
UNIVERSIDADE FEDERAL DE SERGIPE PRÓ-REITORIA DE … · proestro todas as outras ratas foram...
66
UNIVERSIDADE FEDERAL DE SERGIPE PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA MESTRADO EM CIÊNCIAS FISIOLÓGICAS JOSÉ MARCOS MELO DOS SANTOS INFLUÊNCIA DAS VARIAÇÕES HORMONAIS NO CONTATO SOCIAL DE RATAS COM DIFERENTES CONDIÇÕES NOCICEPTIVAS SÃO CRISTÓVÃO 2017
-
Upload
hoangkhanh -
Category
Documents
-
view
213 -
download
0
Transcript of UNIVERSIDADE FEDERAL DE SERGIPE PRÓ-REITORIA DE … · proestro todas as outras ratas foram...

UNIVERSIDADE FEDERAL DE SERGIPE
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
MESTRADO EM CIÊNCIAS FISIOLÓGICAS
JOSÉ MARCOS MELO DOS SANTOS
INFLUÊNCIA DAS VARIAÇÕES HORMONAIS NO
CONTATO SOCIAL DE RATAS COM DIFERENTES
CONDIÇÕES NOCICEPTIVAS
SÃO CRISTÓVÃO
2017

ii
JOSÉ MARCOS MELO DOS SANTOS
INFLUÊNCIA DAS VARIAÇÕES HORMONAIS NO
CONTATO SOCIAL DE RATAS COM DIFERENTES
CONDIÇÕES NOCICEPTIVAS
Dissertação apresentada ao Programa de Pós-
Graduação em Ciências Fisiológicas da
Universidade Federal de Sergipe como
requisito parcial à obtenção ao grau de Mestre
em Ciências Fisiológicas.
Orientador: Prof. Dr. Luís Felipe Souza da
Silva
SÃO CRISTÓVÃO
2017

iii
FICHA CATALOGRÁFICA ELABORADA PELA BIBLIOTECA CENTRAL
UNIVERSIDADE FEDERAL DE SERGIPE

iv
JOSÉ MARCOS MELO DOS SANTOS
INFLUÊNCIA DAS VARIAÇÕES HORMONAIS NO
CONTATO SOCIAL DE RATAS COM DIFERENTES
CONDIÇÕES NOCICEPTIVAS
Dissertação apresentada ao Programa de Pós-
Graduação em Ciências Fisiológicas da
Universidade Federal de Sergipe como
requisito parcial à obtenção ao grau de Mestre
em Ciências Fisiológicas.
______________________________________________________
Prof. Drº. Luis Felipe Souza da Silva- UFS
______________________________________________________
Prof. Drº. Hector Julian Tejada- UFS
______________________________________________________
Prof. Drª. Adriana Gibara Guimarães- UFS

v
“Todo mundo é capaz de dominar uma dor, exceto quem a sente”
William Shakespeare

vi
AGRADECIMENTOS
Agradeço primeiramente a Deus, sem ele nada disso seria possível.
Aos animais por terem doado sua vida em prol de um desenvolvimento científico,
cabe a nós fazer com que isso não tenha sido em vão.
Ao meu orientador, Professor Doutor Luís Felipe, por ter me acolhido, sem nunca
se quer ter me visto anteriormente, ter partido nessa empreitada até então inédita para o
programa e para nós dois, eu na área de biológicas e ele de ter um orientando com uma
inclinação muito forte para as ciências humanas e sociais. Mas não só por isso, também
pela disponibilidade, pelo auxilio, pela orientação e pela amizade. O senhor me inspira
tanto como profissional, como pessoa.
As agências de fomento CAPES e FAPITEC pelos recursos financeiros
disponibilizados para a realização deste projeto.
Ao Programa em Ciências Fisiológicas e aqui eu me refiro a todo o corpo docente
e a toda equipe que coordena esse programa. Agradeço aos professores Daniel Badauê,
José Ronaldo, Enilton Camargo, Julian Tejada por sempre serem muito solícitos no que
se referia as minhas dúvidas, mas também no que se tratava ao meu desempenho como
discente.
A Renivan que nas horas de desespero e de tormenta sempre me recebeu na sua
sala de forma afetuosa e sempre atenciosa tentando assim diminuir ou até mesmo
tamponar os momentos de angustia, também pela sua presteza em sempre auxiliar os
discentes a dirimir suas dúvidas com uma disposição e assertividade louváveis.
A minha mãe, pelo afeto e pelas palavras de apoio nas horas difíceis, pela
confiança, por acreditar em mim e acima de tudo pelo seu amor e compreensão, por isso
eu afirmo mais uma vez, nós chegamos até aqui.
Ao LAFICO e a todos que o compõem, em especial a professora Flávia Teixeira
por ter disponibilizado a estrutura física para que pudéssemos fazer do laboratório nosso
quartel general.
A minha namorada Larissa pela paciência, pela disposição de me ouvir horas a fio
a falar sobre uma temática até então desconhecida para ela, por ter sido inúmeras vezes
minha banca avaliadora hipotética, por ter aguentado todos os períodos de estresse e todas
as dificuldades, por ter entendido os momentos onde foi preciso abdicar de algumas horas
na sua companhia etc.

vii
Ao meu pai, por ter sido compreensivo e sempre solícito com relação aos meus
pedidos, por acreditar no meu potencial e por ter me auxiliado. Ao meu irmão por ter me
auxiliado inúmeras vezes no que se referia a logística do transporte para os experimentos,
mas também pelo incentivo e pela fé em meu potencial.
Aos meus amigos e aqui são muitos, tantos os quais eu acho impossível listar
todos: Primeiramente a Alan Bruno que tem estado comigo desde a pelada com o golzinho
de chinela até os testes de formalina realizados no LAFICO, é isso aí irmão, meu muito
obrigado. Kelly Coutinho por sempre ter me auxiliado, sendo companheira de
experimento e de todas as horas, a José Marcos Meneses meu muito obrigado pela
prestatividade e companheirismo sempre demonstrados. Roas, Clarissa, Amélia, Marcos
Vinicius, José Diogo entre outros, o meu muitíssimo obrigado a todos vocês por serem e
estarem aqui. E a todos os outros amigos que sempre bateram aquele papo terapêutico
no corredor do Departamento de Fisiologia.
Enfim, e no fim agradeço de forma geral a todos os citados e a todos aqueles que
por motivo de esquecimento ou por tentar ser sucinto acabaram não estando aqui, o meu
muito obrigado a todos que direta e indiretamente me ajudaram a chegar até aqui.

viii
RESUMO
INFLUÊNCIA DAS VARIAÇÕES HORMONAIS NO CONTATO SOCIAL DE
RATAS COM DIFERENTES CONDIÇÕES NOCICEPTIVAS. José Marcos Melo
dos Santos, São Cristovão, 2017.
A dor pode ser definida como uma experiência emocional e sensorial desagradável associada a
uma lesão tecidual real ou potencial. Cerca de 70% dos indivíduos que são acometidos por dores
crônicas são do sexo feminino e os hormônios gonadais tem função determinante no que tange o
diformismo e as respostas nociceptivas. Estudos sugerem que roedores são capazes de reconhecer
o componente álgico em co-específicos e despender comportamentos ativos em direção ao
mesmo. O objetivo principal do presente trabalho foi avaliar a influência das variações hormonais
relativas ao ciclo estral no contato social de ratas durante a dor inflamatória e o efeito do contato
social nas respostas nociceptivas de ratas com dor inflamatória induzida por formalina. Foram
utilizadas 120 ratas Wistar com 2 a 3 meses de idade. O projeto foi dividido em dois protocolos
experimentais. O experimento I teve como objetivo a avaliação do comportamento de ratas
residentes durante a exposição a coabitantes com dor inflamatória e foi composto pelos seguintes
grupos e animais experimentais: Ratas residentes que foram submetidas aos animais controle
(CTRL), formalina contato social (FCS) ou salina (SAL). Essa divisão dos grupos se repetiu nas
diferentes fases do ciclo, das ratas residentes que foram denominadas: Residente Estro (ES),
Residente Ovariectomizada (OV) e Residente Proestro (PRO). Excetuando as residentes estro e
proestro todas as outras ratas foram submetidas a cirurgia de ovariectomia. O experimento II foi
composto pelos os grupos: Formalina Contato Social (FCS), e o Formalina Isolado (FI) o qual
recebeu a administração da formalina (1%) na pata posterior esquerda e após foi isolada por 20
minutos. Os animais coabitantes dos grupos FCS/ES, FCS/PRO, FCS/OV e FIS tiveram suas
respostas nociceptivas avaliadas por 40 minutos. Os parâmetros utilizados para avaliação das
respostas nociceptivas foram o tempo de lambida e o número de sacudidas da pata traseira. Todos
os grupos foram compostos por n= 8. No que se refere ao Duração do Contato observou-se
interação entre fase do ciclo e o tratamento (p= 0,0013). Neste sentido as residentes castradas
mostraram duração do contato maiores que as residentes estro (p< 0,05) e as residentes
no proestro (p< 0,001). No experimento II não se observou diferença entre os grupos FCS e FI
nos grupos compostos por animais que tiveram contato com as residentes castradas e no estro. O
grupo composto por animais que haviam tido contato com as residentes proestro mostrou redução
no número de sacudidas o período entre 0 e 30 minutos de observação. Os resultados sugerem
que ratas são capazes de reconhecer o componente álgico em um coabitante e direcionar
comportamentos ativos em pró do mesmo, entretanto a intensidade e a qualidade desse contato
podem variar de acordo com o ciclo. No que tange as respostas nociceptivas observou-se que o
contato social evocado através de comportamentos sociais diretos não foi suficiente para eliciar
analgesia, entretanto comportamentos agressivos por parte da residente eliciaram uma analgesia
de longa duração nas coabitantes que pode indicar a ativação do sistema descendente
antinociceptivo endógeno através do RVM com caráter opioide e sendo induzida por estresse.
Descritores: Fêmeas; Ciclo estral; Empatia; Analgesia; Nocicepção.

ix
ABSTRACT
THE INFLUENCE OF HORMONAL VARIATIONS ON SOCIAL CONTACT
OF FEMALE RATS WITH DIFFERENT NOCICEPTIVE CONDITIONS. José
Marcos Melo dos Santos, São Cristovão, 2017.
Pain can be defined as an unpleasant emotional and sensory experience associated with actual or
potential tissue injury. About 70% of the individuals affected by chronic pain are female and the
gonadal hormones play a decisive role in terms of sex differences and nociceptive responses.
Studies suggest that rodents can recognize pain in co-specifics and expend active behaviors
toward it. The main objective of the present study was to evaluate the influence of hormonal
changes related to the estrous cycle on the social contact of rats during inflammatory pain and the
effect of social contact on the nociceptive responses of female rats with inflammatory pain. Wistar
female rats were used at 2 to 3 months of age. The project was composed of two experimental
protocols. Experiment I evaluate the behavior of resident rats during exposure to cohabitants with
inflammatory pain. It was composed of the following groups and experimental animals: Resident
rats (RES), Control (CTRL), Formalin social contact (FCS) and saline (SAL). The three groups
were repeated in different cycles stages, and three experiments were carried out where the
residents are called Resident Estrous (RES / ES), Resident Ovariectomized (RES / OV) and
Resident Proestrus (RES / PRO) All rats but ES and PRO residents underwent ovariectomy
surgery. Experiment II was composed of the groups: Formalin Social Contact and the Formalin
Isolate, which received the administration of formalin (1%) and was submitted to a 20min
isolation period. After the 20 min of contact or isolation the nociceptive responses were evaluated
for 40 minutes. The parameters used to evaluate the nociceptive responses were the lick time and
the number of flinches. All groups were composed of n = 8. The results showed interaction
between the phase the cycle versus treatment (p = 0.0013) for contact duration. In the case of the
group of OV residents, (P <0.05) and with those residents in proestrus (p <0.001). In the
experiment II, no difference was observed between the FCS and FI groups in groups composed
of animals that had contact with ovariectomized or estrus residents. The group composed of
animals that had contact with the proestrus residents showed analgesia during thirty minutes of
the formalin test between 0 and 30 minutes of observation. The results suggest that rats recognize
pain component in a cohabitant and direct active behaviors towards it, however the intensity and
quality of this contact may vary according to the cycle. Regarding the nociceptive responses, it
was observed that the social contact evoked through direct social behaviors was not enough to
elicit analgesia, however aggressive behaviors on the part of the resident elicited an analgesia that
may be based on the descending system through RVM with opioid character and being induced
by stress.
Keywords: Female, Estrous cycle; Empaty; Analgesia; Nociception.

x
LISTA DE FIGURAS
Figura 1: Oscilação hormonal durante o ciclo, considerando o período estral de 4
dias........................................................................................................................... 22
Figura 2: Rata Wistar na câmara de observação durante o teste de Formalina..........31
Figura 3: Representação esquemática das etapas realizadas com os grupos do
Experimento I ..........................................................................................................34
Figura 4: Representação esquemática das etapas realizadas com os grupos do
Experimento II..........................................................................................................35
Figura 5: Latência, em segundos, para o contato entre o rato residente e os ratos dos
grupos Controle (CTRL) Salina (SAL) ou Formalina (FORM) .............................. 37
Figura 6: Duração, em segundos, do contato das ratas residentes (OVX/ES/PRO)
com os animais dos grupos Controle (CTRL/OVX/ES/PRO), Salina (OVX/
/ES/PRO) e Formalina (FORM/OVX/ES/PRO)...................................................... 38
Figura 7: Duração, em segundos, do contato da rata residente de todos os grupos com
ratas dos grupos Controle (CTRL) Salina (SAL) ou Formalina
(FORM)....................................................................................................................39
Figura 8: Duração, em segundos, do contato do rata residente castrada com ratas dos
grupos Controle (CTRL) Salina (SAL) ou Formalina (FORM)............................. 39
Figura 9: Duração, em segundos, do contato da rata residente estro com ratas dos
grupos Controle (CTRL) Salina (SAL) ou Formalina (FORM)............................. 40
Figura 10: Duração, em segundos, do contato do rata residente proestro com ratas
dos grupos Controle (CTRL) Salina (SAL) ou Formalina (FORM)....................... 41
Figura 11: Duração, em segundos, do contato da rata residente castrada com animais
dos grupos Controle (CTRL), Salina (SAL) e Formalina (FORM) em intervalos de
tempo....................................................................................................................... 42
Figura 12: Duração, em segundos, do contato da rata residente estro com animais dos
grupos Controle (CTRL), Salina (SAL) e Formalina (FORM) em intervalos de
tempo....................................................................................................................... 43
Figura 13: Duração, em segundos, do contato da rata residente proestro com animais
dos grupos Controle (CTRL), Salina (SAL) e Formalina (FORM) em intervalos de
tempo....................................................................................................................... 44
Figura 14: Número de sacudidas da pata dos animais dos grupos Formalina Contato-
social (FC) ou grupo Formalina-isolado (FI) no grupo Residente Estro.................. 45

xi
Figura 15: Número de sacudidas da pata dos animais dos grupos Formalina Contato-
social (FC) ou grupo Formalina-isolado (FI) no grupo Residente Estro....................46
Figura 16: Número de sacudidas da pata dos animais dos grupos Formalina Contato-
social (FC) ou grupo Formalina-isolado (FI) no grupo residente Proestro............... 47
Figura 17: Duração do tempo de lambida, em segundos, dos animais dos grupos
Formalina Contato-social (FCS) ou grupo Formalina-isolado (FI) nas variadas fases
do ciclo..................................................................................................................... 47

xii
LISTA DE ABREVIATURAS E SIGLAS
AI Insúla Bilateral
ANOVA Análise de variância
CCA Córtex cingulado anterior
CDME Corno dorsal da medula espinal
CEPA Comitê de ética em pesquisa animal
CI Córtex insular
CONCEA Conselho Nacional de Controle de Experimentação Animal
CPME Corno posterior da medula espinal
CSS Córtex somatossensorial
CTRL Ratas Controle
CTRL/ES Controle Estro
CTRL/OVX Controle Ovariectomizada
CTRL/PRO Controle Proestro
EPM Erro padrão da média
FCS Formalina Contato Social
FCS/ES Formalina Contato Social Estro
FCS/OVX Formalina Ovariectomizada
FCS/PRO Formalina Contato Social Proestro
FI Formalina Isolado
LC Locus coeruleus
NMR Núcleo magno da rafe
RES Ratas Residentes
RES/ESTRO Residentes Estro
RES/OVX Residentes Ovariectomizadas
RES/PRO Residentes Proestro
RVM Bulbo Rostroventromedial

xiii
SAL Ratas Salinas
SAL/ES Salina Estro
SAL/OVX Salina Ovariectomizada
SAL/PRO Salina Proestro
SCP Substância cinzenta periaquedutal
UFS Universidade Federal de Sergipe

xiv
SUMÁRIO
1.INTRODUÇÃO ......................................................................................................... 15
2. REVISÃO DE LITERATURA ................................................................................ 17
2.1 Dor ........................................................................................................................... 17
2.2 Dor e Dimorfismo Sexual ....................................................................................... 19
2.3 Ciclo Estral .............................................................................................................. 22
2.4 Comportamento Pró-social .................................................................................... 23
2.5 Dimorfismo e Empatia. .......................................................................................... 25
3. OBJETIVOS ............................................................................................................. 28
3.1 Objetivo Geral ......................................................................................................... 28
3.2 Objetivos Específicos .............................................................................................. 28
4. MATERIAL E MÉTODOS: .................................................................................... 29
4.1 Animais .................................................................................................................... 29
4.2 Determinação das fases do Ciclo Estral ................................................................ 29
4.3 Ovariectomia ........................................................................................................... 30
4.4Teste da formalina ................................................................................................... 30
4.5.1 Experimento I: Avaliação do comportamento de ratas residentes durante a
exposição a coabitantes com dor inflamatória. ............................................................... 31
4.5.2 Experimento II: Avaliação do efeito do contato social no comportamento álgico de
ratas submetidos ao teste de formalina. .......................................................................... 35
4.6 Eutanásia dos animais ............................................................................................ 35
4.7 Análise dos resultados ............................................................................................ 35
5. RESULTADOS ......................................................................................................... 37
5..1 Experimento I: Avaliação do comportamento de ratas residentes durante a
exposição a coabitantes com dor inflamatória. .......................................................... 37
5.1.1 Latência para o contato .......................................................................................... 37
5.1.2 Duração de Contato ............................................................................................... 37
5.1.3 Duração do tempo de contato em intervalos ......................................................... 41
5.2 Experimento II: Avaliação do efeito do contato social no comportamento álgico
de ratas submetidos ao teste de formalina. ................................................................ 44
6. DISCUSSÃO ............................................................................................................. 48
7. CONCLUSÃO ........................................................................................................... 56
REFERÊNCIAS ........................................................................................................... 57

xv
ANEXO A- CERTIFICADO COMITÊ DE ÉTICA .................................................... 64

15
1. INTRODUÇÃO
A dor pode ser entendida como uma experiência de caráter desagradável, sendo
este dividido em aspectos sensoriais e psicológicos. A dor está relacionada a uma lesão
que pode ser real ou potencial (MERSKEY e BOGDUK, 1994).
No que se refere ao processamento dos estímulos dolorosos observa-se a
existência de dois componentes: O sensório discriminativo que está envolvido na
localização do ponto acometido pelo estimulo nocivo, nesse componente a informação é
levada através do trato espinotalâmico que através das projeções feitas pelos núcleos
talâmicos laterais chega até o córtex somatossensorial primário e secundário. O afetivo
motivacional que estaria relacionado com os aspectos emocionais que circundam a
mesma, como por exemplo a interpretação que a vivência daquele estimulo doloroso pode
vir a ter, o componente afetivo motivacional tem sua informação transmitida através do
trato espinotalâmico e espinorreticular, chegando até o córtex cingulado anterior (ACC)
e ao córtex insular do sistema límbico (EKMAN et al. , 2008; SEWARDS et al. , 2002;
VOGT et al 2000).
Segundo autores o ACC mostra importante função na modulação das respostas
afetivas e sensoriais da dor, de modo que os neurônios que fazem parte do mesmo se
mostram ativados em respostas a situações de dor infligida contra o indivíduo ou durante
a observação de um coabitante sendo submetido a estimulo doloroso. A inibição da
ativação do ACC está relacionada com a diminuição das respostas de dor em ratos (GU
et al. ,2015; NOGA, 2013; ALOISI et al. ,2002).
No que se refere a dor e dimorfismo sexual observa-se que 70% dos indivíduos
que são acometidos por dores crônicas são do sexo feminino. Os hormônios gonadais,
tem sido descritos como um fator determinante na diferença de sensibilidade dolorosa
(CRAFT, 2008; MANSON, 2010, PEDROSO 2011). Ao investigar as possíveis
diferenças no limiar nociceptivo de mulheres acometidas por cefaleia em diferentes
pontos do ciclo menstrual, observou-se diferença entre as fases do ciclo, assim como
também diferença entre os grupos que tomavam ou não anticoncepcional ( SILVA, 2015).
Em se tratando de animais, não se utiliza o termo ciclo menstrual e sim o ciclo
estral, sendo o mesmo composto por: Metaestro, Diestro, Proestro e Estro, totalizando
quatro fases. O ciclo é caracterizado por diversas flutuações nas concentrações
hormonais, principalmente nos gonadais. Exemplo disso seriam os picos hormonais
encontrados durante o Proestro, assim como os níveis próximos aos parâmetros basais no

16
Estro. Essas variações hormonais podem influenciar comportamentos como os de
ansiedade, estresse, padrão mnemônico entre outros (BIANCHI, 2004; LEAL, 2015).
O comportamento de empatia/pró-social é definido como a capacidade de
compartilhar sentimentos com outrem não sendo necessário que o indivíduo já tenha
passado pela situação em questão (KEUM e SHIN; 2016). A empatia é um construto
amplamente estudado, e existem evidências de que comportamentos pró-empáticos
aconteçam em várias espécies, como por exemplo em aves, roedores, primatas e em
humanos (PRESTON, 2002; LANGFORD et al 2006).
Em se tratando do sexo Bartall e colaboradores (2014) observaram que fêmeas
mostravam comportamento pró-empático de libertar o coabitante com maior velocidade
se comparadas com os machos (BARTAL et al., 2014).Quando compararam homens,
mulheres que haviam sido mães e mulheres que até então nunca tinham tido filhos,
observou-se que independente da condição as mulheres mostravam maior ativação
cortical que os homens o que pode vir corroborar com uma maior sensibilidade do sexo
feminino no que tange o comportamento pró-empático (PROVERBIO, 2006). Além
disso, a ativação das áreas relacionadas aos neurônios espelhos como o ACC e o sistema
límbico se mostrou mais intensa nas mulheres que nos homens (SCHULTE-RHUTER et
al, 2008).
Com base no que foi aludido anteriormente o presente projeto se propõe a avaliar
as possíveis influencias do ciclo estral no contato social entre ratas residentes e com dor
inflamatória. Assim como também se esse contato pode de algum modo influir nas
respostas nociceptivas das mesmas, alterando o padrão álgico induzido pela formalina.
A nossa hipótese no presente trabalho é a de que o contato social é estimulado
pela condição álgica da rata coabitante e varia de acordo com a fase do ciclo estral da rata
residente. Além disso, acreditamos que o contato social induzira hipoalgesia nas ratas
com dor inflamatória. .

17
2. REVISÃO DE LITERATURA
2.1 Dor
A dor pode ser definida como uma experiência emocional e sensorial desagradável
associada a uma lesão tecidual real ou potencial, ou descrita em termos de tal lesão, assim
sendo observa-se um caráter cognitivo e emocional e concomitantemente relacionado a
aspectos culturais e psíquicos (MERSKEY e BOGDUK, 1994).
A detecção dos estímulos nocivos é realizada por nociceptores, os quais
respondem a estímulos mecânicos, térmicos e químicos de grande intensidade. A
transmissão da dor está relacionada a ativação de fibras aferentes do tipo Aδ e C que
conduzem informações a respeito de estímulos nociceptivos da periferia para a medula
espinal. As fibras Aδ estão relacionadas com a transmissão de estímulos de curta duração
e em dores descritas como agudas e são fibras mielinizadas. As fibras do tipo C estão
relacionadas com dores lentas e com uma sensação de ardência e por se tratarem de fibras
amielinizadas tem menor velocidade na transmissão das respostas em comparação as
fibras Aδ. Os neurônios aferentes primários associados a fibras Aδ e C desempenham
três funções principais no que diz respeito a nocicepção. Transdução: Está relacionado a
detecção do estimulo nocivo e a conversão do mesmo em sinais elétricos. Transmissão:
A informação é conduzida da periferia para a medula espinal, onde ocorre as sinapses
com neurônios presentes em lâminas específicas do corno dorsal da medula espinal, mais
precisamente as lâminas I, II e V e destas até áreas supra espinais envolvidas na
modulação da dor. Modulação: este processo representa mudanças que ocorrem no
sistema nervoso em resposta a estimulação nociva (VANDERAH, 2007).
A dor pode ser modulada endogenamente através de um sistema descendente
endógeno localizado no tronco encefálico. Tal sistema supra espinal, tem como principais
estruturas envolvidas a substância cinzenta periaquedutal do mesencéfalo (SCP), o lócus
coerúleos (LC) e o bulbo rostroventromedial (RVM), que inclui o núcleo magno da rafe
(NMR). A SCP, quando ativada, produz efeito analgésico por meio de projeções
excitatórias para o RVM ou para o LC (SILVA e MENESCAL, 2006). Os axônios dos
neurônios do RVM enviam projeções rostrocaudais, via funículo dorsolateral, que
estabelecem sinapses inibitórias com neurônios aferentes primários e nociceptivos de
segunda ordem no corno posterior da medula espinal (CPME), causando inibição da

18
transmissão nociceptiva. Além disso, o RVM contém opioides endógenos que
desempenham um importante papel na desinibição de interneurônios gabaérgicos
promovendo a ativação da via descendente antinociceptiva opioide endógena mediada
pelo RVM (PATEL, 2010; NOBACK et al., 2005).
O processamento cortical dos estímulos nociceptivos pode levar a diferentes
aspectos relacionados a percepção da dor, podendo serem ressaltados o sensorial
discriminativo e o afetivo motivacional. O primeiro deles se refere a capacidade de
localizar o ponto, a escala temporal e a intensidade dos danos efetivos ou potenciais ao
tecido. No que se refere a via do componente sensorial discriminativo, as informações
são levadas através do trato espinotalâmico e processadas no sistema de dor lateral que
se projeta através dos núcleos talâmicos laterais para o córtex somatossensorial primário
e secundário. O componente afetivo motivacional diz respeito aos efeitos que o estimulo
nocivo tem nas emoções e comportamentos, as informações são levadas pelos tratos
espinotalâmicos e espinorreticular, chegando aos núcleos intralaminares e medial do
tálamo e posteriormente ao córtex cingulado anterior (ACC) e ao córtex insular no sistema
límbico (EKMAN et al., 2008; SEWARDS et al., 2002; VOGT et al 2000).
Segundo Noga et al., (2013) a dor e o sofrimento afetivo são funcionalmente e
anatomicamente integrados no córtex cingulado anterior (ACC) e este modula os aspectos
sensoriais e afetivos da dor. Neste sentido, neurônios do ACC são ativados por
estimulação dolorosa e estão envolvidos em respostas à dor afetiva (ALOISI et al., 2002),
e a inibição do ACC, está associada a diminuição da atividade neural no ACC e
consequente diminuição da percepção da dor em ratos (GU et al., 2015).
A relação do ACC com os componentes afetivos da dor foi mostrada em estudo
em humanos que comparou as regiões encefálicas ativadas durante a aplicação de um
estímulo doloroso ou quando os indivíduos viam a pessoa amada recebendo um estímulo
doloroso, fenômeno denominado ´´empatia``. Assim, durante a observação apenas o
sistema associado com os componentes afetivos da dor era ativado (ínsula bilateral
anterior (AI), córtex cingulado anterior rostral (ACC), tronco cerebral, e cerebelo) sem
ativação de áreas relacionadas ao componente sensorial da dor. Assim, este estudo aponta
para relação entre a atividade neural do ACC, interações sociais e a percepção do
componente afetivo da dor (SINGER, et al 2004).
Apesar dos animais não poderem descrever sua dor, são capazes de manifestá-la
por sinais fisiológicos e respostas comportamentais (TEIXEIRA, 2005). Além disso,
roedores podem ainda reconhecer a dor em indivíduos da mesma espécie, de modo que,

19
a coabitação com um animal com dor crônica pode promover uma situação estressante, o
que provoca alterações comportamentais, tais como ansiedade e depressão, e altera as
respostas nociceptivas (SATOSHI et al., 2015).
Assim, em estudo, pesquisadores realizaram testes comportamentais de avaliação
nociceptiva (teste de contorção) e ansiedade (labirinto em cruz elevado e os ensaios de
campo aberto) e mostraram que ratos submetidos a um modelo de dor neuropática, pela
constrição do nervo ciático, induziram efeitos ansiogênicos e hipernocicepção em ratos
coabitantes (BAPTISTA et al 2015).
Em trabalho recente, foi mostrado que ratos são capazes de perceber a ocorrência
de dor em coabitantes e, a partir disso, iniciar comportamentos ativos direcionados ao
animal. Além disso, o contato social reduziu a expressão de comportamentos álgicos
relacionados a dor inflamatória (GONÇALVES, 2016).
2.2 Dor e Dimorfismo Sexual
Em um levantamento dos artigos de pesquisa básica publicados pela revista Pain
usando apenas ratos ou camundongos, no período de 1996 a 2005 foi verificado que 79%
dos artigos publicados testaram apenas machos, apenas 5% usaram ambos os sexos, sem
analisar diferenças sexuais e 3% não relataram os sexos dos sujeitos participantes
(MOGIL e BAILEY, 2010).
Pesquisas epidemiológicas relatam que 70% da população que sofre com dor
crônica é do sexo feminino. Entretanto como fora dito anteriormente os números de
pesquisas em ciências básicas não acompanham os dados clínicos. Pesquisadores
apontam que três motivos justificam esses números: medo de abordar novos referenciais
teóricos ou por temerem não os encontrar não abrem mão das pesquisas com machos,
assim como também a escassa existência de estudos que abordem a nocicepção em fêmeas
o que pode dificultar uma possível comparação entre trabalhos. Outro motivo é a
influência dos hormônios gonadais e a ciclicidade existente nas fêmeas o que por si só já
pode interferir nas pesquisas com cunho de investigação nociceptiva e que pode tornar
necessária uma maior amostra para que exista significância estatística (MOGIL e
BAILEY, 2010).
Em contrapartida observa-se o crescimento do número de estudos experimentais
que analisam a variabilidade de respostas nociceptivas e a diferença entre o efeito de
fármacos analgésicos entre machos e fêmeas. Essas diferenças podem ser atribuídas a

20
diferentes fatores: genéticos, ambientais, sociais e culturais, hormonais
(PEDROSO,2011).
Pesquisas clinicas realizadas tem mostrado que mulheres tendem a relatar maior
prevalência de dor e menor tolerância a mesma. Observa-se que a dor em indivíduos do
sexo feminino tende a ser mais severa, prolongada e recorrente quando em comparação
com os indivíduos do sexo masculino (CRAFT, 2008; MANSON, 2010; PEDROSO,
2011). Entretanto devem ser levados em conta alguns aspectos como qual a modalidade
de estimulo doloroso, o tempo de duração, local de aplicação e alguns fatores como:
aspectos biológicos, psicológicos e sociais, condições patológicas, ciclo estral ou
menstrual, gravidez e menopausa (PEDROSO, 2011).
No que se refere a influência dos hormônios gonadais, tem sido observada uma
tendência dos mesmos a serem vistos como um dos principais responsáveis pelas
diferenças de sensibilidade dolorosa. Um exemplo disso é a maior prevalência e
intensidade de fenômenos dolorosos em mulheres com idade reprodutiva, em decorrência
das flutuações hormonais (CRAFT, 2008; MANSON, 2010; PEDROSO 2011). Ao
investigar as possíveis diferenças no limiar nociceptivo de mulheres acometidas por
cefaleia em diferentes pontos do ciclo menstrual, observou-se diferença entre as fases do
ciclo, assim como também diferença entre os grupos que tomavam ou não
anticoncepcional (ARAÚJO, 2015).
Uma pesquisa utilizando roedores machos e fêmeas, mostrou a existência de uma
diferença no limiar nociceptivo entre os sexos. Nesse estudo onde eram utilizados
modelos de dor inflamatória, observou-se que animais machos se mostraram menos
suscetíveis a sentir dor e concomitantemente se fez necessário utilizar uma maior dose do
agente indutor de resposta inflamatória (formalina ou capsaicina) quando comparado com
fêmeas (BARRET, SMITH, PICKER, 2003).
Pesquisadores apontaram que as mulheres não só são mais acometidas por dores,
como também são mais sensíveis a mesma e menos tolerantes. Outro ponto levantado
pelos autores e que corrobora o que foi citado acima é o fato de que a diferença entre
machos e fêmeas no que tange as respostas nociceptivas é o que diz respeito a
característica do estimulo evocado: químico, térmico, mecânico, sendo relatado que as
maiores diferenças sexuais encontradas são relacionadas a dor por pressão no que se
refere as fêmeas (RILEY et al, 1998; MOGIL; BAILEY,2010; AMANDUSSON e
BLOMQVIST,2013).

21
Diferenças entre dores térmicas relacionada a calor e a mecânica (pressão) se
mostraram existentes, entretanto não se observou aparente diferença entre dores térmicas
provocadas por calor e frio em fêmeas. Em respeito a dor provocada por um agente
químico observou-se uma maior expressão de canais de potencial transiente (TRPV1)
ativados por capsaicina e injeções intramusculares de salina hipertônica e glutamato
(RILEY et al, 1998; YAMAGATA et al 2016).
Estudos apontam que a exposição neonatal a testosterona é necessária para o
desenvolvimento do fenótipo masculino de menor sensibilidade a estímulos nociceptivos,
isso foi bem demonstrado através de estudos com animais que através de indução de dor
inflamatória por carragenina ou por estimulo mecânico apresentaram maior limiar
nociceptivo do que animais machos que haviam sido castrados (LACROIX-FRALISH,
2005; BORZAN, 2006).
Em contrapartida outros trabalhos relatam que a administração de testosterona em
fêmeas, não foi capaz de induzir a mesma antinocicepção que ocorre nos machos. Uma
explicação para isso seria a hipótese da relação entre hormônios gonadais e
neurotransmissores. A analgesia induzida por estresse relacionada aos receptores NMDA
e a analgesia induzida por morfina parecem estar relacionadas à testosterona
(STERNBERG,1996; KRZANOWSKA,2002).
Parece cada vez mais que os machos e as fêmeas podem ter circuitos neurais
altamente diferentes que medeiam a dor. Alguma relação entre os receptores NMDA
(glutamato) e a analgesia induzida por estresse ou por kappa opioides em machos poderia
ser bloqueada através de antagonistas glutamatérgicos, já nas fêmeas isso não ocorre,
mesmo quando tentou-se bloquear essa analgesia (utilizando o mesmo mecanismo) a
mesma continuou a ocorrer (MOGIL e BAILEY,2010).
Em suma quando o assunto é a variada gama de respostas nociceptivas e o
dimorfismo sexual o que se observa é um consenso que relata que o modelo utilizado para
a investigação está intrinsecamente relacionado com as respostas e suas variações. No
que tange as flutuações hormonais pode-se notar uma incongruência da literatura. Alguns
estudos animais mostram que as respostas nociceptivas estão diretamente relacionadas ao
modelo nociceptivo adotado (BARRET, SMITH, PICKER, 2003; MOGIL e
BAILEY,2010; PEDROSO, 2011).

22
2.3 Ciclo Estral
Em se tratando do ciclo estral se mostra de suma importância em um primeiro
momento definir e diferenciar o mesmo do ciclo menstrual. A principal diferença entre
os termos citados anteriormente é no que diz respeito a ritimicidade em que acontecem,
o primeiro deles tem duração de 4 a 5 dias e tendo uma ciclicidade de cerca de 24 horas
(totalizando 4 fases), o segundo tem duração de cerca de 28 dias, em ambos ocorre a
participação do eixo adeno-hipofísario e os principais hormônios envolvidos são o
estrogênio e o progesterona (RODRIGUES, 2005).
No que tange a etimologia da palavra estro, ela deriva da palavra latina “oistros”
e que define o período de desejo sexual da fêmea. Os primeiros trabalhos relacionados ao
ciclo estral, datam de 1917 e 1922 respectivamente, o primeiro estudou a ocorrência dos
ciclos em porcos da Índia (cavia porcellus) e o segundo em camundongos (BIANCHI,
2004; VILELA et al. , 2007).
O ciclo estral é composto por 4 fases, sendo elas: Metaestro, Diestro, Proestro e
Estro. Durante o ciclo estral observa-se uma série de modificações nos parâmetros dos
hormônios gonadais, como por exemplo uma baixa concentração de estrogênio durante o
estro e uma maior concentração durante o proestro, esse apresentando o maior pico de
concentração. Em suma observa-se uma baixa concentração de estrogênio durante
metaestro, ao final do diestro ocorre uma gradativa elevação até o proestro e no período
do estro ocorre o retorno aos parâmetros basais (BIANCHI, 2004; LEAL, 2015).
FIGURA 1- Oscilação hormonal durante o ciclo, considerando o período estral de 4 dias.
Fonte : Leal,2015, p.18.

23
Em estudo que tinha como objetivo avaliar diferenças no nível de ansiedade entre
os diferentes ciclos das fêmeas e os machos através do labirinto em cruz elevado,
observou-se que fêmeas no diestro apresentaram maior perfil do tipo ansioso em relação
aos machos, no que se refere as outras fases do ciclo não se observou diferença (JUNIOR
e MORATO,2002).
Com o objetivo de avaliar possíveis interferências do ciclo estral na resposta
hormonal de estresse agudo, Bianchi (2004) submeteu ratas ovariectomizadas, machos e
ratas com ciclo normalizado a um protocolo de nado forçado, logo após ao experimento
os animais foram eutanasiados e após a dosagem de corticosterona observou-se que
durante a fase do metaestro o grupo controle (não submetido ao estresse agudo)
apresentou uma maior concentração plasmática desse hormônio. Em se tratando dos
grupos tratados observou-se duas coisas: a elevação plasmática de corticosterona em
machos encontrada foi menor quando comparados com as fêmeas, e um aumento abrupto
da concentração do mesmo ocorreu principalmente nas fases do estro e do diestro em
relação aos controles o que pode vir a caracterizar uma maior sensibilidade a agentes
estressores durante essas fases (BIANCHI,2004).
Em outro estudo que objetivava avaliar a influência do ciclo estral no padrão
mnemônico de ratas através do protocolo de reconhecimento de objetivos, os resultados
apontaram que uma maior concentração de estrogênio encontrada durante a fase do
proestro, contribuiu para uma facilitação na evocação da memória (LEAL,2015).
Com base nisso se mostra de suma importância a avaliação do ciclo estral, já que
o mesmo pode influenciar nas diferentes respostas fisiológicas e comportamentais das
fêmeas. Por isso observa-se uma certa resistência por parte da comunidade cientifica para
o trabalho com as mesmas. Entretanto se mostra necessário o trabalho com fêmeas, já que
como fora citado acima observam-se inúmeras diferenças entre respostas
comportamentais e fisiológicas no que tange machos e fêmeas.
2.4 Comportamento Pró-social
A empatia ou o comportamento pró-social vem sendo investigado por muitos
pesquisadores. Especulações apontam que esse tipo de comportamento somente aconteça
em primatas mais evoluídos, outros estudos encontram evidências da existência de

24
comportamentos pró-sociais em ratos e pombos (PRESTON, 2002; LANGFORD et al
2006).
Empatia pode ser definida como a capacidade de compartilhar sentimentos com
os outros, de sentir o que os outros sentem sem necessariamente precisar passar pela
situação, o que para Bandura (1962) pode ser denominada de aprendizagem vicária
(KEUM e SHIN; 2016).
Ainda segundo os autores mencionados anteriormente existe uma certa
dificuldade em definir o termo empatia, isso se deve as suas ramificações e as diversas
subdivisões que buscam explicar a sua origem, sua funcionalidade e quais os circuitos
neurais que estão envolvidos no desenvolvimento dessa habilidade que se mostra de suma
importância para a sobrevivência (KEUM e SHIN; 2016).
No que tange a funcionalidade observa-se a existência do que se define como
contágio emocional, rompendo a barreira da pesquisa animal e adentrando na parte clínica
observa-se isso em estudos de neuroimagem onde indivíduos que observam familiares
sofrerem lesões, tem uma maior ativação cerebral do sistema límbico assim como também
do ACC, região essa relacionada com o componente afetivo motivacional da dor. No dia
a dia observa-se o contagio emocional quando o choro de um recém-nascido elicia o choro
em outros, motiva o estado de alerta dos adultos ou até mesmo o bocejo em adultos
(KEUM e SHIN; 2016).
Atualmente é crescente o número de pesquisas que buscam comprovar a
existência de comportamentos pró-sociais/empáticos em roedores, mais precisamente
comportamentos de contágio emocional (LANGFORD et al. , 2006; CHEN et al. , 2015;
JEON et al. , 2010; AKYAZI e ERASLAN, 2014; KEUM e SHIN; 2016).
Com o objetivo de investigar a existência do comportamento pró-empático
Langford e colaboradores partiram da hipótese de que se a empatia for realmente existente
em ratos a exposição a um co-específico submetido a um estimulo doloroso deve afetar a
resposta do seu par. Primeiramente os animais foram submetidos ao teste de contorção
abdominal e dividido em pares onde ambos os ratos recebiam o ácido acético e em outras
duplas onde somente um sujeito recebia o estimulo. Ambos os ratos foram colocados em
tubos plásticos onde pudessem observar o par. Os ratos em duplas demonstraram maior
resposta nociceptiva que os ratos isolados, mas apenas quando os mesmos eram
habitantes da mesma caixa (LANGFORD et al 2006). Nos últimos anos estudos vem
demonstrando a existência de importante comportamento pró-social entre animais
(DECETY et al. , 2012; SIVASELVACHANDRAN et al. , 2016; KEUM e SHIN, 2016).

25
Neste sentido pesquisas mostraram que ratos apresentam comportamento pró-
social quando percebem que animais da mesma espécie estão em condições de estresse
não doloroso agindo para acabar com esse sofrimento através de ação livre de influência
externa (BARTAL et al., 2014; SATO et al., 2015; MARQUEZ et al 2015).
Comportamento este, que ocorreu independente de aprendizado ou recompensa social.
No que tange a dor, um estudo com ratos, mostrou que esses animais após observarem
seus familiares recebendo estímulo nocivo, apresentavam um aumento na sensibilidade
dolorosa (LANGFORD et al., 2006).
Em se tratando do sexo Bartall e colaboradores (2014) encontraram uma
informação que corrobora o senso comum. No mesmo experimento observou-se que
fêmeas demonstram um comportamento pró-empático de libertar o coabitante com maior
velocidade se comparadas com os machos. Outro ponto importante é o de que a
familiaridade não afetou a ocorrência desses comportamentos, mas afetou a velocidade
com que eles aconteceram. Não obstante encontrou-se que esses ratos respondiam a
coabitantes de cepas diferentes, mas não a animais da mesma espécie que não eram
coabitantes (BARTAL et al., 2014).
No que diz respeito ao controle central da dor associado a respostas sociais, foi
mostrado que respostas de medo social e observacional estão diretamente envolvidas com
ACC e a dor afetiva (JEON et al., 2010). Nesse estudo, ratos ao observarem outras ratas
(manifestantes) recebendo choques repetitivos, apresentaram respostas de medo maiores
quando os manifestantes se relacionavam socialmente com eles. Além disso, a inativação
do ACC e núcleos talâmicos parafascicular ou médio dorsais, que compreendem o
sistema de dor afetiva, reduziram a aprendizagem do medo observacional, enquanto que
as inativações de núcleos talâmicos sensoriais não produziram efeito. Estes resultados
mostram o envolvimento funcional do ACC e do sistema de dor afetiva no medo social e
observacional (JEON et al., 2010).
2.5 Dimorfismo sexual e Empatia
É comum que indivíduos compactuem e dividam de experiencias prazerosas e
desagradáveis, a habilidade de se colocar no lugar do outro é denominada empatia. A
empatia se apresenta como um construto multidimensional e inclui aspectos da
comunicação, da emoção e autoconsciência (DECETY e MORIGUICHI,2007).

26
Quando o assunto são as diferenças sexuais e a empatia observa-se um consenso
entre a literatura e o senso comum. Alguns estudos apontam que se tratando de diferença
sexual as mulheres se mostram muito mais empáticas que os homens e que conseguem
sustentar esse tipo de comportamento por mais tempo (DECETY e MORIGUCHI 2007;
DIMBERG et al 2011; GERY et al 2009; LUO et al 2014).
Em estudo realizado com chimpanzés onde através da aferição das modificações
ocorridas na temperatura da membrana timpânica após a exibição de cenas que continham
estímulos positivos, neutros e negativos (agressões) se observou que mesmo se tratando
de uma medida indireta, mas fidedigna os sujeitos que foram expostos aos estímulos
neutros e positivos não demonstraram alteração, entretanto os sujeitos expostos a cenas
desagradáveis tiveram a temperatura da membrana afetada, o que corrobora a teoria do
contágio emocional (PARR LISA e HOPIKNS 2000).
Em uma pesquisa realizada com homens e mulheres onde os mesmos foram
submetidos a ressonância magnética funcional, onde tinham as suas respostas avaliadas
através de escalas padronizadas de empatia, os indivíduos foram submetidos a expressões
de raiva ou medo e deveriam caracterizar quais as expressões que haviam visto de acordo
com a escala supracitada, denotando assim o comportamento pró-empático do que pode
ser definido como contágio emocional. Observou-se então que a ativação das áreas
relacionadas com os neurônios espelhos como o córtex cingulado anterior, sistema
límbico, se mostrou mais forte nas mulheres que nos homens (SCHULTE-RHUTER et
al, 2008).
Quanto utilizou-se outro método para avaliar os parâmetros fisiológicos
relacionados com o comportamento de empatia, pesquisadores utilizaram a
eletromiografia do musculo zigomático e dividiram os sujeitos em dois grupos : alto
contágio emocional e baixo contágio emocional e observaram que indivíduos que tem
menor contágio emocional tinham dificuldades em diferenciar expressões faciais de raiva
ou felicidade, em se tratando do grupo com alto contágio emocional os indivíduos
conseguiram caracterizar as expressões a que foram expostos (DIMBERG et al, 2011).
Uma grande gama de estudos que tinham como objetivo analisar possíveis
interferências da postura maternal no comportamento pró-empático através de
ressonância magnética apontaram que quando comparadas com mulheres que não haviam
sido mães ou até mesmo com homens, ocorria uma maior ativação na circuitaria pró-
empática nas mulheres que já haviam sido mães. Os resultados encontrados
demonstraram maior ativação nas áreas: córtex orbito frontal, amigdala, cortéx cingulado

27
anterior e substância cinzenta periaquedutal (BARTELS e ZEKI, 2004; LEIBENLUFT
et al., 2004; NITSCHKE et al.,2004; PROVERBIO, 2017).
Em um estudo que tinha como objetivo analisar a diferença neural de ativação
entre mulheres que já haviam sido mães, homens que já haviam sido pais e mulheres que
nunca haviam tido filhos, utilizou-se de um ensaio eletrofisiológico e observou-se que a
ativação cortical das mulheres que já haviam sido mães foi maior que a dos homens, as
mulheres que até então também não tinham sido mães demonstraram maior ativação que
os homens, o que pode vir a sugerir uma maior sensibilidade do sexo feminino
(PROVERBIO, 2006).
Com base nas referências supracitadas observa-se uma consonância da literatura
com o senso comum como já foi mencionado anteriormente. Em pesquisa feita por
Proverbio (2017) onde através de estudos de ressonância magnética foi encontrada uma
maior ativação do que o autor denomina circuitaria pró-empática, os resultados apontaram
para uma maior ativação da mesma por parte dos sujeitos do sexo feminino que
participaram do estudo, de modo que o autor suscita algumas hipóteses para a explicação
do resultado encontrado por ele e que está de acordo com grande parte da literatura : De
acordo com a literatura pode-se explicar esses resultados devido ao perfil biológico
desenvolvido pela mulher através da evolução (cuidadora), necessitando assim ter um
maior poder de distinção entre expressões faciais de alegria ou tristeza, dor ou postura
neutra (PROVERBIO,2017).

28
3. OBJETIVOS
3.1 Objetivo Geral
• Avaliar a influência das variações hormonais de ratas no contato social durante
condições nociceptivas e o efeito do contato social na resposta nociceptiva de ratas
com diferentes condições nociceptivas.
3.2 Objetivos Específicos
• Avaliar o comportamento social de ratas residentes durante a exposição a
coabitantes com diferentes condições nociceptivas.
• Avaliar possíveis influências das variações hormonais do ciclo estral no contato
social de ratas residentes durante a exposição a coabitantes com diferentes condições
nociceptivas.
• Avaliar o efeito do contato social no comportamento álgico de ratas com dor
inflamatória.

29
4. MATERIAL E MÉTODOS:
4.1 Animais:
Neste projeto foram utilizadas 120 ratas Wistar, com 2 a 3 meses de idade,
provenientes do Biotério setorial do Departamento de Fisiologia. Os animais foram
mantidos no biotério do Laboratório de Fisiologia do Comportamento da UFS, em gaiolas
plásticas (41 x 34 x 17,5 cm) com no máximo cinco animais por gaiola. Os mesmos
estavam alojados em ambiente com temperatura controlada entre 22 e 24ºC, e ciclo
claro/escuro de 12h com livre acesso à água e ração. Todos os protocolos experimentais
foram conduzidos em acordo com os princípios éticos estabelecidos pelas Resoluções
Normativas, Decretos e Portarias do Conselho Nacional de Controle de Experimentação
Animal (CONCEA), bem como a Lei Nº 11.794, de 8 de outubro de 2008 (Lei Arouca).
Os experimentos tiveram início após aprovação pelo Comitê de Ética em Pesquisa Animal
(CEPA) da Universidade Federal de Sergipe, sob número de protocolo 52/2016 ( Em
anexo).
4.2 Determinação das fases do Ciclo Estral:
Para avaliar a possível interferência das fases do ciclo estral no comportamento
social das fêmeas, fez-se a análise das células presentes no lavado vaginal obtido através
da técnica do esfregaço vaginal. Assim, as ratas residentes foram submetidas ao lavado e
coleta de conteúdo vaginal diariamente. Para tanto, foi utilizada uma pipeta de plástico
contendo salina e que tinha como objetivo a realização do lavado seguido da coleta do
conteúdo vaginal. Na sequência, o lavado fora colocado em uma lâmina e levado para
análise em um microscópio através das objetivas de 10x e 40x. As diferentes fases foram
identificadas de acordo com a presença e prevalência das células encontradas no lavado.
A coleta do conteúdo vaginal foi sempre feita no mesmo horário durante todo o período,
uma vez que a mudança dos ciclos pode ocorrer durante a manhã, tarde ou noite de um
mesmo dia. Não obstante é importante frisar que a coleta do lavado vaginal ocorreu por
15 dias consecutivos antes dos experimentos com o objetivo de acompanhar as variações
do ciclo estral identificando assim as fases e possibilitando a programação dos
experimentos nas fases escolhidas.

30
4.3 Ovariectomia
Com o objetivo de evitar variações hormonais relacionados ao ciclo estral, foi
realizada a cirurgia de remoção dos ovários de algumas ratas, ovariectomia. Para a
remoção dos ovários, os animais foram anestesiados com solução de ketamina
(100mg/kg) e xilazina (10 mg/kg) por via intraperitoneal (i.p). Após a anestesia foi feita
uma tricotomia da região dorsal, entre a a última costela e a coxa, com o animal
posicionado em decúbito ventral. Na sequência, foi realizada uma incisão longitudinal de
aproximadamente três centímetros na pele e tecido subcutâneo. A parede muscular foi
penetrada até ser obtido acesso a cavidade abdominal. O ovário foi localizado e sua
retirada aconteceu após a ligadura da extremidade da tuba uterina, seccionando-se a
porção situada entre a ligadura e o ovário. Após esse procedimento fez-se uma sutura na
parede muscular e na pele. O procedimento foi realizado bilateralmente. No pós-
operatório administrou-se o petanbiótico (0,1 mg/kg, i.m.) para reduzir as chances de
infecção e o aplanol10 (Flunixinameglumina, 0,1 mg/kg, i.m.) para minimizar a dor pós-
operatória. Optou-se por respeitar o tempo mínimo de 1 mês (30 dias) após o
procedimento cirúrgico para que os animais fossem submetidos aos protocolos
experimentais.
4.4 Teste da formalina
O teste da formalina é um modelo de dor associado à lesão tecidual, no qual se
quantifica a resposta comportamental provocada pela injeção subcutânea de formalina
diluída na pata traseira do animal (DUBUISSON; DENNIS, 1977; MARTINS et al.,
2006). Neste teste é possível avaliar dois tipos diferentes de dor ao longo de um período
prolongado de tempo (RANDOLPH, 1997).
Assim, as respostas comportamentais à formalina possuem duas fases: a primeira
é a fase aguda, que se inicia após a injeção de formalina e se estende pelos primeiros 5
minutos (dor neurogênica ou aguda) e a segunda fase, que é a mais prolongada com
atividade comportamental aumentada, que pode durar até cerca de uma hora. A segunda
fase tem início entre 15 e 30 minutos após a injeção de formalina e está relacionada com
a liberação de vários mediadores pró inflamatórios, como bradicinina, prostaglandinas,
serotonina, entre outros (HUNSKAAR; HOLE, 1987).

31
Nos dois dias que antecederam o teste de formalina, os animais foram colocados
na caixa de observação, com paredes de vidro (30x 17x 12 cm), por 10 minutos para
reconhecimento e habituação a condição experimental. Para realização do teste de
formalina os animais receberam injeções subcutâneas de 50 μL de solução de formalina
a 1% na região dorsal da pata posterior esquerda. Após a injeção de formalina,
dependendo do grupo experimental, as ratas tiveram contato com os animais da sua
colônia ou permaneceram isoladas por 20 minutos. Ao termino desse período de contato
ou isolamento os animais foram alocados na caixa de observação onde tiveram suas
respostas comportamentais avaliadas por 40 minutos. Um espelho foi colocado atrás da
caixa de observação para facilitar a observação dos comportamentos dos animais. As
respostas comportamentais analisadas foram o tempo de lambida e quantidade de
sacudidas da pata.
FIGURA 2- Rata Wistar na câmara de observação durante o teste de formalina
Fonte: Gonçalves, 2016, p 25.
4.5 Protocolo Experimental
4.5.1 Experimento I: Avaliação do comportamento de ratas residentes durante a
exposição a coabitantes com dor inflamatória.
Para a realização do presente experimento optou-se por utilizar a divisão entre
ratas residentes e ratas castradas que tinham como objetivo servir de estimulo para a

32
evocação do comportamento social da rata residente. No que tange as ratas residentes as
mesmas não foram submetidas a ovariectomia, já que um dos objetivos da pesquisa foi o
de analisar a influência das variações hormonais ocorridas no ciclo estral no
comportamento social da rata residente. Para isso usou-se ratas residentes nas seguintes
fases do ciclo: Proestro ( concetrações plasmáticas elevadas de progesterona e estrogênio)
e Estro (concetrações hormonais próximas ao basal), caracterizando assim uma distinção
entre os níveis hormonais nas duas fases. Não obstante fez-se a escolha por um grupo
onde todos os animais fossem castrados, possibilitando a análise dos comportamentos
sociais sem a influência dos hormônios sexuais. Como a densidade de animais é de cinco
por caixa, obteve-se então a seguinte distribuição de animais: uma rata residente, uma
rata controle, uma rata salina, uma rata formalina contato social e uma rata formalina
isolada. As ratas formalina contato social e formalina isolada foram utilizadas também no
experimento II para avaliação de suas respostas nociceptivas. O experimento I foi
composto pelos seguintes distribuição dos animais experimentais: Ratas residentes
(RES), Controle (CTRL), Formalina contato social (FCS) e Salina (SAL). A divisão dos
grupos se repetiu afim de realizar os experimentos nos diferentes ciclos supracitados,
sendo realizadas três baterias de experimentos onde as residentes são denominadas:
Residente Estro (RES/ES), Residente Ovariectomizada (RES/OVX) e Residente Proestro
(RES/PRO), sendo alocados para cada um desse grupo de residentes novos animais que
ainda não haviam sido submetidos aos experimentos. Sendo assim, a distribuição dos
grupos ficou como listado abaixo:
Animais Residentes Estro (RES/ES=8): As ratas foram deixadas nas caixas para
interagir com os animais dos outros grupos. De modo que a mesma residente foi
submetida aos três grupos (CTRL, SAL, FORM).
Grupo Controle (CTRL/ES=8): A rata foi retirada da caixa, e após breve
manipulação/contenção, retornou para caixa onde permaneceu por 20 min com a rata
residente estro. Durante o experimento os animais puderam se movimentar livremente no
interior da caixa e os comportamentos foram filmados para posterior análise.
Grupo Salina (SAL/ES= 8): Após a retirada da caixa o animal recebeu uma injeção
de salina na pata posterior esquerda e retornou a caixa com a residente estro. Durante o
experimento os animais puderam se movimentar livremente no interior da caixa e os
comportamentos foram filmados para posterior análise.
Grupo Formalina (FORM/ES= 8): A rata foi retirada da caixa, e após injeção de
formalina a 1% na pata posterior esquerda, retornou para a caixa, onde permaneceu por

33
20 min com a residente estro. Durante o experimento os animais puderam se movimentar
livremente no interior da caixa e os comportamentos foram filmados para posterior
análise.
Animais Residentes Proestro (RES/PROS=8): Essas ratas foram deixadas nas
caixas para interagir com os animais dos outros grupos. De modo que a mesma residente
foi submetida aos três grupos (CTRL, SAL, FORM).
Grupo Controle (CTRL/PROS=8): A rata foi retirada da caixa, e após breve
manipulação/contenção, retornou para caixa onde permaneceu por 20 min com a rata
residente proestro. Durante o experimento os animais puderam se movimentar livremente
no interior da caixa e os comportamentos foram filmados para posterior análise.
Grupo Salina (SAL/PROS= 8): Após a retirada da caixa o animal recebeu uma
injeção de salina na pata posterior esquerda e retornou a caixa com a residente proestro.
Durante o experimento os animais puderam se movimentar livremente no interior da caixa
e os comportamentos foram filmados para posterior análise.
Grupo Formalina (FORM/PROS= 8): A rata foi retirada da caixa, e após injeção
de formalina a 1% na pata posterior esquerda, retornou para a caixa, onde permaneceu
por 20 min com a residente proestro. Durante o experimento os animais puderam se
movimentar livremente no interior da caixa e os comportamentos foram filmados para
posterior análise.
Animais Residentes Ovariectomizadas (RES/OVX=8): Essas ratas foram
deixadas nas caixas para interagir com os animais dos outros grupos. De modo que a
mesma residente será submetida aos três grupos supracitados (CTRL, SAL, FORM).
Grupo Controle (CTRL/OVX=8): A rata foi retirada da caixa, e após breve
manipulação/contenção, retornou para caixa onde permaneceu por 20 min com a rata
residente ovariectomizada. Durante o experimento os animais puderam se movimentar
livremente no interior da caixa e os comportamentos foram filmados para posterior
análise.
Grupo Salina (SAL/OVX= 8): Após a retirada da caixa o animal recebeu uma
injeção de salina na pata posterior esquerda e retornou a caixa com a residente
ovariectomizada. Durante o experimento os animais puderam se movimentar livremente
no interior da caixa e os comportamentos foram filmados para posterior análise.
Grupo Formalina Contato Social (FORM/OVX= 8): A rata foi retirada da caixa,
e após injeção de formalina a 1% na pata posterior esquerda, retornou para a caixa, onde
permaneceu por 20 min com a residente ovariectomizada. Durante o experimento os

34
animais puderam se movimentar livremente no interior da caixa e os comportamentos
foram filmados para posterior análise.
Foram avaliados como parâmetros comportamentais da residente: o tempo de
latência para o primeiro contato e a duração total do contato.
O tempo de latência pode ser definido como o tempo que a rata residente leva para
direcionar comportamentos ativos para a coespecífica. A duração de contato foi definida
como o tempo que a rata residente passava dispendendo comportamentos e direção a
coabitante, comportamentos esses como: lambedura da região anogenital, montação,
lambedura dorso e da região do fucinho.
Os animais dos grupos controle, formalina e salina constituíram estímulos para
avaliação do comportamento social das ratas residentes.
É importante frisar que com exceção das ratas residentes que não sofreram a
cirurgia para retirada dos ovários (Proestro e Estro), todos os outros grupos foram
compostos por ratas ovariectomizadas.
20 minutos
20 minutos
20 minutos
FIGURA 3- Representação esquemática das etapas realizadas com os grupos do Experimento I.

35
4.5.2 Experimento II: Avaliação do efeito do contato social no comportamento álgico
de ratas submetidas ao teste de formalina.
Nesse experimento participaram os grupos: Grupo Formalina Contato Social
(FCS= 8), o qual participou do experimento I e o Grupo Formalina Isolado (FI= 8) o qual
recebeu a administração da formalina (1%) e logo após foi submetido a um período de
isolamento de 20 minutos. Após o término do experimento I os animais dos grupos
FCS/ES, FCS/PRO, FCS/OVX e FIS tiveram suas respostas nociceptivas avaliadas por
40 minutos. Os parâmetros utilizados para avaliação das respostas nociceptivas foram o
tempo de lambida e o número de sacudidas.
4.6 Eutanásia dos animais
A indução da morte dos animais submetidos aos protocolos experimentais foi
realizada com aplicação de dose excessiva de anestésico.
Para a dose excessiva de anestésico, a anestesia foi induzida por meio da saturação
da atmosfera de uma câmara hermeticamente fechada contendo algodão embebido com o
anestésico inalatório Isoflurano. Não houve contato direto do animal com o algodão.
4.7 Análise dos resultados
Os dados foram expressos em valores médios ± o erro padrão da média (EPM).
Para o tempo de latência os grupos experimentais foram comparados por meio de análise
40 minutos
40 minutos
FIGURA 4- Representação esquemática das etapas realizadas com os grupos do Experimento II.

36
de variância (ANOVA) de duas vias para medidas não repetidas; em se tratando da
duração do contato, a princípio foi feita uma ANOVA de três vias com o objetivo de
verificar a existência de interação entre os fatores tempo, tratamento e ciclo. Além disso,
na ausência de interação realizou-se uma ANOVA de duas vias para medidas repetidas
com as variáveis tempo (fator repetido) e o tratamento (fator independente) ou a
ANOVA de uma via quando a comparação foi feita com base no tempo total. Na
comparação das respostas nociceptivas de número de sacudidas da pata ou tempo de
lambida utilizou-se ANOVA duas vias. Quando necessário avaliar as diferenças entre os
grupos foi utilizado o pós-teste de Bonferroni para duração do contato em intervalos de
tempo, número de sacudidas da pata, tempo de lambida. Os dados foram considerados
estatisticamente significantes para p < 0,05. Para realização das análises foi utilizado o
software SPSS 22.0 e o GraphPad Prism 6.0.
37
5. RESULTADOS
5..1 Experimento I: Avaliação do comportamento de ratas residentes durante a
exposição a coabitantes em diferentes condições nociceptivas.
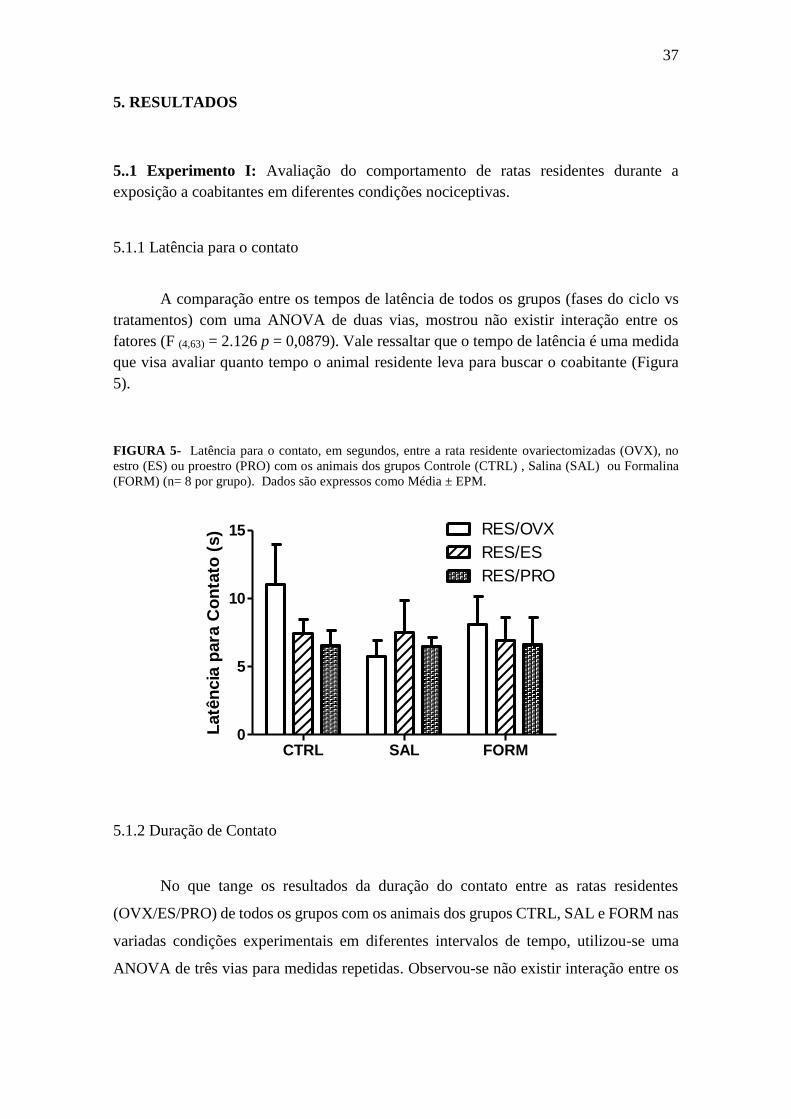
5.1.1 Latência para o contato
A comparação entre os tempos de latência de todos os grupos (fases do ciclo vs
tratamentos) com uma ANOVA de duas vias, mostrou não existir interação entre os
fatores (F (4,63) = 2.126 p = 0,0879). Vale ressaltar que o tempo de latência é uma medida
que visa avaliar quanto tempo o animal residente leva para buscar o coabitante (Figura
5).
FIGURA 5- Latência para o contato, em segundos, entre a rata residente ovariectomizadas (OVX), no
estro (ES) ou proestro (PRO) com os animais dos grupos Controle (CTRL) , Salina (SAL) ou Formalina
(FORM) (n= 8 por grupo). Dados são expressos como Média ± EPM.
CTRL SAL FORM0
5
10
15 RES/OVX
RES/ES
RES/PRO
Latê
ncia
para
Co
nta
to (
s)
5.1.2 Duração de Contato
No que tange os resultados da duração do contato entre as ratas residentes
(OVX/ES/PRO) de todos os grupos com os animais dos grupos CTRL, SAL e FORM nas
variadas condições experimentais em diferentes intervalos de tempo, utilizou-se uma
ANOVA de três vias para medidas repetidas. Observou-se não existir interação entre os

38
fatores Fase do Ciclo, Tratamento e Tempo (F= (12,189) = 0.591 p= 0,848) de modo que se
optou por prosseguir as análises posteriores de modo detalhado e fragmentado (Figura 6).
FIGURA 6- Duração, em segundos, do contato das ratas residentes ovariectomizadas (OVX), no estro (ES)
ou proestro (PRO) com os animais dos grupos Controle (CTRL) , Salina (SAL) ou Formalina (FORM) ao
longo do tempo. (n =8 por grupo). Dados expressos como Média ± EPM. (ANOVA de três vias para
medidas repetidas).
0-5 5-10 10-15 15-200
50
100
150CTRL OVX
CTRL ES
CTRL PRO
SAL OVX
SAL ES
SAL PRO
FORM OVX
FORM ES
FORM PRO
Tempo (min)
Du
ração
do
co
nta
to (
seg
)
A comparação do tempo total de contato entre as fases do ciclo com auxílio de
uma ANOVA de duas vias mostrou haver interação entre as fases do ciclo nos diferentes
tratamentos (F (4,63) = 5,066, p=0,0013). No que diz respeito ao grupo de residentes
ovariectomizadas (RES/OVX) observou-se diferença significativa quando comparadas
com as residentes estro (p < 0,05) e com as residentes no proestro (p< 0,001) somente
quando em contato com ratas FORM. Assim quando em contato com coabitantes com
dor inflamatória, as ratas OVX apresentaram maior duração do contato comparado aos
outros dois grupos. Além disso, considerando a mesma situação experimental, o grupo de
RES/ES mostrou maior duração de contato (p<0,05) que o grupo RES/PRO. Em todos
os outros tratamentos não se observou diferença estatística significativa (Figura 7).

39
FIGURA 7- Duração, em segundos, do contato da rata ovariectomizadas (OVX), no estro (ES) ou proestro
(PRO) com os animais dos grupos Controle (CTRL) , Salina (SAL) ou Formalina (FORM) (n =8 por
grupo). Dados expressos como Média ± EPM. ***p < 0,001, *p < 0,05, #p < 0,05 (ANOVA de duas vias
seguida de pós-test de Bonferroni).
CTRL SAL FORM0
50
100
150
200
250RES/OVX
RES/ES
RES/PRO#
***
*
Tem
po
de c
on
tato
(s)
Na avaliação da duração do contato do grupo RES/OVX nas diferentes condições
álgicas das ratas coabitantes mostrou diferença estatística significativa (F (2,23) = 34,85, p
< 0,0001; ANOVA). O pós teste mostrou que o tempo de contato entre a rata residente e
as coabitantes do grupo formalina foi maior que a duração do contato com as ratas CONT
e SAL, não havendo diferença entre os grupos SAL e CTRL ( Figura 8).
FIGURA 8- Duração, em segundos, do contato da rata residente ovriectomizada com ratas dos grupos
Controle (CTRL) Salina (SAL) ou Formalina (FORM). (n =8 por grupo). Dados expressos como Média ±
EPM. *p<0,05 quando comparado com os demais grupos (ANOVA seguida de pós-test de Bonferroni).
RES/OVX
CONT SAL FORM0
50
100
150
200
250 *
Du
ração
do
co
nta
to (
seg
)

40
No grupo ES a comparação do tempo de contato nos diferentes estímulos
nociceptivos mostrou diferença significativa entre os grupos (F (2,23) = 14,2, p < 0,0001;
ANOVA). Assim, a residente apresentou maior tempo de contato com ratas do grupo
FORM, quando comparado com os grupos CTRL ( p< 0,05), e SAL (p<0,05), não
havendo diferença entre os grupos SAL e CTRL ( Figura 9).
FIGURA 9- Duração, em segundos, do contato do rata residente estro com ratas dos grupos Controle
(CTRL) Salina ( SAL ) ou Formalina (FORM). (n =8 por grupo). Dados expressos como Média ± EPM.
*p<0,001 quando comparado com os grupos CTRL e SAL (ANOVA seguida de pós-test de Bonferroni).
RES/ES
CONT SAL FORM0
50
100
150
200
250
*
Du
ração
do
co
nta
to (
seg
)
A comparação pelo ANOVA da aplicação de diferentes estímulos nociceptivos na
duração do contato de ratas na fase de proestro, mostrou haver diferença significativas
entre os grupos (F (2,23) = 11,08; p = 0,0005). A aplicação do pós teste mostrou que o as
ratas em proestro tiveram maior duração no contato com as ratas com dor inflamatória
(grupo FORM) quando comparadas com as ratas controle, mas não foram diferentes das
do grupo salina. O grupo salina não foi diferente do controle (Figura 10).

41
FIGURA 10- Duração, em segundos, do contato da rata residente proestro com ratas dos grupos Controle
(CTRL) Salina ( SAL ) ou Formalina (FORM). (n =8 por grupo). Dados expressos como Média ± EPM.
*p=0,0005, quando comparado com o CTRL. (ANOVA seguida de pós-test de Bonferroni).
RES/PRO
CONT SAL FORM0
50
100
150
200
250
*
Du
ração
do
co
nta
to (
seg
)
As ratas residentes despenderam uma variada gama de comportamentos sociais
em direção ao animal formalina, sendo observados principalmente: lambedura da pata e
da região anogenital e comportamento de monta (apoiando as patas dianteiras sobre o
dorso do animal lesionado). No grupo CTRL ocorreu excesso de comportamento
exploratório por parte de ambos os animais e no grupo SAL apenas nos primeiros
momentos ocorreram comportamentos de lambedura e de monta.
5.1.3 Duração do tempo de contato em intervalos
No que tange os resultados da duração do contato entre as ratas residentes
ovariectomizadas com os animais dos grupos CTRL, SAL e FORM em diferentes
intervalos de tempo, utilizou-se uma ANOVA de duas vias para medidas repetidas,
seguidas pelo pós teste de Bonferroni. A aplicação do teste estatístico mostrou haver
interação do tempo vs tratamento (F (6,63) = 2,74: p = 0,0198), indicando um efeito do
tratamento ao longo do tempo (Figura 11). O pós- teste de Bonferroni mostrou haver
maior duração do contato dos animais residentes com os animais do grupo FORM, nos
intervalos de 0 -5 (p< 0,0001), 10-15 (p< 0,0001) e 15 – 20 min (p< 0,05), em relação aos
animais do grupo CTRL e SAL. Ao comparar o grupo SAL com o CTRL encontrou-se
diferença apenas nos primeiros cinco minutos 0-5 (p< 0,05; Figura 11).
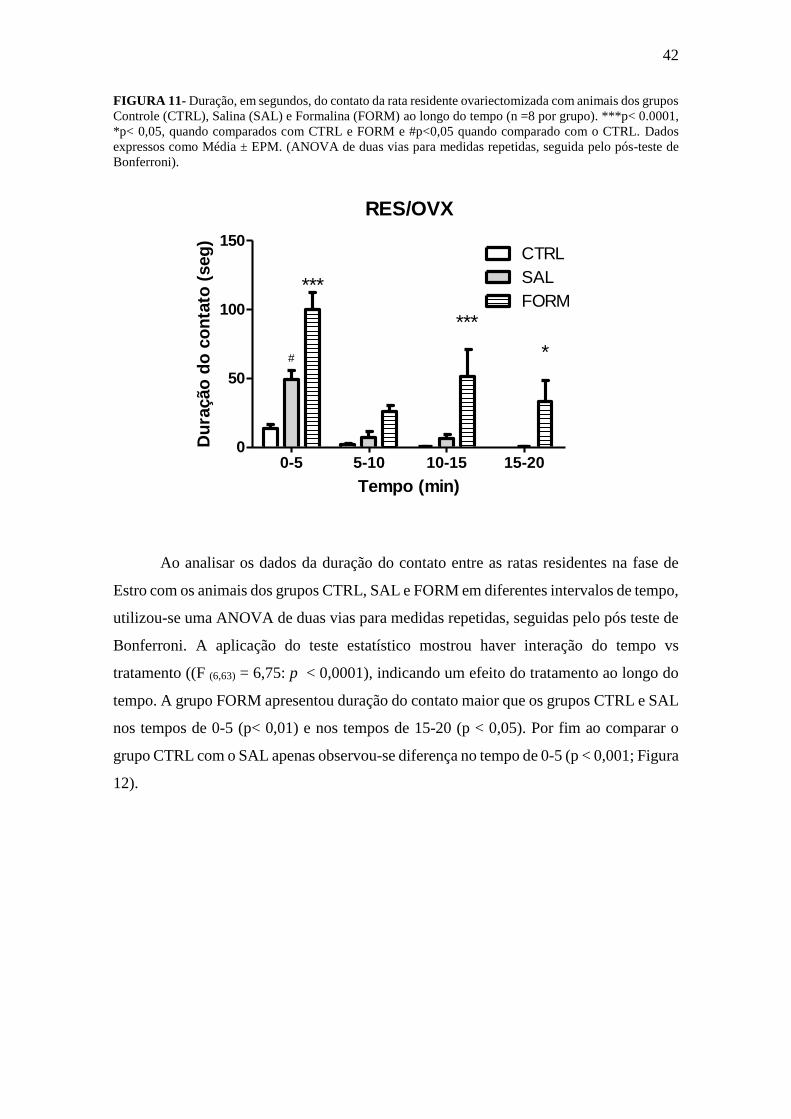
42
FIGURA 11- Duração, em segundos, do contato da rata residente ovariectomizada com animais dos grupos
Controle (CTRL), Salina (SAL) e Formalina (FORM) ao longo do tempo (n =8 por grupo). ***p< 0.0001,
*p< 0,05, quando comparados com CTRL e FORM e #p<0,05 quando comparado com o CTRL. Dados
expressos como Média ± EPM. (ANOVA de duas vias para medidas repetidas, seguida pelo pós-teste de
Bonferroni).
RES/OVX
0-5 5-10 10-15 15-200
50
100
150CTRL
SAL
FORM
#
***
***
*
Tempo (min)
Du
ração
do
co
nta
to (
seg
)
Ao analisar os dados da duração do contato entre as ratas residentes na fase de
Estro com os animais dos grupos CTRL, SAL e FORM em diferentes intervalos de tempo,
utilizou-se uma ANOVA de duas vias para medidas repetidas, seguidas pelo pós teste de
Bonferroni. A aplicação do teste estatístico mostrou haver interação do tempo vs
tratamento ((F (6,63) = 6,75: p < 0,0001), indicando um efeito do tratamento ao longo do
tempo. A grupo FORM apresentou duração do contato maior que os grupos CTRL e SAL
nos tempos de 0-5 (p< 0,01) e nos tempos de 15-20 (p < 0,05). Por fim ao comparar o
grupo CTRL com o SAL apenas observou-se diferença no tempo de 0-5 (p < 0,001; Figura
12).

43
FIGURA 12- Duração, em segundos, do contato da rata residente estro com animais dos grupos Controle
(CTRL), Salina (SAL) e Formalina (FORM) em diferentes intervalos de tempo. (n =8 por grupo).. ###p<
0,001, quando comparado com o CTRL, ***p< 0,01 e p<*0,05 quando comparados com o CTRL e SAL.
Dados expressos como Média ± EPM. (ANOVA de duas vias para medidas repetidas, seguida pelo pós-
teste de Bonferroni).
RES/ES
0-5 5-10 10-15 15-200
50
100
150CTRL
SAL
FORM
###
***
*
Tempo (min)
Du
raç
ão
do
co
nta
to (
se
g)
Os resultados da duração do contato entre as ratas residentes na fase de Proestro
com os animais dos grupos CTRL,SAL e FORM em diferentes intervalos de tempo foram
analisados através de uma ANOVA de duas vias para medidas repetidas, seguidas pelo
pós teste de Bonferroni. A aplicação do teste estatístico mostrou haver interação do tempo
vs tratamento (F (6,63) = 9,23; p < 0,0001), indicando um efeito do tratamento ao longo do
tempo. O grupo FORM apresentou maior duração do contato que o CTRL no tempo de
0-5 (p< 0,001). Por fim ao comparar o grupo CTRL teve menor duração do contato que
grupo SAL no tempo de 0-5 (p < 0,001; Figura 13).

44
FIGURA 13- Duração, em segundos, do contato da rata residente proestro com animais dos grupos
Controle (CTRL), Salina (SAL) e Formalina (FORM) em diferentes intervalos de tempo. (n =8 por grupo).
*** p< 0,001 e ### p<0,001 quando comparados com o CTRL. Dados expressos como Média ± EPM.
(ANOVA de duas vias para medidas repetidas, seguida pelo pós-teste de Bonferroni).
RES/PRO
0-5 5-10 10-15 15-200
50
100
150CTRL
SAL
FORM
###
***
Tempo (min)
Du
raç
ão
do
co
nta
to (
se
g)
Como se pode observar as residentes ovariectomizadas RES/OVX e no período
do ciclo Estro RES/ES, apresentaram comportamentos semelhantes, direcionando uma
gama de comportamentos ativos para o animal formalina nos períodos iniciais e finais 0-
5 e 15-20, em um primeiro momento esses animais exibiam comportamentos de
lambedura da pata lesionada ou da região anogenital, no segundo momento ocorriam
comportamentos de monta, lambedura do dorso e da região da cabeça do coabitante, algo
que apenas ocorreu nos primeiros 0-5 no grupo composto por residentes proestro
RES/PRO . Outro ponto que fica nítido é a existência de uma maior atividade encontrada
entre as residentes para com o animal SAL apenas nos primeiros minutos (0-5) em todos
os grupos de residentes. Em se tratando do animal CTRL, em todos os grupos a residente
apenas exibiu comportamentos de exploração com relação a caixa, não despendendo
comportamentos ativos para com os animais do grupo. No que se refere as residentes no
proestro RES/PRO, observou-se o direcionamento de comportamentos agressivos em
direção a coabitante, algo não observado nas outras fases do ciclo.
5.2 Experimento II: Avaliação do efeito do contato social no comportamento álgico de
ratas submetidos ao teste de formalina.

45
As respostas nociceptivas avaliadas foram o número de sacudidas e tempo de
lambida da pata posterior esquerda. Com relação ao número de sacudidas dos animais
que tiveram contato com a residente ovariectomizada e ao grupo isolado: Após aplicação
da ANOVA de duas vias para medidas repetidas observou-se não haver interação entre
as variáveis tipo de contato e tempo (F(7,98)= 1,571; p = 0,153), indicando não existir efeito
do tipo de contato ao longo do tempo( Figura 14).
FIGURA 14- Número de sacudidas da pata dos animais que receberam formalina na pata e tiveram contato
com ratas ovariectomizadas (RES/OVX) ou ficaram isoladas (ISOL) (n=8 animais por grupo). Dados
expressos como Média ± EPM. (ANOVA de duas vias para medidas repetidas, seguida pelo pós-teste de
Bonferroni).
0-5
5-10
10-1
5
15-2
0
20-2
5
25-3
0
30-3
5
35-4
0
0
50
100
150
200
ISOL
RES/OVX
Tempo (min)
Nú
mero
de S
acu
did
as
Em se tratando do número de sacudidas dos animais que tiveram contato com a
residente estro e ao grupo isolado, após a realização da ANOVA de duas vias para
medidas repetidas observou-se não haver interação entre as variáveis tipo de contato e
tempo (F(7,98)= 0,635; p = 0,725), indicando não existir efeito do tipo de contato ao longo
do tempo ( Figura 15).

46
FIGURA 15- Número de sacudidas da pata dos animais que receberam formalina na pata e tiveram contato
com ratas no estro (RES/ES) ou ficaram isoladas (ISOL). (n=8 animais por grupo). Dados expressos como
Média ± EPM. (ANOVA de duas vias para medidas repetidas, seguida pelo pós-teste de Bonferroni).
0-5
5-10
10-1
5
15-2
0
20-2
5
25-3
0
30-3
5
35-4
0
0
50
100
150
200
RES/ES
ISOL
Tempo (min)
Nú
mero
de S
acu
did
as
Após aplicação da ANOVA de duas vias para medidas repetidas para análise dos
animais que haviam tido contato com a residente proestro e ao grupo isolado observou-
se não haver interação entre as variáveis tipo de contato e tempo (F (7,98)= 1,186; p =
0,318), indicando não existir efeito do tipo de contato ao longo do tempo. Após realização
do pós teste de Bonferroni constatou-se diferença entre os animais nos momentos de: 0-
5 ,10-15,15-20,20-25, 25-30 ocorrendo uma diminuição no número de sacudidas por parte
dos animais que tiveram contato com as ratas no proestro (Figura 16).

47
FIGURA 16- Número de sacudidas da pata dos animais que receberam formalina na pata e tiveram contato
com ratas no proestro (RES/PRO) ou ficaram isoladas (ISOL). (n=8 animais por grupo). Dados expressos
como Média ± EPM. *p<0,05 ** p< 0,01 quando comparado com o grupo ISOL (ANOVA de duas vias
para medidas repetidas, seguida pelo pós-teste de Bonferroni).
0-5
5-10
10-1
5
15-2
0
20-2
5
25-3
0
30-3
5
35-4
0
0
50
100
150
200
RES/PRO
ISOL
***
**** *
Tempo (min)
Nú
mero
de S
acu
did
as
Com relação ao tempo de lambida a aplicação da ANOVA não haver diferença
significativa entre os grupos (F(3,31)= 0,206; p = 0,891). (Figura 17).
FIGURA 17- Duração do tempo de lambida, em segundos, dos animais que receberam formalina na pata
e tiveram contato com ratas ovariectomizadas (RES/OVX), na fase de estro (RES/ES), no proestro
(RES/PRO) ou ficaram isoladas (ISOL) (n=8 animais por grupo). Dados expressos como Média ± EPM.
(ANOVA de duas vias para medidas não repetidas).
ISOL RES/OVX RES/ES RES/PRO0
100
200
300
Tem
po
de L
am
bid
a (
seg
)

48
6. DISCUSSÃO
O presente estudo teve como objetivo avaliar a influência das variações hormonais
no contato social entre ratas residentes e com dor inflamatória, assim como também o
efeito desse nas respostas nociceptivas dessas ratas. Os resultados encontrados apontaram
que o contato social foi modificado pelo tipo de estimulo nociceptivo. Indicando que a
rata residente foi capaz de identificar a condição álgica da coabitante, despendendo para
esta atenção diferenciada quando em comparação ao controle. Não obstante observou-se
que a quantidade e a variedade de comportamentos sociais dispendidos pela residente
variaram de acordo com a fase do ciclo que a mesma se encontrava. Neste sentido, as
residentes ovariectomizadas mostraram maior tempo de contato que as fêmeas no estro e
no proestro, diferença essa ocorrendo também quando se comparou o grupo estro com o
proestro.
Alguns estudos apontam a existência de comportamentos pró-sociais por parte dos
roedores. Esses estudos submetem animais a situações potencialmente estressoras de
caráter doloroso ou não, e tem encontrado como resposta alterações comportamentais
relacionadas a condição em que co-específico se encontra, sejam elas através do aumento
da sensibilidade dolorosa, ou através de ações deliberadas com o intuito de libertar o
coabitante em situações de aprisionamento (MARQUEZ et al. 2015; SATO et al. , 2015;
BARTAL et al. , 2014; LANGFORD et al. , 2006).
Com o objetivo de analisar a existência de um perfil pró-social por parte de
roedores, ratos foram submetidos a condições onde um dos sujeitos tinha acesso a comida,
o outro era submetido a uma situação potencialmente estressora (afogamento). O rato em
situação definida como confortável despendeu uma variedade de comportamentos
direcionados ao co-específico afim de libertá-lo, isso ocorrendo independente de
recompensa ou auto-benefício (SATO, 2015).
Os resultados trazidos por Marquez (2015) mostraram que roedores atuavam de
forma deliberada para fornecer alimento a um co-específico mesmo na ausência de
compensação, apontando a existência de algum mecanismo relacionado a reciprocidade
atuando de forma pró-empática. Assim, esses animas eram submetidos a duas situações
distintas onde poderiam atuar de forma a só permitirem a liberação de alimento para eles
mesmos, ou atuar de forma empática, liberando comida para o co-específico observou-se
assim um maior número de escolhas pró-sociais (MARQUEZ, 2015).

49
A literatura aponta que roedores são capazes de reconhecer estados afetivos em
coabitantes, de modo que esse tipo de comportamento se assemelha ao do tipo empatia
em humanos (KEUM e SHIM, 2016). Atualmente um grande número de pesquisas tem
mostrado a relação entre dor e alterações no comportamento de roedores (LANGFORD
et al., 2006; CHEN et al., 2015; JEON et al., 2010). Em experimento onde os animais
eram expostos a uma situação em que o coabitante era submetido ao aprisionamento, o
animal que poderia agir de forma deliberada, optou por despender uma gama de
comportamentos com o intuito de libertar o co-específico (BARTALL et al 2014).
Em nosso estudo observamos o contato social foi estimulado e determinado pelo
tipo resposta álgica da rata coabitante. Nesse sentido houve a diferenciação entre os tipos
de estímulos nociceptivos, já que se observou que nas ratas que receberam injeção de
salina na pata o contato com a residente ocorreu principalmente nos primeiros 5 minutos
e nas ratas com dor inflamatória (formalina) o contato social foi bifásico, ocorrendo no
primeiro nos cinco minutos iniciais e retornando após quinze minutos de aplicação do
estimulo. Além disso, na ausência de estimulo nocivo o contato social entre a rata
residente e coabitante foi reduzido. Demostrando que a rata residente foi capaz de
distinguir entre as condições sensoriais da coabitante e a partir disso modificar a
expressão comportamental incorporando respostas pró-sociais.
No caso da dor inflamatória vale lembrar que o teste da formalina atua de forma
bifásica e as respostas comportamentais à formalina possuem duas fases: a primeira é a
fase inicial aguda, que se inicia após a injeção de formalina e se estende pelos primeiros
5 minutos (dor neurogênica ou aguda) e a segunda fase, que é a mais prolongada com
atividade comportamental aumentada, que pode durar até cerca de uma hora. A segunda
fase tem início entre 10 e 30 minutos após a injeção de formalina e está relacionada com
a liberação de vários mediadores pró-inflamatórios, como bradicinina, prostaglandinas,
serotonina, entre outros (HUNSKAAR e HOLE,1987). Nossos resultados sugerem que
no caso da formalina as ratas residentes conseguiram perceber alterações no padrão
comportamental da coabitante durante as diferentes fases do teste, despendendo assim
comportamentos sociais de forma bifásica.
Os animais controles aqueles que eram apenas submetidos a manipulação e
retornavam para a caixa eliciaram apenas comportamentos exploratórios nos residentes,
mostrando que a ausência de estímulo doloroso de certo modo não modificou o padrão
de interação na caixa. Os animais do grupo salina evocaram respostas de maior contato
no residente durante os primeiros cinco minutos de teste, essas eram caracterizadas pelo

50
ato de lamber a pata lesionada e a região anogenital da coabitante. Uma explicação para
a evocação do comportamento em relação a SAL seria o fato da lesão aguda provocada
pela injeção.
De modo geral o comportamento de lambedura da pata sugere a possibilidade da
existência de um comportamento pró-empático que pode ser interpretado como uma
tentativa de diminuir o sofrimento do rato machucado. Sendo esta inclusive uma das
respostas comportamentais apresentadas pelos animais ao longo do teste de formalina.
Com base nos resultados não acreditamos que o odor da formalina tenha motivado o
processo de busca pelo coabitante, caso contrário esse contato não ocorreria de forma
bifásica podendo ocorrer somente no início ou durante todo o tempo. Além disso, no
grupo proestro as ratas residentes não apresentaram contato social nos minutos finais.
Alguns autores tendem a classificar os comportamentos de acordo com o seu
direcionamento, subdividindo os seus tipos em: comportamentos sociais de investigação
que podem ser expressos através das ações de cheirar e lamber o coabitante,
comportamentos tidos como diretos que são aqueles relacionados a monta (entendidos
como uma postura de dominância), comportamentos atentos que são os relacionados a
perseguição e a busca pelo outro na caixa e por fim os agressivos caracterizados por
mordidas (MOURA e XAVIER,2010).
Em nosso estudo observou-se assim uma variada gama de comportamentos de
investigação diretos e de atenção para com relação ao coabitante lesionado, como as ações
de cheirar e lamber o coabitante, excessivo deslocamento na caixa sempre em direção ao
coespecifico, comportamento de monta. Em se tratando dos animais controles, apenas
comportamentos do tipo exploratório foram despendidos pela rata residente.
No que tange os animais formalina constatou-se uma maior interação nos
primeiros cinco minutos caracterizada pelas investidas da residente sobre o animal
lesionado, sendo os principais comportamentos observados os de lambedura de pata e da
região anogenital, nos minutos finais de teste que se relacionam a dor inflamatória (15 a
20 min) a residente mostrou comportamento de limpeza do coabitante (lambedura da
região do dorso e da cabeça), o posicionamento das patas sobre o animal e a busca pelo
posicionamento próximo do animal lesionado. Durante os outros quadrantes temporais (5
– 15min) a interação social, assim como também o repertorio comportamental descrito
acima caiu consideravelmente, existindo a predominância apenas de comportamentos
exploratórios em relação a caixa por parte dos dois animais.

51
Em estudo recente do nosso grupo, encontrou-se como resultado que machos são
capazes de reconhecer o componente álgico em coabitantes quando em comparação com
os animais do grupo controle, assim como também direcionar alguns comportamentos
ativos em direção ao co-especifíco (GONÇALVES, 2016).
Como mencionado um dos objetivos do presente estudo foi a análise das possíveis
influencias das variações hormonais do ciclo estral no comportamento social de ratas com
dor inflamatória, para isso fez-se uso de dois grupos de residentes que tinham o seu ciclo
regular: Estro, Proestro e apenas um com ratas ovariectomizadas que não apresentavam
variações hormonais. As ratas residentes dos grupos Estro e Ovarietomizadas mostraram
comportamentos semelhantes na presença de coabitantes nas diferentes condições álgicas.
Por outro lado, as ratas no Proestro (esse caracterizado pela maior taxa hormonal de todo
ciclo) apresentaram pouco interesse na rata coabitante nos últimos cinco minutos. Além
de apresentarem comportamentos agressivos nos primeiros cinco minutos de
experimento.
O ciclo estral é caracterizado por variações nas concentrações dos hormônios
gonadais. Em se tratando desses hormônios observa-se a menor concentração de
estrogênio durante a fase denominada estro, ocorrendo um aumento durante o metaestro
e o seu pico durante a fase denominada proestro. Em se tratando da progesterona, observa-
se um pico durante o metaestro e outro pico durante o proestro. De forma resumida ocorre
um aumento gradativo dos níveis de estrogênio e progesterona durante o proestro (LEAL,
2015).
Aparentemente notou-se a existência do comportamento empático independente
das concentrações hormonais. Contudo durante o proestro os hormônios presentes
causam redução do comportamento empático e favorecimento na expressão de
comportamentos agressivos. Uma hipótese para isso seria a alta concentração de
hormônios gonadais que caracteriza essa fase, assim como também o aumento da
expressão dos receptores para esses hormônios na região do sistema límbico, o que pode
influenciar de algum modo o comportamento da residente no proestro (VARGAS, 2016).
Não obstante como fora citado anteriormente o proestro é marcadamente característico
pelas altas concentrações de estrogênio, esse estando relacionado a liberação do
corticosterona, hormônio esse relacionado a respostas de estresse (FIGUEIREDO, 2006).
É um consenso a existência de receptores específicos para hormônios gonadais
localizados na região do encéfalo, inclusive em regiões que compõem o sistema límbico
como o hipotálamo e a amigdala. Assim, a alta desses hormônios pode estar diretamente

52
ligada a modificações nas respostas comportamentais (FIGUEIREDO 2016; VARGAS
et al 2016; LOVICK, 2014; BARHA e GALEA, 2010).
Segundo Vargas et al (2016) a expressão de receptores de estrogênio em regiões
cerebrais como a amigdala e o núcleo dorsal da rafe, concomitante com as altas
concentraçõe hormonais justificam as mudanças em níveis cognitivos e emocionais das
fêmeas (VARGAS et al 2016).
Em nosso estudoas ratas no estro ou ovariectomizadas mostraram
comportamentos pró sociais como os de lambedura da região anogenital, lambedura do
dorso e da cabeça, assim como busca por uma localização mais próxima do coabitante
com dor. Isso ocorreu durante os primeiros cinco minutos nos animais salina e formalina,
e nos últimos cinco minutos nos animais formalina (repetindo o padrão geral). Entretanto,
na fase de proestro as ratas apresentaram maior expressão de comportamentos sociais nos
primeiros cinco minutos, sem aumento nos últimos cinco minutos de experimento. Além
disso, durante o proestro as ratas residentes apresentaram comportamentos do tipo
agressivos em direção às ratas com dor inflamatória nos últimos cinco minutos. Tais
resultados sugerem que durante o proestro ocorre diminuição dos comportamentos sociais
do tipo empático, provavelmente tal alteração seja mediada por fatores hormonais e/ou
alteração na liberação de neurotransmissores em áreas encefálicas envolvidas com a
modulação de tais comportamentos.
As flutuações hormonais relacionadas ao ciclo estral, influenciam de forma direta
o eixo hipotalâmico- hipofisário, agindo na secreção do hormônio liberador de
corticotrofina (CRH), hormônio adrenocorticotrófico (ACTH) e da corticosterona. O
aumento dos níveis fisiológicos de estradiol e progesterona relacionados com a fase do
proestro, resultam em um aumento na liberação de hormônios como o cortisol
(MARCONDES, TANNO, 2002). Neste sentido, ratas com maiores concentrações de
estrogênio apresentaram maiores concentrações séricas de corticosterona comparadas a
ratas ovariectomizadas (FIGUEIREDO et al., 2006). Assim um aumento nas
concentrações de progesterona, estrogênio e corticosterona, poderiam justificar a
diferença de repertório comportamental apresentado entre os ciclos, aumentando a
sensibilidade a eventos estressores, e a comportamentos do tipo agressivo.
Existe também no Proestro, um aumento da progesterona não somente de origem
folicular, mas também de origem adrenal. Esse aumento é acompanhado por uma
elevação da atividade adrenal, relacionada com o período pré-ovulatório e consequente
elevação da secreção de corticosterona (RODRIGUES, 2005).

53
Atualmente a literatura relata a existência de comportamentos empáticos por parte
de roedores, esses estudos relatam um amplo repertorio comportamental por parte dos
sujeitos em direção ao co-específico em diferentes situações como: percepção de
componentes álgicos em familiares, comportamentos ativos para libertar um companheiro
de caixa ou até mesmo a construção de relações empáticas entre linhagens diferentes de
animais que haviam sido criadas juntas (SIVASELVACHANDRAN et al 2016; KEUM
e SHIN,2016; MOGIL,2012; BARTAL,2014; LANGFORD, 2006.)
Com base no que foi abordado acima pode-se inferir que nas variadas fases do
ciclo estral pode se observar a ocorrência de comportamentos pró-sociais. O mesmo
podendo variar com relação a sua quantidade de acordo com as fases. Em resumo
observou-se uma maior interação entre ratas castradas e ratas no estro quando comparadas
com ratas no proestro, essas ultimas direcionando comportamentos agressivos para a
coespecífica em situação dolorosa.
Em relação ao efeito do contato social na dor inflamatória de ratas, não se
constatou diferença entre os grupos pertencentes as ratas castradas ou ratas estro em
comparação as ratas do grupo sem contato social, apesar do maior tempo de contato
social. Contudo as ratas coabitantes que tiveram contato com as residentes no proestro
apresentaram hipoalgesia de longa duração, nos períodos que sucederam o contato.
Porém, um fator interessante e que pode ser levado em consideração é o que se refere ao
fato de que no grupo proestro onde os comportamentos considerados como pró sociais
foram despendidos somente no início e no restante do tempo se observou comportamentos
agressivos. Uma hipótese para isso seria a ocorrência de uma analgesia por estresse e
modulada pela liberação de opioides endógenos.
A exposição a um predador ou a co-específicos agressivos pode ocasionar lesões
corporais, neste sentido a ocorrência de antinocicepção permite que o animal ameaçado
possa utilizar os comportamentos de luta ou fuga, congelamento, entre outros, sem a
necessidade de interromper os comportamentos defensivos e iniciar respostas
recuperativas direcionadas a injuria (BOLLES E FANSELOW, 1980).
A analgesia por estresse depende da ativação de um circuito envolvendo a
substancia cinzenta periaquedutal, bulbo rostral ventro medial e corno dorsal da medula
espinal. Além dessas estruturas, estudos apontam para o envolvimento do ACC na
analgesia por estresse. A desativação de neurônios do ACC através de estimulação
eletrofisiológica acarretou em diminuição de componentes relacionados ao medo
observacional e respostas nociceptivas (JEON et al 2010). Estudos neuroanatômicos

54
mostraram que o ACC envia projeções para a SCP (FLOYD et al., 2000; PRICE, 2000).
Além disso, a SCP emite projeções para RVM e locus ceruleus (LC) e a estimulação
elétrica ou química do LC libera noradrenalina no CDME produzindo analgesia,
denominada de noradrenérgica. Já o RVM contém opióides endógenos que estão
relacionados com a desinibição de neurônios gabaérgicos e consequentemente ativação
da via descendente inibitória da dor (PATEL, 2010; NOBACK et al., 2005). Assim,
dependendo do circuito descendente ativado podemos observar analgesia do tipo opióide
ou não-opóide (adrenérgica) (GERARD, 2009).
Um dos critérios para definir se a analgesia foi opioide ou não, leva em
consideração sua duração, a analgesia noradrenégica é de caráter rápido totalizando no
máximo 10 minutos e estando relacionada a respostas de luta ou fuga, entretanto a do tipo
opioide tem duração de 40 minutos, sendo observada em situações de congelamento,
imobilidade tônica e em respostas defensivas passivas onde a possibilidade de fuga
inexiste (RODGERS e RANDALL,1987).
As ratas que haviam sido submetidas ao contato com a residente proestro,
mostravam respostas comportamentais de imobilidade quando na presença da residente.
Tais comportamentos suportam a hipóteses da ocorrência de analgesia defensiva após o
contato com ratas em proestro. Contudo a confirmação de tal hipótese dependeria de
estudos com antagonistas opioides, como por exemplo a naloxona.
Entretanto as ratas submetidas às ratas residentes ovariectomizadas ou no estro
não apresentaram redução nas respostas nociceptivas quando comparadas com as ratas
que haviam tido contato com a rata proestro. Uma hipótese para isso é a de que o contato
reduza a percepção do estimulo nociceptivo somente durante o contato, porém, não induz
analgesia. Sendo assim, a redução dos comportamentos nociceptivos ocorre apenas
durante o contato ou por um curto período após e com a interrupção do contato após os
20 minutos o animal com dor inflamatória retoma suas respostas nociceptivas.
O presente trabalho partiu da hipótese de que o contato social entre ratas residentes
e coabitantes em diferentes condições nociceptivas sofre influência das flutuações
hormonais oriundas do ciclo estral. Assim sendo observou-se que nas diferentes
condições experimentais ratas ovariectomizadas e no estro mostraram comportamentos
pró-ativos em direção as co-específicas, entretanto a intensidade desses comportamentos
variou com o tipo de estimulo nocivo. Não obstante as ratas residentes no proestro
apresentaram comportamentos agressivos e não exibiram o padrão comportamental
observado nos grupos citados anteriormente. Assim sendo reitera-se a hipótese de que os

55
hormônios gonadais e as flutuações do mesmo em decorrência do ciclo estral podem
influenciar o contato social de ratas.

56
7. CONCLUSÃO
Com base nos resultados pode se concluir que os comportamentos sociais das ratas
residentes são estimulados pela condição álgica de um co-especifico independente da fase
do ciclo. Entretanto a fase do ciclo determina a duração e padrão comportamental
despendidos em direção ao coabitante. Além disso, se observou que o contato social não
interfere na dor inflamatória das ratas coabitantes, porém comportamentos do tipo
agressivo causam hipoalgesia de longa duração.

57
REFERÊNCIAS
AKYAZI, I.; ERASLAN, E. Transmission of stress between cagemates: a study in
rats. Physiology & behavior, v. 123, p. 114-118, 2014.
ALOISI, A. et al. Effects of the essential oil from citrus lemon in male and female rats
exposed to a persistent painful stimulation. Behav Brain, Vol 136, p. 127-135. 2002.
AMANDUSSON A, BLOMQVIST A. Estrogenic influences in pain processing. Front
Neuroendocrinol, v.34 ,p. 329–349, 2013
ARAÚJO, F. M. Effect of interferential current on fibromyalgia : randomized clinical
trial. 2015. 98 f. Dissertação (Mestrado em Ciências Fisiológicas) - Universidade
Federal de Sergipe, São Cristóvão, 2015
BANDURA, A. Social learning through imitation. 1962.
BAPTISTA, S. D. et al. Mice undergoing neuropathic pain induce anxiogenic-like effects
and hypernociception in cagemates. Behav Pharmacol. Agosto. 2015.
BARHA, C. K.; GALEA, L. AM. Influence of different estrogens on neuroplasticity and
cognition in the hippocampus. Biochimica et Biophysica Acta (BBA)-General
Subjects, v. 1800, n. 10, p. 1056-1067, 2010.
BARON, Sheri; BRUSH, F. Robert. Effects of acute and chronic restraint and estrus cycle
on pituitary-adrenal function in the rat. Hormones and behavior, v. 12, n. 3, p. 218-224,
1979.
BARRETT, A. C.; SMITH, E. S.; PICKER, M. J. Sex-related differences in mechanical
nociception and antinociception produced by μ-and κ-opioid receptor agonists in
rats. European journal of pharmacology, v. 452, n. 2, p. 163-173, 2002.
BARTAL, B. A. I. et al. Pro-social behavior in rats is modulated by social experience.
eLife, Jan. 2014.
BARTELS, A.; ZEKI, S. The neural correlates of maternal and romantic
love. Neuroimage, v. 21, n. 3, p. 1155-1166, 2004.

58
BIANCHI, F. J. et al. Influência do sexo, do ciclo estral e do estrese agudo nas respostas
hormonais e metabólicas em ratos. Dissertação de Mestrado. Universidade Estadual de
Campinas. 2004.
BOLLES, R.C. FANSELOW, M.S. A perceptual-defensive-recuperative model of fear
and pain. The behavioral and brain sciences, v.3, p.291-323, 1980.
BORZAN, J.; FUCHS, P. N. Organizational and activational effects of testosterone on
carrageenan-induced inflammatory pain and morphine analgesia. Neuroscience, v. 143,
n. 3, p. 885-893, 2006.
CHEN, J. et al. Empathy for pain: A novel bio-psychosocial-behavioral laboratory animal
model. Acta physiologica Sinica, v. 67, n. 6, p. 561-570, 2015.
CRAFT, R. M. et al. Dose-and time-dependent estradiol modulation of morphine
antinociception in adult female rats. European Journal of Pain, v. 12, n. 4, p. 472-479,
2008.
DECETY, J. et al. Putting together phylogenetic and ontogenetic perspectives on
empathy. Developmental Cognitive Neuroscience, v. 2, n.1, p. 1-24, Jan. 2012.
DECETY, J.; MORIGUCHI, Y. The empathic brain and its dysfunction in psychiatric
populations: Implications for intervention across different clinical
conditions. BioPsychoSocial Medicine, v. 1, n. 1, p. 22, 2007.
DIMBERG, U.; ANDRÉASSON, P.; THUNBERG, M. Emotional empathy and facial
reactions to facial expressions. Journal of Psychophysiology, 2011.
DINA, O. A. et al. Sex hormones regulate the contribution of PKCε and PKA signalling
in inflammatory pain in the rat. European Journal of Neuroscience, v. 13, n. 12, p.
2227-2233, 2001.
DUBUISSON, D.; DENNIS, S.G. The formalin test: a quantitative study of the analgesic
effects of morphine, meperidine and brain stem stimulation in rats and cats. Pain v.4, n.
2, p. 161-174, 1977.
EKMAN, L. L. Neurociencias: fundamentos para reabilitação/ Laurie Lundy-Ekman;
{tradução Fernando Diniz Mundim... et al} - Rio de Janeiro: Elsevier, p. 103-105, 2008.

59
FIGUEIREDO, H. F. et al. Estrogen potentiates adrenocortical responses to stress in
female rats. American Journal of Physiology-Endocrinology and Metabolism, v. 292,
n. 4, p. E1173-E1182, 2007.
FIGUEIREDO, R. M.de. Diferenças associadas ao ciclo estral na reatividade emocional
de ratas a estímulos incondicionados e condicionados de medo. Tese de Doutorado.
Universidade de São Paulo.2016.
GERARD F. G. Controle rostrocaudal da dor. In NETO, O. A. Dor: Princípios e
práticas. Artmed, Cap.16, págs 227-245, 2009.
GERY, I.et al. Empathy and recognition of facial expressions of emotion in sex offenders,
non-sex offenders and normal controls. Psychiatry research, v. 165, n. 3, p. 252-262,
2009.
GONÇALVES, A. S. B. Alterações no contato social induzidas pela dor reduzem
respostas nociceptivas sem modificar comportamento tipo-ansioso.Dissertação de
mestrado. Universidade Federal de Sergipe. São Cristóvão, 2016.
GU, L.et al. Pain Inhibition by Optogenetic Activation of Specific Anterior Cingulate
Cortical Neurons PLoS ONE v. 10, n.2 Academic Editor: Theodore John Price,
University of Texas at Dallas, UNITED STATES. Fev. 2015.
HUNSKAAR, S. HOLE, K. The formalin test in mice: dissociation between
inflammatory and non-inflammatory pain. Pain, v. 30, n.1, p. 103-14, 1987.
JEON, D. et al. Observational fear learning involves affective pain system and Cav1.2
Ca2+ channels in ACC. Nature Neuroscience ,v.13, n.2504, p. 482–488, Fev. 2010.
JÚNIOR, A. G.; MORATO, S. Influências do ciclo estral sobre o desempenho de ratos
no labirinto em cruz elevado. Interação em Psicologia, v. 6, n. 2, 2002.
KEUM, S; SHIN, H.S. Rodent models for studying empathy. Neurobiology of learning
and memory, v. 135, p. 22-26, 2016.
KRZANOWSKA, E. K. et al. Reversal of sex differences in morphine analgesia elicited
from the ventrolateral periaqueductal gray in rats by neonatal hormone
manipulations. Brain research, v. 929, n. 1, p. 1-9, 2002.

60
LACROIX-FRALISH, M. L. et al. The magnitude of mechanical allodynia in a rodent
model of lumbar radiculopathy is dependent on strain and sex. Spine, v. 30, n. 16, p.
1821-1827, 2005.
LANGFORD, D.J. et al. Social modulation of pain as evidence for empathy in mice.
Science, v. 312, n. 5782 p. 967-70, 30 Jun. 2006.
LEAL, J. C. O. Efeito do estrogênio na memória para objetos em ratas Wistar: estampa
temporal. Dissertação de Mestrado. Universidade Federal do Rio Grande do Norte.
2015.
LEIBENLUFT, E. et al. Mothers' neural activation in response to pictures of their children
and other children. Biological psychiatry, v. 56, n. 4, p. 225-232, 2004.
LI, L. et al. Ablation of estrogen receptor α or β eliminates sex differences in mechanical
pain threshold in normal and inflamed mice. Pain, v. 143, n. 1, p. 37-40, 2009.
LOVICK, T. A. Sex determinants of experimental panic attacks. Neuroscience &
Biobehavioral Reviews, v. 46, p. 465-471, 2014.
LUO, P. et al. Gender differences in affective sharing and self–other distinction during
empathic neural responses to others’ sadness. Brain imaging and behavior, v. 9, n. 2, p.
312-322, 2015.
MANSON, J. E. Pain: sex differences and implications for treatment. Metabolism, v. 59,
p. S16-S20, 2010.
MÁRQUEZ, C. et al. Prosocial choice in rats depends on food-seeking behavior
displayed by recipients. Current Biology, v. 25, n. 13, p. 1736-1745, 2015.
MARTINS, M. A. et al. Formalin injection into knee joints of rats: pharmacologic
characterization of a deep somatic nociceptive model. J Pain. V.7, n.2, p. 100-107, 2006.
MERSKEY H., BOGDUK N. International Association for the Study of Pain.
Classification of chronic pain: descriptions of chronic pain syndromes and definitions of
pain terms. Second ed. Seattle: IASP Press; 1994.

61
MOGIL, J.S.; BAILEY, A. L. Sex and gender differences in pain and analgesia. Prog
Brain Res, v. 186, p. 141-157, 2010.
MOURA,P. J; XAVIER, G.F. Memória de reconhecimento social em ratos. Psicologia
USP, v.21,n.2,p 355-289,2010.
NITSCHKE, J. B. et al. Orbitofrontal cortex tracks positive mood in mothers viewing
pictures of their newborn infants. Neuroimage, v. 21, n. 2, p. 583-592, 2004.
NOBACK, C., et al. (2005). The Human Nervous System: Structure and function.
6ªedição. New Jersey, Humana Press, p. 171-172.
NOGAA, W. E. et al. Metabolite concentrations in the anterior cingulate córtex predict
high neuropathic pain impact after spinal cord injury. Pain, v. 154, n.2, p. 204–212.
Julho, 2013.
PARR, L. A.; HOPKINS, W. D. Brain temperature asymmetries and emotional
perception in chimpanzees, Pan troglodytes. Physiology & behavior, v. 71, n. 3, p. 363-
371, 2000.
PATEL, N. Fisiopatologia da Dor. In: KOPF, A. e PATEL, N. (Ed.). Guia para o
tratamento da dor em contextos de poucos recursos. Seattle, IASP Press, p. 9-13.2010.
PEDROSO, L.B.. Avaliação da possível diferença na sensibilidade dolorosa de ratos
machos e fêmeas e da resposta de cada sexo a crotalfina, um analgésico tipo opióide. Tese
de Doutorado. Universidade de São Paulo. 2011
PRESTON, S.D.; DE WAAL, F.BM. Empathy: Its ultimate and proximate
bases. Behavioral and brain sciences, v. 25, n. 01, p. 1-20, 2002.
PROVERBIO, A. M. et al. Gender and parental status affect the visual cortical response
to infant facial expression. Neuropsychologia, v. 44, n. 14, p. 2987-2999, 2006.
PROVERBIO, A. M. Sex differences in social cognition: The case of face
processing. Journal of Neuroscience Research, v. 95, n. 1-2, p. 222-234, 2017.
RANDOLPH, B. C. Analgesic effectiveness of ketorolac compared to meperidine in the
rat formalin test. Anesth Prog. V.44, n.1 p. 11- 16, 1997.

62
RILEY III, J. L. et al. Sex differences in the perception of noxious experimental stimuli:
a meta-analysis. Pain, v. 74, n. 2, p. 181-187, 1998.
RODGERS, R.J. RANDALL, J.I. Defensive analgesia in rats and mice. Psychol Rec,
v.37.
p.335-347, 1987.
SATO, N. et al. Rats demonstrate helping behavior toward a soaked conspecific. Animal
cognition, v. 18, n. 5, p. 1039-1047, 2015.
SATOSHI, F. et al. Receiving of emotional signal of pain from conspecifics in laboratory
rats. Abril. 2015. DOI: 10,1098
SCHULTE-RÜTHER, M. et al. Gender differences in brain networks supporting
empathy. Neuroimage, v. 42, n. 1, p. 393-403, 2008.
SEWARDS, T. V. et al. The medial pain system: neural representations of the
motivational aspect of pain. Brain Res Bull, v.3, n.3, p. 163-80. Nov. 2002.
SILVA. G.A. Análise do limiar de sensibilidade dolorosa à pressão em mulheres com
cefaleia primária durante as fases do ciclo mestrual. Dissertação de mestrado.
Universidade Federal de Pernambuco, 2015.
SILVA, J. C. et al. Modelos experimentais para avaliação da atividade antinociceptiva de
produtos naturais: uma revisão. Brazilian Journal of Pharmacy, v. 94, p. 18-23, 2013.
SILVA, L. F. S; MENESCAlL, O. Cholinergic modulation of tonic immobility and
nociception in the NRM in the guinea pig. Physiol. Behav.v. 87, n.4, p. 821-827.2006.
SIVASELVACHANDRAN, S. et al. Behavioral and mechanistic insight into rodent
empathy. Neuroscience & Biobehavioral Reviews, 2016.
STERNBERG, W. F. et al. Neonatal testosterone exposure influences neurochemistry of
non-opioid swim stress-induced analgesia in adult mice. Pain, v. 63, n. 3, p. 321-326,
1995.
TANNO, A. P. et al. Estresse, ciclo reprodutivo e sensibilidade cardíaca às
catecolaminas. Brazilian Journal of Pharmaceutical Sciences, v. 38, n. 3, 2002.

63
TEIXEIRA, M. W. Dor em Pequenos Animais. Revista CFMV. Conselho Federal de
Medicina Veterinária, Brasília, v. 34, p. 31-41, jan.-abr. 2005.
VANDERAH, T. W. Pathophysiology of pain. Medical Clinics of North America, v.
91, n. 1, p. 1-12, 2007.
VARGAS, K. G. et al. The functions of estrogen receptor beta in the female brain: A
systematic review. Maturitas, v. 93, p. 41-57, 2016.
VILELA, M. et al. Determinação do ciclo estral em ratas por lavado vaginal. Femina, v.
35, n. 10, p. 667-670, 2012.
VOGT, B. A. et al. The medial pain system, cingulate cortex, and parallel processing of
nociceptive information. Prog Brain Res, v. 122, p. 223-35, 2000.
YAMAGATA, K. et al. Estrogens Exacerbate Nociceptive Pain via Up-Regulation of
TRPV1 and ANO1 in Trigeminal Primary Neurons of Female Rats. Endocrinology, v.
157, n. 11, p. 4309-4317, 2016.
ZSCHAECK, L. L.; WURTMAN, R. J. Brain 3H-catechol synthesis and the vaginal
estrous cycle. Neuroendocrinology, v. 11, n. 3, p. 144-149, 1973.

64
ANEXOS A- CERTIFICADO COMITÊ DE ÉTICA

65


















