
UNIVERSIDADE ESTADUAL DE SANTA CRUZ PROGRAMA DE...
59
UNIVERSIDADE ESTADUAL DE SANTA CRUZ PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUÇÃO VEGETAL DEISY JANINY GONÇALVES SILVA ASPECTOS TAXONÔMICOS E PATOGENICIDADE CRUZADA DE Thielaviopsis aff. basicola ISOLADO DE CENOURA (Daucus carota L.) NO ESTADO DA BAHIA ILHÉUS – BAHIA 2017
Transcript of UNIVERSIDADE ESTADUAL DE SANTA CRUZ PROGRAMA DE...

1
UNIVERSIDADE ESTADUAL DE SANTA CRUZ
PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUÇÃO VEGETAL
DEISY JANINY GONÇALVES SILVA
ASPECTOS TAXONÔMICOS E PATOGENICIDADE CRUZADA DE Thielaviopsis
aff. basicola ISOLADO DE CENOURA (Daucus carota L.) NO ESTADO DA
BAHIA
ILHÉUS – BAHIA 2017

2
DEISY JANINY GONÇALVES SILVA
ASPECTOS TAXONÔMICOS E PATOGENICIDADE CRUZADA DE Thielaviopsis
aff. basicola ISOLADO DE CENOURA (Daucus carota L.) NO ESTADO DA
BAHIA
Dissertação apresentada à Universidade Estadual de Santa Cruz para obtenção do título de Mestre em Produção Vegetal. Linha de Pesquisa: Proteção de Plantas Orientador: Prof. Dr. Jadergudson Pereira Co-orientadora: Profª Drª Edna Dora Martins Newman Luz Co-orientadora: Profª Drª Elizabeth Amélia A. Duarte
ILHÉUS – BAHIA 2017

3
DEISY JANINY GONÇALVES SILVA
ASPECTOS TAXONÔMICOS E PATOGENICIDADE CRUZADA DE Thielaviopsis
aff. basicola ISOLADO DE CENOURA (Daucus carota L.) NO ESTADO DA
BAHIA
Ilhéus, 21/02/2017
_________________________________________
Jadergudson Pereira Engenheiro Agrônomo – Doutor em Biologia de Fungos
UESC (Orientador)
_________________________________________
José Luiz Bezerra Biólogo – Ph.D em Fitopatologia
PPGPV – UESC
_________________________________________
Thiago Alves Santos de Oliveira Engenheiro Agrônomo – Doutor em Fitopatologia
UFRB

4
DEDICATÓRIA
A Deus, pela saúde e oportunidade que foram concedidas a mim para a concretização desse sonho.
Aos meus pais Dália e José Lopes, à minha família e a todos que
fizeram o possível ou intercederam para que este sonho fosse
realizado.

5
Dedico.
AGRADECIMENTOS
“Gratidão não é apenas a mais rica das
virtudes, mas sim a mãe de todas as outras”.
Cícero
A Deus, por sempre iluminar meus caminhos, me concedendo saúde,
coragem, amor, força, e alegrias para a realização deste sonho.
Aos meus pais, pelo apoio, dedicação e incentivo nos momentos de
dificuldade, e por ficarem ao meu lado para a realização desse sonho.
Aos meus irmãos José Victor e Débora, pelo amor, compreensão e alegrias.
Aos meus avós (in memorian), por serem exemplo de vida.
A toda minha família, pelo apoio.
Ao meu Orientador, Dr. Jadergudson Pereira, pela confiança, apoio,
compreensão, ensinamentos, conselhos e pela contribuição inestimável para minha
formação como profissional.
À minha Co-orientadora, Drª. Elizabeth Duarte, pelos ensinamentos,
disponibilidade e importante apoio nas análises moleculares.
À minha Co-orientadora, Drª. Edna Dora, pelo carinho, incentivo e
ensinamentos.
À Universidade Estadual de Santa Cruz, pela oportunidade que tive de cursar
mestrado nesta instituição.
Ao Programa de Pós-graduação em Produção Vegetal e todo o corpo
docente, com o qual tive a oportunidade de conviver durante o curso.
Ao Dr. Célio Kersul e ao Dr. Bonadie, pelas mudas de cacau, graviola e
seringueira cedidas para a realização do presente trabalho.
A Delinho e Zezinho, que me ajudaram com o trabalho pesado.
Aos meus companheiros de laboratório e de curso: Gisele, Francis, Mateus,
Thaís e Thaiana, que me ofereceram ajuda sempre que precisei.
Aos colegas amigos que fiz durante esta etapa de vida.
À equipe do laboratório de biologia molecular da Universidade Federal do
Recôncavo da Bahia, pelo apoio.

6
Á CAPES, pelo apoio financeiro através da concessão da bolsa de estudos,
possibilitando o desenvolvimento desta dissertação.
A todos aqueles que, direta e indiretamente, contribuíram para realização
deste sonho.

7
LISTA DE FIGURAS
CAPÍTULO 1
Figura 1 – Ocorrência de Podridão Negra em raízes de cenoura causada por
Thielaviopsis basicola e Ceratocystis fimbriata* (Cf) no Brasil.............................
10
Figura 2 – Raízes de Daucus carota com sintomas de Podridão Negra
(Thielaviopsis aff. basicola) provenientes de estabelecimento comercial
localizado em Ilhéus, Bahia..................................................................................
11
Figura 3 – Crescimento de Thielaviopsis aff. basicola em diferentes meios de
cultura..............................................................................................................
16
Figura 4 – Estruturas teleomórficas de Thielaviopsis aff. basicola.......................
18
Figura 5 – Estruturas anamórficas de Thielaviopsis aff. basicola.......................
19
Figura 6 – Detalhes de estruturas teleomórficas de Thielaviopsis aff.
basicola.................................................................................................................
20
Figura 7 – Filogenia Bayesiana derivada de análises do conjunto de dados
concatenados (CERF 7F)....................................................................................
25
CAPÍTULO 2
Figura 1 – Incisão no caule de Annona muricata 5 cm acima do solo............. 31
Figura 2 – Inoculação de Thielaviopsis aff. basicola em plantas de
Theobroma cacau, Annona muricata, Capsicum annuum e Passiflora edulis
f. flavicarpa.......................................................................................................
32

8
Figura 3 – Inoculação de Thielaviopsis aff. basicola em plantas de
Lycopersicon esculentum e Hevea brasiliensis...............................................
33
Figura 4 – Inoculação de Thielaviopsis aff. basicola em plantas de
Coriandrum sativum e Petroselinum crispum..................................................
34
Figura 5 – Inoculação de Thielaviopsis aff. basicola em raízes feridas de
Beta vulgaris, Ipomoea batatas e Daucus carota............................................
35
Figura 6 – Plantas de Petroselinum crispum (A, B) e Coriandrum sativum
(C, D) após 45 dias da inoculação com Thielaviopsis aff.
basicola...........................................................................................................
38
Figura 7 – Lesões necróticas em caules de Coriandrum sativum e
Petroselinum crispum inoculadas com Thielaviopsis aff. basicola (A, C) e
estruturas reprodutivas do patógeno (B, D)....................................................
39

9
LISTA DE TABELAS
CAPÍTULO 1
Tabela 1 – Comparação morfológica entre espécies de Thielaviopsis e
Chalaropsis (Complexo Ceratocystis)..............................................................
22
CAPÍTULO 2
Tabela 1 – Inoculação do isolado de Thielaviopsis aff. basicola em plantas
de oito famílias botânicas de importância para o Estado da Bahia.................
37

10
SUMÁRIO
RESUMO............................................................................................................ xii
1. INTRODUÇÃO GERAL.................................................................................
1
2. REVISÃO DE LITERATURA........................................................................
3
3. CAPITULO 1 – Aspectos taxonômicos de Thielaviopsis aff. basicola isolado de cenoura (Daucus carota L.) no Estado da Bahia.......................
7
RESUMO....................................................................................................... 7
3.1 INTRODUÇÃO ........................................................................................ 8
3.2 MATERIAL E MÉTODOS........................................................................ 11
3.2.1 Obtenção e manutenção do isolado de Thielaviopsis aff. basicola....................................................................................................
11
3.2.2 Crescimento do isolado de Thielaviopsis aff. basicola em diferentes meios de cultura.....................................................................
12
3.2.3 Caracterização morfométrica do isolado de Thielaviopsis aff. basicola....................................................................................................
12
3.2.4 Extração de DNA, sequenciamento e análises filogenéticas......... 13
3.2.4.1 Extração do DNA................................................................... 13
3.2.4.2 PCR e sequenciamento........................................................ 13
3.2.4.3 Análises filogenéticas............................................................ 14
3.3 RESULTADOS E DISCUSSÃO............................................................... 15
3.3.1 Crescimento do isolado de Thielaviopsis aff. basicola em diferentes meios de cultura.....................................................................
15
3.3.2 Caracterização morfométrica do isolado de Thielaviopsis aff. basicola....................................................................................................
16
3.3.3 Extração de DNA, sequenciamento e análises filogenéticas......... 23
4. CAPITULO 2 – Patogenicidade cruzada de Thielaviopsis aff. basicola isolado de Daucus carota em algumas plantas de importância econômica para o Estado da Bahia................................................................
26
RESUMO............................................................................................................ 26
4.1 INTRODUÇÃO......................................................................................... 27
4.2 MATERIAL E MÉTODOS........................................................................ 29
4.2.1 Obtenção de inoculo de Thielaviopsis aff. basicola ............................. 29

11
4.2.2 Inoculação de Thielaviopsis aff. basicola em hospedeiros de famílias
diversas........................................................................................................
29
4.3 RESULTADOS E DISCUSSÃO............................................................... 36
5. CONCLUSÕES GERAIS............................................................................... 40
REFERÊNCIAS................................................................................................. 41

12
ASPECTOS TAXONÔMICOS E PATOGENICIDADE CRUZADA DE Thielaviopsis
aff. basicola ISOLADO DE CENOURA (Daucus carota L.) NO ESTADO DA
BAHIA
RESUMO
Thielaviopsis basicola foi descrito pela primeira vez como Torula basicola, parasitando bases de haste de Pisum (ervilhas) e Nemophila auriculata em King's Cliffe, na Inglaterra. Anos depois foi relatado em alguns países da Europa, causando a Podridão negra de raízes em Daucus carota (cenoura). No Brasil, há relatos deste fungo em cenoura no Rio Grande do Sul, Distrito Federal e Maranhão. Objetivou-se, neste trabalho, estudar aspectos taxonômicos de T. aff. basicola isolado de cenoura, uma vez que o mesmo apresentou estruturas reprodutivas sexuadas e sua patogenicidade em plantas cultivadas de importância econômica para o sul da Bahia, como Annona muricata, Beta vulgaris, Coriandrum sativum, Petroselinum crispum, Ipomoea batatas, Hevea brasiliensis, Theobroma cacao, Passiflora edulis f. flavicarpa, Capsicum annuum e Lycopersicon esculentum. Foram testados diferentes meios para o desenvolvimento do fungo (Milho-Ágar, Extrato de Malte-Ágar 2% e Batata-Dextrose-Agar). Os estudos morfológicos foram feitos em microscópio ótico, com lâminas preparadas contendo estruturas reprodutivas. Para a análise em Microscópio Eletrônico de Varredura, amostras do tecido de cenoura com estruturas do fungo foram montadas em “stubs” e revestidas em ouro. Para análises moleculares foram utilizadas as respectivas regiões de genes ITS, VG9-R, Cer-MCM7F, Cer-MCM7R, RPB2-5Fb, RPB2-7Rb. A sequências obtidas foram comparadas com depositadas no GenBank. Em mudas de plantas herbáceas e lenhosas foram induzidos ferimentos, em seguida inoculou-se com uma pipeta 100 μL de uma suspensão de 2,5 × 106 fialoconídios/mL, nas testemunhas 100 μL de água estéril. Em raízes de I. batatas e B. vulgaris foram feitos ferimentos superficiais com o auxílio de um estilete, inoculando-se a suspensão conforme descrito anteriormente. Também foram utilizados discos de micélio de 5 mm de diâmetro a partir de cultura do patógeno e BDA puro para testemunhas. O local de inoculação foi vedado com filme de PVC para evitar a dessecação e entrada de patógenos secundários. O isolado de T. aff. basicola apresenta ascomas peritecióides globosos,100–225 × 175–262,5 μm, pescoço (rostro) longo, 500–725 μm de comprimento × 22,5–30 μm largura na base × 17,5–25 μm largura na extremidade. Apêndices na base do ascoma ausentes. Hifas ostiolares 87,5–187,5 μm comp. Ascos não observados. Ascósporos com formato típico de “chapéu”, 4–6 × 3 μm, que acumulam-se como massa de coloração creme a amarelada na extremidade do pescoço. Aleurioconídios, 10–15 μm × 7–10 μm, observados no micélio basal dos ascomas. Fiálides 57,5–232,5 μm comp. × 5–7,5 μm (L da base), 2,5–5 μm (L da ponta). Fialoconídios unicelulares, 10–18 μm × 4 μm. Aleuroconídios (artroconídios ou clamidósporos) 24–55 μm, em cadeias de 3 a 7 células com dimensões 6–9 × 7–10 μm. Os resultados obtidos por comparação de sequências do isolado de T. aff. basicola utilizando-se as regiões gênicas com aquelas depositadas no GenBank indicam haver dois grupos distintos e que o isolado de T. aff basicola não agrupou
Xii

13
com nenhum, indicando ser este geneticamente distinto. As inoculações mostraram que há alguma especialização do fungo quanto ao hospedeiro, pois o mesmo só foi patogênico à C. sativum e P. crispum, ambas da família Apiaceae.
Palavras chave: Ascomycota. Filogenia. Taxonomia. Cacau. Coentro.

1
1. INTRODUÇÃO GERAL
A cenoura (Daucus carota L.) pertence ao grupo das raízes tuberosas e é
uma hortaliça da família Apiaceae de elevada importância mundial, sendo também
expressiva no Brasil, onde é cultivada em larga escala nas Regiões Sudeste,
Nordeste e Sul. Segundo o CEPEA (2016), a cenoura é a quinta hortaliça mais
cultivada no Brasil e 80% da produção são destinados ao mercado interno. Com
uma produção de cenouras estimada em um milhão de toneladas por ano, o país
possui uma área de cultivo superior a 20 mil hectares. Os principais produtores são
Minas Gerais, São Paulo, Paraná e Bahia. A Bahia de 2014 para 2015 ocupou a
quarta posição com 1.200 ha de área plantada nas safras verão e inverno, onde os
municípios produtores que mais se destacam na atualidade são Lapão, Irecê e João
Dourado (DUMBRA, 2015).
A cultura da cenoura é bastante suscetível a diferentes patógenos, tendo os
fungos como principais agentes etiológicos responsáveis por várias doenças. Dentre
as doenças pós-colheita, a Podridão Negra das Raízes, causada pelo fungo
Thielaviopsis basicola/Ceratocystis, promove perdas significativas (LOPES; REIS
2016).
Thielaviopsis basicola (Berk. & Broome) Ferraris é um fungo de solo,
cosmopolita, polífago entre espécies de importância econômica e não apresenta
estádio sexual (teleomorfo) conhecido. Atualmente está classificado no Filo
Ascomycota, Classe Sordariomycetes, Ordem Microascales, Família
Ceratocystidaceae (INDEX FUNGORUM, 2017). O fungo não penetra diretamente
através da epiderme da planta sadia, que oferece uma forte barreira mecânica,
necessitando de vetor ou ferimentos, sendo que em condições de campo, o fungo
penetra principalmente através de cortes efetuados por ferramentas durante os
tratos culturais, como colheita, poda, desbrota, enxertia etc, mas, também pode
decorrer do ataque de insetos. Produz endoconidióforos que emergem lateralmente
da hifa, formando fiálides hialinas a castanho claro que produzem fialoconídios
hialinos, unicelulares com superfície lisa, dispostos em cadeias de comprimento
variável. Aleurioconidióforos emergem lateralmente a partir da hifa, são septados e
os aleuroconídios são marrons, globosos a piriformes, ocorrendo individualmente ou
em cadeias curtas (ENGELBRECHT; HARRINGTON, 2005). Os clamidósporos são
cilíndricos, multicelulados com paredes espessas e de coloração castanho escuro.

2
Este fungo foi relatado em cenoura em alguns países, como a Austrália,
Brasil, Canadá, Estados Unidos da América, Nova Zelândia e Taiwan, sendo
detectado em D. carota L. subsp. sativus [Hoffm.] Arcang. na Bulgária (PUNJA;
GAYE, 1993; FARR; ROSSMAN, 2017). Além disso, outras espécies de
Thielaviopsis também foram relatadas neste mesmo hospedeiro na Bulgária, como
T. punctulata (Hennebert) A. E. Paulin, T. C. Harr. & McNew e Chalaropsis
thielavioides Peyronel (= T. thielavioides [Peyronel] A. E. Paulin, T. C. Harr. &
McNew) (FARR; ROSSMAN, 2017). No Brasil, T. basicola foi relatado no Rio Grande
do Sul (DALBOSCO et al., 2004), Distrito Federal (INÁCIO et al., 2012) e Maranhão
(SILVA; COSTA, 2013). Entretanto, Carvalho e Carmo (2003) visualizaram
estruturas teleomórficas de Ceratocystis fimbriata Ellis & Halst em raízes de cenoura
com sintomas de Podridão Negra comercializadas no Rio de Janeiro.
Dessa forma, frente à ampla gama de hospedeiros, esse trabalho foi proposto
com o objetivo de caracterizar morfológica e molecularmente um isolado de
Thielaviopsis basicola (complexo Ceratocystis) obtido de raízes de cenoura, uma vez
que apresentou o estádio teleomórfico, fato este não relatado até o momento na
literatura mundial, bem como verificar sua patogenicidade a alguns hospedeiros de
importância econômica para o Estado da Bahia.

3
2. REVISÃO DE LITERATURA
No gênero Thielaviopsis Went são encontradas espécies causadoras de
Podridão Negra das raízes em diversos hospedeiros. Fungos deste gênero são
patógenos de solo, sapróbios, cosmopolitas que afetam grande número de espécies
vegetais de importância econômica no Brasil, podendo causar doença em várias
culturas, como alface (Lactuca sativa L.), abacaxi (Ananas comosus L.), cenoura (D.
carota), cana-de-açucar (Saccharum officinarum L.), coco (Cocos nucifera L.), dendê
(Elaeis guineinsis Jacq.), cacau (Theobroma cacao L.) entre outras (ABBAS, 2007;
GAYED 1972; KOIKE; HENDERSON 1998; PEREG, 2013; WILSON WIJERATNAM
et al., 2005).
Thielaviopsis foi descoberto por Went (1893), baseado em uma única espécie
(Thielaviopsis ethacetica Went) em cana-de-açúcar. Antes da descoberta de
Thielaviopsis, De Seynes (1886) já havia descrito Sporoschisma paradoxum, De
Seynes e Saccardo (1892) consideraram que o fungo pertencia ao gênero Chalara
(Corda) Rabenh, propondo, portanto, a combinação Chalara paradoxa (De Seynes)
Sacc.
Höhnel (1904) também relatou um fungo do endosperma de coco, idêntico a
S. paradoxum, considerando que o mesmo era T. ethacetica, confirmado em
seguida por Went, uma vez que o epíteto paradoxa tinha prioridade sobre
ethacetica. Portanto, Höhnel propôs a combinação Thielaviopsis paradoxa (De
Seynes) Höhn.
Berkeley e Broome (1850) relataram Torula basicola Berk. & Broome
parasitando bases de hastes de ervilhas (Pisum sativum L.) e Nemophila auriculata
(Boraginaceae). Em 1910 Ferraris transferiu To. basicola para Thielaviopsis,
surgindo a combinação Thielaviopsis basicola (Berk. & Broome) Ferraris, colocando-
o como anamorfo de Thielavia basicola Zopf. Porém, em 1925, McCormick mostrou
que não existe ligação teloanamorfo entre Thielavia basicola e T. basicola.
Alguns autores consideraram que nomes de gêneros pleomórficos deveriam
ser baseados em caracteres mais concretos (HENNBERT, 1968; NAG RAJ;

4
KENDRICK, 1975; HUGHES, 1953). Barron (1968) opinou que a distinção entre os
clados Chalara (Corda) Rabenh, Chalaropsis Peyrone e Thielaviopsis é arbitrária e,
devido a isso, colocou Thielaviopsis e Chalaropsis como sinônimos de Chalara.
Thielaviopsis, Chalara, Chaetochalara B. Sutton & Piroz, Hughesiella Bat. & A.F.
Vital e Trichocladium Harz mostram semelhança entre si. Chalara e Chaetochalara
têm um anamorfo monomórfico, Trichocladium, que possui conídios monomórficos e
solitários, Hughesiella possui um único conídio, enquanto Thielaviopsis difere de
Chalaropsis e Hughesiella por possuir artroconídios formados em uma cadeia de 4-8
conídios que permanecem ligados, parecendo um conídio multiseptado (ABBAS et
al., 2007; HENNBERT; SUTTON, 1994).
Nag Raj e Kendrick (1975) tentaram agrupar todos os fungos que possuem o
estado Chalara, mas este é encontrado em alguns Discomycetes, bem como em
Pyrenomycetes (PAULIN; HARRINGTON, 2000) e é, portanto, o produto de uma
evolução convergente.
No decorrer de vários estudos Paulin-Mahady et al. (2002), com base na
análise parcimônica das sequências da região do rDNA nuclear e sequências
parciais da subunidade LSU, colocaram quatro espécies de Chalara como um grupo
monofilético dentro do gênero Ceratocystis Ellis & Halst., são elas Ch. ovoidea Nag
Raj & W. B. Kendr., Ch. thielaviodes (Peyronel) Nag Raj & W. B. Kendr., Ch. populi
Veldeman ex Kiffer & Delon, e Ch. elegans Nag Raj & W. B. Kendr., que produzem
aleurioconídios típicos de Thielaviopsis, para o qual as espécies foram transferidas.
Três espécies, T. ovoidea (Nag Raj & W. B. Kendr.) A. E. Paulin, T. C. Harr. &
McNew, T. populi (Veldeman ex Kiffer & Delon) A. E. Paulin, T. C. Harr. & McNew e
T. thielavioides, foram morfologicamente semelhantes entre si, porém, as
sequências de rDNA mostraram-se distintas. Esses autores discordaram de Nag Raj
e Kendrick (1975) em colocar Thielaviopsis, Chalaropsis e Hughesiella como
sinônimos de Chalara, pois consideraram este último como um gênero distinto de
Thielaviopsis e alteraram sua descrição genérica. Além disso, colocaram
Chalaropsis e Hughesiella como sinônimos de Thielaviopsis. Portanto, a
classificação de T. basicola permaneceu inalterada, como confirmado pelos estudos
de rDNA feitos por Paulin-Mahady et al. (2002).
Segundo Abbas et al. (2007), a taxonomia de T. basicola ainda é bastante
confusa, pois diversos estudos demonstraram propostas de mudança na sua
nomenclatura. Recentemente, com base em análises moleculares, De Beer et al.

5
(2014) colocaram T. basicola em Ceratocystis “incertae sedis”, alegando que este
táxon não foi consistentemente acomodado em algum dos sete maiores clados
formados (Ceratocystis s.s., Chalaropsis, Endoconidiophora Münch, Davidsoniella Z.
W. de Beer, T. A. Duong & M. J. Wingf., Thielaviopsis, Huntiella Z. W. de Beer, T. A.
Duong & M. J. Wingf. e Ambrosiella Brader).
Thielaviopsis basicola é um patógeno de solo, filamentoso, polífago e
cosmopolita entre espécies de importância econômica, causando nas mesmas a
Podridão Negra das Raízes. Não apresenta estágio sexual (teleomorfo) conhecido,
produz aleurioconídios marrons, globosos a piriformes que podem ser simples ou em
cadeias curtas, células conidiogênicas fialídicas simples e tubulares, que seguem
em direção ao ápice e produzem cadeias de conídios primários retangulares ou, em
alguns casos, conídios secundários escuros em forma de barril. Enquanto os
conídios germinam rapidamente, sendo responsáveis pela rápida disseminação do
patógeno, há também clamidósporos (artroconídios), que são considerados esporos
de resistência, e permanecem viáveis em condições adversas, os quais são
estimulados a germinar por exsudatos radiculares de plantas hospedeiras (ABBAS
et al., 2007; DE BEER et al., 2014).
Segundo Teixeira-Yañez (2005), acredita-se que T. basicola foi introduzido no
Brasil através de turfa contaminada importada, utilizada na formulação de substrato
para produção de mudas.
No Brasil T. basicola ocasiona diversas doenças em diferentes culturas. Em
cenoura, a Podridão Negra das raízes foi relatada primeiramente no Rio Grande do
Sul (DALBOSCO et al., 2004), em seguida no Distrito Federal (INÁCIO et al., 2012) e
Maranhão (SILVA; COSTA, 2013). Apesar da observação de Thielaviopsis por esses
autores, nesses locais, Carvalho e Carmo (2003) relataram C. fimbriata como sendo
o agente etiológico da doença em raízes de cenoura comercializadas no Rio de
Janeiro. Além de T. basicola, outras espécies também foram relatadas em cenoura
ocasionando a mesma doença, como T. punctulata e Chalaropsis thielavioides
Peyronel, relatados na Bulgária (FARR; ROSSMAN, 2017).
Recentemente, Silva e Pereira (2016) relataram a ocorrência de T. aff.
basicola em cenoura no Estado da Bahia. Dessa forma, o objetivo deste trabalho foi
caracterizar morfologica e molecularmente um isolado de T. aff. basicola (Complexo

6
Ceratocystis) obtido de cenoura, bem como relatar a ocorrência do morfosexual
deste fungo, ainda não reportado na literatura mundial, e verificar a patogenicidade
deste isolado a algumas espécies vegetais de importância econômica para o Estado
da Bahia.

7
3 CAPÍTULO 1
ASPECTOS TAXONÔMICOS DE Thielaviopsis aff. basicola ISOLADO DE
CENOURA (Daucus carota L.) NO ESTADO DA BAHIA
RESUMO
Thielaviopsis basicola foi descrito pela primeira vez como Torula basicola, parasitando bases de haste de Pisum (ervilhas) e Nemophila auriculata em King's Cliffe, na Inglaterra. Anos depois foi relatado em alguns países da Europa, ocasionando a Podridão negra de raízes em Daucus carota (cenoura). No Brasil, há relatos deste fungo em cenoura no Rio Grande do Sul, Distrito Federal e Maranhão. Objetivou-se, neste trabalho, estudar aspectos taxonômicos de T. basicola isolado de cenoura, uma vez que o mesmo apresentou estruturas reprodutivas sexuadas. Foram testados diferentes meios para o desenvolvimento do fungo (Milho-Ágar, Extrato de Malte-Ágar 2% e Batata-Dextrose-Agar). Os estudos morfológicos foram feitos em microscópio ótico, com lâminas preparadas contendo estruturas reprodutivas. Para a análise em Microscópio Eletrônico de Varredura, amostras do tecido de cenoura com estruturas do fungo foram montadas em “stubs” e revestidas em ouro. Para análises moleculares foram utilizadas as respectivas regiões de genes ITS, VG9-R, Cer-MCM7F, Cer-MCM7R, RPB2-5Fb, RPB2-7Rb. A sequências obtidas foram comparadas com depositadas no GenBank. O isolado de T. aff. basicola apresenta ascomas peritecióides globosos,100–225 × 175–262,5 μm, pescoço (rostro) longo, 500–725 μm de comprimento × 22,5–30 μm largura na base × 17,5–25 μm largura na extremidade. Apêndices na base do ascoma ausentes. Hifas ostiolares 87,5–187,5 μm comp. Ascos não observados. Ascósporos com formato típico de “chapéu”, 4–6 × 3 μm, que acumulam-se como massa de coloração creme a amarelada na extremidade do pescoço. Aleurioconídios, 10–15 μm × 7–10 μm, observados no micélio basal dos ascomas. Fiálides 57,5–232,5 μm comp. × 5–7,5 μm (L da base), 2,5–5 μm (L da ponta). Fialoconídios unicelulares, 10–18 μm × 4 μm. Aleuroconídios (artroconídios ou clamidósporos) 24–55 μm, em cadeias de 3 a 7 células com dimensões 6–9 × 7–10 μm. Os resultados obtidos por comparação de sequências do isolado de T. aff. basicola utilizando-se as regiões gênicas com aquelas depositadas no GenBank indicam haver dois grupos distintos e que o isolado de T. aff basicola não agrupou com nenhum, indicando ser este geneticamente distinto.
Palavras chave: Apiaceae. Ascomycota. Filogenia. Taxonomia.

8
3.1 INTRODUÇÃO
Thielaviopsis basicola (Berk. & Broome) Ferraris (= Chalara elegans Nag Raj
& W. B. Kend.) foi relatado amplamente em todo o mundo em uma vasta gama de
hospedeiros. Primeiramente foi descrito como Torula basicola (Berk. & Broome),
parasitando bases de hastes de ervilhas (Pisum sativuml L.) e “Nemophila
auriculata” em King's Cliff, Inglaterra. Anos depois foi relatado em Daucus carota L.
(cenoura) em alguns países, causando a Podridão Negra das Raízes, como na
Austrália, Brasil, Canadá, Estados Unidos da América, Nova Zelândia e Taiwan,
sendo detectado em D. carota L. subsp. sativus [Hoffm.] Arcang. na Bulgária
(PUNJA; GAYE, 1993; FARR; ROSSMAN, 2017). Além da cenoura, esse patógeno é
responsável por doenças em mais de 140 espécies de plantas, destacando-se alface
(Lactuca sativa L.), citros (Citrus spp.), quiabo (Abelmoschus esculentus L.)
Moench), feijão de corda (Vigna unguiculata L.), fumo (Nicotiana tabacum L.),
amendoim (Arachis hypogaea L.), plantas ornamentais, entre outras (FARR;
ROSSMAN, 2017). Atualmente está classificado no Filo Ascomycota, Classe
Sordariomycetes, Ordem Microascales, Família Ceratocystidaceae (INDEX
FUNGORUM, 2017).
Espécies de Thielaviopsis Went são caracterizadas por apresentar células
conidiogênicas fialídicas simples e tubulares, que seguem em direção ao ápice e
produzem cadeias de conídios retangulares ou, em alguns casos, conídios
secundários escuros em forma de barril. Algumas espécies produzem clamidósporos
simples, asseptados, ou mais complexos e multisseptados, esporos de resistência
que podem sobreviver no solo durante muitos anos, em épocas de condições
climáticas adversas, e são estimulados a germinar por compostos produzidos pelas
raízes de plantas suscetíveis (ABBAS et al., 2007).
Com base apenas na morfologia, a primeira ocorrência de T. basicola no
Brasil, causando Podridão Negra em raízes armazenadas de cenoura, foi feita há
pouco mais de uma década no Rio Grande do Sul (DALBOSCO et al., 2004), em
seguida no Distrito Federal (INÁCIO et al., 2012) e Maranhão (SILVA; COSTA,
2013). Carvalho e Carmo (2003) relataram a Podridão Negra da Cenoura causada
por C. fimbriata, no Rio de Janeiro.

9
Milosavljevic et al. (2015) encontraram na superfície de raízes armazenadas
de cenoura (D. carota L. subsp. sativus [Hoffm.] Arcang.), um vegetal importante
para a Sérvia, o micélio negro do fungo, cujo sintomas manifestaram-se como
manchas negras irregulares que progrediam de tamanho. Após confirmação pelos
Postulados de Koch, juntamente com análises moleculares, comprovaram que o
agente causal da doença era Chalara thielavioides.
Além de T. basicola, outras espécies também foram relatadas em cenoura
ocasionando a mesma doença, como T. punctulata (Hennebert) A. E. Paulin, T.C.
Harr. & McNew e Chalaropsis thielavioides Peyronel (= T. thielavioides [Peyronel]
A.E. Paulin, T.C. Harr. & McNew), relatados na Bulgária (FARR; ROSSMAN, 2017).
Recentemente, Silva e Pereira (2016) relataram a ocorrência de T. aff.
basicola em cenoura na Bahia. A distribuição da ocorrência de T. basicola no Brasil
pode ser visualizada na Figura 1.
O objetivo deste trabalho foi caracterizar morfologica e molecularmente um
isolado de T. basicola (Complexo Ceratocystis) obtido de cenoura, bem como relatar
a ocorrência do morfossexual deste fungo ainda não reportado na literatura.

10
Figura 1. Ocorrência de Podridão Negra em raízes de cenoura causada por
Thielaviopsis basicola e Ceratocystis fimbriata* (Cf) no Brasil. BA =
Bahia; DF = Distrito Federal; MA = Maranhão; RJ = Rio de Janeiro; RS =
Rio Grande do Sul. *Provavelmente T. punctulata.
MA (2013)
RJ (2003) - Cf
RS (2004)
DF (2012) BA (2016)

11
3.2 MATERIAL E MÉTODOS
Os estudos foram conduzidos no Laboratório de Fitopatologia e Nematologia,
casa de vegetação do Programa de Pós-Graduação em Produção Vegetal e no
Centro de Microscopia Eletrônica da UESC, Ilhéus, Bahia. As análises moleculares
foram feitas no Laboratório de Biologia Molecular da Universidade Federal do
Recôncavo da Bahia, Cruz das Almas, Bahia.
3.2.1 Obtenção e manutenção do isolado de Thielaviopsis aff. basicola
O isolado de Thielaviopsis utilizado neste estudo foi obtido de raízes de
cenoura produzidas na região de Jaguaquara, Bahia, apresentando sintomas de
Podridão Negra, sendo estas coletadas em um estabelecimento comercial localizado
no Bairro do Salobrinho, município de Ilhéus, Bahia (Fig. 2).
Figura 2. Raízes de cenoura com sintomas de Podridão Negra (Thielaviopsis aff. basicola) provenientes de estabelecimento comercial localizado no município de Ilhéus, Bahia.

12
Amostras do material foram levadas para o Laboratório de Fitopatologia e
Nematologia para realizar o isolamento indireto do patógeno. Para isso, as raízes
foram lavadas em água corrente e detergente neutro e fragmentos do tecido interno
sob a lesão foram cortados e desinfestados, utilizando-se o protocolo padrão: álcool
70% durante 30 segundos; hipoclorito de sódio 2% durante um minuto; água
destilada estéril por um minuto. Após secagem em papel de filtro esterilizado, os
fragmentos foram colocados em meio de cultura Batata-Dextrose-Ágar (BDA),
contido em placas de Petri de 9 cm de diâmetro, as quais foram mantidas à
temperatura de 25ºC ±1 ºC em incubadora B.O.D. Após 48 horas, discos de BDA
contendo micélio das bordas da colônia foram repicados para obtenção de cultura
pura, a qual foi mantida em tubos de cultura com BDA e pelo método de Castellani
(CASTELLANI, 1939).
3.2.2 Crescimento do isolado de Thielaviopsis aff. basicola em diferentes
meios de cultura
Com a finalidade de selecionar o melhor meio de cultura para o
desenvolvimento do fungo, testaram-se quatro meios: Corn Meal Agar (CMA,
Himedia®), Extrato de Malte-Agar 2% (EMA, Merck®) e o Batata-Dextrose-Agar
(BDA) preparado em laboratório (200 g de batata; 10 g de dextrose Himedia®; 17 g
de ágar Merck®; 1000 ml de água destilada). Todos os meios de cultura foram
preparados considerando os cuidados descritos por Dhingra e Sinclair (1995). Os
meios de cultura foram distribuídos em placas de Petri de 9 cm de diâmetro, para os
quais foi repicada a cultura pura de Thielaviopsis, sendo estas armazenadas a 25 ±
1 ºC em incubadora B.O.D. por 7 a 10 dias.
3.2.3 Caracterização morfométrica do isolado de Thielaviopsis aff. basicola
A caracterização morfológica do isolado de Thielaviopsis foi feita segundo
metodologia descrita por Paulin-Mahady et al. (2002) e De Beer et al. (2014). Para
observação de estruturas reprodutivas (peritécios, ascósporos, aleuroconídios,
fialoconídios, etc.) ao microscópio ótico, foram confeccionadas lâminas contendo

13
lactofenol com e sem azul de algodão. Foram feitas medições de 30 fialoconídios, 30
aleuroconídios, 30 ascósporos, 20 fiálides, 15 aleuroconidióforos e 15 peritécios.
Para obtenção de imagens em Microscópio Eletrônico de Varredura (MEV)
(Quanta 25, EI Company®), amostras do tecido de D. carota com estruturas
reprodutivas do fungo foram montadas em “stubs” de metal cobertos com fita
condutora de carbono, os quais passaram por metalizador (SCD050, Bal-Tec®) para
revestimento em ouro e posterior visualização ao MEV.
3.2.4 Extração de DNA, sequenciamento e análises filogenéticas
3.2.4.1 Extração do DNA
O DNA foi extraído a partir de cultura pura do isolado de Thielaviopsis
(anamorfo e teleomorfo), utilizando-se o kit UltraClean® Microbial DNA Isolation
(MoBio, USA), seguindo as recomendações do fabricante. A integridade e a
quantidade do DNA foram verificadas usando eletroforese em gel de agarose 0,8% e
o Qubit® 2.0 Fluorometer (Invitrogen), respectivamente.
3.2.4.2 PCR e sequenciamento
As reações foram preparadas com os seguintes reagentes e concentrações:
60 ng de DNA de cada amostra (anamorfo e teleomorfo); 1x de tampão da enzima
Taq DNA polimerase; 3,7 mM de MgCl2; 0,6 pmol/μL de dNTPs; 0,4 pmol/μL de cada
primer; 2,5 U de Taq DNA polimerase, com volume final ajustado para 50 μL com
água ultra pura. Os ciclos de amplificações foram realizados no Veriti Thermal Cycler
PCR (Appplied Biosystems), seguindo as condições térmicas estabelecidas para
cada primer estudado. Os produtos amplificados foram visualizados em gel de
agarose 1%, corados com brometo de etídio e visualizados sob luz ultravioleta. Em
seguida, os amplicons foram purificados utilizando o kit Illustra® GFX PCR DNA and
Gel Band Purification (GE Healthcare Life Sciences) para posterior identificação
nucleotídica, utilizando o sequenciador automático ABI-Prism 3500 Genetic Analyzer
(Applied Biosystems), da empresa ACTGene Análises Moleculares LTDA. A edição e

14
montagem das sequências foram realizadas com o programa Sequencher 4.1.4
(Gene Code Corporation).
3.2.4.3 Análises filogenéticas
As sequências de diferentes regiões genéticas foram alinhadas com o auxílio
do software MEGA6®. Foram estudadas três regiões de genes diferentes: região ITS
(nuclear ribosomal DNA, Internal Transcribed Spacers), usando ITS4
(TCCTCCGCTTATTGATATGC) e ITS5 (GGAAGTAAAAGTCGTAACAAGG),
descritos por White et al. (1990); os primers ITS1-F (TCCGTAGGTGAACCTGCGG),
descrito por White et al. (1990); VG9-R (TTACGTCCCTGCCCTTTGTA), descrito por
De Hoog e van den Ende (1998); o componente 7 do complexo de manutenção do
minicromossomo (MCM7), Cer-MCM7F (ACICGIGTITCIGAYGTNAAGCC) e Cer-
MCM7R (TTRGCAACACCAGGRTCACCCAT), designados por De Beer et al.
(2014); a segunda subunidade maior da RNA polimerase II (RPB2) utilizando RPB2-
5Fb (GAYGAYMGWGATCAYTTYGG) e RPB2-7Rb
(CCCATRGCTTGYTTRCCCAT), descritos por Liu et al. (1999).
As regiões genicas estudadas (ITS, ITS-VG9, RPB2, MCM7) foram
analisadas separadamente e, posteriormente, comparadas com análise combinada.
As análises de máxima verossimilhança (ML) e IB Inferência Bayseana com 500
repetições bootstrap foram realizadas utilizando-se o software MEGA6® (Tamura et
al., 2013).
A identificação do anamorfo e teleomorfo do isolado de Thielaviopsis foi feita
a partir da comparação com sequências depositadas no banco de dados GenBank,
utilizando o BLASTn (Basic Local Alignment Search Tool), do National Center for
Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov). As sequências
obtidas neste estudo serão depositadas no GenBank.

15
3.3 RESULTADOS E DISCUSSÃO
3.3.1 Crescimento do isolado de Thielaviopsis aff. basicola em diferentes
meios de cultura
Dentre os meios de cultura testados, o que propiciou maior formação de
estruturas reprodutivas foi o Extrato de Malte-Ágar (EMA), no qual observaram-se
abundantes peritécios após 7 a 10 dias da repicagem, mesmo com a colônia
apresentando somente 2 a 3 cm de diâmetro (Figura 3B). Nos demais meios (BDA e
CMA), foram formados peritécios em menor número no mesmo período, apesar do
diâmetro das colônias ter sido similar. Isto é relevante, tendo em vista a necessidade
de se obter ascósporos e outras estruturas reprodutivas em menor tempo para o
preparo de suspensões.
Yarhood (1981) observou que T. basicola se desenvolve bem no meio
Czapek-Dox Agar e principalmente numa faixa de pH de 8-9,5, com temperatura
menor que 20 ºC.
Dhingra e Sinclair (1995) recomendam os meios Czapek-Dox Agar ou V-8
para produção de clamidósporos de T. basicola, o que ocorre entre 3 a 4 semanas.
Paulin-Mahady et al. (2002), Mbenoun et al. (2014) e De Beer et al. (2014)
utilizaram EMA ou BDA para cultivar isolados de Ceratocystis e Thielaviopsis, mas
não abordaram qual o melhor meio para a produção de peritécios ou outras
estruturas reprodutivas em menor tempo. Diferentemente do observado neste
trabalho, Rodrigues (2016) verificou que, dentre quatro meios testados para a
produção de peritécios de Ceratocystis cacaofunesta Engelbr. & T. C. Harr., o
melhor foi o BDA preparado em laboratório. Tais diferenças podem ocorrer, uma vez
que há variação na concentração de nutrientes nos meios semissintéticos e
espécies, ou isolados de uma mesma espécie, podem responder de forma diferente
aos estímulos nutricionais, mantendo-se outros fatores ambientais controlados
(temperatura, luminosidade, etc). Assim, a composição do meio de cultura determina
a quantidade e qualidade do crescimento micelial e esporulação dos fitopatógenos
(DHINGRA; SINCLAIR, 1995).

16
Figura 3. Crescimento de Thielaviopsis aff. basicola em diferentes meios de cultura. A - Batata-Dextrose-Agar (BDA); B - Extrato de Malte-Agar (EMA), propiciando a formação de peritécios com produção abundante de ascósporos (detalhe); C – Corn Meal Agar (CMA).
3.3.2 Caracterização morfométrica do isolado de Thielaviopsis aff. basicola
Thielaviopsis basicola (Berk. & Broome) Ferraris, Fl. Ital. Crypt., Fungi 1: 233.
1912.
= Chalara elegans Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 111 (1975)
= Torula basicola Berk. & Broome, Ann. Mag. nat. Hist., Ser. 2 5: 461 (1850)
= Trichocladium basicola (Berk. & Broome) J.W. Carmich., in Carmichael, Kendrick, Conners &
Sigler, Genera of Hyphomycetes (Edmonton): 185 (1980)
Ascomas peritecióides globosos, pretos, ostiolados, superficiais ou imersos
no substrato, 100–225 × 175–262,5 μm; pescoços (rostros) longos, retos, pretos,
500–725 μm de comprimento, 22,5–30 μm largura na base e 17,5–25 μm largura na
extremidade. Apêndices na base do ascoma ausentes. Hifas ostiolares (fimbrias)
castanhas claras a hialinas, não septadas, paredes lisas, 87,5–187,5 μm comp.
Ascos não observados. Ascósporos hialinos, unicelulares com formato típico de
“chapéu”, 4–6 × 3 μm, que acumulam-se como massa de coloração creme a
A B C

17
amarelada na extremidade do pescoço (Figuras 4 e 6). Aleurioconídios marrons,
globosos a piriformes, simples ou em cadeias curtas, 10–15 μm × 7–10 μm,
observados no micélio basal dos ascomas.
Fiálides castanhas claras a hialinas, 57,5–232,5 μm comp. × 5–7,5 μm (L da
base), 2,5–5 μm (L da ponta). Fialoconídios unicelulares, hialinos, cilíndricos, 10–18
μm × 4 μm. Aleurioconidióforos hialinos a marrons, 1–4 septos ,18–66 × 3–4 μm;
Aleuroconídios (artroconídios ou clamidósporos) castanho escuros, 24–55 μm, em
cadeias de 3 a 7 células, 6–9 × 7–10 μm (Figura 5).
Espécime examinado: BRASIL. BAHIA: Ilhéus, Salobrinho, Frutaria Claudinete
(14°79'78''S, 39°17'77''W), 15.X.2015, leg. Jad. Pereira, em raízes de Daucus carota
com sintomas de Podridão Negra (TFB 658).
Desde o seu estabelecimento, diversos autores não descreveram ou
observaram o estádio sexuado de T. basicola (BERKELEY; BROOME, 1850;
YARWOOD, 1981; PUNJA; SUN, 2000; PAULIN-MAHADY et al. 2002), fato este
ressaltado por De Beer et al. (2014) na mais recente revisão taxonômica de
Ceratocystis e gêneros próximos. Neste estudo, foram observadas estruturas
sexuadas no isolado de T. aff. basicola utilizado (IB = isolado da Bahia), o que torna
este fato inédito para a ciência e o caracteriza como importante contribuição no
conhecimento do ciclo reprodutivo deste fungo. Assim como T. cerberus, T.
ethacetica, T. paradoxa e T. punctulata, o isolado IB de T. aff. basicola produz
peritécios com rostros longos e ascósporos em forma de chapéu (Figura 4),
característicos de Ceratocystis sensu lato. Na Tabela 1 é possível observar alguns
caracteres taxonômicos de diversas espécies próximas de T. basicola, permitindo
uma comparação adequada das principais diferenças entre estas.
Relatos da ocorrência de T. basicola em cenoura foram feitos tanto no exterior
(PUNJA; GAYE, 1993; FARR; ROSSMAN, 2017) como no Brasil (DALBOSCO et al.,
2004; INÁCIO et al., 2012; SILVA; COSTA, 2013). Carvalho e Carmo (2003) fizeram
o primeiro relato da Podridão Negra em raízes armazenadas de cenoura no Brasil,
entretanto, relataram ser o agente causal Ceratocystis fimbriata, não registrando
formação de estruturas típicas de T. basicola naquela ocasião.

18
Figura 4. Estruturas teleomórficas de Thielaviopsis aff. basicola. A – Ascomas
peritecióides com massa de ascósporos na abertura ostiolar sobre raiz de
Daucus carota; B – Morfologia do ascoma com evidência do rostro longo;
C - Hifas ostiolares (fímbrias) e ascósporos sendo liberados em cadeia.

19
Figura 5. Estruturas anamórficas de Thielaviopsis aff. basicola. A – Aleuroconídios catenulados (artroconídios); B – Fiálide terminal. C – Fialoconídios cilíndricos, unicelulares. D – Aleurioconídios globosos a piriformes, simples.

20
O mais provável é que o fungo estudado por aqueles autores tratava-se de T.
punctulata (= Ceratocystis radicicola), o qual também foi isolado deste hospedeiro na
Austrália (VAN WYK et al., 2009), uma vez que esta espécie também forma
estruturas sexuadas e os aleuroconídios apresentam morfologia semelhante ao
ilustrado por Carvalho e Carmo (2003). Apesar disso, estes autores não
descreveram as medidas das estruturas reprodutivas e nem fizeram análises
moleculares, não permitindo que inferências seguras sobre a verdadeira identidade
daquele fungo sejam feitas com base na publicação.
Quando se compara as características morfológicas do isolado IB com as de
outras espécies causadoras da Podridão Negra em raízes de cenoura, observam-se
diferenças marcantes. O isolado IB forma peritécios no substrato natural e in vitro,
enquanto Chalaropsis thielavioides (= T. thielavioides), por exemplo, não forma,
além do primeiro apresentar dois tipos de aleuroconídios, sendo um formado nas
hifas basais do ascoma (simples) e o outro no estádio assexuado (artroconídios).
Duas diferenças marcantes entre o isolado IB e T. punctulata (= Ceratocystis
radicicola) são a formação de apêndice do ascoma (ausente vs. presente) e o
tamanho dos ascósporos (4–6 × 3 µm vs. 8–15 × 3–4 µm). Ressalta-se, que até o
momento, Ch. thielavioides e T. punctulata não foram relatados em D. carota no
Brasil.
Figura 6: Imagens em Microscópio Eletrônico de Varredura A – Detalhe da base do peritécio; B – Hifas ostiolares com acúmulo de ascósporos de Thielaviopsis aff. basicola.

21
Não se conheciam relatos anteriores da ocorrência de T. basicola em raízes
armazenadas de cenoura na Bahia, sendo o primeiro relato da Podridão Negra neste
estado feito a partir do estudo de Silva e Pereira (2016), ampliando o conhecimento
da distribuição desta doença no território nacional, como mostrado na Figura 1.
Historicamente, a taxonomia de T. basicola tem sido bastante confusa, fato
este bem ilustrado por Abbas et al. (2007), os quais registram a cronologia das
mudanças propostas na nomenclatura deste fungo por diversos autores.
Recentemente, com base em análises moleculares, De Beer et al. (2014) colocaram
T. basicola em Ceratocystis “incertae sedis”, alegando que este táxon não foi
consistentemente acomodado em algum dos sete maiores clados formados
(Ceratocystis s.s., Chalaropsis, Endoconidiophora, Davidsoniella, Thielaviopsis,
Huntiella e Ambrosiella). De fato, ao se observar os agrupamentos filogenéticos
obtidos considerando-se as regiões gênicas LSU, 60S e MCM7, T. basicola ficou
isolado em todas as análises. Algumas considerações sobre o posicionamento
genético do isolado IB são feitas no item 2.3.3.
Harrington e McNew (1997) citam que há variação em espécies de
Ceratocystis quanto à autocompatibilidade e autoesterilidade, o que pode ser
visualizado na Tabela 1, onde espécies de Ceratocystis, Thielaviopsis e gêneros
afins apresentam homo ou heterotalismo. No caso deste estudo, observou-se a
primeira condição, sendo que o isolado IB forma fácil e rapidamente estruturas
sexuadas in vitro, tanto em Extrato de Malte-Agar 2% como Batata-Dextrose-Ágar,
conforme discutido no item 2.3.1. O homotalismo pode proporcionar vantagem
reprodutiva para o fungo e permitir a manutenção de genótipos bem adaptados em
locais de escassez de linhagens compatíveis (PAOLETTI et al., 2007). Paulin-
Mahady et al. (2002) tentaram a produção de peritécios in vitro pareando diferentes
isolados de T. basicola e Ch. thielavioides, mas não obtiveram sucesso. Um dos
aspectos atribuídos pelos autores a este fato é que parece ter havido a perda do
estádio sexuado pelo menos uma vez em Ceratocystis.

22
Tabela 1 – Comparação morfológica entre espécies de Thielaviopsis e Chalaropsis (Complexo Ceratocystis).
Táxon Comp. Sexual
2
Teleomorfo
Peritécio (μm)
Rostro (μm)
Apêndice do ascoma
Aleuroconídios (μm)
Ascósporos (μm)
Conídios primários
(μm)
Conídios secundários
(μm)
Referência
Chalaropsis ovoidea - - - - - 8–16 × 4–10 - 10–21 × 2–4 - Paulin-Mahady et al. (2002)
Ch. populi - - - - - 6.7–12 × 6–9 - 6–18 × 2.2–3.8 - Kiffer and Delon (1983)
Ch. thielavioides - - - - - Simples, 10–20 × 8–
16 - 10-21 x 4-10 Paulin-Mahady et al. (2002)
T. basicola IB1 HO + 100–225
μm × 175–262,5
500–725 Ausente 1) globosos a piriformes, simples ou
em cadeias curtas, 10–15 μm × 7–10 μm;
2) artroconídios em cadeias de 3 a 7
células, 6-9 × 7-10 cada
4–6 × 3 10–18 × 4 Não observados Silva e Pereira (2016)
T. basicola - - - - - Artroconídios em
cadeias de 4 a 8 células, 7–12 × 10–17
- 7-17 × 2,5-4,5 - Ellis (1971)
T. cerberus
HO + 260–340 × 268–348
650–984 Digitado a estelar
Obovoides a subglobosos, simples ou em cadeias, 9–12
× 6–8
7–9 × 3–4 7–12 × 4–5 - Mbenoun et al. (2014)
T. ethacetica HT + 154–215 ×
156–216 672–862 Digitado Obovoides a
subglobosos, simples ou em cadeias, 14–
18 × 8–11
7–9 × 3–4 7–8 × 4–5 8–12 × 6–7 Mbenoun et al. (2014)
T. euricoi - - - - Ausente Obovoides a
subglobosos, simples ou em cadeias, 11–
14 × 8–11
- 9–11 × 4–5 11–16 × 6–7 Mbenoun et al. ( 2014)
T. musarum Indetermi
nada + 300 × 350 1100–
1200 Ausente Obovoides a
subglobosos, simples ou em cadeias, 12–
16 × 6–9
6–11 × 2-3.5 10–12 × 4–5 10–13 × 6–7 Mbenoun et al. (2014)
T. paradoxa HT + 237–317 ×
279–348 1063–1367
Digitado Obovoides a subglobosos, simples ou em cadeias, 10–
16 × 8–12
9–11 × 3–4 10–14 × 4–5 6–7 × 9–13 Mbenoun et al. (2014)
T. punctulata HT + 180–320
diam. 440–980 Digitado Subglobosos,
simples, 13–16 × 10–12
8–15 × 3–4 8–14 × 4–5 4–6 × 10–14 Mbenoun et al. (2014)
1IB = Isolado da Bahia.
2HO = Homotálico; HT = Heterotálico.

23
3.3.3 Extração de DNA, sequenciamento e análises filogenéticas
Na Figura 7, os resultados obtidos por comparação de sequências do isolado
IB de T. aff. basicola (anamorfo e teleomorfo), utilizando-se a região gênica CERF
7F, com algumas existentes no GenBank de táxons de Ceratocystis sensu lato de
diferentes partes do mundo, depositadas por De Beer et al. (2014), nota-se a
presença de dois clados distintos. No primeiro clado encontram-se T. paradoxa, T.
euricoi, T. musarum, T. ethacetica, T. cerberus, T. radicicola, Huntiella oblonga, T.
basicola, Chalaropsis thielavioides e Ch. ovoidea. No segundo clado, encontram-se
C. cacaofunesta e C. platani.
Claramente vê-se que o isolado IB (anamorfo e teleomorfo) não agrupou com
nenhum dos clados, indicando ser este geneticamente distinto. Na revisão de
Ceratocystis sensu lato, De Beer et al. (2014) verificaram que T. basicola não
agrupou com nenhum dos clados formados, o que levou os autores a incluírem este
fungo em Ceratocystis “incertae sedis”. Ressalta-se que as sequências de T.
basicola utilizadas naquele estudo foram obtidas apenas de estruturas anamórficas,
uma vez que os autores enfatizam não haver estádio sexuado conhecido para este
fungo, o que é corroborado por outros autores.
O isolado IB possui a característica de se reproduzir sexuadamente, o que
pode ter sido em virtude de possível cruzamento (hibridação) com alguma espécie
de Ceratocystis. Por ter possivelmente herdado genes que expressam
particularidades genéticas e morfológicas, o isolado IB ficou mais próximo do clado
dois (Figura 7). Como citado anteriormente, Engelbrecht e Harrington (2005)
afirmam a possibilidade de ocorrer hibridação entre espécies de Ceratocystis.
Outro aspecto importante a ser ressaltado é que o isolado IB diferenciou-se
de T. punctulata e Ch. Thielavioides, ambas espécies causadoras da Podridão
Negra em raízes de cenoura, tanto no aspecto morfológico como genético (Tabela 1
e Figura 7, respectivamente). Resultados semelhantes para T. basicola foram
obtidos por Paulin-Mahady et al. (2002).
Estudos moleculares complementares serão feitos para ampliar a
comparação do isolado IB com outras sequências de T. basicola depositadas no
GenBank, uma vez este estudo restringiu-se apenas na utilização de CERF 7F,
marcador utilizado por De Beer et al. (2014). Entretanto, pode-se inferir que o
isolado IB é um novo táxon, o qual será devidamente acomodado e ampliará a lista

24
de espécies causadoras da Podridão Negra da Cenoura. Não há dúvidas que a
descoberta da fase sexuada deste fungo permitirá definir a classificação de T.
basicola, o qual tem sido tão controverso na história micológica mundial.

25
Figura 7. Filogenia Bayesiana derivada de análises do conjunto de dados concatenados (CERF 7F). Ramos representam probabilidades BI (inferência Bayesiana) de 95%; bootstrap com valores iguais ou superiores a 70% a partir de 500 replicas, indicados nos nós os valores ML (Maximum Likelihood).
Clado 1
Clado 2

26
4 CAPÍTULO 2
PATOGENICIDADE CRUZADA DE Thielaviopsis aff. basicola ISOLADO DE
Daucus carota L. EM ALGUMAS PLANTAS DE IMPORTÂNCIA ECONÔMICA
PARA O ESTADO DA BAHIA
RESUMO
Daucus carota é uma hortaliça de elevada importância mundial. No Brasil, é cultivada em larga escala nas Regiões Sudeste, Nordeste e Sul, com produção anual de mais de um milhão de toneladas de raízes. A doença conhecida como Podridão Negra, ocasionada por Thielaviopsis basicola, causa perdas significativas na produção. Objetivou-se neste trabalho estudar a patogenicidade do isolado de T. aff. basicola em plantas cultivadas de importância econômica para o sul da Bahia, como Annona muricata, Beta vulgaris, Coriandrum sativum, Petroselinum crispum, Ipomoea batatas, Hevea brasiliensis, Theobroma cacao, Passiflora edulis f. flavicarpa, Capsicum annuum e Lycopersicon esculentum. Em mudas de plantas herbáceas e lenhosas foram induzidos ferimentos, em seguida inoculou-se com uma pipeta 100 μL de uma suspensão de 2,5 × 106 fialoconídios/mL, nas testemunhas 100 μL de água estéril. Em raízes de I. batatas e B. vulgaris foram feitos ferimentos superficiais com o auxílio de um estilete, inoculando-se a suspensão conforme descrito anteriormente. Também foram utilizados discos de micélio de 5 mm de diâmetro a partir de cultura do patógeno e BDA puro para testemunhas. O local de inoculação foi vedado com filme de PVC para evitar a dessecação e entrada de patógenos secundários. As inoculações mostraram que há alguma especialização do fungo quanto ao hospedeiro, pois o mesmo só foi patogênico à C. sativum e P. crispum, ambas da família Apiaceae.
Palavras chave: Podridão Negra. Coentro. Salsa. Cacau.

27
3.1 INTRODUÇÃO
O gênero Thielaviopsis contempla diversas espécies fúngicas distribuídas em
vários lugares do mundo em uma ampla gama de hospedeiros, incluindo plantas
herbáceas e lenhosas de diferentes famílias (FARR; ROSSMAN 2017).
No Brasil há relatos de poucas espécies pertencentes a este gênero, dentre
elas Thielaviopsis musarum (R. S. Mitchell) Riedl na bananeira, Thielaviopsis
paradoxa na cana-de-açúcar, cacau, coqueiro, dendê e outras espécies de
palmeiras (CARVALHO et al., 2013), e o mais importante, T. basicola em cenoura
(SILVA; COSTA, 2013) e outros hospedeiros.
Thielaviopsis basicola tem uma gama de hospedeiros de mais de 140
espécies, sendo frequentemente encontrado nas famílias Apiaceae, Fabaceae,
Malvaceae, Solanaceae e Cucurbitaceae (FARR; ROSSMAN 2017).
Daucus carota é uma hortaliça da família Apiaceae de elevada importância
mundial. No Brasil, é cultivada em larga escala nas Regiões Sudeste, Nordeste e
Sul, com produção anual de mais de um milhão de toneladas de raízes. Esta planta
é bastante suscetível a diferentes patógenos tendo como os principais agentes
etiológicos os fungos, responsáveis por várias doenças como Podridão de Pré e
Pós-emergência, Queima-das-Folhas, Podridão das Raízes, Tombamento de
Plântulas, entre outras. Dentre as doenças pós-colheita, a Podridão Negra das
Raízes, causada pelo fungo T. basicola, promove perdas significativas.
Abbas et al. (2007) observaram, no Paquistão, raízes de feijão Caupi
(Vigna unguiculata L.) com sintomas de podridão, confirmando ser T. basicola o
agente causal da doença. Pereg (2013) relatou a ocorrência de T. basicola em
plantas de algodão na Austrália com sintomas de crescimento retardado ou lento no
início da estação, em comparação com plantas saudáveis ao redor atrasando a
floração ou maturidade da planta.
A primeira ocorrência de Podridão Negra em raízes armazenadas de cenoura
foi feita há pouco mais de uma década no Rio Grande do Sul (DALBOSCO et al.,
2004), e posteriormente no Distrito Federal (INÁCIO et al., 2012) e Maranhão

28
(SILVA; COSTA, 2013). No Rio de Janeiro, Carvalho e Carmo (2003) atribuíram a
Podridão Negra em raízes de cenoura a C. fimbriata.
Desta forma, objetivo deste trabalho foi estudar a possível patogenicidade de
um isolado de T. aff. basicola a algumas espécies vegetais de importância
econômica para o Estado da Bahia.

29
4.2 MATERIAL E MÉTODOS
Os experimentos foram realizados no Laboratório de Fitopatologia e
Nematologia da UESC, onde o isolado foi cultivado para a obtenção da suspensão
de inóculo e discos de micélio, e na casa de vegetação do Programa de Pós-
Graduação em Produção Vegetal, onde foram cultivadas e inoculadas plantas das
famílias Apiaceae, Annonaceae, Amaranthaceae, Convolvulaceae, Euphorbiaceae,
Malvaceae, Passifloraceae e Solanaceae.
4.2.1 Obtenção de inóculo de Thielaviopsis aff. basicola
O isolado do patógeno foi cultivado em meio Extrato de Malte-Agar (EMA 2%)
contido em placa de Petri que foram mantidas em incubadora a 25 ºC durante 10
dias. Após esse período, foram adicionados 5 mL de água destilada esterilizada
sobre a colônia formada na placa, fazendo-se uma raspagem da superfície micelial
com uma alça de Drigalski e filtração em gaze, para obtenção de uma suspensão de
inóculo (fialoconídios) na concentração de 2,5 × 106 fialoconídios/mL, seguindo o
utilizado por Laia et al. (2000). Também foram utilizados discos de micélio de 5 mm
de diâmetro a partir da cultura do isolado.
4.2.2 Inoculação de Thielaviopsis aff. basicola em hospedeiros de famílias
diversas
Para verificar a patogenicidade cruzada do isolado de T. aff. basicola obtido
de cenoura a outros hospedeiros, inocularam-se plantas das seguintes famílias:
Annonaceae - Annona muricata L. (graviola); Amaranthaceae – Beta vulgaris L.
(beterraba); Apiaceae - Coriandrum sativum L. (coentro) e Petroselinum crispum
(Mill.) Nym. (salsa); Convolvulaceae - Ipomoea batatas [L.] Lam. (batata doce);
Euphorbiaceae - Hevea brasiliensis [Willd. Ex A.Juss.] Müll. Arg. (seringueira);

30
Malvaceae - Theobroma cacao L. (cacau); Passifloraceae - Passiflora edulis f.
flavicarpa O. Deg. (maracujá); Solanaceae - Capsicum annuum L. (pimentão) e
Lycopersicon esculentum Mill. (tomate).
Em mudas de cacau, graviola, maracujá, pimentão e tomate foram feitas
incisões superficiais no caule 5 cm acima do solo, com o auxílio de uma lâmina
desinfestada com álcool 70% (Figura 1). Sobre os ferimentos foram pipetados 100
µL da suspensão do inóculo, vedando-os, logo após, com filme de PVC para evitar a
dessecação e entrada de patógenos secundários (Figuras 2 e 3). Na seringueira,
devido à liberação do látex que dificulta a aplicação da suspensão de inóculo, discos
de micélio de 5 mm de diâmetro foram utilizados para inocular plantas dos clones
CDC 312, IAN 6590, FX 3864, SIAL 893 e PMB1 (Figura 3: C e D). Para cada clone
foram consideradas a testemunha e o tratamento (planta inoculada).
Em mudas de coentro e salsa, por possuírem o caule mais frágil, foram feitos
ferimentos superficiais com auxilio de um estilete desinfestado com álcool 70% e
inoculação com discos de micélio (Figura 4).
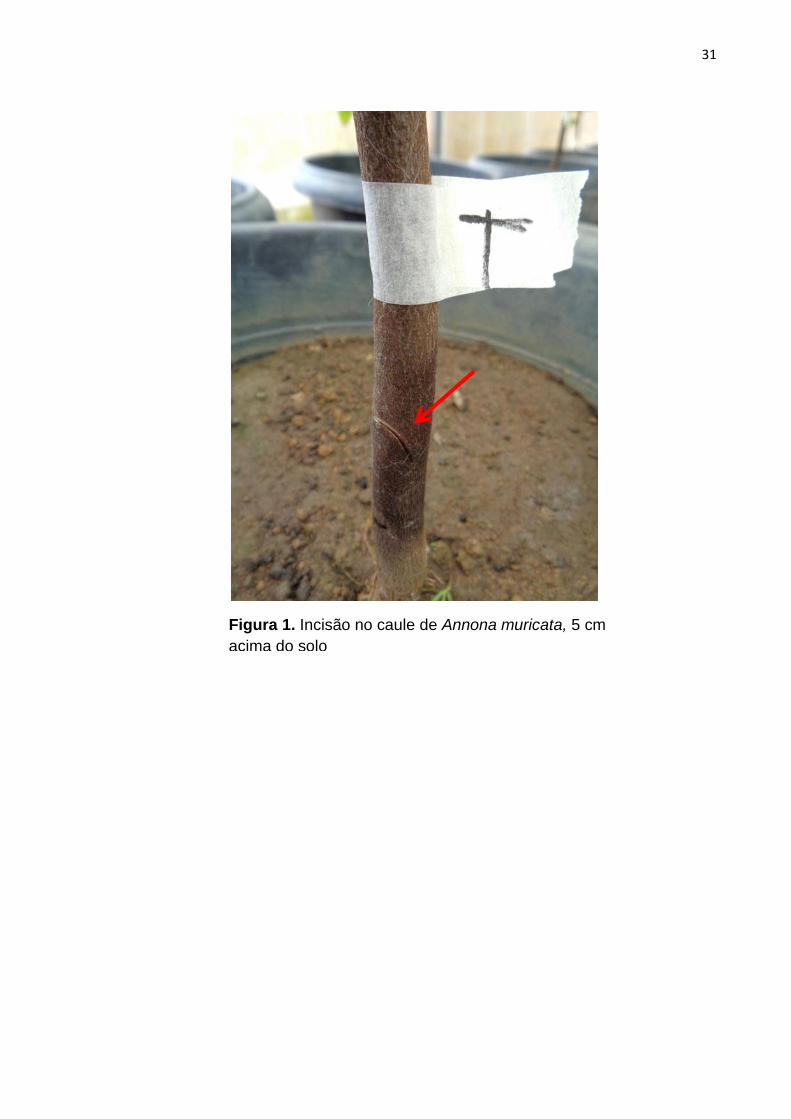
31
Figura 1. Incisão no caule de Annona muricata, 5 cm
acima do solo

32
Figura 2. Inoculação cruzada de Thielaviopsis aff. basicola em plantas de Theobroma cacao, Annona muricata, Capsicum annuum e Passiflora edulis f. flavicarpa. A, C, E e G – inoculadas; B, D, F e H – testemunha.

33
Figura 3. Inoculação de Thielaviopsis aff. basicola em plantas de Lycopersicon esculentum e Hevea brasiliensis. A e C – inoculadas; B e D – testemunha.

34
Figura 4. Inoculação de Thielaviopsis aff. basicola em plantas de Coriandrum sativum e Petroselinum crispum. A – inoculada; B – testemunha; C: detalhe da manutenção da umidade nos locais de inoculação.
Em raízes de batata e beterraba também foram feitos ferimentos superficiais
com o auxílio de um estilete, inoculando-se a suspensão conforme descrito
anteriormente. Raízes sadias de cenoura foram inoculadas paralelamente com a
mesma suspensão para confirmação da patogenicidade do isolado (Figura 5).
Foram utilizadas dez plantas/raízes de cada espécie, sendo cinco
testemunhas positivas (suspensão de inóculo ou disco de micélio) e cinco
testemunhas negativas (água estéril ou disco de meio de cultura EMA 2%).
Realizaram-se observações do terceiro ao 10º dia após a inoculação para verificar a
ocorrência de sintomas em raízes, e do terceiro até o 45º dia após a inoculação em
mudas. Para confirmação da condição patogênica do isolado utilizado, este foi
novamente inoculado em raízes sadias de cenoura e posteriormente reisolado,
cumprindo-se os Postulados de Koch.

35
Figura 5. Inoculação de Thielaviopsis aff. basicola em raízes feridas de Beta vulgaris, Ipomoea batatas e Daucus carota (A, C, E e G – inoculadas; B, D, F e H – testemunha).

36
4.3 RESULTADOS E DISCUSSÃO
No teste de patogenicidade cruzada do Thielaviopsis aff. basicola em
hospedeiros de diferentes famílias, o isolado não foi patogênico às espécies de
graviola, pimentão, seringueira, tomate, maracujá e cacau aos 45 dias após a
inoculação, e à beterraba e batata doce aos 10 dias após a inoculação. Não houve
manifestação de sintomas durante o período de avaliação (Tabela 1). Deste modo, a
ausência de sintomas nesses hospedeiros pode ser em virtude de vários fatores,
como variabilidade genética e especialização por hospedeiro.
O fato do isolado de T. aff. basicola ter sido patogênico ao coentro e à salsa
(Figuras 6 e 7), e não aos demais hospedeiros, mostrou haver especificidade deste
em nível da Família Apiaceae. Isto sinaliza a importância de se entender melhor os
mecanismos de patogenicidade, tendo em vista enorme gama de hospedeiros de T.
basicola em diversas partes do mundo. Punja e Sun (2000) verificaram substancial
variação genética de isolados de T. basicola obtidos de diversas culturas,
especulando que esta variabilidade pode ser em função da adaptação aos
hospedeiros específicos.

37
Tabela 1 – Inoculação do isolado de Thielaviopsis aff. basicola em plantas de oito famílias botânicas de importância para o Estado da Bahia.
Família Espécie Método de Inoculação1
Reação à Inoculação2
Annonaceae Annona muricata SI -
Amaranthaceae Beta vulgaris SI -
Apiaceae Coriandrum sativum DM +
Daucus carota SI +
Petroselinum crispum DM +
Convolvulaceae Ipomoea batatas SI -
Euphorbiaceae Hevea brasiliensis DM -
Malvaceae Theobroma cacao SI -
Passifloraceae Passiflora edulis f. flavicarpa
SI -
Solanaceae Capsicum annuum SI -
Lycopersicon esculentum SI -
1SI = suspensão de inoculo e; DM = disco de micélio.
2+ = presença de sintomas/sinais e; - =
ausência de sintomas/sinais.

38
Figura 6. Plantas de Petroselinum crispum (A, B) e Coriandrum sativum (C, D) após 45 dias da inoculação com Thielaviopsis aff. basicola. A e C – inoculadas; B e D – testemunha.

39
Figura 7. Lesões necróticas em caules de Coriandrum sativum e Petroselinum crispum inoculadas com Thielaviopsis aff. basicola (A, C) e estruturas reprodutivas do patógeno (B, D).

40
4 CONCLUSÕES GERAIS
1. A formação de estruturas reprodutivas de Thielaviopsis aff. basicola deu-se
de forma mais rápida no meio Extrato de Malte-Ágar, observando-se
abundância destas 10 dias após a repicagem.
2. Com base nas características morfológicas, identificou-se isolado do fungo
obtido de Daucus carota como T. aff. basicola, uma vez que este apresenta
estruturas semelhantes a T. basicola mas também se reproduz de forma
sexuada, fato este característico do complexo Ceratocystis.
3. As análises moleculares permitiram obter grupos distintos de Ceratocystis
spp. e Thielaviopsis spp., mas revelou que o isolado de T. aff. basicola, objeto
deste estudo, não alinhou com os mesmos, indicando ser este geneticamente
distinto.
4. O isolado foi patogênico apenas a Coriandrum sativum e Petroselinum
crispum, ambas da família Apiaceae, nas quais observaram-se lesões
necróticas escurecidas, bem como estruturas reprodutivas do patógeno. Esse
resultado indica haver alguma especialização do isolado a espécies desta
família.
5. Estudos moleculares complementares são necessários para ampliar a
comparação de sequências gênicas do isolado estudado com outras
sequências de T. basicola depositadas no GenBank, uma vez este estudo
restringiu-se apenas na utilização de CERF 7F.
6. Pode-se inferir que o isolado estudado é um novo táxon, permitindo definir a
classificação de T. basicola.

41
REFERÊNCIAS
ABBAS, S. Q.; NIAZ, M.; GHAFFAR A.; Thielaviopsis basicola: a potential threat to
agriculture and forestry in Pakistan. Pakistan Journal of Botany, v. 39, n. 3, p.
985–990, 2007.
BARRON, G.L. The genera of Hyphomycetes from soil. The Williams and wilkins
Company, Baltimore, p. 366, 1968.
BERKELEY, M. J.; BROOME, C. E. XL. Notices of Bristish fungi. Ann. Mag. Nat.
Hist., Ser. 2, v. 2, p. 323–379, 1850.
CARVALHO, A. O.; CARMO, M. G. F. Podridão pós-colheita de cenoura causada por
Ceratocystis fimbriata. Fitopatologia Brasileira, v. 28, p. 108, 2003.
CARVALHO, R. R. D. E. et al. WARWICK "In vitro activity of essential oils of Lippia
sidoides and Lippia gracilis and their major chemical components against
Thielaviopsis paradoxa, causal agent of stem bleeding in coconut palms."
Quimica Nova, São Paulo, v. 36, n. 2, p. 241–244, fev. 2013.
CASTELLANI, A. The viability of sme athogenc fung in terile distlled water. Journal
of Tropical Medicine Hygiene, v. 42, p. 225–226, 1939.
CEPEA Disponível em: <http://www.cepea.esalq.usp.br/br> Acesso em: 16 Dez.
2016.
DALBOSCO, M.; EL TASSA, S. O. M.; DUARTE, V. Ocorrência de Podridão Negra,
causada por Chalara elegans, em raízes de cenoura no Rio Grande do Sul.
Fitopatologia Brasileira, v. 29, p. 336, 2004.
DE BEER, Z. W. de; DUONG, T. A.; BARNES, I.; WINGFIELD, B. D.; WINGFIELD,
M. J. Redefining Ceratocystis and allied genera. Studies in Mycology, v. 79, p.
187–219, 2014.

42
DE HOOG, G. S.; VAN DEN ENDE, A. H. G. Molecular diagnostics of clinical strains
of filamentous Basidiomycetes. Mycoses, v. 41, p.183–189, 1998.
DE SEYNES, J. Recherches pour servir a I’Histoire naturelle des vegetaux inferieurs.
Des fistulines. I, F. Savy, v. 1, p. 86, 1886.
DHINGRA, O. D.; SINCLAIR, J. B. Basic plant pathology methods. 2th ed. Boca
Raton, CRC/Lewis Publishers, p. 434, 1995.
DUMBRA, J.G. R. Cenoura – Tempo seco favorece produção em boa parte do ano.
Hortifruti Brasil, ano. 13, n. 141, p. 33, 2015.
ELLIS, M. B Dematiaceous hyphomycetes. Kew, England, CAB, p. 608, 1971.
ENGELBRECHT, C. J.; HARRINGTON, T. C. Intersterility, morphology and
taxonomy of Ceratocystis fimbriata on sweet potato, cacao and sycamore.
Micologia, v. 97, p. 57–69, 2005.
FARR D. F.; ROSSMAN A. Y. Fungal Databases, Systematic Mycology and
Microbiology Laboratory, ARS, USDA. 2017. <http://nt.ars-grin.gov/
fungaldatabases/> Acessado em 10/01/2017.
FERRARIS, T. Flora Italica Cryptogama. Pars. I: Fungi yphales,Tuberculariaceae-
Stilbaceae. Fasc. n. 6, p. 979, 1910.
GAYED, S. K. Host range and persistence of Thielaviopsis basicola in tobacco soils.
Canadian Journal of Plant Science, v. 52, p. 869–873, 1972.
GENBANK. Disponível em: <http://www.ncbi.nlm.nih.gov/genbank/> Acesso em: 16
Jan. 2017.
HARRINGTON, T. C.; MCNEW, D. L. Self-fertility and uni-directional mating-type
switching in Ceratocystis coerulescens, a filamentous ascomycete. Curr.
Genet., v. 32, p. 52–59, 1997.

43
HARRINGTON, T. C.; THORPE, D. J.; ALFENAS, A. C. Genetic variation and
variation in aggressiveness to native and exotic hosts among Brazilian
populations of Ceratocystis fimbriata. Phytopathology, v. 101, n. 5, p. 555–
566, 2011.
HENNEBERT, G.L. Echinobotryum, Wardomyces and Mamaria. Tans. Brit. Mycol.
Soc., v. 51, p. 749–762, 1968.
HENNEBERT, G.L.; SUTTON. B. C. Unitary parameters in conidiogeneses. In:
Ascomycetes Systematic: Problems and Perspectives in the nineties. Plenum,
p. 65–76. 1994.
HOHNEL, F. V. Zur Kenntnis einiger Fadenpilze. Hedwigia, v. 43, 295–299, 1904.
HUGHES, S. J. Conidiophores, Conidia and classification. Canadian Journal
Botany, v. 31, n. 5, 577–659, 1953.
INÁCIO, C. A.; LEMES, G. P.; CAFÉ-FILHO, A. C.; LOPES, C. A. First report of
Thielaviopsis basicola causing black rot on carrot in Central Brazil. Tropical
Plant Pathology, v. 37 (Supl.), p. 752, 2012.
INDEX FUNGORUM. Disponível em: <http://www.indexfungorum.org/
Names/Names.asp> Acesso em: 16 Jan. 2017.
KIFFER, E.; DELON, R. Chalara elegans (Thielaviopsis basicola) and allied species.
II. Validation of two taxa. Mycotaxon, v. 18, n. 1, p. 165–174, 1983.
KOIKE, S. T.; HENDERSON, D. M. Black root rot, caused by Thielaviopsis basicola,
on tomato transplants in California. Plant Disease, v. 82, p. 447, 1998.
LAIA, M. L.; ALFENAS, A. C.; HARRINGTON, T. C. Isolation, detection in soil, and
inoculation of Ceratocystis fimbriata, causal agent of wilting, die-back and
canker in Eucalyptus. Fitopatologia Brasileira, v. 25 (Supl), p. 384, 2000.

44
LIU, Y. J; WHELEN, S.; HALL, B. D. Phylogenetic relationships among ascomycetes:
evidence from an RNA polymerse II subunit. Molecular Biology and
Evolution, v. 16, p. 1799–1808, 1999.
LOPES, C. A.; REIS, A. Doenças da cenoura. Brasília, Embrapa Hortaliças, p. 67,
2016.
MBENOUN, M.; DE BEER, Z. W.; WINGFIELD, M. J. et al. Reconsidering species
boundaries in the Ceratocystis paradoxa complex, including a new species from
oil palm and cacao in Cameroon. Mycologia,v. 106, n. 4, 757–784, 2014.
MCCORMICK, F.A. Perithecia of Thielavia basicola. Zopf. in culture. Bull. Conn.
Agric. Exp. St., p. 269, 1925.
MILOSAVLJEVIC, A.; TRKULJA, N.; POPOVIĆ, T. et al. First report of Thielaviopsis
thielavioides, a causal agent of postharvest blackening on Daucus carota in
Serbia. Plant Disease, v. 99, p. 1274 , 2015.
MONDAL, A. H.; NEHL, D. B.; ALLEN, S. J. First report Thielaviopsis basicola on
soybean in Australia. Australasian Plant Pathology, Queensland, v. 33, n. 3,
p. 451–452, 2004.
NAG RAJ, T.R.; KENDRICK, W. D. A monograph of Chalara and allied genera.
Deptt. Bio. Univ, p. 200, 1975.
PAOLETTI, M.; SEYMOUR, F. A.; ALCOCER, M. J. C.; KAUR, N.; CALVO, A. M.;
ARCHER, D. B.; DYER, P. S. Mating Type and the Genetic Basis of Self-
Fertility in the Model Fungus Aspergillus nidulans. Current Biology, v. 17, p.
1384–1389, 2007.
PAULIN, A. E; HARRINGTON, T. C. Phylogenetic placement of anamorphic species
of Chalara among Ceratocystis and other ascomycetes. Studies in Mycology,
v. 45, p. 169–186, 2000.

45
PAULIN-MAHADY, A. E.; HARRINGTON, T. C.; MCNEW, D. Phylogenetic and
taxonomic evaluation of Chalara, Chalaropsis, and Thielaviopsis anamorphs
associated with Ceratocystis. Mycologia, v. 94, p. 62–72, 2002.
PEREG, L. L. Black root rot of cotton in Australia: the host, the pathogen and disease
management. Crop & Pasture Science, v. 64, p. 1112–1126, 2013.
PUNJA, Z. K; GAYE, M. M. Influence of postharvest handling practices and dip
treatment on development of Black Root Rot on fresh market carrots. Plant
Disease, v. 77, p. 989–995, 1993.
PUNJA, Z. K; SUN, L. J. Morphological and molecular characterization of Chalara
elegans (Thielaviopsis basicola), cause of black root rot on diverse plant
species. Can. J. Bot., v. 77, n. 12, p. 1801–1812, 2000.
RODRIGUES, G. de S. Antagonismo de Trichoderma spp. e leveduras à
Ceratocystis cacaofunesta. 2016. 84 p. Dissertação de Mestrado. Universidade
Estadual de Santa Cruz/PPG em Produção Vegetal, Ilhéus, 2016.
SACCARDO, P. A. Syll. Fung., v. 10, p. 1–964, 1892.
SALA, F. C.; COSTA, C. P.; TEIXEIRA L. D.; FABRI, E. G.; BLAT, S. F. Reação de
cultivares de alface a Thielaviopsis basicola. Horticultura Brasileira, v. 26,
p. 398–400, 2008.
SALA, F. C; COSTA, C. P; TEIXEIRA-YAÑEZLDD; BLAT, S. F. Reação de chicória
a murchadeira (Thielaviopis basicola). Summa Phytopathologica, v. 30, n. 1,
2004.
SILVA, A. M. S.; AKIBA, F.; CARVALHO, A. O.; RIBEIRO, R. L. D.; ARAÚJO, J. S. P.
Podridão negra de raízes de quiabeiro e alface incitada por Thielaviopsis
sp. Fitopatologia Brasileira, Brasília, v. 24, n. 4, p. 328, 1999.

46
SILVA, G. S; COSTA, D. L. M. M. Ocorrência da podridão negra da cenoura no
Estado do Maranhão. Summa Phytopathologica, v. 39, p. 294, 2013.
SILVA, D. J. G.; PEREIRA, J. Ocorrência de Thielaviopsis basicola em Daucus
carota no Estado da Bahia. In: VII Congresso Brasileiro de Micologia, 2016,
Florianópolis-SC. Resumo, Florianópolis: UFSC, 2016.
TEIXEIRA-YAÑEZ, L. D. D. Resistência genética, fungicidas e solarização para o
controle de Thielaviopsis basicola (Berk. & Broome) Ferraris na cultura de
alface (Lactuca sativa L.). 2005. 103 p. Tese de Doutorado. Escola Superior de
Agricultura Luiz de Queiroz, Piracicaba, 2005.
VAN WYK, M. V.; WINGFIELD, B. D.; CLEGG, P. A.; WINGFIELD, M.
J. Ceratocystis larium sp. nov., a new species from Styrax benzoin wounds
associated with incense harvesting in Indonesia. Persoonia, v. 22, p. 75–82,
2009.
WENT, F.A. Die Ananasziekte van het suikerriet. Mededeelingen van het
Proefstation "West-Java" Kagok-Tegal, p. 8, 1893.
WHITE, T. J.; BRUNS, T.; LEE, S. et al. Amplification and direct sequencing of fungal
ribosomal RNA genes for phylogenetics. In: PCR protocols: a guide to methods
and applications (Innis, M.A.; Gelfand, D.H.; Shinsky, T.J.; White, T.J., eds).
Academic Press Inc, New York, pp. 315–322, 1990.
WILSON WIJERATNAM, R. S.; HAWEJULIGE, I. G. N.; ABEYRETNE N.
Postharvest hot water treatment for the control of Thielaviopsis black rot of
pineapple. Postharvest Biology and Technology, v. 36, p. 323–327, 2005.
YARWOOD, C. E. The Occurrence of Chalara elegans. Mycologia, v. 73, p. 524–
530, 1981.


















