
Helena Lúcia Carneiro Santos Diagnóstico diferencial da...
201
Helena Lúcia Carneiro Santos Diagnóstico diferencial da amebíase: aplicação de métodos moleculares para a detecção e diferenciação de espécies de entamoeba spp. Em fezes humanas Orientadores: Dr. José Mauro Peralta/UFRJ Dr. Alexandre Januário da Silva/CDC UNIVERSIDADE FEDERAL DO RIO DE JANEIRO INSTITUTO DE MICROBIOLOGIA PROF. PAULO DE GÓES RIO DE JANEIRO Agosto/2010 Tese de Doutorado apresentada ao Programa de Pós-Graduação em Ciências (Microbiologia), Instituto de Microbiologia Prof. Paulo de Góes, da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Doutor em Ciências Biológicas (Microbiologia)
Transcript of Helena Lúcia Carneiro Santos Diagnóstico diferencial da...

Helena Lúcia Carneiro Santos
Diagnóstico diferencial da amebíase: aplicação de métodos moleculares para a detecção e diferenciação de espécies de
entamoeba spp. Em fezes humanas
Orientadores: Dr. José Mauro Peralta/UFRJ
Dr. Alexandre Januário da Silva/CDC
UNIVERSIDADE FEDERAL DO RIO DE JANEIRO INSTITUTO DE MICROBIOLOGIA
PROF. PAULO DE GÓES
RIO DE JANEIRO Agosto/2010
Tese de Doutorado apresentada ao Programa de Pós-Graduação em Ciências (Microbiologia), Instituto de Microbiologia Prof. Paulo de Góes, da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Doutor em Ciências Biológicas (Microbiologia)

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
Santos, Helena Lúcia Carneiro Diagnóstico diferencial da amebíase: aplicação de métodos moleculares para a detecção e diferenciação de espécies de Entamoeba spp. em fezes humanas/Helena Lúcia Carneiro Santos – Rio de Janeiro, 2010. XXI, 176 Tese Doutorado em Ciências (Microbiologia) Universidade Federal do Rio de Janeiro/Instituto de Microbiologia Prof. Paulo de Góes, 2010. Orientadores: Dr. José Mauro Peralta e Dr. Alexandre Januário da Silva Referências bibliográficas: f 102 1. Entamoeba 2. PCR 3. Diagnóstico molecular 4. Luminex 5. Sequenciamento I. José Mauro Peralta II. UFRJ, Instituto de Microbiologia Prof. Paulo de Góes, Doutorado em Ciências (Microbiologia). III. Diagnóstico diferencial da amebíase: aplicação de métodos Moleculares para a detecção e diferenciação de espécies de Entamoeba spp em fezes humanas.

iii
Helena Lúcia Carneiro Santos
DIAGNÓSTICO DIFERENCIAL DA AMEBÍASE: APLICAÇÃO DE MÉTODOS MOLECULARES PARA DIFERENCIAÇÃO DE ENTAMOEBA SPP.
ENCONTRADAS EM FEZES HUMANAS
Rio de Janeiro, 19 de agosto de 2010
José M. Peralta, Ph.D., Universidade Federal do Rio de Janeiro (Orientador) Maria Aparecida Gomes, Ph.D., Universidade Federal Minas Gerais Adeilton Alves Brandão, Ph.D., Instituto Oswaldo Cruz – FIOCRUZ Marcio Neves Bóia, Ph.D., Instituto Oswaldo Cruz – FIOCRUZ Walter Oelemann, Ph.D., Universidade Federal Rio de Janeiro (Revisor)

iv
O presente trabalho foi realizado no Laboratório de Diagnóstico Imunológico e Molecular de Doenças Infecciosas, Departamento de Imunologia, Instituto de Microbiologia Prof. Paulo de Góes, Centro de Ciências da Saúde (CCS), Universidade Federal do Rio de Janeiro, sob a orientação do Prof José Mauro Peralta e no Laboratório de Parasitologia Molecular do Centers for Disease Control & Prevention (CDC) sob a orientação do Dr. Alexandre J. da Silva.

v
Não devemos nos embriagar pelas grandes alegrias e nem nos deixar abater pelas grandes frustações.
Porque tudo na vida é passageiro!!!
“Autor desconhecido”

vi
AGRADECIMENTOS
A Deus, meu grande amigo, que em todos os momentos me guia pelos melhores caminhos, restabelece minhas esperanças e sonhos, dá vida a minha vida e está sempre presente em meus problemas, necessidades, nos meus desafios e nas minhas alegrias. Tu és maravilhoso.
À minha irmã América, minha maior torcida e pela sempre disposição em me ajudar. Muito, muito obrigada!!!!!
Aos meus mais sinceros agradecimentos ao Dr. José Mauro Peralta pela orientação, pela confiança, pelos conselhos e incentivos nas dificuldades. Obrigada Peralta!
Ao Dr. Alexandre Januário da Silva pela oportunidade de estagiar e me orientar durante onze mEses (doutorado sanduíche) no Centers for Disease Control and Prevention, onde grande parte deste trabalho se realizou. Obrigada pela acolhida, pelos ensinamentos e pela inestimável força durante este período. Você foi dez. Valeu !!!
À Rebecca Bandea pelo acolhimento, carinho e pelas inumeráveis ajudas. Foi muito bom ter convivido com você.
À Giliane Trindade, Isabel Mcauliffe, Marcos, Michael Omandi, pelos bons momentos de convivios e pelas inúmeras ajudas. Acho que eu deveria ter clonado vocês.
Ao grupo do lab. 027, Filomena, Fabíola, Camille, Felipe Piedade, Felipe Cruz, Vânia, Giselle pelo convívio harmonioso. Em especial, a Jacqueline meu muito obrigada por sempre me receber com um sorriso e por ter quebrado meus galhos.
A Drª Lúcia Teixeira por me receber sempre tão bem em seu laboratório e pelo exemplo de profissionalismo. Agradeço, ainda, pela atenção e cuidado que teve comigo durante o período do doutorado sanduíche. Meu muitissímo obrigada.
Ao professor Walter Oelemann por sempre manter as portas de seu laboratório abertas para qualquer ajuda e por ter revisado a tese. O seu coração é infinitamente maior do que o seu tamanho. Muito obrigada!!!
Aos meus amigos do grupo dos surtados da acrilamida: Laurinha, Margareth, Diva, Tiana, Jorge, Mauro “Jr’’, Didio, Bruna, Fadinha e Nathália pelas trocas de ideias, risadas, encontros, por me fazerem sentir sempre “em casa” e tornarem o meu cotidiano muito mais divertido. Espero que a chama da nossa amizade, solidariedade e do bem em comum do nosso grupo permaneça sempre acessa. Valeu!!!!
Ao Carlos, gente muito fina que aturou todas as minhas bagunças no laboratório, pelo apoio técnico realizado nos bastidores, mas crucial para o andamento do trabalho. Você é dez!!!!!
Ao Thiago Guimarães, que se tornou um anjo e está fazendo conexão direta com Deus. Obrigada por ter compartilhado comigo os momentos de estresse e sufoco durante os cultivos das amebas! Você deixou muita saudade.

vii
Ao Setor de Pesquisa Clínica do HUAP por ter autorizado e cedido a utilização do equipamento LUMINEX. E ao Santiago por sempre estar disposto a me ajudar.
À Bruna de Paula Fonseca do LATED de Biomanguinho/FIOCRUZ pela sua simpatia, cordialidade e pela troca de experiência na utilização do equipamento LUMINEX 200. Muito obrigada!!!
À minha chefe e amiga, Maria Conceição do IFF/FIOCRUZ, por ter me concedido a licença integral durante todo o período do doutorado.
Ao CNPq, pelos quatro anos de bolsa (incluindo a bolsa swe).
À coordenação da pós-graduação do Instituto de Microbiologia Prof. Paulo de Góes, na pessoa da Profª Ana Paula Vieira Colombo.
Ao Instituto de Microbiologia Prof. Paulo de Góes, na pessoa da Profª Agnes Marie de Sá Figueiredo.
A todos que ajudaram, de qualquer forma, mas não foram citados aqui por falha minha.
A todos vocês, minha eterna gratidão.
“Cada pessoa que passa em nossa vida, passa sozinha, mas não vai só, nem nos deixa só;
leva um pouco de nós mesmos, deixa um pouco de si mesma”.
Muito obrigada.

viii
Dedico este trabalho aos meus pais Jair Santos (in memorian) e Luzia Carneiro Santos por ter me ensinado a caminhar na estrada da vida.

ix
RESUMO
Helena Lúcia Carneiro Santos
DIAGNÓSTICO DIFERENCIAL DA AMEBÍASE: APLICAÇÃO DE MÉTODOS MOLECULARES PARA A DETECÇÃO E DIFERENCIAÇÃO DE ESPÉCIES DE
ENTAMOEBA SPP. EM FEZES HUMANAS
Orientadores: Dr. José Mauro Peralta e Dr. Alexandre Januário da Silva
Resumo da Tese de Doutorado submetida ao Programa de Pós-Graduação em Ciências (Microbiologia), Instituto de Microbiologia Prof. Paulo de Góes da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários para a obtenção do título de Doutor em Ciências (Microbiologia).
O gênero Entamoeba contém seis espécies que podem ser encontradas nas fezes humanas (E. histolytica, E. dispar, E. poleckii, E. moshkovskii, E. coli e E. hartmanni). Todas as espécies devem ser relatadas quando evidenciadas na amostra clínica. Porém, somente E. histolytica é capaz de causar quadros de amebíase intestinal e extraintestinal, enquanto que as outras espécies são consideradas não patogênicas. E. histolytica, E. dispar, E. moshkovskii, E. poleckii e E. hartmanni são morfologicamente semelhantes, mas podem ser diferenciadas por métodos moleculares. Nos últimos anos, a reação em cadeia da polimerase (PCR) tem sido aplicada na diferenciação entre E. histolytica e E. dispar. Recentemente, houve uma crescente difusão na literatura sobre as técnicas do tipo multiplex incluindo a PCR em tempo real e a PCR seguida da reação de hibridação multianalítica em meio líquido utilizando microesferas fluorescente como suporte sólido associada a citometria de fluxo, sistema Luminex (PCR/Luminex). Estas técnicas visam detectar, simultaneamente, diferentes fragmentos de DNA em uma única amostra clínica e em um único tubo de reação. Este trabalho tem como objetivo aplicar técnicas moleculares inovadoras e do tipo multiplex no diagnóstico diferencial entre as espécies de ameba. Para tal foi utilizado a PCR seguido por sequenciamento, PCR em tempo real e a PCR/LUMINEX. Um total de 45 amostras de fezes positivas para uma ou mais espécies de Entamoeba na microscopia óptica foram identificadas na PCR/sequenciamento. Destas, seis amostras foram identificadas como E. histolytica, 29 como E. dispar, nove como E. hartmanii, uma como E. moshkovskii e cinco como E. coli. As sequências de rDNA das espécies de Entamoeba obtidas neste estudo foram alinhadas e utilizadas no desenho de oligonucleotídeos iniciadores, sondas específicas para cada espécies de Entamoeba e sondas grupo especifica (E. histolytica, E dispar, E. moshkovskii e E. hartmanni). Estes iniciadores e sondas foram empregados na padronização da PCR/Luminex. Na padronização da reação de hibridação multianalítica em meio líquido, utilizando microsferas fluorescente como suporte sólido, associada a citometria de fluxo foram desenhados sete oligonucleotídeos iniciadores e 23 sondas ao todo para as seguintes espécies: E. histolytica, E. dispar, E. hartamanni, E. coli, E. moshkovskii, e sondas grupo especificas. Observamos alto sinal de fluorescência na reação de hibridação em meio líquido, quando utilizamos produto amplificado

x
biotinilado obtido a partir de DNA clonado de E. dispar, E.coli, E. hartmanni e E. moshkovski. No entanto, a sonda para E. histolytica apresentou uma intensidade de fluorescência média. Um total de 34 amostras clínicas foram utilizadas na PCR/Luminex e os resultados mostraram que o teste foi capaz de identificar especificamente E. histoltyica, E. dispar, E hartmanni, E. coli e E. moshkovskii. Estes resultados preliminares sugerem que este método pode ser útil na identificação de amebas entéricas.
Palavras-chave: Amebíase, métodos moleculares, citometria de fluxo empregando microsesferas fluorescentes, sequenciamento, PCR, Entamoeba spp.
Rio de Janeiro Agosto de 2010

xi
ABSTRACT
Helena Lúcia Carnerio Santos
DIFFERENTIAL DIAGNOSIS OF AMEBIASIS: APPLICATION OF MOLECULAR METHODS FOR DETECTION AND DIFFERENTIATION OF
SPECIES OF ENTAMOEBA SPP. IN HUMAN FECES
Advisers: José Mauro Peralta and Alexandre Januário da Silva
Summary of thesis submitted to the Graduate Program in Science (Microbiology), Institute of Microbiology Prof. Paulo de Góes, Federal University of Rio de Janeiro, as part of the requirements for obtaining the title of Doctor of Science (Microbiology).
The genus Entamoeba comprises six species which can be found in human feces (E. histolytica, E. dispar, E. poleckii, E. moshkovskii, E. coli and E. hartmanni). All species should be reported when detected in clinical samples. However, only E. histolytica can cause intestinal amebiasis and extra-intestinal, while other species are considered nonpathogenic. E. histolytica, E dispar, E. moshkovskii, E. poleckii and E. hartmanni are morphologically similar but can be differentiated by molecular methods. In recent years, polymerase chain reaction (PCR) has been applied to differentiate E. histolytica and E. dispar. Recently, there has been a growing spread in the literature on multiplex assays such as real time PCR and PCR followed by multi-analytical hybridization using fluorescent microspheres as solid supports coupled with flow cytometry, Luminex (PCR/Luminex system). These techniques aim to detect different DNA fragments simultaneously in a single clinical specimen and in a single reaction tube. This work aims to apply molecular techniques and innovative multiplex in the differential diagnosis of the species of amoeba. We used the PCR/sequencing, real-time PCR and PCR/System LUMINEX. A total of 45 stool samples positive for one or more species of Entamoeba in optical microscopy were identified by PCR/ sequencing. Of these, six were identified as E. histolytica, 29 as E. dispar, nine as E. hartmanii , one as E. moshkovskii and five as E. coli. The sequences of the rDNA species of Entamoeba obtained in this study were aligned and used to design oligonucleotide primers, probes specific for each species group and Entamoeba (E. histolytica, E. dispar, E. moshkovskii and E. hartmanni) specific probes. These primers and probes were used in the standardization of PCR/Luminex. In the standardization of the hybridization reaction of multi-analysis in liquid medium using fluorescent microspheres as solid support, coupled with flow cytometry were designed seven primers, 23 probes for the following species: E. histolytica, E. dispar, E. hartamanni, E. coli, E. moshkovskii, and group specific probes. A strong fluorescence signal was observed in the reaction of hybridization in liquid medium when using biotinylated amplified product obtained from DNA cloned from E. dispar, E. coli, E. hartmanni and E. moshkovski in PCR. However, the probe for E. histolytica showed mean fluorescence intensity. A total of 34 clinical samples were used in PCR/Luminex and the results showed that the test was able to specifically identify E. histoltyica, E.

xii
hartmanni, E. moshkovskii, E. coli and E. dispar. These preliminary results suggest that this method may be useful in identifying enteric amoebae.
Key Words: Amebiasis, molecular methods, flow cytometry using fluorescent microspheres, sequencing, PCR, Entamoeba spp.
Rio de Janeiro August of 2010

xiii
LISTA DE ABREVIATURAS
% Porcentagem
°C Graus Celsius
µg Micrograma
µl Microlitro
µM Micro Molar
Bp Pares de base
CDC Centers for Diseases Control and Prevention
cm Centímetro
cols Colaboradores
CT “Cycle treshold” (limiar de detecção)
DNA Ácido desoxiribonucleico
dNTP Desoxinucleotídeos
E. hart Entamoeba hartmanni
E. moshki Entamoeba moshkovskii
E. coli Entamoeba coli
E. disp Entamoeba díspar
E. hist Entamoeba histolytica
EDC (N-(3-dimetil aminodipropil)-N’etilcarbodiimida)
EDTA Acido etilenodiamino tetra-acético
ELISA “Enzyme Linked Imunosorbent Assay” (Ensaio imuno-enzimático)
EPF Exame Parasitológico de Fezes
EUA Estados Unidos da América
FAM 6-carboxifluoresceína
Fig Figura
FRET “Fluorescence Ressonance Energy” (Transferência de energia por ressonância de fluorescência)
HBV Vírus da Hepatite B
HCV Vírus da Hepatite C
HEX Hexaclorocarbonilfluoresceína
HIV Vírus da imunodeficiência adquirida
HPV Papiloma Vírus Humano
Kb Kilobases

xiv
kDa Kilodaltons
LB Luria Bertani
LSSP-PCR Low Stringency single primer (PCR em baixa estringência, usando um único iniciador)
M Molar
Mabs Anticorpos monoclonais
MIF Intensidade de fluorescência mediana
ml Mililitro
mM Milimolar
N Número de amostras
NBLAST Nucleotide Basic Local Allignment Search Tool
OMS Organização Mundial da Saúde
PBS Solução Tampão Salina- Fosfato
PCR Polymerase chain reactions (Reação em Cadeia da Polimerase)
PCR/Luminex PCR seguida da reação de hibridação multianalítica em meio líquido utilizando microesferas fluorescente como suporte sólido associada a citometria de fluxo
pH Potencial hidrogeniônico
pmol Picomol
PPS Solução de Precipitação de proteínas
PVA Álcool polivinilico
PVP Polivinilpirilidona
RAPD Randomly Amplified Polimorphic DNA (DNA polimórfico amplificado ao acaso)
RAP-PCR RNA Arbitrarily Primed-PCR (RNA amplificado aleato-riamente)
rDNA DNA ribossômico
RFLP Perfis dos fragmentos obtidos por endonucleases de restrição
Rpm Rotação por minuto
S Segundos
SA-PE Streptavidina-R-Ficoeritrina
SDS Dodecil sulfato de sódio
Spp Espécies
ß-galactosidase Beta-galactosidase

xv
SSU-rDNA Subunidade menor do DNA ribossômico
TaqDNA DNA Polimerase Termoestável
TBE Tris Borato EDTA
TE Tris –EDTA
TMAC Cloreto de tetrametilamonio
Tris Hidroxi-metil-amino-metano
Tris-HCl Tris-ácido clorídrico
tRNA RNA transportador
U Unidades
UFF Universidade Federal Fluminense
V Volts
v/v Volume por volume
x Concentração
Χg Vezes o valor da gravidade

xvi
LISTA DE QUADROS
Quadro 1. Características utilizadas na diferenciação de trofozóitos de amebas encontrados no intestino grosso humano 3
Quadro 2. Características utilizadas na diferenciação de cistos maturos
de amebas encontrados no intestino grosso do humano 3 Quadro 3. Sequência alvo e oligonucleotideos iniciadores utilizado na
PCR convencional descritos na literatura 18

xvii
LISTA DE FIGURAS
Figura 1. Representação esquemática das etapas realizadas neste trabalho 32
Figura 2. Mapa do vetor de clonagem pCR 2.1 TOPO (Invitrogem) 43 Figura 3. Representação esquemática da microesfera de poliestireno
de 5,6 µm recoberta por grupamentos carboxila e da sonda ligada a um espaçador de carbono e um grupamento amina 46
Figura 4. Gel de agarose a 2%. Amplificação de DNA de Entamoeba
spp. usando o par de oligonucleotideo JVF/DSPR2 52 Figura 5. Gel de agararose a 1,5 %. Amplificação utilizando o par de
iniciador JVF/DSRP2 para confirmar a presença do inserto durante o processo da clonagem molecular 57
Figura 6. Análise dos clones obtidos em eletroforese de gel de agarose
2 % através da digestão com a enzima de restrição EcoRI 58 Figura 7. Gel de agarose a 2%. Amplificação de DNA de Entamoeba
spp. 63 Figura 8. Árvore filogenética obtida a partir da análise de sequências
parcial do gene que codifica para o SSU-rRNA das espécies de Entamoeba identificadas neste estudo 119
Figura 9. Representação esquemática do loca de hibridação das
sondas com as suas respectivas sequências alvos 121

xviii
LISTA DE TABELAS
Tabela 1. Frequências das parasitoses intestinais nas amostras proveniente do Serviço de Saúde Pública dos EUA 50
Tabela 2. Frequências das parasitoses intestinais nas amostras proveniente de Ilhéus na Bahia 51
Tabela 3. Resultados obtidos nas amostras de cultura padrão, pela PCR em tempo real e na PCR/Sequenciamento 53
Tabela 4. Resultados obtidos no exame parasitológico de fezes, na PCR em tempo real e na PCR/ Sequenciamento nas amostras de fezes oruindas do Serviço de Saúde Pública da Geórgia, EUA 54
Tabela 5. Resultados obtidos no exame parasitlógico de fezes, na PCR em tempo real e na PCR/Sequenciamento das amostras de fezes de indivíduos moradores de Ilhéus na Bahia, Brasil 56
Tabela 6. Sondas de captura do tipo grupo e espécie específica utilizadas no ensaio multi-analítico em meio líquido na PCR/Luminex 61
Tabela 7. Oligonucleotídeos iniciadores desenhados e utilizados na PCR/Luminex 62
Tabela 8. Novos Oligonucleotídeos iniciadores utilizados na PCR e sondas na reação de hibridação multianalitica 66
Tabela 9. Relação das sondas que apresentaram sinais específicos de fluorescência na PCR/Luminex 70
Tabela 10. Resultados das mif obtidas na PCR/sistema Luminex em 12 amostras clínicas amplificadas com o par de iniciador JVF/Entarev390b 72
Tabela 11. Resultados das MIF obtidas na PCR/Sistema Luminex de oito amostras clínicas amplificadas com o par de iniciador JVF/Enta417A-B 73
Tabela 12. Comparação entre os resultados da PCR/Luminex com a PCR em tempo real, PCR/sequenciamento e com a análise microscópica de amostras clínicas 74
Tabela 13. Resultados da PCR/Luminex e da PCR em tempo real em 15 amostras clínicas 75

xix
LISTA DE GRÁFICOS
Gráfico 1. Resultados dos ensaios de hibridação multianalítica realizada a 46º C/1h com diferentes combinações de sondas, para a detecção e diferenciação de espécies de Entamoeba spp. 64
Gráfico 2. Resultados dos ensaios de hibridação multianalítica
realizada a 41º C/1h com diferentes combinações de sondas, para a detecção e diferenciação de espécies de Entamoeba spp. 67
Gráfico 3. Resultados dos ensaios de hibridação multianalítica
realizada a 43º C/1h com diferentes combinações de sondas, para a detecção e diferenciação de espécies de Entamoeba spp. 68
Gráfico 4. Resultados dos ensaios de hibridação multianalítica
realizada a 46º C/1h com diferentes combinações de sondas, para a detecção e diferenciação de espécies de Entamoeba spp. 68
Gráfico 5 Resultados dos ensaios de hibridação multianalítica
realizada a 48º C/1h com diferentes combinações de sondas, para a detecção e diferenciação de espécies de Entamoeba spp. 69

xx
SUMÁRIO
Lista de Abreviaturas Lista de Quadros Lista de Figuras
xiiixvixvii
Lista de Tabelas xviiiLista de Gráficos xix 1. INTRODUÇÃO 1 2. OBJETIVOS 31 2. 1 Geral 31
2. 2 Específicos 31
3. MATERIAL E MÉTODOS 323. 1 Amostragem 333. 2 Amostras Controles 333. 3 Exame Parasitológico de Fezes (EPF).. 33
3. 3.1 PARATEST® 34 3. 3.2 Método de Richtie modificado 34
3. 4 Extração do DNA 353. 5 PCR em Tempo Real (Sistema TaqMan). 373. 6 PCR/Sequenciamento 38
3. 6.1 Análise do produto amplificado 38 3. 6.2 Purificação do produto amplificado 39 3. 6.3 Reação de sequenciamento 39 3. 6.4 Purificação dos produtos sequenciados 40 3. 6.5 Sequenciamento dos produtos 40 3. 6.6 Análise das sequências 41
3. 7 Análise Filogenética 413. 8 Clonagem Molecular 423. 9 Extração do DNA Plasmidial de E. coli (Minipreparação) 443.10 Reação de hibridação em meio líquido utilizando microesferas
fluorescente como suporte sólido associada a citometria de fluxo, sistema Luminex 443.10.1 Desenho dos oligonucleotídeos utilizados na PCR/Luminex 453.10.2 PCR/Luminex 473.10.3 Ligação das sondas de DNA amino modificadas á superfície
das microesferas carboxiladas 473.10.4 Reação de hibridação multianalítica em meio líquido utilizando
microesferas fluorescente como suporte sólido associada a citometria de fluxo, sistema Luminex® 48
4. RESULTADOS 50
4. 1 Exame parasitológico de fezes 504. 2 PCR/Sequenciamento 514. 3 Clonagem Molecular 574. 4 Análise das Sequências 59

xxi
4. 5 Reação de hibridação multianalítica em meio líquido, utilizandomicroesferas fluorescentes como suporte sólido associado a citometria de fluxo, sistema Luminex 60
5. DISCUSSÃO 76
6. CONCLUSÃO 101 REFERÊNCIAS BIBLIOGRÁFICAS 102
ANEXOS 117Anexo 1. Árvore filogenética obtida a partir da análise da sequência
parcial do gene que codifica a SSU-rRNA das espécies de Entamoeba identificadas neste estudo 118
Anexo 2. Representação esquemática do local de hibridação das sondas com as suas respectivas sequências alvos 120
Anexo 3. Alinhamento das sequências parciais de referências do GenBank utilizando o programa Clustal W inserido no pacote do programa GeneStudio suite 122
Anexo 4. Alinhamento da sequência parcial do gene SSU-rRNA de E. moshkovskii 126
Anexo 5. Regiões polimórficas presente no gene SSU-rRNA de E. hartmannii obtidas a partir do alinhamento das sequências deste estudo 128
Anexo 6. Regiões polimórficas presente no gene SSU-rRNA de E.coli obtidas a partir do alinhamento das sequências deste estudo 131
Anexo 7. Alinhamento múltiplo entre as sequências parciais do gene Ssu-rRNA de Entamoeba obtidas neste estudo 136
Anexo 8. SANTOS, H.L.; BANDEA, R.; MARTINS, L.A. et al. Differential identification of Entamoeba spp. based on the analysis of 18S rRNA. Parasitol Res 106(4): 883-8, 2010 157
Anexo 9. OLIVEIRA, L.M.; SANTOS, H.L.C.; GONÇALVES, M.M.L.; BARRETO, G.M.; PERALTA, J. M. 2010. Evaluation of PCR as an alternative tool for the diagnosis of low-intensity Schistosoma mansoni infection. Diagn Microbiol and Infect Disease, 2010 164

1
1. INTRODUÇÃO
O gênero Entamoeba é composto por protozoários que se locomovem
e incorporam os alimentos através da emissão de pseudópodes. Este gênero
encontra-se largamente distribuído nos hospedeiros vertebrados e pertence ao filo
Sarcomastigophora, família Endamoebidae e classe Lobosea (LEVINE et al.,
1980). Possui um ciclo de vida simples que consiste de um estágio cístico
infectante e um estágio de trofozoíto responsável pela perpetuação do parasito.
Cada espécie do gênero Entamoeba tende a produzir a forma cística que, quando
madura, contém um número definido de núcleos. Baseado nesta característica, o
gênero foi dividido em quatro grupos distintos: amebas com núcleo único, com
quatro núcleos, com oito núcleos e ameba com forma cística desconhecida (REY,
2008).
Várias espécies de amebas que constituem o gênero Entamoeba
podem naturalmente habitar o trato intestinal humano, tais como: Entamoeba coli,
Entamoeba hartmanni, Entamoeba histolytica, Entamoeba histolytica, Entamoeba
polecki e Entamoeba moshkovskii que é considerada como uma ameba de vida
livre, amplamente encontrada em diferentes coleções hídricas. Nota-se ainda, que
E. moshkovskii tem sido encontrada parasitando o intestino humano (ALI et al.,
2003; PARIJA & KHAIRNAR, 2005; FOTEDAR et al., 2007). Por sua vez, E.
polecki é uma espécie encontrada no intestino de porco e macaco, fortuitamente
pode parasitar o intestino humano (DESOWITZ & BARNISH, 1986). As espécies
E. dispar, E. moshkovskii, e E. histolytica são morfologicamente semelhantes na
microscopia óptica, mas podem ser diferenciadas uma das outras por meio de
análise do perfil de isoenzimas, por anticorpos monoclonais, pelo polimorfismo do
tamanho dos fragmentos de restrição e através da técnica da reação em cadeia
da polimerase (PCR – Polymerase Chain Reaction). As formas evolutivas de E.
hartmanni apresentam dimensões menores o que possibilita a sua diferenciação

2
através da análise morfométrica de cistos e trofozoíto. Cistos imaturos de E. coli
apresentam caracterírticas morfológicas bastante semelhante aos cistos maduro
tetranucleados (HECKENDORN et al., 2002).
Sabe-se que Endolimax nana e Iodamoeba butschlii são amebas que
não pertencem ao gênero Entamoeba. Estas espécies colonizam o intestino
humano, vivendo na condição de organismos comensais, aos quais não se
atribuíu, até o momento, ação patogênica. Em 1918, a espécie Dientamoeba
fragilis foi originalmente descrita como uma espécie de ameba. Recentemente,
algumas evidências sugerem que este parasito é um flagelado que pertence
ordem Trichomonadida e ao gênero Dientamoeba (STARK et al., 2006).
Tradicionalmente, o diagnóstico das amebas intestinais inclui métodos
parasitológicos que identificam o parasito através da microscopia óptica. Embora,
largamente utilizado, a diferenciação entre as espécies de amebas pelas análises
morfológicas, de modo geral, não é uma tarefa fácil. Visto que um dos principais
critérios de identificação baseia-se no número de núcleos presente no cisto
maduro, na disposição da cromatina distribuída em torno da membrana nuclear,
na localização e tamanho do cariossoma, no formato do corpo cromatóide quando
presente nos cistos e nas dimensões de ambas formas evolutivas. A seguir, os
quadros mostram um resumo das características morfológicas de trofozoítos e
cistos utilizados na diferenciação entre espécies de amebas que parasitam o
intestino humano.
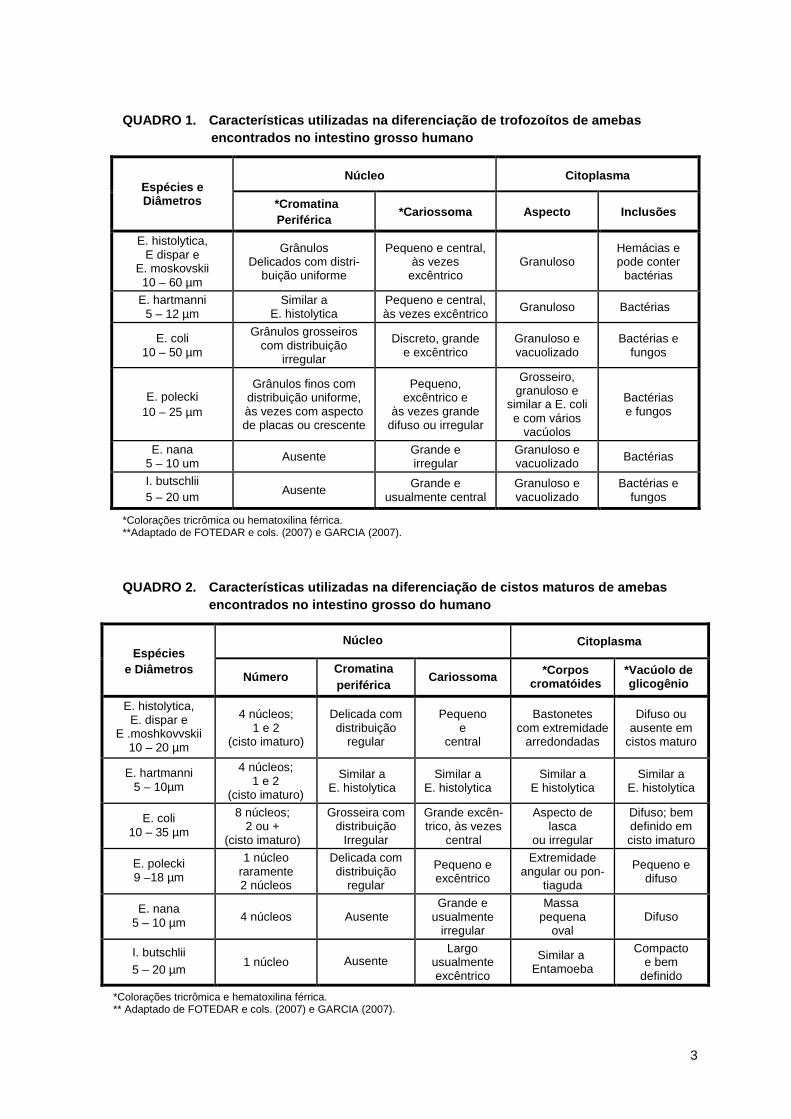
3
QUADRO 1. Características utilizadas na diferenciaç ão de trofozoítos de amebas encontrados no intestino grosso humano
Espécies e Diâmetros
Núcleo Citoplasma
*Cromatina Periférica
*Cariossoma Aspecto Inclusões
E. histolytica, E dispar e
E. moskovskii 10 – 60 µm
Grânulos Delicados com distri-
buição uniforme
Pequeno e central, às vezes
excêntrico Granuloso
Hemácias e pode conter
bactérias
E. hartmanni 5 – 12 µm
Similar a E. histolytica
Pequeno e central, às vezes excêntrico Granuloso Bactérias
E. coli 10 – 50 µm
Grânulos grosseiros com distribuição
irregular
Discreto, grande e excêntrico
Granuloso e vacuolizado
Bactérias e fungos
E. polecki 10 – 25 µm
Grânulos finos com distribuição uniforme, às vezes com aspecto de placas ou crescente
Pequeno, excêntrico e
às vezes grande difuso ou irregular
Grosseiro, granuloso e
similar a E. coli e com vários
vacúolos
Bactérias e fungos
E. nana 5 – 10 um Ausente Grande e
irregular Granuloso e vacuolizado Bactérias
I. butschlii 5 – 20 um
Ausente Grande e usualmente central
Granuloso e vacuolizado
Bactérias e fungos
*Colorações tricrômica ou hematoxilina férrica. **Adaptado de FOTEDAR e cols. (2007) e GARCIA (2007).
QUADRO 2. Características utilizadas na diferenciaç ão de cistos maturos de amebas encontrados no intestino grosso do humano
Espécies e Diâmetros
Núcleo Citoplasma
Número Cromatina periférica
Cariossoma *Corpos cromatóides
*Vacúolo de glicogênio
E. histolytica, E. dispar e
E .moshkovvskii 10 – 20 µm
4 núcleos; 1 e 2
(cisto imaturo)
Delicada com distribuição
regular
Pequeno e
central
Bastonetes com extremidade
arredondadas
Difuso ou ausente em
cistos maturo
E. hartmanni 5 – 10µm
4 núcleos; 1 e 2
(cisto imaturo)
Similar a E. histolytica
Similar a E. histolytica
Similar a E histolytica
Similar a E. histolytica
E. coli 10 – 35 µm
8 núcleos; 2 ou +
(cisto imaturo)
Grosseira com distribuição
Irregular
Grande excên- trico, às vezes
central
Aspecto de lasca
ou irregular
Difuso; bem definido em cisto imaturo
E. polecki 9 –18 µm
1 núcleo raramente 2 núcleos
Delicada com distribuição
regular
Pequeno e excêntrico
Extremidade angular ou pon-
tiaguda
Pequeno e difuso
E. nana 5 – 10 µm 4 núcleos Ausente
Grande e usualmente
irregular
Massa pequena
oval Difuso
I. butschlii 5 – 20 µm
1 núcleo Ausente Largo
usualmente excêntrico
Similar a Entamoeba
Compacto e bem
definido
*Colorações tricrômica e hematoxilina férrica. ** Adaptado de FOTEDAR e cols. (2007) e GARCIA (2007).

4
Vale ressaltar que a presença de características sobrepostas nas
diferentes amebas dificulta a identificação do parasito, sendo necessária a análise
de várias formas evolutivas e de estruturas internas bem preservadas (LEBER &
NOVAK, 2005), o que nem sempre é possível observar em amostras de fezes má
acondicionadas, colhidas sem conservantes e observadas sem métodos de
coloração. Nota-se ainda que, a sutileza da morfologia, a percepção microscópica
aliada ao conhecimento, a experiência profissional, a qualidade óptica e o nível de
parasitemia na amostra fecal são fatores que afetam os resultados e que nem
sempre constituem um processo inteiramente fidedigno.
Existe um consenso geral e conceitual, entre os pesquisadores, de que
a amebíase é uma parasitose humana causada pelo protozoário E. histolytica. Em
1997, a Organização Mundial de Saúde anuiu que E. histolytica e E. dispar eram
duas espécies morfologicamente idênticas à microscopia óptica, formando o
complexo E. histolytica/E. dispar, sendo a primeira patogênica e invasiva no
organismo humano e a segunda se limitando ao lúmen do intestino (WHO, 1997).
Mas, recentemente, vários pesquisadores têm mencionado a inclusão de E.
moshkovskii como parte deste complexo, em decorrência dos casos de infecções
parasitárias concomitantes destas espécies e da elevada frequência de E.
moshkovskii em algumas populações estudadas na região de Dhaka na Índia,
Sidney na Austrália, Bangkok na Tailândia e Tanzânia na África (ALI et al., 2003;
PARIJA & KHAIRNAR, 2005; HAMZAH et al., 2006; FOTEDAR et al., 2007; BECK
et al., 2008). Este achado sugere que esta espécie não é simplesmente uma
espécie de vida livre, mas um parasito próprio da espécie humana. Estes estudos
utilizaram a PCR na diferenciação e detecção de rDNA destas espécies.
Entre as espécies de amebas entéricas, somente E. histolytica é
reconhecidamente capaz de invadir o organismo humano e causar doença em
determinadas condições (TANYUKSEL & PETRI, 2003). As demais espécies são
consideradas como comensais e não invasivas. Porém, a identificação destas

5
espécies é uma condição essencial pela possibilidade de serem confundidas com
E. histolytica. Ademais, o conhecimento epidemiológico das amebas é de suma
importância, já que os dados epidemiológicos contribuem de forma consistente
nas questões e resoluções relacionadas a problemas de saúde pública. Assim
sendo, é necessário realizar o diagnóstico diferencial entre as espécies de
amebas que habitam o intestino humano, a fim de orientar a conduta terapêutica,
evitar o tratamento empírico e obter dados epidemiológicos. Os sintomas da
amebíase, de uma maneira geral, são variados. Em torno de 90%, as infecções
são assintomáticas, constituindo o hospedeiro um vasto reservatório do parasito.
Neste caso, E. histolytica na luz intestinal se comporta como um simples
comensal, não existindo nenhuma evidência de infecção, a não ser a eliminação
de cistos nas fezes. Estes indivíduos, a qualquer momento da sua vida, podem
desenvolver as formas clínicas da doença que variam de colite não disentérica,
disenteria amebiana e amebíase extraintestinal (TANYUKSEL & PETRI, 2003). A
colite não disentérica apresenta sintomas vagos, que muitas vezes passam
despercebidos. Manifesta-se por evacuações diarreicas, podendo conter sangue
ou muco, embora alguns pacientes nem apresentem diarreia. Cólicas e
desconforto abdominal podem surgir e, raramente, observa-se febre. Esta
infecção é caracterizada por períodos silenciosos alternados por manifestações
clínicas (HAQUE et al., 2003; TANYUKSEL & PETRI, 2003). Já na disenteria
amebiana, o quadro clínico geralmente se instala de forma insidiosa, com dor
abdominal, flatulência, diarreia com evacuações muco sanguinolentas e febre
moderada. Pode ocorrer de oito a dez evacuações diárias. As complicações da
amebíase intestinal resultam da confluência das úlceras e necrose do cólon e são
muito variáveis, podendo atingir cerca de 4% dos casos. As mais comuns são:
perfurações intestinais com peritonite, hemorragia, colites pós-disentéricas e
estenoses. O ameboma é encontrado no ceco, geralmente como uma lesão
única, formada por massas granulomatosas que podem ser confundidas com

6
neoplasia (HAQUE et al., 2003). Na amebíase extraintestinal a manifestação
clínica mais comumente encontrada é a necrose liquefativa do fígado (abscesso
hepático). O lobo direito do fígado é frequentemente mais afetado do que o
esquerdo. O parasito provoca um processo inflamatório difuso, degeneração
celular e necrose liquefativa do parênquima, em consequência da ação
enzimática dos trofozoítos. O indivíduo apresenta dor no hipocôndrio direito, febre
intermitente e irregular, hepatomegalia e fraqueza geral. Se houver infecção
bacteriana concomitante ocorre o agravamento do quadro clínico (TANYUKSEL &
PETRI, 2003). De 2% a 20% dos indivíduos sintomáticos evoluem para doença
grave, como: formação de abscesso hepático, perfuração intestinal, peritonite,
ameboma, podendo ainda com menor frequência, desenvolver lesões em outros
órgãos como pulmão, cérebro e região cutânea (AQUINO, 2001; RIGOTHIER et
al., 2002; STANLEY, 2003).
De acordo com as normas estabelecidas pela OMS, o tratamento da
amebíase deverá ser adotado somente nos casos que E. histolytica for
especificamente confirmada devido à possibilidade de se desenvolver resistência
ao Metronidazol (CLARK, 1998; WASSMANN et al., 1999). O tratamento com este
fármaco permanece efetivo no combate à doença, mas as taxas de reinfecção
são altas (CARRERO et al., 2007). Há relatos de pacientes que não respondem
ao tratamento, o que pode ser devido a uma resistência dos trofozoítos expostos
a múltiplas drogas ou ao aparecimento de uma cepa resistente (ABD-ALLA et al.,
2006). Segundo CLARK (1998), para se diferenciar as amebas intestinais e
reduzir o uso desnecessário de agentes antiparasitários, são necessários testes
diagnósticos precisos.
A amebíase é amplamente distribuída no mundo. Cerca de 10% da
população mundial encontra-se infectada por E. histolytica, sendo que 90% dos
indivíduos parasitados não apresentam sintomatologia clínica (STANLEY, 2003).
Estima-se que 50 milhões de pessoas desenvolvem amebíase intestinal, e que

7
10% dos indivíduos com colite amebiana desenvolverão abscesso hepático
(RIGOTHIER et al., 2002). Em uma escala global, as formas graves da amebíase
determinam cerca de 40 a 110 mil mortes por ano. No entender de Ximenez e
cols. (2009), a estimativa da taxa de infecção por E histolytica no mundo pode não
refletir a situação real, visto que, estes dados foram obtidos através da análise
morfológica e/ou sorologia como descrito por Walsh (1986). Segundo Pritt & Clark
(2008) os dados epidemiológicos relatados sobre a amebíase, ao longo dos
últimos anos, foram ambíguos porque foram baseados principalmente na análise
morfológica.
A presença de E. histolytica nos países desenvolvidos está confinada a
agregados populacionais, sendo que os principais grupos acometidos são os
viajantes de áreas endêmicas, imigrantes, homossexuais masculinos e indivíduos
internados em instituições coletivas (asilos, orfanatos e manicômio) (TANYUKSEL
& PETRI, 2003). O contato direto entre o indivíduo infectado por E. histolytica e o
indivíduo sadio certamente constitui a mais importante fonte de infecção em
gregados populacionais com elevado grau de promiscuidade e baixo nível
higiênico. Segundo Evangeloupos e cols. (2000), a detecção e diferenciação entre
E. histolytica e E. dispar é de grande importância, tanto para o diagnóstico, quanto
para os estudos epidemiológicos. Inúmeros estudos têm sido realizados depois do
reconhecimento de E. dispar como uma nova espécie de Entamoeba
morfologicamente semelhante a E. histolytica, visando à distinção entre as
espécies e o melhor entendimento da incidência e prevalência da doença.
Na Grécia, Evangelopoulos, Legakis & Vakalis (2001) realizaram um
estudo epidemiológico para estimar a prevalência de E. histolytica e E. dispar,
através da análise por técnicas de microscopia e de ELISA para detecção de
antígenos e PCR. Os resultados mostraram uma baixa prevalência destes
parasitos na Grécia. A PCR foi o método de escolha para o diagnóstico da
amebíase quando comparado com o teste de ELISA. Posteriormente, Antoniou e

8
cols. (2002) realizaram um estudo epidemiológico utilizando métodos sorológicos,
observando uma soroprevalência de 2,5% na Grécia.
O Japão é um dos poucos países desenvolvidos onde os casos de E.
histolytica são autóctones. Entretanto, no Japão, a amebíase é prevalente
somente em populações restritas: homossexuais masculinos e residentes de
instituições mentais (HAGHIGHI et al., 2002).
Tanyuksel & Petri (2003), relataram que nos países desenvolvidos
como a Itália, Japão e Estados Unidos, a prevalência de E. histolytica na
população de homossexuais masculino está entre 4 a 21%, mas a maioria das
infecções é causada por E. dispar, a qual não requer tratamento.
Em relação a prevalência da amebíase, algumas regiões da África,
Ásia, América Central e do Sul se destacam no cenário mundial, devido aos
baixos níveis sócio-econômicos e higiênico-sanitários da população (TANYUKSEL
& PETRI, 2003).
O México é um dos países do mundo que mais sofre com a
enfermidade amebiana, onde 15% dos casos de disenteria aguda em crianças,
que requerem hospitalização, estão associados a infecção por E. histolytica
(GUTIERREZ, 1986). Palacio-Sanchez e cols. (1997) realizaram um estudo
epidemiológico na cidade de Chihuahua, no México e observaram uma frequência
de 16% quando utilizaram a pesquisa de antígenos de E. histolytica nas fezes.
Na cidade de Bangladesh, Haque e cols. (1997), realizaram um estudo
com 1049 crianças com diarreia e 987 crianças assintomáticas, observando que a
prevalência de E. histolytica e de E. dispar era quase a mesma nas crianças com
diarreia. Entretanto, nas crianças assintomáticas da área rural, a infecção por E.
dispar foi sete vezes mais comum.
Em 2003, Verweij e cols. encontraram alta prevalência do complexo E.
histolytica/E. dispar pelo EPF em amostras fecais oriundas de uma área rural de
Ghana, porém quando utilizaram a “PCR em tempo real”, identificaram somente

9
um caso de E. histolytica e uma alta frequência de E. dispar (VERWEIJ et al, 2003
b). Posteriormente, Kebede e cols. (2004), observaram diferentes percentuais de
prevalência na Etiópia quando compararam seus resultados obtidos pela técnica
de PCR com o do exame parasitológico de fezes.
No Brasil, o número de indivíduos infectados por E. histolytica oscila de
região para região, apresentando percentuais extremamente variáveis. Observa-
se alta prevalência na região norte e baixa prevalência na região nordeste,
sudeste e sul. Na região amazônica, os casos de amebíase intestinal e
extraintestinal são mais frequentes e graves do que nas demais regiões (GOMES
et al., 2000b). Contudo, ainda não existe uma real estimativa da prevalência da
amebíase no território Brasileiro. A presença do Complexo E. histolytica/E. dispar
tem sido relatada em vários estados como Pará (SILVA et al., 2005), Rondônia
(ORLANDI et al., 2001), Pará (MIRANDA et al., 1998), São Paulo (LUDWIG et al.,
1999; MARTINS et al., 2007), Bahia (SANTOS, SANTOS & SOARES, 2007), Rio
Grande do Sul (ASSIS et al., 2003); Santa Catarina (ANDRADE et al., 2008)
Minas Gerais (ROCHA et al., 2000) Piauí (ALVES et al., 2003); Ceará (BRAGA et
al., 1998; 2001) e Rio de Janeiro (SANTOS et al., 2007).
Um estudo realizado em Manaus por Menezes (1986), identificou
vários casos de amebíase hepática e demonstrou a predominância de casos
sintomáticos (65,2%) sobre os assintomáticos. Porém, as formas de colite não
disentérica foram as mais comuns entre os indivíduos sintomáticos. Em 2000,
Póvoa e cols. relataram que 29% das amostras analisadas de indivíduos
moradores da região metropolitana de Belém, Pará, foram positivas para E.
histolytica quando utilizaram a detecção de antígeno nas fezes. Posteriormente,
Silva e cols. (2005) encontraram uma prevalência de 29,35% de indivíduos
infectados por E. histolytica na grande Bélem/Pará, quando empregaram o
conjunto de diagnóstico Entamoeba histolytica test II da TechLab.

10
Por sua vez, Benetton e cols. (2005) encontraram na cidade de
Manaus uma prevalência de 21,5% de indivíduos infectados por E. histolytica/E.
dispar e destes apenas 1,5% foram positivos por E. histolytica.
Em 2001, Braga e cols. realizaram um estudo em uma favela no
Estado de Ceará/Fortaleza, Nordeste do Brasil, através de EPF e detecção de
antígenos nas fezes (ELISA). Observaram que 25% das amostras eram positivas
para o complexo E. histolytica/E. dispar. Neste estudo a prevalência de E.
histolytica foi de 15% quando empregaram a pesquisa de antígeno nas fezes.
Na cidade de Recife, em Pernanbuco, Pinheiro e cols. (2004),
realizaram um estudo sobre a prevalência de E. histolytica, observando somente
a presença de E. dispar quando utilizaram a PCR e a pesquisa de antígeno nas
fezes.
Santos e cols. (2007), utilizaram a técnica de Multiplex-PCR para
diferenciar o complexo E. histolytica/E. dispar descrita previamente por Nunez e
cols. (2001). O teste foi empregado para avaliar a ocorrência e o perfil da
amebíase em duas comunidades do Estado do Rio de Janeiro. Os resultados
mostraram uma discrepância entre o EPF e a Multiplex-PCR, que pode estar
relacionada com diferenças de sensibilidade e especificidade dos métodos. Um
número significativo de amostras fecais que apresentavam o complexo E.
histolytica/E. dispar pelo EPF, foi negativo para as duas espécies na PCR. Assim,
os autores sugeriram que esta discrepância poderia estar refletindo a presença de
outras amebas comensais que não fazem parte do complexo E. histolytica/E.
dispar.
Vale ressaltar que é bem provável que a verdadeira prevalência e
incidência de E. histolytica seja hiperestimada, devido à presença de outras
espécies morfologicamente semelhantes em nosso meio, associada às
dificuldades encontradas na diferenciação destas espécies e à falta de dados
epidemiológicos contundentes.

11
Sabe-se que as prevalências de amebas intestinais comensais em
nosso meio são escassas, principalmente em relação as espécies, E hartmanii e
E. moshkovskii. Infelizmente, os estudos sobre prevalência, em sua maioria,
utilizaram diferentes métodos de diagnóstico, bem como diferentes peculiaridades
locais e características das populações selecionadas, o que dificulta uma análise
comparativa destes dados relativos à frequência de amebas.
O diagnóstico clínico é raramente uma circunstância com margem de
segurança satisfatória nas parasitoses, como sucede na maioria das doenças. Os
indivíduos com amebíase apresentam sintomas não específicos que podem ser
confundidos com os de outras doenças, tais como: criptosporidiose, salmonelose,
shigelose, rotavirose, adenovirose, doença inflamatória intestinal, carcinoma,
colite isquêmica, diverticulite, fazendo com que o diagnóstico laboratorial seja
uma ferramenta vital para confirmar ou descartar a suspeita da infecção por E.
histolytica (TANYUKSEL & PETRI, 2003; PRITT & CLARK, 2008). Para tal fim, é
indispensável um método preciso, sensível, específico para a determinação do
diagnóstico etiológico com adoção de medidas preventivas e de controle da
doença.
O diagnóstico laboratorial da amebíase atualmente pode ser feito
através de uma variedade de técnicas, utilizadas individualmente ou em conjunto,
como o exame parasitológico de fezes (EPF) e métodos moleculares como a
pesquisa de antígenos e de DNA nas fezes (HAQUE et al., 2003; STANLEY,
2003).
O exame parasitológico de fezes é baseado em critérios morfológicos
das formas evolutivas do parasito presente nas fezes. Nas fezes diarreicas
predominam os trofozoítos, sendo recomendada a realização do método direto
em fezes recém emitidas. Já nas fezes pastosas encontramos as formas císticas,
sendo empregados métodos de concentração. Estes métodos são baseados em
dois princípios: a flutuação dos parasitos em solução de alta densidade por

12
centrifugação, como a solução de sulfato de zinco e a sedimentação espontânea
ou por centrifugação.
Os métodos coproscópicos têm sido largamente utilizados na detecção
de amebas intestinais. Todavia, estes métodos não são capazes de diferenciar o
complexo E. histolytica/E. dispar, nem as outras espécies de amebas
morfologicamente semelhantes. Assim, a análise morfológica não é
suficientemente informativa para o diagnóstico preciso das espécies, exceto
quando se evidencia a presença de eritrócitos fagocitados no interior do
trofozoítos, característica esta restrita a E. histolytica (PETRI & SINGH,1999;
STANLEY, 2003). Este achado é extremamente raro em amostras de fezes
diarreicas recém emitidas. Ademais, as formas císticas são eliminadas nas fezes
intermitentemente, podendo o seu número variar de dia para dia. Assim, o exame
parasitológico de fezes realizado em fezes coletadas em dias alternados, dentro
de um período de 10 dias, como geralmente recomendado, aumenta a
sensibilidade do EPF.
É indispensável que o microscopista seja capaz de distinguir as formas
evolutivas de artefatos presente nas fezes, tais como: células epiteliais,
leucócitos, macrófagos, leveduras. Ainda assim, vale ressaltar, que os métodos
de concentração empregados na análise de cistos apresentam uma variação
substancial em relação à sensibilidade quando comparados entre si. Já está bem
estabelecido que o método de flutuação por centrifugação, utilizando sulfato de
zinco a 33% com uma densidade 1,18 (FAUST et al., 1938) é o mais indicado
para o diagnóstico de ameba intestinais (TOBIE et al., 1951; BARRETO, 1962;
REY, 2008). Em geral, na rotina laboratorial para a visualização e identificação
direta de trofozoítos utiliza-se o corante temporário lugol, da mesma forma que
em amostras previamente concentradas. Entretanto, na maioria das vezes, é
necessário utilizar as técnicas de colorações permanentes, tais como: tricrômica

13
ou hematoxilina férrica, pois permite uma melhor visualização das características
morfológicas mencionadas nos quadros 1 e 2.
A caracterização de padrões isoenzimáticos é um método bioquímico
pioneiro para diferenciar E. histolytica de E. dispar e outras espécies de amebas.
Este método é considerado “padrão ouro”, cujo princípio reside na mobilidade
eletroforética das isoenzimas da via glicolítica (hexoquinase, fosfoglicomutase,
malato desidrogenase e fosfoisomerase) de trofozoítos de cultura destas amebas
(SARGEAUNT et al.,1978; ORTNER, et al.,1997). Estas isoenzimas foram
agrupadas em padrões eletroforéticos (zimodemas) capazes de diferenciar as
espécies. O comportamento eletroforético da isoenzima hexoquinase é
considerado marcador de patogenicidade (BLANC & SARGEAUNT, 1991;
ACKERS, 2002). No entanto, a eficácia deste método apresenta certas restrições,
como a necessidade de uma cultura prévia, por um período de 7 a 14 dias, que
não positivam em aproximadamente 30% das amostras de fezes positivas para
cistos (ACKERS, 2002), tornando o método inviável para a rotina laboratorial.
Sendo assim, os cultivos de amebas são de valor para estudos bioquímicos e
imunológicos, produção de antígenos e anticorpos, estudos diferenciais entre
cepas patogênicas e não-patogênicas (zimodemos), triagem de novos
medicamentos in vitro, infecção de animais de laboratório como modelo
experimental em estudos de patogenia e para conhecimento da organização do
parasito em nível ultraestrutural (VISVESVARA & GARCIA, 2002).
A cultura é considerada um método de detecção menos sensível do
que a detecção de antígenos nas fezes e a detecção de anticorpos no soro.
Sabemos que o índice de positividade da cultura varia de acordo com a forma
clínica da doença. Em amostras de indivíduos assintomáticos, a cultura mostrou-
se menos sensível do que a detecção de antígenos nas fezes (ABD-ALLA et al.,
2000). Por sua vez, Sheehan e cols. (1979), relataram que a detecção de
anticorpos anti-E. histolytica no soro de indivíduos com evidências clínicas de

14
amebíase mostrou-se mais sensível que os resultados obtidos pela cultura. Parija
& Rao (1995) recomendaram o uso da cultura no diagnóstico da amebíase, após
ter realizado um estudo de comparação entre a cultura de E. histolytica com o
método direto e de concentração utilizando formol–éter.
Segundo Evangelopoulos e cols. (2000), em amostras com infecção
mista por amebas, não podemos excluir que uma espécie possa suprimir o
crescimento da outra. Ademais, em regiões que apresentam uma alta prevalência
de Blastocystis hominis é frequentemente encontrado uma associação com E.
histolytica e E. dispar no cultivo poliaxênico. O crescimento exacerbado B.
hominis no meio, se sobrepõe muitas vezes o da ameba, impedindo a
manutenção destas em cultura (GONÇALVES et al., 2007).
O diagnóstico definitivo de um processo infeccioso é a demonstração
do patógeno nos tecidos ou fluídos biológicos do hospedeiro. Porém, nem sempre
possível, devido à ausência do agente infeccioso, pela falta de sensibilidade dos
métodos utilizados ou por falhas técnicas (FERREIRA & ÁVILA, 2001).
O diagnóstico imunológico tem sido utilizado para a pesquisa de
antígeno e de anticorpo. Em áreas endêmicas, um dos maiores problemas
encontrados nos métodos imunológicos que detectam anticorpo é a incapacidade
de distinguir entre os indivíduos portadores de infecção recente ou passada, uma
vez que os títulos de anticorpos permanecem elevados por vários meses após a
cura da infecção. Entretanto, a detecção de anticorpo anti-E. histolytica tem valor
diagnóstico nos casos da amebíase extraintestinal, como no abscesso hepático,
onde altos títulos de anticorpos podem ser observados (SANCHEZ-GUILLÉN et
al., 2002).
Os testes sorológicos detectam anticorpos específicos para E.
histolytica em aproximadamente 95% dos pacientes com amebíase
extraintestinal, 70% dos pacientes com amebíase intestinal invasiva e em 5% dos
portadores assintomáticos (PETRI & SINGH, 1999; AQUINO, 2001).

15
Kraoul e cols. (1997) realizaram um estudo comparativo entre testes
sorológicos para a detecção de anticorpos anti-E. histolytica. Nesse estudo
utilizaram 143 amostras de soro, oriundas de indivíduos com quadro de abscesso
hepático amebiano, de indivíduos com outras doenças hepáticas e de indivíduos
saudáveis. Os autores mencionaram que a hemoaglutinação indireta possui uma
sensibilidade de 97,6% e uma especificidade de 97%, enquanto que o teste de
ELISA possui uma sensibilidade de 93% e uma especificidade de 100%.
Os anticorpos monoclonais (MAbs) vêm sendo empregados, ao longo
dos anos, com sucesso no laboratório clínico, através da identificação de
estruturas de diferentes moléculas de superfície celular e de organismos
patogênicos. Representa uma ferramenta, que apresenta especificidade para um
único determinante antigênico. Porém, reações cruzadas podem ser observadas,
uma vez que os micro-organismos apresentam epítopos homólogos em sua
estrutura antigênica. A aplicação de MAbs no imunodiagnóstico da amebíase vem
sendo utilizada na pesquisa de antígenos nas fezes. O conjunto de diagnóstico, E.
histolytica test (TechLab Inc., Blacksburgh, VA) é um teste de ELISA tipo captura,
amplamente utilizado. Este teste utiliza MAbs que reconhecem epítopos
altamente conservados presentes na proteína de adesão GAL/GALNac
encontrada na superfície de trofozoítos de E. histolytica. Segundo o fabricante é
possível detectar no mínimo 1000 trofozoítos em cada amostra analisada
(TELLEZ-SIERRA et al., 1992; GARCIA, SHIMIZU & BERNARD, 2000). A
primeira geração deste teste, o “Entamoeba Test’’, detectava lectina de aderência
GAL/GALNac de E. histolytica e E. dispar. Já o conjunto de diagnóstico Merlin
Optimun Kit (Merlin Diagnostika Bornheim-Hersel, Alemanha) utiliza Mabs que
reconhecem o antígeno de superfície rico em serina. Este teste requer pelo
menos 100 trofozoítos de E. histolytica na amostra analisada. Contudo, apresenta
reação cruzada na presença de alta concentração de antígenos de E. dispar
(MIRELMAN, NUCHAMOWITZ & STOLARSKY, 1997). Outros conjuntos de

16
diagnóstico, Alexon (ProspectT test, Alexon-Trend Ransey MN) e ENZYMEBA
Test utilizam anticorpos policlonais que reconhecem epítopos antigênicos de
E.histolytica e E. dispar (LUACES et al., 1993; MIRELMAN, NUCHAMOWITZ &
STOLARSKY, 1997; PILLAI et al., 1999).
Sabemos que quando os métodos tradicionais não são suficientes ou
definitivos para estabelecer o diagnóstico, os métodos moleculares são utilizados
como um método suplementar por serem eficientes e sensíveis.
Faz pouco mais de duas décadas que as técnicas de biologia
molecular têm sido utilizadas na área de diagnóstico de doenças parasitárias.
Presumivelmente, ainda na era pré-PCR, o uso de sondas específicas de DNA,
utilizada na detecção de E. histolytica pela técnica de “Dot blot” foi uma das
primeiras aplicações no diagnóstico molecular. Garfinkel e cols. (1989)
descreveram as sondas P145 e B133, obtidos a partir de fragmentos de DNA
digerido com enzimas de restrição. Estas sondas hibridizavam com DNA de cepas
de E. histolytica isoladas de pacientes sintomáticos e assintomáticos, sendo
consideradas como uma ferramenta em potencial para o diagnóstico. No mesmo
período, o grupo de pesquisa de Samuelson e cols. (1989), identificaram a
mesma sequência descrita por Garfinkel e a utilizou como sonda, que hibridizava
com DNA de E. histolytica, extraído de cistos concentrados diretamente do
material fecal. Posteriormente, estas sequências foram utilizadas por Bacha e
cols. (1990), num ensaio de hibridação realizado com DNA não amplificado
isolado diretamente de trofozoítos de cultura, oriundos de material fecal. Os
autores concluíram que o ensaio não apresentou sensibilidade suficiente para a
visualização de DNA em amostras clínicas com quantidades diminutas de
parasito.
Pouco tempo depois, surgiram os primeiros trabalhos utilizando a PCR
no diagnóstico de E. histolytica. Em 1991, Tachibana e cols. diferenciaram E
histolytica de isolados de indivíduos assintomático e sintomático através da PCR.

17
Os pares de iniciadores P1, P2, P3 e P4 utilizados codificavam parte do gene de
uma proteína de 30 kDa e o produto amplificado foi visualizado em gel de
agarose. Também, no ano seguinte, Clark e Diamond (1992), utilizaram a técnica
de perfil dos fragmentos obtidos por endonucleases de restrição (RFLP) para
diferenciar produtos amplificados do gene da SSU-rRNA em isolados
provenientes de pacientes sintomáticos e assintomáticos. Posteriormente, estes
autores utilizaram a PCR, tendo como alvo o gene SSU-rRNA para o diagnóstico
diferencial entre E. histolytica e outros protozoários intestinais. No entanto, além
dos iniciadores PsP5/PsP3 específicos para E. histolytica e o NPsp5/NpSP3 para
E. dispar utilizaram oligonucleotídeos iniciadores universais, RD5/RD3. O produto
amplificado utilizando os iniciadores universais foi incubado com enzimas de
restrição, mostrando fragmentos de restrição distintos para E. nana, E coli, E.
histolytica, E. hartmanni, E. moshkovskii e Blastocystis hominis (CLARK &
DIAMOND, 1992). No mesmo ano, Romero e cols. (1992) utilizaram iniciadores
que amplificava as sequências P145 e B 133 descritas por Garfinkel e cols.
(1989), sendo os produtos amplificados detectados através da técnica de “Dot
Blot”, utilizando as sondas P145 e B133 não radioativas. Neste trabalho, a PCR
detectou 42 das 45 amostras positivas para E. histolytica, onde 60% das amostras
eram oriundas de crianças com quadro de disenteria.
O primeiro estudo epidemiológico utilizando a PCR para o diagnóstico
diferencial entre E. histolytica e E. dispar foi o de Acuna-Soto et al. (1993). Neste
trabalho, os autores utilizaram a sequência descrita por Garfinkel e cols. (1989) e
amostras de fezes fixadas com formalina, oriundas de moradores de uma
comunidade rural no México. O produto amplificado foi visualizado em gel de
agarose corado com brometo de etídeo e também pela técnica de “Dot blot”. A
especificidade e sensibilidade da PCR foi de 96% e 98%, respectivamente.
Desde então, a PCR convencional tem sido amplamente utilizada na
detecção e na diferenciação entre E. histolytica e E. dispar ao longo dos últimos

18
anos. Várias publicações utilizando diferentes estratégias de amplificação de
DNA, baseado na PCR convencional, principalmente para o diagnóstico de E.
histolytica, E. dispar e algumas para E. moskovskii estão sumarizadas no quadro
3, onde destaca-se os principais alvos e oligonucleotídeos iniciadores publicados
ultimamente na literatura.
Entre as sequências gênicas descritas e que têm sido alvo da PCR na
diferenciação entre E. histolytica e E. dispar, destacam-se o gene SSU-rDNA e a
sequência repetitiva em tandem, presente em múltiplas cópias no DNA extra-
cromossômica, sendo estimado cerca de 200 cópias no genoma destas espécies
(BHATTACHARYA et al.,1989). Segundo ACKERS (2002), a utilização de
sequência alvo de múltiplas cópias propicia um aumento na sensibilidade da PCR.
Ademais, estudos sobre tipagem gênica no tocante da epidemiológica
molecular de E. histolytica têm sido extensivamente investigado, sendo menos
estudado com relação ao E. dispar. Os principais alvos moleculares destes
estudos são: o gene SREHP que codifica um antígeno de superfície (GHOSH et
al., 2000; HAGHIGHI et al., 2003; ZAKI et al., 2003; RIVERA et al., 2006), o gene
da quitinase (GHOSH et al., 2000; HAGHIGHI, et al., 2003), região intergênica
localizada entre o gene da actina e o da superóxido desmutase (GHOSH et al.,
2000), a região repetitiva no locus 1/2 e 5/6 de microsatélite (ZAKI & CLARK,
2001; ZAKI et al., 2002; HAGHIGHI, et al., 2003; PINHEIRO et al., 2005) e a
sequência repetitiva em tandem que flanqueia os genes do tRNA (ALI, ZAKI &
CLARK, 2005).

19
QUADRO 3. Sequência alvo e oligonucleotídeos inicia dores utilizado na PCR convencional descritos na literatura
Sequência Gene alvo
Tamanho do produto
amplificado Iniciadores Referência(s)
Proteína/ 30 kDa
100 bp 101 bp
374 bp
P11 /P12 (E.h) P13 / P14 ( E.d)
HF/HR (Eh/Ed)
Sanuki et al.,1997; Rivera et al., 1998; Haghighi et al., 2002; Pinheiro et al., 2004 e 2005; Tachibana et al., 1992, 2000 e 2001. Hooshyar et al., 2004
Hemolisina 256 bp Eh6F/Eh6R Zindrou et al., 2001
Actina 300 bp ActF/Act R Freitas et al.,2004
Cisteino protease
242 bp Ehcp6F/ Ehcp6R Freitas et al., 2004
M17 482 pb P1-S17/P1 As20 Tannich & Burchard, 1991; Gomes et al., 1997; 1999; Valle et al., 2000
SSU-rRNA
876 bp
880 bp
135 bp
900 bp
600 pb
1950 bp
Nested-PCR 1076 bp
427 bp/195 bp
260 pb
166 bp 752 bp/580pb
Psp F/PspR (E.h) NPspF/NPspR (Ed)
Eh5/Eh3 e Ed3/Ed5
Eh1/Ed1e Eh2
Eh1/Eh2 (Ed/Eh) EHP1/EHP2 (Eh) EHN1/ENH2(Ed)
Enta 1 /Enta2 Entamoeba sp
RD5/RD3
Entamoeba sp
E1/E2 (Ed/Eh) Eh1-L/Ed-R (Ed/Eh)
Em1/Em2 (Em) Em1/nEm (Em)
Enta F
EhR/EdR/EmR
Clark e Diamond ,1992; Mirelman et al., 1997; Ramos et al., 2000 ; Verweij et al., 2000; Lebbad & Svard, 2005; Moran et al., 2005; Troll et al.,1997; Heckendorn et al., 2002; Gonin & Trude, 2003; Calderaro et al., 2006; Katzwinkel-Wladarch et al., 1994; Haque et al., 1998; Zaman et al.,2000
Verweij et al., 2003 a
Clark & Diamond, 1992; Ramos et al., 2005;
Evangeloupos et al., 2000 e 2001; Pa-glia e Visca, 2004
Ali et al., 2003; Parija e Khairnar 2005; Fotedar et al., 2007; Beck et al., 2008
Hamzah et al., 2006
Sequência repetitiva em tandem localizada no
DNA extracromossômicro
132 bp 96 bp
145 bp 133 bp 145 bp 133 bp
EhP1 /EhP2 (E.h) EdP1/ EdP2 (E.d)
P1/ P2 (Eh) NP1/ NP2 (Ed)
EHP1 / EHP2 (E.h)
EHNP1/EHNP2
Nunez et al., 2003; Santos et al., 2007 Acuna-Soto et al., 1993; Aguirre et al., 1995; Britten et al., 1997; Romero et al., 1992
E.h = E histolytica; E.d = E. dispar; E.m = E. moshkovskii.

20
No Brasil, estudos de variabilidade genética de cepas de E. histolytica
têm sido realizados empregando técnicas do tipo DNA polimórfico amplificado ao
acaso, RAPD (GOMES et al., 2000 a; VALE et al., 2000), RNA amplificado
aleatoriamente, RAP-PCR (VALE et al., 2000), PCR em baixa estringência,
usando um único iniciador (Low Stringency single specific primer, LSSP-PCR)
(GOMES et al., 1997). Um estudo envolvendo cepa de E. dispar foi realizado por
Pinheiro e cols. (2005), onde avaliaram regiões polimórficas localizado no locus
1/2 e 5/6 da região de microsatélite.
A aplicação da PCR no diagnóstico da amebíase ainda é restrita aos
laboratórios de pesquisa ou de referência. Atualmente, ainda encontra-se em fase
de padronização, não havendo até o momento, um marcador molecular
padronizado, nem método de extração e de purificação de DNA, que tenha sido
avaliado em diferentes cenários e utilizado unanimemente para o diagnóstico.
Inicialmente, a PCR apresentou uma alta sensibilidade na detecção e
diferenciação de E. histolytica e E. dispar, quando foram utilizados DNA extraídos
de trofozoítos de cultura. No entanto, estes resultados não foram obtidos quando
o DNA utilizado foi extraído diretamente do material fecal. A amostra fecal é um
material extremamente heterogêneo e rico em substâncias inibidoras para a PCR.
Vale ressaltar que para a PCR ser exequível é necessário ter um método de
extração de DNA simples e rápido, obtendo-se DNA livre de substâncias
inibidoras da PCR. Assim, a extração de DNA de amostras clínicas desempenha
um papel fundamental para a obtenção de resultados satisfatórios da PCR.
Contudo, a extração de DNA das fezes é um processo complexo, em decorrência
da coextração de substâncias inibidoras e da presença dos resíduos
contaminantes, tais como: sais biliares, bilirrubina, etanol, complexos de
polissacarídeos. Essas substâncias interferem na ação da enzima DNA
polimerase ou aderem no DNA, podendo causar a degradação do mesmo
(DEUTER et al, 1995; MONTEIRO et al., 1997; WILSON, 1997; VANDENBERG &

21
VAN OORSCHOT, 2002). A remoção total das substâncias inibidoras nem
sempre é fácil. Porém, é possível verificar a presença destes inibidores na
amostra, introduzindo na reação controles internos de amplificação ou realizando
a amplificação em duplicata ou triplicata com diferentes diluições do DNA extraído
(NECHVATAL et al., 2008). Outra estratégia bastante utilizada é contaminar
(Spiking) a mistura de amplificação com DNA padrão nas amostras que
apresentaram resultados previamente negativos pela PCR (DA SILVA et al., 1997;
OMBROUCK et al., 1997; PELT-VERKUIL, BELKUM & HAYS, 2008b). Com estas
estratégias torna-se possível uma avaliação qualitativa da eficiência da extração.
Recentemente, alguns protocolos comerciais de extração e purificação de DNA
conseguiram melhorar as condições de isolamento de DNA fecal, mas nenhum
método ainda garante o isolamento de DNA livre de substâncias inibidoras.
Com o advento da PCR em tempo real, resultados melhores foram
alcançados na detecção e diferenciação de E. histolytica e E. dispar. Esta técnica
possibilita o monitoramento e a quantificação do fragmento de DNA, utilizando
como detector do produto amplificado, corantes ou sondas marcadas com
fluoróforo, específica para uma região interna da sequência alvo. A quantificação
do fragmento amplificado ocorre à medida que o sinal de fluorescência é emitido
durante a extensão de cada ciclo, sendo diretamente proporcional à quantidade
de produto de PCR. Várias estratégias de monitoramento da amplificação pela
PCR em tempo real têm sido adotadas. O método mais simples é o que utiliza
corante que se intercala na dupla fita de DNA, o SYBR GREEN 1. Já o sistema
TaqMan, emprega sonda que hibridiza especificamente com a sequência alvo.
Durante o processo de polimerização dos nucleotídeos, a sonda é degradada e a
fluorescência liberada é capturada pelo sistema detector e amplificador de sinal.
Existem outros métodos que empregam sondas fluorescentes como a farol
molecular (molecular beacon) e o LightCycler. O farol molecular utiliza uma sonda
que é um DNA de fita simples que forma uma estrutura secundária entre as

22
extremidades 3’ e 5’. Assim, no momento que a sonda encontra o seu alvo,
durante a etapa de amplificação, a mesma assume uma mudança
conformacional, tornando-a capaz de emitir fluorescência. Já o sistema
LightCycler utiliza duas sondas de hibridação que só produzem fluorescência
quando estão hibridizadas juntas na sequência-alvo.
Alguns métodos da PCR em tempo real têm sido empregados na
detecção e diferenciação de E histolytica e E. dispar nos últimos anos.
Blessemann e cols. (2002) amplificaram um fragmento de 310 bp contido no SSU-
rRNA, utilizando o sistema Light Cycler. A PCR foi capaz de detectar menos de
0,1 parasito por grama de fezes. No ano seguinte, Verweij et al. (2003b) utilizaram
o gene SSU-rRNA e a sequência epissômica repetitiva, descrita por Garfinkel et
al. (1989), em dois ensaios de PCR em tempo real, empregando sondas do tipo
TaqMan. Neste trabalho foram utilizadas 192 amostras de fezes obtidas de
indivíduos moradores na região Norte de Gana. Nesta amostragem foi
evidenciada somente uma amostra positiva para E. histolytica, o que revelou um
elevado percentual de E. dispar na população estudada. Posteriormente, Roy e
cols. (2005) empregaram o sistema “molecular beacon” na identificação de E.
histolytica e E. dispar em secreção hepática e em amostras de fezes. Também,
em 2005, Qvarnstrom e cols., em seu trabalho singular, compararam três métodos
de PCR em tempo real, até então publicado na literatura, além de adaptar a PCR
descrita por Clark e Diamond (1992) para um ensaio do tipo PCR em tempo real,
utilizando o corante SYBR-GREEN 1. O limite de detecção e a eficiência de cada
ensaio foi testado utilizando DNA de amostras fecais, amostras artificialmente
contaminada e de trofozoítos de cultura. Os autores concluíram que o método que
apresentou uma maior sensibilidade foi o sistema TaqMan, que amplifica o
segmento gênico do SSU-rDNA, descrito por Verweij et al. (2003 b).
A PCR em tempo real realiza a detecção e a quantificação do DNA de
maneira precisa e com maior sensibilidade. Este método tem mostrado ser mais

23
sensível na detecção simultânea de E. histolytica e E. dispar em amostras fecais,
principalmente em indivíduos com infecção mista, quando comparado com a PCR
convencional (QVARNSTROM et al., 2005), além de ser um método bastante
propício para o laboratório de diagnóstico pela sua característica de eliminar a
análise pós-PCR, diminuindo, assim, seu tempo de execução e, também,
minimizando o risco de contaminação do ambiente do laboratorial com produtos
amplificados.
O emprego da PCR em tempo real em ensaios do tipo multiplex para a
detecção simultânea de E. histolytica, Giardia lamblia e Cryptosporidium parvum
em amostras fecais foram padronizados por Verweij et al. (2004) e Haque et al.
(2007).
Cabe ressaltar que a PCR em tempo real tem sido utilizada como uma
interface experimental na elaboração de teste do tipo multianalítico, que utiliza
uma plataforma de instrumentação que pode ser implementada nos laboratórios
clínicos. Mesmo assim, ainda restam alguns problemas, tendo como a principal
desvantagem da PCR em tempo real em relação a PCR convencional, a sua
limitação quanto ao número de fragmentos de DNA a serem detectados. Os
equipamentos de PCR em tempo real possuem um número limitado de
grupamentos fluorescente, isto ganha um forte impacto quando se pretende
identificar vários alvos simultaneamente.
Outros fatores que limitam a amplificação do DNA, de uma maneira em
geral, é a necessidade prévia de regiões alvos, parcialmente ou totalmente,
sequenciadas. Isto restringe bastante a expansão da PCR para estudo de micro-
organismos com um pequeno número de sequências disponíveis nos bancos
públicos de DNA, GenBank. No entanto, o aumento do número de “projetos
genoma” de vários organismos tem proporcionado a identificação de várias
sequências gênicas, que podem ser utilizadas para o desenvolvimento de testes
de diagnóstico, baseado na PCR. O sequenciamento do genoma total de E.

24
histolytica foi concluído em 2005 (LOFTUS et al. 2005). Já os sequenciamentos
do genoma de E. moshkovskii e E. dispar estão na etapa final de montagem.
Paradoxalmente, somente as sequências que codificma o gene SSU-rRNA de E.
hartmanii, E. poleckii, e E. coli estão disponíveis no GenBank. De fato, os dados
moleculares que fornecem informações importantes para a avaliação da
diversidade inter e intraespecífica de amebas que pertencem ao gênero
Entamoeba são escassos. A única sequência gênica que é comum a todas as
amebas que compõe o gênero Entamoeba, atualmente disponível no GenBank, é
o gene da SSU-rRNA. Isto reduz bastante a expansão de metodologia e
estratégias de diagnóstico para o estudo das espécies de amebas.
Em 1992, Clark e Diamond utilizaram os iniciadores universais,
RD5/RD3 para a amplificação de um segmento do gene SSU-rDNA. O
diagnóstico diferencial entre E. histolytica e outros protozoários intestinais foi
realizado por meio da análise de restrição de fragmentos polimórficos. Somente
uma década após, a mesma estratégia para a diferenciação de espécies de
ameba foi utilizada por Verweij e cols. (2003a). Estes autores, descreveram o
método de hibridação reversa em membrana como intuito de identificar espécies
do gênero Entamoeba. Inicialmente foi empregada a PCR que amplifica parte do
gene do SSU-rDNA das amebas, empregando os iniciadores Entam1 e Entam2
biotinilados. O produto amplificado biotinilado foi hibridizado com sondas ligadas
na membrana de nitrocelulose e a formação dos híbridos foram detectados e
visualizados por meio de adição de substrato quimiluminescente seguida da
autorradiografia.
O DNA ribossômico é uma ferramenta importante e que tem sido
utilizado na avaliação de polimorfismo de micro-organismo de uma maneira em
geral. A diferenciação genética entre as populações de uma espécie constitui o
primeiro estágio da divergência evolutiva. Estas diferenças resultam, na maioria
das vezes, da ação de diferentes ambientes a que cada população está

25
submetida ao longo dos anos sobre a variabilidade preexistente na espécie.
Assim, as diferenças interespécies podem ser avaliadas. Sabemos que o gene
SSU-rDNA está presente em multicópia no DNA extracromossômico e representa
cerca de 200 cópias no genoma de E. histolytica e E. dispar (ALI, CLARK &
PETRI, 2008). Esta sequência possui regiões altamente conservadas e
intercaladas com sequências polimórficas entre o gênero Entamoeba, o que
propicia a sua utilização para desenhos de iniciadores universais ou semi-
universais capazes de amplificar múltiplos produtos, além de sondas do tipo
espécie específica, utilizadas na detecção de sequência-alvos empregadas no
diagnóstico de amebas intestinais.
Uma alternativa utilizada na identificação de espécies de parasitos é
através da reação de sequenciamento de DNA. É uma técnica altamente
fidedigna, porém, trabalhosa, dispendiosa, demorada e que não é prática como
ferramenta de diagnóstico em laboratório clínico. Mas, permite identificar,
caracterizar e avaliar a aplicabilidade das sequências consideradas como uma
potente ferramenta para o diagnóstico. Nos últimos anos, várias tecnologias têm
emergido como uma ferramenta diagnóstica usando como princípio o estilo
multianalítico. Assim, o processo de diagnóstico torna-se rápido, vários patógenos
podem ser detectados simultaneamente, sendo menos dispendioso e decaindo o
tempo de trabalho porque estes métodos podem ser parcialmente automatizados.
A grande vantagem dessa metodologia consiste na capacidade de atender a uma
forte demanda no cenário de diagnóstico por métodos multianalíticos, onde
poucos procedimentos podem diagnosticar uma grande quantidade de micro-
organismos.
Genes de diversos patógenos, dentro do mesmo grupo
taxonomicamente relacionados, podem ser amplificados quando empregamos
iniciadores universais para este grupo. Estes iniciadores são capazes de
amplificar DNA de múltiplos patógenos simultaneamente na amostra clínica.

26
Assim, ensaios do tipo microarranjo podem ser usados para identificar os
produtos amplificados através da reação de hibridação empregando sondas
patógenos-específicas.
Sabemos que a ideia de imobilizar e hibridizar moléculas em suporte
sólido foi primeiro proposta por Denhardt (1966), conduzindo ao desenvolvimento
de metodologias de identificação de sequência no DNA genômico ou oriunda da
clonagem molecular. Posteriormente, o segundo passo foi dado por Southern
(1975), quando demonstrou que poderia ser feita a transferência de DNA do gel
para membranas de nylon. Segundo Dyson (1991), desde as descobertas de
Denhardt e Southern, as técnicas de hibridação em suporte sólidos tornaram-se
gradativamente sofisticadas, principalmente devido a melhor compreensão acerca
dos fatores que influenciam a taxa de hibridação e estabilidade dos híbridos.
No entanto, a tecnologia de microarranjos de DNA foi descrita no final
da década de 80 e tem impulsionado de maneira importante o estudo da
genômica funcional, sendo recentemente aplicada na área de diagnóstico
(PALLACIOS et al., 2007). O ensaio de microarranjos usa o princípio da
complementaridade de bases dos ácidos nucléicos, sendo essa técnica
essencialmente de hibridação de ácidos nucléicos em grande escala. Estudos têm
demonstrado a sua aplicação no diagnóstico de bactérias, vírus e de parasitas.
Wangs, Vora & Stenger (2004) descreveram a utilização deste ensaio na
detecção de E. histolytica, E. dispar, Giárdia lamblia e Cryptosporidium parvum
em amostras fecais. Contudo, a arquitetura dos microarranjos consiste em uma
ordem predefinida de fragmentos de DNA quimicamente ligada à superfície sólida
(lâmina de vidro, membrana etc.). Assim, uma grande quantidade de sondas está
presente nestes arranjos, conferindo a técnica uma maior sensibilidade, devido a
alta densidade das sondas específicas para um único alvo, além de possibilitar a
detecção de diferentes organismos no mesmo ensaio. Estes arranjos pre-
definidos de oligonucleotídeos podem ser aderidos ou sintetizados sobre suportes

27
sólidos que contenham uma superfície positivamente carregada. Este processo é
todo robotizado e requer uma estrutura complexa para a sua execução ou pode
ser terceirizado por empresas especializadas nestas áreas. Isto faz com que esta
tecnologia não seja exponencialmente difundida no cenário do diagnóstico.
No final da década de 70 surgiram os primeiros trabalhos de análise de
imunoensaio, utilizando o suporte sólido associado a citometria de fluxo (HORAN
& WHEELESS, 1977). Desde então, a literatura vem apresentando, de forma
crescente, a utilização de um método de detecção ensaio molecular inovador que
associou a filosofia da citometria de fluxo ao uso de microesferas fluorescentes
(DUNBAR, 2006). Esta tecnologia específica é representada pelo sistema da
empresa Luminex®, que foi a pioneira no desenvolvimento do equipamento e das
microesferas. Esta estratégia apresenta uma amplitude vasta de utilização,
permitindo não somente a realização de imunoensaio, como de hibridação direta
do DNA sobre a superfície de microesferas. Nota-se, ainda, que esta tecnologia
não é controlada por licenças especiais e que aumenta o seu potencial para que
novos ensaios possam ser desenvolvidos.
Este sistema está fundamentado no uso de microesferas de
poliestireno, contendo internamente uma mistura de dois fluorocromos, que
emitem intensidades de fluorescência distintas. A utilização de proporções precisa
desses fluorocromos produziu mais de 100 diferentes microesferas com espectros
de emissão de fluorescências distintos. Assim,cada microesfera apresenta um
sinal de fluorescência único e especifico.
Em cada uma das superfícies das 100 microesferas podem ser
acopladas moléculas de capturas como: anticorpos monoclonais com
especificidade única, peptídeos, antígenos protéicos ou sondas específicas para
uma sequência de DNA. Isto permite a análise simultânea de até 100 analitos
numa única amostra, através de uma simples reação em um único tubo. A ligação
das moléculas de captura, já acopladas a cada microesfera, no seu alvo é

28
detectada pela adição de um conjugado fluorescente. Assim, a microesfera que
tiver os híbridos, produto amplificado biotinilado/sonda no sistema líquido, emitirá
dois tipos de fluorescência quando interceptada pelos dois feixes de laser. O laser
vermelho excita o fluoróforo localizado no interior da microesfera e o laser verde
estimula a emissão de fluorescência do conjugado biotina/estreptavidina-R-
Ficoeritrina (SA-PE) que funciona como uma molécula repórter, localizada
próximo a superfície da microesfera. O sistema óptico classifica as microesferas,
baseado na identidade spectral e detecta a presença ou ausência da emissão de
fluorescência da molécula repórter.
Assim, como a PCR em tempo real, esta tecnologia requer uma
plataforma de instrumentação que contenha um sistema óptico para a excitação e
detecção da emissão da fluorescência, além de um sistema computacional para a
aquisição de dados e análise final da reação. A citometria de fluxo associada ao
uso de microesferas fluorescentes é referida como uma técnica de rápida
execução, fidedigna e que apresenta uma elevada especificidade, merecendo
destaque pelo fato de que a técnica ainda pode ser otimizada para uma ampla
variedade de aplicações, incluindo desde diagnóstico de perfil alérgico, mutações
genéticas (fibrose cística), doenças autoimune, determinação de citosinas e
quimiosinas e até o diagnóstico de doenças infecciosas.
O potencial do uso da citometria de fluxo, utilizando microesferas no
diagnóstico de doenças infecciosas, vem sendo cada vez mais relatado na
literatura. Resultados promissores vêm sendo demonstrados na detecção
simultânea de antígenos do vírus da influenza A e B (YAN et al., 2005) e de
bactérias (DUNBAR et al., 2003). Da mesma forma, tem funcionado para a
detecção de fragmentos de DNA de vírus HIV tipo 1, HCV, HBV (DEFOORT et al.,
2000), HPV (SCHMITT et al., 2006) e de Escherichia coli, Salmonella, Listeria
monocytogenes e Campylobacter jejuni (DUNBAR et al., 2003). Este método tem
mostrado ser igualmente promissor na detecção de fungos do gênero

29
Trichosporon (DIAZ & FELL, 2004), espécies de Ascomicetos (PAGE et al., 2006)
e de Candida (PAGE & KURTZMAN, 2005), onde o diagnóstico ainda é um
desafio (DIAZ, 2007).
Sabemos que a padronização de teste multiplex é de extrema
complexidade, mas não impossível quando utilizamos metodologia como a
desenvolvida pela Luminex. A exemplo disto, Mahony et al. (2007) utilizaram a
técnica para o desenvolvimento de um teste multiplex para a detecção e
diferenciação de 20 diferentes tipos de vírus respiratórios.
E, por fim, tem sido aplicado na diferenciação entre espécies de
protozoários como: Cryptosporidium hominis e C. parvum (BANDYOPADHYAY et
al., 2007) e na detecção e semiquantificação da carga parasitária de quatro
espécies de Plasmodium causadores da malária humana (MCNAMARA et al,
2006).
O emprego desta metodologia no diagnóstico de amebas intestinais foi
incentivado em função dos dados relatados por Bandyopadhyay et al. (2007), que
constataram ser uma técnica prática, acurada e excelente para a identificação de
espécies de Cryptosporidium. Neste trabalho, o limite de detecção da técnica foi
menor que 10 oocistos de Cryptosporidium em 300 µl de fezes e apresentou
100% de sensibilidade.
O sistema de citometria de fluxo, empregando microesferas
fluorescente, já vem sendo utilizado pelas grandes redes de laboratórios clínicos
no Brasil, em decorrência da aplicabilidade da técnica e da disponibilidade de
vários conjuntos de diagnóstico comercializado pela BioRad, Millipore, Qiagen etc.
Portanto, a padronização desta técnica no diagnóstico da amebíase e na
genotipagem de amebas do gênero Entamoeba poderá ser incorporado a rotina
laboratorial futuramente. Isto gera uma nova expectativa e um importante passo
no diagnóstico de parasitoses intestinais, abrindo novos horizontes e a
implementação de um novo método de diagnóstico na área de doenças

30
parasitárias, além de propiciar o domínio de uma metodologia que tende a se
expandir na área de diagnóstico.
No entender de Dunbar (2006), a medida que essa metodologia se
tornar consolidada, muitos outros ensaios serão desenvolvidos e o custo do
equipamento e das microesferas chegarão a um patamar razoável de custo, como
sucedeu na maioria das tecnologias utilizadas atualmente.
Nos últimos anos, têm-se buscado novos métodos de diagnóstico com
o intuito de diferenciar E. histolytica, E. dispar e, mais recentemente, a presença
de outras amebas comensais. De acordo com a literatura e com o exposto acima,
vários argumentos justificam o desenvolvimento de métodos para a diferenciação
das amebas intestinais. Diante destes fatos, nos pareceu importante avaliar um
segmento gênico do DNA ribossômico das amebas, através da PCR, seguida da
reação de sequenciamento. A partir de então, poderemos utilizar esta sequência
no desenvolvimento e na padronização da PCR seguida do ensaio de hibridação
multianalítico, em meio líquido, utilizando microesferas fluorescente como suporte
sólido, associado a citometria de fluxo da plataforma de instrumentação da
Luminex®. Assim, esta nova metodologia será utilizada no diagnóstico diferencial
entre amebas que habitam o intestino humano.

31
2. OBJETIVOS
2.1. Geral
Desenvolver e avaliar a PCR, seguida da reação de hibridação
multianalítica em meio líquido, utilizando microesferas fluorescentes como suporte
sólido associadas à citometria de fluxo no diagnóstico diferencial entre espécies
de Entamoeba presente no intestino humano.
2.2. Específicos
♦ Utilizar a PCR em tempo real para diferenciação das espécies E.
histolytica e E. dispar, em amostras de fezes humanas;
♦ Padronizar e avaliar a técnica de PCR convencional seguida da
reação de sequenciamento, utilizando oligonucleotídeos do tipo universal para o
gene SSU-rRNA na detecção de Entamoeba spp;
♦ Padronizar a técnica multianalítica em meio líquido, utilizando
microesferas fluorescentes como suporte sólido, associada a citometria de fluxo
para detecção e diferenciação de amebas encontradas no intestino humano.

32
3. MATERIAL E MÉTODOS
O presente trabalho foi desenvolvido no Laboratório de Diagnóstico Imunológico e Molecular de Doenças Infecciosas
localizado no Departamento de Imunologia do Instituto de Microbiologia Prof. Paulo de Góes da Universidade Federal do Rio de
Janeiro e no Laboratório de Parasitologia Molecular do Center for Disease Control and Prevention – CDC, Atlanta, USA. Uma
representação explicativa das etapas do trabalho pode ser visualizada na figura 1.
Figura 1. Diagrama explicativo das etapas realizada s neste trabalho
Detecção de Entamoeba spp em fezes
Exame direto Paratest
Método de Ritchie
Amostras Brasil
PCR em tempo real Sistema TaqMan
Verweij et al.
Análise sequências desenhos de inciadores e sondas
PCR/Luminex
PCR/Sequenciamento
Clonagem molecular
Sequenciamento
Purificação de DNA QIAquik PCR purification kit
Amostras do CDC Fast DNA Kit
Métodos moleculares
Amostras do Brasil Protocolo in house Santos et al. (2007)
Extração de DNA
Métodos morfológicos E. P. F.
Amostras CDC
Método de centrifugação Acetato de etila
Coloração Tricrômica

33
3.1 Amostragem
Um total de 45 amostras de fezes foi empregado neste estudo. Destas,
16 foram de pacientes atendidos no Serviço de Saúde Pública dos EUA, enviadas
ao CDC para exame confirmatório, 20 amostras de fezes de indivíduos moradores
do assentamento de Frei Vantuy em São Jorge dos Ilhéu na Bahia, Brasil e nove
amostras de cultura, sendo que destas, cinco de E. histolytica (n=5), três de E.
dispar (n=3) e uma de E. moshkovskii (n=1). Foram utilizadas outras 15 amostras
de DNA oriundas do CDC com o diagnóstico positivo para E. histolytica e/ou E.
dispar. Estas amostras nos foram enviadas sem o conhecimento prévio dos
resultados.
3.2 Amostras Controles de DNA
Produtos de clonagem de DNA de E. histolytica, E. dispar, E.
hartamanii, E. moskhovski e E. coli foram utilizadas como controles. Além disto,
foram utilizados DNA extraído de amostra fecal de Endolimax nana (n=1),
Blastocystis hominis (n=4), Giardia lamblia (n=2), Microsporidium sp (n=1),
Cryptosporidium parvum (n=2) e C. hominis (n= 1).
3.3 Exame Parasitológico de Fezes (EPF)
De cada indivíduo, uma única amostra foi coletada e fracionada em
alíquotas. Essas alíquotas foram mantidas sob refrigeração, sendo então
transportadas de São Jorge dos Ilhéus na Bahia para o Laboratório de
Parasitologia do Hospital Universitário Antônio Pedro, UFF, localizado no
município de Niterói, Rio de Janeiro. O diagnóstico parasitológico das 20
amostras clínicas foi realizado pelo método comercial Paratest® (Diagnostek-
Indústria e Comércio de Produtos Científicos Ltda, SP, Brasil) e pelo método de
Ritchie modificado.

34
3.3.1 PARATEST®
Cerca de um grama de cada amostra fecal foi transferida com o auxílio
de uma espátula para um frasco contendo formol 5% tamponado e,
posteriormente, homogeneizado. Preparou-se uma lâmina para a visualização das
formas parasitárias com uma gota do sedimento fecal acrescido do corante
temporário Lugol, sendo esta submetida à observação por microscopia ótica
(microscópio Nikon Eclipse E20).
3.3.2 Método de Richtie modificado
O frasco contendo o material diluído em formol 5% tamponado foi
homogeneizado e 3,5 ml do homogenato foram transferido para tubo cônico de
plástico. Foi adicionada uma gota de detergente doméstico e 1,5 ml de acetato de
etila. O tubo foi arrolhado e agitado vigorosamente por dez segundos. Após este
período, o tubo foi então destampado e tampado rapidamente para liberar a
pressão e, então, centrifugado a 1500 rpm por dois minutos. O sobrenadante foi
cuidadosamente desprezado e o sedimento ressuspenso com 5,0 ml de água
destilada. O tubo foi centrifugado a 1500 rpm por dois minutos e o sobrenadante
descartado. Após homogeneizar bem o sedimento, uma lâmina foi preparada
contendo uma gota do sedimento e uma gota do corante temporário Lugol para a
visualização das formas parasitárias.
A análise das amostras provenientes do Serviço de Saúde Pública dos
EUA, foi realizada segundo os preceitos do laboratório de parasitologia dos
serviços. De cada individuo, uma única amostra foi coletada e fracionadas em três
alíquotas. A primeira foi conservada em formol a 5%, a segunda em PVA (álcool
polivinílico) e a terceira alíquota, congelada a –20ºC para a extração de DNA. A
análise microscópica das amostras foi feita a partir do sedimento do método de
concentração por centrifugação, usando acetato de etila e formalina, seguido da

35
técnica de coloração tricrômica, de acordo com o protocolo descrito por Garcia
(2007) e www.dpd.cdc.gov/dpdx/diagnosticprocedure, acessado em 10/06/2009.
3.4 Extração do DNA
A extração do DNA nas amostras de fezes proveniente da Bahia foi
realizada segundo o protocolo descrito por Santos et al. (2007). Cerca de 1g de
fezes foi dissolvido em água destilada e filtrada em gaze. O filtrado foi lavado três
vezes por centrifugação a 1.500 rpm (centrífuga de sorologia, Fanem) e o
sedimento ressuspenso em 1,0 ml de água destilada. A uma alíquota de 100 µl
das fezes, adicionou-se 500 µl da solução de tiocianato de guanidina 5M, seguido
por agitação manual por inversão e incubação a temperatura ambiente por 10
minutos. O lisado foi resfriado no gelo por dois minutos e adicionou-se 250 µl de
acetato de amônio à 7,5 M. Após agitação, por várias vezes, do tubo por inversão,
este foi novamente incubado no gelo por 10 minutos. O volume de 500 µl de uma
solução de clorofórmio-álcool isoamílico (24:1 v/v) foi adicionado e após agitação
manual por inversão, centrifugou-se a amostra a 13.800 x g (centrífuga Eppendorf
– 5415 C) por 10 minutos. A fase aquosa foi transferida para um novo tubo de
microcentrífuga e 0,54 volume de isopropanol foi adicionado para precipitação do
DNA. O tubo foi invertido, várias vezes, para misturar as soluções e depois
centrifugado a 3.448 x g por um minuto. Seguiu, então, a lavagem do DNA com
etanol 70% (v/v) por cinco vezes. Após secagem do tubo em estufa a 37°C, o
DNA foi diluído em 50 µl de tampão Tris-EDTA pH 8,0 (TE) e incubado na estufa a
56°C por uma hora. Nenhuma etapa de purificação do DNA extraído foi realizada.
A extração do DNA das amostras de fezes, proveniente do Serviço de
Saúde Pública dos EUA, foi realizada utilizando o conjunto de diagnóstico
FastDNA® (MP Biomedicals Inc, Solon, USA) com ligeiras modificações por Da
Silva et al. (1999). Cerca de 300 a 500 µl de fezes foram diluídas em PBS/EDTA,
centrifugadas a 14000 x g por cinco minutos e lavadas três vezes por

36
centrifugação a 14.000 Χg por cinco minutos. O sedimento foi diluído em um
volume final de 300 µl de PBS/EDTA. Posteriormente, foram adicionados pérolas
de vidros (matrixE), 400 µl de solução de lise (CLS-VF), 200 µl de solução de
precipitação de proteína (PPS) e 20 µl de polivinilpirilidona a 10% (PVP). Após a
etapa de homogeneização e de agitação no desagregador celular (FP120 cell
disruptor instrument) por cinco segundos, centrifugou-se a 16000 x g por cinco
minutos. Foi transferido 600 µl do sobrenadante para um novo tubo de 1,7 ml,
adicionou-se 600 µl da matriz de ligação (Binding Matrix) e, após agitação manual
por inversão, incubou-se o tubo na temperatura ambiente por cinco minutos,
procedendo em seguida uma etapa de centrifugação a 16000 x g por cinco
minutos. O sedimento foi diluído em 500 µl de tampão de lavagem (Sews-M) e
centrifugado a 16000 g por 1 minuto. Novamente, o sedimento foi diluído em 100
µl de tampão de eluição (DES) e incubado por três minutos na temperatura
ambiente, seguida de uma etapa de centrifugação por 16000 x g por um minuto.
O sobrenadante obtido contendo o DNA foi transferido para um novo tubo e
armazenado a 4°C. Após a extração, cada amostra de DNA foi purificada,
utilizando o conjunto de diagnóstico da QIAquik PCR purification kit (QIAGEN Inc.,
Valencia, Calif.), seguindo as instruções do fabricante. Adicionou-se cinco
volumes de tampão PB a um volume de DNA extraído seguida de um etapa de
homogeneização. O material foi transferido para a coluna de sílica Qiaquick e foi
centrifugado a 16000 x g por um minuto. O líquido do tubo coletor foi descartado e
acrescentou-se a coluna 0,75 ml de tampão de lavagem (PE), seguido de duas
etapas de centrifugação a 16000 x g por um minuto. O líquido do tubo coletor foi
desprezado. A coluna foi, então, colocada num microtubo de 1,5 ml, seguida da
adição de 50 µl do tampão de eluição EB (Tris 10mM, pH 8,5). Após a etapa de
incubação na temperatura ambiente por um minuto, a coluna foi centrifugada a
16000 x g por um minuto. O DNA purificado foi armazenado a 4°C.

37
3.5 PCR em Tempo Real (Sistema TaqMan)
A fim de empregar um diagnóstico rápido e específico na diferenciação
do complexo E. histolytica e E. dispar foi utilizado o ensaio de PCR em tempo
real. Este ensaio é baseado na hidrólise de sondas fluorescentes (sonda
TaqMan®), complementar a uma região interna do produto amplificado. Na
extremidade 5’ de cada sonda é adicionada uma molécula fluorescente, no caso,
FAM (6-carboxi-fluoresceína) para E. histolytica e Hex (hexaclorocarbo-
nilfluoresceína) para E. dispar. Na extremidade 3’, uma molécula quencher-BHQ-1
(Black hole quencher-1) capaz de, por um fenômeno físico denominado FRET
(Fluorescence ressonance Energy), absorver a fluorescência emitida pelo
fluoróforo.
Inicialmente, a sonda hibridiza na região complementar da fita de DNA
a ser amplificada. Durante a etapa de polimerização dos nucleotídeos, a sonda
então é clivada pela ação da atividade 5’-3’ exonuclease da enzima Taq DNA
polimerase, levando a separação dos dois fluoróforo, aniquilando o fenômeno
FRET e ocasionando a liberação da fluorescência na reação. À medida que vai
ocorrendo a amplificação, a fluorescência gerada é capturada pelo sistema óptico
do equipamento.
Neste estudo, foi utilizado o par de oligonucleotídeos iniciadores
(Ehd/239F ATTGTCGTGGCATCCTAACTCA e o Ehd/88R GCGGACGGCTCATT
ATAACA) e as sondas histolytica/96T (FAM UCAUUGAAUGAAUUGGCCAUUU–
BHQ1) e dispar/ 96T (HEX-UUACUUACAUAAAUUGGCCACUUUG-BHQ1), que
flanqueiam uma região do gene da Ssu-rRNA de E. histolytica e E. dispar. O
volume final da amplificação foi de 25 µl contendo: Platinum Quantative PCR
Super mix-UDG com ROX (Invitrogen), 0,5 µM do oligonucleotídeo iniciador Ehd-
239F⁄ Ehd-88R e 0,1 µM da sonda espécie-específica, histolytica-96T⁄FAM e
dispar⁄HEX. A amplificação foi feita em 40 ciclos, com a etapa de desnaturação a
95oC/30 seg., anelamento a 60oC/30 seg. e extensão a 72oC/30 seg. A

38
amplificação, detecção e análises de dados foram feitas no termociclador
Mx3000P real time PCR (Stratagene, La Jolla, CA, USA). Esta metodologia é
utilizada na rotina do laboratório de parasitologia do CDC para a diferenciação de
E. histolytica e E. dispar.
3.6 PCR/Sequenciamento
O fragmento a ser amplificado é parte do gene que codifica a
subunidade menor do rRNA das amebas. O par de iniciadores (JVF-5’
GTTGATCCTGCCAGTA TTATATG ‘3 e DSPR2 (5’-CACTATTGGAGCTGGAAT
TAC-3’) gerou um produto de amplificação em torno de 650 bp. O volume final da
amplificação foi de 50 µl, contendo a mistura de reação AmpliTaq Gold (Applied
Biosystems, Foster City, CA), 12 pmoles de cada iniciador e 10 µl do DNA diluído
(1/5). A amplificação foi feita em 40 ciclos, com a etapa de desnaturação a
95oC/30 seg., anelamento a 57oC/30 seg. e extensão a 72oC/30 seg. Antes do
primeiro ciclo, uma fase de predesnaturação a 95°C por cinco minutos, e após o
último ciclo a fase de extensão final a 72°C por se te minutos. Três controles foram
incluídos em todos os experimentos: 1) Todos os reagentes exceto o DNA;
2) DNA de E. histolytica de uma cepa padrão patogênica HM1-IMSS; 3) DNA de
E. dispar de uma cepa isolada de um indivíduo assintomático.
3.6.1 Análise do produto amplificado
Alíquotas de 10 µl do DNA amplificado foram analisadas através da
eletroforese (cuba de eletroforese, modelo HE33/Hoefer Scientific Instruments,
USA) em gel de agarose a 2% (Invitrogen Life Technologies, USA). Como
marcador de peso molecular foi utilizado 5,0 µl do “100 pb ladder” (Invitrogen Life
Technologies, USA) para estimar os tamanhos dos produtos amplificados. O gel a
2% foi preparado em tampão TBE 1 X (1M Tris-borato, 0,01 M EDTA, pH 8,4) e
submetido à tensão constante de 100 V. Após a corrida, o gel foi corado com

39
brometo de etídio na concentração de 0,5 µg/ml por dez minutos e lavado em
água destilada por trinta minutos. A inspeção visual do gel foi realizada no
transiluminador (Sigma Chem. Co., USA, modelo T1201) de luz ultravioleta,
seguida da foto documentação pelo sistema mini Bis Pro, Bio-Imaging Systems
(BioAmerica Inc, USA).
3.6.2 Purificação do produto amplificado
Para a realização desta etapa foi utilizado o conjunto de diagnóstico de
purificação da StrataPrep PCR Purification Kit (Stratagene, La Jolla, CA), segundo
as recomendações do fabricante. Foi adicionado um volume da solução de
ligação ao DNA (DNA binding solution) a um volume de produtos da PCR seguido
da homogeneização. O material foi transferido para uma coluna e centrifugado a
16000 x g por 30 segundos. O líquido do tubo coletor foi descartado e 0,75 ml de
tampão de lavagem foi acrescentado na coluna. A coluna foi centrifugada e o
líquido do tubo coletor foi desprezado. Uma nova etapa de centrifugação foi
realizada e a coluna então foi colocada num microtubo de 1,5 ml, seguida da
adição de 50 µl de água deionizada. O tubo foi incubado na temperatura ambiente
por cinco minutos, seguida de uma etapa de centrifugação na rotação máxima por
trinta segundos. O produto da PCR purificado foi armazenado a 4°C até o
momento do sequenciamento do DNA.
3.6.3 Reação de sequenciamento
Para a realização desta etapa foram utilizados os produtos da PCR
purificado e clones obtidos pelo conjunto de clonagem pCR2.1-TOPO vetor
(Invitrogem, Carlsbad CA). As sequências nucleotídicas do produto amplificado
e/ou clonado foram determinadas pelo método do Dye terminator, utilizando o
conjunto de diagnóstico da ABI PrimTM Dye Terminator Cycle Sequencing (Applied
Biosystems, Foster City, CA). O volume final da reação foi de 20 µl, contendo:

40
1,0 µl do mix dye Terminator, 3,5 µl de tampão, 3,2 pmol de cada iniciador (JVF e
DSPR 2), 1,0 µl DNA e 13,5 µl de água destilada, tanto para os produtos clonados
como para o sequenciamento direto do produto da PCR purificados. As condições
térmicas da reação foram: 1° ciclo de 96ºC/um min, seguido de 25 ciclos de
96ºC/10 seg, 55ºC/cinco seg e 60ºC/4 min.
3.6.4 Purificação dos produtos sequenciados
Os produtos da reação de sequenciamento foram purificados em uma
coluna de gel filtração Sephadex G50 (Sigma Sigma, St Louis, MO), com o intuito
de retirar os nucleotídeos não incorporados e outros componentes da reação. A
coluna de gel filtração foi preparada em uma placa de 96 poços (Multiscreen 96
well filtration plate-Millipore). Adicionou-se a cada poço sephadex G-50 (Sigma
Sigma, St Louis, MO) e 300 µl de água ultrapura e a placa foi mantida por, no
mínimo, três horas a 4ºC. Após a etapa de centrifugação a 910 x g por cinco
minutos, foi adicionado 20 µl do produto da reação de sequenciamento nos
respectivos poços, seguida de uma nova etapa de centrifugação a 910 x g por
quatro minutos. A placa de 96 poços (Costar #6509) de fundo V foi centrifugada a
vácuo (DNA speed Vac) por uma hora e o filtrado seco armazenado a 4ºC e
protegido da luz até o momento da reação de sequenciamento.
3.6.5 Sequenciamento dos produtos
As amostras foram diluídas em 20 µl de formamida (Applied
Biosystems, Foster CA, USA) e adicionadas a uma placa de 96 poços, seguida de
uma etapa de desnaturação no termociclador GeneAmp PCR System 2400
(Applied – Biosystems, CA, USA) a 95ºC por cinco minuto e mantida em banho de
gelo. As amostras foram encaminhadas à análise eletroforética capilar no
sequenciador de DNA automático da AB 3730 (Applied Biosystems, Foster CA,
USA), utilizando capilar de 50 cm, polímero de acrilamida POP7 (Applied

41
Biosystems, Foster, CA USA) e a programação padrão de corrida para estes
parâmetros.
3.6.6 Análise das sequências
As sequências foram obtidas em forma de cromatogramas e analisadas
no programa DNA STAR SeqMAn (DNASTAR Inc., Madison, Wis WI). Assim,
durante a edição das sequências foi verificado que os iniciadores JVF e DSPR2
estavam ancorados e, portanto, amplificando uma região do rDNA, além de
verificar possíveis ambiguidades em decorrência de erros de leitura do
sequenciamento.
Posteriormente, as sequências nucleotídicas consenso editadas foram
alinhadas pelo algoritmo CLUSTAL W (THOMPSON, HIGGINS & GIBBSON,
1994) no pacote do programa GeneStudio suite (GeneStudio, Inc., Suwanee, GA).
Seguida da análise de homologia, utilizando-se o servidor BLAST 2.0 (“Basic
Local Alignement Search Tool”) (ALTSCHUL et al., 1997) do “National Center for
Biotechnology Information” (NCBI) da Biblioteca Nacional de Medicina do NIH
(“National Institute of Health”), Maryland, EUA. Ademais, as sequências do gene
do rRNA do gênero Entamoeba, previamente depositadas no Genbank, cujos
números de acesso: E. histolytica/X64142, E. dispar/Z49256, E.coli/AF149915,
E. coli/AF14 9915, E. hartmanni/AF149907, E. moshkovskii/AF149906, E.
chattoni/AF149913 e E. polecki/AF149912, foram alinhadas com as sequências
obtidas neste estudo com o auxílio do programa pelo algoritmo do CLUSTAL W
(THOMPSON, HIGGINS & GIBBSON, 1994) no pacote do programa GeneStudio
suite (GeneStudio, Inc., Suwanee, GA).
3.7 Análise Filogenética
A análise filogenética foi baseada no alinhamento das sequências
consenso do gene da SSU-rRNA das Entamoeba spp. Assim, o alinhamento das
sequências serviu como base para a construção da árvore filogenética, utilizando

42
o software MEGA versão 4.1 (TAMURA et al., 2007), disponível em http://www.
megasoftware. net/mega4.1.html, sendo que a estimativa da distância utilizada foi
a equação de Jin & Nei (1990) modelo Kimura 2 parâmetros. As árvores
filogenéticas foram construídas empregando o algoritmo de neighbor-joining
(SAITOU & NEI, 1987). Para cada construção, a veracidade dos ramos foi
conferida por analise de “bootstrap” (1.000 repetições).
3.8 Clonagem
A clonagem do fragmento amplificado com os oligonucleotídeos
iniciadores JVF/DSPR2 foi feita, utilizando o conjunto de clonagem pCR2.1 –
TOPO TA Cloning (Invitrogem, Carlsbad, CA), conforme as instruções do
fabricante. O plasmídeo usado como vetor de clonagem foi o pCR® 2.1 TOPO,
cujo esquema se encontra na figura 1. Possui uma marca de resistência aos
antibióticos ampicilina e kanamicina e o gene estrutural lac Z que codifica para a
enzima β-galactosidase, além de conter um fragmento de poli-linker com vários
sítios únicos para enzimas de restrição inserido no gene lac Z.
Para a ligação do fragmento ao vetor de clonagem foram adicionados
3,0 µl do produto amplificado, 1,0 µl da solução de sais (salt solution), 1,0 µl de
água destilada estéril e 1,0 µl do vetor pCR® 2.1-TOPO. A reação foi
homogeneizada e incubada por cinco minutos na temperatura ambiente.
Posteriormente, adicionou-se 2,0 µl da reação de ligação à 200 µl de células
competentes. As células foram homogeneizadas gentilmente por inversão e
incubadas no gelo por cinco minutos e, posteriormente, a 42°C por trinta
segundos. A mistura foi imediatamente recolocada no gelo, 250 µl de meio SOC
foi adicionado no tubo e uma nova etapa de incubação foi realizada a 37°C sobre
agitação (200 rpm) por uma hora. Cerca de 30 µl das células transformadas foram
inoculadas em meio sólido LB (Lúria Bertani) contendo ampicilina 50 ug/ml, X-gal
(5-bromo-4-chloro-3-indolyl-b-D-galactopyranoside) e IPTG (Isopropyl β-D-1-

43
Tiogalactopyranosídeo). As placas foram incubadas na temperatura de 37°C/12
horas. Após a etapa de incubação, cerca de oito colônias que exibiram coloração
branca foram selecionadas aleatoriamente. Para a confirmação da transformação
realizou-se a PCR das colônias selecionadas, utilizando o par de iniciador
JVF/DSPR2. Para isto, parte da colônia foi transferida para um tubo de PCR
contendo 5 µl de água destilada. Os parâmetros da reação da PCR foram os
mesmos empregados no item 3.6. As colônias que foram transformadas com
sucesso foram transferidas para um tubo contendo 50 ml de meio líquido LB com
cloreto de sódio a 5%, sendo adicionado 5,0 µl de ampicilina (50 ug/ml). O meio
de cultura foi incubado a 37°C sob agitação e no di a seguinte centrifugado a
2050 x g por cinco minutos. O sobrenadante foi descartado e o DNA plasmidial
contendo o fragmento de interesse foi purificado.
FIGURA 2. Mapa do vetor de clonagem PCR 2.1 topo (I nvitrogem) sitio de ligação do inserto com timidina nas extremidades 3’, genes de resistência a ampicilina e kanamicina, sistema de seleção de colônias baseado na expresão do gene lac z e sequências universais m13R e T7.

44
3.9. Extração do DNA Plasmidial de E. coli (Minipreparações Plasmidiais )
Para a realização desta etapa foi empregado o conjunto de purificação
QIaprep Spin Miniprep (QiAGEN Inc., Valencia, CA.), seguindo as instruções do
fabricante. O sedimento bacteriano foi ressuspenso em 250 µl do tampão P1,
seguida da adição de 250 µl do tampão P2. O tubo foi homogeneizado
cuidadosamente por inversão por 4 a 6 vezes e adicionado 300 µl do tampão N3.
Uma outra etapa de homogeneização por inversão e centrifugação a 16000 x g
por um minuto foi realizada. O sobrenadante foi adicionado à coluna e
centrifugado a 16000 x g por um minuto. Duas etapas de lavagens da microcoluna
por centrifugação a 16000 x g por um minuto foram realizadas com os tampões
PB e PE. Na coluna foi adicionado 50 µl do tampão de eluição (EB) mantendo a
temperatura ambiente por um minuto e seguida de uma etapa de centrifugação a
14000 x g por um minuto. O DNA plasmidial foi armazenado a 4ºC. Ao término da
purificação, os plasmídeos foram incubados com a enzima Eco RI (100 U/µl), visto
que o vetor clonado possui sítios de restrição para a enzima Eco RI nas regiões
franqueadoras dos insertos. Para tal fim, foram adicionados cerca de um µl do
DNA plasmidial, 1,0 µl de tampão de EcoR1, 0,1 µl de enzima EcoR1 e 7,9 µl de
água destilada, seguida de uma etapa de incubação a 37ºC por no mínimo três
horas. A clivagem do inserto foi analisada em gel de agarose a 2 % corado com
brometo de etídio, utilizando o marcador molecular de 1 kb DNA ladder
(Invitrogem Carlsbad CA) para padrão de comparação.
3.10 Reação de hibridação em meio líquido utilizand o microesferas
fluorescente como suporte sólido associada a citome tria de fluxo,
sistema LUMINEX®
O princípio deste ensaio baseia-se na utilização de microesferas de
poliestireno de 5,6 µm de diâmetro, que desempenham o papel de suporte sólido
na captura de ácidos nucléico, durante a reação de hibridação em um ambiente

45
líquido. Cada microesferas carboxilada possui no seu interior um fluoróforo com
uma identidade espectral única. Assim, a sonda de captura contendo um
grupamento amina é ligada covalentemente à superfície da microesfera. Produtos
da PCR biotinilados ligam-se as sondas já acopladas à superfície da microesfera,
formando um híbrido. Essa interação é detectada utilizando uma molécula
repórter, Estreptavidina-R-ficoeritrina (SA-PE). Assim, as microesferas que
possuem os híbridos, produto amplificado biotinilado/sonda no sistema líquido,
emitirão dois espectros de fluorescência distintos quando interceptadas pelos dois
feixes de lasers do citômetro de fluxo. O laser vermelho excita o fluoróforo
localizado no interior da microesfera e o laser verde estimula a emissão de
fluorescência do conjugado biotina/estreptavidina-R-Ficoeritrina (SA-PE).
As fases que compõe todo o ensaio são: desenhos de iniciadores e
sondas, ligação da sonda na superfície da microesferas, PCR multianalítica e a
reação de hibridação.
3.10.1 Desenho dos oligonucleotídeos utilizados na PCR/Luminex
Como relatado anteriormente no item 3.66, foi realizado o levantamento
das sequências de DNA do rRNA das espécies Entamoeba, descritas e
depositadas no banco de dados (GenBank), cujos números de acesso são:
X64142, Z49256, AF149907, AF149906, AF149915, AF149914, AF149913 e
AF149912. Essas sequências foram alinhadas pelo algoritmo do CLUSTAL W
(THOMPSON, HIGGINS & GIBBSON, 1994) no pacote no programa GeneStudio
suite (GeneStudio, Inc., Suwanee, GA). As regiões conservadas da sequência
consenso, que apresentaram 100% de identidade foram identificadas visualmente
e selecionadas para os desenhos dos iniciadores. O iniciador desenhado para o
ensaio de PCR/sequenciamento, também foi adaptado e utilizado para o teste de
PCR/Luminex. Outros pares de oligonucleotídeos iniciadores que melhor

46
atenderam aos parâmetros recomendados foram selecionados e os iniciadores
reverso foram biotinilados na extremidade 5’.
As regiões que apresentaram polimorfismos entre as sequências foram
selecionadas para os desenhos de sondas complementares do tipo grupo
específica e espécie específica para a parte interna destes segmentos a serem
amplificados. Assim, as sondas desenhadas receberam um grupamento amina
nas extremidades 5’ e um espaçador de carbono (C-6), o que possibilita a sua
ligação na microesfera carboxiladas (figura 3).
FIGURA 3: Representação esquemática da microesfera de poliestireno de 5,6 µm recoberta por grupamentos carboxila e da sonda ligada a um es paçador de carbono e um grupamento amina
As sondas foram sempre posicionadas na fita direta, de maneira que as
suas sequências fossem complementares à fita estendida, a partir do iniciador
reverso, ou seja, à fita biotinilada.
Ademais, verificou-se a possível presença de estrutura secundária nas
sequências alvos, através da ferramenta computacional “DNA folding”, disponível
no site http://mfold.bioinfo.rpi.edu/cgi-bin/dna-form1.cgi, a fim de evitar a
competição entre as estruturas secundárias presentes na sequência amplificada
com a sonda durante a etapa de hibridação.
Microesfera carboxilada
O
OH
NH2 CCCCCC CGATCCAGTTTGATTTAGT
Sonda amino-modificada

47
3.10.2 PCR/Luminex
O fragmento amplificado é parte do gene que codifica a subunidade
menor do rRNA. O par de iniciadores JVF-5’GTTGATCCTGCCAGTATTATATG ‘3
e EntaRev 390 (5’ ATTCCTCGTTATCCGTTAT-Biotina) gerou um produto de
amplificação em torno de 400 bp. O iniciador reverso foi biotinilado na
extremidade 5’. A amplificação foi realizada num volume final de 50 µl, contendo:
tampão 20mM Tris-HCl pH 8,4; 50 mM KCl; 1,5 mM MgCl2; 12 pmoles de cada
iniciador, JVF e EntaRev390; 20 mM de cada nucleotídeos (dNTP) e 1,25
unidades de Taq DNA polimerase (Invitrogen) e 10 µl do DNA extraído diluído. A
amplificação foi feita em 40 ciclos, com a etapa de desnaturação a 95°C/5
minutos, de anelamento a 50°C/30 segundos e de exte nsão a 72°C/1 minuto.
Antes do primeiro ciclo, foi feita uma fase de predesnaturação a 95°C por cinco
minutos, e após o último ciclo, uma fase de extensão final a 72°C por sete
minutos.
Nesta etapa foram testados outros protocolos de amplificação,
utilizando como iniciador direto o JVF, os iniciadores reverso Rev417 e o DSPR2
biotinilados. Foram utilizadas as mesmas concentrações dos reagentes e
condição de amplificação utilizada com o iniciador EntaRev 390, exceto as
temperaturas de anelamento que foram de 55 e 57ºC, respectivamente.
3.10.3 Ligação das sondas de DNA amino modificadas à super fície das
microesferas carboxiladas
Todo o processo de ligação das sondas as microesferas foi realizado
segundo as instruções descritas no manual da LUMINEX®. Cada sonda foi ligada
a um único conjunto de microesferas em uma reação separada. Assim, as sondas
de DNA específicas para E. histolytica, E. dispar, E. hartmanni, E. moshkovskii, E.
coli e E. poleckii foram acopladas covalentemente às diferentes microesferas
carboxiladas codificadas por uma determinada cor (Luminex Corp, Austin, TX),

48
utilizando o processo de acoplamento com carbodiimida. As suspensões de
microesferas não ligadas foram homogeneizadas com auxílio do vortex e
sonicadas por 30 segundos. Foi transferido 200 µl de microesferas não ligadas (5
x 106) para um microtubo de 1,7 ml e o mesmo centrifugado a 10.000 x g por 2
minutos. O sobrenadante foi removido e ao sedimento adicionou-se 50 µl de 0,1
M MES (ácido 2-N-morfolino-etanosulfonico, Sigma, St Louis, MO), pH 4,5. Após
a etapa de sonicagem por 20 segundos foi adicionado 2,0 nM da sonda de
captura e 2,5 µl de 30 mg/ml de EDC (N-(3-dimetil aminodipropil) – N’
etilcarbodiimida) recém preparado, seguida de homogeneização no vortex. Após a
etapa de incubação por 30 minutos na temperatura ambiente, na ausência de luz
e sob homogeneização contínua, foi adicionada novamente a solução de EDC
recém preparada, seguida de uma etapa de incubação. Ao sedimento foi
adicionado 1,0 ml de 0,02% Tween-20. Após centrifugação a 10.000 x g por dois
minutos, o sobrenadante foi removido e o sedimento foi diluído em 1,0 ml de 0,1%
de SDS, seguida de uma etapa de centrifugação a 10.000 x g por dois minutos.
Novamente o sobrenadante foi removido e o sedimento foi diluído em 50 µl de TE
pH = 8,0. As microesferas ligadas as sondas de DNA foram armazenadas a 2-8°C
na ausência de luz.
3.10.4 Reação de hibridação multianalítico em meio líquido utilizando
microesferas fluorescente como suporte sólido assoc iada a
citometria de fluxo (PCR/ Luminex®)
Todo o processo de hibridação foi realizado segundo as instruções
descritas no manual da LUMINEX® com algumas modificações.
Durante o desenvolvimento inicial da técnica, o DNA amplificado,
oriundo de produto de clonagem, foi hibridizado com as sondas acopladas as
microesferas. Posteriormente, o ensaio de hibridação foi realizado com o produto
da PCR de amostras clínicas. Reações contendo combinações diferentes de cada
sonda ligada a microesfera e diferentes volumes do produto amplificado foram

49
avaliadas, de maneira que cada combinação sempre continha um conjunto de
sonda específica para cada espécie.
A reação de hibridação foi baseada na ligação da sonda de captura,
acoplada à microesfera, ao produto amplificado biotinilado e desnaturado. O
volume total da reação foi de 50 µl, contendo 33 µl de diferentes grupos de
microesferas ligada às sondas de captura e 17 µl do produto amplificado, sendo
que no controle negativo (branco) foi adicionado 17 µl de tampão TE. Para o
preparo da mistura das microesferas, um volume calculado entre 1.500-2.500 por
cada grupo de microesferas foi adicionado a 1,5 x do tampão TMAC (cloreto de
tetrametil amônio 4,5 M, Tris-HCl 75 mM, pH 8,0, EDTA 6 mM, pH 8,0 e sarcosil
0,15%)(DUNBAR & JACOBSON, 2007; DIAZ & FELL, 2004). Dessa forma, cada
cavidade da placa de fundo cônico (#6509 Costar, Corning, NY) contém em torno
de 2.000 microesferas específica para cada sequência de DNA Entamoeba
amplificada. A microplaca foi selada e os produtos de PCR foram desnaturados a
95ºC por 5 min, seguido de uma incubação a 46ºC por uma hora no termociclador
(Veriti 96 Well Thermal Cycler/Applied Biosystems). Após a incubação foram
adicionado 25 µl de uma diluição de 1/50 de estreptavidina-R-ficoeritrina pré
aquecida (Molecular Probes, Eugene, OR) na concentração de 1mg/ml. A placa
foi incubada a 46ºC por 5 min e sua leitura foi realizada em placa aquecida à
mesma temperatura na plataforma Luminex 200. Para aquisição dos dados foi
utilizado o programa Xponent Software®, versão 3.0. Em cada ensaio, a amostra
foi testada duas vezes e no mínimo quatro brancos foram acrescentados na
reação. A intensidade de fluorescência mediana (MFI), emitida por 100
microesferas de cada população foi detectada.
Tomando esse protocolo como o padrão e os resultados obtidos como
referência, algumas modificações foram testadas. Avaliou-se o efeito de
diferentes temperaturas de hibridação que variou entre 40ºC até 52ºC, além da
retirada da etapa de lavagem, diferentes diluições do produto amplificado e da
molécula repórter.

50
4. RESULTADOS
4.1 Exame Parasitológico de Fezes
A frequência geral dos enteroparasitos encontrados nas amostras
provenientes do Serviço de Saúde Pública dos EUA está listada na tabela 1.
TABELA 1. Frequências das parasitoses intestinais n as amostras provenientes do Serviço de Saúde Pública dos EUA
Tot
al
Pos
itiva
Neg
ativ
a
Eh
/Ed
E. p
ole
ckii
B. h
om
inis
D. f
rag
ilis
E. h
art
Gia
rdia
E. c
oli
E. n
ana
En
ta s
p
16 14 2 10 2 9 2 3 2 1 2 2
% 94 12 62 12 56 12 18 12 6 12 12
D. fragilis = Dientamoeba fragilis; Eh/Ed = Complexo E. histolytica/E. dispa; Bh = Blastocystis hominis; E. hart = Entamoeba hartmanni; E. coli = Entamoeba coli; E. nana = Endolimax nana; Enta sp = Entamoeba sp.; % = percentual.
Nas 16 amostras relacionadas na tabela acima, foram encontrados
somente protozoários intestinais. Este fato pode ser justificado devido ao método
de concentração empregado, que é mais específico para protozoários intestinais.
Os resultados do EPF estão sumarizados na tabela 4.
A frequência geral dos enteroparasitos encontrados nas amostras
provenientes de indivíduos moradores no assentamento de Frei Vantuy em Ilhéus,
na Bahia, está listada na tabela 2.
Nas 20 amostras relacionadas na tabela acima foram encontrados
protozoários e helmintos. Os resultados do EPF de cada amostra estão
sumarizados na tabela 5.

51
TABELA 2 Frequências das parasitoses intestinais da s amostras provenientes de Ilhéus na Bahia
Tot
al
Pos
itiva
Eh/
Ed
B. h
om
inis
An
cylo
sto
mo
E. n
ana
E.c
oli
A. l
um
bri
coid
es
T. T
rich
iura
S. s
terc
ora
lis
Gia
rdia
20 20 20 9 2 9 12 6 2 2 3
% 100 100 45 10 45 60 30 10 10 15
Eh/Ed = Complexo E. histolytica/E. dispar; B hominis = Blastocystis hominis; A. lumbricoides = Ascaris lumbricoides; T. trichiura = Trichuris trichiura; E. coli = Entamoeba coli; E. nana = Endolimax nana; S. stercoralis = Strongyloides stercoralis; % = Percentual.
4.2 PCR/Sequenciamento
De um total de 45 amostras analisadas, todas (100%) foram
consideradas positivas pela PCR/Sequenciamento para a presença de uma ou
mais espécie de Entamoeba. Dentre as espécies encontradas, seis foram
identificadas como E. histolytica, 29 como E. dispar, nove como E. hartmanii,
cinco como E. coli e uma espécie como E. moshkovskii. Das 45 amostras, 16
foram provenientes do Serviço de Saúde Pública dos EUA, nove foram obtidas de
cultura e 20 eram provenientes de indivíduos moradores do assentamento de Frei
Vantuy em São Jorge dos Ilhéus, na Bahia. Estas amostras foram testadas para a
presença de E. histolytica e E. dispar, utilizando a PCR em tempo real,
empregada na rotina laboratorial da Divisão de Parasitologia do CDC.
Inicialmente, a PCR/Sequenciamento e a PCR em tempo real foram
realizadas com o DNA não diluído e em diferentes diluições (1/5, 1/10 e 1/20). As
diluições das amostras tiveram o propósito de minimizar a ação inibidores, mas
com DNA em concentração suficiente para ser amplificado.
O par de iniciadores, JVF/DSPR2, empregado na PCR/
Sequenciamento pode ser considerado iniciadores do tipo universal, pois

52
amplificou fragmentos de DNA da região parcial do 18S do rDNA de Entamoeba
spp., que variaram entre 622 a 668 pares de bases (pb), dependendo da espécie.
Este par de iniciadores, JVF/DSPR2, foi descrito por Verweij et al. (2003a). A
especificidade da PCR foi testada, utilizando DNA de Endolimax nana,
Blastocystis hominis, Giardia intestinalis, Microsporidium sp, Cryptosporidium
parvum e C. hominis.
A figura 4 ilustra o fragmento amplificado em torno de 650 pb resultante
da PCR de algumas amostras de fezes.
FIGURA 4. Gel de agarose a 2%. Amplificação de DNA de Entamoeba spp. usando o par de iniciador JVF/DSPR2. 1 – Padrão de peso molecular ( 1-100 bp); 2-7 e 10-11 amostras clínicas; 8 e 9 – branco da reação; 12 – c ontrole positivo; 13 – controle negativo
A PCR em tempo real, empregada neste estudo, é capaz de detectar
DNA de E. histolytica e E. dispar nas amostras analisadas. Sendo assim, somente
44 amostras foram avaliadas na PCR em tempo real, visto que a amostra 106 de
cultura era sabidamente positiva para E. moshkovskii. Das 44 amostras, oito
foram negativas pela PCR em tempo real. Nestas oito amostras foram
evidenciadas a presença de E. coli e E. hartmanni pela PCR/Sequenciamento, a
qual é totalmente compatível com o perfil do teste.
600 bp
100 bp

53
Nas nove amostras de cultura padrão identificadas pela
PCR/Sequencia-mento, observamos 100% de concordância com a PCR em
tempo real (tabela 3).
TABELA 3. Resultados obtidos nas amostras de cultur a padrão, pela PCR em tempo real e na PCR/Sequenciamento
Amostra Cultura *PCR em Tempo Real PCR/ Sequenciamento
103/Brasil E.histolytica E.histolytica/CT=22.34 E.histolytica
104/Brasil E.histolytica E.histolytica/CT=20.33 E.histolytica
105/Brasil E.histolytica E.histolytica/CT=21.09 E.histolytica
106/Brasil E. moshkovskii -------- E. moshkovskii*
107/Brasil E.dispar E.dispar/CT=27.35 E. dispar
109/Brasil E.histolytica E.histolytica/CT=20.88 E.histolytica
101/Brasil E.histolytica E.histolytica/CT=21.74 E.histolytica
P2/Brasil E. dispar E. dispar/CT=34.99 E. dispar
03 C/Brasil E. dispar E. dispar/CT=29,76 E. dispar
*PCR em Tempo Real específica para E.dispar e E. histolytica.
A PCR/Sequenciamento detectou a presença de DNA nas 16 amostras
de fezes provenientes do Serviço de Saúde Pública dos EUA (tabela 4). Entre
estas amostras, quatro foram negativas para o complexo E. histolytica/E. dispar e
duas negativas para qualquer parasito pela análise microscópica. Os resultados
da PCR/Sequenciamento nas seis amostras consideradas negativas pela análise
microscópica evidenciaram a presença de DNA de E. coli na amostra 3977/CDC e
de E. hartmanni nas amostras 100/CDC e 3684/CDC. As amostras 9891/CDC,
9893/CDC e 9343 foram positivas para E. dispar por ambas técnicas de PCR.

54
TABELA 4. Resultados obtidos no exame parasitológic o de fezes, na PCR em tempo real e na PCR/Sequenciamento nas amostras de fezes oriundas do Serviço de Saúde Pública dos EUA
Amostra Exame Parasitológico de Fezes
PCR em tempo real
PCR/ Sequenciamento
4093/CDC E.h/E.d;E. Polecki E.histolytica/CT= 30.97 E.histolytica
9893/CDC Bh; D. f; E. hart; Giardia E. dispar/CT= 25,00 E. dispar
9891/CDC Bh; D. f ; Giardia, Enta sp E. dispar/CT= 28,00 E. dispar
9343/CDC negativa E. dispar/CT= 30.97 E. dispar
3162/CDC E.d/E.h ; Bh E. dispar/CT= 25,21 E. dispar
3449/CDC E.d/E.h ; Bh E. dispar/CT= 26,47 E. dispar
3450/CDC E.d/E.h ; Bh E. dispar/CT= 23,52 E. dispar
6553/CDC E.d/E.h ; Bh E. dispar/CT= 20,52 E. dispar
200/CDC E.d/E.h ; Bh E. dispar/CT= 24.9 E. dispar
201/CDC E.d/E.h ; E.hart; E. nana E. dispar/CT= 26,75 E. dispar
33/CDC E.h/E.d ; E. poleckii E.histolytica/CT= 33,28 E.histolytica
3182/CDC E.d/E.h ; E. coli E. dispar/CT= 32,05 E.dispar; E. hart
204/CDC E.d/E.h ; E.hart; E. nana E. dispar/CT=28,90 E. hartmanni
100/CDC B. h Negativa E. hartmanni
3684/CDC B. h Negativa E. hartmanni
3977/CDC Negativa Negativa E. coli
D. f = Dientamoeba fragilis; Eh/Ed = Complexo E. histolytica/E. disp; B.h = Blastocystis hominis; E. Polecki = Entamoeba polecki; E. hart = Entamoeba hartmanni; A.l = Ascaris lumbricoides; T.t = Trichuris trichiura; E. coli = Entamoeba coli; E. nana = Endolimax nana; Anc.= ancilostomideo; Ss = Strongyloides stercoralis; Enta sp = Entamoeba sp.
Quando comparamos os resultados das 16 amostras obtidas na
PCR/Sequenciamento, PCR em tempo real e na análise microscópica
observamos resultados discrepantes em duas amostras. A amostra 3182/CDC foi
identificada como E. dispar pela PCR em tempo real e como E. hartmanni pela
PCR/Sequenciamento. Quando analisamos o eletroferograma desta amostra
gerado pelo sequenciador, observamos a presença de picos bem definidos e a

55
ausência de picos sobrepostos, indicando a presença de uma única sequência na
amostra. A primeira análise microscópica desta amostra não detectou a presença
de E. hartmanni. Diante destes resultados, o produto da PCR desta amostra foi
clonado e oito clones foram sequenciados. Destes, cinco clones foram positivos
para E. hartmanni e três para E. dispar. Sendo assim, foi feita uma reavaliação da
análise microscópica desta amostra que evidenciou a presença de E. hartmanni.
A amostra 204/CDC foi identificada como E. dispar pela PCR em tempo real e
como E. hartmanni pela PCR/Sequenciamento e, também, pela análise
microscópica. O produto da PCR desta amostra foi clonada e 15 clones foram
sequenciados, onde identificamos somente a presença de E. hartmanni. A
PCR/Sequenciamento não detectou a presença de DNA de E. poleckii nas
amostras 33/CDC e 4093/CDC como observado na análise microscópica. A
amostra 4093 foi clonada e oito clones foram sequenciados, sendo evidenciada
somente a presença de sequências de E. histolytica.
Conforme demonstrado na tabela 5, através da PCR/Sequenciamento,
foi detectada a presença de DNA de Entamoeba spp. nas 20 amostras
provenientes de indivíduos moradores no assentamento de Frei Vantuy em Ilhéus,
na Bahia, Brasil. Entre estas amostras, 11 foram positivas para E. dispar, três
positivas para E. coli e duas amostras para E. hartmanni pela PCR/
Sequenciamento. Além destas, três amostras apresentaram infecção mista, sendo
que destas, três foram positivas para E. hartmanni e E. dispar e uma para E.
dispar e E.coli.
As amostras 22 e 26/Brasil apresentaram infecção mista por E.
histolytica e E. dispar na PCR em tempo real. Contudo, a PCR/Sequenciamento
não evidenciou a presença de E. histolytica, mesmo após a clonagem molecular
destas amostras. Por outro lado, entre as 20 amostras, cinco foram negativas na
PCR em tempo real. Destas, duas foram positivas para E. hartmanni e três para
E. coli pela PCR/sequenciamento. A análise microscópica das amostras negativas
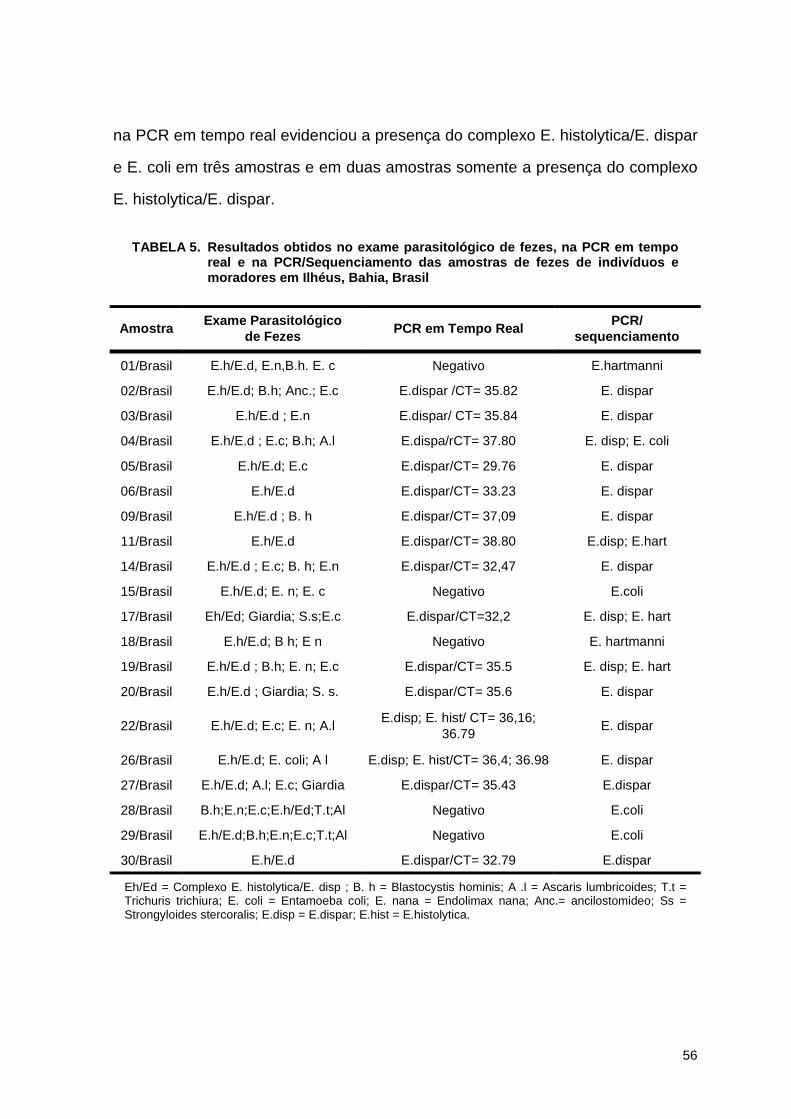
56
na PCR em tempo real evidenciou a presença do complexo E. histolytica/E. dispar
e E. coli em três amostras e em duas amostras somente a presença do complexo
E. histolytica/E. dispar.
TABELA 5. Resultados obtidos no exame parasitológic o de fezes, na PCR em tempo real e na PCR/Sequenciamento das amostras de fezes de indivíduos e moradores em Ilhéus, Bahia, Brasil
Eh/Ed = Complexo E. histolytica/E. disp ; B. h = Blastocystis hominis; A .l = Ascaris lumbricoides; T.t = Trichuris trichiura; E. coli = Entamoeba coli; E. nana = Endolimax nana; Anc.= ancilostomideo; Ss = Strongyloides stercoralis; E.disp = E.dispar; E.hist = E.histolytica.
Amostra Exame Parasitológico
de Fezes PCR em Tempo Real
PCR/ sequenciamento
01/Brasil E.h/E.d, E.n,B.h. E. c Negativo E.hartmanni
02/Brasil E.h/E.d; B.h; Anc.; E.c E.dispar /CT= 35.82 E. dispar
03/Brasil E.h/E.d ; E.n E.dispar/ CT= 35.84 E. dispar
04/Brasil E.h/E.d ; E.c; B.h; A.l E.dispa/rCT= 37.80 E. disp; E. coli
05/Brasil E.h/E.d; E.c E.dispar/CT= 29.76 E. dispar
06/Brasil E.h/E.d E.dispar/CT= 33.23 E. dispar
09/Brasil E.h/E.d ; B. h E.dispar/CT= 37,09 E. dispar
11/Brasil E.h/E.d E.dispar/CT= 38.80 E.disp; E.hart
14/Brasil E.h/E.d ; E.c; B. h; E.n E.dispar/CT= 32,47 E. dispar
15/Brasil E.h/E.d; E. n; E. c Negativo E.coli
17/Brasil Eh/Ed; Giardia; S.s;E.c E.dispar/CT=32,2 E. disp; E. hart
18/Brasil E.h/E.d; B h; E n Negativo E. hartmanni
19/Brasil E.h/E.d ; B.h; E. n; E.c E.dispar/CT= 35.5 E. disp; E. hart
20/Brasil E.h/E.d ; Giardia; S. s. E.dispar/CT= 35.6 E. dispar
22/Brasil E.h/E.d; E.c; E. n; A.l E.disp; E. hist/ CT= 36,16;
36.79 E. dispar
26/Brasil E.h/E.d; E. coli; A l E.disp; E. hist/CT= 36,4; 36.98 E. dispar
27/Brasil E.h/E.d; A.l; E.c; Giardia E.dispar/CT= 35.43 E.dispar
28/Brasil B.h;E.n;E.c;E.h/Ed;T.t;Al Negativo E.coli
29/Brasil E.h/E.d;B.h;E.n;E.c;T.t;Al Negativo E.coli
30/Brasil E.h/E.d E.dispar/CT= 32.79 E.dispar

57
4.3 Clonagem Molecular
Entre as 45 amostras positivas pela PCR/Sequenciamento, 16 tiveram
seu produto amplificado purificado e clonado. A clonagem foi realizada somente
para as amostras que apresentaram sequências muito curtas, baixo sinal de
fluorescência ou a presença de picos sobrepostos nas reações de
sequenciamentos prévios. Além disto, algumas amostras apresentaram um baixo
rendimento na amplificação, apresentando bandas com intensidade tênue como
pode ser observado na figura 3 (canaleta 5, 6, 7 e 11), não sendo possível
sequenciá-las. Assim, nestes casos optamos pela clonagem destas amostras.
Inicialmente, as colônias brancas transformadas, resistentes à
ampicilina e que não degradavam o substrato cromogênico X-gal foram
selecionadas randomicamente. Assim, para confirmar a presença do fragmento
de DNA inserido no vetor de clonagem foi realizada a PCR diretamente da
colônia, utilizando os iniciadores JVF/DSPR2. Como podemos observar na figura
5, a despeito do crescimento de colônias em meio sólido, nem todos os clones
analisados possuíam o fragmento de DNA inserido.
FIGURA 5. Gel de agarose a 1,5%. Amplificação de DN A utilizando o par de iniciador JVF/DSRP2 para confirmar a presença do inserto dura nte o processo da clonagem molecular. 1 e 14: marcador de peso molecu lar; 2 e 4-13: clones selecionados; 3: ausência do inserto, seta b ranca.
100 pb
600 pb

58
Uma vez selecionados, os clones positivos foram expandidos e
purificados. O DNA plasmidial foi digerido com a enzima Eco RI, conforme
descrito em Material e Métodos (item 3.9) para a comprovação da presença do
fragmento de DNA inserido no vetor. Foram considerados positivos os clones que
após a digestão apresentaram no gel de agarose uma banda de
aproximadamente 3,9 Kb, correspondente a do vetor pCR® 2.1 TOPO sem o
inserto e uma outra banda de cerca de 0,65 Kb, correspondente ao fragmento
clonado. Na figura 6 podemos observar o resultado da digestão dos clones, onde
é possível verificar a presença das duas bandas.
FIGURA 6: Análise dos clones obtidos em eletrofore se de gel de agarose 2% através da digestão com a enzima de restrição EcoRI gerando bandas de tamanho esperado para o vetor pCR 2.1 e para o inse rto de aproximadamente 0.65 Kb. (1); Marcador molecular 1 Kb (Invitrogem); 2-12: clones de Entamoeba spp.
Dentre as dezesseis amostras clonadas neste estudo, quatro foram
positivas somente para E. dispar, dois para E. hartmanni, três para E. coli e cinco
apresentaram infecção mista, sendo que destas, quatro foram positivas para E.
hartmanni e E. dispar e uma para E. dispar e E. coli. Além destas, os produtos

59
amplificados das amostras 4093/CDC e 106/Brasil foram clonadas para obtenção
de controles que foram utilizadas na padronização da PCR/Luminex.
4.4 Análise das Sequências
A disponibilidade de dados decorrentes do sequenciamento permitiu
avaliar a diversidade genética entre as espécies de Entamoeba. Assim, todas as
sequências consenso do gene SSU-rRNA obtidas neste estudo foram
comparadas com as sequências SSU-rRNA de Entamoeba, previamente
depositadas no GenBank, utilizando o programa BLAST (http:// www.ncbi.nlm.nih.
gov/blast) para a comprovação da identidade das sequências obtidas. O
alinhamento representado no anexo 7, serviu como base para a análise
filogenética através do programa MEGA versão 2.1, utilizando o modelo de
Kimura 2, parâmetros para a elaboração da matriz de distância e o método de
Neighbor-Joining para a construção da árvore filogenética. Na árvore pode-se
observar uma visível separação das distintas espécies estudadas. Os cinco
grupos formados estão representados na figura 8 (anexo 1) e correspondem,
respectivamente, as espécies de: E. histolytica, E. dispar, E. moshkovskii, E.
hartmanni e E.coli.
Nos alinhamentos múltiplos das sequências oriundas das 45 amostras
estudadas, observamos regiões altamente conservadas intercaladas com áreas
polimórficas. A análise deste alinhamento evidenciou a presença de sítios
polimórficos de substituição do tipo transição, transversão e eventos de inserção
e/ou deleção.
Quando analisamos o alinhamento das seis sequências de E.
histolytica, observamos ausência de polimorfismos intraespecíficos e 100% de
similaridade com a sequência de E. histolytica, depositada no GenBank sob o nº
de acesso X64142. Em contrapartida, a análise comparativa com a outra
sequência de E. histolytica, depositada no Genbank sob o nº ABI97936, revelou

60
alterações em duas bases nucleotídicas do tipo transição (C/T) nas posições 207
e 212, respectivamente (anexo 7). A análise das sequências de E. dispar obtida
demonstrou 100% de homologia entre si. Mas, quando comparamos com a
sequência de E. dispar depositada no GenBank sob o nº de acesso z49256,
observamos um único sítio polimórfico do tipo transição G/A na posição 33 do
alinhamento (anexo 7). Da mesma forma, ao comparar a sequência de E.
moshkovskii (106/Brasil) com a sequência depositada no Genbank sob o nº de
acesso AF149906, observamos onze sítios polimórficos e duas inserções de base
nucleotídicas na posição 486 e 487 (anexo 4). Na análise das sequências de E.
hartmanni foi observado um elevado grau de homologia e a presença de sítios
polimórficos do tipo transição, transversão e sitos de deleção nucleotídica entre si
e em relação à sequência de E. hartmanni depositada no GenBank sob o nº de
acesso A149907 (anexo 5). Por sua vez, o alinhamento das sequências de E. coli
originou dois grupos que mostraram uma alta homologia entre si. As amostras
3977 e 04B apresentaram uma homologia de 100% entre si e com a sequência E.
coli depositada no GenBank sob o nº de acesso AF149915. O restante das
sequências apresentou uma homologia em torno de 100% entre si e com a outra
sequência de E. coli depositada no GenBank sob o nº de acesso AF149914.
Quando comparamos os dois grupos observamos vários sítios polimórficos do tipo
transição e transversão (Anexo 6).
4.5 Reação de hibridação multianalítica em meio líq uido, utilizando
microesferas fluorescentes como suporte sólido asso ciado à
citometria de fluxo, sistema Luminex®
Somente metodologias que empregam sondas possuem um poder
discriminatório que permite o desenvolvimento de teste do tipo multianalítico. Para
isto, sonda espécie e grupo específicas foram desenhados a partir do alinhamento
das sequências de Entamoeba spp. depositadas no GenBank. Os códigos de

61
acesso das sequências utilizadas no alinhamento encontram-se no anexo 3.
Inicialmente foram sintetizadas quatro sondas específicas para E. histolytica, duas
sondas para E. dispar, duas para E. moshkovskii, duas para E. coli e duas sondas
para E. hartmanii. Além destas, duas sondas grupo especificas (E.histolytica, E.
dispar, E. moshkovskii e E.hartmanni) denominadas de EGP1e EGP2, e uma
sonda 275Ehist/Edispar que hibridiza em uma região em comum de E. histolytica
e E. dispar (tabela 6).
TABELA 6. Sondas de captura do tipo grupo e espécie específica utilizadas no ensaio de hibridação multianalitíco em meio líquido na PCR/Lu minex
Espécies Sonda Sequência 5’ ���� 3’ Posição no Gene SSU-rRNA*
E. histolytica histo168 5'CGATCCAGTTTGTATTAGT'3 183-204
E. histolytica hist200 5'TATTAGTACAAAATGGCCAAT'3 224-239
E. histolytica hist242 5'AA TGA ATTGAGAAATGACAT'3 234-256
E. histolytica hist 1 5'TAGTACAAAATGGCCAATT'3 201-227
E. dispar Disp238 5' GTAAGTAAATTGAGAAATGAC'3 231-253
E. dispar Dis186 5'GACGATCCAATTTGTATT'3 180-201
E. moshkovskii Emosh2 5' TA AATACTCTTACGAAATC'3 560-576
E. moshkovskii Emosh1 5'AGACGATCCGGTTTGTAT'3 165-200
E. hartmanni Ehart123 5' ATTAGTAAGTACAAGGAT'3 115-135
E. hartmanni Ehart1 5' ATGAGAATATCTGATCTA'3 326-344
E. coli Ecoli165 5' TGACGGTTTTCACCCCTT'3 162-179
E. coli Ecoli310 5'AGAGATTTTCACAAGTCA'3 305-323
EGP2 EGP2 5'TGAATGATAAAGATAATACT'3 142-161
EGP1 EGP1 5' TACAGGATAGCTTTGTGAAT'3 127-146
E.hist/dispar 275histo/disp 5'TTAGGATGCCACGACAATT'3 268-395
*Conforme o alinhamento (anexo 3).
A etapa de padronização da reação de hibridação multianalítica em
meio líquido foi realizada com o produto amplificado, a partir de DNA clonado de
E. coli, E. dispar, E.moshkovskii, E. histolytica e E. hartmanni. Na reação de
hibridação multianalítica foi utilizado o tampão de hibridação TMAC 1,5 Χ,
contendo volumes iguais das microesferas acopladas às sondas. Essa mistura foi

62
hibridada com o produto amplificado com o iniciador JVF/DSPR2 biotinilado, que
amplifica um fragmento de DNA, que varia entre 622 a 668 pb, de acordo com as
espécies amplificadas. A reação de hibridação foi realizada em temperaturas e
tempos distintos: 40ºC, 42ºC, 43ºC, 46ºC, 48ºC, 50ºC e 52ºC por 30 minutos, 45
minutos e 1 hora. Neste protocolo foi adicionado uma etapa de sedimentação,
onde as placas de hibridação foram centrifugadas a 2230 x g por 3 min e 25 µl do
sobrenadante foram cuidadosamente retirados. Posteriormente, foram
adicionados 75 µl da solução do fluoróforo repórter, estreptavidina-R-ficoeritrina à
1,0 mg/ml (Molecular Probes, Eugene, OR) diluída a 1:120, 1:80 e 1:50 em
tampão TMAC 1Χ, respectivamente.
As hibridação multianalíticas entre os produtos amplificados biotinilados
e as sondas correspondentes (tabela 6), realizadas nas condições mencionadas
acima, não geraram sinais específicos de fluorescências. Os valores de
intensidade de fluorescência mediana (MIF) ficaram muito próximos dos valores
encontrados na reação do branco (reação contendo as microesferas e TE no
lugar do produto da PCR).
Uma vez que os resultados não foram satisfatórios com os iniciadores
JVF/DSPR2, outros iniciadores foram desenhados conforme descritos na tabela 7,
a fim de se obter produtos amplificados de menores tamanhos e, talvez, facilitar
uma melhor ligação entre a sonda com o produto amplificado.
TABELA 7. Oligonucleotídeos iniciadores reversos bi otinilados utilizados na PCR/Luminex
C=Parreamento incorreto com a sequência de E. coli B* = Biotina **De acordo com o alinhamento (anexo 3)
Iniciadores Sequência 5’ 3’
Posição no Gene
SSU-rRNA**
EntaRev390-B* 5` TATTGCCCATTGCTCCTTA3 382 -401
EntaRev390-B* 5` TATTGTCCATTGCTCCTTA ’3 382-401
Rev417A-B* 5' AAAGCTCCTCTCCGATGT’3 410-429

63
Conforme a tabela acima, o par de iniciadores JVF/EntaRev390
amplifica um fragmento que varia entre 378 a 404 pb, dependendo da espécie. O
iniciador EntaRev390 possui um pareamento incorreto (mismatch) de C/T na
sequência de E. coli. Contudo, a eficiência de amplificação com o par de
iniciadores JVF/EntaRev-B não foi afetada, devido ao pareamento incorreto
localizado próximo à região 5’. O par de iniciadores JVF/Rev417A amplifica um
fragmento em torno de 404 a 429 bp, dependendo da espécie a ser amplificada.
A figura 7, ilustra os fragmentos amplificados pelos pares de iniciadores
JVF/EntaRev390-B e JVF/REV417A.
FIGURA 7. Gel de agarose a 2%. Amplificação de DNA de Entamoeba spp. A: produto amplificado com o par de inciador JVF/re v417A; 1 – Padrão de peso molecular (100 bp); 2-3 controles positivo, 4- 7 branco, 8 – negativo, 9–18: amostras clínicas. B: produto amplificado com o par de iniciador JVF/EntaRev390B; 1 – Padrão de peso molecular (100 bp), 2 e 3 – controle positivo, 4-6 – amostras postivas, 7– bran co, 8 e 9 – amostras negativas
A reação de hibridação multianalítica com o produto amplificado pelo
par de iniciador JVF/EntaRev-B e as sondas correspondentes imobilizadas nas
microesferas foram realizadas nas seguintes temperaturas e tempos distintos:
42ºC, 43ºC, 46ºC, 48ºC, 50ºC e 52ºC por 45 minutos e 1 hora. Após a etapa de
lavagem, onde as placas de hibridação foram centrifugadas a 2230 x g por 3 min,
B 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
400 pb
100 bp
A 1 2 3 4 5 6 7 8 9
400 bp
100 pb

64
foram retirados, cuidadosamente, 25 µl do sobrenadante. Posteriormente, foram
adicionadas 75 µl da solução do fluoróforo repórter, estreptavidina-R-ficoeritrina
1,0 mg/ml (Molecular Probes, Eugene, OR) diluída à 1:80 e 1:50 em tampão
TMAC 1Χ, respectivamente. As intensidades de fluorescência (MIF) obtidas com
a temperatura de 46ºC por 1 hora e a diluição 1:50 da SA-PE foram superiores às
demais temperaturas, tempos e diluições do fluoróforo repórter. Nestas
condições, observamos sinais específicos de intensidade de fluorescência
mediana (MIF) entre os produtos amplificados pelo par de iniciador JVF/Enta
Rev390-B e as sondas correspondentes, tendo obtido valores de MIF superior aos
valores encontrados na reação do branco (Gráfico 1). De fato, observamos sinais
de hibridações específicas e com poder discriminatório com as sondas E. hart123
e E.coli165 na detecção de E. hartmanni e E. coli. Estas sondas apresentaram
MIF alta, sendo bastante reprodutíveis. Similarmente, as sondas E. mosh1 e
EGP2 específica para E. histolytica, E dispar, E hartmanni e E. moshkovskii
geraram uma MIF média reprodutível. As sondas de E. dispar e E. histolytica
apresentaram MIFs baixas e não reprodutíveis. Nestes ensaios realizados não
foram observadas reações cruzadas entre os produtos da PCR e as sondas. Os
resultados dos sinais de fluorescência, após a leitura no equipamento Luminex
100, foram corrigidos, subtraindo do valor da MIF das amostras a média da MIF
dos brancos. As MIF corrigidas para cada sonda encontram-se no gráfico 1.
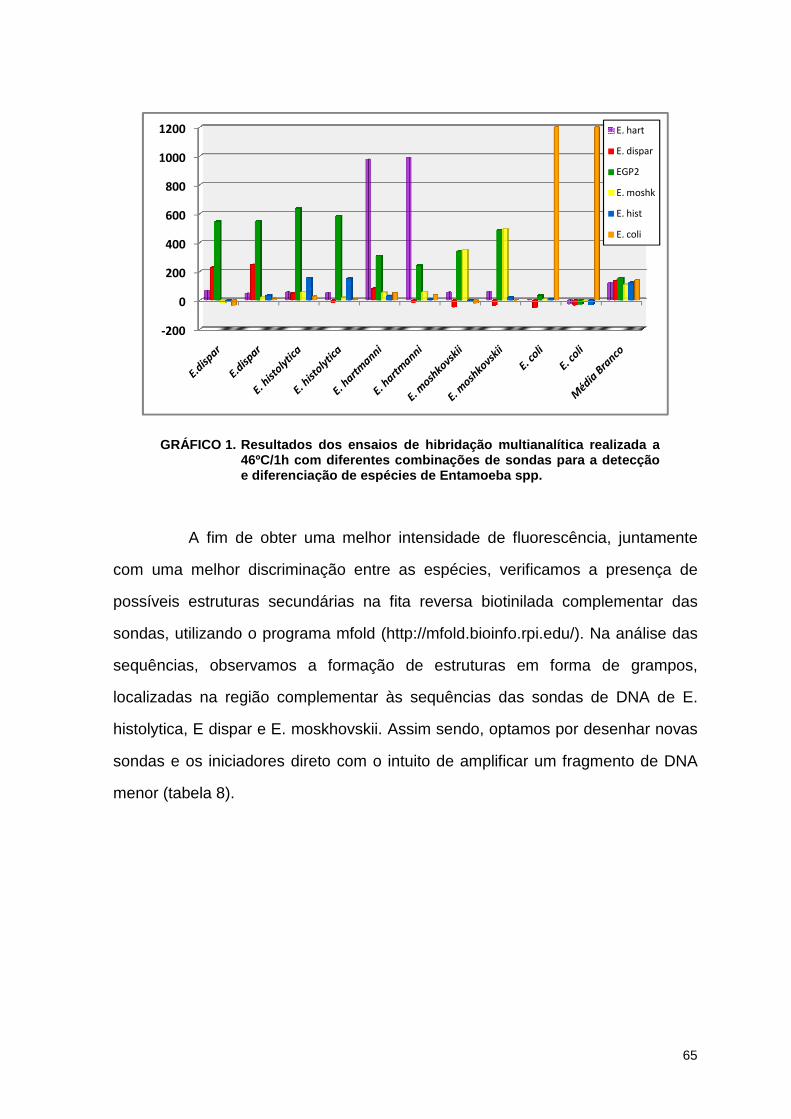
GRÁFICO 1. Resultados dos ensaio46ºC/1h com diferentes combinações de sondas para a detecção e diferenciação de espécies de
A fim de obter uma melhor intensidade
com uma melhor discriminação entre as e
possíveis estruturas secundárias na fita reversa biotinilada complementar
sondas, utilizando o programa mfold (
sequências, observamos a formação de estruturas em forma de
localizadas na região complementar
histolytica, E dispar e E. moskhovskii
sondas e os iniciadores direto
menor (tabela 8).
-200
0
200
400
600
800
1000
1200
Resultados dos ensaio s de hibridação multianalítica realizada a 46ºC/1h com diferentes combinações de sondas para a detecção e diferenciação de espécies de Entamoeba spp .
fim de obter uma melhor intensidade de fluorescência,
com uma melhor discriminação entre as espécies, verificamos a presença de
possíveis estruturas secundárias na fita reversa biotinilada complementar
utilizando o programa mfold (http://mfold.bioinfo.rpi.edu/
observamos a formação de estruturas em forma de
complementar às sequências das sondas de DNA de
E. moskhovskii. Assim sendo, optamos por desenhar novas
sondas e os iniciadores direto com o intuito de amplificar um fragmento de DNA
65
de hibridação multianalítica realizada a 46ºC/1h com diferentes combinações de sondas para a detecção
de fluorescência, juntamente
spécies, verificamos a presença de
possíveis estruturas secundárias na fita reversa biotinilada complementar das
http://mfold.bioinfo.rpi.edu/). Na análise das
observamos a formação de estruturas em forma de grampos,
s sequências das sondas de DNA de E.
por desenhar novas
ragmento de DNA
E. hart
E. dispar
EGP2
E. moshk
E. hist
E. coli

66
TABELA 8. Novos oligonucleotídeos iniciadores utili zados na PCR e sondassondas na reação de hibridação multianalítica
Sondas Sigla Sequências Posição no
gene SSU-rRNA*
E. moshkovskii M1 5’GTATGACAATTGTAGAGC3’ 285-302
E. moshkovskii M2 5’ATGGTATGACAATTGTAGA3’ 282-300
E. moshkovskii M3 5’GACAATGTAGAGCACACAG3’ 286-308
E.histolytica hist116 5’GGTTAGTAAAATACAAGG3’ 116-134
E. dispar Edis1 5'ACGATCCAATTTGTATT'3 188-194
E.dispar Edis2 5’GTTAGAGATTAAGCCAT’3 34-54
E.dispar Edis3 5’TAGAGATTAAGCCATGC’3 36-51
E.dispar Edis4 5' ATGTTAGAGATTAAGCCA3' 31-50
Iniciador direto EntaF1 5’CTGCGGACGGCTCATTATAACAG’3 84-102
Iniciador direto EntaF2 5’ATTATAACAGTAATAGTTTCTTGG’3 97-117
Iniciador direto EntaF3 5’ CGGCTCATTATAACAGTAATAG’3 92-108
Iniciador direto EntaF4 5’GGCTCATTATAACAGTAATAGTTTC’3 91-112
*Conforme alinhamento (anexo 3).
Deve ser ressaltado que os iniciadores diretos Enta 1, 2 , 3 e 4 não
foram avaliados até o presente momento. Contudo, todas as sondas desenhadas
foram testadas na reação de hibridação multianalítica em meio líquido.
A hibridação multianalítica com o produto amplificado pelo par de
iniciadores JVF/EntaRev-B, assim como as sondas correspondentes (tabela 8)
imobilizadas nas microesferas foi realizada nas seguintes temperaturas: 41ºC,
43ºC, 46ºC e 48ºC por 1h, sem a etapa de sedimentação. Após esta etapa, foram
adicionados 25 µl de uma diluição de 1/50 de estreptavidina-R-ficoeritrina pré
aquecida na concentração de 1mg/ml. A placa foi incubada nas respectivas
temperaturas mencionadas acima por 5 min e a sua leitura foi realizada em placa
aquecida na plataforma Luminex 200.

A hibridação
fluorescências para as sondas
coli, e E. dispar. Contudo, fo
de Ehart 123 e o produto da PCR de
sonda hist 116 apresentou uma discreta reaç
dispar e E. moshkovskii.
clonado utilizada neste ensaio
amplificado diluído a 1/100.
leitura no equipamento Luminex 200 foram corrigidos
das amostras, a média da MIF dos brancos
encontram-se no gráfico 2.
GRÁFICO 2. Resultados dos ensaios de hibridação multianalítica realizada a 41ºC/1h com diferentes combinações de sondas para a detecção e diferenciação de espécies de
A hibridação multianalítica
fluorescência para as sondas de
coli, e E. dispar. Entretanto,
-200
0
200
400
600
800
1000
1200
hibridação multianalítica, realizada a 41ºC/1h,
para as sondas de E. histolytica, E. hartmanni, E. moshkovskii, E
. Contudo, foram observadas reações cruzadas entre a
o produto da PCR de E. histolytica, E. moshkovskii
sonda hist 116 apresentou uma discreta reação cruzada com os produtos de
E. moshkovskii. O volume dos produtos amplificados, a partir
clonado utilizada neste ensaio, foi de 17 µl e 5 µl sem diluição e 17 µ
1/100. Os resultados dos sinais de fluores
leitura no equipamento Luminex 200 foram corrigidos, subtraindo do valor da MIF
média da MIF dos brancos. As MIF corrigidas para cada sonda
no gráfico 2.
Resultados dos ensaios de hibridação multianalítica realizada a 41ºC/1h com diferentes combinações de sondas para a detecção e diferenciação de espécies de Entamoeba spp
A hibridação multianalítica, realizada a 43ºC/1h,
para as sondas de E. histolytica, E. hartmanni, E. moshkovskii, E
Entretanto, ainda foram observadas reações cruzadas entre a
67
gerou sinais de
E. histolytica, E. hartmanni, E. moshkovskii, E
reações cruzadas entre as sondas
E. moshkovskii e E. dispar. A
cruzada com os produtos de E.
dos produtos amplificados, a partir de DNA
sem diluição e 17 µl do produto
Os resultados dos sinais de fluorescência, após a
subtraindo do valor da MIF
corrigidas para cada sonda
Resultados dos ensaios de hibridação multianalítica realizada a 41ºC/1h com diferentes combinações de sondas para a
spp
gerou sinais de
E. histolytica, E. hartmanni, E. moshkovskii, E
reações cruzadas entre a
E hartmanii
E dispar
E histolytica
E.moshk
E. coli

-200
0
200
400
600
800
1000
1200
sonda E hart123 com o
moshkovskii. Neste ensaio
clonado utilizado na hibridação
produto amplificado dilu
após a leitura no equipamento Luminex 200 foram
da MIF das amostras a
sonda encontram-se no gráfico 3.
GRÁFICO 3. Resultados dos ensaios de hibridação multianalítica realizada a 43ºC/1h com diferentes combinações de sondas para a detecçã o e diferenciação de espécies de
A hibridação multianalítica, realizada
forneceu sinais de fluorescência satisfatórios para a discriminação entre as cinco
espécies de Entamoeba,
o volume dos produtos amplificados, a partir de DN
hibridação multianalítica, foi
sonda E hart123 com o produto amplificado de E. histolytica
Neste ensaio, o volume dos produtos amplificados, a par
hibridação multianalítica foi de 5 µl sem diluição e 17 µ
uído a 1/100. Os resultados dos sinais de fluorescência,
após a leitura no equipamento Luminex 200 foram corrigidos, subtraindo do valor
das amostras a média da MIF dos brancos. As MIF corrigidas para cada
no gráfico 3.
Resultados dos ensaios de hibridação multianalítica realizada a 43ºC/1h com diferentes combinações de sondas para a detecçã o e diferenciação de espécies de Entamoeba spp
hibridação multianalítica, realizada à temperatura de 46ºC/1h,
forneceu sinais de fluorescência satisfatórios para a discriminação entre as cinco
Entamoeba, não sendo observadas reações cruzadas. Neste ensaio,
os produtos amplificados, a partir de DNA clonado utilizado na
multianalítica, foi o mesmo utilizado a 43ºC/1hora
68
E.hart
E dispar
Ehist
Emoshk
Ecoli
E. histolytica, E. dispar e E.
produtos amplificados, a partir de DNA
sem diluição e 17 µl do
Os resultados dos sinais de fluorescência,
subtraindo do valor
corrigidas para cada
Resultados dos ensaios de hibridação multianalítica realizada a 43ºC/1h com diferentes combinações de sondas para a detecçã o e diferenciação
temperatura de 46ºC/1h,
forneceu sinais de fluorescência satisfatórios para a discriminação entre as cinco
não sendo observadas reações cruzadas. Neste ensaio,
A clonado utilizado na
/1hora. As sondas

específicas para E. hartmanni,
alto sinal de MIF, sendo bastante reprodutíveis. No entanto, a sonda de
histolytica apresentou um v
fluorescência, após a leitura no equipamento Luminex 200 foram corrigidos,
subtraindo do valor da MIF das amostras, a média da MIF dos brancos. As MIF
corrigidas para cada sonda encontram
GRÁFICO 4. Resultados dos ensaiosde hibridação multianalítica realizada a 46ºC/1h com diferentes combinações de sondas para a detecçã o e diferenciação de espécies de
A hibridação multianalítica
fluorescência somente para as sondas de
sendo observadas reações cruzadas.
amplificados, a partir de DN
mesmo utilizado a 43 ºC
-200
0
200
400
600
800
1000
1200
E. hartmanni, E. coli, E. dispar e E. moshkovskii
alto sinal de MIF, sendo bastante reprodutíveis. No entanto, a sonda de
apresentou um valor médio de MIF. Os resultados dos sinais de
fluorescência, após a leitura no equipamento Luminex 200 foram corrigidos,
subtraindo do valor da MIF das amostras, a média da MIF dos brancos. As MIF
corrigidas para cada sonda encontram-se no gráfico 4.
Resultados dos ensaiosde hibridação multianalítica realizada a 46ºC/1h com diferentes combinações de sondas para a detecçã o e diferenciação de espécies de Entamoeba spp
hibridação multianalítica, realizada a 48ºC/1h,
para as sondas de E. dispar, E. coli e E. hartmanni
reações cruzadas. Neste ensaio, o volume d
r de DNA clonado utilizado na hibridação multianalítica
ºC/1h. Os resultados dos sinais de fluorescência, após a
69
moshkovskii apresentaram
alto sinal de MIF, sendo bastante reprodutíveis. No entanto, a sonda de E.
alor médio de MIF. Os resultados dos sinais de
fluorescência, após a leitura no equipamento Luminex 200 foram corrigidos,
subtraindo do valor da MIF das amostras, a média da MIF dos brancos. As MIF
Resultados dos ensaiosde hibridação multianalítica realizada a 46ºC/1h com diferentes combinações de sondas para a detecçã o e diferenciação
gerou sinais de
E. hartmanni, não
o volume dos produtos
multianalítica, foi o
. Os resultados dos sinais de fluorescência, após a
Ehart
dispar
E hist
Emoshko
E coli

leitura no equipamento Luminex 200 foram
das amostras, a média da MIF dos brancos
encontram-se no gráfico
GRÁFICO 5. Resultados dos ensaios de hibridação multianalítica realizada a 48ºC/1h com diferentes combinações de sondas para a detecçã o e diferenciação de espécies de
Diante dos ensaios realizados, as MIF obtidas na temperatura de 46ºC
por 1 hora e na diluição 1
temperaturas, tempos e diluições do fluoróforo repórter, onde obtivemos valores
de MIF superior aos valores encontrados na reação do branco. Nesta temperatura
não observamos reações cruzadas entre as sond
Tanto os produtos amplificados pelo par de iniciador
pelo JVF/Rev417A hibridaram com as sondas correspondentes, imobilizadas nas
microesferas, onde observamos sinais de hibridaç
discriminatório na diferenciação de
e E. histolytica. As sondas que apresentaram os melhores sinais de fluorescência
estão relacionadas na tabela abaixo. Um esquema representativo que mostra o
-200
0
200
400
600
800
1000
1200
leitura no equipamento Luminex 200 foram corrigidos, subtraindo do valor da MIF
média da MIF dos brancos. As MIF corrigidas para cada sonda
se no gráfico 5.
Resultados dos ensaios de hibridação multianalítica realizada a 48ºC/1h com diferentes combinações de sondas para a detecçã o e diferenciação de espécies de Entamoeba spp
Diante dos ensaios realizados, as MIF obtidas na temperatura de 46ºC
diluição 1:50 da SA-PE, foram superiores
temperaturas, tempos e diluições do fluoróforo repórter, onde obtivemos valores
de MIF superior aos valores encontrados na reação do branco. Nesta temperatura
não observamos reações cruzadas entre as sondas e os produtos amplificados.
Tanto os produtos amplificados pelo par de iniciadores JVF/ EntaRev
pelo JVF/Rev417A hibridaram com as sondas correspondentes, imobilizadas nas
microesferas, onde observamos sinais de hibridação específico
discriminatório na diferenciação de E. hartmanni, E. coli, E. dispar, E. moshkovskii
As sondas que apresentaram os melhores sinais de fluorescência
estão relacionadas na tabela abaixo. Um esquema representativo que mostra o
70
subtraindo do valor da MIF
As MIF corrigidas para cada sonda
Resultados dos ensaios de hibridação multianalítica realizada a 48ºC/1h com diferentes combinações de sondas para a detecçã o e diferenciação
Diante dos ensaios realizados, as MIF obtidas na temperatura de 46ºC
PE, foram superiores às demais
temperaturas, tempos e diluições do fluoróforo repórter, onde obtivemos valores
de MIF superior aos valores encontrados na reação do branco. Nesta temperatura
as e os produtos amplificados.
JVF/ EntaRev-B quanto
pelo JVF/Rev417A hibridaram com as sondas correspondentes, imobilizadas nas
ão específicos e com poder
E. coli, E. dispar, E. moshkovskii
As sondas que apresentaram os melhores sinais de fluorescência
estão relacionadas na tabela abaixo. Um esquema representativo que mostra o
Ehart
Edisp
Ehist
Emoskh
Emoshk
E coli

71
local de hibridação das sondas com as suas respectivas sequências alvos
encontra-se no anexo 2.
TABELA 9. Relação das sondas que apresentaram sinai s específicos de fluores-cência na PCR/Luminex
**Conforme o alinhamento das sequências no anexo 3.
A hibridação multianalítica foi avaliada quanto a sua capacidade em
detectar e diferenciar o DNA de Entamoeba em amostras clínicas. As condições
da hibridação multianalítica utilizadas, foram as mesmas preestabelecidas frente
aos produtos amplificados, a partir de DNA clonado. Desta forma, o volume dos
produtos amplificados, foi de 5 µl sem diluição e 17 µl do produto amplificado
diluído a 1/100. Os ensaios foram realizados a 46ºC por 1 hora e a diluição do
fluoróforo repórter, estreptavidina-R-ficoeritrina à 1,0 mg/ml (Molecular Probes,
Eugene, OR) foi de 1:50 da SA-PE em tampão TMAC 1Χ pré aquecido. A placa
foi incubada, novamente, à 46ºC/5 min e a sua leitura foi realizada em placa
aquecida à mesma temperatura na plataforma Luminex 200. Os resultados dos
sinais de fluorescência, após a leitura foram corrigidos, subtraindo do valor da MIF
das amostras a média das MIF dos brancos.
Espécies Sonda Sequência 5’ ���� 3’ Posição no
Gene SSu-rRNA*
E. histolytica hist 116 5’GGTTAGTAAAATACAAGG3’ 116-134
E.dispar Disp238 5'GTAAGTAAATTGAGAAATGAC'3 218-247
E.moshkovskii M3 5’GACAATGTAGAGCACACAG3’ 288-304
E.moshkovskii Emosh1 5'AGACGATCCGGTTTGTAT'3 171-193
E.hartmani Ehart1 5' ATGAGAATATCTGATCTA'3 333-350
E.coli Ecoli310 5'AGAGATTTTCACAAGTCA'3 312-329
EGP2 EGP2 5'TGAATGATAAAGATAATACT'3 148-168

72
As amostras clínicas analisadas pelas PCR/Luminex foram
previamente utilizadas na PCR em tempo real e na PCR/Sequenciamento.
Inicialmente, onze amostras foram amplificadas com o par de iniciadores
JVF/EntaRev390-B. As amostras 2, 3, 5 e 6 foram positivas para E. dispar por
ambas PCRs, assim como as amostras 17 e 19 foram positivas para E. dispar e
E. hartmanni. Por outro lado, as amostras 15 e 28 foram positivas para E. coli,
sendo a amostra 18 positiva para E. hartmanni pela PCR/Sequenciamento. As
amostras Em e Eh são oriundas de cultura. Os resultados da PCR/Luminex
destas amostras encontram-se na tabela 10, onde podemos observar que os
resultados ressaltados na cor amarela representam sinais específicos de
hibridação, as quais foram parcialmente concordantes com a PCR em tempo real
e a PCR/Sequenciamento.
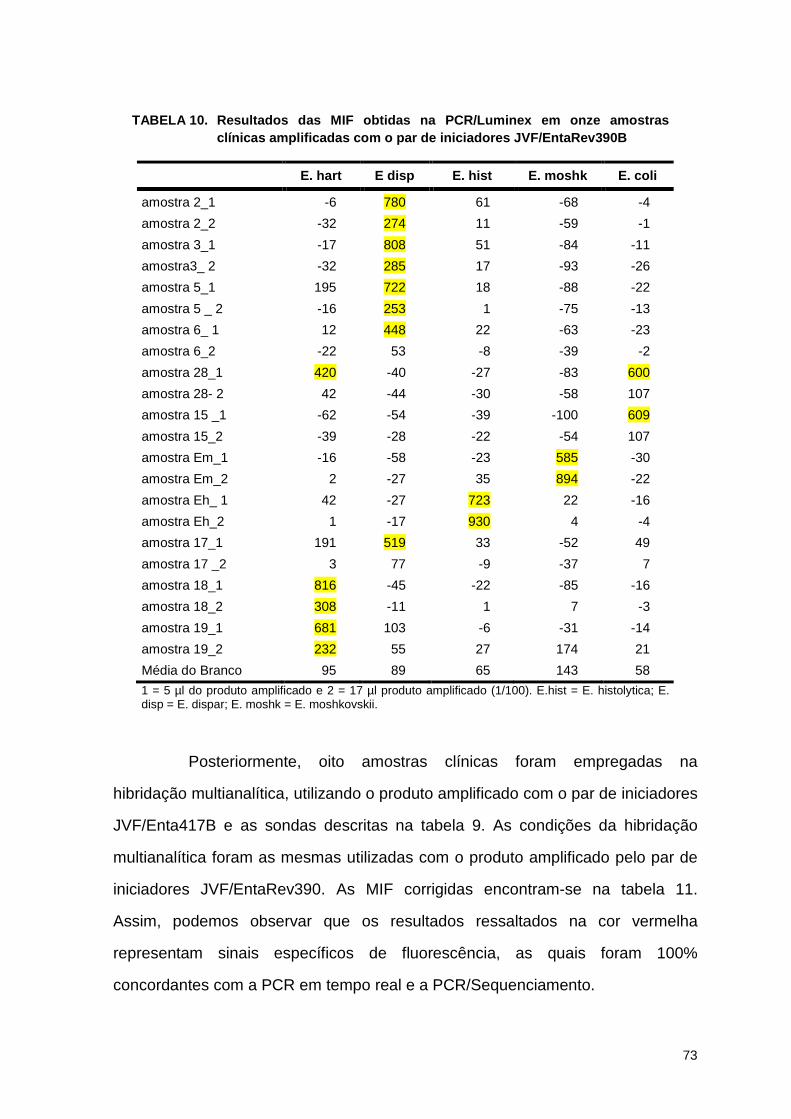
73
TABELA 10. Resultados das MIF obtidas na PCR/Lumine x em onze amostras clínicas amplificadas com o par de iniciadores JVF/ EntaRev390B
E. hart E disp E. hist E. moshk E. coli
amostra 2_1 -6 780 61 -68 -4
amostra 2_2 -32 274 11 -59 -1
amostra 3_1 -17 808 51 -84 -11
amostra3_ 2 -32 285 17 -93 -26
amostra 5_1 195 722 18 -88 -22
amostra 5 _ 2 -16 253 1 -75 -13
amostra 6_ 1 12 448 22 -63 -23
amostra 6_2 -22 53 -8 -39 -2
amostra 28_1 420 -40 -27 -83 600
amostra 28- 2 42 -44 -30 -58 107
amostra 15 _1 -62 -54 -39 -100 609
amostra 15_2 -39 -28 -22 -54 107
amostra Em_1 -16 -58 -23 585 -30
amostra Em_2 2 -27 35 894 -22
amostra Eh_ 1 42 -27 723 22 -16
amostra Eh_2 1 -17 930 4 -4
amostra 17_1 191 519 33 -52 49
amostra 17 _2 3 77 -9 -37 7
amostra 18_1 816 -45 -22 -85 -16
amostra 18_2 308 -11 1 7 -3
amostra 19_1 681 103 -6 -31 -14
amostra 19_2 232 55 27 174 21
Média do Branco 95 89 65 143 58
1 = 5 µl do produto amplificado e 2 = 17 µl produto amplificado (1/100). E.hist = E. histolytica; E. disp = E. dispar; E. moshk = E. moshkovskii.
Posteriormente, oito amostras clínicas foram empregadas na
hibridação multianalítica, utilizando o produto amplificado com o par de iniciadores
JVF/Enta417B e as sondas descritas na tabela 9. As condições da hibridação
multianalítica foram as mesmas utilizadas com o produto amplificado pelo par de
iniciadores JVF/EntaRev390. As MIF corrigidas encontram-se na tabela 11.
Assim, podemos observar que os resultados ressaltados na cor vermelha
representam sinais específicos de fluorescência, as quais foram 100%
concordantes com a PCR em tempo real e a PCR/Sequenciamento.

74
TABELA 11. Resultados das MIF obtidas na PCR/ Lumin ex de oito amostras clínicas amplificadas com o par de iniciadores JVF/ Enta417B
E. dispar E. hist E. moshk E. hart E.coli
02_ 1 -32 -20 -8 -17 691
02_2 286 16 -16 18 -2
03_ 1 1239 63 -31 276 -38
03_ 2 290 20 4 19 2
15_ 1 -30 -26 -24 15 1162
15_2 -6 -8 -12 10 161
17_ 1 738 12 -39 598 113
17 _ 2 171 3 -7 55 24
18_ 1 -18 -24 -32 1831 -34
18_ 2 13 22 10 315 12
19_ 1 216 -16 -39 1.762 -34
19_2 40 10 -4 287 12
28 _ 1 -35 -23 -19 1314 910
28_2 7 3 -3 161 120
Em_ 1 -33 103 1660 556 -36
Em_ 2 6 41 734 25 10
Média/Branco 72 72 78 78 79
1 = 5 µl do produto amplificado e 2 = 17 µl produto amplificado (1/100). E. hist = E. histolytica; E. disp = E. dispar; E. moshk = E. moshkovskii.
Na tabela 11, observamos reação cruzada entre o produto amplificado
da amostra EM-1 e a sonda Ehart 123. Tal fato pode ser devido a uma possível
contaminação entre os poços de reação durante o ensaio, visto que esta mesma
amostra não apresentou reação cruzada quando foi utilizado o produto
amplificado pelos iniciadores JVF/EntaRev (tabela 10).
Os resultados das amostras apresentados nas tabelas 10 e 11 estão
sumarizados na tabela 12 e confrontados com os resultados prévios da análise
microscópica, da PCR em tempo real e da PCR/Sequenciamento. Nesta tabela,
podemos observar 100% de concordância entre os resultados da PCR/Luminex
com os demais testes.

75
TABELA 12. Comparação entre os resultados da PCR/Lu minex com a PCR em tempo real, PCR/Sequenciamento e da análise microscópica de amostras clínicas
Amostras Microscopia* PCR em
tempo real PCR/
sequenciamento PCR/
Luminex
Nº 02/Brasil Eh/Ed; E. coli E.dispar E.dispar E.dispar; E.coli Nº 03/Brasil Eh/Ed E.dispar E.dispar E.dispar
Nº 05/Brasil Eh/Ed; E. coli E.dispar E.dispar E.dispar
Nº 06/Brasil Eh/Ed E.dispar E.dispar E.dispar
Nº 15/Brasil Eh/Ed; E. coli Negativa E.coli E. coli
Nº 17/Brasil Eh/Ed E.dispar E.dispar; E.hart E.dispar/E.hart
Nº 18/Brasil Eh/Ed Negativa E. hatmanni E. hartmanni
Nº 19/Brasil Eh/Ed; E. coli E.dispar E. dispar; E.hart E.dispar; E.hart
Nº 28/Brasil Eh/Ed; E. coli negativa E.coli E. coli; E.hart
*Eh/Ed = Complexo E. histolytica/E. dispar; E. hart = E. hartmanni.
Um grupo de quinze amostras, com resultados desconhecidos no
momento do experimento (“estudo cego”), foi utilizado para avaliar a
PCR/Luminex. Destas amostras, cinco foram negativas na amplificação utilizando
o par de iniciadores JVF/Rev417A. Estas amostras foram gentilmente cedidas
pelo CDC, onde tinha sido previamente realizadao o ensaio de PCR em tempo
real, conforme descrito em material e métodos (item 3.5). Os resultados da
PCR/Luminex foram enviados ao CDC e confrontados com o da PCR em tempo
real, onde observamos 100% de concordância entre os resultados dos testes. As
MIFs corrigidas destas amostras encontram-se na tabela 13.

76
TABELA 13. Resultados da PCR/Luminex e da PCR em te mpo real em 15 amostras clínicas
1 = 5 µl do produto amplificado e 2 = 17 µl produto amplificado (1/100). E. hist = E. histolytica; E. disp = E. dispar; E. moshk = E. moshkovskii.
E.disp E. hist E.moshk E. hart E.coli
PCR em Tempo real
CTL+ Eh_ 1 -41 981 93 140 -89
CTL+Eh_ 2 -60 734 -32 -47 -66
CTL+ Ed_1 1368 54 -61 214 -79
CTL+Ed_ 2 277 -24 -65 -47 -60
CTL+Em_ 1 -101 -82 64 -65 -111
CTL+ Em_ 2 -76 -48 547 -45 -86
CTL+ Ehart_ 1 -82 -87 -80 1844 -85
CTL+ Ehart 2 -70 -67 -68 323 -76
CTL+ Ecoli_ 1 -75 -60 -84 -53 1194
CTL+ Ecoli_ 2 -78 -66 -76 -54 121
Amostra 1cdc_ 1 530 -15 -64 181 -73 E.dispar
Amostra1cdc_ 2 111 -26 -36 -9 -59
Amostra2cdc_ 1 772 -10 -74 728 -76 E.dispar
Amostra2cdc_ 2 142 -26 -40 70 -56
Amostra4 cdc1 -44 1094 135 201 59 E. hist
Amostra4 cdc_ 2 -27 727 22 -2 -16
Amostra 5cdc_ 1 -43 1144 154 185 75 E. hist
Amostra 5 cdc_ 2 -27 714 -13 16 -14
Amostra 6 cdc_ 1 597 -4 -75 578 -80 E.dispar
Amostra 6 cdc_ 2 138 -28 -47 52 -54
Amostra 8 cdc_ 1 -50 1027 101 172 44 E. hist
Amostra 8 cdc_ 2 -51 712 -27 -45 -53
Amostra9 cdc_ 1 -39 1144 117 197 49 E. hist
Amostra 9 cdc_ 2 -7 706 26 -4 17
Amostra10 cdc_ 1 -39 1145 172 255 97 E. hist
Amostra 10 cdc_ 2 -16 728 23 11 -6
Amostra 13 cdc_ 1 487 -23 -78 394 -73 E.dispar
Amostra 13 cdc_ 2 121 -19 -44 34 -43
Amostra 15 cdc_ 1 530 -11 -65 478 -63 E.dispar
Amostra 15 cdc_ 2 148 -13 38 63 -36
Amostra 3 cdc_ neg* -77 -76 -71 -79 -68 Negativa
Amostra 7 cdc_neg* -79 -70 -77 -89 -87 Negativa
Amostra 11 cdc_neg* -85 -69 -86 -94 -93 Negativa
Amostra 12 cdc_ neg* -83 -84 -91 -95 -88 Negativa
amostra 14 cdc_ neg* -75 -69 -81 -78 -71 Negativa
Média do branco 111 101 116 117 125
111 101 116 117 125

77
5. DISCUSSÃO
O exame parasitológico de fezes é baseado em critérios morfológicos
das formas evolutivas de parasitos presentes nas fezes. As técnicas mais
empregadas no diagnóstico de amebas intestinais são: o método direto a fresco, o
método de Lutz ou Hoffman Pons e Janer (HOFFMAN, PONS & JANER 1934),
Faust (FAUST et al., 1938) e o método Ritchie (RITCHIE,1948). Estes métodos
foram desenvolvidos há mais de sete décadas, quando a população humana
apresentava altos níveis de intensidade de infecção. Nestas condições, qualquer
uma dessas técnicas mostrava bom desempenho no diagnóstico, fornecendo
resultados satisfatórios. Entretanto, com o passar dos anos, melhorias nas
condições de saúde e uma maior eficácia dos medicamentos antiparasitários
levou a uma redução da intensidade do parasitismo, resultando na perda da
sensibilidade do EPF. Embora largamente utilizado até a atualidade, o EPF não é
capaz de diferenciar o complexo E. histolytica/E. dispar e E. moshkovskii de E.
histolytica. Ademais, E. hartmanni, E. poleckii e cistos imaturos de E. coli
apresentam diminutas diferenças e/ou características sobrepostas, que, muitas
vezes, são imperceptíveis na análise microscópica, acarretando um diagnóstico
impreciso.
A análise microscópica não é suficientemente informativa para o
diagnóstico preciso das amebas intestinais, visto que o EPF está sujeito a
variáveis que afetam a sua sensibilidade e especificidade. Entretanto, a
ineficiência diagnóstica resulta de diversos fatores, sendo uma parte inerente à
própria técnica, a biologia dos parasitos e a outra referente a fatores humanos,
passíveis de correção. Segundo Petri e cols. (2000), o EPF é uma técnica
ultrapassada que não deveria ser usada para o diagnóstico da colite
amebiana, pois é incapaz de diferenciar E. histolytica de E. dispar e
apresenta uma sensibilidade que varia entre 5 e 60% e uma especificidade

78
entre 10 e 50%. Nota-se, ainda, que o EPF só é capaz de detectar E. histolytica
quando evidenciamos a presença de eritrofagocitose pelos trofozoítos. Este
achado é extremamente raro em fezes diarreicas recém emitidas. No entanto,
existem controvérsias entre os pesquisadores em relação ao real valor do EPF no
diagnóstico da amebíase. Gonzáles-Ruiz e cols. (1994) descreveram que a
eritrofagocitose na amebíase disentérica é 100% específica e que a mesma
prenuncia a infecção invasiva. Entretanto, Doganci, Tanyuksel & Doganci (2004)
mencionaram que a acurácia no diagnóstico da amebíase é essencial e que a
eritrofagocitose por trofozoítos não é um meio adequado de diagnóstico, já que foi
comprovado que E. dispar é capaz de fagocitar eritrócitos in vitro. Os autores
sumarizaram este trabalho, relatando que o EPF não é sensível e nem específico
para o diagnóstico da amebíase. Por sua vez, E. hartmanni pode ser diferenciada
do complexo E. histolytica/E. dispar e de E. moshkovskii por biometria de cistos e
trofozóitos, utilizando-se para tal fim uma ocular micrométrica no microscópio
óptico. Entretanto, as formas císticas podem sofrer retrações e deformação no
material fecal, dificultando o diagnóstico diferencial com E. histolytica.
Para a correta identificação do parasito durante a análise microscópica
é necessário que a morfologia do parasito esteja íntegra, constituindo este um
fator relevante para o diagnóstico, juntamente com a capacitação do
microscopista. Hamzah e cols. (2006) relataram que das 30 amostras fezes
positivas para Entamoeba spp., pela análise microscópica, cinco foram negativas
em três técnicas de PCR que utilizaram iniciadores do tipo gênero e espécie
específicos. Os autores sugeriram que as amostras negativas na PCR foram
identificadas erroneamente como cistos de Entamoeba spp.
De fato, resultados semelhantes vêm sendo observados em relação à
análise microscópica. Em 1999, Ferrer e cols. observaram que somente um
pequeno percentual de técnicos que trabalhavam em laboratórios da província de
Cienfuegos em Cuba eram capacitados para identificar corretamente as amebas

79
no EPF. Neste estudo, 56% dos profissionais foram capazes de identificar os
estágios de pré-cisto, 62% de cisto e somente 36% de trofozoitos. No Brasil, a
baixa reprodutibilidade dos resultados do EPF, principalmente entre diferentes
laboratórios clínicos, tem sido relatada. Em 1997, o Instituto Nacional de
Metrologia, Normalização e Qualidade Industrial (INMETRO) avaliou
criteriosamente a eficiência diagnóstica dos resultados dos exames
parasitológicos, fornecidos por diferentes laboratórios de análises clínicas das
cidades de São Paulo e do Rio de Janeiro. O INMETRO relatou que três entre
quatro laboratórios da cidade de São Paulo e sete entre dez da cidade do Rio de
Janeiro enviaram resultados com um número significativo de erros. O instituto
concluiu que os resultados apresentados pelos laboratórios foram, em sua
maioria, insatisfatórios (INMETRO, 1997).
Neste mesmo ano, após a aceitação de E. dispar como nova espécie, a
OMS anuiu que fossem desenvolvidos métodos de diagnóstico capazes de
diferenciar E. histolytica de E. dispar (WHO, 1997). Assim, diante das
recomendações da OMS, várias pesquisas foram desenvolvidas, baseadas em
métodos moleculares para a detecção de antígeno e de DNA de E. histolytica e E.
dispar nas fezes. Ao longo destes anos, vários estudos vêm utilizando a PCR na
detecção e diferenciação de E. histolytica e E. dispar em amostras de fezes. Para
tal, diferentes fragmentos de DNA-alvo, estratégias de amplificação e protocolos
de extração de DNA têm sido utilizados (ACUNA-SOTO et al.,1993; RIVERA et
al., 1996; AGUIRRE et al., 1997; BRITTEN et al., 1997; MIRELMAN et al., 1997;
TROLL et al., 1997; VERWEIJ et al., 2000; EVANGELOPOUS et al., 2000;
BLESSMANN et al., 2002; GONIN & TRUDEL, 2003; VERWEIJ et al., 2004).
Contudo, a maioria destes estudos não abordou a diferenciação entre
E. histolytica de outras espécies de amebas comensais no diagnóstico da
amebíase, com exceção de E. dispar e, mais recentemente, E. moshkovskii.
Entretanto, a presença de outras espécies de Entamoeba vem sendo aos poucos

80
descrita na literatura, mas ainda não existe uma estimativa da ocorrência destas
espécies. De acordo com Choe e cols. (1996), a presença de E. histolytica na
Coréia é superestimada em decorrência da falta de diferenciação entre E.
histolytica, E. dispar e E. hartmanii. Verweij e cols. (2000) utilizaram a PCR
Colorimétrica (solution hybridization enzyme-linked immunoassay – PCR-SHELA)
na detecção e diferenciação de E. histolytica e E. dispar. Os autores relataram
que cinco amostras positivas para o complexo Eh/Ed foram negativas na PCR.
Diante destes resultados, os autores sugeriram que as amostras negativas na
PCR poderiam estar parasitadas por outras espécies de amebas,
morfologicamente semelhantes. Tais resultados foram observados por Kebede e
cols. (2004), que encontraram uma alta prevalência do complexo E.
histolytica/ E. dispar em amostras de fezes na Etiópia. Das 108 amostras
analisadas contendo o complexo E. histolytica/E. dispar, somente 77 foram
positivas para E. dispar, uma para E. histolytica e as 30 amostras
remanescentes foram negativas na PCR em tempo real. Os autores
concluíram que, provavelmente, outras amebas não patogênicas tinham sido
diagnosticadas erroneamente como complexo E. histolytica/E. dispar. Além
disso, alguns estudos têm demonstrado uma elevada frequência de E.
moshkovskii em algumas populações estudadas na região de Dhaka na Índia, em
Sidney na Austrália, em Bangkok na Tailândia e na Tanzânia na África (ALI et al.,
2003; PARIJA & KHAIRNAR, 2005; HAMZAH et al., 2006; FOTEDAR et al., 2008;
BECK et al., 2008).
Todavia, poucos estudos abordam a aplicação de métodos moleculares
no diagnóstico diferencial entre as amebas intestinais pertencentes ao gênero
Entamoeba. Em 1992, Clark e Diamond utilizaram iniciadores universais na
amplificação de um segmento do gene SSU-rRNA de protozoários intestinais. O
diagnóstico diferencial entre E. histolytica e outros protozoários foi realizado por
meio da análise de restrição de fragmentos polimórficos. Neste estudo, a

81
PCR/RFLP foi capaz de diferenciar E. histolytica de E coli, E. hartmanni, E.
moshkovski, E. nana e Blastocystis hominis nas amostras analisadas.
Posteriormente, Verweij e cols. (2003a) empregaram o método de hibridação
reversa em membrana com o intuito de detectar o DNA ribossômico do gênero
Entamoeba.
Diante da importância do diagnóstico correto das espécies de
Entamoeba que parasitam o intestino humano e dos escassos testes de
diagnóstico disponíveis, buscou-se desenvolver ensaios moleculares do tipo
multianalítico, específicos, sensíveis e semiautomatizados, como a PCR/Luminex
e a PCR/Sequenciamento, para o diagnóstico das amebas intestinais. Assim, o
presente trabalho contribui para esta necessidade, quer seja avaliando um
marcador molecular através da PCR/Sequenciamento, quer avaliando a
PCR/Luminex para a detecção e diferenciação das amebas intestinais do gênero
Entamoeba.
Inicialmente, realizamos o levantamento das sequências de Entamoeba
spp disponíveis no GenBank, com o intento de obter uma sequência gênica que
pudesse ser utilizada como um marcador molecular. Neste levantamento,
observamos que somente o gene SSU-rRNA estava disponível para todas as
espécies de Entamoeba intestinais. Tal fato reduz bastante a expansão de
metodologia e estratégias para o diagnóstico molecular. Por isso, a única
alternativa foi trabalhar com o gene SSU rRNA. Sendo assim, avaliamos parte do
segmento gênico do DNA ribossômico das amebas do gênero Entamoeba através
da PCR/Sequenciamento. Os resultados obtidos neste estudo demostraram que a
sequência do gene da SSU-rRNA pode ser utilizada como alvo da PCR para o
diagnóstico de Entamoeba intestinais. Este gene tem sido utilizado como alvo na
PCR convencional e na PCR em tempo real na detecção e diferenciação entre E.
histolytica e E. dispar.

82
Como apresentado nos resultados, a PCR/Sequenciamento detectou a
presença de uma ou mais espécies Entamoeba nas 45 amostas analisadas. É
importante ressaltarmos que o presente trabalho não foi elaborado para
caracterizar as amostras clínicas. Para isto, deveríamos ter clonado todas as
amostras e sequenciado um número representativo de clones, a fim de detectar
todas as espécies de Entamoeba presente em cada amostra. A clonagem foi
realizada somente paras as amostras que apresentaram sequências
inconclusivas, sendo sequências muito curtas, baixo sinal de fluorescência,
presença de picos sobrepostos ou com um baixo rendimento na amplificação.
Nosso objetivo principal foi avaliar a PCR/Sequenciamento padronizada neste
estudo. Todavia, quando comparamos os resultados da PCR/Sequenciamento
com os da PCR em tempo real e da microscopia, observamos uma discrepância.
Possivelmente, alguns fatores podem ter influenciado na divergência
dos resultados entre os testes. Devemos salientar que a análise microscópica é
susceptível a resultados falso-positivos em decorrência da presença de espécies
morfologicamente similares, características morfológicas sobrepostas entre as
várias espécies de amebas, artefatos presente nas fezes ou pela interpretação
subjetiva do microscopista. Ademais, resultados negativos podem ser observados
devido à eliminação irregular e intermitente das formas evolutivas de Entamoeba,
sendo necessário analisar mais de uma amostra (REY, 2008). Este fatores podem
acarretar um diagnóstico impreciso. Além disto, a amostra fecal é um material
extremamente heterogêneo e rico em substâncias inibidoras para a PCR. A
coextração destes inibidores pode inibir a reação, bloqueando o sítio ativo da
enzima DNA polimerase (ABU Al-SOULD & RADSTROMl, 1998). Componentes
da dieta e da microbiota intestinal, como proteases bacterianas e nucleases, tanto
quanto debris celular podem afetar a amplificação. Estas interferêncais são
extremamente variávies entre diferentes indivíduos, talvez em decorrência dos
hábitos alimentares que possam estar influenciando o tipo e a quantidade de

83
substância inibidora presente no material fecal (HOLLAND et al. 2000; LI,
HULLAR & LAMPE, 2007; NECHVATAL et al., 2008). Assim, a obtenção de DNA
altamente purificado é essencial para que se tenha resultados confiáveis na PCR.
Ademais, a conservação e o tempo de armazenamento da amostra fecal pode
alterar a chance de detecção do parasito (CNOPS & ESBROECK, 2010). Neste
estudo, foram realizadas diluições nas amostras de DNA, com o intuito de
minimizar a ação de substâncias inibidoras nas PCR. Esta estratégia também foi
utilizada por Stacy-Phipps, Mecca & Weiss (1995) e Wilson (1997), que
observaram uma redução dos inibidores fecais quando diluíram as amostras de
DNA. Contudo, resultados negativos, nestes casos, podem ocorrer devido à
presença de DNA-alvo abaixo do limite detectável da PCR.
Neste estudo, foram empregados dois métodos de extração de DNA e
diferentes métodos de análises morfológicas. Assim, criaram-se três tabelas de
acordo com a origem das amostras para facilitar a interpretação dos resultados
em função das estratégias utilizadas no processamento das amostras, na análise
microscópica e na extração de DNA, evitando, assim, que estas variáveis
influenciem ainda mais a análise dos resultados. Como apresentado nos
resultados, na tabela 3, observamos 100% de concordância entre os resultados
da PCR/Sequenciamento e da PCR em tempo real. Em contrapartida, resultados
divergentes em algumas amostras foram observados nas tabela 4 e 5. As
amostras 9893 e 9343 foram negativas pela análise microscópica para o
complexo E. dispar/E. histolytica e positivas para E. dispar por ambas PCR. Além
das citadas acima, as amostras 100, 3684 e 3977 também foram negativas no
EPF, evidenciando a presença de E. hartmanii e E. coli pela PCR/
Sequenciamento. Estes resultados estão de acordo com os resultados obtidos por
Verweij e cols (2003) em estudo realizado em Gana na África. Neste, das 114
amostras negativas na análise microscópica, a PCR em tempo real específica
para E. histolytica e E. dispar foi capaz de amplificar DNA de E. dispar em 87

84
amostras, confirmando, assim, a baixa sensibilidade do EPF. Uma explicação
plausível para os resultados negativos pela análise microscópica seria o número
de amostras analisadas, pois, quando se analisa uma única amostra de fezes,
somente um terço ou metade das espécies parasitárias presentes na massa fecal
são detectadas (GARCIA, 2007). Isto pode ser, pelo menos em parte, devido à
eliminação irregular e intermitente de parasitos pelas fezes, podendo ocorrer
grande variação da quantidade de cistos e trofozoítos no bolo fecal (UNGAR,
YOLKEN & QUINN, 1985; FONTE et al.,1998; GARCIA, 2007). A amostra
3182/CDC foi positiva para E. dispar pela PCR/sequenciamento somente após a
clonagem. Um total de oito clones foram sequenciados e, destes, cinco foram
positivos para E. hartmanii e três para E. dispar. A análise microscópica inicial
desta amostra falhou em detectar a presença de E. hartmanni. Diante destes
resultados foi realizada uma segunda análise microscópica, que evidenciou a
presença de E. hartmanni. Contudo, E. coli não foi detectada pela PCR/
Sequenciamento, nem após a clonagem molecular. Neste caso, é válido
considerar que cistos imaturos de E. coli apresentam uma morfologia similar aos
cistos tetranucleados de amebas ou uma possível distorção na morfologia pode
ter contribuído para tal fato. Para Heckendorn e cols. (2002), existe um risco real
de identificar erroneamente cistos imaturos de E. coli como o complexo E.
histolytica/E. dispar. Na análise microscópica da amostra 33/CDC, foi evidenciada
a presença moderada de cistos do complexo E. histolytica/E. dispar e de raras
formas cisticas de E. poleckii. Contudo, a PCR/Sequenciamento foi positiva
somente para E. histolytica. Para esta amostra, não foi realizada a clonagem. Por
sua vez os produtos amplificados obtidos da amostra 4093/CDC, positiva para E.
poleckii, foram clonados, sendo evidenciada somente a presença de E. histolytica
nos oito clones sequenciados. Da mesma forma, a análise microscópica da
amostra 204/CDC evidenciou a presença moderada de cistos de E. hartmanni e
raras formas císticas do complexo E. dispar/E. histolytica. Esta amostra foi

85
positiva para E. dispar somente na PCR em tempo real. Entretanto, somente E.
hartmanni foi detectada pela PCR/ Sequenciamento, mesmo após o
sequenciamento de oito clones. Devemos levar em consideração que a
PCR/Sequenciamento da amostra 204/CDC foi positiva somente na alíquota
conservada em unifix, o que pode ter ocasionado a redução da sensibilidade da
PCR/Sequenciamento.
É importante salientar que, diante dos resultados obtidos para as
amostras 3182/CDC, 4093/CDC e 204/CDC, podemos sugerir que há uma
quantidade heterogênea de DNA de diferentes espécies de ameba na mesma
amostra. Neste caso, possivelmente houve uma competição entre os DNA alvos e
os iniciadores, o que pode ter proporcionado uma amplificação preferencial na
PCR/Sequenciamento, ou seja, um maior rendimento para a espécie presente em
maior quantidade na reação. Em contrapartida, isto não foi observado na PCR em
tempo real, devido à amplificação de uma única sequência-alvo, não ocorrendo,
assim, uma competição entre diferentes alvos. Para Ali e cols. (2008), vários
fatores podem mascarar a infecção por múltiplos genótipos no caso de uma
espécie ser mais representativa do que a outra na amostra analisada. Os autores
realizaram um ensaio experimental, no qual demonstraram que é possível
detectar o produto amplificado em amostras mistas, quando a espécie minoritária
representa cerca de até 4% do total. Outros autores relataram que, em amostras
mistas, quando os iniciadores são compartilhados em um ensaio do tipo duplex,
uma grande quantidade de DNA de uma espécie presente na PCR pode ofuscar a
amplificação de uma segunda espécie que apresente uma pequena quantidade
de DNA alvo (FOTERDAR et al., 2007; QVARNSTROM et al., 2005). Uma outra
questão a ser discutida diante dos resultados encontrados nas amostras
3182/CDC, 4093/CDC e 204/CDC é a quantidade de clones selecionados e
sequenciados. De fato, não podemos garantir que o número de clones

86
sequenciados foram representativos ou insuficientes para demonstrar a presença
de poucas moléculas DNA-alvo amplificadas.
Diantes destes fatos, pode-se esperar que as PCR, apresentem
rendimentos diferentes e, consequentemente, resultados variados, visto que
mostram faixas de sensibilidade diferentes. Por outro lado, a literatura tem
descrito que a PCR em tempo real é mais sensível que a PCR convencional no
diagnóstico de doenças infecciosas (KLEIN, 2002; ROY et al., 2005; SINGH,
HOUP & PETRI, 2009; STARK et al., 2010). A principal desvantagem da PCR em
tempo real em relação a PCR convencional é a sua limitação quanto ao número
de alvos de DNA a serem detectados (PAUL, SRIVASAVA & BHATTACHARYA,
2007).
Além disto, devemos considerar que estamos avaliando resultados de
duas técnicas que apresentam formatos e sensibilidades diferentes de detecção
do produto amplificado. A PCR em tempo real detecta o produto amplificado à
medida que vai ocorrendo a amplificação por meio de sondas específicas para E.
histolytica e E. dispar, que emitem um sinal de fluorescência. Diferentemente da
PCR convencional, cujo produto é detectado por meio de gel de agarose, corado
com brometo de etídio, que apresenta uma faixa de sensibilidade em torno de 20
ng/ml a 20 µg/ml (PELT-VERKUIL, BELKUM & HAYS, 2008a). Alguns estudos
têm mostrados que a PCR em tempo real é mais sensível do que a PCR
convencional na diferenciação entre duas espécies somente. Roy e cols. (2005)
empregaram a PCR em tempo real, utilizando o sistema ‘’molecular beacon’’ na
identificação e diferenciação entre E. histolytica e E. dispar em amostras de fezes
e de secreção de abscesso hepático. Os autores relataram que a PCR em tempo
real apresentou uma maior sensíbilidade quando comparada com a PCR
convencional. No presente estudo, foi empregada a PCR em tempo real baseada
no sistema Taqman, que amplifica um segmento gênico do SSU-rRNA de E.
histolytica e E. dispar (VERWEIJ et al. 2003b). Este método foi considerado como

87
o melhor utilizado na detecção e diferenciação entre E. histolytica e E. dispar
descrito na literatura (QVARNSTROM et al 2005). Os referidos autores
concluíram que, dentre todos os métodos de PCR em tempo real até então
publicados, o que apresentou uma maior sensibilidade foi o sistema TaqMan, que
amplifica o segmento do SSU-rRNA, descrito por Verweij et al., 2003. Haque e
cols (2007), utilizando a PCR em tempo real para a detecção de E. histolytica,
Giardia e Cryptosporidium spp observaram que a PCR em tempo real
padronizada para amplifcação de um único alvo, foi mais sensível do que ensaio
do tipo multiplex. Sendo assim, estes dados nos sugerem que ensaios de PCR
em tempo real do tipo multiplex padronizados para mais de dois alvos tendem a
perder em sensibilidade. Para Mackay (2004), existe a disponibilidade de um
maior número de fluoroforos que pode ser utilizados na PCR em tempo real.
Apesar disto, ensaios do tipo multiplex não têm sido aplicados ao diagnóstico de
doenças infecciosas numa escala significativa. Segundo Shipley (2008), a última
geração de equipamento de PCR em tempo real possibilita uma maior
combinação de reações do tipo multiplex de maneira mais rápida e utilizando um
maior número de amostras na mesma corrida, quando comparado com os antigos
equipamentos. Contudo, ainda não está claro porque este limite não foi alcançado
na prática.
Com relação às amostras brasileiras, não podemos descartar a
hipótese da ocorrência de um baixo rendimento de DNA extraído. Estas amostras
foram congeladas por mais de um ano, e foi utilizado um método de extração não
comercial sem a etapa de purificação, diferentemente das amostras do CDC
extraídas num curto período de tempo, utilizando um método de extração de DNA
comercial, seguido de uma etapa de purificação. Como relatado na literatura, o
longo tempo de armazenamento de amostras fecais e a presença de inibidores
presentes no DNA extraído afetam adversamente a cinética e a eficiência da PCR
(NECHVATAL et al., 2008; CNOPS & ESBROECK, 2010). Sendo assim, quando

88
comparamos os valores do CT cycle threshold (limiar de detecção) da PCR em
tempo real das amostras brasileiras com as do CDC, observamos uma diferença
bastante acentuada entre os resultados. A média do CT das amostras clínicas
oriundas do Brasil foi de 35, enquanto a média das amostras clínicas do CDC foi
de 27, sendo este resultado compatível com os resultados das amostras de
cultura. Tal fato sugere uma diferença significativa quanto ao rendimento da PCR
em tempo real, de grau de pureza do DNA extraído e da ação de inibidores na
PCR nas amostras brasileiras. Sabemos que na PCR em tempo real quanto maior
a concentração inicial de DNA-alvo, mais precocemente o valor do CT é atingido,
enquanto reações contendo menores concentrações de DNA alvo necesitam de
um maior número de ciclos para que a fluorescência emitida ultrapasse os valores
de background da reação.
Diante do acima exposto, observamos resultados discrepantes entre a
microcópia e os ensaios de PCR para as amostras provenientes de Ilhéus, no
estado da Bahia. A análise microscópica destas amostras não foram submetidas
a nenhum método de coloração, o que reduz a sensibilidade e a especificidade do
teste. De fato, os métodos de coloração evidenciam detalhes morfológicos,
essenciais para a identificação das amebas. E. hartmanni não foi mencionda nos
resultados da análise microscópica porque os cistos e trofozoítos não foram
mensurados, o que justifica os resultados das amostras 1 e 18 que foram
positivas para E. hartmanii na PCR/Sequenciamento e negativas na PCR em
tempo real e na análise microscópica. Portanto, os resultados da PCR em tempo
real destas amostras estão corretos, pois o teste é específico somente para a
detecção e diferenciação de E. histolytica e E. dispar.
Em relação aos resultados divergentes entre a PCR em tempo real e a
PCR/Sequenciamento, uma outra explicação para tal fato seria a qualidade do
DNA utilizado. As amostras brasileiras foram transportadas de Ilhéus na Bahia
para o Rio de Janeiro, resfriadas em gelo e congeladas por mais de um ano até o

89
momento da extração de DNA. Por maiores que fossem os cuidados que tivemos
com o transporte das amostras fecais, persiste a possibilidade de degradação do
DNA, principalmente nas amostras diarreicas. É importante ressaltar que, em
fezes diarreicas predomina a forma de trofozoíto que degenera rapidamente nas
fezes, expondo o DNA à lise por componentes presente no material fecal. É
notorio que os cistos são mais resistente ao processo de degradação e possuem
uma maior quantidade de material genômico, pois cada cisto contêm até oito
núcleos dependendo da espécie, diferentemente do trofozoíto que possuem
apenas um núcleo. Assim, amostras de fezes diarreicas requerem que sejam
congeladas rapidamente com o intento evitar a lise DNA, visto que as formas
prevalentes são sensivieis a degradação. As amostras 15, 28 e 29 são diarreicas
e foram positivas somente para E. coli pela PCR/Sequenciamento. Outro fator
importante que não podemos deixar de mencionar é uma possível degradação do
DNA extraído. Tal fato pode ter ocorrido durante o transporte do DNA na
temperatura ambiente para os EUA. Para algumas amostras contendo poucos
alvos, isto pode ter influenciado a eficiência da PCR. A PCR em tempo real
amplifica um fragmento do DNA ribossômico de E. histolytica e E. dispar de 160
bp e a PCR/Sequenciamento um fragmento em torno de 630 bp. Sabemos que
fragmentos de DNA menores têm maior chance de serem amplificados quando o
DNA sofre degradação. Sendo assim, pode se esperar que as amostras, frente
aos diferentes ensaios de PCR, tenham rendimentos diferentes. As amostras 22 e
26 foram positivas para E. histolytica e E. dispar pela PCR em tempo real. Nestas
amostras a PCR/sequencimento falhou em detectar a presença de E. histolytica.
De fato, como relatado anteriormente, as diferentes concentrações nos DNA-alvos
competindo pelo mesmo oligonucleotideo iniciador e a presença de inibidores
podem afetar adversamente a cinética da PCR. Segundo Flekna e cols. (2007), o
efeito das substâncas inibidoras sobre os resultados da PCR e na variabilidade da
eficiência da extração do DNA são mais pronunciados em amostras com baixo

90
número de alvo da PCR. Portanto, o êxito na amplificação de DNA na PCR
depende da qualidade e da quantidade do DNA-alvo.
A literatura tem relatado diferença entre os resultados de PCR em
relação ao diagnóstico diferencial entre E. histolytica e E. dispar. Verweij e cols.
(2003b), ao utilizarem dois ensaios de PCR em tempo real, na diferenciação e
detecção de E. histolyica e E. dispar em amostras de fezes, verificaram que, de
um total de sete amostras, cinco foram positivas somente na PCR que amplificava
a sequência localizada no DNA epissômico, enquanto que as outras duas
amostras foram positivas somente na PCR que amplificava uma sequência do
gene do SSU-rRNA. Além destas, cinco amostras foram negativas por ambas as
PCR em tempo real. Quando os autores utilizaram iniciadores gênero específico
na PCR seguido do sequenciamento, observaram sequências de E. coli e E.
hartmanni em três das cinco amostras. Assim, mencionaram um provável erro em
diagnosticar cistos pequenos de E. coli e cistos maiores de E. hartmanni como
pertencentes ao complexo E. histolytica/E. dispar durante a análise microscópica.
Resultados similares também foram observados por Hamzah e cols. (2006). Os
autores relataram que 15 amostras contendo o complexo Eh/Ed foram negativas
em dois ensaios de PCR, específicos para E. histolytica, E dispar e E.
moshkovskii. No entanto, quando empregaram a PCR gênero específico
evidenciaram a presença de E. coli e E. hartmanni através da reação de
sequenciamento.
Diante do acima exposto, é importante salientar a possibilidade de
perda de sensibilidade da PCR/Sequenciamento em amplificar o DNA de algumas
amostras do presente trabalho. Tal fato poderia ser amenizado utilizando como
estratégia a Nested-PCR que aumentaria a eficiência da PCR, principalmente nas
amostras contendo poucas moléculas de DNA-alvo .
Neste estudo, foi selecionado o gene SSU-rRNA como região alvo na
padronização das PCRs devido à disponibilidade das sequências. O gene SSU-

91
rRNA é uma região altamente conservada (PELT-VERKUIL, BELKUM & HAYS
2008c). O alto grau de homologia na sequência presente no gene SSU-rRNA
entre as diferentes espécies de Entamoeba permite o seu uso para desenho de
iniciadores específicos que amplificam diversas Entamoeba spp. A região
hipervariável do gene SSU-rRNA discrimina, entre todas as espécies, assim como
entre os múltiplos genótipos em nível de espécie. A variabilidade interespécie
deste gene observada neste estudo já foi demonstrada através da análise dos
padrões de restrição do DNA ribossômico amplificado (CLARK & DIAMOND,
1997a, 1997b) e através de sequenciamento do gene SSU-rRNA de Entamoeba
spp (SILBERMAN et al., 1999).
De fato, a análise dos dados genotípicos gerados pela PCR/
Sequenciamento permitiu avaliar as variações moleculares encontradas no gene
SSu-rRNA de Entamoeba presentes nas amostras oriundas de EUA e de Ilhéus
na Bahia, Brasil. Nestas amostras, observamos que as sequências gênicas foram
bastante similares entre si e com as sequências disponíveis no “GenBank” (anexo
7). Por sua vez, as variações nucleotídicas encontradas nas amostras
mencionadas acima serviram para avaliar a especificidade das sondas
empregadas na detecção e diferenciação de E. histolytica, E. moshkovskii, E.
hartmanni, E. coli e E. dispar.
Um dos objetivos do presente trabalho foi desenvolver a PCR/Luminex
capaz de diferenciar cinco espécies de ameba que parasitam o intestino humano.
Esta técnica tem emergido como uma ferramenta diagnóstica inovadora e
promissora que atende a demanda na área de diagnóstico por métodos
multianalíticos. A PCR/Luminex é uma técnica de hibridação multianalítica em
meio líquido, que emprega sonda de DNA abdsorvida na superfície de
microesferas fluorescentes, associada a citometria de fluxo. As vantagens desta
tecnologia são: rápida aquisição de informação, excelente sensibilidade e
especificidade e a análise simultânea de diferentes DNA-alvo.

92
Devemos salientar que a padronização de teste do tipo multiplex é uma
condição complexa e depende de suporte computacionais e algoritmos bem
definidos. Atualmente, o ensaio de hibridação multianalítica em meio líquido
representa um grande desafio a ser enfrentado durante o seu desenvolvimento.
Infelizmente, a falta de programas computacionais específicos para o desenhos
de sondas de DNA, estratégias que permitam a predição do comportamento de
diversos oligonucleotideos na reação do tipo multiplex e auxiliem a determinação
da temperatura de hibridação, a concentração das sondas e dos produto
biotinilados na reação, torna seu desenvolvimento baseado na experiência
empírica. Segundo Naiser e cols. (2009), apesar do bom conhecimento sobre a
estabilidade do DNA em solução, a predição da afinidade da reação de hibridação
em ensaios de microarranjos permanece sendo realizada de maneira empírica. Já
Diaz (2007) ressaltou que não existe regras específicas para o desenhos de
sondas, verificação da eficiência das sondas, previsão da cinética de hibridização
multianalítica e interação da sonda na sequência alvo, prinicpalmente nas regiões
alvo que possuem estruturas secundárias. Todavia, no entender de Dunbar
(2006), à medida que esta tecnologia se torna consolidada e disseminada muitos
outros ensaios serão desenvolvidos e o custo do equipamento e das microsferas
chegarão ao patamar de razoável de custo, como se sucedeu na maioria das
tecnologias utilizadas atualmente no diagnóstico laboratorial.
Neste trabalho, o desenho das sondas foi realizado de maneira
empírica, baseado em tentativa e erro. As sondas foram desenhadas a partir do
alinhamento das sequências de DNA de Entamoeba spp. disponíveis no
“GenBank” (anexo 3), levando em consideração somente as diferenças de bases
nucleotídicas entre as espécies. Desta forma, 23 sondas de DNA espécie
específicas e duas grupo específicas para o gênero Entamoeba foram
desenhadas. O tamanho das sondas variou entre 18 a 21 bp e estão de acordo
com a literatura.

93
Segundo Diaz (2007), sondas com tamanho maiores que 50
nucleotídeos aumentam a sensibilidade do teste, mas estão sujeitas à formação
de estruturas secundárias, que podem gerar resultados falso-positivos e a perda
da especifidade. Por outro lado, sondas entre 8 e 19 nucleotídeos não permitem
uma condição de estringência ideal devido à baixa temperatura de hibridação e,
consequentemente, a sonda perde em especificidade. Um equilíbrio razoável
entre a especificidade e sensibilidade na hibridação multianalítica em meio líquido
é conseguido quando se utiliza sondas contendo entre 20 e 30 nucleotídeos.
Assim, podemos concluir que o tamanho da sonda na reação de hibridação está
diretamente relacionado com a sensibilidade e inversamente, com a
especificidade do teste (DUNBAR, 2006).
Quando avaliamos o alinhamento das sequências obtidas neste estudo
(anexo 7), observamos que as sondas Ecoli165, M1, M2, Ed2, Ed3 e Ed4
apresentaram, nas suas sequências, bases nucleotídicas discordantes, isto é,
variações nucleotídicas intra-espécie, que afetaram o poder discriminatório e a
especificidade das sondas. Tal fato sugere que deve ser analisado um maior
número de sequências de DNA durante a elaboração de sondas, afim de verificar
possíveis variações intra-espécie. Resultados semelhantes foram salientados por
Verweij e cols (2003), quando descreveram o método de hibridação reversa em
membrana, com o intuito de identificar quaisquer espécies do gênero Entamoeba.
Inicialmente, os autores empregaram iniciadores biotinilados para amplificar parte
do gene do SSU-rRNA das amebas. O produto amplificado foi hibridizado com
sondas ligadas à membrana de nitrocelulose e os resultados da reação de
hibridação foram visualizados por meio de adição de substrato quimiluminescente,
seguida da autoradiografia. No entanto, os autores obtiveram resultados
satisfatórios na detecção simultânea de espécies de Entamoeba, mas, em relação
a E. coli, os resultados indicaram a necessidade de analisar mais sequências para

94
verificar possíveis variações intra-espécie, visto que algumas amostras positivas
pela EPF não foram detectadas pelas sondas utilizadas.
Durante o processo de padronização da PCR/Luminex, optamos por
utilizar produto biotinilado amplificado a partir de DNA clonado, devido à carência
de amostras controle de E. coli e E. hartmanii. A hibridação multianalíticas entre o
produto amplificado biotinilado e as sondas correspondentes foram realizadas em
diferentes temperaturas e tempos, e com diferentes diluições da molécula
repórter. Inicialmente, na hibridação multianalítica, foi utilizado o produto
amplificado pelo par de iniciadores JVF/DSPR2-B, que amplifica um fragmento do
gene do SSU-rDNA de Entamoeba spp. de, aproximadamente, 650 bp. Os
resultados obtidos com este produto e as sondas correspondentes (tabela 6) não
geraram sinais específicos de fluorescência. Os valores da intensidade de
fluorescência mediana (MIF) ficaram muito próximos dos valores encontrados na
reação do branco (reação contendo as microesferas e TE no lugar do produto da
PCR). O insucesso da PCR/Luminex obtido com este fragmento pode ser
explicado devido a sua conformação e/ou à presença de estruturas secundárias
na região de hibridação das sondas. Por sua vez, quando utilizamos o par de
iniciadores JVF/EnatRev390, que amplifica um fragmento em torno de 400 bp,
obtivemos elevados valores de MIF. Estes resultados se repetiram quando
utilizamos o par de inciadores JVF/EntaRev417A que amplifca um fragmento de
DNA em torno de 429 bp. Contudo, o tamanho ideal do produto amplificado
biotinilado a ser utilizado na PCR/Luminex é extremamente controverso na
literatura. Para Dunbar (2006), a utilizacão de fragmento amplificado de até 300
bp minimiza o potencial do impedimento alostérico que afeta a eficiência da
hibridação. Procedendo de maneira semelhante, Fitzgerald e cols. (2007)
utilizaram produtos amplificados entre 100 e 400 bp para minimizar o potencial do
efeito do impedimento alostérico, que afeta a eficiência da hibridação. Por outro
lado, Diaz & Fell (2004) obtiveram sucesso quando utilizaram fragmentos de

95
DNA-alvo maiores que 600 bp. Os autores relataram que uma sequência-alvo de
grande dimensões exibe uma conformação estrutural complexa e que a presença
de várias estruturas secundárias, ao longo da sequência, pode induzir a uma
flexão ou torção do fragmento, afetando a acessibilidade e a eficiência de ligação
da sonda. Este fato sugere que a eficiência da reação de hibridação multianalítica
em meio líquido dependente da sequência e, totalmente, da presença de estrutura
secundária (DIAZ & FELL 2004; DUNBAR, 2006).
No entanto, outros fatores que afetam a especificidade e a
sensibilidade hibridação multianalítica são: temperatura, tempo de hibridação e
quantidade de DNA- alvo adicionada à reação de hibridação (DUNBAR, 2006).
No presente trabalho, observamos que, dentre os volumes de DNA-
alvo utilizados na hibridação multianalítica, os melhores resultados foram obtidos
quando utilizamos 5 µl do produto amplificado biotinilado e 17 µl do diluído a
1/100 nas reações de hibridação. Segundo Dunbar & Jacobson (2007), quando
utilizamos concentrações de produto amplificado além dos níveis de saturação do
ensaio, a eficiência da hibridação pode se reduzir significativamente devido à
competição com a fita complementar biotinilada. Ademais, a renaturação do
produto amplificado da PCR é mais evidente quando o mesmo está presente em
altas concentrações. Assim, é importante determinar uma faixa de concentração
do DNA-alvo que gere uma hibridação eficiente sem sacrificar o poder
discriminatório do ensaio.
É válido considerar que a temperatura e o tempo da reação de
hibridação afeta a estabilidade do híbrido (sonda pareada com a sua sequência
alvo). De fato, a temperatura de hibridação é influenciada pelo conteúdo CG das
sequências de DNA. Todavia, neste estudo, foi utilizado o tampão de hibridação
contendo cloreto de tetrametil amônio (TMAC), que se liga ao DNA nas regiões
contendo bases nucleotídicas do tipo AT (MECHIOR & VON HIPPEL, 1973;
JACOBS et al., 1988). Assim, o uso de TMAC a uma concentração final em torno

96
de 3 a 4 M elimina a influência do conteúdo CG na determinação da Tm dos
híbridos, fazendo com que a hibridação seja dependente do tamanho e não da
composição das bases nucleotídicas (ANDERSON,1995). De fato, o poder de
equalização das temperaturas de dissociação dos pares AT e CG, exercido pelo
uso do TMAC, permite a utilização de diferentes conjuntos de sondas, com
características distintas e sob as mesmas condições de hibridação em ensaios do
tipo multiplex (WOOD et al., 1985). Contudo, pequenas variações da temperatura
na reação de hibridação proporcionam mudanças bastante significativas nos
resultados. Durante a etapa de padronização, observamos que, a temperturas
mais baixas de 41ºC/1h e 43ºC/1 h, houve uma diminuição da especificidade do
teste, gerando resultando falso-positivos e reações cruzadas. Nestas
temperaturas, observamos a presença de reações cruzadas entre a sondas de
Ehart 123 com o produto da PCR de E. histolytica, E. moshkovskii e E. dispar. A
sonda hist 116 apresentou uma discreta reação cruzada com os produtos de E.
dispar e E. moshkovskii somente a 41ºC/1h. Nesta etapa, utilizamos produtos
amplificados a partir de DNA obtido na clonagem molecular. Assim, podemos
afirmar que se tratava de DNA altamente purificado e sem a possibilidade de
coinfecção entre espécies de Entamoeba na amostras amplificadas. Por outro
lado, observamos, a temperatura mais elevada, a 48ºC/1h, existiu uma maior
dificuldade na formação do híbrido, tornando o ensaio pouco eficiente. Nossos
resultados demostraram que a 48 ºC/ 1h a sonda de E. coli manteve uma elevada
MIF e foi observado uma diminuição dos valores da MIF das demais sondas. Já
na temperatura intermediária, a 46 ºC/1h, obtivemos bons resultados onde todas
as cinco sondas geraram sinais de fluorescência satisfatórios, apesar de ainda
persistir uma discreta interação inespecífica entre a sondas de Ehart 123 com o
produto da PCR de E. histolytica. Por fim, a temperatura de 46ºC/1h, que forneceu
os melhores resultados na reação de hibridação, foi adotada para a realização
dos testes das amostras clínicas.

97
É importante lembrarmos que a sensibilidade e especificidade analítica
de um determinado teste diagnóstico não dependem da população ou da
localidade em que o teste será utilizado, pois são características intrínsecas a ele.
Caso este seja realizado conforme padronização, esperam-se resultados
reprodutíveis e constantes. Entretanto, a eficiência do teste está relacionada com
os indivíduos verdadeiramente positivos e verdadeiramente negativos presentes
na população estudada. O conhecimento sobre essas questões é fundamental na
avaliação de um teste que se pretende implantar como diagnóstico. No entanto, é
válido ressaltar que, atualmente, não existe um método de diagnóstico
considerado padrão ouro para a detecção e diferenciação de espécies de
Entamoeba intestinais. Tal fato dificulta a avaliação de novos métodos a serem
desenvolvidos.
Todavia, os resultados obtidos com as 11 amostras clínicas
demonstraram a consistência da PCR/Luminex, uma vez que não houve casos de
discrepância na determinação das espécies. Entretanto, devemos mencionar que
a amostra 28 foi positiva para E. hartmanni somente pelos dois ensaios de
PCR/Luminex (tabela 10 e 11). Lembramos, que durante a etapa de
padronização, não houve indícios de reação cruzada entre a sonda de E.
hartmanni com o produto amplificado de E. coli. Deste modo, podemos sugerir
que os resultados encontrados na amostra 28 representam sinais específicos de
hibridação. Já as amostras 17 e 19 apresentaram sinais de fluorescência com
intensidade bem próxima à leitura média do branco para E. dispar e E. hartmanni,
respectivamente, quando o produto foi amplificado com o par de iniciadores
JFV/EntaRev390B. De fato, melhores resultados foram obtidos com o produto
amplificado com os iniciadores JVF/ Enta417A-B. Contudo, observamos 100% de
concordância entre os resultados da PCR/Sistema Luminex com a PCR em tempo
real e a PCR/Sequenciamento, quando analisamos o somatórios dos resultados
das amostras testadas com ambos os produtos amplificados (tabela 12).

98
Além das amostras citadas acima, um grupo de 15 amostras com
identificação não conhecida foi utilizado na PCR/Luminex. Neste, somente
tivemos acesso aos resultados da PCR em tempo real após a análise pela
PCR/Luminex e verificamos 100% de concordantes entre estes testes.
Até o momento da realização deste estudo, não foi descrito o uso da
PCR/Sistema Luminex na detecção e diferenciação entre espécies de ameba.
Entretanto, esta técnica já vem sendo aplicada para o diagnóstico de uma série
de doenças infecciosas. Em 2007, Bandyopadhyay e cols. empregaram a
PCR/Luminex na diferenciação entre Cryptosporidium hominis e C. parvum. Neste
estudo, o limite de detecção do teste foi inferior a 10 oocistos de Cryptosporidium
spp em 300 µl de fezes, com uma sensibilidade de 100%. Etienne, Kano &
Balajee (2009) utilizaram a PCR/Luminex na identificação de espécies de fungo
do gênero Aspergillus. Os autores relatam que o ensaio foi 100% concordante
com os resultados do sequenciamento. Procedendo da mesma maneira,
Landlinger e cols. (2009) empregaram a PCR/Luminex no diagnóstico de fungos
patogênico de interesse médico. Neste estudo, foi possível identificar, de maneira
rápida e confiável, diferentes espécies de fungos. No mesmo ano, Boving,
Perdersesn & Moller relataram a utilização da PCR/Luminex na detecção de nove
micro-organismos presente no líquido encefalorraquidiano de indivíduos com
suspeita de meningite. Segundo os autores, o ensaio foi sensível e rápido,
permitindo ao médico determinar o tratamento adequado num curto espaço de
tempo. Por sua vez, Gadsby e cols. (2010) utilizaram a PCR em tempo real e a
PCR/Luminex na detecção de um grupo de vírus respiratórios. Neste estudo, os
autores concluíram que a PCR/Luminex apresentou uma boa sensibilidade,
quando comparada com a PCR em tempo real, exceto para as amostras de
adenovírus ou para aquelas que continham uma baixa carga viral. Diante do
acima exposto, observamos que a PCR/Luminex vem sendo utilizada no
diagnóstico de doenças infectoparasitárias, principalmente quando os métodos

99
tradicionais não são suficientes ou definitivos para estabelecer o diagnóstico
correto de maneira rápida e eficaz.
Contudo, assim como sucede com as outras metodologias, a
PCR/Luminex possui variáveis que podem afetar a sua eficácia. Um fator que
pode comprometer a sensibilidade da PCR/Luminex é a natureza multianalítica do
teste, que consiste na análise simultânea de diferentes alvos. Para tanto, é
necessário a utilização da PCR e da reação de hibridação de caráter
multianalítico. Todavia, a utilização de uma PCR multiplex aumenta a
complexidade e pode reduzir a sensibilidade do teste. Para Diaz (2007), a
complexidade da PCR multiplex coloca em risco a sensibilidade da amplificação
de DNA. Principalmente, em amostras clínicas com baixa concentração de DNA-
alvo. Estas amostras necessitam de um sistema de extração altamente eficiente,
para reduzir a presença das substâncias inibidoras da PCR. A adição de
facilitadores da PCR, como BSA, melhora a eficiência da amplificação, assim
como a utilização de DNA polimerase do tipo Tth ou Tfl, que são mais resistentes
à ação de inibidores (ABU AL SOULD & RADSTROM, 1998). Ademais, uma outra
estratégia relatada por DIAZ (2007) é a de aumentar a intensidade do sinal
repórter da PCR/Luminex. Como substitutos do iniciador biotinilado, que
acrescenta uma única molécula de biotina ao alvo, podem ser utilizados
nucleotídeos biotinilados ou dendrimeros de DNA. Segundo Lowe et al. (2004), a
marcação do DNA com dendrímeros resultou em um aumento de dez vezes na
intensidade de fluorescência, permitindo, assim, a detecção de uma única
molécula de DNA por microesferas. Já Das et al. (2006) observaram melhores
resultados na reação de hibridação multianalítica quando utilizaram PCR
assimétrica. Nesta reação, utilizou-se o iniciador reverso quatro vezes mais
concentrado do que o iniciador direto na amplificação. É notório que a
amplificação de DNA, utilizando-se, na PCR, quantidades equimolares de
iniciadores direto e reverso, gera um produto de DNA bifilamentar. Entretanto, se

100
a quantidade do iniciador reverso for excessiva em relação ao direto, a PCR irá
gerar mais produto de DNA monofilamentar do que bifilamentar, ou seja, isto
acontecerá após o consumo do iniciador direto. De fato, tanto o produto
bifilamentar quanto o monofilamentar, após a etapa de desnaturação, participarão
da reação de hibridação.
A PCR/Luminex estudada ainda se encontra em fase de
desenvolvimento. Assim, avaliações na reprodutibilidade intrateste, na
sensibilidade da otimização da PCR/Luminex precisam ser realizadas, bem como
cálculos para se estabelecer a região de corte (cutoff) para a obtenção dos
resultados. Além disto, é necessário, ainda, validar o teste frente a um maior
número de amostras.
As otimizações futuras na PCR/Luminex devem ser concentrada na
etapa de amplificação, testando a adição de BSA na reação e o uso de DNA
polimerase mais resistente à ação de inbidores presentes na amostra fecal. É
preciso também analisar os efeitos da adição de diferentes quantidades do
produto amplificado na reação de hibridização e avaliar outros métodos de
marcação dos produtos da PCR.
PCR/Luminex ainda possui um custo elevado, sendo que a maioria dos
laboratórios de diagnóstico de rotina não possui infraestrutura adequada para sua
implementação. A utilização da PCR/Luminex no Brasil encontra-se em um
estágio inicial de adoção e seu uso ainda é limitado às grandes redes de
laboratórios clínicos, principalmente nos grandes centros urbanos. Todavia, o seu
poder multianalítico, aliado à sua flexibilidade de excluir ou inserir alvos
moleculares ao ensaio, conforme a necessidade epidemiológica em que será
utilizado, é o que torna a técnica vantajosa e atrativa para o diagnóstico clínico. A
implementação de metodologia de ponta no diagnóstico laboratorial, de forma
geral, é dispendiosa. Porém, com o passar do tempo, novas padronizações e

101
reagentes vão sendo testados, reduzindo, assim, gradativamente, os custos, sem
o sacrifício da qualidade até que a metodologia esteja ao alcance de todos.
No entanto, a PCR/Luminex deve ser implementada e o seu custo não
pode ser uma barreira intransponível, quando refletirmos sobre a qualidade do
diagnóstico, o custo do tratamento das consequências adversas da infecção por
E. histolytica e a obtenção de dados epidemiológicos fidedignos. Este novo
método gera uma nova perspectiva e um novo horizonte no diagnóstico de
doenças infectoparasitárias, visto que os resultados alcançados neste estudo
mostraram que a PCR/Luminex é um método exequível, sensível e promissor na
detecção e diferenciação de espécies de Entamoeba intestinais. Ademais, esta
plataforma pode ser estendida e implementada para o diagnóstico de outros
patógenos causadores de doenças infectoparasitárias.

102
6. CONCLUSÃO
Os resultados alcançados neste trabalho permitem concluir:
1) O gene Ssu-rRNA pode ser utlizado como marcador molecular para
o diagnóstico de amebas intestinais;
2) A capacidade da PCR/sequenciamento em efetuar análises
múltiplas é limitada, sendo necessário a clonagem de amostras mistas;
3) A utilização de métodos multianalíticos na diferenciação entre as
espécies de amebas intestinais é importante;
4) A PCR/Luminex padronizada neste estudo mostrou-se exequível e
viável para o diagnóstico da amebíase e para a detecção e diferenciação de
Entamoeba comensais na fezes.;
5) Etapas futuras de otimização e validação da PCR/Luminex serão
necessárias
6) Este trabalho não só permitiu o desenvolvimento de novo teste para
o diagnóstico de Entamoeba spp. intestinais, como proporcionou o treinamento e
conhecimento prático da PCR/Luminex, gerando uma nova perspectiva e um novo
horizonte para o desenvolvimento tecnológico no diagnóstico de doenças
infectoparasitárias.

103
REFERÊNCIAS BIBLIOGRÁFICAS
ABD-ALLA, M.D.; WAHIB, A.A.; RAVDIN, J.I. Comparison of antigen-capture ELISA to stool-culture methods for the detection of asymptomatic Entamoeba species infection in Kafer Daoud, Egypt. Am J Trop Med Hyg. 62(5):579-82, 2000.
ABD-ALLA, M.D. et al. Mucosal immunity to asymptomatic Entamoeba histolytica and Entamoeba dispar infection is associated with a peak intestinal anti-lectin immunoglobulin A antibody response. Infect Immun, 74(7):3897-903, 2006.
ABU AL-SOUD, W. & RADSTROM, P. Capacity of nine thermostable DNA polymerases To mediate DNA amplification in the presence of PCR-inhibiting samples. Appl Environ Microbiol. 64(10):3748-53, 1998.
______. Effects of amplification facilitators on diagnostic PCR in the presence of blood, feces, and meat. J Clin Microbiol. 38(12):4463-70, 2000.
ACKERS, J.P. The diagnostic implications of the separation of Entamoeba histolytica and E. dispar. Journal of Biosciences 27(Suppl. 3): 573–578, 2002.
ACUNA-SOTO, R.J. et al. Application of the polymerase chain reaction to the epidemiology of pathogenic and nonpathogenic Entamoeba histolytica. Am. J. Trop. Med. Hyg. 48(1):58–70, 1993.
AGUIRRE, A. et al. Polymerase chain reaction-solution hybridization enzyme-linked immunoassay (PCR-SHELA) for the differential diagnosis of pathogenic and non-pathogenic Entamoeba histolytica. Trans. R. Soc. Trop. Med. Hyg. 89: 187-188, 1995.
AGUIRRE, A. et al. Diagnosis of Entamoeba histolytica and Entamoeba dispar in clinical specimens by PCR-SHELA. Arch Med Res. 28:282-4,1997.
ALI, I.K. et al. Entamoeba moshkovskii infections in children, Bangladesh. Emerg Infect Dis 9(5): 580-4, 2003.
ALI, I.K.; ZAKI, M. & CLARK, C.G. Use of PCR amplification of tRNA gene-linked short tandem repeats for genotyping Entamoeba histolytica. J Clin Microbiol. 43(12):5842-7, 2005.
ALI, I.K.; CLARK, C.G. & PETRI, W.A. Molecular epidemiology of amebiasis. Infect Genet Evol. 8(5):698-707, 2008.
ALVES, J.R. et al. Intestinal parasite infections in a semiarid area of Northeast Brazil: preliminary findings differ from expected prevalence rates. Cad Saude Publica 19(2): 667-70, 2003.
ALTSCHUL, S.F. et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25:3389-3402, 1997.

104
ANDERSON, M.L. Hybridization Strategy. In: Gene Probe 2, 17-29. Edited by Hames, B.D. & Higgins, J.J. Oxford, New York, 1995.
ANDRADE, F. et al. Parasitoses intestinais em um centro de educação infantil público do município de Blumenau (SC), Brasil, com ênfase em Cryptosporidium spp e outroa parasitos intestinais. Rev Patol Tropical 37(4):332-40, 2008
ANTONIOU, M. et al. Fourteen-year sero- epidemiological study of zoonoses in a Greek village. Am J Trop Med Hyg 66(1): 80-5, 2002.
AQUINO, J.L. Amebíase. In: FERREIRA, A.W. & ÁVILA, S.L.M. Diagnóstico laboratorial das principais doenças infecciosas e auto-imunes. 2a ed.. Rio de Janeiro, Guanabara Koogan, 2001. cap. 21 p. 232- 240.
ASSIS, M. et al. Prevalência de enteroparasitoses em moradores de vilas periféricas de Porto Alegre. RBAC 35(4):215-17, 2003
BANDYOPADHYAY, K. et al. Rapid microsphere assay for identification of cryptosporidium hominis and cryptosporidium parvum in stool and environmental samples. J Clin Microbiol. 45(9):2835-40, 2007.
BARRETO, M.P. Diagnóstico da Amebiase Intestinal pelo exame Parasitológico. Rev. Inst. Med. Trop. São Paulo 2: 305-312, 1962.
BECK, D.L. et al. High prevalence of Entamoeba moshkovskii in a Tanzanian HIV population. Acta Trop 107(1): 48-9, 2008.
BENETTON, M.L. et al. Risk factors for infection by the Entamoeba histolytica/E. dispar complex: an epidemiological study conducted in outpatient clinics in the city of Manaus, Amazon Region, Brazil. Trans R Soc Trop Med Hyg 99(7): 532-40, 2005.
BHATTACHARYA, S. Circular DNA of Entamoeba histolytica encodes ribosomal RNA. J Protozool, 36, (5):455-8, 1989.
BLANC, D. & SARGEAUNT, P.G. Entamoeba histolytica zymodemes: exhibition of gamma and delta bands only of glucose phosphate isomerase and phosphoglucomutase may be influenced by starch content in the medium. Exp Parasitol 72(1): 87-90, 1991.
BLESSMANN, J. et al. Epidemiology of amebiasis in a region of high incidence of amebic liver abscess in central Vietnam. Am J Trop Med Hyg 66(5): 578-83, 2002.
BOVING, M.K.; PEDERSEN, L.N. & MOLLER, J.K. Eight-plex PCR and liquid-array detection of bacterial and viral pathogens in cerebrospinal fluid from patients with suspected meningitis. J Clin Microbiol. 47(4):908-13, 2009.
BRACHA, R. et al. Differentiation of clinical isolates of Entamoeba histolytica by using specific DNA probes. J Clin Microbiol 28(4): 680-4, 1990.

105
BRAGA, L.L. et al. Seropositivity for and intestinal colonization with Entamoeba histolytica and Entamoeba dispar in individuals in northeastern Brazil. J Clin Microbiol 36(10): 3044-5, 1998.
BRAGA, L.L. et al. Entamoeba histolytica and Entamoeba dispar infections as detected by monoclonal antibody in an urban slum in Fortaleza, Northeastern Brazil. Rev Soc Bras Med Trop 34(5): 467-71, 2001.
BRITTEN, D. et al. Detection and differentiation of Entamoeba histolytica and E. dispar using an improved colorimetric polymerase chain reaction method. Arch Med Res 28:279-81, 1997.
CALDERARO, A. et al. Entamoeba histolytica and Entamoeba dispar: comparison of two PCR assays for diagnosis in a non-endemic setting. Trans R Soc Trop Med Hyg. 100(5):450-7, 2006.
CARRERO, J.C. et al. The role of the secretory immune response in the infection by Entamoeba histolytica. Parasite Immunol 29(7): 331-8, 2007.
CHOE, S.C. et al . Differentiation of Korean isolates of Entamoeba histolytica from Entamoeba dispar. Korean J Parasitol. 34(1):15-20, 1996.
CLARK, C.G. & DIAMOND, L.S. Differentiation of pathogenic Entamoeba histolytica from other intestinal protozoa by riboprinting. Arch Med Res 23(2):15-6, 1992.
______. Intraspecific variation and phylogenetic relationships in the genus Entamoeba as revealed by riboprinting. J Eukaryot Microbiol. 44(2):142-54, 1997a.
______. Molecular phylogeny of the genus Entamoeba as revealed by riboprinting. Arch Med Res. 28:69-70, 1997b.
CLARK, C.G. Royal Society of Tropical Medicine and Hygiene meeting at Manson House, London, 19 February 1998. Amoebic disease. Entamoeba dispar, an organism reborn. Trans R Soc Trop Med Hyg 92(4): 361-4, 1998.
CNOPS, L. & ESBROECK, M.V. Freezing of stool samples improves real-time PCR detection of Entamoeba dispar and Entamoeba histolytica. J Microbiol Methods 80(3):310-2, 2010.
DA SILVA, A.J. et al . Fast and reliable extraction of protozoan parasite DNA from fecal specimens. Mol Diagn.; 4(1):57-64, 1999.
DA SILVA, A.J. et al. Diagnosis of Enterocytozoon bieneusi (microsporidia) infections by polymerase chain reaction in stool samples using primers based on the region coding for small-subunit ribosomal RNA. Arch Pathol Lab Med.121(8):874-9, 1997.
DAS, S. et al. DNA probes for the rapid identification of medically important Candida species using a multianalyte profiling system. FEMS Immunol Med Microbiol. 46(2):244-50, 2006.

106
DEFOORT, J.P. et al. Simultaneous detection of multiplex-amplified human immunodeficiency virus type 1 RNA, hepatitis C virus RNA, and hepatitis B virus DNA using a flow cytometer microsphere-based hybridization assay. J Clin Microbiol. 38(3):1066-71, 2000.
DENHARDT, D.T. A membrane-filter technique for the detection of complementary DNA. Biochem Biophys Res Commun, 5(13):641-6, 1966.
DESOWITZ, R.S. & BARNISH, G. Entamoeba polecki and other intestinal protozoa in Papua New Guinea Highland children. Ann Trop Med Parasitol 80(4): 399-402, 1986.
DEUTER, R.S. et al. A method for preparation of fecal DNA suitable for PCR. Nucleic Acids Res, 23(18):3800-1, 1995.
DIAZ, M.R. & FELL, J.W. High-throughput detection of pathogenic yeasts of the genus trichosporon. J Clin Microbiol 42(8): 3696-706, 2004.
DIAZ, M.R. Bead Suspension Arrays for Identifying Fungal Pathogens. Microbe 2(2):74-81, 2007.
DOGANCI, L.; TANYUKSEL, M. & DOGANCI, T. Accurate diagnosis is essential for amebiasis. World J Gastroenterol. 15;10(8):1231, 2004.
DUNBAR, S.A. Quantitative, multiplexed detection of bacterial pathogens: DNA and protein applications of the Luminex LabMAP system. J Microbiol Methods. 53(2):245-52, 2003.
DUNBAR, S.A. Applications of Luminex xMAP technology for rapid, high-throughput multiplexed nucleic acid detection. Clin Chim Acta 363:71-82, 2006.
DUNBAR, S.A. & JACOBSON, J.W. Quantitative, multiplexed detection of Salmonella and other pathogens by Luminex xMAP suspension array. Methods Mol Biol. 394:1-19, 2007.
DYSON, N.J. Immobilization of nucleic acids and hybridization analysis. In Essential Molecular Biology: A Practical Approach, Vol. 2, 111-156. Edited by Brown, T.A., Oxford University Press, Oxford, 1991.
ETIENNE, K.A.; KANO, R. & BALAJEE,S.A. Development and validation of a microsphere-based Luminex assay for rapid identification of clinically relevant aspergilli. J Clin Microbiol. 47(4):1096-100, 2009.
EVANGELOPOULOS, A., SPANAKOS, G. & PATSOULA, E. A nested, multiplex, PCR assay for the simultaneous detection and differentiation of Entamoeba histolytica and Entamoeba dispar in faeces. Ann Trop Med Parasitol 94(3): 233-40, 2000.
EVANGELOPOULOS, A.; LEGAKIS, N. & VAKALIS, N. Microscopy, PCR and ELISA applied to the epidemiology of amoebiasis in Greece. Parasitol Int 50(3): 185-9, 2001.

107
FAUST, E.C. et al. A critical study of clinical laboratory technics for the diagnosis of protozoan cysts and helminth eggs in feces. I. Preliminary communication. Am. J. Trop. Med. 18: 169-183, 1938.
FERREIRA, A.W. & ÁVILA, S.L.M. Sorologia: Importância e Parâmetros. In: Diagnóstico laboratorial das principais doenças infecciosas e auto-imunes, 1-8. Rio de Janeiro, ISBT Brasil, 2001.
FERRER, M.A. et al. Sobrediagnóstico microscópico de amebiasis intestinal: encuesta a técnicos de laboratorio. Rev Cubana Invest Biomed 18(3): 197-202, 1999.
FITZGERALD, C. et al . Multiplex, bead-based suspension array for molecular determination of common Salmonella serogroups. J Clin Microbiol. 45(10):3323-34, 2007.
FLEKNA, G. et al. Real-time PCR method with statistical analysis to compare the potential of DNA isolation methods to remove PCR inhibitors from samples for diagnostic PCR. Mol Cell Probes. 21(4):282-7, 2007.
FONTE, L. et al Overdiagnosis of intestinal amoebiasis associated to serial microscopical examination of faeces. Some precisions on a problem. Mem Inst Oswaldo Cruz 93(6):799-800, 1998.
FOTEDAR, R.D. et al. Laboratory diagnostic techniques for Entamoeba species. Clin Microbiol Rev 20(3): 511-32, 2007.
FOTEDAR, R.D. et al. Entamoeba moshkovskii infections in Sydney, Australia. Eur J Clin Microbiol Infect Dis 27(2): 133-7, 2008.
FREITAS, M.A. et al. A single step duplex PCR to distinguish Entamoeba histolytica from Entamoeba dispar. Parasitology 128( 6): 625-8, 2004.
GADSBY, N.J. et al . Comparison of the Luminex Respiratory Virus Panel fast assay with in-house real-time PCR for respiratory viral infection diagnosis. J Clin Microbiol. 48(6):2213-6, 2010.
GARCIA, L.S.; SHIMIZU, R.Y. & BERNARD, C.N. Detection of Giardia lamblia, Entamoeba histolytica/Entamoeba dispar, and Cryptosporidium parvum antigens in human fecal specimens using the triage parasite panel enzyme immunoassay. J Clin Microbiol 38(9): 3337-40, 2000.
GARCIA, L.S. Intestinal protozoa: Amebae. In: Diagnostic medical parasitology, 28. Edited by Lynne Shore Garcia, ASM Press. Washington, D.C.: ASM Press. 2007.
GARFINKEL, L.I. et al. DNA probes specific for Entamoeba histolytca possessing pathogenic and nonpathogenic zymodemes. Infect Immun 57(3): 926-31, 1989.

108
GHOSH, S. et al. Molecular epidemiology of Entamoeba spp.: evidence of a bottleneck (Demographic sweep) and transcontinental spread of diploid parasites. J Clin Microbiol. 38(10):3815-21, 2000.
GOMES, M.A. et al. LSSP-PCR for characterization of strains of Entamoeba histolytica isolated in Brazil. Parasitology. 114 (Pt 6):517-20, 1997.
GOMES, M.A. et al. An improved method to distinguish Entamoeba histolytica and Entamoeba dispar. Parasitology. 119 (Pt 4):359-62, 1999.
GOMES, M.A. et al. RAPD in the analysis of isolates of Entamoeba histolytica. Acta Trop 25;75(1):71-7, 2000a.
GOMES, M.A. et al. Characterization of Entamoeba histolytica and Entamoeba dispar by biological, biochemical, and molecular parameters. Arch Med Res. 31(4):S249-50, 2000b.
GONCALVES, A.Q. et al. The use of the antifungal agent miconazole as an inhibitor of Blastocystis hominis growth in Entamoeba histolytica/E. dispar cultures. Rev Inst Med Trop Sao Paulo 49(3):201-2, 2007.
GONIN, P. & L. TRUDEL, L. Detection and differentiation of Entamoeba histolytica and Entamoeba dispar isolates in clinical samples by PCR and enzyme-linked immunosorbent assay. J Clin Microbiol 41(1): 237-4, 2003.
GONZALEZ-RUIZ, A. et al. Value of microscopy in the diagnosis of dysentery associated with invasive Entamoeba histolytica. J Clin Pathol. 47(3):236-9,1994.
GUTIERREZ, G. Epidemiology and control of amebiasis in Mexico. Arch Invest Med (Mex) 17(1): 375-83, 1986.
HAGHIGHI, A. et al. Geographic diversity among genotypes of Entamoeba histolytica field isolates. J Clin Microbiol 41(8):3748-56, 2003.
HAGHIGHI, A. et al. Remarkable genetic polymorphism among Entamoeba histolytica isolates from a limited geographic area. J Clin Microbiol 40(11): 4081-90, 2002.
HAMZAH, Z. et al. Differential detection of Entamoeba histolytica, Entamoeba dispar, and Entamoeba moshkovskii by a single-round PCR assay. J Clin Microbiol 44(9): 3196-200, 2006.
HAQUE, R. et al. Entamoeba histolytica and Entamoeba dispar infection in children in Bangladesh. J Infect Dis 175(3): 734-6, 1997.
HAQUE, R.; ALI, I.K.; AKTHER, S. Comparison of PCR, isoenzyme analysis, and antigen detection for diagnosis of Entamoeba histolytica infection. J Clin Microbiol. 36(2):449-52, 1998.
HAQUE, R. et al. Amebiasis. N Engl J Med 348 (16): 1565-73, 2003.

109
HAQUE, R. et al. Multiplex real-time PCR assay for detection of Entamoeba histolytica, Giardia intestinalis, and Cryptosporidium spp. Am J Trop Med Hyg 76(4):713-7, 2007.
HECKENDORN, F. et al. Species-specific field testing of Entamoeba spp. in an area of high endemicity. Trans R Soc Trop Med Hyg 96(5): 521-8, 2002.
HOFFMAN, W.A.; PONS, J.A. & JANER, J.L. The sedimentation concentration method in schistosomiasis mansoni. Puerto Rico J Publ Health Trop Med 9:283-298, 1934.
HOLLAND, J.L. et al. PCR detection of Escherichia coli O157:H7 directly from stools: evaluation of commercial extraction methods for purifying fecal DNA. J Clin Microbiol. 38(11): 4108-13, 2000.
HOOSHYAR, H. et al. The distribution of Entamoeba histolytica and Entamoeba dispar in northern, central, and southern Iran. Parasitol Res 94(2): 96-100, 2004.
HORAN, P.K. & WHEELESS, L.L. Quantitative single cell analysis and sorting. Science 198 (4313):149-57, 1977.
INMETRO. Curso de formação de Auditores Internos de laboratórios Clínicos de Patologia: implantação de Qualidade, Auditoria e Credenciamento. INMETRO, CTLE-04, 1997.
JACOBS, K.A. et al. The thermal stability of oligonucleotide duplexes is sequence independent in tetraalkyl ammonium salt solutions: application to identifying recombinant DNA clones. Nucleic Acids Res. 16(10):4637-50, 1988.
JIN, L. & NEI, M. Limitations of the evolutionary parsimony method of phylogentic analysis. Mol Biol Evol 7:82-102, 1990
KATZWINKEL-WLADARSCH, S. et al.. Direct amplification and differentiation of pathogenic and nonpathogenic Entamoeba histolytica DNA from stool specimens. Am J Trop Med Hyg 51(1): 115-8,1994.
KEBEDE, A. et al. The use of real-time PCR to identify Entamoeba histolytica and E. dispar infections in prisoners and primary-school children in Ethiopia. Ann Trop Med Parasitol 98(1): 43-8, 2004.
KLEIN, D. Quantification using real-time PCR technology: applications and limitations. Trends Mol Med. 8(6):257-60, 2002.
KRAOUL, L. et al. Evaluation of a rapid enzyme immunoassay for diagnosis of hepatic amoebiasis. J Clin Microbiol 35(6): 1530-2, 1997.
LANDLINGER, C. et al. Species-specific identification of a wide range of clinically relevant fungal pathogens by use of Luminex xMAP technology. J Clin Microbiol. 47(4):1063-73, 2009.

110
LEBBAD, M. & SVARD, S.G. PCR differentiation of Entamoeba histolytica and Entamoeba dispar from patients with amoeba infection initially diagnosed by microscopy. Scand J Infect Dis 37(9): 680-5, 2005.
LEBER, A.L. & NOVAK, S.M. Intestinal and urogenital amebae, flagellates, and ciliates. In Manual of clinical microbiology, 1391-1405. Edited by P.R. MURRAY, E.J. BARON, M.A. PFALLER, F.C. TENOVER, AND R.H. YOLKEN. American Society of Microbiology Press, Washington, D.C., 2005.
LEVINE, N.D. et al. A newly revised classification of the protozoa. J Protozool. 27: 37-58, 1980.
LI, F.; HULLAR, M. & LAMPE, J.W. Optimization of terminal restriction fragment polymorphism (TRFLP) analysis of human gut microbiota. J Microbiol Methods. 68(2):303-11, 2007.
LOFTUS, B. et al. The genome of the protist parasite Entamoeba histolytica. Nature, 433(7028): 865-8, 2005.
LOWE, M. et al. Multiplexed, particle-based detection of DNA using flow cytometry with 3DNA dendrimers for signal amplification. Cytometry A. 60(2):135-44, 2004.
LUACES, A.L. et al. A new test for infection by Entamoeba histolytica. Parasitol Today 9(2): 69-71, 1993.
LUDWIG, K.M. et al . Correlation between sanitation conditions and intestinal parasitosis in the population of Assis, State of São Paulo. Rev Soc Bras Med Trop 32(5): 547-55, 1999.
MACKAY, I.M. Real-time PCR in the microbiology laboratory. Clin Microbiol Infect. 10(3):190-212, 2004.
MAHONY, J. et al. Development of a respiratory virus panel test for detection of twenty human respiratory viruses by use of multiplex PCR and a fluid microbead-based assay. J Clin Microbiol. 45(9):2965-70, 2007.
MARTINS, L.P. et al. Frequência de Blastocystis hominis e outras enteroparasitoses em amostras fecais analisadas no laboratório de parasitologia da faculdade de medicina de Marília, SP. Rev Patol Tropical 36(1):47-53, 2007.
MCNAMARA, D.T. et al. Diagnosing infection levels of four human malaria parasite species by a polymerase chain reaction/ligase detection reaction fluorescent microsphere-based assay. Am J Trop Med Hyg 74(3): 413-21, 2006.
MELCHIOR, W.B. & VON HIPPEL, P.H. Alteration of the relative stability of dA-dT and dG-dC base pairs in DNA. Proc Natl Acad Sci USA 70 (2): 298-302, 1973.
MENEZES, L.F. Aspectos da amebíase intestinal e hepática no Hospital Universitário Getúlio Vargas. Manaus. A.M. (Dissertação de mestrado), Departamento de Parasitologia do Instituto de Ciências Biológicas da Universidade federal de Minas Gerais, 1986, 89 p.

111
MIRANDA, R.A.; XAVIER, F.B. & MENEZES, R.C. Intestinal parasitism in a Parakana indigenous community in southwestern Para State, Brazil. Cad Saude Publica 14(3): 507-11, 1998.
MIRELMAN, D.; NUCHAMOWITZ, Y. & STOLARSKY, T. Comparison of use of enzyme-linked immunosorbent assay-based kits and PCR amplification of rRNA genes for simultaneous detection of Entamoeba histolytica and E. dispar. J Clin Microbiol 35(9): 2405-7, 1997.
MONTEIRO, L. et al. Complex polysaccharides as PCR inhibitors in feces: Helicobacter pylori model. J Clin Microbiol, 35(4):995-8, 1997.
MORAN, P. et al. Entamoeba histolytica and/ or Entamoeba dispar: infection frequency in HIV+/AIDS patients in Mexico city. Exp Parasitol. 110(3):331-4, 2005.
NAISER, T. et al. Stability of a surface-bound oligonucleotide duplex inferred from molecular dynamics: a study of single nucleotide defects using DNA microarrays. Phys Rev Lett. 102 (21): 218301-4, 2009.
NECHVATAL, J.M. et al. Fecal collection, ambient preservation, and DNA extraction for PCR amplification of bacterial and human markers from human feces. J Microbiol Methods 72(2): 124-32, 2008.
NUNEZ, Y.O. et al. Multiplex polymerase chain reaction amplification and differentiation of Entamoeba histolytica and Entamoeba dispar DNA from stool samples. Am J Trop Med Hyg 64(5-6): 293-7, 2001.
OMBROUCK, C. et al. Specific PCR assay for direct detection of intestinal microsporidia Enterocytozoon bieneusi and Encephalitozoon intestinalis in fecal specimens from human immunodeficiency virus-infected patients. J Clin Microbiol. 35(3):652-5, 1997.
ORLANDI, P.P. et al. Enteropathogens associated with diarrheal disease in infants of poor urban areas of Porto Velho, Rondonia: a preliminary study. Mem Inst Oswaldo Cruz 96(5): 621-5, 2001.
ORTNER, S. et al. Molecular biology of the hexokinase isoenzyme pattern that distinguishes pathogenic Entamoeba histolytica from nonpathogenic Entamoeba dispar. Mol Biochem Parasitol 86(1): 85-94, 1997.
PAGE, B.T. et al. Rapid identification of ascomycetous yeasts from clinical specimens by a molecular method based on flow cytometry and comparison with identifications from phenotypic assays. J Clin Microbiol. 44(9):3167-71, 2006.
PAGE, B.T. & KURTZMAN, C.P. Rapid identification of Candida species and other clinically important yeast species by flow cytometry. J Clin Microbiol. 43(9):4507-14, 2005.
PAGLIA, M.G. & VISCA, P. An improved PCR-based method for detection and differentiation of Entamoeba histolytica and Entamoeba dispar in formalin-fixed stools. Acta Trop 92(3): 273-7, 2004.

112
PALACIOS, G. et al. Panmicrobial oligonucleotide array for diagnosis of infectious diseases. Emerg Infect Dis. 13(1):73-81, 2007.
PALACIO-SANCHEZ, A. et al. Epidemiological study of amebiasis in Chihuahua, Mexico. Arch Med Res 28: 309-10, 1997.
PARIJA, S.C. & RAO, R.S. Stool culture as a diagnostic aid in the detection of Entamoeba histolytica in the faecal specimens. Indian J Pathol Microbiol. 38(4):359-63, 1995.
PARIJA, S.C. & KHAIRNAR, K. Entamoeba moshkovskii and Entamoeba dispar associated infections in Pondicherry, India. J Health Popul Nutr 23(3): 292-5, 2005.
PAUL, J.; SRIVASTAVA, S. & BHATTACHARYA, S. Molecular methods for diagnosis of Entamoeba histolytica in a clinical setting: an overview. Exp Parasitol. 116(1):35-43, 2007.
PELT-VERKUIL, E.; BELKUM, A. & HAYS, J. Analysis of PCR Amplification Products. In: Principles and Technical Aspects of PCR amplification, 141-182. Edited by Pelt-Verkuil, E.; Belkum, A. & Hays, Springer, 2008a.
______. Ensuring PCR Quality – Laboratory organization, PCR optimization and Controls. In: Principles and Technical Aspects of PCR amplification, 141-182. Edited by Pelt-Verkuil, E.; Belkum, A. & Hays, Springer, 2008 b.
______. The different Types and Varieties of Nucleic Acid Target MoleculesEnsuring. In: Principles and Technical Aspects of PCR amplification, 141-182. Edited by Pelt-Verkuil, E.; Belkum, A. & Hays, Springer, 2008 c.
PETRI, W. . & SINGH, U. Diagnosis and management of amebiasis. Clin Infect Dis 29(5): 1117-25, 1999.
PETRI, W.A. et al. Estimating the impact of amebiasis on health. Parasitol. Today, 16(8):320-321, 2000.
PILLAI, D.R. et al. Entamoeba histolytica and Entamoeba dispar: epidemiology and comparison of diagnostic methods in a setting of nonendemicity. Clin Infect Dis 29(5): 1315-8, 1999.
PINHEIRO, S.M. et al. Determination of the prevalence of Entamoeba histolytica and E. dispar in the pernambuco state of northeastern Brazil by a polymerase chain reaction. Am J Trop Med Hyg 70(2): 221-4, 2004.
PINHEIRO, S.M. et al. Genetic characterization of Entamoeba dispar isolates in Northeast Brazil. Acta Trop 94(1): 35-40, 2005.
POVOA, M.M. et al. Diagnosis of intestinal amebiasis using coproscopic and immunological methods in a population sample in greater metropolitan Belem, Para, Brazil. Cad Saude Publica 16(3):843-6, 2000.

113
PRITT, B.S. & CLARK, C.G. Amebiasis. Mayo Clin Proc 83(10): 1154-9 2008.
QVARNSTROM, Y. et al. Comparison of real-time PCR protocols for differential laboratory diagnosis of amebiasis. J Clin Microbiol 43(11): 5491-7, 2005.
RAMOS, F. et al. Entamoeba histolytica and Entamoeba dispar: prevalence infection in a rural Mexican community. Exp Parasitol. 110(3):327-30, 2005.
RAMOS, F. et al. Prevalence of Entamoeba histolytica and Entamoeba dispar in a highly endemic rural population. Arch Med Res 31(4): 34-5, 2000.
REY, L. Entamoeba histolytica e Amebíase. In: Parasitologia: parasitos e doenças parasitárias do homen nos trópicos ocidentais, 147-180. Editado por Rey, L., Rio de Janeiro, Brasil, 2008.
RIGOTHIER, M.C. et al. Fate of Entamoeba histolytica during establishment of amoebic liver abscess analyzed by quantitative radioimaging and histology. Infect Immun 70(6): 3208-15, 2002.
RITCHIE, L.S. An ether sedimentation technique for routine stool examinations. Bull. U.S. Army Med. Dept., 8: 326, 1948.
RIVERA, W.L. et al, Differentiation of Entamoeba histolytica and E. dispar DNA from cysts present in stool specimens by polymerase chain reaction: its field application in the Philippines. Parasitol Res. 82(7):585-9, 1996.
RIVERA, W.L.; TACHIBANA, H. & KANBARA, H. Application of the polymerase chain reaction PCR) in the epidemiology of Entamoeba histolytica and ntamoeba dispar infections. Tokai J Exp Clin Med 23(6): 413-5, 1998 .
RIVERA, W.L. et al. Prevalence and genetic diversity of Entamoeba histolytica in an institution for the mentally retarded in the Philippines. Parasitol Res. 98(2):106-10, 2006.
ROCHA, R.S. et al. Assessment of schistosomiasis and other intestinal parasitoses in school children of the Bambui municipality, Minas Gerais, Brazil. Rev Soc Bras Med Trop 33(5): 431-6, 2000.
ROMERO, J.L. et al. Use of polymerase chain reaction and nonradioactive DNA probes to diagnose Entamoeba histolytica in clinical samples. Arch Med Res 23(2): 277-9, 1992.
ROY, S. et al. Real-time-PCR assay for diagnosis of Entamoeba histolytica infection. J Clin Microbiol 43(5): 2168-72, 2005.
SAITOU, N. & NEI, M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. Jul; 4(4):406-25, 1987.
SAMUELSON, J. et al. DNA hybridization probe for clinical diagnosis of Entamoeba histolytica. J Clin Microbiol 27(4): 671-6, 1989.

114
SANCHEZ-GUILLEN MDEL, C. et al. Differentiation of Entamoeba histolytica/Entamoeba dispar by PCR and their correlation with humoral and cellular immunity in individuals with clinical variants of amoebiasis. Am J Trop Med Hyg 66(6):731-7, 2002.
SANTOS, H.L. et al. Comparison of multiplex-PCR and antigen detection for differential diagnosis of Entamoeba histolytica. Braz J Infect Dis 11(3): 365-70, 2007.
SANTOS, L.P.; SANTOS, F.L. & SOARES, N.M. Prevalência de parasitoses intestinais em pacientes atendidos no hospital universitário professor Edgar Santos, Salvador, Bahia. Rev Patol Tropical 36(6): 237-46, 2007.
SANUKI, J. et al. Identification of Entamoeba histolytica and E. dispar cysts in stool by polymerase chain reaction. Parasitol Res 83(1): 96-8, 1997.
SARGEAUNT, P.G. et al. The differentiation of invasive and non-invasive Entamoeba histolytica by isoenzyme electrophoresis.Trans R Soc Trop Med Hyg 72(5): 519-21, 1978.
SCHMITT, M. et al. Bead-based multiplex genotyping of human papillomaviruses. J Clin Microbiol. 44(2):504-12, 2006.
SHEEHAN, D.J. et al. Entamoeba histolytica: efficacy of microscopic, cultural, and serological techniques for laboratory diagnosis. J Clin Microbiol.10(2):128-33, 1979.
SHIPLEY, G. Real-Time PCR instrumentation – an overview. In: Real time PCR, 1- 32. Edited by M. Tevfik Dorak , UK, 2008
SILBERMAN, J.D. et al. Phylogeny of the genera Entamoeba and Endolimax as deduced from small-subunit ribosomal RNA sequences. Mol Biol Evol. 16(12):1740-51, 1999.
SILVA, M.C. et al. Determination of Entamoeba histolytica infection in patients from Greater Metropolitan Belem, Para, Brazil, by enzyme-linked immunosorbent assay (ELISA) for antigen detection. Cad Saude Publica 21(3): 969-73, 2005.
SINGH, A.; HOUPT, E. & PETRI, W.A. Rapid Diagnosis of Intestinal Parasitic Protozoa, with a Focus on Entamoeba histolytica. Interdiscip Perspect Infect Dis. 2009:547090-8, 2009.
SOUTHERN, E.M.. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J Mol Biol. 98(3):503-17, 1975.
STACY-PHIPPS, S.; MECCA, J.J. & WEISS, J.B. Multiplex PCR assay and simple preparation method for stool specimens detect enterotoxigenic Escherichia coli DNA during course of infection. J Clin Microbiol. 33(5):1054-9, 1995.
STANLEY, S.L. Amoebiasis. Lancet 361(9362): 1025-34, 2003.

115
STARK, D. et al. Comparison of microscopy, two xenic culture techniques, conventional and real-time PCR for the detection of Dientamoeba fragilis in clinical stool samples. Eur J Clin Microbiol Infect Dis. 29(4):411-6, 2010.
STARK, D.J. et al. Dientamoebiasis: clinical importance and recent advances. Trends Parasitol 22(2): 92-6, 2006.
TACHIBANA, H. et al. Distinguishing pathogenic isolates of Entamoeba histolytica by polymerase chain reaction. J Infect Dis 164(4): 825-6, 1991.
TACHIBANA, H. et al. Detection of pathogenic Entamoeba histolytica DNA in liver abscess fluid by polymerase chain reaction. Int J Parasitol. 22(8):1193-6, 1992.
TACHIBANA, H. et al. Asymptomatic cyst passers of Entamoeba histolytica but not Entamoeba dispar in institutions for the mentally retarded in Japan. Parasitol Int 49(1): 31-5, 2000.
TACHIBANA, H. et al . High prevalence of infection with Entamoeba dispar, but not E. histolytica, in captive macaques. Parasitol Res. 87(1):14-7, 2001.
TAMURA, K. et al. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Molecular Biology and Evolution 24:1596-1599, 2007.
TANNICH, E. & BURCHARD, G.D. Differentiation of pathogenic from nonpathogenic Entamoeba histolytica by restriction fragment analysis of a single gene amplified in vitro. J Clin Microbiol. 29(2):250-5, 1991.
TANYUKSEL, M. & PETRI, W. A. Laboratory diagnosis of amebiasis. Clin Microbiol Rev 16(4): 713-29, 2003.
TELLEZ SIERRA, A. et al. Amebiasis in Nicaragua: class specific serum antibody responses. Arch Med Res 23(2): 261-4,1992.
TOBIE, J.E. et al. The efficiency of the zinc sulfate technic in the detection of intestinal protozoa by successive stool examinations. Am. J. trop. Med., 31:552-560, 1951.
THOMPSON, J.D.; HIGGINS, D.G. & GIBSON, T.J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22:4673-4680, 1994.
TROLL, H. et al. Simple differential detection of Entamoeba histolytica and Entamoeba dispar in fresh stool specimens by sodium acetate-acetic acid-formalin concentration and PCR. J Clin Microbiol 35(7): 1701-5, 1997.
UNGAR, B. P.; YOLKEN, R.H. & QUINN,T.C. Use of Monoclonal Antibody in an Enzyme Immunoassay for the Detection of Entamoeba histolytica in Fecal Specimens. Am. J. Med. Hyg 34(3): 465-472, 1985.

116
VALLE, P.R. et al. Arbitrarily primed PCR fingerprinting of RNA and DNA in Entamoeba histolytica. Rev Inst Med Trop São Paulo. 42(5):249-53, 2000.
VANDENBERG, N. & VAN OORSCHOT, R.A. Extraction of human nuclear DNA from feces samples using the QIAamp DNA Stool Mini Kit. J Forensic Sci, 47(5):993-5, 2002.
VERWEIJ, J.J. et al. Differentiation of Entamoeba histolytica and Entamoeba dispar using PCR-SHELA and comparison of antibody response. Arch Med Res 31(4):44-6, 2000.
VERWEIJ, J.J. et al. Detection and identification of entamoeba species in stool samples by a reverse line hybridization assay. J Clin Microbiol 41(11): 5041-5, 2003(a).
VERWEIJ, J.J. et al. Short communication: Prevalence of Entamoeba histolytica and Entamoeba dispar in northern Ghana. Trop Med Int Health, 8(12):1153-6, 2003 (b).
VERWEIJ, J.J. et al. Simultaneous detection of Entamoeba histolytica, Giardia lamblia, and Cryptosporidium parvum in fecal samples by using multiplex real-time PCR. J Clin Microbiol, 42(3):1220-3, 2004.
VISVESVARA, G.S. & GARCIA, L.S. Culture of protozoan parasites. Clin Microbiol Rev. 15(3):327-8, 2000.
WALSH, J.A. Problems in recognition and diagnosis of amebiasis: estimation of the global magnitude of morbidity and mortality. Rev Infect Dis 8(2): 228-38, 1986.
WANG, Z.; VORA, G.J. & STENGER, D.A. Detection and genotyping of Entamoeba histolytica, Entamoeba dispar, Giardia lamblia, and Cryptosporidium parvum by oligonucleotide microarray. J Clin Microbiol. 42(7):3262-71, 2004.
WASSMANN, C. et al. Metronidazole resistance in the protozoan parasite Entamoeba histolytica is associated with increased expression of iron-containing superoxide dismutase and peroxiredoxin and decreased expression of ferredoxin 1 and flavin reductase. J Biol Chem. 274(37):26051-6, 1999.
WILSON, I.G. Inhibition and facilitation of nucleic acid amplification. Appl Environ Microbiol, 63(10):3741-51, 1997.
WOOD, W.I.; GITSCHIER, J.; LASKY, L.A. et al. Base composition-independent hybridization in tetramethylammonium chloride: a method for oligonucleotide screening of highly complex gene libraries. Proc Natl Acad Sci USA 82(6):1585-8, 1985.
WORLD HEALTH ORGANIZATION. Amoebiasis. Report on the WHO/Pan American Health Organization/UNESCO Expert Consultation Mexico City. Geneva-WHO W Epidemiol Rec 72:97-100, 1997.

117
XIMENEZ, C. et al. Reassessment of the epidemiology of amebiasis: state of the art. Infect Genet Evol 9(6): 1023-32, 2009.
YAN, X. et al. Multiplexed flow cytometric immunoassay for influenza virus detection and differentiation. Anal Chem. 77(23):7673-8, 2005.
ZAKI, M. & CLARK, C.G. Isolation and characterization of polymorphic DNA from Entamoeba histolytica. J Clin Microbiol 39(3):897-905, 2001.
ZAKI, M. et al. Simultaneous differentiation and typing of Entamoeba histolytica and Entamoeba dispar. J Clin Microbiol 40(4):1271-6, 2002.
ZAKI, M. et al. Genotyping of Entamoeba species in South Africa: diversity, stability, and transmission patterns within families. J Infect Dis 187(12):1860-9, 2003.
ZAMAN, S. et al. Direct amplification of Entamoeba histolytica DNA from amoebic liver abscess pus using polymerase chain reaction. Parasitol Res. 86(9):724-8, 2000.
ZINDROU, S. et al. Specific detection of Entamoeba histolytica DNA by hemolysin gene targeted PCR. Acta Trop 78(2): 117-25, 2001.

117
Anexos

118
Anexo 1 Árvore filogenética obtida a partir da análise das
sequências parciais do gene que codifica a SSU-rRNA das espécies de Entamoeba identificadas neste estudo, deduzida pelo algoritimo de Neighbor-Joining com teste
de bootstrap de 1000 réplicas

119
FIGURA 8. Árvore filogenética obtida a partir da an álise de sequências parcial do gene que codifica para o SSU-rRNA das espécies de Entamoeba identificadas neste estudo.
3182/1 3182/3 9343 3449 3162 3450 6553 200 201 9891 9893 03C P2 107 30B 27B 26B 22B/2 20B 19B/1 17B/A 14B/A 11B 09B/A 06B 05B 04B/A 03B 02B z49256 3182/4 AB197936 x64142 4093 CDC 33cdc 103 104 105 109 101 AF149906 106B 204/E 204/F 204/I 204/G 204/D 204/B 204/A 204 17B/3 17B/7 17B/2 17B/6 17B/4 17B/5 19B/5 19B/7 19B/4 19B/2 3684 18B/1 18B/4 18B/5 18B/6 18B/8 3182/7 3182/8 3182/6 3182/5 3182/2 AF149907 01B 100 18B/7 18B/3 19B/6 3977 04B AF149915 AF149914 15B 15B/1 15B/3 15B/4 15B/5 15B/6 15B/8 28B 28B/1 28B/2 28B/3 28B/5 28B/6 28B/8 29B 29B/2 29B/3 29B/5 29B/6 29B/7 29B/8 15B/2 15B/7 Endolimax nana
E. dispar
E. histolytica
E. moshkovskii
E. hartmanni
E. coli

120
Anexo 2 Representação esquemática do local de hibridação da s
sondas com as suas respectivas sequências alvos
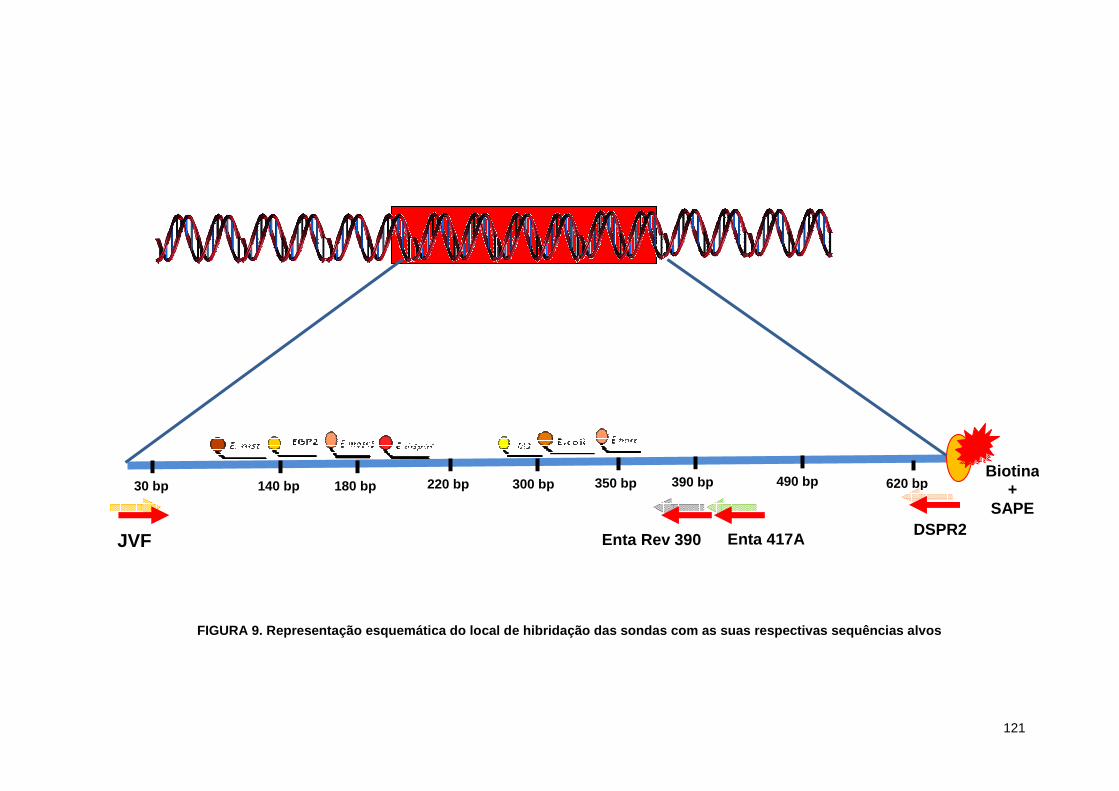
121
FIGURA 9. Representação esquemática do local de hib ridação das sondas com as suas respectivas sequênci as alvos
30 bp 390 bp 350 bp 300 bp 180 bp 220 bp 140 bp 620 bp 490 bp Biotina
+ SAPE
JVF Enta Rev 390 Enta 417A DSPR2

122
Anexo 3 Alinhamento múlitplo das sequências parciais
do gene Ssu-rRNA de Entamoeba spp depositadas no GenBank

123
10 20 3 0 40 50 60 70 80 90 100
....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....|....|....|....|
x64142_E histolytica GTTGATCCTGCCAGTATTATATGCTGATGTTAAAGATTAAGCCATGCATGTGTAAGTATAAAGACCAAGTAGGATGAAACTGCGGACGGCTCATTATAAC
AB197936_E. histolytica GTTGATCCTGCCAGTATTATATGCTGATGTTAAAGATTAAGCCATGCATGTGTAAGTATAAAGACCAAGTAGGATGAAACTGCGGACGGCTCATTATAAC
z49256._E.dispar GTTGATCCTGCCAGTATTATATGCTGATGTTAGAGATTAAGCCATGCATGTGTAAGTATAAAGACCAAGTAGGATGAAACTGCGGACGGCTCATTATAAC
AF149906 E moshkovskii GTTGATCCTGCCAGTATTATATGCTGATGTTAAAGATTAAGCCATGCATGTGTAAGTATAAAGACCAAGTAGGATGAAACTGCGGACGGCTCATTATAAC
AF149907_E. hartmanni GTTGATCCTGCCAGTATTATATGCTGATGTTAAAGATTAAGCCATGCATGTGTAAGTATAAAGTCCAAGAAGGATGAAACTGCGAACGGCTCATTAGAAC
AF149915_E. coli GTTGATCCTGCCAGTATTATATGCTTCTGTCAAAGATTAAGCCATGCATGTCTAAGCACAAAGTCCTAGTATGATGAAGCTGCGAACGGCTCATTACAAC
AF149914_E. coli GTTGATCCTGCCAGTATTATATGCTTCTGTCAAAGATTAAGCCATGCATGTCTAAGCATAAAGTCCTAGTAAGATGAAGCTGCGAACGGCTCATTACAAC
Iniciador JVF 110 120 130 140 150 160 170 180 190 200
....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....|....|....|....|
x64142_E histolytica AGTAATAGTTTCTTTGGTTAGTAAAATACAAGGATAGCTTTGTGAATGATAAAGATAATACTTGA---------------GACGATCCAGTTTG---TAT
AB197936_E. histolytica AGTAATAGTTTCTTTGGTTAGTAAAATACAAGGATAGCTTTGTGAATGATAAAGATAATACTTGA---------------GACGATCCAGTTTG---TAT
z49256._E.dispar AGTAATAGTTTCTTTGGTTAGTAAAGTACAAGGATAGCTTTGTGAATGATAAAGATAATACTTGA---------------GACGATCCAATTTG---TAT
AF149906 E moshkovskii AGTAATAGTTTCTTTGGTTAGTAAAGTACAAGGATAGCTTTGTGAATGATAAAGATAATACTTGA---------------GACGATCCGGTTTG---TAT
AF149907_E. hartmanni AGTTATAGTTTCTTCGATTAGTAA-GTACAAGGATAGCTTTGTGAATGATAAAGATAATACTTGA---------------GACGATCTTGGATG---TAG
AF149915_E. coli AGTTATAATCTTTTTGATGAAGTACGTACAAGGATATCTTTGAGAATGTCAAAGCTAATACTTGACGGTTTTCACCCCTTGTCGAGTCCATTCCCTCTGG
AF149914_E. coli AGTTATAATCTTTTTGATGAAATACGTACAAGGATATCTTTGAGAATGTCAAAGCTAATACTTGACGGGTAACGCCTTCAGTCGAGTGTATTCC-TCCGG
210 220 230 240 250 260 270 280 290 300
....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....|....|....|....|
x64142_E histolytica TAGTACAAAAT------GGCCAAT---TCATTCAA---TGAATTGAGAAATGACATTCTAAGT-GAGTTAGG---------ATGCCACGACAATTGTAGA
AB197936_E. histolytica TAGTACAAAAT------GGCCAAT---TTATTTAA---TGAATTGAGAAATGACATTCTAAGT-GAGTTAGG---------ATGCCACGACAATTGTAGA
z49256._E.dispar TAGTACAAAGT------GGCCAAT---TTATGTAAG--TAAATTGAGAAATGACATTCTAAGT-GAGTTAGG---------ATGCCACGACAATTGTAGA
AF149906 E moshkovskii TAGTACAAGTC------GGCCACT---CTCTTCACG--GGGAGTGCGAA-TGCCATTCTGAATTGAATAAGG---------ATGGTATGACAATTGTAGA
AF149907_E. hartmanni AGATACATTCA------GGCCTTTGCATCATTTAATGGTGTGAAGAGAAAGGATATCCAAAGT-GAAAGTGG---------GTATCATGACAATTGTAGA
AF149915_E. coli GAGTGTAATCGA-TGAGGGCGGG-GGATGCTTCACGTATCCTCTACATATTACCACTTTTTTTTGAATGAGGGTGGAATATATGCCAAGAGAATTGTAGA
AF149914_E. coli GAGTATAATCTACTGAGGAGGGGAGGACCCCCAAGGTCCTTTCAATATATTACCACTTCTTTGTGAATAAGGGTGGATTATATGCCAAGAGAATTGTAGA

124
310 320 330 340 350 360 370 380 390 400
....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....|....|....|....|
x64142_E histolytica ACACACAGTGTTTA-ACAAGTAACCAATGAGAATTTCTGATCTATCAATCAGTTGGTAGTA---TCGAGGACTACCAAGATTATAACGGATAACGAGGAA
AB197936_E. histolytica ACACACAGTGTTTA-ACAAGTAACCAATGAGAATTTCTGATCTATCAATCAGTTGGTAGTA---TCGAGGACTACCAAGATTATAACGGATAACGAGGAA
z49256._E.dispar ACACACAGTGTTTA-ACAAGTAACCAATGAGAATTTCTGATCTATCAATCAGTTGGTAGTA---TCGAGGACTACCAAGATTATAACGGATAACGAGGAA
AF149906 E moshkovskii GCACACAGTGTTTA-ACAAGTAACCAATGAGAATTTCTGATCTATCAATTTGTTGGTAGTA---TCGAGGACTACCAAGATTATAACGGATAACGAGGAA
AF149907_E. hartmanni ATA-GCGATATTTA-ACAAGTAATCGATGAGAATATCTGATCTATCAACTAGTTGGTAGTA---TCGAGGACTACCAAGGTTATAACGGATAACGAGGAA
AF149915_E. coli AATCGAGAGATTTTCACAAGTCATCATTAAGAATATCTGACCTATCAACTGGTTGTTGGTAAGGTAGTGGCTTACCAAGGTGATAACGGGTAACGAGGAA
AF149914_E. coli AATCGAGAGATTTTCACAAGTCATCATTAAGAATATCTGACCTATCAACTGGTTGTTGGTAAGGTAGTGGCTTACCAAGGTGATAACGGGTAACGAGGAA
Iniciador: EntaRev390
410 420 430 440 450 460 470 480 490 500
....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....|....|....|....|
x64142_E histolytica TTGGGGTTCGACATCGGAGAGGGAGCTTTACAGATGGCTACCACTTCTAAGGAAGGCAGCAGGCSCGTAAATTACCCACTTTCGA---ATTGAAGAGGTA
AB197936_E. histolytica TTGGGGTTCGACATCGGAGAGGGAGCTTTACAGATGGCTACCACTTCTAAGGAAGGCAGCAGGCGCGTAAATTACCCACTTTCGA---ATTGAAGAGGTA
z49256._E.dispar TTGGGGTTCGACATCGGAGAGGGAGCTTTACAGATGGCTACCACTTCTAAGGAAGGCAGCAGGCGCGTAAATTACCCACTTTCGA---ATTGAAGAGGTA
AF149906 E moshkovskii TTGGGGTTCGACATCGGAGAGGGAGCTTTACAGATGGCTACCACTTCTACGGAAGGCAGCAGGCGCGTAAATTACCCACTTTCGA---CGTGAAGAGGTA
AF149907_E. hartmanni TTAGGGTTTGACATCGGAGAGGGAGCTTTCAAGATGGCTACCACTTCCAAGGAAGGCAGCAGGCGCGTAAATTACCCAATTTCAACAGATAGAAGAGGTA
AF149915_E. coli TAAGGGTTCGACATCGGAGAGGGAGCTTTAGAGATGGCTACCACTTCTAAGGAAGGCAGCAGGCGCGAAAATTACCCAATCTTAACAAATGAAGGAGGTA
AF149914_E. coli TAAGGGTTCGACATCGGAGAGGGAGCTTTAGAAATGGCTACCACTTCTAAGGAAGGCAGCAGGCGCGAAAATTACCCAATCTTAACAACTGAAGGAGGTA
Iniciador reverso 417A
510 520 530 540 550 560 570 580 590 600
....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....|....|....|....|
x64142_E histolytica GTGACGACACATAACTCTAGAGTTGAGTAAAATCAATTCTTGAAGGAATGAGTAGGAGGTAAATTCTCCTACGAAATCAATTGGAGGGCAAGTCTGGTGC
AB197936_E. histolytica GTGACGACACATAACTCTAGAGTTGAGTAAAATCAATTCTTGAAGGAATGAGTAGGAGGTAAATTCTCCTACGAAATCAATTGGAGGGCAAGTCTGGTGC
z49256._E.dispar GTGACGACACATAACTCTAGAGTTGAGTAAAATCAATTCTTGAAGGAATGAGTAGGAGGTAAATTCTCCTACGAAATCAATTGGAGGGCAAGTCTGGTGC
AF149906 E moshkovskii GTGACGACAAATAACTCTCGAGGTGGTTAACTCCACTTCTTGAAGGAATGAGTAAGAAGTAAATACTCTTACGAAATCAATTGGAGGGCAAGTCTGGTGC
AF149907_E. hartmanni GTGACGACAAATAACTCACGACATGCGTAAG--CATGTCTTGAAGGAATGAGTAAGAAGCAAAGAGTCTTACGAAATCAATTGGAGGGCAAGTCTGGTGC
AF149915_E. coli GTGACGACAATTAACGCTATCCTCGTCTTTTGGCGAGGATCGTCGGAATGATTTCGATTTAAAATATCGAAAGAAAGCAATTGGAGGGCAAGTCTGGTGC
AF149914_E. coli GTGACGACAATTAACGCTTTCCCTCGCTTTACGTGAGGATTGCCGGAATGATCACGACATAAACCATCGTGAGAAAGCGATTGGAGGGCAAGTCTGGTGC

125
610 620 630 640 650 660
....|....|....|....|....|. ...|....|....|....|....|....|....|....|....
x64142_E histolytica CAGCAGCCGCGGTAATTCCAGCTCCAATAGTGTATATTAAAGTTGCTGTGATTAAAACGCTCGTAGTTG
AB197936_E.histolytica CAGCAGCCGCGGTAATTCCAGCTCCAATAGTGTATATTAAAGTTGCTGTGATTAAAACGCTCGTAGTTG
z49256._E.dispar CAGCAGCCGCGGTAATTCCAGCTCCAATAGTGTATATTAAAGTTGCTGTGATTAAAACGCTCGTAGTTG
AF149906 E moshkovskii CAGCAGCCGCGGTAATTCCAGCTCCAATAGTGTATATTAAAGTTGCTGTGATTAAAACGCTCGTAGTTG
AF149907_E. hartmanni CAGCAGCCGCGGTAATTCCAGCTCCAATAGTGTATATTAAAGTTGTTGTGATTAAAACGCTCGTAGTTG
AF149915_E. coli CAGCAGCCGCGGTAATTCCAGCTCCAATAGTGTATTGTAAAGTTGTTGTGATTAAAACGCTCGTAGTTG
AF149914_E. coli CAGCAGCCGCGGTAATTCCAGCTCCAATAGTGTATTGTAAAGTTGTTGTGATTAAAACGCTCGTAGTTG
DSPR2
Leganda:
- Oligonucleotideo iniciador direto e reverso
- As sondas de DNA que apresentaram os melhores sinais de fluorescência na PCR/Luminex

126
Anexo 4 Alinhamento da sequência parcial do gene Ssu-rRNA
de E. moshkovskii obtida neste estudo

127
10 20 30 40 50 60 70 80 90 100 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 106B -TTGATCCTGCCAGTATTATATGCTGATGTTAAAGATTAAGCCATGCATGTGTAAGTATAAAGACCAAGTAGGATGAAACTGCGGACGGCTCATTATAAC AF149906 G................................................................................................... 110 120 130 140 150 160 170 180 190 200 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 106B AGTAATAGTTTCTTTGGTTAGTAAAGTACAAGGATAGCTTTGTGAATGATAAAGATAATACTTGAGACGATCCGGTTTGTATTAGTACAAGTCGGCCACT AF149906 .................................................................................................... 210 220 230 240 250 260 270 280 290 300 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 106B TTCTTAACGGAGAGTGCGAATGCCATCCTGAATTGAACAAGGATGGCACGACAATTGTAGAGCACACAGTGTTTAACAAGTAACCAATGAGAATTTCTGA AF149906 C....C....G...............T..........T........T.T................................................... 310 320 330 340 350 360 370 380 390 400 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 106B TCTATCAATTTGTTGGTAGTATCGAGGACTACCAAGATTATAACGGATAACGAGGAATTGGGGTTCGACATCGGAGAGGGAGCTTTACAGATGGCTACCA AF149906 .................................................................................................... 410 420 430 440 450 460 470 480 490 500 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 106B CTTCTACGGAAGGCAGCAGGCGCGTAAATTACCCACTTTCGACGTGAAGAGGTAGTGACGACAAATAACTCTCGAGGTGGTTTAC--CACTTCTTGAAGG AF149906 ..................................................................................A..TC............. 510 520 530 540 550 560 570 580 590 600 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 106B AATGAGTAAGAAGTAAAATCTCTTACAAAATCAATTGGAGGGCAAGTCTGGTGCCAGCAGCCGCGGTAATTCCAGCTCCAATAGTGTATATTAAAGTTGC AF149906 .................TA.......G......................................................................... 610 620 ....|....|....|....|... 106B TGTGATTAAAACGCTCGTAGTTG AF149906 ....................... ...... - Indica identidade nucleotídica AF149906 – Número de acesso no Genbank da sequência parcial do gene sa SSu-rRNA de Entamoeba moshkovskii

128
Anexo 5 Regiões polimórficas presente no gene SSu-rRNA E.
hartmanni, obtidas a partir do alinhamento das sequências determinadas neste estudo, utilizando o programa Clustal W inserido no pacote do programa
GenStudio

129
210 220 230 240 250 260 270 280 2 90 300 ....|....|....|....|....|....|....|... .|....|....|....|....|....|....|....|....|....|.... |....|....| AF149907 CATCATTTAATGGTGTGAAGAGAAAGGATATCCAAAGTGAAAGTGGGTATCATGACAATTGTAGAATAGCGATATTTAACAAGTAATCGATGAGAATATC 01B_2009010 .....................................................................T.............................. 08Bclone1_2 ......................................................................A............................. 08B_clone3_ .............................................................................G...................... 08B_clone4_ ......................................................................A............................. 08B_clone5_ ......................................................................A............................. 08B_clone6_ ......................................................................A............................. 08B_clone7_ .................................................................................................... 08B_clone8_ ......................................................................A............................. 17Bclone2_2 .....C..G-...............A...........................................AA............................. 17B_clone3_ .......CG-...............A...........................................AA............................. 17B_clone4_ .....C..G-...............A...........................................AA............................. 17B_clone5_ .....C..G-...............A...........................................AA............................. 17B_clone6_ .....C..G-...............A...........................................AA............................. 17B_clone7_ .......CG-...............A...........................................AA............................. 19B_clone2_ .....A.C.T............................................................A............................. 19B_clone4_ .....A.C.T............................................................A............................. 19B_clone5_ .....A................................................................A......G...................... 19B_clone6_ ......----............................................G..............GA......G...................... 19B_clone7_ .....A................................................................A......G...................... 100CDC_2008 ......................................................................A............................. 3182clone2_ ......................................................................A......G...................... 3182clone5_ ......................................................................A......G...................... 3182clone6_ ......................................................................A......G...................... 3182clone7_ ......................................................................A......G...................... 3182clone8_ ......................................................................A......G...................... 3684_CDC_20 ......................................................................A............................. 204_CDC_200 ...T...-...AA....................................................................................... 204_cdc_CLO ...T...-...AA....................................................................................... 204_cdc_CLO ...T...-...AA....................................................................................... 204_cdc_CLO ...T...-...AA....................................................................................... 204_cdc_CLO ...T...-...AA.......G............................................................................... 204_cdc_CLO ...T...-...AA....................................................................................... 204_cdc_CLO ...T...-...AA....................................................................................... 204_cdc_CLO ...T...-...AA.......................................................................................

130
410 420 430 440 450 460 470 480 4 90 500 ....|....|....|....|....|....|....|... .|....|....|....|....|....|....|....|....|....|.... |....|....| AF149907 CCACTTCCAAGGAAGGCAGCAGGCGCGTAAATTACCCAATTTCAACAGATAGAAGAGGTAGTGACGACAAATAACTCACGACATGCGTAAGCATGTCTTG 01B_2009010 .................................................................................................... 08Bclone1_2 .................................................................................................... 08B_clone3_ ...........................................................................................T........ 08B_clone4_ .................................................................................................... 08B_clone5_ .................................................................................................... 08B_clone6_ .................................................................................................... 08B_clone7_ ...........................................................................................T........ 08B_clone8_ .................................................................................................... 17Bclone2_2 .................................................................................TG...........A..... 17B_clone3_ .................................................................................TG...........A..... 17B_clone4_ .................................................................................TG...........A....A 17B_clone5_ .................................................................................TG...........A....A 17B_clone6_ .................................................................................TG...........A..... 17B_clone7_ .................................................................................TG...........A..... 19B_clone2_ .................................................................................................... 19B_clone4_ .................................................................................................... 19B_clone5_ .................................................................................................... 19B_clone6_ ...........................................................................................T........ 19B_clone7_ .................................................................................................... 100CDC_2008 ...........................................................................................T........ 3182clone2_ .................................................................................................... 3182clone5_ .................................................................................................... 3182clone6_ .................................................................................................... 3182clone7_ .................................................................................................... 3182clone8_ .................................................................................................... 3684_CDC_20 .....................................................................................T.............. 204_CDC_200 .................................................................................TG................. 204_cdc_CLO .................................................................................TG................. 204_cdc_CLO .................................................................................TG................. 204_cdc_CLO .................................................................................TG................. 204_cdc_CLO .................................................................................TG................. 204_cdc_CLO ..........................A......................................................TG................. 204_cdc_CLO .................................................................................TG................. 204_cdc_CLO .................................................................................TG................. ...... Indica identidade nucleotídica AF 149907 - Nº de acesso no Gen Bank da sequência parcial do gene da SSu-rRNA de- Entamoeba hartmanni

131
Anexo 6 Regiões polimórficas presente nas sequências do gen e
SSu-rRNA de E. coli obtidas neste estudo
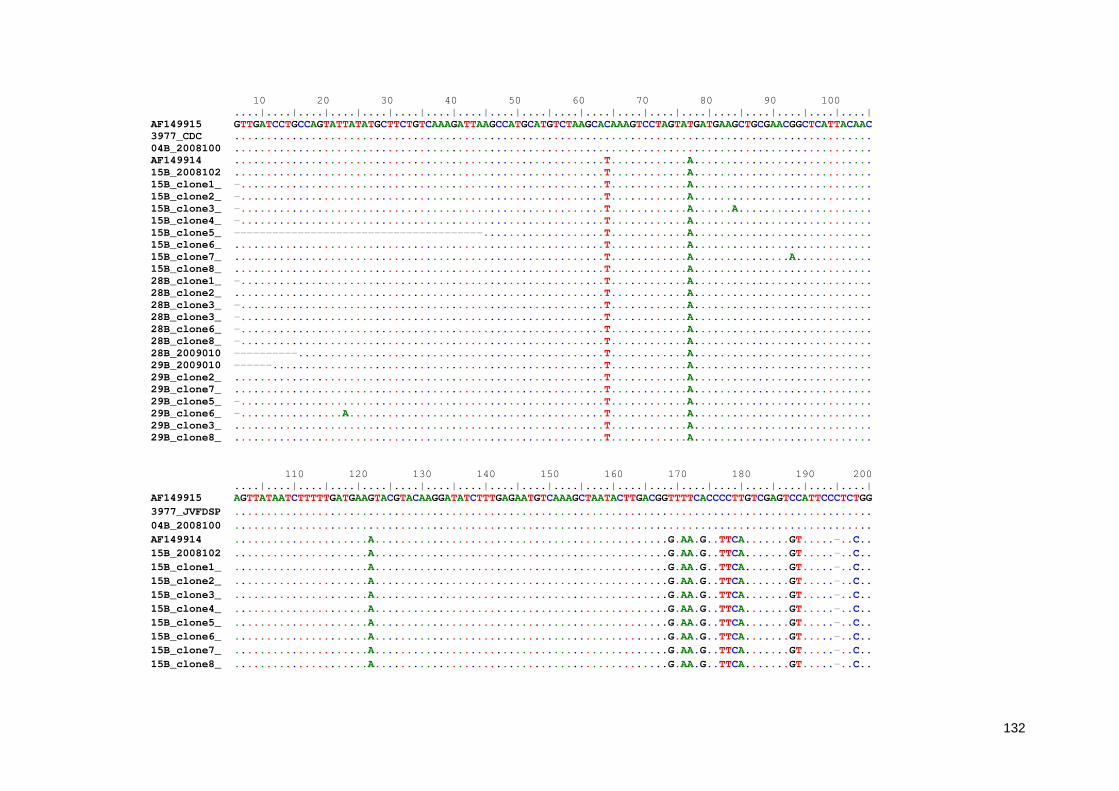
132
10 20 30 40 50 60 70 80 90 100 ....|....|....|....|....|....|....|... .|....|....|....|....|....|....|....|....|....|.... |....|....| AF149915 GTTGATCCTGCCAGTATTATATGCTTCTGTCAAAGATTAAGCCATGCATGTCTAAGCACAAAGTCCTAGTATGATGAAGCTGCGAACGGCTCATTACAAC 3977_CDC .................................................................................................... 04B_2008100 .................................................................................................... AF149914 ..........................................................T............A............................ 15B_2008102 ..........................................................T............A............................ 15B_clone1_ -.........................................................T............A............................ 15B_clone2_ -.........................................................T............A............................ 15B_clone3_ -.........................................................T............A......A..................... 15B_clone4_ -.........................................................T............A............................ 15B_clone5_ ---------------------------------------...................T............A............................ 15B_clone6_ ..........................................................T............A............................ 15B_clone7_ ..........................................................T............A...............A............ 15B_clone8_ ..........................................................T............A............................ 28B_clone1_ -.........................................................T............A............................ 28B_clone2_ ..........................................................T............A............................ 28B_clone3_ -.........................................................T............A............................ 28B_clone3_ -.........................................................T............A............................ 28B_clone6_ -.........................................................T............A............................ 28B_clone8_ -.........................................................T............A............................ 28B_2009010 ----------................................................T............A............................ 29B_2009010 ------....................................................T............A............................ 29B_clone2_ ..........................................................T............A............................ 29B_clone7_ ..........................................................T............A............................ 29B_clone5_ -.........................................................T............A............................ 29B_clone6_ -................A........................................T............A............................ 29B_clone3_ ..........................................................T............A............................ 29B_clone8_ ..........................................................T............A............................ 110 120 130 140 150 160 170 180 1 90 200 ....|....|....|....|....|....|....|... .|....|....|....|....|....|....|....|....|....|.... |....|....| AF149915 AGTTATAATCTTTTTGATGAAGTACGTACAAGGATATCTTTGAGAATGTCAAAGCTAATACTTGACGGTTTTCACCCCTTGTCGAGTCCATTCCCTCTGG 3977_JVFDSP .................................................................................................... 04B_2008100 .................................................................................................... AF149914 .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 15B_2008102 .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 15B_clone1_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 15B_clone2_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 15B_clone3_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 15B_clone4_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 15B_clone5_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 15B_clone6_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 15B_clone7_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 15B_clone8_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C..

133
28B_clone1_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 28B_clone2_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 28B_clone3_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 28B_clone3_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 28B_clone6_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 28B_clone8_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 28B_2009010 .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 29B_2009010 .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 29B_clone2_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 29B_clone7_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 29B_clone5_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 29B_clone6_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 29B_clone3_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C.. 29B_clone8_ .....................A..............................................G.AA.G..TTCA.......GT.....-..C..
210 220 230 240 250 260 270 280 2 90 300 ....|....|....|....|....|....|....|... .|....|....|....|....|....|....|....|....|....|.... |....|....| AF149915 GAGTGTAATCGA-TGAGG-GCGGGGGATGCTTCACGTATCCTCTACATATTACCACTTTTTTTTGAATGAGGGTGGAATATATGCCAAGAGAATTGTAGA 3977_JVFDSP ............-.....-............................................-.................................... 04B_2008100 ............-.....-................................................................................. AF149914 ....A.....T.C.....A.G..A...CC.CCA.G..CCTT..A.T............C...G.....A........T...................... 15B_2008102 ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 15B_clone1_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 15B_clone2_ ...CA.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 15B_clone3_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 15B_clone4_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 15B_clone5_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 15B_clone6_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 15B_clone7_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 15B_clone8_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 28B_clone1_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 28B_clone2_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 28B_clone3_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 28B_clone3_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 28B_clone6_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 28B_clone8_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 28B_2009010 ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 29B_2009010 ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 29B_clone2_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 29B_clone7_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 29B_clone5_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T......................

134
29B_clone6_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 29B_clone3_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 29B_clone8_ ....A.....T.C.....A.G..A....C..-ATG..CCTT..A.T............C...G.....A........T...................... 410 420 430 440 450 460 470 480 4 90 500 ....|....|....|....|....|....|....|... .|....|....|....|....|....|....|....|....|....|.... |....|....| AF149915 TAAGGGTTCGACATCGGAGAGGGAGCTTTAGAGATGGCTACCACTTCTAAGGAAGGCAGCAGGCGCGAAAATTACCCAATCTTAACAAATGAAGGAGGTA 3977_JVFDSP .................................................................................................... 04B_2008100 .................................................................................................... AF149914 ................................A.......................................................C........... 15B_2008102 ................................A.......................................................C........... 15B_clone1_ ................................A.......................................................C........... 15B_clone2_ ................................A.......................................................C........... 15B_clone3_ ................................A.......................................................C........... 15B_clone4_ ................................A.......................................................C........... 15B_clone5_ ................................A.......................................................C........... 15B_clone6_ ................................A.......................................................C........... 15B_clone7_ ................................A.......................................................C........... 15B_clone8_ ................................A.......................................................C........... 28B_clone1_ ................................A.......................................................C........... 28B_clone2_ ................................A.......................................................C........... 28B_clone3_ ................................A.......................................................C........... 28B_clone3_ ................................A.......................................................C........... 28B_clone6_ ................................A.......................................................C........... 28B_clone8_ ................................A.......................................................C........... 28B_2009010 ................................A.......................................................C........... 29B_2009010 ................................A.......................................................C........... 29B_clone2_ ................................A.......................................................C........... 29B_clone7_ ................................A.......................................................C........... 29B_clone5_ ................................A.......................................................C........... 29B_clone6_ ................................A.......................................................C........... 29B_clone3_ ................................A.......................................................C........... 29B_clone8_ ................................A.......................................................C...........
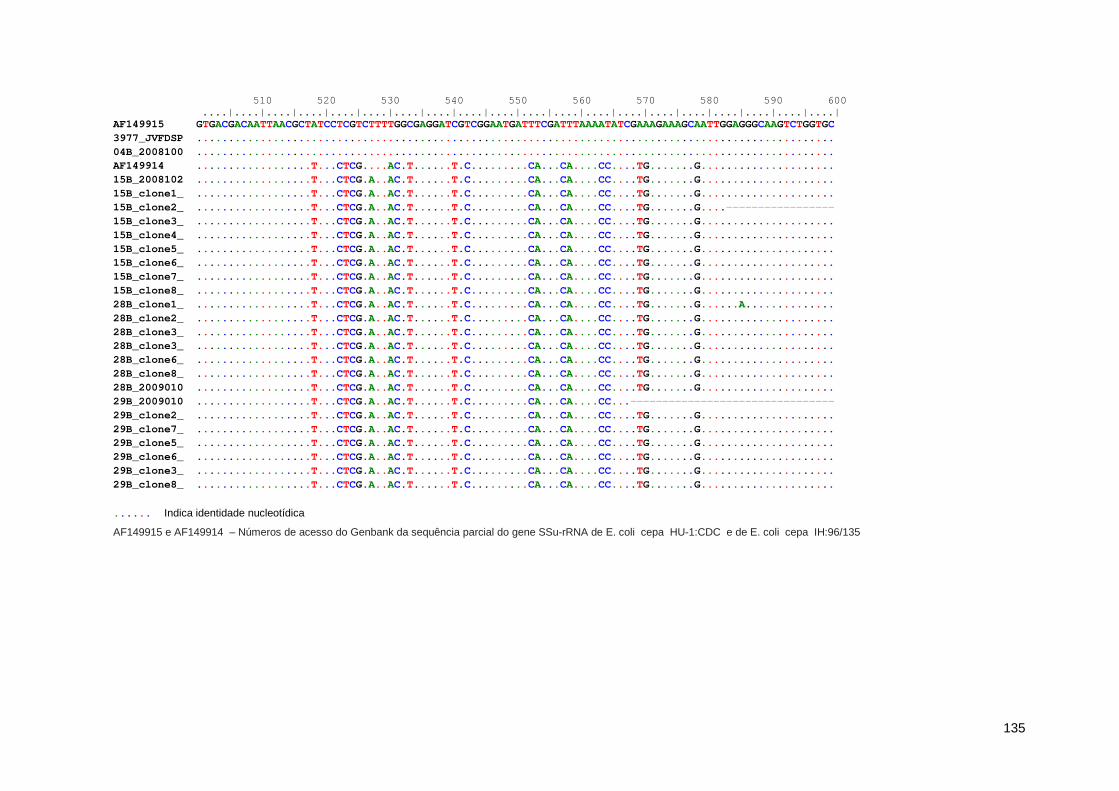
135
510 520 530 540 550 560 570 580 590 600 ....|....|....|....|....|....|....|....|... .|....|....|....|....|....|....|....|....|....|.... |....|
AF149915 GTGACGACAATTAACGCTATCCTCGTCTTTTGGCGAGGATCGTCGGAATGATTTCGATTTAAAATATCGAAAGAAAGCAATTGGAGGGCAAGTCTGGTGC 3977_JVFDSP .................................................................................................... 04B_2008100 .................................................................................................... AF149914 ..................T...CTCG....AC.T......T.C.........CA...CA....CC....TG.......G..................... 15B_2008102 ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 15B_clone1_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 15B_clone2_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G....----------------- 15B_clone3_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 15B_clone4_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 15B_clone5_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 15B_clone6_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 15B_clone7_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 15B_clone8_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 28B_clone1_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G......A.............. 28B_clone2_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 28B_clone3_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 28B_clone3_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 28B_clone6_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 28B_clone8_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 28B_2009010 ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 29B_2009010 ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC...-------------------------------- 29B_clone2_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 29B_clone7_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 29B_clone5_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 29B_clone6_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 29B_clone3_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G..................... 29B_clone8_ ..................T...CTCG.A..AC.T......T.C.........CA...CA....CC....TG.......G.....................
...... Indica identidade nucleotídica
AF149915 e AF149914 – Números de acesso do Genbank da sequência parcial do gene SSu-rRNA de E. coli cepa HU-1:CDC e de E. coli cepa IH:96/135

136
Anexo 7 Alinhamento múltiplo entre as sequências parcias do
gene Ssu-rRNA de Entamoeba spp obtidas neste estudo

137
10 20 30 40 50 60 70 80 90 100 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| x64142 GTTGATCCTGCCAGTATTATATGCTGATGTTAAAGATTAAGCCATGCATGTGTAAGTATAAAGACCAAGTAGGATGAAACTGCGGACGGCTCATTATAAC AB197936 .................................................................................................... 4093_CDC -................................................................................................... 33cdc .................................................................................................... 101 -................................................................................................... 103 .................................................................................................... 104 .................................................................................................... 105 .................................................................................................... 109 .................................................................................................... z49256 ................................G................................................................... 02B -................................................................................................... 03B .................................................................................................... 04B/A .................................................................................................... 05B .................................................................................................... 06B .................................................................................................... 09B/A .................................................................................................... 11B -................................................................................................... 14B/A -................................................................................................... 17B/A ------------------------------------................................................................ 19B/1 -................................................................................................... 20B .................................................................................................... 22B/2 -................................................................................................... 26B -................................................................................................... 27B -................................................................................................... 30B ------------------------------------------.......................................................... 107 .................................................................................................... P2 -................................................................................................... 03C .................................................................................................... 9893 -................................................................................................... 9891 .................................................................................................... 201 -................................................................................................... 200 -................................................................................................... 6553 -................................................................................................... 3450 .................................................................................................... 3162 ----------------------------------------------------------------------------------.................. 3449 .................................................................................................... 9343 .................................................................................................... 3182/1 ---------------..................................................................................... 3182/3 ---------........................................................................................... 3182/4 ---------...........................................................................................
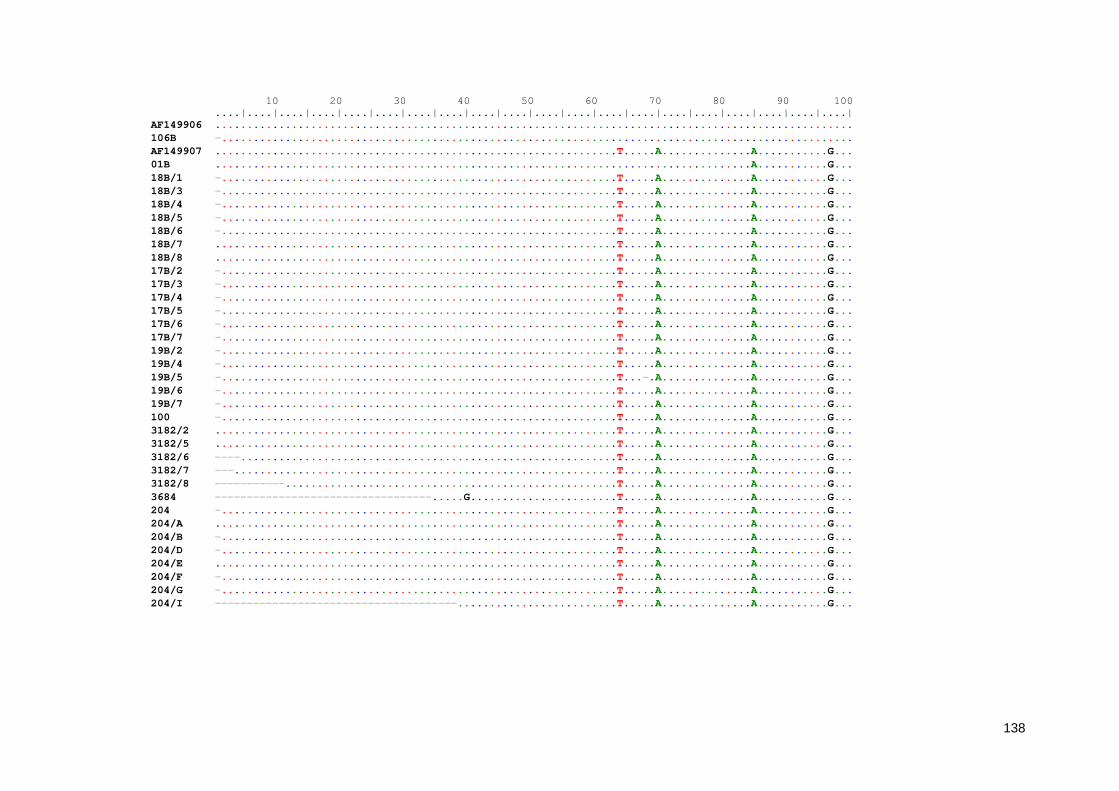
138
10 20 30 40 50 60 70 80 90 100 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| AF149906 .................................................................................................... 106B -................................................................................................... AF149907 ...............................................................T.....A..............A...........G... 01B ....................................................................................A...........G... 18B/1 -..............................................................T.....A..............A...........G... 18B/3 -..............................................................T.....A..............A...........G... 18B/4 -..............................................................T.....A..............A...........G... 18B/5 -..............................................................T.....A..............A...........G... 18B/6 -..............................................................T.....A..............A...........G... 18B/7 ...............................................................T.....A..............A...........G... 18B/8 ...............................................................T.....A..............A...........G... 17B/2 -..............................................................T.....A..............A...........G... 17B/3 -..............................................................T.....A..............A...........G... 17B/4 -..............................................................T.....A..............A...........G... 17B/5 -..............................................................T.....A..............A...........G... 17B/6 -..............................................................T.....A..............A...........G... 17B/7 -..............................................................T.....A..............A...........G... 19B/2 -..............................................................T.....A..............A...........G... 19B/4 -..............................................................T.....A..............A...........G... 19B/5 -..............................................................T...-.A..............A...........G... 19B/6 -..............................................................T.....A..............A...........G... 19B/7 -..............................................................T.....A..............A...........G... 100 -..............................................................T.....A..............A...........G... 3182/2 ...............................................................T.....A..............A...........G... 3182/5 ...............................................................T.....A..............A...........G... 3182/6 ----...........................................................T.....A..............A...........G... 3182/7 ---............................................................T.....A..............A...........G... 3182/8 -----------....................................................T.....A..............A...........G... 3684 ----------------------------------.....G.......................T.....A..............A...........G... 204 -..............................................................T.....A..............A...........G... 204/A ...............................................................T.....A..............A...........G... 204/B -..............................................................T.....A..............A...........G... 204/D -..............................................................T.....A..............A...........G... 204/E ...............................................................T.....A..............A...........G... 204/F -..............................................................T.....A..............A...........G... 204/G -..............................................................T.....A..............A...........G... 204/I --------------------------------------.........................T.....A..............A...........G...

139
10 20 30 40 50 60 70 80 90 100 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| AF149915 .........................TC...C....................C....C.C....T..T....T......G.....A...........C... 3977 .........................TC...C....................C....C.C....T..T....T......G.....A...........C... 04B .........................TC...C....................C....C.C....T..T....T......G.....A...........C... AF149914 .........................TC...C....................C....C......T..T....A......G.....A...........C... 15B .........................TC...C....................C....C......T..T....A......G.....A...........C... 15B/1 -........................TC...C....................C....C......T..T....A......G.....A...........C... 15B/2 -........................TC...C....................C....C......T..T....A......G.....A...........C... 15B/3 -........................TC...C....................C....C......T..T....A............A...........C... 15B/4 -........................TC...C....................C....C......T..T....A......G.....A...........C... 15B/5 ---------------------------------------............C....C......T..T....A......G.....A...........C... 15B/6 .........................TC...C....................C....C......T..T....A......G.....A...........C... 15B/7 .........................TC...C....................C....C......T..T....A......G.....A..A........C... 15B/8 .........................TC...C....................C....C......T..T....A......G.....A...........C... 28B ----------...............TC...C....................C....C......T..T....A......G.....A...........C... 28B/1 -........................TC...C....................C....C......T..T....A......G.....A...........C... 28B/2 .........................TC...C....................C....C......T..T....A......G.....A...........C... 28B/3 -........................TC...C....................C....C......T..T....A......G.....A...........C... 28B/5 -........................TC...C....................C....C......T..T....A......G.....A...........C... 28B/6 -........................TC...C....................C....C......T..T....A......G.....A...........C... 28B/8 -........................TC...C....................C....C......T..T....A......G.....A...........C... 29B ------...................TC...C....................C....C......T..T....A......G.....A...........C... 29B/2 .........................TC...C....................C....C......T..T....A......G.....A...........C... 29B/3 .........................TC...C....................C....C......T..T....A......G.....A...........C... 29B/5 -........................TC...C....................C....C......T..T....A......G.....A...........C... 29B/6 -................A.......TC...C....................C....C......T..T....A......G.....A...........C... 29B/7 .........................TC...C....................C....C......T..T....A......G.....A...........C... 29B/8 .........................TC...C....................C....C......T..T....A......G.....A...........C...

140
110 120 130 140 150 160 170 180 190 200 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| x64142 AGTAATAGTTTCTTTGGTTAGTAAAATACAAGGATAGCTTTGTGAATGATAAAGATAATACTTGA-----GACGATCCAGTTTGTATTAGTACAAAATGG AB197936 .................................................................-----.............................. 4093_CDC .................................................................-----.............................. 33cdc .................................................................-----.............................. 101 .................................................................-----.............................. 103 .................................................................-----.............................. 104 .................................................................-----.............................. 105 .................................................................-----.............................. 109 .................................................................-----.............................. 110 120 130 140 150 160 170 180 190 200 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| z49256 .........................G.......................................-----.........A................G... 02B .........................G.......................................-----.........A................G... 03B .........................G.......................................-----.........A................G... 04B/A .........................G.......................................-----.........A................G... 05B .........................G.......................................-----.........A................G... 06B .........................G.......................................-----.........A................G... 09B/A .........................G.......................................-----.........A................G... 11B .........................G.......................................-----.........A................G... 14B/A .........................G.......................................-----.........A................G... 17B/A .........................G.......................................-----.........A................G... 19B/1 .........................G.......................................-----.........A................G... 20B .........................G.......................................-----.........A................G... 22B/2 .........................G.......................................-----.........A................G... 26B .........................G.......................................-----.........A................G... 27B .........................G.......................................-----.........A................G... 30B .........................G.......................................-----.........A................G... 107 .........................G.......................................-----.........A................G... P2 .........................G.......................................-----.........A................G... 03C .........................G.......................................-----.........A................G... 9893 .........................G.......................................-----.........A................G... 9891 .........................G.......................................-----.........A................G... 201 .........................G.......................................-----.........A................G... 200 .........................G.......................................-----.........A................G... 6553 .........................G.......................................-----.........A................G... 3450 .........................G.......................................-----.........A................G... 3162 .........................G.......................................-----.........A................G... 3449 .........................G.......................................-----.........A................G... 9343 .........................G.......................................-----.........A................G...

141
3182/1 .........................G.......................................-----.........A................G... 3182/3 .........................G.......................................-----.........A................G... 3182/4 .........................G.......................................-----.........A................G... AF149906 .........................G.......................................-----........G................GTC.. 106B .........................G.......................................-----........G................GTC.. AF149907 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 01B ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 18B/1 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 18B/3 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 18B/4 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 18B/5 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 18B/6 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 18B/7 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 18B/8 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 110 120 130 140 150 160 170 180 190 200 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 17B/2 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 17B/3 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 17B/4 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 17B/5 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 17B/6 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 17B/7 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 19B/2 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 19B/4 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 19B/5 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 19B/6 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 19B/7 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 100 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 3182/2 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 3182/5 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 3182/6 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 3182/7 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 3182/8 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 3684 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 204 ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 204/A ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 204/B ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 204/D ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 204/E ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 204/F ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. 204/G ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA..

142
204/I ...T..........C.A.......-G.......................................-----.......TT.GA....GAGA....TTCA.. AF149915 ...T...A.C.T....A.G.AGT.CG..........T.....A.....TC....C..........CGGTTTT.ACC..TTG.C.AG.CCA.T.CCTC... 3977 ...T...A.C.T....A.G.AGT.CG..........T.....A.....TC....C..........CGGTTTT.ACC..TTG.C.AG.CCA.T.CCTC... 04B ...T...A.C.T....A.G.AGT.CG..........T.....A.....TC....C..........CGGTTTT.ACC..TTG.C.AG.CCA.T.CCTC... AF149914 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 15B ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 15B/1 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 15B/2 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 15B/3 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 15B/4 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 15B/5 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 15B/6 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 15B/7 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 15B/8 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 28B ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 28B/1 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 28B/2 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 10 20 30 40 50 60 70 80 90 100 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 28B/3 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC..
28B/5 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 28B/6 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 28B/8 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 29B ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 29B/2 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 29B/3 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 29B/5 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 29B/6 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 29B/7 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 29B/8 ...T...A.C.T....A.G.AAT.CG..........T.....A.....TC....C..........CGGGTA...CCTTCAG.C.AG.GTA.T.C-TCC.. 210 220 230 240 250 260 270 280 290 300 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| x64142 CCAATTCAT-TC-----AATGAATTGAG----AAATGACATTCTAAGT----------------GAGTTAGG---------ATGCCACGACAATTGTAGA AB197936 ......T..-.T-----...........----................----------------........---------................... 4093_CDC .........-..-----...........----................----------------........---------................... 33cdc .........-..-----...........----................----------------........---------................... 101 .........-..-----...........----................----------------........---------................... 103 .........-..-----...........----................----------------........---------................... 104 .........-..-----...........----................----------------........---------................... 105 .........-..-----...........----................----------------........---------...................

143
109 .........-..-----...........----................----------------........---------................... z49256 ......T..G.A-----.G.A.......----................----------------........---------................... 02B ......T..G.A-----.G.A.......----................----------------........---------................... 03B ......T..G.A-----.G.A.......----................----------------........---------................... 04B/A ......T..G.A-----.G.A.......----................----------------........---------................... 05B ......T..G.A-----.G.A.......----................----------------........---------................... 06B ......T..G.A-----.G.A.......----................----------------........---------................... 09B/A ......T..G.A-----.G.A.......----................----------------........---------................... 11B ......T..G.A-----.G.A.......----................----------------........---------................... 14B/A ......T..G.A-----.G.A.......----................----------------........---------................... 17B/A ......T..G.A-----.G.A.......----................----------------........---------................... 19B/1 ......T..G.A-----.G.A.......----................----------------........---------................... 20B ......T..G.A-----.G.A.......----................----------------........---------................... 22B/2 ......T..G.A-----.G.A.......----................----------------........---------................... 26B ......T..G.A-----.G.A.......----................----------------........---------................... 27B ......T..G.A-----.G.A.......----................----------------........---------................... 30B ......T..G.A-----.G.A.......----................----------------........---------................... 210 220 230 240 250 260 270 280 290 300 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 107 ......T..G.A-----.G.A.......----................----------------........---------................... P2 ......T..G.A-----.G.A.......----................----------------........---------................... 03C ......T..G.A-----.G.A.......----................----------------........---------................... 9893 ......T..G.A-----.G.A.......----................----------------........---------................... 9891 ......T..G.A-----.G.A.......----................----------------........---------................... 201 ......T..G.A-----.G.A.......----................----------------........---------................... 200 ......T..G.A-----.G.A.......----................----------------........---------................... 6553 ......T..G.A-----.G.A.......----................----------------........---------................... 3450 ......T..G.A-----.G.A.......----................----------------........---------................... 3162 ......T..G.A-----.G.A.......----................----------------........---------................... 3449 ......T..G.A-----.G.A.......----................----------------........---------................... 9343 ......T..G.A-----.G.A.......----................----------------........---------................... 3182/1 ......T..G.A-----.G.A.......----................----------------........---------................... 3182/3 ......T..G.A-----.G.A.......----................----------------........---------................... 3182/4 ......T..G.A-----.G.A.......----................----------------........---------................... AF149906 ...C.CTC.TCA-----CGG.G.G..C.----..-..C......G.A.T---------------..A.A...---------...GT.T............ 106B ...C..TC.TAA-----CGGAG.G..C.----..-..C...C..G.A.T---------------..ACA...---------...G............... AF149907 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 01B ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 18B/1 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 18B/3 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 18B/4 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............

144
18B/5 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 18B/6 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 18B/7 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 18B/8 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 17B/2 ..TT.G...CA.TTG-TGG..TGAA...----...A..T..C.A....----------------..AAGT..---------G.AT..T............ 17B/3 ..TT.G...CATTCG-TGG..TGAA...----...A..T..C.A....----------------..AAGT..---------G.AT..T............ 17B/4 ..TT.G...CA.TTG-TGG..TGAA...----...A..T..C.A....----------------..AAGT..---------G.AT..T............ 17B/5 ..TT.G...CA.TTG-TGG..TGAA...----...A..T..C.A....----------------..AAGT..---------G.AT..T............ 17B/6 ..TT.G...CA.TTG-TGG..TGAA...----...A..T..C.A....----------------..AAGT..---------G.AT..T............ 17B/7 ..TT.G...CATTCG-TGG..TGAA...----...A..T..C.A....----------------..AAGT..---------G.AT..T............ 19B/2 ..TT.G...CAATCATTGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 19B/4 ..TT.G...CAATCATTGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 19B/5 ..TT.G...CAATTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 19B/6 ..TT.G...CAT----TGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T.G.......... 19B/7 ..TT.G...CAATTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 100 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 3182/2 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 3182/5 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 3182/6 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 210 220 230 240 250 260 270 280 290 300 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 3182/7 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 3182/8 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 3684 ..TT.G...CATTTAATGG..TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 204 ..TT.G...TATT-AAT....TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 204/A ..TT.G...TATT-AAT....TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 204/B ..TT.G...TATT-AAT....TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 204/D ..TT.G...TATT-AAT....TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 204/E ..TT.G...TATT-AAT....TGAA.G.----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 204/F ..TT.G...TATT-AAT....TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 204/G ..TT.G...TATT-AAT....TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ 204/I ..TT.G...TATT-AAT....TGAA...----...G..T..C.A....----------------..AAGT..---------G.AT..T............ AF149915 GAGTG.A..CGA-TGAGG-GCGGGG..TGCTTC.CGT.TCC....CA.ATTACCACTTTTTTTT..A.G...GTGGAATAT......A..G......... 3977 GAGTG.A..CGA-TGAGG-GCGGGG..TGCTTC.CGT.TCC....CA.ATTACCACTTTTTTT-..A.G...GTGGAATAT......A..G......... 04B GAGTG.A..CGA-TGAGG-GCGGGG..TGCTTC.CGT.TCC....CA.ATTACCACTTTTTTTT..A.G...GTGGAATAT......A..G......... AF149914 GAGTA.A..C.ACTGAGG.G.GGAG..CCCCC..GGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 15B GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 15B/1 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 15B/2 GAGCA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 15B/3 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 15B/4 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G.........
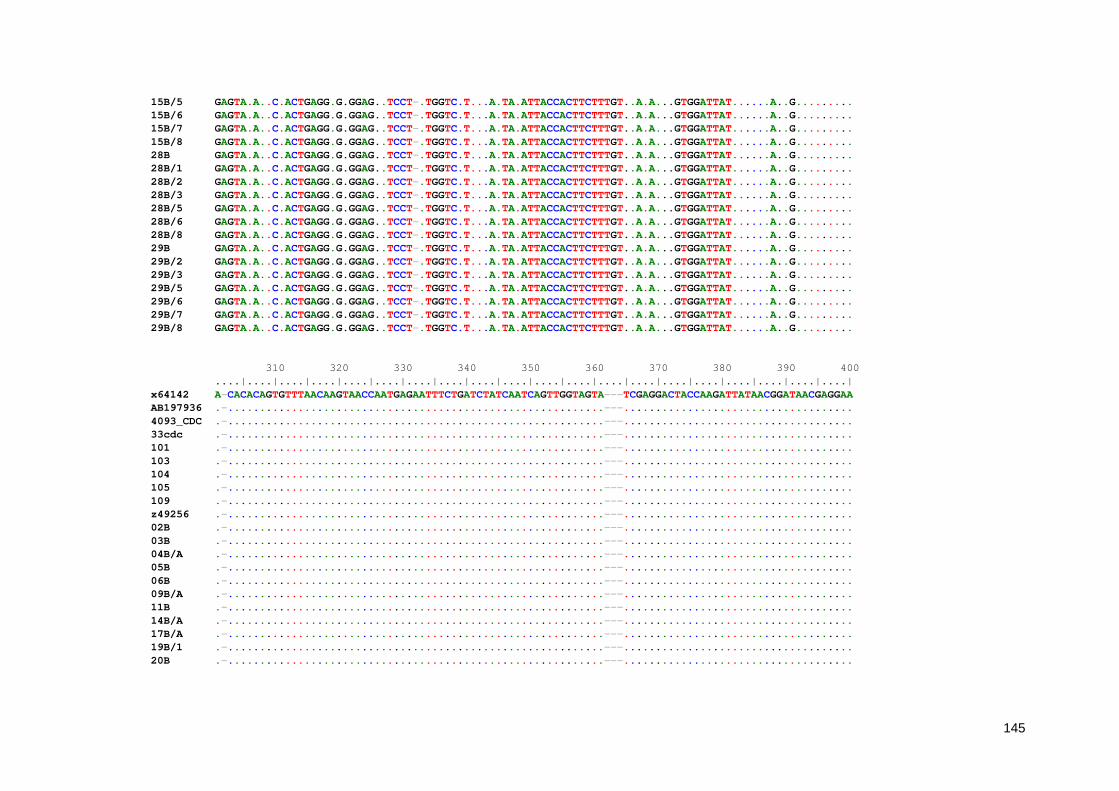
145
15B/5 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 15B/6 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 15B/7 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 15B/8 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 28B GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 28B/1 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 28B/2 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 28B/3 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 28B/5 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 28B/6 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 28B/8 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 29B GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 29B/2 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 29B/3 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 29B/5 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 29B/6 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 29B/7 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 29B/8 GAGTA.A..C.ACTGAGG.G.GGAG..TCCT-.TGGTC.T...A.TA.ATTACCACTTCTTTGT..A.A...GTGGATTAT......A..G......... 310 320 330 340 350 360 370 380 390 400 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| x64142 A-CACACAGTGTTTAACAAGTAACCAATGAGAATTTCTGATCTATCAATCAGTTGGTAGTA---TCGAGGACTACCAAGATTATAACGGATAACGAGGAA AB197936 .-...........................................................---.................................... 4093_CDC .-...........................................................---.................................... 33cdc .-...........................................................---.................................... 101 .-...........................................................---.................................... 103 .-...........................................................---.................................... 104 .-...........................................................---.................................... 105 .-...........................................................---.................................... 109 .-...........................................................---.................................... z49256 .-...........................................................---.................................... 02B .-...........................................................---.................................... 03B .-...........................................................---.................................... 04B/A .-...........................................................---.................................... 05B .-...........................................................---.................................... 06B .-...........................................................---.................................... 09B/A .-...........................................................---.................................... 11B .-...........................................................---.................................... 14B/A .-...........................................................---.................................... 17B/A .-...........................................................---.................................... 19B/1 .-...........................................................---.................................... 20B .-...........................................................---....................................

146
22B/2 .-...........................................................---.................................... 26B .-...........................................................---.................................... 27B .-...........................................................---.................................... 30B .-...........................................................---.................................... 107 .-...........................................................---.................................... P2 .-...........................................................---.................................... 03C .-...........................................................---.................................... 9893 .-...........................................................---.................................... 9891 .-...........................................................---.................................... 201 .-...........................................................---.................................... 200 .-...........................................................---.................................... 6553 .-...........................................................---.................................... 3450 .-...........................................................---.................................... 3162 .-...........................................................---.................................... 3449 .-...........................................................---.................................... 9343 .-...........................................................---.................................... 3182/1 .-...........................................................---.................................... 3182/3 .-...........................................................---.................................... 3182/4 .-...........................................................---.................................... AF149906 G-...............................................TT..........---.................................... 310 320 330 340 350 360 370 380 390 400 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 106B G-...............................................TT..........---.................................... AF149907 .-T.-G.GA.A............T.G........A.............CT...........---...............G.................... 01B .-T.-GTGA.A............T.G........A.............CT...........---...............G.................... 18B/1 .-T.-G..A.A............T.G........A.............CT...........---...............G.................... 18B/3 .-T.-G.GA.A...G........T.G........A.............CT...........---...............G.................... 18B/4 .-T.-G..A.A............T.G........A.............CT...........---...............G.................... 18B/5 .-T.-G..A.A............T.G........A.............CT...........---...............G.................... 18B/6 .-T.-G..A.A............T.G........A.............CT...........---...............G.................... 18B/7 .-T.-G.GA.A............T.G........A.............CT...........---...............G.................... 18B/8 .-T.-G..A.A............T.G........A.............CT...........---...............G.................... 17B/2 .-T.-GA.A.A............T.G........A.............CT...........---...............G.................... 17B/3 .-T.-GA.A.A............T.G........A.............CT...........---...............G.................... 17B/4 .-T.-GA.A.A............T.G........A.............CT...........---...............G.................... 17B/5 .-T.-GA.A.A............T.G........A.............CT...........---...............G.................... 17B/6 .-T.-GA.A.A............T.G........A.............CT...........---...............G.................... 17B/7 .-T.-GA.A.A............T.G........A.............CT...........---...............G.................... 19B/2 .-T.-G..A.A............T.G........A.............CT...........---...............G.................... 19B/4 .-T.-G..A.A............T.G........A.............CT...........---...............G.................... 19B/5 .-T.-G..A.A...G........T.G........A.............CT...........---...............G....................
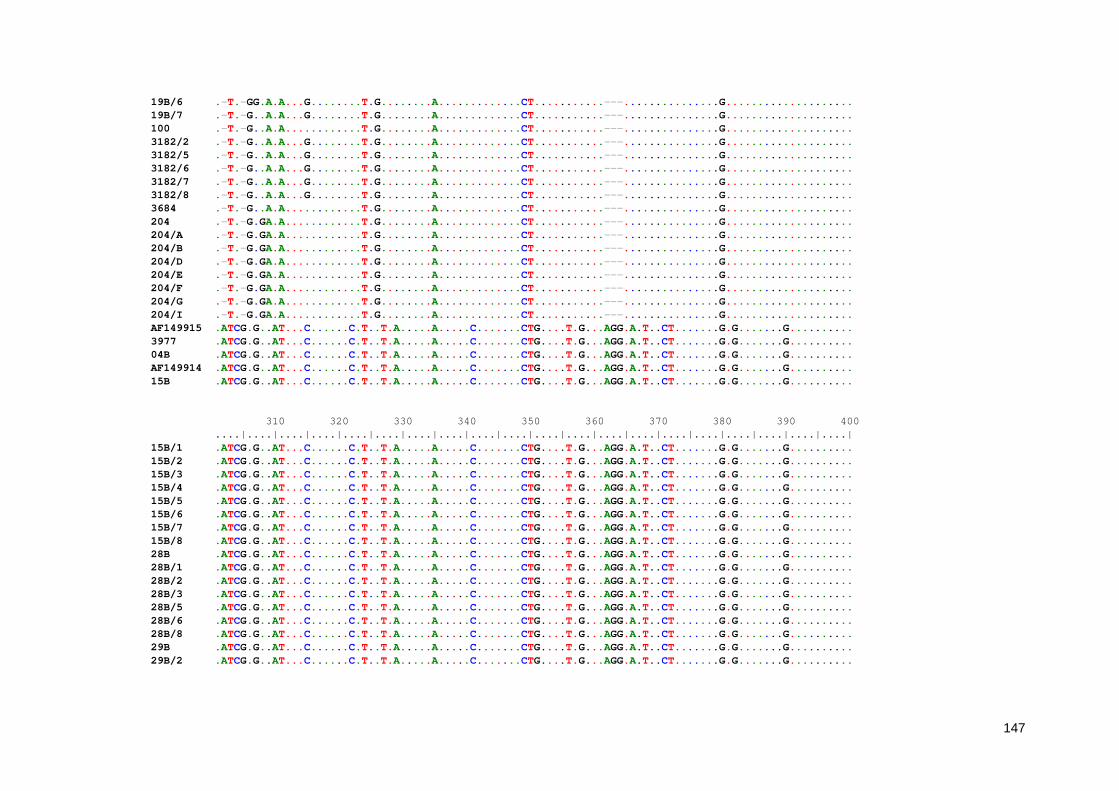
147
19B/6 .-T.-GG.A.A...G........T.G........A.............CT...........---...............G.................... 19B/7 .-T.-G..A.A...G........T.G........A.............CT...........---...............G.................... 100 .-T.-G..A.A............T.G........A.............CT...........---...............G.................... 3182/2 .-T.-G..A.A...G........T.G........A.............CT...........---...............G.................... 3182/5 .-T.-G..A.A...G........T.G........A.............CT...........---...............G.................... 3182/6 .-T.-G..A.A...G........T.G........A.............CT...........---...............G.................... 3182/7 .-T.-G..A.A...G........T.G........A.............CT...........---...............G.................... 3182/8 .-T.-G..A.A...G........T.G........A.............CT...........---...............G.................... 3684 .-T.-G..A.A............T.G........A.............CT...........---...............G.................... 204 .-T.-G.GA.A............T.G........A.............CT...........---...............G.................... 204/A .-T.-G.GA.A............T.G........A.............CT...........---...............G.................... 204/B .-T.-G.GA.A............T.G........A.............CT...........---...............G.................... 204/D .-T.-G.GA.A............T.G........A.............CT...........---...............G.................... 204/E .-T.-G.GA.A............T.G........A.............CT...........---...............G.................... 204/F .-T.-G.GA.A............T.G........A.............CT...........---...............G.................... 204/G .-T.-G.GA.A............T.G........A.............CT...........---...............G.................... 204/I .-T.-G.GA.A............T.G........A.............CT...........---...............G.................... AF149915 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 3977 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 04B .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... AF149914 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 15B .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 310 320 330 340 350 360 370 380 390 400 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 15B/1 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 15B/2 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 15B/3 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 15B/4 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 15B/5 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 15B/6 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 15B/7 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 15B/8 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 28B .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 28B/1 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 28B/2 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 28B/3 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 28B/5 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 28B/6 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 28B/8 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 29B .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 29B/2 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G..........

148
29B/3 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 29B/5 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 29B/6 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 29B/7 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 29B/8 .ATCG.G..AT...C......C.T..T.A.....A.....C.......CTG....T.G...AGG.A.T..CT.......G.G.......G.......... 410 420 430 440 450 460 470 480 490 500 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| x64142 TTGGGGTTCGACATCGGAGAGGGAGCTTTACAGATGGCTACCACTTCTAAGGAAGGCAGCAGGCSCGTAAATTACCCACTTTCGA---ATTGAAGAGGTA AB197936 .....................................................................................---............ 4093_CDC .....................................................................................---............ 33cdc .....................................................................................---............ 101 .....................................................................................---............ 103 .....................................................................................---............ 104 .....................................................................................---............ 105 .....................................................................................---............ 109 .....................................................................................---............ z49256 .....................................................................................---............ 02B .....................................................................................---............ 03B .....................................................................................---............ 04B/A .....................................................................................---............ 05B .....................................................................................---............ 06B .....................................................................................---............ 410 420 430 440 450 460 470 480 490 500 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 09B/A .....................................................................................---............ 11B .....................................................................................---............ 14B/A .....................................................................................---............ 17B/A .....................................................................................---............ 19B/1 .....................................................................................---............ 20B .....................................................................................---............ 22B/2 .....................................................................................---............ 26B .....................................................................................---............ 27B .....................................................................................---............ 30B .....................................................................................---............ 107 .....................................................................................---............ P2 .....................................................................................---............ 03C .....................................................................................---............ 9893 .....................................................................................---............ 9891 .....................................................................................---............

149
201 .....................................................................................---............ 200 .....................................................................................---............ 6553 .....................................................................................---............ 3450 .....................................................................................---............ 3162 .....................................................................................---............ 3449 .....................................................................................---............ 9343 .....................................................................................---............ 3182/1 .....................................................................................---............ 3182/3 .....................................................................................---............ 3182/4 .....................................................................................---............ AF149906 .................................................C...................................---CG.......... 106B .................................................C...................................---CG.......... AF149907 ..A.....T....................CA................C..............................A....A.CAG..A......... 01B ..A.....T....................CA................C..............................A....A.CAG..A......... 18B/1 ..A.....T....................CA................C..............................A....A.CAG..A......... 18B/3 ..A.....T....................CA................C..............................A....A.CAG..A......... 18B/4 ..A.....T....................CA................C..............................A....A.CAG..A......... 18B/5 ..A.....T....................CA................C..............................A....A.CAG..A......... 18B/6 ..A.....T....................CA................C..............................A....A.CAG..A......... 18B/7 ..A.....T....................CA................C..............................A....A.CAG..A......... 18B/8 ..A.....T....................CA................C..............................A....A.CAG..A......... 17B/2 ..A.....T....................CA................C..............................A....A.CAG..A......... 17B/3 ..A.....T....................CA................C..............................A....A.CAG..A......... 17B/4 ..A.....T....................CA................C..............................A....A.CAG..A......... 17B/5 ..A.....T....................CA................C..............................A....A.CAG..A......... 17B/6 ..A.....T....................CA................C..............................A....A.CAG..A......... 17B/7 ..A.....T....................CA................C..............................A....A.CAG..A......... 410 420 430 440 450 460 470 480 490 500 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 19B/2 ..A.....T....................CA................C..............................A....A.CAG..A......... 19B/4 ..A.....T....................CA................C..............................A....A.CAG..A......... 19B/5 ..A.....T....................CA................C..............................A....A.CAG..A......... 19B/6 ..A.....T....................CA................C..............................A....A.CAG..A......... 19B/7 ..A.....T....................CA................C..............................A....A.CAG..A......... 100 ..A.....T....................CA................C..............................A....A.CAG..A......... 3182/2 ..A.....T....................CA................C..............................A....A.CAG..A......... 3182/5 ..A.....T....................CA................C..............................A....A.CAG..A......... 3182/6 ..A.....T....................CA................C..............................A....A.CAG..A......... 3182/7 ..A.....T....................CA................C..............................A....A.CAG..A......... 3182/8 ..A.....T....................CA................C..............................A....A.CAG..A......... 3684 ..A.....T....................CA................C..............................A....A.CAG..A.........

150
204 ..A.....T....................CA................C..............................A....A.CAG..A......... 204/A ..A.....T....................CA................C..............................A....A.CAG..A......... 204/B ..A.....T....................CA................C..............................A....A.CAG..A......... 204/D ..A.....T....................CA................C..............................A....A.CAG..A......... 204/E ..A.....T....................CA................C..............................A....A.CAG..A......... 204/F ..A.....T....................CA................C..................A...........A....A.CAG..A......... 204/G ..A.....T....................CA................C..............................A....A.CAG..A......... 204/I ..A.....T....................CA................C..............................A....A.CAG..A......... AF149915 .AA...........................G....................................A..........A.C.TA.CAA..GA.G...... 3977 .AA...........................G....................................A..........A.C.TA.CAA..GA.G...... 04B .AA...........................G....................................A..........A.C.TA.CAA..GA.G...... AF149914 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 15B .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 15B/1 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 15B/2 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 15B/3 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 15B/4 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 15B/5 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 15B/6 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 15B/7 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 15B/8 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 28B .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 28B/1 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 28B/2 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 28B/3 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 28B/5 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 28B/6 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 28B/8 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 29B .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 29B/2 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 410 420 430 440 450 460 470 480 490 500 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 29B/3 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 29B/5 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 29B/6 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 29B/7 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G...... 29B/8 .AA...........................G.A..................................A..........A.C.TA.CAAC.GA.G......

151
510 520 530 540 550 560 570 580 590 600 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| x64142 GTGACGACACATAACTCTAGAGTTGAGTAAAATCAATTCTTGAAGGAATGAGTAGGAGGTAAATTCTCCTACGAAATCAATTGGAGGGCAAGTCTGGTGC AB197936 .................................................................................................... 4093_CDC .................................................................................................... 33cdc .................................................................................................... 101 .................................................................................................... 103 .................................................................................................... 104 .................................................................................................... 105 .................................................................................................... 109 .................................................................................................... z49256 .................................................................................................... 02B .................................................................................................... 03B .................................................................................................... 04B/A .................................................................................................... 05B .................................................................................................... 06B .................................................................................................... 09B/A .................................................................................................... 11B .................................................................................................... 14B/A .................................................................................................... 17B/A .................................................................................................... 19B/1 .................................................................................................... 20B .................................................................................................... 22B/2 .................................................................................................... 26B .................................................................................................... 27B .................................................................................................... 30B .................................................................................................... 107 .................................................................................................... P2 .................................................................................................... 03C .................................................................................................... 9893 .................................................................................................... 9891 .................................................................................................... 510 520 530 540 550 560 570 580 590 600 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 201 .................................................................................................... 200 .................................................................................................... 6553 .................................................................................................... 3450 .................................................................................................... 3162 .................................................................................................... 3449 .................................................................................................... 9343 ....................................................................................................

152
3182/1 .................................................................................................... 3182/3 .................................................................................................... 3182/4 .................................................................................................... AF149906 .........A........C...G..GT...CTC..C..................A..A......A...T............................... 106B .........A........C...G..GT.T.C--..C..................A..A.....A....T...A........................... AF149907 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 01B .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 18B/1 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 18B/3 .........A.......AC..CA..C....G--T.TG.................A..A.C...GAG..T............................... 18B/4 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 18B/5 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 18B/6 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 18B/7 .........A.......AC..CA..C....G--T.TG.................A..A.C...GAG..T............................... 18B/8 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 17B/2 .........A.......AC..TG..C....G--..TA.................A..A.C...GAG..T............................... 17B/3 .........A.......AC..TG..C....G--..TA.................A..A.C...GAG..T..............----------------- 17B/4 .........A.......AC..TG..C....G--..TA....A............A..A.C...GAG..T............................... 17B/5 .........A.......AC..TG..C....G--..TA....A............A..A.C...GAG..T............................... 17B/6 .........A.......AC..TG..C....G--..TA.................A..A.C...GAG..T............................... 17B/7 .........A.......AC..TG..C....G--..TA.................A..A.C...GAG..T............................... 19B/2 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 19B/4 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 19B/5 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 19B/6 .........A.......AC..CA..C....G--T.TG.................A..A.C...GAG..T............................... 19B/7 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 100 .........A.......AC..CA..C....G--T.TG.................A..A.C...GAG..T............................... 3182/2 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 3182/5 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 3182/6 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 3182/7 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG.T................................ 3182/8 .........A.......AC..CA..C....G--..TG.................A..A.C...GAG..T............................... 3684 .........A.......AC..CA..T....G--..TG.................A..A.C...G------------------------------------ 204 .........A.......AC..TG..C....G--..TG.................A..A.C...GAG..T............................... 204/A .........A.......AC..TG..C....G--..TG.................A..A.C...GAG..T............................... 510 520 530 540 550 560 570 580 590 600 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....|....|....|....|....|.. ..|....| 204/B .........A.......AC..TG..C....G--..TG.................A..A.C...GAG..T............................... 204/D .........A.......AC..TG..C....G--..TG.................A..A.C...GAG..T............................... 204/E .........A.......AC..TG..C....G--..TG.................A..A.C...GAG..T............................... 204/F .........A.......AC..TG..C....G--..TG.................A..A.C...GAG..T............................... 204/G .........A.......AC..TG..C....G--..TG.................A..A.C...GAG..T...............................

153
204/I .........A.......AC..TG..C....G--..TG.................A..A.C...GAG..T............................... AF149915 .........AT....G...TCC.C.TC.TTTGG.G.GGA.C.TC.......T.TC..TT....A.A..GA.A....G....................... 3977 .........AT....G...TCC.C.TC.TTTGG.G.GGA.C.TC.......T.TC..TT....A.A..GA.A....G....................... 04B .........AT....G...TCC.C.TC.TTTGG.G.GGA.C.TC.......T.TC..TT....A.A..GA.A....G....................... AF149914 .........AT....G..TTCCC.CGC.TT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 15B .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 15B/1 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 15B/2 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G....----------------- 15B/3 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 15B/4 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 15B/5 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 15B/6 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 15B/7 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 15B/8 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 28B .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 28B/1 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G......A.............. 28B/2 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 28B/3 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 28B/5 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 28B/6 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 28B/8 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 29B .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..-------------------------------- 29B/2 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 29B/3 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 29B/5 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 29B/6 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 29B/7 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 29B/8 .........AT....G..TTCCC.CGCATT.CGTG.GGA...CC.......TC.C..CA....CCA..G.GA....G.G..................... 610 620 630 640 650 660 670 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....| x64142 CAGCAGCCGCGGTAATTCCAGCTCCAATAGTGTATATTAAAGTTGCTGTGATTAAAACGCTCGTAGTTG- AB197936 .....................................................................- 4093_CDC .....................................................................- 33cdc ....................................................................-- 101 .....................................................................- 103 ................................................................------ 104 .....................................................................- 105 .................................................................----- 109 ..................................................................---- z49256 .....................................................................- 02B ....................................................................--

154
03B .........................--------------------------------------------- 04B/A .....................................................................- 05B .....................................................................- 06B .....................................................................- 09B/A .....................................................................- 11B .....................................................................- 14B/A .....................................................................- 17B/A ...............................--------------------------------------- 19B/1 .....................................................................- 20B .....................................................................- 22B/2 .....................................................................- 26B .....................................--------------------------------- 27B ...........................------------------------------------------- 30B .....................................................................- 107 .....................................................................- P2 .....................................................................- 03C .....................................................................A 9893 .....................................................................- 9891 .....................................................................- 201 .....................................................................- 200 .....................................................................- 6553 .....................................................................- 3450 .....................................................................- 3162 .....................................................................- 3449 .....................................................................- 9343 .....................................................................- 3182/1 .............................................T............------------ 3182/3 ............................................................---------- 3182/4 ...........................................................----------- AF149906 .....................................................................- 106B .....................................................................- 610 620 630 640 650 660 670 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....| AF149907 .............................................T.......................- 01B .............................................T.......................- 18B/1 .............................................T.......................- 18B/3 .............................................T.......................- 18B/4 .............................................T.......................- 18B/5 .............................................T.......................- 18B/6 .............................................T.......................- 18B/7 .............................................T......................--

155
18B/8 .............................................T..................------ 17B/2 .............................................------------------------- 17B/3 ---------------------------------------------------------------------- 17B/4 .............................................------------------------- 17B/5 .............................................T.......................- 17B/6 .............................................T.......................- 17B/7 .............................................T.......................- 19B/2 .............................................T.......................- 19B/4 .............................................T.......................- 19B/5 .............................................T.......................- 19B/6 .............................................T.......................- 19B/7 .............................................T.......................- 100 ................-......................------------------------------- 3182/2 .............................................T.......................- 3182/5 .............................................T.......................- 3182/6 .............................................T.......................- 3182/7 .............................................T................-------- 3182/8 .............................................T................-------- 3684 ---------------------------------------------------------------------- 204 .............................................T.......................- 204/A .............................................T.......................- 204/B .............................................T.......................- 204/D .............................................T.......................- 204/E .............................................T.......................- 204/F .............................................T.......................- 204/G .............................................T.......................- 204/I .............................................T.......................- AF149915 ...................................TG........T.......................- 3977 ...................................TG........T.......................- 04B ...................................TG........T.......................- AF149914 ...................................TG........T.......................- 15B ...................................TG........T.......................- 15B/1 ...................................TG........T.......................- 15B/2 ---------------------------------------------------------------------- 610 620 630 640 650 660 670 ....|....|....|....|....|....|....|....|. ...|....|....|....|....|....| 15B/3 ...................................TG........T.......................- 15B/4 ...................................TG........T.......................- 15B/5 ...................................TG........T......................-- 15B/6 ...................................TG........T......................-- 15B/7 ...................................TG........T.......................-

156
15B/8 ...................................TG........T.......................- 28B ...................................TG........T.................------- 28B/1 ...................................TG........T.......................- 28B/2 ...................................TG........T.......................- 28B/3 ...................................TG........T.......................- 28B/5 ............................------------------------------------------ 28B/6 ...................................----------------------------------- 28B/8 ...................................TG........T.......................- 29B ---------------------------------------------------------------------- 29B/2 ...................................TG........T.......................- 29B/3 ...................................TG........T.......................- 29B/5 ...................................TG........T.......................- 29B/6 ...................................TG........T.......................- 29B/7 ...................................TG........T.......................- 29B/8 ...................................TG........T.......................-

157
ANEXO 8 SANTOS, H.L.; BANDEA, R.; MARTINS, L.A. et al.
Differential identification of Entamoeba spp. based on the analysis of 18S rRNA. Parasitol Res 106(4): 883-8, 2010

158

159

160

161

162

163

164
ANEXO 9
OLIVEIRA, L.M.; SANTOS, H.L.C.; GONÇALVES, M.M.L.; BARRETO, G.M.; PERALTA, J. M. 2010. Evaluation of P CR
as an alternative tool for the diagnosis of low-int ensity Schistosoma mansoni infection. Diagn Microbiol and Infect
Disease, 2010

165
Evaluation of PCR as an additional alternative tool for the diagnosis
of low-intensity Schistosoma mansoni infection
Laura Maria Abreu Oliveira, Helena Lucia Carneiro Santos, Margareth Maria Lessa Gonçalves, Magali Gonçalves Muniz. Barreto*, José Mauro Peralta.
Departamento de Imunologia, Instituto de Microbiologia, Universidade Federal do Rio de Janeiro, Brazil
*Laboratório de Avaliação e Promoção da Saúde Ambiental, Instituto Oswaldo Cruz, Fiocruz, Brazil
Av. Brasil 4365, Pavilhão Lauro Travasso, Rio de Janeiro Cep. 21040350, Tel (fax): 25606474 Running: PCR for schistosomiasis diagnosis Corresponding author: José Mauro Peralta, Departamento de Imunologia, Instituto de Microbiologia Professor Paulo de Góes – CCS, Universidade Federal do Rio de Janeiro. 21941590 Rio de Janeiro, Brazil. [email protected]
ABSTRACT
The aim of the present study was to evaluate polimerase chain reaction (PCR) as an additional tool for diagnosing schistosomiasis in individuals with low-level parasite burden from areas of low endemicity or under occasional risk of infection by Schistosoma mansoni. A total of 102 samples were tested in this study using two PCR assays utilizing distinct primer pairs. One of the primer pairs was targeted to a highly repeated 121-bp sequence of S. mansoni, and the other was targeted to Schistosoma 28S rDNA. The samples were divided into five groups according to parasite burden of the individual as follows: 16 individuals with schistosomiasis excreting less than 10 eggs per gram of feces (epg); 18 individuals excreting higher than 10 epg; 22 individuals with reactive ELISA-IgG against S. mansoni soluble membrane antigen and negative coproscopy; 25 individuals with other intestinal parasites and; 21 individuals with negative parasitological examination. The results obtained with stool samples from individuals with schistosomiasis showed a high sensitivity for PCR as S. mansoni DNA was detected in 91% (31/34) of the samples analyzed. No amplification was observed in three stool samples from individuals excreting below 10 epg. The specificity of the test for both pairs of primers was 100%. In the group of seropositive individuals, S. mansoni DNA was detected in 59% (13/22) of fecal samples, corroborating the serological results. Overall, PCR can be an important tool for detecting S. mansoni infection in individuals excreting few eggs in feces. Moreover, the determination of the infection through the detection of S. mansoni DNA in stool samples from seropositive individuals represents a new means of confirming the results of ELISA-IgG for schistosomiasis. Therefore, studies in this direc-tion should be encouraged and extended.
Keywords: Schistosoma mansoni, schistosomiasis, low parasitic load, diagnosis, PCR.

166
1 Introduction
Schistosomiasis remains an important world health problem, especially in developing
countries, even after the strategy for morbidity control through treatment programs
and preventive measures. In Brazil, Schistosoma mansoni is the only causative
species of schistosomiasis, and morbidity control through chemotherapy has been
successful. In areas where control programs have been applied, the epidemiological
features of schistosomiasis, intensity of infection and morbidity have profoundly
changed. In this respect, an epidemiological situation with a low prevalence and
intensity of infection has reduced the value of traditional diagnostics of
schistosomiasis (KATO et al., 1972) However, the sensitivity of this technique
decreases, particularly in areas where individuals present low-levels of parasitic
burden, mild, absent morbidity, or and have been previously treated. Thus, a
considerable number of schistosomiasis patients can be misdiagnosed. The Kato-
Katz method has also been used as standard for the evaluation of other methods for
diagnosis of schistosomiasis. Low sensitivity in stool examination in individuals with
low infection intensity and doubts about the estimation of cure after therapeutic
intervention affect the development of new tests. How do we diagnose
schistosomiasis in individuals with low intensity infections? New approaches for
improving diagnosis are needed. Antibody-based assays generally have high
sensitivities; however, serology-based analyses are unable to discriminate between
active and past S. mansoni infections. Methods to determine circulating antigens
levels, such as CAA (circulating anodic antigen) and CCA (circulating cathodic
antigen) by capture-ELISA are the most accurate immunodiagnostic test for active S.
mansoni infection. However, these show a similar sensitivity to stool examination for
low infection intensity (DOENHOFF et al., 2004). On the other hand, polimerase
chain reaction (PCR)-based methods have shown high sensitivity and specificity for
detection of parasitic DNA. Recently, investigators have reported the successful
application of PCR to the diagnosis of schistosomiasis. The usefulness of PCR for
the detection of S. mansoni DNA in human feces was first described by Pontes et al.
who used artificial mixtures of eggs with stool that were later evaluated using clinical
samples of patients living in endemic areas (PONTES et al., 2002; 2003). These
authors designed a set of primers to target a tandem repeat sequence described by
Hamburger et al. (1991) that was used to monitor the S. mansoni infestation of water

167
and stool samples (HAMBURGER et al., 1998). Another PCR method based on the
specific amplification of a sequence from the 28S rDNA yielded strong results when
applied to human urine samples, showing diagnostic promise (SANDOVAL et al.,
2006b). Recently, Allam et al. (2009) described a PCR reaction using stool samples
in a region of low-intensity S. mansoni infections. Their work compared the Percoll
technique with PCR and Kato-Katz techniques. In this first moment, we applied
conventional PCR using previously published primers with highly sensitive and
specific detection of S. mansoni (PONTES et al., 2002; SANDOVAL et al., 2006 a) in
regions with infected individuals with low-level parasitic burden. The aim of this study
was to evaluate PCR methods in a region with low intensity infections, taking into
account individuals with serologically-positive samples but negative stool samples.
The PCR approach may constitute an accurate procedure to assess the specificity of
serology and it can be used as an additional test for S. mansoni infection diagnosis.
2 Material and Methods
2.1 Control samples
S. mansoni eggs were extracted from the liver of infected mice. The livers were
minced and homogenized with saline solution in a blender. The liver homogenate
was washed with phosphate-buffered saline solution (PBS) by successive
centrifugation to clarify the supernatant. Subsequently, the egg-containing pellet was
poured in PBS into Petri dishes. Each egg was observed and collected in Petri dishes
using a magnifying glass. A single stool sample, obtained from a healthy donor from
non-endemic area, was collected and aliquots were stored at –20°C. Each aliquot
containing 0.2 gram of stool was spiked with 1, 3, 5, 10 or 25 S. mansoni eggs.
Control samples were prepared by adding eggs in the quantities given to aliquots
containing 100 µl of PBS.
2.2 Fecal Samples
For evaluation of PCR, 102 stool samples obtained from 46 females and 56 males
living in Sumidouro (Rio de Janeiro, Brazil) were collected. Sumidouro is a village
considered a low endemic area because the population has undergone mass
treatment for schistosomiasis. All participants gave their informed consent and the
study was approved by the Ethical Review Board of the Oswaldo Cruz Institute/

168
Fiocruz (No. CAAE-0038.0.011.00-0). Three fresh stool specimens from each
individual were collected in special containers on three consecutive days. Stool
samples were kept frozen at -20oC until DNA extraction. The Kato-Katz method was
used to quantify the egg number in the stools. The number of eggs per gram of feces
(EPG) was calculated through the sum of the S. mansoni eggs found in each slide
prepared by the Kato-Katz method. This number was divided by total number of
slides examined for each person. The result was multiplied by 24 (Katz et al., 1972).
All of the values found in each slide, including zeros, were combined for the sum.
The samples were divided into four groups according to the EPG, as follows:
Group 1 – 16 stool samples from infected individuals excreting below 10 S. mansoni
EPG.
Group 2 – 18 stool samples from infected individuals excreting above 10 S. mansoni
10 EPG.
Group 3 – 22 stool samples from individuals with positive IgG-ELISA serology but
negative results for the detection of S. mansoni eggs in feces.
Group 4 – 46 stool samples from individuals with negative coproscopy and serology
results for S. mansoni infection. Among them, 25 had other intestinal parasites in
their feces including Strongyloides stercoralis (1), Hymenolepis diminuta (2),
hookworms (3), Giardia intestinalis (3), Entamoeba histolytica (4), Iodamoeba
butschlii (4), Entamoeba coli (9), Blastocystis hominis (10) and Endolimax nana (14).
These parasitoses were diagnosed in this study utilizing the technique Hoffman et al.
(1934), except for E. histolytica, which was differentiated from E. dispar by PCR
(SANTOS et al., 2007).
2.3 IgG-ELISA
Serum samples were tested for anti-schistosoma antibodies by IgG-ELISA. Serology
was performed according to the protocol described by Gonçalves et al. (2006). For
acceptable results, the cut-off value had to be in the 0.35-0.45 range.

169
2.4 DNA extraction
The fecal and egg DNA were obtained according protocols previously described by
Pitcher et al. (1989) and Silva et al. (1999), with slight modifications. Approximately 1
g of stool was homogenized in deionized water (1 ml). Suspensions of 300 µl of fecal
homogenate were then resuspended in 1 ml of phosphate buffered saline (PBS),
centrifuged twice at 16,000 × g for 5 min and stored at -20°C. Aliquots of 300 µl were
submitted to mechanical and chemical lysis steps in eppendorf tubes by adding 0,1 g
of glass beads (106 microns) and 0,5 ml of 5 M guanidine isothiocyanate (Promega
Corporation, USA). The tubes were vigorously agitated using a vortex and incubated
at room temperature for 10 minutes. The lysate was cooled on ice for 2 min and 0,25
ml of cold 7,5 M ammonium acetate was added. This mixture was kept on ice for a
further 10 min, and then 0,5 ml chloroform isoamyl-alcohol (24:1 v/v) was added. The
phases were mixed by inversion, centrifuged at 13,800 × g for 10 min and the
aqueous phases were transferred to new eppendorf tubes. Subsequently, 0,43 ml of
2-propanol and 1 µl of 1 µg/µl of glycogenic were added to DNA solution. After
centrifugation at 3,500 x g for 2 min, the sediment was washed five times with 70%
ethanol with successive centrifugation (4,000 x g for 20 min) and dried at 37°C for
24h. The dried sediment was suspended in 50 µl of Tris-EDTA buffer (10mM Tris-
HCL; 1mM EDTA pH 8.0) at 37°C for 1 hour and then s tored at –20°C.
2.5 Polymerase chain reaction (PCR)
Two protocols were followed with some modifications. The first PCR was carried out
according to the protocols detailed by Pontes et al. (2002). A 110-base pair (bp)
product was amplified for DNA sequences highly repeated in the genome of S.
mansoni, using primers (SMEEG1-5’GATCTAACCGACCAACC-3’ and SMEGG2 5’-
ATATTAACGCCCACGCTCTC). PCR reactions were performed in a final volume of
50 µl, containing 20 mM Tris-HCl pH 8.4; 50 mM KCl; 1.5 mM MgCl2; 10 pmoles of
each oligonucleotide primer; 250 µM of each deoxynucleoside triphosphate (dNTPs);
1.25 units of Taq platinum DNA polymerase (Invitrogen Life technologies, USA);
0.1% bovine serum albumin (BSA Sigma Chem. Co., USA) and 2 µl of DNA sample.
PCR cycling parameters consisted of 3 min at 94°C followed by 40 cycles of 30
seconds at 94°C, 30 seconds at 55°C, and 30 seconds at 72°C with a final extension

170
at 72oC for 7 min. The second protocol described by Sandoval et al. (2006a) was
designed for the amplification of the 28S rDNA region from S. mansoni, and was
applied using primers (SmF 5’–GAGATCAAGTGTGACAGTTTTGC-3’ and SmR
5’ACAGTGCGC GCGTCGTAAGC-3’), putatively giving species-specific PCR
products of 350 bp. PCR was performed in a final volume of 50 µL containing 20 mM
of Tris-HCl pH 8.4; 50 mM of KCl; 3 mM of MgCl2; 10 pmoles of each oligonucleotide
primers; 250 µM of each deoxynucleoside triphosphate (dNTPs); 1.25 units of Taq
platinum DNA polymerase (Invitrogen Life Technologies, USA); 0.1% of bovine
serum albumin (BSA Sigma Chem. Co., USA); 2 µl of DNA sample. PCR cycling
parameters consisted of 3 min at 94°C followed by 35 cycles of 30 seconds at 94°C,
20 seconds at 65°C, and 20 seconds at 72°C with a f inal extension at 72°C for 7 min.
In both procedures, the amplified products were analyzed by electrophoresis using
12% polyacrylamide gel after silver staining.
3 Results
In preliminary experiments, the two protocols amplifying different fragments of S.
mansoni using primers SMEEG1/SMEGG2 and SmF/SmR were standardized.
Evaluating the sensitivity of the PCR protocols required samples with known
concentrations of S. mansoni eggs; therefore, a set of standard samples was
prepared based on stool samples spiked with 1, 5, 10, 15 or 25 S. mansoni eggs to
ensure that the detection limit could be reached even for highly sensitive assays. All
these samples were tested and yielded positive results, regardless of the PCR
protocol that was used. The limit of the PCRs on S. mansoni DNA with
SMEEG1/SMEGG2 and SmF/SmR primer pair was one egg. The results were found
to be reproducible, with respect to either of the PCR protocols carried out on the
same DNA samples on different days (data not shown). The specificity of these PCR
tests was demonstrated by the absence of amplification when DNA from healthy
donors and individuals with other parasite infections were added as template for the
PCR reactions. A total of 102 samples were tested in this study using the two PCR
assays, and the results were summarized in Table 1. In these samples the outcome
varied depending on the primer pair used for amplification. Neither protocol amplified
all samples. The results obtained with stool samples from individuals with
schistosomiasis showed a high sensitivity for PCR. S. mansoni DNA was detected in

171
91% (31/34) of the samples analyzed. Amongst 16 infected individuals excreting <10
EPG (group 1), PCR results were positive for 13 (81%). From these, eight samples
were amplified with both PCR assays. Nevertheless, four samples were amplified by
PCR using the SmF/SmR primer pair, which had been negative in previous PCR
using the SMEEG1/SMGG2 primer pair, and one sample was amplified PCR using
the SMEEG1/SMGG2 primer pair, which was negative by another PCR assay. In this
group, both PCR protocols failed to amplify three samples. All 18 samples from
individuals excreting >10 EPG (group 2), were positive by PCR. From these, 14
samples were amplified with both PCRs. The PCR using the SmF/SmR primer pair
failed to amplify two samples, which were positive by another PCR assay. The PCR
using the SMEEG1 /SMEGG2 primers pair were negative in two samples that were
positive by another PCR assay. Out of 22 samples with S. mansoni IgG-ELISA-
positive serology and negative parasitological examination (group 3), 13 tested
positive by PCR. From these, six samples were amplified by both PCR assays,
whereas one sample was amplified using only the SMEEG1/SMGG2 primer pair, and
a further six samples were amplified using SmF/SmR primer pair. No amplification
was obtained in stool samples from healthy individuals or from those that presented
parasites other than S. mansoni (group 4). No internal inhibition control was included
to rule out the presence of inhibitors of DNA amplification. Such inhibitors may stick
to the DNA or inhibit DNA polymerase interfering with the efficiency of the
amplification process. Several methods have been adopted to monitor the action
inhibitors substances in the PCR procedure and to identify false negative results.
Therefore, we verified the action of the inhibitor by spiking the PCR reaction with 10
fg DNA control of S. mansoni. In all samples, amplification of the control DNA
occurred, showing that the previous negative results were not due to action of
inhibiting substances in the DNA sample.
4. Discussion and Conclusion
This work was based on the use of the PCR technique for the diagnosis of low-
intensity S. mansoni infections, including a group of individuals with serologically-
positive samples and S. mansoni egg-negative stool samples. Five groups of
individuals with different profiles of parasite burden were analyzed by to PCR. In
Brazil, some regions that present clusters of asymptomatic individuals harbor a minor

172
parasite burden due to the mass treatment for schistosomiasis in the population. In
this context, conventional parasitological examination has presented low sensitivity
(Gonçalves et al., 2006). PCR techniques have demonstrated their high potential as
a diagnostic tool of several intestinal parasitic diseases, including schistosomiasis
(GIBBONS-MATTHEWS & PRESCOTT, 2003; GONIN & TRUDEL, 2003; PONTES
et al., 2003; VERWEIJ et al., 2003; SUBRUNGRUANG et al., 2004; SANDOVAL et
al., 2006b, GOMES et al., 2006; TEN HOVE et al., 2008; ALLAM et al., 2009;
GOMES et al., 2009). PCR has been used for S. mansoni detection in areas with
medium and heavy infections (PONTES et al., 2003). Recently, Allam et al. (2009)
described a PCR reaction with good results when applied to stool samples from
areas of low intensity of S. mansoni infections. The author compared the efficiency of
the Percoll technique with PCR and Kato-Katz techniques. In our research,
individuals with S. mansoni-positive serology and negative stool examination were
also analyzed. This is the first application of PCR for the diagnosis of S. mansoni
infection in region with a low intensity of infection, taking into account serological
results. We evaluated two target regions of the S. mansoni DNA to improve diagnosis
of the disease. The most important finding in this study was that 59% of patients with
positive serological test and negative parasitological examination were PCR positive.
These data once again confirm the low sensitivity of stool examination for the
diagnosis of schistosomiasis in areas where there is low worm burden. Pairs of
primers from two regions of the S. mansoni DNA were selected according to the
results obtained by other authors. Hamburger et al. (1991) described an S. mansoni
sequence containing a 121-bp tandem repeat that was amplified by the
SMEGG1/SMEEG2 primers. The authors showed that PCR is a sensitive and
specific test for the identification of cercariae in water, which was further used by
Pontes et al. (2002; 2003), in detecting of S. mansoni DNA in human serum and in
the stool of individuals from an endemic area. The minimum detectable DNA quantity
was found to be 1 fg. The authors revealed that simple DNA extraction techniques
and rapid-step PCR allowed amplification of parasitic DNA in fecal samples with a
sensitivity equivalent to the detection of 2.4 EPG, which makes it 10 times more
sensitive than the Kato-Katz technique. Recently, modifications in this technique
were described (GOMES et al., 2009) in order to improve the sensitivity of the test.
Even after such modifications, the PCR missed two parasitological positive cases,
but showed a higher sensitivity (92.9%) than parasitological examination (65.1%) in a

173
population of 67 patients. Another pair of primers, SmF/SmR to the 28S rDNA region
of S. mansoni genome was described by Sandoval et al. (2006a). The authors
showed a sensitivity of 100% when PCR were used to analyze urine samples from
eight infected individuals (SANDOVAL et al., 2006b). Using the combination of the
results of these two pairs of primers, we found 100% sensitivity when stool samples
from patients that excreted more than 10 EPG were analyzed. However, 3 of 18
individuals with less than 10 EPG were PCR-negative for both pairs of primers. The
patients with schistosomiasis included in this study were only identified as positive by
the coproscopy analyses of three fecal samples from different days of collection
during one week. It must be considered that PCR was applied to a minimal part of the
fecal samples after it had been divided to prepare thick smears. We used one aliquot
from one of the three stool samples collected from the same person. We are not sure
if the stool sample that we analyzed was the parasitological positive one.
Investigation to demonstrate that possibility is ongoing. Sandoval et al. (2006b)
suggested that the failure of PCR in detecting patients with schistosomiasis
confirmed by coproscopy could be attributed to “individual specific” inhibitors present
in their samples. PCR sensitivity measured on S. mansoni DNA with the SmF and
SmR primer pairs was shown to be 1.9 pg. Nevertheless, the authors demonstrated
that the inhibitory phenomena lowered from 0.98 to 3.7 pg of the DNA and combined
with DNA losses during extraction procedures, changed the PCR sensitivity to 30 pg.
In our test, a higher detection limit was obtained (data not shown) and when negative
samples were spiked with S. mansoni DNA as a control, amplification was detected
in all samples. The possibility of false-negative cases in PCR is due to the reduced
egg output and uneven distribution in feces. Pontes et al. (2003) reported that
amongst 194 patients studied, no eggs were found in the stool of 16 individuals with
positive PCR results, despite the fact that two patients with eggs in their stool
(diagnosed by Kato-Katz technique) were PCR negative. Similar results were
described by Allam et al. (2009) in which they found seven patients with eggs in the
stools (diagnosed by Percoll technique) and negative PCR results. We did not find
any positive results when S. mansoni negative samples were tested, even when stool
samples from individuals infected with other parasites were analyzed, showing 100%
specificity. Recently, Ten Hove et al. (2008) described a Real Time PCR for
schistomiasis. However, a valid comparison with this method is difficult due to
diversity in their use of DNA targets, DNA isolation, PCR procedure and methods of

174
sample selection. PCR detection of pathogens in stool samples is difficult because of
the presence of PCR inhibitors. An important factor affecting the sensitivity of PCR is
the quality of the extracted template, which must be as pure as possible. Ideally, the
extracted template should not be contaminated with any kind of substances that
inhibit PCR amplification. Despite the vast number of methods described, none allow
isolation of DNA that is totally inhibitor free. Schistosomiasis remains a challenge for
sensitive and specific diagnostic method in those cases of low-intensity infection. A
number of techniques based on parasitological or immunological exams are
available, but some of them lack sensitivity, and others do not have enough
specificity or are expensive to use in public health services. In the future,
technological developments might be able to address the cost of diagnosis. The
search for accurate methods to detect Schistosoma spp infections must be
continued. We demonstrated that PCR possesses an advantage in overcoming
problems with coproscopy and may be able to assist in validating immunodiagnostic
tests. Immunological techniques for the detection of antibodies against Schistosoma
have been shown to have satisfactory sensitivity and specificity, but these techniques
are viewed with criticism because antibodies persist after treatment and cross-
reactivity between Schistosoma and other helminthes can occur. Thus, it is possible
that the results of some serologic tests were not accepted due to the low sensitivity of
the coproscopy for the detection of S mansoni eggs in feces. This could be
demonstrated in the group of individuals with serological tests for S. mansoni
infection with the absence of eggs in coproscopy that were PCR positive in 13
individuals. We conclude that PCR is an alternative test for diagnosing light S.
mansoni infections and can be used to confirm positive serological tests without a
high number of parasitological examination. Therefore, studies in this direction should
be encouraged and extended.
5 References
ALLAM, A.F. et al. Assessing the marginal error in diagnosis and cure of Schistosoma mansoni in areas of low endemicity using Percoll and PCR techniques. Trop Med Int Health 14:316-321, 2009.
DOENHOFF, M.J.; CHIODINI, P.L.; HAMILTON, J.V. Specific and sensitive diagnosis of schistosome infection: can it be done with antibodies? Trends Parasitol 20:35-39, 2004.

175
GIBBONS-MATTHEWS, C.; PRESCOTT, A.M. Intra-isolate variation of Cryptosporidium parvum small subunit ribosomal RNA genes from human hosts in England. Parasitol Res 90:439-44, 2003.
GOMES, A.L.V. et al. Development of a real time polymerase chain reaction for quantitation of Schistosoma mansoni DNA. Mem Inst Oswaldo Cruz 101:133-136, 2006.
GOMES, L.I. et al. Further evaluation of an updapted PCR assay for the detection of Schistosoma mansoni DNA in human stool samples. Mem Inst Oswaldo Cruz 10:1194-96, 2009.
GONÇALVES, M.M.L et al. Immunoassays as an auxiliary tool for the serodiagnosis of Schistosoma mansoni infection in individuals with low intensity of egg elimination. Acta tropica 100:24-30, 2006.
GONIN, P.; TRUDEL, L. Detection and Differentiation of Entamoeba histolytica and Entamoeba dispar isolates in clinical samples by PCR and Enzyme-Linked. Immunosorbent Assay. J Clin Microbiol 41:237-241, 2003.
HAMBURGER, J. et al. Highly repeated hort DNA sequences in the genome of Schistosoma mansoni recognized by a species-specific probe. Mol Bioch Parasitol 44:73-80, 1991.
HAMBURGER, J. et al. Development and laboratory evaluation of a polymerase chain reaction for monitoring Schistosoma mansoni infestation of water. Am J Trop Med Hyg 59:468-473, 1998.
HOFFMAN, W.A.; PONS, J.A.; JANER, J.L. The sedimentation-concentration method in schistosomiasis. Puerto Rico J Publ Hlth 9:281-298, 1934.
KATZ, N.; CHAVES, A.; PELLEGRINO, J. A simple device for quantitative stool thick-smear technique in schistosomiasis mansoni. Rev Inst Med Trop São Paulo 14:397-400, 1972.
PITCHER, D.G.; SAUNDERS, N.A.; OWEN, R.J. Rapid extraction of bacterial genomic DNA with guanidine thiocyanate. Lett Appl Microbiol 4:1513-1521, 1989.
PONTES, L.A.; DIAS-NETO, E.; RABELLO, A. Detection by polymerase chain reaction of Schistosoma mansoni DNA in human serum and feces. Am J Trop Med Hyg 66:157-162, 2002.
PONTES, L.A. et al. Comparison of a polymerase chain reaction and the Kato-Katz technique for diagnosing infection with Schistosoma mansoni. Am J Trop Med Hyg 68:652-656, 2003.
SANTOS, H.L. et al. Comparison of multiplex-PCR and antigen detection for differential diagnosis of Entamoeba histolytica. BrazJ Infect Dis 11:365-70, 2007.
SANDOVAL, N. et al. Schistosoma mansoni: A diagnostic approach to detect acute schistosomiasis infection in a murine model by PCR. Exp Parasitol 114:84-8a, 2006.

176
SANDOVAL, N. et al. A new PCR-based approach for the specific amplification of DNA from different Schistosoma species applicable to human urine samples. Parasitol 133:581-7b, 2006.
SILVA, A.J. et al. Fast and reliable extraction of protozoan parasite DNA from fecal specimens. Mol. Diagn 4:57-64, 1999.
SUBRUNGRUANG, I. et al. Evaluation of DNA extraction and PCR methods for detection of Enterocytozoon bienusi in stool specimens. J Clin Microbiol 42:3490-3494, 2004.
TEN HOVE, R.J. et al. Multiplex real-time PCR for the detection and quantification of Schistosoma mansoni and S. haematobium infection in stool samples collected in northern Senegal. Trans R Soc Trop Med Hyg 102:179-85, 2008.
VERWEIJ, J.J. et al. short communication: Prevalenceof Entamoeba histolytica and Entamoeba dispar in northen Ghana. Trop Med Int Heath 8:1153-56, 2003.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo


















