
Caracterização dos genes de resistência aos … · 1. Valor estimativo de um potencial deficit...
123
Maria João da Silva Costa Caracterização dos genes de resistência aos macrólidos e estreptograminas em Enterococcus sp. isolados de frangos para consumo humano Faculdade de Farmácia da Universidade do Porto Porto 2004
Transcript of Caracterização dos genes de resistência aos … · 1. Valor estimativo de um potencial deficit...

Maria João da Silva Costa
Caracterização dos genes de resistência aos macrólidos e estreptograminas em Enterococcus sp. isolados de frangos
para consumo humano
Faculdade de Farmácia da Universidade do Porto Porto 2004

Maria João da Silva Costa
Caracterização dos genes de resistência aos macrólidos e
estreptograminas em Enterococcus sp. isolados de frangos
para consumo humano
Faculdade de Farmácia da Universidade do Porto Porto 2004

Maria João da Silva Costa
Caracterização dos genes de resistência aos macrólidos e estreptograminas em
Enterococcus sp. isolados de frangos para consumo humano
Dissertação de candidatura ao grau de Mestre em Controlo da Qualidade apresentada à
Faculdade de Farmácia da Universidade do Porto
Orientadora : Professora Doutora Luísa Vieira Peixe
Faculdade de Farmácia da Universidade do Porto
Porto 2004

Agradecimentos
Agradecimentos
É para a Professora Doutora Luísa Vieira Peixe que dirijo, em primeiro lugar, os meus
agradecimentos. A paciência e persistência com que conduziu a orientação científica deste
trabalho, pautou-se, ao longo destes anos, na valorização do meu conhecimento.
Fica a certeza que jamais conseguirei retribuir tal ajuda, amizade e ânimo.
Ao Professor Doutor João Carlos Figueiredo de Sousa agradeço a possibilidade da
realização deste trabalho no Laboratório de Microbiologia da Faculdade de Farmácia da
Universidade do Porto.
À Professora Doutora Beatriz Quinaz, à Professora Doutora Eugenia Pinto, à Professora
Doutora Helena Ferreira, à Professora Doutora Isaura Pinto de Sousa, á Professora Doutora
Maria de Lurdes Basto e à Professora Doutora Nazaré Pestana gostaria de agradecer todo o
incentivo e a simpatia demonstrada ao longo destes anos..
À colega Carla Novais dirijo o meu profundo agradecimento pela forma incansável que
apoiou o meu trabalho laboratorial, que constituiu um suporte fundamental neste sucesso, a
disponibilidade e a amizade demonstrada. Às colegas Elisabete Machado, Patrícia Antunes
e Sandra Quinteira agradeço o apoio nos momento menos bons, o encorajamento, a
amizade e o agradável ambiente de trabalho que sempre proporcionarm.
À D. Alda, à D. Deolinda e à D. Filomena agradeço toda a disponibilidade demonstrada.
Finalmente, porque o sucesso ou o insucesso se mede por tudo quanto tivemos de abrir
mão, é dos meus pais, da minha irmã e avó, que guardo para sempre a compreensão, o
carinho e o apoio que nunca deixaram de me dar.
Aqui fica a minha justificação para a minha ausência e a pouca atenção que lhes dei nos
últimos tempos.
v

Resumo
Resumo
A incidência de bactérias resistentes a antibióticos isoladas de animais de explorações
intensivas ou de carcaças de animais para consumo humano constituem uma preocupação
crescente, em virtude da utilização de antibióticos na produção animal poder seleccionar
bactérias ( comensais ou patogénicas) resistentes a estes agentes antimicrobianos. Os
enterococos fazem parte da flora bacteriana gastrointestinal normal do Homem e dos
animais, podendo ocasionalmente provocar infecções, pelo que o aparecimento de
resistências neste género bacteriano poderão dificultar o seu tratamento. Na produção de
frangos os antibióticos são utilizados em grandes quantidades, não só na profilaxia e
terapia, mas também como promotores de crescimento, não sendo conhecida, em Portugal,
a incidência dos genes de resistência a macrólidos e estreptograminas, nos isolados de
Enterococcus sp. com origem nestas aves.
Neste estudo procedeu-se à caracterização, por PCR, dos genes de resistência aos
macrólidos e estreptograminas existentes nos isolados de Enterococcus sp. e à avaliação da
capacidade de transferência dos genes codificadores de resistência aos antibióticos
referidos, recorrendo a ensaios de conjugação.
Utilizaram-se 388 isolados do género Enterococcus com origem em carcaças de frangos
de dez marcas diferentes, com ampla distribuição a nível nacional,as quais foram
recolhidas entre Setembro de 1999 e Junho de 2001, em talhos da cidade do Porto. Foram
incluídos isolados de Enterococcus faecium, de Enterococcus faecalis, de Enterococcus
gallinarum e de Enterococcus sp., os quais possuíam diferente susceptibilidade à
vancomicina, à eritromicina e à associação quinupristina-dalfopristina.
O gene ermB foi detectado nos isolados de Enterococcus sp. resistentes à eritromicina
(49%), nos isolados com resistência intermédia (4%) e nos isolados sensíveis (5%). Não
foi observada a presença de genes ermA, ermC, nem de mefA. O gene vatD estava apenas
vi

Resumo
presente nos isolados com resistência à associação quinupristina-dalfopristina (2.6%). O
gene vatE foi observado num isolado de E. faecalis resistente à associação quinupristina-
dalfopristina, em isolados de E. faecium resistentes, com comportamento intermédio e em
isolados sensíveis a este férmaco. Em isolados de E. gallinarum não foi possível fazer a
sua observação. Pelo contrário, este gene foi observado em isolados de Enterococcus sp.
com comportamento intermédio e resistente ao antibiótico referido.
Embora não tenha sido possível verificar em todos os enterococos testados a transferência
por conjugação dos genes ermB e vatE, efectuou-se a transferência por conjugação do gene
ermB em dois isolados de E. faecium e em um isolado de E. faecalis, resistentes à
eritromicina., assim como do gene vatE de um isolado de E. faecalis resistente à
associação quinupristina-dalfopristina. De referir a obtenção de um transconjugante que
adquiriu a resistência à eritromicina e à associação quinupristina-dalfopristina por
expressão em simultâneo dos genes ermB e vatE, presentes num E. faecalis testado,
resistente a estes antibióticos.
Os resultados obtidos demonstram a existência de um reservatório de genes de resistência
aos MLS (macrólidos, lincosamidas e estreptograminas) em alimentos para consumo
humano, pelo que o seu contributo para a elevada incidência, observada em Portugal, de
enterococos resistentes aos MLS, em doentes deverá ser objecto de estudos posteriores.
Os resultados obtidos no âmbito desta tese de Mestrado são parte integrante do artigo
intitulado " Insights about recent epidemiology of Enterococcus resistance in the
Portuguese Community " Novais C , Costa M. J., Sousa J.C., Coque T.M., Peixe L.V.,
submetido para publicação.
Vii

Abstract
Abstract
The prevalence of resistant bacteria, isolated from food animals or animal carcasses to
antimicrobials, is of increasing concern, as the use of antimicrobial agents in animal
production may select resistant bacteria (commensal or pathogenic) to these antimicrobials.
As enterococci form part of the normal host flora of both human and animal
gastrointestinal tract, where they seldom cause infections, the emergence of resistant
enterococci may constitute a risk for therapy. In poultry industry, antibiotics are used in
high quantities not only in profilaxy and therapy, but also as antimicrobial growth
promoters in animal feeds. In Portugal it is unknown the prevalence of macrolide and
streptogramin resistant determinants, in isolates of Enterococcus sp, recovered from retail
poultry. In this work we characterized the macrolide and streptogramin resistant genes in
Enterococcus sp. isolates, by PCR, and evaluated the in vitro transfer of the resistance
genes to the previously mentioned antibiotics, by conjugation.
389 Enterococcus sp isolates collected from poultry carcasses, belonging to 10 different
brands, wide distributed in Portugal, were recovered from retail stores in Oporto, from
September 1999 to March 2001. We studied Enterococcus faecium (n=112), Enterococcus
faecalis (n= 121), Enterococcus gallinarum (n=61) and Enterococcus sp. (n=95), which
demonstrated susceptibility to vancomicin, to erithromicyn and to quinupristin-
dalfopristin. ermB resistance determinant was observed in erythromycin-resistant isolates (49%), in
intermediate resistant isolates (4%) and in sensitive isolates (5%). The ermA, ermC and
me/A resistance determinants were not detected in any of the isolates.
The vatD resistance determinant was only observed in the quinupristin-dalfopristin-
resistant enterococci isolates (2.6%) The vatE resistance determinant was observed in the
quinupristin-dalfopristin-resistant E. faecalis isolate, in E. faecium isolates with viii

Abstract
intermediate resistance and in sensitive isolates. The vatE resistance determinant was not
observed in E. gallinarum isolates. On the other hand, this resistance determinant was
observerd in Enterococcus sp. isolates with intermediate resistance and in resistant isolates.
Although it was not possible to transfer the ermB and vatE resistance determinants in all
the tested isolates, we were able to transfer ermB gene in 2 E. faeciwn isolates and in one
E. faecalis isolate erythromycin-resistant.The vatE resistance determinant was transfered
from one E. faecalis isolate quinupristin-dalfopristin-resistant.
We also obtained one transconjugant that acquired resistance to erithomicin and to
quinupristin-dalfopristin, by expressing both ermb and vatE, in one E. faecalis isolate.
As the obtained results show the existence of a reservoir of resistance genes to MLS in
food animals to human consuption, the contribution to the high incidence, reaveled in
Portugal, of MLS-resistant-enterococci, in ill- people, should be a matter of further studies.
The data obtained was included in the paper " Insights about recent epidemiology of
Enterococcus resistance in the Portuguese Community " Novais C , Costa M. J., Sousa
J.C., Coque T.M., Peixe L.V., submitted for publication.
IX

Siglas
Siglas
ABC- ATP-binding cassete
BHI- Brain Heart Infusion
CMI- Concentração Mínima Inibitória
CNS- Staphylococcus coagulase-negativo
FDA- Food and Drug Administration
FEDES A- European Federation of Animal Health Industries
GRE- Enterococcus faecium resistente aos glicopéptidos
LPS- Lipopolissacarídeos MLS- Macrólidos, lincosamidas e estreptograminas
MFS-Major Facilitator Superfamily
MRSA- Staphylococcus aureus resistente à meticilina
NOAH- National Office of Animal Health
PCR- Polimerase Chain Reaction
PFGE- Pulsed-field Gel Electrophoresis
USD A- United States Department of Agriculture
VRE- Enterococcus sp resistente à vancomicina
WHO- World Health Organization
x

índice
r
Indice
1- Introdução , 17
1.1. Utilização de antibióticos em produção animal, 18
1.1.1. Utilização dos antibióticos em veterinária, 18
a) Profilaxia de infecções, 19
b) Terapia de infecções, 19
c) Promoção de crescimento animal, 20
1.2. Consumo de antibióticos, 24
1.2.1. Consumo de antibióticos em animais, 24
1.2.2 .Consumo de antibióticos em medicina humana, 28
1.2.3. Consumo humano versus consumo animal, 29
1.3. Aparecimento e evolução da resistência, 30
1.3.1. Consequências da utilização dos antibióticos, 35
1.3.2. Impacto da utilização de antibióticos em produção animal na resistência
bacteriana de bactérias potencialmente patogénicas para o Homem, 39
1.4. Género Enterococcus, 46
1.4.1. Principais características, 46
1.4.2. Significado clínico, 48
1.5. Macrólidos, lincosamidas e estreptograminas, 49
1.5.1. Modo de acção dos macrólidos, lincosamidas e estreptograminas, 53
1.5.2. Mecanismos de resistência aos macrólidos, lincosamidas e estreptograminas, 54
1.5.2.1. Resistência intrínseca, 55
1.5.2.2. Resistência adquirida, 56
A . Inactivação enzimática do antibiótico, 56
a) Hidrólise de macrólidos e estreptograminas, 56
b) Glicosilação dos macrólidos, 57
XI

índice
c) Fosforilação dos macrólidos, 57
d) Nucleotidiltransferases, 57
e) Acetiltransferases, 58
B. Modificação do alvo, 61
1. Fenótipo constitutivo, 63
2. Fenótipo indutível, 63
C. Sistemas de efluxo do antibiótico, 65
1. Efluxo Tipo-M ou Fenótipo M, 66
2. Efluxo Tipo-MS ou Fenótipo MS, 68
3. Efluxo Tipo Actinomycete, 69
4. Efluxo Tipo mrd , 69
2. Objectivos, 71
3. Material e Métodos, 72
3.1. Origem das estirpes bacterianas, 72
3.2. Isolamento, identificação dos isolados e avaliação da susceptibilidade a agentes
antimicrobianos, 72
3.3. Avaliação de susceptibilidade e identificação dos isolados da colecção
de bactérias, 72
3.4. Detecção de genes de resistência a macrólidos e estreptograminas por PCR, 73
3.4.1. Extracção do DNA genómico das estirpes controlo, 74
3.4.2. Condições de PCR, 75
3.5. Transferência dos determinantes de resistência à eritromicina e às estreptograminas
por conjugação, 79 4. Resultados, 81 4.1. Susceptibilidade a agentes antimicrobianos em Enterococcus sp., 81
XII

Indice
4.2. Distribuição de genes de resistência aos macrólidos e estreptograminas em isolados de
Enterococcus sp., 84
4.2.1. Presença de genes erm, 84
4.2.2. Presença de genes mefA, 87
4.2.3. Presença de genes vatD (satA), 87
4.2.4. Presença de genes vatE (satG), 89
4.2.5. Presença simultânea de genes de resistência, 92
4.3. Distribuição do gene ermB em isolados de Enterococcus sp, com diferentes CMIs
para a vancomicina, 93 4.4. Avaliação da transferência dos determinantes de resistência à eritromicina e às
estreptograminas por conjugação, 95
5. Discussão, 96
6. Conclusão, 110
7. Bibliografia, 112

Lista de tabelas
Lista de tabelas
1. Valor estimativo de um potencial deficit na produção de carne na ausência de promotores de crescimento, 23
2. Uso de antibióticos em diferentes Estados Membros (Kg ), 24
3. Consumo de promotores de crescimento na Dinamarca, em 1995, 25
4. Estimativa de vendas de antibióticos para animais, na União Europeia e Suíça em 1997,26
5. Volumes (toneladas de substância activa) de vendas de antibióticos em diferentes países da União Europeia, em 1997, 27
6. Média anual de consumo de antibióticos pelo Homem e pelos animais, na União Europeia, em 1997, 30
7. Descoberta do antibiótico e desenvolvimento da resistência, 32
8. Percentagem de isolados de Enterococcus e de Streptococcus em frangos com diferentes idades, 37
9. Multiresistência em E. faecium, 40
10. Percentagem de E.faecalis e E. faecium resistentes a diferentes antibióticos, 41
11. Classificação de alguns macrólidos, 50
12. Bactérias onde se encontram genes que provocam a inactivação do antibiótico, 60
13. Bactérias onde se encontram genes codificadores de metilases, 62
14. Bactérias onde se encontram genes que provocam o efluxo do antibiótico, 70
15. Identificação das estirpes controlo das reacções de PCR para detecção dos genes de resistência a macrólidos e estreptograminas, 74
16. Sequência de primers e condições de PCR para os genes de resistência aos macrólidos e estreptograminas, 76
xiv

Lista de tabelas
17. Concentrações dos reagentes usados na reacção de PCR para detecção dos genes
ermA, ermB e ermC, 77
18. Concentrações dos reagentes usados na reacção de PCR para detecção dos genes
mefA, 78
19. Concentrações dos reagentes usados na reacção de PCR para detecção dos vatD
e vatE, 78
20. Distribuição de CMIs para a eritromicina e para a quinupristina-dalfopristina em isolados de Enterococcus sp., 83
21. Distribuição do gene ermB em isolados de Enterococcus sp, com diferentes CMIs para a eritromicina, 85
22. Distribuição do gene vatD (satA) em isolados de Enterococcus sp., com diferentes CMIs para a Quinupristina-dalfopristina, 88
23. Distribuição do gene vatE (satG) em isolados de Enterococcus sp., com diferentes CMIs para a Quinupristina-dalfopristina, 91
24. Número de isolados com os genes vatD e ermB, vatE e ermB e vatD e vatE presentes simultaneamente, 92
25. Distribuição do gene ermB em isolados de Enterococcus sp., com diferentes CMIs para a vancomicina, 94
xv

Lista de figuras
Lista de figuras
Fig. 1 - Estrutura química da eritromicina, 50
Fig. 2- Estrutura química das lincosamidas, 51
Fig. 3- Estrutura química dos grupos A e B das estreptograminas, 52
Fig. 4- Inibição da síntese proteica pelas estreptograminas, 54
Fig. 5- Identificação dos genes ermB por PCR e electroforese em gel de agarose, 84
Fig. 6- Identificação dos genes vafD por PCR e electroforese em gel de agarose, 87
Fig. 7- Identificação dos genes vatE por PCR e electroforese em gel de agarose, 89
xvi

Introdução
1. Introdução
A nível mundial está a ocorrer um aumento no consumo de géneros alimentares
transformados e pré-embalados, preparados por indústrias mais ou menos sofisticadas. Será
necessário duplicar a produção de alimentos de origem animal nos próximos 20 anos,
devido à duplicação previsível da população humana nos próximos 50 anos (Portugal, A.
V., 1998). Como resposta a esta necessidade, surgiram os sistemas intensivos de produção animal,
que vão ao encontro da pressão do mercado. Estas unidades de produção animal,
recorrendo a novas formas de produção, são responsáveis pela obtenção em massa de
alimentos normalizados, os quais devem ser comercializados com a identificação da
origem e a forma como foram obtidos. Como consequência, a produção animal intensiva
deixa de ter características que a vinculam à forma natural, sendo manipulada pelo
Homem, em virtude de se modificarem as actividades fisiológicas normais do animal por
controlo do metabolismo através de agentes externos. Estes sistemas conduzem a ritmos de
produção e à obtenção de lucros superiores, assim como, à elaboração de produtos
diferentes dos obtidos em condições naturais ou em ritmos de produção mais lentos
(Portugal, A. V., 1998 ; Portugal, A. V., 2002).
A conjugação da aplicação de diferentes agentes para o aumento da produção permite a
obtenção de um produto animal de características diferentes das resultantes da aplicação
isolada de cada um deles (Portugal, A. V., 1998 ; Portugal, A. V., 2002).
Em virtude da produção animal ocupar um lugar de destaque na agricultura da União
Europeia (UE), a obtenção de resultados satisfatórios depende de vários factores,
nomeadamente da utilização de alimentos adequados e de boa qualidade para os animais.
No sentido de proteger a saúde pública e animal e o meio ambiente é realizada uma
avaliação da segurança dos aditivos para a alimentação animal, através de procedimentos
17

Introdução
comunitários, antes da sua colocação no mercado. Por esta razão, foi elaborada a Directiva
70/524/CEE do Conselho, de 23 de Novembro de 1970, relativa aos aditivos na
alimentação para animais, que constitui a legislação de base, a qual é sucessivamente
sujeita a grandes modificações (Comunicação do Conselho, Posição Comum (CE)
N°20/2003, adoptada pelo Conselho em 17 de Março de 2003 ; Proposta de Regulamento
do Parlamento Europeu e do Conselho relativa aos aditivos destinados à alimentação
animal, Bruxelas 22/03/02,COM (2002) 153 final, 2002/2003 (COD)).
1.1. Utilização de antibióticos em produção animal
Uma grande variedade de substâncias com actividade antimicrobiana são usadas nos
sistemas de produção animal na profilaxia, na terapia e como promotores de crescimento.
Nos últimos anos a preocupação com a selecção de bactérias resistentes aos antibióticos,
decorrentes da utilização destes agentes na produção animal foi determinante na redução
dos antimicrobianos, autorizados para este fim, na União Europeia.
1.1.1. Utilização dos antibióticos em veterinária
As substâncias usadas com fins profilácticos ou terapêuticos em veterinária pertencem à
mesma classe dos antibióticos utilizados na medicina humana (Schwarz, S. e Chaslus-
Dancla, E., 2001; Smith, J. T., 1993).
Os antibióticos mais usados nos animais para consumo humano pertencem principalmente
a cinco classes - p-lactâmicos, tetraciclinas, aminoglicosídeos, macrólidos e sulfonamidas-
os quais estão sujeitos a legislação (JOCE, n° L 60/16), a qual especifica as substâncias
farmacológicas activas para as quais foram fixados limites máximos de resíduos .
18

Introdução
a) Profilaxia de infecções
O uso profilático dos antibióticos é uma prática comum em veterinária, principalmente em
produções intensivas, onde a densidade dos animais é muito elevada (Mckellar, Q. A.,
1998). O tratamento profilático é um tratamento preventivo, fornecido individualmente,
podendo ser aplicado a um grupo de animais, quando a proporção dos animais que
adoecem durante um período específico de tempo e a probabilidade da maioria, ou de
todos, ficarem infectados é grande. Neste caso dá-se a designação de uso metafilático dos
agentes antimicrobianos (McEwan, S.A. et ai, 2002; Mckellar, Q. A., 1998; Mackinnon, J.
D., 1993; Schwarz, S. et ai, 2001) Esta metodologia permite, igualmente, travar a
disseminação da infecção e prevenir a doença, após o seu diagnóstico num ou mais animais
do grupo (Mackinnon, J. D., 1993; Schwarz, S. et ai, 2001). Este tipo de tratamento
também é efectuado em certos períodos da vida do animal, tal como em cirurgias,
vacinação, transporte e mistura de animais, em virtude de, durante estes períodos, os
animais estarem mais susceptíveis. Sem estes tratamentos preventivos as infecções
ocorreriam com maior frequência e seria necessária a intervenção terapêutica. Apesar
destes benefícios, a aplicação profiláctica dos antibióticos é considerada uma prática
responsável pela selecção de bactérias resistentes, assim como pela disseminação de genes
de resistência (Schwarz, S. et ai, 2001).
b) Terapia de infecções
A terapia envolve o tratamento de um animal ou de um grupo de animais doentes, após o
diagnóstico de uma doença, com um ou mais antibióticos, durante um determinado período
19

Introdução
de tempo e, na maior parte dos países, apenas com prescrição do veterinário (Johnston, A.
M., 1998). A finalidade da terapia antimicrobiana é a de produzir determinadas
concentrações do antibiótico nos tecidos e nos fluídos corporais e manter estes níveis até
que a infecção seja ultrapassada (Johnston, A. M., 1998; Lamb, H. M. et al, 1999).
Os grandes grupos de animais, como por exemplo suínos e aves, podem causar problemas
logísticos para o tratamento com antibióticos (Butaye, P. et ai, 2000; Jackson, E. R., 1993;
Johnston, A. M., 1998). Contudo, os antibióticos com fins profiláticos ou terapêuticos
podem ser administrados através dos alimentos (alimento medicamentoso) ou da água,
podendo, no entanto, ser inevitável o tratamento parental. O alimento de tratamento ou
alimento medicamentoso é preparado de acordo com a duração do tratamento e é
considerado um medicamento (Corpet, D., 1998).
Quando são dados antibióticos através de veículos alimentares, a quantidade de água e de
alimentos consumidos, o peso do animal, a farmacodinâmica e a farmacocinética deste
agente, têm de ser considerados para efectuar o cálculo da taxa de inclusão do antibiótico,
de modo a assegurar um tratamento adequado (Butaye, P. et ai, 2000; Jackson, E. R., 1993,
Johnston, A. M., 1998 ; McEwan, S.A.. et ai, 2002; Smith, T. L. et ai, 1999).
c) Promoção de crescimento animal
Existe um número específico de substâncias utilizadas como promotores de crescimento.
As moléculas e respectivas concentrações encontram-se legisladas, assim como os animais
alvo, a duração e a dosagem em que são fornecidas (McEwan, S.A.. et ai, 2002; Schwarz,
S. et ai, 2001). Os antibióticos autorizados para utilização como promotores de crescimento são a
avilamicina, a salinomicina, o flavofosfolipol e a monensina-sódio (Diário da República I
Série, n° 296 de 27 de Dezembro de 1989; McEwan, S.A. et ai, 2002).
Os antibióticos promotores de crescimento são adicionados às rações animais, em doses
subterapêuticas, durante o crescimento do animal (Aarestrup, F. M., 2000, Aarestrup, F. M.
20

Introdução
et al, 1998, Martel, J.L. et al, 2001; McEwan, S.A. et al, 2002). Por esta razão, os
antibióticos utilizados com esta finalidade não necessitam de prescrição médica. As suas
características como promotores de crescimento são conhecidas desde finais dos anos 40,
tendo esta prática sido imediatamente adoptada por muitas explorações animais (Dunlop,
R. H. et ai, 1998; Finch, R. G. 1998.). Os animais que ingerem promotores de crescimento
necessitam de menor quantidade de alimento para produzirem maior quantidade de carne.
Assim, o índice de consumo - relação entre o peso dos alimentos consumidos e o ganho de
peso do animal - melhora de 3 a 12 %. Para além deste aspecto é de salientar que os
animais crescem mais rapidamente. O ganho médio quotidiano - razão entre o peso total
adquirido pelo animal no final do crescimento e a duração do crescimento, em dias,
aumenta de 3 a 9 % e os lotes são mais homogéneos (Witte, W., 1998).
O efeito dos promotores de crescimento deve-se, provavelmente, à diminuição da flora
intestinal normal e das bactérias patogénicas. Pensa-se que os promotores de crescimento
contribuem para o aumento do peso do animal em virtude de pequenas doses de antibiótico
inibirem o metabolismo da flora intestinal, a qual compete com o hospedeiro para os
nutrientes. O consumo de aminoácidos por estas bactérias diminui, ocorrendo
simultaneamente menor produção de moléculas tóxicas, como por exemplo, amoníaco. A
redução das bactérias patogénicas é um outro factor que permite um maior
desenvolvimento dos animais, dado que a sua presença pode diminuir o crescimento
devido a doença subclínica (Bower, C. K., et ai, 1999; McEwen, S. A. 2002; Wegener, H.
C. étal, 1999). Desta forma, a concentração de antibiótico fornecida tem de ser suficiente para inibir as
bactérias susceptíveis e afectar a composição da flora intestinal. Este efeito inibitório é
confirmado pelo efeito preventivo dos promotores de crescimento contra certas infecções
bacterianas provocadas, por exemplo, por Clostridium perfringens em aves de capoeira e
por Brachyspira hyodysenteriae e Escherichia coli em suínos (Johnston, A. M., 1998;
Wegener, H. C. et ai, 1999).
21

Introdução
Para o National Office of Animal Health (NOAH), o qual representa as companhias
produtoras de promotores de crescimento, os seus benefícios vão para além dos
mencionados. Num comunicado de 9 de Abril de 1999 é referido que os promotores de
crescimento permitem uma maior absorção de nutrientes e de água, diminuindo,
consequentemente, o desperdício devido à excreção de nutrientes, como por exemplo,
fósforo e nitrogénio. Esta será uma grande vantagem pois estes nutrientes ao atingirem os
cursos de água poderão provocar a eutrofização, ou seja, um crescimento excepcional de
algas, esgotando o oxigénio, o que acarreta danos para a comunidade piscícola e para a
microflora. Segundo o NOAH, estima-se que a diminuição do consumo de promotores de
crescimento na Alemanha, França e Reino Unido, levaria a um aumento de 78 mil
toneladas de nitrogénio e de cerca de 15 mil toneladas de fósforo, os quais seriam
libertados para o meio ambiente (National Office of Animal Health (NOAH) Press
Release, 1999).
Desta forma, o NOAH considera que a utilização de promotores de crescimento tem
benefícios para além da produção de carne e de leite, apresentando vantagens para o meio
ambiente e, consequentemente, para o Homem (National Office of Animal Health (NOAH)
Press Release, 1999).
Para além dos factores referidos devem considerar-se as vantagens em termos económicos.
Os promotores de crescimento são baratos, seguros, fáceis de utilizar e tendem a aumentar
o crescimento animal, independentemente do sistema de produção. De acordo com um
estudo publicado (Mackinnon, J. D., 1993), poderia surgir um défice na produção de carne
se não fossem utilizados promotores de crescimento, conforme se pode constatar pela
interpretação dos dados da tabela 1.
22

Introdução
Tabela 1. Valor estimativo de um potencial défice na produção de carne na ausência de promotores de crescimento (Mackinnon, J. D., 1993).
Animais Variação do peso ( Kg )
Suínos - 3.43 Frangos - 0.0065 Vitelos - 6.5
Carne de vaca - 26.1
A distinção entre profiláctico e promotor de crescimento é ainda menos clara do que entre
agente profiláctico e agente terapêutico. Nos Estados Unidos da América, alguns agentes
antimicrobianos podem ser aprovados para uso profiláctico e agente terapêutico, uma vez
que alguns promotores de crescimento podem ajudar a prevenir a doença, mesmo em doses
subterapêuticas. Este é um aspecto da maior importância, porque a administração de
antibióticos, durante um curto intervalo de tempo, pode ser sempre justificada com base na
prevenção da doença (Schwarz, S. et ai, 2001).
No entanto, apesar da utilização dos promotores de crescimento permitir um aumento de
peso entre 1 e 11%, estes benefícios podem não acontecer no caso da utilização de práticas
modernas de produção animal. Verificou-se que estes aditivos são mais eficazes nos
animais jovens do que nos de maior idade e que o ganho médio quotidiano é menor quando
os promotores de crescimento são usados em condições óptimas de higiene. Todavia,
mesmo nestas condições, os resultados são bastante positivos (van den Bogaard, A. E. et
al, 1999). Com o aumento das práticas de higiene e outras medidas para controlar as
doenças, como por exemplo, a vacinação, é levantada a questão sobre os benefícios reais
da utilização de promotores de crescimento. Um estudo dinamarquês revelou que a
remoção dos promotores de crescimento na produção de frangos diminuiu em menos de
1% o seu crescimento e que, apesar de ter ocorrido um aumento de entrites necróticas, não
houve alteração na mortalidade nem diminuição do peso dos frangos (McEwen, S. A. et ai,
2002).
23

Introdução
1.2. Consumo de antibióticos
1.2.1 Consumo de antibióticos em animais
Entre os antibióticos consumidos durante várias décadas, a bacitracina, a espiramicina, a
tilosina e a virginiamicina foram utilizadas quer na terapia, quer como promotores de
crescimento. A avilamicina, o carbadox, o olaquindox, a avoparcina, a flavomicina, a
monensina e a salinomicina estavam aprovados apenas como promotores de crescimento
(Aarestrup, F. M. et ai, 1998b). Em virtude da indústria farmacêutica não ser legalmente
obrigada a fornecer informações relativamente aos volumes de vendas de produtos
utilizados nos animais, os dados disponíveis são escassos (Aarestrup, F. M. et ai, 1998b).
Apenas a Suécia, os Países Baixos, a Dinamarca e a Finlândia fornecem dados passíveis de
utilização (Aarestrup, F. M. et al, 1998b). A Áustria, Bélgica, Espanha, França, Irlanda,
Itália, Luxemburgo, Portugal e Reino Unido fornecem pouca ou nenhuma informação
(Consumer Policy and Consumer Health Protection, 1999; Poyart, C. et al, 1997; Schwarz,
S. et al, 2001; FEDESA, 2004), relativamente às vendas de antibióticos para animais.
A tabela seguinte indica a quantidade de antibióticos usados em Medicina Veterinária, em
alguns países da União Europeia, entre 1994 e 1997. Os dados relativos aos Países Baixos
foram apenas indicados de dois em dois anos e apenas em tratamento de grupo (Report and
Qualitative Risk Assessment by the Committee for Veterinary Medicinal Products, 1999).
Tabela 2. Utilização de antibióticos em diferentes Estados Membros (Kg ) .
1994 1995 1996 1997
Finlândia 17 500 18 197 17 286 16 591 Suécia 30 342 24 569 20 639 19 655
Dinamarca 78 584 40 172 47 454 53 511 Países Baixos 250 200 251500
24

Introdução
No que diz respeito ao volume de antibióticos consumidos por tipo de animal, o " Danish
Plant Directorate " publicou dados, relativos a 1995 ( Tabela 3).
Tabela 3. Consumo de promotores de crescimento na Dinamarca, em 1995 (Aarestrup, F.
M. et ai, 1998b).
Consumo de antibióticos ( Kg de substância activa)
Promotor de crescimento Total Suínos Gado bovino Aves de ca
Carbadox 1.181 1.181 0 0 Olaquindox 16.213 16.213 0 0 Avilamicina 1.665 265 0 1.4 Avoparcina 5.690 2.5 2.09 1.1 Bacitracina 7.910 6 0 610 Flavomicina 48 0 0 48 Monensina 5.007 0 5.007 0 Salinomicina 850 850 0 0 Espiramicina 507 0 0 507 Tilosina 52.275 52.275 0 0 Virginiamicina 2.590 1.5 0 1.09
Estima-se que 60% dos antibióticos utilizados nos animais se destinem a suínos, 20% a
frangos e coelhos, 18% a ruminantes, 1% a peixes e 1% a animais de estimação ( McEwan,
S.A.. et ai, 2002).
Apesar da escassez de dados, verifica-se um decréscimo na venda de antibióticos, quer de
agentes terapêuticos, quer de promotores de crescimento, na maioria dos Países Europeus
(Report and Qualitative Risk Assessment by the Committee for Veterinary Medicinal
Products, 1999).
Em 1997, a Comissão Europeia solicitou à FEDESA (European Federation of Animal
Health Industries) um relatório relativo às quantidades de antibióticos consumidos na
União Europeia. Para a obtenção desses dados foram realizados inquéritos a dezanove
membros desta federação, que representam 60% das vendas na União Europeia e na Suiça. 25

Introdução
Os resultados obtidos encontram-se na tabela 4 (Schwarz, S. et ai, 2001).
Tabela 4. Estimativa de vendas de antibióticos para animais, na União Europeia e Suíça em
1997. * Coccidiostáticos (ionóforos e outros) excluídos (Consumer Policy and Consumer
Health Protection, 1999; McEwan, S.A. et al, 2002).
Antibiótico Quantidade ( Toneladas)
Penicilinas 322 Tetraciclinas 2294 Macrólidos 424 Aminoglicosídeos 154 Fluoroquinolonas 43 Trimetoprim/Sulfonamidas 75 Outros 182
Subtotal Agentes terapêuticos 3494 Promotores de crescimento* 1599
Total 5093
A quantidade de antibióticos (excluindo coccidiostáticos) vendidos em cada país Europeu
encontra-se na tabela 5.
26
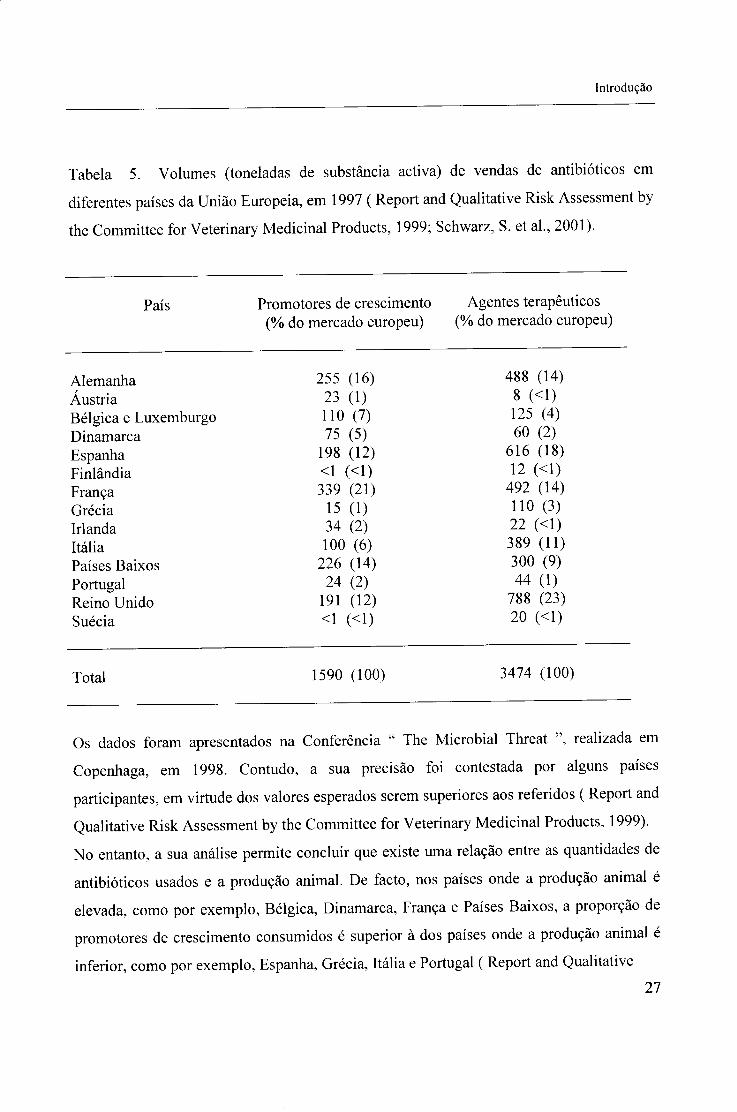
Introdução
Tabela 5. Volumes (toneladas de substância activa) de vendas de antibióticos em
diferentes países da União Europeia, em 1997 ( Report and Qualitative Risk Assessment by
the Committee for Veterinary Medicinal Products, 1999; Schwarz, S. et al., 2001).
País Promotores de crescimento Agentes terapêuticos (% do mercado europeu) (% do mercado europeu)
Alemanha 255 (16) 488 (14) Áustria 23 (1) 8 (<1) Bélgica e Luxemburgo 110 (7) 125 (4) Dinamarca 75 (5) 60 (2) Espanha 198 (12) 616 (18) Finlândia <1 (<1) 12 (<1) França 339 (21) 492 (14) Grécia 15 (1) 110 (3) Irlanda 34 (2) 22 (<1) Itália 100 (6) 389 (11) Países Baixos 226 (14) 300 (9) Portugal 24 (2) 44 (1) Reino Unido 191 (12) 788 (23) Suécia <1 (<1) 20 (<1)
Total 1590 (100) 3474 (100)
Os dados foram apresentados na Conferência " The Microbial Threat ", realizada em
Copenhaga, em 1998. Contudo, a sua precisão foi contestada por alguns países
participantes, em virtude dos valores esperados serem superiores aos referidos ( Report and
Qualitative Risk Assessment by the Committee for Veterinary Medicinal Products, 1999).
No entanto, a sua análise permite concluir que existe uma relação entre as quantidades de
antibióticos usados e a produção animal. De facto, nos países onde a produção animal é
elevada, como por exemplo, Bélgica, Dinamarca, França e Países Baixos, a proporção de
promotores de crescimento consumidos é superior à dos países onde a produção animal é
inferior, como por exemplo, Espanha, Grécia, Itália e Portugal ( Report and Qualitative 27

Introdução
Risk Assessment by the Committee for Veterinary Medicinal Products, 1999).
Apesar dos resultados serem baseados em estimativas, as comparações efectuadas
permitiram classificar os países em grupos, consoante a quantidade de antibióticos que
utilizam com fins terapêuticos e como promotores de crescimento. Os países em que o
consumo é mais elevado são o Reino Unido, a Grécia, a Espanha e os Países Baixos. Os
países que ocupam uma posição intermédia são a Bélgica, a França, a Itália, a Alemanha e
Portugal. Por último, os países onde o consumo é mais baixo são a Suécia, a Dinamarca e a
Finlândia (Schwarz, S. et ai, 2001).
1.2.2. Consumo de antibióticos em medicina humana
Não há muitos dados disponíveis sobre a utilização de antibióticos na maior parte dos
países da União Europeia (Cars, O. et ai, 2001). Alguns estudos revelam que 20% do
consumo total de antibióticos é realizado nos hospitais e os restantes 80% na comunidade.
No entanto, não há um consenso relativamente à quantidade de antibióticos consumidos, na
comunidade e nos hospitais, em virtude de poder haver discrepância entre os diversos
países (Report and Qualitative Risk Assessment by the Committee for Veterinary
Medicinal Products, 1999). Cars e colaboradores (Cars, O. et al, 2001) analisaram dados
sobre as vendas de antibióticos não hospitalares em quinze países da União Europeia, em
1997, de acordo com o sistema de classificação "Anatomic Therapeutic Chemical",
exprimindo o consumo em doses diárias por mil pessoas. Os países com maior consumo
eram França (36,5), Espanha (32,4), Portugal (28,8) e Bélgica (26,7), enquanto que os
Países Baixos (8,9), Dinamarca (11,3), Suécia (13,5) e Alemanha (13,6) apresentavam o
consumo mais baixo (Cars, O. et ai, 2001). Dados semelhantes foram referidos pelo
Comité de Medicina Veterinária ( Report and Qualitative Risk Assessment by the
Committee for Veterinary Medicinal Products, 1999).
No entanto, alguns autores consideram estes estudos pouco significativos em virtude de
não haver uma metodologia consensual para efectuar a comparação entre o consumo de
antibióticos nos diferentes países da União Europeia. Contudo, os dados disponíveis até
1999 permitem verificar que o consumo de antibióticos em medicina humana está a 28

Introdução
aumentar (Report and Qualitative Risk Assessment by the Committee for Veterinary
Medicinal Products, 1999).
Um estudo francês demonstrou um aumento de 3,7% do consumo de antibióticos entre
1981 e 1992. No Reino Unido, o aumento do consumo foi de 5%, entre 1989 e 1991
(Report and Qualitative Risk Assessment by the Committee for Veterinary Medicinal
Products, 1999).
1.2.3. Consumo humano versus consumo animal
É difícil comparar rigorosamente o consumo relativo de antibióticos entre o Homem e os
animais, na União Europeia. Para tal, seria necessário conhecer exactamente a dimensão da
população humana e animal em estudo. Devido à dificuldade de quantificar o consumo de
antibióticos, a maior parte dos dados disponíveis baseia-se em valores monetários
correspondentes às vendas anuais destas substâncias. Estes dados revelam que a quantidade
usada pelo Homem é bastante superior àquela que é usada nos animais. Todavia, o facto
dos antibióticos usados pelo Homem serem mais dispendiosos do que aqueles que são
usados nos animais, não permite uma clara avaliação das quantidades consumidas (Report
and Qualitative Risk Assessment by the Committee for Veterinary Medicinal Products,
1999). Da mesma forma, há parâmetros que são sub ou sobrestimados, em virtude de nem
todos os antibióticos vendidos para consumo humano são de facto consumidos (Schwarz,
S. et ai, 2001). Em 1990, nos Países Baixos, foram utilizados 80 OOOKg de antibióticos (substância activa)
no Homem e 300 OOOKg na medicina veterinária, ou seja, cerca de lOOmg de substâcia
activa /Kg de peso corporal / ano (van den Bogaard, A.E. et al, 2002).
Em termos gerais, a FEDESA forneceu dados que indicam que das 10 493 toneladas de
antibióticos consumidos em 1997, cerca de 52% foram usados em medicina veterinária e
15% na produção animal (Tabela 6) (Millet, A., 1998; Report and Qualitative Risk
Assessment by the Committee for Veterinary Medicinal Products, 1999).
29

Introdução
Tabela 6. Média anual de consumo de antibióticos pelo Homem e pelos animais, na União
Europeia, em 1997 (Millet, A., 1998; Report and Qualitative Risk Assessment by the
Committee for Veterinary Medicinal Products, 1999).
Toneladas de Consumo ingrediente activo com 0,
100 % de pureza
Medicina humana 5400 52
Medicina veterinária 3494 33 (fins terapêuticos)
Aditivos alimentares 1 ,QQ , <-(para animais)
Total 10493 100
1.3. Aparecimento e evolução da resistência
"The war against infectious diseases has been won."
(A guerra contra as doenças infecciosas foi ganha.)
Dr William Stuart, 1969
(Extraído de Persing, D. H., 1993)
Esta frase ilustra o orgulho da Ciência Médica, a qual estava no auge em 1969, devido ao
sucesso no tratamento de doenças infecciosas. Contudo, alguns insucessos nesses
tratamentos, indicaram desde então, que no milénio que se avizinhava, a situação poderia
ser oposta a esta. De facto, recentemente tem-se verificado o aparecimento de um elevado
30

Introdução
número de bactérias multiresistentes, as quais têm provocado um aumento da mortalidade
(Luh, K-T., 2000; Persing, D. H., 1993).
No entanto, pelo facto de terem sido isoladas bactérias resistentes a antibióticos em
glaciares no Canadá, com cerca de dois mil anos, admite-se que os genes e os mecanismos
de resistência já existiriam quando os antibióticos foram introduzidos na clínica (Anglada,
R. R, 1997). A forma como estes genes de resistência apareceram e porque razão levaram tanto tempo a
surgir, gera ainda alguma controvérsia. Alguns destes genes terão sofrido mutações a partir
de genes ancestrais da espécie, tendo posteriormente sido objecto de selecção natural.
Outros poderiam encontrar-se em bactérias ambientais e terem sido transferidos para
espécies animais (Anglada, R. R., 1997).
A existência dos genes de resistência está, igualmente, relacionada com a produção de
agentes antimicrobianos por organismos saprófitos, como por exemplo, Actinomycetes e
Streptomyces. Estes microrganismos possuem genes que codificam resistênca aos agentes
que produzem de forma a evitar a auto-destruição, constituindo uma fonte de genes de
resistência (Anglada, R. R., 1997; Lina, G. et ai, 1999).
Desta forma, pode concluir-se que a resistência aos antibióticos é, provavelmente, tão ou
mais antiga do que a sua síntese e que estes genes de resistência já estariam presentes no
ambiente, como o solo e a água, antes da utilização dos agentes antimicrobianos pelo
Homem (Lina, G. et ai, 1999). Embora com menor frequência do que actualmente, o
contacto entre os agentes antimicrobianos e os microrganismos já acontecia antes da era "
pré-microbiana", o que constituía uma pressão de selecção, levando ao desenvolvimento de
mecanismos de resistência a esses agentes ( Schwarz, S. et ai, 2001).
Contudo, o desenvolvimento de resistência aos antibióticos, a disseminação dos genes de
resistência e o aparecimento de novas estirpes resistentes ocorreu após a utilização
intensiva dos agentes antimicrobianos. Desde a sua introdução, houve um aumento da
prevalência da resistência aos antibióticos em bactéria patogénicas e comensais, no
Homem e nos outros animais (Report and Qualitative Risk Assessment by the Committee
for Veterinary Medicinal Products, 1999).
31

Introdução
Em 1944, alguns anos após ter descoberto a penicilina, Alexander Fleming verificou que
algumas estirpes de Staphylococcus aureus eram capazes de a inactivar. Todavia, só no
final dos anos 60 é que começaram a ser publicados estudos sobre bactérias resistentes a
diferentes antibióticos (van den Bogaard, A. E. et al, 1999). Tal como ocorreu com a
penicilina, alguns anos após a introdução da meticilina na clínica, foram sendo isoladas
em número crescente estirpes de
S. aureus resistentes a esta penicilina semi-sintética. Os doentes infectados por S. aureus
resistentes à meticilina (MRSA) passaram a ser tratados com vancomicina. Da mesma
forma, surgiram outras bactérias resistentes a este antibiótico, como por exemplo,
Enterococcus sp. Esta situação verificou-se, igualmente, para as fluoroquinolonas, que
foram introduzidas na clínica, nos anos oitenta, para tratar, entre outras, as infecções
provocadas por MRSA. No entanto, um ano após a sua introdução na clínica, verificou-se
que várias estirpes de MRSA tinham desenvolvido resistências a este antibiótico. De facto,
a aquisição de resistências aos antibióticos vulgarmente utilizados, tem sido observada
desde a sua introdução na medicina humana e veterinária, tendo-se constatado que cada
vez que um antibiótico é introduzido na clínica, as bactérias adaptam-se, mais ou menos
rapidamente (Tabela 7) (Aarestrup, F. M. et ai, 1998b, Acar, J. e Courvalin, P., 1998;
Fraimow, H.S. e Courvalin, P. 2000; van den Bogaard, A. E., et ai, 2002).
Tabela 7- Descoberta do antibiótico e desenvolvimento da resistência (DND-Dados não
disponíveis) ( Schwarz, S. et ai, 2001).
Antibiótico Descoberta Introdução na clínica
Identificação da Resistência
Penicilina 1940 1943 1940 (Meticilina 1961/5) Estreptogramina 1944 1947 1947,1956
Tetraciclina 1948 1952 1956 Eritromicina 1952 1955 1956 Vancomicina 1956 1972 1987
Ác. Nalidíxico 1960 1962 1966 Gentamicina 1963 1967 1970
Cefalosporinas de 3a geração DND 1980,1985 Fluoroquinolonas 1978 1982 1985
32

Introdução
O rápido aparecimento de estipes resistentes após a introdução de um novo antibiótico na
clínica demonstra a extrema capacidade de adaptação das bactérias a alterações do meio.
Na realidade, os microrganismos devem a sua existência à capacidade de se adaptarem às
mudanças. Uma grave consequência desta capacidade é, sem dúvida, o desenvolvimento de
resistências aos antibióticos ( Anglada, R. R., 1997; Martel, J. L. et ai, 2001; Schwarz, S. et
ai, 2001). Nos últimos anos as bactérias desenvolveram igualmente resistência a substâncias
sintéticas, o que confirma a sua extraordinária capacidade de resistirem às alterações
ambientais e de se desenvolverem na presença de substâncias tóxicas (Schwarz, S. et ai.,
2001).
A frequência da resistência aos antibióticos num microrganismo é determinada por uma
grande variedade de factores epidemiológicos, que facilitam o aparecimento de estirpes
resistentes e que promovem a sua transmissão. A conjugação destes factores é responsável
pela variação da incidência da resistência (Cohen, M. L., 1994). Muitas vezes os
antibióticos são consumidos de uma forma inadequada devido à dificuldade em determinar
se uma infecção é provocada por um vírus ou por uma bactéria. Em caso de dúvida, o
receio de não tratar uma infecção bacteriana leva, por vezes, o médico a prescrever um
antibiótico que não irá ter qualquer efeito se a infecção for provocada por um vírus. A
possibilidade de, em alguns países, se adquirirem facilmente antibióticos sem receita
médica, contribui para o aumento do consumo incorrecto, em doses inadequadas e, muitas
vezes, durante um período de tempo insuficiente. Da mesma forma, as débeis condições
sócio-económicas de alguns países, onde a pobreza e o excesso de população são
predominantes, contribuem para o aparecimento e propagação de bactérias resistentes. Por
outro lado, o aumento do número de viagens e a globalização do mercado dos produtos
alimentares cria condições que levam à rápida propagação de bactérias transmitidas pela
cadeia alimentar (Consumer Policy and Consumer Health Protection, 1999; JOCE, n° C,
407/7; WHO, 2000a; WHO, 2000b).
33

Introdução
A presença de sequências de DNA portadoras de genes de resistência em antibióticos
comercializados, permite, igualmente, que as bactérias tenham um acesso extremamente
facilitado ao reservatório de resistências, que é o ambiente (Boisseau, J., 1993; Corpet, D.
E., 1993).
As características farmacocinéticas das diferentes classes de antibióticos podem favorecer
o desenvolvimento de resistências, assim como a concentração de substância activa, a via
de excreção desse agente e a via de administração. Os antibióticos que são absorvidos em
quantidades reduzidas no tracto gastrointestinal podem exercer uma pressão de selecção
considerável nas bactérias intestinais. Por seu lado, a terapia sistémica ou tópica pode
permitir o desenvolvimento de espécies da flora normal da pele resistentes ao antibiótico e
a sua disseminação. Quanto mais prolongada for a terapia, mais provável é a alteração da
flora normal, a colonização e a superinfecção por microrganismos resistentes (Boisseau, J.,
1993; Corpet, D. E., 1993).
Um outro factor que contribui para o aparecimento de resistências no Homem relaciona-se
com a ingestão de antibióticos através dos alimentos. Estes agentes, quando usados como
promotores de crescimento, podem deixar resíduos na carne ou no leite, os quais, se
conservarem, pelo menos em parte, a sua actividade, podem exercer pressão de selecção e
favorecer o aparecimento de resistências. De um modo geral, a quantidade de resíduos é
praticamente nula quando o antibiótico é administrado como promotor de crescimento, ou
seja, em doses subterapêuticas. No entanto, quando o antibiótico é utilizado com fins
terapêuticos, a dose administrada é mais elevada e, por essa razão, poderá ser absorvido
pelo consumidor. Apesar de existir um intervalo de segurança, legalmente estabelecido, até
que o animal seja abatido, se este não for cumprido, vão permanecer resíduos do
antibiótico no alimento, os quais podem ser transferidos para o consumidor (Boisseau, J.,
1993; Corpet, D. E., 1993; Dayan, A. D., 1993).
No sentido de proteger a saúde pública, nos Estados Unidos da América, a FDA (Food and
Drug Administration), assim como o USDA (United States Department of Agriculture),
cooperam num programa para controlar o uso de antibióticos nos animais, evitando o seu
uso ilegal. Da mesma forma, na União Europeia são tomadas medidas legais no sentido de
proteger a saúde pública, através de legislação que estabelece os limites máximos de
34

Introdução
resíduos de medicamentos veterinários permitidos em alimentos de origem animal e cuja
última alteração data de 4 de Março de 1999. Se estes limites de resíduos forem respeitados
não existe qualquer risco para a saúde pública. O único risco para a saúde verificar-se-ia se
fossem ingeridas grandes quantidades de alimentos, cujos níveis de resíduos fossem muito
superiores aos legais (Corpet, D. E., 1993).
Com efeito, o consumo prolongado de doses subterapêuticas é visto como um dos
principais factores responsáveis pelo desenvolvimento de resistências, exercendo uma
pressão de selecção intensa, permitindo o aumento do número de bactérias resistentes já
existentes na população bacteriana (Consumer Policy and Consumer Health Protection,
1999; JOCE, n° C de 28 de Dezembro de 1998 ; McEwan, S.A.. et al, 2002; Martel, J. L. et
al, 2001; Report and Qualitative Risk Assessment by the Committee for Veterinary
Medicinal Products, 1999; Schwarz, S. et al, 2001).
1.3.1. Consequências da utilização dos antibióticos
A utilização de agentes antimicrobianos na profilaxia, na terapia e como promotores de
crescimento tem como consequência o aparecimento de microrganismos resistentes, não só
entre bactérias patogénicas, mas também entre bactérias da flora endógena normal dos
animais expostos a estas substâncias (dei Campo, R. et ai, 2003; Tollefson, L. et ai, 1997;
van den Bogaard, A. E., et al, 1999; van den Bogaard, A. E., et al, 2002). Da mesma forma,
a utilização de antibióticos na medicina humana contribui para o aumento das resistências
nas bactérias patogénicas e comensais da flora bacteriana no Homem, em virtude dos
antibióticos não actuarem apenas nas bactérias alvo, mas em todas as populações de
bactérias expostas a essas substâncias (Aarts, H. J. M. et ai, 2001; Consumer Policy and
Consumer Health Protection, 1999; van den Bogaard, A. E., et al, 1999). A diminuição
simultânea de susceptibilidade de uma bactéria a três ou mais classes de antibióticos,
através de mecanismos de resistência é designada por multiresistência (Piddock, L. J. V.,
1996).
35

Introdução
Devido à intensa utilização dos agentes antimicrobianos na medicina humana, na medicina
veterinária e como promotores de crescimento, as bactérias vão acumulando genes de
resistência. Desta forma, o progressivo aumento de bactérias resistentes reflète a adaptação
destes organismos a ambientes poluidos por antibióticos. Apesar da maior parte dos
antibióticos utilizados pelo Homem não serem usados nos animais, a resistência cruzada
com antibióticos usados nos animais pode ocorrer, em virtude das moléculas estarem
estruturalmente relacionadas (Piddock, L. J. V., 1996; Report and Qualitative Risk
Assessment by the Committee for Veterinary Medicinal Products, 1999). Está igualmente
demonstrado que um único antibiótico é capaz de seleccionar a resistência a antibióticos
quimicamente não relacionados (Piddock, L. J. V., 1996).
Dado que as bactérias resistentes ocorrem no Homem, nos animais, na vegetação e no meio
ambiente, terá de existir um grande reservatório de genes de resistência acessível a todas as
bactérias, as quais estão expostas a uma pressão de selecção, devido à utilização de
antibióticos nos hospitais, na veterinária e na produção animal (Report and Qualitative
Risk Assessment by the Committee for Veterinary Medicinal Products, 1999).
Tanto no Homem, como nos animais, as bactérias saprófitas são abundantes na pele e nas
mucosas, constituindo a flora bacteriana comensal, a qual se mantém, geralmente,
constante, desde que a saúde do hospedeiro e as condições ambientais permaneçam
estáveis. Cada área colonizada constitui um nicho ecológico particular, onde se verifica um
equilíbrio entre as bactérias e o hospedeiro. A flora bacteriana intestinal é a mais
abundante e possui um papel de extrema importância no equilíbrio da saúde humana e
animal (Levy, S. B., 1998; Nord, C. E., 1993; Report and Qualitative Risk Assessment by
the Committee for Veterinary Medicinal Products, 1999).
Em condições normais, nos omnívoros e carnívoros, a acidez gástrica constitui uma
barreira para os microrganismos ingeridos juntamente com a saliva ou alimentos. Desta
forma, o conteúdo estomacal é geralmente estéril. Pelo contrário, o cólon possui uma flora
complexa (Levy, S. B., 1998; Report and Qualitative Risk Assessment by the Committee
for Veterinary Medicinal Products, 1999).
36

Introdução
A flora intestinal é constituída por mais de 200 espécies de bactérias. Devido ao baixo
potencial redox, os anaeróbios estritos excedem os aeróbios e os anaeróbios facultativos,
na razão de 1:1 000. No Homem, as bactérias anaeróbias mais comuns são Bifibobacterium
(Gram positivo) e Bacteróides (Gram negativo), enquanto que Esherichia coli (Gram
negativo) é o anaeróbio facultativo mais comum. As bactérias do género Enterococcus
(Gram positivo) são os microrganismos anaeróbios facultativos mais abundantes (Levy, S.
B., 1998; Report and Qualitative Risk Assessment by the Committee for Veterinary
Medicinal Products, 1999).
No caso dos frangos, os quais se encontram no âmbito deste estudo, a composição da flora
gastrointestinal varia com a idade. Devriese (Devriese, L. A., 1991) verificou que são
várias as espécies de Enterococcus que se encontram a constituir a flora gastrointestinal
destas aves, assim como algumas espécies do género Streptococcus, contudo as mais
comuns são Enterococcus faecium e Enterococcus faecalis, conforme se pode observar na
tabela 8.
Tabela 8. Percentagem de isolados de Enterococcus sp. e de Streptococcus sp. em frangos
com diferentes idades ( Devriese, L.A., et ai, 1991).
Idade
Espécie 1 dia 3-4 semanas > 12 semanas Enterococcus % % %
faecium 41 46 10 faecalis cecorum
41 0
3 7
10 50
hirae 0 10 1 durans 2 8 0 gallinarum avium
0 0
1 0
0 1
mundii 3 0-5 1 casseliflavus 0 1 0
Streptococcus alactolyticus bovis
2 0
21 0
21 1

Introdução
A flora intestinal desempenha importantes funções fisiológicas no organismo. Nos
herbívoros, a função fundamental é a digestão da celulose e, em alguns casos, constituem
uma fonte de vitaminas e proteínas para o hospedeiro. Em todos os animais, incluindo o
Homem, a microflora afecta a morfologia da mucosa intestinal, participa no metabolismo
dos carbohidratos e participa na produção de vitaminas. A microflora constitui, igualmente,
uma barreira à colonização por bactérias exógenas (Levy, S. B., 1998; Report and
Qualitative Risk Assessment by the Committee for Veterinary Medicinal Products, 1999).
A flora intestinal é determinada geneticamente, mas é influenciada pelos hábitos
alimentares. No entanto, mesmo quando os hábitos alimentares sofrem alterações durante
algum tempo, a sua composição mantém-se estável e a ingestão de bactérias exógenas
através dos alimentos não provoca uma colonização permanente, mantendo-se no intestino
apenas alguns dias ou semanas (Levy, S. B., 1998; Report and Qualitative Risk
Assessment by the Committee for Veterinary Medicinal Products, 1999).
Uma das consequências da utilização de antibióticos é a alteração da constituição da flora
intestinal (Baquero, F., 1997; Boisseau, J., 1993; Corpet, D. E., 1993).
Os antibióticos podem reduzir o número de bactérias e alterar a proporção entre as
diferentes espécies (Baquero, F., 1997; Boisseau, J., 1993; Corpet, D. E., 1993). Esta
modificação conduz à diminuição da resistência à colonização, aumentando o risco de
aquisição de bactérias exógenas resistentes (Huycke, M. M., 1998; Report and Qualitative
Risk Assessment by the Committee for Veterinary Medicinal Products, 1999). Por outro
lado, a morte das bactérias susceptíveis ao antibiótico cria um vazio no intestino, que pode
ser preenchido pelo supercrescimento ou pela aquisição de bactérias e de leveduras,
resistentes ao antibiótico. Quando a resistência à colonização diminui, a dose infecciosa
necessária para provocar doença, também é menor e os animais excretam um maior
número de bactérias e por um período de tempo mais longo, quando comparado com os
animais cuja resistência à colonização não sofreu alteração. Por esta razão, a disseminação
de bactérias resistentes aumenta, assim como o risco de contaminação da carne durante o
abate (van den Bogaard, A. E. et al, 1999).
38

Introdução
1.3.2. Impacto da utilização de antibióticos em produção animal na resistência
bacteriana de bactérias potencialmente patogénicas para o Homem
O facto das bactérias terem acumulado vários genes de resistência a diferentes antibióticos
compromete o tratamento de doenças infecciosas, quer no Homem, quer nos animais,
tornando muito difícil encontrar tratamentos eficazes (Consumer Policy and Consumer
Health Protection, 1999; Jornal Oficial da Comunidades Europeias, N° C, 407/7 de 28 de
Dezembro de 1998).
São vários os exemplos das implicações da resistência na terapia. O primeiro surto bem
documentado envolvendo bactérias multiresistentes relacionou-se com a febre tifóide
provocada por Salmonella typhi, que ocorreu no México em 1970, com mais de 10 000
casos casos confirmados em 1972. A estirpe envolvida possuía genes de resistência ao
cloranfenicol (antibiótico utilizado à época, no combate a esta doença), à ampicilina, à
estreptomicina e à sulfonamida (Consumer Policy and Consumer Health Protection, 1999;
WHO, 1998). Recentemente, têm sido descritas enterobactérias patogénicas como
Salmonella sp., Campylobacter sp. e E. coli resistentes também às fluoroquinolonas
(Consumer Policy and Consumer Health Protection, 1999).
Nos hospitais verificou-se um aumento de infecções provocadas por bactérias Gram-
positivo, particularmente 5". aureus resistentes à meticilina, por Staphylococcus coagulase
negativo e por Enterococcus faecium resistentes à ampicilina e à vancomicina (Firth, N. e
R. A . Skurray, 1998; Luh,K-T., 2000; Report and Qualitative Risk Assessment by the
Committee for Veterinary Medicinal Products, 1999).
Isolados de S. aureus resistentes à meticilina, os quais apresentam também resistência a
todas as penicilinas, cefalosporinas e carbapenemos são, frequentemente, também
resistentes a outros antibióticos, especialmente, macrólidos, quinolonas, aminoglicosídeos,
lincosaminas e trimetropim-sulfametoxazol. Por esta razão, os glicopéptidos, vancomicina
e teicoplanina constituem antibióticos de última escolha para o tratamento de infecções
provocadas por estas estirpes (Report and Qualitative Risk Assessment by the Committee
for Veterinary Medicinal Products, 1999).
39

Introdução
Em Portugal, segundo um estudo realizado em 1994, 47% dos S. aureus eram resistentes à
meticilina, sendo a percentagem de S. epidermidis e de S. haemolyticus superior. De todos,
apenas a vancomicina era activa contra estas estirpes (Cristino, J. M. et ai, 1996). Em
1997, foram detectadas nos Estados Unidos da América, estirpes de S. aureus com um
nível intermédio de resistência à vancomicina (Chang, S. et al, 2003; Report and
Qualitative Risk Assessment by the Committee for Veterinary Medicinal Products, 1999;
Smith, T. L. et al, 1999) e em 2002, o primeiro isolado de S. aureus resistente à
vancomicina por aquisição de van A, o que constitui uma situação alarmante (Chang, S. et
al, 2003; Tenover, F. C. et al, 2004)
Da mesma forma, os Enterococcus sp. tornaram-se uma das mais importantes causas de
infecções nosocomiais. Para além de possuirem resistência intrínseca a muitos antibióticos,
como por exemplo, cefalosporinas, oxacilina, clindamicina e aminoglicosídeos em baixas
concentrações, certas estirpes desenvolveram resistência a todos os antibióticos, incluindo
penicilinas, aminoglicosídeos, em altas concentrações, e glicopéptidos, causando sérios
problemas na clínica humana (Bozdogan, B. et ai, 1999).
Tabela 9 - Multiresistência em E. faecium (Report and Qualitative Risk Assessment by the
Committee for Veterinary Medicinal Products, 1999).
Resistência intrínseca Resistência adquirida
[3-lactâmicos p-lactâmicos (principalmente Aminoglicosídeos cefalosporinas) Fluoroquinolonas Aminoglicosídeos Glicopéptidos(vancomicina e Fluoroquinolonas teicoplanina) Trimetroprim-sulfametoxazol Tetraciclinas Clindamicina Eritromicina
Cloranfenicol Ácido fusídico Nitrofurantoína
40

Introdução
No que diz respeito a Portugal, em 1994, E.faecalis e E.faecium apresentavam resistência
a alguns antibióticos e todas as estirpes resistentes à vancomicina eram também resistentes
a todos os outros antibióticos (Tabela 10).
Tabela 10 - Percentagem de E. faecalis e E. faecium resistentes a diferentes antibióticos
(Cristino, J. M. et ai, 1996).
Bactéria E. faecalis E. faecium Antibiótico
Ampicilina 2% 70%
Estreptomicina 35% 42%
Gentamicina 45% 59%
Vancomicina 1% 9%
Durante os anos 60, o aumento de estirpes de Salmonella resistentes, isoladas em vitelos
para consumo humano, levou à criação no Reino Unido, do Joint Committee on the Use of
Antibiotics in Animal Husbandry and Veterinary Medicine. Num relatório emitido em
1969 esta comissão reconheceu que a administração de antibióticos em doses
subterapêuticas tinha consequências negativas para a saúde humana e animal. Por esta
razão, foram feitas várias recomendações sobre a utilização de antibióticos. Desta forma, a
utilização da penicilina e da tetraciclina como promotores de crescimento, foi proibida.
Estes antibióticos, juntamente com as sulfonamidas, passaram a ser autorizados apenas na
terapia animal e sob prescrição de um médico veterinário. Desde então, os antibióticos
fornecidos aos animais como promotores de crescimento são moléculas diferentes das dos
antibióticos utilizados na terapia humana ( Aarestrup, F. M. et al, 1998b; Consumer Policy
and Consumer Health Protection, 1999; Martel, J.L., 2001; Piddock, L. J. V.,1996;
Schwarz, S. et al, 2001; Witte, W., 1998). 41

Introdução
Estas preocupações levaram a União Europeia a estabelecer uma utilização racional dos
antibióticos, quer como agentes terapêuticos, quer como promotores de crescimento.
Da mesma forma, a World Health Organization (WHO) criou uma comissão para definir
regras na vigilância do desenvolvimento de resistências em bactérias e a sua possível
transferência para o Homem. Para tal, será necessário standardizar a metodologia de
vigilância (Martel J.L., 2001).
Em consequência de tais preocupações, desde os anos 70 têm surgido vários estudos sobre
o desenvolvimento de bactérias multiresistentes (Martel J.L., 2001).
Até que ponto a utilização dos antibióticos na produção animal contribui para o
aparecimento de bactérias resistentes no Homem, permanece em debate há mais de trinta
anos na Europa (Martel, J.L., 2001). Apesar de existirem vários estudos, o conhecimento
científico actual é um pouco ambíguo, não sendo ainda possível identificar todos os
factores que intervêm na selecção e transmissão de resistência. A obtenção de informação
para confirmar e quantificar a relação entre a utilização dos antibióticos e a incidência de
resistência poderá demorar vários anos, pelo que este assunto gera bastante controvérsia
(Consumer Policy and Consumer Health Protection, 1999). Seja qual for a conclusão deste
debate, sabe-se que a vulgarização da utilização dos antibióticos contribuiu para a selecção
de bactérias resistentes aos agentes antimicrobianos. A pressão de selecção exercida pelos
antibióticos contribui para uma rápida aquisição de resistências através de mutações e de
troca de determinantes genéticos, nomeadamente, transposões e integrões. De facto, o
aumento da incidência de bactérias resistente é contemporânea da intensificação da
utilização dos agentes antimicrobianos e é, no início do terceiro milénio, uma ameaça para
a saúde humana e animal ( Hayes, J. R. et ai, 2001; Martel, J.L., 2001).
A associação quinupristina-dalfopristina, tal como a virginiamicina, é uma mistura de duas
pristinamicinas, que actuam sinergicamente (van den Bogaard, A. E. et al, 2002). A
associação quinupristina-dalfopristina, cuja utilização foi aprovada nos EUA em 1999, é
utilizada como agente terapêutico em infecções provocadas por bactérias multiresistentes,
como Staphylococcus aureus resistentes à meticilina e de Enterococcus faecium resistentes
à vancomicina (Hayes, J. R. et ai, 2001; Jensen, L.B., 2002; Solway, S. et ai, 2003).
42

Introdução
Nos EUA a virginiamicina foi utilizada pontualmente no tratamento de infecções
provocadas por Staphylococcus sp. e apenas em alguns estados, mas é utilizada há de mais
de 25 anos, como promotor do crescimento de frangos, tendo sido utilizada também na
Europa, para este fim, durante muitos anos (Solway, S. et ai, 2003; van den Bogaard, A.E.
et al, 2002). No entanto, antes da associação quinupristina-dalfopristina ser utilizada na Europa, na
terapia humana, a resistência a esta associação já era observada em bactérias da flora
intestinal dos frangos, razão pela qual se admitia que tal resistência teria de ser provocada
pela utilização da virginiamicina como promotor de crescimento ( van den Bogaard, A. E.
et ai, 2002). Da mesma forma, admite-se que elevada utilização da virginiamicina na
produção animal poderá ter contribuido para o aparecimento de bactérias Gram-positivo
patogénicas resistentes à associação quinupristina-dalfopristina, em isolados de origem
humana (Hayes, J. R. et ai, 2001; Jensen, L.B., 2002; Simjee, S., 2002a; Simjee, S.,
2002b), o que compromete a sua utilização na clínica (Solway, S. et ai, 2003). Por esta
razão, a utilização da virginiamicina como promotor de crescimento, foi proibida na
Europa em 1999, mas continua a ser utilizada nos EUA (Hayes, J. R. et ai, 2001; Jensen,
L.B., 2002; Regulamento (CE) N° 2821/98 do Conselho, de 17 de Dezembro de 1998;
Simjee, S., 2002a; Simjee, S.2002b; Solway, S. et ai, 2003).
Com a proibição da utilização da virginiamicina na Europa, verificou-se um decréscimo no
número de isolados de Enterococcus faecium resistentes à associação quinupristina-
dalfopristina, com origem em frangos. Com efeito, os estudos realizados na Dinamarca,
onde a virginiamicina não é utilizada desde 1998, demonstraram que em 1997 a incidência
de Enterococcus faecium, isolados de frangos e de suínos, resistentes à virginiamicina era
de 66%, enquanto que em 2000 a incidência era de 34% (Simjee, S., 2002a; Solway, S. et
ai, 2003).
Apesar da incidência da resistência ter diminuido, existiam ainda Enterococcus faecium
resistentes à virginiamicina. Uma possível explicação será o facto de outros agentes
antimicrobianos, como a eritromicina, a tetraciclina e a estreptomicina co-seleccionarem os
isolados resistentes à virginiamicina (Simjee, S., 2002b).
43

Introdução
Uma situação homóloga desta é a que se verifica para a eritomicina e a tilosina. A tilosina
é um antibiótico que foi utilizado durante muito tempo como promotor de crescimento dos
frangos.Vários estudos têm demonstrado que a utilização da tilosina como promotor de
crescimento, está relacionada com o aumento de enterococos resistentes à tilosina na flora
intestinal de frangos e que o número de Enterococcus faecium relativamente aos
enterococos totais também aumentou. Isto deve-se ao facto da eritromicina apresentar
resistência cruzada com a tilosina ( van den Bogaard, A.E. et al, 2002). Por seu lado, os E.
faecium resistentes aos glicopéptidos (GRE) foram detectados pela primeira vez na Europa,
em 1986. Estudos desenvolvidos na Europa admitem que a existência de isolados de GRE
na comunidade se deve à utilização da avoparcina, um glicopéptido utilizado durante mais
de vinte anos, como promotor de crescimento dos frangos e de suínos, estruturalmente
relacionado com a vancomicina. A vancomicina, utilizada em medicina humana desde os
anos cinquenta, possui actividade contra bactérias em fase de crescimento, inibindo a
síntese do peptidoglicano (Anglada, R. R., 1997; Fraise, A. P., 1997; Kirst, H. A. et ai,
1998; Maniatis, A.N. et ai, 2001). Este antibiótico tem ainda a capacidade de inibir a
síntese de RNA e de alterar a permeabilidade da membrana citoplasmática da célula
bacteriana (Anglada, R. R., 1997).
Estudos realizados na Dinamarca e na Noruega permitiram observar que isolados de
enterococos resistentes à vancomicina possuíam resistência cruzada à avoparcina. Desta
forma, admite-se haver uma relação entre a utilização da avoparcina e a selecção de
enterococos resistentes à vancomicina (Khachatourians, G., 1998). Nos Estados Unidos da
América, onde a avoparcina nunca foi utilizada como promotor de crescimento, não foram
detectados Enterococcus sp. resistentes à vancomicina, na flora intestinal dos animais para
consumo. Contudo, o número de doentes hospitalizadoss portadores de Enterococcus sp.
resistentes à vancomicina (VRE) é superior ao observado na Europa. Este facto dever-se-á
à maior utilização dos glicopéptidos nos hospitais dos EUA do que nos hospitais europeus
(Cook, R R, 1997; Fraise, A. P., 1996; Maniatis, A. N. et ai, 2001; McDonald, L. C. et ai,
1997; Shlaes, D. M. e L. B. Rice, 1994; Wegener, H. C. 1998). Desde que, em 1997 a
avoparcina deixou de ser utilizada como promotor de crescimento na Europa ( Directiva
97/6/CE da Comissão de 30 de Janeiro de 1997), estudos realizados em algumas regiões
44

Introdução
alemãs constataram um decréscimo no número de doentes portadores de VRE, de 12% em
1994 para 6% em 1996 e 3% em 1997 ( Sorum, H. e Sunde, M, 2001). Tem sido também
sugerido que a ligação entre os genes de resistência à vancomicina e à eritromicina em
enterococos isolados em animais de produção pode ser importante na co-selecção ou na
manutenção da resistência aos glicopéptidos, quando os macrólidos são usados na terapia
ou como promotores de crescimento, em virtude da resistência simultânea a estes dois
agentes ser frequentemente observada nestes isolados (Lim, J-A. et ai, 2002; Maniatis, A.
N. et ai, 2001; Sorum, H. e Sunde, M, 2001). Na Grécia, o aparecimento de GRE foi mais
tardio do que nos outros países da Europa. Tal facto poderá estar relacionado com uma
menor utilização da avoparcina como promotor de crescimento neste país, relativamente à
sua utilização nos restantes países europeus (Maniatis, A. N. et ai, 2001). Desta forma,
aguardar até que haja confirmação científica para actuar seria perder uma oportunidade de
intervir ( Consumer Policy and Consumer Health Protection. 1999). Por essa razão, o tipo
de moléculas utilizadas como promotores de crescimento e as condições da sua utilização
encontram-se legisladas, sendo especificado os animais alvo, a duração da sua utilização e
a dosagem. Na Europa, os antibióticos (3-lactâmicos e as tetraciclinas não são utilizadas
desde 1975, sendo, no entanto, utilizadas nos EUA (Schwarz, S. et ai, 2001).
Da mesma forma, o Conselho de Ministros da Agricultura decidiu retirar a autorização da
utilização da bacitracina-zinco (um polipeptídeo), da espiramicina e do fosfato de tilosina
(antibióticos do grupo dos macrólidos) e da virginiamicina (uma estreptogramina), como
promotores de crescimento, em Dezembro de 1998 (Regulamento (CE) N°2821/98 do
Conselho, de 17 de Dezembro de 1998). Anteriormente, em 1997, foi retirada a
autorização da utilização da avoparcina (um glicopeptido), como promotor de crescimento
(Directiva 97/6/CE da Comissão de 30 de Janeiro de 1997). Foi igualmente retirada a
autorização da utilização do carbadox e do olaquindox, em Dezembro de 1998, devido aos
possíveis efeitos tóxicos destas substâncias (Regulamento (CE), n° 2788/98, da Comissão
de 22 de Dezembro de 1998; Schwarz, S. et ai, 2001).
Actualmente as moléculas cuja utilização é permitida são o flavofosfolipol, a monensina-
sódio, a salinomicina-sódio e a avilamicina, porque não têm qualquer relação estrutural
45

Introdução
com os agentes antimicrobianos utilizados na medicina humana e veterinária ( Martel, J.L.,
2001; Schwarz, S. et ai, 2001; Regulamento (CE) N° 2821/98 do Conselho de 17 de
Dezembro de 1998).
O flavofosfolipol e a avilamicina têm uma actividade antimicrobiana, enquanto que as
outras duas moléculas são utilizadas devido aos seus efeitos coccidiostáticos (Schwarz, S.
et ai, 2001).
Com estas medidas admite-se que os antibióticos promotores do crescimento animal não
terão como consequência o desenvolvimento de resistência cruzada com os antibióticos
usados em medicina humana (Schwarz, S. et ai, 2001).
Contudo, num parecer do Comité Científico Director, de 28 de Maio de 1999, admite-se
que a utilização, como promotores de crescimento, de agentes antimicrobianos
pertencentes a categorias que são ou que possam vir a ser usados em medicina humana e
veterinária deve ser abolida, devido ao risco de selecção de resistência cruzada aos
medicamentos usados no tratamento de infecções bacterianas (Comunicação do Conselho),
Posição comum (CE) N°20/2003, adoptada pelo Conselho em 17 de Março de 2003).
Num segundo parecer do Comité Científico Director sobre a resistência antimicrobiana,
adoptado em Maio de 2001, foi confirmada a necessidade de planificar e coordenar o
processo de exclusão progressiva destas substâncias. Desta forma, a monensina-sódio, a
salinomicina-sódio, o flavofosfolipol e a avilamicina serão suprimidas a partir de 1 de
Janeiro de 2006 (Proposta de Regulamento do Parlamento Europeu e do Conselho relativa
aos aditivos destinados à alimentação animal, Bruxelas 22/03/02,COM (2002) 153 final,
2002/2003 (COD)).
1.4. Género Enterococcus
1.4.1. Principais características
O género Enterococcus foi descrito pela primeira vez em 1899 como um novo
Streptococcus de origem entérica.
46

Introdução
Os enterococos são cocos Gram positivo cujas células são esféricas ou ovóides com um
diâmetro inferior a 2 um. Estas células podem ocorrer em pares ou em pequenas cadeias
(Lu, H.-Z. et ai, 2002; Fackland, R. R. et ai; Ostrowsky, B. et ai, 1999; Hancock, L. E. e
M. S. Gilmore, 2000). Através da coloração de Gram assemelham-se a Streptococcus,
razão pela qual foram inicialmente classificados como Streptococcus do grupo D
(Ostrowsky, B. et ai, 1999). Contudo, estudos posteriores demonstraram que as bactérias
do género Enterococcus diferem o suficiente das do género Streptococcus , nomeadamente
no que diz respeito à sequência de ácidos nucleícos, de modo a merecerem a sua
classificação num género distinto (Hancock, L. E. e M. S. Gilmore, 2000; Ostrowsky, B.
et ai, 1999). Existem, pelo menos, doze espécies pertencentes ao género Enterococcus. Estão descritas
infecções por E. avium, E. durans, E. raffinosus, E. gallinarum e E. casseliflavus. Contudo,
E. faecalis e E. faecium são as espécies que mais frequentemente provocam infecções
(Ostrowsky, B. et ai, 1999; Joyanes, P et ai, 2000). São organismos anaeróbios facultativos
cuja temperatura óptima de crescimento é de cerca de 35°C, podendo a maior parte das
estirpes crescer entre os 10 e os 45°C (Lu, H.-Z. et ai, 2002 ; Fackland, R. R. et ai).
As bactérias do género Enterococcus podem ser encontrados em quase todos os ambientes,
conseguindo sobreviver nos solos, na água, nos alimentos e nos animais, nomeadamente,
insectos e aves (Hancock, L. E. e M. S. Gilmore, 2000; Lu, H.-Z. et ai, 2002, Fackland, R.
R. et ai). Os enterococos constituem também o componente principal da microflora de
queijos artesanais, produzidos no Sul da Europa. Estas bactérias têm um papel fundamental
na maturação e no desenvolvimento de sabor nos queijos ( Franz, C.M.A.P. et ai, 2001).
Estas bactérias fazem igualmente parte da flora gastrointestinal normal humana e animal
(Lu, H.-Z., 2002; Ostrowsky , B., Eliopoulos, G., 1999; Simjee, S. et ai, 2002). Podem ser
também encontrados na mucosa bucal, na vagina, em úlceras crónicas, em lesões dos
tecidos moles, no tracto hepatobiliar e no sistema nervoso central (Lu, H.-Z., 2002;
Ostrowsky, B., Eliopoulos, G., 1999; Descheemaeker, P. R. M. et ai, 1999; Fackland, R.
R., et ai)
47

Introdução
1.4..2. Significado clínico
As bactérias do género Enterococcus não são patogénicos animais de grande importância.
Contudo, no Homem são responsáveis por várias infecções.( van den Bogaard, A.E. et al,
2002)
E.faecalis é um comensal saprófita que ocorre na flora gastrointestinal humana e de outros
animais, embora se possa também comportar como um patogénico oportunista. Esta
espécie do género Enterococcus domina no que diz respeito às infecções humanas (85%)(
Franz, C.M.A.P. et ai, 2001; Ostrowsky , B. e Eliopoulos, G., 1999), sendo E. faecium
(10%) a outra espécie mais frequentemente isolada (Lu, H.-Z. et ai, 2002; Simjee, S.,
2002a).
Nos últimos anos os enterococos tornaram-se microrganismos patogénicos responsáveis
por um grande número de infecções nosocomials, apesar da sua baixa virulência. A
multiresistência entre estes microrganismos é comum e representa um grave problema no
tratamento de infecções tendo-se tornado, nos últimos anos, importantes patogénicos
nosocomials (Bozdogan, B. et ai, 1999; dei Campo, R., 2003; Donabedian, S.M. et ai,
2003; Eisner, H.-A., et ai. 2000; Franz, C.M.A.P. et ai, 2001; Joyanes, P. et ai, 2002;
Portillo, A. et ai, 2000; Toledo-Arana, A. et ai, 2001; Torell, E. et ai, 1999).
Estes microrganismos são os principais responsáveis por infecções urinárias e da região
abdominal. A bacteremia é o terceiro tipo de infecção mais comum. Estas infecções
ocorrem principalmente nos doentes mais idosos e em doentes imunocomprometidos, com
hospitalização prolongada e sujeitos a uma terapia por antibióticos. Apesar da endocardite
ser uma infecção por Enterococcus sp grave, é menos comum do que a bacteremia
(Bischoff, W. E. et ai, 1999; Fackland, R. R., et ai; Klein, G. et ai, 1998).
As bactérias do género Enterococcus possuem resistência intrínseca a muitos antibióticos,
nomeadamente, às cefalosporinas, penicilinas resistentes às penicilinases,
aminoglicosídeos e aos níveis uilizados na clínica de lincosamidas ( van den Bogaard,
A.E. et ai, 2002). Conseguiram, ainda, ao longo do tempo, adquirir resistência a uma
grande variedade de antibióticos, nomeadamente, resistência de alto nível a
aminoglicosídeos, macrólidos, estreptograminas, penicilina e glicopéptidos (Bozdogan, B.
48

Introdução
et al, 1999; Simjee, S., 2002a ; Singh, K. V.et al, 2002; Toledo-Arana, A. et al, 2001). Por
esta razão os enterococos da flora entérica são indicadores da pressão selectiva a que os
animais de produção estão sujeitos (Aarestrup, F. M. et ai, 1998; Simjee, S., 2002a).
A multiresistência aos antibióticos apresentada por estes microrganismos constitui um
grave problema no tratamento das infecções que provocam, tendo-se tornado um problema
de saúde pública (Donabedian, S.M. et ai, 2003; Portillo, A. et ai, 2000 ).
O aparecimento de Enterococcus sp. como patogénicos nosocomials pode estar
relacionado com o aumento de doentes imunocomprometidos e à sua exposição aos
antibióticos a que as bactérias do género Enterococcus são intrinsecamente resistentes,
como as cefalosporinas de terceira e quarta geração (van den Bogaard, A.E. et al, 2002).
A resistência intrínseca destes microrganismos a muitos antibióticos e a capacidade de
adquirirem resistência a vários antibióticos constitui um grave problema no tratamento de
doentes infectados por estes organismos (Singh, K. V. et ai, 2002).
1.5. Macrólidos, lincosamidas e estreptograminas
Os macrólidos são produzidos por Streptomyces sp e Micromonospora sp. Apresentam
uma estrutura química comum constituída por um anel lactónico, formado por 12, 15 ou 16
átomos de carbono, unidos a açúcares aminados e/ou neutros, razão pela qual apresentam
um elevado peso molecular (Anglada, R. R. 1997; Sousa, J. C. F. et ai, 1991b) (Tabela
11).
49

Introdução
Tabela 11 - Classificação de alguns macrólidos (Anglada, R. R. 1997).
Tamanho do anel lactónico
Antibiótico
14 átomos Claritromicina 15 átomos Eritromicina 16 átomos Oleandomicina
Roxitromicina Azitromicina Espiramicina Josamicina
Tilosina
A eritromicina, descoberta em 1952, é um antibiótico utilizado na terapêutica humana. Este
antibiótico é produzido por Saccharopolyspora erythraea e encontra-se representado na
figura l(Bergogne-Bérézin, E., 1997; Weisblum, B., 1998).
OH-,
a JL
H3C. Y
i o.
OH CH3
Desoamine
CH3
Cladinose
CH3
Fig. 1- Estrutura química da eritromicina A (Weisblum, B., 1998).
Do grupo das lincosamidas fazem parte a lincomicina, isolada em 1962 (Sousa, J. C. F. et
al 1991b) e a clindamicina. Estes antibióticos são apenas utilizados na terapêutica humana
(Lamb, H. M. et al, 1999; Lina, G. et al, 1999). 50

Introdução
A lincomicina é sintetizada por Streptomyces lincolnensis, sendo o seu derivado clorado a
clindamicina (Fig. 2) (Anglada, R. R. 1997).
CH
HO C-R
C — NH - CH
HO
OH SCH
OH
Fig. 2 Estrutura química das lincosamidas. (R = Cl, clindamicina; H, lincomicina)
As estreptograminas são produzidas por Streptomyces spp. São constituídas por uma
mistura de, pelo menos, duas moléculas estruturalmente diferentes: as estreptograminas do
grupo A e as estreptograminas do grupo B (Fig. 3) (Consumer Health Protection. Scientific
Committees. Scientific Committee on Animal Nutrition. 1998a., Consumer Health
Protection. Scientific Committees. Scientific Committee on Animal Nutrition. 1998b;
Jensen, L.B., 2002; Lamb, H. M. et al, 1999, Lina, G. et al, 1999, Pechère, J., 1996). As
estreptograminas do grupo A são macrolactonas polinsaturadas, corn peso molecular 500 e
as estreptograminas do grupo B são hexadepsipéptidos ciclicos, corn peso molecular 800 (
Pechère, J., 1996).
51

Introdução
Estreptogramina do grupo A Estreptogramina do grupo B
Fig 3 - Estrutura química dos grupos A e B das estreptograminas.
A virginiamicina é uma mistura de virginiamicina Ml ( estreptogramina A ) e de
virginiamicina S ( estreptogramina B ) (Jensen, L.B., 2002; Luh, K.-T. et ai, 2000; Simjee,
S. et ai, 2002).
A associação quinupristina-dalfopristina, que surgiu recentemente, é uma estreptogramina
que consiste na mistura de derivados semisintéticos da pristinamicina; a quinupristina
(derivada da pristinamicina IA) é uma estreptogramina do grupo B e a dalfopristina
(derivada da pristinamicina IIA) é uma estreptogramina do grupo A (Fig. 3) (Jensen, L. B.
et ai, 2000, Lamb, H. M. et al, 1999). Estas duas moléculas ligam-se ao ribossoma
bacteriano e, actuando sinergicamente, inibem a síntese proteica e a fase de alongamento
da cadeia peptídica (Solway, S. et ai, 2003).
Esta associação é um antibiótico semi-sintético, análogo à virginiamicina, recentemente
autorizado para uso clínico nos Estados Unidos da América e na Europa, no tratamento de
infecções causadas por patogénicos Gram-positivo multiresistentes, incluindo E. faecium
resistentes à vancomicina (um glicopéptido) e Staphylococcus aureus resistentes à
52

Introdução
meticilina (MRSA). E. faecalis é um resistente intrínseco à associação quinupristina-
dalfopristina (Soltani, M. D. et ai, 2000; Solway, S. et ai; Jensen, L.B., 2002; Soltani, M,
et ai, 2001; Jensen, L.B., 2000; ; Luh, K-T. et ai, 2000).
1.5.1. Modo de acção dos macrólidos, lincosamidas e estreptograminas
Os antibióticos macrólidos, lincosamidas e estreptograminas, apesar de serem
quimicamente distintos, possuem efeitos semelhantes na inibição da síntese proteica
bacteriana (Lamb, H. M. et al, 1999; Lim, J-A. et al, 2002).
As estreptograminas possuem actividade sinérgica. Os componentes do grupo B
(pristinamicina I e virginiamicina S) têm maior actividade na presença dos componentes do
grupo A (pristinamicina II e virginiamicina M). Para além disso, quando cada componente
actua isoladamente tem uma actividade bacteriostática, enquanto que quando os dois
componentes actuam conjuntamente têm actividade bactericida (Ostrowsky, B. et ai, 1999
e Pechère, J., 1996). A união dos componentes do grupo A das estreptograminas ao
ribossoma bacteriano induz neste uma alteração estrutural, aumentando a sua afinidade
pelos componentes do grupo B das estreptograminas (Pechère, J., 1996).
A reacção-chave da síntese proteica é a transpeptidação, a qual é mediada pela
peptidiltransferase, localizada na sub-unidade 50S do ribossoma (Chu, D. T., 1999;
Vannuffel, P. et ai, 1996). A transpeptidação envolve a formação de uma ligação entre a
cadeia peptídica em crescimento (peptidil-tRNA) e o aminoacil-tRNA, as quais estão
ligadas ao local P e ao local A do centro peptidiltransferase, respectivamente (Vannuffel,
P. et ai, 1996). Os macrólidos ligam-se à região 2058-2062 do rRNA 23S, da subunidade 50S do
ribossoma (Chu, D. T., 1999). Esta união provoca o bloqueio da transpeptidação e
provavelmente da translocação. Desta forma é impedido o alongamento da cadeia peptídica
(Vannuffel, P. et ai, 1996). O mecanismo de inibição da síntese proteica pelas estreptograminas é diferente para cada
componente (Fig. 4). A estreptogramina A inibe quer a ligação do peptidil-tRNA ao local
53

Introdução
dador, quer do aminoacil-tRNA ao local receptor da peptidiltransferase (Consumer Health
Protection. Scientific Committees. Scientific Committee on Animal Nutrition. 1998a;
VannuffeLP. etal, 1996).
A estreptogramina B impede a formação da ligação peptídica, provocando a libertação da
cadeia polipeptídica incompleta, do local dador (Consumer Health Protection. Scientific
Committees. Scientific Committee on Animal Nutrition. 1998a e Vannuffel, P. et al, 1996).
Fig 4 - Inibição da síntese proteica pelas estreptograminas
1.5.2. Mecanismos de resistência aos macrólidos, lincosamidas e estreptograminas
As bactérias podem ser resistentes aos macrólidos pelos seguintes mecanismos (Chu, D. T.,
1999; Piddock, L. J. V., 1996):
> Resistência intrínseca
> Resistência adquirida :
• Inactivação enzimática do antibiótico
• Modificação do alvo do antibiótico
• Efluxo activo de antibiótico
54

Introdução
1.5.2.1. Resistência intrínseca
Os antibióticos deste grupo têm de atravessar a parede celular e a membrana citoplasmática
bacteriana de modo a exercerem a sua função a nível dos ribossomas. Estes antibióticos
são moléculas anfifílicas, moderadamente hidrófobas e com peso molecular elevado.
A parede celular das bactérias Gram-positivo, essencialmente constituída por
peptidoglicano e ácidos teicóicos, permite a travessia de compostos de elevado peso
molecular e com diferentes graus de hidrofobia. Pelo contrário, as bactérias Gram-negativo
apresentam na parede celular uma membrana externa constituída principalmente por
fosfolípidos, porinas e lipopolissacarídeos (LPS), a qual funciona como uma barreira de
permeabilidade (Sousa, J. C. F. et ai, 1991).
Nas bactérias Gram-negativo os solutos hidrófilos atravessam a membrana exterior da
parede celular através dos canais de porina. No entanto, devido ao elevado peso molecular
dos macrólidos, os canais de porina são impermeáveis a estes antibióticos. Por esta razão,
muitas estirpes de bactérias Gram-negativo apresentam uma resistência intrínseca à
eritromicina e seus derivados, ao contrário das estirpes de bactérias Gram-positivo (Sousa,
J. C. F. et ai, 1991). Nas estirpes selvagens de Enterobacteriaceae (bactérias Gram-
negativo anaeróbias facultativas) e noutras bactérias Gram-negativo, a penetração dos
macrólidos através da membrana externa não ocorre devido às projecções polissacarídicas
dos LPS, na superfície bacteriana. Desta forma, os solutos hidrófobos não podem difundir
rapidamente através dos canais de porina, quer através dos LPS (Sousa, J. C. F. et ai,
1991).
55

Introdução
1.5.2.2. Resistência adquirida
A - Inactivação enzimática do antibiótico
A inactivação enzimática do antibiótico pode ocorrer por hidrólise, glicosilação, acetilação,
nucleotidilação ou por fosforilação.
a) Hidrólise de macrólidos e estreptograminas
Este processo ocorre por cisão do anel lactónico dos macrólidos mediada pelas enzimas
EreA e EreB, codificadas pelos genes ereA e ereB, respectivamente. Estes genes foram
descritos em Stahpylococcus aureus (Schmitz, F-J. et ai, 2000). Estas enzimas encontram-
se no espaço periplasmático bacteriano, mas são raras as bactérias que as produzem
simultaneamente (Sousa, J. C. F. et ai, 1991). Em E. coli esta resistência verifica-se para a
eritromicina, mas não para os macrólidos com anel lactónico constituído por 16 átomos de
carbono. Nesta espécie bacteriana os genes ereA e ereB encontram-se no plasmídeo
plP1527 (Sousa, J. C. F. et ai 1991; Wondrack, L. et ai, 1996).
Em S. aureus foram descritos genes com actividade semelhante aos genes ere, capazes de
hidrolizar os macrólidos constituídos por 14 ou 16 átomos de carbono no anel lactónico
(Bischoff, W. E. et ai, 1999). Nesta bactéria foi igualmente descrito o gene vgb que
codifica uma lactonase capaz de hidrolizar a quinupristina (estreptogramina B),
inactivando-a (Lamb, H. M. et al, 1999, Lina, G. et al, 1999, Piddock, L. J. V., 1996). Está
igualmente descrito o gene vgbB, igualmente capaz de hidrolizar as estreptograminas B
(Jensen, L. B. et ai, 2000; Jensen, L.B. et ai, 2002; Lina, G. et ai, 1999). Estes genes foram
detectados quer em Staphylococcus sp., quer em Enterococcus sp. (Report and Qualitative
Risk Assessment by the Committee for Veterinary Medicinal Products. 1999; Roberts, M.
C. etal, 1999).
56

Introdução
b) Glicosilação dos macrólidos
Este mecanismo é mediado pelo gene mgt em Streptomyces lividans (Weisblum, B.,
1998). Em S. ambofaciens foi detectada uma glicosiltransferase, codificada pelo gene
gwiA(Chu,D. T., 1999).
c) Fosforilação dos macrólidos
Este processo é mediado, em E. coli, pelos genes mphA, mphB e mphK (Roberts, M. C. et
al, 1999; Weisblum, B., 1998). Dada a sua semelhança, o gene mphK é actualmente
designado por mphA, em virtude das duas proteínas que os codificam, diferirem em apenas
1% dos seus aminoácidos (Roberts, M. C. et al, 1999).
O gene mphA relaciona-se apenas com a fosforilação dos macrólidos com anel lactónico
constituído por 14 átomos de carbono ( eritromicina ), enquanto que o gene mphB se
relaciona com a fosforilação dos macrólidos com anel lactónico constituído quer por 14
quer por 16 átomos de carbono ( espiramicina e tilosina ) (Roberts, M. C. et al, 1999).
Em E. coli BM 2506, isolada na clínica, a elevada resistência aos macrólidos deve-se à
presença da enzima 2'-fosfotransferase II dos macrólidos-MPH ( 2' )( Lina, G. et ai, 1999,
Sousa, J. C. F. et ai, 1991). Esta estirpe possui dois plasmídeos, pTZ 3721(84 Kb) e pTZ
3723 (24 Kb), nos quais se supõe estar localizado o gene mphB, que codifica a enzima
MPH (2'). A transferência destes plasmídeos permite a disseminação do gene mphB, por
conjugação (Roberts, M. C. et al, 1999).
Em Staphylococcus sp. foi descrito o gene mphC, que codifica, igualmente, uma
fosfotransferase (Chu, D. T., 1999; Roberts, M. C. et al, 1999).
d) Nucleotidiltransferases
A resistência às lincosamidas pela produção de nuclotidiltranferases, codificada pelos
genes lin A e lin A', foi detectada em S. haemolyticus e S. aureus, respectivamente. Estas
enzimas modificam o antibiótico, tomando-o ineficaz (Lin, J.A., 2002; Pechére, J. C, 57

Introdução
2001; Roberts, M. C. et al, 1999).
O gene linB, foi detectado em Enterococcus faecium e codifica uma nucleotidiltransferase
das lincosamidas. Esta enzima cataliza a adenilação do grupo hidróxilo na posição 3 [3-(5'-
adenilação)] da lincomicina e clindamicina. Todavia, esta enzima não apresenta qualquer
homologia com as enzimas codificadas pelos genes UnA e /mA' (Lina, G. et ai, 1999;
Roberts, M. C. et ai, 1999).
A expressão do gene UriB foi também observada em E. coli e em S. aureus. Verificou-se a
existência de sequências homólogas do gene linB, em várias estirpes de E. faecium,
indicando a dispersão da resistência entre bactérias desta espécie (Bozdogan, B. et ai,
1999a, Lina, G. et ai, 1999; Roberts, M. C. et al, 1999).
e) Acetiltransferases
Em E. faecium foram detectados os genes sat A e satG, os quais se encontram em
plasmídeos e são codificadores de acetiltransferases, que inactivam a estreptogramina tipo
A, conferindo resistência à associação quinupristina-dalfopristina (Bozdogan, B. et ai,
1999b, Haroche, J.,2000; Jensen, L. B. et ai, 2000; Jensen, L. B. et ai, 2002; Soltani, M. D.
et ai, 2000, Solway, S. et ai, 2003; Wondrack, L. et ai, 1996). Estes genes, foram
inicialmente relacionados com os genes vatA, vatB e vatC, codificadores de
acetiltransferases em Staphylococcus sp. (Haroche, J., 2000; Lina, G. et ai, 1999; Soltani,
M. D. et ai, 2000; Soltani, M., et ai, 2001). Actualmente, os genes sat A e satG recebem a
designação de vatD e vatE, respectivamente. São conhecidas oito variações alélicas do
gene vatE em E. faecium, as quais recebem a designação de vat(E-\) a vat(E-S) (Simjee, S.
et ai , 2002b). Simjee e colaboradores (Simjee, S. et ai, 2002b) admitem ter observado pela
primeira vez variações alélicas do gene vatE em E. faecalis. Contudo, apesar de ser ainda
desconhecida a forma como estas substituições de aminoácidos afectam a actividade da
proteína VatE, supõe-se que não possuem um papael importante na ligação das
estreptograminas, pois não se crê que tenham qualquer efeito ao nível da resistência à
associação quinupristina-dalfopristina (Simjee, S. et ai, 2002b; Soltani, M., et ai, 2001).
58

Introdução
O gene vatA, tal como o gene vatE, modifica a estreptogramina A por adição de um grupo
acetilo (Roberts, M. C. et ai, 1999).
O gene vatD encontra-se distribuído em estirpes do género Enterococcus com origem
humana e não humana. Da mesma forma, foram detectados genes vatE em plasmídeos de
E. faecium, isolados em animais, carnes e em doentes hospitalizados no Reino Unido e
Alemanha (Hammerum, A. M. et ai, 2001; Soltani, M. D. et ai, 2000). Estes genes não
foram detectados em isolados de Staphylococcus sp. (Haroche, J., 2000).
O mecanismo de resistência enzimática, com modificação dos macrólidos, lincosamidas e
estreptograminas está limitado a um pequeno número de isolados clinicos, sendo, por isso,
pouco significativo. Os principais mecanismos são a modificação do alvo do antibiótico e o
efluxo do antibiótico. No entanto, as razões que favorecem o seu aparecimento não são
conhecidas (Weisblum, B., 1998).
Na tabela 12 encontram-se os diferentes genes e os géneros ou espécies onde ocorrem.
59

Introdução
Tabela 12 - Bactérias onde se encontram genes que provocam a inactivação do antibiótico
(Bozdogan B. e R. Leclercq, 1999; Lina, G. et ai, 1999; Roberts, M. C. et ai, 1999; Simjee,
S. et ai, 2002b; Soltani, M. et ai, 2000).
Gene Género/Espécie bacteriana Hidrólise ereA
ereB
Staphylococcus aureus ; Enterobacter ; Escherichia ; Klebsiella ;Citrobacter Staphylococcus aureus ; Esherichia ;Klebsiela ; Proteus
vgbA Staphylococcus aureus ;Enteroc vgbB Staphylococcus aureus
Glicosilação mgt Streptomyces lividans gimA S. ambofaciens
Fosforilação mphA E. coli mphB E. coli mphK E. coli mphC Staphylococcus
Nucleotidiltransferases HnAI lin A ' S. haemolyticus ; S. epi
aureus ; S. hominis HnB Staphylococcus aureus ; E. coli
epidermidis ;Staphylococcus
Acetiltransferases vatA vatB vatC vatD vatE
Enterococcus sp; Staphylococcus S. epidermidis; S. aureus Staphylococcus, S cohnii cohnii E. faecalis E. faecalis
60

Introdução
B. Modificação do alvo
A modificação do alvo é a forma de resistência mais comum na clínica (Wondrack, L. et
ai, 1996).
A modificação do alvo conduziu à resistência cruzada aos macrólidos, lincosamidas e
estreptograminas B (MLSB), aos macrólidos e lincosamida (ML) e aos macrólidos,
cetolidos e estreptograminas A e B (MKS). O primeiro relato de resistência cruzada à
eritromicina data de 1956 (Amezaga, M.R. et ai. 2002; Pechére, J. C, 2001).
Este mecanismo envolve uma alteração conformacional (metilação) pós-transcrição do
rRNA 23S, pela acção da adenina-N-metiltranferase. Esta enzima acrescenta um ou dois
grupos metilo na adenina 2058 no rRNA 23 S, em E. coli, o que diminui ou impede a
ligação dos antibióticos MLS aos ribossomas bacterianos (Sutcliffe, J. et ai, 1996;
Wondrack, L. et ai, 1996).
As metilases que catalizam esta modificação, metilases rRNA 23 S, são designadas pelo
acrónimo Erm (metilase de resistência à eritromicina) (Min. Y-H, et ai, 2003; Weisblum,
B., 1998) e são codificadas pelos genes erm (metilação ribossómica da eritromicina)
(Torres, C, 2002). Estes genes codificam igualmente a resistência às estreptograminas B,
aos macrólidos e às lincosamidas (fenótipo de resistência MLSB), contudo não inactivam
as estreptograminas do grupo A (Jensen, L. B. et ai, 2000; Vannuffel, P. et ai, 1996).
Ao longo dos últimos trinta anos foram descritas adenina-N -metiltransferases em diversos
géneros e espécies, tanto em Gram-negativo como em Gram-positivo, sendo este o
mecanismo de resistência mais frequente aos macrólidos, lincosamidas e estreptograminas
(Roberts, M. C. et al, 1999). Os genes que codificam estas enzimas foram divididos em
várias classes (Tabela 13) (Thomson, C. J., 1999).
61

Introdução
Tabela 13 - Bactérias onde se encontram genes codificadores de metilases (Aarestrup, F.
M, 2000; Amezaga, M. R., 2002; Borgen, K. et ai, 2002; Bozdogan, B. e R. Leclercq,
1999; Lim, J.-A. et ai, 2002; Lina, G. et ai, 1999; Luna, V. A. et ai, 2002; Min. Y.-H. et ai,
2003; Pechére, J. C, 2001; Portillo, A. et ai, 2000; Roberts, M. C. et ai, 1999).
Gene Género/Espécie bacteriana
erm A Actinobacillus, Staphylococcus, Streptococcus, S. hominis, S. haemoliticus,
erm B=ermAM Actinobacillus, Clostridium, Escherichia, Enterococcus, Klebsiella, Neisseria, Staphylococcus, Streptococcus, Wolinella, Bacillus subtilis, Pediococcus, S. haemoliticus
ermBv(ermAMR) E. faecalis erm C Actinobacillus, Bacillus, Eubacterium, Lactobacillus, Neisseria,
Staphylococcus, Streptococcus, Wolinella, S. haemoliticus, S. epidermidis, Enterococcus sp.
erm D Bacillus erm E Streptomyces erm F Actinobacillus, Actinomyces, Bacteroides, Clostridium, Eubacterium,
Fusobacterium, Gardnerella, Haemophilus, Neisseria, Wolinella, Enterococcus sp., Staphylococcus
erm G Porphyromonas, Prevotella, Peptosteptococcus, erm H Selenomonas, Streptococcus, erm I Bacillus, Bacteroides erm M S. epidermidis erm N Streptomyces erm O Streptomyces erm P Clostridium perfringens erm Q Streptomyces; Sreptococus; Clostridium erm R Streptomyces erm S Actinobacillus, Clostridium, Streptococcus, Wolinella; Streptomyces erm T Aeromicrobium; Lactobacillus erm U Streptomyces erm V Lactobacillus; Streptomyces erm W Streptomyces erm X Streptomyces; Corynebacterium erm Y Micromonospora, Corynebacterium, Staphylococcus erm Z Clostridium difficile, E. faecalis erm BC E. coli
62

Introdução
Muitos dos genes erm estão associados a transposões conjugativos ou não conjugativos,
que residem essencialmente no cromossoma, embora alguns tenham sido encontrados em
plasmídeos (Roberts, M. C. et ai, 1999). Os genes erm estão frequentemente associados a
outros genes de resistência, especialmente, a genes de resistência à tetraciclina.
Estes genes podem ser expressos constitutiva ou indutivelmente (Buu-Hoi,A.; Consumer
Health Protection. Scientific Committees. Scientific Committee on Animal Nutrition.
1998b, Sousa, J. C. F. et al, 1991).
1. Fenótipo constitutivo
Uma estirpe com este tipo de fenótipo é resistente a todos os macrólidos, às lincosamidas e
às estreptograminas B. As estreptograminas A não são afectadas, mantendo-se a actividade
sinérgica entre as estreptograminas A e B. A estirpe permanece sensível à pristinamicina
(Buu-Hoi,A.; Lamb, H. M. et al, 1999, Lina, G. et al, 1999, Pechére, J. C , 2001).
2. Fenótipo indutivel
Os determinantes de resistência ermA, ermB e ermC conferem este tipo de resistência por
alteração do sítio alvo do ribossoma (Lina, G. et al, 1999).
Neste caso, a estirpe é resistente à eritromicina e à oleandomicina, mas é sensível aos
outros macrólidos, lincosamidas e estreptograminas. Por esta razão, em presença de
concentrações sub-inibitórias de eritromicina ou de oleandomicina, a resistência MLSB é
induzida e a estirpe torna-se resistente a todos os macrólidos, às lincosamidas e às
estreptograminas B. Por esta razão, torna-se idêntica ao fenótipo constitutivo. Se a estirpe
for cultivada na ausência do antibiótico indutor (eritromicina ou oleandomicina) ocorre
perda de resistência aos outros macrólidos, às lincosamidas e às estreptograminas e a
estirpe volta ao fenótipo induzido inicial (Buu-Hoi,A).
Algumas estirpes podem apresentar resistência induzida pela eritromicina, tornando-se
resistentes isoladamente a este antibiótico. Estas estirpes são sensíveis à oleandomicina e
63

Introdução
aos outros macrólidos, às lincosamidas e às estreptograminas B (Buu-Hoi, A.; Consumer
Health Protection. Scientific Committees. Scientific Committee on Animal Nutrition.
1998b, Pechére, J. C, 2001; Wondrack, L. et al, 1996)
O principal factor que determina se um determinado macrólido induz ou não a resistência é
o seu tamanho. Desta forma, os macrólidos com 15 e especialmente com 14 átomos de
carbono no anel lactónico são indutores poderosos, enquanto que os que possuem 16
átomos de carbono no anel lactónico têm um efeito pouco significativo (Lamb, H. M. et al,
1999; Pechére, J. C, 2001; Wondrack, L. et al, 1996).
Os genes ermA e ermC, determinantes de resistência predominantes na resistência MLSB
em Staphylococcus sp., são mais frequentemente induzidos pelos macrólidos com 14
átomos de carbono no anel lactónico e pela celesticetina (uma lincosamida), do que pelos
macrólidos com 16 átomos de carbono no anel lactónico e lincomicina (uma lincosamida).
No entanto, mutantes de Staphylococcus aureus, seleccionados em laboratório, apresentam
resistência induzida pela lincomicina e pela carbomicina (macrólido com 16 átomos de
carbono no anel lactónico) (Min. Y-H, et ai, 2003; Schmitz, F-J et ai, 2002 ).
Contudo, Schmitz (Schmitz, F.J. et ai, 2002) verificou que os macrólidos com 16 átomos
de carbono no anel lactónico, lincomicina e estreptogramina B, alteram irreversivelmente a
resistência provocada pelo gene ermC de indutível para constitutiva, conferindo resistência
cruzada à eritromicina, à clindamicina e à quinupristina. De facto, recentemente, numa
estirpe clínica de Staphylococcus aureus, foi detectada resistência cruzada indutível à
eritromicina (macrólido com 14 átomos de carbono no anel lactónico), à clindamicina, à
lincomicina e à quinupristina ( Min. Y-H, et ai, 2003; Schmitz, F-J et ai, 2002).
O gene ermD é induzido pela eritromicina e oleandomicina (macrólido com 14 átomos de
carbono no anel lactónico), mas não é induzido pela tilosina, nem pela clindamicina e
lincomicina ( Min. Y-H, et ai, 2003).
O gene ermV, com origem em Streptomyces viridochromogenes é induzido pela
celesticetina (macrólido com 16 átomos de carbono no anel lactónico) e por macrólidos
com 14 átomos de carbono no anel lactónico ( Min. Y-H, et ai, 2003).
64

Introdução
Em Streptomyces fradiae, produtor da tilosina, a transcrição do gene ermS é autoindutível
por este agente ( Min. Y-H, et ai, 2003).
Todos os antibióticos MLSB induzem o gene ermB, o determinante mais comum em
Streptococcus pneumoniae, Streptococcus pyogenes e Enterococcus sp. O gene erm(Bv)
(em que v significa variante), anteriormente designado por erm(AMR), uma variante do
gene ermB, de um isolado clínico de Enterococcus faecalis, é induzido principalmente
pelos macrólidos com 16 átomos de carbono no anel lactónico, josamicina, tilosina e
katasamicina ( Min. Y-H, et ai, 2003).
C. Sistemas de efluxo do antibiótico
Este mecanismo de resistência tornou-se mais conhecido na última década, para um
elevado número de antibióticos. Tal como acontece nas outras células, as paredes celulares
impermeáveis das bactérias têm mecanismos que permitem a entrada de nutrientes e a
eliminação de produtos de metabolismo indesejados. Este efluxo, que é realizado por
transporte activo, também se verifica para os antibióticos. No efluxo de antibióticos estão
envolvidos três componentes: uma bomba no folheto interno da membrana citoplasmática,
uma proteína de canal com lúmen hidrofílico na parede externa e uma proteína que
estabelece a ligação entre o folheto externo e interno da membrana citoplasmática
(Pechére, J. C , 2001).
Os sistemas de efluxo de antibiótico constituem um grave problema de resistência.
Actualmente estão caracterizadas mais de cem proteínas transportadoras que apresentam
um papel importante na expulsão de substâncias tóxicas resultantes do metabolismo
bacteriano (Nikaido, H., 1996; Pechére, J. C , 2001; Roberts, M. C. et al, 1999; Weisblum,
B., 1998).
Em bactérias patogénicas e não patogénicas existem sistemas de efluxo codificados por
genes cromossómicos, sugerindo que estes transportadores não surgiram apenas
recentemente como resposta à terapia por antibióticos (Powell, W. J., 2000).
O efluxo de antibiótico é mediado por transportadores que podem ser distinguidos em
termos funcionais ou com base no tipo de proteínas que os constitui. Em termos funcionais,
65

Introdução
o transporte pode ser mediado por uma bomba de protões ou por adenosina trifosfato
(ATP) (Nikaido, H., 1996; Pechére, J. C, 2001; Roberts, M. C. et ai, 1999; Weisblum, B.,
1998).
Relativamente ao tipo de proteínas, estas podem consistir em proteínas intrínsecas capazes
de excretar um ou vários antibióticos ou em sistemas mais complexos. As diferentes
proteínas são codificadas por uma grande variedade de genes. A função destas proteínas é
bombear um ou mais antibióticos para o exterior da célula, mantendo a sua concentração
intracelular baixa e os ribossomas livres da actividade do antibiótico. Estas bombas podem
mediar o transporte através da membrana citoplasmática ou através da membrana dupla.
No primeiro caso e em Gram negativo, estas bombas deslocam o antibiótico do citoplasma
para o espaço periplasmático (Nikaido, H., 1996, Roberts, M. C. et al, 1999, Weisblum, B.,
1998). Os sistemas de efluxo de macrólidos podem ser agrupados de acordo com a
diversidade de antibióticos ou outras substâncias que transportam (Weisblum, B., 1998):
• Efluxo Tipo-M
• Efluxo Tipo-MS
• Efluxo Tipo-Actinomycetes
• Efluxo Tipo-mrd ou de "largo espectro"
O efluxo Tipo-M e do Tipo-MS são os que apresentam significado clínico (Weisblum, B.,
1998).
66

Introdução
1. Efluxo Tipo-M ou Fenótipo M
As estirpes com este fenótipo são resistentes aos macrólidos de anel lactónico constituído
por 14 e 15 átomos de carbono, mas susceptíveis à clindamicina (uma lincosamida) e
estreptograminas B (Lamb, H. M. et al, 1999; Lim, J-A. et al, 2002; Reinert, R.R., et al,
2003).
Este tipo de resistência deve-se ao gene mefA, originalmente encontrado em Streptococcus
pyogenes, e ao gene meJE, originalmente encontrado em S. pneumoniae. Durante muito
tempo não foi possível distinguir por Polimerase Chain Reaction (PCR) estes dois genes.
Por essa razão, consideraram-se estas duas variantes específicas de determinada espécie.
Desta forma, se o gene mef fosse encontrado em S. pneumoniae assumia-se, geralmente,
que era o gene meJE. Pelo contrário, se fosse encontrado em S. pyogenes assumia-se,
geralmente, que se tratava do gene mefA. No entanto, já foi detectado o gene mefA em
isolados de S. pneumoniae, resistentes a macrólidos (Amezaga, M.R., 2002; Del Grosso,
M. et ai, 2002; Lamb, H. M. et al, 1999; Lim, J-A. et al, 2002; Sutcliffe, J. et al, 1996).
O gene mefA foi, igualmente, detectado em organismos Gram-positivo, como
Corynebacterium spp., Enterococcus sp. e Micrococcus luteus e em organismos Gram-
negativo, como Acinetobacter junni e Neisseria gonorrhoeae (Amezaga, M.R., 2002;
Lamb, H. M. et al, 1999; Lim, J-A. et al, 2002; Luna, V. A. et al, 2002). Por esta reazão,
actualmente estes genes recebem a designação de mefA (Reinert, R.R., et al, 2003).
A actividade das enzimas, codificadas por estes genes, é mediada por uma bomba de
protões (Amezaga, M.R., 2002).
O gene mefA codifica a proteína MefA com homologia com a super-família MFS (Major
Facilitator Superfamily), que efectua o efluxo do antibiótico (Dinh, T. et ai, 1994; Nikaido,
H., 1998; Reinert, R.R., et ai, 2003 ; Roberts, M. C. et ai, 1999). O fenótipo de resistência
M predomina nos Estados Unidos da América, no nordeste da Escócia e no Reino Unido
(Amezaga, M.R., 2002)
Em S. agalactiae foi detectada a presença de um novo gene de efluxo, o gene mreA, que
determina a resistência aos macrólidos de anel lactónico formado por 14, 15 e 16 átomos
67

Introdução
de carbono (Chu, D. T., 1999; Weisblum, B., 1998).
2. Efluxo Tipo-MS ou Fenótipo MS
Este tipo de resistência foi inicialmente identificado em Staphylococcus epidermidis. As
estirpes de bactérias com este fenótipo possuem o gene msrA, que confere resistência à
eritromicina, à oleandomicina e à estreptogramina B, por um mecanismo de efluxo do
antibiótico após indução pela eritromicina (Amezaga, M.R., 2002; Lina, G. et ai, 1999;
Portillo, A. et ai, 2000;).
Este gene foi igualmente identificado em S. pneumoniae. Em Staphylococcus xylosus foi
detectado o gene msrB, relacionado com o gene msrA (Lina, G. et ai, 1999). Em
Enterococcus faecium foi detectado o gene msrC. Estes genes, que codificam as proteínas
transportadoras MsrA, MsrB e MsrC, respectivamente, têm homologia com os
transportadores da família ABC (ATP-binding cassete) (Dinh, T. et ai, 1994; Nikaido, H.,
1998; Roberts, M. C. et ai, 1999, Simjee, S. et ai, 2002b) Por esta razão, alguns autores
admitem que o transportador MS efectua o efluxo de eritromicina e de estreptogramina B,
através da hidrólise de ATP (Chu, D. T., 1999; Weisblum, B., 1998, Wondrack, L. et ai,
1996).
Os genes vgaA e vgaB codificam ATP-" binding proteins", que, provavelmente, estão
envolvidas no efluxo de estreptogramina A (Lina, G. et ai, 1999; Soltani, M. D. et ai,
2000). As proteínas VgaA e VgaB, codificadas por aqueles genes, apresentam homologia
com os transportadores da família ABC (Roberts, M. C. et al, 1999). Estes genes de
resistência às estreptograminas detectados em Enterococcus faecium e Staphylococcus
aureus não foram ainda detectados em Enterococcus faecalis (Bozdogan, B. et ai, 1999b;
Simjee, S. et ai, 2002b).
Em E. faecalis, o gene Isa é determinante no fenótipo de resistência intrínseca LSA
(resistência às lincosamidas e à dalfopristina) e na resistência à associação quinupristina-
dalfopristina. A proteína Lsa, codificada pelo gene Isa, tem semelhanças na sequência de
aminoácidos com os transportadores ABC, incluindo MsrA, VgaA e VgaB, detectados em 68

Introdução
Staphylococcus aureus, (Simjee, S. et al, 2002b), o que sugere que a resistência de E.
faecalis as lincosamidas e à estreptogramina esteja relacionada com o efluxo activo destes
antibióticos. Dado que este gene aparenta ser específico de E. faecalis, Singh e
colaboradores (Singh, K. V. et ai, 2002) admitem que poderá ser utilizado na identificação
de isolados desta espécie. Estes autores demonstraram que o gene Isa é funcional em
isolados de E. faecium e que aumenta os MICs das lincosamidas e dalfopristina e/ou da
quinupristina-dalfopristina, em níveis distintos, consoante a susceptibilidade inicial do
hospedeiro a estes agentes (Singh, K. V. et ai, 2002).
3. Efluxo Tipo Actinomycete
Os genes tlrC de Streptomyces fradiae, os genes car A de Strep, thermotolerans e os genes
srmB de Strep, ambofaciens conferem resistência aos macrólidos constituídos por 16
átomos de carbono no anel lactónico, tal como tilosina, carbomicina e espiramicina,
respectivamente. Foi igualmente detectado em estirpes de Strep, antibioticus, produtoras
de olendomicina, o gene oleB que codifica uma ATPase para os macrólidos com anel
lactónico constituído por 14 átomos de carbono. Não se sabe ainda se os genes oleB
conferem resistência aos macrólidos com anel lactónico constituído por 16 átomos de
carbono (Weisblum, B., 1998).
4. Efluxo Tipo mrd Em E. coli foi detectada uma proteína, MdfA, que reconhece uma grande variedade de
antibióticos, capazes de transportar entre outros a tetraciclina, cloranfenicol, eritromicina,
alguns aminoglicosídeos e fluoroquinolonas. Esta proteína actua através de uma bomba de
protões (Weisblum, B., 1998).
69

Introdução
Tabela 14 - Bactérias onde se encontram genes que provocam o efluxo do antibiótico
(Borgen, K. et ai, 2002; Bozdogan, B. e R. Leclercq, 1999;; Lim, J.-A. et ai, 2002; Lina, G.
et ai, 1999; Luna, V. A. et ai, 2002; Roberts, M. C. et ai, 1999; Singh, K. V. et ai, 2002;
Werner, G. et al, 2001).
Gene Fenótipo M me/A
Género/Espécie bacteriana
Streptococcus pyogenes; Corynebacterium spp; Enterococcus sp; Micrococcus luteus ; Acinetobacter junni ; Neisseria gonorrhoeae S. pneumoniae; S. agalactiae, Staphylococcus aureus Staphylococcus spp., S. haemoliticus, S. intermedius
mre S. agalactiae
Fenótipo MS msrA
msrB msrC
vgaA vgaB
Isa
Staphylococcus epidermidis ; S. pneumoniae; S. haemoliticus; S. hominis; S. aureus; Staphylococcus xylosus Enterococcus faecium
Enterococcus faecium ; Staphylococcus aureus Enterococcus faecium ; Staphylococcus aureus
Enterococcus faecalis ; E. faecium
Efluxo Tipo Actinomycete tlrC carA srm B oleB
Streptomyces fradiae Strep, thermotolerans Strep, ambofaciens Strep, antibioticus
Efluxo Tipo mrd mdfA E. coli
70

Objectivos
Objectivos
Com este trabalho pretendeu-se caracterizar os genes de resistência aos macrólidos e
estreptograminas existentes em isolados do género Enterococcus, com origem em frangos,
recolhidos entre Setembro de 1999 e Junho de 2001, em talhos da cidade do Porto.
Foi também propósito deste estudo avaliar a capacidade de transferência dos genes
codificadores de resistência aos antibióticos supracitados, recorrendo a ensaios de
conjugação.
71

Material e Métodos
3. Material e Métodos
3.1. Origem das estirpes bacterianas
Neste trabalho foram utilizados 388 isolados do género Entervcoccus pertencentes à
colecção do serviço de Microbiologia da Faculdade de Farmácia da Universidade do Porto.
As referidas bactérias, isoladas a partir de 93 lotes de frangos de dez marcas diferentes,
foram recolhidas entre Setembro de 1999 e Junho de 2001, em talhos da cidade do Porto. A
amostragem foi efectuada recolhendo 25g de cada lote de frangos, durante o período de
1999 e 2000. Em 2001, a colheita foi efectuada com zaragatoa, recolhendo-se o material
para amostra na superfície interna das carcaças dos frangos (Novais, C. et ai, 2002).
3.2. Isolamento, identificação dos isolados bacterianos.
Recolheram-se 99 amostras de frango, pertencentes a 93 lotes, tendo sido efectuado um
pré-enriquecimento em água de peptona tamponada sem antibiótico, para as amostras
recolhidas entre 1999 e 2000, ou em meio BHI (Brain Heart Infusion) sem e com
vancomicina (6mg/L), para as amostras de 2001 (Novais, C. et al, 2002).
As suspensões relativas a cada amostra foram plaqueadas em meio M-enterococos ou em
meio de Slanetz-Bartley sem antibiótico e com 6mg/L de vancomicina, 125mg/L de
gentamicina, 1000mg/L de estreptomicina e 500mg/L de canamicina. As placas foram
incubadas a 37°C e examinadas ao fim de 24, 48, 72 e 96 horas (Novais, C. et ai, 2002).
Uma colónia de cada fenótipo de resistência e morfologia foi de seguida congelada em
meio de TSB e Glicerol, a uma temperatura de -72° C (Novais, C. et ai, 2002).
3.3. Avaliação da susceptibilidade e identificação dos isolados da colecção de bactérias
A avaliação da colecção de bactérias da colecção do serviço de Microbiologia da
Faculdade de Farmácia da Universidade do Porto foi efectuada por diluição em agar com 72

Material e Métodos
determinação da Concentração Mínima Inibitória (CMI) para a eritromicina, associação
quinupristina-dalfopristina e para a vancomicina. A avaliação da susceptibilidade aos
antibióticos nos isolados de Enterococcus sp. foi determinada pelo método de diluição em
agar, seguindo as normas NCCLS. Enterococcus faecalis 77F (estirpe cedida pelo Hospital
Rámon e Cajal, Espanha) e Staphylococcus aureus ATCC 29213 foram as estirpes
utilizadas como controlo (Novais, C. et ai, 2002b). Para efectuar a caracterização dos
genes, as bactérias armazenadas foram semeadas em placas de agar de sangue e incubadas
a 32° C, durante 10 a 12 horas. Os isolados foram presumivelmente identificados como
pertencentes ao género Enterococcus recorrendo ao teste de produção da catalase,
coloração de Gram e hidrólise da esculina. A identificação da espécie foi realizada por
PCR Multiplex (Novais, C. et ai, 2002a).
Na totalidade dos isolados que tinham sido identificados procedeu-se à caracterização dos
genes de resistência.
3.4. Detecção de genes de resistência a macrólidos e estreptograminas por PCR
Esta colecção incluía isolados de Enterococcus faecium, de Enterococcus faecalis, de
Enterococcus gallinarum e de Enterococcus sp., tendo-se designado por Enterococcus sp.
todas as outras espécies de Enterococcus, num total de 388 isolados, os quais possuíam
diferente susceptibilidade à vancomicina, à eritromicina e à associação quinupristina-
dalfopristina.
A detecção dos genes ermA, ermB, ermC, me/A, vatD (anteriormente designado por satA)
e vatE (anteriormente designado por satG) foi efectuada em todos os isolados por
Polimerase Chain Reaction, num termociclador (Perkin- Elmer. GeneAmp,
PCRSystem2400).
As estirpes controlo foram gentilmente cedidas por Teresa Coque (Hospital Rámon e
Cajal, Espanha) e encontram-se indicadas na Tabela 16. Os primers foram sistetizados por
Gibco BRL e encontram-se discriminados na tabela 15.
73

Material e Métodos
Tabela 15- Identificação das estirpes controlo usadas nas reacções de PCR para detecção
dos genes de resistência a macrólidos e estreptograminas.
Gene Estirpe controlo
ermA Staphylococcus aureus RN 1389 errriQ Enterococcus faecium H2-4 ermC Staphylococcus aureus RN 4220 mefA Streptococcus pyogenes 02C 1064 vatD Enterococcus faecium H2-4 vatE Enterococcus faecium B306
3.4.1. Extracção do DNA genómico das estirpes controlo
A extracção do DNA genómico das estirpes controlo foi efectuada recorrendo à matriz
InstaGene (Bio-Rad). Para tal suspendeu-se uma colónia da estirpe controlo em 1 ml de
água estéril, centrifugou-se durante 1 minuto entre 10 000 a 12 000 rotações por minuto e
desprezou-se o sobrenadante. De seguida, adicionaram-se 200 \ú de matriz InstaGene ao
"pellet" e incubou-se a 56° C, durante cerca de 30 minutos. Homogeneizou-se durante
aproximadamente 10 segundos. Seguidamente colocou-se, durante 8 minutos, num banho a
100° C. Após o banho homogeneizou-se durante 10 segundos e centrifugou-se, de seguida,
durante 2 a 3 minutos entre 10 000 e 12 000 rotações por minuto. Utilizaram-se 2(il do
sobrenadante para um volume de 25ul na reacção de PCR para detecção dos genes ermA,
ermB e ermC e 2,5|il do sobrenadante na reacção de PCR para detecção dos genes mefA,
vatD e vatE.
74

Material e Métodos
3.4.2. Condições de PCR
A sequência dos primers e as condições das reacções do PCR usadas na detecção dos
genes de resistência aos macrólidos e estreptograminas encontram-se na tabela 16. Nas
tabelas 17, 18 e 19 encontram-se as concentrações dos reagentes e a concentração final
para cada uma das reacções de amplificação dos genes ermA, ermB, ermC, mefA, vatD e
vatE. Em cada reacção de PCR, para a detecção dos genes ermA, ermB e ermC foram
utilizados 2fil, da suspensão bacteriana em estudo, de turvação equivalente a 0,2
MacFarland, e para detecção dos genes mefA, vatD e vatE foram utilizados 2,5fil da
suspensão bacteriana, de turvação equivalente a 0,2 MacFarland, para 25|ll de volume
final. Foi adicionada água ultra-pura até perfazer o volume final de 25|il.
Em todas as reacções de PCR foi incluído um controlo positivo e um controlo negativo.
75
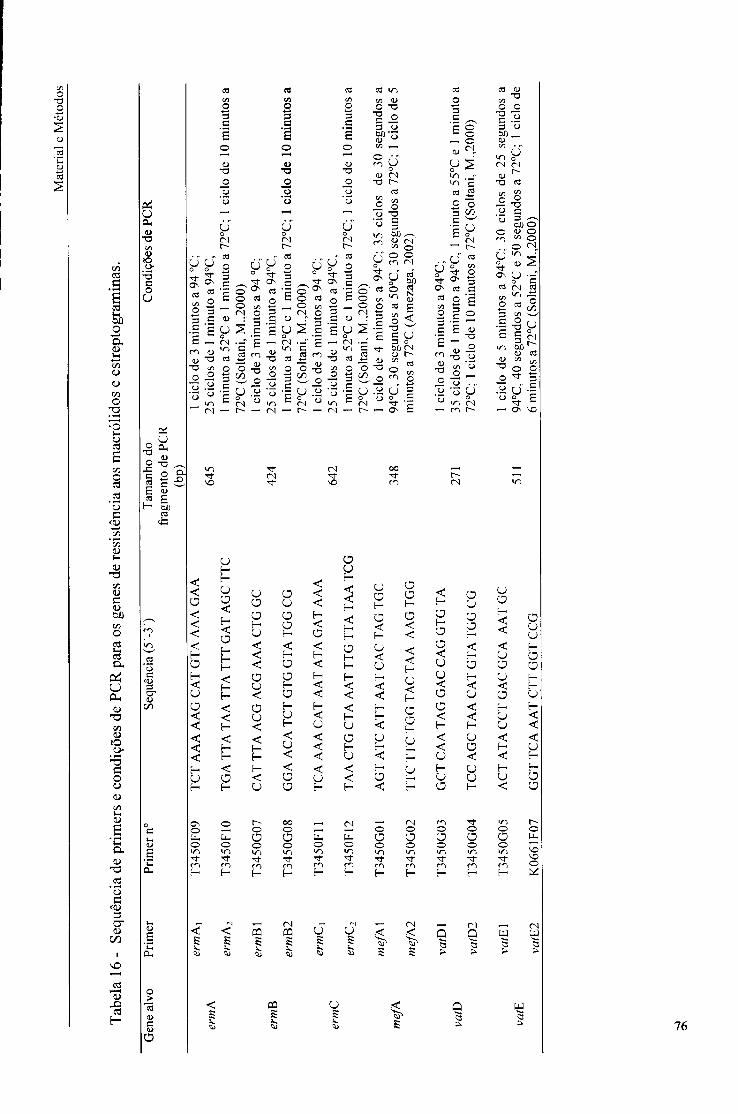
o o o
eu
"C CU
o
05 O Kj
.2 " o a CU I H tu
T3 co <u a CU b û c/1 O RJ s c3 D,
o
a CO cu
IO O -3 d o cu CU cu E G
CU o
CS . —H
CU C
<cu =3 cr cu
E/5
i
_2 "cu
■a H
Pá U CL, cu
a co cu
(O c>
T3 S o U
a!
O "O
c :
CU
E M) ca
U O O o
3 C
T3
U o CN r~ CS
O 3 d
u u
3 g S
o
_o o
' 3
U o CN t~
ca o 3
3 C
CU
a
ca >/i « eu O T3
| O 3 O
t/5 —
o 3 3
i O o , — 1 o
CU CN
ca CO o 3 d
S -
" O on o ° Õ .a _ ^
tN
ca „ O Ol M „
u S o tN • -i n C « ca ca t»
O "5 3 S | u C CN
— r
3 9 C cu E ' g O g — CN
cri ca o 3
cu a
cu a
O o
_ T &<* o ca O 05 CN O
si •E ê ca m
ti
o o r i rca o ^4 í í
ca o 3
E U
u ca F3 y •3 o .a
çu
' 3
r n
O O
^r
o ^
§ g ^ 3 * E 1^ a u o o
u o CN rca CO o
a a 3
O O m CN
y s i n _" i s ca S 2 Õ 3 c£,
CN — r.
. . O ca
ca ca N cu
U <* P
u
« c/3 3 ■Su
o o c 3 bû CN cu t~~ " ca
m O
" IO — CN
r f N
•— r~~ — (N —
CN "0
a U CN rf _ h " 0 > E
3 d
os ca d ca © "S « tt
c
o g o
1 - o ■8 s '
5
_2 "3 . . .2 o O
00 r~ CN
^ CN aa oa S s !» i . ca ÎU
u
< m u s S s V. Ik k cu Cil Î J s 3
ca eu -a
CO O O
l-â 3 u
co ■ *
CN CN eu «
-a ca co s? o S
— T3 .a C ° M O
O U O m « o
o CN U ^ ^
r C3S U • _ o C ca o ) ca CO ^ *
2 « co • E ^ U E = C N
eu ca 73 o o O t 3
S y-S O VO
U o H u
< < a
r
u a <c
< r
H
u a
O U
< < <
r
< <
U a
a a H
< a u
U a
■
< < <
r
u a <c
< r
H
a H u
a o
H < a
< H H
< H
a < <
O H a
a a r
s— < <
o u
> i <
r
u a <c
< r
H < < < o O <
< f —
a < < r
ca r
r
u a <c
< r
H < H b- f- < < < f —
< H u a õ o
r
u a <c
< r
H < a < H U < < f — u a a a
c <cu 3
H < < o a i-
< < r
< u <
u < < u
< r
H a eu
U H <c o < < < u <
o O a U 00 a < a H H < r O
O f—
a < H H < < u O < H H O
O f—
< < U < < r < r U U <
O O f— i- H U <
< < < < < a U u < U < < < H H U < H H u
< a H U < f- H < < U < H u < < r
H < r < < < H U H o H H U O < a U < O r U u U a H f- U o H H <C H O r < a
o o r 00 r i r i r i "* en rc o o o ,_H o o O O o o î _ LL. Ci. a a PH LZ a O a a a u. eu O o o o o o o o o o o ' ' 1 W) V~l IT) '/-> >o m ici cri i n cri cri o 1 ■ *
-* ■<t "tf ^r ^r rr ■ t f ^t <3- rt *õ C r n m m m m Cl m r n m r i m o
OH H r H H H H H f- H r f- U
(N ^ r i _ CN _ c-\
S < 5?
Q û "5 3
^J S 5 > > > >
tu
76

Material e Métodos
Tabela 17 - Concentrações dos reagentes usados na reacção do PCR para detecção dos genes ermA, ermB e ermC
Reagente Concentração
stock Concentração
final
erm Ai 82 (iM 50 pmol
ermA 2 65,2 \M 50 pmol
ermBi 58,2(iM 50 pmol
ermBi 64,4(iM 50 pmol
ermCi 68,4|iM 50 pmol
ermC2 80,2|iM 50 pmol
MgCl2 25 mM 2mM
Tampão lOx l x
Taq polimerase 5U/|ll 0,5 U
DNTP (dATP, dCTP, dGTP,dTTP)
4mM 200 iM
77
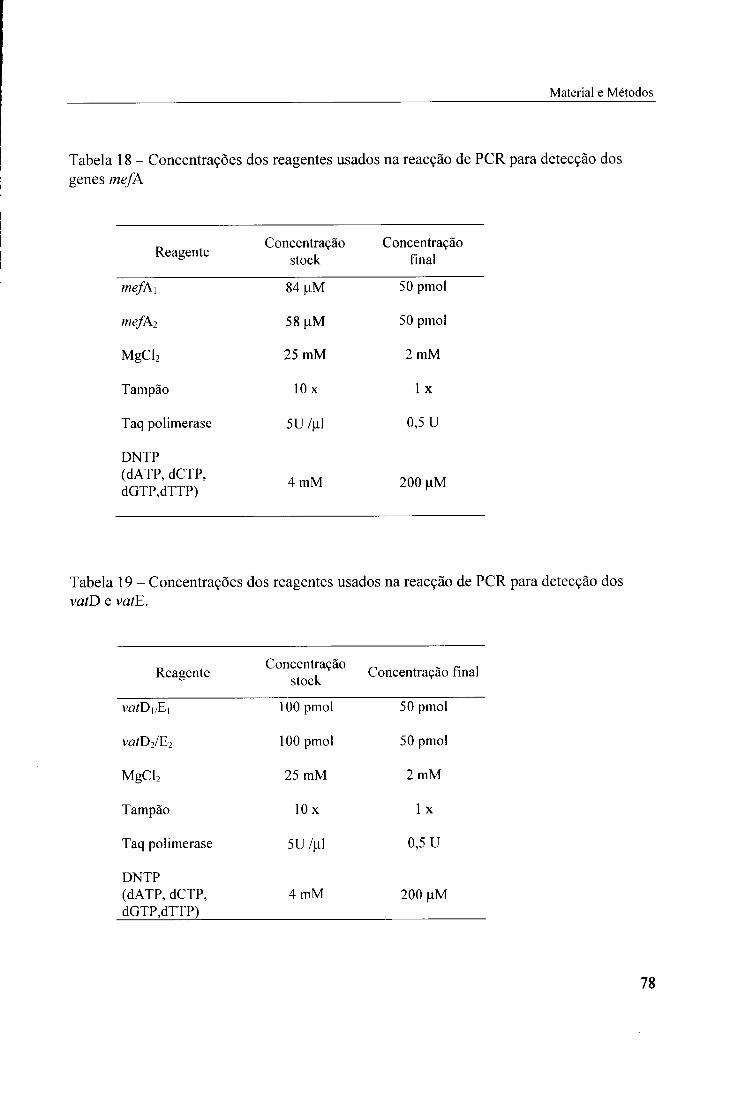
Material e Métodos
Tabela 18 - Concentrações dos reagentes usados na reacção de PCR para detecção dos genes mefA
Concentração Concentração Reagente , , c ,
° stock final me/Ai 84 pM 50 pmol
mefA2 58 pM 50 pmol
MgCl2 25 mM 2mM
Tampão lOx 1 X
Taq polimerase 5U/pl 0,5 U
DNTP (dATP, dCTP, dGTP,dTTP) 4mM 200 pM
Tabela 19 - Concentrações dos reagentes usados na reacção de PCR para detecção dos vatD e vatE.
Concentração „ ~ r- i Reagente , Concentração tinal
vatD\/E\ 100 pmol 50 pmol
vatD2/E2 100 pmol 50 pmol
MgCl2 25 mM 2mM
Tampão lOx 1 X
Taq polimerase 5U/pl 0,5 U
DNTP (dATP, dCTP, dGTP,dTTP)
4 mM 200 pM
78

Material e Métodos
Para avaliar os resultados das reacções de PCR foram realizadas electroforeses em gel de
agarose 1,5% em tampão Tris-acetato EDTA (TAE). Em paralelo realizou-se a
electroforese de um marcador de fragmentos de tamanho conhecido, "lOObp DNA Ladder"
(Promega, Madison, USA). Após a electroforese o gel foi mergulhado em Brometo de
Etídio (0,5 (Ag/ml) e submetido a um transiluminador de radiação ulravioleta (Perkin-
Elmer. GeneAmp, PCRSystem2400).
A aquisição da imagem obtida e a estimativa do tamanho dos fragmentos foi efectuada
através do sistema Kodak Digital Science.
3.5. Transferência dos determinantes genéticos de resistência às estreptograminas e
macrólidos por conjugação
Efectuaram-se ensaios de conjugação nos isolados de E. faecium, E. faecalis, E.
gallinarum e Enterococcus sp. com resistência à eritromicina e à associação quinupristina-
dalfopristina e em isolados de E. faecium com resistência intermédia à associação
quinupristina-dalfopristina e resistência à eritromicina.
Nos ensaios de conjugação foram utilizadas as estirpes receptoras E. faecalis JH2-2 e E.
faecium GE1, as quais são resistentes à rifampicina e ao ácido fusídico, E. faecium H239 e
E. faecalis 21769, como controlos do protocolo e utilizou-se como estirpe controlo a
transconjugante TE 237.1. Sempre que se procedeu à conjugação de isolados de E. faecalis
e E. faecium utilizaram-se como receptoras estirpes da mesma espécie.
Procedeu-se ao isolamento de cada uma das estirpes em meio BHI e colocaram-se as
placas na estufa a 37°C durante 24 horas. Após esse período de crescimento, os isolados
dadores e as estirpes receptoras foram, cultivadas em 5 ml de meio TSB e incubou-se na
estufa a 37°C durante 24 horas .
Posteriormente, prepararam-se placas de meio BHI e colocou-se um filtro de água em cada
uma das placas. Sobre esse filtro colocaram-se 200|ll de meio TSB onde cresceu a bactéria
dadora e sobre esta 20ul de meio TSB onde cresceu a bactéria receptora.
79

Material e Métodos
Homogeneizou-se cuidadosamente e incubou-se na estufa a 37°C durante 24 horas.
O crescimento obtido foi, na totalidade, suspenso em lml de soro fisiológico. De cada uma
das suspensões de soro fisiológico foram retirados IOOJLLI, que foram plaqueados em placas
de BHI contendo ácido fusídico (25ug/ml), rifampicina (100 (i.g/ml) e eritromicina
(8|ig/ml). As placas foram colocadas na estufa a 37°C e foram feitas observações até 72
horas após a incubação. Para os isolados onde se verificou crescimento até às 72 horas,
procedeu-se ao isolamento de uma colónia em placas de BHI, contendo os antibióticos nas
concentrações previamente referidas. Após o crescimento procedeu-se ao isolamento de
uma colónia em meio de Muller-Hinton. De seguida realizaram-se reacções de PCR nas
condições previamente descritas para detecção dos genes ermB e vatE.
80

Resultados
4. Resultados
4.1. Susceptibilidade a agentes antimicrobianos em Enterococcus sp.
Foi analisada uma colecção de isolados de Enterococcus sp. com origem em frangos para
determinar a incidência dos genes de resistência ermA, ermB, ermC, me/Â, vatD e vatE
por reacções de PCR, nas condições específicas, previamente descritas.
Dos isolados de Enterococcus sp. analisados determinou-se ( Carla Novais, comunicação
pessoal) a Concentração Mínima Inibitória à eritromicina, à associação quinupristina-
dalfopristina e vancomicina pelo método de diluição em agar, sendo posteriormente
classificados em sensíveis, intermédias ou resistentes de acordo com as normas
preconizadas pelo NCCLS (NCCLS, 1999). Na tabela 20 encontra-se discriminada a
distribuição das CMIs nos isolados de enterococos analisados, para os antibióticos
testados.
Verificou-se, igualmente, que 14% dos isolados analisados possuíam simultaneamente
resistência à vancomicina (glicopéptido), à associação quinupristina-dalfopristina
(estreptogramina) e à eritromicina (macrólido), 18% possuíam resistência à vancomicina e
à associação quinupristina-dalfopristina, 10% demonstraram ter resistência à vancomicina
e à eritromicina e 1% era resistente à vancomicina e à associação quinupristina-
dalfopristina. Entre os isolados analisados, a resistência simultânea à vancomicina, à
associação quinupristina-dalfopristina e à eritromicina era mais frequente entre os isolados
de E. faecium, assim como a resistência simultânea à associação quinupristina-dalfopristina
e à eritromicina. Verificou-se também a resistência simultânea à vancomicina e à
associação quinupristina-dalfopristina, embora com baixa frequência (4 isolados; 2
isolados de E. faecium e 2 isolados de E. gallinarum).
Constatou-se que dos 131 isolados de Enterococcus faecalis estudados, 35 (26,7 %) eram
sensíveis à eritromicina, 12 (9,1%) apresentavam comportamento intermédio e 84 (64,2%)
eram resistentes a este agente antimicrobiano.
81

Resultados
Dos 138 isolados de Enterococcus faecium estudados, 11 (7,9%) eram sensíveis à
eritromicina, 12 (8,7 %) apresentavam comportamento intermédio e 115 (83,4 %) eram
resistentes a este antibiótico. Dos 61 isolados de Enterococcus gallinarum estudados, 23
(37,7%) eram sensíveis à eritromicina, 5 (8,2 %) apresentavam comportamento intermédio
e 33 (54,1 %) eram resistentes a este antibiótico. Relativamente aos 58 isolados de
Enterococcus sp. analisados verificou-se que 11 (19%) eram sensíveis à eritromicina, 2
(3,4%) apresentavam comportamento intermédio e 45 (77,6%) eram resistentes a este
fármaco.
Relativamente à distribuição de CMIs para a associação quinupristina-dalfopristina, foi-nos
possível constatar que dos 32 isolados de E. faecalis analisados, 2 (6,3%) isolados eram
sensíveis e que 30 (93,7%) isolados eram resistentes a este fármaco. Dos 127 isolados de
E. faecium estudados, 21 (16,5%) eram sensíveis à associação quinupristina-dalfopristina,
24 (18,9%) apresentavam comportamento intermédio e 82 (64,6 %) eram resistentes a este
antibiótico. Dos 54 isolados de E. gallinarum estudados, 4 (7,4%) eram sensíveis à
associação quinupristina-dalfopristina, 11 (20,4 %) apresentavam comportamento
intermédio e 39 (72,2 %) eram resistentes a este antibiótico. Relativamente aos 53 isolados
de Enterococcus sp. analisados verifícou-se que 9 (17%) eram sensíveis à associação
quinupristina-dalfopristina, 9 (17%) apresentavam comportamento intermédio e 35 (66%)
eram resistentes a este fármaco.
82
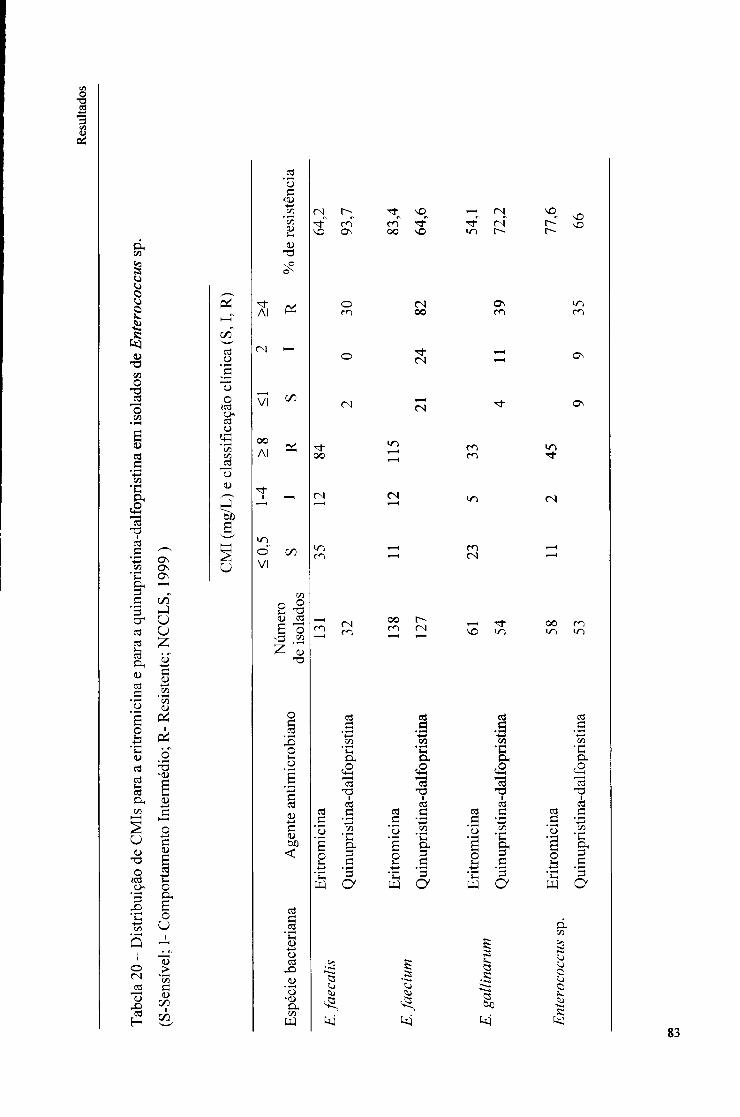
o T3
aí
O CO &5 3 o o o o o 5-Cu
*-* K 1*3 CU
T3 CO
o -d
CS
o to • l"H
s <U cd
.tí "■S3 M
•c D. O
4H
"3 T3 cd
.tí < " ■*
' -M O M o
'd o Q,
r__ 3
r tí O) '3 hJ cr O cd U cd z >-. cd aí CL)
<L> Cd +• tí CO
C/3 o D
Ë tó o s
1
Cd
ò cd '-B Cd
CS
-CU
D. tu ■t»
LO tí *~H *—
S o O 3
(D 1 3
cu
S cd
O o '3 - O
o 0 0 U
•.—I
Q " 1 * ?
CU o > CN '3 -2 tí
cd
Cd
CO
cd o
cu O
ícd % cu S °C0
O) j d "o cu
J "Sb £
o
co
Al
.5 '5 tí
<CU
LO CU
cu -a
Cd
CN «
VI
0 0 Al
CO
Cd
i n o" vi
co
LO
o .2 IH 13 cu cd S o
^ co -a
o c cd
O
cu í—»
tí cu
<
cd tí cd G cu
t—* cu cd
X> .2 ' o cu O LO
CN r-
o
oo
CN
CN 0 0
<N
CN
^i CN CN
i n
O
O
3oo
CN
IO r n
CN
un
CN
ÍH ^ 00
CN
CN
0 0 un
cd .tí
L0 ' C
O.
"cd Ç3 cd
cd H 3 22
cd tí
I * - » LO
•e D,
"3 I
cd tí
cd .tí
CO
•a cd
cd tí S '& cu . 2
cd tí
O
"3 3 tí
g O g 3 s o tí o =3 o 3 o I H tí s tí I H tí I H
«H> +u t—' +-1 U H '3 ' | H '3 °C 3 '£ -Li a UJ O w a W
a Cl
a
cd C
' 3 .22 •**. SH
tí '3
O H LO l>3
S u 'o o
83

Resultados
4.2. Distribuição de genes de resistência aos macrólidos e estreptograminas em
isolados de Enterococcus sp.
4.2.1. Presença de genes erm
bp 1500
500
10 11
Figura 5- Identificação dos genes ermB por PCR e electroforese em gel de agarose
(1-Marcador ; 2-Fragmento de 424 bp correspondente ao gene ermB na estirpe controlo; 3-
5; 6-7; 8-11- Fragmento de 424 bp correspondente ao gene ermB aos isolados 18F5,
18F6, 19F1, 19F2, 19 F5,19F8, 19F9, 19F10, 21F5).
Após a realização das reacções de PCR e de electroforese obtiveram-se bandas com o peso
molecular esperado (424 bp) em 49% dos enterococos analisados, os quais possuíam
resistência à eritromicina (CMI > 8mg/L), em 4 % dos enterococos analisados, os quais
demonstraram comportamento intermédio (lmg/L < CMI < 4mg/L), e em 5 % dos
enterococos analisados, os quais eram sensíveis à eritromicina (CMI < 0,5mg/L),
independentemente da espécie (Tabela 21).
Dos isolados estudados que evidenciavam comportamento sensível à eritromicina (n= 35),
11 (8,4 % do total) isolados possuíam o gene ermB, enquanto que 24 (18,3 % do total)
isolados não apresentavam o referido gene. Dos isolados de E. faecalis com
comportamento intermédio à eritromicina (n= 12), 7 (5,3 % do total) isolados possuíam o
gene ermB , enquanto que em 5 (3,8 % do total) isolados o referido gene não foi detectado.
84
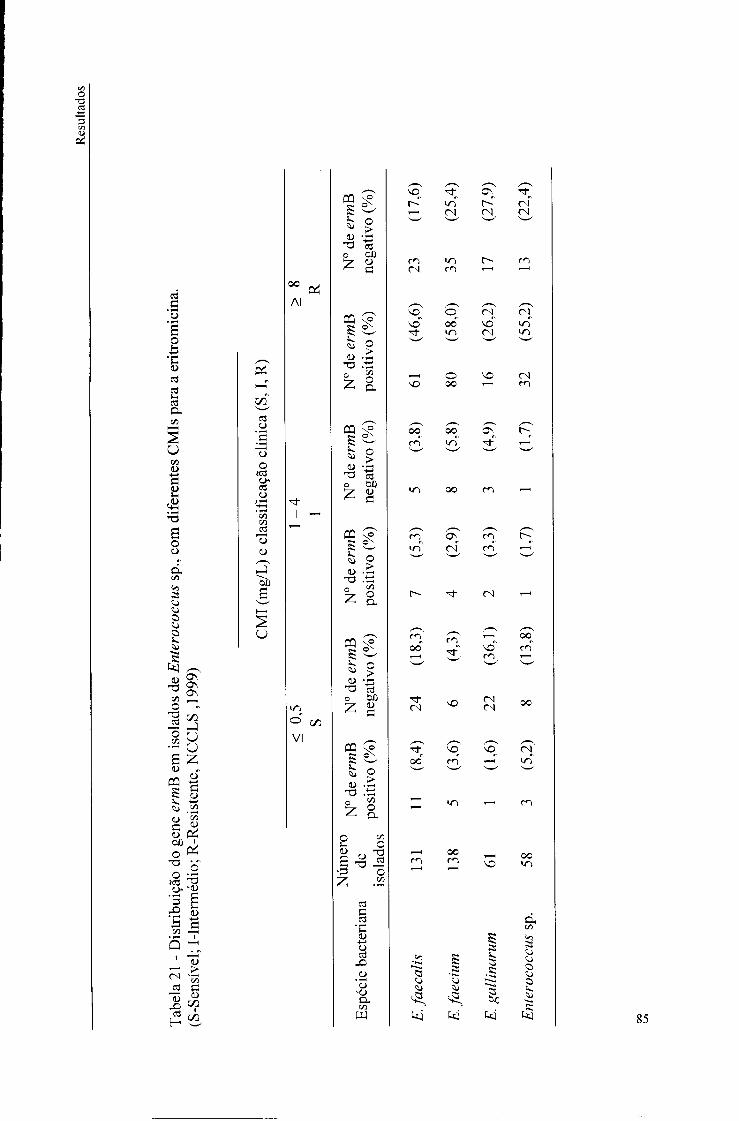
o ca
Pi
C3
d
O
C (D 03 ca i-H ccj
U c/l 1)
S CD SH
C+H
S o o
cfi
S Ca
o Ca o Si
o
° u •SO s z S 0)
<u PS Dû I. O *
O '<D
a s C/3 l—l
CU
c/3 d CU
CO
CN
"03 cci
H
Pi
V3
cu
ê cues CU
• I—I c/3 c/3
-2 "o
CU
~ 5 Û
£
o
00
Al PÍ
*
LO
VI
,^~V CcT ^ o^ Tp t-»" »o r" CN
CN
* > *—' w w s ' cu \ s
13 "S o Dû 7 <u CO IO r m 7 <u CN ro
~ ~ ^
—*. Co C? oT CN ■o CO vo" LO"
<* LO CN LO v _ ca O r > cu .ÏH o .ti ° O Z p.
,—i o VO CN ° O Z p.
vo 00 m
03 ^ °°r,
oo °l r03 ^ rn LO' ^f ^^
^^ ^ ' CU • rt
XJ ta o Dû 7 cu LO oo r i
^ ^ v rn OS rn t > IO r i rn _T
ca o s—' "^ ^^
7, > CD . 1 - j T3 . t i ° O Z cu
t-> ^1- CN —
^^ , N ^ ^—s m ^
v , — i 00 03 ^ oo" en MO c l 03 ^
, — i
en m T —H
v—' ' ' X '
CU ' rt T3 oj o Dû 7 cu ^ d
VO CN CN 00
CQ so Tf vq, vq CN CQ so oo r i r—l LO"
c3 o w
7. > CD .& T3 ."d
^H IO , 1 m
° O Zi o,
O 5S s o
' 3 o m
00 m * — i
VO oo LO
Z .22
cyj
ccj
c .2 cu +-» CU C3
X) CD
'Õ 'CU
O H C/i
0-1
cu ^ 3
hq
ca
ttj k j
O H C/3 to :.
O o ca
—
3 85

Resultados
Nos isolados de E. faecalis resistentes à eritromicina (n= 84), 61 (46,6 % do total) isolados
possuíam o gene ermB, enquanto que 23 (17,6 % do total) isolados não possuíam o
referido gene.
Dos isolados de E. faecium sensíveis a este fármaco (n=l 1), 5 (3,6 % do total) possuíam o
gene ermB, enquanto que 6 (4,3 % do total) dos isolados não o apresentavam. Dos isolados
de E. faecium com comportamento intermédio à eritromicina (n=12), 4 (2,8 % do total)
isolados possuíam o gene ermB, enquanto que em 8 (5,8 % do total) dos isolados o referido
gene não foi detectado. Nos isolados de E. faecium com resistência à eritromicina (n=115),
80 (58 % do total) isolados possuíam o gene ermB e 35 (25,4 % do total) não possuíam o
gene mencionado.
Nos isolados de E. gallinarum sensíveis à eritromicina (n=23), 1 (1,6 % do total) isolado
possuía o gene ermB e 22 (36,1 % do total) não possuíam este gene. Nos isolados de E.
gallinarum com comportamento intermédio à eritromicina (n=5), 2 (3,3 % do total)
isolados possuíam o gene ermB, enquanto que em 3 (4,9 % do total) dos isolados o gene
ermB não foi detectado. Dos isolados de E. gallinarum com resistência a este antibiótico
(n=33), 16 (26,2 % do total) isolados possuíam o gene ermB e 17 (27,9 % do total) não
possuíam o gene referido.
Dos isolados de Enterococcus sp. sensíveis a este antibiótico (n=ll), 3 (5,2 % do total)
possuíam o gene ermB, enquanto que 8 (13,3 % do total) dos isolados não o possuíam. Dos
isolados de Enterococcus sp. que apresentavam comportamento intermédio à eritromicina
(n=2), 1 (1,7 % do total) isolado possuía o gene ermB e 1 (1,7 % do total) isolado não o
possuía. Nos isolados de Enterococcus sp. com resistência à eritromicina (n=45), em 32
(55,2 % do total) isolados foi detectado o gene ermB ao contrário dos 13 (22,4 % do total)
isolados onde este gene não foi detectado.
Em nenhum dos isolados analisados foi detectada a presença dos genes ermA e ermC.
86
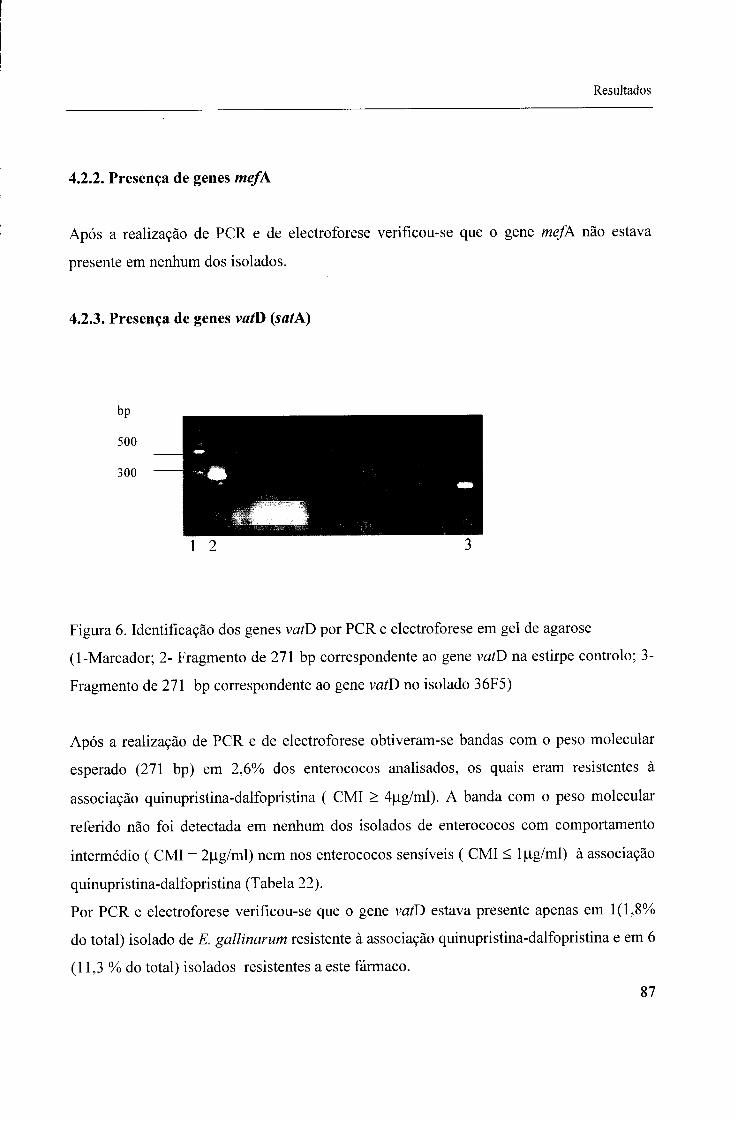
Resultados
4.2.2. Presença de genes mefA
Após a realização de PCR e de electroforese verificou-se que o gene mefA não estava
presente em nenhum dos isolados.
4.2.3. Presença de genes vatD (satA)
Figura 6. Identificação dos genes vatD por PCR e electroforese em gel de agarose
(1-Marcador; 2- Fragmento de 271 bp correspondente ao gene vatD na estirpe controlo; 3-
Fragmento de 271 bp correspondente ao gene vatD no isolado 36F5)
Após a realização de PCR e de electroforese obtiveram-se bandas com o peso molecular
esperado (271 bp) em 2,6% dos enterococos analisados, os quais eram resistentes à
associação quinupristina-dalfopristina ( CMI > 4|0.g/ml). A banda com o peso molecular
referido não foi detectada em nenhum dos isolados de enterococos com comportamento
intermédio ( CMI = 2|ig/ml) nem nos enterococos sensíveis ( CMI < lu.g/ml) à associação
quinupristina-dalfopristina (Tabela 22).
Por PCR e electroforese verificou-se que o gene vatD estava presente apenas em 1(1,8%
do total) isolado de E. gallinarum resistente à associação quinupristina-dalfopristina e em 6
(11,3 % do total) isolados resistentes a este fármaco. 87

o -a
tu
aí B
o*
ê ~? a
I a
Cd
a,
U C/3
CU —
O o d, C/3 &5 3 o o
(D -O
c/3 O
T3 J3 "o
c/3
ON
ON
S W3 CU i—l
õ z a c
p», c/3
« $ <U PS bû '. O *
-73 -
CO I—I
Q 1 cu
<N > cd c
•s H 5S
tó
C/2
Kl O
o o
ícd O cd O
'E/3 C/5
J3 "o
cu
"Sb
u
Al t *
<N ^ H
VI C/3
P £
CU rt T3 td o OJ) 7 cu
P N?
> o T3 . t i ° O
p £ t: "—' P» O
> cu -O
'3 o ao z CU
a
N 3 9s
CU T3
c/3 O O H
P -N?
CU T3
O >
DD CU
B
§ ^ <u . >
T 3 -4-» o c/3
^ O H
CU T 3 C/3
si CU 03
S Õ 3 C/3
cd B cd
O cd
- O cu
' o 'CU O H t/3
w
o' O N
ON. (N
en
<N
CM rn
0 0
■<fr
o"
0 0 rn
O H
m ~
ON oo' o"
tN
-
Vi NO"
CN
2 «*
ON CN)
VO
r-H ° ^
[ >
ON
O O O
O H C/3
C<3 S
o o ti)
tel
88

Resultados
4.2.4. Presença de genes vatE (satG)
Figura 7- Identificação dos genes vatE por PCR e electroforese em gel de agarose
(1-Marcador; 2-Fragmento de 511 bp correspondente ao gene vatE na estirpe controlo; 3-
Fragmento de 511 bp correspondente ao gene vatE no isolado 19 FIO)
Após a realização de PCR e de electroforese obtiveram-se bandas com o peso molecular
esperado (511 bp) em 11 (4,1 %) isolados de enterococos analisados, os quais eram
resistentes (CIM > 4(ig/ml) à associação quinupristina-dalfopristina, em 2 (0,8%) isolados
analisados, os quais demonstraram comportamento intermédio (CIM = 2ug/ml) e em 1( 0,4
%) isolado analisado, o qual era sensível (CIM < lug/ml) à associação quinupristina-
dalfopristina, independentemente da espécie (Tabela 23).
Verificou-se que nos isolados de E. faecalis, o gene vatE estava presente em 1 (3,1% do
total) isolado resistente, mas ausente nos isolados sensíveis e com comportamento
intermédio à associação quinupristina-dalfopristina. Nos isolados de E. faecium o gene
vatE estava presente em 1 (0.8 % do total) isolado sensível e ausente em 20 (15.7 % do
total) isolados sensíveis à associação quinupristina-dalfopristina. O referido gene estava
presente em 1 (0.8 % do total) isolado com resistência intermédia e ausente em 23 (18.1%
do total) dos isolados com resistência intermédia. O gene vatE foi detectado em 7 (5.5 %
do total) isolados resistentes, não tendo sido detectado em 75 (59.1% do total) destes
isolados.
89

Resultados
Relativamente aos isolados de E. gallinarum o gene vatE não estava presente em nenhum
dos isolados analisados.
Nos isolados de Enterococcus sp. o gene vatE encontrava-se em 1 (1.9 % do total) isolado
com comportamento intermédia e em 3 (5.7 % do total) isolados resistentes a este fármaco.
90

o T3
2 3 ÍT>
<D
aí
d a • !H
■ » — »
C/j
•c O H
o(%
) Co o" ON"
I/> r i o
o(%
)
13 o
i
d
o .5S
Z <D o i o o CN
CO
•a O H
0 CN t> m m CO
•a O H
* *
1 Al , s , , , , 1 cn
i n o
un
a 0 w
?" O o3 » S F
3 •o ;g
S o 'c/l O H l/l
M P? Z o ^ O H
1 ' r o m
S c f O * , V / V s—v O
t/D r f C/l
tt 03
a — o " oo o" i n
O ? o *— s—' • •
Si k3 o £ «s "o ? * T3 o
íCd 2, 60 Z u
o CN
^ 0 0
S O cd o
o ( N >—i
o, '35 w ^ o 0 0
o / V
Os » 5 H** © o S "3 CS
w ' ^ s
o i> > o o » .> o o C-T ^ %_»
2 ~5i) o 'c/i o o -
c s—
/
t*3 <L>
T 3 l / l O ^
u <3
vo" I O ^ t--'
c5
"d ov ? O 1—' —'
3 o^ u . > o ° ^ Id c/l ^ ^ ^ Id
ã 2 £ , Ml Z çu
CN o CN
<*t O
" u CO VI o o
CO VI
g z ,—, / s oc , , , ,
^ .? w i°. O ^ o O ■>— i CU ■*"* p , o •ta o ?> o 2 ci o +3 <u 'K o 'c/1
<U K Z 2 O ^ o o
?<A ■ o o O ço
O • " »H O le3 33 '3 1
cu n
• 3 O
CN m
Is-
uri
.-a s Z .22 fí o V I M 03
5 ± 1 à m >
kH D
■ 4 * S [/i C«5
O
<N ' O s
[/i C«5
O
bel
a S
ens 03
X> cu
■S3 ~3
S Ik a .s
[/i C«5
O
bel
a S
ens
o t i "^ H &
o 'CU
O H C/l
5u u « ,
w ^ fei fcj c§ 91

Resultados
4.2.5. Presença simultânea de genes de resistência
Na tabela 24 encontram-se discriminados os isolados que evidenciaram em simultâneo os
genes vatD e ermB, vatE e ermB e vatD e vatE .
Tabela 24 - Isolados com os genes vatD e ermB, vatE e ermB e vatD e vatE presentes simultaneamente.
Número de isolados com presença simultânea de
Espécie bacteriana vatD e ermB vatE e ermB vatD e vatE
E. faecalis 0 E. faecium 0 E. gallinarum 1 Enterococcus sp 3
Os genes vatD e ermB foram observados simultaneamente em um isolado de E. gallinarum
susceptível à associação quinupristina-dalfopristina e em três isolados de Enterococcus sp.
com diferentes comportamentos à associação quinupristina-dalfopristina e à eritromicina.
Verificou-se, igualmente, a presença simultânea dos genes vatE e ermB em um isolado de
E. faecalis, em três isolados de E. faecium e em dois isolados de Enterococcus sp. Todos
os isolados em que se detectou em simultâneo estes genes apresentavam resistência à
eritromicina e comportamento variável à associação quinupristina-dalfopristina.
Os genes vatD e vatE foram observados simultaneamente em dois isolados de
Enterococcus sp.
1 3 0 2
0 0 0 2
92

Resultados
4.3. Distribuição do gene ermB em isolados de Enterococcus sp, com diferentes CMIs
para a vancomicina
O gene ermB foi observado em 68 (51,9 %) isolados de E. faecalis (n = 131), em 45 (32,6
%) isolados de E.faecium (n = 138), em 5 (8,2 %) de E. gallinarum ( n = 61) e em 15 (25,9
%) isolados de Enterococcus sp. ( n - 58) sensíveis à vancomicina. Este gene foi
observado, igualmente, em 14 (23 %) isolados de E. gallinarum ( n = 61) com
comportamento intermédio ao antibiótico referido. Da mesma forma, o gene mencionado
foi observado em 11 ( 8,4 %) isolados de E. faecalis (n = 131), em 43 (31,2 %) isolados de
E. faecium (n = 138) e em 20 (34,5 %) isolados de Enterococcus sp ( n = 58) resistentes a
este fármaco (tabela 25).
93
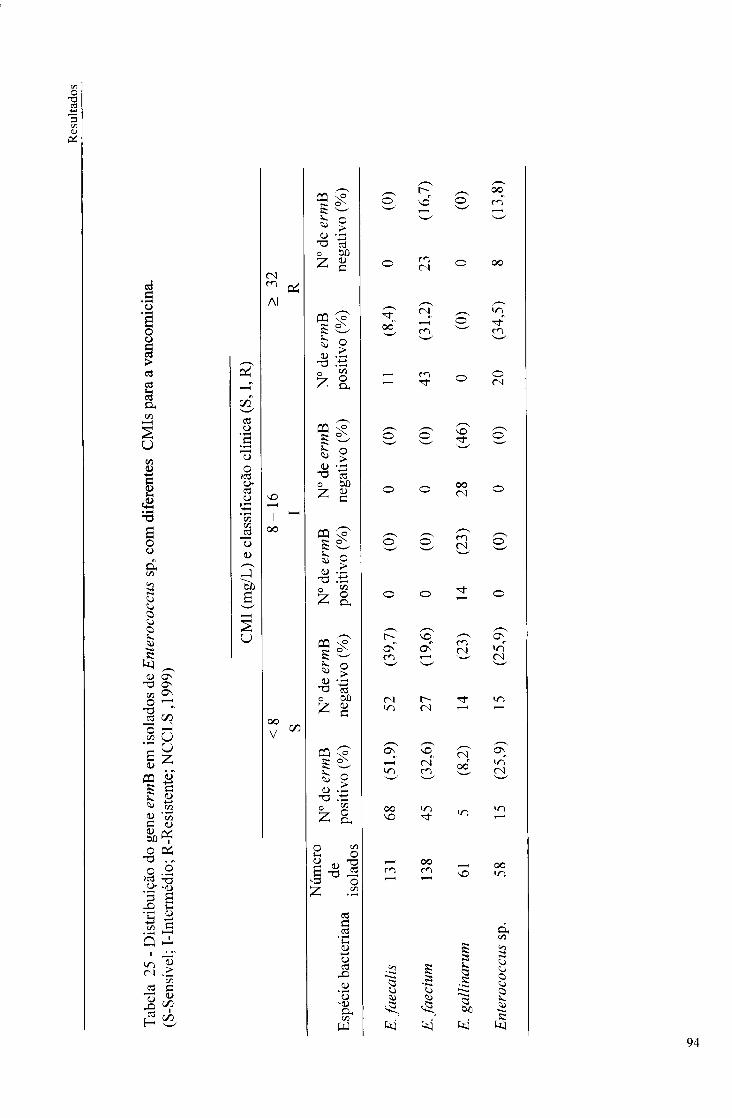
o a ca
CC
si G
'5 S o o
g > ces cd sC3 & en
^ H
s o Kl CU +| S3 CU s
4 H ■ *
-a £ o cu
d c/5 "5 S o CJ o o o s*. cu ~-K t*5 / s cu O N
-o O CO O O ^
-a " _2 C7) "o J CO O
Ï CU
u Ï CU
z m cu
s c <*> cu
CO
cu C/3 a CU CU
I
o cv T3 O o
ÍOCS ■3 O <U
'3 E j O c c/5
cu
1—1
Q 1
i .u I O cu ( N _>
Ë75 JS c 13 CU
C/D ccS
ISl
Cá
x/i cd
o o
ícd o cd cu
'ai CO
"cu CU
U
Al
^O
oo
ífi
m cu O
> cu
-o "S o bO
z cu
oo" CN <s i o
Tf'
v /, cjj O «; > CU . Ï H
-o .-a ° o ^ (O
Tf O o
ffl ^ k. cu o
> cu • r—
T3 3 o ou z cu
m cu CU
-a
O > CO O D.
« £ cu CU
T3
m cu CU
O > M cu G
O O.
O 50
C O
a-si Z .2
ccj
I s. CU *í cu cd
X) <L>
' Õ cu Cleo
W
to <N
Os
o ) I O
i O
oo
5 Cu cu
J3 Uj
Os
SO CN
I O
00
cu cu
Tf
OO CN
t o tN
O ^
to CN
CN oo
r-l ^°
C3 tX3
kj hj
oo CO
O) io ' CN
Os io"
IO ^
00 I O
o, CO
S Cu O cj O
94

Resultados
4.4. Avaliação da transferência dos determinantes de resistência à eritromicina e às
estreptograminas por conjugação
Procedeu-se a ensaios de conjugação em 18 isolados resistentes à eritromicina e que
possuiam o gene ermB e em todos os isolados com mais do que um gene de resistência aos
macrólidos, lincosamidas e estreptograminas, resistentes à eritromicina.
Os ensaios de conjugação demonstraram a transferência, por este processo, dos
determinantes de resistência ermB de duas estirpes dadoras de E. faecium e de uma estirpe
dadora de E. faecalis, resistentes à eritromicina, para as estirpes receptoras da mesma
espécie.
Relativamente ao determinante de resistência vatE observou-se apenas a sua transferência
de uma estirpe de E. faecalis resistente à associação quinupristina-dalfopristina para uma
estirpe receptora da mesma espécie.
No nosso estudo foi possível observar a transferência simultânea dos genes ermB e vatE
entre isolados de E. faecalis resistentes à eritromicina e à associação quinupristina-
dalfopristina. Contudo, não foi possível obter transconjugantes que evidenciassem em
simultâneo os genes ermB e vatE em isolados com comportamento intermédio e sensível à
eritromicina e à associação quinupristina-dalfopristina. Esta impossibilidade de obter
transconjugantes com múltiplos genes de resistência aos macrólidos e estreptograminas foi
também observada quando se procedeu à conjugação com a estipe dadora de Enterococcus
sp. que continha os genes vatD e ermB.
95

Discussão
5. Discussão
Da análise dos resultados constatou-se que a maioria dos isolados de Enterococcus sp.,
com origem em frangos, recolhidos entre 1999 e 2002, apresentavam resistência aos
antibióticos estudados. Dos isolados de Enterococcus sp. analisados, verificou-se que 71%
possuíam resistência à eritromicina e 70% à associação quinupristina-dalfopristina.
Verificou-se, igualmente, que a maior incidência de isolados resistentes à eritromicina
ocorria na espécie E. faecium e que os isolados de E. faecalis apresentavam a maior
incidência de resistência à associação quinupristina-dalfopristina, um facto que parece
estar associado com a presença nesta espécie do gene Isa (Singh, K. V. et ai, 2002). De
salientar ainda que 14% dos isolados de Enterococcus sp. analisados possuíam
simultaneamente resistência à vancomicina (glicopéptido), à associação quinupristina-
dalfopristina (estreptogramina) e à eritromicina (macrólido), 18% possuíam resistência à
vancomicina e à associação quinupristina-dalfopristina, 10% demonstraram ter resistência
à vancomicina e à eritromicina e 1% era resistente à vancomicina e à associação
quinupristina-dalfopristina (Novais, C. et ai, 2002).
Tendo sido estudada a distribuição dos genes de resistência à eritromicina e à associação
quinupristina-dalfopristina em isolados de Enterococcus sp. constatou-se a presença de
genes ermB, vatD e vatE. À semelhança do que foi detectado noutros estudos, com este
trabalho verificou-se que a carne de frango constitui uma fonte de enterococos portadores
de genes de resistência, os quais podem ser transferidos para o Homem, através da cadeia
alimentar (Simjee, S. et ai, 2002). Um aspecto a ter em consideração é o facto das carcaças
de frango pertencerem às dez marcas mais comercializadas em Portugal, tendo por isso
uma ampla distribuição nacional.
O gene ermB estava presente em 57% dos isolados de Enterococcus sp., surgindo em 38%
dos isolados sensíveis à eritromicina, em 45% dos isolados com resistência intermédia e
em 68% dos isolados resistentes a este antibiótico.
96

Discussão
Entre os isolados analisados, a resistência simultânea à vancomicina, à associação
quinupristina-dalfopristina e à eritromicina foi mais frequente entre os isolados de E.
faecium, assim como a resistência simultânea à associação quinupristina-dalfopristina e à
eritromicina. Verificou-se também a resistência simultânea à vancomicina e à associação
quinupristina-dalfopristina, embora com baixa frequência (4 isolados; 2 isolados de E.
faecium e 2 isolados de E. gallinarum).
Por seu lado, o gene vatD foi detectado em 3% dos isolados de Enterococcus sp., tendo
sido observada a sua presença apenas nos isolados que demonstravam resistência à
associação quinupristina-dalfopristina, surgindo em 4% destes isolados. O gene vatE, por
sua vez, surgia em isolados de E. faecalis resistentes à associação quinupristina-
dalfopristina. Este gene foi observado, igualmente, em isolados de E. faecium sensíveis,
com comportamento intermédio e em isolados resistentes à associação quinupristina-
dalfopristina e em isolados de Enterococcus sp. com comportamento intermédio e
resistentes a este fármaco.
Neste trabalho não foi possível observar a presença de genes ermA nos isolados de
Enterococcus sp. analisados. No entanto, Portilho e colaboradores (Portilho A. et ai, 2000)
referem ter observado este gene de resistência em Enterococcus sp., admitindo ter sido a
primeira vez que tal observação foi feita. Jensen e colaboradores, referem ter observado,
igualmente, o gene ermA em isolados de E. faecium, com origem em frangos, mas com
baixa frequência (Jensen L. B. et ai, 2002). Contudo, este gene é mais frequentemente
relacionado com a resistência aos macrólidos em Staphylococcus aureus e em
Staphylococcus coagulase-negativo (Portilho A. et ai, 2000).
Da mesma forma, neste trabalho, não foi possível fazer a observação dos genes ermC em
isolados de Enterococcus sp. Este gene não foi igualmente observado em estudos
realizados por Lim e colaboradores em isolados de Enterococcus sp. com origem em
doentes hospitalizados à semelhança do observado por outros investigadores. No entanto,
tal como os genes ermA, os genes ermC são mais frequentes em Staphylococcus
coagulase-negativo (Lim. J-A. et ai, 2002).
97

Discussão
Neste trabalho, o gene ermB foi detectado em 31.4% dos isolados de E.faecalis sensíveis à
eritromicina, em 41.7% dos isolados com resistência intermédia e em 72.6% dos isolados
resistentes à eritromicina, ou seja, foi detectado em 60% dos isolados de E.faecalis.
Num estudo realizado por Aarestrup e colaboradores (Aarestrup, F. M. et ai, 2000) na
Dinamarca, foi verificado que 44% dos isolados de E. faecalis com origem em frangos,
eram resistentes à eritromicina, frequência inferior à obtida neste trabalho, a qual foi de
64%. No estudo referido de Aarestrup e colaboradores (Aarestrup, F. M. et ai, 2000), o
gene ermB surgia em 64% dos isolados de E. faecalis, com origem em frangos, resistentes
à eritromicina, enquanto que no nosso trabalho o gene mencionado foi observado com uma
incidência superior (72.6%). Por seu lado, neste trabalho, o gene ermB surgiu em 64.5 %
(3.6 % em isolados sensíveis à eritromicina, 2.9% em isolados com comportamento
intermédio e 58% em isolados resistentes) dos isolados de E. faecium. Estes valores são
superiores aos referidos por Aarestrup e colaboradores (Aarestrup, F. M. et ai, 2000), para
a investigação realizada na Dinamarca, onde 74% dos isolados de E. faecium, com origem
em frangos eram resistentes à eritromicina, ocorrendo o gene ermB na totalidade dos
isolados. A incidência da resistência à eritromicina em isolados de E. faecium observada
no nosso estudo e no estudo de Aarestrup e colaboradores (Aarestrup, F. M. et ai, 2000), é
superior à observada por Simjee e colaboradores (Simjee, S. et ai, 2002), num estudo
realizado nos Estados Unidos da América, no qual 13% dos isolados de E. faecium eram
resistentes à eritromicina, surgindo o gene ermB em 30% dos isolados analisados.
Num estudo realizado por Schmitz e colaboradores (Schmitz F-J. et ai, 2000) o gene ermB
foi detectado em 93% dos isolados de E. faecium, procedentes de vários países europeus,
com origem em doentes hospitalizados, resistentes à eritromicina (Schmitz F-J. et ai, 2000.
No estudo referido, o gene ermA foi observado em 4% dos isolados e em 3% dos isolados
foi detectada a presença simultânea do gene ermA e ermB (Schmitz F-J. et ai, 2000),
enquanto que neste estudo não foi observado o gene ermA em nenhum dos isolados.
Neste trabalho, o gene ermB foi detectado em 4.3% dos E. gallinarum sensíveis, em 40%
dos isolados com resistência intermédia e em 48.5% dos isolados resistentes à eritromicina,
ou seja, em 31% (n=19) dos isolados de E. gallinarum. Simjee e colaboradores (Simjee S.
98

Discussão
et al, 2002) detectaram o gene ermB em 100% (n=2) dos isolados de E. gallinarum, com
origem em frangos, resistentes à eritromicina, mas não foi detectado nos isolados sensíveis
(Simjee S. et ai, 2002).
Neste trabalho, o gene ermB foi observado em 27.2% dos isolados de Enterococcus sp
sensíveis à eritromicina, em 58% dos isolados com resistência intermédia e em 71% dos
isolados resistentes.
Os resultados obtidos neste trabalho estão de acordo com os descritos noutras
investigações, tendo-se verificado que o gene ermB era o mais frequentemente encontrado
entre os isolados de Enterococcus sp resistentes analisados.
No entanto, Lim e colaboradores salientam o facto de no fenótipo de resistência MLSB, os
genes demonstrarem especificidade, dado que o gene ermA é mais frequente em S aureus,
o gene ermB em Enterococcus sp e que o gene ermC é mais frequente em Staphylococcus
coagulase-negativo (Lim, J-A et ai, 2002).
De salientar que neste trabalho foi observada a presença de genes ermB em isolados de E.
faecalis, E. faecium, E. gallinarum e Enterococcus sp. sensíveis à eritromicina, à
semelhança do descrito por Del Campo e colaboradores (Del Campo, R. et ai, 2003). Este
fenómeno foi igualmente observado em Streptococcus pneumoniae (Del Campo, R. et al,
2003). Por outro lado, neste trabalho, foram observados isolados de E. faecalis, E. faecium,
E. gallinarum e Enterococcus sp. resistentes aos macrólidos, mas onde o gene ermB não
foi observado. Este fenómeno foi observado por outros autores, que admitem que deverão
existir outros mecanismos de resistência aos macrólidos, para além dos descritos
(Aarestrup, F. M. et ai, 2000; Borgen, K. et ai, 2002; Schmitz F-J. et ai, 2000). O gene
msrC, que está relacionado com uma bomba de efluxo da família ABC, confere resistência
aos macrólidos e estreptograminas (Portillo, A. et ai, 2000). Portillo e colaboradores
(Portillo, A. et ai, 2000) admitem que o gene msrC é um gene intrínseco em E. faecium,
podendo, por esta razão, ser utilizado na identificação desta espécie. Segundo este autor, a
ubiquidade do gene msrC entre os isolados de E. faecium pode indicar que é um gene
ancestral, presente no cromossoma desta espécie bacteriana ou presente num plasmídeo
(Lim, J-A, et ai, 2002; Portillo, A. et ai, 2000).
99

Discussão
Pelo contrário, Werner e colaboradores (Werner, G. et al, 2001) consideram que o gene
msrC não é intrínseco em E. faecium porque não se encontra em todos os isolados desta
espécie bacteriana. No entanto, este autor, à semelhança de Portillo e colaboradores
(Portillo, A. et ai, 2000) admite que o gene msrC não foi adquirido como resposta à
pressão de selecção provocada pela utilização de antibióticos (Portillo, A. et ai, 2000;
Werner, G. et al, 2001). Este gene deverá ter uma função para além do efluxo de
antibióticos, mas a sua expressão tem a consequência referida (Portillo, A. et ai, 2000).
Neste trabalho não foi detectada a presença de genes mefA em nenhum dos isolados
analisados, independentemente de serem sensíveis, de possuírem resistência intermédia ou
de serem resistentes à eritromicina. Esta situação foi também verificada por Portillo e seus
colaboradores ( Portillo A. et ai, 2000). Contudo, Luna e seus colaboradores (Luna, V. A.
et ai, 2002) detectaram a presença do gene mefA em 9% dos isolados de Enterococcus sp.,
Staphylococcus aureus, Streptococcus intermedius e de Streptococcus sp. resistentes aos
macrólidos, num estudo realizado com crianças portuguesas. Estes autores admitem ter
detectado pela primeira vez o gene mefA em isolados de Staphylococcus (Luna, V. A. et ai,
2002). Da mesma forma, Lim e seus colaboradores (Lim, J-A, et ai, 2002) referem a
presença do gene mefA em 2% dos isolados de Enterococcus sp estudados, com origem em
doentes hospitalares (Lim, J-A, et ai, 2002). Luna e seus colaboradores verificaram que o
gene mefA surgia isoladamente, ou juntamente, com o gene ermB em Enterococcus sp. e
em E.faecalis (Luna, V. A. et ai, 2002). Este gene tem sido observado em bactérias Gam-
positivo como S. pyogenes, S. pneumoniae, S. agalactiae, assim como Micrococcus luteus,
Corynebacterium jeikeium, Corynebacterium spp e em "Streptococcus viridans",
demonstrando uma grande distribuição entre as bactérias (Lim, J-A, et ai, 2002).
Num estudo realizado por Lim e colaboradores foi observada a presença simultânea dos
genes mefA e ermA e dos genes mefA e ermC em isolados de Staphylococcus coagulase-
negativo (CNS) e do gene mefA e ermB em isolados de Enterococcus sp., resistentes à
eritromicina( Lim, J-A, et ai, 2002). Os genes ermA, ermB e mefA foram igualmente
observados simultaneamente em isolados de enterococos resistentes à eritromicina (Lim, J-
A, et ai, 2002).
100

Discussão
Portillo e colaboradores, pelo facto de não terem detectado, em estudos realizados, a
presença do gene mefA, admitem que poderá existir uma distribuição geográfica
heterogénea destes genes de efluxo dos macrólidos, em E.faecium (Portillo A. et ai, 2000).
Por seu lado, Del Grosso e colaboradores admitem que o gene mefA não é muito comum
na Europa, entre os isolados de S. pneumoniae resistentes aos macrólidos, sendo, no
entanto, muito frequente nos Estados Unidos da América (Del Grosso, M. et ai, 2002). De
facto, os mecanismos de resistência aos macrólidos mediados pelo gene mefA, são mais
frequentemente encontrados na América do Norte, países do Norte da Europa e no Japão
(Pechère, J.-C, 2001).
Dado que em vários isolados resistentes aos macrólidos não foi observada a presença de
genes mef de genes erm, nem de genes vat, a resistência poderá ser ser devida a outros
genes de resistência, como por exemplo, msrA/B, UnAIA', vga e vgb. Relativamente aos
isolados de E. faecalis, neste trabalho, verificou-se que 6.3% dos isolados eram sensíveis à
associação quinupristina-dalfopristina, 3.1% possuíam resistência intermédia e que 90.6%
possuíam resistência a este fármaco. Contudo, na totalidade dos isolados analisados não foi
detectado, por reacções de PCR, a presença do gene vatD em isolados de E. faecium e de
E. faecalis, à semelhança do que foi observado por Simjee e colaboradores (Simjee, S. et
ai, 2002a; Simjee, S. et ai, 2002b). Hays e seus colaboradores verificaram que 78% dos E.
faecium isolados em frangos, nos contentores utilizados para o seu transporte e nos locais
de produção, eram resistentes à associação quinupristina-dalfopristina, mas não possuíam
igualmente o gene vafD, apesar da frequência de isolados de E. faecium resistentes à
associação quinupristina-dalfopristina, de origem animal, especialmente em frangos, ser
elevada quer na Europa, quer nos Estados Unidos da América, onde o gene vatD nunca foi
observado nestes isolados bacterianos (Simjee, S. et ai, 2002a; Simjee, S. et ai, 2002b;
Hays, J.R. et ai, 2001). Apesar destes resultados Simjee e colaboradores admitem que o
gene vatD predomina na Europa (Simjee, S. et al, 2002a). De facto, este gene foi
observado em isolados de E. faecium resistentes à associação quinupristina-dalfopristina
(21%), com origem animal e em doentes hospitalizados, na Europa (Soltani M. et ai,
2000).
101

Discussão
Ao contrário do observado por Simjee e colaboradores (Simjee, S. et ai, 2002a), no
presente estudo o gene vatD foi detectado em 2.6% dos isolados de E. gallinarum
resistentes à associação quinupristina-dalfopristina. Neste trabalho a incidência do gene
vatD em isolados de Enterococcus sp. foi de 17%, um valor superior ao determinado por
Jensen e seus colaboradores, que verificaram uma incidência de 10 % nos isolados de
Enterococcus sp resistentes à associação quinupristina-dalfopristina, com origem em
frangos (Jensen , L.B. et ai, 2002).
Relativamente ao gene vatE, no nosso estudo foi detectada a sua presença nos isolados de
E. faecalis, o que seria de estranhar em virtude desta espécie possuir resistência intrínseca
às estreptograminas, provavelmente devido a mecanismos de efluxo do antibiótico (Simjee,
S. et ai, 2002b). No entanto, Simjee e seus colaboradores (Simjee S. et ai, 2002b)
publicaram um estudo no qual admitem ter detectado pela primeira vez em E. faecalis o
gene vatE, o qual confere resistência às estreptograminas .
Os mesmos autores (Simjee, S. et ai, 2002a), num estudo realizado nos Estados Unidos da
América, detectaram a presença do gene vatE em 43% dos isolados de E. faecium, com
origem em frangos. Verificaram, igualmente, que apenas 32% dos isolados resistentes à
associação quinupristina-dalfopristina, possuíam o gene vatE (Simjee, S. et ai, 2002a).
Neste trabalho, verificou-se que 64.6% dos isolados de E. faecium possuíam resistência à
associação quinupristina-dalfopristina, mas apenas 8.5% dos isolados resistentes a este
fármaco, possuíam o gene vatE, ou seja, era maior a percentagem de isolados de E.
faecium resistentes à associação quinupristina-dalfopristina, mas o gene vatE estava
presente em menor número de isolados do que no estudo de Simjee e colaboradores
(Simjee, S. et ai, 2002). Estes autores admitem que a incidência do gene vatE em isolados
de E. faecium com baixa resistência à associação quinupristina-dalfopristina (CMI <
32mg/L) é diferente da que se observa na Europa, referindo que tal como os genes vatD, os
genes vatE também não foram observados neste continente (Simjee, S. et ai, 2002a).
Esta situação contrapõe o observado na nossa investigação, uma vez que o gene vatD
surgiu em 5 isolados de Enterococcus sp e em 1 isolado de E. gallinarum resistentes à
associação quinupristina-dalfopristina, mas com CMI inferior a 32mg/L. A mesma situação
foi observada para o gene vatE, dado que este gene foi observado em 1 isolado de 102

Discussão
E. faecium e em 4 isolados de Enterococcus sp., com CMI inferior a 32mg/L. Simjee e
colaboradores (Simjee, S. et ai, 2002a), admitem que em E. faecium resistentes à
associação quinupristina-dalfopristina, os genes vatD e vatE possam estar associados a
plasmídeos. Num estudo realizado, na Dinamarca, por Jensen e colaboradores foi detectada
a presença do gene vatE em 89% dos isolados de E. faecium resistentes à associação
quinupristina-dalfopristina, valor muito superior ao obtido neste trabalho (Jensen , L.B.et
ai, 2002).
Na nossa investigação, à semelhança do observado por Simjee e colaboradores (Simjee, S.
et ai, 2002a) o gene vatE não foi detectado em nenhum dos isolados de E. gallinarum.
Num estudo realizado por Soltani e seus colaboradores antes da utilização da associação
quinupristina-dalfopristina e da proibição da virginiamicina em isolados de diferentes
origens (isolados humanos, porcos e de aves), foi detectada a presença do gene vatE em 50
% dos isolados estudados (Soltani M.et ai, 2000).
Os resultados obtidos em várias investigações levantam sérias preocupações pelo facto de
se observar um aumento da incidência da resistência em isolados de enterococos, a
antibióticos utilizados na clínica para tratamento de infecções. Estas preocupações são
acrescidas pelo facto dos determinantes de resistência se encontrarem em elemento móveis
de DNA, como plasmídeos e transposões, os quais podem ser transferidos a bactérias
patogénicas humanas ou para bactérias da flora gastrointestinal humana, por conjugação ou
transformação (van den Bogaard, A. E. et al, 2002). Estes processos requerem o contacto
próximo entre a bactéria dadora e a bactéria receptora. Este pré-requisito indica que as
bactérias devem partilhar o mesmo ecossistema, como por exemplo, o tracto
gastrointestinal humano, para que a transferência horizontal de DNA possa ocorrer
(Courvalin, P., 1966). A utilização dos antibióticos aumenta a disseminação de bactérias
resistentes devido à pressão de selecção exercida por estes agentes (Tollefson, L. et ai,
1997) A população de enterococos na flora gastrointestinal humana é, provavelmente,
influenciada por vários factores, como por exemplo, a ingestão de enterococos de origem
animal através de alimentos contaminados. Esta ideia é apoiada pelo facto da resistência
aos mesmos antibióticos ter sido observada em isolados de origem humana e animal,
assim como os mesmos genes de resistência (Aarestrup, F. M. et ai, 2000).
103

Discussão
De facto, van den Bogaard e colaboradores observaram a presença de estirpes de E.
faecium resistentes à vancomicina (VRE), com padrões de Pulsed-Field Gel
Electrophoresis (PFGE) semelhante
e com elementos relacionados com o transposão Tnl546, onde se localiza o gene vanA,
que confere resistência à vancomicina, em isolados de um doente hospitalizado e em
suínos, o que apoia a hipótese da transmissão de VRE dos animais para o Homem, através
da cadeia alimentar (Dahl, K. H. et ai, 1999; van den Bogaard et ai, 2002; Lu, H-Z, et ai,
2002). van den Bogaard e colaboradores (van den Bogaard et ai, 2002) admitem que a
transmissão primária ocorre dos animais para o Homem, mas não em sentido contrário. O
mesmo autor, em virtude de não ter sido possível distinguir por PFGE uma estirpe de
Enterococcus sp. resistente à vancomicina isolada num avicultor e em perus da sua unidade
de produção, admite que os enterococos resistentes a este antibiótico, podem colonizar o
tracto gastrointestinal humano (van den Bogaard et ai, 2002). van den Bogaard admite que
a disseminação de enterococos resistentes dos animais para o Homem é mais importante do
que a troca de genes de resistência entre enterococos com origem animal e de origem
humana (van den Bogaard et ai, 2002).
Pelas importantes implicações na saúde humana, o aparecimento e disseminação de GRE
ou de elementos genéticos que conferem resistência aos glicopéptidos, tem sido objecto de
intensos estudos.
Os enterococos resistentes aos glicopéptidos que entram no tracto gastrointestinal humano
ou animal, através da cadeia alimentar ou por contaminação directa, transferem os seus
genes de resistência durante a passagem pelo intestino, não colonizando necessariamente o
hospedeiro (Descheemaeker, P. R. M. et ai, 1999), sendo a transmissão horizontal dos
genes de resistência mais importante do que a disseminação de clones (clonal spread) na
disseminação da resistência à vancomicina (Stobberingh, E. et ai, 1999; Dahl, K. H. et ai,
1999; Borgen, K. et ai, 2002). Esta ideia é partilhada por van den Braak (Bischoff, W. E. et
ai, 1999). A pressão de selecção exercida pelos antibióticos usados pelo Homem e como
promotores de crescimento podem ter estimulado a contaminação cruzada entre os
diferentes hospedeiros (Descheemaeker, P. R. M. et ai, 1999).
104

Discussão
O gene vanA, o qual confere resistência à vancomicina, conforme referido anteriormrnte,
localiza-se no transposão Tnl546 (Borgen, K. et ai, 2002) ou em elementos relacionados e
pode fazer parte do cromossoma em plasmídeos conjugativos ou não conjugativos (Dahl,
K. H. et ai, 1999; Borgen, K. et ai, 2002). Desta forma, devido à localização do gene vanA
num trasposão, a disseminação da resistência mediada por este gene pode ocorrer por
transmissão horizontal dos genes de resistência (Descheemaeker, P. R. M. et ai, 1999;
Robredo, B. et ai, 2000; Stobberingh, E. et ai, 1999)
Num estudo de Aarestrup e colaboradores (Aarestrup, F. M. et ai, 1998a) verificou-se que
nove meses após a proibição da utilização da avoparcina como promotor de crescimento, o
número de animais portadores de E. faecium resistentes à vancomicina, apesar de ter
diminuído ainda se mantinha elevado. Na Alemanha, Itália, Países Baixos e na Dinamarca,
desde que a avoparciana deixou de ser utilizada como promotor de crescimento, ocorreu
um decréscimo de Enterococcus sp. resistentes aos glicopéptidos isolados em frangos
(Portillo, A. et ai, 2000). No entanto, num estudo realizado na Dinamarca, verifícou-se que
a proporção de enterococos resistentes aos glicopéptidos isolados em suínos se manteve
constante após a proibição da utilização da avoparcina e que a sua frequência só baixou
depois da diminuição da utilização da tilosina, um macrólido (Aarestrup, F. M, 2000;
Maniatis, A. N. et ai, 2001). Neste mesmo estudo, verifícou-se, igualmente, que todos os
isolados de enterococos resistentes aos glicopéptidos analisados possuíam resistência à
eritromicina, a qual estava sempre relacionada com a resistência aos glicopéptidos, o que
foi verificado por conjugação (Maniatis, A. N. et ai, 2001; Report of the Scientific
Committee for Animal Nutrition on the Efficacy and Risk for Users of the Therapeutic
Macrolides Antibiotics Tylosin and Spiramycin Used as Feed Additives(opinion expressed
on 05 february 1998. Consumer policy and Consumer Healh Protection). Neste trabalho
constatou-se que 70% dos isolados de Enterococcus sp. resistentes à vancomicina
possuíam o gene ermB, pelo que a utilização de macrólidos em veterinária poderá
contribuir para a manutenção de VRE em frangos..
O gene ermB ocorria em 100% dos isolados de E. faecalis resistentes à vancomicina e em
57% dos isolados sensíveis. Relativamente aos isolados de E. faecium, verifícou-se que o
105

Discussão
gene ermB estava presente em 65% dos isolados resistentes à vancomicina e em 63% dos
isolados sensíveis. Este gene surgia em 1% dos isolados de E. gallinarum com resistência
intermédia à vancomicina e em 26% dos isolados sensíveis. No que diz respeito aos
isolados de Enterococcus sp. observou-se a presença dos genes ermB em 71% dos isolados
resistentes à vancomicina e em 50% dos isolados sensíveis. Admite-se que a ligação entre
os genes de resistência à eritromicina e à vancomicina nos enterococos resistentes aos
glicopéptidos nos animais de produção possa ter um papel importante na co-selecção ou na
manutenção da resistência aos glicopéptidos, quando os macrólidos são utilizados na
terapia animal ou como promotores de crescimento (Aarestrup, F. M., 2000; Maniatis, A.
N. et ai, 2001). Desta forma, a utilização da tilosina como promotor de crescimento terá
levado à manutenção do gene ermB em isolados de E. faecium, conforme demonstrado em
estudos realizados na Dinamarca. Por esta razão, a co-selecção do gene vanA devido à
selecção de genes ermB pela utilização de macrólidos, como por exemplo, a tilosina, tem
sido apontada como uma razão para a manutenção da elevada incidência de VRE
observada na Dinamarca, após a proibição da utilização da avoparcina (Aarestrup, F. M.,
2000; Borgen, K. et ai, 2002). A elevada incidência, observada na Noruega, onde a tilosina
nunca foi utilizada, de enterococos resistentes à vancomicina em isolados resistentes aos
macrólidos com origem em frangos, após a proibição da utilização da avoparcina, poderá
ser devida à utilização da espiramicina (macrólido) na terapia (Borgen, K. et ai, 2002).
Estes estudos demonstram a ligação entre a resistência aos glicopéptidos e à eritromicina,
permitindo a disseminação e manutenção da resistência à vancomicina na ausência da
avoparcina (Borgen, K. et ai, 2002).
A co-transferência da resistência à eritromicina e à vancomicina decorre do facto do gene
ermB e vanA se localizarem no mesmo plasmídeo, tendo sido observada a sua ligação em
vários isolados de E. faecium (Aarestrup, F. M., 2000; Borgen, K. et ai, 2002). O gene
ermB encontra-se junto da terminação 5'do transposão Tnl546, num plasmídeo
conjugativo, o qual é tranferível entre estirpes de E. faecium e, com menor frequência,
entre E. faecium e E. faecalis (Borgen, K. et ai, 2002).
106

Discussão
Por outro lado, estudos realizados em Espanha (Del Campo, R. et at, 2003), onde o
consumo de antibióticos, quer na terapia animal, quer como promotores de crescimento, é
elevado, demonstraram que a incidência da resistência à eritromicina e aos glicopéptidos
entre os enterococos isolados num grupo de manipuladores de alimentos era superior à
observada em indivíduos saudáveis, quatro anos após a proibição da utilização da
avoparcina e da tilosina. Por esta razão, Del Campo e colaboradores, admitem a
possibilidade de transmissão de determinantes de resistência ou de estirpes resistentes entre
os animais e o Homem (Del Campo, R. et at, 2003). Os autores referidos observaram
igualmente que existia uma variedade de mecanismos de resistência nos manipuladores de
alimentos maior do que em indivíduos saudáveis, o que poderá significar que os
manipuladores de alimentos estão expostos a uma maior variedade de enterococos do que
os restantes indivíduos (Del Campo, R. et at, 2003). Estes autores salientam ainda o facto
de terem observado uma única estirpe em indivíduos que trabalhavam em locais diferentes,
o que poderá indicar um potencial risco de disseminação de clones de VRE. Este aspecto
poderá apoiar a hipótese de uma permanência prolongada de certos clones no intestino de
alguns indivíduos, ou de elementos de resistência transferíveis e não pode ser excluída a
hipótese da permanência de microrganismos resistentes nas unidades de produção animal.
Estes autores admitem que os manipuladores de alimentos podem actuar como uma fonte
de transmissão de enterococos resistentes na população humana. Com efeito, vários
estudos levam a considerar os manipuladores de alimentos como um possível elo de
ligação entre os alimentos e o Homem (Del Campo, R. et at, 2003).
Stobbering e colaboradores (Stobbering, E. et ai, 1999) observaram a existência de
isolados de E. faecium resistentes à associação quinupristina-dalfopristina antes desta
associação ter sido introduzida na terapia humana. Admite-se que a utilização, durante
vários anos, da virginiamicina como promotor de crescimento, é responsável pela
resistência observada e pela transferência de enterococos resistentes ou de determinantes
de resistência dos animais para o Homem e pode estar relacionada com a resistência à
associação quinupristina- dalfopristina observada no Homem (Jensen, L. B. et ai, 2002;
Luh, K.-T. et ai, 2000; Stobbering, E. et ai, 1999).
107

Discussão
De facto, a virginiamicina selecciona E. faecium resistentes a este promotor de
crescimento, os quais possuem resistência cruzada com a associação quinupristina-
dalfopristina, surgindo as acetiltransferases VatD e VatE em isolados de enterococos de
diferentes países europeus (Luh, K.-T. et ai, 2000). Neste trabalho foram observados
isolados de E. faecium e de Enterococcus sp onde estavam presentes simultaneamente os
genes ermB e vatE.
Jensen e colaboradores (Jensen, L.B. et ai, 2000) admitem ter observado pela primeira vez
a ligação física entre os genes ermB e vatE, em isolados de E. faecium, na Dinamarca.
Estes autores consideram que estes genes se localizam num elemento genético muito
conservado provavelmente um plasmídeo. Contudo, em estudos realizados nos Estados
Unidos da América esta ligação não foi observada ( Jensen, L.B. et ai, 2000; Simjee, S. et
ai, 2002b).
Neste trabalho foi igualmente observada a presença simultânea dos genes ermB e vatD, em
isolados de E. gallinarum e de Enterococcus sp., assim como de isolados de Enterococcus
sp. que possuíam simultaneamente os genes vatD e vatE. Hamerum e colaboradores, por
seu lado, demonstraram a ligação entre os genes ermB e vatD, em isolados de E. faecium,
tendo efectuado a sua transferência por conjugação (Del Campo, R. et at, 2003). Estes
autores admitem que o gene "cluster" ermB-vatD possa localizar-se num elemento
genético relacionado com o transposão Tnl547ou Tn5385 (Del Grosso, M. et ai, 2002).
Simjee e colaboradores observaram a presença do gene vatE e ermB em isolados de E.
faecalis, o qual como já foi referido é intrinsecamente resistente às estreptograminas, com
origem em frangos (Simjee, S. et ai, 2002b). O facto dos genes ermB e vatE se
encontrarem na mesma estirpe e, em muitos casos, no mesmo plasmídeo, pode ajudar a
explicar a aquisição de vatE por E. faecalis. Este gene pode ter sido co-selecionado devido
à pressão de selecção provocada pela utilização de macrólidos na produção de frangos
(Simjee, S. et ai, 2002b). Por esta razão, um organismo como E. faecalis, que possui
resistência intrínseca às estreptograminas, pode, sob determinadas condições, possuir genes
de resistência como resultado da co-selecção. Esta possibilidade é apoiada pelo facto da
incidência da resistência à virginiamicina observada na Dinamarca em isolados de E.
108

Discussão
faecium, com origem em suínos, ter diminuído após a proibição da utilização deste
promotor de crescimento. Contudo, algum tempo depois verificou-se um aumento da
incidência desta resistência. Apesar de não ser evidente a razão para o aumento desta
incidência, admite-se ter sido uma consequência do aparecimento de isolados que eram
simultaneamente resistentes a vários antibióticos, entre os quais a eritromicina. A
utilização deste antibiótico quer isolada quer conjuntamente com outros antibióticos, pode
co-seleccionar e manter os genes vatE em isolados de E.faecalis (Simjee, S. et ai, 2002b).
Simjee e colaboradores verificaram, recorrendo a ensaios de conjugação, que o gene vatE é
transferível entre isolados de E. faecalis e entre E. faecalis e E. faecium sensíveis à
associação quinupristina-dalfopristina. Esta observação explica a existência de um
reservatório de genes vatE em E. faecalis, os quais podem ser transferidos para E. faecalis
ou E. faecium, podendo, igualmente ser transferidos para Gram-positivo humanos, através
da cadeia alimentar (Simjee, S. et ai, 2002b).
109

Conclusão
6. Conclusão
Os genes encontrados neste estudo, ermB, vatD e vatE, os quais se encontram associados
às resistências aos macrólidos e estreptograminas, apresentam uma incidência elevada e
semelhante ao encontrado em alguns trabalhos realizados por investigadores europeus.
Os nossos resultados corroboram o referido noutros estudos, em que se verifica que a carne
de frango constitui uma fonte de enterococos portadores de genes de resistência, os quais
podem ser transferidos para o Homem, através da cadeia alimentar.
De uma forma semelhante ao demonstrado por outros trabalhos, verificamos que o gene
ermB era o mais frequente entre os isolados de Enterococcus sp. O referido gene, para
além de ter sido detectado em isolados de Enterococcus sp. resistentes e com
comportamento intermédio à eritromicina, foi também observado em sensíveis, facto este
que tinha sido observado em isolados humanos, mas não em isolados de origem animal. A
semelhança do observado por outros investigadores, da análise dos nossos resultados,
verificou-se a ausência do gene ermB em alguns isolados resistentes à eritromicina. Cabe-
nos ainda referir que não nos foi possível observar o gene ermA, nos isolados analisados,
ao contrário de outros investigadores que detectaram a sua presença em isolados de
Enterococcus sp., apesar de ocorrer com baixa frequência. Tal como verificado por outros
investigadores, não nos foi possível observar o gene ermC nos isolados Enterococcus sp
analisados.
Embora o gene mefA tenha sido previamente reportado em isolados de Enterococcus sp.
resistentes aos macrólidos, com origem em crianças portuguesas não foi detectado nos
isolados de frango testados.
Apesar de alguns autores admitirem que a incidência do gene vatD é elevada na Europa, a
nossa investigação não nos permitiu observar a sua presença nos isolados de E. faecium e
de E. faecalis analisados, tendo apenas sido detectado em E. gallinarum e em
Enterococcus sp.
Ao contrário de uma outra investigação, o gene vatE não foi observado nos isolados de E.
faecalis. No presente estudo constatou-se que, apesar da incidência da resistência à 110

Conclusão
associação quinupristina-dalfopristina em E. faecium ser elevada, os genes vatE surgiram
apenas em 8.5% dos isolados. Ao contrário do referido noutros trabalhos, pudemos
observar o gene vatE em isolados de E. faecium sensíveis à associação quinupristina-
daldopristina.
A transferência por conjugação do gene ermB, isoladamente ou em simultâneo com o gene
vatE, verificada nos isolados testados, demonstra a possibilidade de disseminação destes
genes entre enterococos.
111

Bibliografia
7. Bibliografia Aarestrup, F. M. 2000. Characterization of Glycopeptide-Resistant Enterococcus faecium (GRE) from Broilers and Pigs in Denmark : Genetic Evidence that Persistence of GRE in Pig Herds is Associated with Coselection by Resistance to Macrolides. Journal of Clinical Microbiology. 38: 2774 - 2777
Aarestrup, F. M., H. Hasman, L. B. Jensen, M. Moreno, I A. Herrero, L. Dominguez, M. Finn & A. Franklin. 2002. Antimicrobial Resistance among Enterococci from Pigs in Three European Countries. Applied and Environmental Microbiology. 68: 4127-4129
Aarestrup, F. M., Y. Agerso, P. Gerner-Smidt, M. Madsen & L. B. Jensen. 2000. Comparision of antimicrobial resistance phenotypes and resistance genes in Enterococcus faecalis and Enterococcus faecium from humans in the community, broilers and pigs in Denmark. Diagnostic Microbiology and infectious Diseases. 37: 127-137
Aarestrup, F. M., F. Bager, N. E. Jensen, M. Madsen, A. Meyling & H. C. Wegener. 1998a. Resistance to antimicrobial agents used for animal theraphy in pathogenic-, zoonotic- and indicator bacteria isolated from different food animals in Dennmark : a baseline study for the Danish Integrated Antimicrobial Resistance Monitoring Programme (DANMAP). APMIS;106:745 - 770
Aarestrup, F. M., F. Bager, N. E. Jensen, M. Madsen, A. Meyling & H. C. Wegener. 1998b. Surveillance of antimicrobial resistance in bacteria isolated from food animals to antimicrobial growth promoters and related therapeutic agents in Denmark.APMIS;106: 606-622
Aarts, H. J. M., K. S. Boumedine, X. Nesme & A. Cloeckaert .2001. Molecular tools for the characterisation of antibiotic-resistant bacteria. Vet. Res. 32: 363-380
Acar, J. & P. Courvalin .1998. La fin de l'âge d'or des antibiotiques. Il est grand temps d'agir : certaines bactéries échappent à tout les traitements, La Recherche 314
Amezaga, M.R., P. E. Carte, P. Cash & H. Mckenzie. 2002, Molecular Epidemiology of Erytromycin resistance in Streptococcus pneumoniae Isolates from Blood of Noninvasive sites. Journal of Clinical Microbiology. 40: 3313-3318
Anglada, R. R. 1997. Microbiologia Sanitária e Clínica, Ed. Sintesis
Baquero, F. 1997. Gram positive resistance : challenge for the development of new antibiotics, Journal of Antimicrobial Chemotherapy, 39,suppl. A, 1-6
Bergogne-Bérézin, E. 1997. Nouvelles approches pharmacologiques des macrolides. Example de la roxithromycine. La Presse Médicale.26. Suppl.II.
112

Bibliografia
Bischoff, W.E., T. M. Reynolds, G. O. Hall, R. P. Wenzel & M. B. Edmond . 1999. Molecular epidemiology of vancomycin-resistant Enterococcus faecium in a large urban hospital over a 5-year period. Journal of Antimicrobial Chemotherapy. 37:3912-3916
Boerlin, P., A. Wissing, F. M. Aarestrup, J. Frey & J. Nicolet. 2001. Antimicrobial Growth Promoter Ban and Resistance to Macrolides and Vancomycin in Enterococci from Pigs. Journal of Clinical Microbiology. 39: 4193-4195
Boisseau, J. 1993. Basis for the evaluation of the microbiological risks due to veterinary drug residues in food. Veterinary Microbiology. Elsevier Science Publishers. 35 :187-192
Borgen, K., M. Sorum, Y. Wasteson, H. Kruse & H. Oppegaard. 2002. Genetic Linkage Between erm(B) and vanA in Enterococcus hirae of Poultry Origin. Microbial drug Resistance. 8: 363-368
Bower, C. K. & M. A. Daeschel. 1999. Resistance responses of microorganisms in food environments. International Journal of Food Microbiology, 50: 33 - 44
Bozdogan , B. & R. Leclercq. 1999b. Effects of Genes Encoding Resistance to Streptogramins A and B on the Activity of Quinupristin-Dalfopristin against Enterococcus faecium. Antimicrobial Agents and Chemotherapy. 43: 2720-2725
Bozdogan , B., L. Berrezouga, M. Kuo, D. A. Yurek, K. A. Farley, B. J. Stockman & R. Leclercq . 1999a.A New Resistance Gene, linB, Conferring Resistance to Lincosamides by Nucleotidylation in Enterococcus faecium HM1025. Antimicrobial Agents and Chemotherapy, 43: 925 - 929
Bozdogan, B. et al. 1999. Plasmid-Mediated coresistance to Strptogramins and Vancomycin in Enterococcus faecium HM 1032. Antimicrobial Agents and Chemotherapy. 43: 2097-2098
Butaye, P., K. V. Damme, L. A. Devriese, L. V. Damme, M. Baele, S. Lauwers & F. Haesebrouck . 2000. In vitro susceptibility of Enterococcus faecium isolated from food to growth-promoting and therapeutic antibiotics. International Journal of Food Microbiology. 54: 181-187
Buu-Hoi, A .não datado, Cocci a Gram positif et Macrolides-Lincosamides-Streptogramines
Cars, O., S. Môlstad & A. Melander, 2001. Variation in antibiotic use in the European Union.TheLancet.357: 1851-1853
Chang, S. et al. 2003. Infection with Vancomycin-Resistant Staphylococcus aureus Containing the vanA Resistance Gene. The New England Journal of Medicine. 348: 1342-1347
113

Bibliografia
Chu, D. T. 1999. Recent Developments in macrolides and ketolides. Current Opinion in Microbiology. Vol.11: 467-474
Chung, W. O., C. Werckenthin, S. Schwarz & M. C. Roberts. 1999. Host range of the erm F rRNA methylase gene in bacteria of human and animal origin. Journal of Antimicrobial Chemotherapy, 4:5-14
Cohen, M. L.1994. Antimicrobial resistance: prognosis for public health, Trends in microbiology, 2, n° 10
Committee on Drug Use in Food Animals, 1999, The Use of Drugs in Food Animals : Benefits and Risks, Panel on Animal Health, Food Safety and Public Health, National Research Council, National Academy Press, Whashington D.C.
Comunicação do Conselho, Posição comum (CE) N°20/2003, do Conselho de 17 de Março de 2003
Consumer Health Protection. Scientific Committees. Scientific Committee on Animal Nutrition. 1998a. Opinion of Scientific Committee for Animal Nutrition on the immediate and longer - term risk to the value of Streptogramins in Human Medicine posed by the use of Virginiamycin as an animal growth promoter ( produced at the request of the Commission in response to the action taken by Denmark under a safeguard clause to ban virginiamycin as feed addictive) The European Commission. Directorate General XXIV
Consumer Health Protection. Scientific Committees. Scientific Committee on Animal Nutrition. 1998b. Opinion of Scientific Committee for Animal Nutrition on the Efficacy and Risk for Users of the Therapeutics Macrolides Antibiotics Tylosin and Spiramycin Used as Feed Additives. The European Commission. Directorate General XXIV
Consumer Policy and Consumer Health Protection. 1999. Opinion of The Scientific Steering Committee on Antimicrobial Resistance. The European Commission. Directorate General XXIV
Cook, R.1997. Antimicrobial resistance- use in veterinary and human medicine,.Journal of Antimicrobial Chemotherapy.39 : 435-439
Corpet, D. E. 1993. An evaluation of methods to assess the effect of antimicrobial residues on the human gut flora. Veterinary Microbiology. Elsevier Science Publishers. 35:199-212
Corpet, D. 1998. Viande:Après les hormones, les antibiotiques ? Leur usage dans l'alimentation des animaux est controversé. La Recherche 314
Courvalin, P. 1996. Evasion of antibiotic action by bacteria.Journal of Antimicrobial Chemotherapy, 37: 855-869
114

Bibliografia
Cristino, J.M. , et ai .1996. Estudo multicentrico de microrganismos isolados e de resistência aos antimicrobianos em 10 hospitais Portugueses em 1994.Acta Med Port , 9 (4-6) :141-50.
Dahl, K. H., G. S. Simonsen, O. Olsvik, e A. Sundsfjord .1999. Heterogeneity in the van B gene cluster of genomically diverse clinical strains of vancomycin-resistant Enterococci, Antimicrobial Agents and Chemotherapy. 43: 1105-1110
Dayan, A . D. 1993. Allergy to antimicrobial residues in food : assesment of the risk to man. Veterinary Microbiology. Elsevier Science Publishers; 35 :213 - 226
Del Campo, R., P. Ruiz-Garbajosa, M. P. Sánchez-Moreno, F. Baquero, C. Torres, R. Canton & T. M. Croque. 2003. Antimicrobial Resistance in Recent Fecal Enterococci from Healthy Volunteers and Food Handlers in Spain : Genes and Phenotypes. Microbial Drug Resistance. 9:47-60
Del Grosso, M., F. Iannelli, C. Messina, M. Santagati, N. Petrosillo, S. Stefani, G. Pozzi & A. Pantosti. 2002. Macrolide Efflux Genes mef[A) and mefiE) Are Carried by Different Genetic Elements in Streptococcus pnemoniae. Journal of clinical Microbiology. 40: 774-778
Descheemaeker, P.R.M., S. Chapelle, L. A. Devriese, P. Butaye, P. Vandamme & H. Goossens.1999. Comparison of glycopeptide-resistant Enterococcus faecium isolates and glycopeptide resistance genes of human and animal origins. Antimicrobial Agents and Chemotherapy. 43: 2032-2037
Devriese, L.A., J. Hommez, R. Wijfels & F. Haesebrouck, 1991, Composition of enterococcal and streptococcal intestinal flora of poultry. Journal of Applied Bacteriology, 71, 46- 50
Dinh, T., I. T. Paulsen & M. H. Saier, Jr. 1994. A Family of Extracytoplasmatic Proteins That Allow Transport of Large Molecules across the Outer Membrane Of Gram-Negativa Bacteria. Journal of Bavteriology. 176 : 3825-3831
Directiva 70/524/CEE do Conselho, de 23 de Novembro de 1970, Jornal Oficial L270, 14/12/1970, pp.1-17
Directiva 97/6/CE da Comissão de 30 de Janeiro de 1997. Jornal oficial das Comunidades Europeias, N° L 35,05/02/1997, pp. 11-13
115

Bibliografia
Donabedian, S.M., L. A.Thai, M. B. Perri, J. W. Chow, P. Bartlett, R. Jones, K. Joyce, S. Rossiter, K. Gay, J. Johnson, C. Mackinson, E. Debess, J. Madden, F. Angulo & M. J. Zervos. 2003. Molecular Characterization of Gentamicin- Resistant Enterococci in the United States : Evidence of Spread from Animals to humans through Food. Journal of Clinical Microbiology. 41 :1109-1113
Dunlop, R.H.,S.A. McEwen, A . H. Meek, R.C. Clarke,W.D. Black & R.M. Friendship. 1998. Associations among antimicrobial drug treatments and antimicrobial resistance of fecal Escherichia coli of swine on 34 farrow-to-finish farms in Ontario, Canada. Preventive Veterinary Medicine. 34: 283-305
Eisner, H.-A., I. Sobottka, M. Claussen, R. Laufs & R. Wirth. 2000. Virulence Factors of Enterococcus faecalis and Enterococcus faecium blood Culture Isolates. Eur J Clin Microbiol Infect Des. 19: 39-42
Fackland, R. R. , Sahm D. S. & Teixeira, L. M., Manual of Clinical Microbiology, T Ed., ASM Press, Whashington, D.C.
FEDES A, 2004. Dispon+ivel em: http://www.fedesa.be7eng/PublicSite/xtra/dossiers/doss9/. Acedida em 20 de Fevereiro de 2004
Feinman, S. E. Antibiotics in Animal Feed - Drug Resistance Revisited. Routine addition of antibiotics to animal feed raises questions about the spread of antibiotic resistance. ASM News .64: 24 - 30
Fines, M., B. Perichon, P. Reynolds, D. F. Sahm & P. Courvalin. 1999. Van E, a New Type of Acquired Glicopeptide Resistance in Enterococcus faecalis BM 4405. Antimicrobial Agents and Chemotherapy. 43: 2161-2164
Firth, N. & R.A . Skurray.1998. Mobile elements in the evolution and spread of multiple - drug resistance in Staphylococci. Drug Resistance
Fraimow, H. S. & Courvalin P. 2000. Resistance to Glicopeptides in Gram-Positive Pathogens. American Society for Microbiology. Washington D.C.
Fraise, A. P. 1996. The Treatment and Control of Vancomycin Resistant Enterococci Journal of Antimicrobial Chemotherapy. 38: 753 - 756
Fraise, A. P. 1997.Vancomycin Resistant Enterococci in Turkeys and Farmers.The New England Journal of Medicine. 337:1558 - 1559
Franz, C.M.A.P., A. B. Muschill-Silberhorn, N. M. K. Yousif, M. Vancanneyt, J. Swings & W. H. Holzapfel. 2001. Incidence of Virulence Factors and Antibiotic Resistance among Enterococci Isolated from Food, Applied and Environmental Microbiology. 67: 4385-4389
116

Bibliografia
Hammerum, A. M., Flannagan, S. E., Clewell, D. B. & Jensen, L. B.. 2001, Indication of Transposition of a Mobile DNA Element Containing the vat(D) and erm(B) Genes in Enterococcus faecium. Antimicrobial Agents and Chemotherapy. 45: 3223-3225
Haroche, J., J. AUignet, S. Aubert, A. E. van den Bogaard & N. el Solh. 2000 satG, Conferring Resistance to Streptogramin A , Is Widely Distributed in Enterococcus faecium Strains but Not in Staphylococci. Antimicrobial Agents and Chemotherapy, 44:190-191
Hayes, J. R., A.C. Mcintosh, S. Qaiyumi, J. A.Johnson, L. L. English, L. E. Carr, D. D. Wagner & S. W. Joseph. 2001. High-Frequency Recovery of Quinupristin-Dalfopristin-Resistant Enterococcus faecium Isolates from the Poultry Production Environment. Journal of Clinical Microbiology. 39: 2298-2299
Huycke, M. M., D. F. Sahm & M S. Gilmore. 1998. Multiple-Drug Resistant Enterococci : The Nature of the Problem and na Agenda for the Future. Emerging Infectious Diseases.4:239-249
Jackson, E. R. 1993. The proper use and benefits of veterinary antimicrobial agents in practice in cattle. Veterinary Microbiology. Elsevier Science Publishers. 35 :349 - 356
Jensen, L. B., A . M. Hammerum & F. M. Aaretrup.2000. Linkage of vat(E) and erm(B) in Streptogramin-Resistant Enterococcus faecium Isolates from Europe. Antimicrobial Agents and Chemotherapy. 44: 2231-2232
Jensen, L.B. , A. M. Hameerum, F. Bager & F. M. Aarestrup. 2002. Streptogramin Resistance among Enterococcus faecium Isolated from Production Animals in Denmark in 1997. Microbial Drug Resistance. 8: 369-374
Johnston, A . M. 1998. Use of antimicrobial drugs in veterinary practice. British Medical Journal. 317: 665-667
Joyanes, P., Pascual, A., L. Martinez- Martinez, A. Hevia & E. J. Perea. 2002. In Vitro Adherence of Enterococcus faecalis and Enterococcus faecium to Urinary Catheters. Eur J Clin Microbiol Infect Dis, ,19: 124-127
Jornal Oficial da Comunidades Europeias (JOCE), N° C, 407/7 de 28 de Dezembro de 1998
Jornal Oficial das Comunidades Europeias (JOCE), n° L 60/16 de 4 de Março 1999
Jornal Oficial das Comunidades Europeias (JOCE),, 98/C 407/02
Kayser, F. H. 1993. Evolution of resistance in microorganisms of human origin. Veterinary Microbiology. Elsevier Science Publishers. 35: 257 - 267,
117

Bibliografia
Khachatourians, G. 1998. Agricultural use of antibiotics and the evolution and transfer of antibiotic-resistant bacteria.Canadian Medical Association Journal. 159:1129 -1136
Klein, G., A . Pack & G. Reuter.1998. Antibiotic Resistance Patterns of Enterococci and Occurence of Vancomycin-Resistant Enterococci in Raw Minced Beef and Pork in Germany, Appl. Environ. Microbiol.,64:1825-1830
Kirst, H. A., D. G. Thompson & T. I. Nicas. 1998. Historical Yearly Usage of Vancomycin. Antimicrobial Agents and Chemotherapy.42: 1303-1304
Lamb, H. M., D. P. Figgitt & D. Faulds. 1999. Quinupristin/dalfopristin. A Review of its Use in the Management of Serious Gram Positive Infections. Drugs. 58(6):1061-1097
Levy, S. B. 1998. Antibiotic Resistance : an increasing problem ? It always has been but there are things we can do. British Medical Journal. 316:1255 - 1256
Lim, J-A., A.-R. Kwon, S.-K. Kim, Y. Chong, K. Lee & E.-C. Choi. 2002. Prevalence of resistance to macrolide, lincosamide and streptogramin antibiotics in Gram-positive cocci isolated in a Korean hospital. Journal of Antimicrobial Chemotherapy. 49: 489-495
Lina, G., A. Quaglia, M. Reverdy, R. Leclercq, F. Vandenesch & J. Etienne . 1999. Distribution of Genes Encoding Resistance to Macrolides, Lincosamides and Streptogramins among Staphylococci. Antimicrobial Agents and Chemotherapy. 43: 1062-1066
Lopes, H. 1993. Les outils normatifs et réglementaires de la gestion de la qualité. Nadine Fraselle et Hassan Gemei, eds
Lu, H.-Z., X.-H. Weng, H. Li, Y.-K. Pang & Y.-W. Tang, 2002. Enterococcus faecium-Related Outbreak with Molecular Evidence of Transmission from Pigs to Humans. Journal of Clinical Microbiology. 40: 913-917
Luh, K.-T., P.-R. Hsueh, L.-J. Teng, H.-J. Pan, Y.-C. Chen, J.-J. Lu, J.-J. Wu & S.-W. Ho. 2000. Quinupristin-Dalphopristin Resistance among Gram-Positive Bacteria In Taiwan. Antimicrobial Agents and Chemotherapy. 44:,3374-3380
Luna, V. A., Heiken, M., Judge, K, Ulep, C , Kirk, N., V., luis, H., Bernardo, M. & Roberts, M. C. 2002. Distribution of mef(A) in Gram-Positive Bacteria from Healthy Portuguese Children. Antimicrobial Agents and Chemotherapy. 46: 2513-2517
Mackinnon, J. D. 1993. The proper use and benefits of veterinary antimicrobial agents in swine practice. Veterinary Microbiology. Elsevier Science Publishers. 35 :357-367
Maniatis, A.N., S. Pournaras, M. Kanellopoulou, F. Kontos, E. Dimitroulia, E. Papafrangas & A. Tsakris. 2001. Dissemination of Clonaly Unrelated Erytromycin and Glycopeptide-Resistant Enterococcus faecium Isolates in a Terciary Greek Hospital, Journal of Clinical Microbiology. 39: 4571-4574
118

Bibliografia
Martel, J. L. & M. Coudert. 1993. Bacterial resistance monitoring in animals : the French national experiences of surveillance schemes. Veterinary Microbiology. Elsevier Science Publishers. 35 : 321 - 338
Martel, J.L., F. Tardy, P. Sanders & J. Boisseau. 2001. New Trends in regulatory rules and surveillance of antimicrobial resistance in bacteria of animal origin, Vet. Res. 32: 381-392
McDonald, L. C , M.J. Kuehnert, F.C. Tenover & W.R. Jarvis .1997. Vancomycin-resistant Enterococci Outside the Health-Care Setting:Prevalence,Sources and Public Health Implications, Emerging Infectious Diseases.3
McEwan, S.A. & Fedorka-Cray, P., 2002, Antimicrobial Use and Resistance in Animals. Clinical Infectious Diseases. 34: S93-106
McKellar, Q. A .1998. Antimicrobial Resistance : a Veterinary Perspective. Antimicrobials are important for animal welfare but need to be used prudently. British Medical Journal. 317: 610 - 611
Millet, A .1998. Sommet Antimicrobien a Copenhague.Trois cents experts désorientés à la recherche d'un consensus. La Recherche 314
Min, Y-H, J.-H. Jeong, Y.-J. Choi, H.-J. Yun, K. Lee, M.-J. Shim, J.-H. Kwak & E.-C. Choi. 2003. Heterogeneity of Macrolide-Lincosamide-Streptogramin B Resistance Phenotypes in Enterococci. Antimicrobial Agents and Chemotherapy.47: 3415-3420
Murray, B.E. 1998. Diversity among Multidrug-Resistant Enterococci.Emerging Infectious Diseases. 4: 3 7 - 4 7
National Committee for Clinical Laboratory Standards,1999, Performance Standards for Antimicrobial Susceptibility Testing, M-100-S12,vol.22, N°l
National Office of Animal Health ( NOAH ) press release. 1999. Growth promoters environmental benefits. Environmental benefits of dietary enhancing feed additives.http://www.noah.demon.co.uk
Nikaido, H. 1996. Multidrug Efflux Pumps Of Gram Negative Bacteria. Journal of Bacteriology. 178:5853-5859
Nord, C. E. 1993 . The effect of antimicrobial agents on the ecology of the human intestinal microflora . Veterinary Microbiology. Elsevier Science Publishers. 35: 193-197
Novais, C , C. Vital, G. Ribeiro, T. M. Coque & L. V. Peixe, 2002. First characterization of vancomycin-resistant enterococci from a Portuguese hospital. Journal of Antimicrobial Chemotherapy.49: 215-217
119

'
Bibliografia
Novais, C , M. T. Croque, J. C. Sousa, J. Amorim & L. V. Peixe. 2002b. " Vancomycin resistant enterococci from humans and poultry in Portugal : dissemination of Van B phenotype - van A genotype in the community and hospital settings ". 42nd Interscience Conference on Antimicrobial Agents and Chemotherapy (ICAAC). San Diego.
Ostrowsky, B. & G. M. Eliopoulos.1999. Enterococcal Infections : Resistance Mechanisms, Treatment Strategies, and Hospital Issues.14: 62-73
Pechère, J. 1996. Streptogramins. A Unique Class of Antibiotics. 51 :Suppl. 1: 13-19
Pechère, J. C , 2001, Macrolide resistance mechanisms in Gram-positive cocci. International Journal of Antimicrobial Agents. 18: S25-S28
Persing, D. H., 1993, Genotypic Detection of Antimicrobial Resistance, PCR Protocols for Emerging Infectious Diseases, American Society for Microbiology
Piddock, L. J. V. 1996. Does the use of antimicrobial agents in veterinary medicine and animal husbandry select antibiotic-resistent bacteria that infect man and compromise antimicrobial chemotherapy ?. Journal of Antimicrobial Chemotherapy . 38 :l-3
Portillo, A., F. Ruiz-Larrea, M. Zarazaga, A. Alonso J. L. Martinez & C. Torres. 2000. Macrolide Resistance Genes in Enterococcus sp., Antimicrobial Agents and Chemotherapy. 44: 967-971
Portugal, A . V., 1998. Novos desafios que se colocam à exploração pecuária no domínio da Ciência de produção animal, Revista Portuguesa de ciências Veterinárias,.Vol. XCIII, n° 528,171-224
Portugal, A . V., 2002. Sistemas de produção de alimentos de origem animal no futuro.Revista Portuguesa de Ciências Veterinárias, Vol. XCIII, n° 542
Powell, W. J. 2000.Molecular Mechanisms of Antimicrobial Resistance. Technical report ,n°14
Poyart, C , C. Pierre, G. Quesne, B. Pron, P. Berche & P. Trieu-Cuot . 1997. Emergence of vancomycin resistance in the genus Streptococcus : characterization of na van B transferable determinant in Streptococcus bovis, Antimicrobial Agents and Chemotherapy, 41: 24-29
Proposta de Regulamento do Parlamento Europeu e do Conselho relativa aos aditivos destinados à alimentação animal, Bruxelas 22/03/02, COM (2002) 153 final, 2002/2003 (COD)
120

Bibliografia
Regulamento (CE), n° 2788/98, da Comissão de 22 de Dezembro de 1998
Regulamento (CE) N°2821/98 do Conselho, de 17 de Dezembro de 1998. Jornal Oficial das Comunidades Europeias L 351/5 de 29 .12.98
Reinert, R.R., R. Liitticken, A. Bryskier & A. Al-Lhham. 2003. Macrolide-Resistant Streptococcus pneumoniae and Streptococcus pyogenes in the Pediatric Population in Germany during 2000-2001.Antimicrobial Agents and Chemotherapy. 47:489-493
Report and Qualitative Risk Assessment by the Committee for Veterinary Medicinal Products. 1999. The European Agency for the Evaluation of Medicinal Produis. Veterinary Medicines Evaluation Unit. Antibiotic Resistance Associated with Therpeutic Use of Veterinary Medicines. The European comiission. EMEA/CVMP/342/99-corr-Final
Report of the Scientific Committee for Animal Nutrition on the Efficacy and Risk for Users of the Therapeutic Macrolides Antibiotics Tylosin and Spiramycin Used as Feed Additives(opinion expressed on 05 february 1998. Consumer policy and Consumer Healh Protection). The European comiission.
Roberts, M. C , J. Sutcliffe, P. Courvalin, L. B. Jensen, J. Rood & H. Seppala .1999. Nomenclature for Macrolide and Macrolide-Lincosamide-Streptogramin B Resistance Determinants, Antimicrobial Agents and Chemotherapy, 43:2823-2830
Robredo, B., K. V. Singh, F. Baquero, B. E. Murray & C. Torres . 2000. Vancomycin-resistant enterococci isolated from animals and food, International Journal of Food Microbiology, 54,197-204
Schmitz, F.-J., J. Petridou, N. Astfalk, K. Kõhrer, S. Scheuring & S. Schwarz. 2002, Molecular Analysis of Constitutively Expressed erm(C) genes Selected In Vitro by Incubation in the Presence of the Noninducers Quinupristin, Telithromycin, or ABT-773. Microbial Drug Resistance.8:171-177
Schmitz, F-J. R. Sadurski, A. Kray, M. Boos, R. Geisel, K. Kõhrer, J. Verhoel & Ad C. Fluit, 2000, Prevalence of macrolide-resistance genes in Stahpylococcus aureus and Enterococcus faecium isolates from 24 European university hospitals, Journal of Antimicrobial and Chemotherapy. 45. 891-894
Schwarz, S. & E. Chaslus-Dancla. 2001, Use of antimicrobials in veterinary medicine and mechanisms of resistance, Vet. Res., 32,:201-225
Shlaes, D. M., & L. B. Rice .1994. Bacterial resistance to the glycopeptides. Trends in Microbiology. 2: 385-388
121

Bibliografia
Simjee, S., D. G. White, J. Meng, D. D. Wagner. S. Qayumi, S. Zhao, J. R. Hayes & P. F. McDermott. 2002a, Prevalence of streptogramin resistance genes among Enterococcus isolates recovered from retail meats in the Greater Washington DC area, Journal of Antimicrobial Chemotherapy
Simjee, S., D. G. White, D. D. Wagner, J. Meng, S. Qaiyumi, S. Zhao & P. F. McDermott. 2002b. Identification of vat(E) in Enterococcos faecalis Isolates from Retail Poultry and Its Transferability to Enterococcus faecium. Antimicrobial Agents and Chemotherapy. 46: 3823-3828
Singh, K. V., G. M. Weinstock & B. E. Murray. 2002, An Enterococcus faecalis ABC Homologue (Lsa) is Required for the Resistance of This Species to Clindamycin and Quinupristin-Dalfopristin. Antimicrobial Agents and Chemotherapy. 46: 1845-1850
Smith, J. T. & C. S. Lewin. 1993. Mechanisms of antimicrobial resistance and implications for epidemiology. Veterinary Microbiology. Elsevier Science Publishers. 35 :23 3-242
Smith, T. L., M. L. Pearson, K. R. Wilcox, C. Cruz, M. V. Lancaster, B. Robinson-Dunn, F. C. Tenover, M. J. Zervos, J. D. Band, E. White, W. R. Jarvis, for the Glycopeptide-Intermediate Staphylococcus aureus Working Group. 1999. Emergence of Vancomycin Resistance in Staphylococcus aureus, The New England Journal of Medicine, 340: 493-501
Soltani, M., D. Beighton, J. Philpott-Howard & N. Woodford. 2001. Identification of vat(E-3), a novel Gene Encoding Resistance to Quinupristin-Dalphopristin in a Strain of Enterococcus faecium from a Hospital Patient in the United Kingdom. Antimicrobial Agents and Chemotherapy. 45: 645-646
Soltani, M., D. Beighton, J. Philpott-Howard & N. Woodford. 2000. Mechanisms of Resistance to Quinupristin-Dalfopristin among Isolates of Enterococcus faecium from Animals, Raw Meat, and Hospital Patients in Western Europe. Antimicrobial Agents and Chemotherapy. 44: 433-436
Solway, S., L. Vincent, N. Tian, N. Woodford & R. Bendall, 2003, Isolation of streptogramin-resistant Enterococcus faecium from human and non-human sources in a rural community. Journal of Antimicrobial Chemotherapy.
Sorum, H. e Sunde, M., 2001. Resistance to antibiotics in the normal flora of animals.Veterinarian Research. 32:227-241
Sousa, J. C. F. & L. V. N. Prista. 1991. Antibióticos inibidores da síntese proteica. Ordem dos Farmacuticos
122

Bibliografia
Stobberingh, E., A. van den Bogaard, N. London, C. Driessen, J. Top & R. Willems.1999. Enterococci with glycopeptide resistance in turkeys, turkey farmers, turkey slaughterers and (sub)urban residents in the south of the Netherlands: evidence for transmission of vancomycin resistance from animals to humans ?. Antimicrobial Agents and Chemotherapy. 43: 2215-2221
Suppola, J. P., E. Kolho, S. Salmenlinna, E. Tarkka, J. Vuopio-Varkila & M. Vaara. 1999. van A and vanB Incorporate into Endemic Ampicillin-Resistant Vancomycin-Sensitive Enterococcus faecium Strain: Effect on Interpretation of clonality. 37: 3934-3939
Sutcliffe, J., Grebe, T., Tait-Kamradt, A., Wondrack, Lillian. 1996. Detection of Erytromycin-Resistant Determinants by PCR, Antimicrobial Agents and Chemotherapy. 40:,2562-2566
Tenover F. C , L. M. Weigel, P. C. Appelbaum, L. K. McDougal, J. Chaitram, S. McAllister, N. Clark, G. Killgore, C. M. O'Hara, L.Jevitt, J. B. Patel & B. Bozdogan, 2004. Vancomycin-Resistant Staphylococcus aureus Isolate from a Pacient in Pennsylvania. Antimicrobial Agents and Chemotherapy. 48 : 275-280
Thomson, C. J. 1999. The global epidemiology of resistance to ciprofloxacin and the changing nature of antibiotic resistances 10 year perspective.Journal of Antimicrobial Chemotherapy :43,Suppl.A,31 -40
Toledo-Arana, A., J. Valle, C. Solano, M. J. Arrizubieta, C. Cucarella, M. Lamata, B. Amorena, J. Leiva, J. R. Penadés & I. Lasa. 2001. The Enterococcal Surface Protein, Esp, Is Involved in Enterococcus faecalis Biofilm Formation. Applied and Environmental Microbiology. 67: 4538-4545
Tollefson, L., F. J. Angulo & P. J. Fedorka-Cray. 1998. National Surveillance for Antibiotic Resistance in Zoonotic Enteric Pathogens. 4: 141 -150
Tollefson, L., S. F. Altekruse & M. E. Potter. 1997. Therapeutic antibiotics in animal feeds and antibiotic resistance. Rev. Sci.tech.Off.int.Epiz. 16 : 709 - 715
Torell, E., O. Cars, B. Olsson-Liljequist, B. Hoffman, J. Lindbãck, L. G. Burman & The Enterococcal Study Group .1999. Near absence of vancomycin-resistant Enterococci but high carriage rates of quinolone-resistant ampicilin-resitant Enterococci among hospitalized patients and nonhospitalized individuals in Sweden, Journal of Antimicrobial Chemotherapy. 37: 3509-3513
Torres C. 2002. Lectura interpretada del antibiograma de cocos grampositivos. Enferm Infecc Microbiol Clin. 7: 354-364
van den Bogaard, A., E. & E. E. Stobberingh. 1999. Antibiotic Usage in Animals. Impact on Bacterial Resistance and Public Health. 58: 589 - 607
123

Bibliografia
van den Bogaard, A.E., R. Willems, N. London, J. Top & E. E. Stobberingh. 2002. Antibiotic resistance of fecal enterococci in poultry, poultry farmers and poultry slaughterers. Journal of Antimicrobial Chemotherapy. 49:497-505
Vannuffel, P. & C. Cocito.1996. Mechanisms of Action of Streptogramins and Macrolides.Vol.51, Suppl.l: 20 -30
Wegener, H. C. 1998. Historical Yearly Usage of Glycopeptides for Animals and Humans : The American - European Paradox Revisited. Antimicrobial Agents and Chemotherapy, 42: 3049-3049
Wegener, H. C , F.M. Aarestrup, L.B. Jensen , A.M. Hammerum & F. Bager. 1999. Use of antimicrobial growth promoters in food animals and Enterococcus faecium resistance to therapeutic antimicrobial drugs in Europe. Emerging Infectious Diseases. 5
Weisblum, B. 1998. Macrolide Resistance. Drug Resistance Updates. 1: 29-41
Werner, G., B. Hildebrandt, I. Klare & W. Witte. 2000. Linkage of determinants for Streptogramin A, macrolide-lincosamide-Streptogramin B, and chloramphenicol resistance on a conjugative plasmid in Enterococcus faecium and dissemination of this cluster among streptogramin-Resistant Enterococci. Int. J. Med. Microbiol. 290: 543-548
Werner, G., I. Klare & W. Witte. 1999. Large Conjugative vanA Plasmides in Vancomycin-Resistant Enterococcus faecium. Journal of Clinical Microbiology. 37: 2383 -2384
WHO. 2000a. Drug Resistance Threatens to Reverse Medical Progress. Press Release WHO/ 41. http://www.who.int/int-pr-2000/en/pr2000-41.html
WHO. 2000b. Report on Infectious Diseases
Witte, W. 1998. Medical Consequences of Antibiotic Use in Agriculture.The American Association for the Advancement of Science.Vol.279,n° 9553:996997
Wondrack, L., M. Massa, B. V. Yang & J. Sutcliffe, 1996, Clinical Strain of Stahpylococcus aureus Inactivates and Causes Efflux of Macrolides, Antimicrobial agents and Chemotherapy. 40: 992-998
124


![Modulo-8 Deficit Sensorial[1]](https://static.fdocumentos.com/doc/165x107/577ce1381a28ab9e78b50141/modulo-8-deficit-sensorial1.jpg)















