
Rodrigo Barbosa Lima UFPB - AREIA -...
54
UNIVERSIDADE FEDERAL DA PARAÍBA UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO UNIVERSIDADE FEDERAL DO CEARÁ PROGRAMA DE DOUTORADO INTEGRADO EM ZOOTECNIA BALANÇO ELETROLÍTICO EM CODORNAS E MODELOGEM DO CRESCIMENTO DE FRANGAS DE REPOSIÇÃO Rodrigo Barbosa Lima UFPB - AREIA FEVEREIRO – 2012
Transcript of Rodrigo Barbosa Lima UFPB - AREIA -...

UNIVERSIDADE FEDERAL DA PARAÍBA
UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO
UNIVERSIDADE FEDERAL DO CEARÁ
PROGRAMA DE DOUTORADO INTEGRADO EM ZOOTECNIA
BALANÇO ELETROLÍTICO EM CODORNAS E MODELOGEM DO
CRESCIMENTO DE FRANGAS DE REPOSIÇÃO
Rodrigo Barbosa Lima
UFPB - AREIA
FEVEREIRO – 2012

ii
UNIVERSIDADE FEDERAL DA PARAÍBA
UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO
UNIVERSIDADE FEDERAL DO CEARÁ
PROGRAMA DE DOUTORADO INTEGRADO EM ZOOTECNIA
BALANÇO ELETROLÍTICO EM CODORNAS E MODELOGEM DO
CRESCIMENTO DE FRANGAS DE REPOSIÇÃO
Rodrigo Barbosa Lima
Zootecnista
UFPB - AREIA
FEVEREIRO – 2012

iii
RODRIGO BARBOSA LIMA
AVALIAÇÃO BALANÇO ELETROLÍTICO EM CODORNAS E
MODELOGEM DO CRESCIMENTO DE FRANGAS DE REPOSIÇÃO
Tese apresentada ao Programa de
Doutorado Integrado em Zootecnia da
Universidade Federal da Paraíba,
Universidade Federal Rural de
Pernambuco e Universidade Federal do
Ceará como requisito parcial para
obtenção do título de Doutor em
Zootecnia.
Área de Concentração: Produção Animal
Comitê de Orientação:
Prof. Dr. Jose Humberto Vilar da Silva – Orientador Principal
Profª. Drª. Patrícia Emilia Naves Givisiez
Profª. Drª. Terezinha Domiciano Dantas Martins
UFPB - AREIA
FEVEREIRO – 2012

Ficha Catalográfica Elaborada na Seção de Processos Técnicos da
Biblioteca Setorial do CCA, UFPB, Campus II, Areia – PB.
L732b Lima, Rodrigo Barbosa.
Balanço eletrolítico em codornas e modelagem do crescimento de frangas de reposição. / Rodrigo Barbosa Lima. - Areia: UFPB/CCA, 2012.
41 f. : il.
Tese (Doutorado em Zootecnia) - Centro de Ciências Agrárias. Universidade Federal da Paraíba, Areia, 2012.
Bibliografia.
Orientador: José Humberto Vilar da Silva. Co-orientador: Patrícia Emilia Naves Givisiez e Terezinha Domiciano Dantas Martins.
1. Avicultura 2. Aves comerciais – estresse por calor 3. Gompertz 4. Aves – exigências em proteína I. Silva, José Humberto Vilar (Orientador) II.Título.
UFPB/CCA CDU: 636.5(043.2)

v

vi
DADOS CURRICULARES DO AUTOR
RODRIGO BARBOSA LIMA – Filho de José Barbosa Lima e Maria da Glória Pedrosa de
Lima, nasceu no dia 03 de agosto de 1975 na cidade de Recife, no Estado de Pernambuco.
Ingressou no Curso de Zootecnia da Universidade Federal Rural de Pernambuco em 1995,
concluindo o curso no ano 2000. Em 2006 ingressou no Curso de Mestrado em Zootecnia da
Universidade Federal Rural de Pernambuco. Sob a orientação do professor Dr. Carlos Bôa-
Viagem Rabello, trabalhou com avaliação de alimentos para frangos de corte. Recebeu o
título de Mestre em Zootecnia em fevereiro de 2008. Em março ingressou no Doutorado
Integrado em Zootecnia na UFPB, sob a orientação do professor Dr. José Humberto Vilar da
Silva. No ano de 2009 foi bolsista PROCAD na Universidade Estadual Paulista Júlio de
Mesquita Filho/ Jaboticabal sob a tutoria da professora Dr.ª Nilva Kazue Sakomura.

vii
“Tu conservarás em perfeita paz aquele cuja mente está firme em Ti; porque ele confia em
Ti.” Isaías 26:3

viii
A Deus, Meu maior bem, a quem confiei a minha vida, os que me são caros, meus sonhos...
Agradeço.
À minha querida esposa e aos meus filhos Miguel e Aurora, parte dos meus sonhos e da
minha vida.
Aos meus pais, José Barbosa Lima e Maria da Glória Pedrosa de Lima, meus irmãos Diego
Babosa Lima e Bárbara Barbosa Lima, meu sobrinho Lucas e minha cunhada Débora, minha
doce vó, e demais familiares.
Dedico.
Ao povo brasileiro, sobretudo aos outros Severinos e Severinas do meu Nordeste,
Ofereço a força do meu trabalho.

ix
AGRADECIMENTOS
Ao Prof. Dr. José Humberto Vilar da Silva, pela orientação, credibilidade e confiança que
depositou em mim. Agradeço pelas oportunidades. Itens que auxiliaram no meu
desenvolvimento pessoal e profissional.
Ao Profª. Drª. Patrícia Emilia Naves Givisiez, pela orientação, prontidão e atenção
dispensada.
Ao Prof. Dr. Ariosvaldo Nunes de Medeiros, importantíssimo na minha formação, em
nome de quem agradeço a todos os professores e coordenadores do programa de Pós-
Graduação em Zootecnia da UFPB/CCA.
A Profª. Drª. Nilva Kazue Sakomura, com quem aprendi muito sobre planejamento e
disciplina.
Aos professores Dr. Fernando Perazzo, Dr. Edilson Saraiva, Dr. Edgar Pimenta,
Dr. Severino Gonzaga, Dr. Celso Oliveira, Dr.Walter Esfrain, Dr. George Beltrão, Dr.
Marcos Macari, Drª. Elisabeth Gonzalés, pelas orientações.
Agradeço aos membros da banca examinadora: Dr. Fernando Perazzo, Dr. Alex Varela,
Drª. Maria do Carmo M. M. Ludke e Dr. Jorge Ludke.
Agradeço ao Dr. Jorge Vitor Ludke pela colaboração na minha formação.
Ao amigo Marquiliano Moura, Jefferson Siqueira e Jordão Filho pela ajuda.
A meu pai, minha mãe e meus irmãos pela participação direta na execução deste trabalho,
cada um doando aquilo em que são melhores.
A minha doce esposa Denise Fontana Figueiredo Lima, pela companhia e dedicação.
Aos demais colegas e amigos, que tornaram possível a realização deste trabalho, que
Deus os abençoe.
Ao CNPq, pela concessão da bolsa de Doutorado.

x
SUMÁRIO
Página
RESUMO GERAL ..................................................................................... 1
CONSIDERAÇÕES INICIAIS................................................................... 2
CAPÍTULO 1. Influência da temperatura e do balanço eletrolítico sobre o
desempenho de codornas (Coturnix coturnix coturnix). 3
Resumo.............................................................................................. 4
Abstract ............................................................................................. 5
Introdução ......................................................................................... 6
Material e Métodos ........................................................................... 7
Resultados e Discussão...................................................................... 10
Conclusões......................................................................................... 18
Referências Bibliográficas ................................................................ 18
CAPÍTULO 2. Curvas de crescimento e de deposição dos componentes
corporais e estimativas das exigências de proteína bruta para
frangas leves e semipesadas................................... 22
Resumo............................................................................................. . 23
Abstract ........................................................................................... 24
Introdução ........................................................................................ 25
Material e Métodos ........................................................................... 26
Resultados e Discussão....................................................................... 28
Conclusões ......................................................................................... 38
Referências Bibliográficas.................................................................. 38
CONSIDERAÇÕES FINAIS E IMPLICAÇÕES................................ 39

xi
LISTA DE TABELAS
CAPÍTULO 1. Influência da temperatura e do balanço eletrolítico sobre o
desempenho de codornas (Coturnix coturnix coturnix). Página
Tabela 1 Tabela 1. Rações utilizadas nos Experimentos 1 e 2.............................. 08
Tabela 2. Efeito do BE e da temperatura (T) sobre o desempenho codornas
europeias aos 37 dias.......................................................................... 10
Tabela 3. Efeito do BE e da temperatura (T) sobre rendimento de carcaça e o
peso relativo das vísceras de codornas europeias aos 37 dias.............. 10
Tabela 4. Comparação de médias entre ingestão de água e da relação ingestão
de água:consumo de ração para o BE e temperaturas ambientais........ 11
Tabela 5. Efeito da temperatura e do balanço eletrolítico via água de bebida
sobre o desempenho de codornas europeias aos 37 dias...................... 12
Tabela 6. Efeito da temperatura e do balanço eletrolítico via água de bebida
sobre o peso relativo das vísceras de codornas europeias aos 37 dias. 12
Tabela 7. Comparação de médias entre ganho de peso (GP), ingestão de água
(IA), peso relativo do coração (CO) e peso relativo do fígado para o
BE e temperaturas ambientais............................................................. 13
CAPÍTULO 2. Curvas de crescimento e de deposição dos componentes
corporais e estimativas das exigências de proteína bruta para
frangas leves e semipesadas.............................................................. 22
Tabela 1. Composição das dietas do período de cria e recria das frangas leves e
semipesadas........................................................................................ 26
Tabela 2. Estimativa dos parâmetros da equação de Gompertz para peso vivo,
peso corporal depenado e peso de penas de franga leve e
semipesada..................................................................................... 28
Tabela 3. Taxa de crescimento corporal e de penas de frangas leves e
semipesadas.................................................................................... 29
Tabela 4- Estimativa dos parâmetros da equação de Gompertz para proteína,
gordura, matéria mineral e água no corpo vazio de frangas leve e
semipesada....................................................................................... 31
Tabela 5- Taxa de deposição dos componentes corporais de frangas leves e
semipesadas..................................................................................... 32

xii
Tabela 6- Equações alométricas de peso corporal em gordura, água e matéria
mineral em função do peso corporal protéico de frangas leves e
semipesadas.................................................................................... 34
Tabela 7- Estimativa dos parâmetros da equação de Gompertz para proteína na
carcaça das linhagens...................................................... ................ 36
Tabela 8- Equação da proteína corporal (PC) em função do peso de corpo vazio
(PCV)de acordo com a idade das aves.............................................. 36
Tabela 9- Exigências de proteína e consumo de ração em função da fase de
criação de frangas leves e semipesadas.............................................. 37

xiii
LISTA DE FIGURAS
CAPÍTULO 2 - Curvas de crescimento do peso corporal e deposição de
nutrientes, exigências proteicas para frangas leves e semipesadas.
Página
Figura 1. Curvas (a) e taxa de crescimento (b) de peso vivo da franga leve
“Dekalb” e da semipesada “Bovans” em função da idade.................... 30
Figura 2- Curva (a) e taxa de crescimento (b) de peso corporal depenado da
franga leve “Dekalb” e da semipesada “Bovans” em função da
idade................................................................................................... 30
Figura 3- Curva (a) e taxa de crescimento (b) de penas da franga leve “Dekalb”
e da semipesada “Bovans” em função da idade................................... 30
Figura 4- Curva de crescimento (a) e taxa de deposição (b) de proteína no corpo
vazio da franga leve “Dekalb” e da semipesada “Bovans” em função
da idade.............................................................. ................................ 32
Figura 5 - Curva de crescimento (a) e taxa de deposição (b) de gordura no corpo
vazio da franga leve “Dekalb” e da semipesada “Bovans” em função
da idade. ................................................................ ............................ 33
Figura 6- Curva de crescimento (a) e taxa de deposição (b) de matéria mineral
no corpo vazio da franga leve “Dekalb” e da semipesada “Bovans”
em função da idade............................................................... .............. 33
Figura 7- Curva de crescimento (a) e taxa de deposição (b) de água no corpo
vazio da franga leve “Dekalb” e da semipesada “Bovans” em função
da idade........................................................................................ ...... 33
Figura 8- Comportamento da gordura e água no corpo vazio das frangas Dekalb
em função da idade............................................................................. 35

1
BALANÇO ELETROLÍTICO EM CODORNAS E MODELOGEM DO
CRESCIMENTO DE FRANGAS DE REPOSIÇÃO
RESUMO GERAL
O balanço eletrolítico (BE) foi investigado em codornas (Coturnix coturnix coturnix)
criadas sob alta temperatura (34°C) mediante a suplementação de sais via da água ou
ração. Os BE testados na ração foram 0, 120, 240, 360 mEq/ kg em duas temperaturas
(25 e 34ºC). Utilizou-se o bicarbonato de sódio e o cloreto de amônio para obtenção dos
BE nas rações. Os BE avaliado na água de bebida foram 0, 30, 60, 90, 120 mEq/ L.
Utilizou-se apenas o bicarbonato de sódio na água para obtenção dos BE. Os valores de
adição de bicarbonato de sódio na água foram estabelecidos após experimento piloto
para mensurar a tolerância das aves aos tratamentos. Os experimentos foram conduzidos
em câmaras climáticas à temperatura constante. Os ensaios foram conduzidos
paralelamente e ambos tiveram duração de 17 dias (20 a 37 dias de idade). As codornas
aumentaram a ingestão de água a partir do BE de 120 mEq/kg de ração e BE 30 mEq/ L
na água de bebida sem diferir (P > 0,05) dos demais tratamentos. A técnica de abate em
série foi usada para determinar a composição corporal em função da idade para proteína,
lipídio, cinzas e água no corpo depenado de frangas Dekalb White e Bovans Golline de
1 a 16 semanas de idade. Três dietas à base de milho e farelo de soja foram utilizadas
durante o experimento: 1 a 4 semanas, 5 a 11 semanas e 12 a 16 semanas. Relações
alométricas foram utilizadas para descrever as mudanças na composição corporal. Os
dados do peso corporal foram usados para obter os parâmetros do modelo de
crescimento Gompertz. A linhagem Dekalb White (leve) é precoce no crescimento e na
deposição dos componentes corporais e difere na exigência proteica da linhagem
Bovans Golline (semipesada). Apesar disso, tais diferenças têm pouca significado
comercial sem informações adicionais do período de produção. Investigações futuras
sobre a composição corporal das frangas combinadas a fase de postura representam
avanço nessa área de estudo.
Palavras–chave:estresse, bicarbonato de sódio, cloreto de amônio, Gompertz, pré-
postura

2
CONSIDERAÇÕES INICIAIS
Criações de aves comerciais em regiões de clima quente apresentam redução de
desempenho com efeitos negativos sobre o crescimento das aves e qualidade dos
produtos. A adição de sais como bicarbonato de sódio e cloreto de amônio é bem
documentado em dietas para frangos de corte, bem como o balanço eletrolítico (BE) que
proporciona a redução dos efeitos danosos do estresse térmico por calor. Codornas
(Coturnix coturnix coturnix) são originalmente de regiões quentes e pode ter balanço
eletrolítico dietético diferente dos galináceos. Apesar disso pouca atenção tem sido dada
a pesquisas que elucidem os efeitos da elevada temperatura ambiental para codornas em
crescimento. A elaboração de dietas com faixas restritas de balanço eletrolítico
semelhantes às utilizadas com frangos de corte pode não ser apropriada para a
investigação ainda exploratória na criação de codornas. Os resultados demonstraram
que pouco efeito no desempenho das codornas foi alcançado pelos diferentes balanços
eletrolíticos ou do sal. O consumo de água aumentou no BE “nulo” e de 360mEq/kg
para a suplementação de cloreto de amônio e bicarbonato de sódio na ração,
respectivamente. Com relação à cinética de crescimento em função da idade em frangas
das linhagens Dekalb White e Bovans Golline, foram observados diferenças entre as
linhagens na deposição de nutrientes que refletem eficiências diferentes e também
diferentes exigências protéicas para mantença e ganho. Logo, pode-se discutir qual seria
a alternativa mais interessante na formulação de uma ração comercial. Portanto, espera-
se que as informações permitem “manobras” alimentares quer seja na composição da
ração quer seja na diminuição da quantidade de ração fornecida as frangas.

3
CAPÍTULO 1
INFLUÊNCIA DA TEMPERATURA E DO BALANÇO
ELETROLÍTICO SOBRE O DESEMPENHO DE CODORNAS
(Coturnix coturnix coturnix)

4
Influência da temperatura e do balanço eletrolítico sobre o desempenho de
codornas (Coturnix coturnix coturnix)
Resumo
A pesquisa avaliou a eficiência da suplementação de eletrólitos em condições de alta
temperatura ambiental, através da manipulação dos balanços eletrolíticos (BE) da água e
da ração, sobre a sobrevivência e desempenho de codornas europeias. No experimento
1, o delineamento utilizado foi o inteiramente casualizado em esquema fatorial 4 x 2. Os
tratamentos consistiram em quatro valores para o BE na ração (0, 120, 240, 360 mEq/
kg) sob duas temperaturas ambientais (25 e 34ºC). Utilizou-se o bicarbonato de sódio e
o cloreto de amônio para obtenção dos BE nas rações. No experimento 2, as aves foram
distribuídas ao acaso em esquema fatorial 5 x 2. Os tratamentos consistiram em cinco
valores para o BE na água de bebida (0, 30, 60, 90, 120 mEq/ L) sob duas temperaturas
ambientais (25°C e 34°C). Utilizou-se apenas o bicarbonato de sódio na água para
obtenção dos BE. Os experimentos foram conduzidos em câmaras climáticas à
temperatura constante com codornas com idade inicial de 20 dias. Os ensaios foram
conduzidos paralelamente e ambos tiveram duração de 17 dias. As A maioria das
variáveis foram influenciadas pela temperatura em ambos os experimentos. No
Experimento 1, o BE influenciou a ingestão de água e o comprimento do intestino. No
Experimento 2, o BE afetou (p < 0,05) ingestão de água, peso relativo do coração e do
fígado. Para o experimento 1 recomenda-se a o BE de 120 mEq/kg de ração e BE 30
mEq/ L na água de bebida.
Palavras–chave: cloreto de amônio, estresse por calor, bicarbonato de sódio

5
Influence of temperature and electrolyte balance on quails (Coturnix coturnix
coturnix) performance
Abstract
Research evaluated the efficiency of electrolyte supplementation in high environmental
temperature conditions, through manipulation of electrolyte balance (EB) of water and
food on european quails survival and performance. In experiment 1, a completely
randomized design in a factorial scheme 4 x 2 was used. Treatments consisted in four
values for EB in food (0, 120, 240 and 360 mEq/kg) under two environmental
temperatures (25 and 34°C). Sodium bicarbonate and ammonium chloride were used in
order to obtain such EB values in food. In experiment 2, birds were distributed in a
completely randomized design in a factorial scheme 5 x 2. Treatments consisted in five
values for EB in drinking water (0, 30, 60, 90 and 120 mEq/L) under two environmental
temperatures (25 and 34°C). Sodium bicarbonate alone was used in water in order to
reach EB values. Experiments were carried out in climatic chambers at constant
temperature using quails with initial age of 20 days. Most of the variables were
influenced by temperature in both experiments. In experiment 1, EB influenced water
intake, relative weight of heart and liver. In experiment 2, EB affected (p<0.05) water
intake, relative heart and liver weight. For experiment 1, we recommend EB of 120
mEq/kg of ration and EB of 30 mEq/L of drinking water.
Keyword: ammonium chloride, heat stress, sodium bicarbonate

6
Introdução
A alta temperatura ambiental é o principal causador do estresse por calor
(interações com radiação, umidade relativa, velocidade do ar) pode ter efeito adverso
em criações de aves em regiões tropicais. A eficiência de produção nessas regiões
melhora mediante a redução do estresse térmico ou minimização da sensação térmica ou
favorecimento das perdas de calor. A suplementação de eletrólitos via água ou ração é
recomendação bem aceita por pesquisadores e produtores (Smith e Teeter, 1987; Smith,
1994), sobretudo pela praticidade e menor custo. O uso de eletrólitos, sobretudo, de
soluções salinas, atua na prevenção da desidratação e ingestão voluntária de água em
humanos (Wilk et al,1998; Wilk et al,2007), tendo efeitos semelhantes em aves (Macari,
1996).
As concentrações dos eletrólitos e manutenção do equilíbrio ácido-base dos
fluidos corporais (Mongin,1981) que é estudada sob o termo balanço eletrolítico
(BE),variam com a dieta, balanço hídrico e particularmente em aves sob alta
temperatura pelo ofego. Adicionalmente, os valores dietéticos adequados,
provavelmente, são influenciados pela espécie animal, característica produtiva e idade
da ave, além da intensidade e duração do estresse térmico. Experimentos realizados com
frangos de corte, poedeiras comerciais e matrizes de corte, o BE dietético recomendado
varia entre as categorias (Junqueira et al, 2000, Borgatti et al, 2004; Santos et al, 2011) e
temperatura ambiental (Borges, 2006). Codornas originalmente habitavam regiões
quentes (McIlvaine, 2000) e, é provável que sejam mais resistentes às condições de
estresse por calor quando comparadas a frangos e galinhas.
Trabalhos que tratam do BE em codornas não são convencionais na literatura.
Há dúvidas sobre os efeitos do BE em codornas criadas sob altas temperaturas, porque a
extrapolação dos resultados a partir das faixas restritas de BE utilizadas na maioria dos
experimentos com frangos de corte não permitem maiores discussões sobre o assunto
em codornas. Investigando a ingestão de sódio em duas linhagens de frangos de corte
Ross (1979) concluiu que há maior eficiência de utilização do sódio na água de bebida
comparada a ração, portanto não apenas os sais usados, mas a via de suplementação
podem ser fatores relevantes e influenciar os resultados. Tais considerações podem ter
efeitos práticos na criação desta espécie em regiões de clima quente e melhorar a

7
eficiência produtiva da criação. Portanto, objetivou-se com a realização do estudo
avaliar o efeito do balanço eletrolítico na ração ou na água de bebida sobre o
desempenho e sobrevivência de codornas europeias.
Material e métodos
Experimento 1: Foram utilizadas 240 codornas européias de um lote com
machos e fêmeas com 20 dias de idade, distribuídas em delineamento inteiramente
casualizado em esquema fatorial 4 x 2. Os tratamentos consistiram em quatro níveis de
balanço eletrolíticos na ração (0, 120, 240, 360 mEq/ kg) em duas temperaturas
ambientais (34°C e 25°C, ± 2°C), com cinco repetições de seis aves cada.
Os níveis foram obtidos pela adição de bicarbonato de sódio (NaHCO3) e/ou
cloreto de amônio (NH4Cl) à ração basal. O balanço eletrolítico foi calculado como
[Na+ + K +- Cl- ] na ração segundo Mongin (1981): [(%Na +x 104/23) + (%K+ x
104/39,1) – %Cl- x 104/35,5)]. Os valores de BE baixo e alto da ração foram elaborados
para serem superior e inferior ao intervalo normal do equilíbrio de eletrólitos que seria
esperado em dieta prática (Silva e Costa, 2009).
Experimento 2: Trezentos e sessenta codornas europeias de um lote com machos
e fêmeas com 20 dias de idade, distribuídas ao acaso em esquema fatorial 5 x 2. Os
tratamentos consistiram em cinco níveis de balanços eletrolíticos na água (0, 30, 60, 90
e 120 mEq/ L), em duas temperaturas ambientais (34°C e 25°C, ± 2°C), com cinco
repetições com seis aves cada. Em ambos os ensaios as aves foram pesadas
individualmente (peso inicial 126,2 ± 5,4 g) para formação de parcelas homogêneas.Foi
utilizada uma ração com valor “nulo” em mEq/kg, pela adição de 1,18% de NH4Cl à
ração basal. As doses de suplementação de NaHCO3 na água de bebida deionizada
foram estabelecidas após experimento piloto em que se verificou aumento da
mortalidade das aves acima de 120 mEq por inanição. Para as doses de 30, 60, 90 e
120meq / L foram adicionados 2,55, 5,1, 7,65 e 10,2 g de bicarbonato de sódio à água
de bebida, respectivamente. A água foi fornecida em bebedouro plástico para pássaros
(300 mL) fixados nas gaiolas e reabastecidos quatro vezes em 24h.

8
As dietas foram elaboradas com 23% de proteína bruta e 2900 kcal/kg de energia
metabolizável, de acordo com as recomendações de Silva e Costa (2009) conforme
demonstrado na Tabela 1.
Tabela 1. Rações utilizadas nos Experimentos 1 e 2.
BE (1)
Nulo 120 240 360
Ingredientes
Milho 62,3 62,3 62,3 62,3
Farelo de soja 33,3 33,3 33,3 33,3
Inerte 0,281 0,927 1,282 0,26
Fosfato bicálcico 1,01 1,01 1,01 1,01
Calcário 0,84 0,84 0,84 0,84
DL-Metionina 0,249 0,249 0,249 0,249
Sal Comum 0,177 0,177 0,177 0,177
P. Vitamínico(3)
0,1 0,1 0,1 0,1
Clor. de Colina(4)
0,09 0,09 0,09 0,09
L-treonina 0,122 0,122 0,122 0,122
L-lisina HCl(4)
0,112 0,112 0,112 0,112
P. Mineral(5)
0,07 0,07 0,07 0,07
BHT(6)
0,01 0,01 0,01 0,01
NH4Cl 1,179 0,533 0 0
NaHCO3 0,142 0,142 0,32 1,342
Total 100 100 100 100 (1) Ração com BE nulo usada no experimento 2, (2)Conteúdo/kg de mistura vitamínica : Vit. A – 12.000.000 U.I.; Vit. D3 - 3.600.000 U.I.; Vit. B1 – 2.500 mg; Vit.B2 – 8.000 μg; Vit. B6
– 5.000 mg; Ác. pantotênico – 12.000 mg; Biotina – 200 mg; Vit. K3 – 3.000 mg; Ác. Fólico–
1.500 mg; Ác.nicotínico– 40.000 mg; Vit. B12 – 20.000 mcg; Selênio – 150 mg; Veículo, q.s.p. – 1000 g. (3)Foram calculados os teores de cloro oriundos da Lisina HCl e Cloreto de
colina. (4)Conteúdo/kg de mistura mineral: Manganês – 160,0 g; Ferro – 100,0 g; Cobre –
20,0 g; Zinco – 100,0 g; Cobalto – 2,0 g; Iodo - 2,0 g; Veículo q.s.p. – 1000 g. (5) Antioxidante.
As aves foram alojadas em bateria de gaiolas (0,33 x 0,5 x 0,15, cada) verticais,
típicas para codornas, instaladas em ambiente isolado com temperatura controlada
(25ºC e 35ºC). Os animais de cada parcela foram identificados por anilha plástica
colorida por tratamento.
O controle da temperatura de conforto foi feito por condionador de ar (18000
BTUS, Cônsul). O aquecimento do ambiente foi obtido por meio de lâmpadas
(Lâmpada Incandescente Infra-Vermelha para Secagem 250 w Mega Spot 220 v
Empalux IV22514) dispostas a um metro de distância da bateria de gaiolas e a um metro
de altura do chão. Para monitorar o ambiente térmico foram utilizados Termômetro

9
digitais de máxima e mínima INCONTERM e termohigrometro (composto de dois
termômetro escala externa , bulbo seco e úmido para controle da unidade relativa do
ar). A temperatura da água foi medida utilizando termômetro a laser (Termômetro
Digital Infra-Vermelho MINIPA 30~550°C MT-360) em 20% das parcelas
experimentais. As temperaturas medidas pelos termômetros foram registradas
diariamente às 8, 11, 14 e 17 horas.
A ingestão de água e consumo de ração médio foram calculados como
quantidade total por parcela, dividida pelo número de codornas por parcela e os dias de
duração do experimento. O ganho de peso médio foi calculado como ganho de peso
corporal da parcela dividido pelo número de animais de cada parcela. As codornas
foram submetidas a jejum de 12 horas, pesadas e sacrificadas. O rendimento de carcaça
e peso relativo dos órgãos foi mensurado em duas codornas selecionadas considerando-
se o peso médio parcela pela relação entre o peso da carcaça depenada e o peso corporal
do animal em jejum. Os pesos relativos do fígado, moela, intestino e coração foram
calculados em relação ao peso da carcaça.
Foram atendidas as pressuposições de normalidade dos erros estudentizados
(teste de Shapiro-Wilk) e homogeneidade das variâncias (teste Hartley), foram
realizadas as análises de variância. Em caso de significância, foi aplicada a análise de
regressão considerando o ajuste linear e quadrático dos dados conforme o melhor ajuste
do modelo obtido para cada variável, em que o coeficiente de determinação quantifica o
grau de aproximação do modelo às médias. As análises estatísticas foram realizadas
considerando-se um nível de significância de até 5%.
Optou-se pelo desdobramento da interação mesmo quando não for significativa
pelo teste F. Segundo Sampaio (1998), quando o grau de liberdade é superior a dois o
teste F não é conclusivo, portanto comparações entre as médias obtidas foram feitas
para percepção de diferenças importantes.
Os dados de todas as variáveis foram também submetidos à análise canônica
discriminante, visando identificar os parâmetros que permitem discriminar diferenças
quanto ao balanço eletrolítico da água ou ração para codornas criadas em diferentes
temperaturas.

10
Resultados e Discussão
Resultados
Experimento 1. Balanço eletrolítico da ração
A temperatura ambiente influenciou (P<0,01) as variáveis avaliadas no
Experimento 1, com exceção do peso relativo da moela, conforme demonstrado na
Tabela 2.
Tabela 2. Efeito do BE e da temperatura (T) sobre o desempenho codornas
europeias aos 37 dias.
(1) y = 1090,6 - 2,0618x + 0,0059x2; R² = 0,73. (2) y = 56,13+
0,0448x -9E-05x2;R² = 0,66. Médias seguidas por letras iguais não diferem significativamente pelo teste F a 5% de probabilidade. ; *P <
0,05; NS = Não significativo; BE = balanço eletrolítico da ração; T =
temperatura. BExT = Interação dos fatores. GP = Ganho de peso; CR = Consumo de ração; CA = Conversão alimentar; IA = Ingestão de
água.
Tabela 3. Efeito do BE e da temperatura (T) sobre rendimento de carcaça e o peso
relativo das vísceras de codornas europeias aos 37 dias.
BE RC M CO (2)
CI PI
0 76,6 2,67 1,12 56,8 4,89
120 75,7 2,63 1,15 58,2 5,31
240 75,0 2,79 1,09 63,7 5,32
360 75,3 2,78 1,06 59,9 4,74
34 78,3b 2,70 0,89 a 55,85 b 4,26
25 73,0a 2,73 1,32b 63,63a 5,87
BE ns ns ns ** ns
T ** ns ** ** **
BExT ns ns Ns ns ns
CV 4,3 9,3 12,2 7,79 14,81 Médias seguidas por letras iguais não diferem significativamente pelo teste F a
5% de probabilidade. ; *P < 0,05; NS = Não significativo; BE = balanço
eletrolítico da ração; T = temperatura. BExT = Interação dos fatores. C = Rendimento de carcaça; M = Peso relativo da moela; CO = Peso relativo do
coração; FI = Peso relativo do fígado; CI = Comprimento do intestino; PI = Peso relativo do intestino.
BE GP CR (1)
IA CA
0 95,0 353,7 1067,1 3,79
120 96,3 369,6 999,2 3,87
240 96,0 367,4, 866,7 4,1
360 93,1 348,3 1140,3 3,65
34 88,b 305,3b 1316 b 3,48b
25 100,4a 414,2a 721a 4,22a
BE ns ns * ns
T * * * *
BExT ns ns ns ns
% CV 13,4 7,6 15,5 16,6

11
Tabela 4. Comparação de médias entre ingestão de água e da relação ingestão de
água:consumo de ração para o BE e temperaturas ambientais.
A,B = médias seguidas de letras maiúsculas diferentes na coluna diferem entre si pelo
teste SNK (P≤0,05). a,b = médias seguidas de letras minúsculas diferentes na linha
diferem entre si pelo teste “F” (P≤0,05)
As codornas criadas sob temperatura de 34ºC consumiram em média 29,9%
menos ração que àquelas em ambiente a 25ºC. A redução ocorrida no consumo de ração
justifica a diminuição do ganho de peso das aves sob temperatura elevada. A conversão
alimentar também foi influenciada pela temperatura ambiente, sendo que os menores
valores de conversão alimentar foram observados nas aves mantidas no ambiente de
calor. Com relação às características de carcaça, as codornas expostas ao calor
apresentaram maior rendimento de carcaça (Tabela 3), que ocorreu em função do menor
desenvolvimento visceral das aves submetidas à temperatura de 34°C. Mesmo assim, o
comprimento do intestino aumentou com a suplementação de NaHCO3 até atingir o BE
de 246 mEq / kg de ração, diminuindo em seguida. Não houve efeito dos tratamentos
sobre a mortalidade das aves.
Experimento 2. Balanço eletrolítico da água
Os resultados de desempenho das codornas são mostrados na Tabela 5.
Observou-se efeito da interação entre a temperatura ambiente e o balanço eletrolítico
para o ganho de peso e peso relativo do fígado. O BE influenciou a IA e o peso relativo
do coração. A temperatura ambiental de 34°C afetou significativamente todas as
variáveis.
BE IA IA/CR
34°C 25°C 34°C 25°C
0 1369,4aA
764,88bAB
4,67aAB
1,85bAB
120 1323,6aA
674,78bAB
4,05aBC
1,63bAB
240 1141,3aA
592,02bB
3,55aC
1,41bB
360 1428,5aA
852,07bA 5,14
aA 2,06
bA
Efeito ns ns Q Q

12
Tabela 5. Efeito da temperatura e do balanço eletrolítico via água de bebida sobre o
desempenho de codornas européias aos 37 dias.
BE GP CR CA (1)
IA
0 93,08 353,71 3,79 1067,13
30 85,12 351,29 4,17 991,25
60 85,68 346,88 4,12 936,70
90 90,31 377,73 4,24 982,83
120 91,20 364,53 3,90 1173,48
34,0 76,49b 296,59b 3,85 1291,50a
25,0 101,67a 421,07a 4,18 769,06b
BE ns ns ns *
T * * * *
Be*T * ns ns ns
CV 11,223 11,135 14,399 13,831 (1) y = 1079,9- 5,3542x + 0,0503x2; R² = 0,96. (2) y = 2,9629+ 0,0083x -
5E-05x2; R² = 0,77 . Médias seguidas por letras iguais não diferem significativamente pelo teste F a 5% de probabilidade; *P < 0,05; NS =
Não significativo. BE = balanço eletrolítico da água; T = temperatura;
GP= ganho de peso; CR= Consumo de ração em gramas; CA= Conversão alimentar; IA= Ingestão de água em mL;
Tabela 6. Efeito da temperatura e do balanço eletrolítico via água de bebida sobre o
peso relativo das vísceras de codornas européias aos 37 dias.
BE PF C M CO (1)FI CI
0 219,3 76,4 2,29 1,04 2,94 56,8
30 213,0 73,4 2,29 1,02 3,24 64,2
60 214,6 75,0 2,57 1,08 3,17 63,0
90 217,9 74,8 2,55 1,18 3,32 61,6
120 221,5 73,2 2,37 1,09 3,17 61,0
34,0 203,96b 77,29a 2,75ª 1,17a 3,35a 57,16b
25,0 230,57a 71,86b 2,08b 1,00b 2,99b 65,44ª
BE ns ns ns * ns ns
T * * * * * *
Be*T ns ns ns ns * ns CV 5,0 3,9 12,1 10,4 14,5 11,6
(1) y = 2,9629+ 0,0083x -5E-05x2; R² = 0,77 .Médias seguidas por iguais não diferem
significativamente pelo teste F a 5% de probabilidade; *P < 0,05; NS = Não significativo. BE = balanço eletrolítico da água; T = temperatura; RC= Rendimento de carcaça em
percentagem (%); M= Peso relativo da moela (%); CO= Peso relativo do coração (%); CI=
Comprimento do intestino em centímetros; PI= Peso relativo do intestino (%); FI = Peso relativo do intestino (%). Os pesos dos órgãos foram calculados em relação ao peso da
carcaça eviscerada.
O desdobramento dos graus de liberdade para o efeito da interação entre BE e T
das variáveis ganho de peso e peso relativo do fígado de codornas européias obtidos no
experimento 2 é apresentado na Tabela 7.

13
Tabela 7. Comparação de médias entre ganho de peso (GP), ingestão de água (IA), peso
relativo do coração (CO) e peso relativo do fígado para o BE e temperaturas ambientais.
BE GP
(1)IA
(2)CO
(3)FI
34°C 25°C 34°C 25°C 34°C 25°C 34°C 25°C
0 86,7aA 99,5aA 1369,4abA 764,9aB 1,15aA 0,93bB 3,03aA 2,86aA
30 76,3aB 93,9aA 1255,8abA 726,7aB 1,10aA 0,94bB 3,09aA 3,39aA
60 74,4aB 96,9aA 1191,8bA 681,7aB 1,16aA 1,01bB 3,32aA 3,03aA
90 70,2aB 110,4aA 1186,3bA 779,4aB 1,19aA 1,17aA 3,70aA 2,93aB
120 74,8aB 107,6aA 1454,3aA 892,6aB 1,24aA 0,95bB 3,59aA 2,74aB
Efeito Ns ns Q ns Q ns L ns (1) y = 1388,8 – 7,4923x +0,0652x²; R² = 0,9. y = 1,1383- 0,0007x +1E-05x2; R² = 0,88. (3) y = 3,0007 + 0,0058x; R² = 0,86. a,b
= médias seguidas de letras minúsculas diferentes na coluna diferem entre si pelo teste SNK (P≤0,05). A,B = médias seguidas de letras
maiúsculas diferentes na linha diferem entre si pelo teste “F” (P≤0,05). GP= ganho de peso em g; IA= Ingestão de água em mL; CO= Peso relativo do coração (%);FI = Peso relativo do intestino (%).
Os resultados da Tabela 7 para o ganho de peso demonstram que as aves que
receberam o NaHCO3 via água apresentou efeito da interação dos fatores (BE x T). O
efeito da temperatura foi pronunciado sobre o do BE, já que após o desdobramento da
variável não foi encontrada diferença significativa entre as médias para o BE.
O BE influenciou significativamente a ingestão de água pelas codornas criadas
em alta temperatura. A ingestão de água foi semelhante entre os níveis 0, 30 e 120
mEq/ L e superior aos níveis de 60 e 90 mEq/L.
O peso relativo do coração apresentou efeito quadrático em resposta à
suplementação com o NaHCO3 na água de bebida até atingir o ponto máximo de CO
para o BE de 112,5 mEq/L (Tabela 7 ).
Não houve efeito dos tratamentos sobre a mortalidade das codornas
As duas primeiras variáveis canônicas explicaram 94% da variação total, porém
nenhuma delas foi significativa com probabilidade superior a 78%.
Discussão
A alta temperatura ambiental reduz o consumo de ração e o ganho de peso do
animal (Araújo et al., 2007; Welker et al., 2008). A redução no consumo prevê a
diminuição do incremento calórico decorrente da digestão e metabolismo dos nutrientes
(Lesson & Summers, 2001). As codornas sob temperatura de 34°C consumiram 35,65%
menos ração que àquelas criadas em temperatura de 25°C em ambos os experimentos. A

14
redução do ganho de peso das aves alojadas no ambiente de 34°C obedece a um padrão
típico de comportamento de animais em condições de estresse térmico, devido à
demanda aumentada de energia para termoregulação e pela diminuição do consumo de
ração com melhor conversão alimentar (P<0,01). Adicionalmente, o consumo de ração
foi determinante para o efeito sobre o peso do intestino e fígado, sobretudo pelo
consumo de proteína, que foi 26% menor (70 g VS 95 g de proteína bruta) para as aves
criadas em temperatura mais alta. Desse modo, conforme observado em ambos os
experimentos, as aves criadas sob condições de temperaturas ambientais elevadas
tendem a ter os pesos relativos dos órgãos afetados de forma indireta pelo consumo de
ração e água ou diretamente sobre as alterações compensatórias sobre o metabolismo.
Logo o peso relativo dos diferentes órgãos pode obedecer a padrões de desenvolvimento
diferentes de acordo com a finalidade (digestório ou homeostase: excretor (rins),
circulatório e respiratório). Esta dúvida não foi explorada por nenhum dos trabalhos
citados no presente estudo.
O peso relativo do coração apresentou comportamento diferente entre os
experimentos para as mesmas condições ambientais, porque no exp. 2 foi influenciado
pela suplementação com NaHCO3 via água (Tabelas 3 e 6). Infelizmente não foram
investigados dados de hipertensão, medida de parede ou índice cardíaco, mas os
resultados sugerem a interferência da concentração de sódio sobre o peso relativo do
coração dos animais sob altas temperaturas. Há alguns anos foi descoberto que o
coração desempenha importante papel endócrino na excreção do sódio (Bold et al.
(1981) citado por Maack et al. (1985) mediante a ação do hormônio (ANP, atrial
natriuretic peptide), que atua ao mesmo tempo como uma das principais substâncias
endógenas envolvidas no papel anti-hipertrófico do coração (Bubikat et al., 2005). De
fato, no exp. 1 o peso relativo do coração diminuiu sob influencia da temperatura
elevada, mas no exp. 2, o BE (P < 0,05) aumentou o peso relativo do coração, ou seja, o
peso relativo do coração foi influenciado pelos teores de sódio na água de bebida, que
variaram entre 68 a 275mg Na/L. Infelizmente, não foram realizados medida de parede
ou de índice cardíaco que permita-nos analisar os resultados sem especulações. , mas
evidências mostram que o teor de sódio ingerido tem sido correlacionado como uma das
causas de alterações da musculatura cardíaca, como hipertrofia, em experimentos
realizados com ratos em períodos relativamente curtos (Selye & Bajusz, 1959;

15
Lindpaintner & Sen, 1985). Resultado semelhante foi observado por Silva et al. (2006)
em frangas leves e semipesadas, em que, não apenas o peso do coração, mas do fígado,
baço e bursa cloacal aumentaram quando a proporção de sódio foi elevada na ração.
A máxima IA corresponderam às maiores suplementações com os sais à dieta
basal (1,18% de NH4Cl e 1,342% de NaHCO3) independente do balanço eletrolítico da
ração. A utilização destes compostos não está relacionada apenas a promoção da
ingestão de água em condições de estresse por calor. Teeter et al. (1985) demonstrou
que sob estresse por calor tanto a administração do sódio como a administração de
acidificantes na forma de cloreto de amônio, reduzem a gravidade da alcalose
respiratória pelo aumento do pH sanguíneo quando os frangos de corte receberam 0,5%
de NaHCO3 na dieta. Contudo, não foram publicados dados de outras pesquisas
confirmando este resultado.
Na maioria dos trabalhos citados falta qualquer evidência, sobre o efeito do
bicarbonato de sódio ou cloreto de amônio sobre o pH sanguíneo de aves sob estresse
por calor. O CO3 é um produto metabólico que pode obviamente alterar o equilíbrio
ácido-base. Embora para ruminantes a adição de bicarbonato de sódio a ração foi
associado ao aumento no pH sanguíneo (Hart & Doyle 1985; Damir et al., 1990), não
foram encontrados dados que mostrem que isso é possível em frangos.
Quando o “balizador” para medir a eficiência da suplementação de sais, é a
ingestão de água, o uso de fontes de sódio na água de bebida ou ração tem demonstrado
aumento na ingestão voluntária de água pelas aves quando comparado a outros
eletrólitos, desde os primeiros experimentos sobre o assunto (Ross, 1979; Teeter &
Smith, 1986; Mushtaq et al., 2005). Assim, relacionando-se a ingestão de água ao
consumo de sódio para o exp. 1, os teores de sódio variaram de 0,109 a 0,433% da ração
(BE nulo e 360 mEq/kg), sendo o último o que promoveu a maior ingestão de água
pelas aves (Tabela 3). O mesmo aconteceu para o exp. 2 em que os teores de sódio total
variaram de 0,178 e 0,384%. Em estudos realizados com frangos de corte na fase pré-
inicial Maiorka et al. (2004) relataram que 0,45% de sódio na ração promoveu a maior
ingestão de água quando comparado 0,10, 0,22 e 0,34%. Para os tratamentos em que o
bicarbonato de sódio foi diluído na água (exp. 2), a dieta basal contribuiu com 0,109%
de sódio e os teores de sódio na solução eletrolítica variaram de 0,069 a 0,275%. Assim,
embora, o nível mais alto de sódio total no exp. 2 tenha sido inferior ao do exp. 1, o

16
consumo de água foi semelhante. Os resultados indicam que a suplementação de sódio
via água de bebida mostrou-se mais eficiente em promover aumento da ingestão de água
que a suplementação de sódio através da ração. Este resultado confirma o achado de
Ross (1979) que comparou duas formas de suplementação de sódio (dieta basal com
0,095% de sódio) concluiu que as aves utilizaram o sódio mais eficientemente quando a
suplementação foi feita via água de bebida quando comparada à ração. A promoção na
ingestão de água sobre a homeostase térmica baseia-se nas propriedades de calor latente
de vaporização da água líquida e calor específico da água.
A ingestão de soluções salinas na promoção da absorção de nutrientes é pratica
bem conhecida (Currell et al., 2009) e pode justificar o efeito significativo do BE sobre
o comprimento do intestino (Tabela 3). González et al. (1998) verificaram a presença de
sítios específicos de ligação hormônio ANP no intestino de ratos, que atuam como
mediadores da absorção de sódio, glicose e água. Estes resultados foram confirmados
em trabalhos posteriores (Nakamura et al. 2003), e citado em estudo mais recente sobre
o assunto (Yoshikawa et al., 2011).
O uso de cloretos (KCl, NH4Cl) para a indução da ingestão de água tem sido
estudado com aves em condições de estresse por calor (Souza et al., 2005; Souza et al.,
2009), mas aparentemente não dependem apenas da suplementação per se, mas também
da composição da ração, sexo e magnitude e duração do estresse. Em geral, o aumento
nos teores de sódio ingerido (NaHCO3, NaCl) está relacionado com a ativação do
sistema de controle da sede para a manutenção do equilíbrio hídrico.
Pelos trabalhos citados, foi possível concluir que assim como no presente estudo
os resultados das variáveis de desempenho sob condições de estresse térmico,
demonstram elevada variabilidade (Ahmad et al., 2005, Mushtaq et al., 2005 Mushtaq et
al., 2007, Ahmad et al., 2008). Além dos erros experimentais inerentes aos animais e
dietas, fatores como toxidez (Teeter e Smith, 1986; Souza et al., 2005), devem ser
considerados, uma vez que diferentes concentrações (doses) de bicarbonato de sódio ou
cloreto de amônio podem alterar o consumo e a ingestão de água das aves
individualmente. O cloreto de amônio é um acidificante e a sua ingestão crônica pode
ter provocado alterações metabólicas somado a efeitos indiretos como o decorrente da
alteração no nível de cálcio circulante (Dacke et al., 1973) bem como a ingestão de
NaHCO3 (Bottje e Harrison, 1985). Teeter e Smith (1986) concluíram que o NH4Cl é

17
eficiente no combate dos efeitos do estresse por calor, mas níveis de suplementação de
0,5% já apresentam toxidez. Para Souza et al. (2005) tal efeito foi sugerido com 0,4%
de NH4Cl. Portanto, diferenças nas quantidades de cloreto de amônio das dietas
experimentais (exp. 1) podem ter alterado as respostas das aves entre tratamentos
considerando-se que para alcançar os BE, as dietas experimentais não foram formuladas
com os mesmos percentuais de cloreto de amônio. Além disso, a semelhança do
bicarbonato de sódio, o efeito da adição do cloro pode variar com a fonte, já que o
bicarbonato, o íon amônio e sua forma oxidada (amônia) estão envolvidos no equilíbrio
acidobásico (Branton et al., 1986; Ellert 1998; Verlander 2004a, 2004b). . Trabalhos
científicos sobre o uso de sais como prática alimentar para reduzir os efeitos do estresse
por calor, visa apenas à promoção na ingestão de água (Branton et al. 1986; Smith
1994), aspecto que parece não ser o único a ser afetado quando se usa o bicarbonato de
sódio e o cloreto de amônio.
O balanço eletrolítico adequado ou os níveis críticos em dietas para aves em
condições de estresse térmico podem ser influenciados, mesmo considerando-se valores
iguais em mEq/kg. Há dificuldade em comparação dos efeitos e níveis de
suplementação de sais na ração e na água porque, além das respostas à fase de criação
considerada e do tipo de ingrediente, os sais empregados são decisivos nas conclusões.
Mesmo para valores iguais (mEq) precisam-se de diferentes concentrações de K+, Na+
ou Cl-, dependendo da fonte utilizada (KCl, NaCl, NaHCO3), resultando em diferentes
efeitos como pode ser observado nos resultados de Teeter et al. (1986) e Ahmad et al.
(2005). Observa-se que sais como o cloreto de sódio ou potássio fornecem um ânion e
um cátion fortes, sendo a relação (Na+K)/Cl pouco alterada, mesmo em altas
concentrações.
Em conclusão pôde-se constatar que dificilmente a temperatura foi o único
agente desfavorável ou estressor afetando o crescimento das codornas. Em vez disso,
mudanças no padrão dietético e ambiental podem ter influenciado o metabolismo das
aves. As alterações hepáticas podem ilustrar essa hipótese. A tabela 7 mostra que no
exp. 2, o peso relativo do fígado aumentou (P < 0,005) com a suplementação de
bicarbonato de sódio, embora não tenha sido encontrada na literatura consultada relação
direta entre o balanço de eletrólitos e o peso relativo do fígado. O ganho de peso
mínimos, devido ao ambiente com temperatura elevada não foi afetado pelo balanço

18
eletrolítico como a maioria das outras variáveis, mas o BE de 120 mEq/kg na ração e 30
mEq/L na água de bebida é recomendado para aumentar o consumo de água em
codornas européias em crescimento.
Conclusões
Recomenda-se o BE de 120 mEq/ kg de ração (experimento 1) e BE 30 mEq/ L
na água de bebida.
Referências Bibliográficas
Ahmad T, Khalil T, Mushtag T, Mirza MA, Nadeem A, Barabar ME, Ahmad G. Effect
of Potassium Chloride Supplementation in Drinking Water on Broiler
Performance Under Heat Stress Conditions. Poultry Science;87: 1276-1280,
2008.
Ahmad T, Sarwar M, Nisa MU, Haq AU, Hasan ZU. Influence of varying sources of
dietary electrolytes on the performance of broilers reared in a high temperature
environment. Animal Feed Science and Technology;120:277−298, 2005.
Bubikat A, León J. De Windt, Bernd Zetsche, Larissa Fabritz, Heidrun Sickler,
Dominik Eckardt, Axel Gödecke, Hideo A. Baba, and Michaela Kuhn. Local
Atrial Natriuretic Peptide Signaling Prevents Hypertensive Cardiac Hypertrophy
in Endothelial Nitric-oxide Synthase-deficient Mice. The Journal Biological
Chemistry Vol. 280, No. 22, Issue of June 3, pp. 21594 –21599, 2005.
Borgatti LMO, Albuquerque R, Meister NC, Souza LWO, Lima FR, Trindade Neto
MA. Performance of broilers fed diets with different dietary electrolyte balance
under summer conditions. Revista Brasileira de Ciência Avícola;6(3): 153 –
157, 2004.
Dacke CG, X.J Musacchia, Wynn A Volkert, Alexander D Kenny. Cyclical fluctuations
in the levels of blood calcium, pH and pCO2 in Japanese quail. Comparative
Biochemistry and Physiology Part A: Physiology 44(4): 1267-1275, 1973.

19
Damir HA, Scott D, Thomson JK, et al. The effect of a change in blood acid-base on
body composition and mineral retention in growing lambs. Animal Production
51: 527-34,1990.
Jeukendrup AE, Currell K, Clarke J, J Cole and Andrew K Blannin Effect of beverage
glucose and sodium content on fluid delivery . Nutrition & Metabolism
6:9,2009,
Garcia JAD, Incerpi EK. Fatores e mecanismos envolvidos na hipertrofia ventricular
esquerda e o papel anti-hipertrófico do óxido nítrico. Arquivos Brasileiros de
Cardiologia, São Paulo, v. 90, n. 6, 2008..
Gonzalez LV, Vidal NA, Prieto R, et al: Effect of atrial natriuretic peptide on -methyl-
D-glucoside intestinal active uptake in rats. Peptides 19:1249–1253, 1998.
Hart SP, Doyle JJ. Adaptation of early-weaned lambs to high-concentrate diets with
three sources, with or without sodium bicarbonate. Journal Animal Science
61:975-84, 1985.
Junqueira OM, Camargo Filho B, Araújo LF, Araújo CSS, Sakomura NK. Efeitos das
fontes e níveis de sódio, cloro e potássio e da relação (Na + K)/Cl, sobre o
desempenho e características do plasma sangüíneo de poedeiras comerciais.
Revista Brasileira de Zootecinia; 29(4): 1110-1116, 2000.
Lindpaintner K, Sen S. Role of sodium in hypertensive cardiac hypertrophy.
Circulation Research 57: 610-617, 1985.
Maack T, Camargo MJ, Kleinert HD, Laragh JH, Atlas SA. Atrial natriuretic factor:
structure and functional properties. Kidney Int. 27, 607-615. 1985
Macari M. Equilíbrio hídrico em aves. In: Marcos Macari; (Org.). Água na avicultura
industrial. 1 ed. Jaboticabal: FUNEP, p. 27-52, 1996,
Maiorka A, Magro N, Bartels HAS, Kessler AM, Penz Jr AM. Different sodium levels
and electrolyte balances in pre-starter diets for broilers. Revista Brasileira de
Ciência Avícola, 6(3) , 2004.
McIlvaine J (2000). O Biogeography of California Quail (Callipepla californica)
.Disponivel: http://bss.sfsu.edu/holzman/courses/fall00projects/quail.html
Mongin P. Recent advances in dietary ânion-cátion balance: applications in poultry.
Proceedings of the Nutrition Society 40(1):285-294, 1981.

20
Mushtaq T, Mirza M. Aslam, Athar, M., Hooge, D. M., Ahmad, T., Ahmad, G.,
Mushtaq, M. M. H., Noreen, U.Dietary Sodium and Chloride for Twenty-Nine-to
Forty-Two-Day-Old Broiler Chickens at Constant Electrolyte Balance Under
Subtropical Summer Conditions. Journal Applied of Poultry Research 16:161-
170, 2007.
Mushtaq T, Sarwar, M, Nawaz, H, Mirza, MA, Ahmad, T. Effect and interactions of
dietary sodium and chloride on broiler starter performance (hatching to twenty-
eight days of age) under subtropical summer conditions. Poultry Science; 84;
1716-1722, 2005.
Nakamura K. , Takayama, K.a , Nagai, T.a , Maitani, Y. Regional intestinal absorption
of FITC-dextran 4,400 with nanoparticles based on β-sitosterol β-D-glucoside in
rats. Journal of Pharmaceutical Sciences v 92, Issue 2, 1 , p 311-318,2003.
Ross E. The effect of water sodium on the chick requirement for dietary sodium.
Poultry Science 58: 626-630, 1979.
Selye h. & Bajusz E. Effect of various electrolytes upon cardiac and skeletal
musculature. British Journal of Pharmacology; 14: 83-86, 1959.
Silva, JHV, Ribeiro, M.L.G.; Jordão Filho, J., Silva, E.L. O sódio afeta o crescimento
de órgãos dos sistemas circulatório, digestivo e imune de frangas. Revista
AveWorld, n.23, 2006.
Sousa Junior FN, Figueirêdo AV, Lopes JB, Ramos LSN. Bicarbonato de sódio e
cloreto de amonio em rações para frangos de corte sob condições naturais de
extresse térmico. Revista Científica de Produção Animal; 11(1): 23-33, 2009.
Souza BB, Bertechini GA, Teixeira AS, Lima JAF, Freitas RTF. Efeito da
suplementação de cloreto de potássio na dieta sobre o equilíbrio ácido-básico e o
desempenho de frangos de corte no verão. Ciência e Agrotecnologia;26 (6):
1297- 1304, 2002.
Souza Bonifácio Benício de et al . Efeito do nível energético e da suplementação com
cloretos de potássio e de amônia na dieta sobre as respostas fisiológicas e o
desempenho de frangos de corte no verão. Ciência e Agrotecnologia, v. 29, n. 1,
2005.

21
Teeter RG, Smith M. High chronic ambient temperature stress effects on broiler acid-
base balance and their response to supplemantal ammonium chloride potassium
chloride, and potassium carbonate. Poultry Science; 65(9):1777-1781, 1986.
Teeter RG, Smith MO, Owens FN, Arp SC, Sangiah S, Breazile JE. Chronic heat stress
and respiratory alkalosis: occurrence and treatment in broiler chicks. Poultry
Science 64(6):1060-4, 1985.
Verlander JW, Equilibrio ácido-básico.Cunningham JG. TRATADO DE FISIOLOGIA
VETERINÁRIA. Terceira Edição. Editora Guanabara Koogan, p 471-477. 2004b
Verlander JW, Reabsorção de solutos.Cunningham JG. TRATADO DE FISIOLOGIA
VETERINÁRIA. Terceira Edição. Editora Guanabara Koogan, p 452-462. 2004a
Wilk B, Rivera-Brown AM, Bar-Or O. Voluntary drinking and hydration in non-
acclimatized girls exercising in the heat. European Journal of Applied
Physiology 101(6):727-34. 2007.
Wilk B, Kriemler S, Keller H, Bar-Or O.Consistency in preventing voluntary
dehydration in boys who drink a flavored carbohydrate-NaCl beverage during
exercise in the heat. International Journal of Sport Nutrition 8(1):1-9, 1998.
Yoshikawa, T., Inoue, R., Matsumoto, M., Yajima, T., Ushida, K., Iwanaga,
T.Comparative expression of hexose transporters (SGLT1, GLUT1, GLUT2 and
GLUT5) throughout the mouse gastrointestinal tract. Histochemistry and Cell
Biology, 135 (2), pp. 183-194, 2011.

22
CAPÍTULO 2
CURVAS DE CRESCIMENTO DO PESO CORPORAL E
DEPOSIÇÃO DE NUTRIENTES, EXIGÊNCIAS PROTEICAS PARA
FRANGAS LEVES E SEMIPESADAS

23
Curvas de crescimento do peso corporal e deposição de nutrientes, exigências
protéicas para frangas leves e semipesadas
Resumo
Objetivou-se com o estudo ajustar curvas de crescimento e descrever as taxas de
deposição dos componentes corporais e estimar das exigências de proteína de frangas
leves e semipesadas. A técnica de abate em série foi usada para determinar a
composição corporal em proteínas, lipídios, cinzas e água no corpo vazio sem penas de
frangas recebendo ração ad libitum ração de 1 a 16 semanas de idade. Três dietas à base
de milho e farelo de soja foram utilizadas durante o experimento: 1 a 4 semanas, 5 a 11
semanas e 12 a 16 semanas. Relações alométricas foram utilizadas para descrever as
mudanças na composição corporal. No total 1500 frangas divididas em duas linhagens
foram distribuídas em delineamento inteiramente casualizado com 30 repetições de 25
aves. As frangas foram pesadas semanalmente e duas aves por linhagem separadas
como amostra da composição corporal de nutrientes. Essas informações foram usadas
para obter os parâmetros do modelo de crescimento Gompertz. Os coeficiente
alométrico de gordura, água e matéria mineral em função do peso corporal protéico das
linhagens Dekalb e Bovans foram:0114 vs 1,160; 0,811 vs 0,429; 0,930 vs 0,189. A
linhagem leve é precoce no crescimento e na deposição dos componentes corporais. A
exigência em proteína de total (mantença e ganho) para frangas de 1 a 4, 5 a 11 e 12 a
16 semanas de idade são 4,41; 9,814; 12,612 g/d para Dekalb White e 4,82; 8,859;
7,673g/d para Bovans Goldline. Embora algumas das relações alométricas, diferiram
estatisticamente entre as linhagens às 16 semanas, as diferenças têm pouco significado
comercial sem informações adicionais do período de produção. Investigações futuras
sobre a composição corporal das frangas combinadas a fase de postura representam
avanço nessa área de estudo.
Palavras-chave: alométria , composição corporal, crescimento, proteína bruta

24
Growth curves of body weight and nutrient deposition and protein requirements
for light and semi-heavy pullets
Abstract
The aim of this study was to adjust growth curves and to describe the deposition rates of
body components and to estimate protein requirement for light and semi-heavy pullets.
The technique of slaughter in series was used in order to determine body composition in
protein, lipids, ashes and water in the empty, featherless body of pullets receiving ad
libitum ration from 1 to 16 weeks of age. Three diets based on corn and soybean meal
were used during the experiment: 1 to 4 weeks, 5 to 11 weeks and 12 to 16 weeks.
Allometric relations were used to describe the changes in body composition. A total
amount of 1500 pullets, divided into two lines were distributed in a completely
randomized design with 30 replicates of 25 birds. Pullets were weekly weighed and two
birds per line were withdrawn as samples for body composition of nutrients. These
information were used to obtain the parameters for Gompertz growth model. Allometric
coefficient of fat, water and mineral matter in function of protein body weight for
Dekalb and Bovans lines were: 0.114 vs 1.160, 0.811 vs 0.429, 0.930 vs 0.189,
respectively. The light line is more precocious for growth and body components
deposition. Total protein requirement (maintenance and gain) for pullets from 1 to 4, 5
to 11 and 12 to 16 weeks were 4.41; 9.814; 12;612 g/d for Dekalb White and 4.82;
8.859; 7.673g/d for Bovans Goldline. Although some of the allometric relations differed
statistically between the line at 16 weeks, differences has little commercial meaning
without additional information on the production period. Further investigations on
pullets’ body composition combined to laying phase represent an advance in this field
of study.
Keywords: allometry, body composition, growth, crude protein

25
Introdução
Pesquisas com frangas no período de reposição têm sido relegadas a segundo
plano, provavelmente pela aparente falta de lucratividade neste período. Contudo há
relação direta entre composição corporal na fase de reposição e o desempenho durante a
fase de postura. Desse modo, a alimentação adequada na fase de reposição pode ser
convertida em maior massa de ovos por ave.
Em geral, as linhagens modernas de postura são melhoradas para consumirem
menos ração com maior produção de ovos. São, portanto, nutricionalmente mais
exigentes, sob pena de não apresentarem crescimento normal e inferior condição
corporal a idade do primeiro ovo. Estudos anteriores sobre as relações entre o peso
sugerido para ave à fase de postura e a produção de ovos, são pouco conclusivos (Neme
et al., 2006), sem relação direta do crescimento com a produção de ovos. Porém,
determinar modelos de crescimento biologicamente “interpretáveis” pode constituir o
passo inicial dos estudos na estimativa das exigências nutricionais de frangas de
reposição (Hruby et al., 1994, Gous, 2007). Parâmetros estimados pela curva de
Gompertz são capazes descrever as mudanças corporais nas fases de criação de
linhagens de postura (Neme et al., 2006) e frangos de corte (Marcato, 2008).
Modelos de simulação das exigências nutricionais de aminoácidos foram obtidos
para frangos de corte (Hruby et al., 1995; Hruby et al., 1996) e frangas de reposição
(Martin et al., 1994). Tais modelos inter-relacionam fatores biológicos e econômicos,
pelo ajuste da exigência específica do nutriente e o número de dietas por fase de
crescimento. Adicionalmente, o conhecimento das exigências de mantença e ganho das
aves resultam em formulações de rações mais eficientes do ponto de vista nutricional e
econômico.
Com o presente estude prevê-se o ajuste das curvas de crescimento do peso
corporal e deposição de nutrientes, e estimativas das exigências de proteína de frangas
leves e semipesadas (1 a 16 semanas de idade).

26
Material e Métodos
Dois grupos de 750 pintinhas de um dia de idade das linhagens leve (Dekalb
White) e semipesada (Bovans Goldline) foram divididas em 30 repetições de 25 aves
cada, com consumo voluntário de ração e água. Três dietas foram utilizadas em três
períodos sucessivos de quatro semanas cada(1 a 4, 5 a 11 e 12 a 16 semanas). O
fornecimento de nutrientes seguiram as recomendações de Rostagno et al. (2005) e
manuais das linhagens (Granja Planalto, 2005ab) (Tabela 1).
Tabela 1. Composição das dietas do período de cria e recria das frangas leves e
semipesadas.
Ingredientes Fase (semanas)
1 a 4 5 a 11 12 a 16
Milho 49,723 69,270 62,550
Farelo de soja (45%) 35,299 24,290 21,147
Soja extrusada 1,672 0,000 0,000
Calcário 0,831 1,131 4,067
Fosfato bicálcico 2,003 1,883 1,404
Amido 0,400 0,400 0,400
L-Lisina HCl 0,010 0,000 0,007
L-Treonina 0,032 0,000 0,000
Cloreto de colina (70%) 0,100 0,100 0,100
Óleo de soja 4,000 0,000 2,818
Sal 0,289 0,189 0,295
Bicarbonato de sódio 0,000 0,177 0,000
Premix vitamínico(1)
0,100 0,100 0,100
Premix mineral(2)
0,050 0,050 0,050
Promotor de crescimento(3)
0,015 0,015 0,015
Coccidiostático(4)
0,005 0,005 0,005
Antioxidante(5)
0,010 0,010 0,010
Inerte(6)
5,461 2,380 7,032
Total 100,000 100,000 100,000
Composição Química
Proteína bruta (%) 21,000 17,000 15,000
Energia metabolizável (kcal/kg) 2.900 2.900 2.850
Cálcio (%) 0,950 1,000 0,856
Fósforo disponível (%) 0,664 0,450 0,350
Sódio (%) 0,154 0,160 0,150
Metionina+cistina total (%) 0,650 0,560 0,500
Metionina total (%) 0,322 0,275 0,243
Lisina total (%) 1,150 0,848 0,750
Treonina total (%) 0,850 0,660 0,582
Triptofano total (%) 0,267 0,199 0,175 (1)Composição por kg do produto: Vit. A 10.000.000 UI; Vit. D3 2.500.000 UI; Vit. E 6.000 UI; Vit. K
1.600 mg; Vit. B12 11.000 mg; Niacina 25.000 mg; Ácido fólico 400 mg; Ácido pantotênico 10.000 mg; Selênio 300 mg; Antioxidante 30 g; Veículo q.s.p.; (2)Composição por kg do produto: Mg - 150.000 mg;
Zn - 100.000 mg; Fe - 100.000 mg; Cu - 16.000 mg; I -1.500 mg; Veículo q.s.p; (3)Bacitracina de zinco
(150g/t de ração);(4)Coxistac (50g/t de ração); (5)Etoxiquim (100g/t de ração); (6)Caulim.

27
Semanalmente, as aves eram pesadas individualmente, o consumo de ração
mensurado por parcela e dividido pelo numero de aves. As mudanças corporais foram
mensuradas pela comparação entre frangas abatidas no inicio e final de cada fase. Duas
poedeiras de cada grupo eram abatidas (1ª, 2ª, 4ª, 7ª, 11ª, 13ª e 16ª semana) perfazendo
60 repetições por linhagem em cada idade. As frangas selecionadas para o abate
recebiam apenas água a vontade por 24h, em seguida abatidas e pesadas. O peso das
penas foi obtido por diferença após depenagem. As aves foram evisceradas, armazenado
em sacos plásticos e congelado a -20ºC. As amostras foram trituradas em moinho de
carne industrial tipo “Cutter” por três vezes consecutivas e homogeneizadas. Foram
retiradas sub-amostras e, acondicionadas em bandejas de alumínio e posteriormente pré-
secas em estufa de ventilação forçado a 55ºC até peso constante. Antes das análises
laboratoriais, as sub-amostras foram ainda processadas em moinhos tipo “Cutter” e
“Willy” e, trituradas em liquidificador industrial. As metodologias utilizadas para
análises químicas foram as descritas por Silva & Queiroz (2002).
A função de crescimento de Gompertz (1825) foi usada para descrever a
mudança do peso e componentes corporais em função da idade. A equação utilizada
para estimar o peso (P) ou componente estimado da franga (g) no tempo t (dias) foi: Pt
(g) = Pm.exp.{- exp.[-b.(t - T)], em que, Pm o peso corporal ou do componente (g) à
maturidade; b, a taxa de maturidade (por dia); T é a idade em que a taxa de crescimento
é máxima (dias). A taxa de crescimento (TC) e/ou deposição dos pesos e componentes
corporais das frangas foram estimadas pela derivada da função Gompertz (TC =
Pm.b.(exp(-exp(-b.(t-T)))).(exp(-b.(t-T))). As regressões alometricas foram calculadas
usando-se o ln do componente pelo ln da PB, e são usadas para estimar os componentes
corporais em função da PB. Os parâmetros estimados da equação de Gompertz foram
submetidos à análise de variância e as médias foram comparadas pelo teste F (5%).
A exigência de proteína de mantença (PBm) foi obtida conforme o modelo de
Emmans & Fisher (1986) adaptado de Taylor (1970), onde: PBm (g/dia) =
(0,008.PPm0,73).u, em que u = (Pt/PPm), corresponde ao grau de maturidade da
proteína. Para isso, foi utilizada a modificação da função Gompertz, apresentada em
Martin et al. (1994), onde: Pt (kg) = PPm.exp [-exp((LN*(-LN*(PPi÷PPm)))) - (b.t))];
em que: PPt = peso no tempo de proteína corporal após a eclosão; PPm = peso de

28
proteína corporal a maturidade (kg); PPi = peso de proteína inicial (kg); b = taxa de
maturidade da proteína corporal (por dia); e t = idade (dias).
A exigência de ganho de peso foi obtida pela relação da exigência líquida de
ganho pela eficiência de utilização. A exigência líquida foi determinada por meio de
equação de regressão da proteína corporal em função do peso de corpo vazio (parâmetro
“b”, coeficiente de regressão linear). A eficiência de utilização para ganho foi
encontrada segundo a equação: eficiência de utilização = proteína retida ÷ (proteína
ingerida – proteína de mantença).
Resultados e Discussão
Pela equação de Gompertz (Pt (g) = Pm.exp.{- exp.[-b.(t - T)]) o peso a
maturidade foi maior para a Bovans, cuja idade para a taxa de crescimento máxima foi
63 d. O ganho de peso alcançou médias semelhantes para as duas linhagens com 10,7
g/d entre a 1º e 16º semana de idade.
Tabela 2- Estimativa dos parâmetros da equação de Gompertz para peso vivo, peso
corporal depenado e peso de penas de franga leve e semipesada.
Parâmetros PC (g) PD (g) PP (g)
Dekalb Bovans Dekalb Bovans Dekalb Bovans
Pm (g) 1785,90b 1878,75a 1340,718b 1489,213a 135,53b 168,65a
b (d) 0,0210a 0,0198b 0,0255b 0,0285a 0,0232a 0,0236a
T (dias) 58,02b 62,98a 50,60a 46,84b 48,00b 51,08a
R2 0,97 0,96 0,99 0,99 0,96 0,98
PC = Peso corporal; PD = Peso vazio depenado; PP = Peso das penas; Pm = peso à maturidade; b = taxa de maturidade; T = Idade em que a taxa de maturidade é máxima.
As linhagens apresentaram diferentes taxas de crescimento corporal em que as
maiores taxas da linhagem Bovans podem influenciar as exigências protéicas em
relação às Dekalb.
A linhagem Dekalb Branca atingiu e o peso adulto mais cedo que Bovans
Goldline, porém, para o peso corporal vazio depenado e peso das penas o resultado foi
inverso (P≤0,05), com menor tempo de deposição de penas para Dekalb. O peso relativo
das penas, em geral, diminuiu com a idade, representando cerca 7,59% e 8,98% do peso
corporal à maturidade para a Dekalb e Bovans, respectivamente.

29
Semelhantemente, o peso das vísceras acompanha a referida tendência, no
entanto, observou-se o maior peso relativo médio das vísceras para a Dekalb (17,34 vs
11,76% do peso corporal).
Os parâmetros da equação Gompertz (b, T) apresentados na tabela 2, indicam
que as frangas Dekalb White, foram geneticamente melhoradas para atingirem
precocemente o máximo crescimento corporal e de penas que a linhagem Bovans
Goldline. Estas características permitem que a franga leve inicie a postura mais cedo
que a semipesada.
O crescimento do peso vivo das frangas Dekalb foi ligeiramente superior ao das
frangas Bovans até 63 dias de idade (aproximadamente, nove semanas), quando então,
torna a ser menor até o início da postura (Tabela 3).
Tabela 3- Taxa de crescimento corporal e de penas de frangas leves e semipesadas.
Idade (d) PC (g/dia) PD (g/dia) PP(g/dia)
Dekalb Bovans Dekalb Bovans Dekalb Bovans
1 4,528 4,186 3,505 3,902 0,380 0,389
7 5,908 5,448 4,972 5,877 0,552 0,573
14 7,602 7,019 6,837 8,453 0,765 0,814
21 9,263 8,597 8,667 10,980 0,966 1,054
28 10,767 10,074 10,265 13,122 1,128 1,265
35 12,016 11,360 11,486 14,647 1,237 1,425
42 12,948 12,388 12,256 15,459 1,287 1,524
49 13,536 13,121 12,567 15,585 1,283 1,559
56 13,784 13,549 12,464 15,133 1,235 1,539
63 13,724 13,685 12,022 14,249 1,155 1,475
70 13,401 13,559 11,330 13,082 1,056 1,379
77 12,866 13,212 10,471 11,766 0,946 1,262
84 12,175 12,687 9,521 10,405 0,834 1,136
91 11,377 12,029 8,540 9,075 0,726 1,009
98 10,517 11,280 7,573 7,826 0,625 0,885
105 9,631 10,476 6,652 6,686 0,534 0,769
112 8,749 9,649 5,796 5,669 0,453 0,663
O tempo para atingir a maior taxa de maturidade no peso vivo da linhagem
Dekalb White (58 dias) ficou acima do valor obtido com a franga Hy Line W36 (52
dias) e Hisex Branca (55 dias), em trabalho realizado por Neme et al. (2006).
Semelhantemente, para a linhagem semipesada, Bovans Goldline, a taxa de crescimento
máxima ( no ponto de inflexão) foi tardia que para as frangas Hy line Marrom (59 dias)
e Hisex Marrom (59 dias) obtidas no mesmo estudo.
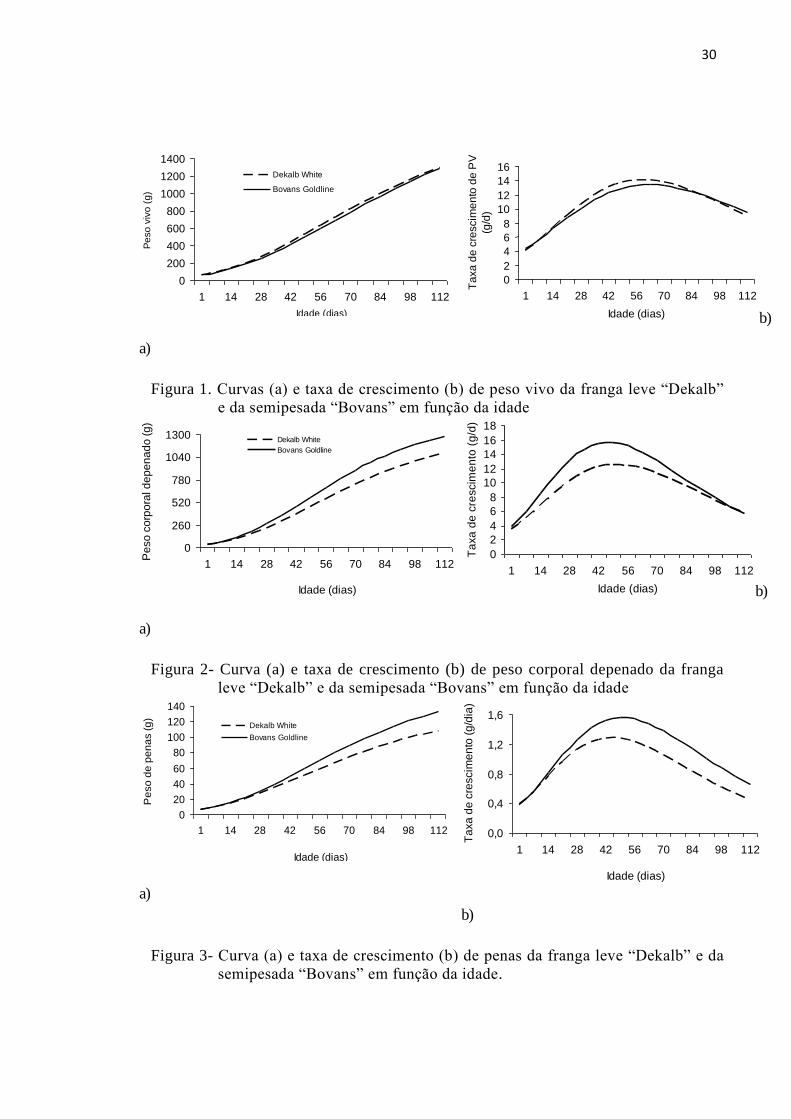
30
0
200
400
600
800
1000
1200
1400
1 14 28 42 56 70 84 98 112
Idade (dias)
Pe
so
viv
o (
g)
Dekalb White
Bovans Goldline
a)
0
2
4
6
8
10
12
14
16
1 14 28 42 56 70 84 98 112
Idade (dias)
Ta
xa
de
cre
scim
en
to d
e P
V
(g/d
)
b)
Figura 1. Curvas (a) e taxa de crescimento (b) de peso vivo da franga leve “Dekalb”
e da semipesada “Bovans” em função da idade
0
260
520
780
1040
1300
1 14 28 42 56 70 84 98 112
Idade (dias)
Pe
so
co
rpo
ral d
ep
en
ad
o (
g)
Dekalb White
Bovans Goldline
a)
0
2
4
6
8
10
12
14
16
18
1 14 28 42 56 70 84 98 112
Idade (dias)
Ta
xa
de
cre
scim
en
to (
g/d
)
b)
Figura 2- Curva (a) e taxa de crescimento (b) de peso corporal depenado da franga
leve “Dekalb” e da semipesada “Bovans” em função da idade
0
20
40
60
80
100
120
140
1 14 28 42 56 70 84 98 112
Idade (dias)
Pe
so
de
pe
na
s (
g)
Dekalb White
Bovans Goldline
a)
0,0
0,4
0,8
1,2
1,6
1 14 28 42 56 70 84 98 112
Idade (dias)
Ta
xa
de
cre
scim
en
to (
g/d
ia)
b)
Figura 3- Curva (a) e taxa de crescimento (b) de penas da franga leve “Dekalb” e da
semipesada “Bovans” em função da idade.

31
A curva de crescimento e a deposição dos componentes corporais diferiram entre as
linhagens (Tabela 4).
Tabela 4- Estimativa dos parâmetros da equação de Gompertz para proteína,
gordura, matéria mineral e água no corpo vazio de frangas leve e
semipesada.
Parâmetro Proteína Gordura Matéria mineral Água
Dekalb Bovans Dekalb Bovans Dekalb Bovans Dekalb Bovans
Pm (g) 231,0b 357,6ª 553,7
b 30155
a 40,5
b 52,0
a 1029,8
b 1160,5
a
b (dia) 0,027ª 0,021b 0,013
a 0,007
b 0,035
a 0,031
a 0,025
b 0,029
a
T (dias) 52,3b 63,3
a 133,8
b 253,6
a 40,8
a 41,9
a 46,1
a 43,6
b
R2 0,95 0,97 0,98 0,98 0,92 0,90 0,99 0,99 Pm = peso a maturidade; b = taxa de maturidade; T = tempo em que a taxa de crescimento é máxima.
Os valores preditos pela equação de Gompertz para a deposição de proteína,
gordura, material mineral e água nas frangas semipesadas (Bovans). Os parâmetros
referentes a deposição de gordura corporal entre as linhagem apresentaram maior
diferenças comparados aos demais nutrientes. O peso em gordura à maturidade foi
544,46% maior na linhagem Bovans que na Dekalb, foi, portanto superestimado. Neme
et al. (2006) encontrou problema semelhante, atribuindo a dificuldade de convergi a
maturidade da gordura por meio da curva Gompertz.
A taxa de maturidade da proteína e matéria mineral nas aves Dekalb foram
superiores às das Bovans, sugerindo que a linhagem Dekalb atinge o desenvolvimento
ósseo e muscular mais cedo que a linhagem Bovans, o que é confirmado pelo menor
tempo. A estimativa dos parâmetros da equação de Gompertz para a deposição de água
corporal coincidiu com a máxima deposição de proteína, explicada pela necessidade de
água para a síntese protéica nas aves.
A deposição dos componentes corporais foi mais precoce nas frangas Dekalb em
relação às frangas Bovans, menos para a água e matéria mineral. Isto confirma a
diferença de crescimento das duas linhagens e de certo modo, justifica o planejamento
alimentar observado a partir de estudos da composição de carcaça (Kwakkel, 1992,
Neme et al., 2006, Marcato, 2008), sendo mais importante que o peso corporal na
preparação do organismo das aves para a produção de ovos (Silva et al., 2000).
A derivada da equação de Gompertz descreveu as taxas de deposição dos
componentes corporais conforme demonstrado na tabela 5 e figuras 4, 5, 6, e 7. As aves
semipesadas “Bovans” depositaram maior quantidade de proteína que as leves

32
“Dekalb”, indicando que, possivelmente, estas aves atingem o início da postura mais
tardiamente. Porque, quanto maior e mais prolongado a deposição protéica, mais tempo
a ave levará para iniciar a postura. Esse resultado é confirmado pelo maior peso vivo e
protéico à maturidade.
Tabela 5- Taxa de deposição dos componentes corporais de frangas leves e semipesadas.
Idade (dias) Proteína (g/d) Gordura (g/d) Cinza (g/d) Água (g/d)
Dekalb Bovans Dekalb Bovans Dekalb Bovans Dekalb Bovans
1 0,480 0,671 0,205 0,281 0,098 0,161 3,540 3,835
7 0,729 0,924 0,268 0,349 0,174 0,247 4,742 5,489
14 1,067 1,256 0,359 0,444 0,280 0,355 6,164 7,515
21 1,417 1,602 0,467 0,556 0,386 0,457 7,458 9,375
28 1,739 1,936 0,594 0,688 0,468 0,535 8,498 10,839
35 1,996 2,234 0,738 0,839 0,516 0,582 9,208 11,774
42 2,169 2,476 0,899 1,012 0,527 0,597 9,568 12,154
49 2,251 2,651 1,073 1,206 0,507 0,584 9,597 12,032
56 2,250 2,754 1,259 1,421 0,467 0,549 9,347 11,510
63 2,178 2,789 1,452 1,656 0,414 0,501 8,882 10,706
70 2,055 2,762 1,649 1,912 0,357 0,446 8,268 9,732
77 1,897 2,683 1,845 2,186 0,301 0,389 7,564 8,681
84 1,719 2,565 2,038 2,477 0,250 0,334 6,820 7,626
91 1,535 2,416 2,223 2,783 0,205 0,283 6,076 6,614
98 1,354 2,249 2,397 3,101 0,166 0,237 5,358 5,678
105 1,182 2,071 2,558 3,428 0,133 0,197 4,686 4,833
112 1,023 1,890 2,702 3,762 0,106 0,163 4,068 4,086
0
50
100
150
200
250
300
1 14 28 42 56 70 84 98 112
Idade (dias)
Pro
teín
a (
g)
Dekalb White
Bovans Goldline
a)
0,0
0,5
1,0
1,5
2,0
2,5
3,0
1 14 28 42 56 70 84 98 112
Idade (dias)
Deposiç
ão d
e p
rote
ína (
g/d
ia)
b)
Figura 4- Curva de crescimento (a) e taxa de deposição (b) de proteína no corpo
vazio da franga leve “Dekalb” e da semipesada “Bovans” em função da
idade.

33
0
50
100
150
200
1 14 28 42 56 70 84 98 112
Idade (dias)
Go
rdu
ra (
g)
Dekalb White
Bovans Goldline
a)
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
1 14 28 42 56 70 84 98 112
Idade (dias)
De
po
siç
ão
de
go
rdu
ra (
g/d
)
b)
Figura 5- Curva de crescimento (a) e taxa de deposição (b) de gordura no corpo
vazio da franga leve “Dekalb” e da semipesada “Bovans” em função da
idade.
0
10
20
30
40
50
1 14 28 42 56 70 84 98 112
Idade (dias)
Ma
téri
a m
ine
ral (g
) Dekalb White
Bovans Goldline
a)
0,0
0,1
0,2
0,3
0,4
0,5
0,6
1 14 28 42 56 70 84 98 112
Idade (dias)
Deposiç
ão d
e m
até
ria m
inera
l
(g/d
ia)
b)
Figura 6- Curva de crescimento (a) e taxa de deposição (b) de matéria mineral no
corpo vazio da franga leve “Dekalb” e da semipesada “Bovans” em função
da idade
0
400
800
1200
1 14 28 42 56 70 84 98 112
Idade (dias)
Ág
ua
(g
)
Dekalb White
Bovans Goldline
a)
3,0
5,0
7,0
9,0
11,0
13,0
1 14 28 42 56 70 84 98 112
Idade (dias)
De
po
siç
ão
de
ág
ua
(g
/dia
)
b)
Figura 7- Curva de crescimento (a) e taxa de deposição (b) de água no corpo vazio da
franga leve “Dekalb” e da semipesada “Bovans” em função da idade .

34
A proteína corporal é o mais confiável indicador do crescimento, porque
desconsidera a variação que os conteúdos de água e gordura causam, devido
especificidades da nutrição e das condições de maio (Hruby et al., 1996). As equações
alométricas do peso de gordura, água e matéria mineral em função do peso protéico
foram discrepantes entre os genótipos avaliados (Tabela 6).
À medida que as frangas ganharam peso em proteína observa-se aumento na
proporção de gordura e pouca redução na proporção de água e matéria mineral corporal
das frangas Bovans em relação às frangas Dekalb. Isto explica a seqüência de deposição
de nutrientes no corpo das frangas, em que a deposição de matéria mineral e água têm a
formação precoce na linhagem Dekalb. O aumento no peso protéico proporciona maior
aumento na deposição de água e matéria mineral para a linhagem Dekalb. Porém para
os menores pesos protéicos, quando as aves são jovens, a maior deposição dos
componentes foi registrada para linhagem Bovans, isto é, essas aves iniciam a vida com
maior proporção de água e matéria mineral que as aves Dekalb.
Tabela 6- Equações alométricas de peso corporal em gordura, água e matéria mineral
em função do peso corporal protéico de frangas leves e semipesadas.
Equação(1)
R2 Coeficiente
alométrico
Frangas Dekalb
Gordura (g) = -1,2609 + 1,142. Proteína corporal 0,97 1,142
Água (g) = 2,4925 + 0,8114. Proteína corporal 0,97 0,811
Matéria mineral (g) = - 1,1977 + 0,9301. Proteína
corporal 0,97 0,930
Frangas Bovans
Gordura (g) = -1,4295 + 1,1601. Proteína corporal 0,98 1,160
Água (g) = 0,8214 + 2,4286. Proteína corporal 0,98 0,429
Matéria mineral (g) = 0,9963 + 0,8908. Proteína corporal 0,98 0,891
As frangas Bovans apresentaram menor coeficiente alométrico de gordura
corporal em relação à proteína corporal, demonstrando que as frangas Dekalb tendem a
depositar mais gordura com o aumento da proporção do peso protéico.

35
De modo geral, a linhagem Dekalb foi precoce no crescimento e deposição de
gordura e matéria mineral corporal, com o aumento do peso que a linhagem Bovans.
Considerando equações alométricas da gordura e água corporal em função do
peso de corpo vazio em escala Ln, observa-se que à medida que as aves ganham peso
corporal, os conteúdos de gordura aumentam enquanto, a água diminui
proporcionalmente (Figura 8).
0
20
40
60
80
100
120
140
1 7 28 49 77 91 112
Idade (dias)
Gord
ura
na c
arc
aça (
g/k
g)
660
680
700
720
740
760
780
800
Água n
a c
arc
aça (
g/k
g)
Gordura
Água
Figura 8- Comportamento da gordura e água no corpo vazio das frangas Dekalb em
função da idade.
O parâmetro que representa o peso de proteína a maturidade das frangas Dekalb
é menor, mas precoce que àquele estimado com frangas Bovans, apesar da taxa de
maturidade não ter sido influenciada (Tabela 7).
O peso da proteína corporal inicial das linhagens Dekalb e Bovans foram
levemente acima das frangas Ross Brown e Amber-Link e ligeiramente inferior aos
valores observados em frangas Hisex estudadas por Martin et al.(1994). O peso da
proteína à maturidade para aves Bovans (0,357 kg) foi semelhante ao obtido para a
linhagem Amber-Link (0,354 kg), mas com taxas de maturidade diferentes (0,0212 vs
0,0178).

36
Tabela 7- Estimativa dos parâmetros da equação de Gompertz para proteína na
carcaça das linhagens.
Parâmetro Dekalb White Bovans Goldline
Proteína corporal inicial (kg) 0,00541 0,00553
Proteína a maturidade (kg) 0,23098b
0,35757a
Taxa de maturidade (por dia) 0,0266a
0,0212b
Tempo de maturidade (dias) 52,30b
63,28a
R2 0,95 0,97
As exigências de proteína bruta para ganho, obtidas por meio da regressão linear
da proteína corporal em função do peso de corpo vazio foram respectivamente: 0,480;
0,696; e 1,108 g proteína/g de ganho, para frangas Dekalb e de 0,456; 0,626 e 0,668 g
proteína/g de ganho para frangas Bovans, de 1 a 4, 5 a 11 e 12 a 16 semanas de idade
(Tabela 8). Estes resultados foram superiores aos valores encontrados por Basaglia et al.
(1998) de 0,406, 0,544 e 0,563 g proteína/g de ganho para frangas leves com 1 a 6, 7 a
12 e 13 a 18 semanas respectivamente. A exigência de proteína líquida para ganho de
ambos os genótipos na pré-postura (12 a 16 semanas) foram as maiores estimativas
dentre as fase estudadas, o que sugere, provavelmente, maior necessidade de proteína
para formação do sistema reprodutivo.
Tabela 8- Equação da proteína corporal (PC) em função do peso de corpo vazio
(PCV)de acordo com a idade das aves.
Fase (sem) Equação R2
ELG,g/d Eficiência
(%)
EGP
(g/d)
Dekalb White
1 a 4 PC = 1,5918+0,96. PCV 0,99 0,096 20 0,480
5 a 11 PC = -20,701+0,195. PCV 0,99 0,195 28 0,696
12-16 PC = -84,862+0,2326. PCV 0,99 0,233 21 1,108
Bovans Goldline
1-4 PC= 0,5236+0,1138. PCV 0,99 0,114 25 0,456
5-11 PC= -21,426+0,1939. PCV 0,99 0,194 31 0,626
12-16 PC= -112,83+0,2543. PCV 0,93 0,254 38 0,668 Exigência Líquida para Ganho; Exigencia de ganho em proteína; Eficiência de utilização = proteína bruta retida ÷ (proteína bruta
ingerida – proteína de mantença).

37
As exigências diárias em proteína de mantença e ganho, o consumo de proteína e
de energia, obtidos a partir das estimativas (Tabela 9) são diferentes entre as linhagens,
independente da fase de criação avaliada. As estimativas de mantença em proteína de 1
a 4, 5 a 11 e 12 a 16 semanas de idade, foram crescentes, tanto para a linhagem leve
como na semipesada. Com o aumento do ganho de peso, maior quantidade de proteína
foi exigida repercutindo na necessidade de maior consumo de ração e de ingestão de
energia, a exceção da fase de 12 a 16 semanas para as frangas Bovans.
Tabela 9- Exigências de proteína e consumo de ração em função da fase de criação de
frangas leves e semipesadas.
Fase (sem) GP (g) Exigência de proteína (g/d)
CR (g/d)(1)
EM(2)
(kcal/d) Mantença Ganho Total
Dekalb White
1 a 4 8,33 0,041 4,0 4,41 19,81 55,47
5 a 11 13,97 0,094 9,72 9,814 57,73 161,64
12 a 16 11,28 0,112 12,50 12,612 84,08 235,42
Bovans Goldline
1 a 4 9,56 0,046 4,36 4,82 21,80 61,04
5 a 11 13,89 0,159 8,70 8,859 52,11 143,11
12 a 16 11,21 0,183 7,49 7,673 51,15 143,22
GP= ganho de peso; CR =consumo de ração; Exigência de proteína de mantença (g/d) = (0,008.PPm0,73).u; PPm =
proteína bruta à maturidade, depois de submetida à equação de Gompertz = 0,357g para frangas bovans e 0,231g para frangas dekalb; u = (PPt÷PPm), onde PPt é o peso potencial da proteína, obtido a partir da derivada de equação.
Exigência de proteína para ganho (g/d): 0,48.GP, 0,696.GP e 1,108.GP para frangas leves; 0,456.GP, 0,626.GP e
0,668.GP para frangas semipesadas 1 a 4, 5 a 11, e 12 a 16 semanas de idade, respectivamente. (1) CR: consumo de ração, considerando 21, 17 e 15% proteína da dieta para 1 a 4, 5 a 11 e 12 a 16 semanas de idade: proteína total ÷ nível
de proteína da dieta;(2)Exigência de energia metabolizável, considerando dieta com 2.800 kcal/kg = CR.2,8.
A exigência para mantença da linhagem semipesada foi maior que a obtida para
a linhagem leve, mas a exigência de ganho foi ligeiramente menor. Os resultados
indicam que as exigências protéicas mudam com o avançar da idade, sugerindo
formulações de rações com concentrações de proteína diferente. A relação EM:PB foi
crescente com o avançar da idade.

38
Conforme os resultados observados neste estudo, ficou demonstrado diferenças
para as curvas de crescimento, taxas de deposição de nutrientes corporais e exigências
em proteína entre frangas leves “Dekalb” e semipesadas “Bovans”. Neste sentido, as
informações sugerem que programas alimentares para a linhagem leve devem ser
diferenciados da linhagem semipesada, para promover o melhor desenvolvimento e
conformação corporal durante a preparação para o ciclo de postura. Contudo, o simples
acompanhamento do peso corporal das aves ao longo do crescimento, provavelmente,
não garante boas condições produtivas. A composição corporal categorizada em
escores no final da recria somados aos dados complementares da fase de produção
podem representar progressos práticos na criação de galinhas poedeiras.
Conclusões
Os dados do crescimento do peso corporal de frangas Dekalb White e Bovans
Goldlina ajustam-se à função Gompertz.
As taxas de crescimento e deposição de nutrientes corporais e das penas são
diferentes entre os dois genótipos. A linhagem leve é mais precoce no crescimento e na
deposição do peso e dos componentes corporais.
A exigência em proteína de total (mantença e ganho) para frangas de 1 a 4, 5 a
11 e 12 a 16 semanas de idade são 4,41; 9,814; 12,612 g/d para Dekalb White e 4,82;
8,859; 7,673g/d para Bovans Goldline.
Referências Bibliográficas
Basaglia R; Sakomura NK; Resende KT. Silva R, Junqueira OM.. Exigência de proteína
para frangas de postura de 1 a 18 semanas de idade. Revista Brasileira de
Zootecnia, v.27, n.3, p.556-563, 1998.
Brody TB, Siegel PB, Cherry JA. Age, body weight and body composition requirements
for the onset of sexual maturity of dwarf and normal chickens. British Poultry
Science, v.25, p.245-252, 1984

39
Emmans GC, Fisher C. Problems in nutritional theory. In: Fisher, C & Booman, K.N.
(Eds). In: Fisher C, Boorman KN. (Ed.) Nutrient Requirements of Poultry and
Nutritional Research. London: Butterworth, 1986. p.9-39. 1986.
Granja Planalto. Guia de manejo de poedeiras bovans goldline: Granja Planalto.
Uberlândia, MG. 37p. 2005a.
Granja Planalto. Guia de manejo de poedeiras Dekald White: Granja Planalto.
Uberlândia, MG. 35p. 2005b.
Gous RM. Methodologies for modelling energy and amino acid responses in poultry.
Revista Brasileira de Zootecnia, 78: 812-821. 2007. Suplemento especial.
Hruby M, Hamre ML, Coon CN. Growth modelling as a tool for predicting amino acid
requirements of Broilers. Journal Applied Poultry Research, 3: 403-415. 1994.
Hurby M, Hamre ML, Coon CN. Predicting amino acid requirements for broilers at
21.11 and 32.21C. Journal Applied Poultry Research, 4:395-401. 1995.
Hruby M, Hamre ML, Coon CN. Non-linear and linear functions in body protein
growth. Journal Applied Poultry Research, 5: 109-115. 1996.
Kwakkel RP, Dekoning FLSM, Verstegen MWA, Hof G. Effect of method and phase of
nutrient restriction during rearing on productive performance of light hybrid
pullets and hens. British Poultry Science, 32:747-761. 1991.
Laird AK, Tyler SA, Barton AD. Dynamics of normal growth. Growth. v. 29, p.233-
248. 1965.
Marcato SM, Sakomura NK, Munari DP, Fernandes JBK, Kawauchi ÍM; Bonato MA
Growth and body nutriente deposition of two broiler comercial genetic lines.
Brazilian Journal of Poultry Science, v.10, n.2, p.117-123, 2008.
Martin PA, Bradford GD, Gous RM. A formal method of determining the dietary amino
acid requirements of laying-type pullets during their growing period. British
Poultry Science, 35: 709-724. 1994.
Neme R, Sakomura NK Fukayama EH, Freitas ER, Fialho FB, Resende KT, Fernandes
JBK. Curvas de crescimento e de deposição dos componentes corporais em aves
de postura de diferentes linhagens. Revista Brasileira de Zootecnia, v.35, n.3,
p.1091-1100, 2006. (Supl.).

40
Rostagno HS, Albino LFT, Donzele JL, et al. Tabelas brasileiras para suínos e aves:
Composição de alimentos e exigências nutricionais. 2ª. ed. Viçosa: UFV; DZO –
Departamento de Zootecnia. 186p. 2005.
Sakomura AE, Rostagno HS. Métodos de pesquisa em nutrição de monogástricos.
Jaboticabal: FCAV/UNESP. 283p. 2007.
Silva JHV, Albino LFT, Rostagno HS. et al. Exigência de lisina para aves de reposição
de 7 a 12 semanas de idade. Revista Brasileira de Zootecnia, 29: 1786-1794.
2000.
Silva DJ, Queiroz AS. Análise de alimentos: métodos químicos e biológicos. 3ª ed.
Viçosa: UFV. 235p. 2002.

41
CONSIDERAÇÕES FINAIS E IMPLICAÇÕES
As pesquisas foram desenvolvidas baseando-se em técnicas conhecidas na criação
animal para codornas de corte e frangas de reposição. Os experimentos com codornas
buscou o aprimoramento de práticas que reduzam efeitos danosos causados pelo
estresse térmico por calor. A manutenção do equilíbrio eletrolítico pela suplementação
de eletrólitos pode ser uma alternativa importante em regiões de clima quente.
Em etapa seguinte foram estimados os parâmetros da função Gompertz para descrição
da cinética do crescimento em frangas Dekalb White e Bovans Goldline têm
implicações zootécnicas na criação comercial. Tais informações são fundamentais para
avanços genéticos e nutricionais. Os parâmetros proporcionaram informações para
estimar a exigência protéica de mantença e ganho das aves. Aparentemente, essas
informações podem ser mais bem aplicadas sob “classificação de escores” para
caracterizar a composição corporal das frangas às 16 semanas em complemento aos
índices zootécnicos durante a fase produtiva. Este conjunto de dados permitirão
manobras na alimentação que representem maior produção com menor custo.


















