
UNIVERSIDADE ESTADUAL DE SANTA CRUZ PROGRAMA DE...
62
UNIVERSIDADE ESTADUAL DE SANTA CRUZ PROGRAMA DE PÓS GRADUAÇÃO EM PRODUÇÃO VEGETAL ANDRESA MUNIZ PEDROSA ANÁLISE FUNCIONAL DOS GENES CsLEA2 E CsNCED3 PARA TOLERÂNCIA À SECA EM CITROS ILHÉUS - BAHIA 2013
Transcript of UNIVERSIDADE ESTADUAL DE SANTA CRUZ PROGRAMA DE...

UNIVERSIDADE ESTADUAL DE SANTA CRUZ
PROGRAMA DE PÓS GRADUAÇÃO EM PRODUÇÃO VEGETAL
ANDRESA MUNIZ PEDROSA
ANÁLISE FUNCIONAL DOS GENES CsLEA2 E CsNCED3 PARA TOLERÂNCIA À
SECA EM CITROS
ILHÉUS - BAHIA
2013

ii
ANDRESA MUNIZ PEDROSA
ANÁLISE FUNCIONAL DOS GENES CsLEA2 E CsNCED3 PARA TOLERÂNCIA À
SECA EM CITROS
Dissertação apresentada para obtenção do
título de mestre em Produção Vegetal à
Universidade Estadual de Santa Cruz.
Área de Concentração: Melhoramento de
Plantas e Biotecnologia.
Orientador: Prof. Márcio Gilberto C. Costa
ILHÉUS - BAHIA
2013

iii
ANDRESA MUNIZ PEDROSA
ANÁLISE FUNCIONAL DOS GENES CsLEA2 E CsNCED3 PARA TOLERÂNCIA À
SECA EM CITROS
Ilhéus, Bahia, 22/02/2013
______________________________
Dr. Márcio Gilberto Cardoso Costa
UESC – Orientador
______________________________
Dr. Carlos Priminho Pirovani
UESC
______________________________
Dr. Abelmon da Silva Gesteira
EMBRAPA CNPMF

iv
DEDICO
Á Deus,
Porque Dele, por Ele e para Ele são todas as coisas.

v
AGRADECIMENTOS
Ao meu bom Deus, por ter me ajudado chegar até aqui. O que direi ao Senhor por
todos os benefícios que me tens feito?
A toda minha família e amigos, pelo apoio e por sempre estar ao meu lado, em
especial ao meu esposo Elisvan, pelas madrugadas que passou comigo na UESC,
pelo amor, carinho e incentivo.
A Luciana Cidade que esteve sempre me ensinando e disposta a me ajudar,
obrigada Lu, sem você seria muito difícil. Agradeço a Cristina, que também foi
essencial nesse trabalho, até na madrugada me acompanhou.
Ao professor Marcio Costa, pela orientação e apoio.
Ao professor Fábio Gomes, pela co-orientação.
Aos colegas do Laboratório de Cultura de Tecido do CBG, pelos momentos de
descontração em meio ao trabalho, pela amizade e auxílio.
Aos professores, colegas e funcionários do Programa de Pós Graduação em
Produção Vegetal, pelos ensinamentos e convivência durante esse período.

vi
SUMÁRIO
RESUMO........................................................................................................ viii
ABSTRACT...................................................................................................... x
LISTA DE FIGURAS ...................................................................................... xi
1 INTRODUÇÃO .............................................................................................. 1
2 OBJETIVOS .................................................................................................. 4
2.1 Geral ............................................................................................................. 4
2.2 Específicos ................................................................................................... 4
3 REVISÃO DE LITERATURA ......................................................................... 5
3.1 Citricultura...................................................................................................... 5
3.2 Respostas das plantas a deficiência hídrica ............................................. 6
3.3 LEA (Late Embryogenesis Abundant)......................................................... 8
3.4 NCED (9-cis-epoxicarotenóide dioxigenase) ............................................. 9
4 MATERIAL E MÉTODOS.............................................................................. 13
4.1 Material vegetal .......................................................................................... 13
4.2 Cultura e manutenção dos isolados de Agrobacterium tumefaciens .... 13
4.3 Alinhamento de múltiplas sequências e análise filogenética ................. 14
4.4 Clonagem molecular dos genes e introdução em vetor de transformação
de plantas ............................................................................................................... 14
4.5 Transformação genética de tabaco, regeneração e aclimatação ........... 17
4.6 Caracterização das plantas transgênicas ................................................ 18
4.6.1 Extração de DNA genômico e análise da inserção dos genes pela técnica de
PCR ......................................................................................................................... 18
4.6.2 Experimento de deficiência hídrica ............................................................... 18

vii
4.6.3 Trocas gasosas ............................................................................................ 19
4.6.4 Potencial hídrico foliar .................................................................................. 19
4.6.5 Potencial mátrico (ψm) do solo ..................................................................... 19
4.6.6 Acúmulo de peróxido de hidrogênio (H2O2) .................................................. 19
4.6.7 Delineamento experimental e análise estatística ......................................... 20
5 RESULTADOS ............................................................................................. 21
5.1 Análise filogenética .................................................................................... 21
5.2 Clonagem molecular das sequências CsLEA2 e CsNCED3 e
transformação de Nicotiana tabacum .................................................................. 22
5.3 Análises fisiológicas ................................................................................... 26
5.3.1 Potencial hídrico foliar .................................................................................. 26
5.3.2 Trocas gasosas foliares ................................................................................. 28
5.4 Acúmulo de peróxido de hidrogênio ......................................................... 33
6 DISCUSSÃO .................................................................................................. 35
7 CONCLUSÕES GERAIS ............................................................................... 40
REFERÊNCIAS ............................................................................................ 41

viii
RESUMO
PEDROSA, Andresa Muniz, M.S., Universidade Estadual de Santa Cruz, Ilhéus, Fevereiro de 2013. Análise funcional dos genes CsLEA2 e CsNCED3 para tolerância a seca em citros. Orientador: Prof. Dr. Márcio Gilberto Cardoso Costa. Co-orientador: Prof. Dr. Fábio Pinto Gomes
O Brasil é um dos maiores produtores de citros do mundo, sendo responsável por
60% da produção mundial de suco de laranja e líder nas exportações deste produto.
Apesar dos citros encontrarem excelentes condições ambientais para seu
crescimento, em algumas regiões produtoras há uma intensificação do período de
estiagem, a exemplo daquela estabelecida na região dos tabuleiros costeiros, no
Nordeste brasileiro. Além disso, a cultura no Brasil é conduzida praticamente sem
irrigação. A citricultura brasileira mostra-se vulnerável devido, principalmente, à
utilização de poucas combinações copa/porta-enxerto, o que vem ocasionando
surtos de pragas e doenças ao longo dos anos. O limoeiro ‘Cravo’ (Citrus limonia
Osbeck) é o principal porta-enxerto da citricultura brasileira, mas devido a sua
vulnerabilidade à Morte Súbita dos Citros (MSC) torna necessário a diversificação
dos portas-enxertos, focada na necessidade de convívio com deficiências hídricas.
O presente estudo teve por objetivo validar a função de genes das famílias LEA
(Late Embryogenesis Abundant) e NCED (9-cis-epoxicarotenóide dioxigenase) de
citros em mecanismos de resposta e tolerância à seca, por meio da superexpressão
em plantas transgênicas de tabaco, visando à futura aplicação em trabalhos de
transformação genética de variedades porta-enxerto de citros. Especificamente, os
genes codificando para uma LEA do grupo 2 (CsLEA2) e uma NCED do grupo 3
(CsNCED3), expressos em bibliotecas de cDNAs de raízes e folhas de citros
submetidos a estresse por deficiência hídrica, foram selecionados para
caracterização. A análise filogenética da sequência predita de aminoácidos
codificada por CsLEA2 confirmou que o mesmo pertence ao grupo 2, das dehidrinas,
enquanto que CsNCED3 é ortologo do gene AtNCED3, envolvido em respostas de
tolerância a seca em Arabidopsis. Linhagens transgênicas de tabaco contendo os
genes CsLEA2 e CsNCED3 foram obtidas e submetidas a 10 dias de estresse
hídrico, pela suspensão de irrigação, e após esse período foram re-irrigadas. O
potencial hídrico foliar reduziu drasticamente em todas as plantas, transgênicas e
plantas controle não-transformadas (NT), ao longo do período de deficiência hídrica,
sendo recuperado após a re-irrigação. As linhagens CsLEA2 e CsNCED3
apresentaram mecanismos de defesa para evitar a perda de água, como redução na
condutância estomática e menor acúmulo de ROS em relação as plantas NT.
Embora as plantas transgênicas apresentaram menor condutância estomática do
que as plantas NT sob condições de estresse, a fotossíntese foi mantida em níveis
similares ao das plantas NT. Após a re-irrigação das plantas, foi observada a
recuperação do potencial hídrico, da condutância estomática e da taxa fotossintética
em todas as plantas analisadas. Conclui-se que os genes CsLEA2 e CsNCED3

ix
contribuem para a estratégia fisiológica de tolerância à desidratação das plantas, por
meio da ativação do mecanismo de fechamento estomático.
Palavras chaves: Deficiência hídrica. Engenharia genética. Agrobacterium
tumefaciens

x
ABSTRACT
PEDROSA, Andresa Muniz, M.S., Universidade Estadual de Santa Cruz, Ilhéus, February 2013. Functional analysis of genes CsLEA2 and CsNCED3 for drought tolerance in citrus. Advisor: Dr. Márcio Gilberto Cardoso Costa. Committee Member: Dr. Fábio Pinto Gomes
Brazil is the major producer of citrus in the world, accounting for 60% of world production of orange juice, and a leader in exports of this product. Although citrus plants find excellent environmental conditions for their growth in Brazil, in some producing regions there is an intensification of the dry season, such as in the region of the coastal plains, in northeastern Brazil. Besides, the citriculture is conducted practically without irrigation in Brazil. Brazilian citriculture is vulnerable, mainly due to the use of fewer rootstock cultivars, which has caused outbreaks by pests and diseases over the years. The rangpur lime (Citrus limonia Osbeck) is the main rootstock used in Brazilian citriculture, but its vulnerability to Citrus Sudden Death (SCD) makes necessary the diversification of rootstocks, focused on the need for interaction with water deficits. This study aimed to validate the function of citrus genes from LEA (Late Embryogenesis Abundant) and NCED (9-cis-epoxicarotenóide dioxygenase) families in mechanisms of response and tolerance to drought, through their overexpression in transgenic tobacco, aiming their future use in genetic transformation of citrus rootstocks. Specifically, genes encoding a Group 2 LEA (CsLEA2) and a Group 3 NCED (CsNCED3), expressed in cDNAs libraries of citrus leaves and roots subjected to water deficit, were selected for characterization. Phylogenetic analysis of the predicted amino acid sequence encoded by CsLEA2 confirmed that it belongs to Group 2 LEA, the group of dehydrins, while CsNCED3 is an ortholog of AtNCED3, involved in mechanismos of response and tolerance to drought in Arabidopsis. Transgenic lines containing the genes CsLEA2 and CsNCED3 were obtained and subjected to 10 days of water stress, by the suspension of irrigation, and after this period they were re-irrigated. The leaf water potential decreased drastically in both transgenic and wild-type (WT) plants over the period of water stress, being recovered after re-watering. The transgenic lines of CsLEA2 and CsNCED3 showed defense mechanisms to prevent water loss, such as reduction in stomatal conductance and low accumulation of ROS as compared to WT. Although the transgenic plants had lower stomatal conductance than the WT plants under stress conditions, the photosynthesis was maintained at similar levels to that of WT plants. After re-irrigation of plants, it was observed recovery of water potential, stomatal conductance and photosynthetic rate in all plants analyzed. It is concluded that the genes CsLEA2 and CsNCED3 contribute to the physiological strategy of drought avoidance , by activating the mechanism of stomatal closure.
Key words: Water deficiency. Genetic Engineering. Agrobacterium tumefaciens

xi
LISTA DE FIGURAS
Figura 1: Via indireta da biossíntese do ácido abscísico (ABA) em plantas. (a): síntese dos carotenóides não-oxigenados nos plastídeos; (b): síntese e clivagem dos carotenóides oxigenados nos plastídeos, constituindo estoque de precursores do ABA; (c): síntese do ABA no citossol. Fonte: Seo e Koshiba (2002) ....................... 11
Figura 2: Esquema da região do T-DNA do vetor binário pCAMBIA1390 modificado (pUESC28), indicando os genes presentes no vetor e o múltiplo sítio de clonagem sob a orientação do promotor CaMV35S ................................................................. 17
Figura 3: Análise filogenética das proteínas da família LEA de Arabidopsis (At) e CsLEA2 (orange1.1g026736m.g) de citros. As seqüências de aminoácidos foram alinhadas utilizando ClustalW e o método Neighbor-Joining. A árvore foi construída com um suporte de bootstrap de 1000 replicações .................................................. 21
Figura 4: Diagnóstico de orientação do gene CsLEA2 inserido do vetor pCambia 1390 modificado para expressão em plantas (oligonucleotídeos LEA 8111-F e NOS-R); M: Marcador molecular 1Kb Plus DNA Ladder; C-: controle negativo; 1-3: colônias 5,6,7 de E. coli transformadas ................................................................... 22
Figura 5: Diagnóstico de orientação do gene CsNCED3. A: Esquema da região do T-DNA do vetor binário pCAMBIA1390 modificado para expressão em plantas, contendo o gene CsNCED3 sob a orientação do promotor CaMV35S, indicando o corte da enzima de restrição XhoI; B: Digestão enzimática do DNA plasmidial. M: Marcador Molecular GeneRulerTM 1kb DNA Ladder Fermentas; 1-3: colônias 1,2,3 de E. coli transformadas ................................................................................................ 23
Figura 6: PCR de colônias de A. tumefaciens transformadas com o produto da ligação do gene CsLEA2 em pCambia 1390 modificado. (A) PCR utilizando os oligonucleotídeos LEA 8111-F E LEA 8111-R; (B) PCR utilizando os oligonucleotídeos do gene hptII-F e hptII-R; M: Marcador Molecular Invitrogen 1kb; C-: controle negativo; 1- 4: amostras das colônias de A.tumefaciens. ..................... 24
Figura 7: PCR de colônias de A. tumefaciens transformadas com o produto da ligação do gene CsNCED3 em pCambia 1390 modificado, utilizando oligonucleotídeos do gene hptII-F e hptII-R. C-: controle negativo; 1- 4: amostras das colônias de A.tumefaciens; M: Marcador Molecular Invitrogen 1kb;. ....................... 24
Figura 8: Etapas na obtenção das plantas transgênicas: explantes foliares transformados com A. tumefaciens (A); obtenção (B), individualização e enraizamento dos brotos (C) e aclimatação das plantas (D). .................................. 25
Figura 9: Amplificação do fragmento do gene hptII em plantas transgênicas de Nicotiana tabacum por PCR. M: Marcador Molecular Invitrogen de 1 kb ; C-: controle negativo ; C+: controle positivo - DNA plasmidial diluído (1:100) de pCAMBIA 1390; (A) 1 a 17: linhagens transgênicas independentes 35S::CsNCED3; (B) 1 a 9 linhagens transgênicas independentes 35S::CsLEA2 ............................................. 26
Figura 10: Potencial hídrico foliar (Ψw) medidos aos dois (controle) e 10 (10DS) dias após a suspensão da irrigação e aos seis dias após a re-irrigação (6DR); A: linhagens CsNCED3; B: linhagens CsLEA2. As colunas são valores médios e as

xii
barras representam o erro padrão da média. ** Diferenças significativas pelo teste t (p< 0,01). *** Diferenças significativas pelo teste t (p< 0,001) ................................. 27
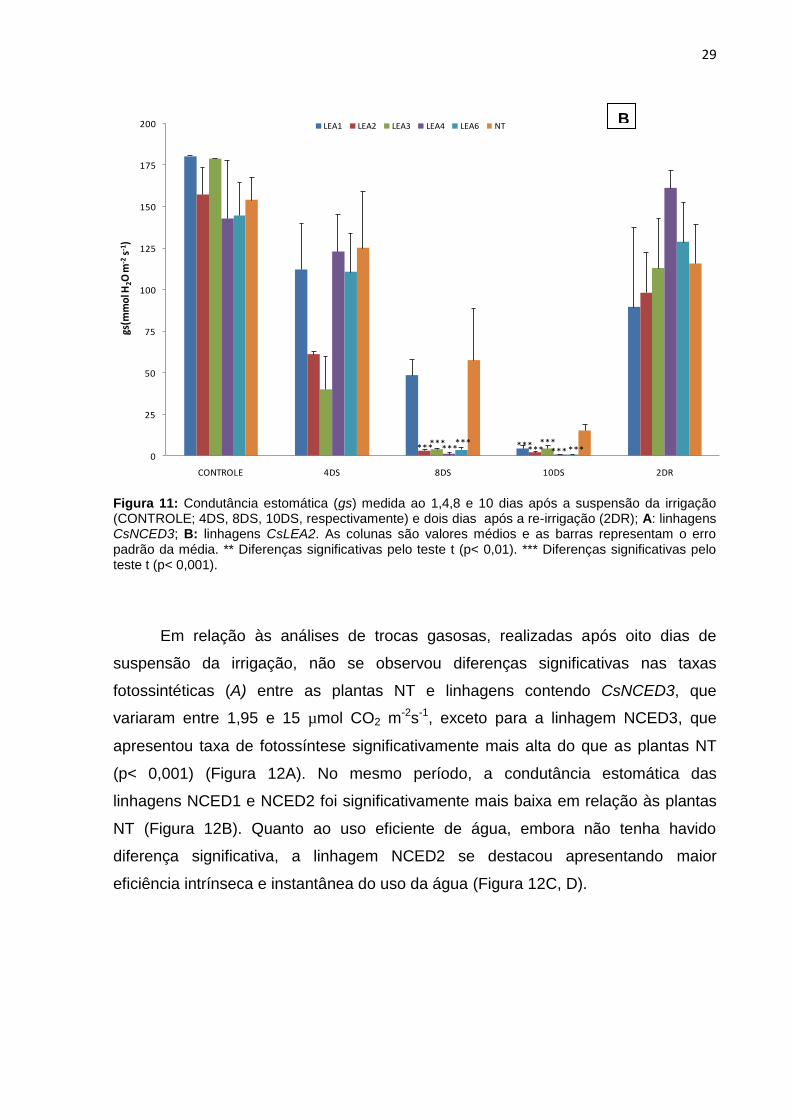
Figura 11: Condutância estomática (gs) medida ao 1,4,8 e 10 dias após a suspensão da irrigação (CONTROLE; 4DS, 8DS, 10DS, respectivamente) e dois dias após a re-irrigação (2DR); A: linhagens CsNCED3; B: linhagens CsLEA2. As colunas são valores médios e as barras representam o erro padrão da média. ** Diferenças significativas pelo teste t (p< 0,01). *** Diferenças significativas pelo teste t (p< 0,001) ............................................................................................................... 29
Figura 12: Trocas gasosas das linhagens CsNCED3 sob oito dias de deficiência hídrica; A - Taxa fotossintética (A) ; B - Condutância estomática (gs); C – Eficiência intrínseca do uso da água (A/gs); D - Eficiência instantanea do uso da água (A/E).......................................................................................................................... 30
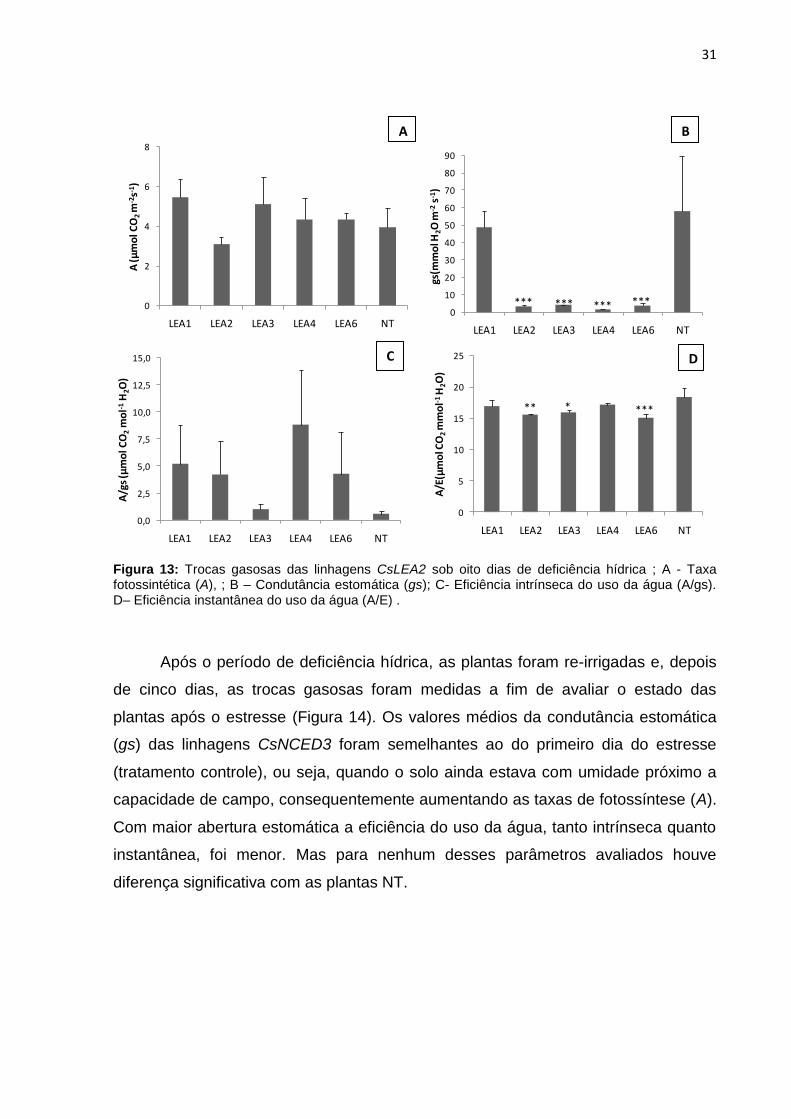
Figura 13: Trocas gasosas das linhagens CsLEA2 sob oito dias de deficiência hídrica ; A - Taxa fotossintética (A), ; B – Condutância estomática (gs); C- Eficiência intrínseca do uso da água (A/gs). D– Eficiência instantanea do uso da água (A/E).......................................................................................................................... 31
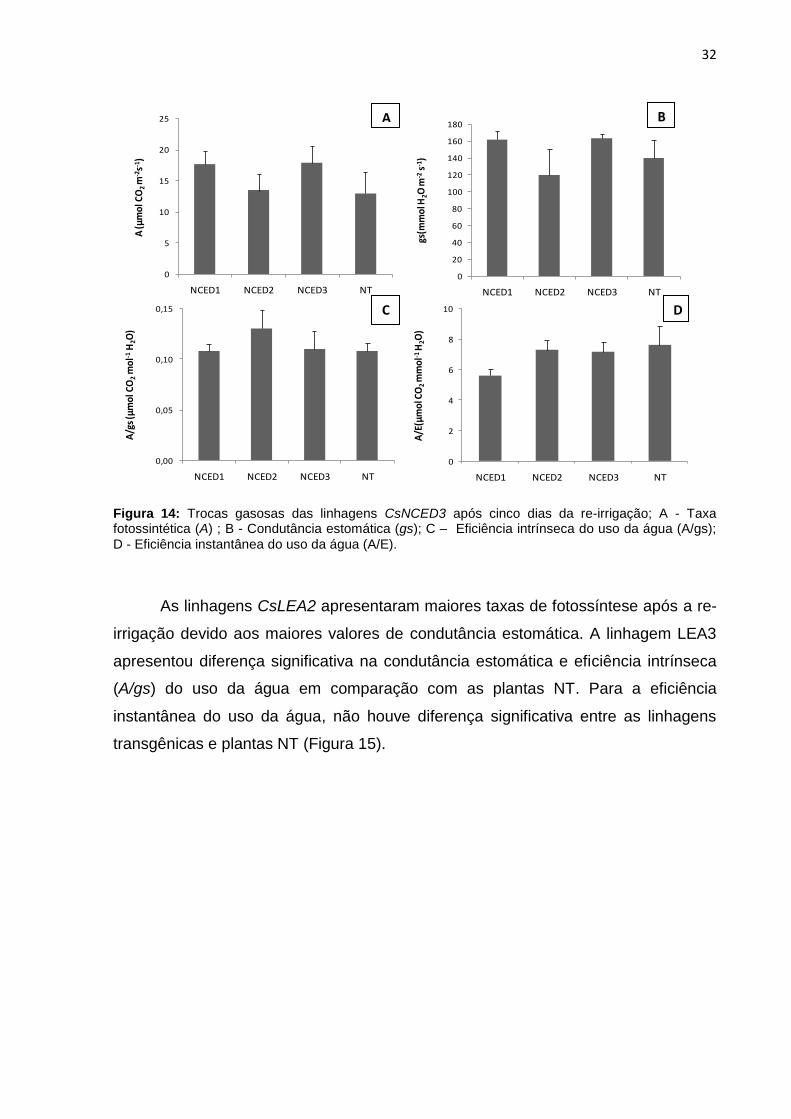
Figura 14: Trocas gasosas das linhagens CsNCED3 após cinco dias da re-irrigação; A - Taxa fotossintética (A) ; B - Condutância estomática (gs); C – Eficiência intrínseca do uso da água (A/gs); D - Eficiência instantanea do uso da água (A/E).......................................................................................................................... 32
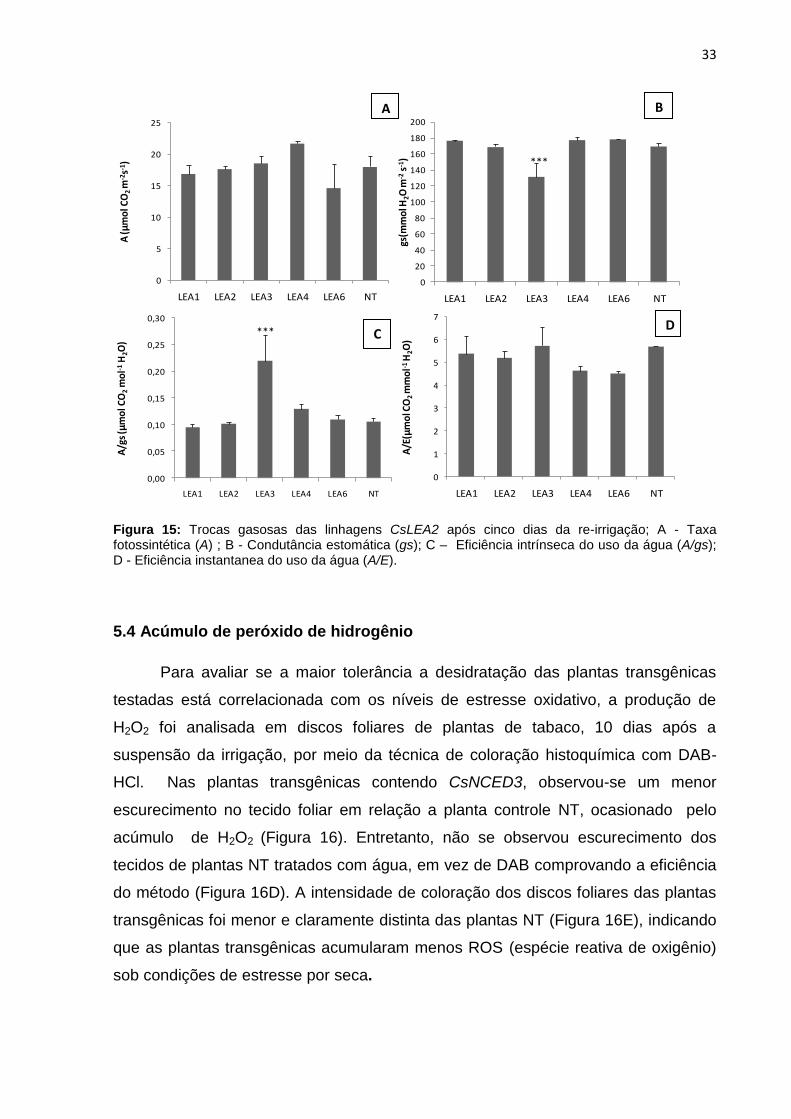
Figura 15: Trocas gasosas das linhagens CsLEA2 após cinco dias da re-irrigação; A - Taxa fotossintética (A) ; B - Condutância estomática (gs); C – Eficiência intrínseca do uso da água (A/gs); D - Eficiência instantanea do uso da água (A/E).......................................................................................................................... 33
Figura 16: Produção de peróxido de hidrogênio em discos foliares de plantas de N. tabacum submetidas à deficiência hídrica, visualizada por meio da coloração com HCL-DAB (1 mg mL-1). A- C: linhagens transgênicas NCED1, NCED2, NCED3 respectivamente submetidas à desidratação; D: planta não transformada fresca infiltrada com água; E: planta não-transformada submetida à desidratação.............................................................................................................. 34
Figura 17: Produção de peróxido de hidrogênio em discos foliares de plantas de N. tabacum submetidas à deficiência hídrica, visualizadapor meio da coloração com HCL-DAB (1 mg mL-1). A- E: linhagens transgênicas LEA1, LEA2, LEA3, LEA4, e LEA6 respectivamente, submetidas à desidratação.; E: planta não transformada infiltrada com água; F: planta não-transformada submetida à desidratação.............................................................................................................. 34

1
1 INTRODUÇÃO
A citricultura é destaque na agroindústria brasileira, sendo responsável por
60% da produção mundial de suco de laranja, com o Brasil liderando as exportações
do produto (MAPA, 2012). Segundo o IBGE, em 2011 a safra da laranja foi de 19,8
milhões de toneladas e a área colhida foi de 777 mil hectares. O cultivo dos citros
está presente em todos estados brasileiros, sendo São Paulo o estado que detém
aproximadamente 77% da produção, e o nordeste é a segunda região produtora
principal, com destaque para a Bahia com 5,1% e Sergipe com 4,1% da produção
nacional (IBGE, 2012).
Apesar dos citros encontrarem excelentes condições ambientais para seu
crescimento e desenvolvimento no estado de São Paulo, há uma intensificação do
período de estiagem do sul para o norte do estado durante os meses de inverno
(SENTELHAS, 2005), o que afeta negativamente a produção. Por outro lado, a
produção citrícola do nordeste brasileiro está estabelecida na região dos tabuleiros
costeiros e os problemas de deficiência hídrica e consequente estresse nas plantas
surgem em função da interação dos horizontes coesos dos solos com o clima
quente, principalmente nos meses de verão (NEVES et al., 2010; SOUZA et al.,
2004). Segundo Donato et al. (2007), o requerimento hídrico para uma boa produção
de frutos em citros é cerca de 900 a 1.200 mm por ano e varia com a demanda
evapotranspirométrica, solo, copa e principalmente com o porta-enxerto utilizado.
Em citros longos períodos de deficiência hídrica reduzem a qualidade e a
produtividade dos frutos.
O limoeiro ‘Cravo’ (Citrus limonia Osbeck) é o principal porta-enxerto da
citricultura brasileira, apresentando características horticulturais favoráveis como
compatibilidade com grande parte das copas cultivadas e alta tolerância à
deficiência hídrica. Entretanto, o uso generalizado de um mesmo porta-enxerto para
todas as cultivares copas, provavelmente não atende às características peculiares
de cada variedade. Além disso, a Morte Súbita dos Citros (MSC), afeta
principalmente plantas enxertadas sobre limoeiro ‘Cravo’ (CANTAGALLO, 2005;
MAGALHÃES FILHO et al., 2008).

2
Os motivos pelos quais o limoeiro ‘Cravo’ se adapta melhor à deficiência
hídrica podem estar relacionados com o crescimento do seu sistema radicular em
condições de estresse hídrico. Magalhães Filho et al. (2008) avaliaram mudas de
laranjeira ‘Valência’ enxertadas sobre limoeiro ‘Cravo’ e também em Poncirus
trifoliata (‘Trifoliata’) e verificaram que a combinação ‘Valência’/ ‘Cravo’ apresentou
sistema radicular mais desenvolvido em comparação com ‘Valência/‘Trifoliata’, tanto
em condições controle (irrigação) como de deficiência hídrica. Esse tipo de resposta
do ‘Cravo’ indica a existência de um mecanismo de adaptação à seca, priorizando o
crescimento das raízes sob deficiência hídrica (MAGALHÃES FILHO et al., 2008).
Dados do genoma funcional de citros obtidos por meio do sequenciamento de
aproximadamente 500.000 ESTs (expressed sequence tags) de Citrus spp. e
Poncirus, foram disponibilizados para acesso pela comunidade científica
(http://harvest.ucr.edu) como parte de um programa (Citrus Genomics Initiative)
objetivando o rápido avanço do conhecimento científico relacionado aos problemas
que limitam a produção citrícola. Por meio de uma busca preliminar neste banco de
dados, foi identificada a existência de diversos genes de resposta e tolerância à
seca, homólogos a Arabidopsis, incluindo aqueles codificando proteínas LEA (Late
Embryogenesis Abundant) e NCED (9-cis-epoxicarotenóide dioxigenase).
As LEAs compreendem um importante grupo de proteínas que normalmente
se acumulam durante os estágios finais da embriogênese, ou em resposta à
desidratação, baixa temperatura, salinidade ou tratamento exógeno de ABA,
indicando a sua capacidade de resposta à desidratação celular (RAMANJULU;
BARTELS, 2002). As proteínas LEA possuem interessantes propriedades funcionais
relacionadas à sua suposta função de estabilizadores celulares sob condições de
estresse. Estudos demonstram que a super expressão de genes da classe LEA
resulta em maior tolerância à desidratação, embora o mecanismo exato ainda seja
desconhecido (HUNDERTMARK; HINCHA, 2008; UMEZAWA et al., 2006).
Evidências genéticas e bioquímicas demonstram que NCED é uma enzima
chave na biossíntese do ácido abscísico (ABA), já que a clivagem oxidativa dos
precursores xantofílicos é fator limitante na rota biossintética deste hormônio (WAN;
LI, 2006). Segundo Guerrini et al. (2005), nos resultados de pesquisa obtidos para
biossíntese de ABA, observa-se que a identificação e isolamento de genes

3
envolvidos na sua produção é extremamente importante, especialmente em relação
ao gene que codifica a enzima NCED, cuja eficiência pode determinar um maior ou
menor grau de resistência de plantas a estresses ambientais. Iuchi et al. (2001)
demonstraram em estudos que a superexpressão de um gene codificando uma
NCED de Arabidopsis, AtNCED3, aumentou a tolerância ao estresse seca em
plantas transgênicas de Arabidopsis.
No presente estudo, os genes CsLEA2 e CsNCED3 foram caracterizados por
meio de análises molecular e funcional em plantas de tabaco. O estudo colaborou
para a ampliação da base de conhecimento potencialmente aplicável ao
melhoramento.

4
2 OBJETIVOS
2.1 Geral
Identificar nas bases de informação genômica de citros genes codificando
proteínas LEA e NCED, candidatos de tolerância à seca, e validar a função dos
mesmos, por meio do desenvolvimento e caracterização de plantas geneticamente
modificadas de Nicotiana tabacum.
2.2 Específicos
Caracterizar a sequência completa de cDNA e aminoácidos codificada pelos
genes de interesse ;
Engenheirar os genes candidatos de tolerância à seca em vetores de
transformação de plantas;
Transformar plantas de tabaco (var. Havana) com os genes candidatos de
tolerância à seca;
Caracterizar as plantas transformadas quanto à integração e expressão dos
genes candidatos e resposta frente a condições simuladas de deficiência
hídrica.

5
3 REVISÃO DE LITERATURA
3.1 Citricultura
As plantas cítricas fazem parte do gênero Citrus, pertencem à família
Rutaceae e são nativas do sudeste do continente asiático. Apesar de seu centro de
origem, as plantas cítricas encontraram no Brasil melhores condições climáticas para
vegetar e produzir, de modo que se expandiram por todo o país, tornando-o um dos
maiores produtores mundiais (DONADIO et al., 2005). Os cítricos utilizados em
plantios comerciais como variedades copas estão, basicamente, distribuídos em seis
grupos: laranjas, tangerinas, limões, limas ácidas, pomelos e outros de menor
importância. O grupo das laranjas doces apresenta maior expressividade, seguido
por tangerinas, limões e limas ácidas (PIO et al., 2005).
A muda cítrica é formada pelo porta-enxerto e copa, sendo o elemento mais
importante na formação do pomar. O uso de um porta-enxerto adequado pode
propiciar frutos de melhor qualidade além de induzir diversas alterações na
variedade copa. Os portas-enxerto influenciam várias características horticulturais e
fitopatólogicas, podendo refletir na aptidão do pomar em relação ao destino da
produção, em função da qualidade da mesma (SCHAFER et al., 2000).
Apesar do impacto na economia brasileira, a citricultura mostra-se vulnerável
devido, principalmente, à utilização de poucas combinações copa/porta-enxerto, o
que vem ocasionando surtos de pragas e doenças ao longo dos anos (SCHINOR et
al., 2006). Diante dessa situação é importante a ampliação da base genética de
variedades de copas e porta-enxertos, e já tem sido uma constante discussão e foco
de trabalho para o aumento da produtividade (MACHADO et al. 2005). Outro
agravante da produtividade dos citros são os estresses abióticos, entre eles, o
estresse por seca, que diminui o crescimento vegetativo, provoca abortamento de
frutos jovens, redução no desenvolvimento e qualidade interna e externa dos frutos,
causando importantes perdas econômicas em pomares (PÉREZ-PÉREZ et al., 2009;
RODRÍGUEZ-GAMIR et al., 2010). Segundo Magalhães Junior (2010), a seleção de
genótipos que apresentem mutações ou recombinações nos genes envolvidos na
tolerância a estresses abióticos é de grande valor, pois se estas diferenças forem
expressas em melhoria, possivelmente esses genes podem ser transferidos para
cultivares comerciais, possibilitando a obtenção de genótipos mais produtivos.

6
3.2 Respostas das plantas a deficiência hídrica
A deficiência hídrica é um dos principais fatores limitantes da produção
vegetal, pois além de comprometer as relações hídricas nas plantas, alterando-lhes
o metabolismo, é um fenômeno que ocorre em grandes extensões de áreas
cultiváveis, sendo frequente e característico do nosso país, com intensidade e
efeitos variáveis no espaço e no tempo. A situação geográfica do território brasileiro
é favorável à ocorrência de episódios de seca. Portanto, este fenômeno deve ser
encarado como um elemento climático de determinada frequência no sentido de que
já ocorreu no passado e ocorrerá no futuro (MENESES et al., 2006; NOGUEIRA et
al., 2001). Mahajan e Tuteja (2005) afirmam que os estresses abióticos causam
perdas no valor de centenas de milhões de dólares a cada ano, devido à redução na
produtividade da cultura e quebra de safra ameaçando a sustentabilidade do setor
agrícola.
As respostas fisiológicas induzidas pela seca incluem o fechamento
estomático, diminuição da atividade fotossintética, ativação da respiração, repressão
do crescimento celular, alteração da elasticidade da parede celular e geração de
metabólitos tóxicos que podem causar a morte da planta. Concomitante, a
reprogramação molecular inclui mudanças extensivas na expressão gênica
causando alterações na maquinaria bioquímica e proteômica (AHUJA et al., 2010).
O ajuste osmótico permite que as plantas mantenham o turgor da folha necessário
para a abertura estomática, sustentando a fotossíntese e o crescimento
(RODRÍGUEZ-GAMIR et al., 2010). As respostas das plantas à seca também são
influenciadas pelo tempo, intensidade, duração e frequência do estresse, bem como
por diversas interações planta-solo-atmosfera (PIERRE et al., 2012). Segundo
Hirayama e Shinozaki (2010), as respostas a estresses abióticos são importantes
para organismos sésseis como as plantas, porque esse tipo de organismo não pode
sobreviver, a menos que eles estejam aptos a resistir às mudanças ambientais.
Estudos envolvendo as relações planta-água têm demonstrado que a maioria
das plantas utiliza uma combinação de duas principais estratégias fisiológicas e
morfológicas em resposta à deficiência hídrica: evitação e tolerância à desidratação
(MORGAN, 1984; YUE et al., 2006). Evitação à desidratação tende a restringir a
perda de água por meio do aumento da resistência estomática e cuticular e de

7
alterações na morfologia e orientação foliar (JONES; CORLETT, 1992). A tolerância
a desidratação, por outro lado, refere-se à capacidade de crescer e permanecer
viável após um ciclo de hidratação e desidratação. Por conseguinte, a tolerância a
desidratação indica a capacidade do tecido para resistir à dessecação. É geralmente
medido como a capacidade da planta para se recuperar do estresse (SHINOZAKI;
YAMAGUCHI-SHINOZAKI, 2007; PIERRE et al., 2012). Os dois mecanismos
primários que contribuem para a tolerância à desidratação são (i) ajuste elástico dos
tecidos (módulo de elasticidade, ) e (ii) ajuste osmótico (SAVÉ et al., 1993;
TOUCHETTE et al., 2007; MUNNS; TESTER, 2008).
Muitas pesquisas têm dado maior atenção ao esclarecimento dos
mecanismos fisiológicos e genéticos relacionados à resistência ou tolerância à
salinidade e à seca, com a identificação de genes importantes na determinação
destas características. Esses genes têm crescido cada vez mais em importância
para a biotecnologia, dada a escassez de água que vem se agravando
progressivamente em termos globais (BENKO-ISEPPON et al., 2011). A
manipulação de genes para proteger e manter as funções celulares ou que mantêm
a estrutura de componentes celulares tem sido o alvo principal de tentativas para
produzir plantas melhoradas tolerantes ao estresse (VALLIYODAN; NGUYEN,
2006). Tem havido muitos esforços para melhorar as culturas em relação à
tolerância à seca e a produtividade sob condições limitantes de água. O
melhoramento da produção das culturas pode ser possível pela incorporação estável
de características pertencente às plantas tolerante à seca. O desenvolvimento de
culturas tolerantes à seca através de uma seleção direta tem sido prejudicado pela
baixa herdabilidade das características tais como produtividade, particularmente sob
seca, e pela grande interação genótipo x ambiente (YUE et al., 2012; AMUDHA;
BALASUBRAMANI, 2011).
Novas tecnologias estão provendo oportunidades para gerar culturas
transgênicas capazes de manter alta produtividade sob estresse e mudanças
ambientais. Com a aplicação de técnicas de biologia molecular, atualmente um
grande número de genes induzíveis por estresses abióticos como a seca, têm sido
isolados e caracterizados e suas funções precisamente caracterizadas em plantas
transgênicas. A estratégia básica da engenharia genética para tolerância à seca é a
introdução de genes que codificam proteínas funcionais ou regulatórias. As

8
proteínas funcionais são aquelas que protegem diretamente as células contra o
estresse, tais como proteínas envolvidas na proteção dos componentes celulares
(LEAs, chaperones), enzimas essenciais para a biossíntese de osmorreguladores e
enzimas detoxificadoras de espécies reativas de oxigênio (ROS). As proteínas
regulatórias são aquelas envolvidas na regulação da expressão dos genes
codificando as proteínas funcionais, tais como fatores de transcrição e proteínas
quinases (AMUDHA; BALASUBRAMANI, 2011; HIRAYAMA; SHINOZAKI, 2010;
REGUERA et al., 2012; UMEZAWA, et al. 2006; SHINOZAKI; YAMAGUCHI-
SHINOZAKI, 2007).
3.3 LEA (Late Embryogenesis Abundant)
As proteínas LEA são formadas durante o período tardio do desenvolvimento
da semente, acompanhado por desidratação. O seu nome está relacionado ao fato
de que as proteínas originalmente descritas são expressas em níveis elevados
durante as fases tardias de desenvolvimento do embrião em sementes. São
compostas principalmente de aminoácidos hidrofílicos ordenados em sequências
repetidas, formando uma estrutura altamente hidrofílica e com estabilidade termal
(HONG –BO, 2005; TUNNACLIFFE;WISE, 2007).
As possíveis funções das proteínas LEA na tolerância seca estão
relacionadas com a manutenção de proteínas de membrana, sequestros de íons,
tampão de hidratação ou ação de chaperonas moleculares, evitando a formação de
agregados de proteínas prejudiciais. A expressão de genes LEA é significativamente
induzida por estresse abióticos, tais como frio, seca e salinidade elevada. Por isso
tem sido postulado que existe uma correlação positiva entre a expressão da LEA e
tolerância a estresse abiótico em plantas. No entanto, a função precisa dessas
proteínas no desenvolvimento da planta e em resposta ao estresse ainda não foi
totalmente esclarecida (REDDY et al., 2012).
A família das proteínas LEA é basicamente dividida em 5 grupos (grupo 1- 5)
na base de domínios específicos e sequências de aminoácidos, de acordo com a
similaridade na composição de peptídeos ou distintivas sequências. Os principais
grupos são 1, 2 e 3, que contêm a maioria das proteínas dessa família. O grupo 1

9
das proteínas LEA está presente principalmente em plantas, o grupo 2, ou
dehidrinas, também é encontrado em algas, e o grupo 3 em nematóides e
procariotos (AMARA et al., 2012). As dehidrinas, também chamadas de D11 ou RAB
(responsiva ao ácido abscísico), são encontradas principalmente em plantas e
acumulam-se tipicamente em tecidos vegetais desidratados, em sementes e, ou em
tecidos submetidos a estresses ambientais, como baixa temperatura, salinidade e
seca. No entanto, como as dehidrinas funcionam nas plantas ainda é uma questão
em aberto e estudos moleculares têm tentado elucidar suas funções. Acredita-se
que elas desenvolvem uma importante função no metabolismo como chaperonas
(ALLAGULOVA et al., 2003; VEERANAGAMALLAIAH et al., 2011; HARA, 2010).
Pesquisas têm demonstrado que genes que codificam para proteínas LEA
conferem tolerância a estresses abióticos em diversas espécies vegetais, como por
exemplo, em amora e arroz superexpressando HVAI, um gene LEA do grupo 3
(CHECKER et al., 2012; BABU et al., 2004), e em tabaco (KIM et al., 2005)
expressando o gene CaLEA6 (Capsicum annuum LEA).
3.4 NCED (9-cis-epoxicarotenóide dioxigenase)
O ácido abscísico (ABA) é um fitohormônio que regula vários processos no
ciclo de vida das plantas. Envolvido nas respostas a estresses ambientais, esse
hormônio também desempenha uma função importante no desenvolvimento e
germinação das sementes. Em plantas sob estresses ambientais, o acúmulo de ABA
desempenha um papel importante em resposta ao estresse, causando mudanças
drásticas no perfil de expressão gênica e processos celulares. Em condição de
estresse osmótico, a ação do ABA ocorre controlando a abertura estomática por
meio da regulação bioquímica dos processos de transporte de íons e água
(HIRAYAMA; SHINOZAKI, 2010; CRAMER et al., 2011; KERBAUY, 2004).
Segundo Pardo (2010), a engenharia de fechamento estomático como um meio para
reduzir a perda de água é uma abordagem atrativa para melhorar o desempenho
das plantas sob limitação de água, a fim de atender a necessidade premente de
culturas em desenvolvimento com maior eficiência no uso da água.

10
Pesquisas recentes revelam que o ABA pode agir em nível molecular, celular
e da planta como um todo, transformando os efeitos exercidos pelo ambiente em
respostas biológicas de proteção, especialmente sob baixa disponibilidade de água,
alta salinidade e temperaturas reduzidas (KERBAUY, 2004). Sob condições de
estresse, especialmente hídrico, a diminuição do teor de água na célula resulta em
alterações no metabolismo e em alguns processos reguladores através da
expressão de genes específicos. O aumento das concentrações de ABA em plantas
durante a exposição ao estresse está relacionado diretamente com o incremento na
expressão de genes, associados à proteção e manutenção do funcionamento
celular. Sendo assim, a ativação da cascata de sinalização do ABA resulta na
melhoria da tolerância a estresse por desidratação (MAGALHÃES JÚNIOR, 2010;
CRAMER et al., 2011).
Segundo Zhu (2002), a função do ABA em plantas sob condição de estresse
está diretamente ligado ao balanço hídrico e tolerância a desidratação celular.
Considerando que a função de balanço hídrico é realizada principalmente através da
regulação das células guardas, a última função está relacionada com a indução dos
genes que codificam as proteínas de tolerância à desidratação em quase todas as
células.
Duas possíveis vias biossintéticas para ABA têm sido sugeridas, a via direta e
indireta. A direta, o farnesil pirofosfato (FPP) origina o ABA diretamente, ou através
do composto intermediário xantoxina. A via indireta inicia-se com o isopentenil-
pirofosfato (IPP) e através de reações enzimáticas ocorridas na rota biossintética
dos carotenóides, forma-se o carotenóide oxigenado violaxantina, catalisada pela
enzima epoxidase da zeaxantina (ZEP), a etapa seguinte consiste na clivagem
oxidativa de 9-cis-violaxantina e/ou 9′-cis-neoxantina para produzir xantoxina,
catalisada pela 9-cis-epoxicarotenóide dioxigenase (NCED), a qual é convertida em
ABA no citossol (Figura 1). Estudos recentes, como a caracterização de mutantes
deficientes ou insensíveis ao ABA, bem como a clonagem de genes, indicam a via
indireta como a mais importante nas plantas (KERBAUY, 2004; SEO;KOSHIBA,
2002).

11
Figura 1: Via indireta da biossíntese do ácido abscísico (ABA) em plantas. (a): síntese dos carotenóides não-oxigenados nos plastídeos; (b): síntese e clivagem dos carotenóides oxigenados nos plastídeos, constituindo estoque de precursores do ABA; (c): síntese do ABA no citossol. Fonte: Seo e Koshiba (2002).
A identificação das NCEDs na via biossintética do ABA proporcionou o
melhor entendimento de como as plantas controlam o nível desse fitohormônio. Os
genes que codificam essas enzimas são ativados por vários tratamentos de estresse
(ZHANG, 2008; HIRAYAMA; SHINOZAKI, 2010). Em todas as espécies de plantas
analisadas, os genes NCED pertencem a uma família multigênica (NAMBARA;
MARION-POLL, 2005). Iuchi et al. (2001) isolaram sete genes NCED (AtNCED1, 2,
3, 4, 5, 6 e 9) do genoma de Arabidopsis e observaram que AtNCED3 tem sua
expressão fortemente induzida pelo estresse por seca, é responsável pela produção
de ABA nessa condição, conferindo maior tolerância a seca nas plantas devido a
redução na abertura estomática causada pelo aumento dos níveis de ABA.
De acordo com Ahrazem et al. (2012), os estudos de expressão gênica
indicam o envolvimento principal das proteínas NCED em respostas a estresse
hídrico em tecidos vegetativos e uma correlação significativa entre expressão de

12
NCED e acúmulo de ABA. Zhang et al. (2008) produziram linhagens transgênicas de
tabaco superexpressando SgNCED1 (S. guianensis), o que resultou no aumento da
biossíntese do ABA, levando a redução da condutância estomática, da taxa
transpiratória, fotossintética e aumento da tolerância à seca. Hwang et al. (2010)
demonstraram que a super-expressão de OsNCED3 em plantas de Arabidopsis
resultou no aumento do acúmulo de ABA, reduziu a perda relativa de água, retardou
a geminação de sementes e conferiu uma maior tolerância à seca em relação à
planta tipo-selvagem. Também em Arabidopsis, Wan e Li (2006) comprovaram que a
tolerância a deficiência hídrica pode ser melhorada pela expressão do gene
AhNCED1 de Arachis hypogaea L. Plantas transgênicas de tabaco (Nicotiana
plumbaginifolia) superexpressando PvNCED1 mostraram um aumento na tolerância
à seca, sendo o mesmo observado em outras espécies, como capim agrostis
(Agrostis palustris L) e feijão caupi (Vigna unguiculata) superexpressando genes
NCED (QIN; ZEEVAART; 2002; ASWATH et al., 2005; IUCHI et al., 2000).

13
4 MATERIAL E MÉTODOS
Os experimentos foram desenvolvidos no laboratório de Cultura de Tecidos
Vegetais, laboratório de Biologia Molecular e casa de vegetação, localizados no
Centro de Biotecnologia e Genética (CBG) da Universidade Estadual de Santa Cruz
(UESC), em Ilhéus-BA.
4.1 Material Vegetal
As plântulas de Nicotiana tabacum var. Havana, de aproximadamente 3
meses de idade, propagadas à partir da germinação in vitro de sementes, foram
utilizadas como material para excisão dos explantes. A espécie N. tabacum foi
empregada no presente trabalho por ser uma planta modelo que apresenta
características desejáveis para o desenvolvimento de estudos de função gênica
(BRASILEIRO; CARNEIRO, 1998).
4.2 Cultura e manutenção dos isolados de Agrobacterium tumefaciens
A estirpe de Agrobacterium tumefaciens EHA 105 foi utilizada nos
experimentos. As culturas foram conservadas em solução de glicerol e meio de
cultura YEP líquido (extrato de levedura, NaCl e peptona), mantido à - 80 ºC. A
partir desse estoque, a bactéria foi plaqueada em meio de cultura YEP sólido
(suplementado com 100 mg L-1 de canamicina e 50 mg L-1 de rifampicina) e
incubada a 28 ºC, por 48 horas. Após esse período, colônias isoladas foram
selecionadas e transferidas para meio de cultura YEP líquido, contendo 100 mg L-1
de canamicina e 50 mg L-1 de rifampicina, e incubado overnight à 28 ºC/200rpm.
Logo após, a cultura foi centrifugada por 15 minutos a 5000 rpm e o precipitado
formado foi ressuspendido em meio de cultura MS para uso em transformação
genética de plantas.

14
4.3 Alinhamento de múltiplas sequências e análise filogenética
A sequência predita de aminoácidos codificada pelo gene CsLEA2,
juntamente com sequências homólogas de LEAs de Arabidopsis thaliana descritas
no trabalho de Hundertmark e Hincha (2008) foram alinhadas para comparação e
análise filogenética. O alinhamento das seqüências de aminoácidos foi efetuado
utilizando o programa ClustalW (http://clustalw.genome.jp/) (THOMPSON et al.,
1994) e a árvore filogenética foi gerada utilizando o método Neighbor-Joining
(SAITOU; NEI, 1987), com a opção de bootstrap (1000 replicações). O
dendrograma foi construído pelo programa MEGA4 (TAMURA et al., 2007). Para
predição de sequências conservadas foi utilizado o programa PredictProtein
(http://cubic.bioc.columbia.edu/predictprotein/). Para o gene CsNCED3 foi utilizado a
análise filogenética realizada por Neves (2011).
4.4 Clonagem molecular dos genes e introdução em vetor de transformação de
plantas
Por meio de uma busca preliminar em bibliotecas de ESTs de citros
disponíveis na base de informação genômica do HarvEST:Citrus
(http://harvest.ucr.edu), identificou-se a presença de diversas sequências com
homologia significativa a genes codificadores de proteínas LEAs e enzimas NCEDs
de Arabidopsis. Dentre as sequências identificadas, selecionou-se para
caracterização nesse estudo aquelas codificando uma LEA do grupo 2 (dehidrinas)
(Unigene C52_8111; http://harvest.ucr.edu) e uma NCED do grupo 3 (Unigene
C52_25167; http://harvest.ucr.edu), por terem sido expressas em bibliotecas de
cDNA de raízes e folhas de citros submetidos a estresse por seca (Tabela 1). No
presente estudo, estas sequências foram nomeadas como Citrus sp. LEA2
(CsLEA2) e NCED3 (CsNCED3), as quais correspondem aos locos
orange1.1g026736m.g (CsLEA2) e orange1.1g007379m.g (CsNCED3) no banco de
informações genômicas de Citrus sinensis, no Phytozome
(http://www.phytozome.net), e aos locos AT1G20450 (LEA2) e AT3G14440 (NCED3)
no banco de informações genômicas de Arabidopsis, no TAIR
(http://www.arabidopsis.org).

15
Tabela 1. Sequência de nucleotídeos dos genes CsLEA2 e CsNCED3
Gene Sequência de nucleotídeos
CsLEA2 CTATAAACGAATTCGGCCATTGTGGCCGGGGGAAGCAAAAATTCCTTAAAGAA
GCTCGAAAACTTACTTTATTGCACATACCACATATCTCTGTGTTCTTGTCCGT
TGATCTGTTGTTAAAAATTGTTGTTTGTGAGTGATAATTGACAAGTATGGCTG
AAGAGATCAAGAAGCAGCAGAAGAGCCATGAGTACGAGCCGAGTGTTGGGACG
GAAGGTGCAGTGGAGACCAAGGATCGCGGGATGCTTGATTTCTTGGGGAAGAA
AGAAGAGGAGAAACCTCAACATCACGATCAGGGGGTGATCGCCACCGAGTTTG
AGAAAGTTCATGTCTCTGAGCCTCAGCCAAAGGTAGAGGAACACAGGAAGGAG
GAGAAAGAGGAAGAGAAGAAGCCCGGTTTCTTAGACAAACTTCACCGATCCAC
CAGCAGCTCTAGCTCTTCAAGCGATGAGGAGGAGGGCGATGATGAGGAAAAGA
AGAAGAAGAAGAAGGAAAAGAAGGGACTGAAGGAGAAGCTCAAGGAGAAAATA
TCAGGTGAGAAAGAAGAGGACACGACAGTCCCTGTCGAGAAACTTGATGATGT
CCATGCTCCCCACCACCAGGAAGAAGCCCATCCTGAAGAAAAGAAAGGATTTC
TCAACAAGATCAAGGAGAAACTACCAGGACAGCAAAAGAAACCTGAAGATCAT
CAGGTCCCTTCTCCTCCTGCTGCTGAGCACCCCACATCAGTCGAGGCTCCTGA
GGCAGAGGCCAAGGAGAAGAAGGGCATATTGGAGAAACTCAAGGAGAAGCTCC
CTGGCTACCACCCAAAGTCTGAGGACGAAAAGGATAAGGACAAAGAAACTGCT
GCCCATTAATTAAGAAGATTAAATTTGTAATGTGTTTGATACAATTACTGTTT
TTGCCGGTCTCCTGTTTGTGTGGTTTGTTCAGTTCTTTCTCCTTTGTAATGCT
TTTTTTTTTTTTTTAAATTTGCCTTTATTACCATTTGATTTTATGCTTAATTT
ATAAGTGCTGTTTATGTACTTGGATGAATAACGTGAAATAGTTTCATTGCTGT
ATGAGTTTTGGTAAACTTTGGTGTGAACGCTGGGTACCACCTGCGTGCTTGTA
AATTGTAATGATGCTTTGATTGCTAAATAAAGCCCCCCCCCCGCCCGGTGCTT
TCTTTTGTTAAAAAAAAAAAAAAAAA
ATG: Códon de iniciação. TAA: Códon de terminação.
Gene Sequência do nucleotídeo
CsNCED3 CTCTGAATAGTCAATACTCAAAAGCACTTAAACGTTAAGCGATTCCCTGAAAA
TCTTACAGTCCTACTTAACTCGCATTACAAGTTTACAACTACTCCCGCTGCTA
CTACTAGAAGTGCTTCTTCTTCAAATACACTTGCGAAAATAGAGAATCAATCC
TCATGGCGGCAGCAACTACTACTTCTTCACTCATAGACTTGGGCTCTTGTAAA
GATTCTTGTTCTTCTCCGTCTTTTTCTTCTTCAAGAGCAAGAATGGCTTTTAG
TTTAAAAAAGCCCACCGAAATCACTTGCTCTCTTCAAACACCTTCCATTCTTC
ATTTCCCCAAGCAGTCCCCAAAATACCCTCCATCACCTGCAGCTTCCCCTCCT
CCATCTTCTCCTCCTCTTACAACTAAGCAGCAACCCAAGGAAAATGACAATTT
CGCCCCTTCAAAATGGAACTTCTTACAAAAAGCAGCAGCTTTAGCCTTGGACG
CAGTCGAAAACGCATTGGTTTCTCAAGAGCGTCAGCATCCTCTCCCCAAAACA
GCCGACCCCACCGTCCAAATCGCCGGAAACTTCTCTCCGGTGCCCGAAAAACC
CGTCTGCCAGAACCTTCCCACTACCGGAAAGGTCCCCGACTGCATTCAAGGCG
TCTACGTCAGAAACGGAGCCAACCCACTTCACGAACCGGTCGCCGGCCACCAT
TTCTTCGACGGAGACGGCATGGTTCATGCAGTCAAATTCAACAAAGGCTCTGT
CAGCTATTCTTGTCGTTTCACTGAAACAAACCGTTTTGTTCAAGAACGTAGCT
TAGGCCACCCCGTATTCCCCAAAGCCATTGGCGAGCTTCACGGCCACACGGGC
ATCGCAAGATTGCTTCTCTTCTACAGCAGAGCGCTCTTCGGTCTCGTTGACCC
CAGCCACGGCACTGGCGTTGCCAACGCCGGCCTTGTTTACTTCAACAACCGTT
TGTTGGCCATGTCTGAAGATGACTTGCCTTATCACGTGCGCGTCACTCCATCC
GGCGACCTCAAAACGGTCGGCCGTTTCGACTTCAGCGGCCAGCTCAAGTCCAC
GATGATAGCTCATCCGAAAGTTGATCCCGTGACGGGTGATTTCTTTGCTCTGA

16
GTTATGACGTTATCAAGAAGCCTTACTTGAAGTACTTTCGGTTTTCGCCCCAA
GGGATCAAGTCTCCGGACGTTGAGATTCCCCTTGAGGAGCCTACAATGATGCA
TGATTTCGCAATAACTGAGAATTTTGTTGTGGTGCCTGACCAGCAAGTGGTGT
TCAAGTTGAATGAGATGATCCGAGGTGGCTCCCCGGTGATTTATGACAAGAAC
AAGGTGTCAAGATTTGGGATTTTGGACAAGAATGCCACCGATGCTTCCAAAAT
GAAGTGGATTGACGCCCCTGATTGCTTCTGTTTCCATCTTTGGAATGCTTGGG
AAGAGCCTGAGAGGGATGAAGTTGTTGTAATTGGGTCCTGTATGACTCCTGCG
GACTCCATTTTCAACGAATGTGATGAGAGTCTGAAGAGTGTTTTGTCCGAAAT
TAGACTCAATCTAAAGACCGGCGAGTCCACTCGCCGCCCCATAATCTCCGAGG
ATGAGCAAGTGAACTTGGAAGCCGGAATGGTGAACAGAAACTTGCTCGGAAGA
AAGACCCGTTTCGCGTACTTAGCCCTCGCCGAGCCATGGCCTAAAGTTTCAGG
TTTTGCCAAAGTTGATCTCTTAACCGGACAAGTAAACAAGTTTATTTACGGAG
ATCAAAGGTACGGGGGCGAGCCTTTGTTCTTTCCGAGAGACCCCAATTCGGAG
AATGAGGATGATGGCTACATTCTCGCTTTTGTACACGATGAAAAGGAGTGGAA
ATCAGAGCTGCAAATTGTTAATGCAATGACTCTGGAGCTTGAAGCTACAGTTA
AGCTTCCATCTCGAGTTCCTTATGGCTTCCACGGGACGTTCATAAGCGCGAAA
GATTTGGCCAAGCAGGCCTAAATTGCAAGAATTAATCAAGAACGATCGAAGCT
TATTAAGGAGAGATTTACCAGAGGGATGCTTGCATACACGTCCCCGGAATCTG
CTCCTCTGTTTTTAGAATATTTTTTTTTTGTCTATTTGGGATTTTACAGTGAG
AGGTTTTGAGAGAGACCAGCTTGTAGCTTGTTGGTGTAGCTAGCATTGGAGCT
CAGCTGGTTTCTGCTTATTCTTTCACTATTGTGTATAATAAATTAAGTACATA
AATTACAACAACATATATATATATATATATATATATATTGTATTTCTATATAC
ATATATATACGCATATGATCATGTCTCTACCTCTACGTATAATTTATCCTCTG
TTTTACTCTGTTCAAAAAAAAAAAAAAA
ATG: Códon de iniciação. TAA: Códon de terminação.
A partir das amostras de cDNA de raiz das plantas de limoeiro Cravo
submetido à deficiência hídrica, cedidas por Neves (2011), foi empregando os
seguintes oligonucleotídeos especificamente desenhados para amplificar a
seqüência dos genes CsLEA2 e CsNCED3:
LEA 8111-F (5’-ATGGCTGAAGAGATCAAGAAGCA-3’),
LEA8111-R (5’-TTAATGGGCAGCAGTTTCTTTGTC-3’),
NCED3-F (5’- ATGGCGGCAGCAACTACTACTT-3’) e
NCED3-R (5’-TTAGGCCTGCTTGGCCAAATC-3’).
Os fragmentos de cDNA amplificados foram clonados no vetor pGEM -T
(Promega, EUA), por meio do sistema de clonagem TA, e utilizados em
transformação de Escherichia coli, estirpe TOP 10. As inserções e a orientação das
sequências no vetor de expressão foram confirmadas por digestão do DNA
plasmidial com endonuclease específica (XhoI) para o gene CsNCED3 e PCR para o
gene CsLEA2, com os oligonucleotídeos LEA 8111-F e NOS-R. As sequências
completas de cDNA dos genes foram retiradas do plasmídeo pGEM-T por meio de

17
digestão com a enzima EcoRI e inseridos no mesmo sítio do vetor de transformação
de plantas pCAMBIA 1390 modificado para expressão em plantas (Figura 2). Esses
vetores foram introduzidos na cepa de Agrobacterium tumefaciens EHA-105 e
utilizados em experimentos de transformação genética de tabaco mediada por
Agrobacterium.
Figura 2: Esquema da região do T-DNA do vetor binário pCAMBIA1390 modificado (pUESC28), indicando os genes presentes no vetor e o sítio múltiplo de clonagem sob a orientação do promotor CaMV35S.
4.5 Transformação genética de tabaco, regeneração e aclimatação
Plantas de tabaco foram transformadas via Agrobacterium tumefaciens com os
genes candidatos de tolerância a seca CsLEA2 e CsNCED3, em construção gênica
em pCambia 1390 modificado, dirigidos pelo promotor CaMV35S, conforme
protocolo descrito por Brasileiro e Carneiro (1998). Os explantes foram obtidos a
partir de folhas de N. tabacum, as quais foram cortadas em quadrados de
aproximadamente 3 cm2 e colocadas em contato com suspensão bacteriana (OD600=
0,5) por 15 minutos a temperatura ambiente sob leve agitação. Os explantes foram
posteriormente lavados com água destilada estéril, secos com papel de filtro e
mantidos durante 48 horas no escuro (período de co-cultivo) em meio de cultura MS
(MURASHIGE; SKOOG, 1962) sólido, sem adição de antibióticos. Após esse
período, os explantes foram transferidos para placas contendo meio seletivo de
regeneração de brotos, com sais e vitaminas de MS, 10 mg L -1 de higromicina e 300
mg L -1 de timentin.
Os brotos com aproximadamente 1,0 cm de tamanho foram excisados e
transferidos para frascos tipo Magenta para o enraizamento, contendo meio de
cultura MS sólido acrescido de 300 mg L -1 timentin. Após o enraizamento dos
brotos, obteve-se quatro clones de cada planta transformada, representando

18
linhagens distintas, por meio de micropropagação. As plantas foram mantidas em
sala de crescimento sob fotoperíodo em 16/8 (luz/escuro) à temperatura de 25±2°C.
Foram selecionadas doze linhagens transformadas de tabaco confirmadas pelo
PCR, sendo seis para cada construção (35S::CsLEA2 e 35S::CsNCED3) e quatro
plantas controle não transformadas (NT). As plantas foram propagadas
vegetativamente, aclimatadas e transferidas para vasos de 20 litros com o substrato
solo, em casa de vegetação.
4.6 Caracterização das plantas transgênicas
4.6.1 Extração de DNA genômico e análise da inserção dos genes pela técnica
de PCR.
Para a confirmação da inserção do transgene nas plantas de tabaco,
extrações de DNA genômico das plantas foram realizadas de acordo com o método
CTAB (DOYLE; DOYLE, 1990), seguidas de PCR utilizando oligonucleotídeos
específicos para as sequências referentes ao gene hptII.
A amplificação foi realizada empregando-se 0,5 μL de DNA (50-100 ng), 0,5
μL de dNTPs (10 mM), 0,5 μL de MgCl2 (25 mM), 1,25 μL de tampão (10x), 1,0 μL
(2,5 μM) dos oligonucleotídeos específicos (F: 5’ CTATTTCTTTGCCCTCGGACGAG
3’ ; R: 5’ ATGAAAAAGCCTGAACTCACCGC 3’) e 0,1 μL de Taq DNA Polimerase
(Fermentas) (5U/μL), em volume de reação de 12 μL. As reações foram
amplificadas em termociclador TC - 412 (Techne, Inglaterra), com uma
desnaturação inicial de 94ºC por 2 minutos, 35 ciclos de 94ºC por 1 minuto, 50ºC por
1 minuto, 72ºC por 2 minutos; e uma extensão final de 72ºC por 10 minutos. O PCR
foi avaliado por eletroforese em gel de agarose (1,0%) corado com brometo de
etídeo (0,5 μg mL-1) e fotografado em luz UV no fotodocumentador Kodak EDAS
290.
4.6.2 Deficiência hídrica
O experimento foi conduzido em casa de vegetação, sendo que por setenta e
um dias as plantas (T0) foram irrigadas, próximo a capacidade de campo. Após esse
período, as plantas foram submetidas à deficiência hídrica pela suspensão total da
irrigação por dez dias, sendo reidratadas posteriormente.

19
4.6.3 Trocas gasosas foliares
No período da deficiência hídrica, a condutância estomática (gs) da folha foi
determinada utilizando o porômetro portátil (SC-1 Leaf Porometer – Decagon
Devices). As medições foram realizadas sob condições de luz e CO2 ambientes, em
dias alternados, entre 09:00 e 11:00 horas, durante o período da deficiência hídrica.
As trocas gasosas foram medidas com um sistema portátil de medição de
fotossíntese Li-6400 (Li-Cor, USA) sob luz saturante artificial de 800 μmol fótons m-
2s-1, de modo a determinar a taxa fotossintética líquida (A) e taxa de transpiração
(E), semanalmente entre 09:00 e 11:00 horas da manhã, em folhas totalmente
expandidas. A eficiência instânea (A/E) e intrínseca (A/gs) de uso da água foi
calculada a partir dos valores obtidos de A, E e gs obtidos.
4.6.4 Potencial hídrico foliar
Para avaliar o potencial hídrico (ψw) foliar, selecionaram-se folhas maduras
da parte mediana das plantas. As determinações foram feitas no período antemanhã
(predawn), com periodicidade semanal, utilizando-se bomba de pressão do tipo
Scholander (m670, Pms Instrument Co., Albany, USA). As leituras foram expressas
em MPa.
4.6.5 Potencial mátrico (ψm) do solo
O potencial mátrico (Ψm) do solo foi estimado por meio da equação derivada
da curva de retenção de água no solo. Para isso, durante o período em que as
plantas foram submetidas à deficiência hídrica, a umidade do solo foi monitorada
através do método gravimétrico no inicio e no final do experimento. As amostras de
solo foram secas em estufa por 48 horas a 104°C.
4.6.6 Acúmulo de peróxido de hidrogênio (H2O2)
Para avaliar o estresse oxidativo, discos foliares foram coletados das plantas
após dez dias da submissão de deficiência hídrica, os quais foram submetidos ao

20
ensaio de coloração histoquímica com DAB-HCl (3,3‘ – diaminobenzidina), conforme
metodologia proposta por THORDAL-CHRISTENSEN et al. (1997). As amostras
foram expostas à infiltração a vácuo com 1 mg mL-1 de DAB-HCl, por 4 horas.
Após esse período, os discos foliares foram mantidos imersos em etanol 70%,
fervidos por 5 horas e posteriormente fotografados em lupa para detecção da
coloração marrom causada pela produção do peróxido de hidrogênio.
4.6.7 Delineamento experimental e análise estatística
O experimento foi conduzido em um delineamento inteiramente casualizado,
com quatro repetições para cada genótipo. Os dados foram submetidos à análise de
variância (ANOVA) e, quando indicado, as médias foram comparadas pelo teste t em
níveis de 5%, 1% e 0.1% de probabilidade.

21
5 RESULTADOS
5.1 Análise filogenética
O alinhamento da sequência predita de aminoácidos codificada pelo gene
CsNCED3 com sequências de outras espécies, realizado por (NEVES, 2011),
demonstrou que o mesmo é um ortólogo do gene AtNCED3 de Aradidopsis. A
sequência codante de CsNCED3 possui 1821 pares de base e a sequencia de
CsLEA2 possui 705 pares de bases. A sequência predita de aminoácidos de
CsLEA2, juntamente com seqüências homólogas de Arabidopsis descrita no
trabalho Hundertmark e Hincha (2008), foram alinhadas para comparação e análise
filogenética (Figura 3). A análise filogenética indicou que CsLEA2 pertence ao grupo
2 das proteínas LEA, também chamado de grupo das dehidrinas, sendo um ortólogo
dos genes At1g20440, At1g20450 e At1g76180 de Arabidopsis.
Figura 3: Análise filogenética das proteínas da família LEA de Arabidopsis (At) e CsLEA2 (orange1.1g026736m.g) de citros. As seqüências de aminoácidos foram alinhadas utilizando ClustalW e o método Neighbor-Joining. A árvore foi construída com um suporte de bootstrap de 1000 replicações.
Dehidrinas

22
5.2 Clonagem molecular das sequências CsLEA2 e CsNCED3 e transformação
de Nicotiana tabacum
As sequências completas de cDNA dos genes CsLEA2 e CsNCED3 foram
clonadas, a partir de amostras de cDNA de raiz de limão Cravo submetidas a seca,
cedidas por Neves (2011), e inseridas no vetor de transformação de plantas
pCAMBIA 1390 modificado, sob o controle do promotor constitutivo CaMV35S. A
inserção e orientação da sequência CsLEA2 no vetor de expressão em plantas foi
diagnosticada por PCR (Figura 4).
Figura 4: Diagnóstico de orientação do gene CsLEA2 inserido do vetor pCambia 1390 modificado para expressão em plantas (oligonucleotídeos LEA 8111-F e NOS-R); M: Marcador Molecular 1Kb Plus DNA Ladder; C-: controle negativo; 1-3: colônias 5,6,7 de E. coli transformadas.
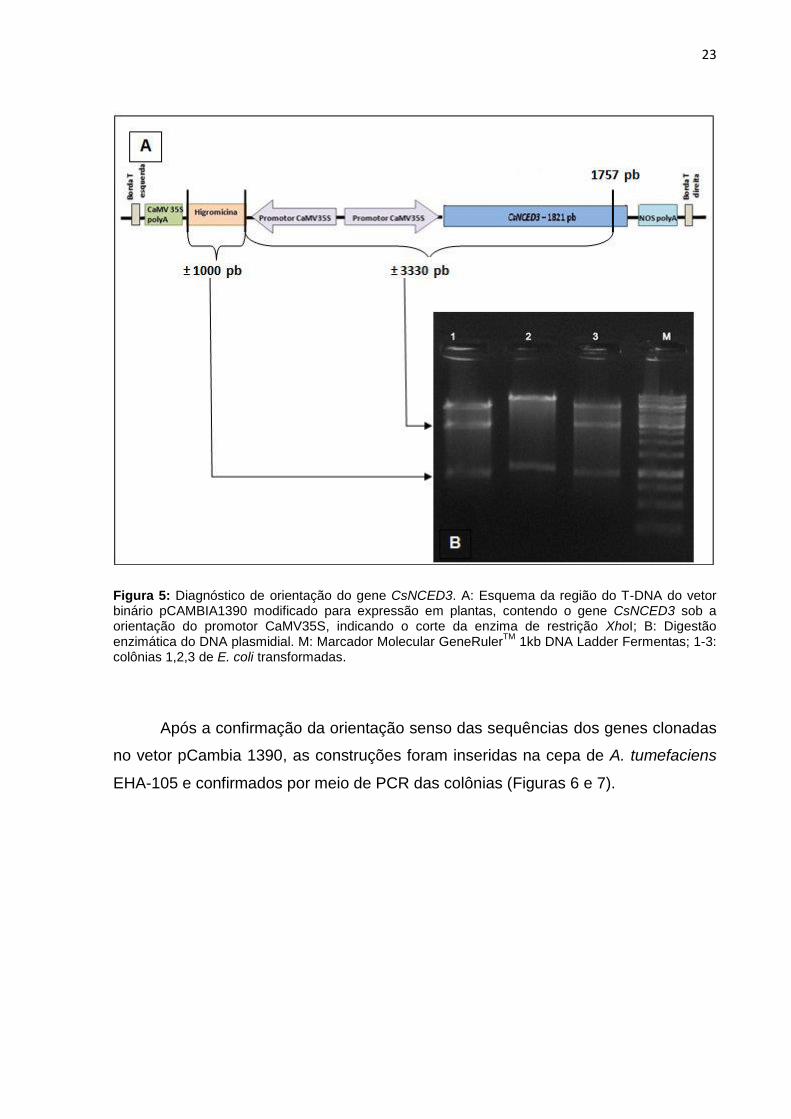
A inserção da sequência de CsNCED3 foi diagnosticada por digestão
enzimática, confirmando a presença e orientação da sequência no referido vetor, já
que a sequencia do CsNCED3 possui sítio de restrição para a enzima XhoI em 1757
pares de bases. Os fragmentos gerados após a digestão enzimática permitiu a
identificação da orientação senso do fragmento inserido (Figura 5).
805 pb
23
Figura 5: Diagnóstico de orientação do gene CsNCED3. A: Esquema da região do T-DNA do vetor binário pCAMBIA1390 modificado para expressão em plantas, contendo o gene CsNCED3 sob a orientação do promotor CaMV35S, indicando o corte da enzima de restrição XhoI; B: Digestão enzimática do DNA plasmidial. M: Marcador Molecular GeneRuler
TM 1kb DNA Ladder Fermentas; 1-3:
colônias 1,2,3 de E. coli transformadas.
Após a confirmação da orientação senso das sequências dos genes clonadas
no vetor pCambia 1390, as construções foram inseridas na cepa de A. tumefaciens
EHA-105 e confirmados por meio de PCR das colônias (Figuras 6 e 7).
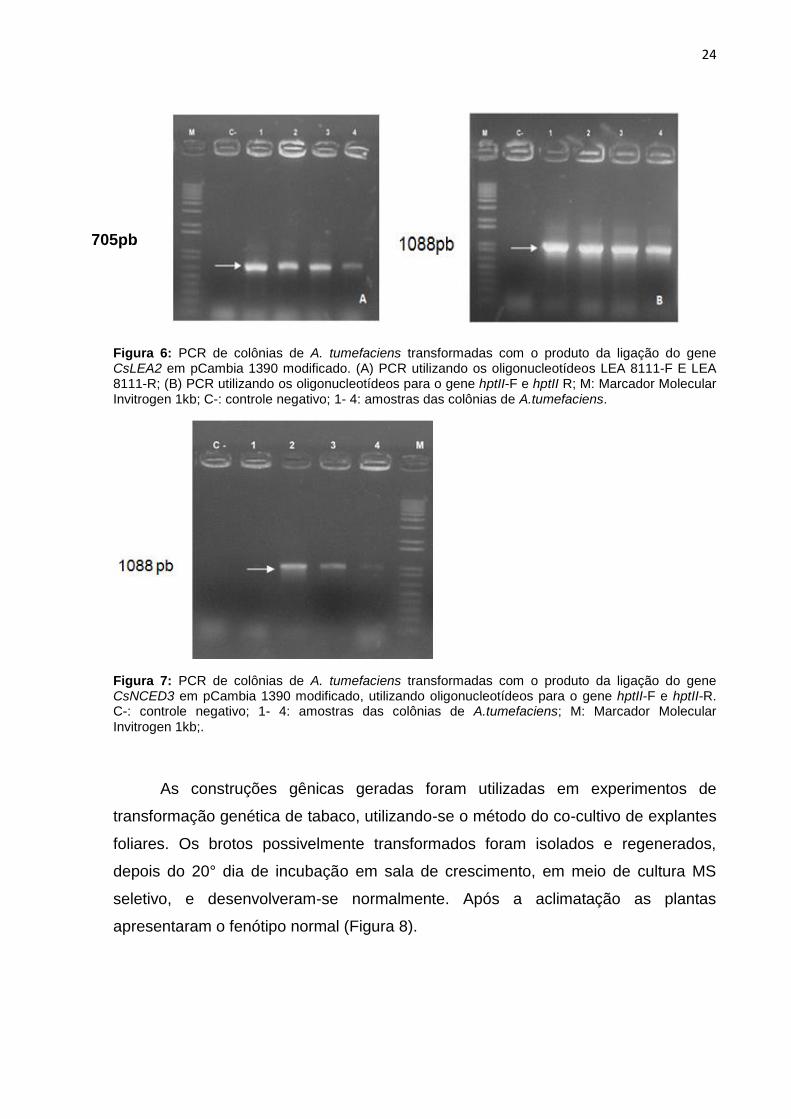
24
Figura 6: PCR de colônias de A. tumefaciens transformadas com o produto da ligação do gene CsLEA2 em pCambia 1390 modificado. (A) PCR utilizando os oligonucleotídeos LEA 8111-F E LEA 8111-R; (B) PCR utilizando os oligonucleotídeos para o gene hptII-F e hptII R; M: Marcador Molecular Invitrogen 1kb; C-: controle negativo; 1- 4: amostras das colônias de A.tumefaciens.
Figura 7: PCR de colônias de A. tumefaciens transformadas com o produto da ligação do gene CsNCED3 em pCambia 1390 modificado, utilizando oligonucleotídeos para o gene hptII-F e hptII-R. C-: controle negativo; 1- 4: amostras das colônias de A.tumefaciens; M: Marcador Molecular
Invitrogen 1kb;.
As construções gênicas geradas foram utilizadas em experimentos de
transformação genética de tabaco, utilizando-se o método do co-cultivo de explantes
foliares. Os brotos possivelmente transformados foram isolados e regenerados,
depois do 20° dia de incubação em sala de crescimento, em meio de cultura MS
seletivo, e desenvolveram-se normalmente. Após a aclimatação as plantas
apresentaram o fenótipo normal (Figura 8).
705pb

25
Figura 8: Etapas na obtenção das plantas transgênicas T0: explantes foliares transformados com A. tumefaciens (A); obtenção (B), individualização e enraizamento dos brotos (C) e aclimatação das
plantas (D).
De acordo com os resultados de PCR, houve inserção dos transgenes em
grande parte dos brotos regenerados, indicando eficiência no método utilizado. Na
reação com o par de oligonucleotídeos correspondente à sequência do gene hptII,
que confere resistência ao antibiótico higromicina, pode-se visualizar a presença de
um fragmento de 1088 pares de bases, conforme o esperado (Figura 9).
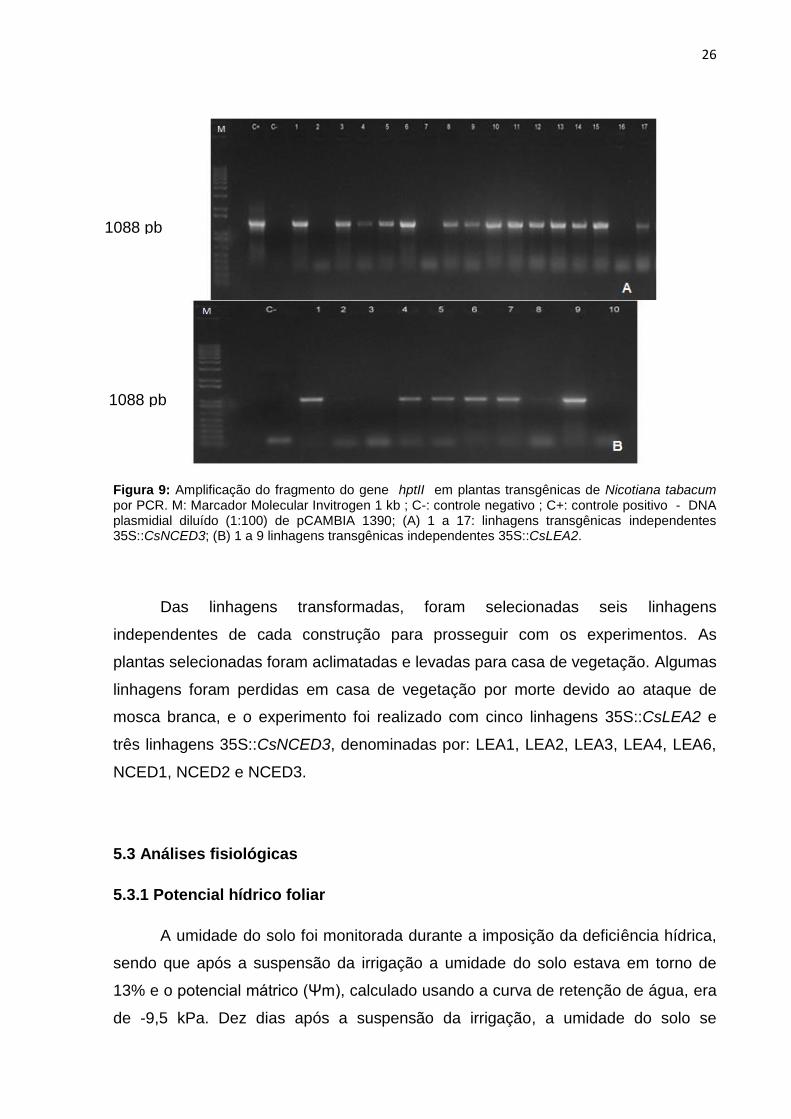
26
nnnnnnn
Figura 9: Amplificação do fragmento do gene hptII em plantas transgênicas de Nicotiana tabacum por PCR. M: Marcador Molecular Invitrogen 1 kb ; C-: controle negativo ; C+: controle positivo - DNA plasmidial diluído (1:100) de pCAMBIA 1390; (A) 1 a 17: linhagens transgênicas independentes 35S::CsNCED3; (B) 1 a 9 linhagens transgênicas independentes 35S::CsLEA2.
Das linhagens transformadas, foram selecionadas seis linhagens
independentes de cada construção para prosseguir com os experimentos. As
plantas selecionadas foram aclimatadas e levadas para casa de vegetação. Algumas
linhagens foram perdidas em casa de vegetação por morte devido ao ataque de
mosca branca, e o experimento foi realizado com cinco linhagens 35S::CsLEA2 e
três linhagens 35S::CsNCED3, denominadas por: LEA1, LEA2, LEA3, LEA4, LEA6,
NCED1, NCED2 e NCED3.
5.3 Análises fisiológicas
5.3.1 Potencial hídrico foliar
A umidade do solo foi monitorada durante a imposição da deficiência hídrica,
sendo que após a suspensão da irrigação a umidade do solo estava em torno de
13% e o potencial mátrico (Ψm), calculado usando a curva de retenção de água, era
de -9,5 kPa. Dez dias após a suspensão da irrigação, a umidade do solo se
1088 pb
1088 pb
27
encontrava em 3,72% e o potencial mátrico (Ψm) de - 3297,0 kPa. As medições de
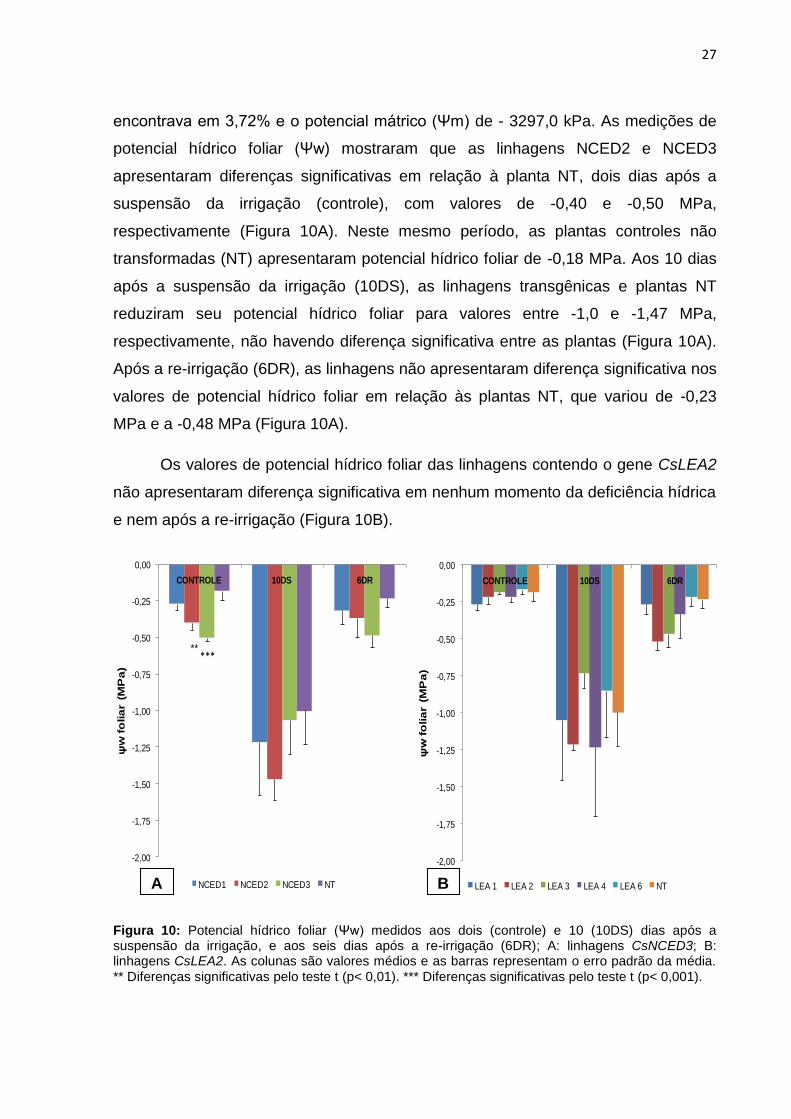
potencial hídrico foliar (Ψw) mostraram que as linhagens NCED2 e NCED3
apresentaram diferenças significativas em relação à planta NT, dois dias após a
suspensão da irrigação (controle), com valores de -0,40 e -0,50 MPa,
respectivamente (Figura 10A). Neste mesmo período, as plantas controles não
transformadas (NT) apresentaram potencial hídrico foliar de -0,18 MPa. Aos 10 dias
após a suspensão da irrigação (10DS), as linhagens transgênicas e plantas NT
reduziram seu potencial hídrico foliar para valores entre -1,0 e -1,47 MPa,
respectivamente, não havendo diferença significativa entre as plantas (Figura 10A).
Após a re-irrigação (6DR), as linhagens não apresentaram diferença significativa nos
valores de potencial hídrico foliar em relação às plantas NT, que variou de -0,23
MPa e a -0,48 MPa (Figura 10A).
Os valores de potencial hídrico foliar das linhagens contendo o gene CsLEA2
não apresentaram diferença significativa em nenhum momento da deficiência hídrica
e nem após a re-irrigação (Figura 10B).
-2,00
-1,75
-1,50
-1,25
-1,00
-0,75
-0,50
-0,25
0,00
CONTROLE 10DS 6DR
ψw
fo
lia
r (M
Pa
)
NCED1 NCED2 NCED3 NT
*****
-2,00
-1,75
-1,50
-1,25
-1,00
-0,75
-0,50
-0,25
0,00
CONTROLE 10DS 6DR
ψw
folia
r (M
Pa
)
LEA 1 LEA 2 LEA 3 LEA 4 LEA 6 NT
Figura 10: Potencial hídrico foliar (Ψw) medidos aos dois (controle) e 10 (10DS) dias após a suspensão da irrigação, e aos seis dias após a re-irrigação (6DR); A: linhagens CsNCED3; B: linhagens CsLEA2. As colunas são valores médios e as barras representam o erro padrão da média.
** Diferenças significativas pelo teste t (p< 0,01). *** Diferenças significativas pelo teste t (p< 0,001).
B A

28
5.3.2 Trocas gasosas foliares
As linhagens contendo o gene CsNCED3 apresentaram uma grande redução
na condutância estomática (gs) a partir do quarto dia após a suspensão da irrigação,
demonstrando que a super-expressão do gene induziu o fechamento estomático
(Figura 11A). As linhagens NCED1 e NCED2 apresentaram diferença significativa
em relação à planta NT, desde os quatro até os 10 dias após a suspensão da
irrigação (Figura 11A). Por outro lado, a linhagem NCED3 apresentou condutância
estomática mais alta do que as demais linhagens transgênicas. Já as plantas
contendo o CsLEA2 (Figura 11B), apresentaram reduções significativas de
condutância estomática em relação ao controle somente depois de oito dias após a
suspensão da irrigação, sendo que a linhagem LEA4 apresentou a menor
condutância (0,83 mmol H2O m-2s-1). É possível observar que ao decorrer do tempo
do estresse, as plantas controle (NT) apresentaram maior condutância estomática,
principalmente nos pontos mais críticos de oito a 10 dias sem irrigação, mostrando o
efeito do gene CsLEA2 na defesa da planta a deficiência hídrica.
0
25
50
75
100
125
150
175
200
CONTROLE 4DS 8DS 10DS 2DR
gs(m
mol
H2O
m-2
s-1)
NCED1 NCED2 NCED3 NT
*****
***
***
******
**
A
29
0
25
50
75
100
125
150
175
200
CONTROLE 4DS 8DS 10DS 2DR
gs(m
mo
l H2O
m-2
s-1)
LEA1 LEA2 LEA3 LEA4 LEA6 NT
************ ****** ***
******
Figura 11: Condutância estomática (gs) medida ao 1,4,8 e 10 dias após a suspensão da irrigação (CONTROLE; 4DS, 8DS, 10DS, respectivamente) e dois dias após a re-irrigação (2DR); A: linhagens CsNCED3; B: linhagens CsLEA2. As colunas são valores médios e as barras representam o erro padrão da média. ** Diferenças significativas pelo teste t (p< 0,01). *** Diferenças significativas pelo teste t (p< 0,001).
Em relação às análises de trocas gasosas, realizadas após oito dias de
suspensão da irrigação, não se observou diferenças significativas nas taxas
fotossintéticas (A) entre as plantas NT e linhagens contendo CsNCED3, que
variaram entre 1,95 e 15 µmol CO2 m-2s-1, exceto para a linhagem NCED3, que
apresentou taxa de fotossíntese significativamente mais alta do que as plantas NT
(p< 0,001) (Figura 12A). No mesmo período, a condutância estomática das
linhagens NCED1 e NCED2 foi significativamente mais baixa em relação às plantas
NT (Figura 12B). Quanto ao uso eficiente de água, embora não tenha havido
diferença significativa, a linhagem NCED2 se destacou apresentando maior
eficiência intrínseca e instantânea do uso da água (Figura 12C, D).
B

30
0
5
10
15
20
25
NCED1 NCED2 NCED3 NT
A (μ
mo
l CO
2 m
-2s-1
)
0,0
0,5
1,0
1,5
2,0
NCED1 NCED2 NCED3 NT
A/g
s (µ
mo
l CO
2m
ol-1
H2O
)
0,0
5,0
10,0
15,0
20,0
25,0
30,0
NCED1 NCED2 NCED3 NT
A/E
(µm
ol C
O2
mm
ol-1
H2O
)
***
0
20
40
60
80
100
120
140
NCED1 NCED2 NCED3 NT
gs(m
mo
l H2O
m-2
s-1)
***
***
Figura 12: Trocas gasosas das linhagens CsNCED3 sob oito dias de deficiência hídrica; A - Taxa fotossintética (A) ; B - Condutância estomática (gs); C – Eficiência intrínseca do uso da água (A/gs);
D - Eficiência instantanea do uso da água (A/E).
Quanto as taxas fotossintéticas (A) das linhagens CsLEA2 (Figura 13), não
houve diferença significativa em relação as plantas NT, variando de 3,1 a 5,5 µmol
CO2 m-2s-1. As linhagens LEA2, LEA3, LEA4 E LEA6 apresentaram valores
significativamente menores de condutância estomática (gs). Entretanto, a eficiência
intrínseca do uso da água (A/gs) não se diferenciou do controle (Figura 13B,C).
Quanto à eficiência instantânea do uso (A/E) de água, as linhagens LEA2, LEA3 E
LEA6 apresentaram diferença significativa em relação às plantas NT, mostrando-se
menos eficientes (Figura 13-D).
A B
C D
31
0
2
4
6
8
LEA1 LEA2 LEA3 LEA4 LEA6 NT
A (μ
mo
l CO
2 m
-2s-1
)
0,0
2,5
5,0
7,5
10,0
12,5
15,0
LEA1 LEA2 LEA3 LEA4 LEA6 NT
A/g
s (µ
mo
l CO
2m
ol-1
H2O
)
0
5
10
15
20
25
LEA1 LEA2 LEA3 LEA4 LEA6 NT
A/E
(µm
ol C
O2
mm
ol-1
H2O
)
0
10
20
30
40
50
60
70
80
90
LEA1 LEA2 LEA3 LEA4 LEA6 NT
gs(m
mo
l H2O
m-2
s-1)
*** ****** ***
** * ***
Figura 13: Trocas gasosas das linhagens CsLEA2 sob oito dias de deficiência hídrica ; A - Taxa fotossintética (A), ; B – Condutância estomática (gs); C- Eficiência intrínseca do uso da água (A/gs). D– Eficiência instantânea do uso da água (A/E) .
Após o período de deficiência hídrica, as plantas foram re-irrigadas e, depois
de cinco dias, as trocas gasosas foram medidas a fim de avaliar o estado das
plantas após o estresse (Figura 14). Os valores médios da condutância estomática
(gs) das linhagens CsNCED3 foram semelhantes ao do primeiro dia do estresse
(tratamento controle), ou seja, quando o solo ainda estava com umidade próximo a
capacidade de campo, consequentemente aumentando as taxas de fotossíntese (A).
Com maior abertura estomática a eficiência do uso da água, tanto intrínseca quanto
instantânea, foi menor. Mas para nenhum desses parâmetros avaliados houve
diferença significativa com as plantas NT.
A
C D
B
32
0
5
10
15
20
25
NCED1 NCED2 NCED3 NT
A (μ
mo
l CO
2 m
-2s-1
)
0
20
40
60
80
100
120
140
160
180
NCED1 NCED2 NCED3 NT
gs(m
mol
H2O
m-2
s-1)
0,00
0,05
0,10
0,15
NCED1 NCED2 NCED3 NT
A/g
s (µm
ol C
O2
mol
-1H
2O)
0
2
4
6
8
10
NCED1 NCED2 NCED3 NT
A/E
(µm
ol C
O2
mm
ol-1
H2O
)
Figura 14: Trocas gasosas das linhagens CsNCED3 após cinco dias da re-irrigação; A - Taxa fotossintética (A) ; B - Condutância estomática (gs); C – Eficiência intrínseca do uso da água (A/gs);
D - Eficiência instantânea do uso da água (A/E).
As linhagens CsLEA2 apresentaram maiores taxas de fotossíntese após a re-
irrigação devido aos maiores valores de condutância estomática. A linhagem LEA3
apresentou diferença significativa na condutância estomática e eficiência intrínseca
(A/gs) do uso da água em comparação com as plantas NT. Para a eficiência
instantânea do uso da água, não houve diferença significativa entre as linhagens
transgênicas e plantas NT (Figura 15).
C D
B A
33
0
5
10
15
20
25
LEA1 LEA2 LEA3 LEA4 LEA6 NT
A (μ
mo
l CO
2 m
-2s-1
)
0
20
40
60
80
100
120
140
160
180
200
LEA1 LEA2 LEA3 LEA4 LEA6 NT
gs(m
mo
l H2O
m-2
s-1) ***
0,00
0,05
0,10
0,15
0,20
0,25
0,30
LEA1 LEA2 LEA3 LEA4 LEA6 NT
A/g
s (µ
mo
l CO
2m
ol-1
H2O
)
0
1
2
3
4
5
6
7
LEA1 LEA2 LEA3 LEA4 LEA6 NT
A/E
(µm
ol C
O2
mm
ol-1
H2O
)
***
Figura 15: Trocas gasosas das linhagens CsLEA2 após cinco dias da re-irrigação; A - Taxa fotossintética (A) ; B - Condutância estomática (gs); C – Eficiência intrínseca do uso da água (A/gs); D - Eficiência instantanea do uso da água (A/E).
5.4 Acúmulo de peróxido de hidrogênio
Para avaliar se a maior tolerância a desidratação das plantas transgênicas
testadas está correlacionada com os níveis de estresse oxidativo, a produção de
H2O2 foi analisada em discos foliares de plantas de tabaco, 10 dias após a
suspensão da irrigação, por meio da técnica de coloração histoquímica com DAB-
HCl. Nas plantas transgênicas contendo CsNCED3, observou-se um menor
escurecimento no tecido foliar em relação a planta controle NT, ocasionado pelo
acúmulo de H2O2 (Figura 16). Entretanto, não se observou escurecimento dos
tecidos de plantas NT tratados com água, em vez de DAB comprovando a eficiência
do método (Figura 16D). A intensidade de coloração dos discos foliares das plantas
transgênicas foi menor e claramente distinta das plantas NT (Figura 16E), indicando
que as plantas transgênicas acumularam menos ROS (espécie reativa de oxigênio)
sob condições de estresse por seca.
A
C D
B

34
Figura 16: Produção de peróxido de hidrogênio em discos foliares de plantas de N. tabacum submetidas à deficiência hídrica, visualizada por meio da coloração com HCL-DAB (1 mg mL-1). A- C: linhagens transgênicas NCED1, NCED2, NCED3 respectivamente submetidas à desidratação; D: planta não transformada fresca infiltrada com água; E: planta não-transformada submetida à desidratação
As plantas transgênicas contendo o gene CsLEA2 (Figura 17), também
apresentaram o mesmo resultado de menor acúmulo de ROS, exceto a linhagem
LEA3 (Figura 17C) que mostrou um nível de escurecimento semelhante a planta
controle.
Figura 17: Produção de peróxido de hidrogênio em discos foliares de N. tabacum submetidas à deficiência hídrica por dez dias, incubado com HCL-DAB (1 mg mL
-1) e produção de H2O2 visualizada
através da coloração com DAB. A- E: linhagens transgênicas LEA1, LEA2, LEA3, LEA4, e LEA6 respectivamente, submetidas à desidratação; F: planta não transformada infiltrada com água; G: planta não-transformada submetida à desidratação.

35
6. DISCUSSÃO
A disponibilidade de água é um fator limitante para o potencial produtivo das
culturas e as estratégias das plantas para lidar com a seca normalmente envolve
uma mistura de mecanismos de prevenção e de tolerância ao estresse. As
respostas das plantas a seca são complexas, envolvendo mudanças adaptativas
e/ou efeitos deletérios. Em condições de campo, essas respostas podem ser
sinergicamente ou antagonicamente modificadas pela superposição de outros
estresses. Portanto, o conhecimento básico dos mecanismos subjacentes ao
funcionamento dos genes induzidos pelo estresse é importante para o
desenvolvimento de plantas transgênicas (CHAVES et al., 2002; MAHAJAN;
TUTEJA, 2005; BARTELS; SALAMINI, 2001). A aplicação da engenharia genética
em culturas, já levou a exemplos de tolerância à seca, melhorando e aumentando o
rendimento em condições de seca (revisado por MANAVALAN; NGUYEN, 2012).
Nesse estudo, membros das famílias LEA (CsLEA2) e NCED (CsNCED3) de
citros, identificados em bibliotecas de cDNA de raízes de plantas de limoeiro ‘Cravo’
submetidas a seca, foram funcionalmente analisados quanto ao envolvimento em
mecanismos de tolerância a seca em plantas de tabaco. CsNCED3 é um ortólogo
do gene AtNCED3 de Arabidopsis, o qual tem sido relacionado com tolerância ao
estresse hídrico (IUCHI et al., 2001). Estudos prévios demonstraram que em plantas
de limão ‘Cravo’ submetidas à deficiência hídrica, a expressão de CsNCED3 foi
altamente induzida nas raízes, sendo correlacionada com aumento na síntese do
ABA (NEVES, 2011). Por outro lado, a análise filogenética de CsLEA2 demonstrou
que este gene pertence ao grupo das dehidrinas (grupo 2). Muitas funções das
dehidrinas têm sido demonstradas em experimentos in vitro, como crioproteção,
prevenção de desnaturação de proteínas, detoxificação de ROS entre outras.
Entretanto não são suficientes para confirmar suas funções fisiológicas, porque se
supõe que as funções in vitro nem sempre são expressas in vivo (HARA, 2010).
O potencial hídrico foliar é um parâmetro de indicação do grau de hidratação
de uma planta, fornecendo um índice relativo do estresse hídrico ao qual a planta
está submetida. O potencial hídrico foliar tem sido usado, principalmente, para o
entendimento dos mecanismos envolvidos e os processos afetados no controle da
deficiência hídrica pela própria planta. Quando determinado no período antemanhã

36
(predawn), fornece uma medida indireta do potencial hídrico do solo, isto porque,
neste período, as perdas de água por evapotranspiração são nulas, ou desprezíveis,
devido à ausência de luz e a alta umidade (próximo a saturação) e, nessa situação,
um equilíbrio no sistema solo:planta:atmosfera é alcançado (TAIZ, ZEIGER; 2004;
MARTINS, et al., 2010). Ao avaliar o potencial hídrico foliar no presente estudo,
observou-se o estado hídrico das plantas no período de deficiência hídrica. O
potencial hídrico foliar reduziu drasticamente em todas as plantas, transgênicas e
NT, ao longo do período de deficiência hídrica, sendo recuperado após a re-
irrigação. Não foram observadas diferenças significativas no potencial hídrico foliar
entre plantas NT e transgênicas nos diferentes tratamentos, exceto para as
linhagens transgênicas contendo CsNCED3 em condições controle. Wang et al.
(2009) avaliou o potencial hídrico das linhagens de Leymus chinensis transformadas
com gene LEA de trigo (TaLEA3), sendo que no tratamento irrigado não houve
diferenças significativas entre as linhagens e a planta NT. Porém, após 49 dias de
estresse hídrico, o potencial hídrico apresentou diferenças pronunciadas entre as
plantas transgênicas e NT.
O fechamento estomático é considerado uma importante linha de defesa da
planta contra a seca, pois a redução na condutância estomática diminui a taxa de
transpiração, evitando assim a desidratação. Porém, pode resultar na redução da
taxa fotossintética (TAIZ; ZEIGER, 2004; MAHAJAN; TUTEJA, 2005). As análises
fisiológicas realizadas nesse trabalho revelaram que quando as linhagens CsLEA2 e
CsNCED3 foram submetidas a deficiência hídrica, o mecanismo de defesa do
fechamento estomático foi ativado na maioria das plantas ao longo do período do
estresse, de modo a evitar maiores perdas de água. Portanto, estes genes
contribuem para a estratégia de evitação à seca das plantas, por meio da ativação
do mecanismo de fechamento estomático. A indução do fechamento estomático em
linhagens transgênicas contendo CsNCED3 pode estar relacionada com a
biossíntese de ABA, fitormônio envolvido no processo hidroativo de fechamento
estomático (TAIZ; ZEIGER, 2004). Um aumento no acúmulo de ABA foi observado
por Iuchi et al. (2001) em plantas transgênicas de Arabidopsis contendo o gene
AtNCED3, que após dezoito dias de estresse por seca apresentaram claramente
maior tolerância à seca do que as plantas do tipo selvagem. Qin e Zeevaart (2002)

37
superexpressaram PvNCED1 em tabaco e também verificaram aumento do nível de
ABA e tolerância a estresse hídrico.
A redução na atividade fotossintética pela deficiência hídrica é devida a vários
eventos coordenados, como a redução da atividade de enzimas fotossintéticas, que
está relacionada com limitações do CO2 imposta pela condutância estomática
(VALLIYODA; NGUYEN, 2006). Sabe-se que o estado o hídrico foliar sempre
interage com a condutância estomática e uma boa correlação entre potencial hídrico
e condutância estomática existe, mesmo sob estresse hídrico (REDDY et al, 2004).
A redução da condutância estomática geralmente implica numa redução das
taxas fotossintéticas. Entretanto, os resultados obtidos no presente estudo
demonstram que, embora as plantas transgênicas apresentassem menor
condutância estomática do que as plantas NT sob condições de estresse, a
fotossíntese foi mantida em níveis similares ao das plantas NT. Estes resultados
indicam que a indução do mecanismo de fechamento estomático nas plantas
transgênicas parece não afetar a produção de biomassa. Resultado similar foi
observado no trabalho de Smeeton (2010), em que linhagens transgênicas de
tomate superexpressando o gene LeNCED1, mesmo com a condutância estomática
reduzida, não apresentaram nenhum efeito na produção biomassa em relação ao
controle não transformado. Esse fato demonstra uma melhor eficiência no uso da
água das linhagens transgênicas, já que a definição básica de eficiência do uso da
água (EUA) é a razão entre a quantidade de biomassa produzida com a quantidade
de água transpirada (JONES, 2004).
Após os dez dias de suspensão da irrigação, as plantas foram reirrigadas,
sendo observada a recuperação do potencial hídrico, da condutância estomática e
da taxa fotossintética em todas as plantas analisadas. Estes dados demonstram que
a superexpressão de CsLEA2 e CsNCED3 não afetou a capacidade de recuperação
das plantas.
As ROS apresentam uma função dupla em plantas, sendo necessárias como
moléculas de sinalização, mas em concentrações elevadas é prejudicial, sendo,
portanto considerado como um sintoma de estresse. As plantas têm de manter a
ROS dentro de um determinado nível necessário para a homeostase celular normal
(HARB et al., 2010). O estresse por seca é acompanhado pela formação de ROS,

38
como O2, H2O2, e OH-. ROS são geradas como um resultado da inibição da
fotossíntese e o predomínio de fotorrespiração. O peróxido de hidrogênio (H2O2) atua
como um sinal para fechamento estomático, aclimatação da folha para alta
irradiância e indução de proteínas de choque térmico (HSPs), mas em altas
concentrações danifica membranas e macromoléculas (NOCTOR et al., 2002;
DESIKAN, et al., 2004). As linhagens transgênicas foram avaliadas quanto à
produção de peróxido de hidrogênio (H2O2), após dez dias sob condições de
deficiência hídrica. Para a maioria das linhagens, tanto de CsNCED3 como de
CsLEA2, o nível de ROS verificado pela intensidade da coloração com DAB foi
menor do que nas plantas NT. As proteínas LEA do tipo dehidrinas também têm
sido implicadas em processo de detoxificação de ROS, o qual funciona no controle e
reparação dos danos causados pelo estresse induzido, resultando em tolerância ao
estresse (MAHAJAN; TUTEJA, 2005). ROYCHOUDHURY et al. (2007) observaram
que o aumento do nível de H2O2 após estresse salino foi comparativamente mais
baixo em todas as linhagens transgênicas de tabaco contendo um gene LEA
(Rab16A) do que na planta tipo selvagem. Em plantas transgênicas de tabaco com
gene PtrMAPK (Poncirus trifoliata L.), quando verificado o nível de ROS nas folhas
coletadas das plantas expostas à seca por 21 dias, as linhagens transgênicas
exibiram claramente coloração menos intensa com DAB em comparação com
plantas tipo selvagem, sugerindo que as linhagens transgênicas acumularam baixos
níveis de ROS sob estresse hídrico (HUANG, et al., 2011). O trabalho de Huang et
al. (2011) também demonstrou que PtrMAPK induz a transcrição de diversos genes
regulatórios e funcionais relacionados a mecanismos de resposta e tolerância a
seca, incluindo NCEDs e LEAS (LEA5, ERD10C, ERD10D).
Em muitos estudos que lidam com as respostas moleculares à seca, o estado
hídrico da planta, condutância foliar e taxa fotossintética geralmente não são
avaliados, o que torna a análise comparativa destes dados uma tarefa difícil de
executar (PINHEIRO; CHAVES, 2011). Entretanto, testes in vitro têm sido realizados
em plantas transgênicas contendo genes das famílias das proteínas LEA e NCED,
com substancias que simulam a deficiência hídrica, tais como PEG (polietileno glicol)
e manitol. Estes estudos têm demonstrado uma correlação da superexpressão de
LEAs e NCEDs com tolerância a deficiência hídrica. Por exemplo, Kim et al. (2005)
submeteram linhagens transgênicas de tabaco contendo o gene CaLEA6 ao

39
tratamento com PEG, o qual atribuiu maior tolerância a desidratação. Em simulação
de seca com PEG, plantas transgênicas de amora (Morus indica L.) com o gene
HVAI demonstraram melhor estabilidade da membrana celular, grande acúmulo de
prolina e rendimento fotossintético comparado com a planta controle não
transformada (CHECKER et al., 2012). Dalal et al. (2009) encontraram resultados
semelhantes a estes citados em linhagens transgênicas de Arabidopsis contendo
BnLEA4-1. A superexpressão do SgNCED em plantas de tabaco em meio contendo
de 0,1 M de manitol conferiu tolerância a desidratação, sendo refletido na melhor
taxa de crescimento em comparação com a planta controle tipo selvagem (ZHANG
et al., 2008). Estes resultados corroboram de alguma forma com os resultados
encontrados no presente trabalho, demonstrando uma maior tolerância das plantas
transgênicas a estresse por deficiência hídrica.

40
7 CONCLUSÕES GERAIS
Os genes candidatos de tolerância a seca de citros, CsLEA2 e CsNCED3,
induziram nas plantas transformadas de N. tabacum mecanismos de defesa contra a
deficiência hídrica, como o fechamento estomático e baixo acúmulo de ROS,
contribuindo para a estratégia fisiológica de evitação à seca das plantas. Esses
genes podem ser empregados em trabalhos de transformação genética de
variedades porta-enxerto de citros, visando à obtenção de plantas tolerantes à seca.

41
REFERÊNCIAS
AHRAZEM, O. et al. Developmental and stress regulation of gene expression for a9-cis-epoxycarotenoid dioxygenase,CstNCED, isolated from Crocus sativus stigmas. Journal of Experimental Botany, v. 63, p. 681–694, 2012.
AHUJA, I.; et al. Plant molecular stress responses face climate change. Trends in Plant Science, v.15, p. 664-674 2010.
ALLAGULOVA, C. R., et al. The plant dehydrins: structure and putative functions. Biochemistry, , v.68, p.945–951, 2003.
AMARA, I. et al. Insights into Maize LEA Proteins: From Proteomics to Functional Approaches. Plant & Cell Physiology. v. 53p. 312–329, 2012.
AMUDHA, J.; BALASUBRAMANI, G. Recent molecular advances to combat abiotic stress tolerance in crop plants. Biotechnology and Molecular Biology Review, v. 6, p. 31-58, 2011.
ASWATH, C. et al. Transgenic Plants of Creeping Bent Grass Harboring the Stress Inducible Gene, 9-cis-epoxycarotenoid dioxygenase, are Highly Tolerant to Drought and NaCl Stress. Plant Growth Regulation, v.47, p. 129-139, 2005.
BABU,C. et al. HVA1, a LEA gene from barley confers dehydration tolerance in transgenic rice (Oryza sativa L.) via cell membrane protection. Plant Science, v.166, p.855–862, 2004.
BARTELS, D.; SALAMINI, F. Desiccation Tolerance in the Resurrection Plant Craterostigma plantagineum . A Contribution to the Study of Drought Tolerance at the Molecular Level. Plant Physiology. v.127, p.1346–1353, 2001.
BENKO-ISEPPON, A. et al. Prospecção de Genes de Resistência à Seca e à Salinidade em Plantas Nativas e Cultivadas. Revista Brasileira de Geografia Física, v. 6, p. 1112-1134, 2011.

42
BRASILEIRO, A. C. M.; CARNEIRO, V. T. C. Manual de Transformação Genética de Plantas. Brasília:Embrapa. 309p. 1998.
CANTAGALLO, F. S. et al. Micropropagação de citrumelo ‗’swingle’ pelo cultivo in vitro de gemas axilares. Revista Brasileira de Fruticultura, v. 27, p. 136-138, 2005.
CHAVES, M.M. et al. How Plants Cope with Water Stress in the Field? Photosynthesis and Growth. Annals of Botany, v.89, p. 907-916, 2002.
CHECKER, V.G.; CHHIBBAR, A.K; KHURANA, P. Stress-inducible expression of barley Hva1 gene in transgenic mulberry displays enhanced tolerance against drought, salinity and cold stress. Transgenic Research, v. 21, p.939–957, 2012.
CRAMER, G.R.; et al. Effects of abiotic stress on plants: a systems biology perspective. BMC Plant Biology, v.11, 2011.
DALAL, M. et al. Abiotic stress and ABA-inducible Group 4 LEA from Brassica napus plays a key role in salt and drought tolerance. Journal of Biotechnology, v.139, p.137–45, 2009.
DESIKAN, R.; et al. ABA, hydrogen peroxide and nitric oxide signaling in stomatal guard cells. Journal Experimental Botany, v.55, p.205–212, 2004.
DONADIO, L.C; MOURÃO FILHO, F.A.A.; MOREIRA,C.S. Centros de origem, distribuição geográfica das plantas cítricas e histórico da citricultura no Brasil. In: MATTOS JUNIOR, D.; NEGRI, J. D.; PIO, R. M.; POMPEU JUNIOR, J. (Ed.). Citros. Campinas: Instituo Agronômico; Fundag, Cap. 1, p. 1-14, 2005.
DONATO, S.L.R. et al. Respostas de combinações de variedades copa e porta-enxerto de citros à deficiência hídrica. Nota Científica. Pesquisa Agropecuária Brasileira, Brasília, v.42, p.1507-1510, 2007.

43
DOYLE, J. J.; DOYLE, J. L. Isolation of plant DNA from fresh tissue. Focus, v.12: p.13-15, 1990.
GUERRINI, I.A; et al. Eucalyptus ESTs involved in the production of 9-cis epoxycarotenoid dioxygenase, a regulatory enzyme of abscisic acid production. Genetics and Molecular Biology, v.28, p.640-643, 2005.
HARA, M. The multifunctionality of dehydrins: An overview. Plant Signaling & Behavior, v.5, p.503–508, 2010.
HARB, A. et al. Molecular and Physiological Analysis of Drought Stress in Arabidopsis Reveals Early Responses Leading to Acclimation in Plant Growth. Plant Physiology, v. 154, p. 1254–1271, 2010.
HIRAYAMA, T.; SHINOZAKI, K. Research on plant abiotic stress responses in the post-genome era: past, present and future. The Plant Journal, v.61, p.1041-1052, 2010.
HONG-BO, S.; ZONG-SUO, L.; MING-AN, S. LEA proteins in higher plants: Structure, function, gene expression and regulation. Colloids and Surfaces B: Biointerfaces, v. 45, p. 131-135, 2005.
HUNDERTMARK, M.; HINCHA, D.K. LEA (Late Embryogenesis Abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genomics, v.9, p 118, 2008.
HUANG, X-S. et al. Cloning and molecular characterization of a mitogen-activated protein kinase gene from Poncirus trifoliata whose ectopic expression confers dehydration/drought tolerance in transgenic tobacco. Journal of Experimental Botany, v.62, p. 5191-5206, 2011.
HWANG, S-G.; et al. Ectopic expression of rice OsNCED3 in Arabidopsis increases ABA level and alters leaf morphology. Plant Science, v. 178, p.12–22, 2010.
IBGE. Instituto Brasileiro de Geografia e Estatística. 2011. Disponível em: http://www.sidra.ibge.gov.br . Acesso em dezembro de 2012.

44
IUCHI, S.; et al. A Stress-Inducible Gene for 9-cis-Epoxycarotenoid Dioxygenase Involved in Abscisic Acid Biosynthesis under Water Stress in Drought-Tolerant Cowpea. Plant Physiology, v. 123, p. 553–562, 2000.
IUCHI, S.; et al. Regulation of drought tolerance by gene manipulation of 9-cis-epoxycarotenoid dioxygenase, a key enzyme in abscisic acid biosynthesis in Arabidopsis. The Plant Journal, v.27, p. 325-333, 2001.
JONES, H.G.; CORLETT, J.E. Current topics in drought physiology. Journal Agriculture Science. v.119, p. 291–29, 1992.
JONES, H. G. What is water use efficiency. In: Water use efficiency in plant biology. 27–41 Edited by Bacon, M. A., Blackwell Publishing, Oxford, 2004.
KERBAUY, G. B. Fisiologia Vegetal. Rio de Janeiro: Guanabara Koogan, 2004, 452p.
KIM, H.-S. et al. Molecular and functional characterization of CaLEA6, the gene for a hydrophobic LEA protein from Capsicum annuum. Gene, v.344, p.115–23, 2005.
MACHADO, A.M. et al. Genética, melhoramento e biotecnologia de citros. In: MATTOS JUNIOR, D.; NEGRI, J. D.; PIO, R. M.; POMPEU JUNIOR, J. (Ed.). Citros. Campinas: Instituo Agronômico; Fundag,Cap. 9, p. 222-277, 2005.
MAHAJAN, S.;TUTEJA, N. Cold, salinity and drought stresses: an overview. Archives of Biochemistry and Biophysics, v.444, p.139-158, 2005.
MAGALHÃES FILHO, J.R. et al. Deficiência hídrica, trocas gasosas e crescimento de raízes em laranjeira ‘Valência’ sobre dois tipos de porta-enxerto. Bragantia, Campinas, v.67, p.75-82, 2008.
MAGALHÃES, A. M. et al. Ácido abscísico e o estresse. Embrapa Clima Temperado. Documentos, 307, p.30, 2010.
MANAVALAN, L.P.; NGUYEN, H.T. Drought tolerance in crops: Physiology to genomics. in: Plant stress physiology, 2012.

45
MAPA. Ministério da Agricultura, Pecuária e Abastecimento. Disponível em: http://www.agricultura.gov.br. Acesso em de 2012.
MARTINS, J.D. et al. Potencial hídrico foliar em milho submetido ao déficit hídrico. Irriga, , v. 15, p. 324-334, 2010.
MENESES,C.H.S.G. et al. Aspectos genéticos e moleculares de plantas submetidas ao déficit hídrico. Revista Brasileira de oleaginosas e fibrosas, v.10, p.1039-1072, 2006.
MORGAN, J.M. Osmoregulation and water stress in higher plants. Annual Review of Plant Physiology, v.35, p. 299–319, 1984.
MUNNS, R.; TESTER, M. Mechanisms of salinity tolerance. Annual Review of Plant Biology, v.59, p. 651–681, 2008.
MURASHIGE, T.; SKOOG, F. A revised medium for rapid growth and bioassay with tobacco tissue culture. Physiologia Plantarum, v. 15, p.473-497, 1962.
NAMBARA, E.; MARION-POLL, M. Abscisic acid biosynthesis and catabolism. Annual Review of Plant Biology, v.56, p.165–185, 2005.
NEVES, M.F. et al. O Retrato da citricultura brasileira. Disponível em: http://www.citrusbr.com.br/download/biblioteca/Apresentacao_Marcos_Fava_evento_valor.pdf. Acesso em: 09 de maio 2011.
NEVES, D. M. Mecanismos fisiológicos e expressão gênica relacionado à seca em porta-enxertos de citros. 2011. 62 f. Dissertação (Mestrado em Genética e Biologia Molecular) - Universidade Estadual de Santa Cruz, Ilhéus, 2011.
NOCTOR, G. et al. Drought and oxidative load in the leaves of C3 plants: a predominant role of photorespiration? Annals of Botany, v.89, p. 841-850, 2002.

46
NOGUEIRA, R.J.M.C. et al. Alterações na resistência à difusão de vapor das folhas e relações hídricas em aceroleiras submetidas a déficit de água. Revista Brasileira de Fisiologia Vegetal, v.13, p.75-87, 2001.
PARDO, J.M. Biotechnology of water and salinity stress tolerance. Current Opinion in Biotechnology, v.21, p.185–196, 2010.
PÉREZ-PÉREZ; J.G. et al. Response to drought and salt stress of lemon ‘Fino 49’ under field conditions: Water relations, osmotic adjustment and gas exchange. Scientia Horticulturae, v.122, p. 83–90, 2009.
PIERRE,C.S. et al. Phenotyping transgenic wheat for drought resistance. Journal of Experimental Botany, v. 63, p. 1799–1808, 2012.
PINHEIRO, C.; CHAVES, M.M. Photosynthesis and drought: can we make metabolic connections from available data? Journal of Experimental Botany, v. 62, p. 869-882, 2011.
PIO, R. M. et al. Variedades copas. In: MATTOS JUNIOR, D.; NEGRI, J. D.; PIO, R. M.; POMPEU JUNIOR, J. (Ed.). Citros. Campinas: Instituo Agronômico; Fundag,Cap. 3, p. 39-60, 2005.
QIN, X.Q; ZEEVAART, J.A.D. Overexpression of a 9-cis-epoxycarotenoid dioxygenase gene in Nicotiana plumbaginifolia increases abscisic acid and phaseic acid levels and enhances drought tolerance. Plant Physiology, v.128, p.544-551, 2002.
RAMANJULU, S.; BARTELS, D. Drought- and desiccation-induced modulation of gene expression in plants. Plant Cell & Environment, v. 25, p.141-151, 2002.
REDDY, A.R; CHAITANYA, V.; VIVEKANANDAN. M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. Journal of Plant Physiology, v.161, p.1189–1202, 2004.
REDDY; P.S. et al. Cloning and molecular characterization of a gene encoding late embryogenesis abundant protein from Pennisetum glaucum: protection against abiotic stresses. Molecular Biology Reports, v. 39, p.7163–7174, 2012.

47
REGUERA, M.; PELEG, Z.; BLUMWALD, E. Targeting metabolic pathways for genetic engineering abiotic stress-tolerance in crops. Biochimica et Biophysica Acta, v.1819, p. 186–194, 2012.
RODRÍGUEZ-GAMIR, J.; et al. Citrus rootstock responses to water stress. Scientia Horticulturae, v.126, p.95–102, 2010.
ROYCHOUDHURY, A. ; ROY, C.; SENGUPTA, D.N. Transgenic tobacco plants overexpressing the heterologous lea gene Rab16A from rice during high salt and water deficit display enhanced tolerance to salinity stress. Plant Cell Reports, v.26, p.1839–1859, 2007.
SAITOU, N.; NEI, M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, v. 4, p.406- 425, 1987.
SAVÉ, R. et al. Changes in leaf osmotic and elastic properties and canopy structure of strawberries under mild water stress. Horticultural Science, v.28, p.925–927, 1993.
SCHÄFER, G. Caracterização molecular, diagnóstico e avaliação de porta-enxertos na citricultura gaúcha. 2000. 81f. Dissertação (Mestrado em Fitotecnia) - Universidade Federal do Rio Grande do Sul, 2000.
SCHINOR, E.H. et al. Organogênese in vitro a partir de diferentes regiões do epicótilo de citrus sp. Revista Brasileira de Fruticultura, v. 28, p. 463-466, 2006.
SENTELHAS, P.C. Agrometeorologia dos citros. In: MATTOS JÚNIOR et al. (Ed.). Citros. Campinas: Instituto Agronômico e Fundag, p. 317-344, 2005.
SEO, M.; KOSHIBA, T. Complex regulation of ABA biosynthesis in plants.TRENDS in Plant Science, v. 7, p. 41-48, 2002.
SHINOZAKI, K.; YAMAGUCHI-SHINOZAKI, K. Gene networks involved in drought stress response and tolerance. Journal of Experimental Botany, v.58, p.221–227, 2007.

48
SMEETON, R.C. THESIS (PhD). An evaluation of the effects of over-production of ABA on whole plant water use, growth and productivity. University of Warwick. January, 2010.
SOUZA, L.D. et al. Avaliação de plantas cítricas, em diferentes profundidades de plantio, em Latossolo Amarelo dos Tabuleiros Costeiros. Revista Brasileira de Fruticultura, v.26, p.241-244, 2004.
SUN, L. et al. Suppression of 9-cis-Epoxycarotenoid Dioxygenase, Which Encodes a Key Enzyme in Abscisic Acid Biosynthesis, Alters Fruit Texture in Transgenic Tomato. Plant Physiology, v.158, p. 283–298, 2012.
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 3 ed. São Paulo: Artmed, 2004. 719p
TAMURA, K.; DUDLEY, J.; NEI, M.; KUMAR, S. MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Molecular Biology and Evolution, v. 24, p. 1596 - 1599. 2007.
THOMPSON, J. D.; HIGGENS, D. G.; GIBSON, T. J.; CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research, v. 22, p. 4673-4680, 1994.
THORDAL-CHRISTENSEN H. et al. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley–powdery mildew interaction. Plant Journal, v.11, p.1187–1194, 1997.
TOUCHETTE, B.W. et al. Drought tolerance versus drought avoidance: a comparison of plant–water relations in herbaceous wetland plants subjected to water withdrawal and repletion. Wetlands, v.27, p.656–667, 2007.
TUNNACLIFFE, A.; WISE, M. J. The continuing conundrum of the LEA proteins. Naturwissenschaften, v. 94, p.791–812, 2007.

49
UMEZAWA,T. et al. Engineering drought tolerance in plants: discovering and tailoring genes to unlock the future. Current Opinion in Biotechnology, v.17, p. 113-122, 2006.
VALLIYODA, B.; NGUYEN H. T. Understanding regulatory networks and engineering for enhanced drought tolerance in plants. Current Opinion in Plant Biology, v.9, p.189–195, 2006.
VEERANAGAMALLAIAH, G.et al. Group 1 and 2 LEA protein expression correlates with a decrease in water stress induced protein aggregation in horsegram during germination and seedling growth. Journal of Plant Physiology, v.168, p.671–677, 2011.
WAN, X-R.; LI, L. Regulation of ABA level and water-stress tolerance of Arabidopsis by ectopic expression of a peanut 9-cis-epoxycarotenoid dioxygenase gene. Biochemical and Biophysical Research Communications, v. 347, p. 1030-1038, 2006.
WANG, W. et al. Enhanced drought tolerance in transgenic Leymus chinensis plants with constitutively expressed wheat TaLEA3. Biotechnology Letters, v.31, p.313–319, 2009.
WANG, W. et al. Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta, v. 218, p. 1–14, 2003.
YUE, B.et al. Genetic basis of drought resistance at reproductive stage in rice: separation of drought tolerance from drought avoidance. Genetics, v.172, p. 1213–1228, 2006.
YUE, et al. Overexpression of the AtLOS5 gene increased abscisic acid level and drought tolerance in transgenic cotton. Journal of Experimental Botany, v. 63, p. 3741-3748, 2012.
ZHANG, Y.et al. Overexpressing SgNCED1 in Tobacco Increases ABA Level, Antioxidant Enzyme Activities, and Stress Tolerance. Journal of Plant Growth Regulation, v. 27, p.151–158, 2008.

50
ZHU, J.K. Salt and drought stress signal transduction in plants. Annual Review Plant Biology, v. 53, p. 247–273, 2002.


















