






Línguas
Páginas
Legal


1
FACULDADES METROPOLITANAS UNIDAS -
UniFMU
MEDICINA VETERINÁRIA
AMANDA OLIVOTTI FERREIRA
ESTUDO DAS MEMBRANAS FETAIS E DA
MORFOLOGIA DO SISTEMA REPRODUTOR
FEMININO DO VEADO-CATINGUEIRO ( MAZAMA
GOUAZOUBIRA) – RELATO DE CASO
SÃO PAULO
2009

2
FACULDADES METROPOLITANAS UNIDAS - UniFMU
MEDICINA VETERINÁRIA
AMANDA OLIVOTTI FERREIRA
Trabalho de Conclusão de Curso
apresentado como exigência parcial para obtenção do título de Médica Veterinária à
Banca Examinadora, sob a orientação do Prof. Dr. João Carlos Borges e supervisor
do Dr. Ricardo Romão Guerra.
São Paulo, 2009

3
AMANDA OLIVOTTI FERREIRA
ESTUDO DAS MEMBRANAS FETAIS E DA MORFOLOGIA DO SISTEMA
REPRODUTOR FEMININO DO VEADO-CATINGUEIRO (MAZAMA
GOUAZOUBIRA) – RELATO DE CASO
Trabalho de Conclusão de Curso apresentado como exigência parcial para obtenção do título de Bacharel junto à Comissão Julgadora da Faculdade Metropolitanas Unidas -UniFMU . Data da aprovação: ___/____/____. Orientador: Prof. Dr. João Carlos Borges ____________________________ Supervisor : Dr. Ricardo Romão Guerra ____________________________ Prof. Dr. Rodolfo Nurmberger Júnior ____________________________

4
RESUMO
O Mazama gouazoubira é uma espécie de cervídeo de porte pequeno, que pode ser
encontrado abundantemente na América do Sul, desde o sul do Uruguai até o norte de
Mato Grosso, no Brasil. Este estudo teve como objetivo descrever as membranas fetais
e o sistema reprodutor feminino dos veados no terço inicial de gestação. Foram
analisadas macroscopicamente e microscopicamente as amostras coletadas. O animal
estudado apresentou uma gestação univitelínea com um embrião medindo 1,3 cm de
CR. No embrião pode ser observado pulmão, coração, fígado, e já apresentava o
crescimento de membros. A placenta apresentou-se oligocotiledonária e o saco
gestacional de uma extremidade a outra media 15 cm, ao abri-lo verificamos a ausência
de placentônios, bem como área de implantação do embrião. A membrana
corioalantoidiana ao microscópio possuía o alantóide bem vascularizado com uma fina
camada de células de núcleo e citoplasma alongados. Na outra face da membrana
visualizamos a tentativa de arranjo de células redondas, de citoplasma escasso e núcleo
grande e arredondado, características de células trofoblásticas. A ausência de
cotilédones foi confirmada na microscopia, sendo a carúncula desprovida de tecido fetal
por entre seus vilos. Envolvendo o embrião visualizou-se o saco amniótico constituído
de duas camadas passíves de separação mecânica, mas que apresentaram a mesma
morfologia macro e microscopicamente, sendo constituídas de células fusiformes, sendo
avascularizada. Não foi observado saco vitelino nesse espécime. O sistema reprodutor
feminino apresentou duas tubas uterinas com 4 cm. O corno úterino era bicornual como
em outros ruminantes. Dentro do útero foi achado 9 carúnculas, este número era
pequeno quando comparado com bovino. A cérvix apresentou-se formada por tecido
conjuntivo frouxo a tecido conjuntivo denso com vilosidades longitudinais como as dos
ruminantes. Concluí-se que o sistema de reprodutor do veado-catingueiro é similar ao
dos outros ruminantes, apresentando algumas diferenças nas membranas fetais, tais
como a falta de saco vitelino, sendo sua placenta classificada como oligocotiledonária
diferentemente dos outros animais ruminantes que possuem placenta cotiledonaria.
Palavras-chave: veado-catingueiro, sistema reprodutor feminino, Mazama
gouazoubira.

5
ABSTRACT
The Mazama gouazoubira is a species of deer from small size, which can be found
abundantly in South America, from the south of Uruguay to the north of Mato Grosso,
Brazil. This study aimed to describe the fetal membranes and reproductive system of
female deer in the initial third of gestation. Were analyzed macroscopically and
microscopically the samples collected. The animal study showed a univitelínea
pregnancy with an embryo measuring 1.3 cm in CR. Embryo can be observed in lung,
heart, liver, and already had the growth of members. The placenta came
oligocotiledonária and the gestational sac from one end to another media 15 cm, open it
to verify the absence of placentônios and area of implantation of the embryo. The
membrane corioalantoidiana the microscope had the allantoic well vascularized with a
thin layer of cells elongated nucleus and cytoplasm. On the other side of the membrane
visualize the attempt to get round cells, scanty cytoplasm and nucleus of large and
round, characteristic of trophoblastic cells. The absence of cotyledons was confirmed on
microscopy, and the caruncle devoid of fetal tissue through its villi. Surrounding the
embryo is viewed the amniotic sac that consists of two layers of mechanical separation,
but had the same macro and microscopic morphology, being composed of spindle cells,
and avascularized. Yolk sac was not observed in specimen. The female reproductive
system showed two oviducts with 4 cm. The uterine horn was bicornual as in other
ruminants. Within the uterus was found caruncle 9, this number was small compared to
beef. The cervix is presented composed of dense connective tissue with villous
longitudinal as ruminants. In conclusion, the reproductive system of the deer Mazama
gouazoubira is similar to other domestic ruminants showing some differences in fetal
membranes such as the lack of yolk sac, being it is placenta classified as
oligocotiledonária, differently of other ruminants which have cotiledonary placenta.
Key words: deer, feminine reproductive system, Mazama gouazoubira

6
INTRODUÇÃO
Os cervídeos (família Cervidae) pertencem à ordem Artiodactyla, que se
caracteriza pela presença de cascos que recobrem totalmente os quatro dedos, sendo que
somente dois deles se apóiam no solo. São ruminantes (subordem Ruminantia),
possuindo quatro compartimentos estomacais (rúmen, retículo, omaso e abomaso),
regurgitando e remastigando várias vezes o seu alimento. Como todos os ruminantes,
apresentam ausência de incisivos superiores, com molares constituídos por quatro
cúspides em forma de meia-lua (selenodontes) (CUBAS et al. 2007).
No Brasil existem oito espécies de cervídeos reconhecidas: Blastocerus
dichotomus, Odocoileus virginiarus, Ozotoceros bezoarticus, Mazama americana,
Mazama bororo, Mazama gouazoubira, Mazama nana e Mazama nemorivaga.
(CUBAS et al. 2007).
Em especial, o veado-catingueiro (Mazama gouazoubira) é uma espécie de porte
pequeno, pesando em média 18kg, raramente excedendo 20kg e altura média de 50cm
na cernelha. A coloração geral dos indivíduos é muito variável, podendo ir do cinza
escuro ao marrom avermelhado, ou até baio. A região ventral é baia, com áreas brancas
na parte inferior da cauda e face interna da orelha. As regiões submandibular e
perioftálmica são mais claras, mas dificilmente chegam a ser brancas. A maioria dos
indivíduos tem uma pinta branca acima dos olhos, que é inexistente em outras espécies.
A orelha é grande e arredondada e os chifres não ramificados, que é uma característica
de todas as espécies pertencentes aos gêneros Mazama e Pudu. Essa é a espécie mais
abundante da América do Sul, ocupando vários tipos de habitat, desde cerrado fechado
até áreas ocupadas pela agricultura. Tem grande poder de adaptação a áreas
modificadas. Ocupa desde o Sul do Uruguai até o Norte do estado do Mato Grosso no
Brasil e desde a cordilheira dos Andes até o Atlântico (CUBAS et al. 2007).
A maioria das espécies brasileiras dá origem a somente um filhote por cria,
sendo raros os casos de gêmeos, com exceção da espécie veado-de-cauda-branca
(Odocoileus virginianus), em que partos gemelares são mais freqüentes que simples. Ao
nascer, eles possuem pintas brancas distribuídas pelo corpo, com, exceção do cervo-do-
pantanal (Blastocerus dichotomus), que nasce com pelagem uniforme, semelhante à do
adulto. As pintas são perdidas na primeira muda de pêlos, entre 2 e 4 meses. (CUBAS et
al. 2007).

7
Os cervídeos neotropicais ainda são pouco estudados quanto a sua taxonomia,
especialmente o gênero Mazama, no qual foram descritas espécies novas nos últimos
anos, como Mazama pandora no México e M. bororo no Brasil. Essa falta de estudo é
gerada pela dificuldade de acesso aos animais, que vivem em sua maioria em florestas.
(CUBAS et al. 2007).
Os cervídeos são poliéstricos estacionais, pelo menos nas espécies de clima
temperado. Sabe-se, que em clima tropical a sazonalidade não é bem definida,
especialmente no gênero Mazama, no qual é citada a ocorrência de cio pós-parto em
veado-mateiro, demonstrando que nessas espécies a reprodução não é sazonal. Em
veado-campeiro é indicada uma certa sazonalidade, pela concentração de nascimentos
na primavera (setembro a novembro) no Brasil e na Argentina, indo até dezembro no
Uruguai. Deve ser enfatizado que há nascimentos por todo o ano, indicando que alguns
machos e fêmeas podem ser férteis por todo esse período. Em cativeiro, cervos-do-
pantanal não apresentam sazonalidade aproximadamente, coincidindo com a gestação
que também é de 8.5 a 9 meses. Nas demais espécies de cervídeos, a duração da
gestação é de aproximadamente sete meses. O ciclo estral em veado-catingueiro é de
aproximadamente 21 dias e, em cervos-do-pantanal, por volta de 24 dias. (CUBAS et
al. 2007).
É sabido que o sistema reprodutor feminino consiste nos ovários, tubas uterinas
(ovidutos) bilaterais, um útero normalmente bicórneo, cérvix, vagina, vestíbulo, vulva e
glândulas associadas. Este é vinculado à produção e transporte de óvulos, ao transporte
dos espermatozóides à fertilização e à acomodação do concepto até o
nascimento (DYCE et al.1997).
As membranas extra-embrionárias são o elo de comunicação entre o feto e a
mãe, que posteriormente irão formar a placenta. Existem quatro tipos de membranas
fetais, o saco coriônico, o alantóide, o âmnio e o saco vitelínico (WOLF et al. 2003).
O constante aumento do número de estradas que fragmenta seu habitat têm sido,
além da caça predatória, uma das principais causas de impacto sobre as populações
naturais. Atropelamentos em estradas têm vitimado muitos veados. Desta forma, a
maioria dos atendimentos à estes animais em centros de atendimentos compreendem
casos de atropelamentos, estando em segundo lugar após os casos de ataques de cães
domésticos (NOWAK, 1991).

8
O objetivo principal deste trabalho é caracterizar a morfologia do sistema
reprodutor do veado-catingueiro, comparando-o com os demais ruminantes devido a
ausência de estudos morfológicos a respeito do trato reprodutor destes nesta espécie. Os
resultados poderão contribuir como fonte de estudo para possíveis intervenções
cirúrgicas e em projetos de reprodução desses animais.
MATERIAIS E MÉTODOS
Animais
Para este estudo, foram coletadas amostras do sistema reprodutor de uma fêmea
adulta de veado-catingueiro (Mazama gouazoubira), que veio a óbito após
atropelamento em rodovia. Devido à gravidade dos traumas por ela sofrido, o mesmo
veio a óbito e foi doado para que estudos morfológicos pudessem ser realizados. Este
animal foi oriundo do SAAS (Serviço de atendimento à Animais Selvagens) da
Universidade Estadual do Centro-Oeste, Guarapuava, Paraná (UNICENTRO). A fêmea
pesava 19.20kg, com 0.92cm de comprimento e 0.60cm de altura de cernelha foi
utilizada. Devido à gravidade dos traumas por ele sofrido, o mesmo veio a óbito e foi
doado para que estudos morfológicos pudessem ser realizados.
Realizou-se uma incisão na linha alba do animal, retirando os órgãos para a
avaliação macroscópica e microscópica do sistema reprodutor feminino. Foram tiradas
fotos com câmera digital Olympus C-315 5.1MP para catalogação e estudo.
Preparações Histológicas e Colorações
Os fragmentos do sistema reprodutor feminino do Mazama gouazoubira foram
tratados em série crescente de etanol (70 a 100%) para desidratação e xilois para
diafanização, utilizando-se procedimento convencional, para posterior inclusão em
Paraplast® (Leica/Germany), confeccionando-se blocos retangulares com base de 2x1
cm.
Foram realizados cortes de 5 µm em micrótomo semi automático para obtenção
das lâminas, as quais foram préviamente silanizadas com 3-aminopropiltrietoxi-silano
(APES) e então desparafinizadas, coradas, analisadas e fotomicrografadas (Microscópio
Olympus BX4160 acoplado a câmera Axio CAM HRc) com o software Zeiss KS 400.

9
Utilizou-se as seguintes colorações: Hematoxilina/Eosina, para caracterização
histológica; tricrômico de Masson, para se diferenciar fibras musculares lisas de
colágeno; picrossírius para corar colágeno e periodic acid Schiff (PAS) para diferenciar
áreas ricas em glicoproteínas.(JUNQUEIRA et al. 2008)
RESULTADOS
Histologia das Membranas Fetais do Mazama gouazoubira
O Mazama gouazoubira apresentou uma gestação univitelínea com um embrião
medindo 1,3 cm de CR (Figura1). No embrião pode ser observado pulmão, coração,
fígado, e já apresentava o crescimento de membros, com separação evidente dos
membros (Figura 2).
FIGURA 1 (e) embrião de Mazama gouazouabira apresentando 1,3cm de CR (Crow-
hump);
FIGURA 2 Fotomicrografia de embrião de Mazama gouazouabira corado com
hematoxilina-eosina (200µm) já evidenciando (f) fígado, (c) coração e (p) pulmão.
10
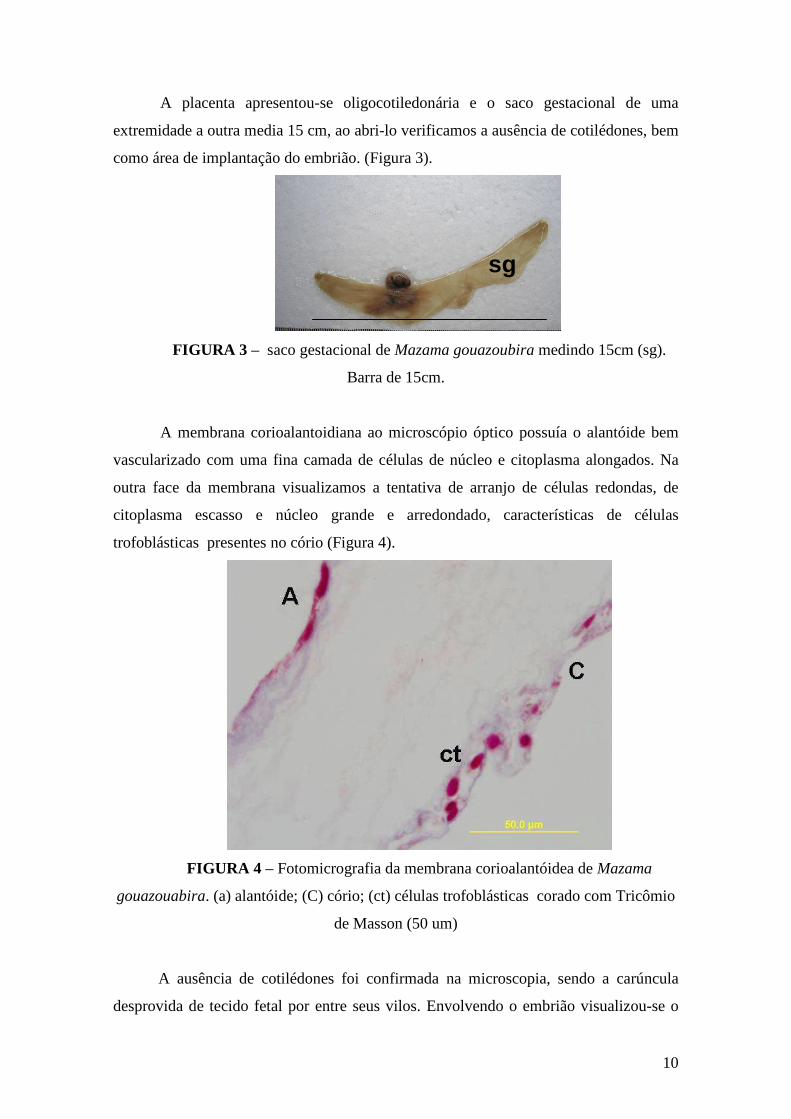
A placenta apresentou-se oligocotiledonária e o saco gestacional de uma
extremidade a outra media 15 cm, ao abri-lo verificamos a ausência de cotilédones, bem
como área de implantação do embrião. (Figura 3).
sgsg
FIGURA 3 – saco gestacional de Mazama gouazoubira medindo 15cm (sg).
Barra de 15cm.
A membrana corioalantoidiana ao microscópio óptico possuía o alantóide bem
vascularizado com uma fina camada de células de núcleo e citoplasma alongados. Na
outra face da membrana visualizamos a tentativa de arranjo de células redondas, de
citoplasma escasso e núcleo grande e arredondado, características de células
trofoblásticas presentes no cório (Figura 4).
FIGURA 4 – Fotomicrografia da membrana corioalantóidea de Mazama
gouazouabira. (a) alantóide; (C) cório; (ct) células trofoblásticas corado com Tricômio
de Masson (50 um)
A ausência de cotilédones foi confirmada na microscopia, sendo a carúncula
desprovida de tecido fetal por entre seus vilos. Envolvendo o embrião visualizou-se o

11
saco amniótico constituído de duas camadas passíves de separação mecânica, mas que
apresentaram a mesma morfologia macro e microscopicamente, sendo constituídas de
células fusiformes, sendo avascularizada (Figura 5). Não foi observado saco vitelino
nesse espécime.
FIGURA 5 – Fotomicrografia de âmnio de Mazama gouazoubira corado com PAS (50µm); (a) âmnio
Anatomia e Histologia do Sistema Reprodutor Feminino do Mazama gouazoubira
O sistema reprodutor feminino apresentou-se com duas tubas uterinas medindo 4
cm cada. O corno úterino era bicornual (direito e esquerdo) como em outros ruminantes
(Figura 6A). Dentro do útero foi encontrado 9 carúnculas, este número era pequeno
quando comparado com o de bovino (Figura 6B). A cérvix era formada por tecido
conjuntivo com vilosidades longitudinais à semelhança dos ruminantes (Figura 7).
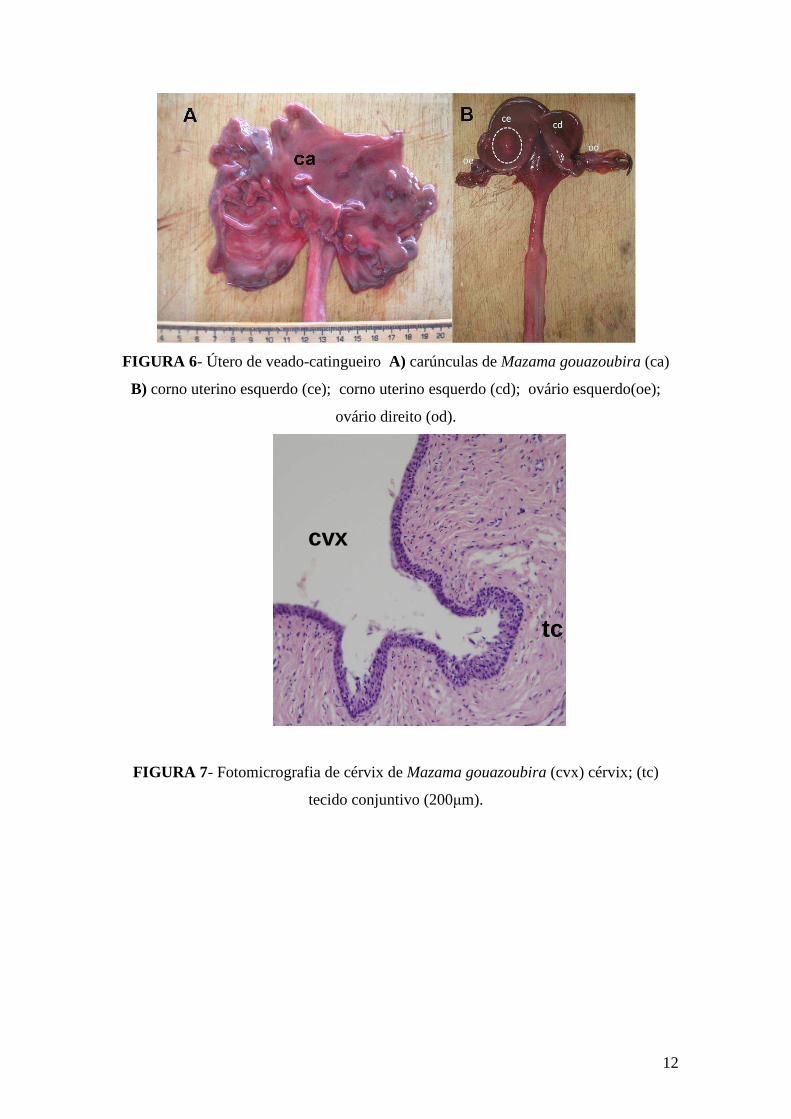
12
FIGURA 6- Útero de veado-catingueiro A) carúnculas de Mazama gouazoubira (ca)
B) corno uterino esquerdo (ce); corno uterino esquerdo (cd); ovário esquerdo(oe);
ovário direito (od).
cvx
tc
cvx
tc
FIGURA 7- Fotomicrografia de cérvix de Mazama gouazoubira (cvx) cérvix; (tc)
tecido conjuntivo (200µm).

13
DISCUSSÃO
Segundo Banks (1982), a placenta dos ruminantes é classificada como
cotiledonária. Neste trabalho podemos observar que a placenta do veado-catingueiro é
do tipo oligocotiledonária, onde o número de cotilédones é menor que em outros
animais, divergindo dos demais ruminantes. Tal fato também foi observado por
Bernirschke (2007), que considera a placenta das diversas espécies de veado
oligocotiledonária, com no máximo 10 cotilédones. A maioria das placentas possuem
entre de 4 a 6 cotilédones. Em contraste com as vacas, carneiros, e outros ruminantes,
que possuem dez vezes mais cotilédones.
As carúnculas compõem a parte materna da placenta, sendo recobertas pelas
membranas fetais que crescem e se expandem para dentro do lúmen uterino, formando
estruturas fetais irregulares, que são conhecidas como cotilédones (SHAFLER et al.
2000). Em contraste com outras espécies Artiodáctilas policotiledonárias (ex. vaca,
carneiros), constatou-se que os cervídeos apresentam poucos cotilédones, possuindo
apenas três carúnculas em cada corno úterino (totalizando seis carúnculas). Bernirschke
(2007).
No presente trabalho o espécime apresentou poucas carúnculas, sendo 9 em sua
totalidade, diferentemente dos bovinos que apresentam cerca de 60 a 90 carúnculas. Não
houve visualização dos cotilédones, apenas das carúnculas, não havendo a formação do
placentoma (PINTO et al. 2008).
De acordo com Banks (1982), o veado-catingueiro apresenta as mesmas
características quanto comparado a outros ruminantes em relação ao grau de
implantação, tendo esta espécie placenta não-decídua (onde os tecidos maternos e fetais
podem estar interdigitados, estando fundidos ou em aposição uns aos outros). Há,
portanto, uma erosão mínima dos tecidos contribuintes. Não havendo perda dos
elementos da mucosa uterina durante o parto.
Percebeu-se que, no caso do veado-catingueiro não havia presença de saco
vitelino, como já descrito por Benirschke (2007), o qual descreve que os veados não
apresentam saco vitelino, diferentemente de outros ruminantes como os bovinos e
bubalinos.

14
Este trabalho vem a confirmar dados encontrados por Banks (1982) e Morini et
al. (2008), os quais observaram que na evolução da gestação a membrana alantoideana
une-se ao cório ficando difícil a percepção das duas separadamente. A extensão do cório
e do alantóide partindo do embrião, também ocorre de maneira concomitante aos outros
ruminantes.
Ao realizarmos dissecações das partes das membranas fetais do veado-
catingueiro podemos visualizar assim como Morini et al. (2008), que o cório formou-se
ao redor do alantóide, envolvendo-o completamente e conseqüentemente envolvendo as
demais membranas. O mesmo fato ocorre com o observado no âmnio, o qual se
apresenta intimamente ligado ao embrião sendo a primeira das membranas fetais, se
considerarmos a ordem embrião-útero.
Tanto em nosso trabalho com o veado-catingueiro, como o realizado por Morini
et al.(2008) em búfalos, não foi possível visualizar vasos macroscopicamente na
membrana. Em ambos estudos observou-se que, o líquido amniótico que é responsável
pela proteção mecânica, sendo este mais denso e viscoso que os demais fluidos
encontrados entre as membranas.
Outra similaridade do veado-catingueiro com outros ruminantes refere-se ao
cório, sendo este constituído por uma camada de células arredondadas, dispostas em
formato linear, igualmente descrito por Banks (1982) e Morini et al. (2008).
Geralmente o cório apresenta uma camada simples de células, mas em algumas
vezes essa membrana pode apresentar mais de uma camada. (BANKS, 1982; MORINI
et al. 2008). Notamos, na maioria de nossas amostras a fusão do cório com a membrana
seguinte, o alantóide, denominado lâmina corioalantóidea, fato também citado em
outras espécies de ruminantes pelos autores supracitados.
Constatamos que, durante a idade gestacional pesquisada não foram observados
placentomas, nem qualquer tipo de conexão entre o endométrio e o cório, porém foram
encontrados pequenos botões que darão origem as vilosidades com o decorrer da
gestação, o que difere do experimento realizado por Hafez (1954) com outros
ruminantes, o qual através de seu estudo histológico, descreveu delicados vilos
aparecendo na superfície da membrana coriônica em oposição às carúnculas maternas.
Acredita-se que provavelmente, essas diferenças entre nossos estudos, ocorreram
devido ao fato dos estudos terem sido realizados em fases gestacionais diferentes. No
período gestacional estudado, não é nítida a presença de vasos provenientes do alantóide

15
na membrana coriônica, porém, percebe-se um contato íntimo entre essas membranas
em diversos locais.
Autores como Kaufmann & Burton (1994) e Morini et al.(2008), relataram em
seus trabalhos sobre a morfologia geral da placenta em diversos mamíferos, que o cório
é um tecido intrinsicamente avascular, sendo vascularizado por vasos do alantóide.
Esses resultados indicam uma semelhança entre búfalos, veados e as demais espécies de
mamíferos.
Foi possível identificar em nosso estudo com o veado-catingueiro, assim como
no estudo realizado por Morini et al. (2008) com búfalos, que a membrana coriônica é
constituída por uma camada simples de células dispostas em formato linear. Fato
também corroborado por Hafez (1954), já descrito anteriormente.
Neste estudo, observou-se que as paredes do alantóide do veado-catingueiro
assemelham-se também com a dos embriões bubalinos relatados por Morini et al.
(2008) onde em ambos estudos constataram aparência fina e translúcida, com um leve
tom esbranquiçado. O que difere do observado por Marshall (1952) e Barone (1986),
que descreveram em outras espécies de ruminantes, essa estrutura apresenta paredes
delgadas e transparentes. Esses resultados indicam que o alantóide do veado-catingueiro
é similar ao de outros ruminantes.
Assis Neto (2005), observou que o alantóide bovino é constituído por uma
delgada camada de células pavimentosas que compõem a parte externa da membrana.
Essas células são sustentadas por uma fina membrana basal e o mesênquima extra-
embrionário. Analisando os resultados deste autor com o nosso, pode-se afirmar que
essa membrana fetal apresenta semelhanças entre essas duas espécies. Assis Neto
(2005), observou no mesênquima dessa membrana em embriões bovinos, vasos e ilhas
eritroblásticas primárias. A similaridade dos resultados indica, mais uma vez, a grande
semelhança entre as membranas fetais dessas duas espécies.
Como descrito por Banks (1982), a cérvix uterina atua como uma válvula para
separar a luz uterina da vagina. Apresenta um epitélio colunar simples com células
caliciformes. A lâmina própria submucosa varia de tecido conjuntivo frouxo a tecido
conjuntivo denso. A camada muscular é bem desenvolvida e rica em fibras elásticas. O
espécime estudado apresentou as mesmas características.

16
CONCLUSÃO
Conclui-se que o sistema reprodutor feminino do Mazama gouazoubira durante
o terço inicial de gestação é similar ao dos outros ruminantes domésticos. Com relação
aos anexos fetais, o presente trabalho demonstrou diferenças em relação a outros
ruminantes como a ausência do saco vitelino, sendo sua placenta classificada como
oligocotiledonária.
REFERÊNCIAS BIBLIOGRÁFICAS
ASSIS, A.C. 2005. Desenvolvimento placentário em bovinos obtidos por gestações naturais e por fecundação in vitro. Tese de Doutorado, Departamento de Cirurgia, Faculdade de Medicina Veterinária e Zootecnia, USP, São Paulo. BANKS, W.J. Histologia Veterinária Aplicada. 2. Ed. São Paulo: Manole, 1992. p.467- 468.
BARONE, R. 1986. Splanchnologie, p.579-605. In: (ed.), Anatomie Comparée dês
Mammifères Domestiques. Tome 2. Vigot Frères, Paris.
BENIRSCHKE, K. Comparative Placentation. 2007. acesso em,
http://placentation.ucsd.edu/indxfs.html
CUBAS, Z.S.; SILVA, J.C.; CATÃO/DIAS, J.L. 2007. Tratado de Anatomia dos
Animais Selvagens.1ed. Editora Roca. Cap.38, p.641.
DYCE, J. M.; W.O.; Wesing, C.I.G. Tratado de anatomia veterinária. 2. Ed. Rio de
Janeiro: Guanabara Koogan, 1997, p. 107-109; 537-538.
HAFEZ, E.S.E.1954. Foetal-maternal attachments in buffalo and camel. Anat. Res. 25 (2):109-115. JUNQUEIRA , L.C.; CARNEIRO,J. Histologia Básica- 11 edição. Rio de Janeiro. Ed. Guanabara, 2008.

17
KAUFMANN P. & BURTON G. 1994. Anatomy and genesis of the placenta, p.441-484. In: Knobil E. & Neill J.D. (ed.), The Physiology of Reproduction. Vol.1. Raven, New York.
PINTO, et.al. Comportamento das células trofoblásticas gigantes na placenta de vacas
Nelore (Bos indicus-Linnaeus, 1758). Revista Brasileira de Reprodução Animal,
2008. 32(2): p. 110-121.
MARSHALL, M.A. 1952. Marshall’s Physiology of Reproduction. A.S. Parkes, London, p.189-211.
MORINI, et al. Caracterização das membranas fetais em búfalas no terço inicial da
gestação. São Paulo, 2008. Pesq. Vet. Bras. 28 (9):437-445, setembro 2008.
NOWAK, K. 1991. Walker’s Mammal of the World , Vol. VI. The Hopkins
University Press, 1101 p.
WOLF, F. B. P. Current topic: The synepitheliochorial placenta of ruminantes:binucleate cell fusions and hormone production. Placenta, London, v. 13, n.2, p.101-113, Mar. 1992.
Top Related