
MUTAGENICIDADE DO CORANTE ALIMENTÍCIO TARTRAZINA NO...
144
i MARIELLY REIS RESENDE MUTAGENICIDADE DO CORANTE ALIMENTÍCIO TARTRAZINA NO ENSAIO Salmonella/MICROSSOMA Limeira 2015
Transcript of MUTAGENICIDADE DO CORANTE ALIMENTÍCIO TARTRAZINA NO...

i
MARIELLY REIS RESENDE
MUTAGENICIDADE DO CORANTE ALIMENTÍCIO
TARTRAZINA NO ENSAIO Salmonella/MICROSSOMA
Limeira
2015

ii

iii
UNIVERSIDADE ESTADUAL DE CAMPINAS
FACULDADE DE TECNOLOGIA
MARIELLY REIS RESENDE
MUTAGENICIDADE DO CORANTE ALIMENTÍCIO TARTRAZINA
NO ENSAIO Salmonella/MICROSSOMA
Dissertação apresentada ao Curso de Mestrado da
Faculdade de Tecnologia da Universidade Estadual de
Campinas, como requisito para a obtenção do título de
Mestra em Tecnologia.
Área de concentração: Tecnologia e Inovação.
ORIENTADORA: Profa. Dra. Nelma De Mello Silva Oliveira
COORIENTADORAS: Profa. Dra. Gisela De Aragão Umbuzeiro
Profa. Dra. Simone Valente Campos
Limeira
2015
Este exemplar corresponde à versão final Dissertação
defendida pela aluna Marielly Reis Resende, e
orientada pela Profa. Dra. Nelma De Mello Silva
Oliveira.

iv

v

vi

vii
Resumo
Embora tenha grande utilidade e diversas aplicações nos setores industriais, há anos
a discussão sobre o potencial genotóxico do corante tartrazina vem sendo abordada, uma vez
que há vários resultados controversos descritos na literatura. É provável que a presença de
impurezas nas amostras possa ser uma das causas do possível potencial mutagênico. Dessa
forma, esse estudo visa avaliar a atividade mutagênica do corante tartrazina com diferentes
graus de pureza e possíveis interferentes presentes nas amostras, utilizando o ensaio
Salmonella/microssoma a partir das linhagens recomendadas pela OECD 471. Os resultados
obtidos demonstraram que o corante tartrazina ≥99 % e o corante tartrazina comercial 90%,
não apresentaram atividade mutagênica para as linhagens TA97a, TA98, TA100, TA1535 e
TA102 demonstrando ausência de impurezas mutagênicas ou que as mesmas estejam em
baixas concentrações nas amostras avaliadas.
Palavras-chave: Aditivos; Corantes alimentícios; Testes de mutagenicidade; Impurezas;
Corante tartrazina.

viii

ix
Abstract
Although very useful and diverse applications in industry, for years the discussion on the
genotoxic potential of the dye tartrazine has been addressed, since there are several
controversial results in the literature. It is likely that the presence of impurities in the samples
may be a cause of the possible mutagenic potential. Thus, this study aims to evaluate the
mutagenic activity of the dye tartrazine with different degrees of purity and possible
interferences present in the samples, using the Salmonella / microsome test from the lines
recommended by the OECD 471. The results showed that the dye tartrazina ≥99% and the
dye tartrazine commercial 90% showed no mutagenic activity for the strains TA97, TA98,
TA100, TA1535 and TA102 showing absence of mutagenic impurities or that they are in low
concentrations in the analyzed samples.
Keywords: Additives; Food dye; Mutagenicity testing; Impurities; Tartrazine dye.

x

xi
SUMÁRIO
1 INTRODUÇÃO ............................................................................................................... 1
2 OBJETIVO ....................................................................................................................... 5
3 REVISÃO BIBLIOGRÁFICA ....................................................................................... 7
3.1 Aditivos Alimentares: Corantes .................................................................................. 7
3.2 Identificação, processo de síntese e impurezas do corante tartrazina.......................... 9
3.3 Mutagenicidade provocadas por impurezas provenientes da síntese dos corantes .. .15
3.4 Toxicocinética ......................................................................................................... 17
3.5 Regulamentações ...................................................................................................... 20
3.6 Estudos de Genotoxicidade ....................................................................................... 23
3.6.1 Lesões no material genético ............................................................................ 23
3.6.2 Ensaios de Salmonella/microssoma ................................................................ 25
3.6.2.1 Características genéticas das linhagens utilizadas no ensaio de
Salmonella/microssoma ........................................................................................... 26
3.6.3 Estudos in vitro de genotoxicidade do corante tartrazina ............................... 28
3.6.4 Estudos in vivo de genotoxicidade do corante tartrazina ............................... 33
3.7 Estudos de carcinogenicidade ............................................................................ 35
3.7.1 Toxicidades reprodutiva e de desenvolvimento ............................................. 36
4 MATERIAIS E MÉTODOS .......................................................................................... 39

xii
4.1 Equipamentos ............................................................................................................ 39
4.2. Vidraria e outros materiais ....................................................................................... 39
4.3 Reagentes .................................................................................................................. 40
4.4 Preparos de Meios de Cultura e Soluções ................................................................ 41
4.5 Amostras de corantes ................................................................................................. 54
4.6 Manutenção das linhagens de Salmonella typhimurium ........................................... 55
4.7 Confirmações das características genéticas das linhagens de Salmonella typhimurium
utilizadas no ensaio .............................................................................................................. 56
4.7.1 Plasmídio pKM101 ........................................................................................ 56
4.7.2 Plasmídio pAQ1 .............................................................................................. 56
4.7.3 Mutação rfa ..................................................................................................... 57
4.7.4 Deleção do uvrB ............................................................................................. 58
4.7.5 Auxotrofia para histidina ............................................................................... 59
4.7.6 Taxa de reversão espontânea .......................................................................... 60
4.8 Protocolo do ensaio de Salmonella/microssoma por incorporação em placas ........... 61
4.9 Controles positivos e negativos ................................................................................... 63
4.10 Critérios de avaliação ................................................................................................ 64
5 RESULTADOS E DISCUSSÃO ................................................................................... 67
5.1 Confirmações das caractetrísticas genéticas ...................................................... 67
5.2 Taxa de reversão espontânea ............................................................................. 70

xiii
5.3 Ensaio de Salmonella/microssoma .................................................................... 71
6 CONCLUSÃO ................................................................................................................. 79
REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................ 81
APÊNDICE ..................................................................................................................... 107

xiv

xv
Dedico este trabalho a toda minha família,
pelo amor, dedicação, carinho, paciência,
e apoio doados a mim.

xvi

xvii
Agradecimentos
À minha orientadora Dra. Nelma De Mello Silva Oliveira pela confiança, paciência,
incentivo e amizade.
Às minhas coorientadoras, Dra. Gisela de Aragão Umbuzeiro e Simone Valente
Campos pelos valiosos ensinamentos, confiança, incentivo, amizade e pelo exemplo de
profissionalismo.
A toda minha família, por ser meu referencial de vida, pelo amor e confiança
incondicional, por sempre acreditar e me apoiar. Amo muito vocês!
Ao meu noivo Luiz Silva de Sousa, por ter sempre me incentivado e encorajado a
lutar pelos meus sonhos, pela confiança e pela paciência nos momentos de ausência. Te amo,
meu amor!
A toda família Amadio, pela amizade, por me acolherem como parte da família e
pelas inesquecíveis confraternizações e deliciosos churrascos.
A toda família Delariva, pela amizade, por sábios conselhos e pelas deliciosas tardes
de domingo. Jamais me esquecerei de vocês.
Às amigas Ádria Caloto de Oliveira, Anjaina Fernandes de Alburqueque e Josiane
Vendemiatti pela simpatia, amizade, apoio dado e ajuda na elaboração do trabalho.
Ao meu amigo Daniel Alexandre Morales, pela ajuda nos ensaios e troca de
informações.
A toda equipe do LEAL e do LACAN pela amizade, troca de conhecimentos e pelos
momentos de distração.
Aos professores Dr. Fábio Kummrow e Dra. Elisangela Franciscon Guimaro Dias,
membros da comissão da banca de defesa, pelas críticas e sugestões que contribuíram para a
melhoria deste trabalho.

xviii
À Karen Mercuri e Fátima Alves da secretaria de pós-graduação pelas dúvidas
solucionadas.
À Coordenação e Aperfeiçoamento Pessoal de Nível Superior, CAPES, pelo apoio,
representado pela bolsa de estudos concedida.
À Rede Mineira de Ensaios Toxicológicos e Farmacológicos da FAPEMIG, pelo
apoio, concedido em forma de material de consumo.

xix
“Um pouco de ciência nos afasta de Deus. Muito, nos aproxima.”
(Louis Pasteur)

xx

xxi
LISTA DE FIGURAS
Figura 1- Processo de síntese do corante tartrazina ............................................................ 10
Figura 2 - Esquema de confirmação da presença de plasmídio das linhagens de Salmonella
typhimurium ........................................................................................................................ 57
Figura 3 - Esquema de confirmação da mutação rfa das linhagens de Salmonella typhimurium
.............................................................................................................................. ...............58
Figura 4 - Esquema de confirmação da deleção do gene uvrB das linhagens de Salmonella
typhimurium ......................................................................................................................... 59
Figura 5 - Esquema de confirmação da auxotrofia para histidina das linhagens de Salmonella
typhimurium ......................................................................................................................... 60
Figura 6 - Esquema do ensaio de Salmonella/microssoma pelo método de incorporação em
placas .................................................................................................................................. 62
Figura 7 - Confirmação da auxotrofia para histidina (A) placas suplementadas com biotina
para a linhagem TA98; (B) placas suplementadas com biotina e histidina para a linhagem
TA98 .................................................................................................................................... 68
Figura 8 - Confirmação da mutação rfa para a linhagem TA98 ......................................... 68
Figura 9 - Confirmação da deleção uvrB (A) para a linhagem TA98; (B) para a linhagem
TA102 .................................................................................................................................. 69
Figura 10 - Confirmação da presença de plasmídio para a linhagem TA98 ....................... 70
Figura 11 - Resultados do ensaio de Salmonella/microssoma do corante tartrazina comercial
Duas Rodas (90%) com as linhagens TA97a, TA98, TA100, TA102 e TA1535, com e sem
ativação metabólica ............................................................................................................. 72

xxii
Figura 12 - Resultados do ensaio de Salmonella/microssoma do corante tartrazina Sigma (≥
99%) com as linhagens TA97a, TA98, TA100, TA102 e TA1535, com e sem ativação
metabólica. ........................................................................................................................... 73

xxiii
LISTA DE TABELAS
Tabela 1 - Identificação e características físicas - químicas do corante tartrazina. ............... 9
Tabela 2 - As especificações de limites máximos de impurezas para síntese do corante
tartrazina .............................................................................................................................. 14
Tabela 3 - Limite máximo de utilização do corante tartrazina em diferentes categorias de
alimentos de acordo com as legislações específica brasileira ............................................ 22
Tabela 4 - Características genética das linhagens de Salmonella typhimurium utilizadas no
ensaio de Samonella/microssoma ........................................................................................ 28
Tabela 5 - Ensaios in vitro de genotoxicidade do corante tartrazina ................................. 30
Tabela 6 - Média histórica da taxa de reversão espontânea das linhagens utilizadas no ensaio
Salmonella/microssoma no LACAN ................................................................................... 61
Tabela 7 - Controles positivos utilizados no ensaio Salmonella/microssoma ..................... 64
Tabela 8 - Taxa de reversão espontânea das linhagens utilizadas no ensaio
Salmonella/microssoma ....................................................................................................... 71

xxiv

xxv
LISTA DE ABREVIATURAS
ANVISA Agência Nacional de Vigilância Sanitária
CPPA Comitê Permanente em Aditivos Alimentares
DNA Ácido desoxirribonucleico
FAO Organização das Nações Unidas para a Alimentação e Agricultura
FAPESP Fundação de Amparo à Pesquisa do Estado de São Paulo
FMN Flavina mononucleotídeo
HCl Ácido clorídrico
HPLC Cromatografia Líquida de Alta Eficiência
IARC Agência Internacional para Pesquisa Sobre o Câncer
LACAN Laboratório Central Analítica
LEAL Laboratório de Ecotoxicologia e Microbiologia Ambiental Prof. Dr. Abílio
Lopes
LPS Lipopolissacarídica
OECD Diretrizes para Ensaios de Produtos Químicos
OMS Organização Mundial de Saúde

xxvi

1
1. INTRODUÇÃO
Aditivos químicos são usados com a finalidade de melhorar a aparência, aroma,
sabor, cor, textura e conservação dos alimentos processados. Por encontrá-los em uma
infinidade de produtos alimentícios, sua ingestão acontece por longos períodos, portanto,
representam potenciais riscos para a saúde (BERZAS et al., 1999, SASAKI et al., 2002).
Dentre estes, estão os corantes que são largamente utilizados para compensar a perda de cores
naturais, que são destruídas durante o processamento e armazenamento de alimentos, além
de proporcionar o aparecimento de novas cores desejadas. Os mesmos podem ser
classificados como corantes naturais e sintéticos (HATHCOCK e RADER, 2003).
Quando comparado com corantes naturais, os sintéticos apresentam várias vantagens,
tais como a alta estabilidade à luz, oxigênio, pH, uniformidade de cor, baixa contaminação
microbiológica, baixo custo de produção, dentre outras (HATHCOCK e RADER, 2003). Os
corantes podem ser classificados de acordo com seu grupamento cromóforo, sendo os mais
relevantes os compostos azóicos, antraquinonas, nitro e quinolinas (GREGORY,1990). Os
mais utilizados são os corantes do tipo azo, que apresentam grupamentos azo (N=N) ligados
a anéis aromáticos (MANSOUR et al., 2007).
Embora tenham grande utilidade e diversas aplicações, algumas dessas substâncias
representam um potencial risco para a saúde humana especialmente se forem excessivamente
consumidas (POLÔNIO, 2002).
Como o uso de corantes alimentícios varia entre os diversos países é necessária uma
regulamentação própria de cada país de acordo com suas necessidades. Sendo observado em
todo o mundo um consenso no uso seguro desses produtos (DALL’AGNOL, 2013). Por esta
razão, dados de segurança, como a ingestão diária aceitável (IDA), com base em estudos
toxicológicos experimentais em animais e clínicos em humanos, têm sido repetidamente
determinados e avaliados pela Organização das Nações Unidas para a Alimentação e
Agricultura (FAO) e a Organização Mundial de Saúde (OMS) (DOWNHAM e COLLINS,
2000).

2
Nesse sentido a toxicologia propõe o estudo desses azocorantes, pela importância da
exposição oral crônica e em baixas concentrações que foram relacionados ao câncer de
bexiga e da porção intestinal do cólon em humanos, a sarcomas esplênicos, hepatocarcinomas
e anomalias nucleares em animais de experimentação e aberrações cromossômicas em células
de mamíferos (PERCY, MOORE e CHIPMAN, 1989; SASAKI, et al., 2002; MANSOUR et
al., 2007).
Estudos indicam que a população infantil constitui o grupo mais vulnerável a ingestão
dessas substâncias, visto que a quantidade ingerida é maior na criança, em relação ao peso
corporal, do que no adulto. Podendo ocorrer alguns efeitos devido à ação direta dos corantes
nas células ou pela formação de metabólitos decorrentes da redução da ligação azo
(POLÔNIO, 2002; SHILS et al., 2003).
Dessa forma, os bioensaios constituem uma ferramenta importante na avaliação de
toxicidade dos corantes. Permitindo assim, fornecer informações sobre o perigo o qual a
população está exposta, uma vez que o ensaio avalia a toxicidade de diversas substâncias
biologicamente ativas (BERZAS et al., 1999).
O ensaio de mutagenicidade com Salmonella typhimurium, também conhecido como
ensaio de Salmonella/microssoma ou ensaio de Ames, é metodologia de triagem mais
utilizada atualmente para identificar substâncias genotóxicas. Utiliza linhagens que possuem
uma mutação específica no gene responsável pela síntese do aminoácido histidina bem como
diferentes alterações nos sistemas de reparo e capacidades metabólicas, permitindo a
detecção de diversos compostos químicos que podem interagir com o ácido
desoxirribonucleico (DNA). A fim de mimetizar a metabolização que ocorreria em
mamíferos, o ensaio emprega um sistema exógeno, fornecido pelas enzimas do citocromo
P450 do fígado de rato, tornando-o capaz de identificar mutagénos que precisam de ativação
metabólica para se tornar ativos (MORTELMANS e ZEIGER, 2000; UMBUZEIRO e
VARGAS, 2003).
As Diretrizes para Ensaios de Produtos Químicos (OECD 471), determinam que pelo
menos cinco linhagens de Salmonella typhimurium devem ser utilizadas no ensaio. A
combinação recomendada é: TA1535, e TA1537 ou TA97a ou TA97, TA98, e TA100, e E.

3
coli WP2 uvrA, ou E. coli WP2 uvrA (pKM101) ou TA102, a fim de detectar um número
maior de mutagénos, uma vez que cada linhagem tem uma característica específica de
detecção.
Por ser um corante azóico, a tartrazina tem despertado uma maior atenção dos
toxicologistas, alergistas e oncologistas diante da possibilidade de causar reações adversas
que variam desde urticária, asma ou até mesmo câncer de cólon intestinal (COMBES e
HAVELAND-SMITH, 1982; BERZAS et al., 1999; YONGLIN et al., 2011; SASAKI et al.,
2002).
Com base nos dados descritos na literatura sobre o corante tartrazina, observa-se que
existe um grande número de ensaios de mutagenicidade com resultados negativos (LÜCK e
RICKERL, 1960, apud EFSA, 2009; BROWN et al., 1978; MUZZAL e COOK, 1979;
CHUNG; FULK e ANDREWS, 1981; BROWN e DIETRICH, 1983; KARPLIUK et al.,
1984, apud EFSA, 2009; ISHIDATE et al., 1984; IZBIRAK et al., 1990, apud EFSA, 2009;
POLLASTRINI et al., 1990, apud EFSA, 2009) e alguns positivos (PATTERSON e
BUTLER, 1982; ISHIDATE et al., 1984; HENSCHLER e WILD, 1985, apud EFSA, 2009;
MUNZNER e WEVER, 1987; PRIVAL et al., 1988; DAS e MUKHEREJEE, 2004;
MPOUNTOUKAS et al., 2010), gerando assim algumas controvérsias sobre o seu potencial
genotóxico, o que vem sendo discutido há anos.
Os corantes não são considerados substâncias químicas puras, podendo conter em sua
formulação mais de 10% de impurezas (EFSA, 2009). Impureza é definida como qualquer
material e/ou substância que afete a pureza de um material de interesse, como por exemplo,
o corante. Estas impurezas podem ser de origens orgânicas ou inorgânicas. As impurezas de
origem inorgânicas podem ser metais pesados e outros materiais empregados durante a
síntese do corante. Já as impurezas orgânicas podem ser provenientes de produto
intermediário ou de degradação (RAO e KIRAM, 2010).
Dessa forma, a identificação, caracterização e os efeitos adversos provenientes a
partir destas impurezas presentes nos lotes dos mesmos, é de importância crucial para garantir
a segurança dos alimentos e medicamentos que contêm em sua formulação esses aditivos,

4
uma vez que muitas dessas impurezas são consideradas mutagênicas e/ou carcinogênicas
(ZIMMER et al., 1980; FDA, 1985).
A EFSA (Autoridade Européia de Segurança Alimentar) propondo esclarecer sobre
essa e outras características, elaborou um parecer científico em 2009, com o intuito de
reavaliar a segurança alimentar do corante tartrazina. Após a análise de banco de dados, o
parecer concluiu que os dados não indicam a necessidade de rever a IDA de 7,5 mg/kg de
peso corporal/dia, uma vez que muitos dos estudos analisados não são esclarecedores e foram
considerados com dados insuficientes. Além disso, o parecer observa que os níveis de
consumos relatados, estão abaixo da IDA estabelecida.
Portanto torna-se necessário a realização de mais estudos sobre o corante tartrazina
para elucidar seu potencial genotóxico por meios de bioensaios como o ensaio de
Salmonella/microssoma, visto que esse pode ser influenciado pelo teor de impurezas presente
no produto.

5
2. OBJETIVO
2.1 Objetivo Geral
Comparar a possível atividade mutagênica do corante alimentício tartrazina grau
analítico como seu respectivo corante comercial, utilizando o ensaio de
Salmonella/microssoma incluindo as linhagens recomendadas pela OECD 471.
2.2 Objetivo Específico
Verificar o comportamento dos dois corantes mediante a aplicação do teste
Salmonella/microssoma para avaliar a mutagenicidade dos corantes em relação ao grau de
pureza de cada um, a fim de verificar a interferência de possíveis contaminantes presentes
nas amostras.

6

7
3. REVISÃO BIBLIOGRÁFICA
3.1 Aditivos Alimentares: Corantes
Os aditivos alimentares são divididos em várias classes de substâncias que podem ser
adicionadas aos alimentos com diversas finalidades. Dentre tais classes encontram-se os
corantes que são substâncias sem valor nutricional, cujo único objetivo é promover a cor e
tornar os alimentos mais atraentes aumentando sua aceitação no mercado consumidor
(POLÔNIO e PERES, 2009). A indústria alimentícia pode ter sofrido um avanço tecnológico
controverso com a utilização destes, enquanto que, para saúde, tais corantes, principalmente
os artificiais não são indicados (SILVA e REED, 2010; CHEESEMAN, 2012). Apesar da
ingestão de corantes não ser indicada por especialistas, a estimativa de produção mundial de
corantes alimentares e pigmentos é aproximadamente 750-800 mil toneladas por ano, dos
quais 26 mil, em média, são consumidos apenas no Brasil (MARMITT; PIROTTAS; STULP,
2010; GOMES et al., 2013).
O controle do uso de corantes alimentares, baseados na IDA, resultou de
investigações dos diversos organismos internacionais (Comitê de Especialistas Aditivos
Alimentares (JECFA), Comitê do Codex sobre Aditivos Alimentares e Contaminantes,
Comissão Científico da Alimentação Humana (CCAH) da União Européia e a Directiva
(94/36/EC) (BESSONOV et al., 2011; GANESAN et al., 2011; GOMES et al., 2013). No
Brasil, a permissão para o uso e determinação dos níveis máximos toleráveis de aditivos
alimentares é de responsabilidade da Agência Nacional de Vigilância Sanitária (ANVISA) -
Ministério da Saúde, por realizar essas atividades de inspeção através do Comitê Permanente
em Aditivos Alimentares (CPAA) (FÁVERO; RIBEIRO; AQUINO, 2011).
No entanto, apesar do controle exigido pelas agências reguladoras, a utilização de
corantes em alimentos ainda levanta uma série de dúvidas quanto à sua toxicidade. Isto é
devido à grande dificuldade em verificar se a quantidade de aditivos utilizados pela indústria
de alimentos está em conformidade com as leis vigentes e seu grau de pureza (POLÔNIO e

8
PERES, 2009). No Brasil não é obrigatório declarar nos rótulos a quantidade de corantes
presentes nos produtos (BRASIL, 2005; GOMES et al., 2013). E muito menos se sabe sobre
o controle de qualidade destes que podem apresentar contaminações por aminas aromáticas
e metais pesados (EFSA, 2009).
Entre os corantes de alimentos, os mais usados pela indústria são os que contêm um
grupo azo (-N = N-), incluindo amarelo crepúsculo, o vermelho bordeaux, e o amarelo
tartrazina. A formação de azocorante se dá por reações de acoplamento, sendo a anilina
composto de partida para a produção dos mesmos enquanto os sais de diazônio obtidos pela
diazotação da anilina podem atacar outro anel aromático como um fenol, ou mesmo, outra
anilina formando o azocorante (MORRISON, 2005).
O corante tartrazina está presente em inúmeros alimentos como cereais, doces,
laticínios, geléias, sorvetes, recheios, licores, sucos em pó, refrigerantes, gelatinas e iogurtes,
além de alguns produtos cosméticos e medicamentos tais como vitaminas, antiácidos e
cápsulas (GOMES et al., 2013). Sendo evidente a abundância de alimentos coloridos que são
altamente atrativos ao público infanto-juvenil, foi realizado um estudo em Hyderabad, na
Índia, com 662 indivíduos em áreas urbanas e 358 em áreas rurais. Foi observado que a
ingestão diária de tartrazina, amarelo crepúsculo e eritrosina em crianças, ultrapassaram os
limites aceitáveis em 104, 284 e 200 %, respectivamente (RAO et al., 2004).
Esses azocorantes entram em contado com o trato gastrointestinal humano que abriga
uma microbiota diversificada composta de pelo menos alguns milhares de espécies. Os
mesmos podem se apresentar tanto insolúveis como solúveis em água e ser reduzidos pelas
bactérias intestinais (FENG, CERNIGLIA, CHEN, 2012). Embora alguns desses corantes
primários não apresentem um potencial cancerígeno e/ou mutagênico, alguns de seus
metabólitos gerados após a sua degradação por atividade de azoredutases, que são
comumente produzidas por bactérias entéricas, se mostram tóxicos e/ou carcinogênico para
os seres humanos (CHUNG et al., 1978; ELHKIM, 2007; FENG, CERNIGLIA, CHEN,
2012).

9
3.2 Identificacão, processo de síntese e impurezas do corante tartrazina
O corante tartrazina também conhecido como INS 102, FD & C Yellow number 5,
CI Food Yellow 4 e E102 é considerado como um aditivo alimentar. A tartrazina apresenta
uma alta solubilidade em meio aquoso, podendo ser identificado por espectrofotometria UV-
VIS no comprimento de onda próximo a 426nm (DIRETIVA 95/45/CE, 1995). Esta característica
ocorre devido à presença de grupos cromóforos, carboxílicos, nitro, hidroxílicos e amino que são
capazes de aumentar a intensidade de absorção específica para um determinado corante (DEL
GIOVINE; BOCCA, 2003).
Segundo Zollinger (1991) a presença de grupos sulfônicos (-SO3H) confere um
caráter ácido ao azocorante, além de torná-lo altamente solúvel em água, uma vez que o
grupo químico é facilmente ionizável, podendo ser encontrados em corantes monoazo
aniônicos. A identificação e características fisico-químicas da tartrazina podem ser
observadas na Tabela 1.
Tabela 1- Identificação e características físico - químicas do corante tartrazina
Nome Usual Amarelo Tartrazina
Nome Químico Sal tri-sódico 5-hidroxi-1-(4-sulfofenil) -4- [(4- sulfofenil) azo] -Pirazole-
3- carboxilato
Sinônimos Tartrazine; FD & C Yellow No. 5, Food Yellow No.4
Classe Monoazo
Fórmula Molecular C16H9N4Na3O=S2
Massa Molar 534, 35781
CAS 1934-21-0
Number Color Index (C.I.) 19140
Absorção Máxima λ max. = 426nm
Kow -10,17
Pka ~9,4
Solubilidade em água 38 mg/L a 2 °C, 200 mg/L a 25 °C
Fonte: (PRADO e GODOY, 2003; U.S. EPA, 2000; PEREZ-URQUIZA e BELTRAN; MARMION, 1991).

10
Durante o processo de síntese da matéria prima e manufatura desses corantes, podem
gerar algumas impurezas, podendo chegar a um total de 10% do composto final (Figura 1).
Algumas destas impurezas, como metais, 4 - aminobifenil, 4 – aminoazobenzeno (ácido
sulfanílico) e benzidina, podem apresentar efeitos carcinogênicos e ou mutagênicos, ou seja,
são capazes de induzirem mutações no DNA bem como gerar a formação de tumores
(ZIMMER et al., 1980; FDA, 1985; CLAXTON, HUGHES e CHUNG, 2001; CHUNG,
CHEN e CLAXTON, 2006). No entando estas podem estar presentes em níveis de segurança
conforme estabelecido pela legislação (FDA, 1985).
Figura 1 – Processo de síntese do corante tartrazina
Fonte: (PRIVAL, PEIPERL, e BELL, 1993).
Apesar de muitos metais serem essenciais ao bom funcionamento do organismo,
outros, no entanto podem ser prejudiciais, causando toxicidade aguda ou crônica,

11
dependendo da sua concentração e a forma disponível no organismo, além do tempo,
frequência da exposição e suscetibilidade do organismo exposto (LINDINO et al., 2008).
A população em geral pode ser exposta a estas substâncias, uma vez que, está presente
em diversos corantes nos mais variados produtos como couro, roupa, brinquedos, além de
alimentos e medicamentos (LANCASTER e LAWRENCE, 1999). Dessa forma, o alto
consumo de corantes pode tornar-se uma fonte de metais tóxicos, se forem consumidos em
concentrações acima do limite permitido pela legislação, mesmo em pequenas quantidades,
podem constituir um sério risco à saúde humana, devido ao seu acúmulo progressivo no
organismo (FRIBERG et al., 1979).
Nesse caso torna-se necessário um rigoroso controle de qualidade durante a
fabricação destes corantes tanto na indústria alimentícia bem como na indústria farmacêutica,
uma vez que o fabricante é responsável pela verificação de possíveis contaminantes presentes
nos mesmos (LINDINO et al., 2008).
O corante tartrazina geralmente é preparado a partir da diazotação do ácido 4 - amino
– benzeno com o ácido nítrico e nitrito de sódio, em seguida o composto diazo é acoplado
com o 4,5- dihidro- 5 - oxo - 1 - (4 - sulfofenil) - 1H - pirazole - 3 - ácido carboxílico ou com
o éster metílico, ou éster etílico, ou um sal de ácido carboxílico, que é isolado e purificado
como o sal de sódio ou cálcio (FDA 21 CRF74.705). Como principais componentes não
corados é possível encontrar cloreto de sódio e/ou sulfato de sódio na sua composição, no
entanto os teores de matérias de corantes totais não devem ser inferiores a 85% (DIRETIVA
95/45/CE, 1995).
De acordo com a Resolução nº 37 de 1977, o preparado de sal a partir do corante é
denominado laca, que consiste em uma combinação com o radical básico de alumínio, cálcio
e/ou sódio, podendo ser comercializados com o nome laca de alumínio, cálcio e/ou sódio de
tartrazina. Essas lacas devem seguir algumas especificações: cloretos e sulfatos como sal de
sódio devem estar em um limite máximo de 2% e matéria inorgânica insolúvel em HCl (ácido
clorídrico) presentes em até 0,5% (BRASIL, 1977). De acordo com a Diretiva 2008/128/CE,
a laca de alumínio de tartrazina não deve conter mais do que 0,5 % de material insolúvel em
HCl e materiais extraíveis com éter não deve exceder mais do que 0,2 % sob condições
neutras. Já o JECFA não dá especificações para lacas de alumínio de tartrazina, porém faz

12
uma referência geral para lacas de alumínios em corantes, determinando que a presença de
cloretos solúveis em água e sulfatos de sais de sódio, matéria insolúvel em ácido clorídrico,
matéria extraível com éter, arsênio e chumbo, não deve conter mais de 2, 0,5, 0,2 %, 3 e
5mg/kg respectivamente (JECFA, 2004).
A benzidina é uma das aminas aromáticas que se origina como uma impureza durante
a oxidação da anilina, um precursor para a síntese do ácido sulfanílico, que por sua vez é
precursor para a síntese do corante tartrazina. Durante a síntese da tartrazina pode ocorrer a
diazotação da benzidina e apenas uma pequena parte permanece como bendizina livre, ou
seja, a maior parte permanece na forma de corantes subsidiários como o corante T diazo
benzidina – pirazolona. Dessa forma após a ingestão da tartrazina pelos mamíferos, se estes
compostos estiverem presentes, por sua vez serão reduzidos no intestino liberando a
benzidina em sua forma livre (PRIVAL, PEIPERL e BELL, 1993; DAVIS e BAILEY, 1993).
Com bases nessas informações Davis e Bailey (1993) avaliaram a presença de
benzidina em 14 amostras do corante tartrazina, onde estes foram quimicamente reduzidos
com ditionito de sódio e as aminas liberadas eram isoladas através de extração com solventes
e posteriormente determinadas por cromatografia líquida de alta eficiência (HPLC). Os
resultados obtidos neste estudo mostram que os lotes comerciais do corante tartrazina
continham benzidina na forma livre e na forma de corante subsidiário em níveis acima
daqueles permitidos pela legislação (≤1 ppb), além disso em 4 das amostras apresentaram
níveis superiores a 100 ppb.
Um total de 53 amostras do corante tartrazina foi avaliado por Prival, Peiperl e Bell
(1993) onde também utilizaram o ditionito de sódio para a redução do corante, com intuito
de converter a benzidina na forma de diazo em benzidina livre, seguida da extração, a
benzidina foi quantificada como corante diazo benzidina-pirazolona T por HPLC. Um total
de 24 amostras que foram positivas para benzidina em níveis que variavam de 7 a 83 ng/g,
haviam sido certificadas pela FDA.
A FDA (1985) limita a benzidina em 1ng/g, apesar de que em alguns estudos o limite
de detecção foI de 3-4 ng/g (LANCASTER e LAWRENCE, 1999). A Tabela 2 apresenta as
especificações estabelecidas pela Directiva 2008/128/CE, JECFA (JECFA, 2006) e FDA

13
(FDA 21 CRF74.705) dos limites máximos de contaminantes que podem estar presentes no
corante tartrazina. No Brasil, o Conselho Nacional de Saúde, aprovou a Resolução CNS/MS
(Brasil, 1988 nº 4) indicando o limite máximo de apenas cinco contaminantes inorgânicos,
como o arsênio, chumbo, cobre, estanho e zinco com limites máximos de 1, 10, 20, 250 e 50
mg/kg do corante, respectivamente (LINDINO et al., 2008). Perante as informações de
acesso público, no Brasil, a legislação determina também os limites máximos tolerados de
contaminantes inorgânicos em diversos alimentos, como refrigerantes, sucos e outros
alimentos (BRASIL,1965). Pode-se notar que há uma divergência de informações e de
critérios no processo de fabricação dos corantes para diversos países, podendo assim, incorrer
num aumento das substâncias classificadas como impurezas presentes no corante e
consequentemente no alimento em que o mesmo é adicionado.
Essas especificações para os aditivos alimentares são elaboradas no intuito de garantir
a qualidade e segurança dos alimentos, incluindo desde o processo de fabricação até o
produto final. Além disso, vale ressaltar que diversas matérias-primas ou os diferentes
métodos de fabricação muitas vezes resultará em diferentes concentrações de pureza
(TEMANORD, 2002).
Portanto durante o processo de síntese de corantes é necessário seguir as boas práticas
de fabricação, seguindo sempre as especificações estabelecidas pela legislação, ou seja, não
se deve envolver compostos que são reconhecidos por apresentarem riscos à saúde, como a
presença de um grande grupo de aminas aromáticas (ANON, 1996; IARC, 2010).
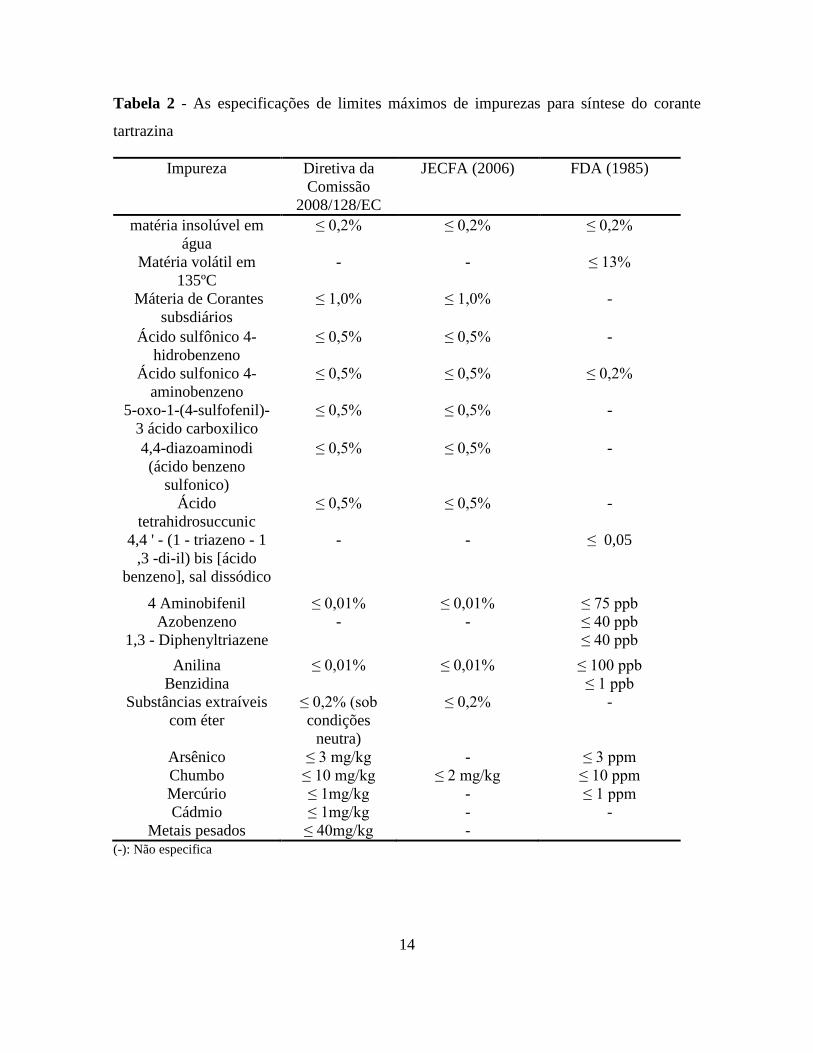
14
Tabela 2 - As especificações de limites máximos de impurezas para síntese do corante
tartrazina
Impureza Diretiva da
Comissão
2008/128/EC
JECFA (2006) FDA (1985)
matéria insolúvel em
água
≤ 0,2% ≤ 0,2% ≤ 0,2%
Matéria volátil em
135ºC
- - ≤ 13%
Máteria de Corantes
subsdiários
≤ 1,0% ≤ 1,0% -
Ácido sulfônico 4-
hidrobenzeno
≤ 0,5% ≤ 0,5% -
Ácido sulfonico 4-
aminobenzeno
≤ 0,5% ≤ 0,5% ≤ 0,2%
5-oxo-1-(4-sulfofenil)-
3 ácido carboxilico
≤ 0,5% ≤ 0,5% -
4,4-diazoaminodi
(ácido benzeno
sulfonico)
≤ 0,5% ≤ 0,5% -
Ácido
tetrahidrosuccunic
≤ 0,5% ≤ 0,5% -
4,4 ' - (1 - triazeno - 1
,3 -di-il) bis [ácido
benzeno], sal dissódico
- - ≤ 0,05
4 Aminobifenil ≤ 0,01% ≤ 0,01% ≤ 75 ppb
Azobenzeno - - ≤ 40 ppb
1,3 - Diphenyltriazene ≤ 40 ppb
Anilina ≤ 0,01% ≤ 0,01% ≤ 100 ppb
Benzidina ≤ 1 ppb
Substâncias extraíveis
com éter
≤ 0,2% (sob
condições
neutra)
≤ 0,2% -
Arsênico ≤ 3 mg/kg - ≤ 3 ppm
Chumbo ≤ 10 mg/kg ≤ 2 mg/kg ≤ 10 ppm
Mercúrio ≤ 1mg/kg - ≤ 1 ppm
Cádmio ≤ 1mg/kg - -
Metais pesados ≤ 40mg/kg - (-): Não especifica

15
3.3 Mutagenicidade provocadas por impurezas provenientes da síntese dos
corantes
Muito dos azocorantes que são análogos da benzidina são considerados mutagênicos
e/ou carcinogênicos, uma vez que através do seu metabolismo os eletrólitos interagem com
o DNA (MATHUR, BHATNAGAR, SHARMA, 2012). Estudos relataram uma alta
incidência de tumores de bexiga (CASE et al, 1954; ZAVON et al, 1973;) de estômago, reto,
pulmão e próstata (UBELIN et al, 1954) em trabalhadores expostos a benzidina e anilina em
fábricas de produtos químicos.
Além disso, vários pesquisadores têm correlacionado um número de aminas
aromáticas carcinogênicas e a capacidade em induzir mutações, quando testadas através do
ensaio de Salmonella/microssoma com e sem combinação de ativação metabólica (AMES et
al., 1972; GARNER et al., 1975; WANG et al., 1975; FERRETTI et al, 1977; LAZEAR e
LOUIE, 1977, CHUNG et al.; 2006; GREGORY, 1981; SHAHIN, 1985; MCCANN et al.,
1975).
Garner (1975) ao avaliar a atividade mutagênica de análogos de benzidina, a 3,3 '-
diclorobenzidina purificada demonstrou que tais substâncias apresentaram uma atividade
mutagênica para a linhagem TA1538 em presença de ativação metabólica e a 3,3’, 5,5'-
tetrametilbenzidina apresentou uma atividade mutagênica tanto em presença quanto em
ausência de S9. De acordo com McCann et al. (1975) a benzidina apresentou um potencial
mutagênico para a linhagem TA1538 em presença de ativação metabólica.
Gregory (1981) avaliou a atividade mutagênica de uma série de compostos derivados
de diazo benzidina e o- tolidina, bem como vários compostos monoazos. Todos os corantes
de benzidina e σ-toluidina testados foram mutagênicos para as linhagens TA98 e TA100,
após sofrerem uma redução química com ditionito de sódio.
Chung et al. (2006) revisaram a mutagenicidade de alguns análogos de benzidina,
demonstrando que muitas destas aminas foram consideradas mutagênicas para as linhagens
TA98 e TA100, em presença de ativação metabólica, segundo estudos realizados por meio

16
do ensaio de Salmonella /microssoma. Algumas aminas com alguns halogênios ou grupos
nitro presentes em sua estrutura apresentaram uma ação direta na mutagenicidade. Porém ao
adicionar um grupamento sulfônico na molécula de benzidina e uma complexação com um
íon metálico gerou uma redução na mutagenicidade.
Em adição, a anilina assim como outras aminas aromáticas são também consideradas
impurezas provenientes do processo de síntese de corantes (EFSA, 2009). Em vários estudos
algumas anilinas foram capazes de causar uma indução cancerígena em bexiga de cães
(BONSER, 1943, apud ZIMMER et al., 1980), em ratos algumas foram consideradas
carcinogênicas como a 2- metil -, 4 - metil -, e 2,4,5 - trimetil - anilina (HOMBURGUER et
al., 1972, RUSSFIELD et al.,1973; MCCANN et al., 1975; ZIMMER et al., 1980), além de
estar associadas ao dano oxidativo no DNA em células de baço de ratos (BOMHARD e
HERBOLD, 2005; MA et al., 2008).
No entanto para alguns autores a anilina apresentou resultados negativos através do
ensaio de Salmonella/microssoma (MCCANN et al., 1975; SUGIMURA et al., 1976;
DEGAWA et al., 1979; SIMON, 1979; MORI et al., 1980) bem como o ácido sulfanílico,
considerado como um de seus metabólicos (MCCANN et al., 1975; DEGAWA et al., 1979;
CHUNG, FULK e ANDREWS, 1981).
Shahin (1985) avaliou as relações de estrutura-atividade, de três nitroanilinas e nove
nitroaminofenols, testados quanto à sua capacidade de induzir mutações em linhagens de
Salmonella typhimurium. Para os compostos m-nitroanilina, 4-nitro-2-aminofenol, o 3-nitro-
6-aminofenol os resultados encontrados foram positivos para as linhagens TA1538 e TA98
em ausência de S9. No entanto os compostos σ-nitroanilina, 2-nitro-3-aminofenol, o 3-nitro-
4-aminofenol, 4-nitro-3-aminofenol, o 3-nitro-2-aminofenol, 2-nitro-6-aminofenol, 2-nitro-
4-aminofenol e 2-nitro-5-aminofenol foram negativos para TA1535, TA100, TA1537,
TA1538 e TA98, tanto na presença quanto em ausência de S9. O autor concluiu que a
atividade mutagênica ou inatividade desses compostos parece ser dependente das posições
dos grupos amino, nitro, e hidroxi nas suas estruturas químicas.
Outro interferente considerado como uma impureza é o composto 4 – aminobifenil.
Estudos tem demonstrado que este apresenta um potencial cancerígeno em camundongos

17
(CLAYSON et al.,1967) e ratos (WALPOLE et al., 1952) além de apresentar um potencial
mutagênico para as linhagens TA100, TA1537 e TA1538 em presença de S9. Seu isômero,
o 2-aminobifenil também foi mutagênico para a linhagem TA100 em presença de S9
(MCCANN et al.,1975). Estes resultados são confirmados por Lazear e Louie (1977) ao
analisar o 4-aminobifenil, bem como o seu sal cloridrato onde observaram uma
mutagenicidade para as linhagens de Salmonella typhimurium TA98 e TA100, com doses
testadas (50 e 100 pg/placa) em presença de ativação metabólica.
3.4 Toxicocinética
Segundo a Agência Internacional para Pesquisa Sobre o Câncer (IARC, 2010) a
ativação metabólica foi aceita como um elemento essencial para explicar a atividade
biológica de uma amina, uma vez que essa atividade está relacionada com a forma de
absorção, hábitos alimentares, tempo de contato, tecido alvo e células específicas além da
susceptibilidade individual ao metabolismo próprio de uma espécie animal.
A exposição a agentes possivelmente tóxicos pode se dar principalmente pelas vias
oral, respiratória e dérmica. Após a exposição oral aos azocorantes, pode ocorrer uma
azoredução numa ampla variedade de espécies de mamíferos, incluindo macacos Rhesus
(RINDE e TROLL, 1975) e seres humanos (FULLER, 1937; SCHRODER e CAMPBELL,
1972; WATABE et al., 1980) por meio de enzimas hepáticas ou por bactérias presentes na
microbiota intestinal sob condições anaeróbicas (WALKER, 1970; GINGELL e WALKER,
1971; SCHELINE, 1973).
Essa azoedução pode gerar como produtos de degradação a formação de
hidroxilaminas, as quais são capazes de causar dano ao DNA. Se forem completamente
reduzidas, as aminas aromáticas (-NH2), podem ser oxidadas novamente a N-
hidroxiderivados por enzimas do sistema citocromo P450. Esses radicais N-hidroxi por sua
vez podem ser acetilados por σ-acetiltransferases gerando íons eletrolíticos, capazes de reagir
com o DNA (BARTSCH, 1981; ARLT et al., 2002; UMBUZEIRO et al., 2005). Dessa forma,

18
a reação metabólica desempenha um importante papel na mutagênese e/ou carcinogênese
destes corantes (CHUNG, 1983; CHUNG e CERNIGLIA, 1992).
O radical nitro (-NO2), apesar de não fazer parte da molécula do corante tartrazina,
pode está presente em outros corantes azóicos, os quais também podem ser reduzidos pelas
nitroredutases bacterianas ou por enzimas presentes no citoplasma de células de mamíferos,
como as xantinas oxidases, ou ainda o citocromo P450, pela via redutiva gerando radicais N-
hidroxi (VOLKER et al., 2004; UMBUZEIRO et al., 2005). As reações de redução com
grupamentos nitro interagem com nitrobenzeno e aminas que são oxidadas principalmente
no fígado onde produzem metabólitos finais como anilinas e estes podem ser distribuídos de
forma diferente. O nitrobenzeno é considerado um causador de adenoma e carcinoma
hepático e anilinas causadoras de sarcoma predominantemente no baço de rato (IARC, 2010).
Com base nos levantamentos históricos, sobre azocorantes, a tartrazina e seu
metabolismo têm sido estudados por vários autores (RYAN e WRIGHT, 1962; JONES et al.,
1964; JECFA, 1966; ROXON et al., 1966, 1967; BERTAGNI et al., 1972; RYAN, 1972;
HONOHAN et al., 1977; KHERA e MUNRO, 1979). Acredita-se que menos de 2% da
tartrazina que é ingerida consegue ser absorvida (MURDOCH et al., 1987; EFSA, 2009).
Estudos têm demonstrado que após a administração intraperitonial ou intravenosa da
tartrazina em diferentes espécies de animais, os compostos resultantes da redução não são
metabolizados no fígado (HONOHAN et al., 1977). No entanto Kuno e Mizutani (2005)
mostraram que a tartrazina não é um substrato para as enzimas CYP2A6 e UDP-
glucuronosiltransferase, através de ensaios utilizando microssomas de fígado bovino para
mimetizar os microssomas hepáticos humanos (HIRANO et al., 2002; HIRANO e
MIZUTANI, 2003; KANOU et al., 2002).
Portanto, as enzimas hepáticas que reduzem ligações azo desempenhariam apenas um
papel menor no metabolismo dos azocorantes. A redução pelas bactérias intestinais seria o
caminho mais provável (JECFA, 1966; BERTAGNI et al., 1972; JONES et al., 1964; RYAN,
1972; CHUNG et al., 1978; KHERA e MUNRO, 1979). A maior parte da tartrazina é
rapidamente metabolizada em aminopirazolone no cólon pela flora intestinal (JECFA, 1966;

19
KHERA e MUNRO, 1979) e degradada a ácido 4 – hidrazinobenzenesulfonico, e em seguida
é reduzida em ácido sulfanílico (RYAN et al, 1969a; JONES et al, 1964).
Segundo Ryan e Wright (1961) após uma administração intravenosa de baixas doses
de tartrazina em ratos, esta não demonstrou ser excretada na bílis. Mais tarde, Ryan e Wright
(1962) comprovaram que o grupamento carboxila presentes na tartrazina foi responsável pelo
bloqueio da excreção biliar. No entanto para Jones et al. (1964) quando administrado por via
oral o corante tartrazina pode ser reduzido in vivo, diferentemente quando administrado por
via intraperitoneal, demonstrando que a flora do trato gastro intestinal é responsável pela
redução desse composto. Após a administração intraperitoneal de uma dose elevada de
tartrazina, a excreção de ácido sulfanílico pode ser explicada através de um ciclo entero-
hepático. A excreção do corante na bílis levaria a uma diminuição no intestino seguido por
uma absorção e subsequente excreção de ácido sulfanílico na urina.
O ácido sulfanílico é o principal metabólito produzido após a redução da tartrazina
pelas bactérias intestinais. Isso sugere que os elétrons extracelulares podem estimular a
redução dos azocorantes. A redução em aminopirazolone e ácido sulfanílico ocorre por meio
da presença de transportadores de életrons como a flavina que são liberadas pelas bactérias
em condições anaeróbicas no cólon (ROXON et al., 1967; CHUNG et al., 1978).
Além disso, em relação a metabolização hepática e renal após a adminstração de
tartazina e carmosina os seus efeitos tornam-se mais evidentes em doses mais elevadas, por
induzirem o estresse oxidativo devido a formação de radicais livres podendo levar a efeitos
tóxicos. O estudo foi realizado administrando-se doses de 15 mg/kg e 500 mg/kg e de 8 mg/kg
e 100 mg/kg de peso corporal dos corantes tartrazina e carmosina, respectivamente, durante
30 dias (AMIN, HAMEID, ELSTTAR, 2010).
Com base nessas informações, pode-se observar que as substâncias como os corantes
são capazes de induzir o estresse oxidativo por formação de radicais livres podendo levar a
efeitos tóxicos. Nesse sentido a realização de estudos com o intuito de avaliar substâncias
indutoras de mutagenicidade devem ser realizados tanto in vitro como in vivo .

20
3.5 Regulamentações
As legislações visam estabelecer padrões para utilização de aditivos alimentares,
incluindo os corantes sintéticos, visto que o emprego dos mesmos tem sido objeto de críticas
em função de possíveis efeitos nocivos à saúde do consumidor. Esses efeitos vão desde
alterações de comportamento em crianças, a asma, urticária, reações alérgicas a alimentos
até apresentação de potencial mutagênico e/ou carcinogênico em alguns casos (RING,
BROCKOWe BEHRENDT, 2001; PRADO e GODOY, 2003; ELHKIM, et al., 2007).
Em virtude de alguns efeitos nocivos e críticas na utilização desses corantes, ocorreu
na Europa e nos EUA um movimento de reavaliação dessas substâncias, com desdobramento
no Brasil e em outros países da América Latina. Diversos órgãos internacionais, entre eles, o
Comitê de Especialistas relacionados FAO/OMS sobre Aditivos Alimentares (JECFA), um
comitê científico internacional de especialistas administrado conjuntamente pela
Organização para a Alimentação e Agricultura das Nações Unidas (FAO) e pela Organização
Mundial da Saúde (OMS) em 1966 e pela Comissão Científica da Alimentação Humana
(CCAH) da União Europeia, em 1975 e 1984 participaram desse processo de reavaliação.
Por meio de vários estudos, esses órgãos estabeleceram uma IDA segura de 7,5 mg/kg de
peso corporal/dia para o corante tartrazina, sendo assim, uma criança de 30 Kg e um adulto
de 60 Kg podem consumir até 225 mg e 450 mg de tartrazina por dia, respectivamente
(JECFA, 1966).
O consumo desse corante acima dos níveis estabelecidos pela legislação pode gerar
desde reações alérgicas (ELHKIM, et al., 2007) como alterações comportamentais atuando
como um catalisador na hiperatividade (TANAKA et al., 2006). Em ratos têm produzido
neurotoxicidade e déficits de aprendizado e memória (YONGLIN et al., 2011).
Devido à grande utilização desses compostos e seu potencial risco para a saúde
humana, os EUA permite a utilização de apenas nove corantes; no Japão segundo a
legislação, somente onze agentes sintéticos são liberados, e na União Europeia, apenas
dezessete, embora a Suécia e a Noruega proibirem o uso desses produtos artificiais em
alimentos (REYES e PRADO, 2001; QUEIJA, et al., 2001; PRADO e GODOY, 2003).

21
No Brasil, as Resoluções n° 382 a 388, de 9 de agosto de 1999, da ANVISA,
regulamentam o uso de onze corantes artificiais, sendo eles: amaranto, vermelho de
eritrosina, vermelho 40, ponceau 4R, amarelo crepúsculo, amarelo tartrazina, azul de
indigotina, azul brilhante, azorrubina, verde rápido e azul patente V. Esses corantes podem
ser utilizados nas indústrias de alimentos e farmacêuticas respeitando os limites máximos de
quantidade permitida para cada um deles e em alimentos específicos. Entre os países do
MERCOSUL, houve necessidade de uma legislação que harmonizasse a utilização desses
corantes alimentares (MERCOSUL Resolução GMC nº 52/98) bem como determinasse
funções de aditivos e seus limites máximos de utilização para todas as categorias de alimentos
(ASSOCIAÇÃO BRASILEIRA DAS INDÚSTRIAS DE ALIMENTAÇÃO, 2001).
O emprego de um novo aditivo alimentar no Brasil apenas é permitido se previamente
for aprovado pela Comissão Nacional de Normas e Padrões para Alimentos (CNNPA do
Ministério da Saúde) e se for utilizado dentro de um limite permitido, garantindo sua
inocuidade (BRASIL, 1969).
O Ministério da Saúde por meio da ANVISA regulamenta, controla e fiscaliza o uso
de aditivos em alimentos e medicamentos, exigindo o cumprimento dos padrões de limites
máximos (Tabela 3) determinados. Uma vez que a utilização de alguns aditivos como os
corantes podem geram uma toxicidade após seu consumo, principalmente se estiverem
presentes impurezas provenientes de sua síntese. Dessa forma, existe uma preocupação dos
órgãos reguladores em relação à rotulagem dos alimentos e medicamentos que contêm como
por exemplo o corante tartrazina em sua formulação. No Brasil é regulamentado pela
ANVISA e nos EUA (FDA) por meio de um Regulamento Federal, os fabricantes de
alimentos e medicamentos que utilizam este corante em sua formulação foram obrigados a
declarar, na lista de ingredientes e bula do medicamento, o nome do corante tartrazina por
extenso (CRF 74.1705; BRASIL, 2002, 2003).

22
Tabela 3 - Limite máximo de utilização do corante tartrazina em diferentes categorias de
alimentos de acordo com as legislações específica brasileira.
Alimentos Limite máximo Resolução (ANVISA)
Balas, confeitos, bombons, chocolates e
similares
0,030 g/100g RDC nº 387, de 5 de agosto de
1999
Cobertura e xarope, recheios e pós para o
preparo de coberturas e xaropes para
produtos de panificação e biscoitos,
produtos de sobremesas, gelados
comestíveis, balas, confeitos, bombons
chocolates e similares
0,050 g/100g
RDC nº 387, de 5 de agosto de
1999
Bebidas não alcoólicas a base de soja 0,01g/100mL RDC nº 25, de 15 de fevereiro de
2005
Bebidas não alcoólicas gaseificadas e
não gaseificadas
0,01g/100mL RDC nº 5, de 15 de janeiro de
2007
Gelados comestíveis 0,015g/100g RDC nº 3, de 15 de janeiro de
2007
Molhos e condimentos 0,05g/100g RDC nº4, 15 de janeiro de 2007
Petiscos 0,02g/100g RDC nº 64, de 16 de setembro
de 2008
Preparações culinárias industriais
(congeladas ou não) desidratadas
0,005g/100g RDC nº 34, de 9 de março de
2001
Sobremesas 0,015g/100g Resolução nº 388, de 05 de agosto
de 1999
Sopas e caldos concentrados/
desidratados
0,005g/100g RDC nº 33, de 09 de março de
2001
Suplementos Vitamínicos e ou de
Minerais (líquidos)
0,01g/100mL RDC nº 24, de 15 de fevereiro de
2005
Suplementos Vitamínicos e ou de
Minerais (sólidos)
0,03g/100g RDC nº 24, de 15 de fevereiro de
2005
Fonte: (ANVISA, 2014)

23
Em função da alta demanda de utilização dos corantes e da sua necessidade de
fiscalização, os métodos de avaliação destes aditivos têm sido rotineiramente requeridos.
Tais determinações visam tanto o controle de qualidade das indústrias alimentícias e
farmacêuticas, quanto à análise destes compostos em órgãos de vigilância e controle da saúde
humana. Na avaliação do TemaNord, 2002, recomendou-se uma reavaliação do corante
tartrazina para a inclusão de dados recém-publicados, em relação aos estudos de
genotoxicidade, toxicidade crônica /carcinogenicidade, reprodutiva e de desenvolvimento
pré-natal.
3.6 Estudos de Genotoxicidade
3.6.1 Lesões no material genético
A molécula de dupla fita de DNA é o local onde está armazenada toda a informação
genética. Estas fitas se ligam através de pontes de hidrogênio com as bases nitrogenadas,
onde uma adenina se liga a uma timina enquanto uma citosina se liga a uma guanina,
compondo uma molécula de DNA. Algumas lesões podem ocorrer na molécula de DNA,
como adições ou deleções de bases nitrogenadas, quebras ou rearranjos cromossômicos.
Dessa forma se a célula não conseguir reparar, ou seja, reconstruir a informação genética
original, a lesão é denominada de mutação, que pode ocorrer de forma espontânea ou
induzida por agentes químicos ou físicos (COMBES, 1992; MORTELMANS e ZEIGER,
2000).
As mutações podem ocorrer em qualquer célula, tanto em células de linhagem
germinativa como em células somáticas. Quando ocorrem em células de linhagem
germinativa, essas podem ser propagadas á geração seguinte, sendo responsáveis por várias
doenças hereditárias assim como em alguns tipos de cânceres (MARON e AMES, 1983;
COMBES, 1992). Porém quando a mutação ocorre em células somáticas, a consequência
mais comum é o desenvolvimento de tumores benignos ou malignos. Além disso, foi

24
proposto que as mutações em células somáticas podem também estar envolvidas na
patogênese de algumas doenças crônicas degenerativas como as cardiovasculares e
neurodegenerativas (VARANDA, 2006),
Uma mutação pode ocorrer nos cromossomos ou nos genes. A mutação
cromossômica se caracteriza quando partes de segmentos de cromossomos, cromossomos
inteiros ou grupamentos de cromossomos se alteram, resultando em rearranjos
cromossômicos anormais (GRIFFITHS et al., 1998a apud FERRAZ, 2008). Dessa maneira
muitas mutações cromossômicas podem conduzir a anomalias no funcionamento da célula e
do organismo (MARON e AMES, 1983).
Na mutação gênica também denominada de mutação de ponto, os eventos de
mutações ocorrem em genes individuais, ou seja, as alterações ocorrem em pares de bases
únicos do DNA ou num pequeno número de bases adjacentes (GRIFFITHS et. al, 2001b apud
FERRAZ, 2008). Estas mutações podem ocorrer através da substituição de pares de bases ou
deslocamento do quadro de leitura do DNA, também chamada de mutação frameshift. A
mutação frameshift ocorre devido a adição ou a perda de uma ou mais bases nitrogenadas do
DNA, levando a uma alteração no quadro de leitura. Quando a leitura da sequência de códons
do ácido ribonucléico mensageiro (mRNA) é “lida” pelo aparelho traducional, esta ocorrerá
de forma incorreta, uma vez que a adição ou deleção de um único par de base do DNA mudará
a matriz de leitura começando a leitura a partir do local que sofreu essa alteração estendendo
até o terminal carboxílico da proteína (GRIFFITHS et. al, 2001b).
Dependendo do gene que sofreu alteração e do tipo de efeito que essa alteração tem
na expressão gênica, isso pode resultar em grandes efeitos, que pode ser desde uma perda
completa da expressão do gene, até à formação de uma proteína variante com propriedades
alteradas (MARON e AMES, 1983).
Com base nessas informações citadas anteriormente há vários sistemas de reparo que
cuidam da remoção de lesões causadas no DNA. Dentre eles, podemos citar o reparo por
excisão, que consiste no reconhecimento da lesão no DNA, corte na fita e excisão da lesão e
de alguns nucleotídeos adjacentes, o que resulta na formação de um novo pedaço de DNA e

25
restauração da dupla hélice original por um processo livre de erros (GREEN, 1978;
HANAWALT, 1979; FRIEDBERG, 1985).
Estes sistemas de reparo são importantes, visto que vários testes de mutagenicidade
se baseiam neles. O ensaio com Salmonella, por exemplo, utiliza linhagens mutantes
deficientes no reparo por excisão através da deleção do gene uvrB, no intuito de aumentar a
capacidade de detecção do ensaio (MARON e AMES, 1983).
3.6.2 Ensaios de Salmonella/microssoma
O ensaio de mutagenicidade com Salmonella typhimurium, também conhecido como
ensaio de Salmonella/microssoma ou teste de Ames, é uma metodologia de triagem mais
utilizada atualmente para detectar substâncias genotóxicas (UMBUZEIRO e VARGAS,
2003). O ensaio foi inicialmente descrito pelo grupo do Dr. Bruce Ames, (AMES,
YAMASAKI, 1971; AMES et al., 1973a; 1973b; MARON, AMES, 1983) e discutido
internacionalmente e revisado por Mortelmans e Zeiger (2000).
Esse teste emprega linhagens de Salmonella typhimurium derivadas da parental LT2,
auxotróficas para o aminoácido histidina, apresentando diferentes mutações no operon deste
aminoácido. Essas linhagens são construídas para detectar mutações do tipo deslocamento
do quadro de leitura ou substituição de pares de base no DNA. São incapazes de crescer em
meio de cultura mínimo sem histidina, a menos que ocorram mutações que restaurem a sua
capacidade de síntese. O número de revertentes é facilmente medido pela contagem de
colônias que crescem em meio mínimo, após a exposição de uma população de células à
substância testada (UMBUZEIRO e VARGAS, 2003).
As linhagens de Salmonella typhimurium empregadas no ensaio são pertencentes ao
grupo das enterobactérias que são capazes de produzir nitroredutases e azoredutases. Alguns
corantes azóicos apenas exibem atividade mutagênica se as ligações azo forem reduzidas
gerando aminas aromáticas, podendo ser mais ou menos mutagênicas quando comparadas ao
corante original, dependendo da estrutura química. Se o corante tiver um ou mais

26
substituintes do grupo nitro, as nitroredutases endógenas irão reduzi-lo às hidroxilaminas ou
a aminas aromáticas (UMBUZEIRO et al., 2005).
Além disso, com a utilização de um sistema exógeno, a partir de uma enzima do
citocromo P450 do fígado de rato, o ensaio é capaz de detectar mutágenos que tem a
necessidade de serem metabolizados para se tornar ativos e interagir com o DNA
(MORTELMANS e ZEIGER, 2000).
3.6.2.1 Características genéticas das linhagens utilizadas no ensaio de
Salmonella/microssoma
Cada uma das linhagens empregadas possui uma mutação específica no gene
responsável pela síntese do aminoácido histidina bem como diferentes alterações nos
sistemas de reparo e capacidades metabólicas. Sendo assim, respondem diferentemente às
variadas classes de compostos químicos. Essas alterações genéticas adicionais são
responsáveis por atribuir uma maior sensibilidade na detecção de diversos mutágenos, tais
como deleção dos genes uvrB que elimina o mecanismo de reparo por excisão, permitindo
que mais lesões no DNA sejam reparadas pelo mecanismo suscetível ao erro, e a mutação
rfa que causa a perda parcial da camada lipopolissacarídica (LPS) da parede bacteriana
tornando-a mais permeável para difusão de moléculas grandes que normalmente não
penetram nas células. A deleção do gene uvrB promoveu a deleção de parte do gene
responsável pela síntese de biotina, tornando-as auxotróficas para biotina (MARON e
AMES; 1983; MORTELMANS e ZEIGER; 2000; UMBUZEIRO e VARGAS, 2003).
Algumas linhagens como a TA98 e YG1041 possuem o plasmídio pKM101 que além
de conferir resistência à ampicilina, contêm os genes mucAB, cuja expressão causa estímulo
ao mecanismo suscetível ao erro (error-prone). A linhagem TA98 apresenta mutação no
gene hisD3052 que codifica para a histidinol desidrogenase, apresentando como ponto
preferencial para a reversão oito resíduos repetitivos de GC e detecta compostos mutagênicos
que causam deslocamento do quadro de leitura do DNA. Apesar de ter a capacidade de

27
reduzir compostos do tipo nitro, ela possui uma menor atividade de nitroredutase. Isso ocorre
devido à deleção de alguns dos genes responsáveis pela produção dessa enzima
(MORTELMANS e ZEIGER; 2000).
Dessa forma, outras linhagens foram desenvolvidas; algumas incapazes de produzir
essa enzima (TA98NR e TA100NR) e outras com alta produção da mesma (YG1021,
YG1026). Já as linhagens YG1041 e YG1042 foram desenvolvidas com alta produção de
ambas as enzimas nitroredutases e o-acetiltransferases. Ensaios empregando essas linhagens
são úteis para o esclarecimento da via de redução do grupo nitro na mutagenicidade de certos
compostos (MARON e AMES; 1983; MORTELMANS e ZEIGER; 2000).
A linhagem TA97a apresenta mutação no gene hisD6610 detectando também
mutágenos que causam deslocamento do quadro de leitura do DNA. Já a linhagem TA102
apresenta uma mutação no gene hisG428, detectando assim mutágenos como formaldeído,
vários hidroperóxidos, bleomicina, fenilidrazina, raios-X, luz UV e agentes cross-link, como
mitomicina C. Já as linhagens TA100 e TA1535 apresentam a mutação hisG46, responsável
por codificar o gene para primeira enzima da biossíntese da histidina (AMES, 1971)
resultando na substituição de uma leucina por uma prolina, tornando assim um marcador para
compostos mutagênicos que causam substituição principalmente nos pares de bases GC
(MARON e AMES, 1983).
Uma combinação dessas linhagens podem auxiliar no papel destas enzimas na
atividade mutagênica de corantes azóicos, tornando assim o ensaio de
Salmonella/microssoma, uma importante ferramenta para contribuir e predizer os possíveis
efeitos desses compostos para a saúde humana após a sua ingestão, uma vez que utilizam
enterobactérias com características metabólicas similares às presentes na microbiota
intestinal de mamíferos (CHADWICK et al., 1992; PORWOLLIK et al., 2001;
UMBUZEIRO et al., 2005; CHUNG, 1992).
Assim, de acordo com OECD 471, as linhagens demonstradas na Tabela 4 são as que
foram utilizadas no presente estudo visando testar a mutagenicidade do corante tartrazina.
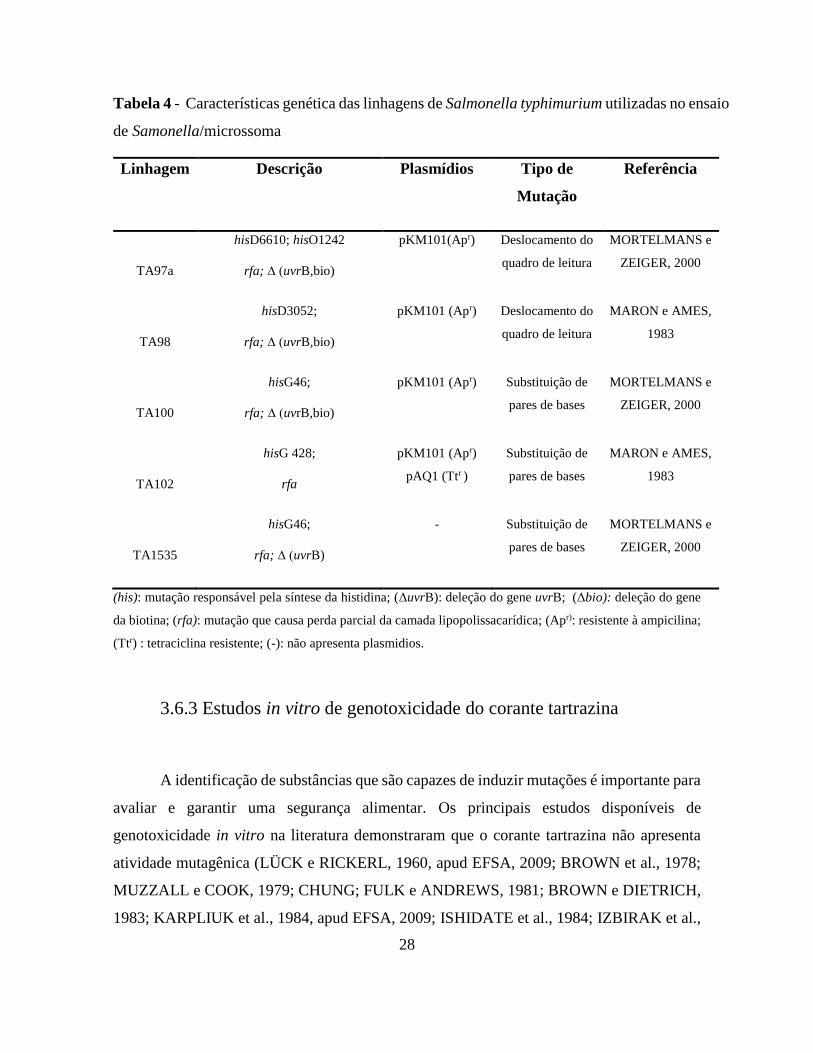
28
Tabela 4 - Características genética das linhagens de Salmonella typhimurium utilizadas no ensaio
de Samonella/microssoma
Linhagem Descrição Plasmídios
Tipo de
Mutação
Referência
TA97a
hisD6610; hisO1242
rfa; ∆ (uvrB,bio)
pKM101(Apr)
Deslocamento do
quadro de leitura
MORTELMANS e
ZEIGER, 2000
TA98
hisD3052;
rfa; ∆ (uvrB,bio)
pKM101 (Apr)
Deslocamento do
quadro de leitura
MARON e AMES,
1983
TA100
hisG46;
rfa; ∆ (uvrB,bio)
pKM101 (Apr)
Substituição de
pares de bases
MORTELMANS e
ZEIGER, 2000
TA102
hisG 428;
rfa
pKM101 (Apr)
pAQ1 (Ttr )
Substituição de
pares de bases
MARON e AMES,
1983
TA1535
hisG46;
rfa; ∆ (uvrB)
- Substituição de
pares de bases
MORTELMANS e
ZEIGER, 2000
(his): mutação responsável pela síntese da histidina; (ΔuvrB): deleção do gene uvrB; (Δbio): deleção do gene
da biotina; (rfa): mutação que causa perda parcial da camada lipopolissacarídica; (Apr): resistente à ampicilina;
(Ttr) : tetraciclina resistente; (-): não apresenta plasmidios.
3.6.3 Estudos in vitro de genotoxicidade do corante tartrazina
A identificação de substâncias que são capazes de induzir mutações é importante para
avaliar e garantir uma segurança alimentar. Os principais estudos disponíveis de
genotoxicidade in vitro na literatura demonstraram que o corante tartrazina não apresenta
atividade mutagênica (LÜCK e RICKERL, 1960, apud EFSA, 2009; BROWN et al., 1978;
MUZZALL e COOK, 1979; CHUNG; FULK e ANDREWS, 1981; BROWN e DIETRICH,
1983; KARPLIUK et al., 1984, apud EFSA, 2009; ISHIDATE et al., 1984; IZBIRAK et al.,

29
1990, apud EFSA, 2009; POLLASTRINI et al., 1990, apud EFSA, 2009), embora alguns
tenham apresentado resultados positivos (PATTERSON e BUTLER, 1982; ISHIDATE et
al., 1984; HENSCHLER e WILD, 1985, apud EFSA, 2009; MUNZNER e WEVER,1987;
PRIVAL et al.,1988; DAS e MUKHEREJEE, 2004; MPOUNTOUKAS et al., 2010) como
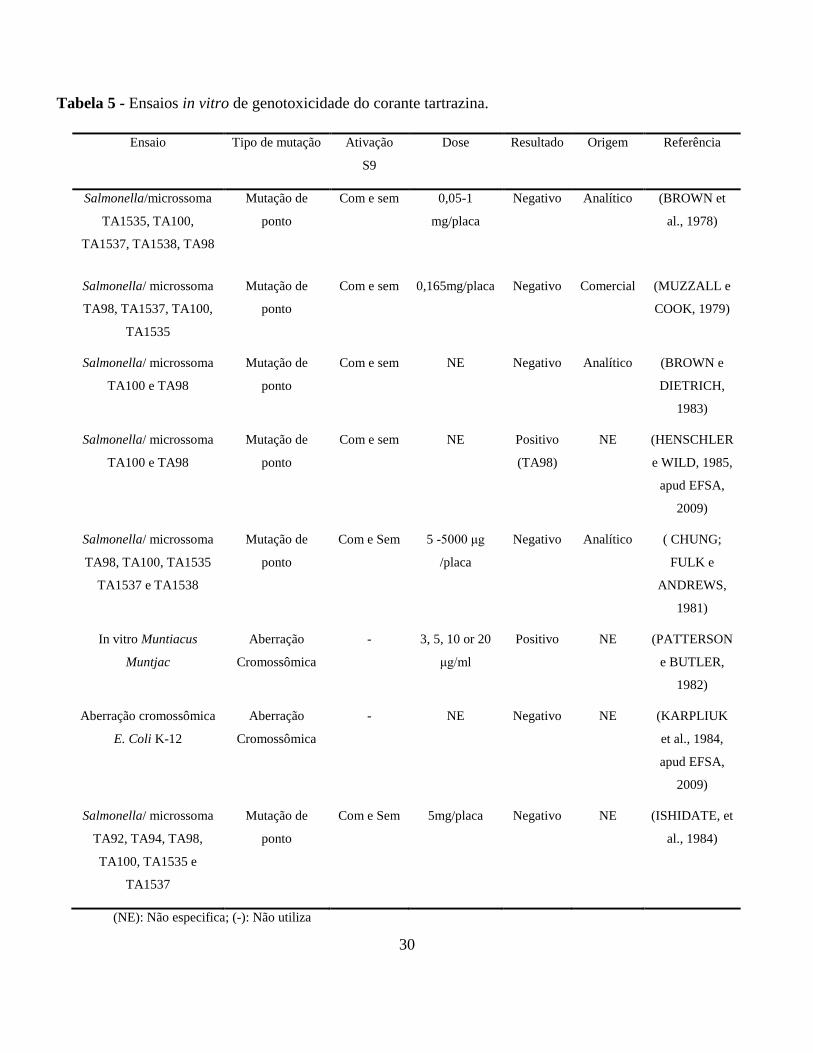
pode ser observados na Tabela 5.
Chung, Fulk e Andrews (1981) realizaram um estudo com dezessete corantes e 16
dos seus metabólitos ou derivados testados no ensaio de mutagenicidade de Salmonella/
microssoma utilizando a metodologia de incorporação em placas, em presença e ausência de
microssomas de fígado de rato (S9), utilizando as linhagens TA1535, TA1537, TA1538,
TA98 e TA100. Os resultados foram positivos com e sem ativação metabólica para 3-
aminopireno, vermelho litol, azul de metileno, amarelo metil, vermelho neutro e vermelho
fenol. Já o 4-amino pirazolone, 2,4- dimetilanilina , N , N - dirmetil - p - fenilenodiamina ,
vermelho de metila e 4 - fenil - azo - 1 – naftilamina, apresentaram resultados positivos
apenas em presença de S9 . Os resultados foram negativos para os azocorantes, tartrazina,
vermelho allura , amaranto, ponceau R, ponceau SX, amarelo crepúsculo em presença e
ausência de S9. Os seguintes metabólitos também apresentaram resultados negativos, como
a anilina, ácido sulfanílico, cloridrato de 1 - amino - 2 - naftol , ácido antranílico, sal
cresidine, pirazolona T, sal de R – amino - ( 1 - amino-2 -naftol-3, 6- sal dissódico
disulfónico), sal de Schaeffer (ácido - 6 - sulfônico 2- naftol, sal de sódio), naphthionate
sódio, sulfanilamida e ácido 4 - Amino-naftalenossulfónico sal sódico (CHUNG, FULK e
ANDREWS, 1981).
A indução de aberração cromossômica foi avaliada após dois diferentes períodos da
cultura celular de fibroblasto de Muntiacus Muntjac expostas em concentrações de 3, 5, 10 e
20 µg/mL de tartrazina. Foi observado um aumento significativo em relação a quebras das
cromátides e cromossomos dicêntricos quando comparados com os grupos controles em
todos os grupos experimentais. Os autores sugerem que novos estudos são necessários para
determinar o potencial mutagênico do corante (PATTERSON e BUTLER, 1982).
30
Tabela 5 - Ensaios in vitro de genotoxicidade do corante tartrazina.
(NE): Não especifica; (-): Não utiliza
Ensaio Tipo de mutação Ativação
S9
Dose Resultado Origem Referência
Salmonella/microssoma
TA1535, TA100,
TA1537, TA1538, TA98
Mutação de
ponto
Com e sem 0,05-1
mg/placa
Negativo Analítico (BROWN et
al., 1978)
Salmonella/ microssoma
TA98, TA1537, TA100,
TA1535
Mutação de
ponto
Com e sem 0,165mg/placa Negativo Comercial (MUZZALL e
COOK, 1979)
Salmonella/ microssoma
TA100 e TA98
Mutação de
ponto
Com e sem NE Negativo Analítico (BROWN e
DIETRICH,
1983)
Salmonella/ microssoma
TA100 e TA98
Mutação de
ponto
Com e sem NE Positivo
(TA98)
NE (HENSCHLER
e WILD, 1985,
apud EFSA,
2009)
Salmonella/ microssoma
TA98, TA100, TA1535
TA1537 e TA1538
Mutação de
ponto
Com e Sem 5 -5000 μg
/placa
Negativo Analítico ( CHUNG;
FULK e
ANDREWS,
1981)
In vitro Muntiacus
Muntjac
Aberração
Cromossômica
- 3, 5, 10 or 20
μg/ml
Positivo NE (PATTERSON
e BUTLER,
1982)
Aberração cromossômica
E. Coli K-12
Aberração
Cromossômica
- NE Negativo NE (KARPLIUK
et al., 1984,
apud EFSA,
2009)
Salmonella/ microssoma
TA92, TA94, TA98,
TA100, TA1535 e
TA1537
Mutação de
ponto
Com e Sem 5mg/placa Negativo NE (ISHIDATE, et
al., 1984)
31
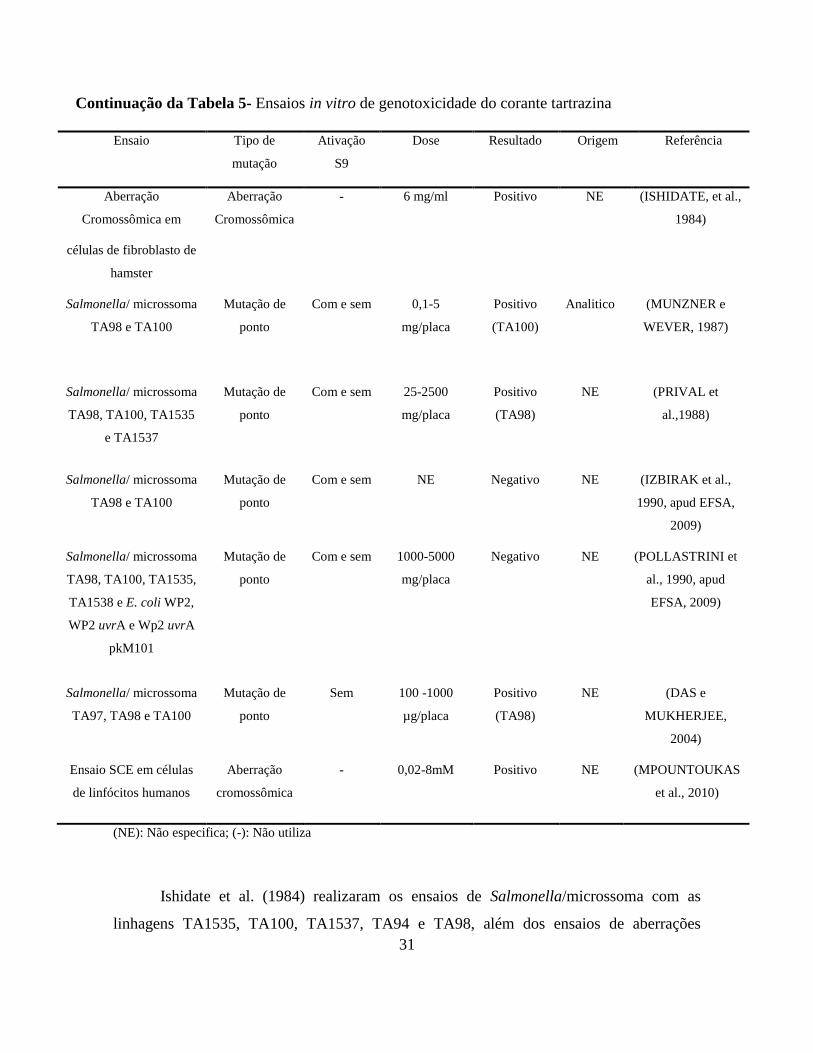
Continuação da Tabela 5- Ensaios in vitro de genotoxicidade do corante tartrazina
Ensaio Tipo de
mutação
Ativação
S9
Dose Resultado Origem Referência
Aberração
Cromossômica em
células de fibroblasto de
hamster
Aberração
Cromossômica
- 6 mg/ml Positivo NE (ISHIDATE, et al.,
1984)
Salmonella/ microssoma
TA98 e TA100
Mutação de
ponto
Com e sem 0,1-5
mg/placa
Positivo
(TA100)
Analitico (MUNZNER e
WEVER, 1987)
Salmonella/ microssoma
TA98, TA100, TA1535
e TA1537
Mutação de
ponto
Com e sem 25-2500
mg/placa
Positivo
(TA98)
NE (PRIVAL et
al.,1988)
Salmonella/ microssoma
TA98 e TA100
Mutação de
ponto
Com e sem NE Negativo NE (IZBIRAK et al.,
1990, apud EFSA,
2009)
Salmonella/ microssoma
TA98, TA100, TA1535,
TA1538 e E. coli WP2,
WP2 uvrA e Wp2 uvrA
pkM101
Mutação de
ponto
Com e sem 1000-5000
mg/placa
Negativo NE (POLLASTRINI et
al., 1990, apud
EFSA, 2009)
Salmonella/ microssoma
TA97, TA98 e TA100
Mutação de
ponto
Sem 100 -1000
µg/placa
Positivo
(TA98)
NE (DAS e
MUKHERJEE,
2004)
Ensaio SCE em células
de linfócitos humanos
Aberração
cromossômica
- 0,02-8mM Positivo NE (MPOUNTOUKAS
et al., 2010)
(NE): Não especifica; (-): Não utiliza
Ishidate et al. (1984) realizaram os ensaios de Salmonella/microssoma com as
linhagens TA1535, TA100, TA1537, TA94 e TA98, além dos ensaios de aberrações

32
cromossômicas in vitro em células de fibroblastos de hamster chinês com 190 aditivos
alimentares sintéticos e 52 aditivos alimentares derivados de fontes naturais. De um total de
200 aditivos testados no ensaio de Salmonella/microssoma 14 apresentaram efeitos positivos
e 54 de um total de 242 foram positivos no teste cromossômico. Em relação ao corante
tartrazina, este apresentou resultados negativos no ensaio de mutagenicidade. Para o ensaio
cromossômico, foi encontrado um pequeno aumento na incidência de células poliplóides
após a exposição das células de fibroblastos de hamster chinês ao corante.
O potencial mutagênico de metabólitos não identificados do corante tartrazina foi
avaliado em amostras de urina de ratos através de ensaios de Salmonella/microssoma com as
linhagens TA98 e TA100, empregando o método de pré - incubação. Após a adição de S9 foi
observado um potencial mutagênico com doses dependentes para a linhagem TA98
(HENSCHLER e WILD, 1985).
Os resultados descritos por Henschler e Wild (1985) citados anteriormente não foram
confirmados por Munzner e Wener (1987) ao administrarem uma dose única de 1,5 g / kg de
peso corporal de tartrazina dissolvida em 2 mL de NaCl a 0,9% em ratos. Uma hora após
essa administração foi coletado bile e por um período de 24horas foi coletado urina e fezes.
Estas amostras foram tratadas e soluções foram preparadas, em seguida foi realizado um
ensaio de mutagenicidade utilizando a metodologia de incorporação em placas com as
linhagens TA98 e TA100 em presença e ausência de ativação metabólica. Os resultados
foram negativos para as amostras de bile e urinas para ambas as linhagens. Porém para as
amostras de fezes em doses de 5 mg/placa em presença de S9 foi observado um aumento
significativo no número de revertentes quando comparadas ao grupo controle (MUNZNER
e WENER, 1987).
Prival et al. (1988) avaliaram a atividade mutagênica de 4 azocorantes (tartrazina,
amarelo crepúsculo, amaranto e vermelho 40) e dos produtos após a sua azoredução,
utilizando as linhagens TA98, TA100, TA1535 e TA1537, por meio do ensaio de
Salmonella/microssoma empregando quatro metodologias diferentes. Os autores não
observaram nenhuma atividade mutagênica para os corantes em doses variando de 0,1 a 10
mg / placa, em ausência e presença de S9, usando a metodologia de incorporação em placas
(1). O corante tartrazina também apresentou resultados negativos quando empregado o ensaio

33
de incorporação em placas utilizando extratos obtidos com éteres provenientes do corante
(2). Já para os outros corantes foi observada uma atividade apenas em doses elevadas. Os
corantes não mostraram atividade mutagênica quando testados com o ensaio de pré-
incubação em presença de flavina mononucleotídeo (FMN) e S9 provenientes de
microssomas de fígado de hamster (3). Porém quando as amostras de corantes foram
reduzidas com ditionito de sódio e em seguida extraída com éter (4), os corantes tartrazina e
amarelo crepúsculo, apresentaram atividade mutagênica para a linhagem TA98 em doses
mais elevadas de 2500 mg/placa na presença de S9. Já os corantes, amaranto e vermelho 40
apresentaram atividade mutagênica em doses menores de 25 mg/placa e 1 mg/placa
respectivamente para a linhagem TA98 em presença de S9, em doses mais altas foi observada
atividade mutagênica para as linhagens TA100 e TA1537.
No entanto, Das e Mukherejee (2004) avaliando o efeito mutagênico do corante
tartrazina por meio do ensaio de Salmonella/microssoma utilizando a metodologia de
incorporação em placas, observaram um aumento significativo no número de colônias
revertentes da linhagem TA98 nas doses de 100 e 250 µg/placa. Além disso, o corante
tartrazina por meio do ensaio (in vitro) em células humanas de sangue periférico apresentou
um efeito tóxico significativo em doses de 4 e 8 mM em relação à qualidade de
cromossomos, de modo que a mancha de diferenciação entre as duas cromatídes não estava
bem clara (MPOUNTOUKAS et al., 2010).
3.6.4 Estudos in vivo de genotoxicidade do corante tartrazina
Os corantes curcumina e tartrazina foram avaliados em células provenientes da
medula óssea de ratos e camundongos após a exposição aguda e crônica. Os resultados
indicam que a tartrazina é mais clastogênica que o corante curcumina, uma vez que resultou
em um aumento significativo de aberrações cromossômicas em doses de 2000 e 1000
mg/kg/peso corporal/dia em exposições aguda e crônica, respectivamente de tartrazina na
alimentação (GIRI et al., 1990, apud EFSA, 2009).

34
Esses resultados também foram observados em células de fibroblasto de hamster
chinês, após uma exposição a 2,5 mg/mL de corante tartrazina por período de 48 horas foi
observado uma incidência de 2 % de células poliploídes e uma incidência de 23% de células
com aberrações cromossômicas estruturais (ISHIDATE et al., 1984).
A atividade citogenética de seis corantes alimentares incluindo tartrazina foram
analisados em camundongos C57B1/ 6, foram administradas por via oral doses diárias de 0,5
e 5,0 mg/kg de tartrazina durante 5 dias. Após o período de tratamento foi realizada a análise
de quinhentas metáfases de células, e uma porcentagem de células com anomalias foram
observadas, porém estas não diferiram dos grupos controle. Para os autores o corante
tartrazina não aumentou a incidência de aberrações cromossômicas em células de medula
óssea de camundongo (DURNEV et al., 1990, apud EFSA, 2009). E segundo o parecer da
EFSA (2009) os tipos de aberrações cromossômicas não foram descritos apresentando
detalhes nem usando a terminologia internacional, indicando uma impossibilidade de
conclusões fundamentadas.
Sasaki et al. (2002) avaliaram a genotoxicidade de 39 produtos que são usados como
aditivos alimentares. Foi realizado o ensaio do cometa em ratos após 3 e 24 horas da
administração de dose máxima de (2000 mg / kg de peso corporal) em células do estômago
glandular , cólon , fígado , rim , bexiga , pulmão, cérebro e da medula óssea. Os resultados
demostraram que os corantes tartrazina, amaranto, allura red e eritrosina foram capazes de
induzir danos ao DNA no estômago, cólon e/ou bexiga e nos órgãos gastrointestinais, a partir
de uma dose de 10 mg/Kg de peso corporal. Entretanto após uma reavaliação pela EFSA
(2009) os autores concluiram que os danos observados no cólon de ratos podem ser
explicados por uma citotoxicidade local do corante.
Das e Mukherejee (2004) avaliaram o corante tartrazina após a administração de
doses de 50, 100 e 200 mg/kg de peso corporal através do ensaio de genotoxicidade por via
intraperitonial em camundongos Swiss. Os resultados demonstraram que o corante não possui
efeito genotóxico, uma vez que a frequência do número de células aberrantes e a quebra
cromossômica não foram significativamente mais elevadas, quando comparado com o grupo
controle. Estes resultados também são confirmados por Poul et al. (2009) através do ensaio
de micronúcleo em células do intestino de roedores. Os autores observaram a ausência de

35
genotoxicidade após a administração oral em doses de até 2000 mg/kg de peso corporal dos
corantes: amaranto, tartrazina, amarelo crepúsculo e metabólitos de aminas aromáticas.
3.7 Estudos de carcinogenicidade
Os resultados de estudos de toxicidade de longo prazo, descritos na avaliação do
JEFCA (1966) demonstram que o corante não apresenta aumento na incidência tumoral em
ratos (WATERMAN e LIGNAC, 1958; WILLHEIM e IVY, 1953; MANNELL et al., 1958
apud JEFCA, 1966) e em cães (DAVIS et al, 1964, apud JEFCA, 1966).
No estudo realizado por Mannell et al. (1958) não foi observado efeitos sobre a
mortalidade, crescimento, histopatologia e incidência tumoral após a administração do
corante tartrazina em dietas de ratos equivalentes a 0, 15, 150, ou 750 mg / kg de peso
corporal/dia por um período de 64 semanas. Neste estudo estabeleceu-se uma ingestão diária
aceitável de 750 mg/Kg de peso corporal/dia baseados no NOAEL (níveis de efeitos não
observados) em um fator 100 de segurança (MANNELL et al.,1958 apud JEFCA, 1966).
Baseando-se neste trabalho e nos citados anteriormente pela JECFA (1966) e
seguindo as avaliações do Comitê Científico da Alimentação Humana (CCAH) em 1975 e
1984, foi estabelecida a ingestão diária aceitável de 7,5 mg/Kg para o corante tartrazina.
Maekawa et al. (1987) não observaram lesões tóxicas após a exposição máxima por
um período de 2 anos em ratos de ambos os sexos nas doses de 0, 500 e 1000 mg/kg de peso
corporal/dia de tartrazina adicionada em água potável. Embora tenha sido observado uma
incidência de mesotelioma e pólipos endometriais em machos e fêmeas respectivamente em
grupos que receberam 500 mg/kg de peso corporal/dia de tartrazina. Para os autores os
mesoteliomas e pólipos endometriais são tumores espontâneos observados frequentemente
nestas linhagens de ratos (F344) e concluíram que o aumento significativo desta incidência
não foi atribuído à administração de tartrazina e o mesmo não foi carcinogênico para a
linhagem de ratos F344.

36
Grupos de camundongos com 42 dias de idade foram tratados com dietas contendo
doses de tartrazina de 0, 714 e 870; 2173 e 2662; 8103 e 9735 mg/kg de peso corporal/dia
(machos e fêmeas respectivamente) a uma exposição máxima por 104 semanas para
avaliação de um estudo crônico de toxicidade e carcinogenicidade. Foi observada uma
incidência de neoplasias e algumas alterações histológicas, porém para os autores as
alterações encontradas não diferiram significativamente entre o controle e os animais
tratados, sugerindo que esses resultados são encontrados normalmente em animais em fase
de envelhecimento (BORZELLECA e HALLAGAN, 1988b).
O uso prolongado do corante tartrazina foi investigado por Moutinho et al. (2007)
através de 45 ratos Wistar, machos, divididos em grupo controle (A) e tratado (B). O grupo
tratado recebeu o corante diariamente em água de beber ad libitum, na dose de 7,5 mg/kg/dia
por um período de dez meses, do desmame aos doze meses de idade. Os resultados
demonstram um aumento significante do número de linfócitos e eosinófilos na mucosa do
antro gástrico. No entanto não foram observadas alterações carcinogênicas em nenhuma das
regiões gástricas com a dose e o tempo utilizados no estudo. O parecer científico elaborado
pela EFSA (2009) considera que essas mudanças observadas pelos autores Moutinho et al.
(2007) não eram relacionadas com a administração do corante tartrazina indicando que os
estudos de carcinogenicidade disponíveis demonstraram que o corante não induz neoplasias
benigna e/ou maligna.
3.7.1 Toxicidade reprodutiva e de desenvolvimento
Levando em consideração que as mutações podem ocorrer em células de linhagem
germinativa podendo ser propagadas a geração seguinte (COMBES, 1992, MARON e
AMES, 1983) torna-se importante a investigação sobre os efeitos de reprodução e
desenvolvimento do feto.
As avaliações da toxidade reprodutiva e de desenvolvimento com corante tartrazina
foram realizados por Collins et al. (1990, 1992) e os resultados demonstraram que não houve
nenhuma alteração relacionada a efeitos teratogênicos após a administração do corante por

37
gavagem em ratos nas doses de 0, 60, 100, 200, 400, 600 e 1000 mg/kg/dia e nem após a
administração do mesmo em água potável nas concentrações de 0, 67, 132, 292, 568 e 1064
mg/kg de peso corporal/dia em ratos.
Borzelleca e Hallagan (1988a) também realizaram um estudo com ratos a partir da
fase uterina onde foi administrado tartrazina na dieta em doses de 48 e 58; 491 e 589; 984 e
1225; 2641 e 3348 mg/kg de peso corporal/dia para machos e fêmeas respectivamente da
geração F0 (ratos 63-70 dias de idade) durante dois meses anteriores de acasalamento. De
forma aleatória após a conclusão da fase uterina foram selecionados dois ratos/sexo/ninhada
para o estudo crônico, sendo estes descendentes da F0, ou seja, os indivíduos da geração F1.
Estes animais foram expostos aos mesmos níveis de dieta dos pais, por um período de 113 e
114 semanas para as doses menores e, nas doses maiores os animais foram expostos por 122
e 125 semanas (machos e fêmeas respectivamente). Uma análise hematológica, bioquímica
e um estudo histopatológico foram realizados regularmente. Para os autores não houve
diferenças entre os grupos controle e os grupos tratados com o corante em relação à
incidência de massas palpáveis. Uma variedade de lesões, incluindo tumores detectados
através dos exames histológicos, alguns dos exames hematológicos e clínicos diferiram
significativamente, porém nenhuma dessas diferenças indica estar relacionado com o
tratamento e pareciam estar comumente relacionados a achados de forma espontânea em
ratos idosos. Com base nesses dados os autores afirmaram que o NOAEL foi de 5,0%
utilizando a dose maior na dieta o equivalente a 2641 e 3348 mg/kg de peso corporal/dia para
ratos machos e fêmeas, respectivamente. O parecer da EFSA, (2009) concorda com esse
NOAEL, uma vez que o peso e mudanças observadas não se fizeram significativas.
Como no estudo anterior, a pesquisa realizada por Tanaka et al. (2006) os autores não
observaram alterações em relação aos parâmetros reprodutivos, após o tratamento de grupos
de 20 ratos (10 animais de cada sexo) com doses de 83, 259, e 773 mg / kg de peso corporal
/ dia do corante tartrazina a partir de cinco semanas de idade da geração F0 a nove semanas
de idade da geração F1. Nos animais da geração F1, não foi observado interferência no
número de animais recém-nascidos, no peso e na relação da proporção entre os sexos. Já no
período da lactação foi observado um índice de sobrevivência significativamente menor nos

38
grupos de doses média e também aumento de peso corporal de animais do sexo masculino,
no grupo de dose mais alta no período de 0 a 21 dias após o nascimento.
Em outro estudo realizado por Tanaka et al. (2008) também não houve indicações de
efeitos adversos relacionados com a tartrazina em relação a reprodução e/ou
desenvolvimento, após o tratamento de ratos com doses de 83, 259 e 773 mg/kg de peso
corporal/dia, do corante tartrazina na dieta, a partir de cinco semanas de idade da geração F0
a nove semanas de idade da geração F2. Os resultados em relação aos ensaios
comportamentais realizados durante o período de lactação apresentaram algumas indicações
de diferenças no desempenho dos animais tratados em comparação com o grupo controle,
apresentando uma coordenação mais acelerada.
No entanto, segundo o parecer elaborado pela EFSA ao rever estes dois estudos
descritos por Tanaka et al. (2006 e 2008) observaram que estes resultados não são
consistentes, e nenhum efeito dose-resposta pode ser observado. O parecer conclui que estes
estudos não demonstraram quaisquer efeitos adversos da tartrazina na reprodução e no
desenvolvimento neurocomportamental. E que uma revisão da IDA com base nestes dados,
portanto, não se justifica (EFSA, 2009).

39
4 MATERIAL E MÉTODOS
4.1 Equipamentos
Agitador de soluções do tipo vortex (Phoenix AP56)
Agitador magnético (Tecnal – TE 0851)
Autoclave vertical (Marconi)
Balança analítica (Marte)
Balança semi-analítica (Shimadzu – AUW220D)
Banho maria (Fiston- Mod. 550)
Banho seco – Dry Block (Marconi – MA4004)
Capela de Segurança Biológica Classe II B2 (Filterflux)
Capela de Segurança Química (Filterflux)
Contador de colônias (Phoenic Luferco – CP600 Plus)
Dispensador automático – Bomba peristáltica (Ominispense® Plus)
Dispenser manual (Eppendorf Varispenser®)
Estufa para esterilização e secagem (Sterilifer)
Estufa Bacteriológica (Marconi-MA032)
Incubadora bacteriológica (Tecnal- TE- 420)
Microondas (Eletrolux - MES27)
Micropipetadores
Refrigerador /Freezer (Consul Pratice 410 duplex)
Ultrapurificador de água (Milliq-Q® - Academic Millipore)
4.2 Vidrarias e outros materiais
Alça de Drigalski
Alça de inoculação
Balão volumétrico
Béqueres
Bico de Busen
Canetas marcadoras para vidro de diferentes cores

40
Criotubos
Estantes para tubos de ensaio
Frascos de vidro com tampa rosqueavél do tipo “Schott”
Frascos para soluções do tipo falcon
Filtros descartáveis para esterilização
Fita de autoclave
Luvas do tipo cirúrgicas
Membrana filtrante 0,45 m (Millipore®)
Óculos de proteção
Provetas
Tubos de ensaio de vidro com tampas
Papel de filtro
Papel alumínio
Parafilme
Placas de Petri descartáveis
Pinças
Ponteiras para micropipetadores
Zaragatoas
4. 3 Reagentes
Acetona PA ACS (CAS: 67-64-1, Qhemis)
Ácido Cítrico monohidratado (CAS: 5949-291, Merck)
Acridina Mutagen ICR 191 (CAS: 17070-45-0, Sigma)
Álcool anidro (Ciclo Farma)
2 aminoantraceno 96% (CAS: 613-13-8, Sigma)
Ampicilina (CAS: 7177- 48-2, Sigma)
Bacto ágar (BD)
Bacto peptona (BD)
β-fosfato de nicotinamida adenina dinucleotídeo ( NADP) (CAS:68999-85B, Sigma)
D- Biotina (CAS: 58-85-5, Sigma)
Dimetilsulfóxido 99,5% (DMSO) (CAS: 67-68-5, Sigma)

41
Caldo nutriente nº2 (Oxoid)
Cloreto de magnésio hexahidratado (CAS: 779-18-6, Sigma)
Cloreto de potássio (CAS: 7447-40-7, Carlo Erba)
Cloreto de sódio (CAS:7647-145, Carlo Erba)
Cristal violeta (CI: 42555, Synth)
Dextrose anidra (D-(+) –Glucose) (CAS: 50-99-7, Sigma)
Fosfato de Potássio dibásico anidro (CAS: 4-11-7758, Sigma)
Fosfato de sódio dibásico anidro (CAS: 7558-79-4, J.T. Baker)
Fosfato de Potássio dibásico anidro (K2HPO4)
Fosfato de sódio monobásico monoidratado (CAS: 1004-21-5, Carlo Erba)
Fosfato de sódio e amônio tetraidratado (CAS: 778313-3, J.T. Baker)
Fração S9 (indutor enzimático Aroclor -1254, Moltox)
D-glicose 6-fosfato (CAS: 3671-99-6, Sigma)
L – Histidina HCl monoidratada (CAS: 5934-29-2, Sigma)
4-Nitroquinolina-1-óxido (4NQO) (CAS: 56-57-5, Sigma)
Sulfato de Magnésio heptahidratado (CAS: 10034-99-8, Sigma)
Tetraciclina cloridrato (CAS: 64-75-5, Sigma)
4. 4 Preparos de Meios de Cultura e Soluções
Os meios de cultura e as soluções utilizadas no ensaio e na confirmação dos genótipos
das linhagens foram preparados conforme Maron e Ames (1983) e Mortelmans e Zeiger
(2000) sendo apresentadas a seguir. Todos os ensaios de mutagenicidade, preparação de
meios e soluções foram realizados no Laboratório Central Analítica (LACAN) da Faculdade
de Tecnologia da Unicamp – Campus em Limeira, São Paulo.
Todos os meios e soluções foram esterilizados em autoclaves a 121 ºC (1 atm) por 20
minutos, exceto os casos que necessitaram de uma esterilização por membrana filtrante. Os
materiais utilizados no preparo dos meios e no ensaio como, ponteiras, dispenser, zaragatoa,

42
frascos do tipo Schott, criotubos e tubos falcon foram acondicionados em local apropriado
envoltos em papel alumínio e esterilizados em autoclave a 121 ºC (1 atm) por 20 minutos.
Os tubos de ensaio com tampa utilizados no ensaio foram armazenados em local apropriado
e envoltos em papel alumínio e submetidos à esterilização em estufa a 180 ºC por 3 horas.
As placas de Petri utilizadas foram adquiridas previamente esterilizadas por radiação γ –
cobalto.
Antecedendo o ensaio de mutagenicidade foi realizado um ensaio de esterilidade
colocando as placas de ágar mínimo e ágar nutriente em estufas a 37 ºC por 12 horas,
descartando as placas que apresentassem qualquer contaminação visual. As amostras do
corante tartrazina foram analisadas quanto à contaminação utilizando placas de ágar
nutriente, previamente ao ensaio.
Ágar Mínimo
Componentes Para 1000 mL
Meio de Vogel Bonner 50x 20 mL
Solução de Glicose 10% 200 mL
Solução de Ágar 780 mL
Em condições assépticas e com o auxílio de um agitador magnético aquecido a (50-60
ºC), os meios foram dissolvidos e em seguida 25 mL foram distribuídos em placa de Petri
estéreis utilizando um dispensador automático. Após a solidificação do meio, as placas foram
invertidas e armazenadas em geladeira a 4 ºC por um período máximo de 14 dias.

43
Meio Vogel Bonner 50x:
Componentes Para 1000 mL
Sulfato de Magnésio heptahidratado 10 g
Ácido Cítrico monohidratado 100 g
Fosfato de Potássio dibásico anidro 500 g
Fosfato de sódio e amônio tetrahidratado 175 g
Água ultrapura q.s.p 1000 mL
Todos os reagentes na ordem acima foram dissolvidos e distribuídos em volumes de
100 mL em frascos âmbar com capacidade de 250 mL. Após a esterilização o meio foi
armazenado em geladeira a 4ºC.
Solução de Glicose 10%
Componentes Para 1000 mL de ágar mínimo
Dextrose Anidra 20 g
Água ultrapura q.s.p. 200 mL
Após a completa dissolução de 20g de dextrose anidra em um frasco contendo 150 mL
de água, o volume final foi ajustado para 200 mL. Após a esterilização a 121ºC por 20
minutos, o meio foi estabilizado em banho-maria a 50-60ºC até o preparo do ágar mínimo.
Para os ensaios com a linhagem TA97a foi utilizada uma solução de glicose a 0,4%, uma vez
que essas linhagens são sensíveis a altas concentrações de glicose.
Solução de Ágar

44
Componentes Para 1000 mL de ágar mínimo
Bacto Ágar 15 g
Água ultrapura 780 mL
15g de bacto ágar foi dissolvido em 780 ml de água em um frasco com tampa, com o
auxílio de um agitador magnético e aquecido a (50-60ºC) até a completa dissolução do meio.
Após a esterilização do meio a 121ºC por 20 minutos, o meio foi estabilizado em banho-
maria a (50-60ºC) até o preparo do ágar mínimo.
Ágar Mínimo com Biotina
O Ágar mínimo com biotina é utilizado nos ensaios para confirmação das características
genéticas das linhagens de Salmonella typhimurium.
Componentes Para1000 mL
Meio de Vogel Bonner 50x 20 mL
Solução de Glicose 10% 200 mL
Solução de Ágar 780 mL
Solução de bitona 0,5 Mm 6,5 mL
Em condições assépticas e com o auxílio de um agitador magnéticoaquecido a (50-60
ºC), os meios foram dissolvidos e em seguida 25mL foram distribuídos em placa de Petri
estéreis utilizando um dispensador automático. Após a solidificação do meio, as placas foram
invertidas e armazenadas em geladeira a 4ºC.
Ágar Mínimo com Biotina/Histidina

45
O Agar mínimo com biotina e histidina, também é utilizado nos ensaios para confirmação
das características genéticas das linhagens de Salmonella typhimurium.
Componentes Para 1000 mL
Meio de Vogel Bonner 50x 20 mL
Solução de Glicose 10% 200 mL
Solução de Ágar 780 mL
Solução de bitona 0,5 Mm 6 mL
Solução de histidina 0,5 % 8 mL
Em condições assépticas e com o auxílio de um agitador magnéticoaquecido a (50-60
ºC), os meios foram dissolvidos e em seguida 25mL foram distribuídos em placa de Petri
estéreis utilizando um dispensador automático. Após a solidificação do meio, as placas foram
invertidas e armazenadas em geladeira a 4 ºC.
Solução de Histidina 0,5 %
A solução de histidina 0,5 % é utilizada no preparo do ágar mínimo com histidina/biotina
para a avaliação da auxotrofia para histidina presentes nas linhagens de Salmonella
typhimurium.
Componentes Para 100 mL
L – Histidina HCl monoidratada 0,5 g
Água ultrapura 100 mL

46
A histidina foi dissolvida em água e volumes de 50mL foram distribuídos em frascos
âmbar e submetidos à esterilização por autoclave a 121 ºC por 20 minutos. Após a
esterilização a solução foi armazenada em geladeira por um período máximo de 45 dias.
Solução de Biotina 0,5mM
A solução de com biotina 0,5 mM é também utilizada no preparo do ágar mínimo com
biotina para a avaliação da auxotrofia para histidina.
Componentes Para 100 mL
D- Biotina (MM = 244,3) 0,012 g
Água ultrapura 100 mL
A biotina foi dissolvida em água e volumes de 50 mL foram distribuídos em frascos
âmbar e submetidos a esterilização por autoclave a 121ºC por 20 minutos. Após a
esterilização a solução foi armazenada em geladeira por um período máximo de 45 dias.
Ágar Nutriente
O ágar nutriente além de ser utilizado para verificar a esterilidade das amostras
antecedendo os ensaios, também é usado para verificar a viabilidade e confirmação de
algumas características genéticas das linhagens utilizadas no ensaio.

47
Componentes Para 1000 mL
Caldo Nutriente Nº2 25 g
Bacto Ágar 15 g
Água ultrapura 1000 mL
Após a homogeneização dos reagentes, o meio foi esterilizado em autoclave a 121ºC por
20 minutos. Em uma câmara de fluxo laminar volumes de 25mL foi distribuído em placas de
Petri esterilizadas previamente com o auxílio de um dispensador automático. Após a
solidificação do meio, as placas foram invertidas e armazenadas em geladeira a 4 ºC. O meio
pode ser utilizado enquanto não apresentar contaminação visual.
Ágar de Superfície (Top ágar)
O meio de ágar de superfície é essencial para o ensaio, uma vez que é fonte do aminoácido
histidina para as linhagens.
Componentes Para 1000 mL
Bacto Ágar 6 g
Cloreto de sódio 6 g
Solução de biotidina 0,5mM 100 mL
Água ultrapura 900 mL
Após a completa homogeneização do bacto ágar e cloreto de sódio foram adicionados
100 mL de solução de biotidina 0,5 mM e volumes de 200 mL foram distribuídos em frascos

48
de 250 mL e autoclavados a 121 ºC por 20 minutos. Após a esterilização os frascos foram
armazenados em geladeira a 4 ºC.
Solução de Biotidina 0,5mM
Esta solução é empregada no top ágar para garantir um excesso de biotina e de histidina
para as linhagens de S.typhimurium usadas no ensaio.
Componentes Para 1000 mL
L – Histidina HCl 0,105 g
D- Biotina 0,122 g
Água ultrapura 1000 mL
Após a completa homogeneização dos reagentes, a solução foi distribuída em volumes
de 100mL em frascos âmbar com capacidade de 250 mL, esterilizada em autoclave a 121 ºC
por 20 minutos e armazenada em geladeira por um período de 60 dias.
Caldo Nutriente
O caldo nutriente é usado para incubar as linhagens antecedendo aos ensaios.
Componentes Para 1000 mL
Caldo Nutriente Nº2 25 g
Água ultrapura 1000 mL

49
Após a dissolução do caldo nutriente em água ultrapura, volumes de 20 mL foram
distribuídos em frascos de vidro com tampa com capacidade de 50 mL e submetidos à
esterilização. Após a esterilização os frascos foram armazenados em geladeira a 4 ºC.
Mistura S9
A mistura de S9 é utilizada no ensaio que necessita de um sistema exógeno, permitindo
identificar mutágenos que precisam de uma ativação metabólica para se tornarem ativos.
Componentes Porcentagem
Água ultrapura estéril 39,5 %
Tampão fosfato 0,2M 50 %
NADP 0,1M 4 %
Glicose-6-fosfato 1M 0,5 %
Solução de Sais 2 %
Fração S9 4 %
Todas as soluções preparadas previamente foram adicionadas assepticamente a um
frasco estéril, homogeneizadas e mantidas a 4ºC. Após o ensaio a solução excedente foi
descartada.

50
Solução de NADP 0,1M
Componentes
NADP 1 g
Água ultrapura estéril 13,06 mL
Em condições assépticas o NADP foi dissolvido em água e esterilizado por membrana
filtrante de 0,45 m. A solução foi transferida para um tubo falcon com capacidade de 25
mL previamente esterilizado e em seguida armazenado em freezer a -20 °C. Uma gota de
solução pronta foi incubada em placa de ágar nutriente por 24h a 37 ºC para controle de
esterilidade.
Solução de Tampão Fosfato 0,2M
Solução A
Componentes Para 100 mL
Fosfato de sódio dibásico anidro 2,84 g
Água ultrapura 100 mL
Solução B
Componentes Para 100 mL
Fosfato de sódio monobásico monohidratado 2,76 g
Água ultrapura 100

51
81 mL da solução A e 19 mL da solução B, foram misturados e o pH ajustado para 7,4,
com a solução A ou B (Solução A para aumentar e solução B para diminuir o pH). Após a
esterilização, a solução foi armazenada frasco de vidro com tampa de rosca e capacidade de
250 mL, em temperatura ambiente.
Glicose - 6 fosfato 0,1M
Componentes Para 10 mL
Glicose 6-fosfato 3,041 g
Água ultrapura estéril 10 mL
Em condições assépticas a glicose-6-fosfato foi dissolvida em água ultrapura, e
esterilizada por membrana filtrante de 0,45 m. Em seguida foi transferida para um tubo
falcon com capacidade de 25 mL previamente esterilizada e armazenada em freezer a -20 °C.
Uma gota de solução pronta foi incubada em placa de ágar nutriente por 24h a 37 ºC para
controle de esterilidade.
Solução de Sais - Cloreto de Magnésio 0,4M e Cloreto de Potássio 1,65M
Componentes Para100 mL
Cloreto de Magnésio 8,13 g
Cloreto de Potássio 12,3 g
Água ultrapura q.s.p. 100 mL

52
Todos os reagentes foram dissolvidos em água ultrapura. Após a esterilização em
autoclave, a solução foi armazenada frasco de vidro com tampa de rosca e capacidade de
250mL em geladeira a 4 ºC.
Fração S9
Componentes
Fração S9 1 frasco
Água ultrapura 2,1 mL
A fração S9 foi reconstituída com 2,1 mL de água ultra pura estéril. Volumes de 200
µL foram distribuídos em criotubos e armazenados em freezer a -20 ºC.
Solução de Ampicilina 8mg/mL
Esta solução é utilizada em meios que servem para confirmar a presença de plasmídio
com marcador de resistência à ampicilina nas linhagens de S.typhimurium
Componentes Para 10 mL
Ampicilina 0,08 g
Água ultrapura 10 mL
Após a homogeneização dos reagentes, a solução foi esterilizada por membrana filtrante
de 0,45 m e transferida para um tubo falcon com capacidade de 25 mL previamente
esterilizado e envolto com papel alumínio, em seguida foi armazenada em geladeira a 4°C.

53
Solução de Tetraciclina 8mg/mL
Usada nos meios que servem para confirmar a presença de plasmídio com marcador de
resistência à tetraciclina nas linhagens de Salmonella typhimurium (pAQ1 na TA102).
Componentes Para10 mL
Tetraciclina cloridrato 0,08 g
Água ultrapura 10 mL
A tetraciclina foi dissolvida em água e em seguida esterilizada por membrana filtrante
0,45 m. Após a esterilização a solução foi transferida para um tubo falcon com capacidade
de 25 mL previamente esterilizado e envolto com papel alumínio, em seguida foi armazenada
em geladeira a 4°C.
Solução de Cristal Violeta a 0,1%
A solução de cristal violeta é utilizada para avaliar as características genéticas, ou seja,
para confirmar a presença da mutação no gene rfa.
Componentes Para 100 mL
Cristal violeta 100 mg
Água ultrapura 100 mL
O cristal violeta foi dissolvido em água ultrapura e transferido para um frasco âmbar com
tampa de rosca e capacidade de 100 mL. A solução foi armazenada em temperatura ambiente.

54
Água de Diluição
Usada para avaliar a viabilidade das linhagens de S.typhimurium.
Componentes Para 1000 mL
Bacto Peptona 1 g
Água ultrapura 1000 mL
Após a dissolução da peptona em água ultrapura, a solução foi distribuída em volumes
de 100 mL em frascos de vidro com capacidade de 250 mL esterilizados e armazenados em
geladeira a 4 ºC.
Solução de Ácido Sulfúrico/Ácido Nítrico
É uma solução utilizada no procedimento de lavagem de tubos.
Solução de ácido sulfúrico a 50%
Componentes Para 1000 mL
Ácido sulfúrico 500 mL
Água destilada 500 mL

55
Solução de ácido nítrico a 50%
Componentes Para 1000 mL
Ácido nítrico 500 mL
Água destilada 500 mL
Adicionou-se 500 mL de água em um balão volumétrico com capacidade de 2000 a
3000 mL e aos poucos o ácido sulfúrico foi sendo adicionado agitando constantemente. A
solução foi esfriada em banho de gelo. O preparo da solução de ácido nítrico foi realizado
adicionando 500 mL de água em um balão volumétrico e aos poucos foi adicionado o volume
de 500 mL de ácido nítrico, agitando constantemente. Em seguida foram adicionados 600
mL de solução de ácido sulfúrico a 50% em um balão volumétrico com capacidade de 2000
a 3000 mL e 100 mL da solução de ácido nítrico a 50 %. O volume final da solução foi
armazenado em frasco de vidro com tampa de rosca e capacidade de 1000 mL e mantida a
temperatura ambiente.
4.5 Amostras de corantes
O corante alimentício tartrazina analítico (≥99%) (CAS Nº 1934-21-0), foi obtido da
empresa Sigma-Aldrich® e o seu respectivo corante comercial (90%) foi adquirido da
empresa Duas Rodas Industrial. As soluções foram preparadas utilizando como solvente água
deionizada estéril.

56
4.6 Manutenção das linhagens de Salmonella typhimurium
As linhagens de Salmonella typhimurium foram armazenadas em freezer a -20 °C, em
criotubos com 180 µL de cultura e 20 µL de dimetilsulfóxido (DMSO) como agente
crioprotetor. As características genéticas de cada linhagem foram avaliadas a cada seis meses,
conforme descrito abaixo. Essas características lhes conferem uma maior sensibilidade na
detecção de diversos tipos de agentes mutagênicos.
Uma alíquota de 100µL de cultura permanente foi inoculada em um frasco contendo
20mL de caldo nutriente e após um período de 12-18 horas overnight a 37 ºC sob agitação
de 150-170 rpm em incubadora bacteriológica tipo shaker, a cultura foi inoculada em placas
de ágar nutriente para o isolamento das colônias.
Após incubação de 24 horas a 37 ºC, em estufa bacteriológica, cinco colônias de cada
linhagem foram escolhidas ao acaso e cada uma foi inoculada em um tubo de ensaio contendo
5mL de caldo nutriente, suplementado com antibiótico correspondente.
4.7 Confirmações das características genéticas das linhagens de Salmonella
typhimurium utilizadas no ensaio
Utilizando uma incubadora bacteriológica tipo shaker cada cultura crescida nos 5mL
de caldo nutriente foi inoculada, com auxílio de zaragatoa previamente esterilizada, em
placas de ágar nutriente para verificação da presença dos plasmídios pKM101 e pAQ1, da
mutação rfa, da deleção de uvrB e auxotrofia para histidina.
4.7.1 Plasmídio pKM101
Para confirmação da existência dos plasmídios pKM101, com o auxílio de uma
zaragatoa foi aplicada uma estria de solução de ampicilina a 8mg/mL em placas de ágar
nutriente. Após secagem desta linha de solução, as linhagens a serem testadas foram estriadas

57
em placas de ágar nutriente sob a linha de antibiótico. As linhagens portadoras desse
plasmídio, que são resistentes à ampicilina, após a incubação a 37 ºC apresentavam
crescimento sobre a linha do antibiótico.
Figura 2- Esquema de confirmação da presença de plasmídio das genéticas das linhagens de
Salmonella typhimurium
(AN): Ágar Nutriente; (amp): ampicilina.
Fonte: (MORALES, 2009).
4.7.2 Plasmídio pAQ1
O procedimento foi o mesmo descrito para a confirmação da presença do plasmídio
pAQ1, porém foi utilizado uma solução de tetraciclina a 8 mg/mL (Figura 2).
4.7.3 Mutação rfa
Para confirmação da mutação rfa, a cultura foi estriada, com o auxílio de uma
zaragatoa, em placas de ágar nutriente (Figura 3). São adicionadas ao centro da placa discos
de papel de filtro de 5 mm de diâmetro embebidos com uma solução de cristal violeta a 0,1%.
A presença dessa mutação confere maior sensibilidade ao cristal violeta, uma vez que essa
mutação causa perda da barreira de lipopolissacarídeos da parede bacteriana, tornando a

58
bactéria mais permeável a moléculas volumosas. Após a incubação observa-se um halo de
inibição de crescimento bacteriano ao redor do disco devido a toxicidade do cristal violeta.
Figura 3 - Esquema de confirmação da mutação rfa das linhagens de Salmonella
typhimurium
(AN): Ágar Nutriente
Fonte: (MORALES, 2009).
4.7.4 Deleção do uvrB
Para confirmação dessa deleção, a cultura foi inoculada em placa de ágar nutriente
(Figura 4). Em seguida metade da placa foi coberta e outra metade exposta à luz ultravioleta
(UV) por 10 segundos. Essa mutação confere às linhagens uma deficiência no mecanismo de
reparo por excisão. Dessa forma as linhagens se tornam sensíveis à radiação ultravioleta. E
após a incubação deve-se observar um crescimento na metade da placa não irradiada e uma
inibição no crescimento na parte irradiada da placa.

59
Figura 4- Esquema de confirmação da deleção do gene uvrB das linhagens de Salmonella
typhimurium
(AN): Ágar Nutriente
Fonte: (MORALES, 2009).
4.7.5 Auxotrofia para histidina
A auxotrofia para a histidina foi avaliada inoculando alíquotas das culturas, por
estriamento, em placas de ágar mínimo contendo biotina e em placa de ágar mínimo contendo
histidina e biotina (Figura 5). Após a incubação deve-se observar ausência de crescimento
nas placas de ágar mínimo contendo biotina e presença de crescimento nas placas de ágar
mínimo contendo histidina e biotina.

60
Figura 5- Esquema de confirmação da auxotrofia para histidina das linhagens de
Salmonella typhimurium
(AM): Ágar Mínimo; (bio): biotina; (his): histidina
Fonte: ( MORALES, 2009).
4.7.6 Taxas de reversão espontânea
Cada linhagem apresenta uma característica especifica em relação a mutação
espontânea. Dessa forma a taxa de reversão espontânea é importante para avaliar a
integridade de cada linhagem a ser utilizada. Cada laboratório tem um intervalo de
crescimento característico de colônias com mutação reversa para cada linhagem que é a que
utilizado como "valores históricos de controle" (MARON e AMES, 1983, MORTELMANS
e ZEIGER, 2000). A média histórica da taxa de reversão das linhagens utilizadas no
laboratório LACAN pode ser observada na Tabela 6. Esta foi avaliada inoculando 100µL da
cultura em 2mL ágar de superfície mantido em banho seco a 45ºC, e verteu-se a mistura após
uma homogeneização sob a placa de ágar mínimo. Após a incubação por um período de 44-
66 horas observou-se e avaliou-se a taxa de reversão espontânea. Uma cultura das 5
analisadas de cada linhagem foi escolhida e utilizadas para o preparo dos ensaios, após a
avaliação das características genéticas.

61
Tabela 6 – Média histórica da taxa de reversão espontânea das linhagens utilizadas no ensaio
Salmonella/microssoma no LACAN
Linhagem
Taxa de Reversão
(LACAN)
TA97a
-S9 140-200
+S9 140-200
TA98 -S9 20-80
+S9 20-80
TA100
-S9 100-170
+S9 100-170
TA102 -S9 100-300
+S9 100-400
TA1535 -S9 5-20
+S9 5-20
4.8 Protocolo do ensaio de Salmonella/microssoma por incorporação em
placas
Foram realizados ensaios com as amostras dos corantes alimentícios tartrazina
analítico (≥99% de pureza - Sigma-Aldrich®) e o corante tartrazina comercial (90% de pureza
- Duas Rodas) com as linhagens TA97a, TA98, TA100, TA102 e TA1535, de acordo com
Mortelmans e Zeiger, 2000, por meio do método de incorporação em placas. A Figura 6
ilustra o protocolo do ensaio de Salmonella/microssoma.
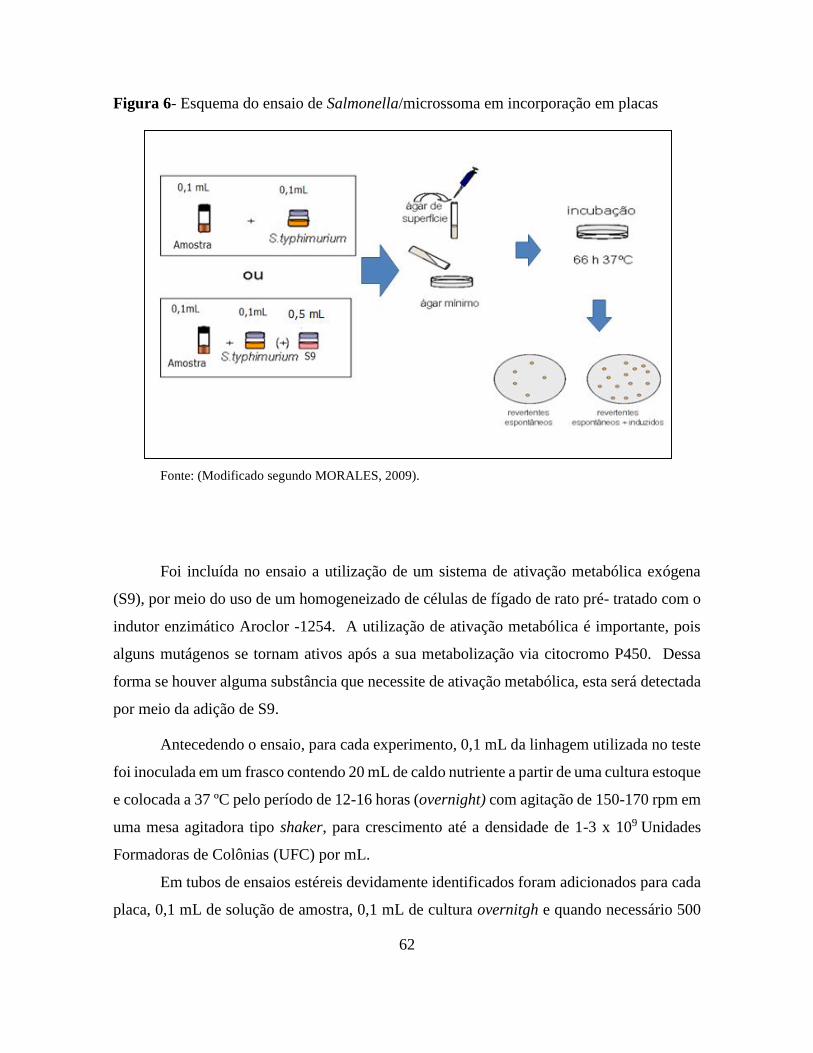
62
Figura 6- Esquema do ensaio de Salmonella/microssoma em incorporação em placas
Fonte: (Modificado segundo MORALES, 2009).
Foi incluída no ensaio a utilização de um sistema de ativação metabólica exógena
(S9), por meio do uso de um homogeneizado de células de fígado de rato pré- tratado com o
indutor enzimático Aroclor -1254. A utilização de ativação metabólica é importante, pois
alguns mutágenos se tornam ativos após a sua metabolização via citocromo P450. Dessa
forma se houver alguma substância que necessite de ativação metabólica, esta será detectada
por meio da adição de S9.
Antecedendo o ensaio, para cada experimento, 0,1 mL da linhagem utilizada no teste
foi inoculada em um frasco contendo 20 mL de caldo nutriente a partir de uma cultura estoque
e colocada a 37 ºC pelo período de 12-16 horas (overnight) com agitação de 150-170 rpm em
uma mesa agitadora tipo shaker, para crescimento até a densidade de 1-3 x 109 Unidades
Formadoras de Colônias (UFC) por mL.
Em tubos de ensaios estéreis devidamente identificados foram adicionados para cada
placa, 0,1 mL de solução de amostra, 0,1 mL de cultura overnitgh e quando necessário 500

63
µL/placa da mistura S9. Em seguida adicionou-se 2 mL de ágar de superfície em cada tubo
e após serem ligeiramente agitados, em um agitador do tipo vórtex, e esse conteúdo foi
transferido para placas com ágar mínimo. As placas foram incubadas por um período de 66
horas a 37 ºC em uma estufa bacteriológica. Após a incubação, a avaliação da citoxicidade
foi realizada por observação visual e as colônias foram contadas manualmente sob um
contador de colônias. As amostras foram analisadas em cinco doses (0; 312,5; 625; 1250;
2500; 5000 µg/ placa) e em duplicata para corante tartrazina analítico e o seu correspondente
corante comercial, juntamente com controles negativos e positivos. Os resultados foram
analisados estatisticamente utilizando o modelo de Bernstein (BERNSTEIN et al., 1982).
4.9 Controles positivos e negativos
Em todos os ensaios foi utilizada água ultrapura Milli-Q estéril como controle
negativo e os controles positivos utilizados em ausência e presença de ativação metabólica
exógena (S9) está descrito na Tabela 7. Todos foram adquiridos da empresa Sigma-Aldrich®
e dissolvidos em dimetilsulfóxido (DMSO). O controle positivo é usado de forma a assegurar
a capacidade de resposta da linhagem e a eficácia do sistema de ativação metabólica,
enquanto o controle negativo é utilizado no intuito de estabelecer o número de revertentes
espontâneos (MARON e AMES, 1983).

64
Tabela 7 - Controles positivos utilizados no ensaio Salmonella/microssoma
Linhagem Controle Positivo Dose
(µg/placa)
TA97a
-S9 9 aminoacridina 50
+S9 2 aminoantraceno (2AA) 2,5
TA98 -S9 4-nitroquinolina-óxido (4NQO) 0,5
+S9 2 aminoantraceno (2AA) 2,5
TA100
-S9 4-nitroquinolina-óxido (4NQO) 0,5
+S9 2 aminoantraceno (2AA) 2,5
TA102 -S9 Mitomicina C 0,5
+S9 2 aminoantraceno (2AA) 2,5
TA1535 -S9 Azida sódica 5,0
+S9 2 aminoantraceno (2AA) 2,5
4.10 Critérios de avaliação
A análise de variância (ANOVA) é aplicada para verificação de diferenças
estatísticas entre o controle negativo e as doses utilizadas, seguida de regressão linear. Os
resultados são apresentados pela razão entre o número de revertentes na placa teste e o
número de revertentes na placa do controle negativo, denominada de razão de
mutagenicidade (RM). A resposta é considerada negativa quando a ANOVA não for
significativa nem for observado um efeito dose-resposta. Porém uma amostra é considerada
positiva quando apresentar, em pelo menos uma das linhagens empregadas, relação dose-
resposta estatisticamente significativa (p ≤ 0,05) entre as concentrações testadas e valores de
revertentes espontâneos, acompanhada da razão de mutagênese maior ou igual a 2, em pelo
menos uma das doses testadas. Esse valor de 2 deve ser apenas usado como guia, pois

65
dependendo da taxa de reversão espontânea da linhagem ele pode ser maior ou menor
(UMBUZEIRO e VARGAS, 2003).
O programa SALANAL, elaborado por Dr. L. Myers do “Integrated Laboratory
Systems”, Carolina do Norte, EUA, foi utilizado para realizar as análises de variância. Para
avaliar a regressão linear foi utilizado um modelo linear, denominado truncagem tipo
Bernstein (BERNSTEIN et al., 1982).

66

67
5. RESULTADOS E DISCUSSÃO
5.1 Confirmações das características genéticas
As características genéticas das linhagens de Salmonella typhimurium foram todas
avaliadas a fim de confirmar a integridade de cada uma das culturas, uma vez que cada uma
delas apresentou uma característica especifica que é responsável por conferir uma maior
sensibilidade na detecção de diversos mutágenos.
Uma vez que a Salmonella typhimurium cresce somente na presença do aminoácido
histidina, foram realizados testes para avaliar a dependência do aminoácido histidina
decorrentes das mutações no operon de histidina (his-), como pode ser observado na Figura
7. As placas suplementadas com biotina e histidina apresentaram um aumento no número de
colônias após a incubação quando comparadas com as placas de ágar mínimo suplementadas
apenas com biotina, em todas as linhagens testadas, confirmando assim a auxotrofia para a
histidina.

68
Figura 7- Confirmação da auxotrofia para histidina (A) placas suplementadas com biotina
para a linhagem TA98; (B) placas suplementadas com biotina e histidina para a linhagem
TA98
(A) (B)
Para a confirmação da mutação rfa, avaliou-se a presença de um halo de inibição
formado após a incubação da placa inoculada com as linhagens testadas e o disco de papel
filtro embebido com cristal violeta, podendo ser comprovado perda parcial da camada de
LPS da membrana externa bacteriana, em todas as linhagens testadas, conforme a Figura 8.
Figura 8 – Confirmação da mutação rfa para a linhagem TA98

69
A deleção do gene uvrB para as linhagens TA97a, TA98, TA100 e TA1535, pode ser
confirmada na Figura 9, uma vez que não apresentam a capacidade de reparo de excisão, esta
pode ser evidenciada pela inibição do crescimento das linhagens quando expostas a luz
ultravioleta. A linhagem TA102 também foi avaliada, porém ela não possui a deleção do
gene uvr, permitindo assim o reparo quando expostas a radiação UV e consequentemente há
o crescimento de colônias após a exposição à luz ultravioleta.
Figura 9- Confirmação da deleção uvrB (A) para a linhagem TA98; (B) para a linhagem
TA102
(A) (B)
Algumas linhagens apresentam plasmídeos que além de conferir uma resistência a
antibióticos, auxilia também na detecção a diferentes mutágenos. Para linhagens TA97a,
TA98 e TA100 foi possível observar na Figura 10, a presença do plasmídeo pKM101 (Apr),
resultado na resistência ao antibiótico ampicilina. Para a linhagem TA102, pode-se confirmar
a presença dos plasmídeos pKM101 (Apr) e pAQ1 (Ttr), conferindo resistência aos
antibióticos ampicilina e tetraciclina respectivamente. Já a linhagem TA1535 não possui
presença de plasmídeo, não sendo resistente aos antibióticos.

70
Figura 10- Confirmação da presença de plasmídeo para a linhagem TA98
5.2 Taxa de Reversão Espontânea
Cada linhagem de Salmonella typhimurium avaliada no presente estudo apresentou
uma taxa de reversão espontânea, conforme a Tabela 8, onde está representado os intervalos
da taxa de reversão espontânea para cada linhagem utilizada no presente estudo, comparando
com as utilizadas como controle no laboratório onde os testes foram realizados. Esta taxa
pode variar em função do solvente utilizado, da concentração histidina no ágar de superfície,
da concentração de S9, ou, ainda, de laboratório para laboratório dependendo das condições
do ensaio (MARON e AMES, 1983; MORTELMANS e ZEIGER, 2000). As taxas de
reversão espontânea encontradas na presente pesquisa estão dentro dos intervalos
demonstrados nos históricos encontrados no Laboratório Central Analítica (LACAN), por
Maron e Ames, 1983; Mortelmans e Zeiger, 2000 e pela Cetesb, 1993.

71
Tabela 8 – Taxa de reversão espontânea das linhagens utilizadas no ensaio
Salmonella/microssoma
Linhagem
Taxa de Reversão
(Ensaio)
TA97a
-S9 150-200
+S9 150-200
TA98 -S9 30-65
+S9 40-75
TA100
-S9 140-160
+S9 130-160
TA102 -S9 140-200
+S9 160-210
TA1535 -S9 12-18
+S9 14-18
5.3 Ensaio de Salmonella/microssoma
Os resultados obtidos demonstraram que os corantes tartrazina comercial (90 %)
(Figura 11) e tartrazina Sigma-Aldrich® (≥ 99 %) (Figura 12) não apresentaram
mutagenicidade para as linhagens TA97a, TA98, TA100, TA102 e TA1535 tanto na presença
quanto ausência de ativação metabólica exógena nas condições do ensaio. Os valores de
razão de mutagenicidade obtidos foram inferiores a 2. Apesar de algumas doses terem
apresentadas um RM em torno de 0,6, ao observar o backgraund não foi detectado sinais de
toxicidade para as linhagens TA97a, TA98, TA100, TA1535. No entanto, para a linhagem

72
TA102 no ensaio com o corante comercial e com o corante analítico, foi observada toxicidade
para a dose de 5000 µg/placa. Os dados brutos estão descritos no Apêndice.
Os ensaios foram realizados com o intuito de avaliar o potencial mutagênico do
corante comercial comparado com o analítico, visando verificar se haveria uma resposta
diferente entre os corantes com grau de pureza variável, uma vez que a presença de impurezas
nas amostras poderiam ser a causa de um potencial mutagênico, concordando com
resultados positivos encontrados (PATTERSON e BUTLER, 1982; ISHIDATE et al., 1984;
HENSCHLER e WILD, 1985, apud EFSA, 2009; MUNZNER e WEVER, 1987; PRIVAL et
al.,1988; DAS e MUKHEREJEE, 2004; MPOUNTOUKAS et al., 2010) para esse corante.
Figura 11- Resultados do ensaio de Salmonella/microssoma do corante tartrazina comercial
Duas Rodas (90%) com as linhagens TA97a, TA98, TA100, TA102 e TA1535, com e sem
ativação metabólica.
0
50
100
150
200
250
300
0 1000 2000 3000 4000 5000
Nº
de
re
vert
en
tes/
pla
ca
Dose (µg/placa)
Tartrazina Comercial
TA98 SEM S9
TA98 COM S9
TA100 SEM S9
TA100 COM S9
TA1535 SEM S9
TA1535 COM S9
TA 102 SEM S9
TA102 COM S9
TA 97A SEM S9
TA 97A COM S9
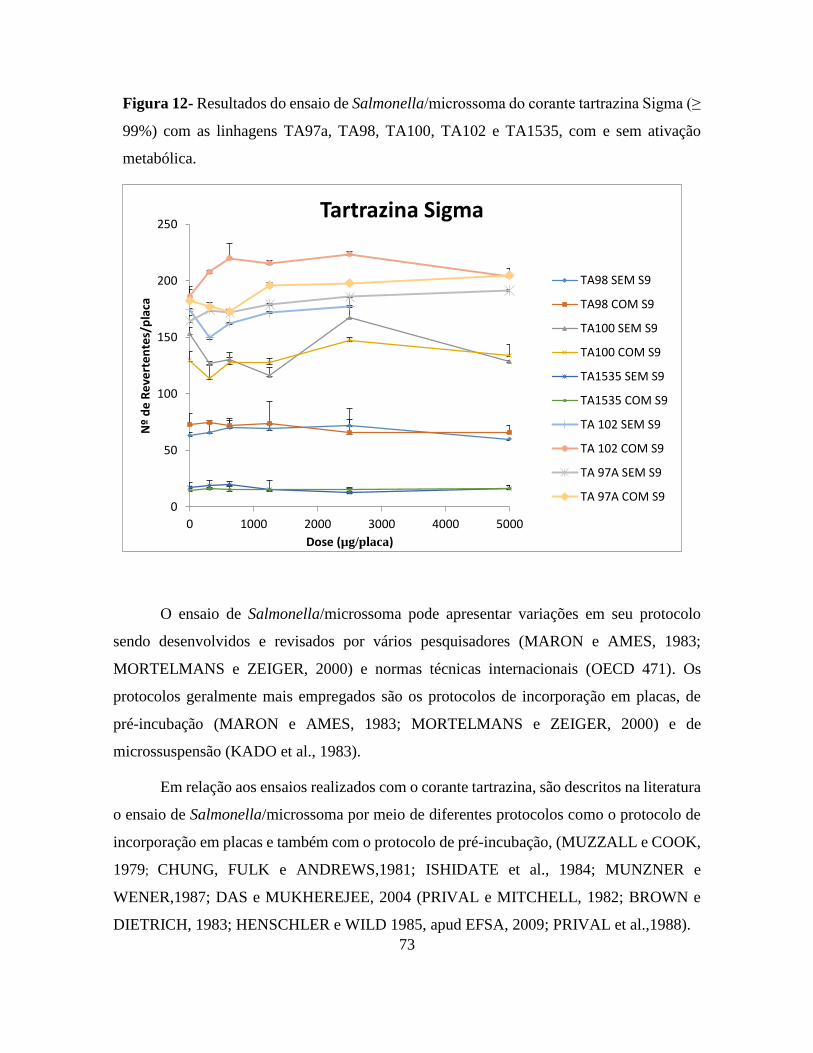
73
Figura 12- Resultados do ensaio de Salmonella/microssoma do corante tartrazina Sigma (≥
99%) com as linhagens TA97a, TA98, TA100, TA102 e TA1535, com e sem ativação
metabólica.
O ensaio de Salmonella/microssoma pode apresentar variações em seu protocolo
sendo desenvolvidos e revisados por vários pesquisadores (MARON e AMES, 1983;
MORTELMANS e ZEIGER, 2000) e normas técnicas internacionais (OECD 471). Os
protocolos geralmente mais empregados são os protocolos de incorporação em placas, de
pré-incubação (MARON e AMES, 1983; MORTELMANS e ZEIGER, 2000) e de
microssuspensão (KADO et al., 1983).
Em relação aos ensaios realizados com o corante tartrazina, são descritos na literatura
o ensaio de Salmonella/microssoma por meio de diferentes protocolos como o protocolo de
incorporação em placas e também com o protocolo de pré-incubação, (MUZZALL e COOK,
1979; CHUNG, FULK e ANDREWS,1981; ISHIDATE et al., 1984; MUNZNER e
WENER,1987; DAS e MUKHEREJEE, 2004 (PRIVAL e MITCHELL, 1982; BROWN e
DIETRICH, 1983; HENSCHLER e WILD 1985, apud EFSA, 2009; PRIVAL et al.,1988).
0
50
100
150
200
250
0 1000 2000 3000 4000 5000
Nº
de
Re
vert
en
tes/
pla
ca
Dose (µg/placa)
Tartrazina Sigma
TA98 SEM S9
TA98 COM S9
TA100 SEM S9
TA100 COM S9
TA1535 SEM S9
TA1535 COM S9
TA 102 SEM S9
TA 102 COM S9
TA 97A SEM S9
TA 97A COM S9

74
Os ensaios realizados pelo método de incorporação em placas com o corante
tartrazina, demostraram resultados controversos (CHUNG, FULK e ANDREWS, 1981;
ISHIDATE et al., 1984; DAS e MUKHEREJEE; 2004), porém para o presente estudo os
resultados não apresentaram mutagenicidade para nehuma das linhagens testadas.
Chung, Fulk e Andrews (1981) obtiveram resultados negativos para o azocorante
tartrazina, por meio do ensaio de mutagenicidade de Salmonella/ microssoma utilizando a
metodologia de incorporação em placas, em presença e ausência de microssomas de fígado
de rato (S9), utilizando as linhagens TA1535, TA1537, TA1538, TA98 e TA100.
Esses resultados também são sustentados por Ishidate et al. (1984) onde realizaram
ensaios de Salmonella/microssoma por meio do protocolo de incorporação em placas com as
linhagens TA1535, TA100, TA1537, TA94 e TA98. De um total de 200 aditivos testados no
ensaio de Salmonella/microssoma 14 apresentaram efeitos positivos. Em relação ao corante
tartrazina, este apresentou resultados negativos no ensaio de mutagenicidade.
Porém, Das e Mukherejee (2004) avaliaram os efeitos mutagênicos por meio do
ensaio de Salmonella/microssoma, empregando o protocolo de incorporação em placas sem
ativação metabólica em concentrações de 10, 100, 250, 500 e 1000 µg/ placa dos corantes
tartrazina e amaranto. Os resultados foram negativos para as linhagens TA97a e TA100,
porém a TA98 apresentou um aumento significativo no número de colônias revertentes
somente nas doses mais baixas para ambos os corantes. Contudo no presente estudo não foi
observado toxicidade e/ou mutagenicidade em todas as doses analisadas para a linhagem
TA98, o que pode estar associado à quantidade de impurezas presente na amostra testada.
No entanto, o presente estudo demonstrou que para a linhagem TA102 o corante
tartrazina comercial apresentou toxicidade em presença de S9, enquanto o corante tartrazina
analítico apresentou toxicidade em ausência de S9 ambos na dosagem de 5000 µg/placa, este
resultado pode ser explicado através da associação de dois fatores: a sensibilidade aumentada
para essa linhagem e as impurezas do corante nessa concentração se tornar significativa,
interferindo no resultado.
Um fator importante que deve ser ressaltado é a redução de azocorantes, uma vez que
pode influenciar nos resultados do ensaio Salmonella/microssoma, ou seja, pode levar a

75
produção de compostos mais ou menos tóxicos e/ou mutagênicos (RAFII et al., 1997) que o
composto original. Alguns estudos têm relatado ausência de atividade mutagênica dos
metabólitos da tartrazina (CHUNG, et al. 1981; EFSA, 2009; HENSCHLER e WILD 1985,
apud EFSA, 2009).
De acordo com Chung, Fulk e Andrews (1981) que avaliaram alguns metabólitos da
tartrazina em presença e ausência de microssomas de fígado de rato (S9), utilizando as
linhagens TA1535, TA1537, TA1538, TA98 e TA100 com doses de até 5000 µg/placa, os
resultados não apresentaram nenhuma atividade mutagênica.
Em contrapartida o parecer elaborado pela EFSA (2009) descreve dois trabalhos onde
foi observado um aumento significativo no número de revertentes quando comparados ao
grupo controle para metabólitos de tartrazina em urina para a linhagem TA98 em presença
de S9 (HENSCHLER e WILD, 1985, apud EFSA, 2009) e em amostras de fezes TA100 em
presença de S9 (MUNZNER e WENER, 1987). Porém para a EFSA (2009) esses estudos
não são relevantes para a avaliação do potencial mutagênico do tartrazina bem como de seus
metabólitos, uma vez que não foi possível identificar os componentes mutagênicos nos
extratos das fezes.
Segundo a OECD, 471, alguns compostos pertencentes à classe química que incluem
cadeia curta, como nitrosaminas, metais, os aldeídos, azo-corantes, compostos diazo, e
alcaloides podem ser detectados de forma mais eficiente usando o método de pré-incubação.
Dessa forma, Prival e Mitchell (1982) elaboraram o protocolo de pré-incubação
utilizando flavina mononucleotídeo (FMN) e a fração S9 de fígado de hamster que possui
uma melhor capacidade para detecção da mutagenicidade de produtos aromáticos
provenientes da redução de alguns azocorantes. No entanto, os resultados descritos por Prival
et al. (1988) não apresentaram uma atividade mutagênica quando testados com o ensaio de
pré-incubação em presença de FMN e fração S9 provenientes de fígado de hamster para o
corante tartrazina em doses de até 2500 mg/placa.
Além disso, outros tipos de ensaios também podem ser realizados para detectar o
potencial mutagênico/genotóxico podendo apresentar os mesmos resultados que o ensaio de
Salmonella/microssoma ou resultados diferentes, isso vai depender do endpoint que será

76
analisado. Dessa forma, outros ensaios foram realizados com o corante tartrazina
apresentando também alguns resultados controversos (PATTERSON e BUTLER, 1982;
ISHIDATE et al., 1984).
A indução de aberração cromossômica foi avaliada após dois diferentes períodos da
cultura celular de fibroblasto de Muntiacus muntjac expostas a concentrações variadas de
tartrazina. Foi observado um aumento significativo em relação a quebras das cromátides e
cromossomos dicêntricos quando comparados com os grupos controles em todos os grupos
experimentais. Os autores sugerem que novos estudos são necessários para determinar o
potencial mutagênico do corante (PATTERSON e BUTLER, 1982).
Ishidate et al. (1984) avaliaram também por meio de ensaios de aberrações
cromossômicas in vitro em células de fibroblastos de hamster chinês com 190 aditivos
alimentares sintéticos e 52 aditivos alimentares derivados de fontes naturais. De um total de
242 aditivos, 54 apresentaram resultados positivos, incluindo o corante tartrazina no ensaio
cromossômico, onde observaram um pequeno aumento na incidência de células poliplóides
após a exposição as células de fibroblastos de hamster chinês ao corante.
Mpountoukas et al. (2010) estudaram o potencial genotóxico, citotóxico e citostático
dos corantes amaranto, eritrosina e tartrazina em células humanas de sangue periférico in
vitro. Nas doses de 1 e 2 mM, houve um aumento significativo em relação ao atraso das
divisões celulares quando comparada ao grupo controle. Em duas concentrações mais
elevadas de tartrazina (4 e 8 mM) foi observado um efeito tóxico significativo sobre a
qualidade de cromossomos de modo que a mancha de diferenciação entre as duas cromatídes
não estava bem clara. Uma explicação para isso seria que a tartrazina foi tóxica durante o
processo de condensação dos cromossomos na mitose.
Além disso, a solubilidade é outro fator que pode interferir nos resultados, visto que
o corante utilizado é altamente solúvel em água devido à presença de grupos sulfônicos (-
SO3H) que confere um caráter ácido ao corante, tornando-o facilmente ionizável
(Zollinger,1991). Dessa forma mesmo com a utilização de um protocolo que auxilie na
azoredução do corante, os produtos gerados após essa azoredução também seriam pouco
absorvidos, devido à sua solubilidade. No presente estudo, pode ser observado que em doses

77
de até 5000 µg/placa do corante tartrazina, as placas apresentam uma coloração amarela,
porém as colônias continuam com a sua coloração característica comparando com as placas
do controle negativo, indicando assim que há uma baixa absorção do corante pelas bactérias
(MURDOCH et al., 1987; EFSA, 2009).
Outro fator que também pode interferir nos resultados é a pureza do corante, ou seja,
quando o corante é obtido dentro dos padrões de qualidade como foi observado em vários
trabalhos, este não apresenta uma atividade mutagênica, embora possam apresentar essa
atividade mutagênica se estiver presente em sua constituição alguns contaminantes, tais como
anilinas, benzidinas, aminas aromáticas, metais pesados entre outros, provenientes da síntese
do próprio corante (ZIMMER et al., 1980; CHUNG; FULK e ANDREWS, 1981; FDA, 1985;
CLAXTON, HUGHES e CHUNG, 2001; CHUNG, CHEN e CLAXTON, 2006).
Dessa forma, Lancaster e Lawrence (1989 e 1991) por meio de um método de HPLC
de pareamento iônico, verificaram a presença de 2 - naftilamina, 4 - aminobifenilo, anilina e
benzidina em amostras comerciais do corante tartrazina utilizadas no Canadá. Após a análise
de sete amostras observaram a presença de benzidina em três delas, em concentrações de até
326 ng/g e todas as amostras continham anilina em concentrações de até 83,2 µg/ g.
Os resultados obtidos por Davis e Bailey (1993) após avaliarem a presença de
benzidina em 14 amostras do corante tartrazina, mostraram que os lotes comerciais do
corante tartrazina continham benzidina na forma livre e na forma de corante subsidiário em
níveis acima daqueles permitidos, 4 das amostras apresentaram níveis superiores a 100 ppb.
Os resultados obtidos por Lancaster e Lawrence (1999) após analisarem 12 amostras
comerciais de tartrazina sugerem que os níveis de contaminantes como a benzidina e anilina
diminuíram após 1990. As amostras de tartrazina antes de 1990 continham níveis de anilina
em 115-182 mg/g e de benzidina 32-38 ng/g. Após o ano de 1990 os níveis de anilina e
benzidina estavam em 0,2-16,6 ng/g e 5 ng/g respectivamente.
Como observado nos trabalhos citados acima pode-se verificar que a presença de
impurezas em diversas amostras de corantes seria uma possível causa das controversas
geradas há anos sobre o potencial genotóxico do corante tartrazina (HENSCHLER e WILD,
1985, apud EFSA, 2009; PATTERSON e BUTLER, 1982; ISHIDATE et al., 1984;

78
MUNZNER e WEVER, 1987; PRIVAL et al., 1988; DAS e MUKHEREJEE, 2004;
MPOUNTOUKAS et al., 2010).
Estes resultados positivos citados na literatura em relação ao potencial mutagênico e
genotóxico do corante tartrazina podem ser explicados pelo fato de que mesmo a utilização
de compostos com alto grau de pureza, ainda há a possibilidade de estarem presentes algumas
impurezas, provenientes de sua síntese, que pode contribuir para a mutagenicidade desse
corante (PRIVAL et al., 1988).
No entanto, os resultados do presente estudo possivelmente se justificam devido aos
relatos de que o corante tartrazina analisado pode não apresentar atividade mutagênica
(EFSA, 2009) principalmente quando não apresentam contaminantes que promovam essa
atividade.
Portanto há necessidade de uma harmozinação das legislações entre os diversos países
predizendo quais são as impurezas que podem fazer parte da composição do corante e os seus
limites máximos permitidos, para que haja um consenso entre os diversos países, visto que
há divergência entre eles nessas informações, com o intuito de sempre proporcionar a
segurança alimentar.

79
6. CONCLUSÃO
Diante dos resultados encontrados no presente estudo, os dois corantes não
apresentaram atividade mutagênica para nenhuma das linhagens utilizadas conforme
recomendações da OECD 471.
Comparando o corante tartrazina analítico com o seu correspondente comercial, os
ensaios realizados demonstraram ausência de atividade mutagênica para as linhagens
testadas, indicando ausência ou baixa concentração de impurezas que promovam esse tipo de
alteração. Entretanto para ambos os corantes pode-se observar uma presença de toxicidade
para a linhagem TA102.
Contudo é importante levar em consideração que os processos de fabricação,
contaminações e a atividade mutagênica podem estar diretamente relacionados com o
controle de qualidade nas indústrias durante a síntese do corante, devido à possibilidade de
contaminação por impurezas.

80

81
REFERÊNCIAS
AMES, B.N.; YAMASAKI, E. The detection of chemical mutagens with enteric bacteria. In:
HOLLANDER, A. (Ed.). Chemical Mutagens: Principles and Methods for their Detection,
Editora New York, Plenum Press, v.1, chap.9, p. 267-282, 1971.
AMES, B.N.; GURNEY, E. G. ; JAMES A. MILLER, J.A.; H. BARTSCH, H. Carcinogens
as frameshift mutagens: metabolites and derivatives of 2-acetylaminofluorene and other
aromatic amine carcinogens (aromatic nitroso carcinogens/carcinogen detection with
Salmonella/DNA intercalation). Proceedings of the National Academy of Sciences of the
United States of America, v. 69, n.11, p. 3128-3132, 1972.
AMES, B.N.; DURSTON, W.E.; YAMASAKI, E.; LEE, F.D. Carcinogens are mutagens: a
simple test system combining liver homogenates for activation and bacteria for detection.
Proceedings of the National Academy of Sciences of the United States of America, v.70,
n.8, p.2281-2285, 1973a.
AMES, B.N.; LEE, F.D.; DURSTON, W.E. An improved bacterial test system for the
detection and classification of mutagens and carcinogens. Proceedings ofthe National
Academy of Sciences of the United States of America, v.70, n.3, p.782-786, 1973b.
AMIN, K.A.; HAMEID II, A.; ELSTTAR, A.H,A. Effect of food azo dyes tartrazine and
carmoisine on biochemical parameters related to renal, hepatic function and oxidative stress
biomarkers in young male rats. Food and Chemical Toxicology, v.48, n.10, p. 2994–2999,
2010.
ANON. Ecological and toxicological association of dyes and pigments manufacturers,
textile chemists and colorist, “German ban of use of certain azo compounds in some
consumer goods: ETAD Information Notice No. 6, v.28, n. 4, p.11, 1996.

82
ARLT, V.M., GLATT, H., MUCKEL, E., PAPEL, U., SORG, B.L., SCHMEISER, H.;
PHILLIPS, D.H. Metabolic activation of the environmental contaminant 3-
nitrobenzanthrone by human acetyltransferases and sulfotransferase. Carcinogenesis, v.23,
n.11, p. 1937–1945, 2002.
ASSOCIAÇÃO BRASILEIRA DAS INDÚSTRIAS DA ALIMENTAÇÃO - ABIA.
Compêndio da Legislação de Alimentos. Consolidação das normas e padrões de alimentos.
Órgão Técnico e Consultivo do Poder Público. Atos do Ministério da Saúde, São Paulo,
v.8, 2001.
BARTSCH, H. Metabolic activation of aromatic amines and azo dyes. IARC Scientific
Publications Series, v. 40, p.13–30, 1981.
BERTAGNI, P., HIROM, P.C., MILLBURN, P., OSIYEMI, F.O., SMITH, R.L.,
TURBERT, H.B., WILLIAMS, R.T. Sex and species differences in the biliary excretion of
tartrazine and lissamine fast yellow in the rat,guinea-pig and rabbit. The influence of sex
hormones on tartrazine excretion in the rat. Journal of Pharmacology and
Pharmacotherapeutics, v. 24, p. 620–624, 1972.
BERNSTEIN, L., KALDOR, J., MCCANN, J., PIKE, M.C. An empirical approach to the
statistical analysis of mutagenesis data from Salmonella test. Mutation Research, v. 97,
p.267–281, 1982.
BERZAS, J.J., FLORES J.R., LLERENA M.J.V.; FARINÄAS R.N. Spectrophotometric
resolution of ternary mixtures of tartrazine, patent blue v and indigo carmine in commercial
products. Analytica Chimica Acta, v. 391, n.3, p. 353-364, 1999.
BESSONOV, V.V.; MALINKIN, A.D.; PEREDERIAEV, O. I.; BOGACHUK,
M.N.; VOLKOVICH, S.V.; MEDVEDEV, I. Development of methods for determining

83
acrylamide in food products by gas-liquid chromatography. Voprosy Pitanni, v.80, n.4,
p.70-83, 2011.
BOMHARD, E.M.; HERBOLD, B.A. Genotoxic activities of aniline and its metabolites and
their relationship to the carcinogenicity of aniline in the spleen of rats. Critical Reviews in
Toxicology, v.35, n.10, p.783-835, 2005.
BONSER, G.M. Epithelial tumours of the bladder in dogs induced by pure β-naphthylamine.
Journal of Pathology and Bacteriology, v. 55, p. 1-6, 1943. In: ZIMMER, D.; MAZUREK,
J.; PETZOLD, G.; BHUYAN, B. K. Bacterial mutagenicity and mammalian cell DNA
damage by several substituted anilines. Mutation Research, v.77, p. 317-326, 1980.
BORZELLECA, J.F.; HALLAGAN J.B. Chronic toxicity/carcinogenicity studies of FD&
C Yellow No.5 (Tartrazine) in rats. Food and Chemical Toxicology, v.26, p.179-187,
1988a.
BORZELLECA, J.F.; HALLAGAN J.B. Chronic toxicity/carcinogenicity studies of FD&C
Yellow no. 5 (Tartrazine) in mice. Food and Chemical Toxicology, v.26, p.189-194, 1988b.
BRASIL. Agência Nacional de Vigilância Sanitária. Portaria nº685, de 27 de agosto de 1998.
Aprova o Regulamento Técnico princípios gerais para o estabelecimento de níveis máximos
de contaminantes químicos em alimentos e seu Anexo: Limites máximos de tolerância para
contaminantes inorgânicos. Diário Oficial da União, Brasília, DF, 1998.
BRASIL. Agência Nacional de Vigilância Sanitária. Decreto nº 50.040, de 24 de Janeiro de
1961 Dispõe sobre as Normas Técnicas Especiais Reguladoras do emprego de aditivos
químicos a alimentos. Diário Oficial da União, Brasília, DF, 1961.
BRASIL. Agência Nacional de Vigilância Sanitária. Decreto nº 55871, de 26 de março de
1965. Modifica o Decreto nº 50.040, de 24 de janeiro de 1961, referente a normas reguladoras

84
do emprego de aditivos para alimentos, alterado pelo Decreto nº 691, de 13 de março de
1962. Diário Oficial da União, Brasília, DF, 1965.
BRASIL. Agência Nacional de Vigilância Sanitária. Decreto-Lei nº 986, de 21 de outubro
de 1969. Institui normas básicas sobre alimentos. Diário Oficial da União, Brasília, DF,
1969.
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução nº 137, de 29 de maio de
2003. Considera Laca o sal preparado a partir do corante incluído na Tabela I, anexa ao
Decreto nº 55871/65, em combinação com o radical básico de alumínio ou cálcio. Diário
Oficial da União, Brasília, DF, 1977.
BRASIL. Agência Nacional de Vigilância Sanitária. Portaria nº 540/97, de 27 de outubro de
1997. Define o Sistema Nacional de Vigilância Sanitária, cria a Agência Nacional de
Vigilância Sanitária, e dá outras providências. Diário Oficial da União, Brasília, DF, 1997.
BRASIL. Agência Nacional de Vigilância Sanitária. Lei nº 9.782, de 26 de Janeiro de 1999.
Define o Sistema Nacional de Vigilância Sanitária, cria a Agência Nacional de Vigilância
Sanitária, e dá outras providências. Diário Oficial da União, Brasília, DF, 1999.
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução nº 387, de 05 de agosto de
1999. Regulamento técnico que aprova o uso de aditivos alimentares, estabelecendo suas
funções e seus limites máximos para a categoria de alimentos 5: balas, confeitos, bombons,
chocolates e similares. Diário Oficial da União, Brasília, DF, 1999.
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução nº 388, de 05 de agosto de
1999. Aprova o Regulamento Técnico que aprova o uso de aditivos alimentares,
estabelecendo suas funções e seus limites máximos para a categoria de alimentos 19 -
sobremesas. Diário Oficial da União, Brasília, DF, 1999.

85
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução nº 33, de 09 de março de
2001 Aprova o Regulamento Técnico que aprova o uso de Aditivos Alimentares,
estabelecendo suas funções e seus limites máximos para a Categoria de Alimentos 12: Sopas
e Caldos. Diário Oficial da União, Brasília, DF, 1999.
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução RDC nº 34, de 9 de março
de 2001. Aprova o Regulamento Técnico que aprova o uso de aditivos alimentares,
estabelecendo suas funções e seus limites máximos para a categoria de alimentos 21:
preparações culinárias industriais. Diário Oficial da União, Brasília, DF, 2001.
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução RDC nº 340, de 13 de
dezembro de 2002. As empresas fabricantes de alimentos que contenham na sua composição
o corante tartrazina (INS 102) devem obrigatoriamente declarar na rotulagem, na lista de
ingredientes, o nome do corante tartrazina por extenso. Diário Oficial da União, Brasília,
DF, 2002.
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução nº 137, de 29 de maio de
2003. Determina que o registro/renovação de registro de medicamentos pertencentes às
classes/princípios ativos relacionados em anexo, só será autorizado se as bulas e embalagens
contiverem a advertência pertinente, conforme relação anexa. Diário Oficial da União,
Brasília, DF, 2003.
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução RDC nº 25, de 15 de
fevereiro de 2005. Aprova o Regulamento técnico que aprova o uso dos aditivos alimentares,
estabelecendo suas funções e limites máximos para a categoria de alimentos: produtos
proteicos - subcategoria: bebidas não alcoólicas a base de soja. Diário Oficial da União,
Brasília, DF, 2005.
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução nº 24, de 15 de fevereiro de
2005. Aprova o Regulamento Técnico para o uso de aditivos alimentares, coadjuvantes de

86
tecnologia, estabelecendo suas funções e limites, e veículos para suplementos vitamínicos e
ou minerais. Diário Oficial da União, Brasília, DF, 2005.
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução RDC nº 2, de 15 de janeiro
de 2007. Aprova o Regulamento Técnico sobre atribuição de aditivos e seus limites máximos
para a categoria de alimentos 16.2: bebidas não alcoólicas, subcategoria 16.2.2: bebidas não
alcoólicas gaseificadas e não gaseificadas. Diário Oficial da União, Brasília, DF, 2007.
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução RDC nº 3, de 15 de janeiro
de 2007. Aprovar o Regulamento Técnico sobre atribuição de aditivos e seus limites
máximos para a categoria de alimentos 3: gelados comestíveis. Diário Oficial da União,
Brasília, DF, 2007.
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução RDC nº 4, de 15 de janeiro
de 2007. Atribuição de aditivos e seus limites máximos para a categoria de alimentos 13:
molhos e condimentos. Diário Oficial da União, Brasília, DF, 2007.
BRASIL. Agência Nacional de Vigilância Sanitária. Resolução RDC nº64, de 16 de setembro
de 2008. Aprova o Regulamento Técnico sobre Atribuição de aditivos e seus limites máximos
para alimentos.Diário Oficial da União, Brasília, DF, 2008.
BRASIL. Agência Nacional de Vigilância Sanitária.http://portal.anvisa.gov.br/ áreas de
atuação/> alimentos> área de interesse> legislação> legislação da área especifica por assunto
> aditivos alimentares e coadjuvantes de tecnologia, acesso 10/05/2014.
BROWN, J., DIETRICH, P. Mutagenicity of selected sulfonated azo dyes in the
Salmonella/microsome assay: use of aerobic and anaerobic activation procedures. Mutation
Research, v. 116, p.305–315, 1983.
BROWN, J.P., ROEHM, G.W.; BROWN, R.J. Mutagenicity testing of certified food colors
and related azo, xanthene and triphenylmethane dyes with the Salmonella/microsome system.
Mutation Research, v. 56, p.249-271, 1978.

87
CASE, R. A. M.; HOSKER, M. E.; MCDONALD, D. B.; PEARSON, J. T. Tumors of the
urinary bladder in workmen engaged in the manufacture and use of certain dyestuff
intermediates in the British Chemical Industry, I , The role of aniline, benzidine, a-
naphthylamine and P-naphthylamine. British Journal of Industrial Medicine, v.11, p. 75,
1954.
CHADWICK, R.W., GEORGE, S.E., CLAXTON, L.D. Role of the gastrointestinal mucosa
and microflora in the bioactivation of dietary and environmental mutagens or carcinogens.
Drug Metabolism Reviews, v.24, p.425-492, 1992.
CHUNG, K.T. The significance of azo-reduction in the mutagenesis and carcinogenesis of
azo dyes. Mutation Research, v. 114, p.269-281, 1983.
CHUNG, K.T., FULK, G.E., EGAN, M. Reduction of azo dyes by intestinal anaerobes.
Applied and Environmental Microbiology, v.35, p.558-562, 1978.
CHUNG, K. T.; FULK, G. E.; ANDREWS, A.W. Mutagenicity testing of some commonly
used dyes. Applied and Environmental Microbiology, v. 42, n.4, p. 641-648, 1981.
CHUNG, K.T.; STEVENS J.R.; CERNIGLIA, C.E. The reduction of dyes by the intestinal
microflora.Critical Reviews in Microbiology, v. 18, n. 3, p. 175-190, 1992.
CHUNG, K. T.; CHEN, S.C.; CLAXTON, L.D. Review of the Salmonella typhimurium
mutagenicity of benzidine, benzidine analogues, and benzidine-based dyes. Mutation
Research, v.12, p. 58–76, 2006.
CLAXTON, L.D.; HUGHES, T.J. CHUNG, K.T. Using base-specific Salmonella tester
strains to characterize the types of mutation induced by benzidine and benzidine congeners
after reductive metabolism. Food and Chemical Toxicology, v.39, n.12, p.1253-1261, 2001.

88
CLAYSON, D. B.; LAWSON, T. A.; PRINGLE, J. A. S. The carcinogenic action of 2-
aminodiphenylene oxide and 4-aminobiphenyl on the bladder and liver of C57XlF mouse.
British Journal of Cancer, v.2, p. 755-757, 1967.
CODE OF FEDERAL REGULATIONS – CRF. Food and Drug Administration, Department
of Health and Human Services, 21CRF74.705, Title 21, v.1, Revised as of April 2013.
COLLINS, T.F.; BLACK, T.N.; BROWN, .LH.; BULHACK, P. Study of the teratogenic
potential of FD & C Yellow No. 5 when given by gavage to rats. Food and Chemical
Toxicology, v.28, n.12, p.821-827, 1990.
COLLINS, T.F.; BLACK, T.N,; O'DONNELL, M.W. JR; BULHACK, P. Study of the
teratogenic potential of FD & C yellow No. 5 when given in drinking-water.
Food and Chemical Toxicology, v.30, n.4, p.263-268, 1992.
COMBES, R.D. On the mutagenicity of tartrazine (FD & C Yellow No 5). Archives
of Toxicology, v.59, p.67-68, 1986.
COMBES, R.D. Genotoxicity testing recent advances and future trends.Chemistry &
Industry, v. 24, p. 950-954, 1992.
COMBES, R.D; HAVELAND-SMITH, R.B. A review of the genotoxicity of food, drug and
cosmetic colours and other azo, triphenylmethane and xanthene dyes. Mutation Research,
v.98, p.101-248, 1982.
CHEESEMAN, M. A. Artificial food color additives and child behavior. Environmental
Health Perspectives, v.20, n.1, p 15-16, 2012.

89
DAS, A.; MUKHERJEE, A. Genotoxicity Testing of the Food Colours Amaranth and
Tartrazine. International Journal of Human Genetics, v.4, n.4, p.277-280, 2004.
DOWNHAM, A.; COLLINS, P. Colouring our food in the last and next millennium.
International Journal of Food Science & Technology, v.35, p.5-22, 2000.
DALL’AGNOL, R. P. SANTOS, J.A.B.; MACHADO, G.J.C.; SILVA, RAFAELA;
SANTOS A.P. Utilização de corantes artificiais em produtos alimentícios no brasil. Anais
do Simpósio Internacional de Inovação Tecnológica-SIMTEC, v. 1, n. 1, p. 26-37, 2013.
DAVIS, V. M.; BAILEY, JR., J.E. Chemical reduction of FD&C Yellow No. 5 to determine
combined benzidine. Journal of Chromatography, v.635, p.160-164, 1993.
DAVIS, K.J.; FITZHUGH, O.G.; NELSON, A.A. Chronic rat and dog toxicity studies on
Tartrazine. Toxicology and Applied Pharmacology, v.6, p. 621-626, 1964. In: JOINT
FAO/WHO EXPERT COMMITTEE ON FOOD ADDITIVES - JECFA. Specifications for
Identity and Purity and Toxicological Evaluation of some Food Colours, FAO Nutrition
Meetings Report Series, WHO Food additives series, n.38B/66.25,Geneva, 1966.
DEGAWA, M.; SHOJL, Y.; MASUKA, Y.; HASTUMOTO, Y. Mutagenicity of metabolites
of carcinogenic aminoazo dyes. Cancer Letters, v.8, p.71-76, 1979.
DEL GIOVINE, L.; BOCCA, A. P. Determination of synthetic dyes in ice cream by capillary
electrophoresis. Food Control, v.14, p.131-135, 2003.
DURNEV, A.D.; ORESHCHENKO, A.V.; KULAKOVA, A.V.; BERESTEN, N.F. Analysis
of cytogenetic activity of food dyes. Voprosy Medit͡sinskoĭ Khimii, v. 41, p.50-53, 1995.In:
EUROPEAN FOOD SAFETY AUTHORITY - EFSA. Scientific Opinion on the re-
evaluation Tartrazine (E 102). EFSA Journal, v.7, n.11, p.1-52, 2009.

90
ELHKIM, M.O.; HERAUD, F.; BEMRAH, N.; GAUCHARD, F.E.; LORINO, T.;
LAMBRE, C.; FREMY, J.M.; JEAN-MICHEL, P. New considerations regarding the risk
assessment on Tartrazine an update toxicological assessment, intolerance reactions and
maximum theoretical daily intake in France. Regulatory Toxicology and Pharmacology,
v. 47, p.308-316, 2007.
EUROPEAN FOOD SAFETY AUTHORITY -EFSA. Opinion of the scientific panel on food
additives, flavourings, processing aids and materials in contact with food on a request from
the Commission to review the toxicology of a number of dyes illegally present in food in the
EU. EFSA Journal, v.263, p.1–71, 2005.
EUROPEAN FOOD SAFETY AUTHORITY - EFSA. Scientific Opinion on the re-
evaluation Tartrazine (E 102). EFSA Journal, v.7, (11):1331, p.1-52, 2009.
EUROPEAN COMMISSION - EC. Commission Directive of the European Parliament and of
the Council 94/36/EC of June 30, 1994. On colours for use in foodstuffs. Official Journal
of the European Communities, L237, 13, 10.09.1994.
EUROPEAN COMMISSION - EC. Commission Directive 2008/128/EC of 22 December
2008 laying down specific purity criteria concerning colours for use in foodstuffs. Official
Journal of the European Communities, L 006, 10.1.2009.
EUROPEAN COMMISSION - EC. Commission Directiva 95/45/CE de 26 de Julho de 1995
que estabelece os critérios de pureza específicos dos corantes que podem ser utilizados nos
gêneros alimentícios. Official Journal of the European Communities, L 226 de 22.9.1995.
FÁVERO, D. M.; RIBEIRO, C. S. G.; AQUINO, A. D. Sulfitos: importância na indústria
alimentícia e seus possíveis malefícios a população. Segurança Alimentar e Nutricional,
v. 18, n. 1, p. 11-20, 2011.

91
FENG, J.; CERNIGLIA, C.E.; CHEN, H.Toxicological significance of azo dye metabolism
by human intestinal microbiota. Frontiers in Bioscience, n.4, p.568-586, 2012.
FERRETTI, J. J.; LU, W.; LIU, H. B. Mutagenicity of benzidine and related compounds
employed in the detection of hemoglobin, American Journal of Clinical Pathology, v.7, p.
526-527, 1977.
FRIEDBERG, E.C. DNA repair. New York, Freeman, 1985.
FRIBERG, L.; KJELLSTROM, T.; NORDBERG, G..; SCATOR, M. Cadmium. In: Friberg,
L. et al., ed. Handbook on the toxicology of metals. Amsterdam, Elsevier, p. 355-381, 1979.
FOOD AND DRUG ADMINISTRATION - FDA. FD&C Yellow No. 5 final rule, removal
of stay. Federal Register, v.50, p. 35774-35783, 1985.
FULLER, A.T. Is p-aminobenzenesulphonamide the active agent in Prontosil therapy?
Lancet, v.232, p.194-198, 1937.
GANESAN, L.; MARGOLLES-CLARK, E.; SONG, Y.; BUCHWALD, P.The food
colorant erythrosine is a promiscuous protein-protein interaction inhibitor. Biochemical
Pharmacology, v.81, n.6, p.810-818, 2011.
GARNER, R. C., WALPOLE, A. C., ROSE, F. L. Testing- of some benzidine analogues for
microsomal activation to bacterial mutagens, Cancer Letters, v.1, p. 39-42, 1975.
GIRI, A.K.; DAS, S.K.; TALUKDER, G.; SHARMA, A. Sister Chromatid exchange and
chromosome aberrations induced by Curcumin and Tartrazine on mammalian cells in vivo.
Cytobios, v.62, n.249, p.111-117, 1990. In: EUROPEAN FOOD SAFETY AUTHORITY -

92
EFSA. Scientific Opinion on the re-evaluation Tartrazine (E 102). EFSA Journal, v.7,
(11):1331, p.1-52, 2009.
GINGELL, R.; WALKER, R. Mechanisms of azo reduction by Streptococcus faecalis, II.
The role of soluble flavins. Xenobiotica, v.1, p.231-239, 1971.
GOMES, K.M.S.; OLIVEIRA, M.V.G.A; CARVALHO, F.R.S.; MENEZES, C.C.; PERON,
A. P. Citotoxicity of food dyes Sunset Yellow (E-110), Bordeaux Red (E-123), and Tatrazine
Yellow (E-102) on Allium cepa L. root meristematic cells. Food Science Technology, v.33,
n.1, p. 218-223, 2013.
GUIDELINE FOR TESTING OF CHEMICALS- OECD/Section 4: Health Effects,
Bacterial Reverse Mutation Test: nº471, 1997.
GREEN, M.L. Mechanisms of bacterial mutagenesis and properties of mutagenesis tester
strains. Archives of Toxicology, 39: 221 – 248, 1978.
GREGORY, A.R.; ELLIOTT, J.; KLUGE, P. Ames testing of Direct Black 38 parallels
carcinogenicity testing. Journal of Applied Toxicology. v.1, n.6, p.308-13, 1981.
GREGORY, P. Classification of dyes by chemical structures. In: Waring, D., Hallas, G.
(Eds.). The Chemistry and Application of Dyes. Plenum Press, New York, 1990.
GRIFFITHS, A.J.F.; GELBART, W.M.; MILLER, J.H.; LEWONTIN, R.C. Mutações
Gênicas. In: Genética Moderna. Rio de Janeiro: Guanabara Koogan, 2001b. cap 7, p.177-
204, 2001. In: FERRAZ, E.R.A. Comparação da mutagenicidade dos azo corante Disperse
Red 1, Disperse Orange 1 e Disperse Red 13 utilizando o teste de mutagenicidade com
Salmonella: USP, 2008. 102 páginas. Dissertação (mestrado). Programa de pós graduação
em Toxicologia, Faculdade de Ciências Farmacêuticas de Ribeirão Preto, Ribeirão Preto,
2008.

93
GRIFFITHS, A.J.F.; MILLER, J.H.; SUSUKI, D.T.; LEWONTIN, R.C.; GELBART, W.M.
Mutação gênica. In: Introdução à Genética. Rio de Janeiro: Guanabara Koogan, 1998a.
cap.7, p.169-194, 1998. In: FERRAZ, E.R.A. Comparação da mutagenicidade dos azos
corantes Disperse Red 1, Disperse Orange 1 e Disperse Red 13 utilizando o teste
de mutagenicidade com Salmonella: USP, 2008. 102 páginas. Dissertação (mestrado).
Programa de pós graduação em Toxicologia, Faculdade de Ciências Farmacêuticas de
Ribeirão Preto, Ribeirão Preto, 2008.
HANAWALT, P.C.; COOPER, P.K.; GANESAN, A.K.; SMITH, C.A. DNA repair in
bacteria and mammalian cells.Annual Reviewof Biochemistry. 48: 783 – 836, 1979.
HATHCOCK, J. N., RADER, J. I. Aditivos, contaminantes e toxinas naturais de alimentos.
In: Shils, M.; Olson, J., Shike, M. Tratado de nutrição moderna na saúde e na doença,
Manole, v.9, 2003.
HENSCHLER, D.; WILD, D. Mutagenic activity in rat urine after feeding with the azodye
Tartrazine. Archives of Toxicology, v.57, p. 214-215, 1985. In: EUROPEAN FOOD
SAFETY AUTHORITY - EFSA. Scientific Opinion on the re-evaluation Tartrazine (E 102).
EFSA Journal, v.7, (11):1331, p.1-52, 2009.
HIRANO, Y., MIZUTANI, T. Study of inhibition of CYP2A6 by some drugs derived from
quinoline. Journal Pharmacy Pharmacology, v.55, p.1667-1672, 2003.
HIRANO, Y.; UEHARA, M.; SAEKI, K.; KATO, T.A.; TAKAHASHI, K.; MIZUTANI, T.
The influence of quinolines on coumarin 7-hydroxylation in bovine liver microsomes and
human CYP2A6. Journal of Health Sciences, v.48, p.118-125, 2002.
HOMBURGER, F.; FRIEDELL, G.H.; WEISBURGER, E.K.; WEISBURGER, J.H.
Carcinogenicity of simple aromatic amine derivatives in mice and rats. Toxicology and
Applied Pharmacology, v. 22, p. 280-281, 1972.

94
HONOHAN, T.; ENDERLIN, F.E.; RYERSON, B.A.; PARKINSON, T.M. Intestinal
absorption of polymeric derivatives of the food dyes sunset yellow and tartrazine in rats.
Xenobiotica, v.7, p.765-774, 1977.
INTERNATIONAL INSTITUTE FOR RESEARCH ON CANCER -IARC. Monographson
the Evaluation of Carcinogenic Risks to Humans: O Some Aromatic Amines, Organic Dyes,
and Related Exposures. International Agency for Research on Cancer, v.99, p.1- 706,
2010.
ISHIDATE, M.; SOFUNI, T.; YOSHIKAWA, K.; HAYASHI, M.; NOHMI, T.; SAWADA,
M.; MATSUKA, A. Primary mutagenicity screening of food additives currently used in
Japan. Food and Chemical Toxicology, v.22, p.623-636, 1984.
IZBIRAK, A.; SUMER, S.; DIRIL, N. Mutagenicity testing of some azo dyes used as food
additives. Mikrobiyol Bulteni, v.24, p.48-56, 1990. In: EUROPEAN FOOD SAFETY
AUTHORITY - EFSA. Scientific Opinion on the re-evaluation Tartrazine (E 102). EFSA
Journal, v.7, (11):1331, p.1-52, 2009.
JOINT FAO/WHO EXPERT COMMITTEE ON FOOD ADDITIVES -
JECFA.Specifications for Identity and Purity and Toxicological Evaluation of some Food
Colours, FAO Nutrition Meetings Report Series, WHO Food additives series,
n.38B/66.25,Geneva, 1966.
JOINT FAO/WHO EXPERT COMMITTEE ON FOOD ADDITIVES - JECFA. 8th Report
of the Joint FAO/WHO Expert Committee on Food Additives. Specifications for identity,
purity, and toxicological evaluation of food colours. WHO Food additives series, v.66.n.25,
p.88-92, 1966.

95
JOINT FAO/WHO EXPERT COMMITTEE ON FOOD ADDITIVES - JECFA. 63rd
meeting of the Joint FAO/WHO Expert Committee on Food Additives. Aluminium Lakes of
Colouring Matters, General Specifications. WHO Food additives series, Geneva, 2004
JOINT FAO/WHO EXPERT COMMITTEE ON FOOD ADDITIVES - JECFA. Evaluation
of certain food contaminants (Sixty-fourth report of the Joint FAO/WHO Expert Committee
on Food Additives). WHO Technical Report Series, n.930, 2006
JOINT FAO/WHO EXPERT COMMITTEE ON FOOD ADDITIVES – JECFA. Laying
down specifications for food additives listed in Annexes II and III to Regulation (EC) nº
1333/2008 of the European Parliament and of the Council. COMMISSION REGULATION
(EU) No 231/2012 of 9 March 2012. Official Journal of the European Union, L 83/1,
22.3.2012.
JONES, R; RYAN, A. J.; WRIGHT, S. E. The Metabolism and Excretion of Tartrazine in
the Rat, Rabbit and Man. Food and Cosmetics Toxicology, v .2, p. 447- 452, 1964.
KANOU, M.; SAEKI, K.; KATO, T.A.; TAKAHASHI, K.; MIZUTANI, T. Study of in vitro
glucuronidation of hydroxyquinolines with bovine liver microsomes. Fundamental and
Clincal Pharmacology, v.16, p. 513-517, 2002.
KARPLIUK, I.A.; VOLKOVA, N.A.; OKUNEVA, L.A.; GOGOL, A.T.; RYBAKOVA,
K.D. Mutagenic effect of the food-coloring agents Tartrazine and indigo carmine. Voprosy
Pitaniia, p.58-61, 1984. In: EUROPEAN FOOD SAFETY AUTHORITY - EFSA. Scientific
Opinion on the re-evaluation Tartrazine (E 102). EFSA Journal, v.7, (11):1331, p.1-52,
2009.
KHERA, K.S., MUNRO, I.C. A review of the specifications and toxicity of synthetic food
colors permitted in Canada. CRC Critical Reviews in Toxicology, v.6, p. 81-133, 1979.

96
KUNO, N.; MIZUTANI, T. Influence of synthetic and natural food dyes on activities of
CYP2A6, UGT1A6, and UGT2B7. Journal of Toxicology and Environmental Health, v.8,
p. 1431-1444, 2005.
Environmental Protection Agency - U.S. EPA 2000. EPI Suite, Version 3.10; KOWWIN
Program, Version 1.66, 2000.
LANCASTER F. E.; LAWRENCE J. F. Determination of total non-sulphonated aromatic
amines in the food colour amaranth by dithionite reduction followed by derivatization and
high-performance liquid chromatography. Food Additives and Contaminants, v.6, p.415-
423, 1989.
LANCASTER F. E.; LAWRENCE J. F. Determination of total non-sulphonated aromatic
amines in tartrazine, Sunset Yellow FCF and allura red by reduction and derivatization
followed by high-performance liquid chromatography. Food Additives and
Contaminants,v.8,n.3, p.249-264, 1991.
LANCASTER, F.E.; LAWRENCE J.F. Determination of benzidine in the food colours
tartrazine and sunset yellow FCF, by reduction and derivatization followed by high-
performance liquid chromatography. Food Additives and Contaminants, v.16, n.9, p.381-
90, 1999.
LAZEAR, E. J., LOUIE, S. C., Mutagenicity of some congeners of benzidine in the
Salmonella typhimurium assay systems, Cancer Letters, v.4, p. 21-25, 1977.
LINDINO, C. A.; GONÇALVES, J. A. C.; SCHREINER, O. G. G.; SCHREINER, J. S.;
FARINA, L. O. Determinação de metais em corantes alimentícios artificiais. Acta
Scientiarum Technology, v.30, n.1, p.93-98, 2008.

97
LÜCK, H.; RICKERL, E. Food additives and mutative effects. VI Report, Z. Lebesmitt.
Untersuch. v. 112, p.157-174, 1960. In: EUROPEAN FOOD SAFETY AUTHORITY -
EFSA. Scientific Opinion on the re-evaluation Tartrazine (E 102). EFSA Journal, v.7, n.11,
p.1-52, 2009.
MA, H.; WANG, J.; ABDEL-RAHMAN, S.Z.; BOOR, P.J.; KHAN, M.F. Oxidative DNA
damage and its repair in rat spleen following subchronic exposure to aniline. Toxicology and
Applied Pharmacology, v.233, n.2, p.247-253, 2008.
MAEKAWA, A.; MATSUOKA, C.; ONODERA, H.; TANIGAWA, H.; FURUTA, K.;
KANNO, J.; JANG, J.J. LacK of Carcinogenecity of Tartrazine (FD & C YELLOW Nº 5)
in the F344 Rat. Food and Chemical Toxicology, v.25, n.12, p.891-896, 1987.
MANSOUR, H.B; CORROLER, D.; BARILLIER,D.; GHEDIRA, K.; CHEKIR, L.;
MOSRATI, R. Evaluation of genotoxicity and pro-oxidant effect of the azo dyes: Acids
yellow 17, violet 7 and orange 52, and of their degradation products by Pseudomonas putida
mt-2. Food and Chemical Toxicology, v .45, p.1670-1677, 2007.
MANNELL, W.A.; GRICE, H.C.; LU, F.C; ALLMARK, M.G.J. Chronic toxicity studies on
food colours. Iv.observations on the toxicity of tartrazine, amaranth and tartrazine in rats.
Journal of Pharmacy and Pharmacology, v.10, p.625-34, 1958. In: JOINT FAO/WHO
EXPERT COMMITTEE ON FOOD ADDITIVES - JECFA.Specifications for Identity and
Purity and Toxicological Evaluation of some Food Colours, FAO Nutrition Meetings
Report Series, WHO Food additives series, n.38B/66.25, Geneva, 1966.
MARMION, D.M. Colorants: Foods, Drugs, and Cosmetics and Medical Devices. Handbook
of U.S. John Wiley & Sons, Inc, v.23,p.1-576, 1991.

98
MARMITT, S.; PIROTTA, L. V.; STÜLP, S. Aplicação de fotólise direta e uv/H2O2 a
efluente sintético contendo diferentes corantes alimentícios. Quimica Nova, v.33, n. 2,
p.384-388, 2010.
MARON, D.M.; AMES, B.N. Revised methods for the Salmonella mutagenicity test.
Mutation Research, v.113, p.173-215, 1983.
MATHUR, N.; BHATNAGAR, P.; SHARMA, P. Review of the Mutagenicity of textile Dye
Products. Universal Journal of Environmental Research and Technology, v.2, p.1-18,
2012.
McCANN, J.; CHOI, E.; YAMASAKI, E.; AMES, B. N. Detection of carcinogens as
mutagens in the Salmonella/microsome test: Assay of 300 chemicals (rapid in vitro
screening/environmental carcinogens and mutagens). Proceedings of the National
Academy of Sciences of the United Stades of America, v.72, n. 12, p. 5135-5139, 1975.
MERCOSUL. Resolução do Grupo Mercado Comum (GMC) nº52 de 1998. Aprova o
Regulamento Técnico sobre “Critérios para determinar funções de aditivos, aditivos e seus
limites máximos para todas as categorias de alimentos”. XXXII GMC – Rio de Janeiro, v.8,
n.12, 1998.
MORI, Y.; NLWA, T.; HORL, T.; TOYOSHT, K. Mutagenicity of 3'-methyl-N, N-
dlmethyl-4-armnoazobenzene metabolites and related compounds. Carcinogenesis, v.1,
p.121-127, 1980.
MORRISON, R.T.; BOYD R. N. Química Orgânica. Trad. M. Alves da Silva. Fundação
Caloustre Gulbenkian, v.14, 2005.
MORTELMANS, K.; ZEIGER, E. The Ames Salmonella/Microsome Mutagenicity Assay.
Mutation Research, v.455, p.29-60, 2000.

99
MOUTINHO, I. L. S.; BERTGES L. C.; ASSIS, R. V. C. Prolonged use of food dye
tartrazine (FD&C yellow n5) and its effects on the gastric mucosa of Wistar rats. Brazilian
Journal Biology, v.67, p. 141-145, 2007.
MPOUNTOUKAS, P.; PANTAZAKI, A.; KOSTARELI, E.; CHRISTODOULOU, P,;
KARELI, D.; POLILIOU, S.; MOURELATOS, C.; LAMBROPOULOU, V.; LIALIARIS,
T. Cytogenetic evaluation and DNA interaction studies of the food colorants amaranth,
erythrosine and tartrazine. Food and Chemical Toxicology. v.48, p.2934-2944, 2010.
MORALES, D.A. Implantação do ensaio de Salmonella/microssoma no laboratório de
Ecotoxicologia Aquática e Limnologia – LEAL: FT-UNICAMP,141 páginas. Trabalho de
conclusão de curso. Programa de Graduação, Faculdade de Tecnologia – FT, Limeira,
2009.
MÜNZNER, R.; WEVER, J. Mutagenic activity of the feces of rats following oral
administration of Tartrazine. Archives Toxicology, v.60, p.328-330, 1987.
MURDOCH, R.D.; POLLOCK, I.; NAEEM, S. Tartrazine induced histamine release in vivo
in normal subjects. Journal of the Royal College of Physicians of London, v.21, n.4, p.
257-261, 1987.
MUZZAL, J.M.; COOK, W.L. Mutagenicity test of dyes used in cosmetics with the
Salmonella/ mammalian microsome test. Mutation Research, v. 6, p.1-8, 1979.
PATTERSON, R.M.; BUTLER, J.S. Tartrazine induced chromosomal aberrations in
mammalian cells. Food and Chemical Toxicology, v.20, n 4, p.461-465, 1982.

100
PERCY, A.J.; MOORE, N.; CHIPMAN, J.K. Formation of nuclear anomalies in rat intestine
by benzidine and its biliary metabolites. Toxicology, v.57, n.2, p. 217-223, 1989.
PEREZ-URQUIZA, M.; BELTRAN J.L. Determination of the dissociation constants of
Sulfonated azo dyes by capillary zone electrophoresis methods.., v.917, p.331-336, 2001.
PORWOLLIK, S.; WONG, R.M.Y.; SIMS, S.H.; SCHAAPER, R.M.; DEMARINI, D.M.;
MCCLELLAND, M. The Delta uvrB mutations in the Ames strains of Salmonella span 15
to 119 genes. Mutation. Research, v.483, p.1-11, 2001.
PRADO, M. A.; GODOY, H. T. Corantes artificiais em alimentos. Alimentos e Nutrição,
Araraquara, v.14, n.2, p. 237-250, 2003.
PRIVAL, M.J.; MITCHELL, V.D. Analysis of a method for testing azo dyes for mutagenic
activity in Salmonella typhimurium in the presence of flavin mononucleotide and hamster
liver S9. Mutation Research, v.97, p.103-116, 1982.
PRIVAL, M.J.; DAVIS, V. M.; PEIPERL, M.D.; SANDRA J.; BELL, S.J. Evaluation of
azo food dyes for mutagenicity and inhibition of mutagenicity by methods using Salmonella
typhimurium. Mutation Research, v. 206, p. 247-259, 1988.
PRIVAL, M.J.; PEIPERL, M.D.; BELL, S.J. Determination of combined benzidine in fd &
c yellow no. 5 (tartrazine), using a highly sensitive analytical method. Food and Chemical
Toxicoloy, v. 31, n. 10, p. 751-758, 1993.
POLLASTRINI, M.T.; BAREA, M.; SALAS, J. Genotoxic study of commercial dyes with
Tartrazine base in S. typhimurium his- and E. coli trp-. Rev. Sanid. Hig. Publica. v.64, p.203-
209, 1990. In: TEMA NORD, 2002. Food additives in Europe 2000; Status of safety
assessments of food additives presently permitted in the EU. TemaNord 560, 53-60, 2002.

101
POUL, M.; JARRY, G.; ELHKIM, M.O.; POUL, J. Lack of genotoxic effect of food dyes
amaranth, sunset yellow and tartrazine and their metabolites in the gut micronucleus assay in
mice. Food and Chemical Toxicology, v.47, p. 443-448, 2009.
POLÔNIO, M. L. T. Aditivos alimentares e saúde infantil. In: ACCIOLY, E.; SAUNDERS,
C.; LACERDA, E. M. Nutrição em Obstetrícia e Pediatria. Cultura Médica, p.511- 527,
2002.
POLÔNIO, M. L. T.; PERES, F. Consumo de aditivos alimentares e efeitos à saúde: desafios
para a saúde pública brasileira. Caderno de Saúde Pública, v.25, n.8, p. 1653-1666, 2009.
QUEIJA, C.; QUEIRÓS, M. A.; RODRIGUES, L. M. A cor dos Alimentos. Quimica,
Boletim da Sociedade Portuguesa de Quimica, v. 80, p. 6-11, 2001.
RAFII, F.; HALL, J.D.; CERNIGLIA, C.E. Mutagenicity of azo dyes used in foods, drugs
and cosmetics before and after reduction by Clostridium species from the human intestinal
tract, Food and Chemical Toxicology, v.35, p.897–901, 1997.
RAO, P.; BHAT, R.V.; SUDERSHAN, R.V.; KRISHNA, T.P.; NAIDU, N. Exposure
assessment to synthetic food colours of a selected population in Hyderabad, India. Food
additives and contaminants, v. 21, n.5, p. 415-421, 2004.
RAO NR, KIRAM S.S.M. Pharmaceutical impurities: an overview. Indian Journal of
Pharmaceutical Education and Research, v. 44, n.33, p. 301-310, 2010.
REYES, F. G. R.; PRADO, M. A. Aditivos e Contaminantes Alimentares. International
Life Science Institute of Brasil, v. 9, n.1, p.5-6, 2001.
RINDE, E.; TROLL, W. Metabolic reduction of benzidine azo dyes to benzidine in the
Rhesus monkey. Journal of the National Cancer Institute, v.55, p.181-182, 1975.

102
RING, J.; BROCKOW, K.,; BEHRENDT, H. Adverse reactions to foods. Journal of
Chromatography B: Biomedical Sciences and Applications, v.756, p.3-10, 2001.
ROXON, J.J.; RYAN, A.J.; WRIGHT, S.E. Reduction of tartrazine by a Proteus species
isolated from rats. Food and Cosmetics Toxicology, v.4, n.4, p. 419-426, 1966.
ROXON, J.J.; RYAN, A.J.; WRIGHT, S.E. Reduction of water-soluble azo dyes by
intestinal bacteria. Food and Cosmetics Toxicology, v.5, n.3, p. 367-369, 1967. .
RUSSFIELD, A.B.; HOMBURGER, F.; WEISBURGER, E.K.; WEISBURGER, J.H.
Further studies on carcinogenicity of environmental chemicals including simple aromatic
amines. Toxicology and Applied Pharmacology, v.25, p.446-447, 1973.
RYAN, A. J.; WRIGHT, S. E. The excretion of some azo dyes in rat bile. Journal of
Pharmacy and Pharmacology, v.13, p.492-495, 1961.
RYAN, A.J.; WRIGHT, S.E. Biliary excretion of some azo dyes related to tartrazine. Nature,
v. 8, p.195-1009, 1962.
RYAN, A.J.; WELLING, P.G.; WRIGHT, S.E. Further studies on the metabolism of
tartrazine and related compounds in the intact rat. Food and Cosmetics Toxicology, v.7, n.4,
p.287-295,1969a.
RYAN, A.J.; WELLING, P.G.; ROXON, J.J. Metabolism of a tartrazine analogue by
intestinal bacteria. Food and Cosmetics Toxicology, v.7n.4, p.297-299, 1969b.
RYAN, A.J. Factors involved in the metabolism of xenobiotics in mammals. Australian
Journal Pharmaceutical Sciences, p. 30-34, 1972.

103
SASAKI, Y.F.; KAWAGUCHI, S.; KAMAYA, A.; OHSHITA, M.; KABASAWA, K.;
IWAMA, K.; TANIGUCHI, K.; TSUDA, S. The comet assay with 8 mouse organs: results
with 39 currently used food additives. Mutation Research, v. 519, p. 103-119, 2002.
SCHELINE, R.R. Metabolism of foreign compounds by gastrointestinal microorganisms.
Pharmacological Reviews, v.25, p. 451-523, 1973.
SCHRODER, H.; CAMPBELL, D.E.S. Absorption, metabolism, and excretion of
salicylazosulfapyridine in man. Clinical Pharmacology & Therapeutics, v.13, p.539-551,
1972.
SHAHIN, M.M. Mutagenic evaluation of nitroanilines and nitroaminophenol in Salmonella
typhimurium, International Journal of Cosmetic Science, v. 7, p. 277–289, 1985.
SHILS, M.; OLSON, J. A.; SHIKE, M.; ROSS, A.C.; CABALERRO, B. Tratado de nutrição
moderna na saúde e na doença. Manole, v. 10, p. 1-2256, 2009.
SILVA, N. O.; REED, E. Estudos sobre corantes artificiais em alimentos:quais os riscos mais
comuns pelo consumo excessivo. Cadernos de Educação, Tecnologia e Sociedade, v. 2, n.
1, p. 10-14, 2010.
SIMON, V.F. In vitro mutagenicity assays of chemical carcinogens and related compounds
with Salmonella typhimurium. Journal of the National Cancer Institute, v. 62, p.893–899,
1979.
SUGIMURA, T.; SATO, S.; NAGAO, M.; YAHAGI, T.; MATSUSHIMA, T.; SEINO, Y.;
TAKEUCHI, M.; KAWACHI, T. Overlapping of carcinogens and mutagens, in:
Fundamentals in Cancer Prevention. International Symposium Princess Takamatsu
Cancer Research Fund, v.6, p. 191–215, 1976.

104
TANAKA, T. Reproductive and neurobehavioural toxicity study of tartrazine administered
to mice in the diet. Food Chemical Toxicology, v.44, p.179-87, 2006.
TANAKA, T.; TAKAHASHI, O.; OISHI, S.; OGATA, A. Effects of Tartrazine on
exploratory behavior in a threegeneration toxicity study in mice. Reproductive Toxicology,
v.26, p.156–163, 2008.
TEMA NORD, 2002. Food additives in Europe 2000; Status of safety assessments of food
additives presently permitted in the EU. TemaNord 560, 53-60, 2002.
UMBUZEIRO, G.A.; HAROLD, F.; WARREN, S.H.H.; KUMMROW, F.; LARRY D.;
CLAXTON, F.D. Mutagenicity evaluation of the commercial product CI Disperse Blue 291
using different protocols of the Salmonella assay. Food and Chemical Toxicology, v.43,
p.49-56, 2005.
UMBUZEIRO, G.A.; VARGAS, V.M.F. Teste de mutagenicidade com Salmonella
typhimurium (Teste de Ames) como indicador de carcinogenicidade em potencial para
mamíferos. In: Mutagênese Ambiental, Eds. RIBEIRO, L.R.; SALVADORI, D.M.F.;
MARQUES, E.K., ULBRA, capítulo 4, p. 81-112, 2003.
UBELIN, F, VON. PLETSCHER, A., Atiologie und prophylaxe gewerblicher tumoren in der
farbstoff-industrie, Schweiz Med Wochenschr Journal, v. 84, p. 917, 1954.
VARANDA, E.A. Atividade mutagênica de plantas medicinais. Revista de Ciências
Farmacêuticas Básica Aplicada. v. 27, n.1,p.1-7, 2006),
VOLKER, M.; ARLT, V.M.; HEWER, A.; SORG,B.L.; SCHMEISER, H.; PJILLIPS, D.H.;
STIBOROVA, M. 3-Aminobenzanthrone, a Human Metabolic of the Environmental
Pollutant 3-Nitrobenzanthone, Forms DNA Adducts after Metabolic Activation by Human

105
and Rat, Liver Microsomes: Evidence for Activation by Cytochrome P450 1A1 and P450
1A2. Chemical Research Toxicology, v.17, p.1092-1101, 2004.
WALKER, R. The metabolism of azo compounds: A review of the literature. Food and
Cosmetic Toxicology, v.8, p.659-676, 1970.
WALPOLE, A. L.; WILLIAMS, M. H. C.; ROBERTS, D. C. The carcinogenic action of 4-
aminodiphenyl and 3: 2'-dimethyl-4-aminodiphenyl. British Journal of Industrial
Medicini, v.9, p. 255-256, 1952.
WANG, C.Y.; MURAOKA, K.; BRYAN, G.T. Mutagenicity of nitrofurans, nitrothiophenes,
nitropyroles , nitroimidazole, aminothiophenes and aminothiazoles in Salmonella
typhimurium, Cancer Research,v. 35, p. 3611-3617, 1975.
WATABE, T.; OZAWA, N.; KOBAYASHI, F.; KURATA, H. Reduction of sulphonated
water-soluble azo dyes by microorganisms from human faeces. Food and Cosmetic
Toxicology, v.18, p.349-352, 1980.
WATERMAN, N. LIGNAC, G.O. The influence of the feeding of a number of food colours
on the occurrence of tumours in mice. Acta Physiologica Pharmacologica Neerlandica, v.
7, p.35-55, 1958. In: JOINT FAO/WHO EXPERT COMMITTEE ON FOOD ADDITIVES
- JECFA.Specifications for Identity and Purity and Toxicological Evaluation of some Food
Colours, FAO Nutrition Meetings Report Series, WHO Food additives series,
n.38B/66.25,Geneva, 1966.
WILLHEIM, R.; IVY, A.C. A preliminary study concerning the possibility of dietary
carcinogenesis. Gastroenterology, v.23, p.1-19, 1953. In: JOINT FAO/WHO EXPERT
COMMITTEE ON FOOD ADDITIVES - JECFA.Specifications for Identity and Purity and
Toxicological Evaluation of some Food Colours, FAO Nutrition Meetings Report Series,
WHO Food additives series, n.38B/66.25,Geneva, 1966.

106
YONGLIN, G.; CHUNMEI L.; JINGYU, S.; HUAXIAN, Y.; XIULIN, A; HAIZHU, J.
Effect of Food Azo Dye Tartrazine on Learning and Memory Functions in Mice and Rats,
and the Possible Mechanisms Involved. Journal of Food Science, v.76, n. 6, 2011.
ZAVON, M.R.; HOEGG, U.; BINGHAM, E. Benzidine exposure as a cause of bladder
tumors. Archives of Environmental Health an International Journal, v. 27 p.1–7, 1973.
ZIMMER, D.; MAZUREK, J.; PETZOLD, G.; BHUYAN, B. K. Bacterial mutagenicity and
mammalian cell DNA damage by several substituted anilines. Mutation Research, v.77, p.
317-326, 1980.
ZOLLINGER, H. Color chemistry: syntheses, properties, and applications of organic dyes
and pigments. WHILEY - VCH, v.3, p. 492,1991.

107
APÊNDICES

108
Apêndice A – Resultados do ensaio de Salmonella/microssoma
TABELA 1- Resultados do Ensaio de Salmonella/microssoma para o corante Tartrazina
comercial 90% com a linhagem TA98 em presença (+S9) e ausência (-S9) de ativação
metabólica.
Tartrazina Comercial 90%
Dose (µg/placa)
TA98-S9 TA98+S9
P1 P2 X DP RM P1 P2 X DP RM
Controle Negativo 27 32 30 3,5 1,0 44 47 46 2,1 1,0
312,5 34 32 33 1,4 1,1 29 34 32 3,5 0,7
625 34 36 35 1,4 1,2 36 34 35 1,4 0,8
1250 36 33 35 2,1 1,2 39 30 35 6,4 0,8
2500 28 33 31 3,5 1,0 42 37 40 3,5 0,9
5000 28 29 29 0,7 1,0 40 35 38 3,5 0,8
Controle Positivo 541 498 520 30,4 17,6 1483 1342 1413 100 31,0
Viabilidade (bactérias/mL) 2,3x109
Controle Negativo (água ultrapura Milli-Q estéril); Controle Positivo (-S9: 4NQO, +S9:2AA); P1 e P2 (Placas 1 e 2); X
(Média); DP (Desvio Padrão); RM (Razão de Mutagenicidade)

109
TABELA 2- Resultados do Ensaio Salmonella/microssoma para o corante Tartrazina Sigma
99% com a linhagem TA98 em presença (+S9) e ausência (-S9) de ativação metabólica.
Tartrazina Sigma 99%
Dose (µg/placa)
TA98-S9 TA98+S9
P1 P2 X DP RM P1 P2 X DP RM
Controle Negativo 65 62 64 2,1 1.0 66 80 73 9,9 1,0
312,5 72 60 66 8,5 1,0 76 73 75 2,1 1,0
625 76 64 70 8,5 1,1 75 69 72 4,2 1,0
1250 52 86 69 24,0 1,1 74 74 74 0 1,0
2500 76 68 72 5,7 1,1 81 50 66 21,9 0,9
5000 60 60 60 0 0,9 62 70 66 5,7 0,9
Controle Positivo 327 410 369 59 5,8 1410 1553 1482 101,1 20,3
Viabilidade (bactérias/mL) 2,5 x 109
Controle Negativo (água ultrapura Milli-Q estéril); Controle Positivo (-S9: 4NQO, +S9:2AA); P1 e P2 (Placas 1 e 2); X
(Média); DP (Desvio Padrão); RM (Razão de Mutagenicidade)

110
TABELA 3- Resultados do Ensaio Salmonella/microssoma para o corante Tartrazina
comercial 90% com a linhagem TA100 em presença (+S9) e ausência (-S9) de ativação
metabólica.
Tartrazina Comercial 90%
Dose (µg/placa)
TA100-S9 TA100+S9
P1 P2 X DP RM P1 P2 X DP RM
Controle Negativo 141 158 150 12 1,0 173 140 157 23,3 1,0
312,5 133 133 133 0 0,9 137 145 141 5,7 0,9
625 124 157 141 23,3 0,9 135 122 129 9,2 0,8
1250 136 130 133 4,2 0,9 148 127 138 14,8 0,9
2500 148 120 134 19,8 0,9 120 122 121 1,4 0,8
5000 149 130 140 13,4 0,9 132 127 130 3,5 0,8
Controle Positivo 4380 4060 4220 226,3 28,2 3976 3870 3923 75 25,1
Viabilidade (bactérias/mL) 2,98 x 109
Controle Negativo (água ultrapura Milli-Q estéril); Controle Positivo (-S9: 4NQO, +S9:2AA); P1 e P2 (Placas 1 e 2); X
(Média); DP (Desvio Padrão); RM (Razão de Mutagenicidade)
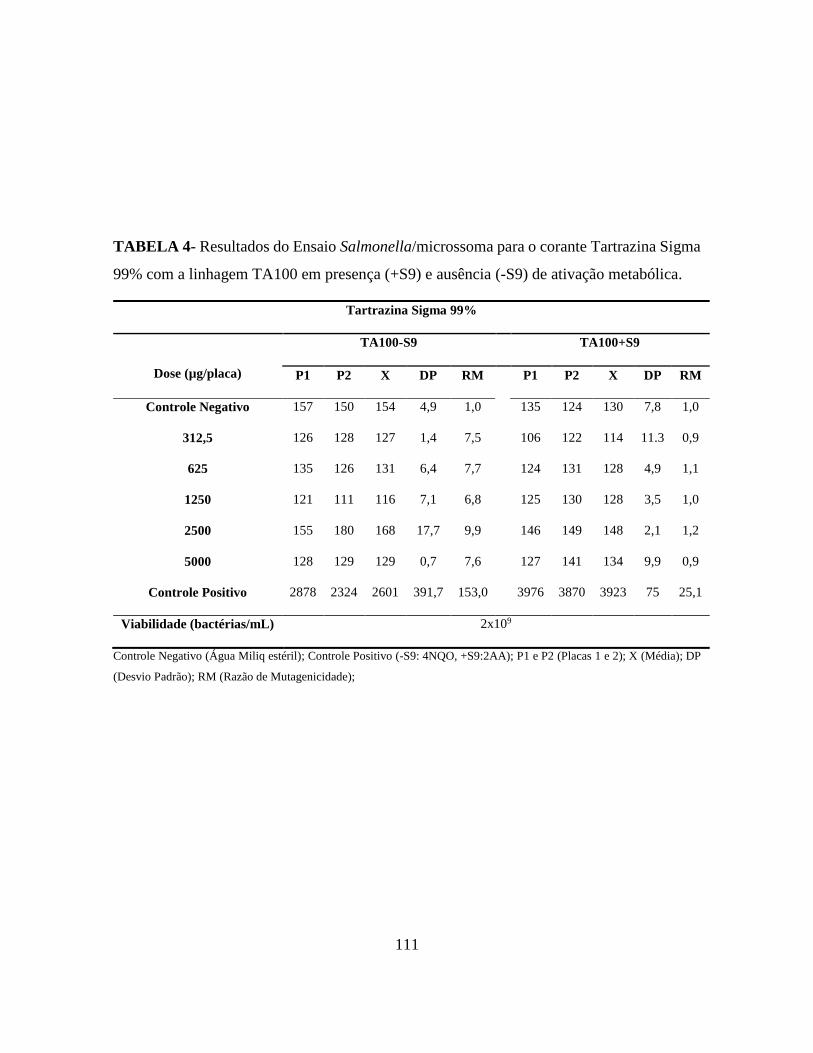
111
TABELA 4- Resultados do Ensaio Salmonella/microssoma para o corante Tartrazina Sigma
99% com a linhagem TA100 em presença (+S9) e ausência (-S9) de ativação metabólica.
Tartrazina Sigma 99%
Dose (µg/placa)
TA100-S9 TA100+S9
P1 P2 X DP RM P1 P2 X DP RM
Controle Negativo 157 150 154 4,9 1,0 135 124 130 7,8 1,0
312,5 126 128 127 1,4 7,5 106 122 114 11.3 0,9
625 135 126 131 6,4 7,7 124 131 128 4,9 1,1
1250 121 111 116 7,1 6,8 125 130 128 3,5 1,0
2500 155 180 168 17,7 9,9 146 149 148 2,1 1,2
5000 128 129 129 0,7 7,6 127 141 134 9,9 0,9
Controle Positivo 2878 2324 2601 391,7 153,0 3976 3870 3923 75 25,1
Viabilidade (bactérias/mL) 2x109
Controle Negativo (Água Miliq estéril); Controle Positivo (-S9: 4NQO, +S9:2AA); P1 e P2 (Placas 1 e 2); X (Média); DP
(Desvio Padrão); RM (Razão de Mutagenicidade);

112
TABELA 5- Resultados do Ensaio Salmonella/microssoma para o corante Tartrazina Comercial
90% com a linhagem TA1535 em presença (+S9) e ausência (-S9) de ativação metabólica.
Tartrazina Comercial90%
Dose (µg/placa)
TA1535-S9 TA1535+S9
P1 P2 X DP RM P1 P2 X DP RM
Controle Negativo 17 15 16 1,4 1,0 21 14 18 4,9 1,0
312,5 22 19 21 2,1 1,3 18 12 15 4,2 0,9
625 21 17 19 2,8 1,2 13 12 13 0,7 0,7
1250 17 20 19 2,1 1,2 13 12 13 0,7 0,7
2500 20 23 22 2,1 1,3 12 10 11 1,4 0,6
5000 16 16 16 0 1,0 10 11 11 0,7 0,6
Controle Positivo 1296 1225 1261 50,2 78,8 206 205 206 1 11,7
Viabilidade (bactérias/mL) 2,5 x109
Controle Negativo (água ultrapura Milli-Q estéril); Controle Positivo (-S9: Azida Sódica, +S9:2AA); P1 e P2 (Placas 1 e
2); X (Média); DP (Desvio Padrão); RM (Razão de Mutagenicidade)
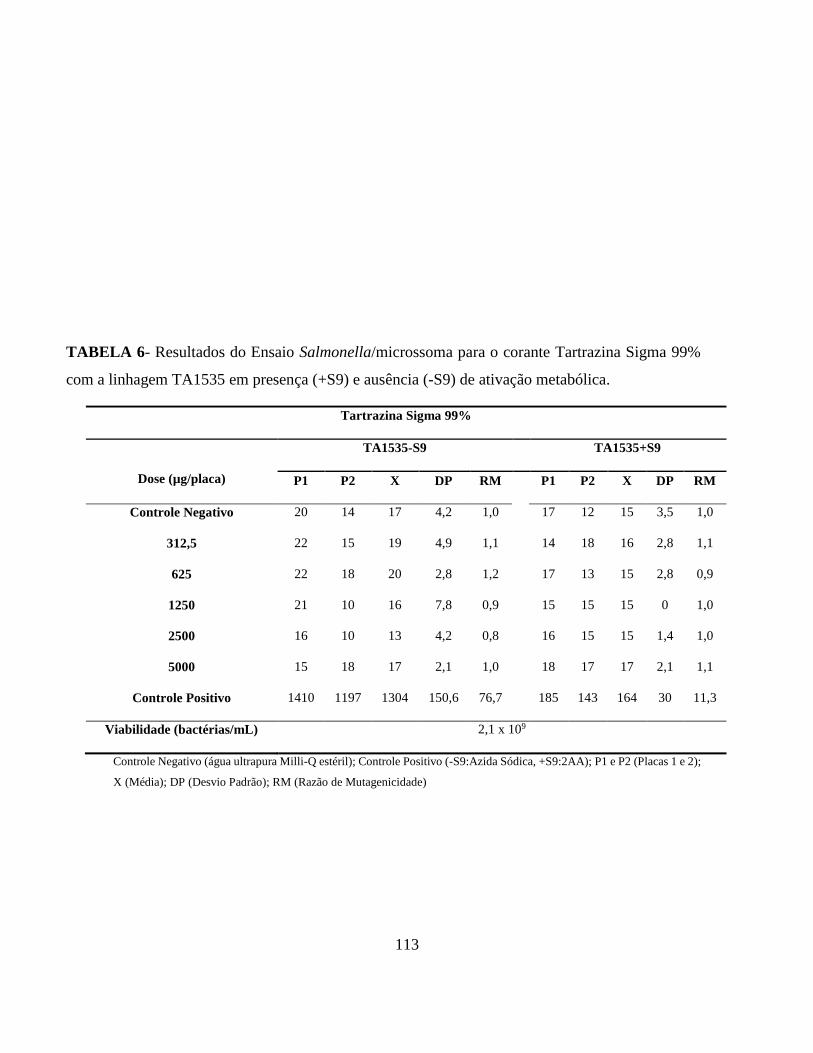
113
TABELA 6- Resultados do Ensaio Salmonella/microssoma para o corante Tartrazina Sigma 99%
com a linhagem TA1535 em presença (+S9) e ausência (-S9) de ativação metabólica.
Tartrazina Sigma 99%
Dose (µg/placa)
TA1535-S9 TA1535+S9
P1 P2 X DP RM P1 P2 X DP RM
Controle Negativo 20 14 17 4,2 1,0 17 12 15 3,5 1,0
312,5 22 15 19 4,9 1,1 14 18 16 2,8 1,1
625 22 18 20 2,8 1,2 17 13 15 2,8 0,9
1250 21 10 16 7,8 0,9 15 15 15 0 1,0
2500 16 10 13 4,2 0,8 16 15 15 1,4 1,0
5000 15 18 17 2,1 1,0 18 17 17 2,1 1,1
Controle Positivo 1410 1197 1304 150,6 76,7 185 143 164 30 11,3
Viabilidade (bactérias/mL) 2,1 x 109
Controle Negativo (água ultrapura Milli-Q estéril); Controle Positivo (-S9:Azida Sódica, +S9:2AA); P1 e P2 (Placas 1 e 2);
X (Média); DP (Desvio Padrão); RM (Razão de Mutagenicidade)

114
TABELA 7- Resultados do Ensaio de Salmonella/microssoma para o corante Tartrazina
comercial 90% com a linhagem TA102 em presença (+S9) e ausência (-S9) de ativação
metabólica.
Amarelo Tartrazina Comercial 90%
Dose (µg/placa)
TA102-S9 TA102+S9
P1 P2 P3 P4 X DP RM P1 P2 P3 P4 X DP RM
Controle
Negativo
153 163 192 152 165 18,7 1,0 223 169 223 174 197 29,8 1,0
312,5 134 154 144 14,1 0,9 215 202 209 9,2 1,1
625 166 172 169 4,2 1,0 221 206 214 10,6 1,1
1250 185 190 188 3,5 1,1 211 226 219 10,6 1,1
2500 196 197 197 0,7 1,2 263 236 250 19,1 1,3
5000 Tox Tox Tox Tox
Controle
Positivo
2569 2431 2500 97,6 15,2 2089 2120 1541 2105 22 11,3
Viabilidade
(bactérias/mL)
3,1 x109
Controle Negativo (água ultrapura Milli-Q estéril); Controle Positivo (-S9: Mitomicina C, +S9:2AA); P1, P2, P3 e P4
(Placas 1,2,3, e 4); X (Média); DP (Desvio Padrão); RM (Razão de Mutagenicidade).
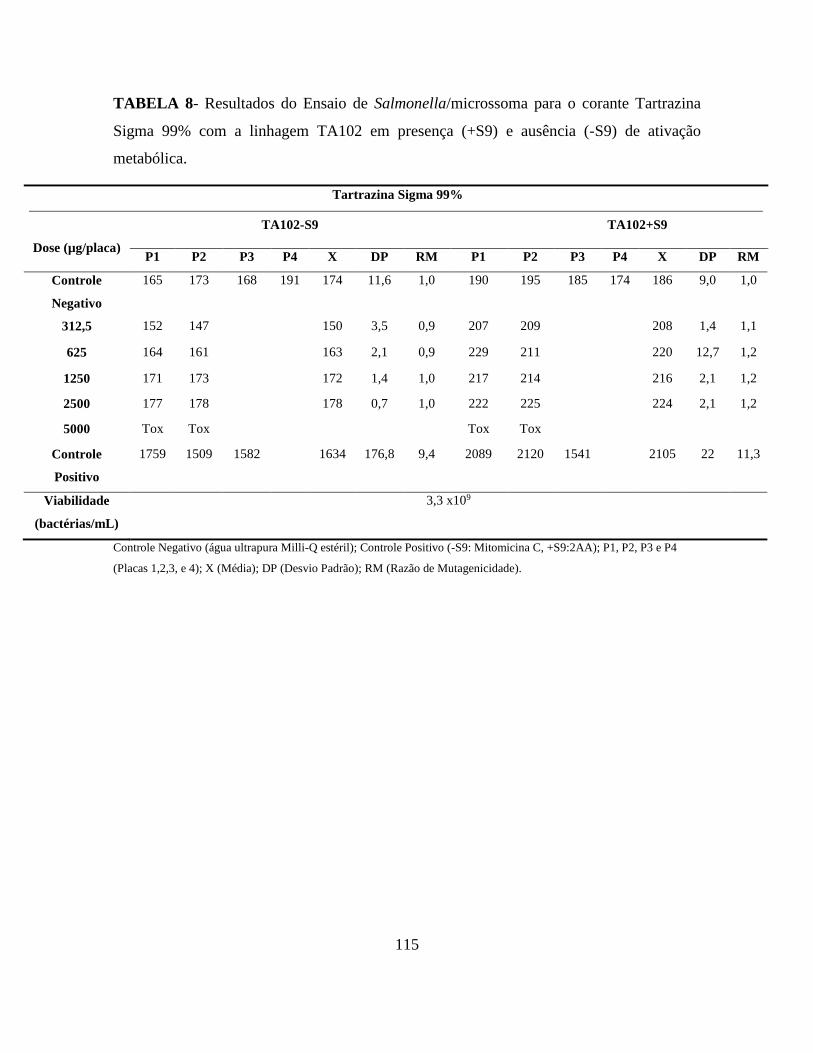
115
TABELA 8- Resultados do Ensaio de Salmonella/microssoma para o corante Tartrazina
Sigma 99% com a linhagem TA102 em presença (+S9) e ausência (-S9) de ativação
metabólica.
Tartrazina Sigma 99%
Dose (µg/placa)
TA102-S9 TA102+S9
P1 P2 P3 P4 X DP RM P1 P2 P3 P4 X DP RM
Controle
Negativo
165 173 168 191 174 11,6 1,0 190 195 185 174 186 9,0 1,0
312,5 152 147 150 3,5 0,9 207 209 208 1,4 1,1
625 164 161 163 2,1 0,9 229 211 220 12,7 1,2
1250 171 173 172 1,4 1,0 217 214 216 2,1 1,2
2500 177 178 178 0,7 1,0 222 225 224 2,1 1,2
5000 Tox Tox Tox Tox
Controle
Positivo
1759 1509 1582 1634 176,8 9,4 2089 2120 1541 2105 22 11,3
Viabilidade
(bactérias/mL)
3,3 x109
Controle Negativo (água ultrapura Milli-Q estéril); Controle Positivo (-S9: Mitomicina C, +S9:2AA); P1, P2, P3 e P4
(Placas 1,2,3, e 4); X (Média); DP (Desvio Padrão); RM (Razão de Mutagenicidade).

116
TABELA 9- Resultados do Ensaio de Salmonella/microssoma para o corante Tartrazina Comercial 90% com
a linhagem TA97a em presença (+S9) e ausência (-S9) de ativação metabólica.
Tartrazina Comercial 90%
Dose
(µg/placa)
TA97a-S9 TA97a+S9
P1 P2 P3 P4 P5 X DP RM P1 P2 P3 P4 P5 X DP RM
Controle
Negativo
184 178 197 181 182 184 7,4 1,0 170 189 175 155 156 169 13,
4
1,0
312,5 187 179 183 5,7 1,0 198 205 202 4,9 1,2
625 172 194 183 15,6 1,0 212 208 210 2,8 1,2
1250 184 176 180 5,7 1,0 214 195 205 13,
4
1,2
2500 185 181 183 2,8 1,0 189 195 192 4,2 1,1
5000 197 189 193 5,7 1,0 207 226 217 13,
4
1,3
Controle
Positivo
665 757 769 730 56,9 4,0 1187 2351 1329 1622 635 9,6
Viabilidade
(bactérias/
mL)
3,1 x 109
Controle Negativo (água ultrapura Milli-Q estéril); Controle Positivo (-S9: 9 aminoacridine, +S9:2AA); P1, P2,P3,P4 e P5
(Placas 1, 2,3,4 e 5); X (Média); DP (Desvio Padrão); RM (Razão de Mutagenicidade)

117
TABELA 10- Resultados do Ensaio de Salmonella/microssoma para o corante Tartrazina Sigma 99% com a
linhagem TA97a em presença (+S9) e ausência (-S9) de ativação metabólica.
Tartrazina Sigma 99%
Dose
(µg/placa)
TA97a-S9 TA97a+S9
P1 P2 P3 P4 P5 X DP RM P1 P2 P3 P4 P5 X DP RM
Controle
Negativo
157 162 165 166 171 164 5,2 1,0 188 174 172 188 190 182 9,
9
1,0
312,5 171 177 174 4,2 1,1 174 180 177 4,2 1,0
625 170 174 172 2,8 1,0 171 174 173 2,1 0,9
1250 180 178 179 1,4 1,1 194 198 196 2,8 1,1
2500 186 186 186 0,0 1,1 197 199 198 1,4 1,1
5000 191 192 192 0,7 1,2 201 209 205 5,7 1,1
Controle
Positivo
1125 1083 1104 29,7 6,7 1955 1792 2342 203
0
283 11,1
Viabilidade
(bactérias/
mL)
3,2 x109
Controle Negativo (água ultrapura Milli-Q estéril); Controle Positivo (-S9: 9 aminoacridine, +S9:2AA); P1, P2, P3, P4 e P5
(Placas 1, 2,3,4 e 5); X (Média); DP (Desvio Padrão); RM (Razão de Mutagenicidade)

118


















