







Línguas
Páginas
Legal


Triptofano hidroxilase Química Bioinorgânica
Catarina Cruz Vaz Patrícia Ferreira de Castro (turma P4)
Licenciatura em Bioquímica 2º ano, 2º semestre Maio de 2013

hTPH1
Na imagem: primeira estrutura de chTPH1, sem substratos ou inibidores presentes no local activo.
Faculdade de Ciências da Universidade do Porto
EC 1.14.16.4
1. Oxidorreductases
Catalisam reacções de oxidação redução.
1.14. Actua sobre dadores acoplados, por incorporação ou redução de moléculas
1.14.16. Com a pteridina reduzida como dador, e incorporação de um átomo.
Coordenação de um catião Fe(III) no centro activo e de um cofactor ou
cofactor análogo.
Outras nomenclaturas • L-triptofano hidroxilase. • Triptofano 5-hidroxilase. • Triptofano 5-monoxigenase

Triptofano hidroxilase
Faculdade de Ciências da Universidade do Porto
SEROTONINA
Importante hormona e neurotransmissor regulador
de variados aspectos psicofisiológicos no cérebro
humano (sono, dor, apetite, humor) e da contracção
do músculo liso.
Função: Catálise do primeiro passo limitante da velocidade da reacção da síntese de
serotonina, um percursor da hormona melatonina.
Diversos distúrbios psiquiátricos (depressão, esquizofrenia, DOC, tendências suicidas) estão
directamente relacionados com anomalias no processo de regulação onde participa a serotonina e
outros neurotransmissores, nomeadamente resultantes de baixos níveis dos mesmos.

Triptofano hidroxilase
A TPH é uma enzima que contém Fe(III), e que catalisa a hidroxilação de
triptofano a 5-hidroxi-triptofano, na presença de O2 e 5,6,7,8 -
tetrahidrobiopterina (BH4) . O BH4 reduz o Fe(III) a Fe(II).
O cofactor enzimático BH4 actua como doador de electrões para a redução
de O2 e para a hidroxilação de Trp.
O O2 e o Trp actuam como substratos.
Faculdade de Ciências da Universidade do Porto

TrpOH Triptofano hidroxilase
A TrpOH participa na hidroxilação do L-Trp na posição 5, o passo limitante na biossíntese da serotonina e o passo inicial da biossíntese da melatonina.
Faculdade de Ciências da Universidade do Porto
O L-triptofano sofre a acção da enzima
triptofano hidroxilase, passando para a
forma de L-5 hidroxitriptofano. Por
conseguinte, ocorre a transformação
dessa última forma em serotonina, pela
acção da L-amina ácida descarboxilase.

Família e proteínas relacionadas
Faculdade de Ciências da Universidade do Porto
Hidroxilases de aminoácidos aromáticos
(AAAH)
PheOH
TyrOH TrpOH
Fenilalanina hidroxilase
Tirosina hidroxilase (domínio de tetramerização)

Família e proteínas relacionadas
Faculdade de Ciências da Universidade do Porto
Hidroxilases de aminoácidos aromáticos
(AAAH)
PheOH
TyrOH TrpOH
5,6,7,8-tetrahidrobiopterina
COFACTOR ENZIMÁTICO Hidroxilação do respectivo
aminoácido aromático

Isoformas: TPH1 e TPH2
As duas isoformas humanas consistem em 444 (hTPH1) e 490 (hTPH2) aminoácidos, respetivamente e têm uma sequência idêntica de 71%. A diferença mais pronunciada entre as isoformas é visto nos domínios de regulação, que é mais longo para a hTPH2 (mais 46 resíduos).
A divisão dos três domínios da TPH1 (hTPH1) e da TPH2 (hTPH2). Faculdade de Ciências da Universidade do Porto
•Expressa principalmente na glândula pineal, e nas partes periféricas do corpo, tais como as células da pele, células cancerosas e na mucosa intestinal.
•Controla a síntese da serotonina na periferia. hTPH1
•Expressa principalmente em neurónios serotoninérgicos do cérebro e intestino;
•Responsável pela síntese da serotonina no sistema nervoso central. hTPH2

Estrutura dos domínios e centro activo Triptofano hidroxilase
Faculdade de Ciências da Universidade do Porto

Estrutura geral
Todas as enzimas pertencentes a esta família contêm três domínios:
• Domínio regulador (N-terminal)
• Domínio catalítico
• Domínio de tetramerização (C-terminal).
Para a enzima triptofano hidroxilase apenas se conhece a estrutura do domínio
catalítico.
Os domínios reguladores e de tetramerização são semelhantes às outras duas enzimas
(tirosina hidroxilase e fenilalanina hidroxilase). Portanto, a caracterização da TPH é
principalmente construída através de estudos sobre as duas outras enzimas desta
família.
Faculdade de Ciências da Universidade do Porto

A estrutura geral do domínio catalítico de hTPH1 com ferro(III) e 7,8-dihidrobiopterina (BH2).
Faculdade de Ciências da Universidade do Porto

Centro activo
A TPH contém Fe(II) no seu centro activo, quando na sua forma activa.
Octaedricamente coordenado com His272, His277, Glu317 numa face
e 3 H2O na outra.
Faculdade de Ciências da Universidade do Porto
Os centros activos residem dentro do domínio catalítico e são responsáveis pela hidroxilação
dos respectivos resíduos aminoácidos aromáticos.
O BH2 é um inibidor competitivo, análogo ao BH4
(compete com o cofactor pelo centro activo da
enzima). Da mesma forma actuam os compostos
análogos ao triptofano p-clorofenilalanina e p-
etinilfenilalanina.

Centro activo
O centro activo, com o Fe(III)
(em verde) coordenado com a
His272,His277, Glu317 e três
H2O (em vermelho).
O BH2, análogo ao cofactor
BH4, é mostrado com átomos
de C a roxo, átomos de O a
vermelho, e átomos de N a
azul.
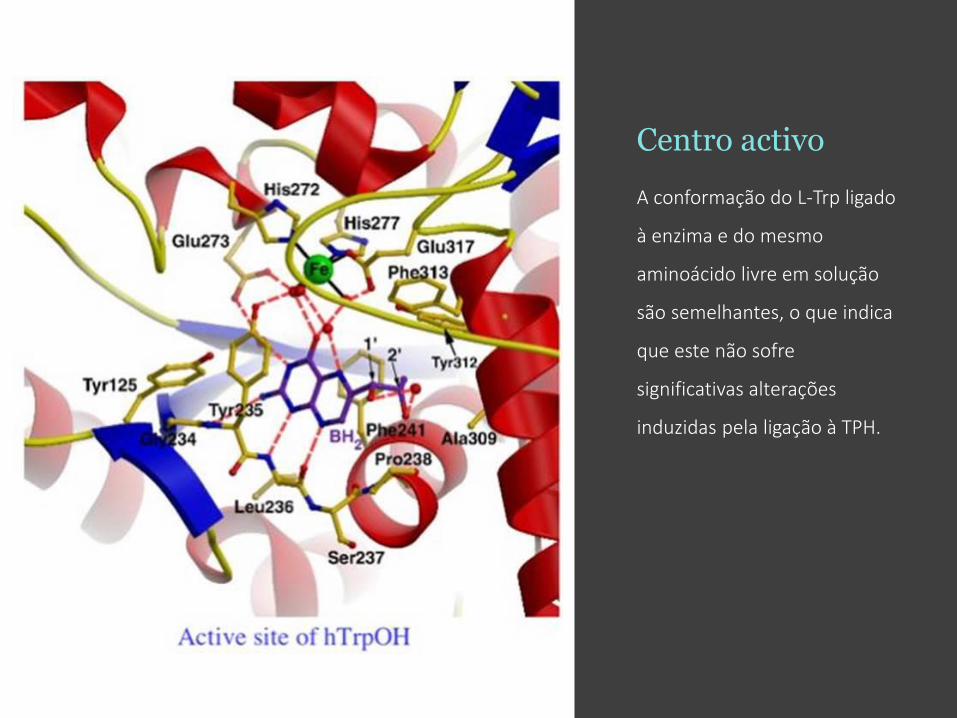
Centro activo
A conformação do L-Trp ligado
à enzima e do mesmo
aminoácido livre em solução
são semelhantes, o que indica
que este não sofre
significativas alterações
induzidas pela ligação à TPH.

Centro activo
A Phe313 interage com o
substrato através de ring
stacking define fortemente a
especificidade da ligação ao
substrato, sendo um resíduo
não conservado entre a TPH e
a PAH.
A conformação do L-Trp ligado
à enzima e do mesmo
aminoácido livre em solução
são semelhantes, o que indica
que este não sofre
significativas alterações
induzidas pela ligação à TPH.
University of Bergen, Noruega

Centro activo
A Phe313 interage com o
substrato através de ring
stacking define fortemente a
especificidade da ligação ao
substrato, sendo um resíduo
não conservado entre a TPH e
a PAH.
A conformação do L-Trp ligado
à enzima e do mesmo
aminoácido livre em solução
são semelhantes, o que indica
que este não sofre
significativas alterações
induzidas pela ligação à TPH.
University of Bergen, Noruega

Centro activo da chTPH1
O ião Fe(III) (a verde)
encontra-se coordenado
octaedricamente por uma
His272, His277, Glu317 e
ainda por 3 H2O .
Faculdade de Ciências da Universidade do Porto

Domínio de tetramerização
Os domínios de tetramerização das
quatro TPH interagem através de
interacções hidrofóbicas, com um
motivo superenrolado. Isto deve-se à
presença de uma repetição 3,4-
hidrofóbica, também presente no
domínio de tetramerização de TPH.
Este domínio é o responsável pela
oligomerização através de interacções
hidrofóbicas num motivo
superenrolado.

Estrutura da TPH com BH4
• Tyr125 a Asp130 • Densidade electrónica alterada
• chTPH1 tem outra conformação.
• Primeira estrutura de chTPH1 sem substratos presentes no centro activo e apenas a estrutura com nº de coordenação seis da hTPH.
Faculdade de Ciências da Universidade do Porto

Catálise pela hTPH Mecanismo, substratos e inibidores
Faculdade de Ciências da Universidade do Porto

Mecanismo da reacção enzimática
Diferentes intermediários de hidroxilação têm sido propostos.
O mecanismo de formação do intermediário Fe(IV)=O não é clara.
Forma activa do ferro nas hidroxilases de aminoácidos aromáticos:
Estado ferroso Fe(II) Fe(III) passa a Fe(II) por acção da BH4.
Fe(II)-peroxibiopterina Espécie de peroxo-BH4 Faculdade de Ciências da Universidade do Porto

• A formação e a reacção seguinte da espécie de peroxo-BH4 não inclui
o ferro, e como a hidroxilação do aminoácido não ocorre sem a
presença de ferro, a espécie de peroxo-BH4 não é um candidato
provável.
• Se a espécie de hidroxilação formada for a Fe(II)-peroxibiopterin, a
quebra da ligação O-O deve ser combinada com a adição do oxigénio
ao aminoácido.
• Nas hidroxilases de aminoácidos aromáticos, pensa-se que a espécie
Fe(IV)=O é formada pela quebra da ligação O-O existente na espécie
intermediária.
Faculdade de Ciências da Universidade do Porto

Coordenação no centro activo
• Ausência de substratos
Os ligandos que se ligam ao ferro são 2 histidinas, um glutamato, e três moléculas de água.
• Ligação dos substratos
Duas das três moléculas de água dissociam-se e o glutamato passa de um ligando monodentado a um ligando bidentado.
Diminuição do número de ligandos de seis para cinco, levando à abertura de um espaço para o O2.
Rearranjo por parte dos ligandos no centro activo quando tanto o aminoácido como o BH4 se
ligam, sendo esta a forma da enzima que reage com o oxigénio.
Faculdade de Ciências da Universidade do Porto
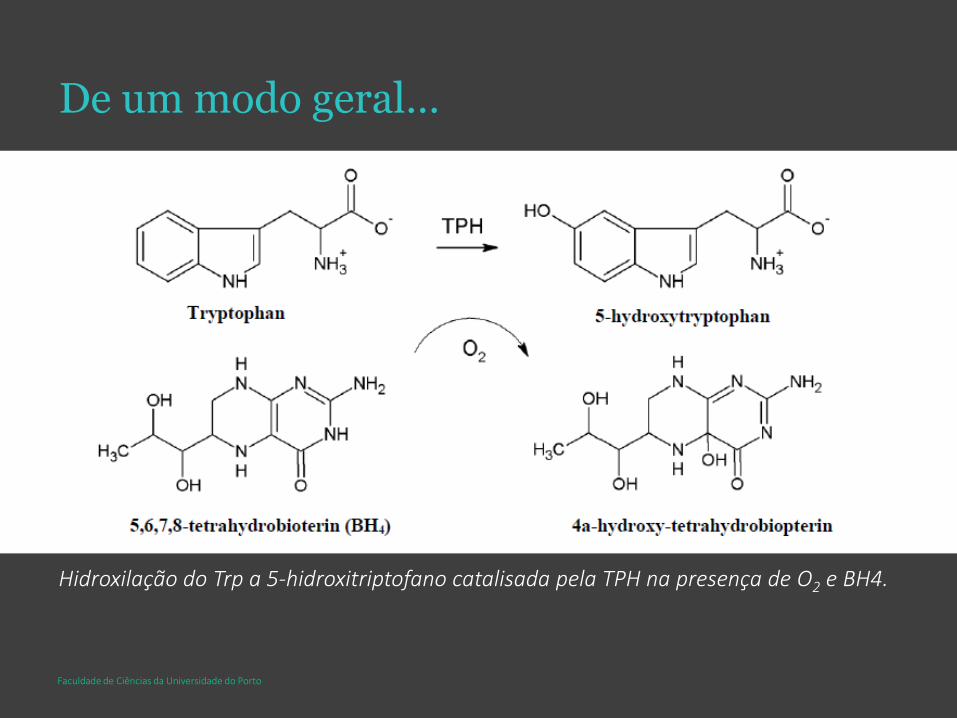
De um modo geral…
Hidroxilação do Trp a 5-hidroxitriptofano catalisada pela TPH na presença de O2 e BH4.
Faculdade de Ciências da Universidade do Porto

1. Reacção entre o sítio activo de ferro, oxigénio e o BH4 para formar um
intermediário reactivo , o Fe(IV)=O.
2. Hidroxilação do aminoácido pela espécie intermediária referida.
O aminoácido é hidroxilado pelo intermediário Fe(IV)=O através de um mecanismo
de substituição eletrofílico.
Estas duas reacções estão bastante acopladas, sendo que por cada molécula de BH4
oxidada, uma molécula de aminoácido é hidroxilada. No entanto, nem sempre ocorre
desta forma, sendo um mecanismo ainda pouco estudado.
Faculdade de Ciências da Universidade do Porto
2 “meias reacções” sequenciais:
De um modo geral…

1. Formação do intermediário de hidroxilação Intermediário: Fe(IV)=O
O mecanismo pelo qual se forma ainda não é bem conhecido
Mecanismo proposto
O Fe(II) liga-se ao O2, é convertido num complexo que vai atacar o C4 da BH4 formando então o intermediário Fe(II)-peroxibiopterina
Quebra da ligação O-O na Fe-peroxibiopterina
Faculdade de Ciências da Universidade do Porto

2. Mecanismo da hidroxilação do L-triptofano Regeneração da BH4
1. Ataque eletrofílico do Fe (IV) = O no átomo de C aromático originando um catião intermediário.
• Durante a formação do intermediário, Fe (IV) = O, BH4 é oxidado a 4-hidroxi-BH4, e é libertado pela enzima.
• A regeneração de BH4 é importante para o fornecimento contínuo de substrato reduzido para o mecanismo catalítico.
Faculdade de Ciências da Universidade do Porto

Reacção catalisada pela TPH e a regeneração da BH4
Faculdade de Ciências da Universidade do Porto
A TPH usa o BH4 como
dador de um electrão na
redução do oxigénio e na
hidroxilação do triptofano.
O 4a-hidroxi-BH4 é
convertido pela enzima
desidratase pterina-4a-
carbinolamina para qBH2.
A qBH2 é então reduzida
para BH4 pela
dihidrobiopteridina
redutase, à custa do NADH.

Inibidores da hTPH
Faculdade de Ciências da Universidade do Porto
BH2 Análogo ao cofactor BH4
p-etinilfenilalanina Análoga ao triptofano
p-clorofenilalanina Análogo ao triptofano

Estudo da triptofano hidroxilase • Dicroísmo Circular (DC) e Dicroísmo Circular Magnético
(DCM) • Cristalografia de raios-X • Mutações no centro activo
Faculdade de Ciências da Universidade do Porto

Dicroísmo Circular
• Estrutura geométrica
Separação entre as orbitais d depende da geometria de coordenação do ferro (teoria de campo de ligando).
Transições d-d difíceis de observar, por apresentarem ε muito baixas (transições proibidas por paridade- Laporte).
Recentemente começaram-se a usar técnicas que provam essas transições: DC e DCM.
Faculdade de Ciências da Universidade do Porto
Transições electrónicas dependentes da coordenação geométrica do Fe(II)
Determinação da energia de transição d-d (separação de orbitais)

Dicroísmo Circular e DC Magnético
Técnicas de Dicroísmo Circular e Dicroísmo Circular Magnético
• Estudo da coordenação do ferro no local ativo da TPH
na ligação do substrato e / ou cofator.
• Usaram-se estas técnicas para estudar a estrutura
geométrica e eletrónica do ferro no local ativo do
domínio catalítico de chTPH2 tendo em conta as
transições eletrónicas d-d.
• Este método pode ser usado para determinar a
coordenação do Fe (II), em enzimas não-heme, como os
AAAHS.
Faculdade de Ciências da Universidade do Porto

Espectros DCM para as 4 combinações de TPH substrato/cofator.
Resultados obtidos por DCM e DC para a TPH
Faculdade de Ciências da Universidade do Porto

Resultados obtidos por DCM e DC para a TPH
Faculdade de Ciências da Universidade do Porto
Espectros obtidos por DC para as 4 combinações de TPH substrato/cofactor

Análise dos resultados obtidos por DC e DCM
• TPH em repouso
Dois picos, que correspondem ao ferro hexacoordenado.
• Ligação do triptofano
A ligação deste sozinho não desencadeia a coordenação do ferro para uma penta coordenação, logo o ferro continua hexacoordenado. Há apenas uma ligeira perturbação na coordenação, mas não chega para haver uma mudança (o que é melhor visualizado pelos espectros de DC).
O pico deslocou-se ligeiramente para uma energia inferior.
Faculdade de Ciências da Universidade do Porto

Análise dos resultados obtidos por DC e DCM • Ligação do BH4
Dois picos observados pelos espectros do DC não evidentes por DCM.
À medida que o espectro de CD mostra claramente dois picos, conclui-se que a ligação de BH4 a TPH não altera a coordenação de ferro de forma significativa uma vez que permanece 6C. Não há grande alteração do pico relativamente à TPH em repouso.
A mudança no espectro é visualizada quando tanto o triptofano e BH4 são adicionados.
Um pico de baixa energia que aparece corresponde à mudança na coordenação do ferro de 6 para 5.
Posição dos picos retirados a partir dos espectros obtidos por DC para as quatro combinações de TPH substrato/cofactor
Faculdade de Ciências da Universidade do Porto

Cristalografia macromolecular de raios-X
• Cristalização por sitting drop vapour diffusion e posterior criopreservação
• Estudo macromolecular a resolução >5 Å permite estudo da estrutura cristalina e ainda do mecanismo reaccional/catalítico
• Incidência de raios X refractados pelos átomos na estrutura cristalina.
Faculdade de Ciências da Universidade do Porto
PADRÃO DE DIFRACÇÃO
Estrutura da biomolécula determinada por análise da intensidade e posição dos pontos de difracção
obtidos
Department of Molecular Biology, The Scripps Research Institute
chTPH1

Mutações no centro activo
Importância dos resíduos do centro activo
para a actividade da enzima.
Glu363 Coordenação com o Fe(II)
Proteína inactiva
Glu363 essencial à catálise
Tyr358 Proteína activa
Importante, mas não essencial à catálise
Faculdade de Ciências da Universidade do Porto

Investigadores e respectivas instituições envolvidas no estudo da TPH1 • Lin Wang, Heidi Erlandsen, Jan Haavik, Per M. Knappskog and Raymond C. Stevens (2002)
“Determinação da estrutura tridimensional da triptofano hidroxilase e o seu papel
na biossíntese de serotonina”
Department of Molecular Biology, The Scripps Research Institute
Department of Biochemistry and Molecular Biology, UniVersity of Bergen
Center of Medical Genetics and Molecular Medicine, UniVersity of Bergen
• Giovanni Cianchetta, Terry Stouch, Wangsheng Yu, Zhi-Cai Shi, Leslie W. Tari,Ronald V. Swanson, Michael J Hunter, Isaac D. Hoffman and Qingyun Liu (2010)
“Mecanismo de inibição da triptofano hidroxilase com a determinação das
estruturas cristalinas”
Department of Medicinal Chemistry, Lexicon Pharmaceuticals
Department of Pharmaceutical Discovery, Lexicon Pharmaceuticals
• Denmark Technical University
Faculdade de Ciências da Universidade do Porto

Bibliografia • M. S. Nielsen, Expression, purification and characterization of tryptophan hydroxylase,
Department of Chemistry, Technical University of Denmark, Kgs. Lyngby, 2007, Ph.D. dissertation
• K. M. Fisher and P. G. Nielsen, Cloning and expression of tryptophan hydroxylase, Department of Chemistry, Technical University of Denmark, Kgs. Lyngby, 2002, B.Sc. thesis
• C. R. Petersen and T. V. Rasmussen, Purification and stabilization of the catalytic domain of tryptophan hydroxylase, Department of Chemistry, Technical University of Denmark, Kgs. Lyngby, 2004, B.Sc. thesis
• M. S. Windahl, C. R. Petersen, H. E. M. Christensen and P. Harris, Crystal structure of tryptophan hydroxylase with bound amino acid substrate, Biochemistry, 47, 2008
• P. F. Fitzpatrick, Mechanism of aromatic amino acid hydroxylation, Biochemistry, 41, 2003, 14083-14091
• Enzyme Structures Database - The European Bioinformatics Institute (parte integrante do European Molecular Biology Laboratory)
http://www.ebi.ac.uk/thornton-srv/databases/enzymes/
EMBL-EBI
Faculdade de Ciências da Universidade do Porto
Top Related