







Línguas
Páginas
Legal


INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA
INFLUÊNCIA COMBINADA DOS RIOS COMO BARREIRA E DO
GRADIENTE DE INUNDAÇÃO NOS PADRÕES BIOGEOGRÁFICOS
DE ANFÍBIOS E RÉPTEIS SQUAMATA NO SUDESTE DA AMAZÔNIA
LEANDRO JOÃO CARNEIRO DE LIMA MORAES
Manaus, AM
Outubro, 2015

LEANDRO JOÃO CARNEIRO DE LIMA MORAES
INFLUÊNCIA COMBINADA DOS RIOS COMO BARREIRA E DO
GRADIENTE DE INUNDAÇÃO NOS PADRÕES BIOGEOGRÁFICOS
DE ANFÍBIOS E RÉPTEIS SQUAMATA NO SUDESTE DA AMAZÔNIA
Orientadora: Dra. Camila Cherem Ribas
Coorientador: Dr. Dante Pavan
Dissertação apresentada ao Instituto Nacional de
Pesquisas da Amazônia como parte dos requisitos
para obtenção do título de Mestre em Biologia
(Ecologia).
Manaus, AM
Outubro, 2015

II
BANCA EXAMINADORA DA DEFESA ORAL PÚBLICA:
Dra. Fernanda de Pinho Werneck
(Instituto Nacional de Pesquisas da Amazônia)
Dr. Fernando Mendonça d’Horta
(Instituto Nacional de Pesquisas da Amazônia)
Dr. Igor Luis Kaefer
(Universidade Federal do Amazonas)
Aprovado por unanimidade

III
M827 Moraes, Leandro João Carneiro de Lima
Influência combinada dos rios como barreira e do gradiente de
inundação nos padrões biogeográficos de anfíbios e répteis
Squamata no sudeste da Amazônia / Leandro João Carneiro de Lima
Moraes. --- Manaus: [s.n.], 2015.
viii, 78 f. : il. color.
Dissertação (Mestrado) --- INPA, Manaus, 2015.
Orientador : Camila Cherem Ribas.
Coorientador : Dante Pavan.
Área de concentração : Ecologia.
1. Rios como barreira. 2. Tipos florestais. 3. Herpetofauna. I.
Título.
CDD 597.6
Sinopse:
Estudei a influência relativa dos rios como barreira e tipos florestais (formados pelo
gradiente de inundação) na geração dos padrões biogeográficos das assembleias de
anfíbios e répteis Squamata da região do Médio Rio Tapajós, sudeste da Amazônia.
Detectei que esses fatores afetam diferencialmente a distribuição das espécies e os
rios são barreiras mais efetivas para determinadas assembleias e grupos funcionais,
permitindo a previsão dos impactos causado pelas grandes hidrelétricas.
Palavras chave: Assembleias, tipos florestais, Rio Tapajós, herpetofauna,
hidrelétricas.

IV
AGRADECIMENTOS
Agradeço aos meus orientadores Camila Ribas e Dante Pavan por despertarem e
apoiarem a ideia deste projeto e pelo aprendizado e disposição constantes.
Aos meus pais Maria Helena e João Carlos e minha irmã Bia pelo apoio desde o
primeiro momento.
À Marina, minha eterna companheira de aventuras, por aceitar o desafio da
mudança e estar presente em cada momento desta caminhada, iniciando a formação da
nossa família, com a Sushi!
Aos amigos que participaram ativamente da nossa vida em Manaus e nos
ajudaram muito: Renata, Felipe, Marcelo, Vanessa, Priscilla, Andrea Vanessa, Rafa.
Ao INPA e ao Programa de Pós-Graduação em Ecologia por toda a estrutura
disponibilizada, pelas disciplinas e também à todos os integrantes das turmas 2013 e
2014 da Ecologia.
À todos os integrantes que passaram pela Equipe de Herpetofauna e pelo
exigente levantamento dos dados para este projeto: Dante, Luis, Cassimiro, Jerriane, Ana
Barbara, Tainá, Daniel, Luanna, Eder, Elizângela, Mauro, Willian, Wilzon, João Paulo,
Michelle, Rafael, Guilherme, Hallana, Luciana, Albedi e Zé Mário. Agradeço também aos
responsáveis por valiosos registros ocasionais de espécies: Gustavo, Ciça, Odgley, Victor,
André Ravetta, Dante Buzzetti, Claudeir, Elizângela, Wilzon, Mauro, Mariel e Eduardo.
Ao ajudantes de campo Edcarlos, Adson ‘Pote’, Sebastião I, II e III, Dill, Carlinhos,
Clayton, Alcindo ‘Kassamba’, Francisco ‘Lorô’, Ronaldo, Waldir, Wesley, Raimundo,
Gerson ‘Marabá’, Antônio, Warlysson ‘Nhonho’, Mariel e Edhimar, pela companhia e
auxílio durante as árduas caminhadas, pelos ensinamentos e por permanecerem firmes
perante as adversidades.
Ao Seu Léo, Dona Maria I, Dona Maria II, Dona Esmeralda (e os outros moradores
da Vila Rayol), Gisleine, Luciana, Dona Francisca, moradores da Vila Machado, Chico
Pires, Duro, Adriano, Vilica, Baixinho, Bodó, irmãos ‘Frank’, Airton, Seu Zé, e também
outros moradores da região do estudo pela ajuda logística no campo, seja abrindo
trilhas, cozinhando ou pilotando os veículos, mas também pelos ensinamentos
constantes.
À CNEC WorleyParsons Engenharia S/A e aos integrantes da empresa pela
possibilidade do estudo, através do Estudo de Impacto Ambiental do AHE São Luiz do

V
Tapajós, construção das trilhas de amostragem, financiamento e logística das campanhas
de amostragem.
Aos integrantes da Força Nacional, Polícia Federal e Polícia Rodoviária Federal
pelo auxílio na segurança durante a realização de algumas campanhas de amostragem.
Ao Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis
(IBAMA) pela concessão da licença de coleta de espécimes na área de estudo.
À Claudene Barros e aos integrantes do Laboratório de Biologia Molecular
GENBIMOL da Universidade Estadual do Maranhão – campus Caxias (UEMA) pelo auxílio
no sequenciamento dos espécimes.
À Fernanda Werneck, Richard Vogt, Ariane Silva e Vinicius Carvalho pelo auxílio e
possibilidade de acesso à Coleção de Anfíbios e Répteis do INPA.
A todos os outros pesquisadores que ajudaram de alguma maneira. Na
identificação de espécies/linhagens: Pedro Ivo Simões, José Cassimiro, Igor Kaefer,
Vinicius Carvalho, Marcelo Gordo, Rafael Fraga, Pedro Peloso, Marinus Hoogmoed, Paulo
Bernarde, Alfredo Santos-Jr, Albertina Lima, T.C. Ávila-Pires, Omar Entiauspe, Diego
Santana, Sarah Mângia, Rafael de Sá, Antoine Fouquet, Renata Amaro, Stefan Lötters,
Thiago Kashi e Willian Duellman. Nas análises estatísticas: Pedro Martins, Juliana
Schietti, Randolpho Dias-Terceiro. Na revisão do inglês: Guilherme Sampaio.
Aos Drs. Sérgio Borges, Luciano Naka, Pedro Ivo Simões, Igor Kaefer, Adrian
Barnett, Albertina Lima, Fernanda Werneck, Fernando d’Horta e Marcelo Gordo e aos
integrantes do grupo de Biogeografia e Evolução de Vertebrados pelas sugestões
valiosas ao projeto, através de discussões, correções do plano, aula de qualificação e
defesa da dissertação.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPQ) pela
concessão da bolsa de mestrado.
E finalmente, às principais estrelas deste projeto, os anfíbios e répteis da região
do Médio Rio Tapajós, pela possibilidade da realização deste estudo e por fomentarem a
nossa curiosidade acadêmica. Espero que este trabalho desperte a necessidade da
conservação da biodiversidade nesta região pouco explorada mas muito ameaçada e
com alto valor biológico.

VI

VII
RESUMO
A histórica busca pelo reconhecimento dos padrões e processos evolutivos de
organismos amazônicos visando reconstruir a história evolutiva desta paisagem megadiversa
levou à diversas hipóteses. Com o avanço nas técnicas de amostragem e análise, é crescente a
utilização de locais inexplorados e organismos pouco recorridos como modelo nesta busca,
como os anfíbios e répteis Squamata. Neste estudo, buscamos entender a influência integrada
de rios como barreira e do gradiente de inundação na geração e manutenção dos padrões
atuais de distribuição das assembleias de anfíbios e répteis Squamata na região do Médio Rio
Tapajós, na Amazônia Oriental, que é pouco conhecida e atualmente ameaçada pela iminente
atividade antrópica. Para isso, detectamos os indivíduos em pontos amostrais localizados em
todas as margens dos grandes rios, Tapajós e Jamanxim, através de busca ativa, armadilhas de
interceptação e queda e encontros ocasionais nos principais períodos do ciclo hidrológico
fluvial. Identificamos os táxons com uma abordagem integrativa morfológica, acústica,
ecológica e molecular, detectamos e testamos os padrões de distribuição com ordenações uni
e multivariadas, regressões lineares e segmentadas e análise de variância. No total, detectamos
92 anfíbios e 101 répteis, 26 e sete, respectivamente, mostraram efeito dos rios como barreira
à sua distribuição, evidenciando o Rio Tapajós como uma possível barreira recente e o Rio
Jamanxim como uma barreira fraca. Foram encontradas diferenças na composição das
assembleias restritas ou não às florestas ripárias e as mais afetadas pelo rio como barreira
foram os anfíbios e répteis não ripários, e os anfíbios ripários de igarapés. Anfíbios pequenos,
diurnos e terrestres e répteis pequenos a médios, diurnos e semi-arborícolas também
demonstram ser especialmente afetados pelos rios como barreira, e devem ser priorizados em
novos estudos com esta temática. A região do Médio Rio Tapajós é evidenciada como uma
zona de contato faunística, limitando a distribuição de linhagens típicas das regiões Oeste e
Leste da Amazônia. A dinâmica fluvial controla os padrões de distribuição na região e pode
mudar com alterações provenientes de ações antrópicas, promovendo ou impedindo a
segregação dos táxons e extinguindo ambientes relevantes, reforçando a necessidade da
preservação desta dinâmica.
Palavras-chave: Assembleias, tipos florestais, Rio Tapajós, herpetofauna, hidrelétricas.

VIII
ABSTRACT
Combined influence of riverine barriers and flooding gradient on biogeographic
patterns of amphibians and squamates in South-eastern Amazonia
The historical search for recognition of evolutionary patterns and processes of
Amazonian organisms aiming to reconstruct the evolutionary history of this mega-diverse
landscape led to several hypotheses. Advances in sampling and analysis methods allowed the
use of unexplored localities and organisms as a model for this search, such as amphibians and
squamates. In this study, we seek to understand the integrated influence of riverine barriers
and the flooding gradient in current assemblages distribution patterns of amphibians and
squamates in the Middle Tapajós River region, eastern Amazonia, which is little known and
threatened by imminent anthropogenic activity. For this, we detect individuals at sampling
units located in all banks of the main rivers, Tapajós and Jamanxim, through active search,
pitfall traps and occasional encounters in all periods of fluvial hydrological cycle. We identify
the taxa with a morphological, acoustic, ecological and molecular integrative approach, detect
and test the distribution patterns with uni- and multivariate ordinations, linear and piecewise
regressions and analysis of variance. In total, we found 92 amphibians and 101 squamates,
and 26 and seven, respectively, showed riverine barrier effect in their distribution, evidencing
the Tapajós River as a possible recent barrier and the Jamanxim River as a weak barrier.
Differences were found in the composition of assemblages restricted or not to riparian forests
and the most affected by the riverine barrier were non-riparian amphibians and squamates,
and small stream riparian amphibians. Small, diurnal terrestrial amphibians and small-
medium, diurnal semi-arboricole squamates prove to be especially affected by riverine
barriers, and should be priorized in further studies with this theme. The Middle Tapajós River
region is evidenced as a faunistic contact zone, limiting the distribution of typical lineages of
West and East Amazonian regions. The fluvial dynamics controls the distribution patterns in
the region and may be affected by changes from anthropogenic activities, promoting or
preventing the taxa segregation and extinguishing relevant environments, highlighting the
importance to preserve this dynamics.
Keywords: Assemblages, forest types, Tapajós Basin, herpetofauna, hydroelectric power
plants.

IX
LISTA DE FIGURAS
Figure 1. Study area. (a) Location in South America, Brazil, Pará and Amazon Basin. (b)
Sampling units, showing the two main rivers and the main interfluves: (M-T) Madeira-
Tapajós; (M-T/J) Madeira-Tapajós/Jamanxim; (T-J) Tapajós-Jamanxim; (T-X) Tapajós-
Xingu and (T/J-X) Tapajós/Jamanxim-Xingu. (c) Scheme of sampling units, with transect (in
black), plots (uniformly distributed in brown and riparian in grey) and pitfall traps (in red). . 32
Figure 2. Direct ordination of the relative abundance of amphibians (a) and squamates (b) in
relation to distance to water gradient. Note that some species increase in abundance or are
restricted to more humid areas (red square). ............................................................................ 33
Figure 3. Linear regression for the three main taxonomical groups studied, between the
NMDS Axis 1 (species composition) and distance to water, divided in modal distances with
three plots agregated: (1) 20-35 m; (2) 36-50 m; (3) 53-62 m;(4) 63-80 m; (5) 82-95 m; (6)
100-150 m; (7) 180-200 m; (8) 250-300 m; (9) 320-380 m; (10) 440- 490 m (11) 500-580 m;
(12) 615-700 m; (13) 715-1120 m; (14) 1200-1400 m; (15) 1700-2140 m. ............................ 34
Figure 4. Direct ordination of presence-absence data for all survey methods revealing a
higher number of amphibians (a) than squamates (b), which are exclusive to one of the main
rivers' banks. Sampling units (letters) and interfluves (acronyms) are on the top. Different
colours correspond to main interfluves and species written in red are significantly (ANOVA,
p≤0.05) more abundant in one interfluve (inside parentheses). ............................................... 35
Figure 5. NMDS ordinations and composition changes between interfluves for amphibian and
squamate assemblages showing the relative riverine barrier strength for these taxa. (a-c)
amphibians; (d-f) lizards and (g-i) snakes. Interfluves: (M-T) Madeira-Tapajós; (M-T/J)
Madeira-Tapajós/Jamanxim; (T-J) Tapajós-Jamanxim; (T-X) Tapajós-Xingu and (T/J-X)
Tapajós/Jamanxim-Xingu. ........................................................................................................ 36
Figure 6. Relative frequency of amphibian (a) and squamate (b) functional groups with
distribution patterns indicating a riverine barrier influence. In black are the proportion of
species which suffer barrier effect and in gray which do not suffer barrier effect. The total
number of species in each category are inside the bars. Micro-habitat: (TE) terrestrial; (FO)
fossorial; (AR) arboricole; (AQ) aquatic; (SR) semi-arboricole and (SA) semi-aquatic. Body
size: (SM) small; (MD) medium and (LA) large. Activity period: (CR) crepuscular/nocturnal;
(DI) diurnal; (NO) nocturnal and (DN) diurnal/nocturnal........................................................ 37

X
Figure 7. NMDS ordinations and composition changes for amphibian and squamate
assemblages associated with distinct forest types, clustered by composition variation in
relation to distance to water, showing the relative riverine barrier strength. (a-c) Non-riparian
amphibians; (d-f) Small stream riparian amphibians; (g-i) Large river riparian amphibians; (j-
l) Non-riparian squamates and (m-o) Riparian squamates. Interfluves: (M-T) Madeira-
Tapajós; (M-T/J) Madeira-Tapajós/Jamanxim; (T-J) Tapajós-Jamanxim; (T-X) Tapajós-
Xingu and (T/J-X) Tapajós/Jamanxim-Xingu. ......................................................................... 38

SUMÁRIO
RESUMO ..................................................................................................................... VII
ABSTRACT .............................................................................................................. VIII
LISTA DE FIGURAS ................................................................................................... IX
INTRODUÇÃO ............................................................................................................... 1
OBJETIVOS ................................................................................................................... 4
CAPÍTULO I ................................................................................................................... 5
ABSTRACT 7
INTRODUCTION 8
MATERIALS AND METHODS 100
RESULTS 122
DISCUSSION 155
ACKNOWLEDGEMENTS 211
REFERENCES 211
SUPPORTING INFORMATION 29
BIOSKETCHES 29
LIST OF FIGURES LEGENDS 30
FIGURES 32
CONCLUSÕES ............................................................................................................. 39
REFERÊNCIAS BIBLIOGRÁFICAS ......................................................................... 61
APÊNDICES ................................................................................................................. 65

1
INTRODUÇÃO
Na floresta Equatorial Amazônica, uma complexa heterogeneidade ambiental e limites
ambientais pouco evidentes são ofuscados pela uniformidade de grandes extensões de terras
baixas cobertas pelas florestas densas (Hoorn et al., 2010). Com o avanço do conhecimento
científico nesta região, resultado do aumento do esforço e padronização das amostragens,
inovações nas análises e exploração de áreas consideradas como lacunas de conhecimento,
diversos estudos demonstram que variações ambientais são diretamente refletidas nas
variações das distribuições de assembleias distintas (Costa e Magnusson, 2010; Magnusson,
2014; Simões et al., 2014).
Com o acesso à esta variação fina em estudos biogeográficos utilizando abordagens
intra ou interespecíficas e focados em uma determinada região, padrões de distribuição
ofuscados ao longo do tempo são evidenciados (e.g. Naka, 2011; Pomara et al., 2013; Boubli
et al., 2015; Leite et al., 2015; Dias-Terceiro et al., 2015). Assim como os principais
processos causadores da diversificação nesta região, que envolvem interações bidirecionais
entre forças históricas e ecológicas (Losos et al., 2013; Beheregaray et al., 2015). O
reconhecimento da história evolutiva dos organismos permite a associação com as principais
teorias biogeográficas reconhecidas acerca da evolução da paisagem amazônica (Leite e
Rogers, 2013; Smith et al., 2014). Com o avanço e a soma deste conhecimento em
determinadas regiões e utilizando diversos organismos como modelo, pode-se compreender
melhor a história evolutiva da bacia como um todo.
Répteis e anfíbios estão entre os organismos com elevado número de linhagens
conhecidas para a Amazônia (Duellman, 1979, 2005). Na porção brasileira do bioma, ocorrem
cerca de 30% do total de anfíbios e 40% do total de répteis do país (Museu Paraense Emílio
Goeldi, 2012; Costa e Bérnils, 2014; Segalla et al., 2014). Porém, a cada ano diversas
espécies são reconhecidas e descritas (e.g. Orrico et al., 2014; Lima et al., 2014) ou possuem
sua área de ocorrência ampliada (e.g. Simões et al., 2011; Dal Vechio et al., 2015). Estas
descobertas indicam que o reconhecimento pleno da diversidade está longe de ser alcançado,
e este é o primeiro passo para a identificação dos seus principais padrões biogeográficos.
Revisões taxonômicas de espécies amazônicas anteriormente consideradas de ampla
distribuição revelam táxons com diferenças fenotípicas sutis, por exemplo, os répteis Plica
plica (Murphy e Jowers, 2013) e Thecadactylus rapicauda (Bergmann e Russell, 2007) e os
anfíbios Trachycephalus resinifictrix (Gordo et al., 2013), Adenomera spp. (Fouquet et al.,

2
2014), Allobates femoralis (Simões et al., 2010), Hypsiboas fasciatus e H. calcaratus
(Caminer e Ron, 2014).
A dificuldade do reconhecimento pleno da diversidade de anfíbios e répteis
amazônicos reside principalmente no fato de muitos táxons apresentarem elevado grau de
conservadorismo fenotípico e polimorfismo intraespecífico, dificultando a definição dos
limites específicos (Fouquet et al., 2014; Funk et al., 2011; Jansen et al., 2011). Para a
identificação dos táxons que compõem estes complexos de espécies são necessárias
abordagens integrativas, com o estudo refinado da morfologia, acústica, ecologia,
comportamento e diversidade molecular (Simões et al., 2013; Nascimento, 2014), já que
mesmo táxons semelhantes morfologicamente podem apresentar grandes divergências
genéticas (Fouquet et al., 2014). Neste cenário de métodos integrativos de identificação e o
aumento da densidade de amostragens na Amazônia, a real diversidade destes grupos na
região amazônica está sendo desvendada, assim como as suas complexas histórias evolutivas.
Dentre as muitas lacunas de conhecimento para estes grupos localizadas na porção
brasileira da Amazônia, a região do Médio Rio Tapajós é uma das mais importantes. Os
levantamentos e estudos de anfíbios e répteis realizados na Bacia do Rio Tapajós são
concentrados no trecho do Baixo Rio Tapajós (Neckel-Oliveira et al., 2000; Azevedo-Ramos
e Gallati, 2001; Ribeiro Jr. et al., 2012), enquanto o conhecimento das regiões do Alto e
Médio Rio Tapajós é incipiente (Lima et al., 2014). A exploração destes locais é
historicamente difícil em virtude destes trechos do rio possuírem pequena representatividade
viária e baixa navegabilidade, devido ao leito rochoso (Azevedo-Ramos e Gallatti, 2001). Esta
característica promove a formação de trechos com corredeiras e cachoeiras, tornando esta
região alvo de projetos recentes que visam a construção de um complexo de usinas
hidrelétricas (Brasil, PR, 2011). Com essa iminente ameaça antrópica e o conhecimento que
variações em pequena escala da composição de assembleias nos diferentes ecossistemas
florestais da Amazônia são relatados (e.g. Menin et al., 2007; Pavan, 2007; Jorge, 2014), o
avanço no conhecimento dos padrões de distribuição dos organismos nesta região e seus
processos causadores são extremamente necessários. Especialmente a influência dos grandes
rios nas assembleias de anfíbios e répteis que habitam as florestas ripárias, tipo florestal mais
ameaçado pelo represamento dos rios (Pavan, 2007; Ferreira et al., 2013). Dessa forma, visa-
se evitar a perda desta informação biológica e a previsão e mitigação dos futuros impactos
ambientais causados por estes empreendimentos.

3
Considerando o avanço do conhecimento para traçar a história evolutiva da Amazônia,
a identificação dos mecanismos geradores da sua megadiversidade, e o incipiente
conhecimento sobre os padrões de distribuição das assembleias de anfíbios e répteis Squamata
no bioma, nós realizamos uma amostragem intensa destes grupos na ameaçada região do
Médio Rio Tapajós. As linhagens foram identificadas através de técnicas integrativas,
buscando evidenciar os principais padrões de distribuição dos organismos e os mecanismos
históricos e ecológicos causadores destes padrões. As questões que pretendemos responder
com esta abordagem foram: se os grandes rios presentes na região atuam de maneira desigual
como barreiras, determinando os limites das distribuições das espécies e se os diferentes tipos
florestais, florestas de terra firme e alagáveis, influenciam os padrões de distribuição das
espécies. Esperamos com este estudo contribuir para o avanço do conhecimento sobre a
biogeografia destes grupos, a história evolutiva da paisagem e a conservação biológica desta
região.

4
OBJETIVOS
Objetivo geral
Determinar como os padrões biogeográficos encontrados para as assembleias de anfíbios e
répteis Squamata, na região do Médio Rio Tapajós, relacionam-se a fatores históricos e
ecológicos.
Objetivos específicos
Avaliar a influência relativa de barreiras geográficas estabelecidas pelos grandes rios
da região (Tapajós e Jamanxim) sobre os padrões de distribuição de assembleias
encontrados.
Avaliar a influência relativa das variações ambientais determinadas pelo tipo florestal
(Florestas de Terra Firme e Inundáveis) sobre os padrões de distribuição de
assembleias encontrados.
Identificar se assembleias de determinados tipos florestais e organismos de
determinados grupos funcionais são mais afetados pelas barreiras geográficas
estabelecidas pelos grandes rios na região.

CAPÍTULO I
Moraes, L.J.C.L.; Pavan, D.; Barros, M.C.; Ribas,
C.C. 2015. Combined influence of riverine barriers
and flooding gradient on biogeographical patterns of
amphibians and squamates in South-eastern
Amazonia. Journal of Biogeography em revisão.

6
Original Article
Distribution patterns of amphibians and squamates in South-eastern Amazonia
Combined influence of riverine barriers and flooding gradient on
biogeographical patterns of amphibians and squamates in South-
eastern Amazonia
LEANDRO JOÃO CARNEIRO DE LIMA MORAES1,5, DANTE PAVAN
2, MARIA CLAUDENE BARROS3
AND CAMILA CHEREM RIBAS4
1 Graduate Program in Ecology, Instituto Nacional de Pesquisas da Amazônia (INPA), Av.
André Araújo, 2936, 69067-375. Manaus, AM, Brazil.
2 Ecosfera Consultoria e Pesquisa em Meio Ambiente LTDA., R. Gioconda Mussolini, 291,
05591-120. São Paulo, SP, Brazil.
3 Graduate Program in Biodiversity, Environment and Health, Universidade Estadual do
Maranhão, Centro de Estudos Superiores de Caxias, Praça Duque de Caxias, S/N, 65604-000.
Caxias, MA, Brazil.
4 Biodiversity Section and Zoological Collections, Instituto Nacional de Pesquisas da
Amazônia (INPA), Av. André Araújo, 2936, 69067-375. Manaus, AM, Brazil
5 Correspondence: Leandro J.C.L. Moraes. E-mail: [email protected]
Word Count
Abstract: 300
Text: 7236

7
ABSTRACT
Aim To investigate how the distribution patterns of amphibian and squamate assemblages in
the Middle Tapajós River region are influenced by the distance to water (flooded and non-
flooded forests) and riverine barriers (Tapajós and Jamanxim rivers) and, considering the
planned hydroelectric dams on both rivers, to discuss the possible impacts on these
assemblages.
Location Middle Tapajós River, South-eastern Amazonia.
Methods We conducted diurnal and nocturnal surveys combining pitfall traps and active
search along both banks of the Tapajós and Jamanxin Rivers. We identified specimens
through an integrative morphological, acoustic, ecological and molecular approach and
evaluated the influence of riverine barriers and distance to water using uni- and multivariate
ordinations, regressions and ANOVA.
Results We found changes in species composition for both groups along the flooding gradient
and differential riverine barrier effects. The rivers restrict the distribution of 33% of
amphibians and 8% of squamates. For amphibians, the main distribution barrier is the Tapajós
River and for squamates both rivers were of similar importance. The assemblages most
affected by riverine barriers were non-riparian amphibians and squamates, as well as riparian
amphibians associated with small streams. The functional groups most affected were small,
diurnal terrestrial amphibians and small-medium, diurnal semi-arboricole squamates.
Main conclusions The Tapajós River is a distributional boundary for lineages from the
western and eastern Amazonia. The fact that many taxa occur on both banks suggests that the
Tapajós is a recent or semi-permeable barrier and that the Jamanxim is an even weaker
barrier. Anthropogenic actions that affect water level, flooding cycles and river flow may
influence these natural patterns and cause changes to the equilibrium of the riverine barrier
effect. Studies seeking to identify these influences should focus on the most affected
functional groups identified here.
Keywords Assemblages, dispersal limitation, Eastern Amazonia, forest types, herpetofauna,
hydropower plants, Tapajós Basin.

8
INTRODUCTION
Current distribution patterns are shaped by multidirectional interactions between
several processes acting at different temporal and spatial scales (Vellend, 2010; Losos et al.,
2013). Historically, several processes have been considered drivers of the megadiversity
currently found in the Amazonian rain forest. At the regional scale, these processes include
Andean orogeny and drainage evolution (Hoorn et al., 2010; Latrubesse et al., 2010),
neotectonism (Rossetti, 2014), and climate changes (Haffer, 1969); whereas at the local scale,
they may include interactions among biotic and abiotic factors or intrinsic ecological
constraints (Cornell & Lawton, 1992). The dynamic equilibrium between historical events
and local ecological conditions may have led to the development and maintenance of current
Amazonian diversity patterns (Tuomisto & Ruokolainen, 1997).
At the regional scale, the fluvial dynamics of the Amazon Basin seem to play an
important role in the origin and maintenance of diversity patterns (Wallace 1852). The
influence of the evolution of large rivers on the segregation and vicariance of sister-taxa and
the distribution patterns of some vertebrates, especially birds (Cracraft, 1985; da Silva et al.,
2005) and monkeys (Boubli et al., 2014), strongly support this hypothesis. Nevertheless,
Amazonian rivers vary in hydromorphological dynamics and evolution (Sioli, 1968) and
therefore exert unequal influence on the evolutionary history of the biota (Gascon et al., 2000;
Bates et al., 2004). In addition, species' life-histories, including functional traits such as size
or microhabitat preference, directly influence their dispersal capacity (Gascon et al., 1998;
Burney & Brumfield, 2009; Smith et al., 2014; Fouquet et al., 2015). For example, riparian or
semi-aquatic species are expected to have greater capacity of dispersing across rivers due to
the increased contact with aquatic surface (Schiesari et al., 2003; Aleixo, 2006; Cadena et al.,
2011).
Species assemblages are also influenced by local ecological factors that are assessed
only through fine-scale studies of distribution patterns. Environmental variables structuring
the assemblages of several Amazonian taxa have already been identified. For example,
topographical characteristics, soil composition and microhabitat variation influence the
distribution of plants (Emílio et al., 2013; Schietti et al., 2013; Zuquin et al., 2014) and
squamates (Pavan, 2007; Fraga et al., 2011; Garda et al., 2012). Distance to streams (Menin,
2011), forest type (Gascon et al., 2000; Pavan, 2007) and leaflitter morphology (Menin et al.,

9
2007) all influence anuran distribution. However, the overall difficulty researchers have faced
in detecting general patterns suggests that the relationship between ecological factors and
distribution patterns is idiosyncratic (Dias-Terceiro et al., 2015).
Amphibians and squamates are among the most diverse Amazonian vertebrate groups,
include several threatened species and play important ecological roles in ecosystem
operations, but few studies have tried to correlate their distribution patterns with evolutionary
processes (Ron, 2000). Studying these groups as a model to investigate the evolutionary
history of the Amazonian landscape may be relevant because they are sensitive to
environmental and climatological changes (Kerby et al., 2010; Mitchell & Janzen, 2010) and
frequently have low individual motility, showing greater vulnerability to these changes and
retaining signals of historical events (Simões et al., 2014). Also, they are generally widely
distributed throughout the basin (Duellman, 1979).
Most of the studies investigating this correlation focus on intraspecific morphological,
acoustic or molecular variation (e.g., Simões et al., 2008; Souza et al., 2013; Fouquet et al.
2014), whereas few address assemblage variations. The studies that do exist show that both
historical and ecological factors help shape species distributions within these groups at
different scales (Ávila-Pires, 1995; Gascon et al., 2000; Ron, 2000; Dias-Terceiro et al.,
2015). The lack of assemblage-level research approaches to investigate these relationships is
caused primarily by the lack of basic knowledge of species distributions that has resulted from
a concentration of studies in a few accessible areas, lack of standardization and short-term
samplings (Azevedo-Ramos & Galatti, 2001). Moreover, incomplete taxonomic knowledge,
can hinder the understanding of real distribution patterns (Fouquet et al., 2007).
Recently, the Amazon basin has been the focus of large developmental projects,
including building dams for hydroelectric energy generation on the primary Amazonian rivers
(Brasil, PR, 2011). These dams will affect the water flow and flooding dynamics of the main
Amazonian rivers and adjacent forests (Nilsson & Berggren, 2000), and it is unknown how
these anthropogenic changes will affect biodiversity. Currently, one of the most threatened
Amazonian rivers is the Tapajós as several ongoing projects aim to build consecutive dams
along this river and its tributaries (Fearnside, 2015).
Here, we aim to understand how the distribution patterns of amphibians and squamates
are influenced by riverine barriers and by distance to water, as a proxy for forest type (upland
or flooded forest) within the poorly known and threatened region of the Middle Tapajós
River. We investigate whether assemblage composition changes with distance from the water

10
and on the opposite banks of the Tapajós and Jamanxim Rivers, the largest rivers that drain
the area. We also ask whether the riverine barrier effect varies in relation to specific
assemblages or functional groups that occur in forests with different flooding regimes.
MATERIALS AND METHODS
The clear-water Tapajós River is one of the main tributaries of the Amazon River. Its
headwaters drain the Precambrian terrain of the Brazilian Shield, which provides low
sediment input (Sioli, 1968). The middle Tapajós is located in a geological contact zone,
including the northern limit of the cratonic Brazilian Shield and the sedimentary plains of the
Amazon River Basin. The dry season occurs between June and August and the rainy season
between November and March. Average annual temperature is 26°C and annual rainfall
varies from 50 to 375 mm (Alvares et al., 2013).
Amphibian and reptile sampling
Sampling was conducted on both banks of the Tapajós and Jamanxin rivers, at 10
sampling units (Fig. 1). Each unit was comprised of one 4 km transect and seven
perpendicular 250 m plots, four of these in upland forests (non-flooded) and three in
periodically flooded riparian forests. The plots follow the contour lines of the terrain. Detailed
descriptions of environmental variation observed in each sampling unit are available in
Appendix S1. The sampling scheme aims to increase standardization and independence
(Magnusson et al., 2005). We sampled both transects and plots during six survey campaigns
from July 2012 to November 2013, covering all phases of the hydrological cycle.
We used complementary methods to register individuals, seeking to maximize
regional diversity sampling and to reduce the effect of sampling bias. Using the active search
method (Heyer, 1994), five researchers searched for individuals along the centre line of trails
at diurnal and nocturnal periods and in all microhabitats accessible visually or through
vocalization (in the case of male breeding anurans). The number of anurans calling at each
point was estimated during sampling. Each plot was sampled for at least 14 days and nights,
and each transect was sampled for six days and nights, totalling more than 340 days of active
search surveys.

11
Pitfall traps (Heyer, 1994) were installed at four random plots, each consisting of three
equidistant parallel rows of five buckets. These traps were checked at 24 h intervals for five
consecutive days during each survey campaign, totalling an effort of 600 trap nights.
Individuals registered through methods other than those described here, or collected by third
parties, were considered casual encounters.
Taxa delimitation and identification
We used an integrative approach to taxa delimitation, combining morphological,
acoustic, ecological and molecular techniques (Padial et al., 2010). The morphological,
acoustic and ecological analyses was based in author’s observations, species descriptions,
taxonomic revisions and field guides, and included comparisons with voucher specimens
deposited in the collection of amphibians and reptiles at the National Institute for Amazonian
Research, including some type series. Molecular species identification was employed for
cryptic species complexes. DNA was extracted from tissue samples and the 16S rRNA gene,
which is one of the standard markers for these groups (Vences et al., 2012), was amplified
through a polymerase chain reaction (PCR) and sequenced in an ABI PRISM 3500 (Life
Technologies) sequencer. Additional sequences from related taxa and outgroups were
obtained from Genbank. Sequences were aligned in CLUSTAL OMEGA (Sievers et al., 2011)
and phylogenetic trees were inferred using Maximum Likelihood in MEGA 5 (Tamura et al.,
2011). Clade support was estimated by bootstrap. See Appendix S2 for details of integrative
species identification.
Data analyses
To detect whether and how environmental variations between distinct forest types
influences assemblage structure, we constructed direct ordinations and dissimilarity matrices
using the qualitative (presence-absence) data from the plot surveys. Dissimilarity was
measured using the Jaccard index, and the dimensionality was reduced using one axis of a
non-metric multidimensional scale (NMDS) (Clarke, 1993). Because some upland plots were
located next to water, we considered forest type variation as a continuous measure based on
proximity of plots to water bodies. Using regional hydrographic maps, we measured the

12
straight-line distance at 90° from the beginning of the plot to the nearest water body.
Correlation between species composition and water proximity (flooding gradient) was
analysed by linear regression. We excluded the species for which we sampled less than 5
individuals because it is more difficult to determine whether they are associated with a
particular forest type.
To evaluate the riverine barrier effect, we constructed dissimilarity matrices using the
entire qualitative data from sampling units in each interfluve (Madeira-Tapajós, Tapajós-
Jamanxim, Madeira-Tapajós/Jamanxim, Tapajós/Jamanxim-Xingu and Tapajós-Xingu).
Dissimilarity was measured using the Jaccard index, and we determined the riverine effects
with NMDS. We used these same methods to investigate how riverine barriers affect
assemblages from distinct forest types based on the qualitative data of all surveys, considering
only the species occurring in each assemblage. For this, we determine the size of riparian
forest through assemblages (NMDS Axis1) breakings in relation to distance to water with
piecewise regressions. We generated ordinations for the assemblages of (1) non-riparian
amphibians, (2) small stream riparian amphibians, (3) large river riparian amphibians, (4)
non-riparian squamates, and (5) riparian squamates. The last assemblage showed no
composition changes across the two riparian forests types. We used one first NMDS axis to
test the assemblage differences in interfluves with an ANOVA and Tukey post-hoc test.
To detect significant differences in species abundance between interfluves we also
used ANOVA and Tukey test. We evaluated the riverine barrier effect for species from
different functional groups based on traits that reflect their life-history: activity period,
microhabitat and body size, comparing the relative number of affected species in each
category. All of the analyses were performed in R (R Development Core Team, 2014).
RESULTS
We registered 14,253 amphibians of 92 species and 3,410 squamates of 101 species (Table
S1). We excluded species recorded at only one site and obtained a final dataset of 79
amphibians and 82 squamates.

13
Flooding gradient
We detected significant changes in species composition associated with proximity to
water for the amphibians and lizards, and weak changes for the snakes (Table 1). The
abundance of some species increased close to a watercourse, whereas some species occurred
exclusively in these areas, including the amphibians Hypsiboas multifasciatus, H.
leucocheilus and Allobates magnussoni and the squamates Bothrops atrox and Uranoscodon
superciliosus (Fig. 2, 3). However, no species occurred exclusively in upland forests, which
was expected because of their varying degrees of dependence on water.
Riverine barriers: widely distributed taxa
Of 79 amphibian species, 49 were detected on all banks of both main rivers. Of these,
33 are also widely distributed in the Amazonia, whereas three are found only south of the
Amazon River, five are restricted to the central portion of the southern Amazonia and one is
distributed in the eastern Amazonia. Nine other species are poorly known or new taxa with
unknown distributions. At least 16 of the widely distributed taxa are species complexes and
intraspecific analyses may reveal restricted lineages (Table S1).
Regarding the squamates, of the 82 species identified, 54 were widely distributed; and
of these, 48 are also widely distributed in the Amazonia. However, at least two are species
complexes, especially snakes of the genus Atractus. One species is restricted to the south of
the Amazon River and one to the central portion of the southern Amazonia. Two species are
distributed in the eastern Amazonia and at least two have uncertain distribution due to
conspicuous habits or taxonomic uncertainty (Table S1).
Riverine barriers: taxa with distributions restricted by rivers
The distribution patterns of 30 amphibian and 27 reptile species indicated some
riverine barrier effect, with species occurrence restricted to one bank of the main river (Fig.
4). We excluded four amphibians and 20 squamates from the analyses as they were already

14
historically registered on opposite margins or are poorly understood, indicating that the
riverine effect on these species could have been due to sampling bias.
Fifteen amphibians are restricted to the western bank of the Tapajós, 10 to the eastern
bank and one to the eastern bank of the Jamanxim. Only the Tapajós represented an effective
barrier, with different species composition on opposite banks. No composition difference was
found on opposite banks of the Jamanxim (Table 1). NMDS ordination reflects this
dissimilarity and shows a large break in composition associated with the Tapajós and no break
associated with the Jamanxim (R²=0.74, stress=0.05) (Fig. 5).
Among the squamates, two species were restricted to the western bank of the Tapajós,
three to the eastern bank and two to the eastern bank of the Jamanxim. The lizard assemblages
on interfluves show some influence from both rivers as barriers. For snakes, no riverine
barrier effect was detected (Table 1). In the NMDS ordination, the Tapajós and Jamanxim
show some influence as geographical boundaries for lizard assemblages (R²=0.82,
stress=0.09) and not significant for snake assemblages (R²=0.49, stress=0.16) (Fig. 5).
Some species were present on both banks of these rivers but had extremely unequal
abundances on opposite banks. For example, we recorded 49 individuals of the lizard Norops
trachyderma on the eastern bank of the Tapajós and only one on the western bank; and for the
frog Rhinella gr. margaritifera1, we identified 88 individuals on the western bank and one on
the eastern bank. In total, six amphibians and six squamates have unequal abundances
between interfluves. Of these, eight are more abundant in the Madeira-Tapajós, two in the
Tapajós-Jamanxim and two in the Tapajós-Xingu (Fig. 4).
The amphibian functional groups most affected by the riverine barrier effect were
small, diurnal and terrestrial species, especially the genera Pristimantis (four taxa), Allobates
and Leptodactylus (three taxa each). The most affected squamate functional groups were
small-medium sized, diurnal and semi-arboricole species, especially lineages of the genus
Gonatodes (two species) (Fig. 6).
Riverine barrier effects on species along the flooding gradient
Because amphibians and squamates are not uniformly distributed in forests with
different degrees of flooding, we tested the riverine barrier effect for different classes of
assemblages. For amphibians, the riverine barrier effect was stronger for non-riparian and

15
riparian assemblages associated with small streams. In the latter case, this result was
influenced by the fact that some species typical of this environment such as Hypsiboas
leucocheilus, H. multifasciatus and Leptodactylus leptodactyloides, are often restricted to one
bank. No significant riverine barrier effect was detected for riparian assemblages associated
with large rivers (Table 1). These effects were visualized in NMDS ordinations: two clusters
were segregated by the Tapajós for non-riparian assemblages (R²=0.84, stress=0.004) and the
small stream riparian assemblages (R²=0.87, stress=0.05), but no clustering occurred for large
river riparian assemblages (R²=0.81, stress=0.06) (Fig. 7).
For squamates, the non-riparian assemblages showed a riverine barrier effect
associated with opposite banks of the Tapajós, whereas the riparian assemblages were
homogeneous (Table 1). In the NMDS ordinations, the Tapajós also segregated two clusters
for the non-riparian assemblages (R²=0.86, stress=0.09) but no clustering occurred for the
riparian assemblages (R²=0.91, stress=0.15) (Fig. 7).
DISCUSSION
We registered several undescribed or recently described taxa in addition to some
distribution extensions, demonstrating the difficulty of characterizing biogeographical
patterns based on current taxonomic knowledge without large sampling efforts to collect
primary data and the use of integrative species identification approaches. Analyses of
amphibian and squamate distribution patterns in the Middle Tapajós River region showed that
both ecological (flooding regimes) and historical (riverine barriers) factors influence the
structure of assemblages, as reported for other Amazonian regions (Gascon et al., 2000; Dias-
Terceiro et al. 2015). Changes in assemblage composition occurred both on opposite banks of
the Tapajós and in forests with different flooding regimes. In addition, some functional
groups are more affected by riverine barriers than others.
Flooding gradient and species assemblages
Several studies have detected changes in assemblage composition in distinct
Amazonian forest ecosystems, e.g., for plants (Wittmann et al., 2004; Schietti et al., 2013),
mammals (Haugaasen & Peres, 2005), birds (Bueno et al., 2012) and amphibians and reptiles

16
(Doan & Arriaga, 2002; Pavan, 2007). In the Middle Tapajós River region, despite the
relatively small development of riparian ecosystems due to riverine channel morphology, we
detected a change in composition between assemblages along the flooding gradient (distance
to water). This was especially the case for amphibians, which have different patterns even
between distinct riparian forests. These differences may be due to species endogenous
processes, such as intrinsic ecological requirements related to reproduction, behaviour and
physiological constraints, which restrict their distributional ranges (Dale & Fortin, 2014).
The high intra- and interspecific homogenization among riparian assemblages
documented for other Amazonian organisms (e.g., Aleixo, 2006; Cadena et al., 2011) was
corroborated, especially for the assemblages associated with large rivers (Igapó forests). This
indicates that dispersal across and along the river is facilitated for these organisms. It has been
suggested that strong water flow may facilitate dispersal by carrying "rafts" of land or
vegetation across or along the rivers (Schiesari et al., 2003; Pavan, 2007).
Contrastingly to literature (e.g. Aleixo, 2006; Cadena et al., 2011), we detected
changes in the composition of small stream riparian amphibian assemblages on opposite
banks of the Tapajós. That difference can be generated by species physiological and
behavioural constraints because individuals remain sedentary in these wetlands and never use
the upland forests and large river riparian forests, thus hindering the exchange of individuals
between banks and may increase morphological and genetic divergence. Analysing the
genetic differences in one amphibian with these characteristics, Mullen et al. (2010) noted
that gene flow can be low even between nearby streams.
Rivers as barriers to dispersal
For amphibians, the Tapajós is the eastern distributional limit of many groups from the
south-western Amazonian highlands, including Hemiphractus scutatus, Dendropsophus
bokermanni and Hypsiboas leucocheilus. Some typical lowland lineages from the Solimões
Basin, such as Ameerega trivittata, Allobates masniger, Scinax cruentommus and
Chiasmocleis bassleri are also registered exclusively on western bank of the Tapajós.
The eastern banks of Tapajós and Jamanxim are the distributional limits for some
amphibians from the Brazilian Shield highlands, such as Adelphobates galactonotus,
Ranitomeya amazonica and Leptodactylus paraensis, and the eastern lowlands, such as
Leptodactylus leptodactyloides and Hypsiboas multifasciatus. Some additional amphibians

17
with distributions bounded by the Tapajós are enigmatic, and it is difficult to infer
biogeographical patterns. These species include: Synapturanus sp., a genus extremely rare
south of the Amazon River (Albertina Lima, com. pess.); Pristimantis gr. lacrimosus3, an
undescribed taxon, and Pristimantis reichlei, an Andean lineage that was registered on only
the eastern bank of the Tapajós.
The Tapajós is also a distributional limit for squamates. Some typical lowland lineages
of the Solimões Basin and highland lineages of the south-western Amazonia that occur
exclusively on the western bank of the Tapajós include Norops tandai (Solimões Basin) and
Gonatodes hasemani (south-western Amazonia highlands). Typical lineages of the Brazilian
Shield highlands occur exclusively on the eastern banks of the Tapajós and Jamanxim, such
as Enyalius leechi, Gonatodes tapajonicus and Thamnodynastes pallidus, or are much more
abundant on this bank, such as Norops trachyderma.
In the studied region, the Tapajós and Jamanxin rivers contribute unequally to the
origin and maintenance of species distribution patterns. The exchange of individuals between
opposite banks of the Tapajós River is higher for riparian than for non-riparian assemblages,
whereas the Jamanxim was not an effective barrier for amphibians and was a weak barrier for
squamates. Previous studies provided discordant results on the effects of primary Amazonian
rivers as barriers to amphibian and reptile dispersal (Ávila-Pires, 1995; Gascon et al., 2000;
Ron, 2000; Dias-Terceiro et al., 2015). For amphibians, the Tapajós appears to be a stronger
barrier than other Western Amazonian rivers, such as the Juruá River (Ron, 2000), whereas
the relatively young Madeira River has impact as a barrier (Dias-Terceiro et al., 2015). The
Tapajós had a weak barrier effect for lizard assemblages and is not historically considered a
barrier for this group (Ávila-Pires, 1995; da Silva Jr. & Sites, 1995). The river had an even
weaker barrier effect on snake assemblages, which was expected as most species are widely
distributed in the Amazonia due to their high dispersal capacity (da Silva Jr. & Sites, 1995),
or may be due to greater difficulty in detecting them, resulting in fewer records and
preventing the patterns definition (Fraga et al., 2014).
A small portion of taxa recorded in the present study had their distributions bounded
by rivers (33% of amphibians and 8% of squamates), suggesting that the Tapajós might be a
recent or semi-permeable barrier. It seems paradoxical that an ancient river located on stable
geological terrain (Hoorn et al., 2010) has a small influence on the structure of species
assemblages; however, recent phylogeographic studies of several vertebrates (e.g., birds
(Ribas et al., 2012, Fernandes et al., 2013), frogs (Jungfer et al., 2013; Simões et al., 2014)

18
and snakes (Nascimento, 2014)) show shallow genetic divergences between intraspecific
lineages associated with opposite banks of Brazilian shield rivers. These results may indicate
that the intrinsic ability of each lineage to cross a river and to establish populations on the
other bank are more relevant than the age, hydromorphology or stability of rivers; or, they
may add to the evidence that Brazilian Shield rivers and the surrounding environments have
gone through important historical changes in the recent past that may have affected
distribution patterns (Rossetti, 2014; Ribas et al., in press.).
Diversification of specific functional groups
Small diurnal terrestrial and semi-terrestrial amphibians with restricted territories, such
as those of the genera Allobates, Pristimantis and Adenomera, are abundant in the forest litter
and have a low dispersal capacity. The combination of dispersal limitation, which has already
been postulated as one major driver of speciation (e.g. for birds, Burney & Brumfield, 2009;
Smith et al., 2014 and anurans, Wollenberg et al., 2011), and potential to use new habitats or
ecological niches, due to the greater independence from water during the reproductive cycle
(Duellman & Trueb, 1994), may promote diversification in these groups.
Small semi-arboricole diurnal Gonatodes lizards are equally abundant in forested areas
and live mainly in tree trunks. High abundance and niche competition may promote
ecological divergence in sympatric species of this genus (Vitt et al., 2000). These biotic
interactions and ecological constraints may increase the riverine barrier effect, accelerating
allopatric diversification (Gutiérrez et al., 2014). Based on dated phylogenies, Gamble et al.,
(2008) already postulated that Amazonian Tertiary orogeny and drainage evolution are major
drivers of diversification in this group.
These amphibian and reptile functional groups are most affected by the emergence of
geographical barriers like large rivers, which segregate populations. Therefore, species of
these genera or functional groups should be the preferred model organisms in the study of the
recent evolution of the Amazon Basin (Simões et al., 2014). However, their recent
diversification (Araújo et al., 2008) creates difficulties in identifying taxonomic units, and
better taxonomic knowledge is essential for further biogeographical studies.

19
Integrated influence of rivers and their floodplains on biodiversity patterns
This study demonstrates the relevance of large rivers as drivers of species diversity
patterns. River dynamics both create distinct environments that allow the establishment of
ecologically distinct assemblages and establish barriers with varying permeabilities for
distinct components of these assemblages, both in current and past times. River dynamics also
create distinct opportunities for fauna exchange between opposite banks as water flow may
increase the longitudinal and transversal passive dispersal of individuals inhabiting riparian
forests but not of those inhabiting upland forests, thereby preventing or promoting lineage
differentiation.
The studied region is a contact zone for amphibians and squamates from the eastern
and western Amazonia. This may be due to rainfall seasonality, temperature and
geomorphological gradients (Sombroek, 2001). The western bank of the Tapajós River is
geomorphically influenced by the Solimões and Amazon sedimentary basins, whereas the
eastern bank is highly influenced by the Brazilian Shield (Sombroek, 2000). This difference is
evident even in the current river morphology, with a sharp curve along the Brazilian Shield
limit. In this region, fauna group components from dense western Amazonian forests,
influenced by the Andes and the sedimentary basin, mix with components from drier eastern
Amazonian forests that are influenced by the Amazonia-Cerrado transition following these
environmental gradients. This mixed influence is evident in the fact that most species
restricted to the western bank of the Tapajós inhabit the Western lowlands (69%), whereas
those restricted to the eastern bank (78%) are typical of the Brazilian Shield.
The river prevents the formation of a continuous gradient in the contact zone of these
two distinct Amazonian regions, acting both as an ecological and physical barrier (Tuomisto
& Ruokolainen, 1997). For this reason, most of the amphibians and squamates that have
shown some riverine barrier effect in this region have no sister-taxa or have unequal
abundances on opposite banks, weakening the effect of this river as a vicariant agent to these
groups and reinforcing the combined action of recent geomorphological changes and various
barrier effects.
Ecological constraints also prevent the establishment of many species detected in this
study within the lower Tapajós River region of the Amazon Sedimentary Basin, where species
richness and composition, especially of amphibian assemblages, change dramatically (see

20
Neckel-Oliveira et al., 2000). Most of the species that occur in this region are typical of
riparian forests associated with large rivers in the middle Tapajós River region.
Considerations about the potential impacts of planned large dams
The imminent construction of hydropower plants, associated with a waterway to
increase navigability in the Tapajós Basin (Fearnside, 2015), will significantly affect
biogeographical patterns, with the potential to cause several environmental impacts at
different scales. After the dams are complete, the rivers on which they are constructed will be
in a permanently flooded state. The natural fast water flow of the rivers will transform into
lentic systems (Nilsson & Berggren, 2000), breaking the rivers’ longitudinal and transversal
functionality (Junk et al., 1989; Vannote et al., 1990) and affecting several organisms that
have eco-physiological constraints.
Based on the results presented here, we predict that these changes will affect mainly
the riparian assemblages, which are distinct from the upland assemblages and include some
restricted species, by eliminating the environments needed to complete the species’ life cycle.
In the short-term, the fauna rescue programme and river filling process may affect the
historical distribution boundaries, mixing components from opposite margins. In the long-
term, the higher water volume and lower energy may increase the potential of these rivers to
become stronger barriers to dispersal (Ward & Stanford, 1995; Hayes & Sewlal, 2004),
thereby affecting both riparian and non-riparian assemblages. This is especially true on the
Jamanxim, which is currently highly permeable to dispersion during the dry season. A
decrease in the rivers’ permeability will prevent species exchanges among regional species
pools, leading to the development of distinct and poorer local diversities on opposite banks
(Brown et al., 2011). Furthermore, the Tapajós Basin is known to harbour distinct flora and
fauna, with many endemic lineages of plants (Ferreira et al., 2013), frogs (Lima et al., 2014),
lizards (Rodrigues, 1980) and birds (Olmos & Pacheco, 2003). Changes in the fluvial
dynamics of this region will certainly impact these lineages, whose ecological constraints and
evolutionary history associated with the natural fluvial dynamics of the region have led to
their current distribution patterns.
Anthropogenic actions can thus affect the role of rivers as current and historical
drivers of biodiversity patterns, disturbing the regional ecosystem as a whole. Because the

21
Amazonian rain forest is intrinsically linked to the largest river basin in the world, preserving
the integrity of its rivers is crucial to the maintenance of the forested ecosystems. We stress
that the biogeographical patterns highlighted here should be part of the discussion of the
feasibility of all large developmental projects that affect Amazonian rivers.
ACKNOWLEDGEMENTS
We thank L.F. Storti, J. Cassimiro, M. Hoffman, T.F.D. Rodrigues, J.M.B. Ghelere, A.B.
Barros, E.S. Brito and all field mates who helped us in data survey. I.L Kaefer, F.P. Werneck,
A.P. Lima, M. Gordo, S.H. Borges, L.N. Naka, P.I. Simões, F.M. d’Horta and A. Barnett
provided valuable suggestions. The specimens were collected under collection permit number
066/2012 provided by Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais
Renováveis (IBAMA). CNEC Worleyparsons Engenharia S.A. provided financial and
logistical support. Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq,
Brazil) provided a Master's degree scholarship to L.J.C.L. Moraes and a productivity
fellowship to C.C. Ribas (307951/2012-0).
REFERENCES
Aleixo, A. (2006) Historical diversification of floodplain forest specialist species in the
Amazon: A case study with two species of the avian genus Xiphorhynchus (Aves:
Dendrocolaptidae). Biological Journal of the Linnean Society, 89, 383-395.
Alvares, C.A., Stape, J.L., Sentelhas, P.C., Gonçalves, J.L.M. & Sparovek, G. (2013)
Koppen's climate classification map for Brazil. Meteorologische Zeitschrift, 22, 711-
728.
Araújo, M.B., Nogués-Bravo, D., Diniz-Filho, A.F., Haywood, A.M., Valdes, P.J. &
Rahbeck, C. (2008) Quaternary climate changes explain diversity among reptiles and
amphibians. Ecography, 31, 8-15.
Ávila-Pires, T.C.S. (1995) Lizards of Brazilian Amazonia (Reptilia: Squamata). Zoologische
Verhandelingen, 299, 1-706.

22
Azevedo-Ramos, C. & Galatti, U. (2002) Patterns of amphibian diversity in Brazilian
Amazonia: conservation implications. Biological Conservation 103, 103-111.
Bates, J.M., Haffer, J. & Grismer, E. (2004) Avian mitochondrial DNA sequence divergence
across a headwater stream of the Rio Tapajós, a major Amazonian river. Journal of
Ornithology, 145, 199-205.
Boubli, J.P., Ribas, C., Lynch Alfaro, J., da Silva, M.N.F., Pinho, G.M., & Farias, I.P. (2014)
Spatial and temporal patterns of diversification on the Amazon: a test of the riverine
hypothesis for all diurnal primates of Rio Negro and Rio Branco in Brazil. Molecular
Phylogenetics and Evolution, 82, 400-412.
Brasil, PR (Presidência da República). (2011) PAC-2 Relatórios. PR, Brasília, DF. Available
at: www.brasil.gov.br.
Brown, B.L., Swan, C.M., Auerbach, D.A., Grant, E.H.C., Hitt, N.P., Maloney, K.O., Patrick,
P. (2011) Metacommunity theory as a multispecies, multiscale framework for studying
the influence of river network structure on riverine communities and ecosystems.
Journal of the North American Benthological Society, 30, 310-327.
Bueno, A.S., Bruno, R.S., Pimentel, T.P., Sanaiotti, T.M. & Magnusson, W.E. (2012) The
width of the riparian habitats for understory birds in an Amazonian forest. Ecological
Applications, 22, 722-734.
Burney, C.W. & Brumfield, R.T. (2009) Ecology predicts levels of genetic differentiation in
neotropical birds. American Naturalist, 174, 358-368.
Cadena, C.D., Gutiérrez-Pinto, N., Dávila, N. & Chesser, R.T. (2011) No population genetic
structure in a widespread aquatic songbird from the Neotropics. Molecular
Phylogenetics and Evolution, 58, 540-545.
Clarke, K.R. (1993) Non-parametric multivariate analyses of changes in community structure.
Australian Journal of Ecology, 18, 117-143.
Cornell, H.V. & Lawton, J.H. (1992) Species interactions, local and regional processes, and
limits to the richness of ecological communities: a theoretical perspective. Journal of
Animal Ecology, 61, 1-12.
Cracraft, J. (1985) Historical biogeography and patterns of differentiation within the South
American avifauna: areas of endemism. Ornithological Monographs, 36, 49-84.
Dale M.R.T. & Fortin M.J. (2014). Spatial analysis: a guide for ecologists. 2nd edn.
Cambridge University Press, Cambridge.

23
Dias-Terceiro, R.G., Kaefer, I.L., de Fraga, R., de Araújo, M.C., Simões, P.I. & Lima, A.P.
(2015) A matter of scale: historical and environmental factors structure anuran
assemblages from the Upper Madeira river, Amazonia. Biotropica, 47, 259-266.
Doan, T.M. & Arriaga, W.A. (2002) Microgeographic variation in species composition of the
herpetofaunal commities of Tambopata region, Peru. Biotropica, 34, 101-117.
Duellman, W.E. (1979) The South American herpetofauna: Its origin, evolution, and
dispersal. Museum of Natural History/University of Kansas, Kansas.
Duellman, W.E. & Trueb L. (1994) Biology of Amphibians. Johns Hopkins University Press,
Baltimore.
Emílio, T., Quesada, C.A., Costa, F. et al. (2013) Soil physical constraints limit palm and tree
basal area in Amazonian forests. Plant Ecology & Diversity, 7, 130204073052004.
Fearnside, P.M. 2015. Amazon Dams and Waterways: Brazil’s Tapajós Basin Plans. Ambio,
44, 426-439.
Ferreira, L.V., Cunha, D.A., Chaves, P.P., Matos, D.C.L. & Parolin, P. 2013. Impacts of
hydroelectric dams on alluvial riparian plant communities in eastern Brazilian
Amazonian. Anais da Academia Brasileira de Ciências, 85, 241-251.
Fernandes, A.M., Gonzalez, J., Wink, M. & Aleixo, A. (2013) Multilocus phylogeography of
the wedge-billed woodcreeper Glyphorynchus spirurus (Aves, Furnariidae) in lowland
Amazonia: widespread cryptic diversity and paraphyly reveal a complex diversification
pattern. Molecular Phylogenetics and Evolution, 66, 270-282.
Fouquet, A., Gilles, A., Vences, M., Marty, C., Blanc, M. & Gemmell, N.J. (2007)
Underestimation of species richness in Neotropical frogs revealed by mtDNA analyses.
PLoSOne, 32, e1109.
Fouquet, A., C.S. Cassini, C.F.B. Haddad, N. Pech & Rodrigues, M.T. (2014) Species
delimitation, patterns of diversification and historical biogeography of the Neotropical
frog genus Adenomera (Anura, Leptodactylidae). Journal of Biogeography, 41, 855-
870.
Fouquet, A., Courtois, E.A., Baudain, D., Jucivaldo Dias Lima, J.D., Souza, S.M., Noonan,
B.P. & Rodrigues, M.T. (2015) The trans-riverine genetic structure of 28 Amazonian
frog species is dependent on life history. Journal of Tropical Ecology, 31, 361-373.
Fraga, R., Lima, A.P. & Magnusson, W.E. (2011) Mesoscale spatial ecology of a tropical
snake assemblage: the width of riparian corridors in Central Amazonia. The
Herpetological Journal, 21, 51-57.

24
Fraga, R., Stow, A.J., Magnusson, W.E. & Lima, A.P. (2014) The Costs of Evaluating
Species Densities and Composition of Snakes to Assess Development Impacts in
Amazonia. PLoS ONE, 9, e105453.
Gascon, C., Lougheed, S.C. & Bogart, J.P. (1998) Patterns of genetic population
differentiation in four species of Amazonian frogs: a test of the Riverine Barrier
Hypothesis. Biotropica, 30, 104–119.
Gascon, C., Malcolm, J.R., Patton, J.L., Silva, M.N.F., Bogart, J.P., Lougheed, S.C., Peres,
C.A., Neckel, S. & Boag, P. (2000) Riverine barriers in the geographic distribution of
Amazonian species. Proceedings of the National Academy of Sciences, 97, 13672-
13677.
Garda, A.A., Wiederhecker, H.C., Gainsbury, A.M., Costa, G.C., Pyron, R.A., Vieira, G.H.C.,
Werneck, F.P. & Colli, G.R. (2012) Microhabitat Variation Explains Local-scale
Distribution of Terrestrial Amazonian Lizards in Rondônia, Western Brazil. Biotropica,
45, 245–252.
Gutiérrez, E.E., Boria, R.A. & Anderson, R.P. (2014) Can biotic interactions cause allopatry?
Niche models, competition, and distributions of South American mouse opossums.
Ecography, 37: 741–753.
Haffer, J. (1969) Speciation in Amazonian forest birds. Science, 165, 131-137.
Haugaasen, T. & Peres, C.A. (2005) Mammal assemblage structure in Amazonian flooded
and unflooded forests. Journal of Tropical Ecology, 21, 133-145.
Hayes F.E. & Sewlal, J.A.N. (2004) The Amazon River as a dispersal barrier to passerine
birds: effects of river width, habitat and taxonomy. Journal of Biogeography, 31, 1809-
1818.
Heyer, W.R., Donnelly, M.A., Mcdiarmid, R.W., Hayek, L.C. & Foster, M.S. (1994)
Measuring and monitoring biological diversity: Standard methods for amphibians.
Smithsonian Institution Press, Washington.
Hoorn, C., Wesselingh, F.P., terSteege, H., Bermudez M.A., Mora A., Sevink J., Sanmartín
I.A., Sanchez-Meseguer, A, Anderson, C.L., Figueiredo, J.P., Jaramillo, C., Riff, D.,
Negri, F.R., Hooghiemstra, H., Lundberg, J., Stadler, T., Särkinen, T. & Antonelli, A.
(2010) Amazonia through time: Andean uplift, climate change, landscape evolution, and
biodiversity. Science, 330, 927-931.
Jungfer, K.H., Faivovich, J., Padial, J.M. et al. (2013) Systematics of spiny-backed treefrogs
(Hylidae: Osteocephalus): an Amazonian puzzle. Zoologica Scripta, 42, 351-380.

25
Junk W.J., P.B. Bayley & Sparks, R.E. (1989) The flood pulse concept in river – floodplain
systems. Canadian Special Publication of Fisheries and Aquatic Sciences, 106, 110-
127.
Kerby, J.L., Richards-Hrdlicka, K.L., Storfer, A. & Skelly, D.K. (2010) An examination of
amphibian sensitivity to environmental contaminants: are amphibians poor canaries?
Ecology Letters, 13, 60-67.
Latrubesse, E.M., Cozzuol, M., Silva-Caminha, S.A.F., Rigsby, C.A., Absy, M.L. &
Jaramillo, C. (2010) The Late Miocene paleogeography of the Amazon Basin and the
evolution of the Amazon river system. Earth-Science Reviews, 99, 99-124.
Lima, A.P., Simões, P.I. & Kaefer, I.L. (2014). A new species of Allobates (Anura:
Aromobatidae) from the Tapajós River basin, Pará State, Brazil. Zootaxa 3889: 355-
387.
Losos, J.B., Arnold, S.J., Bejerano, G., Brodie III, E.D., Hibbett, D., Hoekstra, H.E., Mindell,
D.P., Monteiro, A., Moritz, C., Allen Or, H., Petrov, D.A., Renner, S.A., Ricklefs, R.E.,
Soltis, P.S. & Turner T.L. (2013) Evolutionary biology for the 21st century. PLoS
Biology, 11, e1001466.
Magnusson, W.E., Lima, A.P., Luizão, R., Luizão, F., Costa, F.R.C., de Castilho, C.V. &
Kinupp, V.P. (2005) RAPELD: a modification of the Gentry method for biodiversity
surveys in long-term ecological research sites. Biota Neotropica. 5, 1-5.
Menin, M., Lima, A.P., Magnusson, W.E. & Waldez, F. (2007) Topographic and edaphic
effects on the distribution of terrestrially reproducing anurans in Central Amazonia:
mesoscale spatial patterns. Journal of Tropical Ecology, 23, 539-547.
Menin, M., Waldez, F. & Lima, A.P. (2011) Effects of environmental and spatial factors on
the distribution of anuran species with aquatic reproduction in central Amazonia.
Herpetological Journal, 21, 255-261.
Mitchell N.J. & Janzen, F.J. (2010) Temperature-dependent sex determination and
contemporary climate change. Sexual Development, 4, 129-140.
Mullen, L.B., Woods, H.A., Schwartz, M.K., Sepulveda, A.J., Lowe, W.H. (2010) Scale-
dependent genetic structure of the Idaho giant salamander (Dicamptodon atterimus) in
stream networks. Molecular Ecology, 19, 898-909.
Nascimento, D.S. (2014) Filogenia molecular de serpentes neotropicais do grupo Bothrops
atrox (Linnaeus, 1758) (Viperidae: Crotalinae). Msc. dissertation. Universidade de
Brasília, Brasília.

26
Nilsson C. & Berggren, K. (2000) Alteration of riparian ecosystems caused by river
regulation. BioScience, 50, 783-792.
Olmos, F. & Pacheco, J.F. (2003) Rediscovery of Golden-crowned Manakin Lepidothrix
vilasboasi. Cotinga, 20, 48-50.
Padial, J.M., Miralles, A., de la Riva, I. & M. Vences (2010) The integrative future of
taxonomy. Frontiers in Zoology, 7, 16.
Pavan, D. (2007) Assembléias de répteis e anfíbios do Cerrado ao longo da bacia do rio
Tocantins e o impacto do aproveitamento hidrelétrico da região na sua conservação.
Phd. thesis. Universidade de São Paulo, São Paulo.
R Development Core Team. (2014) R: A language and environment for statistical computing.
R Foundation for Statistical Computing, Vienna, Austria.
Ribas, C.C., Aleixo, A., Nogueira, A.C.R., Miyaki, C.Y. & Cracraft, J. (2012) A
palaeobiogeographic model for biotic diversification within Amazonia over the past
three million years. Proceedings of the Royal Society B: Biological Sciences, 279, 681-
689.
Rodrigues, M.T. 1980. Descrição de uma nova espécie de Gonatodes da Amazônia (Sauria,
Gekkonidae). Papéis Avulsos de Zoologia, 33, 309-314.
Ron, S. (2000) Biogeographic area relationships of lowland Neotropical rainforest based on
raw distributions of vertebrate groups. Biological Journal of Linnean Society, 71, 379-
402.
Rossetti, D.F. (2014) The role of tectonics in the late Quaternary evolution of Brazil’s
Amazonian landscape. Earth Science Reviews, 139, 362-389.
Schiesari, L., Zuanon, J., Azevedo-Ramos, C., Garcia, M., Gordo, M., Messias, M. & Vieira,
E.M. (2003) Macrophyte rafts as dispersal vectors for fishes and amphibians in the
Lower Solimões River, Central Amazon. Journal of Tropical Ecology, 19, 333-336.
Schietti, J., Emilio, T., Rennó, C.D., Drucker, D.P., Costa, F.R.C., Nogueira, A., Baccaro,
F.B., Figueiredo, F., Castilho, C.V., Kinupp, V., Guillaumet, J.L., Garcia, A.R.M. Lima,
A.P. & Magnusson, W.E. (2013) Vertical distance from drainage drives floristic
composition changes in an Amazonian rainforest. Plant Ecology & Diversity, 6, 1-13.
Simões, P.I., Lima, A.P., Magnusson, W.E., Hödl, W. & Amézquita, A. (2008) Acoustic and
morphological differentiation in the frog Allobates femoralis: Relationships with the
upper Madeira River and other potential geological barriers. Biotropica, 40, 607-614.

27
Simões, P.I., Stow, A., Hödl, W., Amézquita, A., Farias, I.P. & Lima, A.P. (2014) The value
of including intraspecific measures of biodiversity in environmental impact surveys is
highlighted by the Amazonian brilliant-thighed frog (Allobates femoralis). Tropical
Conservation Science, 7, 811-828.
da Silva Jr., N.J. & Sites Jr., J.W. (1995) Patterns of diversity of neotropical squamate reptiles
species with emphasis on the Brazilian Amazon and the conservation potential of
indigenous reserves. Conservation Biology, 9, 873-901.
da Silva, J.M.D., Rylands, A.B. & da Fonseca, G.A.B. (2005) The fate of the Amazonian
areas of endemism. Conservation Biology, 19, 689-694.
Sioli, H. (1968) Hydrochemistry and geology in the Brazilian Amazon region. Amazoniana,
1, 267-277.
Smith, B.T., McCormack, J.E., Cuervo, A.M., Hickerson, M.J., Aleixo, A., Cadena, C.D.,
Pérez-Emán, J., Burney, C.W., Xie, X., Harvey, M.G., Faircloth, B.C., Glenn, T.C.,
Derryberry, E.P., Prejean, J., Fields, S. & Brumfield, R.T. (2014) The drivers of
tropical speciation. Nature, 515, 406-409.
Sombroek, W.G. (2000) Amazon landforms and soils in relation to biological diversity. Acta
Amazonica, 30, 81-100.
Sombroek, W. (2001) Spatial and temporal patterns of Amazon rainfall - Consequences for
the planning of agricultural occupation and the protection of primary forests. Ambio, 30,
388-396.
Souza, S.M., Rodrigues, M.T. & Cohn-Haft, M. (2013) Are amazonia rivers biogeographic
barriers for lizards? A study on the geographic variation of the spectacled lizard
Leposoma osvaldoi Ávila-Pires (Squamata, Gymnophthalmidae). Journal of
Herpetology, 47, 511-519.
Tuomisto, H. & Ruokolainen, K. (1997) The role of ecological knowledge in explaining
biogeography and biodiversity in Amazonia. Biodiversity and Conservation, 6, 347-357
Vannote, R.L., Minshal, G.W., Cummins, K.W., Sedell, J.R. & Cushing, C.E. (1980) The
River Continuum Concept. Canadian Journal of Fisheries and Aquatic Sciences, 37,
130-137
Vellend, M. (2010) Conceptual synthesis in community ecology. The Quarterly Review of
Biology, 85, 183-206.

28
Vences, M., Nagy, Z.T., Sonet, G. & Verheyen, E. (2012) DNA barcoding amphibians and
reptiles. In: Kress, W.J., & D. L. Erickson (eds.): DNA Barcodes: Methods and
Protocols. Springer Protocols Methods in Molecular Biology, 858, 79-107.
Vitt, L.J., Souza, R.A., Sartorius, S.S., Ávila-Pires, T.C.S. & Espósito, M.C. (2000)
Comparative ecology of sympatric Gonatodes (Squamata: Gekkonidae) in the western
Amazon of Brazil. Copeia, 2000, 83-95.
Wakeley, J. & Aliacar, N. (2001) Gene genealogies in a metapopulation. Genetics 159, 893-
905.
Wallace, A.R. (1852) On the monkeys of the Amazon. Proceedings of the Zoological Society
of London, 20, 107-110.
Ward J.V. & Stanford, J.A. (1995) The serial discontinuity concept: extending the model to
floodplain rivers. Regulated rivers: Research & Management, 10, 159-168.
Wittmann, F., Junk, W.J. & Piedade, M.T.F. (2004) The várzea forests in Amazonia: flooding
and the highly dynamic geomorphology interact with natural forest succession. Forest
Ecology and Management, 196, 199-212.
Wollenberg, K.C., Vieites, D.R., Glaw, F. & Vences, M. (2011) Speciation in little: the role
of range and body size in the diversification of Malagasy mantellid frogs. BMC
Evolutionary Biology, 11, 217.
Zuquim, G., Tuomisto, H., Jones, M.M., Prado, J., Figueiredo, F.O.G., Moulatlet, G.M.,
Costa, F.R.C., Quesada, C.A. & Emilio T. (2014). Predicting environmental gradients
with fern species composition in Brazilian Amazonia. Journal of Vegetation Science,
25, 1195-1207.

29
Table 1. Summary statistical results of distance to water (linear regression) and riverine barriers effects (ANOVA)
to changes in species composition of distinct taxa and assemblages, and the main rivers differential barrier
strengths (Tukey test). Italicized results are significant (≤0.05). Interfluve barriers: (Tapajós 1) Madeira-Tapajós
vs. Tapajós-Xingu; (Tapajós 2) Madeira-Tapajós vs. Tapajós-Jamanxim; (Jamanxim 1) Madeira-
Tapajós/Jamanxim vs. Tapajós/Jamanxim-Xingu; (Jamanxim 2) Tapajós-Jamanxim vs. Tapajós/Jamanxim-Xingu.
Taxa Groups and
assemblages
Distance to
water
Riverine
barrier
Tapajós
1
Tapajós
2
Jamanxim
1
Jamanxim
2
F1,13 p F4,18 p p p p p
Am
ph
ibia
ns
Total 34.9 0.001 7.6 <0.001 0.001 0.005 0.2 0.9
Non-riparians - - 9.2 <0.001 <0.001 0.003 0.09 0.9
Small streams
riparians - - 3.9 0.01 0.02 0.03 0.8 0.8
Large rivers
riparians - - 0.9 0.4 - - - -
Sq
uam
ate
s Lizards 12.5 0.003 7.3 0.001 0.02 0.6 0.004 0.09
Snakes 3.7 0.07 0.7 0.6 - - - -
Non-riparians - - 5.8 0.003 0.006 0.02 0.2 0.09
Riparians - - 0.6 0.6 - - - -
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online version of this article:
Appendix S1. Forest types description.
Appendix S2. Integrative taxa identification.
Table S1. List of registered species.
BIOSKETCHES
Leandro J.C.L. Moraes is a graduate student in Ecology at the National Institute of
Amazonian Research (INPA), and this study is part of his Msc. Dissertation. He is interested
in biogeographical patterns and integrated evolutionary history of amphibians/reptiles and
landscape in Neotropical region.
Author’s contributions: L.J.C.L.M., D.P. and C.C.R. conceived the idea. L.J.C.L.M. and D.P.
collected the data. M.C.B. performed molecular sequencing. L.J.C.L.M, D.P. and C.C.R.

30
analyzed the data. L.J.C.L.M. led the writing and all authors contributed to writing and
reviewing the manuscript.
Editor: Şerban Procheş
LIST OF FIGURES LEGENDS
Figure 1. Study area. (a) Location in South America, Brazil, Pará and Amazon Basin. (b)
Sampling units, showing the two main rivers and the main interfluves: (M-T) Madeira-
Tapajós; (M-T/J) Madeira-Tapajós/Jamanxim; (T-J) Tapajós-Jamanxim; (T-X) Tapajós-
Xingu and (T/J-X) Tapajós/Jamanxim-Xingu. (c) Scheme of sampling units, with transect (in
black), plots (uniformly distributed in brown and riparian in grey) and pitfall traps (in red).
Figure 2. Direct ordination of the relative abundance of amphibians (a) and squamates (b) in
relation to distance to water gradient. Note that some species increase in abundance or are
restricted to more humid areas (red square).
Figure 3. Linear regression for the three main taxonomical groups studied, between the
NMDS Axis 1 (species composition) and distance to water, divided in modal distances with
three plots agregated: (1) 20-35 m; (2) 36-50 m; (3) 53-62 m;(4) 63-80 m; (5) 82-95 m; (6)
100-150 m; (7) 180-200 m; (8) 250-300 m; (9) 320-380 m; (10) 440- 490 m (11) 500-580 m;
(12) 615-700 m; (13) 715-1120 m; (14) 1200-1400 m; (15) 1700-2140 m.
Figure 4. Direct ordination of presence-absence data for all survey methods revealing a
higher number of amphibians (a) than squamates (b), which are exclusive to one of the main
rivers' banks. Sampling units (letters) and interfluves (acronyms) are on the top. Different
colours correspond to main interfluves and species written in red are significantly (ANOVA,
p≤0.05) more abundant in one interfluve (inside parentheses).
Figure 5. NMDS ordinations and composition changes between interfluves for amphibian and
squamate assemblages showing the relative riverine barrier strength for these taxa. (a-c)
amphibians; (d-f) lizards and (g-i) snakes. Interfluves: (M-T) Madeira-Tapajós; (M-T/J)
Madeira-Tapajós/Jamanxim; (T-J) Tapajós-Jamanxim; (T-X) Tapajós-Xingu and (T/J-X)
Tapajós/Jamanxim-Xingu.
Figure 6. Relative frequency of amphibian (a) and squamate (b) functional groups with
distribution patterns indicating a riverine barrier influence. In black are the proportion of
species which suffer barrier effect and in gray which do not suffer barrier effect. The total

31
number of species in each category are inside the bars. Micro-habitat: (TE) terrestrial; (FO)
fossorial; (AR) arboricole; (AQ) aquatic; (SR) semi-arboricole and (SA) semi-aquatic. Body
size: (SM) small; (MD) medium and (LA) large. Activity period: (CR) crepuscular/nocturnal;
(DI) diurnal; (NO) nocturnal and (DN) diurnal/nocturnal.
Figure 7. NMDS ordinations and composition changes for amphibian and squamate
assemblages associated with distinct forest types, clustered by composition variation in
relation to distance to water, showing the relative riverine barrier strength. (a-c) Non-riparian
amphibians; (d-f) Small stream riparian amphibians; (g-i) Large river riparian amphibians; (j-
l) Non-riparian squamates and (m-o) Riparian squamates. Interfluves: (M-T) Madeira-
Tapajós; (M-T/J) Madeira-Tapajós/Jamanxim; (T-J) Tapajós-Jamanxim; (T-X) Tapajós-
Xingu and (T/J-X) Tapajós/Jamanxim-Xingu.
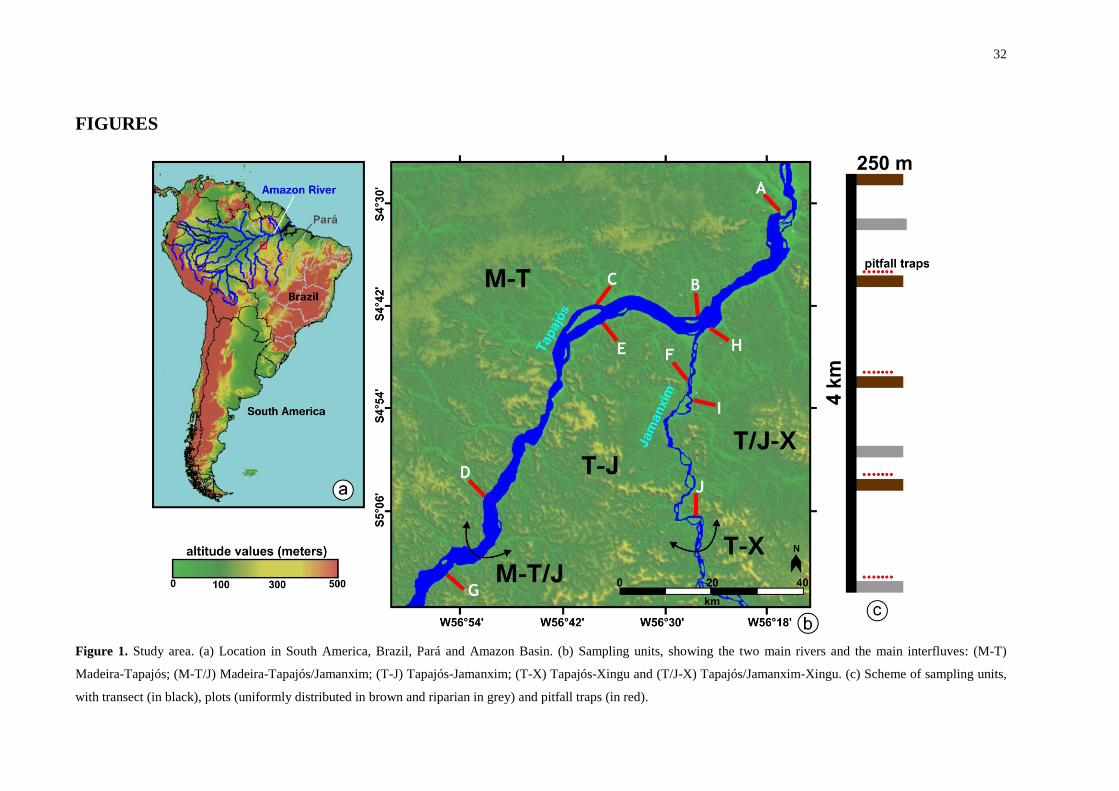
32
FIGURES
Figure 1. Study area. (a) Location in South America, Brazil, Pará and Amazon Basin. (b) Sampling units, showing the two main rivers and the main interfluves: (M-T)
Madeira-Tapajós; (M-T/J) Madeira-Tapajós/Jamanxim; (T-J) Tapajós-Jamanxim; (T-X) Tapajós-Xingu and (T/J-X) Tapajós/Jamanxim-Xingu. (c) Scheme of sampling units,
with transect (in black), plots (uniformly distributed in brown and riparian in grey) and pitfall traps (in red).

33
Figure 2. Direct ordination of the relative abundance of amphibians (a) and squamates (b) in relation to distance to water gradient. Note that some species increase in
abundance or are restricted to more humid areas (red square).

34
Figure 3. Linear regression for the three main taxonomical groups studied, between the NMDS Axis 1 (species composition) and distance to water, divided in modal distances
with three plots agregated: (1) 20-35 m; (2) 36-50 m; (3) 53-62 m;(4) 63-80 m; (5) 82-95 m; (6) 100-150 m; (7) 180-200 m; (8) 250-300 m; (9) 320-380 m; (10) 440- 490 m
(11) 500-580 m; (12) 615-700 m; (13) 715-1120 m; (14) 1200-1400 m; (15) 1700-2140 m.
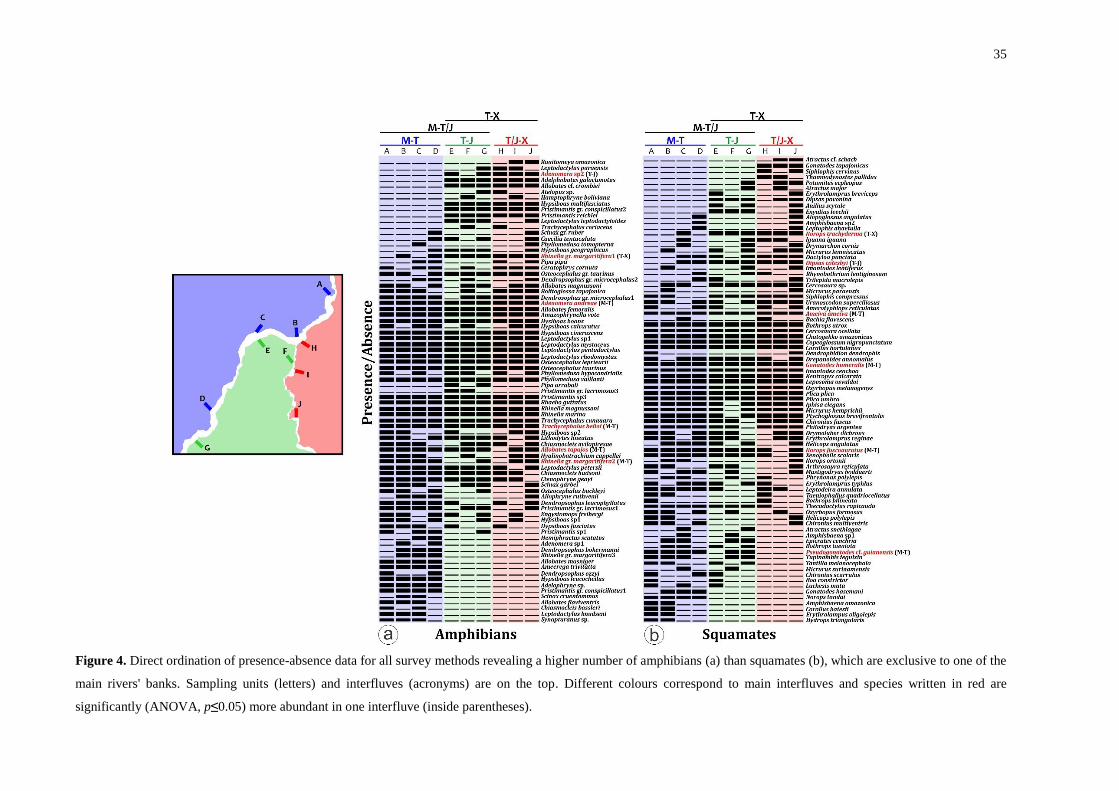
35
Figure 4. Direct ordination of presence-absence data for all survey methods revealing a higher number of amphibians (a) than squamates (b), which are exclusive to one of the
main rivers' banks. Sampling units (letters) and interfluves (acronyms) are on the top. Different colours correspond to main interfluves and species written in red are
significantly (ANOVA, p≤0.05) more abundant in one interfluve (inside parentheses).

36
Figure 5. NMDS ordinations and composition changes between interfluves for amphibian and squamate assemblages showing the relative riverine barrier strength for these
taxa. (a-c) amphibians; (d-f) lizards and (g-i) snakes. Interfluves: (M-T) Madeira-Tapajós; (M-T/J) Madeira-Tapajós/Jamanxim; (T-J) Tapajós-Jamanxim; (T-X) Tapajós-
Xingu and (T/J-X) Tapajós/Jamanxim-Xingu.

37
Figure 6. Relative frequency of amphibian (a) and squamate (b) functional groups with distribution patterns indicating a riverine barrier influence. In black are the proportion
of species which suffer barrier effect and in gray which do not suffer barrier effect. The total number of species in each category are inside the bars. Micro-habitat: (TE)
terrestrial; (FO) fossorial; (AR) arboricole; (AQ) aquatic; (SR) semi-arboricole and (SA) semi-aquatic. Body size: (SM) small; (MD) medium and (LA) large. Activity period:
(CR) crepuscular/nocturnal; (DI) diurnal; (NO) nocturnal and (DN) diurnal/nocturnal.
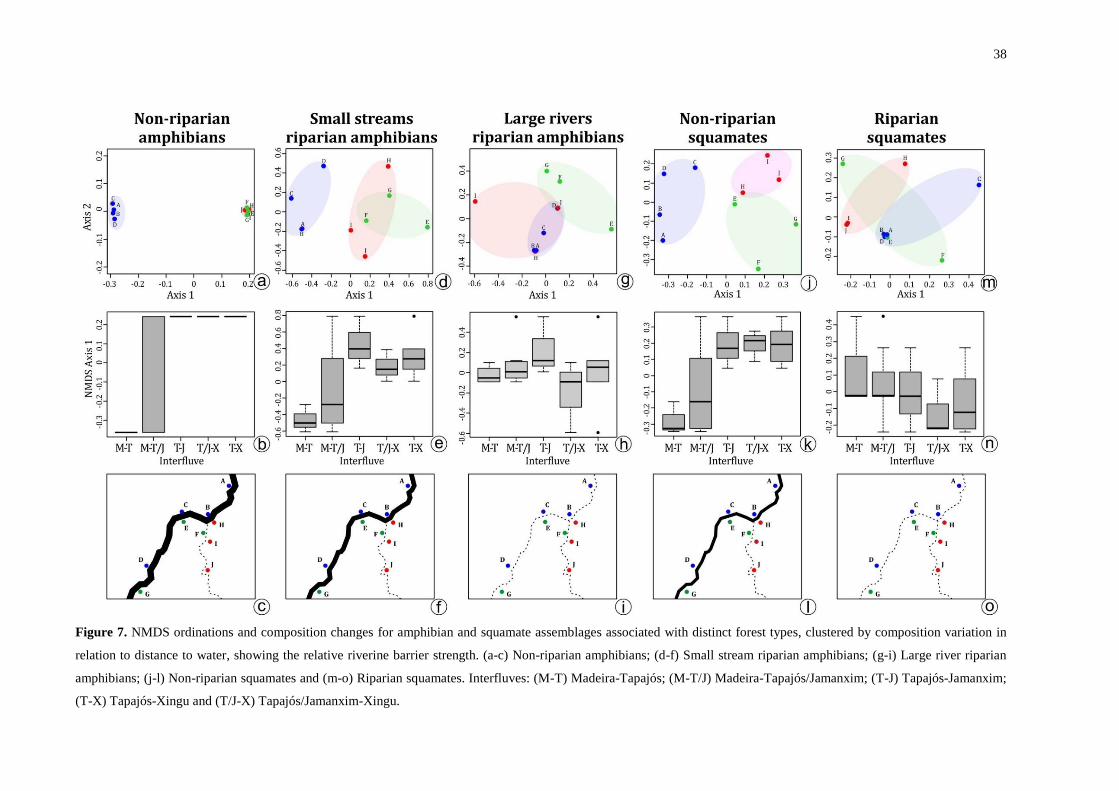
38
Figure 7. NMDS ordinations and composition changes for amphibian and squamate assemblages associated with distinct forest types, clustered by composition variation in
relation to distance to water, showing the relative riverine barrier strength. (a-c) Non-riparian amphibians; (d-f) Small stream riparian amphibians; (g-i) Large river riparian
amphibians; (j-l) Non-riparian squamates and (m-o) Riparian squamates. Interfluves: (M-T) Madeira-Tapajós; (M-T/J) Madeira-Tapajós/Jamanxim; (T-J) Tapajós-Jamanxim;
(T-X) Tapajós-Xingu and (T/J-X) Tapajós/Jamanxim-Xingu.

39
CONCLUSÕES
O Médio Rio Tapajós e o Rio Jamanxim possuem contribuição desigual na geração
dos padrões de distribuição das assembleias de anfíbios e répteis Squamata nesta
região.
O Médio Rio Tapajós é uma barreira mais efetiva para os anfíbios, enquanto o Rio
Jamanxim é uma barreira fraca, sendo mais efetiva para os répteis Squamata.
As assembleias de anfíbios e répteis Squamata divergem em relação aos tipos
florestais presentes na região (florestas ripárias e de terra firme).
As assembleias que sofrem efeito dos grandes rios como barreira são os anfíbios e
répteis Squamata não ripários, e os anfíbios ripários de igarapés. Os anfíbios ripários
de igapó e os répteis ripários não sofrem este efeito, indicando uma maior dispersão
entre margens. No entanto, para os répteis, a detecção dos padrões reais pode ser
influenciada pela baixa detectabilidade das espécies, especialmente de serpentes.
Os anfíbios pequenos, diurnos e terrestes e os répteis pequenos a médios, diurnos e
semi-arborícolas são os grupos funcionais especialmente afetados pelos grandes rios
como barreira nesta região, especialmente os gêneros de anfíbios Allobates,
Adenomera, Pristimantis e Leptodactylus e de répteis Gonatodes. Esta característica é
conferida pela baixa capacidade de dispersão destas espécies, evidenciando esses
organismos como bons modelo em novos estudos biogeográficos.
Os gradientes ambientais na região do Médio Rio Tapajós geram uma zona de contato
faunística, onde assembleias provenientes dos ambientes à Oeste e Leste da Amazônia
se encontram. O Rio Tapajós funciona como uma barreira ecológica e física para essas
faunas distintas, devido à diferentes restrições ecológicas.
Alterações antrópicas na região, como a construção de usinas hidrelétricas, podem
alterar os efeitos naturais dos grandes rios como mecanismos geradores dos padrões de
diversidade, assim como extinguir linhagens endêmicas e ambientes relevantes para o
ciclo de vida das espécies, especialmente as florestas ripárias. A preservação da
integridade fluvial é tão importante quanto a preservação dos ecossistemas florestais.

40
SUPPORTING INFORMATION
Appendix S1. Forest types description.
We briefly described the principal forest types on study area based in observations along the sampling surveys and literature records. The
fluvial dynamic of Tapajós River led to the development and maintenance of two principal forest types: riparian and non-riparian forests. The
riparian forests can be further divided into two main types, those that occur on the banks of the large rivers (Igapó) and those occurring along the
banks of small streams within the forest (Palm swamp forests):
Non-riparian forest Riparian forest Riparian forest
Upland forests Large rivers riparian forests Small streams riparian forests
Name in Portuguese Terra firme. Igapó. Açaizal.
Location in study
area1
Predominant forest type in the region,
homogeneously distributed among interfluves.
Mainly on western margin of the Tapajós
River. Rarer on eastern margin and in the
banks of the Jamanxim River.
Homogeneously distributed among interfluves,
following the course of small streams.
Forest type origin1
In the dry land of plateaus and slopes the
forests do not suffer the direct effect of
riverine flood pulse, creating this forest type.
The maintenance of natural patterns of their
assemblages actively depends on the integrity
of adjacent flooded forests, as many species
use them during its life cycle.
Flooding period of Tapajós and Jamanxim
rivers during the rainy season is essential for
nutrients retention in ecosystem and
maintenance of high biomass in their banks as
these are quickly exported downstream due to
the rapid water flow. The interaction of this
flood pulse, rapid water flow and low
sediments input create that distinct forest type.
The water flow is lower and often interrupted by
fall of organic material from the marginal forest,
generating a muddy environment dominated by
adapted palms.
Flood pulse
oscillation1,4
Do not suffers. Sazonal, one pulse by year and predictable. Higher, frequent and unpredictable, suffering
greater effect of local rainfall.
Productivity2 Lower. Higher. Higher.
Canopy1,3 Tall, non-uniform. Tall, uniform or non-uniform Short, usually uniform.
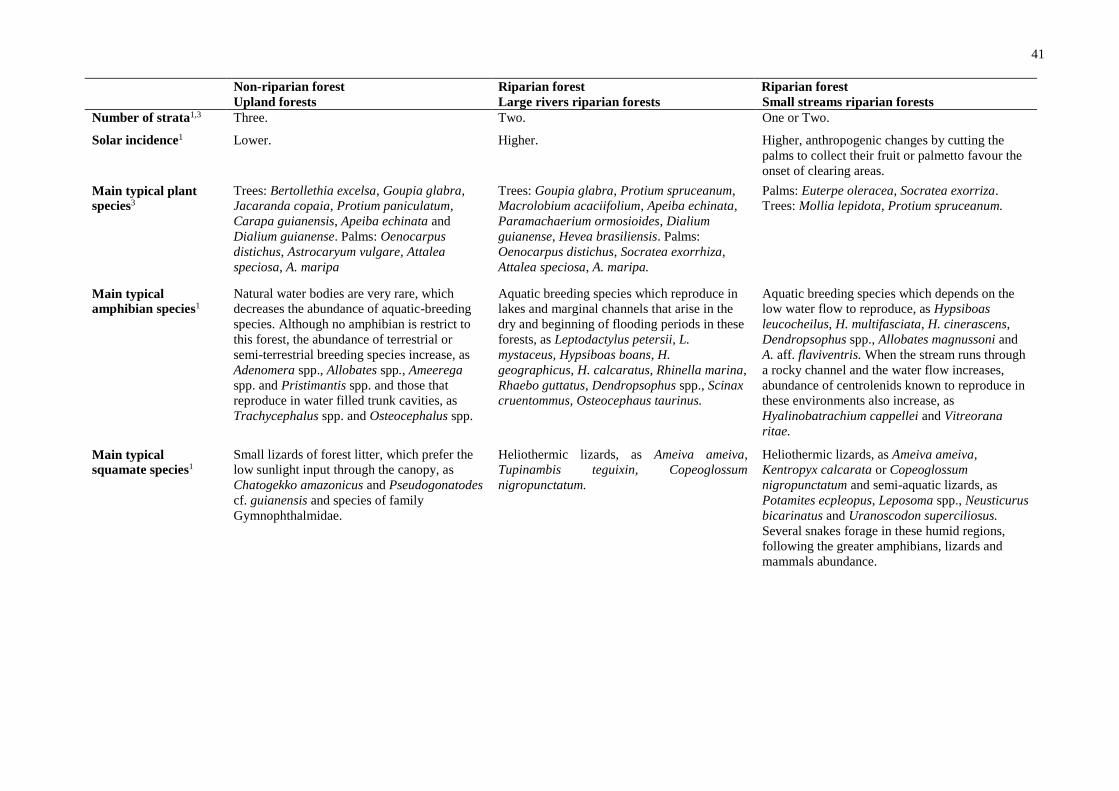
41
Non-riparian forest Riparian forest Riparian forest
Upland forests Large rivers riparian forests Small streams riparian forests
Number of strata1,3 Three. Two. One or Two.
Solar incidence1 Lower. Higher. Higher, anthropogenic changes by cutting the
palms to collect their fruit or palmetto favour the
onset of clearing areas.
Main typical plant
species3
Trees: Bertollethia excelsa, Goupia glabra,
Jacaranda copaia, Protium paniculatum,
Carapa guianensis, Apeiba echinata and
Dialium guianense. Palms: Oenocarpus
distichus, Astrocaryum vulgare, Attalea
speciosa, A. maripa
Trees: Goupia glabra, Protium spruceanum,
Macrolobium acaciifolium, Apeiba echinata,
Paramachaerium ormosioides, Dialium
guianense, Hevea brasiliensis. Palms:
Oenocarpus distichus, Socratea exorrhiza,
Attalea speciosa, A. maripa.
Palms: Euterpe oleracea, Socratea exorriza.
Trees: Mollia lepidota, Protium spruceanum.
Main typical
amphibian species1
Natural water bodies are very rare, which
decreases the abundance of aquatic-breeding
species. Although no amphibian is restrict to
this forest, the abundance of terrestrial or
semi-terrestrial breeding species increase, as
Adenomera spp., Allobates spp., Ameerega
spp. and Pristimantis spp. and those that
reproduce in water filled trunk cavities, as
Trachycephalus spp. and Osteocephalus spp.
Aquatic breeding species which reproduce in
lakes and marginal channels that arise in the
dry and beginning of flooding periods in these
forests, as Leptodactylus petersii, L.
mystaceus, Hypsiboas boans, H.
geographicus, H. calcaratus, Rhinella marina,
Rhaebo guttatus, Dendropsophus spp., Scinax
cruentommus, Osteocephaus taurinus.
Aquatic breeding species which depends on the
low water flow to reproduce, as Hypsiboas
leucocheilus, H. multifasciata, H. cinerascens,
Dendropsophus spp., Allobates magnussoni and
A. aff. flaviventris. When the stream runs through
a rocky channel and the water flow increases,
abundance of centrolenids known to reproduce in
these environments also increase, as
Hyalinobatrachium cappellei and Vitreorana
ritae.
Main typical
squamate species1
Small lizards of forest litter, which prefer the
low sunlight input through the canopy, as
Chatogekko amazonicus and Pseudogonatodes
cf. guianensis and species of family
Gymnophthalmidae.
Heliothermic lizards, as Ameiva ameiva,
Tupinambis teguixin, Copeoglossum
nigropunctatum.
Heliothermic lizards, as Ameiva ameiva,
Kentropyx calcarata or Copeoglossum
nigropunctatum and semi-aquatic lizards, as
Potamites ecpleopus, Leposoma spp., Neusticurus
bicarinatus and Uranoscodon superciliosus.
Several snakes forage in these humid regions,
following the greater amphibians, lizards and
mammals abundance.

42
Non-riparian forest Riparian forest Riparian forest
Upland forests Large rivers riparian forests Small streams riparian forests
Images
Photo by Elizângela Brito
Photo by Leandro Moraes
Photo by Leandro Moraes
Photo by Dante Pavan
Photo by Dante Pavan
Photo by Dante Pavan
Photo by Dante Pavan
Photo by Marina Maximiano
Photo by Leandro Moraes

43
Non-riparian forest Riparian forest Riparian forest
Upland forests Large rivers riparian forests Small streams riparian forests
Photo by Dante Pavan
Photo by Leandro Moraes
1 Author’s, personal observations; 2 Furch, 1997; 3 Domingues, 2014, 4 Junk et al., 1989.

44
Appendix S2. Integrative taxa identification.
We adopt the evolutionary species concept (Wiley, 1978) and the integrative approach
of taxa identification was realized through several steps in order to delimit units for
biogeographical analyses (Padial et al., 2010). The first step performed was the morphological
delimitation and taxa were delimited by morphometric and colouration patterns analyses.
Simultaneously, we performed acoustic analyses, by comparison of sonograms and
spectrograms for amphibians, as well as environmental analyses, such as preferred habitat of
the species and behavioral observations. These steps filtered cryptic groups located in taxa
delimited by morphological approach. The identification of taxa delimited through these steps
was performed with original descriptions, field guides and taxonomic revisions, as well as
comparisons with voucher specimens deposited in the collection of amphibians and reptiles at
the National Institute for Amazonian Research. Several researchers and taxonomic experts
also assisted in identifying the taxa as Pedro Ivo Simões (PUC-RS), José Cassimiro da Silva
Junior (USP), Igor Luis Kaefer (UFAM), Vinicius Tadeu de Carvalho (UFAM), Jerriane
Oliveira Gomes (MPEG), Marcelo Gordo (UFAM), Miguel Trefaut Urbano Rodrigues (USP),
Rafael de Fraga (INPA), Pedro Luis Vieira Peloso (AMNH), Marinus Steven Hoogmoed
(MPEG), Paulo Sérgio Bernarde (UFAC), Alfredo Pedroso dos Santos-Jr (PUC-RS),
Albertina Pimentel Lima (INPA), Teresa Cristina Ávila-Pires (MPEG), Omar Machado
Entiauspe (IFSUL), Diego José Santana Silva (UFMS), Sarah Mângia Barros (UFPB), Rafael
O. de Sá (U. Richmond), Renata C. Amaro (USP), Antoine Fouquet (CNRS Guyane), Stefan
Lötters (U. Trier), Thiago de Carvalho Kashi (UFU) e Willian Edward Duellman (U. Kansas).
If the taxonomic delimitation after analyzing all the data of the previous steps still
remained confusing and irresolute, we performed molecular species identification. Among
amphibians, molecular data was obtained for the following genera and species groups:
Caecilia sp., Allobates sp., Amazophrynella sp., Rhinella gr. margaritifera, Rhinella gr.
marina, Pristimantis gr. lacrimosus, Pristimantis gr. conspicillatus, Pristimantis aff.
ockendeni, Hemiphractus sp., Dendropsophus gr. microcephalus, Hypsiboas gr.
albopunctatus, Hypsiboas aff. boans, Hypsiboas aff. geographicus, Osteocephalus sp.,
Phyllomedusa gr. hypocondrialis, Scinax gr. ruber, Scinax gr. rostratus, Trachycephalus sp.,
Adenomera sp., Leptodactylus gr. melanonotus, Leptodactylus gr. pentadactylus e
Leptodactylus aff. mystaceus. Among squamates molecular data was used for the following
genera and species groups: Chironius sp., Norops gr. chrysolepis, Plica sp. and Cercosaura

45
sp. In total, we obtained sequences of the mtDNA gene coding for the 16S rRNA for 132
individuals. Total DNA was isolated using the phenol-chloroform protocol (Sambrook &
Russel, 2001). Amplifications of mtDNA 16S gene region were performed using the primers:
L1987-5'GCCTCGCCTGTTTACCAAAAAC3' and H2609-5'CCGGTCTGAACTCAGATC
ACGT3' (Palumbi et al., 1991). Sequencing was performed using the Big Dye Terminator
sequencing kit (Applied Biosystems) and an automatic DNA sequencer ABI PRISM 3500
(Life Technologies). Sequences from closely related taxa were downloaded from GenBank.
Alignment was performed in CLUSTAL OMEGA (Sievers et al., 2011) and corrected manually,
and phylogenetic trees were constructed using maximum likelihood in MEGA 5 (Tamura et al.,
2011). Clade support was estimated by bootstrap (1000 replicates).
To determine to which lineage within a species complex each individual belong we
evaluated bootstrap support and p-distances. The molecular identification was circularly
compared with the morphological, acoustic and ecological delimitation, increasing the
reliability of distribution patterns See Fig. S1 for an example of this integrative identification,
for the genus Allobates.

46
Figure S1. Example of integrative taxa identification for genus Allobates. (a) taxa are delimited by comparative morphological (a1), acoustic (a2) and ecological (a3) data. In (a3),
green dots indicate species that occur preferentially in upland forests and blue dots species of small streams riparian forests. (b) Phylogenetic tree of 16S mtDNA used for molecular
delimitation, diferent colours represent distinct lineages with elevated clade support, and taxa recorded in the study are highlighted, located inserted in some of these lineages. These
delimitations approaches are cyclically compared, defining the seven taxonomic units for biogeographical analyses.
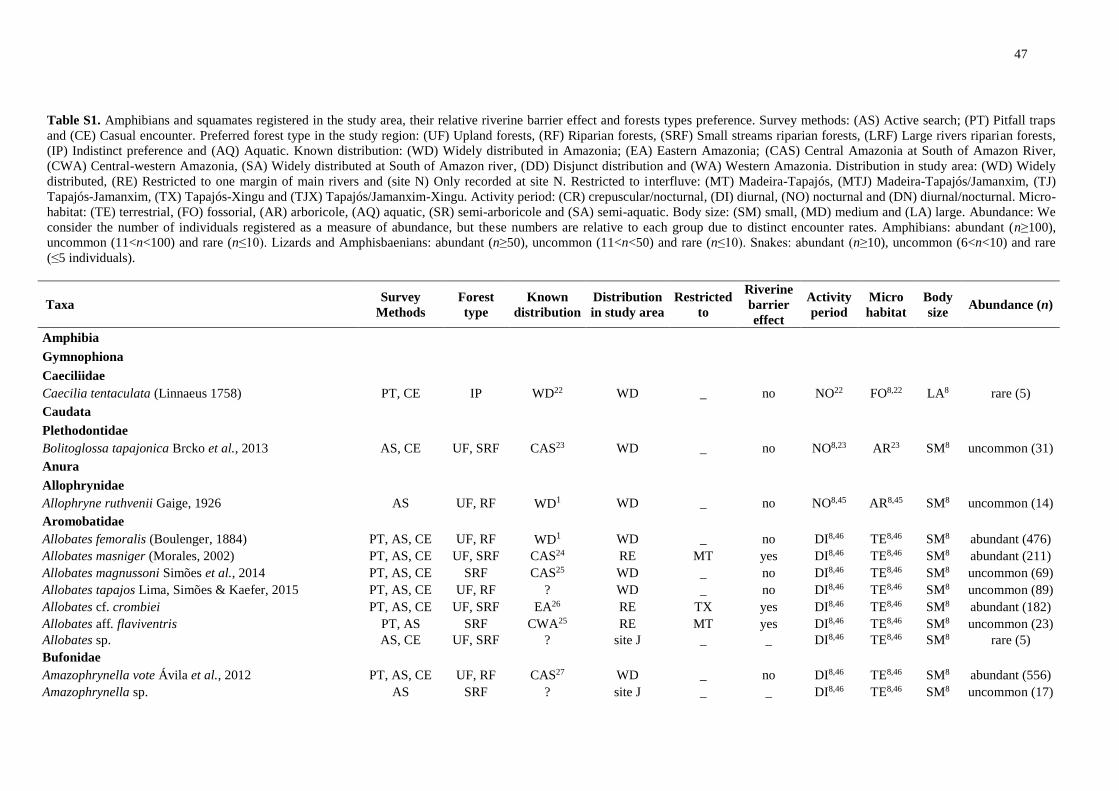
47
Table S1. Amphibians and squamates registered in the study area, their relative riverine barrier effect and forests types preference. Survey methods: (AS) Active search; (PT) Pitfall traps
and (CE) Casual encounter. Preferred forest type in the study region: (UF) Upland forests, (RF) Riparian forests, (SRF) Small streams riparian forests, (LRF) Large rivers riparian forests,
(IP) Indistinct preference and (AQ) Aquatic. Known distribution: (WD) Widely distributed in Amazonia; (EA) Eastern Amazonia; (CAS) Central Amazonia at South of Amazon River,
(CWA) Central-western Amazonia, (SA) Widely distributed at South of Amazon river, (DD) Disjunct distribution and (WA) Western Amazonia. Distribution in study area: (WD) Widely
distributed, (RE) Restricted to one margin of main rivers and (site N) Only recorded at site N. Restricted to interfluve: (MT) Madeira-Tapajós, (MTJ) Madeira-Tapajós/Jamanxim, (TJ)
Tapajós-Jamanxim, (TX) Tapajós-Xingu and (TJX) Tapajós/Jamanxim-Xingu. Activity period: (CR) crepuscular/nocturnal, (DI) diurnal, (NO) nocturnal and (DN) diurnal/nocturnal. Micro-
habitat: (TE) terrestrial, (FO) fossorial, (AR) arboricole, (AQ) aquatic, (SR) semi-arboricole and (SA) semi-aquatic. Body size: (SM) small, (MD) medium and (LA) large. Abundance: We
consider the number of individuals registered as a measure of abundance, but these numbers are relative to each group due to distinct encounter rates. Amphibians: abundant (n≥100),
uncommon (11<n<100) and rare (n≤10). Lizards and Amphisbaenians: abundant (n≥50), uncommon (11<n<50) and rare (n≤10). Snakes: abundant (n≥10), uncommon (6<n<10) and rare
(≤5 individuals).
Taxa Survey
Methods
Forest
type
Known
distribution
Distribution
in study area
Restricted
to
Riverine
barrier
effect
Activity
period
Micro
habitat
Body
size Abundance (n)
Amphibia
Gymnophiona
Caeciliidae
Caecilia tentaculata (Linnaeus 1758) PT, CE IP WD22 WD _ no NO22 FO8,22 LA8 rare (5)
Caudata
Plethodontidae
Bolitoglossa tapajonica Brcko et al., 2013 AS, CE UF, SRF CAS23 WD _ no NO8,23 AR23 SM8 uncommon (31)
Anura
Allophrynidae
Allophryne ruthvenii Gaige, 1926 AS UF, RF WD1 WD _ no NO8,45 AR8,45 SM8 uncommon (14)
Aromobatidae
Allobates femoralis (Boulenger, 1884) PT, AS, CE UF, RF WD1 WD _ no DI8,46 TE8,46 SM8 abundant (476)
Allobates masniger (Morales, 2002) PT, AS, CE UF, SRF CAS24 RE MT yes DI8,46 TE8,46 SM8 abundant (211)
Allobates magnussoni Simões et al., 2014 PT, AS, CE SRF CAS25 WD _ no DI8,46 TE8,46 SM8 uncommon (69)
Allobates tapajos Lima, Simões & Kaefer, 2015 PT, AS, CE UF, RF ? WD _ no DI8,46 TE8,46 SM8 uncommon (89)
Allobates cf. crombiei PT, AS, CE UF, SRF EA26 RE TX yes DI8,46 TE8,46 SM8 abundant (182)
Allobates aff. flaviventris PT, AS SRF CWA25 RE MT yes DI8,46 TE8,46 SM8 uncommon (23)
Allobates sp. AS, CE UF, SRF ? site J _ _ DI8,46 TE8,46 SM8 rare (5)
Bufonidae
Amazophrynella vote Ávila et al., 2012 PT, AS, CE UF, RF CAS27 WD _ no DI8,46 TE8,46 SM8 abundant (556)
Amazophrynella sp. AS SRF ? site J _ _ DI8,46 TE8,46 SM8 uncommon (17)
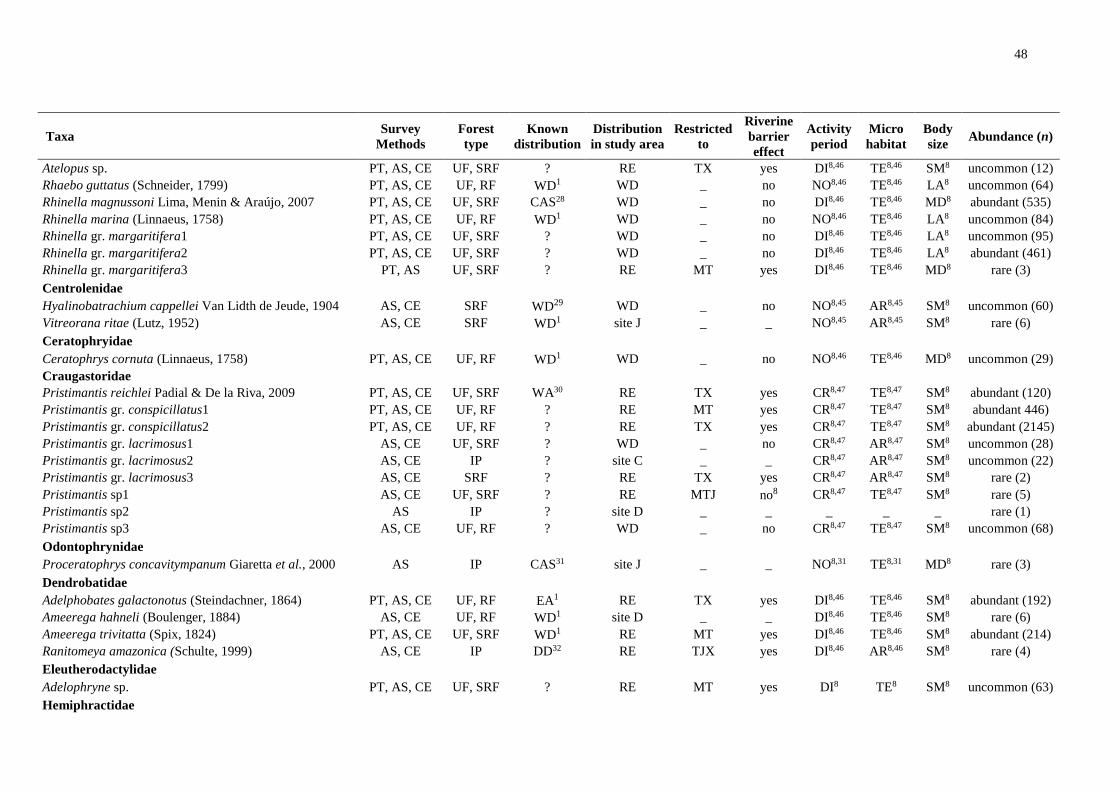
48
Taxa Survey
Methods
Forest
type
Known
distribution
Distribution
in study area
Restricted
to
Riverine
barrier
effect
Activity
period
Micro
habitat
Body
size Abundance (n)
Atelopus sp. PT, AS, CE UF, SRF ? RE TX yes DI8,46 TE8,46 SM8 uncommon (12)
Rhaebo guttatus (Schneider, 1799) PT, AS, CE UF, RF WD1 WD _ no NO8,46 TE8,46 LA8 uncommon (64)
Rhinella magnussoni Lima, Menin & Araújo, 2007 PT, AS, CE UF, SRF CAS28 WD _ no DI8,46 TE8,46 MD8 abundant (535)
Rhinella marina (Linnaeus, 1758) PT, AS, CE UF, RF WD1 WD _ no NO8,46 TE8,46 LA8 uncommon (84)
Rhinella gr. margaritifera1 PT, AS, CE UF, SRF ? WD _ no DI8,46 TE8,46 LA8 uncommon (95)
Rhinella gr. margaritifera2 PT, AS, CE UF, SRF ? WD _ no DI8,46 TE8,46 LA8 abundant (461)
Rhinella gr. margaritifera3 PT, AS UF, SRF ? RE MT yes DI8,46 TE8,46 MD8 rare (3)
Centrolenidae
Hyalinobatrachium cappellei Van Lidth de Jeude, 1904 AS, CE SRF WD29 WD _ no NO8,45 AR8,45 SM8 uncommon (60)
Vitreorana ritae (Lutz, 1952) AS, CE SRF WD1 site J _ _ NO8,45 AR8,45 SM8 rare (6)
Ceratophryidae
Ceratophrys cornuta (Linnaeus, 1758) PT, AS, CE UF, RF WD1 WD _ no NO8,46 TE8,46 MD8 uncommon (29)
Craugastoridae
Pristimantis reichlei Padial & De la Riva, 2009 PT, AS, CE UF, SRF WA30 RE TX yes CR8,47 TE8,47 SM8 abundant (120)
Pristimantis gr. conspicillatus1 PT, AS, CE UF, RF ? RE MT yes CR8,47 TE8,47 SM8 abundant 446)
Pristimantis gr. conspicillatus2 PT, AS, CE UF, RF ? RE TX yes CR8,47 TE8,47 SM8 abundant (2145)
Pristimantis gr. lacrimosus1 AS, CE UF, SRF ? WD _ no CR8,47 AR8,47 SM8 uncommon (28)
Pristimantis gr. lacrimosus2 AS, CE IP ? site C _ _ CR8,47 AR8,47 SM8 uncommon (22)
Pristimantis gr. lacrimosus3 AS, CE SRF ? RE TX yes CR8,47 AR8,47 SM8 rare (2)
Pristimantis sp1 AS, CE UF, SRF ? RE MTJ no8 CR8,47 TE8,47 SM8 rare (5)
Pristimantis sp2 AS IP ? site D _ _ _ _ _ rare (1)
Pristimantis sp3 AS, CE UF, RF ? WD _ no CR8,47 TE8,47 SM8 uncommon (68)
Odontophrynidae
Proceratophrys concavitympanum Giaretta et al., 2000 AS IP CAS31 site J _ _ NO8,31 TE8,31 MD8 rare (3)
Dendrobatidae
Adelphobates galactonotus (Steindachner, 1864) PT, AS, CE UF, RF EA1 RE TX yes DI8,46 TE8,46 SM8 abundant (192)
Ameerega hahneli (Boulenger, 1884) AS, CE UF, RF WD1 site D _ _ DI8,46 TE8,46 SM8 rare (6)
Ameerega trivitatta (Spix, 1824) PT, AS, CE UF, SRF WD1 RE MT yes DI8,46 TE8,46 SM8 abundant (214)
Ranitomeya amazonica (Schulte, 1999) AS, CE IP DD32 RE TJX yes DI8,46 AR8,46 SM8 rare (4)
Eleutherodactylidae
Adelophryne sp. PT, AS, CE UF, SRF ? RE MT yes DI8 TE8 SM8 uncommon (63)
Hemiphractidae

49
Taxa Survey
Methods
Forest
type
Known
distribution
Distribution
in study area
Restricted
to
Riverine
barrier
effect
Activity
period
Micro
habitat
Body
size Abundance (n)
Hemiphractus scutatus (Spix, 1824) AS, CE UF, SRF WA1 RE MT yes NO8,46 TE8,46 MD8 rare (3)
Hylidae
Cruziohyla craspedopus (Funkhouser, 1957) CE IP WD33 site B _ _ NO8,46 AR8,46 MD8 rare (2)
Dendropsophus bokermanni (Goin, 1960) AS, CE LRF WA1 RE MT yes NO8,46 AR8,46 SM8 uncommon (16)
Dendropsophus leucophyllatus (Beireis, 1783) AS, CE RF WD1 WD _ no NO8,46 AR8,46 SM8 uncommon (27)
Dendropsophus minutus (Peters, 1872) AS, CE IP WD1 site C _ _ _ _ _ rare (1)
Dendropsophus ozzyi Orrico et al., 2014 AS, CE SRF CAS34 RE MT yes NO8,46 AR8,46 SM8 rare (2)
Dendropsophus gr. microcephalus1 AS, CE RF ? WD _ no NO8,46 AR8,46 SM8 abundant (187)
Dendropsophus gr. microcephalus2 AS, CE RF ? WD _ no NO8,46 AR8,46 SM8 uncommon (63)
Dendropsophus gr. microcephalus3 AS IP ? site C _ _ _ _ _ rare (1)
Dryaderces sp. AS IP ? site E _ _ _ _ _ rare (1)
Hypsiboas boans (Linnaeus, 1758) AS, CE LRF WD1 WD _ no NO8,46 AR8,46 LA8 abundant (170)
Hypsiboas calcaratus (Troschel, 1848) AS, CE LRF WD1 WD _ no NO8,46 AR8,46 MD8 uncommon (13)
Hypsiboas cinerascens (Spix, 1824) AS, CE RF WD1 WD _ no NO8,46 AR8,46 SM8 abundant (318)
Hypsiboas fasciatus (Günther, 1858) AS, CE LRF WD1 WD _ no NO8,46 AR8,46 SM8 uncommon (47)
Hypsiboas geographicus (Spix, 1824) AS, CE LRF WD1 WD _ no NO8,46 AR8,46 MD8 rare (6)
Hypsiboas leucocheilus (Caramaschi & Niemeyer, 2003) AS, CE SRF CAS35 RE MT yes NO8,46 AR8,46 MD8 uncommon (71)
Hypsiboas multifasciatus (Günther, 1859) AS, CE SRF EA1 RE TX yes NO8,46 AR8,46 MD8 abundant (208)
Hypsiboas sp1 AS, CE SRF ? WD _ no NO8,46 AR8,46 MD8 uncommon (27)
Hypsiboas sp2 AS, CE SRF ? WD _ no NO8,46 AR8,46 MD8 uncommon (14)
Osteocephalus buckleyi (Boulenger, 1882) AS, CE SRF WD1 WD _ no NO8,46 AR8,46 SM8 uncommon (41)
Osteocephalus leprieurii(Duméril and Bibron, 1841) AS, CE UF, RF WD1 WD _ no NO8,46 AR8,46 SM8 abundant (156)
Osteocephalus taurinus Steindachner, 1862 AS, CE UF, RF WD36 WD _ no NO8,46 AR8,46 LA8 abundant (287)
Osteocephalus gr. taurinus AS, CE UF, RF WD36 WD _ no NO8,46 AR8,46 MD8 abundant (805)
Phyllomedusa hypochondrialis (Daudin, 1800) AS, CE UF, RF EA37 WD _ no NO8,46 AR8,46 SM8 uncommon (86)
Phyllomedusa tomopterna (Cope, 1868) CE UF, RF WD1 WD _ no NO8,46 AR8,46 SM8 rare (2)
Phyllomedusa vaillanti Boulenger, 1882 AS, CE UF, RF WD1 WD _ no NO8,46 AR8,46 MD8 abundant (101)
Scinax cruentommus (Duellman, 1972) AS, CE LRF WA1 RE MT yes NO8,46 AR8,46 SM8 uncommon (28)
Scinax garbei (Miranda-Ribeiro, 1926) AS, CE UF, RF WD1 WD _ no NO8,46 AR8,46 SM8 uncommon (12)
Scinax gr. ruber CE LRF WD1 WD _ no NO8,46 AR8,46 SM8 rare (8)
Trachycephalus coriaceus (Peters, 1867) AS UF WD1 RE TX no1 NO8,46 AR8,46 LA8 rare (2)
Trachycephalus cunauaru Gordo et al., 2013 AS, CE UF WD38 WD _ no NO8,46 AR8,38 LA8 abundant (398)

50
Taxa Survey
Methods
Forest
type
Known
distribution
Distribution
in study area
Restricted
to
Riverine
barrier
effect
Activity
period
Micro
habitat
Body
size Abundance (n)
Trachycephalus helioi Nunes et al., 2013 AS, CE UF CAS39 WD _ no NO8,46 AR8,39 LA8 abundant (303)
Leptodactylidae
Adenomera andreae (Müller, 1923) PT, AS, CE UF, RF WD41 WD _ no DI8,41 TE8,41 SM8 abundant (2822)
Adenomera sp1 PT, AS, CE UF, SRF CAS41 RE MT yes DI8,41 TE8,41 SM8 rare (3)
Adenomera sp2 PT, AS, CE UF, SRF EA41 RE TX yes DI8,41 TE8,41 SM8 abundant (164)
Engystomops freibergi (Donoso-Barros, 1969) PT, AS, CE UF, RF SA40 WD _ no NO8,40 TE8,40 SM8 abundant (583)
Leptodactylus knudseni Heyer, 1972 AS UF WA42 RE MT yes NO8,42 TE8,42 LA8 rare (3)
Leptodactylus leptodactyloides (Andersson, 1945) AS,CE SRF DD42 RE TX yes NO8,42 TE8,42 SM8 uncommon (28)
Leptodactylus mystaceus (Spix, 1824) PT, AS, CE UF, RF WD42 WD _ no NO8,42 TE8,42 SM8 abundant (110)
Leptodactylus paraensis Heyer, 2005 PT, AS, CE UF EA42 RE TX yes NO8,42 TE8,42 LA8 rare (4)
Leptodactylus pentadactylus (Laurenti, 1768) PT, AS, CE RF WD42 WD _ no NO8,42 TE8,42 LA8 abundant (152)
Leptodactylus petersii (Steindachner, 1864) PT, AS, CE RF WD42 WD _ no NO8,42 TE8,42 SM8 uncommon (56)
Leptodactylus rhodomystax Boulenger, 1884 PT, AS, CE UF, RF WD42 WD _ no NO8,42 TE8,42 MD8 uncommon (79)
Leptodactylus stenodema Jiménez de la Espada, 1875 CE IP WA42 site G _ _ _ _ _ rare (1)
Leptodactylus sp1 PT, AS, CE UF ? WD _ no NO8 TE8 LA8 abundant (173)
Leptodactylus sp2 AS IP ? site J _ _ _ _ _ rare (1)
Lithodytes lineatus (Schneider, 1799) PT, AS, CE UF WD1 WD _ no NO8,46 TE8,46 MD8 uncommon (49)
Microhylidae
Chiasmocleis avilapiresae Peloso and Sturaro, 2008 PT, AS, CE UF, RF SA43 WD _ no NO8,43 FO8,43 SM8 uncommon (34)
Chiasmocleis bassleri Dunn, 1949 PT, AS UF, RF WA43 RE MT yes NO8,43 FO8,43 SM8 rare (5)
Chiasmocleis hudsoni Parker, 1940 PT, AS, CE UF, RF WD43 WD _ no NO8,43 FO8,43 SM8 uncommon (63)
Ctenophryne geayi Mocquard, 1904 PT, AS UF, RF WD44 WD _ no NO8,46 FO8,46 MD8 abundant (142)
Hamptophryne boliviana (Parker, 1927) PT, AS UF, RF WD1 RE TJX no1 NO8,46 FO8,46 SM8 rare (3)
Synapturanus sp. AS UF ? RE MT yes NO8,46 FO8,46 SM8 uncommon (11)
Pipidae
Pipa arrabali Izecksohn, 1976 PT, AS, CE AQ DD1 RE TJ no1 NO8,46 AQ8,46 SM8 rare (8)
Pipa pipa (Linnaeus, 1758) AS, CE AQ WD1 WD _ no NO8,46 AQ8,46 LA8 rare (8)
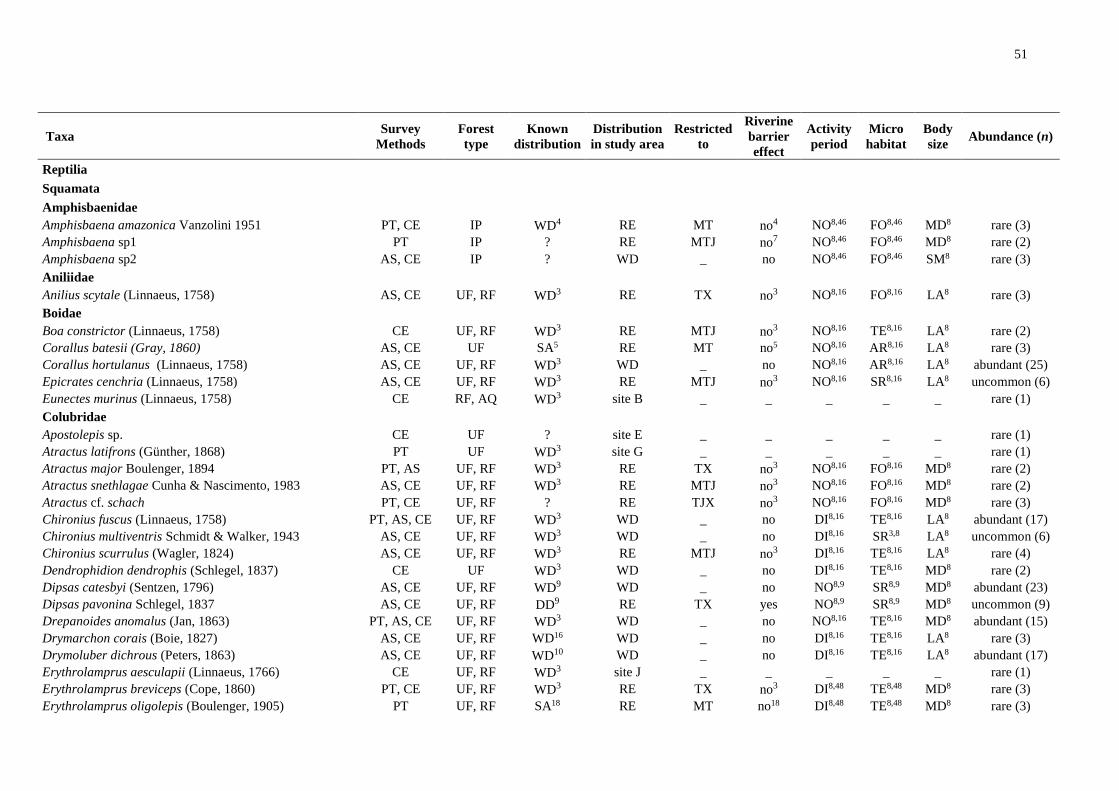
51
Taxa Survey
Methods
Forest
type
Known
distribution
Distribution
in study area
Restricted
to
Riverine
barrier
effect
Activity
period
Micro
habitat
Body
size Abundance (n)
Reptilia
Squamata
Amphisbaenidae
Amphisbaena amazonica Vanzolini 1951 PT, CE IP WD4 RE MT no4 NO8,46 FO8,46 MD8 rare (3)
Amphisbaena sp1 PT IP ? RE MTJ no7 NO8,46 FO8,46 MD8 rare (2)
Amphisbaena sp2 AS, CE IP ? WD _ no NO8,46 FO8,46 SM8 rare (3)
Aniliidae
Anilius scytale (Linnaeus, 1758) AS, CE UF, RF WD3 RE TX no3 NO8,16 FO8,16 LA8 rare (3)
Boidae
Boa constrictor (Linnaeus, 1758) CE UF, RF WD3 RE MTJ no3 NO8,16 TE8,16 LA8 rare (2)
Corallus batesii (Gray, 1860) AS, CE UF SA5 RE MT no5 NO8,16 AR8,16 LA8 rare (3)
Corallus hortulanus (Linnaeus, 1758) AS, CE UF, RF WD3 WD _ no NO8,16 AR8,16 LA8 abundant (25)
Epicrates cenchria (Linnaeus, 1758) AS, CE UF, RF WD3 RE MTJ no3 NO8,16 SR8,16 LA8 uncommon (6)
Eunectes murinus (Linnaeus, 1758) CE RF, AQ WD3 site B _ _ _ _ _ rare (1)
Colubridae
Apostolepis sp. CE UF ? site E _ _ _ _ _ rare (1)
Atractus latifrons (Günther, 1868) PT UF WD3 site G _ _ _ _ _ rare (1)
Atractus major Boulenger, 1894 PT, AS UF, RF WD3 RE TX no3 NO8,16 FO8,16 MD8 rare (2)
Atractus snethlagae Cunha & Nascimento, 1983 AS, CE UF, RF WD3 RE MTJ no3 NO8,16 FO8,16 MD8 rare (2)
Atractus cf. schach PT, CE UF, RF ? RE TJX no3 NO8,16 FO8,16 MD8 rare (3)
Chironius fuscus (Linnaeus, 1758) PT, AS, CE UF, RF WD3 WD _ no DI8,16 TE8,16 LA8 abundant (17)
Chironius multiventris Schmidt & Walker, 1943 AS, CE UF, RF WD3 WD _ no DI8,16 SR3,8 LA8 uncommon (6)
Chironius scurrulus (Wagler, 1824) AS, CE UF, RF WD3 RE MTJ no3 DI8,16 TE8,16 LA8 rare (4)
Dendrophidion dendrophis (Schlegel, 1837) CE UF WD3 WD _ no DI8,16 TE8,16 MD8 rare (2)
Dipsas catesbyi (Sentzen, 1796) AS, CE UF, RF WD9 WD _ no NO8,9 SR8,9 MD8 abundant (23)
Dipsas pavonina Schlegel, 1837 AS, CE UF, RF DD9 RE TX yes NO8,9 SR8,9 MD8 uncommon (9)
Drepanoides anomalus (Jan, 1863) PT, AS, CE UF, RF WD3 WD _ no NO8,16 TE8,16 MD8 abundant (15)
Drymarchon corais (Boie, 1827) AS, CE UF, RF WD16 WD _ no DI8,16 TE8,16 LA8 rare (3)
Drymoluber dichrous (Peters, 1863) AS, CE UF, RF WD10 WD _ no DI8,16 TE8,16 LA8 abundant (17)
Erythrolamprus aesculapii (Linnaeus, 1766) CE UF, RF WD3 site J _ _ _ _ _ rare (1)
Erythrolamprus breviceps (Cope, 1860) PT, CE UF, RF WD3 RE TX no3 DI8,48 TE8,48 MD8 rare (3)
Erythrolamprus oligolepis (Boulenger, 1905) PT UF, RF SA18 RE MT no18 DI8,48 TE8,48 MD8 rare (3)

52
Taxa Survey
Methods
Forest
type
Known
distribution
Distribution
in study area
Restricted
to
Riverine
barrier
effect
Activity
period
Micro
habitat
Body
size Abundance (n)
Erythrolamprus reginae (Wagler, 1824) PT, AS, CE UF, RF WD3 WD _ no DI8,48 TE8,48 MD8 abundant (17)
Erythrolamprus typhlus (Linnaeus, 1758) PT, AS, CE UF, RF WD3 WD _ no DI8,48 TE8,48 MD8 abundant (14)
Helicops angulatus (Linnaeus, 1758) AS, CE RF, AQ WD3 WD _ no DN8,16 AQ8,16 MD8 abundant (15)
Helicops polylepis Günther, 1861 AS, CE RF, AQ WD16 WD _ no DN8,16 AQ8,16 MD8 rare (3)
Hydrodynastes bicinctus (Herrmann, 1804) CE RF, AQ WD16 site B _ _ DI8,16 AQ8,16 LA8 rare (2)
Hydrops martii (Wagler, 1824) CE AQ WD3 site B _ _ NO8,16 AQ8,16 LA8 rare (2)
Hydrops triangularis (Wagler, 1824) CE AQ WD12 RE MT no3 NO8,12 AQ8,12 MD8 rare (2)
Imantodes cenchoa (Linnaeus, 1758) AS, CE UF, RF WD3 WD _ no NO8,16 AR8,16 MD8 abundant (38)
Imantodes lentiferus (Cope, 1894) AS, CE UF, RF WD16 WD _ no NO8,16 AR8,16 MD8 rare (4)
Leptodeira annulata (Linnaeus, 1758) PT, AS, CE UF, RF WD3 WD _ no NO8,16 SR8,16 MD8 abundant (16)
Leptophis ahaetulla (Linnaeus, 1758) CE UF, RF WD3 WD _ no DI8,16 SR8,16 MD8 rare (2)
Mastigodryas boddaerti (Sentzen, 1796) AS, CE UF, RF WD3 WD _ no DI8,16 TE8,16 MD8 uncommon (6)
Oxyrhopus formosus (Wied, 1820) PT, AS, CE UF, RF WD16 WD _ no NO8,16 SR8,16 MD8 abundant (11)
Oxyrhopus melanogenys (Tschudi, 1845) PT, AS, CE RF WD16 WD _ no NO8,16 TE8,16 MD8 abundant (19)
Oxyrhopus petolarius Reuss, 1834 AS UF, RF WD16 site C _ _ _ _ _ rare (1)
Philodryas argentea (Daudin, 1803) AS, CE UF, RF WD3 WD _ no DI8,16 SR8,16 MD8 abundant (19)
Pseudoboa coronata Schneider, 1801 PT IP WD3 site I _ _ _ _ _ rare (1)
Phrynonax polylepis (Peters, 1867) AS, CE UF, RF WD3 WD _ no DI8,16 TE8,16 LA8 rare (5)
Rhynobothrium lentiginosum (Scopoli, 1785) AS, CE UF, RF WD11 WD _ no NO8,16 TE8,16 LA8 rare (4)
Siphlophis cervinus (Laurenti, 1768) AS, CE UF, RF WD3 RE TJX no3 NO8,16 SR8,16 MD8 rare (2)
Siphlophis compressus (Daudin, 1803) PT, AS, CE UF, RF WD3 WD _ no NO8,16 SR8,16 MD8 abundant (27)
Siphlophis worontzowi (Prado, 1940) CE RF WD17 site B _ _ _ _ _ rare (1)
Spilotes pullatus (Linnaeus, 1758) CE UF, RF WD3 site J _ _ _ _ _ rare (1)
Spilotes sulphureus (Wagler, 1824) CE UF WD3 site E _ _ _ _ _ rare (1)
Taeniophallus brevirostris (Peters, 1863) PT UF WD3 site B _ _ _ _ _ rare (1)
Taeniophallus quadriocellatus Santos-Jr et al., 2008 PT, AS UF, RF EA19 WD _ no DN8,19 TE8,19 MD8 rare (3)
Taeniophallus sp. PT UF, RF ? site A _ _ DN8,19 TE8,19 MD8 rare (2)
Tantilla melanocephala (Linnaeus, 1758) PT, AS, CE UF, RF WD3 RE MTJ no3 DN8,16 TE8,16 MD8 uncommon (7)
Thamnodynastes cf. pallidus (Linnaeus, 1758) AS,CE UF, SRF EA16 RE TJX yes NO8,16 SR8,16 MD8 uncommon (9)
Xenopholis scalaris (Wucherer, 1861) PT, AS, CE UF, RF WD3 WD _ no NO8,16 TE8,16 SM8 abundant (17)
Dactyloidae
Dactyloa phyllorhina (Myers & Carvalho, 1945) PT IP CAS2 site I _ _ _ _ _ rare (1)
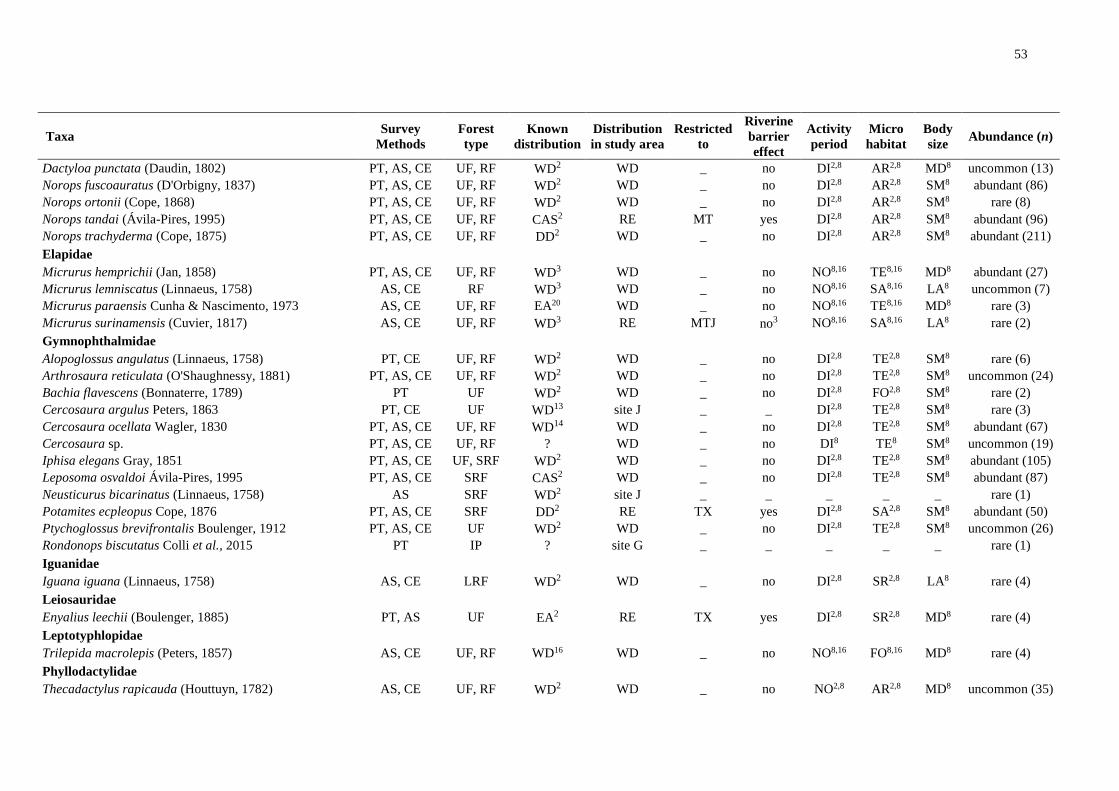
53
Taxa Survey
Methods
Forest
type
Known
distribution
Distribution
in study area
Restricted
to
Riverine
barrier
effect
Activity
period
Micro
habitat
Body
size Abundance (n)
Dactyloa punctata (Daudin, 1802) PT, AS, CE UF, RF WD2 WD _ no DI2,8 AR2,8 MD8 uncommon (13)
Norops fuscoauratus (D'Orbigny, 1837) PT, AS, CE UF, RF WD2 WD _ no DI2,8 AR2,8 SM8 abundant (86)
Norops ortonii (Cope, 1868) PT, AS, CE UF, RF WD2 WD _ no DI2,8 AR2,8 SM8 rare (8)
Norops tandai (Ávila-Pires, 1995) PT, AS, CE UF, RF CAS2 RE MT yes DI2,8 AR2,8 SM8 abundant (96)
Norops trachyderma (Cope, 1875) PT, AS, CE UF, RF DD2 WD _ no DI2,8 AR2,8 SM8 abundant (211)
Elapidae
Micrurus hemprichii (Jan, 1858) PT, AS, CE UF, RF WD3 WD _ no NO8,16 TE8,16 MD8 abundant (27)
Micrurus lemniscatus (Linnaeus, 1758) AS, CE RF WD3 WD _ no NO8,16 SA8,16 LA8 uncommon (7)
Micrurus paraensis Cunha & Nascimento, 1973 AS, CE UF, RF EA20 WD _ no NO8,16 TE8,16 MD8 rare (3)
Micrurus surinamensis (Cuvier, 1817) AS, CE UF, RF WD3 RE MTJ no3 NO8,16 SA8,16 LA8 rare (2)
Gymnophthalmidae
Alopoglossus angulatus (Linnaeus, 1758) PT, CE UF, RF WD2 WD _ no DI2,8 TE2,8 SM8 rare (6)
Arthrosaura reticulata (O'Shaughnessy, 1881) PT, AS, CE UF, RF WD2 WD _ no DI2,8 TE2,8 SM8 uncommon (24)
Bachia flavescens (Bonnaterre, 1789) PT UF WD2 WD _ no DI2,8 FO2,8 SM8 rare (2)
Cercosaura argulus Peters, 1863 PT, CE UF WD13 site J _ _ DI2,8 TE2,8 SM8 rare (3)
Cercosaura ocellata Wagler, 1830 PT, AS, CE UF, RF WD14 WD _ no DI2,8 TE2,8 SM8 abundant (67)
Cercosaura sp. PT, AS, CE UF, RF ? WD _ no DI8 TE8 SM8 uncommon (19)
Iphisa elegans Gray, 1851 PT, AS, CE UF, SRF WD2 WD _ no DI2,8 TE2,8 SM8 abundant (105)
Leposoma osvaldoi Ávila-Pires, 1995 PT, AS, CE SRF CAS2 WD _ no DI2,8 TE2,8 SM8 abundant (87)
Neusticurus bicarinatus (Linnaeus, 1758) AS SRF WD2 site J _ _ _ _ _ rare (1)
Potamites ecpleopus Cope, 1876 PT, AS, CE SRF DD2 RE TX yes DI2,8 SA2,8 SM8 abundant (50)
Ptychoglossus brevifrontalis Boulenger, 1912 PT, AS, CE UF WD2 WD _ no DI2,8 TE2,8 SM8 uncommon (26)
Rondonops biscutatus Colli et al., 2015 PT IP ? site G _ _ _ _ _ rare (1)
Iguanidae
Iguana iguana (Linnaeus, 1758) AS, CE LRF WD2 WD _ no DI2,8 SR2,8 LA8 rare (4)
Leiosauridae
Enyalius leechii (Boulenger, 1885) PT, AS UF EA2 RE TX yes DI2,8 SR2,8 MD8 rare (4)
Leptotyphlopidae
Trilepida macrolepis (Peters, 1857) AS, CE UF, RF WD16 WD _ no NO8,16 FO8,16 MD8 rare (4)
Phyllodactylidae
Thecadactylus rapicauda (Houttuyn, 1782) AS, CE UF, RF WD2 WD _ no NO2,8 AR2,8 MD8 uncommon (35)

54
Taxa Survey
Methods
Forest
type
Known
distribution
Distribution
in study area
Restricted
to
Riverine
barrier
effect
Activity
period
Micro
habitat
Body
size Abundance (n)
Polychrotidae
Polychrus marmoratus (Linnaeus, 1758) AS IP WD2 site J _ _ _ _ _ rare (1)
Scincidae
Copeoglossum nigropunctatum (Spix, 1825) PT, AS, CE UF, RF WD2 WD _ no DI2,8 SR2,8 SM8 abundant (121)
Sphaerodactylidae
Chatogekko amazonicus (Andersson, 1918) PT, AS, CE UF WD2 WD _ no DI2,8 TE2,8 SM8 abundant (170)
Gonatodes hasemani Griffin, 1917 PT, AS, CE UF, RF WA2 RE MT yes DI2,8 SR2,8 SM8 rare (4)
Gonatodes humeralis (Guichenot, 1855) PT, AS, CE UF, RF WD2 WD _ no DI2,8 SR2,8 SM8 abundant (449)
Gonatodes tapajonicus Rodrigues, 1980 PT, AS, CE UF, SRF CAS2 RE TJX yes DI2,8 SR2,8 SM8 abundant (118)
Pseudogonatodes cf. guianensis Parker, 1935 PT, AS UF WD2 RE MTJ no2 DI2,8 TE2,8 SM8 uncommon (14)
Teiidae
Ameiva ameiva (Linnaeus, 1758) PT, AS, CE UF, RF WD2 WD _ no DI2,8 TE2,8 MD8 abundant (429)
Kentropyx calcarata Spix, 1825 PT, AS, CE UF, RF WD2 WD _ no DI2,8 TE2,8 MD8 abundant (593)
Tupinambis longilineus Ávila-Pires, 1995 AS IP CAS15 site E _ _ DI2,8 TE2,8 LA8 rare (2)
Tupinambis teguixin (Linnaeus, 1758) PT, AS, CE LRF WD2 RE MTJ no2 DI2,8 TE2,8 LA8 rare (9)
Tropiduridae
Plica plica (Linnaeus, 1758) PT, AS, CE UF, RF WD2 WD _ no DI2,8 AR2,8 MD8 abundant (51)
Plica umbra (Linnaeus, 1758) PT, AS, CE UF, RF WD2 WD _ no DI2,8 AR2,8 MD8 uncommon (39)
Uranoscodon superciliosus (Linnaeus, 1758) PT, AS, CE RF WD2 WD _ no DI2,8 AR2,8 MD8 uncommon (26)
Typhlopidae
Amerotyphlops reticulatus (Linnaeus, 1758) PT, CE UF WD3 WD _ no NO8,16 FO8,16 MD8 uncommon (6)
Viperidae
Bothrops atrox (Linnaeus, 1758) AS, CE RF WD3 WD _ no NO8,16 TE8,16 LA8 abundant (57)
Bothrops bilineata (Wied, 1821) AS, CE UF SA21 WD _ no NO8,16 AR8,16 MD8 rare (3)
Bothrops taeniata (Wagler, 1824) AS, CE UF SA6 RE MTJ no6 NO8,16 SR8,16 LA8 rare (4)
Lachesis muta (Linnaeus, 1766) AS, CE UF WD3 RE MTJ no3 NO8,16 TE8,16 LA8 rare (3)
References: 1 Frost, 2015; 2 Ávila-Pires, 1995; 3 Fraga et al., 2013; 4 Gans, 2005; 5 Henderson et al., 2009; 6 Souza et al., 2013; 7 Jerriane Gomes, pers. comm.; 8 Authors, personal
observations; 9 Lima & Prudente, 2009; 10 Caldeira-Costa et al., 2013; 11 Arruda et al., 2015; 12 Albuquerque & de Lema, 2008; 13 Freitas et al., 2013; 14 Sales et al., 2014; 15 Caldeira-Costa
et al., 2008; 16 Wallach et al., 2014; 17 Dal Vechio et al., 2015; 18 França et al., 2013; 19 Santos Jr. et al., 2008; 20 Feitosa et al., 2007; 21 Bernarde et al., 2011; 22 Maciel & Hoogmoed, 2011; 23 Brcko et al., 2013; 24 Tsuji-Nishikido et al., 2012; 25 Lima et al., 2014; 26 Lima et al., 2012; 27 Ávila et al., 2012; 28 Lima et al., 2007; 29 Noronha et al., 2012; 30 Padial & De la Riva, 2009;

55
31 Bernardo et al., 2012; 32 Brown et al., 2011; 33 Bitar et al., 2015; 34 Orrico et al., 2014; 35 Pansonato et al., 2011; 36 Jungfer et al., 2013; 37 Bruschi et al., 2013; 38 Gordo et al., 2013; 39
Nunes et al., 2013; 40 Funk et al., 2008; 41 Fouquet et al., 2014; 42 Sá et al., 2014; 43 Peloso et al., 2014; 44 Freitas et al., 2014, 45 Guayasamin et al., 2009, 46 Pough et al., 2015, 47 Lynch &
Duellmann, 1997, 48Dixon, 1989.

56
REFERENCES
Albuquerque, N.R. & de Lema, T. (2008) Taxonomic revision of the Neotropical water snake
Hydrops triangularis (Serpentes, Colubridae). Zootaxa, 1685, 55-66.
Arruda, L.A.G, Carvalho, M.A. & Kawashita-Ribeiro, R.A. (2015) New records of the
Amazon banded snake Rhinobothryum lentiginosum (Serpentes: Colubridae) from Mato
Grosso State, Brazil, with natural history notes. Salamandra, 51, 199-205.
Ávila, R.W., Carvalho, V.T., Gordo, M., Kawashita-Ribeiro R.A. & Morais, D.H. (2012) A
new species of Amazophrynella (Anura: Bufonidae) from southern Amazonia. Zootaxa,
3484, 65-74.
Ávila-Pires, T.C.S. (1995) Lizards of Brazilian Amazonia (Reptilia: Squamata). Zoologische
Verhandelingen, 299, 1-706.
Bernarde, P.S., Costa, H.C., Machado R.A. & São-Pedro, V.A. (2011) Bothriopsis bilineata
bilineata (Wied, 1821) (Serpentes: Viperidae): New records in the states of Amazonas,
Mato Grosso and Rondônia, northern Brazil. Check List, 7, 343-347.
Bernardo, P.H., Matiazzi, W. & Guerra-Fuentes, A. (2012) Distribution extention and
distribution map of Chiasmocleis jimi Caramaschi and Cruz 2002 (Amphibia: Anura:
Microhylidae) and Proceratophrys concavitympanum Giaretta, Bernarde and Kokubum,
2000 (Amphibia: Anura: Cycloramphidae). Check List, 8, 152-154.
Bitar, Y.O.C, Silva, K.R.A., Filho, H.F.S. & Pinheiro, L.C. (2015) Amphibia, Anura, Hylidae,
Cruziohyla craspedopus (Funkhouser, 1957): distribution extension and first record
from the state of Pará, Brazil. Check List, 11, 1-2.
Brcko, I.C., Hoogmoed, M.S. & Neckel-Oliveira, S. (2013) Taxonomy and distribution of the
salamander genus Bolitoglossa Duméril, Bibron & Duméril, 1854 (Amphibia, Caudata,
Plethodontidae) in Brazilian Amazonia. Zootaxa 3686, 401-431.
Bruschi, D.P., Busin, C.S., Toledo, L.F., Vasconcellos, G.A., Strussmann, C., Weber, L.N.,
Lima, A.P., Lima, J.D. & Recco-Pimentel, S.M. (2013) Evaluation of the taxonomic
status of populations assigned to Phyllomedusa hypochondrialis (Anura, Hylidae,
Phyllomedusinae) based on molecular, chromosomal, and morphological approach.
BMC Genetics, 14, 1-14.
Brown, J.L., Twomey, E., Amézquita A., de Souza, M.B., Caldwell, J.P., Lötters, S., von
May, R., Melo-Sampaio, P.R., Mejía-Vargas, D., Pérez-Peña, P.E., Pepper, M.,
Poelman, E.H., Sanchez-Rodriguez, M. & Summers, K. (2011) A taxonomic revision of

57
the Neotropical poison frog genus Ranitomeya (Amphibia: Dendrobatidae). Zootaxa,
3083, 1-120.
Caldeira-Costa, H., São Pedro, V.A., Péres Jr, A.K. & Feio, R.N. (2008) Reptilia, Squamata,
Teiidae, Tupinambis longilineus: Distribution extension. Check List, 4, 267-268.
Caldeira-Costa, H., Moura, M.R. & Feio, R.N. (2013) Taxonomic revision of Drymoluber
Amaral, 1930 (Serpentes: Colubridae). Zootaxa 3716, 349-394.
Dal Vechio, F., Junior, M.T., Neto, A.M. & Rodrigues, M.T. (2015) On the snake Siphlophis
worontzowi (Prado, 1940): notes on its distribution, diet and morphological data.
Checklist, 11, 1-5
Dixon, J.R. (1989) A key and checklist to the neotropical snake genus Liophis, with country
list and maps. Smithsonian Herpetological Information Service, 79, 1-40.
Domingues, M.J. (2014) Flora da AID. In: Estudo de impacto ambiental – EIA, AHE São
Luiz do Tapajós. CNEC WorleyParsons. São Paulo, SP.
Feitosa, D.T., Prudente, A.L.C. & Lima, A.C. (2007): Redescription and variation of
Micrurus paraensis Cunha & Nascimento 1973 (Serpentes, Elapidae). Zootaxa, 1470,
35-45.
Fouquet, A., Cassini, C.S., Haddad, C.F.B., Pech, N. & Rodrigues, M.T. (2014) Species
delimitation, patterns of diversification and historical biogeography of the Neotropical
frog genus Adenomera (Anura, Leptodactylidae). Journal of Biogeography, 41, 855-
870.
Fraga, R. de, Lima, A.P., Prudente, A.L.C. & Magnusson, W.E. (2013) Guide to the snakes of
the Manaus region – Central Amazonia. Editora INPA, Manaus.
França, D.P.F, Freitas, M.A., Bernarde, P.S. & Uhlig, V.M. (2013) Erythrolamprus oligolepis
(Boulenger, 1905) (Serpentes: Dipsadidae): First record for the state of Acre, Brazil.
Check List, 9, 668–669.
Freitas, M.A., Oliveira e Sousa, S., Vieira, R.S., Farias, T. & Moura, G.J.B. (2013) First
record of Cercosaura argulus (Peters, 1863) (Squamata: Gymnophthalmidae) for the
state of Maranhão, Brazil. Check List, 9,1541-1542
Freitas, M.A., Dias, I.R., Farias, T., Oliveira e Sousa, S., Vieira, R.S., de Moura, G.J.B. &
Uhlig, V. (2014) First record of Ctenophryne geayi Mocquard, 1904 (Amphibia: Anura)
for the state of Maranhão, Brazil. Check List, 10, 585-587.
Furch, K. (1997) Chemistry of várzea and igapó soils and nutrient inventory of their
floodplain forests. In: Junk, W.E. (Ed.) The Central Amazon Floodplain. Berlin
Heidelberg: Springer-Verlag.

58
Funk W.C., Angulo A., Caldwell J.P., Ryan M.J. & Cannatella D.C. (2008) Comparison of
morphology and calls of two cryptic species of Physalaemus (Anura: Leiuperidae).
Herpetologica, 64, 290-304.
Frost, D.R. (2015) Amphibian Species of the World: an Online Reference. Version 6.0.
Electronic Database accessible at http://research.amnh.org/herpetology/amphibia/
index.html. American Museum of Natural History, New York, USA. Acess 06 Jun
2015.
Gans, C. (2005) Checklist and bibliography of the amphisbaenia of the world. Bulletin of the
American Museum of Natural History, 289, 1-130.
Gordo, M., Toledo, L.F., Suárez, P., Kawashita-Ribeiro, R.A., Ávila, R.W., Morais, D.H. &
Nunes, I. (2013). A New Species of Milk Frog of the Genus Trachycephalus tschudi
(Anura, Hylidae) from the Amazonian Rainforest. Herpetologica, 69, 466-479.
Guayasamin, J.M., Castroviejo-Fisher, S., Trueb, L., Ayarzagüena, J., Rada, M. & Vilà, C.
(2009) Phylogenetic systematics of Glassfrogs (Amphibia: Centrolenidae) and their
sister taxon Allophryne ruthveni. Zootaxa, 2100, 1-97.
Henderson, R.W., Passos, P. & Feitosa, D. (2009) Geographic variation in the Emerald
Treeboa, Corallus caninus (Squamata: Boidae). Copeia, 2009, 572-582.
Jungfer, K.H., Faivovich, J., Padial, J.M., Castroviejo-Fisher, S., Lyra, M.L., Berneck, B.M.,
Iglesias, P.P., Kok, P.J.R., MacCulloch, R.D., Rodrigues, M.T., Verdade, V.K, Torres-
Gastello, C.P., Chaparro, J.C., Valdujo, P.H., Reichle, S., Moravec, J., Gvoždík, V.,
Gagliardi-Urrutia, G., Ernst, R., De la Riva, I., Means, D.B., Lima, A.P., Señaris, J.C.,
Wheeler, W.C. & Haddad, C.F.B. (2013) Systematics of spiny-backed treefrogs
(Hylidae: Osteocephalus): an Amazonian puzzle. Zoologica Scripta, 42, 351-380.
Junk W.J., P.B. Bayley & Sparks, R.E. (1989) The flood pulse concept in river – floodplain
systems. Canadian Special Publication of Fisheries and Aquatic Sciences, 106, 110-
127.
Lima, A.P., Menin, M. & Araújo, M.C. (2007) A new species of Rhinella (Anura: Bufonidae)
from Brazilian Amazon. Zootaxa, 1663, 1-15.
Lima, A.C. & Prudente, A.L.C. (2009) Morphological variation and systematics of Dipsas
catesbyi (Sentzen, 1796) and Dipsas pavonina Schlegel, 1837 (Serpentes: Dipsadinae).
Zootaxa, 2203, 31-48.
Lima, A.P., Erdtmann, L.K. & Amézquita, A. (2012) Advertisement call and colour in life of
Allobates crombiei (Morales) “2000” [2002] (Anura: Aromobatidae) from the type
Locality (Cachoeira do Espelho), Xingu River, Brazil. Zootaxa, 3475, 86-88.

59
Lima, A.P., Simões, P.I. & Kaefer, I.L. (2014) A new species of Allobates (Anura:
Aromobatidae) from the Tapajós River basin, Pará State, Brazil. Zootaxa, 3889, 355-
387.
Lynch, J.D. & Duellman, W.E. (1997) Frogs of the genus Eleutherodactylus in western
Ecuador. Systematics, ecology, and biogeography. Special Publication. Natural History
Museum, University of Kansas, 23, 1-236.
Maciel, A.O. & Hoogmoed, M.S. (2011) Taxonomy and distribution of caecilian amphibians
(Gymnophiona) of Brazilian Amazonia, with a key to their identification. Zootaxa,
2984, 1-53.
Noronha, J.C., Rodrigues, D.J., Barros, A.B. & Almeida, E.J. (2012) New record and
distribution map of Hyalinobatrachium cappellei (van Lidth de Jeude 1904) (Anura:
Centrolenidae). Herpetology Notes, 5, 467-468.
Nunes, I., Suárez, P., Gordo, M. & Pombal, J.P. (2013) A second species of Trachycephalus
Tschudi (Anura: Hylidae) with a single vocal sac from the Brazilian Amazon. Copeia,
2013, 634-640.
Orrico, V.G.D., Peloso, P.L.V., Sturaro, M.J., Da Silva-Filho, H.F., Neckel-Olivera, S.,
Gordo, M., Faivovich, J. & Haddad, C.F.B. (2014) A new "Bat-voiced" species of
Dendropsophus Fitzinger, 1843 (Anura, Hylidae) from the Amazon Basin, Brazil.
Zootaxa, 3881, 341-361.
Padial, J.M. & De la Riva, I. (2009) Integrative taxonomy reveals cryptic Amazonian species
of Pristimantis (Anura: Strabomantidae). Zoological Journal of the Linnean Society,
155: 97–122.
Palumbi, S.R., Martin, A., Romano, S., Mcmillan, W.O., Stice, L. & Grabowski, G. (1991)
The Simple Fool's Guide to PCR. University of Hawaii, Honolulu.
Pansonato, A., Ávila, R.W., Kawashita-Ribeiro, R.A. & Morais, D.H. (2011) Advertisement
call and new distribution records of Hypsiboas leucocheilus (Anura: Hylidae).
Salamandra, 47, 55-58.
Peloso, P.L.V., Sturaro, M.J., Forlani M.C., Gaucher, P., Motta, A.P. & Wheeler, W.C. (2014)
Phylogeny, taxonomic revision, and character evolution of the genera Chiasmocleis and
Syncope (Anura, Microhylidae) in Amazonia, with descriptions of three new species.
Bulletin of the American Museum of Natural History, 136, 1-96.
Sá, R.O., Grant, T., Camargo, A. Heyer, W.R., Ponssa, M.L. & Stanley, E. (2014)
Systematics of the Neotropical genus Leptodactylus Fitzinger, 1826 (Anura:

60
Leptodactylidae): Phylogeny, the relevance of non-molecular evidence, and species
accounts. South American Journal of Herpetology, 9, 1-128.
Sales, R.F.D., Jorge J.S., Meira-Ribeiro, M. & Freire, E.M.X. (2014) New record of
Cercosaura ocellata Wagler, 1830 (Squamata, Gymnophthalmidae) in northeastern
Brazil, with a distribution map for the species in South America. Check List 10, 1531-
1534.
Sambrook, J. & Russel D.W. (2001) Molecular Cloning: a Laboratory Manual. 3rd edn. Cold
Spring Harbor Laboratory Press, New York.
Santos Jr., A.P., Di-Bernardo, M., & Lema, T. (2008) New Species of the Taeniophallus
occipitalis Group (Serpentes, Colubridae) from Eastern Amazonia, Brazil. Journal of
Herpetology, 42, 419-426.
Sievers, F., Wilm, A., Dineen, D., Gibson, T.J., Karplus, K., Li, W., Lopez, R., McWilliam,
H., Remmert, M., Söding, J., Thompson, J.D. & Higgins, D.G. (2011) Fast, scalable
generation of high-quality protein multiple sequence alignments using Clustal Omega.
Molecular Systems Biology, 7, 539.
Souza, J.D.R., Venâncio, N.M., Freitas, M.A., Souza, M.B. & Veríssimo, D. (2013) First
record of Bothrops taeniatus Wagler, 1824 (Reptilia: Viperidae) for the state of Acre,
Brazil. Check List, 9, 430-431.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M. & Kumar, S. (2011) MEGA5:
Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary
Distance, and Maximum Parsimony Methods. Molecular Biology and Evolution, 28,
2731-2739.
Tsuji-Nishikido, B.M., Kaefer, I.L., Freitas, F.C., Menin, M. & Lima, A.P. (2012) Significant
but not diagnostic: differentiation through morphology and calls in the Amazonian frogs
Allobates nidicola and A. masniger. Herpetological Journal, 22, 105–114.
Wallach, V., Williams, K.L. & Boundy, J. (2014) Snakes of the World: a Catalogue of Living
and Extinct Species. CRC Press, Boca Raton, 1209 pp.
Wiley, E.O. (1978) The evolutionary species concept reconsidered. Systematic Zoology, 27,
17–26.

61
REFERÊNCIAS BIBLIOGRÁFICAS
Azevedo-Ramos, C.; Galatti, U. 2002. Patterns of amphibian diversity in Brazilian Amazonia:
conservation implications. Biological Conservation, 103: 103-111.
Beheregaray, L.B.; Cooke, G.M.; Chao, N.L.; Landguth, E.L. 2015. Ecological speciation in
the tropics: insights from comparative genetic studies in Amazonia. Frontiers in
Genetics, 5: 1-19.
Boubli, J.P.; Ribas, C.; Lynch Alfaro, J.; da Silva, M.N.F.; Pinho, G.M.; Farias, I.P. 2014.
Spatial and temporal patterns of diversification on the Amazon: a test of the riverine
hypothesis for all diurnal primates of Rio Negro and Rio Branco in Brazil. Molecular
Phylogenetics and Evolution, 82: 400-412.
Brasil, PR (Presidência da República). 2011. PAC-2 Relatórios. PR, Brasília, DF. Disponível
em: http://www.brasil.gov.br.
Caminer, M.; Ron, S.R. 2014. Systematics of treefrogs of the Hypsiboas calcaratus and
Hypsiboas fasciatus species complex (Anura, Hylidae) with the description of four new
species. ZooKeys, 370: 1-68.
Costa, F.R.C.; Magnusson, W.E. 2010. The need for large-scale, integrated studies of
biodiversity - the experience of the program for biodiversity research in brazilian
amazonia. Natureza & Conservação, 8: 3-12.
Costa, H.C.; Bérnils, R.S. 2014. Répteis brasileiros: Lista de espécies. Disponível em
<http://www.sbherpetologia.org.br>. Acesso em 06 jun 2015.
Dal Vechio, F.; Junior, M.T.; Neto, A.M.; Rodrigues, M.T. 2015. On the snake Siphlophis
worontzowi (Prado, 1940): notes on its distribution, diet and morphological data.
Checklist, 11(1).
Duellman, W.E. 1979. The South American herpetofauna: Its origin, evolution, and dispersal.
v.7. Kansas: Museum of Natural History/University of Kansas. 485 p.
Duellman, W.E. 2005. Cusco Amazônico. The Lives of amphibians and reptiles in an
Amazonian Rainforest. Comstock Publishing Associates, Cornell University Press,
Ithaca, New York. 488pp.
Fouquet, A.; C.S. Cassini, C.S.; Haddad, C.F.B.; Pech, N.; Rodrigues, M.T. 2014. Species
delimitation, patterns of diversification and historical biogeography of the Neotropical
frog genus Adenomera (Anura, Leptodactylidae). Journal of Biogeography, 41(5): 855-
870.

62
Funk, W.C.; Caminer, M.; Ron, S.R. 2011. High levels of cryptic species diversity uncovered
in Amazonian frogs. Proceedings of the Royal Society B: Biological Sciences, 279:
1806-1814.
Gordo, M.; Toledo, L.F.; Suárez, P.; Kawashita-Ribeiro, R.A.; Ávila, R.W., Morais, D.H.;
Nunes, I. 2013. A new species of Milk Frog of the genus Trachycephalus Tschudi
(Anura, Hylidae) from the Amazonian rainforest. Herpetologica, 69: 466-479.
Hoorn, C.; Wesselingh, F.P.; terSteege, H.; Bermudez M.A.; Mora A.; Sevink J.; Sanmartín
I.A.; Sanchez-Meseguer, A; Anderson, C.L.; Figueiredo, J.P.; Jaramillo, C.; Riff, D.;
Negri, F. R.; Hooghiemstra, H.; Lundberg, J.; Stadler, T.; Särkinen, T.; Antonelli, A.
2010. Amazonia through time: Andean uplift, climate change, landscape evolution, and
biodiversity. Science, 330: 927-931.
Jansen, M.; Bloch, R.; Schulze, A.; Pfenninger, M. 2011. Integrative inventory of Bolivia’s
lowland anurans reveals hidden diversity. Zoologica Scripta, 40: 567-583.
Jorge, R.F. 2014. Fatores determinantes dos padrões de distribuição e densidade de Allobates
sumtuosus e Atelopus spumarius em duas bacias de drenagem em 64 Km² de floresta de
terra-firme na Amazônia central. Dissertação de Mestrado. Instituto Nacional de
Pesquisas da Amazônia, Manaus, Amazonas.
Leite, Y.L.R.; Kok, P.R.J.; Weksler, M. 2015. Evolutionary affinities of the ‘Lost World’
mouse suggest a late Pliocene connection between the Guiana and Brazilian shields.
Journal of Biogeography, 42: 706–715.
Leite, R.N.; Rogers, D.S. 2013. Revisiting Amazonian phylogeography: insights into
diversification hypotheses and novel perspectives. Organisms Diversity and Evolution,
13(4): 639-664.
Lima, A.P.; Simões, P.I.; Kaefer, I.L. 2014. A new species of Allobates (Anura:
Aromobatidae) from the Tapajós River basin, Pará State, Brazil. Zootaxa, 3889: 355-
387.
Losos, J.B.; Arnold, S.J.; Bejerano, G.; Brodie III, E.D.; Hibbett, D.; Hoekstra, H.E.; Mindell,
D.P.; Monteiro, A.; Moritz, C.; Allen Or, H.; Petrov, D.A.; Renner, S.A.; Ricklefs, R.E.;
Soltis, P.S.; Turner, T.L. 2013 Evolutionary biology for the 21st century. PLoS Biology,
11: e1001466.
Magnusson, W.E. 2014. Uncertainty and the design of in-situ biodiversity-monitoring
programs. Nature Conservation, 8: 77-94.

63
Menin, M.; Lima, A.P.; Magnusson, W.E.; Waldez, F. 2007. Topographic and edaphic effects
on the distribution of terrestrially reproducing anurans in Central Amazonia: mesoscale
spatial patterns. Journal of Tropical Ecology, 23: 539-547.
Murphy J.C.; Jowers, M.J. 2013. Treerunners, cryptic lizards of the Plica plica group
(Squamata, Sauria, Tropiduridae) of northern South America. ZooKeys, 355: 49-77.
Museu Paraense Emílio Goeldi, 2012. Censo da Biodiversidade da Amazônia Brasileira.
Disponível em < http://www.museu-goeldi.br/censo/>. Acesso em 06 jun 2015.
Nascimento, D.S. 2014. Filogenia molecular de serpentes neotropicais do grupo Bothrops
atrox (Linnaeus, 1758) (Viperidae: Crotalinae). Dissertação de Mestrado. Universidade
de Brasília, Brasília.
Naka, L.N. 2011. Avian distribution patterns in the Guiana Shield: implications for the
delimitation of Amazonian areas of endemism. Journal of Biogeography, 38: 681-696.
Neckel-Oliveira, S.; Magnusson, W.E.; Lima, A.P. 2000. Diversity and distribution of frogs
in an Amazonian savanna in Brazil. Amphibia-Reptilia, 21(3):317-326.
Orrico, V.G.D.; Peloso, P.L.V.; Sturaro, M.; da Silva, J.; Filho, H.F.; Neckel-Oliveira, S.;
Gordo, M.; Faivovich, J.; Haddad, C.F.B. 2014. A new “Bat-Voiced” species of
Dendropsophus Fitzinger, 1843 (Anura, Hylidae) from the Amazon Basin, Brazil.
Zootaxa, 3881: 341-361.
Pavan, D. 2007. Assembléias de répteis e anfíbios do Cerrado ao longo da bacia do rio
Tocantins e o impacto do aproveitamento hidrelétrico da região na sua conservação.
Tese de doutorado. Universidade de São Paulo, São Paulo.
Pomara, L.Y.; Ruokolainen, K.; Young, K.R. 2013. Avian species composition across the
Amazon river: the roles of dispersal limitation and environmental heterogeneity.
Journal of Biogeography, 41: 784-796.
Ribeiro Jr., J.W.; Lima, A.P.; Magnusson, W.E. 2012. The effect of riparian zones on species
diversity of frogs in Amazonian forests. Copeia, 2012(3):375-381.
Segalla, M.V.; Caramaschi, U.; Cruz, C.A.G; Grant, T.; Haddad, C.F.B.; Langone, J.A.;
Garcia, P.C.A. 2014. Brazilian Amphibians: List of Species. Disponível em
<http://www.sbherpetologia.org.br>. Acesso em 06 jun 2015.
Simões, P.I.; Lima, A.P.; Faria, I.P. 2010. The description of a cryptic species related to the
pan-Amazonian frog Allobates femoralis (Boulenger 1883) (Anura: Aromobatidae).
Zootaxa, 2406: 1-18
Simões, P.I.; Kaefer, I.L.; Lima, I.P. 2011. The first record of the rare microhylid Altigius
alios Wild, 1995 in Brazil. Herpetology Notes, 4: 141-142.

64
Simões, P.I.; Kaefer, I.L.; Farias, I.P.; Lima, A.P. 2013. An integrative appraisal of the
diagnosis and distribution of Allobates sumtuosus (Morales, 2002) (Anura,
Aromobatidae). Zootaxa, 3746: 401-421.
Simões, P.I.; Stow, A.; Hödl, W.; Amézquita, A.; Farias, I.P.; Lima, A.P. 2014. The value of
including intraspecific measures of biodiversity in environmental impact surveys is
highlighted by the Amazonian brilliant-thighed frog (Allobates femoralis). Tropical
Conservation Science, 7(4): 811-828.
Smith, B.T.; McCormack, J.E.; Cuervo, A.M.; Hickerson, M.J.; Aleixo, A.; Cadena, C.D.;
Pérez-Emán, J.; Burney, C.W.; Xie, X.; Harvey, M.G.; Faircloth, B.C.; Glenn, T.C.;
Derryberry, E.P.; Prejean, J.; Fields, S.; Brumfield, R.T. 2014. The drivers of tropical
speciation. Nature, 515, 406-409.

65
APÊNDICES
Ata da Aula de Qualificação

66
Ata da Defesa da Dissertação
Top Related