
UNIVERSIDADE BANDEIRANTE DE SÃO PAULO · Figura 2 Aspecto macroscópico de neoplasia mamária...
71
UNIVERSIDADE BANDEIRANTE ANHANGUERA Programa de Pós-Graduação em Farmácia RODRIGO CASEMIRO PINTO MONTEIRO ESTUDO HISTOLÓGICO E EPIGENÉTICO DE TECIDOS NORMAIS E TUMORAIS DA GLÂNDULA MAMÁRIA CANINA São Paulo 2013 RODRIGO CASEMIRO PINTO MONTEIRO
-
Upload
nguyentuyen -
Category
Documents
-
view
213 -
download
0
Transcript of UNIVERSIDADE BANDEIRANTE DE SÃO PAULO · Figura 2 Aspecto macroscópico de neoplasia mamária...

UNIVERSIDADE BANDEIRANTE ANHANGUERA
Programa de Pós-Graduação em Farmácia
RODRIGO CASEMIRO PINTO MONTEIRO
ESTUDO HISTOLÓGICO E EPIGENÉTICO DE TECIDOS NORMAIS E TUMORAIS DA GLÂNDULA MAMÁRIA CANINA
São Paulo 2013
RODRIGO CASEMIRO PINTO MONTEIRO

ESTUDO HISTOLÓGICO E EPIGENÉTICO DE TECIDOS NORMAIS E TUMORAIS DA GLÂNDULA MAMÁRIA CANINA
São Paulo 2013
Dissertação apresentada para
defesa de Tese em Farmácia pelo
Programa de Pós-Graduação em
Farmácia da Universidade
Bandeirante Anhanguera.
Orientadora: Profa. Dra. Susana
Nogueira Diniz

RODRIGO CASEMIRO PINTO MONTEIRO
ESTUDO HISTOLÓGICO E EPIGENÉTICO DE TECIDOS NORMAIS E TUMORAIS DA GLÂNDULA MAMÁRIA CANINA
São Paulo, 12 de abril de 2013.
Banca Examinadora
__________________________________________ Profa.Dra. Susana Nogueira Diniz
Orientadora
__________________________________________ Profa.Dra.Sílvia Regina Kleeb
Examinadora
___________________________________________ Prof.Dra. Márcia Regina Machado
Examinadora
Dissertação apresentada para
obtenção do título de Mestre em
Farmácia pelo Programa de Pós-
Graduação em Farmácia da
Universidade Bandeirante
Anhanguera.
Orientadora: Profa. Dra. Susana
Nogueira Diniz

AGRADECIMENTOS
À professora e orientadora Dra. Susana Nogueira Diniz pelo
enorme empenho e dedicação na elaboração deste trabalho e paciência em passar
seus ensinamentos.
Ao Prof. Dr. Paulo Pardi, por suas aulas que tanto nos ajudaram
no ponto de vista filosófico, e pelos conhecimentos passados.
A todos os professores colaboradores do Programa de Mestrado
em Farmácia da Universidade Bandeirante de São Paulo pela dedicação e carinho.
Aos colegas do Hospital Veterinário da Universidade
Bandeirante de São Paulo pela atenção e ajuda prestadas.
À professora Dra. Renata Afonso Sobral pelos seus
ensinamentos passados durante o período de graduação em Medicina Veterinária e
pela sua contribuição na participação no exame de qualificação.
Aos meus familiares pela atenção e paciência neste período de
elaboração da tese.
Aos meus colegas de mestrado pelo companheirismo e
solidariedade ao decorrer desta jornada.
Ao professor e amigo Marcelo Contieri pela ajuda e dedicação
na parte histopatológica.
A professora Sílvia Regina Kleeb pelos ensinamentos passado
no período da graduação e pelo aceite do convite para a defesa da tese.
Aos professores Luis Marques e Márcia Regina pelo
ensinamentos passados.
E a todos aqueles que contribuíram, de maneira direta ou
indireta para a elaboração deste trabalho.

RESUMO
As neoplasias mamárias em cadelas vêm ganhando grande destaque na casuística
da clínica veterinária, uma vez que os animais estão vivendo cada vez mais. A idade
de aparecimento dos tumores varia de 4 a 12 anos e está associado com fatores
hormonais e de envelhecimento, assim como no ser humano. O diagnóstico é
clínico, devendo ser realizados exames complementares para pesquisa de
metástases e o tratamento se baseia na remoção das mamas acometidas por
cirurgia e castração. O diagnóstico definitivo é obtido por exame histopatológico.
Neste trabalho foram realizados estudos histopatológicos e epigenéticos de
amostras tumorais e amostras de mamas livres de tumores. Na histopatologia além
da coloração padrão HE (hematoxilina-eosina),foram realizadas as técnicas de PAS
(ácido periódico de Shiff) e Tricômio de Masson, com o objetivo de avaliar a
presença de glicoproteínas e tecido conjuntivo nas amostras. A classificação das
amostras tumorais baseada na histopatologia demonstrou a prevalência de 100% de
carcinomas distribuídos em variados graus de malignidade. Foi comprovado
queexiste correlação estatística entre tumores malignose a presença de
glicoproteínas e açúcares no tecido estudado e escassez de tecido conjuntivo, mais
especificamente de fibras colágenas. Em relação ao estudoepigenético por MSAP-
PCR (Methilations Sensitive Arbitraly Primed- PCR), foi demonstradoalteração do
padrão de metilação de fragmentos gênicos provenientes de tecidos normais e
tumorais. Essa alteração pôde ser observada tanto por um padrão de metilação em
amostras tumorais e hipometilação em tecido normal (banda de 340pb). Ou pela
presença de fragmentos hipometilados em tumor e metilados em tecido normal
(banda de 600pb), que pode representar representar novas regiões do DNA tumoral
com padrão de hipometilação. Foram identificadas regiões genômicas que
apresentaram padrão de metilação alterado entre os subtipos tumorais. Foi mostrado
um fragmento de aproximadamente 400pb que se apresentou metilado emamostra
de tumor sólido e desmetilado nas demais amostras tumorais, do tipo tubular e
papilífero. Observou-se também uma banda de 390 pb que estava metilada em
amostras tubulares complexas de grau II e desmetilada em amostras de tumor
papilífero. Todos esses fragmentos representam potenciais marcadores específicos
de subtipos de tumores de mama canino, fato este que corrobora com a
necessidade do estudo mais aprofundado do padrão molecular das neoplasias

mamárias caninas, com objetivo de encontrar possíveis marcadores de significância
clínica.
Palavras-chave: neoplasia, mamária, cadela, histopatologia, epigenética, DNA, carcinoma
ABSTRACT
The mammary neoplasms is gaining great prominence in the veterinary clinic , since
the animal sare living increasingly. The age of tumor`s onset varies from 4 to12 years
old and is associated with hormonal factor sand aging, (just like)in humans. The
diagnosis is based on clinical signals and must be undertaken for metastasis
research and treatment involves surgery to remove the breasts involved and
neutering. The definitive diagnosis is obtained by histopathology. In this case, breast
tumor and tumor-free samples were in histopathology terms and molecular biology
were evaluated. In histopathology HE staining(hematoxylin-eosin), PAS(periodic
acid-Schiff) and Tricômio of Masson colorations were performed, in order toassess
the presence of glycoproteins and tissue in tumors .The tumor samples classification
showed the prevalence of 100% carcinomas distributed invarying degrees of
malignancy. It was observed that the more undifferentiated tumors, there are
prevalence of glycoproteins and scarcity of tissue, but specifically collagen. To this
observation we took into account the PAS staining and Tricômio of Masson,
respectively. All normal and tumor samples genomic DNA were demonstrated intact
with high molecular weight bands, a fact that confers reliability to this study.The
methylation profile of the samples was determined after digestion of DNA by
restriction enzymes,PCR (Polymerase Chain Reaction) and polyacrylamide gel.It was
observed that there are significant differences in the methylation status of tumor
tissues and tumor-free tissue. These changes could be observed both by a
methylation pattern in tumor samples and hypomethylation in normal tissue (band of
340pb) or by the presence of fragments hipometilated, but methylated in the tumor
and in normal tissue (Band 600pb), which may represent new represent regions of
DNA hypomethylation of tumor pattern. Genomic regions were identified andreveled
altered methylation pattern between tumor subtypes. It was shown a fragment of

approximately 400pb which appeared methylated in tumor sample and demethylated
in other solid tumor samples, tubular and papillary type. We also observed a band of
390 bp in samples that was methylated and demethylated in tubular complex II in
samples of papillary tumors. All these fragments represent potential markers of
specific subtypes of canine mammary tumors, a fact that confirms the need for further
study of the molecular pattern of canine mammary tumors, in order to find possible
markers of clinical significance.
Key Words: neoplasms, mammary, bitch, histopathology, epigenetics, DNA, carcinoma

LISTA DE ILUSTRAÇÕES
Figura 1 Conformação e estrutura dos ácinos mamários 16 Quadro 1 Tipos histológicos adotados no Consenso de 2011 23 Figura 2 Aspecto macroscópico de neoplasia mamária canina. 33 Figura 3 Fragmento representativo de neoplasia mamária, para realização
do estudo epigenético 33
Gráfico 1 Classificação citopatológica das amostras tumorais obtidas 38 Gráfico 2 Distribuição dos tumores mamários avaliados segundo a
classificação histológica 40
Quadro 2 Tabela comparativa entre diagnóstico citológico (coluna2) e histopatológico(coluna 3) de 11 amostras estudadas representadas (coluna 1), sendo que na histopatologia utilizou-se a coloração padrão hematoxilina-eosina (HE)
41
Figura 4 Tecido glandular controle-normal (HE 40X) 42 Figura 5 Tecido glandular controle-normal (Tricrômio de Masson 40X) 43 Figura 6 Tecido glandular controle-normal (PAS 40X) 43 Figura 7 Tecido glandular com alto índice de proliferação (HE 40/100X) 44 Figura 8 Tecido conjuntivo escasso e com pouca produção de colágeno
(Tricrômio de Masson 40/100X) 44
Figura 9 Tecido conjuntivo de sustentação é delicado e apresenta de grande quantidade de açúcares (PAS 40/100X)
45
Quadro 3 Dados obtidos das amostras tumorais, de acordo com o numero de identificação do animal
46
Quadro 4 Determinação da concentração do DNA em ng/ul 47 Tabela 1 Amostras tumorais – Coloração de PAS 48 Tabela 2 Amostras tumorais – Coloração de MASSON 49 Tabela 3 Tecido mamário normal x Adenocarcinomas – Coloraçao de PAS 50 Tabela 4 Tecido mamário normal x Adenocarcinomas – Coloração de
MASSON 50
Tabela 5 Dados de leitura espectrofomêtrica do DNA genômico obtido de tecidos normais (N) e tumorais (T) da mama de cadelas
53
Figura 10 Eletroforese em gel de Agarose. Amostras realizadas 55 Figura 11 Banda de 600 pb metilada em amostra normal e desmetilada em
tecido tumoral 56
Figura 12 Bandas de 400 pb indicando metilação em carcinoma sólido e desmetilação nas demais amostras tumorais tubulares e papilíferas
57
Figura 13 Banda de 390 pb metilada em amostras de carcinomas tubulares complexos e desmetilada em carcinomas papilíferos
58

LISTA DE ABREVIATURAS E SIGLAS
ABS ABSORBÂNCIA
CEUA Comitê de Ética no Uso de Animais
CpG ILHA CpG
DNA Ácido dexorribonucléico
DnmtsDNA Metil-transferases
DNTP DEXORRIBONUCLEOTÍDEO 5-FOSFATO
EDTA ÁCIDO ETILENODIAMINOTETRACÉTICO
HE Hematoxilina-eosina
Kb QUILOBASE
LBMGF Laboratório de Biologia Molecular e Genômica Funcional
MIN MINUTOS
MASSON Tricômio de Masson
C GRAUS CELSIUS
MSAP-PCR METILAÇÃO SENSÍVEL COM PRIMERS ALEATÓRIOS -PCR
OMS Organização Mundial de Saúde
OSH Ováriosalpingohisterectomia
ON OVERNIGHT
PAS ÁCIDO PERIÓDICO DE SHIFF
Pb PARES DE BASES
PH POTENCIAL HIDROGENIÔNICO
RE Receptores de estrogênio
RP Receptores de progesterona
SDS
SEG SEGUNDOS
TNM Tumor primário, linfonodos e metástase a distância
TRIS-HCL TRI(HIDROXIMETIL) AMINOMETANO – ÁCIDO CLORÍDRICO

SUMÁRIO
AGRADECIMENTOS...................................................................................................4 RESUMO.....................................................................................................................5 ABSTRACT..................................................................................................................6 LISTA DE ILUSTRAÇÕES...........................................................................................7
LISTA DE ABREVIATURAS E SIGLAS.......................................................................9 SUMÁRIO..................................................................................................................10 1. INTRODUÇÃO.......................................................................................................12 2 REVISÃO DE LITERATURA...................................................................................15 2.1 Características citológicas e histológicas da glândula mamária canina..............15 2.2.Câncer de mama em cadelas..............................................................................16 2.3.Características histológicas do tumor de mama em cadelas...............................19 2.4.Epigenética e câncer............................................................................................24 2.5 Marcadores moleculares das neoplasias mamárias caninas...............................27 3. JUSTIFICATIVA.....................................................................................................30 4. OBJETIVOS...........................................................................................................31 4.1. Geral....................................................................................................................31 4.2. Específicos..........................................................................................................31 5. MATERIAL E MÉTODOS.......................................................................................32 5.1. Ética.....................................................................................................................32 5.2 Material.................................................................................................................32 5.2.1 Coleta do material.............................................................................................32 5.2.2 Transporte do material......................................................................................33 6. METODOLOGIA-histologia....................................................................................34 6.1. Técnica da parafina ............................................................................................34 6.1.1 Colheita.............................................................................................................34 6.1.2. Fixação.............................................................................................................34 6.1.3.Desidratação e diafanização.............................................................................35 6.1.4. Inclusão............................................................................................................35 6.1.5. Corte.................................................................................................................35 6.1.6. Coloração.........................................................................................................35 6.1.6.1. Hematoxilina-eosina (HE)........................................................................... 36 6.1.6.2. Tricômio de Masson......................................................................................36 6.1.6.3. Coloração Ácido Periódico de Shiff (PAS)....................................................36 6.2 Extração de DNA genômico.................................................................................37 7. RESULTADOS E DISCUSSÃO.............................................................................37 7.1 Citopalotogia........................................................................................................38 7.2 Histopatologia- Coloração Hematoxilina-eosina (HE)..........................................39 7.3 Estudo epigenético...............................................................................................52 7.3.1 Quantificação e avaliação da integridade de DNA genômico............................52 7.3.2 Avaliação do perfil de metilação por MSAP-PCR..............................................55 8. CONCLUSÃO.........................................................................................................59 9. REFERÊNCIAS BIBLIOGRÁFICAS.......................................................................61 10. ANEXO 1 - Termos de consentimento livre e esclarecido (TCLE).......................67

11. ANEXO 2 -Parecer de aprovação CEUA - UNIBAN......................................................................................................................69 12. ANEXO 3 - Fotos do pré-cirúrgico das pacientes portadorasde neoplasiasmamárias do estudo...................................................................................................70 12. ANEXO 4 - Fotos das peças enviadas para estudo histopatológico (coluna B) e para o estudo epigenética (coluna A).........................................................................71

12
1. INTRODUÇÃO
A importância do estudo das neoplasias mamárias em cadelas vem
aumentando devido à frequência com que casos deste tipo surgem na clínica de
animais de companhia e devido as semelhanças histológicas que têm com os
tumores de mama em mulheres, podendo representar um modelo de estudos
translacionais para prognóstico, diagnóstico e terapia antitumoral(DALECK;
FONSECA, 2000). Este tipo de neoplasia representa 25 a 50% dos tumores caninos,
sendo que a metade são considerados malignos. São detectados em animais acima
de 5 anos, podendo em menor proporção acometer fêmeas com menor faixa etária.
Não existe uma predisposição racial, no entanto, apresenta maior risco nas raças
puras, com maior semelhança genética (DALECK, et al, 1998).
A cadela possui 5 pares de glândulas, as quais podem apresentar diversos
tipos de tumorações. A maneira definitiva de se obter o diagnóstico é pela análise
histopatológica de biópsia excisional. Caso haja suspeita de invasão ou aumento de
tamanho de linfonodos regionais, recomenda-se sua remoção e a realização de
análises histopatológicas. O tratamento cirúrgico é a terapia de escolha para todas
as cadelas com neoplasias mamárias, exceto para carcinomas inflamatórios e
neoplasias com metástase a distância. A técnica cirúrgica dependerá da localização
dos tumores nas glândulas. Na prática veterinária, alguns fatores são considerados
prognósticos, tais como tipo histológico, grau de invasão e de diferenciação nuclear,
tamanho do tumor, presença ou ausência de ulceração e presença ou ausência de
metástases a distância ou em linfonodos regionais (QUEIROGA; LOPES, 2002).
Deve-se ressaltar a importância da realização da castração precoce na
prevenção de neoplasias mamárias em cadelas, uma vez que caso seja feita antes
do primeiro cio reduz quase a zero o percentual de câncer de mama quando estiver
em idade adulta(QUEIROGA; LOPES, 2002).
No entanto, na medicina humana, têm se atentado a outros fatores
prognósticos e preditivos de resposta a terapia antitumoral, enfatizando o estudo de
fatores moleculares. Consideram-se fator prognóstico as características clínicas,
patológicas e biológicas de pacientes com câncer e seus tumores, que prevêem o
resultado clínico, numa situação sem tratamento (ZUCCARI, et al, 2088).
Em cadelas, os maiores fatores preditivos para metástase, tais como,
comprometimento de linfonodo ou grau histológico do tumor e seus histotipos,
falham em classificar os tumores mamários de acordo com seu comportamento

13
clínico. Com isso, a avaliação de marcadores moleculares de prognóstico apresenta-
se como uma ferramenta mais útil e conclusiva. As informações como expressão de
receptores hormonais ou mesmo características proliferativas são definitivas na
escolha de um esquema terapêutico correto (ZUCCARI, et al, 2008).
É importante ressaltar que existe uma grande similaridade entre genes
humanos e caninos, comprovando que é possível estudar mecanismos genéticos em
cães que servirá de modelo comparativo a seres humanos ( SANTOS, et al, 2010).
Os potenciais fatores prognósticos para carcinoma mamário em mulheres
podem ser agrupados em quatro gerações. Na primeira geração estariam os fatores
clássicos, já conhecidos como o estágio TNM (tumor primário, linfonodos e
metástase a distância). O estágio TNM é utilizado para realizar o estadiamento
tumoral, levando-se em conta o tamanho do tumor primário, a invasão dos
linfonodos adjacentes por células neoplásicas e a existência de metástases à
distância. A segunda geração inclui marcadores de reconhecida validade, como os
receptores de estrogênio (RE) e progesterona (RP) e marcadores de expressão
celular (Ki-67 e PCNA). Na terceira geração estão os marcadores genéticos, como
oncogenes, genes mutados ou não expressos e proteases. A quarta geração é
constituída pelos chamados preditores de metástase órgão-específicos (ZUCCARI,
et al, 2008; Eisemberg e Koifman, 2001; Terzian et al, 2007).
O estudo das mudanças na função gênica mantidos durante a mitose, que
não podem ser explicadas pela mudança na sequência do DNA é denominado de
epigenética. Os mecanismos epigenéticos de controle de expressão gênica são
fenômenos que, por impor um padrão de expressão gênica específico e herdado
pela progênie sem alterar a sequência de nucleotídeos no DNA, seriam capazes de
promover a especialização de células e suas linhagens, dando origem ao fenótipo
durante o desenvolvimento. O epigenoma é composto pela cromatina e pela
modificação covalente do DNA pela metilação. A metilação do DNA, uma
característica de genomas eucarióticos, é a transferência de um grupo metil pelas
DNA metiltransferases (Dnmts) ao carbono 5´ da citosina nos dinucleotídeosCpG.
Diversos estudos têm demonstrado que os mecanismos epigenéticos, incluindo a
metilação do DNA, são dinâmicos e podem ser alterados durante a vida por múltiplos
fatores fisiológicos, patológicos, nutricionais e ambientais.
O distúrbio de qualquer mecanismo epigenético pode alterar o balanço entre
conformação de cromatina ativa ou silenciada, resultando em um estado

14
transcricional alterado. Por exemplo, no câncer, o padrão de metilação do DNA é
invertido, havendo hipermetilação de promotores ricos em CG e hipometilação geral
do genoma, contribuindo para uma instabilidade genômica. Este processo alterado
de metilação não é aleatório e a hipermetilação específica de genes (por exemplo:
gene supressor de tumor) parece estar associada com a tumorigênese. Estudos
demonstram uma correlação positiva entre o grau de hipometilação e o grau de
malignidade crescente, sugerindo que a hipometilação pode ser um biomarcador útil
com significado prognóstico em tumores mamários (BANCROFT et al, 1996).
A classificação dos tumores, com critérios histogenéticos e de acordo com o
grau de diferenciação das células, tem grande utilidade na aplicação prática
(BANCROFT et al, 1996). Com base nestas informações, a proposta desse estudo é
avaliar as características histológicas de tumores de mama de cadelas para a
classificação de tipos tumorais. E, realizar uma análise global do epigenoma de
tecidos normais e tumorais, com o objetivo de se investigar alterações de padrões de
metilação do DNA na célula tumoral e buscar marcadores moleculares que possam
ser utilizados juntamente com a análise histológica para a classificação de diferentes
tipos tumorais.

15
2 REVISÃO DE LITERATURA
2.1. Características citológicas e histológicas da glândula mamária canina
As glândulas mamárias caninas são caracterizadas como estruturas
especializadas da pele, originadas embriologicamente da invaginação de brotos
ectodérmicos para o interior do mesoderma subjacente. Histologicamente são
classificadas como glândulas túbulo-alveolares compostas, que consistem de
unidades secretoras formadas por lóbulos separados por septos de tecido conjuntivo
(Botelho, 2009).
Na espécie canina (Canis familiaris), as glândulas mamárias
desenvolvem-se como botões epiteliais que crescem a partir de espessamentos
lineares, formando as cristas mamárias. Essas cristas desenvolvem-se em cinco
pares nas regiões torácica, abdominal e inguinal (DYCE et al., 2004). A proliferação
do mesênquima que circunda o botão eleva um mamilo sobre a superfície corpórea.
Um ou mais brotos epidérmicos surgem no botão mamário, no tecido conjuntivo, e
começam a formar canais por ocasião do nascimento. Cada broto constituirá o
sistema de ductos (DYCE et al., 2004). Esses ductos se abrem em ductos
interlobulares, formando dúctulos nos septos do tecido conjuntivo, os quais
convergem para formar os ductos lactíferos que drenam os lobos da glândula (DYCE
et al, 2004.) O epitélio de revestimento dos ductos é formado por uma dupla
camada de células epiteliais cúbicas ou cilíndricas baixas e à medida que vão se
ramificando para formar dúctulos, o epitélio transforma-se em simples cúbico ou
cilíndrico (BANKS, 1991). As células epiteliais ductais são responsáveis pela
secreção de proteínas lácteas e lipídios durante a lactação (BANKS, 1991).
Os ductos, assim como os alvéolos, são circundados por células
mioepiteliais contráteis, que se contraem sob a influência da ocitocina, secretando o
leite (FRANDSON, 1979). Além do tecido epitelial, ainda há a presença de
fibroblastos e células que compõem e sustentam o parênquima mamário. Estas
células estromais liberam fatores de crecimento, diretamente ligados à inibição e à
proliferação das células epiteliais (ZUCCARI et al., 2001).

16
Figura 1.(HE/40X):Fotomicrografia de tecido glandular mamário em aumento de 40X. A estrutura é composta por lóbulo pequeno e bem delimitado por tecido conjuntivo denso (setas). As estruturas glandulares são delicadas e apresentam epitélio monomorfico e em monocamada (Arquivo pessoal, 2012) 2.2. O câncer de mama em cadelas
A prevalência das neoplasias mamárias em cadelas está aumentando
consideravelmente nos últimos anos. A crescente incidência de doenças neoplásicas
na espécie canina tem várias razões, entre elas está o aumento da expectativa de
vida observado nestes animais. Fatores como nutrição com dietas balanceadas,
prevenção de doenças infecciosas com vacinação, métodos precisos de diagnóstico
e avanços na área terapêutica contribuem para o aumento da longevidade dos cães
(HATORE, 2011).
Os tumores mamários caninos constituem cerca de 50% de todos os
tumores que afetam as fêmeas desta espécie, embora a incidência mostre tendência
de diminuição, uma vez que a prática de ováriosalpingohisterectomia (OSH) em
fêmeas jovens é cada vez mais comum (QUEIROGA E LOPES, 2002). Para Hatore,
50% dos tumores mamários em cadelas são malignos, acomentendo em maior
proporção fêmeas entre 10 e 11 anos de idade, sendo mais raras em animais com
menos de 5 anos de vida. No entanto a idade está diretamente relacionada ao porte
do animal no que diz respeito ao aparecimento de neoplasias mamárias. Animais de
porte grande podem apresentar as alterações neoplásicas mais cedo que animais de

17
porte pequeno, pois possuem expectativa de vida menor. Para De Nardi, et al 2002,
o percentual de neoplasias malignas em relação ao número total de neoplasias
mamárias é de 68%.
Não existe uma predisposição racial evidente, embora algumas raças
sejam apontadas por alguns autores nas quais a incidência é maior. Em Beagles e
Boxer existem relatos com menor risco de apresentarem tumores de mama
(QUEIROGA e LOPES, 2002).A determinação do risco em relação a raça pode
variar de acordo com a região geográfica, como por exemplo no Japão, onde foi
relatada a incidência de 25% de neoplasias mamárias em cães de raça pequena e
58,5% de neoplasias malignas em cães de grande porte (FILHO, 2010).
Entre os tumores de mama malignos, a grande maioria se constitui de
carcinomas, sendo o tipo histológico um fator prognóstico importante para estes
animais. Entre as cadelas com neoplasias benignas, 26% delas desenvolvem
tumores em outras glândulas mamárias ao decorrer da vida. Cadelas com vários
nódulos podem apresentar tumores malignos e benignos concomitantemente
(OLIVEIRA et al., 2003).
Em relação ao tamanho dos tumores mamários, a realização de estudos
multicêntricos contribui muito para o estudo da epidemiologia deste tipo de câncer,
uma vez que pode-se observar que em países em desenvolvimento, geralmente
observam-se tumores relativamente maiores que em países desenvolvidos. Fato
este é suportado pela alta conscientização da castração precoce e do controle da
natalidade dos animais nos países ricos. Outro fator a ser levado em consideração
seria o poder aquisitivo dos proprietários em poder tratar do seu animal logo que
percebe a neoplasia. No estudo de Oliveira et al em 2003, foi observado que no
Brasil mais de 70% dos casos as neoplasias mamárias ultrapassavam 5 cm. No
entanto, segundo Itoh em 2005, no Japão, este número era de menos de 30 %.
De acordo com Daleck (1998), 40% dos tumores mamários são
carcinomas, 50% são tumores mistos e 10% são de outro tipo histológico. Afirma
ainda que a hiperplasia cística ocorre frequentemente em cadelas e está associada
a uma alteração pré-neoplásica. No desenvolvimento destas neoplasias são
envolvidos fatores genéticos, ambientais e hormonais. O papel dos hormônios
sexuais na origem destas neoplasias está extensamente estudado e estabelecido.
Este fato é suportado pelo aumento pronunciado da incidência dos tumores quando
a castração é efetuada após o segundo ciclo estral do animal. Quando as cadelas

18
são castradas antes do 1º cio o risco de desenvolverem tumores de mama é 0,05%.
Se a castração é efetuada após o 1º cio, o risco é de 8%, aumentando para 26%, se
a castração for efetuada após o 2º ciclo éstrico. Após instalado o tumor mamário, a
remoção dos ovários não interfere na prevenção do câncer de mama, apenas como
prevenção de outras afecções reprodutivas (QUEIROGA e LOPES, 2002).
As neoplasias presentes nos 3 pares de mamas craniais, correspondem a
cerca de 5,9% dos casos, enquanto que nas 3 mamas caudais correspondem a
51,8%. Tumores em todas as mamas ocorreram em 36,5% dos casos. A terceira
mama esteve envolvida em 5,9% dos casos. Nódulos tumorais múltiplos foram
identificados em 77,6% dos casos e tumores presentes nas 2 cadeias mamárias
concomitantemente 55,3% dos casos (OLIVEIRA et al, 2003).
Dentre as cadelas que apresentam tumores de mama, 48% apresentam ou já
apresentaram pseudociese, e o percentual de cadelas portadoras de neoplasia
mamária que tiveram filhotes é praticamente o mesmo de cadelas que nunca
pariram (OLIVEIRA, et al, 2003)
A administração de progestágenos foi relatada em 44,4% das cadelas dos
tumores mamários. Destas, 72% foram acometidas por neoplasias malignas.
Tumores benignos também são relatados em cadelas que receberam progestágenos
com objetivos anticoncepcionais (OLIVEIRA, et al, 2003).
As hipóteses mais citadas sobre a etiologia dos tumores mamários referem-
se à obesidade e a atividade hormonal. Os autores citados na sequência são
unânimes em afirmar que alguns tumores mamários são hormônio-dependentes
(ZUCCARI, et al, 2001; HATORE, 2011; SANTIN et al, 2009).
Em relação ao potencial metastático, foi relatado que em cerca de 15% dos
casos de tumores malignos mamáriosapresentaram metástase pulmonar
(OLIVEIRA, et al, 2003).
Segundo Filho et al, (2010) em estudo retrospectivo de 19 anos de rotina de
atendimento hospitalar, nos últimos 5 anos houve um aumento significativa da
ocorrência de tumores mamários em cadelas. O tipo histológico prevalente neste
estudo foi o carcinoma simples e a ocorrência mais frequente de metástase é para
linfonodos regionais, linfonodos mediastínicos e pulmões.
O diagnóstico das neoplasias mamárias caninas é realizado através de
exame físico minucioso, palpação de linfonodos regionais e verificação do estado
geral do animal, associados ao histórico clínico. Para pesquisa de metástases são

19
realizadas radiografias de tórax em 3 incidências e ultrassonografia abdominal
(CASSALI, et al, 2011; CIRILLO, 2008; SALIBA, et al, 2010).
O tratamento cirúrgico é o de escolha, podendo ser seguido por tratamento
quimioterápico após cirurgia, com exceção dos carcinomas inflamatórios, cuja
indicação é apenas tratamento quimioterápico. A escolha da técnica cirúrgica
depende da conduta do cirurgião, no entanto podem ser realizadas mastectomias
parciais ou totais, unilaterais ou bilaterais (CASSALI, et al 2011).
O tratamento pós cirúrgico envolve o uso de quimioterápicos, cuja escolha
depende do tipo histológico. Os quimioterápicos mais utilizados na medicina
veterinária são a doxorrubicina associada com ciclofosfamida e vincristina ou o uso
de cisplatina ou carboplatina como drogas únicas (CASSALI, et al 2011; SALIBA, et
al, 2010). No entanto, a utilização de mais de um fármaco associado a outro, com
mecanismos de ação diferentes (poliquimioterapia) tem demonstrado maior eficácia
(CIRILLO, 2008).
Os quimioterápicos têm como alvo células em intensa atividade proliferativa,
inclusive células normais que estão em constante mitose, como epitélio intestinal,
medula óssea e folículos pilosos. Por estes motivos, mielossupressão, sinais
gastrointestinais e perda de pelo são os efeitos colaterais mais comuns (CIRILLO,
2008). O tempo de tratamento varia de acordo com o protocolo escolhido, sendo que
a média de duração está entre 3 a 6 meses (CASSALI, et al 2011; SALIBA, et al,
2010). Após este período o animal entra nos segmentos oncológicos, consistindo de
1 consulta a cada 3 meses no primeiro ano após o término do tratamento, e 1
consulta a cada 6 meses no segundo ano pós tratamento (CASSALI, et al, 2011)
2.3 Características histológicas do tumor de mama de cadelas
Os tumores malignos de mama são classificados conforme a origem do
tecido envolvido (de origem epitelial ou mesenquimal). Histopatologicamente, os
tumores malignos da mama são classificados de acordo com a Organização Mundial
da Saúde (OMS) da seguinte forma:
1-Tumores de origem epitelial (carcinoma):
- não invasivo (in situ) ocorrem quando as células epiteliais não
ultrapassam a membrana basal;
- invasivos – quando as células epiteliais rompem a membrana basal e invadem
os tecidos adjacentes;

20
2- Tumores de origem conjuntiva (sarcomas);
3- Tumores mistos – tumores que se originam de ambos os tecidos, tanto do
tecido epitelial quanto do tecido conjuntivo, e são chamados de
carcinossarcomas;
Quase todas as malignidades da mama são carcinomas, de modo que tipos
como carcinoma de células escamosas, tumores filóides, linfomas e sarcomas
representam menos do que cinco por cento do total. Desses tumores, sarcomas são
os menos frequentes.
Os tumores prevalentes nas glândulas mamárias caninas são o adenoma,
carcinoma, carcinomas sólidos e tumores mistos da glândula mamária. Os
adenomas mamários, tumores benignos, ocorrem em 2 tipos histológicos sendo os
intra-ductais e lobulares. Os adenomas intraductais, têm origem nas células epiteliais
dos grandes ductos mamários e dos ductulosinterlobulares, geralmente possuem
ductos císticos, podem ser solitários ou múltiplos, com coloração branca ao corte. Ao
microscópio óptico são caracterizados por massas bem organizadas, revestidas por
uma camada simples de células epiteliais cuboides. As células revestem longas
projeções papilares de um estroma fibrovascular delicado, ou surgem como túbulos
alongados circundados por tecido fibrovascular, que, em conjunto, formam uma
massa polipoide (JONES, 2000).
Os adenomas lobulares, também chamados de adenomas acinares ou
tubulares, são neoplasias epiteliais benignas com origem nos ácinos mamários ou
nos pequenos ductulosintra-lobulares. Macroscopicamente são bem circunscritos e
sólidos, mas podem conter cistos disseminados. Ao microscópio óptico observa-se
células neoplásicas cuboides dispostas em ácinos compactados bem diferenciados
em dimensões variáveis ou em túbulos alongados, podendo apresentar
ramificações. Ao lúmen podem apresentar secreção rica em proteína. Possuem
margem bem definida e tecidos adjacentes podem se apresentar comprimidos
devido à expansão do tumor (JONES, 2000; ZUCCARI et al, 2001).
Os carcinomas são neoplasias epiteliais malignas originadas de células
epiteliais dos ductos mamários maiores, ductos interlobulares, ductos intralobulares
e epitélio secretório dos ácinos (alvéolos). Geralmente ocorrem em 3 padrões de
proliferação, sendo acinar, tubular e papilar, mas pode-se observar mais de um
padrão em uma mesma neoplasia. Além disso, o tecido pode estar acompanhado

21
por uma proliferação mioepitelial circundada por matriz condromucinosa, fato este
que faz o carcinoma ser denominado de simples ou complexo, respectivamente. São
observadas áreas de invasão no estroma e sistema linfático em todos os tipos
histológicos de adenocarcinomas, podendo haver até oclusão dos vasos linfáticos
por células tumorais (JONES, 2000).
Os carcinomas papilares têm origem no epitélio de grandes dutos e dutos
mamários intralobulares e crescem no interior de seus lúmens, sendo denominados
de carcinomas papilares intraductais. Diferem dos adenomas papilares por presença
de maior pleomorfismo celular, sendo as células dispostas em várias camadas,
muitas vezes empilhadas sobre projeções papilares. O prognóstico é melhor nos
casos em que o tumor fica confinado no lúmen do ducto comparado a tumores com
invasão de tecidos circunjacentes. Ocorre invasão do estroma e do sistema linfático,
podendo haver oclusão linfática por êmbolos tumorais. Quando está presente uma
proliferação mioepitelial, o tumor passa a ser chamado de carcinoma papilar
complexo e carcinoma papilar simples na ausência desta característica. Caso os
túbulos neoplásicos estejam císticos, a neoplasia é chamada de
cistoadenocarcinoma papilar ( JONES, 2000).
Os carcinomas tubulares também têm origem nos dutos da glândula mamária,
mas o padrão de crescimento é tubular e não papilar. Constituem a grande maioria
dos tumores de mama da espécie canina. Ao microscópio óptico, consistem de
túbulos de diâmetro uniforme, colabados ou desobstruídos, revestidos por uma
camada simples ou por várias camadas de células epiteliais cuboides. Em alguns
casos, os túbulos podem estar dilatados. Podem ser denominados de simples ou
complexos, dependendo da ausência ou presença de tecido mioepitelial. Também
possuem a característica de invasão do sistema linfático e do estroma (JONES,
2000).
Os carcinomas lobulares ou acinares possuem semelhanças com o tipo
histológico anterior, no entanto as células são arranjadas em ácinos distintos e não
em túbulos. Ambos os padrões podem existir em uma mesma neoplasia. As
características de malignidade são as mesmas dos tumores malignos citados
anteriormente (JONES, 2000).
Os chamados carcinomas infiltrativos da glândula mamária podem ser
papilares ou tubulares, mas possuem uma proliferação fibroblástica significativa, que
se diferencia em um tecido conjuntivo colagenoso denso e maturo. Esta proliferação

22
representa uma resposta desmoplásica pelo tecido conjuntivo do hospedeiro à
invasão por células neoplásicas. Seria uma tentativa de isolar as células invasoras.
As células neoplásicas, encontradas em ninhos, estão incrustradas em tecido
conjuntivo colagenoso. Portanto, neste tipo histológico a presença e identificação do
colágeno é de suma importância para o diagnóstico preciso (JONES, 2000;
ZUCCARI et al., 2001).
Os carcinomas sólidos microscopicamente consistem de nódulos sólidos ou
folhetos de células epiteliais neoplásicas sem evidências de padrão tubular, acinar
ou papilar. Frequentemente invadem sistema linfático e estroma adjacente. Os
carcinomas anaplásicos são neoplasias das glândulas mamárias caninas,
constituídas por células extremamente pleomórficas, podendo ser redondas,
poliédricas e fusiformes. Possuem núcleos mais hipercromáticos, pleomórficos com
aspecto polipoide. Estes tumores são altamente invasivos e com alto potencial de
metástases (JONES, 2000).
Os tumores mistos da glândula mamária são comuns na espécie canina. Ao
microscópio óptico apresentam uma proliferação neoplásica das células epiteliais e
mioepiteliais com diferenciação dessas células em ilhas de tecido cartilaginoso ou
ósseo. A diferenciação entre tumores mistos benignos de malignos pode ser difícil,
mas critérios de invasibilidade de sistema linfático e estroma são levados em
consideração para o diagnóstico, além das características neoplásicas das células
envolvidas (CASSALI, et al, 2011).
Em 2011 foi publicado no Brazilian Journal of Veterinary Pathology o Documento
de Consenso para o Diagnóstico, Prognóstico e Tratamento dos Tumores Mamários
Caninos, o qual estabelece uma modificação na classificação do tumores mamários
caninos, que, até então, eram classificados de acordo com características
morfológicas celulares e nucleares. Recentemente os pesquisadores propõem a
graduação das neoplasias mamárias caninas levando-se em conta o índice de
Nottingham, o qual já é utilizado na medicina humana.
O Índice de Nottingham leva em conta muitos critérios objetivos para
classificação da neoplasia, como a formação de estruturas tubulares no tecido
neoplásico e número de mitoses atípicas encontradas por campo de 40x. Todos
estes dados levam a obtenção de um índice, que já classifica as neoplasias
mamárias em mulheres mais objetivamente (CASSALI, et al 2011).

23
Quadro 1: Tipos histológicos adotados no Consenso de 2011, todos adaptados da
classificação já estabelecida da OMS e AFIP, de 1999:
Lesões epiteliais não neoplásicas Hiperplasia epithelial Hiperplasia ductal Hiperplasia lobular Adenose Lesão de células colunares Alteração de células colunares Hiperplasia de células colunares Lesões atípicas de células colunares Tumores benignos Adenoma Adenoma complexo Adenoma basalóide Fibroadenoma Tumor misto benigno Papiloma ductal Tumores malignos Carcinomas Carcinomas in situ Carcinoma ductalin situ Carcinoma lobular in situ Tumor misto-carcinoma Carcinoma complexo Carcinoma papilar Carcinoma tubular Carcinoma sólido Tipos especiais de carcinomas Carcinoma micropapilar Carcinoma lobular invasivo Carcinoma lobular pleomórfico Carcinoma secretório Carcinoma mucinoso Carcinoma rico em lipídeos Carcinoma de células escamosas Carcinoma anaplásico Neoplasias mamárias com diferenciação sebácea Sarcomas Fibrossarcoma Osteossarcoma Carcinossarcoma Tumor misto-sarcomas Outros sarcomas Condrossarcoma puro Lipossarcoma Hemangiossarcoma
Adaptado de CASSALI, et al 2011.

24
2.4 Epigenética e câncer
Epigenética é o estudo da transmissão da memória celular através de mitose,
que não é baseada na sequência do DNA (BRONNER et al., 2010). Modificações
epigenéticas influenciam o padrão de expressão gênica e oferecem uma única
assinatura de um estado de diferenciação celular (NG et al., 2008). A metilação do
DNA é a modificação epigenética mais extensivamente estudada em mamíferos. Ela
fornece um mecanismo de silenciamento estável do gene, desempenhando um
papel importante na regulação da expressão dos genes e da estrutura da cromatina
(SHARMA et al., 2010). A metilação do DNA desempenha seus efeitos de regulação
da transcrição genica diretamente, bloqueando o acesso de fatores de transcrição
chave para os sítios de ligação ao DNA, ou indiretamente através do controle
epigenético do seu acesso ao DNA por meio de alterações na estrutura da cromatina
(SINGAL et al., 1997; BEDNARIK et al., 1991 apud CHAPPELL et al., 2006).
O silenciamento gênico ao nível de cromatina é necessário para a vida de
organismos eucarióticos e é particularmente importante para a orquestração dos
principais processos biológicos, incluindo diferenciação, imprinting e silenciamento
de grandes domínios cromossômicos (JONES et al., 2007). Estudos de como os
genes são regulados e os mecanismos que estão envolvidos neste processo são de
grande importância para a compreensão dos processos normais e estados de
doença (COSTA, 2010; MULAS, 2005).
Diversas enzimas e vias bioquímicas que participam no estabelecimento dos
padrões epigenéticos têm sido identificadas. A metilação do DNA é mediada pelas
DNA metiltransferases (DNMT1, 3a e 3b) na presença da S-adenosil-L-metionina,
doadora de grupos metil. DNMT3a e DNMT3b foram descritas como
metiltransferasesde novo que catalisam a metilação do DNA em sequências
anteriormente não metiladas. DNMT1 é uma metiltransferase de manutenção que
tem como alvo preferencial o DNA hemimetilado, gerado após a replicação de uma
sequência metilada. Dessa forma, a DNMT1 assegura que os padrões de metilação
do DNA possam ser transmitidos para a próxima geração
O câncer é uma doença que resulta do acúmulo e interação de alterações
genéticas e epigenéticas (ROUNTREE et al., 2001). O epigenoma do câncer é
caracterizado por mudanças na metilação do DNA, bem como perfis de expressão
alterado de enzimas modificadoras da cromatina (SHARMA et al., 2010), composto

25
por desmetilação global e hipermetilação do promotor (TING et al., 2006). Sugere-se
que a hipometilação leva à ativação de proto-oncogenes, tais como h-Ras e c-Myc
(COSTELLO et al., 2001 apud ATTWOOD et al., 2002). Estudos demonstram uma
correlação positiva entre o grau de hipometilação e o grau de malignidade crescente,
sugerindo que a hipometilação pode ser um biomarcador útil com significado
prognóstico. Enquanto testes mostram uma diminuição global da metilação em todo
o genoma em muitos tumores, eles não fornecem informações sobre onde a
hipometilação ocorre (SOARES et al. apud PLASS et al., 2002).
Tem sido documentado que o silenciamento do gene supressor de tumor por
hipermetilação do promotor contribui no processo de transformação maligna
(ESTELLER, 2002 apud EL-HAMID et al., 2010). O aumento da metilação das ilhas
CpG é freqüentemente acompanhada por uma diminuição no nível global de
metilação do DNA e isto pode contribuir para a diminuição da instabilidade do
cromossomo (CHEN et al., 1998 apud JONES, 1999). A hipermetilação de ilhas CpG
normalmente não metiladas nas regiões promotoras de genes-chave para o câncer,
incluindo as ciclinas e proteínas relacionadas com a regulação do ciclo celular, por
exemplo os genes p16, p15, E-caderina, VHL e hMLH1, correlaciona-se com a perda
da sua transcrição em tumores humanos (HERMAN et al., 1998 ; ESTELLER et al.,
1998 apud ESTELLER et al. 1998).
Em tumores mamários caninos parece existir uma correlação entre perda de
expressão das E-caderinas (moléculas de adesão celular) e outros indicadores
conhecidos de prognóstico ruim, tais como tamanho do tumor, ulceração, tipo
histológico e metástases em linfonodos. Isto indica que a perda de expressão das E-
caderinas é um fator prognóstico importante ( PIEKARZ et al, 2008).
A hipometilação do DNA deve-se, principalmente, à perda da metilação em
sequências repetitivas, como as repetições do DNA ribossomal, repetições satélites
ou centroméricas, que se encontram altamente metiladas em tecidos normais, mas
que perdem a metilação em tumores. Os mecanismos desta perda de metilação do
DNA não estão claros, mas pode ocorrer por um mecanismo passivo que envolve a
replicação do DNA e a inibição da DNMT1, a metiltransferase de manutenção do
DNA ( PIEKARZ et al, 2008). A hipometilação de sequências repetitivas coincide
com mudanças na cromatina, provocando instabilidade genômica, uma das
características dos cânceres e que pode ser o evento iniciador da tumorigênese,

26
como demonstrado em modelos de camundongos com expressão diminuída de
DNMT1.
A hipermetilação do DNA em sequências ricas em CpG denominadas de ilhas de
CpG, normalmente não metiladas, é um dos mecanismos pelo qual as alterações
epigenéticas contribuem para a tumorigênese. Foi demonstrado que, em muitos
genes relacionados ao câncer e em genes supressores de tumor, as modificações
epigenéticas, tendo como alvo preferencial a região promotora de genes, resultam
em silenciamento (ESTELLER et al., 1998 apud ESTELLER et al. 1998). Além disso,
a hipermetilação das ilhas CpG pode preceder o processo neoplásico e aumentar
continuamente de lesões iniciais pré-neoplásicas até o câncer invasivo. Isto foi
demonstrado em vários cânceres, incluindo câncer de mama, cólon, próstata,
pulmão, esôfago, cervical, gástrico e síndrome mielodisplásica e pode ser
recapitulado em modelos animais para a progressão do câncer (BAYLIN,2005;
KOPELOVICH, 2003; LEE, 2010).
A terapia epigenética, que tem como objetivo a utilização de drogas para a
correção das alterações epigenéticas, é uma nova e atraente área para o
desenvolvimento de novas estratégias terapêuticas para a prevenção e tratamento
de várias doenças, incluindo o câncer (DUCASSE; BROWN 2006). No câncer de
mama, por exemplo, a ausência/perda da expressão de ERα está ligada à
insensibilidade à terapia antiestrogênica e a possibilidade de sensibilizar estes
tumores ao tamoxifeno pela indução da expressão de ERα utilizando-se HDIs e/ou
agentes desmetilantes é atualmente um dos objetivos dos novos tratamentos(GAMA;
SCHMITT, 2008; UVA, 2009; RIVERA, 2011).
Do ponto de vista clinico, a epigenética oferece uma via promissora, isto porque,
diferentemente das alterações genéticas como as mutações e deleções, por
exemplo, as alterações epigenéticas são potencialmente reversíveis (JAENISCH;
BIRD, 2003). Genes silenciados por metilação podem ser desmetilados e os
complexos de histonas podem tornar-se transcricionalmente ativos, através da
acetilação e metilação das histonas, via nutrientes, drogas ou outras intervenções na
dieta, fatores que representam uma oportunidade de desenvolvimento de novas
terapias quimiopreventivas e terapêuticas.
Todos esses estudos mostram um papel crucial dos mecanismos epigenéticos,
principalmente a metilação do DNA, no desenvolvimento tumoral e destaca a

27
importancia de se modular esses mecanismos como terapias alternativas para o
câncer (COSTA, 2010)
A identificação das modificações epigenéticas que implicam na expressão gênica
é um passo muito importante para que haja um entendimento de todos os processos
normais e patológicos que ocorrem em uma pessoa ou animal. Alguns
medicamentos são produzidos baseados nas alterações epigenéticas relatadas, tais
como inibidores de DNA metil-transferases e inibidores de modificação nas histonas.
No entanto estes medicamentos não são específicos e seus efeitos colaterais são os
maiores problema relacionados a seu uso (COSTA, 2010)
2.5 Marcadores moleculares das neoplasias mamárias caninas
Os estudos sobre marcadores moleculares em neoplasias mamárias na medicina
veterinária vêm ganhando cada vez mais espaço, uma vez que a busca por um
prognóstico mais acurado e por um tratamento mais personalizado cresce a cada
dia.
Os receptores hormonais (receptores de estrógeno-ER e receptores de
progesterona-PR), COX-2, marcador de angiogênese ( CD31), marcador de índice
proliferativo (MIB-1), fator de crescimento epidermal (EGF), moléculas de adesão (E-
caderinas) e p53 são exemplos de marcadores importantes para fins prognósticos
avaliados em neoplasias mamárias de cadelas, através da imunohistoquímica (
CASSALI et al, 2011).
Em relação ao número de receptores hormonais, existe uma relação proporcional
inversa com a capacidade proliferativa das neoplasias mamárias. Quando mais
agressivo o tumor, menor será a expressão de receptores hormonais do
mesmo.(CASSALI, et al, 2011). Os receptores de estrógeno e de progesterona
atualmente são encarados como marcadores preditivos de resposta a terapia
hormonal (ZUCCARI, et al, 2008).
Em cadelas, a utilização de anticorpos contra os receptores hormonais não tem
alcançado sucesso, pois não houve reação cruzada dos anticorpos utilizados da
espécie humana para cadelas, inviabilizando estudo comparativo. Como não houve
expressão destes marcadores na espécie canina, podemos utilizar a história clínica
do paciente para realizar a OSH no momento da exérese tumoral, com intuito de que
este animal não receba mais estímulo hormonal (ZUCCARI, et al, 2008).

28
O marcador de proliferação celular (MIB-1) é muito expresso nos tumores
malignos pouco diferenciados, sendo que nos mesmos a expressão de receptores
de progesterona é baixa (CASSALI, et al 2011).
O documento de consenso ainda comenta sobre a ciclooxigenase-2 (COX-2) é
um importante marcador tanto para cadelas quanto para mulheres, sendo expresso
em maior proporção em tumores com prognóstico ruim. Está associado com alta
capacidade proliferativas das células neoplásicas, decréscimo no índice apoptótico,
possibilidade de metastatização e neovascularização (CASSALI, et al, 2011).
As moléculas de adesão celular, principalmente as e-caderinas, têm expressão
reduzida conforme o grau de diferenciação da neoplasia diminui, revelando alta
capacidade proliferativa da mesma (CASSALI, et al, 2011). Alguns estudos
comprovam que a e caderina é um potente supressor de invasão nos tumores
mamários e a perda de sua expressão tem relação com um prognóstico ruim
(ZUCCARI, et al, 2008). Alta expressão da mesma indica que a adesão entre as
células está preservada, e deste modo, o risco de metástases à distância é
considerado baixo (ZUCCARI, et al, 2008).
Na década de 70, demonstrou-se que o estresse térmico induzia a expressão
de genes, permitindo a síntese de grande quantidade de proteínas chamadas de
‘’proteínas de estresse”. A expressão destas proteínas no câncer está associada às
condições estressantes das células neoplásicas, tais como baixa concentração de
nutrientes, oferta de oxigênio reduzida e espaço limitado para crescimento.Estas
proteínas ainda estão estudo, mas já estão classificadas como potenciais
marcadores de resposta a terapia (CARVALHO, 2006)
Além disso, mutações do gene p53 são frequentes em tumores mamários de
cadelas, e a expressão da proteína p53 modificada está associada com altas taxas
de invasão e progressão tumoral (CASSALI, et al. 2011)
O gene p53 é um gene supressor de tumor, codificando uma fosfoproteína
nuclear com atividade supressora de tumor. A mesma está presente em condições
normais e tem a função de regulação da proliferação celular e apoptose. Com a
mutação do gene p53, a proteína defeituosa codificada se acumula na célula, sendo
eliminada de forma mais lenta, fato este que permite sua detecção por imuno-
histoquímica (ZUCCARI, et al, 2008).
Nos carcinomas mamários de mulheres, a expressão do p53 está associada a
manifestações mais agressivas e pior prognóstico. A mesma lesão pode ainda

29
expressar c-erb-B2, que é também um gene localizado no cromossomo 17 em
humanos. Ambos são expressos em lesões de alto grau (ZUCCARI, et al, 2008)
O c-erb-B2 é um oncogene cuja expressão está associada com aumento da
atividade proliferativa celular. Na espécie canina é descrita uma superexpressão
deste gene, correlacionada com diagnóstico histopatológico de malignidade mas não
com a presença de invasão local ou doença metastática regional (ZUCCARI et al,
2008).
Em relação aos marcadores de proliferação celular, os mais estudados na
medicina veterinária são o Ki-67 e o PCNA. O Ki-67 é um antígeno nuclear presente
nas fases ativas do ciclo celular. Valores aumentados do mesmo têm correlação
positiva com metástase e baixa taxa de sobrevivência sem a doença (ZUCCARI, et
al, 2008). O PCNA (antígeno nuclear de proliferação celular) funciona como um
cofator da DNA polimerase delta, está presente em todas as fases do ciclo celular,
em maior proporção na fase S. Seus níveis podem-se manter elevados quando
induzidos por fatores de crescimento, como resultados de danos no DNA, fato este
que faz que o PCNA não pode ser considerado um bom marcador (ZUCCARI, et al,
2008).
As alterações moleculares que são observadas em células cancerosas estão
diretamente relacionadas com o processo de divisão e morte celular. A morte celular
programada, chamada de apoptose, tem um papel importante na determinação do
crescimento tumoral. Para iniciar o processo de apoptose algumas proteases são de
suma importância, tais como a caspase-3. Estudos mostram que a marcação da
mesma possa ser usada no controle da resposta aos tratamentos quimioterápicos, e
que a diminuição de sua expressão possa indicar um mecanismo de sobrevivência
da célula tumoral (ZUCCARI, et al, 2008).
Pela técnica de imunoistoquímica, pode-se determinar o fator de crescimento
endotelial vascular (VEGF), que é uma proteína que estimula a migração e
proliferação de células endoteliais vasculares, podendo aumentar as chances de
metástase, pois induz neovascularização, estimula proliferação das células
neoplásicas e induz produção colagenase e outras enzimas de degradação.O VEGF
é considerado um marcador de angiogênese (FELICIANO, et al, 2012).
Outro marcador que vem ganhando certa importância no estudo de câncer de
mama em cadelas é o maspin, que é um marcador muito sensível , cujos estudos
sugerem ter ação supressora de tumor, com ação indutora de adesão celular entre

30
membrana basal e matriz extracelular. Segundo Zuccari, et al, 2009, foi demonstrada
uma perda de expressão de maspin nas células mamárias malignas de cães quando
comparadas a um pool de tecido mamário normal de cadelas, analisado por PCR em
tempo real. No entanto, existe uma escassez de estudo sobre o maspin na medicina
veterinária
3. JUSTIFICATIVA
O câncer está incluído entre as entidades patológicas que mais acometem
a mama feminina. Segundo a Organização Mundial de Saúde (2010), o câncer de
mama é a segunda causa mais comum de óbitos em mulheres no mundo inteiro. A
cada ano, cresce o número de mulheres acometidas pelo câncer de mama e
também as pesquisas sobre a biologia desse tumor, incluídas aquelas que utilizam
modelos animais, principalmente os tumores de mama de cadelas. O estilo de vida
da sociedade moderna contribui para aumentar a exposição da população a alguns
fatores ambientais, nutricionais, químicos e hormonais, potencialmetne
carcinogênicos. As semelhanças entre os tumores de mama humana e de cadelas,
tais como, a morfologia, a presença de receptores de estrógenos, os órgãos-alvo de
metástases, a evolução clínica e a hereditariedade, motiva o interesse no estudo da
patologia comparada, a partir do uso desses modelos animais. Estudos
comparativos são importantes para o conhecimento da biologia do crescimento e da
organização funcional normal dos tecidos de órgãos humanos e de animais, bem
como de tumores, que poderão produzir conhecimento importante para diagnóstico,
tratamento, prognóstico e prevenção das neoplasias malignas.
O câncer é uma doença que resulta do acúmulo e interação de alterações
genéticas e epigenéticas, afetando a expressão gênica. O epigenoma do câncer
écaracterizado pormudanças na metilação do DNA, bem como perfis deexpressão
alterado deenzimasmodificadoras dacromatina, composto por desmetilação global e
hipermetilação do promotor. Sugere-se que a hipometilação leva à ativação de proto-
oncogenes. Estudos demonstram uma correlação positiva entre o grau de
hipometilação e o grau de malignidade crescente, sugerindo que a hipometilação
pode ser um biomarcador útil com significado prognóstico.
A classificação dos tumores, com critérios histogenéticos e de acordo com o
grau de diferenciação das células, tem grande utilidade na aplicação prática

31
(BANCROFT et al, 1996). Assim, a proposta desse estudo é avaliar as
características histológicas de tumores de mama de cadelas para a classificação de
tipos tumorais. E, realizar uma análise global do epigenoma de células normais e
células neoplásicas, com o objetivo de se investigar alterações de padrões de
metilação do DNA na célula tumoral e buscar marcadores moleculares que possam
ser utilizados juntamente com a análise histológica para a classificação de diferentes
tipos tumorais.
4. OBJETIVOS
4.1 Objetivo Geral
Estudar alterações histológicas estromais e epigenéticas associadas com
diferentes tipos tumorais provenientes da glândula mamária de cadelas.
4.2 Objetivos específicos:
- Analisar os cortes histológicos de tecido normal e tumoral;
- Avaliar a presença de glicoproteínas e colágeno nos cortes histológicos através de
coloração PAS e Tricômio de Masson, além da coloração padrão Hematoxilina-
Eosina (HE)
- Estabelecer a metodologia de obtenção de DNA gênomico de fragmentos de
tecidos normais e tumorais de mama de cadelas, no laboratório LBMGF ( Biologia
Molecular e Genômica Funcional) da UNIBAN;
- Comparar o padrão de metilação do DNA na célula normal e tumoral por MSAP-
PCR.

32
5. MATERIAL E MÉTODOS
5.1 Ética
Para esta pesquisa, foi criado um banco de DNA de amostras de tumores de
mama de cadelas de animais operados no centro cirúrgico do Hospital Veterinário da
UNIBAN/Anhanguera. Os dados clínicos e anátomo-patológicos foram coletados e
depositados em um banco de dados para futura correlação com os dados
epigenéticos.
Embora a remoção cirúrgica do tumor seja um procedimento muito invasivo, é
o principal tratamento indicado para esse tipo de neoplasia. Desta forma, nenhum
procedimento além do convencional será realizado no animal para uso exclusivo
dessa pesquisa. Os responsáveis pelos animais serão informados sobre a pesquisa
e permitindo a utilização do material, deverão assinar o termo de consentimento livre
e esclarecido (ANEXO 1).
Este presente estudo possui aprovação no Comitê de Ética e Experimentação
Animal (CEUA) da Universidade Bandeirante Anhanguera (UNIBAN) sob o registro
275/2012 (ANEXO 2).
5.2 Material
5.2.1 Coleta do material
A despeito dos diversos subtipos histológicos dos tumores caninos,
reconhecidos, foram utilizadas neoplasias genericamente classificadas como
carcinomas. As amostras de parênquima mamário canino, normal e tumoral foram
obtidas por cirurgias realizadas no Hospital Veterinário da Universidade Bandeirante
Anhanguera, sob a responsabilidade do cirurgião veterinário Rodrigo Monteiro.
Momentos antes do início da cirurgia, eram realizadas as punções aspirativas das
formações para estudo citológico, realizava-se a punção com agulha hipodérmica
20x5,5 mm e fazia-se esfregaço em lâminas de microscopia. As lâminas eram
coradas pelo métido de coloração panótico e avaliadas sob microscopia óptica em
aumento de 40 e 100 vezes. Foram coletados 10 fragmentos de mamas normais e

33
10 fragmentos de tumores malignos, que variavam entre 1 a 10 cm de diâmetro de
cadelas acima de 10 anos de idade, todas inteiras, sem raça definida (Figura
2;Figura 3).Foram escolhidos animais que nunca receberam aplicação de
anticoncepcionais. As técnicas cirúrgicas empregadas foram as de mastectomia
unilateral total ou mastectomia bilateral total, dependendo de cada caso.
Figura 2: Aspecto macroscópico de neoplasia mamária canina.
Fonte: Arquivo pessoal, 2012
Figura 3: Fragmento representativo de neoplasia mamária, para realização do
estudo epigenético Fonte: Arquivo pessoal, 2012
5.2.2 Transporte do material

34
Para cada animal operado, obtiveram-se amostras normais e tumorais. Para cada
uma foram gerados 2 fragmentos, sendo 1 enviado para estudo histológico
acondicionado em formaldeído a 10% e outro colocado no tampão de lise para
estudo epigenético.
Após remoção cirúrgica, os fragmentos para análise molecular foram transportados
para o Laboratório de Biologia Molecular e Genômica Funcional da UNIBAN, dentro
de frascos estéreis contendo 4 ml de tampão de lise ( 40 ul Tris-HCl + 400ul EDTA +
200ul SDS 10% + 3360ul de água bidestilada + 4 ul de Proteinase K), onde foram
colocadas em banho maria a 55º C por 72 horas. As peças cirúrgicas extraídas para
estudo histopatológico foram colocadas em formol 10% na proporção de 1:30 em
coletores apropriados até serem encaminhadas ao setor de patologia do Hospital
Veterinário da UNIBAN, onde após 10 dias seria emitido o laudo histopatológico das
amostras enviadas para constatação e diagnóstico histopatológico dos tecidos
enviados
6 METODOLOGIA – histologia
6.1 Técnica da parafina
Esta técnica é um procedimento simples e confiável que permite inferências
da situação in vivo. É o método mais utilizado para a preparação das lâminas
histológicas para estudo histopatológico para utilização em microscopia óptica.
6.1.1 Colheita
Feita a colheita do material para histopatologia, o mesmo deve ser
rapidamente colocado no fixador, visando inativar as enzimas por desnaturação
protéica responsáveis pelo processo de autólise pós-morte.
6.1.2 Fixação
Após serem coletadas as amostras para histopatologia, as mesmas foram
colocadas em fixação ao formaldeído 10%, que é constituída em 10 volumes de
formalina comercial (40% deformaldeído em água) e 90 volumes de solução tampão
fosfato.

35
Com a fixação, previne-se a autólise pós-morte, facilita-se o corte dos
tecidos por tornarem mais endurecidos e estabilizam-se os componentes estruturais
em uma condição mais próxima da que havia in vivo (BANKS, 1991).
A imersão no fixador ocorreu imediatamente sua remoção do organismo.
A proporção ideal do volume do fixador e do volume do tecido deve ser de 30:1. O
tempo de fixação com o formaldeído gira em torno de 24 horas (BANKS, 1991).
6.1.3 Desidratação e Diafanização
Antes da inclusão da parafina para realização do corte histológico, procedeu-
se a retirada de cerca de 70 a 75% de água presente nos tecidos. Após a fixação, o
tecido é submetido a intensa lavagem com água e submetido a desidratação. Muitos
agentes desidratantes foram utilizados, sendo o de escolha o etanol.
6.1.4 Inclusão
Esta etapa permitirá que o espécime seja cortado fino o suficiente para que
possa ser estudada em microscópio óptico. Após a diafanização, os tecidos são
mergulhados em parafina fundida em uma estufa a 60º C. Devido ao calor o xilol
evapora, e os espaços ocupados por eles serão ocupados pela parafina. Em
seguida coloca-se o tecido em um recipiente retangular contendo um pouco de
parafina fundida e deixa-se solidificar a temperatura ambiente, formando-se um
bloco de parafina com o tecido no seu interior (JUNQUEIRA & CARNEIRO, 1999).
6.1.5 Corte
Após o endurecimento da parafina, os moldes foram removidos e os blocos
aparados para expor o tecido incluído. O bloco foi levado ao aparelho chamado
micrótomo rotativo de parafina MR2 e fatias finas (5 a 7 micrômetros) são removidas
da superfície de corte. O processo de corte é associado a uma leve compressão,
sendo as fitas colocadas para flutuar em banho de água quente, sendo
posteriormente recolhidas nas lâminas. (BANKS, 1991 ; JUNQUEIRA & CARNEIRO,
1999).
6.1.6 Coloração
O uso dos corantes facilita o estudo de componentes celulares e teciduais
por permitir uma adequada visualização dos mesmos em microscopia óptica. Alguns

36
deles são muito seletivos, sendo muito específicos para alguns componentes
celulares ou teciduais. Outros são mais seletivos para materiais extra-celulares.
Existem os corantes que não são seletivos, como a hematoxilina (H) e a eosina(E),
pois coram de uma forma geral os componentes celulares e teciduais. (BANKS,
1991). Após a fixação, as lâminas passarão pelos métodos de coloração:
6.1.6.1 Hematoxilina-eosina (HE)
A hematoxilina, corante básico, é aplicada primeiramente. Confere cor
púrpura-azulada aos componentes como cromatina e o ergastoplasma. A eosina,
corante ácido, confere cor rosa ou vermelha aos componentes celulares básicos
como citoplasma e produtos extra-celulares (BANKS, 1991).
6.1.6.2 Tricrômico de Masson
A coloração de Masson ou tricômico de Masson é um método de coloração
que visa ressaltar fibras colágenas na cor azul, tecido muscular e citoplasma em
vermelho. A presença de tecido conjuntivo, principalmente de fibras colágenas, foi
classificada histologicamente em pouco intensa, moderada e intensa.
6.1.6.3 Coloração Ácido Periódico de Shiff (PAS)
Esta coloração identifica a presença de glicogênio e glicoproteínas no citoplasma
celular. As células tumorais podem expressar uma glicoproteína na membrana
plasmática, denominada glicoproteína-P. Tal fato confere à célula neoplásica
habilidade reduzida em acumular fármacos anticâncer, sendo esta a principal causa
de falência dos tratamentos quimioterápicos (Andrade, et al, 2008) . Esta coloração
confere tonalidade avermelhada ao tecido em estudo. A presença de açúcares no
tecido analisado foi caracterizada histologicamente em pouco intensa, moderada e
intensa.

37
6.2 Metodologia -Extração de DNA genômico
Fragmentos dos tecidos serão digeridos com um volume da enzima
proteinase K (20mg/mL) de 1 ul para cada ml de tampão contendo Tris-HCl 10mM
pH 8.0, EDTA 50mM pH 8.0, SDS 0,5%, a 550C, por 72 horas. Após a lise das
células, o DNA genômico foi extraído pela metodologia de fenol/clorofórmio,
quantificado em espectrofotômetro e armazenado a -200C até o uso.
"Methylation sensitive arbitrarily primed-PCR” (MSAP-PCR)
O DNA genômico foi fragmentado com a enzima de restrição MseI
(Biolabs) e digerido com enzimas que compartilham do mesmo sítio de
reconhecimento (isoesquizomeros), mas mostram uma sensibilidade diferencial à
metilação do DNA (MspI e HpaII). A digestão incubada “overnight” (ON) a 37°C foi
amplificada em reações de PCR contendo MgCl2 (1.5mM), dNTP (1.25mM), 20uM de
primer aleatório (MLG-2 ou MGE-2 ou MGF-0) e 0.5U de TaqDNAPolimerase por 5
min. a 95° e 35 ciclos de 95° por 45 seg., 40° por 1 min., 72° por 1 min., e extensão
a 72º por 7 min. O produto da amplificação foi submetido a eletroforese em gel de
poliacrilamida que será revelado com uma solução contendo 0.6g de AgNO3. O perfil
de bandas gerado foi comparado entre tecido normal e tumoral, e, bandas que
possuírem um padrão de metilação diferencial nessa comparação serão os
potenciais marcadores moleculares de tumores de glândula mamária de cadelas.
7 RESULTADOS E DISCUSSÃO
Neste estudo foram obtidas 11 amostras de animais portadores de
neoplasias mamárias, que foram subdividos em11 amostras tumorais e 11 amostras
de tecidos mamários normais (ANEXO 4) .Em todas as amostras tumorais foi
realizada punção aspirativa por agulha fina, e biópsia de fragmento tumoral para fins
de comparação do diagnóstico citológico com o histopatológico.

38
7.1 - Citopatologia
Considerando os resultados obtidos pela citopatologia, foi mostrado que
dos 11 casos estudados 82% foram classificadas como carcinomas, 9% como
adenomas e 9% com resultado inconclusivo, demonstrando uma alta sensibilidade
diagnóstica, de cerca de 82%(Gráfico 1), e especificidade igual a zero.
Gráfico 1: Classificação citopatológica das amostras tumorais obtidas:
Segundo Zuccari, Santana e Rocha, 2001, o diagnóstico citológico possui
grande correlação com o diagnóstico definitivo histopatológico, fato este que confere
confiabilidade na realização da citologia aspirativa no pré-cirúrgico. Neste estudo
82% das amostras tumorais coletadas por citologia aspirativa por agulha fina tiveram
seu diagnóstico citológico de carcinomas, enquanto que 9% tiveram resultados
inconclusivos e 9% tiveram resultados positivos para adenoma.
No entanto, segundo Magalhães, et al, 2001, o diagnóstico por citologia
aspirativa por agulha fina não confere resultados confiáveis um vez que pode existir
grande variação de tumores e interferências como ulceração e inflamação, devendo
este método ser substituído por citologia esfoliativa. Neste trabalho, o único caso
com diagnóstico citológico inconclusivo constava de uma neoplasia ulcerada, com
Classificação citopatológica das amostras tumorais obtidas
Carcinoma
Adenoma
Inconclusiva 82%
9%
9%

39
extensas áreas de necrose e presença de infecção bacteriana local (ANEXO 3 B),
fato este que se correlaciona com o estudo de Magalhães, et al, 2001.
A citologia apresentou-se como técnica útil, especialmente quando se
observa sinais de malignidade nas células, como pleomorfismo, nucléolo evidente,
cromatina fortemente corada,entre outros. No entanto, quando não observados
estes sinais pode deixar indefinido o quadro, sendo fundamental estudo
histopatológico. Além disso, é um exame rápido de ser realizado, não necessitando
de anestesia do animal para sua realização e que muitas vezes revela um
diagnóstico preliminar. (REIS, F.R, et al, 2010).
7.2 - Histopatologia (Coloração Hematoxilina-eosina - HE)
Na análise dos resultados histopatológicos foram considerados os seguintes
critérios e achados histológicos para a graduação das neoplasias em carcinomas de
grau I, II e III, de acordo com a graduação preconizada pela Organização Mundial de
Saúde (OMS) /AFIP – Armed Forces InstituteofPathology (1999).
Carcinomas Grau I: Presença de estratificação do tecido glândulas
mamário podendo ou não ocupar toda a luz tubular. Tecido conjuntivo
bem delimitado nas margens. Ausência de alterações em estruturas
vasculares. Alto grau de atipia nuclear, presença de mais de 1 nucléolo
por célula.
Carcinoma Grau II: Aumento da atipia celular, presença de elevado
pleomorfismo nuclear e citoplasmático, elevado número de mitoses,
células neoplásicas com estruturas glandulares pouco delimitadas por
tecido conjuntivo, com caracteristicas de invasão aos tecidos
adjacentes.
Carcinoma Grau III: Presenca de todos os achados descritos
anteriormente, adicionado a invasão de estruturas vasculares,
sanguíneas ou linfaticas por células neoplásicas. Presenca de tecidos
linfoides adjacentes com infiltrado neoplásico
Todos podendo variar de simples a complexo, sendo que neste último
observam-se matrizes cartilaginosas no componente mesenquimal. Para fins de
análise de dados, os carcinomas foram agrupados em simples ou complexos e a
graduação de 1 a 3.

40
A análise histopatológica pela coloração de hematoxilina-eosina (HE)
mostrou que 36,4% das amostras foram classificadas como carcinomas mamários
simples grau I, 27,2% carcinomas mamários simples grau II, 18,2 % carcinomas
mamários simples grau III e 18,2% carcinomas mamários complexos grau I (gráfico
2). A alta prevalência de carcinomas simples também foi relatada por outros estudos
(DALECK et al. 1998, ZUCCARI et al. 2001, OLIVEIRA et al.2003).
Gráfico 2.: Distribuição dos tumores mamários avaliados segundo a
classificação histológica
Segundo Hatore, 2011, em um estudo constando de 21 cadelas
portadoras de neoplasias mamárias, mais de 76% dos casos revelaram carcinoma
mamário, reforçando a elevada prevalência deste tipo tumoral. No presente estudo,
100% dos casos revelaram carcinomas mamários, de variados graus. Esta diferença
pode ser explicada pelo fato de que neste trabalho todas as cadelas que
participaram do estudo eram consideradas idosas (acima de 10 anos e no estudo
anterior de Hatore, das 21 cadelas, 5 eram consideradas adultas e 16 idosas. No
entanto, deve-se considerar o porte do animal estudado. Animais de raças
pequenas, como poodles, yorkshires têm expectativa de vida maior que um cão de
raça grande, como um rottweiler por exemplo. Isto deve ser levado em consideração
nos estudos. Neste trabalho todos os animais eram sem raça definida, mas
possuíam porte médio, com peso entre 10 a 25 kg.
A comparação realizada entre o diagnóstico citológico e histopatológico
as amostras neoplásicas pode ser mostrada no Quadro 1.Na coluna 1 está
Classificação histopatológica das amostras tumorais obtidas
Carcinoma mamário simples Grau I
Carcinoma mamário simples Grau II
Carcinoma mamário simples Grau III
Carcinoma complexo Grau I
36,4%
27,3%
18,2%
18,2%

41
representado o número de prontuário de cada amostra. Na coluna 2 a classificação
baseada na análise citopatológica e na coluna 3 a classificação baseada na análise
histológica.
Quadro 2: Comparação entre diagnóstico citológico e histopatológico de amostras
tumorais avaliadas nesse estudo e baseada na coloração padrão hematoxilina-
eosina (HE)
Número do prontuário Diagnóstico citológico Diagnóstico histopatológico
14.217 Carcinoma mamário Carcinoma papilíferosimples II
13.864 Carcinoma mamário Carcinoma
sólidopoucodiferenciado
14.170 Carcinoma mamário Carcinoma túbulo
papilíferosimples III
14.110 Carcinoma mamário Carcinoma tubular complexo I
13.943 Carcinoma mamário Carcinoma tubular simples I
14.055 Carcinoma mamário Carcinoma tubular complexo I
14.134 Adenoma mamário Carcinoma túbulo papilífero
simples III
14.289 Carcinoma mamário Carcinoma túbulo
papilíferosimples II
13.957 Inconclusivo Carcinoma papilíferosimples I
13.464 Carcinoma mamário Carcinoma tubular simples I
14.179 Carcinoma mamário Carcinoma tubular simples I
Os critérios de malignidade mais observados foram atipia e pleomorfismo
celular. Já o padrão nuclear se mostrou decisivo no diagnóstico, visto que a
presença de um ou mais nucléolos e o padrão irregular da cromatina só podem estar
presentes em tecidos neoplásicos. No entanto, o componente inflamatório, quando
presente no esfregaço, se mostrou um fator negativo para o diagnóstico preciso das
neoplasias mamárias. A única amostra 13.957 (Quadro 1) cujo resultado da
avaliação citológica foi inconclusivo contava com grande ulceração e áreas de
necrose pelo tecido, conforme pode ser evidenciado no ANEXO 4 , figura B.
A avaliação citológica pode revelar falsos positivos, uma vez que a
amostra coletada pode não condizer com o processo neoplásico do tecido como um

42
todo. Fato este pode ser evidenciado na amostra 14.134 ( Quadro 1). Neste caso a
punção ocorreu na periferia da lesão.
A avaliação citológica é de grande valia para diagnósticos precoce, mas
no entanto deve ser sempre confirmada com a histopatologia convencional.
A título de comparação, em todas as amostras de mamas aparentemente
normais foram realizadas as colorações histológicas hematoxilina-eosina (HE)
(Figura 4), Tricômio de Masson (MASSON) (Figura 5) e ácido periódico de shiff
(PAS) (Figura 6). Para todas as amostras consideradas normais coletadas durante o
procedimento cirúrgico o exame histopatológico revelou tecido mamário cuja
estrutura é composta por lóbulo pequeno e bem delimitado por tecido conjuntivo
denso. As estruturas glandulares são delicadas e apresentam epitélio monomórfico e
em monocamada, caracterizando um tecido normal.
Figura 4: (HE/40X): Fotomicrografia em aumento de 40X de tecido glandular controle-normal. Percebe-se que a estrutura e composta por lóbulo pequeno e bem delimitado por tecido conjuntivo denso (setas). As estruturas glandulares são delicadas e apresentam epitélio monomorfico e em monocamada

43
Figura 5 (Tricrômio de Masson /40X): Fotomicrografia em aumento de 40X
tecido glandular controle-normal. Tecido conjuntivo perilobular com marcação
intensa azulada (setas).
Figura 6 (PAS/40X): Fotomicrografia em aumento de 40X tecido glandular controle-normal. Marcação pouco intensa de açucares em rosa (setas) em tecido conjuntivo
Em todas as amostras tumorais foram realizadas as colorações
histológicas hematoxilina-eosina (HE) (Figura 7), ácido periódico de shiff (PAS)
(Figura 8) e Tricômio de Masson (MASSON) (Figura 9). Através destas colorações

44
especiais (MASSON e PAS) foi avaliada a presença de tecido conjuntivo, em
especial colágeno e de açúcares, respectivamente, nas amostras tumorais.
Observou-se que nas neoplasias malignas existe marcação de moderada a intensa
para açúcares e de pouco intensa a moderada de colágeno, variando de acordo com
cada caso.
Figura 7- (HE/40X): Fotomicrografia em aumento de 40X de tecido glandular com alto índice de proliferação com característica maciça não permitindo a visualização do tecido conjuntivo (seta) (A). (HE 100X): lóbulos glandulares com difícil distinção. Células neoplásicas com núcleo grande dando uma característica mais basofílica (seta) ao tecido na histologia. Carcinoma papilar simples grau I (B).
Figura 8 (Tricrômio de Masson/40X): Fotomicrografia em aumento de 40X de tecido conjuntivo escasso e com pouca produção de colágeno,marcação pouco intensa (seta). Somente é possível caracterizá-lo na periferia do lóbulo (A) . (Tricrômio de Masson/100X): Fotomicrografia em aumento de 100X de tecido conjuntivo delicado e com moderada quantidade de colágeno (seta).Carcinoma papilar simples grau I (B).
A B
A B

45
Figura 9 (PAS/40X): Fotomicrografia em aumento de 40X. Observamos que tecido conjuntivo perilobular apresenta pouca eosinofilia (A). (PAS/100X): Fotomicrografia em aumento de 100X o tecido conjuntivo de sustentação é delicado e apresenta de grande quantidade de açúcares caracterizados pela intensa eosinofilia (seta).Carcinoma papilar simples Grau 1 (B) Os dados obtidos de todas as colorações das amostras normais estão
agrupados no quadro a seguir (Quadro 2), seguindo o critério adotado de marcação
pouco intensa, moderada e intensa, tanto para tecido conjuntivo, principalmente
colágeno, quanto para açúcares.

46
Quadro 3: Dados obtidos das amostras normais, de acordo com o numero de identificação do animal (numero do prontuário, histopatológico, colorações de PAS e MASSON)
Número do
prontuário
Diagnóstico
histopatológico
(HE)
PAS MASSON
14.217 Tecido mamário normal
POUCO
INTENSA
INTENSA
13.864 Tecido mamário normal
POUCO
INTENSA
INTENSA
14.170 Tecido mamário normal
POUCO
INTENSA
INTENSA
14.110 Tecido mamário normal
POUCO
INTENSA
INTENSA
13.943 Tecido mamário normal
POUCO
INTENSA
MODERADA
14.055 Tecido mamário normal
POUCO
INTENSA
INTENSA
14.134 Tecido mamário normal
POUCO
INTENSA
INTENSA
14.289 Tecido mamário normal
POUCO
INTENSA
INTENSA
13.957 Tecido mamário normal
POUCO
INTENSA
INTENSA
13.464 Tecido mamário normal
POUCO
INTENSA
INTENSA
14.179 Tecido mamário
normal
POUCO
INTENSA
MODERADA
Os dados obtidos de todas as colorações das amostras tumorais estão
agrupados no quadro a seguir, seguindo o critério adotado de marcação pouco

47
intensa, moderada e intensa , tanto para tecido conjuntivo, principalmente colágeno ,
quanto para açúcares.
Quadro 4: Dados obtidos das amostras tumorais, de acordo com o numero de identificação do animal (numero do prontuário, diagnostico citológico, histopatológico, colorações de PAS e MASSON)
Número do
prontuário-
Número da
amostra
Diagnóstico
citológico
Diagnóstico
histopatológico (HE)
PAS MASSON
14.217 –T10 Carcinoma
mamário
Carcinoma
papilíferosimples II
INTENSA POUCO
INTENSA
13.864 –T9 Carcinoma
mamário
Carcinoma
sólidopoucodiferenciado
INTENSA POUCO
INTENSA
14.170 -T7 Carcinoma
mamário
Carcinoma túbulo
papilíferosimples III
MODERADA POUCO
INTENSA
14.110-T6 Carcinoma
mamário
Carcinoma tubular
complexo I
INTENSA POUCO
INTENSA
13.943-T3 Carcinoma
mamário
Carcinoma tubular
simples I
INTENSA MODERADA
14.055-T1 Carcinoma
mamário
Carcinoma tubular
complexo I
INTENSA POUCO
INTENSA
14.134-T2 Adenoma
mamário
Carcinoma túbulo
papilífero simples III
INTENSA POUCO
INTENSA
14.289-T5 Carcinoma
mamário
Carcinoma túbulo
papilífero simples II
INTENSA MODERADA
13.957-T8 Inconclusivo Carcinoma
papilíferosimples I
INTENSA POUCO
INTENSA
13.464-T11 Carcinoma
mamário
Carcinoma tubular
simples I
INTENSA MODERADA
14.179-T4 Carcinoma
mamário
Carcinoma tubular
simples I
INTENSA MODERADA

48
Notou-se que as células de uma glândula mamária normal possuem intensa
marcação para tecido conjuntivo e colágeno e marcação pouco intensa para
açúcares segundo as colorações especiais de MASSON e PAS, respectivamente.
Nos carcinomas observamos nitidamente o contrário, fato este que pode ser
explicado pelo hipermetabolismo e intensa produção de açucares pela célula
neoplásica.
Para verificação da existência de correlação entre as colorações de PAS e de
MASSON relacionadas com os subtipos tumorais, foi utilizado o teste estatístico do
Qui-Quadrado (X2), uma vez que as variáveis são qualitativas. As 22 amostras,
sendo 11 normais e 11 tumorais foram avaliadas quanto à marcação pelos corantes
de PAS e MASSON.
Na tabela 1 foram colocadas nas linhas a classificação histológica do tumor e
nas colunas a intensidade de marcação pelo corante de PAS (pouco intensa,
moderada e intensa), e o número de amostras correspondentes:
Tabela 1.Amostras tumorais – Coloração de PAS
PAS INTENSA MODERADA POUCO INTENSA
Carcinoma tubular simples I
3 0 0
Carcinoma papilifero simples II
1 0 0
Carcinoma sólido
1 0 0
Carcinoma túbulo-papilifero simples III
1 1 0
Carcinoma tubular complexo I
2 0 0
Carcinoma papilifero simples I
1 0 0
Carcinoma túbulo papilifero simples II
1 0 0
P=0,55 Não existe diferença estatística
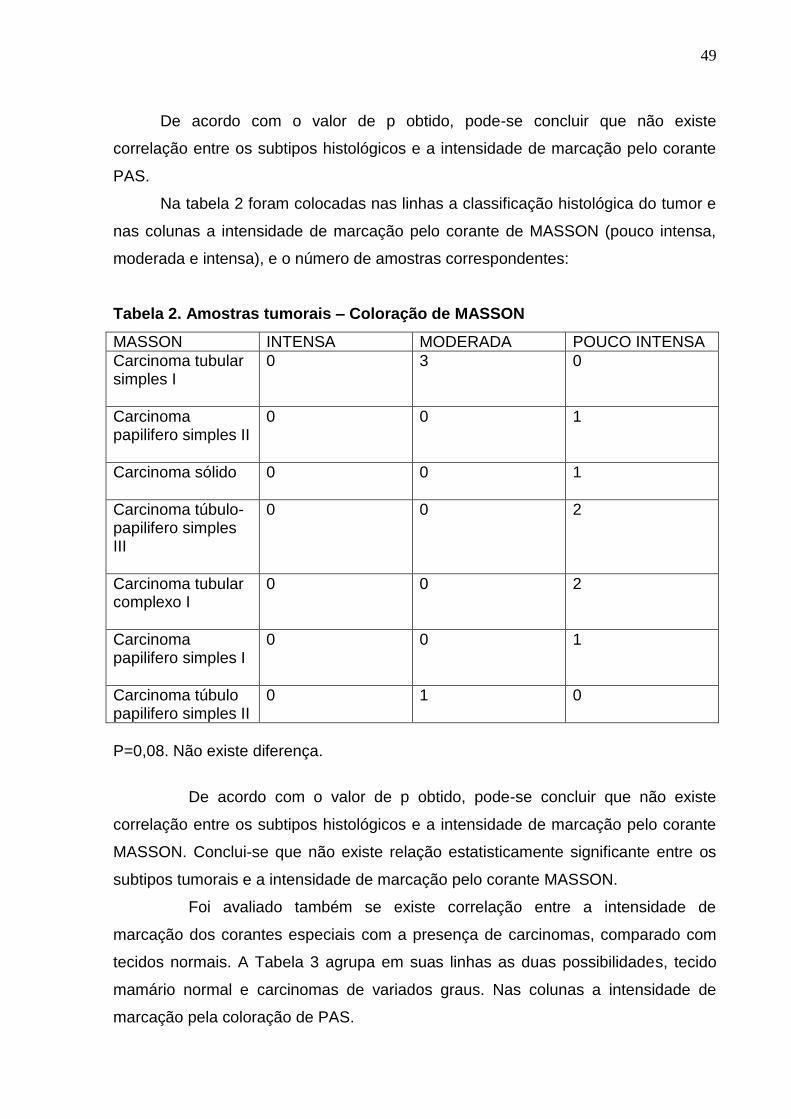
49
De acordo com o valor de p obtido, pode-se concluir que não existe
correlação entre os subtipos histológicos e a intensidade de marcação pelo corante
PAS.
Na tabela 2 foram colocadas nas linhas a classificação histológica do tumor e
nas colunas a intensidade de marcação pelo corante de MASSON (pouco intensa,
moderada e intensa), e o número de amostras correspondentes:
Tabela 2. Amostras tumorais – Coloração de MASSON
MASSON INTENSA MODERADA POUCO INTENSA
Carcinoma tubular simples I
0 3 0
Carcinoma papilifero simples II
0 0 1
Carcinoma sólido
0 0 1
Carcinoma túbulo-papilifero simples III
0 0 2
Carcinoma tubular complexo I
0 0 2
Carcinoma papilifero simples I
0 0 1
Carcinoma túbulo papilifero simples II
0 1 0
P=0,08. Não existe diferença.
De acordo com o valor de p obtido, pode-se concluir que não existe
correlação entre os subtipos histológicos e a intensidade de marcação pelo corante
MASSON. Conclui-se que não existe relação estatisticamente significante entre os
subtipos tumorais e a intensidade de marcação pelo corante MASSON.
Foi avaliado também se existe correlação entre a intensidade de
marcação dos corantes especiais com a presença de carcinomas, comparado com
tecidos normais. A Tabela 3 agrupa em suas linhas as duas possibilidades, tecido
mamário normal e carcinomas de variados graus. Nas colunas a intensidade de
marcação pela coloração de PAS.

50
Tabela 3. Tecido mamário normal x Carcinomas – Coloração de PAS
POUCO INTENSA MODERADA INTENSA
Carcinomas 0 1 10
Tecido mamário
normal
11 0 0
Feito teste Qui-quadrado: p < 0,0001
Através do valor de p obtido pelo teste do qui-quadrado, nota-se que há
forte associação entre as variáveis. O carcinoma está associado à coloração intensa
de PAS, revelando a presença de glicoproteínas e glicogênio desta célula
modificada. Em tecido mamário normal, a coloração em questão mostrou-se com
pouca intensidade de marcação.
Tabela 4. Tecido mamário normal x Carcinomas – Coloração de MASSON
POUCO INTENSA MODERADA INTENSA
Carcinomas 7 4 0
Tecido mamário
normal
0 2 9
Feito teste Qui-quadrado: p < 0,0002
Através do p valor obtido, nota-se que existe associação entre as variáveis. O
tecido tumoral está associado à marcação pouco intensade MASSON, uma vez que
ocorre perda da estrutura tecidual normal da glândula mamária.
De uma maneira peculiar, as células cancerosas produzem energia
preferencialmente pela glicólise anaeróbia em detrimento da fosforilação oxidativa
mitocondrial (WARBURG, 1966; REITZER, 1979; ROSSIGNOL, 2004 ).
A fosforilação oxidativa fornece energia para o citoplasma e a glicólise
anaeróbia para o núcleo. O núcleo parece ser o compartimento mais susceptível à
deficiência de ATP, não somente nas células normais como nas células malignas. A
perda de ATP no núcleo de células malignas possui conseqüências drásticas

51
provocando o impedimento de cruciais funções celulares como a transcrição e a
replicação do DNA.
As células cancerosas apresentam uma grande variedade de estados de
diferenciação, indo de células altamente diferenciadas, em relação a célula da qual o
tumor se originou, com uma glicólise anaeróbia normal e uma baixa taxa de
crescimento; até células altamente indiferenciadas com alta glicólise anaeróbia e
rápida velocidade de crescimento (BAGGETTO, 1992).
A intensidade da marcação de açucares nos tecidos neoplásicos pode ser
explicada pela alta taxa metabólica das células tumorais, principalmente pela alta
capacidade de metabolismo anaeróbico.
As células cancerosas são metabolicamente adaptadas para o rápido
crescimento e proliferação em condições de pH ácido e baixas tensões de oxigênio,
condições nas quais as células normais pouco cresceriam ou mesmo não
conseguiriam sobreviver (GRIFFITHS, 2001)
Em 2004, Isidoro e Cuezva, analisaram marcadores da glicólise e da
mitocôndria em outros tipos de câncer: adenocarcinoma de mama, estômago e
próstata; carcinoma de pulmão e carcinoma epidermóide de esôfago. Quando
comparado com os tecidos normais correspondentes, os autores encontraram
significante diferença na expressão dos marcadores glicolíticos e mitocondriais em
todos esses tipos de câncer. A conseqüência metabólica do impedimento
mitocondrial é o desvio de produção de ATP celular via glicólise anaeróbia.
Segundo Martins, Tamaso e Guerra 2002, em um estudo retrospectivo sobre
a matriz extracelular de carcinomas mamários caninos, quanto maior for a
malignidade neoplásica menor será a presença de tecido conjuntivo estrutural
permeando as células neoplásicas, uma vez que as mesmas possuem alta taxa de
crescimento e o microambiente tecidual com ph extremamente ácido é prejudicial ao
crescimento de matriz conjuntiva, principalmente colágeno.
Em humanos a presença de glicogênio e demais açúcares no tecido
neoplásico mamário está diretamente relacionada com o alto grau de malignidade
(MARKOPOULOS, 2008)
Em um estudo histoquímico de tumores testiculares de cães, foi verificado
presença do componente colágeno de forma desorganizada ao redor do processo
neoplásico, enquanto que ausência do mesmo por entre as células neoplásicas
(STAUT, et al 2008).Estes dados também são observados neste trabalho, ficando

52
evidente a escassez de tecido conjuntivo por entre as células de um carcinoma
mamário.
O estado de hipermetabolismo ou catabolismo persistente é comum em
estados avançados do câncer, isto explica a rapidez das células neoplásicas
malignas em captar glicose como fonte de energia. O aumento desta captação está
relacionado com o grau de malignidade e poder de invasão celular.
A célula tumoral tem como principal substrato energético a glicose,
consumindo de 10 a 50 vezes mais em relação ás células normais. Dependendo do
tamanho do tumor e do estágio da doença a produção da glicose é maior. O
consumo excessivo de glicose pelo tumor aumenta a produção de glicose hepática a
partir do lactato e de aminoácidos musculares do hospedeiro, podendo levar o
paciente à caquexia ( GIBNEY, 2007; DA SILVA, 2005).
Com estas análises pode-se afirmar que existem diferenças significativas de
intensidade de coloração entre tecido mamário normal e tecido mamário neoplásico.
No entanto, as mudanças de intensidade de marcação do tecido tumoral não têm
correlação com o subtipo histológico. Desta forma, passamos a avaliar marcadores
moleculares que pudessem ser utilizados para classificação dos subtipos de tumores
encontrados nessa casuística.
7.3 ESTUDO EPIGENÉTICO
7.3.1 - Quantificação e avaliação da integridade de DNA genômico
Nesse estudo foi padronizada a lise dos fragmentos de tecidos normais e
tumorais com proteinase K em tampão de lise. Após a lise das células, o DNA
genômico foi extraídopelo método fenol-clorofórmio, pois apesar de ser um método
antigo e tradicional, é o método que obtemos um grau maior de pureza do DNA
obtido. Após, foi realizada a leitura espectrofotométrica em absorbância de 260nm e
calculada a concentração do DNA em ng/ml. Para o cálculo da mesma, utilizamos a
seguinte fórmula: Concentração DNA (ng/ul) = Abs(260nm) X 50 (fator de correção
para DNA) X 40 (diluição). Este método se mostrou muito eficaz e forneceu amostras
de DNA íntegros de alta qualidade (Figura 13 – gel de agarose)

53
Neste trabalho foram obtidos o DNA genômico das amostras a seguir, sendo
que para cada paciente, foram analisadas 2 amostras, sendo uma tumoral (T) e a
outra normal (N). Os números de prontuário dos animais estão representados em
conjunto com a nomenclatura das amostras:
Tabela 5 – Dados de leitura espectrofomêtrica do DNA genômico obtido de tecidos
normais (N) e tumorais (T) da mama de cadelas.
Amostras-
n.prontuário
Diagnóstico
histopatológico
Abs 260
nm
Abs
280
nm
Concentração
ng/ul
N1- 14.055 Tecido mamário normal 0,9 0,533 1800
T1- 14. 055 Carcinoma tubular
complexo I
0,107 0,021 214
N2- 14.134 Tecido mamário normal 0,023 0,009 46
T2- 14.134 Carcinoma túbulo
papilífero simples III
0,319 0,143 638
N3- 13.943 Tecido mamário normal 0,083 0,104 166
T3- 13.943 Carcinoma tubular
simples I
0,5 0,243 1000
N4- 14.179 Tecido mamário normal 0,48 0,369 960
T4- 14.179 Carcinoma tubular
simples I
0,464 0,292 928
N5- 14.289 Tecido mamário normal 0,065 0,251 130
T5- 14.289 Carcinoma túbulo
papilífero simples II
0,166 0,052 332
N6-14.110 Tecido mamário normal Erro
técnica
T6-14.110 Carcinoma tubular
complexo I
Erro
técnica
N7-14.170 Tecido mamário normal 0,824 0,399 1648
T7-14.170 Carcinoma túbulo
papilíferosimples III
0,879 0,474 1758
N8-13.957 Tecido mamário normal 0,572 0,219 1144
T8-13.957 Carcinoma 0,796 0,385 1592

54
papilíferosimples I
N9-13.864 Tecido mamário normal 0,248 0,099 496
T9-13.864 Carcinoma
sólidopoucodiferenciado
0,901 0,560 1802
N10-14.217 Tecido mamário normal 0,458 0,201 916
T10-14.217 Carcinoma
papilíferosimples II
0,688 0,310 1375
Determinação da concentração do DNA em ng/ul, utilizando-se a fórmula:
Concentração DNA (ng/ul) = Abs(260nm) X 50 (fator de correção) X 40
(diluição), onde T = tecido tumoral e N= tecido normal
A análise da integridade dessas amostras pode ser visualizada por
eletroforese em gel de agarose corado com brometo de etídio (Figura 10). Pode se
observar bandas de alto peso molecular (acima de 23 kb, setas amarelas Figura 10)
demonstrando a presença de DNA de excelente qualidade para prosseguimento dos
estudos moleculares.
Figura 10: Eletroforese em gel de Agarose corado com brometo de etídio. As
amostras de DNA genomico (setas brancas) foram aplicadas nas canaletas de 1-4,
gel 1 e 2-6 gel 02).Bandas de 23 Kb (setas amarelas).
T3 N1 N5 N2 PM T 2 N3 T5 T1 PM T4
Gel 01 Gel 02

55
7.3.2 - Avaliação do perfil de metilação por MSAP-PCR
Padrões de metilação do DNA foram gerados pela técnica de MSAP-PCR
utilizando primer aleatório MGE-2 e eletroforese em gel de poliacrilamida corado
pela prata. Utilizamos a enzima MseIcujo sítio de reconhecimento está representado
no DNA genômico humano com uma alta frequência. A geração desseperfilde
bandas permitiu a diminuição da complexidade da análise,embora a exploração de
mais regiões genômicas diferencialmente metiladas seja restringida pelo tratamento
com essa enzima.
As enzimas HpaII e MspI são isoesquizômeros, ou seja, reconhecem a
mesmasequência de DNA. Desta forma, os fragmentos gerados por MseI que
apresentam sitio para estas enzimas, devem necessariamente ser digeridos com
MspI (enzima não sensível a metilação). Esta cliva o fragmento independentemente
de estar ou não metilado. Se os fragmentos estiverem metilados, a enzima HpaI
(sensível a metilação), não irá digerir o DNA, mas se o fragmento estiver
desmetilado essa enzima reconhecerá o sitio e irá clivar o DNA, não gerando bandas
após a amplificação do DNA por PCR. Desta forma, através da análise do padrão de
bandas geradas é determinado o perfil de metilação de diversos fragmentos gênicos.
O perfil de bandas gerado por MSAP-PCR foi comparado entre amostras
normais (N) e os subtipos tumorais (T), de acordo com a classificação histológica
convencional, conforme mostra tabela 5.
Neste estudo foi demonstrado alteração do padrão de metilação de
fragmentos gênicos provenientes de tecidos normais e tumorais. Essa alteração
pôde ser observada tanto por um padrão de metilação em amostras tumorais e
hipometilação em tecido normal (banda de 340pb). Ou pela presença de fragmentos
hipometilados em tumor e metilados em tecido normal (banda de 600pb – Figura 11).

56
Eventos de alteração no padrão demetilação do DNA em células tumorais já é
bem conhecido em diversos tipos de neoplasias malignas. Tem sido mostrado que o
padrão de metilação do DNA é invertido, havendo hipermetilação de promotores
ricos em CG e hipometilação geral do genoma, contribuindo para uma instabilidade
genômica. Ainda, este processo alterado de metilação não é aleatório e a
hipermetilação específica de genes (por exemplo: genes supressor de tumor) parece
estar associada com a tumorigênese, facilitando o processo de invasão e metástase
(OLIVEIRA, et al, 2010) (COSTA, 2010). Os estudos sobre marcadores
moleculares em neoplasias mamárias na medicina veterinária vêm ganhando cada
vez mais espaço, uma vez que a busca por um prognóstico mais acurado e por um
tratamento mais personalizado cresce a cada dia.
Neste trabalho foram identificadas regiões genômicas que apresentaram
padrão de metilação alterado entre os subtipos tumorais. Foi mostrado um fragmento
de aproximadamente 400pb que se apresentou metilado na amostra de tumor sólido
(T9) e desmetiladas nas demais amostras tumorais, do tipo tubular (T3e T4) e
papilífero (T8) (Figura 12).
Figura 11: Banda de 600pbmetilada em amostras normal e desmetilada no tecido
tumoral
250 pb
500 pb

57
Observou-se também uma banda de 390 pb que estava metilada em
amostras tubulares complexas II (animal 3 e 4) e desmetilada em amostras de
tumores papilíferos (animal 8). Todos esses fragmentos representam potenciais
marcadores específicos de subtipos de tumores de mama canino e, futuros estudos
para sua identificação através de sequenciamento devem ser realizados.
Figura 12 – Bandas de 400pb, indicando metilação em carcinoma sólido e
desmetilação nas demais amostras tumorais tubulares e papilíferas

58
Figura 13 = Banda de 390pbmetilada em amostras de carcinomas tubulares
complexos e desmetilados em carcinomas papilíferos
O fenótipo dos tumores mamários em humanos está intimamente ligado com
o padrão de metilação encontrado nas ilhas CPG. Foi comprovado que os tumores
mamários em seres humanos possuem diferente padrão de metilação das ilhas CpG
(PARK, 2012). Em um estudo comparativo entre o perfil de metilação e fenótipo
tumoral, Baeet al em 2004 demonstrou que o gene BRCA1 mostrou-se com um perfil
de hipermetilação em carcinomas mucinosos, cujas células neoplásicas estão
depositadas sobre muco, comparado com carcinomas ductais em seres humanos
(BAE, et al, 2004). Em um outro estudo, a metilação aberrante do DNA foi associada
com os subtipos tumorais classificados de acordo com critérios clinico-patológicos de
câncer de mama em humanos. A expressão de bcl-2 e de receptores hormonais esta
associada com o grau de metilação do gene SFRP1 (JEONG, et al 2013). É possível
que as mesmas alterações aconteçam em cadelas, uma vez que existe grande
similaridade entre os tipos celulares dos tumores mamários caninos e humanos.
Esses estudos demonstram a identificação de possíveis marcadores de
subtipos tumorais com padrão de hipermetilação e hipometilação. Além disso, os
estudos mostram uma diminuição global da metilação em todo o genoma em muitos
tumores, entretanto, eles não fornecem informações sobre onde a hipometilação
ocorre (SOARES et al. apud PLASS et al., 2002). Os fragmentos genicos
observados neste trabalho podem representar novas regiões do DNA tumoral com

59
padrão de hipometilação. Foi observada uma banda de 600pb desmetilada em todos
os subtipos tumorais (tubular, papilífero e sólido) avaliados. Outra banda de 400pb
também apresentou padrão de hipometilação em tumores papilíferos e tubulares,
mas não em sólido. E uma banda de 390pbapresentou-se desmetiladasomente em
tumorpapilífero.
Estudos posteriores de identificação desses marcadores se fazem
necessários para caracterizar marcadores moleculares de classificação de subtipos
tumorias de tumores mamários caninos e identificar novas regiões hipometiladas no
DNA genômico da célula tumoral.
8 CONCLUSÃO
O presente estudo demonstrou que existe uma concordância entre os
diagnósticos citológico e histopatológico na ordem de 82%., de acordo com a
literatura. Em amostras tumorais, à coloração HE, nota-se perda da estrutura
tecidual lobular mamária, perda de organização tecidual, estratificação do tecido,
pleomorfismo celular, condizentes com carcinomas.
Com as colorações especiais de PAS e MASSON, observa-se diferença
significativa entre amostras normais e tumorais. Em tecido normal existe marcação
intensa para MASSON e pouco intensa para PAS. Em tecido tumoral existe
marcação intensa para PAS e pouco intensa para MASSON. No entanto a
intensidade de coloração tanto para PAS quanto para MASSON, não tem
correlaçãocom os subtipos histológicos estudados.
O estudo epigenético revelou alterações no perfil de metilaçãodos tecidos
normais e tumorais da glândula mamária canina. Foram constatadas bandas com
padrão de metilação específico de subtipos tumorais. Padrão metilado em tumor
sólido e em tumores tubulares complexo I. Epadrão desmetilado em tumor papilífero,
fato este que reforça ainda mais a necessidade do prosseguimento dos estudos,
afim de identificar genes com perfil de metilação alterado, e com isso, buscar
marcadores moleculares para a espécie canina.

60
9 REFERÊNCIAS BIBLIOGRÁFICAS
ANDRADE, A L. et al. Avaliação imunoistoquímica da expressão da
glicoproteína-P em tumores de mama de cadelas tratadas com prednisona.
Veterinaria e Zootecnia.v.15,n.1, p.75-84, 2008
BAE, et al. Hypermethylation in histologically distinct classes of breast cancer.Department of Pathology, Yeungnam University College of Medicine. Clinical Cancer Research. 2004 Sep 15;10(18 Pt 1):5998-6005.
BAGGETTO,LG. Deviant energetic metabolism of glycolytic cancer cells.
Biochimie. 74,959-974,1992
BANKS, W. Histologia veterinária aplicada.1998.2ª edição, São Paulo;Manole
BAYLIN, S B. DNA methylation and gene silencing in cancer. Nat. Clin. Pract.
Oncology. 2005; 1:4-11.
BOTELHO, C F. Estudo histopatológico, imunohistoquímico e de cultura de
células de tecidos normais e cancerosos da mama humana e canina.Dissertação
de mestrado. Universidade Federal de Viçosa, 2009.
CARVALHO, T.B; Neoplasia mamária em cadelas: Caracterização
histopatológica e expressão de proteínas de estresse (HSP 72).Tese de
Mestrado.Universidade Federal de Viçosa. Minas Gerais, 2006.
CASSALI, et al. Consensus for the Diagnosis, Prognosis and Treatment of
Canine Mammary Tumors.Braz. Journal of Vet. Pathology, 2011, 4(2), 153-180.
CIRILLO, J. V; Tratamento quimioterápico das neoplasias mamárias em cadelas
e gatas. Rev. Inst. Ciências da Saúde. 2008:26(3); p-325-327

61
COSTA, F. F; Epigenomics in cancer management. Cancer Management and
Research. 2010:2, p.255-265.
CUEZVA, J.M.; et al.. The bioenergetic signature of cancer : a marker of tumor progression. Cancer Res. 62, 6674-6681, 2002 DA SILVA, M. P. N; Síndrome da anorexia-caquexia em portadores de câncer. Revista Brasileira de Cancerologia 2006; 52(1): 59-77
DALECK, C et al. Aspectos clínico e cirúrgicos do tumor mamário canino.Ciência
Rural, Santa Maria, v.28. n.1, p.95-100, 1998.
DE NARDI, A B et al. Prevalência de neoplasias e modalidades de tratamento em
cães, atendidos no hospital veterinário da Universidade Federal do Paraná.
Archives Of Veterinary Science. V.7, n.2, p.15-26, 2002.
DUCASSE, M; BROWN, MA. Epigenetic aberrations and cancer. Mol Cancer
2006;5:60
DYCE, K. M.; SACK, W. O. WENSING, C. J. Tratado de anatomia veterinária
2004. Rio de Janeiro, Guanabara Koogan
EINSEMBERG, A L A; KOIFMAN, S. Câncer de mama:marcadores tumorais
(revisão de literatura).RevBras Cancerologia 2001; 47 (4):377-388
FELICIANO, M.A.R; et al. Estudo clínico, histopatológico e imunoistoquímico de
neoplasias mamárias em cadelas. Arq. Bras. Med. Vet. Zootec. v.64. n.5.
p.1094-1100, 2012.
FILHO,J C et al. Estudo retrospectivo de 1647 tumores mamários em cães.Pesq.
Vet Bras. 30(2): 177-185, 2010
FILHO, J.C. O. Estudo retrospectivo de 1647 tumores mamários em cães.
Dissertaçao de Mestrado.Santa Maria, 2010.

62
FRANDSOM, R. D.Anatomia das glândulas mamárias. In:
FrandsomRD.Anatomia e fisiologia dos animais domésticos 1979;Rio de
Janeiro; Guanabara Koogan
FONSECA, C S; DALECK, C R. Neoplasias mamárias em cadelas: influência
hormonal e efeitos da ovário-histerectomia como terapia adjuvante.Ciência
Rural, Santa Maria, v.30, n.4,p.731-735, 2000
GAMA, A; ALVES, A; SCHMITT F. Identification of molecular phenotypes in
canine mammary carcinomas with clinical implications: application of the human
classification. VirchowsArch. 2008;453(2):123-32
GIBNEY, M. J. et al. Nutrição clinica. Rio deJaneiro: Guanabara Koogan. 314-
343, 2007 p
GRIFFTHS, J.R.. Causes and consequenses of hypoxia and acidity in tumour microenvironments . Bioessays, 23:295-296, 2001
HATORE, T et al. Incidência de carcinoma nas neoplasias mamárias de cadelas
atendidas no Hospital Veterinário da Cesumar.VIIEPCC-
CESUMAR.Maringá,2011
ISODORO, A.et al. Alteration of the bioenergetic phenotype of mitochondria is a hallmark of breast, gastric, lung and oesophageal cancer. Biochem J. 378(17-20) 2004
ITOH, T. et al. Clinicopathological survey of 101 canine mammary gland tumors:
differences between small-breed dogs and others. Journal of Veterinary
Medical Science, Tokyo, v.67, n.3, p.345-347, 2005
JAENISCH, R; BIRD A. Epigenetic regulation of gene expression: how the
genome integrates intrinsic and environmental signals. Nat Genet, 2003; 33
:245–54.

63
JEONG, et al. Low methylation levels of the SFRP1 gene are associated with the basal-like subtype of breast cancer. Oncology Report. Department of Surgery, College of Medicine, Catholic University of Daegu, Daegu 705-718, Republic of Korea, 2013.
JONES, C . T; HUNT, L; KING, N. Patologia Veterinária.2000.6a edição, São
Paulo; Manole
JUNQUEIRA, L. C. &CARNEIRO. Histologia Básica . 9ª edição, Rio de Janeiro;
Guanabara Koogan.
KOPELOVICH L; CROWELL. J A; FAY, J R. The epigenome as a target for
cancer chemoprevention. J Natl Cancer Inst 2003; 95: 1747-57.
PARK, SY, et al. Distinct patterns of promoter CpG island methylation of breast cancer subtypes are associated with stem cell phenotypes. Mod Pathology.Feb;25(2):185-96
WARBURG, O.. Oxygen, the creator of differentiation. In: Current aspects of biochemical energetics ( eds. N.O. Kaplan and E. Kennedy) pp.103-109. London-New York: Academic Press 1966 KOWATA, C. H.; BENEDETTI, G. V.; TRAVAGLIA, T.; ARAÚJO, E. J. A. Fisiopatologia da caquexia no câncer: uma revisão. Arq.Ciênc. Saúde UNIPAR, Umuarama, v. 13, n. 3, p. 267-272, set/dez. 2009
LEE, J S; FACKLER, M J; LEE, J H; CHOI, C; PARK, M H; YOON, J H; ZHANG,
Z; SUKUMAR, S. Basal-like breast cancer displays distinct patterns of promoter
methylation. CancerBiol. Theraphy. 2010; 9(12):1017-24.
MAGALHÃES, et al.Estudo comparativo entre citopatologia e histopatologia no
diagnóstico de neoplasias caninas. Pesq. Vet. Bras. 21(1):23-32, jan./mar. 2001

64
MARKOPOULOS, C. Glycogen-rich clear cell carcinoma of the breast-case
report.World Journal of Surgical Oncology, 2008.
MARTINS, A.M.C.R.P.F.; TAMASO, E.; GUERRA, J.L. Retrospective review and systematic study of mammary tumors in dogs and characteristics of the extracellular matrix Braz. J. Vet. Res. Anim. Science., São Paulo, v.39, n.1, p. 38-42, 2002 MULAS, J M; MILLÁN, Y; DIOS, R. A prospective analysis of
munohistochemically determined estrogen receptor alpha and progesterone
receptor expression and host and tumor factors as predictors of disease-free
period in mammary tumors of the dog. Vet. Pathology. 2005; 42(2):200-12.
OLIVEIRA,L O et al. Aspectos epidemiológicos da neoplasia mamária canina.
Acta Scientiae Veterinariae.31 (2): 105-110, 2003.
PARK, SY, et al. Distinct patterns of promoter CpG island methylation of breast cancer subtypes are associated with stem cell phenotypes. Mod.Pathology.Feb;25(2):185-96
PELETEIRO, M C. Tumores mamários na cadela e na gata. Rev Port C Vet,
1994; LXXXIX (59) 10-29.
PIEKARZ, C H et al.Expressão das caderinas nos tumores mamários em
cadelas. ArchivesofVeterinary Science, v.13, n.1, p.13-21, 2008.
QUEIROGA, F; LOPES, C. Tumores mamários caninos – Novas perspectivas.
Congresso de CiênciasVeterinárias, 2002, SPCV, 10-12 Out, p.183-190.
REITZER, L.; WICE, B.; KENNEL, D. Evidence that glutamine, not sugar, is the major energy source for cultured Hela cells. J.Biol. Chem., 254:2669-2676, 1979
REIS, F.R, et al. Indícios sobre a correlação entre diferentes métodos
diagnósticos em casos de tumor de mama em cadelas. Revista Eletrônica Novo
Enfoque, ano 2010, v. 09, n. 09, p. 14 – 31

65
RIVERA, P; VON EULER, H. Molecular biológica
laspectsoncanineandhumanmammarytumors.Vet Pathology. 2011;48(1):132-46.
ROSSIGNOL, R.; GILKERSON, R.; AGGELER, R.; YAMAGATA, K.; REMINGTON, S.J.; CAPALDI, R.A.. Energy Substrate Mitochondrial Structure and Oxidative Capacity in Cancer Cells. CancerResearch 64, 985-993, 2004.
SALIBA, R. et al. Tratamento quimioterápico de adenocarcinomas mamário em
cadelas.FEMM/FIO-Ourinhos. São Paulo, 2011.
SANTOS, I L V et al. Construção de um novo par de primers para análises do
éxon 2 do proto-oncogene p 21 em cadelas.X Jornada de ensino, pesquisa e
extensão – JEPEX 2010, UFRPE.Recife, 2010
TERZIAN, A C et al. Avaliação da caspase-3 e KI-67 como marcadores
prognósticos nas neoplasias mamárias em cadelas. Braz. Vet. Res. Anim.
Science.São Paulo, v.44, n.2, p.96-102, 2007
UVA P; AURISICCHIO, L; WATTERS, J; LOBODA, A; KULKARNI, A; CASTLE,
J; RINALDIS, E. Comparative expression pathway analysis of human and canine
mammary tumors. BMC Genomics 2009; 10:135
YUAN, J. et al. Aberrant methylation and silencing of ARHI, an imprinted tumor
suppressor gene in which the function is lost in breast cancers. Department of
Experimental Therapeutics, The University of Texas, M. D. Anderson
CancerCenter,Cancer Research.Jul 15;63(14):4174-80 Houston, Texas 77030,
USA, 2003
ZUCCARI, D A; SANTANA, A E; ROCHA, N S. Correlação entre biópsia
aspirativa por agulha fina e a histologia no diagnóstico de tumores mamários de
cadelas. Brazilian Journal of Veterinary Research and Animal Science 2001;
3: 1-8.

66
ZUCCARI, D A et al. Fatores prognósticos e preditivos nas neoplasias mamárias-
importância dos marcadores imuno-histoquímicos nas espécies humana e
canina- estudo comparativo.Arq. Ciências da Saúde. Out/Dez 2008,15(4): 189-
98
ZUCCARI, D A et al. The maspin expression. In canine mammary tumors: An
imunohistochemical and molecular study. PesqVet Bras. 29(2):167-173,2009

67
10 ANEXO 1 Termo de Consentimento Livre e Esclarecido (TCLE) do proprietário ou responsável pelo animal TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO (modelo)
Título da Pesquisa: “Avaliação de padrões de metilação de DNA alterados em
neoplasias mamárias de cadelas”
Nome do (a) Pesquisador (a) Responsável: .Susana Nogueira Diniz. RG 1.15.615.
CPF 685656666-20. Uniban, Rua Maria Cândida 1813 - Vila Guilherme, São Paulo
Fone: 2967-9147
Nome dos demais participantes: Rodrigo Casemiro Pinto Monteiro. RG 29.213.092-
2.CPF 316.053.768-41. Uniban, Rua Maria Cândida 1813 - Vila Guilherme, São
Paulo Fone: 2967-9147
1. Natureza da pesquisa: o Sr. (sra.) está sendo convidada (o) a autorizar a
participação de seu(s) animal(is) nesta pesquisa que tem como finalidade avaliar
alterações de mecanismos epigenéticos no DNA da célula tumoral para buscar
biomarcadores de diferentes tipos tumorais.
2. Identificação do(s) animal(is): (identificar espécie, sexo, raça, quantidade,
nome ou número de registro (se for o caso).
3. Envolvimento na pesquisa: ao participar deste estudo o Sr. (Sra.) permitirá ao
(a) pesquisador (a) aproveite do material biológico que seria descartado para
extração do DNA da célula tumoral da mama de sua cadela. O Sr. (Sra.) tem a
liberdade de se recusar a participar e ainda se recusar a continuar participando
em qualquer fase da pesquisa, sem qualquer prejuízo para o seu animal. Sempre
que quiser poderá pedir mais informações sobre a pesquisa através do telefone
do (a) pesquisador e, se necessário, através do telefone da Comissão de Ética
no Uso de Animais (CEUA).
4. Riscos e desconforto: a participação nesta pesquisa não traz complicações
legais. Os termos de concordância com a realização do procedimento cirúrgico e

68
os riscos que este procedimento confere estão anexados e o Sr. (Sra.) também
deverar assinar. Os procedimentos adotados nesta pesquisa obedecem à
Resolução 879, de 15 de fevereiro de 2008.e à Lei Federal 11794, de 08 de
outubro de 2008.
5. Confidencialidade: todas as informações coletadas neste estudo são
estritamente confidenciais. Somente os pesquisadores terão conhecimento dos
dados.
6. Benefícios: esperamos que este estudo traga informações importantes sobre as
alterações epigenéticas em diferentes tipos tumorais, e, devido às semelhanças
com os tumores de mama na mulher,o conhecimento que será construído a partir
desta pesquisa poderá trazer constribuições importantes para diagnóstico,
tratamento, prognóstico e prevenção das neoplasias mamarias malignas.
7. Pagamento: o Sr. (Sra.) não terá nenhum tipo de despesa para participar desta
pesquisa, bem como, nada será pago por sua participação. Somente o
procedimento cirúrgico será custeado pelo Sr. (Sra.)Após estes esclarecimentos,
solicitamos o seu consentimento de forma livre para participar desta pesquisa.
Tendo em vista os itens acima apresentados, eu, de forma livre e esclarecida, manifesto meu consentimento em participar da pesquisa. ___________________________ Nome do Proprietário (CPF/RG) ______________________________ Assinatura do Proprietário __________________________________ Assinatura do Pesquisador Data: _____________________. TELEFONES Pesquisador: Susana Nogueira Diniz – 89758844 Rodrigo Monteiro – 8444-5050 CEUA: 2967-9022__________

69
11 ANEXO 2 – PARECER CEUA

70
12 ANEXO 3 – FOTOS DO PRÉ-CIRÚRGICO DAS PACIENTES PORTADORAS DE NEOPLASIAS MAMÁRIAS DO ESTUDO
B A
C D
E F

71
13 ANEXO 4: FOTOS DAS PEÇAS PÓS-CIRÚRGICAS ENVIADAS PARA EXAME HISTOPATOLÓGICO (COLUNA B) E DAS AMOSTRAS PARA ESTUDO EPIGENÉTICO (COLUNA A)
A B


















