
AVALIAÇÃO DA GENOTOXICIDADE DO CLORETO DE …livros01.livrosgratis.com.br/cp144726.pdf · Figura...
102
SERVIÇO PÚBLICO FEDERAL UNIVERSIDADE FEDERAL DO PARÁ INSTITUTO DE CIÊNCIAS BIOLÓGICAS PROGRAMA DE PÓS-GRADUAÇÃO EM NEUROCIÊNCIAS E BIOLOGIA CELULAR AVALIAÇÃO DA GENOTOXICIDADE DO CLORETO DE METILMERCÚRIO EM DUAS ESPÉCIES DE PEIXES NEOTROPICAIS CARLOS ALBERTO MACHADO DA ROCHA BELÉM - PA 2009
Transcript of AVALIAÇÃO DA GENOTOXICIDADE DO CLORETO DE …livros01.livrosgratis.com.br/cp144726.pdf · Figura...

SERVIÇO PÚBLICO FEDERAL
UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM NEUROCIÊNCIAS E BIOLOGIA CELULAR
AVALIAÇÃO DA GENOTOXICIDADE DO CLORETO DE METILMERCÚRIO EM DUAS ESPÉCIES DE
PEIXES NEOTROPICAIS
CARLOS ALBERTO MACHADO DA ROCHA
BELÉM - PA 2009

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

CARLOS ALBERTO MACHADO DA ROCHA
AVALIAÇÃO DA GENOTOXICIDADE DO CLORETO DE METILMERCÚRIO EM DUAS ESPÉCIES DE
PEIXES NEOTROPICAIS
Tese apresentada ao Programa de Pós-Graduação em Neurociências e Biologia Celular, do Instituto de Ciências Biológicas da Universidade Federal do Pará, como requisito para obtenção do grau de Doutor em Neurociências e Biologia Celular. Orientador: Prof. Dr. Rommel Mario Rodriguèz Burbano Co-orientadora: Profa. Dra. Marta Margarete Cestari
BELÉM - PA 2009

CARLOS ALBERTO MACHADO DA ROCHA
AVALIAÇÃO DA GENOTOXICIDADE DO CLORETO DE METILMERCÚRIO EM DUAS ESPÉCIES DE PEIXES
NEOTROPICAIS
Tese apresentada ao Programa de Pós-Graduação em Neurociências e Biologia Celular, do Instituto de Ciências Biológicas da Universidade Federal do Pará, como requisito para obtenção do grau de Doutor em Neurociências e Biologia Celular.
Data de aprovação: 16 de junho de 2009. Orientador:
Prof. Dr. Rommel Mario Rodríguez Burbano - Orientad or Instituto de Ciências Biológicas, UFPA
Avaliadores:
Prof. Dr. Edivaldo Herculano Correa de Oliveira Instituto de Ciências Biológicas, UFPA Prof. Dr. Edmar Tavares da Costa Instituto de Ciências Biológicas, UFPA Profa. Dra. Fabíola Raquel Tenório Oliveira Universidade do Estado do Pará – UEPA Prof. Dr. Wallace Gomes Leal Instituto de Ciências Biológicas, UFPA
Belém - PA 2009

Aos meus pais Círio Rocha, in memoriam, e
Corina, pessoas que me motivaram por superar
seus limites;
Aos meus filhos Carlos Júnior, Carla Fernanda e
Carlinho, na esperança de que lhes sirva de
motivação.

AGRADECIMENTOS
Ao meu orientador, Prof. Dr. Rommel Burbano, primeiramente por sua
amizade. No que se refere a esta orientação, agradeço pela valiosa ajuda e por ter
acreditado que eu poderia conseguir mesmo vindo de outra área e com um projeto
que se desviava um pouco do perfil do laboratório. Valeu mesmo tio!
À minha co-orientadora, Profa. Marta Margarete Cestari, pela receptividade na
Universidade Federal do Paraná, minha iniciação nos testes de genotoxicidade em
peixes e pelas numerosas sugestões e recomendações para a tese. Também
agradeço ao Marcos Ferraro, que me acompanhou no laboratório da UFPR.
Ao Bruno Coêlho do Laboratório de Onocologia Experimental da Universidade
Federal do Ceará, pela acolhida em Fortaleza e grande ajuda na execução do
ensaio cometa.
Aos amigos do Laboratório de Citogenética Humana da UFPA que
contribuíram de alguma forma para a realização desta tese: André, Dani, Adriana,
Patrícia e, particularmente, ao Marcelo, colaborador neste projeto e quase mais um
orientador nestes anos de doutorado.
Aos amigos do Laboratório e parceria de GDP: Presidente Plínio, Diego,
Helem, Luiz, Leopoldo, Gabi, Priscila, Amanda, Tati, Leomá, João, Gal e Rodrigo.
Meu agradecimento também à Glorita e tomara não tenha esquecido ninguém.
Aos colaboradores no IFPA: Lorena Cunha, meu braço direito neste trabalho;
Henrique Pinheiro, o estagiário “da hora”. Também agradeço ao André (primeiro
estagiário), Seu Cinésio e Vitor.
À Pós-Gradução de Neurociências e Biologia Celular, pela qualidade do curso
e pela organização. Aos funcionários do Instituto de Ciências Biológicas que, de
algum modo, foram úteis na execução desta tese.
A toda a minha família, pela torcida e confiança. À Aldair, por sua paciência,
amor, incentivo e pelas horas doadas de nosso convívio.
A todos meus sinceros agradecimentos.

SUMÁRIO
1. INTRODUÇÃO 1
1.1 CONSIDERAÇÕES GERAIS 1
1.2 MERCÚRIO: OCORRÊNCIA E CICLO BIOGEOQUÍMICO 4
1.2.1 Metilação do mercúrio inorgânico 7
1.3 BIOACUMULAÇÃO E BIOMAGNIFICAÇÃO 9
1.4 MERCÚRIO E METILMERCÚRIO EM SISTEMAS HUMANOS 11
1.5 IMPACTOS AMBIENTAIS E À SAÚDE HUMANA
RELACIONADOS A EMISSÕES DE MERCÚRIO
12
1.5.1 O mal de Minamata 13
1.5.2 Acidentes com fungicidas 15
1.5.3 A questão do garimpo de ouro na Amazônia 16
1.6 PEIXES DA BACIA DO TAPAJÓS 18
1.7 ANÁLISES DE GENOTOXICIDADE 19
1.7.1 Estudos de efeitos genotóxicos de metais pesa dos em peixes 25
1.8 JUSTIFICATIVA 29
2. OBJETIVOS 30
2.1 OBJETIVO GERAL 30
2.2 OBJETIVOS ESPECÍFICOS 30
3. MATERIAIS E MÉTODOS 31
3.1 MATERIAIS 31
3.1.1 Organismos utilizados como bioindicadores 31
3.1.2 Coleta e aclimatação dos animais 33
3.2 MÉTODOS 34
3.2.1 Tratamento dos animais e obtenção do sangue p ara os
bioensaios
34
3.2.2 Ensaio Cometa 35
3.2.3 Teste do Micronúcleo 37
3.2.4 Bioensaios com Colossoma macropomum 38
3.2.5 Bioensaios com Aequidens tetramerus 39
3.2.6 Análises estatísticas 40
4. RESULTADOS 41
4.1 RESULTADOS DOS EXPERIMENTOS COM C. MACROPOMUM 41

4.2 RESULTADOS DOS EXPERIMENTOS COM A. TETRAMERUS 46
5. DISCUSSÃO 51
5.1 USO DE ERITRÓCITOS PERIFÉRICOS 52
5.2 MAIOR SENSIBILIDADE DO ENSAIO COMETA 53
5.3 TESTE DO MICRONÚCLEO 54
5.4 MAIOR ÍNDICE DE DANOS NO ACARÁ 56
6. CONCLUSÕES 57
7. REFERÊNCIAS BIBLIOGRÁFICAS 58
ANEXOS: TABELAS DE RESULTADOS
APÊNDICE 1: Artigo “Evaluation of Genotoxic Effect s of
Xenobiotics in Fishes Using Comet Assay – A Review”
APÊNDICE 2: Artigo “The Micronucleus Assay in Fis h
Species as an Important Tool for Xenobiotic Exposur e Risk
Assessment – A Brief Review and an Example Using
Neotropical Fish Exposed To Methylmercury”

LISTA DE FIGURAS E TABELAS
Figura 1 Horizontes do solo. Figura 2 Origem e ciclo global do mercúrio no ambiente. Figura 3 Metilação do mercúrio inorgânico. Figura 4 Bioacumulação e Biomagnificação do metilmercúrio. Tabela 1 Recentes estudos de genotoxicidade de metais pesados em
diversas espécies de peixes. Figura 5 Espécime de Colossoma macropomum. Figura 6 Espécime de Aequidens tetramerus. Figura 7 Aquários no Laboratório de Biologia Aquática do IFPA. Figura 8 Esquema resumido dos bioensaios. Figura 9 Resumo das sucessivas etapas que integram o Ensaio cometa. Figura 10 Visão de aquários no Laboratório de Biologia Aquática do IFPA,
por ocasião da exposição de C. macropomum ao metilmercúrio. Figura 11 Freqüência de nucleóides por classe de dano. Figura 12 Resultados do teste de Mann-Whitney obtidos no BioEstat 5.0
com dados do Ensaio cometa em C. macropomum. Figura 13 Gráfico representativo dos escores (por 100 células) do Ensaio
cometa em C. macropomum. Figura 14 Eritrócitos de C. macropomum corados com Giemsa, em
ampliação de 1.000X. A seta indica um micronúcleo. Figura 15 Resultado do teste do qui-quadrado obtido no BioEstat 5.0, com
dados do Teste do micronúcleo em C. macropomum. Tabela 2 Médias e desvios de micronúcleos (MN) e alterações
morfológicas nucleares (AMN) de C. macropomum. Figura 16 Freqüências de micronúcleos + alterações morfológicas
nucleares (por 4.000 eritrócitos) em C. macropomum. Figura 17 Ensaio cometa em A. tetramerus. Comparação do índice de
danos nos grupos controle (C) e tratados com MeHg 2mg.L-1 nos dois tempos de exposição.
Figura 18 Freqüência de nucleóides por classe de dano. Figura 19 Resultados do teste de Kruskal-Wallis obtidos no BioEstat 5.0
com dados do Ensaio cometa em A. tetramerus. Figura 20 Ensaio cometa em A. tetramerus. Comparação do índice de
danos ao DNA nos grupos controle (C) e tratados com MeHg 2mg.L-1 nos dois tempos de exposição.
Figura 21 Resultado do teste do qui-quadrado obtido no BioEstat 5.0, com dados do Teste do micronúcleo em A. tetramerus.
Tabela 3 Médias e desvios de micronúcleos (MN) e alterações morfológicas nucleares (AMN) de A. tetramerus.
Figura 22 Representação das freqüências de micronúcleos + alterações morfológicas nucleares (por 4.000 eritrócitos) em A. tetramerus.
Tabela 4 Efeitos genotóxicos e mutagênicos do MeHg 2mg.L-1. Comparação entre os resultados nas duas espécies de peixes.

LISTA DE ABREVIATURAS E SIGLAS
AMN Alterações morfológicas nucleares
DNPM Departamento Nacional de Produção Mineral
EDTA Ácido etilenodiamino tetra-acético
g Grama
Hg Mercúrio
IARC International Agency for Research on Câncer
IFPA Instituto Federal de Educação, Ciência e Tecnologia do Pará
kg Quilograma
L Litro
LMP Low melting point (baixo ponto de fusão)
mA Miliampere
MeHg Metilmercúrio
mg Miligrama
mL Mililitro
mM Milimolar
MN Micronúcleos
ng Nanograma
nm Nanômetro
OMS Organização Mundial de Saúde
PM Peso molecular
ppm Parte por milhão
SCGE Single cell gel electrophoresis
SNC Sistema Nervoso Central
UFPA Universidade Federal do Pará
UFRA Universidade Federal Rural da Amazônia
V Volt
WHO World Health Organization
µg Micrograma
µL Microlitro

RESUMO
A preocupação com a contaminação ambiental por compostos contendo
metais pesados é decorrente, além da alta toxicidade, de sua capacidade de
bioacumulação e potencialidade de induzir danos ao material genético. O mercúrio é
um metal xenobiótico, contaminante ambiental. A biotransformação do mercúrio
inorgânico em metilmercúrio nos ambientes aquáticos é um processo bem conhecido
e favorece a contaminação de algas, moluscos, crustáceos, peixes e, finalmente, o
homem, nos níveis mais elevados das cadeias alimentares. O objetivo deste estudo
foi determinar as alterações genotóxicas induzidas pelo cloreto de metilmercúrio
(CH3HgCl) em duas espécies de peixes comuns na bacia do rio Tapajós (Colossoma
macropomum e Aequidens tetramerus) e que, no ambiente natural, ocupam
diferentes níveis tróficos. Os peixes das duas espécies foram mantidas sob as
mesmas condições de manejo e, após exposição ao CH3HgCl por via hídrica,
tiveram o sangue coletado e utilizado em dois bioensaios: o Ensaio cometa e o Teste
do micronúcleo. Nossos resultados demonstraram que o CH3HgCl pode ser
altamente genotóxico. O Ensaio cometa apresentou diferenças mais significativas
que o Teste do micronúcleo para ambas as espécies em relação aos seus
respectivos controles. Ao compararmos os resultados entre as duas espécies
estudadas, percebemos níveis bem mais elevados de danos ao material genético da
espécie carnívora – A. tetramerus. Além disso, o nível de danos aumentou com o
tempo de exposição. Tais resultados, entretanto, não dependeram de suas posições
nas cadeias alimentares em ambiente natural, mas de características específicas
que determinam sua sensibilidade ao xenobionte ou sua susceptibilidade à
bioacumulação, ou ambas.
Palavras-chave : Metilmercúrio, Genotoxicidade, Colossoma, Aequidens, Ensaio
cometa, Teste do micronúcleo.

ABSTRACT
Concerns regarding environmental contamination caused by heavy metal
compounds is due their high toxicity and ability to bioaccumulate and cause DNA
damage. Mercury is a xenobiotic metal, being a environmental contaminant itself.
The process that transforms inorganic mercury in methylmercury in aquatic
environment is well known and favors algae, mollusks, crustaceans and fishes
contamination, as well as mankind on higher levels of the food chain. This study
aimed to evaluate genotoxic alterations induced by methylmercury chlorite (CH3HgCl)
in two fish species commons in Tapajós river bay (Colossoma macropomum and
Aequidens tetramerus) which, in their natural habitat, occupy different trophic levels.
Specimens of both species were kept under similar management conditions and,
after fishes have been exposed to MeHg via water, their blood was collected under
laboratory conditions and used in two bioassays: Comet Assay and Micronucleus
test. Our data showed that CH3HgCl can be highly genotoxic. Comet assay showed
to be more sensitive than the Micronucleus test for both species. When comparing
data between the two studied species, we noticed a higher DNA damage index in the
carnivore specie, A. tetramerus. Furthermore, DNA damage levels increased with
longer exposure to MeHg. Our result, however, were not consequence of their
position on the trophic chain in their natural environment, but were probably due to
specific features in those species that determine their sensibility to xenobiotics or its
susceptibility to bioaccumulation, or both.
Key words: Methylmercury, Genotoxicity, Colossoma, Aequidens, Comet assay,
Micronucleus test.

1. INTRODUÇÃO
1.1 CONSIDERAÇÕES GERAIS
O mercúrio é um metal xenobiótico e corresponde a um dos contaminantes
órgãos-específicos mais largamente difundidos e deletérios (Silva-Pereira et al.,
2005). O nome mercúrio é uma homenagem ao planeta Mercúrio. Já na mitologia
romana, Mercúrio é o mensageiro dos deuses, filho do deus Júpiter e de Maia, a filha
do titã Atlante. Mercúrio era também o deus dos mercadores e do comércio e tinha
muitos dos atributos do deus grego Hermes (Azevedo, 2003). Os gregos chamaram
o metal de Hidrargiro (hidro = elemento de composição com sentido de água;
argyros = prata). Já os romanos transcreveram essa expressão para o latim como
Hidrargyrum.
Os primeiros estudos sobre a problemática da presença do mercúrio em
vários compartimentos dos ecossistemas na Bacia Amazônica, os quais datam cerca
de duas décadas, reconhecem duas principais fontes como responsáveis por essa
contaminação: os garimpos de ouro e a agricultura do tipo corte e queima (Sampaio
da Silva et al., 2006). Em outras palavras, o rejeito de mercúrio no sistema aquático,
proveniente de operações de mineração e pela lixiviação de solos após
desflorestamentos, é considerado a maior contribuição para a contaminação na
Bacia Amazônica (Amorin et al., 2000).
Um levantamento realizado na década de 1980, pelo Departamento Nacional
de Produção Mineral – DNPM, em várias áreas de garimpo do Pará, Mato Grosso e
Goiás, detectou uma relação de cerca de dois gramas de mercúrio para cada grama
de ouro. Portanto, se considerarmos que a produção real (a oficial mais a não
computada) de ouro por garimpagem na região de 1980 a 1988 foi próxima de 900
toneladas, tivemos um lançamento de mercúrio na ordem de 1.800 toneladas ao
ambiente (Rocha & Canto-Lopes, 2002).
Realmente a liberação de mercúrio das atividades de garimpos de ouro
contribuiu para aumentar localmente suas concentrações, mas para toda a
Amazônia essas cargas seriam quase insignificantes, considerando-se os níveis
elevados em solos distantes das áreas de garimpo. Um processo razoável que deve
explicar essas observações no solo é que horizontes B funcionam como uma
esponja que acumula mercúrio durante um período de tempo geológico, liberando-o
de volta ao ciclismo por ocasião de erosões e incêndios florestais (Wasserman et al.,
2003).

Figura 1 – Horizontes do solo. Desenho: Nonato Reis (2009). (O) horizonte orgânico; (A) horizonte rico em atividade biológica, participando da humificação da matéria e da ciclagem de nutrientes; (B) horizonte iluvial, caracterizado por maior concentração de argila, com menor teor de matéria orgânica, bastante afetado pelos processos de formação do solo e que recebe, por infiltração da água, a influência das camadas superficiais; (C) material de origem (rocha-mãe) em decomposição (Andrade et al., 2004).
O metilmercúrio é um composto classificado como grupo 2B, pela IARC
(International Agency for Research on Câncer), indicando-o como possível
carcinogênico para humanos (Hallenbeck, 1993) e se acumula em extensão muito
maior quando comparado às outras formas do mercúrio, apesar de todas as formas
poderem se acumular (Azevedo, 2003).
Estudos têm mostrado que o mercúrio metálico liberado para a atmosfera ou
em sistemas aquáticos pode ser oxidado em Hg2+ e posteriormente convertido a
metilmercúrio por diversos mecanismos, principalmente por meio de reações
mediadas por bactérias. A lipossolubilidade de compostos orgânicos de mercúrio
contribui para os seus maiores níveis encontrados nos predadores, devido à
bioamplificação ao longo da cadeia alimentar (Barkay et al., 2003). O consumo de
organismos aquáticos contaminados com o metilmercúrio é a principal via de
exposição humana.

O peixe é um componente importante da dieta saudável. Alguns dos benefícios à
saúde do peixe incluem: riqueza em proteína, gordura insaturada e ácidos graxos
ômega 3 (David, 2009) e pobreza em gordura saturada. Os peixes são a principal
fonte de alimentação da população amazônida, e podem apresentar elevadas
concentrações de mercúrio, principalmente metilmercúrio, constituindo-se em
importante fonte de exposição humana ao metal (WHO, 1990).

1.2 MERCÚRIO: OCORRÊNCIA E CICLO BIOGEOQUÍMICO
O mercúrio é um metal pesado de aspecto argênteo, inodoro, cujo símbolo Hg
deriva do latim hydrargyrum, que, normalmente, é encontrado em três estados de
oxidação: 0, +1 e +2. Ele é um elemento químico de transição, de caráter metálico,
com elevado peso molecular (PM = 80) e alta densidade. Por esse motivo é
classificado como um metal pesado, que em sua forma elementar se apresenta
como um líquido denso, prateado, nas condições normais de temperatura e pressão
(Schroeder & Munthe, 1998).
Ocorre normalmente, em pequenas concentrações, nos vários
compartimentos da natureza: hidrosfera, litosfera, atmosfera e biosfera. Entre esses
compartimentos há um contínuo fluxo de mercúrio (Figura 2). Raramente é
encontrado como elemento livre na natureza, estando amplamente distribuído, em
baixas concentrações, por toda a crosta terrestre (Azevedo, 2003). O aporte
antrópico ocorre através de indústrias que queimam combustíveis fósseis, produção
eletrolítica de cloro-soda, produção de acetaldeído, incineradores de lixo, polpa de
papel, tintas, pesticidas, fungicidas, lâmpadas de vapor de mercúrio, baterias,
produtos odontológicos, amalgamação de mercúrio em extração de ouro, entre
outros (Micaroni, 2000).
Figura 2 – Origem e ciclo global do mercúrio no ambiente. Desenho: Nonato Reis (2009).

Por apresentar uma espécie química estável na atmosfera como sua forma
volátil, o vapor de mercúrio (Hg0) pode ser transportado em escala global, afetando
áreas remotas naturais longe de fontes pontuais de contaminação (Lacerda & Malm,
2008).
No ciclo atmosférico, também chamado ciclo global, ocorre a circulação de
espécies voláteis de mercúrio, como o dimetilmercúrio, (CH3)2Hg, e o mercúrio
elementar, Hg0, liberadas de fontes naturais e antropogênicas. Pouco se conhece a
respeito das reações da fase gasosa dessas espécies na atmosfera, mas tudo indica
que ocorre um processo de oxidação fotoquímica, através do qual as formas pouco
solúveis em água sejam convertidas em espécies mais solúveis (WHO, 1990).
Nos sedimentos do fundo dos rios as concentrações de mercúrio observadas
são relativamente baixas, demonstrando que o meio aquático não é uma via de
transporte importante para este elemento (Lechler et al., 2000). Entretanto,
particulados ricos em Hg2+ são transportados para o sedimento, onde o metal pode
ser metilado por bactérias sulfato-redutoras (Bisinoti & Jardim, 2004).
O mercúrio inorgânico apresenta uma toxicidade relativamente baixa, mas a
ação bacteriana organifica este elemento formando o metilmercúrio, que foi o grande
“vilão” dos principais acidentes de contaminação humana (Wasserman et al., 2001).
Para o íon Hg2+, a maioria dos ânions, principalmente óxidos, sulfetos e
cloretos, formam compostos de natureza covalente, e existem em solução quase
totalmente não dissociados, como moléculas discretas (Magalhães, 1995).
O metilmercúrio por ser lipossolúvel é muito bem absorvido pelas membranas
biológicas em geral, assim como pelos tratos digestivos dos animais em
praticamente todas as cadeias alimentares (Lacerda & Malm, 2008).
Percebe-se, assim, que a distribuição das diversas espécies de mercúrio que
entram no sistema aquático é regulada por processos físicos, químicos e biológicos,
os quais ocorrem nas interfaces ar/água e água/sedimento. A conversão entre estas
diferentes formas é a base do complexo padrão de distribuição do mercúrio no ciclo
e de seu enriquecimento biológico (Micaroni, 2000).

A complexidade dos ecossistemas amazônicos e o conhecimento limitado do
ciclo biogeoquímico do mercúrio em florestas tropicais explicam as dificuldades
enfrentadas pela comunidade científica em avaliar o impacto da contaminação de
mercúrio. Assim, o conhecimento atual sobre o transporte, transformação e destino
do mercúrio nos meios aquáticos e terrestres do sistema amazônico é ainda
relativamente superficial. Esses problemas afetam o potencial de exposição das
populações locais e, conseqüentemente, o risco envolvido (Hacon et al., 2008).

1.2.1 Metilação do mercúrio inorgânico
A alquilação é a notável transformação que o mercúrio inorgânico pode sofrer,
levando à formação do metil ou do dimetilmercúrio com as interconversões
correspondentes, sendo de fundamental importância para a avaliação da
contaminação local por mercúrio (Tena, 1981).
Nas camadas superiores do sedimento, que são biologicamente ativas, o
mercúrio bivalente é, em parte, metilado por bactérias bênticas a metilmercúrio e
depois a dimetilmercúrio, o que eleva sua capacidade para vencer membranas
biológicas (Figura 3). Nas camadas inferiores do sedimento o mercúrio é inativo,
principalmente sob a forma de sulfeto de mercúrio. Já o metilmercúrio irá se integrar
nas cadeias tróficas ou, se as condições de pH forem apropriadas, dará origem ao
dimetilmercúrio, o qual por ser menos solúvel e volátil passará à atmosfera e será
recolhido nas águas das chuvas. Se estas forem ácidas, o dimetilmercúrio irá se
transformar no metilmercúrio, retornando ao meio aquático e, assim, completando o
ciclo (Azevedo, 2003).
Figura 3 – Metilação do mercúrio inorgânico. Elaboração: Carlos Rocha e Nonato Reis (2009).

A metilação do mercúrio depende de bactérias e outros microrganismos que
tendem a prosperar em condições baixas de oxigênio dissolvido, como na interface
sedimento-água ou nos tapetes de algas. Inúmeros fatores ambientais influenciam
as taxas de metilação de mercúrio bem como a reação reversa, conhecida como
desmetilação. Estes fatores incluem a temperatura, o carbono orgânico dissolvido, a
salinidade, a acidez (pH), as condições de oxidação-redução, bem como a forma e a
concentração de enxofre na água e sedimentos (Alpers & Hunerlach, 2000).

1.3 BIOACUMULAÇÃO E BIOMAGNIFICAÇÃO
Como mencionado anteriormente, o mercúrio inorgânico, ao sofrer o processo
de alquilação, ganha lipossolubilidade, condição indispensável para o fácil transporte
através das membranas celulares, e, assim, pode se depositar – bioacumulação –
nos tecidos de um organismo. Tal fenômeno se dá em invertebrados, peixes e
mamíferos, podendo ocorrer em plantas aquáticas. As espécies inorgânicas do metal
(incluindo-se o mercúrio elementar) e o metilmercúrio têm capacidade de reagir com
ligantes intracelulares, o que explica o alto grau de acumulação desses compostos
(WHO, 1989).
Sempre que um organismo contaminado por mercúrio ocupa um nível inferior
em uma cadeia trófica, seu predador absorverá aquele mercúrio orgânico, mas
revelará concentrações comparativamente aumentadas – biomagnificação. Esse
termo se refere à acumulação de alguns metais pesados (e algumas outras
substâncias) por níveis tróficos sucessivos. Em estudo que avaliava a presença de
Hg em diversos tipos de peixes, em ambiente contaminado, observou-se que o metal
se concentrava mais intensamente à medida em que se crescia na cadeia alimentar:
os peixes vegetarianos apresentavam 6,64 ppm; os peixes que se alimentavam de
invertebrados, 12,4 ppm; os onívoros, 26,6 ppm; e os piscívoros, 40,2 ppm (Boening,
2000).
O homem torna-se um dos principais afetados pela intoxicação mercurial
quando usufrui do rio poluído para diversas atividades, como a alimentação, pois os
peixes que apresentam os maiores níveis de MeHg são os mais consumidos pelo
homem, como o tucunaré e a traíra (Lebel et al., 1998).
Muitos fatores no ambiente aquático influenciam a metilação do mercúrio e,
conseqüentemente, sua biomagnificação. A bioacumulação e a biomagnificação
representam processos extremamente complexos e envolvem biogeoquímica e
interações ecológicas. Como resultado, o grau de biomagnificação de mercúrio no
peixe é difícil de ser prognosticado, embora a acumulação/magnificação possa ser
facilmente observada. Desse modo, o mercúrio é transferido e concentrado por
intermédio de vários níveis da cadeia alimentar. As cadeias alimentares aquáticas
tendem a ter mais níveis tróficos do que as terrestres e, conseqüentemente, a
biomagnificação aquática alcança níveis elevados (UNEP, 2002).

Figura 4 – Bioacumulação e Biomagnificação do metilmercúrio. Elaboração: Carlos Rocha e Nonato Reis (2009).

1.4 MERCÚRIO E METILMERCÚRIO EM SISTEMAS HUMANOS
Evidências em diversas fontes indicam que a exposição crônica a baixas concentrações
de metais pesados, incluindo o mercúrio, resulta em disfunções imunológicas (Pollard & Hultman,
1997). Um estudo de Ben-Ozer et al. (2000) aponta a apoptose de linfócitos e monócitos induzida
por mercúrio, como tendo um papel importante na disfunção imune causada por este metal.
Contudo, vários resultados são contraditórios; por exemplo, alguns autores não
observaram alterações imunológicas já registradas por outros (Barregård et al., 1997). Isso deve
ser decorrente das diferenças metodológicas, genéticas e de características da exposição:
intensidade, tempo, higiene, alimentação, etc (Medrado-Faria, 2003).
O sistema nervoso central é o alvo principal do MeHg, onde afeta, principalmente, áreas
específicas do cérebro e cerebelo. A intoxicação por MeHg se caracteriza por ataxia (perda da
coordenação dos movimentos voluntários), disartria (problemas na articulação das palavras),
parestesia (perda da sensibilidade nas extremidades das mãos e pés e em torno da boca), visão
de túnel (constrição do campo visual) e perda da audição. Uma contaminação severa pode
causar cegueira, coma e morte. O período médio de latência varia, freqüentemente, de 16 a 38
dias (Bahia, 1997). As alterações na função visual (redução do campo de visão) estão entre os
mais freqüentes sinais relacionados à intoxicação por metilmercúrio (WHO, 1990).
A síndrome de Hunter-Russel é uma doença provocada pelo MeHg, cujos sintomas são
disfunção do sistema nervoso, ataxia e diminuição do campo visual. No entanto, vários outros
sintomas podem ser causados, dependendo do grau de exposição (Bisinoti & Jardim, 2004).
A neurotoxicidade do metilmercúrio é bem evidente no desenvolvimento embrionário e
fetal, sendo a exposição intra-uterina muito eficaz. No caso de Minamata, onde gestantes
expostas ao metilmercúrio através da ingestão de peixes contaminados, os efeitos neurológicos
foram bem evidentes, comprovados por casos de microcefalia e retardo mental (WHO, 1990).
De acordo com Nascimento et al. (2008), a neurotoxicidade induzida por metilmercúrio
(MeHg) aumenta a formação de radicais reativos e acelera as reações de radicais livres. O
estresse oxidativo no SNC pode produzir danos pela interação de diversos mecanismos,
incluindo danos mitocondriais, com aumento de Ca++ livre no compartimento intracelular, ativação
e inibição de enzimas, liberação de aminoácidos excitatórios, expressão de metalotioneínas e
disrupção de microtúbulos.
1.5 IMPACTOS AMBIENTAIS E À SAÚDE HUMANA RELACIONADOS A
EMISSÕES DE MERCÚRIO
Desde o surgimento das sociedades humanas, focos localizados de
contaminação ambiental ocorreram ao longo dos séculos. Porém, nunca na escala

global que testemunhamos hoje. Praticamente, todos os ambientes do planeta
encontram-se sob graus variados de contaminação (Lacerda & Malm, 2008).
Segundo os mesmos autores, de modo geral podemos dividir os poluentes
que mais causam danos aos ecossistemas em dois grandes grupos. O primeiro
inclui substâncias presentes nos efluentes de grandes áreas urbanas, especialmente
associadas à disposição imprópria de resíduos sólidos (lixo) e ao tratamento
inadequado ou inexistente de esgoto sanitário. Os ambientes aquáticos, como rios,
estuários e áreas costeiras, são os mais afetados pelos contaminantes gerados por
essas fontes. O segundo grupo, composto pelos poluentes de origem industrial e da
mineração, inclui substâncias tóxicas, como metais, gases de efeito estufa e
poluentes orgânicos, especialmente aqueles gerados pela queima de petróleo. Ao
contrário dos contaminantes do primeiro grupo, cujo efeito é geralmente local ou, no
máximo, regional, esses têm o poder de afetar o ambiente em escala global.
A contaminação humana pelo mercúrio presente na água e nos alimentos é
de extrema importância, haja vista ter sido responsável por trágicos episódios de
intoxicação, como os ocorridos no Japão, nas décadas de 50 e 60, e no Iraque na
década de 70 (Almeida et al., 2006).

1.5.1 O mal de Minamata
O exemplo mais notório de um envenenamento maciço por mercúrio ocorreu
na baía de Minamata, Japão, entre os anos de 1930 e 1960. A Chisso Corporation
de Tokyo (maior indústria de plásticos do Japão) despejou o mercúrio na Baía de
Minamata. O MeHg alcançou altas concentrações nos peixes, cerca de 40 µg/g, uma
quantidade enorme quando comparada à recomendada pela FDA dos EUA de não
consumir peixe com concentração de MeHg acima de 1,5 µg/g (Schwartz, 2005), ou
ao limite máximo permitido pela legislação vigente no Brasil, que é de 0,5 µgHg/g
(Vieira et al., 2005).
Segundo Magalhães (1995), os níveis de mercúrio nas águas próximas à
descarga eram de 2000 µg/L, sendo que o máximo permitido poderia ser de 1,6 a
3,6 µg/L. Os peixes, base da dieta alimentar da população da região de Minamata,
atingiam uma concentração de mercúrio muito acima do máximo admitido, que
atualmente é de 0,5 µg/g.
A contaminação provocada pelo mercúrio na Baía de Minamata, a partir de
1932 passou a chamar atenção da opinião pública mundial para o perigo do uso
indiscriminado desse metal, principalmente pelos óbitos registrados no início da
década de 1950. Porém, somente 24 anos depois do início da produção da Chisso,
1956, constatou-se que a doença era resultado da ingestão de metal pesado sem,
no entanto, identificar qual era o metal. Ainda na década de 50 o médico Hosakawa
constatou que o envenenamento era provocado por mercúrio. Somente em 1968, no
entanto, é que o governo japonês determinou que a Universidade de Kumamoto
pesquisasse o assunto; e os resultados vieram confirmar a constatação feita por
Hosakawa (Rocha & Canto-Lopes, 2002).
As vilas em torno da baía eram economicamente dependentes da pesca, de
modo que o peixe fazia parte da alimentação regular dos aldeões. Por não terem
sido informados sobre os perigos dos altos níveis de MeHg, cerca de 200.000
pessoas foram contaminadas (Schwartz, 2005). Centenas destas morreram e, com o
passar do tempo, milhares passaram a ser vítimas de uma doença crônica,
debilitante do sistema nervoso, conhecida hoje como Mal de Minamata. Nos
indivíduos expostos ao metilmercúrio que apresentaram intoxicação ocorreu
neuropatia, que começava por sensação de formigamento das extremidades distais,
dos lábios e da língua, disartria, surdez, cegueira e espasticidade. Surgiram,
também, tremores e movimentos involuntários, deficiência mental, infecções

recorrentes, coma e morte, no período entre 26 dias e quatro anos após o contato
(Karland et al., 1960)
Ainda segundo Karland et al. (1960), nas crianças evidenciou-se síndrome
neurológica de descorticação, e as que foram expostas ao metilmercúrio na fase
fetal nasceram com síndrome de paralisia cerebral. Autópsias de pessoas e animais
(pássaros e gatos) intoxicados e mortos mostraram grande destruição do cerebelo,
do córtex e dos gânglios cerebrais basais.

1.5.2 Acidentes com fungicidas
Na década de 70, no Iraque, Paquistão, Gana e Guatemala ocorreram vários
casos de contaminação de agricultores e seus familiares, que utilizavam grãos, para
confecção de pão caseiro, tratados com fungicidas a base de metil e etilmercúrio. No
caso particular do Iraque, mais de 6.900 pessoas foram hospitalizadas e pelo menos
459 morreram. Em 1969, nos Estados Unidos, a intoxicação resultou da ingestão de
carne de porco alimentado com grãos tratados com fungicidas organomercuriais
(Barce, 2006).
Com base nos estudos epidemiológicos dos casos de Minamata e de
envenenamento por ingestão de grãos contaminados com fungicida mercurial no
Iraque, a Organização Mundial de Saúde (WHO, 1990) estabeleceu as
recomendações da concentração máxima de mercúrio no peixe segura para a
saúde. De acordo com a OMS, peixes contendo até 300 ng/g (partes por bilhão)
podem ser consumidos diariamente, enquanto peixes contendo entre 300 e 500 ng/g
deverão ser consumidos eventualmente, enquanto que peixes contendo acima do
limite de 500 ng/g não deverão ser consumidos. Grupos de risco como as mulheres
grávidas e crianças devem estar mais atentos devido à
sua elevada sensibilidade em relação à intoxicação do mercúrio.

1.5.3 A questão do garimpo de ouro na Amazônia
A poluição ambiental pelo mercúrio, resultante da utilização desse metal na
atividade garimpeira de ouro, é um exemplo importante de ameaça à saúde de
populações da Amazônia decorrente da ocupação acelerada dessa região pelo
homem nas últimas décadas (Silveira et al., 2004).
No início da década de 1970, com o aumento da produção do ouro, o
interesse da sociedade capitalista voltou-se para os garimpos, os quais foram
parcialmente mecanizados, com a introdução de retroescavadeiras, pás mecânicas,
bombas de água potentes, uso de explosivos, entre outros. A transformação desses
ambientes foi expressa pelo desmatamento, pela poluição dos rios com o mercúrio
advindo do processamento do ouro, assoreamento e desvios dos cursos dos rios, e
aparecimento de gigantescas crateras, à medida que a frente garimpeira avançou
(Couto, 1990).
O crescimento acentuado da produção brasileira de ouro nas décadas de
1970 e 1980 colaborou para uma alteração marcante do padrão de morbidade na
região amazônica (Santos et al., 1995).
Dada a elevada contribuição do ouro do garimpo para a produção total, bem
como a notória dependência da garimpagem em relação à mão-de-obra, pode-se
desde logo concluir que o crescimento do produto implicou intensa mobilização de
contingentes humanos. É difícil precisar o número de garimpeiros existentes no
Brasil naquele período. A única fonte oficial que existe é um cadastro realizado pelo
Departamento Nacional de Produção Mineral em 1990. Este cadastro mostrou um
número, provavelmente subestimado, de 419.920 trabalhadores, distribuídos
principalmente no Estado do Pará (52,7%), sendo seguido por Mato Grosso,
Rondônia e Goiás (Santos et al., 1995).
A poluição da Amazônia em termos de mercúrio representa grave problema
ambiental. De acordo com Meech et al. (1997), 70 a 170 toneladas são lançadas
anualmente no meio ambiente pelas atividades informais de mineração de ouro,
além dos incêndios, em que a vegetação queimada constitui uma fonte primária de
emissões de Hg. Essa grande quantidade do metal sofre metilação, acumulando-se
em peixes da cadeia alimentar.
A oxidação do metal, que permite sua melhor dissolução para posterior
metilação, garantindo sua estabilidade em ambientes aquáticos, foi avaliada por

análises termodinâmicas e eletroquímicas. Evidenciou-se que, na presença de
ácidos orgânicos, há formação de complexos orgânico-Hg. Embora a metilação
dessas espécies solúveis de mercúrio não seja totalmente elucidada, a formação de
tais complexos em águas de rios deve contribuir para aumentar a disponibilidade do
Hg ali presente como contaminante (Meech et al., 1997).
As comunidades ribeirinhas e grupos indígenas são os principais afetados
pela contaminação de mercúrio. Estudos demonstraram que em algumas regiões a
concentração de MeHg nos peixes está abaixo do limite recomendado para consumo
pela legislação brasileira, porém o consumo contínuo de peixes e em grande
quantidade pelas comunidades indígenas pode tornar-se um importante fator de
risco de toxicidade crônica (Lebel et al., 1998).
A região Oeste do Estado do Pará apresenta um histórico de intensa atividade
garimpeira arcaica com grande derramamento de mercúrio no ambiente. Os níveis
do mercúrio detectados por diferentes trabalhos em populações expostas da bacia
do Rio Tapajós foram elevados em certas comunidades (São Luís do Tapajós,
Barreiras e Rainha), no entanto, apresentando uma diminuição gradual com o passar
dos anos (Sá et al., 2006).
O risco de intoxicação pela ingestão de peixe contaminado depende de
alguns fatores, sendo que o teor de mercúrio, a quantidade ingerida e a freqüência
de ingestão são determinantes para a acumulação de metilmercúrio no organismo
humano. Cuidados especiais, no entanto, devem ser tomados com grupos de risco,
como gestantes, pós-gestantes e recém-nascidos (Yallouz et al., 2001).
Segundo Harada et al. (2001), o aparecimento, na Amazônia, de
manifestações clínicas semelhantes às do Mal de Minamata, deverá seguir a regra
da evolução da contaminação ambiental, em cinco estágios: 1) contaminação
ambiental pelo mercúrio inorgânico; 2) transformação do mercúrio inorgânico em
mercúrio orgânico (biometilação); 3) acúmulo do mercúrio orgânico na cadeia
alimentar aquática; 4) acúmulo do organo-metal pelo homem através da ingestão de
peixes; 5) aparecimento dos sinais e sintomas do Mal de Minamata.

1.6 PEIXES DA BACIA DO TAPAJÓS
A população ribeirinha do Tapajós, Estado do Pará, encontra no pescado sua
principal fonte de alimentação. Nos últimos anos, alguns estudos têm demonstrado
que peixes da região podem apresentar teores de mercúrio acima do recomendável
para o consumo humano, tornando essa população grupo de risco de contaminação
ambiental pelo metal (Pinheiro et al., 2000), dados nem sempre confirmados por
outros autores.
A Bacia do Tapajós é uma das regiões mais estudadas da Amazônia, sendo
vários os trabalhos encontrados sobre determinação dos teores de Hg total em
peixes. Para esta região os peixes mais comumente analisados são dourado, jaú,
piraíba, mandubé, cachorro, traíra, apapá, pescada, tucunaré, filhote, pirarucu,
acará, aruanã, pacu, surubim, matrinxã, jaraqui, sarda, jiju, jacundá, tambaqui e
aracu. As concentrações de Hg total para estas espécies em área de mineração
variaram de 0,02 até 2,75 mg/kg e ficaram abaixo de 0,1 mg/kg para peixes
coletados em rios não contaminados (Malm et al., 1997).
Vieira et al. (2005) quantificaram o teor de mercúrio total em peixes
provenientes de Itaituba-Pará, através de análise por espectrofotometria de
absorção atômica com amalgamação em lâminas de ouro dos teores de mercúrio
em amostras de filé de peixes. A concentração média de mercúrio total em
Colossoma macrapomum (tambaqui) foi de 0,28±0,08µg/g, enquanto que em Cichla
sp. (tucunaré) foi de 0,35±0,09µg/g. Os dados indicam que os teores médios do
metal nas espécies analisadas, encontram-se abaixo do limite máximo permitido
pela legislação vigente, isto é 0,5 µgHg/g. Entretanto, em dois exemplares, um de
cada espécie, correspondente a 4,25% das amostras, os valores superaram tal
limite, caracterizando, assim, a possível exposição humana ao metal.
A dinâmica específica dos ambientes aquáticos da região, em função do
processo de inundação sazonal, pode causar modificações na alimentação dos
peixes e/ou facilitar o desenvolvimento de condições favoráveis à produção do
MeHg. De acordo com o estudo de Roulet et al. (2000), realizado no baixo Tapajós,
os espaços adjacentes aos cursos d’água constituem sítios importantes para a
produção do MeHg. Nesses sítios, a produção e a acumulação do MeHg é
estreitamente relacionada à inundação e à degradação da matéria orgânica
(Guimarães et al., 2000).

1.7 ANÁLISES DE GENOTOXICIDADE
Um aspecto comum a todos os metais pesados é uma forte afinidade por
enxofre e sua interação com sistemas biológicos é geralmente por meio da ligação
com grupamentos sulfidrila (–SH) de proteínas. Resíduos cisteinil têm papéis
essenciais na função de muitas enzimas, particularmente aquelas envolvidas em
hidrólises e reações do tipo redox, como a glutationa redutase (Bilitewski & Turner,
2000).
O mercúrio, como alguns outros metais e certos compostos organomercuriais,
tem demonstrado propriedades mutagênicas. Essa toxicidade ocorre sobre a
tubulina, a subunidade estrutural dos microtúbulos das células, os quais estão
envolvidos na organização citoplasmática e constituem as fibras do fuso, interferindo
na polimerização e provocando contração dos cromossomos na metáfase, retardo
na divisão do centrômero e movimento anafásico reduzido (Cassidy & Furr, 1978;
Thier et al., 2003).
Análises de genotoxicidade são aquelas que levam em consideração a
vulnerabilidade do material genético (DNA) a agressões impostas pelo ambiente.
Tais avaliações dividem-se basicamente em análises mutagênicas, carcinogênicas e
teratogênicas (Ferreira, 2004). De acordo com a mesma autora, dentre as análises
mutagênicas mais comuns realizadas com organismos aquáticos, podem-se
destacar: teste de aberrações cromossômicas, teste de trocas entre cromátides-
irmãs, teste dos micronúcleos, deformidades citoplasmáticas e nucleares, e ensaio
cometa.
O teste de aberrações cromossômicas consiste na avaliação de alterações
provocadas por xenobióticos, em uma ou ambas as cromátides e que resultam de
quebras apenas ou quebras seguidas de soldadura. As quebras resultam na
diminuição do tamanho do cromossomo e são identificadas por um fragmento
deslocado, não alinhado com o resto da cromátide ou que apresenta
descontinuidade em relação ao resto do cromossomo, maior que a largura da própria
cromátide (Rabelo-Gay, 1991).
A respeito do teste das trocas entre cromátides-irmãs (TCI), Varela-Garcia
(1991) fez um relato sobre o histórico, execução e viabilidade. Como descrito por
esse autor, as TCI são manifestações citológicas de quebras que ocorrem no

mesmo lócus das duas cromátides de um cromossomo, seguidas de intercâmbio e
reparo. Para a coloração assimétrica das cromátides-irmãs podem ser utilizados
fluorocromos (por exemplo, alaranjado de acridina), que requerem análise ao
microscópio de epifluorescência, ou corantes como Giemsa.
O teste do micronúcleo desenvolvido por Schmid (1975) usando células da
medula óssea de mamíferos tem sido aplicado extensivamente para testar a
genotoxicidade de compostos químicos. Excelentes resultados têm sido obtidos com
esse teste em invertebrados, peixes e anfíbios, no monitoramento de áreas
contaminadas (Campana et al., 2003).
Os micronúcleos (MN) provêm de fragmentos cromossômicos acêntricos ou
cromossomos inteiros que se atrasaram e não foram incorporados ao núcleo
principal da célula-filha durante a divisão celular. São considerados micronúcleos
clássicos aquelas estruturas esféricas ou ovais, bem delimitadas, de mesma
refringência que o núcleo principal, não ligadas a este. No caso específico dos
peixes, devido ao tamanho normalmente reduzido dos cromossomos, os
micronúcleos variam na faixa de 1/30 a 1/10 do tamanho do núcleo normal (Al-Sabti
& Metcalfe, 1995; Ayllon & Garcia-Vazquez, 2000).
Pelo teste dos micronúcleos podem-se detectar compostos que interferem na
formação do fuso mitótico, alterando a distribuição eqüitativa dos cromossomos
durante a divisão celular, ou ao nível de proteínas diretamente envolvidas na
segregação cromossômica, com a vantagem de ser mais rápido que a análise de
aberrações cromossômicas (Al-Sabti & Metcalfe, 1995). Assim, este teste detecta
tanto eventos aneugênicos (alterações no número de cromossomos do genoma,
devido a erros na distribuição destes durante o processo de divisão celular), quanto
eventos clastogênicos (quebras que produzem alterações na estrutura dos
cromossomos).
Uma das vantagens é que pode ser aplicado em qualquer população de
células em proliferação sem depender do cariótipo envolvido. Devido aos peixes
terem um grande número de cromossomos, e muitas vezes de pequeno tamanho, as
análises das metáfases para avaliação de aberrações cromossômicas são
dificultadas, enquanto que o estudo de micronúcleos é fácil e possível de ser
realizada em eritrócitos, devido ao fato destes serem nucleados (Hayashi et al.,
1998).

Micronúcleos resultam de lesões no DNA ou cromossomos, ou em nível de
proteínas direta ou indiretamente envolvidas na segregação cromossômica (como a
tubulina, por exemplo). A formação de micronúcleos depende da perda de
fragmentos cromossômicos ou de cromossomos inteiros, e requer divisão mitótica ou
meiótica (Kirsch-Volders et al., 2003).
Sabe-se que perdas cromossômicas e a não segregação de cromossomos
(não disjunção), são importantes eventos no câncer e que eles são causados por
defeitos no fuso, centrômero ou como uma conseqüência da não-condensação da
estrutura cromossômica antes da metáfase (Fenech, 2000).
As vantagens do teste do micronúcleo incluem, além da simplicidade e
rapidez: (1) MN pode ser observado durante o ciclo celular, e o número de células
contáveis é ilimitado; (2) A contagem pode ser feita por qualquer pessoa com pouco
treinamento em citogenética; (3) não é necessário um cariótipo favorável; (4) o MN
formado persiste pelo menos até a próxima intérfase; (5) não é necessário nenhum
reagente para bloquear o fuso (Heddle et al., 1983).
É possível fazer a contagem de MN por programas computacionais; como o
Cytoscan, que é um programa de captura de imagens que não requer a contínua
transferência de informação digital do microscópio para o computador, podendo
escanear uma área de 10cm2 por lâmina em 5 minutos (Al-Sabti & Metcalfe, 1995).
Carrasco et al. (1990) sugerem que se utilize, além da contagem dos MN,
também a contagem de alterações morfológicas nucleares. Estas alterações foram
descritas e classificadas como:
(1) Blebbed: núcleos com uma pequena evaginação da membrana nuclear,
parecendo conter eucromatina ou heterocromatina (mais escuro). Os tamanhos
destas evaginações situam-se na faixa de pequenas protuberâncias até estruturas
completamente circunscritas, semelhantes aos micronúcleos, mas ainda ligadas ao
núcleo principal.
(2) Lobed: núcleos com evaginações mais largas do que as descritas para
Blebbed. Sua estrutura não é tão definida como a anterior. Alguns núcleos
apresentam várias destas estruturas.
(3) Vacuolated: núcleos que apresentam uma região que lembra os vacúolos
no seu interior. Estes “vacúolos” apresentam-se destituídos de qualquer material
visível no seu interior.

(4) Notched: núcleos que apresentam um corte bem definido em sua forma.
Geralmente com uma profundidade apreciável no núcleo. Estes cortes parecem não
possuir nenhum material nuclear e parecem ser delimitados pelo envelope nuclear.
As deformidades citoplasmáticas e nucleares são produzidas em resposta
a xenobióticos e consideradas como manifestações quantificáveis de genotoxicidade
em organismos aquáticos. Muitas vezes essas estruturas observadas nas células do
sangue periférico de vertebrados aquáticos, exibindo DNA extranuclear, podem ser
confundidas com micronúcleos. Tais alterações caracterizam-se por fragmentações
e vacuolização de eritrócitos, irregularidades na carioteca, eritrócitos exibindo
aglomerados de cromatina e aumento dos espaços de intercromatina (Ferreira,
2004).
O ensaio cometa , também conhecido como SCGE (single cell gel
electrophoresis), é uma técnica rápida e sensível de análise e quantificação de
danos ao DNA de células individuais. Assim, é usada na pesquisa de câncer,
avaliação de genotoxicidade e efetividade da quimioprevenção. A imagem que é
obtida lembra um “cometa” com cabeça e cauda distintas. Esta técnica foi
desenvolvida por Östling & Johansson em 1984, porém Singh et al. (1988) a
modificaram, como o Ensaio Cometa Alcalino.
A técnica do ensaio cometa consiste em células individualizadas,
colocadas em agarose sobre uma lâmina de microscopia, lisadas, submetidas
a eletroforese e coradas com brometo de etídio. O resultado é observado ao
microscópio, sendo que as células sem DNA danificado apresentam um
núcleo redondo, enquanto o DNA resultante de quebras simples ou múltiplas
migra para fora do núcleo, formando uma cauda, parecendo um cometa.
Como a cabeça é composta de DNA intacto e a cauda consiste de DNA danificado
ou quebrado, a extensão da migração do DNA reflete a extensão dos danos
(Fairbairn et al., 1995).
O papel da lise no ensaio cometa é o de remover os conteúdos
celulares, com exceção do material nuclear. O DNA permanece bem
condensado devido à presença de uma pequena quantidade de proteínas

não-histônicas. Porém, quando colocado na solução de eletroforese, a qual
possui um pH > 13, a espiralização do DNA começa a relaxar a partir de
pontos de quebra da fita. Permitindo desta maneira que os mesmos sejam
revelados pela eletroforese na seqüência do teste (Yendle et al., 1997; Collins
et al., 1997).
O ensaio cometa é habitualmente realizado com eritrócitos, pois estes são
facilmente obtidos por métodos não destrutivos e não necessitam do passo adicional
de isolamento, porém outros tecidos também têm sido testados, pois os efeitos de
genotoxicidade de contaminantes podem ser muitas vezes tecido-específicos. Os
tecidos mais pesquisados, além do sanguíneo, são do fígado, por se tratar do
principal órgão do metabolismo, das brânquias, devido ao seu contínuo contato com
a fase aquosa e do rim, tecido produtor de sangue em peixes (Belpaeme et al.,
1998).
Ao contrário de outros tipos de ensaio como os testes de micronúcleo, de
aberrações cromossômicas ou de trocas de cromátides irmãs, que necessitam de
células em proliferação para sua viabilidade, o ensaio cometa não necessita desta
condição, podendo ser utilizado em, virtualmente, qualquer tipo de célula. Também
não pode ser esquecido que o tipo de dano observado pelo ensaio é possivelmente
reversível, o que já foi observado por vários autores em estudos de monitoramento
ambiental (Pandrangi et al., 1995).
As vantagens dessa técnica incluem além da sensibilidade na
detecção de dano no DNA: (a) a coleta de dados em nível de células
individuais; (b) o uso de um número pequeno de células para a análise; (c) a
possibilidade de aplicação em qualquer população de células eucarióticas
isoladas (Tice & Vasquez, 1999).
Uma das dificuldades do uso do teste cometa em trabalhos
ambientais, é a comparação dos resultados com diferentes métodos de
quantificação do dano. Atualmente utiliza-se tanto tamanho da cauda com
relação ao núcleo quanto porcentagem de fragmentação da cauda, ou ambos
(Lee & Steinert, 2002).
Várias são as metodologias empregadas para avaliar a extensão do
dano ocasionado ao DNA. Uma das medidas utilizadas na avaliação deste

dano é feita pela relação entre o raio do núcleo e a extensão das caudas
formadas pelo DNA em migração (classificados como classe 0 = nenhum
dano, até classe 4 = máximo dano). Esta análise tanto pode ser feita
visualmente como através de softwares especiais (Ferraro et al., 2004).
Durante o curso de doutoramento reunimos numerosas publicações
abordando o uso do Ensaio cometa na avaliação de efeitos genotóxicos em peixes,
o que nos permitiu a elaboração de uma breve revisão intitulada “Evaluation of
Genotoxic Effects of Xenobiotics in Fishes Using Comet Assay – A Review”, a qual
foi publicada no periódico Reviews in Fisheries Science 17(2): 170–173, 2009. O
referido artigo encontra-se em um apêndice ao final deste trabalho.

1.7.1 Estudos de efeitos genotóxicos de metais pes ados em peixes
Para melhor compreender os efeitos de xenobiontes nos ecossistemas, são
realizados bioensaios com a utilização de biomarcadores. Os bioensaios auxiliam no
conhecimento das reações biológicas de um xenobionte em um dado ser vivo dentro
de uma cadeia trófica. Este conhecimento permite comparações com dados obtidos
diretamente do ambiente, ou ainda deduzir se a concentração do agente genotóxico
utilizado pode ou não provocar alterações no DNA ou patologias nos tecidos e
órgãos (Lopes-Poleza, 2004).
Pode-se detectar se o agente genotóxico está interagindo com o material
genético pelo aparecimento de adutos em proteínas (albumina e hemoglobina) e no
DNA, assim como pelas trocas entre cromátides-irmãs, refletindo uma exposição
primária. Um resultado positivo a este nível, entretanto, não indica necessariamente
conseqüências adversas, já que parte do dano genotóxico primário pode ser
reversível (Benito, 2002).
O uso de bioensaios permite estudar os efeitos tóxicos de determinados
contaminantes de forma isolada ou associados, minimizando a influência das
variáveis ambientais. Os resultados obtidos através de bioensaios não podem ser
transferidos diretamente para o ambiente, mas auxiliam no fornecimento de uma
base de dados visando um melhor entendimento dos fatores que estão interferindo
na saúde dos organismos e/ou alterando as condições do próprio ambiente
(Ramsdorf, 2007). A maioria dos estudos que avaliam a toxicidade de poluentes aos
organismos aquáticos e seus sistemas têm investigado exposições a tóxicos
individuais (Bagdonas & Vosyliené, 2006).
Um número considerável de estudos dos efeitos genotóxicos e mutagênicos
de metais pesados e seus compostos inorgânicos e orgânicos em peixes foi
publicado nos últimos 10 anos, tanto provenientes de trabalhos de biomonitoramento
in situ, quanto de bioensaios. Alguns são apresentados na Tabela 1.

Tabela 1 – Recentes estudos de genotoxicidade de metais pesados em diversas espécies de peixes.
Organismos Tipos de Célula Técnicas Metais Contaminantes Referências
Anguilla Anguilla Células de Rim Teste do Micronúcleo Cádmio e Mercúrio Sanchez-Galan et al., 2001.
Hoplias malabaricus Eritrócitos Aberrações Cromossômicas e Ensaio
Cometa Chumbo Cestari et al., 2004.
Hoplias malabaricus Eritrócitos Aberrações Cromossômicas Ensaio Cometa e Teste do Micronúcleo
Chumbo Ferraro et al., 2004.
Hoplias malabaricus Eritrócitos Aberrações Cromossômicas Ensaio Cometa e Teste do Micronúcleo
Mercúrio Lopes-Poleza, 2004.
Danio rerio Eritrócitos Citometria de Fluxo e Teste do
Micronúcleo Urânio Barillet et al., 2005.
Cyprinus carpio, Carassius gibelio e Corydoras paleatus
Eritrócitos e Células de Brânquias e de Fígado
Teste do Micronúcleo Cobre e Cádmio Cavas et al., 2005.
Prochilodus nigricans, Mylossoma duriventris e Hoplias malabaricus
Eritrócitos Teste do Micronúcleo Mercúrio Porto et al., 2005.
Genidens genidens, Cathorops spixii, Sciadeichtys luniscutis, Haemulon steindachneri e Micropogonias furnieri
Eritrócitos Teste do Micronúcleo Mercúrio Ramos et al., 2005.
Oncorhynchus mykiss Eritrócitos Teste do Micronúcleo Cobre e Zinco Bagdonas & Vosyliené, 2006.
Oreochromis niloticus Eritrócitos Ensaio Cometa e Teste do
Micronúcleo Cromo Matsumoto et al., 2006.
Clarias gariepinus, Alburnus orontis e Mugil cephalus
Eritrócitos Teste do Micronúcleo Cobre, Cádmio, Níquel e
Chumbo Ergene et al., 2007.
Oncorhynchus mykiss Eritrócitos Teste do Micronúcleo Cobre, Zinco, Chumbo, Níquel,
Cromo e Manganês Andreikënaitë et al., 2007.
Carassius auratus auratus Eritrócitos e Células
Epiteliais de Brânquias e Nadadeiras
Teste do Micronúcleo Mercúrio e Chumbo Cavas, 2008
Chondrostoma nasus e Barbus capito pectoralis
Eritrócitos Teste do Micronúcleo Cobre, Zinco, Cádmio, Cobalto
e Chumbo Koca et al., 2008.

Sanchez-Galan et al. (2001) utilizaram as análises de teor de cádmio e de
mercúrio no fígado e o Teste do Micronúcleo em células renais da enguia Anguilla
anguilla de sítios de rios com diferentes níveis de poluição com esses metais
pesados. Apesar do teor de cádmio no fígado apresentar associação com o nível de
cádmio dos sedimentos, as outras análises (teor de mercúrio no fígado,
micronúcleos) não apresentaram resultados significativos.
Lopes-Poleza (2004) avaliou o efeito genotóxico do metilmercúrio por via
trófica na traíra Hoplias malabaricus, um peixe predador de hábito alimentar
piscívoro. As análises foram feitas através das freqüências de aberrações
cromossômicas (rim anterior), de micronúcleos e alterações na forma dos núcleos e
do Ensaio Cometa em eritrócitos de sangue periférico. Neste estudo, apenas foram
observadas diferenças significativas entre os grupos controle e contaminado no
Teste de Aberrações Cromossômicas.
Porto et al. (2005) utilizaram o Teste do Micronúcleo em três espécies de
peixes Characiformes de diferentes hábitos alimentares para avaliar efeitos
genotóxicos da poluição por mercúrio. Obtiveram distintas freqüências de
micronúcleos nos peixes coletados em dois rios da Bacia Amazônica: rio Madeira
(área poluída) e rio Solimões (área não poluída). Também observaram que a
freqüência média de MN na espécie piscívora foi cerca de cinco vezes maior que
nas espécies detritívora e onívora.
Ramos et al. (2005) avaliaram a contaminação por mercúrio em tecido
muscular, sangue total, plasma e hemácias de cinco espécies de peixes (Genidens
genidens, Cathorops spixii, Sciadeichtys luniscutis, Haemulon steindachneri e
Micropogonias furnieri) da Baía do Ribeira, município de Angra dos Reis (RJ, Brasil).
Um dos objetivos foi avaliar efeitos genotóxicos por meio do teste do micronúcleo e
núcleo bilobado em eritrócitos, comparando com resultados anteriores em
espécimes de Netuma barba da Baía de Guanabara. Foi verificado que tanto a
freqüência de micronúcleos quanto a de núcleos bilobados eram estatisticamente
diferentes nas duas áreas, sugerindo que os peixes da Baía de Guanabara
apresentaram uma freqüência maior de efeitos cromossômicos por estarem
expostos a diversos tipos de contaminantes, inclusive o mercúrio.
Cavas (2008) utilizou o Teste do Micronúcleo, sob coloração por alaranjado
de acridina, para avaliação in vivo da genotoxicidade do cloreto de mercúrio e do

acetato de chumbo em diferentes células de Carassius auratus auratus. As
freqüências de MN nos três tipos de células foram elevadas nos peixes expostos
tanto ao cloreto de mercúrio quanto ao acetato de chumbo. Por outro lado, as
freqüências de MN, tanto no grupo controle quanto no contaminado, foram maiores
em células de brânquias que nas outras células testadas. O estudo também
confirmou que a coloração por alaranjado de acridina é conveniente para o estudo
de micronúcleos in vivo em peixes.

1.8 JUSTIFICATIVA
O trabalho proposto se justifica ao considerarmos que a forma mais tóxica do
mercúrio, o metilmercúrio (MeHg), é um composto orgânico com grande estabilidade
química, o que aliado à sua alta afinidade por lipídios (membranas biológicas) o
conduz a um trânsito preferencial e estável pela biota. O MeHg, por outro lado,
apresenta o fenômeno de biomagmificação de forma mais eficiente do que qualquer
outro poluente metálico e este fenômeno é muito melhor documentado em sistemas
aquáticos onde, dependendo do grau de poluição ambiental, podem ser
contaminados organismos de vários níveis tróficos (WHO, 1989; WHO, 1990; Lebel
et al., 1998; Boening, 2000; UNEP, 2002; Azevedo, 2003; Barkay et al., 2003;
Lacerda & Malm, 2008).
A ingestão pelo homem representa a passagem do MeHg a um elo mais
elevado na cadeia alimentar, o que pode representar conseqüências dramáticas à
saúde de populações humanas com hábitos alimentares restritos, que incluem
basicamente peixes e outros organismos aquáticos contaminados. Além disso,
particularmente na região do rio Tapajós, diversas comunidades humanas têm
exposição confirmada ao metilmercúrio (MeHg), exibindo, algumas vezes, índices
acima dos limites de segurança (WHO, 1990; Malm et al., 1997; Lebel et al., 1998;
Pinheiro et al., 2000; Harada et al., 2001; Silveira et al., 2004; Schwartz, 2005; Vieira
et al., 2005; Almeida et al., 2006; Sá et al., 2006).
Na literatura encontramos um grande número de trabalhos sobre
biomagnificação de metais pesados, como o mercúrio, o que lhe garante uma
condição de conhecimento relativamente bem documentado. Constatamos também
a carência de trabalhos que, como este, avaliem as respostas genotóxicas de peixes
isentos da influência de suas posições tróficas.
Além disso, os peixes freqüentemente respondem aos tóxicos de forma
semelhante aos vertebrados superiores, podendo ser usados em bioensaios para
exposição aos xenobiontes que têm potencial de causar efeitos teratogênicos e
cancerígenos em humanos (Al-Sabti & Metcalfe, 1995; Lopes-Poleza, 2004).
Finalmente, devido à ainda escassa informação sobre genotoxicidade em espécies
nativas, trabalhos como este se fazem necessários como contribuição na geração de
bancos de dados mais consistentes (Cestari et al., 2004).

2. OBJETIVOS
2.1 OBJETIVO GERAL
Avaliar as alterações genotóxicas induzidas pelo cloreto de metilmercúrio
(CH3HgCl), por via hídrica em peixes que ocupam diferentes níveis da cadeia trófica
no rio Tapajós, porém com o intuito de demonstrar possíveis diferenças específicas,
independentemente de suas relações tróficas no ambiente natural. As avaliações
foram feitas por meio de bioensaios, utilizando o Ensaio cometa e o Teste do
micronúcleo.
2.2 OBJETIVOS ESPECÍFICOS
Os seguintes objetivos específicos foram delineados:
• Determinar o dano no DNA através do Ensaio Cometa em núcleos de
eritrócitos de sangue periférico de Colossoma macropomum e
Aequidens tetramerus;
• Determinar a freqüência de Micronúcleos Písceos (MNP) e de
alterações morfológicas nucleares em eritrócitos de sangue periférico
de Colossoma macropomum e Aequidens tetramerus;
• Comprovar a provável diferença entre efeitos genotóxicos primários e
efeitos mutagênicos do cloreto de metilmercúrio nos peixes estudados;
• Verificar se existe diferença significativa de respostas entre as espécies
Colossoma macropomum e Aequidens tetramerus, quanto aos efeitos
genotóxicos e mutagênicos do MeHg.

3. MATERIAIS E MÉTODOS
3.1 MATERIAIS
3.1.1 Organismos utilizados como bioindicadores
Foram utilizados peixes de duas espécies neotropicais (Colossoma
macropomum e Aequidens tetramerus) que ocorrem naturalmente nas bacias da
Amazônia e que ocupam diferentes nichos tróficos. Essas espécies foram escolhidas
por serem de fácil adaptação e manutenção nas condições laboratoriais:
Colossoma macropomum (Cuvier, 1818) – o tambaqui (Figura 5), de hábito
onívoro, nativo da Bacia Amazônica, Orinoco e seus rios afluentes, como o Tapajós.
C. macropomum, pertencente à família Characidae, constitui-se num dos maiores
peixes de valor econômico da região Amazônica. É o segundo maior peixe de
escamas e o maior Characiforme do rio Solimões/Amazonas que, no ambiente
natural, pode chegar até 100 cm de comprimento e 30 kg de peso (Araújo-Lima &
Goulding, 1998). Por possuir carne bastante apreciada pela população local e por
apresentar certo declínio na captura em ambiente natural, o tambaqui é a principal
espécie cultivada na região Norte, destacando-se na piscicultura de seis dos sete
estados da região (Val et al., 2000). Estudos citogenéticos demonstram que C.
macropomum apresenta um número diplóide 2n = 54, com o cariótipo composto de
20 metacêntricos e 34 submetacêntricos, e um número fundamental (NF) de 108
(Almeida–Toledo et al., 1987; Nirchio et al., 2003).
Aequidens tetramerus (Heckel, 1840) – o acará sela (Figura 6), de hábito
carnívoro (mais precisamente piscívoro) é amplamente distribuído na bacia do Rio
Amazonas, incluindo rios do Peru, Colômbia, Equador, Brasil e Bolívia. Também
ocorre no Rio Tocantins, Rio Parnaíba, na Guiana Francesa, Suriname, Guiana, e na
bacia do Rio Orinoco. A análise cariotípica realizada em A. tetramerus revelou a
presença de 48 cromossomos, fórmula cariotípica 8M-SM+40ST-A (Ribeiro, 2007).

Figura 5 – Espécime de Colossoma macropomum. Foto: O autor (2008).
Figura 6 – Espécime de Aequidens tetramerus. Foto: O autor (2008).
1 cm
1 cm

3.1.2 Coleta e aclimatação dos animais
Os tambaquis utilizados nos bioensaios foram fornecidos pela Estação de
Piscicultura da Universidade Federal Rural da Amazônia (UFRA), no município de
Castanhal, a 70 km de Belém; os acarás foram doados pelo professor Augusto
César Paes de Souza, de seu cultivo particular em Belém.
Os peixes, de ambas as espécies, foram mantidos em aquários (Figura 7),
sob manejo, no Laboratório de Biologia Aquática do Instituto Federal de Educação,
Ciência e Tecnologia do Pará (IFPA), em Belém – PA.
Figura 7 - Visão geral dos aquários no Laboratório de Biologia Aquática do IFPA, onde ocorreram os períodos de aclimatação e exposição dos animais. Foto: O autor (2008).
Os aquários funcionaram em sistema semi-estático, com limpeza e troca de
1/3 da água a cada 48 horas. Durante este período os peixes foram alimentados
diariamente, com ração comercial (28% de proteína bruta). Estas condições foram
mantidas durante o período de aclimatação. Após a contaminação, apenas foi
suspensa a troca de água. Os peixes foram mantidos em água desclorificada,
temperatura controlada (em torno de 28ºC), aeração constante e periodicidade
luminosa (12/12 h) por um período de aclimatação de 30 dias.
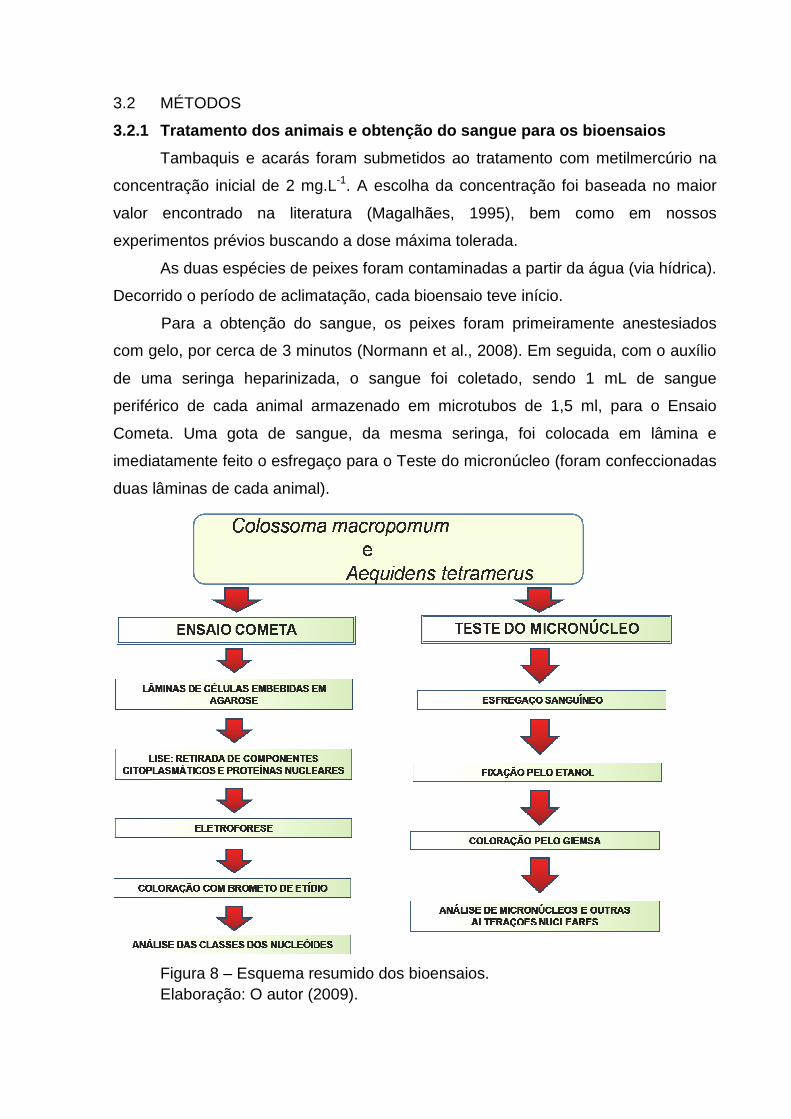
3.2 MÉTODOS
3.2.1 Tratamento dos animais e obtenção do sangue p ara os bioensaios
Tambaquis e acarás foram submetidos ao tratamento com metilmercúrio na
concentração inicial de 2 mg.L-1. A escolha da concentração foi baseada no maior
valor encontrado na literatura (Magalhães, 1995), bem como em nossos
experimentos prévios buscando a dose máxima tolerada.
As duas espécies de peixes foram contaminadas a partir da água (via hídrica).
Decorrido o período de aclimatação, cada bioensaio teve início.
Para a obtenção do sangue, os peixes foram primeiramente anestesiados
com gelo, por cerca de 3 minutos (Normann et al., 2008). Em seguida, com o auxílio
de uma seringa heparinizada, o sangue foi coletado, sendo 1 mL de sangue
periférico de cada animal armazenado em microtubos de 1,5 ml, para o Ensaio
Cometa. Uma gota de sangue, da mesma seringa, foi colocada em lâmina e
imediatamente feito o esfregaço para o Teste do micronúcleo (foram confeccionadas
duas lâminas de cada animal).
Figura 8 – Esquema resumido dos bioensaios. Elaboração: O autor (2009).

3.2.2 Ensaio Cometa
Para o Ensaio cometa usamos o procedimento de Singh et al. (1988), com
pequenas modificações (Hartmann & Speit, 1997). Foram preparadas duas lâminas
de cada animal. Utilizamos lâminas previamente cobertas com agarose normal
(Gibco). Do sangue coletado, 10 µL foram diluídos em 1 mL de soro bovino fetal.
Desta solução, foram coletados 10 µL e misturados com 120 µL de gel de agarose
com baixo ponto de fusão (“low melting point” – LMP), previamente preparada e
levemente aquecida (37ºC). As lâminas foram então incubadas em tampão de lise
(2,5 M NaCl, 100 mM EDTA, 10 mM Tris, 10% DMSO, 1% TRITON X-100), pH 10,
por 1 hora, na ausência de luz, para remoção das proteínas celulares.
Posteriormente, as lâminas passaram pelo processo em tampão de
desnaturação (NaOH 30 mM, 1 mM EDTA) pH > 13, pré-resfriado entre 4-8 ºC e
incubadas de maneira a cobrir as lâminas, por 10 minutos. A eletroforese foi feita em
uma cuba horizontal e transcorreu a 25 V e 300 mA, por um período de 20 minutos.
Todas as etapas anteriores foram realizadas sob luz amarela ou no escuro para
prevenir danos adicionais ao DNA. Após a eletroforese, as lâminas foram removidas,
neutralizadas (Tris 0,4 M, por 10 minutos), fixadas em etanol absoluto e coradas
com 20 µL de 20 µL/mL de brometo de etídio.
Figura 9 – Resumo das sucessivas etapas que integram o Ensaio cometa. Elaboração: O autor (2009).

Para a visualização dos danos do DNA, as lâminas foram observadas em
aumento 400X, usando-se microscópio de epifluorescência equipado com filtro de
excitação de 515–560 nm e um filtro de barreira de 590 nm. Foram analisadas 100
células (nucleóides) por lâmina, estipulando-se previamente, de acordo com Collins
et al. (2001), cinco Classes de Danos. A avaliação foi visual, segundo o
comprimento da cauda nas classes e para a análise estatística foram atribuídos
valores numéricos (Ranques) de 0 a 4 para cada uma das classes, respectivamente.
As células foram ranqueadas visualmente, de acordo com a intensidade relativa de
fluorescência do DNA na cauda (de 0 = incólumes, sem DNA na cauda; a 4 =
maximamente danificadas, mais de 80% de DNA na cauda). O escore total para 100
células (entre 0 e 400) foi obtido multiplicando o número de nucleóides de cada
classe pelo ranque da classe de dano. Assim, o índice de dano (ID) foi indicado
usando a seguinte fórmula:
ID = (0 x n0) + (1 x n1) + (2 x n2) + (3 x n3) + (4 x n4),
onde n = número de células em cada classe analisada. (Collins et al., 1995; Silva et
al., 2000; Collins et al., 2001). Finalmente, o escore total por espécime corresponde
à média dos escores de suas duas lâminas e representa, então, a freqüência de
quebras no seu DNA.
Em nosso estudo, o Ensaio cometa foi realizado no ano de 2008, no
Laboratório de Oncologia Experimental, do Departamento de Fisiologia e
Farmacologia da Universidade Federal do Ceará.
[

3.2.3. Teste do Micronúcleo
Para a análise dos Micronúcleos (MN) foi empregada a técnica descrita por
Heddle (1973), com algumas modificações. Assim, uma vez coletado o sangue do
peixe, imediatamente foi pingada uma gota em lâmina bem limpa; com o auxílio de
uma lamínula, foi feito o esfregaço, que secou ao ar, em temperatura ambiente.
Após a secagem, fez-se a fixação em etanol absoluto por 10 minutos e a
lâmina foi corada com solução de Giemsa 10%, diluída em tampão fosfato a pH 6,8,
por 10 minutos e lavada em água corrente. Somente foram consideradas as células
com a membrana citoplasmática intacta. As laminas para o escore de MN foram
observadas ao microscópio sob ampliação de 1000X, no Laboratório de Citogenética
Humana e Genética Toxicológica da Universidade Federal do Pará. Foram
analisados, em teste cego, 2.000 eritrócitos por lâmina e, assim, 4.000 por animal.
Em nosso estudo os MN foram definidos como corpos citoplasmáticos
esféricos ou ovais, não conectados ao núcleo principal, com diâmetro de 1/30 – 1/10
do núcleo maior e no mesmo plano óptico deste (Al-Sabti & Metcalfe, 1995; Ayllon e
Garcia-Vazquez, 2000). Três outras anormalidades nucleares foram também
consideradas: brotos, lobos e invaginações (Ayllon & Garcia-Vazquez, 2000;
Bolognesi et al., 2006).
]

3.2.4 Biensaios com Colossoma macropomum
Os espécimes de C. macropomum foram mantidos sob densidade de três
indivíduos por aquário de 30 L (Figura 10). Para esses bioensaios foram utilizados
18 indivíduos, os quais constituíram dois grupos de nove, sendo um grupo exposto
ao metilmercúrio 2 mg.L-1 e o outro, não exposto, utilizado como controle. Após 120
horas (cinco dias) de exposição, os tambaquis de ambos os grupos foram
anestesiados e sacrificados e o sangue coletado para os bioensaios.
Figura 10 – Visão de aquários no Laboratório de Biologia Aquática do IFPA, por ocasião da exposição de C. macropomum ao metilmercúrio. Foto: O autor (2008).
No Ensaio cometa, as lâminas foram observadas em microscópio de
epifluorescência usando-se aumento de 400X. Foram analisados 100 nucleóides por
lâmina e, assim, 200 por indivíduo. No total, foram averiguados 3.600 nucleóides
neste bioensaio.
O Teste de micronúcleos foi realizado pela análise de 4.000 eritrócitos por
animal, incluindo aquelas com micronúcleos típicos e alterações na forma do núcleo.
No total, foram averiguados 72.000 eritrócitos neste bioensaio.

3.2.5 Bioensaios com Aequidens tetramerus
Nos bioensaios com A. tetramerus um protocolo diferente foi adotado. Devido
à sua agressividade, a manutenção foi feita em aquários individuais de 15 L até
coleta do sangue.
Foram utilizados 33 espécimes, divididos em três grupos de 11: um grupo
controle (C) e dois grupos expostos (A) e (B). O metilmercúrio foi adicionado à água
até a concentração planejada (2 mg.L-1) e a obtenção do sangue para as análises
feita após 24 h e 120 h, respectivamente.
No Ensaio cometa, as lâminas foram observadas em microscópio de
epifluorescência usando-se aumento de 400X. Foram analisados 100 nucleóides por
lâmina e, assim, 200 por indivíduo. No total, foram averiguados 6.600 nucleóides
neste bioensaio.
As laminas para o escore de MN foram observadas ao microscópio usando
ampliação de 1000X. Foram analisados 2.000 eritrócitos por lâmina e, assim, 4.000
por animal. No total, foram averiguados 132.000 eritrócitos neste bioensaio.
[

3.2.6 Análises estatísticas
Para comparar diferenças entre grupos controle e tratados, os resultados
foram avaliados por meio de três testes não paramétricos: teste de Mann-Whitney,
teste de Kuskal-Wallis e teste do Qui-quadrado (χ2). Para isso, usamos o programa
BioEstat 5.0 (Ayres et al., 2007) e o nível mínimo de significância considerado nas
análises foi de 0,05.
As análises estatísticas dos resultados obtidos no Ensaio Cometa foram
efetuadas com os testes de Mann-Whitney (teste U) e de Kuskal-Wallis, de acordo
com Ferraro (2003), Ferraro et al. (2004), Rigonato et al. (2005), Benincá (2006),
Bücker et al. (2006) e Vanzella (2006), por se tratar de dados não paramétricos e
independentes.
O teste U baseia-se no seguinte raciocínio: se na amostra A os valores são,
em geral, menores do que na amostra B, quando se ordenam do menor ao maior os
valores das duas amostras juntas, os postos ocupados pelos indivíduos da amostra
A serão, em geral, menores do que os ocupados pelos da amostra B.
conseqüentemente, o posto médio em A será também menor do que o posto médio
em B. Uma diferença estatisticamente significativa entre os dois postos médios
estará indicando que a população A tem valor de tendência central menor do que a
população B. O teste de Kuskal-Wallis é uma generalização do teste de Mann-
Whitney e serve, portanto, para se compararem duas ou mais populações (Cellegari-
Jacques, 2003).
Este tipo de teste estatístico nos permitiu comparar os grupos
simultaneamente. Os escores utilizados nos testes foram estimados por 100 células.
Assim, o escore máximo possível nos experimentos foi de 400 (dano máximo) e o
escore mínimo de zero (nenhum dano).
Aos resultados do Teste do Micronúcleo foi aplicado o teste do Qui-quadrado,
de acordo com Ferraro (2003), Ferraro et al. (2004), Lopes-Poleza (2004), Banasik
et al. (2005) e Cavas et al. (2005). Como não existe hipótese para o esperado, o qui-
quadrado foi obtido através de tabelas de contingência.

4. RESULTADOS
4.1 RESULTADOS DOS EXPERIMENTOS COM C. MACROPOMUM
No Ensaio cometa, a avaliação foi feita analisando-se visualmente 100
nucleóides por lâmina, 200 por espécime, os quais foram distribuídos em cinco
classes, de acordo com a intensidade da cauda. Procedeu-se o somatório do
número de células identificadas em cada classe e a multiplicação pelo valor da
classe. Em seguida, o escore total por indivíduo foi obtido pela média dos escores
de suas duas lâminas, representando, portanto, o nível de danos no seu DNA.
No gráfico da Figura 11 estão representados os resultados encontrados na
análise do Ensaio Cometa, com as informações sobre as classes de danos. Foram
analisados 3.600 nucleóides, 1.800 de cada grupo. Do total, 1.380 nucleóides do
grupo controle e 112 do grupo contaminado não apresentaram danos (classe 0). No
caso dos nucleóides com cauda (células com danos), verificou-se maior freqüência
da classe 1 tanto no grupo controle quanto no tratado.
Ensaio cometa em C. macropomum
0
100
200
300
400
500
600
700
800
classes de danos
frequ
ênci
a/90
0 cé
lula
s
Série1 Série2
Figura 11 – Freqüência de nucleóides por classe de dano.
Os resultados observados foram tratados estatisticamente com o teste de
Mann-Whitney e a diferença mostrou-se significativa entre os grupos controle e
contaminado (Figuras 12 e 13).
Controle Tratado
0 1 2 3 4

Figura 12 – Resultados do teste de Mann-Whitney obtidos no BioEstat 5.0 (Ayres et al., 2007) mostrando diferença significativa entre os grupos controle e tratado no Ensaio cometa em C. macropomum (p < 0,0005).
Ensaio Cometa: Controle x Exposição ao MeHg
Figura 13 – Gráfico representativo dos escores obtidos (por 100 células) em C. macropomum. Ensaio cometa para os animais tratados e os do grupo controle.
Esc
ores
das
cla
sses
de
nucl
eóid
es /
100
célu
las

No Teste do Micronúcleo, foram analisadas 4.000 células por espécime. Do
total de 72.000 células, o número de micronúcleos típicos (Figura 14) foi bastante
reduzido, o que dificultaria bastante a análise estatística. Entretanto, de acordo com
nossa proposta, os micronúcleos foram incluídos com as alterações morfológicas
nucleares, que apareceram com maior freqüência.
Figura 14 - Eritrócitos de C. macropomum corados com Giemsa, em ampliação de 1.000X. A seta indica um micronúcleo.
Das 72.000 células analisadas, uma metade foi do grupo controle e a outra do
grupo exposto ao xenobiótico. No primeiro grupo, 307 células se apresentaram
alteradas (micronúcleos típicos + alterações morfológicas nucleares); no grupo
exposto foram 367 células alteradas. Esses resultados foram tratados
estatisticamente com o teste do qui-quadrado através da montagem de uma tabela
de contingência (2 x 2). A hipótese H0 (a proporção de células alteradas é
independente da exposição ao metilmercúrio) foi testada e rejeitada, devido à
significativa diferença entre os grupos controle e tratado (Figura 15), onde p < 0,05.

Figura 15 – Resultado do teste do qui-quadrado obtido no BioEstat 5.0 (Ayres et al., 2007), demonstrando que houve diferença significativa entre a ocorrência de células alteradas nos grupos controle e exposto ao metilmercúrio (p < 0,05).
Os dados são também apresentados na Tabela 2 e no gráfico da Figura 16,
onde estão disponibilizadas as médias e os desvios padrões de micronúcleos e
alterações morfológicas nucleares.
Tabela 2 – Médias e desvios de micronúcleos (MN) e alterações morfológicas nucleares (AMN) de C. macropomum expostos ao metilmercúrio 2 mg.L-1.
MÉDIA E DESVIO PADRÃO DE MN E AMN / 4000 CÉLULAS
MN AMN MN + AMN
Controle 9,66 ± 5,02 24,44 ± 9,48 34,11 ± 13,84
MeHg /120 h 8,44 ± 7,42 32,33 ± 6,61 40,77 ± 12,19

Figura 16 – Representação das freqüências de micronúcleos + alterações morfológicas nucleares (por 4.000 eritrócitos) em C. macropomum.
Ainda durante o curso de doutoramento reunimos numerosas publicações
abordando o uso do Teste do micronúcleo em peixes, na avaliação de riscos da
exposição a xenobióticos. A esta breve revisão, acrescentamos nossos resultados
com espécimes de C. macropomum expostos ao metilmercúrio e produzimos o
artigo “The Micronucleus Assay in Fish Species as an Important Tool for Xenobiotic
Exposure Risk Assessment – A Brief Review and an Example Using Neotropical Fish
Exposed To Methylmercury”, que foi publicado no periódico Reviews in Fisheries
Science 17(4):478–484, 2009. O referido artigo encontra-se em um apêndice ao final
deste trabalho.
Núm
ero
de c
élul
as a
ltera
das
/ 4.0
00 c
élul
as
Teste do Micronúcleo: Controle x Exposição ao MeHg

4.2 RESULTADOS DOS EXPERIMENTOS COM A. TETRAMERUS
No Ensaio cometa, a avaliação foi feita analisando-se visualmente 100
nucleóides por lâmina, 200 por espécime. Após sua distribuição em classes, de
acordo com a intensidade da cauda, procedeu-se o somatório do número de células
identificadas em cada classe e a multiplicação pelo valor da classe. O escore total
por indivíduo foi então obtido pela média dos escores de suas duas lâminas,
representando o nível de danos no seu DNA.
Verificou-se um aumento muito significativo de nucleóides com cauda nos
eritrócitos de peixes tratados com MeHg em relação aos do grupo controle não
tratado (Figura 17).
Comparativo
C 24h 120h0
50
100
150
200
250
300
350
Exposição
Índi
ce d
e da
no
Figura 17 – Ensaio cometa em A. tetramerus. Comparação do índice de danos nos grupos controle (C) e tratados com MeHg 2mg.L-1 nos dois tempos de exposição. Gráfico obtido no programa GraphPad.Prism.5.
No gráfico da Figura 18 estão representados os resultados encontrados na
análise do Ensaio Cometa, com as informações sobre as classes de danos. Foram
analisados 6.600 nucleóides, 2.200 de cada grupo. Do total, 1.976 nucleóides do
grupo controle e 58 do grupo exposto por 24h não apresentaram danos, porém
todos os nucleóides do grupo exposto por 120h apresentaram algum nível de dano.
No caso dos nucleóides com cauda (células com danos), verificou-se maior
freqüência da classe 1 no grupo C (controle), da classe 2 no grupo A (exposto por
24h) e da classe 3 no grupo B (exposto por 120h).

Ensaio cometa em A. tetramerus
0
200
400
600
800
1000
1200
classes de danos
frequ
ênci
a/11
00 c
élul
as
Série1 Série2 Série3
Figura 18 – Freqüência de nucleóides por classe de dano.
Os resultados observados foram tratados estatisticamente com o teste de
Kruskal-Wallis (Figuras 19 e 20) e a diferença mostrou-se altamente significativa
entre os grupos controle e tratados (p < 0,0001). A opção por esse teste estatístico
decorreu principalmente de sua semelhança operacional com o teste de Mann-
Whitney e da sua capacidade de comparar os três grupos simultaneamente.
Figura 19 – Resultados do teste de Kruskal-Wallis obtidos no BioEstat 5.0 (Ayres et al., 2007) para os escores do Ensaio cometa em A. tetramerus. Na tabela, temos: 1 = grupo C, 2 = grupo A e 3 = grupo B.
Controle
0 1 2 3 4
MeHg 24 h MeHg 120 h

Como se pode notar, houve diferença altamente significativa entre o grupo
controle (C) e cada um dos grupos tratados, porém entre estes (A e B) a diferença
no índice de danos não chegou a ser significativa. Os escores médios e desvios em
cada grupo podem ser observados no gráfico da Figura 20.
Ensaio Cometa: Controle x Exposição ao MeHg
Figura 20 – Ensaio cometa em A. tetramerus. Comparação do índice de danos ao DNA nos grupos controle (C) e tratados com MeHg 2mg.L-1 nos dois tempos de exposição.
Foram analisadas 132.000 células de Aequidens tetramerus no Teste do
micronúcleo, 44.000 em cada grupo. No grupo controle, 314 células se
apresentaram alteradas; no grupo exposto ao MeHg por 24h foram 553 células
alteradas e no grupo exposto por 120h foram 824 células alteradas. Os resultados
foram tratados estatisticamente com o teste do qui-quadrado através da montagem
de uma tabela de contingência (3 x 2). A hipótese H0 (a proporção de células
alteradas é independente das exposições ao metilmercúrio) foi testada e rejeitada,
devido à diferença altamente significativa entre os grupos controle e tratado, onde p
< 0,0001 (Figura 21).
Esc
ores
das
cla
sses
de
nucl
eóid
es /
100
célu
las
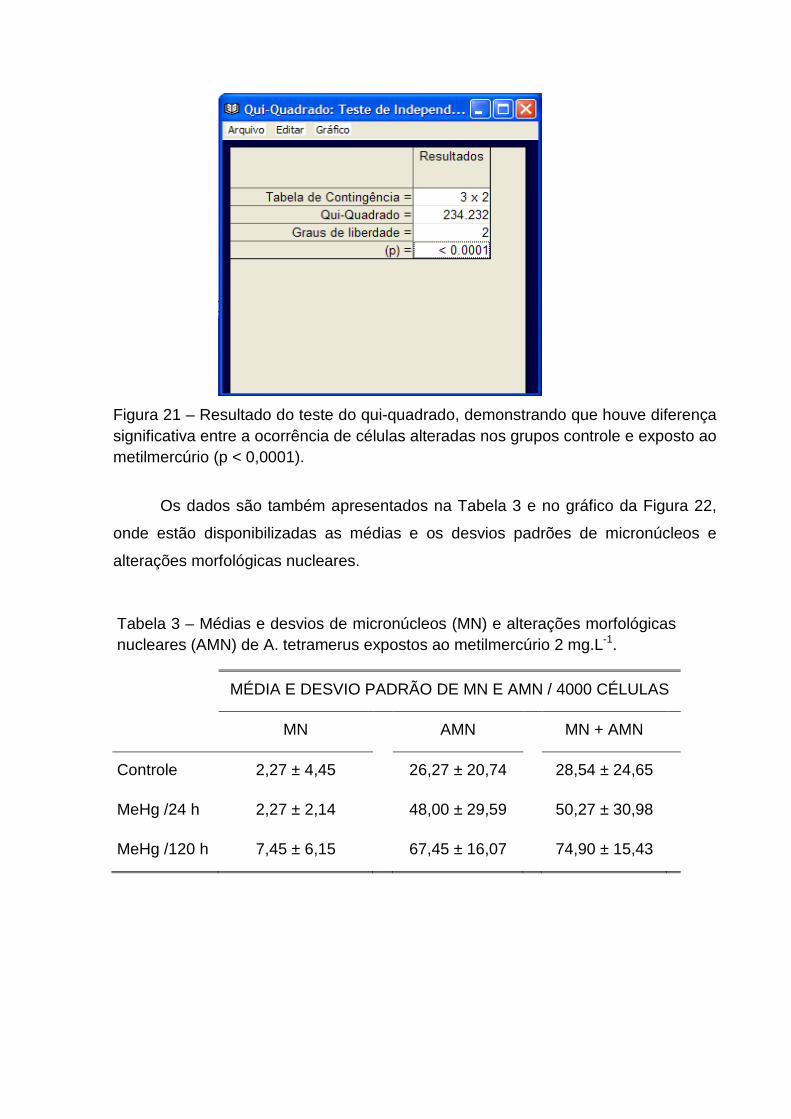
Figura 21 – Resultado do teste do qui-quadrado, demonstrando que houve diferença significativa entre a ocorrência de células alteradas nos grupos controle e exposto ao metilmercúrio (p < 0,0001).
Os dados são também apresentados na Tabela 3 e no gráfico da Figura 22,
onde estão disponibilizadas as médias e os desvios padrões de micronúcleos e
alterações morfológicas nucleares.
Tabela 3 – Médias e desvios de micronúcleos (MN) e alterações morfológicas nucleares (AMN) de A. tetramerus expostos ao metilmercúrio 2 mg.L-1.
MÉDIA E DESVIO PADRÃO DE MN E AMN / 4000 CÉLULAS
MN AMN MN + AMN
Controle 2,27 ± 4,45 26,27 ± 20,74 28,54 ± 24,65
MeHg /24 h 2,27 ± 2,14 48,00 ± 29,59 50,27 ± 30,98
MeHg /120 h 7,45 ± 6,15 67,45 ± 16,07 74,90 ± 15,43

Figura 22 – Representação das freqüências de micronúcleos + alterações morfológicas nucleares (por 4.000 eritrócitos) em A. tetramerus.
Núm
ero
de c
élul
as a
ltera
das
/ 4.0
00 c
élul
as

5. DISCUSSÃO
O uso de diferentes ensaios e sistemas biológicos garante uma triagem mais
acurada dos efeitos causados por contaminantes no ambiente aquático. Nesse
sentido, vários biomarcadores estão sendo desenvolvidos e dentre eles, os
biomarcadores genéticos vêm alcançando grande aceitação (Ramsdorf, 2007).
O desenvolvimento de novas metodologias e aplicação de técnicas mais
refinadas para a avaliação da genotoxicidade em ambientes aquáticos são objeto de
muitos estudos científicos (Van der Oost et al., 2003). Em nosso estudo, o Ensaio
cometa e o Teste do micronúcleo foram utilizados para avaliar os efeitos do
metilmercúrio em Colossoma macropomum e Aequidens tetramerus, espécies de
distribuição relativamente ampla na região neotropical.
Os resultados obtidos no presente trabalho mostraram, primeiramente, que os
peixes, neste caso C. macropomum e A. tetramerus, podem representar um bom
modelo experimental, sendo considerados ótimos indicadores de efeitos genotóxicos
de xenobiontes, pelo menos se o poluente for mercurial. Por outro lado, a
bioacumulação de compostos organometálicos pode ser comparada à do homem,
pois tanto em um quanto no outro, tais compostos têm capacidade de reagir com
ligantes intracelulares.
Em nosso estudo, C. macropomum e A. tetramerus foram expostos ao
metilmercúrio na concentração de 2 mg.L-1, o maior valor encontrado na literatura
(Magalhães, 1995) para algumas áreas da baía de Minamata (Japão) nas décadas
de 50 e 60. Neste sentido, as chances de observação de efeitos genotóxicos eram
praticamente totais. Assim, nosso principal foco de análise direcionava-se às
diferenças de resultados entre os testes de genotoxicidade empregados e entre as
espécies usadas nos bioensaios.

5.1 USO DE ERITRÓCITOS PERIFÉRICOS
Testes de genotoxicidade em peixes são habitualmente realizados com
eritrócitos, pois estes são nucleados (Hayashi et al., 1998), facilmente obtidos por
métodos não destrutivos e não necessitam do passo adicional de isolamento, porém
outros tecidos também têm sido testados, pois os efeitos de genotoxicidade de
contaminantes podem ser muitas vezes tecido-específicos. Os tecidos mais
pesquisados, além do sanguíneo, são do fígado, do rim (Belpaeme et al., 1998),
além das brânquias (Cavas et al., 2005; Cavas, 2008).
Apesar de alguns estudos com MN demonstrarem maior sensibilidade das
células epiteliais de brânquias em relação aos eritrócitos, como Hayashi et al. (1998)
e Cavas (2008), o uso de células do sangue periférico tem predominado pela sua
facilidade técnica, possível obtenção sem necessariamente sacrificar os animais da
amostra e, principalmente, pela possibilidade de comparações reais dos resultados
com um maior número de publicações na área (Cestari et al., 2004; Ferraro et al.,
2004; Lopes-Poleza, 2004; Barillet et al., 2005; Porto et al., 2005; Ramos et al.,
2005; Bagdonas & Vosyliené, 2006; Matsumoto et al., 2006; Ergene et al., 2007;
Andreikënaitë et al., 2007; Koca et al., 2008; Ferraro, 2009).

5.2 MAIOR SENSIBILIDADE DO ENSAIO COMETA
As diferenças de respostas nos grupos controle e contaminados, para ambas
as espécies, foram significativas tanto no Ensaio cometa quanto no Teste do
micronúcleo, porém com maiores índices de danos no primeiro. Provavelmente esta
diferença relaciona-se aos eficientes mecanismos de reparo do DNA e à seleção
que elimina grande número de células danificadas. Muitos outros estudos de
genotoxicidade têm obtido resultados semelhantes, ou seja, com dados mais
significativos no Ensaio cometa (Benincá, 2006; Bücker et al., 2006; Vanzella, 2006).
A alta porcentagem de células apoptóticas foi descrita em peixes marinhos
expostos a efluente de refinaria de petróleo (Frenzilli et al. 2004,). Este fenômeno
ocorre quando os danos celulares são intensos e os mecanismos de defesa não
conseguem repará-los. Em conseqüência, o número de células micronucleadas
disponíveis para a análise, também decresceria.
Embora nos Ensaios Cometa e dos Micronúcleos Písceos sejam analisados
eritrócitos circulantes, a natureza do dano detectado é diferente. No Ensaio cometa,
analisam-se danos que envolvem segmentos muito curtos na molécula de DNA,
enquanto que no Teste do micronúcleo os danos detectáveis são, em termos de
escala, de natureza muito maior (Ferraro, 2003).

5.3 TESTE DO MICRONÚCLEO
Em nossa pesquisa de micronúcleos e outras anormalidades nucleares,
analisamos 4.000 eritrócitos por espécime, porém há na literatura uma grande
divergência quanto ao número de células que devem ser utilizadas: 1.000 (Ayllon
and Garcia-Vazquez, 2000; Gravato and Santos, 2002; Russo et al., 2004); 1.500
(Cavas, 2008); 2.000 (Palhares and Grisolia, 2002; Ramsdorf, 2007; Normann et al.,
2008, Ferraro, 2009; Kirschbaum et al., 2009); 3.000 (Galindo and Moreira, 2009;
Grisolia et al., 2005; Grisolia et al., 2009); 4.000 (Metcalfe, 1988; Ferraro et al., 2004;
Bolognesi et al., 2006); 5.000 (Porto et al, 2005; Andreikënaitë et al., 2007). De
acordo com Bolognesi et al. (2006), a grande variabilidade interindividual associada
à baixa freqüência basal deste biomarcador confirma a necessidade de avaliação de
um número consistente de células (em torno de 4000) em um adequado número de
animais.
Tal como em alguns estudos anteriores (Ferraro, 2003; Ferraro et al., 2004;
Bolognesi et al., 2006; Andreikënaitë et al., 2007; Grisolia et al., 2009), no presente
trabalho também houve dificuldades em observar a presença de micronúcleos mas
nem tanto de alterações morfonucleares. A freqüência espontânea de micronúcleos
típicos é geralmente muito baixa (Ferraro et al., 2004). No nosso caso, MN foram
pouco freqüentes em todos os grupos. Quando as amostras foram consideradas
como um todo, reunindo MN típicos e outras alterações nucleares, o teste do qui-
quadrado apresentou diferença significativa entre os grupos controle e contaminado.
Do mesmo modo que em Ferraro (2003), algumas das alterações
morfológicas nucleares descritas em trabalhos anteriores foram encontradas e
fotografadas no presente estudo. Embora, muitas destas pudessem ser facilmente
identificadas dentro de um determinado tipo, outras mostraram-se dúbias quanto à
sua caracterização e, por este motivo, todas as alterações morfológicas nucleares
foram computadas em conjunto.
Vanzella (2006), trabalhando com Prochilodus lineatus, concluiu que em
testes de micronúcleos a exposição de 24h é o melhor tempo experimental para
detectar alterações, pois neste intervalo de tempo as lesões celulares ocorridas não
foram reparadas. Nossos resultados com A. tetramerus expostos ao MeHg por 24h e
120h, entretanto, não corroboram tal conclusão, pois verificamos aumento no índice
de danos em função do tempo de exposição.

Ayllon & Garcia-Vazquez (2000), trabalhando com as espécies Phoxinus
phoxinus e Poecilia latipinna, verificaram que agentes reconhecidamente
genotóxicos, com forte ação clastogênica tanto in vivo como in vitro, podem agir de
maneiras diferentes. Enquanto a colchicina e a mitomicina C induziram aumento na
freqüência de micronúcleos e de anormalidades nucleares, a ciclofosfamida induziu
apenas aumento de anormalidades, não levando à formação de micronúcleos.
Esses resultados indicam que alguns compostos podem agir formando
anormalidades nucleares, porém sem a formação de micronúcleos. Dessa forma,
mais estudos são necessários para se conhecer melhor o mecanismo de formação
das anormalidades nucleares, para que possa ser determinada mais precisamente a
ação genotóxica dos contaminantes.
Por outro lado, se considerarmos que alterações morfológicas nucleares são
indicadores de citotoxicidade e micronúcleos típicos são indicadores de
genotoxicidade, como proposto por Cavas et al. (2005), nossos resultados indicam
que os danos promovidos pelo metilmercúrio nos eritrócitos de peixes são tão
citotóxicos quanto genotóxicos.

5.4 MAIOR ÍNDICE DE DANOS NO ACARÁ
Por fim, como pode ser visto na Tabela 4, os resultados encontrados indicam
que nas condições empregadas em nosso estudo, o metilmercúrio mostrou-se
altamente genotóxico nas espécies estudadas, o que foi demonstrado pelos
elevados escores obtidos no Ensaio cometa, com A. tetramerus (acará)
apresentando níveis mais acentuados de danos em comparação com C.
macropomum (tambaqui). Neste caso tal diferença não pode ser atribuída ao
fenômeno de biomagnificação, uma vez que ambos apresentaram baixo nível de
danos no grupo controle e foram expostos ao xenobionte por via hídrica. Os efeitos
clastogênicos e aneugênicos avaliados no Teste do micronúcleo não foram tão
marcantes quanto as lesões do DNA detectadas no Ensaio cometa, mas foram
significativas nas duas espécies e novamente mais acentuadas em A. tetramerus.
Tabela 4 – Efeitos genotóxicos e mutagênicos do MeHg 2mg.L-1. Comparação entre os resultados nas duas espécies usadas nos bioensaios.
Colossoma macropomum Aequidens tetramerus
MÉDIAS DOS ESCORES NO ENSAIO COMETA
Controle 27,28 10,18
MeHg 24 h – 243,00
MeHg 120 h 140,83 303,13
MÉDIAS DE FREQUÊNCIAS DE MN + AMN
Controle 0,85% 0,71%
MeHg 24 h – 1,25%
MeHg 120 h 1,02% 1,87%
Percebe-se, portanto, que além da influência do nível trófico e
conseqüentemente do fenômeno de biomagnificação nas possíveis agressões de
contaminantes no ambiente aquático, existem variações interespecíficas que podem
resultar de diferenças na sensibilidade entre as espécies ou na susceptibilidade à
bioacumulação, ou ambas.

6. CONCLUSÕES
a) O metilmercúrio apresenta acentuados efeitos genotóxicos em peixes na
concentração de 2mg.L-1, o que foi demonstrado pelas diferenças significativas entre
grupos controles e tratados;
b) As espécies neotropicais Colossoma macropomum e Aequidens tetramerus
funcionam como bons modelos para a análise de exposições a produtos à base de
mercúrio;
c) Os dois testes de genotoxicidade, Ensaio cometa e Teste do micronúcleo,
tiveram sua eficiência confirmada na avaliação de danos ao material genético de
peixes por contaminantes ambientais;
d) O nível de danos detectados pelo Ensaio cometa foi maior que pelo Teste
do micronúcleo, o que pode ser considerado normal, haja vista que o primeiro teste
avalia danos primários ao DNA, muitos dos quais não são passados às futuras
gerações de células e, então, não podem ser detectados pelo segundo teste;
e) A espécie carnívora Aequidens tetramerus apresentou maior índice de
danos em ambos os testes, quando comparada à espécie onívora Colossoma
macropomum. Entretanto, neste caso, a diferença não deve ser atribuída ao seu
comportamento trófico no ambiente natural, uma vez que suas condições de manejo
foram as mesmas e a contaminação exclusivamente por via hídrica.
f) O nível de danos genotóxicos provocados por metais pesados, como o
mercúrio, depende da associação de diferentes fatores. Além da já bem
documentada magnificação trófica, devem também ser consideradas, para cada
espécie, sua sensibilidade ao contaminante e sua susceptibilidade à bioacumulação
do mesmo.

7. REFERÊNCIAS BIBLIOGRÁFICAS
ALMEIDA SS, VIEIRA JLF, PINHEIRO MCN, FREITAS-JÚNIOR JAB, RIBEIRO
DRG, AZEVEDO AA, FERREIRA MN (1987) Concentrações de Mercúrio Total em
Peixes Consumidos pela População Amazônida. III Encontro da ANPPAS, Brasília-
DF.
ALMEIDA-TOLEDO LF, FORESTI F, TOLEDO-FILHO SA, BERNARDINO G,
FERRARI VA e ALCANTARA RCG (1987) Cytogenetic studies in C. mitrei, C.
macropomum and their interspecific hybrid. In: Tiews K (ed) Selection, Hybridization
and Genetic Engineering in Aquaculture. Berlin Heenemann Verlagsgesellshaft mb II
v. 1, pp 189-195.
ALPERS CN e HUNERLACH MP (2000) Mercury Contamination from Historic Gold
Mining in California. USGS Fact Sheet. Disponível em:
<http://ca.water.usgs.gov/mercury/fs06100.pdf> Acesso em 08 jun. 2009.
AL-SABTI K e METCALFE CD (1995) Fish micronuclei for assessing genotoxicity in
water. Mutat. Res. 343: 121–135.
AMORIM MIM, MERGLER D, BAHIA MO, DUBEAU H, MIRANDA DC, LEBEL J,
BURBANO RR e LUCOTTE M (2000) Cytogenetic damage related to low levels of
methylmercury contamination in the Brazilian Amazon. An. Acad. Bras. Ci., 72(4):
497-507.
ANDRADE G, ALBINO UB e NOGUEIRA MA (2004) Meio Ambiente. In: BORÉM A
(ed) Biotecnologia e Meio Ambiente. Editora Folha de Viçosa, Viçosa, MG, pp 21-49.
ANDREIKËNAITË L, BARSIENË J e VOSYLIENË (2007) Studies of micronuclei and
other nuclear abnormalities in blood of rainbow trout (Oncorhynchus mykiss) treated
with heavy metal mixture and road maintenance salts. Acta Zoologica Lituanica,
17(3): 213-219.

ARAÚJO-LIMA C e GOULDING M (1998) Os frutos do tambaqui: Ecologia,
conservação e cultivo na Amazônia. Tefé, AM: Sociedade Civil Mamirauá; Brasília:
CNPQ, 186p.
AZEVEDO FA (2003) Toxicologia do mercúrio. Rima Editora, São Paulo, SP, 292p.
AYLLON F e GARCIA–VAZQUEZ E (2000) Induction of micronuclei and other
nuclear abnormalities in European minnow Phoxinus phoxinus and mollie Poecilia
latipinna: an assessment of the fish micronucleus test. Mutation Research – Genetic
Toxicology and Environmental Mutagenesis, 467: 177–186.
AYRES M, AYRES-JUNIOR M, AYRES D e SANTOS A (2007) Bioestat 5.0:
Statistical Applications in Biological Science and Medicine. Belém, PA: Sociedade
Civil Mamirauá; Brasília: CNPq, Brazil. 364p.
BAGDONAS E e VOSYLIENÉ MZ (2006) A study of toxicity and genotoxicity of
copper, zinc and their mixture to rainbow trout (Oncorhynchus mykiss). Biologija 1:
8–13.
BAHIA MO (1997) Le potentiel génotoxique du mercure: mutations HPRT- et effects
cytogénétiques. Dissertação de Mestrado, Montreal, Canada: Université du Quebéc
à Montreal, 119p.
BARCE MS (2006) Poluição em Rondônia. Revista Eletrônica de Ciências 32.
<http://www.cdcc.sc.usp.br/ciencia/artigos/art_32/aprendendo4.html>
BARKAY T, MILLER MS e SUMMERS AO. (2003) Bacterial mercury resistance from
atoms to ecosystems. FEMS Microbiol Rev, 27:355-84.
BARREGÅRD L, ENESTROM S, LJUNGHUSEN O, WIESLANDER J e HULTMAN P
(1997) A study of autoantibodies and circulating immune complexes in mercury-
exposed chloralkali workers. Int Arch Occup Environ Health 70:101-106.

BELPAEME K, COOREMAN K e KIRSCH-VOLDERS M (1998) Development and
validation of the in vivo alkaline comet assay for detecting genomic damage in
marine flatfish. Mutation Research, 415(3): 167-184.
BENINCÁ C (2006) Biomonitoramento das lagoas estuarinas do Camacho –
Jaguaruna (SC) e Santa Marta - Laguna (SC); utilizando Geophagus brasiliensis
(Cichlidae). Dissertação de Mestrado. Universidade Federal do Paraná.
BENITO SP (2002) Biomonitarización citogenética de cuatro poblaciones agrícolas
europeas, expuestas a plaguicidas, mediante el ensayo de micronúcleos. Tesis
Doctoral. Universitat Autònoma de Barcelona, España.
BEN-OZER EY, ROSENSPIRE AJ, McCABE Jr MJ, WORTH RG, KINDZELSKII AL,
WARRA NS e PETTY HR (2000) Mercury Chloride Damages Cellular DNA by a Non-
Apoptotic Mechanism. Mutat. Res. 470(1): 19-27.
BILITEWSKI U e TURNER APF (eds.) (2000) Biosensors for environmental
monitoring. Harwood Academic Publishers, 421p.
BISINOTI MC e JARDIM WF (2004) O comportamento do metilmercúrio (MetilHg) no
ambiente. Química Nova, 27(4): 593-600.
BOENING DW (2000) Ecological effects, transport and fate of mercury: a general
review. Chemosphere, v. 40, pp. 1335-1351.
BÜCKER A, CARVALHO W e ALVES-GOMES J (2006) Evaluation of mutagenicity
and genotoxicity in Eingenmannia virescens (Teleostei: Gymnotiformes) exposed to
benzene. Acta Amazônica, 36: 357–230 (2006).
CALLEGARI-JACQUES SM. Bioestatística: princípios e aplicações. Editora Artmed,
Porto Alegre, RS, 255p.

CAMPANA MA, PANZERI AM, MORENO VJ e DULOUT FN (2003) Micronuclei
induction in Rana catesbeiana tadpoles by the pyrethroid insecticide lambda-
cyhalothrin. Genetics and Molecular Biology 26:99-103.
CARRASCO, K. R.; TILBURY, K. L. e MYERS, M.S. (1990). Assessment of the
piscine micronucleus test as na in situ biological indicator of chemical contaminant
effects. Can. J. Fish. Sci., Ottawa, 47: 2123-2136.
CASSIDY DR e FURR A. (1978) Toxicity of inorganic and organic mercury
compounds in animals. In: OEHEM FW (Ed.). Toxicity of heavy metals in the
environment. New York: Marcel Dekker. v. 1, p.303-330.
CAVAS T, GARANKO NN e ARKHIPCHUK VV (2005) Induction of micronuclei and
binuclei in blood, gill and liver cells of fishes subchronically exposed to cadmium
chloride and copper sulphate. Food and Chemical Toxicology 43: 569–574.
CAVAS T (2008) In vivo genotoxicity of mercury chloride and lead acetate:
Micronucleus test on acridine orange stained fish cells. Food and Chemical
Toxicology 46: 352–358.
CESTARI MM, LEMOS PMM, RIBEIRO CAO, COSTA JRMA, PELLETIER E,
FERRARO MVM MANTOVANI MS e FENOCCHIO S (2004) Genetic damage
induced by trophic doses of lead in the neotropical fish Hoplias malabaricus as
revealed by the comet assay and chromosomal aberrations. Genetics and Molecular
Biology 27:270-274.
COLLINS AR DUSINSKÁ M e HORSKÁ A (2001) Detection of alkylation damage in
human lymphocyte DNA with the comet assay. Acta Biochim. Pol., 48: 611–614.
COUTO RCS (1990) Condições de saúde nos garimpos de ouro da Amazônia.
Seminário Nacional: Riscos e conseqüências do uso do mercúrio, Rio de Janeiro.

DAVID IMB (2009) Salmão selvagem e de cativeiro: benefícios, riscos e
sustentabilidade. Artigos Médicos, Associação Brasileira de Nutrologia. Disponível
em <www.abran.org.br/inf_artigos/002.doc> Acesso em: 10 jun. 2009.
ERGENE S, CAVAS T, CELIK A, KÖLELI N, KAYA F e KARAHAN A (2007)
Monitoring of nuclear abnormalities in peripheral erythrocytes of three fish species
from the Goksu Delta (Turkey): genotoxic damage in relation to water pollution.
Ecotoxicology 16: 385-391.
FAIRBAIRN DW, OLIVE PL e O’NEILL KL (1995) The Comet assay: a
comprehensive review. Mutation Research, 339: 37-59.
FENECH M (2000) The in vitro micronucleus technique. Mutation Research, 455: 81-
95.
FERRARO MV (2003) Avaliação do efeito mutagênico do tributilestanho (TBT) e do
chumbo inorgânico (Pb II) em Hoplias malabaricus (Pisces) através dos ensaios:
cometa, micronúcleo e de aberrações cromossômicas. Dissertação de Mestrado.
Universidade Federal do Paraná.
FERRARO MV, FENOCCHIO AS, MANTOVANI MS, CESTARI MM e RIBEIRO CAO
(2004) Mutagenic effects of tributyltin (TBT) and inorganic lead (PbII) on the fish H.
malabaricus as evaluated using the Comet Assay, Piscine Micronucleus and
Chromosome Aberrations tests. Genetics and Molecular Biology, 27(1): 103-107.
FERRARO MV (2009) Avaliação de três espécies de peixes – Rhamdia quelen,
Cyprinus carpio e Astyanax bimaculatus, como potenciais bioindicadores em
sistemas hídricos através dos ensaios: cometa e dos micronúcleos. Tese de
Doutorado. Universidade Federal do Paraná.
FERREIRA CM (2004) Análises complementares obtidas a partir de testes de
toxicidade aquática. In: RIZANI-PAIVA MJT, TAKEMOTO RM e LIZAMA MAP (eds)
Sanidade de organismos aquáticos. Livraria Varela Editora, São Paulo, pp 273-284.

FRENZILLI G, SCARELLI V, DEL BARGA I, NIGRO M, FÖRLIN L, BOLOGNESI C e
STURVE J. (2004) DNA in eelpout (Zoarces viviparous) from Göteborg harbour,
Mutat. Res. 552: 187-195.
GRISOLIA CK, RIVERO CLG, STARLING FLRM, SILVA ICR, BARBOSA AC e
DOREA JG (2009) Profile of micronucleus frequencies and DNA damage in different
species of fish in a eutrophic tropical lake. Genetics and Molecular Biology, 32(1):
138-143.
GUIMARÃES J-RD, ROULET M e LUCOTTE M. (2000) Mercury methylation
potentials along a lake-forest transect in the Tapajós floodplain, Brazilian Amazon:
seasonal and vertical variations. Sci. Tot. Environment, 261: 91-98.
HACON S, BARROCAS PRG, VASCONCELLOS ACS, BARCELLOS C,
WASSERMAN JC, CAMPOS RC, RIBEIRO C e AZEVEDO-CARLONI FB (2008) An
overview of mercury contamination research in the Amazon basin with an emphasis
on Brazil. Cad. Saúde Pública, vol.24, n.7, pp. 1479-1492.
HARADA M, NAKANISHI J, YASADA E, PINHEIRO MC, OIKAWA T de ASSIS
GUIMARÃES G, da SILVA CARDOSO B, KIZAKI T e OHNO H (2001) Mercury
pollution in the Tapajós River basin, Amazon: mercury level of head hair and health
effects. Environmental International, 27(4): 285-290.
HARTMANN A e SPEIT G (1997) The contribution of cytotoxicity to effects seen in
the alkaline comet assay. Toxicol. Lett. 90: 183-188.
HAYASHI M, UEDA T, UYENO K, WADA K, KINAE N, SAOTOME K, TANAKA N.
TAKAI A, SASAKI YF, ASANO N, SOFUNI T e OJIMA Y (1998) Development of
genotoxicity assays systems that use aquatic organisms. Mutation Research, 399 (2):
125-133.
HEDDLE JA (1973) A rapid in vivo test for chromossomal damage. Mutation
Research. v. 18, p. 187–190.

HALLENBECK, WH 1993. Quantitative risk assessment for environmental and
occupational health. Lewis Publishers. INC.224pp.International Agency Research
Cancer (IARC). 1993. Lyon, IARC Monographs, 58: 119.
http:/www.monographs.iarc.fr/htdocs/monografs/vol58/mono58-2.htm
KARLAND LT, FARO SH e SIEDLER H (1960) Minamata disease. Wed. Neurol.,
vol.1, p.370-395.
KOCA S, KOCA YB, YILDIZ S e GÜRCÜ B (2008) Genotoxic and Histopathological
Effects of Water Pollution on Two Fish Species, Barbus capito pectoralis and
Chondrostoma nasus in the Büyük Menderes River, Turkey. Biol Trace Elem Res
122: 276–291.
LACERDA LD e MALM O (2008) Contaminação por mercúrio em ecossistemas
aquáticos: uma análise das áreas críticas. Estudos Avançados 22(63): 173-190.
LEBEL J, MERGLER D, BRANCHES F, LUCOTTE M, AMORIM M, LARRIBE F e
DOLBEC J. (1998) Neurotoxic Effects of Low-Level Methylmercury Contamination in
the Amazoniam Basin. Environmental Research, 79: 20-32.
LECHLER, PJ MILLER JR, LACERDA LD, VINSON D, BOZONGO J-C, LYONS, WB
e WARWICK JJ (2000) Elevated mercury concentrations in soils, sediments, water,
and fish of the Madeira River basin, Brazilian Amazon: a function of natural
enrichments? Science of the Total Environment, 260(1-3): 87-96.
LOPES-POLEZA SCG (2004) Avaliação do efeito do metilmercúrio (CH3Hg+) em
Hoplias malabaricus através da freqüência de Aberrações Cromossômicas e dos
ensaios Cometa e Micronúcleo. Dissertação de Mestrado. Universidade Federal do
Paraná.
MAGALHÃES MEA (1995) Estudo cinético de dissolução/oxidação do mercúrio
metálico em solução aquosa de cloreto de sódio. Tese de Doutorado. Universidade
Estadual de Campinas.

MALM O, GUIMARÃES JRD, CASTRO MB, BASTOS WR, VIANA JP, BRANCHES
FJP, SILVEIRA EG, PFEIFFER WC (1997) Follow-up of mercury levels in fish,
human hair and urine in the Madeira and Tapajos basins, Amazon, Brazil. Water, Air,
Soil Pollut. 97(1-2): 45-51.
MATSUMOTO ST, MANTOVANI MS, MALAGUTTII MIA, DIAS AL, FONSECA IC e
Marin-Morales MA. (2006) Genotoxicity and mutagenicity of water contaminated with
tannery effluents, as evaluated by the micronucleus test and comet assay using the
fish Oreochromis niloticus and chromosome aberrations in onion root-tips. Genetics
and Molecular Biology, 29, 1, 148-158.
MEDRADO-FARIA MA (2003) Mercuralismo metálico crônico ocupacional. Rev.
Saúde Pública, 37(1):116-127.
MEECH JA, VEIGA MM e TROMANS D (1997) Emissions and stability of mercury in
the Amazon. Canadian Metallurgical Quaterly, 36, 4, 231-239.
MICARONI R, BUENO M e JARDIN W (2000) Compostos de Mercúrio: revisão de
métodos de determinação, tratamento e descarte. Química Nova, 23: 487-495.
NASCIMENTO JLM, OLIVEIRA KRM, CRESPO-LOPEZ ME, MACCHI BM, MAUÉS
LAL, PINHEIRO MCN, SILVEIRA LCL e HERCULANO AM (2008) Methylmercury
neurotoxicity & antioxidant defenses. Indian J Med Res 128: 373-382.
NIRCHIO M, FENOCCHIO AS, SWARÇA AC, PÉREZ JE, GRANADO A, ESTRADA
A e RON E (2003) Cytogenetic characterization of hybrids offspring between
Colossoma macropomum (Cuvier, 1818) and Piaractus brachypomus (Cuvier, 1817)
from Caicara del Orinoco, Venezuela. Caryologia 56: 405-411.
NORMANN CABM, MOREIRA JCF e CARDOSO VV (2008) Micronuclei in red blood
cells of armored catfish Hypostomus plecotomus exposed to potassium Dichromate.
African Journal of Biotechnology 7(7): 893-896.

NORTH CAROLINA DIVISION OF POLLUTION PREVENTION AND
ENVIRONMENTAL ASSISTANCE (DPPEA). Mercury. Disponível em:
<http://www.p2pays.org/mercury/health.asp>. Acesso em: 25 nov. 2005.
PANDRANGI R, PETRAS M, RALPH S e VRZOC M (1995) Alkaline single cell
(comet): assay and genotoxicity monitoring using bullhead and carp. Environmental
and Molecular Mutagenesis, 26: 345-356.
PINHEIRO MCN, GUIMARÃES GA, NAKANISHI J, VIEIRA JL, QUARESMA M,
CARDOSO B e AMORAS W. (2000) Avaliação da contaminação mercurial mediante
análise do teor de Hg total em amostras de cabelo em comunidades ribeirinhas do
Tapajós, Pará, Brasil. Rev. Socied. Bras. Med. Trop., 33, 2, 181-184.
POLLARD KM e HULTMAN P (1997) Effects of Mercury on the Immune System.
Metal Ions Biol. Syst. 34: 421-440.
RABELO-GAY MN (1991) Teste de metáfases da medula óssea. In: RABELO-GAY
MN, RODRIGUES MAR, MONTELEONE-NETO R (eds) Mutagênese,
Carcinogênese e Teratogênese: métodos e critérios de avaliação. São Paulo,
Sociedade Brasileira de Genética, pp: 77-83.
RAMOS AS, CASTILHOS ZC e RODRIGUES APC (2005) Avaliação de risco
ecológico e de parâmetros citogenéticos em diversas espécies de peixes da Baía da
Ribeira, Angra dos Reis, RJ – Brasil. XIII Jornada de Iniciação Científica – CETEM.
RAMSDORF W (2007). Utilização de duas espécies de Astyanax (Astyanax sp. B e
A. altiparanae) como bioindicadores de região contaminada por agrotóxico (Fazenda
Cangüiri – UFPR). Dissertação de Mestrado, Universidade Federal do Paraná.
RIBEIRO HB (2007) Estrutura e evolução cariotípica de peixes ciclídeos Sul
Americanos. Dissertação de Mestrado. Universidade Estadual Paulista –
Botucatu/SP, Brasil.

RIGONATO J, MANTOVANI MS e JORDÃO BQ (2005) Comet assay comparison of
different Corbicula fluminea (Mollusca) tissues for the detection of genotoxicity.
Genetics and Molecular Biology, 28(3): 464-468.
ROCHA CAM e CANTO-LOPES LO (2002) Ambiente: ecologia para a escola básica.
Distribel Editora, Belém, PA, 117p.
ROULET M, LUCOTTE M e GUIMARÃES J-RD (2000) Methylmercury in the water,
seston and epiphyton of an Amazonian River and foodplains, Tapajós River, Brazil.
Sci. Total Environ., 261: 43-59.
SÁ AL, HERCULANO AM, PINHEIRO MC, SILVEIRA LCL, NASCIMENTO JLM e
CRESPO-LÓPEZ ME (2006). Exposição humana ao mercúrio na região oeste do
estado do Pará. Revista Paraense de Medicina V.20 (1): 19-25.
SAMPAIO DA SILVA D, LUCOTTE M, ROULET M, POIRIER H, MERGLER D e
CROSSA M (2006) Mercúrio nos peixes do rio Tapajós, Amazônia brasileira.
INTERFACEHS – Revista de Gestão Integrada em Saúde do Trabalho e Meio
Ambiente - v.1, n.1.
SANCHEZ-GALAN S, LINDE AR, AYLLON F e GARCIA-VAZQUEZ E (2001)
Induction of micronuclei in eel (Anguilla anguilla L.) by heavy metals. Ecotoxicol-
Environ-Saf. 49(2): 139-143.
SANTOS EO, LOUREIRO ECB, JESUS IM, BRABO E, SILVA RSU, SOARES MCP,
CÂMARA VM, SOUZA MRS e BRANCHES F (1995) Diagnóstico das Condições de
Saúde de uma Comunidade Garimpeira na Região do Rio Tapajós, Itaituba, Pará,
Brasil, 1992. Cad. Saúde Pública vol.11 no.2 Rio de Janeiro.
SCHMID W (1975) The micronucleus test. Mutat Res 31:9-15.
SCHROEDERK WH e MUNTHE J (1998) Atmospheric Mercury - An overview. Atm.
Env., 32(5): 809-822.

SCHWARTZ M (2005) Bioaccumulation of Mercury in Sport Fish from the San
Francisco Bay. Mercury. Disponível em: <http://www.des.ucdavis.edu/
courses/ESP10/10D_drafts/Kuspa.pdf>. Acesso em: 31 jan. 2006.
SILVA-PEREIRA LC, CARDOSO PCS, LEITE DS, BAHIA MO, BASTOS WR, SMITH
MAC e BURBANO RR (2005) Cytotoxicity and genotoxicity of low doses of mercury
chloride and methylmercury chloride on human lymphocytes in vitro. Brazilian Journal
of Medical and Biological Research 38: 901-907.
SILVEIRA LCL, VENTURA DF e PINHEIRO MCN (2004) Toxicidade mercurial -
avaliação do sistema visual em indivíduos expostos a níveis tóxicos de mercúrio.
Ciência e Cultura V.56 (1): 36-38.
SINGH NP, McCOY MT, TICE RR e SCHNEIDER EL (1988) A simple technique for
quantification of low levels of DNA damage in individual cells. Exp. Cell Res. 175:
184-191.
TENA G (1981) Los problemas gerados por el mercurio en España. In: Simposio
sobre ambiente y salud: Mercurio y Ecología, 5. Boletim de la Academia Nacional de
Medicina de Buenos Aires.
THIER R, BONACKER D, STOIBER T, BÖHM KJ, WANG M, UNGER E, BOLT HM e
DEGEN G (2003) Interaction of metal salts with cytoskeletal motor protein systems.
Toxicology letters vol. 140-41: 75-81.
UNEP Chemicals Mercury Program (2002) Disponível em:
<http://www.chem.unep.ch/mercury/WG-meeting1-documents.htm> Acesso em 16
mar. 2006.
VAL A, ROLIM P e TABELO H (2000) Situação atual da aqüicultura na Região Norte.
In: Valenti W, Poli C, Pereira J e Borghetti J [eds.] Aqüicultura no Brasil: Bases para
seu Desenvolvimento Sustentável. CNPq, Brasília, p.247-266.

VAN Der OOST R, BEYER J e Vermeulen N (2003) Fish bioaccumulation and
biomarkers in environmental risk assessment: a review. Environ. Toxicol. Pharmacol.
13: 57-149.
VANZELLA TP (2006) Efeitos genotóxicos e mutagênicos da fração solúvel do óleo
diesel em uma espécie de peixe neotropical. Dissertação de Mestrado, Universidade
Estadual de Londrina.
VARELA-GARCIA M (1991) Teste de trocas entre cromátides-irmãs (TCI). In:
RABELO-GAY MN, RODRIGUES MAR, MONTELEONE-NETO R (eds) Mutagênese,
Carcinogênese e Teratogênese: métodos e critérios de avaliação. São Paulo,
Sociedade Brasileira de Genética, pp: 123-141.
VIEIRA JLF, PINHEIRO MCN, RIBEIRO DRG, AZEVEDO AA e FERREIRA MN
(2005) Mercúrio total em peixes provenientes da cidade de Itaituba, Pará. Revista
Paraense de Medicina 19(2): 21-24.
WASSERMAN JC, HACON S, WASSERMAN MA. (2001) O ciclo do mercúrio no
ambiente amazônico. Mundo & Vida 2(1/2): 46-53.
WASSERMAN JC, HACON S, WASSERMAN MA (2003) Biogeochemistry of
mercury in the Amazonian environment. Ambio 32:336-42.
WHO. WORLD HEALTH ORGANIZATION (1989) Mercury – environmental aspects.
Geneva. Environmental Health Criteria 86. 115p.
WHO. WORLD HEALTH ORGANIZATION (1990) Mercury – environmental aspects.
Geneva. Environmental Health Criteria 101. 144p.
YALLOUZ A, CAMPOS RC e LOUZADA A (2001) Níveis de mercúrio em atum sólido
enlatado comercializado na cidade do Rio de Janeiro. Ciênc. Tecnol. Aliment.,
Campinas, 21(1): 1-4

ANEXOS
TABELAS DE RESULTADOS

Número de nucleóides de eritrócitos analisados, normais e alterados, com os respectivos danos, em C. macropomum não contaminados (controle) e
contaminados com metilmercúrio 2 mg.L-1.
Número de células Classes/100 células Escores
Analisadas Alteradas 0 1 2 3 4 Controle
C1 200 56 72 22.5 5 0.5 0 34
C2 200 52 74 22 2.5 1.5 0 31.5
C3 200 70 65 27 8 0 0 43
C4 200 39 80.5 16 3 0.5 0 23.5
C5 200 60 70 24.5 5.5 0 0 35.5
C6 200 46 77 18 4 0.5 0.5 29.5
C7 200 28 86 14 0 0 0 14
C8 200 16 92 8 0 0 0 8
C9 200 53 73.5 26.5 0 0 0 26.5
Total 1800 420 690 178.5 28 3 0.5 X= 27.28
Tratado
Hg1 200 200 0 24 22.5 38 15.5 245
Hg2 200 181 9.5 53 30.5 7 0 135
Hg3 200 169 15.5 60.5 20 3.5 0.5 113
Hg4 200 195 2.5 70 20 5.5 2 134.5
Hg5 200 198 1 60.5 31 5 2.5 147.5
Hg6 200 189 5.5 73 18 3.5 0 119.5
Hg7 200 183 8.5 70.5 17 3.5 0.5 117
Hg8 200 183 8.5 77 13 1.5 0 107.5
Hg9 200 190 5 55 31 4.5 4.5 148.5
Total 1800 1688 56 543.5 203 72 25.5 X= 140.83
C= controle; X= média; Hg= tratado.

Número de células com núcleos normais e alterados em Colossoma macropomum (tambaqui) nos grupos controle e exposto.
Células Normais
Células
Alteradas TOTAL Alt Morf MN
Controle C1 3972 23 5 4000 C2 3975 20 5 4000 C3 3954 32 14 4000 C4 3965 23 12 4000 C5 3960 30 10 4000 C6 3947 39 14 4000 C7 3969 21 10 4000 C8 3957 27 16 4000 C9 3994 5 1 4000
Total 35693 220 87 36000
MeHg 2mg/L 120h Hg20 3965 30 5 4000 Hg21 3955 35 10 4000 Hg22 3957 36 7 4000 Hg23 3971 26 3 4000 Hg24 3937 45 18 4000 Hg25 3944 33 23 4000 Hg26 3963 36 1 4000 Hg27 3970 24 6 4000 Hg28 3971 26 3 4000 Total 35633 291 76 36000

Número de nucleóides de eritrócitos analisados, normais e alterados, com os respectivos danos, em A. tetramerus não contaminados (controle) e contaminados
com metilmercúrio 2 mg.L-1. Número de células Classes/100 células
Escores Analisadas Alteradas 0 1 2 3 4
Controle
C1 200 22 89 11 0 0 0 11
C2 200 15 92.5 7.5 0 0 0 7.5
C3 200 20 90 10 0 0 0 10
C4 200 18 91 9 0 0 0 9
C5 200 18 91 9 0 0 0 9
C6 200 14 93 7 0 0 0 7
C7 200 32 84 16 0 0 0 16
C8 200 28 86 14 0 0 0 14
C9 200 18 91 9 0 0 0 9
C10 200 19 90.5 9.5 0 0 0 9.5
C11 200 20 90 10 0 0 0 10
Total 2200 224 988 112 0 0 0 X = 10.18
MeHg 2mg/L 24h
Hg14 200 190 5 14 31.5 35 14.5 240
Hg15 200 200 0 8.5 34 40 17.5 266.5
Hg18 200 200 0 9.5 29.5 50 11 262.5
Hg19 200 200 0 15.5 39.5 39.5 5.5 235
Hg20 200 200 0 5 46.5 46.5 13.5 268.5
Hg21 200 183 8.5 15.5 31.5 25.5 19 231
Hg22 200 200 0 18 38.5 34.5 9 234.5
Hg23 200 177 11.5 17 35.5 24 12 208
Hg24 200 200 0 20 39.5 30 10.5 231
Hg38 200 197 1.5 9.5 33.5 33.5 22 265
Hg39 200 195 2.5 15.5 40 32.5 9.5 231
Total 2200 2142 29 148 399.5 391 144 X = 243
MeHg 2mg/L 120h
Hg25 200 200 0 0 23.5 51 25.5 302
Hg26 200 200 0 0 21.5 48.5 30 308.5
Hg27 200 200 0 0 29 46 25 296
Hg28 200 200 0 0 27 45.5 27.5 300.5
Hg29 200 200 0 0 22.5 51 26.5 304
Hg30 200 200 0 0 21 53 26 305
Hg31 200 200 0 0 23 50 27 304
Hg32 200 200 0 0 23 48.5 28.5 305.5
Hg33 200 200 0 0 26.5 47 26.5 300
Hg34 200 200 0 0 19 56 25 306
Hg35 200 200 0 0 21.5 54 24.5 303
Total 2200 2200 0 0 257.5 550.5 292 X = 303.13
C= controle; X= média; Hg= tratado

Número de células com núcleos normais e alterados em Aequidens tetramerus (acará) nos grupos controle e exposto.
Células Normais
Células
Alteradas TOTAL Alt Morf MN
Controle 1 3990 10 0 4000 2 3991 9 0 4000 3 3982 18 0 4000 4 3950 43 7 4000 5 3986 14 0 4000 6 3977 20 3 4000 7 3977 22 1 4000 8 3910 76 14 4000 9 3962 38 0 4000
10 3965 35 0 4000 11 3996 04 0 4000
Total 43686 289 25 44000
MeHg 2mg/L 24h 14 3975 21 4 4000 15 3980 18 2 4000 18 3976 24 0 4000 19 3960 40 0 4000 20 3986 13 1 4000 21 3965 35 0 4000 22 3915 83 2 4000 23 3937 61 2 4000 24 3917 79 4 4000 38 3943 54 3 4000 39 3893 100 7 4000
Total 43540 528 25 44000
MeHg 2mg/L 120h 25 3926 60 14 4000 26 3928 56 16 4000 27 3903 91 6 4000 28 3917 71 12 4000 29 3942 50 8 4000 30 3918 66 16 4000 31 3936 59 5 4000 32 3953 47 0 4000 33 3927 70 3 4000 34 3925 73 2 4000 35 3901 99 0 4000
Total 43176 742 82 44000

APÊNDICES

APÊNDICE 1:
Artigo* “Evaluation of Genotoxic Effects of Xenobio tics in Fishes Using Comet Assay – A Review”
*Publicado em Reviews in Fisheries Science 17(2):170–173, 2009.





APÊNDICE 2:
Artigo* “The Micronucleus Assay in Fish Species as an Important Tool for Xenobiotic Exposure Risk Assessment—A Brief Review and an Example Using
Neotropical Fish Exposed To Methylmercury”
*Publicado em Reviews in Fisheries Science 17(4):478–484, 2009.








Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo


















