W, Thompson R, Alcantara L,
Carvalho EM. “HTLV-1 decreases Th2 type of immune response in patients with
strongyloidiasis”. Parasite Immunol. 2001;23(9):503-7.
Porto AF, Oliveira Filho J, Neva FA, Orge G, Alcantara L, Gam A, Carvalho
EM. “Influence of human T-cell lymphocytotropic virus type 1 infection on serologic
and skin tests for strongyloidiasis”. Am J Trop Med Hyg. 2001;65(5):610-3.
Porto MA, Muniz A, Oliveira Junior J, Carvalho EM. “Clinical and
immunological consequences of the association between HTLV-1 and
strongyloidiasis”.

Roselle GA, Mendenhall CL, Chedid A, Moritz E, Gartside P. “Alcohol
modulation of immune function: clinical and experimental data. Veterans Affairs
Cooperative Study Groups 119 and 275”. Alcohol Clin Exp Res. 1995;19(3):551-
4.
Rossi CL, Takahashi EEH, Partel CD, Teodoro LGVL, Silva LJ. “Total serum
IgE and parasite-specific IgG and IgA antibodies in human strongyloidiasis”. Rev
Inst Med Trop São Paulo. 1993;35(4):361-65.
Salazar SA, Berk SH, Howe D, Berk Sl. “Ivermectin vs thiabendazole in the
treatment of strongyloidiasis”. Infect Med. 1994;50-59.
Sato Y, Shiroma Y, Kiyuna S, Toma H, Kobayashi J. “Reduced efficacy of
chemotherapy might accumulate concurrent HTLV-1 infection among
strongyloidiasis patients in Okinawa, Japan”. Trans R Soc Trop Med Hyg.
1994;88(1):59.
Sato Y, Kobayashi J, Shiroma Y. “Serodiagnosis of strongyloidiasis. The
application and significance”. Rev Inst Med Trop Sao Paulo. 1995;37(1):35-41.
Sato Y, Kobayashi J, Toma H, Shiroma Y. “Efficacy of stool examination for
detection of Strongyloides infection”. Am J Trop Med Hyg. 1995;53(3):248-50.
Schad GA, Hellman ME, Muncey DW. “Strongyloides stercoralis:
hyperinfection in immunosuppressed dogs”. Exp Parasitol. 1984;57(3):287-96.
Schad GA. “Cyclosporine may eliminate the threat of overwhelming
storngyloidiais in immunosuppressed patients”. J Infect Dis. 1986;57(3):287-96.
Schad GA, Aikens LM, Smith G. “Strongyloides stercoralis: is there a
canonical migratory route through the host?” J Parasitol. 1989;75(5):740-9.
Schad GA, Smith G, Megyeri Z, Bhopale VM, Niamatali S, Maze R.
“Strongyloides stercoralis: an initial autoinfective burst amplifies primary infection”.
Am J Trop Med Hyg. 1993;48:716-25.
Shiwaku K, Chigusa Y, kadosaka T, Kaneto K. “Factors influencing
development of free-living generations os Strongyloides stercoralis”. Parasitology.
1988;97:129-38.
Siddiqui AA, Berk SL. “Diagnosis of Strongyloides stercoralis infection”. Clin
Infect Dis. 2001;33(7):1040-7.

Silva LP, Barcelos IS, Passos-Lima AB, Espindola FS, Campos DM, Costa-
Cruz JM. “Western blotting using Strongyloides ratti antigen for the detection of IgG
antibodies as confirmatory test in human strongyloidiasis”. Mem Inst Oswaldo
Cruz. 2003;98(5):687-91.
Steven WM, Kumar SN, Stewart GL, Seelig LL Jr. “The effects of ethanol
consumption on the expression of immunity to Trichinella spiralis in rats”. Alcohol
Clin Exp Res. 1990;14(1):87-91.
Szabo G. “Consequences of alcohol consumption on host defence”.
Alcohol Alcohol. 1999;34(6):830-41.
Takafuji S, Tadokoro K, Ito K, Dahinden CA. “Degranulation from human
eosinophils stimulated with C3a and C5a. Int Arch Allergy Immunol”. 1994;104
Suppl 1(1):27-9.
Terry SI, Blattner W, Neva FA, Murphy CP, Murphy EL, Golden MHN,
Bundy DAP, Hanchard B, Robinson R. “Coincidental HTLV-1 infection influences
outcome of treatment of strongyloidiasis”. West Indian Med J. 1989:38(suppl
1):36.
Terashima A, Alvarez H, Tello R, Infante R, Freedman DO, Gotuzzo E.
“Treatment failure in intestinal strongyloidiasis: an indicator of HTLV-I infection”.
Int J Infect Dis. 2002;6(1):28-30.
Toma H, Sato Y, Shiroma Y, Kobayashi J, Shimabukuro I, Takara M.
“Comparative studies on the efficacy of three anthelminthics on treatment of
human strongyloidiasis in Okinawa, Japan”. Southeast Asian J Trop Med Public
Health. 2000;31(1):147-51.
Trione N, Corti M, Castello T, Abuin JC, Bellegarde E. “Disseminated
infection due to strongyloides stercoralis in AIDS patients. A report of 2 cases”.
Acta Gastroenterol Latinoam. 2001;31(5):399-402.
van der Feltz M, Slee PHThJ, van Hees PAM, Tersmette M. “Strongyloides
stercoralis: how to diagnose best?” The Netherlands J of Medicine. 1999;
55:128-31.

Vannachone B, Kobayashi J, Nambanya S, Manivong K, Inthakone S, Sato
Y. An epidemiological survey on intestinal parasite infection in Khammouane
Province, Lao PDR, with special reference to Strongyloides infection. Southeast
Asian J Trop Med Public Health. 1998;29(4):717-22.
Vieira ARM, Carvalho FG, Pagani MB, Gonçalves AJ. “Infecção por
múltiplos microorganismos oportunistas em hospedeiro com imunodeficiência
adquirida. Relato de um caso fatal”. 1985. Arq Bras Med, 59(2):55-60.
Viney ME, Brown M, Omoding NE, Bailey JW, Gardner MP, Roberts E,
Morgan D, Elliott AM, Whitworth JA. “Why does HIV infection not lead to
disseminated strongyloidiasis?” J Infect Dis. 2004;15;190(12):2175-80.
Waltenbaugh C, Vasquez K, Peterson JD. “Alcohol consumption alters
antigen-specific Th1 responses: mechanisms of deficit and repair”. Alcohol Clin
Exp Res. 1998;22(5 Suppl):220S-223S.

Zago-Gomes MP, Aikawa KF, Perazzio SF, Goncalves CS, Pereira FE.
“Prevalence of intestinal nematodes in alcoholic patients”. Rev Soc Bras Med
Trop. 2002;35(6):571-4.

ANEXO I
TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO Venho por meio deste solicitar a permissão para usar os dados contidos no seu prontuário do Hospital Universitário Cassiano Antônio Moraes nas consultas realizadas no ambulatório de atendimento ao alcoolista e ambulatório de fígado , no trabalho denominado “ Strongyloides stercoralis e Alcoolismo”. Todas as informações obtidas no seu prontuário serão analisadas em conjunto com aquelas obtidas com outros pacientes, resguardando, desta forma, a confidencialidade, mesmo se a pesquisa vier a ser publicada em revista científica. Cada participante deste projeto deverá fazer três amostras de exame parasitológico das fezes, que será realizado no laboratório do Núcleo de Doenças Infecciosas ou no Laboratório de Análises Clínicas do HUCAM. Sua participação é isenta de despesas. Você tem o direito de recusar participar deste estudo, sem penalidades ou prejuízos no seu atendimento neste Serviço. ___________________________________ Assinatura do paciente/representante legal ____________________________________ Assinatura da testemunha Declaro que obtive de forma apropriada e voluntária o Consentimento Livre e Esclarecido deste paciente ou representante legal para participação neste estudo. _________________________________ Dra Carla Couzi Marques HUCAM ( Casa 3 – Núcleo de Doenças Infecciosas- 33357319 / 33357210 )

ANEXO II
Strongyloides stercoralis Nome: Idade: Cor: Sexo: ( ) M ( ) F Prontuário: Estado conjugal: Endereço: Telefone: Rede de esgoto: ( ) S ( ) N Água encanada: ( ) S ( ) N Ocupação ( ofício exercido ): Andar descalço: ( ) S ( ) N Grau de instrução: Uso etanol: ( ) sim ( ) não Cortisol: Tempo de uso:................................ Quantidade estimada:............................ Data da última ingestão:....................... EPF: S. stercoralis ( ) sim ( ) não Outros:............................................. Hemograma: FA: GGT: TGO: TGP: HbsAg: Anti-VHC: Albumina: Cirrose: ( ) sim ( ) não Etiologia: ( ) álcool ( ) B ( ) C ( ) outros

ANEXO III
Tabela 1- Prevalência de S. stercoralis em pacientes alcoolistas crônicos atendidos no Programa
de Atendimento ao Alcoolista do HUCAM e controles não alcoolistas, nos quais os exames
parasitológicos das fezes foram feitos pelo método de Baermann, no ano de 2003.
Variáveis Alcoolistas
(N=190)
Controles
(N= 111)
OR* (IC 95%) p**
Todos os casos
S. stercoralis
Positivo
Negativo
Outros nematóides
Positivo
Negativo
Sexo masculino
S. stercoralis
Positivo
Negativo
Outros nematóides
Positivo
Negativo
Sexo feminino
S. stercoralis
Positivo
Negativo
Outros nematóides
Positivo
Negativo
48 (25,2%)
142 (74,7%)
22 (11,5%)
168 (88,4%)
45 (26,3%)
126 (73,6%)
20 (11,6%)
151 (88,3%)
3 (15,7%)
16 (84,2%)
2 (10,5%)
17 (89,4%)
10 (9%)
101 (90,9%)
13 (11,7%)
98 (88,2%)
4 (8%)
46 (92%)
6 (12%)
44 (88%)
6 (9,8%)
55 (90,1%)
7 (11,4%)
54 (88,5%)
3,41 (1,58-7,57)
0,99 (0,40-1,49)
4,11 (1,32-14,2)
0,91 (0,34-2,89)
1,72 (0,50-9,11)
0,91 (0,72-5,14)
0,000
N.S.
0,010
N.S.
N.S.
N.S.
*OR (razão de chances); **χ² e Teste de Fisher quando n<5; N.S. (não significativa)

ANEXO IV
Tabela 2- Prevalência de S. stercoralis em 156 pacientes alcoolistas atendidos no Programa de
Atendimento ao Alcoolista do HUCAM e nos controles, nos quais o exame parasitológico de fezes
foi feito pelo método de sedimentação.
Variáveis Álcool (N=156) Controle
(N=591)
OR*(IC 95%) p**
Todos os casos
S.stercoralis
Positivo
Negativo
Outros nematóides
Positivo
Negativo
Sexo masculino
S. stercoralis
Positivo
Negativo
Outros nematóides
Positivo
Negativo
Sexo feminino
S. stercoralis
Positivo
Negativo
Outros nematóides
Positivo
Negativo
27 (17,3%)
129 (82,6%)
7 (4,4%)
149 (95,5%)
27 (19,1%)
114 (80,8%)
6 (4,2%)
135 (95,7%)
0 (0%)
15 (100%)
1 (6,6%)
14 (93,3%)
26 (4,3%)
565 (95,6%)
42 (7,1%)
549 (92,8%)
22 (4,7%)
443 (95,2%)
31 (6,6%)
434 (93,3%)
4 (3,2%)
121 (96,8%)
11 (8,8%)
114 (91,2%)
4,55 (2,48 – 8,36)
0,95 (0,36 – 2,39)
4,21 (2,23 –7,97)
0,62 (0,28 – 1,60)
N.D.
N.D.
0,000
N.S.
0,000
N.S.
N.S.
N.S.
*OR (razão de chances); **χ² e Teste de Fisher quando n<5; N.S. (não significativa); N.D. (não
determinada)


ANEXO VI
Tabela 4- Prevalência de S. stercoralis em pacientes alcoolistas crônicos e controles atendidos na
Unidade de Saúde Feu Rosa (Município de Serra), nos quais o exame parasitológico foi feito pelo
método de sedimentação.
Variáveis Alcoolistas
(N=49)
Controle (N=129) OR* (IC 95%) p**
Todos os casos
S. stercoralis
Positivo
Negativo
Outros nematóides
Positivo
Negativo
Sexo masculino
S. stercoralis
Positivo
Negativo
Outros nematóides
Positivo
Negativo
Sexo feminino
S. stercoralis
Positivo
Negativo
Outros nematóides
Positivo
Negativo
6 (12,2%)
43 (87,7%)
6 (12,2%)
43 (87,7%)
6 (13%)
40 (86,9%)
6 (13%)
40 (86,9%)
0 (0%)
3 (100%)
0 (0%)
3 (100%)
0 (0%)
129 (100%)
10 (7,7%)
119 (92,2%)
0 (0%)
98 (100%)
7 (7,1%)
91 (92,8%)
0 (0%)
31 (100%)
3 (9,6%)
28 (90,3%)
indefinido
1,66 (0,50- 5,36)
indefinido
1,95 (0,54-7,01)
indefinido
indefinido
0,001
N.S.
0,001
N.S.
N.S.
N.S.
*OR (razão de chances); **χ² e Teste de Fisher quando n<5; N.S.(não significativa)

ANEXO VII
Tabela 5- Relação entre a quantidade de etanol ingerida e a prevalência do exame de fezes
positivo para S. stercoralis. Dados tomados de 548 alcoolistas crônicos nos quais o exame
parasitológico foi feito pelo método de sedimentação.
Quantidade média diária
de etanol ingerida (g)
Strongyloides stercoralis
Positivo Negativo
(IC a 95%)
48-150
151 – 300
301 – 450
451 – 600
>600
Todos
17
30
24
11
8
90
124
197
104
25
8
458
11,9 (6,6-17,2)
13,2 (8,7-16,6)
22,6 (15,3-29,8)
30,5 (15,6-45,7)
50 (26,4-70,4)
19,6 (16,3-22,9)
Freqüência de S. stercoralis em 591 controles, não alcoolistas: 4,3%(IC 95%:2,7 –6,1)

ANEXO VIII
Tabela 6- Prevalência de exame positivo para S. stercoralis em 385 alcoolistas crônicos sem
cirrose e em 76 com cirrose hepática
Grupos S. stercoralis
Positivo Negativo
p
Alcoolistas sem cirrose (N=385)
Alcoolistas com cirrose (N=76)
Total = 461
76 (19,7%)
9 (11,8%)
309 (80,2%)
67 (88,1%)
N.S.
N.S. (não significativa)

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo