
ACTIVIDADE DE MYOTIS MYOTIS (CHIROPTERA) DURANTE … · cada vez que eu tinha que ir sozinha para...
51
UNIVERSIDADE DE LISBOA FACULDADE DE CIÊNCIAS DEPARTAMENTO DE BIOLOGIA ANIMAL ACTIVIDADE DE MYOTIS MYOTIS (CHIROPTERA) DURANTE O INVERNO: INFLUÊNCIA DA ABUNDÂNCIA ALIMENTAR ANA MARGARIDA PONTES DA SILVA AUGUSTO Mestrado em Biologia da Conservação 2008
Transcript of ACTIVIDADE DE MYOTIS MYOTIS (CHIROPTERA) DURANTE … · cada vez que eu tinha que ir sozinha para...

UNIVERSIDADE DE LISBOA FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
ACTIVIDADE DE MYOTIS MYOTIS
(CHIROPTERA) DURANTE O INVERNO: INFLUÊNCIA DA ABUNDÂNCIA ALIMENTAR
ANA MARGARIDA PONTES DA SILVA AUGUSTO Mestrado em Biologia da Conservação
2008

UNIVERSIDADE DE LISBOA FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
ACTIVIDADE DE MYOTIS MYOTIS
(CHIROPTERA) DURANTE O INVERNO: INFLUÊNCIA DA ABUNDÂNCIA ALIMENTAR
ANA MARGARIDA PONTES DA SILVA AUGUSTO
Mestrado em Biologia da Conservação
Dissertação orientada pelo Prof. Doutor Jorge M. Palmeirim
(DBA/CBA - FCUL)
2008

iii
Agradecimentos
A minha sincera gratidão a todos aqueles que cruzaram o meu caminho durante a
realização desta tese, e que duma maneira ou de outra, contribuíram para que este
trabalho fosse possível. Em especial agradeço:
Ao meu orientador Prof. Doutor Jorge M. Palmeirim por me ter permitido
conhecer o fantástico mundo dos morcegos.
À minha orientadora “não-oficial” Ana Rainho, por todas as preciosas dicas,
sugestões, rabugices e afins. Nunca te poderei agradecer o suficiente...
Aos responsáveis pela Herdade dos Machados, Herdade dos Lameirões e Mina
da Preguiça pela autorização de acesso aos terrenos. Em particular ao “Quim-Zé”,
responsável pelo terreno dos Montejuntos (Herdade dos Machados), por se preocupar
cada vez que eu tinha que ir sozinha para perto dos touros.
Ao Israel Silva (CBA) e à Sónia Ferreira (CIBIO) pelo apoio na identificação
dos desgraçados dos insectos.
Ao Ricardo Correia, Ewelina Rogozińska e Miguel Augusto por toda a ajuda que
me foi prestada na triagem das amostras de insectos no laboratório.
À Ana Rainho, Sofia Lourenço e Teresa Santos pelas sugestões durante a escrita
da tese.
À Mónica Almeida por todas as dicas e ajuda com as “picuices”; mas
principalmente pelas imensas horas de companhia. Também à Nina e à Daisy pelos
bocadinhos de descontracção que me proporcionaram.

iv
À D. Antónia Patinho, uma senhora muito querida que me “adoptou” durante o
trabalho de campo, um grande obrigado por ter sempre um sorriso e uma palavra amiga,
mesmo quando eu espezinhava a sua casa toda com aquela famosa lama alentejana, que
se agarra a tudo.
A todos os morcególogos pelas palavras de incentivo que me deram. Em
particular à Luísa Rodrigues, à Sofia Lourenço e ao Hugo Rebelo, por me tornarem
numa “viciada” em trabalho de campo com morcegos.
A todos os meus amigos, agradeço que me desculpem por ter “desaparecido”
durante tanto tempo.
À minha mãe por me apoiar incondicionalmente, ajudando-me a nunca desistir
dos meus sonhos…”Resiste e vencerás!”
Ao Ricardo…se começar a agradecer tudo o que fizeste por mim, ocupo o
espaço todo dedicado a esta tese...por isso digo apenas: Obrigada por seres quem és!

v
Resumo
Em regiões temperadas, os morcegos passam a maior parte do Inverno em
hibernação, mas este estado é frequentemente interrompido, por razões que ainda não
estão bem compreendidas. Uma das explicações propostas para estas interrupções é a
necessidade de se alimentarem. Assim, o objectivo deste trabalho foi determinar a
importância da actividade de caça durante o Inverno numa região Mediterrânica,
caracterizada por Invernos amenos. Neste sentido, (i) verificou-se se os morcegos
revelaram actividade fora do abrigo, que possa estar associada a actividade de caça
durante o Inverno; (ii) verificou-se se os excrementos produzidos durante o Inverno
continham peças de artrópodes; (iii) comparou-se a abundância de presas durante o
Inverno com dados de outras estações do ano; e (iv) avaliou-se a disponibilidade de
presas em diferentes usos do solo e densidades de vegetação. O estudo foi conduzido no
sul de Portugal, tendo como população modelo, uma colónia de Myotis myotis.
Os morcegos saíram frequentemente dos abrigos durante os meses de Inverno, e
nestes meses houve produção de excrementos contendo peças de artrópodes,
confirmando que se alimentam frequentemente durante este período. A quantificação da
abundância de presas, nos usos de solo predominantes da área amostrada, demonstrou:
(a) a existência de menor abundância de presas durante o Inverno, que no Outono e na
Primavera, e que (b) as pastagens, forragens e cereais são os tipos de uso do solo que
apresentam maior abundância de presas potenciais para M. myotis. A avaliação da
disponibilidade das presas revelou, no entanto, que é nestes usos do solo, com coberto
mais denso ao nível do estrato herbáceo, que M. myotis revela menor sucesso de
captura.
Os resultados sugerem que os Invernos amenos da região de estudo, possibilitam
a existência de presas activas durante este período, permitindo a alimentação de M.
myotis, embora o alimento não seja abundante nesta época do ano. Assim, como a
alimentação de Inverno pode ser essencial para a sobrevivência dos morcegos
Mediterrânicos, deve ter-se em conta esta necessidade, aquando da gestão das áreas
envolventes aos principais abrigos.
Palavras-chave: Myotis myotis, Inverno, dieta, artrópodes do solo, Mediterrâneo

vi
Abstract
In the temperate zones, bats spend most of the winter in hibernation, but they
frequently interrupt it for reasons that are not always understood. One of the reasons
suggested relates this winter activity with a need to feed. This way, the objective of this
work was to determine if foraging was a relevant activity during winter in a
Mediterranean region, characterized by mild winters. To do this, we (i) checked if bats
frequently made potential foraging trips during winter, (ii) verified if the fecal pellets
produced in the winter contained arthropod pieces, (iii) compared the abundance of prey
during the winter with data of other seasons, and (iv) evaluated the availability of prey
in different land uses and vegetation densities. This study was conducted in the south of
Portugal, using a colony of Myotis myotis as a model population.
Bats often made potential foraging flights throughout the winter, and they
produced pellets containing prey, which confirms that they are often foraging. The
quantification of the abundance of prey in the major types of the land cover,
demonstrated that (a) there is significantly less prey during the winter than in autumn
and spring, and that (b) grasslands and cereal fields have the greatest abundance of prey.
Data from prey availability analysis showed nevertheless that bat prey capture
efficiency declines with ground density cover, with grasslands among the land uses
where prey is less available to bats.
The results suggest that the mild winters of the study region, allow the existence
of active bat prey during this period, making winter foraging to M. myotis possible,
despite the fact that food is not abundant at this time of the year. As winter feeding may
be important for the survival of Mediterranean bats, management around key colonies
should take into consideration the winter foraging needs of bats.
Keywords: Myotis myotis, winter, diet, ground arthropods, Mediterranean

vii
Índice
Agradecimentos ............................................................................................................... iii
Resumo ............................................................................................................................. v
Abstract............................................................................................................................ vi
Índice de figuras .............................................................................................................. ix
1. Introdução e Objectivos................................................................................................ 1
1.1. Os Morcegos em Portugal...................................................................................... 1
1.2. O Inverno em regiões temperadas ......................................................................... 1
1.3. Os morcegos alimentam-se durante o Inverno, em Portugal? ............................... 3
1.4. Abundância e disponibilidade de presas ................................................................ 4
1.5. Objectivos .............................................................................................................. 5
2. Material e Métodos....................................................................................................... 6
2.1. Espécie estudada .................................................................................................... 6
2.2. Área de estudo ....................................................................................................... 6
2.3. Recolha de dados ................................................................................................... 8
2.3.1. Actividade dos morcegos durante o Inverno................................................... 8
2.3.2. Recolha de excrementos de morcegos .......................................................... 10
2.3.3. Abundância sazonal de presas de morcegos ................................................. 11
Amostragem com armadilhas do tipo pitfall ....................................................... 11
Contagem de cantos de Orthoptera..................................................................... 13
2.3.4. Disponibilidade de presas em diferentes densidades de vegetação............... 13
Amostragem da densidade da vegetação............................................................. 14
Experiências em cativeiro ................................................................................... 14
2.4. Análise estatística ................................................................................................ 15
3. Resultados................................................................................................................... 16

viii
3.1. Actividade dos morcegos durante o Inverno ....................................................... 16
3.2. Composição da dieta ............................................................................................ 17
3.3. Abundância sazonal de presas de morcegos ........................................................ 18
3.3.1. Amostragem com armadilhas do tipo pitfall ................................................. 18
Comparação entre consumo e abundância de presas ......................................... 20
Abundância de presas potenciais nas pitfalls ..................................................... 21
3.3.2. Contagem de cantos de Orthoptera ............................................................... 24
3.4. Disponibilidade de presas em diferentes densidades de vegetação ..................... 25
3.4.1. Amostragem da densidade da vegetação....................................................... 25
3.4.2. Experiências em cativeiro ............................................................................. 26
4. Discussão.................................................................................................................... 28
4.1. Actividade dos morcegos durante o Inverno ....................................................... 28
4.2. Composição da dieta ............................................................................................ 28
4.3. Abundância sazonal de presas de morcegos ........................................................ 30
4.4. Disponibilidade de presas em diferentes densidades de vegetação ..................... 32
5. Bibliografia................................................................................................................. 33
ANEXO I........................................................................................................................ 40
ANEXO II ...................................................................................................................... 41

ix
Índice de figuras
Figura 1 – Área de estudo................................................................................................. 7
Figura 2 – Espectrograma de M. myotis. .......................................................................... 9
Figura 3 – Localização das armadilhas de pitfall e pontos de escuta realizados na área de
estudo.............................................................................................................................. 11
Figura 4 – Número médio de passagens de M. myotis registadas à saída do abrigo, por
mês.................................................................................................................................. 16
Figura 5 – Percentagem (%) de ocorrência dos artrópodes identificados na dieta de M.
myotis entre Outubro de 2007 e Abril de 2008............................................................... 17
Figura 6 – Abundância relativa de artrópodes capturados nas pitfalls. .......................... 19
Figura 7 – Frequência de ocorrência de artrópodes capturados nas pitfalls e nos
excrementos. ................................................................................................................... 20
Figura 8 – Número médio de presas potenciais capturadas nas pitfalls nos diferentes
usos do solo. ................................................................................................................... 21
Figura 9 – Variação do número médio de presas capturadas nas pitfalls por uso do solo.
........................................................................................................................................ 22
Figura 10 – Variação mensal das principais presas de M. myotis capturadas nas pitfalls,
considerando todos os biótopos amostrados................................................................... 23
Figura 11 – Número médio de cantos de Gryllidae e Gryllotalpidae gravados por noite.
........................................................................................................................................ 24
Figura 12 – Relação entre a abundância de presas e a densidade da vegetação nos
diferentes usos do solo e microrelevos. (Gira = Girassol, Oli = Olival, Past = Pastagem,
Cere = Cereal, Forr = Forragem; B = baixio, C = cabeço)............................................. 25
Figura 13 – Tempo médio utilizado pelos morcegos na captura das presas em diferentes
densidades do coberto (hh:mm:ss). ................................................................................ 26
Figura 14 – Taxa de sucesso de captura das presas em diferentes densidades do coberto.
........................................................................................................................................ 27

Introdução
1
1. Introdução e Objectivos
1.1. Os Morcegos em Portugal
Em Portugal, quase metade da fauna de mamíferos pertence ao grupo dos
quirópteros. Existem cerca de 26 espécies de morcegos no país, 12 das quais com
estatuto de ameaça (Cabral et al. 2006). Este facto atribui a Portugal responsabilidades
na conservação destas espécies.
Os morcegos cavernícolas estão entre as espécies mais ameaçadas em Portugal.
Os indivíduos destas espécies são extremamente dependentes do seu abrigo, como
protecção diurna, bem como da área envolvente ao abrigo para se alimentarem. Por
estas razões, a conservação deste grupo passa não só por proteger as espécies
directamente, mas principalmente por preservar os seus abrigos e áreas de caça
(Palmeirim & Rodrigues 1992).
Apesar de os morcegos não serem bem encarados pela sociedade, desempenham
um papel fundamental no funcionamento dos ecossistemas naturais e humanizados
(Rainho et al. 1998). Em Portugal, este grupo é quase exclusivamente insectívoro,
comendo diariamente mais de metade do seu peso em insectos durante os meses quentes
do ano. A sua importância no controle de pragas agrícolas e combate a vectores de
doenças, conferem aos morcegos bastante relevância a nível económico (Rainho et al.
1998). Esta razão, associado ao seu papel no equilíbrio do ecossistema, realça a
importância da sua conservação.
1.2. O Inverno em regiões temperadas
O conhecimento da ecologia dos morcegos, principalmente durante os períodos
em que se encontram mais vulneráveis, é essencial no delineamento de uma estratégia
de conservação adequada a estas espécies (Lawton 1996, Wallington et al. 2005). Nas
regiões temperadas, o Inverno é uma época desfavorável à maioria dos seres vivos,
sendo durante este período que a taxa de mortalidade é, regra geral, mais elevada
(Storey & Storey 1996, Stamou et al. 2004). A escassez de alimento e as maiores
exigências energéticas devido às baixas temperaturas levaram diversas espécies a
adoptar estratégias fisiológicas e comportamentais para superar esta época (Park et al.
2000).

Introdução
2
Entre estas espécies, encontram-se os morcegos, que podem hibernar (Park et al.
2000, Zukal et al. 2005, Wojciechowski et al. 2007), migrar e/ou acumular reservas
energéticas durante os períodos mais favoráveis (Cryan & Veilleux 2007).
Segundo Wojciechowski et al. (2007), existem dois factores determinantes na
capacidade dos morcegos de sobreviverem durante a hibernação: (a) a sua condição
corporal no início da hibernação, e (b) a temperatura ambiente – quanto mais baixa a
temperatura, maior é o estado de torpor, garantindo menos gasto de reservas
energéticas.
No sul da Europa, nomeadamente em Portugal, o período de hibernação dura
cerca de dois meses, sendo mais curto do que nos países do Norte, devido ao clima
mediterrânico desta região (Rodrigues et al. 2003). As temperaturas observadas durante
o Inverno em algumas regiões de Portugal, poderão impedir que algumas espécies de
morcegos atinjam uma temperatura corporal suficientemente baixa para hibernar por
períodos superiores a alguns dias ou semanas (Rodrigues et al. 2003). Nesta situação,
surge a necessidade de adoptar uma estratégia, que permita compensar os gastos
energéticos relacionados com a manutenção da temperatura corporal. Esta estratégia
poderá passar pela reposição das reservas energéticas, por via da alimentação. No
entanto, tratando-se de espécies insectívoras, a abundância e disponibilidade alimentar
neste período poderá ser crítica, dado que a abundância de insectos e outros artrópodes é
também bastante afectada pela temperatura (Stamou et al. 2004). Assim, dando-se o
caso de não existirem condições quer para a hibernação, quer para a alimentação
durante este período, a sobrevivência dos morcegos poderá ser seriamente afectada.

Introdução
3
1.3. Os morcegos alimentam-se durante o Inverno, em Portugal?
A questão da alimentação dos morcegos durante o período de hibernação traz
associada uma problemática com alguma controvérsia, já que certos autores defendem
que alguns morcegos se alimentam durante esta fase do seu ciclo anual, enquanto outros
afirmam o contrário.
É sabido que os morcegos têm períodos de actividade durante a hibernação
(Arlettaz 1999, Kanuch et al. 2005, Lausen et al. 2006, Dunbar et al. 2007) durante os
quais os morcegos podem sair do seu abrigo, eventualmente para beber e/ou se
alimentar, ou até mesmo para mudar de abrigo (Whitaker et al. 1997, Park et al. 2000,
Wojciechowski et al. 2007). Esta actividade nos meses de Inverno parece ocorrer
apenas em dias de temperatura mais elevada e vento reduzido, facto que poderá estar
associado a uma maior disponibilidade de presas, o que potenciará a actividade de caça
(Park et al. 2000, Celuch & Kanuch 2005). No entanto, a relação entre a actividade dos
morcegos e a sua alimentação durante o Inverno é geralmente feita com base em
métodos indirectos (e.g. análise de excrementos). Por vezes, até mesmo a produção de
excrementos durante o Inverno poderá não ser um bom indício de actividade alimentar,
já que foi comprovado que durante este período, há produção de excrementos
resultantes de um processo de acumulação de sucos gástricos e de resíduos no tubo
digestivo dos morcegos (Dunbar et al. 2007). Assim sendo, estes indivíduos poderão
não se ter alimentado nos meses de Inverno, apesar de ter havido produção de
excrementos.
Torna-se então importante determinar a natureza destas incursões ao exterior.
Tentar compreender se o facto de os morcegos interromperem a hibernação, poderá
estar relacionado com a necessidade de se alimentarem.
Independente desta problemática é o facto de que os morcegos perdem peso
durante o Inverno, mesmo em Portugal (Zahn et al. 2007). Fica, no entanto, por
esclarecer se esta perda de peso se prende com (a) a não alimentação durante o Inverno,
(b) a impossibilidade de se alimentar devidos a factores climáticos, ou (c) a reduzida
abundância e/ou disponibilidade de presas.

Introdução
4
1.4. Abundância e disponibilidade de presas
Se for confirmada a necessidade de os morcegos se alimentarem durante o
Inverno, torna-se essencial garantir que há alimento disponível. Para tal, é fundamental
a compreensão de quais os usos do solo que poderão beneficiar a alimentação desta
espécie, de forma a melhor definir propostas de gestão da paisagem envolvente aos
principais abrigos de hibernação desta espécie.
A intensificação da exploração agrícola que se tem verificado desde os meados
do século XIX, resultou numa diminuição da biodiversidade, (Pfiffner & Luka 2000,
Zalidis et al. 2002, Donald 2004, Pywell et al. 2005). Esta diminuição da biodiversidade
poderá limitar a disponibilidade de alimento para muitas das espécies de morcegos
dependentes destes agro-ecossistemas, pelo facto de serem tão dependentes das zonas
adjacentes aos seus abrigos.
É reconhecido que a homogeneização das culturas afecta principalmente os
artrópodes do solo (Benton et al. 2002, Cole et al. 2002), grupo que constitui grande
parte da dieta de muitas espécies ameaçadas de morcegos (Arlettaz 1999, Ransome
2000). O estudo dos habitats onde estes artrópodes se encontram em maior abundância
durante as fases mais cruciais do ciclo anual dos morcegos, é fundamental na definição
de estratégias para a sua conservação (Safi & Kerth 2004). No entanto, a abundância das
presas pode não reflectir a sua disponibilidade para um determinado predador (Siemers
& Güttinger 2006). Assim, para predadores que se alimentem de artrópodes no solo, as
características do coberto vegetal (e.g. densidade da vegetação) poderá ser limitante na
detectabilidade e/ou capturabilidade de uma presa; desta forma o uso do solo poderá ser
mais ou menos limitante no sucesso de captura dos artrópodes aí presentes. É
fundamental saber quais os habitats em que há maior abundância de presas, mas
também relevante averiguar a disponibilidade das mesmas.
Com um melhor entendimento da utilização dos recursos pelas espécies que
dependem destes agro-sistemas, nomeadamente os morcegos, poderão extrapolar-se
medidas de gestão da paisagem agrícola de forma a melhor proteger todos os grupos de
animais que ocorrem nestes habitats.

Introdução
5
1.5. Objectivos
Este trabalho, tem como objectivo responder às questões: Os morcegos
alimentam-se durante o Inverno, numa região temperada? Poderão a abundância e a
disponibilidade de presas durante o Inverno ser limitativas para a sua alimentação?
Para responder a estas questões, seleccionou-se o Morcego-rato-grande, Myotis myotis
(Borkhausen, 1797) como espécie de estudo. Assim para esta espécie, verificou-se se:
(i) os morcegos fizeram incursões ao exterior do abrigo durante o Inverno;
(ii) os excrementos produzidos durante o Inverno, continham peças de
artrópodes;
(iii) a abundância de presas durante o Inverno é idêntica à de outras estações
do ano;
(iv) a capacidade de M. myotis capturar as presas é diferente em densidades
distintas de vegetação.

Material e Métodos
6
2. Material e Métodos
2.1. Espécie estudada
O Morcego-rato-grande, Myotis myotis (Borkhausen, 1797) é uma espécie
colonial que se distribui por toda a zona Euroasiática Ocidental, desde a Península
Ibérica até à Ucrânia, Turquia, Israel, Líbano e Síria (Mitchell-Jones et al. 1999). Em
Portugal é relativamente frequente no Norte e no Centro, surgindo esporadicamente no
Algarve (Palmeirim et al. 1999). Esta espécie utiliza geralmente abrigos subterrâneos
para colónias de criação no país, ao contrário da tendência para utilizar edifícios como
abrigos de criação na Europa Central (Rodrigues et al. 2003). No que diz respeito à
hibernação, conhecem-se poucos abrigos subterrâneos com colónias desta espécie em
Portugal (Palmeirim & Rodrigues 1992).
A destruição dos abrigos e a degradação dos biótopos de alimentação são
consideradas as principais causas de declínio de M. myotis na Europa, sendo estes
também considerados como factores de ameaça para esta espécie em Portugal. Estes
factores, associados ao facto de ser uma espécie dependente de abrigos subterrâneos, faz
com que esta espécie seja classificada com o estatuto de Vulnerável segundo o Livro
Vermelho dos Vertebrados de Portugal (Cabral et al. 2006).
Dados referentes à dieta de M. myotis em Portugal, mostram que esta espécie se
alimenta quase exclusivamente de artrópodes do solo, preferencialmente indivíduos da
Família Carabidae, Gryllidae e da Classe Arachnida (Pereira et al. 2002). Esta espécie
caça em áreas abertas e de vegetação rasteira, onde pode capturar livremente artrópodes
do solo (Arlettaz 1999, Zahn et al. 2005). Quando caça, recorre à utilização de ultra-
sons, bem como a pistas sonoras para detectar as suas presas no meio da vegetação
(Russo et al. 2007).
2.2. Área de estudo
Este estudo incidiu sobre uma das principais colónias de M. myotis em Portugal,
cujo abrigo se localiza no concelho de Moura, distrito de Beja (Fig.1). Este abrigo
consiste numa mina desactivada de grande extensão, com uma grande diversidade
climática no seu interior (Palmeirim & Rodrigues 1992). Este local abriga uma grande
diversidade de espécies, podendo ser encontradas um total de 11 espécies de morcegos

Material e Métodos
7
cavernícolas ao longo de todo o ano (ICNB 2007). Alberga ainda uma colónia de
criação que, para além de acolher cerca de 2000 indivíduos de M. myotis, acolhe ainda
cerca de 5000 Morcego-de-peluche Miniopterus schreibersii e ainda algumas centenas
de Morcego-de-ferradura-mourisco Rhinolophus mehelyi. Esta diversidade de espécies e
o tamanho da colónia de criação são únicas em relação aos outros abrigos do país, o que
realça a sua importância a nível nacional. Este abrigo, bem como parte da área
amostrada, estão integrados na Rede Natura 2000 (vd.
http://europa.eu.int/comm./environment/nature/home.htm).
Geomorfologicamente, a área circundante ao abrigo, é dominada por planícies,
embora se encontrem zonas com relevo acentuado. Destaca-se um sistema de serras que
inclui Savos (356m), Adiça (476m), Ficalho (518m) e Preguiça (326m). O clima
tipicamente mediterrânico desta região caracteriza-se por verões quentes e secos, com
temperaturas por vezes excedendo os 40ºC e baixa precipitação (precipitação média:
520mm/ano; temperatura média anual: 17ºC). As linhas de água pertencem à bacia do
rio Guadiana. Por consequência do substrato geológico (essencialmente calcário) e do
clima, todos os cursos e água são de regime temporário e torrencial.
Figura 1 – Área de estudo.

Material e Métodos
8
Esta região tem sofrido uma forte influência humana, tendo sido a paisagem
progressivamente alterada ao longo dos séculos. Os principais usos do solo da região
reflectem esta intervenção humana, sendo predominantemente resultantes de actividades
agro-silvo-pastoris. Neste estudo foram considerados os seis tipos de uso do solo mais
representativos da zona: cevada (cultura da gramínea cerealífera Hordeum vulgare),
pastagem (área onde a vegetação natural é usado para alimentação de gado), forragem
(área semelhante a uma pastagem, mas enriquecida com gramíneas da Família Poaceae),
montado (área onde o coberto arbóreo é constituído predominantemente por azinho
(Quercus rotundifolia) onde, nalguns casos, o sub-coberto é aproveitado para
pastagem), olival (plantação de Olea europaea) e trigo (cultura da gramínea Triticum
spp.).
2.3. Recolha de dados
2.3.1. Actividade dos morcegos durante o Inverno
Os morcegos insectívoros utilizam um sistema de ecolocalização para sua
orientação e captura de presas. Este sistema consiste na emissão contínua de pulsos
ultra-sonoros que podem ser detectados recorrendo a um detector de ultra-sons. Este
tipo de aparelho converte ultra-sons em sons audíveis ao ouvido humano e permite o
registo das vocalizações dos morcegos para posterior análise. Este é na generalidade um
bom método de monitorização de morcegos (Ahlén & BaagØe 1999) que, pelo facto de
permitir registos à distância, não interfere com a sua actividade.
A actividade de M. myotis à saída do abrigo foi avaliada entre os meses de
Dezembro de 2007 e Abril de 2008, com o auxílio de um detector de ultra-sons
(Pettersson D1000X). As passagens de indivíduos (vocalizações detectadas) foram
contabilizadas em média 4 dias por mês, ao fim do dia. Em cada dia de amostragem, as
contagens duraram uma hora após a saída do primeiro indivíduo da espécie M. myotis,
de forma a evitar a contagem de indivíduos que regressavam ao abrigo. Cada vez que
era detectada a presença de um morcego procedia-se ao registo da vocalização. Este
método está sujeito a erros, já que (1) é dependente da rapidez do operador do detector;
(2) podem não ter sido gravados vocalizações de indivíduos que não tenham emitido
som durante a saída do abrigo; e (3) poderão ter sido incluídos nas contagens morcegos
que regressaram ao abrigo ou circulavam no local durante o período de saída. No
entanto, dado que a amostragem foi realizada sempre pelo mesmo observador,
Material e Métodos
9
considera-se que estas fontes de erro se mantiveram aproximadamente constantes
durante todo o período de amostragem, não influenciando assim as comparações.
As vocalizações detectadas durante a amostragem foram gravadas em suporte
digital (formato WAVE, taxa de amostragem de 96 kHz) sem qualquer tipo de
transformação (High Frequency). Posteriormente, as vocalizações gravadas foram
analisadas recorrendo a um programa de análise de som – BatSound Pro – Sound
Analysis v3.31 (Pettersson Elektronic AB. Uppsala). Este é um sistema para
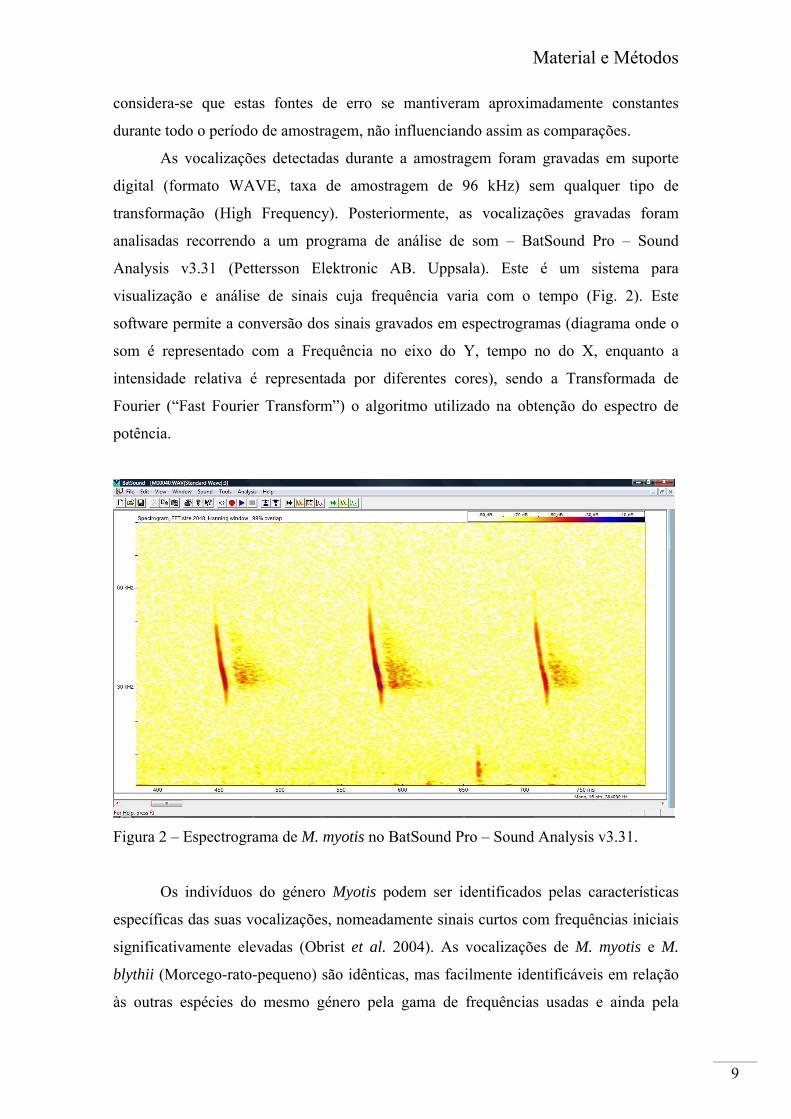
visualização e análise de sinais cuja frequência varia com o tempo (Fig. 2). Este
software permite a conversão dos sinais gravados em espectrogramas (diagrama onde o
som é representado com a Frequência no eixo do Y, tempo no do X, enquanto a
intensidade relativa é representada por diferentes cores), sendo a Transformada de
Fourier (“Fast Fourier Transform”) o algoritmo utilizado na obtenção do espectro de
potência.
Figura 2 – Espectrograma de M. myotis no BatSound Pro – Sound Analysis v3.31.
Os indivíduos do género Myotis podem ser identificados pelas características
específicas das suas vocalizações, nomeadamente sinais curtos com frequências iniciais
significativamente elevadas (Obrist et al. 2004). As vocalizações de M. myotis e M.
blythii (Morcego-rato-pequeno) são idênticas, mas facilmente identificáveis em relação
às outras espécies do mesmo género pela gama de frequências usadas e ainda pela

Material e Métodos
10
frequência de emissão a meio da duração do pulso (≈35kHZ) (Russo & Jones 2002). A
semelhança entre as vocalizações de M. myotis e M. blythii impede a sua distinção. No
entanto, M. blythii é uma espécie rara em Portugal e poderá surgir neste abrigo apenas
esporadicamente e em número muito baixo, pelo que a probabilidade de enviesamento
dos resultados será reduzida, ou até mesmo nula.
2.3.2. Recolha de excrementos de morcegos
De forma a caracterizar a dieta de M. myotis durante o período de Outono,
Inverno e Primavera, procedeu-se à recolha mensal de excrementos no interior do
abrigo, entre Outubro de 2007 e Abril de 2008.
Para recolha dos excrementos, foram colocadas placas de madeira, no interior do
abrigo, por baixo da colónia e/ou de indivíduos isolados. Não foi, assim, realizada
qualquer captura ou manuseio dos indivíduos, reduzindo deste modo a perturbação da
colónia. As visitas ao abrigo foram realizadas sob licença emitida pelo ICNB (Licença
nº 19 / 2008 / RECOL).
As amostragens tiveram duração de 10 dias por mês; no final de cada período
foram seleccionados aleatoriamente 30 dos excrementos encontrados sobre as placas de
madeira. Apesar de o abrigo ser utilizado por várias espécies de morcegos, foi possível
distinguir os excrementos de M. myotis dos das restantes espécies devido às diferenças
de tamanho, cor e textura. Após recolha, os excrementos foram secos e analisados em
laboratório. Cada excremento foi amolecido em álcool a 70% e triado com o auxílio de
uma lupa binocular (45x). As peças de artrópodes encontradas foram montadas numa
lâmina. Sempre que possível os itens encontrados nos excrementos foram identificados
até à Família, usando como base algumas publicações sobre a morfologia destes grupos
(McAney et al. 1991, Whitaker 1988) e ainda por comparação com exemplares de uma
colecção de referência elaborada a partir de artrópodes capturados na região.
Apesar de muitas das vezes a identificação apenas ser fiável até ao nível da
Ordem ou Família, esta é uma técnica considerada fiável para a classificação da dieta de
mamíferos insectívoros (Jordan 2005), tendo sido também utilizado em morcegos
(Arlettaz et al. 2000, Almenar et al. 2008, Bontadina et al. 2008).
Material e Métodos
11
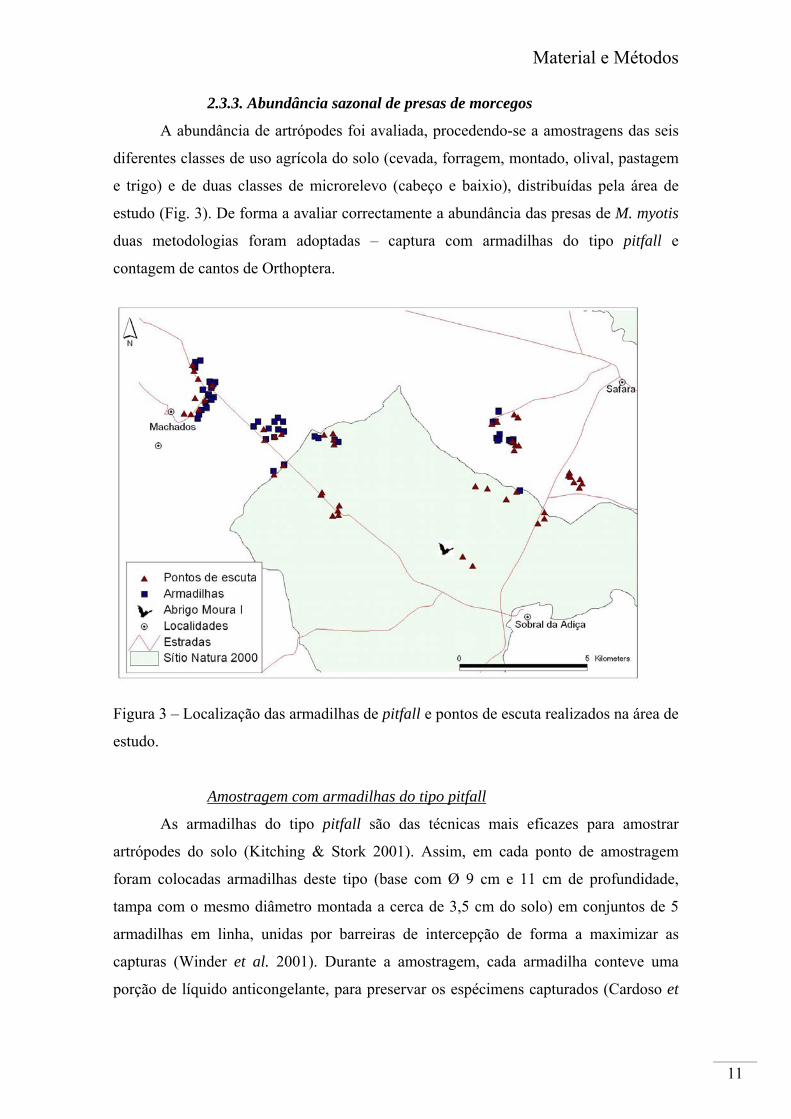
2.3.3. Abundância sazonal de presas de morcegos
A abundância de artrópodes foi avaliada, procedendo-se a amostragens das seis
diferentes classes de uso agrícola do solo (cevada, forragem, montado, olival, pastagem
e trigo) e de duas classes de microrelevo (cabeço e baixio), distribuídas pela área de
estudo (Fig. 3). De forma a avaliar correctamente a abundância das presas de M. myotis
duas metodologias foram adoptadas – captura com armadilhas do tipo pitfall e
contagem de cantos de Orthoptera.
Figura 3 – Localização das armadilhas de pitfall e pontos de escuta realizados na área de
estudo.
Amostragem com armadilhas do tipo pitfall
As armadilhas do tipo pitfall são das técnicas mais eficazes para amostrar
artrópodes do solo (Kitching & Stork 2001). Assim, em cada ponto de amostragem
foram colocadas armadilhas deste tipo (base com Ø 9 cm e 11 cm de profundidade,
tampa com o mesmo diâmetro montada a cerca de 3,5 cm do solo) em conjuntos de 5
armadilhas em linha, unidas por barreiras de intercepção de forma a maximizar as
capturas (Winder et al. 2001). Durante a amostragem, cada armadilha conteve uma
porção de líquido anticongelante, para preservar os espécimens capturados (Cardoso et

Material e Métodos
12
al. 2007). O período de amostragem foi de 10 dias por mês, entre Maio de 2007 e Abril
de 2008 (com excepção do mês de Agosto), mantendo-se as armadilhas fechadas nos
intervalos entre amostragens. No total foram usadas 240 armadilhas por mês entre
Setembro de 2007 e Abril de 2008. Entre Maio de 2007 e Julho de 2007 não foi
amostrado o uso do solo Montado, tendo sido utilizadas apenas 200 armadilhas pitfall
por mês.
As amostras recolhidas em cada armadilha foram mantidas em recipientes
separados e conservadas em líquido anticongelante até à sua triagem em laboratório.
Durante o processo de triagem, foram considerados todos os artrópodes com
comprimento total superior a 5 mm. A utilização deste valor resultou de uma medida
precaucionária, já que apesar de estudos anteriores (e.g. Arlettaz & Perrin 1995)
indicarem um comprimento total de aproximadamente 15 mm para as presas de M.
myotis durante a Primavera e Verão, não há dados referentes às presas capturadas por
esta espécie durante o Inverno.
Durante a triagem, cada exemplar da Classe Insecta foi identificado até à
Ordem, enquanto os restantes artrópodes, foram identificados apenas até à Classe,
recorrendo a chaves de identificação como Chinery (1993) e de Quartau & Luna de
Carvalho (1998). Os exemplares de Orthoptera capturados foram ainda identificados até
à Família. Os exemplares da Ordem Coleoptera e Família Gryllidae (Orthoptera) foram
agrupados em morfotipos. Para cada morfotipo foi recolhido um exemplar de forma a
criar uma colecção de referência. Os restantes artrópodes capturados nas armadilhas
foram conservados em álcool a 70%. Posteriormente, a colecção de referência foi
identificada com o máximo de especificidade possível (sempre que possível foi
identificado até à espécie, e os restantes até à Família) por um especialista (vd.
Agradecimentos). Um exemplar de cada morfotipo foi de seguida desmembrado, sendo
as suas principais peças montadas numa preparação definitiva, de forma a ser possível a
comparação com os itens encontrados nos excrementos de morcego (vd. Secção 2.3.2.).
Foram excluídas da análise todas as amostras de armadilhas que revelaram um
número anormal de capturas devido a problemas durante o período de amostragem –
captura de vertebrados, armadilhas secas ou inundadas, armadilhas pisadas pelo gado ou
destruídas por máquinas agrícolas. Em resultado desta triagem, das 2520 amostras
recolhidas nas pitfalls durante o ciclo anual, apenas se usou informação de 2215
amostras.

Material e Métodos
13
Contagem de cantos de Orthoptera
Em simultâneo com a armadilhagem, foi realizada a contagem de cantos de
Orthoptera, com o auxílio de um detector de ultra-sons (Pettersson D1000X; vd. Secção
2.3.1.) nas diferentes classes de uso agrícola do solo e microrelevo. A implementação
desta metodologia deve-se ao facto de as pitfalls terem uma eficiência reduzida nas
capturas de insectos da Família Orthoptera (Gardiner et al. 2005), que inclui algumas
das presas preferenciais de M. myotis (Pereira et al. 2002).
Assim, foram realizados pontos de escuta, onde durante 2 minutos foram
gravados todos os cantos de Orthoptera, de forma a, posteriormente, se poder
contabilizar os cantos de grilos (Gryllidae) e de ralos (Gryllotalpa). Em cada ponto de
escuta, dividiu-se o espaço por 6 secções (gravação durante 10 segundos em cada
secção, repetido por duas vezes), formando um ângulo de 180º, de forma a facilitar a
localização dos indivíduos na análise posterior.
Esta metodologia foi utilizada em média 5 noites por mês entre Setembro 2007 e
Abril 2008. Em cada mês foram realizadas gravações em 12 pontos de escuta. Excepção
foi feita nos meses de Dezembro a Fevereiro, em que, face ao flagrante decréscimo no
número de cantos, houve uma redução do número de pontos de escuta (10 em
Dezembro e 8 em Janeiro e Fevereiro).
As gravações foram analisadas posteriormente usando o programa de análise de
sons BatSound Pro (v3.31 Pettersson Elektronik AB, vd. Secção 2.3.1), e identificadas
por comparação com registos de cantos previamente identificados ou com os registos de
Ragge & Reynolds (1998).
2.3.4. Disponibilidade de presas em diferentes densidades de vegetação
Com o objectivo de determinar o modo como a densidade do coberto vegetal
pode afectar a disponibilidade de presas para M. myotis, foi amostrada a densidade da
vegetação nos principais usos do solo da área de estudo, e foram realizadas experiências
de curta duração com morcegos em cativeiro (licença de detenção emitida pelo ICNB a
25 de Março de 2008).

Material e Métodos
14
Amostragem da densidade da vegetação
A amostragem da densidade da vegetação em diferentes tipos de uso do solo e
micro-relevos (cabeço e baixio) foi realizada em Junho de 2008, usando o método de
intercepção de pontos (vd. Damgaard 2008 para mais detalhes). Neste método, por cada
amostragem, um arame é colocado perpendicularmente ao solo a distâncias regulares
(neste caso em intervalos de 10 cm até completar 1 m), sendo contabilizadas as vezes
que o arame intercepta, ou não, a vegetação herbácea. Foram realizados 15 replicados
em cada classe.
Experiências em cativeiro
As experiências em cativeiro consistiram na análise do comportamento de
indivíduos de M. myotis quando confrontados com a possibilidade de capturar uma
presa (Acheta domesticus) em tabuleiros com 3 densidades distintas do coberto,
nomeadamente Ralo [0 a 33% de coberto], Intermédio [34 a 67%] e Denso [68 a 100%].
Para tal, recorreu-se a 8 machos de M. myotis (mantidos em cativeiro, 2 de cada
vez durante cerca de uma semana) durante os meses de Maio e Junho de 2008. As
experiências decorreram em laboratório, onde os morcegos eram mantidos durante o dia
dentro de uma caixa-abrigo desenhada para o efeito (Lollar & Schmidt-French 2002) e
durante a noite eram libertados dentro de uma tenda (LxWxH 4x2x3m) onde o seu
comportamento de caça era permanentemente acompanhado com recurso a 4 câmaras
de infra-vermelhos (Cameras: 690IRBW - B/W IR Camera). As imagens simultâneas
das câmaras foram gravadas usando para tal uma placa de captura de vídeo (Geovision-
GV650). Este registo permitiu a posterior análise dos comportamentos.
No início de cada experiência, um dos morcegos foi colocado num pouso
localizado em frente a um dos tabuleiros (1,10m x 70m) com vegetação. Cada um dos
tabuleiros tinha disponível uma presa. A duração das experiências terminava com a
captura da presa ou, no caso de insucesso na captura, findos 30 min. Este processo foi
repetido usando sucessivamente os três tabuleiros com os diferentes tipos de coberto. Os
morcegos tiveram água sempre disponível quer na caixa abrigo, quer na tenda durante
as experiências.
Da posterior análise das filmagens, foram retirados vários parâmetros, mas
tivemos em conta apenas: (1) tempo que demorou desde que pousou no tabuleiro até
que capturou a presa; (2) captura vs. não captura a presa.

Material e Métodos
15
2.4. Análise estatística
Salvo onde especificado, os resultados serão apresentados em termos de médias
e proporções relativas de ocorrência. Estes valores e os seus respectivos intervalos de
confiança a 95% foram calculados usando o programa Microsoft Excel 2007 (Microsoft
Corporation). Este programa foi também utilizado na aplicação de um teste-T
emparelhado, para comparação do comportamento de caça dos morcegos face a
diferentes densidades da vegetação (Sokal & Rohlf 1997).
Na análise do número médio de artrópodes capturados em cada uso do solo em
estudo, e a respectiva variação sazonal, recorreu-se a uma análise de variância
(ANOVA) e ao teste de Fisher a posteriori, para inferir a existência de diferenças no
número de artrópodes capturados entre os meses amostrados (Sokal & Rohlf 1997). O
teste foi realizado com recurso ao programa STATISTICA 7.0 - Data Analysis Software
System (StatSoft, Inc. 2004).
Em todas as análises o nível de significância considerado foi de α = 0,05.

Resultados
16
3. Resultados
3.1. Actividade dos morcegos durante o Inverno
Entre os meses de Dezembro de 2007 e Abril de 2008 foram realizados 21 dias
de observação, em que foram gravadas um total de 5974 passagens de M. myotis.
Dezembro foi o mês em que foi registado um menor número de passagens de M.
myotis (média de 38 passagens registadas) à saída do abrigo. Abril foi o mês em que foi
registado um maior número de passagens (média de 454 passagens registadas). No
decorrer dos meses amostrados houve um aumento do número de passagens de M.
myotis registadas, com excepção do mês de Março, provavelmente devido ao mau
tempo que se fez sentir durante os dias de amostragem. (Fig. 4)
0
100
200
300
400
500
600
Dezembro Janeiro Fevereiro Março Abril
Nº de
passagens
Mês
Variação mensal da actividade
Figura 4 – Número médio de passagens (± intervalos confiança a 95%) de M. myotis
registadas à saída do abrigo, por mês.

Resultados
17
3.2. Composição da dieta
Foram identificadas sete taxa de presas nos 210 dejectos analisados, totalizando
390 artrópodes. Do total de artrópodes identificados, 87,9% das ocorrências são
pertencentes aos taxa: Gryllidae (Orthoptera) – 13%, Chilopoda – 8%, Arachnida – 6%,
Carabidae (Coleoptera) – 18% e outros Coleoptera – 43%. Devido à inexistência dos
principais caracteres diagnosticantes das Famílias pertencentes a Arachnida (fieiras do
abdómen e/ou conjuntos de olhos no cefalotórax) e Chilopoda (segmentos das antenas,
posição dos ocelos, número de segmentos do tórax e/ou morfologia do tergitos do tórax)
não foi possível identificar as famílias pertencentes a estes grupos (Roberts 1995,
Edgecombe & Giribet 2007). A classe Hymenoptera foi representada por 0,3% das
ocorrências, enquanto outros artrópodes não identificados ocorreram em 12%.
Ocasionalmente foram também encontrados pedaços de vegetação e parasitas no guano
analisado.
Apesar das presas potenciais serem as mesmas ao longo da amostragem, a
proporção em que ocorrem na dieta varia entre as distintas épocas do ano (Fig. 5).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Outubro Novembro Dezembro Janeiro Fevereiro Março Abril
% de ocorrência
Mês
Dieta de M. myotis
Hymenoptera
Outros
Arachnida
Chilopoda
Carabidae
outros Coleoptera
Gryllidae
Figura 5 – Percentagem (%) de ocorrência dos artrópodes identificados na dieta de M.
myotis entre Outubro de 2007 e Abril de 2008.

Resultados
18
A Ordem Coleoptera, de um modo geral, predominou na dieta de M. myotis nos
meses amostrados, tendo a sua percentagem variado entre 40% e 80%. As restantes
Ordens apresentaram no geral, percentagens menos relevantes. No entanto, é de realçar
a importância de Chilopoda no mês de Março (20%) e de Gryllidae nos meses de Março
e Abril (20% em ambos os meses).
3.3. Abundância sazonal de presas de morcegos
3.3.1. Amostragem com armadilhas do tipo pitfall
O total de capturas nas pitfalls foi de 89081 artrópodes (distribuídos pelo total de
2215 amostras recolhidas nas armadilhas pitfall entre Maio de 2007 e Abril de 2008)
com tamanho corporal maior que 5mm.
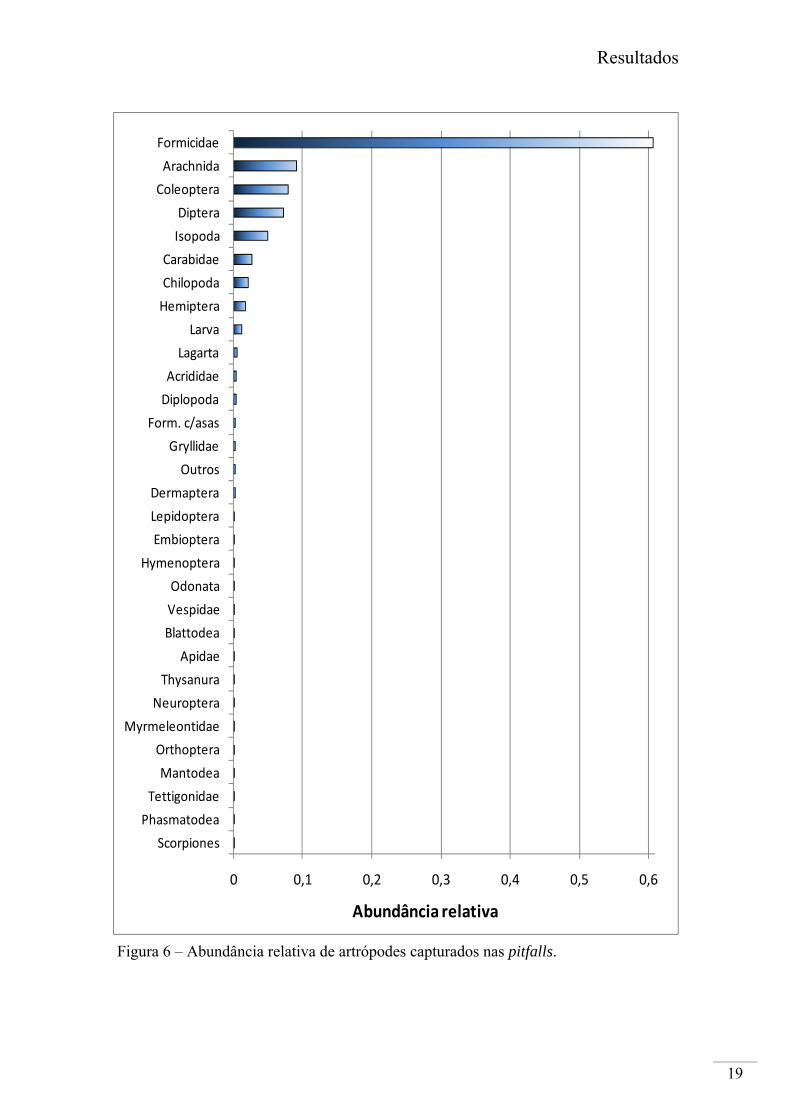
Os taxa mais abundantes, por ordem decrescente foram Formicidae, Arachnida,
Coleoptera (excluindo Carabidae) e Diptera (Fig. 6). Todos eles foram capturados em
maior abundância no mês de Maio.
A Família Formicidae foi o grupo mais abundante em todos os biótopos
amostrados, em especial na Pastagem e no microrelevo de baixio. A Classe Arachnida
ocorreu em maior abundância na Forragem, com maior incidência no baixio. Coleoptera
teve maior taxa de captura no Trigo, em particular no microrelevo de baixio. No caso da
Ordem Diptera ocorreu em abundância muito semelhante nos biótopos Forragem,
Pastagem e Trigo, e mais uma vez em maior número no baixio.
No total dos 6 usos do solo amostrados foram identificados 31 grupos,
pertencentes a 20 Ordens, representados por indivíduos que ultrapassavam os 5 mm de
comprimento corporal (ANEXO I). Os Coleoptera foram identificados com maior
especificidade possível, tendo sido identificados 65 morfotipos diferentes, enquanto os
Gryllidae identificados correspondem a 9 espécies diferentes (ANEXO II).
Resultados
19
0 0,1 0,2 0,3 0,4 0,5 0,6
Scorpiones
Phasmatodea
Tettigonidae
Mantodea
Orthoptera
Myrmeleontidae
Neuroptera
Thysanura
Apidae
Blattodea
Vespidae
Odonata
Hymenoptera
Embioptera
Lepidoptera
Dermaptera
Outros
Gryllidae
Form. c/asas
Diplopoda
Acrididae
Lagarta
Larva
Hemiptera
Chilopoda
Carabidae
Isopoda
Diptera
Coleoptera
Arachnida
Formicidae
Abundância relativa
Figura 6 – Abundância relativa de artrópodes capturados nas pitfalls.

Resultados
20
0 0,2 0,4 0,6
Outros
Formicidae
Diptera
Isopoda
Hemiptera
Acrididae
Diplopoda
Arachnida
Chilopoda
Gryllidae
Carabidae
Coleoptera
Frequência de ocorrência
Dieta
Pitfalls
Comparação entre consumo e abundância de presas
Comparando a frequência de ocorrência de artrópodes nas armadilhas com a sua
frequência de ocorrência nos excrementos, verificou-se que existem diferenças na
abundância dos diferentes grupos de artrópodes. Formicidae, Diptera, Arachnida e
Coleoptera (à excepção da Família Carabidae) foram os grupos mais abundantes nas
pitfalls. Os grupos mais abundantes nos excrementos foram: outros Coleoptera,
Carabidae, Gryllidae, Chilopoda e Arachnida (Fig. 7). Estes últimos taxa constituem as
presas preferenciais de M. myotis.
Figura 7 – Frequência de ocorrência de artrópodes (± intervalos confiança a 95%)
capturados nas pitfalls e nos excrementos.
Resultados
21
Abundância de presas potenciais nas pitfalls
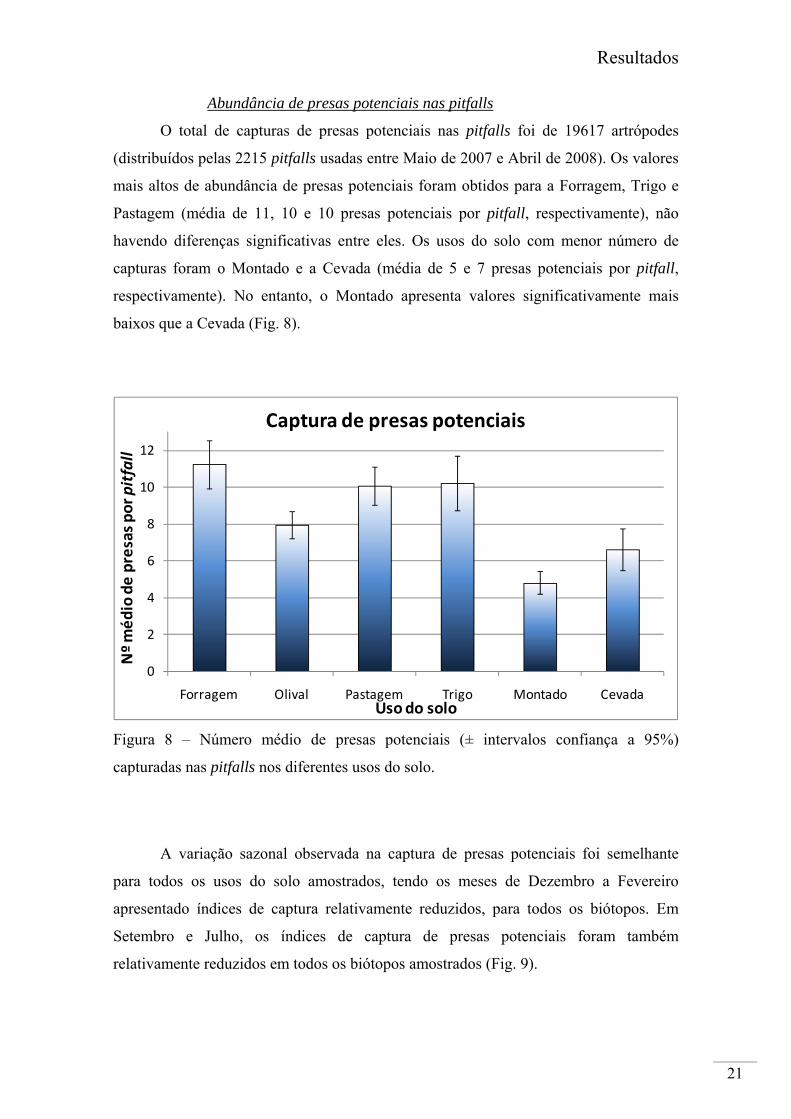
O total de capturas de presas potenciais nas pitfalls foi de 19617 artrópodes
(distribuídos pelas 2215 pitfalls usadas entre Maio de 2007 e Abril de 2008). Os valores
mais altos de abundância de presas potenciais foram obtidos para a Forragem, Trigo e
Pastagem (média de 11, 10 e 10 presas potenciais por pitfall, respectivamente), não
havendo diferenças significativas entre eles. Os usos do solo com menor número de
capturas foram o Montado e a Cevada (média de 5 e 7 presas potenciais por pitfall,
respectivamente). No entanto, o Montado apresenta valores significativamente mais
baixos que a Cevada (Fig. 8).
0
2
4
6
8
10
12
Forragem Olival Pastagem Trigo Montado Cevada
Nº méd
io de presas por pitfall
Uso do solo
Captura de presas potenciais
Figura 8 – Número médio de presas potenciais (± intervalos confiança a 95%)
capturadas nas pitfalls nos diferentes usos do solo.
A variação sazonal observada na captura de presas potenciais foi semelhante
para todos os usos do solo amostrados, tendo os meses de Dezembro a Fevereiro
apresentado índices de captura relativamente reduzidos, para todos os biótopos. Em
Setembro e Julho, os índices de captura de presas potenciais foram também
relativamente reduzidos em todos os biótopos amostrados (Fig. 9).
Resultados
22
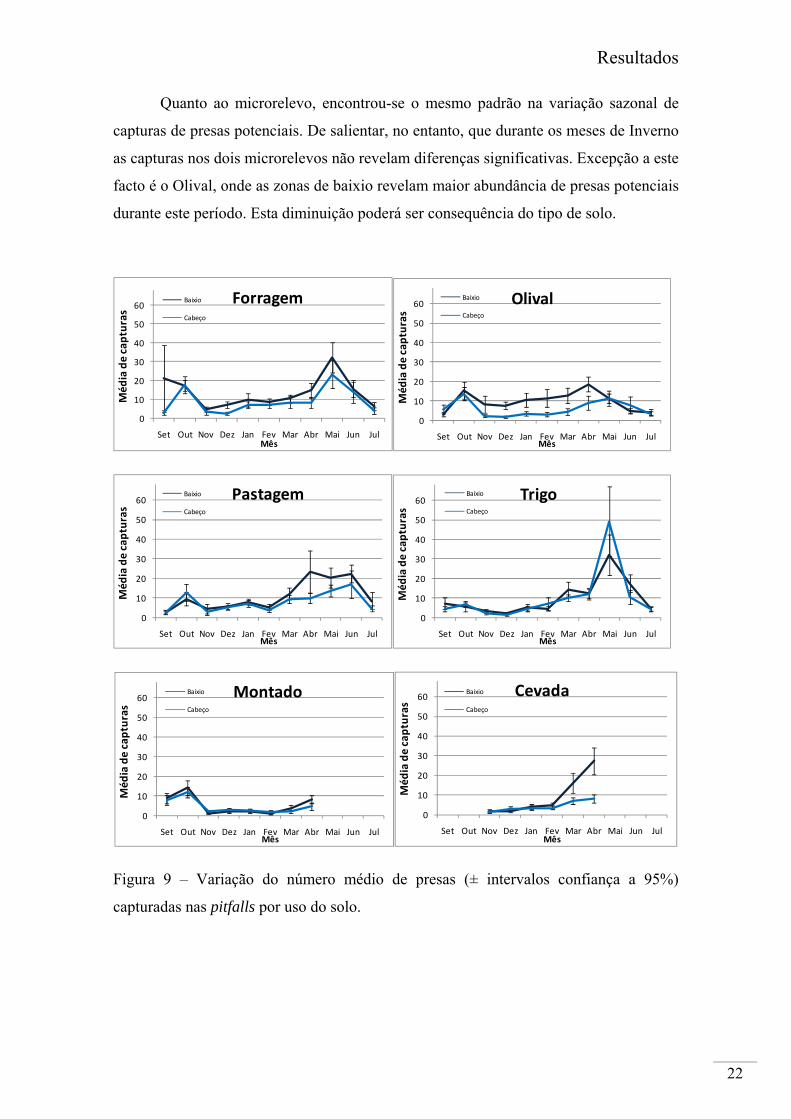
Quanto ao microrelevo, encontrou-se o mesmo padrão na variação sazonal de
capturas de presas potenciais. De salientar, no entanto, que durante os meses de Inverno
as capturas nos dois microrelevos não revelam diferenças significativas. Excepção a este
facto é o Olival, onde as zonas de baixio revelam maior abundância de presas potenciais
durante este período. Esta diminuição poderá ser consequência do tipo de solo.
0
10
20
30
40
50
60
Set Out Nov Dez Jan Fev Mar Abr Mai Jun Jul
Méd
ia de capturas
Mês
ForragemBaixio
Cabeço
0
10
20
30
40
50
60
Set Out Nov Dez Jan Fev Mar Abr Mai Jun Jul
Méd
ia de capturas
Mês
OlivalBaixio
Cabeço
0
10
20
30
40
50
60
Set Out Nov Dez Jan Fev Mar Abr Mai Jun Jul
Méd
ia de capturas
Mês
PastagemBaixio
Cabeço
0
10
20
30
40
50
60
Set Out Nov Dez Jan Fev Mar Abr Mai Jun Jul
Méd
ia de capturas
Mês
TrigoBaixio
Cabeço
0
10
20
30
40
50
60
Set Out Nov Dez Jan Fev Mar Abr Mai Jun Jul
Méd
ia de capturas
Mês
MontadoBaixio
Cabeço
0
10
20
30
40
50
60
Set Out Nov Dez Jan Fev Mar Abr Mai Jun Jul
Méd
ia de capturas
Mês
CevadaBaixio
Cabeço
Figura 9 – Variação do número médio de presas (± intervalos confiança a 95%)
capturadas nas pitfalls por uso do solo.
Resultados
23
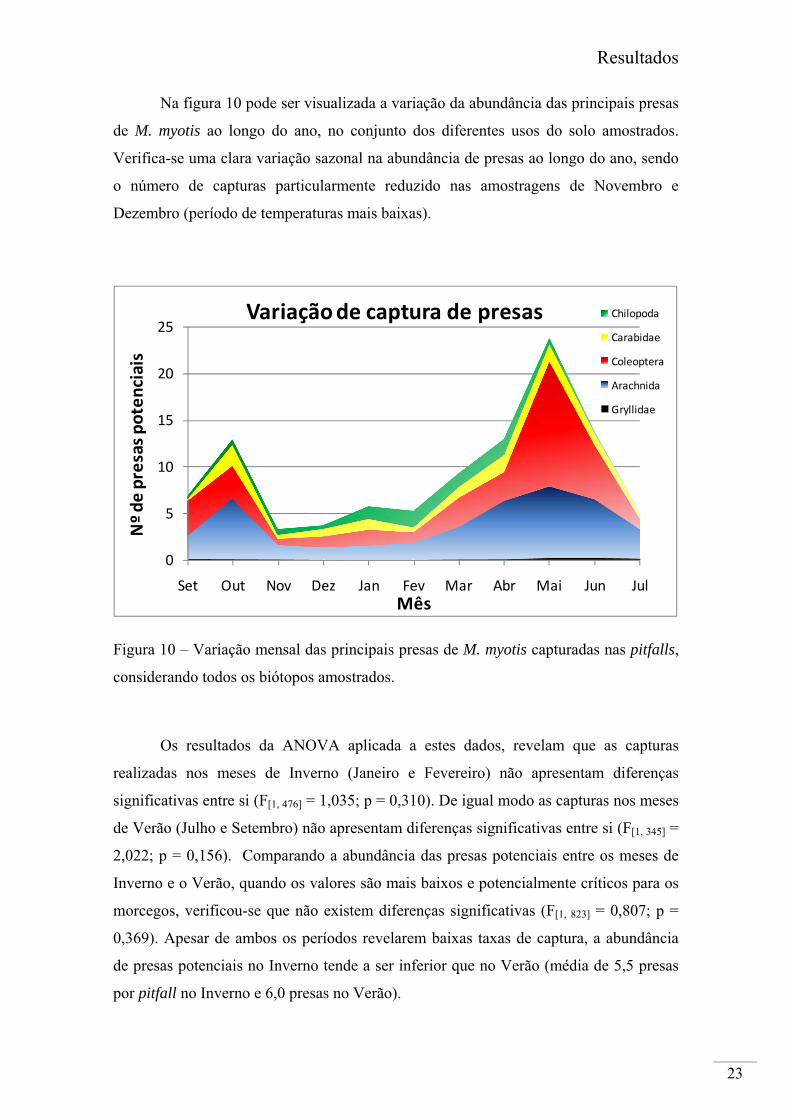
Na figura 10 pode ser visualizada a variação da abundância das principais presas
de M. myotis ao longo do ano, no conjunto dos diferentes usos do solo amostrados.
Verifica-se uma clara variação sazonal na abundância de presas ao longo do ano, sendo
o número de capturas particularmente reduzido nas amostragens de Novembro e
Dezembro (período de temperaturas mais baixas).
0
5
10
15
20
25
Set Out Nov Dez Jan Fev Mar Abr Mai Jun Jul
Nº de
presas po
tenciais
Mês
Variação de captura de presas Chilopoda
Carabidae
Coleoptera
Arachnida
Gryllidae
Figura 10 – Variação mensal das principais presas de M. myotis capturadas nas pitfalls,
considerando todos os biótopos amostrados.
Os resultados da ANOVA aplicada a estes dados, revelam que as capturas
realizadas nos meses de Inverno (Janeiro e Fevereiro) não apresentam diferenças
significativas entre si (F[1, 476] = 1,035; p = 0,310). De igual modo as capturas nos meses
de Verão (Julho e Setembro) não apresentam diferenças significativas entre si (F[1, 345] =
2,022; p = 0,156). Comparando a abundância das presas potenciais entre os meses de
Inverno e o Verão, quando os valores são mais baixos e potencialmente críticos para os
morcegos, verificou-se que não existem diferenças significativas (F[1, 823] = 0,807; p =
0,369). Apesar de ambos os períodos revelarem baixas taxas de captura, a abundância
de presas potenciais no Inverno tende a ser inferior que no Verão (média de 5,5 presas
por pitfall no Inverno e 6,0 presas no Verão).

Resultados
24
3.3.2. Contagem de cantos de Orthoptera
No caso de Gryllidae, a amostragem por pitfalls foi algo deficiente, facto que
dificulta a avaliação da variação da sua abundância. Numa tentativa de compensar esta
deficiência de amostragem, foram realizadas gravações dos cantos deste grupo.
No total, foram gravados 1333 cantos de Gryllidae e 140 cantos de
Gryllotalpidae. Os diferentes usos do solo e microrelevos foram amostrados, num total
de 494 gravações (Fig. 11). Nesta análise nota-se um padrão semelhante às capturas de
presas potenciais nas pitfalls, tendo sido registado um menor número de cantos nos
meses de Novembro a Fevereiro.
0
1
2
3
4
5
6
7
8
Setembro Outubro Novembro Dezembro Janeiro Fevereiro Março Abril
Nº de
cantos po
r no
ite
Mês
Cantos de Orthoptera
Gryllidae
Gryllotalpidae
Figura 11 – Número médio de cantos de Gryllidae e Gryllotalpidae (± intervalos
confiança a 95%) gravados por noite.
Resultados
25
3.4. Disponibilidade de presas em diferentes densidades de vegetação
3.4.1. Amostragem da densidade da vegetação
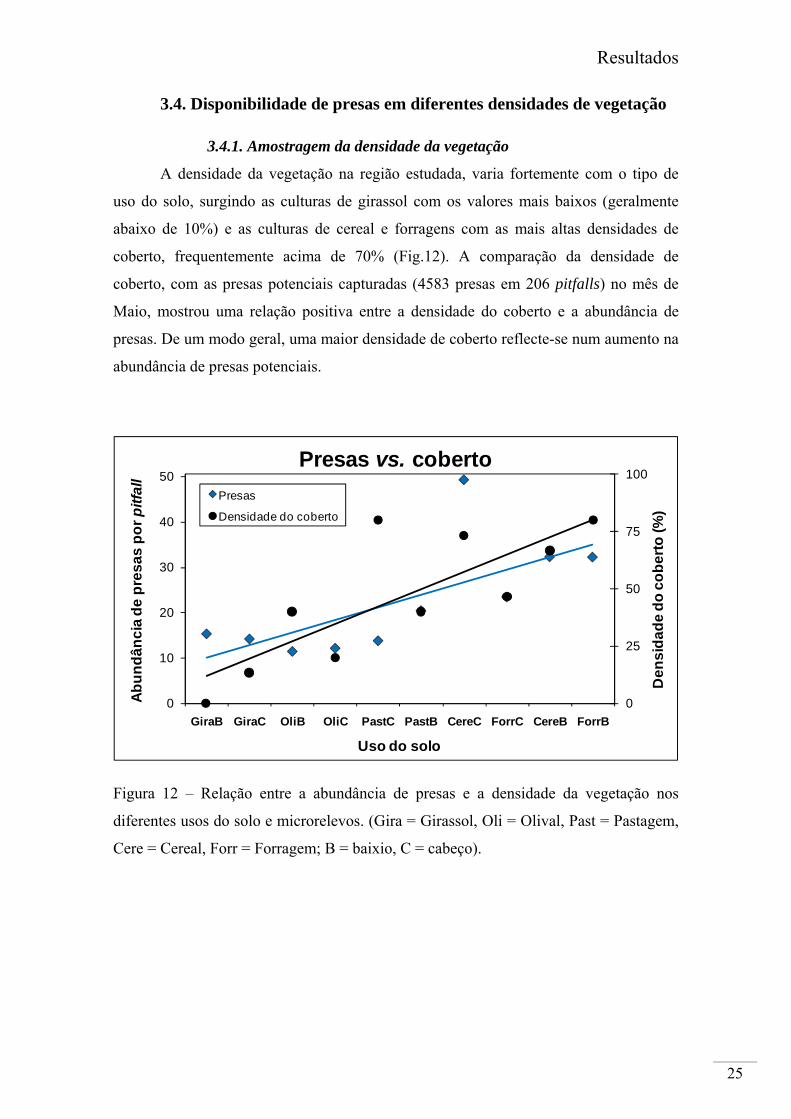
A densidade da vegetação na região estudada, varia fortemente com o tipo de
uso do solo, surgindo as culturas de girassol com os valores mais baixos (geralmente
abaixo de 10%) e as culturas de cereal e forragens com as mais altas densidades de
coberto, frequentemente acima de 70% (Fig.12). A comparação da densidade de
coberto, com as presas potenciais capturadas (4583 presas em 206 pitfalls) no mês de
Maio, mostrou uma relação positiva entre a densidade do coberto e a abundância de
presas. De um modo geral, uma maior densidade de coberto reflecte-se num aumento na
abundância de presas potenciais.
0
25
50
75
100
0
10
20
30
40
50
GiraB GiraC OliB OliC PastC PastB CereC ForrC CereB ForrB
Den
sida
de d
o co
berto
(%)
Abun
dânc
ia d
e pr
esas
por
pitf
all
Uso do solo
Presas vs. coberto Presas
Densidade do coberto
Figura 12 – Relação entre a abundância de presas e a densidade da vegetação nos
diferentes usos do solo e microrelevos. (Gira = Girassol, Oli = Olival, Past = Pastagem,
Cere = Cereal, Forr = Forragem; B = baixio, C = cabeço).

Resultados
26
3.4.2. Experiências em cativeiro
A influência da densidade da vegetação no sucesso de captura de presas por M.
myotis foi testada através dos tempos médios e taxas de captura de presas.
Os tempos médios de captura foram calculados a partir de dados de 204 réplicas,
em que os 8 indivíduos amostrados, capturaram com sucesso a presa em diferentes
densidades da vegetação. A captura de presas na vegetação rala demorou em média
1min 2seg. Este intervalo de tempo aumentou, no entanto, significativamente quando o
morcego teve de capturar a presa em condições de vegetação intermédia (tempo médio
de 4min 51seg.; T[Ralo vs. Intermédio] = 2,6853; g.l. = 7; p = 0,0313) e em condições de
vegetação densa (tempo médio de 11min 44seg.; T[Intermédio vs. Denso] = 3,7232; g.l. = 7 p =
0,0074 e T[Ralo vs. Denso] = 6,0358; g.l. = 7; p = 0,0005 (Fig. 13)).
00:00:00
00:02:53
00:05:46
00:08:38
00:11:31
00:14:24
00:17:17
Ralo Intermédio Denso
Tempo
de captura
Densidade do coberto
Captura de presas
Figura 13 – Tempo médio utilizado pelos morcegos na captura das presas (± intervalos
confiança a 95%) em diferentes densidades do coberto (hh:mm:ss).
Resultados
27
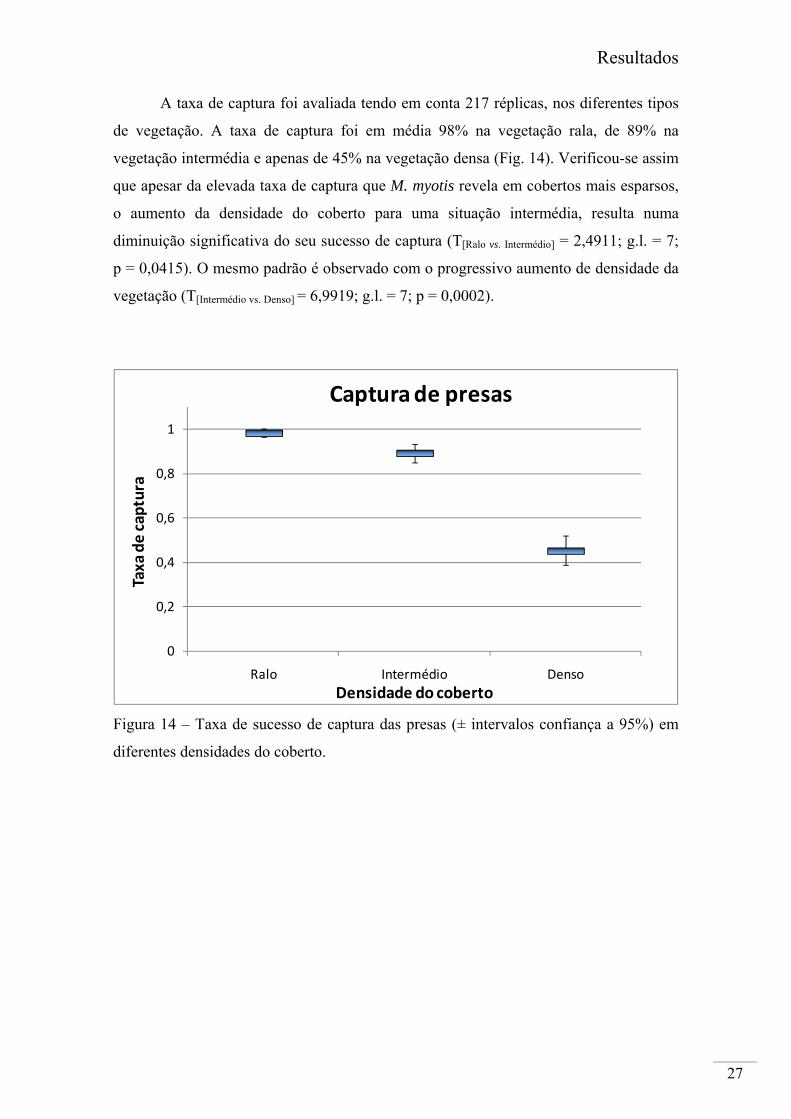
A taxa de captura foi avaliada tendo em conta 217 réplicas, nos diferentes tipos
de vegetação. A taxa de captura foi em média 98% na vegetação rala, de 89% na
vegetação intermédia e apenas de 45% na vegetação densa (Fig. 14). Verificou-se assim
que apesar da elevada taxa de captura que M. myotis revela em cobertos mais esparsos,
o aumento da densidade do coberto para uma situação intermédia, resulta numa
diminuição significativa do seu sucesso de captura (T[Ralo vs. Intermédio] = 2,4911; g.l. = 7;
p = 0,0415). O mesmo padrão é observado com o progressivo aumento de densidade da
vegetação (T[Intermédio vs. Denso] = 6,9919; g.l. = 7; p = 0,0002).
0
0,2
0,4
0,6
0,8
1
Ralo Intermédio Denso
Taxa de captura
Densidade do coberto
Captura de presas
Figura 14 – Taxa de sucesso de captura das presas (± intervalos confiança a 95%) em
diferentes densidades do coberto.

Discussão
28
4. Discussão
4.1. Actividade dos morcegos durante o Inverno
Apesar do escasso conhecimento acerca dos abrigos de hibernação de M. myotis
no nosso país (Palmeirim & Rodrigues 1992), alguns indivíduos permaneceram no
abrigo em estudo durante o Inverno (individualmente ou em pequenos grupos),
permitindo o registo da sua actividade durante este período.
Com base nos registos obtidos, foi possível confirmar que pelo menos uma parte
dos indivíduos da espécie M. myotis está activa e sai regularmente do abrigo durante as
noites dos meses de Inverno. No entanto, o facto de se encontrarem em locais de difícil
acesso no interior do abrigo, dificulta a contabilização do número de M. myotis que
utilizaram o abrigo durante estes meses, impedindo a determinação da proporção de
indivíduos que está activa e sai do abrigo.
A baixa taxa de passagens registadas durante os meses de Dezembro e Janeiro,
poderá dever-se apenas a uma eventual redução no número de indivíduos presentes no
abrigo. O número de passagens contabilizado revela um progressivo aumento da
actividade desde Dezembro até Abril. Este padrão pode, no entanto, resultar de factores
como (1) o aumento do número de indivíduos dentro do abrigo com o aproximar do
período de criação (Março/Abril), ou (2) o aumento da abundância de alimento,
resultado da melhoria das condições climáticas. Os dados obtidos não permitem
esclarecer como estes factores influenciam os resultados, mas é de salientar que durante
o mês de Março houve uma redução da actividade contabilizada, provavelmente
associada a condições meteorológicas adversas nos dias de amostragem.
A análise detalhada da variação do número de indivíduos dentro do abrigo e o
esclarecimento de como os factores meteorológicos afectam e/ou limitam a actividade
dos morcegos, será fundamental para melhor responder a este tópico, devendo por isso
ser abordados em trabalhos futuros.
4.2. Composição da dieta
Em todos os excrementos recolhidos foram encontradas peças de artrópodes. O
tipo e condição das peças encontradas sugerem que, aquando da recolha dos
excrementos, os indivíduos se tinham alimentado recentemente. Ou seja, as peças

Discussão
29
encontradas eram numerosas e estavam rígidas, contrariando a ideia de que possam ser
restos de alimentação anterior ao início da hibernação.
Confirmada a ocorrência de actividade de caça durante o Inverno, verificou-se
também que a composição da dieta durante este período não revela grandes diferenças
em relação a outros períodos do ano. Ao longo dos meses amostrados, a maioria da
dieta foi sempre constituída pelos mesmos grupos, embora com pequenas variações.
Nos meses de Inverno foi maioritariamente constituída por Carabidae e outros
Coleoptera. A partir de Fevereiro observou-se um aumento no consumo de Gryllidae,
que deverá estar associado a um aumento das temperaturas e da abundância de grilos.
A composição da dieta de M. myotis durante o Inverno, não difere muito da
descrita por Pereira et al. (2002) e Zahn et al. (2007), para a Primavera e Verão. No
entanto, dado que a área de estudo é a mesma, são de realçar três diferenças em relação
aos resultados de Pereira et al. (2002): (a) uma maior proporção de Chilopoda, (b) uma
menor proporção de Aracnidae, e (c) a inexistência de Gryllotalpidae na dieta de M.
myotis durante o Inverno.
A elevada proporção de Chilopoda observada na dieta durante o Inverno parece
reflectir uma maior abundância deste artrópode durante esta estação (vd. Fig. 10). No
entanto, os resultados apresentados por Zahn et al. (2007) revelam também uma elevada
proporção de Chilopoda na dieta noutras alturas do ano. Sendo uma presa com uma
biomassa considerável, M. myotis poderá explorar oportunisticamente este grupo,
aproveitando os seus picos de abundância. Este tipo de exploração dos recursos,
justifica também as diferenças observadas no consumo de Arachnida. Novamente, a
reduzida proporção de aranhas na dieta está associada a uma baixa abundância no meio,
padrão aliás também observado por Zahn et al. (2007).
No que se refere ao uso de Gryllotalpidae como presa, ambos os estudos
anteriores confirmam que os ralos são de facto capturados por M. myotis nesta região,
tendo sido também capturados nas armadilhas pitfall (Pereira et al. 2002, Zahn et al.
2007). No presente estudo, o grupo Gryllotalpidae não foi detectado nem nos
excrementos, nem nas armadilhas pitfall. A sua presença foi apenas confirmada através
das contagens dos cantos de Orthoptera, que revelaram também uma abundância muito
reduzida deste grupo (vd. Fig. 11). A inexistência de contagens deste tipo em anos
anteriores impossibilita a comparação, mas não será de excluir a hipótese de que a
abundância deste grupo (juntamente com os grilos) esteja em declínio, como é aliás
referido por alguns agricultores da região.

Discussão
30
4.3. Abundância sazonal de presas de morcegos
Embora os taxa mais capturados nas armadilhas do tipo pitfall tenham sido
Formicidae, Arachnida, Coleoptera e Diptera, nem todos foram detectados nos
excrementos. Assim, apesar da abundância de Formicidae e Diptera ser bastante elevada
(em relação a Gryllidae e Chilopoda), M. myotis não parece alimentar-se destes grupos.
Apesar de existirem espécies de Formicidae com tamanho corporal maior que 15mm,
estes não apresentam uma biomassa corporal elevada, o que pode torná-las menos
conspícuas e desejáveis. Podem, no entanto, ser simplesmente rejeitadas, devido ao
facto de possuírem ácido fórmico que lhes poderá conferir um mau sabor, pouco
atractivo para o predador (Glendinning 2007).
Coleoptera, Gryllidae, Arachnida e Chilopoda apresentam frequentemente
elevada biomassa corporal. M. myotis provavelmente é um predador selectivo em
relação ao tamanho/biomassa da presa, não fazendo distinção por um taxa específico, tal
como foi sugerido para outros morcegos insectívoros (Vaughan 1997). Com efeito,
sendo M. myotis uma das maiores espécies de morcegos da fauna de Portugal, este
poderá facilmente caçar e manusear presas de elevado porte sem grande dispêndio
energético, como já referido por Arlettaz (1996). Em contrapartida, caçar várias presas
de tamanho menor, implicaria um gasto energético adicional para esta espécie. Para tal,
seriam necessários vários processos de detecção, captura e manipulação da presa. Deste
modo, não seria compensatório a captura de várias presas pequenas em relação a presas
maiores e mais conspícuas (Blanckenhorn 2000). Outros estudos indicam ainda que
presas de tamanho menor produzem menos ruído quando se movimentam na vegetação,
o que poderá não ser detectado ao ouvido do predador (Siemers & Güttinger 2006).
Outro factor de “escolha” das presas a capturar, é a actividade das mesmas,
factor que pode influenciar a sua detectabilidade (Schaub et al. 2008). De facto os
coleópteros da Família Carabidae, algumas Famílias de Arachnida e Chilopoda, são
activos predadores nocturnos (Foelix 1996, Lang et al. 1999, Sabo & Power 2002,
Edgecombe & Giribet 2007), tornando-se mais fáceis de detectar devido ao seu
movimento. Também os Gryllidae estão maioritariamente activos durante a noite,
principalmente as fêmeas, como resposta aos cantos dos machos (Ragge & Renolds
1998). Assim, o período de actividade levará também a que alguns taxa aparentemente
abundantes, não sejam encontrados nos excrementos. Pelo facto de terem hábitos

Discussão
31
essencialmente diurnos, mantêm-se imóveis durante a noite, resultando numa maior
dificuldade de detecção por parte de M. myotis.
Sabendo então quais as presas preferenciais de M. myotis, foi possível visualizar
o panorama da abundância dessas presas ao longo do ano e nos diferentes usos do solo
amostrados.
No que se refere ao uso do solo, as forragens, os trigos e pastagens revelaram as
abundâncias mais elevadas, facto que lhes poderá conferir alguma importância como
potenciais áreas de alimentação para M. myotis durante o Inverno. De um modo geral,
as zonas de baixo relevo (“baixio”) revelaram uma maior abundância de presas
potenciais. Este facto poderá dever-se a características do solo, como a qualidade (maior
disponibilidade de matéria orgânica) e humidade (associada à escorrência das águas).
Ao longo dos meses amostrados (tanto nas capturas com pitfalls como nas
gravações de cantos de Orthoptera), nota-se um padrão semelhante em todos os
biótopos, com a captura de muito menos presas entre Novembro e Fevereiro do que nos
restantes meses. Assim, apesar de haver presas potenciais activas nos meses de Inverno,
o seu número é relativamente reduzido. No entanto, os resultados da ANOVA para a
comparação entre a abundância de presas no Inverno com a verificada no Verão,
revelou que não existem diferenças significativas entre as duas estações. É assumido
que o Verão é provavelmente uma época crítica para os M. myotis Mediterrânicos em
resultado desta baixa abundância de presas, facto que se reflecte na sua condição
corporal (Zahn et al. 2007). Por outro lado, é no Inverno (nomeadamente em Fevereiro)
que a condição de M. myotis se apresenta mais baixa ao longo do ano, facto
provavelmente associado aos elevados dispêndios energéticos na manutenção da
temperatura corporal e à escassez de alimento neste período (Zahn et al. 2007). Deste
modo, o Inverno representará provavelmente também uma época crítica para a
sobrevivência de M. myotis.
Por estes motivos, a alimentação no Inverno poderá não ser energeticamente
vantajosa quando comparada com a hibernação. Este facto leva-nos a pensar que a
reduzida taxa de hibernação pode estar relacionada com as condições no interior dos
abrigos (temperatura, pressão barométrica, etc.). No entanto, as razões pelas quais M.
myotis se encontra activo no Inverno nas regiões Mediterrâncias não são ainda
totalmente compreendidas, situação que deverá ser alvo de estudos futuros. De qualquer
modo, e tendo em conta que M. myotis está activo e se alimenta, torna-se importante

Discussão
32
existir uma gestão dos habitats adjacentes às colónias-chave, de forma a garantir suas
necessidades alimentares de Inverno.
4.4. Disponibilidade de presas em diferentes densidades de vegetação
A densidade da vegetação revelou-se como um factor fulcral no sucesso de
captura de M. myotis. Esta espécie, que tipicamente captura insectos do solo, revelou
uma maior facilidade de caça em terrenos com densidades de vegetação mais baixas.
Apesar de ser capaz de capturar presas noutras densidades da vegetação, poderá ver o
seu sucesso de captura bastante reduzido em usos do solo onde o coberto seja muito
denso.
Extrapolando a situação das diferentes densidades da vegetação testadas em
laboratório, com a realidade existente no terreno, verificou-se que os usos do solo com
baixa densidade de coberto herbáceo e consequentemente bons locais de caça para M.
myotis, são geralmente pobres em abundância de presas. Pelo contrário as forragens e os
campos de cereais, usos do solo onde a vegetação herbácea é em geral muito densa,
dificultando a captura das presas, são as áreas onde a abundância de presas potenciais é
maior. Por forma a maximizar a relação ganho/gasto de energia, calcula-se que M.
myotis, utilize os usos do solo onde a densidade da vegetação seja intermédia, de forma
a ter maior sucesso de captura. As pastagens surgem assim entre os melhores candidatos
como áreas de caça para esta espécie. Dependendo do nível de pastoreio, poderão ter um
coberto com densidade intermédia a baixa, podendo simultaneamente manter uma
abundância relativamente elevada de presas potenciais (vd. Fig. 12).
Sendo estes tipos de habitat fundamentais na conservação desta espécie, propõe-
se que o pastoreio seja mantido, de forma a controlar o crescimento da vegetação.

Bibliografia
33
5. Bibliografia
Ahlén, I. & BaagØe, H. J. 1999. Use of ultrasound detectors for bat studies in Europe:
experiences from field identification, surveys, and monitoring. Acta Chiropterologica 1:
137-150.
Almenar, D., Aihartza, J., Goiti, U., Salsamendi, E. & Garin, I. 2008. Diet and prey
selection in the trawling long-fingered bat. Journal of Zoology 274: 340-348.
Arlettaz, R. & Perrin, N. 1995. The trophic niches of sympatric sibling Myotis myotis
and Myotis blythii: do Mouse-eared bats select prey? In Ecology, evolution and
behaviour of bats (Racey, P. A. & Swift, S. M.) (Eds.). Symposia of the Zoological
Society of London, 67. Clarendon Press, Oxford, xxii.
Arlettaz, R. 1996. Feeding behavior and foraging strategy of free-living mouse-eared
bats, Myotis myotis and Myotis blythii. Animal Behaviour 51: 1-11.
Arlettaz, R. 1999. Habitat selection as a major resource partitioning mechanism between
the two sympatric sibling bat species Myotis myotis and Myotis blythii. Journal of
Animal Ecology 68: 460-471.
Arlettaz, R., Godat, S. & Meyer, H. 2000. Competition for food by expanding pipistrelle
bat populations (Pipistrellus pipistrellus) might contribute to the decline of lesser
horseshoe bat (Rhinolophus hiposideros). Biological Conservation 93: 55-60.
Benton, T. G., Bryant, D. M., Cole, L., & Crick, H. Q. P. 2002. Linking agricultural
practice to insect and bird populations: a historical study over three decades. Journal of
Applied Ecology 39: 673-687.
Blanckenhorn, W. U. 2000. The evolution of body size: what keeps organism small?
The Quarterly Review of Biology 75: 385-407.

Bibliografia
34
Bontadina, F., Schmied, S. F., Beck, A. & Arlettaz, R. 2008. Changes in prey
abundance unlikely to explain the demography of a critically endangered Central
European bat. Journal of Applied Ecology 45: 641-648.
Cabral, M. J. (coord.), Almeida, P. R., Dellinger, T., Ferrand de Almeida, N., Oliveira,
M. E., Palmeirim, J. M., Queiroz, A. I., Rogado, L. & Santos-Reis, M. (Eds.) 2006.
Livro Vermelho dos Vertebrados de Portugal. 2ª ed. Instituto da Conservação da
Natureza / Assírio & Alvim. Lisboa.
Cardoso, P., Silva, I., Oliveira, N. G. & Serrano, A. R. 2007. Seasonality of spiders
(Araneae) in Mediterranean ecosystems and its implications in the optimum sampling
period. Ecological Entomology 32: 516-526.
Celuch, M. & Kanuch, P. 2005. Winter activity and roosts of the noctule (Nyctalus
noctula) in an urban area (Central Slovakia). Lynx (Praha) 36: 39-45.
Chinery, M. 1993. Insects of Britain and Northern Europe. 3ª ed. Harper Collins.
London.
Cole, L. J., McCracken, D. I., Dennis, P., Downie, I. S., Griffin, A. L., Foster, G. N.,
Murphy, K. J. & Waterhouse, T. 2002. Relationships between agricultural management
and ecological groups of ground beetles (Coleoptera: Carabidae) on Scottish farmland.
Agriculture, Ecosystems and Environment 93: 323-336.
Cryan, P. M. & J. P. Veilleux. 2007. Migration and use of autumn, winter, and spring
roosts by tree bats. In Lacki, M. J., Kurta, A. & Hayes, J. P. (Eds.). Conservation and
management of bats in forests. John Hopkins University Press. Baltimore, Maryland.
Pp. 153–175.
Damgaard, C. 2008. Modelling pin-point plant cover data along an environmental
gradient. Ecological Modelling 214: 404-410.
Donald, P. F. 2004. Biodiversity Impacts of Some Agricultural Commodity Production
Systems. Conservation Biology 18: 17-37.

Bibliografia
35
Dunbar, M., Whitaker, J. O. & Robbins, L. W. 2007. Winter feeding by bats in
Missouri. Acta Chiropterologica 9: 305-322.
Edgecombe, G. D. & Giribet, G. 2007. Evolutionary Biology of Centipedes
(Myriapoda: Chilopoda). Annual Review of Entomology 52: 151-170.
Foelix, R. F. 1996. Biology of spiders. Oxford University Press. Oxford.
Gardiner, T., Hill, J. & Chesmore, D. 2005. Review of the methods frequently used to
estimate the abundance of Orthoptera in grassland ecosystems. Journal of Insect
Conservation 9: 151-173.
Glendinning, J. I. 2007. How do predators cope with chemically defended food? The
Biological Bulletin 213: 252–266.
ICNB 2007. Base de observações de quirópteros. Informação disponibilizada a Outubro
2008.
Jordan, M. J. R. 2005. Dietary analysis for mammals and birds: a review of field
techniques and animal-management applications. International Zoo Yearbook 39: 108-
116.
Kanuch, P., Janecková, K. & Kristín, A. 2005. Winter diet of the noctule bat Nyctalus
noctula. Folia Zoologica 54: 53-60.
Kitching, R. L., Li, D. & Stork, N. E. 2001. Assessing biodiversity “sampling
packages”: how similar are arthropod assemblages in different tropical rainforests?
Biodiversity and Conservation 10: 793-813.
Lang, A., Filser, J. & Henschel, J. R. 1999. Predation by ground beetles and wolf
spiders on herbivorous insects in a maize crop. Agriculture, Ecosystems & Environment
72: 189-199.

Bibliografia
36
Lausen, C. L. & Barclay, R. M. R. 2006. Winter bat activity in the Canadian prairies.
Canadian Journal of Zoology 84: 1079-1086.
Lawton, J. 1996. Corncrake pie and prediction in ecology. Oikos 76: 3-4.
Lollar, A. & Schmidt-French, B. 2002. Captive Care and Medical Reference for the
Rehabilitation of Insectivorous Bats. Bat World Publication. Mineral Wells, TX.
Mcaney, C., Shiel, C., Sullivan, C. & Fairley, J. 1991. The analysis of bat droppings.
Occasional Publication of the Mammal Society 14: 1-48.
Mitchell-Jones, A. J., Amori, G., Bogdanowicz, W., Kryštufek, B., Reijnders, P. J. H.,
Spitzenberger, F., Thissen, J. B. M., Vohralik, V. & Zima, J. 1999. The atlas of
European mammals. 1ª ed. Poyser-Academic Press. London.
Obrist, M. K., Boesch, R. & Flückinger, P. F. 2004. Variability in echolocation call
design of 26 Swiss bat species: consequences, limits and options for automated field
identification with a synergetic pattern recognition approach. Mammalia 68: 307-322.
Palmeirim, J. M. & Rodrigues, L. 1992. Plano Nacional de Conservação dos Morcegos
Cavernícolas. Estudos de Biologia e Conservação da Natureza 8, Serviço Nacional de
Parques Reservas e Conservação da Natureza. Lisboa.
Palmeirim, J.M., Rodrigues, L., Rainho, A. & Ramos, M. J. 1999. Chiroptera. In:
Mamíferos terrestres de Portugal Continental, Açores e Madeira. Instituto da
Conservação da Natureza & Centro de Biologia Ambiental (Ed.). Lisboa. Pp. 41-95.
Park, K. J., Jones, G. & Ransome, R. D. 2000. Torpor, arousal and activity of
hibernating Greater Horseshoe Bats (Rhinolophus ferrumequinum). Functional Ecology
14: 580-588.
Pereira, M. J., Rebelo, H., Rainho, A. & Palmeirim, J. M. 2002. Prey selection by
Myotis myotis (Vespertilionidae) in a Mediterranean region. Acta Chiropterologica 4:
183-193.

Bibliografia
37
Pfiffner, L. & Luka, H. 2000. Overwintering of arthropods in soils of arable fields and
adjacent semi-natural habitats. Agriculture, Ecosystems & Environment 78: 215-222.
Pywell, R. F., James, K. L., Herbert, I., Meek, W. R., Carvell, C., Bell, D. & Sparks, T.
H. 2005. Determinants of overwintering habitat quality for beetles and spiders on arable
farmland. Biological Conservation 123: 79-90.
Quartau, J. A. & Carvalho, E. L. 1998. Contribuição para o melhor conhecimento dos
insectos em Portugal: chaves para a determinação das ordens. Publicações Avulsas 2ª
série 5, Museu Bocage. Lisboa.
Ragge, D. R. & Renolds, W. J. 1998. The songs of the Gasshoppers and Crickets of
Western Europe. Harley Books. England.
Rainho, A., Rodrigues, L., Bicho, S., Franco, C. & Palmeirim, J. 1998. Morcegos das
Áreas Protegidas Portuguesas I. Estudos de Biologia e Conservação da Natureza 26,
Instituto da Conservação da Natureza. Lisboa.
Ransome, R. D. 2000. Monitoring diets and population changes of greater horseshoe
bats in Gloucestershire and Somerset. English Nature Research Reports 341: 55.
Roberts, M. J. 1995. Spiders of Britain & Northern Europe. HarperCollins Publishers.
London.
Rodrigues, L., Zahn, A., Rainho, A. & Palmeirim, J. M. 2003. Contrasting the roosting
behaviour and phenology of an insectivorous bat (Myotis myotis) in its southern and
northern distribution ranges. Mammalia 67: 321-335.
Russo, D. & Jones, G. 2002. Identification of twenty-two bat species (Mammalia:
Chiroptera) from Italy by analysis of time-expanded recordings of echolocation calls.
Journal of Zoology 258: 91-103.

Bibliografia
38
Russo, D., Jones, G. & Arlettaz, R. 2007 Echolocation and passive listening by foraging
mouse-eared bats Myotis myotis and M. blythii. Journal of Experimental Biology 210:
166-176.
Sabo, J. L. & Power, M. E. 2002. River-watershed exchange: effects of riverine
subsidies on riparian lizards and their terrestrial prey. Ecology 83: 1860-1869.
Safi, K. & Kerth, G. 2004. A Comparative Analysis of Specialization and Extinction
Risk in Temperate-Zone Bats. Conservation Biology 18: 1293-1303.
Schaub, A., Ostwald, J. & Siemers, B. M. 2008. Foraging bats avoid noise. The Journal
of Experimental Biology 211: 3174-3180.
Siemers, B. M. & Güttinger, R. 2006 Prey conspicuousness can explain apparent prey
selectivity. Current Biology 16: 157-159.
Sokal, R. R. & Rohlf, F. J. 1997. Biometry: the principles and practice of statistics in
biological research. 3ª ed. Freeman & Company. New York.
Stamou, G. P., Stamou, G. V., Papatheodorou, E. M., Argyropoulou, M. D. & Tzafestas,
S. G. 2004. Population dynamics and life history tactics of arthropods from
Mediterranean-type ecosystems. Oikos 104: 98-108.
Storey, K. B. & Storey, J. M. 1996. Natural freezing survival in animals. Annual Review
of Ecology and Systematics 27: 365-386.
Vaughan, N. 1997. The diets of British bats (Chiroptera). Mammal Review 27: 77–94.
Wallington, T. J., Hobbs, R. J. & Moore, S. A. 2005. Implications of current ecological
thinking for biodiversity conservation: a review of the salient issues. Ecology and
Society 10: 15.

Bibliografia
39
Whitaker, J. O. 1988. Food habits analysis of insectivorous bats. In Kunz, T. H. (Ed.)
Ecological and behavioral methods for the study of bats. Smithsonian Institution Press.
Washington. Pp. 171–190.
Whitaker, J. O., Rose, R. K. & Padgett, T. M. 1997. Food of the Red Bat Lasiurus
borealis in winter in the great dismal swamp, North Carolina and Virginia. American
Midland Naturalist 137: 408-411.
Winder, L., Holland, J. M., Perry, J. N., Woolley, C. & Alexander, C. J. 2001. The use
of barrier-connected pitfall trapping for sampling predatory beetles and spiders.
Entomologia Experimentalis et Applicata 98: 249-258.
Wojciechowski, M. S., Jefimow, M. & Tegowska, E. 2007. Environmental conditions,
rather than season, determine torpor use and temperature selection in large mouse-eared
bats (Myotis myotis). Comparative Biochemistry and Physiology - Part A 147: 828-840.
Zahn, A., Haselbach, H. & Güttinger, R. 2005. Foraging activity of central European
Myotis myotis in a landscape dominated by spruce monocultures. Mammalian Biology
70: 265-270.
Zahn, A., Rodrigues, L., Rainho, A. & Palmeirim, J. M. 2007. Critical times of year for
Myotis myotis, a temperate zone bat: roles of climate and food resources. Acta
Chiropterologica 9: 115-125
Zalidis, G., Stamatiadis, S., Takavakoglou, V., Eskridge, K. & Misopolinos, N. 2002.
Impacts of agricultural practices on soil and water quality in the Mediterranean region
and proposed assessment methodology. Agriculture, Ecosystems & Environment 88:
137-146.
Zukal, J., Berkova, H. & Rehak, Z. 2005. Activity and shelter selection by Myotis
myotis and Rhinolophus hipposideros hibernating in the Katerinska cave (Czech
Republic). Mammalian Biology 70: 271-281.
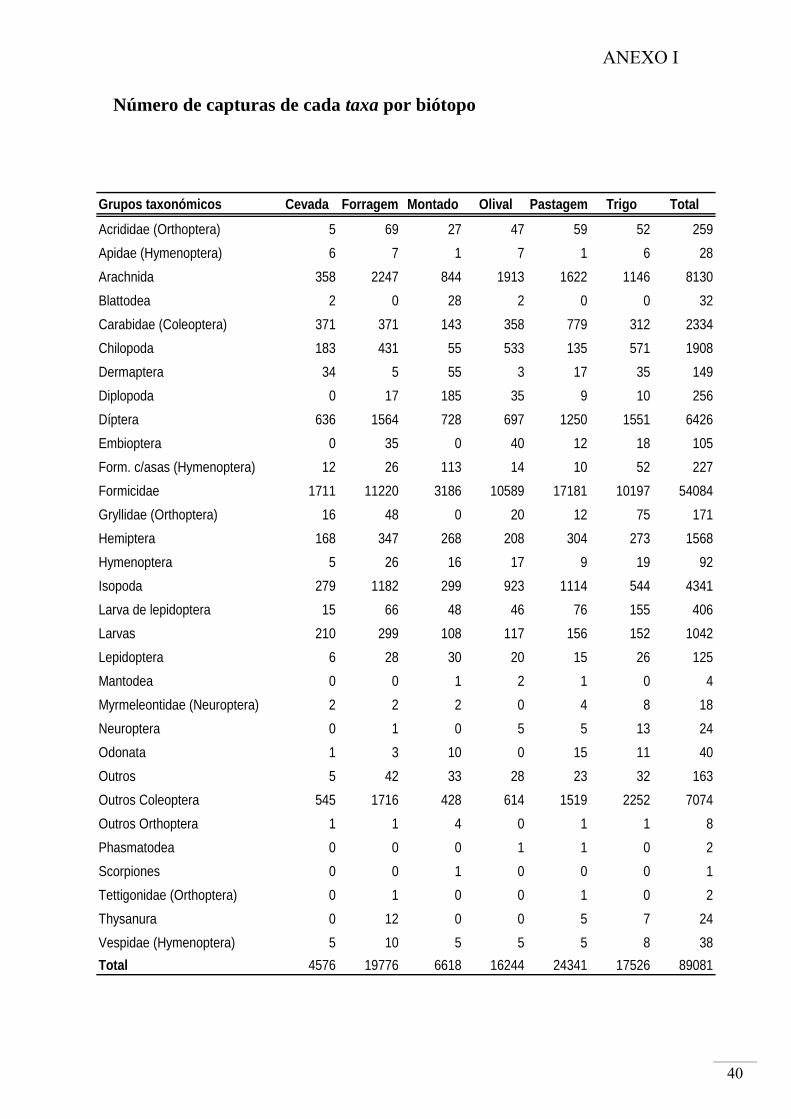
ANEXO I
40
Grupos taxonómicos Cevada Forragem Montado Olival Pastagem Trigo TotalAcrididae (Orthoptera) 5 69 27 47 59 52 259Apidae (Hymenoptera) 6 7 1 7 1 6 28Arachnida 358 2247 844 1913 1622 1146 8130Blattodea 2 0 28 2 0 0 32Carabidae (Coleoptera) 371 371 143 358 779 312 2334Chilopoda 183 431 55 533 135 571 1908Dermaptera 34 5 55 3 17 35 149Diplopoda 0 17 185 35 9 10 256Díptera 636 1564 728 697 1250 1551 6426Embioptera 0 35 0 40 12 18 105Form. c/asas (Hymenoptera) 12 26 113 14 10 52 227Formicidae 1711 11220 3186 10589 17181 10197 54084Gryllidae (Orthoptera) 16 48 0 20 12 75 171Hemiptera 168 347 268 208 304 273 1568Hymenoptera 5 26 16 17 9 19 92Isopoda 279 1182 299 923 1114 544 4341Larva de lepidoptera 15 66 48 46 76 155 406Larvas 210 299 108 117 156 152 1042Lepidoptera 6 28 30 20 15 26 125Mantodea 0 0 1 2 1 0 4Myrmeleontidae (Neuroptera) 2 2 2 0 4 8 18Neuroptera 0 1 0 5 5 13 24Odonata 1 3 10 0 15 11 40Outros 5 42 33 28 23 32 163Outros Coleoptera 545 1716 428 614 1519 2252 7074Outros Orthoptera 1 1 4 0 1 1 8Phasmatodea 0 0 0 1 1 0 2Scorpiones 0 0 1 0 0 0 1Tettigonidae (Orthoptera) 0 1 0 0 1 0 2Thysanura 0 12 0 0 5 7 24Vespidae (Hymenoptera) 5 10 5 5 5 8 38Total 4576 19776 6618 16244 24341 17526 89081
Número de capturas de cada taxa por biótopo

ANEXO II
41
Diversidade de Coleoptera capturados nas pitfalls (superiores a 5mm)
Alleculidae: Heliotaurus ruficollisBuprestidae: Capnodis sp.CantharidaeCarabidae: Amara aeneaCarabidae: Brachinus cf. bellicosusCarabidae: Carabus (Macrothorax) rugosusCarabidae: Carabus (Mesocarabus) lusitanicusCarabidae: Carabus (Rhabdotocarabus) melancholicusCarabidae: Carabus sp. (larva)Carabidae: Carterus interceptusCarabidae: Celia sp.Carabidae: Chlaeniellus olivieriCarabidae: Dinodes decipiensCarabidae: Harpalus cf. attenuatusCarabidae: Harpalus distinguendusCarabidae: Licinus punctatulusCarabidae: Notiophilus geminatusCarabidae: Olisthopus sp.Carabidae: Ophonus opacusCarabidae: Ophonus subquadratusCarabidae: Parophonus sp.Carabidae: Percus politusCarabidae: Poecilus purpurascensCarabidae: Steropus globosusCarabidae: Trichochlaenius chrysocephalusCarabidae: Zabrus ignavusCerambycidaeChrysomelidaeChrysomelidae: CassidiniChrysomelidae: Chrysomela sp.Chrysomelidae: Timarcha sp.CoccinellidaeCoccinellidae: Coccinella septempunctataCoccinellidae: Hyperaspis sp.CurculionidaeCurculionidae: Brachycerus sp.DasytidaeDermestidae: Dermestes sp.Elateridae: Athous sp.Geotrupidae: Thorectes sp.Geotrupidae: Typhaeus typhoeusHydrophilidae: Helophorus sp.Lagriidae: Lagria sp.Meloidae

ANEXO II
42
Scarabaeidae: AphodiinaeScarabaeidae: Cetoniinae: Protaetia sp.Scarabaeidae: Copris sp. (macho)Scarabaeidae: Melolonthidae: Chasmatopterus sp.Scarabaeidae: MelolonthinaeScarabaeidae: Scarabaeinae: Onthophagus sp.Silphidae: Silpha sp.Staphylinidae: Lobopaederus meridionalisStaphylinidae: Ocypus obscuroaeneusStaphylinidae: Ocypus olensTenebrionidae: Asida sp.Tenebrionidae: Belopus sp.Tenebrionidae: Blaps sp.Tenebrionidae: Cossyphus sp.Tenebrionidae: Crypticus sp.Tenebrionidae: Glabrasida sp.Tenebrionidae: Gonocephalum sp.Tenebrionidae: Misolampus sp.Tenebrionidae: Scaurus cf. punctatusTenebrionidae: Tentyria sp.Trogidae: Trox sp.
Diversidade de Gryllidae capturados nas pitfalls (superiores a 5mm)
Acheta hispanicusEumodicogryllus bordigalensis Eumodicogryllus theryiGryllomorpha uclensis Gryllus bimaculatus Melanogryllus desertusOecanthus pellucensSciobia lusitanica Stenonemobius gracilis


















