
Prescrição de Medicamentos Jeniffer Barni Balconista de Farmácia.

0
WALDEMAR ALVES DA SILVA NETO
Tomada de decisão individual e aprendizado em Dinoponera quadriceps
(Ponerinae, Hymenoptera) forrageando em ambientes dinâmicos.
Dissertação apresentada à
Universidade Federal do Rio
Grande do Norte para obtenção
do título de Mestre em
Psicobiologia.
NATAL / RN
2014

1
WALDEMAR ALVES DA SILVA NETO
Tomada de decisão individual e aprendizado em Dinoponera quadriceps
(Ponerinae, Hymenoptera) forrageando em ambientes dinâmicos.
Dissertação apresentada à
Universidade Federal do Rio
Grande do Norte para obtenção
do título de Mestre em
Psicobiologia.
Orientador: Arrilton Araújo
NATAL / RN
2014

2
Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de
Biociências
Silva Neto, Waldemar Alves da.
Tomada de decisão individual e aprendizado em Dinoponera quadríceps (Ponerinae, Hymenoptera)
forrageando em ambientes dinâmicos / Waldemar Alves da Silva Neto. – Natal, RN, 2014.
39 f.: il.
Orientador: Prof. Dr. Arrilton Araújo.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências.
Programa de Pós-Graduação em Psicobiologia.
1. Dinoponera quadríceps. – Dissertação. 2. Aprendizado. – Dissertação. 3. Decisões de
forrageamento – Dissertação. I. Araújo, Arrilton. II. Universidade Federal do Rio Grande do Norte. III.
Título.
RN/UF/BSE-CB CDU 595.796

3
Título: Tomada de decisão individual e aprendizado em formigas Ponerinae
Dinoponera quadriceps forrageando em ambientes dinâmicos.
Autor: Waldemar Alves da Silva Neto
Data da defesa: 26 de agosto de 2014 – 14:30
Banca Examinadora
____________________________________________
Prof. Arrilton Araújo (Orientador)
Universidade Federal do Rio Grande do Norte
____________________________________________
Prof. Nicolas Chaline
Universidade de São Paulo
____________________________________________
Prof. Felipe Nalon de Castro
Universidade Potiguar
NATAL / RN
2014

4
“O desgosto e alegria dependem mais do que somos do que daquilo que nos
acontece.” (Multatuli)

5
AGRADECIMENTOS
Agradeço a minha família pelo eterno apoio às minhas escolhas. Cada
um é um grande exemplo e fonte de inspiração.
A minha namorada, Cíntia, pelo constante e fundamental apoio na
realização desse trabalho. Pela imensa paciência e compreensão ao longo da
realização desse trabalho. Agradeço imensamente também pela leitura, revisão
e correções sugeridas.
A Arrilton, pelas orientações, seja em questões acadêmicas, seja na vida
de maneira geral. Todas são de grande importância. Agradeço também pela
confiança depositada e contínuo suporte oferecido sempre que possível.
A Wall, pela colaboração e ensinamentos estatísticos ao longo desse
trabalho. Também pela paciência, afinal, aluno que não sabe estatística
costuma dar muito trabalho e “perturbar” bastante, hehehe.
A Dina e Jeniffer, que me apresentaram, ensinaram e me ajudaram a
trabalhar com a “Tocandira”. Dos primeiros contatos até hoje já se vão sete
anos (Nísia Floresta – 2008.1).
A recém doutora (uhul!), Priscila Fernandes, pela leitura e sugestões
feitas para melhorias no texto final.
Aos meus amigos da Biologia, em especial Bruno e Jessica, pelos quase
dez anos de companhia, conversas, angústias e sonhos compartilhados.
A todos do Laboratório de Biologia Comportamental (LBC), os pós-
graduandos, IC´s, voluntários, frequentadores outros, visitantes ou aqueles que
somente passaram rapidamente, mas contribuíram de alguma maneira com o
grupo (LBC).

6
A professora Fátima Arruda, pela participação na minha qualificação e
importante contribuição no projeto, assim como pela experiência passada a
todos nós alunos. Ah sim, também agradeço bastante pelo empréstimo do
notebook para que eu pudesse usar nas minhas observações. Parece besteira,
mas me ajudou bastante, hehe.
E finalmente a todos que dividem ou dividiram comigo momentos felizes.
A vida ultrapassa qualquer conhecimento.

7
RESUMO
No presente trabalho, avaliamos o efeito da distância do alimento,
sucesso de captura e tamanho do alimento e taxa de recompensa nas decisões
de forrageio tomadas por formigas da espécie Dinoponera quadriceps.
Também investigamos, medindo o tempo de permanência em cada área, a
influência do aprendizado no desempenho das operárias ao longo de
sucessivas viagens. Foram simulados quatro cenários. Cada operária realizou
10 viagens em cada cenário. Cenário 1: operárias sempre encontravam
alimento de alta qualidade; Cenário 2: operárias encontravam alimento de alta
qualidade em somente 50% das viagens; Cenário 3: operárias encontravam
alimento de alta e baixa qualidade com probabilidades de ocorrência de 0,5
para cada tamanho de alimento. Cenário 4: operárias tinham três
possibilidades, encontrar alimento de alta qualidade (33%), encontrar alimento
de baixa qualidade (33%) e não encontrar alimento. Em todos os cenários,
havia duas rotas possíveis de exploração, uma com 300 cm e outra com 600
cm de comprimento. A pesquisa mostrou que operárias da espécie D.
quadríceps tendem a retornar ao mesmo local onde o alimento foi encontrado
na viagem anterior, não importando a distância, tamanho do alimento ou taxa
de recompensa. Nos casos de viagens sem captura, operárias eram mais
propensas a trocar de área em busca de alimento. No entanto, no cenário 4
essa decisão de “troca” foi menos evidente, possivelmente pela maior dinâmica
do cenário. Resultados também indicaram um processo de aprendizado das
rotas de exploração assim como das condições das áreas de exploração. Com
a repetição das viagens, forrageadoras reduziram o tempo de busca nas áreas
nas viagens que não capturavam alimento e rapidamente trocavam de área.

8
ABSTRACT
When searching for food, animals often make decisions of where to go,
how long to stay in a foraging area and whether or not to return to the last
visited spot. These decisions can be enhanced by cognitive traits and adjusted
based on previous experience. In social insects such as ants, foraging
efficiency have an impact on both individual and colony level. The present study
investigated, in the laboratory, the effect of distance from food, capture success
and food size, and reward rate on decisions of where to forage in Dinoponera
quadriceps, a ponerine ant that forage solitarily and individually make their
foraging decisions. We also investigated the influence of learning on the
performance of workers over successive trips searching for food by measuring
the patch residence time in each foraging trip. Four scenarios were created
differing in food reward rates, food size offered and distances colony-food site.
Our work has shown that as a rule-of-thumb, workers of D. quadriceps return to
the place where a prey item was found on the previous trip, regardless of
distance, food size and reward rate. When ants did not capture preys, they were
more likely to change path to search for food. However, in one of the scenarios,
this decision to switch paths when unsuccessful was less evident, possibly due
to the greater variation of possible outcomes ants could experience in this
scenario and cognitive constraints of D. quadriceps to predict variations of food
distribution. Our results also indicated a learning process of routes of
exploration as well as the food site conditions for exploration. After repeated
trips, foragers reduced the patch residence time in areas that they did not
capture food and quickly changed of foraging area, increasing their foraging
efficiency.

9
SUMÁRIO
INTRODUÇÃO .........................................................................................................................10
OBJETIVOS .............................................................................................................................16
1º Objetivo: ...........................................................................................................................16
2º Objetivo: ...........................................................................................................................16
HIPÓTESES E PREDIÇÕES .................................................................................................16
MATERIAIS E MÉTODOS .................................................................................................17
Manutenção das colônias ...............................................................................................17
Aparato experimental ......................................................................................................17
Procedimento experimental ...........................................................................................18
Análise dos dados ...........................................................................................................21
RESULTADOS .....................................................................................................................22
Decisões de forrageio .....................................................................................................22
Tempo de permanência na rota.....................................................................................25
DISCUSSÃO/CONCLUSÃO ..............................................................................................27
AGRADECIMENTOS ..........................................................................................................33
BIBLIOGRAFIA ....................................................................................................................33

10
INTRODUÇÃO
O sucesso ecológico de uma espécie depende da sua habilidade em
ajustar as estratégias de forrageio para maximizar a obtenção de alimento
gastando o mínimo de energia possível. As estratégias de forrageamento
envolvem duas questões fundamentais: a escolha da dieta, p.e., qual item
alimentar consumir, e a exploração de rotas, p.e., quando deixar uma rota. Por
isso, quando forrageando, supõe-se que o animal faz decisões particulares que
se acredita ser ideal em termos de taxa de consumo de energia (Stephens &
Krebs, 1971).
A perspectiva Darwiniana diz que uma forte pressão seletiva favorece
indivíduos capazes de perceber mudanças temporais e espaciais na
disponibilidade de alimento, pois essa habilidade poderá implicar em fortes
impactos positivos no desempenho desses animais (Krebs 1978). Esses
ajustes de estratégias diários são influenciados principalmente por variáveis
limitantes como disponibilidade e distribuição do alimento e as necessidades
nutricionais do animal (Giraldeau & Caraco 2000). Segundo Caraco e Real
(1986), a variação dos recursos quase sempre influencia o comportamento
alimentar dos animais. Portanto, podem ser considerados no momento da
decisão de qual estratégia adotar, fatores como o tempo para encontrar
alimento e a qualidade energética do mesmo, assim como os riscos inerentes
ao processo de busca pelo alimento, como predação, desidratação, e perda de
tempo e energia com deslocamentos aleatórios.
Teorias de otimização do forrageamento têm sido desenvolvidas para
estudar diversos animais. Charnov (1976) propôs o teorema do valor marginal,

11
que prediz que um predador deverá deixar uma área assim que a taxa marginal
de captura se torna inferior à média da mesma. Essa taxa declina
continuamente quanto mais o predador permanece e explora a área chegando
um momento em que todo o alimento foi consumido e nenhuma energia pode
mais ser adquirida. O teorema do valor marginal tem sido aplicado na
compreensão das estratégias ótimas em comportamento animal (Krebs &
Kacelnik 1991).
No caso das formigas, as estratégias de forrageio das diversas espécies
têm sido coerentes com as predições de outro modelo de otimização: a teoria
do ponto central (Orians & Pearson 1979) que considera que o tamanho da
carga não interfere na velocidade nem tempo de viagem e prevê que com o
aumento da distância do alimento do ninho, os indivíduos terão que selecionar
alimentos mais ricos em termos energéticos, evidenciando os custos potenciais
associados à ocupação de um único ninho. Além disso, é conhecido que o
investimento realizado por uma colônia através de suas estratégias está
diretamente relacionado à habilidade e eficiência de cada operária, às
variações ambientais, e a fatores como distância da comida ao ninho e
densidade demográfica (Gordon 1999).
Devigne e Detrain (2000) demonstraram que formigas da espécie
Messor barbarus otimizam o forrageio sendo mais seletiva na captura do
alimento com o aumento da distância do ninho. Resultado semelhante foi
encontrado em Pogonomyrmex barbatus por Davidson (1978) em laboratório.
Também Holder Bailey e Polis (1987) e Taylor (1978) perceberam que
operárias das espécies Pogonomyrmex californicus e Solenopsis geminata,
respectivamente, coletam preferencialmente sementes com maior retorno

12
energético indicando uma otimização do forrageio. Observando Solenopsis
invicta, Martin e Vinson (2008) perceberam que a espécie carregava maior
quantidade de alimento quando esse era coletado há grandes distâncias e
menor quantidade de alimento quando a busca era feita mais próximo do ninho.
No entanto, no estudo Willott et. al. (2000) com Messor bouvieri, não foi
encontrada seletividade por sementes maiores quando houve aumento das
distâncias percorridas, resultado contrário às predições da Teoria do
Forrageamento Central. Em Pogonomyrmex occidentalis, um indicativo de
otimização com relação à distância foi percebido por Crist e MacMahon (1992)
somente quando os animais ultrapassavam as distâncias limites de forrageio.
Não foi visto também correlação entre tamanho do alimento e distância
percorrida no trabalho de Rissing e Pollock (1984) com a espécie Veromessor
(= Messor) pergandei.
Em 1999, Holway e Case em experimentos com Linepithema humile,
demonstrou que essa espécie reduz os custos de forrageio reduzindo as
distâncias de busca por alimento, o número de viagens e o tempo de exposição
a predadores. Isso é feito através da realocação dos ninhos (espécie
polidômica), das operárias, larvas e pulpas dentro dos ninhos. Entretanto, essa
capacidade de re-alocação em resposta a distribuição de recursos não ocorre
em todas as espécies polidômicas.
Estudos (Caraco 1980, Pulliam & Millikan 1982) argumentam que a
sensibilidade ao risco também pode ajudar a explicar o comportamento de
forrageio social. A sensibilidade ao risco caracteriza-se quando o animal
modifica sua estratégia alimentar em função de uma ou mais variáveis. A
principal variável identificada nos estudos tem sido o saldo energético do

13
animal. Entretanto qualquer aspecto fisiológico (ex: período reprodutivo ou
cuidado da prole) ou ambiental (ex: estiagem, predador, distância, etc.) que
preveja corretamente a demanda energética futura poderá ser utilizado como
um fator para a modificação da estratégia alimentar. O animal é propenso ao
risco quando, dentre duas ou mais possibilidades de forrageio, adota uma
estratégia que implicará em algum tipo de risco tais como não encontrar o
alimento, ficar exposto por maior tempo e ser predado durante a busca ou
consumir um determinado recurso que ao final não compense energeticamente
os custos da procura e manipulação do mesmo. No caso da escolha de uma
estratégia que envolva os mesmos riscos da estratégia anterior, mas em
probabilidades menores ou nulas, o animal será considerado avesso ao risco.
Em animais sociais, essa permuta de estratégia poderá ocorrer considerando o
saldo energético da colônia e não dos animais que a compõe (Cartar 1991).
Decidir por estratégias ótimas quanto à obtenção de alimento,
considerando ainda os riscos dessas estratégias requer um aprendizado e
memorização da distribuição de alimento no ambiente para que o animal seja
capaz de fazer as escolhas apropriadas. Isso implica em três processos
cognitivos: percepção, em que o animal coleta e armazena uma informação
sobre o ambiente na memória; manipulação, quando o animal processa as
várias informações armazenadas; por último a representação, quando é
formada a “imagem” do ambiente e baseado nela o animal poderá realizar as
decisões. Portanto as teorias de otimização podem ser refinadas quando são
levados em conta os processos cognitivos de cada animal (Krebs & Davies
1997). Nos estudos de Nonacs e Dill em 1988 e 1990, operárias de Lasius
pallitarsis preferiram a rota de menor risco de predação quando as

14
recompensas eram iguais e Myrmica incompleta respondeu às variações de
risco e recursos em laboratório. Observando abelhas, Cartar e Dill (1990),
perceberam que as operárias onde as reservas do ninho se exauriam,
mudavam para o comportamento de propensão ao risco. Já Waddington et. al.
(1981), e Real et. al. (1981), em estudos diferentes com abelhas, tentaram
induzir o comportamento de propensão ao risco privando os ninhos de
alimento. Porém, esses animais permaneceram avessos ao risco. Apesar de
diferentes resultados na literatura, grande parte dos estudos (Kacelnik &
Bateson 1996) tem sugerido uma preferência parcial entre as estratégias.
Os insetos sociais têm sido utilizados como um sistema modelo para o
estudo do comportamento de forrageio. Essa importância se dá principalmente
pela complexa e modulada organização social (Oster & Wilson 1978; Traniello
1989; Holldobler & Wilson 1990). No forrageamento social, há uma
dependência recíproca das recompensas e a eficiência de forrageio depende
tanto das estratégias de cada animal como dos outros indivíduos da colônia.
Formigas, cupins e algumas vespas e abelhas constituem o grupo de
insetos eusociais devido a três características comuns: cooperação no cuidado
dos jovens, divisão reprodutiva do trabalho e sobreposição de pelo menos duas
gerações capazes de contribuir para o trabalho da colônia (Wilson 1971). Junto
com abelhas e vespas, as formigas constituem a ordem Hymenoptera e
formam uma família distinta, Formicidae, por possuir uma diferença estrutural:
a presença do pecíolo, uma cintura longa entre o tórax e abdômen. Dentro
dessa única família, existe a subfamília das Ponerinae. É encontrada em todo o
mundo, porém, predominantemente nos ambientes tropicais. São em grande
parte, predadoras (Caetano et. al. 2002), mas também se alimentam de

15
sementes, néctar floral, secreções de homópteros e larvas de lepidópteras. O
modo de forrageio pode ser solitário, buscando alimento de forma individual e
independente uma da outra ou em grupo, podendo ser especialistas ou
generalistas.
Entre as Ponerinae, está o gênero Dinoponera Roger (1861), que possui
formigas que atingem ou até ultrapassam os 3 cm de comprimento sendo por
isso uma das maiores formigas conhecidas. Estão distribuídas apenas na
América do Sul, mais especificamente: sudeste da Colômbia, leste do Peru,
Bolívia e Paraguai, nordeste da Argentina e todo o Brasil. Já a espécie
Dinoponera quadriceps Kempf (1971), é endêmica do nordeste brasileiro.
Possuem proporções pouco menores que a Dinoponera gigantea e se difere da
Dinoponera mutica pelo abdômen menos brilhante e um pecíolo em formato
peculiar: uma extremidade anterior marcante e estreitamente arredondada e
extremidade posterior amplamente arredondada (Kempf 1971).
O comportamento de forrageio de Dinoponera quadriceps, descrito por
Araújo & Rodrigues (2006), é solitário e sem recrutamento. As operárias saem
da colônia e já iniciam a busca de alimento de forma lenta. Ao encontrar o
alimento, retornam em maior velocidade para a colônia. Comportamento
semelhante foi visto também em Dinoponera gigantea (Fourcassié et. al. 1999).
Estudos de tomada de decisão durante o forrageio, levando em conta
características ecológicas assim como as capacidades e limitações cognitivas
dos animais, são de grande relevância visto que a procura por alimento é uma
atividade fundamental para o sucesso ecológicos dos animais. Nesse sentido,
experimentos que buscam investigar essas questões têm sido importantes para
avanços na área da Ecologia Comportamental.

16
OBJETIVOS
1º Objetivo:
Avaliar a influência do sucesso de captura e tamanho da presa, da
distância do alimento e da taxa de recompensas nas decisões de permanência
ou troca de área de forrageio em Dinoponera quadriceps.
2º Objetivo:
Avaliar o efeito do aprendizado no desempenho de forrageio das
operárias de Dinoponera quadriceps.
HIPÓTESES E PREDIÇÕES
● 1ª Hipótese: a captura de alimento influenciará o retorno das
operárias ao local visitado na viagem anterior.
○ Predição: forrageadoras irão retornar diretamente ao local
anterior onde capturaram uma presa; E, nos casos de não
captura, irão procurar alimento em outra área.
● 2ª Hipótese: durante o forrageio, com a repetição das visitas aos
locais de oferta de alimento, as operárias modificarão o tempo de
resposta para permanecer ou trocar de área de forrageio.
○ Predição: operárias irão aumentar a eficiência de forrageio
através da redução do tempo de permanência nas rotas quando não
capturam presas.

17
MATERIAIS E MÉTODOS
Manutenção das colônias
Nós utilizamos quatro colônias de Dinoponera quadriceps que foram
coletadas nos arredores da Universidade Federal do Rio Grande do Norte, Rio
Grande do Norte, Brasil. As colônias “C”, “E”, “G” e “H” continham,
respectivamente, 15, 9, 30 e 28 animais (média ± dp 19,7 ± 5,3). No laboratório
as colônias foram colocadas em caixas de plástico (30 X 15 X 10 cm) que eram
mantidas dentro de arenas (100 X 50 X 20 cm) construídas de madeira e
revestidas de fórmica para evitar que as formigas escapassem da arena. Todas
as formigas foram marcadas individualmente com uma etiqueta plástica colada
na região do tórax utilizando uma cola a base de cianoacrilato (Corbara et al.
1986).
A temperatura foi mantida entre 25 ºC e 30 ºC, umidade relativa do ar
em 70% e foto período de 12 horas claro/ 12 horas escuro. Nós alimentávamos
as formigas com água três vezes por semana e Tenebrio molitor (larva e
inseto). As arenas eram molhadas regularmente para manutenção da umidade.
Aparato experimental
O aparato experimental (Fig. 1) consistia de uma arena (150 x 50 X 20
cm) conectada a dois tubos de plástico de comprimentos distintos, sendo um
tubo com 300 centímetros (rota curta) e o outro com 600 centímetros (rota
longa). Ambos os tubos tinham nove centímetros de diâmetro. No final de cada
tubo foi colocado um placa de petri (35 x 7 mm) onde colocávamos o alimento
(Fig. 1). Cada tubo com sua respectiva placa de petri foi considerado um área
de forrageio.

18
Procedimento experimental
Nós observamos uma operária por dia. Outras operárias ativas que
saiam do ninho durante o experimento eram removidas e colocas numa caixa
plástica preta. Antes de cada dia de observação, as colônias eram privadas de
alimento. No entanto elas podiam explorar livremente toda arena e as duas
áreas de alimentação. Nós observamos somente operárias que tinham visitado
as duas áreas ao menos uma vez durante os dois dias de livre exploração.
Dessa maneira as forrageadoras poderiam conhecer e se acostumar com o
aparato experimental. Nós estimamos o peso das presas (minhocas) usadas no
experimento medindo o comprimento desses animais. Minhocas de 10±2 mm
(média ± dp, N=26), correspondem ao peso aproximado de 0.49±0.10 mg
(média ± dp, N=26), enquanto as minhocas com 21±3 mm (média ± dp, N=31),
correspondiam ao peso de 101±28 mg (média ± dp, N=31). Portanto, uma
presa tinha aproximadamento duas vezes o peso de outra e por isso foi
considerada o dobro mais rica em termos energéticos quando comparada com
as minhocas de tamanhos menores. A partir de agora, iremos nos referir a
essas presas como “presa grande” e “presa pequena”.
Figura 1: Aparato experimental: dois tubos de plásticos transparentes conectados a arena
princiapal. Dessa maneira era possível acompanhar a formiga por todo o trajeto.

19
Foram testadas 21 forrageadoras, que realizaram 10 viagens de
forrageio em cada dos seguintes cenários:
Cenário 1: forrageadoras poderiam experienciar uma condição por viagem ao
chegarem no final dos tubos: uma minhoca grande. Taxa de recompensa:
100%.
Cenário 2: forrageadoras poderiam experienciar duas condições ao chegarem
no final dos tubos: ou uma presa grande ou nenhuma presa. Taxa de
recompensa: 50%.
Cenário 3: forrageadoras poderiam experienciar duas condições ao chegarem
no final dos tubos: ou uma presa grande ou uma pequena. Taxa de
recompensa: 100%, sendo 50% para cada tamanho de alimento.
Cenário 4: forrageadoras poderiam experienciar três condições ao chegarem
no final dos tubos: ou uma presa grande, ou uma presa pequena ou nenhuma
presa. Taxa de recompensa: 66%, sendo 33% para cada tamanho de alimento
e 33% para nenhum alimento. Cenário 4 foi criado para ser o mais próximo
possível do cenário natural em termos de sucesso de forrageio. Em ambientes
naturais, formigas podem ou encontrar um alimento de alto conteúdo
energético, ou um alimento de baixo conteúdo energético ou não encontrar
alimento algum.
Embora os cenários pareçam redundantes, as diferenças nesles foram
criadas para avaliar os efeitos de cada variável de forma mais significante. Por
exemplo, no cenário 1 e 2, a qualidade do alimento e as distâncias são as
mesmas, porém a taxa de recompensa é diferente nos dois cenários (100% x
50%). No cenário 1 e no 3 as formigas tinham as mesmas taxas de
recompensas e distâncias a percorrer. Entretanto a qualidade do alimento

20
ofertado era diferente entre os cenários (presa grande X presa pequena). Além
disso, o método estatístico utilizado (abaixo) nos permitiu combinar essas
varáveis e compará-las de forma mais precisa.
As condições que as formigas encontravam no final dos tubos foram
randomicamente distribuídas nas 10 viagens que cada operária realiza em
cada cenário. O alimento era recolocado ou removido enquanto o animal
estava fora dos tubos. Portanto, as forrageadoras não notavam qualquer
movimentação nas placas de petri. Nenhuma operária era observada
consecutivamente nos cenários para evitar qualquer influência de experiências
recentes nas operárias durante a realização do cenário seguinte. Todas as
forrageadoras tinham pelo menos um mês de intervalo entre os cenários.
Estudos definiram a memória de longo-prazo em insetos entre 24 e 48 horas
(Dreier et al 2007; Haehnel & Menzel 2012). Além disso, uma vez que ambas
as rotas foram construídas na mesma direção, a fidelidade direcional
apresentada por esse espécime não um problema. No início de cada dia de
observação, nós colocávamos uma presa em ambas as placas de petri para
estimular o forrageio em algum animal que esteja explorando as áreas. O
experimento era iniciado assim que alguma operária capturava a presa
ofertada nas placas de petri. Nas próximas 10 viagens realizadas por essa
formiga, nós coletávamos os seguintes dados: 1) rota escolhida (300 cm X 600
cm); 2) tempo de viagem; 3) se a operária captura ou não a presa; 4) tamanho
da presa; 5) a decisão tomada na viagem seguinte (retorno X troca de rota).
Nós consideramos a decisão “retorno” a rota visitada anteriormente como
“zero” (0) e a decisão “troca” derota como “um” (1). O tempo foi medido na
escala dos minutos. Após cada dia de obsevação nós tínhamos 10 decisões

21
tomads por uma operária. No final de cada cenário nós tínhamos 210 decisões.
E por fim, 840 decisões ao término do trabalho.
Nós também medimos o tempo gasto pelas forrageadoras para sair de
cada tubo após a chegada na placa de petri (tempo de permanência na área).
Nas viagens que as operárias não capturavam alimento, se elas gradualmente
reduzissem o tempo gasto nos tubos, isso poderia ser uma evidência de que os
animais aprendem baseado em experiências passadas, o que elas podem
encontrar em cada placa assim como as rotas que estão explorando.
Análise dos dados
Para verificar os efeitos de cada fator nas decisões de retornar ao ponto
anteriormente visitado ou trocar de rota, nós realizamos o teste de Equações
Estimativas Generalizadas (EEG). Nós consideramos como efeitos fixos o
cenário (taxa de recompensa), a distância, o sucesso de captura e tamanho do
alimento. Incluimos também as interações desses fatores. Como efeitos
randômicos nós consideramos o indivíduo e a colônia. Além disso, nós
incluímos o tempo total de viagem como covariável.
Realizamos o EEG também para verificar o efeito das viagens
consecutivas (experiência) e do sucesso de captura nas viagens, incluindo o
efeito de interação dessas duas variáveis no tempo de permanência das
formigas nos tubos de forrageio. Essas mesmas análises foram feitas para
cada cenário individualmente e não houve diferença da análise geral incluindo
todos os cenários juntos. Por esse motivo escolhemos descrever a análise que
engloba os cenários. Para ambos os EEGs, foi utilizado a matriz de correção
não-estruturada e o teste post hoc foi o seqüencial de Sidak. O nível de

22
significância adotado foi de 5% para todos os testes estatísticos realizados
nesse trabalho.
RESULTADOS
Decisões de forrageio
Nós investigamos os efeitos principais e as interações das variáveis:
cenário, distância e sucesso de captura e tamanho do alimento nas decisões
tomadas durante o forrageio. Nós observamos um efeito das interações dos
três fatores nas “decisões” (F1, 4 = 253.29, P < 0.001). Esse modelo foi
controlado pela covariável “tempo total”. (Tabela 1).
Tabela1: Efeitos principais e de interação nas tomadas de decisões individuais em D.
quadriceps.
Main and interaction effects
Type III
Wald Chi-Square DF P
(Intercept) 4240.05 1 <0.001
Scenarios 318.63 3 <0.001
Distance 11.59 1 0.001
Capture success 805.56 2 <0.001
Scenarios * Distance 75.16 3 <0.001
Scenarios * Capture success 1536.74 4 <0.001
Distance * Capture success 80.85 2 <0.001
Scenarios * Distance * Capture success 253.29 4 <0.001
Total trip time 29.69 1 <0.001
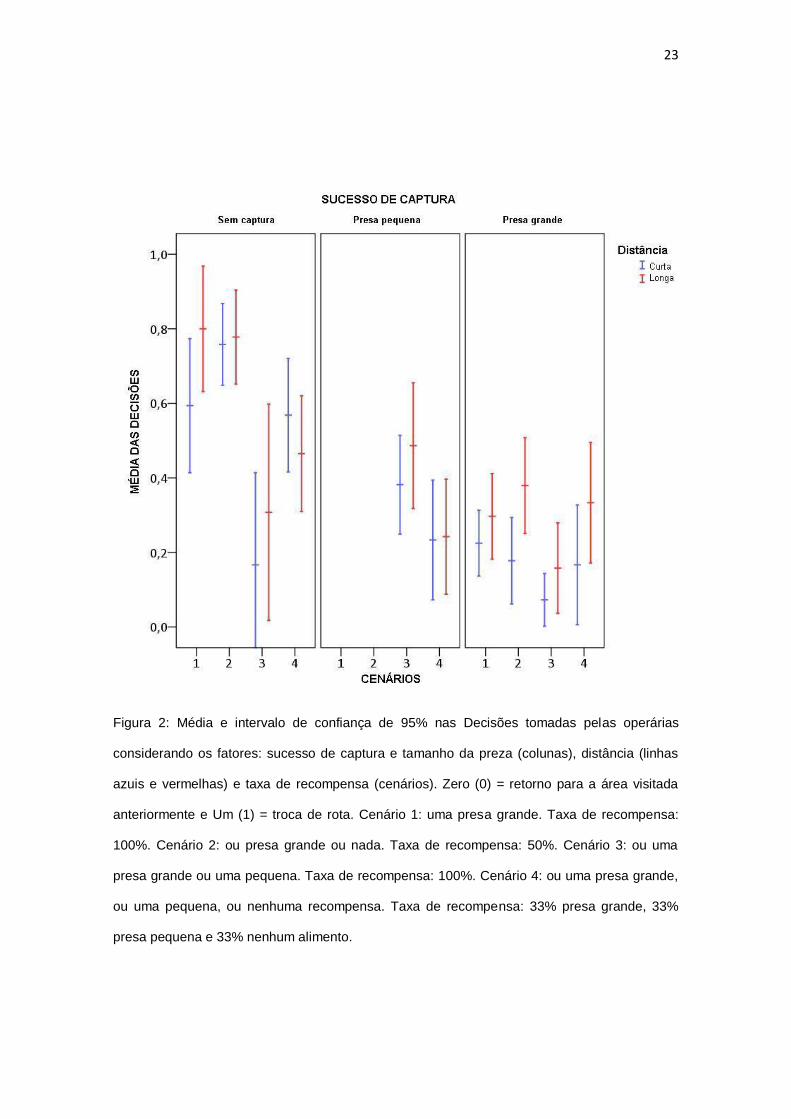
23
Figura 2: Média e intervalo de confiança de 95% nas Decisões tomadas pelas operárias
considerando os fatores: sucesso de captura e tamanho da preza (colunas), distância (linhas
azuis e vermelhas) e taxa de recompensa (cenários). Zero (0) = retorno para a área visitada
anteriormente e Um (1) = troca de rota. Cenário 1: uma presa grande. Taxa de recompensa:
100%. Cenário 2: ou presa grande ou nada. Taxa de recompensa: 50%. Cenário 3: ou uma
presa grande ou uma pequena. Taxa de recompensa: 100%. Cenário 4: ou uma presa grande,
ou uma pequena, ou nenhuma recompensa. Taxa de recompensa: 33% presa grande, 33%
presa pequena e 33% nenhum alimento.

24
Comparações entre sucesso de captura, considerando cada distância e
cenário, mostraram algumas diferenças (Fig. 2). Para cenário 1 e ambas as
distâncias, nós observamos diferenças entre “não capturar” (média rota
curta=0.59, média rota longa=0.80) e “captura de presa grande” (rota
curta=0.22, rota longa=0.30) (P < 0.002). Para cenário 2, nós também
observamos diferenças entre “não capturtar” (rota curta=0.76, rota longa=0.78)
e “capturar presa grande” (rota curta=0.18, rota longa=0.38), para ambas as
distâncias (P < 0.002). Em suma, para ambos os cenários e distâncias, as
formigas trocaram derota mais quando elas não capturaram alimento,
comparado aos momentos em que elas capturaram “presas grandes”. Para o
cenário 4 e rotas curtas, os indivíduos trocaram de rota mais quando não
capturavam presas (média=0.57) do que quando capturavam presas grandes
(média=0.17) (P=0.004). Esse resultado é similar ao padrão observado nos
cenários 1 e 2. Nenhuma outra diferença foi encontrada entre “sucesso de
captura”, considerando cada cenário e cada distância (P > 0.088).
Adcionalmente, comparações entre cenários, levando em conta sucesso
de captura e distância, indicaram algumas diferenças (Fig. 2). Em casos de
“não captura” em rotas curtas, nós observamos que no cenário 1 (média=0.59),
2 (média=0.76), e 4 (média=0.57) as operárias trocaram de rota mais do que no
cenário 3 (média=0.17) (P < 0.001). Padrão semelhante foi visto para rota
longa, mas nesse caso somente no cenário 1 (média=0.80) as formigas
trocaram mais de rota que no cenário 3 (média=0.31) (P = 0.001).
No cenário 3 na rota curta, os animais trocaram mais quando eles
capturavam presas pequenas (média=0.38), comparado quando eles
capturavam presas grandes (média=0.07) (P < 0.001). Note que o tamanho do

25
alimento teve um efeito somente neste cenário e em rotas curtas. Note também
que a influência ocorreu na mesma direção comportamental (“retorno após
captura”). Nenhuma outra diferença foi observada quando consideramos
presas pequenas e grandes em todos os cenários para as duas rotas (P >
0.479).
Finalmente, comparações entre rota curta e longa, em cada cenário e
sucesso de captura também não mostrou diferenças significativas (P > 0.273)
(Fig. 2).
Tempo de permanência na rota
Foram investigados nesse trabalho os efeitos principais e de interação
dos fatores no tempo de permanência das operárias nas rotas de forrageio.
Nós observamos um efeito com a interação das variáveis (F1, 16 = 81553024.35,
P < 0.001) (Tabela 2).
Tabela 2: Efeitos princiapais e de interação no tempo de saída dos tubos ao longo das 10
viagens em D. quadriceps.
Main and interaction effects
Type III
Wald Chi-Square DF P
(Intercept) 156.42 1 <0.001
Capture success 22.00 2 <0.001
Trip experience 36.11 9 <0.001
Capture success * Trip experience 81553024.35 16 <0.001

26
Figura 3: média do tempo de permanência na rota em cada viagem quando as forrageadoras
capturavam presas grandes (linha vermelha), quando capturavam presas pequenas (linha
verde) e quando não capturavam alimento (linha azul).
Nós comparamos o tempo de permanência nas rotas entre os três
diferentes “sucessos de captura” (grande X pequeno X nada) (Fig. 3).
Analisando as viagens que resultaram em nenhuma captura, foi visto que as
formigas permaneciam mais tempo na rota na viagem 1, (média=0.12) do que
na viagem 9 (média=0.04) e 10 (média=0.04) (P < 0.045). Quando capturavam
presas pequenas, as formigas retornaram mais rapidamente para o ninho nas
viagens 1 (média=0.02) e 2 (média= 0.01) comparados a viagem 7

27
(média=0.05) (P < 0.046). Nenhuma diferença foi encontrada na análise das
viagens que os animais capturavam presas grandes (P > 0.730).
Por fim, nós comparamos o tempo de permanência nas rotas entre os
sucessos de capturas para cada viagem (Fig. 3). Observamos que as fomigas
gastam mais tempo para sair das rotas na viagem 1 e 2 quando não capturam
presa (média=0.12 e 0.12), comparado quando elas capturam presas
pequenas (média=0.02 e 0.01) ou presas grandes (média=0.02 e 0.03) (P <
0.001 e P < 0.026). Nenhuma outra diferença foi encontrada entre os sucessos
de captura nas viagens 3 até 10 (P > 0.112).
DISCUSSÃO/CONCLUSÃO
Esse trabalho demonstrou a forte influência do sucesso de captura na
tomada de decisão das forrageadoras na viagem subsequente. Em todos os
cenários, quando as formigas capturaram presas grandes, elas tendiam a
retornar para o mesmo local onde elas foram durante a viagem anterior. Esse
padrão de decisão foi também observado quando as operárias capturavam
presas pequenas. Essa tendência a retornar a áreas onde itens alimentares
foram encontrados na viagem anterior tem sido descrita em outros estudos com
espécimes de formigas que forrageiam de forma solitária (Wehner et al. 1983;
Fresneau 1985; Fourcassie & Traniello 1994; Breed et al. 1996; Mercier &
Lenoir 1999; Schultheiss & Cheng 2012). Nos casos onde as formigas não
capturavam presas, nós observamos uma alta variação nas respostas. No
cenário 1 (recompensa constante), as operárias trocavam de rota quando elas
não encontravam alimento. Essa recusa de alimento pode ter sido
conseqüência de uma possível saciação de proteínas (principal componente de

28
insetos). De fato, nós observamos que 66% das operárias (N=14) recusaram
alimento (57 de 210 viagens), mas elas mantiveram a exploração das rotas.
Estudos anteriores descrevem diferentes efeitos do tipo de comida no
comportamento alimentar individual das formigas. Por exemplo, a estrutura das
rotas de buscas e a duração da busca em Formica schaufussi são
principalmente influenciadas pelo tipo de alimento. Operárias persistiram mais
no alimento quando esses eram carboidratos, comparados por alimentos
protéicos (Traniello et al. 1992). Formigas do deserto Cataglyphis bicolor
persistiram menos na busca em uma determinada área de sucesso anteior se
elas tivessem capturado uma mosca nessa última viagem, comparado a
quando elas capturavam um pedaço de queijo (lipídeos) (Schimd-Hempel
1984). Não obstante, formigas da espécie Lasius Níger ingerem
significativamente mais gotas de açúcar do que gotas de compostos protéicos
(Portha et al. 2003).
No cenário 2, a tendência para trocar de rotas após uma viagem não
bem sucessiva se deu pelo fato de não haver a presença de alimento, condição
essa que ocorreu em 50% das viagens de cada operária. Forrageadoras não
persistentes se beneficiam quando estão buscando alimentos que estão
distribuídos aleatóriamente no ambiente (Schimd-Hempel 1984), fato esse
bastante evidente no cenário 2 (50% de recompensa). Variações de curto
prazo na distribuição de recursos no ambiente podem ser superadas pelas
formigas através de uma frequente amostragem do ambiente para detecção de
alimento (Stephens & Krebs 1986). De fato, Dinoponera quadriceps se alimenta
principalmente de artrópodes (Araujo & Rodrigues 2006), um tipo de recurso
que está disperso de forma imprevisível no ambiente natural. Embora fidelidade

29
direcional seja bem descrito como uma das estratégias de forrageio de
formigas (Traniello 1989), a revisitação a uma área onde o indivíduo não
encontrou alimento na viagem anterior foi raramente observada em campo.
Operárias de Melophorus bagoti treinadas com recompensa constante por dois
dias retornaram para o mesmo local mesmo após a remoção do alimentador
(Schultheiss & Cheng 2012). Além disso, Breed et al. (1996) estudando
operárias de Paraponera clavata em ambiente natural, observaram que após
duas recompensas consecutivas alguns indivíduos (10 de 21) retornaram ao
local de alimentação algumas vezes seguidas mesmo sem mais a presença de
qualquer alimento. Este pode ter sido o caso de D. quadriceps forrageando no
cenário 4 quando elas não capturavam presas na rota longa. Nesse cenário, a
decisão de vistar novamente uma placa de petri mesmo após uma viagem mal
sucedida aparente ser um efeito de visitas bem sucedidas consecutivas que
ocorreram anteriormente dentro das 10 viagens de cada operária. Uma
segunda explicação, mas que não exclui a primeira é a de que os indivíduos
estavam mais confusos com relação à distribuição do alimento. Vale lembrar
que o cenário 4 foi realmente criado para ser o mais variado possível, similar
aos padrões naturais. Os indivíduos poderiam encontrar três situações ao
chegar ao final de cada tupo: obter uma presa de tamanho grande; obter uma
presa de tamanho pequeno ou não obter presa alguma. Fourcassié & Traniello
(1993), observando o forrageio individual de Formica schaufussi, mostrou que
essa espécie não é capaz de prever distribuição de alimento em ambientes
dinâmicos. Animais podem erroneamente avaliar o tempo e energia investido
no forrageio devido às limitações cognitivas, e, portanto se tornam
forrageadores “sub-ótimos” (Janetos & Cole 1981).

30
A tendência de revisitar um local anterior mesmo após uma viagem mal
sucedida foi também vista no cenário 3 em ambas as rotas (longa e curta).
Uma pergunta que surge é porque formigas agiram diferentes quando não
capturam alimento no cenário 1 do cenário 3. Duas possíveis explicações são
levantadas: primeiro, a freqüência de revisitação no cenário 3 permaneceu
muito similar ao cenário 1 (N=19; N=18; respectivamente). Entretanto, quando
analisamos a frequência de viagens que as operárias não capturaram presas
nós encontramos uma diferença notável (cenário 3, N=25; cenário 1, N=57).
Uma vez que ambos os cenários tinham taxas de recompensas de 100%, essa
flutuação não tinha como ser controlada. Então, a média de “retornos após
insucesso” aparenta ter sido subestimada. Outro questionamento importante é
porque as formigas diferiram quanto a freqüência de viagens mal sucedidas no
cenário 1 e 3, se o alimento era oferecido constantemente nesses cenários. A
resposta surge quando analisamos o tamanho do alimento que foi oferecido no
cenário 1 e 3 e o quão rápido as colônias ficam saciadas durante o
experimento. No cenário 1, formigas tinham presas grandes disponíveis
durante todas as 10 viagens realizadas, enquanto que no cenário 3 dois
tamanhos de presas foram oferecidas aleatoriamente. Independente da
quantidade de alimento que cada colônia nessecita, esse motante pode ser
alcançado em menos viagens no cenário 1, comparado com cenário 3. Isso
resulta, portanto, numa diferença no número de viagens onde não houve
captura de alimento mesmo com a presa sendo oferecida. A segunda possível
razão para esse padrão de resposta “aparentemente” contrastante entre
cenários 1 e 3 quando as formigas não capturavam alimento é que 94% (18 de
19) dos “retornos após um insucesso” no cenário 3 e 66% (12 de 18) no

31
cenário 1 foram realizados por somente 5 operárias, que pertenciam as
menores colônias do experimento (“C” e “E”). Considerando o tamanho
reduzido das colônias “E” e “C”, que significa menor demanda energégica, e
que por isso elas suprem suas necessidades mais rapidamente que as outras
duas colônias (“G” e “H”), nós hipotetizamos que essas cinco formigas estavam
forrageando de maneira avessa ao risco, tentando evitar os riscos inerentes a
exploração de ambientes dinâmicas como mortalidade, gastos energéticos
desnecessários e tempo excessivo. Operárias de Lasius pallitarsis respondem
ao risco e a variações na qualidade do alimento enquanto forrageiam.
Forrageadoras reduziram o risco de mortalidade buscando alimento de maneira
aversiva ao risco quando os ganhos de alimento da colônia decresceram
(Nonacs & Dill 1990). Gómez & Espadaler (1998) sugeriu que estratégias
avessas ou tendenciosas ao risco em formigas Aphaenogaster senilis podem
ser um reflexo das diferentes idades de cada animal. O comportamento de
forrageio de abelhas é afetado por tanto as necessidades energéticas das
colônias (Cartar & Dill 1990) como pelo padrão espaço/temporal de distribuição
dos recursos (Wadington et al 1981). Abelhas também se comportam avessas
ao risco quando forrageando néctar. Vespas (Vespula maculifrons)
apresentaram sensibildiade ao risco quando forrageando néctar (Real 1981).
Entretanto elas foram menos avessas ao risco do que abelhas quando o
alimento era néctar visto que esse tipo de alimento não é a principal fonte de
energia para reprodução dessa espécie. Ao invés disso, o crescimento de
colônias de vespas depende principalmente de uma dieta carnívora. (Real &
Caraco 1986).

32
Com relação às decisões tomadas após cada viagem, o fator “distância”
foi o que teve menor efeito nos nossos experimentos. Nenhuma diferença foi
observada quando comparamos cada situação entre as rotas curta e longa. A
distância criada em laboratório foi menor que a distância média percorridas por
D. quadriceps em ambiente natural (24.67 m) (Azevedo et al. 2014). Embora
nós não tenhamos encontrado efeito significativo da distância na busca pelo
alimento, vários estudos apontam para um efeito nesse comportamento das
formigas (Traniello 1989). Portanto. Futuros trabalhos realizados em laboratório
com essa espécime para avaliar questões relacionadas a distância devem levar
em cosideração maiores áreas de exploração e maiores variações de
recompensas.
Nossos dados sugerem que as decisões individuais tomadas durante a
busca por alimento são melhoradas por capacidades cognitivas. Durante
viagens mal sucedidas, operárias apresentaram um tempo de permanência
maior nas primeiras viagens. No entanto, com a repetição das viagens, as
forrageadoras foram gradualmente reduzindo o tempo de permanência nas
rotas nos casos em que não capturavam alimento, até o ponto que o tempo de
busca por alimento nas rotas era o mesmo, independente se as forrageadoras
capturavam ou não uma presa. Isso é um indicativo que esses animais
aprendem e memorizam pequenos atributos das áreas de forrageio. Com isso,
operárias podiam ajustar o tempo e energia investida na busca por alimento em
determinado lugar baseado em experiências passadas. Decisões mais rápidas
significam menos tempo e energia gasta em áreas de forrageio indesejadas, e,
portanto, uma maior eficiência de forrageio. Franz & Wcislo (2003) demonstrou
que forrageadoras da espécie Ectatomma aumentam sua eficiência de

33
forrageio realizando ajustes temporais e direcionais baseados experiências
anteriores positivas. Além disso, Schwarz & Cheng (2011) mostraram que a
formiga do deserto australiana Melophorus bagoti não somente discrimina
estímulos visuais associados a recompensas como também aprende pistas
visuais durante o retorno ao ninho.
Em suma, nós demonstramos com esse estudo que o principal fator de
influência nas decisões de forrageio de Dinoponera quadriceps é o sucesso de
captura da busca anterior. Como regra geral, formigas tendem a retornar a um
determinado ponto caso elas tenham tido sucesso nessa viagem anterior, e, no
caso de uma viagem mal sucedida, trocar de área de busca e começar uma
nova exploração. Demonstramos também a importância de traços cognitivos no
aumento da eficiência de forrageio através da redução do esforço de busca em
locais de baixo retorno energético.
AGRADECIMENTOS
Agradecemos a Universidade Federal do Rio Grande do Norte pelo
apoio logístico e estrutural assim como ao Conselho Nacional de
Desenvolvimento Científico e Tecnológico (CNPq) (WASN - Apoio
#132139/2012-0; AA - Apoio #303466/2012-0 e #307418/2009-0).
BIBLIOGRAFIA
Araujo, A. & Rodrigues, Z. 2006. Foraging behavior of the queenless ant
Dinoponera quadriceps Santschi (Hymenoptera: Formicidae). Neotropical
Entomology, 35, 159–164.

34
Azevedo, D. L., Medeiros, J. C. & Araújo, A. 2014. Adjustments in the time,
distance, and direction of foraging in Dinoponera quadriceps Workers. Journal
of Insect Behaviour, 27, 177-191.
Beckers, R., Goss, S., Deneubourg, J. L. & Pasteels, J. M. 1989. Colony
size, communication and ant foraging strategy. Psyche, 96, 239-256.
Breed, M. D., Fewell, J. H., Moore, A. J. & Williams, K. R. 1987. Graded
recruitment in a ponerine ant. Behavioral Ecology and Sociobiology, 20, 407-
411.
Breed, M. D., Stierstofer, C., Furness, E. D., Jeral, J. M. & Fewell, J. H.
1996. Individual constancy of local search strategies in the giant tropical ant,
Paraponera clavata (Hymenoptera: Formicidae). Journal of Insect Behavior, 9,
673-682.
Caetano, F. H., Jaffé, K. & Zara, J. 2002. Formigas: biologia e anatomia. Ed.
Araras: Topázio.
Caraco, T. 1980. On foraging time allocation in a stochastic environment.
Ecology, 61, 119-28.
Caraco, T. & Real, L. 1986. Risk and foraging in stochastic environments.
Annual Review of Ecology and Systematics, 17, 371-390.
Cartar, R. V. & Dill, L. M. 1990. Colony energy requirements affect the foraging
currency of bumble bees. Behavioral Ecology and Sociobiology, 27, 337-383.
Cartar, R. V. 1991. A test of risk-sensitive foraging in wild bumble bees.
Ecology, 72, 888-895.
Charnov, E. L. 1976. Optimal Foraging, the Marginal Value Theorem.
Theoretical Population Biology, 9, 129-136.

35
Corbara, B., Fresneau, D., Lachaud, J. P., Leclerc, Y. & Goodall, G. 1986.
An automated photographic technique for behavioural investigations of social
insects. Behavioural Process, 13, 237–249.
Davidson, D. W. 1978. Experimental tests of the optimal diet in two social
insects. Behavioral Ecology and Sociobiology, 4, 35-41.
Deneubourg, J. L., Fresneau, D., Goss, S., Lachaud, J. P. & Pasteels, J. M.
1987. A simple model to simulate the organization of individual foraging in
Neoponera apicalis. In Chemistry and Biology of Social Insects, Eds J. Eder &
H. Rembold. Verlag Peperny, Munich, 527-528.
Devigne, C. & Detrain, C. 2006. How does food distance influence foraging in
the ant Lasius niger: the importance of home-range marking. Insectes Sociaux,
53, 46-55.
Dreier, S., van Zweden, J. & D´Ettorre, P. 2007. Long-term memory of
individual identity in ant queens. Biology Letters, 3, 459-462. DOI:
10.1098/rsbl.2007.0224.
Fewell, J. H., Harrison, J. F., Stiller, T. M., & Breed, M. D. 1992. A cost
benefit analysis of distance effects on foraging and recruitment in the giant
tropical ant, Paraponera clavata. Oecologia, 92, 542-547.
Franz, N. M. & Wcislo, W. T. 2003. Foraging behavior in two species of
Ectatomma (Formicidae: Ponerinae): individual learning of orientation and
timing. Journal of Insect Behavior, 16, 381-410.
Fresneau, D. 1985. Individual foraging and path fidelity in a ponerine ant.
Insectes Sociaux, 32, 109-116.

36
Fourcassié, V. & Traniello, J. F. A. 1993. Effects of experience on food-
searching behavior in the ant Formica schaufussi (Hymenoptera: Formicidae).
Journal of Insect Behavior, 6, 287-299.
Fourcassié, V. & Traniello, J. F. A. 1994. Food searching behaviour in the ant
Formica schaufussi (Hymenoptera, Formicidae): response of naïve foragers to
protein and carbohydrate food. Animal Behaviour, 48, 69-79.
Fourcassié V., Henriques, A. & Fontella, C. 1999. Route fidelity and spatial
orientation in the ant Dinoponera gigantea (Hymenoptera, Formicidae) in a
primary forest: a preliminary study. Sociobiology, 34, 505-524.
Giraldeau, L. A. & Caraco, T. 2000. Social Foraging Theory. pp.3-31.
Princeton: Princeton University Press.
Gómez, C. & Espadaler, X. 1998. Aphaenogaster senilis Mayr (Hymenoptera,
Formicidae): a possible parasite in the myrmecochory of Euphorbia characias
(Euphorbiaceae). Sociobiology, 32, 441-450.
Gordon, D. M. 1999. Ants at work: How an insect society is organized. New
York, The Free Press Ed., 182p.
Gordon, D. M. & Keller, L. 2009. The live of ants. Oxford University Press.
252p.
Haehnel, M. & Menzel, R. 2012. Long-term memory and response
generalization in mushroom body extrinsic neurons in the honeybee Apis
mellifera. The Journal of Experimental Biology, 215, 559-565.
doi:10.1242/jeb.059626.
Holder-Bailey, K., & Polis, G. A. 1987. Optimal and central place foraging
theory applied to a desert harvester ant, Pogonomyrmex californicus.
Oecologia,

37
72, 440-48.
Holldobler, B. & Wilson, E. O. 1990. The Ants. Harvard Univisty Press,
Cambridge, MA.
Holway, D. A. & Case, T. J. 1999. Mechanisms of dispersed central-place
foraging in polydomous colonies of the Argentine ant. Animal Behaviour, 59,
433-441.
Kalcenik, A. & Bateson, M. 1996. Risky theories: the effects of variance on
foraging decisions. American Zoologist, 36, 402–434.
Kempf, W.W. 1971. A preliminary review of the Ponerine ant genus Dinoponera
Roger (Hymenoptera, Formicidae). Studia Entomologica, 14, 369-394.
Krebs, J. R. 1978. Optimal foraging: decision rules for predators. Ch. 2 in: J. R.
Krebs & N. B. Davies (eds.). Behavioural Ecology: and evolutionary approach.
Blackwell Scientific Publications, Oxford.
Krebs, J. R. & Davies, N. B. 1997. Behavioural Ecology: an Evolutionary
approach. Blackwell Scientific Publications, Oxford.
Krebs, J. R. & Kacelnik, A. C. 1991. Individual decisions and the distribution of
predators in a patchy environment. II. The influence of travel costs and structure
of the environment. Journal of Animal Ecology, 60, 205- 225.
Janetos, A. C. & Cole, B. J. 1981. Imperfectly optimal animals. Behavioral
Ecology and Sociobiology, 9, 203-209.
Johnson, R. A. 1991. Learning, memory, and foraging efficiency in two species
of desert seed-harvester ants. Ecology, 72, 1408-1419.
Latty, T. & Beekman, M. 2013. Keeping track of changes: the performance of
ant colonies in dynamic environments. Animal Behaviour, 85, 637-643.

38
Levings, S. C. & Franks, N. R. 1982. Patterns of nest dispersion in a tropical
ground ant community. Ecology, 63(2), 338-344.
Martin, J. B. & Vinson, S. B. 2008. The influence of travel distance on sugar
loading and water balance in the central place foraging ant Solenopsis invicta.
Insectes Sociaux, 55, 129-136.
Mercier, J. L. & Lenoir, A. 1999. Individual flexibility and choice of foraging
strategy in Polyrhachis laboriosa F. Smith (Hymenoptera, Formicidae). Insectes
Sociaux, 46, 267-272.
McNair, J. M. 1982. Optimal giving-up times and marginal value theorem.
Journal theoretical Biology, 119, 511-529.
McNamara, J. M. & Houston, A. I. 1985. Optimal foraging and learning.
Journal theoretical Biology, 117, 231-249.
Nonacs, P. & L. M. Dill. 1988. Foraging responses of the ant Lasius pallitarsis
to food sources with associated mortality risk. Insectes Sociaux, 35, 293-303.
Nonacs, P. & Dill, L. M. 1990. Mortality risk vs. food quality trade-offs in a
common currency: ant patch preferences. Ecology, 71, 1886-1892.
Oster, G. F. & Wilson, E. O. 1978. Caste and Ecology in the Social Insects.
Princeton, NJ: Princeton University Press.
Orians, G.H. & Pearson, N. E. 1979. On the theory of central place foraging. In
Analysis of Ecological System (Horn, D.J, Mitchel, R.D e Stairs, G.R. eds). Ohio
State University Press, Columbus.
Pelé, M. & Sueur, C. 2013. Decision-making theories: linking the disparate research
areas of individual and collective cognition. Animal Cognition, DOI: 10.1007/s10071-
013-0631-1.

39
Portha, S., Deneubourg, J. L. & Detrain, C. 2004. How food type and brood
influence foraging decisions of Lasius niger scouts. Animal Behaviour, 68, 115-
122.
Pyke, G. H. 1984. Optimal Foraging Theory: a critical review. Annual Review of
Ecology and Systematics, 15, 523-575.
Pullian, H. R. & Millikan, G. C. 1982. Social organization in the non-
reproductive season. In: Avian Biology, 6, ed. D. S. Farmer, J. R. King, pp. 169-
97. New York: Academic Samuel.
Real, L. A. 1981. Uncertainty and pollinator-plant interactions: The foraging
behavior of bees and wasps on artificial flowers. Ecology, 62, 20-26.
Real, L. 1986. Risk and foraging in stochastic environments. Annual Reviews of
Ecology and Systematics, 17, 371-390.
Schatz, B., Lachaud, J. P., & Beugnon, G. 1996. Polyethism within hunters of
the ponerine ant, Ectatomma ruidum Roger (Formicidae, Ponerinae). Insectes
Sociaux, 43, 111–118.
Rissing, S. W. & Pollock, G. B. 1984. Worker size variability and foraging
efficiency in Veromessor pergandei (Hymenoptera: Formicidae). Behavioral
Ecology and Sociobiology, 15, 121-26.
Schmid-Hempel, P. 1984. Individually different foraging methods in the desert
ant Cataglyphis bicolor (Hymenoptera, Formicidae). Behavioral Ecology and
Sociobiology, 14, 263-271.
Schultheiss, P. & Cheng, K. 2012. Finding food: outbound searching behavior
in the Australian desert ant Melophorus bagoti. Behavioral Ecology, 24, 128-
135. DOI:10.1093/beheco/ars143.

40
Schwarz, S. & Cheng, K. 2011. Visual discrimination, sequential learning and
memory retrieval in the Australian desert ant Melophorus bagoti. Animal
Cognition, 6, 861-870. DOI: 10.1007/s10071-011-0419-0.
Stephens, D. W. & Krebs, J. R. 1986. Foraging Theory. Princeton, NJ:
Princeton University Press.
Taylor. F. 1976. Foraging behavior in ants: experiments with two species of
myrmecine ants. Behavioral Ecology and Sociobiology, 2, 147-168.
Traniello, J. F. A. 1989. Foraging strategies of ants. Annual Review of
Entomology, 34, 191-210.
Traniello, J. F. A., Kozol, A. J & Fournier, M. A. 1992. Resource-related
spatial patterns of search in the ant Formica schaufussi: a field study. Psyche,
99, 87-93.
Waddington, K. D., Allen, T. & Heinrich, B. 1981. Floral preferences of
bumblebees (Bombus edwardsii) in relation to intermittent versus continuous
rewards. Animal Behaviour, 29, 779-784.
Wehner, R., Harkness, R.D. & Schmid-Hempel, P. 1983. Foraging strategies
in individually searching ants, Cataglyphis bicolor (Hymenoptera: Formicidae).
Akademie derWissenschaften und der Literatur, Mainz, Mathematisch-
Naturwissenschaftliche Klasse (ed. by M. Lindauer), 1–79, two maps, Fischer,
Stuttgart.
Willot, S. J., Compton, S. G. & Incoll, L. D. 2000. Foraging, food selection and
worker size in the seed harvesting ant Messor bouvieri, Oecologia, 125, 35-44.
Wilson, E. O. 1971. The Insect Societies. pp. 1-6. Londres: Belknap Press of
Harvard University Press.


















